Перекрестная ссылка на родственные заявки
Данная заявка заявляет приоритет патентных заявок США № 62/107824 (поданной 26 января 2015 года), 62/008229 (поданной 5 июня 2014 года; рассматривается), и 62/004571 (поданной 29 мая 2014 года, рассматривается), каждая из которых включена в настоящее описание ссылкой во всей полноте.
Ссылка на перечень последовательностей
Эта заявка включает один или несколько перечней последовательностей в соответствии с 37 CFR 1,821 и далее, которые раскрыты в машиночитаемом носителе (Название файла: 1301_0119PCT_Sequence_Listing_ST25.txt, созданный 18 мая 2015 года, и имеющий размер 416,408 байт), этот файл включен в данное описание ссылкой во всей полноте.
Область техники, к которой относится изобретение
Настоящее изобретение относится к триспецифичным связывающим молекулам, представляющим собой многоцепочечные полипептидные молекулы, которые обладают тремя связывающими доменами и, таким образом, способны опосредовать скоординированное связывание с тремя эпитопами. Триспецифичная связывающая молекула предпочтительно отличается тем, обладает связывающими доменами, которые позволяют ей иммуноспецифично связываются с: (1) эпитопом первого антигена злокачественной опухоли (2) эпитопом второго антигена злокачественной опухоли, и (3) эпитопом молекулы, которая экспрессируется на поверхности эффекторных клеток иммунной системы, и, таким образом, способным локализовать эффекторную клетку иммунной системы к клетке, которая экспрессирует антиген злокачественной опухоли, с тем чтобы тем самым облегчить уничтожение такой злокачественной опухолевой клетки.
Уровень техники
I. Иммунная система млекопитающих
Иммунная система млекопитающих служит защитой от различных состояний, в том числе, например, травмы, инфекции и новообразования. Эффективность, с которой у людей и других млекопитающих развивается иммунологический ответ на патогены, чужеродные вещества и антигены злокачественной опухоли опирается на две характеристики: изысканную специфичность иммунного ответа на распознавание антигена, и иммунологическую память, которая позволяет быстрее и более энергично отвечать при повторной активации тем же антигеном (Portolés P. et al. (2009) “The TCR/CD3 Complex: Opening the Gate to Successful Vaccination,” Current Pharmaceutical Design 15:3290-3300; Guy, C.S. et al. (2009) “Organization of Proximal Signal Initiation at the TCR:CD3 Complex,” Immunol Rev. 232(1):7-21).
Иммунная система млекопитающих опосредуется двумя отдельными, но взаимосвязанными системами: клеточной и гуморальной иммунными системами. Вообще говоря, гуморальная система опосредуется растворимыми продуктами (антитела или иммуноглобулины), которые имеют возможность объединяться с и нейтрализовать продукты, признанные системой как инородные организму. В противоположность этому, клеточная иммунная система предполагает мобилизацию определенных клеток, называемых "Т-клетки", которые выполняют различные терапевтические роли. Т-клетки представляют собой лимфоциты, которые происходят из тимуса и циркулируют между тканями, лимфатической системой и системой кровообращения. В ответ на присутствие и распознавание инородных структур (антигенов), Т-клетки становятся "активированными", для инициации иммунного ответа. Во многих случаях эти чужеродные антигены экспрессируются на клетках-хозяевах, в результате неоплазии или инфекции. Хотя Т-клетки сами не секретируют антитела, они, как правило, требуются для секреции антитела вторым классом лимфоцитов, В-клетками (которые происходят из костного мозга). Чрезвычайно важно то, что Т-клетки проявляют необычайную иммунологическую специфичность таким образом, что способны различать один антиген от другого). Два типа Т-клеток, "хелперные Т-клетки" и "цитотоксические Т-клетки", имеют особое значение.
Т-хелперы характеризуются экспрессией гликопротеина CD4 (т.е. они являются "CD4+"). CD4+ Т-клетки являются основными организаторами большинства иммунных и аутоиммунных реакций млекопитающих (Dong, С. et al. (2003) “Immune Regulation by Novel Costimulatory Molecules,” Immunolog. Res. 28(1):39-48). Было обнаружено, что активация CD4+ Т-клеток опосредована костимуляторными взаимодействиями между антигеном: молекулой главного комплекса гистосовместимости класса II (МНС II), которая развернута на поверхности антигенпредставляющей клетки (такой как В-клетка, макрофаг или дендритная клетка) и комплексом из двух молекул, Т-клеточного рецептора ("TCR") и лиганда рецептора клеточной поверхности CD3, которые развернуты на поверхности наивных CD4+ Т-клеток. Активированные Т-хелперы способны пролиферировать в клетки Th1, которые способны опосредовать воспалительный ответ на клетки-мишени.
Цитотоксические Т-клетки характеризуются экспрессией CD8 (т.е. они являются "CD8+", и CD3+). Было обнаружено, что активация CD8+ Т-клеток опосредуется через костимуляторные взаимодействия антигеном: молекулой главного комплекса гистосовместимости класса II (МНС I), которая развернута на поверхности клетки-мишени, и комплекса CD8 и Т-клеточного рецептора, которые развернуты на поверхности CD8+ T-клетки. В отличие от молекул MHC II, которые экспрессируются только определенными клетками иммунной системы, молекулы MHC I экспрессируются очень широко. Таким образом, цитотоксические Т-клетки способны связываться с широким спектром типов клеток. Активированные цитотоксические Т-клетки опосредуют гибель клеток, высвобождая цитотоксины перфорин, гранзимы и гранулизин. Благодаря действию перфорина гранзимы входят в цитоплазму клетки-мишени и их серин-протеазная функция запускает каскад каспаз, который представляет собой ряд цистеиновых протеаз, что в конечном итоге приводит к апоптозу (запрограммированной гибели клеток) клеток-мишеней.
Т-клеточный рецептор ("TCR") представляет собой ковалентно связанный гетеродимер альфа и бета цепей ("TCRαβ"). Эти цепи представляют собой мембранные полипептиды класса I длиной 259 (α) и 296 (β) аминокислот. Молекула CD3 представляет собой Т-клеточный корецептор, состоящий из пяти различных полипептидных цепей (CD3 γ цепь, a CD3 δ цепь, две CD3 ε цепи и две дзета-цепи). Отдельные полипептидные цепи объединяясь образуют комплекс из трех димеров (εγ, εδ, ζζ) (Wucherpfennig, K.W. et al. (2010) “Structural Biology Of The T Cell Receptor: Insights into Receptor Assembly, Ligand Recognition, и Initiation of Signaling,” Cold Spring Harb. Perspect. Biol. 2(4):a005140; pages 1-14; Chetty, R. et al. (1994) “CD3: Structure, Function And The Role Of Immunostaining In Clinical Practice,” J. Pathol. 173:303-307; Guy, C.S. et al. (2009) “Organization of Proximal Signal Initiation at the TCR:CD3 Complex,” Immunol Rev. 232(1):7-21; Call, M.E. et al. (2007) “Common Themes In The Assembly And Architecture Of Activating Immune Receptors,” Nat. Rev. Immunol. 7:841-850; Weiss, A. (1993) “T Cell Antigen Receptor Signal Transduction: A Tale Of Tails And Cytoplasmic Protein-Tyrosine Kinases,” Cell 73:209-212). CD3-комплекс ассоциируется с TCR для образования сигнала активации в Т-лимфоцитах. В отсутствие CD3 TCR не собирается должным образом и деградирует (Thomas, S. et al. (2010) “Molecular Immunology Lessons From Therapeutic T Cell Receptor Gene Transfer,” Immunology 129(2):170–177). CD3 обнаруживается связанным с мембранами всех зрелых Т-клеток, и практически ни с одним другим типом клеток (см., Janeway, C.A. et al. (2005) In: Immunobiology: The Immune System In Health And Disease,” 6th ed. Garland Science Publishing, NY, pp. 214-216; Sun, Z. J. et al. (2001) “Mechanisms Contributing To T Cell Receptor Signaling And Assembly Revealed By The Solution Structure Of An Ectodomain Fragment Of The CD3ε:γ Heterodimer,” Cell 105(7):913-923; Kuhns, M.S. et al. (2006) “Deconstructing The Form And Function Of The TCR/CD3 Complex,” Immunity. 2006 Feb;24(2):133-139).
Комплекс TCR и CD3, вместе CD3 ζ цепью дзета цепью (также известной как Т3 дзета цепь Т-клеточного рецептора или CD247) составляют TCR-комплекс (van der Merwe, P.A. etc. (epub Dec. 3, 2010) “Mechanisms For T Cell Receptor Triggering,” Nat. Rev. Immunol. 11:47-55; Wucherpfennig, K.W. et al. (2010) “Structural Biology of the T cell Receptor: Insights into Receptor Assembly, Ligand Recognition, и Initiation of Signaling,” Cold Spring Harb. Perspect. Biol. 2:a005140). Комплекс имеет особое значение, так как он содержит большое количество (десять) активационных тирозинсодержащих мотивов иммунорецепторов (ITAM).
Два взаимодействия необходимы для активации Т-клеток (Viglietta, В. et al. (2007) “Modulating Co-Stimulation,” Neurotherapeutics 4:666-675; Korman, A.J. et al. (2007) “Checkpoint Blockade in Cancer Immunotherapy,” Adv. Immunol. 90:297-339). В первом взаимодействии, клетка должна демонстрировать релевантный антиген, связанный с главным комплексом гистосовместимости, так чтобы он мог связаться с Т-клеточным рецептором ("TCR") наивного Т-лимфоцита. Во втором взаимодействии лиганд клетки должен связываться с корецептором Т-лимфоцита (Dong, C. et al. (2003) “Immune Regulation by Novel Costimulatory Molecules,” Immunolog. Res. 28(1):39-48; Lindley, P.S. et al. (2009) “The Clinical Utility Of Inhibiting CD28-Mediated Costimulation,” Immunol. Rev. 229:307-321). Т-клетки, испытывающие оба стимулирующих сигнала, затем способны реагировать на цитокины (такие как интерлейкин-2 и интерлейкин-12). При отсутствии обоих костимуляторующих сигналов во время взаимодействия TCR, Т-клетки входят функционально неуправляемое состояние, называемое клональная толерантность (Khawli, L.A. et al. (2008) “Cytokine, Chemokine, и Co-Stimulatory Fusion Proteins for the Immunotherapy of Solid Tumors,” Exper. Pharmacol. 181:291-328). В патологических состояниях, Т-клетки являются ключевыми игроками различных орган-специфичных аутоиммунных заболеваний, таких как диабет типа I, ревматоидный артрит и рассеянный склероз (Dong, C. et al. (2003) “Immune Regulation by Novel Costimulatory Molecules,” Immunolog. Res. 28(1):39-48).
Необходимость двух сигналов для активации Т-клеток, приводящей к адаптивному иммунному ответу, как полагают, обеспечивает механизм для избегания ответов на аутоантигены, которые могут присутствовать на антигенпредставляющей клетках в тех местах в системе, где они могут быть распознаны Т-клеткой. Там, где контакт с Т-клетки с клеткой приводит к образованию только одного из двух необходимых сигналов, Т-клетки не активируются и адаптивный иммунный ответ не происходит.
II. Антитела и другие эпитопсвязывающие молекулы
A. Антитела
"Антитела" представляют собой иммуноглобулиновые молекулы, способные специфически связываться с мишенью, например углеводом, полинуклеотидом, липидом, полипептидом, и т.п. По меньшей мере, один сайт распознавания антигена расположен в вариабельном домене иммуноглобулиновой молекулы. При использовании в данном описании термин охватывает не только интактные поликлональные или моноклональные антитела, верблюдизированные антитела, одноцепочечные антитела и антиидиотипические (анти-Id) антитела (включая, например, анти-Id и анти-анти-Id антитела к антителам по настоящему изобретению), и их мутанты, встречающиеся в природе варианты, слитые белки, содержащие часть антитела с сайтом распознавания антигена требуемой специфичности, гуманизированные антитела и химерные антитела, но также любую другую модифицированную конфигурацию иммуноглобулиновой молекулы, которая содержит участок распознавания антигена требуемой специфичности. В данной заявке, нумерация аминокислотных остатков легких и тяжелых цепей антител соответствует индексу EU по Kabat et al. (1992) Sequences of Proteins of Immunological Interest, National Institutes of Health Publication № 91-3242. При использовании в данном описании "антигенсвязывающий фрагмент антитела" представляет собой часть антитела, имеющую по меньшей мере, один участок распознавания антигена. При использовании в данном описании термин охватывает фрагменты (например, фрагменты Fab, Fab ', F (аb')2, Fv), биспецифическые Fv, связанные дисульфидными связями (sdFv), интратела, и одноцепочечные молекулы (например, ScFv). В частности, антитела включают иммуноглобулиновые молекулы и иммунологически активные фрагменты иммуноглобулиновых молекул, т.е. молекулы, которые содержат антигенсвязывающий участок. Иммуноглобулиновые молекулы могут быть любого типа (например, IgG, IgE, IgM, IgD, IgA и IgY), класса (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2) или подкласса.
Естественные антитела (такие как IgG-антитела), состоят из двух легких цепей, образующих комплекс с двумя тяжелыми цепями. Каждая легкая цепь содержит вариабельный домен (VL) и константный домен (CL). Каждая тяжелая цепь содержит вариабельный домен (VH), три константных домена (СН1, СН2 и СН3), и шарнирный домен, расположенный между CH1- и CH2-доменами. Основная структурная единица природных иммуноглобулинов (например, IgG), таким образом, является тетрамером, имеющим две легкие цепи и две тяжелые цепи, как правило, экспрессируемым в виде гликопротеина около 150000 Да. Аминоконцевая ("N") часть каждой цепи включает вариабельную область примерно от 100 до 110 или более аминокислот, в первую очередь, ответственных за распознавание антигена. Карбоксиконцевая ("С") часть каждой цепи определяет константную область, при этом легкие цепи имеют один константный домен, а тяжелые цепи, как правило, имеют три константных домена и шарнирную область. Таким образом, структура легких цепей молекулы IgG представляет собой н-VL-CL-C, а структура тяжелых цепей IgG представляет собой n-VH-CH1-H-CH2-CH3-c (где Н представляет собой шарнирную область, а n и c представляют собой, соответственно, N-конец и С-конец полипептида).
Способность интактного, немодифицированного антитела (например, антитела IgG) к связыванию с эпитопом антигена зависит от наличия вариабельных доменов на легких и тяжелых цепях иммуноглобулина (т.е. VL-домен и VH-домен, соответственно). Взаимодействие легкой цепи и тяжелой цепи антитела, и, в частности, взаимодействие ее VL и VH-доменов, образует один из эпитопсвязывающих участков антитела. Вариабельные области IgG молекулы состоят из гипервариабельных участков (CDR), которые содержат остатки, контактирующие с эпитопом, и не-CDR сегменты, называемые каркасными сегментами (FR), которые в целом поддерживают структуру и определяют позиционирование CDR-петель таким образом, чтобы осуществлять такой контакт (хотя определенные каркасные остатки могут также связываться с антигеном). Таким образом, VL- и VH-домены имеют структуру n-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4-c. Полипептиды, которые являются (или могут служить в качестве) первого, второго и третьего CDR легкой цепи антитела в данном описании соответственно обозначены как домен CDRL1, домен CDRL2, и домен CDRL3. Аналогичным образом, полипептиды, которые являются (или могут служить в качестве) первого, второго и третьего CDR тяжелой цепи антитела в данном описании, соответственно, обозначены домен CDRH1, домен CDRH2, и домен CDRH3. Таким образом, термины домен CDRL1, домен CDRL2, домен CDRL3, домен CDRH1, домен CDRH2, и домен CDRH3 направлены на полипептиды, которые при введении в белок, приводят к тому, что белок становится способным связывать специфический эпитоп вне зависимости от того, является ли такой белок антителом, имеющим легкие и тяжелые цепи, или диателом, или одноцепочечной связывающей молекулой (например, ScFv, BiTe и т.д.), или белком другого типа. В отличие от таких антител конструкт ScFv содержит VL- и VH-домен антитела, находящихся в одной полипептидной цепи, в которой домены отделены гибким линкером достаточной длины так, чтобы позволить самосборку двух доменов в функциональный участок связывания эпитопа. Если самосборка VL- и VH-доменов становится невозможной из-за недостаточной длины линкера (менее чем около 12 аминокислотных остатков), два из ScFv-конструктов могут взаимодействовать друг с другом с другом с образованием двухвалентной молекулы, в которой VL одной цепи ассоциирован с VH другой (рассмотрено в Marvin et al. (2005) “Recombinant Approaches To IgG-Like Bispecific Antibodies,” Acta Pharmacol. Sin. 26:649-658).
В дополнение к их известным применениям в диагностике, антитела, как было показано, могут быть полезными в качестве терапевтических агентов. За последние несколько десятилетий наблюдается возрождение интереса к терапевтическому потенциалу антител, и антитела стали одним из ведущих классов биотехнологически получаемых лекарственных средств (Chan, CE et al.(2009) “The Use Of Antibodies In The Treatment Of Infectious Diseases,” Singapore Med. J. 50(7):663-666). Около 200 лекарственных средств на основе антител были одобрены для использования или находятся в стадии разработки.
Термин "моноклональное антитело" относится к гомогенной популяции антител, где моноклональное антитело состоит из аминокислот (встречающихся в природе и не встречающиеся в природе), которые участвуют в избирательном связывании антигена. Моноклональные антитела являются высокоспецифичными, направленными против одного эпитопа (или антигенного сайта). Термин "моноклональное антитело" охватывает не только интактные моноклональные антитела и полноразмерные моноклональные антитела, но также их фрагменты (такие как Fab, Fab', F(аb')2 Fv), одиночные цепи (ScFv), их мутанты, слитые белки, содержащие часть антитела, гуманизированные моноклональные антитела, химерные моноклональные антитела и любые другие модифицированные конфигурации иммуноглобулиновой молекулы, которые содержат участок распознавания антигена требуемой специфичности и имеют способность связываться с антигеном. Термин не должен быть ограничен в отношении источника антитела или способа, в котором оно получено (например, с помощью гибридом, фагового отбора, рекомбинантной экспрессии, трансгенных животных и т.д.). Термин включает целые иммуноглобулины, и фрагменты, и т.д. описанные выше в соответствии с определением "антитела". Способы получения моноклональных антител известны в данной области техники. Один из способов, который может быть использован, представляет собой способ Kohler, G. et al. (1975) “Continuous Cultures Of Fused Cells Secreting Antibody Of Predefined Specificity,” Nature 256:495-497 или его модификации. Как правило, моноклональные антитела получают в мышах, крысах и кроликах. Эти антитела получают путем иммунизации животного иммуногенным количеством клеток, клеточных экстрактов, или белковых препаратов, которые содержат желаемый эпитоп. Иммуноген может представлять собой, без ограничения перечисленным, первичные клетки, культивированные клеточные линии, раковые клетки, белки, пептиды, нуклеиновые кислоты, или ткани. Клетки, используемые для иммунизации, могут быть выращены в течение определенного периода времени (например, по меньшей мере, 24 часа) до их применения в качестве иммуногена. Клетки могут быть использованы в качестве иммуногенов сами по себе или в сочетании с неденатурирующим адъювантом, таким как RIBI (см., например, Jennings, V.M. (1995) “Review of Selected Adjuvants Used in Antibody Production,” ILAR J. 37(3):119-125).
В общем, клетки должны быть сохранены и предпочтительно являться жизнеспособными при использовании в качестве иммуногенов. Неповрежденные клетки позволяют иммунизированным животным лучше детектировать антигены, чем разрушенные клетки. Применение денатурирующих или жестких адъювантов, например, адъюванта Фрейнда, может привести к разрушению клеток и, следовательно, не рекомендуется. Иммуноген может быть введен несколько раз через определенные промежутки времени, например, дважды в неделю, или еженедельно, или может быть введен таким образом, чтобы поддерживать жизнеспособность в животном (например, в тканевом рекомбинанте). В ином случае, существующие моноклональные антитела и любые другие эквивалентные антитела, которые являются иммуноспецифичными для требуемого патогенного эпитопа могут быть секвенированы и получены рекомбинантно любыми способами, известными в данной области техники. В одном воплощении такое антитело секвенируют и полинуклеотидную последовательность затем клонируют в вектор для экспрессии или размножения. Последовательность, кодирующая представляющее интерес антитело, может быть сохранена в векторе в клетке-хозяине, и клетка-хозяин может быть затем размножена и заморожена для дальнейшего использования. Полинуклеотидная последовательность таких антител может быть использована для генетических манипуляций для создания химерного антитела, гуманизированного антитела, или канинизированного антитела, или для улучшения аффинности или других характеристик антитела. Термин "гуманизированные" антитела относятся к химерной молекуле, как правило, полученной с использованием рекомбинантных способов, имеющей участок связывания антигена, полученного из иммуноглобулина нечеловеческого вида, и остальную иммуноглобулиновую структуру молекулы, основанную на структуре и/или последовательности, иммуноглобулина человека. Полинуклеотидная последовательность вариабельных доменов таких антител может быть использована для генетических манипуляций для получения таких производных и улучшения аффинности или других характеристик таких антител. Общий принцип гуманизации антитела включает сохранение основной последовательности антигенсвязывающей части антитела с заменой остальной части нечеловеческого антитела на последовательности человеческого антитела. Есть четыре основных стадии для гуманизации моноклонального антитела. К ним относятся: (1) определение нуклеотидной и прогнозируемой аминокислотной последовательности исходных вариабельных доменов легкой и тяжелой цепей антитела (2) конструирование гуманизированного антитела или канинизированного антитела, т.е. определение того, каркасный участок какого антитела будет использоваться в ходе процессов гуманизации или канинизации (3) методологии/методы фактической гуманизации или канинизации и (4) трансфекция и экспрессия гуманизированного антитела. См., например, патенты США № 4816567, 5807715; 5866692; и 6331415.
Эпитопсвязывающий домен таких антител может включать либо полные вариабельные домены, слитые на константные домены или только гипервариабельные участки (CDR), привитые на соответствующие каркасные области в вариабельных доменах. Антиген-связывающие участки могут быть дикого типа или модифицированные с помощью одной или нескольких аминокислотных замен. Это исключает константную область как иммуноген у людей, но возможность иммунного ответа на инородные вариабельные области остается (LoBuglio, AF et al. (1989) “Mouse/Human Chimeric Monoclonal Antibody In Man: Kinetics And Immune Response,” Proc. Natl. Acad. Sci. (U.S.A.) 86:4220-4224). Другой подход ориентирован не только на обеспечении константных областей из человека, но и модификации вариабельных областей, и на то, чтобы изменить их как можно ближе к человеческой форме. Известно, что вариабельные области как тяжелой, так и легкой цепей содержат три гипервариабельных участка (CDR), которые изменяются в ответ на антигены, о которых идет речь, и определяют способность связывания, между четырьмя каркасными областями (FRS), которые относительно консервативны в заданном виде и которые предполагаемо обеспечивают поддержку для CDR. Когда нечеловеческие антитела получают к конкретному антигену, вариабельные участки могут быть "видоизменены" или "гуманизированы" прививкой CDR, полученных из нечеловеческого антитела на FR, присутствующих в модифицируемом человеческом антителе. Применение такого подхода к различным антителам сообщалось Sato, K. et al. (1993) Cancer Res 53:851-856. Riechmann, L. et al. (1988) “Reshaping Human Antibodies for Therapy,” Nature 332:323-327; Verhoeyen, M. et al. (1988) “Reshaping Human Antibodies: Grafting An Antilysozyme Activity,” Science 239:1534-1536; Kettleborough, C. A. et al. (1991) “Humanization Of A Mouse Monoclonal Antibody By CDR-Grafting: The Importance Of Framework Residues On Loop Conformation,” Protein Engineering 4:773-3783; Maeda, H. et al. (1991) “Construction Of Reshaped Human Antibodies With HIV-Neutralizing Activity,” Human Antibodies Hybridoma 2:124-134; Gorman, S. D. et al. (1991) “Reshaping A Therapeutic CD4 Antibody,” Proc. Natl. Acad. Sci. (U.S.A.) 88:4181-4185; Tempest, P.R. et al. (1991) “Reshaping A Human Monoclonal Antibody To Inhibit Human Respiratory Syncytial Virus Infection in vivo,” Bio/Technology 9:266-271; Co, M. S. et al. (1991) “Humanized Antibodies For Antiviral Therapy,” Proc. Natl. Acad. Sci. (U.S.A.) 88:2869-2873; Carter, P. et al. (1992) “Humanization Of An Anti-p185her2 Antibody For Human Cancer Therapy,” Proc. Natl. Acad. Sci. (U.S.A.) 89:4285-4289; и Co, M.S. et al. (1992) “Chimeric And Humanized Antibodies With Specificity For The CD33 Antigen,” J. Immunol. 148:1149-1154. В некоторых воплощениях гуманизированные антитела сохраняют все последовательности CDR (например, гуманизированное антитело мыши, которое содержит все шесть CDR из антител мыши). В других воплощениях, гуманизированные антитела имеют один или несколько CDR (один, два, три, четыре, пять, или шесть раз), которые различаются по последовательности относительно исходного антитела.
Был описан ряд "гуманизированных" молекул антител, содержащих сайт связывания антигена, полученного из нечеловеческого иммуноглобулина, в том числе химерные антитела, имеющие V-участки грызуна или модифицированные V-участки грызуна и связанные с ними гипервариабельные участки (CDR), слитые с константными человеческими доменами (см., например, Winter et al. (1991) “Man-made Antibodies,” Nature 349:293-299; Lobuglio et al. (1989) “Mouse/Human Chimeric Monoclonal Antibody In Man: Kinetics And Immune Response,” Proc. Natl. Acad. Sci. (U.S.A.) 86:4220-4224 (1989), Shaw et al. (1987) “Characterization Of A Mouse/Human Chimeric Monoclonal Antibody (17-1A) To A Colon Cancer Tumor-Associated Antigen,” J. Immunol. 138:4534-4538, и Brown et al. (1987) “Tumor-Specific Genetically Engineered Murine/Human Chimeric Monoclonal Antibody,” Cancer Res. 47:3577-3583). Другие источники описывают CDR грызунов, привитые к человеческому поддерживающему каркасному участку (FR) до слияния с соответствующим константным доменом человеческого антитела (см., например, Riechmann L. et al. (1988) “Reshaping Human Antibodies for Therapy,” Nature 332:323-327; Verhoeyen, M. et al. (1988) “Reshaping Human Antibodies: Grafting An Antilysozyme Activity,” Science 239:1534-1536; и Jones et al. (1986) “Replacing The Complementarity-Determining Regions In A Human Antibody With Those From A Mouse,” Nature 321:522-525). Другой источник описывает CDR грызуна, поддерживаемые рекомбинантно винированными каркасными участками грызунов. См., например, публ. Европейского патента № 519596. Эти "гуманизированные" молекулы предназначены для минимизации нежелательного иммунологического ответа по отношению к анти-человеческим антительным молекулам грызунов, что ограничивает продолжительность и эффективность терапевтического применения этих веществ в человеческих реципиентах. Другие способы гуманизации антител, которые также могут быть использованы, описаны Daugherty et al. (1991) “Polymerase Chain Reaction Facilitates The Cloning, CDR-Grafting, And Rapid Expression Of A Murine Monoclonal Antibody Directed Against The CD18 Component Of Leukocyte Integrins,” Nucl. Acids Res. 19:2471-2476 и в пат. США № 6,180,377; 6,054,297; 5,997,867 и 5,866,692.
B. Биспецифичные антитела, мультиспецифичные диатела и диатела DART™
Природные антитела способны связываться только с одним видом эпитопа (т.е. они являются "моно-специфичными"), хотя они могут быть в состоянии связать несколько копий этого вида (т.е. они могут демонстрировать бивалентность или мультивалентность). Было разработано широкое разнообразие форматов рекомбинантных биспецифичных антител (см., например, публ. РСТ № WO 2008/003116, WO 2009/132876, WO 2008/003103, WO 2007/146968, WO 2007/146968, WO 2009/018386, WO 2012/009544, WO 2013/070565), большинство из которых используют линкерные пептиды, либо для слияния антительного ядра (IgA, IgD, IgE, IgG или IgM) с дополнительным связывающим белком (например, scFv, VL VH, и т.д.), или в пределах антительного ядра, либо для слияния множественных антительных частей или для слияния (например, два Fab-фрагменты или ScFv) с доменом, содействующим гетеродимеризации, таким как домен CH2-CH3 или альтернативные полипептиды (WO 2005/070966, WO 2006/107786A WO 2006/107617A, WO 2007/046893). Как правило, такие подходы предусматривают компромиссы и уступки. Например, публ. РСТ № В WO 2013/174873, WO 2011/133886 и WO 2010/136172 раскрывают, что применение линкеров может вызвать проблемы в терапевтических планах, и сообщает, что триспецифичные антитела, в которых домены CL и СН1 переключаются из их соответствующих природных позиций, а VL- и VH-домены диверсифицируются (WO 2008/027236, WO 2010/108127) для того, чтобы дать им возможность связываться с более чем одним дополнительным антигеном. Таким образом, молекулы, описанные в этих документах, обменивают специфическое связывание на способность связывать дополнительные виды антигенов. Публ. РСТ № WO 2013/163427 и WO 2013/119903 раскрывают модификацию CH2-домена, для включения слитого белкового аддукта, содержащего связывающий домен. В документе отмечается, что CH2 домен вероятно, играет минимальную роль в опосредовании эффекторной функции. Публ. РСТ № В WO 2010/028797, WO WO2010028796 и 2010/028795 раскрывают рекомбинантные антитела, у которых Fc-домены были заменены дополнительными VL- и VH-доменами для получения трехвалентных связывающих молекул. Публ. РСТ № WO 2003/025018 и WO2003012069 раскрывают рекомбинантных диатела, чьи отдельные цепи содержат домены ScFv. Публ. РСТ № WO 2013/006544 раскрывает поливалентные молекулы Fab, которые синтезируют в виде единой полипептидной цепи и затем подвергают протеолизу с получением гетеродимерных структур. Таким образом, молекулы, описанные в этих документах обменивают все или некоторые способности опосредовать эффекторные функции на способность связывать дополнительные виды антигенов. Публ. РСТ № WO 2014/022540, WO 2013/003652, WO 2012/162583, WO 2012/156430, WO 2011/086091, WO 2007/075270, WO 1998/002463, WO 1992/022583 и WO 1991/003493 раскрывают добавление дополнительных связывающих доменов или функциональных групп с антителом или частью антитела (например, добавление диатела к легкой цепи антитела, либо добавления дополнительных VL- и VH-доменов к легким и тяжелым цепям антитела, либо добавление гетерологичного слитого белка или добавление в виде цепи множественных доменов Fab друг за другом). Таким образом, молекулы, описанные в этих документах обменивают нативную структуру антитела на способность связывать дополнительные виды антигенов.
В данной области дополнительно отмечена возможность продуцировать диател, которые отличаются от таких естественных антител по способности связывать два или большее количество различных видов эпитопов (например, демонстрировать биспецифичность или мультиспецифичность в дополнение к бивалентности или мультивалентности) (см., например, Holliger et al. (1993) “’Diabodies’: Small Bivalent And Bispecific Antibody Fragments,” Proc. Natl. Acad. Sci. (U.S.A.) 90:6444-6448; US 2004/0058400 (Hollinger et al.); US 2004/0220388 (Mertens et al.); Alt et al. (1999) FEBS Lett. 454(1-2):90-94; Lu, D. et al. (2005) “A Fully Human Recombinant IgG-Like Bispecific Antibody To Both The Epidermal Growth Factor Receptor And The Insulin-Like Growth Factor Receptor For Enhanced Antitumor Activity,” J. Biol. Chem. 280(20):19665-19672; WO 02/02781 (Mertens et al.); Olafsen, T. et al. (2004) “Covalent Disulfide-Linked Anti-CEA Diabody Allows Site-Specific Conjugation And Radiolabeling For Tumor Targeting Applications,” Protein Eng Des Sel. 17(1):21-27; Wu, A. et al. (2001) “Multimerization Of A Chimeric Anti-CD20 Single-chain Fv-Fv Fusion Protein Is Mediated Through Variable Domain Exchange,” Protein Engineering 14(2):1025-1033; Asano et al. (2004) “A Diabody For Cancer Immunotherapy And Its Functional Enhancement By Fusion Of Human Fc Domain,” Abstract 3P-683, J. Biochem. 76(8):992; Takemura, S. et al. (2000) “Construction Of A Diabody (Small Recombinant Bispecific Antibody) Using A Refolding System,” Protein Eng. 13(8):583-588; Baeuerle, P.A. et al. (2009) “Bispecific T-Cell Engaging Antibodies For Cancer Therapy,” Cancer Res. 69(12):4941-4944).
Конструкция диатела основывается на структуре одноцепочечных фрагментов вариабельных доменов (ScFv). Такие молекулы получают путем связывания вариабельных доменов легкой и/или тяжелой цепи друг с другом через короткий связующий пептид. Bird et al. (1988) (“Single-Chain Antigen-Binding Proteins,” Science 242:423-426) описывает пример связывающих пептидов, которые перекрывают приблизительно 3,5 нм между карбоксильным концом одного вариабельного домена и амино-концом другого вариабельного домена. Линкеры с другими последовательностями были разработаны и использованы (Bird et al.(1988) “Single-Chain Antigen-Binding Proteins,” Science 242:423-426). Линкеры в свою очередь могут быть изменены для дополнительных функций, таких как прикрепление лекарственных средств или прикрепления к твердым носителям. Одноцепочечные варианты могут быть получены либо рекомбинантно, либо синтетически. Для синтетического производства ScFv может быть использован автоматизированный синтезатор. Для рекомбинантного получения ScFv подходящая плазмида, содержащая полинуклеотид, который кодирует ScFv, может быть введена в подходящую клетку-хозяина, либо эукариотическую, такую как дрожжи, клетка растений, насекомых или млекопитающих, либо прокариотическую, такую как E.coli. Полинуклеотиды, кодирующие представляющий интерес ScFv, можно получить с помощью обычных манипуляций, таких как лигирование полинуклеотидов. Полученный ScFv может быть выделен с использованием стандартных методов очистки белков, известных в данной области техники.
Пат. США 7585952 и пат. публ. США № 2010-0173978 относятся к молекулам ScFv, которые иммуноспецифичны к ErbB2. Были описаны биспецифические активаторы Т-клеток (Bi-specific T cell engagers, “BiTEs”), тип молекулы ScFv (WO 05/061547; Baeuerle, Р et al. (2008) “BiTE: A New Class Of Antibodies That Recruit T Cells,” Drugs of the Future 33: 137-147; Bargou, et al. 2008) “Tumor Regression in Cancer Patients by Very Low Doses of a T Cell-Engaging Antibody,” Science 321: 974-977). Такие молекулы состоят из одной молекулы полипептидной цепи, имеющей два антигенсвязывающих домена, один из которых иммуноспецифично связывается с эпитопом CD3, а второй из которых иммуноспецифично связывается с антигеном, присутствующим на поверхности клетки-мишени.
Предоставление немоноспецифичных диател обеспечивает существенное преимущество: способность к колигировать и колокализовать клетки, которые экспрессируют различные эпитопы. Бивалентные диатела, таким образом, имеют широкие применения, включая терапию и иммунодиагностику. Бивалентность допускает большую гибкость при проектировании и конструировании диатела в различных применениях, обеспечивая усиленную авидность к мультимерным антигенам, перекрестную связь различных антигенов, и направленное нацеливание на специфические клеточные типы, на основании наличия обоих антигенов-мишеней. Из-за их более высокой валентности, низкой скорости диссоциации и быстрого выведения из кровотока (для диатела малого размера, на уровне или ниже ~50 кДа), молекулы диатела, известные в данной области также показали конкретное применение в области визуализации опухолей (Fitzgerald et al. (1997) “Improved Tumour Targeting By Disulphide Stabilized Diabodies Expressed In Pichia pastoris,” Protein Eng. 10:1221). Особое значение имеет совместное лигирование разных клеток, например, перекрестное сшивание цитотоксических Т-клеток к опухолевыми клетками (Staerz et al. (1985) “Hybrid Antibodies Can Target Sites For Attack By T Cells,” Nature 314:628-631, and Holliger et al. (1996) “Specific Killing Of Lymphoma Cells By Cytotoxic T-Cells Mediated By A Bispecific Diabody,” Protein Eng. 9:299-305).
Двухвалентные эпитопсвязывающие домены могут быть направлены на поверхностную детерминанту любой иммунной эффекторной клетки, такой как CD3, CD16, CD32, CD64 и т.п., которые экспрессируются на Т-лимфоцитах, естественных киллерах (NK) или других мононуклеарных клетках. Во многих исследованиях, связывание диатела с детерминантами эффекторных клеток, например, Fc-рецепторами γ (FcγR), также было обнаружено, как активирующее эффекторные клетки (Холлигер et al. (1996) “Specific Killing Of Lymphoma Cells By Cytotoxic T-Cells Mediated By A Bispecific Diabody,” Protein Eng. 9:299-305; Holliger et al. (1999) “Carcinoembryonic Antigen (CEA)-Specific T-cell Activation In Colon Carcinoma Induced By Anti-CD3 x Anti-CEA Bispecific Diabodies And B7 x Anti-CEA Bispecific Fusion Proteins,” Cancer Res. 59:2909-2916; WO 2006/113665; WO 2008/157379; WO 2010/080538; WO 2012/018687; WO 2012/162068). Как правило, активация эффекторных клеток инициируется связыванием антиген-связанного антитела с эффекторной клеткой через взаимодействие Fc-FcγR; Таким образом, в связи с этим, двухвалентные молекулы могут проявлять lg-подобную функциональность независимо от того, содержат ли они Fc-домен (например, на основе любого функционального эффекторного анализа, известного в данной области, или приведенного в качестве примера в данном документе (например, ADCC-анализ)). Перекрестной сшивкой опухолевых и эффекторных клеток диатело не только помещает эффекторные клетки в непосредственную близость к опухолевым клеткам, но и приводит к эффективному уничтожению опухоли (см., например, Као et al. (2003) “Bispecific Antibody Conjugates In Therapeutics,” Adv. Drug. Deliv. Rev. 55:171-197).
Например, пат. США № 6171586, относится к производству биспецифичных антител путем протеолитического расщепления двух антител для получения их F(аb')2-фрагментов, восстановления таких фрагментов в условиях, которые предотвращают образование межмолекулярных дисульфидных связей, а затем смешивания фрагментов для получения биспецифичных антител). Пат. США № 6551592; 6994853 и 8277806 и публ. РСТ № WO 2012/156430, WO 2002/020039, WO 2000/018806 и WO 1998/003670 касаются получения триспецифичных антител, способных одновременно связываться с Т-клетками и другими антигенами на опухолевой клетке, и, через Fc-часть биспецифичного антитела, с Fc-рецептором клеток, обладающих таким рецептором. Публ. РСТ № WO 2000/018806, WO 1998/003670 и WO 2006/072152 касаются производства триспецифичных антител, способных одновременно связываться с Т-клетками и другими антигенами. Публ. пат. США № 2008-0057054 раскрывает биспецифичные конъюгаты, специфичные для связывающего элемента против олигомеров бета-амилоида и связывающего элемента против трансмембранного белка теленцефалина. Публ. пат. США № 2010-0291112 касается биспецифичных и триспецифичных одноцепочечных молекул Fv, которые специфически связываются с одним (или двумя) опухолевым антигеном(ами) и антигеном эффекторных клеток (например, CD3, CD16 CD32, CD64 и т.д.).
В публ. РСТ № WO 1999/042597 и WO 1998/006749 раскрывают производные антител, которые включают домены связывания главного комплекса гистосовместимости человека, с или без связанных связывающих пептидов MHC. Публ. РСТ № WO 02/072141 касается мультиспецифичных связывающих молекул, чьи скорости ассоциации (скорости, с которыми они связываются с молекулами-мишенями) и скорости диссоциации (скорости, с которыми они высвобождают молекулы-мишени) отличаются тем, чтобы преимущественно связываются с одной мишенью по сравнению с их связыванием с другой такой молекулой-мишенью. Триспецифичные молекулы, например, молекулы, имеющие одновалентную первую часть, которая представляет собой анти-CD3 или анти-CD28-антитело, и вторую часть, содержащую двухвалентную группу демонстрирующую иммунную функцию, которая иммуноспецифично связывается с одним или несколькими целевыми лигандами на целевой пораженной клетке или иммунной клетке.
Пат. США N 7695936 и патентная публикация 2007/0196363 относятся к биспецифичным антителам, которые образуются из тяжелых цепей двух антител, одно из которых обладают выступающей частью, сконструированной на его тяжелой цепи, а второе из которых обладает комплементарным углублением, сконстуированным на его тяжелой цепи. Присутствие таких дополнительных "выпуклостей" и "углублений", как сообщается, преимущественно образует биспецифичные гетероантитела (имеющие одну тяжелую цепь каждого такого антитела) по сравнению с моноспецифичными гомоантителами, которые содержат две тяжелые цепи одного и того же антитела. Предложены различные биспецифичные гетероантитела, в том числе и те, которые являются иммуноспецифическими для CD3 и антигена опухолевых клеток. Также предложены различные триспецифичные гетероантитела, в том числе такие, которые являются иммуноспецифичными для CD3, CD8 и CD37 (трансмембранный белок, экспрессируемый преимущественно на В-клетках, которые участвуют в регулировании пролиферации Т-клеток (Robak, T. et al. (2014) “Anti-CD37 Antibodies For Chronic Lymphocytic Leukemia,” Expert Opin. Biol. Ther. 14 (5): 651-661), однако, никакого механизма для их производства и раскрытия их структуры не предусмотрено.
Публ. РСТ WO2012-162561 касается биспецифических, тетравалентных связывающих молекул, которые включают два полипептида, каждый из которых состоит из двух двухвалентные структур, разделенных промежуточным CH2-CH3 доменом. Этом документ также относится к четырехвалентным связывающим молекулам, состоящим из четырех полипептидных цепей, в которых две из полипептидных цепей содержат вариабельные домены легкой и тяжелой цепей для двух антигенов, и в котором остальные две полипептидные цепи содержат комплементарные вариабельные тяжелые и вариабельные легкие домены для антигенов и концево CH2-CH3 домен. Биспецифические, тетравалентные связывающие молекулы образуются через ассоциацию их соответствующих CH2-CH3 доменов. В конструкции из четырех полипептидных цепей, "легкие" цепи не ковалентно связаны с тяжелыми цепями, что приводит к нестабильности (см., Lu, D. et al. (2005) “A Fully Human Recombinant IgG-Iike Bispecific Antibody To Both The Epidermal Growth Factor Receptor And The Insulin-Like Growth Factor Receptor For Enhanced Antitumor Activity,” J. Biol. Chem. 280(20):19665-19672). В этом документе описан третий конструкт, в котором цепи изменяются для того, чтобы обеспечить такую ковалентную связь, но за счет устранения их биспецифичности (т.е. молекулы являются моно-специфичными). Раскрыты молекулы, имеющие специфичность к CD2, CD3, CD4, CD8, CD161, рецептору хемокинов, CD95, CCR5 и т.д. Биспецифичные молекулы, способные связываться как CD3, так и с CD8, не раскрывается.
Тем не менее, вышеуказанные преимущества подразумевают заметную стоимость. Формирование таких не-моноспецифичных диател требует успешной сборки двух или более отдельных и различных полипептидов (т.е. такое образование требует, чтобы диатела формировались через гетеродимеризацию различных видов полипептидной цепи). Это отличается от моноспецифичных диател, которые образуются через гомодимеризацию идентичных полипептидных цепей. Поэтому должно быть предусмотрено, по меньшей мере, два разнородных полипептида (например, два вида полипептидов), для того, чтобы сформировать немоноспецифическое диатело, и потому что из-за гомодимеризации таких полипептидов получаются неактивные молекулы (Takemura S. et al. (2000) “Construction Of A Diabody (Small Recombinant Bispecific Antibody) Using A Refolding System,” Protein Eng. 13(8):583-588), производство таких полипептидов должно быть выполнено таким образом, чтобы предотвратить образование ковалентной связи между полипептидами одного вида (Takemura, S. et al. (2000) “Construction Of A Diabody (Small Recombinant Bispecific Antibody) Using A Refolding System,” Protein Eng. 13(8):583-588). В данной области, таким образом, сообщается о нековалентной ассоциации таких полипептидов (см., например, Olafsen et al. (2004) “Covalent Disulfide-Linked Anti-CEA Diabody Allows Site-Specific Conjugation And Radiolabeling For Tumor Targeting Applications,” Prot. Engr. Des. Sel. 17:21-27; Asano et al. (2004) “A Diabody For Cancer Immunotherapy And Its Functional Enhancement By Fusion Of Human Fc Domain,” Abstract 3P-683, J. Biochem. 76(8):992; Takemura, S. et al. (2000) “Construction Of A Diabody (Small Recombinant Bispecific Antibody) Using A Refolding System,” Protein Eng. 13(8):583-588; Lu, D. et al. (2005) “A Fully Human Recombinant IgG-Like Bispecific Antibody To Both The Epidermal Growth Factor Receptor And The Insulin-Like Growth Factor Receptor For Enhanced Antitumor Activity,” J. Biol. Chem. 280(20):19665-19672).
Тем не менее, в данной области признано, что биспецифичные диатела, состоящие из нековалентно связанных полипептидов, неустойчивы и легко распадаются на нефункциональные мономеры (см., например, Lu, D. et al. (2005) “A Fully Human Recombinant IgG-Like Bispecific Antibody To Both The Epidermal Growth Factor Receptor And The Insulin-Like Growth Factor Receptor For Enhanced Antitumor Activity,” J. Biol. Chem. 280(20):19665-19672).
Перед лицом этой проблемы, в данной области удалось создать стабильные, ковалентно связанные гетеродимерные немоноспецифичные диатела, названные DART™ (см., например, публ. пат. США № 2013-0295121; 2010-0174053 и 2009-0060910; публ. Европейского патента № ЕР 2714079, ЕР 2601216, ЕР 2376109, ЕР 2158221 и публ. РСТ № WO 2012/162068, WO 2012/018687, WO 2010/080538, и Moore, P.A. et al. (2011) “Application Of Dual Affinity Retargeting Molecules To Achieve Optimal Redirected T-Cell Killing Of B-Cell Lymphoma,” Blood 117(17):4542-4551; Veri, M.C. et al. (2010) “Therapeutic Control Of B Cell Activation Via Recruitment Of Fcgamma Receptor IIb (CD32B) Inhibitory Function With A Novel Bispecific Antibody Scaffold,” Arthritis Rheum. 62(7):1933-1943; Johnson, S. et al. (2010) “Effector Cell Recruitment With Novel Fv-Based Dual-Affinity Re-Targeting Protein Leads To Potent Tumor Cytolysis And in vivo B-Cell Depletion,” J. Mol. Biol. 399(3):436-449). Такие диатела содержат два или более ковалентно комплексированных полипептидов и включают конструирование одного или несколько остатков цистеина в каждом из используемых видов полипептидов, которые позволяют образовать дисульфидные связи и тем самым ковалентно связать две полипептидные цепи. Например, добавление остатка цистеина на C-конце таких конструкций, как было показано, обеспечивает дисульфидную связь между полипептидными цепями, стабилизацию получаемого гетеродимера, не мешая свойствам связывания двухвалентной молекулы.
Есть множество воплощений DART™. Каждый из двух полипептидов в простейшем воплощении DART™ состоит из трех доменов (фигура 1). Первый полипептид включает: (i) первый домен, который содержит связывающую область вариабельного домена легкой цепи первого иммуноглобулина (VL1), (ii) второй домен, который содержит связывающую область вариабельного домена тяжелой цепи второй иммуноглобулин (VH2), и (iii), третий домен, который содержит остаток цистеина (или цистеинсодержащий домен) и домен, содействующий гетеродимеризации, который служит для содействия гетеродимеризации со второй полипептидной цепью. Остаток цистеина (или цистеинсодержащий домен) третьего домена служит для содействия ковалентному связыванию первой полипептидной цепи со второй полипептидной цепью диатела. Второй полипептид содержит: (i) добавочный первый домен (VL2-содержащий домен), (ii) добавочный второй домен (VH1-содержащий домен) и (iii), третий домен, содержащий остаток цистеина (или цистеинсодержащий домен) и, необязательно, дополнительный домен, содействующий гетеродимеризации, который образует комплекс с доменом, содействующим гетеродимеризации, первой полипептидной цепи, с тем чтобы содействовать гетеродимеризации с первой полипептидной цепью. Остаток цистеина (или цистеинсодержащий домен) третьего домена второй полипептидной цепи служит для содействия ковалентному связыванию второй полипептидной цепи с первой полипептидной цепью диатела. Такие молекулы являются стабильными, сильнодействующими и обладают способностью связывать одновременно два или более антигенов. Они способны содействовать перенаправленному опосредованному Т-клетками уничтожению клеток, экспрессирующих антигены-мишени.
В одном воплощении третьи домены первого и второго полипептидов содержат остаток цистеина, который служит для связывания полипептидов через дисульфидную связь. Третий домен одного или обоих полипептидов могут дополнительно обладать последовательностью СН2-СН3-домена, так чтобы комплекс полипептидов диатела образовал Fc-домен, который способен связываться с Fc-рецептором клеток (таких, как В-лимфоциты, дендритные клетки, естественные клетки-киллеры, макрофаги, нейтрофилы, эозинофилы, базофилы и тучные клетки) (фигуры 2А-2В).
Было описано множество вариаций таких молекул (см., например, публ. пат. США № 2013-0295121; 2010-0174053 и 2009-0060910; публ. Европейского патента № ЕР 2714079, ЕР 2601216, ЕР 2376109, ЕР 2158221 и публ. РСТ № WO 2012/162068, WO 2012/018687, WO 2010/080538). Эти Fc-несущие DART могут включать три полипептидные цепи (пример на фигуре 2В). Первая полипептидная цепь такого диатела содержит три домена: (i) а VL1-содержащий домен, (ii) а VH2-содержащий домен и (iii) домен, содержащий остаток цистеина (или цистеинсодержащий домен) и домен, способствующий гетеродимеризации, и (iv) остаток цистеин (или цистеинсодержащий домен и СН3-СН2-домен. Вторая полипептидная цепь такого DART™ содержит: (i) а VL2-содержащий домен, (ii) а VH1-содержащий домен и (iii) домен, содержащий остаток цистеина (или цистеинсодержащий домен) и домен, способствующий гетеродимеризации, который способствует гетеродимеризации с первой полипептидной цепью. Остаток цистеина (или цистеинсодержащий домен) третьего домена второй полипептидной цепи служит для содействия ковалентному связыванию второй полипептидной цепи с первой полипептидной цепью диатела. Третий полипептид такого DART™ содержит остаток цистеина (или цистеинсодержащий домен) и CH2-CH3-домен. Таким образом, первая и вторая полипептидные цепи такого DART™ ассоциированы вместе для того, чтобы сформировать сайт связывания VL1/VH1, который способен связываться с эпитопом, и сайт связывания VL2/VH2, который способен связываться со вторым эпитопом. Первый и второй полипептиды связаны друг с другом через дисульфидную связь с участием остатков цистеина в пределах их соответствующих третьих доменов. Следует отметить, что первая и третья полипептидные цепи образуют комплекс друг с другом с образованием Fc-домена, который стабилизирован через дисульфидную связь. Такие диатела обладают повышенной активностью. Такие Fc-несущие DART™ могут иметь любую из двух ориентаций (таблица 1).
Таблица 1
ориентация
ориентация
Еще более сложные диатела DART™, названные lg-DART™ (фигуры 3A-3B) и диатела Fc-DART™ (фигура 3C) были описаны (WO 2012/018687). Fc-DART™ имеют четыре полипептидные цепи. Первая и третья полипептидные цепи такого диатела содержат три домена: (i) а VL1-содержащий домен, (ii) а VH2-содержащий домен и (iii), домен, содержащую последовательность CH2-CH3. Второй и четвертый полипептид с Fc-DART™ содержат: (i) а VL2-содержащий домен, (ii) а VH1-содержащий домен и (iii) домен, способствующий гетеродимеризации и ковалентному связыванию с первой полипептидной цепью Fc-DART™. Третья и четвертая и первая и вторая полипептидные цепи могут быть одинаковыми или различными, с тем, чтобы сделать возможным четырехвалентное связывание, которое является либо моноспецифичным, биспецифичным или тетраспецифичным. Такие более сложные молекулы DART™ обладают также цистеинсодержащими доменами, функция которых заключается в образовании ковалентно связанного комплекса. Диатела Fc-DART™ содержат домены CH1 и CL.
Альтернативные конструкции известны в данной области техники для применений, в которых желательной является четырехвалентная молекула, но включение Fc не требуется в том числе, без ограничения перечисленным, четырехвалентные тандемные антитела, которые также называют "TandAb" (см., например, публ. пат. США № 2005-0079170, 2007-0031436, 2010-0099853, 2013-0189263 2011-020667, публ. Европейского патента № EP 1078004, ЕР 2371866, ЕР 2361936 и ЕР 1293514; публ. РСТ № WO 1999/057150, WO 2003/025018, и WO 2013/013700), которые образуются при гомодимеризации двух идентичных цепей, каждая из которых несет домены VH1, VL2, VH2 и VL2.
Тем не менее, несмотря на все предыдущие достижения, остается потребность в композициях, которые могли бы обеспечить улучшенную терапевтическую ценность для пациентов, страдающих от злокачественных опухолей или других заболеваний и состояний. Настоящее изобретение направлено на решение этой и других целей.
Раскрытие изобретения
Настоящее изобретение относится к триспецифичным связывающим молекулам, которые являются полипептидными молекулами из нескольких цепей, которые обладают тремя связывающими доменами и, таким образом, способны опосредовать скоординированное связывание с тремя эпитопами. Триспецифичная связывающая молекула предпочтительно отличается тем, что обладает связывающими доменами, которые позволяют ей иммуноспецифично связываться с: (1) эпитопом первого антигена злокачественной опухоли (2) эпитопом второго антигена злокачественной опухоли, и (3) эпитоп молекулы, которая экспрессируется на поверхности эффекторных клеток иммунной системы, и, таким образом, способными локализовать эффекторную клетку иммунной системы к клетке, которая экспрессирует антиген злокачественной опухоли, с тем, чтобы тем самым облегчить уничтожение такой злокачественной опухолевой клетки.
Более подробно, настоящее изобретение относится к триспецифической связывающей молекуле, способной иммуноспецифично связываться с тремя разными эпитопами, где указанные эпитопы представляют собой эпитоп I, эпитоп II, и эпитоп III, где два из трех эпитопов являются эпитопами антигена злокачественной опухоли (антигены), а третий указанный эпитоп представляет собой эпитоп антигена эффекторной клетки.
В частности, изобретение относится к воплощению такой триспецифичной связывающей молекулы, которая включает четыре различных полипептидных цепи, ковалентно объединенных в комплекс вместе, и содержит:
(i), антигенсвязывающий домен I, который способен иммуноспецифично связываться с эпитопом I, присутствующим на первом антигене, и антигенсвязывающий домен II, который способен иммуноспецифично связываться с эпитопом II на втором антигене, где антигенсвязывающий домен I и антигенсвязывающий домен II оба являются связывающими доменами по типу диатела;
(ii), антигенсвязывающий домен III, который способен иммуноспецифично связываться с настоящего эпитопом III на третьем антигене; и
(iii) Fc-домен, который формируется за счет объединения в комплекс двух CH2-CH3-доменов друг с другом;
где один из числа эпитопа I, эпитопа II или эпитопа III представляет собой эпитоп антигена эффекторной клетки, второй из числа эпитопа I, эпитопа II или эпитопа III представляет собой эпитоп первого антигена злокачественной опухоли, а третий из числа эпитопа I, эпитопа II или эпитопа III представляет собой эпитоп второго антигена злокачественной опухоли, и где антигенсвязывающие домены I, II и III связывающих молекул опосредуют скоординированное связывание эффекторной клетки иммунной системы, экспрессирующей антиген эффекторных клеток, и злокачественной опухолевой клетки, экспрессирующей первый и второй антигены злокачественных клеток.
В частности, изобретение относится к воплощению таких триспецифичных связывающих молекул, где Fc-домен способен связываться с Fc-рецептором, расположенном на поверхности клетки.
Кроме того, изобретение относится воплощению таких триспецифичных связывающих молекул, в котором антиген эффекторных клеток расположен на поверхности эффекторной клетки и где антигены злокачественной опухоли расположены на поверхности злокачественной опухолевой клетки, и в котором иммуноспецифического связывания достаточно для колокализации антигена эффекторных клеток и антигенов злокачественной опухоли, тем самым облегчая активацию эффекторных клеток против злокачественной опухолевой клетки.
Кроме того, изобретение относится к воплощению таких триспецифичных связывающих молекул, в котором антиген эффекторных клеток выбран из группы, состоящей из: CD2, CD3, CD16, CD19, CD20, CD22, CD32B, CD64 В-клеточного рецептора (BCR), Т-клеточного рецептора (TCR), и рецептора NKG2D.
Кроме того, изобретение относится к воплощению таких триспецифичных связывающих молекул, в котором первый и второй антигены злокачественной опухоли независимо друг от друга выбраны из группы, состоящей из: антигена рака толстой кишки 19,9; муцина рака желудка; антигена 4.2; гликопротеина A33 (gpA33); ADAM-9; антигена рака желудка AH6; ALCAM; антигена злокачественных лимфоцитов человека APO-1; антигена злокачественной опухоли В1; B7-H3; бета-катенина; группы крови ALeb/Ley; антигена 38.13 лимфомы Беркитта, антигена аденокарциномы толстой кишки C14; антигена рак яичников СА125; карбоксипептидазы М; CD5; CD19; CD20; CD22; CD23; CD25; CD27; CD30; CD33; CD36; CD45; CD46; CD52; CD79a/CD79b; CD103; CD317; CDK4; карциноэмбрионального антигена (СЕА); CEACAM5; CEACAM6; CO17-1A; СО-43 (группа крови Leb); СО-514 (группа крови Leа); СТА-1; CTLA4; цитокератина 8; антигена D1.1; Антигена D1 56-22; DR5; серии E1 (группа крови B); EGFR (рецептора эпидермального фактора роста); рецептора эфрина А2 (EphA2); ErbB1; ErbB3; ErbB4; GAGE-1; GAGE-2; GD2/GD3/GM2; антигена аденокарциномы легкого F3; антигена FC10.2; G49, ганглиозида GD2; ганглиозида GD3; ганглиозида GM2; ганглиозида GM3; GD2; GD3; Gică 19-9; GM2; gp100; антигена Т-клеточного лейкоза человека gp37; антигена меланомы gp75; gpA33; антигена HER2 (p185 HER2); антигена жировых шариков молока человека (HMFG); вируса папилломы человека-E6/вируса папилломы человека-E7; высокомолекулярного антигена меланомы (ВММ-MAA); I антигена (антигена дифференцировки) I (Ma); Интегрина альфа-V-бета-6 интегрина β6 (ITGB6); Интерлейкина-13; Рецептора α2 (IL13Rα2); JAM-3; KID3; KID31; KS 1/4 антигена пан-карциномы; антигенов карциномы легкого человека L6 и L20; LEA; LUCA-2; М1: 22: 25: 8; M18; M39; MAGE-1; MAGE-3; MART; MUC-1; MUM-1; Myl; N-ацетилглюкозаминтрансферазы; неогликопротеина; NS-10; OFA-1; OFA-2; Онкостатина M; p15; меланома-ассоциированного антигена р97; полиморфного эпителиального муцина (PEM); антигена полиморфного эпителиального муцина (РЕМА); PIPA; простат-специфического антигена (ПСА); простат-специфического мембранного антигена (PSMA); фосфата простатической кислоты; R24; ROR1; сфинголипидов; SSEA-1; SSEA-3; SSEA-4; STN; пептида, полученного из Т-клеточного рецептора; T5A7; TAG-72; TL5 (группа крови A); TNF-α-рецептора; TNF-β-рецептора; TNF-γ-рецептора; TRA-1-85 (группа крови Н); рецептора трансферрина; опухолеспецифичного трансплантационного антигена (TSTA), карциноэмбрионального антигена-альфа-фетопротеина (AFP); VEGF; VEGFR, VEP8; VEP9; VIM-D5; и Y-гаптена, Leу.
Кроме того, изобретение относится к воплощению таких триспецифичных связывающих молекул, в котором первый и второй антигены злокачественной опухоли выбирают из группы, состоящей из: CD2, CD317, CEACAM5, CEACAM6, DR5, EphA2, gpA33, Her2, В7-Н3; EGF, EGFR, VEGF и VEGFR.
Кроме того, изобретение относится к воплощению таких триспецифичных связывающих молекул, в котором связывающий домен III не по типу диатела включает связывающий домен Fab-типа (VLIII/VHIII), который способен иммуноспецифично связываться с эпитопом III, где молекула включает:
(А) первую полипептидную цепь, которая включает по направлению от N-конца к С-концу:
(1) вариабельный домен легкой цепи иммуноглобулина, способного связываться с первым из трех эпитопов (VLI);
(2) вариабельный домен тяжелой цепи иммуноглобулина, способного связываться со вторым из трех эпитопов (VHII);
(3) домен, содействующий гетеродимеризации; и
(4) СН2- и СН3-домены IgG;
(В) вторую полипептидную цепь, которая включает по направлению от N-конца к С-концу:
(1) вариабельный домен легкой цепи иммуноглобулина, способного связываться с вторым из трех эпитопов (VHII);
(2) вариабельный домен тяжелой цепи иммуноглобулина, способного связываться с первым из трех эпитопов (VHI); и
(3) комплементарный домен, содействующий гетеродимеризации;
(С) третью полипептидную цепь, которая включает по направлению от N-конца к С-концу:
(1) вариабельный домен тяжелой цепи иммуноглобулина, способного связываться с третьим из трех эпитопов (VHIII); и
(2) СН1-домен, шарнирный домен и СН2-СН3-домен IgG;
и
(D), четвертую полипептидную цепь, которая включает по направлению от N-конца к С-концу:
(1) вариабельный домен легкой цепи иммуноглобулина, способного связываться с третьим из трех эпитопов (VLIII); и
(2) константный домен легкой цепи (CL);
в которой:
(i) VLI и VHI домены ассоциируются с образованием домена, способного связывать первый эпитоп;
(ii) VLII и VHII домены ассоциируются с образованием домена, способного связывать второй эпитоп;
(iii) VLIII и VHIII-домены ассоциируются с образованием домена, способного связывать третий эпитоп;
(iv) СН2-СН3-домен первой полипептидной цепи и СН2-СН3-домен третьей полипептидной цепи ассоциируются с образованием Fc-домена;
(v) первая и вторая полипептидные цепи ковалентно связаны друг с другом;
(vi), первая и третья полипептидные цепи ковалентно связаны друг с другом; и
(vii), третья и четвертая полипептидные цепи ковалентно связаны друг с другом.
Кроме того, изобретение относится к воплощению таких триспецифичных связывающих молекул, в которых:
(A) домен, содействующий гетеродимеризации является E-спиральным, а комплементарный домен, содействующий гетеродимеризации является K-спиральным; или
(B) домен, содействующий гетеродимеризации является K-спиральным, а комплементарный домен, содействующий гетеродимеризации является Е-спиральным.
Кроме того, изобретение относится к воплощению таких триспецифичных связывающих молекул, в которых:
(А) СН2-СН3-домены первой и третьей полипептидных цепей каждый имеют последовательность SEQ ID NO: 1, таким образом, что Fc-домен, образованный при их объединении проявляет нормальную FcγR-опосредованную эффекторную функцию; или
(В) СН2-СН3-домен первой и третьей полипептидных цепей содержат, по меньшей мере, одну аминокислотную замену, по отношению к последовательности SEQ ID NO: 1, таким образом, что Fc-домен, образованный при их объединении демонстрирует видоизмененную FcγR-опосредованную эффекторную функцию.
Кроме того, изобретение относится к воплощению таких триспецифичных связывающих молекул, в котором замещение, по меньшей мере, одной аминокислоты, включает, по меньшей мере, одну аминокислотную замену, выбранную из группы, состоящей из: L235V, F243L, R292P, Y300L, V305I и P396L, где нумерация та же, что и в индексе EU по Kabat.
Кроме того, изобретение относится к воплощению таких триспецифичных связывающих молекул, в котором замещение, по меньшей мере, одной аминокислоты включает:
(А), по меньшей мере, одну замену, выбранную из группы, состоящей из F243L, R292P, Y300L, V305I и P396L;
(В) по меньшей мере, две замены, выбранные из группы, состоящей из:
(1) F243L и P396L;
(2) F243L и R292P; и
(3) R292P и V305I;
(С) по меньшей мере, три замены, выбранные из группы, состоящей из:
(1) F243L, R292P и Y300L;
(2) F243L, R292P и V305I;
(3) F243L, R292P и P396L; и
(4) R292P, V305I и P396L;
(D), по меньшей мере, четыре замены, выбранные из группы, состоящей из:
(1) F243L, R292P, Y300L и P396L; и
(2) F243L, R292P, V305I и P396L;
или
(Е), по меньшей мере, пять замен, выбранных из группы, состоящей из:
(1) F243L, R292P, Y300L, V305I и P396L; и
(2) L235V, F243L, R292P, Y300L и P396L.
Кроме того, изобретение относится к воплощению таких триспецифичных связывающих молекул, в котором СН2-СН3-домен первой и третьей полипептидных цепей отличаются друг от друга и имеют аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 52 и SEQ ID NO: 53.
Кроме того, изобретение относится к воплощению таких триспецифичных связывающих молекул, в котором:
(А) эпитоп I, эпитоп II и эпитоп III представляют собой, соответственно, эпитоп первого антигена злокачественной опухоли эпитоп второго антигена злокачественной опухоли и эпитоп антигена эффекторных клеток;
(B) эпитоп I, эпитоп II и эпитоп III представляют собой, соответственно, эпитоп первого антигена злокачественной опухоли эпитоп антигена эффекторных клеток и эпитоп второго антигена злокачественной опухоли;
(С) эпитоп I, эпитоп II и эпитоп III представляют собой, соответственно, эпитоп второго антигена злокачественной опухоли, эпитоп первого антигена злокачественной опухоли, и эпитоп антигена эффекторных клеток;
(D) эпитоп I, эпитоп II и эпитоп III представляют собой, соответственно, эпитоп второго антигена злокачественной опухоли, эпитоп антигена эффекторных клеток и эпитоп первого антигена злокачественной опухоли;
(Е) эпитоп I, эпитоп II и эпитоп III представляют собой, соответственно, эпитоп антигена эффекторных клеток, эпитоп первого антигена злокачественной опухоли и эпитоп второго антигена злокачественной опухоли;
и
(F) эпитоп I, эпитоп II и эпитоп III представляют собой, соответственно, эпитоп антигена эффекторных клеток эпитоп второго антигена злокачественной опухоли, и эпитоп первого антигена злокачественной опухоли.
Кроме того, изобретение относится к воплощению таких триспецифичных связывающих молекул, в котором:
(А) эпитоп антигена эффекторной клетки представляет собой CD2-эпитоп, распознаваемый антителом Lo-CD2a;
(В) эпитоп антигена эффекторной клетки представляет собой CD3-эпитоп, распознаваемый антителом OKT3, M291, YTH12.5, анти-CD3 mAb 1 или анти-CD3 mAb 2;
(С) эпитоп антигена эффекторной клетки представляет собой CD16 эпитоп, распознаваемый антителом 3G8 или А9;
(D), эпитоп антигена эффекторной клетки представляет собой CD19-эпитоп, распознаваемый антителом MD1342, MEDI-551, блинатумомаб или HD37;
(Е) эпитоп антигена эффекторной клетки CD20-эпитоп, распознаваемый антителом ритуксимаб, ибритумомаб, афатумомаб и тозитумомаб;
(F), эпитоп антигена эффекторной клетки представляет собой CD22 эпитоп, распознаваемый антителом эпратузумаб;
(G), эпитоп антигена эффекторной клетки представляет собой CD32B эпитоп, распознаваемый антителом CD32B mAb 1;
(Н) эпитоп антигена эффекторной клетки представляет собой CD64-эпитоп, распознаваемый антителом CD64 mAb 1;
(i) эпитоп антигена эффекторной клетки представляет собой BCR/CD79 эпитоп, распознаваемый антителом CD79 mAb 1;
(J) эпитоп антигена эффекторной клетки представляет собой TCR эпитоп, распознаваемый антителом BMA 031;
или
(К) эпитоп антигена эффекторной клетки представляет собой эпитоп рецептора NKG2D, распознаваемый антителом KYK-2.0.
Кроме того, изобретение относится к фармацевтической композиции, которая содержит любую из описанных выше триспецифичных связывающих молекул, и фармацевтически приемлемый носитель, наполнитель или разбавитель.
Кроме того, изобретение относится к воплощению фармацевтической композиции или любых таких триспецифичных связывающих молекулах, в котором триспецифичная связывающая молекула используется при лечении онкологических заболеваний.
Кроме того, изобретение относится к воплощению таких фармацевтических композиций или таких триспецифичных связывающих молекул, где злокачественная опухоль характеризуется наличием злокачественных опухолевых клеток, выбранных из группы, состоящей из клеток: опухоли надпочечников, СПИД-ассоциированных злокачественных новообразований, альвеолярной саркомы мягких тканей, астроцитарной опухоли, рака мочевого пузыря, рака кости, рака головного и спинного мозга, метастатической опухоли головного мозга, рака молочной железы, опухолей каротидного гломуса, рака шейки матки, хондросаркомы, хордомы, хроматофобного почечноклеточного рака, светлоклеточного рака, рака толстой кишки, колоректальный рака, кожной доброкачественной фиброзной гистиоцитомы, десмопластической мелкокруглоклеточной опухоли, эпендимомы, опухоли Юинга, внескелетной миксоидной хондросаркомы, несовершенного костного фиброгенеза, фиброзной дисплазии кости, рака желчного пузыря или желчных протоков, рака желудка, гестационной трофобластической болезни, герминомы, рака головы и шеи, гепатоцеллюлярной карциномы, опухоли островковых клеток, саркомы Капоши, рака почки, лейкоза, липомы/ доброкачественной липоматозной опухоли, липосаркомы/злокачественной липоматозной опухоли, рака печени, лимфомы, рака легкого, медуллобластомы, меланомы, менингиомы, множественных эндокринных неоплазий, множественной миеломы, миелодиспластического синдрома, нейробластомы, нейроэндокринной опухоли, рака яичника, поджелудочной железы, папиллярной карциномы щитовидной железы, опухоли паращитовидной железы, педиатрического рака, опухоли оболочки периферического нерва, феохромоцитомы, опухоли гипофиза, рака предстательной железы, задней увеальной меланомы, редкого гематологического заболевания, почечного метастатического рака, палочковидной опухоли, рабдомиосаркомы, саркомы, рака кожи, саркомы мягких тканей, плоскоклеточного рака, рака желудка, синовиальной саркомы, рака яичек, карциномы тимуса, тимомы, метастатического рака щитовидной железы, и рака матки.
Кроме того, изобретение относится к воплощению таких фармацевтических композиций или таких триспецифичных связывающих молекул, где злокачественная опухоль представляет собой колоректальный рак, гепатоцеллюлярный рак, глиому, рак почки, рак молочной железы, множественную миелому, рак мочевого пузыря, нейробластому; саркому, неходжкинскую лимфому, немелкоклеточный рак легкого, рак яичников, рак поджелудочной железы или рак прямой кишки.
Кроме того, изобретение относится к воплощению таких фармацевтических композиций или таких триспецифичных связывающих молекул, где злокачественная опухоль представляет собой острый миелоидный лейкоз (AML), хронический миелоидный лейкоз (CML), острый В-лимфобластный лейкоз (B-ALL), хронический лимфоцитарный лейкоз (CLL), волосатоклеточный лейкоз (HCL), бластные новообразования плазмацитоидных дендритных клеток (BPDCN), неходжкинскую лимфому (NHL), включая мантийноклеточную лимфому (MCL), и мелкоклеточную лимфоцитарную лимфому (SLL), лимфому Ходжкина, системный мастоцитоз или лимфому Беркитта.
Краткое описание чертежей
Фигуры 1A-1B демонстрируют схематическое представление областей диатела DART™. На фигуре 1А показано схематическое представление доменов основного диатела DART™. фигура 1В представляет собой схему ковалентно связанного диатела, состоящего из двух полипептидных цепей, каждая из которых имеет домен, содействующий гетеродимеризации, VL и VH-домены, которые распознают один и тот же эпитоп, показаны с использованием одинакового затенения.
Фигуры 2A-2B обеспечивают схематическое изображение ковалентно связанных диател, состоящих из двух полипептидных цепей, каждая из которых имеет СН2- и СН3-домен (фигура 2А) или в которых только одна имеет СН2- и СН3-домен (фигура 2В), так, что связанные цепи образуют Fc-домен, который включает весь или часть встречающегося в природе Fc-домена. VL и VH-домены, которые распознают один и тот же эпитоп, показаны с использованием одинакового затенения.
Фигуры 3A-3C обеспечивают схематическое изображение, демонстрирующее четырехвалентные диатела, состоящие из двух пар полипептидных цепей. Пары являются различными, что дает биспецифичную молекулу, которая двухвалентна по отношению к каждому из двух эпитопов, из которых один эпитоп DR5, а другой эпитоп молекулы, присутствующей на поверхности эффекторной клетки. Один полипептид из каждой пары обладает СН2- и СН3-доменом, так что соответствующие цепи образуют Fc-домен, который включает все или часть встречающегося в природе Fc-домена. VL и VH-домены, которые распознают один тот же эпитоп, показаны с использованием одинакового затенения. Только одна пара эпитопов (показаны одинаковой штриховкой) способна связываться с DR5. На фигуре 3А показано Ig-диатело. На фигуре 3В показано Ig-диатело, которое содержит E-спиральный и K-спиральный домены, способствующие гетеродимеризации. Фигура 3C демонстрирует диатела Fc-DART™, которые содержат антительные CH1- и CL-домены. Обозначение "VL1" и "VH1" обозначают соответственно вариабельный домен легкой цепи и вариабельный домен тяжелой цепи, которые связывают "первый" эпитоп. Аналогичным образом, обозначение "VL2" и "VH2" обозначают соответственно, вариабельный домен легкой цепи и вариабельный домен тяжелой цепи, которые связывают "второй" эпитоп.
Фигуры 4A-4G обеспечивают графическое представление доменов предпочтительных триспецифичных связывающих молекул по настоящему изобретению. Фигуры схематично иллюстрируют порядок и ориентацию доменов воплощений предпочтительных триспецифичных связывающих молекул по настоящему изобретению. Фигуры 4А, 4В и 4G иллюстрируют воплощения, в которых триспецифичная связывающая молекула состоит из четырех полипептидных цепей. Фигуры 4C, 4D, 4E и 4F иллюстрируют воплощения, в которых связывающая молекула состоит из трех полипептидных цепей. Молекула может обладать шарнирным и/или CL-доменами (фигуры 4А, 4В, 4С, 4Е) или может содержать альтернативный линкерный пептид (фигуры 4D, 4F, 4G).
Фигуры 5A-5E обеспечивают графическое представление о доменах альтернативного воплощения триспецифичных связывающих молекул по настоящему изобретению, в котором домен, связывающий эффекторные клетки, состоит из связывающего домена по типу рецептора эффекторной клетки, а не связывающего домена по типу диатела или связывающего домена Fab-типа. Фигуры 5А и 5В иллюстрируют воплощения, в которых триспецифичная связывающая молекула состоит из четырех полипептидных цепей. Фигуры 5С и 5E иллюстрируют воплощение, в котором связывающая молекула состоит из трех полипептидных цепей. Фигура 5D иллюстрирует воплощение, в котором связывающая молекула состоит из пяти полипептидных цепей. Молекула может обладать шарнирным и/или CL-доменами или может содержать альтернативные линкертные пептиды.
Фигура 6 демонстрирует способность анти-DR5 человека моноклональных антител и DR5 mAb 1 и DR5 mAb 2 связываться с DR5 человека и к DR5 макаки.
Фигура 7, панели AH, демонстрирует кинетику связывания DR5 mAb 1 (панели А и Е), DR5 mAb 2 (панели B и F), DR5 mAb 3 (панели C и G) и DR5 mAb 4 (панели D и H) для DR5 человека (панели AD) и DR5 макаки (панели EH).
Фигура 8 демонстрирует неожиданное превосходство DR5 mAb 1 и DR5 mAb 2. Превосходство оценивали путем сравнения способности к DR5 х CD3 диател, имеющих VL- и VH-домены из DR5 mAb 1, DR5 mAb 2, DR5 mAb 3 или DR5 mAb 4, опосредовать цитотоксичность A549 аденокарциномных человеческих альвеолярных базальных эпителиальных опухолевых клеток.
Фигуры 9А-9С демонстрируют синергическое повышение связывания клетки-мишени, которое достигается, когда оба из двух доменов, связывающих антигены злокачественной опухоли триспецифичные связывающие молекулы по настоящему изобретению способны связываться с клеткой-мишенью. Фигура 9А демонстрирует связывание, получаемое когда триспецифичные молекулы: EphA2 mAb 1 х CD3 mAb 2 х DR5 mAb 1; EphA2 mAb 1 х CD3 mAb 2 х gpA33 mAb 1; и gpA33 mAb 1 х CD3 mAb 2 х DR5 mAb 1 инкубируют в присутствии EphA2-экспрессирующих клеток СНО. Фигура 9 В демонстрирует связывание, которое получается, когда такие триспецифичные молекулы инкубируют в присутствии DR5-экспрессирующих клеток СНО. Фигура 9С демонстрирует связывание, которое получается, когда такие триспецифичные молекулы инкубируют в присутствии DU145 клеток предстательной железы человека, которые экспрессируют EphA2 и DR5, но не gpA33.
Фигуры 10А-10С демонстрируют синергическое повышение цитотоксисности по отношению к клетке-мишени, которая достигается, когда оба из двух доменов, связывающих антигены злокачественной опухоли триспецифичные связывающие молекулы по настоящему изобретению способны связываться с клеткой-мишенью. Фигура 10A показывает процент цитотоксичности, полученный путем инкубирования триспецифичных молекул: EphA2 mAb 1 х CD3 mAb 2 х DR5 mAb 1; EphA2 mAb 1 х CD3 mAb 2 х gpA33 mAb 1; и gpA33 mAb 1 х CD3 mAb 2 х DR5 mAb 1 в присутствии EphA2-экспрессирующих клеток СНО и цитотоксичных лимфоцитов. Фигура 10B демонстрирует процент цитотоксичности, полученной, когда такие триспецифичные молекулы инкубируют в присутствии DR5-экспрессирующих клеток СНО и цитотоксических лимфоцитов. Фигура 10С демонстрирует цитотоксичность, полученную, когда такие триспецифичные молекулы инкубируют в присутствии DU145 клеток предстательной железы человека и цитотоксических лимфоцитов. Клетки DU145 экспрессируют EphA2 и DR5, но не gpA33. Цитотоксичность измеряли по увеличению люминесценции, вызванной выделением люциферазы после лизиса клеток.
Осуществление изобретения
Настоящее изобретение относится к триспецифичным связывающим молекулам, которые являются полипептидными молекулами из нескольких цепей, и которые обладают тремя связывающими доменами и, таким образом, способны опосредовать скоординированное связывание с тремя эпитопами. Триспецифичная связывающая молекула предпочтительно отличается тем, что обладает связывающими доменами, которые позволяют ей иммуноспецифично связываться с: (1) эпитопом первого антигена злокачественной опухоли (2) эпитопом второго антигена злокачественной опухоли, и (3) эпитопом молекулы, которая экспрессируется на поверхности эффекторных клеток иммунной системы, и, таким образом, способную локализовать эффекторную клетку иммунной системы к клетке, которая экспрессирует антиген злокачественной опухоли, с тем, чтобы тем самым облегчить уничтожение такой злокачественной опухолевой клетки.
Триспецифичные связывающие молекулы по настоящему изобретению могут включать эпитопсвязывающие домены гуманизированных, химерных или канинизированных производных вышеописанных антител, например, DR5 mAb 1 или DR5 mAb 2.
I. Общие методы и общие определения
Практика настоящего изобретения будет использовать, если не указано иное, обычные методы молекулярной биологии (в том числе рекомбинантные методы), микробиологии, клеточной биологии, биохимии и иммунологии, которые известны специалистам в данной области техники. Такие методы подробно описаны в литературе, такой как Molecular Cloning: A Laboratory Manual, Third Edition (Sambrook et al. Eds., 2001) Cold Spring Harbor Press, Cold Spring Harbor, NY; Oligonucleotide Synthesis: Methods and Applications (Methods in Molecular Biology), Herdewijn, P., Ed., Humana Press, Totowa, NJ; Oligonucleotide Synthesis (Gait, M.J., Ed., 1984); Methods in Molecular Biology, Humana Press, Totowa, NJ; Cell Biology: A Laboratory Notebook (Cellis, J.E., Ed., 1998) Academic Press, New York, NY; Animal Cell Culture (Freshney, R.I., Ed., 1987); Introduction to Cell and Tissue Culture (Mather, J.P. and Roberts, P.E., Eds., 1998) Plenum Press, New York, NY; Cell and Tissue Culture: Laboratory Procedures (Doyle, A. et al., Eds., 1993-8) John Wiley and Sons, Hoboken, NJ; Methods in Enzymology (Academic Press, Inc.) New York, NY; Weir’s Handbook of Experimental Immunology (Herzenberg, L.A. et al. Eds. 1997) Wiley-Blackwell Publishers, New York, NY; Gene Transfer Vectors for Mammalian Cells (Miller, J.M. et al. Eds., 1987) Cold Spring Harbor Press, Cold Spring Harbor, NY; Current Protocols in Molecular Biology (Ausubel, F.M. et al., Eds., 1987) Greene Pub. Associates, New York, NY; PCR: The Polymerase Chain Reaction, (Mullis, K. et al., Eds., 1994) Birkhäuser, Boston MA; Current Protocols in Immunology (Coligan, J.E. et al., eds., 1991) John Wiley and Sons, Hoboken, NJ; Short Protocols in Molecular Biology (John Wiley and Sons, 1999) Hoboken, NJ; Immunobiology 7 (Janeway, C.A. et al. 2007) Garland Science, London, UK; Antibodies (P. Finch, 1997) Stride Publications, Devoran, UK; Antibodies: A Practical Approach (D. Catty., ed., 1989) Oxford University Press, USA, New York NY); Monoclonal Antibodies: A Practical Approach (Shepherd, P. et al. Eds., 2000) Oxford University Press, USA, New York NY; Using Antibodies: A Laboratory Manual (Harlow, E. et al. Eds., 1998) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY; The Antibodies (Zanetti, M. et al. Eds. 1995) Harwood Academic Publishers, London, UK); и DeVita, Hellman, and Rosenberg's Cancer: Principles & Practice of Oncology, Eighth Edition, DeVita, V. et al. Eds. 2008, Lippincott Williams & Wilkins, Philadelphia, PA.
II. Предпочтительные триспецифические связывающие молекулы по настоящему изобретению
A. Способности к связыванию
Предпочтительные триспецифичные связывающие молекулы по настоящему изобретению, способны координационно и одновременно связываться с тремя различными эпитопами. Предпочтительные триспецифичные связывающие молекулы по настоящему изобретению включают:
(i), "связывающий домен I", который способен иммуноспецифично связываться с "эпитопом I", присутствующем на первом антигене, и "связывающий домен II", который способен иммуноспецифично связываться с "эпитопом II", присутствующем на втором антигене, где указанный связывающий домен I и указанный связывающий домен II оба являются "связывающими доменами по типу диатела";
(ii), A "связывающий домен III", который способен иммуноспецифично связываться с "эпитопом III" присутствующим на третьем антигене; и
(iii) Fc-домен, который формируется за счет объединения в комплекс двух CH2-CH3 доменов друг с другом;
где:
(А) один из числа эпитопа I, эпитопа II или эпитопа III представляет собой эпитоп первого" антигена злокачественной опухоли" антигена злокачественной опухоли;
(B) второй из числа эпитопа I, эпитопа II или эпитопа III представляет собой эпитоп второго антигена злокачественной опухоли; и
(С) третий из числа эпитопа I, эпитопа II или эпитопа III представляет собой эпитоп молекулы экспрессируются на поверхности эффекторных клеток иммунной системы ("антиген эффекторных клеток");
и где связывающие домены I, II и III связывающих молекул опосредуют скоординированное связывание эффекторных клеток иммунной системы и клеток, экспрессирующих и первый и второй антигены злокачественной опухоли, таким образом, совместно локализуя такие клетки.
Эпитопсвязывающие домены диатела могут быть также направлены на поверхностную детерминанту В-клетки, такую как CD19, CD20, CD22, CD30, CD37, CD40 и CD74 (Moore, PA et al. (2011) “Application Of Dual Affinity Retargeting Molecules To Achieve Optimal Redirected T-Cell Killing Of B-Cell Lymphoma,” Blood 117(17):4542-4551; Cheson, B.D. et al. (2008) “Monoclonal Antibody Therapy For B-Cell Non-Hodgkin’s Lymphoma,” N. Engl. J. Med. 359(6):613-626; Castillo, J. et al. (2008) “Newer monoclonal antibodies for hematological malignancies,” Exp. Hematol. 36(7):755-768. Во многих исследованиях, также было обнаружено, что связывание диатела с детерминантами эффекторных клеток, например, Fc-рецепторами γ (FcγR), активирует эффекторную клетку (Holliger et al. (1996) “Specific Killing Of Lymphoma Cells By Cytotoxic T-Cells Mediated By A Bi-specific Diabody,” Protein Eng. 9:299-305; Holliger et al. (1999) “Carcinoembryonic Antigen (CEA)-Specific T-Cell Activation In Colon Carcinoma Induced By Anti-CD3 x Anti-CEA Bi-specific Diabodies And B7 x Anti-CEA Bi-specific Fusion Proteins,” Cancer Res. 59:2909-2916; WO 2006/113665; WO 2008/157379; WO 2010/080538; WO 2012/018687; WO 2012/162068). Как правило, активация эффекторных клеток инициируется антителом, связанным с антигеном с эффекторной клеток через взаимодействие Fc-FcγR; таким образом, в связи с этим, молекулы диател могут проявлять lg-подобную функциональность независимо от того, содержат ли они Fc-домен (например, анализируемый в любом анализе эффекторной функции, известном в данной области, или приведенном для примера в данном документе (например, ADCC-анализ)). Перекрестной сшивкой опухолевых и эффекторных клеток, диатело не только доставляет эффекторные клетки в непосредственную близость к опухолевым клеткам, но и приводит к эффективному уничтожению опухоли (см., например, Као et al. (2003) “Bi-specific Antibody Conjugates In Therapeutics,” Adv. Drug. Deliv. Rev. 55:171-197).
Хотя такие триспецифичные связывающие молекулы являются особенно предпочтительными, изобретение, кроме того в частности, предусматривает триспецифичные связывающие молекулы, которые содержат любую комбинацию связывающих доменов, достаточную для получения молекулы, имеющей три специфичности связывания, две из которых являются обязательными для специфичности, направленные против антигенов злокачественной опухоли, и одна из них демонстрирует специфичность связывания направленную против антигена эффекторной клетки. Так, например, изобретение предусматривает: триспецифичную связывающую молекулу, которая включает три связывающих домена Fab-типа, триспецифичную связывающую молекулу, которая включает один двухвалентный, биспецифичный домен антитела (образованный, например, путем объединения в комплекс двух различных легких цепей и двух различных тяжелых цепей) и один связывающий домен Fab-типа, триспецифичную связывающую молекулу, которая включает два двухвалентных, биспецифичных домена антител (образованных, например, путем объединения в комплекс четырех различных легких цепей и двух различных тяжелых цепей), но в котором один из доменов антитела оказался неактивным, и т.д.
Термины "полипептид", "полипептидная цепь" и "пептид" используются в данном документе взаимозаменяемо для обозначения полимеров аминокислот любой длины, но в особенности, длина которых превышает 3, 5, 10, 15, 20 или 25 аминокислотных остатков, в которых два, а более предпочтительно все, аминокислотные остатки соединены посредством амидной (пептидной) связи (-NH-C(O)-). Однако полимер может быть линейным или разветвленным, он может содержать модифицированные аминокислоты, и он может быть прерван неаминокислотами. Термины также включают аминокислотный полимер, который был изменен естественным образом или путем вмешательства; например, образованием дисульфидной связи, гликозилированием, липидированием, ацетилированием, фосфорилированием, или любыми другими манипуляциями или модификациями, такими как конъюгация с метящим компонентом. Также включены в определение, например, полипептиды, содержащие один или несколько аналогов аминокислот (включающие, например, неприродные аминокислоты и т.п.), и другие модификации, известные в данной области техники. Полипептиды по настоящему изобретению могут находиться в виде отдельных цепей или в виде собранных в комплекс цепей.
"Связывающий домен не относящийся к типу диатела" является эпитопсвязывающим доменом диатела, и особенно, диатела DART®. Термины "диатело" и "диатело DART®" обсуждались выше, и относятся к молекуле, которая содержит, по меньшей мере, две полипептидные цепи, которые предпочтительно собираются в комплекс друг с другом через ковалентное взаимодействие с образованием по меньшей мере, двух эпитопсвязывающих участков, которые могут распознать одинаковые или различные эпитопы. Каждая из двух полипептидных цепей диатела или диатела DART® включает вариабельный участок легкой цепи иммуноглобулина и вариабельный участок тяжелой цепи иммуноглобулина, но эти участки не взаимодействуют с образованием эпитопсвязывающего участка (т.е. они не являются взаимно "комплементарными"), Скорее, вариабельный участок тяжелой цепи иммуноглобулина цепи одного (например, первого) диатела, или диатела DART®, взаимодействует с вариабельным участком легкой цепи иммуноглобулином, выделенным из другого (например, второго) диатела, или диатела DART®, полипептидной цепи с образованием эпитопсвязывающего участка. Точно так же, полипептидные цепи вариабельного участка легкой цепи иммуноглобулина одного (например, первого) диатела, или диатела DART®, взаимодействуют с вариабельным участком тяжелой цепи диатела, выделенным из другого (например, второго) диатела, или диатела DART®, полипептидной цепи с образованием эпитопсвязывающего участка. Молекулы диатела DART® раскрыты в публ. пат. США № 2013-0295121; 2010-0174053 и 2009-0060910; В публ. Европейского патента № ЕР 2714079; EP 2601216; EP 2376109; ЕР 2158221 и публ. публикациях РСТ № WO 2012/162068; WO 2012/018687; WO 2010/080538; WO 2006/113665, WO 2008/157379 и Moore, P.A. et al. (2011) “Application Of Dual Affinity Retargeting Molecules To Achieve Optimal Redirected T-Cell Killing Of B-Cell Lymphoma,” Blood 117(17):4542-4551; Veri, M.C. et al. (2010) “Therapeutic Control Of B Cell Activation Via Recruitment Of Fcgamma Receptor IIb (CD32B) Inhibitory Function With A Novel Bi-specific Antibody Scaffold,” Arthritis Rheum. 62(7):1933-1943; и Johnson, S. et al. (2010) “Effector Cell Recruitment With Novel Fv-Based Dual-Affinity Re-Targeting Protein Leads To Potent Tumor Cytolysis And in vivo B-Cell Depletion,” J. Mol. Biol. 399(3):436-449.
Связывающий домен III предпочтительно представляет собой связывающий домен не относящийся к типу диатела, что означает, что связывающий домен III не имеет структуру-связывающего домена по типу диатела. Предпочтительно, связывающий домен III представляет собой связывающий домен не относящийся к типу диатела, который представляет собой связывающий домен Fab-типа или связывающий домен по типу рецептора эффекторных клеток. Таким образом, в одном из воплощений приведенных в качестве примера на фигурах 4А-4G, связывающий домен III представляет собой связывающий домен Fab-типа. Фигуры 5A-5E иллюстрируют воплощение, в котором связывающий домен III является связывающим доменом по типу рецептора эффекторной клетки. При использовании в данном описании термин "связывающий домен Fab-типа" относится к эпитопсвязывающему домену, который образован взаимодействием VL-домена легкой цепи иммуноглобулина и дополняющим VH-доменом тяжелой цепи иммуноглобулина. Связывающие домены Fab-типа отличаются от связывающего домена по типу диатела в том, что две полипептидные цепи, которые образуют связывающий домен Fab-типа, включают только один эпитопсвязывающий домен, в то время как две полипептидные цепи, которые образуют связывающий домен не относящийся к типу диатела, содержат, по меньшей мере, два эпитопсвязывающих домена. Таким образом, при использовании в данном документе связывающие домены FAB-типа отличаются от связывающего домена по типу диатела. Когда связывающий домен представляет собой связывающий домен Fab-типа или связывающий домен не относящийся к типу диатела, то он будет включать VL-домен и VH-домен, которые могут быть расположены на одной или на разных полипептидных цепях. Отбор таких VL- и VH-доменов координируется таким образом, что домены образовали эпитопсвязывающий домен. При использовании в данном описании термин "связывающий домен по типу рецептора эффекторной клетки" относится к эпитопсвязывающему домену, который образуется в результате взаимодействия вариабельного домена альфа-цепи Т-клеточного рецептора, и вариабельного домена бета-цепи Т-клеточного рецептора. Такие рецепторы распознают пептиды, отображаемые в контексте MHC и, таким образом, способные распознавать внутриклеточные эпитопы.
Триспецифичные связывающие молекулы по настоящему изобретению, таким образом, отличаются от четырехвалентных связывающих молекул, таких, как те, что получаются димеризацией бивалентного антитела, и предпочтительно обладающего тремя, но не четырьмя связывающими доменами. Как будет описано ниже, триспецифичные молекулы по настоящему изобретению, могут обладать дополнительными связывающими доменами (например, альбумин-связывающим доменом, FcγR-связывающим доменом, и т.д.). Такие дополнительные связывающие домены не предназначены для того, чтобы считаться или подсчитываться как один из трех связывающих доменов из триспецифичной связывающей молекулы по настоящему изобретению.
При использовании в данном описании термины "объединение" или "ассоциирование" в отношении полипептидов (например, одного полипептида диатела с другим, легкая цепь иммуноглобулина к тяжелой цепи иммуноглобулина, одного СН2-СН3-домена с другим СН2-СН3-доменом, и т.д.) предназначены для обозначения нековалентного комбинирования полипептидов. Термины "комплексы" или "образование комплекса" предназначены для обозначения ковалентного комбинирования полипептидов.
При использовании в данном описании, связывающие домены триспецифической связывающей молекулы настоящего изобретения упоминаются как опосредующие "координированное связывание", если по меньшей мере, два из ее связывающих доменов и предпочтительно все его связывающие домены, способны одновременно быть связанными с их соответствующими распознаваемыми эпитопами или связываемым лигандом. Такое связывание может быть одновременным. Тем не менее, один аспект настоящего изобретения относится к модификации скорости "ассоциации" и/или "диссоциации", с которыми такие связывающие домены связываются с их распознаваемыми эпитопами. При использовании в данном описании "скорость ассоциации" связывания является мерой аффинности, с которой такие связывающие домены распознают и инициируют связывание с их распознаваемыми эпитопами. В противоположность этому "скорость диссоциации" связывания является мерой степени устойчивости комплекса связывающего домена: эпитопа. Скорости "ассоциации" и/или "диссоциации" связывания могут быть изменены путем изменения аминокислотной последовательности CDR из связывающего домена. Как будет описано ниже, независимо от каких-либо модификаций CDR, степень скоординированного связывания молекул по настоящему изобретению может быть модулирована путем изменения конфигурации из их связывающего домена, так чтобы конкретный связывающий домен (т.е. VLx/VHx домен) присутствовал как связывающий домен III или в качестве внутреннего или внешнего связывающего домена по типу диатела по отношению к связывающему домену III (более подробно описано ниже).
Эти скорости ассоциации и диссоциации связывающих доменов триспецифических связывающих молекул по настоящему изобретению можно легко измерить с помощью методов, хорошо известных в данной области, например, путем анализа Biacore® (Jason-Moller, L. et al.(2006) “Overview Of Biacore Systems And Their Applications,” Curr. Protoc. Protein Sci. Chapter 19:Unit 19.13; Swanson, S.J. (2005) “Characterization Of An Immune Response,” Dev. Biol. (Basel). 122:95-101; Buijs, J. et al. (2005) “SPR-MS In Functional Proteomics,” Brief Funct. Genomic Proteomic. 4(1):39-47; Karlsson, R. et al. (2004) “SPR For Molecular Interaction Analysis: A Review Of Emerging Application Areas,” J. Mol. Recognit. 17(3):151-161; Van Regenmortel, M.H. (2003) “Improving The Quality Of BIACORE-Based Affinity Measurements,” Dev. Biol. (Basel) 112:141-151; Malmqvist, M. (1999) “BIACORE: An Affinity Biosensor System For Characterization Of Biomolecular Interactions,” Biochem. Soc. Trans. 27(2):335-340; Malmqvist, M. et al. (1997) “Biomolecular Interaction Analysis: Affinity Biosensor Technologies For Functional Analysis Of Proteins,” Curr. Opin. Chem. Biol. 1(3):378-383; Fivash, M. et al. (1998) “Biacore For Macromolecular Interaction,” Curr. Opin. Biotechnol. 9(1):97-101; Malmborg, A.C. et al. (1995) “Biacore As A Tool In Antibody Engineering,” J. Immunol. Methods. 183(1):7-13). Эти скорости ассоциации и диссоциации связывающих доменов триспецифических связывающих молекул по настоящему изобретению могут быть легко изменены путем случайного или направленного мутагенеза молекул нуклеиновых кислот, которые кодируют такие связывающие домены, с последующим рутинным скринингом извлекаемых молекул нуклеиновых кислот по их способности кодировать мутантные белки, которые обладают такой измененной кинетикой связывания.
Связующие домены триспецифичных связывающих молекул по настоящему изобретению связываются с эпитопами "иммуноспецифическим" образом. При использовании в данном описании антитело, диатело или другая эпитопсвязывающая молекула упоминается как "иммуноспецифично" связывающая область другой молекулы (т.е. эпитоп), если она вступает в реакцию или ассоциируется чаще, быстрее, с большей продолжительностью и/или с большей аффинностью с этим эпитопом по сравнению с альтернативными эпитопами. Например, антитело, которое связывается иммуноспецифично с вирусным эпитопом, представляет собой антитело, которое связывает этот вирусный эпитоп с большей аффинностью, авидностью, более легко, и/или с большей длительностью, чем оно иммуноспецифично связывается с другими вирусными эпитопами или невирусными эпитопами. Также следует понимать, прочитав это определение, что, например, антитело (или фрагмент, или эпитоп), что иммуноспецифично связывается с первой мишенью, может или не может специфически или преимущественно связываются со вторым мишенью. Таким образом, "специфическое связывание" не обязательно требует (хотя она может включать) исключительное связывание. Как правило, но не обязательно, ссылка на связывание означает "специфическое" связывание. Две молекулы называются способными связываться друг с другом в "физиоспецифическим" образом, если такое связывание демонстрирует специфичность, с которой рецепторы связывают с их соответствующими лигандами.
Функциональность антител может быть повышена за счет генерации мультиспецифичных молекул на основе антител, которые могут одновременно связывать два отдельных и различных антигена (или различные эпитопы одного и того же антигена) и/или за счет генерации на основе антительной молекулы, обладающей более высокой валентностью (т.е. более двух участки связывания) к тому же эпитопу и/или антигену.
Таким образом, в наиболее простом воплощении, предпочтительные связывающие молекулы по настоящему изобретению, по меньшей мере, триспецифичны. Важно отметить, что такие молекулы имеют, по меньшей мере, три участка, которые способны связывать антиген: "внешний" связывающий домен не относящийся к типу диатела, который находится на противоположной стороне от связывающего домена III, "внутренний" связывающий домен не относящийся к типу диатела, который расположен ближе к связывающему домену III, и связывающий домен III сам по себе. Положения таких доменов соответственно обозначены как "участок A" участок B "и" C "участок С (фигуры 4A-4G; фигуры 5A-5E).
Триспецифичные связывающие молекулы по настоящему изобретению, способны координационно связываться с тремя различными эпитопами, потому что содержат три связывающих домена. Два из связывающих доменов таких молекул, способны связываться с эпитопами "антигенов злокачественных опухолей", таким образом, что молекула способна связываться с двумя различными антигенами злокачественной опухоли. Третий связывающий домен таких молекул способен связываться с эпитопом молекулы, экспрессируемой на поверхности эффекторной клетки иммунной системы (т.е. "антиген эффекторных клеток"). Таким образом, триспецифичные связывающие молекулы по настоящему изобретению способны опосредовать одновременного скоординированное связывание со злокачественной опухолевой клеткой, экспрессирующей два антигена злокачественной опухолевой клетки, и эффекторной клетки иммунной системы, экспрессирующей антиген эффекторных клеток. Эпитопы, распознаваемые триспецифичными связывающими молекулами по настоящему изобретению, могут быть непрерывными или прерывистыми (например, конформационными).
Первый и второй антигены злокачественной опухоли, которые связаны с доменами, связывающими антигены злокачественной опухоли, триспецифичных связывающих молекул по настоящему изобретению, могут быть выбраны из любой молекулы, которая характерно присутствует на поверхности злокачественной опухолевой клетки. Один из аспектов настоящего изобретения относится к способности нацеливаться на "антиген злокачественной опухоли с низкой экспрессией" (т.е. антиген злокачественной опухоли, который может быть экспрессирован на злокачественной опухолевой клетке на слишком низком уровне для того, чтобы получить моноспецифичную связывающую молекулу, которая обеспечит эффективную терапию рака). В отличие от таких моноспецифичных связывающих молекул, триспецифичные связывающие молекулы по настоящему изобретению путем ориентации двух антигенов злокачественной опухоли вместо одного, демонстрируют синергическую и кооперативную повышенную связывающую авидность, которая может компенсировать низкую аффинность связывания и, таким образом, могут быть с успехом использованы для нацеливания на виды злокачественных опухолей, которые характеризуются даже антигенами злокачественной опухоли с низкой экспрессией. Второй аспект настоящего изобретения относится к способности нацеливать на "антиген злокачественной опухоли с низкой специфичностью" (т.е. антиген злокачественной опухоли, который может быть экспрессирован в нормальной клетке в дополнение к тому, что он экспрессируется на злокачественной опухолевой клетке). Триспецифичные связывающие молекулы по настоящему изобретению, обеспечивая синергетическую и кооперативную повышенную авидность связывания к двум антигенам злокачественной опухоли, демонстрирует более высокую авидность связывания даже для антигена злокачественной опухоли с низкой специфичностью и, таким образом, обеспечивает средство для лечения онкологических заболеваний, которые характеризуются такими антигенами злокачественной опухоли. Таким образом, триспецифичные связывающие молекулы по настоящему изобретению могут быть использованы для обеспечения противоопухолевой терапии даже в тех случаях, когда один или оба целевых антигенов злокачественной опухоли неэффективны сама по себе для обеспечения такой терапии.
Например, CD32B (рецептор FcγRIIB) широко экспрессируется на гемопоэтических клетках, в том числе моноцитах, макрофагах, В-клетках, NK-клетках, нейтрофилах, тучных клетках и тромбоцитах. После связывания с Fc-доменом IgG, CD32B ингибирует иммунную систему хозяина, тем самым угнетая постоянный иммунный ответ. Хотя такое ингибирование является желательным, чтобы помочь хозяину оправиться от воспалительных реакций, он служит для обострения иммунодефицитов объектов, страдающих от злокачественной опухоли или инфекционного заболевания. Антитела, которые связываются с CD32B, с тем, чтобы блокировать связывание молекул Fc IgG, служат для предотвращения такого ингибирования и, таким образом, полезны в качестве адъюнкт-молекул при лечении злокачественных опухолей и инфекционных заболеваний (Veri, M.C. et al. (2007) “Monoclonal Antibodies Capable Of Discriminating The Human Inhibitory Fcgamma-Receptor IIB (CD32B) From The Activating Fcgamma-Receptor IIA (CD32A): Biochemical, Biological And Functional Characterization,” Immunology 121(3):392-404). К сожалению, CD32B также экспрессируется в синусоидальных эндотелиальных клетках печени ("клетки LSE") (Shahani, T. et al. (2014) “Human Liver Sinusoidal Endothelial Cells But Not Hepatocytes Contain Factor VIII,” J. Thromb. Haemost. 12(1):36-42; Géraud, C. et al. (2013) “Endothelial Transdifferentiation In Hepatocellular Carcinoma: Loss Of Stabilin-2 Expression In Peri-Tumourous Liver Correlates With Increased Survival,” Liver Int. 33(9):1428-1440; Takabe, Y. et al. (2012) “Immunomagnetic Exclusion Of E-Cadherin-Positive Hepatoblasts In Fetal Mouse Liver Cell Cultures Impairs Morphogenesis And Gene Expression Of Sinusoidal Endothelial Cells,” J. Anat. 221(3):229-239). Таким образом, антитела, которые связывают CD32B, атакуют клетки LSE. Тем не менее, за счет формирования триспецифичных связывающих молекул по настоящему изобретению, которые связываются с CD32B и с антигенами (т.е. первым и вторым антигенами злокачественной опухоли), которые не экспрессируются на клетках LSE, либо экспрессируются на низких уровнях такими клетками (т.е. сниженная экспрессия гена антигена(ов) злокачественной опухоли), или экспрессируется с низкой специфичностью на злокачественных опухолевых клетках и таких клетках LSE (т.е. антиген(ы) злокачественной опухоли с низкой специфичностью), настоящее изобретение относится к композициям и способам, которые будут использованы для угнетения CD32B-опосредованной иммунной системы торможения.
B. Примеры доменов, связывающих антигены злокачественной опухоли
Примеры подходящих антигенов злокачественных опухолей включают: 19,9, обнаруженный при раке толстой кишки, муцины рака желудка; 4.2; A33 (антиген колоректального рака; Almqvist, Y. 2006, Nucl Med Biol. Nov;33(8):991-998); ADAM-9 (публ. пат. США № 2006/0172350; публикация РСТ WO 06/084075); AH6, обнаруженный при раке желудка; ALCAM (публ. РСТ WO 03/093443); APO-1 (злокачественный антиген лимфоцитов человека) (Trauth et al. (1989) “Monoclonal Antibody-Mediated Tumor Regression By Induction Of Apoptosis,” Science 245:301-304); B1 (Egloff, A.M. et al. 2006, Cancer Res. 66(1):6-9); BAGE (Bodey, B. 2002 Expert Opin Biol Ther. 2(6):577-84); B7-H3; бета-катенин (Prange W. et al. 2003 J Pathol. 201 (2): 250-9); группа крови ALeb/Ley обнаруженный при ободочной аденокарциноме; антиген лимфомы Беркитта -38,13, C14, обнаруженный при аденокарциноме толстой кишки; CA125 (антиген карциномы яичника) (Bast, R.C. Jr. et al. 2005 Int J Gynecol Cancer 15 Suppl 3:274-81 ; Yu et al. (1991) “Coexpression Of Different Antigenic Markers On Moieties That Bear CA 125 Determinants,” Cancer Res. 51(2):468-475); Карбоксипептидаза M (United States Patent Publication No. 2006/0166291); CD5 (Calin, G.A. et al. 2006 Semin Oncol. 33(2):167-73; CD19 (Ghetie et al. (1994) “Anti-CD19 Inhibits The Growth Of Human B-Cell Tumor Lines In Vitro And Of Daudi Cells In SCID Mice By Inducing Cell Cycle Arrest,” Blood 83:1329-1336; Troussard, X. et al. 1998 Hematol Cell Ther. 40(4):139-48); CD20 (Thomas, D.A. et al. 2006 Hematol Oncol Clin North Am. 20(5):1125-36); CD22 (Kreitman, R.J. 2006 AAPS J. 18;8(3):E532-51); CD23 (Rosati, S. et al. 2005 Curr Top Microbiol Immunol. 5;294:91-107); CD25 (Troussard, X. et al. 1998 Hematol Cell Ther. 40(4):139-48); CD27 (Bataille, R. 2006 Haematologica 91(9):1234-40); CD28 (Bataille, R. 2006 Haematologica 91(9):1234-40); CD33 (Sgouros et al. (1993) “Modeling And Dosimetry Of Monoclonal Antibody M195 (Anti-CD33) In Acute Myelogenous Leukemia,” J. Nucl. Med. 34:422-430); CD36 (Ge, Y. 2005 Lab Hematol. 11(1):31-7); CD40/CD154 (Messmer, D. et al. 2005 Ann N Y Acad Sci. 1062:51-60); CD45 (Jurcic, J.G. 2005 Curr Oncol Rep. 7(5):339-46); CD56 (Bataille, R. 2006 Haematologica 91(9):1234-40); CD46 (пат. США № 7148038; публ. PCT№ WO 03/032814); CD79a/CD79b (Troussard, X. et al. 1998 Hematol Cell Ther. 40(4):139-48; Chu, P.G. et al. 2001 Appl Immunohistochem Mol Morphol. 9(2):97-106); CD103 (Troussard, X. et al. 1998 Hematol Cell Ther. 40(4):139-48); CDK4 (Lee, Y.M. et al. 2006 Cell Cycle 5(18):2110-4); CEA (карциноэмбриональный антиген) (Foon et al. (1995) “Immune Response To The Carcinoembryonic Antigen In Patients Treated With An Anti-Idiotype Antibody Vaccine,” J. Clin. Invest. 96(1):334-42); CEA (карциноэмбриональный антиген; Mathelin, C. 2006 Gynecol Obstet Fertil. 34(7-8):638-46; Tellez-Avila, F.I. et al. 2005 Rev Invest Clin. 57(6):814-9); CO17-1A (Ragnhammar et al. (1993) “Effect Of Monoclonal Antibody 17-1A And GM-CSF In Patients With Advanced Colorectal Carcinoma-Long-Lasting, Complete Remissions Can Be Induced,” Int. J. Cancer 53:751-758); CO-43 (группа крови Leb); CO-514 (группа крови Lea) обнаруженный при аденокарциноме; CTA-1; CTLA4 (Peggs, K.S. et al. 2006 Curr Opin Immunol. 18 (2): 206-13); Цитокератин 8 (публ. РСТ WO 03/024191); D1.1; D1 56-22; DR5 (Abdulghani, J. et al.(2010) “TRAIL Receptor Signaling And Therapeutics,” Expert Opin. Ther. Targets 14(10):1091-1108; Andera, L. (2009) “Signaling Activated By The Death Receptors Of The TNFR Family,” Biomed. Pap. Med. Fac. Univ. Palacky Olomouc Czech. Repub. 153(3):173-180; Carlo-Stella, C. et al. (2007) “Targeting TRAIL Agonistic Receptors for Cancer Therapy,” Clin, Cancer 13(8):2313-2317; Chaudhari, B.R. et al. (2006) “Following the TRAIL to Apoptosis,” Immunologic Res. 35(3):249-262); E1 серия (группа крови B), обнаруженный при раке поджелудочной железы; EGFR (рецептор эпидермального фактора роста) (Adenis, A. et al. 2003 Bull Cancer. 90 Spec No: S228-32); рецепторы эфрина (и, в частности, EphA2 (Пат. США № 7569672; публ. РСТ № WO 06/084226); Erb (ErbB1; ErbB3, ErbB4; Zhou, H. et al.2002 Oncogene 21(57):8732-8740; Rimon, E. et al. 2004 Int J Oncol. 24(5):1325-1338); GAGE (GAGE-1; GAGE-2; Akcakanat, A. et al. 2006 Int J Cancer. 118(1):123-128); GD2/GD3/GM2 (Livingston, P.O. et al. 2005 Cancer Immunol Immunother. 54 (10): 1018-1025); F3, обнаруженный при аденокарциноме легкого; FC10.2 обнаруженный в клетках эмбриональной карциномы и аденокарциноме желудка; G49, ганглиозиды GD2 (Saleh et al. (1993) “Generation Of A Human Anti-Idiotypic Antibody That Mimics The GD2 Antigen,” J.Immunol., 151, 3390-3398); ганглиозид GD3 (Shitara et al. (1993) “A Mouse/Human Chimeric Anti-(Ganglioside GD3) Antibody With Enhanced Antitumor Activities,” Cancer Immunol. Immunother. 36:373-380); ганглиозид GM2 (Livingston et al. (1994) “Improved Survival In Stage III Melanoma Patients With GM2 Antibodies: A Randomized Trial Of Adjuvant Vaccination With GM2 Ganglioside,” J. Clin. Oncol. 12:1036-1044); ганглиозид GM3 (Hoon et al. (1993) “Molecular Cloning Of A Human Monoclonal Antibody Reactive To Ganglioside GM3 Antigen On Human Cancers,” Cancer Res. 53:5244-5250); GD2; GD3; GICA 19-9 (Herlyn et al. (1982) “Monoclonal Antibody Detection Of A Circulating Tumor-Associated Antigen. I. Presence Of Antigen In Sera Of Patients With Colorectal, Gastric, And Pancreatic Carcinoma,” J. Clin. Immunol. 2:135-140); GM2; gp100 (Lotem, M. et al. 2006 J Immunother. 29(6):616-27); Gp37 (антиген Т-клеточного лейкоза человека) (Bhattacharya-Chatterjee et al. (1988) “Idiotype Vaccines Against Human T Cell Leukemia. II. Generation And Characterization Of A Monoclonal Idiotype Cascade (Ab1, Ab2, and Ab3),” J. Immunol. 141:1398-1403); gp75 (антиген меланомы) (Vijayasardahl et al. (1990) “The Melanoma Antigen Gp75 Is The Human Homologue Of The Mouse B (Brown) Locus Gene Product,” J. Exp. Med. 171(4):1375-1380); gpA33; антиген HER2 (p185HER2) (Kumar, Pal S et al. 2006 Semin Oncol. 33(4):386-91); антиген В-лимфомы человека CD20 (Reff et al. (1994) “Depletion Of B Cells In Vivo By A Chimeric Mouse Human Monoclonal Antibody To CD20,” Blood 83:435-445); антиген жировых шариков молока человека; папиломавирус человека-E6/папиломавирус человека-E7 (DiMaio, D. et al. 2006 Adv Virus Res. 66:125-59; HMW-MAA (high высокомолекулярный антиген меланомы) (Natali et al. (1987) “Immunohistochemical Detection Of Antigen In Human Primary And Metastatic Melanomas By The Monoclonal Antibody 140.240 And Its Possible Prognostic Significance,” Cancer 59:55-63; Mittelman et al. (1990) “Active Specific Immunotherapy In Patients With Melanoma. A Clinical Trial With Mouse Antiidiotypic Monoclonal Antibodies Elicited With Syngeneic Anti-High-Molecular-Weight-Melanoma-Associated Antigen Monoclonal Antibodies,” J. Clin. Invest. 86:2136-2144); I антиген (антиген дифференцировки) (Feizi (1985) “Demonstration By Monoclonal Antibodies That Carbohydrate Structures Of Glycoproteins And Glycolipids Are Onco-Developmental Antigens,” Nature 314:53-57) такой, как I(Ma) обнаруженный при аденокарциноме желудка; интегрин альфа-V-бета-6 (публ. PCT № WO 03/087340); JAM-3 (публ. PCT № WO 06/084078); KID3 (публ. PCT № WO 05/028498); KID31 (публ. PCT № WO 06/076584); KS 1/4 антиген пан-карциномы (Perez et al. (1989) “Isolation And Characterization Of A cDNA Encoding The Ks1/4 Epithelial Carcinoma Marker,” J. Immunol. 142:3662-3667; Möller et al. (1991) “Bi-specific-Monoclonal-Antibody-Directed Lysis Of Ovarian Carcinoma Cells By Activated Human T Lymphocytes,” Cancer Immunol. Immunother. 33(4):210-216; Ragupathi, G. 2005 Cancer Treat Res. 123:157-80); L6 and L20 (антигены карциномы легкого человека) (Hellström et al. (1986) “Monoclonal Mouse Antibodies Raised Against Human Lung Carcinoma,” Cancer Res. 46:3917-3923); LEA; LUCA-2 (United States Patent Publication No. 2006/0172349; PCT Publication No. WO 06/083852); M1:22:25:8; M18; M39; MAGE (MAGE-1; MAGE-3; (Bodey, B. 2002 Expert Opin Biol Ther. 2(6):577-84); MART (Kounalakis, N. et al. 2005 Curr Oncol Rep. 7(5):377-82; MUC-1 (Mathelin, C. 2006 Gynecol Obstet Fertil. 34(7-8):638-46); MUM-1 (Castelli, C. et al. 2000 J Cell Physiol. 182(3):323-31); Myl; N-ацетиглюкозаминилтрансфераза (Dennis, J.W. 1999 Biochim Biophys Acta. 6;1473(1):21-34); неогликопротеин; NS-10 обнаруженный в аденокарциномах; OFA-1; OFA-2; онкостатин M (рецептор онкостатина бета) (United States Patent No. 7,572,896; PCT Publication No. WO 06/084092); p15 (Gil, J. et al. 2006 Nat Rev Mol Cell Biol. 7 (9): 667-77); p97 (меланома-ассоциированный антиген) (Estin et al. (1989) “Transfected Mouse Melanoma Lines That Express Various Levels Of Human Melanoma-Associated Antigen p97,” J. Natl. Cancer Instit. 81 (6): 445-454); PEM (полиморфный эпителиальный муцин) (Hilkens et al. (1992) “Cell Membrane-Associated Mucins And Their Adhesion-Modulating Property,” Trends in Biochem. Sci. 17: 359-363); РЕМА (антиген полиморфного эпителиального муцина); PIPA (пат. США № 7405061; публ. РСТ WO 04/043239); ПСА (простат-специфический антиген) (Henttu et al. (1989) “cDNA Coding For The Entire Human Prostate Specific Antigen Shows High Homologies To The Human Tissue Kallikrein Genes,” Biochem. Biophys. Res. Comm. 10(2):903-910; Israeli et al. (1993) “Molecular Cloning Of A Complementary DNA Encoding A Prostate-Specific Membrane Antigen,” Cancer Res. 53:227-230; Cracco, C.M. et al. 2005 Minerva Urol Nefrol. 57 (4): 301-11); PSMA (простата-специфический мембранный антиген) (Ragupathi, G. 2005 Cancer Treat Res. 123:157-180); фосфат простатической кислоты (Tailor et al. (1990) “Nucleotide Sequence Of Human Prostatic Acid Phosphatase Determined From A Full-Length cDNA Clone,” Nucl. Acids Res. 18 (16): 4928); R 24, обнаруженный при меланоме; ROR1 (Пат. США № 5843749); сфинголипиды; SSEA-1; SSEA-3; SSEA-4; STN (Holmberg, LA 2001 Expert Opin Biol Ther.1(5):881-91); пептид, полученный из Т-клеточного рецептора, из Т-клеточной лимфомы кожи (see Edelson (1998) “Cutaneous T-Cell Lymphoma: A Model For Selective Immunotherapy,” Cancer J Sci Am. 4:62-71); T5A7 обнаруженный в миелоидных клетках; TAG-72 (Yokota et al. (1992) “Rapid Tumor Penetration Of A Single-Chain Fv And Comparison With Other Immunoglobulin Forms,” Cancer Res. 52:3402-3408); TL5 (blood group A); TNF-рецептор (TNF-α рецептор, TNF-β рецептор; TNF-γ рецептор (van Horssen, R. et al. 2006 Oncologist. 11(4):397-408; Gardnerova, M. et al. 2000 Curr Drug Targets. 1 (4): 327-64); TRA-1-85 (группа крови Н); рецептор трансферрина (Пат. США № 7572895; публ. РСТ WO 05/121179); TSTA (опухольспецифический трансплантационный антиген), такие как вирусно-индуцированные опухолевые антигены, включая Т-антиген опухолевых ДНК-вирусов и антигены оболочки опухолевых РНК-вирусов, онкофетальный антиген альфа-фетопротеина, такой как СЕА толстой кишки, онкофетальный антиген опухоли мочевого пузыря (Hellström et al. (1985) “Monoclonal Antibodies To Cell Surface Antigens Shared By Chemically Induced Mouse Bladder Carcinomas,” Cancer. Res. 45:2210-2188); VEGF-рецептор (O’Dwyer. PJ 2006 Oncologist.11 (9): 992-998); VEP8; VEP9; VIM-D5; и Y гаптен, Leу, обнаруженные в клетках эмбриональной карциномы.
1. Campath-1 (CD52)-связывающий домен (алемтузумаб)
Аминокислотная последовательность VL-домена гуманизированного анти-CD52 антитела "Алемтузумаб" (SEQ ID NO: 205), показана ниже (остатки CDR выделены подчеркиванием):
DIQMTQSPSS LSASVGDRVT ITCKASQNID KYLNWYQQKP GKAPKLLIYN TNNLQTGVPS RFSGSGSGTD FTFTISSLQP EDIATYYCLQ HISRPRTFGQ GTKVEIKR
Аминокислотная последовательность VH-домена гуманизированного анти-CD52 антитела "Алемтузумаб" (SEQ ID NO: 206), показана ниже (остатки CDR выделены подчеркиванием):
QVQLQESGPG LVRPSQTLSL TCTVSGFTFT DFYMNWVRQP PGRGLEWIGF IRDKAKGYTT EYNPSVKGRV TMLVDTSKNQ FSLRLSSVTA ADTAVYYCAR EGHTAAPFDY WGQGSLVTVS S
2. CD317 (BMST2)-связывающие домены
CD317 (также известный как Антиген 2 стромальных клеток костного мозга; BMST) избыточно экспрессируется на различных злокачественных опухолевых клетках, выделенных из молочной железы, легких, почек, матки и кожи (Kawai S. et al.(2008) “Interferon-α enhances CD317 expression and the antitumor activity of anti-CD317 monoclonal antibody in renal cell carcinoma xenograft models,” Cancer Science 99(12):2461-2466; Cai, D. et al. (2009) “Up-Regulation Of Bone Marrow Stromal Protein 2 (BST2) In Breast Cancer With Bone Metastasis,” BMC Cancer 9:102, pp. 1-10; Wang, W. et al. (2009) HM1.24 (CD317) Is A Novel Target Against Lung Cancer For Immunotherapy Using Anti-HM1.24 Antibody,” Cancer Immunology, Immunotherapy 58(6):967-976; Wang, W. et al. (2009) “Chimeric And Humanized Anti-HM1.24 Antibodies Mediate Antibody-Dependent Cellular Cytotoxicity Against Lung Cancer Cells. Lung Cancer,” 63(1):23-31; Sayeed, A. et al. (2013) “Aberrant Regulation Of The BST2 (Tetherin) Promoter Enhances Cell Proliferation And Apoptosis Evasion In High Grade Breast Cancer Cells,” PLoS ONE 8(6)e67191, pp. 1-10; Yi, E.H. et al. (2013) “BST-2 Is A Potential Activator Of Invasion And Migration In Tamoxifen-Resistant Breast Cancer Cells,” Biochem. Biophys. Res. Commun. 435(4):685-690; Staudinger, M. (2014) “The Novel Immunotoxin HM1.24-ETA' Induces Apoptosis In Multiple Myeloma Cells,” Blood Cancer J. 13;4:e219, pp. 1-11). Антитела, которые иммуноспецифично связываются с CD317, являются коммерчески доступными (Novus Biologicals LLC; BioLegend, Inc.; см. также пат. США № 8,834,876, который ссылается на депозит тяжелых и легких цепей НМ1.24 как FERM BP-5644 и FERM BP-5646; см. также пат. США № 8394374). Аминокислотная последовательность VL-домена анти-CD317 антитела "НМ1.24" (SEQ ID NO: 302), показана ниже (остатки CDR выделены подчеркиванием):
DIVMTQSHKF MSTSVGDRVS ITCKKASQDV NTAVAWYQQK PGQSPKLLIY SASNRYTGVP DRITGSGSGT DFTFTISSVQ AEDLALTTCQ QHYSTPFTFG SGTKLEIK
Аминокислотная последовательность VH-домена антитела "НМ1.24" анти-CD317 (SEQ ID NO: 303), показана ниже (остатки CDR выделены подчеркиванием):
QVQLQQSGAE LARPGASVKL SCKASGYTFT PYWMQWVKQR PGQGLEWIGS IFPGDGDTRY SQKFKGKATL TADKSSSTAY MQLSILAFED SAVYYCARGL RRGGYYFDYW GQGTTLTVSS
3. CEACAM5- и CEACAM6-связывающие домены
Было установлено, что молекулы клеточной адгезии, связанные с карциноэмбриональным антигеном 5 (CEACAM5) и 6 (CEACAM6), связаны с различными типами злокачественных опухолей, включая медуллярный рак щитовидной железы, колоректальный рак, рак поджелудочной железы, гепатоцеллюлярную карциному, рак желудка, рак легкого, рак головы и шеи, рак мочевого пузыря, рак предстательной железы, рак матки, рак эндометрия, рак молочной железы, неоплазии кроветворной системы, лейкоз и рака яичников (публ. РСТ № WO 2011/034660), и, в частности колоректальный рак, рак желудочно-кишечного тракта, рак поджелудочной железы, немелкоклеточный рак легкого (NSCL), карциномы молочной железы, щитовидной железы, желудка, яичников и матки (Zheng, C. et al.(2011) “A Novel Anti-CEACAM5 Monoclonal Antibody, CC4, Suppresses Colorectal Tumor Growth and Enhances NK Cells-Mediated Tumor Immunity,” PLoS One 6(6):e21146, pp. 1-11).
Было установлено, что CEACAM5 избыточно экспрессируется в 90% случаев желудочно-кишечного рака, рака толстой и прямой кишки и рака поджелудочной железы, 70% клеток немелкоклеточного рака легкого и 50% случаев рака молочной железы (Thompson, JA et al.(1991) “Carcinoembryonic Antigen Gene Family: Molecular Biology And Clinical Perspectives,” J. Clin. Lab. Anal. 5:344-366).
Суперэкспрессированная молекула клеточной адгезии, связанная с карциноэмбриональным антигеном 6 (CEACAM6), играет важную роль в инвазии и метастазирования различных злокачественных опухолей человека, в том числе медуллярного рака щитовидной железы, колоректального рака, рака поджелудочной железы, гепатоцеллюлярной карциномы, рака желудка, рака легких, рака головы и шеи, рака мочевого пузыря, рака предстательной железы, рака матки, рака эндометрия, рака молочной железы, неоплазий кроветворной системы, лейкоза и рака яичников (Публ. РСТ № WO 2011/034660, Deng, X. et al.(2014) “Expression Profiling Of CEACAM6 Associated With The Tumorigenesis And Progression In Gastric Adenocarcinoma,” Genet. Mol. Res. 13(3):7686-7697; Cameron, S. et al. (2012) “Focal Overexpression Of CEACAM6 Contributes To Enhanced Tumourigenesis In Head And Neck Cancer Via Suppression Of Apoptosis,” Mol. Cancer 11:74, pp. 1-11; Chapin, C. et al. (2012) “Distribution And Surfactant Association Of Carcinoembryonic Cell Adhesion Molecule 6 In Human Lung,” Amer. J. Physiol. Lung Cell. Mol. Physiol. 302(2):L216-L25; Riley, C.J. et al. (2009) “Design And Activity Of A Murine And Humanized Anti-CEACAM6 Single-Chain Variable Fragment In The Treatment Of Pancreatic Cancer,” Cancer Res. 69(5):1933-1940; Lewis-Wambi, J.S. et al. (2008) “Overexpression Of CEACAM6 Promotes Migration And Invasion Of Oestrogen-Deprived Breast Cancer Cells,” Eur. J. Cancer 44(12):1770-1779; Blumenthal, R.D. et al. (2007) “Expression Patterns Of CEACAM5 And CEACAM6 In Primary And Metastatic Cancers,” BMC Cancer. 7:2, pp. 1-15). Антитела, которые иммуноспецифично связываются с CEACAM5 и CEACAM6 являются коммерчески доступными (Santa Cruz Biotechnology, Inc., Novus Biologicals LLC; Abnova Corporation). Аминокислотная последовательность VL-домена гуманизированного анти-CEACAM5/анти-CEACAM6 антитела 16C3 (ЕР 2585476) (SEQ ID NO: 304), показана ниже (остатки CDR выделены подчеркиванием):
DIQMTQSPSS LSASVGDRVT ITCGASENIY GALNWYQRKP GKSPKLLIWG ASNLADGMPS RFSGSGSGRQ YTLTISSLQP EDVATYYCQN VLSSPYTFGG GTKLEIK
Аминокислотная последовательность VH-домена гуманизированного анти-CEACAM5/анти-CEACAM6 антитела 16C3 (ЕР 2585476) (SEQ ID NO: 305), показана ниже (остатки CDR выделены подчеркиванием):
QVQLQQSGPE VVRPGVSVKI SCKGSGYTFT DYAMHWVKQS HAKSLEWIGL ISTYSGDTKY NQNFKGKATM TVDKSASTAY MELSSLRSED TAVYYCARGD YSGSRYWFAY WGQGTLVTVS S
Аминокислотная последовательность VL-домена гуманизированного анти-CEACAM5/CEACAM6 антитела hMN15 (WO 2011/034660) (SEQ ID NO: 306), показана ниже (остатки CDR выделены подчеркиванием):
DIQLTQSPSS LSASVGDRVT MTCSASSRVS YIHWYQQKPG KAPKRWIYGT STLASGVPAR FSGSGSGTDF TFTISSLQPE DIATYYCQQW SYNPPTFGQG TKVEIKR
Аминокислотная последовательность VH-домена гуманизированного анти-CEACAM5/CEACAM6 антитела hMN15 (WO 2011/034660) (SEQ ID NO: 307), показана ниже (остатки CDR выделены подчеркиванием):
QVQLVESGGG VVQPGRSLRL SCSSSGFALT DYYMSWVRQA PGKGLEWLGF IANKANGHTT DYSPSVKGRF TISRDNSKNT LFLQMDSLRP EDTGVYFCAR DMGIRWNFDV WGQGTPVTVS S
4. DR5-связывающие домены
DR5 является предпочтительным Антигеном злокачественной опухоли по настоящему изобретению. Предпочтительные связывающие молекулы против DR5 человека по настоящему изобретению обладают VL и/или VH-доменами мышиных анти-DR5 человека моноклональных антител "DR5 mAb 1" и/или "DR5 mAb 2", и более предпочтительно имеют 1, 2 или все 3 из CDR в VL-домене и/или 1, 2 или все 3 CDR, используемые в VH-домене таких моноклональных антител против DR5 человека. В ином случае, любое моноклональное антитело против DR5 человека может быть использовано, в частности: дрозитумаб (обозначенный в настоящем описании как "DR5 mAb 3"), конатимумаб (обозначенный в настоящем описании как "DR5 mAb 4"), тигатумумаб (обозначенный в настоящем описании как "DR5 mAb 5"), LBY135-1 (обозначенный в настоящем описании как "DR5 mAb 6"), LBY135-2 (обозначенный в настоящем описании как "DR5 mAb 7") и KMTR2 (обозначенный в настоящем описании как "DR5 mAb 8").
a. антитело DR5 mAb 1 против DR5 человека
DR5 потенциально полезен при лечении широкого спектра онкологических заболеваний (примером являются колоректальный рак, гепатоцеллюлярный рак, глиома, рак почки, рак молочной железы, множественная миелома, рак мочевого пузыря, нейробластома; саркома, неходжкинская лимфома, немелкоклеточный рак легкого, рак яичников, рак поджелудочной железы и рака прямой кишки. Аминокислотная последовательность предшественника DR5 человека (NCBI Последовательность NP_003833.4) (SEQ ID NO: 2):
MEQRGQNAPA ASGARKRHGP GPREARGARP GLRVPKTLVL VVAAVLLLVS AESALITQQD LAPQQRVAPQ QKRSSPSEGL CPPGHHISED GRDCISCKYG QDYSTHWNDL LFCLRCTRCD SGEVELSPCT TTRNTVCQCE EGTFREEDSP EMCRKCRTGC PRGMVKVGDC TPWSDIECVH KESGTKHSGE APAVEETVTS SPGTPASPCS LSGIIIGVTV AAVVLIVAVF VCKSLLWKKV LPYLKGICSG GGGDPERVDR SSQRPGAEDN VLNEIVSILQ PTQVPEQEME VQEPAEPTGV NMLSPGESEH LLEPAEAERS QRRRLLVPAN EGDPTETLRQ CFDDFADLVP FDSWEPLMRK LGLMDNEIKV AKAEAAGHRD TLYTMLIKWV NKTGRDASVH TLLDALETLG ERLAKQKIED HLLSSGKFMY LEGNADSAMS
Аминокислотная последовательность VL-домена DR5 mAb 1 (SEQ ID NO: 3), показана ниже (остатки CDR выделены подчеркиванием):
DIVLTQSPAS LAVSLGQRAT ISCRASKSVS SSGYSYMHWY QQKPGQPPKV LIFLSSNLDS GVPARFSGSG SGTDFTLNIH PVEDGDAATY YCQHSRDLPP TFGGGTKLEI K
CDRL1 из DR5 mAb 1 (SEQ ID NO: 4): RASKSVSSSGYSYMH
CDRL2 из DR5 mAb 1 (SEQ ID NO: 5): LSSNLDS
CDRL3 DR5 mAb 1 (SEQ ID NO: 6): QHSRDLPPT
VL-домен DR5 mAb 1 предпочтительно кодируется полинуклеотидом (SEQ ID NO: 7), имеющим последовательность, показанную ниже (полинуклеотиды, кодирующие CDR, показаны подчеркиванием):
gacattgtgc tgacacagtc tcctgcttcc ttagctgtat ctctcgggca gagggccacc atctcatgca gggccagcaa aagtgtcagt tcctctggct atagttatat gcactggtac caacagaaac caggacagcc acccaaagtc ctcatctttc tttcatccaa cctagattct ggggtccctg ccaggttcag tggcagtggg tctgggacag acttcaccct caacatccat cctgtggagg atggggatgc tgcaacctat tactgtcagc acagtaggga tcttcctccg acgttcggtg gaggcaccaa gctggaaatc aaa
Аминокислотная последовательность VH-домена DR5 mAb 1 (SEQ ID NO: 8) показана ниже (остатки CDR выделены подчеркиванием). С-концевая аминокислота может быть замещена аланином для облегчения субклонирования этого VH-домена.
EVKFLESGGG LVQPGGSLKL SCVASGFDFS RYWMSWVRQA PGKGLEWIGE INPDSNTINY TPSLKDKFII SRDNAKNTLY LQMTKVRSED TALYYCTRRA YYGNPAWFAY WGQGTLVTVSS
CDRН1 из DR5 mAb 1 (SEQ ID NO: 9): GFDFSRYWMS
CDRН2 из DR5 mAb 1 (SEQ ID NO: 10): EINPDSNTINYTPSLKD
CDRН3 из DR5 mAb 1 (SEQ ID NO: 11): RAYYGNPAWFAY
VH-домен DR5 mAb 1 предпочтительно кодируется полинуклеотидом (SEQ ID NO: 12), имеющим последовательность, показанную ниже (полинуклеотиды, кодирующие CDR, показаны подчеркиванием):
gaggtgaagt ttctcgagtc tggaggtggc ctggtgcagc ctggaggatc cctgaaactc tcctgtgtag cctcaggatt cgattttagt agatactgga tgagttgggt ccggcaggct ccagggaaag ggctagaatg gattggagaa attaatccag atagcaatac gataaactat acgccatctc taaaggataa attcatcatc tccagagaca acgccaaaaa tacgctgtat ctgcaaatga ccaaagtgag atctgaggac acagcccttt attattgtac aagaagggcc tactatggta acccggcctg gtttgcttac tggggccaag ggactctggt cactgtctct tcc
b. антитело DR5 mAb 2 против DR5 человека
(1) Мышиное антитело mAb 2 против DR5 человека
Аминокислотная последовательность VL-домена DR5 mAb 2 (SEQ ID NO: 13), показана ниже (остатки CDR выделены подчеркиванием):
DIVMTQSHKF MSTSVGDRVS ITCKASQDVN TAVAWYQQKP GQSPKLLIYW ASTRHTGVPD RFTGSGSGTD YTLTIKSVQA EDLTLYYCQQ HYITPWTFGG GTKLEIK
CDRL1 из DR5 mAb 2 (SEQ ID NO: 14): KASQDVNTAVA
CDRL2 из DR5 mAb 2 (SEQ ID NO: 15): WASTRHT
CDRL3 из DR5 mAb 2 (SEQ ID NO: 16): QQHYITPWT
VL-домен DR5 mAb 2 предпочтительно кодируется полинуклеотидом (SEQ ID NO: 17), имеющим последовательность, показанную ниже (полинуклеотиды, кодирующие CDR, показаны подчеркиванием):
gacattgtga tgacccagtc tcacaaattc atgtccactt cagtaggaga cagggtcagc atcacctgca aggccagtca ggatgtgaat actgctgtag cctggtatca acaaaaacca gggcaatctc ctaaactact gatttactgg gcatccaccc ggcacactgg agtccctgat cgcttcacag gcagtggatc tgggacagat tatacactca ccatcaaaag tgtgcaggct gaagacctga cactttatta ctgtcagcaa cactatatca ctccgtggac gttcggtgga ggcaccaagc tggaaatcaaa
Аминокислотная последовательность VH-домена DR5 mAb 2 (SEQ ID NO: 18), показана ниже (остатки CDR выделены подчеркиванием):
KVQLQQSGAE LVKPGASVKL SCKASGYTFT EYILHWVKQK SGQGLEWIGW FYPGNNNIKY NEKFKDKATL TADKSSSTVY MELSRLTSED SAVYFCARHE QGPGYFDYWG QGTTLTVSS
CDRН1 из DR5 mAb 2 (SEQ ID NO: 19): GYTFTEYILH
CDRН2 из DR5 mAb 2 (SEQ ID NO: 20): WFYPGNNNIKYNEKFKD
CDRН3 из DR5 mAb 2 (SEQ ID NO: 21): HEQGPGYFDY
VH-домен DR5 mAb 2 предпочтительно кодируется полинуклеотидом (SEQ ID NO: 22), имеющим последовательность, показанную ниже (полинуклеотиды, кодирующие CDR, показаны подчеркиванием):
aaggtccagc tgcagcagtc tggagctgaa ctggtgaaac ccggggcatc agtgaagctg tcctgcaagg cttctgggta caccttcact gagtatattt tacactgggt aaagcagaag tctggacagg gtcttgagtg gattgggtgg ttttatcctg gaaataataa tataaagtac aatgagaaat tcaaggacaa ggccacactg actgcggaca aatcctccag cacagtctat atggaactta gtagattgac atctgaagac tctgcggtct atttctgtgc aagacacgaa caaggaccag gttactttga ctactggggc caaggcacca ctctcacagt ctcctcc
(2) Гуманизированное DR5 mAb 2 ("hDR5 mAb 2")
Описанные выше мышиные антитела против DR5 человека mAb 2 были гумманизированы для того, чтобы продемонстрировать способность гуманизации антитела против DR5 человека с тем, чтобы уменьшить его антигенность при введении в организм человека-реципиента. Гуманизация дала четыре гуманизированных VL-домена обозначенных в настоящем документе как "hDR5 mAb 2 VL-2", "hDR5 mAb 2 VL-3", "hDR5 mAb 2 VL-4" и "hDR5 mAb 2 VL-5", и один гуманизированный VH-домен, обозначенный в настоящем описании как "hDR5 mAb 2 VH-2". Любой из гуманизированных VL-доменов могут быть соединены с гуманизированным VH-доменом. Соответственно, любое антитело, содержащее один из гуманизированных VL-доменов, спаренных с гуманизированным VH-доменом, называется обобщенно как "hDR5 mAb 2" и конкретную комбинацию гуманизированных VL/VH-доменов обозначают ссылкой на VL-домен.
Аминокислотная последовательность VL-домена hDR5 mAb 2 VL-2 (SEQ ID NO: 23), показана ниже (остатки CDR выделены подчеркиванием):
DIQMTQSPSF LSASVGDRVT ITCKASQDVN TAVAWYQQKP GKAPKLLIYW ASTRHTGVPS RFSGSGSGTD FTLTISSLQP EDVATYYCQQ HYITPWTFGG GTKLEIK
hDR5 mAb 2 VL-2 предпочтительно кодируется полинуклеотидом (SEQ ID NO: 24), имеющим последовательность, показанную ниже:
gatattcaga tgacccagag tccctcattt ctgtccgcct ccgtcggtga ccgcgtgact attacttgta aagcttctca ggatgtcaac accgccgtgg cttggtacca gcagaagccc ggtaaagcac ctaagctgct gatctattgg gccagcactc ggcacaccgg agtcccatct aggttctctg gcagtggatc agggacagac tttaccctga caattagctc cctgcagccc gaggatgtgg ctacttacta ttgtcagcag cactacatca ctccttggac cttcggcggg ggcacaaaac tggaaatcaa a
Аминокислотная последовательность VL-домена hDR5 mAb 2 VL-3 (SEQ ID NO: 25), показана ниже (остатки CDR выделены подчеркиванием):
DIQMTQSPSF LSASVGDRVT ITCRASQDVN TAVAWYQQKP GKAPKLLIYW ASTRHTGVPD RFSGSGSGTD FTLTISSLQP EDVATYYCQQ HYITPWTFGG GTKLEIK
hDR5 mAb 2 VL-3 предпочтительно кодируется полинуклеотидом (SEQ ID NO: 26), имеющим последовательность, показанную ниже:
gatattcaga tgacccagag tccctcattt ctgtccgcct ccgtcggtga ccgcgtgact attacttgtc gggcttctca ggatgtcaac accgccgtgg cttggtacca gcagaagccc ggtaaagcac ctaagctgct gatctattgg gccagcactc ggcacaccgg agtcccagat aggttctctg gcagtggatc agggacagac tttaccctga caattagctc cctgcagccc gaggatgtgg ctacttacta ttgtcagcag cactacatca ctccttggac cttcggcggg ggcacaaaac tggaaatcaa a
Аминокислотная последовательность VL-домена hDR5 mAb 2 VL-4 (SEQ ID NO: 27), показана ниже (остатки CDR выделены подчеркиванием):
DIQMTQSPSF LSASVGDRVT ITCRASQDVN TAVAWYQQKP GKAPKLLIYW ASTRHTGVPS RFSGSGSGTD FTLTISSLQP EDIATYYCQQ HYITPWTFGG GTKLEIK
hDR5 mAb 2 VL-4 предпочтительно кодируется полинуклеотидом (SEQ ID NO: 28), имеющий последовательность, показанную ниже:
gatattcaga tgacccagag tccctcattt ctgtccgcct ccgtcggtga ccgcgtgact attacttgtc gggcttctca ggatgtcaac accgccgtgg cttggtacca gcagaagccc ggtaaagcac ctaagctgct gatctattgg gccagcactc ggcacaccgg agtcccatct aggttctctg gcagtggatc agggacagac tttaccctga caattagctc cctgcagcca gaggatatcg ctacatacta ttgtcagcag cactacatca ctccttggac cttcggcggg ggcacaaaac tggaaatcaa a
Аминокислотная последовательность VL-домена hDR5 mAb 2 VL-5 (SEQ ID NO: 29), показана ниже (остатки CDR выделены подчеркиванием):
DIQMTQSPSF LSASVGDRVT ITCRASQDVN TAVAWYQQKP GKAPKLLIYW ASTRHTGVPD RFSGSGSGTD FTLTISSLQP EDIATYYCQQ HYITPWTFGG GTKLEIK
hDR5 mAb 2 VL-5 предпочтительно кодируется полинуклеотидом (SEQ ID NO: 30), имеющим последовательность, показанную ниже:
gatattcaga tgacccagag tccctcattt ctgtccgcct ccgtcggtga ccgcgtgact attacttgtc gggcttctca ggatgtcaac accgccgtgg cttggtacca gcagaagccc ggtaaagcac ctaagctgct gatctattgg gccagcactc ggcacaccgg agtcccagat aggttctctg gcagtggatc agggacagac tttaccctga caattagctc cctgcagccc gaggatatcg ctacttacta ttgtcagcag cactacatca ctccttggac cttcggcggg ggcacaaaac tggaaatcaa a
Аминокислотная последовательность VH-домена hDR5 mAb 2 VH-2 (SEQ ID NO: 31), показана ниже (остатки CDR выделены подчеркиванием):
QVQLVQSGAE VKKPGASVKV SCKASGYTFT EYILHWVRQA PGQGLEWMGW FYPGNNNIKY NEKFKDRVTI TADKSTSTVY MELSSLRSED TAVYYCARHE QGPGYFDYWG QGTLVTVSS
hDR5 mAb 2 VH-2 предпочтительно кодируется полинуклеотидом (SEQ ID NO: 32), имеющим последовательность, показанную ниже:
caggtccagc tggtgcagag tggggcagag gtgaaaaagc caggggcatc agtgaaagtg tcttgtaaag catcaggtta tacatttact gagtacatcc tgcactgggt gcgacaggca ccaggacagg gactggaatg gatggggtgg ttctaccctg gcaacaacaa cattaagtac aacgagaagt ttaaagaccg ggtgaccatc acagcggata agtctaccag tacagtctat atggagctga gctccctgag aagcgaagac accgccgtct actattgcgc tcgccacgaa cagggtccag gttactttga ttattggggg cagggaactc tggtcacagt cagctcc
CDR1 из VL-домена hDR5 mAb 2 VL-3, hDR5 mAb 2 VL-4 и hDR5 mAb VL-5 имеет аминокислотную последовательность: RASQDVNTAVA (SEQ ID NO: 196).
c. Дрозитумаб ("DR5 mAb 3")
Аминокислотная последовательность VL-домена дрозитумаба ("DR5 mAb 3") (SEQ ID NO: 54), показана ниже (остатки CDR выделены подчеркиванием):
SELTQDPAVS VALGQTVRIT CSGDSLRSYY ASWYQQKPG QAPVLVIYGA NNRPSGIPDR FSGSSSGNTA SLTITGAQAE DEADYYCNSA DSSGNHVVFG GGTKLTVLG
CDRL1 из DR5 mAb 3 (SEQ ID NO: 55): SGDSLRSYYAS
CDRL2 из DR5 mAb 3 (SEQ ID NO: 56): GANNRPS
CDRL3 из DR5 mAb 3 (SEQ ID NO: 57): NSADSSGNHVV
Аминокислотная последовательность VH-домена дрозитумаба ("DR5 mAb 3") (SEQ ID NO: 58), показана ниже (остатки CDR выделены подчеркиванием):
EVQLVQSGGG VERPGGSLRL SCAASGFTFD DYAMSWVRQA PGKGLEWVSG INWQGGSTGY ADSVKGRVTI SRDNAKNSLY LQMNSLRAED TAVYYCAKIL GAGRGWYFDY WGKGTTVTVS S
CDRН1 из DR5 mAb 3 (SEQ ID NO: 59): GFTFDDYAMS
CDRН2 из DR5 mAb 3 (SEQ ID NO: 60): INWQGGSTGYADSVKG
CDRН3 из DR5 mAb 3 (SEQ ID NO: 61): ILGAGRGWYFDY
d. Конатумумаб ("DR5 mAb 4")
Аминокислотная последовательность VL-домена Конатумумаба ("DR5 mAb 4") (SEQ ID NO: 62), показана ниже (остатки CDR выделены подчеркиванием):
EIVLTQSPGT LSLSPGERAT LSCRASQGIS RSYLAWYQQK PGQAPSLLIY GASSRATGIP DRFSGSGSGT DFTLTISRLE PEDFAVYYCQ QFGSSPWTFG QGTKVEIK
CDRL1 из DR5 mAb 4 (SEQ ID NO: 63): RASQGISRSYLA
CDRL2 из DR5 mAb 4 (SEQ ID NO: 64): GASSRAT
CDRL3 из DR5 mAb 4 (SEQ ID NO: 65): QQFGSSPWT
Аминокислотная последовательность VH-домена Конатумумаба ("DR5 mAb 4") (SEQ ID NO: 66), показана ниже (остатки CDR выделены подчеркиванием):
QVQLQESGPG LVKPSQTLSL TCTVS GGSIS SGDYFWS WIR QLPGKGLEWI G HIHNSGTTY YNPSLKS RVT ISVDTSKKQF SLRLSSVTAA DTAVYYCAR D RGGDYYYGMD V WGQGTTVTV С. С.
CDRН1 из DR5 mAb 4 (SEQ ID NO: 67): GGSISSGDYFWS
CDRН2 из DR5 mAb 4 (SEQ ID NO: 68): HIHNSGTTYYNPSLKS
CDRН3 из DR5 mAb 4 (SEQ ID NO: 69): DRGGDYYYGMDV
e. Тигатумумаб ("DR5 mAb 5")
Аминокислотная последовательность VL-домена тигатумумаба ("DR5 mAb 5") (SEQ ID NO: 70), показана ниже (остатки CDR выделены подчеркиванием):
DIQMTQSPSS LSASVGDRVT ITCKASQDVG TAVAWYQQKP GKAPKLLIYW ASTRHTGVPS RFSGSGSGTD FTLTISSLQP EDFATYYCQQ YSSYRTFGQG TKVEIK
CDRL1 из DR5 mAb 5 (SEQ ID NO: 71): KASQDVGTAVA
CDRL2 из DR5 mAb 5 (SEQ ID NO: 72): WASTRHT
CDRL3 из DR5 mAb 5 (SEQ ID NO: 73): QQYSSYRT
Аминокислотная последовательность VH-домена тигатумумаба ("DR5 mAb 5") (SEQ ID NO: 74), показана ниже (остатки CDR выделены подчеркиванием):
EVQLVESGGG LVQPGGSLRL SCAASGFTFS SYVMSWVRQA PGKGLEWVAT ISSGGSYTYY PDSVKGRFTI SRDNAKNTLY LQMNSLRAED TAVYYCARRG DSMITTDYWG QGTLVTVSS
CDRН1 из DR5 mAb 5 (SEQ ID NO: 75): GFTFSSYVMS
CDRН2 из DR5 mAb 5 (SEQ ID NO: 76): TISSGGSYTYYPDSVKG
CDRН3 из DR5 mAb 5 (SEQ ID NO: 77): RGDSMITTDY
f. LBY135-1 (“DR5 mAb 6”)
Аминокислотная последовательность VL-домена LBY135-1 ("DR5 mAb 6") (SEQ ID NO: 78), показана ниже (остатки CDR выделены подчеркиванием):
DIAMTQSHKF MSTLVGDRVS ITCKASQDVN TAIAWYQQKP GQSPKLLIYW ASTRHTGVPD RFYGSGSGTD YTLTISSMEA EDAATYYCQQ WSSNPLTFGA GTKLELKRA
CDRL1 из DR5 mAb 6 (SEQ ID NO: 79): QDVNTAIA
CDRL2 из DR5 mAb 6 (SEQ ID NO: 80): WASTRHT
CDRL3 из DR5 mAb 6 (SEQ ID NO: 81): QQWSSNPLT
Аминокислотная последовательность VH-домена LBY135-1 ("DR5 mAb 6") (SEQ ID NO: 82), показана ниже (остатки CDR выделены подчеркиванием):
KVQLQQSGAE LVKPGASVKL SCKAS GYTFT DYTIH WVKQR SGQGLEWIG W FYPGGGYIKY NEKFKD RATL TADKSSNTVY MELSRLTSEG SAVYFCAR HE EGIYFDY WGQ GTTLTVSS
CDRН1 из DR5 mAb 6 (SEQ ID NO: 83): GYTFTDYTIH
CDRН2 из DR5 mAb 6 (SEQ ID NO: 84): WFYPGGGYIKYNEKFKD
CDRН3 из DR5 mAb 6 (SEQ ID NO: 85): HEEGIYFDY
g. LBY135-2 ("DR5 mAb 7")
Аминокислотная последовательность VL-домена LBY135-2 ("DR5 mAb 7") (SEQ ID NO: 86), показана ниже (остатки CDR выделены подчеркиванием):
DIVMTQSHKF MSTSVGDRVS ITCKASQDVN TAIAWYQQKP GQSPKLLIYW ASTRHTGVPD RFTGSGSGTD YTLTISSVQA EDLALYYCQQ HYTTPFTFGS GTKL
CDRL1 из DR5 mAb 7 (SEQ ID NO: 87): KASQDVNTAIA
CDRL2 из DR5 mAb 7 (SEQ ID NO: 88): WASTRHT
CDRL3 из DR5 mAb 7 (SEQ ID NO: 89): QQHYTTPFT
Аминокислотная последовательность VH-домена LBY135-2 ("DR5 mAb 7") (SEQ ID NO: 90), показана ниже (остатки CDR выделены подчеркиванием):
KVQLQQSGAE LVKPGASVKL SCKAS GYTFT DYTIH WVKQR SGQGLEWIG W FYPGGGYIKY NEKFKD RATL TADKSSNTVY MELSRLTSED SAVYFCAR HE EGIYFDY WGQ GTTLTVSS
CDRН1 из DR5 mAb 7 (SEQ ID NO: 91): GYTFTDYTIH
CDRН2 из DR5 mAb 7 (SEQ ID NO: 92): WFYPGGGYIKYNEKFKD
CDRН3 из DR5 mAb 7 (SEQ ID NO: 93): HEEGIYFDY
час KMTR2 ("DR5 mAb 8")
Аминокислотная последовательность VL-домена KMTR2 ("DR5 mAb 8") (SEQ ID NO: 94), показана ниже (остатки CDR выделены подчеркиванием):
EIVLTQSPAT LSLSPGERAT LSC RASQSVS Syla WYQQKP GQAPRLLIY D ASNRAT GIPA RFSGSGSGTD FTLTISSLEP EDFAVYYC QQ RSNWPLT FGG GTKVEIKR
CDRL1 из DR5 mAb 8 (SEQ ID NO: 95): RASQSVSSYLA
CDRL2 из DR5 mAb 8 (SEQ ID NO: 96): DASNRAT
CDRL3 DR5 mAb 8 (SEQ ID NO: 97): QQRSNWPLT
Аминокислотная последовательность VH-домена KMTR2 ("DR5 mAb 8") (SEQ ID NO: 98), показана ниже (остатки CDR выделены подчеркиванием):
QVQLVQSGAE MKKPGASVKV SCKTSGYTFT NYKINWVRQA PGQGLEWMGW MNPDTDSTGY PQKFQGRVTM TRNTSISTAY MELSSLRSED TAVYYCARSY GSGSYYRDYY YGMDVWGQGT TVTVSS
CDRН1 из DR5 mAb 8 (SEQ ID NO: 99): GYTFTNYKIN
CDRН2 из DR5 mAb 8 (SEQ ID NO: 100): WMNPDTDSTGYPQKFQG
CDRН3 из DR5 mAb 8 (SEQ ID NO: 101): SYGSGSYYRDYYYGMDV
5. EphA2-связывающие домены
Рецепторная тирозинкиназа, рецептор эфрина А 2 (EphA2) является предпочтительным антигеном злокачественной опухоли по настоящему изобретению. EphA2 обычно экспрессируется в участках межклеточного контакта во взрослых эпителиальных тканях, однако, недавние исследования показали, что он также избыточно экспрессируется в различных типах эпителиальных карцином, с наибольшим уровнем экспрессии EphA2, наблюдаемым в метастатических поражениях. Высокие уровни экспрессии EphA2 были найдены в широком диапазоне видов злокачественных опухолей и в многочисленных линий опухолевых клеток, включая рак простаты, рак молочной железы, немелкоклеточный рак легкого и меланому (Xu, J. et al.(2014) “High Epha2 Protein Expression In Renal Cell Carcinoma Is Associated With A Poor Disease Outcome,” Oncol. Lett. Август 2014; 8 (2): 687-692; Мяо, Б. et al.(2014) “EphA2 is a Mediator of Vemurafenib Resistance and a Novel Therapeutic Target in Melanoma,” Cancer Discov. pii: CD-14-0295. EphA2, похоже, не является просто маркером для злокачественной опухоли, а, скорее, по- видимому, постоянно избыточно экспрессируется и функционально изменен в многочисленных человеческих злокачественных опухолевых клетках (Chen, P. et al.(2014) “Epha2 Enhances The Proliferation And Invasion Ability Of Lncap Prostate Cancer Cells,” Oncol. Lett. 8 (1): 41-46).
В частности, изобретение предусматривает выбор EphA2 как антигена злокачественной опухоли, и применение анти-EphA2 антител для получения домена связывания с антигеном злокачественной опухоли триспецифичных связывающих молекул по настоящему изобретению. Примеры анти-EphA2 антитела включают "EphA2 mAb 1", "EphA2 mAb 2" и "EphA2 mAb 3".
а. EphA2 mAb 1
Аминокислотная последовательность VL-домена предпочтительного антитела против человеческого EphA2 ("EphA2 mAb 1") (SEQ ID NO: 153), показана ниже (остатки CDR выделены подчеркиванием):
DIQMTQTTSS LSASLGDRIT ISCRASQDIS NYLNWYQQKP DGTVKLLIYY TSRLHSGVPS RFSGSGSGTD YSLTISNLEQ EDIATYFCQQ GYTLYTFGGG TKLEIK
CDRL1 из EphA2 mAb 1 (SEQ ID NO: 154): RASQDISNYLN
CDRL2 из EphA2 mAb 1 (SEQ ID NO: 155): YTSRLHS
CDRL3 EphA2 mAb 1 (SEQ ID NO: 156): QQGYTLYT
VL Домен EphA2 mAb 1 предпочтительно кодируется полинуклеотидом (SEQ ID NO: 157), имеющим последовательность, показанную ниже (полинуклеотиды, кодирующие CDR, показаны подчеркиванием):
gatatccaga tgacacagac tacatcctcc ctgtctgcct ctctgggaga cagaatcacc atcagttgca gggcaagtca ggacattagc aattatttaa actggtatca gcagaaacca gatggaactg ttaaactcct gatctactac acatcaagat tacactcagg agtcccatca aggttcagtg gcagtgggtc tggaacagat tattctctca ccattagcaa cctggagcaa gaagatattg ccacttactt ttgccaacag ggttatacgc tgtacacgtt cggagggggg accaagctgg aaataaaa
Аминокислотная последовательность VH-домена EphA2 mAb 1 (SEQ ID NO: 158), показана ниже (остатки CDR выделены подчеркиванием):
QVQLKESGPG LVAPSQSLSI TCTVSGFSLS RYSVHWVRQP PGKGLEWLGM IWGGGSTDYN SALKSRLSIS KDNSKSQVFL KMNSLQTDDT AMYYCARKHG NYYTMDYWGQ GTSVTVSS
CDRН1 из EphA2 mAb 1 (SEQ ID NO: 159): GFSLSRYSVH
CDRН2 из EphA2 mAb 1 (SEQ ID NO: 160): MIWGGGSTDYNSALKS
CDRН3 из EphA2 mAb 1 (SEQ ID NO: 161): KHGNYYTMDY
VH-домен EphA2 mAb 1 предпочтительно кодируется полинуклеотидом (SEQ ID NO: 162), имеющим последовательность, показанную ниже (полинуклеотиды, кодирующие CDR, показаны подчеркиванием):
caggtgcagc tgaaggagtc aggacctggc ctggtggcac cctcacagag cctgtccatc acatgcactg tctctgggtt ctcattatcc agatatagtg tacactgggt tcgccagcct ccaggaaagg gtctggagtg gctgggaatg atatggggtg gtggaagcac agactataat tcagctctca aatccagact gagtatcagc aaggacaact ccaagagcca agttttctta aaaatgaaca gtctgcaaac tgatgacaca gccatgtact actgtgccag aaaacatggt aactactata ctatggacta ctggggtcaa ggaacctcag tcaccgtctc ctcc
b. EphA2 mAb 2
Аминокислотная последовательность VL-домена второго предпочтительного антитела против человеческого EphA2 ("EphA2 mAb 2") (SEQ ID NO: 163), показана ниже (остатки CDR выделены подчеркиванием):
DVVMTQTPLS LPVSLGDQAS ISCRSSQSLV HSSGNTYLHW YLQKPGQSPK LLIYKVSNRF SGVPDRFSGS GSGTDFTLKI SRVEAEDLGV YFCSQSTHVP TFGSGTKLEI K
CDRL1 из EphA2 mAb 2 (SEQ ID NO: 164): RSSQSLVHSSGNTYLH
CDRL2 из EphA2 mAb 2 (SEQ ID NO: 165): KVSNRFS
CDRL3 из EphA2 mAb 2 (SEQ ID NO: 166): SQSTHVPT
VL Домен EphA2 mAb 2 предпочтительно кодируется полинуклеотидом (SEQ ID NO: 318), имеющим последовательность, показанную ниже (полинуклеотиды, кодирующие CDR, показаны подчеркиванием):
gatgttgtga tgacccaaac tccactctcc ctgcctgtca gtcttggaga tcaagcctcc atctcttgca gatctagtca gagccttgta cacagtagtg gaaacaccta tttacattgg tacctgcaga agccaggcca gtctccaaag ctcctgatct acaaagtttc caaccgattt tctggggtcc cagacaggtt cagtggcagt ggatcaggga cagatttcac actcaagatc agcagagtgg aggctgagga tctgggagtt tatttctgct ctcaaagtac acatgttccc acgttcggct cggggacaaa gttggaaata aaa
Аминокислотная последовательность VH-домена EphA2 mAb 2 (SEQ ID NO: 167), показана ниже (остатки CDR выделены подчеркиванием):
QIQLVQSGPE LKKPGETVKI SCKASGFTFT NYGMNWVKQA PGKGLKWMGW INTYIGEPTY ADDFKGRFVF SLETSASTAY LQINNLKNED MATYFCAREL GPYYFDYWGQ GTTLTVSS
CDRН1 из EphA2 mAb 2 (SEQ ID NO: 168): GFTFTNYGMN
CDRН2 из EphA2 mAb 2 (SEQ ID NO: 169): WINTYIGEPTYADDFKG
CDRН3 из EphA2 mAb 2 (SEQ ID NO: 170): ELGPYYFDY
VH-домен EphA2 mAb 2 предпочтительно кодируется полинуклеотидом (SEQ ID NO: 171), имеющим последовательность, показанную ниже (полинуклеотиды, кодирующие CDR, показаны подчеркиванием):
cagatccagt tggtgcagtc tggacctgag ctgaagaagc ctggagagac agtcaagatc tcctgcaagg cttctgggtt taccttcaca aactatggaa tgaactgggt gaagcaggct ccaggaaagg gtttaaagtg gatgggctgg ataaacacct atattggaga gccgacatat gctgatgact tcaagggacg gtttgtcttc tctttggaaa cctctgccag cactgcctat ttgcagatca acaacctcaa aaatgaggac atggccacat atttctgtgc aagagaactg ggaccatact actttgacta ctggggccaa ggcaccactc tcacagtctc ctcc
c. EphA2 mAb 3
Аминокислотная последовательность VL-домена другого предпочтительного антитела к человеческому EphA2 ("EphA2 mAb 3") (SEQ ID NO: 172), показана ниже (остатки CDR выделены подчеркиванием):
DIVLTQSHRS MSTSVGDRVN ITCKASQDVT TAVAWYQQKP GQSPKLLIFW ASTRHAGVPD RFTGSGSGTD FTLTISSVQA GDLALYYCQQ HYSTPYTFGG GTKLEIK
CDRL1 из EphA2 mAb 3 (SEQ ID NO: 173): KASQDVTTAVA
CDRL2 из EphA2 mAb 3 (SEQ ID NO: 174): WASTRHA
CDRL3 из EphA2 mAb 3 (SEQ ID NO: 175): QQHYSTPYT
VL Домен EphA2 mAb 3 предпочтительно кодируется полинуклеотидом (SEQ ID NO: 176), имеющим последовательность, показанную ниже (полинуклеотиды, кодирующие CDR, показаны подчеркиванием):
gacattgtgc tgacccagtc tcacagatcc atgtccacat cagtaggaga cagggtcaac atcacctgca aggccagtca ggatgtgact actgctgtag cctggtatca acaaaaacca gggcaatctc ctaaattact gattttctgg gcatccaccc ggcacgctgg agtccctgat cgcttcacag gcagtggatc tgggacagat tttactctca ccatcagcag tgtgcaggct ggagacctgg cactttatta ctgtcaacaa cattatagca caccgtacac attcggaggg gggaccaagc tggaaataaa a
Аминокислотная последовательность VH-домена EphA2 mAb 3 (SEQ ID NO: 177), показана ниже (остатки CDR выделены подчеркиванием):
EVQLVESGGG SVKPGGSLKL SCAASGFTFT DHYMYWVRQT PEKRLEWVAT ISDGGSFTSY PDSVKGRFTI SRDIAKNNLY LQMSSLKSED TAMYYCTRDE SDRPFPYWGQ GTLVTVSS
CDRН1 из EphA2 mAb 3 (SEQ ID NO: 178): GFTFTDHYMY
CDRН2 из EphA2 mAb 3 (SEQ ID NO: 179): TISDGGSFTSYPDSVKG
CDRН3 из EphA2 mAb 3 (SEQ ID NO: 180): DESDRPFPY
VH-домен EphA2 mAb 3 предпочтительно кодируется полинуклеотидом (SEQ ID NO: 319), имеющим последовательность, показанную ниже (полинуклеотиды, кодирующие CDR, показаны подчеркиванием):
gaagtgcagc tggtggagtc tgggggaggc tcagtgaagc ctggagggtc cctgaaactc tcctgtgcag cctctggatt cactttcact gaccattaca tgtattgggt tcgccagact ccggaaaaga ggctggagtg ggtcgcaacc attagtgatg gcggtagttt cacctcctat ccagacagtg tgaaggggcg attcaccatc tccagagaca ttgccaagaa caacctgtac ctccaaatga gcagtctgaa gtctgaggac acagccatgt attactgtac aagagatgag agcgataggc cgtttcctta ctggggccaa gggactctgg tcactgtctc ctcc
6. gpA33-связывающие домены
gpA33 также является предпочтительным антигеном злокачественной опухоли по настоящему изобретению. Колоректальный рак является одним из наиболее распространенных злокачественных заболеваний западного мира и является ведущей причиной смерти от злокачественных новообразований (Silverberg, E. et al.(1989) “Cancer Statistics, 1989,” CA Cancer J Clin. 39 (1): 3-20). Одной из потенциально полезных мишеней для рака толстой кишки является 43 кДа трансмембранный гликопротеин A33 (gpA33), который экспрессируется в> 95% всех колоректальных карцином (Heath, JK et al.(1997) “The Human A33 Antigen Is A Transmembrane Glycoprotein And A Novel Member Of The Immunoglobulin Superfamily,” Proc. Natl. Acad. Sci. (U.S.A.) 94(2):469-474; Ritter, G. et al. (1997) “Characterization Of Posttranslational Modifications Of Human A33 Antigen, A Novel Palmitoylated Surface Glycoprotein Of Human Gastrointestinal Epithelium,” Biochem. Biophys. Res. Commun. 236(3):682-686; Wong, N.A. et al. (2006) “EpCAM and gpA33 Are Markers Of Barrett's Metaplasia,” J. Clin. Pathol. 59 (3): 260-263). gpA33 был впервые обнаружен по возникновению моноклональных мышиных антител против клеточной линии ASPC1, полученной из карциномы поджелудочной железы человека.
В частности, изобретение предусматривает выбор gpA33 как антигена злокачественной опухоли, и применение анти-gpA33 антител для того, чтобы обеспечить домен, связывающий антиген злокачественной опухоли в триспецифичных связывающих молекулах по настоящему изобретению. Примерное анти-gpA33 антитело представляет собой "gpA33 mAb 1".
Аминокислотная последовательность VL-домена предпочтительного антитела против человеческого gpA33 ("gpA33 mAb 1") (SEQ ID NO: 181), показана ниже (остатки CDR выделены подчеркиванием):
DIQLTQSPSF LSASVGDRVT ITC SARSSIS FMY WYQQKPG KAPKLLIY DT SNLAS GVPSR FSGSGSGTEF TLTISSLEAE DAATYYC QQW SSYPLT FGQG TKLEIK
CDRL1 из gpA33 mAb 1 (SEQ ID NO: 182): SARSSISFMY
CDRL2 из gpA33 mAb 1 (SEQ ID NO: 183): DTSNLAS
CDRL3 gpA33 mAb 1 (SEQ ID NO: 184): QQWSSYPLT
VL Домен gpA33 mAb 1 предпочтительно кодируется полинуклеотидом (SEQ ID NO: 185), имеющим последовательность, показанную ниже (полинуклеотиды, кодирующие CDR, показаны подчеркиванием):
gacattcagc tgactcagtc cccctctttt ctgtccgcat ccgtcggaga tcgagtgact attacttgct ctgctaggtc ctcaatcagc ttcatgtact ggtatcagca gaagcccggc aaagcaccta agctgctgat ctacgacaca agcaacctgg cctccggggt gccatctcgg ttctctggca gtgggtcagg aactgagttt accctgacaa ttagctccct ggaggctgaa gatgccgcta cctactattg ccagcagtgg agcagctatc ctctgacctt cggacagggg actaaactgg aaatcaag
Аминокислотная последовательность VH-домена gpA33 mAb 1 (SEQ ID NO: 186), показана ниже (остатки CDR выделены подчеркиванием):
QVQLVQSGAE VKKPGASVKV SCKASGYTFT GSWMNWVRQA PGQGLEWIGR IYPGDGETNY NGKFKDRVTI TADKSTSTAY MELSSLRSED TAVYYCARIY GNNVYFDVWG QGTTVTVSS
CDRН1 из gpA33 mAb 1 (SEQ ID NO: 187): GYTFTGSWMN
CDRН2 из gpA33 mAb 1 (SEQ ID NO: 188): RIYPGDGETNYNGKFKD
CDRН3 из gpA33 mAb 1 (SEQ ID NO: 189): IYGNNVYFDV
VH-домен gpA33 mAb 1 предпочтительно кодируется полинуклеотидом (SEQ ID NO: 190), имеющим последовательность, показанную ниже (полинуклеотиды, кодирующие CDR, показаны подчеркиванием):
caggtccagc tggtccagag cggggccgaa gtcaaaaaac ccggagcaag cgtgaaggtc tcctgcaaag catcaggcta tacatttaca ggcagctgga tgaactgggt gaggcaggct ccaggacagg gactggagtg gatcgggcgc atctaccctg gagacggcga aactaactat aatggaaagt tcaaagaccg agtgaccatc acagccgata agtctactag taccgcctac atggagctga gctccctgcg gtctgaagat accgccgtct actattgcgc tagaatttac ggaaacaatg tctattttga cgtgtggggg cagggaacaa ctgtgactgt ctcctcc
7. HER2-связывающие домены
Изобретение также, в частности, предполагает выбор Her2 как антигена злокачественной опухоли, и применение анти-HER2 антител для того, чтобы обеспечить домен, связывающий антиген злокачественной опухоли из триспецифичных связывающих молекул по настоящему изобретению. Примерные антитела против HER2 включают "Her2 mAb 1" и трастузумаб.
а. Her2 mAb 1
Аминокислотная последовательность VL-домена анти- HER2 антитела "Her2 mAb 1" (SEQ ID NO: 191), показана ниже (остатки CDR выделены подчеркиванием):
DIQMTQSPSS LSASVGDRVT ITCRASQDVN TAVAWYQQKP GKAPKLLIYS ASFLESGVPS RFSGSRSGTD FTLTISSLQP EDFATYYCQQ HYTTPPTFGQ GTKVEIKRT
Аминокислотная последовательность VH-домена анти-HER2 антитела "Her2 mAb 1" (SEQ ID NO: 192), показана ниже (остатки CDR выделены подчеркиванием):
QVQLQQSGPE LVKPGASLKL SCTASGFNIK DTYIHWVKQR PEQGLEWIGR IYPTNGYTRY DPKFQDKATI TADTSSNTAY LQVSRLTSED TAVYYCSRWG GDGFYAMDYW GQGASVTVSS
b. Трастузумаб
Аминокислотная последовательность VL-домена гуманизированного анти-HER2 антитела "трастузумаб" (SEQ ID NO: 193), показана ниже (остатки CDR выделены подчеркиванием):
DIQMTQSPSS LSASVGDRVT ITCRASQDVN TAVAWYQQKP GKAPKLLIYS ASFLYSGVPS RFSGSRSGTD FTLTISSLQP EDFATYYCQQ HYTTPPTFGQ GTKVEIKR
Аминокислотная последовательность в домене VH гуманизированного анти-HER2 антитела "трастузумаб" (SEQ ID NO: 194), показана ниже (остатки CDR выделены подчеркиванием):
EVQLVESGGG LVQPGGSLRL SCAASGFNIK DTYIHWVRQA PGKGLEWVAR IYPTNGYTRY ADSVKGRFTI SADTSKNTAY LQMNSLRAED TAVYYCSRWG GDGFYAMDYW GQGTLVTVSS
8. B7-H3-связывающие домены
В дополнение к экспрессии в клетках нейробластомы, также известно, что человеческий В7-Н3 экспрессируется в различных других злокачественных опухолевых клетках (например, клетках рака желудка, яичников и немелкоклеточного рака легкого). Экспрессия белка В7-Н3 была обнаружена иммуногистологически в опухолевых клеточных линиях (Chapoval А. et al. (2001) “B7-H3: A Costimulatory Molecule For T Cell Activation and IFN-γ Production,” Nature Immunol. 2: 269-274; Saatian, В. et al. (2004) “Expression Of Genes For B7-H3 And Other T Cell Ligands By Nasal Epithelial Cells During Differentiation And Activation,” Amer. J. Physiol. Lung Cell. Mol. Physiol. 287: L217-L225; Castriconi et al. (2004) “Identification Of 4Ig-B7-H3 As A Neuroblastoma-Associated Molecule That Exerts A Protective Role From An NK Cell-Mediated Lysis,” Proc. Natl. Acad. Sci. (США) 101 (34): 12640-12645); Вс, M. et al. (2002) “Characterization of Mouse and Human B7-H3 Genes,” J. Immunol. 168: 6294-6297). Экспрессия мРНК была обнаружена в сердце, почках, яичках, легких, печени, поджелудочной железе, предстательной железе, толстой кишке и остеобластах (Collins, M. et al. (2005) “The B7 Family Of Immune-Regulatory Ligands,” Genome Biol. 6:223.1-223.7). На белковом уровне, В7-Н3 встречается в человеческой печени, легких, мочевом пузыре, яичке, предстательной железе, молочной железе, плаценте и лимфоидных органах (Hofmeyer, K. et al. (2008) “The Contrasting Role Of B7-H3,” Proc. Natl. Acad. Sci. (U.S.A.) 105 (30): 10277-10278).
Изобретение также в особенности предусматривает выбор B7-H3 как антигена злокачественной опухоли, и применение анти-В7-Н3 антитела для обеспечения домена, связывающего антиген злокачественной опухоли, из триспецифичных связывающих молекул по настоящему изобретению. Примерные анти-В7-Н3 антитела включают, "В7-Н3 mAb 1", "В7-Н3 mAb 2" и "В7-Н3 mAb 3".
a. B7-H3 mAb 1
Аминокислотная последовательность VL-домена анти-В7-Н3 антитела "В7-Н3 mAb 1" (SEQ ID NO: 195), показана ниже (остатки CDR выделены подчеркиванием):
DIAMTQSQKF MSTSVGDRVS VTCKASQNVD TNVAWYQQKP GQSPKALIYS ASYRYSGVPD RFTGSGSGTD FTLTINNVQS EDLAEYFCQQ YNNYPFTFGS GTKLEIK
Аминокислотная последовательность VH-домена анти-В7-Н3 антитела "В7-Н3 mAb 1" (SEQ ID NO: 196), показана ниже (остатки CDR выделены подчеркиванием):
DVQLVESGGG LVQPGGSRKL SCAASGFTFS SFGMHWVRQA PEKGLEWVAY ISSDSSAIYY ADTVKGRFTI SRDNPKNTLF LQMTSLRSED TAMYYCGRGR ENIYYGSRLD YWGQGTTLTV SS
b. B7-H3 mAb 2
Аминокислотная последовательность VL-домена анти-В7-Н3 антитела "В7-Н3 mAb 2" (SEQ ID NO: 197), показана ниже (остатки CDR выделены подчеркиванием):
DIQMTQTTSS LSASLGDRVT ISCRASQDIS NYLNWYQQKP DGTVKLLIYY TSRLHSGVPS RFSGSGSGTD YSLTIDNLEQ EDIATYFCQQ GNTLPPTFGG GTKLEIK
Аминокислотная последовательность VH-домена анти-В7-Н3 антитела "В7-Н3 mAb 2" (SEQ ID NO: 198), показана ниже (остатки CDR выделены подчеркиванием):
QVQLQQSGAE LARPGASVKL SCKASGYTFT SYWMQWVKQR PGQGLEWIGT IYPGDGDTRY TQKFKGKATL TADKSSSTAY MQLSSLASED SAVYYCARRG IPRLWYFDVW GAGTTVTVSS
c. B7-H3 mAb 3
Аминокислотная последовательность VL-домена анти-В7-Н3 антитела "В7-Н3 mAb 3" (SEQ ID NO: 199), показана ниже (остатки CDR выделены подчеркиванием):
DIQMTQSPAS LSVSVGETVT ITCRASESIY SYLAWYQQKQ GKSPQLLVYN TKTLPEGVPS RFSGSGSGTQ FSLKINSLQP EDFGRYYCQH HYGTPPWTFG GGTNLEIK
Аминокислотная последовательность VH-домена анти-В7-Н3 антитела "В7-Н3 mAb 3" (SEQ ID NO: 200), показана ниже (остатки CDR выделены подчеркиванием):
EVQQVESGGD LVKPGGSLKL SCAASGFTFS SYGMSWVRQT PDKRLEWVAT INSGGSNTYY PDSLKGRFTI SRDNAKNTLY LQMRSLKSED TAMYYCARHD GGAMDYWGQG TSVTVSS
9. Домены связывающие EGF рецептор (цетуксимаб)
Аминокислотная последовательность VL-домена химерного анти-EGFR антитела "цетуксимаб" (SEQ ID NO: 201), показана ниже (остатки CDR выделены подчеркиванием):
DILLTQSPVI LSVSPGERVS FSCRASQSIG TNIHWYQQRT NGSPRLLIKY ASESISGIPS RFSGSGSGTD FTLSINSVES EDIADYYCQQ NNNWPTTFGA GTKLELKR
Аминокислотная последовательность VH-домена химерного анти-EGFR антитело "цетуксимаб" (SEQ ID NO: 202), показана ниже (остатки CDR выделены подчеркиванием):
QVQLKQSGPG LVQPSQSLSI TCTVSGFSLT NYGVHWVRQS PGKGLEWLGV IWSGGNTDYN TPFTSRLSIN KDNSKSQVFF KMNSLQSNDT AIYYCARALT YYDYEFAYWG QGTLVTVSA
Панитумумаб (например, Vectibix®, Amgen) является альтернативным EGF-рецептор-связывающим антителом, которое может быть использовано в соответствии с настоящим изобретением.
10. VEGF-связывающие домены (бевацизумаб)
Аминокислотная последовательность VL-домена гуманизированного анти-VEGF-антитело "бевацизумаб" (SEQ ID NO: 203), показана ниже (остатки CDR выделены подчеркиванием):
DIQMTQSPSS LSASVGDRVT ITCSASQDIS NYLNWYQQKP GKAPKVLIYF TSSLHSGVPS RFSGSGSGTD FTLTISSLQP EDFATYYCQQ YSTVPWTFGQ GTKVEIKR
Аминокислотная последовательность VH-домена гуманизированного анти-VEGF-антитело "бевацизумаб" (SEQ ID NO: 204), показана ниже (остатки CDR выделены подчеркиванием):
EVQLVESGGG LVQPGGSLRL SCAASGYTFT NYGMNWVRQA PGKGLEWVGW INTYTGEPTY AADFKRRFTF SLDTSKSTAY LQMNSLRAED TAVYYCAKYP HYYGSSHWYF DVWGQGTLVT VSS
11. 5T4-связывающие домены
Карциноэмбриональный белок, 5T4, представляет собой ассоциированный с опухолью белок, представленный на клеточной мембране многих карцином, в том числе почек, толстой кишки, простаты, легких, рак и острого лимфобластного лейкоза (см., Boghaert, Е. Р. et al.(2008) “The Oncofetal Protein, 5T4, Is A Suitable Target For Antibody-Guided Anti-Cancer Chemotherapy With Calicheamicin,” Int. J. Oncol. 32(1):221-234; Eisen, T. et al. (2014) “Naptumomab Estafenatox: Targeted Immunotherapy with a Novel Immunotoxin,” Curr. Oncol. Rep. 16:370, pp. 1-6). Аминокислотная последовательность вариабельного домена легкой цепи примерного анти-5T4 антитела ("5T4 mAb 1") приведена ниже (остатки CDR выделены подчеркиванием): (SEQ ID NO: 308):
DIQMTQSPSS LSASVGDRVT ITCRASQGIS NYLAWFQQKP GKAPKSLIYR ANRLQSGVPS RFSGSGSGTD FTLTISSLQP EDVATYYCLQ YDDFPWTFGQ GTKLEIK
Аминокислотная последовательность вариабельного домена тяжелой цепи такого примерного анти-5T4 антитела показана ниже (остатки CDR выделены подчеркиванием): (SEQ ID NO: 309):
QVQLVQSGAE VKKPGASVKV SCKASGYTFT SFWMHWVRQA PGQGLEWMGR IDPNRGGTEY NEKAKSRVTM TADKSTSTAY MELSSLRSED TAVYYCAGGN PYYPMDYWGQ GTTVTVSS
Аминокислотная последовательность вариабельного домена легкой цепи второго примерного анти-5T4 антитела ("5T4 mAb 2") приведена ниже (остатки CDR выделены подчеркиванием): (SEQ ID NO: 310):
DVLMTQTPLS LPVSLGDQAS ISCRSSQSIV YSNGNTYLEW YLQKPGQSPK LLIYKVSNRF SGVPDRFSGS GSGTDFTLKI SRVEAEDLGV YYCFQGSHVP FTFGSGTKLE IK
Аминокислотная последовательность вариабельного домена тяжелой цепи такого второго примерного антитела анти-5T4 показана ниже (остатки CDR выделены подчеркиванием) (SEQ ID NO: 311):
QVQLQQPGAE LVKPGASVKM SCKASGYTFT SYWITWVKQR PGQGLEWIGD IYPGSGRANY NEKFKSKATL TVDTSSSTAY MQLSSLTSED SAVYNCARYG PLFTTVVDPN SYAMDYWGQG TSVTVSS
12. IL13Rб2-связывающие домены
Рецептор α2 Интерлейкина-13 (IL13Rα2) избыточно экспрессируется в различных злокачественных опухолях, включая глиобластому, колоректальный рак, рак шейки матки, рак поджелудочной железы, множественную меланому, остеосаркому, лейкоз, лимфому, рак предстательной железыи рак легких (публ. РСТ № WO 2008/146911;
Brown, C.E. et al. (2013) “Glioma IL13Rα2 Is Associated With Mesenchymal Signature Gene Expression And Poor Patient Prognosis,” PLoS One. 18;8(10):e77769; Barderas, R. et al. (2012) “High Expression Of IL-13 Receptor Α2 In Colorectal Cancer Is Associated With Invasion, Liver Metastasis, And Poor Prognosis,” Cancer Res. 72(11):2780-2790; Kasaian, M.T. et al. (2011) “IL-13 Antibodies Influence IL-13 Clearance In Humans By Modulating Scavenger Activity Of IL-13Rα2,” J. Immunol. 187 (1): 561-569; Bozinov, О. et al. (2010) “Decreasing Expression Of The Interleukin-13 Receptor IL-13Ralpha2 In Treated Recurrent Malignant Gliomas,” Neurol. Med. Chir. (Tokyo) 50(8):617-621; Fujisawa, T. et al. (2009) “A novel role of interleukin-13 receptor alpha2 in pancreatic cancer invasion and metastasis,” Cancer Res. 69(22):8678-8685). Антитела, которые иммуноспецифично связываются с IL13Rα2, являются коммерчески доступными (Abnova Corporation, Biorbyt, LifeSpan Biosciences, United States Biologicals; см также публ. РСТ WO 2008/146911). Аминокислотная последовательность вариабельного домена легкой цепи примерного антитела против IL13Rα2 ("hu08", публ. РСТ WO 2014/072888), показана ниже (остатки CDR выделены подчеркиванием): (SEQ ID NO: 308):
DIQMTQSPSS LSASVGDRVT ITCKASQDVG TAVAWYQQKP GKAPKLLIYS ASYRSTGVPS RFSGSGSGTD FTLTISSLQP EDFATYYCQH HYSAPWTFGG GTKVEIK
Аминокислотная последовательность вариабельного домена тяжелой цепи такого примерного антитела против IL13Rα2 ("hu08", публ. РСТ WO 2014/072888), показана ниже (остатки CDR выделены подчеркиванием): (SEQ ID NO: 309):
EVQLVESGGG LVQPGGSLRL SCAASGFTFS RNGMSWVRQA PGKGLEWVAT VSSGGSYIYY ADSVKGRFTI SRDNAKNSLY LQMNSLRAED TAVYYCARQG TTALATRFFD VWGQGTLVTV SS
13. Домены связывающие интегрин бета 6
Интегрин β6 (ITGB6) является подтипом интегринов, который экспрессируется исключительно на поверхности эпителиальных клеток, и является рецептором для белков внеклеточного матрикса (ECM). ITGB6 специфически экспрессируется в опухолевых тканях (таких, как рак толстой кишки, предстательной железы, почки), но его, как правило, невозможно обнаружить в здоровой эпителиальной ткани (Liang, B. et al. (2014) “Integrinβ6-targeted Immunoliposomes Mediate Tumor Specific Drug Delivery and Enhance Therapeutic Efficacy in Colon Carcinoma,” Clin. Cancer Res. Dec 30. pii: clincanres.1194.2014). Моноклональные антитела, которые иммуноспецифично связываются с ITGB6, являются коммерчески доступными (например, MAB2075Z клон R6G9, EMD Millipore; см также, Weinacker, А. et al.(1994) “Role Of The Integrin Alpha V Beta 6 In Cell Attachment To Fibronectin. Heterologous Expression Of Intact And Secreted Forms Of The Receptor,” J. Biol. Chem. 269:6940-6948). Анти-ITGB6 моноклональные антитела 3G9 и 8G6, и их варианты описаны в публ. РСТ № В WO 03/100033 и WO 2007/008712.
Аминокислотная последовательность вариабельного домена легкой цепи примерного гуманизированного анти-ITGB6 антитела (полученного из антитела 3G9, в публ. РСТ WO 2007/008712), показана ниже (остатки CDR выделены подчеркиванием): (SEQ ID NO: 312):
EIVLTQSPAT LSLSPGERAT LSCSASSSVS SSYLYWYQQK PGQAPRLLIY STSNLASGIP ARFSGSGSGT GFTLTISSLE PEDFAVYYCH QWSTYPPTFG GGTKVEIK
Аминокислотная последовательность вариабельного домена тяжелой цепи такого примерного гуманизированного анти-ITGB6 антитела (полученного из антитела 3G9, в публ. РСТ WO 2007/008712), показана ниже (остатки CDR выделены подчеркиванием): (SEQ ID NO: 313):
EVQLVESGGG LVQPGGSLRL SCAASGFTFS RYWMSWVRQA PGKGLEWVAS ISSGGRMYYP FTVKGRFTIS RDNAKNSLYL QMNSLRAEDT AVYYCARGSI YDGYYVFPYW GQGTLVTVSS
Аминокислотная последовательность вариабельного домена легкой цепи примерного анти-ITGB6 антитела (полученного из антитела 8G6, в публ. РСТ WO 2007/008712), показана ниже (остатки CDR выделены подчеркиванием): (SEQ ID NO: 314):
EIVLTQSPAT LSLSPGERAT LSCRASQSVS TSSYSYMYWY QQKPGQAPRL LIYYASNLES GIPARFSGSG SGTDFTLTIS SLEPEDFAVY YCQHNWEIPF TFGGGTKVEI K
Аминокислотная последовательность вариабельного домена тяжелой цепи такого примерного анти-ITGB6 антитела (полученного из антитела 8G6, в публ. РСТ WO 2007/008712), показана ниже (остатки CDR выделены подчеркиванием): (SEQ ID NO: 315):
QVQLVQSGAEVKKPGASVKVSCKASGYTFTDYAMHWVRQAPGQGLEWMGVISTYYGNTNYNQKFKGRVTMTRDTSISTAYMELSRLRSDDTAVYYCARGGLRRGDRPSLQYAMDYWGQGTLVTVSS
14. Дополнительные домены, связывающие антигены злокачественных опухолей
Дополнительные антитела против антигенов злокачественных опухолей, которые могут быть использованы в соответствии с настоящим изобретением, включают следующие коммерчески доступные антитела: брентуксимаб (например, Adcetris®), которыое связывается с CD30; гемтузумаб (например, Mylotarg®, Wyeth), которое связывается с CD33; и ипилимумаб (например, Yervoy®), которое связывается с CTLA-4.
C. Примерные домены связывания с эффекторной клеткой
Антитела, которые способны связываться с эффекторными клетками иммунной системы, могут быть использованы для обеспечения доменов связывания с эффекторными клетками триспецифических связывающих молекул по настоящему изобретению. Особенно подходящими являются антитела, которые связываются с CD2, CD3, CD16, CD19, CD20, CD22, CD64, CD32B В-клеточным рецептором (BCR), T-клеточным рецептором (TCR) и рецептором NKG2D.
1. CD2-связывающие домены
CD2 представляет собой молекулу клеточной адгезии, которая находится на поверхности Т-клеток и естественных клеток-киллеров (NK). CD2 усиливает клеточную цитотоксичность NK, возможно, в качестве промотора формирования нанотрубок NK-клеток (Mace, Е. М. et al.(2014) “Cell Biological Steps And Checkpoints In Accessing NK Cell Cytotoxicity,” Immunol. Cell. Biol. 92 (3): 245-255; Комерси, CJ et al. (2012) “CD2 Promotes Human Natural Killer Cell Membrane Nanotube Formation,” PLoS One 7(10):e47664:1-12). Аминокислотная последовательность VL-домена анти-CD2-антитела (Lo-CD2a; АТСС №: 11423) представляет собой (SEQ ID NO: 102) (остатки CDR выделены подчеркиванием):
DVVLTQTPPT LLATIGQSVS ISCRSSQSLL HSSGNTYLNW LLQRTGQSPQ PLIYLVSKLE SGVPNRFSGS GSGTDFTLKI SGVEAEDLGV YYCMQFTHYP YTFGAGTKLE LK
Аминокислотная последовательность VH-домена анти-CD2-антитело (Lo-CD2a; АТСС №: 11423) представляет собой (SEQ ID NO: 103) (остатки CDR выделены подчеркиванием):
EVQLQQSGPE LQRPGASVKL SCKASGYIFT EYYMYWVKQR PKQGLELVGR IDPEDGSIDY VEKFKKKATL TADTSSNTAY MQLSSLTSED TATYFCARGK FNYRFAYWGQ GTLVTVSS
2. CD3-связывающие домены
В предпочтительном воплощении второй эпитоп, который связывается триспецифичными связывающими молекулами по настоящему изобретению будет эпитопом CD3. CD3 представляет собой Т-клеточный корецептор, который состоит из четырех отдельных цепей (Wucherpfennig, K.W. et al. (2010) “Structural Biology Of The T-Cell Receptor: Insights Into Receptor Assembly, Ligand Recognition, And Initiation Of Signaling,” Cold Spring Harb. Perspect. Biol. 2(4):a005140; pages 1-14). У млекопитающих комплекс содержит цепь CD3γ, цепь CD3δ и две цепи CD3ε. Эти цепи связываются с молекулой, известной как Т-клеточный рецептор (TCR), для создания сигнала активации в Т-лимфоцитах. В отсутствие CD3, TCR не собираться должным образом и деградирует (Thomas, S. et al. (2010) “Molecular Immunology Lessons From Therapeutic T-Cell Receptor Gene Transfer,” Immunology 129(2):170–177). CD3 обнаруживается связанным с мембранами всех зрелых Т-клеток, и практически ни с одним другим типом клеток (см. Janeway, C.A. et al. (2005) In: Immunobiology: The Immune System In Health And Disease,” 6th ed. Garland Science Publishing, NY, pp. 214-216; Sun, Z. J. et al. (2001) “Mechanisms Contributing To T Cell Receptor Signaling And Assembly Revealed By The Solution Structure Of An Ectodomain Fragment Of The CD3ε:γ Heterodimer,” Cell 105(7):913-923; Kuhns, M.S. et al. (2006) “Deconstructing The Form And Function Of The TCR/CD3 Complex,” Immunity. 2006 Feb;24(2):133-139).
Как будет описано ниже, для того, чтобы проиллюстрировать данное изобретение, получали биспецифичные античеловеческий CD3 х античеловеческий DR5-связывающие молекулы. Антитело против CD3 человека, используемое для таких конструкций обозначается в данном документе как "CD3 mAb 2". Аминокислотная последовательность VL-домена CD3 mAb 2 (SEQ ID NO: 104), показана ниже (остатки CDR выделены подчеркиванием):
QAVVTQEPSL TVSPGGTVTL TCRSSTGAVT TSNYANWVQQ KPGQAPRGLI GGTNKRAPWT PARFSGSLLG GKAALTITGA QAEDEADYYC ALWYSNLWVF GGGTKLTVLG
CDRL1 из CD3 mAb 2 (SEQ ID NO: 105): RSSTGAVTTSNYAN
CDRL2 из CD3 mAb 2 (SEQ ID NO: 106): GTNKRAP
CDRL3 из CD3 mAb 2 (SEQ ID NO: 107): ALWYSNLWV
Аминокислотная последовательность VH-домена CD3 mAb 2 (SEQ ID NO: 108), показана ниже (остатки CDR выделены подчеркиванием):
EVQLVESGGG LVQPGGSLRL SCAASGFTFS TYAMNWVRQA PGKGLEWVGR IRSKYNNYAT YYADSVKDRF TISRDDSKNS LYLQMNSLKT EDTAVYYCVR HGNFGNSYVS WFAYWGQGTL VTVSS
CDRН1 из CD3 mAb 2 (SEQ ID NO: 109): TYAMN
CDRН2 из CD3 mAb 2 (SEQ ID NO: 110): RIRSKYNNYATYYADSVK D
CDRН3 из CD3 mAb 2 (SEQ ID NO: 111): HGNFGNSYVSWFAY
В некоторых CD3-конструкциях был использован вариант VH-домена для CD3 mAb 2. Вариант VH-домена несет замену D65G, и таким образом, имеет аминокислотную последовательность, показанную ниже (SEQ ID NO: 112) (остатки CDR выделены подчеркиванием):
EVQLVESGGG LVQPGGSLRL SCAASGFTFS TYAMNWVRQA PGKGLEWVGR IRSKYNNYAT YYADSVKGRF TISRDDSKNS LYLQMNSLKT EDTAVYYCVR HGNFGNSYVS WFAYWGQGTL VTVSS
Замещение приводит к тому, что CDRH2 имеет аминокислотную последовательность (SEQ ID NO:113) RIRSKYNNYATYYADSVKG. Замещенная позиция (D65G) показана двойным подчеркиванием.
Второе анти-CD3 антитело, используемое в данном описании, представляет собой антитело муромонаб-CD3 "ОКТ3" (Xu et al. (2000) “In Vitro Characterization Of Five Humanized OKT3 Effector Function Variant Antibodies,” Cell. Immunol. 200:16-26); Norman, D.J. (1995) “Mechanisms Of Action And Overview Of OKT3,” Ther. Drug Monit. 17 (6): 615-620; Canafax, DM et al. (1987) “Monoclonal Antilymphocyte Antibody (OKT3) Treatment Of Acute Renal Allograft Rejection,” Pharmacotherapy 7(4):121-124; Swinnen, L.J. et al. (1993) “OKT3 Monoclonal Antibodies Induce Interleukin-6 And Interleukin-10: A Possible Cause Of Lymphoproliferative Disorders Associated With Transplantation,” Curr. Opin. Nephrol. Hypertens. 2 (4): 670-678). Аминокислотная последовательность VL-домена ОКТ3 (SEQ ID NO: 114), показана ниже (остатки CDR выделены подчеркиванием):
QIVLTQSPAI MSASPGEKVT MTCSASSSVS YMNWYQQKSG TSPKRWIYDT SKLASGVPAH FRGSGSGTSY SLTISGMEAE DAATYYCQQW SSNPFTFGSG TKLEINR
Аминокислотная последовательность VH-домена ОКТ3 (SEQ ID NO: 115) показана ниже (остатки CDR выделены подчеркиванием):
QVQLQQSGAE LARPGASVKM SCKASGYTFT RYTMHWVKQR PGQGLEWIGY INPSRGYTNY NQKFKDKATL TTDKSSSTAY MQLSSLTSED SAVYYCARYY DDHYCLDYWG QGTTLTVSS
3. CD16-связывающие домены
CD16 представляет собой рецептор FcγRIIIA. CD16 экспрессируется нейтрофилами, эозинофилами, естественных киллерами (NK) и тканевыми макрофагами, которые связывают агрегированный но не мономерный человеческий IgG (Peltz, GA et al. (1989) “Human Fc Gamma RIII: Cloning, Expression, And Identification Of The Chromosomal Locus Of Two Fc Receptors For IgG,” Proc. Natl. Acad. Sci. (U.S.A.) 86 (3): 1013-1017; Bachanova, В. et al. (2014) “NK Cells In Therapy Of Cancer,” Crit. Rev. Oncog. 19(1-2):133-141; Miller, J.S. (2013) “Therapeutic Applications: Natural Killer Cells In The Clinic,” Hematology Am. Soc. Hematol. Educ. Program. 2013: 247-253; Youinou, P. et al. (2002) “Pathogenic Effects Of Anti-Fc Gamma Receptor IIIB (CD16) On Polymorphonuclear Neutrophils In Non-Organ-Specific Autoimmune Diseases,” Autoimmun Rev. 1(1-2):13-19; Peipp, M. et al. (2002) “Bi-specific Antibodies Targeting Cancer Cells,” Biochem. Soc. Trans. 30(4):507-511).
Аминокислотная последовательность вариабельного домена легкой цепи анти-CD16 антитела 3G8 представляет собой (SEQ ID NO: 116) (остатки CDR выделены подчеркиванием):
DTVLTQSPAS LAVSLGQRAT ISCKASQSVD FDGDSFMNWY QQKPGQPPKL LIYTTSNLES GIPARFSASG SGTDFTLNIH PVEEEDTATY YCQQSNEDPY TFGGGTKLEI K
Аминокислотная последовательность вариабельного домена тяжелой цепи анти-CD16 антитело 3G8 представляет собой (SEQ ID NO: 117) (остатки CDR выделены подчеркиванием):
QVTLKESGPG ILQPSQTLSL TCSFSGFSLR TSGMGVGWIR QPSGKGLEWL AHIWWDDDKR YNPALKSRLT ISKDTSSNQV FLKIASVDTA DTATYYCAQI NPAWFAYWGQ GTLVTVSA
Аминокислотная последовательность вариабельного домена легкой цепи анти-CD16 антитела А9 представляет собой (SEQ ID NO: 118) (остатки CDR выделены подчеркиванием):
DIQAVVTQES ALTTSPGETV TLTCRSNTGT VTTSNYANWV QEKPDHLFTG LIGHTNNRAP GVPARFSGSL IGDKAALTIT GAQTEDEAIY FCALWYNNHW VFGGGTKLTVL
Аминокислотная последовательность вариабельного домена тяжелой цепи анти-CD16 антитела А9 представляет собой (SEQ ID NO: 119) (остатки CDR выделены подчеркиванием):
QVQLQQSGAE LVRPGTSVKI SCKASGYTFT NYWLGWVKQR PGHGLEWIGD IYPGGGYTNY NEKFKGKATV TADTSSRTAY VQVRSLTSED SAVYFCARSA SWYFDVWGAR TTVTVSS
4. CD19-связывающие домены
CD19-антиген представляет собой трансмембранный гликопротеин I типа, принадлежащий к суперсемейству иммуноглобулинов Ig.CD19 экспрессируется на фолликулярных дендритных клетках и В-клетках. Считается, что он является пан-В-клеточным маркером, экспрессируемым в ходе развития В-клеток, но с более высокой экспрессией в зрелых клетках по сравнению с незрелыми В-клетками (Raufi А. et al. (2013) “Targeting CD19 In B-Cell Lymphoma: Emerging Role Of SAR3419,” Cancer Manag. Res. 5:225-233). Многие CD19-антитела были описаны (например, MD1342, Medi-551 и т.д.). (Mei, H.E. et al. (2012) “Rationale Of Anti-CD19 Immunotherapy: An Option To Target Autoreactive Plasma Cells In Autoimmunity,” Arthritis Res. Ther. 14(Suppl 5):S1:1-16). Анти-CD19 связывающая молекула "блинатумомаб" раскрыта в ЕР 2186527.
Аминокислотная последовательность VL-домена предпочтительного анти-CD19 антитела (HD37) представляет собой (SEQ ID NO: 120) (остатки CDR выделены подчеркиванием):
DILITQSPKS MSMSVGERVT LTCKASENVV TYVSWYQQKP EQSPKLLIYG ASNRYTGVPD RFTGSGSATD FTLTISSVQA EDLADYHCGQ GYSYPYTFGG GTKLEIKR
Аминокислотная последовательность VH-домена анти-CD19 антитела HD37 представляет собой (SEQ ID NO: 121) (остатки CDR выделены подчеркиванием):
QVQLQQSGAE LVRPGSSVKI SCKASGYAFS SYWMNWVKQR PGQGLEWIGQ IWPGDGDTNY NGKFKGKATL TADESSSTAY MQLSSLASED SAVYFCARRE TTTVGRYYYA MDYWGQGTSV TVSS
5. CD20-связывающие домены
CD20 представляет собой антиген, специфичный для В-клеточной дифференцировки, который экспрессируется на зрелых В-клетках и в большинстве В-клеточных неходжкинских лимфом, но не на ранних клетках-предшественниках В-клеток или поздних зрелых клетках плазмы (Maloney, DG (2012) “Anti-CD20 Antibody Therapy for B-Cell Lymphomas,” N. Engl. J. Med. 366:2008-2016). Ритуксимаб является примерным анти-CD20-антителом человека. Аминокислотная последовательность VL-домена химерного анти-CD20 антитела (ритуксимаб) представляет собой (SEQ ID NO: 122) (остатки CDR выделены подчеркиванием):
QIVLSQSPAI LSASPGEKVT MTCRASSSVS YIHWFQQKPG SSPKPWIYAT SNLASGVPVR FSGSGSGTSY SLTISRVEAE DAATYYCQQW TSNPPTFGGG TKLEIKR
Аминокислотная последовательность VH-домена анти-CD20-антитела (ритуксимаб) представляет собой (SEQ ID NO: 123) (остатки CDR выделены подчеркиванием):
QVQLQQPGAE LVKPGASVKM SCKASGYTFT SYNMHWVKQT PGRGLEWIGA IYPGNGDTSY NQKFKGKATL TADKSSSTAY MQLSSLTSED SAVYYCARST YYGGDWYFNV WGAGTTVTVS A
Альтернативные анти-CD20-антитела, которые могут быть использованы в соответствии с настоящим изобретением, включают следующие коммерчески доступные антитела: ибритумомаб (например, Zevalin®, Spectrum Pharmaceuticals, Inc.), офатумумаб (например, Arzerra®, SmithKlineGlaxo) и тозитумомаб (например, Bexxar®, GlaxoSmithKline).
6. CD22-связывающие домены
CD22 представляет собой сахар-связывающий трансмембранный белок, обнаруженный на поверхности зрелых В-клеток и в меньшей степени на некоторых незрелых В-клетках (WO 2011/032633; Poe, JC et al.(2012) “CD22 And Siglec-G In B Cell Function And Tolerance,” Trends Immunol. 33(8):413-420; Chen, W.C. et al. (2012) “Targeting B Lymphoma With Nanoparticles Bearing Glycan Ligands Of CD22,” Leuk. Lymphoma 53(2):208-210; Walker, J.A. (2008) “CD22: An Inhibitory Enigma,” Immunology 123(3):314-325; Coleman, M. et al. (2003) “Epratuzumab: Targeting B-Cell Malignancies Through CD22,” Clin. Cancer Res. 9 (10 Pt 2):3991S-3994S).
Аминокислотная последовательность VL-домена анти-CD22 антитела (эпратузумаб) представляет собой (SEQ ID NO: 124) (остатки CDR выделены подчеркиванием):
DIQLTQSPSS LSASVGDRVT MSCKSSQSVL YSANHKNYLA WYQQKPGKAP KLLIYWASTR ESGVPSRFSG SGSGTDFTFT ISSLQPEDIA TYYCHQYLSS WTFGGGTKVQ IKR
Аминокислотная последовательность VH-домена анти-CD22 антитела (эпратузумаб) представляет собой (SEQ ID NO: 125) (остатки CDR выделены подчеркиванием):
QVQLVQSGAE VKKPGSSVKV SCKASGYTFT SYWLHWVRQA PGQGLEWIGY INPRNDYTEY NQNFKDKATI TADESTNTAY MELSSLRSED TAFYFCARRD ITTFYWGQGT TVTVSS
7. CD32B-связывающие домены
Предпочтительная последовательность для VL-домена-антитела, которое связывается с человеческим CD32B, представляет собой CD32B mAb 1 (SEQ ID NO: 126) (остатки CDR выделены подчеркиванием):
DIQMTQSPSS LLAALGERVS LTCRASQEIS GYLSWLQQKP DGTIKRLIYA ASTLDSGVPK RFSGSESGSD YSLTISSLES EDFADYYCLQ YFSYPLTFGA GTKLELK
Предпочтительная последовательность для VH-домена из антитела CD32B mAb 1, которое связывается с человеческим CD32B, представляет собой (SEQ ID NO: 127) (остатки CDR выделены подчеркиванием):
EVKLEESGGG LVQPGGSMKL SCEASGFTFS DAWMDWVRQS PEKGLEWVAE IRNKAKNHAT YYAESVIGRF TISRDDSKSS VYLQMNSLRA EDTGIYYCGA LGLDYWGQGT TLTVSS
8. CD64-связывающие домены
CD64 представляет собой рецептор FcγRI и экспрессируется на моноцитах/макрофагах, дендритных клетках и активированных гранулоцитах. Экспрессия может быть положительно ативирована стимуляцией IFN-гамма. CD64 связывается с иммунным комплексом IgG. CD64 играет роль в захвате антигена, фагоцитозе IgG/антигенных комплексов, и антитело-зависимой клеточной цитотоксичности (WO 2006/002438).
Предпочтительная последовательность для VL-домена антитела, которое связывается с человеческим CD64, представляет собой CD64 mAb 1 (SEQ ID NO: 128) (остатки CDR выделены подчеркиванием):
EIVLTQSPAT LSLSPGERAT LSCRASQSVS SYLAWYQQKP GQAPRLLIYD ASSRATGIPA RFGGSGSGGT DFTLTISSLE PEDFAVYYCQ LRSNWPPYTF GQGTKLEIK
Предпочтительная последовательность для VH-домена антитела, которое связывается с человеческим CD64, представляет собой (SEQ ID NO: 129) (остатки CDR выделены подчеркиванием):
QVQLVESGGG VVQPGRSLRL SCAASGFIFS GYGMHWVRQA PGKGLEWVTV IWYDGSNKYY ADSVKGRFTI SRDNSKNTLY LQMNSLRAED TAVYYCARDT GDRFFDYWGQ GTLVTVSS
9. BCR/CD79-связывающие домены
BCR состоит из мембранного иммуноглобулина, который вместе с нековалентно связанными альфа и бета субъединицами CD79 ("CD79a" и "CD79b" соответственно), образует комплекс BCR. CD79a и CD79b представляют собой субъединицы передачи сигнала, которые содержат консервативный активационный тирозинсодержащий мотив иммунорецепторов ("ITAM"), необходимый для передачи сигнала (Dylke, J. et al. (2007) “Role Of The Extracellular And Transmembrane Domain Of Ig-Alpha/Beta In Assembly Of The B Cell Antigen Receptor (BCR),” Immunol. Lett. 112 (1): 47-57; Cambier, JC (1995) “New Nomenclature For The Reth Motif (or ARH1/TAM/ARAM/YXXL),” Immunol. Today 16:110). Агрегация комплекса BCR поливалентным антигеном инициирует трансфосфорилирование CD79a и CD79b ITAM и активацию рецептор-ассоциированных киназ (DeFranco, AL (1997) “The Complexity Of Signaling Pathways Activated By The BCR,” Curr. Opin. Immunol. 9:296-308; Kurosaki, T. (1997) “Molecular Mechanisms In B Cell Antigen Receptor Signaling,” Curr. Opin. Immunol. 9:309-318; Kim, K.M. et al. (1993) “Signalling Function Of The B-Cell Antigen Receptors,” Immun. Rev. 132: 125-146). Фосфорилированные ITAM рекрутируют дополнительные эффекторы, такие как PI3K, PLC-гамма и представителей пути Ras/MAPK. Эти сигнальные события ответственны как за пролиферацию В-клеток, так и за повышенную экспрессии маркеров активации (например, MHCII и CD86), которые требуются для первичных В-клеток для их последующего взаимодействия с Т-хелперами ("TH").
Предпочтительная последовательность для VL-домена антитела, которое связывается с человеческим В-клеточным рецептором (CD79) представляет собой CD79 mAb 1 (SEQ ID NO: 130) (остатки CDR выделены подчеркиванием):
DVVMTQTPLT LSVNIGQPAS ISCKSSQSLL DTDGKTYLNW LLQRPQGSPN RLIYLVSKLD SGVPDRFTGS GSGTDFTLKI SRVEAEDLGI YYCWQGTHFP LTFGAGTKLE LK
Предпочтительная последовательность для VH-домена CD79 mAb 1 антитела, которое связывается с человеческим B-клеточным рецептором (CD79) представляет собой (SEQ ID NO: 131) (остатки CDR выделены подчеркиванием):
QVQLQQPGAE LVRPGASVKL SCKASGYTFT SYWMNWVKQR PGQGLEWIGM VDPSDSETHY NQMFKDKATL TVDKSSSTAY MQLSSLTSED SAVYYCARAM GYWGQGTSVT VSS
10. Домены, связывающиеся с Т-клеточным рецептором
В альтернативном воплощении второй эпитоп, который связывается триспецифичными связывающими молекулами по настоящему изобретению будет представлять собой эпитоп Т-клеточного рецептора (TCR). Т-клеточный рецептор изначально экспрессируется CD4+ или CD8+ Т-клетками, и позволяет таким клеткам распознавать антигенные пептиды, которые связаны и презентированы белками MHC класса I или класса II антигенпредставляющих клеток. Распознавание комплекса pMHC (пептид-МНС) с помощью TCR инициирует распространение клеточного иммунного ответа, что приводит к продукции цитокинов и лизису антигенпредставляющей клетки (см., например, Armstrong, KМ et al.(2008) “Conformational Changes And Flexibility In T-Cell Receptor Recognition Of Peptide–MHC Complexes,” Biochem. J. 415 (Pt 2): 183-196; Willemsen, Р. (2008) “Selection Of Human Antibody Fragments Directed Against Tumor T-Cell Epitopes For Adoptive T-Cell Therapy,” Cytometry A. 73(11):1093-1099; Beier, K.C. et al. (2007) “Master Switches Of T-Cell Activation And Differentiation,” Eur. Respir. J. 29: 804-812; Mallone, R. et al. (2005) “Targeting T Lymphocytes For Immune Monitoring And Intervention In Autoimmune Diabetes,” Am. J. Ther. 12 (6): 534-550). CD3 представляет собой рецептор, который связывается с TCR (Thomas S. et al. (2010) “Molecular Immunology Lessons From Therapeutic T-Cell Receptor Gene Transfer,” Immunology 129(2):170-177; Guy, C.S. et al. (2009) “Organization Of Proximal Signal Initiation At The TCR:CD3 Complex,” Immunol. Rev. 232(1):7-21; St. Clair, E.W. (Epub 2009 Oct 12) “Novel Targeted Therapies For Autoimmunity,” Curr. Opin. Immunol. 21(6):648-657; Baeuerle, P.A. et al. (Epub 2009 Jun 9) “Bi-specific T-Cell Engaging Antibodies For Cancer Therapy,” Cancer Res. 69(12):4941-4944; Smith-Garvin, J.E. et al. (2009) “T Cell Activation,” Annu. Rev. Immunol. 27:591-619; Renders, L. et al. (2003) “Engineered CD3 Antibodies For Immunosuppression,” Clin. Exp. Immunol. 133(3):307-309).
Антитела, которые специфически связываются с Т-клеточным рецептором, включают анти-TCR-антитела BMA 031 (EP 0403156; Kurrle R. et al. (1989) “BMA 031 – A TCR-Specific Monoclonal Antibody For Clinical Application,” Transplant Proc. 21(1 Pt 1):1017-1019; Nashan, B. et al. (1987) “Fine Specificity Of A Panel Of Antibodies Against The TCR/CD3 Complex,” Transplant Proc. 19(5):4270-4272; Shearman, C.W. et al. (1991) “Construction, Expression, And Biologic Activity Of Murine/Human Chimeric Antibodies With Specificity For The Human α/β T Cell,” J. Immunol. 146(3):928-935; Shearman, C.W. et al. (1991) “Construction, Expression And Characterization of Humanized Antibodies Directed Against The Human α/β T Cell Receptor,” J. Immunol. 147(12):4366-4373).
Аминокислотная последовательность VL-домена анти-TCR антитела BMA 031 (SEQ ID NO: 132) (остатки CDR выделены подчеркиванием):
EIVLTQSPAT LSLSPGERAT LSCSATSSVS YMHWYQQKPG KAPKRWIYDT SKLASGVPSR FSGSGSGTEF TLTISSLQPE DFATYYCQQW SSNPLTFGQG TKLEIK
Аминокислотная последовательность VH-домена анти-TCR антитело BMA 031 (SEQ ID NO: 133) (остатки CDR выделены подчеркиванием):
QVQLVQSGAE VKKPGASVKV SCKASGYKFT SYVMHWVRQA PGQGLEWIGY INPYNDVTKY NEKFKGRVTI TADKSTSTAY LQMNSLRSED TAVHYCARGS YYDYDGFVYW GQGTLVTVSS
11. Домены связывающие рецептор NKG2D
В альтернативном воплощении второй эпитоп, который связывается триспецифичными связывающими молекулами по настоящему изобретению будет представлять собой эпитоп рецептора NKG2D. Рецептор NKG2D экспрессируется на всех человеческих (и других видов млекопитающих) естественных киллерах (Bauer, S. et al.(1999) “Activation Of NK Cells And T Cells By NKG2D, A Receptor For Stress-Inducible MICA,” Science 285(5428):727-729; Jamieson, A.M. et al. (2002) “The Role Of The NKG2D Immunoreceptor In Immune Cell Activation And Natural Killing,” Immunity 17(1):19-29), а также на всех CD8+ T-клетках (Groh, V. et al. (2001) “Costimulation Of CD8αβ T Cells By NKG2D Via Engagement By MIC Induced On Virus-Infected Cells,” Nat. Immunol. 2 (3): 255-260; Джемисон, А. М. et al. (2002) “The Role Of The NKG2D Immunoreceptor In Immune Cell Activation And Natural Killing,” Immunity 17(1):19-29). Такие связующие лиганды, и особенно те, которые не экспрессируются на нормальных клетках, включают молекулу гистосовместимости 60 (H60), продукт раннего индуцируемого ретиноевой кислотой гена-1 (RAE-1), и мышиный UL16-связывающий белковоподобный транскрипт 1 (mult1) (Raulet DH (2003) “Roles Of The NKG2D Immunoreceptor And Its Ligands,” Nature Rev. Immunol. 3: 781-790; Coudert, JD et al. (2005) “Altered NKG2D Function In NK Cells Induced By Chronic Exposure To Altered NKG2D Ligand-Expressing Tumor Cells,” Blood 106:1711-1717). Антитела, которые специфически связываются с рецептором NKG2D, включают KYK-2.0(Kwong, KY et al. (2008) “Generation, Affinity Maturation, And Characterization Of A Human Anti-Human NKG2D Monoclonal Antibody With Dual Antagonistic And Agonistic Activity” J. Mol. Biol. 384:1143-1156; и PCT/US09/54911).
Аминокислотная последовательность VL-домена анти-NKG2D антитела KYK-1.0 представляет собой (SEQ ID NO: 134) (остатки CDR выделены подчеркиванием):
QPVLTQPSSV SVAPGETARI PCGGDDIETK SVHWYQQKPG QAPVLVIYDD DDRPSGIPER FFGSNSGNTA TLSISRVEAG DEADYYCQVW DDNNDEWVFG GGTQLTVL
Аминокислотная последовательность VH-домена анти-NKG2D антитела KYK-1.0 представляет собой (SEQ ID NO: 135) (остатки CDR выделены подчеркиванием):
EVQLVESGGG VVQPGGSLRL SCAASGFTFS SYGMHWVRQA PGKGLEWVAF IRYDGSNKYY ADSVKGRFTI SRDNSKNTKY LQMNSLRAED TAVYYCAKDR FGYYLDYWGQ GTLVTVSS
Аминокислотная последовательность VL-домена анти-NKG2D антитела KYK-2.0 представляет собой (SEQ ID NO: 136) (остатки CDR выделены подчеркиванием):
QSALTQPASV SGSPGQSITI SCSGSSSNIG NNAVNWYQQL PGKAPKLLIY YDDLLPSGVS DRFSGSKSGT SAFLAISGLQ SEDEADYYCA AWDDSLNGPV FGGGTKLTVL
Аминокислотная последовательность VH-домена анти-NKG2D антитела KYK-2.0 представляет собой (SEQ ID NO: 137) (остатки CDR выделены подчеркиванием):
QVQLVESGGG LVKPGGSLRL SCAASGFTFS SYGMHWVRQA PGKGLEWVAF IRYDGSNKYY ADSVKGRFTI SRDNSKNTLY LQMNSLRAED TAVYYCAKDR GLGDGTYFDY WGQGTTVTVS S
D. Предпочтительные триспецифичные связывающие молекулы по настоящему изобретению
1. Предпочтительный Fc-домены
CH2 и CH3 домены двух тяжелых цепей взаимодействуют с образованием Fc-домена, который представляет собой домен, который распознается клеточными Fc-рецепторами (FcγRs). При использовании в данном описании термин "Fc-домен" используется для определения C-концевой области тяжелой цепи IgG. Аминокислотная последовательность СН2-СН3 домена примерного IgG1 человека представляет собой (SEQ ID NO: 1):
│CH2 ◊
APELLGGPS VFLFPPKPKD TLMISRTPEV TCVVVDVSHE DPEVKFNWYV
231 240 250 260 270
DGVEVHNAKT
280
⇓CH2│CH3◊
KPREEQYNST YRVVSVLTVL HQDWLNGKEY KCKVSNKALP APIEKTISKA
290 300 310 320 330
KGQPREPQVY
340
TLPPSREEMT KNQVSLTCLV KGFYPSDIAV EWESNGQPEN NYKTTPPVLD
350 360 370 380 390
SDGSFFLYSK
400
⇓CH3│
LTVDKSRWQQ GNVFSCSVMH EALHNHYTQK SLSLSPGK
410 420 430 440
В настоящем описании нумерация остатков в тяжелой цепи IgG является нумерацией индекса EU по Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, NH1, MD (1991), специально включенного в настоящее описание ссылкой. "Индекс EU по Kabat" относится к нумерации человеческого антитела IgG1 EU. Аминокислоты из вариабельных областей зрелых тяжелых и легких цепей иммуноглобулинов обозначены положением аминокислоты в цепи. В Kabat описаны многочисленные аминокислотные антительные последовательности, идентифицирована консенсусная аминокислотная последовательность для каждой подгруппы, и присвоен номер остатка до каждой аминокислоты. Схема нумерации Kabat расширяется на антитела, не включенные в этот компендиум, путем выравнивания первичной структуры исследуемого антитела с одной из консенсусных последовательностей списка Kabat путем ссылки на консервативные аминокислоты. Этот способ назначения номеров остаткам стал стандартом в этой области, и легко идентифицирует аминокислоты в эквивалентных положениях различных антител, в том числе химерных или гуманизированных вариантах. Например, аминокислота в положении 50 легкой цепи антитела человека занимает эквивалентное положение по отношению к аминокислоте в положении 50 легкой цепи антитела мыши.
Хотя границы могут слегка отличаться, домен СН2 Fc-домена человеческого IgG обычно простирается от аминокислоты 231 до аминокислоты 341 человеческого IgG в соответствии с системой нумерации Kabat. Домен СН3 человеческого IgG обычно простирается от аминокислоты 342 до 447 в соответствии с системой нумерации Kabat. "Шарнирный участок" или "шарнирный домен", как правило, определяется как простирающийся от Glu216 до Pro230 IgG1 человека.
Полиморфизм наблюдали при нескольких различных положениях в пределах константных участков антитела (например, Fc-положения, в том числе, без ограничения перечисленным, положения 270, 272, 312, 315, 356, и 358, согласно нумерации индекса EU, как это указано в Kabat), и, таким образом, существуют небольшие различия между представленной последовательностью и последовательностями в известном уровне техники. Полиморфные формы иммуноглобулинов человека были хорошо охарактеризованы. В настоящее время, известны 18 Gm-аллотипов: G1m (1, 2, 3, 17) или G1m (a, x, f, z), G2m (23) или G2m (n), G3m (5, 6, 10, 11, 13, 14, 15, 16, 21, 24, 26, 27, 28) или G3m (b1, c3, b3, b0, b3, b4, s, t, g1, c5, u, v, g5) (Lefranc, et al., The human IgG subclasses: molecular analysis of structure, function and regulation. Pergamon, Oxford, pp. 43-78 (1990); Lefranc, G. et al., 1979, Hum. Genet.: 50, 199-211). Специально предусмотрено, что антитела, предлагаемые в настоящем изобретении, могут быть включены в любой аллотип, изоаллотип или гаплотип любого гена иммуноглобулина, и не ограничиваются аллотипом, изоаллотипом или гаплотипом последовательностей, представленных в данном документе.
Активирующие и ингибирующие сигналы трансдуцируются через Fc-рецепторы (FcγRs) после их лигирования с Fc-доменом. Эти диаметрально противоположные функции являются результатом структурных различий между различными изоформами рецепторов. Два различных домена в пределах цитоплазматических сигнальных доменов рецептора, названные активационные тирозинсодержащие мотивы иммунорецепторов (ITAM) или ингибирующие тирозинсодержащие мотивы иммунорецепторов (ITIMS) предназначены для различных ответов. Рекрутирование различных цитоплазматических ферментов к этим структурам диктует исход FcγR-опосредованных клеточных реакций. ITAM-содержащие FcγR-комплексы включают FcγRI, FcγRIIA, FcγRIIIA, в то время как ITIM-содержащие комплексы включают только FcγRIIB. Человеческие нейтрофилы экспрессируют ген FcγRIIA. Кластеризация FcγRIIA с помощью иммунных комплексов или перекрестного связывания специфичными антителами служит для агрегации ITAM вместе с рецептор-ассоциированными киназами, которые облегчают фосфорилирование ITAM. Фосфорилирование ITAM служит стыковочным участком для Syk-киназы, активация которой приводит к активации нисходящих субстратов (например, PI3K). Активация клеток приводит к высвобождению провоспалительных медиаторов. Ген FcγRIIB экспрессируется в В-лимфоцитах; его внеклеточный домен на 96% идентичен FcγRIIA и связывает IgG комплексы неотличимым образом. Присутствие ITIM в цитоплазматическом домене FcγRIIB определяет этот ингибирующую подкласс FcγR. Недавно была раскрыта молекулярная основа этого ингибирования. При совместном лигировали вместе с активирующим FcγR, ITIM в FcγRIIB фосфорилируется и привлекает SH2-домен инозитполифосфат 5'-фосфатазы (SHIP), которая гидролизует мессенджер фосфоинозитол, высвобождаемый в результате активации ITAM-содержащей FcγR- опосредованной тирозинкиназы, вследствие чего предотвращается приток внутриклеточного Са++. Таким образом, перекрестная сшивка FcγRIIB гасит активирующий ответ лигирования FcγR и ингибирует клеточную респонсивность. Таким образом, прерывается активация В-клеток, пролиферация В-леток и секреция антител.
Fc-домен связывающих молекул по настоящему изобретению, может представлять собой либо полный Fc-домен (например, полный Fc-домен IgG) или только фрагмент полного Fc-домена. Несмотря на то, Fc-домен биспецифичных моновалентных Fc-диател по настоящему изобретению, могут обладать способностью связываться с одним или несколькими Fc-рецепторами (например, FcγR), более предпочтительно, если такой Fc-домен будет вызывать измененное связывание с FcγRIA (CD64), FcγRIIA (CD32A), FcγRIIB (CD32B), FcγRIIIA (CD16a) или FcγRIIIB (CD16b) (по отношению к связыванию, демонстрируемому Fc-доменом дикого типа) или существенно исключать возможность такого Fc-домена связываться с ингибиторным рецептором(ами). Таким образом, Fc-домен из Fc-домен-содержащих диател по настоящему изобретению может включать часть и весь СН2-домен и/или часть или весь СН3-домен полного Fc-домена, или может включать последовательность вариантного СН2 и/или вариантного СН3 (которая может включать, например, одну или более вставок и/или одну или более делеций относительно СН2 или СН3-доменов полного Fc-домена). Такие Fc-домены могут содержать части не относящиеся к Fc-полипептиду, или могут содержать части, неестественного полного Fc-домена, или могут содержать неестественные ориентации СН2 и/или СН3-доменов (такие как, например, два CH2-домена или два СН3-домена, или в направлении от N-конца к С-концу, СН3-домен, связанный с CH2-доменом и т.д.).
Модификации Fc-домена, идентифицированные как изменяющие эффекторную функцию, известны в данной области техники, в том числе и модификации, которые увеличивают его связывание с активирующими рецепторами (например, FcγRIIA (CD16A) и уменьшающие связывание с ингибирующими рецепторами (например, FcγRIIB (CD32B) (см., например, Stavenhagen, J.B. et al. (2007) “Fc Optimization Of Therapeutic Antibodies Enhances Their Ability To Kill Tumor Cells In Vitro And Controls Tumor Expansion In Vivo Via Low-Affinity Activating Fcgamma Receptors,” Cancer Res. 57(18):8882-8890).
В частности, предпочтительно, чтобы полипептидные цепи СН2-СН3-доменов Fc-домен-содержащих диател по настоящему изобретению демонстрировали снижение (или, по существу отсутствие) связывания с FcγRIA (CD64), FcγRIIA (CD32A), FcγRIIB (CD32B), FcγRIIIA (CD16a) или FcγRIIIB (CD16b) (по отношению к связыванию, демонстрируемому диким типом Fc-домена (SEQ ID NO: 1). Fc-варианты и мутантные формы, способные опосредовать такие измененные связывания, описаны выше. В предпочтительном воплощении СН2-СН3-домен первой и/или третьей полипептидных цепей таких диател включает любые 1, 2, 3, 4, 5, 6, или 7 из числа замен: L234A, L235A, F243L, R292P, Y300L, V305I и P396L. Примерные варианты Fc-доменов IgG1 человека с ограниченной способностью к связыванию с CD32B и/или увеличенным связыванием с CD16A содержат замены F243L, R292P, Y300L, V305I или P296L. Эти аминокислотные замены могут присутствовать в Fc-домене IgG1 человека в любой комбинации. В одном из воплощений вариант Fc-домена IgG1 человека содержит замены F243L, R292P и Y300L. В другом воплощении изобретения вариант Fc IgG1, домен человека содержит замены F243L, R292P, Y300L, V305I и P296L. В одном воплощении CH2-CH3 домена первой и/или третьей полипептидных цепей таких диател включают любую 1, 2, или 3, из числа замен: L234A, L235A, N297G, N297Q. В другом воплощении изобретения вариант Fc-домена IgG1 человека содержит замену N297Q, замены L234A и L235A или замену D265A, так как эти мутации отменяют связывание с FcR. В ином случае используется СН2-СН3-домен, который от природы демонстрирует уменьшение (или фактическое отсутствие) связывания с FcγRIIIA (CD16a), и/или пониженную эффекторную функцию (по отношению к связыванию, демонстрируемого Fc-доменом IgG1 дикого типа (SEQ ID NO: 1)). В конкретном воплощении изобретения Fc-домен-содержащие диатела по настоящему изобретению включают Fc-домен IgG2 или Fc-домен IgG4. Там, где согласно настоящему изобретению используется Fc-домен IgG4, также включено введение стабилизирующей мутации, такой как S228P, согласно нумерации по индексу EU, как указано в Kabat (Lu et al., (2008) “The Effect Of A Point Mutation On The Stability Of Igg4 As Monitored By Analytical Ultracentrifugation,” J Pharmaceutical Sciences 97:960-969) для того, чтобы уменьшить частоту обмена цепей. Другие стабилизирующие мутации, известные в данной области могут быть введены в Fc-домен IgG4 (Peters, P et al., (2012) “Engineering an Improved IgG4 Molecule with Reduced Disulfide Bond Heterogeneity and Increased Fab Domain Thermal Stability”, J. Biol. Chem., 287:24525-24533; пат. публ. PCT № WO 2008/145142). Так как замены N297A, L234A, L235A и D265A отменяют эффекторную функцию, в обстоятельствах, при которых эффекторная функция является желательной, будет предпочтительно не использовать эти замены.
СН2- и/или СН3-домены таких полипептидных цепей, не обязательно должны быть идентичными в определенной последовательности, и, преимущественно модифицируются для того, чтобы способствовать образованию комплекса между двумя полипептидными цепями. Например, аминокислотная замена (предпочтительно замена аминокислотой, содержащей объемную боковую группу, образующую "выпуклость", например, триптофан) может быть введена в СН2- или СН3-домен таким образом, чтобы стерические помехи предотвращали взаимодействие с доменом со схожей мутацией, и помогала спаривание с доменом, в который была введена комплементарная или вмещающая мутация, т.е. "углубление" (например, замена глицином). Такие наборы мутаций могут быть сконструированы в любых двух полипептидах триспецифичных связывающих молекул. Методы белковой инженерии, способствующие гетеродимеризации относительно гомодимеризации, хорошо известны в данной области техники, в частности, в отношении инженерии иммуноглобулин-подобных молекул, и охватываются в данном описании (см., например, Ridgway et al. (1996) “‘Knobs-Into-Holes’ Engineering Of Antibody CH3 Domains For Heavy Chain Heterodimerization,” Protein Engr. 9:617-621, Atwell et al. (1997) “Stable Heterodimers From Remodeling The Domain Interface Of A Homodimer Using A Phage Display Library,” J. Mol. Biol. 270: 26-35, и Се et al. (2005) “A New Format Of Bi-specific Antibody: Highly Efficient Heterodimerization, Expression And Tumor Cell Lysis,” J. Immunol. Methods 296:95-101; каждый из которых включен в описание данного документа в ссылкой во всей полноте). Предпочтительно "выпуклость" спроектирована в СН2-СН3-доменах первой полипептидной цепи, а "углубление" спроектировано в CH2-CH3 домене другой CH2-CH3-содержащей полипептидной цепи. Таким образом, "выпуклость" поможет помочь в предотвращении гомодимеризации первой полипептидной цепь через ее СН2 и/или СН3-домены. Полипептидная цепь CH2-CH3 с "углублением" будет гетеродимеризоваться с полипептидной цепью CH2-CH3 с "выпуклостью", и гомодимеризоваться с собой. Предпочтительная выпуклость создается путем изменения нативного Fc-домена IgG модификацией T366W. Предпочтительное углубление создается путем модификации нативного Fc-домена IgG модификациями T366S, L368A и Y407V. Для облегчения очистки гомодимера полипептидной цепи с "углублением" из окончательных триспецифичных связывающих молекул, сайт связывания с протеином А из СН2- и СН3-доменов Fc-домена с "углублением" предпочтительно мутируют путем замены аминокислоты в положении 435 (H435R). Таким образом, гомодимер Fc-домена с "углублением" не будет связываться с протеином А, в то время как искомая триспецифическая связывающаяся молекула сохранит свою способность связываться с протеином А с помощью сайта связывания с протеином А на первой полипептидной цепи.
Предпочтительная последовательность для СН2- и СН3-доменов первой полипептидной цепи Fc-домен-содержащих диатела по настоящему изобретению будет иметь последовательность с "выпуклостью" (SEQ ID NO: 52):
APEAAGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVKFNWYVD GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLWCLVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE ALHNHYTQKS LSLSPGK
Предпочтительная последовательность для СН2- и СН3-доменов второй полипептиднуюой цепи Fc-домен-содержащего диатела по настоящему изобретению, имеющего две полипептидные цепи (или третью полипептидную цепь Fc-домен-содержащего диатела, имеющего три полипептидных цепи) будет иметь последовательность с "углублением" (SEQ ID NO: 53):
APEAAGGPSV FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVKFNWYVD GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA PIEKTISKAK GQPREPQVYT LPPSREEMTK NQVSLSCAVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS DGSFFLVSKL TVDKSRWQQG NVFSCSVMHE ALHNRYTQKS LSLSPGK
Как будет отмечено, СН2-СН3-домены из SEQ ID NO: 52 и SEQ ID NO: 53, включают замену в положении 234 на аланин и в 235 на аланин, и, таким образом, образуют Fc-домен демонстрирующий снижение (или по существу отсутствие) связывания с FcγRIA (CD64), FcγRIIA (CD32A), FcγRIIB (CD32B), FcγRIIIA (CD16a) или FcγRIIIB (CD16b) (относительно связывания, демонстрируемого Fc-доменом дикого типа (SEQ ID NO: 1).
Предпочтительно, если первая полипептидная цепь будет иметь "несущую выпуклость" последовательность СН2-СН3, как, например, SEQ ID NO: 52. Однако, как будет понятно, CH2-CH3 домен с "углублением" (например, SEQ ID NO: 53), можно было бы использовать в первой полипептидной цепи, в таком случае, CH2-CH3 домен с "выпуклостью" (например, SEQ ID NO: 52), будет использоваться во второй полипептидной цепи Fc-домен-содержащего диатела по настоящему изобретению, имеющего две полипептидные цепи (или третью полипептидную цепь Fc-домен-содержащего диатела, имеющего три полипептидных цепи).
2. Предпочтительная первая полипептидная цепь
Первая полипептидная цепь предпочтительной связывающей молекулы по настоящему изобретению будет содержать вариабельный домен легкой цепи, способный связываться с эпитопом I (VLI), вариабельный домен тяжелой цепи, способный связываться с эпитопом II (VHII), домен, способствующий гетеродимеризации и СН3-СН2-домен.
Поскольку вариабельный домен легкой цепи и вариабельный домен тяжелой цепи первого полипептида направлены на различные эпитопы, они не могут связать друг с другом, образуя связывающий домен, который способен связывать либо эпитоп I, либо эпитоп II. Вариабельный домен легкой цепи и вариабельный домен тяжелой цепи первого полипептида разнесены друг от друга с помощью промежуточного пептидного линкера, который является достаточно коротким, чтобы по существу предотвратить ассоциацию этих доменов. Примерный линкер, называемый "линкер 1", имеет последовательность (SEQ ID NO: 33): GGGSGGGG.
Вариабельный домен тяжелой цепи первого полипептида и домен, способствующий гетеродимеризации, из этого полипептида предпочтительно разнесены друг от друга с помощью промежуточного линкерного пептида, который содержит 1, 2, 3 или более остатков цистеина. Предпочтительный цистеинсодержащий спейсерный пептид ("линкер 2") имеет последовательность, которая представляет собой SEQ ID NO: 34: GGCGGG.
Линкеры, которые могут быть использованы для связывания СН2-СН3-домена с полипептидной цепью молекул по настоящему изобретению, включают: ASTKG (SEQ ID NO:47), DKTHTCPPCP (SEQ ID NO:48), LEPKSS (SEQ ID NO:49) и APSSSPME (SEQ ID NO:50), APSSS (SEQ ID NO:152) и GGG или GCG. SEQ ID NO:49, могут быть использованы вместо GGG или GCG для облегчения клонирования. Кроме того, за последовательностью SEQ ID NO: 49, может непосредственно следовать последовательность SEQ ID NO: 47 для того, чтобы сформировать альтернативный линкер (LEPKSSDKTHTCPPCP; SEQ ID NO: 51).
Домен, содействующий гетеродимеризации, первого полипептида и домен, содействующий гетеродимеризации, второго полипептида выбираются скоординированно. Домены отличаются друг от друга и предназначены для того, чтобы связываться друг с другом с тем, чтобы содействовать объединению первой и второй полипептидных цепей. Например, один из доменов, содействующих гетеродимеризации, будет разработан так, чтобы он имел отрицательный заряд при рН 7, в то время как другой из второй полипептидной цепи, будут сконструирован так, чтобы иметь положительный заряд при рН = 7. Наличие таких заряженных доменов способствует связи между первым и вторым полипептидами, и, таким образом, способствует гетеродимеризации. Несущественно, который из доменов, содействующих гетеродимеризации, предлагается в каждой из цепей, при условии, что домены, используемые в первой и второй полипептидных цепях, отличаются так, чтобы способствовать гетеродимеризации между этими цепями.
Домены, содействующие гетеродимеризации, могут быть доменами, CL и СН1 IgG или могут представлять собой пептид, имеющий аминокислотную последовательность GVEPKSC (SEQ ID NO: 35) или VEPKSC (SEQ ID NO: 36), полученные из шарнирного домена человеческого IgG, а вместо домена CL, можно применять С-концевые 6 аминокислот человеческой легкой цепи каппа, GFNRGEC (SEQ ID NO: 37) или FNRGEC (SEQ ID NO: 38).
Более предпочтительно, однако, если домены, способствующие гетеродимеризации, таких диател образуются из одного, двух, трех или четырех тандемно повторяющихся доменов спиральных доменов противоположного заряда, которые включают последовательность, по меньшей мере, шести, по меньшей мере, семи, по меньшей мере, восьми заряженных аминокислотных остатков (Apostolovic, B. et al. (2008) “pH-Sensitivity of the E3/K3 Heterodimeric Coiled Coil,” Biomacromolecules 9:3173–3180; Arndt, K.M. et al. (2001) “Helix-stabilized Fv (hsFv) Antibody Fragments: Substituting the Constant Domains of a Fab Fragment for a Heterodimeric Coiled-coil Domain,” J. Molec. Biol. 312:221-228; Arndt, K.M. et al. (2002) “Comparison of In Vivo Selection and Rational Design of Heterodimeric Coiled Coils,” Structure 10:1235-1248; Boucher, C. et al. (2010) “Protein Detection By Western Blot Via Coiled–Coil Interactions,” Analytical Biochemistry 399:138-140; Cachia, P.J. et al. (2004) “Synthetic Peptide Vaccine Development: Measurement Of Polyclonal Antibody Affinity And Cross-Reactivity Using A New Peptide Capture And Release System For Surface Plasmon Resonance Spectroscopy,” J. Mol. Recognit. 17: 540-557; De Crescenzo, GD et al.(2003) “Real-Time Monitoring of the Interactions of Two-Stranded de novo Designed Coiled-Coils: Effect of Chain Length on the Kinetic and Thermodynamic Constants of Binding,” Biochemistry 42:1754-1763; Fernandez-Rodriquez, J. et al. (2012) “Induced Heterodimerization And Purification Of Two Target Proteins By A Synthetic Coiled-Coil Tag,” Protein Science 21:511-519; Ghosh, T.S. et al. (2009) “End-To-End And End-To-Middle Interhelical Interactions: New Classes Of Interacting Helix Pairs In Protein Structures,” Acta Crystallographica D65:1032-1041; Grigoryan, G. et al. (2008) “Structural Specificity In Coiled-Coil Interactions,” Curr. Opin. Struc. Biol. 18: 477-483; Litowski, JR et al.(2002) “Designing Heterodimeric Two-Stranded α-Helical Coiled-Coils: The Effects Of Hydrophobicity And α-Helical Propensity On Protein Folding, Stability, And Specificity,” J. Biol. Chem. 277:37272-37279; Steinkruger, J.D. et al. (2012) “The d′--d--d′ Vertical Triad is Less Discriminating Than the a′--a--a′ Vertical Triad in the Antiparallel Coiled-coil Dimer Motif,” J. Amer. Chem. Soc. 134 (5): 2626-2633; Straussman, R. et al. (2007) “Kinking the Coiled Coil – Negatively Charged Residues at the Coiled-coil Interface,” J. Molec. Biol. 366: 1232-1242; Tripet, В. et al.(2002) “Kinetic Analysis of the Interactions between Troponin C and the C-terminal Troponin I Regulatory Region and Validation of a New Peptide Delivery/Capture System used for Surface Plasmon Resonance,” J. Molec. Biol. 323: 345-362; Woolfson, DN (2005) “The Design Of Coiled-Coil Structures And Assemblies,” Adv. Prot. Chem. 70:79-112; Zeng, Y. et al. (2008) “A Ligand-Pseudoreceptor System Based On de novo Designed Peptides For The Generation Of Adenoviral Vectors With Altered Tropism,” J. Gene Med. 10:355-367).
Такие повторяющиеся спиральные домены могут быть точные повторами или могут иметь замены. Например, домен, способствующий гетеродимеризации, первой полипептидной цепи, может содержать последовательность из восьми отрицательно заряженных аминокислотных остатков, а домен, содействующий гетеродимеризации, второй полипептидной цепи, может содержать последовательность из восьми отрицательно заряженных аминокислотных остатков. Несущественно, какая из спиралей предусмотрена на первой или второй полипептидной цепи, при условии, что спираль с противоположным зарядом используется для другой полипептидной цепи. Положительно заряженными аминокислотами могут быть собой лизин, аргинин, гистидин и т.п. и/или отрицательно заряженными аминокислотами может быть глутаминовая кислота, аспарагиновая кислота и т.п. Положительно заряженная аминокислота, предпочтительно представляет собой лизин и/или отрицательно заряженная аминокислота, предпочтительно представляет собой глутаминовую кислоту. Вполне возможно, что только один домен, содействующий гетеродимеризации, будет использоваться (так как такой домен будет препятствовать гомодимеризации и тем самым способствовать гетеродимеризации), тем не менее, предпочтительно, если и первая и вторая полипептидные цепи диатела настоящего изобретения содержат домены, способствующие гетеродимеризации.
В предпочтительном варианте один из доменов, содействующих гетеродимеризации, будет состоять из четырех тандемных спиральных доменов "E-спираль" (SEQ ID NO:39: EVAALEK-EVAALEK-EVAALEK-EVAALEK), чьи остатки глутамата образуют отрицательный заряд при рН 7, в то время как другой из доменов, содействующих гетеродимеризации, будет включать четыре тандемных домена "К-спираль" (SEQ ID NO:40: KVAALKE-KVAALKE-KVAALKE-KVAALKE), чьи остатки лизина будут образовывать положительный заряд при рН = 7. Присутствие таких заряженных доменов способствует связи между первым и вторым полипептидами, и, таким образом, способствует гетеродимеризации. Особенно предпочтительным является домен, содействующий гетеродимеризации, в котором один из четырех тандемных спиральных доменов "E-спираль" SEQ ID NO: 39 был изменена для включения остатка цистеина: E EVAACEK-EVAALEK-EVAALEK-EVAALEK (SEQ ID NO:41). Подобным же образом, особенно предпочтительным является домен, содействующий гетеродимеризации, в котором один из четырех тандемных спиральных доменом "К-спираль" SEQ ID NO: 40 была модифицирован для включения остатка цистеина: KVAACKE-KVAALKE-KVAALKE-KVAALKE (SEQ ID NO:42).
Как раскрыто в WO 2012/018687, для того, чтобы улучшить фармакокинетические свойства диател in vivo, диатело может быть модифицировано для включения полипептидной части сывороточного связывающего белка на одном или нескольких концах диатела. Наиболее предпочтительно, если такая полипептидная часть сывороточного связывающего белка будет установлена на С-конце диатела. Альбумин является наиболее распространенным белком в плазме и имеет период полужизни 19 дней в организме человека. Альбумин обладает несколькими участками связывания малых молекул, которые позволяют ему нековалентно связываться с другими белками и тем самым увеличивать их периоды полужизни в сыворотке. Альбумин-связывающий домен 3 (ABD3) белка G из штамма Streptococcus G148 состоит из 46 аминокислотных остатков, образующих устойчивый трехспиральный пучок и имеет широкую специфичность связывания альбуминов (Johansson, MU et al. (2002) “Structure, Specificity, And Mode Of Interaction For Bacterial Albumin-Binding Modules,” J. Biol. Chem. 277(10):8114-8120. Таким образом, особенно предпочтительная полипептидная часть белка для связывания сыворотки для улучшения фармакокинетических свойств диатела in vivo представляет собой Альбумин-связывающий домен (ABD) стрептококкового протеина G, а более предпочтительно, альбумин-связывающий домен 3 (ABD3) протеина G штамма Streptococcus G148 (SEQ ID NO: 43): LAEAKVLANR ELDKYGVSDY YKNLIDNAKS AEGVKALIDE ILAALP.
Как раскрыто в WO 2012/162068 (включен в данный документ ссылкой), "деиммунизированные" варианты SEQ ID NO: 43 обладают способностью ослаблять или устранять связывание с МНС класса II. На основе результатов комбинаторных мутаций, следующие комбинации замен считаются предпочтительными заменами для формирования такого деиммунизированного альбумин-связывающего домена: 66S/70S + 71A; 66S/70S + 79а; 64A/65A/71A + 66S; 64A/65A/71A + 66D; 64A/65A/71A + 66E; 64A/65A/79A + 66S; 64A/65A/79A + 66D; 64A/65A/79A + 66E. Вариантные ABD имеют модификации L64A, I65A и D79A или модификации N66S, T70S и D79A. Вариантный деиммунизированный ABD, имеющий аминокислотную последовательность:
LAEAKVLANR ELDKYGVSDY YKNA64A65NNAKT VEGVKALIA79E ILAALP (SEQ ID NO: 44),
или аминокислотную последовательность:
LAEAKVLANR ELDKYGVSDY YKNLIS66NAKS70 VEGVKALIA79E ILAALP (SEQ ID NO: 45),
является предпочтительным в качестве деиммунизиронных альбумин-связывающих доменов, демонстрирующих по существу связывание дикого типа, при этом обеспечивая ослабленное связывание с МНС класса II. Таким образом, первая полипептидная цепь такого диатела, имеющего альбумин-связывающий домен, содержит третий линкер (Линкер 3) предпочтительно расположенный на С-конце домена E-спирали (или K-спирали) такой полипептидной цепи, таким образом, чтобы располагаться между доменом Е-спирали (или K-спирали) и Альбумин-связывающим доменом (который предпочтительно представляет собой деиммунизированный альбумин-связывающий домен). Предпочтительная последовательность для такого линкера 3 представляет собой последовательность SEQ ID NO: 46: GGGS.
Таким образом, резюмируя, предпочтительная первая полипептидная цепь предпочтительной триспецифичной связывающей молекулы по настоящему изобретению будет содержать домены и линкеры: (VLI-домен) ─ (линкер 1) ─ (VHII-домен) ─ (линкер 2) ─ (домен E-спирали, способствующий гетеродимеризации) ─ (Линкер 3) ─ (CH2-CH3-домен, несущий выпуклость).
3. Альтернативная первая полипептидная цепь
В одном из воплощений ориентации описанных выше доменов будут находиться в направлении от N-конца к С-концу. Настоящее изобретение, однако, предусматривает также их изменение, в котором ориентации доменов первой полипептидной цепи следующие: NH2-(CH3-CH2-домен, несущий выпуклость)-(VLI-домен) ─ (линкер 1) ─ (VHII-домен) ─ (Линкер 2) ─ (домен Е-спирали, способствующий гетеродимеризации). Предпочтительно, если присутствует цистеинсодержащий пептид на N-конце такого CH2-CH3-домена. Последовательность примерного пептида представляет собой последовательность (SEQ ID NO: 48): DKTHTCPPCP. Предпочтительно в этом воплощении СН3-домен отстоит от VLI-домена на дистанцию промежуточного пептидного линкера (Линкер 4), например, линкера, имеющего аминокислотную последовательность (SEQ ID NO: 152): APSSS, а более предпочтительно-аминокислотную последовательность (SEQ ID NO: 50): APSSSPME.
4. Предпочтительная вторая полипептидная цепь
Вторая полипептидная цепь таких предпочтительных триспецифичных связывающих молекул будет включать, в направлении от N-конца к С-концу, вариабельный домен легкой цепи, способный связываться с эпитопом II (VLII), вариабельный домен тяжелой цепи, способный связываться к эпитопом I (VHI), и домен, содействующий гетеродимеризации.
Поскольку вариабельный домен легкой цепи и вариабельный домен тяжелой цепи второго полипептида направлены на различные эпитопы, они не могут связать друг с другом, образуя связывающий домен, который способен связывать либо эпитоп I, либо эпитоп II. Вариабельный домен легкой цепи и вариабельный домен тяжелой цепи второго полипептида разнесены друг от друга с помощью промежуточного пептидного линкера, который является достаточно коротким, чтобы по существу предотвратить ассоциацию этих доменов. "Линкер 1", имеющий последовательность (SEQ ID NO: 33): GGGSGGGG является примерным линкером для этой цели.
Как и в случае первой полипептидной цепи, вариабельный домен тяжелой цепи второго полипептида и домен, содействующий гетеродимеризации, этого полипептида предпочтительно разнесены друг от друга с помощью промежуточного линкерного пептида, который содержит 1, 2, 3 или более остатков цистеина. "Линкер 2", имеющий последовательность (SEQ ID NO: 34) GGCGGG является примерным линкером для этой цели. Такие остатки цистеина могут образовывать дисульфидные связи с остатками цистеина в цистеинсодержащем спейсерном пептиде, отделяющем вариабельный домен тяжелой цепи первого полипептида и домен, содействующий гетеродимеризации, этого полипептида. Таким образом, первый и второй полипептиды связывающих молекул по настоящему изобретению, ковалентно связаны друг с другом.
Как уже обсуждалось выше, домен, способствующий гетеродимеризации, на второй полипептидной цепи выбирают таким образом, чтобы он координировался с доменом, способствующим гетеродимеризации, на первой полипептидной цепи. Таким образом, в предпочтительном воплощении изобретения домен, способствующий гетеродимеризации, первой полипептидной цепи является либо Доменом с "К-спиралью" (SEQ ID NO: 40) либо доменом с "Е-спиралью" (SEQ ID NO: 39). Если цистеинсодержащая Е-спираль (SEQ ID NO: 41) используется в первой полипептидной цепи, то цистеинсодержащая K-спираль (SEQ ID NO: 42), предпочтительно применяется во второй полипептидной цепи. И наоборот, если цистеинсодержащая K-спираль (SEQ ID NO: 42) используется в первой полипептидной цепи, то цистеинсодержащая Е-спираль (SEQ ID NO: 41), предпочтительно применяется во второй полипептидной цепи. Так как первая полипептидная цепь будет предпочтительно обладать доменом "E-спирали", то вторая полипептидная цепь предпочтительно содержит домен "K-спирали".
Поскольку первая и вторая полипептидные цепи являются полипептидными цепями диатела, то они способны связываться вместе с образованием домен I-связывающего домена (VLА/VHА), который распознает и иммуноспецифично связывается с эпитопом, I, а домен II-связывающий домен (VLB/VHB), который распознает и иммуноспецифично связывается с эпитопом II.
Таким образом, резюмируя, предпочтительная вторая полипептидная цепь предпочтительной связывающей молекулы по настоящему изобретению будет содержать домены и линкеры: (VLII домен) ─ (линкер 1) ─ (VHI домен) ─ (линкер 2) ─ (домен К-спирали, содействующий гетеродимеризации).
5. Предпочтительная третья полипептидная цепь
Третья полипептидная цепь предпочтительной связывающей молекулы по настоящему изобретению представляет собой полипептид, который включает, в направлении от N-конца к С-концу, связывающий домен, дополнительный CH1-шарнирный домен и СН2-СН3-домен. Связывающий домен третьей полипептидной цепи предпочтительной связывающей молекулы по настоящему изобретению, может представлять собой вариабельный домен тяжелой цепи, способной связываться с эпитопом III (VH III), и в этом случае, четвертая полипептидная цепь предпочтительной связывающей молекулы по настоящему изобретению (обсуждается ниже), представляет собой полипептид, который содержит вариабельный домен легкой цепи, способный связываться с эпитопом III (VLIII), таким образом, что связывающий домен способен иммуноспецифически связываться с антигеном, обладающим эпитопом III. В ином случае, связывающий домен третьей полипептидной цепи предпочтительных связывающих молекул по настоящему изобретению может включать связывающий домен по типу рецептора эффекторной клетки, и, в этом случае, четвертая полипептидная цепь предпочтительных связывающих молекул по настоящему изобретению (обсуждается ниже) представляет собой полипептид, который включает комплементарный связывающий домен по типу рецептора эффекторной клетки, таким образом, что взаимодействие двух полипептидных цепей образует связывающий домен, способный физиоспецифически связывать молекулу, присутствующую на поверхности эффекторных клеток. Третья полипетидная цепь может быть выделена из естественных антител. В ином случае она может быть сконструирована рекомбинантным способом. Примерный СН1-домен представляет собой СН1-домен IgG1 человека, имеющий аминокислотную последовательность (SEQ ID NO: 207):
ASTKGPSVFP LAPSSKSTSG GTAALGCLVK DYFPEPVTVS WNSGALTSGV HTFPAVLQSS GLYSLSSVVT VPSSSLGTQT YICNVNHKPS NTKVDKKV
Вариант СН1-домена человеческого IgG1 последовательности SEQ ID NO: 207 представляет собой (SEQ ID NO: 208):
ASTKGPSVFP LAPSSKSTSG GTAALGCLVK DYFPEPVTVS WNSGALTSGV HTFPAVLQSS GLYSLSSVVT VPSSSLGTQT YICNVNHKPS NTKVDKRV
Примерный шарнирный домен представляет собой шарнирный домен IgG1 человека, имеющий аминокислотную последовательность (SEQ ID NO: 209): EPKSCDKTHTCPPCP. Как будет понятно, примерный шарнирный домен содержит несколько остатков цистеина (Elkabetz et al.(2005) “Cysteines In CH1 Underlie Retention Of Unassembled Ig Heavy Chains,” J. Biol. Chem. 280:14402-14412), которые могут участвовать в ковалентной связи между цепями.
Хотя может быть использован СН2-СН3-домен дикого типа, предпочтительно, как описано выше, использовать модифицированный CH2-CH3-домен, который способствует гетеродимеризации с СН3-СН2-доменом первой полипептидной цепи.
Предпочтительно, следовательно, если СН2-СН3-домен третьей полипептидной цепи будет СН2-СН3-доменом с "углублением", чья аминокислотная последовательность является комплементарной СН2-СН3-домену с "выпуклостью" (SEQ ID NO: 52) реализуемом в первом полипептиде. Как уже обсуждалось выше, CH2-CH3-домен с "углублением" предпочтительно должен содержать замену в положении 435 (H435R) для удаления участка связывания с протеином А. Примерный СН2-СН3-домен CH2-CH3-домен с "углублением" с заменой H435R для третьего полипептида представляет собой SEQ ID NO: 53.
Как будет понятно, СН2-СН3-домен с "выпуклостью" (например, SEQ ID NO: 52), можно использовать в третьей полипептидной цепи, и в этом случае СН2-СН3-домен с "углублением" (например, SEQ ID NO: 53) будет использоваться в первой полипептидной цепи.
В воплощении, в котором третья (и четвертая) полипептидные цепи предпочтительных триспецифичных связывающих молекул по настоящему изобретению, каждая содержит полипептидную цепь связывающего домена по типу рецептора эффекторной клетки, способы получения таких связывающих доменов по типу рецептора эффекторной клетки хорошо известны (например, US2012/0294874A1).
Таким образом, резюмируя, третья полипептидная цепь из предпочтительных связывающих молекул настоящего изобретения включает домены и линкеры:
(VHIII-домен) ─ (необязательно СН1-домен) ─ (необязательно шарнирный домен) ─ (CH2-CH3 домен с "углублением"), или (домен, связывающий Т-клеточный рецептор; его первый или второй полипептид) ─ (необязательно CH1-домен) ─ (необязательно шарнирный домен) ─ (CH2-CH3 домен с "углублением").
6. Предпочтительная четвертая полипептидная цепь
Четвертая полипептидная цепь предпочтительных триспецифичных связывающих молекул по настоящему изобретению, представляет собой либо полипептид связывающего домена по типу рецептора эффекторной клетки, (в котором третий и четвертый полипептиды образуют лиганд для рецептора, обнаруживаемого на поверхности эффекторной клетки, или более предпочтительно, вариабельный домен тяжелой цепи указанного выше антитела, который иммуноспецифично связывается с эпитопом III или который комплементарен связывающему домену третьей полипептидной цепи.
Таким образом, если третий и четвертый полипептиды образуют связывающий домен Fab-типа, такая четвертая полипептидная цепь содержит в направлении от N-конца к С-концу, вариабельный домен легкой цепи, способный связываться с эпитопом III (VL III), и Домен, содействующий ковалентному связыванию с третьей полипептидной цепью или связывающий домен и домен, содействующий ковалентному связыванию с третьей полипептидной цепью. Такой домен может быть CL-доменом, или его цистеинсодержащей частью, например, (SEQ ID NO: 38) FNRGEC или линкером, таким как линкер 2 (имеющим последовательность (SEQ ID NO: 34) GGCGGG. Примерный цистеинсодержащий пептид, который образует дисульфидные связи с таким линкером 2 содержит аминокислотную последовательность VEPKSC (SEQ ID NO: 36) или шарнирный домен.
Четвертая полипептидная цепь может быть выделена из природных антител. В ином случае он может быть сконструирована рекомбинантным способом. Предпочтительный CL-домен представляет собой CL каппа домен IgG1 человека, имеющий аминокислотную последовательность (SEQ ID NO: 210):
RTVAAPSVFI FPPSDEQLKS GTASVVCLLN NFYPREAKVQ WKVDNALQSG NSQESVTEQD SKDSTYSLSS TLTLSKADYE KHKVYACEVT HQGLSSPVTK SFNRGEC
В ином случае, примерный CL-домен представляет собой CL Лямбда2 домен IgG1 человека, имеющий аминокислотную последовательность (SEQ ID NO: 211):
QPKAAPSVTL FPPSSEELQA NKATLVCLIS DFYPGAVTVA WKADSSPVKA GVETTPSKQS NNKYAASSYL SLTPEQWKSH RSYSCQVTHE GSTVEKTVAP TECS
Как можно заметить, CL-домен или другой цистеинсодержащий домен, четвертой полипептидной цепи содержит остатки цистеина. Такие остатки цистеина способны ковалентно связываться с остатками цистеина СН1-домена третьей полипептидной цепи, чтобы, таким образом, ковалентно образовать комплекс третьей и четвертой полипептидных цепей связывающих молекул настоящего изобретения друг с другом. Таким образом, третья и четвертая полипептидной цепи ковалентно связаны друг с другом.
Кроме того, остатки цистеина CH2-CH3-домена первой полипептидной цепи могут образовывать дисульфидные связи с остатками цистеина в CH2-CH3-домене третьей полипептидной цепи. Таким образом, первый и третий полипептидные цепи ковалентно связаны друг с другом.
E. Вариантные Fc-домены
В традиционной иммунной функции взаимодействие комплексов антиген-антитело с клетками иммунной системы дает широкий спектр ответов, начиная от эффекторных функций, таких как антитело-зависимая цитотоксичность, дегрануляция тучных клеток и фагоцитоз, и заканчивая иммуномодулирующими сигналами, такими как регуляция пролиферации лимфоцитов и секреция антител. Все эти взаимодействия инициируются через связывание Fc-домена антител или иммунных комплексов со специализированными рецепторами клеточной поверхности на кроветворных клетках. Разнообразие клеточных реакций, вызываемых антителами и иммунными комплексами, является результатом структурной неоднородности трех Fc-рецепторов: FcγRI (CD64), Fc RII (CD32) и FcγRIII (CD16). FcγRI (CD64), FcγRIIA (CD32A) и FcγRIII (CD16) являются активизирующими (т.е. усиливающими иммунную систему) рецепторами; FcγRIIB (CD32B) является ингибирующим (т.е. ослабляющим иммунную систему) рецептором. Аминокислотная последовательность примерного Fc-домена IgG1 (SEQ ID NO: 1) приведена выше.
Модификация Fc-домена, как правило, приводит к измененному фенотипу, например, измененному периоду полужизни в сыворотке, измененной стабильности, измененной восприимчивости к клеточным ферментам или к измененной эффекторной функции. Может оказаться желательным модифицировать антитело по изобретению в отношении эффекторной функции, с тем чтобы повысить эффективность антитела при лечении, например, злокачественной опухоли. Снижение или устранение эффекторной функции является желательным в некоторых случаях, например в случае антител, механизм действия которых включает блокирование или антагонизм, но не уничтожение клеток, несущих антиген-мишень. Увеличение эффекторной функции, как правило, желательно, когда она направлена по отношению к нежелательным клеткам, таким как опухолевые и чужеродные клетки, в которых FcγRs экспрессируется на низком уровне, например, опухоль-специфичные В-клетки с низким уровнем FcγRIIB (например, при неходжкинской лимфоме, CLL и лимфоме Беркитта). В указанных воплощениях молекулы по изобретению с предусмотренной или измененной активностью эффекторной функции могут быть использованы для лечения и/или профилактики заболевания, расстройства или инфекции, при которых желательна улучшенная эффективность активность эффекторной функции.
В некоторых воплощениях настоящего изобретения триспецифичные связывающие молекулы по настоящему изобретению содержат Fc-домен, который обладает одной или несколькими модификациями (например, заменами, делециями или вставками) относительно аминокислотной последовательности Fc-домена дикого типа (SEQ ID NO: NO: 1), которые уменьшают, таким образом, аффинность и авидность Fc-доменамолекулы по изобретению к одному или нескольким рецепторам FcγR. В других воплощениях молекулы по изобретению включают Fc-домен, который обладает одной или несколькими модификациями аминокислот Fc-домена дикого типа, которые увеличивают аффинность и авидность Fc-домена и, таким образом, молекулы по изобретению, к одному или нескольким рецепторам FcγR. В других воплощениях молекулы включают вариантный Fc-домен, в котором указанный вариант придает или опосредует повышенную ADCC-активность и/или увеличение связывания с FcγRIIA, относительно молекулы, не содержащей Fc-домен или содержащей Fc-домен дикого типа. В альтернативных воплощениях молекулы включают вариантный Fc-домен, в котором указанный вариант придает или опосредует снижение ADCC-активности (или другой эффекторной функции) и/или увеличение связывания с FcγRIIB, относительно молекулы, не содержащей Fc-домен или содержащей Fc-домен дикого типа. В некоторых воплощениях настоящее изобретение включает триспецифичные связывающие молекулы, содержащий вариантный Fc-домен, где вариантной Fc-домен не демонстрирует детектируемого связывания с каким-либо FcγR, относительно сопоставимой молекулы, содержащей Fc-домен дикого типа. В других воплощениях настоящее изобретение включает триспецифичные связывающие молекулы, содержащие вариантный Fc-домен, где вариантной Fc-домен связывается только с одним FcγR, предпочтительно одним из числа FcγRIIA, FcγRIIB или FcγRIIIA. Любая такая повышенная аффинность и/или авидность предпочтительно оценивается с помощью измерения in vitro степени детектируемого связывания с FcγR или FcγR-связанной активности в клетках, которые экспрессируют низкие уровни FcγR, когда активность связывания родительской молекулы (без модифицированного Fc-домена) не может быть обнаружена в клетках, или в клетках, которые экспрессируют не являющиеся FcγR-рецептором целевые антигены при плотности 30 000-20 000 молекул/клетку, при плотности 20000-10000 молекул/клетку, при плотности 10000-5000 молекул/клетку, при плотности 5000-1000 молекул/клетку при плотности 1000-200 молекул/клетку или при плотности 200 молекул/клетку или менее (но, по меньшей мере, 10, 50, 100 или 150 молекул/клетку).
Триспецифичные связывающие молекулы по настоящему изобретению могут включать измененные аффинности по отношению к активирующему и/или ингибирующему рецептору Fcγ. В одном из воплощений триспецифичная связывающая молекула содержит вариант Fc-домена, который имеет повышенную аффинность к FcγRIIB и пониженную аффинность к FcγRIIIA и/или FcγRIIA, по сравнению с сопоставимой молекулой с Fc-доменом дикого типа. В другом воплощении триспецифичная связывающая молекула по настоящему изобретению включает вариантный Fc-домен, который имеет пониженную аффинность к FcγRIIB и повышенную аффинность к FcγRIIIA и/или FcγRIIA, по сравнению с сопоставимой молекулой с Fc-доменом дикого типа. В еще одном воплощении триспецифичные связывающие молекулы по настоящему изобретению включают вариант Fc-домена, который имеет пониженную аффинность к FcγRIIB и пониженную аффинность к FcγRIIIA и/или FcγRIIA, по сравнению с сопоставимой молекулой с Fc-доменом дикого типа. В еще одном воплощении триспецифичные связывающие молекулы по настоящему изобретению включают вариантный Fc-домен, который имеет неизмененную аффинность к FcγRIIB и пониженную (или повышенную) аффинность к FcγRIIIA и/или FcγRIIA, по сравнению с сопоставимой молекулой с Fc-доменом дикого типа.
В некоторых воплощениях настоящего изобретения триспецифичные связывающие молекулы по настоящему изобретению включают вариантный Fc-домен, имеющий измененную аффинность к FcγRIIIA и/или FcγRIIA, так что иммуноглобулин имеет повышенную эффекторную функцию, например, антитело-зависимую клеточно-опосредованную цитотоксичность. Не ограничивающие примеры функций эффекторных клеток включают антитело-зависимую клеточно-опосредованную цитотоксичность (ADCC), антитело-зависимый фагоцитоз, фагоцитоз, опсонизацию, опсонофагоцитоз, связывание клеток, розеткообразование, связывание C1q, и комплемент-зависимую клеточно-опосредованную цитотоксичность.
В предпочтительном воплощении изменение аффинности или эффекторной функции, по меньшей мере, в 2 раза, предпочтительно, по меньшей мере, в 4 раза, по меньшей мере, в 5 раз, по меньшей мере, в 6 раз, по меньшей мере, в 7 раз, по меньшей мере, 8 раз, по меньшей мере, в 9 раз, по меньшей мере, в 10 раз, по меньшей мере, в 50 раз, или, по меньшей мере, в 100 раз, по сравнению с сопоставимой молекулой, содержащей Fc-домен дикого типа. В других воплощениях настоящего изобретения вариантный Fc-домен иммуноспецифично связывает один или несколько FcR, по меньшей мере, на 65%, предпочтительно, по меньшей мере, 70%, 75%, 80%, 85%, 90%, 95%, 100%, 125%, 150%, 175%, 200%, 225%, или 250% большей аффинностью по отношению к молекуле, содержащей Fc-домена дикого типа. Такие измерения могут быть анализами in vivo или in vitro, и в предпочтительном воплощении анализами in vitro, такими как тИФА или поверхностный плазмонный резонанс.
В других воплощениях настоящего изобретения триспецифичные связывающие молекулы по настоящему изобретению включают вариантный Fc-домен, где указанный вариант выполняет роль агониста, по меньшей мере, одной активности FcγR-рецептора, или выполняет роль антагониста, по меньшей мере, одной активность FcγR-рецептора. В предпочтительном воплощении молекулы включают вариант, который является антагонистом одной или нескольких активностей FcγRIIB, например, сигнала, опосредуемого В-клеточным рецептором, активации В-клеток, пролиферации В-клеток, образования антител, внутриклеточного притока кальция в В-клетках, прогрессии клеточного цикла, FcγRIIB-опосредованного ингибирования сигнального пути FcεRI, фосфорилирования FcγRIIB, рекрутирования SHIP, фосфорилирования SHIP и ассоциации с Shc или активности одной или нескольких молекул, расположенных далее по сигнальному пути (например, MAP киназы, JNK, p38 или Akt) при передаче сигнала по пути FcγRIIB. В другом воплощении триспецифичные связывающие молекулы по настоящему изобретению включают вариант, который выполняет роль агониста одной или нескольких активностей FcεRI, например, активации тучных клеток, мобилизации кальция, дегрануляции, выработки цитокинов или высвобождения серотонина.
В некоторых воплощениях молекулы содержат Fc-домен, включающий участки из двух или более изотипов IgG (например, IgG1, IgG2, IgG3 и IgG4). Различные IgG изотипы демонстрируют различные физические и функциональные свойства, в том числе период полужизни в сыворотке, фиксация комплемента, аффинность связывания FcγR и активность эффекторной функции (например, ADCC, CDC и т.п.) из-за различий в аминокислотных последовательностей их шарнирных и/или Fc-доменов, например, как описано в Flesch and Neppert (1999) J. Clin. Lab. Anal. 14:141-156; Chappel et al. (1993) J. Biol. Chem. 33:25124-25131; Chappel et al. (1991) Proc. Natl. Acad. Sci. (U.S.A.) 88:9036-9040; или Brüggemann et al. (1987) J. Exp. Med 166:1351-1361. Этот тип вариантного Fc-домена может быть использован отдельно или в сочетании с аминокислотной модификацией для того, чтобы влиять на Fc-опосредованную эффекторную функцию и/или активность связывания. В комбинации, аминокислотная модификация и шарнирный/Fc-домен IgG могут демонстрировать аналогичные функциональные возможности (например, повышенную аффинность к FcγRIIA) и могут действовать аддитивно или, более предпочтительно, синергично для того, чтобы изменить эффекторную функциональность в молекуле согласно изобретению, по отношению к молекуле по изобретению, содержащей Fc-домен дикого типа. В других воплощениях аминокислотная модификация и Fc-домен IgG могут демонстрировать противоположные функциональные возможности (например, повышенную и пониженную аффинности к FcγRIIA, соответственно), и может действовать, чтобы выборочно смягчить или уменьшить определенную функциональность в молекуле согласно изобретению, по отношению к молекуле по настоящему изобретению, не содержащей Fc-домен или содержащей Fc-домен дикого типа того же изотипа.
В предпочтительном конкретном воплощении триспецифичные связывающие молекулы по настоящему изобретению включают вариантный Fc-домен, который содержит, по меньшей мере, одну аминокислотную модификацию по сравнению с Fc-доменом дикого типа, так, что указанная молекула обладает измененной аффинностью к FcR, при условии, что указанный вариантный Fc-домен не имеет замену в положениях, благодаря которым осуществляется прямой контакт с FcγR, согласно кристаллографическому и структурному анализу взаимодействий Fc-FcR, таких как те, что описаны Sondermann et al.(2000) Nature 406:267-73. Примеры положений в пределах Fc-домена, которые благодаря которым осуществляется прямой контакт с FcγR, представляют собой аминокислотные остатки 234-239 (шарнирная область), аминокислотные остатки 265-269 (B/C петля), аминокислотные остатки 297-299 (C'/E петля), и аминокислотные остатки 327-332 (F/G петля). В некоторых воплощениях молекулы по изобретению включают вариантные Fc-домены, которые содержат модификацию, по меньшей мере, одного остатка, который не осуществляет прямой контакт с FcγR, согласно структурному и кристаллографическому анализу, например, не находится в пределах участка связывания Fc-FcγR.
Вариантные Fc-домены хорошо известны в данной области, и любой известный вариант Fc может быть использован в настоящем изобретении для придания или изменения эффекторной функции, демонстрируемой молекулой по изобретению, содержащей Fc-домен (или его часть) в качестве функционально анализируемого, например, в NK-зависимом или макрофаг-зависимом анализе. Например, варианты Fc-домена, идентифицированные как изменяющие эффекторную функцию, раскрыты в the Antibody Engineering Technology Art, и любой подходящий вариант, раскрытый в этом документе, может быть использован в настоящих молекулах.
В некоторых воплощениях триспецифичные связывающие молекулы по настоящему изобретению включают вариантный Fc-домен, имеющий одну или несколько аминокислотных модификаций в одном или нескольких участках так, что модификация(и) изменяют (по сравнению с Fc-доменом дикого типа) Соотношение величин аффинности вариантного Fc-домена с активирующим FcγR (например, FcγRIIA или FcγRIIIA) относительно ингибирующего FcγR (например, FcγRIIB):

Соотношение аффинностей = (изменение дикого типа к варианту в аффинности к FcgRактивирущий)/ (изменение дикого типа к варианту в аффинности к FcgRингибирующий)
Особенно предпочтительными являются триспецифичные связывающие молекулы по настоящему изобретению, которые обладают вариантным Fc-доменом (относительно Fc-домена дикого типа), в котором вариантный Fc имеет соотношение аффинности больше 1. Такие молекулы имеют особое применение в обеспечении терапевтического или профилактического лечения заболевания, расстройства или инфекции, или облегчения их симптомов, при которых требуется улучшенная эффективность эффекторной функции клеток (например, ADCC), опосредуемая FcγR, например, злокачественная опухоль или инфекционное заболевание. В отличие от этого, вариант Fc, имеющий соотношение аффинности менее 1 опосредует пониженную эффективность функции эффекторных клеток. Таблица 1 перечисляет примеры одинарных, двойных, тройных, четверных и пятерных мутаций по соотношению их аффинностей больше или меньше 1.
Таблица 1. Примерные одиночные и множественные мутации, перечисленные по соотношению аффинностей
D270E
R292G
R292P
F243L & Y300L
F243L & P396L
D270E & P396L
R292P & Y300L
R292P & V305I
R292P & P396L
Y300L & P396L
P396L & Q419H
F243L, R292P & Y300L
F243L, R292P & V305I
F243L, R292P & P396L
F243L, Y300L & P396L
P247L, D270E и N421K
R255L, D270E и P396L
D270E, G316D & R416G
D270E, K392T & P396L
D270E, P396L & Q419H
V284M, R292L & K370N
R292P, Y300L & P396L
L235I, F243L, R292P & Y300L
L235Q, F243L, R292P & Y300L
F243L, P247L, D270E и N421K
F243L, R255L, D270E и P396L
F243L, D270E, G316D и R416G
F243L, D270E, K392T & P396L
F243L, D270E, P396L & Q419H
F243L, R292P, Y300L, и P396L
F243L, R292P, V305I & P396L
P247L, D270E, Y300L & N421K
R255L, D270E, R292G & P396L
R255L, D270E, Y300L & P396L
D270E, G316D, P396L & R416G
L235P, F243L, R292P, Y300L & P396L
F243L, R292P, V305I, Y300L & P396L
P396L
P247L & N421K
R255L & P396L
R292P & V305I
K392T & P396L
P396L & Q419H
В конкретном воплощении в вариантных Fc-доменах, любые аминокислотные модификации (например, замены) в любом из положений 235, 240, 241, 243, 244, 247, 262, 263, 269, 298, 328, или 330, и предпочтительно в одном или нескольких из следующих остатков: A240, I240, L241, L243, H244, N298, I328 или V330. В другом конкретном воплощении, в вариантных Fc-доменах, любые аминокислотные модификации (например, замены) в любом из положений 268, 269, 270, 272, 276, 278, 283, 285, 286, 289, 292, 293, 301, 303, 305, 307, 309, 331, 333, 334, 335, 337, 338, 340, 360, 373, 376, 416, 419, 430, 434, 435, 437, 438 или 439,и предпочтительно в одном или нескольких из следующих остатков: H280, Q280, Y280, G290, S290, T290, Y290, N294, K295, P296, D298, N298, P298, V298, I300 или L300.
В предпочтительном воплощении, в вариантных Fc-доменах, которые связывают FcγR с измененной аффинностью, любые аминокислотные модификации (например, замены) в любом из положений 255, 256, 258, 267, 268, 269, 270, 272, 276, 278, 280, 283, 285, 286, 289, 290, 292, 293, 294, 295, 296, 298, 300, 301, 303, 305, 307, 309, 312, 320, 322, 326, 329, 330, 332, 331, 333, 334, 335, 337, 338, 339, 340, 359, 360, 373, 376, 416, 419, 430, 434, 435, 437, 438 или 439. Предпочтительно, если вариантный Fc-домен имеет любой из следующих остатков: A256, N268, Q272, D286, Q286, S286, A290, S290, A298, M301, A312, E320, M320, Q320, R320, E322, A326, D326, E326, N326, S326, K330, T339, A333, A334, E334, H334, L334, M334, Q334, V334, K335, Q335, A359, A360 или A430.
В другом воплощении в вариантных Fc-доменах, которые связывают FcγR (с помощью своего Fc-домена) с пониженной аффинностью, любые аминокислотные модификации (например, замены) в любом из положений 252, 254, 265, 268, 269, 270, 278, 289, 292, 293, 294, 295, 296, 298, 300, 301, 303, 322, 324, 327, 329, 333, 335, 338, 340, 373, 376, 382, 388, 389, 414, 416, 419, 434, 435, 437, 438 или 439.
В другом воплощении, в вариантных Fc-доменах, которые связывают FcγR (с помощью своего Fc-домена) с повышенной аффинностью, любые аминокислотные модификации (например, замены) в любом из положений 280, 283, 285, 286, 290, 294, 295, 298, 300, 301, 305, 307, 309, 312, 315, 331, 333, 334, 337, 340, 360, 378, 398 или 430. В другом воплощении, в вариантных Fc-доменах, которые связывают FcγRIIA с повышенной аффинностью, любой из следующих остатков: A255, A256, A258, A267, A268, N268, A272, Q272, A276, A280, A283, A285, A286, D286, Q286, S286, A290, S290, M301, E320, M320, Q320, R320, E322, A326, D326, E326, S326, K330, A331, Q335, A337 или A430.
Предпочтительные варианты включают одну или несколько модификаций в любом из положений 228, 230, 231, 232, 233, 234, 235, 239, 240, 241, 243, 244, 245, 247, 262, 263, 264, 265, 266, 271, 273, 275, 281, 284, 291, 296, 297, 298, 299, 302, 304, 305, 313, 323, 325, 326, 328, 330 или 332.
Особенно предпочтительные варианты включают одну или несколько модификаций, выбранных из групп A-AI:
Еще более предпочтительные варианты включают одну или несколько модификаций, выбранных из группы 1-105:
В одном воплощении мультивалентная DR5-связывающая молекула по изобретению будет включать вариантный Fc-домен, имеющий, по меньшей мере, одну модификацию в Fc-домене. В некоторых воплощениях вариантный Fc-домен содержит, по меньшей мере, одну замену, выбранную из группы, состоящей из L235V, F243L, R292P, Y300L, V305I и P396L, в котором указанная нумерация представлена по индексу EU, как в Kabat.
В конкретном воплощении изобретения вариантный Fc-домен содержит:
(А), по меньшей мере, одну замену, выбранную из группы, состоящей из F243L, R292P, Y300L, V305I и P396L;
(В) по меньшей мере, две замены, выбранные из группы, состоящей из:
(1) F243L и P396L;
(2) F243L и R292P; и
(3) R292P и V305I;
(С) по меньшей мере, три замены, выбранные из группы, состоящей из:
(1) F243L, R292P и Y300L;
(2) F243L, R292P и V305I;
(3) F243L, R292P и P396L; и
(4) R292P, V305I и P396L;
(D), по меньшей мере, четыре замены, выбранные из группы, состоящей из:
(1) F243L, R292P, Y300L и P396L; и
(2) F243L, R292P, V305I и P396L; или
(Е), по меньшей мере, пять замен, выбранных из группы, состоящей из:
(1) F243L, R292P, Y300L, V305I и P396L; и
(2) L235V, F243L, R292P, Y300L и P396L.
В другом конкретном воплощении вариантный Fc-домен содержит замены:
(A) F243L, R292P и Y300L;
(B) L235V, F243L, R292P, Y300L и P396L; или
(C) F243L, R292P, Y300L, V305I и P396L.
В других воплощениях настоящее изобретение охватывает применение любого вариантного Fc, известного в данной области, такого как описанные в Jefferis, BJ et al. (2002) “Interaction Sites On Human IgG-Fc For FcgammaR: Current Models,” Immunol. Lett. 82:57-65; Presta, L.G. et al. (2002) “Engineering Therapeutic Antibodies For Improved Function,” Biochem. Soc. Trans. 30:487-90; Idusogie, E.E. et al. (2001) “Engineered Antibodies With Increased Activity To Recruit Complement,” J. Immunol. 166:2571-75; Shields, R.L. et al. (2001) “High Resolution Mapping Of The Binding Site On Human IgG1 For Fc Gamma RI, Fc Gamma RII, Fc Gamma RIII, And FcRn And Design Of IgG1 Variants With Improved Binding To The Fc gamma R,” J. Biol. Chem. 276:6591-6604; Idusogie, E.E. et al. (2000) “Mapping Of The C1q Binding Site On Rituxan, A Chimeric Antibody With A Human IgG Fc,” J. Immunol. 164:4178-84; Reddy, M.P. et al. (2000) “Elimination Of Fc Receptor-Dependent Effector Functions Of A Modified IgG4 Monoclonal Antibody To Human CD4,” J. Immunol. 164:1925-1933; Xu, D. et al. (2000) “In Vitro Characterization of Five Humanized OKT3 Effector Function Variant Antibodies,” Cell. Immunol. 200:16-26; Armour, K.L. et al. (1999) “Recombinant human IgG Molecules Lacking Fcgamma Receptor I Binding And Monocyte Triggering Activities,” Eur. J. Immunol. 29:2613-24; Jefferis, R. et al. (1996) “Modulation Of Fc(Gamma)R And Human Complement Activation By IgG3-Core Oligosaccharide Interactions,” Immunol. Lett. 54:101-04; Lund, J. et al. (1996) “Multiple Interactions Of IgG With Its Core Oligosaccharide Can Modulate Recognition By Complement And Human Fc Gamma Receptor I And Influence The Synthesis Of Its Oligosaccharide Chains,” J. Immunol. 157:4963-4969; Hutchins et al. (1995) “Improved Biodistribution, Tumor Targeting, And Reduced Immunogenicity In Mice With A Gamma 4 Variant Of Campath-1H,” Proc. Natl. Acad. Sci. (U.S.A.) 92:11980-84; Jefferis, R. et al. (1995) “Recognition Sites On Human IgG For Fc Gamma Receptors: The Role Of Glycosylation,” Immunol. Lett. 44:111-17; Lund, J. et al. (1995) “Oligosaccharide-Protein Interactions In IgG Can Modulate Recognition By Fc Gamma Receptors,” FASEB J. 9:115-19; Alegre, M.L. et al. (1994) “A Non-Activating "Humanized" Anti-CD3 Monoclonal Antibody Retains Immunosuppressive Properties In Vivo,” Transplantation 57:1537-1543; Lund et al. (1992) “Multiple Binding Sites On The CH2 Domain Of IgG For Mouse Fc Gamma R11,” Mol. Immunol. 29: 53-59; Lund et al. (1991) “Human Fc Gamma RI And Fc Gamma RII Interact With Distinct But Overlapping Sites On Human IgG,” J. Immunol. 147:2657-2662; Duncan, A.R. et al. (1988) “Localization Of The Binding Site For The Human High-Affinity Fc Receptor On IgG,” Nature 332:563-564; US Patent Nos. 5624821; 5885573; 6194551; 7276586; и 7317091; и PCT Publications WO 00/42072 и PCT WO 99/58572.
В некоторых воплощениях молекулы по изобретению дополнительно содержат один или несколько сайтов гликозилирования, так что один или нескольких углеводных остатков ковалентно присоединены к молекуле. Предпочтительно молекулы согласно изобретению с одним или несколькими сайтами гликозилирования, и/или одним или несколькими модификациями в Fc-домене придают или обладают повышенной антителоопосредуемый эффекторной функцией, например, повышенной ADCC-активностью, по сравнению с исходным антителом. В некоторых воплощениях настоящее изобретение дополнительно включает молекулы, включающие одну или несколько аминокислотных модификаций, которые, как известно, прямо или косвенно взаимодействуют с углеводной частью антитела, в том числе, без ограничения перечисленным, аминокислотных модификаций в положениях 241, 243, 244, 245, 245, 249, 256, 258, 260, 262, 264, 265, 296, 299, и 301. Аминокислоты, которые прямо или косвенно взаимодействуют с углеводным фрагментом антитела, известны в данной области, см., например, Jefferis et al., 1995 Immunology Letters, 44: 111-7, которая включена в настоящее описание ссылкой во всей полноте.
В другом воплощении настоящее изобретение охватывает молекулы, которые были модифицированы путем введения одного или нескольких сайтов гликозилирования, в один или несколько участков молекул, предпочтительно, без изменения функциональности молекул, например, активности связывания с антигеном-мишенью или с FcγR. Сайты гликозилирования, могут быть введены в вариабельную и/или константную области молекулы по изобретению. При использовании в данном описании термин "сайты гликозилирования" включают любую конкретную аминокислотную последовательность в антителе, к которой олигосахариды (т.е. углеводы, содержащие два или более простых сахара, связанных друг с другом) специфически и ковалентного присоединены. Олигосахаридные боковые цепи, как правило, связаны с основной цепью антитела либо через N-, либо через O-связь. N-связанное гликозилирование относится к присоединению олигосахаридных фрагментов к боковой цепи остатка аспарагина. O-связанное гликозилирование относится к присоединению олигосахаридных фрагментов к гидроксиаминокислоте, например, серину, треонину. Молекулы по изобретению могут содержать один или несколько сайтов гликозилирования включая участки для N-связанного и О-связанного гликозилирования. Любой сайт гликозилирования для N-связанного или О-связанного гликозилирования, известный в данной области техники, может быть использован в соответствии с настоящим изобретением. Примерный сайт N-связанного гликозилирования, который является полезным в соответствии со способами по настоящему изобретению, представляет собой аминокислотную последовательность: Asn-X-Thr/Ser, где Х может быть любой аминокислотой, и Thr/Ser обозначает треонин или серин. Такой сайт или сайты могут быть введены в молекулу настоящего изобретения с помощью способов, хорошо известных в области техники, к которой относится настоящее изобретение (см., например, In vitro Mutagenesis, Recombinant DNA: A Short Course, J. D. Watson, et al. W.H. Freeman and Company, New York, 1983, глава 8, стр. 106-116, которая включена в данное описание ссылкой во всей полноте. Примерный способ введения сайта гликозилирования в молекулу по изобретению может включать: модификацию или мутацию аминокислотной последовательности молекулы таким образом, чтобы получить требуемую последовательность Asn-X-Thr/Ser.
В некоторых воплощениях изобретение относится к способам модификации содержания углеводов молекулы по изобретению путем добавления или удаления сайта гликозилирования. Способы модификации содержания углеводов антител (и молекул, содержащих домены антитела), хорошо известны в данной области техники и охватываются настоящим изобретением, см., например, пат. США № 6218149; EP 0 359 096 B1; публ. США № 2002/0028486; WO 03/035835; публ. США № 2003/0115614; пат. США № 6218149; пат. США № 6472511; все из которых включены в настоящее описание ссылкой во всей полноте. В других воплощениях изобретение относится к способам модификации содержания углеводов молекулы по изобретению путем удаления одной или нескольких эндогенных углеводных групп молекулы. В конкретном воплощении настоящее изобретение включает перенос сайта гликозилирования Fc-домена антитела, путем изменения положения, примыкающего к 297. В конкретном воплощении настоящее изобретение охватывает модификации положения 296, с тем чтобы положение 296, а не положение 297 было гликозилированным.
Эффекторная функция также может быть модифицирована с помощью известных методов, например, путем введения одного или нескольких остатков цистеина в Fc-домен, что позволит создать межцепочечную дисульфидную связь в этой области, в результате чего образуется гомодимерное антитело, которое может иметь улучшенную способность к интернализации и/или повышенному комплемент-опосредованному уничтожению клеток и ADCC (Caron, PC et al.(1992) “Engineered Humanized Dimeric Forms Of IgG Are More Effective Antibodies,” J. Exp. Med. 176:1191-1195; Shopes, B. (1992) “A Genetically Engineered Human IgG Mutant With Enhanced Cytolytic Activity,” J. Immunol. 148(9):2918-2922. Гомодимерные антитела с повышенной противоопухолевой активностью также могут быть получены с использованием гетеробифункциональных сшивающих агентов, как описано в Wolff, EA et al.(1993) “Monoclonal Antibody Homodimers: Enhanced Antitumor Activity In Nude Mice,” Cancer Research 53:2560-2565. В ином случае, может быть сконструировано антитело, которое имеет двойные Fc-домены и поэтому может усилить комплементный лизис и способности к ADCC (Stevenson, G.T. et al. (1989) “A Chimeric Antibody With Dual Fc Domains (bisFabFc) Prepared By Manipulations At The IgG Hinge,” Anti-Cancer Drug Design 3:219-230).
III. Примеры триспецифичных связывающих молекул
F. gpA33 mAb 1 х CD3 mAb 2 х DR5 mAb 1
Была сконструирована примерная триспецифичная связывающая молекула, состоящая из четырех полипептидных цепей. Триспецифичная связывающая молекула включает VL- и VH-домены из gpA33 mAb 1, VL- и VH-домены из антитела CD3 mAb 2, и VL- и VH-домены из DR5 mAb 1, и был обозначена соответствующим образом "gpA33 mAb 1 х CD3 mAb 2 х DR5 mAb 1". Аминокислотная последовательность первой полипептидной цепи этой триспецифичной связывающей молекулы представляет собой (SEQ ID NO: 212):
DIQLTQSPSF LSASVGDRVT ITCSARSSIS FMYWYQQKPG KAPKLLIYDT SNLASGVPSR FSGSGSGTEF TLTISSLEAE DAATYYCQQW SSYPLTFGQG TKLEIKGGGS GGGGEVQLVE SGGGLVQPGG SLRLSCAASG FTFSTYAMNW VRQAPGKGLE WVGRIRSKYN NYATYYADSV KGRFTISRDD SKNSLYLQMN SLKTEDTAVY YCVRHGNFGN SYVSWFAYWG QGTLVTVSSG GCGGGEVAAL EKEVAALEKE VAALEKEVAA LEKGGGDKTH TCPPCPAPEA AGGPSVFLFP PKPKDTLMIS RTPEVTCVVV DVSHEDPEVK FNWYVDGVEV HNAKTKPREE QYNSTYRVVS VLTVLHQDWL NGKEYKCKVS NKALPAPIEK TISKAKGQPR EPQVYTLPPS REEMTKNQVS LWCLVKGFYP SDIAVEWESN GQPENNYKTT PPVLDSDGSF FLYSKLTVDK SRWQQGNVFS CSVMHEALHN HYTQKSLSLS
PGK
В SEQ ID NO: 212, аминокислотные остатки 1-106 соответствуют аминокислотной последовательности VL-домена gpA33 mAb 1 (SEQ ID NO: 181), остатки 107-114 соответствуют промежуточному спейсерному пептиду GGGSGGGG (линкер 1) (SEQ ID NO: 33), остатки 115-239 соответствуют аминокислотной последовательности VH-домена CD3 mAb 2, имеющего замену D65G (SEQ ID NO: 112), остатки 240-245 соответствуют линкеру GGCGGG (SEQ ID NO: NO: 34), остатки 246-273 соответствуют домену с Е-спиралью (SEQ ID NO: 39), остатки 274-276 представляют собой линкер GGG, остатки 277-286 представляют собой линкер DKTHTCPPCP (SEQ ID NO: 48), а остатки 287-503 представляют собой СН2-СН3-домен, несущий "выпуклость" (SEQ ID NO: 52).
Полинуклеотид, который кодируется SEQ ID NO: 212 представляет собой SEQ ID NO: 213:
gacattcagc tgactcagtc cccctctttt ctgtccgcat ccgtcggaga tcgagtgact attacttgct ctgctaggtc ctcaatcagc ttcatgtact ggtatcagca gaagcccggc aaagcaccta agctgctgat ctacgacaca agcaacctgg cctccggggt gccatctcgg ttctctggca gtgggtcagg aactgagttt accctgacaa ttagctccct ggaggctgaa gatgccgcta cctactattg ccagcagtgg agcagctatc ctctgacctt cggacagggg actaaactgg aaatcaaggg tggaggatcc ggcggcggag gcgaggtgca gctggtggag tctgggggag gcttggtcca gcctggaggg tccctgagac tctcctgtgc agcctctgga ttcaccttca gcacatacgc tatgaattgg gtccgccagg ctccagggaa ggggctggag tgggttggaa ggatcaggtc caagtacaac aattatgcaa cctactatgc cgactctgtg aagggtagat tcaccatctc aagagatgat tcaaagaact cactgtatct gcaaatgaac agcctgaaaa ccgaggacac ggccgtgtat tactgtgtga gacacggtaa cttcggcaat tcttacgtgt cttggtttgc ttattgggga caggggacac tggtgactgt gtcttccgga ggatgtggcg gtggagaagt ggccgcactg gagaaagagg ttgctgcttt ggagaaggag gtcgctgcac ttgaaaagga ggtcgcagcc ctggagaaag gcggcgggga caaaactcac acatgcccac cgtgcccagc acctgaagcc gcggggggac cgtcagtctt cctcttcccc ccaaaaccca aggacaccct catgatctcc cggacccctg aggtcacatg cgtggtggtg gacgtgagcc acgaagaccc tgaggtcaag ttcaactggt acgtggacgg cgtggaggtg cataatgcca agacaaagcc gcgggaggag cagtacaaca gcacgtaccg tgtggtcagc gtcctcaccg tcctgcacca ggactggctg aatggcaagg agtacaagtg caaggtctcc aacaaagccc tcccagcccc catcgagaaa accatctcca aagccaaagg gcagccccga gaaccacagg tgtacaccct gcccccatcc cgggaggaga tgaccaagaa ccaggtcagc ctgtggtgcc tggtcaaagg cttctatccc agcgacatcg ccgtggagtg ggagagcaat gggcagccgg agaacaacta caagaccacg cctcccgtgc tggactccga cggctccttc ttcctctaca gcaagctcac cgtggacaag agcaggtggc agcaggggaa cgtcttctca tgctccgtga tgcatgaggc tctgcacaac cactacacgc agaagagcct ctccctgtct ccgggtaaa
Аминокислотная последовательность второй полипептидной цепи gpA33 mAb 1 х CD3 mAb 2 х DR5 mAb 1 (SEQ ID NO: 214):
QAVVTQEPSL TVSPGGTVTL TCRSSTGAVT TSNYANWVQQ KPGQAPRGLI GGTNKRAPWT PARFSGSLLG GKAALTITGA QAEDEADYYC ALWYSNLWVF GGGTKLTVLG GGGSGGGGQV QLVQSGAEVK KPGASVKVSC KASGYTFTGS WMNWVRQAPG QGLEWIGRIY PGDGETNYNG KFKDRVTITA DKSTSTAYME LSSLRSEDTA VYYCARIYGN NVYFDVWGQG TTVTVSSGGC GGGKVAALKE KVAALKEKVA ALKEKVAALK E
В SEQ ID NO: 214 аминокислотные остатки 1-110 соответствуют аминокислотной последовательности VL-домена CD3 mAb 2 (SEQ ID NO: 104), остатки 111-118 соответствуют промежуточному спейсерному пептиду GGGSGGGG (линкер 1) (SEQ ID NO: 33), остатки 119-237 соответствуют аминокислотной последовательности VH-домена gpA33 mAb 1 (SEQ ID NO: 186), остатки 238-243 соответствуют линкеру GGCGGG (SEQ ID NO: 34), а остатки 244-271 представляют собой домен К-спирали (SEQ ID NO: 40).
Полинуклеотид, который кодируется SEQ ID NO: 214, представляет собой (SEQ ID NO: 215):
caggctgtgg tgactcagga gccttcactg accgtgtccc caggcggaac tgtgaccctg acatgcagat ccagcacagg cgcagtgacc acatctaact acgccaattg ggtgcagcag aagccaggac aggcaccaag gggcctgatc gggggtacaa acaaaagggc tccctggacc cctgcacggt tttctggaag tctgctgggc ggaaaggccg ctctgactat taccggggca caggccgagg acgaagccga ttactattgt gctctgtggt atagcaatct gtgggtgttc gggggtggca caaaactgac tgtgctggga gggggtggat ccggcggagg tggacaggtc cagctggtcc agagcggggc cgaagtcaaa aaacccggag caagcgtgaa ggtctcctgc aaagcatcag gctatacatt tacaggcagc tggatgaact gggtgaggca ggctccagga cagggactgg agtggatcgg gcgcatctac cctggagacg gcgaaactaa ctataatgga aagttcaaag accgagtgac catcacagcc gataagtcta ctagtaccgc ctacatggag ctgagctccc tgcggtctga agataccgcc gtctactatt gcgctagaat ttacggaaac aatgtctatt ttgacgtgtg ggggcaggga acaactgtga ctgtctcctc cggaggatgt ggcggtggaa aagtggccgc actgaaggag aaagttgctg ctttgaaaga gaaggtcgcc gcacttaagg aaaaggtcgc agccctgaaa gag
Аминокислотная последовательность третьей полипептидной цепи из gpA33 mAb 1 х CD3 mAb 2 х DR5 mAb 1 (SEQ ID NO: 216):
EVKFLESGGG LVQPGGSLKL SCVASGFDFS RYWMSWVRQA PGKGLEWIGE INPDSNTINY TPSLKDKFII SRDNAKNTLY LQMTKVRSED TALYYCTRRA YYGNPAWFAY WGQGTLVTVS SASTKGPSVF PLAPSSKSTS GGTAALGCLV KDYFPEPVTV SWNSGALTSG VHTFPAVLQS SGLYSLSSVV TVPSSSLGTQ TYICNVNHKP SNTKVDKRVE PKSCDKTHTC PPCPAPEAAG GPSVFLFPPK PKDTLMISRT PEVTCVVVDV SHEDPEVKFN WYVDGVEVHN AKTKPREEQY NSTYRVVSVL TVLHQDWLNG KEYKCKVSNK ALPAPIEKTI SKAKGQPREP QVYTLPPSRE EMTKNQVSLS CAVKGFYPSD IAVEWESNGQ PENNYKTTPP VLDSDGSFFL VSKLTVDKSR WQQGNVFSCS VMHEALHNRY TQKSLSLSPG
K
В SEQ ID NO: 216, аминокислотные остатки 1-121 соответствуют аминокислотной последовательности VH-домена DR5 mAb 1 (SEQ ID NO: 8), остатки 122-219 соответствуют измененному СН1-домену (SEQ ID NO: 208), остатки 220-234 соответствуют линкеру (SEQ ID NO: 209), а остатки 235-451 соответствуют CH2-CH3-домену с "углублением" (SEQ ID NO: 53).
Полинуклеотид, который кодируется SEQ ID NO: 216, представляет собой (SEQ ID NO: 217):
gaggtgaagt ttctcgagtc tggaggtggc ctggtgcagc ctggaggatc cctgaaactc tcctgtgtag cctcaggatt cgattttagt agatactgga tgagttgggt ccggcaggct ccagggaaag ggctagaatg gattggagaa attaatccag atagcaatac gataaactat acgccatctc taaaggataa attcatcatc tccagagaca acgccaaaaa tacgctgtat ctgcaaatga ccaaagtgag atctgaggac acagcccttt attattgtac aagaagggcc tactatggta acccggcctg gtttgcttac tggggccaag ggactctggt cactgtctct tccgcctcca ccaagggccc atcggtcttc cccctggcac cctcctccaa gagcacctct gggggcacag cggccctggg ctgcctggtc aaggactact tccccgaacc ggtgacggtg tcgtggaact caggcgccct gaccagcggc gtgcacacct tcccggctgt cctacagtcc tcaggactct actccctcag cagcgtggtg accgtgccct ccagcagctt gggcacccag acctacatct gcaacgtgaa tcacaagccc agcaacacca aggtggacaa gagagttgag cccaaatctt gtgacaaaac tcacacatgc ccaccgtgcc cagcacctga agccgcgggg ggaccgtcag tcttcctctt ccccccaaaa cccaaggaca ccctcatgat ctcccggacc cctgaggtca catgcgtggt ggtggacgtg agccacgaag accctgaggt caagttcaac tggtacgtgg acggcgtgga ggtgcataat gccaagacaa agccgcggga ggagcagtac aacagcacgt accgtgtggt cagcgtcctc accgtcctgc accaggactg gctgaatggc aaggagtaca agtgcaaggt ctccaacaaa gccctcccag cccccatcga gaaaaccatc tccaaagcca aagggcagcc ccgagaacca caggtgtaca ccctgccccc atcccgggag gagatgacca agaaccaggt cagcctgagt tgcgcagtca aaggcttcta tcccagcgac atcgccgtgg agtgggagag caatgggcag ccggagaaca actacaagac cacgcctccc gtgctggact ccgacggctc cttcttcctc gtcagcaagc tcaccgtgga caagagcagg tggcagcagg ggaacgtctt ctcatgctcc gtgatgcatg aggctctgca caaccgctac acgcagaaga gcctctccct gtctccgggt aaa
Аминокислотная последовательность четвертой полипептидной цепи из gpA33 mAb 1 х CD3 mAb 2 х DR5 mAb 1 (SEQ ID NO: 218):
DIVLTQSPAS LAVSLGQRAT ISCRASKSVS SSGYSYMHWY QQKPGQPPKV LIFLSSNLDS GVPARFSGSG SGTDFTLNIH PVEDGDAATY YCQHSRDLPP TFGGGTKLEI KRTVAAPSVF IFPPSDEQLK SGTASVVCLL NNFYPREAKV QWKVDNALQS GNSQESVTEQ DSKDSTYSLS STLTLSKADY EKHKVYACEV THQGLSSPVT KSFNRGEC
В SEQ ID NO: 218, аминокислотные остатки 1-111 соответствуют аминокислотной последовательности VL-домена DR5 mAb 1 (SEQ ID NO: 3), а остатки 112-218 соответствуют CL Каппа домену (SEQ ID NO: 210).
Полинуклеотид, который кодируется SEQ ID NO: 218, представляет собой (SEQ ID NO: 219):
gacattgtgc tgacacagtc tcctgcttcc ttagctgtat ctctcgggca gagggccacc atctcatgca gggccagcaa aagtgtcagt tcctctggct atagttatat gcactggtac caacagaaac caggacagcc acccaaagtc ctcatctttc tttcatccaa cctagattct ggggtccctg ccaggttcag tggcagtggg tctgggacag acttcaccct caacatccat cctgtggagg atggggatgc tgcaacctat tactgtcagc acagtaggga tcttcctccg acgttcggtg gaggcaccaa gctggaaatc aaacgtacgg tggctgcacc atcggtcttc atcttcccgc catctgatga gcagttgaaa tctggaactg cctctgttgt gtgcctgctg aataacttct atcccagaga ggccaaagta cagtggaagg tggataacgc cctccaatcg ggtaactccc aggagagtgt cacagagcag gacagcaagg acagcaccta cagcctcagc agcaccctga cgctgagcaa agcagactac gagaaacaca aagtctacgc ctgcgaagtc acccatcagg gcctgagctc gcccgtcaca aagagcttca acaggggaga gtgt
G. gpA33 mAb 1 х CD3 mAb 2 х DR5 mAb 2
Была сконструирована вторая примерная триспецифичная связывающая молекула, состоящая из четырех полипептидных цепей. Триспецифическая связывающаяся молекула содержит VL- и VH-домены из gpA33 mAb 1, VL- и VH-домены из антитела CD3 mAb 2 и VL- и VH-домены из DR5 mAb 2, и обозначается соответствующим образом как "gpA33 mAb 1 х CD3 mAb 2 х DR5 mAb 2". Аминокислотная последовательность первой полипептидной цепи данной триспецифичной связывающей молекулы представляет собой (SEQ ID NO: 220):
DIQLTQSPSF LSASVGDRVT ITCSARSSIS FMYWYQQKPG KAPKLLIYDT SNLASGVPSR FSGSGSGTEF TLTISSLEAE DAATYYCQQW SSYPLTFGQG TKLEIKGGGS GGGGEVQLVE SGGGLVQPGG SLRLSCAASG FTFSTYAMNW VRQAPGKGLE WVGRIRSKYN NYATYYADSV KGRFTISRDD SKNSLYLQMN SLKTEDTAVY YCVRHGNFGN SYVSWFAYWG QGTLVTVSSG GCGGGEVAAL EKEVAALEKE VAALEKEVAA LEKGGGDKTH TCPPCPAPEA AGGPSVFLFP PKPKDTLMIS RTPEVTCVVV DVSHEDPEVK FNWYVDGVEV HNAKTKPREE QYNSTYRVVS VLTVLHQDWL NGKEYKCKVS NKALPAPIEK TISKAKGQPR EPQVYTLPPS REEMTKNQVS LWCLVKGFYP SDIAVEWESN GQPENNYKTT PPVLDSDGSF FLYSKLTVDK SRWQQGNVFS CSVMHEALHN HYTQKSLSLS PGK
В SEQ ID NO: 220, аминокислотные остатки 1-106 соответствуют аминокислотной последовательности VL-домена gpA33 mAb 1 (SEQ ID NO: 181), остатки 107-114 соответствуют промежуточному спейсерному пептиду GGGSGGGG (линкер 1) (SEQ ID NO: 33), остатки 115-239 соответствуют аминокислотной последовательности VH-домена CD3 mAb 2, содержащего замену D65G (SEQ ID NO: 112), остатки 240-245 соответствуют линкеру GGCGGG (SEQ ID NO: NO: 34), остатки 246-273 соответствуют домену с Е-спиралью (SEQ ID NO: 39), остатки 274-276 представляют собой линкер GGG, остатки 277-286 представляют собой линкер DKTHTCPPCP (SEQ ID NO: 48), а остатки 287-503 представляют собой СН2-СН3-домен, несущий "выпуклость" (SEQ ID NO: 52).
Полинуклеотид, который кодируется SEQ ID NO: 220 представляет собой (SEQ ID NO: 221):
gacattcagc tgactcagtc cccctctttt ctgtccgcat ccgtcggaga tcgagtgact attacttgct ctgctaggtc ctcaatcagc ttcatgtact ggtatcagca gaagcccggc aaagcaccta agctgctgat ctacgacaca agcaacctgg cctccggggt gccatctcgg ttctctggca gtgggtcagg aactgagttt accctgacaa ttagctccct ggaggctgaa gatgccgcta cctactattg ccagcagtgg agcagctatc ctctgacctt cggacagggg actaaactgg aaatcaaggg tggaggatcc ggcggcggag gcgaggtgca gctggtggag tctgggggag gcttggtcca gcctggaggg tccctgagac tctcctgtgc agcctctgga ttcaccttca gcacatacgc tatgaattgg gtccgccagg ctccagggaa ggggctggag tgggttggaa ggatcaggtc caagtacaac aattatgcaa cctactatgc cgactctgtg aagggtagat tcaccatctc aagagatgat tcaaagaact cactgtatct gcaaatgaac agcctgaaaa ccgaggacac ggccgtgtat tactgtgtga gacacggtaa cttcggcaat tcttacgtgt cttggtttgc ttattgggga caggggacac tggtgactgt gtcttccgga ggatgtggcg gtggagaagt ggccgcactg gagaaagagg ttgctgcttt ggagaaggag gtcgctgcac ttgaaaagga ggtcgcagcc ctggagaaag gcggcgggga caaaactcac acatgcccac cgtgcccagc acctgaagcc gcggggggac cgtcagtctt cctcttcccc ccaaaaccca aggacaccct catgatctcc cggacccctg aggtcacatg cgtggtggtg gacgtgagcc acgaagaccc tgaggtcaag ttcaactggt acgtggacgg cgtggaggtg cataatgcca agacaaagcc gcgggaggag cagtacaaca gcacgtaccg tgtggtcagc gtcctcaccg tcctgcacca ggactggctg aatggcaagg agtacaagtg caaggtctcc aacaaagccc tcccagcccc catcgagaaa accatctcca aagccaaagg gcagccccga gaaccacagg tgtacaccct gcccccatcc cgggaggaga tgaccaagaa ccaggtcagc ctgtggtgcc tggtcaaagg cttctatccc agcgacatcg ccgtggagtg ggagagcaat gggcagccgg agaacaacta caagaccacg cctcccgtgc tggactccga cggctccttc ttcctctaca gcaagctcac cgtggacaag agcaggtggc agcaggggaa cgtcttctca tgctccgtga tgcatgaggc tctgcacaac cactacacgc agaagagcct ctccctgtct ccgggtaaa
Аминокислотная последовательность второй полипептидной цепи gpA33 mAb 1 х CD3 mAb 2 х DR5 mAb 2 (SEQ ID NO: 222):
QAVVTQEPSL TVSPGGTVTL TCRSSTGAVT TSNYANWVQQ KPGQAPRGLI GGTNKRAPWT PARFSGSLLG GKAALTITGA QAEDEADYYC ALWYSNLWVF GGGTKLTVLG GGGSGGGGQV QLVQSGAEVK KPGASVKVSC KASGYTFTGS WMNWVRQAPG QGLEWIGRIY PGDGETNYNG KFKDRVTITA DKSTSTAYME LSSLRSEDTA VYYCARIYGN NVYFDVWGQG TTVTVSSGGC GGGKVAALKE KVAALKEKVA ALKEKVAALK E
В SEQ ID NO: 222 аминокислотные остатки 1-110 соответствуют аминокислотной последовательности VL-домена CD3 mAb 2 (SEQ ID NO: 104), остатки 111-118 соответствуют промежуточному спейсерному пептиду GGGSGGGG (линкер 1) (SEQ ID NO: 33), остатки 119-237 соответствуют аминокислотной последовательности VH-домена gpA33 mAb 1 (SEQ ID NO: 186), остатки 238-243 соответствуют линкеру GGCGGG (SEQ ID NO: 34) а остатки 244-271 представляют собой домен с K-спиралью (SEQ ID NO: 40).
Полинуклеотид, который кодируется SEQ ID NO: 222 представляет собой (SEQ ID NO: 223):
caggctgtgg tgactcagga gccttcactg accgtgtccc caggcggaac tgtgaccctg acatgcagat ccagcacagg cgcagtgacc acatctaact acgccaattg ggtgcagcag aagccaggac aggcaccaag gggcctgatc gggggtacaa acaaaagggc tccctggacc cctgcacggt tttctggaag tctgctgggc ggaaaggccg ctctgactat taccggggca caggccgagg acgaagccga ttactattgt gctctgtggt atagcaatct gtgggtgttc gggggtggca caaaactgac tgtgctggga gggggtggat ccggcggagg tggacaggtc cagctggtcc agagcggggc cgaagtcaaa aaacccggag caagcgtgaa ggtctcctgc aaagcatcag gctatacatt tacaggcagc tggatgaact gggtgaggca ggctccagga cagggactgg agtggatcgg gcgcatctac cctggagacg gcgaaactaa ctataatgga aagttcaaag accgagtgac catcacagcc gataagtcta ctagtaccgc ctacatggag ctgagctccc tgcggtctga agataccgcc gtctactatt gcgctagaat ttacggaaac aatgtctatt ttgacgtgtg ggggcaggga acaactgtga ctgtctcctc cggaggatgt ggcggtggaa aagtggccgc actgaaggag aaagttgctg ctttgaaaga gaaggtcgcc gcacttaagg aaaaggtcgc agccctgaaa gag
Аминокислотная последовательность третьей полипептидной цепи из gpA33 mAb 1 х CD3 mAb 2 х DR5 mAb 2 (SEQ ID NO: 224):
KVQLQQSGAE LVKPGASVKL SCKASGYTFT EYILHWVKQK SGQGLEWIGW FYPGNNNIKY NEKFKDKATL TADKSSSTVY MELSRLTSED SAVYFCARHE QGPGYFDYWG QGTTLTVSSA STKGPSVFPL APSSKSTSGG TAALGCLVKD YFPEPVTVSW NSGALTSGVH TFPAVLQSSG LYSLSSVVTV PSSSLGTQTY ICNVNHKPSN TKVDKRVEPK SCDKTHTCPP CPAPEAAGGP SVFLFPPKPK DTLMISRTPE VTCVVVDVSH EDPEVKFNWY VDGVEVHNAK TKPREEQYNS TYRVVSVLTV LHQDWLNGKE YKCKVSNKAL PAPIEKTISK AKGQPREPQV YTLPPSREEM TKNQVSLSCA VKGFYPSDIA VEWESNGQPE NNYKTTPPVL DSDGSFFLVS KLTVDKSRWQ QGNVFSCSVM HEALHNRYTQ KSLSLSPGK
В SEQ ID NO: 224 аминокислотные остатки 1-119 соответствуют аминокислотной последовательности VH-домена DR5 mAb 2 (SEQ ID NO: 18), остатки 120-217 соответствуют модифицированному СН1-домену (SEQ ID NO: 208), остатки 218-232 соответствуют линкеру (SEQ ID NO: 209), а остатки 233-449 соответствуют CH2-CH3-домену, несущему "углубление" (SEQ ID NO: 53).
Полинуклеотид, который кодируется SEQ ID NO: 224 представляет собой (SEQ ID NO: 225):
aaggtccagc tgcagcagtc tggagctgaa ctggtgaaac ccggggcatc agtgaagctg tcctgcaagg cttctgggta caccttcact gagtatattt tacactgggt aaagcagaag tctggacagg gtcttgagtg gattgggtgg ttttatcctg gaaataataa tataaagtac aatgagaaat tcaaggacaa ggccacactg actgcggaca aatcctccag cacagtctat atggaactta gtagattgac atctgaagac tctgcggtct atttctgtgc aagacacgaa caaggaccag gttactttga ctactggggc caaggcacca ctctcacagt ctcctccgcc tccaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagagagt tgagcccaaa tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaagccgc ggggggaccg tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag gtcacatgcg tggtggtgga cgtgagccac gaagaccctg aggtcaagtt caactggtac gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggaggagatg accaagaacc aggtcagcct gagttgcgca gtcaaaggct tctatcccag cgacatcgcc gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg gactccgacg gctccttctt cctcgtcagc aagctcaccg tggacaagag caggtggcag caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaaccg ctacacgcag aagagcctct ccctgtctcc gggtaaa
Аминокислотная последовательность четвертой полипептидной цепи gpA33 mAb 1 х CD3 mAb 2 х DR5 mAb 2 (SEQ ID NO: 226):
DIVMTQSHKF MSTSVGDRVS ITCKASQDVN TAVAWYQQKP GQSPKLLIYW ASTRHTGVPD RFTGSGSGTD YTLTIKSVQA EDLTLYYCQQ HYITPWTFGG GTKLEIKRTV AAPSVFIFPP SDEQLKSGTA SVVCLLNNFY PREAKVQWKV DNALQSGNSQ ESVTEQDSKD STYSLSSTLT LSKADYEKHK VYACEVTHQG LSSPVTKSFN RGEC
В SEQ ID NO: 226 аминокислотные остатки 1-107 соответствуют аминокислотной последовательности VL-домена DR5 mAb 2 (SEQ ID NO: 13), а остатки 108-214 соответствуют CL каппа домену (SEQ ID NO: 210).
Полинуклеотид, который кодируется SEQ ID NO: 226 представляет собой (SEQ ID NO: 227):
gacattgtga tgacccagtc tcacaaattc atgtccactt cagtaggaga cagggtcagc atcacctgca aggccagtca ggatgtgaat actgctgtag cctggtatca acaaaaacca gggcaatctc ctaaactact gatttactgg gcatccaccc ggcacactgg agtccctgat cgcttcacag gcagtggatc tgggacagat tatacactca ccatcaaaag tgtgcaggct gaagacctga cactttatta ctgtcagcaa cactatatca ctccgtggac gttcggtgga ggcaccaagc tggaaatcaa acgtacggtg gctgcaccat cggtcttcat cttcccgcca tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gt
H. EphA2 mAb 1 х CD3 mAb 2 х DR5 mAb 1
Была сконструирована дополнительная примерная триспецифичная связывающая молекула, состоящая из четырех полипептидных цепей. Триспецифичная связывающая молекула содержит VL- и VH-домены из EphA2 mAb 1, VL- и VH-домены из антитела CD3 mAb, 2 и VL- и VH-домены из DR5 mAb 1, и обозначается соответствующим образом "EphA2 mAb 1 х CD3 mAb 2 х DR5 mAb 1". Аминокислотная последовательность первой полипептидной цепи этой триспецифичной связывающей молекулы представляет собой (SEQ ID NO: 228):
DIQMTQTTSS LSASLGDRIT ISCRASQDIS NYLNWYQQKP DGTVKLLIYY TSRLHSGVPS RFSGSGSGTD YSLTISNLEQ EDIATYFCQQ GYTLYTFGGG TKLEIKGGGS GGGGEVQLVE SGGGLVQPGG SLRLSCAASG FTFSTYAMNW VRQAPGKGLE WVGRIRSKYN NYATYYADSV KGRFTISRDD SKNSLYLQMN SLKTEDTAVY YCVRHGNFGN SYVSWFAYWG QGTLVTVSSG GCGGGEVAAL EKEVAALEKE VAALEKEVAA LEKGGGDKTH TCPPCPAPEA AGGPSVFLFP PKPKDTLMIS RTPEVTCVVV DVSHEDPEVK FNWYVDGVEV HNAKTKPREE QYNSTYRVVS VLTVLHQDWL NGKEYKCKVS NKALPAPIEK TISKAKGQPR EPQVYTLPPS REEMTKNQVS LWCLVKGFYP SDIAVEWESN GQPENNYKTT PPVLDSDGSF FLYSKLTVDK SRWQQGNVFS CSVMHEALHN HYTQKSLSLS PGK
В SEQ ID NO: 228, аминокислотные остатки 1-106 соответствуют аминокислотной последовательности VL-домена EphA2 mAb 1 (SEQ ID NO: 153), остатки 107-114 соответствуют промежуточному спейсерному пептиду GGGSGGGG (линкер 1) (SEQ ID NO: 33), остатки 115-239 соответствуют аминокислотной последовательности VH-домена CD3 mAb 2, содержащего замену D65G (SEQ ID NO: 112), остатки 240-245 соответствуют линкеру GGCGGG (SEQ ID NO: NO: 34), остатки 246-273 соответствуют домену с Е-спиралью (SEQ ID NO: 39), остатки 274-276 представляют собой линкер GGG, остатки 277-286 представляют собой линкер DKTHTCPPCP (SEQ ID NO: 48), а остатки 287-503 представляют собой СН2-СН3-домен, несущий "выпуклость" (SEQ ID NO: 52).
Полинуклеотид, который кодируется SEQ ID NO: 228 представляет собой (SEQ ID NO: 229):
gatatccaga tgacacagac tacatcctcc ctgtctgcct ctctgggaga cagaatcacc atcagttgca gggcaagtca ggacattagc aattatttaa actggtatca gcagaaacca gatggaactg ttaaactcct gatctactac acatcaagat tacactcagg agtcccatca aggttcagtg gcagtgggtc tggaacagat tattctctca ccattagcaa cctggagcaa gaagatattg ccacttactt ttgccaacag ggttatacgc tgtacacgtt cggagggggg accaagctgg aaataaaagg tggaggatcc ggcggcggag gcgaggtgca gctggtggag tctgggggag gcttggtcca gcctggaggg tccctgagac tctcctgtgc agcctctgga ttcaccttca gcacatacgc tatgaattgg gtccgccagg ctccagggaa ggggctggag tgggttggaa ggatcaggtc caagtacaac aattatgcaa cctactatgc cgactctgtg aagggtagat tcaccatctc aagagatgat tcaaagaact cactgtatct gcaaatgaac agcctgaaaa ccgaggacac ggccgtgtat tactgtgtga gacacggtaa cttcggcaat tcttacgtgt cttggtttgc ttattgggga caggggacac tggtgactgt gtcttccgga ggatgtggcg gtggagaagt ggccgcactg gagaaagagg ttgctgcttt ggagaaggag gtcgctgcac ttgaaaagga ggtcgcagcc ctggagaaag gcggcgggga caaaactcac acatgcccac cgtgcccagc acctgaagcc gcggggggac cgtcagtctt cctcttcccc ccaaaaccca aggacaccct catgatctcc cggacccctg aggtcacatg cgtggtggtg gacgtgagcc acgaagaccc tgaggtcaag ttcaactggt acgtggacgg cgtggaggtg cataatgcca agacaaagcc gcgggaggag cagtacaaca gcacgtaccg tgtggtcagc gtcctcaccg tcctgcacca ggactggctg aatggcaagg agtacaagtg caaggtctcc aacaaagccc tcccagcccc catcgagaaa accatctcca aagccaaagg gcagccccga gaaccacagg tgtacaccct gcccccatcc cgggaggaga tgaccaagaa ccaggtcagc ctgtggtgcc tggtcaaagg cttctatccc agcgacatcg ccgtggagtg ggagagcaat gggcagccgg agaacaacta caagaccacg cctcccgtgc tggactccga cggctccttc ttcctctaca gcaagctcac cgtggacaag agcaggtggc agcaggggaa cgtcttctca tgctccgtga tgcatgaggc tctgcacaac cactacacgc agaagagcct ctccctgtct ccgggtaa
Аминокислотная последовательность второй полипептидной цепи EphA2 mAb 1 х CD3 mAb 2 х DR5 mAb 1 (SEQ ID NO: 230):
QAVVTQEPSL TVSPGGTVTL TCRSSTGAVT TSNYANWVQQ KPGQAPRGLI GGTNKRAPWT PARFSGSLLG GKAALTITGA QAEDEADYYC ALWYSNLWVF GGGTKLTVLG GGGSGGGGQV QLKESGPGLV APSQSLSITC TVSGFSLSRY SVHWVRQPPG KGLEWLGMIW GGGSTDYNSA LKSRLSISKD NSKSQVFLKM NSLQTDDTAM YYCARKHGNY YTMDYWGQGT SVTVSSGGCG GGKVAALKEK VAALKEKVAA LKEKVAALKE
В SEQ ID NO: 230, аминокислотные остатки 1-110 соответствуют аминокислотной последовательности VL-домена CD3 mAb 2 (SEQ ID NO: 104), остатки 111-118 соответствуют промежуточному спейсерному пептиду GGGSGGGG (линкер 1) (SEQ ID NO: 33), остатки 119-236 соответствуют аминокислотной последовательности VH-домена EphA2 mAb 1 (SEQ ID NO: 158), остатки 237-242 соответствуют линкеру GGCGGG (SEQ ID NO: 34) а остатки 243-270 представляют собой домен K-спирали (SEQ ID NO: 40).
Полинуклеотид, который кодируется SEQ ID NO: 230 представляет собой (SEQ ID NO: 231):
caggctgtgg tgactcagga gccttcactg accgtgtccc caggcggaac tgtgaccctg acatgcagat ccagcacagg cgcagtgacc acatctaact acgccaattg ggtgcagcag aagccaggac aggcaccaag gggcctgatc gggggtacaa acaaaagggc tccctggacc cctgcacggt tttctggaag tctgctgggc ggaaaggccg ctctgactat taccggggca caggccgagg acgaagccga ttactattgt gctctgtggt atagcaatct gtgggtgttc gggggtggca caaaactgac tgtgctggga gggggtggat ccggcggagg tggacaggtg cagctgaagg agtcaggacc tggcctggtg gcaccctcac agagcctgtc catcacatgc actgtctctg ggttctcatt atccagatat agtgtacact gggttcgcca gcctccagga aagggtctgg agtggctggg aatgatatgg ggtggtggaa gcacagacta taattcagct ctcaaatcca gactgagtat cagcaaggac aactccaaga gccaagtttt cttaaaaatg aacagtctgc aaactgatga cacagccatg tactactgtg ccagaaaaca tggtaactac tatactatgg actactgggg tcaaggaacc tcagtcaccg tctcctccgg aggatgtggc ggtggaaaag tggccgcact gaaggagaaa gttgctgctt tgaaagagaa ggtcgccgca cttaaggaaa aggtcgcagc cctgaaagag
Аминокислотная последовательность третьей полипептидной цепи EphA2 mAb 1 х CD3 mAb 2 х DR5 mAb 1 (SEQ ID NO: 232):
EVKFLESGGG LVQPGGSLKL SCVASGFDFS RYWMSWVRQA PGKGLEWIGE INPDSNTINY TPSLKDKFII SRDNAKNTLY LQMTKVRSED TALYYCTRRA YYGNPAWFAY WGQGTLVTVS SASTKGPSVF PLAPSSKSTS GGTAALGCLV KDYFPEPVTV SWNSGALTSG VHTFPAVLQS SGLYSLSSVV TVPSSSLGTQ TYICNVNHKP SNTKVDKRVE PKSCDKTHTC PPCPAPEAAG GPSVFLFPPK PKDTLMISRT PEVTCVVVDV SHEDPEVKFN WYVDGVEVHN AKTKPREEQY NSTYRVVSVL TVLHQDWLNG KEYKCKVSNK ALPAPIEKTI SKAKGQPREP QVYTLPPSRE EMTKNQVSLS CAVKGFYPSD IAVEWESNGQ PENNYKTTPP VLDSDGSFFL VSKLTVDKSR WQQGNVFSCS VMHEALHNRY TQKSLSLSPG
K
В SEQ ID NO: 232, аминокислотные остатки 1-121 соответствуют аминокислотной последовательности VH-домена DR5 mAb 1 (SEQ ID NO: 8), остатки 122-219 соответствуют измененному СН1-домену (SEQ ID NO: 208), остатки 220-234 соответствуют линкеру (SEQ ID NO: 209), а остатки 235-451 соответствуют CH2-CH3-домену, несущему "выпуклость" (SEQ ID NO: 53).
Полинуклеотид, который кодируется SEQ ID NO: 232 представляет собой (SEQ ID NO: 233):
gaggtgaagt ttctcgagtc tggaggtggc ctggtgcagc ctggaggatc cctgaaactc tcctgtgtag cctcaggatt cgattttagt agatactgga tgagttgggt ccggcaggct ccagggaaag ggctagaatg gattggagaa attaatccag atagcaatac gataaactat acgccatctc taaaggataa attcatcatc tccagagaca acgccaaaaa tacgctgtat ctgcaaatga ccaaagtgag atctgaggac acagcccttt attattgtac aagaagggcc tactatggta acccggcctg gtttgcttac tggggccaag ggactctggt cactgtctct tccgcctcca ccaagggccc atcggtcttc cccctggcac cctcctccaa gagcacctct gggggcacag cggccctggg ctgcctggtc aaggactact tccccgaacc ggtgacggtg tcgtggaact caggcgccct gaccagcggc gtgcacacct tcccggctgt cctacagtcc tcaggactct actccctcag cagcgtggtg accgtgccct ccagcagctt gggcacccag acctacatct gcaacgtgaa tcacaagccc agcaacacca aggtggacaa gagagttgag cccaaatctt gtgacaaaac tcacacatgc ccaccgtgcc cagcacctga agccgcgggg ggaccgtcag tcttcctctt ccccccaaaa cccaaggaca ccctcatgat ctcccggacc cctgaggtca catgcgtggt ggtggacgtg agccacgaag accctgaggt caagttcaac tggtacgtgg acggcgtgga ggtgcataat gccaagacaa agccgcggga ggagcagtac aacagcacgt accgtgtggt cagcgtcctc accgtcctgc accaggactg gctgaatggc aaggagtaca agtgcaaggt ctccaacaaa gccctcccag cccccatcga gaaaaccatc tccaaagcca aagggcagcc ccgagaacca caggtgtaca ccctgccccc atcccgggag gagatgacca agaaccaggt cagcctgagt tgcgcagtca aaggcttcta tcccagcgac atcgccgtgg agtgggagag caatgggcag ccggagaaca actacaagac cacgcctccc gtgctggact ccgacggctc cttcttcctc gtcagcaagc tcaccgtgga caagagcagg tggcagcagg ggaacgtctt ctcatgctcc gtgatgcatg aggctctgca caaccgctac acgcagaaga gcctctccct gtctccgggt aaa
Аминокислотная последовательность четвертой полипептидной цепи EphA2 mAb 1 х CD3 mAb 2 х DR5 mAb 1 (SEQ ID NO: 234):
DIVLTQSPAS LAVSLGQRAT ISCRASKSVS SSGYSYMHWY QQKPGQPPKV LIFLSSNLDS GVPARFSGSG SGTDFTLNIH PVEDGDAATY YCQHSRDLPP TFGGGTKLEI KRTVAAPSVF IFPPSDEQLK SGTASVVCLL NNFYPREAKV QWKVDNALQS GNSQESVTEQ DSKDSTYSLS STLTLSKADY EKHKVYACEV THQGLSSPVT KSFNRGEC
В SEQ ID NO: 234, аминокислотные остатки 1-111 соответствуют аминокислотной последовательности VL-домена DR5 mAb 1 (SEQ ID NO: 3), а остатки 112-218 соответствуют CL Каппа домену (SEQ ID NO: 210).
Полинуклеотид, который кодируется SEQ ID NO: 234 представляет собой (SEQ ID NO: 235):
gacattgtgc tgacacagtc tcctgcttcc ttagctgtat ctctcgggca gagggccacc atctcatgca gggccagcaa aagtgtcagt tcctctggct atagttatat gcactggtac caacagaaac caggacagcc acccaaagtc ctcatctttc tttcatccaa cctagattct ggggtccctg ccaggttcag tggcagtggg tctgggacag acttcaccct caacatccat cctgtggagg atggggatgc tgcaacctat tactgtcagc acagtaggga tcttcctccg acgttcggtg gaggcaccaa gctggaaatc aaacgtacgg tggctgcacc atcggtcttc atcttcccgc catctgatga gcagttgaaa tctggaactg cctctgttgt gtgcctgctg aataacttct atcccagaga ggccaaagta cagtggaagg tggataacgc cctccaatcg ggtaactccc aggagagtgt cacagagcag gacagcaagg acagcaccta cagcctcagc agcaccctga cgctgagcaa agcagactac gagaaacaca aagtctacgc ctgcgaagtc acccatcagg gcctgagctc gcccgtcaca aagagcttca acaggggaga gtgt
I. EphA2 mAb 2 х CD3 mAb 2 х DR5 mAb 1
Была сконструирована дополнительная примерная приметриспецифичная связывающая молекула, состоящая из четырех полипептидных цепей. Триспецифичная связывающая молекула содержит VL- и VH-домены из EphA2 mAb 2, VL- и VH-домены антитела CD3 mAb 2 и VL- и VH-домены из DR5 mAb 1, и обозначается соответствующим образом "EphA2 mAb 2 х CD3 mAb 2 х DR5 mAb 1". Аминокислотная последовательность первой полипептидной цепи этой триспецифичной связывающей молекулы представляет собой (SEQ ID NO: 236):
DVVMTQTPLS LPVSLGDQAS ISCRSSQSLV HSSGNTYLHW YLQKPGQSPK LLIYKVSNRF SGVPDRFSGS GSGTDFTLKI SRVEAEDLGV YFCSQSTHVP TFGSGTKLEI KGGGSGGGGE VQLVESGGGL VQPGGSLRLS CAASGFTFST YAMNWVRQAP GKGLEWVGRI RSKYNNYATY YADSVKGRFT ISRDDSKNSL YLQMNSLKTE DTAVYYCVRH GNFGNSYVSW FAYWGQGTLV TVSSGGCGGG EVAALEKEVA ALEKEVAALE KEVAALEKGG GDKTHTCPPC PAPEAAGGPS VFLFPPKPKD TLMISRTPEV TCVVVDVSHE DPEVKFNWYV DGVEVHNAKT KPREEQYNST YRVVSVLTVL HQDWLNGKEY KCKVSNKALP APIEKTISKA KGQPREPQVY TLPPSREEMT KNQVSLWCLV KGFYPSDIAV EWESNGQPEN NYKTTPPVLD SDGSFFLYSK LTVDKSRWQQ GNVFSCSVMH EALHNHYTQK SLSLSPGK
В SEQ ID NO: 236, аминокислотные остатки 1-111 соответствуют аминокислотной последовательности VL-домена EphA2 mAb 2 (SEQ ID NO: 163), остатки 112-119 соответствуют промежуточному спейсерному пептиду GGGSGGGG (линкер 1) (SEQ ID NO: 33), остатки 120-244 соответствуют аминокислотной последовательности VH-домена CD3 mAb 2, содержащего D65G замену (SEQ ID NO: 112), остатки 245-250 соответствуют GGCGGG линкеру (SEQ ID NO: NO: 34), остатки 251-278 соответствуют домену Е-спирали (SEQ ID NO: 39), остатки 279-281 представляют собой линкер GGG, остатки 282-291 представляют собой линкер DKTHTCPPCP (SEQ ID NO: 48), а остатки 292-508 представляют собой СН2-СН3-домен, несущий выпуклость (SEQ ID NO: 52).
Полинуклеотид, который кодируется SEQ ID NO: 236, представляет собой (SEQ ID NO: 237):
gatgttgtga tgacccaaac tccactctcc ctgcctgtca gtcttggaga tcaagcctcc atctcttgca gatctagtca gagccttgta cacagtagtg gaaacaccta tttacattgg tacctgcaga agccaggcca gtctccaaag ctcctgatct acaaagtttc caaccgattt tctggggtcc cagacaggtt cagtggcagt ggatcaggga cagatttcac actcaagatc agcagagtgg aggctgagga tctgggagtt tatttctgct ctcaaagtac acatgttccc acgttcggct cggggacaaa gttggaaata aaaggtggag gatccggcgg cggaggcgag gtgcagctgg tggagtctgg gggaggcttg gtccagcctg gagggtccct gagactctcc tgtgcagcct ctggattcac cttcagcaca tacgctatga attgggtccg ccaggctcca gggaaggggc tggagtgggt tggaaggatc aggtccaagt acaacaatta tgcaacctac tatgccgact ctgtgaaggg tagattcacc atctcaagag atgattcaaa gaactcactg tatctgcaaa tgaacagcct gaaaaccgag gacacggccg tgtattactg tgtgagacac ggtaacttcg gcaattctta cgtgtcttgg tttgcttatt ggggacaggg gacactggtg actgtgtctt ccggaggatg tggcggtgga gaagtggccg cactggagaa agaggttgct gctttggaga aggaggtcgc tgcacttgaa aaggaggtcg cagccctgga gaaaggcggc ggggacaaaa ctcacacatg cccaccgtgc ccagcacctg aagccgcggg gggaccgtca gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc acatgcgtgg tggtggacgt gagccacgaa gaccctgagg tcaagttcaa ctggtacgtg gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga ggagatgacc aagaaccagg tcagcctgtg gtgcctggtc aaaggcttct atcccagcga catcgccgtg gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag agcctctccc tgtctccggg taaa
Аминокислотная последовательность второй полипептидной цепи EphA2 mAb 2 х CD3 mAb 2 х DR5 mAb 1 представляет собой (SEQ ID NO: 238):
QAVVTQEPSL TVSPGGTVTL TCRSSTGAVT TSNYANWVQQ KPGQAPRGLI GGTNKRAPWT PARFSGSLLG GKAALTITGA QAEDEADYYC ALWYSNLWVF GGGTKLTVLG GGGSGGGGQI QLVQSGPELK KPGETVKISC KASGFTFTNY GMNWVKQAPG KGLKWMGWIN TYIGEPTYAD DFKGRFVFSL ETSASTAYLQ INNLKNEDMA TYFCARELGP YYFDYWGQGT TLTVSSGGCG GGKVAALKEK VAALKEKVAA LKEKVAALKE
В SEQ ID NO: 238, аминокислотные остатки 1-110 соответствуют аминокислотной последовательности VL-домена CD3 mAb 2 (SEQ ID NO: 104), остатки 111-118 соответствуют промежуточному спейсерному пептиду GGGSGGGG (линкер 1) (SEQ ID NO: 33), остатки 119-236 соответствуют аминокислотной последовательности VH-домена EphA2 mAb 2 (SEQ ID NO: 167), остатки 237-242 соответствуют линкеру GGCGGG (SEQ ID NO: 34) а остатки 243-270 представляют собой домен K-спирали (SEQ ID NO: 40).
Полинуклеотид, который кодируется SEQ ID NO: 238, представляет собой (SEQ ID NO: 239):
caggctgtgg tgactcagga gccttcactg accgtgtccc caggcggaac tgtgaccctg acatgcagat ccagcacagg cgcagtgacc acatctaact acgccaattg ggtgcagcag aagccaggac aggcaccaag gggcctgatc gggggtacaa acaaaagggc tccctggacc cctgcacggt tttctggaag tctgctgggc ggaaaggccg ctctgactat taccggggca caggccgagg acgaagccga ttactattgt gctctgtggt atagcaatct gtgggtgttc gggggtggca caaaactgac tgtgctggga gggggtggat ccggcggagg tggacagatc cagttggtgc agtctggacc tgagctgaag aagcctggag agacagtcaa gatctcctgc aaggcttctg ggtttacctt cacaaactat ggaatgaact gggtgaagca ggctccagga aagggtttaa agtggatggg ctggataaac acctatattg gagagccgac atatgctgat gacttcaagg gacggtttgt cttctctttg gaaacctctg ccagcactgc ctatttgcag atcaacaacc tcaaaaatga ggacatggcc acatatttct gtgcaagaga actgggacca tactactttg actactgggg ccaaggcacc actctcacag tctcctccgg aggatgtggc ggtggaaaag tggccgcact gaaggagaaa gttgctgctt tgaaagagaa ggtcgccgca cttaaggaaa aggtcgcagc cctgaaagag
Аминокислотная последовательность третьей полипептидной цепи EphA2 mAb 2 х CD3 mAb 2 х DR5 mAb 1 представляет собой (SEQ ID NO: 240):
EVKFLESGGG LVQPGGSLKL SCVASGFDFS RYWMSWVRQA PGKGLEWIGE INPDSNTINY TPSLKDKFII SRDNAKNTLY LQMTKVRSED TALYYCTRRA YYGNPAWFAY WGQGTLVTVS SASTKGPSVF PLAPSSKSTS GGTAALGCLV KDYFPEPVTV SWNSGALTSG VHTFPAVLQS SGLYSLSSVV TVPSSSLGTQ TYICNVNHKP SNTKVDKRVE PKSCDKTHTC PPCPAPEAAG GPSVFLFPPK PKDTLMISRT PEVTCVVVDV SHEDPEVKFN WYVDGVEVHN AKTKPREEQY NSTYRVVSVL TVLHQDWLNG KEYKCKVSNK ALPAPIEKTI SKAKGQPREP QVYTLPPSRE EMTKNQVSLS CAVKGFYPSD IAVEWESNGQ PENNYKTTPP VLDSDGSFFL VSKLTVDKSR WQQGNVFSCS VMHEALHNRY TQKSLSLSPG K
В SEQ ID NO: 240, аминокислотные остатки 1-121 соответствуют аминокислотной последовательности VH-домена DR5 mAb 1 (SEQ ID NO: 8), остатки 122-219 соответствуют модифицированному СН1-домену (SEQ ID NO: 208), остатки 220-234 соответствуют линкеру (SEQ ID NO: 209), а остатки 235-451 соответствуют "несущему углубление" CH2-CH3-домену (SEQ ID NO: 53).
Полинуклеотид, который кодируется SEQ ID NO: 240, представляет собой (SEQ ID NO: 241):
gaggtgaagt ttctcgagtc tggaggtggc ctggtgcagc ctggaggatc cctgaaactc tcctgtgtag cctcaggatt cgattttagt agatactgga tgagttgggt ccggcaggct ccagggaaag ggctagaatg gattggagaa attaatccag atagcaatac gataaactat acgccatctc taaaggataa attcatcatc tccagagaca acgccaaaaa tacgctgtat ctgcaaatga ccaaagtgag atctgaggac acagcccttt attattgtac aagaagggcc tactatggta acccggcctg gtttgcttac tggggccaag ggactctggt cactgtctct tccgcctcca ccaagggccc atcggtcttc cccctggcac cctcctccaa gagcacctct gggggcacag cggccctggg ctgcctggtc aaggactact tccccgaacc ggtgacggtg tcgtggaact caggcgccct gaccagcggc gtgcacacct tcccggctgt cctacagtcc tcaggactct actccctcag cagcgtggtg accgtgccct ccagcagctt gggcacccag acctacatct gcaacgtgaa tcacaagccc agcaacacca aggtggacaa gagagttgag cccaaatctt gtgacaaaac tcacacatgc ccaccgtgcc cagcacctga agccgcgggg ggaccgtcag tcttcctctt ccccccaaaa cccaaggaca ccctcatgat ctcccggacc cctgaggtca catgcgtggt ggtggacgtg agccacgaag accctgaggt caagttcaac tggtacgtgg acggcgtgga ggtgcataat gccaagacaa agccgcggga ggagcagtac aacagcacgt accgtgtggt cagcgtcctc accgtcctgc accaggactg gctgaatggc aaggagtaca agtgcaaggt ctccaacaaa gccctcccag cccccatcga gaaaaccatc tccaaagcca aagggcagcc ccgagaacca caggtgtaca ccctgccccc atcccgggag gagatgacca agaaccaggt cagcctgagt tgcgcagtca aaggcttcta tcccagcgac atcgccgtgg agtgggagag caatgggcag ccggagaaca actacaagac cacgcctccc gtgctggact ccgacggctc cttcttcctc gtcagcaagc tcaccgtgga caagagcagg tggcagcagg ggaacgtctt ctcatgctcc gtgatgcatg aggctctgca caaccgctac acgcagaaga gcctctccct gtctccgggt aaa
Аминокислотная последовательность четвертой полипептидной цепи EphA2 mAb 2 х CD3 mAb 2 х DR5 mAb 1 представляет собой (SEQ ID NO: 242):
DIVLTQSPAS LAVSLGQRAT ISCRASKSVS SSGYSYMHWY QQKPGQPPKV LIFLSSNLDS GVPARFSGSG SGTDFTLNIH PVEDGDAATY YCQHSRDLPP TFGGGTKLEI KRTVAAPSVF IFPPSDEQLK SGTASVVCLL NNFYPREAKV QWKVDNALQS GNSQESVTEQ DSKDSTYSLS STLTLSKADY EKHKVYACEV THQGLSSPVT KSFNRGEC
В SEQ ID NO: 242, аминокислотные остатки 1-111 соответствуют аминокислотной последовательности VL-домена DR5 mAb 1 (SEQ ID NO: 3), а остатки 112-218 соответствуют домену CL Каппа (SEQ ID NO: 210).
Полинуклеотид, который кодируется SEQ ID NO: 242, представляет собой (SEQ ID NO: 243):
gacattgtgc tgacacagtc tcctgcttcc ttagctgtat ctctcgggca gagggccacc atctcatgca gggccagcaa aagtgtcagt tcctctggct atagttatat gcactggtac caacagaaac caggacagcc acccaaagtc ctcatctttc tttcatccaa cctagattct ggggtccctg ccaggttcag tggcagtggg tctgggacag acttcaccct caacatccat cctgtggagg atggggatgc tgcaacctat tactgtcagc acagtaggga tcttcctccg acgttcggtg gaggcaccaa gctggaaatc aaacgtacgg tggctgcacc atcggtcttc atcttcccgc catctgatga gcagttgaaa tctggaactg cctctgttgt gtgcctgctg aataacttct atcccagaga ggccaaagta cagtggaagg tggataacgc cctccaatcg ggtaactccc aggagagtgt cacagagcag gacagcaagg acagcaccta cagcctcagc agcaccctga cgctgagcaa agcagactac gagaaacaca aagtctacgc ctgcgaagtc acccatcagg gcctgagctc gcccgtcaca aagagcttca acaggggaga gtgt
J. EphA2 mAb 3 х CD3 mAb 2 х DR5 mAb 1
Была сконструирована дополнительная примерная триметриспецифичная связывающая молекула, состоящая из четырех полипептидных цепей. Триспецифичная связывающая молекула содержит VL- и VH-домены из EphA2 mAb 3, VL- и VH-домены антитела CD3 mAb, 2 и VL- и VH-домены из DR5 mAb 1, и обозначается соответствующим образом "EphA2 mAb 3 х CD3 mAb 2 х DR5 mAb 1". Аминокислотная последовательность первой полипептидной цепи этой триспецифичной связывающей молекулы представляет собой (SEQ ID NO: 244):
DIVLTQSHRS MSTSVGDRVN ITCKASQDVT TAVAWYQQKP GQSPKLLIFW ASTRHAGVPD RFTGSGSGTD FTLTISSVQA GDLALYYCQQ HYSTPYTFGG GTKLEIKGGG SGGGGEVQLV ESGGGLVQPG GSLRLSCAAS GFTFSTYAMN WVRQAPGKGL EWVGRIRSKY NNYATYYADS VKGRFTISRD DSKNSLYLQM NSLKTEDTAV YYCVRHGNFG NSYVSWFAYW GQGTLVTVSS GGCGGGEVAA LEKEVAALEK EVAALEKEVA ALEKGGGDKT HTCPPCPAPE AAGGPSVFLF PPKPKDTLMI SRTPEVTCVV VDVSHEDPEV KFNWYVDGVE VHNAKTKPRE EQYNSTYRVV SVLTVLHQDW LNGKEYKCKV SNKALPAPIE KTISKAKGQP REPQVYTLPP SREEMTKNQV SLWCLVKGFY PSDIAVEWES NGQPENNYKT TPPVLDSDGS FFLYSKLTVD KSRWQQGNVF SCSVMHEALH NHYTQKSLSL SPGK
В SEQ ID NO: 244, аминокислотные остатки 1-107 соответствуют аминокислотной последовательности VL-домена EphA2 mAb 3 (SEQ ID NO: 172), остатки 108-115 соответствуют промежуточному спейсерному пептиду GGGSGGGG (линкер 1) (SEQ ID NO: 33), остатки 116-240 соответствуют аминокислотной последовательности VH-домена CD3 mAb 2, содержащего D65G замену (SEQ ID NO: 112), остатки 241-246 соответствуют GGCGGG линкеру (SEQ ID NO: NO: 34), остатки 247-274 соответствуют домену с Е-спиралью (SEQ ID NO: 39), остатки 275-277 представляют собой линкер GGG, остатки 278-287 представляют собой линкер DKTHTCPPCP (SEQ ID NO: 48), а остатки 288-504 представляют собой "несущий выпуклость" СН2-СН3-домен (SEQ ID NO: 52).
Полинуклеотид, который кодируется SEQ ID NO: 244, представляет собой (SEQ ID NO: 245):
gacattgtgc tgacccagtc tcacagatcc atgtccacat cagtaggaga cagggtcaac atcacctgca aggccagtca ggatgtgact actgctgtag cctggtatca acaaaaacca gggcaatctc ctaaattact gattttctgg gcatccaccc ggcacgctgg agtccctgat cgcttcacag gcagtggatc tgggacagat tttactctca ccatcagcag tgtgcaggct ggagacctgg cactttatta ctgtcaacaa cattatagca caccgtacac attcggaggg gggaccaagc tggaaataaa aggtggagga tccggcggcg gaggcgaggt gcagctggtg gagtctgggg gaggcttggt ccagcctgga gggtccctga gactctcctg tgcagcctct ggattcacct tcagcacata cgctatgaat tgggtccgcc aggctccagg gaaggggctg gagtgggttg gaaggatcag gtccaagtac aacaattatg caacctacta tgccgactct gtgaagggta gattcaccat ctcaagagat gattcaaaga actcactgta tctgcaaatg aacagcctga aaaccgagga cacggccgtg tattactgtg tgagacacgg taacttcggc aattcttacg tgtcttggtt tgcttattgg ggacagggga cactggtgac tgtgtcttcc ggaggatgtg gcggtggaga agtggccgca ctggagaaag aggttgctgc tttggagaag gaggtcgctg cacttgaaaa ggaggtcgca gccctggaga aaggcggcgg ggacaaaact cacacatgcc caccgtgccc agcacctgaa gccgcggggg gaccgtcagt cttcctcttc cccccaaaac ccaaggacac cctcatgatc tcccggaccc ctgaggtcac atgcgtggtg gtggacgtga gccacgaaga ccctgaggtc aagttcaact ggtacgtgga cggcgtggag gtgcataatg ccaagacaaa gccgcgggag gagcagtaca acagcacgta ccgtgtggtc agcgtcctca ccgtcctgca ccaggactgg ctgaatggca aggagtacaa gtgcaaggtc tccaacaaag ccctcccagc ccccatcgag aaaaccatct ccaaagccaa agggcagccc cgagaaccac aggtgtacac cctgccccca tcccgggagg agatgaccaa gaaccaggtc agcctgtggt gcctggtcaa aggcttctat cccagcgaca tcgccgtgga gtgggagagc aatgggcagc cggagaacaa ctacaagacc acgcctcccg tgctggactc cgacggctcc ttcttcctct acagcaagct caccgtggac aagagcaggt ggcagcaggg gaacgtcttc tcatgctccg tgatgcatga ggctctgcac aaccactaca cgcagaagag cctctccctg tctccgggta aa
Аминокислотная последовательность второй полипептидной цепи EphA2 mAb 3 х CD3 mAb 2 х DR5 mAb 1 представляет собой (SEQ ID NO: 246):
QAVVTQEPSL TVSPGGTVTL TCRSSTGAVT TSNYANWVQQ KPGQAPRGLI GGTNKRAPWT PARFSGSLLG GKAALTITGA QAEDEADYYC ALWYSNLWVF GGGTKLTVLG GGGSGGGGEV QLVESGGGSV KPGGSLKLSC AASGFTFTDH YMYWVRQTPE KRLEWVATIS DGGSFTSYPD SVKGRFTISR DIAKNNLYLQ MSSLKSEDTA MYYCTRDESD RPFPYWGQGT LVTVSSGGCG GGKVAALKEK VAALKEKVAA LKEKVAALKE
В SEQ ID NO: 246, аминокислотные остатки 1-110 соответствуют аминокислотной последовательности VL-домена CD3 mAb 2 (SEQ ID NO: 104), остатки 111-118 соответствуют промежуточному спейсерному пептиду GGGSGGGG (линкер 1) (SEQ ID NO: 33), остатки 119-236 соответствуют аминокислотной последовательности VH-домена EphA2 mAb 3 (SEQ ID NO: 177), остатки 237-242 соответствуют линкеру GGCGGG (SEQ ID NO: 34) а остатки 243-270 представляют собой домен K-спирали (SEQ ID NO: 40).
Полинуклеотид, который кодируется SEQ ID NO: 246, представляет собой (SEQ ID NO: 247):
caggctgtgg tgactcagga gccttcactg accgtgtccc caggcggaac tgtgaccctg acatgcagat ccagcacagg cgcagtgacc acatctaact acgccaattg ggtgcagcag aagccaggac aggcaccaag gggcctgatc gggggtacaa acaaaagggc tccctggacc cctgcacggt tttctggaag tctgctgggc ggaaaggccg ctctgactat taccggggca caggccgagg acgaagccga ttactattgt gctctgtggt atagcaatct gtgggtgttc gggggtggca caaaactgac tgtgctggga gggggtggat ccggcggagg tggagaagtg cagctggtgg agtctggggg aggctcagtg aagcctggag ggtccctgaa actctcctgt gcagcctctg gattcacttt cactgaccat tacatgtatt gggttcgcca gactccggaa aagaggctgg agtgggtcgc aaccattagt gatggcggta gtttcacctc ctatccagac agtgtgaagg ggcgattcac catctccaga gacattgcca agaacaacct gtacctccaa atgagcagtc tgaagtctga ggacacagcc atgtattact gtacaagaga tgagagcgat aggccgtttc cttactgggg ccaagggact ctggtcactg tctcctccgg aggatgtggc ggtggaaaag tggccgcact gaaggagaaa gttgctgctt tgaaagagaa ggtcgccgca cttaaggaaa aggtcgcagc cctgaaagag
Аминокислотная последовательность третьей полипептидной цепи EphA2 mAb 3 х CD3 mAb 2 х DR5 mAb 1 представляет собой (SEQ ID NO: 248):
EVKFLESGGG LVQPGGSLKL SCVASGFDFS RYWMSWVRQA PGKGLEWIGE INPDSNTINY TPSLKDKFII SRDNAKNTLY LQMTKVRSED TALYYCTRRA YYGNPAWFAY WGQGTLVTVS SASTKGPSVF PLAPSSKSTS GGTAALGCLV KDYFPEPVTV SWNSGALTSG VHTFPAVLQS SGLYSLSSVV TVPSSSLGTQ TYICNVNHKP SNTKVDKRVE PKSCDKTHTC PPCPAPEAAG GPSVFLFPPK PKDTLMISRT PEVTCVVVDV SHEDPEVKFN WYVDGVEVHN AKTKPREEQY NSTYRVVSVL TVLHQDWLNG KEYKCKVSNK ALPAPIEKTI SKAKGQPREP QVYTLPPSRE EMTKNQVSLS CAVKGFYPSD IAVEWESNGQ PENNYKTTPP VLDSDGSFFL VSKLTVDKSR WQQGNVFSCS VMHEALHNRY TQKSLSLSPG K
В SEQ ID NO: 248, аминокислотные остатки 1-121 соответствуют аминокислотной последовательности VH-домена DR5 mAb 1 (SEQ ID NO: 8), остатки 122-219 соответствуют модифицированному СН1-домену (SEQ ID NO: 208), остатки 220-234 соответствуют линкеру (SEQ ID NO: 209), а остатки 235-451 соответствуют "несущему углубление" CH2-CH3-домену (SEQ ID NO: 53).
Полинуклеотид, который кодируется SEQ ID NO: 248, представляет собой (SEQ ID NO: 249):
gaggtgaagt ttctcgagtc tggaggtggc ctggtgcagc ctggaggatc cctgaaactc tcctgtgtag cctcaggatt cgattttagt agatactgga tgagttgggt ccggcaggct ccagggaaag ggctagaatg gattggagaa attaatccag atagcaatac gataaactat acgccatctc taaaggataa attcatcatc tccagagaca acgccaaaaa tacgctgtat ctgcaaatga ccaaagtgag atctgaggac acagcccttt attattgtac aagaagggcc tactatggta acccggcctg gtttgcttac tggggccaag ggactctggt cactgtctct tccgcctcca ccaagggccc atcggtcttc cccctggcac cctcctccaa gagcacctct gggggcacag cggccctggg ctgcctggtc aaggactact tccccgaacc ggtgacggtg tcgtggaact caggcgccct gaccagcggc gtgcacacct tcccggctgt cctacagtcc tcaggactct actccctcag cagcgtggtg accgtgccct ccagcagctt gggcacccag acctacatct gcaacgtgaa tcacaagccc agcaacacca aggtggacaa gagagttgag cccaaatctt gtgacaaaac tcacacatgc ccaccgtgcc cagcacctga agccgcgggg ggaccgtcag tcttcctctt ccccccaaaa cccaaggaca ccctcatgat ctcccggacc cctgaggtca catgcgtggt ggtggacgtg agccacgaag accctgaggt caagttcaac tggtacgtgg acggcgtgga ggtgcataat gccaagacaa agccgcggga ggagcagtac aacagcacgt accgtgtggt cagcgtcctc accgtcctgc accaggactg gctgaatggc aaggagtaca agtgcaaggt ctccaacaaa gccctcccag cccccatcga gaaaaccatc tccaaagcca aagggcagcc ccgagaacca caggtgtaca ccctgccccc atcccgggag gagatgacca agaaccaggt cagcctgagt tgcgcagtca aaggcttcta tcccagcgac atcgccgtgg agtgggagag caatgggcag ccggagaaca actacaagac cacgcctccc gtgctggact ccgacggctc cttcttcctc gtcagcaagc tcaccgtgga caagagcagg tggcagcagg ggaacgtctt ctcatgctcc gtgatgcatg aggctctgca caaccgctac acgcagaaga gcctctccct gtctccgggt aaa
Аминокислотная последовательность четвертой полипептидной цепи EphA2 mAb 3 х CD3 mAb 2 х DR5 mAb 1 (SEQ ID NO: 250):
DIVLTQSPAS LAVSLGQRAT ISCRASKSVS SSGYSYMHWY QQKPGQPPKV LIFLSSNLDS GVPARFSGSG SGTDFTLNIH PVEDGDAATY YCQHSRDLPP TFGGGTKLEI KRTVAAPSVF IFPPSDEQLK SGTASVVCLL NNFYPREAKV QWKVDNALQS GNSQESVTEQ DSKDSTYSLS STLTLSKADY EKHKVYACEV THQGLSSPVT KSFNRGEC
В SEQ ID NO: 250, аминокислотные остатки 1-111 соответствуют аминокислотной последовательности VL-домена DR5 mAb 1 (SEQ ID NO: 3), а остатки 112-218 соответствуют домену CL Каппа (SEQ ID NO: 210).
Полинуклеотид, который кодируется SEQ ID NO: 250 представляет собой (SEQ ID NO: 251):
gacattgtgc tgacacagtc tcctgcttcc ttagctgtat ctctcgggca gagggccacc atctcatgca gggccagcaa aagtgtcagt tcctctggct atagttatat gcactggtac caacagaaac caggacagcc acccaaagtc ctcatctttc tttcatccaa cctagattct ggggtccctg ccaggttcag tggcagtggg tctgggacag acttcaccct caacatccat cctgtggagg atggggatgc tgcaacctat tactgtcagc acagtaggga tcttcctccg acgttcggtg gaggcaccaa gctggaaatc aaacgtacgg tggctgcacc atcggtcttc atcttcccgc catctgatga gcagttgaaa tctggaactg cctctgttgt gtgcctgctg aataacttct atcccagaga ggccaaagta cagtggaagg tggataacgc cctccaatcg ggtaactccc aggagagtgt cacagagcag gacagcaagg acagcaccta cagcctcagc agcaccctga cgctgagcaa agcagactac gagaaacaca aagtctacgc ctgcgaagtc acccatcagg gcctgagctc gcccgtcaca aagagcttca acaggggaga gtgt
Хотя примерные триспецифичные связывающие молекулы, описанные выше, включают три CDR легкой цепи (VL) и три CDR тяжелой цепи (VH) для каждого связывающего домена, будет понятно, что изобретение также включает триспецифичные связывающие молекулы, которые содержат:
(1) по меньшей мере, одну из CDR VL-домена любого такого связывающего домена;
(2) по меньшей мере, две из CDR VL-домена любого такого связывающего домена;
(3) три CDR VL-домена любого такого связывающего домена;
(4) по меньшей мере, одну из CDR VH-домена любого такого связывающего домена;
(5) по меньшей мере, две из CDR VH-домена любого такого связывающего домена;
(6) три CDR VH-домена любого такого связывающего домена;
(7) по меньшей мере, одну из CDR VL-домена любого такого связывающего домена и, по меньшей мере, одну из CDR VH-домена этого связывающего домена;
(8) по меньшей мере, две из CDR VL-домена любого такого связывающего домена и, по меньшей мере, две из CDR VH-домена этого связывающего домена;
(9) три CDR VL-домена любого такого связывающего домена и три CDR VH-домена этого связывающего домена;
(10) VL-домен любого такого связывающего домена;
(11) VH-домена любого такого связывающего домена; или
(12) VL и VH-домены любого такого связывающего домена.
K. gpA33 mAb 1 х CD3 mAb 2 х EphA2 mAb 1
Триспецифичная связывающая молекула состоит из четырех полипептидных цепей включает VL- и VH-домены из gpA33 mAb 1, VL- и VH-домены антитела CD3 mAb, 2 и VL- и VH-домены из EphA2 mAb 1. Триспецифическая связывающаяся молекула был соответствующим образом обозначена "gpA33 mAb 1 х CD3 mAb 2 х EphA2 mAb 1". Аминокислотная последовательность первой полипептидной цепи этой триспецифичной связывающей молекулы представляют собой (SEQ ID NO: 252):
DIQLTQSPSF LSASVGDRVT ITCSARSSIS FMYWYQQKPG KAPKLLIYDT SNLASGVPSR FSGSGSGTEF TLTISSLEAE DAATYYCQQW SSYPLTFGQG TKLEIKGGGS GGGGEVQLVE SGGGLVQPGG SLRLSCAASG FTFSTYAMNW VRQAPGKGLE WVGRIRSKYN NYATYYADSV KGRFTISRDD SKNSLYLQMN SLKTEDTAVY YCVRHGNFGN SYVSWFAYWG QGTLVTVSSG GCGGGEVAAL EKEVAALEKE VAALEKEVAA LEKGGGDKTH TCPPCPAPEA AGGPSVFLFP PKPKDTLMIS RTPEVTCVVV DVSHEDPEVK FNWYVDGVEV HNAKTKPREE QYNSTYRVVS VLTVLHQDWL NGKEYKCKVS NKALPAPIEK TISKAKGQPR EPQVYTLPPS REEMTKNQVS LWCLVKGFYP SDIAVEWESN GQPENNYKTT PPVLDSDGSF FLYSKLTVDK SRWQQGNVFS CSVMHEALHN HYTQKSLSLS PGK
В SEQ ID NO: 252, аминокислотные остатки 1-106 соответствуют аминокислотной последовательности VL-домена gpA33 mAb 1 (SEQ ID NO: 181), остатки 107-114 соответствуют промежуточному спейсерному пептиду GGGSGGGG (линкер 1) (SEQ ID NO: 33), остатки 115-239 соответствуют аминокислотной последовательности VH-домена CD3 mAb 2, содержащего D65G замену (SEQ ID NO: 112), остатки 240-245 соответствуют GGCGGG линкер (SEQ ID NO: NO: 34), остатки 246-273 соответствуют домену с Е-спиралью (SEQ ID NO: 39), остатки 274-276 представляют собой линкер GGG, остатки 277-286 представляют собой линкер DKTHTCPPCP (SEQ ID NO: 48), а остатки 287-503 представляют собой "несущий выпуклость" СН2-СН3-домен (SEQ ID NO: 52).
Полинуклеотид, который кодируется SEQ ID NO: 252, представляет собой (SEQ ID NO: 253):
gacattcagc tgactcagtc cccctctttt ctgtccgcat ccgtcggaga tcgagtgact attacttgct ctgctaggtc ctcaatcagc ttcatgtact ggtatcagca gaagcccggc aaagcaccta agctgctgat ctacgacaca agcaacctgg cctccggggt gccatctcgg ttctctggca gtgggtcagg aactgagttt accctgacaa ttagctccct ggaggctgaa gatgccgcta cctactattg ccagcagtgg agcagctatc ctctgacctt cggacagggg actaaactgg aaatcaaggg tggaggatcc ggcggcggag gcgaggtgca gctggtggag tctgggggag gcttggtcca gcctggaggg tccctgagac tctcctgtgc agcctctgga ttcaccttca gcacatacgc tatgaattgg gtccgccagg ctccagggaa ggggctggag tgggttggaa ggatcaggtc caagtacaac aattatgcaa cctactatgc cgactctgtg aagggtagat tcaccatctc aagagatgat tcaaagaact cactgtatct gcaaatgaac agcctgaaaa ccgaggacac ggccgtgtat tactgtgtga gacacggtaa cttcggcaat tcttacgtgt cttggtttgc ttattgggga caggggacac tggtgactgt gtcttccgga ggatgtggcg gtggagaagt ggccgcactg gagaaagagg ttgctgcttt ggagaaggag gtcgctgcac ttgaaaagga ggtcgcagcc ctggagaaag gcggcgggga caaaactcac acatgcccac cgtgcccagc acctgaagcc gcggggggac cgtcagtctt cctcttcccc ccaaaaccca aggacaccct catgatctcc cggacccctg aggtcacatg cgtggtggtg gacgtgagcc acgaagaccc tgaggtcaag ttcaactggt acgtggacgg cgtggaggtg cataatgcca agacaaagcc gcgggaggag cagtacaaca gcacgtaccg tgtggtcagc gtcctcaccg tcctgcacca ggactggctg aatggcaagg agtacaagtg caaggtctcc aacaaagccc tcccagcccc catcgagaaa accatctcca aagccaaagg gcagccccga gaaccacagg tgtacaccct gcccccatcc cgggaggaga tgaccaagaa ccaggtcagc ctgtggtgcc tggtcaaagg cttctatccc agcgacatcg ccgtggagtg ggagagcaat gggcagccgg agaacaacta caagaccacg cctcccgtgc tggactccga cggctccttc ttcctctaca gcaagctcac cgtggacaag agcaggtggc agcaggggaa cgtcttctca tgctccgtga tgcatgaggc tctgcacaac cactacacgc agaagagcct ctccctgtct ccgggtaaa
Аминокислотная последовательность второй полипептидной цепи gpA33 mAb 1 х CD3 mAb 2 х EphA2 mAb 1 (SEQ ID NO: 254):
QAVVTQEPSL TVSPGGTVTL TCRSSTGAVT TSNYANWVQQ KPGQAPRGLI GGTNKRAPWT PARFSGSLLG GKAALTITGA QAEDEADYYC ALWYSNLWVF GGGTKLTVLG GGGSGGGGQV QLVQSGAEVK KPGASVKVSC KASGYTFTGS WMNWVRQAPG QGLEWIGRIY PGDGETNYNG KFKDRVTITA DKSTSTAYME LSSLRSEDTA VYYCARIYGN NVYFDVWGQG TTVTVSSGGC GGGKVAALKE KVAALKEKVA ALKEKVAALK E
В SEQ ID NO: 254, аминокислотные остатки 1-110 соответствуют аминокислотной последовательности VL-домена CD3 mAb 2 (SEQ ID NO: 104), остатки 111-118 соответствуют промежуточному спейсерному пептиду GGGSGGGG (линкер 1) (SEQ ID NO: 33), остатки 119-237 соответствуют аминокислотной последовательности VH-домена gpA33 mAb 1 (SEQ ID NO: 186), остатки 238-243 соответствуют линкера GGCGGG (SEQ ID NO: 34) а остатки 244-271 представляют собой домен K-спирали (SEQ ID NO: 40).
Полинуклеотид, который кодируется SEQ ID NO: 254, представляет собой (SEQ ID NO: 255):
caggctgtgg tgactcagga gccttcactg accgtgtccc caggcggaac tgtgaccctg acatgcagat ccagcacagg cgcagtgacc acatctaact acgccaattg ggtgcagcag aagccaggac aggcaccaag gggcctgatc gggggtacaa acaaaagggc tccctggacc cctgcacggt tttctggaag tctgctgggc ggaaaggccg ctctgactat taccggggca caggccgagg acgaagccga ttactattgt gctctgtggt atagcaatct gtgggtgttc gggggtggca caaaactgac tgtgctggga gggggtggat ccggcggagg tggacaggtc cagctggtcc agagcggggc cgaagtcaaa aaacccggag caagcgtgaa ggtctcctgc aaagcatcag gctatacatt tacaggcagc tggatgaact gggtgaggca ggctccagga cagggactgg agtggatcgg gcgcatctac cctggagacg gcgaaactaa ctataatgga aagttcaaag accgagtgac catcacagcc gataagtcta ctagtaccgc ctacatggag ctgagctccc tgcggtctga agataccgcc gtctactatt gcgctagaat ttacggaaac aatgtctatt ttgacgtgtg ggggcaggga acaactgtga ctgtctcctc cggaggatgt ggcggtggaa aagtggccgc actgaaggag aaagttgctg ctttgaaaga gaaggtcgcc gcacttaagg aaaaggtcgc agccctgaaa gag
Аминокислотная последовательность третьей полипептидной цепи из gpA33 mAb 1 х CD3 mAb 2 х EphA2 mAb 1 (SEQ ID NO: 256):
QVQLKESGPG LVAPSQSLSI TCTVSGFSLS RYSVHWVRQP PGKGLEWLGM IWGGGSTDYN SALKSRLSIS KDNSKSQVFL KMNSLQTDDT AMYYCARKHG NYYTMDYWGQ GTSVTVSSAS TKGPSVFPLA PSSKSTSGGT AALGCLVKDY FPEPVTVSWN SGALTSGVHT FPAVLQSSGL YSLSSVVTVP SSSLGTQTYI CNVNHKPSNT KVDKRVEPKS CDKTHTCPPC PAPEAAGGPS VFLFPPKPKD TLMISRTPEV TCVVVDVSHE DPEVKFNWYV DGVEVHNAKT KPREEQYNST YRVVSVLTVL HQDWLNGKEY KCKVSNKALP APIEKTISKA KGQPREPQVY TLPPSREEMT KNQVSLSCAV KGFYPSDIAV EWESNGQPEN NYKTTPPVLD SDGSFFLVSK LTVDKSRWQQ GNVFSCSVMH EALHNRYTQK SLSLSPGK
В SEQ ID NO: 256, аминокислотные остатки 1-118 соответствуют аминокислотной последовательности VH-домена EphA2 mAb 1 (SEQ ID NO: 158), остатки 119-216 соответствуют модифицированному СН1-домену (SEQ ID NO: 208), остатки 217-231 соответствуют линкеру (SEQ ID NO: 209), а остатки 232-448 соответствуют "несущему углубление" CH2-CH3-домен (SEQ ID NO: 53).
Полинуклеотид, который кодируется SEQ ID NO: 256 представляет собой (SEQ ID NO: 257):
caggtgcagc tgaaggagtc aggacctggc ctggtggcac cctcacagag cctgtccatc acatgcactg tctctgggtt ctcattatcc agatatagtg tacactgggt tcgccagcct ccaggaaagg gtctggagtg gctgggaatg atatggggtg gtggaagcac agactataat tcagctctca aatccagact gagtatcagc aaggacaact ccaagagcca agttttctta aaaatgaaca gtctgcaaac tgatgacaca gccatgtact actgtgccag aaaacatggt aactactata ctatggacta ctggggtcaa ggaacctcag tcaccgtctc ctccgcctcc accaagggcc catcggtctt ccccctggca ccctcctcca agagcacctc tgggggcaca gcggccctgg gctgcctggt caaggactac ttccccgaac cggtgacggt gtcgtggaac tcaggcgccc tgaccagcgg cgtgcacacc ttcccggctg tcctacagtc ctcaggactc tactccctca gcagcgtggt gaccgtgccc tccagcagct tgggcaccca gacctacatc tgcaacgtga atcacaagcc cagcaacacc aaggtggaca agagagttga gcccaaatct tgtgacaaaa ctcacacatg cccaccgtgc ccagcacctg aagccgcggg gggaccgtca gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc acatgcgtgg tggtggacgt gagccacgaa gaccctgagg tcaagttcaa ctggtacgtg gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga ggagatgacc aagaaccagg tcagcctgag ttgcgcagtc aaaggcttct atcccagcga catcgccgtg gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac tccgacggct ccttcttcct cgtcagcaag ctcaccgtgg acaagagcag gtggcagcag gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccgcta cacgcagaag agcctctccc tgtctccggg taaa
Аминокислотная последовательность четвертой полипептидной цепи gpA33 mAb 1 х CD3 mAb 2 х EphA2 mAb 1 (SEQ ID NO: 258):
DIQMTQTTSS LSASLGDRIT ISCRASQDIS NYLNWYQQKP DGTVKLLIYY TSRLHSGVPS RFSGSGSGTD YSLTISNLEQ EDIATYFCQQ GYTLYTFGGG TKLEIKRTVA APSVFIFPPS DEQLKSGTAS VVCLLNNFYP REAKVQWKVD NALQSGNSQE SVTEQDSKDS TYSLSSTLTL SKADYEKHKV YACEVTHQGL SSPVTKSFNR GEC
В SEQ ID NO: 258, аминокислотные остатки 1-106 соответствуют аминокислотной последовательности VL-домена EphA2 mAb 1 (SEQ ID NO: 153), а остатки 107-213 соответствуют CL каппа домен (SEQ ID NO: 210).
Полинуклеотид, который кодируется SEQ ID NO: 258, представляет собой (SEQ ID NO: 259):
gatatccaga tgacacagac tacatcctcc ctgtctgcct ctctgggaga cagaatcacc atcagttgca gggcaagtca ggacattagc aattatttaa actggtatca gcagaaacca gatggaactg ttaaactcct gatctactac acatcaagat tacactcagg agtcccatca aggttcagtg gcagtgggtc tggaacagat tattctctca ccattagcaa cctggagcaa gaagatattg ccacttactt ttgccaacag ggttatacgc tgtacacgtt cggagggggg accaagctgg aaataaaacg tacggtggct gcaccatcgg tcttcatctt cccgccatct gatgagcagt tgaaatctgg aactgcctct gttgtgtgcc tgctgaataa cttctatccc agagaggcca aagtacagtg gaaggtggat aacgccctcc aatcgggtaa ctcccaggag agtgtcacag agcaggacag caaggacagc acctacagcc tcagcagcac cctgacgctg agcaaagcag actacgagaa acacaaagtc tacgcctgcg aagtcaccca tcagggcctg agctcgcccg tcacaaagag cttcaacagg ggagagtgt
L. gpA33 mAb 1 х CD3 mAb 2 х EphA2 mAb 2
Была сконструирована триспецифичная связывающая молекула, которая состоит из четырех полипептидных цепей, которая включает VL- и VH-домены из gpA33 mAb 1, VL- и VH-домены антитела CD3 mAb, 2 и VL- и VH-домены из EphA2 mAb 2. Триспецифическая связывающаяся молекула был соответствующим образом обозначен "gpA33 mAb 1 х CD3 mAb 2 х EphA2 mAb 2". Аминокислотная последовательность первой полипептидной цепи этой триспецифичной связывающей молекулы является (SEQ ID NO: 260):
DIQLTQSPSF LSASVGDRVT ITCSARSSIS FMYWYQQKPG KAPKLLIYDT SNLASGVPSR FSGSGSGTEF TLTISSLEAE DAATYYCQQW SSYPLTFGQG TKLEIKGGGS GGGGEVQLVE SGGGLVQPGG SLRLSCAASG FTFSTYAMNW VRQAPGKGLE WVGRIRSKYN NYATYYADSV KGRFTISRDD SKNSLYLQMN SLKTEDTAVY YCVRHGNFGN SYVSWFAYWG QGTLVTVSSG GCGGGEVAAL EKEVAALEKE VAALEKEVAA LEKGGGDKTH TCPPCPAPEA AGGPSVFLFP PKPKDTLMIS RTPEVTCVVV DVSHEDPEVK FNWYVDGVEV HNAKTKPREE QYNSTYRVVS VLTVLHQDWL NGKEYKCKVS NKALPAPIEK TISKAKGQPR EPQVYTLPPS REEMTKNQVS LWCLVKGFYP SDIAVEWESN GQPENNYKTT PPVLDSDGSF FLYSKLTVDK SRWQQGNVFS CSVMHEALHN HYTQKSLSLS PGK
В SEQ ID NO: 260, аминокислотные остатки 1-106 соответствуют аминокислотной последовательности VL-домена gpA33 mAb 1 (SEQ ID NO: 181), остатки 107-114 соответствуют промежуточному спейсерному пептиду GGGSGGGG (линкер 1) (SEQ ID NO: 33), остатки 115-239 соответствуют аминокислотной последовательности VH-домена CD3 mAb 2, содержащего D65G замену (SEQ ID NO: 112), остатки 240-245 соответствуют GGCGGG линкер (SEQ ID NO: NO: 34), остатки 246-273 соответствуют домену с Е-спиралью (SEQ ID NO: 39), остатки 274-276 представляют собой линкер GGG, остатки 277-286 представляют собой линкер DKTHTCPPCP (SEQ ID NO: 48), а остатки 287-503 представляют собой "несущий выпуклость" СН2-СН3-домен (SEQ ID NO: 52).
Полинуклеотид, который кодируется SEQ ID NO: 260 представляет собой (SEQ ID NO: 261):
gacattcagc tgactcagtc cccctctttt ctgtccgcat ccgtcggaga tcgagtgact attacttgct ctgctaggtc ctcaatcagc ttcatgtact ggtatcagca gaagcccggc aaagcaccta agctgctgat ctacgacaca agcaacctgg cctccggggt gccatctcgg ttctctggca gtgggtcagg aactgagttt accctgacaa ttagctccct ggaggctgaa gatgccgcta cctactattg ccagcagtgg agcagctatc ctctgacctt cggacagggg actaaactgg aaatcaaggg tggaggatcc ggcggcggag gcgaggtgca gctggtggag tctgggggag gcttggtcca gcctggaggg tccctgagac tctcctgtgc agcctctgga ttcaccttca gcacatacgc tatgaattgg gtccgccagg ctccagggaa ggggctggag tgggttggaa ggatcaggtc caagtacaac aattatgcaa cctactatgc cgactctgtg aagggtagat tcaccatctc aagagatgat tcaaagaact cactgtatct gcaaatgaac agcctgaaaa ccgaggacac ggccgtgtat tactgtgtga gacacggtaa cttcggcaat tcttacgtgt cttggtttgc ttattgggga caggggacac tggtgactgt gtcttccgga ggatgtggcg gtggagaagt ggccgcactg gagaaagagg ttgctgcttt ggagaaggag gtcgctgcac ttgaaaagga ggtcgcagcc ctggagaaag gcggcgggga caaaactcac acatgcccac cgtgcccagc acctgaagcc gcggggggac cgtcagtctt cctcttcccc ccaaaaccca aggacaccct catgatctcc cggacccctg aggtcacatg cgtggtggtg gacgtgagcc acgaagaccc tgaggtcaag ttcaactggt acgtggacgg cgtggaggtg cataatgcca agacaaagcc gcgggaggag cagtacaaca gcacgtaccg tgtggtcagc gtcctcaccg tcctgcacca ggactggctg aatggcaagg agtacaagtg caaggtctcc aacaaagccc tcccagcccc catcgagaaa accatctcca aagccaaagg gcagccccga gaaccacagg tgtacaccct gcccccatcc cgggaggaga tgaccaagaa ccaggtcagc ctgtggtgcc tggtcaaagg cttctatccc agcgacatcg ccgtggagtg ggagagcaat gggcagccgg agaacaacta caagaccacg cctcccgtgc tggactccga cggctccttc ttcctctaca gcaagctcac cgtggacaag agcaggtggc agcaggggaa cgtcttctca tgctccgtga tgcatgaggc tctgcacaac cactacacgc agaagagcct ctccctgtct ccgggtaaa
Аминокислотная последовательность второй полипептидной цепи gpA33 mAb 1 х CD3 mAb 2 х EphA2 mAb 2 (SEQ ID NO: 262):
QAVVTQEPSL TVSPGGTVTL TCRSSTGAVT TSNYANWVQQ KPGQAPRGLI GGTNKRAPWT PARFSGSLLG GKAALTITGA QAEDEADYYC ALWYSNLWVF GGGTKLTVLG GGGSGGGGQV QLVQSGAEVK KPGASVKVSC KASGYTFTGS WMNWVRQAPG QGLEWIGRIY PGDGETNYNG KFKDRVTITA DKSTSTAYME LSSLRSEDTA VYYCARIYGN NVYFDVWGQG TTVTVSSGGC GGGKVAALKE KVAALKEKVA ALKEKVAALK E
В SEQ ID NO: 262, аминокислотные остатки 1-110 соответствуют аминокислотной последовательности VL-домена CD3 mAb 2 (SEQ ID NO: 104), остатки 111-118 соответствуют промежуточному спейсерному пептиду GGGSGGGG (линкер 1) (SEQ ID NO: 33), остатки 119-237 соответствуют аминокислотной последовательности VH-домена gpA33 mAb 1 (SEQ ID NO: 186), остатки 238-243 соответствуют линкера GGCGGG (SEQ ID NO: 34) а остатки 244-271 представляют собой домен K-спирали (SEQ ID NO: 40).
Полинуклеотид, который кодируется SEQ ID NO: 262, представляет собой (SEQ ID NO: 263):
caggctgtgg tgactcagga gccttcactg accgtgtccc caggcggaac tgtgaccctg acatgcagat ccagcacagg cgcagtgacc acatctaact acgccaattg ggtgcagcag aagccaggac aggcaccaag gggcctgatc gggggtacaa acaaaagggc tccctggacc cctgcacggt tttctggaag tctgctgggc ggaaaggccg ctctgactat taccggggca caggccgagg acgaagccga ttactattgt gctctgtggt atagcaatct gtgggtgttc gggggtggca caaaactgac tgtgctggga gggggtggat ccggcggagg tggacaggtc cagctggtcc agagcggggc cgaagtcaaa aaacccggag caagcgtgaa ggtctcctgc aaagcatcag gctatacatt tacaggcagc tggatgaact gggtgaggca ggctccagga cagggactgg agtggatcgg gcgcatctac cctggagacg gcgaaactaa ctataatgga aagttcaaag accgagtgac catcacagcc gataagtcta ctagtaccgc ctacatggag ctgagctccc tgcggtctga agataccgcc gtctactatt gcgctagaat ttacggaaac aatgtctatt ttgacgtgtg ggggcaggga acaactgtga ctgtctcctc cggaggatgt ggcggtggaa aagtggccgc actgaaggag aaagttgctg ctttgaaaga gaaggtcgcc gcacttaagg aaaaggtcgc agccctgaaa gag
Аминокислотная последовательность третьей полипептидной цепи gpA33 mAb 1 х CD3 mAb 2 х EphA2 mAb 2 представляет собой (SEQ ID NO: 264):
QIQLVQSGPE LKKPGETVKI SCKASGFTFT NYGMNWVKQA PGKGLKWMGW INTYIGEPTY ADDFKGRFVF SLETSASTAY LQINNLKNED MATYFCAREL GPYYFDYWGQ GTTLTVSSAS TKGPSVFPLA PSSKSTSGGT AALGCLVKDY FPEPVTVSWN SGALTSGVHT FPAVLQSSGL YSLSSVVTVP SSSLGTQTYI CNVNHKPSNT KVDKRVEPKS CDKTHTCPPC PAPEAAGGPS VFLFPPKPKD TLMISRTPEV TCVVVDVSHE DPEVKFNWYV DGVEVHNAKT KPREEQYNST YRVVSVLTVL HQDWLNGKEY KCKVSNKALP APIEKTISKA KGQPREPQVY TLPPSREEMT KNQVSLSCAV KGFYPSDIAV EWESNGQPEN NYKTTPPVLD SDGSFFLVSK LTVDKSRWQQ GNVFSCSVMH EALHNRYTQK SLSLSPGK
В SEQ ID NO: 264, аминокислотные остатки 1-118 соответствуют аминокислотной последовательности VH-домена EphA2 mAb 2 (SEQ ID NO: 167), остатки 119-216 соответствуют модифицированному СН1-домену (SEQ ID NO: 208), остатки 217-231 соответствуют линкеру (SEQ ID NO: 209), а остатки 232-448 соответствуют "несущему углубление" CH2-CH3-домену (SEQ ID NO: 53).
Полинуклеотид, который кодируется SEQ ID NO: 264, представляет собой (SEQ ID NO: 265):
cagatccagt tggtgcagtc tggacctgag ctgaagaagc ctggagagac agtcaagatc tcctgcaagg cttctgggtt taccttcaca aactatggaa tgaactgggt gaagcaggct ccaggaaagg gtttaaagtg gatgggctgg ataaacacct atattggaga gccgacatat gctgatgact tcaagggacg gtttgtcttc tctttggaaa cctctgccag cactgcctat ttgcagatca acaacctcaa aaatgaggac atggccacat atttctgtgc aagagaactg ggaccatact actttgacta ctggggccaa ggcaccactc tcacagtctc ctccgcctcc accaagggcc catcggtctt ccccctggca ccctcctcca agagcacctc tgggggcaca gcggccctgg gctgcctggt caaggactac ttccccgaac cggtgacggt gtcgtggaac tcaggcgccc tgaccagcgg cgtgcacacc ttcccggctg tcctacagtc ctcaggactc tactccctca gcagcgtggt gaccgtgccc tccagcagct tgggcaccca gacctacatc tgcaacgtga atcacaagcc cagcaacacc aaggtggaca agagagttga gcccaaatct tgtgacaaaa ctcacacatg cccaccgtgc ccagcacctg aagccgcggg gggaccgtca gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc acatgcgtgg tggtggacgt gagccacgaa gaccctgagg tcaagttcaa ctggtacgtg gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga ggagatgacc aagaaccagg tcagcctgag ttgcgcagtc aaaggcttct atcccagcga catcgccgtg gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac tccgacggct ccttcttcct cgtcagcaag ctcaccgtgg acaagagcag gtggcagcag gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccgcta cacgcagaag agcctctccc tgtctccggg taaa
Аминокислотная последовательность четвертой полипептидной цепи gpA33 mAb 1 х CD3 mAb 2 х EphA2 mAb 2 представляет собой (SEQ ID NO: 266):
DVVMTQTPLS LPVSLGDQAS ISCRSSQSLV HSSGNTYLHW YLQKPGQSPK LLIYKVSNRF SGVPDRFSGS GSGTDFTLKI SRVEAEDLGV YFCSQSTHVP TFGSGTKLEI KRTVAAPSVF IFPPSDEQLK SGTASVVCLL NNFYPREAKV QWKVDNALQS GNSQESVTEQ DSKDSTYSLS STLTLSKADY EKHKVYACEV THQGLSSPVT KSFNRGEC
В SEQ ID NO: 266, аминокислотные остатки 1-111 соответствуют аминокислотной последовательности VL-домена EphA2 mAb 1 (SEQ ID NO: 163), а остатки 112-218 соответствуют домену CL каппа (SEQ ID NO: 210).
Полинуклеотид, который кодируется SEQ ID NO: 266, представляет собой (SEQ ID NO: 267):
gatgttgtga tgacccaaac tccactctcc ctgcctgtca gtcttggaga tcaagcctcc atctcttgca gatctagtca gagccttgta cacagtagtg gaaacaccta tttacattgg tacctgcaga agccaggcca gtctccaaag ctcctgatct acaaagtttc caaccgattt tctggggtcc cagacaggtt cagtggcagt ggatcaggga cagatttcac actcaagatc agcagagtgg aggctgagga tctgggagtt tatttctgct ctcaaagtac acatgttccc acgttcggct cggggacaaa gttggaaata aaacgtacgg tggctgcacc atcggtcttc atcttcccgc catctgatga gcagttgaaa tctggaactg cctctgttgt gtgcctgctg aataacttct atcccagaga ggccaaagta cagtggaagg tggataacgc cctccaatcg ggtaactccc aggagagtgt cacagagcag gacagcaagg acagcaccta cagcctcagc agcaccctga cgctgagcaa agcagactac gagaaacaca aagtctacgc ctgcgaagtc acccatcagg gcctgagctc gcccgtcaca aagagcttca acaggggaga gtgt
М. gpA33 mAb 1 х CD3 mAb 2 х EphA2 mAb 3
Конструировали триспецифичную связывающую молекулу, состоящую из четырех полипептидных цепей, которая содержит VL- и VH-домены из gpA33 mAb 1, VL- и VH-домены антитела CD3 mAb 2 и VL- и VH-домены из EphA2 mAb 3. Триспецифичную связывающую молекулу назвали соответствующим образом "gpA33 mAb 1 х CD3 mAb 2 х EphA2 mAb 3". Аминокислотная последовательность первой полипептидной цепи этой триспецифичной связывающей молекулы представляет собой (SEQ ID NO: 268):
DIQLTQSPSF LSASVGDRVT ITCSARSSIS FMYWYQQKPG KAPKLLIYDT SNLASGVPSR FSGSGSGTEF TLTISSLEAE DAATYYCQQW SSYPLTFGQG TKLEIKGGGS GGGGEVQLVE SGGGLVQPGG SLRLSCAASG FTFSTYAMNW VRQAPGKGLE WVGRIRSKYN NYATYYADSV KGRFTISRDD SKNSLYLQMN SLKTEDTAVY YCVRHGNFGN SYVSWFAYWG QGTLVTVSSG GCGGGEVAAL EKEVAALEKE VAALEKEVAA LEKGGGDKTH TCPPCPAPEA AGGPSVFLFP PKPKDTLMIS RTPEVTCVVV DVSHEDPEVK FNWYVDGVEV HNAKTKPREE QYNSTYRVVS VLTVLHQDWL NGKEYKCKVS NKALPAPIEK TISKAKGQPR EPQVYTLPPS REEMTKNQVS LWCLVKGFYP SDIAVEWESN GQPENNYKTT PPVLDSDGSF FLYSKLTVDK SRWQQGNVFS CSVMHEALHN HYTQKSLSLS PGK
В SEQ ID NO: 268, аминокислотные остатки 1-106 соответствуют аминокислотной последовательности VL-домена gpA33 mAb 1 (SEQ ID NO: 181), остатки 107-114 соответствуют промежуточному спейсерному пептиду GGGSGGGG (линкер 1) (SEQ ID NO: 33), остатки 115-239 соответствуют аминокислотной последовательности VH-домена CD3 mAb 2, содержащего D65G замену (SEQ ID NO: 112), остатки 240-245 соответствуют GGCGGG линкеру (SEQ ID NO: NO: 34), остатки 246-273 соответствуют домену с Е-спиралью (SEQ ID NO: 39), остатки 274-276 представляют собой линкер GGG, остатки 277-286 представляют собой линкер DKTHTCPPCP (SEQ ID NO: 48), а остатки 287-503 представляют собой "несущий выпуклость" СН2-СН3-домен (SEQ ID NO: 52).
Полинуклеотид, который кодируется SEQ ID NO: 268, представляет собой (SEQ ID NO: 269):
gacattcagc tgactcagtc cccctctttt ctgtccgcat ccgtcggaga tcgagtgact attacttgct ctgctaggtc ctcaatcagc ttcatgtact ggtatcagca gaagcccggc aaagcaccta agctgctgat ctacgacaca agcaacctgg cctccggggt gccatctcgg ttctctggca gtgggtcagg aactgagttt accctgacaa ttagctccct ggaggctgaa gatgccgcta cctactattg ccagcagtgg agcagctatc ctctgacctt cggacagggg actaaactgg aaatcaaggg tggaggatcc ggcggcggag gcgaggtgca gctggtggag tctgggggag gcttggtcca gcctggaggg tccctgagac tctcctgtgc agcctctgga ttcaccttca gcacatacgc tatgaattgg gtccgccagg ctccagggaa ggggctggag tgggttggaa ggatcaggtc caagtacaac aattatgcaa cctactatgc cgactctgtg aagggtagat tcaccatctc aagagatgat tcaaagaact cactgtatct gcaaatgaac agcctgaaaa ccgaggacac ggccgtgtat tactgtgtga gacacggtaa cttcggcaat tcttacgtgt cttggtttgc ttattgggga caggggacac tggtgactgt gtcttccgga ggatgtggcg gtggagaagt ggccgcactg gagaaagagg ttgctgcttt ggagaaggag gtcgctgcac ttgaaaagga ggtcgcagcc ctggagaaag gcggcgggga caaaactcac acatgcccac cgtgcccagc acctgaagcc gcggggggac cgtcagtctt cctcttcccc ccaaaaccca aggacaccct catgatctcc cggacccctg aggtcacatg cgtggtggtg gacgtgagcc acgaagaccc tgaggtcaag ttcaactggt acgtggacgg cgtggaggtg cataatgcca agacaaagcc gcgggaggag cagtacaaca gcacgtaccg tgtggtcagc gtcctcaccg tcctgcacca ggactggctg aatggcaagg agtacaagtg caaggtctcc aacaaagccc tcccagcccc catcgagaaa accatctcca aagccaaagg gcagccccga gaaccacagg tgtacaccct gcccccatcc cgggaggaga tgaccaagaa ccaggtcagc ctgtggtgcc tggtcaaagg cttctatccc agcgacatcg ccgtggagtg ggagagcaat gggcagccgg agaacaacta caagaccacg cctcccgtgc tggactccga cggctccttc ttcctctaca gcaagctcac cgtggacaag agcaggtggc agcaggggaa cgtcttctca tgctccgtga tgcatgaggc tctgcacaac cactacacgc agaagagcct ctccctgtct ccgggtaaa
Аминокислотная последовательность второй полипептидной цепи gpA33 mAb 1 х CD3 mAb 2 х EphA2 mAb 3 (SEQ ID NO: 270):
QAVVTQEPSL TVSPGGTVTL TCRSSTGAVT TSNYANWVQQ KPGQAPRGLI GGTNKRAPWT PARFSGSLLG GKAALTITGA QAEDEADYYC ALWYSNLWVF GGGTKLTVLG GGGSGGGGQV QLVQSGAEVK KPGASVKVSC KASGYTFTGS WMNWVRQAPG QGLEWIGRIY PGDGETNYNG KFKDRVTITA DKSTSTAYME LSSLRSEDTA VYYCARIYGN NVYFDVWGQG TTVTVSSGGC GGGKVAALKE KVAALKEKVA ALKEKVAALK E
В SEQ ID NO: 270, аминокислотные остатки 1-110 соответствуют аминокислотной последовательности VL-домена CD3 mAb 2 (SEQ ID NO: 104), остатки 111-118 соответствуют промежуточному спейсерному пептиду GGGSGGGG (линкер 1) (SEQ ID NO: 33), остатки 119-237 соответствуют аминокислотной последовательности VH-домена gpA33 mAb 1 (SEQ ID NO: 186), остатки 238-243 соответствуют линкеру GGCGGG (SEQ ID NO: 34) а остатки 244-271 представляют собой домен K-спирали (SEQ ID NO: 40).
Полинуклеотид, который кодируется SEQ ID NO: 270, представляет собой (SEQ ID NO: 271):
caggctgtgg tgactcagga gccttcactg accgtgtccc caggcggaac tgtgaccctg acatgcagat ccagcacagg cgcagtgacc acatctaact acgccaattg ggtgcagcag aagccaggac aggcaccaag gggcctgatc gggggtacaa acaaaagggc tccctggacc cctgcacggt tttctggaag tctgctgggc ggaaaggccg ctctgactat taccggggca caggccgagg acgaagccga ttactattgt gctctgtggt atagcaatct gtgggtgttc gggggtggca caaaactgac tgtgctggga gggggtggat ccggcggagg tggacaggtc cagctggtcc agagcggggc cgaagtcaaa aaacccggag caagcgtgaa ggtctcctgc aaagcatcag gctatacatt tacaggcagc tggatgaact gggtgaggca ggctccagga cagggactgg agtggatcgg gcgcatctac cctggagacg gcgaaactaa ctataatgga aagttcaaag accgagtgac catcacagcc gataagtcta ctagtaccgc ctacatggag ctgagctccc tgcggtctga agataccgcc gtctactatt gcgctagaat ttacggaaac aatgtctatt ttgacgtgtg ggggcaggga acaactgtga ctgtctcctc cggaggatgt ggcggtggaa aagtggccgc actgaaggag aaagttgctg ctttgaaaga gaaggtcgcc gcacttaagg aaaaggtcgc agccctgaaa gag
Аминокислотная последовательность третьей полипептидной цепи из gpA33 mAb 1 х CD3 mAb 2 х EphA2 mAb 3 (SEQ ID NO: 272):
EVQLVESGGG SVKPGGSLKL SCAASGFTFT DHYMYWVRQT PEKRLEWVAT ISDGGSFTSY PDSVKGRFTI SRDIAKNNLY LQMSSLKSED TAMYYCTRDE SDRPFPYWGQ GTLVTVSSAS TKGPSVFPLA PSSKSTSGGT AALGCLVKDY FPEPVTVSWN SGALTSGVHT FPAVLQSSGL YSLSSVVTVP SSSLGTQTYI CNVNHKPSNT KVDKRVEPKS CDKTHTCPPC PAPEAAGGPS VFLFPPKPKD TLMISRTPEV TCVVVDVSHE DPEVKFNWYV DGVEVHNAKT KPREEQYNST YRVVSVLTVL HQDWLNGKEY KCKVSNKALP APIEKTISKA KGQPREPQVY TLPPSREEMT KNQVSLSCAV KGFYPSDIAV EWESNGQPEN NYKTTPPVLD SDGSFFLVSK LTVDKSRWQQ GNVFSCSVMH EALHNRYTQK SLSLSPGK
В SEQ ID NO: 272, аминокислотные остатки 1-118 соответствуют аминокислотной последовательности VH-домена EphA2 mAb 3 (SEQ ID NO: 177), остатки 119-216 соответствуют модифицированному СН1-домену (SEQ ID NO: 208), остатки 217-231 соответствуют линкеру (SEQ ID NO: 209), а остатки 232-448 соответствуют CH2-CH3-домену, "несущему углубление" (SEQ ID NO: 53).
Полинуклеотид, который кодируется SEQ ID NO: 272, представляет собой (SEQ ID NO: 273):
gaagtgcagc tggtggagtc tgggggaggc tcagtgaagc ctggagggtc cctgaaactc tcctgtgcag cctctggatt cactttcact gaccattaca tgtattgggt tcgccagact ccggaaaaga ggctggagtg ggtcgcaacc attagtgatg gcggtagttt cacctcctat ccagacagtg tgaaggggcg attcaccatc tccagagaca ttgccaagaa caacctgtac ctccaaatga gcagtctgaa gtctgaggac acagccatgt attactgtac aagagatgag agcgataggc cgtttcctta ctggggccaa gggactctgg tcactgtctc ctccgcctcc accaagggcc catcggtctt ccccctggca ccctcctcca agagcacctc tgggggcaca gcggccctgg gctgcctggt caaggactac ttccccgaac cggtgacggt gtcgtggaac tcaggcgccc tgaccagcgg cgtgcacacc ttcccggctg tcctacagtc ctcaggactc tactccctca gcagcgtggt gaccgtgccc tccagcagct tgggcaccca gacctacatc tgcaacgtga atcacaagcc cagcaacacc aaggtggaca agagagttga gcccaaatct tgtgacaaaa ctcacacatg cccaccgtgc ccagcacctg aagccgcggg gggaccgtca gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc acatgcgtgg tggtggacgt gagccacgaa gaccctgagg tcaagttcaa ctggtacgtg gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga ggagatgacc aagaaccagg tcagcctgag ttgcgcagtc aaaggcttct atcccagcga catcgccgtg gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac tccgacggct ccttcttcct cgtcagcaag ctcaccgtgg acaagagcag gtggcagcag gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccgcta cacgcagaag agcctctccc tgtctccggg taaa
Аминокислотная последовательность четвертого полипептидной цепи gpA33 mAb 1 х CD3 mAb 2 х EphA2 mAb 3 (SEQ ID NO: 274):
DIVLTQSHRS MSTSVGDRVN ITCKASQDVT TAVAWYQQKP GQSPKLLIFW ASTRHAGVPD RFTGSGSGTD FTLTISSVQA GDLALYYCQQ HYSTPYTFGG GTKLEIKRTV AAPSVFIFPP SDEQLKSGTA SVVCLLNNFY PREAKVQWKV DNALQSGNSQ ESVTEQDSKD STYSLSSTLT LSKADYEKHK VYACEVTHQG LSSPVTKSFN RGEC
В SEQ ID NO: 274, аминокислотные остатки 1-107 соответствуют аминокислотной последовательности VL-домена EphA2 mAb 3 (SEQ ID NO: 172), а остатки 108-214 соответствуют домену CL каппа (SEQ ID NO: 210).
Полинуклеотид, который кодируется SEQ ID NO: 274, представляет собой (SEQ ID NO: 275):
gacattgtgc tgacccagtc tcacagatcc atgtccacat cagtaggaga cagggtcaac atcacctgca aggccagtca ggatgtgact actgctgtag cctggtatca acaaaaacca gggcaatctc ctaaattact gattttctgg gcatccaccc ggcacgctgg agtccctgat cgcttcacag gcagtggatc tgggacagat tttactctca ccatcagcag tgtgcaggct ggagacctgg cactttatta ctgtcaacaa cattatagca caccgtacac attcggaggg gggaccaagc tggaaataaa acgtacggtg gctgcaccat cggtcttcat cttcccgcca tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gt
N. EphA2 mAb 1 х CD3 mAb 2 х gpA33 mAb 1
Конструировали альтернативную триспецифичную связывающую молекулу EphA2/CD3/gpA33. Молекула состоит из четырех полипептидных цепей, и включает VL- и VH-домены из EphA2 mAb 1, VL- и VH-домены антитела CD3 mAb, 2 и VL- и VH-домены из gpA33 mAb 1. Молекула была названа "EphA2 mAb 1 х CD3 mAb 2 х gpA33 mAb 1". Аминокислотная последовательность первой полипептидной цепи этой триспецифичной связывающей молекулы представляет собой (SEQ ID NO: 276):
DIQMTQTTSS LSASLGDRIT ISCRASQDIS NYLNWYQQKP DGTVKLLIYY TSRLHSGVPS RFSGSGSGTD YSLTISNLEQ EDIATYFCQQ GYTLYTFGGG TKLEIKGGGS GGGGEVQLVE SGGGLVQPGG SLRLSCAASG FTFSTYAMNW VRQAPGKGLE WVGRIRSKYN NYATYYADSV KGRFTISRDD SKNSLYLQMN SLKTEDTAVY YCVRHGNFGN SYVSWFAYWG QGTLVTVSSG GCGGGEVAAL EKEVAALEKE VAALEKEVAA LEKGGGDKTH TCPPCPAPEA AGGPSVFLFP PKPKDTLMIS RTPEVTCVVV DVSHEDPEVK FNWYVDGVEV HNAKTKPREE QYNSTYRVVS VLTVLHQDWL NGKEYKCKVS NKALPAPIEK TISKAKGQPR EPQVYTLPPS REEMTKNQVS LWCLVKGFYP SDIAVEWESN GQPENNYKTT PPVLDSDGSF FLYSKLTVDK SRWQQGNVFS CSVMHEALHN HYTQKSLSLS PGK
В SEQ ID NO: 276, аминокислотные остатки 1-106 соответствуют аминокислотной последовательности VL-домена EphA2 mAb 1 (SEQ ID NO: 153), остатки 107-114 соответствуют промежуточному спейсерному пептиду GGGSGGGG (линкер 1) (SEQ ID NO: 33), остатки 115-239 соответствуют аминокислотной последовательности VH-домена CD3 mAb 2, содержащего D65G замену (SEQ ID NO: 112), остатки 240-245 соответствуют GGCGGG линкеру (SEQ ID NO: NO: 34), остатки 246-273 соответствуют домену с Е-спиралью (SEQ ID NO: 39), остатки 274-276 представляют собой линкер GGG, остатки 277-286 представляют собой линкер DKTHTCPPCP (SEQ ID NO: 48), а остатки 287-503 СН2-СН3-домен, представляют собой "несущий выпуклость" (SEQ ID NO: 52).
Полинуклеотид, который кодируется SEQ ID NO: 276, представляет собой (SEQ ID NO: 277):
gatatccaga tgacacagac tacatcctcc ctgtctgcct ctctgggaga cagaatcacc atcagttgca gggcaagtca ggacattagc aattatttaa actggtatca gcagaaacca gatggaactg ttaaactcct gatctactac acatcaagat tacactcagg agtcccatca aggttcagtg gcagtgggtc tggaacagat tattctctca ccattagcaa cctggagcaa gaagatattg ccacttactt ttgccaacag ggttatacgc tgtacacgtt cggagggggg accaagctgg aaataaaagg tggaggatcc ggcggcggag gcgaggtgca gctggtggag tctgggggag gcttggtcca gcctggaggg tccctgagac tctcctgtgc agcctctgga ttcaccttca gcacatacgc tatgaattgg gtccgccagg ctccagggaa ggggctggag tgggttggaa ggatcaggtc caagtacaac aattatgcaa cctactatgc cgactctgtg aagggtagat tcaccatctc aagagatgat tcaaagaact cactgtatct gcaaatgaac agcctgaaaa ccgaggacac ggccgtgtat tactgtgtga gacacggtaa cttcggcaat tcttacgtgt cttggtttgc ttattgggga caggggacac tggtgactgt gtcttccgga ggatgtggcg gtggagaagt ggccgcactg gagaaagagg ttgctgcttt ggagaaggag gtcgctgcac ttgaaaagga ggtcgcagcc ctggagaaag gcggcgggga caaaactcac acatgcccac cgtgcccagc acctgaagcc gcggggggac cgtcagtctt cctcttcccc ccaaaaccca aggacaccct catgatctcc cggacccctg aggtcacatg cgtggtggtg gacgtgagcc acgaagaccc tgaggtcaag ttcaactggt acgtggacgg cgtggaggtg cataatgcca agacaaagcc gcgggaggag cagtacaaca gcacgtaccg tgtggtcagc gtcctcaccg tcctgcacca ggactggctg aatggcaagg agtacaagtg caaggtctcc aacaaagccc tcccagcccc catcgagaaa accatctcca aagccaaagg gcagccccga gaaccacagg tgtacaccct gcccccatcc cgggaggaga tgaccaagaa ccaggtcagc ctgtggtgcc tggtcaaagg cttctatccc agcgacatcg ccgtggagtg ggagagcaat gggcagccgg agaacaacta caagaccacg cctcccgtgc tggactccga cggctccttc ttcctctaca gcaagctcac cgtggacaag agcaggtggc agcaggggaa cgtcttctca tgctccgtga tgcatgaggc tctgcacaac cactacacgc agaagagcct ctccctgtct ccgggtaaa
Аминокислотная последовательность второй полипептидной цепи EphA2 mAb 1 х CD3 mAb 2 х gpA33 mAb 1 (SEQ ID NO: 278):
QAVVTQEPSL TVSPGGTVTL TCRSSTGAVT TSNYANWVQQ KPGQAPRGLI GGTNKRAPWT PARFSGSLLG GKAALTITGA QAEDEADYYC ALWYSNLWVF GGGTKLTVLG GGGSGGGGQV QLKESGPGLV APSQSLSITC TVSGFSLSRY SVHWVRQPPG KGLEWLGMIW GGGSTDYNSA LKSRLSISKD NSKSQVFLKM NSLQTDDTAM YYCARKHGNY YTMDYWGQGT SVTVSSGGCG GGKVAALKEK VAALKEKVAA LKEKVAALKE
В SEQ ID NO: 278, аминокислотные остатки 1-110 соответствуют аминокислотной последовательности VL-домена CD3 mAb 2 (SEQ ID NO: 104), остатки 111-118 соответствуют промежуточному спейсерному пептиду GGGSGGGG (линкер 1) (SEQ ID NO: 33), остатки 119-236 соответствуют аминокислотной последовательности VH-домена EphA2 mAb 1 (SEQ ID NO: 158), остатки 237-242 соответствуют линкеру GGCGGG (SEQ ID NO: 34) а остатки 243-270 представляют собой домен K-спирали (SEQ ID NO: 40).
Полинуклеотид, который кодируется SEQ ID NO: 278, представляет собой (SEQ ID NO: 279):
caggctgtgg tgactcagga gccttcactg accgtgtccc caggcggaac tgtgaccctg acatgcagat ccagcacagg cgcagtgacc acatctaact acgccaattg ggtgcagcag aagccaggac aggcaccaag gggcctgatc gggggtacaa acaaaagggc tccctggacc cctgcacggt tttctggaag tctgctgggc ggaaaggccg ctctgactat taccggggca caggccgagg acgaagccga ttactattgt gctctgtggt atagcaatct gtgggtgttc gggggtggca caaaactgac tgtgctggga gggggtggat ccggcggagg tggacaggtg cagctgaagg agtcaggacc tggcctggtg gcaccctcac agagcctgtc catcacatgc actgtctctg ggttctcatt atccagatat agtgtacact gggttcgcca gcctccagga aagggtctgg agtggctggg aatgatatgg ggtggtggaa gcacagacta taattcagct ctcaaatcca gactgagtat cagcaaggac aactccaaga gccaagtttt cttaaaaatg aacagtctgc aaactgatga cacagccatg tactactgtg ccagaaaaca tggtaactac tatactatgg actactgggg tcaaggaacc tcagtcaccg tctcctccgg aggatgtggc ggtggaaaag tggccgcact gaaggagaaa gttgctgctt tgaaagagaa ggtcgccgca cttaaggaaa aggtcgcagc cctgaaagag
Аминокислотная последовательность третьей полипептидной цепи из EphA2 mAb 1 х CD3 mAb 2 х gpA33 mAb 1 (SEQ ID NO: 280):
QVQLVQSGAE VKKPGASVKV SCKASGYTFT GSWMNWVRQA PGQGLEWIGR IYPGDGETNY NGKFKDRVTI TADKSTSTAY MELSSLRSED TAVYYCARIY GNNVYFDVWG QGTTVTVSSA STKGPSVFPL APSSKSTSGG TAALGCLVKD YFPEPVTVSW NSGALTSGVH TFPAVLQSSG LYSLSSVVTV PSSSLGTQTY ICNVNHKPSN TKVDKRVEPK SCDKTHTCPP CPAPEAAGGP SVFLFPPKPK DTLMISRTPE VTCVVVDVSH EDPEVKFNWY VDGVEVHNAK TKPREEQYNS TYRVVSVLTV LHQDWLNGKE YKCKVSNKAL PAPIEKTISK AKGQPREPQV YTLPPSREEM TKNQVSLSCA VKGFYPSDIA VEWESNGQPE NNYKTTPPVL DSDGSFFLVS KLTVDKSRWQ QGNVFSCSVM HEALHNRYTQ KSLSLSPGK
В SEQ ID NO: 280, аминокислотные остатки 1-119 соответствуют аминокислотной последовательности VH-домена gpA33 mAb 1 (SEQ ID NO: 186), остатки 120-217 соответствуют модифицированному СН1-домену (SEQ ID NO: 208), остатки 218-232 соответствуют линкеру (SEQ ID NO: 209), а остатки 233-449 соответствуют "несущему углубление" CH2-CH3-домен (SEQ ID NO: 53).
Полинуклеотид, который кодируется SEQ ID NO: 280, представляет собой (SEQ ID NO: 281):
caggtccagc tggtccagag cggggccgaa gtcaaaaaac ccggagcaag cgtgaaggtc tcctgcaaag catcaggcta tacatttaca ggcagctgga tgaactgggt gaggcaggct ccaggacagg gactggagtg gatcgggcgc atctaccctg gagacggcga aactaactat aatggaaagt tcaaagaccg agtgaccatc acagccgata agtctactag taccgcctac atggagctga gctccctgcg gtctgaagat accgccgtct actattgcgc tagaatttac ggaaacaatg tctattttga cgtgtggggg cagggaacaa ctgtgactgt ctcctccgcc tccaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagagagt tgagcccaaa tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaagccgc ggggggaccg tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag gtcacatgcg tggtggtgga cgtgagccac gaagaccctg aggtcaagtt caactggtac gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggaggagatg accaagaacc aggtcagcct gagttgcgca gtcaaaggct tctatcccag cgacatcgcc gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg gactccgacg gctccttctt cctcgtcagc aagctcaccg tggacaagag caggtggcag caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaaccg ctacacgcag aagagcctct ccctgtctcc gggtaaa
Аминокислотная последовательность четвертой полипептидной цепи EphA2 mAb 1 х CD3 mAb 2 х gpA33 mAb 1 (SEQ ID NO: 282):
DIQLTQSPSF LSASVGDRVT ITCSARSSIS FMYWYQQKPG KAPKLLIYDT SNLASGVPSR FSGSGSGTEF TLTISSLEAE DAATYYCQQW SSYPLTFGQG TKLEIKRTVA APSVFIFPPS DEQLKSGTAS VVCLLNNFYP REAKVQWKVD NALQSGNSQE SVTEQDSKDS TYSLSSTLTL SKADYEKHKV YACEVTHQGL SSPVTKSFNR GEC
В SEQ ID NO: 282, аминокислотные остатки 1-106 соответствуют аминокислотной последовательности VL-домена gpA33 mAb 1 (SEQ ID NO: 181), а остатки 107-213 соответствуют домену CL каппа (SEQ ID NO: 210).
Полинуклеотид, который кодируется SEQ ID NO: 282, представляет собой (SEQ ID NO: 283):
gacattcagc tgactcagtc cccctctttt ctgtccgcat ccgtcggaga tcgagtgact attacttgct ctgctaggtc ctcaatcagc ttcatgtact ggtatcagca gaagcccggc aaagcaccta agctgctgat ctacgacaca agcaacctgg cctccggggt gccatctcgg ttctctggca gtgggtcagg aactgagttt accctgacaa ttagctccct ggaggctgaa gatgccgcta cctactattg ccagcagtgg agcagctatc ctctgacctt cggacagggg actaaactgg aaatcaagcg tacggtggct gcaccatcgg tcttcatctt cccgccatct gatgagcagt tgaaatctgg aactgcctct gttgtgtgcc tgctgaataa cttctatccc agagaggcca aagtacagtg gaaggtggat aacgccctcc aatcgggtaa ctcccaggag agtgtcacag agcaggacag caaggacagc acctacagcc tcagcagcac cctgacgctg agcaaagcag actacgagaa acacaaagtc tacgcctgcg aagtcaccca tcagggcctg agctcgcccg tcacaaagag cttcaacagg ggagagtgt
О. EphA2 mAb 2 х CD3 mAb 2 х gpA33 mAb 1
Конструировали вторую альтернативную EphA2/CD3/gpA33 триспецифичную связывающую молекулу. Молекула состоит из четырех полипептидных цепей, и включает VL- и VH-домены из EphA2 mAb 2, VL- и VH-домены антитела CD3 mAb, 2 и VL- и VH-домены из gpA33 mAb 1. Молекула была обозначена как "EphA2 mAb 2 х CD3 mAb 2 х gpA33 mAb 1". Аминокислотная последовательность первой полипептидной цепи этой триспецифичной связывающей молекулы представляет собой (SEQ ID NO: 284):
DVVMTQTPLS LPVSLGDQAS ISCRSSQSLV HSSGNTYLHW YLQKPGQSPK LLIYKVSNRF SGVPDRFSGS GSGTDFTLKI SRVEAEDLGV YFCSQSTHVP TFGSGTKLEI KGGGSGGGGE VQLVESGGGL VQPGGSLRLS CAASGFTFST YAMNWVRQAP GKGLEWVGRI RSKYNNYATY YADSVKGRFT ISRDDSKNSL YLQMNSLKTE DTAVYYCVRH GNFGNSYVSW FAYWGQGTLV TVSSGGCGGG EVAALEKEVA ALEKEVAALE KEVAALEKGG GDKTHTCPPC PAPEAAGGPS VFLFPPKPKD TLMISRTPEV TCVVVDVSHE DPEVKFNWYV DGVEVHNAKT KPREEQYNST YRVVSVLTVL HQDWLNGKEY KCKVSNKALP APIEKTISKA KGQPREPQVY TLPPSREEMT KNQVSLWCLV KGFYPSDIAV EWESNGQPEN NYKTTPPVLD SDGSFFLYSK LTVDKSRWQQ GNVFSCSVMH EALHNHYTQK SLSLSPGK
В SEQ ID NO: 284, аминокислотные остатки 1-111 соответствуют аминокислотной последовательности VL-домена EphA2 mAb 2 (SEQ ID NO: 163), остатки 112-119 соответствуют промежуточному спейсерному пептиду GGGSGGGG (линкер 1) (SEQ ID NO: 33), остатки 120-244 соответствуют аминокислотной последовательности VH-домена CD3 mAb 2, содержащего D65G замену (SEQ ID NO: 112), остатки 245-250 соответствуют GGCGGG линкеру (SEQ ID NO: NO: 34), остатки 251-278 соответствуют домену с Е-спиралью (SEQ ID NO: 39), остатки 279-281 представляют собой линкер GGG, остатки 282-291 представляют собой линкер DKTHTCPPCP (SEQ ID NO: 48), а остатки 292-508 представляют собой "несущий выпуклость" СН2-СН3-домен (SEQ ID NO: 52).
Полинуклеотид, который кодируется SEQ ID NO: 284, представляет собой (SEQ ID NO: 285):
gatgttgtga tgacccaaac tccactctcc ctgcctgtca gtcttggaga tcaagcctcc atctcttgca gatctagtca gagccttgta cacagtagtg gaaacaccta tttacattgg tacctgcaga agccaggcca gtctccaaag ctcctgatct acaaagtttc caaccgattt tctggggtcc cagacaggtt cagtggcagt ggatcaggga cagatttcac actcaagatc agcagagtgg aggctgagga tctgggagtt tatttctgct ctcaaagtac acatgttccc acgttcggct cggggacaaa gttggaaata aaaggtggag gatccggcgg cggaggcgag gtgcagctgg tggagtctgg gggaggcttg gtccagcctg gagggtccct gagactctcc tgtgcagcct ctggattcac cttcagcaca tacgctatga attgggtccg ccaggctcca gggaaggggc tggagtgggt tggaaggatc aggtccaagt acaacaatta tgcaacctac tatgccgact ctgtgaaggg tagattcacc atctcaagag atgattcaaa gaactcactg tatctgcaaa tgaacagcct gaaaaccgag gacacggccg tgtattactg tgtgagacac ggtaacttcg gcaattctta cgtgtcttgg tttgcttatt ggggacaggg gacactggtg actgtgtctt ccggaggatg tggcggtgga gaagtggccg cactggagaa agaggttgct gctttggaga aggaggtcgc tgcacttgaa aaggaggtcg cagccctgga gaaaggcggc ggggacaaaa ctcacacatg cccaccgtgc ccagcacctg aagccgcggg gggaccgtca gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc acatgcgtgg tggtggacgt gagccacgaa gaccctgagg tcaagttcaa ctggtacgtg gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga ggagatgacc aagaaccagg tcagcctgtg gtgcctggtc aaaggcttct atcccagcga catcgccgtg gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag agcctctccc tgtctccggg taaa
Аминокислотная последовательность второй полипептидной цепи EphA2 mAb 2 х CD3 mAb 2 х gpA33 mAb 1 (SEQ ID NO: 286):
QAVVTQEPSL TVSPGGTVTL TCRSSTGAVT TSNYANWVQQ KPGQAPRGLI GGTNKRAPWT PARFSGSLLG GKAALTITGA QAEDEADYYC ALWYSNLWVF GGGTKLTVLG GGGSGGGGQI QLVQSGPELK KPGETVKISC KASGFTFTNY GMNWVKQAPG KGLKWMGWIN TYIGEPTYAD DFKGRFVFSL ETSASTAYLQ INNLKNEDMA TYFCARELGP YYFDYWGQGT TLTVSSGGCG GGKVAALKEK VAALKEKVAA LKEKVAALKE
В SEQ ID NO: 286, аминокислотные остатки 1-110 соответствуют аминокислотной последовательности VL-домена CD3 mAb 2 (SEQ ID NO: 104), остатки 111-118 соответствуют промежуточному спейсерному пептиду GGGSGGGG (Линкер 1) (SEQ ID NO: 33), остатки 119-236 соответствуют аминокислотной последовательности VH-домена EphA2 mAb 2 (SEQ ID NO: 167), остатки 237-242 соответствуют линкеру GGCGGG (SEQ ID NO: 34) а остатки 243-270 представляют собой домен K-спирали (SEQ ID NO: 40).
Полинуклеотид, который кодируется SEQ ID NO: 286, представляет собой (SEQ ID NO: 287):
caggctgtgg tgactcagga gccttcactg accgtgtccc caggcggaac tgtgaccctg acatgcagat ccagcacagg cgcagtgacc acatctaact acgccaattg ggtgcagcag aagccaggac aggcaccaag gggcctgatc gggggtacaa acaaaagggc tccctggacc cctgcacggt tttctggaag tctgctgggc ggaaaggccg ctctgactat taccggggca caggccgagg acgaagccga ttactattgt gctctgtggt atagcaatct gtgggtgttc gggggtggca caaaactgac tgtgctggga gggggtggat ccggcggagg tggacagatc cagttggtgc agtctggacc tgagctgaag aagcctggag agacagtcaa gatctcctgc aaggcttctg ggtttacctt cacaaactat ggaatgaact gggtgaagca ggctccagga aagggtttaa agtggatggg ctggataaac acctatattg gagagccgac atatgctgat gacttcaagg gacggtttgt cttctctttg gaaacctctg ccagcactgc ctatttgcag atcaacaacc tcaaaaatga ggacatggcc acatatttct gtgcaagaga actgggacca tactactttg actactgggg ccaaggcacc actctcacag tctcctccgg aggatgtggc ggtggaaaag tggccgcact gaaggagaaa gttgctgctt tgaaagagaa ggtcgccgca cttaaggaaa aggtcgcagc cctgaaagag
Аминокислотная последовательность третьей полипептидной цепи EphA2 mAb 2 х CD3 mAb 2 х gpA33 mAb 1 (SEQ ID NO: 288):
QVQLVQSGAE VKKPGASVKV SCKASGYTFT GSWMNWVRQA PGQGLEWIGR IYPGDGETNY NGKFKDRVTI TADKSTSTAY MELSSLRSED TAVYYCARIY GNNVYFDVWG QGTTVTVSSA STKGPSVFPL APSSKSTSGG TAALGCLVKD YFPEPVTVSW NSGALTSGVH TFPAVLQSSG LYSLSSVVTV PSSSLGTQTY ICNVNHKPSN TKVDKRVEPK SCDKTHTCPP CPAPEAAGGP SVFLFPPKPK DTLMISRTPE VTCVVVDVSH EDPEVKFNWY VDGVEVHNAK TKPREEQYNS TYRVVSVLTV LHQDWLNGKE YKCKVSNKAL PAPIEKTISK AKGQPREPQV YTLPPSREEM TKNQVSLSCA VKGFYPSDIA VEWESNGQPE NNYKTTPPVL DSDGSFFLVS KLTVDKSRWQ QGNVFSCSVM HEALHNRYTQ KSLSLSPGK
В SEQ ID NO: 288, аминокислотные остатки 1-119 соответствуют аминокислотной последовательности VH-домена gpA33 mAb 1 (SEQ ID NO: 186), остатки 120-217 соответствуют модифицированному СН1-домену (SEQ ID NO: 208), остатки 218-232 соответствуют линкеру (SEQ ID NO: 209), а остатки 233-449 соответствуют "несущему углубление" CH2-CH3-домену (SEQ ID NO: 53).
Полинуклеотид, который кодируется SEQ ID NO: 288, представляет собой (SEQ ID NO: 289):
caggtccagc tggtccagag cggggccgaa gtcaaaaaac ccggagcaag cgtgaaggtc tcctgcaaag catcaggcta tacatttaca ggcagctgga tgaactgggt gaggcaggct ccaggacagg gactggagtg gatcgggcgc atctaccctg gagacggcga aactaactat aatggaaagt tcaaagaccg agtgaccatc acagccgata agtctactag taccgcctac atggagctga gctccctgcg gtctgaagat accgccgtct actattgcgc tagaatttac ggaaacaatg tctattttga cgtgtggggg cagggaacaa ctgtgactgt ctcctccgcc tccaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagagagt tgagcccaaa tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaagccgc ggggggaccg tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag gtcacatgcg tggtggtgga cgtgagccac gaagaccctg aggtcaagtt caactggtac gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggaggagatg accaagaacc aggtcagcct gagttgcgca gtcaaaggct tctatcccag cgacatcgcc gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg gactccgacg gctccttctt cctcgtcagc aagctcaccg tggacaagag caggtggcag caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaaccg ctacacgcag aagagcctct ccctgtctcc gggtaaa
Аминокислотная последовательность четвертой полипептидной цепи EphA2 mAb 2 х CD3 mAb 2 х gpA33 mAb 1 (SEQ ID NO: 290):
DIQLTQSPSF LSASVGDRVT ITCSARSSIS FMYWYQQKPG KAPKLLIYDT SNLASGVPSR FSGSGSGTEF TLTISSLEAE DAATYYCQQW SSYPLTFGQG TKLEIKRTVA APSVFIFPPS DEQLKSGTAS VVCLLNNFYP REAKVQWKVD NALQSGNSQE SVTEQDSKDS TYSLSSTLTL SKADYEKHKV YACEVTHQGL SSPVTKSFNR GEC
В SEQ ID NO: 290, аминокислотные остатки 1-106 соответствуют аминокислотной последовательности VL-домена gpA33 mAb 1 (SEQ ID NO: 181), а остатки 107-213 соответствуют CL каппа домен (SEQ ID NO: 210).
Полинуклеотид, который кодируется SEQ ID NO: 290, представляет собой (SEQ ID NO: 291):
gacattcagc tgactcagtc cccctctttt ctgtccgcat ccgtcggaga tcgagtgact attacttgct ctgctaggtc ctcaatcagc ttcatgtact ggtatcagca gaagcccggc aaagcaccta agctgctgat ctacgacaca agcaacctgg cctccggggt gccatctcgg ttctctggca gtgggtcagg aactgagttt accctgacaa ttagctccct ggaggctgaa gatgccgcta cctactattg ccagcagtgg agcagctatc ctctgacctt cggacagggg actaaactgg aaatcaagcg tacggtggct gcaccatcgg tcttcatctt cccgccatct gatgagcagt tgaaatctgg aactgcctct gttgtgtgcc tgctgaataa cttctatccc agagaggcca aagtacagtg gaaggtggat aacgccctcc aatcgggtaa ctcccaggag agtgtcacag agcaggacag caaggacagc acctacagcc tcagcagcac cctgacgctg agcaaagcag actacgagaa acacaaagtc tacgcctgcg aagtcaccca tcagggcctg agctcgcccg tcacaaagag cttcaacagg ggagagtgt
P. EphA2 mAb 3 х CD3 mAb 2 х gpA33 mAb 1
Конструировали третью альтернативную EphA2/CD3/gpA33 триспецифичную связывающую молекулу. Молекула состоит из четырех полипептидных цепей, и включает VL- и VH-домены из EphA2 mAb 3, VL- и VH-домены антитела CD3 mAb, 2 и VL- и VH-домены из gpA33 mAb 1. Молекула была обозначена "EphA2 mAb 3 х CD3 mAb 2 х gpA33 mAb 1". Аминокислотная последовательность первой полипептидной цепи этой триспецифичной связывающей молекулы является (SEQ ID NO: 292):
DIVLTQSHRS MSTSVGDRVN ITCKASQDVT TAVAWYQQKP GQSPKLLIFW ASTRHAGVPD RFTGSGSGTD FTLTISSVQA GDLALYYCQQ HYSTPYTFGG GTKLEIKGGG SGGGGEVQLV ESGGGLVQPG GSLRLSCAAS GFTFSTYAMN WVRQAPGKGL EWVGRIRSKY NNYATYYADS VKGRFTISRD DSKNSLYLQM NSLKTEDTAV YYCVRHGNFG NSYVSWFAYW GQGTLVTVSS GGCGGGEVAA LEKEVAALEK EVAALEKEVA ALEKGGGDKT HTCPPCPAPE AAGGPSVFLF PPKPKDTLMI SRTPEVTCVV VDVSHEDPEV KFNWYVDGVE VHNAKTKPRE EQYNSTYRVV SVLTVLHQDW LNGKEYKCKV SNKALPAPIE KTISKAKGQP REPQVYTLPP SREEMTKNQV SLWCLVKGFY PSDIAVEWES NGQPENNYKT TPPVLDSDGS FFLYSKLTVD KSRWQQGNVF SCSVMHEALH NHYTQKSLSL SPGK
В SEQ ID NO: 292, аминокислотные остатки 1-107 соответствуют аминокислотной последовательности VL-домена EphA2 mAb 3 (SEQ ID NO: 172), остатки 108-115 соответствуют промежуточному спейсерному пептиду GGGSGGGG (линкер 1) (SEQ ID NO: 33), остатки 116-240 соответствуют аминокислотной последовательности VH-домена CD3 mAb 2, содержащего D65G замену (SEQ ID NO: 112), остатки 241-246 соответствуют GGCGGG линкер (SEQ ID NO: NO: 34), остатки 247-274 соответствуют домену с Е-спиралью (SEQ ID NO: 39), остатки 275-277 представляют собой линкер GGG, остатки 278-287 представляют собой линкер DKTHTCPPCP (SEQ ID NO: 48), а остатки 288-504 представляют собой "несущий выпуклость" СН2-СН3-домен (SEQ ID NO: 52).
Полинуклеотид, который кодируется SEQ ID NO: 292, представляет собой (SEQ ID NO: 293):
gacattgtgc tgacccagtc tcacagatcc atgtccacat cagtaggaga cagggtcaac atcacctgca aggccagtca ggatgtgact actgctgtag cctggtatca acaaaaacca gggcaatctc ctaaattact gattttctgg gcatccaccc ggcacgctgg agtccctgat cgcttcacag gcagtggatc tgggacagat tttactctca ccatcagcag tgtgcaggct ggagacctgg cactttatta ctgtcaacaa cattatagca caccgtacac attcggaggg gggaccaagc tggaaataaa aggtggagga tccggcggcg gaggcgaggt gcagctggtg gagtctgggg gaggcttggt ccagcctgga gggtccctga gactctcctg tgcagcctct ggattcacct tcagcacata cgctatgaat tgggtccgcc aggctccagg gaaggggctg gagtgggttg gaaggatcag gtccaagtac aacaattatg caacctacta tgccgactct gtgaagggta gattcaccat ctcaagagat gattcaaaga actcactgta tctgcaaatg aacagcctga aaaccgagga cacggccgtg tattactgtg tgagacacgg taacttcggc aattcttacg tgtcttggtt tgcttattgg ggacagggga cactggtgac tgtgtcttcc ggaggatgtg gcggtggaga agtggccgca ctggagaaag aggttgctgc tttggagaag gaggtcgctg cacttgaaaa ggaggtcgca gccctggaga aaggcggcgg ggacaaaact cacacatgcc caccgtgccc agcacctgaa gccgcggggg gaccgtcagt cttcctcttc cccccaaaac ccaaggacac cctcatgatc tcccggaccc ctgaggtcac atgcgtggtg gtggacgtga gccacgaaga ccctgaggtc aagttcaact ggtacgtgga cggcgtggag gtgcataatg ccaagacaaa gccgcgggag gagcagtaca acagcacgta ccgtgtggtc agcgtcctca ccgtcctgca ccaggactgg ctgaatggca aggagtacaa gtgcaaggtc tccaacaaag ccctcccagc ccccatcgag aaaaccatct ccaaagccaa agggcagccc cgagaaccac aggtgtacac cctgccccca tcccgggagg agatgaccaa gaaccaggtc agcctgtggt gcctggtcaa aggcttctat cccagcgaca tcgccgtgga gtgggagagc aatgggcagc cggagaacaa ctacaagacc acgcctcccg tgctggactc cgacggctcc ttcttcctct acagcaagct caccgtggac aagagcaggt ggcagcaggg gaacgtcttc tcatgctccg tgatgcatga ggctctgcac aaccactaca cgcagaagag cctctccctg tctccgggta аа
Аминокислотная последовательность второй полипептидной цепи EphA2 mAb 3 х CD3 mAb 2 х gpA33 mAb 1 (SEQ ID NO: 294):
QAVVTQEPSL TVSPGGTVTL TCRSSTGAVT TSNYANWVQQ KPGQAPRGLI GGTNKRAPWT PARFSGSLLG GKAALTITGA QAEDEADYYC ALWYSNLWVF GGGTKLTVLG GGGSGGGGEV QLVESGGGSV KPGGSLKLSC AASGFTFTDH YMYWVRQTPE KRLEWVATIS DGGSFTSYPD SVKGRFTISR DIAKNNLYLQ MSSLKSEDTA MYYCTRDESD RPFPYWGQGT LVTVSSGGCG GGKVAALKEK VAALKEKVAA LKEKVAALKE
В SEQ ID NO: 294, аминокислотные остатки 1-110 соответствуют аминокислотной последовательности VL-домена CD3 mAb 3 (SEQ ID NO: 104), остатки 111-118 соответствуют промежуточному спейсерному пептиду GGGSGGGG (линкер 1) (SEQ ID NO: 33), остатки 119-236 соответствуют аминокислотной последовательности VH-домена EphA2 mAb 3 (SEQ ID NO: 177), остатки 237-242 соответствуют линкера GGCGGG (SEQ ID NO: 34) а остатки 243-270 представляют собой домен K-спирали (SEQ ID NO: 40).
Полинуклеотид, который кодируется SEQ ID NO: 294, представляет собой (SEQ ID NO: 295):
caggctgtgg tgactcagga gccttcactg accgtgtccc caggcggaac tgtgaccctg acatgcagat ccagcacagg cgcagtgacc acatctaact acgccaattg ggtgcagcag aagccaggac aggcaccaag gggcctgatc gggggtacaa acaaaagggc tccctggacc cctgcacggt tttctggaag tctgctgggc ggaaaggccg ctctgactat taccggggca caggccgagg acgaagccga ttactattgt gctctgtggt atagcaatct gtgggtgttc gggggtggca caaaactgac tgtgctggga gggggtggat ccggcggagg tggagaagtg cagctggtgg agtctggggg aggctcagtg aagcctggag ggtccctgaa actctcctgt gcagcctctg gattcacttt cactgaccat tacatgtatt gggttcgcca gactccggaa aagaggctgg agtgggtcgc aaccattagt gatggcggta gtttcacctc ctatccagac agtgtgaagg ggcgattcac catctccaga gacattgcca agaacaacct gtacctccaa atgagcagtc tgaagtctga ggacacagcc atgtattact gtacaagaga tgagagcgat aggccgtttc cttactgggg ccaagggact ctggtcactg tctcctccgg aggatgtggc ggtggaaaag tggccgcact gaaggagaaa gttgctgctt tgaaagagaa ggtcgccgca cttaaggaaa aggtcgcagc cctgaaagag
Аминокислотная последовательность третьей полипептидной цепи из EphA2 mAb 3 х CD3 mAb 2 х gpA33 mAb 1 (SEQ ID NO: 296):
QVQLVQSGAE VKKPGASVKV SCKASGYTFT GSWMNWVRQA PGQGLEWIGR IYPGDGETNY NGKFKDRVTI TADKSTSTAY MELSSLRSED TAVYYCARIY GNNVYFDVWG QGTTVTVSSA STKGPSVFPL APSSKSTSGG TAALGCLVKD YFPEPVTVSW NSGALTSGVH TFPAVLQSSG LYSLSSVVTV PSSSLGTQTY ICNVNHKPSN TKVDKRVEPK SCDKTHTCPP CPAPEAAGGP SVFLFPPKPK DTLMISRTPE VTCVVVDVSH EDPEVKFNWY VDGVEVHNAK TKPREEQYNS TYRVVSVLTV LHQDWLNGKE YKCKVSNKAL PAPIEKTISK AKGQPREPQV YTLPPSREEM TKNQVSLSCA VKGFYPSDIA VEWESNGQPE NNYKTTPPVL DSDGSFFLVS KLTVDKSRWQ QGNVFSCSVM HEALHNRYTQ KSLSLSPGK
В SEQ ID NO: 296, аминокислотные остатки 1-119 соответствуют аминокислотной последовательности VH-домена gpA33 mAb 1 (SEQ ID NO: 186), остатки 120-217 соответствуют модифицированному СН1-домену (SEQ ID NO: 208), остатки 218-232 соответствуют линкеру (SEQ ID NO: 209), а остатки 233-449 соответствуют "несущему углубление" CH2-CH3-домен (SEQ ID NO: 53).
Полинуклеотид, который кодируется SEQ ID NO: 296, представляет собой (SEQ ID NO: 297):
caggtccagc tggtccagag cggggccgaa gtcaaaaaac ccggagcaag cgtgaaggtc tcctgcaaag catcaggcta tacatttaca ggcagctgga tgaactgggt gaggcaggct ccaggacagg gactggagtg gatcgggcgc atctaccctg gagacggcga aactaactat aatggaaagt tcaaagaccg agtgaccatc acagccgata agtctactag taccgcctac atggagctga gctccctgcg gtctgaagat accgccgtct actattgcgc tagaatttac ggaaacaatg tctattttga cgtgtggggg cagggaacaa ctgtgactgt ctcctccgcc tccaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagagagt tgagcccaaa tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaagccgc ggggggaccg tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag gtcacatgcg tggtggtgga cgtgagccac gaagaccctg aggtcaagtt caactggtac gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggaggagatg accaagaacc aggtcagcct gagttgcgca gtcaaaggct tctatcccag cgacatcgcc gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg gactccgacg gctccttctt cctcgtcagc aagctcaccg tggacaagag caggtggcag caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaaccg ctacacgcag aagagcctct ccctgtctcc gggtaaa
Аминокислотная последовательность четвертого полипептидной цепи EphA2 mAb 3 х CD3 mAb 2 х gpA33 mAb 1 (SEQ ID NO: 298):
DIQLTQSPSF LSASVGDRVT ITCSARSSIS FMYWYQQKPG KAPKLLIYDT SNLASGVPSR FSGSGSGTEF TLTISSLEAE DAATYYCQQW SSYPLTFGQG TKLEIKRTVA APSVFIFPPS DEQLKSGTAS VVCLLNNFYP REAKVQWKVD NALQSGNSQE SVTEQDSKDS TYSLSSTLTL SKADYEKHKV YACEVTHQGL SSPVTKSFNR GEC
В SEQ ID NO: 298, аминокислотные остатки 1-106 соответствуют аминокислотной последовательности VL-домена gpA33 mAb 1 (SEQ ID NO: 181), а остатки 107-213 соответствуют домену CL каппа (SEQ ID NO: 210).
Полинуклеотид, который кодируется SEQ ID NO: 298, представляет собой (SEQ ID NO: 299):
gacattcagc tgactcagtc cccctctttt ctgtccgcat ccgtcggaga tcgagtgact attacttgct ctgctaggtc ctcaatcagc ttcatgtact ggtatcagca gaagcccggc aaagcaccta agctgctgat ctacgacaca agcaacctgg cctccggggt gccatctcgg ttctctggca gtgggtcagg aactgagttt accctgacaa ttagctccct ggaggctgaa gatgccgcta cctactattg ccagcagtgg agcagctatc ctctgacctt cggacagggg actaaactgg aaatcaagcg tacggtggct gcaccatcgg tcttcatctt cccgccatct gatgagcagt tgaaatctgg aactgcctct gttgtgtgcc tgctgaataa cttctatccc agagaggcca aagtacagtg gaaggtggat aacgccctcc aatcgggtaa ctcccaggag agtgtcacag agcaggacag caaggacagc acctacagcc tcagcagcac cctgacgctg agcaaagcag actacgagaa acacaaagtc tacgcctgcg aagtcaccca tcagggcctg agctcgcccg tcacaaagag cttcaacagg ggagagtgt
IV. Эталонные Антитела и диатела
Для помощи в описании характеристик триспецифичных связывающих молекул по настоящему изобретению были сконструированы следующие эталонные диатела.
Q. диатело DR5 mAb 1 х CD3 mAb 2
Конструировали примерное биспецифичное диатело, состоящее из двух полипептидных цепей, содержащих VL- и VH-домены антитела DR5 mAb 1 к человеческому DR5 и VL- и VH-домены из CD3 mAb 2. Диатело было названо "диатело DR5 mAb 1 х CD3 mAb 2". Аминокислотная последовательность первой полипептидной цепи этого диатела представляет собой (SEQ ID NO: 140):
DIVLTQSPAS LAVSLGQRAT ISCRASKSVS SSGYSYMHWY QQKPGQPPKV LIFLSSNLDS GVPARFSGSG SGTDFTLNIH PVEDGDAATY YCQHSRDLPP TFGGGTKLEI KGGGSGGGGE VQLVESGGGL VQPGGSLRLS CAASGFTFST YAMNWVRQAP GKGLEWVGRI RSKYNNYATY YADSVKGRFT ISRDDSKNSL YLQMNSLKTE DTAVYYCVRH GNFGNSYVSW FAYWGQGTLV TVSSASTKGE VAACEKEVAA LEKEVAALEK EVAALEK
В SEQ ID NO: 140, аминокислотные остатки 1-111 соответствуют аминокислотной последовательности VL-домена DR5 mAb 1 (SEQ ID NO: 3), остатки 112-119 соответствуют промежуточному спейсерному пептиду GGGSGGGG (Линкер 1) (SEQ ID NO: 33), остатки 120-244 соответствуют аминокислотной последовательности VH-домена CD3 mAb 2, содержащего замену D65G (SEQ ID NO: 112), остатки 245-249 соответствуют ASTKG линкеру (SEQ ID NO: NO: 47) а остатки 250-277 соответствуют цистеинсодержащему домену E-спирали (SEQ ID NO: 41). Полинуклеотид, который кодируется SEQ ID NO: 140, представляет собой SEQ ID NO: 141:
gacattgtgc tgacacagtc tcctgcttcc ttagctgtat ctctcgggca gagggccacc atctcatgca gggccagcaa aagtgtcagt tcctctggct atagttatat gcactggtac caacagaaac caggacagcc acccaaagtc ctcatctttc tttcatccaa cctagattct ggggtccctg ccaggttcag tggcagtggg tctgggacag acttcaccct caacatccat cctgtggagg atggggatgc tgcaacctat tactgtcagc acagtaggga tcttcctccg acgttcggtg gaggcaccaa gctggaaatc aaaggaggcg gatccggcgg cggaggcgag gtgcagctgg tggagtctgg gggaggcttg gtccagcctg gagggtccct gagactctcc tgtgcagcct ctggattcac cttcagcaca tacgctatga attgggtccg ccaggctcca gggaaggggc tggagtgggt tggaaggatc aggtccaagt acaacaatta tgcaacctac tatgccgact ctgtgaaggg tagattcacc atctcaagag atgattcaaa gaactcactg tatctgcaaa tgaacagcct gaaaaccgag gacacggccg tgtattactg tgtgagacac ggtaacttcg gcaattctta cgtgtcttgg tttgcttatt ggggacaggg gacactggtg actgtgtctt ccgcctccac caagggcgaa gtggccgcat gtgagaaaga ggttgctgct ttggagaagg aggtcgctgc acttgaaaag gaggtcgcag ccctggagaa
Аминокислотная последовательность второй полипептидной цепи диатела DR5 mAb 1 х CD3 mAb 2 представляет собой (SEQ ID NO: 142):
QAVVTQEPSL TVSPGGTVTL TCRSSTGAVT TSNYANWVQQ KPGQAPRGLI GGTNKRAPWT PARFSGSLLG GKAALTITGA QAEDEADYYC ALWYSNLWVF GGGTKLTVLG GGGSGGGGEV KFLESGGGLV QPGGSLKLSC VASGFDFSRY WMSWVRQAPG KGLEWIGEIN PDSNTINYTP SLKDKFIISR DNAKNTLYLQ MTKVRSEDTA LYYCTRRAYY GNPAWFAYWG QGTLVTVSAA STKGKVAACK EKVAALKEKV AALKEKVAAL KE
В SEQ ID NO: 142, аминокислотные остатки 1-110 соответствуют аминокислотной последовательности VL-домена CD3 mAb 2 (SEQ ID NO: 104), остатки 111-118 соответствуют промежуточному спейсерному пептиду GGGSGGGG (линкер 1) (SEQ ID NO: 33), остатки 119-239 соответствуют аминокислотной последовательности VH-домена DR5 mAb 1 (SEQ ID NO: 8), за исключением того, что с-концевой остаток серина последовательности SEQ ID NO: 8 был заменен остатком аланина), остатки 240-244 соответствуют ASTKG линкеру (SEQ ID NO: 47), а остатки 245-272 соответствуют цистеинсодержащему домену К-спирали (SEQ ID NO: 42). Полинуклеотид, который кодируется SEQ ID NO: 142, представляет собой SEQ ID NO: 143:
caggctgtgg tgactcagga gccttcactg accgtgtccc caggcggaac tgtgaccctg acatgcagat ccagcacagg cgcagtgacc acatctaact acgccaattg ggtgcagcag aagccaggac aggcaccaag gggcctgatc gggggtacaa acaaaagggc tccctggacc cctgcacggt tttctggaag tctgctgggc ggaaaggccg ctctgactat taccggggca caggccgagg acgaagccga ttactattgt gctctgtggt atagcaatct gtgggtgttc gggggtggca caaaactgac tgtgctggga ggtggtggat ccggcggcgg aggcgaggtg aagtttctcg agtctggagg tggcctggtg cagcctggag gatccctgaa actctcctgt gtagcctcag gattcgattt tagtagatac tggatgagtt gggtccggca ggctccaggg aaagggctag aatggattgg agaaattaat ccagatagca atacgataaa ctatacgcca tctctaaagg ataaattcat catctccaga gacaacgcca aaaatacgct gtatctgcaa atgaccaaag tgagatctga ggacacagcc ctttattatt gtacaagaag ggcctactat ggtaacccgg cctggtttgc ttactggggc caagggactc tggtcactgt ctctgcagcc tccaccaagg gcaaagtggc cgcatgtaag gagaaagttg ctgctttgaa agagaaggtc gccgcactta aggaaaaggt cgcagccctg aaagag
R. Диатело DR5 mAb 2 х CD3 mAb 2
Конструировали примерное биспецифичное диатело, состоящее из двух полипептидных цепей, содержащих VL- и VH-домены антитела DR5 mAb 2 к DR5 и VL- и VH-домены из CD3 mAb 2. Диатело назвали "диатело DR5 mAb 2 х CD3 mAb 2". Аминокислотная последовательность первой полипептидной цепи этого диатела представляет собой (SEQ ID NO: 144):
DIVMTQSHKF MSTSVGDRVS ITCKASQDVN TAVAWYQQKP GQSPKLLIYW ASTRHTGVPD RFTGSGSGTD YTLTIKSVQA EDLTLYYCQQ HYITPWTFGG GTKLEIKGGG SGGGGEVQLV ESGGGLVQPG GSLRLSCAAS GFTFSTYAMN WVRQAPGKGL EWVGRIRSKY NNYATYYADS VKGRFTISRD DSKNSLYLQM NSLKTEDTAV YYCVRHGNFG NSYVSWFAYW GQGTLVTVSS ASTKGEVAAC EKEVAALEKE VAALEKEVAA LEK
В SEQ ID NO: 144, аминокислотные остатки 1-107 соответствуют аминокислотной последовательности VL-домена DR5 mAb 2 (SEQ ID NO: 13), остатки 108-115 соответствуют промежуточному спейсерному пептиду (линкер 1) (SEQ ID NO: 33), остатки 116-240 соответствуют аминокислотной последовательности VH-домена CD3 mAb 2, содержащего замену D65G (SEQ ID NO: 112), остатки 241-245 соответствуют ASTKG линкеру (SEQ ID NO: 47) а остатки 246-273 соответствуют цистеинсодержащему домену E-спирали (SEQ ID NO: 41). Полинуклеотид, который кодируется SEQ ID NO: 144, представляет собой SEQ ID NO: 145:
gacattgtga tgacccagtc tcacaaattc atgtccactt cagtaggaga cagggtcagc atcacctgca aggccagtca ggatgtgaat actgctgtag cctggtatca acaaaaacca gggcaatctc ctaaactact gatttactgg gcatccaccc ggcacactgg agtccctgat cgcttcacag gcagtggatc tgggacagat tatacactca ccatcaaaag tgtgcaggct gaagacctga cactttatta ctgtcagcaa cactatatca ctccgtggac gttcggtgga ggcaccaagc tggaaatcaa aggaggcgga tccggcggcg gaggcgaggt gcagctggtg gagtctgggg gaggcttggt ccagcctgga gggtccctga gactctcctg tgcagcctct ggattcacct tcagcacata cgctatgaat tgggtccgcc aggctccagg gaaggggctg gagtgggttg gaaggatcag gtccaagtac aacaattatg caacctacta tgccgactct gtgaagggta gattcaccat ctcaagagat gattcaaaga actcactgta tctgcaaatg aacagcctga aaaccgagga cacggccgtg tattactgtg tgagacacgg taacttcggc aattcttacg tgtcttggtt tgcttattgg ggacagggga cactggtgac tgtgtcttcc gcctccacca agggcgaagt ggccgcatgt gagaaagagg ttgctgcttt ggagaaggag gtcgctgcac ttgaaaagga ggtcgcagcc ctggagaaa
Аминокислотная последовательность второй полипептидной цепи диатела DR5 mAb 2 х CD3 mAb 2 представляет собой (SEQ ID NO: 146):
QAVVTQEPSL TVSPGGTVTL TCRSSTGAVT TSNYANWVQQ KPGQAPRGLI GGTNKRAPWT PARFSGSLLG GKAALTITGA QAEDEADYYC ALWYSNLWVF GGGTKLTVLG GGGSGGGGKV QLQQSGAELV KPGASVKLSC KASGYTFTEY ILHWVKQKSG QGLEWIGWFY PGNNNIKYNE KFKDKATLTA DKSSSTVYME LSRLTSEDSA VYFCARHEQG PGYFDYWGQG TTLTVSSAST KGKVAACKEK VAALKEKVAA LKEKVAALKE
В SEQ ID NO: 146, аминокислотные остатки 1-110 соответствуют аминокислотной последовательности VL-домена CD3 mAb 2 (SEQ ID NO: 104), остатки 111-118 соответствуют промежуточному спейсерному пептиду GGGSGGGG (линкер 1) (SEQ ID NO: 33), остатки 119-237 соответствуют аминокислотной последовательности VH-домена DR5 mAb 2 (SEQ ID NO: 18), остатки 238-242 соответствуют ASTKG линкеру (SEQ ID NO: 47), а остатки 243-270 соответствуют цистеинсодержащему домену K-спирали (SEQ ID NO: 42). Полинуклеотид, который кодируется SEQ ID NO: 146, представляет собой последовательность SEQ ID NO: 147:
caggctgtgg tgactcagga gccttcactg accgtgtccc caggcggaac tgtgaccctg acatgcagat ccagcacagg cgcagtgacc acatctaact acgccaattg ggtgcagcag aagccaggac aggcaccaag gggcctgatc gggggtacaa acaaaagggc tccctggacc cctgcacggt tttctggaag tctgctgggc ggaaaggccg ctctgactat taccggggca caggccgagg acgaagccga ttactattgt gctctgtggt atagcaatct gtgggtgttc gggggtggca caaaactgac tgtgctggga gggggtggat ccggcggcgg aggcaaggtc cagctgcagc agtctggagc tgaactggtg aaacccgggg catcagtgaa gctgtcctgc aaggcttctg ggtacacctt cactgagtat attttacact gggtaaagca gaagtctgga cagggtcttg agtggattgg gtggttttat cctggaaata ataatataaa gtacaatgag aaattcaagg acaaggccac actgactgcg gacaaatcct ccagcacagt ctatatggaa cttagtagat tgacatctga agactctgcg gtctatttct gtgcaagaca cgaacaagga ccaggttact ttgactactg gggccaaggc accactctca cagtctcctc cgcctccacc aagggcaaag tggccgcatg taaggagaaa gttgctgctt tgaaagagaa ggtcgccgca cttaaggaaa aggtcgcagc cctgaaagag
S. диатело DR5 mAb 3 х CD3 mAb 2
Конструировали примерное биспецифичное диатело, состоящее из двух полипептидных цепей, содержащих VL- и VH-домены антитела DR5 mAb 3 к DR5 и VL- и VH-домены из CD3 mAb 2. Аминокислотная последовательность первой полипептидной цепи диатела содержала последовательность (SEQ ID NO: 148) (остатки CDR выделены подчеркиванием):
SELTQDPAVS VALGQTVRIT CSGDSLRSYY ASWYQQKPGQ APVLVIYGAN NRPSGIPDRF SGSSSGNTAS LTITGAQAED EADYYCNSAD SSGNHVVFGG GTKLTVLGGG GSGGGGEVQL VESGGGLVQP GGSLRLSCAA SGFTFSTYAM NWVRQAPGKG LEWVGRIRSK YNNYATYYAD SVKGRFTISR DDSKNSLYLQ MNSLKTEDTA VYYCVRHGNF GNSYVSWFAY WGQGTLVTVS SASTKGEVAA CEKEVAALEK EVAALEKEVA ALEK
В SEQ ID NO: 148, аминокислотные остатки 1-108 соответствуют VL-домену DR5 mAb 3 (SEQ ID NO: 54), остатки 109-116 соответствуют промежуточному спейсерному пептиду GGGSGGGG (линкер 1) (SEQ ID NO: 33), остатки 117-241 соответствуют аминокислотной последовательности VH-домена CD3 mAb 2, содержащего замену D65G (SEQ ID NO: 112), остатки 242-246 соответствуют ASTKG линкеру (SEQ ID NO: 47), а остатки 247-275 соответствуют цистеинсодержащему домену K-спирали (SEQ ID NO: 42).
Аминокислотная последовательность второй полипептидной цепи, диатела содержала последовательность (SEQ ID NO: 149) (остатки CDR выделены подчеркиванием):
QAVVTQEPSL TVSPGGTVTL TCRSSTGAVT TSNYANWVQQ KPGQAPRGLI GGTNKRAPWT PARFSGSLLG GKAALTITGA QAEDEADYYC ALWYSNLWVF GGGTKLTVLG GGGSGGGGEV QLVQSGGGVE RPGGSLRLSC AASGFTFDDY AMSWVRQAPG KGLEWVSGIN WQGGSTGYAD SVKGRVTISR DNAKNSLYLQ MNSLRAEDTA VYYCAKILGA GRGWYFDYWG KGTTVTVSSA STKGKVAACK EKVAALKEKV AALKEKVAAL KE
В SEQ ID NO: 149, аминокислотные остатки 1-110 соответствуют VL-домену CD3 mAb 2 (SEQ ID NO: 104), остатки 111-118 соответствуют промежуточному спейсерному пептиду GGGSGGGG (линкер 1) (SEQ ID NO: 33), остатки 119-239 соответствуют аминокислотной последовательности VH-домена DR5 mAb 3 (SEQ ID NO: 58), остатки 240-244 соответствуют ASTKG линкеру (SEQ ID NO: 47), а остатки 245- 272 соответствуют цистеинсодержащему домену K-спирали (SEQ ID NO: 42).
T. диатело DR5 mAb 4 х CD3 mAb 2
Конструировали примерное биспецифичное диатело, состоящее из двух полипептидных цепей, содержащих VL- и VH-домены антитела DR5 mAb 4 к человеческому DR5 и VL- и VH-домены из CD3 mAb 2. Аминокислотная последовательность первой полипептидной цепи диатела содержала последовательность (SEQ ID NO: 150) (остатки CDR выделены подчеркиванием):
EIVLTQSPGT LSLSPGERAT LSCRASQGIS RSYLAWYQQK PGQAPSLLIY GASSRATGIP DRFSGSGSGT DFTLTISRLE PEDFAVYYCQ QFGSSPWTFG QGTKVEIKGG GSGGGGEVQL VESGGGLVQP GGSLRLSCAA SGFTFSTYAM NWVRQAPGKG LEWVGRIRSK YNNYATYYAD SVKGRFTISR DDSKNSLYLQ MNSLKTEDTA VYYCVRHGNF GNSYVSWFAY WGQGTLVTVS SASTKGEVAA CEKEVAALEK EVAALEKEVA ALEK
В SEQ ID NO: 150, аминокислотные остатки 1-108 соответствуют VL-домену DR5 mAb 4 (SEQ ID NO: 62), остатки 109-116 соответствуют промежуточному спейсерному пептиду GGGSGGGG (линкер 1) (SEQ ID NO: 33), остатки 117-241 соответствуют аминокислотной последовательности VH-домена CD3 mAb 2, содержащего замену D65G (SEQ ID NO: 112), остатки 242-246 соответствуют ASTKG линкеру (SEQ ID NO: 47), а остатки 247-275 соответствуют цистеинсодержащему домену E-спирали (SEQ ID NO: 41).
Аминокислотная последовательность второй полипептидной цепи диатела содержала последовательность (SEQ ID NO: 151) (остатки CDR выделены подчеркиванием):
QAVVTQEPSL TVSPGGTVTL TCRSSTGAVT TSNYANWVQQ KPGQAPRGLI GGTNKRAPWT PARFSGSLLG GKAALTITGA QAEDEADYYC ALWYSNLWVF GGGTKLTVLG GGGSGGGGQV QLQESGPGLV KPSQTLSLTC TVSGGSISSG DYFWSWIRQL PGKGLEWIGH IHNSGTTYYN PSLKSRVTIS VDTSKKQFSL RLSSVTAADT AVYYCARDRG GDYYYGMDVW GQGTTVTVSS ASTKGKVAAC KEKVAALKEK VAALKEKVAA LKE
В SEQ ID NO: 151, аминокислотные остатки 1-110 соответствуют VL-домену CD3 mAb 2 (SEQ ID NO: 104), остатки 111-118 соответствуют промежуточному спейсерному пептиду GGGSGGGG (линкер 1) (SEQ ID NO: 33), остатки 119-240 соответствуют аминокислотной последовательности VH-домена DR5 mAb 4 (SEQ ID NO: 66), остатки 241-245 соответствуют ASTKG линкеру (SEQ ID NO: 47), а остатки 246- 273 соответствуют цистеинсодержащему домену K-спирали (SEQ ID NO: 42).
U. Эталонное диатело gpA33 х CD3 mAb 2
Для дальнейшей иллюстрации биспецифичных триспецифичных связывающих молекул по настоящему изобретению, конструировали диатело, состоящее из двух полипептидных цепей, с использованием доменов VL и VH из gpA33 mAb 1 и CD3 mAb 2. Аминокислотная последовательность первой полипептидной цепи диатела содержала последовательность (SEQ ID NO: 316) (остатки CDR выделены подчеркиванием):
DIQLTQSPSF LSASVGDRVT ITCSARSSIS FMYWYQQKPG KAPKLLIYDT SNLASGVPSR FSGSGSGTEF TLTISSLEAE DAATYYCQQW SSYPLTFGQG TKLEIKGGGS GGGGEVQLVE SGGGLVQPGG SLRLSCAASG FTFSTYAMNW VRQAPGKGLE WVGRIRSKYN NYATYYADSV KGRFTISRDD SKNSLYLQMN SLKTEDTAVY YCVRHGNFGN SYVSWFAYWG QGTLVTVSSA STKGEVAACE KEVAALEKEV AALEKEVAAL EK
В SEQ ID NO: 316, аминокислотные остатки 1-106 соответствуют VL-домену gpA33 mAb 1 (SEQ ID NO: 181), остатки 107-114 соответствуют промежуточному спейсерному пептиду GGGSGGGG (линкер 1) (SEQ ID NO: 33), остатки 115-239 соответствуют аминокислотной последовательности VH-домена CD3 mAb 2, содержащего замену D65G (SEQ ID NO: 112), остатки 240-244 соответствуют ASTKG линкеру (SEQ ID NO: 47), а остатки 245-272 соответствуют цистеинсодержащему домену E-спирали (SEQ ID NO: 41).
Аминокислотная последовательность второй полипептидной цепи диатела содержала последовательность (SEQ ID NO: 317) (остатки CDR выделены подчеркиванием):
QAVVTQEPSL TVSPGGTVTL TCRSSTGAVT TSNYANWVQQ KPGQAPRGLI GGTNKRAPWT PARFSGSLLG GKAALTITGA QAEDEADYYC ALWYSNLWVF GGGTKLTVLG GGGSGGGGQV QLVQSGAEVK KPGASVKVSC KASGYTFTGS WMNWVRQAPG QGLEWIGRIY PGDGETNYNG KFKDRVTITA DKSTSTAYME LSSLRSEDTA VYYCARIYGN NVYFDVWGQG TTVTVSSAST KGKVAACKEK VAALKEKVAA LKEKVAALKE
В SEQ ID NO: 317, аминокислотные остатки 1-110 соответствуют VL-домену CD3 mAb 2 (SEQ ID NO: 104), остатки 111-118 соответствуют промежуточному спейсерному пептиду GGGSGGGG (линкер 1) (SEQ ID NO: 33), остатки 119-237 соответствуют аминокислотной последовательности VH-домена gpA33 mAb 1 (SEQ ID NO: 186), остатки 238-242 соответствуют ASTKG линкеру (SEQ ID NO: 47), а остатки 243- 270 соответствуют цистеинсодержащему домену K-спирали (SEQ ID NO: 42).
V. Эталонное антитело к флуоресцеину
Антитело 4-4-20 к флуоресцеину (Gruber, M. et al. (1994) “Efficient Tumor Cell Lysis Mediated By A Bi-specific Single Chain Antibody Expressed In Escherichia coli,” J. Immunol. 152(11):5368-5374; Bedzyk, W.D. et al. (1989) “Comparison Of Variable Region Primary Structures Within An Anti-Fluorescein Idiotype Family,” J. Biol. Chem. 264(3): 1565-1569) использовали в контрольных диателах. Аминокислотные последовательности вариабельных доменов легкой и тяжелой цепей антитела 4-4-20 к флуоресцеину представлены далее:
Аминокислотная последовательность вариабельного домена легкой цепи антитела 4-4-20 к флуоресцеину (SEQ ID NO: 138) (CDR остатки подчеркнуты):
DVVMTQTPFS LPVSLGDQAS ISCRSSQSLV HSNGNTYLRW YLQKPGQSPK VLIYKVSNRF SGVPDRFSGS GSGTDFTLKI SRVEAEDLGV YFCSQSTHVP WTFGGGTKLE IK
Аминокислотная последовательность вариабельного домена тяжелой цепи антитела 4-4-20 к флуоресцеину(SEQ ID NO: 139) (CDR остатки подчеркнуты):
EVKLDETGGG LVQPGRPMKL SCVASGFTFS DYWMNWVRQS PEKGLEWVAQ IRNKPYNYET YYSDSVKGRF TISRDDSKSS VYLQMNNLRV EDMGIYYCTG SYYGMDYWGQ GTSVTVSS
V. Способы получения
Триспецифичные связывающие молекулы по настоящему изобретению, могут быть созданы из полинуклеотидов и/или последовательностей антител, которые иммуноспецифичны к DR5, целевому антигену злокачественной опухоли, и целевой эффекторной клетке, способами, известными в данной области техники, например, синтетически или рекомбинантно. Один из способов получения таких пептидных агонистов, антагонистов и модуляторов включает химический синтез полипептида с последующей обработкой при окислительных условиях, подходящих для получения нативной конформации, т.е. корректных дисульфидных связей. Это может быть достигнуто с помощью методик, хорошо известных специалистам в данной области техники (см., например, Kelley, R. F. et al. (1990) в: Genetic Engineering Principles and Methods, Setlow, J.K. Ed., Plenum Press, N.Y., vol. 12, pp 1-19; Stewart, J.M et al. (1984) Solid Phase Peptide Synthesis, Pierce Chemical Co., Rockford, IL; см. также Патенты США No. 4105603; 3972859; 3842067; и 3862925).
Полипептиды по изобретению могут быть получены стандартным способом с использованием твердофазного пептидного синтеза (Merrifield, B. (1986) “Solid Phase Synthesis,” Science 232(4748):341-347; Houghten, R.A. (1985) “General Method For The Rapid Solid-Phase Synthesis Of Large Numbers Of Peptides: Specificity Of Antigen-Antibody Interaction At The Level Of Individual Amino Acids,” Proc. Natl. Acad. Sci. (U.S.A.) 82(15):5131-5135; Ganesan, A. (2006) “Solid-Phase Synthesis In The Twenty-First Century,” Mini Rev. Med. Chem. 6(1):3-10).
Еще в одном альтернативном варианте, подходящие антитела, содержащие один или несколько CDR из целевого антитела к DR5, антитела к антигену злокачественной опухоли или антитела к антигену эффекторных клеток, могут быть получены с помощью коммерчески доступных мышей, которые были сконструированы для экспрессии конкретных иммуноглобулиновых белков человека. Трансгенные животные, которые предназначены для получения более целесообразного (например, с использованием полностью человеческих антител) или более сильного иммунного ответа, также могут быть использованы для получения гуманизированных или человеческих антител. Примерами такой технологии являются Xenomouse ™ (Abgenix, Inc., Fremont, CA) и HuMAb-Mouse ® и TC ™ Mouse (оба из Medarex, Inc., Принстон, Нью-Джерси).
В ином случае, антитела могут быть получены рекомбинантным способом и экспрессированы с использованием любого способа, известного в данной области. Антитела могут быть сделаны рекомбинантным способом сначала путем выделения антител, сделанных из животных-хозяев, с получением последовательности гена, и с использованием генной последовательности для экспрессии антитела рекомбинантно в клетках-хозяевах (например, клетках СНО). Другой метод, который может быть использован, это экспрессия последовательности антитела в растениях (например, в табаке) или в трансгенном молоке. Были раскрыты подходящие методы экспрессии рекомбинантных антител в растениях или молоке (см., например, Peeters et al. (2001) “Production Of Antibodies And Antibody Fragments In Plants,” Vaccine 19:2756; Lonberg, N. et al. (1995) “Human Antibodies From Transgenic Mice,” Int. Rev. Immunol 13:65-93; и Pollock et al. (1999) “Transgenic Milk As A Method For The Production Of Recombinant Antibodies,” J. Immunol Methods 231:147-157). Подходящие методы получения производных антител, например, гуманизированных, одноцепочечных и т.д., известны в данной области техники. В другом альтернативном варианте, антитела могут быть получены рекомбинантным способом с помощью технологии фагового дисплея (см., например, пат. США № 5565332;. 5580717; 5733743; 6265150; и Winter, G. et al. (1994) “Making Antibodies By Phage Display Technology,” Annu. Rev. Immunol. 12.433-455).
Антитела или белок, представляющие интерес, могут быть подвергнуты секвенированию с помощью деградации Эдмана, которая хорошо известен специалистам в данной области техники. Информация о пептиде, полученная по результатам масс-спектрометрии или деградации по Эдману, может быть использована для разработки зондов или праймеров, которые используются для клонирования белка, представляющего интерес.
Альтернативный метод клонирования белка, представляющего интерес, представляет собой "пэннинг" с использованием очищенного DR5, и/или целевого антигена злокачественной опухоли, и/или молекулы, экспрессированной на поверхности целевых эффекторных клеток (или части любых таких молекул), для клетки, экспрессирующей антитело или белок, представляющий интерес, который содержит одну или несколько CDR с тем, чтобы быть способным связываться с DR5 или с таким целевым антигеном злокачественной опухоли или молекулы эффекторной клетки. Процедура "пэннинга" может быть осуществлена путем получения библиотеки кДНК из тканей или клеток, которые экспрессируют DR5, сверхэкспрессии кДНК во втором типе клеток, и скринингом трансфецированных клеток второго типа клеток на предмет специфичного связывания с DR5 в присутствии или в отсутствии известного антитела, способного связываться с такой молекулой (например, DR5, mAb 1 или DR5 mAb 2 в случае пэннинга для новых антител к DR5 и т.д.). Подробное описание методов, используемых в клонировании генов млекопитающих, кодирующих белки клеточной поверхности с помощью "пэннинга" можно найти в данной области (см., например, Aruffo, A. et al.(1987) “Molecular Cloning Of A CD28 cDNA By A High-Efficiency COS Cell Expression System,” Proc. Natl. Acad. Sci.(U.S.A.) 84:8573-8577 и Stephan, J. et al. (1999) “Selective Cloning Of Cell Surface Proteins Involved In Organ Development: Epithelial Glycoprotein Is Involved In Normal Epithelial Differentiation,” Endocrinol. 140: 5841-5854).
Векторы, содержащие полинуклеотиды, представляющие интерес, могут быть введены в клетку-хозяин с помощью любого из целого ряда соответствующих средств, в том числе электропорации, трансфекции с использованием хлорида кальция, хлорида рубидия, фосфат кальция, ДЭАЭ-декстрана или других веществ; бомбардировки микрочастицами; липофекции; и инфекции (например, если вектор представляет собой инфекционный агент, такой как вирус коровьей оспы). Выбор внедряющих векторов или полинуклеотидов часто будет зависеть от особенностей клетки-хозяина.
Любые клетки-хозяева, способные сверхэкспрессировать гетерологичные ДНК, можно использовать для целей выделения генов, кодирующих антитело, полипептид или белок, представляющий интерес. Не ограничивающие примеры подходящих клеток-хозяев млекопитающих включают, без ограничения перечисленным, COS, HeLa и клетки СНО. Предпочтительно, клетки-хозяева экспрессируют кДНК на уровне около в 5 раз выше, более предпочтительно в 10 раз выше, еще более предпочтительно в 20 раз выше, чем у соответствующего эндогенного антитела или белка, представляющего интерес, если он присутствует, в клетках-хозяевах. Скрининг клеток-хозяев на предмет специфичного связывания с DR5 осуществляется с помощью иммунологического анализа или FACS. Клетка, сверхэкспрессирующая антитело или белок, представляющий интерес, может быть идентифицирована.
Изобретение включает полипептиды, содержащие аминокислотную последовательность антител по настоящему изобретению. Полипептиды данного изобретения могут быть получены способами, известными в данной области техники. Полипептиды могут быть получены путем протеолитической или другой деградации антител, с помощью рекомбинантных методов (т.е. в виде одиночных или слитых полипептидов), как описано выше, или путем химического синтеза. Полипептиды антител, особенно более короткие полипептиды вплоть до около 50 аминокислот, удобно получать с помощью химического синтеза. Методы химического синтеза известны в данной области техники и коммерчески доступны. Например, полипептид антитела к DR5 может быть получен с помощью автоматизированного синтезатора полипептидов использованием твердофазного метода.
Изобретение включает модификации любых таких антител (или любых из их полипептидных фрагментов, которые связываются с DR5, антигеном злокачественной опухоли или эффекторных клеток, в зависимости от обстоятельств), и агонисты, антагонисты и модуляторы таких молекул, в том числе функционально эквивалентные антитела и слитые полипептиды, которые не оказывают существенного влияния на свойства таких молекул, и варианты, которые обладают повышенной или пониженной активностью. Модификация полипептидов является обычной практикой в данной области и не должна быть подробно описываться в настоящем документе. Примеры модифицированных полипептидов включают полипептиды с консервативными заменами аминокислотных остатков, одной или несколькими делециями или вставками аминокислот, которые не оказывают существенного вредного изменения функциональной активности, или использование химических аналогов. Аминокислотные остатки, которые могут консервативно заменять друг друга, включают, без ограничения перечисленным: глицин/аланин; серин/треонин; валин/изолейцин/лейцин; аспарагин/глютамин; аспарагиновая кислота/глутаминовая кислота; лизин/аргинин; и фенилаланин/тирозин. Эти полипептиды также включают гликозилированные и негликозилированные полипептиды, и полипептиды с другими посттрансляционными модификациями, такие как, например, гликозилирование с использованием различных сахаров, ацетилирование и фосфорилирование. Предпочтительно, если аминокислотные замены будет консервативными, если замещенные аминокислоты будут обладать одинаковыми химическими свойствами с исходными аминокислотами. Такие консервативные замены известны в данной области, и примеры приведены выше. Аминокислотные модификации могут варьировать от изменения или модификации одной или нескольких аминокислот до полной перестройки области, такой как вариабельная область. Изменения в вариабельной области могут изменять аффинность связывания и/или специфичность. Другие способы модификации включают использование методов конденсации, известных в данной области техники, включая, но не ограничиваясь ими, ферментные средства, окислительный замещения и комплексообразование. Модификации могут быть использованы, например, для присоединения меток для иммунологического анализа, как например, присоединение радиоактивных компонентов для радиоиммунологического анализа. Модифицированные полипептиды получают с использованием установленных процедур в данной области и могут быть подвергнуты скринингу с использованием стандартных анализов, известных в данной области техники.
Изобретение охватывает слитые белки, содержащие один или более из полипептидов по настоящему изобретению. В одном воплощении предлагается слитый полипептид, при условии, что он содержит вариабельный домен тяжелой цепи, тяжелую цепь, или как легкую, так и тяжелую цепь. В другом воплощении слитый полипептид содержит константную область гетерологичного иммуноглобулина. В еще одном воплощении слитый полипептид содержит вариабельную область легкой цепи и вариабельную область тяжелой цепи антитела, полученного из общедоступной гибридомы. Для целей настоящего изобретения, слитый белок антитела содержит один или несколько полипептидных доменов, которые специфично связываются с DR5, антигеном злокачественной опухоли или эффекторной клетки (в зависимости от обстоятельств), и другую аминокислотную последовательность, с которой он не соединен в нативной молекуле, например, с гетерологичной последовательностью или гомологичными последовательностями из другой области.
VI. Применения триспецифичных связывающих молекул по настоящему изобретению
Триспецифичные связывающие молекулы по настоящему изобретению обеспечивают общую терапию злокачественных опухолей. Злокачественные опухоли, которые могут быть подвергнуты лечению с помощью таких молекул, включают злокачественные опухоли, характеризующиеся наличием злокачественной опухолевой клетки, выбранной из группы, состоящей из клеток: опухоли надпочечников, СПИД-ассоциированной злокачественной опухоли, альвеолярной саркомы мягких тканей, опухоли астроцитов, рака мочевого пузыря, рака кости, рака мозга и спинного мозга, метастатической опухоли мозга, рака груди, опухоли каротидной железы, рака шейки матки, хондросаркомы, хордомы, хромофобного почечно-клеточного рака, светлоклеточного рака, рака толстой кишки, колоректального рака, кожной доброкачественной фиброзной гистиоцитомы, десмопластической мелкокруглоклеточной опухоли, эпендимомы, опухоли Юинга, внескелетной миксоидной хондросаркомы, несовершенного костного фиброгенеза, фиброзной дисплазии костей, рака желчного пузыря или желчных протоков, рака желудка, гестационной трофобластической болезни, опухоли половых клеток, рака головы и шеи, гепатоцеллюлярной карциномы, опухоли островковых клеток, саркомы Капоши, рака почки, лейкозов, липомы/доброкачественной липоматозной опухоли, липосаркомы/злокачественной липоматозной опухоли, рака печени, лимфомы, рака легкого, медуллобластомы, меланомы, менингиомы, множественных эндокринных неоплазий, множественной миеломы, миелодиспластического синдрома, нейробластомы, нейроэндокринной опухоли, рака яичника, поджелудочной железы, папиллярного рака щитовидной железы, паратиреоидной опухоли, педиатрического рака, опухоли периферической нервной оболочки, феохромоцитомы, опухоли гипофиза, рака простаты, задней увеальной меланомы, редкого гематологического заболевания, почечного метастатического рака, палочковидной опухоли, рабдомиосаркомы, саркомы, рака кожи, саркомы мягких тканей, плоскоклеточного рака, рака желудка, синовиальной саркомы, тестикулярного рака, карциномы тимуса, тимомы, метастатического рака щитовидной железы и рака матки. Триспецифичные связывающие молекулы по настоящему изобретению могут быть использованы при лечении колоректального рака, печеночно-клеточного рака, глиомы, рак почки, рак молочной железы, множественной миеломы, рака мочевого пузыря, нейробластомы; саркомы, неходжкинской лимфомы, немелкоклеточного рака легкого, рака яичников, рака поджелудочной железы и рака прямой кишки.
Триспецифичные связывающие молекулы по настоящему изобретению содействуют терапии злокачественной опухоли, которая обеспечивается антителом к антигену злокачественной опухоли, характерному для клеток-мишеней, будучи дополнительно способным связываться с молекулами DR5, выстроенных на поверхности таких опухолевых клеток. Полезность данного изобретения, в частности, рассматривается в условиях, в которых плотность антигена злокачественной опухоли является низкой, или когда кинетика связывания антитела против антигена злокачественной опухоли является неоптимальной (или недостаточной) для содействия клинически достаточному терапевтическому ответу. В таких случаях способность молекул по настоящему изобретению связываться как с антигеном злокачественной опухоли, так и с DR5 опухолевых клеток обеспечивает повышенное связывание (посредством авидности), что является достаточным для содействия клинически достаточному терапевтическому ответу. Кроме того, также с помощью обладания связывающим доменом, способным связываться с молекулой на поверхности эффекторных клеток иммунной системы, триспецифичные связывающие молекулы по настоящему изобретению позволяют совместную локализацию таких клеток иммунной системы с опухолевыми клетками, таким образом, содействуя цитотоксическому ответу против опухолевых клеток через перенаправленное уничтожение.
Как показано в таблице 2, триспецифичные связывающие молекулы по настоящему изобретению, которые содержат конкретные комбинации доменов связывания с антигеном злокачественной опухоли, обладают предпочтительной применимостью при лечении некоторых злокачественных опухолей.
Таблица 2
Дополнительно к их применимости в терапии, триспецифичные связывающие молекулы по настоящему изобретению могут быть детектируемо помечены и могут использоваться в диагностике злокачественной опухоли или в визуализации опухолей и опухолевых клеток.
VII. Фармацевтические композиции
Композиции по изобретению включают объемные лекарственные композиции, полезные при изготовлении фармацевтических композиций (например, не очищенные или нестерильные композиции) и фармацевтические композиции (например, композиции, которые пригодны для введения объекту или пациенту), которые могут быть использованы в приготовление лекарственных форм. Такие композиции содержат профилактически или терапевтически эффективное количество триспецифичных связывающих молекул по настоящему изобретению или комбинацию таких агентов и фармацевтически приемлемый носитель. Предпочтительно, композиции по изобретению содержат профилактически или терапевтически эффективное количество триспецифичных связывающих молекул по настоящему изобретению и фармацевтически приемлемый носитель. В частности, изобретение охватывает такие фармацевтические композиции, в которых триспецифичная связывающая молекула содержит DR5-связывающим домен:
(1) DR5 mAb 1 антитела;
(2) DR5 mAb 2 антитела;
(3) DR5 mAb 3 антитела;
(4) DR5 mAb 4 антитела;
(5) DR5 mAb 5 антитела;
(6) DR5 mAb 6 антитела;
(7) DR5 mAb 7 антитела; или
(8) DR5 mAb 8 антитела
(или гуманизированного производного любого из таких антител).
Кроме того, изобретение, в частности, охватывает такие фармацевтические композиции, в которых триспецифичная связывающая молекула содержит домен связывания антигена злокачественной опухоли, который:
(А) связывается с эпитопом EphA2, особенно, если триспецифичная связывающая молекула содержит домен связывания антигена злокачественной опухоли из EphA2 mAb 1, EphA2 mAb 2 или EphA2 mAb 3 или их гуманизированного или химерного варианта; или
(В) связывается с эпитопом gpA33, особенно, если триспецифичная связывающая молекула имеет домен связывания антигена злокачественной опухоли из gpA33 mAb 1 или гуманизированного или химерного вариант; или
(С) связывается с эпитопом Her2, особенно, если триспецифичная связывающая молекула содержит домен связывания антигена злокачественной опухоли HER2 mAb 1 или трастузумаб или их гуманизированного или химерного варианта; или
(D), связывается с эпитопом B7-H3, особенно, если триспецифичная связывающая молекула имеет домен связывания антигена злокачественной опухоли В7-Н3 mAb 1, В7-Н3 mAb 2 или В7-Н3 mAb 3, или их гуманизированного или химерного варианта.
Кроме того, изобретение, в частности, охватывает такие фармацевтические композиции, в которых триспецифичная связывающая молекула содержит домен связывания эффекторных клеток, который связывается с CD2, CD3, CD17, CD20, CD22, CD32B, CD64, BCR/CD79, Т-клеточным рецептором или с NKG2D-рецептором. Кроме того, изобретение, в частности, охватывает такие фармацевтические композиции, в которых триспецифичная связывающая молекула содержит домен связывания эффекторных клеток антитела: Lo-CD2a, CD3 mAb 2, ОКТ3, 3G8, A9, HD37, ритуксимаб, эпратузумаб, CD32B mAb 1, CD64 mAb 1, CD79 mAb 1, BMA 031, KYK-1,0, или KYK-2.0.
Изобретение конкретно рассматривает триспецифичные связывающие молекулы, фармацевтические композиции, которые содержат такую связывающую молекулу и применения таких триспецифичных связывающих молекул, в которых:
(1) к DR5-связывающий домен представляет собой DR5-связывающий домен любого антитела к DR5;
(2) домен связывания злокачественной опухоли представляет собой любой из антигенов злокачественной опухоли, описанных в данном документе;
и
(3) домены связывания эффекторных клеток связываются с любым из CD2, CD3, CD17, CD20, CD22, CD64, CD32B, BCR/CD79, Т-клеточного рецептора или NKG2D-рецептора.
Кроме того, изобретение, в частности, предусматривает триспецифичные связывающие молекулы, фармацевтические композиции, которые содержат такую связывающую молекулу и применения таких триспецифичных связывающих молекул, в которых:
(1) DR5-связывающий домен представляет собой DR5-связывающий домен любого антитела к DR5;
(2) домен связывания злокачественной опухоли представляет собой любой из следующих: EphA1, gpA33, Her2 или В7-Н3;
и
(3) домены связывания эффекторных клеток связываются с любым из CD2, CD3, CD17, CD20, CD22, CD64, CD32B, BCR/CD79, Т-клеточного рецептора или NKG2D-рецептора.
В частности, изобретение охватывает каждую из триспецифичных связывающих молекул, а также фармацевтические композиции, которые содержат такую связывающую молекулу и применения таких триспецифичных связывающих молекул, в которых:
(1) DR5-связывающий домен представляет собой DR5-связывающий домен любого из: DR5 mAb 1 антитела, DR5, mAb 2 антитела, DR5 mAb 3 антитела, DR5 mAb 4 антитела, DR5 mAb 5 антитела, DR5, mAb 6 антитела, DR5 mAb 7 антитела, или к DR5 mAb 8 антитела;
(2) домен, связывающий антиген злокачественной опухоли, представляет собой связывающий домен любого из: EphA2 mAb 1, EphA2 mAb 2, EphA2 mAb 3, gpA33 mAb 1, Her2 mAb 1, трастузумаб, B7-H3 mAb 1, B7-H3 mAb 2 или В7-Н3 mAb 3;
и
(3) домен связывания эффекторных клеток представляет собой связывающий домен любого из: Lo-CD2a, CD3 mAb 2, ОКТ3, 3G8, A9, HD37, ритуксимаб, эпратузумаб, CD32B mAb 1, CD64 mAb 1, CD79 mAb 1, BMA 031, KYK-1,0, или KYK-2.0.
Поскольку приведены 8 DR5-связывающих доменов антител, 9 доменов связывания антигенов злокачественной опухоли из антител и 14 доменов связывания эффекторных клеток из антител, то такая перспектива охватывает все (8 х 9 х 14 =) 1008 комбинаций таких доменов связывания.
Изобретение также охватывает такие фармацевтические композиции, которые дополнительно включают второе терапевтическое антитело (например, опухоль-специфичное моноклональное антитело), которое является специфичным для конкретного антигена злокачественной опухоли, и фармацевтически приемлемый носитель.
В конкретном воплощении термин «фармацевтически приемлемый» означает одобренный регулирующим органом федерального правительства или правительства штата или перечисленные в фармакопее США или другой общепризнанной фармакопее для применения на животных и более конкретно у людей. Термин "носитель" относится к разбавителю, адъюванту (например, адъюванту Фрейнда (полный и неполный), наполнителю или носителю, с которым вводят терапевтическое средство. Такие фармацевтические носители могут быть стерильными жидкостями, такими как вода и масла, в том числе, полученные из нефти, животного, растительного или синтетического происхождения, такие как арахисовое масло, соевое масло, минеральное масло, кунжутное масло и тому подобное. Вода является предпочтительным носителем, если фармацевтическую композицию вводят внутривенно. Солевые растворы и водные растворы декстрозы и глицерини могут быть использованы в качестве жидких носителей, в частности, для растворов для инъекций. Подходящие фармацевтические наполнители включают крахмал, глюкозу, лактозу, сахарозу, желатин, солод, рис, муку, мел, силикагель, стеарат натрия, моностеарат глицерина, тальк, хлорид натрия, сухое обезжиренное молоко, глицерин, пропилен, гликоль, воду, этанол и тому подобное. Композиция, если целесообразно, могут также содержать незначительные количества смачивающих или эмульгирующих агентов или рН-буферные агенты. Эти композиции могут принимать форму растворов, суспензий, эмульсий, таблеток, пилюль, капсул, порошков, препаратов с замедленным высвобождением и тому подобное.
Как правило, ингредиенты композиций по изобретению поставляются либо отдельно, либо в смеси друг с другом в виде стандартной лекарственной формы, например, в виде сухого лиофилизированного порошка или безводного концентрата в герметично закрытом контейнере, таком как ампула или саше, с указанием количества активного агента, когда композиция предназначена для введения путем инфузии, она может быть налита в инфузионный флакон, содержащий стерильную воду фармацевтической чистоты или физиологический раствор. Когда композицию вводят путем инъекции, может обеспечиваться ампула стерильной воды для инъекций или физиологического раствора, так что ингредиенты могут быть смешаны перед введением.
Композиции по изобретению могут быть приготовлены в виде нейтральной или солевой формы. Фармацевтически приемлемые соли включают, без ограничения перечисленным, соли, образованные с анионами, такие как соли, полученные из соляной, фосфорной, уксусной, щавелевой, винной кислот и т.д., и соли, образованные с катионами, такие как соли, полученные из натрия, калия, аммония, кальция, трехвалентного железа, гидроксиды, изопропиламин, триэтиламин, 2-этиламино этанол, гистидин, прокаин и т.д.
В изобретении также предлагается фармацевтическая упаковка или набор, содержащие один или несколько контейнеров, заполненных триспецифичными связывающими молекулами по настоящему изобретению (и более предпочтительно, любой из описанных специфичных связывающих молекул или из проиллюстрированных выше). Кроме того, один или более других профилактических или терапевтических агентов, пригодных для лечения заболевания, также могут быть включены в фармацевтическую упаковку или набор. В изобретении также предлагается фармацевтическая упаковка или набор, включающий один или несколько контейнеров, заполненных одним или несколькими ингредиентами фармацевтических композиций по изобретению. Необязательно к такому контейнеру(ам) может быть привязано уведомление в форме, предписанной правительственным агентством, регулирующим производство, применение или продажу фармацевтических или биологических продуктов, которое отражает одобрение органом производства, применения или продажи для введения человеку.
В настоящем изобретении предлагаются наборы, которые могут быть использованы в описанных выше способах. Набор может содержать любую из триспецифичных связывающих молекул по настоящему изобретению. Набор может дополнительно содержать один или более других профилактических и/или терапевтических агентов, пригодных для лечения злокачественной опухоли, в одном или нескольких контейнерах; и/или набор может дополнительно содержать одно или несколько цитотоксических антител, которые связываются с одним или несколькими антигенами злокачественной опухоли, ассоциированными с злокачественной опухолью. В некоторых воплощениях, другой профилактический или терапевтический агент представляет собой химиотерапевтическое средство. В других воплощениях, профилактический или терапевтический агент представляет собой биологическое или гормональное терапевтическое средство.
VIII. Методы введения
Композиции по настоящему изобретению могут быть получены для лечения, профилактики и облегчения одного или более симптомов, ассоциированных с заболеванием, расстройством или инфекцией, путем введения объекту эффективного количества слитого белка или конъюгированной молекулы по изобретению, или фармацевтической композиции, содержащей слитый белок или конъюгированную молекулу по изобретению. В предпочтительном аспекте, такие композиции по существу чистые (т.е. по существу, свободны от веществ, которые ограничивают ее эффект или производят нежелательные побочные эффекты). В конкретном воплощении объект представляет собой животное, предпочтительно млекопитающее, такое как млекопитающее не являющееся приматом (например, бычьи, лошадиные, кошачьи, собачьи, грызуны и т.д.) или примат (например, обезьяны, такие как, макака, человек и т.д.). В предпочтительном воплощении объектом является человек.
Известны различные системы доставки и они могут быть использованы для введения композиций по изобретению, например, инкапсуляция в липосомы, микрочастицы, микрокапсулы, рекомбинантные клетки, способные экспрессировать антитело или слитый белок, рецептор-опосредованный эндоцитоз (см., например, Wu et al. (1987) “Receptor-Mediated In Vitro Gene Transformation By A Soluble DNA Carrier System,” J. Biol. Chem. 262: 4429-4432), конструкция нуклеиновой кислоты в качестве части ретровирусного или другого вектора и т.д.
Способы введения молекулы по изобретению включают, без ограничения перечисленным, парентеральное введение (например, внутрикожное, внутримышечное, внутрибрюшинное, внутривенное и подкожное), эпидуральное, и через слизистую оболочку (например, интраназальный и пероральный пути). В конкретном воплощении триспецифичные связывающие молекулы по настоящему изобретению вводят внутримышечно, внутривенно или подкожно. Композиции могут быть введены любым удобным способом, например, путем инфузии или болюсной инъекции, путем абсорбции через эпителиальные или слизисто-кожные накладки (например, через слизистую оболочку полости рта, прямой кишки и кишечника и т.д.) и могут быть введены вместе с другими биологически активными агентами. Введение может быть системным или локальным. Кроме того, также может быть использовано легочное введение, например, путем использования ингалятора или распылителя и препарата с распылительным агентом. См., например, пат. США № 6019968. 5985320; 5985309; 5934272; 5874064; 5855913; 5290540; и 4880078; и публ. РСТ WO № 92/19244; WO 97/32572; WO 97/44013; WO 98/31346; и WO 99/66903, каждый из которых включен в настоящее описание в ссылкой в полном объеме.
Изобретение также предусматривает, что триспецифичные связывающие молекулы по настоящему изобретению упаковывают в герметично закрытом контейнере, таком как ампула или саше, с указанием количества молекул. В одном воплощении такие молекулы поставляются в виде сухого стерилизованного лиофилизированного порошка или безводного концентрата в герметично закрытом контейнере, и они могут быть восстановлены, например, водой или физиологическим раствором до нужной концентрации для введения объекту. Предпочтительно, триспецифичные связывающие молекулы по настоящему изобретению поставляются в виде сухого стерильного лиофилизированного порошка в герметично закрытом контейнере, при стандартной дозе, составляющей, по меньшей мере, 5 мкг, более предпочтительно, по меньшей мере, 10 мкг, по меньшей мере, 15 мкг, по меньшей мере, 25 мкг, по меньшей мере, 50 мкг, по меньшей мере, 100 мкг, или, по меньшей мере, 200 мкг.
Лиофилизированные триспецифичные связывающие молекулы по настоящему изобретению, следует хранить при температуре от 2 до 8°C в их первоначальном контейнере, и молекулы должны быть введены в течение 12 часов, предпочтительно в течение 6 часов, в течение 5 часов, в течение 3-х часов, или в пределах 1 часа после восстановления. В альтернативном воплощении, такие молекулы поставляются в жидком виде, в герметично закрытом контейнере, с указанием количества и концентрации молекулы, слитого белка или конъюгированной молекулы. Предпочтительно, такие триспецифичные связывающие молекулы, когда они предусмотрены в жидком виде, поставляются в герметично закрытом контейнере, в котором молекулы присутствуют в концентрации, составляющей не менее 1 мкг/мл, более предпочтительно, по меньшей мере, 2,5 мкг/мл, по меньшей мере, 5 мкг/мл, по меньшей мере, 10 мкг/мл, по меньшей мере, 50 мкг/мл, или, по меньшей мере, 100 мкг/мл.
Количество композиции по изобретению, которое будет эффективным при лечении, профилактике или облегчении одного или более симптомов, ассоциированных с расстройством, может быть определено стандартными клиническими методами. Точная доза, используемая в композиции, также будет зависеть от пути введения и тяжести состояния, и должна быть определена в соответствии с оценкой лечащего врача и согласно обстоятельствам каждого пациента. Эффективные дозы могут быть экстраполированы из кривых доза-ответ, полученных in vitro или на животных модельных тестовых системах.
Для одновалентных диател триспецифичных связывающих молекул, охваченных изобретением, доза, вводимая пациенту, предпочтительно определяется в расчете на массу тела (кг) объекта-реципиента. Вводимая доза обычно составляет от, по меньшей мере, около 0,3 нг/кг в день до около 0,9 нг/кг в день, по меньшей мере, от около 1 нг/кг в день до около 3 нг/кг в день, по меньшей мере, около 3 нг/кг в день до около 9 нг/кг в день, по меньшей мере, около 10 нг/кг в день до около 30 нг/кг в день, по меньшей мере, около 30 нг/кг в день до около 90 нг/кг в день, от, по меньшей мере, около 100 нг/кг в день до около 300 нг/кг в день, по меньшей мере, около 200 нг/кг в день до около 600 нг/кг в день, по меньшей мере, около 300 нг/кг в день до около 900 нг/кг в день, по меньшей мере, около 400 нг/кг в день до около 800 нг/кг в день, по меньшей мере, около 500 нг/кг в день до около 1000 нг/кг в день, по меньшей мере, около 600 нг/кг в день до около 1000 нг/кг в день, по меньшей мере, около 700 нг/кг в день до около 1000 нг/кг в день, по меньшей мере, около 800 нг/кг в день до около 1000 нг/кг в день, по меньшей мере, около 900 нг/кг в день до около 1000 нг/кг в день, или, по меньшей мере, около 1000 нг/кг в день.
В другом воплощении пациенту вводят схему лечения, включающую одну или несколько доз такого профилактически или терапевтически эффективного количества триспецифичной связывающей молекулы по настоящему изобретению, где схему лечения вводят в течение 2 дней, 3 дней, 4 дней, 5 дней, 6 дней или 7 дней. В некоторых воплощениях, схема лечения включает периодическое введение дозы профилактически или терапевтически эффективного количества триспецифичных связывающих молекул по настоящему изобретению (например, введение дозы в 1 день, 2-й день, 3-й день и 4-й день данной недели, и отсутствие введения дозы профилактически или терапевтически эффективного количества триспецифичной связывающей молекулы.
В частности изобретением охвачено введение таких триспецифичных связывающих молекул, которые содержат любую из конкретных комбинаций DR5-связывающих доменов, доменов, связывающие антигены злокачественной опухоли, и доменов, связывающих эффекторные клетки, описанных выше, на 5-й день, 6-й день и 7-й день той же недели). Как правило, проводят 1, 2, 3, 4, 5 или более курсов лечения. Каждый курс может представлять собой ту же схему или другую.
В другом воплощении, введенной дозы увеличивается в первой четверти, первой половине или в первых двух третях или трех четвертях схемы (например, в течение первой, второй или третьей схем из 4 курсов лечения) до достижения ежедневного профилактически или терапевтически эффективное количества триспецифичной связывающей молекулы. В таблице 3 приведены 5 примеров различных режимов дозирования, описанных выше, для типичного курса лечения.
Таблица 3
Дозировка и частота введения триспецифичной связывающей молекулы по настоящему изобретению, могут быть уменьшены или изменены путем увеличения поглощения и проницаемости тканей для молекулы путем модификаций, таких как, например, липидирование.
Дозировка триспецифичных связывающих молекулу по изобретению, вводимая пациенту, может быть рассчитана для использования в качестве единственного терапевтического агента. В ином случае, молекула может быть использована в сочетании с другими терапевтическими композициями, и доза, вводимая пациенту, будет ниже, чем когда указанные молекулы используют в качестве единственного терапевтического агента.
Фармацевтические композиции по настоящему изобретению могут быть введены местно в область, нуждающуюся в лечении; это может быть достигнуто, например, а не в качестве ограничения, местной инфузии, инъекцией, или с помощью имплантата, причем указанный имплантат будет из пористого, непористого или желатинового материала, включая мембраны, такие как силастические мембраны, или волокна. Предпочтительно, чтобы при введении молекулы согласно изобретению, соблюдалась осторожность, чтобы использовались материалы, на которые молекула не абсорбируется.
Композиции по изобретению могут быть доставлены в визикуле, конкретно, в липосоме (See Langer (1990) “New Methods Of Drug Delivery,” Science 249:1527-1533); Treat et al., in Liposomes in the Therapy of Infectious Disease and Cancer, Lopez-Berestein and Fidler (eds.), Liss, New York, pp. 353- 365 (1989); Lopez-Berestein, ibid., pp. 3 17-327).
Композиции по изобретению могут быть доставлены в систему с контролируемым высвобождением или с замедленным высвобождением. Любой метод, известный специалисту в данной области, может быть использован для получения композиций с замедленным высвобождением, включающих одну или несколько из триспецифичных связывающих молекул по изобретению. См., например, пат. США № 4526938; публикацию РСТ WO 91/05548; публ. РСТ WO 96/20698; Ning et al. (1996) “Intratumoral Radioimmunotheraphy Of A Human Colon Cancer Xenograft Using A Sustained-Release Gel,” Radiotherapy & Oncology 39:179-189, Song et al. (1995) “Antibody Mediated Lung Targeting Of Long-Circulating Emulsions,” PDA Journal of Pharmaceutical Science & Technology 50:372-397; Cleek et al. (1997) “Biodegradable Polymeric Carriers For A bFGF Antibody For Cardiovascular Application,” Pro. Int’l. Symp. Control. Rel. Bioact. Mater. 24:853-854; и Lam et al. (1997) “Microencapsulation Of Recombinant Humanized Monoclonal Antibody For Local Delivery,” Proc. Int’l. Symp. Control Rel. Bioact. Mater. 24: 759- 760, каждая из которых включена в настоящее описание в ссылкой в полном объеме. В одном воплощении может быть использована помпа в системе с контролируемым высвобождением (см Langer, выше; Sefton, (1987) “Implantable Pumps,” CRC Crit. Rev. Biomed. Eng. 14:201-240; Buchwald et al. (1980) “Long-Term, Continuous Intravenous Heparin Administration By An Implantable Infusion Pump In Ambulatory Patients With Recurrent Venous Thrombosis,” Surgery 88:507-516; и Saudek et al. (1989) “A Preliminary Trial Of The Programmable Implantable Medication System For Insulin Delivery,” N. Engl. J. Med. 321: 574-579). В другом воплощении могут быть использованы полимерные материалы для достижения контролируемого высвобождения молекул (см например, Medical Applications of Controlled Release, Langer and Wise (eds.), CRC Pres., Boca Raton, Florida (1974); Controlled Drug Bioavailability, Drug Product Design and Performance, Smolen and Ball (eds.), Wiley, New York (1984); Levy et al. (1985) “Inhibition Of Calcification Of Bioprosthetic Heart Valves By Local Controlled-Release Diphosphonate,” Science 228:190-192; During et al. (1989) “Controlled Release Of Dopamine From A Polymeric Brain Implant: In Vivo Characterization,” Ann. Neurol. 25:351-356; Howard et al. (1989) “Intracerebral Drug Delivery In Rats With Lesion-Induced Memory Deficits,” J. Neurosurg. 7 (1): 105-112); пат. США № 5679377; пат. США № 5916597; пат. США № 5912015; пат. США № 5989463; пат. США № 5128326; Публ. РСТ № WO 99/15154; и публ. РСТ WO 99/20253). Примеры полимеров, используемых в композициях с замедленным высвобождением, включают, без ограничения перечисленным, поли (2-гидрокси-этил-метакрилат), поли (метилметакрилат), поли (акриловая кислота), поли (этилен-со-винилацетат), поли (метакриловая кислота), полигликолиды (ПЛГ), полиангидриды, поли (N-винилпирролидон), поли (виниловый спирт), полиакриламид, поли (этиленгликоль), полилактиды (PLA), поли (лактид-со-гликолиды) (PLGA), и полиортоэфиры. Система с контролируемым высвобождением может быть размещена в непосредственной близости от терапевтической мишени (например, в легких), при этом требуется только часть системной дозы (см., например, Goodson, in Medical Applications of Controlled Release, выше, vol. 2, pp. 115-138 (1984)). Полимерные композиции, применяемые в качестве имплантатов с контролируемым высвобождением, могут быть использованы в соответствии с Dunn et al. (см. U.S. 5945155). Данный метод основан на терапевтическом эффекте in situ контролируемого высвобождения биоактивных веществ из полимерной системы. Имплантация, как правило, может иметь место в любой точке внутри тела пациента, нуждающегося в терапевтическом лечении. Может быть использована неполимерная система замедленной доставки, в результате чего неполимерный имплантат в теле объекта используется в качестве системы доставки лекарственного средства. После имплантации в теле, органический растворитель имплантата будет рассеиваться, диспергироваться или вытекать из композиции в окружающую тканевую жидкость, а неполимерный материал будет постепенно коагулировать или осаждаться с образованием твердой микропористой матрицы (см. US 5888533).
Системы с контролируемым высвобождением обсуждаются в обзоре Langer (1990, “New Methods Of Drug Delivery,” Science 249:1527-1533). Любой метод, известный специалисту в данной области, может быть использован для получения композиций с замедленным высвобождением, включающих один или более терапевтических агентов по изобретению. См., например, пат. США № 4526938; Международную публикацию № WO 91/05548 и WO 96/20698. Ning et al. (1996) “Intratumoral Radioimmunotheraphy Of A Human Colon Cancer Xenograft Using A Sustained-Release Gel,” Radiotherapy & Oncology 39:179-189, Song et al. (1995) “Antibody Mediated Lung Targeting Of Long-Circulating Emulsions,” PDA Journal of Pharmaceutical Science & Technology 50:372-397; Cleek et al. (1997) “Biodegradable Polymeric Carriers For A bFGF Antibody For Cardiovascular Application,” Pro. Int’l. Symp. Control. Rel. Bioact. Mater. 24: 853-854; и Lam et al. (1997) “Microencapsulation Of Recombinant Humanized Monoclonal Antibody For Local Delivery,” Proc. Int’l. Symp. Control Rel. Bioact. Mater. 24: 759-760, каждый из которых включен в настоящее описание ссылкой в полном объеме.
В случае, если композиция по изобретению представляет собой нуклеиновую кислоту, кодирующую триспецифичную связывающую молекулу по настоящему изобретению, нуклеиновая кислота может быть введена in vivo, чтобы способствовать экспрессии кодируемой триспецифической связывающей молекулы, путем создания ее конструкции в виде части соответствующего вектора для экспрессии нуклеиновых кислот, и введения его таким образом, что он становится внутриклеточным, например, путем использования ретровирусного вектора (см. пат. США 4980286), или путем прямой инъекции, или при помощи бомбардировки микрочастицами (например, генной пушки; Biolistic, Dupont), или покрытия липидами или рецепторами клеточной поверхности, или трансфицирующими агентами, либо путем введения его вместе с гомеобоксподобным пептидом, который, как известно, проникает в ядро (см., например, Joliot et al. (1991) “Antennapedia Homeobox Peptide Regulates Neural Morphogenesis,” Proc. Natl. Acad. Sci. (U.S.A.) 88: 1864-1868) и т.д. В ином случае, нуклеиновая кислота может быть введена внутриклеточно и включена в ДНК клетки-хозяина для экспрессии с помощью гомологичной рекомбинации.
Лечение субъекта терапевтически или профилактически эффективным количеством триспецифичной связывающей молекулы по настоящему изобретению может включать один курс или, предпочтительно, может включать серию курсов. В предпочтительном примере субъект подвергается лечению таким диателом один раз в неделю в течение от около 1 до 10 недель, предпочтительно от 2 до 8 недель, более предпочтительно, от около 3 до 7 недель, и еще более предпочтительно в течение около 4, 5 или 6 недель. Фармацевтические композиции по изобретению может вводиться один раз в день, два раза в день, или три раза в день. В ином случае, фармацевтические композиции могут вводиться один раз в неделю, два раза в неделю, раз в две недели, раз в месяц, один раз в шесть недель, один раз в два месяца, два раза в год или один раз в год. Следует также понимать, что эффективная доза молекул, используемых для лечения, может увеличиваться или уменьшаться в течение конкретного курса лечения.
Примеры
Теперь, после того, как в целом изобретение описано, то же самое будет легче понять посредством ссылки на следующие примеры, которые приведены в качестве иллюстрации и не предназначены для ограничения объема настоящего изобретения, если не указано иное. Специалистам в данной области техники будет очевидно, что многочисленные модификации как материалов, так и методов могут быть осуществлены на практике, не отходя от сущности и объема настоящего изобретения.
W. Пример 1. Описание характеристик моноклональных антител DR5 mAb 1 и DR5 mAb 2 против DR5 человека
Два моноклональных антитела выделили как способные иммуноспецифично связываться с человеческим DR5, и назвали "DR5 mAb 1" и "DR5 mAb 2". Как обсуждалось выше, было обнаружено, что CDR этих антител отличаются. Для того, чтобы определить, связываются ли антитела с различными эпитопами DR5, получали человеческий слитый белок DR5-Fc и наносили его в виде покрытия на иммобилизуемую поверхность. DR5 mAb 1 (1 мкг/мл) биотинилировали и инкубировали либо с контрольным IgG, либо с DR5 mAb 2 (10 мкг/мл), и оценивали способность антитела IgG или DR5 mAb 2 конкурировать за связывание (с человеческим DR5-Fc слитым белком) с DR5 mAb 1 путем измерения количества иммобилизованного биотинилированного антитела. Кроме того, оценивали способность IgG или антитела DR5 mAb 1 конкурировать за связывание с биотинилированным DR5 mAb 2. Результаты этого эксперимента представлены в таблице 4.
Таблица 4
Результаты этого эксперимента показывают, что биотинилированное антитело способно связываться с белком DR5 даже в присутствии избыточных количеств небиотинилированного антитела. Таким образом, результаты показывают, что DR5 mAb 1 и DR5, mAb 2 связываются с различными эпитопами DR5.
Для того чтобы дополнительно описать характеристики антител DR5 mAb 1 и DR mAb 2, оценивали их способность блокировать связывание между DR5 и лигандом TRAIL. Таким образом, биотинилированное DR5 mAb 1, биотинилированное DR5 mAb 2 или биотинилированный слитый белок DR5-Fc (каждый по 2 мкг/мл) по отдельности инкубировали с иммобилизованным слитым белком DR5-Fc (1 мкг/мл) в присутствии либо буфера или меченного гистидином TRAIL (20 мкг/мл). Оценивали количество иммобилизованного биотинилированного антитела. Результаты этого эксперимента представлены в таблице 5.
Таблица 5
Результаты демонстрируют, что количество DR5 mAb 1 или DR5 mAb 2, связанного с иммобилизованным DR5-Fc, не зависело от присутствия меченного гистидином TRAIL, что свидетельствует о том, что ни DR5 mAb 1, ни DR5 mAb 2 не блокируют сайт связывания лиганда TRAIL и DR5. Кроме того, никакое антитело не было способно связываться с меченным гистидином лигандом TRAIL.
X. Пример 2. Видовая специфичность моноклональных антител DR5 mAb 1 и DR5 mAb 2 к DR5 человека
Для того чтобы оценить видовую специфичность моноклональных антител DR5 mAb 1 и DR5, mAb 2 против DR5 человека, способность антител связываться с DR5 человека сравнивали с их способностью связываться с DR5 макаки (Macaca fascicularis). Результаты этого эксперимента показаны на фигуре 6. Результаты демонстрируют, что оба антитела способны связываться с DR5 макаки, но, что каждый из них проявляют более высокую аффинность связывания в отношении человеческого DR5.
Кинетику связывания исследовали с использованием Biacore анализа, как показано на фигуре 7. Биспецифичные диатела DR5 х CD3 инкубировали с His-меченым DR5, и кинетику связывания определяли с помощью анализа Biacore. Применяли диатела DR5 mAb 1 х CD3 mAb 2 (фигура 7, панели А и Е), DR5 mAb 2 х CD3 mAb 2 (фигура 7, панели B и F), DR5 mAb 3 х CD3 mAb 2 (фигура 7, панели С и G), и DR5, mAb 4 х CD3 Mab, 2 (фигура 7, панели D и Н). Фигура 7, панели A-D показаны результаты для DR5 человека. Фигура 7, панели E-H показаны результаты для DR5 макаки. Рассчитанные ka, kd и KD представлены в таблице 6.
Таблица 6
Результаты показывают, что DR5 mAb 1 и DR5 mAb 2 демонстрируют измененные кинетики связывания по сравнению с эталонными антителами DR5 mAb 3 и DR5 mAb 4.
Y. Пример 3. Неожиданное преимущество DR5 mAb 1 и DR5 mAb 2
Способность к DR5-связывающих молекул DR5 mAb 1 и DR5, mAb 2 по настоящему изобретению опосредовать цитотоксичность сравнивали с такой способностью эталонных анти-DR5 антител: DR5 mAb 3 и DR5 mAb 4. Для того чтобы осуществить такое сравнение, получали биспецифичное DR5 х CD3 диатело, содержащее VL и VH-домены этих антител и VL и VH-домены CD3 mAb 2. Полученные диатела: DR5 mAb 1 х CD3 mAb 2; DR5 mAb 2 х CD3 mAb 2; DR5 mAb 3 х CD3 mAb 2; и DR5 mAb 4 х CD3 mAb 2.
Применяемое контрольное диатело содержало VL- и VH-домены антитела 4-4-20 к флуоресцеину (соответственно, SEQ ID NO: 138 и 139) и VL- и VH-домены из CD3 mAb 2 (соответственно, SEQ ID NO: 102 и 108), и было предназначено в качестве контрольного диатела анти-флуоресцеин х анти-CD3 "4-4-20 х CD3 mAb 2". Диатело состояло из двух полипептидных цепей. Первая полипептидная цепь диатела содержала аминокислотную последовательность (SEQ ID NO: 300) (CDR показаны подчеркиванием):
DVVMTQTPFS LPVSLGDQAS ISCRSSQSLV HSNGNTYLRW YLQKPGQSPK VLIYKVSNRF SGVPDRFSGS GSGTDFTLKI SRVEAEDLGV YFCSQSTHVP WTFGGGTKLE IKGGGSGGGG EVQLVESGGG LVQPGGSLRL SCAASGFTFN TYAMNWVRQA PGKGLEWVAR IRSKYNNYAT YYADSVKDRF TISRDDSKNS LYLQMNSLKT EDTAVYYCVR HGNFGNSYVS WFAYWGQGTL VTVSSGGCGG GEVAALEKEV AALEKEVAAL EKEVAALEK
В SEQ ID NO: 300, аминокислотные остатки 1-112 соответствуют VL-домену анти-флуоресцеин-антитела 4-4-20 (SEQ ID NO: 138), остатки 113-120 соответствуют промежуточному спейсерному пептиду GGGSGGGG (Линкер 1) (SEQ ID NO: 33), остатки 121-245 соответствуют VH-домену CD3 mAb 2 (SEQ ID NO: 108), остатки 246-251 представляют собой цистеинсодержащий спейсерный пептид (GGCGGG) (SEQ ID NO: 34), и остатки 252-280 соответствуют домену Е-спирали (SEQ ID NO: 39).
Вторая полипептидная цепь диатела содержала аминокислотную последовательность (SEQ ID NO: 301) (CDR показаны подчеркиванием):
QAVVTQEPSL TVSPGGTVTL TCRSSTGAVT TSNYANWVQQ KPGQAPRGLI GGTNKRAPWT PARFSGSLLG GKAALTITGA QAEDEADYYC ALWYSNLWVF GGGTKLTVLG GGGSGGGGEV KLDETGGGLV QPGRPMKLSC VASGFTFSDY WMNWVRQSPE KGLEWVAQIR NKPYNYETYY SDSVKGRFTI SRDDSKSSVY LQMNNLRVED MGIYYCTGSY YGMDYWGQGT SVTVSSGGCG GGKVAALKEK VAALKEKVAA LKEKVAALKE
В SEQ ID NO: 301, аминокислотные остатки 1-110 соответствуют VL-домену CD3 mAb 2 (SEQ ID NO: 104), остатки 111-118 соответствуют промежуточному спейсерному пептиду GGGSGGGG (Линкер 1) (SEQ ID NO: 33), остатки 119-236 соответствуют VH-домену антитела 4-4-20 к флуоресцеину (SEQ ID NO: 139), остатки 237-242 представляют собой цистеинсодержащий спейсерный пептид (GGCGGG) (SEQ ID NO: 34), и остатки 243-270 соответствуют домену K-спирали (SEQ ID NO: 40).
Опухолевые клетки-мишени инкубировали с одним из этих диател или с контрольным (диателом 4-4-20 х CD3 mAb 2) в присутствии мононуклеаров периферической крови (РВМС) и альвеолярных базальных эпителиальных клеток А549 аденокарциномы человека в течение 24 часов при соотношении эффекторных клеток к клеткам-мишеням 20: 1. Процент цитотоксичности определяли путем измерения высвобождения лактатдегидрогеназы (ЛДГ) в среду поврежденными клетками.
Результаты этого исследования представлены на фигуре 8. Аналогичные результаты были получены с использованием злокачественных опухолевых клеток SKMES легкого человека, злокачественных опухолевых клеток DU145 предстательной железы человека, злокачественных клеток A375 меланомы человека, HER2-сверхэкспрессирующих клеток SKBR3 карциномы молочной железы и клеток JIMT карциномы молочной железы человека. Результаты выявили, что VL- и VH-домены из DR5 mAb 1 и DR5 mAb 2 значительно и неожиданно более эффективны в отношении индукции цитотоксичности, чем у эталонного DR5 mAb.
Z. Пример 4. Триспецифичные связывающие молекулы опосредуют координированное и одновременное связывание с клетками-мишенями
Изучали способность триспецифичных связывающих молекул по настоящему изобретению связываться с клетками-мишенями. Применяемые триспецифичные молекулы: EphA2 mAb 1 х CD3 mAb 2 х DR5 mAb 1; EphA2 mAb 1 х CD3 mAb 2 х gpA33 mAb 1; и gpA33 mAb 1 х CD3 mAb 2 х DR5 mAb 1. Как показано на фигуре 9А, было обнаружено, что такие триспецифичные связывающие молекулы, которые содержат домен EphA2 связывания с антигеном злокачественной опухоли, способны связываться с EphA2-экспрессирующими клетками-мишенями CHO. Как показано на фигуре 9В, было обнаружено, что такие триспецифичные связывающие молекулы, которые содержат домен DR5 связывания с антигеном злокачественной опухоли, способны связываться с DR5-экспрессирующими клетками-мишенями CHO. Как показано на фигуре 9С, было обнаружено, что такие триспецифичные связывающие молекулы, которые содержат домен EphA2 связывания с антигеном злокачественной опухоли или домен DR5 связывания с антигеном злокачественной опухоли, способны связываться с клетками DU145. Клетки DU145 представляют собой человеческие клеточные линии клеток простаты, которые экспрессируют оба, EphA2 и DR5, но не экспрессируют gpA33. Вышеописанное эталонное gpA33 mAb 1 х CD3 mAb 2 диатело использовали в качестве контроля.
Важно отметить, что данные демонстрируют, что, когда оба из двух доменов связывания с антигеном злокачественной опухоли триспецифичной связывающей молекулы по настоящему изобретению способны связываться с клеткой-мишенью, то такое двойное связывание ассоциировано с синергетическим (например, 5- 25 кратным) усилением связывания мишени.
AA. Пример 5. Триспецифичные связывающие молекулы опосредуют цитотоксичность связанных клеток-мишеней
Изучали способность триспецифичных связывающих молекул по настоящему изобретению опосредовать цитотоксичность связанных клеток-мишеней в присутствии цитотоксических лимфоцитов. Применяемые триспецифичные молекулы: EphA2 mAb 1 х CD3 mAb 2 х DR5 mAb 1; EphA2 mAb 1 х CD3 mAb 2 х gpA33 mAb 1; и gpA33 mAb 1 х CD3 mAb 2 х DR5 mAb 1. Вышеописанное эталонное диатело gpA33 mAb 1 х CD3 mAb 2 и диатело 4-4-20 х CD3 mAb 2 использовали в качестве контроля.
Как показано на фигуре 10А, те триспецифичные связывающие молекулы, которые содержат домен связывания с антигеном злокачественной опухоли EphA2 и, таким образом, способны связываться с EphA2-экспрессирующими клетками СНО, были способны опосредовать цитотоксичность на таких клетках в присутствии цитотоксических лимфоцитов. Как показано на фигуре 10В, было обнаружено, что такие триспецифичные связывающие молекулы, которые включают домен связывания с антигеном злокачественной опухоли DR5 и, таким образом, способны связываться с DR5-экспрессирующими клетками СНО, способны опосредовать цитотоксичность на DR5-экспрессирующих клетках-мишенях CHO в присутствии цитотоксических лимфоцитов.
Как показано на фигуре 10С, такие триспецифичные связывающие молекулы, которые содержат домен EphA2 связывания с антигеном злокачественной опухоли или домен DR5 связывания с антигеном злокачественной опухоли и, таким образом, способны связываться с клетками DU145, были способны опосредовать цитотоксичность таких клеток в присутствии цитотоксических лимфоцитов. Важно отметить, что данные демонстрируют, что, когда оба из двух доменов связывания с антигеном злокачественной опухоли триспецифичной связывающей молекулы по настоящему изобретению способны связываться с клеткой-мишенью, то такое двойное связывание ассоциировано с синергическим усилением связывания мишени. Таким образом, EphA2 mAb 1 х CD3 mAb 2 х DR5 mAb 1, которые способны связываться с обоими EphA2 и DR5, опосредовали существенно больше цитотоксичности, чем EphA2 mAb 1 х CD3 mAb 2 х gpA33 mAb 1 или gpA33 mAb 1 х CD3 mAb 2 х DR5 mAb 1, которые были способны связываться только с молекулой EphA2 или DR5 клеток DU146 (поскольку такие клетки лишены gpA33).
В связи с этим, при приблизительно ЕС50 для EphA2 mAb 1 х CD3 mAb 2 х DR5 mAb 1 не наблюдается никакого ответа цитотоксических лимфоцитов ни для EphA2 mAb 1 х CD3 mAb 2 х gpA33 mAb 1, ни для gpA33 mAb 1 х CD3 mAb 2 х DR5 mAb 1. Приблизительно при EC90 для EphA2 mAb 1 х CD3 mAb 2 х DR5 mAb 1, EphA2 mAb 1 х CD3 mAb 2 х gpA33 mAb 1 демонстрирует только EC15, а gpA33 mAb 1 х CD3 mAb 2 х DR5 mAb 1 демонстрирует полное отсутствие ответа цитотоксических лимфоцитов.
Все публикации и патенты, упомянутые в данном описании, включены в ссылкой в той же степени, как если бы каждая отдельная публикация или патентная заявка была конкретно и отдельно указана для включения ссылкой в полном объеме. В то время как изобретение было описано в связи с его конкретными воплощениями, следует понимать, что возможны дополнительные модификации, и подразумевается, что данная заявка охватывает любые вариации, применения или адаптации изобретения, следующие, в целом, принципам изобретения и включающие такие отклонения от настоящего описания, которые соответствуют известной или обычной практике в данной области, к которой относится изобретение, и которые могут быть применены к существенным признакам, описанным выше в данном документе.
| название | год | авторы | номер документа |
|---|---|---|---|
| БИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СПОСОБНЫЕ СВЯЗЫВАТЬ CD137 И ОПУХОЛЕВЫЕ АНТИГЕНЫ, И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2805648C2 |
| ВАРИАНТНЫЕ СВЯЗЫВАЮЩИЕ CD3 ДОМЕНЫ И ИХ ПРИМЕНЕНИЕ В КОМБИНИРОВАННОЙ ТЕРАПИИ ПРИ ЛЕЧЕНИИ ЗАБОЛЕВАНИЙ | 2019 |
|
RU2810222C2 |
| МОЛЕКУЛЫ, СВЯЗЫВАЮЩИЕ ADAM9, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2783619C2 |
| ТРИФУНКЦИОНАЛЬНАЯ АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА | 2015 |
|
RU2753882C2 |
| СВЯЗЫВАЮЩИЕ IL-1 БЕЛКИ | 2011 |
|
RU2615173C2 |
| НОВОЕ PSMA-СВЯЗЫВАЮЩЕЕ АНТИТЕЛО И ЕГО ПРИМЕНЕНИЯ | 2017 |
|
RU2762704C2 |
| БИСПЕЦИФИЧНЫЕ МОЛЕКУЛЫ, ИММУНОРЕАКТИВНЫЕ С ИММУННЫМИ ЭФФЕКТОРНЫМИ КЛЕТКАМИ, ЭКСПРЕССИРУЮЩИМИ АКТИВИРУЮЩИЙ РЕЦЕПТОР | 2014 |
|
RU2721707C2 |
| БИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ АГЕНТЫ С МЕЖВИДОВОЙ СПЕЦИФИЧНОСТЬЮ | 2008 |
|
RU2535992C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С HLA-A2/WT1 | 2018 |
|
RU2815176C2 |
| АНТИТЕЛА, СОДЕРЖАЩИЕ ТОЛЬКО ТЯЖЕЛЫЕ ЦЕПИ, КОТОРЫЕ СВЯЗЫВАЮТСЯ С CD19 | 2019 |
|
RU2824170C2 |
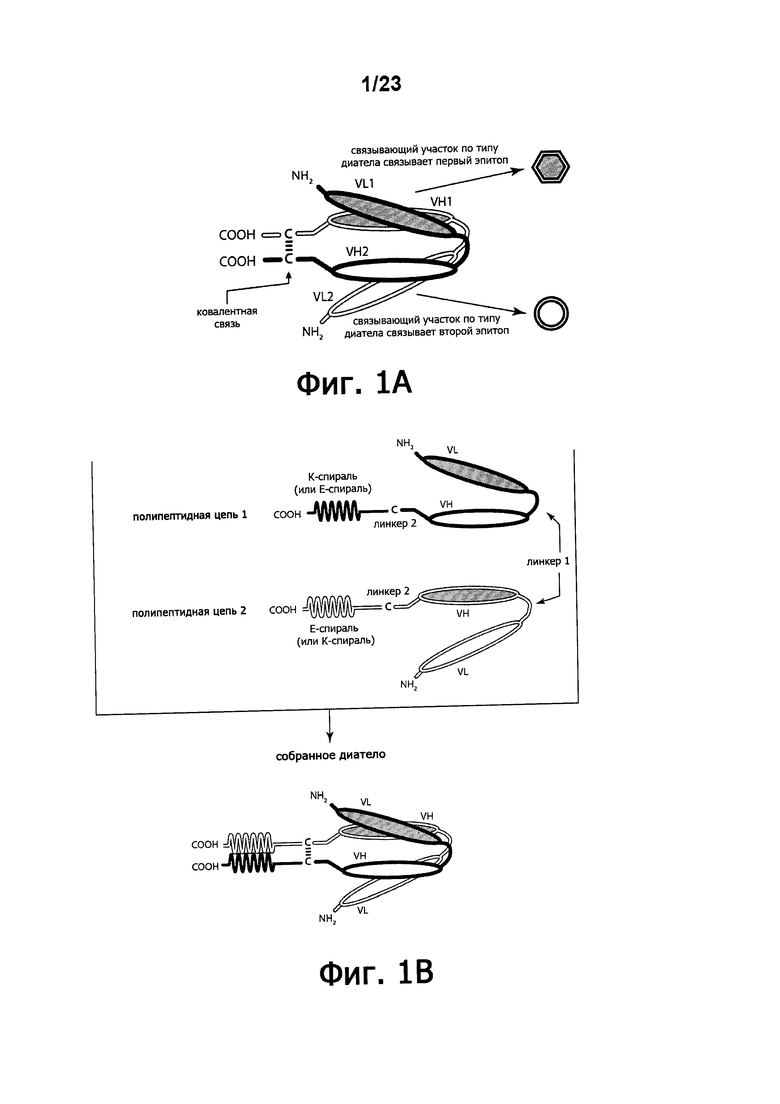
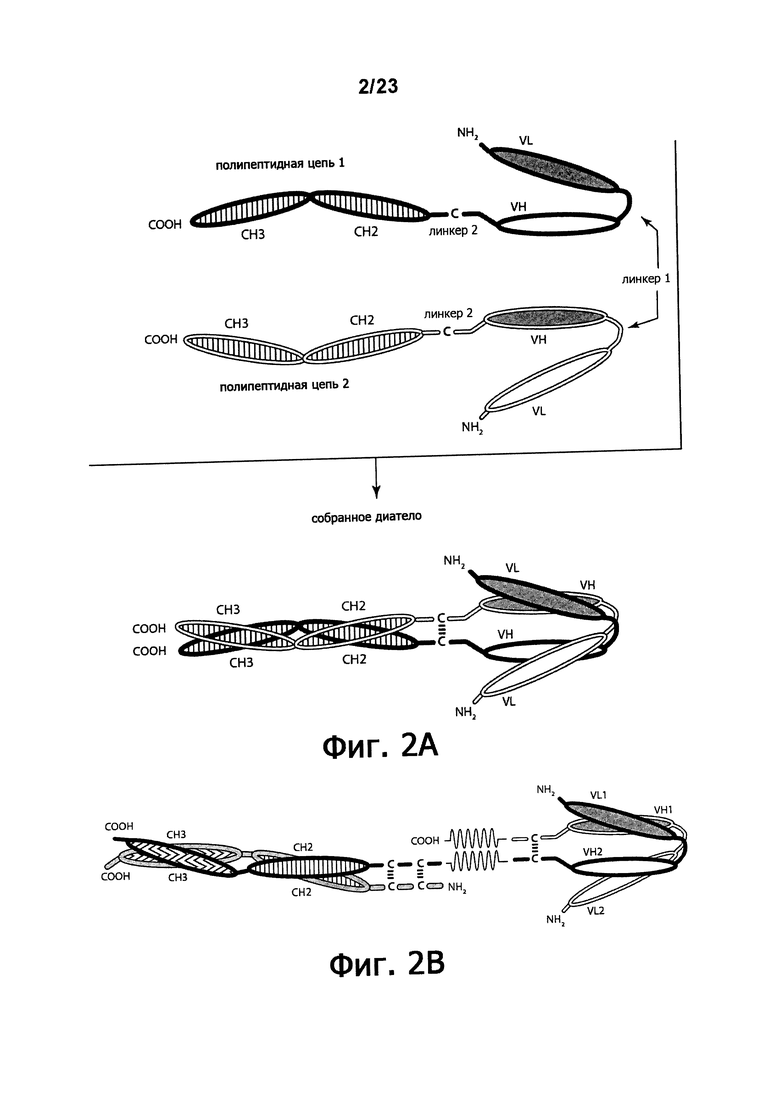
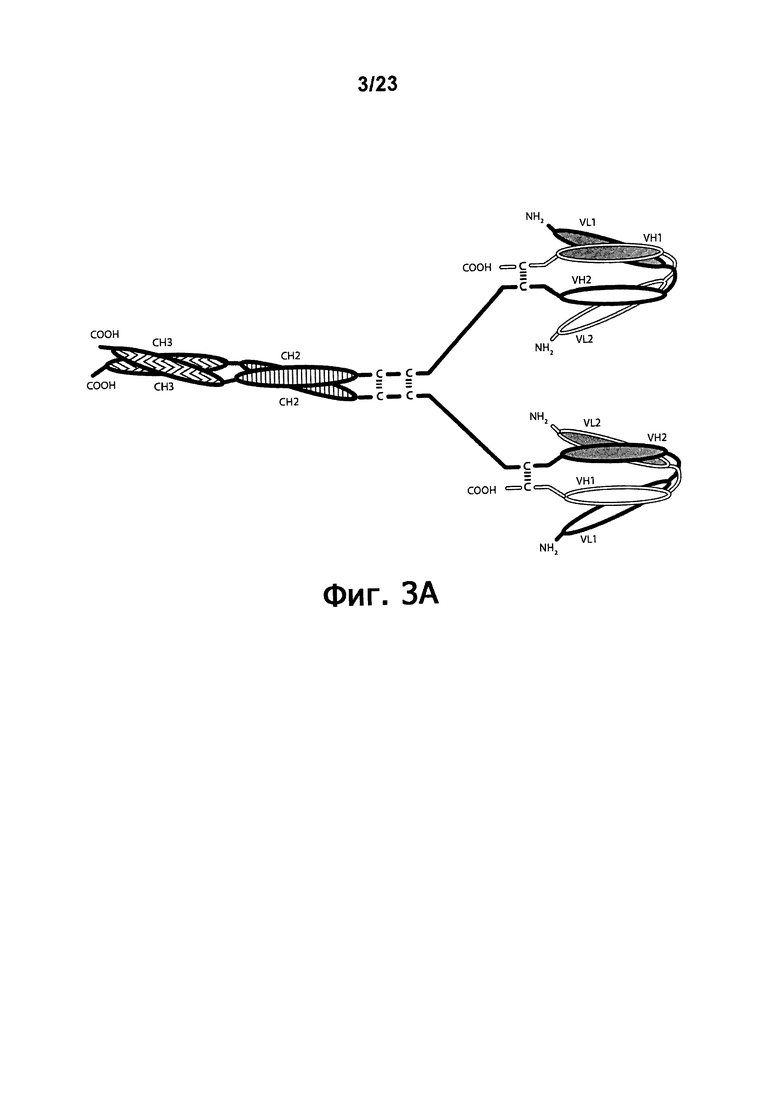
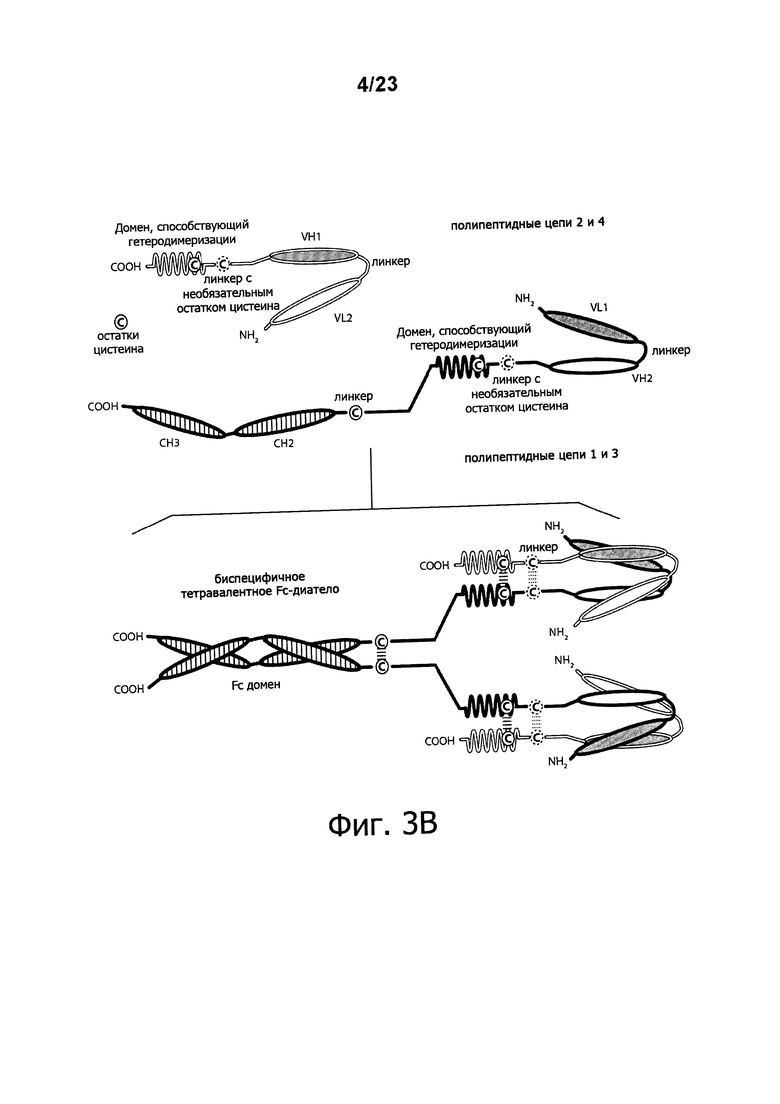
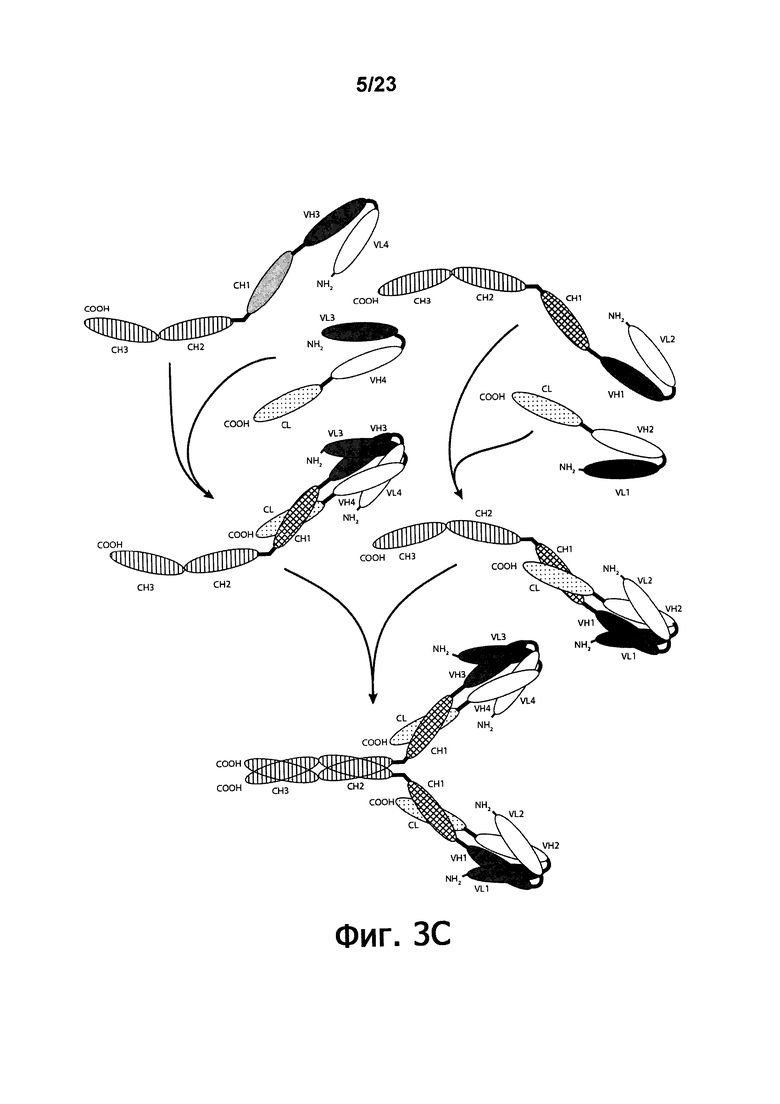
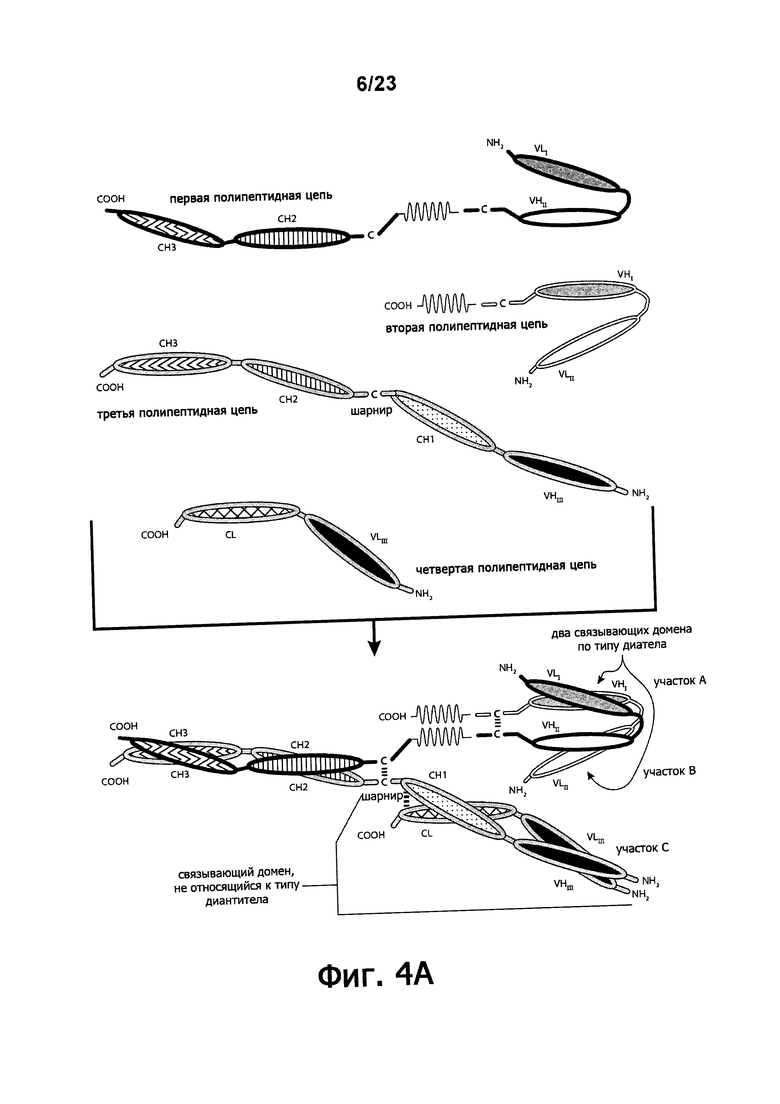
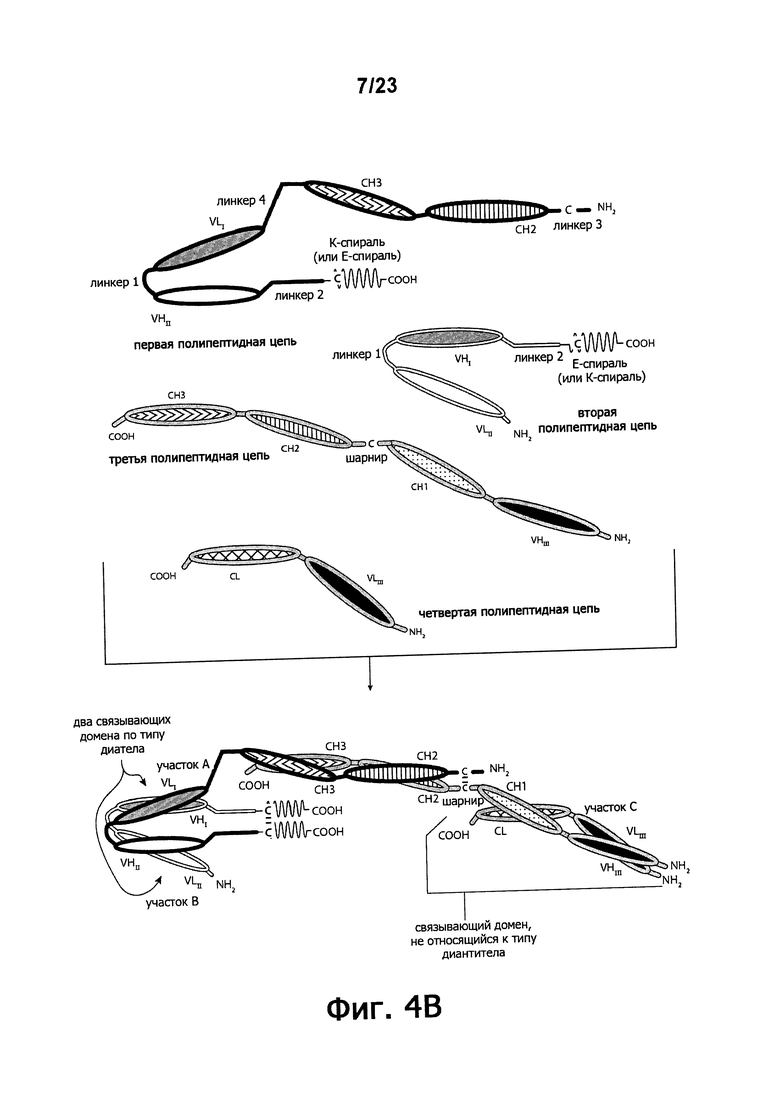
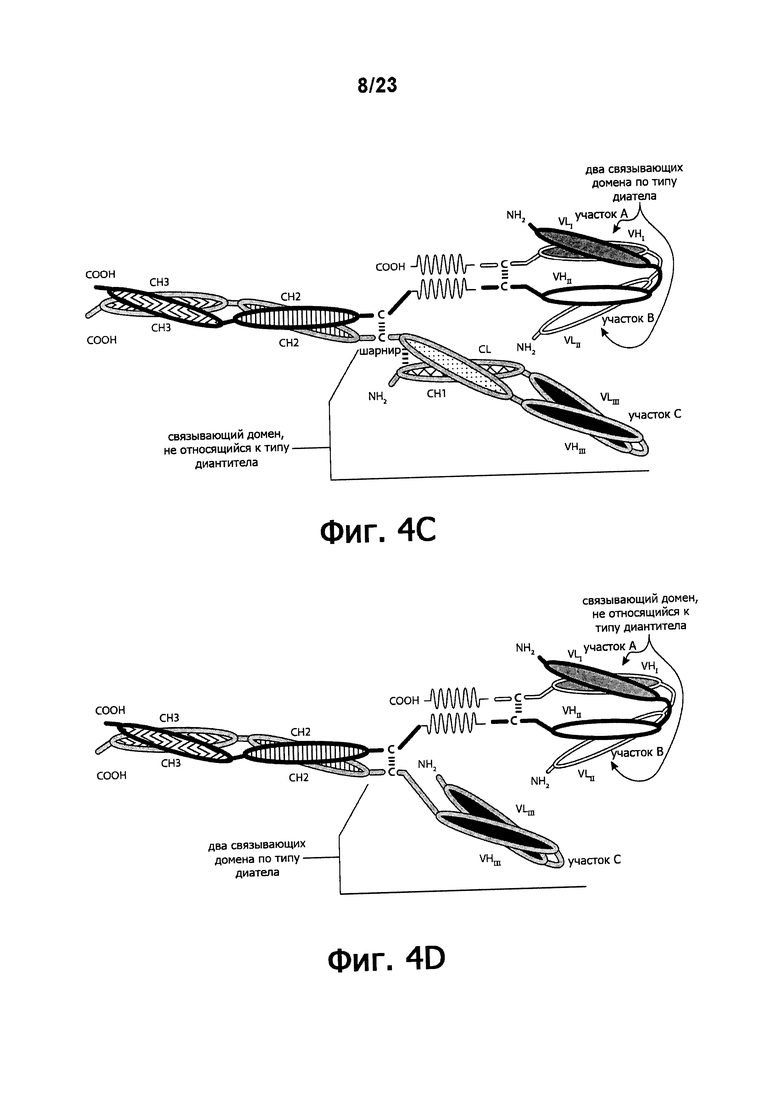
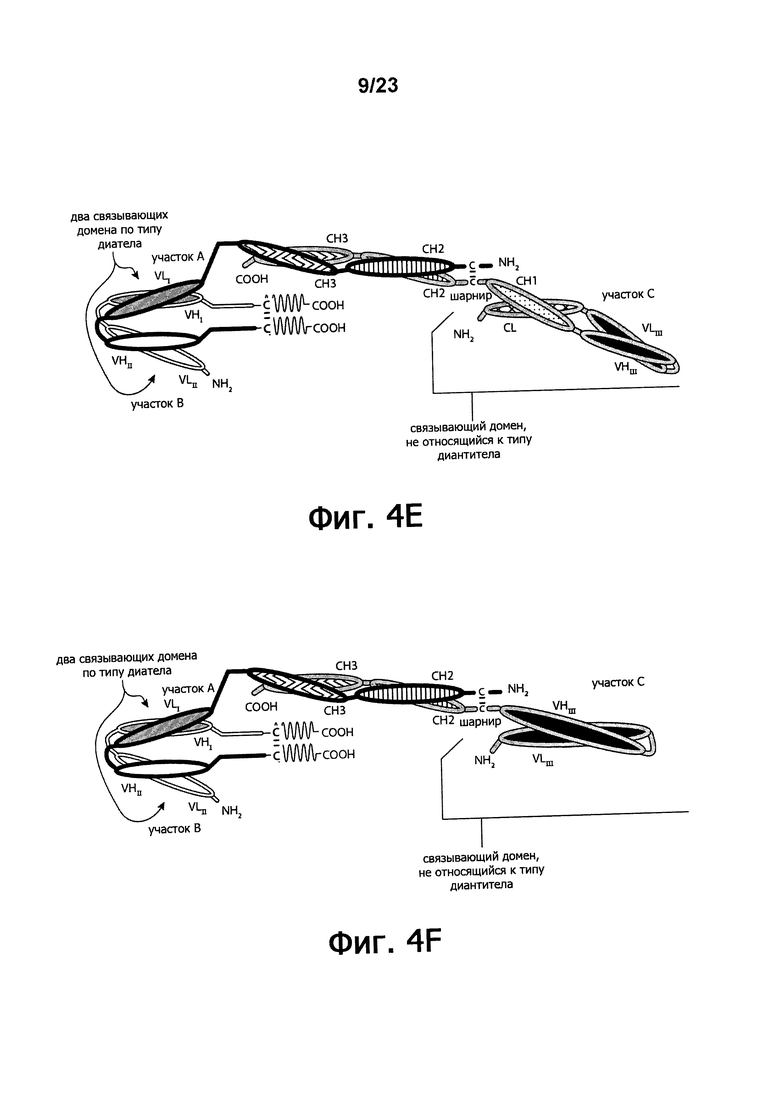
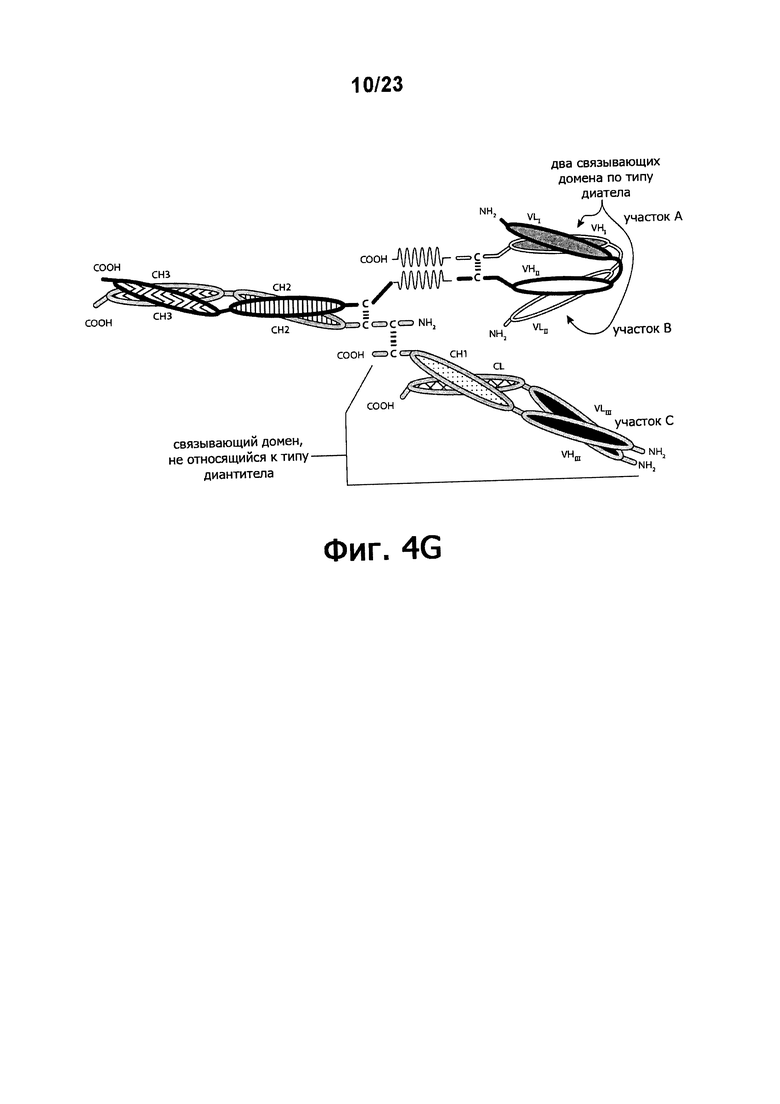
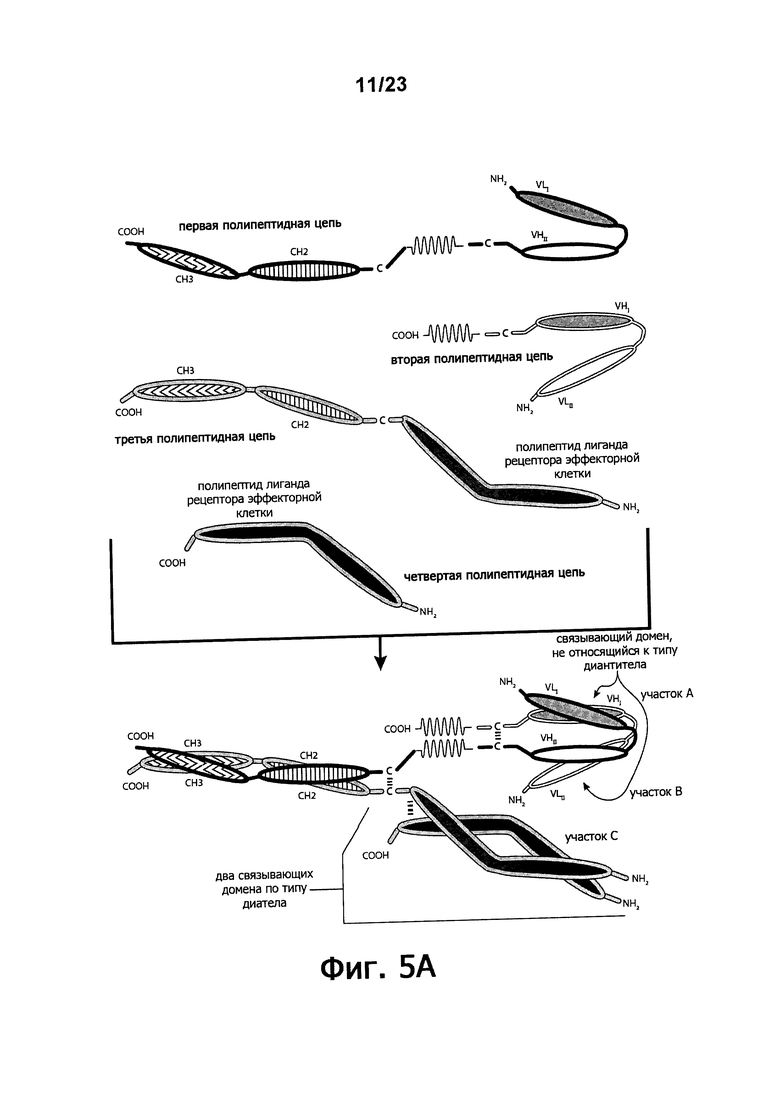
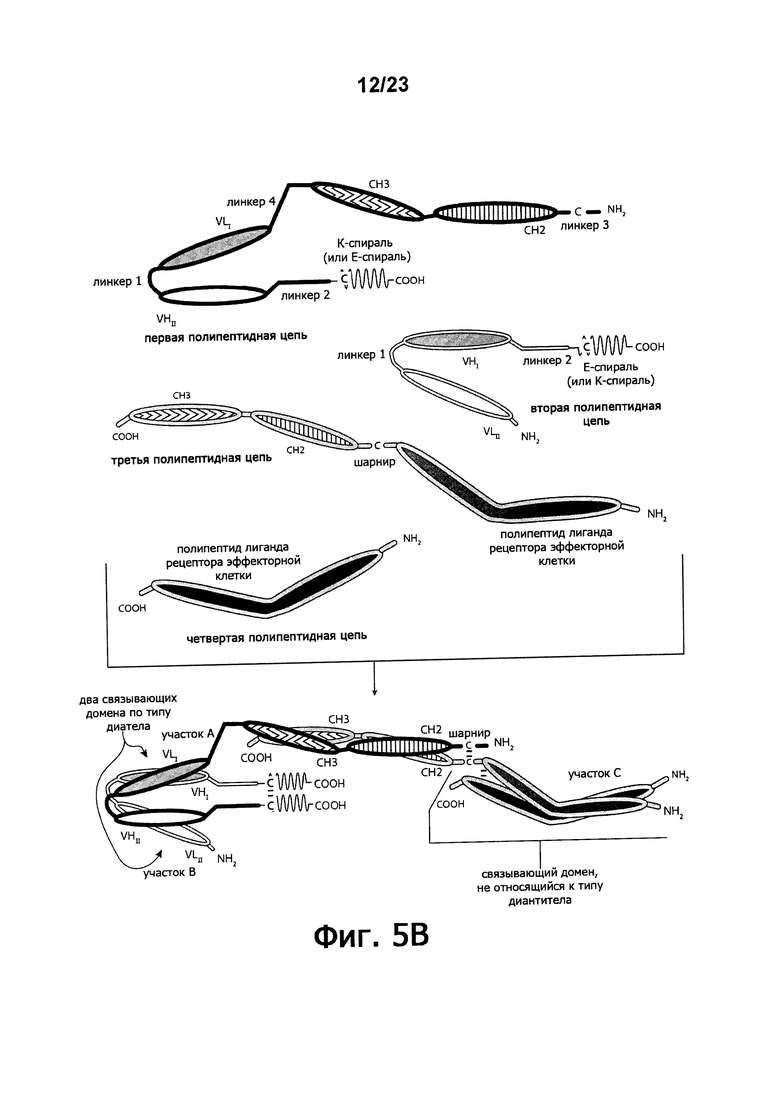

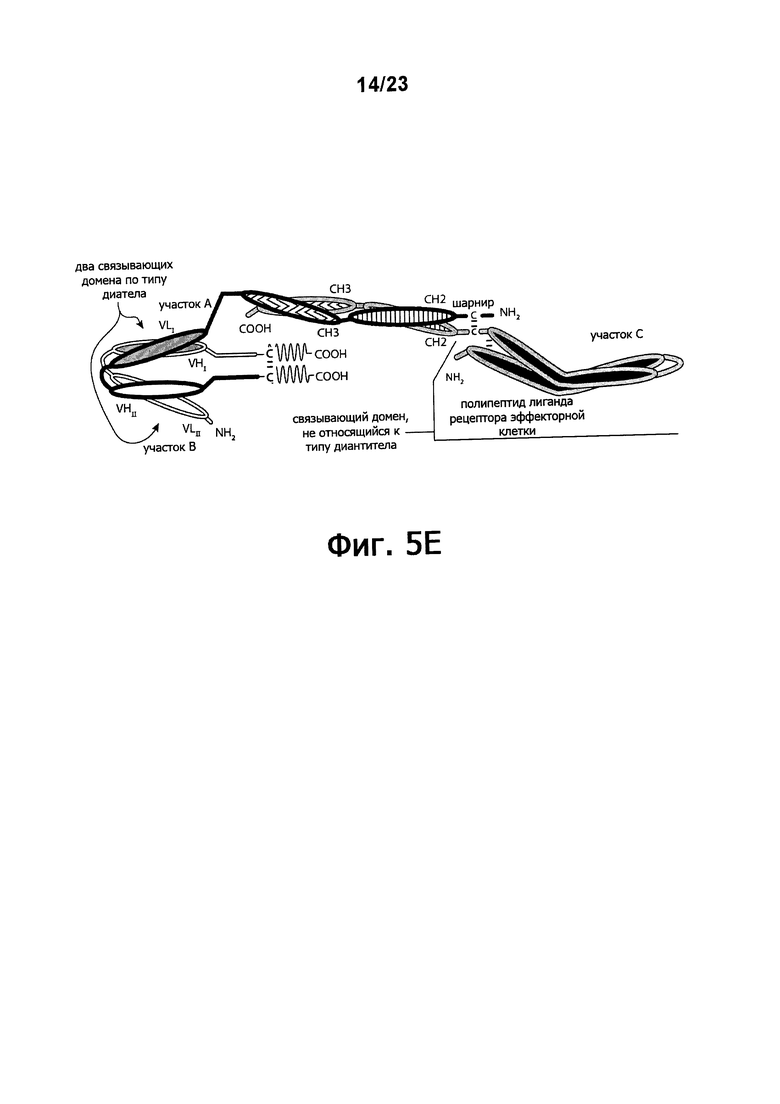
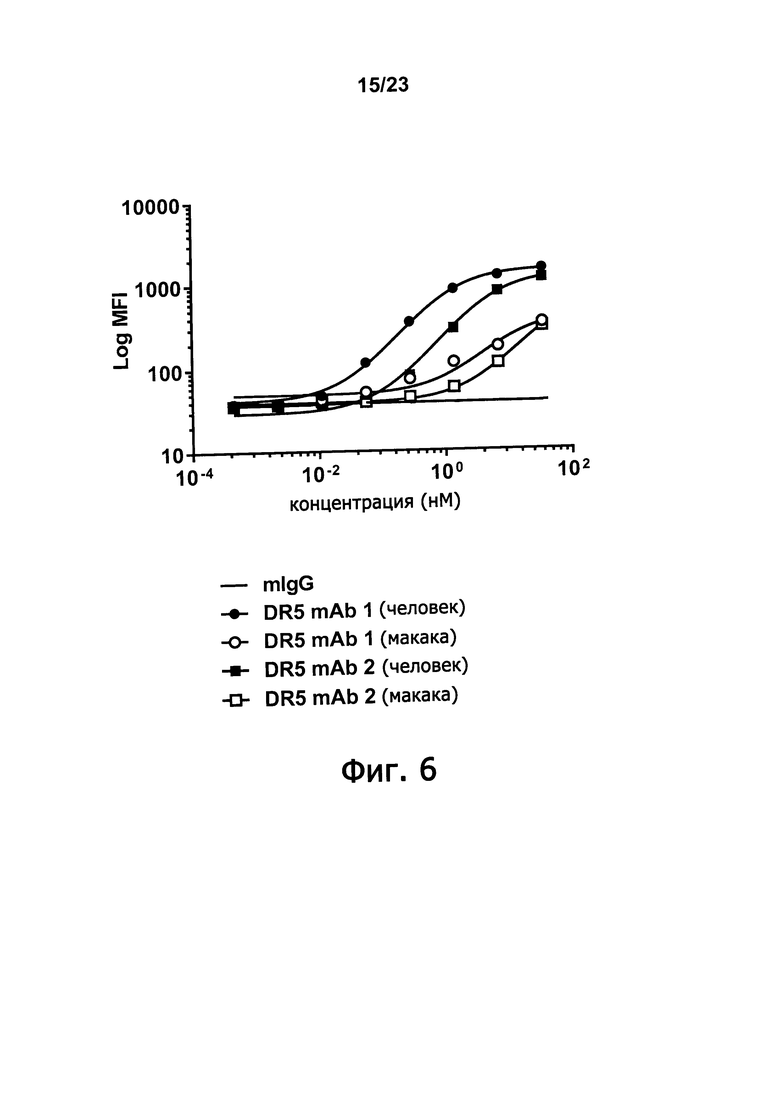
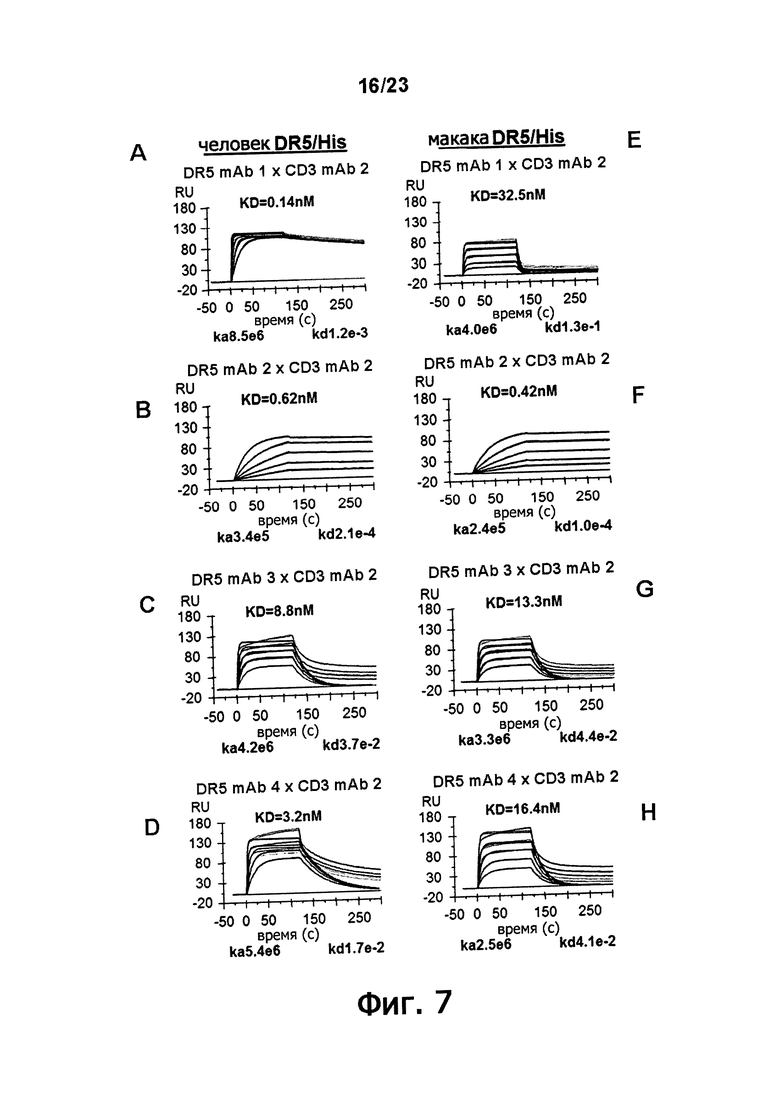
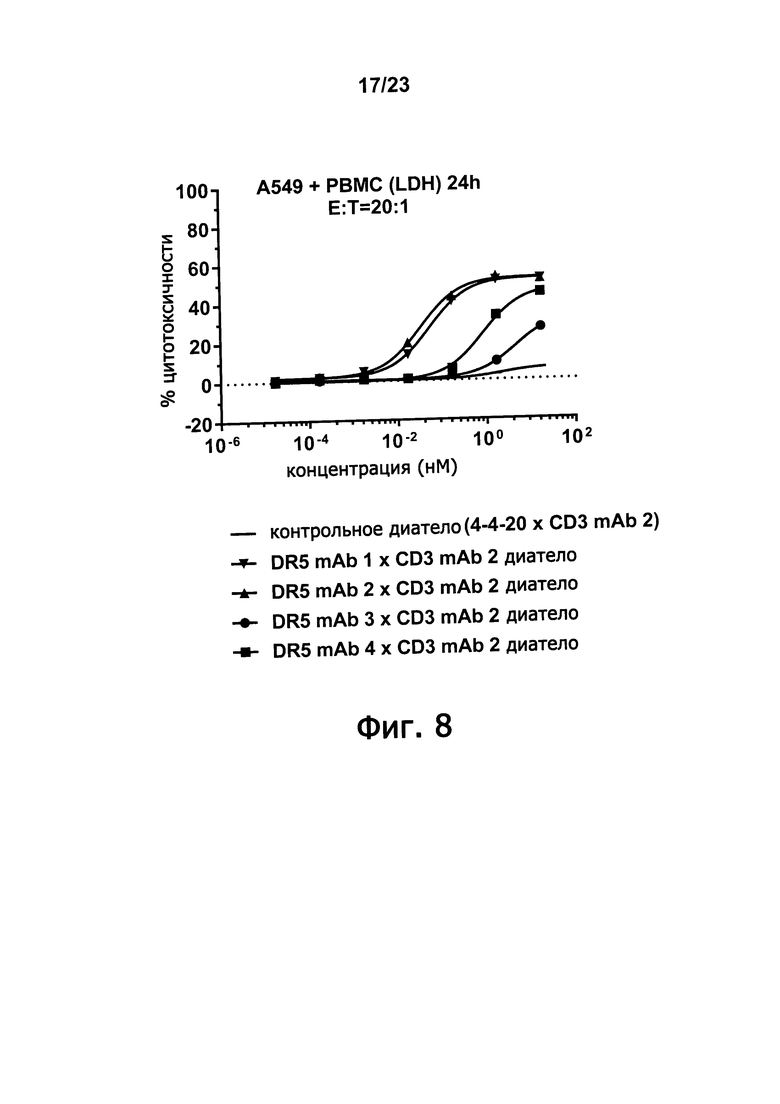
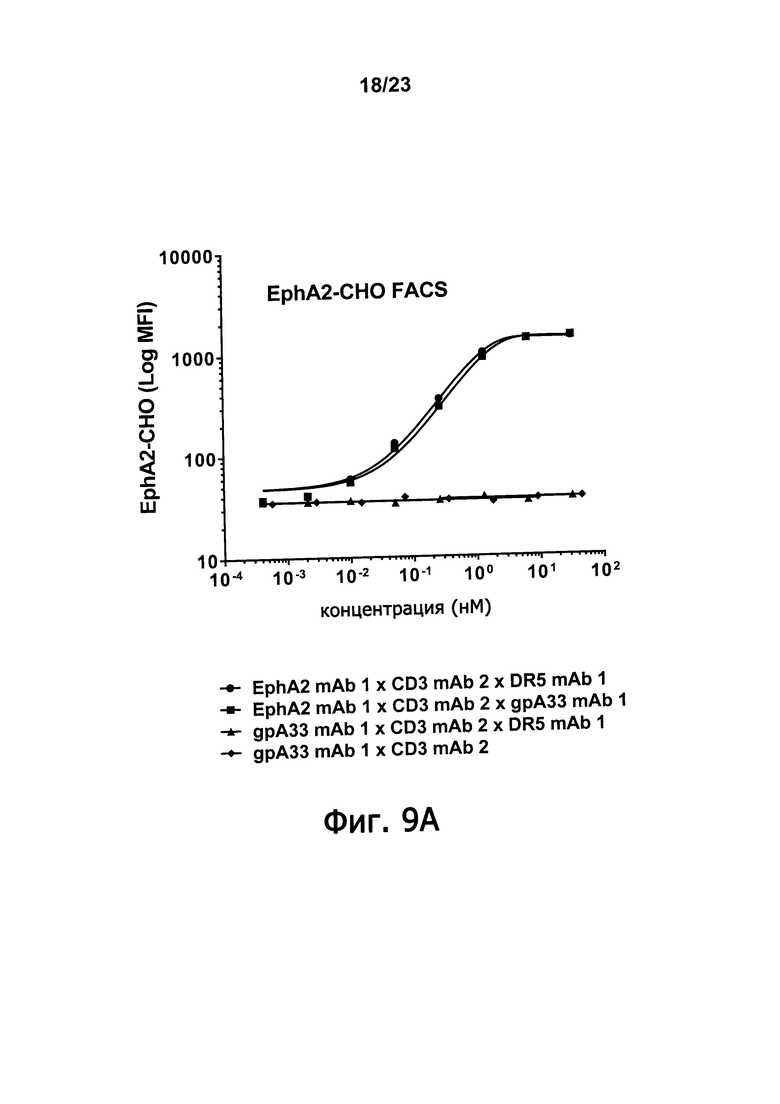


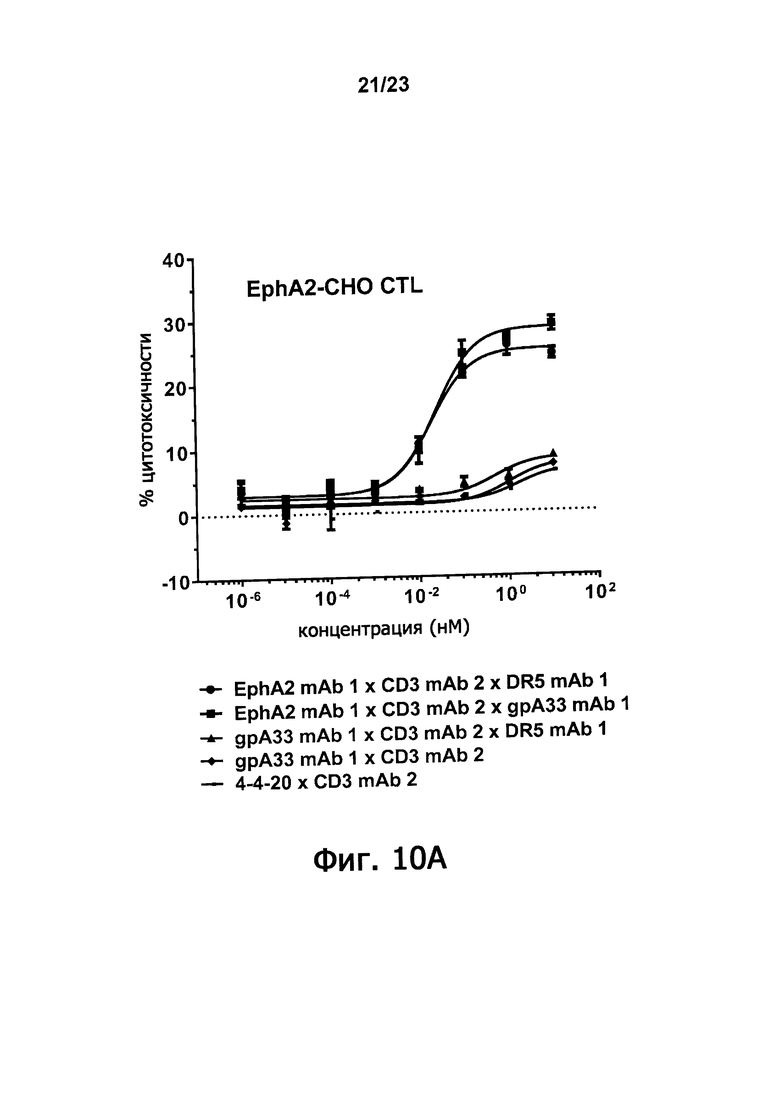
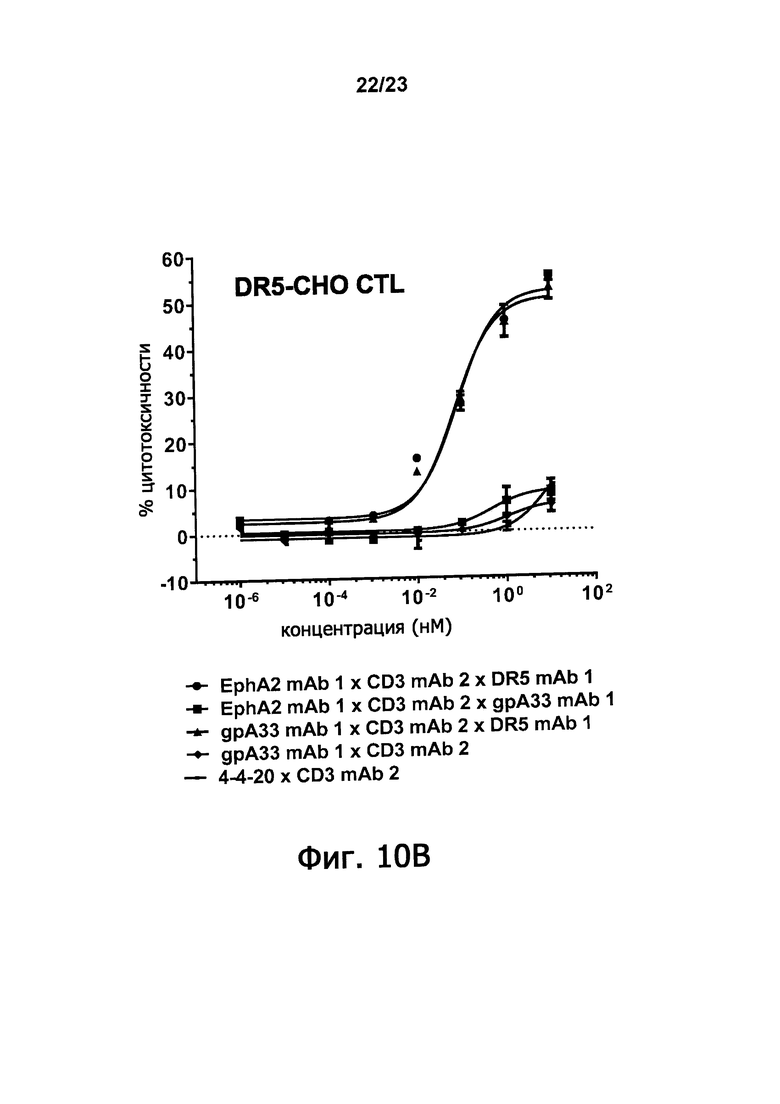
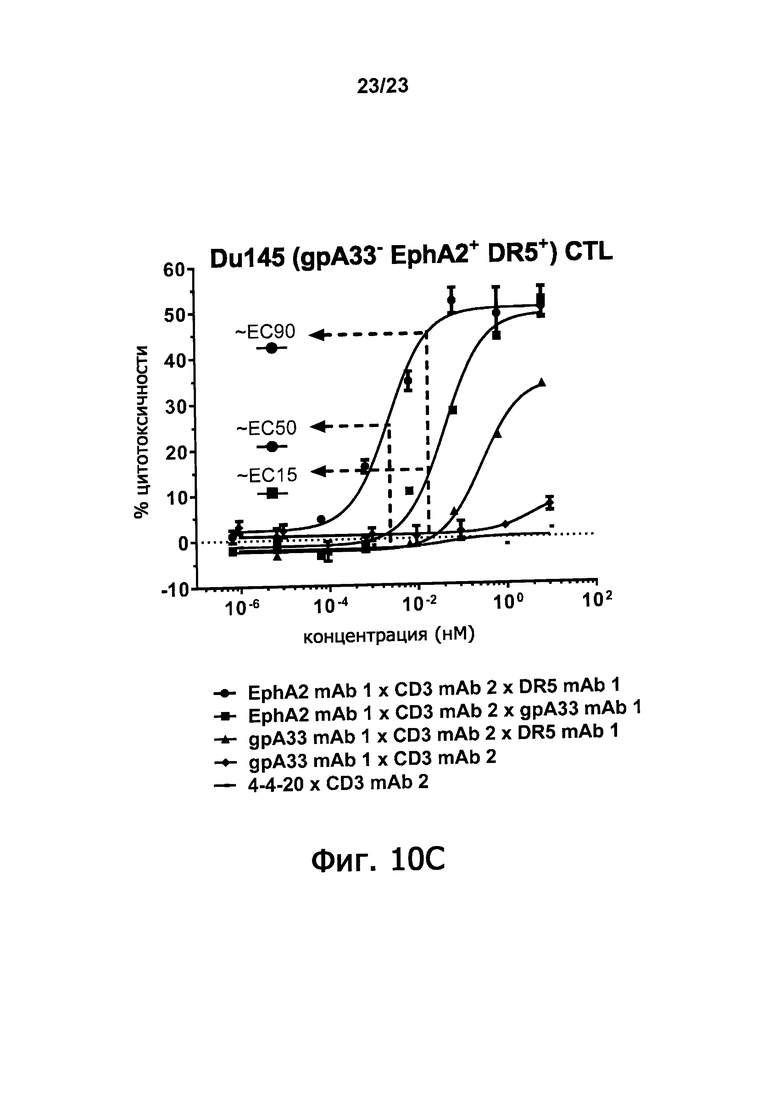
Изобретение относится к триспецифичным связывающим молекулам, которые представляют собой мультицепочечные полипептидные молекулы, которые обладают тремя связывающими доменами и, таким образом, способны опосредовать скоординированное связывание с тремя эпитопами. Триспецифичная связывающая молекула предпочтительно отличается тем, что обладает связывающими доменами, которые позволяют ей иммуноспецифично связываться с: (1) эпитопом первого антигена злокачественной опухоли, (2) эпитопом второго антигена злокачественной опухоли, и (3) эпитопом молекулы, которая экспрессируется на поверхности эффекторных клеток иммунной системы, и, таким образом, способна локализовать эффекторную клетку иммунной системы с клеткой, которая экспрессирует антиген злокачественной опухоли, с тем, чтобы тем самым облегчить уничтожение такой злокачественной опухолевой клетки. 5 н. и 21 з.п. ф-лы, 10 ил., 6 табл., 5 пр.
1. Триспецифичная связывающая молекула, способная иммуноспецифично связываться с тремя различными эпитопами, содержащая:
(I) четыре различные полипептидные цепи, ковалентно связанные вместе;
(II) антигенсвязывающий домен I, который способен иммуноспецифично связываться с эпитопом I, присутствующим на первом антигене, и антигенсвязывающий домен II, который способен иммуноспецифично связываться с эпитопом II, присутствующим на втором антигене;
(III), антигенсвязывающий домен III, который способен иммуноспецифично связываться с эпитопом III, присутствующим на третьем антигене; и
(IV) Fc-домен, где
(А) один из эпитопа I, эпитопа II или эпитопа III представляет собой эпитоп антигена эффекторной клетки, второй из эпитопа I, эпитопа II или эпитопа III представляет собой эпитоп первого опухолевого антигена, а третий из эпитопа I, эпитопа II или эпитопа III представляет собой эпитоп второго опухолевого антигена;
(B) первая полипептидная цепь содержит в направлении от N-конца к C-концу:
(домен VLI) - (линкер 1) - (домен VHII) - (линкер 2) - (домен, промотирующий формирование гетеродимера) - (линкер 3) - (домен CH2-CH3); или
(домен CH2-CH3) - (линкер 4) - (домен VLI) - (линкер 1) - (домен VHII) - (линкер 2) - (домен, промотирующий формирование гетеродимера); или
(цистеинсодержащий домен) - (домен CH2-CH3) - (линкер 4) - (домен VLI) - (линкер 1) - (домен VHII) - (линкер 2) - (домен, промотирующий формирование гетеродимера);
(C) вторая полипептидная цепь содержит в направлении от N-конца к C-концу:
(домен VLII) - (линкер 1) - (домен VHI) - (линкер 2) - (домен, промотирующий формирование гетеродимера);
(D) третья полипептидная цепь содержит в направлении от N-конца к C-концу:
(домен VHIII) - (цистеинсодержащий домен) - (домен CH2-CH3);
(E) четвертая полипептидная цепь содержит в направлении от N-конца к C-концу:
(домен VLIII) - (цистеинсодержащий домен);
(F) домен VLI представляет собой вариабельный домен легкой цепи иммуноглобулина, способного связываться с эпитопом I, домен VHI представляет собой вариабельный домен тяжелой цепи иммуноглобулина, способного связываться с эпитопом I, домен VLII представляет собой вариабельный домен легкой цепи иммуноглобулина, способного связываться с эпитопом II, домен VHII представляет собой вариабельный домен тяжелой цепи иммуноглобулина, способного связываться с эпитопом II, домен VLIII представляет собой вариабельный домен легкой цепи иммуноглобулина, способного связываться с эпитопом III, и домен VHIII представляет собой вариабельный домен тяжелой цепи иммуноглобулина, способного связываться с эпитопом III; и
(G) домен VLI и домен VHI ассоциируются с образованием антигенсвязывающего домена I, домен VLII и домен VHII ассоциируются с образованием антигенсвязывающего домена II, домен VLIII и домен VHIII ассоциируются с образованием антигенсвязывающего домена III и домен CH2-CH3 первой полипептидной цепи и домен CH2-CH3 третьей полипептидной цепи ассоциируются с образованием домена Fc, и
(H) указанный линкер 1 содержит последовательность SEQ ID NO: 33;
указанный линкер 2 содержит последовательность SEQ ID NO: 34 или 47;
указанный домен, промотирующий формирование гетеродимера, на первой полипептидной цепи представляет собой E-спиральный домен, а указанный домен, промотирующий формирование гетеродимера, на второй полипептидной цепи представляет собой K-спиральный домен, или указанный домен, промотирующий формирование гетеродимера, на первой полипептидной цепи представляет собой K-спиральный домен, а указанный домен, промотирующий формирование гетеродимера, на второй полипептидной цепи представляют собой E-спиральный домен, причем указанный E-спиральный домен независимо содержит последовательность SEQ ID NO: 39 или 41, а указанный K-спиральный домен независимо содержит последовательность SEQ ID NO: 40 или 42;
указанный линкер 3 содержит последовательность SEQ ID NO: 46, 47, 48, 49, 50, 51, 152, или GCG, или GGG; или
указанный цистеинсодержащий домен независимо содержит последовательность SEQ ID NO: 34, 36, 38, 48, 210 или 211; или
имеет место комбинация вышесказанного.
2. Триспецифичная связывающая молекула по п. 1, где указанная третья полипептидная цепь содержит домен CH1, и необязательно указанная четвертая полипептидная цепь содержит домен CL.
3. Триспецифичная связывающая молекула по любому из пп. 1 или 2, где (i) первая полипептидная цепь и вторая полипептидная цепь ковалентно связаны друг с другом; (ii) первая полипептидная цепь и третья полипептидная цепь ковалентно связаны друг с другом; и (iii) третья полипептидная цепь и четвертая полипептидная цепь ковалентно связаны друг с другом.
4. Триспецифичная связывающая молекула, способная иммуноспецифически связываться с тремя различными эпитопами, содержащая:
(I) три разные полипептидные цепи, ковалентно связанные вместе;
(II) антигенсвязывающий домен I, который способен иммуноспецифически связываться с эпитопом I, присутствующим на первом антигене, и антигенсвязывающий домен II, который способен иммуноспецифически связываться с эпитопом II, присутствующим на втором антигене;
(III) антигенсвязывающий домен III, который способен иммуноспецифически связываться с эпитопом III, присутствующим на третьем антигене; и
(IV) домен Fc, где:
(A) один из эпитопа I, эпитопа II или эпитопа III представляет собой эпитоп антигена эффекторной клетки, второй из эпитопа I, эпитопа II или эпитопа III представляет собой эпитоп первого ракового антигена и третий из эпитопа I, эпитопа II или эпитоп III представляет собой эпитоп второго ракового антигена;
(B) первая полипептидная цепь содержит в направлении от N-конца к C-концу:
(домен VLI) - (линкер 1) - (домен VHII) - (линкер 2) - (домен, промотирующий формирование гетеродимера) - (линкер 3) - (домен CH2-CH3); или
(домен CH2-CH3) - (линкер 4) - (домен VLI) - (линкер 1) - (домен VHII) - (линкер 2) - (домен, промотирующий формирование гетеродимера); или
(цистеинсодержащий домен) - (домен CH2-CH3) - (линкер 4) - (домен VLI) - (линкер 1) - (домен VHII) - (линкер 2) - (домен, промотирующий формирование гетеродимера);
(C) вторая полипептидная цепь содержит в направлении от N-конца к C-концу:
(домен VLII) - (линкер 1) - (домен VHI) - (линкер 2) - (домен, промотирующий формирование гетеродимера);
(D) третья полипептидная цепь содержит в направлении от N-конца к C-концу:
(домен VLIII) - (гибкий линкер) - (домен VHIII) - (цистеинсодержащий домен) - (домен CH2-CH3); или
(цистеинсодержащий домен 1) - (домен VLIII) - (гибкий линкер) - (домен VHIII) - (цистеинсодержащий домен 2) - (домен CH2-CH3);
(E) домен VLI представляет собой вариабельный домен легкой цепи иммуноглобулина, способного связываться с эпитопом I, домен VHI представляет собой вариабельный домен тяжелой цепи иммуноглобулина, способного связываться с эпитопом I, домен VLII представляет собой вариабельный домен легкой цепи иммуноглобулина, способного связываться с эпитопом II, домен VHII представляет собой вариабельный домен тяжелой цепи иммуноглобулина, способного связываться с эпитопом II, домен VLIII представляет собой вариабельный домен легкой цепи иммуноглобулина, способного связываться с эпитопом III, и домен VHIII представляет собой вариабельный домен тяжелой цепи иммуноглобулина, способного связываться с эпитопом III; и
(F) домен VLI и домен VHI ассоциируются с образованием антигенсвязывающего домена I, домен VLII и домен VHII ассоциируются с образованием антигенсвязывающего домена II, домен VLIII и домен VHIII ассоциируются с образованием антигенсвязывающего домена III, и домен CH2-CH3 первой полипептидной цепи и домен CH2-CH3 третьей полипептидной цепи ассоциируются с образованием домена Fc, и
(H) указанный линкер 1 содержит последовательность SEQ ID NO: 33;
указанный линкер 2 содержит последовательность SEQ ID NO: 34 или 47;
указанный домен, промотирующий формирование гетеродимера, на первой полипептидной цепи представляет собой E-спиральный домен, а указанный домен, промотирующий формирование гетеродимера, на второй полипептидной цепи представляет собой K-спиральный домен, или указанный домен, промотирующий формирование гетеродимера, на первой полипептидной цепи представляет собой K-спиральный домен, а указанный домен, промотирующий формирование гетеродимера, на второй полипептидной цепи представляют собой E-спиральный домен, причем указанный E-спиральный домен независимо содержит последовательность SEQ ID NO: 39 или 41, а указанный K-спиральный домен независимо содержит последовательность SEQ ID NO: 40 или 42;
указанный линкер 3 содержит последовательность SEQ ID NO: 46, 47, 48, 49, 50, 51, 152, или GCG, или GGG; или
указанный цистеинсодержащий домен независимо содержит последовательность SEQ ID NO: 34, 36, 38, 48, 210 или 211; или
имеет место комбинация вышесказанного.
5. Триспецифичная связывающая молекула по п.4, в которой третья полипептидная цепь содержит домен CL, и необязательно третья полипептидная цепь содержит домен CH1.
6. Триспецифичная связывающая молекула по любому из пп.1-5, в которой:
домен CH2-CH3 первой полипептидной цепи представляет собой несущий выступ домен CH2-CH3, а домен CH2-CH3 третьей полипептидной цепи представляет собой несущий впадину домен CH2-CH3; или
домен CH2-CH3 третьей полипептидной цепи представляет собой домен CH2-CH3, несущий выступ, а домен CH2-CH3 первой полипептидной цепи представляет собой домен CH2-CH3, несущий впадину.
7. Трехспецифичная связывающая молекула по п.6, в которой домен CH2-CH3, несущий выступ, содержит последовательность SEQ ID NO: 52, а несущий впадину домен CH2-CH3 содержит последовательность SEQ ID NO: 53.
8. Триспецифичная связывающая молекула по любому из пп.1-7, в которой домен CH2-CH3 первой полипептидной цепи и третьей полипептидной цепи содержит по меньшей мере одну аминокислотную замену относительно последовательности SEQ ID NO: 1, и домен Fc, образованный посредством их ассоциации, демонстрирует измененную FcγR-опосредованную эффекторную функцию.
9. Триспецифичная связывающая молекула по п.8, в которой домен CH2-CH3 первой полипептидной цепи и третьей полипептидной цепи включает:
(А) одну замену, выбранную из группы, состоящей из:
F243L, R292P, Y300L, V305I и P396L;
(Β) две замены, выбранные из группы, состоящей из:
(1) F243L и P396L;
(2) F243L и R292P; и
(3) R292P и V305I;
(С) три замены, выбранные из группы, состоящей из:
(1) F243L, R292P и Y300L;
(2) F243L, R292P и V305I;
(3) F243L, R292P и P396L; и
(4) R292P, V305I и P396L;
(D) четыре замены, выбранные из группы, состоящей из:
(1) F243L, R292P, Y300L и P396L; и
(2) F243L, R292P, V305I и P396L; или
(Ε) пять замен, выбранных из группы, состоящей из:
(1) F243L, R292P, Y300L, V305I и P396L; и
(2) L235V, F243L, R292P, Y300L и P396L.
10. Триспецифичная связывающая молекула по любому из пп.2, 3 и 5-9, в которой:
домен CH1 содержит последовательность SEQ ID NO: 207 или 208; а также
домен CL содержит последовательность SEQ ID NO: 210 или 211.
11. Триспецифичная связывающая молекула по п.10, в которой
E-спиральный домен SEQ ID NO: 41 или K-спиральный домен SEQ ID NO: 42 соседствует с линкером 2, когда линкер 2 содержит последовательность SEQ ID NO: 47;
E-спиральный домен SEQ ID NO: 39 или K-спиральный домен SEQ ID NO: 40 соседствует с линкером 2, когда линкер 2 содержит последовательность SEQ ID NO: 34; или
домен E-катушки SEQ ID NO: 41 или домен K-катушки SEQ ID NO: 42 соседствует с линкером 2, когда линкер 2 содержит последовательность SEQ ID NO: 34.
12. Триспецифичная связывающая молекула по любому из пп.1-11, в которой:
(A) эпитоп I, эпитоп II и эпитоп III являются, соответственно, эпитопом
первого ракового антигена, эпитопом второго ракового антигена и
эпитопом антигена эффекторной клетки;
(Β) эпитоп I, эпитоп II и эпитоп III являются, соответственно, эпитопом
первого ракового антигена, эпитопом антигена эффекторной клетки и
эпитопом второго ракового антигена;
(C) эпитоп I, эпитоп II и эпитоп III являются, соответственно, эпитопом
второго ракового антигена, эпитопом первого ракового антигена и
эпитопом антигена эффекторной клетки;
(D) эпитоп I, эпитоп II и эпитоп III являются, соответственно, эпитопом
второго ракового антигена, эпитопом антигена эффекторной клетки и
эпитопом первого ракового антигена;
(Ε) эпитоп I, эпитоп II и эпитоп III являются, соответственно, эпитопом
антигена эффекторной клетки, эпитопом первого ракового антигена, и
эпитопом второго ракового антигена; или
(F) эпитоп I, эпитоп II и эпитоп III являются, соответственно, эпитопом
антигена эффекторной клетки, эпитопом второго ракового антигена и
эпитопом первого ракового антигена.
13. Триспецифичная связывающая молекула по любому из пп. 1-12, где антиген эффекторной клетки выбран из группы, состоящей из: CD2, CD3, CD16, CD19, CD20, CD22, CD32B, CD64, B-клеточного рецептора (BCR), Т-клеточного рецептора (TCR) и рецептора NKG2D.
14. Триспецифичная связывающая молекула по любому из пп.1-13, в которой
(A) эпитоп антигена эффекторной клетки представляет собой эпитоп CD2, распознаваемый антителом Lo-CD2a;
(B) эпитоп антигена эффекторной клетки представляет собой эпитоп CD3, распознаваемый антителом OKT3, M291, YTH12.5, анти-CD3 mAb 1 или анти-CD3mAb 2;
(C) эпитоп антигена эффекторной клетки представляет собой эпитоп CD16, распознаваемый антителом 3G8 или A9;
(D) эпитоп антигена эффекторной клетки представляет собой эпитоп CD19, распознаваемый антителом MD1342, MEDI-551, блинатумомабом или HD37;
(E) эпитоп антигена эффекторной клетки представляет собой эпитоп CD20, распознаваемый антителами к ритуксимабу, ибритумомабу, офатумумабу и тозитумомабу;
(F) эпитоп антигена эффекторной клетки представляет собой эпитоп CD22, распознаваемый антителом эпратузумаб;
(G) эпитоп антигена эффекторной клетки представляет собой эпитоп CD32B, распознаваемый антителом CD32B mAb 1;
(H) эпитоп антигена эффекторной клетки представляет собой эпитоп CD64, распознаваемый антителом CD64 mAb 1;
(I) эпитоп антигена эффекторной клетки представляет собой эпитоп BCR/CD79, распознаваемый антителом CD79 mAb 1;
(J) эпитоп антигена эффекторной клетки представляет собой эпитоп TCR, распознаваемый антителом BMA 031;
(K) эпитоп антигена эффекторной клетки представляет собой эпитоп рецептора NKG2D, распознаваемый антителом KYK-2.0; или
(L) один из эпитопа I, эпитопа II или эпитопа III представляет собой эпитоп антигена эффекторной клетки, и антигенсвязывающий домен, который способен иммуноспецифически связываться с эпитопом, включает шесть CDR SEQ ID NO: 102 и 103; 104 и 108; 104 и 112; 114 и 115; 116 и 117; 118 и 119; 120 и 121; 122 и 123; 124 и 125; 126 и 127; 128 и 129; 130 и 131; 132 и 133; 134 и 135; или 136 и 137; или
вариабельный домен легкой цепи SEQ ID NO: 102 и вариабельный домен тяжелой цепи SEQ ID NO: 103; вариабельный домен легкой цепи SEQ ID NO: 104 и вариабельный домен тяжелой цепи SEQ ID NO: 108; вариабельный домен легкой цепи SEQ ID NO: 104 и вариабельный домен тяжелой цепи SEQ ID NO: 112; вариабельный домен легкой цепи SEQ ID NO: 114 и вариабельный домен тяжелой цепи SEQ ID NO: 115; вариабельный домен легкой цепи SEQ ID NO: 116 и вариабельный домен тяжелой цепи SEQ ID NO: 117; вариабельный домен легкой цепи SEQ ID NO: 118 и вариабельный домен тяжелой цепи SEQ ID NO: 119; вариабельный домен легкой цепи SEQ ID NO: 120 и вариабельный домен тяжелой цепи SEQ ID NO: 121; вариабельный домен легкой цепи SEQ ID NO: 122 и вариабельный домен тяжелой цепи SEQ ID NO: 123; вариабельный домен легкой цепи SEQ ID NO: 124 и вариабельный домен тяжелой цепи SEQ ID NO: 125; вариабельный домен легкой цепи SEQ ID NO: 126 и вариабельный домен тяжелой цепи SEQ ID NO: 127; вариабельный домен легкой цепи SEQ ID NO: 128 и вариабельный домен тяжелой цепи SEQ ID NO: 129; вариабельный домен легкой цепи SEQ ID NO: 130 и вариабельный домен тяжелой цепи SEQ ID NO: 131; вариабельный домен легкой цепи SEQ ID NO: 132 и вариабельный домен тяжелой цепи SEQ ID NO: 133; вариабельный домен легкой цепи SEQ ID NO: 134 и вариабельный домен тяжелой цепи SEQ ID NO: 135; или вариабельный домен легкой цепи SEQ ID NO: 136 и вариабельный домен тяжелой цепи SEQ ID NO: 137.
15. Триспецифичная связывающая молекула по любому из пп. 1-14, где первый опухолевый антиген и второй опухолевый антиген независимо выбраны из группы, состоящей из: антигена 19,9 злокачественной опухоли ободочной кишки; муцина злокачественной опухоли желудочно-кишечного тракта; антигена 4.2; гликопротеина A33 (gpA33); ADAM-9; антигена AH6 злокачественной опухоли желудочно-кишечного тракта; ALCAM; антигена APO-1 злокачественных лимфоцитов человека; антигена В1 злокачественной опухоли; B7-H3; бета-катенина; антигена ALeb/Ley группы крови; антигена 38.13 лимфомы Беркитта, антигена C14 аденокарциномы ободочной кишки; антигена СА125 карциномы яичников; Карбоксипептидазы М; CD5; CD19; CD20; CD22; CD23; CD25; CD27; CD30; CD33; CD36; CD45; CD46; CD52; CD79a/CD79b; CD103; CD317; CDK4; канцероэмбрионального антигена (СЕА); CEACAM5; CEACAM6; CO17-1A; СО-43 (антиген группы крови Leb); СО-514 (антиген группы крови Leа); СТА-1; CTLA4; Цитокератина 8; антигена D1.1; Антигена D1 56-22; DR5; серии E1 (группа крови B); EGFR (рецептор эпидермального фактора роста); рецептора эфрина А2 (EphA2); ErbB1; ErbB3; ErbB4; GAGE-1; GAGE-2; GD2/GD3/GM2; антигена F3 аденокарциномы легкого; антигена FC10.2; G49, ганглиозида GD2; ганглиозида GD3; ганглиозида GM2; ганглиозида GM3; GD2; GD3; GICA 19-9; GM2; gp100; Т-клеточного антигена Gp37 лейкоза человека; антигена gp75 меланомы; gpA33; HER2 антигена (p185 HER2); антиген жировых глобул человеческого молока (HMFG); вирус папилломы человека-E6/вирус папилломы человека-E7; антигена меланомы высокой молекулярной массы (ВММ-MAA); I антигена (дифференцировочный антиген) I (Ma); Интегрина альфа-V-бета-6 Интегрина β6 (ITGB6); α2 Рецептора Интерлейкина-13 (IL13Rα); JAM-3; KID3; KID31; антигена KS 1/4 пан-карциномы; антигенов L6 и L20 карциномы легкого человека; LEA; LUCA-2; М1: 22: 25: 8; M18; M39; MAGE-1; MAGE-3; MART; MUC-1; MUM-1; Myl; N-ацетилглюкозаминилтрансферазы; неогликопротеина; NS-10; OFA-1; OFA-2; Онкостатина M; p15; меланома-ассоциированного антигена р97; полиморфного эпителиального муцина (PEM); антигена полиморфного эпителиального муцина (РЕМА); PIPA; простата-специфического антигена (PSA); простата-специфического мембранного антигена (PSMA); фосфатна простатической кислоты; R24; ROR1; сфинголипидов; SSEA-1; SSEA-3; SSEA-4; sTn; пептида, полученного из Т-клеточного рецептора; T5A7; TAG-72; TL5 (группа крови A); TNF-α-рецептора; TNF-β-рецептора; TNF-γ-рецептора; TRA-1-85 (группа крови Н); рецептора трансферрина; опухолеспецифичного антигена трансплантации (TSTA), канцероэмбрионального антигена-альфа-фетопротеина (AFP); VEGF, VEGFR; VEP8; VEP9; VIM-D5; и Y гаптена, Leу и 5Т4.
16. Триспецифичная связывающаяся молекула по п.15, где один из эпитопа I, эпитопа II или эпитопа III представляет собой эпитоп ракового антигена, и антигенсвязывающий домен, который способен иммуноспецифически связываться с эпитопом, включает шесть CDR: SEQ ID NO: 3 и 8; 13 и 18; 23 и 31; 25 и 31; 27 и 31; 29 и 31; 54 и 58; 62 и 66; 70 и 74; 78 и 82; 86 и 90; 94 и 98; 153 и 158; 163 и 167; 172 и 177; 181 и 186; 191 и 192; 193 и 194; 195 и 196; 197 и 198; 199 и 200; 201 и 202; 203 и 204; 205 и 206; 302 и 303; 304 и 305; 306 и 307; 308 и 309; 310 и 311; 312 и 313; 314 и 315; или 321 и 322; или
вариабельный домен легкой цепи SEQ ID NO: 3 и вариабельный домен тяжелой цепи SEQ ID NO: 8; вариабельный домен легкой цепи SEQ ID NO: 13 и вариабельный домен тяжелой цепи SEQ ID NO: 18; вариабельный домен легкой цепи SEQ ID NO: 23 и вариабельный домен тяжелой цепи SEQ ID NO: 31; вариабельный домен легкой цепи SEQ ID NO: 25 и вариабельный домен тяжелой цепи SEQ ID NO: 31; вариабельный домен легкой цепи SEQ ID NO: 27 и вариабельный домен тяжелой цепи SEQ ID NO: 31; вариабельный домен легкой цепи SEQ ID NO: 29 и вариабельный домен тяжелой цепи SEQ ID NO: 31; вариабельный домен легкой цепи SEQ ID NO: 54 и вариабельный домен тяжелой цепи SEQ ID NO: 58; вариабельный домен легкой цепи SEQ ID NO: 62 и вариабельный домен тяжелой цепи SEQ ID NO: 66; вариабельный домен легкой цепи SEQ ID NO: 70 и вариабельный домен тяжелой цепи SEQ ID NO: 74; вариабельный домен легкой цепи SEQ ID NO: 78 и вариабельный домен тяжелой цепи SEQ ID NO: 82; вариабельный домен легкой цепи SEQ ID NO: 86 и вариабельный домен тяжелой цепи SEQ ID NO: 90; вариабельный домен легкой цепи SEQ ID NO: 94 и вариабельный домен тяжелой цепи SEQ ID NO: 98; вариабельный домен легкой цепи SEQ ID NO: 153 и вариабельный домен тяжелой цепи SEQ ID NO: 158; вариабельный домен легкой цепи SEQ ID NO: 163 и вариабельный домен тяжелой цепи SEQ ID NO: 167; вариабельный домен легкой цепи SEQ ID NO: 172 и вариабельный домен тяжелой цепи SEQ ID NO: 177; вариабельный домен легкой цепи SEQ ID NO: 181 и вариабельный домен тяжелой цепи SEQ ID NO: 186; вариабельный домен легкой цепи SEQ ID NO: 191 и вариабельный домен тяжелой цепи SEQ ID NO: 192; вариабельный домен легкой цепи SEQ ID NO: 193 и вариабельный домен тяжелой цепи SEQ ID NO: 194; вариабельный домен легкой цепи SEQ ID NO: 195 и вариабельный домен тяжелой цепи SEQ ID NO: 196; вариабельный домен легкой цепи SEQ ID NO: 197 и вариабельный домен тяжелой цепи SEQ ID NO: 198; вариабельный домен легкой цепи SEQ ID NO: 199 и вариабельный домен тяжелой цепи SEQ ID NO: 200; вариабельный домен легкой цепи SEQ ID NO: 201 и вариабельный домен тяжелой цепи SEQ ID NO: 202; вариабельный домен легкой цепи SEQ ID NO: 203 и вариабельный домен тяжелой цепи SEQ ID NO: 204; вариабельный домен легкой цепи SEQ ID NO: 205 и вариабельный домен тяжелой цепи SEQ ID NO: 206; вариабельный домен легкой цепи SEQ ID NO: 302 и вариабельный домен тяжелой цепи SEQ ID NO: 303; вариабельный домен легкой цепи SEQ ID NO: 304 и вариабельный домен тяжелой цепи SEQ ID NO: 305; вариабельный домен легкой цепи SEQ ID NO: 306 и вариабельный домен тяжелой цепи SEQ ID NO: 307; вариабельный домен легкой цепи SEQ ID NO: 308 и вариабельный домен тяжелой цепи SEQ ID NO: 309; вариабельный домен легкой цепи SEQ ID NO: 310 и вариабельный домен тяжелой цепи SEQ ID NO: 311; вариабельный домен легкой цепи SEQ ID NO: 312 и вариабельный домен тяжелой цепи SEQ ID NO: 313; вариабельный домен легкой цепи SEQ ID NO: 314 и вариабельный домен тяжелой цепи SEQ ID NO: 315; или вариабельный домен легкой цепи SEQ ID NO: 321 и вариабельный домен тяжелой цепи SEQ ID NO: 322.
17. Композиция нуклеиновой кислоты, содержащая полинуклеотиды, которые кодируют полипептидные цепи триспецифичной связывающей молекулы по любому из пп.1-16.
18. Фармацевтическая композиция, которая содержит триспецифичную связывающую молекулу по любому из пп. 1-16 и фармацевтически приемлемый носитель, вспомогательное вещество или разбавитель.
19. Триспецифичная связывающая молекула по любому из пп. 1-16, где указанную молекулу используют как медикамент.
20. Фармацевтическая композиция по п. 18, где указанную композицию используют как медикамент.
21. Применение триспецифичной связывающей молекулы по любому из пп. 1-16 для лечения рака.
22. Применение по п.21, где указанная злокачественная опухоль характеризуется наличием злокачественных опухолевых клеток, выбранных из группы, состоящей из клеток: опухоли надпочечников, СПИД-ассоциированной злокачественной опухоли, альвеолярной саркомы мягких тканей, опухоли астроцитов, рака мочевого пузыря, рака кости, рака мозга и спинного мозга, метастатической опухоли мозга, рака груди, опухоли каротидной железы, рака шейки матки, хондросаркомы, хордомы, хромофобного почечно-клеточного рака, светлоклеточного рака, рака толстой кишки, колоректального рака, кожной доброкачественной фиброзной гистиоцитомы, десмопластической мелкокруглоклеточной опухоли, эпендимомы, опухоли Юинга, внескелетной миксоидной хондросаркомы, несовершенного костного фиброгенеза, фиброзной дисплазии костей, рака желчного пузыря или желчных протоков, рака желудка, гестационной трофобластической болезни, опухоли половых клеток, рака головы и шеи, гепатоцеллюлярной карциномы, опухоли островковых клеток, саркомы Капоши, рака почки, лейкоза, липомы/доброкачественной липоматозной опухоли, липосаркомы/злокачественной липоматозной опухоли, рака печени, лимфомы, рака легкого, медуллобластомы, меланомы, менингиомы, множественных эндокринных неоплазий, множественной миеломы, миелодиспластического синдрома, нейробластомы, нейроэндокринной опухоли, рака яичника, рака поджелудочной железы, папиллярного рака щитовидной железы, паратиреоидной опухоли, педиатрического рака, опухоли периферической нервной оболочки, феохромоцитомы, опухоли гипофиза, рака простаты, задней увеальной меланомы, редкого гематологического заболевания, почечного метастатического рака, палочковидной опухоли, рабдомиосаркомы, саркомы, рака кожи, саркомы мягких тканей, плоскоклеточного рака, рака желудка, синовиальной саркомы, тестикулярного рака, карциномы тимуса, тимомы, метастатического рака щитовидной железы и рака матки.
23. Применение по п.21, где указанная злокачественная опухоль представляет собой колоректальный рак, гепатоцеллюлярный рак, глиому, рак почки, рак молочной железы, множественную миелому, рак мочевого пузыря, нейробластому; саркому, неходжкинскую лимфому, немелкоклеточный рак легкого, рак яичников, рак поджелудочной железы или рак прямой кишки, острый миелоидный лейкоз (AML), хронический миелолейкоз (CML), острый лимфобластный лейкоз В (B-ALL), хронический лимфоцитарный лейкоз (CLL), волосатоклеточный лейкоз (HCL), бластное неопластическое образование плазмоцитодных дендритных клеток (BPDCN), неходжкинские лимфомы (NHL), включая мантийноклеточный лейкоз (MCL), и мелкоклеточную лимфоцитарную лимфому (SLL), лимфому Ходжкина, системный мастоцитоз или лимфому Беркитта.
24. Применение фармацевтической композиции по п. 18 для лечения рака.
25. Применение по п. 24, где указанная злокачественная опухоль характеризуется наличием злокачественных опухолевых клеток, выбранных из группы, состоящей из клеток: опухоли надпочечников, СПИД-ассоциированной злокачественной опухоли, альвеолярной саркомы мягких тканей, опухоли астроцитов, рака мочевого пузыря, рака кости, рака мозга и спинного мозга, метастатической опухоли мозга, рака груди, опухоли каротидной железы, рака шейки матки, хондросаркомы, хордомы, хромофобного почечно-клеточного рака, светлоклеточного рака, рака толстой кишки, колоректального рака, кожной доброкачественной фиброзной гистиоцитомы, десмопластической мелкокруглоклеточной опухоли, эпендимомы, опухоли Юинга, внескелетной миксоидной хондросаркомы, несовершенного костного фиброгенеза, фиброзной дисплазии костей, рака желчного пузыря или желчных протоков, рака желудка, гестационной трофобластической болезни, опухоли половых клеток, рака головы и шеи, гепатоцеллюлярной карциномы, опухоли островковых клеток, саркомы Капоши, рака почки, лейкоза, липомы/доброкачественной липоматозной опухоли, липосаркомы/злокачественной липоматозной опухоли, рака печени, лимфомы, рака легкого, медуллобластомы, меланомы, менингиомы, множественных эндокринных неоплазий, множественной миеломы, миелодиспластического синдрома, нейробластомы, нейроэндокринной опухоли, рака яичника, рака поджелудочной железы, папиллярного рака щитовидной железы, паратиреоидной опухоли, педиатрического рака, опухоли периферической нервной оболочки, феохромоцитомы, опухоли гипофиза, рака простаты, задней увеальной меланомы, редкого гематологического заболевания, почечного метастатического рака, палочковидной опухоли, рабдомиосаркомы, саркомы, рака кожи, саркомы мягких тканей, плоскоклеточного рака, рака желудка, синовиальной саркомы, тестикулярного рака, карциномы тимуса, тимомы, метастатического рака щитовидной железы и рака матки.
26. Применение по п.24, где указанная злокачественная опухоль представляет собой колоректальный рак, гепатоцеллюлярный рак, глиому, рак почки, рак молочной железы, множественную миелому, рак мочевого пузыря, нейробластому, саркому, неходжкинскую лимфому, немелкоклеточный рак легкого, рак яичников, рак поджелудочной железы или рак прямой кишки, острый миелоидный лейкоз (AML), хронический миелолейкоз (CML), острый лимфобластный лейкоз В (B-ALL), хронический лимфоцитарный лейкоз (CLL), волосатоклеточный лейкоз (HCL), бластное неопластическое образование плазмоцитодных дендритных клеток (BPDCN), неходжкинские лимфомы (NHL), включая мантийноклеточный лейкоз (MCL), и мелкоклеточную лимфоцитарную лимфому (SLL), лимфому Ходжкина, системный мастоцитоз или лимфому Беркитта.
| US 2010291112 A1, 18.11.2010 | |||
| WO 2012018687 A1, 09.02.2012 | |||
| US 6994853 B1, 07.02.2006 | |||
| ALBRECHT H | |||
| ET AL: "Recombinant Antibodies: From the Laboratory to the Clinic", CANCER BIOTHERAPY & RADIOPHARMACEUTICALS, 2006, vol | |||
| Выбрасывающий ячеистый аппарат для рядовых сеялок | 1922 |
|
SU21A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| ПЕРЕДВИЖНАЯ ДИАГРАММА ДЛЯ СРАВНЕНИЯ ЦЕННОСТИ РАЗЛИЧНЫХ ПРОДУКТОВ ПО ИХ КАЛОРИЙНОСТИ | 1919 |
|
SU285A1 |
| RU 94028282 A, 20.07.1996. | |||

