Предпосылки изобретения
Область изобретения
Настоящее изобретение касается в целом области молекулярной биологии. Более конкретно настоящее изобретение касается молекулярного клонирования и характеристики генов, кодирующих гомологичные белки молекулярной массой 28 кДа, у Ehrlichia canis и мультигенного локуса, кодирующего гомологичные белки массой 28 кДа у Ehrlichia canis, а также их использования.
Описание уровня техники
Эрлихиоз собак, также известный как тропическая панцитопения собак, - переносимый клещами риккетсиоз домашних собак, впервые описанный в Африке в 1935 году и в США в 1963 году (Donatiеn & Lestoquard, 1935; Ewing, 1963). Заболевание стало лучше диагностируемым после эпизоотической вспышки, произошедшей у армейских собак США в ходе Вьетнамской войны (Walker et al., 1970).
Агентом-возбудителем эрлихиоза собак является Ehrlichia canis - небольшая грам-отрицательная облигатно внутриклеточная бактерия, которая проявляет тропизм в отношении моноядерных фагоцитов (Nyindo et al., 1971) и переносится обыкновенным собачьим клещом Rhipicephalus sanguineus (Groves et al., 1975). Развитие эрлихиоза собак подразделяется на три стадии - острую, субклиническую и хроническую. Острая стадия характеризуется лихорадкой, анорексией, депрессией, лимфаденопатией и мягкой тромбоцитопенией (Troy & Forrester, 1990). Обычно собаки выздоравливают после острой стадии, но становятся постоянно инфицированными носителями возбудителя без клинических симптомов заболевания в течение месяцев и даже лет (Harrus et al., 1998). Хроническая стадия, развивающаяся в некоторых случаях, характеризуется тромбоцитопенией, гиперглобулинемией, анорексией, истощением и кровотечениями, в частности носовым кровотечением, с последующей смертью (Troy & Forrester, 1990).
Молекулярно-таксономический анализ, основанный на параметрах гена 163-рРНК, определил, что E.canis и E.chaffeensis, являющаяся возбудителем моноцитарного эрлихиоза человека (МЭХ), близкородственны (Anderson et al., 1991; Andersоn et al., 1992; Dawson et al., 1991; Chen et al., 1994). Сообщалось о существенной перекрестной реактивности антигенов с молекулярными массами 64, 47, 40, 30, 29 и 23 кДа у E.canis и E.chaffeensis (Chen et al., 1994; Chen et al., 1997; Rikihisa et al., 1994; Rikihisa et al., 1992). Анализ методом иммуноблоттинга иммунореактивных антигенов из сывороток человека и собаки, взятых в период выздоровления, позволил идентифицировать многочисленные иммунодоминантные белки E.canis, включая белок с молекулярной массой 30 кДа (Chen et al., 1997). Кроме того, белок массой 30 кДа E.canis был описан как основной иммунодоминантный антиген, распознаваемый ранним иммунным ответом, который по антигенным свойствам отличается от белка массой 30 кДа E.chaffeensis (Rikihisa et al., 1992; Rikihisa et al., 1994). Другие иммунодоминантные белки E.canis с молекулярными массами в диапазоне 20-30 кДа также были идентифицированы (Brouqui et al., 1992; Nyindo et al., 1991; Chen et al., 1994; Chen et al., 1997).
Недавно клонирование и секвенирование мультигенного семейства (оmр-1), кодирующего белки с массой 23-28 кДа, было описано у E.chaffeensis (Ohashi et al., 1998). Был клонирован ген иммунодоминантного белка внешней мембраны массой 28 кДа (р28) у E.chaffeensis, гомологичный гену map-1 Cowdria ruminantium. Мыши, иммунизованные рекомбинантным белком Р28, оказывались защищенными против инфицирования гомологичным штаммом в соответствии с данными ПЦР-анализа периферической крови через 5 дней после инфекции (Ohashi et al., 1998). Также было сообщено о молекулярном клонировании двух сходных, но неидентичных тандемно организованных генов белков массой 28 кДа у E.canis, гомологичных генному семейству оmр-1 E.chaffeensis и гену mар-1 С.ruminantium (Reddy et al., 1998).
Для уровня техники характерно отсутствие данных по клонированию и характеристике новых генов гомологичных иммунореактивных белков массой 28 кДа у Ehrlichia canis и единого мультигенного локуса, включающего гены гомологичных белков массой 28 кДа. Кроме того, в известном уровне техники отсутствуют данные по рекомбинантным белкам, кодируемым такими иммунореактивными генами у Ehrlichia canis. Настоящее изобретение заполняет этот давно существующий пробел и назревшую потребность данной области техники.
Сущность изобретения
Настоящее изобретение описывает молекулярное клонирование, секвенирование, охарактеризование и экспрессию генов гомологичных зрелых иммунореактивных белков массой 28 кДа у Ehrlichia canis (обозначенных Еса28-1, ECa28SA3 и ECa28SA2), a также идентификацию единого локуса (5592 пары нуклеотидов (п.н.)), включающего пять генов белков массой 28 кДа Ehrlichia canis (ECa28SAl, ECa28SA2, ECa28SA3, Еса28-1 и ЕСа28-2). Сравнения с E.chaffeensis и между генами белков массой 28 кДа Е.canis показали, что ЕСа28-1 проявляет наиболее высокий уровень гомологии аминокислотных последовательностей с мультигенным семейством omp-1 E.chaffeensis и высококонсервативен среди изолятов Е.canis. Пять белков массой 28 кДа, как предсказывается, включают сигнальные сегменты, отщепление которых дает зрелые белки, а уровень аминокислотной гомологии находится в диапазоне 51-72%. Анализ межгенных спейсеров выявил гипотетические промоторные участки для каждого гена: это подтверждает, что данные гены могут быть экспрессированы независимо и дифференцированно. Размер некодирующих межгенных спейсеров составляет от 299 до 355 п.н., и они гомологичны на 48-71%.
В одном осуществлении настоящее изобретение относится к последовательностям ДНК, кодирующим иммунореактивный белок массой 30 кДа Ehrlichia canis. Предпочтительно белок имеет аминокислотную последовательность, выбранную из группы, которая включает SEQ ID NO: 2, SEQ ID NO: 4 и SEQ ID NO: 6, а ген имеет нуклеотидную последовательность, выбранную из группы, которая включает SEQ ID NO: 1, SEQ ID NO: 3 и SEQ ID NO: 5, и входит в состав полиморфного мультигенного семейства. Как правило, белок включает N-концевую сигнальную последовательность, которая отщепляется в ходе посттрансляционного процессинга, в результате чего образуется зрелый белок массой 28 кДа. Еще более предпочтительно ДНК, кодирующие белки массой 28 кДа, входят в состав единого мультигенного локуса, размер которого составляет 5592 п.н. и который кодирует все пять гомологичных белков массой 28 кДа у Ehrlichia canis.
В другом осуществлении настоящее изобретение относится к экспрессирующему вектору, включающему ген, кодирующий иммунореактивный белок массой 28 кДа Ehrlichia canis, способный экспрессировать ген по внесении вектора в клетку.
Еще в одном осуществлении настоящее изобретение относится к рекомбинантному белку, включающему аминокислотную последовательность, выбранную из группы, которая включает SEQ ID NO: 2, SEQ ID NO: 4 и SEQ ID NO: 6. Предпочтительно аминокислотная последовательность кодируется последовательностью нуклеиновой кислоты, выбранной из группы, которая включает SEQ ID NO: I, SEQ ID NO: 3 и SEQ ID NO: 5. Предпочтительно рекомбинантный белок включает четыре вариабельных расположенных на клеточной поверхности участка, являющихся гидрофильными и антигенными. Рекомбинантный белок можно использовать в качестве антигена.
Еще в одном осуществлении настоящее изобретение относится к способу получения рекомбинантного белка, включающему стадии получения вектора, который включает экспрессирующий участок, включающий последовательность, которая кодирует аминокислотную последовательность, выбранную из группы, которая включает SEQ ID NO: 2, SEQ ID NO: 4 и SEQ ID NO: 6, функционально присоединенный к промотору; трансфекцию клетки вектором; и культивирование клетки в условиях, эффективных для экспрессии экспрессирующего участка.
Также настоящее изобретение может быть описано в некоторых осуществлениях как способ подавления инфекции Ehrlichia canis у субъекта, включающий стадии: идентификации субъекта, для которого предполагается воздействие или инфекция Ehrlichia canis; и введение композиции, содержащей антиген массой 28 кДа Ehrlichia canis, в количестве, эффективном для подавления инфекции Ehrlichia canis. Подавление может происходить любыми путями, такими как, например, стимуляция у субъекта гуморального или клеточного иммунных ответов, или за счет других путей, таких как подавление нормальной функции антигена массой 28 кДа, или даже конкуренция с антигеном за взаимодействие с некоторым агентом в организме субъекта.
Другие и последующие аспекты, свойства и преимущества настоящего изобретения будут ясны из нижеследующего описания рассматриваемых предпочтительных осуществлении изобретения, приводимых в связи с целью его раскрытия.
Краткое описание чертежей
С учетом того, что материал, в котором понятны указанные выше свойства, преимущества и объекты настоящего изобретения, а также и другое, что становится ясным, изложен и может быть понят в деталях, более подробное описание изобретения, краткое содержание которого было дано выше, может быть выполнено путем ссылки на некоторые его осуществления, которые проиллюстрированы на прилагающихся чертежах. Эти чертежи образуют часть данной заявки. Необходимо отметить, однако, что прилагающиеся чертежи иллюстрируют предпочтительные осуществления настоящего изобретения и, следовательно, не призваны ограничить его объем.
На фигуре 1 показаны нуклеотидная последовательность (SEQ ID NO: 1) и расшифрованная аминокислотная последовательность (SEQ ID NO: 2) гена ЕСа28-1, включая фланкирующие 5’- и 3’-некодирующие последовательности. Старт-кодон ATG и стоп-кодон ТАА выделены жирным шрифтом, а 23-аминокислотная лидерная сигнальная последовательность подчеркнута.
На фигуре 2 показаны данные электрофореза в ДСН-ПААГ экспрессированного рекомбинантного химерного белка ЕСа28-1-тиоредоксина с молекулярной массой 50 кДа (дорожка 1, стрелка) и контрольного тиоредоксина массой 16 кДа (дорожка 2, стрелка), а также соответствующий иммуноблот рекомбинантного химерного белка ЕСа28-1-тиоредоксина, распознанного антисывороткой собаки, находящейся на стадии выздоровления от заражения E.canis (дорожка 3). Контрольный тиоредоксин антисывороткой к E.canis не выявлялся (не показано).
На фигуре 3 показано сопоставление аминокислотных последовательностей белка ЕСа28-1 (SEQ ID NO: 2) и ECa28SA2 (частичная последовательность: SEQ ID NO: 7) и ECa28SAl (SEQ ID NO: 8), P28 E.chaffeensis (SEQ ID NO: 9), семейства ОМР-1 E.chaffeensis (SEQ ID NO: 10-14) и MAP-1 С.ruminantium (SEQ ID NO: 15). Аминокислотная последовательность ЕСа28-1 представлена как консенсусная последовательность. Не показаны аминокислоты, идентичные аминокислотам ЕСа28-1 - они даны точками. Отличающиеся аминокислоты показаны соответствующими однобуквенными обозначениями. Пробелы, внесенные для соблюдения максимального соответствия аминокислотных последовательностей, даны черточками. Вариабельные сегменты подчеркнуты и обозначены (VR1, VR2, VR3 и VR4). Стрелки указывают на предполагаемый сигнал сайта пептидазного отщепления сигнального пептида.
На фигуре 4 показаны филогенетические отношения ЕСа28-1 с ЕСа28SА2 (частичная последовательность) и ECa28SAl E.canis, шестью членами мультигенного семейства omp-1 E.chaffeensis и mар-1 C.ruminantium по расшифрованным аминокислотным последовательностям с использованием конструирования несбалансированного дерева. Длина каждой пары ветвей соответствует расстоянию между аминокислотными последовательностями в парах. Масштабная линейка определяет расстояние между последовательностями.
На фигуре 5 показаны данные Саузерн-блоттинга геномной ДНК E.canis, полностью расщепленной шестью отдельными рестриктазами и гибридизованной на помеченный дигоксигенином зонд ЕСа28-1 (дорожки 2-7); помеченные дигоксигенином маркеры молекулярной массы (дорожки 1 и 8).
На фигуре 6 отображено сравнение характеристик предсказанных белков ЕСа28-1 (штамм Jake) и Р28 E.chaffeensis (штамм Arkansas). Поверхностная вероятность предсказывает поверхностные остатки с использованием гексапептидного окна. Поверхностным остатком является остаток с превышением 2,0 нм2 по площади поверхности, доступной для воды. Гексапептид, характеризующийся величиной более 1, рассматривался как поверхностный участок. Антигенный индекс предсказывает потенциальные антигенные детерминанты. Участки, характеризующиеся величиной выше 0, являются потенциальными антигенными детерминантами. Т-клеточный мотив локализует потенциальные Т-клеточные антигенные детерминанты с использованием мотива из 5 аминокислот: остаток 1 - глицин или полярный, остаток 2 - гидрофобный, остаток 3 - гидрофобный, остаток 4 - гидрофобный или пролин и остаток 5 - полярный или глицин. Линейка указывает на положения аминокислот.
На фигуре 7 показаны последовательности нуклеиновых кислот и расшифрованные аминокислотные последовательности генов белков массой 28 кДа E.canis - ECa28SA2 (нуклеотиды 1-849: SEQ ID NO: 3; аминокислотная последовательность: SEQ ID NO: 4) и ECa28SA3 (нуклеотиды 1195-2031: SEQ ID NO: 5; аминокислотная последовательность SEQ ID NO: 6), включая межгенные некодирующие последовательности (NC2, нуклеотиды 850-1194: SEQ ID NO: 31). Старт-кодон ATG и стоп-кодоны выделены жирным шрифтом.
На фигуре 8 схематически показаны пять генов локуса белков массой 28 кДа E.canis (5592 п.н.), что указывает на геномную ориентацию и межгенные некодирующие участки (28NC1-4). Гены белков массой 28 кДа, показанные в локусе 1 и 2 (затенены), были описаны ранее (McBride et al., 1999; Reddy et al., 1998; Ohashi et al., 1998). Была определена полная последовательность ECaSA2 и гена нового белка массой 28 кДа, обозначенного ECa28SA3 (без затенения). Некодирующие межгенные участки (28NC2-3) между ECaSA2, ECa28SA3 и ЕСа28-1 были определены полностью путем соединения ранее несвязывавшихся локусов 1 и 2.
На фигуре 9 показаны филогенетические отношения пяти генов белков массой 28 кДа E.canis по параметрам аминокислотных последовательностей с использованием конструирования несбалансированного дерева. Длина каждой пары ветвей соответствует расстоянию между парами аминокислот. Масштабная линейка определяет расстояние между последовательностями.
На фигуре 10 показано сопоставление последовательностей нуклеиновых кислот некодирующего межгенного спейсера генов белков массой 28 кДа E.canis (SEQ ID NO: 30-33). Непоказанные нуклеиновые кислоты, обозначенные точкой (.), идентичны некодирующему участку 1 (28NC1). Дивергенция показана с использованием соответствующих однобуквенных обозначений. Пробелы, внесенные для максимально информативного сопоставления аминокислотных последовательностей, обозначены черточкой (-). Предполагаемые участки промоторов транскрипции (-10 и -35) и сайты рибосомного связывания (RBS) обведены.
Подробное описание изобретения
Настоящее изобретение относится к клонированию, секвенированию и экспрессии гомологичных генов, кодирующих белок Ehrlichia canis с молекулярной массой 30 килодальтон (кДа). Также был проведен сравнительный молекулярный анализ гомологичных генов у семи изолятов A.canis и мультигенного семейства omp-1 E.chaffeensis. Идентифицированы два новых гена, кодирующих белки массой 28 кДа, - ЕСа28-1 и ECa28SA3. ЕСа28-1 имеет 834-нуклеотидную открытую рамку считывания, кодирующую белок из 278 аминокислот (SEQ ID NO: 2) с расчетной молекулярной массой 30,5 кДа. Была идентифицирована N-концевая сигнальная последовательность: это подтверждает, что белок характеризуется посттрансляционной модификацией в зрелый белок массой 27,7 кДа. ECa28SA3 имеет открытую рамку из 840 п.н., которая кодирует белок из 280 аминокислот (SEQ ID NO: 6).
С использованием ПЦР для амплификации генов белков массой 28 кДа E.canis ранее несеквенированный участок ECa28SA2 был определен полностью. Анализ последовательности ECa28SA2 показал наличие 849-нуклеотидной открытой рамки считывания, кодирующей белок из 283 аминокислот (SEQ ID NO: 4). В результате ПЦР-амплификации с использованием праймеров, специфичных для межгенных некодирующих участков генов белков массой 28 кДа, были соединены два ранее разделявшихся локуса с идентификацией единого локуса (5592 п.н.), включающего все пять генов белков массой 28 кДа. Пять белков массой 28 кДа, как было предсказано, включают сигнальные сегменты, обусловливающие образование зрелых белков, а уровень их аминокислотной гомологии составляет 51-72%. Анализ межгенных участков (спейсеров) указал на предполагаемые промоторные участки для каждого гена: это подтверждает то, что эти гены могут экспрессироваться независимо и дифференцированно. Размер межгенных некодирующих участков (28NCl-4) - в пределах от 299 до 355 п.н. при уровне гомологии 48-71%.
Настоящее изобретение относится к двум новым гомологичным генам белков массой 28 кДа у Ehrlichia canis - Еса28-1 и ECa28SA3 - и на полную последовательность ранее частично секвенированного ECa28SA2. Также заявляется мультигенный локус, кодирующий все пять гомологичных внешнемембранных белков массой 28 кДа Ehrlichia canis.
В одном осуществлении настоящее изобретение относится к последовательности ДНК, кодирующей иммунореактивный белок массой 30 кДа Ehrlichia canis. Предпочтительно белок характеризуется аминокислотной последовательностью, выбранной из группы, которая включает SEQ ID NO: 2, SEQ ID NO: 4 и SEQ ID NO: 6, а ген характеризуется нуклеотидной последовательностью, выбранной из группы, которая включает SEQ ID NO: 1, SEQ ID NO: 3 и SEQ ID NO: 5, и входит в состав полиморфного мультигенного семейства. Более предпочтительно белок имеет N-концевую сигнальную последовательность, которая отщепляется в ходе посттрансляционного процессинга, в результате чего образуется зрелый белок массой 28 кДа. Еще предпочтительнее ДНК, кодирующие белки массой 28 кДа, входят в состав единого мультигенного локуса, размер которого составляет 5592 п.н. и который кодирует все пять гомологичных белков массой 28 кДа у Ehrlichia canis.
В другом осуществлении настоящее изобретение относится к экспрессирующему вектору, включающему ген, который кодирует иммунореактивный белок массой 28 кДа Ehrlichia canis, и способный экспрессировать ген после внесения вектора в клетку.
Еще в одном осуществлении настоящее изобретение относится к рекомбинантному белку, включающему аминокислотную последовательность, выбранную из группы, которая включает SEQ ID NO: 2, SEQ ID NO: 4 и SEQ ID NO: 6. Предпочтительно аминокислотная последовательность кодируется последовательностью нуклеиновой кислоты, выбранной из группы, которая включает SEQ ID NO: 1, SEQ ID NO: 3 и SEQ ID NO: 5. Предпочтительно рекомбинантный белок включает четыре вариабельных сегмента, которые находятся на поверхности клетки и являются гидрофильными и антигенными. Еще предпочтительнее рекомбинантный белок является антигеном.
Еще в одном осуществлении настоящее изобретение относится к способу выработки рекомбинантного белка, включающему стадии формирования вектора, который включает экпрессирующий участок, включающий последовательность, которая кодирует аминокислотную последовательность, выбранную из группы, которая включает SEQ ID NO: 2, SEQ ID NO: 4 и SEQ ID NO: 6, функционально присоединенный к промотору; трансфекцию вектором клетки; и культивирование данной клетки в условиях, эффективных по экспрессии экспрессирующего участка.
Также настоящее изобретение может быть описано некоторыми вариантами в виде способа подавления инфекции Ehrlichia canis у субъекта, включающего стадии: идентификации субъекта, предположительно находящегося под влиянием или инфицированного Ehrlichia canis; и введение композиции, содержащей антиген массой 28 кДа Ehrlichia canis в количестве, эффективном по подавлению инфекции Ehrlichia canis. Подавление может происходить за счет любого пути, например за счет стимуляции у субъекта гуморального или клеточного иммунных ответов, или за счет других путей, таких как подавление нормальной функции антигена массой 28 кДа или даже конкуренция с антигеном за взаимодействие с некоторым агентом в организме субъекта.
В соответствии с настоящим изобретением могут быть применены стандартные методы молекулярной биологии, микробиологии и технологий рекомбинантных ДНК, известных специалистам в данной области техники. Такие методы полностью описаны в научной литературе: см., например, Maniatis, Fritsch & Sambrook, "Molecular Cloning: A Laboratory Manual" (1982); "DNA Cloning: A Practical Approach", Volumes I & II (D.N.Glover, ed., 1985); "Oligonucleotide Synthesis" (M.J.Gait, ed., 1984); "Nucleic Acid Hybridization" [B.D.Hames & S.L.Higgins, eds. (1985)]; "Transcription and Translation" [B.D.Hames & S.L.Higgins, eds. (1984)]; "Animal Cell Culture" [R.I.Freshney, ed. (1986)]; "Immobilized Cells and Enzymes" [IRL Press (1986)]; B.Perbal, "A Practical Guide to Molecular Cloning" (1984).
Так, по мере появления в данном тексте следующие термины должны быть определены в соответствии с указанным ниже.
“Репликон” обозначает любой генетический элемент (например, плазмиду, хромосому, вирус), который функционирует в качестве автономной единицы процесса репликации ДНК in vivo: т.е. способен реплицироваться под своим собственным контролем.
“Вектор” обозначает репликон, такой как плазмида, фаг или космида, к которой может быть присоединен другой сегмент ДНК таким образом, чтобы обеспечить репликацию присоединенного сегмента.
“Молекула ДНК” обозначает полимерную форму дезоксирибонуклеотидов (аденин, гуанин, тимин или цитозин) либо в одноцепочечной форме, либо в виде двухцепочечной спирали. Этот термин обозначает только первичную и вторичную структуру молекулы и не ограничивает ее по какой-либо конкретной четвертичной форме. Таким образом, данный термин включает двухцепочечную ДНК, обнаруживаемую, помимо прочего, в линейных молекулах ДНК (например, рестрикционных фрагментах, вирусах, плазмидах и хромосомах). При обсуждении структуры в данном тексте в соответствии с принятой нормой дается только последовательность в направлении от 5’ к 3’ в виде нетранскрибируемой (т.е. кодирующей) цепи ДНК (т.е. цепи, имеющей такую же последовательность, как и у мРНК).
“Кодирующая последовательность” ДНК обозначает двухцепочечную последовательность ДНК, которая транскрибируется и транслируется в полипептид in vivo при попадании под контроль подходящих регуляторных последовательностей. Границы кодирующей последовательности определяются старт-кодоном в 5’- конце (N-конце) и стоп-кодоном в 3’-конце (С-конце). Кодирующая последовательность может включать, тем самым не ограничиваясь, прокариотические последовательности, кДНК с эукариотических мРНК, геномные последовательности ДНК эукариот (например, млекопитающих) и даже синтетические последовательности ДНК. Сигнал полиаденилирования и сайт терминации транскрипции обычно должны быть расположены с 3’-стороны от кодирующей последовательности.
Контролирующие транскрипцию и трансляцию последовательности являются регуляторными последовательностями ДНК, такими как промоторы, энхансеры, сигналы полиаденилирования, терминаторы и подобное, которые обеспечивают экспрессию кодирующей последовательности в клетке-хозяине.
“Промоторная последовательность” обозначает ДНК-регуляторный участок, способный связывать РНК-полимеразу в клетке и инициировать транскрипцию расположенной ниже (в направлении 3’) кодирующей последовательности. Для целей определения в настоящем изобретении промоторная последовательность связана по ее 3’-концу с сайтом инициации транскрипции и продолжается вверх (в направлении 5’) так, чтобы включить минимальное число нуклеотидов или элементов, необходимых для инициации транскрипции на уровнях, отчетливо превышающий фоновый уровень. В пределах промоторной последовательности можно найти сайт инициации транскрипции, а также домены белкового связывания (консенсусные последовательности), отвечающие за связывание РНК-полимеразы. Часто, но не всегда, промоторы у эукариот включают боксы ТАТА и боксы CAT. Прокариртические промоторы включают последовательности Шайна-Дальгарно в дополнение к консенсусным последовательностям в положениях -10 и -35.
“Контролирующая экспрессию последовательность” обозначает последовательность ДНК, которая контролирует и регулирует транскрипцию и трансляцию другой последовательности ДНК. Кодирующая последовательность находится “под контролем” последовательностей-регуляторов транскрипции и трансляции в клетке тогда, когда РНК-полимера за транскрибирует кодирующую последовательность в мРНК, которая затем транслируется в белок, кодируемый кодирующей последовательностью.
“Сигнальная последовательность” может быть включена рядом с кодирующей последовательностью. Данная последовательность кодирует сигнальный сегмент, находящийся в N-концевой части полипептида и обеспечивающий клетке-хозяину перенесение полипептида на клеточную поверхность или секрецию полипептида в окружающую среду, причем данный сигнальный сегмент отщепляется клеткой-хозяином перед тем, как белок покидает клетку. Сигнальные последовательности могут быть обнаружены в связи с различными белками, нативными для прокариот и эукариот.
Термин “олигонуклеотид”, по использованию в данном тексте обозначающий зонд по настоящему изобретению, определяет молекулу, состоящую из двух или большего числа рибонуклеотидов, предпочтительно из более чем трех. Его точная длина будет зависеть от многих факторов, которые, в свою очередь, зависят от окончательной функции и использования олигонуклеотида.
Термин “праймер” по использованию в данном тексте обозначает олигонуклеотид, как встречающийся в естественных условиях в виде очищенного рестрикционного фрагмента, так и полученный синтетически, который способен выполнять роль сайта инициации синтеза при попадании в условия, в которых запускается синтез достраиваемого с праймера продукта, комплементарного цепи нуклеиновой кислоты (матрице), т.е. в присутствии нуклеотидов и индуцирующего агента, такого как ДНК-полимераза, и при подходящих температуре и рН. Праймеp может быть либо одноцепочечным, либо двухцепочечным и должен быть достаточно длинным для того, чтобы затравлять синтез желательного достроенного продукта в присутствии индуцирующего агента. Точная длина праймера будет зависеть от многих факторов, включая температуру, источник праймера и используемый метод. Например, для диагностических применений в зависимости от сложности последовательности-мишени олигонуклеотидный праймер обычно состоит из 15-25 или большего числа нуклеотидов, хотя он может включать и меньшее число нуклеотидов.
Праймеры в настоящей заявке выбирают как “существенно” комплементарные разным цепям конкретной последовательности-мишени ДНК. Это означает, что праймеры должны быть комплементарны настолько, чтобы гибридизовать с соответствующими им цепями. Следовательно, последовательность праймера необязательно должна отражать точную последовательность матрицы. Например, некомплементарный нуклеотидный фрагмент может быть присоединен к 5’-концу праймера, а остальная часть последовательности праймера будет комплементарна данной цепи. Как альтернатива, некомплементарные нуклеотиды или более длинные последовательности могут быть диспергированы по праймеру, лишь бы эта последовательность праймера была по существу комплементарной последовательности или гибридизовала бы с ней, тем самым образуя матрицу для синтеза достроенного продукта.
Клетка тогда считается “трансформированной” экзогенной или гетерологичной ДНК, когда такая ДНК была внесена внутрь клетки. Трансформирующая ДНК может интегрироваться (путем ковалентного связывания) или не интегрироваться в геном клетки. У прокариот, дрожжей и клеток млекопитающих, например, трансформирующая ДНК может поддерживаться в виде эписомного элемента, такого как плазмида. В случае эукариотических клеток стабильно трансформированной клеткой является клетка, в которой трансформирующая ДНК становится интегрированной в хромосому таким образом, чтобы она передавалась дочерним клеткам в результате хромосомной репликации. Такая стабильность отражается способностью эукариотической клетки формировать клеточные линии или клоны, включающие популяцию дочерних клеток, несущих трансформирующую ДНК. “Клоном” является популяция клеток, образующаяся от единственной клетки или предка в результате митозов. “Клеточная линия” - это клон первичной клетки, который способен стабильно расти in vitro в течение многих поколений.
Две последовательности ДНК “по существу гомологичны” тогда, когда по крайней мере примерно 75% (предпочтительно по крайней мере примерно 80% и наиболее предпочтительно по крайней мере примерно 90% или 95%) нуклеотидов совпадают на определенном участке последовательностей ДНК. Последовательности, которые по существу гомологичны, могут быть идентифицированы путем сравнения последовательностей с использованием стандартных компьютерных программ на материале последовательностей из баз данных или с помощью метода Саузерн-блоттинга, например, в жестких условиях гибридизации, определенных для данной конкретной системы. Определение подходящих условий гибридизации известно специалистам в данной области техники: см., например, Maniatis et al., цит. выше; "DNA Cloning", Vol.I & II, цит. выше; "Nucleic Acid Hybridization", цит. выше.
“Гетерологичный” участок в ДНК-конструкции - это идентифицируемый сегмент ДНК в большей молекуле ДНК, для которого неизвестна связь с данной более крупной молекулой в природе. Таким образом, когда гетерологичный участок кодирует ген млекопитающего, данный ген обычно фланкирован ДНК, которая не фланкирует ДНК генома млекопитающего в геноме организма-источника. В другом примере кодирующей последовательностью является конструкция, в которой сама кодирующая последовательность в природе не обнаруживается (например, кДНК, в которой геномная кодирующая последовательность включает интроны, или синтетические последовательности, включающие кодоны, отличающиеся от таковых в нативном гене). Аллельные варианты или естественные мутационные события в связи с данным определением не образуют гетерологичного сегмента ДНК.
Метками, наиболее обычно используемыми для данных исследований, являются радиоактивные элементы, ферменты, химические агенты, которые флуоресцируют при облучении ультрафиолетом, и другие. Известен ряд флуоресцентных материалов, которые можно использовать в качестве меток. Ими являются, например, флуоресцеин, родамин, аурамин, техасский красный, АМСА голубой и люциферовый желтый. Конкретным детекционным материалом является антикроличье антитело, генерированное у коз и соединенное с флуоресцеином через изотиоцианат.
Также белки могут быть помечены радиоактивным изотопом или ферментом. Радиоактивная метка может быть выявлена с помощью любой из доступных в настоящее время процедур. Предпочтительный изотоп можно выбрать из 3H,14С, 32Р, 35S, 36Cl, 51Cr, 57Co, 58Co, 59Fe, 90Y, 125I, 131I и 186Re.
Также применимы ферментные метки: они могут быть выявлены с помощью любого из существующих колориметрических, спектрофотометрических, флуороспектрофотометрических, амперометрических или газометрических методов. Фермент соединяют с отобранной частицей с помощью реакции с промежуточной молекулой, такой как карбодиимиды, диизоцианаты, глутаровый альдегид и подобное. Известны и применимы многие ферменты, которые можно использовать в данных процедурах. Предпочтительными являются пероксидаза, β-глюкуронидаза, β-D-глюкозидаза, β-D-галактозидаза, уреаза, глюкооксидаза с пероксидазой и щелочная фосфатаза. В качестве примера можно сослаться на патенты США № 3654090, 3850752 и 4016043 в связи с изложенными в них альтернативными материалами и способами мечения.
По использованию в данном тексте термин “хозяин” призван включить не только прокариот, но также и эукариот, таких как клетки дрожжей, растений и животных. Рекомбинантную молекулу ДНК или ген, которые кодируют иммунореактивный белок массой 28 кДа Ehrlichia canis по настоящему изобретению, можно использовать для трансформации хозяина с использованием любого из методов, хорошо известных специалистам в данной области техники. Наиболее предпочтительным является использование вектора, включающего кодирующие последовательности гена, кодирующего иммунореактивный белок массой 28 кДа Ehrlichia canis по настоящему изобретению, для целей трансформации прокариот.
Прокариотическими хозяевами могут являться E.coli, S.typhimurium, Serratia marcescens и Bacillus subtilis. Эукариотическими хозяевами являются дрожжи, такие как Pichia pastoris, клетки млекопитающих и клетки насекомых.
В целом, экспрессирующие векторы, включающие промоторные последовательности, которые обеспечивают эффективную транскрипцию встроенного фрагмента ДНК, используют согласованно с хозяином. Обычно экспрессирующий вектор включает сайт начала репликации, промотор (промоторы), терминатор (терминаторы), а также конкретные гены, которые способны обеспечивать фенотипический отбор в трансформированных клетках. Трансформированные хозяева могут быть ферментированы и прокультивированы в соответствии со способами, известными в данной области техники в связи с достижением оптимального роста клеток.
Настоящее изобретение включает по существу чистую ДНК, кодирующую иммунореактивный белок массой 28 кДа Ehrlichia canis, цепь которой будет гибридизовать при высокой жесткости с зондом, включающим последовательность из по крайней мере 15 расположенных подряд нуклеотидов из SEQ ID NO: 1 или SEQ ID NO: 3, или SEQ ID NO: 5. Белок, кодируемый ДНК по настоящему изобретению, может характеризоваться по крайней мере 80%-ным уровнем идентичности последовательности (предпочтительно 85%, более предпочтительно 90% и наиболее предпочтительно 95%) с аминокислотами, перечисленными в SEQ ID NO: 2 или SEQ ID NO: 4, или SEQ ID NO: 6. Более предпочтительно ДНК включает кодирующую последовательность нуклеотидов SEQ ID NO: 1 или SEQ ID NO: 3, или SEQ ID NO: 5, или вырожденный вариант такой последовательности.
Зонд, с которым ДНК по настоящему изобретению гибридизует, предпочтительно состоит из по крайней мере 20 расположенных подряд нуклеотидов, более предпочтительно 40 нуклеотидов, даже более предпочтительно 50 нуклеотидов и наиболее предпочтительно 100 нуклеотидов или больше (вплоть до полного размера) из кодирующей последовательности нуклеотидов, перечисленных в SEQ ID NO: 1, или SEQ ID NO: 3, или SEQ ID NO: 5, или их комплементов. Такой зонд применим для выявления экспрессии иммунореактивного белка массой 28 кДа Ehrlichia canis в клетке человека с помощью способа, включающего стадии (а) контакта мРНК, полученной из клетки, с помеченным гибридизационным зондом; и (b) выявление гибридизации зонда с мРНК.
Настоящее изобретение также относится к существенно чистой ДНК, включающей последовательность по крайней мере из 15 расположенных подряд нуклеотидов (предпочтительно 20, более предпочтительно 30, даже более предпочтительно 50 и наиболее предпочтительно все) из участка нуклеотидов, перечисленных в SEQ ID NO: 1, или SEQ ID NO: 3, или SEQ ID NO: 5.
Под “высокой жесткостью” понимаются условия гибридизации ДНК и промывки, отличающиеся высокой температурой и низкой солевой концентрацией, например условиями промывки при 65°С при концентрации солей примерно 0,1×SSC или их функциональных эквивалентов. Например, условия высокой жесткости могут включать гибридизацию при примерно 42°С в присутствии примерно 50% формамида; первую промывку при примерно 65°С примерно в 2× SSC, содержащем 1% ДСН; с последующей второй промывкой при примерно 65°С примерно в 0,1× SSC.
Под “по существу чистой ДНК” подразумевается ДНК, которая перестала быть частью среды, в которой ДНК встречается в нативных условиях, в результате отделения (частичной или полной очистки) от некоторых или всех молекул этой среды или в результате изменения последовательностей, которые фланкируют заявленную ДНК. Следовательно, данный термин охватывает, например, рекомбинантную ДНК, которая встроена в вектор, в автономно реплицирующиеся плазмиду или вирус или в геномную ДНК прокариотического или эукариотического организма; или которая существует в виде отдельной молекулы (например, кДНК или фрагмент геномной или кДНК, полученный в полимеразной цепной реакции (ПЦР) или при расщеплении рестрикционными эндонуклеазами) независимо от других последовательностей. Также он охватывает рекомбинантную ДНК, которая является частью химерного гена, кодирующего дополнительную пептидную последовательность, например химерный белок. Также охватывается рекомбинантная ДНК, которая включает часть нуклеотидов, перечисленных в SEQ ID NO: 1, или SEQ ID NO: 3, или SEQ ID NO: 5, которая кодирует альтернативный сплайсинговый вариант гена, кодирующего иммунореактивный белок массой 28 кДа Ehrlichia canis.
ДНК может характеризоваться по крайней мере примерно 70%-ной идентичностью последовательности по сравнению с кодирующей последовательностью нуклеотидов, перечисленных SEQ ID NO: 1, или SEQ ID NO: 3, или SEQ ID NO: 5, предпочтительно по крайней мере 75%-ной (например, по крайней мере 80%-ной), а наиболее предпочтительно по крайней мере 90%-ной. Идентичность двух последовательностей является прямой функцией числа совпадающих или идентичных положений. Когда положение мономера в обеих последовательностях занято одним и тем же мономером, например, если данное положение занято аденином в двух молекулах ДНК, то они идентичны по этому положению. Например, если 7 положений в последовательности из 10 нуклеотидов идентичны соответствующим положениям во второй 10-нуклеотидной последовательности, то эти две последовательности характеризуются 70%-ной идентичностью последовательностей. Длина сравниваемых последовательностей в целом должна составлять по крайней мере 50 нуклеотидов, предпочтительно по крайней мере 60 нуклеотидов, более предпочтительно по крайней мере 75 нуклеотидов и наиболее предпочтительно 100 нуклеотидов. Идентичность последовательностей обычно определяют с использованием компьютерной программы для анализа последовательностей (например, пакет программ для анализа последовательностей от Genetics Computer Group, University Wisconsin Biotechnology Center, 1710 University av., Madison, WI 53705).
Настоящее изобретение относится к вектору, включающему кодирующую последовательность ДНК гена, который кодирует иммунореактивный белок массой 28 кДа Ehrlichia canis, где указанный вектор способен реплицироваться в хозяине, который включает в функциональной связи: (а) сайт начала репликации; (b) промотор и (с) последовательность ДНК, кодирующую упомянутый белок. Предпочтительно вектор по настоящему изобретению включает часть последовательности ДНК, показанной в SEQ ID NO: 1, или SEQ ID NO: 3, или SEQ ID NO: 5.
“Вектор” может быть определен как реплицирующаяся конструкция нуклеиновой кислоты, например плазмида или вирусная нуклеиновая кислота. Векторы могут быть использованы для амплификации и/или экспрессии нуклеиновой кислоты, кодирующей иммунореактивный белок массой 28 кДа Ehrlichia canis. Экспрессирующий вектор является способной к репликации конструкцией, в которой последовательность нуклеиновой кислоты, кодирующей полипептид, функционально присоединена к подходящим контрольным последовательностям, способным обеспечивать экспрессию полипептида в клетке. Потребность в таких регуляторных последовательностях будет варьироваться в зависимости от выбранной клетки и выбранного способа трансформации. В целом, регуляторные последовательности включают транскрипционный промотор и/или энхансер, подходящие сайты связывания мРНК на рибосомах и последовательности, которые контролируют терминацию транскрипции и трансляции. Способы, которые хорошо известны специалистам в данной области техники, могут быть использованы для конструирования экспрессирующих векторов, включающих подходящие транскрипционные и трансляционные регуляторные элементы: см., например, методы, описанные у Sambrook et al., 1989, "Molecular Cloning: A Laboratory Manual" (2nd ed.). Cold Spring Harbor Press, NY. Ген и его транскрипционные регуляторные последовательности определяются как “функционально присоединенные” тогда, когда транскрипционные регуляторные последовательности эффективно контролируют транскрипцию гена. Векторы по настоящему изобретению включают, тем самым не ограничиваясь, плазмидные векторы и вирусные векторы. Предпочтительными вирусными векторами по настоящему изобретению являются те векторы, которые происходят от ретровирусов, аденовирусов, аденоассоциированного вируса, вируса SV40 или герпесвирусов.
Под “существенно чистым белком” понимается белок, который был отделен от по крайней мере некоторых из тех компонентов, которые сопровождают его в нативных условиях. Обычно белок является существенно чистым, если он по крайней мере на 60% по массе свободен от белков и других встречающихся в естественных условиях органических молекул, с которыми он нативно связан in vivo. Предпочтительно чистота препарата составляет по крайней мере 75%, более предпочтительно по крайней мере 90% и наиболее предпочтительно по крайней мере 99% по массе. Существенно чистый иммунореактивный белок массой 28 кДа Ehrlichia canis может быть получен, например, путем экстракции из естественного источника; путем экспрессии рекомбинантной нуклеиновой кислоты, кодирующей иммунореактивный белок массой 28 кДа Ehrlichia canis; или путем химического синтеза белка. Чистота может быть определена с помощью любого подходящего метода, например, хроматографии на колонках, такой как иммуноаффинная хроматография с использованием антитела, специфичного в отношении иммунореактивного белка массой 28 кДа Ehrlichia canis, электрофореза в полиакриламидном геле или ВЭЖХ-анализа. Белок является существенно свободным от компонентов, связанных с ним в нативных условиях, когда он отделен по крайней мере от части тех загрязняющих компонентов, которые сопровождают его в естественном состоянии. Таким образом, белок, который химически синтезирован или выработан в клеточной системе, отличающейся от клетки, от которой он происходит в естественных условиях, по определению будет существенно свободным от компонентов, связанных с ним в естественных условиях. Следовательно, существенно чистые белки включают эукариотические белки, синтезированные в E.coli, других прокариотах или любом ином организме, в котором они в естественных условиях не встречаются.
В дополнение к существенно полноразмерным белкам настоящее изобретение также представляет фрагменты (например, фрагменты со свойствами антигенов) иммунореактивного белка массой 28 кДа Ehrlichia canis (SEQ ID NO: 2 или SEQ ID NO: 4, или SEQ ID NO: 6). По использованию в данном тексте “фрагмент” в приложении к полипептиду будет обозначать по крайней мере 10 остатков, обычнее по крайней мере 20 остатков и предпочтительнее по крайней мере 30 (например, 50) остатков в длину, но меньше полной, интактной последовательности. Фрагменты иммунореактивного белка массой 28 кДа Ehrlichia canis могут быть получены с помощью методов, известных специалистам в данной области техники, например с помощью каталитического расщепления нативного или рекомбинантного иммунореактивного белка массой 28 кДа Ehrlichia canis, с помощью технологий рекомбинантных ДНК с использованием экспрессирующего вектора, который кодирует определенный фрагмент иммунореактивного белка массой 28 кДа Ehrlichia canis, или с помощью химического синтеза. Способность фрагмента-"кандидата" проявлять свойство иммунореактивного белка массой 28 кДа Ehrlichia canis (например, свойства связывания с антителом, специфичным для иммунореактивного белка массой 28 кДа Ehrlichia canis) может быть оценена с помощью описанных в данном тексте способов. Очищенный иммунореактивный белок массой 28 кДа Ehrlichia canis или антигенные фрагменты иммунореактивного белка массой 28 кДа Ehrlichia canis могут быть использованы для получения новых антител или для анализа существующих антител (например, в качестве позитивных контролей в диагностическом тесте) согласно стандартным прописям, известным специалистам в данной области техники. Настоящим изобретением охватываются поликлональные антисыворотки, полученные с использованием иммунореактивного белка массой 28 кДа Ehrlichia canis или фрагмента иммунореактивного белка массой 28 кДа Ehrlichia canis в качестве иммуногена в отношении, например, кроликов. Используют стандартные протоколы получения моноклонального и поликлонального антитела, известные специалистам в данной области техники. Моноклональные антитела, полученные с помощью такой процедуры, могут быть протестированы по способности идентифицировать рекомбинантные клоны кДНК Ehrlichia canis и отличать их от известных клонов кДНК.
Далее настоящее изобретение относится к фрагментам иммунореактивного белка массой 28 кДа Ehrlichia canis, которые кодируются по крайней мере участками SEQ ID NO: 1, или SEQ ID NO: 3, или SEQ ID NO: 5, например, продуктами альтернативного сплайсинга мРНК или альтернативных событий процессинга белков или продуктами, в которых участок последовательности был делетирован. Фрагмент или интактный иммунореактивный белок массой 28 кДа Ehrlichia canis может быть ковалентно соединен с другим полипептидом, который, например, выполняет роль метки, лиганда или средства для повышения антигенности.
Выражение “фармацевтически приемлемый” обозначает молекулярные компоненты и композиции, которые не вызывают аллергической или сходной нежелательной реакции при введении человеку. Препарат водной композиции, который содержит белок в качестве активного компонента, хорошо известен в данной области техники. Обычно такие композиции приготавливают в виде инъецируемых жидких либо растворов, либо суспензий: также могут быть приготовлены твердые формы, подходящие для превращения в раствор или суспензию перед инъекцией. Также препарат можно эмульгировать.
Белок может быть включен в состав композиции нейтральным или в форме соли. Фармацевтически приемлемые соли включают кислотно-добавочные соли (образуемые свободными аминогруппами белков), которые образуют неорганические кислоты, такие как, например, соляная или фосфорная кислоты, или такие органические кислоты, как уксусная, щавелевая, тартаровая, миндальная кислоты и подобное. Соли, образованные свободными карбоксильными группами, также могут быть получены с помощью неорганических оснований, таких как, например, гидроксиды натрия, калия, аммония, кальция или железа, и таких органических оснований, как изопропиламин, триметиламин, гистидин, прокаин и подобное.
После приготовления композиции растворы могут быть введены путем, соответствующим форме дозы, и в таком количестве, которое является терапевтически эффективным. Препараты легко могут быть введены в формах стандартных доз, таких как растворы для инъекций.
Для парентерального введения водного раствора, например, раствор должен быть подходящим образом забуферен, если необходимо, и жидкий разбавитель делают изотоническим с помощью достаточного количества соли или глюкозы. Такие конкретные водные растворы, в частности, подходят для внутривенного, внутримышечного, подкожного и внутрибрюшинного введения. В этой связи стерильные водные среды, которые можно использовать, хорошо известны специалистам в данной области техники в свете раскрытия настоящего изобретения. Например, одна доза может быть растворена в 1 мл изотонического раствора NaCl и либо добавлена к 1000 мл гиподермолитической жидкости, либо инъецирована в предполагаемом сайте инфузии (см., например, "Remington’s Pharmaceutical Sciences", 15th Edition, pp.1035-1038, pp.1570-1580). Некоторые изменения доз будут необходимыми в зависимости от состояния субъекта, лечение которого проводят. В любом случае лицо, ответственное за введение, сможет определить подходящую дозу для отдельного пациента.
Как хорошо известно в данной области техники, данный полипептид может варьироваться по своей иммуногенности. Часто бывает, следовательно, необходимым сочетать иммуноген (например, полипептид по настоящему изобретению) с носителем. Примером и предпочтительными носителями являются гемоцианин слизня (KLH) и человеческий альбумин. Другими носителями могут быть различные лимфокины и адъюванты, такие как IL2, IL4, IL8 и другие.
Средства для соединения полипептида с белком-носителем хорошо известны в данной области техники и включают глутаровый альдегид, сложный эфир m-малеимидобензоил-N-гидроксисукцинимида, карбодиимид и бис-биазотизированный бензидин. Также должно быть понятно, что пептид может быть соединен с белком методами генетической инженерии, которые хорошо известны в данной области техники.
Также в данной области техники хорошо известно, что иммуногенность конкретного иммуногена может быть усилена путем использования неспецифических стимуляторов иммунного ответа, называемых адъювантами. Примерами и предпочтительными адъювантами являются полные BCG, Detox (RIBI Immunochem Research Inc.), ISCOMS и гидроксид алюминия (Superphos, Biosector).
По использованию в данном тексте термин “комплемент” обозначает цепь нуклеиновой кислоты, которая будет гибридизовать с последовательностью первой нуклеиновой кислоты с образованием двухцепочечной молекулы при жестких условиях. Жесткими условиями являются те условия, которые обеспечивают гибридизацию между последовательностями двух нуклеиновых кислот, имеющих высокую степень гомологичности, но исключают гибридизацию случайных последовательностей. Например, гибридизация при низкой температуре и/или высокой ионной силе определяется как низкая жесткость, а гибридизация при высокой температуре и/или низкой ионной силе определяется как высокая жесткость. Понятно, что температура и ионная сила для желаемого уровня жесткости будут применяться, исходя из конкретных размеров зондов, размеров и нуклеотидного состава последовательностей и присутствия формамида в гибридизационной смеси.
По использованию в данном тексте термин “искусственно преобразованная” или “рекомбинантная” клетка призван обозначить клетку, в которую был внесен рекомбинантный ген, такой как ген, кодирующий антиген Ehrlichia chaffeensis. Следовательно, искусственно преобразованные клетки отличаются от нативных клеток, которые не включают внесенного рекомбинантного гена. Искусственно преобразованные клетки, таким образом, являются клетками, несущими ген или гены, внесенные “руками человека”. Внесенные рекомбинантные гены будут находиться в форме либо кДНК гена, либо копии геномного гена, либо будут включать гены, расположенные рядом с промотором, который в естественных условиях не связан с конкретным вносимым геном. Кроме того, рекомбинантный ген может быть интегрирован в геном хозяина или может находиться в векторе, или в бактериальном геноме, которым была трансфицирована клетка-хозяин.
Нижеследующие примеры даны для цели иллюстрирования различных вариантов настоящего изобретения и не призваны в чем-либо ограничить настоящее изобретение.
Пример 1
Эрлихии и очистка
Ehrlichia canis (штамм Florida и изоляты Demon, DJ, Jake и Fuzzy) были предоставлены Д-ром Edward Breitschwerdt (College of Veterinary Medicine, North Carolina Sate University, Raleigh, NC). E.canis (штамм Louisiana) была предоставлена Д-ром Richard E. Corstvet (School of Veterinary Medicine, Louisiana State University, Baton Rouge, LA) и E.canis (штамм Oklahoma) была предоставлена Д-ром Jacqueline Dawson (Centers of Disease Control & Prevention, Atlanta, GA). Воспроизводство эрлихий осуществляли в клетках линии DH82 в культуральной среде DMEM, дополненной 10% плодной телячьей сыворотки и 2 мМ L-глутамина при 37°С. Внутриклеточный рост в клетках DH82 контролировали по присутствию морул E.canis, для чего применяли методы цитологического окрашивания. Клетки собирали тогда, когда 100% клеток оказывались инфицированы эрлихиями, и затем пеллетировали в центрифуге при 17000 g в течение 20 минут. Клеточные сгустки разрушали в соникаторе Braun-Sonic 2000 дважды при 40 Вт в течение 30 с на льду. Эрлихий очищали в соответствии с описанным ранее (Weiss et al., 1975). Лизат загружали в прерывистый градиент ренографина - 42%-36%-30% - и центрифугировали при 80000 g в течение 1 часа. Тяжелые и легкие бэнды, содержащие эрлихий, собирали и промывали сахарозо-фосфатно-глутаматным буфером (СФГ, 218 мМ сахарозы, 3,8 мМ КН2РО4, 7,2 мМ К2НРО4, 4,9 мМ глутамата, рН 7,0) и пеллетировали центрифугированием.
Пример 2
Приготовление нуклеиновых кислот
Геномную ДНК Ehrlichia canis приготавливали путем ресуспендирования очищенных с помощью ренографина эрлихий в 600 мкл 10 мМ буфера Трис-НСl (рН 7,5) с 1% додецилсульфатом натрия (ДСН, маc/об) и 100 нг/мл протеиназы-К в соответствии с описанным ранее (McBride et al., 1996). Эту смесь инкубировали в течение 1 часа при 56°С, и нуклеиновые кислоты экстрагировали дважды смесью фенола/хлороформа/изоамилового спирта (24:24:1). ДНК пеллетировали методом преципитации в абсолютном этаноле, один раз промывали 70%-ным этанолом, высушивали и ресуспендировали в 10 мМ Трис (рН 7,5). Плазмидную ДНК очищали с использованием набора High Pure Plasmid Isolation Kit (Boehringer Mannheim, Indianapolis, IN), а ПЦР-продукты очищали с использованием набора QIAquick PCR Purification Kit (Qiagen, Santa Clarita, CA).
Пример 3
ПЦР-амплификация генов белков массой 28 кДа E.canis
Участки гена ЕСа28-1 E.canis, отобранные для амплификации методом ПЦР, выбирали на основе гомологии, выявленной (>90%) в консенсусной последовательности, полученной с помощью сопоставления в программе Jotun-Hein последовательностей генов р28 Е.chaffeensis и map-1 Cowdria ruminantium. Прямой праймер 793 (5’-GСАGGАGСТGТТGGТТАСТС-3’) (SEQ ID NO: 16) и обратный праймер 1330 (5’-CCTTCCTCCAAGTTCTATGCC-3’) (SEQ ID NO: 17) соответствовали нуклеотидам 313-332 и 823-843 MAP-1 С.ruminantium и нуклеотидам 307-326 и 834-814 Р28 E.chaffeensis. ДНК E.canis (изолят из Северной Каролайны -Jake) амплифицировали с помощью праймеров 793 и 1330 с динамикой термополимеразного цикла - 2 минуты при 95°С и 30 циклов по 30 секунд при 95°С, 1 минуте при 62°С, 2 минуты при 72°С с последующей достройкой при 72°С в течение 10 минут и хранением при 4°С. ПЦР-продукты анализировали в 1%-ном агарозном геле. Этот ПЦР-амплифицированный продукт секвенировали напрямую с использованием праймеров 793 и 1330.
Праймеры, специфичные для гена ECa28SA2, - сконструированный 46f (5’-ATATACTTCCTACCTAATGTCTCA-3’, SEQ ID NO: 18) и праймер 1330 (SEQ ID NO: 17) - использовали для амплификации намеченного участка. Амплифицированный продукт очищали в геле и клонировали в состав клонирующего вектора ТА (Invitrogen, Santa Clarita, CA). Клон секвенировали в двух направлениях с использованием праймеров: обратного М13 из вектора, 46f, ECa28SA2 (5’-AGTGCAGAGTCTTCGGTTTC-3’, SEQ ID NO: 19), ECa5.3 (5’-GTTACTTGCGGAGGACAT-3’, SEQ ID NO: 20). ДНК амплифицировали в термополимеразном цикле: 2 минуты при 95°С и 30 циклов по 30 секунд при 95°С, 1 минута при 48°С, 1 минута при 72°С с последующей достройкой при 72°С в течение 10 минут и хранением при 4°С.
Пример 4
Секвенирование неизвестных 5’- и 3’-сегментов гена ЕСа28-1
Полноразмерную последовательность ЕСа28-1 определяли с использованием набора Universal Genome Walker Kit (Clontech, Palo Alto, CA) в соответствии с прописью, предоставляемой производителем. Геномную ДНК E.canis (изолят Jake) расщепляли полностью пятью рестриктазами (DraI, EcoRV, PvuII, ScaI, StuI), которые образуют в ДНК “тупые концы”. Предлагаемый набором адаптер (AP1) лигировали на каждый из концов ДНК E.canis. Геномные библиотеки использовали в качестве матриц для обнаружения неизвестной последовательности ДНК гена ЕСа28-1 с помощью ПЦР с использованием праймера, комплементарного известной части последовательности ЕСа28-1, и праймера, специфичного для адаптера AP1. Праймеры, специфичные для ЕСа28-1, использованные для “прогулок” по геному, конструировали по известной последовательности ДНК, полученной в результате ПЦР-амплификации ЕСа28-1 с праймерами 793 (SEQ ID NO: 16) и 1330 (SEQ ID NO: 17). Праймеры 394 (5’-GCATTTCCACAGGATCATAGGTAA-3’; нуклеотиды 687-710, SEQ ID NO: 21) и 394С (5’-TTACCTATGATCCTGTGGAAATGC-3’; нуклеотиды 710-687, SEQ ID NO: 22) использовали в сочетании с предоставляемым AP1 для амплификации неизвестных 5’- и 3’-сегментов гена ЕСа28-1 с помощью ПЦР. ПЦР-продукт, соответствующий 5’-сегменту гена ЕСа28-1, амплифицированный с праймерами 394С и AP1 (2000 п.н.), секвенировали в одном направлении с праймером 793С (5’-GAGTAACCAACAGCTCCTGC-3’, SEQ ID NO: 23). ПЦР-продукт, соответствующий 3’-сегменту гена ЕСа28-1, амплифицированному с праймерами 394 и AP1 (580 п.н.), секвенировали в двух направлениях в теми же праймерами. Секвенировали некодирующие участки 5’- и 3’-сегментов, соседствующих с открытой кодирующей рамкой, и праймеры EC28ОM-F (5’-TCTACTTTGCACTTCCACTATTGT-3’, SEQ ID NO: 24) и EC28ОM-R (5’-АТТСТТТТGССАСТАТТТТТСТТТ-3’, SEQ ID NO: 25), комплементарные этим сегментам, конструировали с целью амплификации полноразмерного гена ЕСа28-1.
Пример 5
Секвенирование изолятов E.canis
ДНК была секвенирована с помощью ДНК-секвенатора ABI Prism 377 (Perkin-Elmer Applied Biosystems, Foster City, CA). Полноразмерные гены ЕСа28-1 семи изолятов E.canis (четыре из Северной Каролайны и по одному из Оклахомы, Флориды и Луизианы) амплифицировали с помощью ПЦР с праймерами EC28ОM-F (SEQ ID NO: 24) и EC28ОM-R (SEQ ID NO: 25) в термополимеразном цикле: 5 минут при 95°С и 30 циклов по 30 секунд при 95°С, 1 минута при 62°С и 2 минуты при 72°С с достройкой при 72°С в течение 10 минут. Полученные ПЦР-продукты секвенировали в обоих направлениях с использованием тех же праймеров.
Пример 6
Клонирование и экспрессия ЕСа28-1 E.canis
Полноразмерный ген ЕСа28-1 E.canis ПЦР-амплифицировали с праймерами EC28ОM-F и EC28ОM-R и клонировали в клонирующий вектор pCR2.1-TOPO ТА с целью получения желательной группы сайтов расщепления рестриктазами (Invitrogen, Carlsbad, CA). Вставку вырезали из состава pCR2.1-TOPO рестриктазой BstXl и лигировали в состав pcDNA3.1 - эукариотического экспрессирующого вектора (Invitrogen, Carlsbad, CA), обозначенного pcDNA3.1/ЕС28, для последующего изучения. Плазмиду pcDNA3.1/ЕС28 амплифицировали и ген вырезали путем обработки двумя рестриктазами KpnI/XbaI и непосредственно лигировали в прокариотический экспрессирующий вектор pThioHis (Invitrogen, Carlsbad, CA). Клон (обозначенный pThioHis/EC28) вырабатывал рекомбинантный химерный белок тиоредоксина в Escherichia coli BL21. Рекомбинантный химерный белок подвергали грубой очистке в нерастворимой фазе с помощью центрифугирования. Контрольный химерный белок тиоредоксина очищали из растворимых клеточных лизатов в нативных условиях с использованием вращающихся колонок Ni-NTA (Qiagen, Santa Clarita, CA).
Пример 7
Анализ методом Вестерн-иммуноблоттинга
Рекомбинантный химерный белок ЕСа28-1 E.canis подвергали электрофорезу в ДСН-полиакриламидном геле (ДСН-ПААГ) в градиентных гелях 4-15% с Трис-НСl (Bio-Rad, Hercules, CA) и переносили на чистую нитроцеллюлозу (Schleicher & Schuell, Keene, NH) с использованием метода полусухого переноса клеток (Bio-Rad, Hercules, CA). Мембрану инкубировали с антисывороткой от выздоравливающей собаки, инфицированной E.canis, в разведении 1:5000, в течение 1 часа, промывали и затем инкубировали с антисобачьим IgG (H&L) вторичным антителом, очищенным аффинным способом, соединенным с щелочной фосфатазой, в разведении 1:1000 в течение 1 часа (Kirkegaard & Perry Laboratories, Gaithersburg, MD). Связанное антитело визуализовали с использованием в качестве субстрата 5-бром-4-хлор-3-индолилфосфат/нитроголубого тетразолия (BCIP/NBT) (Kirkegaard & Perry Laboratories, Gaithersburg, MD).
Пример 8
Анализ методом Саузерн-блоттинга
Для установления того, присутствуют ли в геноме E.canis множественные гены, гомологичные ЕСа28-1, геномный анализ методом Саузерн-блоттинга осуществляли с использованием стандартной процедуры (Sambrook et al., 1989). Геномную ДНК E.canis расщепляли полностью каждой из рестриктаз BanII, EcoRV, HaeII, KpnI и SpeI, которые не имеют сайтов в пределах гена ЕСа28-1, а также рестриктазой AseI, которая разрезает ген ЕСа28-1 по нуклеотидам 34, 43 и 656. Зонд получали с помощью ПЦР-амплификации с праймерами EC28ОM-F и EC28ОM-R и помеченными дигоксигенином (DIG) дезоксирибонуклеотидтрифосфатами (dNTP) (Boehringer Mannheim, Indianapolis, IN) и расщепляли его рестриктазой AseI. Расщепленный зонд (566 п.н.) разделяли электрофорезом в агарозном геле, очищали в геле и затем использовали для гибридизации. Полностью расщепленную геномную ДНК E.canis подвергали электрофорезу, переносили на нейлоновую мембрану (Boehringer Mannheim, Indianapolis, IN) и гибридизовали при 40°С в течение 16 часов с DIG-помеченным зондом гена ЕСа28-1 в буфере DIG Easy Hyb в соответствии с прописью производителя (Boehringer Mannheim, Indianapolis, IN). Связанный зонд детектировали с антидигоксигениновым антителом, соединенным с щелочной фосфатазой, и люминесцирующим субстратом (Boehringer Mannheim, Indianapolis, IN) и экспонировали на специальную фотопленку BioMax (Eastman Kodak, Rochester, NY).
Пример 9
Анализ и сравнение последовательностей
Последовательности ДНК р28 Е.chaffeensis и map-1 С.ruminantium были получены от National Center Biotechnology Information (NCBI) (сайт в Интернете http://www.ncbi.nlm.nih.gov/Entrez). Нуклеотидные и расшифрованные аминокислотные последовательности, а также белковый и филогенетический анализ проводили с помощью программы LASERGENE (DNASTAR Inc., Madison, WI). Анализ посттрансляционного процессинга осуществляли с помощью метода McGeoch & von Heijne поиска сигнальной последовательности с использованием программы PSORT (McGeoch, 1985; von Heijne, 1986) (сайт в Интернете: PRIVATE HREF - http://www.imcb.osaka-u.ac.jp/nakai/form.htm; MACROBUTTON HtmlResAnchor http://www.imcb.osaka-u.ac.jp/nakai/form. htm).
Депозитарные номера в GenBabk для нуклеотидных и аминокислотных последовательностей генов ЕСа28-1 E.canis, описанных в настоящем исследовании, таковы: Jake - AF082744; Louisiana - AF082745; Oklahoma - AF082746; Demon - AF082747; DJ - AF082748; Fuzzy - AF082749; Florida - AF082750.
Анализ последовательностей ЕСа28-1 от семи разных штаммов E.canis проводили с праймерами, сконструированными с целью амплификации полного гена. Проведенный анализ показал, что последовательность данного гена была консервативна среди изолятов, происходящих из Северной Каролайны (четыре), Луизианы, Флориды и Оклахомы.
Пример 10
ПЦР-амплификация, клонирование, секвенирование и экспрессия ЕСа28-1
Сопоставление нуклеотидных последовательностей р28 Е.chaffeensis и map-1 Cowdria ruminantium с помощью программы Jotun-Hein позволила получить консенсусную последовательность с сегментами высокого уровня гомологии (>90%). Эти гомологичные сегменты (нуклеотиды 313-332 и 823-843 map-1 С.ruminantium; нуклеотиды 307-326 и 814-834 р28 Е.chaffeensis) выбрали в качестве сайтов для отжига праймеров в ПЦР-амплификации. ПЦР-амплификацию ЕСа28-1 E.canis и гена р28 Е.chaffeensis проводили с использованием праймеров 793 и 1330 с получением в результате ПЦР-продукта длиной 518 п.н. Нуклеотидная последовательность ПЦР-продукта E.canis была определена путем прямого секвенирования продукта с праймерами 793 и 1330. Анализ последовательности выявил открытую рамку считывания, кодирующую белок из 170 аминокислот, а сопоставление последовательности 518-п.н., полученной в результате ПЦР-амплификации у E.canis, с последовательностью ДНК р28 E.chaffeensis указало на уровень сходства более 70%: это указывает на гомологичность данных генов. ПЦР с адаптером с праймерами 394 и 793С проводили с целью определения 5’- и 3’-сегментов последовательности полноразмерного гена. Праймер 394 дал четыре ПЦР-продукта (3 т.п.н., 2 т.п.н., 1 т.п.н. и 0,8 т.п.н.): продукт длиной 0,8 т.п.н. секвенировали в двух направлениях с использованием праймеров 394 и AP1. Расшифрованная последовательность перекрывалась с 3’-концом 518-п.н. продукта, удлиняя открытую рамку считывания на 12 п.н. в сторону стоп-кодона. Также были секвенированы дополнительные 625 п.н. некодирующей последовательности в 3’-конце гена ЕСа28-1. Праймер 394С использовали для амплификации 5’-конца гена ЕСа28-1 наряду с предоставляемым праймером AP1. Амплификация с данными праймерами дала в результате три ПЦР-продукта (3,3 т.п.н., 3 т.п.н. и 2 т.п.н.). Фрагмент длиной 2 т.п.н. секвенировали в одном направлении по праймеру 793С. Последовательность дала предполагаемый старт-кодон гена ЕСа28-1 и полную открытую рамку считывания из 834 п.н., кодирующую белок, состоящий из 278 аминокислот. Были получены дополнительные 144 п.н. считываемой последовательности в 5’-нетранслируемом сегменте гена ЕСа28-1. Праймеры EC28ОM-F и EC28ОM-R были сконструированы по комплементарным некодирующим сегментам, фланкирующим ген ЕСа28-1.
ПЦР-продукт, амплифицированный с этими праймерами, был секвенирован напрямую с теми же праймерами. Полная последовательность ДНК (SEQ ID NO: 1) для гена ЕСа28-1 E.canis показана на фиг.1. ПЦР-фрагмент ЕСа28-1, амплифицированный с этими праймерами, включал полноразмерную открытую рамку считывания и 17 дополнительных аминокислот по 5’-некодирующему праймерному сегменту. Ген был напрямую субклонирован в состав экспрессирующего вектора pThioHis, и этой конструкцией трансформировали клетки E.coli (BL21). Экспрессированный химерный белок ЕСа28-1-тиоредоксин оказался нерастворимым. Экспрессированный белок включал дополнительные 114 аминокислот, связанных с тиоредоксином, 5 аминокислот сайта энтерокиназного распознавания и 32 аминокислоты сайта множественного клонирования и 5’-некодирующего праймерного сегмента на N-конце. Антисыворотка выздоравливающей собаки, инфицированной E.canis, распознавала экспрессированный рекомбинантный белок, но не взаимодействовала с контрольным тиоредоксином (фиг.2).
Пример 11
Гомология последовательностей
Последовательности нуклеиновой кислоты ЕСа28-1 (834 п.н.) и генов семейства omp-1 E.chaffeensis, включая сигнальные последовательности (ЕСа28-1, omp-1A, В, С, D, Е и F), сопоставляли с использованием метода Clustal с целью анализа гомологии между этими генами (само сопоставление не показано). Гомология нуклеиновых кислот была в равной степени консервативна (68,9%) между ЕСа28-1 и р28 E.chaffeensis, и omp-1F. Гены других предполагаемых внешнемембранных белков из семейства omp-1 E.chaffeensis, omp-1D (68,2%), omp-1E (66,7%), omp-1C (64,1%), map-1 Cowdria ruminantium (61,8%), ген белка-1 массой 28 кДа E.canis (60%) и ген белка-2 массой 28 кДа (частично) (59,5%) также были гомологичны ЕСа28-1. Наименьшая гомология нуклеотидных последовательностей (45,1%) отмечена между omp-1B E.chaffeensis и ЕСа28-1.
Сопоставление предсказанных аминокислотных последовательностей ЕСа28-1 (SEQ ID NO: 2) и Р28 E.chaffeensis позволило выявить аминокислотные замены, приводящие к появлению четырех вариабельных сегментов (VR). Были идентифицированы замены или делеции в аминокислотной последовательности и расположение вариабельных сегментов в ЕСа28-1 и у членов семейства ОМР-1 E.chaffeensis (фиг.3). Сравнение аминокислот, включая сигнальный сегмент, показало, что ЕСа28-1 проявляет наибольший уровень гомологии с OMP-1F (68%) из семейства ОМР-1 E.chaffeensis, далее с Р28 E.chaffeensis (65,5%), ОМР-1Е (65,1%), OMP-1D (62,9%), ОМР-1С (62,9%), MAP-1 Cowdria ruminantium (59,4%), белком-1 массой 28 кДа E.canis (55,6%) и белком-2 массой 28 кДа (частичным) (53,6%), и ОМР-1В (43,2%). Филогенетические реконструкции на основе аминокислотных последовательностей показывают, что ЕСа28-1 и MAP-1 C.ruminantium, белки ОМР-1 E. chaffeensis и белки 1 и 2 (частичный) массой 28 кДа E.canis являются родственными (фиг.4).
Пример 12
Предсказанные поверхностная вероятность и иммунореактивность
Анализ ЕСа28-1 E.canis по профилям гидропатичности и гидрофильности позволил предсказать участки, расположенные на поверхности молекулы ЕСа28-1 (фиг.6). Восемь основных расположенных на поверхности участков, составленных 3-9 аминокислотами, были идентифицированы в ЕСа28-1 и оказались сходными с профилем поверхностных участков у Р28 E.chaffeensis (фиг.6). Пять наиболее крупных поверхностных участков ЕСа28-1 располагались в N-концевой части белка. Расположенные на поверхности гидрофильные участки были обнаружены во всех четырех вариабельных сегментах ЕСа28-1. Десять Т-клеточных мотивов были предсказаны в ЕСа28-1 с использованием метода Ротбарда-Тейлора (Rothbard & Taylor, 1988), а высокой уровень антигенности ЕСа28-1 был предсказан с помощью алгоритма оценки антигенности по Джеймсону-Вольфу (фиг.6) (Jameson & Wolf, 1988). Сходство антигенности и Т-клеточных мотивов было обнаружено между ЕСа28-1 и Р28 E.chaffeensis.
Пример 13
Выявление гомологичных геномных копий гена ЕСа28-1
Геномный анализ методом Саузерн-блоттинга ДНК E.canis, полностью и по отдельности расщепленной рестриктазами BanII, EcoRV, HaeII, KpnI, SpeI, для которых отсутствуют сайты распознавания в пределах гена ЕСа28-1, а также рестриктазой AseI, для которой имеются рестрикционные сайты в нуклеотидах 34, 43 и 656, показал присутствие по крайней мере трех гомологичных копий гена ЕСа28-1 (фиг.5). Хотя ЕСа28-1 включает внутренний сайт распознавания рестриктазой AseI, DIG-помеченный зонд, использованный в эксперименте по гибридизации, помечал участок гена в пределах единственного ДНК-фрагмента, полученного в результате обработки AseI этого гена. Расщепление рестриктазой AseI привело к образованию 3 бэндов (длиной приблизительно 566 п.н., 850 п.н. и 3 т.п.н.), которые гибридизовали с ДНК-зондом ЕСа28-1, что указывает на присутствие множественных генов, гомологичных ЕСа28-1, в геноме. Расщепление рестриктазами EcoRV и SpeI давало два бэнда, которые гибридизовали с зондом гена ЕСа28-1.
Пример 14
Идентификация локуса генов белков массой 28 кДа
Специфичные праймеры, обозначенный ЕСаSА3-2 (5’-CTAGGATTAGGTTATAGTATAAGTT-3’, SEQ ID NO: 26), соответствующий участкам в пределах ECa28SA3, и праймер 793С (SEQ ID NO: 23), который соединяется с участком ЕСа28-1, использовали для амплификации межгенного спейсера между генами SA3 и ЕСа28-1. Продукт длиной 800 п.н. был секвенирован с использованием тех же праймеров. ДНК амплифицировали в термополимеразной схеме: 2 минуты при 95°С и 30 циклов по 30 секунд при 95°С, 1 минута при 50°С, 1 минута при 72°С с последующей достройкой при 72°С в течение 10 минут и хранением при 4°С.
Пример 15
ПЦР-амплификация генов белков массой 28 кДа и идентификация мультигенного локуса
С целью специфичной амплификации возможных неизвестных генов, расположенных вниз от ECa28SA2, для амплификации использовали специфичный для ECa28SA2 праймер 46f и праймер 1330, который соответствует консервативному сегменту с 3’-конца гена ЕСа28-1. По этим праймерам был амплифицирован ПЦР-продукт длиной 2 т.п.н., в котором имелись две открытые рамки считывания. Первая открытая рамка считывания включала известный участок гена - ECaSA2 - и ранее несеквенированный 3’-участок гена. Вниз от ECaSA2 был найден дополнительный неидентичный, но гомологичный ген белка массой 28 кДа - его обозначили ECa28SA3. Два известных локуса соединили с помощью амплификации с праймером SA3-2, специфичным для 3’-конца гена ECa28SA3, который использовали в сочетании с обратным праймером 793С, который соединяется с 5’-концом ЕСа28-1. Амплифицировали ПЦР-продукт длиной 800 п.н., который включал 3’-конец ECa28SA3, межгенный спейсер между ECa28SA3 и ЕСа28-1 (28NC3) и 5’-конец ЕСа28-1, соединяющий ранее раздельные локусы (фиг.8). Открытая рамка считывания длиной 849 п.н. ECa28SA2 кодирует белок из 283 аминокислот, а ECa28SA3 имеет открытую рамку считывания из 840 п.н., кодирующую белок из 280 аминокислот. Межгенный некодирующий участок между ECa28SA3 и ЕСа28-1 имел в длину 345 п.н. (фиг.7 и 8).
Пример 16
Нуклеотидная и аминокислотная гомология
Нуклеотидные и аминокислотные последовательности всех пяти генов белков массой 28 кДа E.canis были сопоставлены с использованием метода Clustal с целью анализа гомологии между генами. Гомология нуклеиновых кислот варьировалась в пределах 58-75%, а аналогичная гомология аминокислотных последовательностей составила 67-72% при сравнении генов, кодирующих белки массой 28 кДа E.canis (фиг.9).
Пример 17
Участки контролирующих транскрипцию промоторов
Межгенные участки между генами белков массой 28 кДа анализировали на присутствие промоторных последовательностей путем сравнения с консенсусными промоторными участками Escherichia coli и промотором E.chaffeensis (Yu et al., 1997; McClure, 1985).
Предполагаемые промоторные последовательности, включая участки RBS, -10 и -35, были идентифицированы в четырех межгенных последовательностях, соответствующих генам ECa28SA2, ECa28SA3, ECa28-1 и ЕСа28-2 (фиг.10). Некодирующий участок выше гена ECa28SA1 неизвестен и не был исследован.
Пример 18
N-концевая сигнальная последовательность
Анализ аминокислотных последовательностей показал, что полноразмерный ECa28-1 E.canis имеет расшифрованную молекулярную массу 30,5 кДа, а полноразмерный ECa28SA3 имеет расшифрованную молекулярную массу 30,7 кДа. Оба белка включают предсказанный N-концевой сигнальный сегмент из 23 аминокислот (MNCKKILITTALMSLMYYAPSIS, SEQ ID NO: 27), который сходен с предсказанным для Р28 E.chaffeensis (MNYKKILITSALISLISSLPGVSFS, SEQ ID NO: 28) и белков семейства ОМР-1 (Yu et al., 1998; Ohashi et al., 1998b). Предпочтительный сайт расщепления сигнальными пептидазами (SIS; серин-Х-серин) (Oliver, 1985) найден в аминокислотах 21, 22 и 23 у ECa28-1. Также присутствовал дополнительный сайт предполагаемого расщепления по аминокислотному положению 25 (MNCKKILITTALISLMYSIPSISSFS, SEQ ID NO: 29), идентичный предсказанному сайту расщепления в Р28 E.chaffeensis (SFS), который обусловливал образование зрелого белка ЕСа28-1 с предсказанной молекулярной массой 27,7 кДа. Сайт сигнального расщепления ранее опубликованной частичной последовательности ECa28SA2 предполагается на 30-й аминокислоте. Однако анализ сигнальной последовательности предсказал, что ЕСа28SА1 имел неотщепляемую сигнальную последовательность.
Краткое изложение
Белки со сходными молекулярными массами были идентифицированы и клонированы у нескольких агентов-риккетсий, включая E.canis, E.chaffeensis и С.ruminantium (Reddy et al., 1998; Jongejan et al., 1993; Ohashi et al., 1998). Ранее были описаны единственный локус у Ehrlichia chaffeensis с 6 гомологичными генами р28 и два локуса у E.canis, каждый из которых включает по несколько гомологичных генов белков массой 28 кДа.
Настоящее изобретение показало клонирование, экспрессию и охарактеризование генов, кодирующих зрелый белок массой 28 кДа E.canis, которые гомологичны мультигенному семейству отр-1 E.chaffeensis и гену mар-1 С.ruminantium. Были идентифицированы два новых гена белков массой 28 кДа - ЕСа28-1 и ЕСа28SА3. Другой ген белка массой 28 кДа E.canis - SCa28SA2, который частично был секвенирован ранее (Reddy et al., 1998), - был секвенирован полностью в настоящем изобретении. Также заявляется идентификация и охарактеризование единого локуса E.canis, включающего все пять генов белков массой 28 кДа Е.canis.
Белок массой 28 кДа E.canis гомологичен семейству ОМР-1 E.chaffeensis и белку MAP-1 С.ruminantium. Наиболее гомологичные белки массой 28 кДа E.canis (ECa28SA3, ECa28-1 и ЕСа28-2) выстроены в локусе тандемным образом. Гомология этих белков варьировалась от 67,5% до 72,3%. Уровень дивергенции этих белков массой 28 кДа составил от 27,3% до 38,6%. Белки массой 28 кДа E.canis ECa28SAl и ECa28SA2 характеризовались наименьшей гомологичностью при уровне гомологии в диапазоне от 50,9% до 59,4% и при дивергенции от 53,3% до 69,9%. Различия между генами приурочены в основном к четырем гипервариабельным участкам: это подтверждает, что данные участки расположены на поверхности и находятся под прессом отбора с участием иммунной системы. Был описан консерватизм ECa28-1 у семи изолятов E.canis (McBride et al., 1999): это подтверждает, что E.canis может характеризоваться клональным воспроизводством в Северной Америке. Напротив, было сообщено о существенном уровне дивергенции белков р28 у изолятов E.chaffeensis (Yu et al., 1998).
Все белки массой 28 кДа у E.canis предположительно претерпевают посттрансляционный процессинг, превращаясь из белка массой 30 кДа в зрелый белок массой 28 кДа. Недавно была идентифицирована сигнальная последовательность в Р28 E.chaffeensis (Yu et al., 1998), а секвенирование N-концевых аминокислот подтвердило, что этот белок подвержен посттрансляционному процессингу, заключающемуся в отщеплении сигнальной последовательности с образованием зрелого белка (Ohashi et al., 1998). Лидерные последовательности в составе OMP-1F и ОМР-1Е также предположительно являются лидерными сигнальными пептидами (Ohashi et al., 1998). Сигнальные последовательности, идентифицированные в составе OMP-1F, ОМР-1Е И Р28 у E.chaffeensis, гомологичны лидерной последовательности белка массой 28 кДа E.canis. Промоторные последовательности генов р28 не были установлены экспериментальным путем, однако предполагаемые промоторные участки были идентифицированы с помощью сравнения консенсусных последовательностей промоторных сегментов RBS, -10 и -35 у E.coli и у других эрлихий (Yu et al., 1997; McClure, 1985). Такие промоторные последовательности должны обеспечивать гену потенциальные транскрипцию и трансляцию: это подтверждает, что данные гены могут быть в хозяине экспрессированы по-разному. Персистенция инфекции у собак может быть связана с дифференцированной экспрессией генов р28, приводящей к антигенным преобразованиям in vivo, что позволяет возбудителю избегать иммунного ответа.
Гены белков массой 28 кДа E.canis, как было установлено, характеризуются гомологией нуклеотидных и аминокислотных последовательностей с генами семейства omp-1 E.chaffeensis и с геном mар-1 С.ruminantium. В ранее проведенных исследованиях был идентифицирован белок массой 30 кДа у E.canis, который взаимодействует с антисывороткой к E.chaffeensis, полученной от выздоравливающих собак, но считается, что по антигенным свойствам они неодинаковы (Rikihisa et al., 1994). Данные сравнения аминокислотных замен в четырех вариабельных участках белков массой 28 кДа E.canis подтверждают такую возможность. Наряду с этими данными также подтверждается, что аминокислоты, ответственные за антигенные различия между Р28 E.canis и E.chaffeensis, находятся в этих вариабельных участках и легко доступны для иммунной системы. Было сообщено, что иммунореактивные пептиды располагались в вариабельных сегментах белков массой 28 кДа С.ruminantium, E.chaffeensis и E.canis (Ready et al., 1998). Анализ Р28 E.canis и E.chaffeensis показал, что все вариабельные сегменты имеют предсказанные аминокислоты, расположенные на поверхности молекулы. Анализ на собаках показал отсутствие перекрестной реактивности между E.canis и E.chaffeensis (Dawson & Ewing, 1992). Это наблюдение может иметь отношение к антигенной дифференцировке вариабельных сегментов в составе Р28, равно как и в других иммунологически важных антигенах данных видов эрлихий. В другом исследовании было обнаружено, что человеческая антисыворотка стадии выздоровления у пациентов, инфицированных E.chaffeensis, распознает белок (белки) массой 29/28 кДа E.chaffeensis, а также реагирует с гомологичными белками E.canis (Chen et al., 1997). Гомологичные и перекрестно реагирующие эпитопы белков массой 28 кДа E.canis и Р28 E.chaffeensis предположительно распознаются иммунной системой.
Белки массой 28 кДа E.canis могут быть важными иммунореактивными антигенами. В некоторых сообщениях было показано, что антиген массой 30 кДа E.canis проявляет интенсивную иммунореактивность (Rikihisa et al., 1994; Rikihisa et al., 1992). Антитела в составе антисывороток стадии выздоровления, полученных от человека и домашней собаки, характеризуются существенной реактивностью в отношении белков Е.chaffeensis и E.canis в данном размерном диапазоне: это подтверждает, что они могут быть важными иммунореактивными антигенами (Rikihisa et al., 1994; Chen et al., 1994; Chen et al., 1997). Кроме того, антитела к белкам массой 30, 24 и 21 кДа образовывались на ранних этапах формирования иммунного ответа на E.canis (Rikihisa et al., 1994; Rikihisa et al., 1992): это подтверждает, что данные белки могут быть особенно важны для иммунных ответов в острой стадии заболевания. Недавно семейство гомологичных генов, кодирующих внешнемембранные белки с молекулярными массами 28 кДа, было идентифицировано у Е.chaffeensis, а у мышей, иммунизованных рекомбинантным Р28 E.chaffeensis, по-видимому, развивался иммунитет против гомологичной инфекции (Ohashi et al., 1998). Белок Р28 E.chaffeensis, как было показано, присутствует на внешней мембране, а по данным иммуноэлектронной микроскопии Р28 был картирован на поверхности данного организма: следовательно, это подтвердило, что он может выполнять роль адгезионного белка (Ohashi et al., 1998). Вполне вероятно, что белки массой 28 кДа E.canis, идентифицированные в настоящем исследовании, имеют такую же локализацию и возможно выполняют сходные функции.
Сравнение ЕСа28-1 у разных штаммов E.canis показало, что ген, по-видимому, полностью консервативен. Исследования с привлечением E.chaffeensis дали иммунологические и молекулярные доказательства дивергенции ЕСа28-1. Пациенты, инфицированные E.chaffeensis, проявили варьирующуюся иммунореактивность в отношении белков массой 29/28 кДа: это подтверждает их антигенную дифференцировку (Chen et al., 1997). Недавно было получено молекулярное подтверждение антигенной неоднородности гена р28 у E.chaffeensis (Yu et al., 1998). Сравнение пяти изолятов E.chaffeensis показало, что два изолята (Sapulpa и St.Vincent) идентичны на 100%, но три других изолята (Arkansas, Jax и 91НЕ17) дивергировали на уровне аминокислотных последовательностей по меньшей мере на 13,4%. Консерватизм ЕСа28-1 подтверждает, что штаммы E.canis, обнаруживаемые в США, могут быть генетически идентичными, т.е. белок массой 28 кДа E.canis является привлекательным материалом для создания вакцин против эрлихиоза собак в США. Дальнейший анализ изолятов E.canis из-за пределов США может предоставить информацию, касающуюся происхождения и эволюции E.canis. Консерватизм белка массой 28 кДа делает его важным потенциальным кандидатом для проведения эффективной серодиагностики эрлихиоза собак.
Роль множественных гомологичных генов в данный момент неизвестна: однако персистенция возбудителя E.canis в организме собак предположительно может быть связана с антигенной изменчивостью, обусловливаемой варьирующейся экспрессией генов гомологичных белков массой 28 кДа, что тем самым позволяет E.canis избегать активности иммунной системы. Изменчивость генов msp-3 A.marginale отчасти ответственна за изменчивость белка MSP-3, что приводит к персистенции возбудителя (Alleman et al., 1997). Исследования экспрессии генов белков массой 28 кДа E.canis у собак с острой и хронической инфекцией этим возбудителем дадут информацию о роли семейства генов белков массой 28 кДа в персистенции возбудителя.
Любые патенты или публикации, упоминавшиеся в данной заявке, соответствуют уровню специалистов в той области техники, к которой относится настоящее изобретение. Эти патенты и публикации включены здесь для сведения в виде библиографических ссылок в той же степени, в которой каждая отдельная публикация призвана быть включенной для сведения в виде библиографической ссылки.
Для специалиста в данной области техники будет вполне очевидно, что настоящее изобретение может быть легко применено для осуществления объектов, объема и преимуществ, упоминавшихся выше, а также тех, которые им присущи. Настоящие примеры наряду с методами, процедурами, обработками, молекулами и конкретными соединениями, описанными в данном тексте, являются репрезентативными для предпочтительных вариантов, являются примерами и не призваны в чем-либо ограничить объем настоящего изобретения. Изменения в нем и другие пути использования будут ясны специалистам в данной области техники, которые охватываются сутью настоящего изобретения, определяемой объемом формулы изобретения.
Список последовательностей приведен в конце описания
Источники информации
AllemanA.R., et al., (1997) Infect Immun 65: 156-163.
Anderson B.E., et al., (1991) J Clin Microbiol 29: 2838-2842.
Anderson B.E., et al., (1992) Int J Syst Bacteriol 42: 299-302.
Brouqui P., et al., (1992) J Clin Microbiol 30: 1062-1066.
Chen S.M., et al., (1997) Clin Diag Lab Immunol 4: 731-735.
Chen S.M., et al., (1994) Am J Trop Med Hyg 50: 52-58.
Dawson J.E., et al., (1992) Am J Vet Res 53: 1322-1327.
Dawson J.E., et al., (1991) J Infect Dis 163: 564-567.
Donation, et al., (1935) Bull Soc Pathol Exot 28: 418-9.
Ewing, (1963) J Am Vet Med Assoc 143: 503-6.
Groves M.G., et al., (1975) Am J Vet Res 36: 937-940.
Harrus S., et al., (1998) J Clin Microbiol 36: 73-76.
Jameson В.A., et al., (1988) CABIOS 4: 181-186.
Jongejan F., et al., (1993) Rev Elev Med Vet Pays Trop 46: 145-152.
McBride J.W., et al., (1996) J Vet Diag Invest 8: 441-447.
McBride, et al., (1999) Clin Diagn Lab Immunol.: (In press).
McClure, (1985) Ann Rev Biochem 54: 171-204.
McGeoch D.J. (1985) Virus Res 3: 271-286.
Nyindo M., et al., (1991) Am J Vet Res 52: 1225-1230.
Nyindo, et al., (1971) Am J Vet Res 32: 1651-58.
Ohashi, et al., (1998) Infect Immun 66: 132-9.
Ohashi, et al., (1998) J Clin Microb 36: 2671-80.
Reddy, et al., (1998) Biochem Biophys Res Comm 247: 636-43.
Rikihisa, et al., (1994) J Clin Microbiol 32: 2107-12.
Rothbard J.B., et al., (1988) The EMBO J7: 93-100.
Sambrook J., et al., (1989) In Molecular Cloning: A Laboratory Manual. Cold Spring Harbor: Cold Spring Harbor Press.
Troy G.C., et al., (1990) Canine ehrlichiosis. In Infectious diseases of the dog and cat. Green C.E. (ed). Philidelphia: W.B. Sauders Co. von Heijne, (1986) Nucl Acids Res 14: 4683-90.
Walker, et al., (1970) J Am Vet Med Assoc 157: 43-55.
Weiss E., et al., (1975) Appil Microbiol 30: 456-463.
Yu, et al., (1997) Gene 184: 149-154.
Yu, et al., (1998) J. Clin. Microbiol. (In press).


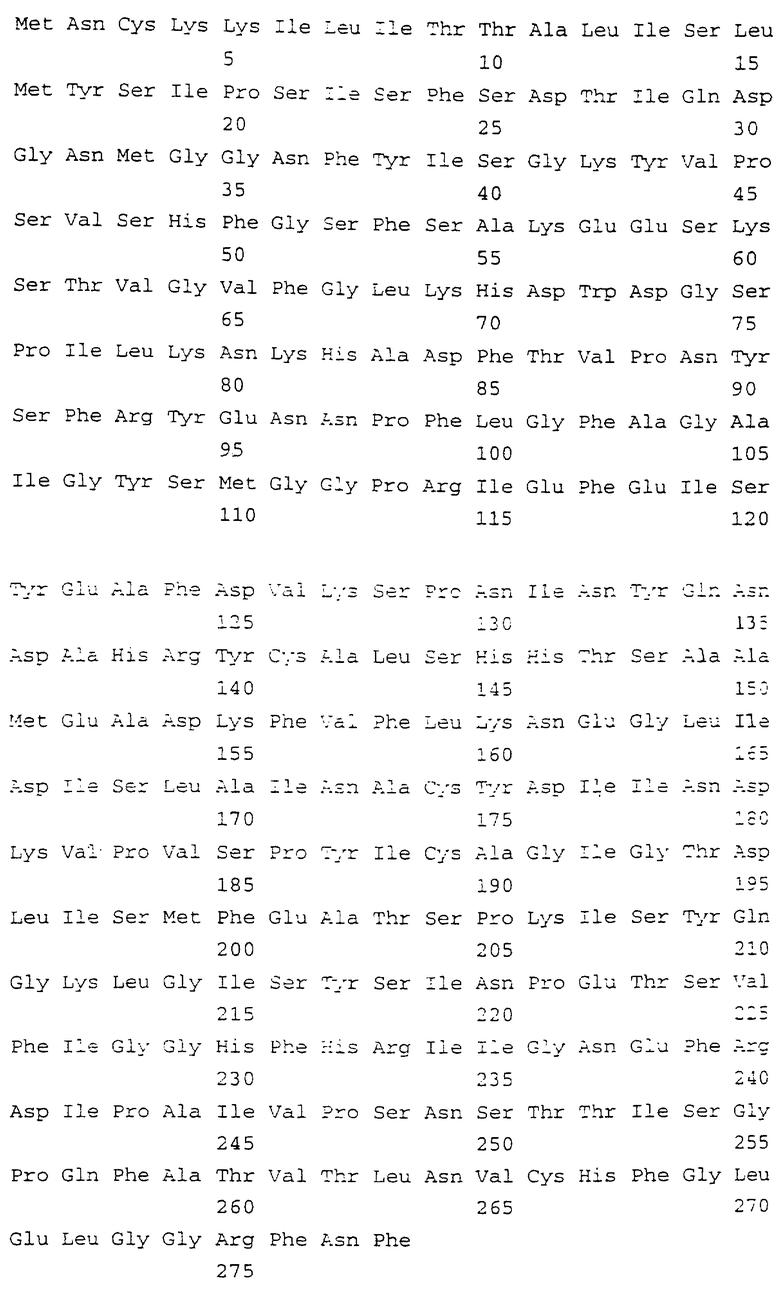



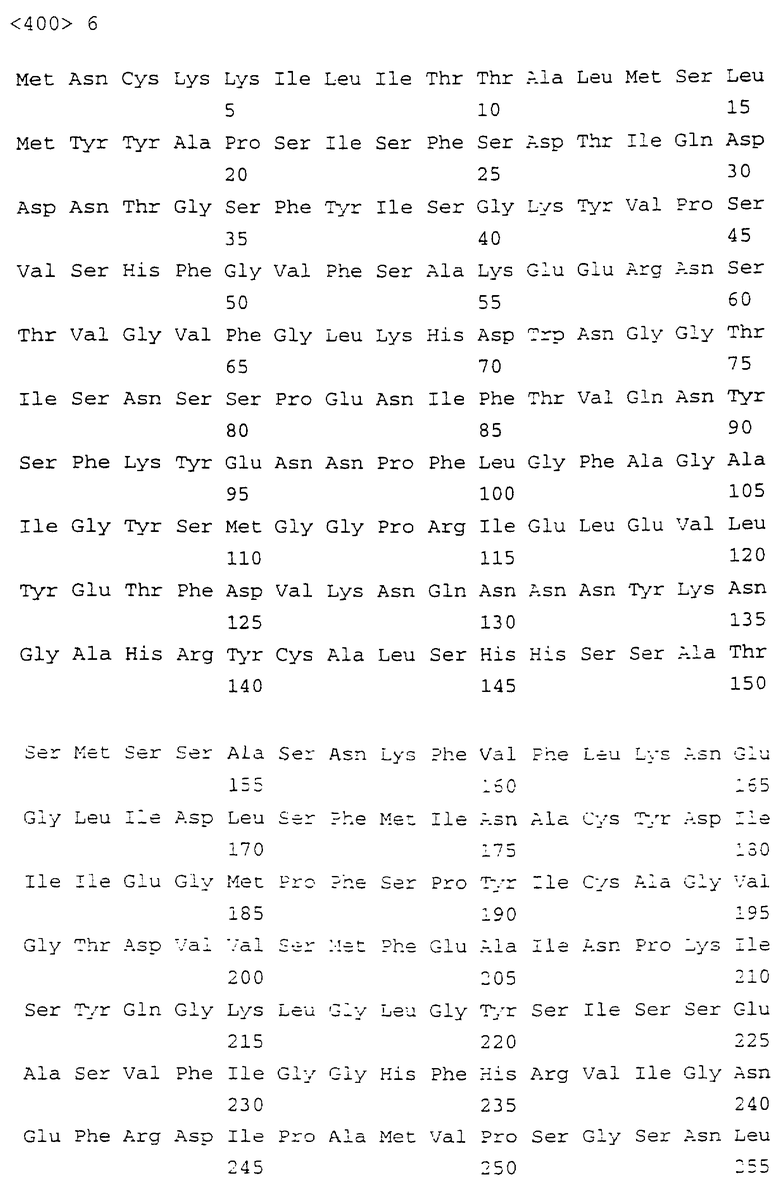





















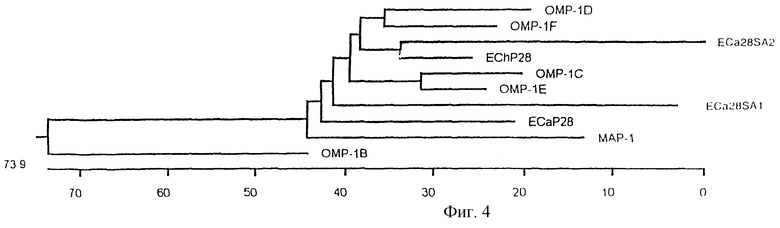
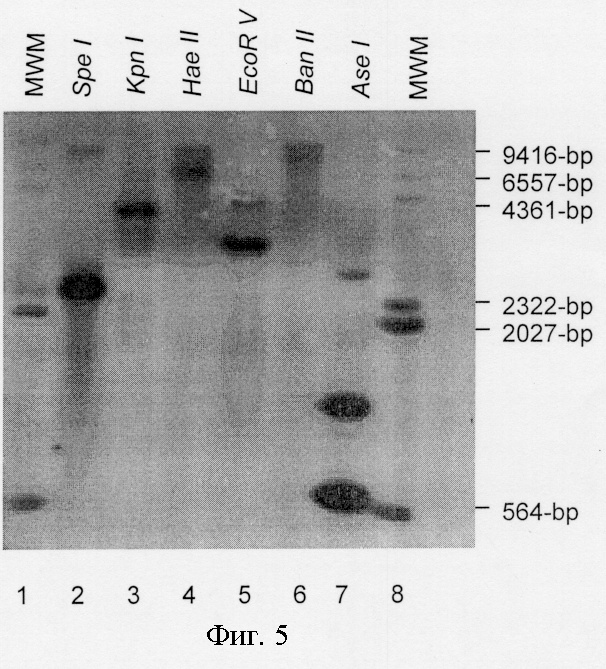
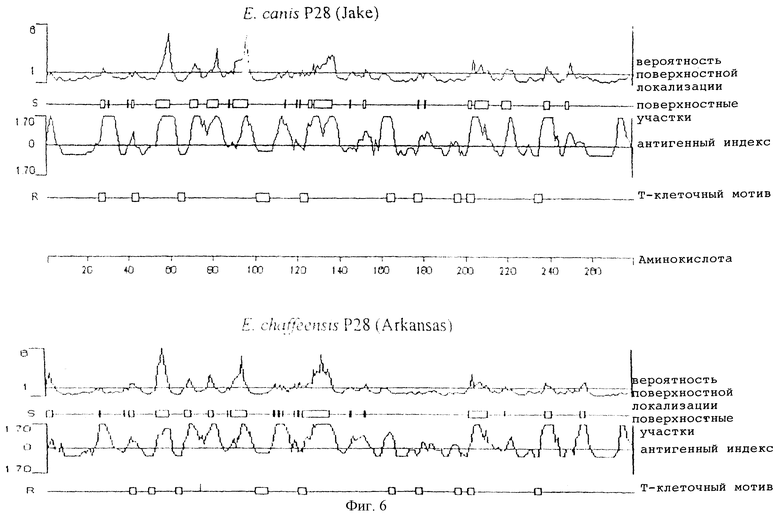


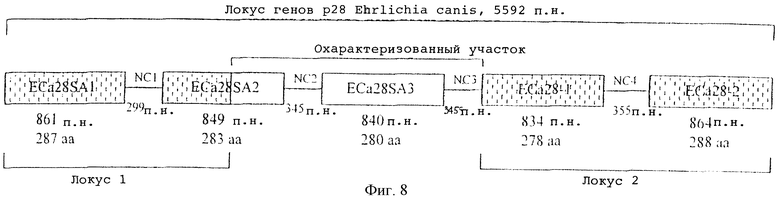
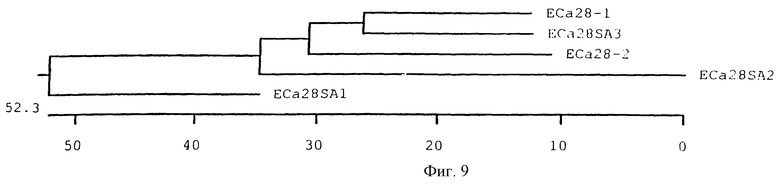

Изобретение относится к области молекулярной биологии. Предложена последовательность ДНК, кодирующая белок Ehrlichia canis с молекулярной массой 30 килодальтон. Также предложены вектор для получения белка, содержащий последовательность ДНК, рекомбинантный белок для получения антител, выбранный из группы, состоящей из SEQ ID NO:4 и 6, и способ его получения. Рекомбинантный белок Ehrlichia canis с молекулярной массой 30 килодальтон иммунореактивен к сыворотке против Ehrlichia canis. Предложенная группа изобретений может быть использована для создания вакцин против эрлихиоза собак. 4 с. и 8 з.п. ф-лы, 10 ил.

| REDDY G.R | |||
| et al | |||
| Видоизменение прибора с двумя приемами для рассматривания проекционные увеличенных и удаленных от зрителя стереограмм | 1919 |
|
SU28A1 |
| Biochem | |||
| Biophys | |||
| Res | |||
| Commun | |||
| Способ и аппарат для получения гидразобензола или его гомологов | 1922 |
|
SU1998A1 |
| Солесос | 1922 |
|
SU29A1 |
| CHEN S.M | |||
| et al | |||
| Western immunoblotting analysis of the antibody responses of patients with human monocytotropic ehrlichiosis to different strains of Ehrlichia chaffeensis and Ehrlichia canis | |||
| Clin | |||
| Diagn | |||
| Lab | |||
| Immunol | |||
| Электрическое сопротивление для нагревательных приборов и нагревательный элемент для этих приборов | 1922 |
|
SU1997A1 |

