ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Данная заявка имеет приоритет по предварительной заявке США № 61/263526, зарегистрированной 23-го ноября 2009 г., которая, таким образом, полностью включена в настоящее описание, и по предварительной заявке США № 61/263530, зарегистрированной 23-го ноября 2009 г., которая, таким образом, полностью включена в настоящее описание.
ВКЛЮЧЕНИЕ СПИСКА ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Список последовательностей, который содержится в файле с названием «56887-0001_seqlisting.txt», размер которого составляет 19,6 килобайт (размер, измеренный в Microsoft Windows®), и который был создан 12-го ноября 2010 г., регистрируется при этом путем электронной подачи и, таким образом, полностью включается в настоящий документ путем ссылки.
ОБЛАСТЬ ТЕХНИКИ
Изобретение относится к области разведения растений, их исследований и сельского хозяйства. Более конкретно, изобретение относится к трансгенному событию MON 87427 маиса и нуклеотидным молекулам, растениям, частям растения, семенам растения, клеткам растения, сельскохозяйственным продуктам и способам, связанным с трансгенным событием MON 87427 маиса. Оно также относится к прогнозу развития метелок маиса и использования этого в способах скрещивания растений, исследований и сельском хозяйстве, и полученным с помощью этого гибридным семенам маиса.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Зерновые культуры, имеющие новые желаемые признаки, полезны для скрещивания растений, исследований и сельского хозяйства. Такие культуры можно получать с использованием биотехнологических способов. Однако получение и отбор подходящего для коммерческих целей трансгенного события может требовать проведения интенсивных исследований, анализа и характеристики большого числа событий трансформации индивидуальных растений для отбора события, имеющего как желаемый признак, так и оптимальные фенотипические и агротехнические характеристики, необходимые для его соответствия коммерческим и сельскохозяйственным целям. Для данного процесса отбора события часто необходимы теплицы и полевые испытания с большим числом событий на протяжении многих лет, во множестве мест и при различных условиях, для того чтобы можно было собрать достоверное количество агротехнических, фенотипических и молекулярных данных. Полученные в результате данные и наблюдения должны быть затем проанализированы командами ученых и агрономов с целью отбора коммерчески пригодного события. Изобретение относится к такому коммерчески пригодному событию, результатом которого является новый желаемый признак у маиса.
Точное определение репродуктивной зрелости маиса также полезно для скрещивания растений, исследований и сельскохозяйственных целей, например, при получении гибридных семян маиса. Средства, обычно используемые в данной области для прогнозирования и оценки стадий роста и развития маиса, включают шкалы, например, V-стадии, которые основаны на вегетативных характеристиках, и градусо-единицы роста (GDU), которые основаны на числе градусо-дней роста. Однако оба этих подхода дают оценку стадии развития метелки, которая сильно варьирует среди генотипов маиса. Поэтому, использование указанных измерений может привести к пропуску оптимально эффективного времени в случае схем обработки, для которых стадия развития является важным фактором. Изобретение относится к Относительной шкале развития, основанной на развитии метелки, согласованной по генотипам, которая полезна для мониторинга и прогноза развития метелки у растений маиса с различными генотипами.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Изобретение относится к рекомбинантной молекуле ДНК, включающей в себя молекулу нуклеиновой кислоты, содержащую нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-8. Изобретение также относится к рекомбинантной молекуле ДНК, образованной соединением встроенной гетерологичной молекулы нуклеиновой кислоты и геномной ДНК растения маиса, клетки растения или семени. Изобретение также относится к рекомбинантной молекуле ДНК, полученной из трансгенного события MON 87427 маиса, репрезентативный образец семян которого был помещен в Американскую коллекцию типовых культур (АТСС®) под номером доступа № PTA-7899. Изобретение также относится к рекомбинантной молекуле ДНК, которая представляет собой ампликон диагностический в отношении присутствия ДНК, полученной из трансгенного события MON 87427 маиса. Изобретение также относится к рекомбинантной молекуле ДНК, которая находится в растении маиса, клетке растения, семени, потомстве растения, части растения или товарном продукте, полученном из трансгенного события MON 87427 маиса.
Изобретение также относится к молекуле ДНК, включающей в себя молекулу нуклеиновой кислоты, имеющую нуклеотидную последовательность с достаточной длиной непрерывной нуклеотидной последовательности из SEQ ID NO: 10 для функционирования в качестве зонда ДНК, которая гибридизуется при жестких условиях с молекулой ДНК, включающей в себя нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-10, и не гибридизуется при жестких условиях с молекулой ДНК, не включающей в себя нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-10.
Изобретение также относится к паре молекул ДНК, состоящей из первой молекулы ДНК и второй молекулы ДНК, отличающейся от первой молекулы ДНК, причем каждая из первой и второй молекул ДНК содержит молекулу нуклеиновой кислоты, имеющую нуклеотидную последовательность с достаточной длиной непрерывной нуклеотидной последовательности из SEQ ID NO: 10 для функционирования в качестве ДНК-праймеров при совместном использовании в реакции амплификации с ДНК, полученной из события MON 87427, чтобы получить ампликон, диагностический в отношении ДНК трансгенного события MON 87427 маиса в образце.
Изобретение также относится к способу детекции присутствия молекулы ДНК, полученной из MON 87427, в образце посредством контакта образца с зондом ДНК, подвергание указанного образца и указанного зонда ДНК жестким условиям гибридизации и детекции гибридизации зонда ДНК с молекулой ДНК в образце, причем гибридизация зонда ДНК с молекулой ДНК указывает на присутствие молекулы ДНК, полученной из трансгенного события MON 87427 маиса, в образце.
Изобретение также относится к способу детекции присутствия молекулы ДНК, полученной из трансгенного события MON 87427 маиса, в образце посредством контакта образца с парой молекул ДНК, проведения реакции амплификации, достаточной для получения ампликона ДНК, содержащего последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-10, и детекции присутствия ДНК-ампликона в реакции, причем присутствие ДНК-ампликона в реакции указывает на присутствие молекулы ДНК, полученной из MON 87427, в образце.
Изобретение также относится к набору для детекции ДНК, включающему в себя по меньшей мере одну молекулу ДНК, содержащую нуклеотидную последовательность с достаточной длиной непрерывной нуклеотидной последовательности из SEQ ID NO: 10 для функционирования в качестве ДНК-праймера или пробы, специфичных для детекции присутствия ДНК, полученной из трансгенного события MON 87427 маиса, причем детекция ДНК является диагностической в отношении присутствия ДНК трансгенного события MON 87427 маиса в образце.
Изобретение также относится к рекомбинантному растению маиса, его семени, клетке или части, содержащим молекулу нуклеиновой кислоты, имеющую нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-10. Изобретение также относится к рекомбинантному растению маиса, семени, клетке или части растения, имеющим ткане-избрирательную устойчивость к обработке гербицидом глифосат. Изобретение также относится к рекомбинантному растению маиса, семени, клетке или части растения, геном которых продуцирует ампликон, содержащий молекулу ДНК, выбранную из группы, состоящей из SEQ ID NO: 1-10, при тестировании в способе ДНК-амплификации.
Изобретение также относится к растению или семени маиса, причем растение или семя маиса получены из трансгенного события MON 87427 маиса. Изобретение также относится к растению или семени маиса, причем растение или семя маиса являются гибридом, имеющим по меньшей мере одно родительское растение, полученное из трансгенного события MON 87427 маиса.
Изобретение также относится к неживому растительному материалу, содержащему рекомбинантную молекулу ДНК, выбранную из группы, состоящей из SEQ ID NO: 1-10.
Изобретение также относится к микроорганизму, содержащему молекулу нуклеиновой кислоты, имеющую нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-10.
Изобретение также относится к микроорганизму, который является растительной клеткой.
Изобретение также относится к продукту потребления, получаемому из трансгенного события MON 87427 маиса и содержащему молекулу нуклеиновой кислоты, имеющую нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-10, причем детекция нуклеотидной последовательности в образце, полученном из продукта потребления, определяет то, что продукт потребления получен из трансгенного события 87427 маиса. Изобретение также относится к продукту потребления, выбранному из группы, состоящей из целых или переработанных семян, питания для животных, масла, муки крупного помола, муки, хлопьев, отрубей, биомассы и топливных продуктов. Изобретение также относится к способу изготовления продукта потребления путем получения растения маиса или его части, включающих трансгенное событие MON 87427 маиса, и изготовления продукта потребления маиса из растения маиса или его части.
Изобретение также относится к способу контроля сорняков в поле путем высаживания растений MON 87427 в поле и применения эффективной дозы гербицида глифосата для контроля сорняков в поле без поражений растений трансгенного события MON 87427 маиса. Изобретение также относится к способу контроля сорняков в поле, в котором эффективная доза гербицида глифосата составляет от примерно 0,1 фунта до примерно 4 фунтов на акр.
Изобретение также относится к способу получения растения маиса, которое устойчиво к применению гербицида глифосата в результате полового скрещивания растения трансгенного события MON 87427 маиса, содержащего молекулу нуклеиновой кислоты, содержащую нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-10, со вторым растением маиса, таким образом получая семена, сбора семян, полученных в результате скрещивания, выращивания семян для получения множества растений-потомков, обработки растений-потомков глифосатом и отбора растения-потомка, которое устойчиво к глифосату. Изобретение также относится к способу получения растения маиса, которое устойчиво к применению гербицида глифосата самоопылением растения трансгенного события MON 87427 маиса, содержащего молекулу нуклеиновой кислоты, содержащую нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-10, в результате чего получаются семена, сбором семян, полученных после самоопыления, выращиванием семян для получения множества растений-потомков, обработкой растений-потомков глифосатом и отбором растения-потомка, которое устойчиво к глифосату.
Изобретение также относится к способу получения гибридного семени маиса посредством высаживания семени трансгенного события MON 87427 маиса на местность, выращивания растения маиса из семени, обработки растения эффективной дозой гербицида глифосата до образования пыльцы, для того чтобы сделать мужские соцветия стерильными, не повреждая растение, опыления растения пыльцой со второго родительского растения и сбора семян от растения, причем семя является гибридным семенем маиса, полученным скрещиванием растений трансгенного события MON 87427 маиса со вторым родительским растением. Изобретение также относится к способу получения гибридного семени маиса, причем эффективная доза гербицида глифосата составляет от примерно 0,1 фунта до примерно 4 фунтов на акр. Изобретение также относится к способу получения гибридного семени маиса, дополнительно включающему в себя высаживание семени второго родительского растения на местность и выращивание растения маиса из второго родительского растения. Изобретение также относится к способу получения гибридного семени маиса, причем второе родительское растение является устойчивым к глифосату.
Изобретение также относится к способу прогнозирования времени развития метелки маиса посредством выбора диапазона на Относительной шкале развития, причем диапазон указывает на созревание до желаемой стадии развития метелки. Изобретение также относится к способу прогнозирования времени развития метелки маиса, причем желаемой стадией развития метелки является стадия развития, оптимальная для репродуктивного скрещивания, стерилизации метелки, удаления метелки и/или проведения модулирующей развитие обработки на растении маиса. Изобретение также относится к способу прогнозирования времени развития метелки маиса, в котором определенной стадией развития цветков, используемой для создания Относительной шкалы развития, является выделение пыльцы примерно 50 процентами популяции растений маиса, и в которой диапазон составляет от примерно 0,62 и примерно до 0,75 на Относительной шкале развития. Изобретение также относится к способу прогнозирования времени развития метелки маиса, дополнительно включающему в себя проведение модулирующей развитие обработки на растении маиса на желаемой стадии развития метелки.
Изобретение также относится к способу получение гибридного семени маиса путем высаживания семени маиса для первого родительского растения на местность, выращивания первого родительского растения из семени маиса, определения времени развития метелки для первого родительского растения путем выбора диапазона, который указывает созревание до желаемой стадии развития метелки на Относительной шкале развития, использования определения времени развития метелки для своевременного проведения модулирующей развитие обработки первого родительского растения, таким образом предупреждая самоопыление первого родительского растения, проведения модулирующей развитие обработки первого родительского растения, оплодотворения первого родительского растения пыльцой второго родительского растения и сбора семян первого родительского растения, причем семя является гибридным семенем маиса, полученным скрещиванием первого родительского растения со вторым родительским растением. Изобретение также относится к гибридному семени маиса, полученному с использованием данного способа. Изобретение также относится к способу получения гибридного семени маиса, причем модулирующей развитие обработкой является обработка глифосатом, а первое родительское растение имеет тканеизбирательную устойчивость к глифосату. Изобретение также относится к способу получения гибридного семени маиса, причем первым родительским растением является растение трансгенного события MON 87427 маиса. Изобретение также относится к способу получения гибридного семени маиса, причем второе родительское растение является устойчивым к глифосату.
Нижеследующие и другие аспекты изобретения станут более очевидны из следующие подробного описания.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг. 1 проиллюстрирована организация трансгенного события MON 87427 маиса. На фигуре [A1], [A2] и [A3] соответствуют относительной позиции соответственно SEQ ID NO: 1, SEQ ID NO: 3 и SEQ ID NO: 5, которые охватывают геномную ДНК маиса, фланкирующую 5’-конец трансгенной вставки, и 5’-участок ДНК трансгенной вставки; [B1], [B2] и [B3] соответствуют относительной позиции соответственно SEQ ID NO: 2, SEQ ID NO: 4 и SEQ ID NO: 6, которые охватывают геномную ДНК маиса, фланкирующую 3’-конец трансгенной вставки и 3’-участок ДНК трансгенной вставки; [C] соответствует относительной позиции SEQ ID NO: 7, которая включает геномную ДНК маиса, фланкирующую 5’-конец трансгенной вставки и участок 5’-конца трансгенной вставки; [D] соответствует относительной позиции SEQ ID NO: 8, которая включает геномную ДНК маиса, фланкирующую 3’-конец трансгенной вставки и участок 3’-конца трансгенной вставки; [E] соответствует относительной позиции SEQ ID NO: 9 и различным элементам в трансгенной вставке; а [F] представляет непрерывную последовательность MON 87427, приведенную в виде SEQ ID NO: 10 и включающую SEQ ID NO: 1-9.
На фигуре 2 показана урожайность гибридов MON 87427 при скрещивании с событием маиса NK603 и двухкратной за сезон обработке глифосатом с помощью распыления в количестве 2,25 фунтов (1,02 кг) на акр (4047 м2) на каждое распыление.
На фигуре 3 проиллюстрированы стадии развития метелки, используемые в создании Относительной шкалы развития. Приблизительный размер показан между квадратными скобками. На фигуре Vg представляет собой меристему на вегетативной стадии; Т0 представляет собой переход от вегетативной к репродуктивной стадии; Т1 представляет собой видимую репродуктивную точку роста (0,9 мм); Т2 представляет собой видимые боковые примордии (1,8 мм); Т3 представляет собой видимые примордии колосков (4,1 мм); Т4 представляет собой удлинение центральной оси и боковой оси (12,9 мм); Т5 представляет собой начало дифференцировки пыльников (41,0 мм); Т6 представляет собой начало дифференцировки пыльцы (175 мм); и Т7 представляет собой раскрытие пыльников и выделение пыльцы (285,0 мм).
На фигуре 4 проиллюстрирован разброс размера метелок для трех генотипов маиса на двух стадиях развития (V8 и V10).
На фигуре 5 проиллюстрирована корреляция между требованиями GDU для стадии Т5 и этими требованиями к Р50% и более определенно показана линия регрессии, полученная с использованием корреляции между требованиями GDU к Т5 и к Р50%. Каждая точка представляет инбредную линию, усредненную по местоположению.
На фигуре 6 проиллюстрирован пример того, как Относительная шкала развития выявляет оптимальное окно эффективности химического агента для получения стерильных метелок маиса, измеряемой риском выбрасывания пыльника (АЕ-риском (%)), которое находится между 0,62 и 0,75 на Относительной шкале развития, где выполняются 62-75% общих требований GDU для достижения Р50, и в которой АЕ-риск минимизирован между инбредными линиями и группами спелости. Каждая точка представляет собой средние значения для 1 участка или двух рядов в общем из 32 растений. N=620.
На фигуре 7 проиллюстрированы Т-стадии как функция от GDU (A) и Относительной шкалы развития (В). Каждая линия регрессии представляет разную инбредную линию.
На фигуре 8 проиллюстрирован процент риска выбрасывания пыльников (ось y), измеренный на разных стадиях выметывания пестичных столбиков (ось x) для блоков MON 87427 и CMS.
На фигуре 9 проиллюстрирована генетическая чистота и чистота признака гибридных семян, полученных на MON 87427 с использованием системы гибридизации Roundup® (RHS) и с помощью CMS-системы, с 95%-ым уровнем достоверности. Черная линия на графике представляет желаемые стандарты качества для генетической чистоты и чистоты признака семян, соответственно.
КРАТКОЕ ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
SEQ ID NO: 1 является 20-нуклеотидной последовательностью, представляющей 5’-область соединения геномной ДНК маиса и встроенной трансгенной экспрессионной кассеты.
SEQ ID NO: 2 является 20-нуклеотидной последовательностью, представляющей 3’-область соединения геномной ДНК маиса и встроенной трансгенной экспрессионной кассеты.
SEQ ID NO: 3 является 60-нуклеотидной последовательностью, представляющей 5’-область соединения геномной ДНК маиса и встроенной трансгенной экспрессионной кассеты.
SEQ ID NO: 4 является 60-нуклеотидной последовательностью, представляющей 3’-область соединения геномной ДНК маиса и встроенной трансгенной экспрессионной кассеты.
SEQ ID NO: 5 является 100-нуклеотидной последовательностью, представляющей 5’-область соединения геномной ДНК маиса и встроенной трансгенной экспрессионной кассеты.
SEQ ID NO: 6 является 100-нуклеотидной последовательностью, представляющей 3’-область соединения геномной ДНК маиса и встроенной трансгенной экспрессионной кассеты.
SEQ ID NO: 7 является 5’-последовательностью, фланкирующей встроенную ДНК MON 87427 до области и включая область трансгенной ДНК-вставки.
SEQ ID NO: 8 является 3’-последовательностью, фланкирующей встроенную ДНК в MON 87427 до области и включая область трансгенной ДНК-вставки.
SEQ ID NO: 9 является последовательностью, полностью встроенной в геномную ДНК маиса и содержащей экспрессионную ДНК-кассету.
SEQ ID NO: 10 является нуклеотидной последовательностью, представляющей контиг 5’-последовательности, фланкирующей встроенную ДНК MON 87427 (SEQ ID NO: 7), последовательности, полностью встроенной в геномную ДНК маиса и содержащей экспрессионную кассету (SEQ ID NO: 9), и 3’-последовательности, фланкирующей встроенную ДНК MON 87427 (SEQ ID NO: 8), и включает SEQ ID NO: 1-6.
SEQ ID NO: 11 является праймером 1 (SQ20052) специфичного к трансгену метода анализа события, используемого для идентификации MON 87427. ПЦР-ампликон, получаемый в результате метода анализа TAQMAN® (PE Applied Biosystems, Foster City, Calif.) с использованием комбинации праймеров SEQ ID NO: 11 и SEQ ID NO: 12, является положительным результатом на присутствие события MON 87427.
SEQ ID NO: 12 является праймером 1 (SQ20053) специфичного к трансгену метода анализа события, используемого для идентификации MON 87427.
SEQ ID NO: 13 является 6FAM-пробой (PB10016) специфичного к трансгену метода анализа события, используемого для идентификации MON 87427. Эта проба представляет собой 6FAM™-меченный синтетический олигонуклеотид. Высвобождение флуоресцентного сигнала в реакции амплификации с использованием праймеров SEQ ID NO: 11-12 в комбинации с 6FAM™-меченной пробой является диагностическим критерием события MON 87427 в методе анализа TAQMAN®.
SEQ ID NO: 14 является внутренним контрольным праймером 1 (SQ1241) специфичного к трансгену метода анализа события.
SEQ ID NO: 15 является внутренним контрольным праймером 1 (SQ1242) специфичного к трансгену метода анализа события.
SEQ ID NO: 16 является внутренней контрольной VIC-пробой (PB0084) специфичного к трансгену метода анализа события.
SEQ ID NO: 17 является праймером 1 (SQ12763) специфичного к событию метода анализа события, используемого для идентификации MON 87427. ПЦР-ампликон, получаемый в результате метода анализа TAQMAN® (PE Applied Biosystems, Foster City, Calif.) с использованием комбинации праймеров SEQ ID NO: 17 и SEQ ID NO: 18, является положительным результатом на присутствие события MON 87427.
SEQ ID NO: 18 является праймером 1 (SQ12886) специфичного к событию метода анализа события, используемого для идентификации MON 87427.
SEQ ID NO: 19 является 6FAM-пробой (PB4352) специфичного к событию метода анализа события, используемого для идентификации MON 87427.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Следующие определения и способы приведены для лучшего описания изобретения и в качестве руководства по применению изобретения на практике для средних специалистов в данной области. Если не указано иное, то термины следует понимать в соответствии с их обычным применением средними специалистами в соответствующей области.
Используемый в настоящем описании термин «маис» означает «кукурузу» или Zea mays и включает все сорта растений, которые могут быть скрещены с маисом, включая дикие виды маиса, а также растения, принадлежащие роду Zea, которые позволяют межвидовое скрещивание.
«Глифосат» относится к N-фосфонометилглицину, который представляет собой гербицид, являющийся ингбитором енолпирувилшикимат-3-фосфат синтазы (EPSPS). Глифосат нарушает синтез ароматических аминокислот, ингибируя EPSPS. Глифосат доступен в продаже как гербицид Roundup® (Monsanto Company, St. Louis, Mo.).
Изобретение относится к трансгенному событию MON 87427 маиса (также именуемому в настоящем документе MON 87427). Используемый в настоящем документе термин «событие» относится к продукту, созданному в результате встраивания трансгенной молекулы нуклеиновой кислоты в геном растения, то есть в результате трансформации растения для получения трансгенного растения. Поэтому, «событие» получают в результате действий человека: (i) трансформации растительной клетки в лаборатории молекулой нуклеиновой кислоты, которая включает представляющий интерес трансген, то есть встраивания в геном растительной клетки конструкции или молекулы нуклеиновой кислоты, (ii) регенерации популяции трансгенных растений, получающихся в результате встраивания молекулы нуклеиновой кислоты в геном растения, и (iii) отбора конкретного растения, отличающегося встраиванием молекулы нуклеиновой кислоты в определенное местоположение в геноме растения. Таким образом, событие можно однозначно и определенно описать последовательностью нуклеиновой кислоты, представляющей по меньшей мере часть непрерывной молекулы ДНК, которая получается в событии в результате встраивания молекулы нуклеиновой кислоты в конкретное местоположение генома растения, и которая включает участок геномной ДНК самого растения, который фланкирует (примыкает) и физически соединен с встроенной молекулой ДНК, и встроенную молекулу нуклеиновой кислоты. Событие является рекомбинантным, полученным в результате действия человека и не найдено в нетрансгенных растениях.
Термин «событие», поэтому, относится к исходному трансформированному растению («трансформанту»), который включает молекулу нуклеиновой кислоты, встроенную в конкретное местоположение в геноме растения. Термин «событие» также относится ко всему потомству от трансформанта, которое включает молекулу нуклеиновой кислоты, встроенную в конкретное положение в геноме растения. Такое потомство является соответственно трансгенным и содержит событие. Потомство можно получить любыми средствами, включая самоопыление, скрещивание с другим растением, которое содержит такой же или другой трансген, и/или скрещивание с нетрансгенным растением, например, растением другого сорта. Даже после многих поколений в любом растении, именуемом потомством растения MON 87427, будут присутствовать встроенная ДНК и фланкирующая ДНК из исходного трансформированного растения, и их будет легко идентифицировать.
Термин «событие» также относится к непрерывной молекуле ДНК, созданной в исходном трансформанте (содержащей встроенную ДНК и и фланкирующую геномную ДНК маиса, непосредственно примыкающую к краям встроенной ДНК) или к любой молекуле ДНК, содержащей эту последовательность нуклеиновой кислоты. Непрерывная молекула ДНК была создана в результате встраивания трансгенной молекулы нуклеиновой кислоты в геном растения, то есть в результате действия трансформации, и является специфичной и уникальной для конкретного события. Положение встроенной ДНК в MON 87427 относительно окружающей геномной ДНК растения маиса, поэтому, является специфичным и уникальным для MON 87427. Эта молекула ДНК также является неотъемлемой частью хромосомы маиса MON 87427, и как таковая является стационарной в растении и может наследоваться потомством.
Растения трансгенного события MON 87427 маиса проявляют приемлемую с коммерческой точки зрения тканеизбирательную устойчивость к глифосату. В MON 87427 вегетативные части маиса и женские репродуктивные части маиса являются устойчивыми к глифосату, но ключевые мужские репродуктивные ткани маиса, критичные для развития пыльцы маиса, не устойчивы к глифосату. Поэтому, обработанные глифосатом растения MON 87427 можно использовать в качестве женского родительского растения при получении гибридных семян.
Используемый в настоящем документе, термин «рекомбинантный» относится к неприродной ДНК и/или белку, и/или организму, которые в норме не обнаружены в природе и были созданы в результате вмешательства человека, то есть руками человека. В результате такого вмешательства человека может быть получена молекула ДНК, и/или растение или семя. В контексте настоящего документа «рекомбинантная молекула ДНК» является молекулой ДНК, содержащей комбинацию молекул ДНК, которые обычно не встречаются вместе, и является результатом вмешательства человека, например, молекулой ДНК, в которой содержится комбинация по меньшей мере двух молекул ДНК, гетерологичных друг другу, и/или молекулой ДНК, которая синтезирована искусственно и имеет нуклеотидную последовательность, отличающуюся от нуклеотидной последовательности, обычно присутствующей в природе, и/или молекулой ДНК, которая содержит молекулу нуклеиновой кислоты, искусственно встроенную в геномную ДНК клетки-хозяина и ассоциированную фланкирующую ДНК генома клетки-хозяина. Примером рекомбинантной молекулы ДНК является молекула ДНК, содержащая по меньшей мере одну последовательность, выбранную из SEQ ID NO: 1-10. В контексте настоящего документа «рекомбинантное растение» является растением, которое в норме не существует в природе, является результатом вмешательства человека, и содержит трансген и/или гетерологичную молекулу ДНК, включенную в его геном. В результате такого геномного изменения, рекомбинантное растение очевидно отличается от родственного растения дикого типа. Примером рекомбинантного растения является растение трансгенного события 87427 маиса.
Используемый в настоящем документе термин «трансген» относится к молекуле нуклеиновой кислоты, искусственно встроенной в геном организма в результате вмешательства человека. Такой трансген может быть гетерологичным для клетки-хозяина. Термин «трансгенный» относится к содержанию трансгена, например, «трансгенное растение» относится к растению, содержащему трансген, то есть, молекулу нуклеиновой кислоты, искусственно встроенную в геном организма в результате вмешательства человека. Используемый в настоящем документе термин «гетерологичный» относится к первой молекуле, обычно не найденной в природе в комбинации со второй молекулой. Например, молекула может быть получена из первого биологического вида и встроена в геном второго вида. Поэтому, молекула будет гетерологичной молекулой, то есть гетерологичной для организма и искусственно встроенной в геном организма.
Используемый в настоящем документе термин «химерный» относится к одной молекуле ДНК, полученной слиянием первой молекулы ДНК со второй молекулой ДНК, где ни первая, ни вторая молекулы ДНК не находятся обычно в такой конфигурации, то есть, слитыми друг с другом. Химерная молекула ДНК, поэтому, является новой молекулой ДНК, иным образом не присутствующей в природе. Примером химерной молекулы ДНК является молекула ДНК, содержащая по меньшей мере одну последовательность, выбранную из SEQ ID NO: 1-10.
Изобретение относится к молекулам ДНК и их соответствующим нуклеотидным последовательностям. Используемый в настоящем документе термин «ДНК», «молекула ДНК», «молекула нуклеиновой кислоты» относится к двухцепочечной молекуле ДНК геномного или синтетического происхождения, то есть полимеру из дезоксинуклеотидных оснований или нуклеотидной молекуле, читаемой с 5′ (верхнего) конца до 3′ (нижнего) конца. Используемый в настоящем документе термин «последовательность ДНК» или «нуклеотидная последовательность» относится к нуклеотидной последовательности молекулы ДНК. Используемой в настоящем документе номенклатурой является требуемая разделом 37 Свода нормативных документов США, § 1.822 и указанная в таблицах в WIPO Standard ST.25 (1998), приложение 2, таблицы 1 и 3. Как принято и в настоящем документе, нуклеотидные последовательности по изобретению, такие как приведенные как SEQ ID NO: 1-10 и их фрагменты, приведены относительно только одной цепи из двух комплементарных цепей нуклеотидной последовательности, но подразумевается, что комплементарные последовательности (то есть последовательности комплементарной цепи) также именуемые в данной области обратными комплементарными последовательностями, входят в объем изобретения и, определенно предполагается, что они входят в объем заявленного объекта изобретения. Поэтому, в контексте настоящего документа ссылки на SEQ ID NO: 1-10 и их фрагменты включают последовательность и относятся к последовательности комплементарной цепи и ее фрагментов.
Используемый в настоящем описании термин «фрагмент» относится к участку или неполной меньшей части целого. Например, фрагменты SEQ ID NO: 10 включали бы последовательности, которые составляют по меньшей мере 10 нуклеотидов, по меньшей мере 20 нуклеотидов или по меньшей мере 50 нуклеотидов полной последовательности SEQ ID NO: 10.
Нуклеотидная последовательность, соответствующая полной нуклеотидной последовательности встроенной трансгенной ДНК и достаточно большим сегментам геномной ДНК маиса, фланкирующим оба конца встроенной трансгенной ДНК, приведена в настоящем документе как SEQ ID NO: 10. Частью данной последовательности является встроенная трансгенная ДНК (также именуемая в настоящем документе как трансгенная вставка или встроенная ДНК), приведенная как SEQ ID NO: 9. Нуклеотидная последовательность геномной ДНК маиса, физически соединенная фосфодиэфирной связью и, поэтому, фланкирущая 5′-конец встроенной трансгенной ДНК и содержащая 10 нт трансгенной встроенной ДНК, изложена в SEQ ID NO: 7. Нуклеотидная последовательность геномной ДНК маиса, физически соединенная фосфодиэфирной связью и, поэтому, фланкирующая 3′-конец встроенной трансгенной ДНК и содержащая 10 нт трансгенной встроенной ДНК, изложена в SEQ ID NO: 8.
MON 87427 дополнительно содержит две области, именуемые «соединениями». «Соединение» представляет собой участок, в котором один конец встроенной трансгенной ДНК встроен в геномную ДНК и соединен с ней. Соединение охватывает, то есть перекрывает, участок встроенной трансгенной ДНК и примыкающей фланкирующей геномной ДНК, и как таковое включает в себя точку соединения этих двух элементов в виде одной непрерывной молекулы. Одно соединение находится на 5’-конце встроенной трансгенной ДНК, а другое на 3’-конце встроенной трансгенной ДНК, которые именуют в настоящем документе как 5′- и 3′-соединения, соответственно. «Последовательность соединения» или «область соединения» относятся к последовательности ДНК и/или соответствующей молекуле ДНК соединения. Специалист в данной области может подобрать последовательности соединений в MON 87427, используя SEQ ID NO: 10. Примеры последовательностей соединений в MON 87427 приведены как SEQ ID NO: 1-6. SEQ ID NO: 1 представляет собой 20-нуклеотидную последовательность перекрывающую соединение между геномной ДНК маиса и 5’-концом ДНК трансгенной вставки; SEQ ID NO: 3 представляет собой 60-нуклеотидную последовательность перекрывающую соединение между геномной ДНК маиса и 5’-концом ДНК трансгенной вставки; SEQ ID NO: 5 представляет собой 100-нуклеотидную последовательность перекрывающую соединение между геномной ДНК маиса и 5’-концом ДНК трансгенной вставки. SEQ ID NO: 2 представляет собой 20-нуклеотидную последовательность перекрывающую соединение между геномной ДНК маиса и 3’-концом ДНК трансгенной вставки; SEQ ID NO: 4 представляет собой 60-нуклеотидную последовательность перекрывающую соединение между геномной ДНК маиса и 3’-концом ДНК трансгенной вставки; SEQ ID NO: 6 представляет собой 100-нуклеотидную последовательность перекрывающую соединение между геномной ДНК маиса и 3’-концом ДНК трансгенной вставки. На фигуре 1 проиллюстрировано физическое расположение SEQ ID NO: 1-10 в направлении от 5′ до 3′. Любой сегмент ДНК, полученный из трансгенного MON 87427, который включает SEQ ID NO: 1-6, входит в объем изобретения. Изобретение, таким образом, относится к молекуле ДНК, которая содержит по меньшей мере одну из нуклеотидных последовательностей, указанных в SEQ ID NO: 1-6.
Последовательности соединений события MON 87427 присутствуют как части генома растения, семени или клетки трансгенного события MON 87427 маиса. Идентификация любой одной или нескольких из SEQ ID NO: 1-6 в образце, полученном из растения маиса, семени или части растения, определяет, что ДНК получена из MON 87427 и является диагностическим признаком присутствия в образце ДНК из MON 87427.
Изобретение относится к примерам молекул ДНК, которые можно использовать либо в качестве праймеров, либо в качестве проб для диагностики присутствия ДНК, полученной из растения события MON 87427, в образце. Такие праймеры или пробы специфичны в отношении последовательности нуклеиновой кислоты-мишени и, как таковые, их можно использовать для идентификации последовательностей нуклеиновой кислоты MON 87427 способами по изобретению, описанными в настоящем документе.
«Праймер» является молекулой нуклеиновой кислоты, которая подобрана для использования в способах отжига или гибридизации, которые включают температурную амплификацию. Пару праймеров можно использовать с матричной ДНК, такой как образец геномной ДНК маиса, в температурной амплификации, такой как полимеразная цепная реакция (ПЦР), для получения ампликона, причем ампликон, полученной в такой реакции, будет иметь последовательность ДНК, соответствующую последовательности матричной ДНК, расположенной между двумя участками гибридизации праймеров на матрице. В контексте настоящего документа «ампликон» является ДНК, которая синтезирована с использованием методик амплификации. Ампликоны по изобретению имеют последовательность, содержащую одну или несколько из SEQ ID NO: 1-10 или их фрагменты. Праймер обычно подбирают для гибридизации с комплементарной цепью ДНК-мишени с образованием гибрида между праймером и цепью ДНК-мишени, и присутствие праймера является точкой узнавания полимеразой для начала удлинения праймера (то есть полимеризации дополнительных нуклеотидов в удлиняющуюся нуклеотидную молекулу) с использованием в качестве матрицы цепи ДНК-мишени. Предполагается, что пары праймеров, используемые в изобретении, относятся к использованию двух праймеров, связывающихся с противоположными цепями двухцепочечного нуклеотидного сегмента для амплификации линейного нуклеотидного сегмента между позициями, являющихся мишенями связывания индивидуальных членов пары праймеров. Примеры последовательностей праймеров приведены как SEQ ID NO: 11-12, SEQ ID NO: 14-15 и SEQ ID NO: 17-18. Пара праймеров SEQ ID NO: 14-15 и пара праймеров SEQ ID NO: 17-18, каждая имеет первую молекулу ДНК и вторую молекулу ДНК (которая отличается от первой молекулы ДНК), причем обе молекулы имеют достаточный непрерывный нуклеотидный участок для функционирования в качестве праймеров, которые при совместном использовании в реакции амплификации ДНК с матрицей ДНК, полученной из MON 87427, дают ампликон, который является диагностическим критерием присутствия в образце ДНК из MON 87427.
«Проба» представляет собой молекулу нуклеиновой кислоты, комплементарную цепи нуклеиновой кислоты-мишени, для использования в способах отжига или гибридизации. Пробы по изобретению включают не только дезоксирибонуклеиновые или рибонуклеиновые кислоты, но также полиамиды и другие материалы проб, которые специфично связывают последовательность ДНК-мишени, и детекция такого связывания может быть полезна в диагностике, выделении, определении или подтверждении присутствия последовательности ДНК-мишени в конкретном образце. Проба может быть соединена с обычной детектируемой меткой или репортерной молекулой, например, радиоактивным изотопом, лигандом, хемилюминесцентным агентом или ферментом. Примеры последовательностей, которые можно использовать в качестве проб для детекции MON 87427, являются SEQ ID NO: 1-2, SEQ ID NO: 13, SEQ ID NO: 16, SEQ ID NO: 19.
Способы подбора и использования праймеров и проб хорошо известны в данной области и описаны, например, Joseph Sambrook, Molecular Cloning: A Laboratory Manual, Third Edition, Cold Spring Harbor Laboratory Press (2001) и Current Protocols in Molecular Biology, Wiley-Blackwell. Специалист в данной области может легко подобрать молекулы ДНК, содержащие фрагменты SEQ ID NO: 1-10 и пригодные в качестве праймеров и проб для детекции MON 87427, для использования в качестве проб в гибридизационных способах детекции, например, в способе Саузерн-блота.
Молекулы ДНК и соответствующие нуклеотидные последовательности, приведенные в настоящем документе, поэтому, пригодны, среди прочего, для идентификации MON 87427, отбора сортов или гибридов растений, включающих в себя MON 87427, детекции присутствия ДНК, полученной из трансгенного MON 87427, в образце и мониторинга образцов на присутствие и/или отсутствие MON 87427 или частей растения, полученных из MON 87427.
Изобретение относится к растениям маиса, потомству, семенам, клеткам растения, частям растения и продуктам потребления. Эти растения, потомство, семена, клетки растения, части растения и продукты потребления содержат детектируемое количество нуклеотида по изобретению, то есть, такого как молекула нуклеиновой кислоты, содержащая по меньшей мере одну из последовательностей, приведенных как SEQ ID NO: 1-10. Растения, потомство, семена, клетки растения и части растения по изобретению могут также содержать один или несколько дополнительных трансгенов. Таким трансгеном может быть любая нуклеотидная последовательность, кодирующая белок, или молекула РНК, придающие желаемый признак, включая, но не ограниченный этим, повышенную устойчивость к насекомым, повышенную эффективность использования воды, повышенную урожайность, повышенную устойчивость к засухе, повышенное качество семян, повышенные питательные качества и/или повышенную устойчивость к гербицидам, причем желаемый признак измеряют относительно растения маиса, не имеющего такой дополнительный трансген.
Изобретение относится к растениям маиса, потомству, семенам, клеткам растения и частям растения, а также листьям, полученным от растения трансгенного события MON 87427 маиса. Репрезентативный образец семян MON 87427 депонирован в соответствии с Будапештским договором с целью воспроизведения изобретения. Хранилищем для помещения депозита является Американская коллекция типовых культур (АТСС), имеющая адрес 10801 University Boulevard, Manassas, Va. USA, Zip Code 20110. В хранилище АТСС семенам события MON 87427 был присвоен номер доступа PTA-7899.
Изобретение относится к микроорганизму, содержащему молекулу ДНК, имеющую SEQ ID NO: 1-10, присутствующие в его геноме. Примером такого микроорганизма является клетка трансгенного растения. Микроорганизм, такой как растительная клетка по изобретению, можно использовать во многих промышленных применениях, включая без ограничений: (i) применение в качестве исследовательского средства для научного поиска или промышленных исследований; (ii) применение в культуре для получения эндогенных или рекомбинантных углеводных, липидных, нуклеотидных или белковых продуктов или низкомолекулярных соединений, которые могут быть полезны для последующего научного исследования или в качестве промышленных продуктов; и (iii) применение в современных методиках культивирования тканей растений для получения трансгенных растений или культур тканей растений, которые затем можно использовать для сельскохозяйственных исследований или производства. При использовании микроорганизмов, таких как клетки трансгенных растений, применяют современные микробиологические методики и вмешательство человека для получения искусственного уникального микроорганизма. В этом способе рекомбинантную ДНК встраивают в геном растительной клетки для создания трансгенной растительной клетки, которая отличается от и является уникальной среди природных растительных клеток. Эту трансгенную растительную клетку затем можно культивировать примерно также как и бактериальные и дрожжевые клетки с использованием современных микробиологических методик, и она может существовать в недифференцированнм, одноклеточном состоянии. Новая генетическая композиция и фенотип растительной клетки являются техническим эффектом, созданным в результате встраивания гетерологичной ДНК в геном клетки. Другим аспектом изобретения является способ использования микроорганизма по изобретению. Способы использования микроорганизма по изобретению, такого как трансгенные растительные клетки, включают (i) способы получения трансгенных клеток в результате встраивания рекомбинантной ДНК в геном клетки и затем использования этой клетки для получения дополнительных клеток, обладающих такой же гетерологичной ДНК; (ii) способы культивирования клеток, которые содержат рекомбинантную ДНК, с использованием современных микробиологических методик; (iii) способы получения и очистки эндогенных или рекомбинантных углеводных, липидных, нуклеотидных или белковых продуктов из культивируемых клеток; и (iv) способы использования современных методик культивирования тканей растений с трансгенным растительными клетками для получения трансгенных растений или культур тканей трансгенных растений.
Растения по изобретению могут передавать ДНК события, включая трансгенную вставку, потомству. В контексте настоящего документа «потомство» включает любое растение, семя, клетку растения и/или регенерируемую часть растения, содержащие ДНК события, полученную от растения-предшественника, и/или молекулу нуклеиновой кислоты, имеющую по меньшей мере одну из последовательностей, приведенных как SEQ ID NO: 1-10. Растения, потомство и семена могут быть гомозиготными или гетерозиготными по трансгену. Потомство можно выращивать из семян, продуцируемых растением MON 87427, и/или из семян, продуцируемых растением, оплодотворенным пыльцой от растения MON 87427. Растения по изобретению можно получить самоопылением или перекрестным опылением, и/или их можно использовать в способах самоопыления или перекрестного опыления. Поэтому, в одном варианте осуществления изобретения растение MON 87427 можно перекрестно опылять пыльцой от другого растения маиса для получения гибридного потомства. Способы скрещивания, подходящие для растений маиса MON 87427, известны в данной области.
Изобретение относится к части растения, которая получена из MON 87427. В контексте настоящего документа «часть растения» относится к любой части растения, в которой содержится материал, полученный из растения MON 87427. Части растения включают, но не ограничены этим, пыльцу, семязачаток, стручок, цветок, ткань корня или стебля, волокна и листья. Части растения могут быть живыми, неживыми, регенерируемыми и/или нерегенерируемыми.
Изобретение относится к продукту потребления, который получают из трансгенного события MON 87427 маиса и включает в себя молекулу нуклеиновой кислоты, имеющую нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-10. В контексте настоящего документа «продукт потребления» относится к любой композиции или продукту, в котором содержится материал, полученный из растения, семени, растительной клетки или части растения MON 87427. Продукты потребления могут быть живыми или неживыми. Неживые продукты потребления включают, но не ограничены этим неживые семена и зерна; переработанные семена, части семян и части растения; дегидратированную ткань растения, замороженную ткань растения и переработанную ткань растения; семена и части растения, переработанные для питания животных для потребления наземными и/или водными животными, масло, муку крупного помола, муку, хлопья, отруби, клетчатку, молоко, сыр, бумагу, сливки, вино и любой другой продукт питания для потребления человеком; и биомассу и топливные продукты. Живые продукты потребления включают, но не ограничены этим, семена и клетки растения. Трансгенное событие MON 87427 маиса, поэтому, можно использовать для изготовления любого продукта потребления, обычно получаемого из маиса. Продукт потребления, который получен из MON 87427, может содержать детектируемое количество специфичной и уникальной ДНК, соответствующей MON 87427, и, более конкретно, может содержать детектируемое количество молекулы нуклеиновой кислоты, имеющей по меньшей мере одну из последовательностей, приведенных как SEQ ID NO: 1-10. Детекция одной или нескольких из этих последовательностей в образце продукта потребления, полученного из, изготовленного из, состоящего из или содержащего растение кукурузы, семя кукурузы, клетку растения кукурузы или часть растения кукурузы, является заключающей и определяющей присутствие биологического материала, полученного из события MON 87427 кукурузы, в таком продукте потребления, и детекцию такой молекулы нуклеиновой кислоты можно использовать для определения содержания и/или источника продукта потребления. Можно использовать любой стандартный способ детекции молекул нуклеиновой кислоты, включая способы детекции, раскрытые в настоящем документе.
Растения, потомство, семена, растительные клетки, части растения и продукты потребления по изобретению, поэтому, можно, использовать, помимо прочего, для выращивания растений с целью получения семян и/или частей растения MON 87427 для сельскохозяйственных нужд, получения потомства MON 87427 для скрещивания растений и исследовательских целей, использования с микробиологическими методиками для промышленного и исследовательского применения, и продажи потребителям.
Изобретение относится к способам контроля сорняков с использованием гербицида глифосата и MON 87427. Приведен способ контроля сорняков в поле, который состоит из высаживания сортовых или гибридных растений MON 87427 в поле и применения гербицидно-эффективной дозы глифосата на поле для контроля сорняков на поле без поражения растений MON 87427. Такое применение гербицида глифосата можно осуществить до всхода семян, то есть в любое время после посева семян MON 87427 и перед всходом растений MON 87427, или после всхода семян, то есть в любое время после всхода растений MON 87427. Гербицидно эффективная доза глифосата при применении на поле для контроля сорняков должна находиться в диапазоне от примерно 0,1 фунта до такого количества, как примерно 4 фунта на акр в течение вегетационного периода. В течение вегетационного периода глифосата можно применять многократно, например, два раза (например, до посадки и после всходов или до всходов и после всходов) или три раза (например, до посадки, до всходов и после всходов).
Изобретение относится к способам получения толерантного к гербициду растения трансгенного события MON 87427 маиса. Потомство, полученное с помощью данных способов, может быть сортовыми или гибридными растениями; оно может быть выращено из семян, продуцированных растением MON 87427 и/или из семян, продуцированных растением, оплодотворенным пыльцой от растения MON 87427; и оно может быть гомозиготым или гетерозиготным по трансгену. Потомство можно впоследствии самоопылять или перекрестно опылять.
Растение маиса, которое устойчиво к применению гербицида глифосата, можно получить в результате полового скрещивания растения MON 87427, содержащего молекулу нуклеиновой кислоты, содержащую по меньшей мере одну из последовательностей, приведенных как SEQ ID NO: 1-10, с другим растением маиса, и, в результате получения семян, которые затем собирают и выращивают из них растения-потомство. Это потомство можно затем обработать гербицидом глифосат, и потомство, устойчивое к гербициду глифосат можно отобрать. В альтернативном варианте, эти растения-потомство можно проанализировать, используя диагностические способы для отбора растений-потомства, которые содержат ДНК MON 87427.
В применении на практике способов по изобретению стадию полового скрещивания одного растения с другим растением, то есть перекрестного опыления, можно выполнить или обеспечить в результате вмешательства человека, например: ручного сбора пыльцы одного растения и контактирования этой пыльцы со столбиком или рыльцем второго растения и затем необязательно предупреждения дополнительного оплодотворения оплодотворенного растения; ручного и/или автоматического удаления (например, путем удаления метелок), разрушения (например, путем использования химических агентов) или закрывания тычинок или пыльников растения, так чтобы предупредить природное самоопыление, и для оплодотворения должно произойти перекрестное опыление; размещения человеком опыляющих насекомых в позиции для «прямого опыления» (например, путем помещения пчелиных роев в плодовые питомники или поля или путем помещения растений под сетку с опыляющими насекомыми); открытия или удаления человеком частей цветка для помещения или контакта чужеродной пыльцы на столбик или рыльце (например, в маисе, который в природе имеет цветки, препятствующие или предупреждающие перекрестное опыление, что делает их природно облигатными самоопыляющимися растениями в отсутствие вмешательства человека); избирательного помещения растений в определенную область (например, преднамеренной посадки растений на расстоянии, позволяющем опыление); и/или применения химических соединений для ускорения цветения или усиления восприимчивости (рыльца к пыльце).
При применении на практике способов по изобретению стадию полового оплодотворения растения маиса в результате самоопыления можно выполнить или обеспечить путем вмешательства человека, например: ручного сбора пыльцы одного растения и контактирования этой пыльцы со столбиком или рыльцем того же растения и затем необязательно предупреждения дополнительного оплодотворения оплодотворенного растения; ручного и/или автоматического удаления (например, путем удаления метелок), разрушения (например, путем использования химических агентов) или закрывания тычинок или пыльников растения, так чтобы предупредить природное самоопыление, и для оплодотворения должно произойти ручное самоопыление; размещения человеком опыляющих насекомых в позиции для «прямого опыления» (например, путем помещения индивидуального растения под сетку с опыляющими насекомыми); проведения манипуляций человеком с репродуктивными частями растения для обеспечения возможности или усиления самоопыления; избирательного помещения растений в определенную область (например, преднамеренной посадки других растений на расстоянии, превышающем расстояние, позволяющее опыление); и/или применения химических соединений для ускорения цветения или усиления восприимчивости (рыльца к пыльце).
Изобретение относится к растениям и способам, полезным в продукции гибридных семян маиса.
Растение маиса имеет раздельные мужские и женские цветущие части. Метелка является мужской структурой, а початок является женской цветочной структурой растения. Стадия цветения у маиса включает выделение пыльцы и выметывание пестичных столбиков. Пыльца маиса может оплодотворять то же растение (самоопыление) или другое растение (перекрестное опыление). Если мужские структуры растения не удалены до выделения пыльцы, то растение маиса самоопыляется в некоторой степени. Для продукции гибридных семян женские структуры первого растения маиса перекрестно опыляют пыльцой от второго растения маиса. Поэтому, для эффективного получения гибридных семян необходимо, чтобы пыльца самого растения не могла самооплодотворять растение. Способы увеличения продукции гибридных семян маиса, приведенные в настоящем документе, включают в себя выращивание на местности семени или растения, содержащих MON 87427 и одно или несколько других растений маиса. Растения события MON 87427 затем обрабатывают глифосатом до образования пыльцы, таким образом делая мужские части растения события MON 87427 стерильными и неспособными к самоопылению. Растения события MON 87427 затем опыляют пыльцой от другого растения маиса, используя любой из способов, описанных в настоящем документе. Другие растения маиса могут быть устойчивыми или неустойчивыми к глифосату. Затем собирают семена маиса от растений события MON 87427, причем семена, собранные от обработанных растений MON 87427 имеют больший выход гибридных семян маиса (то есть более высокий процент собираемых гибридных семян или большую чистоту гибридных семян) относительно семян маиса, собранных от необработанных растений события MON 87427 или от других растений маиса в таких же условиях. Семена маиса, собранные от необработанных растений события MON 87427 при таких же условиях, будут иметь более высокий процент негибридных семян (то есть, инбредных семян, полученных в результате самоопыления) и, поэтому, меньший выход гибридных семян маиса.
Растения и способы по изобретению можно также использовать для скрещивания маиса с использованием способов, известных в данной области, включая использование способов, описанных в патенте США № 7314970, который, таким образом, включен в настоящий документ путем ссылки, и патентной публикации США № 20090165166, которая, таким образом, включена в настоящий документ путем ссылки.
Растения, потомство и семена, охваченные этими способами и полученные с использованием этих способов, будут отличаться от других растений маиса. Например, растения MON 87427, потомство и семена по изобретению являются трансгенными и рекомбинантными, и, как таковые, созданы в результате вмешательства человека и содержат детектируемое количество молекулы нуклеиновой кислоты, имеющей по меньшей мере одну из последовательностей, приведенных как SEQ ID NO: 1-10.
Способы по изобретению, поэтому, полезны среди прочего для контроля сорняков на поле при выращивании растений для получения семян и/или частей растений MON 87427 для сельскохозяйственных или исследовательских целей, отбора потомства MON 87427 для скрещивания растений или исследовательских целей и получения растений-потомства и семян MON 87427.
Растения, потомство, семена, клетки растения, части растения и продукты потребления по изобретению можно оценить по композиции ДНК, экспрессии генов и/или экспрессии белков. Такую оценку можно осуществить, используя стандартные способы, такие как ПЦР, Нозерн-блоттинг, Саузерн-анализ, Вестерн-блоттинг, иммунопреципитация и ELISA, или используя способы детекции и/или наборы для детекции, приведенные в настоящем документе.
Изобретение относится к способам детекции в образце присутствия материала, специфичного для MON 87427. Присутствие молекулы нуклеиновой кислоты по изобретению можно детектировать, используя пробы и праймеры по изобретению с любым способом детекции нуклеиновых кислот, используемым в данной области, таким как полимеразная цепная реакция (ПЦР) или гибридизация ДНК. Один способ обеспечивает приведение образца ДНК в контакт с парой праймеров, которые способны продуцировать ампликон на ДНК события MON, проведение реакции амплификации и, таким образом, получение ДНК-ампликона, содержащего по меньшей мере одну из нуклеотидных последовательностей, приведенных как SEQ ID NO: 1-10, и затем детекцию присутствия или отсутствия молекул ампликона и, необязательно, подтверждение присутствия в последовательности ампликона последовательности, содержащей по меньшей мере одну из последовательностей, приведенных как SEQ ID NO: 1-10. Присутствие такого ампликона является определяющим и/или диагностическим в отношении присутствия ДНК, специфичной для MON 87427, и, таким образом, присутствия биологического материала MON 87427 в образце. Другой способ относится к приведению образца ДНК в контакт с зондом ДНК, помещению пробы и образца ДНК в жесткие условия гибридизации и, Затем детекции гибридизации между пробой ДНК образца-мишени. Детекция гибридизации является диагностическим признаком присутствия ДНК, специфичной для MON 87427, в образце ДНК. Амплификацию нуклеиновых кислот, гибридизацию нуклеиновых кислот и секвенирование ДНК можно выполнить любым из способов, известных в данной области. Одним примером методик, которые можно использовать в практическом осуществлении данного изобретения, является TAQMAN® (PE Applied Biosystems, Foster City, Calif.).
Последовательность гетерологичной вставки ДНК, последовательности соединений или фланкирующие последовательности из MON 87427 (с репрезентативными образцами семян, депонированным как ATCC PTA-7899) можно подтвердить (и исправить при необходимости) путем амплификации таких последовательностей на матрице из события с использованием праймеров, полученных из последовательностей, приведенных в настоящем документе, с последующим стандартным ДНК-секвенированием ампликона или клонированной ДНК.
Изобретение относится к наборам для детекции ДНК. Варианты таких наборов также можно разработать с использованием композиций и способов, раскрытых в настоящем документе, и способов, хорошо известных в области детекции ДНК. Наборы для детекции ДНК можно использовать для идентификации ДНК MON 87427 DNA в образце и можно применять для способов скрещивания растений маиса, содержащих ДНК MON 87427. Наборы могут содержать ДНК-праймеры или пробы, которые аналогичны или комплементарны SEQ ID NO: 1-10 или их фрагментам.
Наборы и способы детекции по изобретению, поэтому, можно использовать среди прочего для идентификации MON 87427, отбора сортов или гибридов растений, содержащих MON 87427, детекции присутствия ДНК, полученной из трансгенного MON 87427, в образце и мониторинга образов на присутствие и/или отсутствие MON 87427 или частей растения, полученных из MON 87427.
Изобретение относится к Относительной шкале развития, пригодной для мониторинга и/или определения репродуктивного развития у маиса. Эта новая Относительная шкала развития решает проблему различий в развитии и репродуктивном созревании среди различных сортов и инбредных линий маиса, обеспечивая временную шкалу, которая выражает стадии развития метелки относительно цветения. Относительная шкала развития сокращает наблюдаемые между генотипами отличия в развитии и росте метелки. Развитие метелки в различные стадии созревания проиллюстрированы на фигуре 3.
Развитие маиса часто определяют по шкале стадий, основанной на вегетационных событиях, обычно известных как V-стадии. Эти стадии определяют в соответствии с самым верхним листом, у которого виден листовой узел. VE соответствует прорастанию, V1 соответствует первому листу, V2 соответствует второму листу, V3 соответствует третьему листу, V(n) соответствует n-му листу. VT происходит, когда видно последнее разветвление метелки, но до выметывания пестичных столбиков. При оценке стадий для поля маиса, каждую конкретную V-стадию указывают, только когда 50 процентов или больше растений на поле имеют эту или более высокую стадию. Однако применение этой вегетационной шкалы для определения репродуктивной зрелости может осложняться тем фактом, что вегетативное развитие не обязательно коррелирует с репродуктивным развитием для всех генотипов. Кроме того, не у всех инбредных линий вырастает одинаковое число листьев, агрономы не всегда последовательны в своей оценке, и первые развивающиеся листья начинают вянуть достаточно рано в сезон, так что если листья не помечены правильно в течение ранних стадий, то позднее становится очень трудно правильно идентифицировать V-стадии.
Другим распространенным средством прогноза и оценки стадий роста и развития маиса являются градусо-единицы роста (GDU). Основополагающим фактором роста и развития маиса является тепло. Тепло обычно измеряют в одной точке времени и выражают в виде температуры, но его также можно измерить в течение отрезка времени и выражать в тепловых единицах. Эти тепловые единицы обычно именуют GDU. GDU можно определить, как разницу между средней дневной температурой и выбранной базовой температурой с наложенными некоторыми ограничениями. GDU вычисляют, используя следующее уравнение:
Градусо-единицы роста={(H+L)/2}−B
в котором Н является дневной максимальной температурой (но не выше 86°F, L является дневным минимумом (но не ниже 50°F, а B является базовой температурой 50°F (10°С). Поскольку рост маиса сильно замедляется при температурах выше 86°F или ниже 50°F, то на максимальную и минимальную дневные температуры, используемые в формуле, наложены ограничения. Нижний порог дневной температуры также предупреждает получение отрицательных значений. Поэтому, если дневной максимум превышает 86°F, то дневную максимальную температуру, используемую в GDU-формуле, указывают как 86°F. В свою очередь если дневной минимум падает ниже 50°F, то дневную минимальную температуру, используемую в GDU-формуле, указывают как 50°F. Если дневная максимальная температура не превышает 50°F, то для этого дня не регистрируют GDU. Максимальная величина GDU, которую растение маиса может набрать в день, составляет 36, а минимальная равняется 0. Оценку созревания растений маиса определяют по сумме дневных величин GDU за определенный промежуток времени. Временным периодом, который используют большинство производителей семян маиса, является время от точки посадки до физиологической спелости или точки, в которой заполнение зерен практически завершено. В большинстве штатов США накопленные данные GDU сохраняются для большинства географических районов и доступны от Crop Reporting Service (Службы информации об урожаях USDA) или от State Extension Services (Служб по распространению опыта в штатах). Кроме того, инструмент получения информации о GDU в конкретном местоположении также описан в патенте США № 6967656, который, таким образом, полностью включен в настоящий документ путем ссылки. Как и в случае V-стадий, изменения GDU могут значительно варьировать относительно стадии развития метелки среди генотипов и не могут являться надежным способом прогноза развития метелки.
В контексте настоящего документа «Относительная шкала развития» определена как шкала, созданная делением GDU на данную стадию развития метелки на GDU, требуемую для достижения определенной стадии выделения пыльцы. Затем конструируют линию регрессии с этой информацией для каждого генотипа или инбредного сорта. Относительную шкалу развития можно сконструировать, используя способы, описанные в настоящем документе, и она основана на корреляции между требованиями GDU, необходимыми для достижения некоторой стадии развития цветков кукурузы относительно данной стадии развития метелки. Как таковую, Относительную шкалу развития можно использовать для прогноза развития метелки у маиса для различных генотипов и инбредных сортов, и ее можно использовать в качестве альтернативы использованию V-стадий или GDU в скрещивании растений и агротехнических способах.
В контексте настоящего изобретения «стадия развития цветков» определена согласно степени, до которой популяция растений выделяет пыльцу, именуемую Р-стадией. Стадию развития цветков выражают как Рх, где Р обозначает «пыльцу», а «х» указывает на процент растений в популяции, которые выделяют пыльцу. Относительная шкала развития по изобретению основана на регрессии, полученной делением GDU на данной стадии развития метелки на число GDU, требуемое для достижения конкретной стадии выделения пыльцы. Это соотношение выражено следующей формулой:
Относительная шкала развития = (GDU для Tn/GDU для Px)
где «GDU для Tn» является количеством GDU (градусо-единиц роста), требуемым для достижения некоторой стадии развития метелки, где n может варьировать от 0 до 7, и где «GDU для Px» является количеством GDU, требуемым для достижения некоторой стадии развития цветка или Р-стадии, в которой х может варьировать от 0 до 100 (примером является р50, определяемая как 50% растений на поле начали выделять пыльцу).
Регрессия может быть основана на корреляционном взаимодействии между любыми стадией развития метелки и стадией развития цветка или GDU-требованиями для Р-стадии. Такое корреляционное взаимодействие выражается делением GDU, требуемых для достижения некоторой стадии развития цветков или Р-стадии. В одном варианте осуществления изобретения стадия развития цветков или Р-стадия для регрессии является Р50, в которой 50% популяции растений маиса выделяют пыльцу. В другом варианте осуществления изобретения стадия развития цветков или Р-стадия выделения пыльцы для вычисления регрессии может составлять примерно от 1% до 100%, включая примерно 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 и 99%. Стадия развития метелки для регрессии может находиться между Т0 и Т7, такая как T0, T1, T2, T3, T4, T5, T6 и T7. Вне зависимости от стадии развития метелки и стадии развития цветков, выбранных для создания регрессии, Относительную шкалу развития можно получить построением графика зависимости GDU, требуемых для достижения определенной стадии выделения пыльцы, относительно числа GDU, требуемого для достижения данной стадии развития метелки. Этот аспект проиллюстрирован на фиг. 5.
Используемый в настоящем документе термин «определяющий» относится к действию измерения, определения, оценки, расчета, мониторинга и/или прогноза. Например, «определение развития метелки» в контексте настоящего документа включает измерение текущей стадии развития метелки, мониторинг прогресса развития метелки и/или прогноз начала следующей стадии развития метелки.
Поэтому, изобретение относится к способу получения Относительной шкалы развития, включающему в себя измерение градусо-единиц роста, требуемых для достижения популяцией растений маиса определенной стадии развития метелки; измерение градусо-единиц роста, требуемых для достижения указанной популяцией растений маиса определенной стадии развития цветков; и создание линии регрессии делением указанных измеренных градусо-единиц роста, требуемых для достижения указанной популяцией растений маиса указанной определенной стадии развития метелки, на указанные измеренные градусо-единицы роста, требуемые для достижения указанной популяцией растений маиса указанной определенной стадии развития цветков. Стадию измерения можно повторить по меньшей мере для двух популяций растений маиса. Стадию измерения можно повторить для множества стадий развития метелки и/или множества стадий развития цветков. Определенной стадией развития цветков может быть выделение пыльцы примерно 50 процентами популяции растений маиса.
Изобретение также относится к способу определения оптимального диапазона в пределах Относительной шкалы развития для схемы обработки, сцепленной с развитием метелки, таким образом позволяющему определить оптимальное время схемы обработки, в которой стадия развития является важным фактором. Примером такой обработки является применение однократной схемы обработки стандартным химическим агентом для максимальной эффективности в обеспечении стерильности метелок маиса среди различных родительских генотипов при получении гибридных семян маиса безотносительно генотипа или группы спелости.
Используемый в настоящем документе термин «гибридное семя» является семенем, получаемым перекрестным опылением двух растений. Растения, выращенные из гибридных семян, могут иметь улучшенные агротехнические характеристики, такие как большая урожайность, большая однородность и/или устойчивость к заболеваниям. Гибридные семена не являются чисто линейными, то есть семя, полученное в результате самооплодотворения гибридного растения (растения, выращенного из гибридного семени), не обязательно даст в следующем поколении идентичное гибридное растение. Поэтому, новые гибридные семена необходимо получать из линейных родительских растений для каждой посадки. Поскольку большинство зерновых растений имеют как мужские, так и женские органы гибридное семя можно получать, только предупреждая самоопыление женского родительского растения и позволяя или способствуя опылению желательной пыльцой. Существует множество способов предупреждения самоопыления женского родительского растения, причем в одном способе самоопыление предупреждают механическим удалением органа, продуцирующего пыльцу, перед выделением пыльцы. Коммерческое получение гибридных семян маиса (маис, Zea mays) обычно включает в себя посадку желаемых мужских и женских родительских линий, обычно отдельными рядами или блоками на выделенном поле, обработку женских родительских растений для предупреждения поллинации, гарантии опыления женских родительских растений только пыльцой предназначенных мужских растений и сбора гибридных семян только у женских родительских растений. Гибридные семена могут быть результатом однократного скрещивания (например, первым поколением при скрещивании между двумя инбредными линиями), модифицированного однократного скрещивания (например, первым поколением при скрещивании между двумя инбредными линиями, одна или другая из которых могли быть немного модифицированы с применением близкородственного скрещивания), двойного скрещивания (например, первого поколения скрещивания между двумя однократными скрещиваниями), тройного скрещивания (например, первого поколения скрещивания между однократным скрещиванием и инбредной линией), линейного скрещивания (например, первым поколением скрещивания между инбредной линией и сорт-популяцией, или первым поколением скрещивания между однократным скрещиванием и сорт-популяцией) или является сорт-популяцией (например, популяцией растений, отобранных по стандарту, который может иметь вариации, но обладает характеристиками, по которым сорт можно отличить от других сортов).
При получении гибридных семян, продукцию и/или выделение пыльцы женским родительским растением можно предупредить, для того чтобы способствовать опылению женского родительского растения только предназначенным мужским родительским растением и, таким образом, получить гибридные семена. Этого можно достичь любым способом или средством, известным опытным специалистам в данной области, включая без ограничений, ручное удаление (или удаление руками) метелок, механическое удаление метелок, использование генетических средств контроля опыления и/или использование химического агента. Любые из них можно объединять или использовать индивидуально. Удаление метелок можно осуществлять вручную или руками, и обычно его проводит человек, обрывая метелки у растения маиса, обычно отделяя метелки. При механическом или машинном удалении метелок обычно используют машину для удаления метелок, называемую «косилкой», которая движется по рядам маиса и срезает верхнюю часть растений. Затем через несколько дней машина «вытягиватель» движется по рядам маиса и обрывает метелки у растений, захватывая их между двумя валками, движущимися с высокой скоростью. Машины для удаления метелок, пригодные в практическом применении способов по настоящему изобретению, включают эти устройства, установленные на машины с высоким клиренсом. Косилкой может быть вращающееся лезвие или вращающейся нож, который режет в различных плоскостях от горизонтальной до вертикальной, регулируемый по высоте, для срезания или отрезания верхушки растения маиса, включая метелку. Вытягиватель может представлять собой два небольших колеса или валка, регулируемых по высоте, которые вращаются в противоположных направлениях и захватывают метелки и верхние листья, вытягивая их вверх, аналогично ручному удалению метелок. Вытягиватели и косилки можно использовать отдельно или совместно и/или в комбинации с другими способами удаления метелок. Временное окно для удаления метелок обычно является наиболее критичным и трудным в организации периодом при продукции гибридных семян маиса. В данной области также используют химические агенты и/или генетические средства для предупреждения образования жизнеспособной пыльцы или выделения пыльцы.
Изобретение относится к способу определения времени удаления метелок путем выбора диапазона на Относительной шкале развития, причем выбранный диапазон указывает на созревание до желаемой стадии развития метелки. Желаемой стадией развития метелки является стадия с Т0 до Т7, например, стадия Т5. Стадией развития метелки, представляющей особый интерес, является стадия развития метелки, оптимальная для репродуктивного скрещивания, стадия развития метелки, оптимальная для стерилизации метелок или удаления метелок, и/или стадия, оптимальная для проведения модулирующей развитие обработки на растении маиса. При создании и использовании Относительной шкалы развития определенной стадией развития цветков, используемой для создания Относительной шкалы развития, может быть выделение пыльцы примерно 50 процентами популяции растений маиса. Примером диапазона на Относительной шкале развития, который можно использовать со способом по изобретению, является диапазон от примерно 0,62 и до примерно 0,75 на Относительной шкале развития.
Определение времени развития метелки может быть полезно для агротехнических способов, включая планирование и/или стандартизацию способов практического применения, которые связаны с развитием растений. Примеры включают: способы, требующие применения химического агента по времени, такие как применение гербицида, фунгицида, фертилайзера и/или регулятора роста для инбредных растений, имеющих противоположные группы спелости; способы, требующие мониторинга, прогнозирования и/или регулирования развития метелки, такие как мониторинг мужских инбредных растений по раннему развитию метелки, которое может привести к снижению выделения пыльцы, и обеспечение соответствующей обработки для влияния на развитие метелки; способы, требующие применение гормона и/или регулятора роста по времени для коррекции нарушений и/или для получения желаемого агротехнического результата; и/или любые способы, требующие применения модулирующей развитие обработки на растения маиса на указанной желательной стадии развития метелки. Изобретение можно использовать для полевого планирования и/или исследовательской работы, например, для прогнозирования требуемого объема работы, связанной с удалением метелок или развитием растений; для ожидания требований, связанных с развитием метелок; для определения того, как стресс влияет на развитие метелок; и/или для применения в скрининге и оценке признаков, и/или инбредных растений или гибридов при наложении стресса в определенные стадии развития, определенные с помощью прогнозирования стадий развития метелки.
Изобретение относится к способам, которые можно использовать для определения времени, когда обработка, модулирующая развитие, является оптимально эффективной, с использованием Относительной шкалы развития. Используемый в настоящем документе термин «модулирующая развитие обработка» относится к применению по меньшей мере одного физического воздействия и/или химического агента, которые влияют на развитие растения. Развитие растения включает, но не ограничено этим, развитие цветков, развитие корней, развитие листьев, развитие стеблей, развитие метелок, развитие репродуктивных органов, развитие гамет, развитие пыльцы, развитие семян и/или развитие любой другой части растения. Модулирующая обработка может вызывать остановку, замедление, предупреждение, задержку или усиление развития. Модулирующей развитие обработкой может быть физическое воздействие, такое как удаление метелок, обжигание (использование горелки для обжигания верхушек мужских растений как средства замедления созревания) и/или обдирание, истирание, соскабливание, надсекание, отрезание, прокалывание, ультразвуковая обработка, отсоединение, разрушение, удаление, измельчение, обрезка и/или укрывание любой части растения. Обработкой, модулирующей развитие, может быть химический агент, такой как природные или синтетические соединения. Химические агенты, которые могут быть полезны в качестве модулирующей развитие обработки включают регуляторы роста растений, ингибиторы регуляторов роста растений, растительные гормоны, ингибиторы растительных гормонов, стимуляторы роста растений, замедлители роста растений, фунгициды, инсектициды, гербициды, ауксины, антиауксины, цитокинины, дефолианты, ингибиторы этилена, соединения, способствующие высвобождению этилена, гиббереллины, морфактины и гаметоциды. Примером физического воздействия для применения в способах по изобретению является удаление метелок и/или обжигание. Примером химического агента для использования в способах по изобретению является гербицид глифосат. Модулирующую развитие обработку можно применять на любой стадии развития растения маиса, например, когда стадия развития метелки соответствует диапазону от 0,62 до 0,75 по Относительной шкале развития, который включает в себя стадию развития метелки Т5. Способность идентифицировать оптимально эффективный период для применения обработки, модулирующей развитие метелки, с использованием приведенной Относительной шкалы развития является одним аспектом изобретения. В данной области известно множество обработок, модулирующих развитие метелки, включая химические агенты, способные предупреждать развитие пыльцы или выделение пыльцы, которые были бы полезны в применении на практике способов по изобретению.
Изобретение можно применять для получения гибридных семян с использованием способов по изобретению. Изобретение относится к способам, при помощи которых первое родительское растение маиса скрещивают с вторым родительским растением маиса, причем продукция пыльцы первым родительским растением маиса ингибирована в результате применения модулирующей развитие обработки. Способы по изобретению можно использовать для определения стадии развития и/или времени, когда применение модулирующей развитие обработки будет максимально эффективным. Изобретение может быть особенно полезным в способах, приведенных в патенте США № 6762344 и публикации патентной заявки США № 2009/0165166. В одном варианте осуществления изобретение представляет собой гибридное семя, полученное с использованием способов по изобретению, включая растения и части растения, выращенного из гибридного семени и продукты потребления, произведенные из них.
Изобретение относится к способу применения растений трансгенного события MON 87427 маиса для продукции гибридных семян, причем глифосат используют в качестве обработки, модулирующей развитие метелки, у растений MON 87427 для предупреждения образования пыльцы. Это обусловлено возможностью предупреждать выделение пыльцы женскими родительскими линиями, содержащими MON 87427, своевременным применением глифосата, таким образом, предупреждая самоопыление. Если применять глифосат слишком рано относительно развития метелок, то мужские репродуктивные тельца могут недостаточно развиться, чтобы обработка была полностью эффективной. Если применять глифосат слишком поздно относительно развития метелки, то может уже происходить выбрасывание пыльников, и обработка глифосатом может быть не способна предупредить развитие пыльцы. Поэтому, время применения глифосата относительно развития метелки важно для гарантии максимальной эффективности и, поэтому, максимальной чистоты получаемых гибридных семян. В одном варианте осуществления изобретения оптимальное время применения глифосата для этого способа можно идентифицировать, определяя время развития метелки растений MON 87427 с использованием Относительной шкалы развития и выбора диапазона на Относительной шкале развития, который указывает на достижение желаемой стадии развития метелки. Это определение времени развития метелки затем можно использовать для идентификации времени применения глифосата в качестве модулирующей развитие обработки на растении MON 87427, таким образом, предупреждая самооплодотворение и увеличивая продукцию гибридных семян. Способы по изобретению выявляют, что развитие метелки в диапазоне от 0,62 до 0,75 по Относительной шкале развития или Т5, является оптимальным диапазоном для применения глифосата на растениях MON 87427 для предупреждения образования пыльцы. Поэтому, в системе гибридизации Roundup® для любой данной родительской женской линии любой данной группы спелости оптимальное время будет прогнозироваться умножением GDU, требуемых для достижения P50 для этой родительской женской линии на любое значение в пределах этого диапазона. Когда результат этого вычисления будет равен GDU данного вегетационного сезона, то будет выяснено оптимальное время применения глифосата. Относительные шкалы развития можно получать с использованием других показателей выделения пыльцы, и других показателей Т-шкалы, которые можно будет применять, не выходя за приделы объема изобретения.
Еще один аспект изобретения относится к способу, который полезен для определения стадии развития метелки, при которой репродуктивное скрещивание является оптимальным. Аналогично тому, как различия в развитии по генотипам препятствуют достоверному прогнозированию оптимальных модулирующих обработок, основанных только на V-стадиях или GDU, определение времени для перекрестного опыления с помощью Относительной шкалы развития также может быть эффективным. Простое исследование с использованием Относительной шкалы развития может выявить, что перекрестное опыление является оптимальным в некотором диапазоне на этой шкале. Кроме того, зная GDU, требуемые для Р50 данного генотипа, можно будет точно определить время достижения диапазона, не полагаясь на недостоверные вегетативные показатели или не проводя времязатратную физическую оценку развития метелки. Описания способов скрещивания, которые обычно используются для различных признаков и зерновых, можно найти в одном из нескольких справочников

При применении на практике способов по изобретению одно или оба родительских растений маиса могут содержать один или несколько желаемых признаков, представляющих агротехнический интерес. Например, родительское растение маиса MON 87427 можно использовать в продукции гибридных семян для скрещивания со вторым родительским растением маиса, которое содержит по меньшей мере один ген и/или признак, представляющие агротехнический интерес. В этом варианте осуществления изобретения Относительную шкалу развития можно использовать для мониторинга и/или определения репродуктивной стадии развития родительского растения маиса MON 87427 для точного определения времени обработки родительского растения маиса MON 87427 глифосатом до образования пыльцы, таким образом предупреждая самооплодотворение. Растения события MON 87427 затем опылялись бы вторым родительским растением маиса, и гибридные семена маиса собирали бы от растений события MON 87427, причем семена, собранные от обработанных растений MON 87427 имели больший выход гибридных семян маиса (то есть, более высокое процентное содержание собираемых гибридных семян или более высокую чистоту гибридных семян), чем семена маиса, собранные от необработанных или обработанных в несоответствующее время растений события MON 87427 или от другого(их) растения(й) маиса в таких же условиях.
Признаки и гены, представляющие агротехнический интерес, хорошо известны в данной области и включают, но не ограничены этим, например, гены устойчивости к гербицидам, мужской стерильности, повышения урожайности, контроля насекомых, устойчивости к грибковым заболеваниям, устойчивости к вирусам, устойчивости к нематодам, устойчивости к бактериальным заболеваниям, роста и развития растений, продукции крахмала, модифицированной и/или высокой продукции масла, модифицированного содержания жирных кислот, продукции большого количества белка, созревания фруктов, увеличенной питательности для животных и/или человека, биополимеров, устойчивости к стрессу, вызываемому окружающей средой, гены фармацевтических пептидов и секретируемых пептидов, гены, признаков, улучшающих переработку, гены повышенной перевариваемости, низкого содержания раффинозы, промышленной продукции ферментов, улучшенного аромата, фиксации азота, продукции гибридных семян, продукции клетчатки и/или продукции биотоплива. Примерами растений, имеющих один или несколько желаемых признаков являются зарегистрированные в Службе по контролю за здоровьем животных и растений (APHIS) Департамента сельского хозяйства США как устойчивые к гербицидам (например, события маиса MON 88017, NK603, DAS-68416-4, HCEM485, DP-098140-6, P-356043-5, MIR604, 59122, TC-6275, Line 1507, MON 802, T14 и/или T25), как устойчивые к насекомым (например, события маиса MON 863, MON 809, MON 810, MON 89034, MON 88017, MON 802, MIR-162, TC-6275, DBT418, B16, TC-1507, DAS 59122-7, MIR604 и/или MON 80100), и/или имеющие другие желаемые признаки (например, события маиса LY038, MON 87460 и/или 3272) (полный список и описание таких признаков доступен на веб-сайте Правительства США, http://www.aphis.usda.gov/brs/not_reg.html).
В применении на практике способов по изобретению можно использовать инбредную линию, сорт или гибрид (или любой другой генотип). Например, элитной инбредной линией является линия растений маиса, которая получена скрещиванием и отбором по высочайшим агротехническим показателям. Генотипы могут быть трансформированы и/или использованы в способах скрещивания, чтобы включить в них ген, представляющий агротехнический интерес, такой как ген устойчивости к глифосату, и события можно отбирать по их пригодности в качестве женского или мужского родительского растения в системе продукции гибридных семян.
Элитные генотипы маиса для использования в практическом применении изобретения включают, но не ограничены этим: CI9805 (патентная публикация США № 20030093826); LH321 (патентная публикация США № 20030106086); H0I002 (патентная публикация США № 20030154524); HOI001 (патентная публикация США № 20030172416); 5750 (патентная публикация США № 20030177541); G0502 (патентная публикация США № 20030177543); G1102 (патентная публикация США № 20030177544); HX879 (патентная публикация США № 20040068771); 6803 (патентная публикация США № 20040088767); 5020 (патентная публикация США № 20040088768); G3001 (патентная публикация США № 20040098768); LH268 (патентная публикация США № 20040111770); LH311 (патентная публикация США № 20040111771); LH306 (патентная публикация США № 20040111772); LH351 (патентная публикация США № 20040111773); LHE323 (патентная публикация США № 20040111774); 402A (патентная публикация США № 20040123352); 366C (патентная публикация США № 20040139491); NP2315 (патентная публикация США № 20040143866); PH0GC (патентная публикация США № 20040194170); SE8505 (патентная публикация США № 20050015834); D201 (патентная публикация США № 20050028236); BE1146BMR (патентная публикация США № 20050076402); PHCAM (патентная публикация США № 20050114944); PHCK5 (патентная публикация США № 20050114945); PHC77 (патентная публикация США № 20050114951); PHCND (патентная публикация США № 20050114952); PHCMV (патентная публикация США № 20050114953); PHB00 (патентная публикация США № 20050114955); PHCER (патентная публикация США № 20050114956); PHCJP (патентная публикация США № 20050120437); PHADA (патентная публикация США № 20050120439); PHB8V (патентная публикация США № 20050120443); 6XN442 (патентная публикация США № 20050125864); 4XP811 (патентная публикация США № 20050125865); PHCCW (патентная публикация США № 20050125866); MN7224 (патентная публикация США №20050132433); BE9514 (патентная публикация США № 20050132449); PHCA5 (патентная публикация США № 20050138697); PHCPR (патентная публикация США № 20050144687); PHAR1 (патентная публикация США № 20050144688); PHACV (патентная публикация США № 20050144689); PHEHG (патентная публикация США № 20050144690); NP2391 (патентная публикация США № 20050160487); PH8WD (патентная публикация США № 20050172367); D501 (патентная публикация США № 20050177894); D601 (патентная публикация США № 20050177896); D603 (патентная публикация США № 20050177904); PHCEG (патентная публикация США № 20050223443); W16090 (патентная публикация США № 20050273876); M10138 (патентная публикация США №. 20050273877); N61060 (патентная публикация США № 20050273878); NP2460 (патентная публикация США № 20060048243); BS112 (патентная публикация США № 20060070146); PHDWA (патентная публикация США № 20060107393); PH8JV (патентная публикация США № 20060107394); PHEWW (патентная публикация США № 20060107398); PHEDR (патентная публикация США № 20060107399); PHE67 (патентная публикация США № 20060107400); PHE72 (патентная публикация США № 20060107408); PHF1J (патентная публикация США № 20060107410); PHE35 (патентная публикация США № 20060107412); PHEHR (патентная публикация США № 20060107415); PHDPP (патентная публикация США № 20060107416); PHEHC (патентная публикация США № 20060107418); PHANF (патентная публикация США № 20060107419); PHC78 (патентная публикация США № 20060107420); PH8T0 (патентная публикация США № 20060107421); PHDRW (патентная публикация США № 20060107422); PHEGV (патентная публикация США № 20060107423); PHEBA (патентная публикация США № 20060107426); PHENE (патентная публикация США № 20060112463); PHEJW (патентная публикация США № 20060112464); PHAPT (патентная публикация США № 20060112465); PHCND (патентная публикация США № 20060130188); PHCEG (патентная публикация США № 20060130189); PHADA (патентная публикация США № 20060130190); PHEED (патентная публикация США № 20060143744); PHHB (патентная публикация США № 5633427); LH262 (патент США № 5633428); LH227 (патент США № 5633429); LH226 (патент США № 5639941); LH235 (патент США № 5639942); LH234 (патент США № 5639943); PHDP0 (патент США № 5639946); PH06N (патент США № 5675066); LH177 (патент США № 5684227); PH24E (патент США № 5689034); PHP38 (патент США № 5708189); ASG06 (патент США № 5714671); CG00685 (патент США № 5723721); PHND1 (патент США № 5723722); PH44A (патент США № 5723723); ZS01591 (патент США № 5723724); ZS01101 (патент США № 5723725); ZS01452 (патент США № 5723726); ZS01429 (патент США № 5723727); ZS01819 (патент США № 5723728); ZS01250 (патент США № 5723729); ZS01595 (патент США № 5723730); CG3ND97 (патент США № 5728923); NP938(934) (патент США № 5728924); PHNG2 (патент США № 5731491); CG5NA58 (патент США № 5731502); NP948 (патент США № 5731503); LH236 (патент США № 5731504); CG00766 (патент США № 5731506); PHOAA (патент США № 5750829); PH15A (патент США № 5750830); PH25A (патент США № 5750831); PH44G (патент США № 5750832); PH0CD (патент США № 6084160); ASG25 (патент США № 6084161); 86ISI15 (патент США № 6084162); BE4547 (патент США № 6084163); PH21T (патент США № 6091007); 01DHD16 (патент США № 6096952); PH224 (патент США № 6096953); ASG26 (патент США № 6103958); ASG28 (патент США № 6103959); PH0V0 (патент США № 6107550); 90LCL6 (патент США № 6111171); 22DHD11 (патент США № 6111172); ASG17 (патент США № 6114606); AR5253bm3 (патент США № 6114609); ASG27 (патент США № 6114610); WDHQ2 (патент США № 6114611); PH3GR (патент США № 6114613); PH1NF (патент США № 6118051); PH0JG (патент США № 6118053); PH189 (патент США № 6118054); PH12J (патент США № 6118055); PH1EM (патент США № 6118056); 90DJD28 (патент США № 6121519); PH12C (патент США № 6121520); PH55C (патент США № 6121522); PH3EV (патент США № 6121523); ZS4199 (патент США № 6121525); PH2V7 (патент США № 6124529); PH4TF (патент США № 6124530); PH3KP (патент США № 6124531); PH2MW (патент США № 6124532); PH2N0 (патент США № 6124533); PH1K2 (патент США № 6124534; PH226 (Патент США № 6124535); PH2VJ (Патент США № 6127609); PH1M8 (Патент США № 6127610); WQCD10 (Патент США № 6130369); PH1B8 (патент США № 6130370); 17DHD5 (патент США № 6133512); PH0WD (патент США № 6133513); PH3GK (патент США № 6133514); PH2VK (патент США № 6137036); PH1MD (патент США № 6137037); SM4603 (патент США № 6137038); PH04G (патент США № 6140562); NP2151 (патент США № 6140563); PH5DR (патент США № 6727413); LH254 (патент США № 6730833); PH5WB (патент США № 6730834); PH7CH (патент США № 6730835); PH54M (патент США № 6730836); PH726 (патент США № 6730837); PH48V (патент США № 6734348); PH3PV (патент США № 6737566); PH77V (патент США № 6740795); PH7JB (патент США № 6740796); NP2316 (патент США № 6740797); PH70R (патент США № 6740798); RAA1 (патент США № 6747194); VMM1 (патент США № 6747195); PH3RC (патент США № 6747196); MNI1 (патент США № 6753465); 5750 (патент США № 6756527); PH6KW (патент США № 6756528); PH951 (патент США № 6756530); PH6ME (патент США № 6759578); NP2171 (патент США № 6759579); PH87H (патент США № 6759580); PH26N (патент США № 6765132); RII1 (патент США № 6765133); PH9AH (патент США № 6770802); PH51H (патент США № 6774289); PH94T (патент США № 6774290); PH7AB (патент США № 6777599); PH5FW (патент США № 6781042); PH75K (патент США № 6781043); KW7606 (патент США № 6784348); PH8CW (патент США № 6784349); PH8PG (патент США № 6784350); RBOl (патент США № 6797869); 9SM990 (патент США № 6803509); PH5TG (патент США № 6806408); 1501150 (патент США № 6806409); 1390186 (патент США № 6806410); PH6JM (патент США № 6809240); KW4636 (патент США № 6809243); 1363128 (патент США № 6809244); LH246 (патент США № 6812386); 2JK221 (патент США № 6812387); PHN46 (патент США № 5567861); ZS0223 (патент США № 5569813); phajo (патент США № 5569816); PHJJ3 (патент США № 5569817); phap8 (патент США № 5569818); PHPP8 (патент США № 5569819); ZS1284 (патент США № 5569820); PHT11 (патент США № 5569821); phte4 (патент США № 5569822); ZS0114 (патент США № 5569826); 7054 (патент США № 5576473); ZS0560 (патент США № 5585533); ZS0853 (патент США № 5585534; ZS1791 (патент США № 5585539); ZS1513 (патент США № 5585541); ZS1679 (патент США № 5589606); ZS1022 (патент США № 5602314); ZS1202 (патент США № 5602315); ZS1783 (патент США № 5602316); PHDG1 (патент США № 5602318); PHKV1 (патент США № 5608138); PH05F (патент США № 5608139); PH38B (патент США № 5608140); PH42B (патент США № 5618987); PHDD6 (патент США № 5625129); ZS0541 (патент США № 5625131); PH08B (патент США № 5625132); PHOC7 (патент США № 5625133); LH233 (патент США № 5625135); ASG05 (патент США № 5723731); LH281 (патент США № 5723739); PHBF0 (патент США № 5728919); CG5NA01 (патент США № 5728922); AR5651bm3 (патент США № 5977458); LH266 (патент США № 5977459); LH303 (патент США № 5977460); LH301 (патент США № 5981855); 4SQ601 (патент США № 5986182); PH1TB (патент США № 5986184); PH24D (патент США № 5986185); LH229 (патент США № 5986186); LH277 (патент США № 5986187); PH1CN (патент США № 5990393); LH261 (патент США № 5990394); W1498A (патент США № 5990395); WQDS2 (патент США № 5994631); NL085B (патент США № 5998710); PH09E (патент США № 5998711); 5 LH284 (патент США № 6015944); PH1B5 (патент США № 6020543); PH1CA (патент США № 6025547); 70LDL5 (патент США № 6031160); GM9215 (патент США № 6031161); 90LDI1 (патент США № 6031162); 90LDC2 (патент США № 6034304); 90QDD1 (патент США № 6034305); R398D (патент США № 6034306); RDBQ2 (патент США № 6037531); HX621 (патент США № 6040506); HX622 (патент США № 6040507); 01HG12 (патент США № 6040508); HX740 (патент США № 6043416); 79314N1 (патент США № 6043417); 17INI20 (патент США № 6043418); 17DHD7 (патент США № 6046387); 83INI8 (патент США № 6046388); 83InI14 (патент США № 6046389); 01INL1 (патент США № 6046390); LH286 (патент США № 6049030); ASG29 (патент США № 6054640); ASG07 (патент США № 6060649); QH111 (патент США № 6069303); 09DSQ1 (патент США № 6072108); JCRNR113 (патент США № 6072109); NP2029 (патент США № 6072110); ASG09 (патент США № 6077996); PHOWE (патент США № 6077997); 86AQV2 (патент США № 6077999); PH1GG (патент США № 6080919); RPK7346 (патент США № 6506965); NP2044BT (патент США № 6573438); PH8W4 (патент США № 6600095); M42618 (патент США № 6617500); MV7100 (патент США № 6624345); 3JP286 (патент США № 6627800); BE4207 (патент США № 6632986); CI9805 (патент США № 6632987); JCR503 (патент США № 6635808); NR401 (патент США № 6635809); 4VP500 (патент США № 6635810); 7SH385 (патент США № 6642440); KW4773 (патент США № 6642441); NP2073 (патент США № 6646187); PSA104 (патент США № 6646188); 5XH755 (патент США № 6653536); 1445-008-1 (патент США № 6653537); NP2015 (патент США № 6657109); 7SH383 (патент США № 6660916); LH310 (патент США № 6664451); I880S (патент США № 6670531); RR728-18 (патент США № 6677509); LH320 (патент США № 6683234); 11084BM (патент США № 6686519); W60028 (патент США № 6686520); PH1GD (патент США № 6693231); LH295 (патент США № 6693232); PH1BC (патент США № 6700041); PH4V6 (патент США № 6706954); NP2276 (патент США № 6706955); NP2222 (патент США № 6710233); Ph0R8 (патент США № 6717036); PH581 (патент США № 6717037); PH6WR (патент США № 6717038); PH5HK (патент США № 6717039); PH5W4 (патент США № 6717040); PH0KT (патент США № 6720486); PH4GP (патент США № 6720487); PHJ8R (патент США № 6723900); NP2052 (патент США № 6723901); PH7CP (патент США № 6723902); PH6WG 15 (патент США № 6723903); PH54H (патент США № 6727412); 4P33339 (патент США № 5489744); PHKM5 (патент США № 5491286); LH225 (патент США № 5491293); LH185 (патент США № 5491294); LH176 (патент США № 5491296); PHW06 (патент США № 5495065); LH252 (патент США № 5495067); LH231 (патент США № 5495068); PHTE4 (патент США № 5495069); PHP38 (патент США № 5506367); PHN82 (патент США № 5506368); PHTD5 (патент США № 5527986); 899 (патент США № 5530181); PHAP1 (патент США № 5530184); PHKW3 (патент США № 5534661); CG00653 (патент США № 5536900); PHRD6 (патент США № 5541352); PHK46 (патент США № 5543575); PHBG4 (патент США № 5545809); LH189 (патент США № 5545811); PHNJ2 (патент США № 5545812); PHRF5 (патент США № 5545813); PHFR8 (патент США № 5545814); PHN18 (патент США № 5557034); PHTP9 (патент США № 5557038); PH54B (патент США № 5563320); PHGF5 (патент США № 5563321); PHAG6 (патент США № 5563322); PHAP9 (патент США № 5563323); PHBE2 (патент США № 5563325); ZS0510 (патент США № 5563327); PHAAO (патент США № 5602317); LH273 (патент США № 5880348); 7571 (патент США № 5880349); LH237 (патент США № 5880350); PH0B4 (патент США № 5889188); FEBS (патент США № 5902922); 8F286 (патент США № 5905191); 3AZA1 (патент США № 5910625); 91DFA-5 (патент США № 5910635); ASG20 (патент США № 5910636); ZS03940 (патент США № 5912420); 91ISI6 (патент США № 5912421); MF1113B (патент США № 5914452); PH03D (патент США № 5917125); PHDN7 (патент США № 5917134); 01DIB2 (патент США № 5920003); 82DHB1 (патент США № 5922935); 8M222 (патент США № 5922936); PHMJ2 (патент США № 5929313); SBB1 (патент США № 5932787); 86ISI3 (патент США № 5932788); ZS01231 (патент США № 5936144); 87DIA4 (патент США № 5936145); 79310J2 (патент США № 5936146); PH1GC (патент США № 5936148); 01DHD10 (патент США № 5939606); PH2CB (патент США № 5939607); PH080 (патент США № 5939608); PH14T (патент США № 5942670); PH185 (патент США № 5942671); PH 19V (патент США № 5948957); ZS09247 (патент США № 5952551); CRAUGSH2W-89 (патент США № 5952552); 91DHA1 (патент США № 5962770); LH300 (патент США № 5965798); 91ISI4 (патент США № 5965799); 79103A1 (патент США № 5969212); ASG22 (патент США № 5969220); 82IUH1 (патент США № 5969221); (патент США № 5969222); LH302 (патент США № 5973238); LH265 (патент США № 5973239); PHFW4 (патент США № 5977451); 01IBH10(патент США № 5977452); 91CSI-1 (патент США № 5977453); WKBC5 (патент США № 5977455); PH1M7 (патент США № 5977456); R327H (патент США № 6399860); FR2108 (патент США № 6407320); FR3383 (патент США № 6410830); IT302 (патент США № 6414227); FR3303 (патент США № 6414228); 9034 (патент США № 6420634); G1500 (патент США № 6420635); FR3311 (патент США № 6420636); 1389972 (патент США № 6420637); PH77C (патент США № 6423888); IT201 (патент США № 6426451); G3000 (патент США № 6426453); 94INK1B (патент США № 6429363); PH3HH (патент США № 6433259); 6TR512 (патент США № 6433260); 89AHD12 (патент США № 6433261); 1889291 (патент США № 6433262); 2070BT (патент США № 6437223); 3323 (патент США № 6437224); G1900 (патент США № 6441279); 16IUL6 (патент США № 6441280); 7RN401 (патент США № 6444881); UBB3 (патент США № 6444882); 6077 (патент США № 6444883); 1014738 (патент США № 6444884); TDC1 (патент США № 6452074); GF6151 (патент США № 6452075); 7180 (патент США № 6452076); WQDS7 (патент США № 6455764); X532Y (патент США № 6459021); 1465837 (патент США № 6459022); 1784S (патент США № 6469232); LH176Bt810 (патент США № 6469233); 6RC172 (патент США № 6469234); 3327 (патент США № 6469235); 7SH382 (патент США № 6476298); 1181664 (патент США № 6476299); NP2010 (патент США № 6483014); FR3361 (патент США № 6483015); 1778S (патент США № 6486386); 1362697 (патент США № 6492581); RPK7250 (патент США № 6506964); и 6RT321 (патент США № 6911588).
Используемый в настоящем документе термин «включающий» означает «включающий, но не ограниченный этим».
Следующие примеры включены для демонстрации примеров некоторых предпочтительных вариантов осуществления изобретения. Опытным специалистам в данной области следует понимать, что методики, раскрытые в нижеследующих примерах, представляют подходы, которые, как обнаружили авторы изобретения, функционируют хорошо в применении изобретения на практике, и, поэтому, могут рассматриваться в качестве составляющих примеров предпочтительных вариантов осуществления изобретения на практике. Однако опытным специалистам в данной области в свете настоящего раскрытия изобретения должно быть ясно, что можно осуществить множество изменений в конкретных вариантах осуществления, которые раскрыты в описании, и все еще получить подобный или аналогичный результат, не отступая от сути и объема изобретения.
ПРИМЕРЫ
Пример 1: трансформация маиса и отбор события MON 87427
Растение маиса MON 87427 получали Agrobacterium-опосредованной трансформацией маиса. Клетки маиса были трансформированы и регенерированы в целые растения маиса, и из популяции растений были отобраны отдельные растения, которые показывали сохранность трансгенной экспрессионной кассеты и устойчивость к глифосату. Из этой популяции было отобрано и охарактеризовано растение события MON 87427 маиса.
Трансгенное устойчивое к глифосату растение маиса MON 87427 было получено посредством Agrobacterium-опосредованной трансформации незрелых зародышей маиса с использованием трансформирующего вектора pMON58401. Трансгенная вставка и экспрессионная кассета из MON 87427 включает в себя промотор и лидер из гена 35S-РНК вируса мозаики цветной капусты (CaMV), содержащий дуплицированную энхансерную область (P-e35S); функционально связанные с ДНК-лидером из первого интрона гена белка теплового шока 70 (I-HSP70) маиса; функционально связанным с молекулой ДНК, кодирующей N-концевой пептид-переносчик в хлоропласты из гена shkG, кодирующего EPSPS из Arabidopsis thaliana (Ts-CTP2); функционально связанной с молекулой ДНК из гена aroA из штамма CP4 агробактерий и кодирующего белок CP4 EPSPS; функционально связанной с молекулой ДНК 3′-UTR из гена нопалин-синтетазы (T-NOS) из Agrobacterium tumefaciens.
Клетки маиса можно трансформировать различными способами. Например, для получения трансгенного растения маиса, содержащего кассету для экспрессии в растениях по изобретению, можно использовать следующий способ. Жидкие культуры Agrobacterium tumefaciens, содержащие кассету для экспрессии в растениях, инициировали из законсервированных в глицерине культур или с чашки со свежими штрихами бактерий и выращивали в течение ночи при 26-28°C с качанием (приблизительно 150 оборотов в минуту, об/мин) до середины логарифмической фазы роста в жидкой среде LB, рН 7,0, содержащей 50 мг/л (миллиграммов на литр) канамицина и либо 50 мг/мл стрептомицина, либо 50 мг/л спектиномицина, и 25 мг/л хлорамфеникола с 200 мкМ ацетосирингоном (AS). Клетки агробактерий ресуспендировали в среде для заражения (жидкая СМ4С, описанная в таблице 8 патента США № 6573361), и плотность клеток регулировали таким образом, что ресуспендированные клетки имели оптическую плотность 1 при измерении в спектрофотометре при длине волны 660 нм (то есть, OD660). Свежевыделенные незрелые зародыши заражали агробактериями и совместно культивировали 2-3 дня в темноте при 23°C. Затем зародыши помещали в замедляющую среду (N6 1-100-12, описана в таблице 1 патента США № 5424412) с добавленными карбенициллином (500 мг/л, Sigma-Aldrich, St Louis, Mo.) и 20 мкМ AgNO3 и инкубировали при 28°C в течение 4-5 дней. Все последующие культуры содержали при данной температуре.
Зародышей переносили в первую среду для отбора (N61-0-12, описана в таблице 1 патента США № 5424412) с добавленными карбенициллином (500 мг/л) и 1,0 мМ глифосатом. Через две недели выжившие ткани переносили во вторую среду для отбора (N61-0-12) с добавленными карбенициллином (500 мг/л) и 1,0 мМ глифосатом. Выживший каллюс субкультивировали каждые 2 недели примерно 3 раза на 1,0 мМ глифосате. После идентификации устойчивых к глифосату тканей, ткань наращивали для регенерации. Для регенерации каллюсные тккани переносили в регенерационную среду (MSOD, описана в таблице 1 патента США № 5424412) с добавленной 0,1 мМ абсцизовой кислотой (ABA; Sigma-Aldrich, St Louis, Mo.) и инкубировали в течение двух недель. Регенерирующиеся каллюсы переносили в среду с высоким содержанием сахарозы и инкубировали в течение двух недель. Ростки переносили в среду MSOD (без АВА) в культуральном сосуде и инкубировали в течение двух недель. Растения с корнями и нормальными фенотипическими характеристиками отбирали и переносили в почву для роста и дальнейшей оценки. Растения R0, созданные с помощью вышеуказанной трансформации, переносили в почву для роста и последующего самоопыления для получения семян R1. Растения отбирали комбинацией аналитических методик, включая TaqMan, ПЦР-анализ и опрыскивание гербицидом.
Событие MON 87427 отбирали из 45 индивидуальных трансгенных событий на основе многолетних анализов, демонстрирующих превосходные фенотипические и молекулярные характеристики события и его желательную ассоциацию с гаплотипом (хромосома 9, 60 cM). Процесс отбора для события MON 87427 начинали с трансформированных растений маиса, представляющих 45 событий R0. Их опрыскивали глифосатом (64 унции/акр (1,81 кг/4047 м2) на стадии V7) и затем оценивали по вегетативной устойчивости и стерильности после опрыскивания глифосатом. Из исходных 45 событий, 35 событий R0 проявляли вегетативную устойчивость к глифосату и мужскую стерильность после опрыскивания тестируемой концентрацией глифосата в указанное время. Проводили дополнительную характеристику растений этих 35 событий R0 с помощью молекулярного анализа. 35 событий R0 были молекулярно охарактеризованы с использованием ПЦР-анализа Taqman® и Саузерн-блот анализа. Из 35 проанализированных событий 29 были отобраны для дальнейшей работы и тестирования в поле. 29 событий R1 были проанализированы в полевых испытаниях на полевую эффективность и урожайность. Кроме этого, были проведены дополнительные молекулярные характеристики, включая геномную характеристику и характеристику экспрессии белка. Данные были проанализированы, и исходя из всестороннего анализ растений R1 и результатов полевых испытаний были отобраны три лидирующих события и выдвинуты для полевого тестирования R2. Последующий анализ и тестирование этих трех лидирующих событий привели к отбору события MON 87427.
Дополнительный полевой скрининг включал обработку события MON 87427 глифосатом в количестве 32 унции/акр (907 г/4047 м2) (Roundup Ultra®, Monsanto Co., St. Louis, Mo.) на вегетативной стадии роста 4 (V4) и вегетативной стадии роста (V10). Подсчет положительного и отрицательного ответа растений проводили после опрыскивания на стадии V4. Кроме того, у обработанных растений проводили оценку хлороза и нарушений развития через 10-14 дней после обработки (DAT) как на стадии V4, так и на стадии V10. Степень стерильности метелок в момент цветения также оценивали для растений, опрысканных на стадии V4 и V10.
Пример 2: характеристика последовательностей ДНК MON 87427
ДНК, встроенная в геном растения маиса MON 87427, и фланкирующая геномная последовательность были подробно охарактеризованы методами молекулярного анализа. Эти методы анализа включали: секвенирование трансгенной вставки ДНК и геномной ДНК, фланкирующей трансгенную вставку, определение числа трансгенных вставок (числа сайтов интеграции в геноме маиса), определение числа копий (числа копий трансгенной ДНК в одном локусе), анализ целостности встроенной генной кассеты, анализ геномной ДНК, фланкирующей вставку, и ассоциацию вставки с гаплотипными областями генома маиса.
Последовательности, фланкирующие трансгенную вставку ДНК в MON 87427 были определены с использованием методик ПЦР. Геномную ДНК растений выделяли из ткани растений трансгенной линии, выращенных в стандартных условиях теплицы. Ткань растения заливали жидким азотом и растирали в порошок, используя ступку и пестик. ДНК выделяли, используя набор для выделения геномной ДНК Nucleon™ PhytoPure™ Genomic DNA extraction kit (RPN8511, Amersham, Piscataway, N.J.) в соответствии с протоколом изготовителя. После конечной стадии осаждения ДНК ресуспендировали в 0,5 мл ТЕ-буфера (10 мМ Tris-HCl pH 8,0, 1 мМ EDTA). Опытный специалист в данной области может модифицировать этот способ для выделения ДНК из любой ткани маиса, включая, без ограничений, семена. Аликвоту ДНК расщепляли рестрикционными эндонуклеазами, исходя из рестрикционного анализа трансгенной ДНК. После лигирования рестрикционных фрагментов самих на себя проводили ПЦР с использованием праймеров, подобранных на основе последовательности трансгенной ДНК, которые будут амплифицировать последовательности, простирающиеся от 5′- и 3′-концов трансгенной ДНК. Продукты ПЦР разделяли электрофорезом в агарозном геле и очищали с использованием набора QIAGEN gel purification kit (Qiagen, Valencia, Calif.). Полученные ДНК-продукты секвенировали напрямую с использованием стандартных протоколов секвенирования ДНК. 5′-фланкирующая последовательность, которая переходит в правую граничную (RB) последовательность трансгенной ДНК экспрессионной кассеты представлена как SEQ ID NO: 7. 3′-фланкирующая последовательность, которая переходит в левую граничную (LB) последовательность трансгенной ДНК экспрессионной кассеты представлена как SEQ ID NO: 8. Последовательность, полностью встроенная в геномную ДНК маиса и содержащая ДНК экспрессионной кассеты, представлена как SEQ ID NO: 9.
Последовательности выделенных молекул ДНК сравнивали с последовательностью трансгенной ДНК для идентификации фланкирующей последовательности и совместно выделенного фрагмента трансгенной ДНК. Подтверждение присутствия экспрессионной кассеты достигали с помощью ПЦР с праймерами, подобранными, исходя из выведенных данных фланкирующей последовательности и известной последовательности трансгенной ДНК. Последовательность дикого типа, соответствующая той же области, в которую была встроена трансгенная ДНК в трансформированной линии, была выделена с использованием праймеров, подобранных из фланкирующих последовательностей в MON 87427. ПЦР-реакции проводили с использованием системы амплификации Elongase® (Invitrogen, Carlsbad, Calif.). Фланкирующие последовательности ДНК в MON 87427 и последовательность дикого типа LH198 анализировали относительно множества нуклеотидных и белковых баз данных. Эту информацию использовали для исследования взаимодействия трансгена с растительным геномом и для поиска консервативности сайта встраивания.
Пример 3: способы, пригодные для идентификации ДНК MON 87427 в образце
В этом примере описаны способы, пригодные для идентификации ДНК события MON 87427 в образце. Фланкирующую(ие) последовательность(и), геномную последовательность дикого типа маиса и/или трансгенную последовательность можно использовать для подбора праймеров и проб для использования в таких способах. В анализ можно включить (или можно не включать) внутренние контрольные праймер(ы) и пробу(ы).
Описаны способы температурной амплификации TAQMAN® с детекцией по конечной точке для идентификации события MON 87427 (специфичный к событию анализ) и/или синтетического гена CP4-EPSPS (также именуемого CP4-Zm) (специфичный к трансгену анализ) события MON 87427 в образце. Фланкирующая(ие) последовательность(и) события, геномная последовательность дикого типа маиса и/или трансгенная последовательность были использованы для подбора праймеров и проб для использования в этих анализах (таблица 1). ДНК-праймеры, используемые в специфичном к событию анализе, являются праймерами SQ12763 (SEQ ID NO: 17) и SQ12886 (SEQ ID NO: 18) с 6-FAM™-меченной пробой PB4352 (SEQ ID NO: 19). ДНК-праймеры, используемые в специфичном к трансгену анализе, являются праймерами SQ20052 (SEQ ID NO: 11) и SQ20053 (SEQ ID NO: 12) с 6-FAM™-меченной пробой PB10016 (SEQ ID NO: 13). 6-FAM™ является флуоресцентным красителем от Applied Biosystems (Foster City, Calif.), присоединенным к зонду ДНК. Контроль для данного анализа должен включать положительный контроль из ДНК маиса, содержащей событие MON 87427, отрицательный контроль из нетрансгенного маиса и отрицательный контроль, который не содержит ДНК-матрицу.
Кроме того, необязательный контроль для ПЦР может включать праймеры внутреннего контроля и пробу внутреннего контроля, специфичные к однокопийному гену в геноме маиса. Опытный специалист в данной области будет знать, как подобрать праймеры, специфичные к однокопийному гену в геноме маиса. ДНК-праймерами, используемыми в специфичном к трансгену анализе в качестве внутренних контролей являются праймеры SQ1241 (SEQ ID NO: 14) и SQ1242 (SEQ ID NO: 15) с VIC TAMRA-меченной пробой PB0084 (SEQ ID NO: 16). Праймеры и пробу внутреннего контроля для специфичного к трансгену анализа можно использовать на необязательных стадиях 5-6 (ниже). В специфичном к событию анализе не использовали внутренних контролей.
Праймеры и пробы
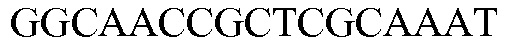



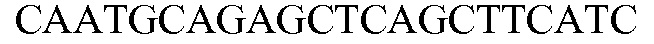



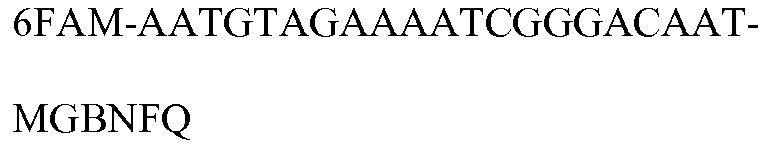
Примеры условий, подходящих для способов TAQMAN® с детекцией по конечной точке, являются следующими:
Стадия 1: воду (18 МОм) подводят до конечного объема 10 мкл.
Стадия 2: добавить 5,0 мкл 2х универсальной «мастер»-смеси (все 4 дезоксинуклеотида, фермент, буфер) до 1х конечной концентрации.
Стадия 3: добавить 0,5 мкл смеси специфичного к событию праймера 1 (SEQ ID NO: 17) и специфичного к событию праймера 2 (SEQ ID NO: 18) (ресуспендированных в воде (18 МОм) до концентрации 20 мкМ для каждого праймера) до 1,0 мкМ конечной концентрации (например, в микроцентрифужной пробирке чтобы получить 500 мкл с конечной концентрацией 20 мкМ следует добавить: 100 мкл специфичного к событию праймера 1 (SEQ ID NO: 17) с концентрацией 100 мкМ; 100 мкл специфичного к событию праймера 2 (SEQ ID NO: 18) с концентрацией 100 мкМ; 300 мкл воды (18 МОм)).
Стадия 4: добавить 0,2 мкл специфичной к событию 6-FAM™ MGB-пробы (SEQ ID NO: 19) (ресуспендированной в воде (18 МОм) до концентрации 10 мкМ) до конечной концентрации 0,2 мкМ.
Стадия 5 (необязательная): добавить 0,5 мкл смеси праймера 1 внутреннего контроля и праймера 2 внутреннего контроля (ресуспендированных в воде (18 МОм) до концентрации каждого праймера 20 мкМ) до конечной концентрации 1,0 мкМ.
Стадия 6 (необязательная): добавить 0,2 мкл VIC™-пробы внутреннего контроля (ресуспендированной в воде (18 МОм) до концентрации 10 мкМ) до конечной концентрации 0,2 мкМ.
Стадия 7: добавить 3,0 мкл выделенной ДНК (матрицы) для каждого образца с каждым одним из следующего, включая: 1. образцы листьев для анализа; 2. отрицательный контроль (нетрансгенную ДНК); 3. отрицательный контроль - воду (нет матрицы); 4. положительный контроль - ДНК MON 87427.
Стадия 8: условия термоциклера были следующими: один цикл при 50°C в течение 2 минут; один цикл при 95°C в течение 10 минут; десять циклов (95°C в течение 15 секунд, затем 64°C в течение 1 минуты с −1°C/цикл); тридцать циклов (95°C в течение 15 секунд, затем 54°C в течение 1 минуты); конечный цикл при 10°C.
Эти анализы оптимизированы для использования либо с Applied Biosystems GeneAmp® PCR System 9700 (операции на максимальной скорости) или термоциклером MJ Research DNA Engine PTC-225. Другие способы и аппараты, известные опытным специалистам в данной области, которые продуцируют ампликоны, идентифицирующие ДНК события MON 87427, входят в объем данной области.
SEQ ID NO: 11 и SEQ ID NO: 12 или SEQ ID NO: 17 и SEQ ID NO: 18, каждые являются примером пары молекул ДНК (пары праймеров), состоящих из первой молекулы ДНК и второй молекулы ДНК, отличающейся от первой молекулы ДНК, причем указанные каждая первая и вторая молекулы ДНК содержат молекулу нуклеиновой кислоты с достаточной длиной непрерывной нуклеотидной последовательности из SEQ ID NO: 10 для функционирования в качестве ДНК-праймеров при совместном использовании в реакции амплификации с ДНК, полученной из события MON 87427, чтобы получить диагностический ампликон для ДНК MON 87427 в образце. Эти праймеры можно использовать в других способах на основе полимеразной цепной реакции (ПЦР) для детекции события.
Пример 4: использование события MON 87427 для продукции гибридных семян
В следующем примере описано как можно использовать MON 87427 для скрещивания маиса, включая использование способов, описанных в патентной публикации США № 20090165166 и/или патенте США № 7314970.
При получении гибридных семян растения маиса, содержащие MON 87427, высаживают на местности, такой как открытое поле. Другие родительские растения маиса могут присутствовать (или могут не присутствовать) в той же местности. Для контроля сорняков в ходе получения семян и на промышленных полях можно применять глифосат на растениях маиса, содержащих MON 87427, на вегетативных стадиях, как указано на товарных этикетках к агротехническому продукту Roundup®, в таких же количествах, как используемые для событий Roundup Ready® NK603 и MON 88017 маиса. Для продукции гибридных семян две обработки глифосата, начиная непосредственно перед стадиями и в течение стадий развития метелки (приблизительно стадии вегетативного роста маиса в диапазоне от V8 до V13), применяли у растений MON 87427 для получения мужского стерильного фенотипа посредством тканеизбирательной устойчивости к глифосату. В системе продукции гибридных семян маиса растения MON 87427 после применения глифосата во время развития метелки будут иметь мужскую стерильность и, поэтому, будут легко опыляться пыльцой от других растений-доноров пыльцы (мужских), давая в результате живые гибридные семена маиса, несущие ген тканеизбирательной устойчивости к глифосату. Растения-доноры пыльцы могут присутствовать, а могут и не присутствовать, в той же местности. Опыление можно осуществить любыми средствами, известными в данной области, включая размещение растений на близком расстоянии или ручное опыление. Только обработки глифосатом, проведенные в точное время, начиная непосредственно перед стадиями и/или в течение стадий развития метелки (приблизительно стадии вегетативного роста маиса в диапазоне от V8 до V13) будут давать мужской стерильный фенотип посредством тканеизбирательной устойчивости к глифосату у MON 87427. Глифосат является системным гербицидом, который легко перемещается по флоэме у растений. Как только глифосат оказывается во флоэме, он перемещается в области с высокой меристемной активностью, следуя типичному распределению источников и стоков. Развитие пыльцы у растений маиса занимает приблизительно четыре недели. Ранние стадии роста метелки начинаются приблизительно на вегетативной стадии V9 маиса, поэтому, обработка глифосатом, проведенная приблизительно на этой стадии и в это время позволяет максимальное перемещение глифосата в мужские репродуктивные ткани. Обработки глифосатом, проведенные в течение ранних вегетативных стадий в соответствии с временем применения, указанным на товарной этикетке существующего агротехнического продукта Roundup® для контроля сорняков, не влияют на продукцию пыльцы растениями MON 87427, поскольку чувствительные мужские репродуктивные ткани не развиваются активно в это время. В условия обработки глифосатом можно внести модификации, которые известны специалистам в области применения гербицидов, и входят в объем изобретения.
MON 87427 при скрещивании с другим устойчивым к глифосату событием маиса, таким как событие NK603 маиса (патент США № 6825400), для получения гибридных семян не показывают потерю урожайности по сравнению с урожайностью стандартного гибрида NK603 (см. фиг. 2). Поле гибридных растений маиса обрабатывали двумя последовательными опрыскиваниями глифосатом в количестве 2,25 фунтов a.e./акр, каждое из которых применяли для контроля сорняков, и не наблюдалось никакой разницы в повреждении или мужской фертильности между различными гибридами события MON 87427 и гибридом NK603. Это иллюстрирует то, что гибридные растения F1, полученные в результате скрещивания события MON 87427, являются полностью устойчивыми к глифосату при его использовании для контроля сорняков.
Пример 5: измерение стадий развития метелки
Стадии развития метелки проиллюстрированы на фиг. 3, с приблизительными размерами в миллиметрах, показанных между скобками. На фигуре Vg представляет собой меристему на вегетативной стадии; T0 является переходом от вегетативной к репродуктивной стадии; Т1 представляет собой видимую репродуктивную точку роста (0,9 мм); Т2 представляет собой видимые боковые примордии (1,8 мм); Т3 представляет собой видимые примордии колосков (4,1 мм); Т4 представляет собой удлинение центральной оси и боковой оси (12,9 мм); Т5 представляет собой начало дифференцировки пыльников (41,0 мм); Т6 представляет собой начало дифференцировки пыльцы (175 мм); и Т7 представляет собой раскрытие пыльников и выделение пыльцы (285,0 мм). Стадию развития метелки для данного растения измеряли, исследуя метелку на разных стадиях созревания. Используя скальпель и маленький пинцет под препаративным микроскопом, меристему метелки отсекали от развивающихся листьев. Затем меристему надрезали скальпелем в ее основании и оценивали в соответствии со стадиями развития метелки (показанными на фиг. 3) под микроскопом.
Пример 6: стадии вегетативного развития (V-стадия) относительно стадий развития метелки
В этом примере продемонстрировано, что сухой вес метелки, длина метелки и стадия развития метелки значительно варьируют по генотипам при измерении относительно вегетативных стадий растения и вегетативного роста растения.
Было высажено десять генотипов: инбредные линии LH198, LH287, O1DKD2, 19HGZI, 17IDI6 и гибриды DKC 44-46, DKC 47-10, DKC 52-40, DKC 58-80, DKC 63-81. Гибриды были выбраны таким образом, чтобы они представляли генотипы, относящиеся к различным вариантам развития. Исследование проводили в городах Farmer City (IL), Kearney (NE) и Williamsburg (IA). Десять генотипов высаживали с использованием конусной сеялки и прореживали до конечной плотности насаждений 38000 растений на акр (4047 м2). Длина участка составляла 20 футов (6 м) с промежутками в 3 фута (0,9 м) через 4 ряда, для того чтобы гарантировать достаточное число растений для всех обработок и снизить граничные эффекты. Данные собирали для регистрации наблюдений как вегетативного развития растений, так и развития метелки, относительно стадий развития метелки.
Явные различия в развитии метелки наблюдались по генотипам для идентичных соответствующих вегетативных стадий (V-стадий). Например, различия в размере метелки между генотипами были очевидны для каждой V-стадии. Средняя длина метелки на стадии V8 составляла 7 мм для LH198, 40,2 мм для LH287 и 47,8 мм для DKC44-46 (фиг. 4). Этот диапазон в длине метелки на стадии V8 представляет собой 7-кратную разницу между генотипами. На стадии V10 средняя длина метелки для этих трех генотипов соответственно составляла 70,1 мм, 148,2 мм и 277,3 мм (фиг. 4). Это давало практически 4-кратную разницу между генотипами. Генетическая вариабельность в росте метелки относительно V-стадий также очевидна при исследовании накопления сухого веса метелок.
В дополнительном исследовании использовали семьдесят две инбредных линии для охвата широкого диапазона спелости. Эти инбредные линии были сгруппированы в 6 групп спелости для упрощения процесса препарирования (таблица 2). Были выбраны инбредные линии без характерных признаков для того чтобы избежать сложности проведения препарирования метелок на контролируемых полях. Этот эксперимент отражает данные, собранные на четырех различных местоположениях полей: Williamsburg (IA), Waterman (IL), Farmer city (IL) и Constantine (MI). Растения выращивали на четырехрядных участках длиной 20 футов (6 м). Целевая конечная популяция составляла 38000 растений на акр (4047 м2). Подсчет конечных насаждений документировали на стадии V3. Помечали пятый и десятый листья трех репрезентативных растений на участок для того чтобы отслеживать V-стадии; все листья подсчитывали, включая колеоптили. Три репрезентативных растения из каждой группы отбирали в качестве образцов на 60, 70 и 80% средних градусо-единиц роста для цветения (определяемого как приблизительно половина метелок в группе выделяют пыльцу, представляемое как Р50), и проводили препарирование метелок, как описано ранее.
Растения выбирали из средних рядов, чтобы избежать растений из граничных рядов. В момент первого отбора образцов, растения на соответствующей стадии развития были помечены для использования для второго и третьего отбора образцов. Даты каждого препарирования вместе с V-стадией на момент отбора образцов документировали. Стадию развития метелки идентифицировали по Относительной шкале развития, описанной выше. Участки продолжали мониторить на протяжении цветения и регистрировали дату Р50. Вегетативная стадия варьировала среди инбредных линий относительно стадий развития метелки. Например, V-стадия для Т5 (начало дифференцировки пыльников) варьировала в диапазоне более чем 6,5 листьев среди инбредных линий, которые на этот момент имели от 7 до 14 листьев. Это составляло приблизительно 64% от общего среднего значения 10,3 листьев для достижения данной стадии.
Пример 7: средние GDU относительно стадий развития метелки и цветков
Этот пример демонстрирует, что средние GDU, требуемые для достижения данной стадии развития метелки или стадии развития цветков могут значительно варьировать среди генотипов, и что GDU, поэтому, не является достоверным средством прогнозирования развития метелки. Данные были получены из участков на поле и инбредных растений, описанных выше. Проводили мониторинг температуры каждый час от начала посадки и до цветения, используя метеорологические станции Onset, и данные использовали для вычисления дневных кумулятивных GDU, следуя традиционному способу (то есть усреднению дневных максимальной и минимальной температур). Данные препарирования образцов, указывающие на стадию развития метелки, откладывали относительно требований GDU. Обычно ожидается, что инбредные линии, у которых развивается большее число листьев, имеют более высокие требования GDU для достижения определенной стадии развития метелки. Однако описанная выше вариабельность, наблюдаемая в V-стадиях для Т5, не объясняет всю вариабельность, наблюдаемую в GDU для Т5. Это позволяет предположить, что филлохрон, представляющий собой время между развертыванием двух следующих листов, может варьировать между инбредными линиями. Результаты этого исследования показали, что GDU для достижения стадии Т5 варьировали более чем на 400 тепловых или градусо-единиц роста среди инбредных линий; то есть разброс составлял примерно 40% от общего среднего.
Наблюдалась сильная корреляция между требованиями GDU для Р50 и данной стадией развития метелки среди инбредных сортов. Используя данные от полевых участков и инбредных растений, регистрировали средние требования GDU для Р50 среди инбредных растений и сравнивали с данными стадиями развития метелки. Требования GDU для Р50 варьировали от 1283 до 1645 единиц, от инбредной линии наиболее быстрой спелости до инбредной линии наиболее поздней спелости; то есть разница в GDU составляла немного больше 360 единиц. Было найдено, что эта разница коррелирует с разницей в средних требованиях GDU для стадии Т5 в пределах инбредных линий (фиг. 5).
Пример 8: конструирование Относительной шкалы развития
В этом примере продемонстрировано конструирование стандартизированной шкалы для мониторинга и/или предупреждения развития метелки. Эта Относительная шкала развития успешно стандартизирует стадии развития метелки по инбредным линиям.
Принимая во внимание сильную корреляцию между требованиями GDU для достижения Р50 на данной стадии развития метелки среди инбредных сортов, описанную выше, была создана Относительная шкала развития. Она была вычислена путем выражения роста метелки для каждого генотипа относительно температурного времени для выделения пыльцы, как указано ниже:
Относительная шкала развития = (GDU для Tn/GDU для Px)
Использовали данные от полевых участков и инбредных растений, описанные выше. Величину GDU на данной стадии развития метелки (GDU для Tn) делили на число GDU, которое, как известно, требуется для достижения конкретной стадии выделения пыльцы (GDU для Px), которая в данном случае являлась Р50 для некоторого генотипа. Это согласовало различия в стадии развития метелки по генотипам. Например, требования GDU для каждой Т5 были довольно постоянными на Относительной шкале развития для всех инбредных линий и варьировали только от 69 до 75% GDU, требуемых для P50. Вариабельность в линиях регрессии требований GDU различных генотипов относительно стадии развития метелки по сравнению с более согласованными линиями регрессии тех же генотипов с использованием стандартизированной Т-шкалы использовали для оценки стандартизации шкалы вне зависимости от группы спелости (фиг. 7).
Пример 9: прогнозирование оптимального времени для модулирующей развитие обработки
В этом примере продемонстрировано применение Относительной шкалы развития для определения оптимального времени для схемы опрыскивания химическим агентом для того чтобы достичь полной стерильности метелок маиса. В этом примере химическим агентом был гербицид глифосат Roundup®, используемый в комбинации с растениями маиса MON 87427 в системе гибридизации Roundup® (RHS). Оптимальное время опрыскивания было согласовано с фактической стадией развития метелки, полная стерильность метелок маиса достигалась в результате только одной эффективной дозы Roundup®.
Тридцать два инбредных исходных линии, содержащие событие MON 87427, были отобраны для исследования и сгруппированы в две группы спелости. Инбредные растения высаживали рядами по 20 футов с промежутками по три фута. Расстояние между растениями в ряду составляло 30 дюймов. Ряды высаживали таким образом, чтобы были четыре ряда женских «тестируемых» растений, за которыми следовали два ряда мужских опыляющих растений. Для данного исследования отбирали трансгенные события, которые имели устойчивость вегетативных и женских репродуктивных тканей к глифосату, но не имели устойчивости к глифосату у мужских репродуктивных тканей (то есть, тканеизбирательную устойчивость к глифосату). Мужские опыляющие растения также имели устойчивость мужских репродуктивных тканей к глифосату, в то время как женские растения-реципиенты были чувствительны к пыльце мужских растений после обработки глифосатом. Обработку опрыскиванием делили на блоки и разбивали на две подгруппы исходя из группы спелости инбредной линии. Сразу после каждого опрыскивания отбирали три репрезентативных растения с каждого участка и препарировали на поле. Записывали длину метелки, стадию развития метелки, дату и GDU в момент опрыскивания. Обработки опрыскиванием (SS1) Roundup PowerMAX™ с объемом воды 15 галлонов/акр применяли однократно на группе спелости, соответствующей каждой обработке (Trt), с использованием опрыскивателя с высоким клиренсом. Обработки опрыскиванием для индукции стерильности применяли в диапазоне от 50% до 80% GDU, требуемых для достижения Р50 (усредненной в пределах группы спелости инбредных линий), как показано в таблице 3, в которой WC=«контроль сорняков»; Trt 1 SS1=50% GDU для P50; Trt 2 SS1=57,5% GDU для P50; Trt 3 SS1=65% GDU для P50; Trt 4 SS1=72,5% GDU для P50; Trt 5 SS1=80% GDU для P50.
После всех обработок опрыскиванием проводили оценку стерильности/способности к оплодотворению метелок путем оценки выбрасывания пыльников и выделения пыльцы относительно выметывания пестичных столбиков. Эти оценки проводили, когда каждый участок находился на определенной стадии развития: 10% растений входят в стадию выметывания пестичных столбиков (S10); 50% растений входят в стадию выметывания пестичных столбиков (S50); 90% растений входят в стадию выметывания пестичных столбиков (S90); 3 дня после даты S90 (S90+3); и 6 дней после даты S90 (S90+6). У растений наблюдали выбрасывание пыльников (АЕ), а стерильность измеряли, используя индекс риска выбрасывания пыльников (AE-риск), который является взвешенным средним, объединяющим процент растений на участке, показывающих выбрасывание пыльников, с интенсивностью явления. Например, слабораскрытой (LP) является метелка с 10 или меньшим числом выброшенных пыльников. Среднераскрытой (МР) является метелка, у которой >11 и до 25% пыльников выброшены. Сильнораскрытой (НР) является метелка, у которой выброшено >25% пыльников. Как показано на фиг. 6, относительная шкала развития выявляет оптимальное окно эффективности химического агента для получения стерильности метелок маиса между 0,62 и 0,75, в котором минимизирован АЕ-риск по инбредным линиям и группам спелости. Это исследование подтверждает эффективность Относительной шкалы развития в качестве средства для предоставления рекомендаций по опрыскиванию для применения системы гибридизации Roundup® на инбредных линиях. Она особенно полезна с MON 87427.
Пример 10: способ получения гибридных семян с увеличенной чистотой семян
Способы получения гибридных семян и получаемую в результате чистоту семян измеряли с использованием двадцати четырех пилотных блоков получения, расположенных в Kearney, Nebr.; Williamsburg, Iowa; Waterman, Ill.; Farmer City, Ill.; и Constantine, Mich. В каждом месте высаживали по четыре блока MON 87427 и по два блока растений с цитоплазматической стерильностью (CMS). Блоки MON 87427 состояли из 01DKD2MON87427-MON89034 (женские растения) × 80IDM2MON88017 (мужские растения), а блоки CMS состояли из 01DKD2NK603B-CMS (женские растения») × 80IDM2MON88017 (мужские растения).
Растения высаживали в соотношении женских к мужским растениям 4:1 на 30-дюймовых рядах. Каждый экспериментальный блок по размеру состоял из 10 панелей 100-150 футов в длину. Блоки были окружены 30 футами (12 рядами) мужских растений по сторонам, а также спереди и сзади. Блоки были удалены по меньшей мере на 200 футов от других потенциальных источников пыльцы и отделены друг от друга 45 футами. Для исследования высаживали популяцию из 40000 и 38000 растений для орошаемых и неорошаемых земель, соответственно. Женские и мужские ряды высаживали одновременно. Мужские ряды обжигали на стадии роста V3 для замедления роста и задержки выделения пыльцы на мужских растениях путем чередующегося обжигания секций рядов длиной 20 футов. Пропановые горелки устанавливали позади трактора и располагали над мужскими рядами и чередовали обжигание и отсутствие обжигания примерно каждые 20 футов. При посадке использовали инсектициды для минимизации вариабельности вследствие заражения насекомыми. Мужские ряды уничтожали после опыления. Все блоки опрыскивали Roundup PowerMax™ в количестве 0,75 фунтов a.e./акр примерно на стадии V3 для контроля сорняков. Кроме того, блоки MON 87427 опрыскивали Roundup PowerMax™ два раза по 0,75 фунтов a.e./акр при 825 и 975 градусо-единиц роста (GDU) от посадки. Объем опрыскивания оставался постоянным - 15 галлонов на акр (GPA).
Стерильность метелок оценивали, наблюдая за растениями через день от появления метелок и до 6 дней после выметывания пестичных столбиков (P90+6 дней). При возникновении прорыва (выделения пыльцы), отдельные растения дополнительно делили на категории как растения с низким выделением пыльцы (LP; менее 10 выброшенных пыльников), средним выделением пыльцы (МР; от 11 пыльников и до 25% площади поверхности метелки с выброшенными пыльниками) или высоким выделением пыльцы (НР; более 25% площади поверхности метелки с выброшенными пыльниками). Затем вычисляли риск выбрасывания пыльника, как:
AE-риск (%) = ([(LP×0,25)+(MP×0,5)+(HP×1,0)]/ число растений)×100
После достижения физиологической спелости и содержания влаги в зернах примерно 30-35%, вручную собирали составной образец из 100 початков на блок, следуя заранее определенной схеме отбора образцов, для представления всех панелей в блоке. Образцы высушивали и взвешивали для определения конечной урожайности. Первый набор образцов обрабатывали вручную и отсылали на анализ качества (оценку холодного проращивания и теплого проращивания с использованием двух повторов по 100 семян). Второй набор образцов посылали на анализ генетической чистоты (анализ однонуклеотидного полиморфизма (SNP)) и анализ чистоты признака (ELISA на специфичный маркер мужского растения). Третий набор образцов использовали для документации распределения размеров семян. Урожай на пилотных блоках продукции семян собирали комбайном и доводили до содержания влажности 15%, чтобы определить урожайность в бушелях на акр (25,40 кг/4047 м2).
В общем оба блока, CMS и MON 87427, превосходили стандарты стерильности метелок и чистоты семян маиса. После выброса пыльников риск был значительно ниже желаемого стандартного показателя 0,5% даже через 6 дней после того как у 90% популяции женских растений произошло выпадения пестичных столбиков (фиг. 8). Практически никаких прорывов не было задокументировано на блоках MON 87427, но немного больший процент прорывов на последних стадиях выпадения пестичных столбиков наблюдался на блоках CMS. Как женские, так и мужские родительские растения маиса для CMS и MON 87427 тестировали на генетическую чистоту, и результаты показали 100%-ю чистоту. Высокий уровень стерильности метелок маиса родительских растений как MON 87427, так и CMS, давал высокий уровень генетической чистоты и чистоты признака у гибридных семян, полученных в результате данных испытаний (фиг.9). Между MON 87427 и CMS не было статистически достоверной разницы по чистоте признака, но между MON 87427 и CMS была измерена статистически достоверная разница (при p<0,05) для генетической чистоты. Уровень генетической чистоты гибридных семян, полученных с использованием MON 87427 и системы гибридизации Roundup® (RHS) составлял 98,7%, что было достоверно выше уровня генетической чистоты гибридных семян, полученных с использованием CMS (98,0%). В результате женские родительские растения MON 87427 продуцировали примерно на 0,2% меньше событий самоопыления и на 0,5% меньше событий опыления других растений по сравнению с родительскими растениями CMS-системы, демонстрируя, что использование MON 87427 с системой гибридизации Roundup® (RHS) можно применять для увеличения чистоты семян маиса при продукции гибридных семян маиса.
Репрезентативный образец семян MON 87427, раскрытых выше и изложенных в формуле изобретения, депонирован в соответствии с Будапештским договором в Американской коллекции типовых культур (АТСС), 10801 University Boulevard, Manassas, Va., 20110. Номером доступа АТСС для этого депозита является PTA-7899. Депозит будет храниться в хранилище в течение 30 лет или в течение 5 лет после последнего запроса, или в течение эффективного времени действия патента, любого наиболее длинного периода времени из вышеперечисленных, и в течение этого периода при необходимости будет производиться замена депозита.
Имея проиллюстрированные и описанные принципы изобретения опытным специалистам в данной области должно быть ясно, что изобретение можно модифицировать по структуре и деталям, не выходя за рамки данных принципов. Авторы изобретения заявляют права на все модификации, которые входят в суть и объем приложенной формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ТРАНСГЕННОЕ СОБЫТИЕ MON 87427 МАИСА И ОТНОСИТЕЛЬНАЯ ШКАЛА РАЗВИТИЯ | 2010 |
|
RU2764586C2 |
| ОБЪЕКТ КУКУРУЗЫ MON 87411 | 2013 |
|
RU2648155C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ИЗБИРАТЕЛЬНОЙ РЕГУЛЯЦИИ ЭКСПРЕССИИ БЕЛКА | 2012 |
|
RU2667424C2 |
| ТРАНСГЕННЫЙ ОБЪЕКТ СОИ MON 87708 И СПОСОБЫ ЕГО ПРИМЕНЕНИЯ | 2010 |
|
RU2624025C2 |
| КОМПОЗИЦИИ ДЛЯ ИНДУКЦИИ ГАПЛОИДИИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2771141C2 |
| ГЕНЫ, КОНСТРУКЦИИ И ОБЪЕКТ DP-202216-6 МАИСА | 2019 |
|
RU2833371C2 |
| ЛИНИИ ТРАНСГЕННОЙ СОИ, ГЕНЕТИЧЕСКОЕ СОБЫТИЕ 8264.42.32.1, УСТОЙЧИВОЕ К ГЕРБИЦИДАМ С ПАКЕТИРОВАННЫМИ ГЕНАМИ НА ЕГО ОСНОВЕ, И ИХ ДЕТЕКТИРОВАНИЕ | 2012 |
|
RU2636021C2 |
| ГЕН-ВОССТАНОВИТЕЛЬ RF4 ДЛЯ ЦИТОПЛАЗМАТИЧЕСКОЙ МУЖСКОЙ СТЕРИЛЬНОСТИ (CMS) C-ТИПА КУКУРУЗЫ, МОЛЕКУЛЯРНЫЕ МАРКЕРЫ И ИХ ПРИМЕНЕНИЕ | 2011 |
|
RU2603005C2 |
| СПОСОБЫ БОРЬБЫ С СОРНЯКАМИ С ИСПОЛЬЗОВАНИЕМ РАСТЕНИЙ AAD-1 И ПРЕДПОСЕВНОГО И/ИЛИ ДОВСХОДОВОГО ВНЕСЕНИЯ ГЕРБИЦИДОВ | 2012 |
|
RU2610664C2 |
| ГЕНЕТИЧЕСКАЯ МОДИФИКАЦИЯ MON89788 СОИ И СПОСОБЫ ЕЕ ОПРЕДЕЛЕНИЯ | 2006 |
|
RU2411720C2 |
















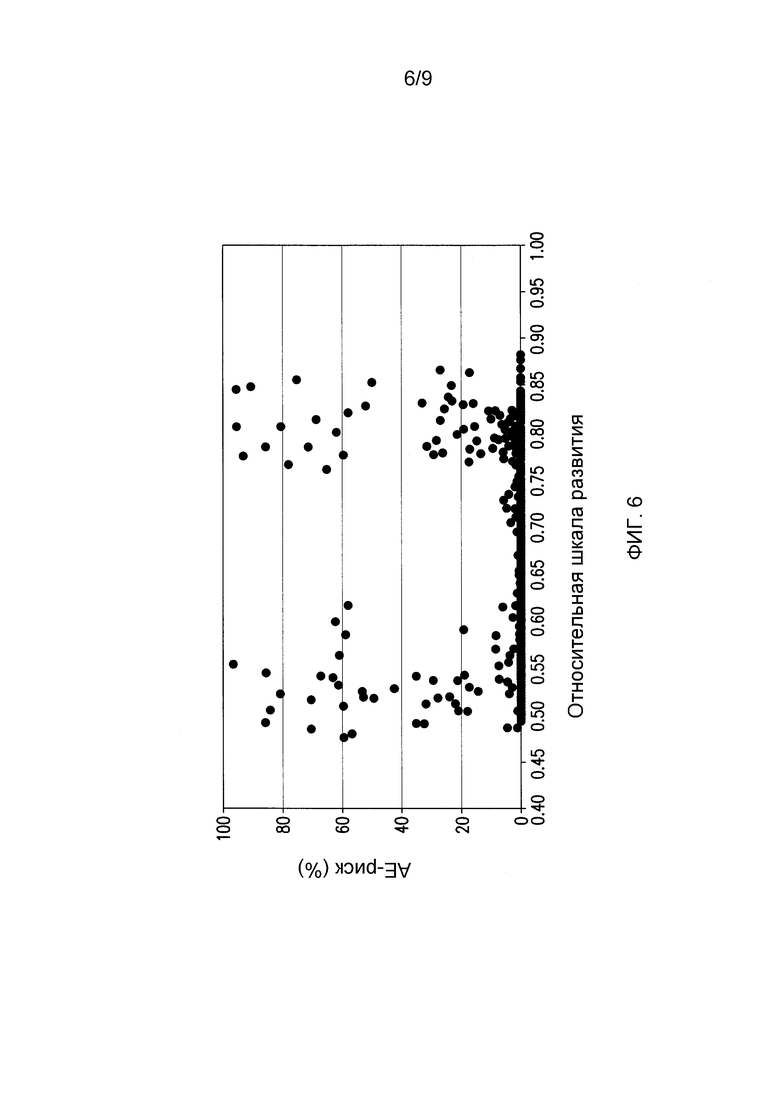


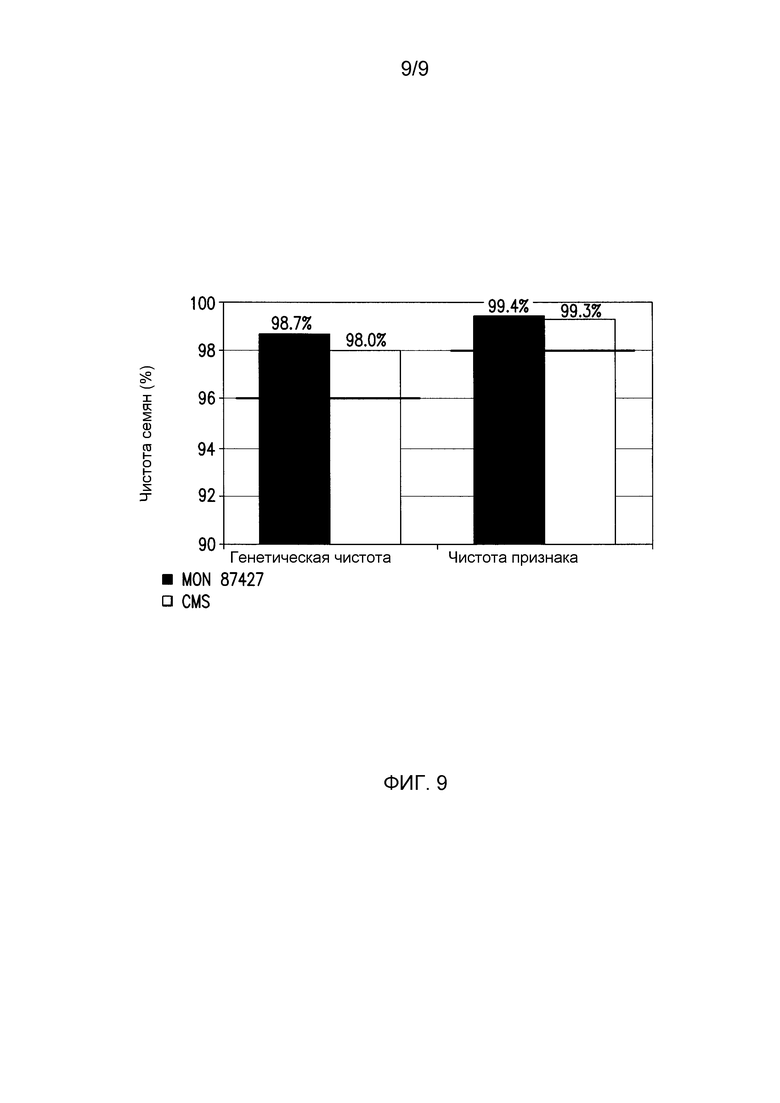
Изобретение относится к области биохимии, в частности к рекомбинантной молекуле ДНК, указывающей на присутствие трансгенного события, где репрезентативный образец семени, содержащий указанное трансгенное событие, депонирован как АТСС РТА-7899. Также раскрыт способ детекции присутствия молекулы ДНК из вышеуказанного трансгенного события в образце, который осуществляется путем использования зонда или пары молекул ДНК. Изобретение также относится к рекомбинантному растению маиса, устойчивому к глифосату, а также к способу борьбы с сорняками с его использованием, части указанного растения, его клетки и семени. Также раскрыт продукт потребления, содержащий вышеуказанное трансгенное событие, а также способ его изготовления. Изобретение позволяет эффективно получать растение маиса, устойчивое к глифосату. 12 н. и 16 з.п. ф-лы, 9 ил., 3 табл., 10 пр.
1. Рекомбинантная молекула ДНК, включающая нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-8 и 10, указывающая на присутствие трансгенного события, где репрезентативный образец семени, содержащий указанное трансгенное событие, депонирован как АТСС РТА-7899.
2. Рекомбинантная молекула ДНК по п. 1, причем указанная рекомбинантная молекула ДНК образована соединением встроенной гетерологичной молекулы нуклеиновой кислоты и геномной ДНК растения, клетки растения или семени маиса.
3. Рекомбинантная молекула ДНК по п. 2, причем указанная молекула ДНК представляет собой диагностический ампликон.
4. Рекомбинантная молекула ДНК по п. 1, причем указанная молекула ДНК находится в растении, клетке растения, семени, потомстве растения, части растения маиса или продукте потребления, содержащих трансгенное событие, где репрезентативный образец семени, содержащий указанное трансгенное событие, депонирован как АТСС РТА-7899.
5. Зонд ДНК, включающий молекулу нуклеиновой кислоты, имеющую нуклеотидную последовательность с достаточной длиной непрерывной нуклеотидной последовательности из SEQ ID NO: 10 для функционирования в качестве зонда ДНК, указывающего на присутствие трансгенного события, где репрезентативный образец семени, содержащий указанное трансгенное событие, депонирован как АТСС РТА-7899, где указанный зонд гибридизуется при жестких условиях с молекулой ДНК, включающей нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-8 и 10, и не гибридизуется при жестких условиях с молекулой ДНК, не включающей нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-8 и 10.
6. Пара молекул ДНК, состоящая из первой молекулы ДНК и второй молекулы ДНК, отличающейся от первой молекулы ДНК, причем каждая из первой и второй молекул ДНК содержит молекулу нуклеиновой кислоты, имеющую нуклеотидную последовательность с достаточной длиной непрерывной нуклеотидной последовательности из SEQ ID NO: 10 для функционирования в качестве ДНК-праймеров при совместном использовании в реакции амплификации с ДНК, полученной из трансгеного события где репрезентативный образец семени, содержащий указанное трансгенное событие, депонирован как АТСС РТА-7899, для получения ампликона, диагностического в отношении ДНК указанного трансгенного события маиса в образце.
7. Способ детекции присутствия молекулы ДНК из трансгенного события, где репрезентативный образец семени, содержащий указанное трансгенное событие, депонирован как АТСС РТА-7899, в образце, причем указанный способ включает в себя:
a. приведение образца в контакт с зондом ДНК по п. 5;
b. подвергание указанного образца и указанного зонда ДНК жестким условиям гибридизации; и
c. детекцию гибридизации указанного зонда ДНК с молекулой ДНК в указанном образце, причем гибридизация указанного зонда ДНК с указанной молекулой ДНК указывает на присутствие молекулы ДНК, полученной из указанного трансгенного события маиса, в указанном образце.
8. Способ детекции присутствия молекулы ДНК, полученной из трансгенного события маиса, где репрезентативный образец семени, содержащий указанное трансгенное событие, депонирован как АТСС РТА-7899, причем указанный способ включает:
a. приведение образца в контакт с парой молекул ДНК по п. 6;
b. проведение реакции амплификации, достаточной для получения ампликона ДНК, содержащего последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-10; и
c. детекцию присутствия указанного ДНК-ампликона в указанной реакции, причем присутствие указанного ДНК-ампликона в указанной реакции указывает на присутствие молекулы ДНК, полученной из трансгенного события, где репрезентативный образец семени, содержащий указанное трансгенное событие, депонирован как АТСС РТА-7899, в указанном образце.
9. Рекомбинантное растение маиса, содержащее молекулу нуклеиновой кислоты, имеющую нуклеотидную последовательность SEQ ID NO: 10, где указанное растение маиса устойчиво к глифосату.
10. Рекомбинантное растение маиса по п. 9, причем указанное растение маиса имеет тканеизбирательную устойчивость к обработке гербицидом глифосатом.
11. Рекомбинантное растение маиса по п. 9, геном которого продуцирует ампликон, содержащий молекулу ДНК, выбранную из группы, состоящей из SEQ ID NO: 1-10, при тестировании в способе ДНК-амплификации.
12. Часть растения маиса, содержащая молекулу нуклеиновой кислоты, имеющую нуклеотидную последовательность SEQ ID NO: 10, где указанная часть растения маиса устойчива к глифосату.
13. Часть растения маиса по п. 12, причем указанная часть растения маиса имеет тканеизбирательную устойчивость к обработке гербицидом глифосат.
14. Часть растения маиса по п. 12, геном которого продуцирует ампликон, содержащий молекулу ДНК, выбранную из группы, состоящей из SEQ ID NO: 1-10, при тестировании в способе ДНК-амплификации.
15. Рекомбинантное семя маиса, содержащее молекулу нуклеиновой кислоты, имеющую нуклеотидную последовательность SEQ ID NO: 10, где указанное семя маиса устойчиво к глифосату.
16. Рекомбинантное семя маиса по п. 15, причем указанное семя маиса имеет тканеизбирательную устойчивость к обработке гербицидом глифосат.
17. Рекомбинантное семя маиса по п. 15, геном которого продуцирует ампликон, содержащий молекулу ДНК, выбранную из группы, состоящей из SEQ ID NO: 1-10, при тестировании в способе ДНК-амплификации.
18. Рекомбинантная клетка маиса, содержащая молекулу нуклеиновой кислоты, имеющую нуклеотидную последовательность SEQ ID NO: 10, где указанная клетка маиса устойчива к глифосату.
19. Рекомбинантная клетка маиса по п. 18, причем указанная клетка маиса имеет тканеизбирательную устойчивость к обработке гербицидом глифосат.
20. Рекомбинантная клетка маиса по п. 18, геном которой продуцирует ампликон, содержащий молекулу ДНК, выбранную из группы, состоящей из SEQ ID NO: 1-10, при тестировании в способе ДНК-амплификации.
21. Растение маиса, причем указанное растение маиса содержит нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-8 или 10, указывающую на наличие трансгенного события и сообщаемое тем самым наличие устойчивости к глифосату, где репрезентативный образец семени, содержащий указанное трансгенное событие, депонирован как АТСС РТА-7899 и где указанное растение устойчиво к глифосату.
22. Семя маиса, причем указанное семя маиса содержит нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-8 или 10, указывающую на наличие трансгенного события и сообщаемое тем самым наличие устойчивости к глифосату, где репрезентативный образец семени, содержащий указанное трансгенное событие, депонирован как АТСС РТА-7899 и где указанное семя устойчиво к глифосату.
23. Продукт для потребления человеком, полученный из трансгенного растения маиса, содержащего нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-10, и указывающую на наличие трансгенного события, где репрезентативный образец семени, содержащий указанное трансгенное событие, депонирован как АТСС РТА-7899.
24. Продукт для потребления животным, полученный из трансгенного растения маиса, содержащего нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-10, и указывающую на наличие трансгенного события, где репрезентативный образец семени, содержащий указанное трансгенное событие, депонирован как АТСС РТА-7899.
25. Способ изготовления продукта для потребления по п. 23 или 24, причем указанный способ включает:
a. получение растения маиса или его части, включающих нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-10, и указывающую на наличие трансгенного события, где репрезентативный образец семени, содержащий указанное трансгенное событие, депонирован как АТСС РТА-7899; и
b. изготовление продукта для потребления из растения маиса или его части.
26. Способ борьбы с сорняками в поле, включающий высаживание трансгенных растений маиса по любому из пп. 9-11, 21 в поле и применение эффективной дозы гербицида глифосата для борьбы с сорняками на указанном поле без поражения указанных трансгенных растений маиса.
27. Способ по п. 26, причем указанная эффективная доза гербицида глифосата составляет от примерно 0,1 фунта до примерно 4 фунтов на акр.
28. Рекомбинантная молекула ДНК, включающая нуклеотидную последовательность SEQ ID NO: 10, указывающая на присутствие трансгенного события, где репрезентативный образец семени, содержащего указанное событие, депонирован как АТСС РТА-7899.
| WO 2009085982 A1, 09.07.2009 | |||
| Способ получения нефтяного кокса | 1981 |
|
SU975778A1 |
| US 2007130645 A1, 07.06.2007 | |||
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННОГО РАСТЕНИЯ КУКУРУЗЫ | 1991 |
|
RU2114911C1 |

