Изобретение относится к биотехнологии, конкретно - биофармакологии, к областям получения и производства терапевтических и вакцинных препаратов, технологий выделения и очистки рекомбинантных белков, создания изделий медицинского назначения, удовлетворяющих требованиям апирогенности, и касается способа получения рекомбинантной фосфатазы LpxE, способной удалять фосфатные группы липида А бактериального экзотоксина, что приводит к снижению токсичности медицинских препаратов, полученных из бактериальной биомассы.
Основными сферами применения предлагаемого способа получения бактериальной фосфатазы LpxE являются биомедицинские исследования, фармацевтика, разработка и создание производственных технологий получения медицинских лекарственных препаратов (в первую очередь производство вакцин), лабораторные биологические и медицинские исследования, получение апирогенных препаратов рекомбинантных белков в бактериальных системах экспрессии.
Фосфатаза LpxE принадлежит к большому семейству липидных фосфатаз, обнаруживающихся как у про-, так и у эукариотических организмов (Sciorra V.A., Morris A.J. // Biochim. Biophys. Acta - 2002. - V. 1582. - P. 45-51; Brindley D.N., Waggoner D.W. // J. Biol. Chem. - 1998. - V. 273. - P. 24281-24284). Ферменты этого семейства второго типа не задействуют в качестве ко-фактора бивалетные металлы и обнаруживают в последовательности общий тройной мотив KXXXXXXRP-PSGH-SRXXXXXIIXXXD, который задействован в связывании и гидролизе субстрата (Stukey J., Carman, G.M. // Protein Sci. - 1997. - V. 6. - P. 469-472). Большинство этих ферментов представляет собой мембранные белки с шестью трансмембранными доменами. Субстратами для данных ферментов являются различные липиды, в том числе фосфатидилглицеролфосфат, фосфатидная кислота, липофосфатидная кислота, липид А бактериального эндотоксина и др.
Липид А представляет собой гидрофобный компонент в структуре бактериального эндотоксина, ответственный за его токсичность и реактогенность. В результате его взаимодействия с комплексом TLR4/MD2 на иммунных клетках млекопитающих осуществляется выброс клетками цитокинов, в частности фактора некроза опухолей (TNF-α) и интерлейкина-6 (IL-6), активация на антиген-презентирующих клетках молекул 2 класса комплекса гистосовместимости II и Th-1-зависимого иммунного ответа (Baker P.J., Hraba Т., Taylor C.E., Myers K.R., Takayama K., Qureshi Ν., Stuetz P., Kusumoto S., Hasegawa A. // Infect. Immun. - 1992. - V. 60. - P. 2694-2701; Lu Y.C., Yeh W.C., Ohashi P.S. // Cytokine. - 2008. - V.42. - P. 145-151; Moingeon P., Haensler J., Lindberg A. // Vaccine. - 2001. - V.19. - P. 4363-4372). Удаление бактериального эндотоксина для нейтрализации токсических эффектов является принципиальным и необходимым требованием как с точки зрения российских, так и с точки зрения международных стандартов при получении любых медицинских препаратов для инъекций (XII издание Государственной фармакопеи Российской Федерации; Международная фармакопея ВОЗ, документ QAS/11.452 FINAL, июль 2012 г.), однако часто остается технически трудноосуществимым и затратным процессом (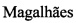 P.O., Lopes A.M., Mazzola P.G., Rangel-Yagui C, Penna T.C., Pessoa A. Jr. // J. Pharm. Pharm. Sci. - 2007. - V. 10. - P. 388-404).
P.O., Lopes A.M., Mazzola P.G., Rangel-Yagui C, Penna T.C., Pessoa A. Jr. // J. Pharm. Pharm. Sci. - 2007. - V. 10. - P. 388-404).
Фермент LpxE, способный отщеплять фосфатную группу в первом положении липида А, был впервые обнаружен у грамположительных почвенных азотофиксирующих бактерий R. leguminosarum (Karbarz M.J., Kalb S.R., Cotter R.J., Raetz C.R.H. // J. Biol. Chem. - 2003. - V. 278. - P. 39269-39279), позже структурно гомологичные ферменты со сходной активностью были выявлены у F. novicida (Wang X., Karbarz M.J., McGrath, S.C., Cotter, R.J., Raetz, C.R.H. // J. Biol. Chem. - 2004. - 279. - P. 49470-49478), H. pylori (Tran A.X., Karbarz M.J., Wang X., Raetz C.R.H., McGrath S.C., Cotter R.J., Trent M.S. // J. Biol. Chem. - 2004. - V. 279. - P. 55780-55791). Результатом гидролиза 1'-фосфата под действием фосфатазы LpxE является образование производного липида А, 4'-монофосфорил-липида A (MPLA). Липид А дикого типа, содержащий две фосфатные группы, активирует сигнальные пути TLR4-TRAM-TRIF и TLR4-Mal-MyD88 (Park B.S., Song D.H., Kim H.M., Choi B.S., Lee H., Lee J.O. // Nature. - 2009. - V.458. - P. 1191-1195; Gaekwad J., Zhang Y., Zhang W., Reeves J., Wolfert M.A., Boons G.J. // J. Biol. Chem. - 2010. - V.285. - P. 29375-29386). MPLA, реактогенность которого существенно снижена по сравнению с липидом дикого типа, является значительно более слабым лигандом. Он индуцирует структурные перестройки комплекса TLR4/MD-2 и избирательно активирует сигнальный путь TLR4-TRAM-TRIF, что приводит к секреции значительно более низких уровней цитокинов (Mata-Haro V., Cekic С., Martin M., Chilton P.M., Casella C.R., Mitchell T.C. // Science. - 2007. - V. 316. - P. 1628-1632). Ранее было продемонстрировано, что монофосфат-производные липида А сохраняют его адъювантные качества, обладая существенно сниженной токсичностью (Qureshi Ν., Takayama K., Ribi Ε. // J. Biol. Chem. - 1982. - V. 257. - P. 11808-11815; Ulrich J.T., Myers K.R. // Vaccine Design. - 1995. - P. 495-524; Persing D.H., Coler R.N., Lacy M.J., Johnson D.Α., Baldridge J.R., Hershberg R.M., Reed S.G. // Trends Microbiol. - 2002. - V.10. - P. S32-S37). В силу этих свойств некоторые варианты MPLA были одобрены в качестве адъювантов для создания вакцин (Kundi M. // Expert Rev. // Vaccines. - 2007. - V.6. - P. 133-140). Таким образом, обработка рекомбинантной фосфатазой LpxE является перспективной стратегией для снижения уровня активного бактериального эндотоксина в препаратах белков и других соединений, выделенных из бактериальной биомассы, решающей проблему борьбы с токсичностью и пирогенностью продукта во многих биотехнологических процессах, в первую очередь, при производстве вакцинных препаратов, для которых адъювантные свойства MPLA могут играть положительную роль.
Получение фосфатаз группы LpxE в гетерологичных системах в Е. coli ранее осуществлялось в исследовательских целях. Сравнение результатов анализа активности гомологичных фосфатаз LpxE H. pylori, R. leguminesarium и F. novicida показывает, что наиболее активной из них является LpxE F. novicida (Tran A.X., Karbarz M.J., Wang X., Raetz C.R., McGrath S.C., Cotter R.J., Trent M.S. // J. Biol. Chem. - 2004. - V.279. - P. 55780-55791; Wang X., Karbarz M.J., McGrath S.C., Cotter R.J., Raetz C.R. // J. Biol. Chem. - 2004. - V. 279. - P. 49470-49478). Фермент LpxE практически 100% консервативен в рамках рода бактерий Francisella и имеет 78% структурной гомологии с ферментом R. leguminesarium. В большинстве исследований, проведенных с рекомбинантными фосфатазами LpxE, очистка фермента не проводилась.
Известен способ получения LpxE в Е. coli с целью создания рекомбинантного штамма, дефицитного по продукции эндотоксина, не предусматривавший очистки фермента, включающий генно-инженерную интеграцию экспрессионной конструкции, продуцирующей LpxE, в бактериальную клетку с последующей геномной экспрессией фермента (патент US 20130230555).
Известен способ продукции LpxE F. novicida в Ε. coli, также не предполагавший очистки фермента, с помощью которого было продемонстрировано, что экспрессия этой фосфатазы в составе рекомбинантной плазмиды в Е. coli приводит к дефосфорилированию липида A (Ingram В.О., Masoudi Α., R. H. Raetz C.R.H. // Biochemistry. - 2010. - V. 49. - P. 8325-8337).
Известен по существу наиболее близкий к заявленному изобретению и единственный описанный для рекомбинантных фосфатаз группы LpxE способ получения фосфатазы LpxE R. leguminesarium в Ε. coli (Karbarz M.J., Six D.A., Raetz C.R. // J. Biol. Chem. - 2009. - V.284. - P. 414-425). Данный способ предполагает очистку гомологичного варианта фермента в виде фьюжн-белка с полигистидиновой последовательностью, однако в аналитическом варианте. Продукция белка в работе не оценивается, однако на основании приведенных иллюстраций можно предположить, что уровень продукции фермента невысок, и значительная часть белка находится в нерастворимой форме.
Изобретение решает задачу создания способа получения больших количеств ферментативно активной рекомбинантной липидной фосфатазы LpxE F. novicida, специфически удаляющей фосфат в первом положении липида А бактериального эндотоксина с целью использования для нужд биотехнологии и фармакопейного производства препаратов, полученных из бактериального сырья.
Технический результат изобретения достигается за счет оптимизации последовательности ДНК, кодирующей фосфатазу LpxE F. novicida, для достижения максимально эффективной трансляции белка фермента в Е. coli.
Также технический результат изобретения достигается за счет конструирования векторной плазмиды, содержащей промоторную последовательность ДНК бактериофага Т7 и последовательности ДНК, кодирующие мальтоза-связывающий белок (МВР), сайт расщепления протеазой фактором Ха, шесть гистидинов для очистки белка металл-хелатной хроматографией и оптимизированную для трансляции в Е. coli последовательность ДНК, кодирующую фосфатазу LpxE F. novicida.
Также технический результат изобретения достигается за счет N-концевой химеризации пептидной последовательности фосфатазы LpxE F. novicida с мальтоза-связывающим белком, способствующим повышенной растворимости синтезируемого белкового продукта и его очистке на смоле с амилозой.
Также технический результат изобретения достигается за счет разделения химеризованных последовательностей сайтом расщепления протеазой фактором Ха для протеолитического удаления мальтоза-связывающего белка с N-конца полученного рекомбинантного белкового продукта.
Также технический результат достигается продукцией рекомбинантного химерного белка MBP-LpxE в бактериях Е. coli штамма BL21(DE3) при температуре 25°С, что обеспечивает накопление рекомбинантного белка в бактериальной цитоплазме преимущественно в растворенном состоянии.
Также технический результат изобретения достигается за счет высокоэффективной процедуры лизиса бактериальных клеток, предусматривающей обработку бактерий детергентом Triton Х-100 в концентрации 4%, и последующего присутствия Triton Х-100 при очистке белка в хроматографических рабочих растворах в концетрации не менее 0,1%, что способствует повышению активности фермента и сохранению его в растворенном состоянии.
Также технический результат изобретения достигается благодаря схеме хроматографической очистки, включающей этапы аффинной хроматографии на смоле с амилозой, металл-хелатной, ионнообменной и гель-фильтрационной хроматографии с конечным переводом фермента в буфер со значением рН 6,5.
Техническим результатом изобретения является создание эффективного способа продукции и очистки рекомбинантной фосфатазы LpxE F. novicida с высоким выходом, применимого для получения больших количеств активного фермента, пригодного для использования в биотехнологических процессах.
Отличием предлагаемого способа является получение фосфатазы на основе синтетической последовательности ДНК, оптимизированной для продукции белка в Е. coli. Адаптация последовательности ДНК, кодирующей целевой белок, для продукции в Е. coli проводилась путем компьютерного анализа и включала замену кодонов нативного гена на кодоны, наиболее часто встречающиеся в белках с высоким уровнем продукции у организма-хозяина, анализ и совершенствование стабильности считываемой мРНК, удаление чрезмерной структурированности мРНК, удаление гомополимерных участков мРНК.
В предлагаемом техническом решении выщепление мальтоза-связывающего белка из состава рекомбинантного полипептида с восстановлением первичной структуры фосфатазы LpxE, близкой к нативной, обеспечивается за счет введенного в состав рекомбинантного полипептида сайта расщепления протеазой фактором Ха, имеющего аминокислотную последовательность I-(E/D)-G-R. В указанной последовательности данная протеаза вносит разрыв полипептидной цепи после радикала аргинина. Поскольку необходимым требованием для многих биотехнологических процессов может оказаться удаление фермента фосфатазы из раствора после проведения дефосфорилирования, на N-конце рекомбинантного белка фосфатазы сохраняется короткая аминокислотная последовательность шести гистидинов, не влияющая на активность фермента, но позволяющая удалить его из раствора адсорбцией на металл-хелатной смоле.
Предлагаемое техническое решение предусматривает проведение нескольких этапов хроматографической очистки, включая аффинную хроматографию на смоле с амилозой и металл-хелатную хроматографию, а также анионообменную и гель-фильтрационную хроматографии, обеспечивающих получение фермента высокой чистоты (до 98% по данным денситометрии) в конечном буферном растворе со значением рН 6,5, пригодном для анализа активности и хранения белка.
Существенным достоинством предлагаемого способа получения рекомбинантной фосфатазы LpxE F. novicida является получение высокого выхода белка фосфатазы (0,4 г с литра бактериальной культуры) в растворимой форме за счет его химеризации с мальтоза-связывающим белком и понижения температуры культивации продуцента E. coli при синтезе белка LpxE. Известно, что мальтоза-связывающий белок способствует повышению растворимости химеризованных с ним рекомбинантных белков (Kapust R.B., Waugh D.S. // Protein Sci. - 1999. - V. 8. - P. 1668-1674).
Изобретение осуществляют следующим образом:
Конструируют рекомбинантную плазмидную ДНК pETLpxE на основе коммерчески доступной плазмиды pET22b(+), в которую вводят нуклеотидную последовательность, кодирующую мальтоза-связывающий белок, полученную прямой ПЦР с генома Е. coli с использованием праймеров, MBP_FNde и MBP_RNheXho (Фиг. 2), содержащих сайты рестрикции эндонуклеаз NdeI (праймер MBP_FNde), NheI и XhoI (праймер MBP_RNheXho). Полученную ДНК расщепляют эндонуклеазами рестрикции NdeI и XhoI и затем лигируют с расщепленной по соответствующим сайтам векторной плазмидной ДНК pET22b(+). Единичные клоны, содержащие плазмиду pET22b(+) со встроенным фрагментом мальтоза-связывающего белка получают электротрансформацией клеток штамма Е. coli DH12S.
Конструируют последовательность ДНК, кодирующую фермент фосфатазу LpxE F. novicida, оптимизированную для продукции белка в Е. coli, и синтезируют данную последовательность при помощи ПЦР с использованием перекрывающихся праймеров. На 5'-конец синтезированной последовательности при помощи ПЦР вводят фрагмент ДНК, кодирующий сайт для протеазы фактора Ха (праймер Lpx_FXa) и последовательность шести гистидинов (праймер Lpx_HIS6/Nhe), на 3'-конец последовательности при помощи ПЦР вводят стоп-кодон и сайт эндонуклеазы рестрикции XhoI (праймер Lpx_RXho). Полученный ПЦР-продукт расщепляют по сайтам эндонуклеаз рестрикции NheI и XhoI и клонируют в расщепленную по тем же сайтам векторную плазмиду, содержащую ДНК, кодирующую мальтоза-связывающий белок. Получают единичные клоны, содержащие заданную последовательность ДНК.
Плазмидой pETLpxE трансформируют клетки штамма Е. coli BL21(DE3) и получают единичные колонии продуцента белка фосфатазы BL-LpxE. На основании аналитической экспрессии выделяют клоны-суперпродуценты белка. Проводят препаративную экспрессию белка клонов-суперпродуцентов. Для этого бактериальные клоны выращивают в течение ночи в среде 2xYT, содержащей ампициллин и 1% глюкозы, 10 мл ночной культуры помещают в 1 литр свежей среды, содержащей 0,1% глюкозы и ампициллин, и наращивают культуру до достижения 1 единицы оптической плотности. Культуру охлаждают до 25°С и индуцируют синтез белка добавлением 1 мМ ИПТГ. Экспрессию белка проводят при 25°С в течение 6 часов. Бактериальную биомассу собирают центрифугированием и хранят при -70°С.
Выделение белка MBP-LpxE из осветленного лизата культуры клеток проводят на колонке со смолой с иммобилизированной амилозой, элюируют белок с колонки буфером, содержащим мальтозу. Выделенный белок MBP-LpxE расщепляют протеазой фактором Ха, после чего отщепленную фосфатазу отделяют с применением металл-хелатной хроматографии. Доочищают полученный фермент анионообменной хроматографией с последующей гель-фильтрацией. На каждом этапе контроль степени очистки проводят с применением электрофореза в ПААГ.
Выход рекомбинантного белка фосфатазы LpxE составляет 0,4 г с литра культуры.
Активность фосфатазы определяют с использованием в качестве субстрата липополисахарида Е. coli (Sigma-Aldrich, США) по накоплению неорганического фосфата в тесте с малахитовым зеленым. Пирогенные свойства бактериальных липополисахаридов оценивают по подъему температуры в тесте на кроликах.
Изобретение иллюстрируют следующие графические материалы:
Фиг. 1. Синтетическая последовательность ДНК, оптимизированная для эффективной продукции фосфатазы LpxE в клетках Е. coli. А - последовательность ДНК LpxE, В - аминокислотная последовательность LpxE. С - полная последовательность плазмиды pETLpxE.
Фиг. 2. Последовательности праймеров, использованных для получения экспрессионной конструкции для продукции фосфатазы LpxE в клетках Е. coli.
Фиг. 3. Карта экспрессионной конструкции, использованной для продукции фосфатазы LpxE в клетках Е. coli.
Фиг. 4. Результаты выделения и очистки белка рекомбинантной фосфатазы LpxE из бактериальной биомассы.
Дорожка 1 - маркер молекулярной массы SM671 (Fermentas, США); Дорожка 2 - рекомбинантный белок MBPLpxE после очистки аффинной хроматографией на сорбенте с амилозой; Дорожка 3 - протеаза фактор Ха; Дорожка 4 - рекомбинантный белок MBPLpxE, расщепленный протеазой фактором Ха; Дорожка 5 - очищенный препарат рекомбинантного белка фосфатазы LpxE.
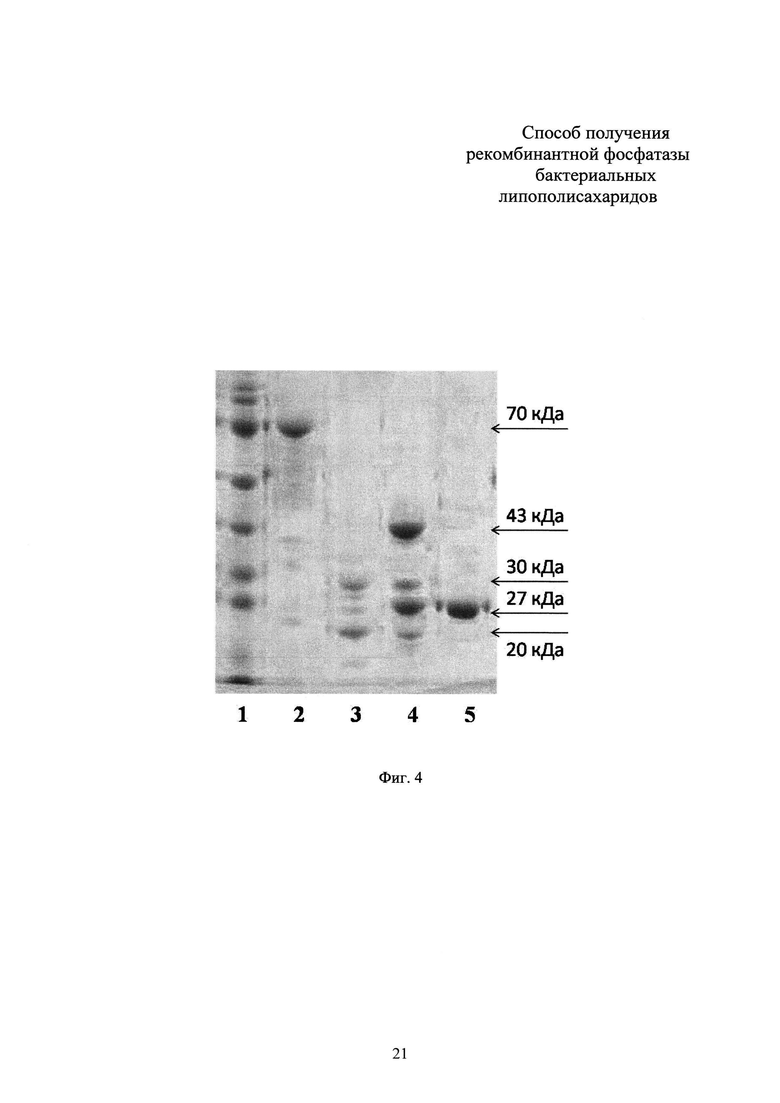
Фиг. 5. Фосфатазная активность рекомбинантного фермента фосфатазы LpxE F. novicida.
График А. Результаты измерения оптической плотности для фосфатного стандарта (калибровочная кривая). Результаты представлены как среднее арифметическое для трех лунок (повторностей). Пунктирной линией обозначена линия тренда.
График В. Результаты измерения оптической плотности в анализе фосфатазной активности рекомбинантного фермента LpxE в отношении липополисахалида Е. coli. Результаты представлены как среднее арифметическое для трех лунок (повторностей). Пунктирной линией обозначена линия тренда.
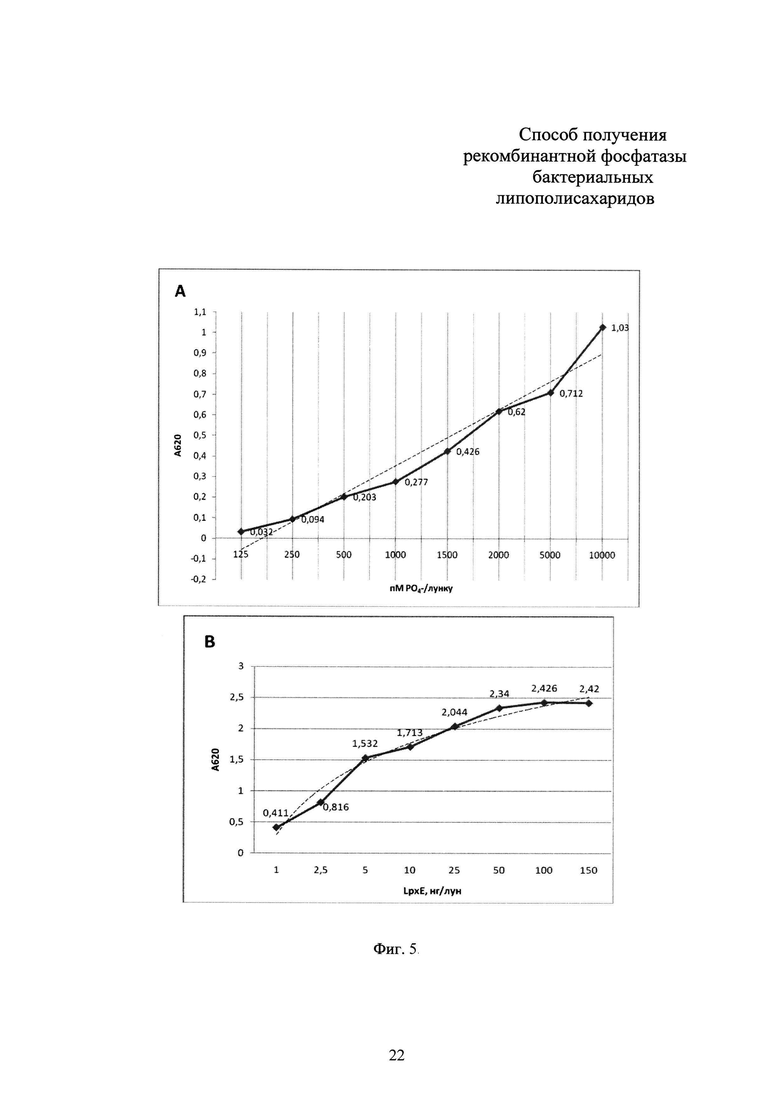
Фиг 6. Результаты теста на пирогенность препарата липополисахалида Е. coli, обработанного рекомбинантной фосфатазой LpxE, на лабораторных животных.
Для лучшего понимания сущности изобретения ниже следуют примеры его конкретного выполнения.
Пример 1. Создание экспрессионной конструкции для продукции белка фосфатазы LpxE F. novicida в Ε. coli.
Конструируют рекомбинантную плазмидную ДНК pETLpxE на основе коммерчески доступной плазмиды pET22b(+) (Novagen, США). Последовательность, кодирующую мальтоза-связывающий белок получают прямым ПЦР с генома Е. coli с использованием праймеров, MBP_FNde и MBP_RNhe, представленных на Фиг. 2. Праймеры, используемые для ПЦР-амплификации, содержат сайты эндонуклеаз рестрикции NdeI (N-конец гена, праймер MBP_FNde), NheI и XhoI (С-конец гена, праймер МВР_RNheXho). Полученную ДНК расщепляют эндонуклеазами рестрикции NdeI и XhoI и затем лигируют с расщепленной по соответствующим сайтам и очищенной элюцией из агарозного геля векторной плазмидной ДНК pET22b(+). Единичные клоны, содержащие плазмиду pET22b(+) со встроенным фрагментом мальтоза-связывающего белка получают электротрансформацией клеток штамма Е. coli DH12S (Thermo Fisher Scientific, США). Клоны, содержащие инсерт длиной 1200 п. н., идентифицируют при помощи ПЦР с праймеров T7forward и T7reverse (Sigma-Aldrich, США). Выделяют плазмидную ДНК и верифицируют корректность встроенной последовательности методом капиллярного секвенирования.
С помощью компьютерного анализа конструируют последовательность ДНК, кодирующую фермент фосфатазу LpxE F. novicida, оптимизированную для продукции белка в Е. coli и синтезируют данную последовательность при помощи ПЦР с перекрывающихся праймеров. На 5'-конец синтезированной последовательности при помощи двух этапов ПЦР с использованием праймеров Lpx_FXa и Lpx_HIS6/Nhe вводят фрагмент ДНК, кодирующий сайт для протеазы фактора Ха и последовательность шести гистидинов, на 3'-конец последовательности при помощи ПЦР с использованием праймера Lpx_RXho вводят стоп-кодон и сайт эндонуклеазы рестрикции XhoI. Полученный ПЦР-продукт расщепляют по сайтам эндонуклеаз рестрикции NheI и XhoI и клонируют в расщепленную по тем же сайтам векторную плазмиду, содержащую ДНК, кодирующую мальтоза-связывающий белок. Единичные клоны, содержащие инсерт длиной 800 п. н. идентифицируют при помощи ПЦР с праймеров T7forward и T7reverse. Выделяют ДНК плазмиды pETLpxE и верифицируют корректность встроенной последовательности ДНК и проведенного конструирования методом капиллярного секвенирования.
Пример 2. Получение продуцентов и продукция белка фосфатазы LpxE F. novicida в Ε. coli.
Штамм-продуцент Ε. coli BL-LpxE получают электротрансформацией компетентных клеток Е. coli BL21(DE3) плазмидой pETLpxE. Проводят аналитическую экспрессию единичных клонов BL21(DE3), несущие плазмиду pETLpxE. Для этого едничные колонии суперпродуцента E. coli BL-LpxE выращивают в течение ночи при 37°С в 5 мл среды 2xYT с добавлением 1% глюкозы и 50 мкг/мл ампициллина, засевают 100 мкл ночной культуры в 10 мл среды 2xΥΤ, содержащей 0.1% глюкозы и 50 мкг/мл ампициллина и выращивают при 37°С в течение 3 часов. Выросшие культуры охлаждают до 25°С, добавляют ИПТГ до 1 мМ и проводят экспрессию 6 часов при температуре 25°С. Бактериальные клетки собирают центрифугированием при 5000 об/мин 10 минут и анализируют уровень экспрессии белка фосфатазы LpxE денатурирующим электрофорезом в 12% полиакриламидном геле.
Для проведения электрофореза равные аликвоты клеток разных клонов центрифугируют при 5000 об/мин 5 минут, осажденные клетки растворяют в 100 мкл лизирующего буфера с красителем бромфеноловым синим, обрабатывают 20 секунд ультразвуком, нагревают 3 минуты при 100°С и наносят на гель. После прохождения электрофореза гель окрашивают кумасси R-250 по стандартной методике и сканируют с помощью денситометра Typhoon FLA9500 (GE Healthcare, США). Клоны, обнаруживающие наибольший выход белка (образование на геле яркой полосы с молекулярной массой 27 кДа) являются суперпродуцентами, их выращивают на богатой среде (2xYT) в течение ночи, добавляют глицерин до 15% и замораживают на -70°С для хранения.
Для препаративной экспрессии штаммы-суперпродуценты Е. coli BL-LpxE выращивают в течение ночи при 37°С с добавлением 1% глюкозы и 50 мкг/мл ампициллина на среде 2xYT, помещают 10 мл ночной культуры в 1000 мл среды 2xΥΤ, содержащей 0.1% глюкозы и 50 мкг/мл ампициллина и выращивают при 37°С до достижения 1 единицы оптической плотности. Выросшую культуру охлаждают до 25°С, добавляют ИПТГ до 1 мМ и проводят экспрессию 6 часов при 25°С. Далее клетки собирают центрифугированием при 5000 об/мин 10 минут и хранят в виде замороженных осадков при температуре -70°С до этапа выделения белка.
Пример 3. Выделение и очистка белка фосфатазы LpxE из бактериальной биомассы.
Все этапы выделения белка фосфатазы проводят при температуре +4°С. 50 г биомассы суспендируют в 200 мл раствора, содержащего 20 мМ трис-HCl рН 7,5, 200 мМ хлорида натрия и ингибитор протеаз Complete (Roche Life Sciences, США), а затем добавляют к суспензии Triton Х-100 до концентрации 0,5%. Проводят обработку полученного лизата ДНКазой и РНКазой в концентрации каждого из ферментов 10 мкг/мл, а затем добавляют Triton Х-100 до концентрации 4% и хлорид натрия до концентрации 0,3 М. Полученный клеточный лизат центрифугируют в течение 1 часа при 18000 об/мин.
Осветленный лизат наносят на колонку со смолой с иммобилизованной амилозой (Amylose Resin, NEB, США), промывают колонку десятью объемами буфера, содержащего 20 мМ трис-HCl рН 7,5, 100 мМ хлорида натрия, 0,1% Triton Х-100 и элюируют белок тем же буфером, содержащим 10 мМ мальтозы (Фиг. 4, дорожка 2).
Очищенный химерный белок MBP-LpxE расщепляют протеазой фактором Ха (NEB, США). Протеазу добавляют к раствору белка в соотношении 1/100 (w/w) и инкубируют в течение 8 часов при температуре 10°С в присутствии 1 мМ хлорида кальция. Расщепление белка контролируют денатурирующим электрофорезом в 12% ПААГ (Фиг. 4, дорожки 3-4).
Отщепленную фосфатазу отделяют от фактора Ха и продуктов гидролиза металл-хелатной хроматографией на колонке cOmplete™ His-Tag Purification Resin (Roche Life Sciences) в буфере, содержащем 20 мМ трис-HCl рН 7,5, 100 мМ хлорида натрия, 0,1% Triton Х-100. Элюцию белка с колонки проводят буфером, содержащим 20 мМ трис-HCl рН 7,5, 20 мМ хлорида натрия, 0,1% Triton Х-100, 50 мМ имидазола.
Анионообменную хроматографию проводят на колонке Q-Sepharose (GE Healthcare). Белок наносят на колонку в буфере, содержащем 20 мМ трис-HCl рН 7,5, 20 мМ хлорида натрия, 0,1% Triton Х-100 и элюируют градиентом хлорида натрия на том же буфере от 20 до 500 мМ в течение 40 минут. Фракции, содержащие белок фосфатазы, идентифицируют электрофорезом в 12% ПААГ, концентрируют при помощи центрифужных концентраторов Amicon Ultra-15 (Millipore, США) и наносят на гель-фильтрационную колонку Superdex 200 10/300 GL, уравновешенную буфером, содержащим 20 мМ MES рН 6,5, 100 мМ хлорида натрия, 0,1% Triton Х-100 (Фиг. 4, дорожка 5). Фракции, содержащие целевой белок, определяют электрофоретически, объединяют, концентрируют и измеряют концентрацию белка. Белок рекомбинантной фосфатазы LpxE хранят в аликвотах при температуре -70°С в присутствии 30% глицерина. Выход рекомбинантного белка фосфатазы LpxE составляет 0,4 г с литра культуры.
Пример 4. Анализ активности рекомбинантного фермента фосфатазы LpxE F. novicida.
Активность фосфатазы LpxE анализируют постановкой теста с малахитовым зеленым в отношении липополисахарида Е. coli (Sigma-Aldrich, США). Для этого готовят 10% стоковый раствор малахитового зеленого с добавлением 0,01% детергента Твин-20. Готовый сток хранят не более 48 часов.
Готовят контрольные разведения фосфатного стандарта KH2PO4 в буфере 20 мМ MES рН 6,5, 100 мМ хлорида натрия, 0,1% Triton Х-100 с добавлением 1 мМ дитиотреитола с содержанием фосфата: 100 нМ/мл; 75 нМ/мл; 50 нМ/мл; 25 нМ/мл; 12,5 нМ/мл; 6,25 нМ/мл.
Готовят разведения рекомбинантной фосфатазы LpxE F. novicida в буферном растворе состава 20 мМ MES рН 6,5, 100 мМ хлорида натрия, 0,1% Triton Х-100 с содержанием фермента: 1 нг/мкл; 2,5 нг/мкл; 5 нг/мкл; 10 нг/мкл; 25 нг/мкл; 50 нг/мкл; 100 нг/мкл; 150 нг/мкл.
В стерильный 96-луночный планшет вносят разведения фосфатного стандарта в объеме 20 мкл на лунку. В качестве отрицательного контроля используют буфер 20 мМ MES рН 6,5, 100 мМ хлорида натрия, 0,1% Triton Х-100 с добавлением 1 мМ дитиотреитола в объеме 20 мкл в лунке. В лунки для тестирования фосфатазной активности вносят по 4 мкл стерильной деионизованной воды, 4 мкл буфера 100 мМ MES, рН 6,5, 500 мМ хлорида натрия, 0,5% Triton Х-100, 2 мкл 100 мМ дитиотреитола, 9 мкл раствора липополисахарида Е. coli с концентрацией 5 мг/мл и по 1 мкл соответствующего разведения фосфатазы (50-1000 нг/лунку). Все точки анализируют не менее чем в трех повторностях.
Выдерживают планшет при температуре 37°С в течение 10 минут с перемешиванием на орбитальном шейкере. По окончании инкубации приливают в каждую лунку по 30 мкл стокового раствора малахитового зеленого. Выдерживают планшет при комнатной температуре в течение 40 минут, после чего считывают показатели оптической плотности на спектрофотометре при длине волны 620 нм.
При помощи контрольных разведений фосфатного стандарта строят калибровочную кривую зависимости оптической плотности от количественного содержания фосфата, откладывая в сетке координат средние для трех повторностей значения оптической плотности. Аналогично строят график зависимости оптической плотности от концентрации фермента фосфатазы в тестируемых лунках. Сравнивая контрольную кривую с тестовой, экстраполяцией получают данные о количественном содержании свободных фосфатных групп в тестируемых лунках с липополисахаридом, вычисляют среднее значение для трех повторностей. На основании имеющихся данных вычисляют удельную активность фосфатазы: количество фосфатных групп, отщепляющихся в течение одной минуты действием 1 нМ фосфатазы (пМ PO4-/мин × нМ). Удельная активность рекомбинантной фосфатазы LpxE F. novicida в отношении липополисахарида Е. coli составляет не менее 7400 пМ PO4-/мин × нМ.
Результаты тестирования активности рекомбинантной фосфатазы LpxE F. novicida в отношении липополисахарида Е. coli представлены на Фиг. 5.
Пример 5. Определение пирогенности бактериальных липополисахаридов, обработанных рекомбинантным ферментом фосфатазой LpxE F. novicida, на примере липополисахарида Е. coli.
Тестирование пирогенности препаратов с использованием лабораторных животных проводят в соответствии со стандартной методикой, описанной в Фармакопейной статье США (USP29-NF24, глава 151). Для проведения теста используют здоровых животных, а именно кроликов породы советская шиншилла (филиал «Андреевка» ГУ Научный центр биомедицинских технологий Отделения медицинских наук РАН). Животных содержат раздельно, при температуре окружающей среды 20-23°С, в покое, без раздражающих факторов, при постоянных условиях. До проведения теста животных выдерживают не менее 7 дней, повторяя все этапы описанного ниже протокола, с внутривенным введением стерильного физиологического раствора. Каждое животное используют для тестирования не чаще, чем раз в 48 часов, а при подъеме температуры тела животного на 0,6°С и выше при проведении одной итерации протокола - не чаще чем раз в две недели. В течение эксперимента (3 ч) животных не допускают к корму и ограничивают питье. Для измерения температуры тела кролика в эксперименте используют медицинский термометр с точностью ±0,1°С. Измерение температуры тела проводят ректально. Не ранее, чем за 30 минут до начала теста делают контрольные замеры температуры у каждого животного. Температура тела каждого животного внутри одной экспериментальной группы не должна отличаться более чем на 1°С по отношению к температуре тела других животных группы.
Для приготовления растворов и всех манипуляций используют апирогенные посуду и расходные материалы. При необходимости стерилизуют посуду при температуре не менее 250°С в течение часа для достижения апирогенности.
В качестве раствора для разведения препаратов используют стерильный раствор 0,9% натрия хлорида (физиологический раствор для инъекций). Этот же раствор используют в качестве контроля сравнения в тесте. Все растворы, вводимые внутривенно, предварительно нагревают до 37±0,2°С.
Используют шесть групп животных по три кролика в каждой - четыре контрольных и две экспериментальных. Каждому животному в ушную вену вводят исследуемый раствор из расчета 10 мл раствора на 1 кг веса животного, раствор вводят медленно, но не более чем за 10 минут.
Схема эксперимента:
1) Контрольная группа, которой вводят 0,9% раствор хлорида натрия;
2) Контрольная группа, которой вводят раствор липополисахарида Е. coli с концентрацией 0,1 мкг/мл в 0,9% растворе хлорида натрия;
3) Экспериментальная группа, которой вводят раствор липополисахарида Е. coli с концентрацией 0,1 мкг/мл в 0,9% растворе хлорида натрия, предварительно проинкубированный с рекомбинантным ферментом фосфатазой LpxE F. novicida в концентрации 500 нг/мл.
Инкубацию липополисахаридов с рекомбинантной фосфатазой LpxE проводят в течение часа при комнатной температуре в буфере состава 20 мМ MES рН 6,5, 100 мМ хлорида натрия, 0,1% Triton Х-100 с добавлением 1 мМ дитиотреитола. По окончании инкубации раствор наносят на гель-фильтрационную колонку Superdex 200 10/300 GL, уравновешенную 0,9% раствором хлорида натрия. Фракции, содержащие липополисахарид, определяют электрофоретически с окрашиванием геля нитратом серебра.
Концентрацию липополисахарида определяют фенол-сернокислым методом в планшетной модификации (Masuko T., Minami Α., Iwasaki Ν., Majima T., Nishimura S.I., Lee Y.С. // Anal. Biochem. - 2005. - Vol. 339, Ν 1. - P. 69-72).
После введения исследуемых растворов измерение температуры тела проводят каждые 30 минут в промежуток времени 1-3 часа от введения дозы исследуемого препарата.
Учет результатов проводят по истечении 3 часов от введения препарата внутривенно. При изменении температуры тела животных не более, чем на 0,5°С по отношению к исходным значениям, препарат считается апирогенным. При увеличении температуры тела животных в группе более чем на 0,5°С, в экспериментальную группу вводят еще 5 животных, после чего повторяют эксперимент по истечении 2 недель. При повышении температуры тела животных более чем на 0,5°С у трех животных из восьми или менее, подсчитывают суммарное повышение температуры по отношению к исходным значениям. Если суммарные значения повышения температуры тела животных в группе не превышают 3,3°С, препарат считается апирогенным. При повышении температуры тела животных на 0,6°С и более у четырех - восьми животных группы препарат признается пирогенным.
Результаты тестирования на апирогенность липополисахарида Е. coli, обработанного рекомбинантной фосфатазой LpxE, представлены в таблице (Фиг. 6).




| название | год | авторы | номер документа |
|---|---|---|---|
| Способ получения рекомбинантного экзопротеина А Pseudomonas aeruginosa | 2016 |
|
RU2636346C1 |
| Способ получения рекомбинантного пептидогликан-ассоциированного липопротеина (PAL) Legionella pneumophila | 2016 |
|
RU2634385C1 |
| ПОЛИНУКЛЕОТИД, КОДИРУЮЩИЙ МУТАНТНУЮ РЕКОМБИНАНТНУЮ IgA1 ПРОТЕАЗУ Neisseria meningitidis СЕРОГРУППЫ В, РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, СОДЕРЖАЩАЯ УКАЗАННЫЙ ПОЛИНУКЛЕОТИД, КЛЕТКА-ХОЗЯИН, СОДЕРЖАЩАЯ УКАЗАННУЮ ПЛАЗМИДНУЮ ДНК, РЕКОМБИНАНТНАЯ IgA1 ПРОТЕАЗА Neisseria memingitidis СЕРОГРУППЫ В, СПОСОБ ПОЛУЧЕНИЯ ЗРЕЛОЙ ФОРМЫ IgA1 ПРОТЕАЗЫ | 2011 |
|
RU2486243C1 |
| Рекомбинантный белок, обладающий протективным действием в отношении менингококков (варианты), полинуклеотид, кодирующий рекомбинантный белок, рекомбинантная плазмидная ДНК, содержащая указанный полинуклеотид, клетка-хозяин, содержащая указанную рекомбинантную плазмидную ДНК, способ получения рекомбинантного белка | 2017 |
|
RU2701964C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pPHOFus, КОДИРУЮЩАЯ СУБСТРАТ ЛЕТАЛЬНОГО ФАКТОРА СИБИРСКОЙ ЯЗВЫ, СЛИТЫЙ СО ЩЕЛОЧНОЙ ФОСФАТАЗОЙ Escherichia coli, И ШТАММ Escherichia coli BL-PHOFus, ПРОДУЦИРУЮЩИЙ БЕЛОК - СУБСТРАТ ЛЕТАЛЬНОГО ФАКТОРА СИБИРСКОЙ ЯЗВЫ В СОСТАВЕ ЩЕЛОЧНОЙ ФОСФАТАЗЫ Escherichia coli | 2009 |
|
RU2416637C1 |
| ШТАММ Escherichia coli BL21(DE3)Gold/pETCYPopti - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО ЦИКЛОФИЛИНА А ЧЕЛОВЕКА | 2015 |
|
RU2603283C1 |
| ПЛАЗМИДА 40Ph, ОПРЕДЕЛЯЮЩАЯ СИНТЕЗ ЩЕЛОЧНОЙ ФОСФАТАЗЫ CmAP, ШТАММ E.coli rosetta(DE3)/40Ph - ПРОДУЦЕНТ ХИМЕРНОГО БЕЛКА, ВКЛЮЧАЮЩЕГО АМИНОКИСЛОТНУЮ ПОСЛЕДОВАТЕЛЬНОСТЬ РЕКОМБИНАНТНОЙ ЩЕЛОЧНОЙ ФОСФАТАЗЫ CmAP, И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2010 |
|
RU2447151C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА, ШТАММ Escherichia coli, ХИМЕРНЫЙ БЕЛОК И ИХ ПРИМЕНЕНИЕ | 2010 |
|
RU2422524C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pD4spGBD, ШТАММ Escherichia coli - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО БЕЛКА D4-GBD, РЕКОМБИНАНТНЫЙ БЕЛОК D4-GBD И СПОСОБ ЕГО ПОЛУЧЕНИЯ, СПОСОБ ИССЛЕДОВАНИЯ СВЯЗЫВАНИЯ БЕЛКА D4-GBD С АНТИТЕЛАМИ СЫВОРОТОК БОЛЬНЫХ, СПОСОБ ПОЛУЧЕНИЯ СПЕЦИФИЧЕСКИХ АНТИТЕЛ К БЕЛКУ D4-GBD | 2008 |
|
RU2401305C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, КОДИРУЮЩАЯ СИНТЕЗ РЕКОМБИНАНТНОГО БЕЛКА TUL4spCBD, ШТАММ ESCHERICHIA COLI M15 [pREP4, pTUL4spCBD] - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО БЕЛКА TUL4spCBD, РЕКОМБИНАНТНЫЙ БЕЛОК TUL4spCBD И СПОСОБ ЕГО ПОЛУЧЕНИЯ, СПОСОБ ПОЛУЧЕНИЯ СПЕЦИФИЧЕСКИХ АНТИТЕЛ К БЕЛКУ TUL4spCBD | 2004 |
|
RU2270249C1 |






Изобретение относится к области биохимии, генной инженерии и биотехнологии, в частности к способу получения рекомбинантной фосфатазы бактериальных липополисахаридов. Данный способ включает синтез оптимизированной для трансляции в E. coli последовательности ДНК, кодирующей белок фосфатазы LpxE F. novicida. Указанная последовательность ДНК представлена на Фиг.1А. Настоящий способ предусматривает последующее конструирование экспрессионного вектора, кодирующего химерный белок-предшественник с N-концевой химеризацией. Указанный белок-предшественник состоит из последовательности мальтоза-связывающего белка, сайта узнавания протеазой фактором Xa, шести гистидинов и фосфатазы LpxE F. novicida. Способ также предполагает получение рекомбинантного белка-предшественника продукцией в E. coli и его выделение и очистку. Указанный способ предусматривает последующее выщепление мальтоза-связывающего белка из состава полипептида и доочистку белка фосфатазы LpxE. Настоящее изобретение позволяет получать рекомбинантную фосфатазу LpxE F. novicida с высоким выходом. 6 ил., 5 пр.
Способ получения рекомбинантной фосфатазы бактериальных липополисахаридов, включающий синтез оптимизированной для трансляции в Е. coli последовательности ДНК, кодирующей белок фосфатазы LpxE F. novicida, имеющей нуклеотидную последовательность, представленную на Фиг. 1А., конструирование экспрессионного плазмидного вектора, кодирующего химерный белок-предшественник с N-концевой химеризацией, составленного пептидными последовательностями мальтоза-связывающего белка, сайта узнавания протеазой фактором Ха, шести гистидинов и фосфатазы LpxE F. novicida, получение рекомбинантного белка-предшественника продукцией при 25°С в Е. coli, выделение белка-предшественника из растворимой фракции клеточного лизата в присутствии 4% Triton Х-100, очистку белка-предшественника в присутствии 0,1% Triton Х-100 с помощью аффинной хроматографии на смоле с амилозой, выщепление мальтоза-связывающего белка из состава полипептида обработкой протеазой фактором Ха и последующую доочистку белка фосфатазы металл-хелатной хроматографией, анионообменной и гель-фильтрационной хроматографией в присутствии 0,1% Triton X-100 с выходом активного белка в количестве не менее 0,4 г с литра бактериальной культуры.
| INGRAM BO ET AL, Escherichia coli mutants that synthesize dephosphorylated lipid A molecules, BIOCHEMISTRY, 2010 | |||
| US 8945587 B2, 03.02.2015 | |||
| ЦИМБАЛИСТОВА МВ, Фосфатазная активность бактерий рода Francisella, автореферат диссертации, 1998. |

