Область техники, к которой относится настоящее изобретение
Настоящее изобретение относится к мультиспецифическому антителу, обладающему специфичностью к IL-13 человека, IL-17A и/или IL-17F человека. Кроме того, изобретение относится к способам получения мультиспецифического антитела и к его терапевтическому применению для лечения атопического дерматита и других заболеваний.
Предшествующий уровень техники настоящего изобретения
Атопический дерматит (AD), также известный как атопическая экзема, представляет собой воспалительное заболевание, который приводит к сильному зуду, покраснению, отеку, намоканию и растрескиванию кожи, которая со временем часто утолщается.
С начала двадцатого века многие воспалительные заболевания слизистых оболочек стали более распространенными; атопический дерматит является классическим примером такого заболевания. В настоящее время он поражает 15-30% детей и 2-10% взрослых в развитых странах, а в Соединенных Штатах за последние тридцать-сорок лет он почти утроился. Более 15 миллионов взрослых и детей в США страдают атопическим дерматитом.
Лечение, используемое для AD, включает системные иммунодепрессанты, такие как циклоспорин, метотрексат, гамма-интерферон, микофенолат, мофетил и азатиоприн. для контроля зуда (жжения) можно использовать антидепрессанты и налтрексон. В 2016 году для экземы легкой и средней степени был одобрен крисаборол, ингибитор фосфодиэстеразы-4, а в 2017 году для лечения экземы средней и тяжелой степени был одобрен дупилумаб, моноклональное антитело - антагонист IL-4Rα.
Из-за ограничений существующих лекарственных средств существует большая потребность в улучшении лечения атопического дерматита.
В WO2013/102042A2 (Abbvie) описаны связывающие белки с двойной специфичностью, направленные против IL-13 и IL-17, и их потенциальное применение при лечении обширного списка заболеваний. Связывающие белки не получили клинического развития.
В WO2015/127405A2 (Genentech) описаны биспецифические антитела против IL-13/IL-17 и способы их применения для лечения астмы средней и тяжелой степени и/или эозинофильной астмы. В фазе I клинических испытаний BITS7201A был связан с высокой частотой образования антител против лекарственного средства (ADA) и был исключен из клинической разработки.
Сущность настоящего изобретения
В настоящем изобретении представлено усовершенствованное мультиспецифическое антитело, способное связывать IL-13 человека, IL-17A человека и/или IL-17F человека.
Антитела согласно настоящему изобретению имеют улучшенные свойства по сравнению с доступными в настоящее время антителами, например, более низкую иммуногенность и/или более хороший фармакокинетический профиль. Кроме того, антитела согласно настоящему изобретению можно сконструировать так, чтобы их можно было более эффективно очищать с использованием улучшенного способа очистки, который включает меньше стадий, чем доступные в настоящее время способы, и является эффективным с точки зрения затрат и времени в промышленном масштабе. Следовательно, антитела согласно настоящему изобретению могут иметь улучшенную технологичность.
В изобретении дополнительно представлены:
Выделенный полинуклеотид, кодирующий мультиспецифическое антитело.
Вектор экспрессии, несущий полинуклеотид.
Клетка-хозяин, содержащая вектор.
Способ получения мультиспецифического антитела, включающий культивирование клетки-хозяина и выделение полученного антитела.
Фармацевтическая композиция, содержащая мультиспецифическое антитело.
Мультиспецифическое антитело или фармацевтическая композиция для использования в способе лечения человека или животного путем терапии.
Способ лечения или профилактики атопического дерматита, хронической экземы рук, микрополипоза или полипоза носа, пищевой аллергии или эозинофильного эзофагита, включающий введение терапевтически эффективного количества мультиспецифического антитела или фармацевтической композиции пациенту.
Краткое описание чертежей
Фиг. 1.
Выравнивания с гуманизацией Ab650.
Выравнивания последовательностей V-области крысиного антитела (донор) с последовательностями V-области зародышевой линии человека (акцептор) вместе с сконструированными гуманизированными последовательностями.
(A) трансплантат 650 легкой цепи:
650=последовательность вариабельной области легкой цепи крысы.
650gL8=гуманизированный трансплантат вариабельной области легкой цепи 650 с использованием зародышевой линии человека IGKV1-39 в качестве акцепторного каркаса.
CDR показаны жирным шрифтом/подчеркиванием.
Донорские остатки показаны жирным шрифтом/курсивом и выделены: I58 и Y71.
(B) Трансплантат тяжелой цепи 650:
650=последовательность вариабельной области тяжелой цепи крысы.
650gH9=гуманизированным трансплантат вариабельной области тяжелой цепи 650 с использованием IGHV1-69 зародышевой линии человека в качестве акцепторного каркаса.
CDR показаны жирным шрифтом/подчеркиванием.
Донорские остатки показаны жирным шрифтом/курсивом и выделены: A67, F69 и V71.
Фиг. 2.
Аминокислотные последовательности и последовательности ДНК.
Фиг. 3.
Очистка мультиспецифического антитела IL-13/IL-17AF.
(A) Анализ BEH200 SEC-UPLC очищенного мультиспецифического антитела, обнаружение с помощью FLR.
(B) Образцы белка, выделенные с помощью Tris-glycine SDS-PAGE в невосстанавливающих (полоса 1) или восстанавливающих (полоса 2) условиях. гель окрашивали быстрым красителем Кумасси и обесцвечивали в dH2O. Белковые маркеры Mark 12 (Life Technologies) использовали в качестве стандартов (M). Молекулярные массы (MW) измеряли в килодальтонах ( кДа).
Фиг. 4.
Ингибирования передачи сигналов STAT6 с помощью мультиспецифического антитела IL-13/IL-17AF.
Фиг. 5.
(A) Ингибирование выработки IL-6 с помощью мультиспецифического антитела IL-13/IL-17AF в ответ на IL-17A человека или яванского макака в комбинации с TNF-α. (i) IL-17A человека. (ii) IL-17A яванского макака.
(B) Ингибирование выработки IL-6 с помощью мультиспецифического антитела IL-13/IL-17AF в ответ на IL-17F человека или яванского макака в комбинации с TNF-α. (i) IL-17F человека. (ii) IL-17F яванского макака.
Фиг. 6.
Одновременная нейтрализация IL-13, IL-17A и IL-17F с помощью мультиспецифического антитела IL-13/IL-17AF в биоанализе высвобождения IL-13/IL-17AF.
Обозначения:
кружок=антитело против IL-13/IL-17AF
квадрат=антитело против IL-17A
треугольник вершиной вверх=антитело против IL-17F
треугольник вершиной вниз=антитело против IL-13
Фиг. 7.
Схематичное изображение мультиспецифического антитела IL-13/IL-17AF согласно настоящему изобретению.
Подробное раскрытие настоящего изобретения
IL-13
IL-13 представляет собой короткоцепочечный цитокин, обладающий 25% идентичностью последовательности с IL-4. Он содержит примерно 132 аминокислоты, образующих вторичную структуру из четырех спиралей, охватывающих остатки 10-21 (спираль A), 43-52 (спираль B), 61-69 (спираль C) и 92-110 (спираль D) наряду с двумя β цепями, охватывающими остатки 33-36 и 87-90. Была предложена структура решения IL-13, раскрывающая прогнозируемую конформацию четырехспирального пучка вверх-вверх-вниз-вниз, которая также наблюдается для IL-4. (Eise нМesser 2001).
IL-13 человека представляет собой гликопротеин 17 кДа и вырабатывается активированными T-клетками линии Th2, хотя Th0 и Th1 CD4+ T-клетки, CD8+ T-клетки и некоторые не T-клеточные популяции, такие как тучные клетки, также вырабатывают IL-13. Функции IL-13 включают переключение изотипа иммуноглобулина на IgE в B-клетках человека и подавление выработки воспалительных цитокинов и у людей, и мышей.
IL-13 связывается со своими рецепторами клеточной поверхности, IL-13R-альфа1 и IL-13R-альфа2. IL-13R-альфа1 взаимодействует с IL-13 с низким сродством (KD~10 нМ) с последующим рекрутированием IL-4R-альфа для образования гетеродимерного рецепторного комплекса передачи сигналов с высоким сродством (KD~0,4 нМ).
Комплекс IL-4R/IL-13R-альфа1 экспрессируется на многих типах клеток, таких как B-клетки, моноциты/макрофаги, дендритные клетки, эозинофилы, базофилы, фибробласты, эндотелиальные клетки, эпителиальные клетки дыхательных путей и гладкомышечные клетки дыхательных путей. Лигирование рецепторного комплекса IL-13R-альфа/IL-4R приводит к активации множества путей передачи сигнала, включая пути передачи сигнала и активатора транскрипции 6 (STAT6) и субстрата инсулинового рецептора 2 (IRS2).
Одна только цепь IL-13R-альфа2 имеет высокое сродство с IL-13 (KD~0,25-0,4 нМ). Она функционирует и как рецептор-приманка, который отрицательно регулирует связывание IL-13 и как сигнальный рецептор, который индуцирует синтез TGF-β и фиброз посредством пути AP-1 в макрофагах и возможно, в других типах клеток.
IL-13 вовлечен в патогенез многих заболеваний человека, и были разработаны терапевтические стратегии ингибирования или противодействия активности IL-13. В частности, искали антитела, которые связывают и нейтрализуют IL-13 в качестве средства ингибирования активности IL-13. Однако в данной области существует потребность в подходящих и/или улучшенных антителах, способных связывать IL-13, особенно IL-13 человека, и в частности, антитела, которые способны нейтрализовать IL-13 человека.
В настоящем изобретении представлено новое семейство связывающих белков, антитела с привитыми CDR, гуманизированные антитела и их фрагменты, способные связывать IL-13 человека, связывать с высоким сродством и связывать и нейтрализовать IL-13 человека.
Антитела, которые ингибируют активность IL-13, могут действовать посредством некоторых возможных механизмов действия. Группа 1 представляет антитело, которое связывается с IL-13 человека и предотвращает связывание IL-13Rα1 и в результате также блокирует связывание IL-4R. Антитела группы 1 могут также предотвращать связывание IL-13 с IL-13Rα2. Группа 2 представляет антитело, которое связывает hIL-13 таким образом, чтобы оно обеспечивало связывание с IL-13Rα1, но предотвращало рекрутирование IL-4R в комплекс. Авторы отбирали антитела, которые действовали посредством группы 1.
В одном варианте осуществления мультиспецифическое антитело связывается с IL-13 человека и предотвращает связывание IL-13Rα1.
В одном варианте осуществления мультиспецифическое антитело связывается с IL-13 человека и предотвращает связывание IL-13Rα2.
В одном варианте осуществления мультиспецифическое антитело связывается с IL-13 человека и предотвращает связывание IL-13Rα1 и IL-13Rα2.
В одном варианте осуществления мультиспецифическое антитело связывается с IL-13 человека с KD <100 пМ.
IL-17
Семейство IL-17 цитокинов состоит из 6 элементов, основанных на схожести структуры, с молекулярной массой 23-36 кДа и димерной структурой. Основной элемент IL-17A (часто называемый в литературе просто IL-17) имеет 16-50% идентичность аминокислотных последовательностей с другими элементами: IL-17B, IL-17C, IL-17D, IL-17E (также известными как IL-25) и IL-17F. IL-17A и IL-17F имеют общую наибольшую гомологию (50%) и связываются с одним и тем же рецепторным комплексом, таким образом было замечено, что эти 2 цитокина имеют общую биологическую активность. Кроме того, IL-17A и IL-17F существуют не только в виде гомодимеров, но также в виде гетеродимера IL-17A/F. IL-17E (IL-25) имеет наименьшую схожесть с IL-17A. Значимым и актуальным для биологической активности IL-17A и IL-17F является тот факт, что они имеют одинаковый рецепторный комплекс IL-17RA/IL-17RC, где IL-17A имеет наибольшее сродство с IL-17RA, тогда как IL-17F более сильно связывается с IL-17RC. Другим членом семейства, использующим IL-17RA, является IL-17E, который передает сигналы посредством рецепторного комплекса IL-17RA/IL-17RB.
IL-17A и IL-17F вырабатывает Th17 подмножество CD4+ T-клеток. Кроме того, другие подмножества T-клеток вырабатывают IL-17A и IL-17F, включая цитотоксичные CD8+ T-клетки (Tc17), gdT клетки и NK T-клетки. Сообщалось, что другие популяции клеток, секретирующие IL-17A, включают нейтрофилы, моноциты, NK-клетки, клетки, подобные индукторам лимфоидной ткани (LTi-like), кишечные клетки Панета и даже B-клетки и тучные клетки. Кроме того, сообщалось, что IL-17F секретируют эпителиальные клетки.
Типы клеток, которые реагируют на цитокины IL-17, отражает экспрессия различных рецепторов. IL-17RA повсеместно экспрессируется, с особенно высокими уровнями в кроветворных тканях, тогда как IL-17RC более сильно экспрессируется в неиммунных клетках суставов, печени, почек, щитовидной железы и предстательной железы. Эта разная экспрессия может объяснить различия биологической активности IL-17A и IL-17F, так как клетки, экспрессирующие высокие уровни IL-17RC, могут быть более чувствительными к IL-17F, тогда как клетки с более высокой экспрессией IL-17RA, чем IL-17RC, могут более легко реагировать на IL-17A. Конкретные типы клеток, которые являются чувствительными к IL-17A и F, включают фибробласты, эпителиальные клетки, кератиноциты, синовиоциты и эндотелиальные клетки, где также сообщалось, что IL-17A действуют на T и B-клетки и макрофаги.
Мультиспецифическое антитело согласно настоящему изобретению способно связываться с IL-17A и/или IL-17F человека. Таким образом, антитело может связываться с гомодимером IL-17A, гомодимером IL-17F и/или гетеродимером IL-17AF.
В одном варианте осуществления мультиспецифическое антитело связывается с IL-17A человека. В одном варианте осуществления мультиспецифическое антитело связывается с IL-17F человека. В одном варианте осуществления мультиспецифическое антитело связывается с IL-17A и IL-17F человека.
В одном варианте осуществления мультиспецифическое антитело связывается с IL-17С человека с KD <50 пМ. В одном варианте осуществления мультиспецифическое антитело связывается с IL-17С человека с KD <25 пМ. В одном варианте осуществления мультиспецифическое антитело связывается с IL-17С человека с KD <10 пМ.
В одном варианте осуществления мультиспецифическое антитело связывается с IL-17F человека с KD <200 пМ. В одном варианте осуществления мультиспецифическое антитело связывается с IL-17F человека с KD <100 пМ.
Альбумин
Высокая специфичность и сродство антител делают их идеальным диагностическим и терапевтическим средствами, особенно для модулирования межбелковых взаимодействий. Однако антитела могут страдать от повышенной скорости клиренса из сыворотки, особенно, когда у них отсутствует Fc-домен, который обеспечивает длительное время жизни in vivo (Medasan et al. 1997, J. Immunol. 158:2211-2217).
Средства улучшения периода полужизни антител известны. Один подход состоит в конъюгации фрагмента с молекулами полимеров. Таким образом, короткий период полувыведения фрагментов Fab’, F(ab’)2 из кровотока у животных был улучшен путем конъюгации с полиэтиленгликолем (PEG; см., например, WO98/25791, WO99/64460 и WO98/37200). Другой подход состоит в модификации фрагмента антитела путем конъюгации со средством, которое взаимодействует с рецептором FcRn (см., например, WO97/34631). Еще один подход увеличения периода полужизни состоит в использовании полипептидов, которые связывают сывороточный альбумин (см., например, Smith et al. 2001, Bioconjugate Chem. 12:750-756; EP0486525; US6267964; WO04/001064; WO02/076489; и WO01/45746).
Сывороточный альбумин является распространенным белком как в сосудистых, так и внесосудистых компартментах с периодом полужизни у человека приблизительно 19 дней (Peters, 1985, Adv Protein Chem. 37:161-245). Он похож на период полужизни IgG1, который составляет приблизительно 21 день (Waldeman & Strober, 1969, Progr. Allergy, 13:1-110).
Были описаны одиночные вариабельные домены, связывающие сывороточный альбумин, наряду с их использованием в качестве конъюгатов для повышения периода полужизни лекарственных средств, включая лекарственные средства NCE (химическое соединение), белки и пептиды, см., например, Holt et al. Protein Engineering, Design & Selection, vol 21, 5, pp283-288, WO04003019, WO2008/096158, WO05118642, WO2006/0591056 и WO2011/006915. Другие антитела против сывороточного альбумина и их использование в мультиспецифических форматах антител были описаны в WO2009/040562, WO2010/035012 и WO2011/086091. В частности, авторы ранее описали антитело против альбумина с улучшенной гуманизацией в WO2013/068571.
Мультиспецифические антитела согласно настоящему изобретению можно сконструировать для связывания с сывороточным альбумином человека для увеличения их периода полужизни в сыворотке in vivo, что приводит к улучшению фармакокинетического профиля.
Антитела
Антитела для использования в контексте настоящего раскрытия включают целые антитела и их функционально активные фрагменты, то есть молекулы, которые специфично связываются с IL-13, IL-17A и/или IL-17F, также называемые антигенсвязывающие фрагменты. Признаки, описанные в данном документе относительно антител, также применимы к фрагментам антитела, если из контекста не следует иное.
Целые антитела, также известные как «иммуноглобулины (Ig)», обычно относятся к интактным или полноразмерным антителам, то есть содержащим элементы двух тяжелых цепей и двух легких цепей, соединенных между собой дисульфидными связями, которые в совокупности образуют характерную Y-образную трехмерную структуру. Классические природные целые антитела являются моноспецифическими в том смысле, что они связывают один тип антигена и бивалентными в том смысле, что они имеют два независимых антигенсвязывающих домена. Термины «интактное антитело», «полноразмерное антитело» и «целое антитело» использованы взаимозаменяемо для обозначения моноспецифического бивалентного антитела, имеющего конструкцию, похожую на нативную структуру антитела, включая Fc-область согласно настоящему изобретению.
Каждая легкая цепь состоит из вариабельной области легкой цепи (сокращенно обозначенной в данном документе VL) и константной области легкой цепи (CL). Каждая тяжелая цепь состоит из вариабельной области тяжелой цепи (сокращенно обозначенной в данном документе VH) и константной области тяжелой цепи (CH), состоящей из трех константных доменов CH1, CH2 и CH3 или четырех константных доменов CH1, CH2, CH3 и CH4 в зависимости от класса Ig. «Класс» Ig или антитела относится к типу константной области и включает IgA, IgD, IgE, IgG и IgM, и некоторые из них могут быть дополнительно разделены на подклассы, например, IgG1, IgG2, IgG3, IgG4. Константные области антител могут опосредовать связывание иммуноглобулина с тканями или факторами хозяина, включая различные клетки иммунной системы (например, эффекторные клетки) и первый компонент (Clq) классической системы комплемента.
Области VH и VL антитела согласно настоящему изобретению могут быть дополнительно подразделены на области гипервариабельности (или «гипервариабельные области»), определяющие распознавание антигена, называемые определяющими комплементарность областями (CDR), чередующимися с областями, которые являются более структурно консервативными, называемыми каркасными областями (FR). Каждая VH и VL состоит из трех CDR и четырех FR, расположенных от аминоконца к карбоксиконцу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. CDR и FR вместе образуют вариабельную область. По соглашению CDR в вариабельной области тяжелой цепи антитела или его антигенсвязывающего фрагмента обозначают CDR-H1, CDR-H2 и CDR-H3, а в вариабельных областях легкой цепи CDR-L1, CDR-L2 и CDR-L3. Их нумеруют последовательно в направлении от N-конца к C-концу каждой цепи.
CDR обычно нумеруют в соответствии с системой, разработанной Kabat et al. Эта система изложена в Kabat et al. 1991, в Sequences of Proteins of Immunological Interest, US Department of Health and Human Services, NIH, USA (далее «Kabat et al. (См. выше)»). Эта система нумерации использована в настоящем описании, если не указано иное.
Обозначения остатков по Kabat не всегда прямо соответствуют линейной нумерации аминокислотных остатков. реальная линейная аминокислотная последовательность может содержать меньше или дополнительные аминокислоты, чем по строгой нумерации согласно Kabat, что соответствует укорочению или вставке в структурный компонент, будь то каркас или определяющая комплементарность область, структуры главного вариабельного домена. Правильную нумерацию остатков согласно Kabat можно определить для заданного антитела путем выравнивания остатков гомологии в последовательности антитела с последовательностью со «стандартной» нумерацией согласно Kabat.
CDR вариабельного домена тяжелой цепи находятся в остатках 31-35 (CDR-H1), остатках 50-65 (CDR-H2) и остатках 95-102 (CDR-H3) в соответствии с нумерацией по системе Kabat. Однако в соответствии с Chothia (Chothia, C. and Lesk, A.M. J. Mol. Biol., 196, 901-917 (1987)) петля, эквивалентная CDR-H1, проходит от остатка 26 до остатка 32. Таким образом, если не указано иное, ‘CDR-H1’ в рамках настоящего изобретения относится к остаткам 26-35, как описано комбинацией нумерации по системе Kabat и определением топологической петли Chothia.
CDR вариабельного домена легкой цепи находятся в остатках 24-34 (CDR-L1) остатки 50-56 (CDR-L2) и остатках 89-97 (CDR-L3) в соответствии с нумерацией по системе Kabat.
В дополнение к петлям CDR между CDR-2 (CDR-L2 или CDR-H2) и CDR-3 (CDR-L3 или CDR-H3) существует четвертая петля, которая образована каркасом 3 (FR3). Нумерация по системе Kabat определяет каркас 3 как позиции 66-94 в тяжелой цепи и позиции 57-88 в легкой цепи.
На основе выравнивания последовательностей различных элементов семейства иммуноглобулинов были предложены схемы нумерации, описанные, например, в Kabat et al. 1991 и Dondelinger et al. 2018, Frontiers in Immunology, Vol 9, article 2278.
Термин «константный домен (домены)», «константная область» согласно настоящему изобретению использованы взаимозаменяемо для обозначения домена (доменов) антитела, который находится за пределами вариабельных областей. Константные домены идентичны во всех антителах одного и того же изотипа, но отличаются от одного изотипа к другому. Обычно константная область тяжелой цепи образована от N к C концу CH1-шарниром -CH2-CH3-необязательно CH4, содержащим три или четыре константных домена.
При наличии, константные домены молекулы антитела согласно настоящему изобретению можно выбрать с учетом предполагаемой функции молекулы антитела и в частности эффекторных функций, которые могут потребоваться. Например, константными доменами могут быть домены IgA, IgD, IgE, IgG или IgM человека. В частности, когда молекула антитела предназначена для терапевтических вариантов применения, и требуются эффекторные функции антитела, можно использовать константные домены IgG человека, особенно изотипы IgG1 и IgG3. Альтернативно, когда молекула антитела предназначена для терапевтических целей и эффекторные функции антитела не требуются, можно использовать изотипы IgG2 и IgG4. Должно быть понятно, что также можно использовать варианты последовательностей этих константных доменов. Например, можно использовать молекулы IgG4, в которых серин в позиции 241 (номер соответствует нумерации по системе Kabat) заменен на пролин, как описано у Angal et al. (Angal et al., 1993. A single amino acid substitution abolishes the heterogeneity of chimeric mouse/human (IgG4) antibody as observed during SDS-PAGE analysis Mol Immunol 30, 105-108), которые в данном документе называются IgG4P.
«Fc», «Fc-фрагмент», «Fc-домен» и «Fc-область» использованы взаимозаменяемо для обозначения C-концевой области антитела, содержащей константную область антитела за исключением первого константного домена иммуноглобулина. Таким образом, Fc относится к последним двум константным доменам CH2 и CH3 IgA, IgD и IgG или последним трем константным доменам IgE и IgM и гибкому шарнирному N-концу этих доменов. Fc-область тяжелой цепи IgG1 человек определена в данном документе, как содержащая остатки C226 на своем карбоксильном конце, где нумерация соответствует индексу ЕС согласно Kabat. В контексте IgG1 человека нижний шарнир относится к позициям 226-236, домен CH2 относится к позициям 237-340, а домен CH2 относится к позициям 341-447 в соответствии с индексом ЕС согласно Kabat. Соответствующую Fc-область других иммуноглобулинов можно идентифицировать с помощью выравнивания последовательностей.
В контексте настоящего раскрытия, при наличии, константная область или Fc-область может быть природной согласно определению выше или же может быть модифицирована различными способами, при условии, что она содержит функциональный связывающий FcR домен и предпочтительно функциональный связывающий FcRn домен. Предпочтительно, модифицированная константная область или Fc-область приводит к улучшению функциональности и/или фармакокинетики. Модификации могут включать делецию определенных участков фрагмента Fc. Модификации могут дополнительно включать различные аминокислотные замены, способные повлиять на биологические свойства антитела. также могут иметься мутации для увеличения связывания FcRn и таким образом, периода полужизни in vivo. Модификации могут дополнительно включать модификация профиля гликозилирования антитела. Природный Fc-фрагмент гликозилирован в домене CH2 с наличием в каждой из двух тяжелых цепей N-гликана, связанного с остатком аспарагина в позиции 297 (Asn297). В контексте настоящего раскрытия антитело может быть гликомодифицировано, то есть сконструировано таким образом, чтобы оно имело определенный профиль гликозилирования, который, например, приводит к улучшенным свойствам, например, к улучшенной эффекторной функции или улучшенному периода полужизни в сыворотке.
Антитела, описанные в данном документе, являются выделенными. «Выделенное» антитело представляет собой антитело, которое было выделено (например, с помощью средств очистки) из компонента его природной среды.
Термин «антитело» охватывает моновалентные антитела, то есть антитела, содержащие только один антигенсвязывающий домен (например, неполные антитела, содержащие соединенные друг с другом полноразмерную тяжелую цепь и полноразмерную легкую цепь, также называемые «полуантитело») и мультивалентные антитела, то есть антитела, содержащие более одного антигенсвязывающего домена.
Термин «антитело» согласно изобретению также охватывает антигенсвязывающие фрагменты антител. Антигенсвязывающие фрагменты антител включают одноцепочечные антитела (например, scFv и dsscfv), Fab, Fab’, F(ab’)2, Fv, однодоменные антитела или нанотела (например, VH или VL или VHH или VNAR). Другие фрагменты антитела для использования в настоящем изобретении включают фрагменты Fab и Fab’, описанные в международных заявках на патент WO2011/117648, WO2005/003169, WO2005/003170 и WO2005/003171.
Способы создания получения этих фрагментов антитела хорошо известны в данной области (см., например, Verma et al. 1998, Journal of Immunological Methods, 216, 165-181).
Термин «фрагмент Fab» согласно настоящему изобретению относится к фрагменту антитела, содержащему фрагмент легкой цепи, содержащий VL (вариабельный легкий) домен и константный домен легкой цепи (CL) и VH (вариабельный тяжелый) домен и первый константный домен (CH1) тяжелой цепи.
Обычный «фрагмент Fab’» содержит пару тяжелой и легкой цепей, в которой тяжелая цепь содержит вариабельную область VH, константный домен CH1 и природную или модифицированную шарнирную область, а легкая цепь содержит вариабельную область VL и константный домен CL. Димеры Fab’ согласно настоящему раскрытию создают F(ab’)2, где, например, димеризация может происходить через шарнир.
Термин «однодоменное антитело» согласно настоящему изобретению относится к фрагменту антитела, состоящему из единственного мономерного вариабельного домена антитела. Примеры однодоменных антител включают VH или VL или VHH или V-NAR.
Термин «Fv» относится к двум вариабельным доменам, например объединенным вариабельным доменам, таким как родственная пара или вариабельные домены с созревшим сродством, то есть пара VH и VL.
«Одноцепочечный вариабельный фрагмент» или «scFv» в рамках настоящего изобретения относится к одноцепочечному вариабельному фрагменту, содержащему или состоящему из вариабельного домена тяжелой цепи (VH) и вариабельного домена легкой цепи (VL), который стабилизирован пептидным линкером между вариабельными доменами VH и VL. Вариабельные домены VH и VL могут быть в любой подходящей ориентации, например C-конец VH может быть связан с N-концом VL, или C-конец VL может быть связан с N-концом VH.
«Стабилизированный дисульфидным мостиком одноцепочечный вариабельный фрагмент» или «dsscFv» в рамках настоящего изобретения относится к одноцепочечному вариабельному фрагменту, который стабилизирован пептидным линкером между вариабельными доменами VH и VL, а также содержит междоменную дисульфидную связь между VH и VL. (см., например, Weatherill et al. Protein Engineering, Design & Selection, 25 (321-329) 2012, WO2007109254.
«Стабилизированный дисульфидным мостиком вариабельный фрагмент» или «dsFv» в рамках настоящего изобретения относится к одноцепочечному вариабельному фрагменту, который не содержит пептидного линкера между вариабельными доменами VH и VL и вместо этого стабилизирован междоменной дисульфидной связью между VH и VL.
В одном варианте осуществления мультиспецифическое антитело согласно настоящему изобретению представляет собой антагонистическое антитело. Согласно настоящему изобретению термин «антагонистическое антитело» описывает антитело, которое способно ингибировать или нейтрализовать биологическую сигнальную активность одного или нескольких антигенов, например путем блокирования связывания или восстанавливающего связывания IL-13, IL-17A и/или IL-17F с их рецепторами.
Антитела для использования в настоящем изобретении могут представлять собой без ограничения моноклональные, гуманизированные, полностью человеческие или химерные антитела.
Моноклональные антитела можно получать с помощью любого способа, известного в данной области, такого как метод гибридомы (Kohler & Milstein, 1975, Nature, 256:495-497), метод триомы, метод B-клеточной гибридомы человека (Kozbor et al. 1983, Immunology Today, 4:72) и метод EBV-гибридомы (Cole et al. Monoclonal Antibodies and Cancer Therapy, pp77-96, Alan R Liss, Inc. 1985).
Антитела также можно создавать с использованием методов получения антител из отдельных лимфоцитов путем клонирования и экспрессии кДНК вариабельной области иммуноглобулина, полученной из отдельных лимфоцитов, выбранных для получения специфических антител, например, с помощью способов, описанных в Babcook, J. et al. 1996, Proc. Natl. Acad. Sci. USA 93(15):7843-7848l; WO92/02551; WO2004/051268 и международной заявке на патент номер WO2004/106377.
Скрининг антител можно проводить с использованием анализов для измерения связывания с антигеном и/или анализов для измерения способности блокировать связывание антигена с одним или несколькими из его рецепторов. Примером анализа связывания является ELISA, например, с использованием белка слияния IL-13, который иммобилизован на планшетах, и использованием конъюгированного вторичного антитела для обнаружения антитела против IL-13, связанного с IL-13. Примером анализа блокирования является анализ на основе проточной цитометрии измерения блокирования связывания белка лиганда IL-13 с IL-13R. для определения количества связывания белка лиганда IL-13 с IL-13R используют флуоресцентно меченное вторичное антитело.
Гуманизированные антитела (которые включают антитела с привитой CDR) представляют собой молекулы антител, имеющие одну или несколько определяющих комплементарность областей (CDR) от видов, отличных от человека, и каркасную область из молекулы иммуноглобулина человека (см., например, US 5585089; WO91/09967). Должно быть понятно, что может быть необходимо переносить только определяющие специфичность остатки CDR, а не всю CDR (см., например, Kashmiri et al. 2005, Methods, 36, 25-34). Гуманизированные антитела могут необязательно дополнительного содержать один или несколько каркасных остатков, полученных от видов, отличных от человека, у которых были получены CDR.
Химерные антитела состоят из элементов, полученных от двух разных видов, так что элемент сохраняет характеристики видов, у которых он получен. Обычно химерное антитело будет содержать вариабельную область от одних видов, например мыши, крысы, кролик и тому подобное, а константную область от других видов, таких как человек.
Антитела также можно создавать с использованием различных методов фаговых дисплеев, известных в данной области, и они включают антитела, раскрытые у Brinkman et al. (в J. Immunol. Methods, 1995, 182: 41-50), Ames et al. (J. Immunol. Methods, 1995, 184:177-186), Kettleborough et al. (Eur. J. Immunol. 1994, 24:952-958), Persic et al. (Gene, 1997 187 9-18), Burton et al. (Advances in Immunology, 1994, 57:191-280) and WO 90/02809; WO 91/10737; WO 92/01047; WO 92/18619; WO 93/11236; WO 95/15982; WO 95/20401; и US 5698426; 5223409; 5403484; 5580717; 5427908; 5750753; 5821047; 5571698; 5427908; 5516637; 5780225; 5658727; 5733743 и 5969108.
Полностью человеческие антитела представляют собой такие антитела, в которых вариабельные области и константные области (где имеются) и тяжелой и легкой цепей все имеют человеческое происхождение или по существу идентичны последовательностям человеческого происхождения, но не обязательно из одного и того же антитела. Примеры полностью человеческих антител могут включать антитела, полученные, например, методами фаговых дисплеев, описанных выше, и антитела, вырабатываемые мышами, в которых гены вариабельной и необязательно константной области мышиного иммуноглобулина были заменены их человеческими аналогами, например, как описано в общих чертах в EP 0546073, US 5545806, US 5569825, US 5625126, US 5633425, US 5661016, US 5770429, EP 0438474 и EP 0463151.
Мультиспецифические антитела
Антитело согласно настоящему изобретению представляет собой мультиспецифическое антитело. «Мультиспецифическое антитело» в рамках настоящего изобретения относится к антителу, описанному в данном документе, которое имеет по меньшей мере два связывающих домена, то есть два или более связывающих домена, например два или три связывающих домена, где по меньшей мере два связывающих домена независимо связывают два разных антигена или два разных эпитопы на одном и том же антигене. Мультиспецифические антитела обычно являются моновалентными для каждой специфичности (антигена). Мультиспецифические антитела, описанные в данном документе, включают в себя моновалентные и мультивалентные, например, бивалентные, трехвалентные, тетравалентные мультиспецифические антитела.
Паратоп представляет собой область антитела, которая распознает и связывается с антигеном. Антитело согласно изобретению может представлять собой мультипаратопное антитело. «Мультипаратопное антитело» в рамках настоящего изобретения относится к антителу, описанному в данном документе, которое содержит два или более разных паратопа, которые взаимодействуют с разными эпитопами либо из одного и того же антигена, либо из двух разных антигенов. Мультипаратопные антитела, описанные в данном документе, могут быть бипаратопными, трипаратопными, тетрапаратопными.
«Антигенсвязывающий домен» в рамках настоящего изобретения относится к участку антитела, который содержит часть или целиком один или несколько вариабельных доменов, например часть или целиком пару вариабельных доменов VH и VL, которые специфично взаимодействуют с антигеном-мишенью. Связывающий домен может содержать однодоменное антитело. В одном варианте осуществления каждый связывающий домен является моновалентным. Предпочтительно каждый связывающий домен содержит не более одной VH и один VL.
«Специфично» в рамках настоящего изобретения относится к связывающему домену, который распознает только антиген, к которому он специфичен, или к связывающему домену, который имеет значительно более высокое сродство связывания с антигеном, к которому он специфичен, по сравнению со сродством с антигенами, с которыми он неспецифичен. Сродство связывания можно измерить с помощью стандартных анализов, таких как поверхностный плазмонный резонанс, например BIAcore.
Было создано множество форматов мультиспецифических антител. были предложены различные классификации, но мультиспецифические форматы IgG антител обычно включают биспецифический IgG, дополненный IgG, мультиспецифические (например, биспецифические) фрагменты антитела, мультиспецифические (например, биспецифические) белки слияния и мультиспецифические (например, биспецифические) конъюгаты антител, как описано, например, в Spiess et al. Alternative molecular formats and therapeutic applications for bispecific antibodies. Mol Immunol. 67(2015):95-106.
Методы получения биспецифических антител включают без ограничения метод CrossMab (Klein et al. Engineering therapeutic bispecific antibodies using CrossMab technology, Methods 154 (2019) 21-31) метод Knobs-in-holes (например, WO1996027011, WO1998050431), метод DuoBody (например, WO2011131746), метод Azymetric (например, WO2012058768). Дополнительные методы получения биспецифических антител были описаны, например, в Godar et al. 2018, Therapeutic bispecific antibody formats: a patent applications review (1994-2017), Expert Opinion on Therapeutic Patents, 28:3, 251-276. Биспецифические антитела включают, в частности, антитела CrossMab, DAF (два в одном) DAF (четыре в одном), DutaMab, DT-lgG, Knobs-in-holes common LC, Knobs-in-holes assembly, Charge pair, Fab-arm exchange, SEEDbody, Triomab, LUZ-Y, Fcab, κλ-body и orthogonal Fab.
Дополненный IgG классически включает в себя полноразмерный IgG, сконструированный путем добавления дополнительного антигенсвязывающего домена или антигенсвязывающего фрагмента к N- и/или C-концу тяжелой и/или легкой цепи IgG. Примеры таких дополнительных антигенсвязывающих фрагментов включают антитела sdAb (например VH или VL) Fv, scFv, dsscFv, Fab, scFav. Дополненные форматы IgG антител включают, в частности, DVD-IgG, lgG(H)-scFv, scFv-(H)lgG, lgG(L)-scFv, scFv-(L)lgG, lgG(L, H)-Fv, lgG(H)-V, V(H)-lgG, lgC(L)-V, V(L)-lgG, KIH IgG-scFab, 2scFv-lgG, lgG-2scFv, scFv4-lg, Zybody и DVI-IgG (четыре в одном), например, как описано в Spiess et al. Alternative molecular formats and therapeutic applications for bispecific antibodies. Mol Immunol. 67(2015):95-106.
Мультиспецифических фрагменты антитела включают нанотело, нанотело-HAS, BiTEs, диатело, DART, TandAb, scDiabody, sc-Diabody-CH3, Diabody-CH3, Triple Body, Miniantibody; Minibody, Tri Bi minibody, scFv-CH3 KIH, Fab-scFv, scFv-CH-CL-scFv, F(ab')2, F(ab')2-scFV3, scFv-KIH, Fab-scFv-Fc, Tetravalent HCAb, scDiabody-Fc, Diabody-Fc, Tandem scFv-Fc; и интратело, как описано, например, в Spiess et al. Alternative molecular formats and therapeutic applications for bispecific antibodies. Mol Immunol. 67(2015):95-106.
Мультиспецифические белки слияния включают Dock и Lock, ImmTAC, HSAbody, scDiabody-HAS и Tandem scFv-Токсин.
Мультиспецифические конъюгаты антител включают IgG-lgG; Cov-X-Body; и scFv1-PEG-scFv2.
Дополнительные мультиспецифические форматы антител были описаны, например, в Brinkmann и Kontermann, The making of bispecific antibodies, mAbs, 9:2, 182-212 (2017), в частности, на Фиг. 2, например, tandem scFv, triplebody, Fab-VHH, taFv-Fc, scFv4-Ig, scFv2-Fcab, scFv4-IgG. Bibodies, tribodies и способы их получения раскрыты, например, в WO99/37791.
В изобретении представлено мультиспецифическое антитело, которое связывает IL-13 человека, IL-17A человека и/или IL-17F человека.
В одном варианте осуществления мультиспецифическое антитело содержит антигенсвязывающий участок, который связывается с IL-13 человека, где участок связывания IL-13 содержит вариабельную область легкой цепи, содержащую последовательность SEQ ID NO:15 для CDR-L1, последовательность SEQ ID NO:16 для CDR-L2, и последовательность SEQ ID NO:17 для CDR-L3.
В одном варианте осуществления мультиспецифическое антитело содержит антигенсвязывающий участок, который связывается с IL-13 человека, где участок связывания IL-13 содержит вариабельную область тяжелой цепи, содержащую последовательность SEQ ID NO:18 для CDR-H1, последовательность SEQ ID NO:19 для CDR-H2, и последовательность SEQ ID NO:20 для CDR-H3.
В одном варианте осуществления участок связывания IL-13 содержит вариабельную область легкой цепи, содержащую последовательность SEQ ID NO:27.
В одном варианте осуществления участок связывания IL-13 содержит вариабельную область тяжелой цепи, содержащую последовательность SEQ ID NO:28.
В одном варианте осуществления участок связывания IL-13 содержит вариабельную область легкой цепи, содержащую последовательность SEQ ID NO:31.
В одном варианте осуществления участок связывания IL-13 содержит вариабельную область тяжелой цепи, содержащую последовательность SEQ ID NO:32.
В одном варианте осуществления мультиспецифическое антитело содержит антигенсвязывающий участок, который связывается с IL-17A и IL-17F человека, содержащий:
вариабельную область легкой цепи, содержащую последовательность SEQ ID NO:1 для CDR-L1, последовательность SEQ ID NO:2 для CDR-L2, и последовательность SEQ ID NO:3 для CDR-L3.
В одном варианте осуществления мультиспецифическое антитело содержит антигенсвязывающий участок, который связывается с IL-17A и IL-17F человека, содержащий:
вариабельную область тяжелой цепи, содержащую последовательность SEQ ID NO:4 для CDR-H1, последовательность SEQ ID NO:5 для CDR-H2, и последовательность SEQ ID NO:6 для CDR-H3.
В одном варианте осуществления антигенсвязывающий участок, который связывается с IL-17A и IL-17F человека, содержит вариабельную область легкой цепи, содержащую последовательность SEQ ID NO:7.
В одном варианте осуществления антигенсвязывающий участок, который связывается с IL-17A и IL-17F человека, содержит вариабельную область тяжелой цепи, содержащую последовательность SEQ ID NO:9.
В одном варианте осуществления в мультиспецифическом антителе отсутствует Fc-домен, а период полужизни обеспечивается антигенсвязывающим участком, который связывается с сывороточным альбумином.
В одном варианте осуществления мультиспецифическое антитело содержит последовательность SEQ ID NO:57 или SEQ ID NO: 59.
В одном варианте осуществления мультиспецифическое антитело содержит последовательность SEQ ID NO:61 или SEQ ID NO: 63.
В одном варианте осуществления мультиспецифическое антитело содержит последовательность SEQ ID NO:59, и последовательность SEQ ID NO: 63.
В одном варианте осуществления мультиспецифическое антитело содержит или состоит из:
полипептидной цепи формулы (I):
VH-CH1-(CH2)s-(CH3)t-X-(V1)p; и
полипептидной цепи формулы (II):
VL-CL-Y-V2;
где:
VH представляет вариабельный домен тяжелой цепи;
CH1 представляет домен 1 константной области тяжелой цепи;
CH2 представляет домен 2 константной области тяжелой цепи;
CH3 представляет домен 3 константной области тяжелой цепи;
X представляет связь или линкер;
V1 представляет dsscFv, dsFv или scFv;
VL представляет вариабельный домен легкой цепи;
CL представляет домен из константной области легкой цепи, такой как Ckappa;
Y представляет связь или линкер;
V2 представляет dsscFv, dsFv или scFv;
p составляет 0 или 1;
s составляет 0 или 1;
t составляет 0 или 1;
где, когда p составляет 0, X отсутствует, а когда q составляет 0, Y отсутствует; и
при этом полипептидная цепь формулы (I) содержит домен связывания белка А; и
при этом полипептидная цепь формулы (II) не связывает белок A.
В одном варианте осуществления, когда s составляет 0, а t составляет 0, мультиспецифическое антитело согласно настоящему раскрытию представлено в виде димера тяжелой и легкой цепи:
формулы (I) и (II), соответственно, где участок VH-CH1 вместе с участком VL-CL образуют функциональный фрагмент Fab или Fab’.
В одном варианте осуществления, когда s составляет 1, а t составляет 1, мультиспецифическое антитело согласно настоящему раскрытию представлено в виде димера двух тяжелых цепей и двух легких цепей:
формулы (I) и (II), соответственно, где две тяжелые цепи соединены с помощью межцепочечного взаимодействия, особенно на уровне CH2-CH3, и при этом участок VH-CH1 каждой тяжелой цепи вместе с участком VL-CL каждой легкой цепи, образуют функциональный фрагмент Fab или Fab’. В таком варианте осуществления два участка VH-CH1- CH2- CH3 вместе с двумя участками VL-CL образуют функциональное полноразмерное антитело. В таком варианте осуществления полноразмерное антитело может содержать функциональную Fc-область.
VH представляет вариабельный домен тяжелой цепи. В одном варианте осуществления VH является гуманизированным. В одном варианте осуществления VH является полностью человеческим.
VL представляет вариабельный домен легкой цепи. В одном варианте осуществления VL является гуманизированным. В одном варианте осуществления VL является полностью человеческим.
Обычно, VH и VL вместе образуют антигенсвязывающий домен. В одном варианте осуществления VH и VL образуют родственную пару.
«Родственная пара» в рамках настоящего изобретения относится к паре вариабельных доменов из одного антитела, которые было получено in vivo, то есть к природному соединению вариабельных доменов, выделенных у хозяина. Следовательно, родственной парой является пара VH и VL. В одном примере родственная пара совместно связывает антиген.
«Вариабельная область» или «вариабельный домен» в рамках настоящего изобретения относится к области в цепи антитела, содержащей CDR и каркас, в частности подходящий каркас.
Вариабельные области для использования в настоящем раскрытии обычно получают из антитела, которые можно получить с помощью любого способа, известного в данной области.
«Полученный из» в рамках настоящего изобретения относится к тому, что используемая последовательность или последовательность очень похожая на используемую последовательность получена из исходного генетического материала, такого как легкая или тяжелая цепь антитела.
«Очень похожая» в рамках настоящего изобретения относится к аминокислотной последовательности, которая по всей своей длине является сходной на 95% или более, например сходной на 96, 97, 98 или 99%.
Вариабельные области для использования в настоящем изобретении, как описано в данном документе выше для VH и VL, могут быть из любого подходящего источника и могут быть, например, полностью человеческими или гуманизированными.
В одном варианте осуществления связывающий домен, образованный VH и VL, является специфическими к первому антигену.
В одном варианте осуществления связывающий домен V1 является специфическим ко второму антигену.
В одном варианте осуществления связывающий домен V2 является специфическим к третьему антигену.
В одном варианте осуществления каждый из VH-VL, V1 и V2, который имеется, раздельно связывает свой соответствующий антиген.
В одном варианте осуществления домен CH1 представляет собой природный домен 1 из тяжелой цепи антитела или его производное. В одном варианте осуществления домен CH2 представляет собой природный домен 2 из тяжелой цепи антитела или его производное. В одном варианте осуществления домен CH2 представляет собой природный домен 3 из тяжелой цепи антитела или его производное.
В одном варианте осуществления фрагмент CL в легкой цепи представляет собой константную последовательность каппа или ее производное. В одном варианте осуществления фрагмент CL в легкой цепи представляет собой константную последовательность лямбда или ее производное.
Производное природного домена в рамках настоящего изобретения относится к ситуации, когда по меньшей мере одна аминокислота в природной последовательности была заменена или удалена, например, для оптимизации свойств домена, например, путем устранения нежелательных свойств, но при этом сохраняется/сохраняются отличительный признак (признаки) домена. В одном варианте осуществления производное природного домена содержит два, три, четыре, пять, шесть, семь, восемь, десять, одиннадцать или двенадцать аминокислотных замен или делеций по сравнению с природной последовательностью.
В одном варианте осуществления в функциональном фрагменте Fab или Fab’ имеется одна или несколько природных или сконструированных межцепочечных (то есть между легкой и тяжелой цепями) дисульфидных связей.
В одном варианте осуществления «природная» дисульфидная связь присутствует между CH1 и CL в полипептидных цепях формулы (I) и (II).
Когда CL домен получен либо из Каппа, либо из Лямбда, природной позицией для образующего связь цистеина является 214 в cКаппа и cЛямбда человека (нумерации согласно Kabat 4th edition 1987).
Точная локализация образующего дисульфидную связь цистеина в CH1 зависит от реально используемого конкретного домена. Таким образом, например, в гамма-1 человека природное положение дисульфидной связи находится в позиции 233 (нумерации согласно Kabat 4th edition 1987). известна позиция образующего связь цистеина для других изотипов человека, таких как гамма 2, 3, 4, IgM и IgD, например, позиция 127 для IgM, IgE, IgG2, IgG3, IgG4 человека и 128 тяжелой цепи IgD и IgA2B человека.
Необязательно, может быть дисульфидная связь между VH и VL полипептидов формулы I и II.
В одном варианте осуществления мультиспецифическое антитело согласно раскрытию имеет дисульфидную связь в позиции, эквивалентной или соответствующей природной позиции между CH1 и CL.
В одном варианте осуществления константная область, содержащая CH1, и константная область, такая как CL, имеет дисульфидную связь, которая находится в неприродной позиции. Ее можно сконструировать в молекуле путем введения цистеина (цистеинов) в аминокислотную цепь в нужной позиции или позициях. Эта неприродная дисульфидная связь имеется в дополнение или в качестве альтернативы природной дисульфидной связи, имеющейся между CH1 и CL. Цистеин (цистеины) в природной позиции могут быть заменены такой аминокислотой, как серин, который неспособен образовать дисульфидный мостик.
Введение сконструированных цистеинов можно проводить с использованием любого способа, известного в данной области. Эти способы включают без ограничения мутагенез с перекрывающимися удлинениями при ПЦР, сайтнаправленный мутагенез или кассетный мутагенез (см, в общем Sambrook et al., Molecular Cloning, A Laboratory Manual, Cold Spring Harbour Laboratory Press, Cold Spring Harbour, NY, 1989; Ausbel et al., Current Protocols in Molecular Biology, Greene Publishing & Wiley-Interscience, NY, 1993). Наборы для сайтнаправленного мутагенеза доступны на рынке, например, набор QuikChange® для сайтнаправленного мутагенеза (Stratagene, La Jolla, CA). Кассетный мутагенез можно проводить на основе Wells et al., 1985, Gene, 34:315-323. Альтернативно, мутанты можно получать с помощью полного синтеза генов путем отжига, лигирования и ПЦР-амплификации и клонирования перекрывающихся олигонуклеотидов.
В одном варианте осуществления дисульфидная связь между CH1 и CL полностью отсутствует, например, межцепочечные цистеины могут быть заменены другой аминокислотой, такой как серин. Таким образом, в одном варианте осуществления в функциональном фрагменте Fab молекулы нет межцепочечных дисульфидных связей. В таких раскрытиях как WO2005/003170, включенных в данный документ посредством ссылки, описано, как обеспечить фрагменты Fab без межцепочечной дисульфидной связи.
Примеры форматов антител для использования в настоящем изобретении включают дополненный IgG и дополненный Fab, где целый фрагмент IgG или Fab, соответственно, конструируют путем добавления по меньшей мере одного дополнительного антигенсвязывающего домена (например одного, двух, трех или четырех дополнительных антигенсвязывающих доменов), например однодоменного антитела (такого как VH или VL, или VHH), scFv, dsscFv, dsFv на N- и/или C-конец легкой цепи указанного IgG или Fab и необязательно в тяжелую цепь указанного IgG или Fab, например как описано в WO2009/040562, WO2010035012, WO2011/030107, WO2011/061492, WO2011/061246 и WO2011/086091, которые все включены в данный документ посредством ссылки. дополненный IgG, включая полноразмерный IgG, сконструированный путем добавления dsFv на C-конец легкой цепи (и необязательно в тяжелую цепь) IgG, был впервые раскрыт в WO2015/197789, включенной в данный документ посредством ссылки.
Предпочтительный формат антитела для использования в настоящем изобретении содержит Fab, связанный с двумя scFv или dsscFv, где каждый scFv или dsscFv связывает одну и ту же или разные мишени (например, один scFv или dsscFv связывает терапевтическую мишень, и один scFv или dsscFv, который увеличивает период полужизни за счет связывания, например, альбумина). Такие фрагменты антитела описаны в публикации международной заявки на патент № WO2015/197772, которая полностью включена в данный документ посредством ссылки и особенно в отношении обсуждения фрагментов антител.
V1 представляет dsscFv, dsFv или scFv.
V2 представляет dsscFv, dsFv или scFv.
В одном варианте осуществления, когда V1 и/или V2 представляют собой dsFv или dsscFv, дисульфидная связь между вариабельными доменами VH и VL из V1 и/или V2 находится между двумя остатками, перечисленными ниже (если из контекста не следует иное, в списке ниже использована нумерация согласно Kabat). Всякий раз, когда ссылка сделана на нумерацию согласно Kabat, соответствующей ссылкой является Kabat et al. 1991 (5th edition, Bethesda, Md.) в Sequences of Proteins of Immunological Interest, US Department of Health and Human Services, NIH, USA.
В одном варианте осуществления дисульфидная связь находится в позиции, выбранной из группы, содержащей:
VH37+VL95C см., например, Protein Scienc 6, 781-788 Zhu et al (1997);
VH44+VL100 см., например; Weatherill et al. Protein Engineering, Design & Selection, 25 (321-329) 2012);
VH44+VL105 см., например, J Biochem. 118, 825-831 Luo et al (1995);
VH45+VL87 см., например, Protein Scienc 6, 781-788 Zhu et al (1997);
VH55+VL101 см., например, FEBS Letters 377 135-139 Young et al (1995);
VH100+VL50 см., например, Biochemistry 29 1362-1367 Glockshuber et al (1990);
VH100b+VL49; см., например, Biochemistry 29 1362-1367 Glockshuber et al (1990);
VH98+VL 46; см., например, Protein Scienc 6, 781-788 Zhu et al (1997);
VH101+VL46; см., например, Protein Scienc 6, 781-788 Zhu et al (1997);
VH105+VL43 см., например; Proc. Natl. Acad. Sci. USA Vol. 90 pp,7538-7542 Brinkmann et al (1993); или Proteins 19, 35-47 Jung et al (1994)
VH106+VL57 см., например, FEBS Letters 377 135-139 Young et al (1995),
и соответствующей ей позиции в паре вариабельных областей, расположенных в молекуле.
В одном варианте осуществления дисульфидная связь образована между позициями VH44 и VL100.
Пары аминокислот, перечисленных выше, находятся в позициях, благоприятных для замены цистеинами, так чтобы можно было образовать дисульфидные связи. Цистеины можно сконструировать в этих нужных позициях с помощью известных методов. В одном варианте осуществления, следовательно, сконструированный цистеин согласно настоящему раскрытию относится к ситуации, когда природный остаток в заданной позиции аминокислоты был заменен остатком цистеина.
Введение сконструированных цистеинов можно проводить с использованием любого способа, известного в данной области. Эти способы включают без ограничения мутагенез с перекрывающимися удлинениями при ПЦР, сайтнаправленный мутагенез или кассетный мутагенез (см, в общем Sambrook et al., Molecular Cloning, A Laboratory Manual, Cold Spring Harbour Laboratory Press, Cold Spring Harbour, NY, 1989; Ausbel et al., Current Protocols in Molecular Biology, Greene Publishing & Wiley-Interscience, NY, 1993). Наборы для сайтнаправленного мутагенеза доступны на рынке, например, набор для сайтнаправленного мутагенеза QuikChange® (Stratagen, La Jolla, CA). Кассетный мутагенез можно проводить на основе Wells et al., 1985, Gene, 34:315-323. Альтернативно, мутанты можно получать с помощью полного синтеза генов путем отжига, лигирования и ПЦР-амплификации и клонирования перекрывающихся олигонуклеотидов.
Соответственно, в одном варианте осуществления, когда V1 и/или V2 представляют собой dsFv или dsscFv, вариабельные домены VH и VL из V1 и/или вариабельные домены VH и VL из V2, могут быть связаны дисульфидной связью между двумя остатками цистеина, где позицию пары остатков цистеина выбирают из группы, состоящей из: VH37 и VL95, VH44 и VL100, VH44 и VL105, VH45 и VL87, VH100 и VL50, VH100b и VL49, VH98 и VL46, VH101 и VL46, VH105 и VL43 и VH106 и VL57.
В одном варианте осуществления, когда V1 и/или V2 представляют собой dsFv или dsscFv, вариабельные домены VH и VL из V1 и/или вариабельные домены VH и VL из V2 могут быть связаны дисульфидной связью между двумя остатками цистеина, один в VH и один в VL, которые находятся за пределами CDR, где позицию пары остатков цистеина выбирают из группы, состоящей из VH37 и VL95, VH44 и VL100, VH44 и VL105, VH45 и VL87, VH100 и VL50, VH98 и VL46, VH105 и VL43 и VH106 и VL57.
В одном варианте осуществления, когда V1 представляет собой dsFv или dsscFv, вариабельные домены VH и VL из V1 связаны дисульфидной связью между двумя сконструированными остатками цистеина, один в позиции VH44, а другой в VL100. В одном варианте осуществления, когда V2 представляет собой dsFv или dsscFv, вариабельные домены VH и VL из V2 связаны дисульфидной связью между двумя сконструированными остатками цистеина, один в позиции VH44, а другой в VL100.
В одном варианте осуществления, когда V1 представляет собой dsscFv, dsFv или scFv, домен VH из V1 присоединен к X.
В одном варианте осуществления, когда V1 представляет собой dsscFv, dsFv или scFv, домен VL из V1 присоединен к X.
В одном варианте осуществления, когда V2 представляет собой dsscFv, dsFv или scFv, домен VH из V2 присоединен к Y.
В одном варианте осуществления, когда V2 представляет собой dsscFv, dsFv или scFv, домен VL из V2 присоединен к Y.
Специалисту будет понятно, что когда V1 и/или V2 представляет dsFv, мультиспецифическое антитело будет содержать третий полипептид, кодирующий соответствующий свободный домен VH или VL, который не присоединен к X или Y. Когда V1 и V2 представляют собой dsFv, то «свободный вариабельный домен» (то есть домен, связанный посредством дисульфидной связи с остальной частью полипептида) будет общим для обеих цепей. Таким образом, тогда как реальный вариабельный домен, слитый или связанный посредством X или Y с полипептидом, может отличаться в каждой полипептидной цепи, свободные вариабельные домены, соединенные с ним, обычно будут идентичны друг другу.
В некоторых вариантах осуществления p составляет 1. В некоторых вариантах осуществления p составляет 0.
В некоторых вариантах осуществления s составляет 1. В некоторых вариантах осуществления s составляет 0.
В некоторых вариантах осуществления t составляет 1. В некоторых вариантах осуществления t составляет 0.
В некоторых вариантах осуществления s составляет 1, а t составляет 1. В некоторых вариантах осуществления s составляет 0, а t составляет 0.
В одном варианте осуществления p составляет 1, q составляет 1, r составляет 0, s составляет 0, а t составляет 0, при этом и V1 и V2 представляют dsscFv. Таким образом, в одном аспекте представлено мультиспецифическое антитело, которое связывает IL-13 человека, IL-17A человека и/или IL-17F человека, содержащее или состоящее из:
a) полипептидной цепи формулы (Ia):
VH-CH1-X-V1; и
b) полипептидной цепи формулы (IIa):
VL-CL-Y-V2;
где:
VH представляет вариабельный домен тяжелой цепи;
CH1 представляет домен 1 константной области тяжелой цепи;
X представляет связь или линкер;
Y представляет связь или линкер;
V1 представляет scFv, dsscFv или dsFv;
VL представляет вариабельный домен легкой цепи;
CL представляет домен из константной области легкой цепи, такой как Cкаппа;
V2 представляет scFv, dsscFv или dsFv;
где по меньшей мере один из V1 или V2 представляет собой dsscFv или dsFv;
при этом полипептидная цепь формулы (Ia) содержит домен связывания белка А; и
при этом полипептидная цепь формулы (IIa) не связывает белок A.
В таком варианте осуществления V2 не связывает белок A, то есть scFv, dsscFv или dsFv из V2 не содержит домен связывания белка А. В одном варианте осуществления V2, то есть scFv, dsscFv или dsFv из V2, содержит домен VH1. В другом варианте осуществления V2, то есть scFv, dsscFv или dsFv из V2, содержит домен VH3, который не связывает белок A. В одном варианте осуществления V2, то есть scFv, dsscFv или dsFv из V2, содержит домен VH2. В одном варианте осуществления V2, то есть scFv, dsscFv или dsFv из V2, содержит домен VH4. В одном варианте осуществления V2, то есть scFv, dsscFv или dsFv из V2, содержит домен VH5. В одном варианте осуществления V2, то есть scFv, dsscFv или dsFv из V2, содержит домен VH6. В одном варианте осуществления полипептидная цепь формулы (Ia) содержит только один домен связывания белка А, присутствующий в VH или V1. В одном варианте осуществления полипептидная цепь формулы (Ia) содержит только один домен связывания белка А, присутствующий в V1. В другом варианте осуществления полипептидная цепь формулы (Ia) содержит два домена связывания белка А, присутствующих в VH и V1, соответственно.
В другом варианте осуществления p составляет 0, q составляет 1, r составляет 0, s составляет 1, t составляет 1, а V2 представляет собой dsscFv. Таким образом, в одном аспекте представлено мультиспецифическое антитело, которое связывает IL-13 человека, IL-17A человека и/или IL-17F человека и содержит или состоит из:
a) полипептидной цепи формулы (Ib):
VH-CH1- CH2 -CH3; и
b) полипептидной цепи формулы (IIb):
VL-CL-Y-V2;
где:
VH представляет вариабельный домен тяжелой цепи;
CH1 представляет домен 1 константной области тяжелой цепи;
CH2 представляет домен 2 константной области тяжелой цепи;
CH3 представляет домен 3 константной области тяжелой цепи;
Y представляет связь или линкер;
VL представляет вариабельный домен легкой цепи;
CL представляет домен из константной области легкой цепи, такой как Cкаппа;
V2 представляет dsscFv;
при этом полипептидная цепь формулы (Ib) содержит домен связывания белка А; и
при этом полипептидная цепь формулы (IIb) не связывает белок A.
В таком варианте осуществления V2 не связывает белок A, то есть dsscFv из V2 не содержит домен связывания белка А. В одном варианте осуществления V2, то есть dsscFv из V2, содержит домен VH1. В другом варианте осуществления V2, то есть dsscFv из V2, содержит домен VH3, который не связывает белок A. В одном варианте осуществления полипептидная цепь формулы (Ib) содержит только один домен связывания белка А, присутствующий в VH или CH2-CH3. В другом варианте осуществления полипептидная цепь формулы (Ib) содержит два домена связывания белка А, присутствующих в VH и CH2-CH3, соответственно.
В другом варианте осуществления p составляет 0, q составляет 1, r составляет 0, s составляет 1, t составляет 1, а V2 представляет собой dsFv. Таким образом, в одном аспекте представлено мультиспецифическое антитело, которое связывает IL-13 человека, IL-17A человека и/или IL-17F человека, содержащее или состоящее из:
a) полипептидной цепи формулы (Ic):
VH-CH1- CH2 -CH3; и
b) полипептидной цепи формулы (IIc):
VL-CL-Y-V2;
где:
VH представляет вариабельный домен тяжелой цепи;
CH1 представляет домен 1 константной области тяжелой цепи;
CH2 представляет домен 2 константной области тяжелой цепи;
CH3 представляет домен 3 константной области тяжелой цепи;
Y представляет связь или линкер;
VL представляет вариабельный домен легкой цепи;
CL представляет домен из константной области легкой цепи, такой как Cкаппа;
V2 представляет dsFv;
при этом полипептидная цепь формулы (Ic) содержит домен связывания белка А; и
при этом полипептидная цепь формулы (IIc) не связывает белок A.
В таком варианте осуществления V2, то есть dsFv из V2, не связывает белок A. В одном варианте осуществления полипептидная цепь формулы (Ic) содержит только один домен связывания белка А, присутствующий в VH или CH2-CH3. В другом варианте осуществления полипептидная цепь формулы (Ic) содержит два домена связывания белка А, присутствующих в VH и CH2-CH3, соответственно.
В одном варианте осуществления мультиспецифического антитела согласно изобретению
VL и VH содержат антигенсвязывающий участок, который связывается с IL-17A человека и/или IL-17F человека,
V1 содержит антигенсвязывающий участок, который связывается с сывороточным альбумином человека, а
V2 содержит антигенсвязывающий участок, который связывается с IL-13 человека.
В одном варианте осуществления VL содержит последовательность SEQ ID NO:1 для CDR-L1, последовательность SEQ ID NO:2 для CDR-L2, и последовательность SEQ ID NO:3 для CDR-L3; VH содержит последовательность SEQ ID NO:4 для CDR-H1, последовательность SEQ ID NO:5 для CDR-H2, и последовательность SEQ ID NO:6 для CDR-H3.
В одном варианте осуществления V1 содержит вариабельную область легкой цепи, содержащую последовательность SEQ ID NO:39 для CDR-L1, последовательность SEQ ID NO:40 для CDR-L2, и последовательность SEQ ID NO:41 для CDR-L3; и вариабельную область тяжелой цепи, содержащую последовательность SEQ ID NO:42 для CDR-H1, последовательность SEQ ID NO:43 для CDR-H2, и последовательность SEQ ID NO:44 для CDR-H3.
В одном варианте осуществления V2 содержит вариабельную область легкой цепи, содержащую последовательность SEQ ID NO:15 для CDR-L1, последовательность SEQ ID NO:16 для CDR-L2, и последовательность SEQ ID NO:17 для CDR-L3; и вариабельную область тяжелой цепи, содержащую последовательность SEQ ID NO:18 для CDR-H1, последовательность SEQ ID NO:19 для CDR-H2, и последовательность SEQ ID NO:20 для CDR-H3;
В одном варианте осуществления VL содержит последовательность SEQ ID NO:7, а VH содержит последовательность SEQ ID NO:9.
В одном варианте осуществления V1 содержит вариабельную область легкой цепи, содержащую последовательность SEQ ID NO:45, и вариабельную область тяжелой цепи, содержащую последовательность SEQ ID NO:46.
В одном варианте осуществления V1 содержит вариабельную область легкой цепи, содержащую последовательность SEQ ID NO:49, и вариабельную область тяжелой цепи, содержащую последовательность SEQ ID NO:50.
В одном варианте осуществления вариабельная область легкой цепи и вариабельная область тяжелой цепи V1 соединены линкером, где указанный линкер содержит последовательность SEQ ID NO:68.
В одном варианте осуществления V1 представляет собой scFv, содержащий последовательность SEQ ID NO:53, или dsscFv, содержащий последовательность SEQ ID NO:55.
В одном варианте осуществления V2 содержит вариабельную область легкой цепи, содержащую последовательность SEQ ID NO:27, и вариабельную область тяжелой цепи, содержащую последовательность SEQ ID NO:28.
В одном варианте осуществления V2 содержит вариабельную область легкой цепи, содержащую последовательность SEQ ID NO:31, и вариабельную область тяжелой цепи, содержащую последовательность SEQ ID NO:32.
В одном варианте осуществления вариабельная область легкой цепи и вариабельная область тяжелой цепи V2 соединены линкером, где указанный линкер содержит последовательность SEQ ID NO:66.
В одном варианте осуществления V2 представляет собой scFv, содержащий последовательность SEQ ID NO:35, или dsscFv, содержащий последовательность SEQ ID NO:37.
В одном варианте осуществления X представляет собой линкер, содержащий последовательность SEQ ID NO:67.
В одном варианте осуществления Y представляет собой линкер, содержащий последовательность SEQ ID NO:65.
В одном варианте осуществления полипептидная цепь формулы (Ia) содержит последовательность SEQ ID NO:57 или SEQ ID NO: 59.
В одном варианте осуществления полипептидная цепь формулы (IIa) содержит последовательность SEQ ID NO:61 или SEQ ID NO: 63.
В одном варианте осуществления полипептидная цепь формулы (Ia) содержит последовательность SEQ ID NO:59, а полипептидная цепь формулы (IIa) содержит последовательность SEQ ID NO: 63.
Должно быть понятно, что в последовательностях, представленных в настоящем изобретении, можно сделать одну или несколько аминокислотных замен, дополнений и/или делеций без значительного изменения способности антитела связываться с антигеном и нейтрализовать его биологическую активность. Эффект любых аминокислотных замен, дополнений и/или делеций может быть легко протестирован специалистом в данной области, например с использованием способов, описанных в данном документе, в частности, способов, показанных в примерах, для определения связывания антигена и ингибирования биологической активности.
Соответственно, в настоящем изобретении представлено мультиспецифическое антитело, содержащее CDR с последовательностями, приведенными в SEQ ID NO:1, 2, 3, 4, 5, 6, 15, 16, 17, 18, 19, 20, 39, 40, 41, 42, 43 и 44, в которых одна или несколько аминокислот в одной или более CDR были заменены другой аминокислотой, например, аналогичной аминокислотой согласно определению в настоящем изобретении ниже.
«Идентичность» согласно настоящему изобретению означает, что в любой определенной позиции в выровненных последовательностях аминокислотный остаток является идентичным в последовательностях. «Схожесть» согласно настоящему изобретению означает, что в любой определенной позиции в выровненных последовательностях аминокислотный остаток относится к аналогичному типу в последовательностях. Например, лейцин может быть заменен на изолейцин или валин. Другие аминокислоты, которые часто можно заменять друг на друга, включают без ограничения:
- фенилаланин, тирозин и триптофан (аминокислоты, имеющие ароматические боковые цепи);
- лизин, аргинин и гистидин (аминокислоты, имеющие основные боковые цепи);
- аспартат и глутамат (аминокислоты, имеющие кислые боковые цепи);
- аспарагин и глутамин (аминокислоты, имеющие амидные боковые цепи); и
- цистеин и метионин (аминокислоты, имеющие серосодержащие боковые цепи).
Степень идентичности и схожести можно легко рассчитать (Computational Molecular Biology, Lesk, A.M., ed., Oxford University Press, New York, 1988; Biocomputing. Informatics and Genome Projects, Smith, D.W., ed., Academic Press, New York, 1993; Computer Analysis of Sequence Data, Part 1, Griffin, A.M., and Griffin, H.G., eds., Humana Press, New Jersey, 1994; Sequence Analysis in Molecular Biology, von Heinje, G., Academic Press, 1987, Sequence Analysis Primer, Gribskov, M. and Devereux, J., eds., M Stockton Press, New York, 1991, программное обеспечение BLAST™, поставляемое NCBI (Altschul, S.F. et al., 1990, J. Mol. Biol. 215:403-410; Gish, W. & States, D.J. 1993, Nature Genet. 3:266-272. Madden, T.L. et al., 1996, Meth. Enzymol. 266:131-141; Altschul, S.F. et al., 1997, Nucleic Acids Res. 25:3389-3402; Zhang, J. & Madden, T.L. 1997, Genome Res. 7:649-656).
В одном варианте осуществления CDR мультиспецифического антитела содержат последовательности, которые имеют по меньшей мере 70%, 80%, 90%, 95% или 98% идентичность или схожесть с последовательностями, приведенными в SEQ ID NO:1, 2, 3, 4, 5, 6, 15, 16, 17, 18, 19, 20, 39, 40, 41, 42, 43 и 44.
В одном варианте осуществления VL содержит последовательность, которая имеет по меньшей мере 70%, 80%, 90%, 95% или 98% идентичность или схожесть с последовательностью, приведенной в SEQ ID NO:7, а VH содержит последовательность, которая имеет по меньшей мере 70%, 80%, 90%, 95% или 98% идентичность или схожесть с последовательностью, приведенной в SEQ ID NO:9.
В одном варианте осуществления V1 содержит вариабельную область легкой цепи, содержащую последовательность, которая имеет по меньшей мере 70%, 80%, 90%, 95% или 98% идентичность или схожесть с последовательностью, приведенной в SEQ ID NO:45, и/или вариабельную область тяжелой цепи, содержащую последовательность, которая имеет по меньшей мере 70%, 80%, 90%, 95% или 98% идентичность или схожесть с последовательностью, приведенной в SEQ ID NO:46.
В одном варианте осуществления V1 содержит вариабельную область легкой цепи, содержащую последовательность, которая имеет по меньшей мере 70%, 80%, 90%, 95% или 98% идентичность или схожесть с последовательностью, приведенной в SEQ ID NO:49, и/или вариабельную область тяжелой цепи, содержащую последовательность, которая имеет по меньшей мере 70%, 80%, 90%, 95% или 98% идентичность или схожесть с последовательностью, приведенной в SEQ ID NO:50.
В одном варианте осуществления вариабельная область легкой цепи и вариабельная область тяжелой цепи V1 соединены линкером, где указанный линкер содержит последовательность, которая имеет по меньшей мере 70%, 80%, 90%, 95% или 98% идентичность или схожесть с последовательностью, приведенной в SEQ ID NO:68.
В одном варианте осуществления V1 представляет собой scFv, содержащий последовательность, которая имеет по меньшей мере 70%, 80%, 90%, 95% или 98% идентичность или схожесть с последовательностью, приведенной в SEQ ID NO:53, или dsscFv, содержащий последовательность, которая имеет по меньшей мере 70%, 80%, 90%, 95% или 98% идентичность или схожесть с последовательностью, приведенной в SEQ ID NO:55.
В одном варианте осуществления V2 содержит вариабельную область легкой цепи, содержащую последовательность, которая имеет по меньшей мере 70%, 80%, 90%, 95% или 98% идентичность или схожесть с последовательностью, приведенной в SEQ ID NO:27, и/или вариабельную область тяжелой цепи, содержащую последовательность, которая имеет по меньшей мере 70%, 80%, 90%, 95% или 98% идентичность или схожесть с последовательностью, приведенной в SEQ ID NO:28.
В одном варианте осуществления V2 содержит вариабельную область легкой цепи, содержащую последовательность, которая имеет по меньшей мере 70%, 80%, 90%, 95% или 98% идентичность или схожесть с последовательностью, приведенной в SEQ ID NO:31, и/или вариабельную область тяжелой цепи, содержащую последовательность, которая имеет по меньшей мере 70%, 80%, 90%, 95% или 98% идентичность или схожесть с последовательностью, приведенной в SEQ ID NO:32.
В одном варианте осуществления вариабельная область легкой цепи и вариабельная область тяжелой цепи V2 соединены линкером, где указанный линкер содержит последовательность, которая имеет по меньшей мере 70%, 80%, 90%, 95% или 98% идентичность или схожесть с последовательностью, приведенной в SEQ ID NO:66.
В одном варианте осуществления V2 представляет собой scFv, содержащий последовательность, которая имеет по меньшей мере 70%, 80%, 90%, 95% или 98% идентичность или схожесть с последовательностью, приведенной в SEQ ID NO:35, или dsscFv, содержащий последовательность, которая имеет по меньшей мере 70%, 80%, 90%, 95% или 98% идентичность или схожесть с последовательностью, приведенной в SEQ ID NO:37.
В одном варианте осуществления X представляет собой линкер, содержащий последовательность, которая имеет по меньшей мере 70%, 80%, 90%, 95% или 98% идентичность или схожесть с последовательностью, приведенной в SEQ ID NO:67.
В одном варианте осуществления Y представляет собой линкер, содержащий последовательность, которая имеет по меньшей мере 70%, 80%, 90%, 95% или 98% идентичность или схожесть с последовательностью, приведенной в SEQ ID NO:65.
В одном варианте осуществления полипептидная цепь формулы (Ia) содержит последовательность, которая имеет по меньшей мере 70%, 80%, 90%, 95% или 98% идентичность или схожесть с последовательностью, приведенной в SEQ ID NO:57 или SEQ ID NO: 59.
В одном варианте осуществления полипептидная цепь формулы (IIa) содержит последовательность, которая имеет по меньшей мере 70%, 80%, 90%, 95% или 98% идентичность или схожесть с последовательностью, приведенной в SEQ ID NO:61 или SEQ ID NO: 63.
В одном варианте осуществления полипептидная цепь формулы (Ia) содержит последовательность, которая имеет по меньшей мере 70%, 80%, 90%, 95% или 98% идентичность или схожесть с последовательностью, приведенной в SEQ ID NO:59, а полипептидная цепь формулы (IIa) содержит последовательность, которая имеет по меньшей мере 70%, 80%, 90%, 95% или 98% идентичность или схожесть с последовательностью, приведенной в SEQ ID NO: 63.
Эпитоп
Эпитоп представляет собой область антигена, которую связывает антитело. Эпитопы можно определять, как структурные или функциональные. Функциональные эпитопы обычно составляют подмножество структурных эпитопов и имеют те остатки, которые прямо вносят вклад в сродство взаимодействия. Эпитопы могут также быть конформационными, то есть состоящими из нелинейных аминокислот. В определенных вариантах осуществления эпитопы могут содержать детерминанты, которые представляют собой химически активные поверхностные группировки молекул, таких как аминокислоты, боковые цепи сахаров, фосфорильные группы или сульфонильные группы, а в определенных вариантах осуществления могут иметь конкретные характеристики трехмерной структуры и/или конкретные характеристики заряда.
Можно легко определить, связывается ли антитело с тем же эпитопом, что и эталонное антитело, или конкурирует ли оно за связывание с ним с использованием стандартных способов, известных в данной области. Например, для определения, связывается ли тестируемое антитело с тем же эпитопом, что и эталонное антитело согласно изобретению, обеспечивают связывание эталонного антитела с белком или пептидом в условиях насыщения. Затем оценивают способность тестируемого антитела связываться с белком или пептидом. если тестируемое антитело способно связываться с белком или пептидом после связывания с эталонным антителом при насыщении, можно сделать вывод, что тестируемое антитело связывается с другим эпитопом, чем эталонное антитело. С другой стороны, если тестируемое антитело не способно связываться с белком или пептидом после связывания с эталонным антителом при насыщении, то тестируемое антитело может связываться с тем же эпитопом, что и эпитоп, связанный эталонным антителом согласно изобретению.
Для определения, конкурирует ли антитело за связывание с эталонным антителом, описанный выше метод связывания выполняют в двух ориентациях. В первой ориентации обеспечивают связывание эталонного антитела с белком/пептидом в условиях насыщения с последующей оценкой связывания тестируемого антитела с молекулой белка/пептида. Во второй ориентации обеспечивают связывание тестируемого антитела с белком/пептидом в условиях насыщения с последующей оценкой связывания эталонного антитела с белком/пептидом. Если в обеих ориентациях только первое (насыщенное) антитело способно связываться с белком/пептидом, то делают вывод, что тестируемое антитело и эталонное антитело конкурируют за связывание с белком/пептидом. Как будет понятно специалисту, антитело, которое конкурирует за связывание с эталонным антителом, не обязательно может связываться с идентичным эпитопом, как эталонное антитело, но может пространственно блокировать связывание эталонного антитела путем связывания с перекрывающимся или соседним эпитопом.
Два антитела связываются с одним и тем же или с перекрывающимся эпитоп, если каждое конкуретно ингибирует (блокирует) связывание другого с антигеном. то есть 1-, 5-, 10-, 20- или 100-кратное превышение одного антитела ингибирует связывание другого по меньшей мере на 50%, 75%, 90% или даже 99% при измерении в анализе конкурентного связывания (см., например, Junghans et al. Cancer Res, 1990:50:1495-1502). Альтернативно, два антитела имеют один и тот же эпитоп, если по существу все аминокислотные мутации в антигене, которые уменьшают или устраняют связывание одного антитела, уменьшают или устраняют связывание другого. Два антитела имеют перекрывающиеся эпитопы, если некоторые аминокислотные мутации, которые уменьшают или устраняют связывание одного антитела, уменьшают или устраняют связывание другого.
Затем можно выполнить дополнительные стандартные эксперименты (например, анализы мутаций и связывания пептидов) для подтверждения, в действительности ли наблюдаемое отсутствие связывания тестируемого антитела обусловлено связыванием с тем же эпитопом, что и эталонное антитело, или же за отсутствие наблюдаемого связывания отвечает стерическое блокирование (или другое явление). Эксперименты этого типа можно проводить с использованием ELISA, RIA, поверхностного плазмонного резонанса, проточной цитометрии или любого другого количественного или качественного анализа связывания антитела, доступного в данной области.
Антитела могут конкурировать за связывание с IL-17A или IL-17F или связываются с тем же эпитопом, что и мультиспецифическое антитело, которое содержит комбинацию последовательностей CDR-L1/CDR-L2/CDR-L3/CDR-H1/CDR-H2/CDR-H3 из SEQ ID NO: 1/2/3/4/5/6.
Антитела могут конкурировать за связывание с IL-13 или связываются с тем же эпитопом, что и мультиспецифическое антитело, которое содержит комбинацию последовательностей CDR-L1/CDR-L2/CDR-L3/CDR-H1/CDR-H2/CDR-H3 из SEQ ID NO: 15/16/17/18/19/20.
Антитела могут конкурировать за связывание с сывороточным альбумином или связываются с тем же эпитопом, что и мультиспецифическое антитело, которое содержит комбинацию последовательностей CDR-L1/CDR-L2/CDR-L3/CDR-H1/CDR-H2/CDR-H3 из SEQ ID NO: 39/40/41/42/43/44.
Эффекторные молекулы
Если нужно мультиспецифическое антитело для использования в настоящем изобретении может быть конъюгировано с одной или несколькими эффекторными молекулами. Должно быть понятно, что эффекторная молекула может включать в себя единственную эффекторную молекулу или две или более таких молекул связанных так, чтобы образовать единый фрагмент, который может связываться с антителами согласно настоящему изобретению. Когда нужно получить фрагмент антитела, связанный с эффекторной молекулой, его можно получить с помощью стандартных химических методов или методов рекомбинантной ДНК, в которых фрагмент антитела связывают либо прямо, либо посредством связывающего средства с эффекторной молекулой. Методы конъюгации таких эффекторных молекул с антителами хорошо известны в данной области (см. Hellstrom et al., Controlled Drug Delivery, 2nd Ed., Robinson et al., eds., 1987, pp. 623-53; Thorpe et al., 1982, Immunol. Rev., 62:119-58 and Dubowchik et al., 1999, Pharmacology and Therapeutics, 83, 67-123). Конкретные химические методы включают, например, те, что описаны в WO 93/06231, WO 92/22583, WO 89/00195, WO 89/01476 и WO 03031581. Альтернативно, когда эффекторная молекула представляет собой белок или полипептид, связь может быть обеспечена с использованием методов рекомбинантной ДНК, например, описанных в WO 86/01533 и EP 0392745.
Термин эффекторная молекула согласно настоящему изобретению включает, например, противоопухолевые средства, лекарственные средства, токсины, биологически активные белки, например ферменты, другие антитела или фрагменты антитела, синтетические или природные полимеры, нуклеиновые кислоты и их фрагменты, например, ДНК, РНК и их фрагменты, радионуклиды, особенно радиоактивный йод, радиоизотопы, хелатные металлы, наночастицы и репортерные группы, такие как флуоресцентные соединения или соединения, которые могут быть обнаружены с помощью спектроскопии ЯМР или ЭПР.
Примеры эффекторных молекул могут включать цитотоксины или цитотоксичные средства, включая любое средство, которое является вредным для клеток (например, убивает их). Примеры включают комбрестатины, доластатины, эпотилоны, стауроспорин, майтанзиноиды, спонгистатины, ризоксин, галихондрины, роридины, гемиастерлины, таксол, цитохалазин B, грамицидин D, бромид этидия, эметин, митомицин, этопозид, тенопозид, винкристин, винбластин, колхицин, доксорубицин, даунорубицин, дигидроксиантрациндион, митоксантрон, митрамицин, актиномицин D, 1-дегидротестостерон, глюкокортикоиды, прокаин, тетракаин, лидокаин, пропранолол и пуромицин и их аналоги или гомологи.
Эффекторные молекулы также включают без ограничения антиметаболиты (например, метотрексат, 6-меркаптопурин, 6-тиогуанин, цитарабин, 5-фторурацил декарбазин), алкилирующие средства (например, мехлорэтамин, тиоэпа хлорамбуцил, мелфалан, кармустин (BSNU) и ломустин (CCNU), циклотосфамид, бусульфан, дибромоманнитол, стрептозотоцин, митомицин C и цис-дихлордиамин платины (II) (DDP) цисплатин), антрациклины (например, даунорубицин (ранее дауномицин) и доксорубицин), антибиотики (например, дактиномицин (ранее актиномицин), блеомицин, митрамицин, антрамицин (AMC), калихеамицины или дуокармицины) и антимитотические средства (например, винкристин и винбластин).
Другие эффекторные молекулы могут включать хелатные радионуклиды, такие как 111In и 90Y, Lu177, висмут213, калифорний252, иридий192 и вольфрам188/рений188; или лекарственные средства, такие как без ограничения алкилфосфохолины, ингибиторы топоизомеразы I, таксоиды и сурамин.
Другие эффекторные молекулы включают белки, пептиды и ферменты. Представляющие интерес ферменты включают без ограничения протеолитические ферменты, гидролазы, лиазы, изомеразы, трансферазы. Представляющие интерес белки, полипептиды и пептиды включают без ограничения иммуноглобулины, токсины, такие как абрин, рицин A, экзотоксин синегнойной палочки или дифтерийный токсин, белок, такой как инсулин, фактор некроза опухоли, α-интерферон, β-интерферон, фактор роста нервов, фактор роста тромбоцитов или тканевой активатор плазминогена, тромботическое средство или антиангиогенное средство, например, ангиостатин или эндостатин, или модификатор биологического ответа, такой как лимфокин, интерлейкин-1 (IL-1), интерлейкин-2 (IL-2), гранулоцитарно-макрофагальный колониестимулирующий фактор (GM-CSF), гранулоцитарный колониестимулирующий фактор (G-CSF), фактор роста нервов (NGF) или другой фактор роста и иммуноглобулины.
Другие эффекторные молекулы могут включать выявляемые вещества, полезные, например, для диагностики. Примеры выявляемых веществ включают различные ферменты, простетические группы, флуоресцентные материалы, люминесцентные материалы, биолюминесцентные материалы, радионуклиды, излучающие позитроны металлы (для использования в позитронно-эмиссионной томографии) и ионы нерадиоактивных парамагнитных металлов. См. в целом патент США № 4741900 для ионов металлов, которые можно конъюгировать с антителами для использования в качестве диагностики. Подходящие ферменты включают пероксидазу хрена, щелочную фосфатазу, бета-галактозидазу или ацетилхолинэстеразу; подходящие простетические группы включают стрептавидин, авидин и биотин; подходящие флуоресцентные материалы включают умбеллиферон, флуоресцеин, флуоресцеинизотиоцианат, родамин, дихлортриазиниламин флуоресцеин, дансилхлорид и фикоэритрин; подходящие люминесцентные материалы включают люминол; подходящие биолюминесцентные материалы включают люциферазу, люциферин и экворин; и подходящие радиоактивные нуклиды включают 125I, 131I, 111In и 99Tc.
В другом пример эффекторная молекула может увеличивать период полужизни антитела in vivo и/или уменьшать иммуногенность антитела и/или усиливать доставку антитела через эпителиальный барьер в иммунную систему. Примеры подходящих эффекторных молекул этого типа включают полимеры, альбумин, связывающие альбумин белки или связывающие альбумин соединения, такие как те, что описаны в WO 05/117984.
Когда эффекторная молекула представляет собой полимер, она может, как правило, быть синтетическим или природным полимером, например, необязательно замещенным полиалкиленовым, полиалкениленовым или полиоксиалкиленовым полимером с прямой или разветвленной цепью, или разветвленным или неразветвленным полисахаридом, например гомо- или гетерополисахаридом.
Конкретные необязательные заместители, которые могут присутствовать в вышеупомянутых синтетических полимерах, включают один или несколько гидроксильных, метильных или метоксигрупп.
Конкретные примеры синтетических полимеров включают необязательно замещенный поли(этиленгликоль) поли(пропиленгликоль) поли(виниловый спирт) с прямой или разветвленной цепью или их производные, особенно необязательно замещенный поли(этиленгликоль), такой как метоксиполи(этиленгликоль) или его производные.
Конкретные природные полимеры включают лактозу, амилозу, декстран, гликоген или их производные.
Термин «производные» согласно настоящему изобретению включает реакционноспособные производные, например тиол-селективные реакционноспособные группы, такие как малеимиды и тому подобное. Реакционноспособная группа может быть связана непосредственно или через линкерный сегмент с полимером. Следует понимать, что остаток такой группы в некоторых случаях образует часть продукта в качестве связующей группы между фрагментом антитела и полимером.
Размер полимера можно менять по желанию, но обычно он находится в диапазоне средней молекулярной массы от 500 до 50000 Да, например, от 5000 до 40000 Да, например, от 20000 до 40000 Да. Размер полимера можно, в частности, выбирать на основе предполагаемого использования продукта, например способности локализоваться в определенных тканях, таких как опухоли, или увеличивать период полувыведения из кровотока (для обзора см. Chapman, 2002, Advanced Drug Delivery Reviews, 54, 531-545). Таким образом, например, когда продукт предназначен для выхода из кровообращения и проникновения в ткани, может быть предпочтительно использовать полимер с низкой молекулярной массой, например, с молекулярной массой около 5000 Да. Для применения, когда продукт остается в кровотоке, может быть предпочтительно использовать полимер с более высокой молекулярной массой, например, имеющий молекулярную массу в диапазоне от 20000 Да до 40000 Да.
Подходящие полимеры включают полиалкиленовый полимер, такой как поли(этиленгликоль) или особенно, метоксиполи(этиленгликоль) или его производное и особенно с молекулярной массой в диапазоне от приблизительно 15000 Да до приблизительно 40000 Да.
В одном примере антитела для применения в настоящем изобретении присоединены к молекулам полиэтиленгликоля (PEG). В одном конкретном примере антитело представляет собой фрагмент антитела, и молекулы PEG могут быть присоединены через любую доступную боковую цепь аминокислоты или концевую аминокислотную функциональную группу, расположенную во фрагменте антитела, например любую свободную амино-, имино-, тиоловую, гидроксильную или карбоксильную группу. Такие аминокислоты могут встречаться в природе во фрагменте антитела или могут быть встроены во фрагмент с использованием методов рекомбинантной ДНК (см., например, US 5219996; US 5667425; WO 98/25971). В одном примере молекула антитела согласно настоящему изобретению представляет собой модифицированный Fab-фрагмент, где модификация представляет собой добавление к С-концу его тяжелой цепи одной или нескольких аминокислот для обеспечения возможности присоединения эффекторной молекулы. Соответственно, дополнительные аминокислоты образуют модифицированную шарнирную область, содержащую один или несколько остатков цистеина, к которым может быть присоединена эффекторная молекула. Можно использовать несколько сайтов для присоединения двух или более молекул PEG.
Подходящим образом молекулы PEG могут быть ковалентно связаны через тиоловую группу по меньшей мере одного остатка цистеина, расположенного во фрагменте антитела. Каждая молекула полимера, присоединенная к модифицированному фрагменту антитела, может быть ковалентно связана с атомом серы остатка цистеина, находящегося во фрагменте. Ковалентная связь обычно представляет собой дисульфидную связь или, в частности, связь сера-углерод. Если в качестве точки присоединения использована тиоловая группа, можно использовать соответствующим образом активированные эффекторные молекулы, например, тиол-селективные производные, такие как малеимиды и производные цистеина. Активированный полимер можно использовать в качестве исходного материала при получении модифицированных полимером фрагментов антител, как описано выше. Активированный полимер может представлять собой любой полимер, содержащий реакционноспособную тиоловую группу, такую как α-галогенкарбоновая кислота или сложный эфир, например йодацетамид, имид, например, малеимид, винилсульфон или дисульфид. Такие исходные материалы могут быть получены коммерчески (например, от Nektar, ранее Shearwater Polymers Inc., Huntsville, AL, USA) или могут быть получены из коммерчески доступных исходных материалов с использованием обычных химических процедур. Конкретные молекулы PEG включают 20К метокси-PEG-амин (получаемый от Nektar, ранее Shearwater; Rapp Polymere и SunBio) и M-PEG-SPA (получаемый от Nektar, ранее Shearwater).
В одном варианте осуществления антитело представляет собой модифицированный Fab-фрагмент или diFab, который пегилирован, то есть имеет ковалентно присоединенный к нему PEG (поли(этиленгликоль)), например, в соответствии со способом, раскрытым в ЕР 0948544 или ЕР 1090037 [см. также «Poly(ethyleneglycol) Chemistry, Biotechnical and Biomedical Applications», 1992, J. Milton Harris (ed), Plenum Press, New York, «Poly(ethyleneglycol) Chemistry and Biological Applications», 1997, J. Milton Harris and S. Zalipsky (eds), American Chemical Society, Washington DC and «Bioconjugation Protein Coupling Techniques for the Biomedical Sciences», 1998, M. Aslam and A. Dent, Grove Publishers, New York; Chapman, A. 2002, Advanced Drug Delivery Reviews 2002, 54:531-545]. В одном примере PEG присоединен к цистеину в шарнирной области. В одном примере Fab-фрагмент, модифицированный PEG, имеет малеимидную группу, ковалентно связанную с одной тиольной группой в модифицированной шарнирной области. Остаток лизина может быть ковалентно связан с малеимидной группой, и к каждой из аминогрупп остатка лизина может быть присоединен полимер метоксиполи(этиленгликоля), имеющий молекулярную массу приблизительно 20000 Да. Таким образом, общая молекулярная масса PEG, присоединенного к Fab-фрагменту, может составлять приблизительно 40000 Да.
В одном варианте осуществления мультиспецифическое антитело не присоединено к эффекторной молекуле.
Полинуклеотиды/Векторы/Клетки-хозяева
В настоящем изобретении также представлен выделенный полинуклеотид, кодирующий полипептидную цепь молекулы мультиспецифического антитела IL-13/IL-17AF.
Вариантный полинуклеотид может содержать 1, 2, 3, 4, 5, до 10, до 20, до 30, до 40, до 50, до 75 или более замен и/или делеций нуклеиновой кислоты из последовательностей, приведенных в перечне последовательностей. Обычно, вариант имеет 1-20, 1-50, 1-75 или 1-100 замен и/или делеций.
Подходящие варианты могут быть по меньшей мере приблизительно на 70% гомологичны полинуклеотиду любой из последовательностей нуклеиновой кислоты, раскрытых в данном документе, обычно гомологичны ему по меньшей мере приблизительно на 80 или 90% и более предпочтительно по меньшей мере приблизительно на 95%, 97% или 99%. Варианты могут сохранять по меньшей мере приблизительно 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичности. Варианты обычно сохраняют приблизительно 60% - приблизительно 99% идентичности, приблизительно 80% - приблизительно 99% идентичности, приблизительно 90% - приблизительно 99% идентичности или приблизительно 95% - приблизительно 99% идентичности. Гомология и идентичность на этих уровнях обычно присутствуют по меньшей мере относительно кодирующих областей полинуклеотидов. Способы измерения гомологии хорошо известны в данной области, и специалистам в данной области будет понятно, что в настоящем контексте гомологию рассчитывают на основе идентичности нуклеиновой кислоты. Такая гомология может существовать на протяжении области по меньшей мере приблизительно 15, по меньшей мере приблизительно 30, например по меньшей мере приблизительно 40, 60, 100, 200 или более смежных нуклеотидов (в зависимости от длины). Такая гомология может существовать по всей длине немодифицированной полинуклеотидной последовательности.
Гомолог может отличаться от последовательности соответствующего полинуклеотида менее чем приблизительно на 3, 5, 10, 15, 20 или более мутаций (каждой из которых может быть замена, делеция или вставка). Например, гомолог может отличаться на 3-50 мутаций, часто на 3-20 мутаций. Эти мутации можно измерить на протяжении области по меньшей мере 30, например по меньшей мере приблизительно 40, 60 или 100 или более смежных нуклеотидов гомолога.
Последовательность ДНК согласно настоящему изобретению может содержать синтетическую ДНК, например полученную путем химической обработки, кДНК, геномную ДНК или любую их комбинацию.
Специалистам в данной области хорошо известны общие способы, с помощью которых можно конструировать векторы, способы трансфекции и способы культивирования. В этом отношении ссылка сделана на «Current Protocols in Molecular Biology», 1999, F. M. Ausubel (ed), Wiley Interscience, New York и the Maniatis Manual, выпущенный Cold Spring Harbor Publishing.
Также предложена клетка-хозяин, содержащая один или несколько векторов клонирования или экспрессии, содержащих одну или несколько последовательностей ДНК, кодирующих мультиспецифическое антитело IL-13/IL-17AF. Можно использовать любую подходящую систему клеток-хозяев/векторов для экспрессии последовательностей ДНК, кодирующих мультиспецифическое антитело IL-13/IL-17AF. Можно использовать бактериальные, например E. Coli, и другие микробные системы, или также можно использовать эукариотические системы экспрессии клеток-хозяев, например млекопитающих. Подходящие клетки-хозяева млекопитающих включают клетки CHO.
В настоящем изобретении также представлен способ получения мультиспецифического антитела IL-13/IL-17AF, включающий культивирование клетки-хозяина, содержащей вектор, в условиях, подходящих для экспрессии белка из ДНК, кодирующей мультиспецифическое антитело IL-13/IL-17AF, и выделение мультиспецифического антитела IL-13/IL-17AF.
Получение мультиспецифических антител
Существует ряд подходов для получения мультиспецифических, особенно биспецифических антител. У Morrison et al (Coloma и Morrison 1997, Nat Biotechnol. 15, 159-163) описано слияние одноцепочечных вариабельных фрагментов (scFv) с целыми антителами, например IgG. У Schoonjans et al. 2000, JouРНКl of Immunology, 165, 7050-7057 описано слияние scFv с фрагментами Fab антитела. В WO2015/197772 описано слияние дисульфидного стабилизированного scFv (dsscFv) с фрагментами Fab.
Стандартные подходы, описанные в предшествующем уровне техники, предусматривают экспрессию в клетке-хозяине по меньшей мере двух полипептидов, каждый из которых кодирует тяжелую цепь (HC) или легкую цепь (LC) целого антитела или его антигенсвязывающего фрагмента, например Fab, с которым дополнительный антигенсвязывающий фрагмент антитела может быть слит в N- и/или C- концевой позиции тяжелой цепи и/или легкой цепи. При попытке рекомбинантного получения таких мультиспецифических антител путем экспрессии двух (одной легкой цепи и одной тяжелой цепи для образования дополненного Fab) или четырех полипептидов (двух легких цепей и двух тяжелых цепей для образования дополненного IgG) обычно требуется избыточная экспрессия легкой цепи по сравнению с тяжелой цепью для обеспечения надлежащей укладки тяжелой цепи при сборке с соответствующей легкой цепью. В частности, предотвращают укладку самого CH1 (домена 1 константной области тяжелой цепи) белками BIP, которые могут быть вытеснены соответствующей LC; следовательно, правильная укладка CH1/HC зависит от доступности его соответствующей LC (Lee et al. 1999, Molecular Biology of the Cell, Vol. 10, 2209-2219).
Авторы наблюдали, что эти способы экспрессии мультиспецифических антител могут приводить к получению избытка легкой цепи по сравнению с тяжелой цепью, что сохраняется при сборе клеток-хозяев и что избыток легкой цепи имеет тенденцию к образованию димерных комплексов (или «LC димеров»), которые присутствуют в виде побочного продукта технологического процесса с нужным мультиспецифическим антителом, особенно мономерным, и, таким образом, к необходимости очистки.
Важно, что техническая проблема, связанная с образованием димеров легких цепей при слиянии на N- и/или C-конце с дополнительным антигенсвязывающим фрагментом (фрагментами), до сих пор не была идентифицирована, а общепринятые аналитические способы не имеют возможности обнаружения и количественного определения этих дополненных димеров LC среди гетерогенных продуктов технологического процесса. Это может приводить к значительной систематической ошибке при оценке количества продуктов с использованием стандартных аналитических способов.
Таким образом, существует необходимость в улучшении мультиспецифических антител и способов их получения, которые обеспечивают легкое и эффективное выделение и удаление дополненных димеров LC на самых ранних стадиях технологического процесса и, таким образом, повышают выход представляющего интерес белка для использования в терапии, который представляет мультиспецифическое антитело, в частности в его мономерной форме.
Мультиспецифические антитела согласно настоящему изобретению были сконструированы, чтобы предоставить улучшенные мультиспецифические антитела с эквивалентной функциональностью и стабильностью при одновременном увеличении выхода материала «мультиспецифических антител», особенно мономерных, полученных после очистки, особенно после одностадийной очистки, включающей хроматографию по сродству на белок А.
Предпочтительно, мультиспецифические антитела согласно настоящему раскрытию можно очищать более эффективно с помощью способа очистки, более совершенного по сравнению со способами, общепринятыми в предшествующем уровне техники, особенно в том, что улучшенный способ предусматривает меньше стадий, и является эффективным с точки зрения затрат и времени в промышленном масштабе. В частности, мультиспецифические антитела согласно настоящему раскрытию дают максимальное количество представляющих интерес белков (то есть правильнаый формат мультиспецифического антитела), полученных после способа одностадийной очистки, предусматривающего хроматографию по сродству на белок А, где очистка представляющих интерес мультиспецифических антител и удаление дополненных димеров LC происходит одновременно. Предпочтительно, способы получения и очистки мультиспецифических антител согласно настоящему раскрытию не требуют дополнительной стадии очистки для захвата избытка свободных, несвязанных легких цепей, особенно дополненных димеров LC.
Белок A
Белок А представляет собой поверхностный белок 42 кДа, первоначально обнаруженный в клеточной стенке бактерий Staphylococcus aureus. Белок А широко используется для обнаружения, количественного определения и очистки иммуноглобулинов. Сообщалось, что белок А связывает часть Fab, полученную из антител семейства VH3, и гамма-область Fc в части константной области IgG (между доменами CH2 и CH3). Кристаллическая структура комплекса, образованного белком А и Fab, описана, например, в Graille et al., 2000, PNAS, 97(10): 5399-5404. В контексте настоящего изобретения белок А включает природный белок А и любой его вариант или производное в той мере, в какой вариант или производное белка А сохраняет свою способность связывать домены VH3 и/или гамма-домены Fc.
Полипептидная цепь формулы (I) согласно настоящему изобретению содержит домен связывания белка А. В одном варианте осуществления полипептидная цепь формулы (I) содержит один, два или три домена связывания белка А.
«Домен связывания белка А» в рамках настоящего изобретения относится к связывающему домену, который конкретно связывается с белком A. Домен связывания белка А может относиться к домену VH3 или участку домена VH3, который связывает белок A, то есть который содержит интерфейс связывания белка А. Участок домена VH3, который связывает белок А, не содержит CDR домена VH3, то есть интерфейс связывания белка А VH3 не включает в себя CDR; следовательно, будет понятно, что домен связывания белка А не конкурирует с антигенсвязывающим доменом, раскрытым в настоящей заявке.
В одном варианте осуществления полипептидная цепь формулы (I) содержит домен связывания белка А, который присутствует в VH и/или CH2-CH3 и/или V1. В одном варианте осуществления полипептидная цепь формулы (I) содержит один, два или три домена связывания белка А, который имеется/имеются в VH и/или CH2-CH3 и/или V1. В одном варианте осуществления полипептидная цепь формулы (I) содержит только один домен связывания белка А, который присутствует в VH или V1. В одном варианте осуществления s составляет 0, t составляет 0, а полипептидная цепь формулы (I) содержит только один домен связывания белка А, который присутствует в VH или V1. В одном варианте осуществления полипептидная цепь формулы (I) содержит только один домен связывания белка А, который присутствует в VH. В одном варианте осуществления s составляет 0, t составляет 0, p составляет 0, а полипептидная цепь формулы (I) содержит только один домен связывания белка А, который присутствует в VH. В одном варианте осуществления полипептидная цепь формулы (I) содержит только один домен связывания белка А, который присутствует в V1. В одном варианте осуществления s составляет 0, t составляет 0, p составляет 1, а полипептидная цепь формулы (I) содержит только один домен связывания белка А, который присутствует в V1.
В одном варианте осуществления полипептидная цепь формулы (I) содержит два домена связывания белка А. В одном варианте осуществления полипептидная цепь формулы (I) содержит два домена связывания белка А, которые имеются в VH и CH2-CH3, соответственно. В другом варианте осуществления полипептидная цепь формулы (I) содержит два домена связывания белка А, которые имеются в VH и V1, соответственно. В другом варианте осуществления полипептидная цепь формулы (I) содержит два домена связывания белка А, которые имеются в CH2-CH3 и V1, соответственно.
В одном варианте осуществления полипептидная цепь формулы (I) содержит три домена связывания белка А, каждый из которых присутствует в VH, CH2-CH3 и V1.
Природные белок А может взаимодействовать, в частности, с гамма-областью Fc, в части константной области IgG. Более конкретно, белок А может взаимодействовать со связывающим доменом между CH2 и CH3. В одном варианте осуществления, когда s составляет 1, t составляет 1, и CH2 и CH3 представляют собой природные домены класса IgG.
В некоторых вариантах осуществления домен (домены) связывания белка А содержит (содержат) или состоит (состоят) из домена VH3 или его варианта, который связывает белок A. В некоторых вариантах осуществления домен (домены) связывания белка А содержит (содержат) или состоит (состоят) из природного домена VH3. В некоторых вариантах осуществления вариант домена VH3, который связывает белок А, представляет собой вариант природного домена VH3, где указанный природный домен VH3 неспособен связывать белок A.
Полипептидная цепь формулы (II) согласно настоящему раскрытию не связывает белок A. В одном варианте осуществления связывающий домен V2 не связывает белок A.
В некоторых вариантах осуществления V2 содержит или состоит из VH1 и/или VH2, и/или VH4, и/или VH5, и/или VH6 и не содержит домен VH3. В некоторых вариантах осуществления V2 содержит или состоит из домена VH3 или его варианта, который не связывает белок A. В некоторых вариантах осуществления V2 содержит или состоит из природного домена VH3, не способного связывать белок A. В некоторых вариантах осуществления вариант домен VH3, который не связывает белок, представляет собой вариант природного VH3, где указанный природный домен VH3 способен связывать белок A.
Гены зародышевой линии VH3 человека и домены VH3 (или каркасы) хорошо охарактеризованы. Многие природные домены VH3 обладают способностью связывать белок А, но определенные природные домены VH3 не обладают способностью связывать белок (см. Roben et al. 1995, J Immunol.;154(12):6437-6445).
Домен VH3 для использования в настоящем изобретении может быть получен несколькими способами. В одном варианте осуществления домен VH3 для использования в настоящем изобретении представляет собой природный домен VH3, выбранный по его способности или неспособности связывать белок А, в зависимости от его положения в полипептиде (I) и/или (II) согласно настоящему изобретению. Например, может быть получена панель антител против представляющего интерес антигена путем иммунизации животного, отличного от человека, затем гуманизирована, а гуманизированные антитела могут быть подвергнуты скринингу и отобраны на основе их способности или неспособности связывать белок А через гуманизированный домен VH3, например, против колонки проведения хроматографии белка А по сродству. Альтернативно, технологии отображения (например, фаговый дисплей, дрожжевой дисплей, рибосомный дисплей, бактериальный дисплей, дисплей на клеточной поверхности млекопитающих, дисплей мРНК, дисплей ДНК) можно использовать для скрининга библиотек антител и отбора антител, содержащих домен VH3, который связывается, в частности, через интерфейс связывания белка А, который не содержит CDR или не связывает белок А.
Альтернативно, домен VH3 для использования в настоящем изобретении представляет собой вариант природного VH3. В одном варианте осуществления вариант VH3 содержит последовательность природного VH3, способного связывать белок А, и дополнительно содержит по меньшей мере одну аминокислотную мутацию, которая устраняет его способность связывать белок А. В одном варианте осуществления вариант VH3, который связывает белок А, содержит последовательность природного VH3, неспособного связывать белок А, и дополнительно содержит по меньшей мере одну аминокислотную мутацию. В таком варианте осуществления мутация (мутации) отвечает (отвечают) за приобретение доменом VH3 способности связывать белок А, то есть мутация (мутации) вносит вклад (вклады) в образование домена связывания белка А, который не существует в природе.
В одном варианте осуществления вариант VH3 содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 аминокислотных мутаций. В одном варианте осуществления вариант VH3 содержит мутацию в позиции 15, 17, 19, 57, 59, 64, 65, 66, 68, 70, 81 или 82 на VH3, нумерации в соответствии с Kabat и как описано например в Graille et al. 2000, PNAS, 97(10): 5399-5404). Мутацией может быть замена, делеция или вставка. В одном варианте осуществления вариант VH3 содержит замену в позиции 15, 17, 19, 57, 59, 64, 65, 66, 68, 70, 81 или 82 на VH3, нумерации в соответствии с Kabat.
Природный VH1, VH2, VH4, VH5 и VH6 не связывает белок A. В одном варианте осуществления домен VH, который не связывает белок А, представляет собой VH1. В одном варианте осуществления домен VH, который не связывает белок А, представляет собой VH2. В одном варианте осуществления домен VH, который не связывает белок А, представляет собой VH4. В одном варианте осуществления домен VH, который не связывает белок А, представляет собой VH5. В одном варианте осуществления домен VH, который не связывает белок А, представляет собой VH6.
Фармацевтические Композиции, Дозировки и Схемы лечения
мультиспецифическое антитело согласно изобретению может быть представлено в фармацевтической композиции. Фармацевтическая композиция обычно стерильна и обычно содержит фармацевтически приемлемый носитель и/или вспомогательное средство. Фармацевтическая композиция согласно настоящему изобретению может дополнительно содержать фармацевтически приемлемое вспомогательное средство и/или носитель.
Согласно настоящему изобретению «фармацевтически приемлемый носитель» включает в себя любой и все растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые средства, изотонические и замедляющие абсорбцию средства и тому подобное, которые являются физиологически совместимыми. Носитель может быть подходящим для парентерального, например, внутривенного, внутримышечного, внутрикожного, внутриглазного, внутрибрюшинного, подкожного, спинального или других парентеральных путей введения, например путем инъекции или инфузии. Альтернативно, носитель может быть подходящим для непарентерального введения, такого как местный, эпидермальный или чрезслизистый путь введения. Носитель может быть подходящим для перорального введения. В зависимости от пути введения модулятор может быть покрыт материалом для защиты соединения от действия кислот и других природных условий, которое могут инактивировать соединение.
Фармацевтические композиции согласно изобретению могут содержать одну или несколько фармацевтически приемлемых солей. «Фармацевтически приемлемая соль» относится к соли, которая сохраняет нужную биологическую активность исходного соединения и не придает никакого ненужного токсикологического эффекта. Примеры таких солей включают соли присоединения кислот и соли присоединения оснований.
Фармацевтически приемлемые носители включают в себя водные носители или разбавители. Примеры подходящих водных носителей, которое можно использовать в фармацевтической композиции согласно изобретению, включают воду, забуференную воду и солевой раствор. Примеры других носителей включают этанол, полиолы (такие как глицерин, пропиленгликоль, полиэтиленгликоль и тому подобное) и их подходящие смеси, растительные масла, такие как оливковое масло и инъецируемые органические сложные эфиры, такие как этилолеат. Во многих случаях будет желательно включить в композицию изотонические средства, например, сахары, полиспирты, такие как маннит, сорбит или хлорид натрия.
Терапевтические композиции обычно должны быть стерильными и стабильными в условиях изготовления и хранения. Композиция может быть составлена в виде раствора, микроэмульсии, липосомы или другой упорядоченной структуры, подходящей для высокой концентрации лекарственного средства.
Фармацевтическая композиции согласно изобретению может содержать дополнительные активные ингредиенты.
Также в объем настоящего изобретения входят наборы, содержащие антитела или модулирующие средства согласно изобретению и инструкция для применения. Набор может дополнительно содержать один или несколько дополнительных реагентов, таких как дополнительное терапевтическое или профилактическое средство, обсуждавшееся выше.
Модуляторы и/или антитела согласно изобретению или лекарственные формы или их композиции можно вводить для профилактического и/или терапевтического лечения.
В терапевтических целях соединения вводят субъекту, уже имеющему заболевание или состояние, как описано выше, в количестве, достаточном для исцеления, ослабления или частичного купирования состояния или одного или нескольких его симптомов. Такое терапевтическое лечение может приводить к снижению тяжести симптомов заболевания или увеличения частоты или продолжительности бессимптомных периодов. Количество, достаточное для достижения этого, определяется как «терапевтически эффективное количество».
В профилактических целях лекарственные формы вводят субъекту с риском заболевания или состояния, как описано выше, в количестве, достаточном для предотвращения или уменьшения последующих результатов состояния или одного или нескольких его симптомов. Количество, достаточное для достижения этого, определяется ка «профилактически эффективное количество». Эффективные количества для каждой цели будут зависеть от тяжести заболевания или поражения, а также от массы и общего состояния субъекта.
Субъектом для введения может быть человек или не являющееся человеком животное. Термин «не являющееся человеком животное» включает всех позвоночных, например, млекопитающих и не млекопитающих, таких как не являющиеся человеком приматы, овцы, собаки, кошки, лошади, коровы, куры, амфибии, рептилии и так далее. Типичным является введение людям.
Антитело/модулятор или фармацевтическую композицию согласно изобретению можно вводить одним или несколькими путями введения с использованием одного или нескольких из множества способов, известных в данной области. Как будет понятно специалистам, путь и/или способ введения будут варьироваться в зависимости от желаемых результатов. Примеры путей введения соединений или фармацевтических композиций согласно изобретению включают внутривенный, внутримышечный, внутрикожный, внутриглазной, внутрибрюшинный, подкожный, спинальный или другие парентеральные способы введения, например, путем инъекции или инфузии. Фраза «парентеральное введение» согласно настоящему изобретению означает способы введения, отличные от энтерального и местного введения, обычно путем инъекции. Альтернативно, антитело/модулирующее средство или фармацевтическую композицию согласно изобретению можно вводить непарентеральным путем, таким как местный, эпидермальный или чрезслизистый способ введения. Антитело/модулирующее средство или фармацевтическая композиция согласно изобретению могут быть предназначены для перорального введения.
Подходящая доза антитела/модулирующего средства или фармацевтической композиции согласно настоящему изобретению может быть определена квалифицированным практикующим врачом. Фактические уровни дозировки активных ингредиентов в фармацевтических композициях согласно настоящему изобретению могут варьироваться для получения количества активного ингредиента, эффективного для достижения желаемого терапевтического ответа для конкретного пациента, композиции и способа введения, без токсичности для пациента. Выбранный уровень дозы будет зависеть от множества фармакокинетических факторов, включая активность конкретных используемых композиций согласно настоящему изобретению, способ введения, время введения, скорость выведения конкретного используемого соединения, продолжительность лечения, другие лекарственные средства, соединения и/или материалы, используемые в сочетании с конкретными применяемыми композициями, возраст, пол, вес, состояние, общее состояние здоровья и предшествующая история болезни пациента, проходящего лечение, и подобные факторы, хорошо известные в области медицины.
Подходящая доза может находиться, например, в диапазоне от примерно 0,01 мкг/кг до примерно 1000 мг/кг массы тела, обычно от примерно 0,1 мкг/кг до примерно 100 мг/кг массы тела пациента, подлежащего лечению. Например, подходящая доза может составлять от примерно 1 мкг/кг до примерно 10 мг/кг массы тела в сутки или от примерно 10 мкг/кг до примерно 5 мг/кг массы тела в сутки.
Схемы лечения могут быть скорректированы для обеспечения оптимального желаемого ответа (например, терапевтического ответа). Например, можно вводить разовую дозу, можно вводить несколько разделенных доз в течение времени, или дозу можно пропорционально уменьшать или увеличивать в зависимости от остроты терапевтической ситуации. Единичная дозированная форма согласно настоящему изобретению относится к физически дискретным единицам, подходящим в качестве единичных доз для субъектов, подлежащих лечению; каждая единица содержит заранее определенное количество активного соединения, рассчитанное для получения желаемого терапевтического эффекта в сочетании с требуемым фармацевтическим носителем.
Введение может быть однократным или многократным. Многократные дозы можно вводить одним и тем же или разными путями и в одно и то же или разные места. В качестве альтернативы дозы можно вводить в виде препарата с замедленным высвобождением, и в этом случае требуется менее частое введение. Дозировку и частоту можно менять в зависимости от периода полувыведения антагониста у пациента и желаемой продолжительности лечения.
Как упоминалось выше, модуляторы/антитела или фармацевтические композиции согласно изобретению можно вводить совместно с одним или несколькими другими терапевтическими средствами.
Комбинированное введение двух или более средств может быть обеспечено рядом различных способов. Оба препарата можно вводить вместе в одной композиции, или их можно вводить в виде отдельных композиций как часть комбинированной терапии. Например, одно можно вводить до, после или одновременно с другим.
Терапевтические показания
Антитела согласно настоящему изобретению можно использовать для лечения, профилактики или облегчения любого состояния, связанного с активностью IL-13 и/или IL-17A и/или IL-17F; например, любого состояния, которое полностью или частично является результатом передачи сигнала через рецептор IL-13, IL-17A и/или IL-17F.
Заболевания включают такие заболевания как первичный и метастатический рак, в том числе карциному молочной железы, толстой кишки, прямой кишки, легких, ротоглотки, гортаноглотки, пищевода, желудка, поджелудочной железы, печени, желчного пузыря и желчных протоков, тонкой кишки, мочевыводящих путей (включая почки, мочевой пузырь и уротелий), женских половых путей (включая шейку матки, матку и яичники, а также хориокарциному и гестационную трофобластическую болезнь), мужских половых путей (включая предстательную железу, семенные пузырьки, яички и опухоли зародышевых клеток), железы внутренней секреции (включая щитовидную железу, надпочечники и гипофиз), и кожи, а также гемангиомы, меланомы, саркомы (в том числе возникающие из костей и мягких тканей, а также сарком Капоши), опухоли головного мозга, нервов, глаз и мозговых оболочек (в том числе астроцитомы, глиомы, глиобластомы, ретинобластомы, невромы, нейробластомы, шванномы и менингиомы), солидные опухоли, возникающие в результате злокачественных новообразований кроветворной системы, таких как лейкемии, и лимфомы (как ходжкинские, так и неходжкинские лимфомы), ревматоидный артрит, остеоартроз, ювенильный хронический артрит, септический артрит, артрит Лайма, псориатический артрит, реактивный артрит, спондилоартропатия, системная красная волчанка, язвенный колит, воспалительные заболевания кишечника, инсулинозависимый сахарный диабет, тиреоидит, аллергические заболевания, псориаз, склеродермический дерматит, трансплантат против хозяина, отторжение трансплантата органов, острое или хроническое иммунное заболевание, связанное с трансплантацией органов, саркоидоз, атеросклероз, диссеминированное внутрисосудистое свертывание крови, болезнь Кавасаки, болезнь Грейвса, нефротический синдром, синдром хронической усталости, гранулематоз Вегенера, пурпура Геноха-Шенлейна, микроскопический васкулит почек, хронический активный гепатит, увеит, септический шок, синдром токсического шока, септический синдром, кахексия, инфекционные заболевания, паразитарные заболевания, синдром приобретенного иммунодефицита, острый поперечный миелит, хорея Хантингтона, болезнь Паркинсона, болезнь Альцгеймера, инсульт, первичный билиарный цирроз, гемолитическая анемия, злокачественные новообразования, сердечная недостаточность, болезнь Аддисона, спорадическая, полигландулярная недостаточность I типа и полигландулярная недостаточность II типа, синдром Шмидта, острый респираторный дистресс-синдром во взрослом возрасте, алопеция, очаговая алопеция, артропатия, болезнь Рейтера, псориатическая артропатия, артропатия при язвенном колите, энтеропатический синовит, хламидиоз, артропатия, связанная с присутствием Yersinia и Salmonella, атероматозное заболевание/атеросклероз, атопическая аллергия, аутоиммунная буллезная болезнь, пузырчатка обыкновенная, листовидная пузырчатка, пемфигоид, болезнь линейного IgA, аутоиммунная гемолитическая анемия, гемолитическая анемия с положительной пробой Кумбса, приобретенная пернициозная анемия, ювенильная пернициозная анемия, миалгический энцефалит/Royal Free Disease, хронический кожно-слизистый кандидоз, гигантоклеточный артериит, первичный склерозирующий гепатит, криптогенный аутоиммунный гепатит, заболевания, связанные с приобретенным иммунодефицитом, гепатит В, гепатит С, неклассифицируемый вариабельный иммунодефицит неклассифицируемая вариабельная гипогаммаглобулинемия), дилатационная кардиомиопатия, женское бесплодие, угасание функции яичников, преждевременное угасание функции яичников, фиброзное заболевание легких, криптогенный фиброзирующий альвеолит, поствоспалительное интерстициальное заболевание легких, интерстициальный пневмонит, заболевание соединительной ткани, ассоциированное с интерстициальным заболеванием легких, смешанное заболевание соединительной ткани, ассоциированное с заболеванием легких, интерстициальное заболевание легких, связанное с системным склерозом, интерстициальное заболевание легких, связанное с ревматоидным артритом, заболевание легких, связанное с системной красной волчанкой, заболевание легких, связанное с дерматомиозитом/полимиозитом, заболевание легких, связанное с болезнью Шегрена, заболевание легких, связанное с анкилозирующим спондилитом, диффузное васкулитное заболевание легких, заболевание легких, связанное с гемосидерозом, медикаментозное интерстициальное заболевание легких, фиброз, лучевой фиброз, облитерирующий бронхиолит, хроническая эозинофильная пневмония, лимфоцитарно-инфильтративное заболевание легких, постинфекционное интерстициальное заболевание легких, подагрический артрит, аутоиммунный гепатит, аутоиммунный гепатит 1 типа (классический аутоиммунный или волчаночный гепатит), аутоиммунный гепатит 2 типа (гепатит с антителами против LKM), аутоиммунно-опосредованная гипогликемия, резистентность к инсулину типа В с акантокератодермией, гипопаратиреоз, острое иммунное заболевание, связанное с трансплантацией органов, хроническое иммунное заболевание, связанное с трансплантацией органов, остеоартроз, первичный склерозирующий холангит, псориаз 1 типа, псориаз 2 типа, идиопатическая лейкопения, аутоиммунная нейтропения, заболевания почек без дополнительного уточнения (БДУ), гломерулонефриты, микроскопический васулит почек, болезнь Лайма, дискоидная красная волчанка, идиопатическое мужское бесплодие или БДУ, аутоиммунитет спермы, рассеянный склероз (все подтипы), симпатическая офтальмия, легочная гипертензия, вторичная по отношению к заболеванию соединительной ткани, синдром Гудпасчера, легочные проявления узелкового полиартериита, острая ревматическая лихорадка, ревматоидный спондилит, болезнь Стилла, системный склероз, синдром Сьергрена, болезнь/артериит Такаясу, аутоиммунная тромбоцитопения, идиопатическая тромбоцитопения, аутоиммунное заболевание щитовидной железы, гипертиреоз, зобный аутоиммунный гипотиреоз (болезнь Хашимото), атрофический аутоиммунный гипотиреоз, первичная микседема, факогенный увеит, первичный васкулит, витилиго, острое заболевание печени, хронические заболевания печени, алкогольный цирроз, алкогольное поражение печени, холеосатит, идиосинкразическое заболевания печени, лекарственный гепатит, неалкогольный стеатогепатит, аллергия, инфекция стрептококков группы В (GBS), психические расстройства, депрессия, шизофрения, заболевания, опосредованные типом Th2 и Th1, острая и хроническая боль, различные формы боли, рак, рак легких, рак молочной железы, рак желудка, рак мочевого пузыря, рак толстой кишки, рак поджелудочной железы, рак яичников, рак предстательной железы, рак прямой кишки, злокачественные новообразования кроветворной системы, лейкемия, лимфома, абеталипопротеемия, акроцианоз, острые и хронические паразитарные или инфекционные процессы, острый лейкоз, острый лимфобластный лейкоз (ALL), острый миелоидный лейкоз (AML), острая или хроническая бактериальная инфекция, острый панкреатит, острая почечная недостаточность, аденокарциномы, предсердные экстрасистолы, комплекс СПИД-деменция, алкогольный гепатит, аллергический конъюнктивит, аллергический контактный дерматит, аллергический ринит (включая сезонный аллергический ринит), неаллергический ринит, отторжение аллотрансплантата, дефицит альфа-I-антитрипсина, боковой амиотрофический склероз, анемия, стенокардия, дегенерация клеток переднего рога спинного мозга, анти-CD3 терапия, антифосфолипидный синдром, реакции гиперчувствительности к антирецепторам, аневризмы аорты и периферических аневризм, расслоение аорты, артериальная гипертензия, артериосклероз, артериовенозная фистула, атаксия, фибрилляция предсердий (устойчивая или пароксизмальная), трепетание предсердий, атриовентрикулярная блокада, В-клеточная лимфома, отторжение костного трансплантата, отторжение трансплантата костного мозга (BMT), блокада ножек пучка Гиса, лимфома Беркитта, ожоги, нарушения сердечного ритма, синдром оглушенного миокарда, опухоли сердца, кардиомиопатия, воспалительная реакция на сердечнолегочное шунтирование, отторжение хрящевого трансплантата, дегенерация коры мозжечка, мозжечковые расстройства, хаотическая или мультифокальная предсердная тахикардия, заболевания, связанные с химиотерапией, хронический миелоцитарный лейкоз (CML), хронический алкоголизм, хронические воспалительные патологии, хронический лимфолейкоз (CLL), хроническая обструктивная болезнь легких (COPD), хроническая интоксикация салицилатами, колоректальная карцинома, застойная сердечная недостаточность, конъюнктивит, контактный дерматит, легочное сердце, ишемическая болезнь сердца, болезнь Крейтцфельдта-Якоба, сепсис с отрицательным результатом посева, кистозный фиброз, расстройства, связанные с цитокиновой терапией, деменция боксеров, демиелинизирующие заболевания, геморрагическая лихорадка денге, дерматит, дерматологические заболевания, диабет, сахарный диабет, диабетическая атеросклеротическая болезнь, диффузная болезнь с тельцами Леви, дилатационная застойная кардиомиопатия, поражение базальных ганглиев, синдром Дауна в среднем возрасте, лекарственные двигательные расстройства, вызванные препаратами, блокирующими дофаминовые рецепторы ЦНС, чувствительность к лекарственным препаратам, экзема, энцефаломиелит, эндокардит, эндокринопатия, эпиглоттит, вирусная инфекция Эпштейна-Барр, эритромелалгия, экстрапирамидные и мозжечковые расстройства, семейный гематофагоцитарный лимфогистиоцитоз, отторжение имплантата эмбрионального тимуса, атаксия Фридрейха, функциональные нарушения периферических артерий, грибковый сепсис, газовая гангрена, язва желудка, гломерулярный нефрит, отторжение трансплантата любого органа или ткани, грамотрицательный сепсис, грамположительный сепсис, гранулемы, вызванные внутриклеточными микроорганизмами, волосатоклеточный лейкоз, болезнь Галлервордена-Шпатца, тиреоидит Хашимото, сенная лихорадка, отторжение трансплантата сердца, гемахроматоз, гемодиализ, гемолитико-уремический синдром/тромболитическая тромбоцитопеническая пурпура, геморрагия, гепатит А, аритмии пучка Гиса, ВИЧ-инфекция/ВИЧ-нейропатия, болезнь Ходжкина, гиперкинетические двигательные расстройства, реакции гиперчувствительности, гиперчувствительный пневмонит, гипертония, гипокинетические двигательные расстройства, оценка гипоталамо-гипофизарно-надпочечниковой оси, идиопатическая болезнь Аддисона, идиопатический фиброз легких, цитотоксичность, опосредованная антителами, астения, детская спинальная мышечная атрофия, воспаление аорты, грипп А, воздействие ионизирующего излучения, иридоциклит/увеит/неврит зрительного нерва, ишемически-реперфузионное повреждение, ишемический инсульт, ювенильный ревматоидный артрит, ювенильная спинальная мышечная атрофия , саркома Капоши, отторжение почечного трансплантата, легионеллез, лейшманиоз, проказа, поражение кортикоспинальной системы, жировой отек, отторжение трансплантата печени, лимфедема, малярия, злокачественная лимфома, злокачественный гистиоцитоз, злокачественная меланома, менингит, менингококкемия, мигрень при метаболическом синдроме, идиопатическая мигрень, митохондриальное мультисистемное нарушение, смешанное заболевание соединительной ткани, моноклональная гаммапатия, множественная миелома, множественные системные дегенерации (Мензеля, Дежерина-Томаса, Шай-Драгера и Мачадо-Джозефа), инфицирование Mycobacterium avium intracellulare, инфицирование Mycobacterium tuberculosis, миелодипластический синдром, инфаркт миокарда, ишемические нарушения миокарда, карцинома носоглотки, хронические заболевания легких у новорожденных, нефрит, нефроз, нейродегенеративные заболевания, нейрогенные мышечные атрофии, нейтропеническая лихорадка, неходжкинская лимфома, окклюзия брюшной аорты и ее ветвей, окклюзионные поражения артерий, терапия okt3, орхит/эпидидимит, орхит/процедуры реверсивной вазэктомии, органомегалия, остеопороз, отторжение трансплантата поджелудочной железы, карцинома поджелудочной железы, паранеопластический синдром/злокачественная гиперкальциемия, отторжение трансплантата паращитовидной железы, воспалительные заболевания органов малого таза, круглогодичный ринит, заболевание перикарда, периферическое атеросклеротическое заболевание, заболевания периферических сосудов, перитонит, пернициозная анемия, пневмоцистная пневмония, пневмония, синдром POEMS (синдром полинейропатии, органомегалии, эндокринопатии, моноклональной гаммапатии и изменения кожи), постперфузионный синдром, постгемодиализный синдром, посткардиотомный синдром после инфаркта миокарда, преэклампсия, прогрессирующий надъядерный паралич, первичная легочная гипертензия, лучевая терапия, феномен и болезнь Рейно, болезнь Рейно, болезнь Рефсума, тахикардия с регулярными узкими комплексами QRS, реноваскулярная гипертензия, реперфузионное поражение, рестриктивная кардиомиопатия, саркомы, сенильная хорея, сенильная деменция с тельцами Леви, серонегативные артропатии, шок, серповидно-клеточная анемия, отторжение кожного аллотрансплантата, синдром кожных изменений, отторжение трансплантата тонкой кишки, солидные опухоли, специфические аритмии, спинальная атаксия, спиноцеребеллярные дегенерации, стрептококковый миозит, структурные поражения мозжечка, подострый склерозирующий панэнцефалит, обмороки, сифилис сердечно-сосудистой системы, системная анафалактика, синдром системной воспалительной реакции, ювенильный идиопатический артрит с системным началом, Т-клеточная или FAB ALL телеангиэктазия, облитерирующий тромбангиит, тромбоцитопения, интоксикация, трансплантаты, травма/кровоизлияние, реакции гиперчувствительности III типа, гиперчувствительность IV типа, нестабильная стенокардия, уремия, уросепсис, пороки клапанов сердца, варикозное расширение вен, васкулит венозные заболевания, венозный тромбоз, фибрилляция желудочков, вирусные и грибковые инфекции, вирусный энцефалит/асептический менингит, вирус-ассоциированный гемафагоцитарный синдром, синдром Вернике-Корсакова, болезнь Вильсона, отторжение ксенотрансплантата какого-либо органа или ткани, острые коронарные синдромы, острый идиопатический полиневрит, острая воспалительная демиелинизирующая полирадикулоневропатия, острая ишемия, болезнь Стилла во взрослом возрасте, анафилаксия, синдром антифосфолипидных антител, апластическая анемия, атопическая экзема, атопический дерматит, аутоиммунный дерматит, аутоиммунное заболевание, связанное со стрептококковой инфекцией, аутоиммунная энтеропатия, аутоиммунная потеря слуха, аутоиммунный лимфопролиферативный синдром (ALPS), аутоиммунный миокардит, аутоиммунное преждевременное угасание функции яичников, блефарит, бронхоэктазы, буллезный пемфигоид, сердечно-сосудистые заболевания, катастрофический антифосфолипидный синдром, глютеновая болезнь, шейный спондилез, хроническая ишемия, рубцовый пемфигоид, клинически изолированный синдром (cis) с риском рассеянного склероза, начало психического расстройства в детском возрасте, дакриоцистит, дерматомиозит, диабетическая ретинопатия, грыжи межпозвоночных дисков, пролапс диска, лекарственная иммунная гемолитическая анемия, эндометриоз, эндофтальмит, эписклерит, многоформная эритема, большая форма мультиформной эритемы, гестационный пемфигоид, синдром Гийена-Барре (GBS), синдром Хьюза, идиопатическая болезнь Паркинсона, идиопатическая интерстициальная пневмония, IgE-опосредованная аллергия, иммунная гемолитическая анемия, миозит с тельцами включения, инфекционное воспалительное заболевание глаз, воспалительное демиелинизирующее заболевание, воспалительное заболевание сердца, воспалительное заболевание почек, IPF/UIP, ирит, кератит, сухой кератоюнтивит, болезнь Куссмауля или болезнь Куссмауля-Мейера, паралич Ландри, гистиоцитоз из клеток Лангерганса, сетчатое ливедо, дегенерация желтого пятна, микроскопический полиангиит, болезнь Бехтерева, поражение двигательных нейронов, пемфигоид слизистых оболочек, полиорганная недостаточность, тяжелая миастения, миелодиспластический синдром, миокардит, поражение нервных корешков, нейропатия, гепатит ни-А-ни-В, неврит зрительного нерва, остеолиз, полиартикулярный ювенильный ревматоидный артрит JRA, окклюзионная болезнь периферических артерий (PAOD), заболевание периферических сосудов (PVD), заболевание периферических артерий (PAD), флебит, узелковый полиартериит (или узелковый периартериит), полихондрит, полиоз, полиартикулярный JRA, синдром полиэндокринной недостаточности, полимиозит, ревматическая полимиалгия (PMR), первичный паркинсонизм, простатит, чистая эритроцитарная аплазия, первичная недостаточность надпочечников, рецидивирующий нейромиелит зрительного нерва, рестеноз, ревмокардит, sapho (синовит, акне, пустулез, гиперостоз и остеит), вторичный амилоидоз, шоковое легкое, склерит, радикулит, вторичная недостаточность надпочечников, заболевания соединительной ткани, связанные с силиконом, дерматоз Снеддона-Уилкинсона, анкилозирующий спондилит, синдром Стивенса-Джонсона (SJS), височный артериит, токсоплазматический ретинит, токсический эпидермальный некролиз, поперечный миелит, TRAPS (рецептор фактора некроза опухоли), аллергическая реакция 1 типа, диабет 2 типа, крапивница, обычная интерстициальная пневмония (UIP), васкулит, весенний конъюнктивит, вирусный ретинит, синдром Фогта-Коянаги-Харада (синдром VKH), влажная дегенерация желтого пятна, заживление ран, аспирин-чувствительная астма, атопическая астма, хроническая экзема рук, аллергический бронхолегочный аспергиллез, глютеновая болезнь, синдром Чарга-Стросса (узелковый периартериит плюс атопия), синдром эозинофильной миалгии, гиперэозинофильный синдром, отечные реакции, включая эпизодический ангионевротический отек, гельминтозы, онхоцеркальный дерматит, эозинофил-ассоциированные заболевания желудочно-кишечного тракта, эозинофильный эзофагит, эозинофильный гастрит, эозинофильный гастроэнтерит, эозинофильный энтерит, эозинофильный колит, микрополипоз и полипоз носа, пищевая аллергия, непереносимость аспирина и обструктивное апноэ во сне, хроническая астма, болезнь Крона и эндомиокардиальный фиброз, рак (например, глиобластома (например, мультиформная глиобластома), неходжкинская лимфома (NHL)), фиброз, воспалительное заболевание кишечника, легочный фиброз (включая идиопатический легочный фиброз (IPF) и легочный фиброз, вторичный по отношению к склерозу), COPD и печеночный фиброз.
Мультиспецифические антитела согласно настоящему изобретению могут быть особенно полезные для лечения или профилактики атопического дерматита, хронической экземы рук, микрополипоза или полипоза носа, пищевой аллергии или эозинофильного эзофагита. Таким образом, в одном варианте осуществления мультиспецифическое антитело или фармацевтическая композиция согласно настоящему изобретению представлена для использования в способе лечения человека или животного путем терапии. В одном варианте осуществления мультиспецифическое антитело или фармацевтическая композиция представлена для использования в способе лечения атопического дерматита, хронической экземы рук, микрополипоза или полипоза носа, пищевой аллергии или эозинофильного эзофагита. В одном варианте осуществления в изобретении представлен способ лечения или профилактики атопического дерматита, хронической экземы рук, микрополипоза или полипоза носа, пищевой аллергии или эозинофильного эзофагита, включающий введение пациенту терапевтически эффективного количества мультиспецифического антитела или фармацевтической композиции.
Следующий Примеры иллюстрируют изобретение.
ПРИМЕРЫ
ПРИМЕР 1. Получение и отбор терапевтического антитела CA650 против IL-13
Крыс иммунизировали либо очищенным IL-13 человека (Peprotech), либо крысиными фибробластами, экспрессирующими IL-13 человека (экспрессирующими примерно 1 мкг/мл в супернатанте клеточной культуры), либо в некоторых случаях их комбинацией. После 3-6 инъекций животных умерщвляли, и собирали РВМС, селезенку, костный мозг и лимфатические узлы. Отслеживали связывание сыворотки с IL-13 человека в ELISA, а также способность нейтрализовать hIL-13 в анализе репортерных клеток HEK-293 IL-13R-STAT-6 (анализ HEK-Blue, Invivogen).
Получали культуры В-клеток, и супернатанты сначала подвергали скринингу на их способность связывать hIL-13 в анализе на основе шариков в анализе FMAT Applied Biosystems. Это был гомогенный анализ с использованием биотинилированного IL-13 человека, нанесенного на шарики стрептавидина, и конъюгата козьего антикрысиного Fc-Cy5 в качестве средства для выявления. Положительные результаты этого анализа затем переносили в анализ репортерных клеток HEK-293 IL-13R-STAT-6 (анализ HEK-Blue, Invivogen) для идентификации нейтрализаторов. Нейтрализующие супернатанты затем анализировали в Biacore для оценки степени диссоциации, а также для характеристики механизма действия нейтрализации. Нейтрализация была классифицирована либо как группа 1, либо как группа 2. Группа 1 представляет собой антитело, которое связывается с человеческим IL-13 и предотвращает связывание IL-13Rα1, и в результате также блокирует связывание IL-4R. Антитела группы 1 также могут предотвращать связывание IL-13 с IL-13Rα2. Группа 2 представляет собой антитело, которое связывает hIL-13 таким образом, что позволяет связываться с IL-13Rα1, но предотвращает рекрутирование IL-4R в комплекс. Авторы выбирали антитела, которые действовали в группе 1.
Примерно 7500 специфических к IL-13 положительных результатов были идентифицированы при первичном скрининге FMAT из 27 экспериментов SLAM на 100 планшетах. 800 лунок продемонстрировали нейтрализацию в анализе HEK-blue. 170 лунок имели желаемые профили Biacore, то есть антитела группы 1 со скоростью диссоциации < 5x 10-4 с-1. Была предпринята попытка клонирования вариабельной области из этих 170 лунок, и в 160 успешно были получены флуоресцирующие фокусы. В 100 лунках были получены пары генов вариабельной области тяжелой и легкой цепи после обратной транскрипции (RT)-PCR. Эти гены V-области клонировали в виде полноразмерных антител мышиного IgG1 и повторно экспрессировали в системе временной экспрессии HEK-293. Анализ последовательности показал, что существует 27 уникальных семейств антител против IL-13 человека. Затем эти рекомбинантные антитела повторно тестировали на их способность блокировать рекомбинантный hIL-13 (полученный из E.coli и полученный от млекопитающих), рекомбинантный вариант hIL-13 (R130Q) (полученный из E.coli), природный дикий тип и вариантный hIL-13. 13 (полученный от донора человека) и IL-13 яванского макака (полученный от млекопитающего) в клеточном анализе. Рекомбинантные антитела также тестировали на их способность связывать вариантный IL-13 человека (R130Q) и IL-13 яванского макака в Biacore. После получения этой характеристики были отобраны семейства антител, отвечающие критериям авторов, то есть антитела менее 100 пМ с минимальным снижением эффективности и сродства для всех препаратов IL-13 человека и яванского макака.
На основании нейтрализующей активности, сродства и содержания донора в гуманизированных трансплантатах (см. Ниже) для дальнейшей разработки был выбран гуманизированный СА650.
ПРИМЕР 2. Гуманизация Антитела CA650
Антитело 650 гуманизировали путем прививки CDR из V-области крысы на каркасы V-области зародышевого антитела человека. Для восстановления активности антитела в гуманизированной последовательности также сохраняли ряд каркасных остатков из V-области крысы. Эти остатки были отобраны с использованием протокола, описанного Adair et al. (1991) (Humanised antibodies. WO91/09967). Выравнивания последовательностей V-области крысиного антитела (донора) с последовательностями V-области зародышевой линии человека (акцептора) показаны на фиг. 1 вместе со сконструированными гуманизированными последовательностями. (Фиг.1(A) трансплантат легкой цепи 650 и Фиг.1(B) трансплантат тяжелой цепи 650). CDR, привитые от донора к акцепторной последовательности, соответствуют определениям Kabat (Kabat et al., 1987), за исключением CDR-H1, где использовано комбинированное определение Chothia/Kabat (см. Adair et al., 1991 Humanised antibodies. WO91/0996).
Гены, кодирующие исходные последовательности V-области, были спроектированы и сконструированы с помощью подхода автоматизированного синтеза Entelechon GmbH и модифицированы для получения привитых версий gL8 и gH9 с помощью мутагенеза, направленного на олигонуклеотиды. Последовательность gL8 субклонировали в вектор экспрессии pVhCK легкой цепи человека UCB Celltech, который содержит ДНК, кодирующую константную область C-Kappa человека (аллотип Km3). Последовательность gH9 субклонировали в pVhg1Fab, который содержит ДНК, кодирующую константную область тяжелой цепи гамма-1 CH1 человека.
V-область IGKV1-39 плюс J-область JK2 человека (International Immunogenetics Information System® IMGT, http://www.imgt.org) выбрали в качестве акцептора для CDR легкой цепи антитела 650. Каркасные остатки легкой цепи в трансплантате gL8 все взяты из гена зародышевой линии человека, за исключением остатков 58 и 71 (нумерация по Kabat), где были сохранены донорные остатки изолейцина (I58) и тирозина (Y71) соответственно. Сохранение остатков I58 и Y71 было необходимо для полной эффективности гуманизированного антитела.
V-область IGHV1-69 плюс J-область JH4 человека (IMGT, http://www.imgt.org) была выбрана в качестве акцептора для CDR тяжелой цепи антитела 650. Все каркасные остатки тяжелой цепи в трансплантатах gH9 взяты из гена зародышевой линии человека за исключением остатков 67, 69 и 71 (нумерация по Kabat), в которых были сохранены донорные остатки аланина (A67), фенилаланина (F69) и валина (V71) соответственно. Сохранение остатков A67, F69 и V71 необходимо для полной активности гуманизированного антитела. Остаток глутамина в позиии 1 каркаса человека был заменен глутаминовой кислотой (Е1), чтобы обеспечить экспрессию и очистку гомогенного продукта: широко сообщается о превращении глутамина в пироглутамат на N-конце антител и фрагментов антител. Окончательно выбранные вариабельные последовательности трансплантата gL8 и gH9 показаны на фиг.1(А) и фиг.1(В), соответственно.
Последовательности аминокислот и ДНК, кодирующие CDR, тяжелые и легкие вариабельные области, форматы scFv и dsscFV антитела 650, показаны на фиг. 2.
ПРИМЕР 3. Получение антитела 496.g3 против IL-17AF
Получение антитела CA028_00496.g3 (также называемого в данном документе антителом 496.g3) против IL-17A человека и IL-17F человека ранее было описано в WO 2012/095662. Антитело связывает IL-17A, IL-17F человека и гетеродимер IL-17A/F с pM-сродством. Аминокислотные и ДНК-последовательности, кодирующие CDR, тяжелые и легкие вариабельные области, а также легкие и тяжелые цепи Fab-формата антитела 496.g3, показаны на фиг. 2. Константные области Fab 496.g3 (связывающие IL-17A/F) содержали константную область C-каппа человека (аллотип K1m3) и константную область гамма-1 CH1 человека и шарнир (аллотип G1m17).
ПРИМЕР 4. Получение of против человек альбумин антитело 645
Получение антитела 645 против альбумина человека было ранее описано в WO 2013/068571. Последовательности аминокислот и ДНК, кодирующие CDR, вариабельные области тяжелой и легкой цепей, форматы scFv и dsscFV антитела 645, показаны на фиг. 2.
ПРИМЕР 5. Мультиспецифическое антитело IL-13/IL-17AF - конструирование транзиентных плазмид и экспрессия в клетках
Были сконструированы мультиспецифические антитела с V-областью анти-IL-17AF (496.g3), фиксированной в положении Fab; V-область антиальбумина (645gL4gH5) и IL-13 (1539gL8gH9) переформатировали в дисульфид-связанный scFv в ориентации HL (dsHL) и присоединили к С-концам соответствующих константных областей тяжелой и легкой цепей Fab через 11-аминокислотные линкеры, богатые глицином и серином. (фиг. 7). Последовательности аминокислот и ДНК, кодирующие полноразмерную тяжелую цепь и легкую цепь мультиспецифического антитела, показаны на фиг. 2.
Гены легкой цепи и тяжелой цепи независимо клонировали в векторы экспрессии млекопитающих для временной экспрессии под управлением промотора hCMV. Обе плазмиды в равных соотношениях трансфицировали в клеточную линию CHO-S XE (UCB) с использованием коммерческого набора для транзиентной экспрессии ExpiFectamine ExpiCHO (Thermo Scientific). Культуры инкубировали в роллерных флаконах Corning с вентилируемыми крышками при 37°C, 8,0% CO2, 190 об/мин. Через 18-22 часа в культуры подавали соответствующие объемы энхансера CHO и питательные вещества для метода HiTiter, как это предусмотрено производителем. Культуры реинкубировали при 32°C, 8,0% CO2, 190 об/мин в течение дополнительных 10-12 дней. Супернатант собирали посредством центрифугирования при 4000 об/мин в течение 1 ч при 4°C перед стерилизацией фильтрованием через фильтр 0,45 мкм, а затем через фильтр 0,2 мкм. Титры экспрессии количественно определяли с помощью Protein G HPLC с использованием колонки GE HiTrap Protein G объемом 1 мл (GE Healthcare) и своих стандартов Fab. Титр экспрессии показан в таблице 1.
Таблица 1: Титры экспрессия из транзиентной экспрессии в клеточной линии S-XE CHO
ПРИМЕР 6. Мультиспецифическое антитело IL-13/IL-17AF - разработка клеточной линии млекопитающих.
Для демонстрации стабильной экспрессии мультиспецифического антитела IL-13/IL-17AF была создана стабильно экспрессирующая клеточная линия млекопитающих. Линию клеток CHO трансфицировали вектором, содержащим 496.g3 Fab, 1539gH9gL8 dsscFv HL (LC, INS0025609), 645gH5gL4 dsscFv HL (HC, INS0025306) и селективный маркер. Линии клеток клонировали и оценивали на соответствие подходящему производственному процессу. Для оценки качества и количества белка и обеспечения выбора оптимальной клеточной линии клеточную линию оценивали в мелкомасштабной модели производственного биореактора с периодической подачей. Выбирали клеточную линию CHO, экспрессирующую мультиспецифические антитела IL-13/IL-17AF в количестве более 1,8 г/л и более 75% мономера.
ПРИМЕР 7. Способ очистки мультиспецифического антитела IL-13/IL-17AF
Белок мультиспецифического антитела очищали на стадии захвата нативного белка А с последующей завершающей стадией препаративной эксклюзионной хроматографии. Очищенные супернатанты после стандартной транзиентной экспрессии CHO загружали в колонку MabSelect (GE Healthcare) с временем контакта 5 минут и промывали буфером для связывания (20 мМ Hepes pH 7,4+150 мМ NaCl). Связанный материал элюировали с помощью ступенчатого элюирования 0,1 М цитрата натрия, рН 3,1, и нейтрализовали 2 М Трис/HCl, рН 8,5, и количественно определяли по поглощению при 280 нм.
Эксклюзионную хроматографию (SE-UPLC) использовали для определения степени чистоты элюированного продукта. Антитело (~2 мкг) загружали в колонку BEH200, 200 Å, 1,7 мкм, ID 4,6 мм x 300 мм (Waters ACQUITY) и хроматографировали с изократическим градиентом 0,2 М фосфата, рН 7, при 0,35 мл/мин. Непрерывное обнаружение осуществляли по поглощению при 280 нм и многоканальному флуоресцентному (FLR) детектору (Waters). Было обнаружено, что элюированное мультиспецифическое антитело представляет собой 72% мономера.
Нейтрализованные образцы концентрировали с использованием концентратора Amicon Ultra-15 (мембрана с отсечкой по молекулярной массе 10 кДа) и центрифугировали при 4000×g в бакет-роторе. Концентрированные образцы наносили на колонку XK16/60 Superdex200 (GE Healthcare), уравновешенную в PBS, pH 7,4, и хроматографировали с изократическим градиентом PBS, pH 7,4, со скоростью 1 мл/мин. Фракции собирали и анализировали с помощью эксклюзионной хроматографии на колонке BEH200, 200Å, 1,7 мкм, 4,6 мм ID x 300 мм (Aquity) и хроматографировали с изократическим градиентом 0,2 М фосфата, pH 7, при 0,35 мл/мин с детектированием попоглощению при 280 нм и с помощью многоканального флуоресцентного (FLR) детектора (Waters). Выбранные мономерные фракции объединяли, подвергали стерильной фильтрации с размером пор 0,22 мкм, а конечные образцы анализировали на концентрацию с помощью сканирования A280 на DropSense96 (Trinean). Уровень эндотоксина был менее 1,0 ЕЭ/мг по оценке с помощью портативной тестовой системы EndoSafe® от Charles River с тестовыми картриджами Limulus Amebocyte Lysate (LAL).
Состояние мономера конечного мультиспецифического антитела определяли эксклюзионной хроматографией на колонке BEH200, 200 Å, 1,7 мкм, 4,6 мм ID x 300 мм (Aquity) и хроматографировали с изократическим градиентом 0,2 М фосфата, рН 7 при 0,35 мл/мин с детектированием по поглощению при 280 нм и с помощью многоканального флуоресцентного (FLR) детектора (Waters). Было обнаружено, что конечное мультиспецифическое антитело является >99% мономерным, как показано на фиг. 3(A).
Для анализа с помощью электрофореза в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE) образцы готовили путем добавления буфера для образцов 4х Novex NuPAGE LDS (Life Technologies) и либо средства для восстановления образца 10х NuPAGE (Life Technologies), либо 100 мМ N-этилмалеимида (Sigma-Aldrich) до ~ 5 мкг очищенного белка и нагревали до 100°С в течение 3 мин. Образцы загружали в 10-луночный планшет с Novex 4-20% Трис-глицин 1,0 мМ SDS-полиакриламидного геля (Life Technologies) и разделяли при постоянном напряжении 225 В в течение 40 мин в рабочем буфере Трис-глицин SDS (Life Technologies). В качестве стандартов использовали стандарты белков широкого диапазона Novex Mark12 (Life Technologies). Гель окрашивали красителем Coomassie Quick Stain (Generon) и обесцвечивали в дистиллированной воде.
При невосстанавливающем SDS-PAGE мультиспецифическое антитело с теоретической молекулярной массой (MW) ~100 кДа мигрировало до ~120 кДа. Когда белок мультиспецифического антитела был восстановлен, обе цепи мигрировали со скоростью подвижности, приближающейся к их соответствующим теоретическим MW, тяжелой цепи (HC) ~ 52 кДа и легкой цепи (LC) ~ 51 кДа. Дополнительные полосы на невосстановленном геле при ~45-50 кДа представляют собой «свободные» LC и HC без дисульфидной связи в части Fab молекулы, они не мигрируют в то же положение, что и LC и HC на полосе 2, так как они полностью не восстановены. (фиг. 3(B)).
ПРИМЕР 8. Связывание Антигена молекулой мультиспецифического антитела IL-13/IL-17AF.
(i) Сродство Связывания Антигена
Кинетику связывания IL-13, IL-17A, AF, F и альбумина человека и яванского макака оценивали с помощью поверхностного плазмонного резонанса (Biacore T200).
Козье антитело против IgG человека, специфичное к фрагменту F(ab’)2 (Jackson ImmunoResearch), иммобилизовали на сенсорном чипе CM5 с помощью химического связывания амина до уровня приблизительно 5000 RU. Каждый цикл анализа состоял из захвата молекулы мультиспецифического антитела IL-13/IL-17AF/альбумина на поверхность анти-F(ab')2, введения аналита (при 25°C со скоростью потока 30 мкл в минуту) с последующей регенерацией поверхности. Аналиты человека и яванского макака вводили в 2-кратных серийных разведениях в рабочем буфере HBS-EP+ (GE Healthcare) в концентрациях от 10 нМ до 0,3125 нМ для IL-13, от 5 нМ до 0,156 нМ для IL-17A, AF, F и от 100 нМ до 3,125. НМ для альбумина. Антигены были приготовлены собственными силами за исключением IL-13 человека (R&D Systems), IL-13 яванского макака (Sinobiologicals) и сывороточного альбумина человека (Jackson ImmunoResearch). Ввод холостой пробы буфера был предусмотрен, чтобы вычесть шум прибора и дрейф.
Кинетические параметры определяли с использованием модели связывания 1:1 с использованием программного обеспечения Biacore T200 Evaluation (версия 3.0) и суммировали в таблицах 2(1) и 2(2). Если скорость диссоциации (KD) была измерена как менее 1,0×10-5, она была зафиксирована на уровне 1,0×10-5 (предел обнаружения прибора Biacore T200, определенный производителем GE Healthcare) для расчета сродства (KD).
Результаты показали, что мультиспецифическое антитело эффективно связывалось с IL-17A, IL-17AF, IL-17F, IL-13 и альбумином человека и яванского макака.
Таблица 2(1): Кинетические Константы связывания мультиспецифического антитела IL-13/IL-17AF с антигенами человека
Результаты отображают среднее значение из указанного количества определений.
Таблица 2(2): Кинетические Константы связывания мультиспецифического антитела IL-13/IL-17AF с антигенами яванского макака.
Результаты отображают среднее значение из указанного количества определений.
(ii) Одновременное связывание антигенов
Поверхностный плазмонный резонанс использовали для демонстрации того, что IL-17A, IL-13 и альбумин могут одновременно связываться с мультиспецифическим антителом IL-13/IL-17AF с использованием форм белков человека или яванского макака.
Формат метода для оценки одновременного связывания аналитов с мультиспецифическим антителом IL-13/IL-17AF заключался в захвате образца антитела иммобилизованным антителом, специфичным к фрагменту F(ab’)2 IgG человека. IL-13, IL-17A и альбумин человека или яванского макака затем вводили по отдельности или в смешанном растворе всех трех аналитов (по 30 мкл/мин в течение 300 с) поверх захваченного мультиспецифического антитела IL-13/IL-17AF (конечная концентрации 30 нМ IL-13, 15 нМ IL-17А, 150 нМ альбумина).
В конце каждого цикла поверхность регенерировали при скорости потока 10 мкл/мин, используя введение в течение 60 с 50 мМ HCl, затем введение в течение 30 с 5 мМ NaOH и итоговое введение в течение 60 с 50 мМ HCl.
Определяли реакцию связывания каждого антигена при инъекции по отдельности, и сумму индивидуальных ответов сравнивали с реакцией связывания при инъекции смеси всех трех антигенов.
Средний ответ связывания смеси IL-17A, IL-13 и альбумина человека с мультиспецифическим антителом IL-13/IL-17AF составлял 100% от суммы индивидуальных ответов на связывание (суммировано в таблице 3), что указывает на то, что мультиспецифическое антитело IL-13/IL-17AF было способно одновременно и независимо связывать каждый антиген.
Таблица 3: Одновременное Связывание мультиспецифического антитела IL-13/IL-17AF с IL-17A, IL-13 и Альбумином человека
Среднее значение ответа на связывание смеси IL-17A яванского макака, IL-13 и альбумина с мультиспецифическим антителом IL-13/IL-17AF составляло 97% суммы индивидуальных ответов на связывание (приведено в таблице 4), что указывает, что мультиспецифическое антитело IL-13/IL-17AF способно одновременно и независимо связывать каждый антиген.
Таблица 4: Одновременное Связывание мультиспецифического антитела IL-13/IL-17AF с IL-17A, IL-13 и Альбумином яванского макака
ПРИМЕР 9. Нейтрализация IL-13 с помощью мультиспецифического антитела IL-13/IL-17AF.
Активность мультиспецифического антитела IL-13/IL-17AF по нейтрализации IL-13 оценивали с использованием анализа репортерной клеточной линии HEK 293 Human IL4/IL-13 SEAP. Секрецию SEAP измеряли при активации пути STAT6 для оценки ответов IL-13.
Репортерные клетки IL4/IL-13 SEAP человека HEK 293 (# hkb-il413) были получены от Invivogen, San Diego. Культивирование, замораживание и обслуживание клеточной линии осуществляли в соответствии с протоколом производителя.
Рекомбинантный человеческий IL-13 был получен от R&D Systems, Minneapolis, MN. (# 213-ILB)
Клетки высевали таким образом, чтобы при стимуляции в 96-луночных планшетах с плоским дном клетки были примерно на 80% конфлюэнтны.
Клетки обрабатывали в двух повторностях антителами, предварительно инкубированными с 250 пг/мл IL-13 в течение 30 минут при 37°C, 5% CO2, 100% влажности перед добавлением к клеткам на 24 часа.
Через 24 часа пипеткой отбирали 20 мкл супернатанта после стимуляции клеток и добавляли к 180 мкл Quanti-blue #rep-qbs (Invivogen, San Diego) в соответствии с инструкциями производителя.
Анализ проводили до тех пор, пока не появлялся видимый градиент изменения цвета, и считывали оптическую плотность с использованием спектрофотометра при 620 нм. IC50 рассчитывали методом нелинейной регрессии с использованием призмы Graphpad (San Diego, CA). На фиг. 4 показан репрезентативный график % ингибирования передачи сигнала STAT6 мультиспецифическим антителом IL-13/IL-17AF.
Таблица 5: IC50 значение для ингибирования передачи сигналов STAT6 с помощью мультиспецифического антитела IL-13/IL-17AF.
ПРИМЕР 10. Нейтрализующее действие мультиспецифического антитела IL-13/IL-17AF на ответы IL-6 дермальных фибробластов человека на IL-17A и IL-17F человека и яванского макака.
Цель этого исследования заключалась в определении нейтрализующей способности мультиспецифического антитела IL-13/IL-17AF против IL-17A и IL-17F человека и яванского макака в системе первичных клеток человека. Провоспалительный ответ индуцируется, когда IL-17 присутствует в комбинации с другими цитокинами, такими как TNF-α. Таким образом, этот синергетический эффект был использован для проведения анализа высвобождения IL-6 из первичных нормальных фибробластов кожи новорожденных (nHDF), стимулированных IL-17 и TNF-α.
В этом анализе измеряли способность мультиспецифического антитела IL-13/IL-17AF ингибировать индуцированное IL-17 высвобождение IL-6 из nHDF. В частности, nHDF стимулировали IL-17A человека или яванского макака (50 пМ) или IL-17F (25000 пМ) в сочетании с TNF-α (25 пМ) в присутствии титрования мультиспецифического антитела IL-13/IL-17AF (диапазон концентраций от 5000 пМ до 0,25 пМ для исследования IL-17A; от 500000 до 25 пМ для исследования IL-17F). Затем полученный ответ IL-6 измеряли с использованием гомогенного FRET с временным разрешением (HTRF).
Клетки nHDF (Sigma #106-05n) культивировали в полных средах (DMEM+10% FCS+2 мМ L-глутамина) и содержали в колбах для тканевых культур с использованием стандартных методик. Клетки собирали из колбы для тканевых культур с использованием TrypLE (Invitrogen #12605036). TrypLE нейтрализовали, используя полную среду (45 мл), и клетки центрифугировали при 300×g в течение 3 минут. Клетки ресуспендировали в полных средах (3-5 мл), подсчитывали и доводили до концентрации 3,125×104 клеток/мл перед добавлением в 384-луночный планшет для анализа (Corning #3701) по 40 мкл/лунку. Клетки инкубировали в течение трех часов при 37°C/5% CO2 для прилипания к планшету. Мультиспецифическое антитело IL-13/IL-17AF серийно разводили в полных средах в 384-луночном планшете для разведения (Greiner #781281) до конечного диапазона концентраций от 5000 пМ до 0,25 пМ для оценки против IL-17A и от 500000 пМ до 25 пМ для оценки против IL-17F. Смеси TNF-α и цитокина IL-17 готовили в полных средах до конечных концентраций TNF-α 25 мкМ с IL-17A человека или яванского макака 50 пМ или IL-17F 25000 пМ. Затем 30 мкл/лунку этих растворов добавляли в 384-луночный планшет для реагентов (Greiner #781281). 10 мкл из планшета для серийных разведений мультиспецифических антител IL-13/IL-17AF затем переносили в планшет для реагентов, содержащий 30 мкл разведенных цитокинов. Затем мультиспецифическое антитело IL-13/IL-17AF инкубировали со смесью цитокинов в течение одного часа при 37°C/5% CO2. После инкубации 10 мкл переносили из планшета с реагентами в планшет для анализа, содержащий клетки. Затем планшет для анализа инкубировали в течение 18 часов ± 2 часа при 37°C/5% CO2. После завершения инкубации криптат европия и антитела Alexa 665 из набора Cisbio IL-6 HTRF (Cisbio #62IL6PEB) разводили в буфере для восстановления и смешивали 1:1; согласно вкладышу в наборе. Затем 10 мкл/лунку этой смеси антител добавляли в белый малообъемный 384-луночный планшет HTRF (Greiner #784075). Затем супернатант из планшета для анализа переносили в планшет HTRF в количестве 10 мкл на лунку. Затем планшет HTRF инкубировали при комнатной температуре в течение 2 часов при осторожном встряхивании. Планшеты HTRF затем считывали на планшетном считывателе Synergy Neo 2 в соответствии с инструкциями производителя, измеряя флуоресценцию при считываниях 330/620 нм и 330/665 нм. Затем рассчитывали значения отношения, используя следующее уравнение (330/665 нм, деленное на 330/620 нм) × 10000, и использовали для определения относительного процентного ингибирования по сравнению с контрольными лунками, используя Microsoft Excel. Подбор кривой 4PL и расчет значений IC50 выполняли с помощью GraphPad Prism 7.0.
Показанные результаты представляют собой среднее значение (+/- SEM) трех независимых экспериментов. Значение IC50 мультиспецифического антитела IL-13/IL-17AF рассчитывали как 42 пМ для IL-17A человека и 49 пМ для IL-17A яванского макака. Значение IC50 мультиспецифического антитела IL-13/IL-17AF было рассчитано как 28030 пМ для IL-17F человека и 34320 пМ для IL-17F яванского макака. (фиг. 5).
Таблица 6: Сводные данные о потенциале, эффективности и значениях коэффициента Хилла для мультиспецифического антитела IL-13/IL-17AF.
Значения были рассчитаны из трех независимых экспериментов.
ПРИМЕР 11. Одновременная нейтрализация IL-13, IL-17A и IL-17F с помощью мультиспецифического антитела IL-13/IL-17AF в биоанализе высвобождения IL-13/IL-17AF
Цель этого анализа заключалась в оценке способности мультиспецифического антитела IL-13/IL-17AF одновременно нейтрализовать IL-13, IL-17A и IL-17F в системе первичных клеток. Когда NHEK обрабатывают IL-13 или IL-17A, или IL-17F по отдельности, они индуцируют секрецию CXCL1, хемокина, ответственного за рекрутирование клеток в очаги воспаления. В контексте атопического дерматита есть доказательства того, что CXCL1 играет роль в сенсибилизации нейронов, так что они имеют более низкий порог возбуждения. Это наблюдение может иметь отношение к зуду, который испытывают пациенты (Yang T.B. and Kim BS 2019; «Pruritus in allergy and immunology" J Allergy Clin Immunol 144(2): 353-360).
NHEK (PromoCell, Heidelberg) хранили, культивировали и использовали в соответствии с протоколом производителя. Клетки высевали таким образом, чтобы они были на 100% конфлюэнтны при стимуляции в 48-луночных планшетах с плоским дном. Клетки предварительно инкубировали с увеличивающимися концентрациями мультиспецифического антитела против IL-13, против IL-17A, против IL-17F или IL-13/IL-17AF соответственно в течение 30 минут перед обработкой 100 нг/мл IL-13 и 100 нг/мл IL-17А, 1 мкг/мл IL-17F в течение 72 часов. По прошествии определенного периода времени собирали 50 мкл бесклеточного супернатанта для количественного определения концентрации CXCL1 с помощью ELISA в соответствии с протоколом производителя (R&D Systems). IC50 каждой экспериментальной группы рассчитывали с помощью нелинейной регрессии с использованием призмы Graphpad (San Diego, CA).
Результаты показали, что мультиспецифическое антитело IL-13/IL-17AF одновременно нейтрализует активность IL-13, IL-17A и IL-17F. (фиг. 6). При нормализации количества сайтов связывания, доступных в анализе, мультиспецифическое антитело IL-13/IL-17AF (81,3%) было более эффективным, чем антитело против IL-17A (52,7%), против IL-17F (0,7%) или против IL-13 (48,8%) при ингибировании высвобождения CXCL1. Повышение концентрации только антитела против IL-17A, против IL-17F или против IL-13 не улучшало достигнутое максимальное ингибирование. Результаты подчеркивают преимущество одновременного ингибирования IL-13, IL-17A и IL-17F по сравнению с нейтрализацией одного цитокина.
ПРИМЕР 12. Сравнение мультиспецифического антитела IL-13/IL-17 с антителами IL-13/IL-17 предшествующего уровня техники
Введение
Пример мультиспецифического антитела IL-13/IL-17AF согласно настоящему изобретению показан на фиг. 7. Оно содержит домен Fab с двойной специфичностью в отношении IL-17A и IL-17F, связаный с 2 доменами scFv, один из которых специфичен для IL-13, а другой для альбумина. домен против альбумин обеспечивает мультиспецифическому антителу увеличенный период полужизни.
Биспецифические антитела, которые связываются с IL-13 и IL-17, ранее были описаны Abbvie (WO2013/102042A2) и Genentech (WO2015/127405A2). Однако мало что известно о том, как они связываются и в какой степени они связываются с IL-13, IL-17A и IL-17F. Чтобы сравнить эти свойства, авторы получили антитела, описанные в предшествующем уровне техники, и исследовали их поведение при связывании по следующим характеристикам:
Сродство с IL-13
Взаимодействие молекулы с IL-13 и IL-13Rα1
Сродство с IL-17A
Сродство с IL-17F
Получение антител сравнения
BITS7201A (Genentech) сконструировали с использованием последовательностей, описанных в примере 6 WO2015/127405A2.
DVD2166 и DVD2174 (Abbvie) сконструировали с использованием последовательностей, описанных в WO2013/102042A2, Таблица 6 и Таблица 7. Эти молекулы были выбраны на основе сообщений об активности отдельных плечей биспецифических антител против IL-13 и IL-17, являющихся почти эквивалентными или эквивалентными их соответствующему исходному антителу (WO2013/102042A2, пример 4, стр. 83, параграф 0195).
Конструкции ДНК трансфицировали в клетки CHO-SXE с использованием протокола высоких титров системы трансфекции ExpiCHO (ThermoFisher Scientific). После сбора клеточные культуры центрифугировали не менее часа при 4000 об/мин, и супернатанты очищали с помощью фильтрования с использованием фильтров Stericup 0,22 мкМ.
Очистка DVD2166+DVD2174
Белки DVD-IgG очищали путем нанесения очищенных супернатантов на колонку MabSelect Sure объемом 10 мл и промывали PBS, pH 7,4, для 3 объемов колонки (CV). Белки элюировали с колонки с помощью ступенчатого элюирования 0,1 М цитрата натрия, рН 3,6, и нейтрализовали 2 М трис-HCl, рН 8,5. Мономерные белки выделяли путем нанесения на колонку HiLoad 16×60 Superdex 200 пг (Sigma), уравновешенную PBS, pH 7,4. Фракции, содержащие мономерные белки, объединяли, стерильно фильтровали и хранили при 4°С.
Очистка BITS7201A
Исходные белки knob and hole очищали путем нанесения очищенных супернатантов на колонку MabSelect Sure объемом 10 мл и промывали 3 CVs PBS, pH 7,4. Белки элюировали с колонки 0,1 М цитратом натрия, рН 3,6. Для нейтрализации и стабилизации белка образцы разводили 1:1 1 М буфера аргинин/сукцинат, рН 8,7. Затем исходные антитела наносили на колонку HiLoad 26×60 Superdex 200 пг (Sigma), уравновешенную 0,15 М ацетата натрия, 0,5 М аргининового буфера, рН 8,5. Затем биспецифический материал получали путем смешивания исходных антител в соотношении 1:1 в присутствии 5 мМ цистеамина и инкубации в течение ночи при комнатной температуре. Вторую стадию препаративной гель-фильтрации проводили для удаления любых высокомолекулярных частиц из обменного биспецифического материала путем нанесения на колонку HiLoad 26×60 Superdex 200 пг, уравновешенную PBS, pH 7,4. Фракции, содержащие мономерные биспецифические белки, объединяли, стерильно фильтровали и хранили при 4°С.
Сравнение характеристик связывания
Кинетику связывания IL-13, IL-17A и IL-17F человека, связывающегося с мультиспецифическим антителом IL-13/IL-17AF, называемым «UCBXXXX», и антителами предшествующего уровня техники, оценивали с помощью поверхностного плазмонного резонанса (Biacore T200). и напрямую сравнивали в рамках одного эксперимента. Кроме того, поверхностный плазмонный резонанс использовали для оценки того, приводит ли связывание мультиспецифического антитела IL-13/IL-17AF UCCBXXXX или молекул сравнения с IL-13 к блокированию взаимодействия IL-13 с рецепторами IL-13.
Козье антитело против IgG человека, специфичное к фрагменту F(ab’)2 (Jackson ImmunoResearch), иммобилизовали на сенсорном чипе CM5 с помощью химического связывания амина до уровня приблизительно 5000 RU. Для оценки сродства каждый цикл анализа состоял из захвата мультиспецифического антитела IL-13/IL-17AF или биспецифической молекулы сравнения на поверхность анти-F(ab’)2 (от 50 до 100 RU), введения аналита (при 25°C при скорости потока 30 мкл в минуту в течение 180 с), после чего отслеживали диссоциацию в течение 1200 с для IL-13 и IL-17A и 600 с для IL-17F. В конце каждого цикла поверхность регенерировали при скорости потока 10 мкл/мин, используя введение в течение 60 с 50 мМ HCl, затем введение в течение 30 с 5 мМ NaOH и итоговое введение в течение 60 с 50 мМ HCl. Аналиты вводили в 2-кратных серийных разведениях в рабочем буфере HBS-EP+ (GE Healthcare) в концентрациях от 10 нМ до 0,3125 нМ для IL-13 и от 5 нМ до 0,156 нМ для IL-17A и IL-17F. IL-17A и IL-17F были получены самостоятельно, тогда как IL-13 человека был получен от R&D Systems. Ввод холостой пробы буфера был предусмотрен, чтобы вычесть шум прибора и дрейф.
Кинетические параметры определяли с использованием модели связывания 1:1 с использованием программного обеспечения Biacore T200 Evaluation (версия 3.0). Результаты обобщены в таблице 7.
Таблица 7: Сравнение кинетических констант для связывания с IL-13, IL-17A и IL-17F.
Чтобы оценить блокирование рецептора IL-13Rα1, каждую молекулу антитела захватывали на поверхности козьего анти-человеческого F(ab')2 IgG (приблизительно до 50-100 RU) с последующим введением IL-13 (25 нМ в течение 180 с при 10 мкл/мин) и введением IL-13Rα1 (R&D Systems, 100 нМ в течение 300 с при 10 мкл/мин). Контрольные введения как IL-13, так и IL-13Rα1 были предусмотрены, чтобы вычесть любой дрейф или фоновые ответы. Как показано в таблице 8, UCBXXXX был способен блокировать взаимодействие IL-13 с IL-13Rα1, тогда как молекулы BITS7210A, DVD2166 и DVD 2174 не блокировали.
Таблица 8: Блокирование рецептора IL-13Rα1 с помощью мультиспецифического антитела IL-13/IL-17AF и молекул сравнения.
Был проведен дополнительный эксперимент для оценки способности мультиспецифического антитела IL-13/IL-17AF блокировать связывание с IL-13Rα1 и IL-13Rα2. В этом эксперименте около 260 RU UCBXXXX было захвачено иммобилизованным мышиным антителом против CH1 человека (собственный UCB), после чего вводили IL-13 (25 нМ в течение 180 с при 10 мкл/мин), затем либо IL-13Rα1, либо IL-13Rα2 (R&D Systems, 100 нМ на 300 с при 10 мкл/мин). Контрольные введения IL-13, IL-13Rα1 и IL-13Rα2 были предусмотрены для вычитания любого дрейфа или фоновых ответов. Результаты показали, что UCXXXX способен блокировать взаимодействие IL-13 как с IL-13Ra1, так и с IL-13Ra2. (таблица 9).
Таблица 9: Блокирование рецептора IL-13Rα1 и IL-13Rα2 с помощью мультиспецифического антитела IL-13/IL-17AF.
Обсуждение
Сравнительное исследование показало, что мультиспецифическое антитело IL-13/IL-17AF UCBXXXX, биспецифическое антитело BITS7210A и антитела с двумя вариабельными доменами DVD2166 и DVD2174 способны связываться с IL-13 с высоким сродством. Однако характеристики связывающих взаимодействий были очень разными. UCXXXX был способен блокировать взаимодействие IL-13 с IL-13-Rα1, тогда как BITS7210A, DVD2166 и DVD2174 не блокировали.
Сравнительное исследование также продемонстрировало, что мультиспецифическое антитело UCXXXX, биспецифическое антитело BITS7210A и антитела с двумя вариабельными доменами DVD2166 и DVD2174 способны связываться с IL-17. Однако опять же характеристики связывающих взаимодействий были очень разными. Сродство связывания UCBXXXX с IL-17A было значительно выше, чем у BITS7210A и антител против DVD. UCXXXX и BITS7210A с одинаковым сродством связывались с IL-17F, тогда как антитела к DVD вообще не были способны связываться с IL-17F.
Взаимодействие между IL-13 и IL-13Rα1 в присутствии этих антител важно в контексте снижения потенциальной иммуногенности. Имеющиеся данные свидетельствуют о том, что иммуногенность, особенно образование антител к лекарственным средствам (ADA), следует тщательно учитывать при изучении потенциальных новых терапевтических средств с биспецифическими антителами. Основной движущей силой образования ADA является взаимодействие и последующая интернализация терапевтического антитела с антигенами-мишенями, экспрессированными на клеточной поверхности (Schellekens, H., 2002; Clin Ther. 24(11):1720-40). Интернализованные терапевтические комплексы антитело/антиген-мишень проходят через различные внутриклеточные компартменты и могут либо возвращаться обратно на клеточную поверхность, либо подвергаться деградации (St Pierre et al, 2011); что может привести к презентации пептида антигенпрезентирующими молекулами и образованию ADA.
В биспецифическом антителе BITS7201A часть молекулы IL-13 F(ab) такая же, как у антитела против IL-13, лебрикизумаба (см. Пример 6 WO2015/127405A2). Считается, что лебрикизумаб связывается с IL-13 в месте, которое позволяет IL-13 связываться с его рецепторами IL-13Rα1 и IL-13Rα2, но блокирует взаимодействие с рецептором IL-4Rα (Popovic et al., 2017; J Mol. Biol., 429(2):208-19). Этот конкретный тип взаимодействия может способствовать интернализации комплекса антитело/антиген-мишень/рецептор через IL-13Rα2 и, таким образом, увеличить вероятность иммуногенного ответа. В фазе I клинических испытаний BITS7201A был связан с высокой частотой образования антилекарственных антител (ADA) и был исключен из клинической разработки.
Точно так же антитела с двумя вариабельными доменами DVD2166 и DVD2174 не были способны блокировать взаимодействие IL-13 с IL-13-Rα1.
Чтобы снизить потенциальный риск иммуногенности, UCBXXXX был специально разработан таким образом, чтобы при взаимодействии с соответствующими антигенами-мишенями IL-13, IL-17A и IL-17F предотвращать их взаимодействие с рецепторами на клетках, что снижает вероятность интернализации, деградации и вероятности образования ADA. Заболеваемость ADA с UCBXXXX у людей будет точно известна только после того, как будут доступны клинические данные.
Данные, полученные выше, показывают, что мультиспецифическое антитело IL-13/IL-17AF UCCBXXXX обладает правильными свойствами, чтобы стать эффективным терапевтическим антителом IL-13/IL-17, с повышенной эффективностью и меньшим риском иммуногенности.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> UCB Biopharma SRL
<120> Мультиспецифическое антитело со специфичностью
связывания IL-13 и IL-17 человека
<130> PF0208-WO-PCT
<150> GB 1919061.0
<151> 2019-12-20
<160> 68
<170> PatentIn version 3.5
<210> 1
<211> 11
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 1
Arg Ala Asp Glu Ser Val Arg Thr Leu Met His
1 5 10
<210> 2
<211> 7
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 2
Leu Val Ser Asn Ser Glu Ile
1 5
<210> 3
<211> 9
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 3
Gln Gln Thr Trp Ser Asp Pro Trp Thr
1 5
<210> 4
<211> 10
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 4
Gly Phe Thr Phe Ser Asp Tyr Asn Met Ala
1 5 10
<210> 5
<211> 17
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 5
Thr Ile Thr Tyr Glu Gly Arg Asn Thr Tyr Tyr Arg Asp Ser Val Lys
1 5 10 15
Gly
<210> 6
<211> 16
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 6
Pro Pro Gln Tyr Tyr Glu Gly Ser Ile Tyr Arg Leu Trp Phe Ala His
1 5 10 15
<210> 7
<211> 107
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 7
Ala Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Asp Glu Ser Val Arg Thr Leu
20 25 30
Met His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Leu Val Ser Asn Ser Glu Ile Gly Val Pro Asp Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Arg Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Trp Ser Asp Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 8
<211> 321
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 8
gcaatccagc tcacccagag tccaagcagt ctctccgcca gcgtaggcga ccgtgtgact 60
attacctgta gagcggacga gtcggtcagg actctcatgc actggtatca acagaagcct 120
ggtaaagctc ctaaactgct catctatctg gtgtccaact cggagatagg tgtgccagat 180
cggtttagtg ggtctggttc aggcactgat ttcagactga ccatatcatc tctacagcca 240
gaggacttcg ccacatatta ctgtcagcaa acctggagtg acccgtggac tttcggccag 300
ggcactaaag tagaaattaa a 321
<210> 9
<211> 125
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 9
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Asn Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Thr Tyr Glu Gly Arg Asn Thr Tyr Tyr Arg Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ser Pro Pro Gln Tyr Tyr Glu Gly Ser Ile Tyr Arg Leu Trp Phe
100 105 110
Ala His Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 10
<211> 375
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 10
gaagttcagc tggtcgagtc tggaggtggc cttgtccaac ctggagggag cctgcgtctc 60
tcttgtgcag caagcggatt cacgttttct gattacaata tggcttgggt tagacaggca 120
ccgggtaagg gccttgaatg ggttgcgacg attacatacg aaggcagaaa tacctattac 180
agggactcag taaaagggcg gtttaccata agccgagata atgctaaaaa cagtctgtat 240
ttgcaaatga acagcctacg agctgaagac actgccgtgt attactgcgc gagtccacct 300
cagtattatg aaggatcaat ctatcgcctc tggttcgcac attggggaca ggggaccctt 360
gtgacagtct cgagt 375
<210> 11
<211> 214
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 11
Ala Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Asp Glu Ser Val Arg Thr Leu
20 25 30
Met His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Leu Val Ser Asn Ser Glu Ile Gly Val Pro Asp Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Arg Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Trp Ser Asp Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 12
<211> 642
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 12
gcaatccagc tcacccagag tccaagcagt ctctccgcca gcgtaggcga ccgtgtgact 60
attacctgta gagcggacga gtcggtcagg actctcatgc actggtatca acagaagcct 120
ggtaaagctc ctaaactgct catctatctg gtgtccaact cggagatagg tgtgccagat 180
cggtttagtg ggtctggttc aggcactgat ttcagactga ccatatcatc tctacagcca 240
gaggacttcg ccacatatta ctgtcagcaa acctggagtg acccgtggac tttcggccag 300
ggcactaaag tagaaattaa acgtacggtg gccgctccct ccgtgttcat cttcccaccc 360
tccgacgagc agctgaagtc cggcaccgcc tccgtcgtgt gcctgctgaa caacttctac 420
ccccgcgagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg caactcccag 480
gaatccgtca ccgagcagga ctccaaggac agcacctact ccctgtcctc caccctgacc 540
ctgtccaagg ccgactacga gaagcacaag gtgtacgcct gcgaagtgac ccaccagggc 600
ctgtccagcc ccgtgaccaa gtccttcaac cggggcgagt gc 642
<210> 13
<211> 228
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 13
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Asn Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Thr Tyr Glu Gly Arg Asn Thr Tyr Tyr Arg Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ser Pro Pro Gln Tyr Tyr Glu Gly Ser Ile Tyr Arg Leu Trp Phe
100 105 110
Ala His Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
210 215 220
Pro Lys Ser Cys
225
<210> 14
<211> 684
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 14
gaagttcagc tggtcgagtc tggaggtggc cttgtccaac ctggagggag cctgcgtctc 60
tcttgtgcag caagcggatt cacgttttct gattacaata tggcttgggt tagacaggca 120
ccgggtaagg gccttgaatg ggttgcgacg attacatacg aaggcagaaa tacctattac 180
agggactcag taaaagggcg gtttaccata agccgagata atgctaaaaa cagtctgtat 240
ttgcaaatga acagcctacg agctgaagac actgccgtgt attactgcgc gagtccacct 300
cagtattatg aaggatcaat ctatcgcctc tggttcgcac attggggaca ggggaccctt 360
gtgacagtct cgagtgcgtc cacaaagggc ccatcggtct tccccctggc accctcctcc 420
aagagcacct ctgggggcac agcggccctg ggctgcctgg tcaaggacta cttccccgaa 480
ccagtgacgg tgtcgtggaa ctcaggtgcc ctgaccagcg gcgttcacac cttcccggct 540
gtcctacagt cttcaggact ctactccctg agcagcgtgg tgaccgtgcc ctccagcagc 600
ttgggcaccc agacctacat ctgcaacgtg aatcacaagc ccagcaacac caaggtcgat 660
aagaaagttg agcccaaatc ttgt 684
<210> 15
<211> 11
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 15
Lys Ala Ser Gln Asn Ile Asn Glu Asn Leu Asp
1 5 10
<210> 16
<211> 7
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 16
Tyr Thr Asp Ile Leu Gln Thr
1 5
<210> 17
<211> 8
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 17
Tyr Gln Tyr Tyr Ser Gly Tyr Thr
1 5
<210> 18
<211> 10
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 18
Gly Tyr Ser Phe Thr Ser Tyr Tyr Ile His
1 5 10
<210> 19
<211> 17
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 19
Arg Ile Gly Pro Gly Ser Gly Asp Ile Asn Tyr Asn Glu Lys Phe Lys
1 5 10 15
Gly
<210> 20
<211> 7
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 20
Phe His Tyr Asp Gly Ala Asp
1 5
<210> 21
<211> 106
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 21
Asp Ile Gln Met Thr Gln Ser Pro Pro Val Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Leu Ser Cys Lys Ala Ser Gln Asn Ile Asn Glu Asn
20 25 30
Leu Asp Trp Tyr His Gln Lys His Gly Glu Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Asp Ile Leu Gln Thr Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ala Thr Tyr Tyr Cys Tyr Gln Tyr Tyr Ser Gly Tyr Thr
85 90 95
Phe Gly Pro Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 22
<211> 318
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 22
gacatccaga tgacccagtc tcctccagtc ctgtctgcat ctgtgggaga cagagtcact 60
ctcagttgca aagcaagtca gaatattaat gagaacttag actggtatca tcaaaagcat 120
ggcgaagctc caaaactcct gatatattat acagacattt tgcaaacggg catcccatca 180
aggttcagtg gcagtggatc tggtacagat tacacactca ccatcagcag cctgcagcct 240
gaagatgttg ccacatatta ctgctatcag tattacagtg ggtacacgtt tggacctggg 300
accaagctgg aaataaaa 318
<210> 23
<211> 116
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 23
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Ile Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Arg Ile Gly Pro Gly Ser Gly Asp Ile Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Lys Ala Thr Phe Thr Val Asp Lys Tyr Phe Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Ser Pro Glu Asp Thr Ala Val Phe Tyr Cys
85 90 95
Ala Arg Phe His Tyr Asp Gly Ala Asp Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 24
<211> 348
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 24
caggtacaac tgcagcagtc tggagctgag ttggtgaagc ctgggtcttc agtgaagatg 60
tcctgcaagg cttctggcta cagtttcacc agctactaca tacactggat aaagcagagg 120
cctggacagg gccttgagtg gattgggcgt attggtcctg gaagtggaga tattaattac 180
aatgagaagt tcaagggcaa ggccacattt actgtggaca aatatttcag cacagcctac 240
atgcaactca gcagcctgtc acctgaggac actgcggtct tttactgtgc aagatttcac 300
tatgatgggg ctgactgggg ccaaggcact ctggtcacag tctcgagc 348
<210> 25
<211> 107
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 25
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Ser Thr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 26
<211> 113
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 26
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser
100 105 110
Ser
<210> 27
<211> 106
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 27
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asn Ile Asn Glu Asn
20 25 30
Leu Asp Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Asp Ile Leu Gln Thr Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Tyr Gln Tyr Tyr Ser Gly Tyr Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 28
<211> 116
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 28
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Gly Pro Gly Ser Gly Asp Ile Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Ala Thr Phe Thr Val Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe His Tyr Asp Gly Ala Asp Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 29
<211> 318
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 29
gacatccaga tgacccagtc cccctcctcc ctgtccgcct ccgtgggcga cagggtgacc 60
atcacctgca aggcctccca gaacatcaac gagaacctgg actggtacca gcagaagccc 120
ggcaaggccc ccaagctgct gatctactac accgacatcc tgcagaccgg catcccctcc 180
aggttctccg gctccggctc cggcaccgac tacaccctga ccatctcctc cctgcagccc 240
gaggacttcg ccacctacta ctgctaccag tactactccg gctacacctt cggccagggc 300
accaagctgg agatcaag 318
<210> 30
<211> 348
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 30
gaggtgcagc tggtgcagtc cggcgccgag gtgaagaagc ccggctcctc cgtgaaggtg 60
tcctgcaagg cctccggcta ctccttcacc tcctactaca tccactgggt gaggcaggcc 120
cccggccagg gcctggagtg gatgggcagg atcggccccg gctccggcga catcaactac 180
aacgagaagt tcaagggcag ggccaccttc accgtggaca agtccacctc caccgcctac 240
atggagctgt cctccctgag gtccgaggac accgccgtgt actactgcgc caggttccac 300
tacgacggcg ccgactgggg ccagggcacc ctggtgaccg tctcgagc 348
<210> 31
<211> 106
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 31
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Lys Ala Ser Gln Asn Ile Asn Glu Asn
20 25 30
Leu Asp Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Asp Ile Leu Gln Thr Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Tyr Gln Tyr Tyr Ser Gly Tyr Thr
85 90 95
Phe Gly Cys Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 32
<211> 116
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 32
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Cys Leu Glu Trp Met
35 40 45
Gly Arg Ile Gly Pro Gly Ser Gly Asp Ile Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Ala Thr Phe Thr Val Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe His Tyr Asp Gly Ala Asp Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 33
<211> 318
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 33
gacatccaga tgacccagtc cccctcctcc ctgtccgcct ccgtgggcga cagggtgacc 60
atcacctgca aggcctccca gaacatcaac gagaacctgg actggtacca gcagaagccc 120
ggcaaggccc ccaagctgct gatctactac accgacatcc tgcagaccgg catcccctcc 180
aggttctccg gctccggctc cggcaccgac tacaccctga ccatctcctc cctgcagccc 240
gaggacttcg ccacctacta ctgctaccag tactactccg gctacacctt cggctgcggc 300
accaagctgg agatcaag 318
<210> 34
<211> 348
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 34
gaggtgcagc tggtgcagtc cggcgccgag gtgaagaagc ccggctcctc cgtgaaggtg 60
tcctgcaagg cctccggcta ctccttcacc tcctactaca tccactgggt gaggcaggcc 120
cccggccagt gcctggagtg gatgggcagg atcggccccg gctccggcga catcaactac 180
aacgagaagt tcaagggcag ggccaccttc accgtggaca agtccacctc caccgcctac 240
atggagctgt cctccctgag gtccgaggac accgccgtgt actactgcgc caggttccac 300
tacgacggcg ccgactgggg ccagggcacc ctggtgaccg tgtcctcc 348
<210> 35
<211> 242
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 35
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Arg Ile Gly Pro Gly Ser Gly Asp Ile Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Ala Thr Phe Thr Val Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe His Tyr Asp Gly Ala Asp Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser Pro
130 135 140
Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Lys
145 150 155 160
Ala Ser Gln Asn Ile Asn Glu Asn Leu Asp Trp Tyr Gln Gln Lys Pro
165 170 175
Gly Lys Ala Pro Lys Leu Leu Ile Tyr Tyr Thr Asp Ile Leu Gln Thr
180 185 190
Gly Ile Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Thr
195 200 205
Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
210 215 220
Tyr Gln Tyr Tyr Ser Gly Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu
225 230 235 240
Ile Lys
<210> 36
<211> 726
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 36
gaggtgcagc tggtgcagtc cggcgccgag gtgaagaagc ccggctcctc cgtgaaggtg 60
tcctgcaagg cctccggcta ctccttcacc tcctactaca tccactgggt gaggcaggcc 120
cccggccagg gcctggagtg gatgggcagg atcggccccg gctccggcga catcaactac 180
aacgagaagt tcaagggcag ggccaccttc accgtggaca agtccacctc caccgcctac 240
atggagctgt cctccctgag gtccgaggac accgccgtgt actactgcgc caggttccac 300
tacgacggcg ccgactgggg ccagggcacc ctggtgaccg tgtcctccgg aggtggcggt 360
tctggcggtg gcggttccgg tggcggtgga tcgggaggtg gcggttctga catccagatg 420
acccagtccc cctcctccct gtccgcctcc gtgggcgaca gggtgaccat cacctgcaag 480
gcctcccaga acatcaacga gaacctggac tggtaccagc agaagcccgg caaggccccc 540
aagctgctga tctactacac cgacatcctg cagaccggca tcccctccag gttctccggc 600
tccggctccg gcaccgacta caccctgacc atctcctccc tgcagcccga ggacttcgcc 660
acctactact gctaccagta ctactccggc tacaccttcg gccagggcac caagctggag 720
atcaag 726
<210> 37
<211> 242
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 37
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Ser Tyr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Cys Leu Glu Trp Met
35 40 45
Gly Arg Ile Gly Pro Gly Ser Gly Asp Ile Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Arg Ala Thr Phe Thr Val Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Phe His Tyr Asp Gly Ala Asp Trp Gly Gln Gly Thr Leu Val
100 105 110
Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser Pro
130 135 140
Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Lys
145 150 155 160
Ala Ser Gln Asn Ile Asn Glu Asn Leu Asp Trp Tyr Gln Gln Lys Pro
165 170 175
Gly Lys Ala Pro Lys Leu Leu Ile Tyr Tyr Thr Asp Ile Leu Gln Thr
180 185 190
Gly Ile Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr Thr
195 200 205
Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys
210 215 220
Tyr Gln Tyr Tyr Ser Gly Tyr Thr Phe Gly Cys Gly Thr Lys Leu Glu
225 230 235 240
Ile Lys
<210> 38
<211> 726
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 38
gaggtgcagc tggtgcagtc cggcgccgag gtgaagaagc ccggctcctc cgtgaaggtg 60
tcctgcaagg cctccggcta ctccttcacc tcctactaca tccactgggt gaggcaggcc 120
cccggccagt gcctggagtg gatgggcagg atcggccccg gctccggcga catcaactac 180
aacgagaagt tcaagggcag ggccaccttc accgtggaca agtccacctc caccgcctac 240
atggagctgt cctccctgag gtccgaggac accgccgtgt actactgcgc caggttccac 300
tacgacggcg ccgactgggg ccagggcacc ctggtgaccg tgtcctccgg aggtggcggt 360
tctggcggtg gcggttccgg tggcggtgga tcgggaggtg gcggttctga catccagatg 420
acccagtccc cctcctccct gtccgcctcc gtgggcgaca gggtgaccat cacctgcaag 480
gcctcccaga acatcaacga gaacctggac tggtaccagc agaagcccgg caaggccccc 540
aagctgctga tctactacac cgacatcctg cagaccggca tcccctccag gttctccggc 600
tccggctccg gcaccgacta caccctgacc atctcctccc tgcagcccga ggacttcgcc 660
acctactact gctaccagta ctactccggc tacaccttcg gctgcggcac caagctggag 720
atcaag 726
<210> 39
<211> 12
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 39
Gln Ser Ser Pro Ser Val Trp Ser Asn Phe Leu Ser
1 5 10
<210> 40
<211> 7
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 40
Glu Ala Ser Lys Leu Thr Ser
1 5
<210> 41
<211> 11
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 41
Gly Gly Gly Tyr Ser Ser Ile Ser Asp Thr Thr
1 5 10
<210> 42
<211> 10
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 42
Gly Ile Asp Leu Ser Asn Tyr Ala Ile Asn
1 5 10
<210> 43
<211> 16
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 43
Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys Gly
1 5 10 15
<210> 44
<211> 13
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 44
Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu
1 5 10
<210> 45
<211> 110
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 45
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ser Ser Pro Ser Val Trp Ser Asn
20 25 30
Phe Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Glu Ala Ser Lys Leu Thr Ser Gly Val Pro Ser Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln
65 70 75 80
Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gly Gly Gly Tyr Ser Ser Ile
85 90 95
Ser Asp Thr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 46
<211> 121
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 46
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 47
<211> 330
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 47
gatattcaga tgacgcaatc accttcgagc gtatccgcct cggtgggaga cagggtgaca 60
atcacttgtc agtcatcccc ctcagtctgg agcaactttt tgtcatggta tcagcagaag 120
cccggaaagg ctccgaaatt gctgatctac gaggcatcga agttgacgag cggtgtacca 180
agcagattct ccggttcggg gtcgggaact gacttcaccc ttacgatctc atcgctgcag 240
ccggaggatt ttgcgaccta ctactgtggg ggtgggtatt cgtcgatttc cgacacaaca 300
ttcgggggcg gcacgaaagt ggaaatcaag 330
<210> 48
<211> 363
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 48
gaagtgcagt tgctggagtc aggtggaggg ctggtgcagc ccggaggatc gctgcggttg 60
tcatgcgcgg tgtccggtat tgatttgtcc aattacgcca tcaattgggt acgccaagcg 120
ccagggaagg gccttgagtg gattggcatc atctgggcgt cggggacgac cttttatgct 180
acttgggcca aaggaagatt cacaatctcc cgagacaact cgaagaacac cgtgtatctt 240
caaatgaact cgctcagggc cgaggacacg gcggtctact actgtgcacg gacagtgccg 300
ggttattcaa cggcacctta ctttgatctt tggggccagg ggaccctcgt gactgtctca 360
agt 363
<210> 49
<211> 110
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 49
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ser Ser Pro Ser Val Trp Ser Asn
20 25 30
Phe Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Glu Ala Ser Lys Leu Thr Ser Gly Val Pro Ser Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln
65 70 75 80
Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gly Gly Gly Tyr Ser Ser Ile
85 90 95
Ser Asp Thr Thr Phe Gly Cys Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 50
<211> 121
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 50
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 51
<211> 330
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 51
gatattcaga tgacgcaatc accttcgagc gtatccgcct cggtgggaga cagggtgaca 60
atcacttgtc agtcatcccc ctcagtctgg agcaactttt tgtcatggta tcagcagaag 120
cccggaaagg ctccgaaatt gctgatctac gaggcatcga agttgacgag cggtgtacca 180
agcagattct ccggttcggg gtcgggaact gacttcaccc ttacgatctc atcgctgcag 240
ccggaggatt ttgcgaccta ctactgtggg ggtgggtatt cgtcgatttc cgacacaaca 300
ttcgggtgcg gcacgaaagt ggaaatcaag 330
<210> 52
<211> 363
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 52
gaagtgcagt tgctggagtc aggtggaggg ctggtgcagc ccggaggatc gctgcggttg 60
tcatgcgcgg tgtccggtat tgatttgtcc aattacgcca tcaattgggt acgccaagcg 120
ccagggaagt gccttgagtg gattggcatc atctgggcgt cggggacgac cttttatgct 180
acttgggcca aaggaagatt cacaatctcc cgagacaact cgaagaacac cgtgtatctt 240
caaatgaact cgctcagggc cgaggacacg gcggtctact actgtgcacg gacagtgccg 300
ggttattcaa cggcacctta ctttgatctt tggggccagg ggaccctcgt gactgtctca 360
agt 363
<210> 53
<211> 251
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 53
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln
130 135 140
Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly Asp Arg Val
145 150 155 160
Thr Ile Thr Cys Gln Ser Ser Pro Ser Val Trp Ser Asn Phe Leu Ser
165 170 175
Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Glu
180 185 190
Ala Ser Lys Leu Thr Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly
195 200 205
Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp
210 215 220
Phe Ala Thr Tyr Tyr Cys Gly Gly Gly Tyr Ser Ser Ile Ser Asp Thr
225 230 235 240
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
245 250
<210> 54
<211> 753
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 54
gaagtgcagt tgctggagtc aggtggaggg ctggtgcagc ccggaggatc gctgcggttg 60
tcatgcgcgg tgtccggtat tgatttgtcc aattacgcca tcaattgggt acgccaagcg 120
ccagggaagg gccttgagtg gattggcatc atctgggcgt cggggacgac cttttatgct 180
acttgggcca aaggaagatt cacaatctcc cgagacaact cgaagaacac cgtgtatctt 240
caaatgaact cgctcagggc cgaggacacg gcggtctact actgtgcacg gacagtgccg 300
ggttattcaa cggcacctta ctttgatctt tggggccagg ggaccctcgt gactgtctca 360
agtggaggtg gcggttctgg cggtggcggt tccggtggcg gtggatcggg aggtggcggt 420
tctgatattc agatgacgca atcaccttcg agcgtatccg cctcggtggg agacagggtg 480
acaatcactt gtcagtcatc cccctcagtc tggagcaact ttttgtcatg gtatcagcag 540
aagcccggaa aggctccgaa attgctgatc tacgaggcat cgaagttgac gagcggtgta 600
ccaagcagat tctccggttc ggggtcggga actgacttca cccttacgat ctcatcgctg 660
cagccggagg attttgcgac ctactactgt gggggtgggt attcgtcgat ttccgacaca 720
acattcgggg gcggcacgaa agtggaaatc aag 753
<210> 55
<211> 251
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 55
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr
20 25 30
Ala Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Ile
35 40 45
Gly Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
115 120 125
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln
130 135 140
Met Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly Asp Arg Val
145 150 155 160
Thr Ile Thr Cys Gln Ser Ser Pro Ser Val Trp Ser Asn Phe Leu Ser
165 170 175
Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Glu
180 185 190
Ala Ser Lys Leu Thr Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly
195 200 205
Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp
210 215 220
Phe Ala Thr Tyr Tyr Cys Gly Gly Gly Tyr Ser Ser Ile Ser Asp Thr
225 230 235 240
Thr Phe Gly Cys Gly Thr Lys Val Glu Ile Lys
245 250
<210> 56
<211> 753
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 56
gaagtgcagt tgctggagtc aggtggaggg ctggtgcagc ccggaggatc gctgcggttg 60
tcatgcgcgg tgtccggtat tgatttgtcc aattacgcca tcaattgggt acgccaagcg 120
ccagggaagt gccttgagtg gattggcatc atctgggcgt cggggacgac cttttatgct 180
acttgggcca aaggaagatt cacaatctcc cgagacaact cgaagaacac cgtgtatctt 240
caaatgaact cgctcagggc cgaggacacg gcggtctact actgtgcacg gacagtgccg 300
ggttattcaa cggcacctta ctttgatctt tggggccagg ggaccctcgt gactgtctca 360
agtggaggtg gcggttctgg cggtggcggt tccggtggcg gtggatcggg aggtggcggt 420
tctgatattc agatgacgca atcaccttcg agcgtatccg cctcggtggg agacagggtg 480
acaatcactt gtcagtcatc cccctcagtc tggagcaact ttttgtcatg gtatcagcag 540
aagcccggaa aggctccgaa attgctgatc tacgaggcat cgaagttgac gagcggtgta 600
ccaagcagat tctccggttc ggggtcggga actgacttca cccttacgat ctcatcgctg 660
cagccggagg attttgcgac ctactactgt gggggtgggt attcgtcgat ttccgacaca 720
acattcgggt gcggcacgaa agtggaaatc aag 753
<210> 57
<211> 492
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 57
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Asn Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Thr Tyr Glu Gly Arg Asn Thr Tyr Tyr Arg Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ser Pro Pro Gln Tyr Tyr Glu Gly Ser Ile Tyr Arg Leu Trp Phe
100 105 110
Ala His Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
210 215 220
Pro Lys Ser Cys Ser Gly Gly Gly Gly Thr Gly Gly Gly Gly Ser Glu
225 230 235 240
Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser
245 250 255
Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr Ala
260 265 270
Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly
275 280 285
Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys Gly
290 295 300
Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu Gln
305 310 315 320
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg
325 330 335
Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly Gln
340 345 350
Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly
355 360 365
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln Met
370 375 380
Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly Asp Arg Val Thr
385 390 395 400
Ile Thr Cys Gln Ser Ser Pro Ser Val Trp Ser Asn Phe Leu Ser Trp
405 410 415
Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Glu Ala
420 425 430
Ser Lys Leu Thr Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser
435 440 445
Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe
450 455 460
Ala Thr Tyr Tyr Cys Gly Gly Gly Tyr Ser Ser Ile Ser Asp Thr Thr
465 470 475 480
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys Arg Thr
485 490
<210> 58
<211> 1476
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 58
gaagttcagc tggtcgagtc tggaggtggc cttgtccaac ctggagggag cctgcgtctc 60
tcttgtgcag caagcggatt cacgttttct gattacaata tggcttgggt tagacaggca 120
ccgggtaagg gccttgaatg ggttgcgacg attacatacg aaggcagaaa tacctattac 180
agggactcag taaaagggcg gtttaccata agccgagata atgctaaaaa cagtctgtat 240
ttgcaaatga acagcctacg agctgaagac actgccgtgt attactgcgc gagtccacct 300
cagtattatg aaggatcaat ctatcgcctc tggttcgcac attggggaca ggggaccctt 360
gtgacagtct cgagtgcgtc cacaaagggc ccatcggtct tccccctggc accctcctcc 420
aagagcacct ctgggggcac agcggccctg ggctgcctgg tcaaggacta cttccccgaa 480
ccagtgacgg tgtcgtggaa ctcaggtgcc ctgaccagcg gcgttcacac cttcccggct 540
gtcctacagt cttcaggact ctactccctg agcagcgtgg tgaccgtgcc ctccagcagc 600
ttgggcaccc agacctacat ctgcaacgtg aatcacaagc ccagcaacac caaggtcgat 660
aagaaagttg agcccaaatc ttgtagcggt ggcggtggca ccggaggtgg cggttcagaa 720
gtgcagttgc tggagtcagg tggagggctg gtgcagcccg gaggatcgct gcggttgtca 780
tgcgcggtgt ccggtattga tttgtccaat tacgccatca attgggtacg ccaagcgcca 840
gggaagggcc ttgagtggat tggcatcatc tgggcgtcgg ggacgacctt ttatgctact 900
tgggccaaag gaagattcac aatctcccga gacaactcga agaacaccgt gtatcttcaa 960
atgaactcgc tcagggccga ggacacggcg gtctactact gtgcacggac agtgccgggt 1020
tattcaacgg caccttactt tgatctttgg ggccagggga ccctcgtgac tgtctcaagt 1080
ggaggtggcg gttctggcgg tggcggttcc ggtggcggtg gatcgggagg tggcggttct 1140
gatattcaga tgacgcaatc accttcgagc gtatccgcct cggtgggaga cagggtgaca 1200
atcacttgtc agtcatcccc ctcagtctgg agcaactttt tgtcatggta tcagcagaag 1260
cccggaaagg ctccgaaatt gctgatctac gaggcatcga agttgacgag cggtgtacca 1320
agcagattct ccggttcggg gtcgggaact gacttcaccc ttacgatctc atcgctgcag 1380
ccggaggatt ttgcgaccta ctactgtggg ggtgggtatt cgtcgatttc cgacacaaca 1440
ttcgggggcg gcacgaaagt ggaaatcaag cgtacc 1476
<210> 59
<211> 492
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 59
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Asn Met Ala Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Thr Tyr Glu Gly Arg Asn Thr Tyr Tyr Arg Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ser Pro Pro Gln Tyr Tyr Glu Gly Ser Ile Tyr Arg Leu Trp Phe
100 105 110
Ala His Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr
115 120 125
Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser
130 135 140
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
145 150 155 160
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
165 170 175
Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser
180 185 190
Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys
195 200 205
Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu
210 215 220
Pro Lys Ser Cys Ser Gly Gly Gly Gly Thr Gly Gly Gly Gly Ser Glu
225 230 235 240
Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly Ser
245 250 255
Leu Arg Leu Ser Cys Ala Val Ser Gly Ile Asp Leu Ser Asn Tyr Ala
260 265 270
Ile Asn Trp Val Arg Gln Ala Pro Gly Lys Cys Leu Glu Trp Ile Gly
275 280 285
Ile Ile Trp Ala Ser Gly Thr Thr Phe Tyr Ala Thr Trp Ala Lys Gly
290 295 300
Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu Gln
305 310 315 320
Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala Arg
325 330 335
Thr Val Pro Gly Tyr Ser Thr Ala Pro Tyr Phe Asp Leu Trp Gly Gln
340 345 350
Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly
355 360 365
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln Met
370 375 380
Thr Gln Ser Pro Ser Ser Val Ser Ala Ser Val Gly Asp Arg Val Thr
385 390 395 400
Ile Thr Cys Gln Ser Ser Pro Ser Val Trp Ser Asn Phe Leu Ser Trp
405 410 415
Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Glu Ala
420 425 430
Ser Lys Leu Thr Ser Gly Val Pro Ser Arg Phe Ser Gly Ser Gly Ser
435 440 445
Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe
450 455 460
Ala Thr Tyr Tyr Cys Gly Gly Gly Tyr Ser Ser Ile Ser Asp Thr Thr
465 470 475 480
Phe Gly Cys Gly Thr Lys Val Glu Ile Lys Arg Thr
485 490
<210> 60
<211> 1476
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 60
gaagttcagc tggtcgagtc tggaggtggc cttgtccaac ctggagggag cctgcgtctc 60
tcttgtgcag caagcggatt cacgttttct gattacaata tggcttgggt tagacaggca 120
ccgggtaagg gccttgaatg ggttgcgacg attacatacg aaggcagaaa tacctattac 180
agggactcag taaaagggcg gtttaccata agccgagata atgctaaaaa cagtctgtat 240
ttgcaaatga acagcctacg agctgaagac actgccgtgt attactgcgc gagtccacct 300
cagtattatg aaggatcaat ctatcgcctc tggttcgcac attggggaca ggggaccctt 360
gtgacagtct cgagtgcgtc cacaaagggc ccatcggtct tccccctggc accctcctcc 420
aagagcacct ctgggggcac agcggccctg ggctgcctgg tcaaggacta cttccccgaa 480
ccagtgacgg tgtcgtggaa ctcaggtgcc ctgaccagcg gcgttcacac cttcccggct 540
gtcctacagt cttcaggact ctactccctg agcagcgtgg tgaccgtgcc ctccagcagc 600
ttgggcaccc agacctacat ctgcaacgtg aatcacaagc ccagcaacac caaggtcgat 660
aagaaagttg agcccaaatc ttgtagcggt ggcggtggca ccggaggtgg cggttcagaa 720
gtgcagttgc tggagtcagg tggagggctg gtgcagcccg gaggatcgct gcggttgtca 780
tgcgcggtgt ccggtattga tttgtccaat tacgccatca attgggtacg ccaagcgcca 840
gggaagtgcc ttgagtggat tggcatcatc tgggcgtcgg ggacgacctt ttatgctact 900
tgggccaaag gaagattcac aatctcccga gacaactcga agaacaccgt gtatcttcaa 960
atgaactcgc tcagggccga ggacacggcg gtctactact gtgcacggac agtgccgggt 1020
tattcaacgg caccttactt tgatctttgg ggccagggga ccctcgtgac tgtctcaagt 1080
ggaggtggcg gttctggcgg tggcggttcc ggtggcggtg gatcgggagg tggcggttct 1140
gatattcaga tgacgcaatc accttcgagc gtatccgcct cggtgggaga cagggtgaca 1200
atcacttgtc agtcatcccc ctcagtctgg agcaactttt tgtcatggta tcagcagaag 1260
cccggaaagg ctccgaaatt gctgatctac gaggcatcga agttgacgag cggtgtacca 1320
agcagattct ccggttcggg gtcgggaact gacttcaccc ttacgatctc atcgctgcag 1380
ccggaggatt ttgcgaccta ctactgtggg ggtgggtatt cgtcgatttc cgacacaaca 1440
ttcgggtgcg gcacgaaagt ggaaatcaag cgtacc 1476
<210> 61
<211> 469
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 61
Ala Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Asp Glu Ser Val Arg Thr Leu
20 25 30
Met His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Leu Val Ser Asn Ser Glu Ile Gly Val Pro Asp Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Arg Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Trp Ser Asp Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
210 215 220
Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly
225 230 235 240
Ser Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Ser
245 250 255
Tyr Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp
260 265 270
Met Gly Arg Ile Gly Pro Gly Ser Gly Asp Ile Asn Tyr Asn Glu Lys
275 280 285
Phe Lys Gly Arg Ala Thr Phe Thr Val Asp Lys Ser Thr Ser Thr Ala
290 295 300
Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr
305 310 315 320
Cys Ala Arg Phe His Tyr Asp Gly Ala Asp Trp Gly Gln Gly Thr Leu
325 330 335
Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
340 345 350
Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser
355 360 365
Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys
370 375 380
Lys Ala Ser Gln Asn Ile Asn Glu Asn Leu Asp Trp Tyr Gln Gln Lys
385 390 395 400
Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Tyr Thr Asp Ile Leu Gln
405 410 415
Thr Gly Ile Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr
420 425 430
Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr
435 440 445
Cys Tyr Gln Tyr Tyr Ser Gly Tyr Thr Phe Gly Gln Gly Thr Lys Leu
450 455 460
Glu Ile Lys Arg Thr
465
<210> 62
<211> 1407
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 62
gcaatccagc tcacccagag tccaagcagt ctctccgcca gcgtaggcga ccgtgtgact 60
attacctgta gagcggacga gtcggtcagg actctcatgc actggtatca acagaagcct 120
ggtaaagctc ctaaactgct catctatctg gtgtccaact cggagatagg tgtgccagat 180
cggtttagtg ggtctggttc aggcactgat ttcagactga ccatatcatc tctacagcca 240
gaggacttcg ccacatatta ctgtcagcaa acctggagtg acccgtggac tttcggccag 300
ggcactaaag tagaaattaa acgtacggtg gccgctccct ccgtgttcat cttcccaccc 360
tccgacgagc agctgaagtc cggcaccgcc tccgtcgtgt gcctgctgaa caacttctac 420
ccccgcgagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg caactcccag 480
gaatccgtca ccgagcagga ctccaaggac agcacctact ccctgtcctc caccctgacc 540
ctgtccaagg ccgactacga gaagcacaag gtgtacgcct gcgaagtgac ccaccagggc 600
ctgtccagcc ccgtgaccaa gtccttcaac cggggcgagt gcagcggtgg cggtggctcc 660
ggaggtggcg gttcagaggt gcagctggtg cagtccggcg ccgaggtgaa gaagcccggc 720
tcctccgtga aggtgtcctg caaggcctcc ggctactcct tcacctccta ctacatccac 780
tgggtgaggc aggcccccgg ccagggcctg gagtggatgg gcaggatcgg ccccggctcc 840
ggcgacatca actacaacga gaagttcaag ggcagggcca ccttcaccgt ggacaagtcc 900
acctccaccg cctacatgga gctgtcctcc ctgaggtccg aggacaccgc cgtgtactac 960
tgcgccaggt tccactacga cggcgccgac tggggccagg gcaccctggt gaccgtgtcc 1020
tccggaggtg gcggttctgg cggtggcggt tccggtggcg gtggatcggg aggtggcggt 1080
tctgacatcc agatgaccca gtccccctcc tccctgtccg cctccgtggg cgacagggtg 1140
accatcacct gcaaggcctc ccagaacatc aacgagaacc tggactggta ccagcagaag 1200
cccggcaagg cccccaagct gctgatctac tacaccgaca tcctgcagac cggcatcccc 1260
tccaggttct ccggctccgg ctccggcacc gactacaccc tgaccatctc ctccctgcag 1320
cccgaggact tcgccaccta ctactgctac cagtactact ccggctacac cttcggccag 1380
ggcaccaagc tggagatcaa gcgtacc 1407
<210> 63
<211> 469
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 63
Ala Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Asp Glu Ser Val Arg Thr Leu
20 25 30
Met His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Leu Val Ser Asn Ser Glu Ile Gly Val Pro Asp Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Arg Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Thr Trp Ser Asp Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
210 215 220
Ser Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly
225 230 235 240
Ser Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Ser
245 250 255
Tyr Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Cys Leu Glu Trp
260 265 270
Met Gly Arg Ile Gly Pro Gly Ser Gly Asp Ile Asn Tyr Asn Glu Lys
275 280 285
Phe Lys Gly Arg Ala Thr Phe Thr Val Asp Lys Ser Thr Ser Thr Ala
290 295 300
Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr
305 310 315 320
Cys Ala Arg Phe His Tyr Asp Gly Ala Asp Trp Gly Gln Gly Thr Leu
325 330 335
Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
340 345 350
Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Gln Met Thr Gln Ser
355 360 365
Pro Ser Ser Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys
370 375 380
Lys Ala Ser Gln Asn Ile Asn Glu Asn Leu Asp Trp Tyr Gln Gln Lys
385 390 395 400
Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr Tyr Thr Asp Ile Leu Gln
405 410 415
Thr Gly Ile Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Tyr
420 425 430
Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr
435 440 445
Cys Tyr Gln Tyr Tyr Ser Gly Tyr Thr Phe Gly Cys Gly Thr Lys Leu
450 455 460
Glu Ile Lys Arg Thr
465
<210> 64
<211> 1407
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 64
gcaatccagc tcacccagag tccaagcagt ctctccgcca gcgtaggcga ccgtgtgact 60
attacctgta gagcggacga gtcggtcagg actctcatgc actggtatca acagaagcct 120
ggtaaagctc ctaaactgct catctatctg gtgtccaact cggagatagg tgtgccagat 180
cggtttagtg ggtctggttc aggcactgat ttcagactga ccatatcatc tctacagcca 240
gaggacttcg ccacatatta ctgtcagcaa acctggagtg acccgtggac tttcggccag 300
ggcactaaag tagaaattaa acgtacggtg gccgctccct ccgtgttcat cttcccaccc 360
tccgacgagc agctgaagtc cggcaccgcc tccgtcgtgt gcctgctgaa caacttctac 420
ccccgcgagg ccaaggtgca gtggaaggtg gacaacgccc tgcagtccgg caactcccag 480
gaatccgtca ccgagcagga ctccaaggac agcacctact ccctgtcctc caccctgacc 540
ctgtccaagg ccgactacga gaagcacaag gtgtacgcct gcgaagtgac ccaccagggc 600
ctgtccagcc ccgtgaccaa gtccttcaac cggggcgagt gcagcggtgg cggtggctcc 660
ggaggtggcg gttcagaggt gcagctggtg cagtccggcg ccgaggtgaa gaagcccggc 720
tcctccgtga aggtgtcctg caaggcctcc ggctactcct tcacctccta ctacatccac 780
tgggtgaggc aggcccccgg ccagtgcctg gagtggatgg gcaggatcgg ccccggctcc 840
ggcgacatca actacaacga gaagttcaag ggcagggcca ccttcaccgt ggacaagtcc 900
acctccaccg cctacatgga gctgtcctcc ctgaggtccg aggacaccgc cgtgtactac 960
tgcgccaggt tccactacga cggcgccgac tggggccagg gcaccctggt gaccgtgtcc 1020
tccggaggtg gcggttctgg cggtggcggt tccggtggcg gtggatcggg aggtggcggt 1080
tctgacatcc agatgaccca gtccccctcc tccctgtccg cctccgtggg cgacagggtg 1140
accatcacct gcaaggcctc ccagaacatc aacgagaacc tggactggta ccagcagaag 1200
cccggcaagg cccccaagct gctgatctac tacaccgaca tcctgcagac cggcatcccc 1260
tccaggttct ccggctccgg ctccggcacc gactacaccc tgaccatctc ctccctgcag 1320
cccgaggact tcgccaccta ctactgctac cagtactact ccggctacac cttcggctgc 1380
ggcaccaagc tggagatcaa gcgtacc 1407
<210> 65
<211> 11
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 65
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 66
<211> 20
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 66
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser
20
<210> 67
<211> 11
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 67
Ser Gly Gly Gly Gly Thr Gly Gly Gly Gly Ser
1 5 10
<210> 68
<211> 20
<212> PRT
<213> Искусственная Последовательность
<220>
<223> рекомбинантная последовательность
<400> 68
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser
20
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Анти-PD-L1 и IL-2 цитокины | 2017 |
|
RU2769282C2 |
| АНТИ-IL-22R-АНТИТЕЛА | 2017 |
|
RU2758721C2 |
| КОМПОЗИЦИИ IL-12, НАЦЕЛЕННЫЕ НА EDB | 2019 |
|
RU2758143C1 |
| IL-18-СВЯЗЫВАЮЩИЕ БЕЛОК (IL-18BP) И АНТИТЕЛА ПРИ ВОСПАЛИТЕЛЬНЫХ ЗАБОЛЕВАНИЯХ | 2016 |
|
RU2755691C2 |
| АНТИ-IL-5 АНТИТЕЛА | 2017 |
|
RU2758008C2 |
| IL-18-СВЯЗЫВАЮЩИЙ БЕЛОК (IL-18BP) И АНТИТЕЛА ПРИ ВОСПАЛИТЕЛЬНЫХ ЗАБОЛЕВАНИЯХ | 2016 |
|
RU2810206C1 |
| АНТИТЕЛО К IL-13RA2 И ЕГО ПРИМЕНЕНИЕ | 2018 |
|
RU2756623C2 |
| АНТИТЕЛО ПРОТИВ IL-17A И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2816204C2 |
| АНТИТЕЛО, СВЯЗЫВАЮЩЕЕ ЧЕЛОВЕЧЕСКИЙ IL-4R, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2019 |
|
RU2779649C1 |
| АНТИТЕЛО ПРОТИВ IL-5, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ЕГО МЕДИЦИНСКОЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2772716C2 |













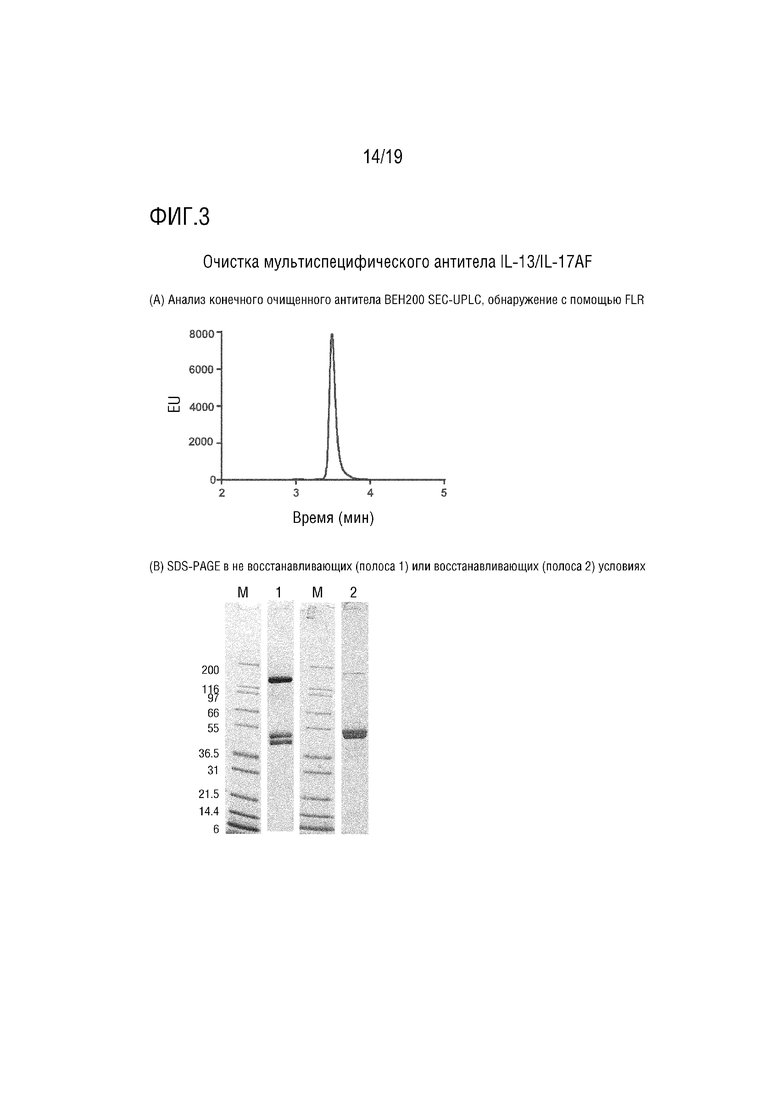
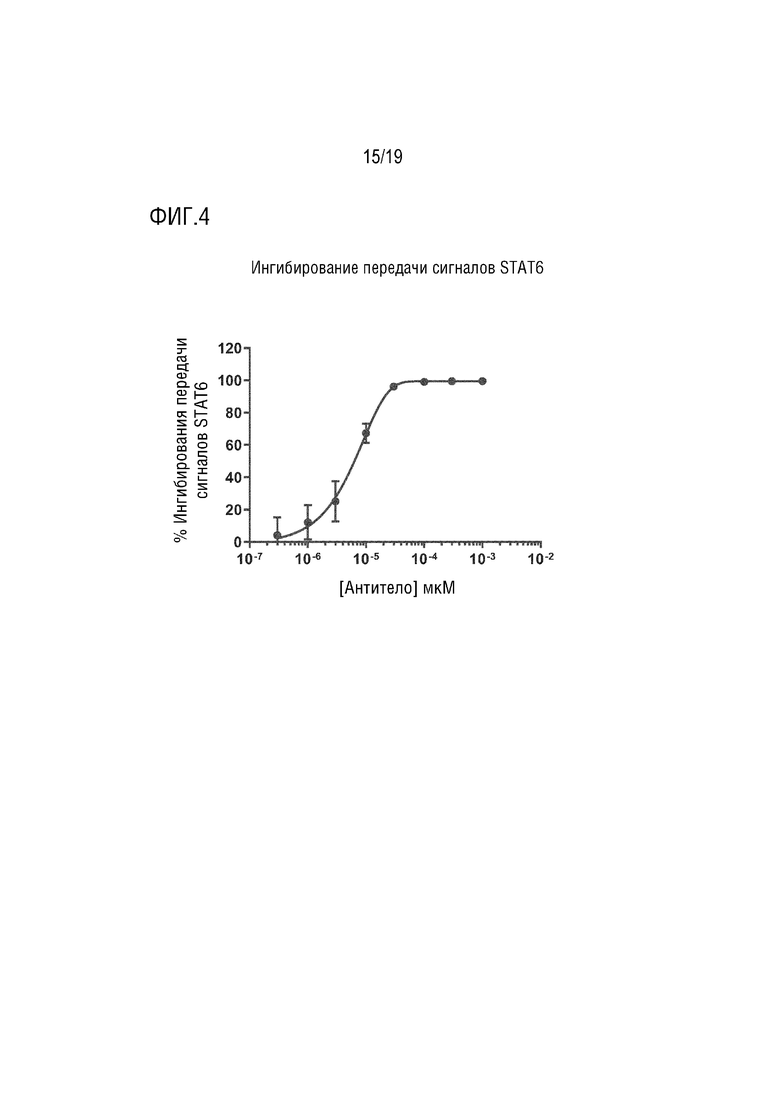
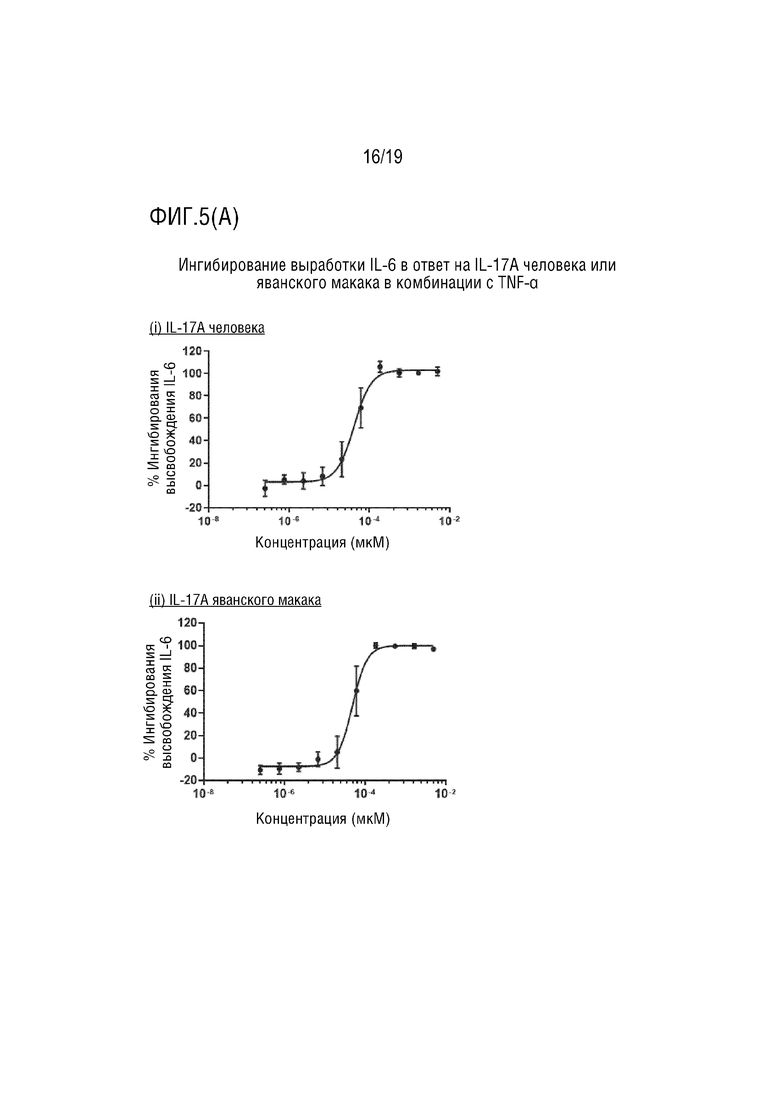
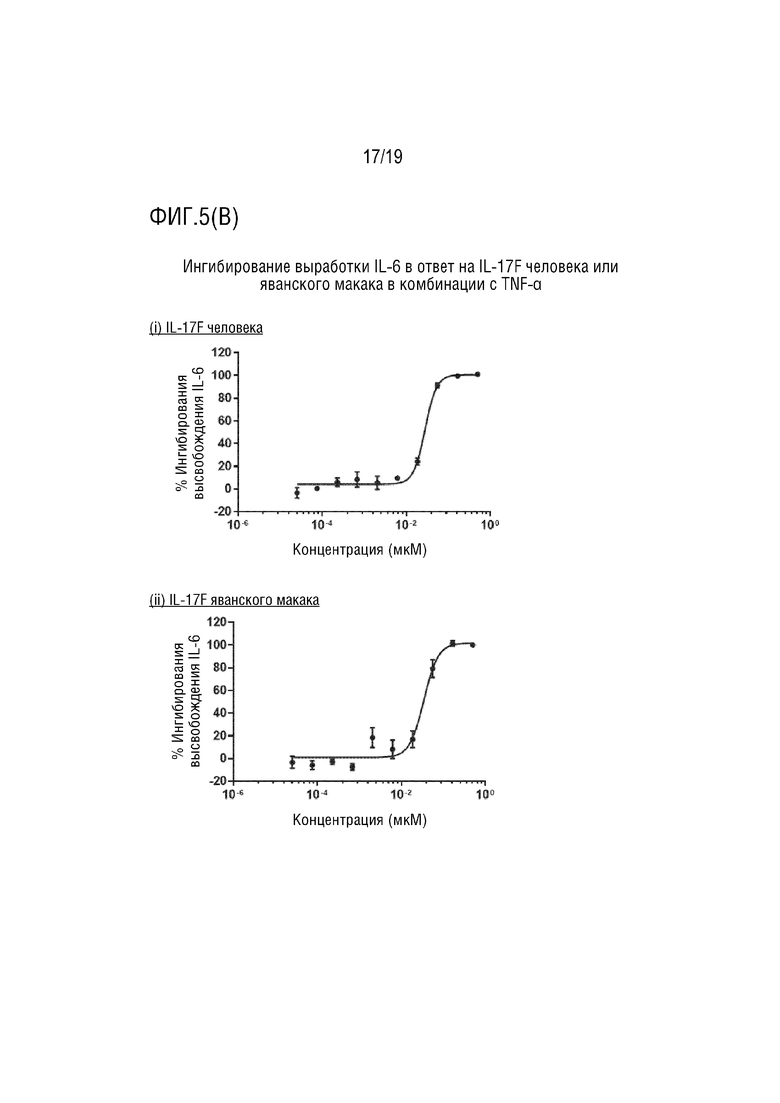
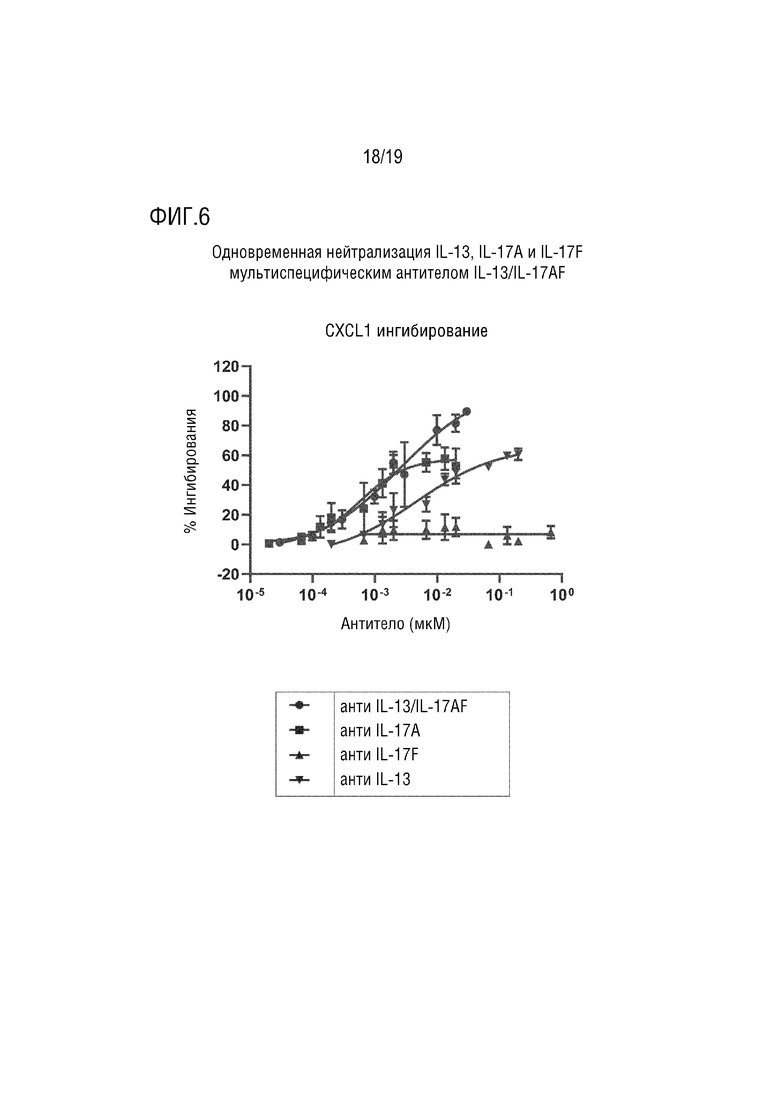

Изобретение относится к области биотехнологии, в частности к мультиспецифическому антителу, которое связывается с IL-13 человека, IL-17A человека и IL-17F, способу его получения, а также к композиции, его содержащей. Также раскрыты выделенный полинуклеотид, кодирующий вышеуказанное антитело, а также вектор и клетка, его содержащие. Изобретение эффективно для лечения или профилактики атопического дерматита, хронической экземы рук, микрополипоза или полипоза носа, пищевой аллергии или эозинофильного эзофагита. 7 н. и 18 з.п. ф-лы, 7 ил., 9 табл., 12 пр.
1. Мультиспецифическое антитело, которое связывается с IL-13 человека, IL-17A человека и IL-17F человека, содержащее IL-13-связывающий участок, который содержит:
вариабельную область легкой цепи, содержащую
CDR-L1 с последовательностью SEQ ID NO:15,
CDR-L2 с последовательностью SEQ ID NO:16 и
CDR-L3 с последовательностью SEQ ID NO:17, и
вариабельную область тяжелой цепи, содержащую
CDR-H1 с последовательностью SEQ ID NO:18,
CDR-H2 с последовательностью SEQ ID NO:19, и
CDR-H3 с последовательностью SEQ ID NO:20,
где указанное антитело содержит антигенсвязывающий участок, который связывается с IL-17A человека и IL-17F человека, который содержит:
вариабельную область легкой цепи, содержащую
CDR-L1 с последовательностью SEQ ID NO:1,
CDR-L2 с последовательностью SEQ ID NO:2 и
CDR-L3 с последовательностью SEQ ID NO:3, и
вариабельную область тяжелой цепи, содержащую
CDR-H1 с последовательностью SEQ ID NO:4,
CDR-H2 с последовательностью SEQ ID NO:5, и
CDR-H3 с последовательностью SEQ ID NO:6,
где указанное антитело содержит:
a) полипептидную цепь формулы (Ia):
VH-CH1-X-V1; и
b) полипептидную цепь формулы (IIa):
VL-CL-Y-V2;
где:
VH представляет вариабельный домен тяжелой цепи;
CH1 представляет домен 1 константной области тяжелой цепи;
X представляет собой связь или линкер;
Y представляет собой связь или линкер;
V1 представляет собой scFv, dsscFv или dsFv;
VL представляет вариабельный домен легкой цепи;
CL представляет домен из константной области легкой цепи, такой как Cкаппа;
V2 представляет собой scFv, dsscFv или dsFv;
где полипептидная цепь формулы (Ia) содержит домен связывания белка А; и
где полипептидная цепь формулы (IIa) не связывается с белком A;
где
VL и VH содержат антигенсвязывающий участок, который связывается с IL-17A человека и IL17F человека,
V2 содержит антигенсвязывающий участок, который связывается с IL-13 человека, и
V1 содержит антигенсвязывающий сайт, который связывается с сывороточным альбумином человека;
где
VL содержит
CDR-L1 с последовательностью SEQ ID NO:1,
CDR-L2 с последовательностью SEQ ID NO:2 и
CDR-L3 с последовательностью SEQ ID NO:3, и
VH содержит
CDR-H1 с последовательностью SEQ ID NO:4,
CDR-H2 с последовательностью SEQ ID NO:5 и
CDR-H3 с последовательностью SEQ ID NO:6;
где
V1 содержит
вариабельную область легкой цепи, содержащую
CDR-L1 с последовательностью SEQ ID NO:39,
CDR-L2 с последовательностью SEQ ID NO:40 и
CDR-L3 с последовательностью SEQ ID NO:41, и
вариабельную область тяжелой цепи, содержащую
CDR-H1 с последовательностью SEQ ID NO:42,
CDR-H2 с последовательностью SEQ ID NO:43 и
CDR-H3 с последовательностью SEQ ID NO:44; и
где
V2 содержит
вариабельную область легкой цепи, содержащую
CDR-L1 с последовательностью SEQ ID NO:15,
CDR-L2 с последовательностью SEQ ID NO:16 и
CDR-L3 с последовательностью SEQ ID NO:17, и
вариабельную область тяжелой цепи, содержащую
CDR-H1 с последовательностью SEQ ID NO:18,
CDR-H2 с последовательностью SEQ ID NO:19 и
CDR-H3 с последовательностью SEQ ID NO:20.
2. Мультиспецифическое антитело по п. 1, где IL-13-связывающий участок содержит
вариабельную область легкой цепи, содержащую последовательность SEQ ID NO:27, и
вариабельную область тяжелой цепи, содержащую последовательность SEQ ID NO:28.
3. Мультиспецифическое антитело по п. 1, где IL-13-связывающий участок содержит
вариабельную область легкой цепи, содержащую последовательность SEQ ID NO:31, и
вариабельную область тяжелой цепи, содержащую последовательность SEQ ID NO:32.
4. Мультиспецифическое антитело по любому из пп. 1-3, где антигенсвязывающий участок, который связывается с IL-17A человека и IL-17F человека, содержит
вариабельную область легкой цепи, содержащую последовательность SEQ ID NO:7, и
вариабельную область тяжелой цепи, содержащую последовательность SEQ ID NO:9.
5. Мультиспецифическое антитело по любому из пп. 1-4, где
VL содержит последовательность SEQ ID NO:7, и
VH содержит последовательность SEQ ID NO:9.
6. Мультиспецифическое антитело по любому из пп. 1-5, где V2 содержит
вариабельную область легкой цепи, содержащую последовательность SEQ ID NO:27, и
вариабельную область тяжелой цепи, содержащую последовательность SEQ ID NO:28.
7. Мультиспецифическое антитело по любому из пп. 1-5, где V2 содержит
вариабельную область легкой цепи, содержащую последовательность SEQ ID NO:31, и
вариабельную область тяжелой цепи, содержащую последовательность SEQ ID NO:32.
8. Мультиспецифическое антитело по любому из пп. 1-7, где V1 содержит
вариабельную область легкой цепи, содержащую последовательность SEQ ID NO:45, и
вариабельную область тяжелой цепи, содержащую последовательность SEQ ID NO:46.
9. Мультиспецифическое антитело по любому из пп. 1-8, где V1 содержит
вариабельную область легкой цепи, содержащую последовательность SEQ ID NO:49, и
вариабельную область тяжелой цепи, содержащую последовательность SEQ ID NO:50.
10. Мультиспецифическое антитело по любому из пп. 1-8,
где вариабельная область легкой цепи и вариабельная область тяжелой цепи V2 соединены линкером,
где указанный линкер содержит последовательность SEQ ID NO:66.
11. Мультиспецифическое антитело по п. 10, где V2 представляет собой
scFv, содержащий последовательность SEQ ID NO:35, или
dsscFv, содержащий последовательность SEQ ID NO:37.
12. Мультиспецифическое антитело по любому из пп. 1-11,
где вариабельная область легкой цепи и вариабельная область тяжелой цепи V1 соединены линкером,
где указанный линкер содержит последовательность SEQ ID NO:68.
13. Мультиспецифическое антитело по п. 12,
где V1 представляет собой
scFv, содержащий последовательность SEQ ID NO:53, или
dsscFv, содержащий последовательность SEQ ID NO:55.
14. Мультиспецифическое антитело по любому из пп. 1-13, где Y представляет собой линкер, содержащий последовательность SEQ ID NO:65.
15. Мультиспецифическое антитело по любому из пп. 1-14, где X представляет собой линкер, содержащий последовательность SEQ ID NO:67.
16. Мультиспецифическое антитело по любому предыдущему пункту, содержащее последовательность SEQ ID NO:57 или SEQ ID NO: 59.
17. Мультиспецифическое антитело по любому предыдущему пункту, содержащее последовательность SEQ ID NO:61 или SEQ ID NO: 63.
18. Мультиспецифическое антитело по любому предыдущему пункту, содержащее последовательность SEQ ID NO:59, и последовательность SEQ ID NO: 63.
19. Выделенный полинуклеотид, кодирующий мультиспецифическое антитело по любому из пп. 1-18,
где указанный полинуклеотид представляет собой последовательность ДНК для экспрессиии указанного мультиспецифического антитела.
20. Вектор экспрессии, несущий полинуклеотид по п. 19 для экспрессиии указанного мультиспецифического антитела.
21. Клетка-хозяин для экспрессиии указанного мультиспецифического антитела, где клетка содержит вектор по п. 20.
22. Способ получения мультиспецифического антитела по любому из пп. 1-15, включающий
культивирование клетки-хозяина по п. 21 в условиях, обеспечивающих получение антитела, и
выделение полученного антитела.
23. Способ по п. 22, включающий стадию очистки белка А.
24. Фармацевтическая композиция для лечения или профилактики атопического дерматита, хронической экземы рук, микрополипоза или полипоза носа, пищевой аллергии или эозинофильного эзофагита, где композиция содержит антитело по любому из пп. 1-18 и фармацевтически приемлемое вспомогательное средство и/или носитель.
25. Способ лечения или профилактики атопического дерматита, хронической экземы рук, микрополипоза или полипоза носа, пищевой аллергии или эозинофильного эзофагита, включающий введение терапевтически эффективного количества мультиспецифического антитела по пп. 1-21 или фармацевтической композиции по п. 24 пациенту.
| WO2012095662 A1, 19.07.2012 | |||
| WO2013102042 A2, 04.07.2013 | |||
| TRACY L | |||
| STATON et al., A phase I, randomized, observer-blinded, single and multiple ascending-dose study to investigate the safety, pharmacokinetics, and immunogenicity of BITS7201A, a bispecific antibody targeting IL-13 and IL17, in healthy volunteers, BMC Pulmonary Medicine, 2019 Jan 7, |

