Область техники изобретения
Данное изобретение относится главным образом к области молекулярной биологии. Точнее, данное изобретение имеет отношение к молекулярному клонированию и характеристике генов гомологичных белков Ehrlichia canis в 28-кДа, мультигенному локусу, кодирующему гомологичные белки Ehrlichia canis в 28-кДа, и их применению.
Описание родственной области
Эрлихиоз собак, также известный как тропическая пангемоцитопения собак, представляет собой переносящееся клещами риккетсионное заболевание собак, впервые описанное в Африке в 1935 году и Соединенных Штатах в 1963 году (Donatien and Lestoquard, 1935; Ewing, 1963). Заболевание стало лучше распознаваться после его эпизоотической вспышки, имевшей место среди американских военных собак во время войны во Вьетнаме (Walker et al., 1970).
Этиологическим агентом эрлихиоза собак является Ehrlichia canis, небольшая грамотрицательная, облигатная внутриклеточная бактерия, которая проявляет тропизм в отношении мононуклеарных фагоцитов (Nyindo et al., 1971) и переносится клещом коричневых собак, Rhipicephalus sanguineus (Groves et al., 1975). При развитии эрлихиоза собак возможно проявление трех форм - острой, бессимптомной и хронической. Острая форма характеризуется лихорадкой, анорексией, депрессией, лимфаденопатией и небольшой тромбоцитопенией (Troy and Forrester, 1990). Обычно после острой формы собаки выздоравливают, но остаются персистентно инфицированными носителями возбудителя без клинических признаков заболевания в течение месяцев и даже лет (Harrus et al., 1998). В некоторых случаях развивается хроническая форма, которая проявляется тромбоцитопенией, гиперглобулинемией, анорексией, истощением и кровотечением, в частности носовым кровотечением, с последующей смертью (Troy and Forrester, 1990).
Регуляция поверхностной антигенности может быть важным фактором механизма установления таких персистентных инфекций у хозяина. Хотя патогенез заболевания недостаточно выяснен, мультигенные семейства, описанные у представителей родственных родов Ehrlichia, Anasplasma и Cowadria, могут быть вовлечены в изменчивость экспрессии главного поверхностного антигена, что помогает, таким образом, избежать иммунного контроля. Anaplasma marginale, микроорганизм, близко родственный Е. canis, проявляет изменчивость генов основного поверхностного белка-3 (msp-3), что приводит к антигенному полиморфизму среди штаммов (Alleman et al., 1997).
Молекулярный таксономический анализ на основании гена рРНК 16S показал, что Е. canis и E.chaffeensis, этиологические возбудители моноцитарного эрлихиоза человека (НМЕ), являются близкородственными (Anderson et al., 1991; Anderson et al., 1992; Dawson et al., 1991; Chen et al., 1994). Описана значительная перекрестная реактивность антигенов 64, 47, 40, 30, 29 и 23-кДА между Е. canis и E.chaffeensis (Chen et al., 1994; Chen et al., 1997; Rikihisa et al., 1994; Rikihisa et al., 1992). Анализ иммунореактивных антигенов посредством иммуноблота с использованием сыворотки человека и собаки, взятой в период выздоровления, позволил идентифицировать многочисленные иммунодоминантные белки Е. canis, включая белок в 30-кДа (Chen et al., 1997). Кроме того, белок Е. canis в 30-кДа был описан как главный иммунодоминантный антиген, рано распознаваемый в иммунном ответе, который отличается по антигенным свойствам от белка Е. chaffeensis в 30-кДа (Rikihisa et al., 1992; Rikihisa et al., 1994). Также были идентифицированы другие иммунодоминантные белки Е. canis с молекулярными массами, колеблющимися от 20 до 30-кДа (Brouqui et al., 1992; Nyindo et al., 1991; Chen et al., 1994; Chen et al., 1997).
Гомологичные иммунодоминантные белки 28-32-кДа, кодируемые мультигенными семействами, были описаны у родственных микроорганизмов, включая Е. chaffeensis и Cowdria ruminantium (Sulsona et al., 1999; Ohashi et al., 1998a; Reddy et al., 1998). Недавно были описаны свойства 21-членного мультигенного семейства, кодирующего белки Е. chaffeensis в 23-28-кДа (Yu et al., 2000). Белки внешней мембраны E.chaffeensis в 28-кДа экспонированы на поверхности и содержат три основные гипервариабельные области (Ohashi et al., 1998a). Оказалось, что рекомбинантный Р28 E.chaffeensis обеспечивает защиту против введенной соответствующей инфекции у мышей, а антисыворотка, полученная против рекомбинантного белка, перекрестно реагирует с белком E.canis в 30-кДа (Ohashi et al., 1998a). Описано многообразие гена р28 среди изолятов E.chaffeensis (Yu et al., 1999a), а исследования с использованием моноклональных антител, кроме того, выявили многообразие в экспрессированных белках Р28 (Yu et al., 1993). Наоборот, описана полная консервативность генов р28 у географически различных изолятов E.canis, и высказано предположение, что в Северной Америке E.canis могут быть консервативными (McBride et al., 1999, 2000).
Предыдущий уровень исследований оказался несовершенным из-за отсутствия данных по клонированию и характеристике новых генов гомологичных иммунореактивных белков Ehrlichia canis в 28-кДа и единого мультигенного локуса, содержащего гены гомологичных белков 28-кДа. Кроме того, предыдущий уровень исследований является несовершенным из-за отсутствия рекомбинантных белков таких иммунореактивных генов Ehrlichia canis. Данное изобретение восполняет эту давнишнюю потребность в данной области.
КРАТКОЕ ИЗЛОЖЕНИЕ ИЗОБРЕТЕНИЯ
Некоторые аспекты согласно изобретению связаны с описанием молекулярного клонирования, секвенирования, свойств и экспрессии генов гомологичных зрелых иммунореактивных белков Ehrlichia canis в 28-кДа (обозначеных р28-1, -2, -3, -5, -6, -7, -9) и идентификацией единого локуса (10677-пар оснований), содержащего девять генов белков Ehrlichia canis в 28-кДа (р28-1 - р28-9). Восемь генов р28 локализованы на одной цепи ДНК, а один ген р28 обнаружен на комплементарной цепи. Гомология нуклеиновых кислот среди девяти представителей генов р28 составляла 37-75%, а гомология аминокислотных последовательностей колебалась от 28 до 72%.
Один из аспектов согласно изобретению связан с последовательностями ДНК, кодирующими иммунореактивный белок Ehrlichia canis в 30-кДа. Предпочтительно, белок имеет аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID No. 2, 4, 6, 40, 42, 44, 46, а ген имеет последовательность нуклеиновых кислот, выбранную из группы, состоящей из SEQ ID No. 1, 3, 5, 39, 41, 43, 45, и является представителем полиморфного, многочисленного семейства генов. Обычно белок имеет N-концевую сигнальную последовательность, которая может отщепляться в результате посттрансляционного процессинга, что приводит к образованию зрелого белка 28-кДа. Кроме того, гены, кодирующие белки 28-кДа, предпочтительно находятся в одном мультигенном локусе, который имеет размер 10677 пар оснований и кодирует девять гомологичных белков Ehrlichia canis в 28-кДа.
В другом аспекте данное изобретение связано с экспрессирующим вектором, содержащим ген, кодирующий иммунореактивный белок Ehrlichia canis в 28-кДа, и способным экспрессировать ген, когда вектор вводят в клетку.
Еще в одном аспекте согласно изобретению представлен рекомбинантный белок, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID No. 2, 4, 6, 40, 42, 44 и 46. Предпочтительно, если аминокислотная последовательность кодируется последовательностью нуклеиновых кислот, выбранной из группы, состоящей из SEQ ID No. 1, 3, 5, 39, 41, 43, 45. Предпочтительно, чтобы рекомбинантный белок содержал четыре вариабельные области, которые могли бы быть гидрофильными, антигенными и экспонированными на поверхности. Рекомбинантный белок может быть использован как антиген.
В другом аспекте согласно изобретению представлен способ получения рекомбинантного белка, включающий в себя стадии получения вектора, который содержит экспрессирующую область, содержащую последовательность, которая кодирует аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID No. 2, 4, 6, 40, 42, 44 и 46, прочно связанную с промотором; трансфекции вектора в клетку; и культивирования клетки в условиях, эффективных для экспрессии экспрессирующей области.
Изобретение также может быть описано в отдельных аспектах как способ ингибирования инфекции Ehrlichia canis у субъекта, включающий в себя стадии идентификации субъекта до воздействия Ehrlichia canis или субъекта, подозреваемого в том, что он подвергся воздействию Ehrlichia canis или инфицирован Ehrlichia canis; и введения композиции, содержащей антиген Ehrlichia canis в 28-кДа в количестве, эффективном для ингибирования инфекции Ehrlichia canis. Ингибирование можно осуществлять любыми способами, такими, например, как стимуляция гуморального или клеточного иммунных ответов субъекта, или посредством других способов, таких как ингибирование нормальной функции антигена 28-кДа или даже конкурирования с антигеном за взаимодействие с каким-либо агентом в организме субъекта.
Другие и дальнейшие аспекты, особенности и преимущества согласно изобретению станут очевидными из следующего описания предпочтительных аспектов изобретения, представленных с целью объяснения.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Для того чтобы основной предмет обсуждения, в котором проясняются вышеизложенные особенности, преимущества и цели изобретения, а также другие вопросы, был бы установлен и понят в деталях, более подробные описания изобретения, кратко суммированные выше, могут быть представлены посредством упоминаний определенных его аспектов, которые проиллюстрированы прилагаемыми чертежами. Эти чертежи составляют часть описания. Однако следует отметить, что прилагаемые чертежи иллюстрируют предпочтительные аспекты изобретения и поэтому не предназначены для его ограничения.
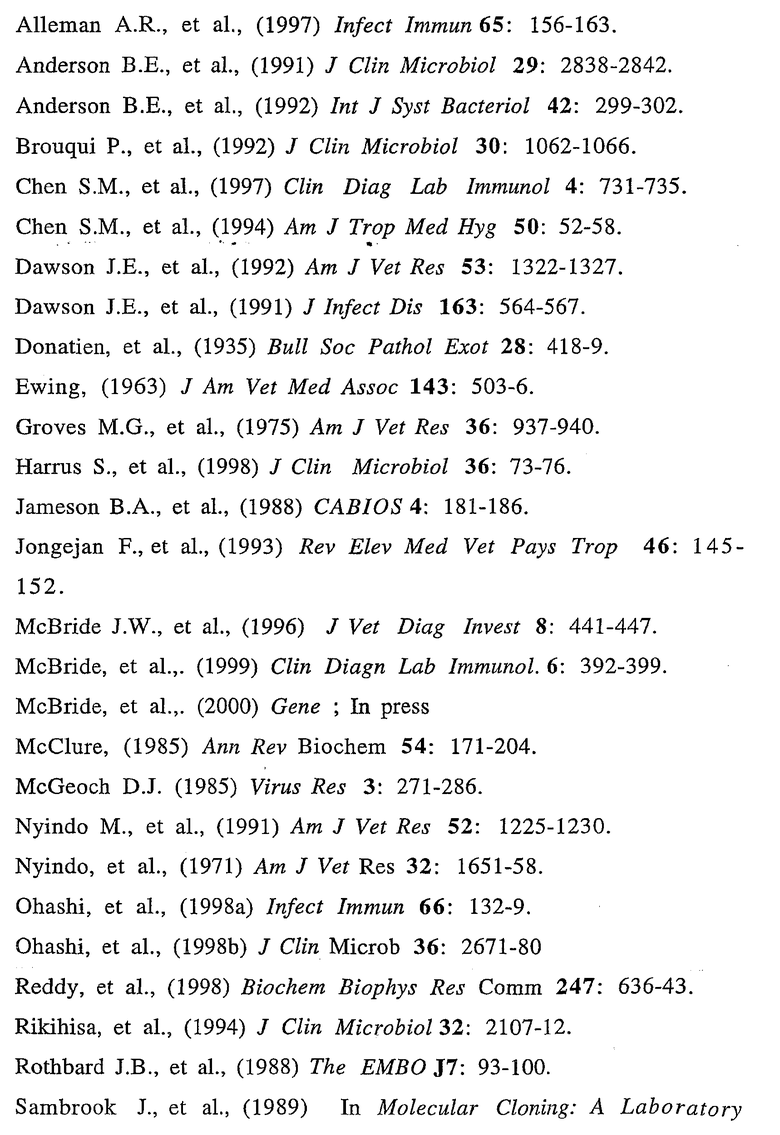
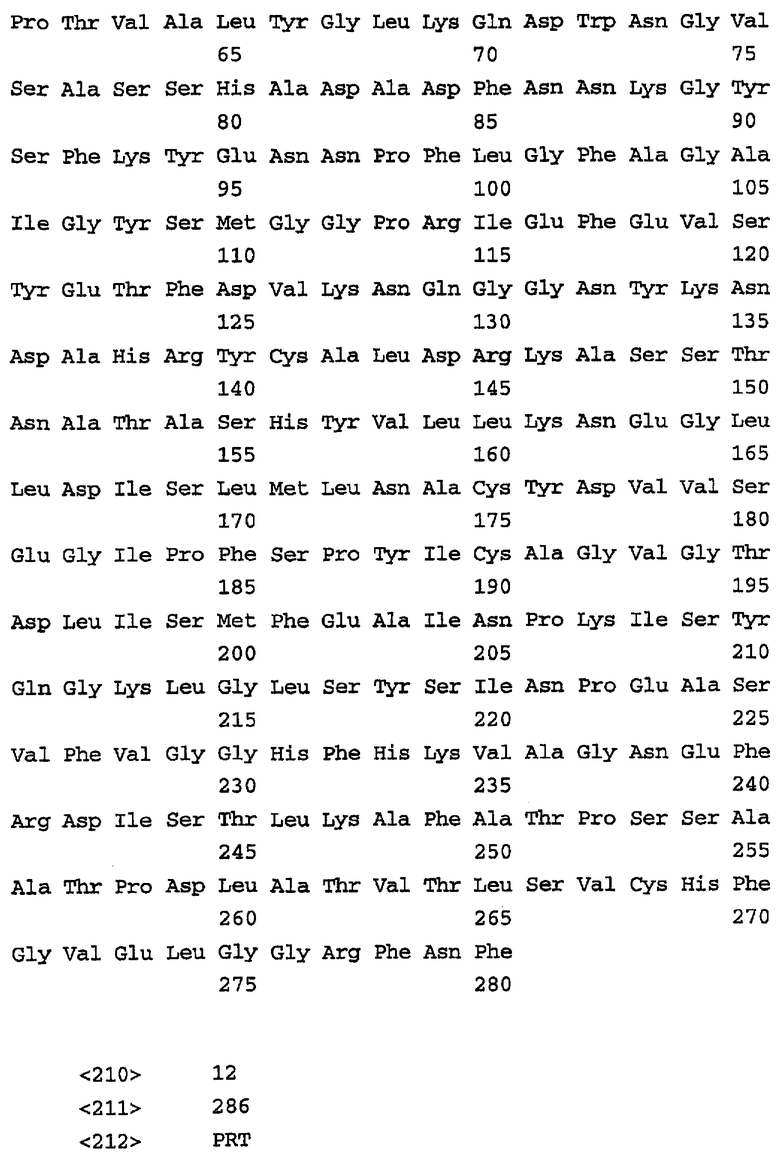
На фиг.1 представлена последовательность нуклеиновых кислот (SEQ ID No. 1) и выведенная аминокислотная последовательность (SEQ ID No. 2) гена р28-7, включая смежные 5' и 3' некодирующие последовательности. Инициирующий кодон ATG и терминирующий кодон ТАА отмечены жирным шрифтом, а 23-членная аминокислотная лидерная сигнальная последовательность подчеркнута.
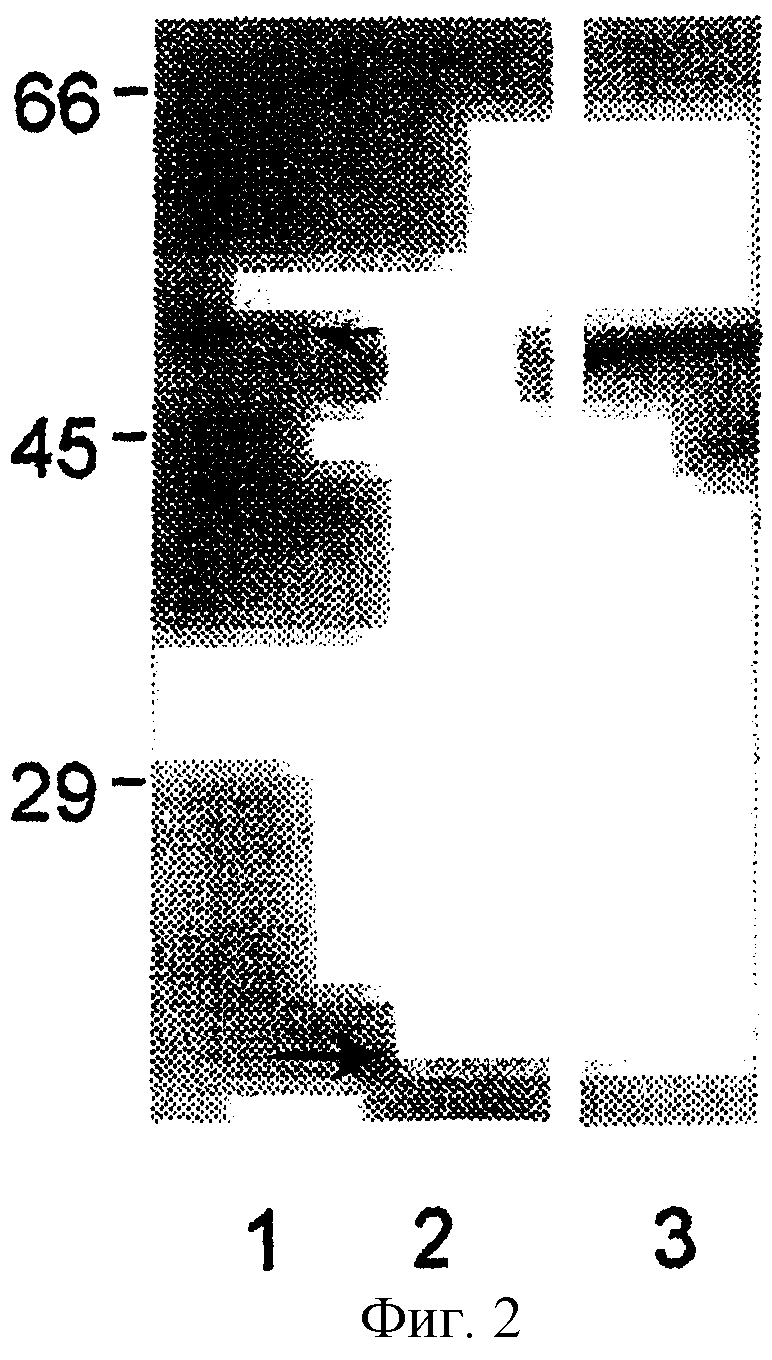
На фиг.2 показан ЭФ в ПААГ в присутствии ДДС экспрессированного рекомбинантного слитого с р28-7-тиоредоксином белка в 50 кДа (дорожка 1, стрелка) и контроля тиоредоксина 16-кДа (дорожка 2, стрелка) и соответствующий иммуноблот рекомбинантного слитого с р28-7-тиоредоксином белка, распознаваемого анти-Е. canis сывороткой собаки, полученной в период выздоровления (дорожка 3). Тиоредоксиновый контроль не детектировался антисывороткой Е. canis (не показано).
На фиг.3 представлен сравнительный анализ аминокислотных последовательностей белка р28-7 (ЕCа28-1, SEQ ID No. 2), белка р28-5 (ЕCа28SA2, частичная последовательность, SEQ ID No. 7), белка р28-4 (ЕCа28SA1, SEQ ID No. 8), E.chaffeensis P28 (SEQ ID No.9), OMP-1 семейства E.chaffeensis (SEQ ID Nos: 10-14) и белка MAP-1 C.ruminantium (SEQ ID No. 15). Аминокислотная последовательность р28-7 представлена как консенсусная последовательность. Не представленные аминокислоты идентичны р28-7 и изображены точками. Дивергентные аминокислоты отмечены посредством соответствующей однобуквенной аббревиатуры. Промежутки, введенные для максимального выравнивания аминокислотных последовательностей, указаны в виде тире. Вариабельные области подчеркнуты и обозначены (VR1, VR2, VR3 и VR4). Стрелки указывают на предсказанный участок расщепления сигнальной пептидазой на сигнальном пептиде.
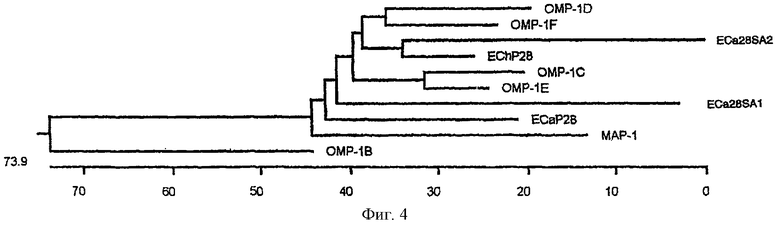
На фиг.4 схематически изображено филогенетическое родство р28-7 (ЕСа28-1), р28-5 (ЕСа28SA2, частичная последовательность), р28-4 (ЕСа28SA1) E.canis, представителей многочисленного семейства генов omp-1 E.chaffeensis, и белка map-1 C.rumanintium с помощью несбалансированной конструкции древа. Длина каждой пары ветвей отражает расстояние между аминокислотной последовательностью пар. Линейка дает возможность оценить расстояние между последовательностями.
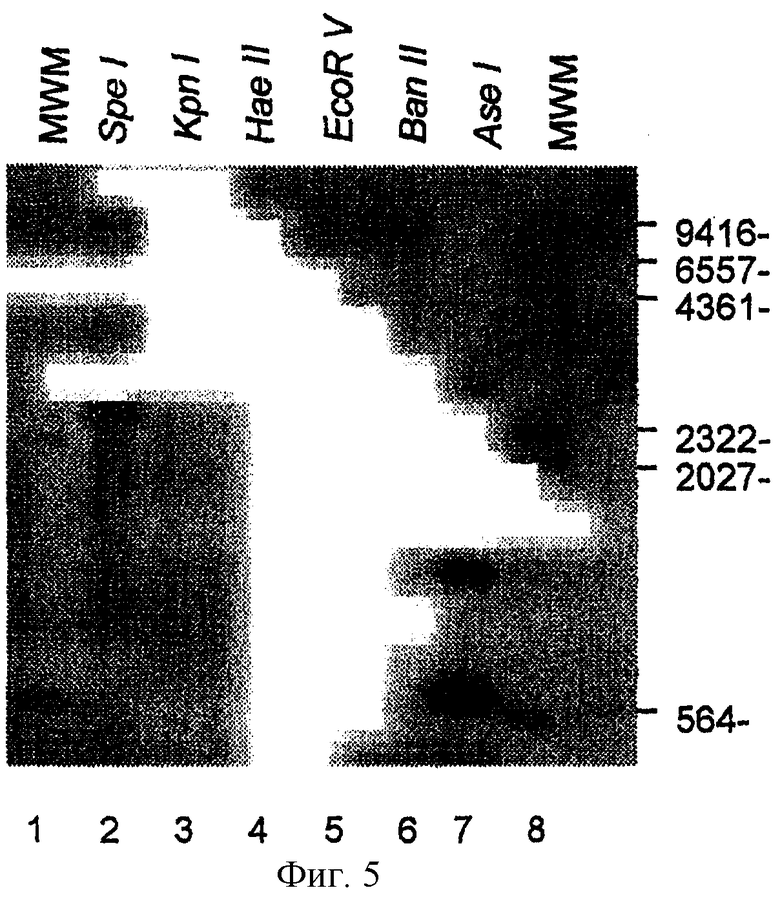
На фиг.5 представлены данные блоттинг-анализа по Саузерну геномной ДНК E.canis, полностью расщепленной шестью индивидуальными рестрицирующими ферментами и гибридизованной с DIG-меченным зондом р28-7 (дорожки 2-7); DIG-меченные метчики молекулярных масс (дорожки 1 и 8).
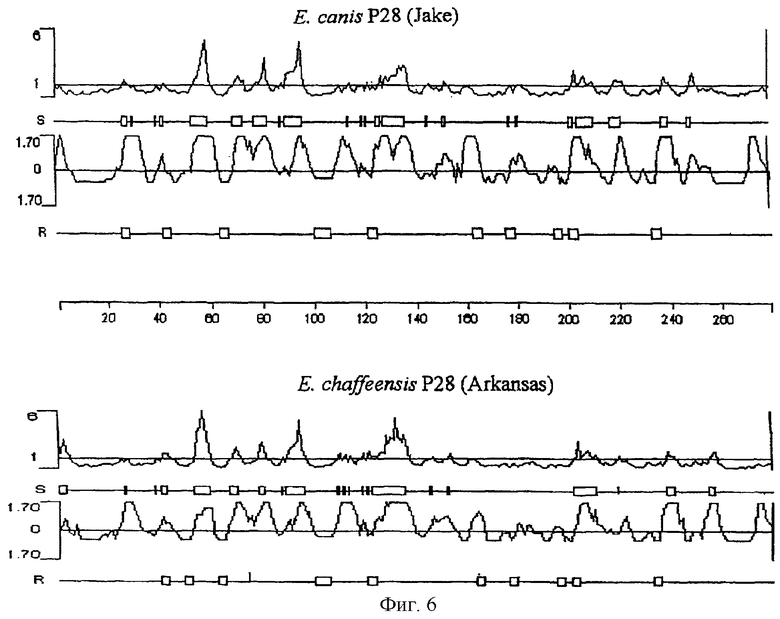
На фиг.6 представлены данные сравнения предсказанных свойств белков р28-7 (ЕCа28-1, штамм Jake) E.canis и Р28 E.chaffeensis (штамм Arkansas). Поверхностная вероятность предсказывает поверхностные остатки посредством использования окна гексапептида. Поверхностным остатком является любой остаток с >2,0 нм2 водной, доступной области поверхности. Гексапептид со значением больше 1 рассматривали как поверхностную область. Антигенный индекс предсказывает антигенные детерминанты. Области со значением выше нуля являются возможными антигенными детерминантами. Т-клеточный мотив определяет местоположение возможных Т-клеточных антигенных детерминант по мотиву из 5 аминокислот с 1 - глициновым или полярным остатком, 2 -гидрофобным остатком, 3 - гидрофобным остатком, 4 - гидрофобным или пролиновым остатком и 5 - полярным или глициновым остатком. Линейка указывает положение аминокислот.
На фиг.7-1 и 7-2 представлены последовательности нуклеиновых кислот и выведенные аминокислотные последовательности генов белков E.canis в 28-кДа: р28-5 (нуклеотид 1-849: SEQ ID No. 3; аминокислотная последовательность: SEQ ID No. 4) и р28-6 (нуклеотид 1195-2031: SEQ ID No. 5; аминокислотная последовательность: SEQ ID No. 6), включая межгенные некодирующие последовательности (NC2, нуклеотид 850-1194: SEQ ID No. 31). Инициирующий кодон ATG и терминирующие кодоны обозначены жирным шрифтом.
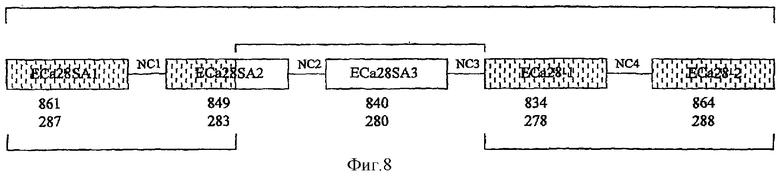
На фиг.8 представлено схематическое изображение локуса генов (5,592 Kb, содержащего пять генов) белков E.canis в 28-кДа, демонстрирующее генную ориентацию и межгенные некодирующие области (28NC1-4). Описаны гены белков 28-кДа, показанные в локусе 1 и 2 (затененные) (McBride et al., 1999; Reddy et al., 1998; Ohashi et al., 1998). Секвенирована полная последовательность р28-5 и последовательность нового гена белка 28-кДа, обозначенного р28-6. Некодирующими межгенными областями (28NC2-3) между р28-5, р28-6 и р28-7 завершено соединение ранее не связанных локусов 1 и 2.
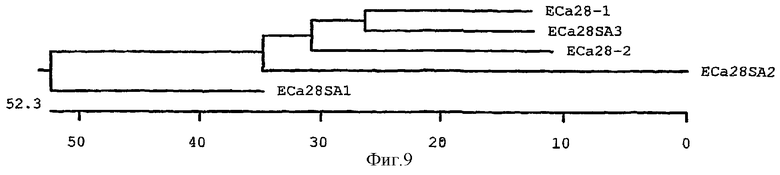
На фиг.9 схематически изображено филогенетическое родство гена р28-4 (EСa28SA1), p28-5 (EСa28SA2), p28-6 (EСa28SA3), p28-7 (EСa28-1) и p28-8 (EСa28-2) белков E.canis в 28-кДа на основании аминокислотных последовательностей с использованием неуравновешенной конструкции древа. Длина каждой пары ветвей изображает расстояние между аминокислотными парами. Линейка измеряет расстояние между последовательностями.
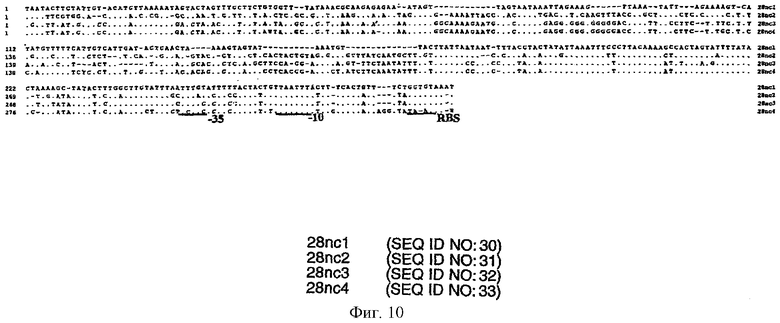
На фиг.10 представлены данные сравнительного анализа межгенных некодирующих последовательностей нуклеиновых кислот (SEQ ID Nos 30-33) генов белков E.canis в 28-кДа. Не представленные нуклеиновые кислоты, обозначенные точками (.), идентичны некодирующей области 1 (28NC1). Дивергенция обозначена с помощью соответствующей однобуквенной аббревиатуры. Введенные для максимального выравнивания аминокислотных последовательностей промежутки указаны в виде тире (-). Выделены предполагаемые транскрипционные промоторные области (-10 и -35) и рибосомный связывающий участок (RBS).
На фиг.11 представлено схематическое изображение локуса (10677-пар оснований) девяти генов р28 E.canis, демонстрирующее геномную ориентацию и межгенные некодирующие области. Гены р28 (р28-1, 2, 3, 9) (незатененные) идентифицированы в примере 8. Затененные гены р28 были идентифицированы ранее и обозначены следующим образом: р28-4. р30а (Ohashi et al., 1998b) и ORF1 (Reddy et al., 1998); р28-5 и р28-6 (McBride, et al., 2000); p28-7, p28 (McBride et al., 1999) и р30 (Ohashi et al., 1998b); и р28-8, р30-1 (Ohashi et al., 1998b).
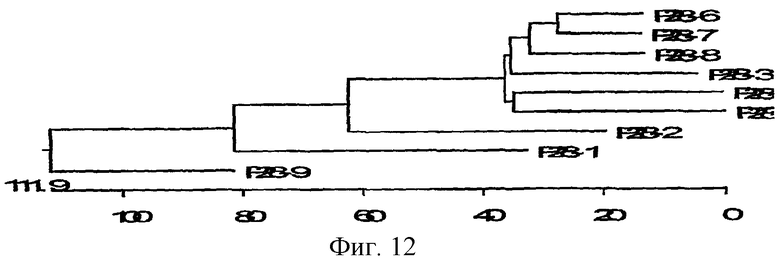
На фиг.12схематически изображено филогенетическое родство р28-1 - р28-9 E.canis на основании аминокислотных последовательностей. Длина каждой пары ветвей отражает расстояние между аминокислотными парами. Линейка измеряет процент расхождения между последовательностями.
На фиг.13представлены последовательность нуклеиновых кислот (SEQ ID No. 39) и выведенная аминокислотная последовательность (SEQ ID No. 40) гена р28-1 E.canis.
На фиг.14представлены последовательность нуклеиновых кислот (SEQ ID No. 41) и выведенная (на ее основании) аминокислотная последовательность (SEQ ID No. 42) гена р28-2 E.canis.
На фиг.15представлены последовательность нуклеиновых кислот (SEQ ID No. 43) и выведенная аминокислотная последовательность (SEQ ID No. 44) гена р28-3 E.canis.
На фиг.16представлены последовательность нуклеиновых кислот (SEQ ID No. 45) и выведенная аминокислотная последовательность (SEQ ID No. 46) гена р28-9 E.canis.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В данном изобретении описаны клонирование, секвенирование и экспрессия гомологичных генов, кодирующих белок в 30-килодальтон (кДа) Ehrlichia canis. Также проведен сравнительный молекулярный анализ гомологичных генов семи изолятов Е. canis и мультигенного семейства omp-1 E.chaffeensis. Некоторые новые гены белка 28-кДа идентифицированы следующим образом:
Р28-7 (ЕСа28-1) имеет открытую рамку считывания из 834-пар оснований, кодирующую белок из 278 аминокислот (SEQ ID No. 2) с предсказанной молекулярной массой в 30,5-кДа. Идентифицирована N-концевая сигнальная последовательность, указывающая, что белок подвергается посттрансляционной модификации в зрелый белок в 27,7-кДа.
Р28-6 (ЕСа28SA3) имеет открытую рамку считывания из 840-пар оснований, кодирующую белок из 280 аминокислот (SEQ ID No. 6).
Используя PCR (ПЦР) для амплификации генов белков E.canis в 28-кДа, завершили определение последовательности ранее несеквенированной области р28-5 (ЕСа28SA2). При анализе последовательности р28-5 обнаружена открытая рамка считывания, 849 пар оснований, кодирующая белок из 283 аминокислот (SEQ ID No. 4).
ПЦР-амплификация с использованием праймеров, специфичных для межгенных некодирующих областей генов белков 28-кДа, приводила к определению первичной структуры областей, соединяющих два ранее отдельных локуса, и тем самым идентифицировали единый локус (5592-kb), содержащий пять генов (р28-4, -5, -6, -7 и -8) белков 28-кДа. Было предсказано, что пять белков 28-кДа имеют сигнальные пептиды, удаление которых приводит к образованию зрелых белков, и имеют аминокислотную гомологию, колеблющуюся от 51 до 72%. Анализ межгенных областей выявил гипотетические промоторные области для каждого гена, что позволяет предположить, что эти гены могут независимо и дифференциально экспрессироваться. Величина межгенных некодирующих областей (28NC1-4) колеблется от 299 до 355 пар оснований, а гомология составляет 48-71%.
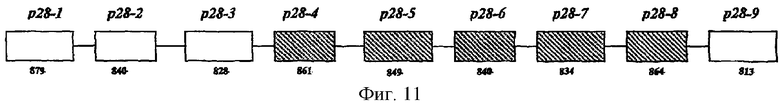
Кроме того, были секвенированы ранее неизвестные области ДНК в обратном и прямом направлении вышеупомянутого локуса, из пяти последовательно расположенных генов р28, и идентифицированы р28-1, -2, -3 и -9. Следовательно, в данном изобретении идентифицирован локус из девяти генов р28 E.canis, охватывающий 10677 пар оснований.
Данное изобретение, среди прочего, нацелено на изучение генов гомологичных белков р28 у Ehrlichia canis, р28-1, -2, -3, -6, -7 и р28-9 и определение полной последовательности частично секвенированного ранее р28-5. Также открытием является мультигенный локус, кодирующий девять гомологичных белков 28-кДа внешней мембраны Ehrlichia canis. Восемь из генов р28 локализованы на одной цепи ДНК, а один ген р28 обнаружен на комплементарной цепи. Гомология нуклеиновых кислот среди представителей девяти генов р28 составляет от 37 до 75%, а аминокислотная гомология колеблется от 28 до 72%.
Согласно данному изобретению специалисты в данной области могут применять общепринятые методы молекулярной биологии, микробиологии и методы рекомбинантных ДНК. Такие методы описаны в литературе в полном объеме. См., например, Maniatis, Fritsch & Sambrook, "Molecular Cloning: A Laboratory Manual (1982); "DNA Cloning: A Practical Approach," Volumes I and II (D.N. Glover ed. 1985); "Oligonucleotide Synthesis" (M.J. Gait ed. 1984); "Nucleic Acid Hybridization" [B.D. Hames & S.J. Higgins eds. (1985)]; "Transcription and Translation" [B.D. Hames & S.J. Higgins eds. (1984)]; "Animal Cell Culture" [R.I. Freshney, ed. (1986)]; "Immobilized Cells and Enzymes" [IRL Press, (1986)]; B. Perbal, "A Practical Guide to Molecural Cloning" (1984).
В изобретении предусмотрено получение по существу чистой ДНК, кодирующей иммунореактивный белок 28-кДа Ehrlichia canis. Белок, кодированный ДНК согласно изобретению, может иметь по меньшей мере 80% идентичности последовательности (предпочтительно 85%, более предпочтительно 90% и наиболее предпочтительно 95%) с аминокислотами, приведенными в SEQ ID No. 2, 4, 6, 40, 42, 44 или 46. Более предпочтительно, когда ДНК содержит кодирующую последовательность нуклеотидов SEQ ID No. 1, 3, 5, 39, 41, 43, 45 или вырожденный вариант такой последовательности.
В данной области хорошо известно, что аминокислотная последовательность белка определяется нуклеотидной последовательностью ДНК, которая кодирует белок. Вследствие вырожденности генетического кода (то есть большинство аминокислот кодируется более чем одним триплетом (кодоном) отдельные аминокислоты или полипептид могут кодироваться различными нуклеотидными последовательностями. Таким образом, полинуклеотидные последовательности рассматриваемого изобретения также содержат упомянутые вырожденные последовательности, которые кодируют полипептид согласно изобретению или его фрагмент или вариант.
Данное изобретение также включает в себя по существу чистую ДНК, содержащую последовательность по меньшей мере из 15 последовательных нуклеотидов (предпочтительно из 20, более предпочтительно из 30, еще более предпочтительно из 50 и наиболее предпочтительно из всех) из области нуклеотидов, приведенных в SEQ ID No. 1, 3, 5, 39, 41, 43 или 45.
Под выражением «по существу чистая ДНК» подразумевают ДНК, которая не является частью среды, в которой ДНК естественно встречается, благодаря отделению (частичная или полная очистка) от некоторых или всех молекул такой среды или благодаря перестройке последовательностей, которые фланкируют названную ДНК. Поэтому термин включает в себя, например, рекомбинантную ДНК, которую вводят в вектор, в автономно реплицирующуюся плазмиду или вирус, или в геномную ДНК прокариота или эукариота; или ДНК, которая существует как отдельная молекула (например, кДНК или фрагмент геномной или кДНК, полученный в результате полимеразно-цепной реакции (PCR; ПЦР) или расщепляется рестриктирующими эндонуклеазами), не зависящая от других последовательностей. Термин также включает в себя рекомбинантную ДНК, которая является частью гибридного гена, кодирующего дополнительную полипептидную последовательность, например слитый белок. Также включенной в данное изобретение является рекомбинантная ДНК, которая заключает в себе часть нуклеотидов, приведенных в SEQ ID No. 1, 3, 5, 39, 41, 43 или 45, которая кодирует иммунореактивный белок Ehrlichia canis в 28-кДа.
ДНК должна иметь по меньшей мере 70% идентичности последовательности с кодирующей последовательностью нуклеотидов, представленных в SEQ ID No. 1, 3, 5, 39, 41, 43 или 45, предпочтительно по меньшей мере 75% (например, по меньшей мере 80%); и наиболее предпочтительно по меньшей мере 90% идентичности. Идентичность между двумя последовательностями является прямой функцией числа соответствующих или идентичных положений. Если положение субъединицы в обеих из двух последовательностей занято такой же мономерной субъединицей, например, если данное положение занято аденином в каждой из двух молекул ДНК, тогда они идентичны по данному положению. Например, если 7 положений в последовательности из 10 нуклеотидов по длине оказываются идентичными соответствующим положениям во второй 10-нуклеотидной последовательности, тогда две последовательности имеют 70% идентичности последовательности. Обычно длина сравниваемых последовательностей составляет по меньшей мере 50 нуклеотидов, предпочтительно по меньшей мере 60 нуклеотидов, более предпочтительно 75 нуклеотидов и наиболее предпочтительно 100 нуклеотидов. Обычно идентичность последовательностей определяют, используя программное обеспечение - анализ последовательности (например, пакет программного обеспечения - анализ последовательности, Генетической компьютерной группы Биотехнологического центра Университета Висконсина, 1710 University Avenue, Madison, WI 53705).
В данном изобретении также рассматривается вектор, содержащий кодирующую последовательность ДНК, которая содержит ген, кодирующий иммунореактивный белок 28-кДа Ehrlichia canis, и названный вектор способен реплицироваться у хозяина, который содержит в оперативном сцеплении: а) ориджин репликации; b) промотор; и с) последовательность ДНК, кодирующую названный белок. Предпочтительно, вектор согласно изобретению содержит часть последовательности ДНК, представленной в SEQ ID No. 1, 3, 5, 39, 41, 43 или 45.
«Вектор» может быть определен как реплицируемая конструкция нуклеиновых кислот, например плазмидная или вирусная нуклеиновая кислота. Можно использовать векторы, чтобы амплифицировать и/или экспрессировать нуклеиновую кислоту, кодирующую иммунореактивный белок 28-кДа Ehrlichia canis. Экспрессирующий вектор является реплицируемой конструкцией, в которой последовательность нуклеиновых кислот, кодирующая полипептид, является прочно связанной с соответствующими контрольными последовательностями, способными эффективно экспрессировать полипептид в клетке. Потребность в таких контрольных последовательностях изменяется в зависимости от избранной клетки и выбранного способа трансформации. Обычно контрольные последовательности включают в себя транскрипционный промотор и/или энхансер, соответствующие рибосомные связывающие участки мРНК и последовательности, которые контролируют терминацию транскрипции и трансляции. Способы, которые хорошо известны специалистам в данной области, можно применять для конструирования экспрессирующих векторов, содержащих соответствующие транскрипционный и трансляционный контрольные сигналы. См., например, способы, описанные Sambrook et al.,1989, Molecular Cloning: A Laboratory Manual (2nd Ed.), Cold Spring Harbor Press, N.Y. Ген и его транскрипционные контрольные последовательности определены как прочно связанные, если транскрипционные контрольные последовательности эффективно контролируют транскрипцию гена. Не ограничиваясь этим, векторы согласно изобретению включают в себя плазмидные векторы и вирусные векторы. Предпочтительными вирусными векторами в данном изобретении являются векторы, происходящие из ретровирусов, аденовируса, адено-ассоциированного вируса, вируса SV40 или вирусов герпеса.
Вообще экспрессирующие векторы, содержащие промоторные последовательности, которые способствуют эффективной транскрипции инсерционного фрагмента ДНК, используют в связи с хозяином. Используемый в описании термин «хозяин» предназначен для того, чтобы охватить не только прокариот, но также и эукариот, таких как дрожжи, растения и животные клетки. Рекомбинантную молекулу ДНК или ген, который кодирует иммунореактивный белок Ehrlichia canis в 28-кДа согласно изобретению, можно использовать, чтобы трансформировать хозяина, применяя любой из способов, обычно известных специалистам в данной области. Особо предпочтительным является применение вектора, содержащего кодирующую последовательность гена, кодирующего иммунореактивный белок 28-кДА Ehrlichia canis согласно изобретению для целей прокариотической трансформации.
Прокариотические хозяева могут включать в себя E.coli, S. typhimurium, Serratia marcescens и Bacillus subtilis. Эукариотические хозяева включают в себя дрожжи, такие как Pichia pastoris, клетки млекопитающих и клетки насекомых. Для достижения оптимального клеточного роста трансформированных хозяев можно ферментировать и культивировать способами, известными в данной области.
Используемый в описании термин «инженерная», или «рекомбинантная», клетка относится к клетке, в которую введен рекомбинантный ген, такой как ген, кодирующий антиген Ehrlichia canis. Поэтому инженерные клетки отличаются от естественно встречающихся клеток, которые не содержат рекомбинантно введенного гена. Таким образом, инженерными клетками являются клетки, содержащие ген или гены, введенные рукой человека. Рекомбинантно введенные гены или будут находиться в виде кДНК-гена, копии геномного гена, или будут включать гены, позиционно смежные с промотором, по природе не ассоциированным с особым введенным геном. Кроме того, рекомбинантный ген может быть интегрирован в геном хозяина или может быть помещен в вектор или в бактериальный геном, трансфицированный в клетку хозяина.
В данном изобретении также получены по существу чистые иммунореактивные белки 28-30 кДа E.canis, которые содержат аминокислотные последовательности, приведенные, например, в SEQ ID No. 2, 4, 6, 40, 42, 44 или 46.
Выражение «по существу чистый белок» означает белок, который был отделен по меньшей мере от нескольких из тех компонентов, которые сопутствуют ему по природе. Обычно белок считают по существу чистым, когда он оказывается по меньшей мере на 60% по массе свободным от белков и других природно встречающихся органических молекул, с которыми он естественно связан in vivo. Предпочтительно чистота препарата составляет по меньшей мере 75%, более предпочтительно по меньшей мере 90% и наиболее предпочтительно 99% по массе. По существу чистый иммунореактивный белок 28-кДа Ehrlichia canis может быть получен, например, путем экстракции из природного источника; экспрессии рекомбинантной нуклеиновой кислоты, кодирующей иммунореактивный белок 28-кДа Ehrlichia canis; или посредством химического синтеза белка. Чистоту можно определять любым подходящим способом, например методом колоночной хроматографии, такой как иммуноаффинная хроматография с использованием антител, специфичных в отношении иммунореактивного белка 28-кДа Ehrlichia canis, электрофореза в полиакриламидном геле или анализа посредством ВЭЖХ. Белок является по существу свободным от естественно ассоциированных компонентов, если он отделен по меньшей мере от некоторых из тех примесей, которые сопутствуют ему в исходном состоянии. Таким образом, белок, который химически синтезирован или получен в клеточной системе, отличающейся от клетки, из которой он происходит по природе, будет, по определению, по существу свободным от его естественно ассоциированных компонентов. Соответственно, по существу чистые белки включают в себя эукариотические белки, синтезированные в E.coli, других прокариотах или любом другом организме, в котором они естественно не встречаются.
Кроме полноразмерных белков, в изобретении также рассматриваются фрагменты (например, антигенные фрагменты) иммунореактивного белка 28-кДа Ehrlichia canis (SEQ ID No. 2, 4, 6, 40, 42, 44 или 46). Термин «фрагмент» относится к полипептиду, который обычно составляет по меньшей мере 10 остатков, чаще по меньшей мере 20 остатков и более предпочтительно 30 (например, 50) остатков по длине, но меньше полной интактной последовательности. Фрагменты иммунореактивного белка 28-кДа Ehrlichia canis можно получать способами, известными специалистам в данной области, например, в результате ферментативного переваривания природно встречающегося или рекомбинантного иммунореактивного белка 28-кДа Ehrlichia canis, с помощью методов рекомбинантной ДНК, используя экспрессирующий вектор, который кодирует определенный фрагмент иммунореактивного белка 28-кДа Ehrlichia canis, или посредством химического синтеза. Способность предполагаемого фрагмента проявлять свойства иммунореактивного белка 28-кДа Ehrlichia canis (например, связывание с антителом, специфическим в отношении иммунореактивного белка 28-кДа Ehrlichia canis) можно оценивать способам, представленными в описании.
Очищенный иммунореактивный белок 28-кДа Ehrlichia canis или антигенный фрагмент иммунореактивного белка 28-кДа Ehrlichia canis может быть использован для образования новых антител или для тестирования существующих антител (например, как позитивные контроли в диагностическом исследовании), применяя стандартные протоколы, известные специалистам в данной области.
Как хорошо известно в данной области, данный полипептид может изменяться по его иммуногенности. Поэтому часто оказывается необходимым связать иммуноген (например, полипептид согласно изобретению) с носителем. Типичными и предпочтительными носителями являются гемоцианин моллюска (KLH) и сывороточный альбумин человека. Средства для соединения полипептида с белком-носителем хорошо известны в данной области и включают в себя глутаральдегид, сложный эфир м-малеимидобензоил-N-гидроксисукцинимида, карбодиимид и бис-биазотизированный бензидин. Также понятно, что пептид может быть соединен с белком генно-инженерными способами, которые хорошо известны в данной области.
Также в данной области хорошо известно, что иммуногенность в отношении определенного иммуногена может быть повышена путем применения неспецифических стимуляторов иммунного ответа, известных как адъюванты. Типичные и предпочтительные адъюванты включают в себя полный BCG, Detox, (RIBI, Immunochem Research Inc.) ISCOMS и адъювант - гидроокись алюминия (Superphos, Biosector).
В данное изобретение включены поликлональные антисыворотки, полученные, например, у кроликов при использовании иммунореактивного белка 28-кДА Ehrlichia canis или фрагмента иммунореактивного белка 28-кДА Ehrlichia canis в качестве иммуногена. Применяют стандартные протоколы для получения моноклональных и поликлональных антител, известные специалистам в данной области. Моноклональные антитела, полученные согласно упомянутому способу, следует проверить на их способность идентифицировать рекомбинантные клоны кДНК Ehrlichia canis и отличать их от известных клонов кДНК.
В изобретении рассматриваются не только интактные моноклональные антитела, но также и иммунологически активный фрагмент антител, например фрагмент Fab или (Fab)2; инженерную молекулу одиночной цепи Fv; или химерную молекулу, например антитело, которое содержит связывающую специфичность одного антитела, например мышиной природы, и оставшиеся части другого антитела, например, человека.
В одном аспекте, антитело или его фрагмент может быть связано с токсином или с детектируемой меткой, например радиоактивной меткой, нерадиоактивной изотопной меткой, флуоресцентной меткой, хемилюминисцентной меткой, парамагнитной меткой, ферментной меткой или колориметрической меткой. Специалистам в данной области хорошо известны перечисленные метки и другие подходящие метки, которые можно применять в соответствии с данным изобретением. Связывание упомянутых меток с антителами или их фрагментами может быть осуществлено при использовании стандартных способов, хорошо известных специалистам в данной области.
Также предполагают, что фармацевтические композиции можно получать, используя новые белки согласно изобретению. В таком случае фармацевтическая композиция включает в себя новый активный препарат(ы) согласно изобретению и фармацевтически приемлемый носитель. Субъект, являющийся обычным специалистом в данной области, сможет легко определить, без чрезмерных исследований, соответствующие дозировки и способы введения активного компонента согласно изобретению.
Фраза «фармацевтически приемлемый» относится к молекулярным частицам или композициям, которые не вызывают аллергическую или подобную неблагоприятную реакцию при введении субъекту. Препарат водной композиции, который содержит белок в качестве активного ингредиента, полностью изучен в данной области. Обычно, такие композиции получают в виде инъецируемой формы, или в виде жидких растворов или суспензий; также могут быть приготовлены твердые формы, подходящие для растворения или суспендирования в жидкости перед инъекцией. Препарат также может быть эмульгированным.
Для композиции белок можно готовить в нейтральной или солевой форме. Фармацевтически приемлемые соли включают в себя аддитивные соли кислот (образованные свободными аминогруппами белка) и соли, образованные неорганическими кислотами, например, такими как хлористоводородная или фосфорная кислоты, или такими органическими кислотами, как уксусная, щавелевая, винная, миндальная и тому подобные. Соли, образованные свободными карбоксильными группами, также можно получать с неорганическими основаниями, например, такими как гидроокиси натрия, калия, аммония, кальция или железа, и такими органическими основаниями, как изопропиламин, триметиламин, гистидин, прокаин и тому подобные.
При приготовлении композиции растворы следует вводить способом, совместимым с дозированным препаратом и в таком количестве, которое является терапевтически эффективным. Препараты легко вводят в целый ряд дозированных форм, таких как инъецируемые растворы.
Например, для парентерального введения в водном растворе раствор должен быть соответственно забуферен, если необходимо, а жидкий разбавитель, прежде всего, сделан изотоническим с достаточным количеством соли и глюкозы. Такие специальные водные растворы особенно подходят для внутривенного, внутримышечного, подкожного и внутрибрюшинного введения. В этой связи в свете согласно изобретению стерильные водные среды, которые можно применять, станут известными специалистам в данной области. Например, одну дозу можно растворить в 1 мл изотонического раствора NaCl и либо добавить к 1000 мл жидкости для подкожного введения либо ввести с помощью инфузии в предполагаемый участок (смотри, например, «Remington's Pharmaceutical Sciences» 15th Edition, pages 1035-1038 и 1570-1580). Некоторые изменения доз неизбежно будут требоваться в зависимости от состояния субъекта, которого лечат.
В одном из аспектов данного изобретения предусмотрены последовательности ДНК, кодирующие иммунореактивный белок 30-кДа Ehrlichia canis. Предпочтительно, белок должен иметь аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID No. 2, 4, 6, 40, 42, 44, 46, а ген должен иметь последовательность нуклеиновых кислот, выбранную из группы, состоящей из SEQ ID No. 1, 3, 5, 39, 41, 43, 45, и быть представителем полиморфного мультигенного семейства. Более предпочтительно, белок имеет N-концевую сигнальную последовательность, которая удаляется в результате посттрансляционного процессинга, что приводит к образованию зрелого белка 28-кДа. Еще более предпочтительно, ДНК, кодирующие белки 28-кДа, располагаются в одном мультигенном локусе, который имеет размер 10677 пар оснований и кодирует девять гомологичных белков 28-кДа Ehrlichia canis.
В другом аспекте данного изобретения представлен экспрессирующий вектор, содержащий ген, кодирующий иммунореактивный белок 28-кДа Ehrlichia canis, и способный экспрессировать ген, когда вектор вводят в клетку.
В еще одном аспекте данного изобретения предусмотрен рекомбинантный белок, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID No. 2, 4, 6, 40, 42, 44, 46. Предпочтительно, аминокислотная последовательность кодируется последовательностью нуклеиновых кислот, выбранной из группы, состоящей из SEQ ID No. 1, 3, 5, 39, 41, 43, 45. Более предпочтительно, рекомбинантный белок содержит четыре вариабельные области, которые являются поверхностно расположенными, гидрофильными и антигенными. Еще более предпочтительно, рекомбинантный белок представляет собой антиген.
В следующем аспекте данного изобретения представлен способ получения рекомбинантного белка, включающий в себя стадии получения вектора, который содержит экспрессирующиую область, заключающую в себе последовательность, кодирующую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID No. 2, 4, 6, 40, 42, 44, 46, функционально связанную с промотором; трансфекцию вектора в клетку; и культивирование клетки в условиях, эффективных для экспрессии экспрессирующей области.
Также в некоторых аспектах изобретения может быть описан способ ингибирования инфекции Ehrlichia canis у субъекта, включающий в себя стадии: идентификации субъекта, подозреваемого в том, что он подвергся воздействию или инфицирован Ehrlichia canis; и введения композиции, содержащей антиген 28-кДа Ehrlichia canis в количестве эффективном, чтобы ингибировать инфекцию Ehrlichia canis. Ингибирование может осуществляться любыми способами, такими как стимуляция гуморальных и клеточных ответов субъектов, или другими способами, такими как ингибирование нормальной функции антигена 28-кДа, или даже конкуренции с антигеном за взаимодействие с каким-то агентом в организме субъекта.
Следующие примеры представлены с целью иллюстрирования различных аспектов изобретения и не предназначены для ограничения изобретения каким-либо образом.
ПРИМЕР 1
Секвенирование неизвестных 5'-
и 3'-областей гена ECa28-1 (p28-7)
Эрлихии и очишеннные Ehrlichia canis (штамм Florida и изоляты Demon, DJ, Jake и Fuzzy) были предоставлены Dr. Edward Breitschwerdt (College of Veterinary Medicine, North Carolina State University, Raleigh, NC). E.canis (штамм Louisiana) предоставлен Dr. Richard E.Corstvet (School of Veterinary Medicine, Louisiana State University, Baton Rouge, LA) и E.canis (штамм Oklahoma) предоставлен Dr. Jacqueline Dawson (Центры контроля и предупреждения заболеваний, Atlanta, GA). Размножение эрлихий осуществляли в клетках DH82 с DMEM, дополненной 10% бычьей телячьей сывороткой и 2мМ L-глутамином при 37°С. Внутриклеточный рост в клетках DH82 контролировали присутствием морул E.canis, используя методы цитологического окрашивания. Когда 100% клеток было инфицировано эрлихиями, клетки собирали, а затем осаждали центрифугированием при 17000 х g в течение 20 минут. Клеточные осадки дважды разрушали в ультразвуковом гомогенизаторе 2000 Braun-Sonic при 40В в течение 30 секунд на льду. Эрлихии очищали, как было описано ранее (Weiss et al., 1975). Лизат наслаивали в прерывистые градиенты ренографина 42%-36%-30% и центрифугировали при 80000 х g в течение 1 часа. Собирали тяжелые и легкие зоны, содержащие эрлихии, промывали сахароза-фосфат-глутаматным буфером (SPG, 218 мМ сахарозы, 3,8 мМ КН2РО4, 7,2 мМ К2НРО4, 4,9 мМ глутамата, рН 7,0) и осаждали центрифугированием.
Получение нуклеиновых кислот. Геномную ДНК Ehrlichia canis получали повторным суспендированием очищенных ренографином эрлихий в 600 мкл 10 мМ трис-HCl буфере (рН 7,5) с 1% додецилсульфатом натрия (SDS, мас./об.) и 100 нг/мл протеиназы К, как описано ранее (McBride et al., 1996). Полученную смесь инкубировали в течение 1 часа при 56°С и нуклеиновые кислоты дважды экстрагировали смесью фенол/хлороформ/изоамиловый спирт (24:24:1). ДНК осаждали абсолютным спиртом, промывали один раз 70% этанолом, высушивали и повторно суспендировали в 10 мМ трисе (рН 7,5). Плазмидную ДНК очищали, используя набор для выделения высокоочищенной плазмиды (Boehringer Mannheim, Indianapolis, IN), а продукты ПЦР очищали, используя набор для QIA-быстрой ПЦР-очистки (Qiagen, Santa Clarita, CA).
Клонирование гена ECa28-1 (p28-7) Полномерную последовательность гена р28-7 определяли, используя универсальный набор Genome Walker Kit (CLONTECH, Palo Alto, CA), в соответствии с протоколом, прилагаемым к руководству. Геномную ДНК E.canis (изолят Jake) полностью расщепляли пятью рестрикционными ферментами (DraI, EcoRV, PvuII, ScaI, StuI), чтобы получить ДНК с «затупленными концами». Внесенный в набор адаптер (АР1) лигировал каждый конец ДНК E.canis. Геномные библиотеки использовали как темплаты, чтобы выявить неизвестную последовательность ДНК гена р28-7 с помощью ПЦР, используя праймер, комплементарный к известной части последовательности р28-7, и праймер, специфичный для адаптера АР1. Специфичные для р28-7 праймеры, использованные для «прогулки» по геному, конструировали из известной последовательности ДНК, полученной при амплификации ПЦР р28-7 с праймерами 793 (SEQ ID No. 16) и 1330 (SEQ ID No. 17). Праймеры 394 (5'-GCATTTCCACAGGATCATAGGTAA-3'; нуклеотиды 687-710, SEQ ID No.21) и 394С (5'-TTACCTATGATCCTGT GGAAATGC-; нуклеотиды 710-687, SEQ ID No. 22) использовали вместе с внесенным праймером АР1, чтобы амплифицировать неизвестные 5' и 3' области гена р28-7 с помощью ПЦР. Продукт ПЦР, соответствующий 5' области гена р28-7, амплифицированной с праймерами 394С и АР1 (2000 пар оснований), секвенировали однонаправленно с праймером 793С (5'-GAGTA ACCAACAGCTCCTGC-3', SEQ ID No. 23). Продукт ПЦР, соответствующий 3' области гена р28-7, амплифицированный с праймерами 394 и АР1 (580 пар оснований), секвенировали двунаправленно с теми же праймерами. Некодирующие области на 5' и 3' областях, смежных с открытой рамкой считывания, секвенировали и выбирали праймеры EC28OM-F (5'-TCTACTTTGCACTTCC ACTATTGT-3', SEQ ID No. 24) и EC28OM-R (5'-ATTCTTTTGCCACTATTT TTCTTT-3', SEQ ID No. 25), комплементарные к этим областям, чтобы амплифицировать полный ген р28-7.
Секвенирование ДНК ДНК секвенировали ДНК-секвенатором ABI Prism 377 (Perkin-Elmer Applied Biosystems, Foster City, CA). Целые гены р28-7 из семи изолятов E.canis (четыре из Северной Каролины и по одному из Оклахомы, Флориды и Луизианы) амплифицировали посредством ПЦР с праймерами EC28OM-F (SEQ ID No. 24) и EC28OM-R (SEQ ID No. 25) c температурным профилем цикла 95°С в течение 5 минут и 30 циклов 95°С в течение 30 секунд, 62°С в течение 1 минуты и 72°С в течение 2 минут с последующим продолжением при 72°С в течение 10 минут. Полученные продукты ПЦР двунаправленно секвенировали с теми же самыми праймерами.
ПРИМЕР 2
ПЦР-амплификация, клонирование, секвенирование и экспрессия гена ЕСа28-1 (р28-7) E.canis
Экспрессирующие векторы Целый ген р28-7 амплифицировали посредством ПЦР с праймерами ЕС28ОМ-F и EC28OM-R и клонировали в клонирующий вектор pCR2.1-TOPO TA, чтобы получить требуемый набор участков расщепления ферментами рестрикции (Invitrogen, Carlsbad, CA). Вставку вырезали из pCR2.1-TOPO с BstX 1 и лигировали в эукариотический экспрессирующий вектор рсДНК 3.1 (Invitrogen, Carlsbad, CA), обозначенный рсДНК3.1/ЕС28 для последующих исследований. Плазмиду рсДНК3.1/ЕС28 амплифицировали, а ген вырезали посредством двойного расщепления KpnI-XbaI и двунаправленно сшивали в прокариотический экспрессирующий вектор pThioHis (Invitrogen, Carlsbad, CA). Клон (обозначеный pThioHis/ЕС28) продуцировал рекомбинантный белок, слитый с тиоредоксином, в Escherichia coli BL21. Рекомбинантный слитый белок грубо очищали в нерастворимую фазу центрифугированием. Контрольный слитый с тиоредоксином белок очищали из растворимых клеточных лизатов в нативных условиях, используя спин-колонки никель-NTA (Qiahen, Santa Clarita, CA).
Вестерн-блоттинг-анализ Рекомбинантный слитый белок р28-7 E.canis подвергали электрофорезу в полиакриламидном геле в присутствии ДДС (SDS) (SDS-PAGE) в градиентных гелях 4-15% в трис-HCl (Bio-Rad, Hercules, CA) и переносили на чистую нитроцеллюлозу (Schleicher & Schuell, Keene, NH), используя полусухую ячейку переноса (Bio-Rad, Hercules, CA). Мембрану инкубировали с антисывороткой, полученной у инфицированной E.canis собаки в период выздоровления, разбавленной 1:5000 в течение 1 часа, промывали, а затем инкубировали с анти-собачьими IgG (H & L), конъюгированными с щелочной фосфатазой аффинно-очищенными вторичными антителами, в разведении 1:1000 в течение 1 часа (Kirkegaard & Perry Laboratories, Gaithersburg, MD). Связанные антитела визуализировали с помощью субстрата 5-бром-4-хлор-3-индолилфосфат/тетразолий нитроголубой (BCIP/NBT) (Kirkegaard & Perry Laboratories, Gaithersburg, MD).
Блоттинг-анализ по Саузерну Чтобы установить, существует ли гомология многочисленных генов с геном р28-7 в геноме E.canis, проводили блоттинг-анализ по Саузерну, используя стандартный метод (Sambrook et al., 1989). Геномную ДНК E.canis полностью расщепляли каждым из ферментов рестрикции BanII, EcoRV, HaeII, KpnI и SpeI, которые не разрезали внутри гена р28-7, и AseI, который расщеплял р28-7 у нуклеотидов 34, 43 и 656. Зонд получали ПЦР-амплификацией с праймерами ЕС28ОМ-F и ЕС28ОМ-R и дигоксигенином (DIG)-меченным дезоксинуклеотидтрифосфатом (dNTPs) (Boehringer Mannheim, Indianopolis, IN) и расщепляли AseI. Расщепленный зонд (566 пар оснований) разделяли посредством гель-электрофореза в агарозе, очищали на геле и затем использовали для гибридизации. Полностью переваренную геномную ДНК E.canis подвергали электрофорезу и переносили на нейлоновую мембрану (Boehringer Mannheim, Indianopolis, IN) и гибридизовали при 40°С в течение 16 часов с DIG-меченным зондом гена р28-7 буфере DIG Easy Hyb, в соответствии с протоколом инструкции (Boehringer Mannheim, Indianopolis, IN). Связанный зонд определяли с помощью конъюгированных с щелочной фосфатазой анти-DIG-антител и люминисцентного субстрата (Boehringer Mannheim, Indianopolis, IN) и экспонировали на изобразительную пленку BioMax (Eastman Kodak, Rochester, NY).
Анализ и сравнение последовательностей ДНКпоследовательностей р28 E.chaffeensis и map-1 C.ruminantium получали из Национального Центра биотехнологической информации (NCBI). Анализ нуклеотидной и выведенной аминокислотной последовательностей и белковый и филогенетический анализы проводили с программным обеспечением LASERGENE (DNASTAR, Inc., Madison, WI). Анализ посттрансляционного процессинга осуществляли по способу McGeoch и von Heijne для распознавания сигнальной последовательности, используя программу PSORT (McGeoch, 1985; von Heijne, 1986).
Анализ последовательностей р28-7 из семи различных штаммов E.canis проводили с праймерами, предназначенными для амплификации целого гена. Анализ показал, что последовательность этого гена оказалась консервативной в числе изолятов из Северной Каролины (четыре), Луизианы, Флориды и Оклахомы.
Результаты
Сравнительный анализ последовательностей нуклеиновых кислот из р28 E.chaffeensis и map-1 Cowdria ruminantium с использованием алгоритма Jotun-Hein установил согласованную последовательность с областями высокой гомологии (>90%). Обнаруженные гомологичные области (нуклеотиды 313-332 и 823-843 из map-1 C.ruminantium; 307-326 и 814-834 р28 E.chaffeensis) таргетировали как участки отжига праймера для ПЦР-амплификации. ПЦР-амплификацию гена р28-7 E.canis осуществляли с праймерами 793 (5'-GCAGGAGCTGTTGGTTACTC-3') (SEQ ID No. 16) и 1330 (5'-CCTTCCTCCAAGTTCTATGCC-3') (SEQ ID No. 17), получая в результате продукт ПЦР 518 пар оснований. ДНК E.canis амплифицировали с праймерами 793 и 1330 с температурным профилем цикла 95°С в течение 2 минут и 30 циклов 95°С в течение 30 секунд, 62°С в течение 1 минуты, 72°С в течение 2 минут с последующим продолжением 72°С в течение 10 минут и выдержкой при 4°С. Последовательность нуклеиновых кислот продукта ПЦР E.canis получали секвенированием продукта непосредственно с праймерами 793 и 1330.
Анализ последовательности выявил открытую рамку считывания, кодирующую белок из 170 аминокислот, а сравнительный анализ последовательности из 518 пар оснований, полученной посредством ПЦР-амплификации E.canis с последовательностью ДНК гена р28 E.chaffeensis, выявил сходство более 70%, указывая, что гены являются гомологичными.
Адаптерную ПЦР с праймерами 394 и 793С проводили, чтобы обнаружить 5' и 3' сегменты последовательности целого гена. Праймер 394 образовывал четыре продукта ПЦР (3-т.п.н., 2-т.п.н., 1-т.п.н. и 0,8-т.п.н.) и продукт 0,8 пар оснований секвенировали двунаправленно, используя праймеры 394 и АР1. Выведенная последовательность частично совпадала с 3' концом продукта 518-пар оснований, простираясь на 12 пар оснований открытой рамки считывания к терминирующему кодону. Также секвенировали дополнительные 625 пар оснований некодирующей последовательности на 3' конце гена р28-7.
Праймер 394С использовали, чтобы амплифицировать 5' конец гена р28-7, с добавленным праймером АР1. Амплификация с названными праймерами давала в результате три продукта ПЦР (3,3-т.п.н., 3-т.п.н. и 2-т.п.н.). Фрагмент 2-т.п.н. однонаправленно секвенировали с праймером 793С. Последовательность дала предполагаемый инициирующий кодон гена р28-7 и завершила открытую рамку считывания 834 пар оснований, кодирующую белок из 278 аминокислот. Образованы дополнительные 144 пар оснований считываемой последовательности в 5' некодирующей области гена р28-7. Праймеры ЕС28ОМ-F и EC28OM-R выбирали из комплементарных некодирующих областей, смежных с геном р28-7.
Амплифицированный с названными праймерами продукт ПЦР секвенировали непосредственно с теми же праймерами. Полная последовательность ДНК для гена р28-7 E.canis (SEQ ID No. 1) представлена на фиг.1. Фрагмент ПЦР р28-7, амплифицированный с названными праймерами, содержал целую открытую рамку считывания и 17 дополнительных аминокислот из 5' некодирующей области праймера. Ген направленно субклонировали в экспрессирующий вектор pThioHis, а E.coli (BL21) трансформировали этой конструкцией. Экспрессированный р28-7-тиоредоксин-слитый белок оказался нерастворимым. Экспрессированный белок имел дополнительные 114 аминокислот, ассоциированные с тиоредоксином, 5 аминокислот для энтерокиназного участка распознавания и 32 аминокислоты из участка многократного клонирования и 5' некодирующей области праймера на N-конце. Антисыворотка от инфицированной E.canis собаки, полученная во время периода выздоровления, распознавала экспрессированный рекомбинантный слитый белок, но не реагировала с тиоредоксиновым контролем (фиг.2).
ПРИМЕР 3
Гомология последовательностей гена р28-7 E.canis
Последовательность нуклеиновых кислот семейства генов р28-7 E.canis (834 пар оснований) и omp-1 E.chaffeensis, включая сигнальные последовательности (р28-7, omp-1A, B, C, D, E и F), выравнивали, используя способ Клустала (Clustal), чтобы установить гомологию между перечисленными генами (сравнительный анализ не представлен). Гомология нуклеиновых кислот сохранялась в равной степени (68,9%) между р28-7 E.canis, р28 E.chaffeensis и omp-1F. Другие предполагаемые гены белков внешней мембраны у семейства omp-1 E.chaffeensis, omp-1D (68,2%), omp-1E (66,7%), omp-1C (64,1%), map-1 Cowdria ruminantium (61,8%), гена 1 белка E.canis в 28-кДа (60%) и гена 2 белка 28-кДа (частичная) (59,5%) также оказались гомологичными с р28-7. OMP-1В E.chaffeensis характеризовался наименьшей гомологией нуклеиновых кислот (45,1%) с р28-7 E.canis.
Сравнительный анализ предсказанных аминокислотных последовательностей р28-7 E.canis. (SEQ ID No. 2) и р28 E.chaffeensis выявил аминокислотные замещения, приводящие к четырем вариабельным областям (VR). Идентифицированы замещения или делеции в аминокислотной последовательности и локализациях вариабельных областей Р28-7 E.canis и семейства ОМР-1 Е. chaffeensis (фиг.3). Сравнение аминокислотных последовательностей, включая сигнальный пептид, показало, что р28-7 E.canis разделяет наибольшую гомологию с OMP-1F (68%) семейства OMP-1 E.chaffeensis, за которыми следуют Р28 E.chaffeensis (65,5%), ОМР-1Е (65,1%), OMP-1D (62,9%), OMP-1C (62,9%), MAP-1 Cowdria ruminantium (59,4%), белок 1 28-кДа E.canis (55,6%) и белок 2 28-кДа (частичная) (53,6%) и ОМР-1В (43,2%). Анализ филогенетического родства на основании аминокислотных последовательностей показал, что Р28-7 E.canis и MAP-1 C.ruminantium, белки ОМР-1 E.chaffeensis и белки 1 и 2 (частичная) 28-кДа Е. canis являются родственными (фиг.4).
ПРИМЕР 4
Предсказанная поверхностная вероятность и иммунореактивность Р28-7 E.canis
Анализ Р28-7 E.canis с использованием профилей гидрообработки и гидрофильности предсказал поверхностно-экспонированные области на Р28-7 (фиг.6). Восемь основных поверхностно-экспонированных областей, состоящих из 3-9 аминокислот, идентифицировали на Р28-7 E.canis и установили сходство с профилем поверхностно-экспонированных областей на Р28 E.chaffeensis (фиг.6). Пять более крупных поверхностно-экспонированных областей на Р28-7 E.canis локализованы в N-концевой области белка. Поверхностно-экспонированные гидрофильные области обнаружены во всех четырех вариабельных областях Р28-7 E.canis. Десять Т-клеточных мотивов было предсказано в Р28-7 при использовании алгоритма Ротбарда-Тейлора (Rothbard-Taylor) (Rothbard and Taylor, 1988), а высокая антигенность Р28-7 E.canis предсказана по алгоритму антигенности Джеймсона-Вольфа (фиг.6) (Jameson and Wolf, 1988). Наблюдали сходство в антигенности и Т-клеточных мотивах между Р28-7 E.canis и Р28 E.chaffeensis.
ПРИМЕР 5
Определение гомологичных геномных копий гена р28-7 E.canis
Геномный блоттинг-анализ по Саузерну ДНК E.canis, независимо полностью расщепленной рестрицирующими ферментами BanII, EcoRV, HaeII, KpnI, SpeI, которые не имеют эндонуклеазных участков рестрикции в гене р28-7, и AseI, который имеет внутренние эндонуклеазные участки рестрикции по нуклеотидам 34, 43 и 656, показал наличие по меньшей мере трех гомологичных копий гена р28-7 (фиг.5). Хотя р28-7 E.canis имеет внутренние участки рестрикции AseI, DIG-меченный зонд, использованный в гибридизационном эксперименте, таргетировал область гена внутри единственного фрагмента ДНК, образованного при расщеплении гена посредством AseI. Расщепление AseI давало 3 зоны (приблизительно 566 пар оснований, 850 пар оснований и 3-т.п.н.), которые гибридизовали с ДНК-зондом р28-7, указывая на присутствие в геноме многочисленных генов, гомологичных р28-7. При расщеплении посредством EcoRV и SpeI образовывались две зоны, которые гибридизовали с зондом гена р28-7.
ПРИМЕР 6
ПЦР-Амплификация генов ЕСа28SA2 (p28-5), ECa28SA3 (p28-6) E.canis и идентификация мультигенного локуса
Для того что специфически амплифицировать возможные неизвестные гены прямого направления ECa28SA2 (p28-5), для амплификации использовали праймер 46f, специфический для р28-5 (5'-ATATACTTCCTACCTAATGTCTCA-3', SEQ ID No. 18), и праймер 1330 (SEQ ID No. 17), который таргетирует консервативную область на 3'конце гена р28-7. Амплифицированный продукт очищали на геле и клонировали в клонирующий вектор ТА (Invitrogen, Santa Clarita, CA). Клон двунаправленно секвенировали с праймерами: М13 обратный от вектора, 46f, ECa28SA2 (5'-AGTGCAGAGTCTTCGGTTTC-3', SEQ ID No. 19), EСa5.3 (5'-GTTACTTGCGGAGGACAT-3', SEQ ID No. 20). Амплифицировали ДНК с температурным профилем цикла: 95°С в течение 2 минут и 30 циклов 95°С в течение 30 секунд, 48°С в течение 1 минуты, 72°Св течение 1 минуты с последующим продолжением 72°Св течение 10 минут и выдержкой 4°С.
Продукт ПЦР 2-т.п.н. амплифицировали с упомянутыми праймерами, которые содержали 2 открытые рамки считывания. Первая открытая рамка считывания включала в себя известную область гена р28-5 и ранее не секвенированную 3' часть гена. В прямом направлении от р28-5 обнаружен неидентичный, но гомологичный ген белка 28-кДа и обозначен ECa28SA3 (р28-6).
Специфические праймеры, обозначенные EСaSA3-2 (5'-CTAGGATTA GGTTATAGTATAAGTT-3", SEQ ID No. 26), соответствующие областям в р28-6, и праймер 793С (SEQ ID No. 23), который присоединяется к области с р28-7, использовали, чтобы амплифицировать межгенную область между геном р28-6 и р28-7. Амплифицировали ДНК с температурным профилем цикла: 95°С в течение 2 минут и 30 циклов 95°С в течение 30 секунд, 50°С в течение 1 минуты, 72°Св течение 1 минуты с последующим продолжением 72°Св течение 10 минут и выдержкой 4°С.
Амплифицировали продукт ПЦР 800 пар оснований, который содержал 3' конец р28-6, межгенную область между р28-6 и р28-7 (28NC3) и 5' конец р28-7, соединяющий ранее отдельные локусы (фиг.8). Открытая рамка считывания 849 пар оснований р28-5 кодирует белок из 283 аминокислот, а р28-6 имеет открытую рамку считывания 840 пар оснований, кодирующую белок из 280 аминокислот. Межгенная некодирующая область между р28-6 и р28-7 составляла 345 пар оснований по длине (фиг.7 и 8).
ПРИМЕР 7
Гомология нуклеиновых кислот и аминокислот белков р28-4, р28-5, р28-6, р28-7 и р28-8 E.canis
Нуклеиновые и аминокислотные последовательности всех пяти генов белков E.canis в 28-кДа выравнивали, используя способ Клустала (Clustal), чтобы исследовать гомологию между этими генами. Гомология нуклеиновых кислот составляла от 58 до 75%, и подобную аминокислотную гомологию, составляющую от 67 до 72%, наблюдали среди представителей генов белков E.canis в 28-кДа (фиг.9).
Транскрипционные промоторные области Анализировали межгенные области между генами белков 28-кДа относительно промоторных последовательностей посредством сравнения согласованных промоторных областей Escherichia coli и промотора из E.chaffeensis (Yu et al., 1997; McClure, 1985). Предполагаемые промоторные последовательности, включающие области RBS, -10 и -35, идентифицировали в 4 межгенных последовательностях, соответствующих генам р28-5, р28-6, р28-7 и р28-8 (ЕСа28-2) (фиг.10). Некодирующая область в обратном направлении р28-4 (ECa28SA1) неизвестна и не исследовалась.
N-концевая сигнальная последовательность Анализ аминокислотной последовательности показал, что целый р28-7 E.canis имеет выведенную молекулярную массу 30,5-кДа, а выведенная молекулярная масса целого р28-6 составляет 30,7-кДа. Оба белка имеют предсказанный N-концевой сигнальный пептид из 23 аминокислот (MNCKKILITTALMSLMYYAPSIS, SEQ ID No. 27), который подобен пептидам, предсказанным для Р28 E.chaffeensis (MNYKKILITSALISLISSLPGV SFS, SEQ ID No. 28) и ОМР-1 белкового семейства (Yu et al., 1999a; Ohashi et al., 1998b).
Предпочтительный участок гидролиза для сигнальных пептидаз (SIS; Ser-X-Ser) (Oliver, 1985) установлен по аминокислотам 21, 22 и 23 р28-7. Также обнаружен дополнительный предполагаемый участок расщепления в положении аминокислоты 25 (MNCKKILITTALISLMYSIPSISSFS, SEQ ID No. 29), идентичный предсказанному участку расщепления Р28 E.chaffeensis (SFS) и который должен обеспечить образование зрелого р28-7 с предсказанной молекулярной массой 27,7-кДа. Участок гидролиза сигнальной последовательности ранее описанной частичной последовательности р28-5 предсказан по аминокислоте 30. Однако анализ сигнальной последовательности показал, что р28-4 имеет нерасщепляемую сигнальную последовательность.
Резюме
Идентифицированы белки с похожей молекулярной массой и клонированы из многочисленных риккетсионых агентов, включающих в себя E.canis, E.chaffeensis и C.ruminantium (Reddy et al., 1998; Jongejan et al., 1993; Ohashi et al., 1998). Ранее были описаны один локус в Ehrlichia chaffeensis c 6 гомологичными генами р28 и 2 локуса в E.canis, каждый содержащий несколько гомологичных генов белков 28-кДа.
В данном изобретении продемонстрировали клонирование, экспрессию и свойства генов, кодирующих зрелые белки E.canis в 28-кДа, которые являются гомологичными мультигенному семейству omp-1 E.chaffeensis и гену map-1 C.ruminantium. Идентифицированы два новых гена белков 28-кДа, р28-7 и р28-6. В данном изобретении полностью секвенировали другой ген белков E.canis в 28-кДа, р28-5, частично секвенированный ранее (Reddy et al., 1998). Также осуществили идентификацию и характеристику единственного локуса в E.canis, содержащего пять генов белков E.canis в 28-кДа (р28-4, р28-5, р28-6, р28-7 и р28-8).
Белки E.canis в 28-кДа являются гомологичными семейству ОМР-1 E.chaffeensis и белку МАР-1 C.ruminantium. Наиболее гомологичные белки E.canis в 28-кДа (р28-6, р28-7 и р28-8) последовательно расположены в локусе. Гомология названных белков колебалась от 67,5% до 72,3%. Дивергенция среди этих белков в 28-кДа составляла от 27,3% до 38,6%. Белки E.canis в 28-кДа, р28-4 и р28-5 оказались наименее гомологичными с гомологией, составляющей от 50,9% до 59,4%, и дивергенцией от 53,3% до 69,9%. Различия между генами относятся, главным образом, к четырем гипервариабельным областям и наводят на мысль, что упомянутые области являются экспонированными на поверхности и подвергаются избирательному воздействию иммунной системы. Описан консерватизм р28-7 в числе семи изолятов E.canis (McBride et al., 1999), позволяя предположить, что E.canis может быть клональным в Северной Америке. Наоборот, описано значительное многообразие р28 среди изолятов E.chaffeensis (Yu et al., 1999a).
Оказалось, что все из белков E.canis в 28-кДа в результате посттрансляционных модификаций превращаются из белка 30-кДа в зрелый белок в 28-кДа. Недавно была идентифицирована сигнальная последовательность Р28 E.chaffeensis (Yu et al., 1999a), а секвенирование N-концевой аминокислотной последовательности подтвердило, что белок подвергается посттрансляционной модификации, что приводит к отщеплению сигнальной последовательности с образованием зрелого белка (Ohashi et al., 1998). Высказано предположение, что лидерные последовательности OMP-1F и OMP-1E также имеют лидерные сигнальные пептиды (Ohashi et al., 1998). Сигнальные последовательности, идентифицированные на OMP-1F и OMP-1E E.chaffeensis и р28, являются гомологичными лидерной последовательности белка E.canis в 28-кДа. Промоторные последовательности для генов р28 экспериментально не были обнаружены, но предполагаемые промоторные области идентифицированы в результате сравнения с согласованными последовательностями промоторных областей RBS, -10 и -35 E.coli и других эрлихий (Yu et al., 1997; McClure, 1985). Такие промоторные последовательности позволяют каждому гену потенциально быть транскрибированным и транслированным, указывая, что эти гены могут дифференциально экспрессироваться у хозяина. Персистентность инфекции у собак может быть связана с дифференциальной экспрессией генов р28, приводя к антигенным изменениям in vivo, таким образом предоставляя микроорганизму возможность уклониться от иммунного ответа.
Обнаружено, что гены белков E.canis в 28-кДа проявляют гомологию последовательности нуклеиновых кислот и аминокислотной последовательности с семейством генов omp-1 E.chaffeensis и геном map-1 C.ruminantium. Предшествующие исследования идентифицировали белок 30-кДА E.canis, который реагирует с антисывороткой против E.chaffeensis, полученной во время периода выздоровления, но, как полагают, отличается по антигенным свойствам (Rikihisa et al., 1994). Выводы, сделанные на основании сравнения аминокислотных замещений в четырех вариабельных областях белков E.canis в 28-кДа, подтвердили эту возможность. В то же время сделанные выводы также позволяют предположить, что аминокислоты, ответственные за антигенные различия между Р28 E.canis и E.chaffeensis, локализованы в упомянутых вариабельных областях и легко доступны иммунной системе.
Описано, что иммунореактивные пептиды локализованы в вариабельных областях белков C.ruminantium, E.chaffeensis и E.canis в 28-кДа (Reddy et al., 1998). Анализ Р28 E.canis и E.chaffeensis показал, что все из вариабельных областей имеют предсказанные поверхностно расположенные аминокислоты. Исследование на собаках продемонстрировало отсутствие перекрестного иммунитета между E.canis и E.chaffeensis (Dawson and Ewing, 1992). Приведенный результат наблюдений может быть связан с антигенными различиями в вариабельных областях Р28, а также в других иммунологически важных антигенах этих видов эрлихий. В другом исследовании было показано, что антисыворотка человека, полученная от инфицированных E.chaffeensis пациентов во время периода выздоровления, распознавала белок(и) 29/28-кДа E.chaffeensis, а также взаимодействовала с гомологичными белками E.canis (Chen et al., 1997). Оказалось, что гомологичные и дающие перекрестную реакцию эпитопы на белке 28-кДа E.canis и Р28 E.chaffeensis распознаются иммунной системой.
Белки E.canis в 28-кДа могут быть важными иммунопротективными антигенами. В нескольких сообщениях было продемонстрировано, что антиген E.canis в 30-кДа проявляет сильную иммунореактивность (Rikihisa et al., 1994; Rikihisa et al., 1992). Антитела к антисыворотке, полученной от человека и собак во время периода выздоровления, реагировали с белками этой величины из E.chaffeensis и E.canis, позволяя предположить, что они могут быть важными иммунопротективными антигенами (Rikihisa et al., 1994; Chen et al., 1994; Chen et al., 1997). Кроме того, антитела к белкам 30, 24 и 21-кДа рано вырабатывались в иммуном ответе на E.canis (Rikihisa et al., 1994; Rikihisa et al., 1992), указывая, что упомянутые белки могут быть особенно важными в иммунных ответах при острой форме заболевания. Недавно, семейство гомологичных генов, кодирующих белки внешней мембраны с молекулярными массами 28-кДа, были идентифицированы у E.chaffeensis, и оказалось, что у мышей, иммунизированных рекомбинантным Р28 E.chaffeensis, вырабатывался иммунитет против заражения гомологами (Ohashi et al., 1998). Показано, что Р28 E.chaffeensis присутствует во внешней мембране, а иммуноэлектронная микроскопия локализовала Р28 на поверхности микроорганизма, и таким образом позволяя предположить, что он может функционировать как адгезин (Ohashi et al., 1998). Возможно, что белки E.canis в 28-кДа, идентифицированные в описываемом исследовании, имеют такую же локализацию и, возможно, выполняют подобную функцию.
Сравнение р28-7 из различных штаммов Е. canis показало, что ген, очевидно, является полностью консервативным. Исследования, выполненные на E.chaffeensis, продемонстрировали иммунологические и молекулярные доказательства многообразия. Пациенты, инфицированные E.chaffeensis, характеризуются изменчивой иммунореактивностью в отношении белков 29/28-кДа, что указывает на антигенное многообразие (Chen et al., 1997). Недавно на молекулярном уровне были получены данные, которые являются подтверждением антигенного разнообразия в гене р28 из Е. chaffeensis (Yu et al., 1999a). Сравнение пяти изолятов Е. chaffeensis выявило, что два изолята (Sapulpa and St. Vincent) имели 100% идентичность, но три других (Arkansas, Jax, 91HE17) отличались до 13,4% на аминокислотном уровне. Консерватизм р28-7 E.canis позволяет предположить, что штаммы E.canis, обнаруженные в Соединенных Штатах, могут быть генетически идентичными и, таким образом, белок E.canis в 28-кДа является привлекательным кандидатом на вакцину относительно эрлихиоза собак в Соединенных Штатах. Дальнейший анализ изолятов E.canis за пределами Соединенных Штатов может обеспечить информацией относительно происхождения и эволюции E.canis. Консерватизм белка 28-кДа делает его важным возможным кандидатом для достоверной серодиагностики эрлихиоза собак.
В настоящее время роль многочисленных гомологичных генов не известна; однако устойчивость инфекций E.canis у собак предположительно может быть связана с антигенной изменчивостью, обусловленной изменчивой экспрессией гомологичных генов белка 28-кДа, таким образом предоставляя возможность E.canis уклоняться от иммунного контроля. Изменчивость генов msp-3 у A. marginale частично является ответственной за изменчивость в белке MSP-3, приводя в результате к устойчивым инфекциям (Alleman et al., 1997). Исследования экспрессии генов белков 28-кДа посредством E.canis у остро и хронически инфицированных собак обеспечат понимание роли семейства генов белков 28-кДа в персистенции инфекции.
ПРИМЕР 8
Идентификация генов р28-1, р28-2, р28-3 и р28-9 E.canis
Секвенировали неизвестные области ДНК в обратном и прямом направлении локуса, из пяти последовательно расположенных генов р28, описанных выше, с помощью выбранных специфических генных праймеров для р28-1 (Еса28-75С) и р28-5 (ЕСа28-5-818f), чтобы пройти генный локус в двух направлениях. Чтобы установить неизвестную последовательность, многократные скрининги генов локуса проводили следующим образом: 1,9 т.п.н. в прямом направлении 5-генного локуса амплифицировали и секвенировали, используя праймеры р28-5-818f-(5'TTA AAC ATA TGC CAC TTC GCA CTA-3', SEQ ID No. 34), получая ампликон 900 пар оснований, и 1191 (5'-TAT GAT CGT GTA AAA TTG CTG TGA GTA T-3', SEQ ID No. 35), получая ампликон 1-т.п.н. 3,67 т.п.н. ДНК в прямом направлении локуса из пяти генов и амплифицировали, секвенировали с праймерами ЕСа28-75С (5'TAC TGG CAC GTG CTG GAC TA-3', SEQ ID No. 36), получая ампликон 1,6-т.п.н.; ЕСа5'-1600 (5'-CAC CAA TAA ATG GAG AGA CTT C-3', SEQ ID No. 37) c образованием ампликона 1,6-т.п.н.; и 3125 (5'-AAT CCA TCA TTT CTC ATT ACA GTG TG-3', SEQ ID No. 38), получая ампликон 800 пар оснований. Локус из девяти тандемно расположенных генов, состоящих из четырех новых генов р28 и пяти генов р28, описанных выше, обозначали р28-1 до р28-9 (фиг.11).
Проводили сравнение последовательностей нуклеиновых кислот и аминокислотных последовательностей генов р28 E.canis, используя способ Клустала (Clustal), чтобы исследовать гомологию между названными генами. Гомология названных белков составляла от 67,5% до 75%, а дивергенция между этими белками Р28 составляла 26,9%-38%. Белки Р28 E.canis, Р28-1, Р28-2 и Р28-9 по меньшей мере оказались гомологичными с другими генами р28, с гомологией от 37% до 49% и с дивергенцией 53-77%. Гомология нуклеиновых кислот девяти генов р28 составляла от 28 до 72%. Филогенетическое родство р28 E.canis, на основании аминокислотных последовательностей, представлено на фиг.12.
Нуклеотидные последовательности и номера депозитов. Номером депозита GenBank для последовательностей нуклеиновых кислот и аминокислотных последовательностей локуса девяти генов р28 E.canis (штамм Jake) является AF082744. Первоначально указанный номер депозита предназначался для р28-7, но был модернизирован для последовательности локуса девяти генов р28, который включает р28-7. Номерами депозитов GenBank для последовательностей нуклеиновых кислот и аминокислотных последовательностей р28-7 в других изолятах E.canis, описанных в данном исследовании, являются: Louisiana, AF082745; Oklahoma, AF082746; Demon, AF082747; DJ, AF082748; Fuzzy, AF082749; Florida, AF082750.
Многочисленные зоны в области 28-килодальтон были выявлены посредством иммуноблотов в сыворотках инфицированных E.canis собак, взятых во время периода выздоровления (Rikihisa et al., 1994), а экспрессия множества белков р28 может быть объяснением полученных данных. Блоттинг-анализы по Саузерну указывают, что другие гены р28, кроме пяти представителей рассматриваемого локуса, присутствуют в геноме (McBride et al., 1999; Ohashi et al., 1998b).
В данном исследовании идентифицирован единственный генный локус, содержащий девять последовательно расположенных генов р28 E.canis, кодирующих гомологичные, но неидентичные гены р28. Локус из девяти генов включает в себя четыре новых гена р28 (фиг.13-16) и пять тандемно расположенных генов р28, которые описаны выше. Восемь из генов р28 локализованы на одной цепи ДНК, а один ген р28 обнаружен на комплементарной цепи. Гомология нуклеиновых кислот среди девяти представителей генов р28 составляла 37-75%, а аминокислотная гомология колебалась от 28 до 72%.
Обнаружено, что р28 E.canis являются близкородственными белкам 28-килодальтон других видов, таких как E.chaffeensis, к которым они относятся (McBride et al., 2000). Различия среди белков обнаружены главным образом в некоторых основных гипервариабельных областях и указывают, что упомянутые области являются поверхностно экспонированными и подвергаются избирательному воздействию иммунной системы (McBride et al., 2000).
В числе семи географически разных изолятов описана консервативность гена р28 (р28-7) E.canis (McBride et al., 1999), позволяя предположить, что E.canis может быть высококонсервативным в Северной Америке. Аналогично, гликопротеин E.canis в 120-кДа также оказался консервативным среди изолятов в Соединенных Штатах (Yu et al., 1997). Наоборот, гены как 120-кДа, так и белка 28-кДа E.chaffeensis оказались дивергентными среди изолятов (Yu et al., 1999a; Chen et al., 1997). Оказалось, разнообразие генов белков E.chaffeensis в 28-кДа является результатом точечных мутаций в гипервариабельных областях, возможно вследствие избирательного иммунного воздействия (Yu et al., 1999a). Представленные данные позволяют предположить, что E.canis был привнесен в Северную Америку относительно недавно, и это может быть объяснением консерватизма, который наблюдали среди изолятов. Консерватизм генов р28 в изолятах E.canis может обеспечить благоприятную возможность для разработки вакцин и серодиагностических антигенов, которые особенно эффективны для предупреждения заболеваний и серодиагностики. Смесь Р28 может обеспечить наиболее надежный серодиагностический тест, однако описано, что серологическая диагностика с одним Р28 можно использовать для иммунодиагностики (Ohashi et al., 1998b; McBride et al., 1999).
Следующие ссылки цитируются в описании.

Любые патенты или публикации, упомянутые в данном описании, указывают на профессиональный уровнь специалистов в данной области, которые имеют отношение к изобретению. Упомянутые патенты и публикации введены в описание путем ссылки в такой степени, как если бы каждая индивидуальная публикация была индивидуально представлена для включения путем ссылки.
Специалист в данной области легко оценит, что данное изобретение удобно адаптировать, чтобы осуществить цели и получить упомянутые результаты и преимущества, а также выполнить собственные задачи. Данные примеры наряду со способами, процедурами, обработками, молекулами и специфическими соединениями, представленными в описании, в настоящее время являются типичными предпочтительными аспектами, являются образцовыми и не предназначены для ограничений рамок изобретения. Изменения аспектов и других применений, которые потребуются специалистами в данной области, будут соответствовать сути изобретения, определенной рамками формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК, КОДИРУЮЩАЯ БЕЛОК EHRLICHIA CANIS С МОЛЕКУЛЯРНОЙ МАССОЙ 30 КИЛОДАЛЬТОН, ВЕКТОР, РЕКОМБИНАНТНЫЙ БЕЛОК И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 1999 |
|
RU2237716C2 |
| ИММУНОДОМИНАНТНЫЙ АНТИГЕННЫЙ БЕЛОК МАССОЙ 120 КДА И ГЕН EHRLICHIA CANIS | 1999 |
|
RU2232814C2 |
| СОСУДИСТЫЙ БЕЛОК - 1 АДГЕЗИИ, ОБЛАДАЮЩИЙ АМИНООКСИДАЗНОЙ АКТИВНОСТЬЮ | 1998 |
|
RU2204838C2 |
| ВАКЦИНЫ ПРОТИВ EHRLICHIA И ИММУНОГЕННЫЕ КОМПОЗИЦИИ | 2020 |
|
RU2824881C2 |
| ПЕСТИЦИДНЫЙ БЕЛОК (ВАРИАНТЫ) | 2000 |
|
RU2267535C2 |
| БЕЛОК СЛИЯНИЯ, СОДЕРЖАЩИЙ ТИМУСНЫЙ СТРОМАЛЬНЫЙ ЛИМФОПОЭТИЧЕСКИЙ БЕЛОК, ВАКЦИНА, СОДЕРЖАЩАЯ ЭКСПРЕССИОННЫЙ ВЕКТОР ДЛЯ ЭКСПРЕССИИ БЕЛКА, И ВАКЦИНА, СОДЕРЖАЩАЯ УКАЗАННЫЙ ВЕКТОР ДЛЯ ЭКСПРЕССИИ | 2012 |
|
RU2617957C2 |
| ПОЛИПЕПТИД ОЖИРЕНИЯ (ОВ)(ВАРИАНТЫ), ЕГО АНАЛОГ (ВАРИАНТЫ) И СЛИТЫЙ БЕЛОК (ВАРИАНТЫ), ИЗОЛИРОВАННАЯ МОЛЕКУЛА НУКЛЕИНОВОЙ КИСЛОТЫ, МОЛЕКУЛА ДНК, РЕКОМБИНАНТНЫЙ ВЕКТОР КЛОНИРОВАНИЯ, РЕКОМБИНАНТНЫЙ ВЕКТОР ЭКСПРЕССИИ, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, МОНОКЛОНАЛЬНОЕ И ПОЛИКЛОНАЛЬНОЕ АНТИТЕЛО | 1995 |
|
RU2273645C9 |
| СОБАЧИЙ ТИМУСНЫЙ СТРОМАЛЬНЫЙ ЛИМФОПОЭТИЧЕСКИЙ БЕЛОК И ЕГО ПРИМЕНЕНИЕ | 2007 |
|
RU2457217C2 |
| СПОСОБ СНИЖЕНИЯ СТЕПЕНИ ГЛИКОЗИЛИРОВАНИЯ БЕЛКОВ, СПОСОБЫ И БЕЛКИ | 2010 |
|
RU2575607C2 |
| ОЧИЩЕННЫЙ ИММУНОГЕННЫЙ БЕЛОК, ЕГО ФРАГМЕНТЫ И ПРОИЗВОДНЫЕ | 1999 |
|
RU2304585C2 |











































Изобретение относится к области молекулярной биологии и ветеринарии. Предложена изолированная последовательность ДНК (варианты), кодирующая белок Ehrlichia canis в 30-килодальтон. Кроме того, предложены вектор, содержащий такую последовательность; рекомбинантный белок Ehrlichia canis в 28 килодальтон, кодируемый этой последовательностью; клетка-хозяин, содержащая ее; способ получения белка; антитело иммунореактивное в отношении этого белка; и способ ингибирования инфекции Ehrlichia canis у субъекта. Рекомбинантный белок Ehrlichia canis в 28 килодальтон является иммунореактивным в отношении сыворотки против Ehrlichia canis. Изобретение может использоваться для разработки вакцин и серодиагностикумов, которые особенно эффективны для предупреждения заболеваний и серодиагностики. 7 н. и 12 з.п. ф-лы, 17 ил.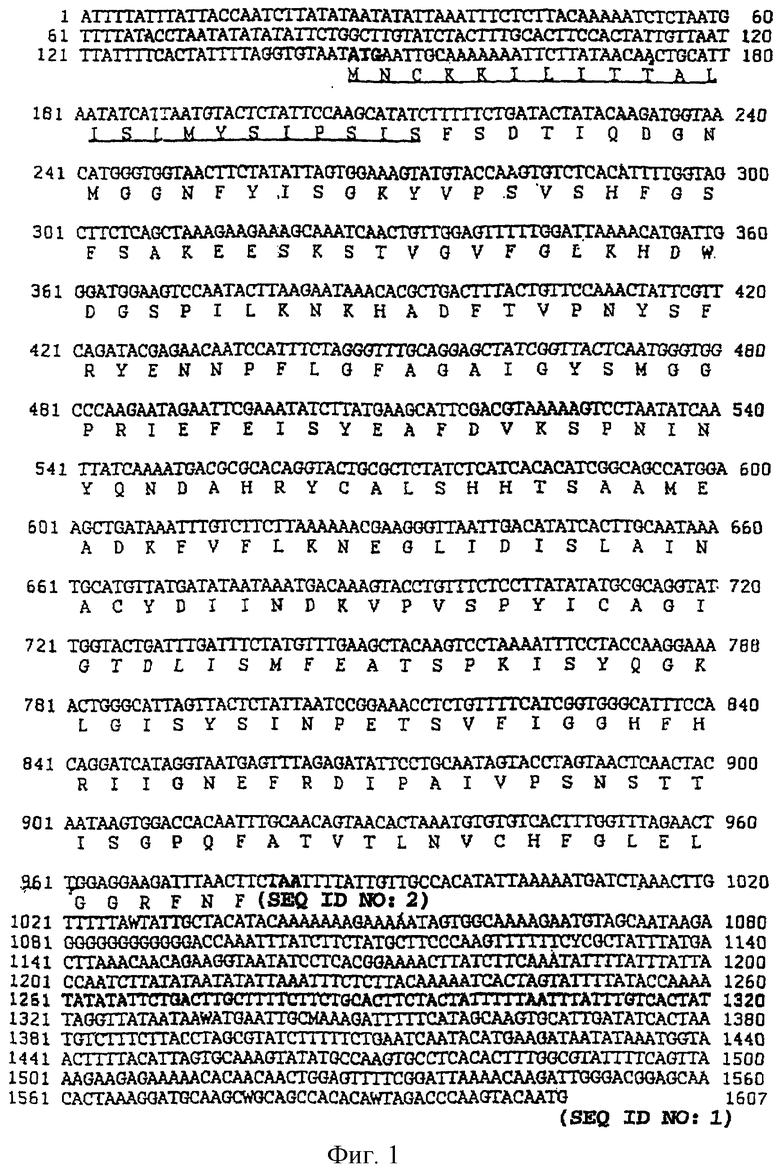
получения вектора, который содержит экспрессирующую область, включающую в себя последовательность, кодирующую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID No.40, 42, 44 и 46, оперативно связанную с промотором; трансфекции указанного вектора в клетку; и культивирования указанной клетки в условиях, эффективных для экспрессии указанной экспрессирующей области.
| УКЛОНОМЕР | 1932 |
|
SU32745A1 |
| REDDY G.R | |||
| et al | |||
| Видоизменение прибора с двумя приемами для рассматривания проекционные увеличенных и удаленных от зрителя стереограмм | 1919 |
|
SU28A1 |
| Biochem | |||
| Biophys | |||
| Res | |||
| Commun | |||
| Способ и аппарат для получения гидразобензола или его гомологов | 1922 |
|
SU1998A1 |

