Область техники
Изобретение относится к биотехнологии, а именно к разработке оптимизированных геннотерапевтических препаратов с высокой стабильностью и пролонгированной экспрессией трансгена vegf.
Современный уровень техники
Геннотерапевтические препараты
Все большее значение в современной медицине приобретают геннотерапевтичекие препараты - группа лекарственных средств, действующим веществом которых являются генные конструкции - нуклеиновые кислоты, содержащие один или несколько генов, кодирующих терапевтические белки.
К настоящему времени в мире уже зарегистрированы и внедрены в клиническую практику 5 таких препаратов, а несколько сотен находятся на стадиях экспериментальных и клинических исследований. Общее количество клинических исследований в рамках генной терапии с 1989 года превысило 1900 [1]. Один из 5 зарегистрированных геннотерапевтических препаратов - «Неоваскулген» (ОАО «Институт Стволовых Клеток Человека») - разработан нами и внедрен в клиническую практику на территории России (РУ № ЛП-000671 от 28.09.11) и Украины (РУ №899/13300200000 от 25.01.2013).
Таким образом, в условиях развития биотехнологии сформировался принципиально новый класс фармацевтических препаратов -геннотерапевтические лекарственные средства.
К наиболее безопасным вариантам генных конструкций, несущих трансген, относится плазмидная ДНК. Именно плазмидная ДНК с геном vegf является действующим веществом упомянутого выше геннотерапевтического препарата «Неоваскулген». Главной проблемой плазмидной ДНК как вектора для доставки трансгена в клетки-мишени абсолютным большинством исследователей признается низкая эффективность трансфекции: лишь 1-2% от общего количества молекул генных конструкций поступает в клетки и/или экспрессируется ими [2]. При этом, количество молекул поступившей в клетку плазмидной ДНК и общее число трансфицированных клеток являются одними из основных факторов, напрямую определяющих уровень продукции терапевтического белка и, соответственно, биологический эффект лекарственного средства. В этой связи, в условиях развития геннотерапевтических препаратов как нового фармакологического класса высоко актуальной является разработка методов, направленных на увеличение продукции терапевтического белка, кодируемого генной конструкцией при снижении количества вводимого в организм препарата или повышении эффективности действия. Это особенно важно для плазмидной ДНК, эффективность трансфекции которой крайне низкая.
Повышение общей концентрации терапевтического белка может быть обеспечено посредством двух основных подходов. Первый - наиболее очевидный - состоит в увеличении уровня трансфекции генных конструкций. Иными словами, чем большее количество плазмидной ДНК поступит в клетки-мишени, тем большая концентрация кодируемого ей белка будет продуцирована. В рамках данного подхода предложен целый ряд физических и химических методов. Однако физические методы увеличения уровня трансфекции требуют применения специального оборудования и не всегда безопасны и выполнимы в клинической практике. Химические же методы требуют изменения состава геннотерапевтического препарата, что требует дополнительных ресурсов для производства, а также может быть сопряжено с негативными эффектами добавленных компонентов.
Второй - альтернативный подход - заключается в продлении времени жизни продукта транскрипции введенной генетической конструкции - мРНК, что приводит к увеличению количества циклов трансляции и, соответственно, повышению результирующего количества терапевтического белка за счет его пролонгированной продукции. Фундаментальные основы данного подхода были сформированы за счет накопления сведений о регуляции времени жизни мРНК. Так, описан целый ряд регуляторных молекул, вовлеченных в модуляцию стабильности мРНК, такие как РНК-связывающие протеины и регуляторные РНК (микроРНК, длинный некодирующие РНК). Важно, что воздействие указанных веществ на мРНК опосредуется через связывание с ее 3'-нетранслируемой областью (3'UTR), содержащей специальные последовательности - дестабиизирующие элементы, в частности, аденин-уридин-обогащенный элемент (AU-rich element). Ряд веществ, такие как AU-rich element RNA-binding protein 1 (AUF1), tristetraprolin (TTP), KH-type splicing regulatory protein (KSRP), связываясь со специфическими сайтами 3'UTR индуцируют деградацию мРНК, тогда как другие, например polyadenylate-binding protein-interacting protein 2 (PAIP2), наоборот, обеспечивают стабилизацию мРНК [3, 4].
Координация процессов деградации и стабилизации мРНК имеет важное значение для нормального функционирования клеток, а некоторые патологические состояния воспалительного или онкологического генеза, по мнению ряда авторов, могут быть связаны с посттранскрипционной дисрегуляцией, приводящей либо к недостаточной, либо к избыточной продукции факторов роста, онкогенов и других биологически активных веществ [4]. В этой связи, последовательность нуклеотидов 3'UTR имеет принципиальное значение для трансдукции регуляторных влияний, эффектором которых является мРНК, и детерминации времени жизни мРНК в целом.
Анализируя данные современного уровня техники, становится очевидным, что изменение последовательности нуклеотидов 3'UTR, особенно дестабилизирующих элементов, может обеспечить продление жизни мРНК. Более того, разработаны методы сайт-направленного мутагенеза, позволяющие технически выполнить необходимые изменения в генной конструкции и, соответственно, мРНК как продукта ее транскрипции. Однако до сих пор нет точных, исчерпывающих и систематизированных данных о том, какие варианты последовательности 3'UTR, какие ее минорные сайты или их комбинации за какие аспекты функционирования мРНК отвечают [3]. Отчасти это связно с тем, что количество изоформ мРНК, отличающихся последовательностью нуклеотидов 3'UTR, варьирует в широких пределах, в том числе из-за альтернативных сплайсинга и полиаденилирования. При этом, качественный и количественный состав дестабилизирующих элементов 3'UTR определяется также и кодирующей областью нуклеиновой кислоты. Все это, в сочетании с тем фактом, что 3'UTR определяет не только время жизни мРНК, но и другие составляющие ее «физиологии», как транспорт из ядра клетки, цитоплазматическая локализация, эффективность трансляции и т.д., предопределяет сложности и, во многом, непредсказуемость моделирования времени жизни мРНК через изменение последовательности 3'UTR, что может выражаться как в увеличении времени жизни транскрипта, так и в снижении.
Несмотря на все трудности и недостаточную изученность вопроса детерминации, или «программирования», времени жизни мРНК через изменение последовательности 3'UTR, предпринимаются попытки эмпирически подобрать такие варианты изменений в соответствующей области конкретных вариантов генных конструкций, которые увеличили бы стабильность мРНК без негативного влияния на ее функционирование.
В частности, описан способ увеличения продукции трансгена за счет замены последовательности AU-rich элемента, представленной AUUUA, на другие варианты и их комбинации, ограниченные следующими: AUGUA, AUAUA, GUGUG, AGGGA, GAGAG [5]. Однако, указанная последовательность дестабилизирующего элемента в 3'UTR характерна далеко не для всех генов (описана большинством исследователей для G-CSF), а, с другой стороны - ей не исчерпывается перечень дестабилизирующих элементов, в связи с чем, продление жизни мРНК может быль нивелировано.
Другими авторами разработана специальная последовательность 3'UTR гена эритропоэтина, обеспечивающая пролонгированную продукцию трансгена, входящего в состав плазмидной ДНК. Последовательность строго конкретизирована и имеет длину 100 нуклеотидов [6].
Описаны также способы элиминации специфических последовательностей 3'UTR, ответственных за связывание с различными микроРНК, индуцирующими деградацию мРНК [7].
Однако большинство предлагаемых изменений последовательности 3'UTR относятся к протяженным делециям или заменам, что неизбежно сопряжено с риском возникновения негативных эффектов на метаболизм мРНК. Кроме того, предлагаемые решения крайне зависимы от кодирующей области гена, поэтому одни неприменимы, другие недостаточно эффективны для гена vegf и продления жизни продукта его транскрипции.
Стоящие же перед нами задачи сводились к необходимости увеличить продукцию VEGF таргетными клетками (на примере клеток человека линии HEK293, мультипотентных мезенхимных стромальных клеток, фибробластов человека и пр.), в которые проникла плазмидная ДНК, за счет увеличения продолжительности жизни специфической мРНК указанного гена с помощью оптимизиции его 3 'UTR.
VEGF и его биологическая роль
VEGF - семейство биологически активных белков, впервые выделенных J. Folkman с соавт. в 1971 [8], которые считаются одними из основных ауто- и паракринных факторов регуляции васкуло-, ангио- (VEGF-А, В; PIGF) и лимфогенеза (VEGF-C, D); вырабатываются клетками всех тканей организма, включая эпителиальные.
В постнатальном периоде развития человека наибольшее влияние на формирование кровеносных сосудов оказывает VEGF-A (изоформы 121, 145, 148, 165, 183, 189, 206) [9]. Обнаружены три типа рецепторов VEGF. 1 и 2 тип вовлечены в ангиогенез, 3 - в образование лимфатических сосудов. При этом, рецептор 1 типа обладает большей афинностью к VEGF, однако его тиразинкиназная активность гораздо ниже, чем у рецептора 2 типа, что расценивается как один из регуляторных механизмов, предотвращающих избыточную активность VEGF. Соответственно, именно через рецептор 2 типа в норме реализуются эффекты VEGF [10, 11]. После взаимодействия VEGF со специфическим рецептором 2 типа происходит аутофосфорилирование его внутриклеточных тирозиновых сайтов (Y951, 1054, 1059, 1175, 1214) киназных и карбокситерминального доменов [11], которые, в свою очередь, активируют ряд внутриклеточных белков, таких как фосфолипазы Cγ, Cβ3, адаптерные белки SRK, NCK, SHB, SCK и др., являющиеся первыми звеньями сложных каскадов трансдукции сигналов, изменяющих морфофункциональное состояние клеток-мишеней (главным образом, эндотелиальных). В частности, фосфолипаза Cγ гидролизирует мембранный фосфолипид PIP2 с образованием диацилглицерола и инозитол-1,4,5-трифосфата, увеличивающего внутриклеточное содержание кальция, которые вместе активируют протеинкиназу С, которая, в свою очередь, запускает последовательную активацию сигнального пути RAS-ERK, приводящего к индукции митоза. В результате, повышается пролиферативная активность эндотелиальных клеток [10]. Фосфолипаза Cβ3 участвует в полимеризации актина и формировании стресс-фибрилл, обеспечивающих миграцию и двигательную активность клеток, в целом [12]. Через активацию сигнального пути «фосфоинозитид-3-киназа - протеинкиназа В» (PI3K/AKT) VEGF блокирует апоптоз, ингибируя каспазы 3, 7 и 9, тем самым повышая выживаемость клеток. Кроме того, ось PI3K/AKT с помощью ионов кальция модулирует работу эндотелиальной NO-синтазы, что сопровождается увеличением продукции NO и повышением проницаемости сосудов, что является необходимым звеном в ангиогенезе (рис. 3) [13, 14]. Таким образом, VEGF через специфический рецептор 2 типа индуцирует активацию, миграцию, пролиферацию и дифференцировку эндотелиоцитов и их клеток-предшественниц, повышение выживаемости клеток, что в сочетании с модуляцией межклеточных взаимодействий и повышением проницаемости сосудов служит необходимыми условиями формирования капилляро-подобных структур с последующим ремоделированием в «зрелые» сосуды [10-16].
Учитывая роль VEGF как ключевого ангиогенного фактора, различные варианты кодирующего его гена используются для создания генных конструкций, показанных для лечения пациентов с заболеваниями сердечнососудистой системы ишемического генеза [17, 18]. Кроме того, неспецифический ангиогенный эффект таких геннотерапевтических препаратов может оказать положительное влияние при других патологических состояниях, требующих активации репаративного процесса: травмы периферических нервов [19], синдром диабетической стопы [20], боковой амиотрофический склероз [21], повреждения костей скелета [22, 23] и др.
Что касается костной ткани, то как в случае первичного, так и вторичного остеогистогенеза именно сосуды, прорастающие в рыхлую волокнистую соединительную или хрящевую ткани, создают необходимые условия для дифференцировки резидентных клеток в остеобластическом направлении, а также для миграции камбиальных резервов (периваскулярно и с кровотоком). Помимо опосредованного через ангиогенез влияния, VEGF оказывает и прямое стимулирующее действие на клетки остеобластического дифферона, которые не только продуцируют VEGF [24], но и экспрессируют его рецепторы 1 и 2 типов как в эмбриогенезе [25], так и в постнатальном периоде развития [26]. Показано, что под воздействием VEGF пролиферация камбиальных клеток костной ткани значительно увеличивается (до 70%), а также активируется миграция остеогенных клеток по градиенту концентрации VEGF [27-29].
В последние годы, помимо канонического - рецепторного - механизма действия VEGF, появились данные о принципиально ином механизме, который обозначают как «интракринный». Подтверждением этому стали результаты исследований, продемонстрировавших, что прогениторные клетки, коммитированные в остеобластическом направлении (экспрессирующие Osx), синтезируют VEGF не только «на экспорт», но и для обеспечения собственной дифференцировки в остеобластическом направлении [30].
Таким образом, VEGF обладает широким спектром действия на клетки эндотелиальной и мезенхимальных клеточных линий. Основной его биологический эффект связан с индукцией формирования кровеносных и лимфатических сосудов, однако, по всей видимости, для VEGF характерны и другие механизмы прямого действия на клетки различных дифферонов через рецепторные и интракринные механизмы.
В этой связи, плазмидная ДНК с геном vegf, характеризующаяся пролонгированной экспрессией трансгена, стала бы эффективным инструментом в разработке лекарственных препаратов и ген-активированных медицинских изделий, предназначенных не только для лечения пациентов с заболеваниями сердечно-сосудистой системы, но и другой патологией, при которой локальное повышение уровня VEGF в affected area обеспечило бы усиление репаративного процесса.
Перечень иллюстраций:
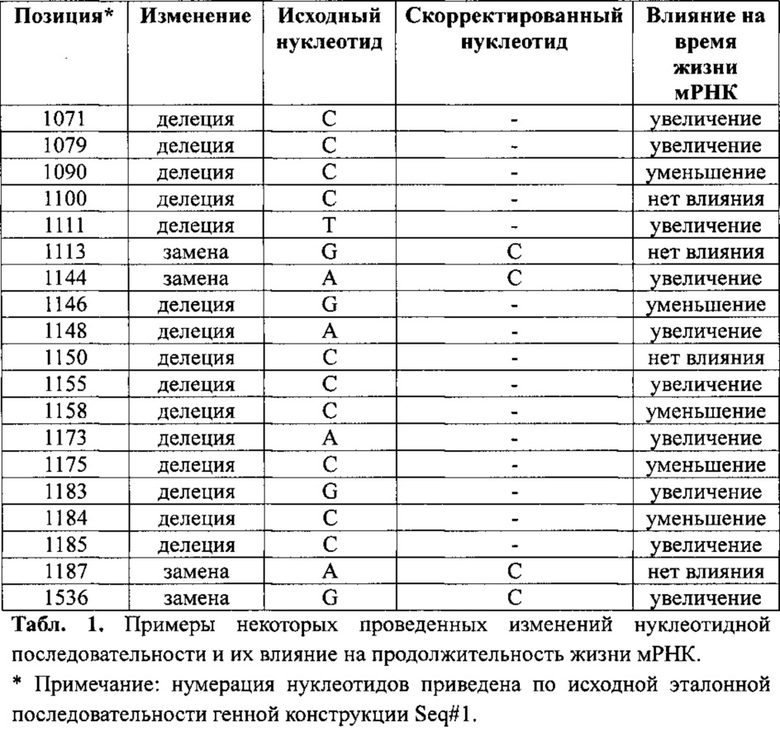
Рис. 1. Динамика продукции (накопления) мРНК культурами клеток, трансфицированными различными вариантами генных конструкций: 1 - Seq#1 (исходная генная конструкция), 2 - Seq#2, 3 - Seq#3, 4 - Seq#4, 5 - Seq#5, 6 - Seq#6, 7 - Seq#7, 8 - Seq#8, 9 - Seq#9, 10 - Seq#10, 11 - Seq#11
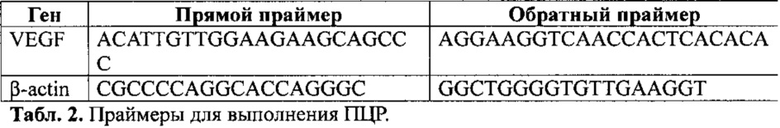
Рис. 2. Динамика продукции (накопления) мРНК гена vegf трансфицированной культурой клеток: А - исходная генная конструкция (Seq#1); В - оптимизированная генная конструкция SEQ#11. Результаты представлены как кратное увеличение по сравнению с уровнем продукции мРНК эндогенного VEGF.
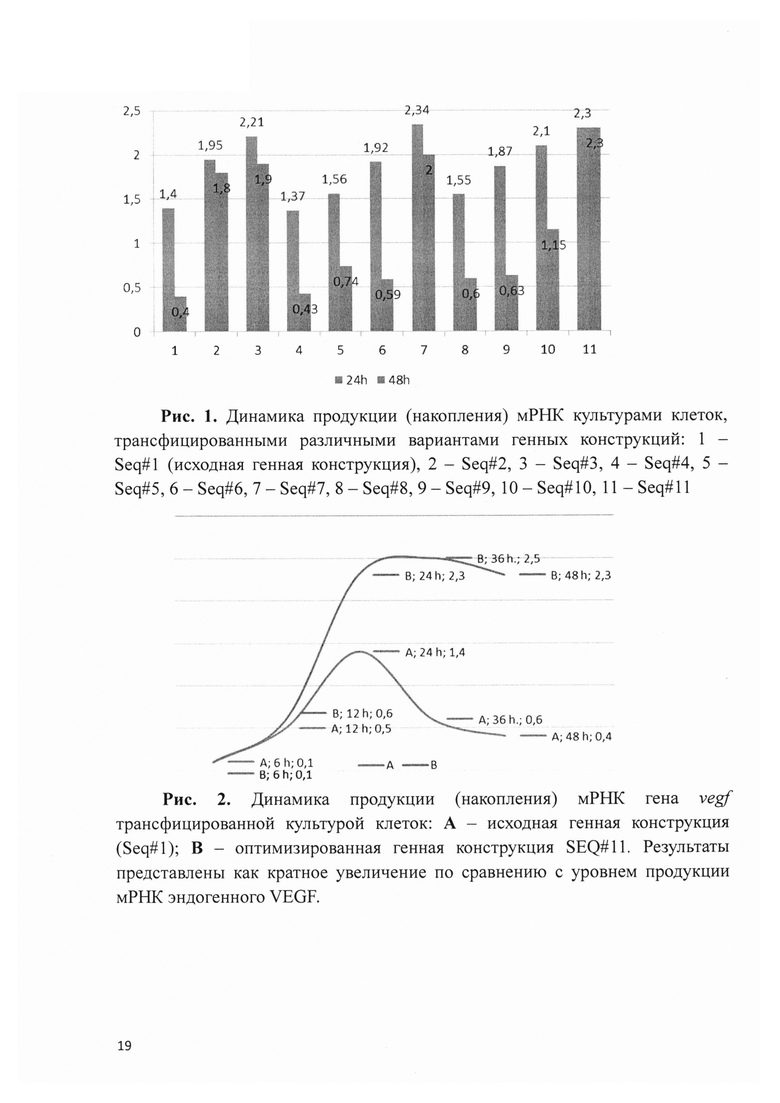
Рис. 3. Динамика продукции мРНК гена vegf трансфицированной культурой клеток: А - эндогенная продукция (культура нетрансфицированных клеток); В - исходная генная конструкция (Seq#1); С - оптимизированная генная конструкция Seq#11.
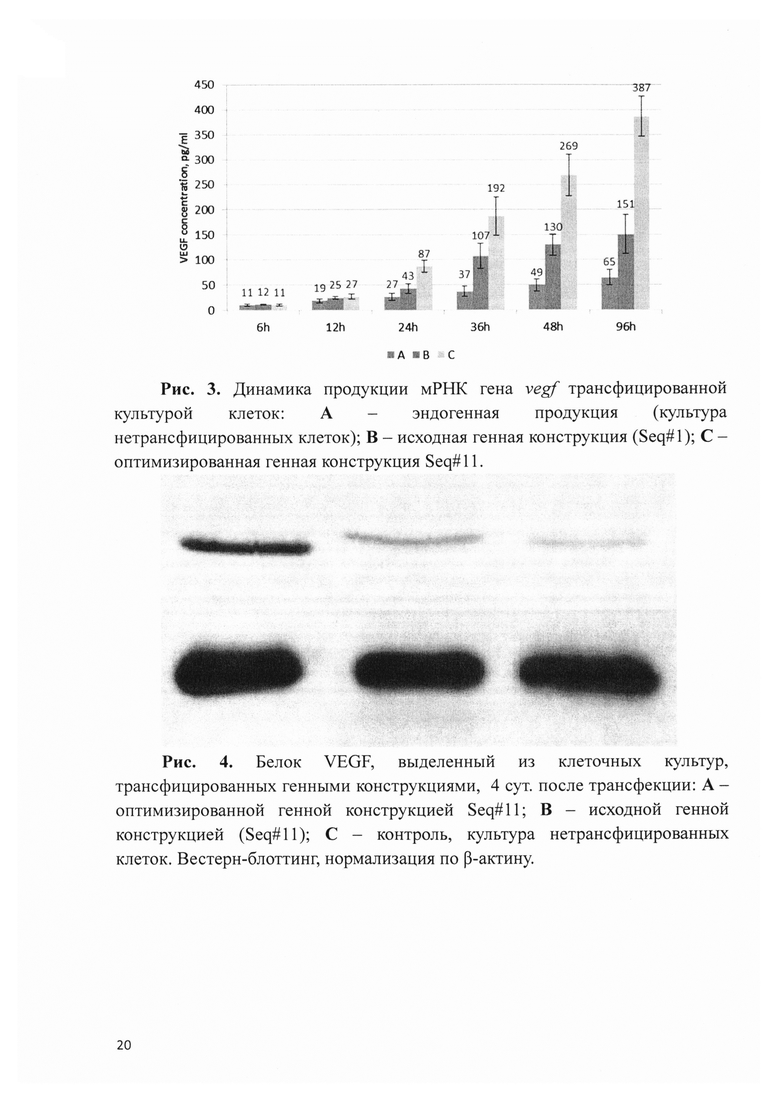
Рис. 4. Белок VEGF, выделенный из клеточных культур, трансфицированных генными конструкциями, 4 сут после трансфекции: А - оптимизированной генной конструкцией Seq#11; В - исходной генной конструкцией (Seq#11); С - контроль, культура нетрансфицированных клеток. Вестерн-блоттинг, нормализация по β-актину.
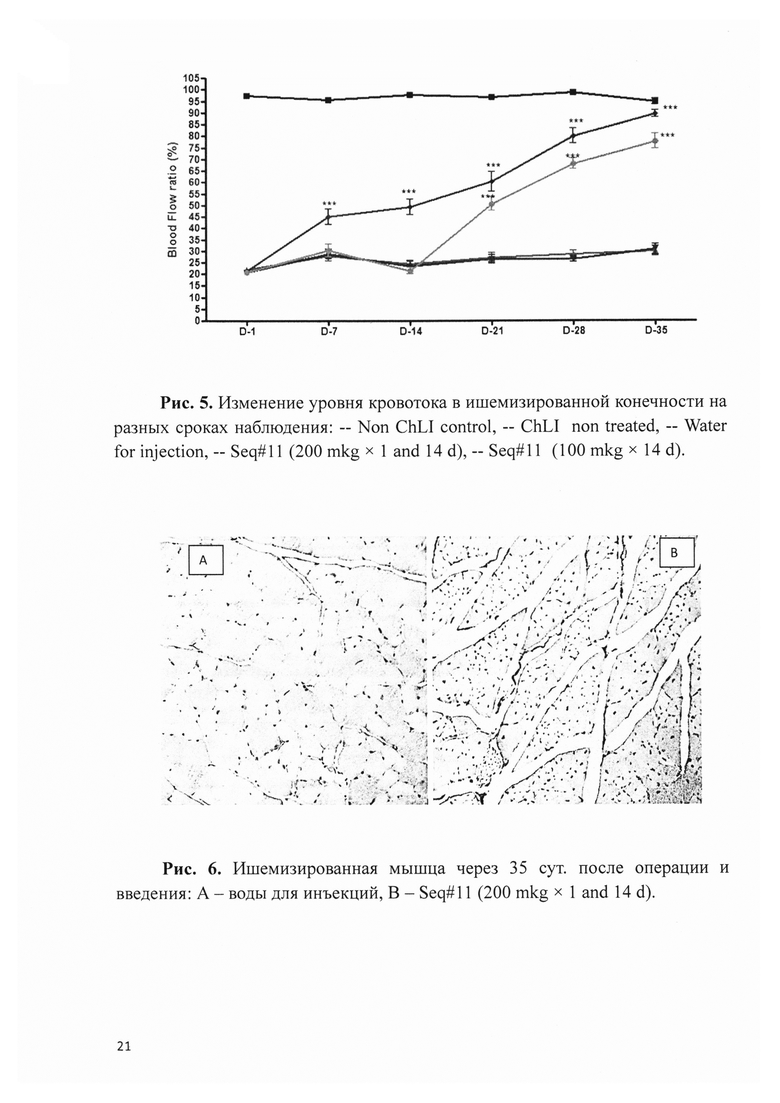
Рис. 5. Изменение уровня кровотока в ишемизированной конечности на разных сроках наблюдения: - Non ChLI control, - ChLI non treated, - Water for injection, - Seq#11 (200 mkg × 1 and 14 d), - Seq#11 (100 mkg × 14 d).
Рис. 6. Ишемизированная мышца через 35 сут после операции и введения: А - воды для инъекций, В - Seq#11 (200 mkg × 1 and 14 d).
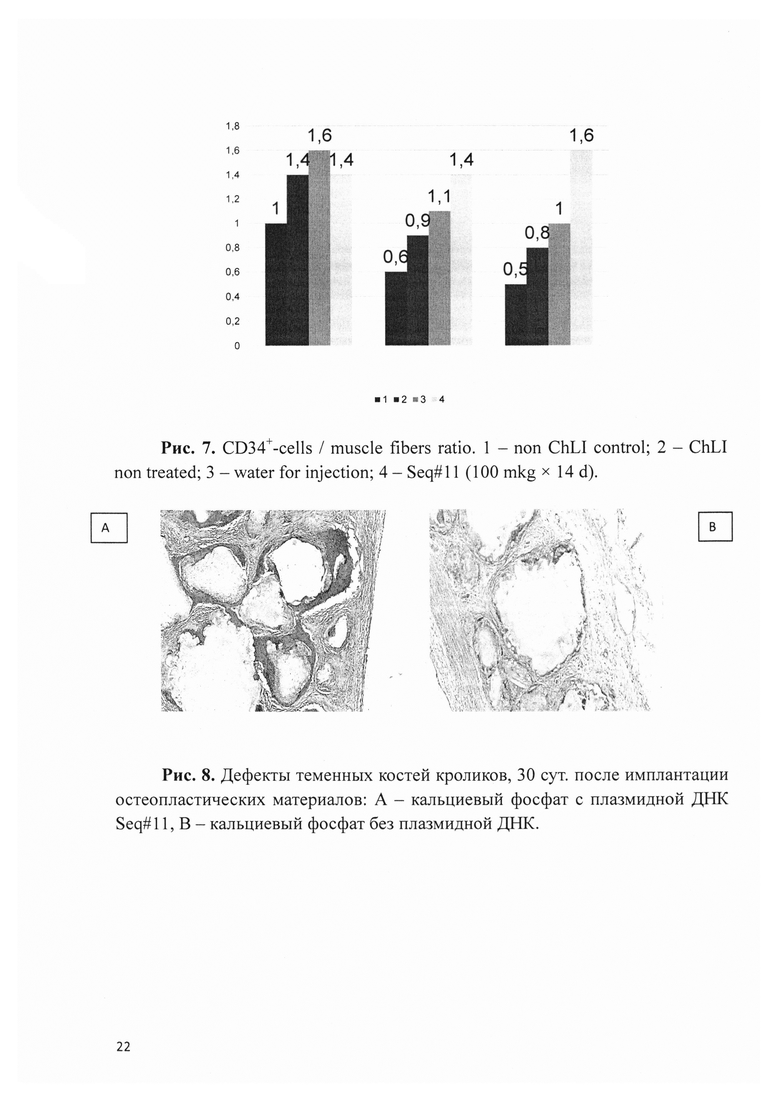
Рис. 7. CD34+-cells / muscle fibers ratio. 1 - non ChLI control; 2 - ChLI non treated; 3 - water for injection; 4 - Seq#11 (100 mkg × 14 d).
Рис. 8. Дефекты теменных костей кроликов, 30 сут после имплантации остеопластических материалов: А - кальциевый фосфат с плазмидной ДНК Seq#11, В - кальциевый фосфат без плазмидной ДНК.
Рис. 9. Карта оптимизированной последовательности Seq#11.
Краткое описание настоящего изобретения
С использованием методов сайт-направленного мутагенеза и количественного ПНР анализа молекул РНК в клетке нами были проанализированы конструкции, несущие различные изменения последовательности нуклеотидов 3'UTR терапевтического гена vegf, по сравнению с исходной генной конструкцией (Seq#1, разработана нами ранее). Примеры влияния некоторых изменений приведены в табл.1. Как видно из приведенных данных, некоторые изменения нуклеотидной последовательности (делеции или замены) сказывались на продолжительности жизни положительным образом (например делеция С 1079), другие не оказывали никакого влияния (например, делеция С 1100) или приводили к уменьшению жизни мРНК (например, делеция С 1090). При этом разница в продолжительности жизни мРНК гена vegf в клетке в крайних случаях (делеция С 1079 - увеличение; делеция С 1090 - уменьшение) составляла в среднем более 6 ч. при средней жизни мРНК 6 ч.
На втором этапе исследования все изменения, оказывающие положительное влияние, были использованы для создания ряда вариантов генных конструкций, несущих от четырех до десяти "положительных" мутаций в различных комбинациях, т.е. отличающихся последовательностью нуклеотидов на отрезке 1070-1600 (Seq#2-11).
На основании полученных данных нами была выбрана и синтезирована оригинальная 3'UTR концевая часть гена vegf, которая в совокупности с кодирующей частью гена обеспечивала увеличение продолжительности жизни транскрипта гена на 70% (SEQ#11), в которую были сведены некоторые мутации, оказавшие в совокупности наибольшее положительное влияние, а именно: делеция С 1079, делеция Τ 1111, замена А на С 1144, делеция А 1148, делеция С 1155, делеция А 1173, делеция С 1185, замена G на С 1536.
Полученный вариант генетической конструкции характеризуется максимальным временем жизни мРНК и, соответственно, наибольшей общей продукцией белка VEGF, превышающей таковую, характерную для исходной генных конструкций (Seq#1).
На основе разработанной оптимизированной генной конструкции и адъювантов, обеспечивающих криопротекцию, стабилизацию рН и получение изотонического раствора для инъекций, были созданы фармацевтические композиции, обладающие выраженной ангиогенной активностью.
Разработанные фармацевтические композиции предназначена для применения в случае заболеваний и патологических состояний, в лечении которых требуется стимуляция ангиогенеза (ишемия тканей) или репаративной регенерации тканей, которая может быть осуществлена через активацию ангиогенеза (восстановление целостности периферических нервов, костей скелета и др.)
Для производства разработанных фармацевтических композиций создан бактериальный (Е.colli) штамм-продуцент генной конструкции Seq#11, депонированный во Всероссийской коллекции микроорганизмов в соответствии с Будапештским Договором по международному депонированию микроорганизмов с целью патентования. Номер депонирования штамма VKM B-2967D. Справку о Международном депонировании штамма прилагаем.
Подробное описание настоящего изобретения
Сайт-направленный мутагенез 3'UTR гена vegf был выполнен с использованием QuikChange Lightning Site-Directed Mutagenesis Kit (Agilent Technologies, США), согласно инструкции производителя.
В результате сайт-направленного мутагенеза исходной плазмидной ДНК (Seq#1), нами были получены различные варианты изменений в 3 'UTR гена vegf. Полученные варианты плазмидной ДНК с геном vegf отличающиеся последовательностью 3'UTR, были использованы для трансфекции клеток линии HEK293 с последующим определением динамики накопления продукта экспрессии генной конструкции - мРНК гена vegf.
Для этого, клетки HEK293 в количестве 2×105 помещали в лунки 6-луночного планшета с культуральной средой DMEM/F-12 с добавлением 10% FBS. После прикрепления клеток в культуральную среду вносили 10 мкг одного из полученных вариантов плазмидной ДНК (в 100 мкл воды для инъекций). В качестве негативного контроля использовали 100 мкл воды для инъекций без плазмидной ДНК, что позволяло оценить уровень продукции мРНК эндогенного vegf. Для определения уровня экспрессии гена vegf методом РТ-ПЦР из клеток через 6, 12, 24, 36 и 48 ч. после трансфекции выделяли общую РНК на колонках с помощью набора PureLink RNA Mini Kit (Invitrogen, США), согласно инструкции производителя. Кратко, осажденные клетки тщательно лизировали в объеме 350 мкл лизирующего буфера в присутствии 1% β-меркаптоэтанола. Добавляли равный объем 70% этанола, вортексировали и переносили на колонки. Центрифугировали 1 мин при 12,4 тыс.об/мин. Сливали отцентрифугированную жидкость. Добавляли на мембрану 600 мкл промывочного раствора 1, центрифугировали 15 сек при 12,4 тыс.об/мин, убирали жидкость. Добавляли на мембрану 500 мкл промывочного раствора 2, центрифугировали при тех же условиях, убирали жидкость. Повторяли отмывку с промывочным раствором 2. Переносили колонки в новые пробирки и центрифугировали 1 мин. при 12,4 тыс.об/мин. Переносили колонки в пробирки для сбора РНК. Непосредственно на мембрану вносили 15 мкл воды, без РНКаз, инкубировали 1 мин. при комнатной температуре, центрифугировали 90 сек при 12,4 тыс.об/мин. До дальнейшего применения пробы хранились при температуре -80°C. Далее проводили синтез первой цепи кДНК с помощью реактивов компании Promega (США). Уровень экспрессии таргетных генов (в молях) определяли по сравнению с экспрессией гена домашнего хозяйства β-актина с помощью набора для проведение ПЦР в реальном времени в присутствии SYBR Green I и референсного красителя ROX (Синтол, Россия) На рис 1. приведен пример одной из нуклеотидных замен, обеспечивающих увеличение жизни мРНК. Постановка реакции обратной транскрипции:
1) на льду: к 10 мкл РНК добавляли 2,5 мкл Random Hexamer, инкубировали 5 мин. при 70°C;
2) убирали лед;
3) осаждали капли;
4) приготавливали смесь для каждого образца:
- 5 мкл буфера для обратной транскриптазы MMLV 5Х
- 1,25 дНТП
- 1,25 RNase Inhibitor
- 1 мкл MMLV Tранскриптазы
- 4 мкл H2O
5) добавляли по 12,5 мкл смеси в каждую пробирку, инкубировали 1 ч. при 37°C;
6) останавливали реакции инкубацией при 75°C в течение 5 мин. Все образцы кДНК хранились при температуре -80°C.
Для выполнении ПЦР в реальном времени использовали прямые и обратные праймеры к vegf и β-actin (табл.2): Реакционную смесь для ПЦР в реальном времени в присутствии SYBR Green I применяли согласно инструкции производителя. Измерения проводили на приборе CFX90 Touch Real-time PCR Detection System.
В результате проведенных исследований были выявлены мутации, положительно влияющие на время жизни мРНК (делеция С 1071, делеция С 1079, делеция Τ 1111, замена А на С 1144, делеция А 1148, делеция С 1155, делеция А 1173, делеция G 1183, делеция С 1185, замена G на С 1536 и др.). Указанные изменения в различных комбинациях из 4-10 мутаций были сведены в синтетические конструкты, включающие кодирующую часть гена vegf и 3'UTR, оптимизированные по отношению к гену vegf. Основные из полученных вариантов представлены Seq#2-11. Чтобы выбрать наиболее оптимальный вариант из разработанных генных конструкций, было выполнено сравнительное исследование, в ходе которого определяли динамику накопления мРНК трансфицированными клетками с помощью ПЦР-РТ, как указано выше. Результаты основных десяти генных конструкций в виде кратности увеличения концентрации мРНК на сроках 24 и 48 ч после трансфекции представлены на рис. 1.
Как следует из графика, в некоторых случаях комбинации нескольких мутаций, каждая из которых в отдельности оказывала положительное влияние на время жизни мРНК, не сопровождались значимым увеличением накопления мРНК, по сравнению с исходной генной конструкцией (Seq#1). Среди генных конструкций, характеризовавшихся увеличением времени жизни мРНК, отсутствие снижения концентрации определялось только в случае Seq#11, тогда как в остальных случаях уровень мРНК снижался через 48 ч. после трансфекции по сравнению с показателем на сроке 24 ч. В этой связи, именно данная генная конструкция была выбрана как наиболее оптимальная и подвергнута дальнейшим сравнительным исследованиям.
На рис. 2. приведен результат ПЦР-РТ конструкции Seq#11 в сравнении с неоптимизированной последовательностью Seq#1. На приведенном графике видно, что уже через 12 ч. уровень мРНК гена VEGF оптимизированной конструкции превышал значение параметра в контроле, где для трансфекции была использована исходная плазмидная ДНК. Важно, что динамика прироста концентрации мРНК в обеих группах совпадала с пиком на отметке в 24 ч. после трансфекции. Однако только в случае исходной генной конструкции наблюдалось постепенное снижение уровня мРНК, обусловленное быстрой биодеградацией молекул, тогда как в случае оптимизированной конструкции (Seq#11) определялась фаза «плато» длительностью не менее 6 ч. с последующим плавным снижением концентрации мРНК. При этом на крайней временной точке разница между группами по данному показателю составила 190%.
Разработанная плазмидная ДНК, несущая ген vegf, была исследована in vitro для количественной оценки продукции белка VEGF клетками HEK273. При этом, концентрация терапевтического белка определялась в культуральной среде с помощью ELISA через 6, 12, 24, 48, 72 и 96 ч. после трансфекции клеток. Оказалось, что накопленная концентрация терапевтического белка в культуральной среде клеток, трансфицированных генной конструкцией, значительно превышала значение параметра в контроле, в том числе в группах с альтернативными вариантами разработанных генных конструкций, характеризовавшихся неоптимальной последовательностью 3'UTR гена vegf и стандартным временем жизни мРНК трансгена (рис. 3). Важно, что максимальный прирост концентрации VEGF в культуральной среде обеих групп трансфицированных клеток (увеличение в 2,3-2,4 раза) наблюдался через 36 часов после начала эксперимента, что соответствовало данным ПЦР-РТ с пиковой концентрацией мРНК генных конструкций на временной точке 24 часа. В дальнейшем, если концентрация белка VEGF в среде трансфицированных оптимизированной генной конструкцией клеток продолжала нарастать высокими темпами, то в группе с исходной плазмидной ДНК прирост существенно замедлялся, незначительно превышая прирост в контрольной группе (культура без трансфекции).
Определение накопленной концентрации белка VEGF в культуральной среде позволяет судить о динамике его продукции клетками и количественном приросте на каждом контрольном сроке, однако не позволяет оценить внутриклеточную концентрацию белка на каждой из временных точек. При этом особенно важным в комплексной оценке времени жизни мРНК генных конструкций является определение уровня белка VEGF в клетках на крайних сроках наблюдения. В этой связи, клетки после 4 сут культивирования были подвергнуты Вестерн-блоттинг анализу по стандартной методике с использованием антител к белку VEGF. Концентрация белка оказалась наибольшей в лизате клеток, трансфицированных оптимизированной генной конструкцией Seq#11 (рис. 4). При этом уровень белка в случае клеток, трансфицированных исходной конструкцией, на данном сроке наблюдения незначительно превышал эндогенную концентрацию (контроль).
Более того, кондиционированная среда клеток, трансфицированных оптимизированными генными конструкциями, характеризовалась статистически значимо большей ангиогеннной активностью, определенной в стандартном тесте HUVEC tube formation assay.
Сопоставляя результаты проведенных исследований, можно заключить, что мРНК оптимизированной генной конструкции (Seq#11) характеризовалась большим временем жизни, что привело к более длительной продукции высоких концентраций терапевтического белка VEGF вплоть до крайних сроков наблюдения. Исходная же генная конструкция характеризовалась меньшей стабильностью мРНК, коротким и менее выраженным биологическим эффектом.
Таким образом, основной технический результат настоящего изобретения состоит в увеличении общей продукции терапевтического белка за счет увеличения времени жизни мРНК, достигнутой посредством выявления оптимальных изменений последовательности 3'UTR гена vegf. Неожиданным стал тот факт, что замена всего одного нуклеотида в целевой области оказала столь выраженный эффект на продолжительность жизни мРНК и, соответственно, продукцию терапевтического белка. Во всех известных работах в данной области исследователи выполняли другие изменения, с вовлечением гораздо большего количества нуклеотидов. Учитывая недостаточную изученность функционального предназначения каждого сайта в 3'UTR мРНК, наиболее благоприятными с точки зрения безопасности и нивелирования влияния на другие аспекты метаболизма мРНК являются именно минимальные, точечные изменения, в случае, если они позволяют добиться нужного эффекта. Важно отметить, что длина последовательности 3'UTR гена vegf составляет не менее 250 нуклеотидов, следовательно, общее количество только единичных, точечных мутаций в виде замены одного нуклеотида на другой составляет не менее 750. Принимая во внимание другие возможные варианты изменений (делеции, дупликации, инсерции), а также с учетом задействования более одного сайта, количество вариантов генных конструкций, отличающихся 3'UTR гена vegf составляет сотни тысяч. Таким образом, выявление нескольких вариантов изменений последовательности, приводящих к увеличению продолжительности жизним мРНК без негативного влияния на другие аспекты ее функционирования равносильно выигрышу в лотерею.
Учитывая опыт в разработке геннотерапевтических препаратов были созданы фармацевтические композиции на основе разработанной оптимизированных генных конструкций и фармакологически приемлемых вспомогательных веществ, представленных, по меньшей мере, одним криопротектантом, обладающим свойствами наполнителя, и стабилизатором рН, в эффективных количествах обеспечивающих получение изотонического раствора плазмидной ДНК для инъекций.
Состав фармацевтической композиции, обеспечивающий сохранение свойств оптимизированной плазмидной ДНК, представляет собой:
- плазмидная ДНК от 0,1 до 10 мг/мл, предпочтительно от 0,5 до 4 мг/мл, наболее предпочтительно от 0,8 до 1,2 мг/мл.
- глюкоза (декстроза) от 200 до 400 мМ, предпочтительно от 250 до 350 мМ, наболее предпочтительно от 280 мМ до 320 мМ.
- фосфат натрия (смесь тризамещенного, двузамещенного и однозамещенного фосфатов натрия) в концентрации от 3 до 30 мМ, предпочтительно от 5 до 20 мМ, наболее предпочтительно от 8 до 12 мМ.
рН раствора от 7,0 до 9,0, предпочтительно от 7,2 до 8,5, наиболее предпочтительно от 7,4 до 8,2.
Состав одного из вариантов фармацевтической композиции следующий:
- плазмидная ДНК по Seq#11 - 1,2 мг;
- декстрозы моногидрат - 60 мг;
- натрия гидрофосфата додекагидрат - 3,94 мг;
- натрия дигидрофосфата дигидрат - 0,16 мг.
Данный вариант были использован в исследованиях, описанных в примерах.
Разработанные генные конструкции и фармацевтические композиции на их основе были исследованы in vivo в экспериментальных моделях, воспроизводящих основные патофизиологические и патоморфологические симптомы, характерные для заболеваний, являющихся известными показаниями к применению плазмидной ДНК с геном vegf.
Пример 1. Модель хронической ишемии нижних конечностей.
Исследование было выполнено на иммунодефицитных мышах (n=90), которым для моделирования хронической ишемии нижней конечности (ХИНК) выполнялось пересечение правой бедренной артерии в паховой области. Введение фармацевтической композиции на основе плазмидной ДНК по Seq#11 выполнялось в двух вариантах: 100 мкг однократно на 14-е сут и по 200 мкг двукратно с на 1-е и 14-е сут - в проксимальный и дистальный участок послеоперационной раны. Контролем служили три группы животных: здоровые, оперированные но без введения каких-либо веществ; оперированные с введением воды для инъекций. Животных выводили из эксперимента через 7, 21, 35 сут после операции, конечности подвергали гистологическому исследованию с определением количества сосудов и отношения числа эндотелиоцитов к мышечным волокнам. На сроках 1, 7, 14, 21, 28 и 35 сут выполняли лазерную сканирующую доплерометрию для определения уровня кровотока (blood flow perfusion rate) в ишемизированной и здоровой конечности.
Оказалось, что независимо от дозы и режима введения, уровень кровотока в ишемизированной конечности увеличивался только в экспериментальных группах через 7 сут после введение разработанного геннотерапевтического препарата. Уровень кровотока увеличивался, достигая к крайнему сроку наблюдения значений, близких к параметру здоровых животных. В контрольных группах оперированных животных уровень кровотока оставался на низком уровне без положительной динамики (рис. 5).
Функциональные результаты были подтверждены данными гистологического исследования. В экспериментальных группах было выявлено большее количество сосудов (рис. 6), а также большее отношение CD34+-клеток (эндотелиоцитов) к мышечным волокнам (рис. 7).
Пример 2. Модель костного дефекта критического размера. Разработанная генная конструкция по Seq#11 была объединена с матриксом из кальциевого фосфата по ранее разработанному протоколу:
1. Подготовка носителя:
а) отмывка (инкубирование в 0,5 M фосфатного буфера в объеме 1 мл при температуре 37°C при постоянном встряхивании в течение 12 ч.);
б) уравновешивание (обработка 10 мМ фосфатного буфера в объеме 1 мл при температуре 37°C при постоянном встряхивании, 3 раза по 10 мин.);
в) высушивание (инкубирование при температуре 37°C до полного высыхания - 3 ч.).
2. Нанесение нуклеиновой кислоты (инкубирование с раствором плазмидной ДНК по Seq#11 в 10 мМ фосфатном буфере в концентрации 1 мкг/мкл при температуре 37°C и постоянном встряхивании в течение 12 ч.).
3. Обработка полученного комплекса «носитель-генная конструкция»:
а) отмывка (обработка 5 мМ раствором фосфата в объеме 1 мл 3 раза);
б) высушивание (инкубирование при температуре 37°C до полного высыхания - 3 ч.).
Полученный ген-активированный костный графт был исследован в ортотопических условиях. Исследование было выполнено на кроликах породы Шиншилла (n=15). Каждому животному выполнялись два одинаковых симметричных полнослойных дефекта обеих теменных костей, диаметром по 10 мм, которые являются «критическими» для кроликов, так как естественный восстановительный процесс, без каких-либо оптимизирующих влияний не завершается полной консолидацией. В дефекты правых теменных костей имплантировали ген-активированный остеопластический материал (экспериментальная группа), состоящий из кальциевого фосфата и плазмидной ДНК Seq#11, в дефекты левых теменных костей - носитель без плазмидной ДНК (контрольная группа). Животных выводили из эксперимента через 30, 60 и 90 сут, результаты оценивали с использованием компьютерной томографии и гистологических методов.
В связи с исходно высокой плотностью выбранного носителя (около 1800 HU) и длительностью его биорезорбции (более 6 мес.) объективно оценить признаки репаративного остеогенеза в сравнительном аспекте не удалось. Однако по данным гистологического исследования, только в случае применения ген-активированного остеопластического материала признаки остеогенеза наблюдались уже через 30 сут после операции в центральной части дефекта (рис. 8).
Как видно на рис. 8 гранулы материла, расположенные в центральной части дефекта, выполненного рыхлой волокнистой соединительной тканью, являлись источником репаративного остеогенеза, тогда как в контроле кальциевый фосфат без плазмидной ДНК был окружен только рыхлой волокнистой соединительной тканью без признаков образования костной ткани. Полученные данные свидетельствуют о том, что введение плазмидной ДНК Seq#11 в область костного дефекта критических размеров привело к выраженной индукции репаративного остеогенеза, что может быть обусловлено как опосредованным через ангиогенез действием, так и прямым влиянием VEGF на клетки мезенхимальной линии.
Таким образом, представленные выше примеры иллюстрируют тот факт, что разработанная оптимизированная генная конструкция и фармацевтическая композиция на ее основе обладают выраженной ангиогенной активностью, что позволяет рассчитывать на эффективность ее использования в лечении ишемических заболеваний сердечно-сосудистой системы. Кроме того, оптимизированная генная конструкция с геном vegf может оказаться эффективной в лечении других патологических состояний, требующих активации репаративного процесса, таких как: повреждения кожи и опорно-двигательного аппарата, травмы периферических нервов, синдром диабетической стопы, боковой амиотрофический склероз.
Литература
1. Gene therapy clinical trials worldwide, http://www.abedia.com/wiley/years.php.
2. Grigorian A.S., Schevchenko K.G. Some possible molecular mechanisms of VEGF encoding plasmids functioning. Cell. Tanspl. Tiss. Engin. 2011. VI (3): 24-8.
3. Baboo S., Cook P.R. "Dark matter" worlds of unstable RNA and protein. Nucleus 2014; 5(4): 281-6.
4. Vislovukh Α., Vargas T.R., Polesskaya A. et al. Role of 3'-untranslated region translational control in cancer development, diagnostics and treatment. World J. Biol. Chem. 2014; 5(1): 40-57.
5. Malter J. Method to increase regulatory molecule production. US patent №5,587,300; 24.12.1996.
6. Sytkowski A.J., Grodberg J. Erythropoietin DNA having modified 5' and 3' sequences and its use to prepare EPO therapeutics. US patent №6,153,407; 28.11.2000
7. Mauro VP, Chappell S.A., Zhou W. et al. Reengineering mRNA primary structure for enhanced protein production. US patent application №2012/0053333 Al; 01.03.2012.
8. Folkman J., Merler E., Abernathy C. et al. Isolation of a tumor factor responsible for angiogenesis. J. Exp. Med. 1971; 133(2): 275-88.
9. Goel H.L., Mercurio A.M. VEGF targets the tumour cell. Nat. Rev. Cancer. 2013; 13(12): 871-82.
1 O. Koch S., Claesson-Welsh L. Signal transduction by vascular endothelial growth factor receptors. Cold Spring Harb. Perspect. Med. 2012; 2(7): a006502.
11. Matsumoto T., Bohman S., Dixelius J. et al. VEGF receptor-2 Y951 signaling and a role for the adapter molecule TSAd in tumor angiogenesis. EMBO J. 2005; 24(13): 2342-53.
12. Bhattacharya R., Kwon J., Li X. et al. Distinct role of PLCbeta3 in VEGF-mediated directional migration and vascular sprouting. J. Cell Sci. 2009; 122(Pt 7): 1025-34.
13. Coultas L., Chawengsaksophak K., Rossant J. Endothelial cells and VEGF in vascular development. Nature 2005; 438(7070): 937-45.
H. Olsson A.K., Dimberg Α., Kreuger J. et al. VEGF receptor signalling - in control of vascular function. Nat. Rev. Mol. Cell Biol. 2006; 7(5): 359-71.
15. Ma X.N., Li Q.P., Feng Z.C. Research progress in cytokines and signaling pathways for promoting pulmonary angiogenesis and vascular development. Zhongguo Dang Dai Er Ke Za Zhi. 2013; 15(9): 800-5.
16. Арутюнян И.В., Кананыхина Е.Ю., Макаров A.B. Роль рецепторов VEGF-А165 в ангиогенезе. Клеточная трансплантология и тканевая инженерия 2013; VIII(l): 12-8.
17. Cherviakov Iu.A., Staroverov I.N., Nersesian E.G. et al. The opportunities for genie therapy of chronic obliterating diseases of lower limbs arteries. Khirurgiia (Mosk). 2014; (4): 40-5.
18. Willyard C. Limb-saving medicines sought to prevent amputation. Nature Medicine 2012; 18(3): 328.
19. Hillenbrand M., Holzbach T., Matiasek K. et al. Vascular endothelial growth factor gene therapy improves nerve regeneration in a model of obstetric brachial plexus palsy. Neurol Res. 2015; 37(3): 197-203.
20. Kessler JA. Vascular endothelial growth factor gene transfer for diabetic polyneuropathy. Ann Neurol. 2009; 65(4): 362-4.
21.Zavalishin I.A., Bochkov N.R, Suslina Z.A. et al. Gene therapy of amyotrophic lateral sclerosis.Булл Exp Biol Med. 2008; 145(4): 483-6.
22. Zhao D.M., Yang J.R, Wu S.Q. et al. Effect of vascular endothelial growth factor 165 gene transfection on repair of bone defect: experiment with rabbits. Zhonghua Yi Xue Za Zhi. 2007; 87(25): 1778-82.
23. Geiger F., Bertram H., Berger I. et al. Vascular endothelial growth factor gene-activated matrix (VEGF165-GAM) enhances osteogenesis and angiogenesis in large segmental bone defects. J. Bone Miner. Res. 2005; 20(11): 2028-35.
24. Neve Α., Cantatore F.P., Corrado A. et al. In vitro and in vivo angiogenic activity of osteoarthritic and osteoporotic osteoblasts is modulated by VEGF and vitamin D3 treatment. Regul. Pept. 2013; 184: 81-4.
25. Marini M., Sarchielli E., Toce M. et al. Expression and localization of VEGF receptors in human fetal skeletal tissues. Histol. Histopathol. 2012; 27(12): 1579-87.
26. Tombran-Tink J., Barnstable C.J. Osteoblasts and osteoclasts express PEDF, VEGF-A isoforms, and VEGF receptors: possible mediators of angiogenesis and matrix remodeling in the bone. Biochem. Biophys. Res. Commun. 2004; 316(2): 573-9.
27. Mayr-Wohlfart U., Waltenberger J., Hausser H. et al. Vascular endothelial growth factor stimulates chemotactic migration of primary human osteoblasts. Bone 2002; 30(3): 472-7.
28. D'Alimonte I., Nargi E., Mastrangelo F. et al. Vascular endothelial growth factor enhances in vitro proliferation and osteogenic differentiation of human dental pulp stem cells. J. Biol. Regul. Homeost. Agents. 2011; 25(1): 57-69.
29. Yang Y.Q., Tan Y.Y., Wong R. et al. The role of vascular endothelial growth factor in ossification. Int. J. Oral Sci. 2012; 4(2): 64-8.
30. Berendsen A.D., Olsen B.R. How vascular endothelial growth factor-Α (VEGF) regulates differentiation of mesenchymal stem cells. J. Histochem. Cytochem. 2014; 62(2): 103-8.
















Изобретение относится к области биохимии. Предложен способ продления времени жизни мРНК трансгена vegf в клетке млекопитающего, трансфицированной генной конструкцией. Проводят точечные делеции в 3'некодирующей области гена фактора роста эндотелия сосудов (vegf), при этом делетированный нуклеотид не заменяется либо заменяется на цитозин в точках замещения гуанина или аденина. Затем определяют время жизни мРНК при каждой делеции. Сведения анализируют и конструируют генную конструкцию, содержащую 3'некодирующую область с совокупностью одиночных делеций и/или замен на цитозин, показавших наилучший результат по продлению жизни мРНК. Кроме того, предложены варианты плазмидной ДНК, сконструированной для осуществления указанного способа, и мРНК трансгена vegf, полученная в результате транскрипции плазмидной ДНК. Также изобретение относится к применению плазмидной ДНК, в том числе в составе фармацевтической композиции, для регенерации тканей организма. Изобретение позволяет увеличить общую продукцию терапевтического белка vegf за счет увеличения времени жизни мРНК, достигнутой посредством введения оптимальных изменений в последовательность 3'UTR гена vegf. 6 н. и 2 з.п. ф-лы, 9 ил., 2 табл., 2 пр.
1. Способ продления времени жизни мРНК трансгена vegf в клетке млекопитающего, трансфицированной генной конструкцией, предусматривающий проведение точечных делеций в 3'некодирующей области гена фактора роста эндотелия сосудов (vegf), при котором делетированный нуклеотид не заменяется либо заменяется на цитозин в точках замещения гуанина или аденина, затем определяется время жизни мРНК при каждой делеций, сведения анализируются и конструируется генная конструкция, содержащая 3'некодирующую область с совокупностью одиночных делеций и/или замен на цитозин, показавших наилучший результат по продлению жизни мРНК.
2. Способ по п.1, где трансфицированная генная конструкция позволяет увеличить количество кодируемого трансгеном белка.
3. Способ по п.1, где 3'-некодирующая область мРНК является некодирующей областью гена, выбранного из: vegf121, vegf145, vegf165, vegf185 и других возможных изоформ гена фактора роста эндотелия сосудов.
4. Плазмидная ДНК гена фактора роста эндотелия сосудов, выбранного из изоформ: 121, 145, 165, 185 или других, содержащая в 3'-некодирующей области на отрезке 1070-1600 нуклеотидов одну или несколько делеций и/или замен одиночных нуклеотидов, представленных на Seq#2-10, сконструированная для осуществления способа по п.1.
5. Плазмидная ДНК гена фактора роста эндотелия сосудов vegf 165, представленная на Seq#11, сконструированная для осуществления способа по п.1.
6. мРНК трансгена vegf в клетке млекопитающего, трансфицированной плазмидной ДНК по п.4 или 5, полученная способом по п.1.
7. Фармацевтическая композиция для регенерации соединительной, мышечной, нервной тканей, включающая плазмидную ДНК по п.4 или 5 и, по меньшей мере, один криопротектант и стабилизатор рН.
8. Применение плазмидной ДНК по п.5 в регенерации соединительной, нервной, мышечной, костной тканей, тканей сердечно-сосудистой системы.
| US 7709450 B2, 04.05.2010 | |||
| ЧЕРЕНКОВА Е.Е | |||
| и др | |||
| Конструирование экспрессионных векторов, кодирующих про-ангиогенные, нейротрофические и нейропротекторные факторы для генной и генно-клеточной терапии | |||
| МЕЖДУНАРОДНЫЙ ЖУРНАЛ ПРИКЛАДНЫХ И ФУНДАМЕНТАЛЬНЫХ ИССЛЕДОВАНИЙ, 2013, N11, стр.57-58 | |||
| СПОСОБ ПОЛУЧЕНИЯ ЛЕКАРСТВЕННОГО ПРЕПАРАТА ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫХ КЛЕТОК | 2009 |
|
RU2431669C2 |
| ШВАЛЬБ П.Г | |||
| и др | |||
| Эффективность и безопасность применения препарата "Неоваскулген" в комплексной терапии пациентов с хронической ишемией нижних конечностей (IIB-III фаза клинических испытаний) | |||
| Клеточная трансплантология и тканевая инженерия, 2011, Том 6, N3, стр.76-83. | |||

