Предшествующий уровень техники
Распространенность заболеваний и патологических состояний, сопровождающихся или приводящих к повреждениям тканей и органов человека, чрезвычайно велика. Основными причинами повреждений костей и скелетной мускулатуры, кожи, подкожной жировой клетчатки, слизистых оболочек, сосудов, периферических нервов, внутренних органов являются травмы, онкологические, воспалительные, дегенеративно-дистрофические заболевания, врожденные аномалии развития и деформации, а также первые этапы их хирургического лечения.
Травмы занимают ведущее место в структуре причин формирования повреждений тканей и органов. По данным ВОЗ, ежегодное количество пострадавших только в результате дорожно-транспортных происшествий составляет 30-50 млн чел. [1]. В России, по данным ГИБДД, около 230 тыс человек ежегодно получают травмы в результате ДТП [2]. В части онкологических заболеваний, по прогнозам ВОЗ, ежегодное количество вновь диагностированных случаев увеличится с 14 млн (в 2012 г. ) до 22 млн к 2020 г. [3].
В этой связи, ежегодное количество пациентов, нуждающихся в реконструктивно-восстановительных операциях, чрезвычайно велико. Эффективное лечение большинства из них может быть достигнуто с применением стандартных методов и средств, однако у значительной части пациентов не удается добиться действительно успешных результатов, восстановления исходного уровня качества жизни. Это обусловлено тем фактом, что при выраженных (объемных, протяженных) повреждениях тканей и органов, при наличии сопутствующей патологии и факторов риска, отягощающих клиническую ситуацию, в зоне дефекта количество камбиальных клеток и биологически активных веществ, индуцирующих репаративную регенерацию тканей, минимизировано и не позволяет реализовать полное гистотипическое восстановление. Эффективное лечение в таких случаях может быть обеспечено только теми методами и средствами, которые воздействуют на репаративную регенерацию тканей, восполняют утраченные структуры, камбиальный резерв, привносят факторы, регулирующие восстановительный процесс и т.д. Фактически, понимание этого положения и привело к формированию так называемой «регенеративной медицины» [4].
В рамках регенеративной медицины ведутся активные исследования и разработки новых биофармацевтических препаратов (генных [5], клеточных [6]), медицинских изделий и технологий, способных оказать выраженное влияние на репаративную регенерацию тканей, восстановить утраченную структуру и функцию поврежденных органов [7] и позволяющих выполнить их частичную и даже полную замену (персонализированные технологии с применением трехмерной печати) [8].
В регенеративной медицине сформирован арсенал инструментов, относящихся к клеточным, генным и постгеномным технологиям. Живые клетки, факторы роста, генные конструкции отдельно или в различных комбинациях как без дополнительных элементов, так и в составе различных матриксов-носителей разрабатываются и оцениваются на предмет безопасности и эффективности в восстановлении поврежденных тканей и органов [8, 9].
В качестве матриксов-носителей используются любые материалы, главным образом, биорезорбируемые, не содержащие в своем составе биологически активные компоненты, стандартизированные по качественным и количественным показателям. Основными материалами являются аллогенные и ксеногенные матриксы различных технологий обработки (децеллюляризация, деминерализация, депротеинизация и т.п.) [10]; неорганические соединения (β-трикальция фосфат [11], октакальциевый фосфат (ОКФ) [12], натуральный или синтетический гидроксиапатит [13], силикаты [14] и т.п.); синтетические (PLGA, агароза и т.д.) [15] и натуральные (коллаген, хитозан, желатин, альгинат натрия) органические вещества, полимеры и соли органических кислот [16], а также композитные изделия из вышеперечисленных материалов. Матриксы-носители могут быть твердыми веществами, гелями (в том числе гидрогелями), аэрогелями [17, 18].
Большинство матриксов без биологически активных компонентов оказывают лишь оптимизирующее влияние на репаративную регенерацию тканей, но не способны ее активировать и поддержать на высоком уровне до полного гистотипического заживления. Иными словами, они выполняют только механическую функцию (восполнение зоны дефекта, направление формирующихся тканей со стороны краев дефекта к центру), а также способны доставить биологически активные вещества, предварительно внесенные в материал [9]. Основными направлениями совершенствования матриксов-носителей являются оптимизация состава (в том числе за счет комбинации нескольких веществ), модификация поверхности как в виде обработки веществами, улучшающими прикрепление биологически активных компонентов (клеток, белков, генных конструкций) [19], так и в форме изменения макро- и микрорельефа поверхности, наноструктурирования, что позволяет модулировать функцию клеток, совмещенных с матриксов до введения in vivo, и (или) клеток реципиентного ложа [20, 21].
Нашей исследовательской группой ранее был разработан биокомпозит, состоящий из матрикса-носителя, плазмидной ДНК, несущей гены vegf и (или) sdf, и клеток, вовлеченных в репаративную регенерацию тканей [22]. Другие исследователи остановили свой выбор на сложной четырехкомпонентной системе, представленной матриксом-носителем, клетками, генными конструкциями и факторами роста [23]. Основной предпосылкой для разработки таких биокомпозитов стала неудовлетворенность результатами двухкомпонентных систем: «матрикс-носитель + клетки», «матрикс-носитель + факторы роста», «матрикс-носитель + генные конструкции».
Тканеинженерные материалы
Данная группа материалов представляет собой изделия, состоящие из двух основных компонентов: биорезорбируемый носитель и живые (ауто- или аллогенные) клетки. Основная идея подхода состоит в восполнении утраченных камбиальных резервов и повышении концентрации факторов, индуцирующих репаративный процесс, в области введения материала. Трансплантированные клетки в реципиентном ложе в случае сохранения жизнеспособности могут оказать благоприятный терапевтический эффект за счет двух механизмов действия: непосредственный - дифференцировка в специализированные клетки поврежденных тканей (показано для аутогенных клеток [24]), а также опросредованный - паракринный эффект - регуляция морфофункциональной активности других клеток за счет продукции биологически активных веществ - факторов локальной регуляции гистогенеза. Важно, что, по мнению многих авторов, именно паракринная активность клеток тканеинженерных материалов является основным их механизмом действия [25].
Клетки, использующиеся для создания тканеинженерных материалов, могут подвергаться технологиям культивирования или использоваться непосредственно после выделения из тканевого источника. К основным применяющимся для создания тканеинженерных материалов видам культивированных клеток относятся мультипотентные мезенхимальные стромальные клетки (ММСК) [26, 27], остеогенные клетки и остеобласты [28, 29], эндотелиоциты [30, 31], фибробласты, эпителиоциты [32], стволовые клетки с индуцированной плюрипотентностью [33] и другие виды малодифференцированных и специализированных клеток отдельно и в различных комбинациях. К числу некультивированных клеточных популяций относятся клетки костного мозга («гетерогенная смесь» ММСК, фибробластов, эндотелиальных прогениторных клеток, гемопоэтических стволовых и дефинитивных клеток гемопоэтической линии и др.) [34] и стромально-васкулярная фракция жировой ткани (СВФ ЖТ) (ММСК, эндотелиоциты и эндотелиальные прогенеторные клетки, лейомиоциты, фибробласты, преадипоциты и иммунокомпетентные клетки) [35].
Изготовление тканеинженерных материалов с аллогенными клетками может быть выполнено с использованием принципиально иных технологий, предполагающих сберегательную обработку донорских тканей с сохранением компонентов межклеточного матрикса и живых клеток [36].
Несмотря на то что некоторые тканеинженерные изделия уже зарегистрированы и разрешены для применения в рутинной клинической практике («Osteocel plus» (NuVasive, США) (2005), «Trinity Evolution)) (Orthofix, США) (2009), «AlloStem» (AlloSource) (2011), «Cellentra VCBM» (BioMet, США) (2012), «OvationOS» (Osiris Therapeutics, США) (2013), BioSeed-Oral Bone» (BioTissue Technologies, Германия) (2001)), данный подход имеет ряд недостатков:
- недостаточная эффективность в случае протяженных (объемных) дефектов тканей и органов из-за гибели большинства клеток, входящих в состав тканеинженерного материала непосредственно после трансплантации - тканеинженерные изделия без префабрикации аваскулярны, а клетки требуют активного кровоснабжения, которое критически минимизировано в зоне повреждения;
- высокая себестоимость и сложность технологического процесса (клеточного сервиса) создания тканеинженерного материала, который должен осуществляться в соответствии со стандартами GMP и GTP;
- невозможность организации полноценного серийного производства наиболее эффективных - персонализированных (содержащих аутогенные клетки) - тканеинженерных изделий;
- особые условия хранения, не всегда являющиеся доступными для лечебно-профилактических учреждений (например, температура ниже -80°C);
- сохраняющиеся сложности правового регулирования и регистрации медицинских изделий, содержащих живые клетки.
Материалы с факторами роста
К данной группе медицинских изделий относятся материалы, состоящие из матрикса-носителя и факторов роста (одного или нескольких), обеспечивающих регуляцию репаративной регенерации тканей. Данный технологический подход является наиболее успешным в аспекте «клинической трансляции». Уже зарегистрированы и разрешены для клинического применения такие изделия как «Emdogain» (Straumann, Германия) - материал с эмалевыми матричными протеинами (1997); «ОР-1» (Stryker Biotech, США) - с рекомбинантным ВМР-7 (2001); «Infuse» (Medtronic, США) (2002, 2004, 2007) - с рекомбинантным ВМР-2; «GEM21S», «Augment bone graft» (BioMimetic Therapeutics Inc., США) - с рекомбинантным PDGF-BB (2005, 2009), «i-Factor Putty» (Cerapedics, США) - с белком P-15 (лиганд для интегринов α2β1, экспрессирующихся клетками остеобластического дифферона) (2008) и др.
Передовые исследования и разработки материалов с факторами роста направлены на два основных аспекта. Первый предполагает комбинацию нескольких факторов, в том числе неспецифических (ангиогенных) и специфических (селективно влияющих на определенный механизм в общем процессе репаративной регенерации какой-либо ткани), в одном изделии. Например, иммобилизация VEGF и ВМР-2 на одном носителе [37]. Второй - обеспечение пролонгированного контролируемого высвобождения терапевтических белков из структуры матрикса, в частности, за счет управления динамикой биодеградации гидрогелевых матриксов [38] или инкапсулирования молекул факторов роста в микросферы из органических полимеров [39]. Некоторые авторы за счет специальных технологий (например, сайт направленный мутагенез) изменяют структуру факторов роста, объединяют несколько в один, создавая «мутантные» молекулы, обладающие большей эффективностью в активации репаративного процесса. Например, P. Kasten с со авт. (2010) модифицировали фактор роста и дифференцировки-5 (growth-and-differentiation factor-5, GDF-5), добавив в его последовательность сайты ВМР-2, ответственные за связывание со специфическими рецепторами. В результате полученная молекула GDF-5 приобрела свойства, характерные для ВМР-2 [40].
Однако двухкомпонентные биокомпозиты, состоящие из матрикса-носителя и факторов роста, также имеют ряд недостатков, ограничивающих их эффективность. Во-первых, молекулы белка в условиях операционной раны (экссудация, высокая активность протеолитических ферментов) подвергаются быстрой биодеградации, являются короткоживущими и короткодистантными, что не позволяет материалу в полной мере проявить индуцирующее действие. Во-вторых, количество терапевтического белка в материале ограничено, а его действие даже при условии контролируемого и пролонгированного высвобождения - кратковременное и «грубое». Иными словами, та меньшая часть молекул белка, которая отделилась от носителя, сохранила свою биологическую активность и достигла клеток-мишеней - вступит во взаимодействие со специфическими рецепторами на их поверхности и вызовет биологический эффект. При этом быстро произойдет инактивация рецепторов вместе с лигандом как компенсаторно-адаптационный механизм, предохраняющий клетки от избыточной стимуляции, биологическое действие фактора роста прекратится, а его количество иссякнет. Этот аспект механизма действия таких материалов оказывает особенно негативное влияние на их эффективность при отсутствии заданной кинетики высвобождения факторов роста (что характерно для большинства изделий данной группы). Непосредственно после имплантации происходит последовательная смена ранних фаз воспалительного процесса (альтерация, затем экссудация), при которых высвободившиеся из материала факторы роста - регуляторы репаративной регенерации - еще не требуются. Более того, на столь ранней (первой) стадии репаративного процесса для факторов роста из состава имплантированного материала может не оказаться достаточного рецепторного поля, поскольку основными клетками в зоне повреждения будут иммунокомпетентные - нейтрофилы и макрофаги. Указанных недостатков теоретически лишен третий альтернативный подход.
Ген-активированные материалы
Данная группа материалов представляет собой комплекс «матрикс-носитель + генные конструкции (кодирующие нуклеиновые кислоты)», компоненты которого объединены различными методами: за счет технологий «химического связывания» [41], использования вспомогательных веществ (например, гелевых биополимеров) [42], непосредственного включения нуклеиновых кислот в состав носителей на этапе синтеза матрикса и т.д. Развитие ген-активированных материалов напрямую связано с достижениями генной терапии в целом, в рамках которой генные конструкции разрабатываются как действующие вещества генно-терапевтических лекарственных препаратов.
В механизме остеоиндуктивного действия ген-активированного материала можно выделить два последовательных этапа: неспецифический и специфический. Первый состоит в высвобождении нуклеиновых кислот из структуры носителя после имплантации в зону дефекта, поступлении в клетки реципиентного ложа и экспрессии в них. Фактически, он является одинаковым для любых генных конструкций, а вариабельность в части поступления в клетку обусловлена лишь системами доставки трансгена. Второй состоит в специфическом действии регуляторной молекулы белковой природы, кодируемой трансгеном и продуцируемой трансфицированными клетками, которые работают как «биореакторы» терапевтического белка, синтезируя его в течение контролируемого периода времени. В отличие от изделий, содержащих факторы роста, основной компонент ген-активированных остеопластических материалов действует «мягко». Иными словами, попадание трансгена в ядро клетки-мишени не принуждает ее к облигатной экспрессии терапевтического белка. Клетка сохраняет свое нормальное функциональное состояние и реакцию на стимулы микроокружения, в связи с чем при отсутствии потребности в терапевтическом белке в конкретный промежуток времени она за счет внутриклеточных посттранскрипционных механизмов регуляции полужизни мРНК способна снизить уровень мРНК трансгена и, тем самым, предотвратить продукцию белка [43]. Кроме того, даже без заданной кинетики высвобождения генных конструкций, их поступление в клетки реципиентного ложа, последующая экспрессии и начало пролонгированной продукции терапевтического белка требует времени. В этой связи, пик секреции белка трансфицированными клетками придется уже на вторую стадию репаративного процесса, после завершения воспалительной. Такой механизм действия генных конструкций значительно повышает эффективность ген-активированного материала по сравнению с изделиями, содержащими факторы роста [44].
Любые генные конструкции состоят из терапевтического гена (в виде кДНК или РНК) и системы его внутриклеточной доставки (вектор). Векторы разделяются на две основные группы: вирусные и невирусные. В первом случае трансген заключается в частице ретро-, ленти-, аденовируса или адено-ассоциированного вируса и др., а во втором - в плазмиду - кольцевую молекулу нуклеиновой кислоты, содержащую ряд вспомогательных последовательностей, обеспечивающих экспрессию трансгена. Вирусная и невирусная системы доставки отличаются по эффективности трансфекции. В первом случае до 40% и более генных конструкций поступают в клетки-мишени, а во втором - показатель не превышает 1-2% в виду размеров и отрицательного заряда «голой» плазмидной ДНК. Предложены подходы (физических, химических), обеспечивающие повышение эффективности трансфекции плазмидной ДНК до 8-10% [45].
Ряд вирусных векторов (ретро-, лентивирусные и др.) встраиваются в геном. Иными словами, экспрессия трансгена, который они переносят, длительная, в то время как остальные, в том числе плазмидная ДНК, в геном не интегрируются, поэтому экспрессируются временно, в течение 10-14 сут. Ретро- и лентивирусные векторы чаще применяются в генно-клеточном подходе, когда культура клеток трансфицируется ex vivo и после этого совмещается с матриксом-носителем [46].
Все ген-активированные материалы могут быть разделены по технологическому варианту совмещения матрикса и генных конструкций, а также по составу биологически активного компонента: природе вектора и трансгена, количеству трансгенов или различных генных конструкций в составе одного изделия. Однако главным дифференциальным признаком, обусловливающим различия в биологическом действии ген-активированных материалов, является именно трансген. Наиболее часто применяющимися при создании ген-активированных материалов трансгенами являются последовательности нуклеотидов, кодирующие основные факторы роста и транскрипционные факторы, вовлеченные в положительную регуляцию репаративного процесса - индукцию миграции, пролиферации, дифференцировки клеток, синтеза ими компонентов гистотипического межклеточного матрикса и биологически активных веществ [9]. Кроме того, в качестве трансгенов могут быть использованы последовательности нуклеотидов, кодирующие белки рецепторов клеток (увеличение рецепторного поля увеличивает восприимчивость клеток реципиентного ложа к факторам микроокружения и веществам, введенным извне), цитокины (интерлейкины, фактор некроза опухолей-α и др.), гормоны и гормоноподобные вещества, что может улучшить регенерацию тканей.
Однако разработанные к настоящему времени ген-активированные материалы и технологии их создания имеют ряд специфических недостатков.
Во-первых, невирусные генные конструкции, входящие в состав ген-активированных материалов, характеризуются крайне низкой эффективностью трансфекции, а использование физических (электрофорез, ультразвук и т.д.) или химических (совмещение с катионными белками, липосомы и т.д.) методов увеличения уровня поступления биологически активного компонента в клетки не всегда возможно. В этой связи, исследователи вынуждены использовать высокие дозы генных конструкций, чтобы увеличить количество акцептировавших их клеток и добиться желаемого биологического эффекта. Увеличение дозы закономерно влечет к повышению себестоимости изделия, а также может скомпрометировать его безопасность. Кроме того, технологии создания ген-активированных материалов, основанные на формировании химических связей между матриксом-носителем и нуклеиновыми кислотами, далеко не всегда позволяют увеличить дозу генных конструкций, так как такое увеличение требует изменения состава матрикса.
Во-вторых, начало экспрессии трансгена после имплантации материала in vivo у большинства ген-активированных материалов является поздним - приходится на вторую стадию репаративного процесса, т.е. после завершения воспалительной стадии (через 2 недели после введения in vivo), а максимальный уровень продукции трансгена - на еще более поздние сроки, что зависит от кинетики высвобождение генных конструкций из матрикса-носителя. Однако инициировать более раннюю экспрессию генных конструкции и обеспечить смещение сроков достижения максимального уровня продукции терапевтического белка «влево» стандартные ген-активированные материалы не способны. Для этого разрабатываются различные способы оптимизации ген-активированных материалов, направленные на изменение и «программирование» кинетики высвобождения биологически активного компонента из структуры матрикса-носителя. Наиболее известными являются методы инкапсуляции молекул нуклеиновых кислот с последующим связыванием комплексов с матриксом-носителем за счет чувствительных к определенным факторам (температура, pH, ультразвук, специфические ферменты) посредников. Постепенное разрушение вещества, обеспечивающего связь микрокапсул или иных комплексов, содержащих нуклеиновую кислоту, с матриксом-носителем обеспечивает контролируемое высвобождение генных конструкций [47].
В-третьих, как было отмечено выше, в случаях, когда объединение матрикса-носителя и нуклеиновых кислот обеспечивается за счет химического взаимодействия указанных компонентов, точность дозы генных конструкций практически невозможно проконтролировать. Этот недостаток особенно характерен для твердых материалов, выбранных в качестве матриксов-носителей. В частности, известна способность материалов, содержащих соединения кальция, удерживать молекулы нуклеиновых кислот за счет комплексообразования, реализуемого, вероятно, по механизму донорно-акцепторной (координативной) связи. Данный механизм используется в хроматографических колонках при очистке нуклеиновых кислот [48]. Этот же принцип экстраполирован на создание ген-активированных материалов с использованием матриксов, содержащих фосфаты кальция [49]. Однако образцы разных серий изготовления и даже в пределах одной серии могут существенно отличаться по количеству связываемых ими нуклеиновых кислот, а среднее количество невозможно прогнозировать и контролировать - оно вычисляется экспериментальным путем при объединении каждого варианта матрикса с нуклеиновой кислотой.
В-четвертых, важнейшим условием для внедрения в клиническую практику любого имплантируемого медицинского изделия, к которым относятся и ген-активированные материалы, является стерильность. На заключительном этапе производственной технологии подавляющего большинства медицинских изделий предусмотрена стерилизация. Однако все методы стерилизации направлены на элиминацию в том числе и нуклеиновых кислот, которые в случае ген-активированных материалов являются важнейшим, отличительным компонентом изделия, обеспечивающим его главный механизм действия и терапевтический эффект. В случае геннотерапевтических препаратов (генные конструкции + матрикс- носитель), таких как «Неоваскулген» (ПАО «Институт Стволовых Клеток Человека»), в виду отсутствия в их составе матрикса-носителя, реализуема стерильная микрофильтрация. Однако проблема изготовления стерильного ген-активированного материала до настоящего времени не решена. В каждом частном случае разработчики вынуждены искать удобоваримые варианты.
Два первых из вышеуказанных недостатков предопределяют менее выраженную эффективность ген-активированных материалов в регенерации тканей по сравнению с теоретически возможной. Две последние проблемы в большей степени обусловливают экономические издержки и являются препятствиями для внедрения ген-активированных материалов в качестве медицинских изделий в клиническую практику.
Учитывая вышеизложенное, наши усилия были направлены на исследования и разработки по созданию оптимизированных ген-активированных материалов, в той или иной степени лишенных указанных выше недостатков.
Перечень иллюстраций:
Рис. 1. Регенерат в центральной зоне дефекта теменной кости кролика, 30 сут после имплантации материалов: А - обычный ген-активированный материал; Б - оптимизированный ген-активированный материал с двумя фракциями нуклеиновых кислот. Окраска: гематоксилин и эозин. Ув.: ×40.
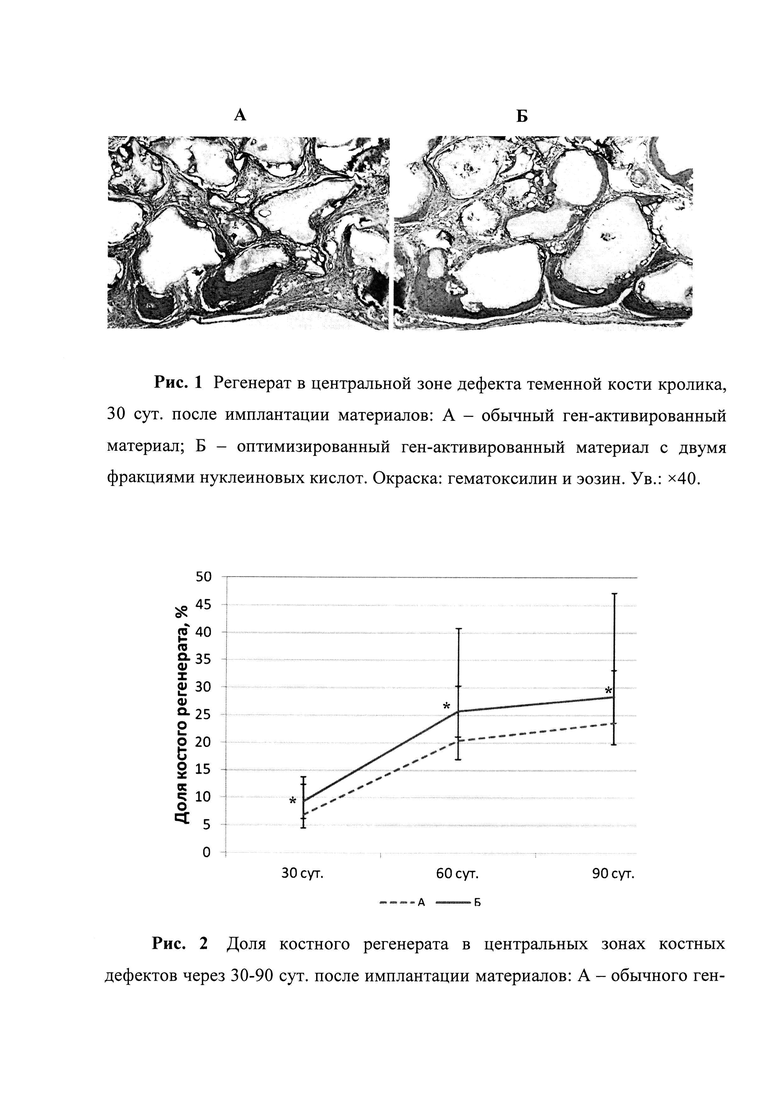
Рис. 2. Доля костного регенерата в центральных зонах костных дефектов через 30-90 сут после имплантации материалов: А - обычного ген-активированного материала; Б - оптимизированного ген-активированного материала с двумя фракциями нуклеиновых кислот.
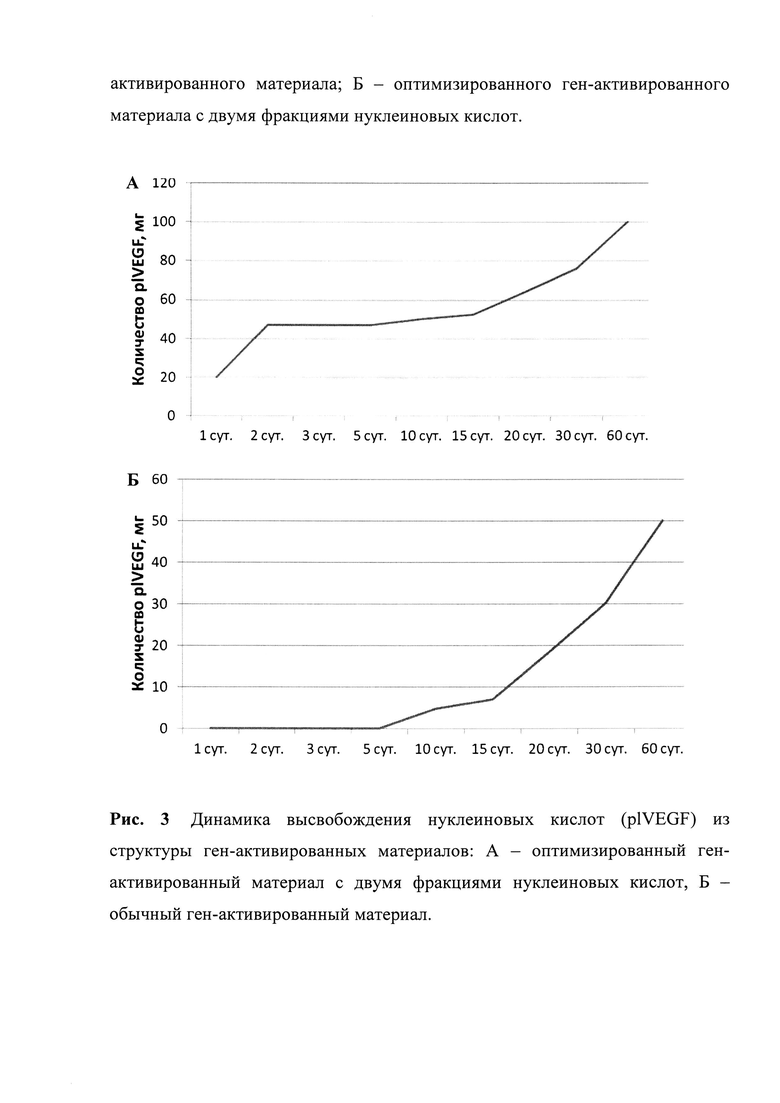
Рис. 3. Исследование выполняли в стандартных модельных условиях без эвакуации/деструкции выделившихся в раствор нуклеиновых кислот, т.е. определяли накопленную концентрацию в растворе.
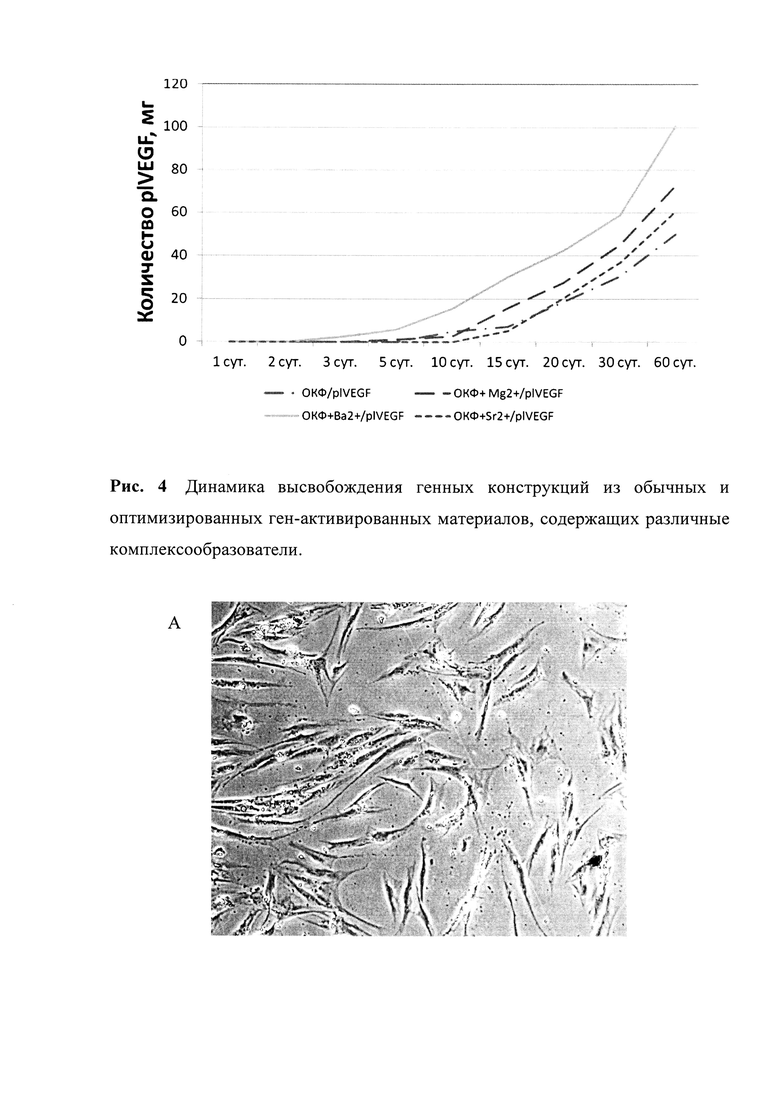
Рис. 4. Динамика высвобождения генных конструкций из обычных и оптимизированных ген-активированных материалов, содержащих различные комплексообразователи.
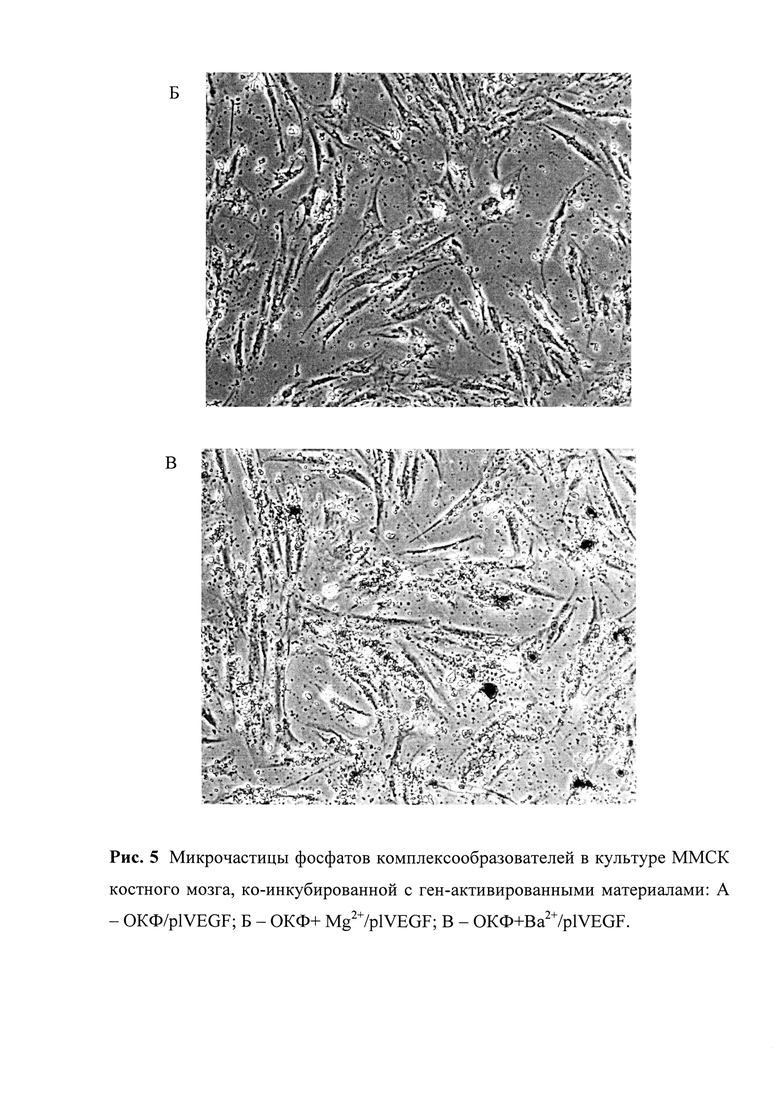
Рис. 5. Микрочастицы фосфатов-комплексообразователей в культуре ММСК костного мозга, ко-инкубированной с ген-активированными материалами: А - ОКФ/plVEGF; Б - ОКФ+ Mg2+/plVEGF; В -ОКФ+Ba2+/plVEGF.
Подробное описание изобретения
Задачей настоящего изобретения была оптимизация ген-активированных материалов за счет:
- повышения минимальной дозы нуклеиновых кислот в составе ген-активированного материала;
- обеспечения стандартизации состава ген-активированного материала за счет точного воспроизведения выбранной дозы нуклеиновых кислот в каждом образце изделия каждой серии;
- коррекции динамики высвобождения генных конструкций из состава матрикса-носителя в соответствии со стадийностью репаративного процесса;
- создания стерильного ген-активированного материала, включающего твердый матрикс-носитель.
Технический результат изобретения состоит в разработке способа получения оптимизированного твердого ген-активированного материала, включающего создание матрикса-носителя, связавшего как минимум одну молекулу нуклеиновой кислоты, с последующим размещением на его поверхности еще как минимум одной дополнительной молекулы нуклеиновой кислоты любым физическим методом, способа получения твердого матрикса-носителя, пригодного для создания оптимизированного твердого ген-активированного материала, самого оптимизированного, в том числе стерильного, твердого ген-активированного материала для регенерации тканей. В контексте настоящего исследования под термином «оптимизированный ген-активированный материал» понимается изделие, состоящее из матрикса-носителя и нуклеиновых кислот, отличающееся тем, что, по меньшей мере, два из четырех вышеперечисленных направлений оптимизации были успешно реализованы при создании такого материала.
Оптимизированные ген-активированные материалы с двумя фракциями нуклеиновых кислот
Первый способ касается ген-активированных материалов, в составе которых как минимум часть матрикса-носителя представлена твердым веществом, элементы которого способны к транзиторному удержанию (связыванию) молекул нуклеиновых кислот. Перечень таких матриксов включает, но не ограничивается следующими группами: гидроксиапатит, фосфаты кальция, костные матриксы различных технологий обработки и др.
Как было отмечено выше, в таких материалах до настоящего времени невозможно было предсказать и контролировать дозу генных конструкций, а также не были разработаны эффективные способы повышения этой дозы. Каждый вариант материала из вышеуказанных групп отдельно или в составе комплекса с какими-либо другими дополнительными компонентами (коллагеном, желатином, альгинатами, агарозой, полимерами органических кислот и др.) способен связать некоторое количество нуклеиновых кислот, которое может существенно различаться между образцами матриксов-носителей одного и того же состава, даже одной серии.
Нам удалось разработать первый способ создания оптимизированных ген-активированных материалов, в составе которых нуклеиновые кислоты были разделены на две фракции. Первая - связанная с матриксом-носителем, вероятно, за счет комплексообразования с соединениями кальция. Вторая - несвязанная, размещенная на поверхности твердого матрикса-носителя фракция нуклеиновых кислот.
Разработанный способ включает нижеприведенные этапы.
1. Создание первичного комплекса «матрикс-носитель - нуклеиновая кислота» в следующих вариантах: а) синтез матрикса-носителя, способного связать на своей поверхности нуклеиновую кислоту, с последующим объединением с как минимум одной молекулой нуклеиновой кислоты; б) добавление как минимум одной молекулы нуклеиновой кислоты к исходным материалам в процессе синтеза матрикса-носителя (выполнимо для материалов, имеющих на одном из этапов изготовления жидкую или гелевую формы). Указанные два технологических варианта создания первичного комплекса не являются взаимоисключающими. В частности, после реализации варианта «б», полученный матрикс-носитель с генными конструкциями может быть подвергнут объединению с дополнительным количеством нуклеиновых кислот, согласно варианту «а».
2. Размещение на поверхности полученного первичного комплекса «матрикс-носитель - нуклеиновая кислота» еще как минимум одной дополнительной молекулы нуклеиновой кислоты любым физическим методом, позволяющим осуществить такое размещение без образования химической связи между указанным матриксом-носителем и дополнительной нуклеиновой кислотой. Данный этап является критически значимой отличительной особенностью разработанного способа, так как позволяет повысить общую дозу нуклеиновых кислот до необходимого уровня.
Реализация второго этапа осуществима только в случае твердых матриксов-носителей. Увеличение эффективности и количества нуклеиновых кислот, размещенных на поверхности твердых материалов без образования химических связей с ними, закономерно, зависит от общей площади поверхности, ее макро- и микрорельефа, гидрофильности, сорбционных свойств матрикса-носителя и иных факторов. В этой связи за счет модификации указанных факторов возможно увеличивать количество размещенных на поверхности матрикса-носителя генных конструкций.
Для реализации второго этапа наиболее приемлемым является использование нуклеиновых кислот в водном растворе (вода, различные фосфатные или иные буферные растворы). Для более эффективного размещения несвязанной фракции нуклеиновых кислот на поверхности матриксов-носителей в растворы нуклеиновых кислот могут быть добавлены фармацевтически приемлемые вспомогательные вещества, такие как глюкоза, декстроза, натрия гидрофосфата додекагидрат, натрия дигидрофосфата дигидрат и любые другие, корректирующие заряд поверхности матриксов-носителей, нуклеиновых кислот, функционализирующие поверхность и предназначенные для удержания на ней молекулы, а также обеспечивающие первичное размещение нуклеиновых кислот для последующей фиксации на поверхности под воздействием физических методов без образования химических связей. Кроме того, фармацевтически приемлемые вещества обеспечивают стабилизацию нуклеиновых кислот, что увеличивает сроки хранения.
Перечень таких физических методов представлен, но не ограничен следующими: сушка при любых удобоваримых температуре, влажности, газовом составе среды, давлении, времени экспозиции; воздействие низких температур, ультразвука, магнитных и электрических полей и т.д.
Разрабатывая данный способ, мы рассчитывали, главным образом, на точность дозирования и повышение минимально возможной концентрации нуклеиновых кислот в составе ген-активированного материала.
Для исследований в качестве матриксов-носителей были отобраны следующие 10 наименований твердых изделий: ксеногенный деминерализированный костный матрикс - «Биоматрикс» (Конектбиофарм, Россия); аллогенный деминерализированный костный матрикс - «Перфоост» (Тканевый банк ЦИТО им. Н.Н. Приорова, Москва), «Остеоматрикс» (Конектбиофарм, Россия), «Аллоплант» (Всероссийский центр глазной и пластической хирургии, Уфа); ксеногенный депротеинизированный костный матрикс - «Bio-oss spongiosa» (Geistlich, Швейцария); композитный материал из синтетического гидроксиапатита и ксеногенного коллагена - «Колапол-КПЗ» (НПО Полистом, Россия); β-трикальция фосфат - «Cerasorb» (Curasan, Германия), «Chronos» (Synthes, Швейцария), «Трикафор» (БиоНова, Россия); октакальциевый фосфат (ОКФ, Институт металлургии и материаловедения РАН им. А.А. Байкова, Москва).
В качестве генных конструкций была использована высокоочищенная сверхскрученная плазмидная ДНК с геном, кодирующим VEGF (pCMV-VEGF165). Указанная генная конструкция является действующим веществом лекарственного препарата «Неоваскулген» (ПАО «Институт Стволовых Клеток Человека», РУ ЛП-000671 от 28.09.11), содержащего в качестве адъювантов декстрозы моногидрат - 60,0 мг (НД 42-11395-07), натрия гидрофосфата додекагидрат - 3,94 мг (ГОСТ 4172-76), натрия дигидрофосфата дигидрат - 0,160 мг (ГОСТ 245-76). Препарат выпускается в форме лиофилизированного порошка белого цвета в стерильных стеклянных флаконах с содержанием плазмидной ДНК 1,2 мг и показан для лечения пациентов с хронической ишемией нижних конечностей 2а-3 ст по классификации Фонтейна в модификации А.В. Покровского.
Совмещение вышеуказанных носителей и генных конструкций выполнялось по разработанному оригинальному протоколу, описанному в патенте РФ №2519326, и частично модифицированному. Содержание основных этапов лабораторного протокола:
1) Отмывка носителей. Образцы стандартной массы (100 мг) инкубировали в 0,5 М фосфатном буфере в объеме 1 мл при 37°С при постоянном встряхивании в течение 10 ч.
2) Уравновешивание. Образцы отмывали 10 мМ фосфатным буфером в объеме 1 мл при 37°С и постоянном встряхивании 4 раза по 10 мин.
3) Высушивание. Образцы выдерживали при 37°С до полного высыхания в течение 10 ч.
4) Совмещение носителей и генных конструкций. Раствор плазмидной ДНК (в концентрации 1 мкг/мкл) наносили в объеме 0,5 мл на образцы носителей. Материалы инкубировали при 37°С и постоянном встряхивании в течение 10 ч.
5) Промывка. Образцы отмывали 5 мМ фосфатным буфером в объеме 1 мл 3 раза, что позволяло убрать из раствора не связавшуюся с матриксом-носителем плазмидную ДНК.
6) Высушивание. Образцы выдерживали при 37°С до полного высыхания в течение 10 ч.
Для каждого материала выполнялось 4 повторения. Результаты - количество связавшейся с носителем плазмидной ДНК - оценивались после ее диссоциации (обработка 0,5 М раствором фосфатного буфера в объеме 200-600 мкл при 37°С и постоянном встряхивании в течение 10 мин) с определением концентрации нуклеиновых кислот в растворе с помощью флуориметра Qubit 2.0 (Invitrogen, США).
Оказалось, что все выбранные для исследования материалы были способны связать некоторое количество нуклеиновых кислот. По всей видимости, связь реализовывалась за счет комплексообразования между соединениями кальция, входящими в состав матриксов, и плазмидной ДНК. Однако различные материалы даже в пределах оной и той же группы (β-трикальция фосфат, деминерализованный костный матрикс) существенно различались по количеству нуклеиновых кислот, которое они удерживали (табл. 1). Это полностью согласуется с вышеуказанными особенностями и недостатками разрабатываемых к настоящему времени твердых ген-активированных материалов.
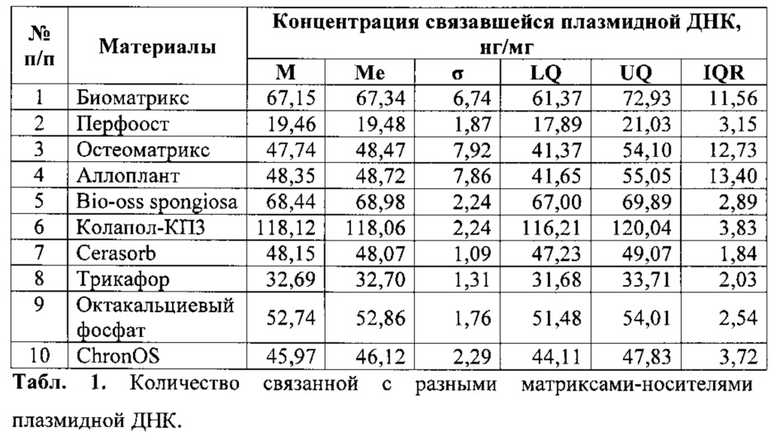

На втором этапе данного исследования для повышения уровня нуклеиновых кислот в составе ген-активированного материала и точного их дозирования в количествах 100, 300 и 500 мкг на 1,0 г указанных в табл.1 материалов, технология изготовления была модифицирована:
- совмещение матриксов-носителей с генными конструкциями выполнялось сразу в растворе с целевыми концентрациями нуклеиновых кислот (100, 300 и 500 мкг на 1,0 г матрикса-носителя), раствор также содержал 10 мМ фосфатного буфера и 60,0 мг декстрозы моногидрата, инкубирование выполнялось в течение 4 ч;
- этап последующей промывки был отменен - сразу после инкубации материалы не извлекались из раствора и помещались в лиофилизатор;
- лиофилизация выполняла по различным протоколам (варьировались основные параметры) продолжительностью до 4 сут, однако результаты существенно не отличались.
После выполнения указанного способа лиофилизированные ген-активированные материалы подвергались инкубации сначала с 0,9% раствором NaCl с определением количества несвязанной фракции плазмидной ДНК в растворе, а затем после отмывки образцов материалов 0,9% раствором NaCl до промывных вод с нулевым содержанием плазмидной ДНК и сушки при температуре 37°С, инкубировались с 0,5М раствором фосфатного буфера для диссоциации связанной фракции плазмидной ДНК.
Было установлено, что общая доза плазмидной ДНК во всех случаях точно соответствовала заданному уровню - 100, 300 или 500 мкг на 1,0 г. Количество связанной фракции нуклеиновых кислот было аналогичным таковому, показанному для использованных матриксов-носителей ранее и представленных в табл. 1. Количество же несвязанной фракции, закономерно, было равным разнице между целевой концентрацией и уровнем связанной фракции.
Таким образом, разработанный способ позволил решить две из четырех вышеуказанных задач: повышение минимальной дозы и точность дозирования. В нашем исследовании для размещения несвязанной фракции нуклеиновых кислот на поверхности твердого матрикса-носителя, предварительно связавшего максимально возможное количество нуклеиновых кислот, была использована лиофилизация. При этом является очевидным, что для этого могут быть использованы и другие физические методы: экспозиция при высоких или низких температурах различной продолжительности, ультразвуковые и электрофоретические методы и др.
Однако технический результат разработанного способа и полученных с помощью него оптимизированных ген-активированных материалов не ограничился только лишь точностью дозы и ее повышением до необходимого уровня. Неожиданные результаты были получены при дальнейших исследованиях разработанных оптимизированных ген-активированных материалов в ортотопических условиях in vivo.
Исследования in vivo были выполнены на кроликах-самцах породы Шиншилла, массой 2,0-2,5 кг с соблюдением международных правил гуманного обращения с лабораторными животными. Каждому животному выполнялись два одинаковых симметричных полнослойных дефекта обеих теменных костей, диаметром по 10 мм, которые являются «критическими» для кроликов, так как естественный восстановительный процесс, без каких-либо оптимизирующих влияний не завершается замещением дефекта новообразованной костной тканью. Ранее нами уже были получены и опубликованы данные о влиянии обычных ген-активированных материалов, т.е. содержащих только одну - связанную - фракцию нуклеиновых кислот, на регенерацию костной ткани в данной стандартной экспериментальной модели. В качестве матриксов-носителей в тех ранних исследованиях были использованы три материала из табл.1, с которыми связалось наибольшее количество плазмидной ДНК с геном vegf при минимальной вариативности данного показателя (Колапол-КП3, Bio-oss spongiosa, ОКФ) [41, 49]. Однако при сравнении результатов с данными, полученными при оценке в такой же экспериментальной модели оптимизированных ген-активированных материалов, состоящих из аналогичных матриксов-носителей, но отличающихся наличием сразу двух фракций плазмидной ДНК с геном vegf, была выявлена существенная разница. Оказалось, что оптимизированные ген-активированные материалы приводили к формированию статистически значимо большему объему костного регенерата в центральной зоне костного дефекта, т.е. там, где именно фрагменты имплантированного материала, а не края теменной кости являются источниками репаративного остеогенеза.
*различия между группами статистически значимы, р<0,05.
Вероятно, большая эффективность разработанных оптимизированных ген-активированных материалов была обусловлена сразу двумя причинами. Во-первых, исходно большей дозой генных конструкций в составе изделия (100, 300 и 500 мкг на 1,0 г матрикса-носителя). Однако если принять во внимание фундаментальные данные о динамике и стадиях репаративного процесса, мы нашли и другую вероятную причину. Как было указано выше, начало и достижение максимального уровня экспрессии трансгена после имплантации материала in vivo у обычных ген-активированных материалов несколько запаздывает относительно реализации стадий репаративного процесса. При этом камбиальные клетки, а также фибробласты и их предшественники мигрируют в зону повреждения гораздо раньше - уже в пролиферативную фазу воспаления, т.е. в первую стадию репаративного процесса (5-6 сут после имплантации in vivo). Таким образом, в ранние сроки после введения материала в зоне повреждения уже имеется и пул клеток-реципиентов генных конструкций (префибробласты, фибробласты, макрофаги), и популяция клеток-мишеней для терапевтического белка, кодируемого генной конструкцией (клетки основных дифферонов поврежденных тканей, эндотелиоциты и их предшественники). В этой связи, несвязанная фракция оптимизированного ген-активированного материала практически сразу после имплантации изделия высвобождается с поверхности матрикса-носителя, создавая первую нагрузочную дозу, которая в свою очередь обеспечивает ранний пик экспрессии трансгена, приходящийся уже на пролиферативную стадию воспаления. Вторая фракция, связанная с матриксом-носителем, высвобождается постепенно, в том числе по мере его биодеградации, создавая второй, но уже пролонгированный максимальный уровень продукции терапевтического белка во вторую - регенеративную - стадию восстановительно процесса. Таким образом, оптимизированный ген-активированный материал, содержащий две фракции нуклеиновых кислот, начинает оказывать свое биологически активное действия в зоне повреждения тканей гораздо раньше обычного ген-активированного материала, что и позволяет первому ускорить репаративную регенерацию тканей.
Независимо от вида генных конструкций, предназначенных для размещения на поверхности матриксов-носителей, состава растворов, содержащих эти нуклеиновые кислоты, состава вспомогательных веществ, физических методов, обеспечивающих транзиторную фиксацию генных конструкций на поверхности матриксов-носителей, а также модификаций свойств поверхности матриксов-носителей отличительной особенностью разработанного способа и изделий, изготовленных с его помощью, является отсутствие химической связи между второй фракцией нуклеиновых кислот и матриксом-носителем при наличии связанной с ним первой фракции нуклеиновых кислот. Наличие сразу двух фракций - связанной и несвязанной - является главным отличием разработанных изделий от других известных ген-активированных материалов [50].
Пример №1
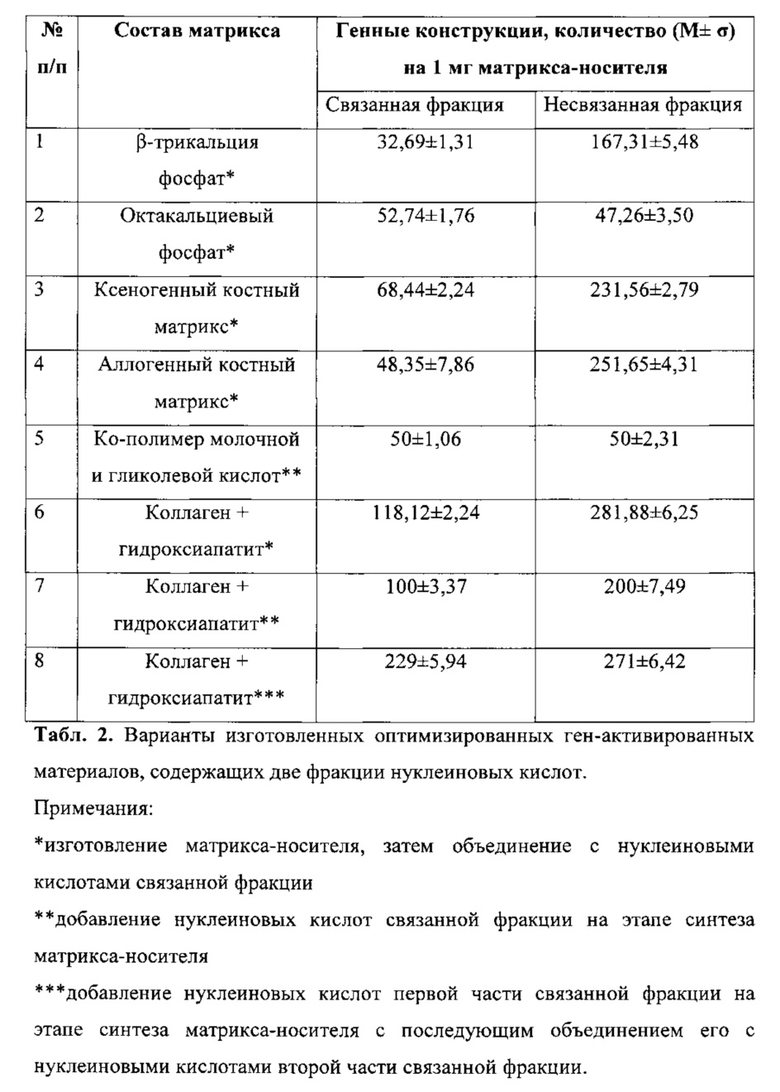
С использованием разработанного способа мы изготовили ряд вариантов оптимизированных ген-активированных материалов, содержащих две фракции нуклеиновых кислот (связанную и несвязанную) (табл. 2).
Разработанные оптимизированные ген-активированные материалы были исследованы in vitro на предмет построения кривой высвобождения генных конструкций. Результат, типичный для всех полученных нами ген-активированных материалов с двумя фракциями нуклеиновых кислот, в сравнении с обычными ген-активированными материалами представлен на Рис. 3. Исследование выполняли в стандартных модельных условиях без эвакуации/деструкции выделившихся в раствор нуклеиновых кислот, т.е. определяли накопленную концентрацию в растворе.
Важно, что разработанный способ позволяет использовать для создания оптимизированных ген-активированных материалов любые твердые матриксы-носители и любые нуклеиновые кислоты, что свидетельствует об универсальности подхода и предопределяет значительные перспективы для внедрения в клиническую практику для разных показаний (восстановление тканей и органов опорно-двигательного аппарата, кожи, слизистых оболочек, органов сердечно-сосудистой системы, периферических нервов, полых и паренхиматозных органов и т.п.).
За счет применения различных вариантов генных конструкций, в том числе кодирующих различные трансгены, можно изготавливать оптимизированные ген-активированные материалы, фракции нуклеиновых кислот, в которых различны по составу вектора и (или) трансгена(ов). В частности, мы использовали для создания изделий, указанных в табл. 2, не только плазмидную ДНК, несущую ген vegf, но и плазмидную ДНК, несущую гены других белков: фактора стромальных клеток-1 (SDF-1), основного фактора роста фибробластов (bFGF), нейрального фактора роста (NGF), эпидермального фактора роста (EGF), костных морфогенетических белков (BMP), интерлейкинов (IL) и др.), в том числе комбинации генов нескольких белков (SDF-VEGF; BMP-VEGF и т.д.) и в различных сочетаниях.
Для создания оптимизированных ген-активированных материалов могут быть использованы и другие генные конструкции, кодирующие любые другие белки, в том числе цитокины, мембранные и цитоплазматические рецепторы, транскрипционные факторы и т.п. в зависимости от биологического эффекта, который необходимо достичь за счет применения ген-активированного материала. Выбор определяется также необходимой динамикой развития биологического эффекта.
Например, мы изготовили оптимизированный ген-активированный материал, состоящий из ОКФ, связанной фракции нуклеиновых кислот в виде плазминой ДНК с геном vegf и несвязанной фракции в виде двухкассетной плазмидной ДНК с генами vegf и sdf. Идеология такого выбора была продиктована необходимостью привлечь в зону костного дефекта в максимально ранние сроки после имплантации значительное количество камбиальных резервов (клеток-предшественниц остеобластов и эндотелиоцитов), в том числе из системных источников. За эту часть механизма действия отвечала несвязанная фракция двухкассетной плазмидной ДНК, так как SDF является известным индуктором хоуминга клеток. На стадии же регенерации более важным является воздействие на уже привлеченные в зону дефекта клетки-предшественницы, в частности активизация ангиогенеза. Этот биологический эффект был реализован за счет связанной фракции генных конструкций - плазминой ДНК с геном vegf, - которая постепенно высвобождалась из структуры матрикса-носителя, в том числе по мере его биодеградации, и реализовывала свой механизм действия.
Указанный пример является лишь частным вариантов успешного использования разработанного способа создания оптимизированных ген-активированных материалов. Варьирование составом двух фракций нуклеиновых кислот может позволить добиться комплексных, сложных биологических эффектов, различающихся как по времени реализации за счет разницы в динамике высвобождения двух фракций нуклеиновых кислот, так и по свой природе за счет возможности использовать различные генные конструкции в составе двух фракций.
Таким образом, оптимизированные ген-активированные материалы, содержащие две фракции нуклеиновых кислот (связанную и несвязанную), характеризуются не только точностью дозы и возможностью ее повысить до необходимого уровня, но и позволяют осуществить более тонкое и сложное влияние на репаративный процесс, что недостижимо при использовании обычных ген-активированных материалов.
Оптимизированные ген-активированные материалы, содержащие функционализированный матрикс-носитель
Другое направление из разработанных нами, позволяющее получить оптимизированные ген-активированные материалы, состоит в повышении минимальной дозы и точности дозирования связанной фракции нуклеиновых кислот за счет модификации матрикса-носителя.
Как было указано выше, твердые материалы, содержащие соединения кальция, способны связывать некоторое количество нуклеиновых кислот за счет комплексообразования, реализуемого, возможно, по механизму донорно-акцепторной (координативной) связи. Выявление этого эффекта, который нашел свое основное применение в хроматографии, привело к попыткам модификации состава гидроксиапатита хроматографических колонок другими металлами, такими как стронций [51]. Кроме того, известны прецеденты модификации матриксов из фосфатов кальция другими металлами, такими как магний, для изменения физико-химических свойств материала и оптимизации их использования в качестве матриксов-носителей для клеток в составе тканей инженерных изделий [52].
Однако до настоящего времени не были разработаны способы модификации материала, содержащего соединения кальция, комплексообразователями для последующего связывания с нуклеиновыми кислотами и создания ген-активированного материала.
Разработанный нами способ включает обработку исходного материала, из которого планируется синтезировать матрикс-носитель, раствором соли(ей) других металлов-комплексообразователей (в частности, стронция, бария, магния) с инициацией химической реакции, приводящей к получению материала, содержащего как соединения кальция, так и соединения другого металла-комплексобразователя. Полученный таким образом матрикс-носителя подвергается объединению с нуклеиновыми кислотами с получением оптимизированного ген-активированного материала.
В исходный материал одновременной или последовательной обработкой растворами солей разных металлов могут быть внесены несколько различных комплексообразователей в различных соотношениях.
Повышение минимальной дозы генных конструкций и точность дозирования обеспечиваются варьированием рассчитанного экспериментальным путем количества, состава и соотношения внесенных в состав матрикса-носителя комплексообразователей.
Пример 2
С использованием разработанного способа из трикальциевого фосфата (ТКФ) (Ca3(PO4)2) с атомным соотношением элементов Ca/P 1,5±0,1 был синтезирован материал из октакальциевого фосфата (ОКФ), в котором от 1 до 3% Ca были заменены на Mg2+, Sr2+ и Ba2+ в том числе в различных соотношениях.
Особенностью данного частного варианта изготовления оптимизированных ген-активированных материалов являлось гармоничное встраивание этапа обработки исходного материала раствором комплексообразователя в известный этап синтеза ОКФ [53]. Иными словами функционализация матрикса-носителя была осуществлена в процессе его синтеза, а не в качестве дополнительного этапа. В зависимости от природы материала возможен любой из вариантов.
Модификацию ТКФ осуществляли в специальных буферных растворах А и Б. Для получения раствора А готовили водный 1,5 М раствор ацетата натрия и 0,15±0,02 М глутаминовой аминокислоты и доводили его ортофосфорной кислотой до значения pH 5,5±0,1 раствора [53]. Ацетат натрия служит буфером. Для получения раствора Б готовили водный 1,5 М раствор ацетата натрия и ацетата магния, стронция или бария со значением pH 8,7±0,1 раствора. Раствор А предназначен для перевода ТКФ в дикальцийфосфат дигидрат (ДКФД). Процесс ведут при 35±1°С. Соотношение матрикса-носителя ТКФ и раствора А составляет 1:100. Процесс трансформации ТКФ в ДКФД полностью завершается к 7 сут. При проведении процесса при температуре выше 35±1°С вместе с ДКФД начинает образовываться дикальцийфосфат (ДКФ), который является цитотоксичным. При температуре менее 30°С реакция перехода замедляется, так что даже через месяц выдержки в растворе достигается значение менее 50% ДКФД. Далее полученные матриксы-носители отмывали в дистиллированной воде до значения pH не ниже 6,5, высушивали при температуре 35±1°С в течение суток.
Раствор Б служит для трансформации ДКФД в ОКФ. Химическую обработку необходимо осуществлять также при 35±1°С. Соотношение ДКФД и раствора Б составляет 1:100. Процесс трансформации ДКФД в ОКФ протекает в течение 7 сут. Именно в данном процессе, одномоментно с переходом ДКФД в ОКФ, реализуется частичное катионное замещение Mg, Sr2+, Ba2+на Ca2+, т.е. создание положительнозаряженных комплексов в процессе формирования структуры апатита (ДКФД→ОКФ), что является также отличительным признаком данной технологии от описанной в [53]. При замещении 1-3% ионов кальция на Mg2+, Sr2+ или Ba2+происходит образование однофазного замещенного ОКФ. Повышение содержания ионов металлов в растворе приводит к ингибированию процесса гидролиза ДКФД в ОКФ, что обусловлено влиянием ионов металлов на процессы "осаждения - растворения" ДКФД.
Важно, что данный способ реализуем при физиологических температурах (не превышающих 40°С), что очень существенно при синтезе и функционализации таких матриксов-носителей, как ОКФ (термически нестабильное соединение).
Модифицированные комплексообразователями матриксы-носители были объединены с нуклеиновыми кислотами (в частности, плазмидной ДНК с геном vegf) по описанному в патенте РФ №2519326 и частично модифицированному способу, изложенному выше.
Способ создания оптимизированных ген-активированных материалов, содержащих функционализированный комплексообразователями матрикс-носитель, позволил разработать изделия из ОКФ с минимальными концентрациями генных конструкций, превышающими в 1,2-2,3 количество связанных со стандартным ОКФ нуклеиновых кислот (табл.1, 2).
Однако неожиданные данные были получены при построении кривой высвобождения генных конструкций из полученных таким образом оптимизированных ген-активированных материалов и при оценке эффективности трансфекции культур клеток in vitro.
Оказалось, что функционализация матрикса-носителя (в частности, ОКФ) комплексообразователями (в частности, магнием, барием и стронцием) приводила к изменению динамики высвобождения генных конструкций из состава оптимизированных ген-активированных материалов, по сравнению со стандартными, содержащими обычный ОКФ и те же нуклеиновые кислоты (рис. 4).
Вероятной причиной является различная сила связи в комплексах соединений металлов с молекулами нуклеиновых кислот. Подтверждение данной гипотезы требует проведения дополнительных исследований, которые позволят в различных экспериментальных системах in vitro и in vivo выявить динамику высвобождения генных конструкций и в дальнейшем программировать ее за счет варьирования составом, количеством и соотношением комплексообразователей в составе матрикса-носителя.
Другим неожиданным техническим результатом разработанного способа стало повышение эффективности трансфекции культур клеток in vitro, ко-инкубированных с оптимизированными ген-активированными материалами, содержащими функционализированный матрикс-носитель из фосфатов кальция. Широко известным является кальций-фосфатный метод оптимизации трансфекции, основанный на применении хлорида кальция и растворов фосфатных буферов, которые формируют преципитаты фосфатов кальция, связывающие молекулы генных конструкций и поступающие в клетки за счет эндоцитоза [54]. Более того, уже выдвигались предположения, что ген-активированные материалы, состоящие из фосфатов кальция и плазмидной ДНК обладают более высокой эффективностью трансфекции [55]. Однако до настоящего времени не было данных, свидетельствующих о повышении уровня трансфекции за счет модификации кальций-фосфатного матрикса-носителя другими комплексообразователями. В наших же исследованиях такой эффект был детектирован. В эксперименте in vitro мультипотентные мезенхимальные стромальные клетки были ко-инкубированы с обычными (ОКФ/plVEGF) и оптимизированными ген-активированными материалами (ОКФ + Mg2+/plVEGF; ОКФ+Sr2+/plVEGF; ОКФ+Ba2+/plVEGF; ОКФ+Mg2+Ba2+/plVEGF), помещенными во вставки «transwell»-системы (мембрана с величиной пор 3 мкм). Иными словами, клетки и материалы были разделены мембраной, проницаемой для культуральной среды. С помощью ПЦР-РТ и ИФА была выявлена в 1,5-2 раза большая эффективность трансфекции оптимизированных ген-активированных материалов. Причиной такого эффекта стали, по всей видимости, обнаруженные на светооптическом уровне в зоне с клетками микрочастицы материалов, расположенные как вне-, так и внутриклеточно (рис. 5). По всей видимости, эти микрочастицы диаметром менее 3 мкм представляли собой преципитаты фосфатов кальция и фосфатов других комплексообразователей (магния, бария, стронция), которые связывали молекулы генных конструкций и поступали в клетки. Аналогичные на светооптическом уровне частицы были обнаружены и в случае обычных ген-активированных материалов из ОКФ. Однако повышение уровня трансфекции в случае оптимизированных изделий могло быть связано с рядом причин: более высокий уровень поступления преципитатов других комплексообразователей внутрь клеток, иная сила связи с молекулами генных конструкций, наличие протективных свойств у микрочастиц фосфатов других металлов, защищающих генные конструкции от разрушения ферментами клетки. Безусловно, гипотетические причины повышенной эффективности трансфекции требуют дальнейших исследований.
Важно отметить, что разработанный способ создания оптимизированных ген-активированных материалов из функционализированного комплексообразователями матрикса-носителя и генных конструкций может быть использован в сочетании с первым из описанных в настоящем изобретении способов. Иными словами к полученному по второму способу ген-активированному материалу может быть добавлена несвязанная фракция нуклеиновых кислот. В этом случае оптимизация ген-активированного материала будет достигнута сразу по двум направлениям: и функционализация матрикса-носителя с увеличением дозы связанной фракции нуклеиновых кислот, и внесение несвязанной фракции генных конструкций. Представляется очевидным, что такой комплексный подход позволит создать еще более эффективные изделия, объединяющие в себе преимущества каждого из подходов.
Стерильное медицинское изделие из оптимизированных ген-активированных материалов
Все ген-активированные материалы, находящиеся на различных стадиях экспериментальных исследований в РФ и за рубежом, являются прототипами или основными составляющими медицинских изделий. Любое медицинское изделие, предназначенное для имплантации в организм реципиента, должно быть стерильным. Однако все методы стерилизации направлены на уничтожение всех видов микроорганизмов и их компонентов, включая нуклеиновые кислоты как класс органических веществ. Однако нуклеиновые кислоты являются биологически активным отличительным компонентом ген-активированных материалов, обеспечивающих их специфическое действие. В этой связи, разработка стерильного медицинского изделия из ген-активированного материала является крайне сложной задачей.
В ходе проведенных исследований разработали технологию, позволяющую создать медицинские изделия из оптимизированных ген-активированных материалов по любому (или обоим сразу) из вышеприведенных способов, но в стерильном виде.
Разработанный способ включает следующие этапы:
1) создание исходных компонентов: стерильного раствора высокоочищенных нуклеиновых кислот (плазмидная ДНК с геном vegf, с геном sdf, другими генами и их комбинациями) и твердого матрикса-носителя;
2) обработка полученного матрикса-носителя в растворе фосфатного буфера, последующая стерилизация любым удобоваримым методом (автоклавирование, ионизирующее излучение и т.д.);
3) внесение нуклеиновых кислот в растворе фосфатного буфера с фармацевтически приемлемыми вспомогательными веществами, обеспечивающими стабильность нуклеиновых кислот, в стерильную емкость с заданным количеством матрикса-носителя в соотношении не менее 100 нг нуклеиновых кислот на 1 мг матрикса-носителя, инкубация в течение не менее 4 ч при оптимальной температуре (чаще - 37°С);
4) лиофильная сушка матрикса-носителя, связавшего часть добавленных нуклеиновых кислот, в растворе с нуклеиновыми кислотами, в качестве которого может быть использован оставшийся в емкости раствор или другой раствор с нуклеиновыми кислотами.
В качестве стерильной емкости на третьем этапе разработанного способа может быть использована емкость, в которой изготовленное медицинское изделие в дальнейшем будет поставляться потребителям (флакон, шприц-дозатор и т.д.). Более того, расфасовка в указанные емкости может быть выполнена еще на втором этапе с последующей стерилизацией уже в них. Все манипуляции в ходе третьего этапа должны быть выполнены в стерильных условиях (помещения классов А или Б).
Лиофилизация в рамках четвертого этапа должна осуществляться в стерильных условиях по любому удобоваримому протоколу. По завершению лиофилизации крышки стерильных емкостей с полученными медицинскими изделиями должны быть плотно закрыты (в оптимальном варианте - поддоном лиофилизатора сразу по завершению процесса). Закрытые емкости с оптимизированными ген-активированными материалами могут быть переданы в стерильных условиях в зону упаковки (материалы индивидуальной упаковки и маркировки предварительно стерилизуются), если требуется сохранить и наружную поверхность флаконов стерильной.
Согласно разработанному способу были изготовлены несколько вариантов медицинских изделий из ОКФ или депротеинизированного костного матрикса и плазмидных ДНК, кодирующих различные гены (vegf, sdf и др.). Результаты контрольных исследований показали абсолютную стерильность медицинских изделий.
Литература
1. Дорожно-транспортные травмы. Информационный бюллетень, май 2016. http://www.who.int/mediacentre/factsheets/fs358/ru/.
2. Сведения о показателях безопасности дорожного движения. http://www.gibdd.ru/stat.
3. Рак. Информационный бюллетень №297, февраль 2015 г. http://www.who.int/mediacentre/factsheets/fs297/ru/.
4. Suh Н. Tissue restoration, tissue engineering and regenerative medicine. Yonsei Med J. 2000 Dec; 41(6): 681-4.
5. Deev RV, Bozo IY, Mzhavanadze ND, Voronov DA, Gavrilenko AV, Chervyakov YV, Staroverov IN, Kalinin RE, Shvalb PG, Isaev AA. pCMV-vegf165 Intramuscular Gene Transfer is an Effective Method of Treatment for Patients With Chronic Lower Limb Ischemia. J Cardiovasc Pharmacol Ther. 2015 Sep; 20(5): 473-82.
6. Bang OY, Lee JS, Lee PH, Lee G. Autologous mesenchymal stem cell transplantation in stroke patients. Ann Neurol. 2005 Jun; 57(6): 874-82.
7. Tatara AM, Mikos AG. Tissue Engineering in Orthopaedics. J Bone Joint Surg Am. 2016 Jul 6; 98(13): 1132-9.
8. Sadri-Ardekani H, Atala A. Regenerative medicine. Methods. 2016 Apr 15; 99: 1-2.
9. Deev R.V., Drobyshev A.Y., Bozo I.Y., Isaev A.A. Ordinary and Activated Bone Grafts: Applied Classification and the Main Features. Biomed Res Int. 2015; 2015: 365050.
10. Yu Y, Alkhawaji A, Ding Y, Mei J. Decellularized scaffolds in regenerative medicine. Oncotarget. 2016 Jul 29. doi: 10.18632/oncotarget.10945. [Epub ahead of print].
11. Wang Z., Guo Z., Bai H. et al. Clinical evaluation of β-TCP in the treatment of lacunar bone defects: a prospective, randomized controlled study. Mater. Sci. Eng. С Mater. Biol. Appl. 2013; 33(4): 1894-9.
12. Komlev V.S., Barinov S.M., Bozo I.I. et al. Bioceramics composed of octacalcium phosphate demonstrate enhanced biological behaviour. ACS Appl. Mater. Interfaces 2014; 6(19): 16610-20.
13. Zakaria S.M., Sharif Zein S.H., Othman M.R. et al. Nanophase hydroxyapatite as a biomaterial in advanced hard tissue engineering: a review. Tissue Eng. Part В Rev. 2013; 19(5): 431-41.
14. Wang S., Wang X., Draenert F.G. et al. Bioactive and biodegradable silica biomaterial for bone regeneration. Bone 2014; 67: 292-304.
15.  Lanao R.P., Jonker A.M., Wolke J.G. et al. Physicochemical properties and applications of poly(lactic-co-glycolic acid) for use in bone regeneration. Tissue Eng. Part В Rev. 2013; 19(4): 380-90.
Lanao R.P., Jonker A.M., Wolke J.G. et al. Physicochemical properties and applications of poly(lactic-co-glycolic acid) for use in bone regeneration. Tissue Eng. Part В Rev. 2013; 19(4): 380-90.
16. Li X., Wang X., Miao Y., Yang G. et al. Guided bone regeneration at a dehiscence-type defect using chitosan/collagen membranes in dogs. Zhonghua Kou Qiang Yi Xue Za Zhi. 2014; 49(4): 204-9.
17. Sabri F, Gerth D, Tamula GR et al. Novel technique for repair of severed peripheral nerves in rats using polyurea crosslinked silica aerogel scaffold. J Invest Surg. 2014; 27(5): 294-303.
18. De Cicco F, Russo P, Reverchon E et al. Prilling and supercritical drying: A successful duo to produce core-shell polysaccharide aerogel beads for wound healing. Carbohydr Polym. 2016; 147: 482-9.
19. He L, Tang S, Prabhakaran MP, Liao S, Tian L, Zhang Y, Xue W, Ramakrishna S. Surface modification of PLLA nano-scaffolds with laminin multilayer by LbL assembly for enhancing neurite outgrowth. Macromol Biosci. 2013 Nov; 13(11): 1601-9.
20. Motamedian SR, Hosseinpour S, Ahsaie MG, Khojasteh A. Smart scaffolds in bone tissue engineering: A systematic review of literature. World J Stem Cells. 2015 Apr 26; 7(3): 657-68.
21. Jahani H, Jalilian FA, Wu CY, et al. Controlled surface morphology and hydrophilicity of polycaprolactone toward selective differentiation of mesenchymal stem cells to neural like cells. J Biomed Mater Res A. 2015 May; 103(5): 1875-81.
22. Патент биокомпозит.
23. Huang YC, Kaigler D, Rice KG et al. Combined angiogenic and osteogenic factor delivery enhances bone marrow stromal cell-driven bone regeneration. J Bone Miner Res. 2005 May; 20(5): 848-57.
24. Kan I., Melamed E., Offen D. Integral therapeutic potential of bone marrow mesenchymal stem cells. Curr. Drug Targets. 2005; 6(1): 31-41.
25. Knight M.N., Hankenson K.D. Mesenchymal Stem Cells in Bone Regeneration. Adv. Wound Care (New Rochelle). 2013; 2(6): 306-16.
26. Zorin V.L., Komlev V.S., Zorina A.I. et al. Octacalcium phosphate ceramics combined with gingiva-derived stromal cells for engineered functional bone grafts. Biomed. Mater. 2014; 9(5): 055005.
27. Деев P.B., Исаев A.A., Кочиш А.Ю. и др. Пути развития клеточных технологий в костной хирургии. Травматология и ортопедия России, 2008; 1(47): 65-75.
28. Liu X., Zhang G., Hou С.et al. Vascularized bone tissue formation induced by fiber-reinforced scaffolds cultured with osteoblasts and endothelial cells. Biomed. Res. Int. 2013; 2013: 854917.
29. Kim B.S., Kim J.S., Lee J. Improvements of osteoblast adhesion, proliferation, and differentiation in vitro via fibrin network formation in collagen sponge scaffold. J. Biomed. Mater. Res. A. 2013; 101(9): 2661-6.
30.  Y.M., Cheng L.M., Pei G.X. et al. Experimental study of repairing femoral bone defects with nHA/RHLC/PLA scaffold composite with endothelial cells and osteoblasts in canines. Zhonghua Yi Xue Za Zhi. 2013; 93(17): 1335-40.
Y.M., Cheng L.M., Pei G.X. et al. Experimental study of repairing femoral bone defects with nHA/RHLC/PLA scaffold composite with endothelial cells and osteoblasts in canines. Zhonghua Yi Xue Za Zhi. 2013; 93(17): 1335-40.
31. Shim J.B., Ankeny R.F., Kim H. et al. A study of a three-dimensional PLGA sponge containing natural polymers co-cultured with endothelial and mesenchymal stem cells as a tissue engineering scaffold. Biomed. Mater. 2014; 9(4): 045015.
32. Andreev DIu, Abramova NV, Blinova MI, Pinaev GP. Efficacy of dermoplasty and the dermal equivalent in treatment of vast leg ulcers of mixed genesis. Vestn Khir Im I I Grek. 2013; 172(1): 104-7.
33. Illich D.J., Demir N., 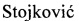 M. et al. Concise review: induced pluripotent stem cells and lineage reprogramming: prospects for boneregeneration. Stem Cells 2011; 29(4): 555-63.
M. et al. Concise review: induced pluripotent stem cells and lineage reprogramming: prospects for boneregeneration. Stem Cells 2011; 29(4): 555-63.
34. Pelegrine A.A., Aloise A.C., Zimmermann A. et al. Repair of critical-size bone defects using bone marrow stromal cells: a histomorphometric study in rabbit calvaria. Part I: use of fresh bone marrow or bone marrow mononuclear fraction. Clin. Oral. Implants Res. 2014; 25(5): 567-72.
35. Kim B.S., Kim J.S., Lee J. Improvements of osteoblast adhesion, proliferation, and differentiation in vitro via fibrin network formation in collagen sponge scaffold. J. Biomed. Mater. Res A. 2013; 101(9): 2661-6.
36. Eastlack RK, Garfin SR, Brown CR, Meyer SC. Osteocel Plus cellular allograft in anterior cervical discectomy and fusion: evaluation of clinical and radiographic outcomes from a prospective multicenter study. Spine (Phila Pa 1976). 2014 Oct 15; 39(22): E1331-7.
37. Zhang W., Zhu C., Wu Y. et al. VEGF and BMP-2 promote bone regeneration by facilitating bone marrow stem cell homing and differentiation. Eur. Cell Mater. 2014; 27: 1-11.
38. Holloway J.L., Ma H., Rai R. et al. Modulating hydrogel crosslink density and degradation to control bone morphogenetic protein delivery and in vivo bone formation. J. Control. Release. 2014; 191: 63-70.
39. Chang P.C., Dovban A.S., Lim L.P. et al. Dual delivery of PDGF and simvastatin to accelerate periodontal regeneration in vivo. Biomaterials 2013; 34(38): 9990-7.
40. Kleinschmidt K., Ploeger F., Nickel J. et al. Enhanced reconstruction of long bone architecture by a growth factor mutant combining positive features of GDF-5 and BMP-2. Biomaterials 2013; 34(24): 5926-36.
41. Деев P.B., Дробышев А.Ю., Бозо И.Я. и др. Создание и оценка биологического действия ген-активированного остеопластического материала, несущего ген VEGF человека. Клеточная трансплантология и тканевая инженерия 2013; VIII (3): 78-85.
42. Wegman F., Bijenhof A., Schuijff L. et al. Osteogenic differentiation as a result of BMP-2 plasmid DNA based gene therapy in vitro and in vivo. Eur. Cell Mater. 2011; 21: 230-42.
43. Baboo S., Cook P.R. "Dark matter" worlds of unstable RNA and protein. Nucleus 2014 Jun 20; 5(4) [Epub ahead of print].
44. Evans C.H. Gene delivery to bone. Adv. Drug Deliv. Rev. 2012; 64(12): 1331-40.
45. Григорян A.C., Шевченко К.Г. Возможные молекулярные механизмы функционирования плазмидных конструкций, содержащих ген VEGF. Клеточная трансплантология и тканевая инженерия, 2011; VI(3): 24-8.
46. Rose Т., Peng Н., Usas A. et al. Ex-vivo gene therapy with BMP-4 for critically sized defects and enhancement of fracture healing in an osteoporotic animal model. Unfallchirurg 2005; 108(1): 25-34.
47. Lu H, Lv L, Dai Y, Wu G, Zhao H, Zhang F. Porous chitosan scaffolds with embedded hyaluronic acid/chitosan/plasmid-DNA nanoparticles encoding TGF-β1 induce DNA controlled release, transfected chondrocytes, and promoted cell proliferation. PLoS One. 2013 Jul 23; 8(7): e69950.
48. Giovannini R, Freitag R. Comparison of different types of ceramic hydroxyapatite for the chromatographic separation of plasmid DNA and a recombinant anti-rhesus D antibody. Bioseparation. 2000; 9(6): 359-68.
49. Бозо И.Я., Деев P.B., Дробышев А.Ю. и др. Эффективность ген-активированного остеопластического материала на основе октакальциевого фосфата и плазмидной ДНК с геном vegf в восполнении «критических» костных дефектов. Вестник травматологии и ортопедии им. Н.Н. Приорова. 2015; 1: 35-42.
50. Гоулдстейн С.А. Способы переноса генов in vivo для заживления ран. Патент №2170104 от 10.07.2001.
51. Kawasaki T, Niikura M, Takahashi S, Kobayashi W. Strontium-phosphate hydroxyapatite high-performance liquid chromatography. Biochem Int. 1987 Dec; 15(6): 1137-49.
52. Zhang J, Ma X, Lin D, Shi H, Yuan Y, Tang W, Zhou H, Guo H, Qian J, Liu C. Magnesium modification of a calcium phosphate cement alters bone marrow stromal cell behavior via an integrin-mediated mechanism. Biomaterials 2015 Jun; 53: 251-64.
53. «Способ получения керамики на основе октакальциевого фосфата». Дата международной подачи: 31.08.2016. Номер международной заявки: PCT/RU2016/000587.
54. Graham FL, van der Eb AJ. Transformation of rat cells by DNA of human adenovirus 5. Virology. 1973 Aug; 54(2): 536-9.
55. Keeney M., van den Beucken J.J., van der Kraan P.M. et al. The ability of a collagen/calcium phosphate scaffold to act as its own vector for gene delivery and to promote bone formation via transfection with VEGF(165). Biomaterials 2010; 31(10): 2893-902.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ СОЗДАНИЯ ПЕРСОНАЛИЗИРОВАННОГО ГЕН-АКТИВИРОВАННОГО ИМПЛАНТАТА ДЛЯ РЕГЕНЕРАЦИИ КОСТНОЙ ТКАНИ | 2015 |
|
RU2597786C2 |
| БИОКОМПОЗИТ ДЛЯ ОБЕСПЕЧЕНИЯ ВОССТАНОВИТЕЛЬНЫХ ПРОЦЕССОВ ПОСЛЕ ПОВРЕЖДЕНИЯ У МЛЕКОПИТАЮЩЕГО, СПОСОБ ЕГО ПОЛУЧЕНИЯ (ВАРИАНТЫ) И ПРИМЕНЕНИЯ | 2011 |
|
RU2519326C2 |
| Оптимизированная нуклеотидная последовательность и фармацевтическая композиция на ее основе с пролонгированной экспрессией трансгена vegf | 2015 |
|
RU2612497C2 |
| Способ получения керамических гранул для регенерации костной ткани | 2018 |
|
RU2695342C1 |
| Способ стимуляции репаративного ангиогенеза и регенерации соединительной ткани, при её повреждении, методом генной терапии с использованием видоспецифичных генов белковых факторов vegf и fgf2, в ветеринарии, и генетическая конструкция для реализации заявленного способа | 2015 |
|
RU2614665C1 |
| Способ стимуляции репаративного ангиогенеза и регенерации кожного покрова собак при его повреждении методом генной терапии с использованием видоспецифичных генов белковых факторов vegf и fgf2 в ветеринарии и генетическая конструкция для реализации заявленного способа | 2019 |
|
RU2719513C1 |
| НОСИТЕЛЬ ДЛЯ НАПРАВЛЕННОЙ ДОСТАВКИ НУКЛЕИНОВЫХ КИСЛОТ В КЛЕТКИ, ЭКСПРЕССИРУЮЩИЕ РЕЦЕПТОР CXCR4 | 2012 |
|
RU2522810C1 |
| Способ получения плазмидной ДНК в составе полимерных наносфер для доставки | 2022 |
|
RU2817116C1 |
| Способ изготовления средства для клеточно-опосредованной генной терапии и средство для клеточно-опосредованной генной терапии | 2019 |
|
RU2716013C2 |
| РЕКОМБИНАНТНЫЙ ГЕН, КОДИРУЮЩИЙ БЕЛОК HBD-EPO, РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PL610, СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО БЕЛКА HBD-EPO, РЕКОМБИНАНТНЫЙ БЕЛОК HBD-EPO, КОМПОЗИЦИЯ ДЛЯ СПЕЦИФИЧЕСКОЙ ИНДУКЦИИ РЕГЕНЕРАЦИИ КОСТНОЙ ТКАНИ, СПОСОБ СПЕЦИФИЧЕСКОЙ ИНДУКЦИИ РЕГЕНЕРАЦИИ КОСТНОЙ ТКАНИ | 2017 |
|
RU2664192C1 |
Изобретение относится к биотехнологии. Описан способ получения оптимизированного твердого ген-активированного материала, включающего создание матрикса-носителя, связавшего как минимум одну молекулу нуклеиновой кислоты, с последующим размещением на его поверхности еще как минимум одной дополнительной молекулы нуклеиновой кислоты любым физическим методом, позволяющим осуществить такое размещение без образования химической связи между указанным матриксом-носителем и дополнительной нуклеиновой кислотой. Также представлен способ получения твердого матрикса-носителя, используемого в описанном способе, и предназначенного для связывания как минимум одной молекулы нуклеиновой кислоты, включающий введение в состав твердого матрикса-носителя на этапе его синтеза любого комплексообразователя за счет обработки исходного материала раствором, содержащим соль металла-комплексообразователя, способного связать как минимум одну молекулу нуклеиновой кислоты. Также представлены продукты, полученные указанными способами. Изобретение позволяет получать оптимизированные остеопластические материалы, отличающиеся точной дозой нуклеиновых кислот, более эффективной динамикой высвобождения генных конструкций из структуры матрикса-носителя и повышенным уровнем трансфекции клеток реципиентного ложа биологически активным веществом. 4 н. и 1 з.п. ф-лы, 5 ил., 2 табл., 2 пр.
1. Способ получения оптимизированного твердого ген-активированного материала, включающего создание матрикса-носителя, связавшего как минимум одну молекулу нуклеиновой кислоты, с последующим размещением на его поверхности еще как минимум одной дополнительной молекулы нуклеиновой кислоты любым физическим методом, позволяющим осуществить такое размещение без образования химической связи между указанным матриксом-носителем и дополнительной нуклеиновой кислотой.
2. Способ по п. 1, отличающийся тем, что матрикс-носитель, предварительно подготовленный для связывания как минимум одной молекулы нуклеиновой кислоты, подвергается стерилизации, с последующим связыванием как минимум одной молекулы нуклеиновой кислоты, затем лиофилизации в растворе с еще как минимум одной молекулой нуклеиновой кислоты.
3. Способ получения твердого матрикса-носителя, используемого в способе по п. 1 и предназначенного для связывания как минимум одной молекулы нуклеиновой кислоты, включающий введение в состав твердого матрикса-носителя на этапе его синтеза любого комплексообразователя за счет обработки исходного материала раствором, содержащим соль металла-комплексообразователя, способного связать как минимум одну молекулу нуклеиновой кислоты.
4. Оптимизированный твердый ген-активированный материал для регенерации тканей, изготовленный способом по п. 1, характеризующийся началом высвобождения в первые сутки после имплантации оптимизированного ген-активированного материала как минимум одной дополнительной молекулы нуклеиновой кислоты, размещенной любым физическим методом на поверхности твердого матрикса-носителя, с последующим пролонгированным высвобождением нуклеиновой кислоты, связанной с твердым матриксом-носителем.
5. Стерильный оптимизированный твердый ген-активированный материал для регенерации тканей, изготовленный способом по п. 1, характеризующийся началом высвобождения в первые сутки после имплантации оптимизированного ген-активированного материала как минимум одной дополнительной молекулы нуклеиновой кислоты, размещенной любым физическим методом на поверхности твердого матрикса-носителя, с последующим пролонгированным высвобождением нуклеиновой кислоты, связанной с твердым матриксом-носителем.
| ДЕЕВ Р | |||
| В | |||
| Генные технологии в костной реконструкции | |||
| Пластическая хирургия и косметология | |||
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| DEEV R | |||
| V.Ordinary and activated bone grafts: applied classification and the main features | |||
| BioMed research international, 2015 | |||
| RAFTERY R | |||
| M.Development of a gene-activated scaffold platform for tissue engineering applications using chitosan-pDNA nanoparticles on collagen-based scaffolds | |||
| Journal of Controlled Release, 2015, 210, 84-94 | |||
| MEHTA M | |||
| Biomaterial delivery of morphogens to mimic the natural healing cascade in bone | |||
| Advanced drug delivery reviews, 2012, 64(12), с | |||
| Приспособление для ослабления силы удара при столкновениях поездов | 1923 |
|
SU1257A1 |
| БИОКОМПОЗИТ ДЛЯ ОБЕСПЕЧЕНИЯ ВОССТАНОВИТЕЛЬНЫХ ПРОЦЕССОВ ПОСЛЕ ПОВРЕЖДЕНИЯ У МЛЕКОПИТАЮЩЕГО, СПОСОБ ЕГО ПОЛУЧЕНИЯ (ВАРИАНТЫ) И ПРИМЕНЕНИЯ | 2011 |
|
RU2519326C2 |

