Перекрестная ссылка на родственные заявки
Настоящая заявка не является предварительной заявкой и испрашивает приоритет согласно 35 U.S.С. §119(е) предварительной заявки США с серийным номером 60/875, поданной 14 декабря 2006, содержание которой включено здесь путем ссылки во всей полноте
Область техники
Настоящее изобретение относится к собачьему тимусному стромальному лимфопоэтическому белку (собачьему ТСЛБ), молекулам нуклеиновым кислот, векторам и клеткам хозяина, кодирующим собачий ТСЛБ, и способам получения и применения собачьего ТСЛБ.
Уровень техники
Животные, включая человека, которые страдают от реагин-опосредованных заболеваний, таких как атопические заболевания, наследственно предрасположены к развитию прямых аллергических реакций, включающих в себя иммуноглобулины Е (антитела IgE). Множественные генетические факторы вносят свой вклад в проявление результирующего фенотипа, который наблюдается у таких животных. Прямая гиперчувствительность, наблюдаемая при атопических заболеваниях, происходит в результате воздействия специфических аллергенов, таких как клещ домашней пыли (Dermatophagoides pteronyssinus), пыльцы, плесени и перхоти. Неудивительно, что индивидуумы, имеющие атопическое заболевание, наиболее вероятно страдают от астмы, атопического дерматита, также как и от остальных заболеваний, обусловленных высвобождением эндогенного IgE.
Атопические заболевания, такие как аллергический дерматит, астма и тому подобные, также появляются среди собачьих видов, включающих в себя домашних собак. У таких собак обычно начинают проявляться признаки атопии в возрасте от 1 до 3 лет. Из-за наследственной природы заболевания некоторые породы, включающие в себя золотистого ретривера, большинство терьеров, ирландского сеттера, лхасского апсо, далматинов, бульдогов и староанглийских овчарок, несут в себе большую тенденцию к атопии, несмотря на то, что известно, что другие виды собак, включая смешанные породы, также страдают от этого. Частота случаев по крайней мере одного частного вида атопии, атопического дерматита, значительно увеличивается схожим образом как у человека, так и у собак.
Собаки с атопией обычно начинают тереть, лизать, жевать, кусать или царапать свои лапы, морду, уши, подмышки или область паха, что будет приводить к потере волос, покраснению и утолщениям кожи. В некоторых отдельных случаях состояния кожи сочетаются и вызывают зуд у животного, в то время, как только одна аллергия не могла бы привести к такому зуду. Такие ухудшающиеся проблемы могут быть обусловлены аллергенами, передающимися воздушным путем (пыльца и т.д.), аллергенами в пище и аллергенами от паразитов (блохи и т.д.). Бактериальные и/или дрожжевые инфекции кожи могут также усиливать чувство зуда.
Одним простым средством для облегчения раздражающих симптомов атопии является избежание аллергенов-возбудителей. К сожалению, такое избежание обычно невозможно. Прежде ветеринары-практики лечили атопический дерматит у собак применением оральных антигистаминных средств, оральных или топических противовоспалительных агентов класса кортикостероидов, других супрессоров иммунной системы, таких как циклоспорин или такролимус, производные жирных кислот, и аллерген-специфическая иммунотерапия (которая требует введения определенного антигена). Однако ни один из этих видов лечения не работает во всех случаях. Более того, такие виды лечения дороги и/или вызывают значительные побочные эффекты. Таким образом, существует давняя нужда в более безопасных, более эффективных и более экономичных подходах к лечению или подавлению симптомов атопического дерматита у собак. Иммунная система млекопитающих основана на серии сложных клеточных взаимодействий, которая называется «иммунная сеть». Многие из иммунных ответов вращаются вокруг взаимодействий подобно сетевым взаимодействиям лимфоцитов, макрофагов, гранулоцитов и других клеток с растворимыми белками, которые называются цитокинами и играют критическую роль в опосредовании/контроле/регулировании этих клеточных взаимодействий. Таким образом, цитокины и клетки иммунной системы служат для запуска специфических физиологических механизмов или путей, ведущих к различным воспалительным заболеваниям.
Аллергическое воспаление является результатом сложного иммунного каскада, который ведет Т клетки к продуцированию нерегулируемых ТН2-цитокинов, таких как IL-4, IL-5 и IL-13. Эти цитокины, в свою очередь, вызывают бронхиальную гиперактивность, продуцирование IgE, эозинофилию и продуцирование слизи (см., например, Busse and Lemanske, Jr (2001) N. Engl. J. Med. 344:350-62; Holgate (2000) Br. Med. J. 320:231-234, and Renauld (2001) J/Clin Pathol. 54:577-589).
Тимусный стромальный лимфопоетический белок (ТСЛБ) является IL-7 подобным цитокином, который был первоначально открыт у мышей как фактор, поддерживающий: (i) продуцирование in vitro иммуноглобулина IgM на поверхности В-клеток и (ii) пролиферацию В и Т клеток (Friend et al., 1994 Exp Hematology 22:321-328, см. также Levin et al., 199, J. Immunol. 162:677-683). Как известно, ТСЛБ связывает клеточный рецептор, содержащий в себе IL-7R-альфа субъединицу и уникальную субъединицу рецептора, называемую ТСЛБ-R. Это взаимодействие запускает передачу сигнала посредством активации STAT или экспрессией тимусного и регулируемого активацией хемокина (TARC) в гемопоэтических клетках, таких как миелоидные клетки, такие как моноциты, или дендритные клетки (см., например, патент США №6890734, включен здесь в качестве ссылки).
ТСЛБ также может играть значительную роль у мышей в патогенезе аллергических заболеваний, таких как атопический дерматит и астма. Например, трансгенные мыши, у которых в коже была специфически индуцирована экспрессия гена ТСЛБ, обладают иммунологическим и клиническими признаками атопического дерматита, такими как экзематозные повреждения, содержащие воспалительные кожные клеточные инфильтраты, резкое увеличение количества TH2-CD4*T клеток, экспрессирующих кожные рецепторы для наведения, и увеличенный уровень IgE в сыворотке. Более того, легкие мышей, экспрессирующих легочный специфический трансген ТСЛБ обладают иммунологическими и клиническими признаками астмы, включающими крупную инфильтрацию лейкоцитов, гиперплазию бокаловидных клеток, субэпителиальный фиброз, увеличение цитокинов в Т-хелперах 2 типа и увеличение уровня IgE.
Sims et al. получили последовательность кДНК мышиного ТСЛБ, использующуюся в экспрессионном клонировании, но не имели возможности клонировать человеческий гомолог в пробы для гибридизации, основанные на мышином ТСЛБ (Sims et al. 200. J Exp Med, 192:671-680). В последствии человеческий гомолог был идентифицирован посредством детального EST анализа. Человеческий нуклеотид, соответствующий ТСЛБ, как было обнаружено, имеет только 43% гомологии с соответствующим нуклеотидом у мышей. Таким образом, остается необходимость в обеспечении новых и более практических способов лечения атопических заболеваний, включающих атопический дерматит и связанные с ним клинические проявления, у собак. Более того, необходимо выделить факторы, которые вовлечены в иммунный каскад, который ведет к атопическим заболеваниям у собак, что могло бы привести к развитию таких способов лечения.
Цитирование любой ссылки здесь не должно быть истолковано как принятие этой ссылки в качестве предшествующего уровня техники для настоящей заявки.
Краткое описание изобретения
Настоящее изобретение предоставляет новые и более применимые на практике способы лечения атопических заболеваний у собак, включающих в себя атопический дерматит и связанные с ним клинические проявления. Соответственно, настоящее изобретение предоставляет новые выделенные и/или рекомбинантные тимусные стромальные лимфопоэтические белки (ТСЛБ), которые вовлечены в иммунный каскад, который ведет к атопическим заболеваниям. Настоящее изобретение далее предоставляет антигенные фрагменты таких белков ТСЛБ. В частном аспекте настоящего изобретения белок ТСЛБ является собачьим ТСЛБ.
Таким образом, настоящее изобретение предоставляет белок ТСЛБ, содержащий в себе аминокислотную последовательность, которая на 80% или более идентична аминокислотной последовательности SEQ ID NO:2, за исключением аминокислотного остатка 28 сигнальной последовательности, который, при назначении белка собаке, служит в качестве вакцины, антитела, которые связывают собачий ТСЛБ белок, содержащие в себе аминокислотную последовательность SEQ ID NO:2, распознаются в конечной собачьей сыворотке, полученной из вакцинированной собаки. В сходном варианте выполнения белок ТСЛБ содержит в себе аминокислотную последовательность, которая на 80% или более идентична аминокислотной последовательности SEQ ID NO:2, за исключением 28 аминокислотного остатка сигнальной последовательности; и она является перекрестно-реагирующей с антителом к собачьему ТСЛБ белку, содержащему аминокислоту из SEQ ID NO:2.
Настоящее изобретение далее предоставляет белок ТСЛБ, содержащий аминокислотную последовательность, которая на 80% или более идентична аминокислотной последовательности SEQ ID NO:2 (за исключением 28 аминокислотного остатка сигнальной последовательности), которая связывается моноклональным антителом, специфичным к эпитопу собачьего белка ТСЛБ.
В более частном варианте выполнения белок ТСЛБ содержит аминокислотную последовательность, которая на 90% или более идентична аминокислоной последовательности SEQ ID NO:2, за исключением 28 аминокислотного остатка сигнальной последовательности. В другом варианте выполнения белок ТСЛБ содержит аминокислотную последовательность, которая на 95% или более идентична аминокислоной последовательности SEQ ID NO:2, за исключением 28 аминокислотного остатка сигнальной последовательности.
В частном варианте выполнения настоящего изобретения белок ТСЛБ является собачьим белком ТСЛБ, который содержит в себе аминокислотную последовательность SEQ ID NO:2. В другом варианте выполнения белок ТСЛБ является зрелым собачьим белком ТСЛБ, который содержит аминокислотные остатки 29-155 из SEQ ID NO:2.
Также в настоящем изобретении предоставляются антигенные фрагменты белков ТСЛБ. Такие антигенные фрагменты включают в себя те, которые содержат один или более эпитопов, определенных индивидуально по аминокислотным последовательностям SEQ ID NO:8-101. В частном варианте выполнения антигенный фрагмент в настоящем изобретении содержит в себе один или более эпитопов, которые содержат в себе аминокислотные последовательности SEQ ID NO:30, 31, 32, и/или 34. В другом варианте выполнения изобретения антигенные фрагменты могут иметь аминокислотную последовательность внутри перекрывания аминокислотных последовательностей SEQ ID NO:30, 31, 32, и/или 34, т.е. NPPDCLARIERLTLHRIRGCAS (SEQ ID NO:118). В частном варианте выполнения изобретения антигенный фрагмент собачьего белка ТСЛБ способен связывать моноклональное антитело к человеческому ТСЛБ. Размер антигенных фрагментов аминокислотной последовательности NPPDCLARIERLTLHRIRGCAS (SEQ ID NO:118) может лежать в приближенном интервале от 5 до 21 аминокислотных остатков.
Также предоставляются вакцины, которые могут включать в себя эффективное количество любого белка ТСЛБ настоящего изобретения, один или более антигенных фрагментов этих белков, или комбинации таких полноразмерных белков и одного или более таких фрагментов. В одном варианте выполнения белок ТСЛБ является собачьим ТСЛБ, который содержит в себе аминокислотную последовательность SEQ ID NO:2. В частном варианте выполнения изобретения вакцина содержит один или более антигенных фрагментов собачьего ТСЛБ белка, который содержит в себе от 5 до 22 смежных аминокислот из аминокислотных остатков 71-92 из SEQ ID NO:2 (определенная здесь как SEQ ID NO:118). Примеры таких антигенных фрагментов включают в себя эпитопы, показанные здесь, которые содержат аминокислотные последовательности SEQ ID NO:30, SEQ ID NO:31, SEQ ID NO:32. SEQ ID NO:33 или SEQ ID NO:34. все вакцины настоящего изобретения могут далее содержать в себе фармацевтически приемлемый адъювант.
Вакцина настоящего изобретения может быть использована в способе получения антител к собачьему ТСЛБ. Один из таких способов содержит в себе иммунизацию млекопитающего эффективным количеством вакцины. Этот способ необязательно включает в себя способ регуляции активности ТСЛБ по его афферентным путям у собак и/или способ лечения или предотвращения аллергических симптомов у собак с атопией, который включает в себя иммунизацию собаки эффективным количеством вакцины. Улучшенные аллергические симптомы могут включать в себя аллергический дерматит, астму и тому подобное.
Вакцина настоящего изобретения может назначаться посредством внутримышечных инъекций, подкожных инъекций, внутривенных инъекций, внутрикожных инъекций, перорального применения, интраназального применения, скарификации и их комбинаций.
Настоящее изобретение далее предоставляет молекулу нуклеиновой кислоты, кодирующую белок ТСЛБ настоящего изобретения или его антигенный фрагмент. В одном таком варианте выполнения молекула нуклеиновой кислоты кодирует аминокислотную последовательность SEQ ID NO:2. В частном варианте выполнения этого типа молекула нуклеиновой кислоты содержит нуклеотидную последовательность SEQ ID NO:1. Фрагменты нуклеотидной последовательности SEQ ID NO:1 приблизительно из 18 смежных нуклеотидов, приблизительно из 24 смежных нуклеотидов, приблизительно из 36 смежных нуклеотидов, приблизительно из 45 смежных нуклеотидов, приблизительно из 66 смежных нуклеотидов или больше также являются частью настоящего изобретения. Нуклеиновые кислоты приблизительно из 18 нуклеотидов, приблизительно 24 нуклеотидов, приблизительно 36 нуклеотидов, приблизительно 45 нуклеотидов, приблизительно 66 нуклеотидов или больше, включая в себя нуклеиновые кислоты, кодирующие полноразмерные белки ТСЛБ, которые гибридизуются с SEQ ID NO:1 в условиях точной гибридизации, также предоставляются настоящим изобретением. Все молекулы нуклеиновых кислот и их фрагменты настоящего изобретения могут далее содержать в себе гетерологичные нуклеотидные последовательности.
Настоящее изобретение также предоставляет вектор для экспрессии, который включает в себя ранее отмеченные молекулы нуклеиновых кислот и/или их фрагменты. В дополнение настоящее изобретение предоставляет клетки хозяина, которые содержат в себе такие вектора для экспрессии. Клетка хозяина необязательно либо прокариотическая, либо эукариотическая. В одном варианте выполнения прокариотической клеткой хозяина является Esherichia coli. В частном варианте выполнения этого типа клетка хозяина является E.coli BL21(DE3)/pLysS, которая содержит ген Т7 РНК полимеразы под контролем изопропил-β-тиогалактопиранозид (ИПТГ)-индуцибельного промотора lacUV5.
Настоящее изобретение далее предоставляет рекомбинантные вирусные вектора и/или пустые ДНК-вектора, содержащие одну из вышеупомянутых молекул нуклеиновых кислот, кодирующих собачий ТСЛБ, например SEQ ID NO:1, и/или фрагменты их. Такие вектора могут быть использованы, например, в вакцинах, подходящих для назначения собакам с атопическим дерматитом.
Настоящее изобретение также предоставляет способы получения белка ТСЛБ настоящего изобретения. Один из таких способов содержит в себе культивирование клетки хозяина настоящего изобретения в подходящей культуральной среде. Этот способ может далее включать в себя стадию выделения и/или очистки белка ТСЛБ из культивируемой клетки хозяина или культуральной среды. Конечный выделенный и/или очищенный белок ТСЛБ также является частью настоящего изобретения.
Антитела к ТСЛБ, полученные из гибридомной системы при действии вакцины настоящего изобретения, также являются частью настоящего изобретения. В одном из вариантов выполнения такого типа используется гибридомная система млекопитающих. В частном варианте выполнения антитела выделяют и/или очищают. Антитела могут быть либо поликлональными, либо моноклональными. Согласно изобретению, моноклональное антитело, полученное не от собаки, может быть необязательно сконструировано «особаченным» для того, чтобы проявлять минимальный антигенный эффект при введении собаке. В определенных предпочтительных вариантах выполнения связывающие домены любого антитела согласно изобретению необязательно преобразуются в связывающие фрагменты меньшего размера, чем первоначальное антитело, например, посредством расщепления, и/или рекомбинантные Fv, Fab и F(ab’)2 связывающие белки. Производные от антител терапевтические белки, которые обладают уникальными структурными и функциональными свойствами встречающихся в природе антител с тяжелыми с цепями (например, NANOBODIES®) также включены в изобретение. В дополнение, заменители антител, которые обладают высокой аффинностью к ТСЛБ и низкой иммуногенностью (например, авимерные белки, сконструированные из связывающих участков рецептора к ТСЛБ) также включены в настоящее изобретение. Изобретенные авимерные антитела к собачьему ТСЛБ могут активно использоваться при лечении аллергических симптомов у собак с атопией посредством применения эффективного количества такого антитела к собачьему ТСЛБ.
Настоящее изобретение также предоставляет вакцину, содержащую эффективное количество иммуногена не ТСЛБ природы в сочетании с эффективным количеством белка ТСЛБ настоящего изобретения, одним или более антигенных фрагментов этих белков, или сочетания полноразмерного белка с одним или более таким фрагментом. В частном варианте выполнения этого типа, белок ТСЛБ является собачьим белком ТСЛБ. В более частном варианте выполнения, собачий белок ТСЛБ содержит аминокислотную последовательность SEQ ID NO:2.
Настоящее изобретение дополнительно предоставляет диагностические способы с использованием собачьего белка ТСЛБ, его фрагменты и/или антитела, полученные к собачьему ТСЛБ и его фрагментам. В одном варианте выполнения настоящее изобретение предоставляет способ диагностики атопического дерматита у собак, содержащий в себе полученный образец эпидермы от собаки и детектирующий присутствие собачьего белка ТСЛБ в образце эпидермы. Эти и другие аспекты настоящего изобретения будут лучше поняты при помощи ссылок на следующие иллюстрации и детальное описание.
Краткое описание чертежей
Фиг.1 иллюстрирует анализ белка из эукариотических клеток - системы для синтеза свободного белка, экспрессирующего собачий белок ТСЛБ - способом SDS электрофореза в полиакриламидном геле (SDS-PAGE). Дорожка 1: Стандартный белок; Дорожка 2: Общий белок; Дорожка 3: Растворимый белок; Дорожка 4: Нерастворимый белок. Полосы белка ТСЛБ помечены стрелками.
Фиг.2А иллюстрирует анализ белка из эукариотических клеток - системы для синтеза свободного белка, экспрессирующего собачий белок ТСЛБ - способом Вестерн блоттинга. Белки были проинкубированы с антителами Anti-His (С Term)/AP (Invitrogen). Дорожка 1: Стандартный белок; Дорожка 2: Полный белок; Дорожка 3: Растворимый белок; Дорожка 4: Нерастворимый белок. Собачий белок ТСЛБ был обнаружен в общем белке и в нерастворимом белке (как помечено стрелками).
Фиг.2В иллюстрирует анализ белка из эукариотических клеток - системы для синтеза свободного белка, экспрессирующего собачий белок ТСЛБ - способом Вестерн блоттинга. Белки были проинкубированы с крысиными моноклональными антителами, специфическими к человеческому ТСЛБ. Дорожка 1: Стандартный белок; Дорожка 2: Полный белок; Дорожка 3: Растворимый белок; Дорожка 4: Нерастворимый белок. Собачий белок ТСЛБ был обнаружен в общем белке и в нерастворимом белке (как помечено стрелками).
Фиг.3А иллюстрирует экспрессию и очистку ТСЛБ из клеток хозяина E.coli и показывает полосу 61 кДа, которая присутствует в растворимой фракции E.coli, которая представляет собой слияние собачьего белка ТСЛБ, белка GST и концевого пептида из 6 остатков гистидина. «М» обозначет стандартный белок (один и тот же на Фиг.3A-3D). Дорожка 1 и дорожка 2 являются растворимыми фракциями E.coli B121(DE3)pLysS, содержащую плазмиду 1265-93В без и с индукцией ИПТГ, соответственно. Стрелки показывают полосу слияния белка GST-ТСЛБ-His (один и тот же на Фиг.3A-3D).
Фиг.3В показывает, что меченый белок слияния GST-ТСЛБ-His может быть очищен смолой на основе глутатион Сефарозы 4В. Дорожки с 1 по 3 показывают различные фракции после элюции на глутатион Сефарозе 4В.
Фиг.3С показывает, что белок слияния на дорожке В может быть в дальнейшем очищен при использовании смолы Ni-NTA. Этот рисунок иллюстрирует повторную очистку белка слияния GST-ТСЛБ-His на смоле Ni-NTA после очистки на глутатион Сефарозе 4В. Дорожка 1 обозначает элюат, дорожка 2 обозначает элюирование смолы Ni-NTA.
Фиг.3D иллюстрирует анализ белка слияния GST-ТСЛБ-His с помощью Вестерн блоттинга и подтверждает, что белок слияния распознается антителом к GST (GE Health Care Cat No.27457701).
Фиг.4 иллюстрирует окрашивание FITC участка запечатанной в парафин части поврежденной кожи, полученной от собаки 10197 с диагнозом «атопический дерматит». Этот участок был проинкубирован с кроличьими поликлональными антителами к человеческому ТСЛБ и реакция визуализировалась с помощью Стрептавидин-FITC (Флуорескаин изотиоцианат). Интенсивность флуоресценции (светлые области) определяли связывание кроличьего поликлонального антитела к человеческому ТСЛБ с ТСЛБ, представленному в ткани.
Фиг.5А иллюстрирует окрашивание FITC участка запечатанной в парафин части поврежденной кожи, полученной от собаки 10197 с диагнозом «атопический дерматит». В этой части наблюдается диффузное окрашивание (темные области) эпидермальной области кожи, помеченной крысиным моноклональным антителом к человеческому ТСЛБ.
Фиг.5В иллюстрирует контрольный участок. Этот участок был из запечатанной в парафин части поврежденной кожи, полученной от собаки 10197 с диагнозом «атопический дерматит», которая обрабатывалась только контролем в виде фосфатного буфера.
Фиг.6 иллюстрирует картирование эпитопов собачьего белка ТСЛБ с помощью крысиного монокпонального антитела к человеческому ТСЛБ. Пики, вызывающие особенный интерес - это пики от эпитопов под номером 22-26 (SEQ ID NO:29-33). Эпитопы 22-26 были также проинкубированы с N-концевым производным (пик 55 и выше) для того, чтобы подтвердить, что связывающиеся эпитопы не требуют N-концевого остатка.
Фиг.7 иллюстрирует сравнение собаки (SEQ ID NO:32) и человеческого аналога эпитопа 25 (SEQ ID NO:3) пептидной последовательности ТСЛБ.
Фиг.8А иллюстрирует последовательность ДНК гена собачьего ТСЛБ (SEQ ID NO:1).
Фиг.8В иллюстрирует предсказанный полипептид ТСЛБ, экспрессирующийся с последовательности ДНК, показанной на Фиг.8А (SEQ ID NO:2). Звездочка помечает N-конец первоначальной сигнальной последовательности (остатки 1-28) и подчеркнутые остатки 71-92 (SEQ ID NO:118), представляющие область, на которой были определены перекрывающиеся эпитопы.
Подробное описание изобретения
Атопический дерматит (АД) - это Th2-опосредованное аллергическое воспалительное заболевание. Это заболевание проявляется множеством схожих клинических признаков у человека и у собаки. Вероятно, что иммунопатогенез АД у собак схож с АД человека в отношении типа клеток и цитокинов, вовлеченных в повреждение кожи.
Связывание лиганда TARC (CCL22) с рецептором 4 хемокина СС (CCR4), который селективно экспрессируется на ТН2-лимфоцитах, вызывает селективную миграцию этих клеток к аллергическим повреждениям. Сообщалось, что TARC и его рецептор CCR4 регулируются по эфферентным путям при повреждениях кожи у собак с АД. Так как ТСЛБ является сильным индуктором TARC у человека, было предположено, что ТСЛБ может присутствовать при повреждениях у собак с АД. Антитела, полученные к человеческому ТСЛБ, были поэтому исследованы на поврежденной коже от собак с АД. Иммуногистохимия таких образцов кожи подтвердила присутствие антигена, реакционно-способного по отношению к антителам к человеческому ТСЛБ при повреждениях, как показано на Фиг.4. Однако задача по определению собачьего гомолога генов, кодирующих мышиный и человеческий ТСЛБ, как было доказано, особенно тяжело из-за высокой степени расхождения последовательностей нуклеиновых кислот и аминокислотных последовательностей ТСЛБ у млекопитающих, как обнаружено здесь.
Иммунизация домашней собаки ТСЛБ настоящего изобретения и/или одним или более его антигенных фрагментов должна служить для уменьшения уровней активности эндогенного ТСЛБ и поэтому сдерживать, снимать или предотвращать один или более симптомов атопии, таких, которые проявляются при астме и/или при атопическом дерматите, у иммунизированных собак. В дополнение, собачий белок ТСЛБ может быть использован в качестве иммуногена для расщепления антител к собачьему ТСЛБ для использования в качестве исследовательского или диагностического реагента у домашних собак или других видов млекопитающих. В качестве альтернативы, в частных примерах, собачий белок ТСЛБ и/или нуклеиновые кислоты, которые кодируют собачий ТСЛБ, могут служить для регулирования элементов эфферентных путей иммунной системы у собак с ослабленным иммунитетом, например, посредством активации STAT или экспрессии TARC, например, гемопоэтическими клетками.
Для того, чтобы более полно понять настоящее изобретение, предлагаются следующие определения.
Использование уникальных терминов для удобства в описании ни в коей мере не ограничивается. Так, например, ссылка на состав, содержащий в себе «полипептид», включает в себя ссылку на один или более таких полипептидов. Используемый здесь термин «приблизительно» используется попеременно с термином «примерно» и обозначает, что величина находится внутри интервала с 20% отклонением от определенной величины, т.е., пептид, содержащий «приблизительно» 50 аминокислотных остатков, может содержать от 40 до 60 аминокислотных остатков.
Термин «связывающий состав» относится к молекулам, которые специфично связываются с собачьим ТСЛБ, например, во взаимодействии антиген-антитело. Специфичность может быть более или менее включающей, например, специфичной для частного варианта выполнения, или для группы родственных вариантов выполнения, например, собачий ТСЛБ и/или собачьи антитела.
Используемый здесь термин «собака» включает всех домашних собак, Canis lupus familiaris или Canis familiahs, если не указано иное.
Используемый здесь термин «полипептид» используется попеременно с термином «белок» и «пептид» и обозначает полимер, содержащий в себе 2 или более аминокислот, связанных между собой пептидными связями. Термин «полипептид», используемый здесь, включает в себя значительный фрагмент или сегмент и содержит ряд аминокислотных остатков примерно по крайней мере из 8 аминокислот, в основном примерно из 12 аминокислот, обычно по крайней мере 16 аминокислот, предпочтительней по крайней мере 20 аминокислот, и в особенно предпочтительных вариантах выполнения по крайней мере 30 или более аминокислот, например, 35, 40, 45, 50 и т.д. Такие фрагменты могут нести концы, которые начинаются и/или заканчиваются фактически на всех позициях, например, начало на остатках 1, 2, 3 и конец на, например, 155, 154, 153 и т.д., во всех практических комбинациях.
Необязательно, у полипептида может не хватать определенных аминокислотных остатков, которые кодируются геном или мРНК. Например, ген или молекула мРНК может кодировать последовательность аминокислотных остатков на N-конце полипептида (т.е. сигнальную последовательность), которая потом отщепляется и поэтому может не являться частью конечного белка.
Используемая здесь аминокислотная последовательность на 100% является «гомологичной» второй аминокислотной последовательности, если две аминокислотные последовательности идентичны и/или отличаются только нейтральными или консервативными заменами, как определено ниже. Соответственно аминокислотная последовательность приблизительно на 80% является «гомологичной» второй аминокислотной последовательности, если приблизительно 80% двух аминокислотных последовательностей идентичны и/или различаются только по нейтральным или консервативным заменам.
Функционально эквивалентные аминокислотные остатки часто могут замещаться остатками в пределах последовательности, приводя к консервативным аминокислотным заменам. Такие изменения определяют термин «консервативная замена», используемый здесь. Например, один или более аминокислотные остатки в пределах последовательности могут замещаться другой аминокислотой схожей полярности, которая действует как функциональный эквивалент, приводя к «молчащим» изменениям. Замены аминокислот в пределах последовательности могут быть выбраны из других членов того класса, к которым принадлежит аминокислота. Например, неполярные (гидрофобные) аминокислоты включают в себя аланин, лейцин, изолейцин, валин, пролин, фенилаланин, триптофан и тирозин. Полярные нейтральные аминокислоты включают в себя глицин, серин, треонин, цистеин, тирозин, аспарагин и глутамин. Положительно заряженные (основные) аминокислоты включают в себя аргинин, лизин и гистидин. Отрицательно заряженные (кислотные) аминокислоты включают в себя аспартамовую кислоту и глутаминовую кислоту. Не ожидается, что такие изменения повлияют на молекулярный вес, как определено с помощью электрофореза в полиакриламидном геле или по изоэлектрической точке.
Особенно предпочтительными консервативными заменами являются: лизин на аргинин и наоборот, что в любом случае обеспечивает положительный заряд; глутаминовую кислоту на аспарагиновую и наоборот, что в любом случае обеспечивает отрицательный заряд; серин на треонин, который в любом случае обеспечивает свободную ОН-группу; и глутамин на аспарагин, что в любом случае обеспечивает свободную аминогруппу. Аминокислоты также могут быть замещены в пределах следующих сходных групп: (1) пролин, аланин, глицин, серин и треонин; (2) глутамин, аспарагин, глутаминовая кислота и аспарагиновая кислота; (3) гистидин, лизин и аргинин; (4) цистеин; (5) валин, лейцин, изолейцин, метионин; и (6) фенилаланин, тирозин и триптофан.
В родственном варианте выполнения две высоко гомологичные последовательности ДНК могут быть идентифицированы по собственной гомологии, или по гомологии аминокислот, которые они кодируют. Такое сравнение последовательностей может быть представлено с использованием стандартного пакета программ, доступного в банках данных таких последовательностей. В частном варианте выполнения две высоко гомологичные последовательности ДНК кодируют аминокислотные последовательности, имеющие приблизительно 80% идентичности, более предпочтительно приблизительно 90% идентичности и еще более предпочтительно приблизительно 95% идентичности. В более частном случае две высоко гомологичные аминокислотные последовательности имеют приблизительно 80% идентичности, более предпочтительно приблизительно 90% идентичности и еще более предпочтительно приблизительно 95% идентичности.
Как используется здесь, процент идентичности белковой и ДНК последовательности может быть определен при помощи пакета программ, такого как MacVector v9, который доступен коммерчески у фирмы Accelrys (Burlington, Massachusetts) и алгоритм Clustal W для параметров по умолчанию для сопоставления и параметров по умолчанию для идентичности. Смотри, например, Thompson, et al, 1994, Nucleic Acids Res. 22:4673-4680. Clustal W - бесплатно загружаемая программа для платформ dos, Macintosh и Unix, например, с EMBLI, Европейского Института Биоинформатики. Настоящая ссылка для скачивания найдена на http://www.ebi.ac.uk/clustalw/. Эти и другие доступные программы могут быть также использованы для определения сходства последовательностей, используя такие же или аналогичные параметры по умолчанию.
«Полинуклеотид» или «молекула нуклеиновой кислоты» является молекулой, содержащей в себе нуклеотиды, включающая в себя без ограничений РНК, кДНК, геномную ДНК и даже синтетические последовательности ДНК. Эти термины также предназначены для рассмотрения молекул нуклеиновых кислот, которые включают в себя какие-либо из известных в науке аналогов ДНК и РНК.
Настоящее изобретение предоставляет нуклеиновые кислоты, которые гибридизуются с нуклеотидными последовательностями, кодирующими белок ТСЛБ настоящего изобретения. Молекула нуклеиновой кислоты способна к гибридизации с другой молекулой нуклеиновой кислоты, такой как кДНК, геномная ДНК или РНК, когда одноцепочечная форма молекулы нуклеиновой кислоты может отжигаться с другой молекулой нуклеиновой кислоты при соответствующих значениях температуры и ионной силы раствора (см. Sambrook and Russel Molecular Cloning: A Laboratory Manual. 3rd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, L.I, (2000)).
Очень строгие условия гибридизации соответствуют, например, самой высокой температуре Tm, 50% формамида, 5Х или 6XSSC. При гибридизации необходимо, чтобы две молекулы нуклеиновых кислот содержали в себе комплементарные последовательности, хотя, в зависимости от строгости гибридизации, возможны неспаренные участки (мисмэтчи) между основаниями. Соответствующая строгость гибридизации нуклеиновых кислот зависит от длины нуклеиновых кислот и степени комплементарности, разновидности которой хорошо известны в данной области. Чем степень сходства или гомологии между двумя нуклеотидными последовательностями, тем больше значение Tm, для гибридов нуклеиновых кислот, имеющих такие последовательности. Относительная стабильность (соответствующая более высокой Tm) гибридизации нуклеиновых кислот уменьшается в следующем порядке: РНК:РНК, ДНК.-РНК, ДНК:ДНК. Для гибридов длиной более 100 нуклеотидов было получено уравнение для вычисления Tm (Sambrook and Russel Molecular Cloning: A Laboratory Manual. 3rd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, L.I, (2000)). Для гибридизации с более короткими молекулами нуклеиновых кислот, т.е., олигонуклеотидами, участок с неспаренными основаниями (мистмэтчами) становится более важным и длина олигонуклеотида определяет его специфичность.
Предпочтительно минимальная длина молекулы нуклеиновой кислоты, способной к гибридизации, составляет по крайней мере приблизительно 12 нуклеотидов; более предпочтительно по крайней мере приблизительно 18 нуклеотидов; еще более предпочтительно длина составляет по крайней мере приблизительно 24 нуклеотида, и наиболее предпочтительный вариант - по крайней мере 36 нуклеотидов. В специфическом варианте выполнения, к термину «стандартные условия гибридизации» относится Tm 55°С, и условия использования, установленные ранее. В другом специфическом варианте выполнения, строгие условия обозначают Tm 65°С для условий гибридизации и отмывки, соответственно.
«Кодирующая последовательность» ДНК или «последовательность, кодирующая» частный белок или пептид, - это последовательность ДНК, которая транскрибируется и транслируется в полипептид in vitro и in vivo под контролем соответствующих регуляторных элементов.
Границы кодирующей последовательности определяются стартовым кодоном на 5’-конце и стоп-кодоном на 3’-конце. Кодирующая последовательность может включать в себя, но без ограничений, прокариотические последовательности, кДНК с мРНК эукариот, последовательности геномной ДНК от эукариот (например, млекопитающих) и даже синтетические последовательности ДНК. Последовательность терминации транскрипции обычно будет располагаться на 3’-конце кодирующей последовательности.
Термин «реально связанный» относится к расположению элементов, где описанные компоненты располагаются так, чтобы выполнять свои обычные функции. Так, контрольные элементы, реально связанные с кодирующей последовательностью, способны влиять на экспрессию кодирующей последовательности. Необходимо, чтобы контрольные элементы не располагались по соседству с кодирующей последовательностью все время, пока они действуют, управляя их экспрессией. Так, например, промежуточные, еще нетранслированные, транскрибированные последовательности могут присутствовать между промотором и кодирующей последовательностью, и промотор может пока рассматриваться как «реально связанный» с кодирующей последовательностью.
Термин «гетерологичная нуклеотидная последовательность», используемый здесь, обозначает нуклеотидную последовательность, которая добавляется к нуклеотидной последовательности настоящего изобретения рекомбинационными способами с образованием молекулы нуклеиновой кислоты, которая не является в действительности сформированной в природе. Такие молекулы нуклеиновой кислоты могут кодировать белки слияния (т.е. химерные). Так, гетерологичная нуклеотидная последовательность может кодировать пептиды и/или белки, которые обладают регуляторными и/или структурными свойствами. В другом таком варианте выполнения гетерологичные нуклеотидные последовательности могут кодировать белок или пептид, который функционирует как средство обнаружения белка или пептида, закодированного нуклеотидной последовательностью настоящего изобретения после того, как экспрессируется молекула рекомбинантной нуклеиновой кислоты. В другом варианте выполнения гетерологичная нуклеотидная последовательность может функционировать как средство для обнаружения нуклеотидной последовательности настоящего изобретения. Гетерологичная нуклеотидная последовательность может содержать в себе некодирующие последовательность, включающие в себя сайты рестрикции, сайты регуляции, промоторы и тому подобное.
Используемый здесь термин «белок слияния» или «пептид слияния» используется попеременно и заключает в себе «химерные белки и/или химерные пептиды» и слияние «интеиновых белков/пептидов». Белок слияния содержит в себе по крайней мере один участок собачьего белка ТСЛБ настоящего изобретения, соединенный пептидной связью с по крайней мере участком другого белка, например, несобачьего белка ТСЛБ, и/или содержит в себе комбинацию двух или более не смежных участков собачьего белка ТСЛБ, например, эпитопов, которые обычно не встречаются в смежном последовательном порядке в собачьем полипептиде ТСЛБ (например, белке слияния из 10 аминокислотных остатков, который состоит из аминокислотных остатков 71-75 и 101-105 из SEQ ID NO:2, соединенных пептидной связью). В предпочтительных вариантах выполнения изобретения участок(-ки) собачьего белка ТСЛБ являются функциональными, например, обладают своей антигенностью. Белки слияния также могут содержать в себе белок-маркер или белок, который способствует выделению и/или очистке (например, концевая метка FLAG, см. примеры ниже) и/или антигенности собачьего белка ТСЛБ настоящего изобретения.
Последовательности несобачьего ТСЛБ могут находиться на N- или С-конце у последовательностей собачьего ТСЛБ.
Рекомбинантная молекула ДНК, кодирующая белок слияния настоящего изобретения, например, может содержать в себе последовательность, кодирующую по крайней мере участок несобачьего белка ТСЛБ, объединенного рамкой считывания с кодирующей последовательностью собачьего белка ТСЛБ, и в дальнейшем может кодировать сайт расщепления для специфической протеазы, например, тромбина или Фактора Ха, предпочтительно на или рядом с местом соединения последовательности собачьего ТСЛБ и несобачьего ТСЛБ. В специфическом варианте выполнения изобретения белок слияния экспрессируется прокариотическими клетками. Такой белок слияния может быть использован для выделения собачьего ТСЛБ настоящего изобретения посредством использования колонки для аффинной хроматографии, специфической для белка и/или концевой метки, «пришитой» к собачьему ТСЛБ (см. примеры ниже).
Очищенный собачий белок ТСЛБ, например, может быть отделен от белка слияния посредством использования протеолитических ферментов и сайта расщепления, на который, например, ссылались выше.
«Вектор» или «вектор для репликации» - репликон, такой как плазмида, вирус, фаг или космида, к которому может быть прикреплен или вставлен другой сегмент ДНК, так, что это приведет к репликации прикрепленного сегмента. Термин также включает в себя репликон, который включает в себя вставленный или прикрепленный сегмент ДНК, представляющий интерес.
Векторы, которые могут использоваться в этом изобретении, включают в себя бактериальные плазмиды, вирусы, бактериофаги, фрагменты ДНК для вставки и другие переносчики, которые могут осуществлять интеграцию нуклеиновой кислоты в геном хозяина. Плазмиды являются наиболее часто используемым вектором, но все другие векторы, которые обладают эквивалентными функциями и известны или становятся известны в науке и технике, подходят для использования в настоящем изобретении (см., например, Pouwels et al., Cloning Vectors: A Laboratory Manual, 1985, Supplement, Elsevier, N.Y.; Rodriguez et al. (eds.), Vectors: A Survey of Molecular Cloning Vectors and Their Uses, 1988, Buttersworth, Boston, MA).
Вставка ДНК, кодирующей собачий белок ТСЛБ настоящего изобретения, в вектор легко протекает, когда концы молекулы ДНК и вектора содержат в себе сопоставимые сайты рестрикции. Если это невозможно сделать, то может быть необходимо модифицировать концы ДНК и вектора путем обрезания одноцепочечной ДНК с помощью эндонуклеаз рестрикции, так, чтобы сформировать тупые концы, или достигнуть такого же результата путем достройки одноцепочечной ДНК с помощью подходящей ДНК-полимеразы. В качестве альтернативного варианта, желаемые сайты можно получить, например, путем лигирования нуклеотидных последовательностей (линкеров) на концы. Такие линкеры могут содержать в себе специфические олигонуклеотидные последовательности, которые определяют желаемые сайты рестрикции. Сайты рестрикции могут быть также получены путем использования полимеразной цепной реакции (ПЦР) (см., например, Saiki et al., Science 239:487 (1988)). Расщепленный вектор и фрагменты ДНК могут также быть модифицированы, если это требуется, посредством гомополимерного удлинения концов молекулы. Рекомбинантные вектора для экспрессии, используемые в этом изобретении, обычно представляют собой самореплицирующиеся конструкты ДНК или РНК, содержащие в себе нуклеиновые кислоты, кодирующие собачий белок ТСЛБ настоящего изобретения и/или его антигенный фрагмент, обычно реально связанный с подходящими контрольными генетическими элементами, которые способны регулировать экспрессию нуклеиновых кислот в совместимых клетках хозяина. Контрольные генетические элементы могут включать в себя систему прокариотического промотора или систему эукариотического промотора для контроля экспрессии, и обычно включают в себя транскрипционный промотор, необязательный оператор для контроля начала транскрипции, энхансеры транскрипции для увеличения уровня экспрессии мРНК, последовательность, которая кодирует подходящий сайт связывания рибосомы, и последовательности, которые терминируют транскрипцию и трансляцию. Вектора для экспрессии могут также содержать точку начала репликации, которая позволяет вектору реплицироваться независимо от клетки хозяина.
Экспрессия нуклеиновых кислот, кодирующих собачий белок ТСЛБ по изобретению, может быть проведена с помощью обычных способов как в прокариотических, так и в эукариотических клетках.
«Клеткой хозяина» является клетка, которая содержит или способна содержать и экспрессировать экзогенную молекулу нуклеиновой кислоты, временно или постоянно. Клетка была «трансформирована» посредством экзогенного ДНК введением такой экзогенной ДНК за клеточную мембрану. Экзогенная ДНК может включаться или не включаться (ковалентно связываться) в хромосомную ДНК, изменяя геном клетки. Например, в прокариотах и дрожжах экзогенная ДНК может включаться в эписомальный элемент, такой как плазмида. Что касается эукариотических клеткок, то стабильная трансформированная клетка - это такая клетка, в которой экзогенная ДНК была включена в хромосому так, что она наследуется дочерними клетками путем репликации хромосом. Эта стабильность демонстрируется способностью эукариотических клеток к образованию клеточных линий или клонов, содержащихся в популяции дочерних клеток, содержащих экзогенную ДНК.
Прокариоты включают в себя и грамм-отрицательные, и грамм-положительные организмы, например, Е.coli и В.subtilis. Эукариоты включают в себя клеточные линии, полученные из ткани и клеток животных, как от животных, не относящихся к млекопитающим, например, насекомых, так и от птиц, и от млекопитающих, например, человека, приматов, грызунов.
Прокариотические системы «хозяин - вектор» включают в себя большое разнообразие векторов для множества различных видов. Вектора для амплификации ДНК включают в себя pBR322 и множество его производных, или pET42b(+)вектор для экспрессии (Novagen).
Обычно используемые Прокариотические последовательности контроля экспрессии включают в себя промоторы, включая промоторы - производные от систем β-лактамазного и пастозного промоторов (Chang et al., Nature, 198:1056 (177)), например, серии pUC, системы триптофанового промотора (trp) (Goeddel et al., Nucleic Acids Res. 8:4057 (1980)), например, pBR322-trp, системы промотора λ PL (Shimatake et al., Nature, 292:128 (181)), промоторов λ рР или pR (pOTS), арабинозо-индуцибельных промоторов (Invitrogen), tac промоторов (De Boer et al., Proc. Natl. Acad. Sci. USA 292:128 (1983)), промоторов ipp (pIN-серия); или гибридных промоторов таких, как ptac (pDR540). Многочисленные другие вектора для экспрессии, содержащие такие контрольные последовательности, также известны в этой области и доступны коммерчески (см., например Brosius et al., "Expression Vectors Employing Lambda-, trp-, lac- and lpp-derived Promoters", Rodrigues and Denhardt (eds.) Vectors: A Survey of Molecular Cloning Vectors and Their Uses, 1988, Buttersworth, Boston, pp.205-236).
В общем, эквивалентные подходящим E.coli вектора, которые могут быть использованы в других прокариотах, также могут быть использованы для экспрессии белков ТСЛБ настоящего изобретения.
Дрожжи, также как и культуры клеток высших эукариот, также рассматриваются в качестве хозяев для получения рекомбинантного собачьего белка ТСЛБ по изобретению и/или антител к собачьему ТСЛБ и/или фрагментам этих антител. Хотя могут использоваться культуры клеток любых эукариот, включая в себя бакуловирусные системы для экспрессии в насекомых, предпочтительными являются млекопитающие. Трансформация или трансфекция и размножение таких клеток становится рутинной процедурой. Примерами используемых клеточных линий являются клетки HeLa, клеточные линии яичников китайского хомячка (СНО), клеточные линии почки детенышей крысы (BRK), линии клеток насекомых (например, SF9), клеточные линии птиц (например, DF-11), клетки почки крупного рогатого скота Мадин-Дарби (MDBK), клетки почки собаки Мадин-Дарби (MDCK), клетки Vero, HEK-23 клеточные линии и клетки обезьяны (COS).
Вектора для экспрессии в таких клетках обычно включают в себя, например, точку начала репликации, промотор, сайт инициации трансляции, сайты сплайсинга РНК (при использовании геномной ДНК), сайт полиаденилирования и терминации транскрипции. Эти вектора обычно содержат ген селекции или ген амплификации. Подходящими векторами для экспрессии могут быть плазмиды, вирусы или ретровирусы, несущие производные от промотора, например, из таких источников, как аденовирус, SV40, парвовирус, вирус для вакцины или цитомегаловирус. Примерам подходящих векторов для экспрессии включают в себя pCR®3.1, pCDNA-1, pCD (Okayama et al., Mol Cell Biol. 5:1136 (1985)), pMdneo Poly-A (Thomas et al., Cell 5:503 (1987)), pUC19, pREP8, pSVSPORT и их производные, и бакуловирусные вектора, такие как рАС373 и рАС610.
После экспрессии собачий белок ТСЛБ по изобретению может быть очищен согласно стандартным процедурам в этой области, включая переосаждение сульфатом аммония, аффинную хроматографию, колоночную хроматографию и тому подобное (см., в основном, R. Scorpes. Protein Purification, Springer-Verlag, N.Y. (1982)). Достаточно чистые смеси по крайней мере 90-95% гомогенности являются предпочтительными, а 98-100% или более гомогенности наиболее предпочтительны для использовании в фармакологии. Очистка может быть частичная или до желаемой гомогенности. Если собачий ТСЛБ необходимо использовать терапевтически, белок должен быть достаточно чистым от эндотоксина. Стратегии избирательной очистки ТСЛБ после экспрессии на аффинной колонке с антителами к ТСЛБ или рецепторами к ТСЛБ доступны для получения высокоочищенного собачьего белка ТСЛБ.
Способы очистки хорошо известны в этой области. Например, нуклеиновые кислоты могут быть очищены переосаждением, хроматографией, ультрацентрифугированием и пр. Белки и полипептиды, также как и пептиды, могут быть очищены различными способами, включающими в себя без ограничения препаративный дисковый гель-электрофорез, изоэлектрическая фокусировка, ВЭЖХ, ВЭЖХ с обращенной фазой, гель-фильтрация, ионообменная и заместительная хроматография, переосаждение и высаливающая хроматография, экстракция и противоточное распределение. По ряду причин предпочтительно получать полипептид в рекомбинантной системе, в которой белок содержит дополнительную концевую метку, которая способствует очистке, таких, например, но без ограничений, как полигистидиновая последовательность или последовательность, которая специфически связывается с антителом, такая как FLAG® и GST. Полипептид затем может быть выделен из неочищенного лизата клеток хозяина при помощи хроматографии с подходящей матрицей для твердой фазы. В качестве альтернативного варианта как агенты для очистки могут быть использованы антитела или их связывающие фрагменты к полипептиду.
Растворитель и электролиты обычно находятся в биологически допустимом буфере, того типа, который используется для сохранения биологической активности, и обычно в приближении содержат физиологический растворитель на основе воды. Обычно растворитель имеет нейтральный рН, обычно между 5 и 10, и предпочтительней всего около 7.5. в некоторых случаях могут быть добавлены один или более детергентов, обычно мягкие и неденатурирующие, например, CHS (гемисукцинат холестерила) или CHAPS (3-(3-холамидопропил)диметиламмоний-)1-пропан сульфонат, или достаточно малая концентрация, чтобы избежать значительного разрушения структурных или физиологических свойств белка. В других примерах может быть использован жесткий детергент для достижения значительной денатурации.
В качестве альтернативного варианта, функциональные гетерологичные белки из E.coli или других бактерий могут быть выделены из телец включения посредством солюбилизации с использованием жестких денатурирующих агентов и последующего рефолдинга. Известные в этой области денатурирующие агенты включают в себя, например, мочевину, тиоцианат калия, гуанидин хлорид (GuHCl), иодат калия, иодид натрия комбинации из них. Предпочтительно, GuHCl используется как уменьшающий агент, например, в концентрации приблизительно от 6 до 8 М, при щелочных условиях, например, рН приблизительно 8. Необязательно используется другой уменьшающий агент, дитиотреитол (DTT), либо отдельно, либо в комбинации с GuHCl. При использовании DTT интервал концентраций просто путем примера составляет приблизительно от 50 мМ до 0.5 мМ DTT. В течение этапа солюбилизации, хорошо известного в этой области, уменьшающий агент должен присутствовать для разделения или денатурации дисульфидных связей. Один буфер для примера содержит, 0.1 М Tris рН 8.0, 8 М гуанидин, 2 мМ EDTA и 0.3 М DTE (дитиоэритритол).
Ренатурация обычно сопровождается разведением (например, в 100 раз) денатурированного и уменьшенного белка в буфере для рефолдинга в присутствии окисляющего агента. Может быть использован любой известный в этой области подходящий окисляющий агент, если это позволяет протекать правильному рефолдингу с высоким выходом. Например, окисление и рефолдинг могут быть обеспечены низкомолекулярными реагентами тиола в уменьшенной и окисленной форме, как описано у Saxena et al., 1970, Biochemistry 9:5015-5021, включено здесь в качестве ссылки, и особенно как описывалось у Buchner, et al., выше. Ренатурация обычно сопровождается разбавлением (например, в 100 раз) денатурированного и уменьшенного белка в буфере для рефолдинга. Один из примерных буферов для рефолдинга представляет собой Tris HCI 100 мМ рН 10.0, 25 мМ EDTA, NaCl 0.1 М, GSSG 551 мг/л, 0.5 М аргинин. GSSG является окисленной формой глутатиона.
Размер и структура полипептида должны в основном быть в существенно стабильном состоянии и обычно не в денатурированной состоянии. Полипептид может быть связан с другими полипептидами в квадратичную структуру, например, для того, чтобы предоставить растворимость, или с липидами или детергентами.
Значительно чистый, например, в отношении белка, обычно обозначает, что белок не содержит других загрязняющих белков, нуклеиновых кислот или других биологических объектов, полученных из первоначального организма. Чистота может быть проанализирована при помощи стандартных способов, обычно по весу, и должна составлять по крайней мере 40% чистоты, в основном приблизительно 50% чистоты, часто по крайней мере 60% чистоты, типично по крайней мере приблизительно 80% чистоты, предпочтительно по крайней мере приблизительно 90% чистоты и в наиболее предпочтительных вариантах выполнения по крайней мере 95% чистоты. Часто добавляются носители или наполнители. Чистота может быть оценена при помощи хроматографии, гель-электрофореза, иммуноанализа, анализа состава, биологического анализа и других способов, известных в этой области. Из функциональных аспектов выделенный собачий белок ТСЛБ, согласно изобретению является достаточно отделенным от остальных белков, включая предшественника собачьего белка ТСЛБ и/или зрелый собачий белок ТСЛБ, так что он способен вызывать иммунный ответ, который является специфическим для собачьего белка ТСЛБ.
Растворимость полипептида или фрагмента зависит от окружения и от полипептида. Многие параметры влияют на растворимость полипептида, включая температуру, электролитическое окружение, размер и молекулярные характеристики полипептида, и природу растворителя. Обычно температура, при которой используется полипептид, лежит в интервале от 4 до 65°С. Обычно температура больше, чем 18°С. Для диагностических целей температура обычно комнатная или теплее, но меньше, чем температура денатурации компонентов теста. Для терапевтических целей температура обычно равна температуре тела, обычно от 36 до 40°С (например, 39°С для собаки), хотя при определенных условиях температура может быть повышена или понижена in situ или in vitro.
Используемый здесь термин «антигенный фрагмент» по отношению к определенному белку является фрагментом этого белка (включая большие фрагменты, в которых пропущено только одна аминокислота из полноразмерного белка), который является антигенным, т.е. способным к специфическому взаимодействию с антиген-распознающей молекулой иммунной системы, такой как иммуноглобулин (антитело) или Т-клеточный антигенный рецептор. Например, антигенный фрагмент собачьего ТСЛБ настоящего изобретения является фрагментом антигенного собачьего ТСЛБ. Такие фрагменты не иммуногены сами по себе, т.е., способны вызывать иммунный ответ в отсутствие переносчика, так что они могут быть использованы для получения антитела к белку ТСЛБ после конъюгирования фрагмента с молекулой переносчика для иммунизации. Предпочтительно, однако, чтобы антигенный фрагмент настоящего изобретения являлся иммунодоминантным для антитела и/или для распознавания Т-клеточными рецепторами.
В частном варианте выполнения антигенный фрагмент собачьего белка ТСЛБ содержит от 5 до 150 аминокислотных остатков. В одном частном варианте выполнения антигенный фрагмент собачьего ТСЛБ содержит более, чем 120 аминокислотных остатков. В другом варианте выполнения антигенный фрагмент собачьего ТСЛБ содержит от 10 до 120 аминокислотных остатков. Еще в одном варианте выполнения антигенный фрагмент собачьего ТСЛБ содержит от 20 до 100 аминокислотных остатков. Еще в одном варианте выполнения антигенный фрагмент собачьего ТСЛБ содержит от 25 до 75 аминокислотных остатков.
Антигенный фрагмент собачьего ТСЛБ может быть получен из рекомбинантного источника, из белка, полученного из природного источника, или с помощью химического синтеза. Более того, антигенный фрагмент может быть получен при последующем протеолитическом расщеплении собачьего ТСЛБ или его фрагмента посредством рекомбинационной экспрессии или, как альтернативный вариант, он может быть получен de novo, например, посредством пептидного синтеза.
Вакцины
Настоящее изобретение далее предоставляет вакцины, которые включают в себя эффективное количество белка ТСЛБ настоящего изобретения, один или более его антигенных фрагментов, или комбинации полноразмерного белка и одного или более таких фрагментов. Например, собачий белок ТСЛБ и/или его фрагменты, такие как например пронумерованные в таблице 2 ниже, могут быть включены в совместимую с белками или пептидами вакцину любого состава. Вакцины такого состава хорошо известны в этой области и могут, необязательно, включать в себя, например, физиологически совместимые буфера и солевые растворы и тому подобное, также как и фармакологически допустимые адъюванты, такие как CARBOPOL® или Emulsigen®.
Смесь для вакцинирования может быть использована для назначения эндогенных антител к ТСЛБ собаке, которая нуждается в этом, например, для того, чтобы лечить клинические проявления заболевания или расстройства, которые отвечают за регуляцию активности афферентных путей ТСЛБ у собак. В качестве альтернативного варианта или в сочетании с этим, вакцина настоящего изобретения может также быть использована для того, чтобы выявлять антисыворотку для скрининга и/или идентификации собачьего ТСЛБ, например, в качестве вспомогательного агента в ките для идентификации собак с гиперэкспрессией ТСЛБ.
Пептиды из ТСЛБ, такие как показано в таблице 2 ниже, и их варианты, могут быть использованы в качестве иммуногенов, как индивидуально, так и в различных сочетаниях. Такие пептиды могут быть необязательно связаны друг с другом и/или с более крупными белками, известными как носители, как с помощью химических способов, так и с помощью рекомбинантных ДНК. Действие носителей направлено на увеличение распознавания пептида клеткой животного-хозяина как мишени для иммунного ответа и увеличения иммуногенности пептидов ТСЛБ. Отдельные носители известны в этой области и включают в себя столбнячный анатоксин или нетоксичный C-концевой фрагмент столбнячного анатоксина, дифтерийный анатоксин, РПоР белок, гемоцианин лимфы улитки, бета-галактозидаза, gD белок из вируса BHV-1, G белок вируса бешенства, F белок вируса собачьей чумы и синтетические носители, полученные в результате полимеризации известных «универсальных» эпитопов Т-клетки.
Пептиды ТСЛБ, используемые как иммуногены, могут быть выбраны из таблицы 2, и их варианты, при использовании известных алгоритмов, которые оценивают свойства, такие как доступность поверхности у природного белка ТСЛБ, гидрофильность, мобильность атомов и антигенность. Эпитопы на пептидах, указанных в таблице 2, и их варианты могут также быть выбраны на основании их реакционной способности при взаимодействии с поликлональными или моноклональными антителами, которые реагируют с нативными белками ТСЛБ и особенно те антитела, которые способны нейтрализовать биоактивность ТСЛБ. Такие антигены могут включать в себя синтетические пептиды, полученные из последовательностей, которые здесь обнаружены, используя стандартный протокол пептидного синтеза и/или в качестве альтернативного варианта, могут быть фрагментами, полученными из рекомбинантного или природного белка ТСЛБ.
Фармакологически допустимые адъюванты настоящего изобретения могут быть получены из любого источника, включая природные, рекомбинантные и/или синтезированные химически, и т.д. примеры химических соединений, используемых как адъюванты, включают в себя, но без ограничений, соединения алюминия, преобразующиеся или не преобразующиеся в ходе обмена веществ, блокполимеры, ISCOM (иммуностулирующие комплексы), витамины и минералы (включающие в себя, но без ограничений: витамин Е, витамин А, селен и витамин В12) и Quil А (сапонины), полный адъювант Freund, полимеры акриловой кислоты поперечно сшивались с полиалкенильными эфирами или дивинилгликолем, как продается под торговой маркой CARBOPOL® (например, CARBOPOL® 941) и единообразно диспергировались в виде капель масла микронного размера в воде (например, как продается под торговой маркой Emulsigen®). Дополнительные примеры адъювантов, на которые иногда ссылались как на специфические иммуностимуляторы, включают в себя компоненты клеточной стенки бактерий и грибов (например, липополисахариды, липопротеины, гликопротеины, мурамилпептиды, бета-1,3/1,6-глюканы), различные сложные углеводороды, полученные из растений (например, глюканы, ацеманнаны), различные белки и пептиды, полученные из животных (например, гормоны, цитокины, ко-стимулирующие факторы) и новые нуклеиновые кислоты, полученные из вирусов и других источников (например, двуцепочечная РНК, CpG). В дополнение, любое количество комбинаций вышеупомянутых соединений может обеспечивать адъювантный эффект, и поэтому может формировать адъювант настоящего изобретения.
Вакцины настоящего изобретения могут назначаться любым путем, включая внутримышечное введение, подкожное введение, внутривенное введение, внутрикожное введение, пероральное назначение, интраназальное назначение и их комбинации.
Антитела
Настоящее изобретение также включает в себя поликлональные и моноклональные (mAb) антитела, которые специфически связываются с собачьим белком ТСЛБ по изобретению. Используемый здесь термин «антитело» подразумевает под собой иммуноглобулин и/или его фрагменты. Встречающийся в природе иммуноглобулин состоит из одного или более полипептидов, закодированных существенно в генах иммуноглобулинов. Распознанные гены иммуноглобулинов включают в себя гены каппа, лямбда, альфа, гамма, дельта, эпсилон и мю константных областей также как и множество генов вариабельных участков иммуноглобулина. Антитело или антитела согласно настоящему изобретению также включают в себя фраменты антител, т.е. антиген-связывающие фрагменты, например, сконструированные одноцепочечные Fv, Fab и F(ab’)2 связывающие белки (например, Huston et al., Proc. Natl. Acad. Sci. USA, 85, 5879-5883 (1988), Bird et al., Science, 242, 423-426 (1988), включенные сюда в качестве ссылок), также как и бифункциональные гибридные антитела (например, Lanzavecchia et al., Eur. J. Immunol. 17, 105 (1987)). См., в основном, Hood et al., Immunology, Benjamin, N.Y., 2nd ed. (1984), Harlow and Lane, Antibodies. A Laboratory Manual, Cold Spring Harbor Laboratory (1988), Hunkapiller and Hood, Nature, 323, 15-16 (1986), все включены сюда в качестве ссылок.
Например, сыворотка, полученная от животных, иммунизированных собачьим белком ТСЛБ по изобретению, при использовании стандартных способов, могут быть использованы напрямую или фракция IgG может быть выделена из сыворотки при использовании стандартных способов, таких как плазмофорез или адсорбционная хроматография со специфическими к IgG адсорбентами, такими как иммобилизованный белок А или белок G. В качестве альтернативы, могут быть приготовлены моноклональные антитела и из таких mAb необязательно получены антиген-связывающие фрагменты или рекомбинантные связывающие белки. Такие mAb или их фрагменты могут необязательно быть гуманизированы или «особачены» при помощи известных в данной области способов или их прямой модификации.
Используемый здесь термин «эпитоп-специфичное» собачье антитело к ТСЛБ - это антитело, которое взаимодействует с фрагментом собачьего ТСЛБ, который содержит в себе одну или более из пяти последующих аминокислотных последовательностей: SEQ ID NO:30, SEQ ID NO:31, SEQ ID NO:32, SEQ ID NO:33 или SEQ ID NO:34; и которые в дальнейшем связывают белок, имея аминокислотную последовательность SEQ ID NO:2, и/или белок, имеющий аминокислотную последовательность SEQ ID NO:2, за исключением сигнальной последовательности 28 аминокислотного остатка. В частном варианте выполнения эпитоп-специфичное антитело к собачьему ТСЛБ является моноклональным антителом.
Гибридомы, продуцирующие mAb, которые селективно связывают собачий белок ТСЛБ по изобретению, получаются с использованием хорошо известных способов. Обычно процесс включает в себя слияние иммортализованной клеточной линии с В-лимфоцитом, который продуцирует желаемое антитело. В качестве альтернативы, могут быть использованы способы, не включающие в себя слияние, для получения иммортализованных антитело-продуцирующих клеток, например, вирус-индуцированная трансформация (Casali et al., Science 234:476 (1986)). Иммортализованные клеточные линии являются обычно трансформированными клетками млекопитающих, в частности, клетки миеломы грызунов, крупного рогатого скота и человека. Наиболее часто используются крысиные или мышиные клетки миеломы в целях удобства и доступности.
Способы для получения анитело-продуцирующих лимфоцитов млекопитающих, иньецированных антигенов, хорошо известны. Обычно используются лимфоциты периферической крови, если используются клетки человеческого происхождения, или клетки селезенки или лимфоузлов при использовании антител от клеток млекопитающего, но не человека. Животное-хозяин иньецируется повторяющимися дозами очищенного антигена (клетки человека сенсибилизируются in vitro) и животное получает возможность продуцировать желаемые антитело-продуцирующие клетки перед тем, как их нарастят для слияния с использованием иммортализованной линии клеток. Способы для слияния также хорошо известны в этой области, и, в основном, включают в себя смешивание клеток с агентом слияния, таким как полиэтиленгликоль.
Гибридомы выбираются путем стандартных процедур, таких как HAT (гипоксантин-аминоптерин-тимидин) селекция. Те, которые секретируют желаемое антитело, отбираются при использовании стандартных иммунных тестов, таких как Вестерн блоттинг, ELISA (иммуноферментный анализ), RIA (радиоиммуноанализ) и тому подобное. Антитела восстанавливаются из среды при использовании стандартных протоколов очистки белков (Tijssen, Practice and Theory of Enzyme Immunoassays (Elsevier, Amsterdam, 1985)).
Доступно множество ссылок для обеспечения руководства для применения вышеуказанных способов (Kohler et al., Hybridoma Techniques (Cold Spring Harbor Laboratory, New York, 1980), Tijssen, Practice and Theory of Enzyme Immunoassays (Elsevier, Amsterdam, 1985), Campbell, Monoclonal Antibody Technologies (Elsevier, Amsterdam, 1984), Hurrel, Monoclonal Hybridoma Antibodies: Techniques and Applications (CRC Press, Boca Raton, FL, 1982)). Моноклональные антитела могут также продуцироваться с использование хорошо известных фаговых библиотек (см., например, Huse et al., Science 246:1275 (1989); Ward et al., Nature, 341:544 (1989)).
Антитела, получаемые таким образом, поликлональные или моноклональные, могут быть использованы, например, в иммобилизованной форме, связанной с твердой подложкой хорошо известными способами для очистки собачьего белка ТСЛБ с помощью иммуноаффиннной хроматографии.
Антитела к собачьему белку ТСЛБ также могут быть использованы, меченые или немеченые стандартными способами, как основа для иммунных тестов для детекции или определения количества собачьего белка ТСЛБ. В частности используемая метка зависит от типа иммунного теста. Примерами меток могут служить, но без ограничений, радиоактивные метки, такие как 32P, 125I, 3H и 14С; флуоресцентные метки, такие как флуорескаин и его производные, родамин и его производные, дансил и умбелиферон; хемилюминесцентные метки, такие как люциферин и 2,3-дигидрофталазинедионы; и ферменты, такие как пероксидаза хрена, щелочная фосфатаза, лизоцим и глюкоза-6-фосфат дегидрогеназа.
Антитела могу быть помечены такими метками с помощью известных способов. Например, «сшивающие» агенты, такие как альдегиды, карбодиимиды, дималеимиды, имидаты, сукцинимиды, бис-диазобензадин и тому подобное могут быть использованы для мечения антител флуоресцентными, хемилюминесцентными или ферментными метками. Основные способы, включенные в этот процесс, хорошо известны в данной области и описаны, например, в Immunoassay: A Practical Guide, 1987, Chan (Ed.), Academic Press, Inc., Orlando, FL Такие иммунные тесты могут проводиться, например, на фракциях, полученных в ходе очистки рецепторов.
Антитела настоящего изобретения могут также быть использованы для идентификации определенных клонов кДНК, экспрессирующих собачий белок ТСЛБ в системах для клонирования и экспрессии. Нейтрализующие антитела, специфичные для лиганд-связывающего сайта рецептора, также могут быть использованы как антагонисты (ингибиторы) для того, чтобы блокировать или ослаблять функцию собачьего белка ТСЛБ. Такие нейтрализующие антитела могут быть легко идентифицированы посредством обычных экспериментов.
Антагонизм активности собачьего белка ТСЛБ может сопровождаться использованием целых молекул антител или хорошо известных антиген-связывающих фрагментов, таких как Fv, Fab и F(ab’)2 фрагменты. Определение таких фрагментов может быть найдено в описанном выше или, например, в Klein, Immunology (John Wiley, New York, 1982), Parham, Chapter 12, Weir, ed. Immunochemistry, 4th Ed. (Blackwell Scientific Publishers, Oxford, 1986). Применение и получение фрагментов антител также было описано, например, Fab фрагментов (Tijssen, Practice and Theory of Enzyme Immunoassays (Elsevier, Amsterdam, 1985), Fv фрагментов (Hochman et al., Biochemistry 12:1130 (1973); Sharon et al., Biochemistry 15:1591 (1976); Ehrich et al., U.S. Patent No. 4355023) и полной молекулы антител (Auditore-Hargreaves, U.S. Patent No. 4470925). Способы получения рекомбинантных Fv фрагментов, основанные на последовательностях тяжелых и легких цепей вариабельной области известного антитела были описаны далее, например, Moore et al. (патент США No. 4642334) и Pluckthun (Biotechnology 9:545 (1991)). В качестве альтернативы они могут быть синтезированы химически с помощью стандартных способов.
Настоящее изобретение также охватывает анти-идиотипические антитела, как поликлональные, так и моноклональные, которые продуцируются при использовании описанных выше антител в качестве антигенов. Эти антитела являются полезными, так как они могут имитировать структуры лигандов. Антитела несобачьего происхождения или из несобачьих гибридомных систем могут быть необязательно сконструированы для того, чтобы переводить их в существенно неантигенную форму при введении собаке, т.е. могут быть «особачены». Процесс модификации моноклонального антитела от животного для того, чтобы перевести его в менее иммуногенную форму для терапевтического использования у человека (гуманизация) активно разрабатывался и описан в ряде публикаций (например, Antibody Engineering: A practical Guide. Carl А. K. Borrebaeck ed. W.H. Freeman and Company, 1992; Reichman, L. et al., "Reshaping human antibodies for therapy", Nature 332:323-327 (1988)). В качестве альтернативы моноклональные антитела несобачьего происхождения, например, мышиные моноклональные антитела, модифицируются в химерные с собачьими антителами или их последовательностями для того, чтобы получить антитела менее иммуногенные для реципиента, чем стандартные мышиные антитела. См, например, Патент США No. 5593861, "Dog-Mouse Heterohybridoma and Gene Fragment Coding for Constant Region of Canine Immunoglobulins", который включен сюда в качестве ссылки.
В дополнение, Wasserman and Capra (Biochem. 16:3160 (1977)) определили аминокислотную последовательность тяжелой цепи вариабельных областей как собачьего IgM, так и собачьего IgA. Эти исследователи далее определили аминокислотную последовательность каппа легкой цепи собачьего IgA (Wasserman and Capra, Immunochem. 15:303 (1978)). McCumber and Capra (Mol. Immunol. 16:565 (1979)) определили полную аминокислотную последовательность собачьей мю цепи. Tang et al., (Vet. Immunology Immunopathology 80:259 (2001)) определили одну последовательность кДНК гамма цепи собачьих IgG-A и четыре белковые последовательности гамма цепи собачьих IgG-A. Tang et al., выше, далее описывают ПЦР-амплификацию кДНК из библиотеки кДНК селезенки собаки с вырожденными праймерами, сконструированными на консервативные области IgG человека, мыши, свиньи и крупного рогатого скота. Более того, Krah, et al. (U.S. Publication No. 20040181039, опубликовано 16 сентября 2004 и включено сюда в качестве ссылки) описывает детально один из процессов «особачивания» несобачьих антител.
Выделение собачьего гена ТСЛБ
А. Первоначальные попытки
Первоначальные попытки идентификации собачьего ТСЛБ были основаны на сопоставлении клонированных последовательностей кДНК ТСЛБ человека и мыши с последовательностями кДНК ТСЛБ крысы, шимпанзе и макаки резуса, которые собраны в BLAT (публичная геномная база данных Университета Калифорнии, Санта Круз). ТСЛБ шимпанзе был на 100% гомологичен ТСЛБ человека, в то время как ТСЛБ макаки резуса был гомологичен ТСЛБ человека на 90% (12 из 151 аминокислотных остатков не совпадали) в зрелом состоянии. Однако последовательности белка и кДНК ТСЛБ человека и приматов сильно расходятся с последовательностями ТСЛБ мыши. Последовательности кДНК ТСЛБ человека и мыши имеют только 43% гомологии, что не позволяет проводить кпонирование посредством нестрогой межвидовой гибридизации между этими видами. Более того, последовательность ТСЛБ крысы показала 39 изменений из 121 аминокислотных остатков в зрелом белке по сравнению с мышиным ТСЛБ, что показывает, что даже у близкородственных видов грызунов последовательности ТСЛБ значительно различаются.
К сожалению, как определено здесь, было доказано, что последовательность собачьего ТСЛБ расходится со всеми мышиными последовательностями и последовательностями у приматов. Поэтому получение собачьего ТСЛБ посредством нестрогой межвидовой гибридизации было признано неуспешным. Действительно, праймеры, сконструированные для попытки клонирования ортолога собачьего ТСЛБ методом «гнездовой» ПЦР, используя последовательности человека, мыши, крысы и обезьяны не идентифицировали ни одной полосы, соответствующей собачьему ТСЛБ.
В. Успешное выделение гена собачьего ТСЛБ
При поиске собачьих последовательностей в доступной геномной базе данных (производной от полной первоначальной геномной последовательности, доступ открыт Университетом Калифорнии, Санта Круз) с последовательностью ТСЛБ человека привело к частичной идентификации 1 и 4 экзонов собачьего ТСЛБ. Вкратце, были идентифицированы некоторые удачные попытки поиска значительной гомологии последовательностей в первоначальном поиске (см. удачные попытки («хиты») 1-6 ниже). Эти последовательности были собраны и использованы в качестве запросов для расширения и сбора частичных последовательностей собачьего ТСЛБ в электронном виде.
Хит 1
Счет = 60.8 бит (146), Ожидание = 1е-08
Идентичность = 33/58 (56%), Позитивные позиции = 39/58 (57%), Пропуски = 1/58 (1%)

SEQ ID NO:102: ТСЛБ человека
SEQ ID NO:103; >gi36323560gbAACN010632090.1 Canis familiaris ctg19866851299046, полная длина первоначальной геномной последовательности = 1007
Хит 2
Счет = 59.7 бит (143), Ожидание = 3е-08
Идентичность = 30/42 (71%), Позитивные позиции = 33/42 (78%), Сдвиг рамки = -1

SEQ ID NO:104: ТСЛБ человека
SEQ ID NO:105; >gi 36314527gbAACN010674832.1 Canis familiaris ctg19866851282529, полная длина первоначальной геномной последовательности = 963
Хит 3
Счет = 42.0 бит (97), Ожидание = 0.006

SEQ ID NO:106: ТСЛБ человека
SEQ ID NO:107; >gi36442813gbAACN011084208.11 Canis familiaris ctg19866851499233, полная длина первоначальной геномной последовательности = 370
Хит 4
Счет = 38.9 бит (89), Ожидание = 0.047
Идентичность = 15/32 (46%), Позитивные позиции = 22/32 (78%), Сдвиг рамки = +2

SEQ ID NO:108: ТСЛБ человека
SEQ ID NO:109; >gi36211043gbAACN010354273.1 Canis familiaris ctg19866851087147, полная длина первоначальной геномной последовательности = 1369
Хит 5
Счет = 42.0 бит (97), Ожидание = 0.006 Идентичность = 21/44 (47%), Позитивные позиции = 27/44 (78%), Сдвиг рамки = -2

Последовательность:
SEQ ID NO:110: ТСЛБ человека
SEQ ID NO:111; >gi36211043gbAACN010354273.1 Canis familiaris ctg19866851087147, полная длина первоначальной геномной последовательности = 1369
Хит 6
Счет = 38.9 бит (89), Ожидание = 0.047
Идентичность = 15/32 (46%), Позитивные позиции = 22/32 (78%), Сдвиг рамки = +2

SEQ ID NO:108: ТСЛБ человека
SEQ ID NO:109; >gi36211043gbAACN010354273.1 Canis familiaris ctg19866851087147, полная длина первоначальной геномной последовательности = 1369.
Сравнение этой электронно сконструированной последовательности с ТСЛБ человека, обезьяны, крысы и мыши показало границы консервативных экзонов/интронов и существенную идентичность, ведущую к идентификации этой последовательности как части собачьего ортолога ТСЛБ. Праймеры для ПЦР были впоследствии сконструированы на основе этого открытия и использованы для амплификации пропущенных сегментов гена. С помощью продублированной «гнездовой» ПЦР были получены два частичных перекрывающихся клона из библиотеки кДНК активированных периферических мононуклеарных клеток крови (РВМС) собаки. Дополнительные попытки для обнаружения полной последовательности кДНК собачьего ТСЛБ посредством «гнездовой» ПЦР или попытки расширить последовательности по направлению от 5’ к 3’ концу были неудачными. Однако повторяющимися раундами поиска по базе данных с использованием расширенной информации о последовательности, полученной от этих клонов по данным о первоначальной геномной последовательности собаки (Университет Калифорнии, Санта Круз) в сочетании с ручным сбором черновых последовательностей ДНК из этой библиотеки привели к электронному набору полноразмерной кДНК собачьего ТСЛБ. Физический клон этой последовательности кДНК был потом синтезирован in vitro с использованием синтезатора ДНК. В заключение, используя текущие и установленные в данной области методики молекулярного клонирования не было возможным получить последовательность собачьего ТСЛБ непосредственно от последовательностей человека, мыши, крысы или обезьяны. Только точные повторяющиеся поиски по базе данных с использованием накопленных генов ТСЛБ человека, мыши, крысы и NHP, с использованием приближений о границах экзонов/интронов и идентичности последовательности из геномной базы данных, в сочетании с методиками молекулярного клонирования с помощью ПЦР, привели к идентификации гена, кодирующего собачий ТСЛБ.
При получении собачий ТСЛБ показал 58 изменений из 132 по сравнению с аминокислотной последовательностью зрелого белка ТСЛБ человека (61% идентичности) и 83 изменений из 129 по сравнению с аминокислотной последовательностью зрелого белка ТСЛБ мыши (33% идентичности) (см. ниже).
Сравнение последовательностей у зрелого белка ТСЛБ Canis familiaris и человека
ТСЛБ_СР (Canis familiaris) Длина 141 (1…141)
ТСЛБ JH (человек) Длина 145 (1…145)
Счет = 167 бит (423), Ожидание = 1е-40
Идентичность = 85/139 (61%), Positives = 101/139 (72%)

Последовательность:
121
SEQ ID NO:114: Canine familiaris ТСЛБ
SEQ ID NO:115: TSLP человека
Сравнение последовательностей у зрелого белка ТСЛБ Canis familiaris и мыши
ТСЛБ_СР Длина 141 (1…141)
ТСЛБ_М Длина 136 (1…136)
Счет = 72.0 бит (175), Ожидание = 7е-12
Идентичность = 46/138 (33%), Позитивные позиции = 67/138 (48%), Пропуски = 8/138(5%)

SEQ ID NO:116: Canine familiaris ТСЛБ
SEQ ID NO:117: ТСЛБ мыши
Таким образом, при преодолении ранее отмеченных трудностей, настоящее изобретение предоставляет последовательности ДНК, кодирующие собачий ТСЛБ и закодированный собачий белок ТСЛБ. Собачий белок ТСЛБ и определенные его фрагменты являются хорошими антигенами, т.е. иммуногенами, для продуцируемых антител к различным эпитопам белка, как к линейным, так и к конформационным. ДНК, кодирующая собачий ТСЛБ, также важна в обеспечении векторов и клеток хозяина для получения белка ТСЛБ для иммунизации и/или в качестве объекта для исследования, также как и для получения вакцин, основанных на ДНК, для получения антител к ТСЛБ, либо в форме «голой» ДНК, либо в форме плазмиды, либо в форме вирусного вектора, подходящего для экспрессии ТСЛБ клетками вакцинированного животного.
Полученная таким образом последовательность гена собачьего ТСЛБ проиллюстрирована ФИГ.8А (SEQ ID NO:1), и предсказанный экспрессированный белок ТСЛБ проиллюстрирован ФИГ. 8 В (SEQ ID NO:2). Остатки 1-28 представляют собой сигнальную последовательность, а остатки 29-155 представляют зрелый белок.
Исследование для идентификации гомологичных ТСЛБ белков
Настоящее изобретение также обеспечивает белки ТСЛБ, которые содержат в себе аминокислотную последовательность SEQ ID NO:2, за исключением сигнальной последовательности из 28 аминокислотных остатков, которая при применении у собак в качестве вакцины, вызывает продукцию антител, которые связываются с собачьим белком ТСЛБ, содержащим аминокислотную последовательность SEQ ID NO:2. предоставляются также антигенные фрагменты такого белка ТСЛБ.
Действительно, одним из вариантов демонстрации того, что предполагаемый белок ТСЛБ - это белок ТСЛБ настоящего изобретения, является проверка того, может ли такой белок продуцировать антитела, которые связываются с собачьим ТСЛБ, содержащим аминокислотную последовательность SEQ ID NO:2. Одним из таких способов является вакцинация (т.е. инъекция) собак различными дозами, лежащими в интервале от 5 до 500 мкг предполагаемого антигена ТСЛБ-GST. Такие антигены могут быть приготовлены на основе адъюванта, содержащего гидроксид алюминия, такого как Rehydrogel. Собакам вакцина затем вводится внутримышечно три раза: в день 0, день 21 и день 42. Образцы сыворотки собирают от вакцинированных и контрольных (неиммунизированных) собак в день 0, 21, 42 и 63.
Индукция антител у собак, вакцинированных антигенами, можно оценить с помощью ИФА в соответствии со следующим протоколом: собачий белок ТСЛБ, содержащий в себе аминокислотную последовательность SEQ ID NO:2, растворялся в концентрации 5 мкг/мл в покрывающем буфере (бикарбонат натрия рН 9.0) и раскапывался 100 мкл/лунка в 96-луночном планшете (Pierce). Планшеты инкубировали при 4°С в течение ночи. Затем планшеты три раза промывали фосфатно-солевым буфером, содержащим 0.05% Tween-20 (PBST). Затем в каждую лунку добавляли по 200 мкл блокирующего буфера (2% снятого молока в PBST) и планшеты инкубировали при комнатной температуре 60 мин. Планшеты затем промывали три раза PBST. Затем добавляли 100 мкл/лунка раствора антисыворотки тестируемой собаки в воде 1:100 в верхний ряд соответствующих лунок. Образцы сыворотки затем разбавляли в 10 раз до соответствующей позиции планшета. После инкубации планшетов при комнатной температуре в течение 60 мин планшеты промывали три раза PBST.
Затем в каждую лунку добавляли раствор 1:20000 пероксидазы хрена, конъюгированной с козьими антителами к собачьему IgG (Bethyl Laboratories). Затем планшеты инкубировали при комнатной температуре в течение 60 мин. Затем планшеты промывали три раза PBST и добавляли в каждую лунку 100 мкл субстрата ТМВ (3,3’, 5,5’ тетраметилбензидин, Sigma Chemical Co., St. Louis, МО). Давали развиться качественной реакции в течение 10-120 мин при комнатной температуре перед тем, как остановить реакцию с помощью добавления 50 мкл на лунку 0.18 М серной кислоты.
Оптическая плотность (OD) во всех лунках определяли при длине волны 450 нм, используя ELISA reader для планшетов (Thermo Max; Molecular Devices, Sunnyvale, CA). Образцы сыворотки, полученные от собак, вакцинированных предполагаемыми антигенами ТСЛБ, считаются детектируемыми и, следовательно, антигены идентифицируются как белки ТСЛБ настоящего изобретения, когда анализ показывал значения OD, равные или больше, чем в три раза, чем фоновые, полученные от образцов сыворотки от собак перед иммунизацией. Подобным образом, титры родственных антител к антигенам ТСЛБ могут быть определены на основании наибольшего разведения сыворотки, дающего значения OD, равные или больше, чем в три раза, чем фоновые, полученные от образцов сыворотки от собак перед иммунизацией этими антигенами.
Антитела к специфическим эпитопам собачьего белка ТСЛБ
Антитела могут быть получены к различным эпитопам собачьих белков ТСЛБ, включая в себя виды, полиморфы или аллельные варианты и их фрагменты, как во встречающейся в природе форме, так и в рекомбинантных вариантах.
Дополнительно, антитела могут быть получены к собачьим ТСЛБ, либо в своих активных формах, либо в неактивных, включая нативный или денатурированный вариант. Также рассматриваются антиидиотипические антитела.
Антитела, включающие в себя связывающие фрагменты и варианты с одиночной цепью, к определенным заранее антигенам, могут быть получены посредством иммунизации животных собачьим ТСЛБ и/или его фрагментами, совместно с стандартными в данной области адъювантами и/или конъюгированы с иммуногенными белками. Иммунизированные таким образом животные могут являться собаками, которых иммунизировали для того, чтобы понизить активность собачьего ТСЛБ.
Подходящего хозяина, например, инбредную линию мышей таких, как Balb/c, иммунизируют выбранным белком, обычно используя стандартный адъювант и стандартный протокол иммунизации мышей (см., Harlow and Lane, Id, выше). Адъювант может назначаться целевому животному до, в сочетании или после применения вакцины.
В качестве альтернативы, синтетический пептид, полученный от последовательностей, описанных здесь, и конъюгированный с белком носителем, может служить в качестве иммуногена. Поликлональные сыворотки собирают и титруют белком-иммуногеном при иммунном анализе, например, твердофазном иммунном анализе с иммуногеном, иммобилизованном на твердой подложке. Поликлональные антисыворотки с титром 1*104 или более, отбирают и тестируют по их перекрестной реактивности по отношению к другим членам семейства IL-7, например, мышиный IL-7, используя конкурентный иммунный анализ, как было описано в Harlow and Lane, Id, выше, стр.570-573. Предпочтительно, по крайней мере, один из остальных членов семейства IL-7 использовался в этом определении, например, IL-7 приматов. Члены семейства IL-7 могут быть получены как рекомбинантные белки и выделены с помощью стандартных молекулярно-биологических и белково-химических способов, как описано здесь. Иммунные анализы в виде конкурентного связывания могут быть использованы для определения перекрестной реактивности. Например, белок SEQ ID NO:2 может быть иммобилизован на твердой подложке. Белки, используемые в тесте, конкурируют за связывание антисыворотки с иммобилизованным антигеном. Способность конкурировать вышеуказанных белков за связывание антисыворотки с иммобилизованным белком сравнивается с белком, содержащим аминокислотную последовательность SEQ ID NO:2. Процент перекрестной реактивности вышеуказанных белков вычисляют с помощью стандартных расчетов. Те антисыворотки, которые имеют менее чем 10% перекрестной реактивности с каждым из белков, перечисленных выше, отбирают и группируют. Перекрестно реактивные антитела затем удаляют из сгруппированных сывороток с помощью иммуноабсорбции с вышеуказанными белками.
Иммуноабсорбированные и сгруппированные антисыворотки затем используют в конкурентном связывающем иммуноанализе как описано выше, для того, чтобы сравнить второй белок с иммуногенным белком (например, IL-7-подобный белок из SEQ ID NO:2). Для того, чтобы сделать это сравнение, два белка тестируют в широком диапазоне концентраций и определяется требуемое количество каждого белка, способное на 50% ингибировать связывание антисывороток. Если требуемое количество второго белка меньше, чем удвоенное количество белка из выбранного белка или белков, чем требуется, говорят затем, что второй белок специфически связывается с антителом, полученным при действии иммуногена. Антитела настоящего изобретения могут также использоваться в диагностических целях. Как ловушка или не-нейтрализующие антитела, с ними может быть проведен скрининг способности связывать антигены без ингибирования связывания рецептора. Как нейтрализующие антитела, они могут использоваться в анализе конкурентного связывания. Они также будут использоваться при определении и количественной оценке собачьего белка ТСЛБ или его рецептора (см., например, Chan (ed. 1987) Immunology: A Practical Guide. Academic Press, Orlando, Fla.; Price and Newman (eds. 1991) Principles and Practice of Immunoassay. Stockton Press, N.Y.; and Ngo (ed. 1988) Nonisotopic Immunoassay. Plenum Press, N. Y). Перекрестная абсорбция, расщепление или другие средства обеспечивают получение определенной селективности, например, уникальные признаки вида или встречающиеся у других видов. Это может быть основой для тестов для определения различных групп антигенов.
Далее, антитела, включающие в себя антиген-связывающие фрагменты настоящего изобретения, могут быть потенциальными антагонистами, которые связываются с антигеном и ингибируют функциональное связывание, например, с рецептором, который может вызывать биологический ответ. Далее, эти антитела могут быть конъюгированы с лекарствами или другими терапевтическими агентами, либо напрямую, либо не напрямую посредством линкера, и могут влиять на нацеливание лекарственных средств.
Синтетический пептид, произведенный от последовательности, описанной здесь и конъюгированный с белком носителем, может использоваться в качестве иммуногена. В любом случае фрагменты антигена могут быть присоединены к другим веществам, в частности, полипептидам, объединенных или ковалентно связанных, для использования в качестве иммуногенов. Антиген и его фрагменты могут быть объединены или ковалентно сшиты с различными иммуногенами, такими как гемоцианин лимфы улитки, бычий сывороточный альбумин, столбнячный анатоксин и т.д. см. Microbiology. Hoeber Medical Division, Harper and Row, 1969; Landsteiner (1962) Specificity of Serological Reactions. Dover Publications, New York; Williams, et al. (1967) Methods in Immunology and Immunochemistrv, vol. 1, Academic Press, New York; и Harlow and Lane (1988) Antibodies: A Laboratory Manual. CSH Press, NY, для описания способов приготовления поликлональных антисывороток.
В некоторых случаях желательно получить моноклональные антитела от различных млекопитающих, таких как мыши, грызуны, приматы, человек и т.д. Описание способов для приготовления таких моноклональных антител можно найти, например, в Stites, et al. (eds.) Basic and Clinical Immunology (4th ed.), Lange Medical Publications, Los Altos, Calif, в ссылках, цитируемых здесь (Harlow and Lane (1988) Antibodies: A Laboratory Manual. CSH Press; Coding (1986) Monoclonal Antibodies: Principles and Practice (2d ed.), Academic Press, New York) и особенно в Kohler and Milstein (1975) in Nature 256:495-497, которые описывают один способ получения моноклональных антител.
Другие подходящие способы включают в себя in vitro представление лимфоцитов антигенным полипептидам для выбора библиотеки антител в фагах или похожих векторах (см. Huse, et al. (1989) "Generation of a Large Combinatorial Library of the Immunoglobulin Repertoire in Phage Lambda," Science 246:1275-1281; и Ward, et al. (1989) Nature 341:544-546.]). Полипептиды и антитела настоящего изобретения могут быть использованы как с, так и без модификации, включающую в себя получение химерных антител, «особачивание» или гуманизацию антител.
Часто полипептиды и антитела настоящего изобретения метят соединениями, которые обеспечивают детектируемый сигнал. Такое присоединение может производиться как ковалентно, так и нековалентно. Известно большое количество меток и способов конъюгирования и о них многое сообщается и в научной, и в патентной литературе. Подходящие метки включают в себя радионуклиды, ферменты, субстраты, кофакторы, ингибиторы, флуоресцентные участки, хемилюминесцентные участки, магнитные частицы и тому подобное. Патенты, раскрывающие использование таких меток, включают в себя патенты США №№3,817,837; 3,850,752; 3,939,350; 3,996,345; 4,277,437; 4,275,149; и 4,366,241. Также рекомбинантные или химерные иммуноглобулины могут быть получены по Cabilly, патенту США No. 4,816,567; Moore, et al., патенту США No. 4,642,334; и Queen, et al. (1989) Proc. NaVI Acad. Sci. USA 86:10029-10033; или получены в трансгенных мышах, см. Mendez, et al. (1997) Nature Genetics Vx 146-156. Эти ссылки включены здесь в виде ссылок.
Антитела настоящего изобретения могут также использоваться для аффинной хроматографии при выделении белков. Колонки могут быть приготовлены, когда антитела связаны с твердой подложкой (см. Wilchek et al. (1984) Meth. Enzymol. 104:3-55). В качестве альтернативы, антитела, связанные с твердой подложкой, могут быть использованы для очистки соответствующих антител.
Антитела, полученные к каждому собачьему ТСЛБ, могут также быть использованы для получения антиидиотипических антител. Эти антитела будут использоваться для детекции или диагностики различных иммунологических состояний, связанных с экспрессией соответствующего антигена.
Ингибирование РНК
Интерференция с РНК, кодирующей собачий ТСЛБ в клетках, продуцирующих собачий ТСЛБ, является дополнительным средством ингибирования биологической активности ТСЛБ и, следовательно, лечения ряда связанных с ТСЛБ заболеваний, таких как атопический дерматит. Для этой цели химически синтезированные или клонированные с помощью подходящего вектора для доставки, такого как плазмида или вирусный вектор, двухцепочечные молекулы РНК могут вводиться в клетку, активно продуцирующую мРНК ТСЛБ, с целью понижения уровня эндогенной мРНК, кодирующей ТСЛБ. После попадания в клетку таких молекул РНК (в случае экзогенно доставленных молекул или транскрипции РНК, которая следует за введением плазмиды или вирусного вектора в желаемые клетки), они процессируются посредством расщепляющей активности рибонуклеазы типа III на короткие нуклеотидные фрагменты, которые называют миРНК. Эти фрагменты миРНК затем включаются в содержащий нуклеазу мультипротеиновый комплекс, который называется RISC (РНК-индуцирующий «молчащий» комплекс), который приходит в активное состояние в результате раскручивания дуплекса миРНК при действии РНК-хеликазы. Потом одноцепочечные фрагменты миРНК направляют комплекс RISC к целевой мРНК, которая затем расщепляется и деградирует при действии эндонуклеазной активности RISC.
В более частном случае плазмиды, содержащие ген ТСЛБ или его фрагменты, клонируют в любой из ряда коммерчески доступных эукариотических плазмид, где транскрипция гена ТСЛБ или его фрагментов управляется соответствующим промотором, например, CMV или SV40. Очищенная плазмидная ДНК (1-100 мкг) затем иньецируется в место повреждения на коже или в области, окружающие повреждение кожи, характерное для атонического дерматита. Введение плазмидной ДНК может затем повторяться с частотой, необходимой для достижения значительного снижения мРНК ТСЛБ. Это снижение можно оценить получением биопсии кожи с пораженных участков и определения уровня мРНК ТСЛБ такими способами, как количественная ПЦР. Следующие примеры осуществления настоящего изобретения служат для обеспечения дальнейшего понимания изобретения, но не означают ни в коей мере ограничение объема изобретения.
ПРИМЕРЫ
Пример 1
ДНК и белковая последовательности собачьего ТСЛБ
Собачий ген, экспрессирующий собачий ТСЛБ, был идентифицирован с помощью способа последовательных итераций с использованием данных, найденных в электронных базах данных, и молекулярно-биологических способов, как описывается детально выше.
Результаты
Последовательность гена собачьего ТСЛБ иллюстрирует ФИГ.8А (SEQ ID NO:1) и предсказанный экспрессированный белок ТСЛБ проиллюстрирован ФИГ.8В (SEQ ID NO:2). Остатки 1-28 представляют собой сигнальную последовательность, а остатки 29-155 представляют зрелый белок.
Пример 2
Клонирование и экспрессия собачьего ТСЛБ
ДНК, кодирующая собачий ТСЛБ, была идентифицирована, как описано здесь, и клонирована в донорский вектор pDONR221 (Invitrogen Gateway System) стандартными способами в этой области. Отбор гена и клонирование его в донорский вектор было проведено исследовательской организацией, работающей по контракту, которая называется DNA 2.0. и привело к конструкции плазмиды, названной pDONR221.G03276, которая содержала в себе идентифицированный геномный ген собачьего ТСЛБ. ДНК, кодирующая зрелый (т.е. без сигнальной последовательности) собачий белок ТСЛБ была амплифицирована с помощью ПЦР из pDONR221.G03276 с использованием праймеров, которые содержали V сайты Nco I и EcoR, соответственно.
Праймеры
#1: 5’ AATAATCCATGGCATACAATTTCATTGACTGTGAC-3’ (SEQ ID NO:4); и #2: 5’-AAAATAGATATCTGAAATGCGACTGAAACGACG-3’ (SEQ ID NO:5).
После получения Nco I и EcoR V, ПЦР-продукты были вставлены в сайты Nco I и Sma I вектора pIVEX 1.3 WG (Roche Applied Sciences, Cat# 3728803). Это привело к конструкции плазмиды, содержащей ген, который кодирует зрелый собачий ТСЛБ, соединенный с шестью остатками гистидина на С-конце ("His6 хвост"). Плазмида, содержащая правильную последовательность вставки, была названа 1265-93.D. Плазмида 1265-93.D была использована для экспрессии ТСЛБ в RTS Proteomaster Instrument согласно рекомендациям производителя (Roche Applied Sciences, Cat# 3064859). Как показано на ФИГ.1, полоса в @16 кДА очевидна в дорожках 2 и 4 (стрелки). Анализ с помощью Вестерн-блоттинга (ФИГ.2А и 2В) показывает, что эта полоса специфически реагирует с антителом к «His хвосту» (ФИГ.2А) и с крысиным моноклональным антителом, специфичным к ТСЛБ человека (ФИГ.2В).
Пример 3
Получение собачьего ТСЛБ из клеток хозяина
Для того, чтобы экспрессировать рекомбинантный белок ТСЛБ в E.coli, нуклеотидная последовательность, кодирующая кТСЛБ (т.е. ТСЛБ без нуклеотидов, кодирующих сигнальную последовательность) была амплифицирована с помощью ПЦР с использованием плазмиды 1265-66С в качестве образца совместно с прямым и обратным праймерами, содержащими сайты NcoI и Hind III, соответственно.
Прямой праймер
5’-AATAATCCATGGCATACAATTTCATTGACTGTGAC-3’ (SEQ ID NO:6) Обратный праймер
5’-ACATAAAAGCTTTGAAATGCGACTGAAACGACG-3’ (SEQ ID NO:7)
После растворения Nco I и Hind III ПЦР-продукты вставляли в сайты NcoI/HindIII вектора для экспрессии pET42b(+) (Novagen). В результате этого процесса получали плазмиду, которая кодировала зрелый белок кТСЛБ, соединенный с GST-хвостом на N-конце и 6-His хвосте на С-конце. Плазмида, содержащая корректные последовательности вставки, была названа 1265-93В. Экспрессия белка слияния GST-TSLP-His проводилась в Е.coli BL21(DE3)/pLysS, которая содержала ген Т7 РНК-полимеразы под контролем изопропил-(бета)-D-тиогалактопиранозидного (ИПТГ)-индуцируемого промотора lacUV5. Клетки E.coli, несущие плазмиду 1265-93В, выращивали при 30°С до O.D. 600 из 0.6 и затем экспрессию белка индуцировали посредством добавления 0.5 мМ ИПТГ и далее инкубировали при температуре при 30°С в течение 2 часов. SDS-электрофорез в полиакриламидном геле выявил полосу белка (стрелка) с корректной массой (~61 кДа), присутствующего в растворимой фракции Е.coli (ФИГ.3А). Анализ с помощью Вестерн-блоттинга показал, что экспрессируемый белок реагирует с антителом к GST (Фиг.3D). Белок GST-TSLP-His может быть очищен с помощью смолы Глутатион Сефароза 4В (ФИГ.3В). После дополнительной очистки на смоле Ni-NTA большее количество белка GST-TSLP содержится в элюате с колонки (ФИГ.3С).
Пример 4
Иммунофлуоресцентная детекция собачьего ТСЛБ
Экспрессию собачьего белка ТСЛБ в коже собаки и тканях миндалин определяли посредством иммуногистохимии (ИГХ), используя поликпональные антитела кролика к белку ТСЛБ человека. Иммуногистохимию проводили на образцах ткани в парафиновых блоках, полученных от здоровых собак, инъецированных солевым раствором также как и от собак с диагнозом, включающим в себя атопический дерматит, эритему в мультиформе, кожную красную волчанку и синаптический буллезный эпидермолиз. Дополнительно, экспрессия белка ТСЛБ была определена в замороженном образце ткани миндалины, полученной от двух собак. Процедура для определения экспрессии ТСЛБ способом ИГХ была следующей:
I. Подготовка образцов
1. Парафиновые блоки с заключенными в них образцами кожи были разрезаны на образцы толщиной 5-7 микрон и прикреплены к подложке, обработанной поли-L-лизином для того, чтобы вызвать адгезию.
2. Секции депарафинизировали с помощью ксилена и регидратировали с помощью серии этанольных растворов.
3. Восстановление антигена проводили в цитратном буфере [10 мМ цитрата натрия, содержащего Tween-20 в концентрации 0.5 мл/л в течение 25 мин с использованием лабораторной микроволновой установки для достижения 99-100°С]. Это процесс, который восстанавливает антигенность секций ткани, которые закрываются в течение процесса заливания в парафин.
II. Иммуноокрашивание:
1. Секции инкубировали в 10% нормальной ослиной сыворотке, разведенной в фосфатном буфере (PBS), на 1 час при комнатной температуре для неспецифического связывания с антителом.
2. Избыток сыворотки осторожно удаляли и секции обрабатывали кроличьим антителом (1:100), разведенном в PBS затем инкубировали либо при комнатной температуре в течение 1 часа или в течение ночи при 100°С во влажной атмосфере.
3. Секции затем промывали дважды в течение 5 мин в PBS, затем осторожно встряхивали.
4. Избыток PBS затем осторожно удаляли и секции обрабатывали биотинилированными ослиными антителами к IgG кролика, разведенными 1:5000 в PBS в течение 30 мин при комнатной температуре во влажной атмосфере.
5. Секции затем дважды промывали PBS, осторожно встряхивая.
6. Избыток PBS удаляли и секции инкубировали в течение 30 мин при комнатной температуре с конъюгатом Стрептавидин-флуорескаин изозотиоцианат (Стрептавидин-FITC) в PBS при концентрации 5 микрограмм/мл.
7. Секции затем промывали дважды в течение 5 мин в PBS, при осторожном встряхивании.
8. Секции затем окрашивали второй раз гемотоксилином в течение 2-3 мин.
9. Секции затем исследовали под флуоресцентным микроскопом.
10. Подходящие изображения фотографировали.
11. Контроли эксперимента включали в себя пропуск первичных антител к ТСЛБ, или замещение первичных антител к ТСЛБ нормальными антителами кролика.
Таблица 1 внизу объединяет результаты экспериментов с ИГХ.
Экспрессия ТСЛБ была отмечена в 80% образцов кожной ткани от собак с диагнозом АД, но только в 20% нормальных кожных тканей, в которые произвели инъекцию солевого раствора. ТСЛБ также был обнаружен в 66% и 100% ткани собак с эритемой в мультиформе, и собак с генетическим заболеванием кожи, синаптическим буллезным эпидермолизом, соответственно. Не наблюдалось экспрессии белка ТСЛБ в кожной ткани собак с кожной красной волчанкой. В образцах кожной ткани, залитым парафином, экспрессия ТСЛБ была отмечена в потовых железах. Экспрессия ТСЛБ в замороженной ткани миндалины была обнаружена в наслоенном чешуйчатом эпителии и была связана со слюнными железами. Пример положительного ИГХ окрашивания образцов ткани собак показан на ФИГ.4, которая представляет образцы кожной ткани, запаянные в парафин, от собак с диагнозом «атопический дерматит».
Пример 6
Иммунопероксидазная детекция собачьего ТСЛБ
Экспрессия собачьего белка ТСЛБ в блоках, запаянных в парафин и приготовленных из кожи собак с диагнозом «атопический дерматит» была также определена иммуногистохимически при использовании окрашивания иммунопероксидазой в качестве способа детекции. В этом способе эпитоп-специфичное моноклональное антитело к белку ТСЛБ человека было использовано в качестве первичного антитела. Процедура определения экспрессии ТСЛБ с помощью окрашивания иммунопероксидазой была следующей:
Специальные реагенты:
Нормальная сыворотка новорожденных телят: #N-4762 Sigma
Запаянная в парафин ткань
Первичные антитела: крысиное монокпональное антитело IgG2a к ТСЛБ человека
Вторичные антитела: кроличьи биотинилированные антитела к IgG человека: ВА-4000 Vector Lab, Burlingame, CA.
Реагент для детекции: Стрептавидин-HRP: #43-8323 Zymed Labs. San Francisco, CA
Набор на основе АЕС: Biogenex #HK129-5K San Ramon, CA
1. Отбирали образцы 4-6 мкл.
2. Высушивали на воздухе 10 мин при комнатной температуре.
3. 10 мин фиксировали ацетоном.
4. Промывали в PBS (0.01 фосфатно-солевой буфет) 3 мин.
5. Ингибировали с помощью инкубации в 0.3% пероксиде водорода с 0.1% азидом натрия в течение 7-10 мин.
6. Промывали в течение 5 мин PBS.
7. Блокировали секции с помощью 1% нормальной сыворотки новорожденных телят в течение 20 мин во влажной камере.
8. Удаляли со срезов жидкость и действовали первичными антителами в разведении 1:100 в течение 2 часов при комнатной температуре.
9. Промывали 5 мин.
10. Действовали вторичными антителами (кроличьи антитела к крысиному IgG в разведении 1:400) в течение 30 мин во влажной камере при комнатной температуре.
11. Промывали 5 мин.
12. Удаляли жидкость и действовали реагентом для детекции (Стрептавидин-HRP в разведении 1:400) в течение 30 мин при комнатной температуре.
13. Промывали 2Х5 мин.
14. Действовали АЕС 2.5 мин. Выравнивали в соответствии с желаемой интенсивностью окраски и фоновым окрашиванием.
15. Окрашивали вторично при помощи Гематоксилина и переносили на подложку. Выборка из кожной ткани от собак с АД была исследована с помощью ИГХ с использованием эпитопа специфичного собачьего антитела к ТСЛБ. Результаты, представленные на ФИГ.5, показывают, что это антитело реагирует с молекулой, которая делит антигенные эпитопы с ТСЛБ человека. Режимы окрашивания образцов кожи собак с АД были строгими в областях хронического воспаления, где эпидермис утолщается. Эта модель совпадает с тем, что известно о локализации экспрессии ТСЛБ при повреждениях кожи у человека и далее предполагает, что распознанная молекула при повреждениях кожи у собак - это собачий ТСЛБ. Не наблюдалось окрашивания при обработке PBS или при использовании другого крысиного моноклонального тела к другому белку (белок из лимфоцитов).
Пример 7
Картирование эпитопов собачьего ТСЛБ
Для того, чтобы идентифицировать эпитопы в собачьем ТСЛБ, которые полезны для включения в вакцину, способную нейтрализовать активность ТСЛБ, была синтезирована выборка перекрывающихся пептидов, основанных на последовательности собачьего белка ТСЛ, и исследована на наличие способности реагировать, нейтрализуя моноклональное тело к ТСЛБ человека. С этой целью была синтезирована выборка перекрывающихся пептидов, из 15 аминокислотных остатков каждый и заканчивающихся 2 аминокислотами для образования шпильки, фирмой MIMOTOPES (Minneapolis, MN). Последовательности этих пептидов перечислены в Таблице 2. Пептиды 1-57 были синтезированы с аминированным концом в конфигурации NH2-ПЕПТИД-ШПИЛЬКА. Пептиды 58-94 (дубликаты родственных пептидов 1-37) были синтезированы с ацетилированным концом в конфигурации АЦЕТИЛ-ПЕПТИД-ШПИЛЬКА.
Шпильки, несущие пептид, перечисленные в Таблице 2, были исследованы с помощью ИФА в соответствии с методиками, рекомендованными производителем (Mimotopes, Minneapolis, MN). Как показано на ФИГ. 6, пептид 25 (эпитоп 25) с аминокислотной последовательностью NH2-ARIERLTLHRIRGCA (SEQ ID NO:32) имел самую большую реакционную способность по отношению к моноклональному антителу РАВ100. Сравнение последовательности этого пептида с соответствующей предполагаемой последовательностью пептида ТСЛБ человека показано на ФИГ.7.
Настоящее изобретение не должно быть ограничено рамками конкретных вариантов выполнения, описанных здесь. Действительно, различные модификации изобретения в дополнение к описанным здесь будут очевидны для специалистов в данной области техники исходя из предшествующего описания. Такие модификации планируется включить в рамки приложенной формулы изобретения.
Далее необходимо понять, что все размеры оснований или размеры аминокислот, все значения молекулярных весов или молекулярных масс, данные для нуклеиновых кислот или полипептидов, являются приближенными, и предлагаются для описания.
Здесь цитируются различные публикации, описания которых включены здесь в виде полной ссылки.


























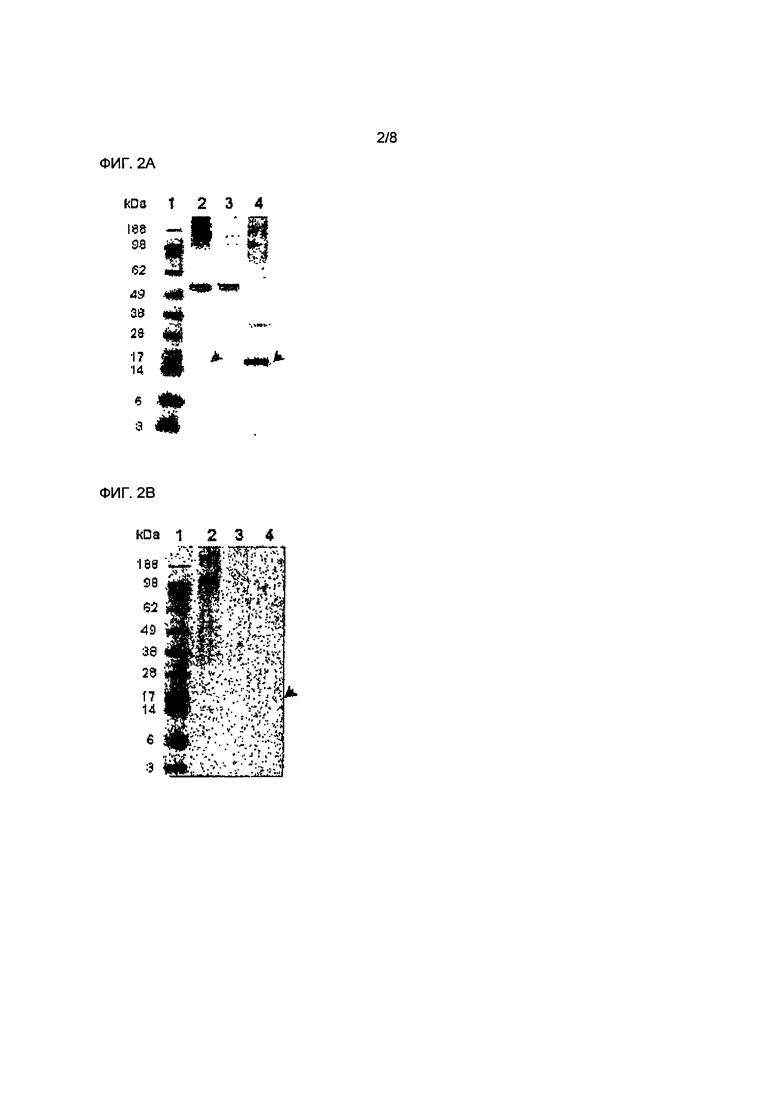



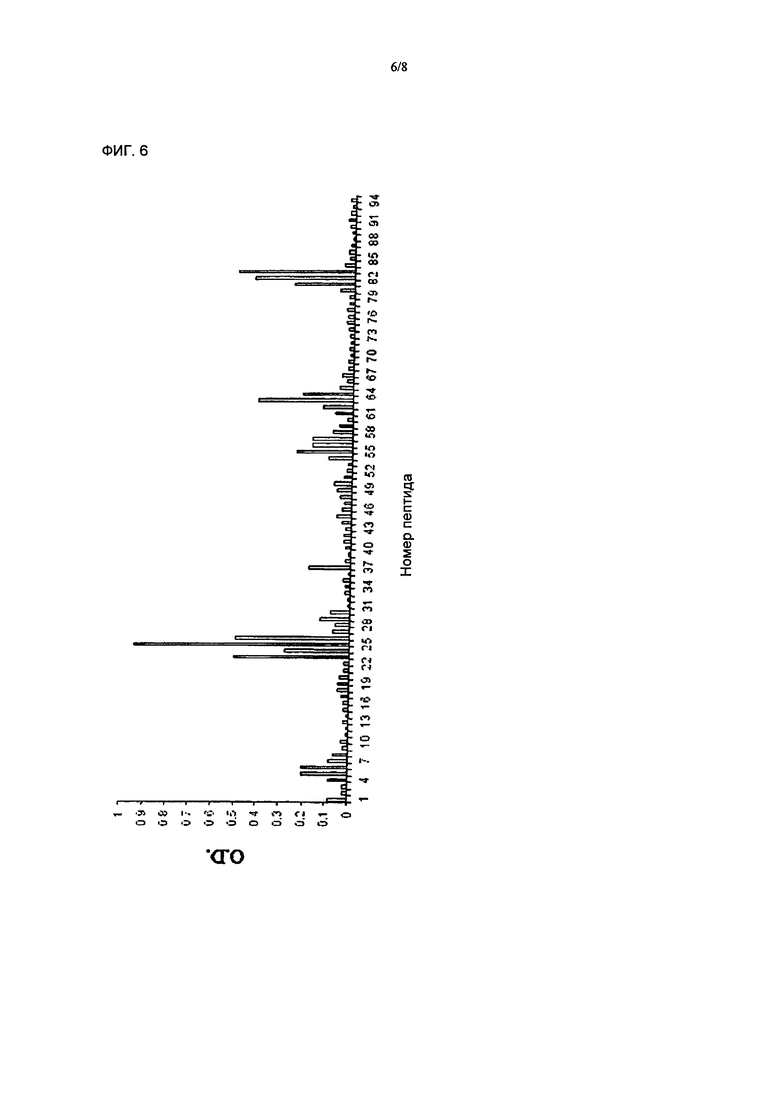


Настоящее изобретение относится к области биотехнологии и может быть использовано для получения антитела против собачьего тимусного стромального лимфопоэтического белка (ТСЛБ). Антитело против ТСЛБ получают путем иммунизации млекопитающего ТСЛБ или его антигенным фрагментом, включающим антителосвязывающий эпитоп собачьего ТСЛБ. Изобретение позволяет получить блокирующую пролиферацию антитела против собачьего ТСЛБ. 2 з.п. ф-лы, 8 ил., 2 табл., 7 пр.
1. Выделенное антитело против собачьего тимусного стромального лимфопоэтического белка (ТСЛБ), полученное путем иммунизации млекопитающего ТСЛБ или его антигенным фрагментом, где указанный ТСЛБ имеет аминокислотную последовательность, которая имеет 90% или более идентичность с аминокислотной последовательностью SEQ ID NO: 2, за исключением сигнальной последовательности из 28 аминокислотных остатков, где указанный ТСЛБ или его антигенный фрагмент включают антителосвязывающий эпитоп собачьего ТСЛБ.
2. Антитело против собачьего ТСЛБ по п. 1, где антитело представляет собой моноклональное антитело.
3. Антитело против собачьего ТСЛБ по п. 2, где моноклональное антитело является канинизированным (caninized).
| Междуэтажное перекрытие из заранее изготовленных железобетонных | 1932 |
|
SU29581A1 |
| LIU Y.J., Thymic stromal lymphopoietin: master switch for allergic inflammation, J | |||
| Exp | |||
| Med., 2006, v.203, n.2,p.269-273 | |||
| АНТИИДИОТИПИЧЕСКОЕ АНТИТЕЛО К АНТИТЕЛАМ, ИНГИБИРУЮЩИМ СВЯЗЫВАНИЕ ИММУНОГЛОБУЛИНА С ЕГО ВЫСОКОАФФИННЫМ РЕЦЕПТОРОМ | 2000 |
|
RU2253655C2 |

