ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] По настоящей заявке испрашивается приоритет временной заявки на патент США 61/504102, поданной 1 июля 2011 г., которая включена в настоящее описание в качестве ссылки в полном объеме.
ВКЛЮЧЕНИЕ СПИСКА ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0002] Список последовательностей содержится в файле под названием «MONS294WO.txt» размером 41 килобайт (согласно оценке Microsoft Windows®), который был создан 29 июня 2012 г., представлен в электронном виде и включен в настоящее описание посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ
[0003] Изобретение в целом относится к области сельского хозяйства, растениеводству и молекулярной биологии. В частности, изобретение относится к способам и композициям для избирательной супрессии экспрессии рекомбинантного белка в репродуктивной ткани мужских особей трансгенных растений и их применению.
УРОВЕНЬ ТЕХНИКИ
[0004] Гибридные семена, т.е. семена, полученные способом гибридизации или перекрестного опыления близкородственных растений, могут быть выращены в потомственные гибридные растения, обладающие требуемой комбинацией признаков, отсутствующих у любого из родительских растений. Гибридные растения могут демонстрировать лучшие агрономические характеристики, включая улучшение размера растений, урожайность, питательную композицию, устойчивость к заболеваниям, толерантность к гербицидам, толерантность к стрессу, климатическую адаптацию и другие желательные признаки. Эффективное получение гибридных семян требует отсутствия самоопыления растения собственной пыльцой.
[0005] При получении гибридных семян у женской особи родительского растения можно предотвратить образование пыльцы и/или сбрасывание пыльцы для облегчения перекрестного опыления женской особи, а не для самоопыления. Такое предотвращение может быть достигнуто, например, удалением вручную содержащих пыльцу структур (например, удаления соцветия метелки у кукурузы ручным или механическим способом), использованием генетических способов контроля опыления (например, цитоплазматическая стерильность мужских особей, ядерная мужская стерильность) и/или использованием химического препарата.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0006] Изобретение относится в целом к способам избирательной супрессии экспрессии рекомбинантного белка в мужской репродуктивной ткани трансгенных растений, конструкциям рекомбинантной ДНК, пригодным в таких способах, а также трансгенным растениям, клеткам и семенам, содержащим такие конструкции рекомбинантной ДНК. Конструкции рекомбинантной ДНК и трансгенные растения, клетки и семена, содержащие такие конструкции, обеспечивают значительно улучшенный способ использования гербицидов для индукции мужской стерильности у трансгенных растений для получения гибридных семян.
[0007] В одном аспекте изобретение предоставляет конструкцию рекомбинантной ДНК, включающую кодирующую белок последовательность, которая кодирует рекомбинантный белок и специфический для мужской ткани элемент миРНК (мтс-миРНК), функционально связанный с кодирующей белок последовательностью. В одном варианте воплощения изобретения элемент мтс-миРНК включен в 3’-нетранслируемый участок кодирующей белок последовательности. В другом варианте воплощения изобретения элемент мтс-миРНК расположен между кодирующей белок последовательностью и последовательностью полиаденилирования, которая является частью 3’-нетранслируемого участка. В другом варианте воплощения изобретения элемент мтс-миРНК включает по меньшей мере одну последовательность мтс-миРНК. В другом варианте воплощения изобретения элемент мтс-миРНК включает по меньшей мере одну последовательность мтс-миРНК, выбранную из группы, состоящей из SEQ ID NO: 1-56 и 105-149. В другом варианте воплощения изобретения элемент мтс-миРНК выбран из группы, состоящей из SEQ ID NO: 57-94 и 96-104. В другом варианте воплощения изобретения экспрессия рекомбинантного белка в трансгенном растении придает растению по меньшей мере вегетативную толерантность к гербициду. В другом варианте воплощения изобретения рекомбинантный белок является толерантным к глифосату EPSPS.
[0008] Другой аспект изобретения предоставляет способ получения конструкции рекомбинантной ДНК, включая идентификацию элемента мтс-миРНК, включая по меньшей мере одну последовательность мтс-миРНК и функционально связанный элемент мтс-миРНК с кодирующей белок последовательностью, например, последовательностью ДНК, кодирующей рекомбинантный белок. В одном варианте воплощения изобретения элемент мтс-миРНК включает по меньшей мере одну последовательность мтс-миРНК, выбранную из группы, состоящей из SEQ ID NO: 1-56 и 105-149, или по меньшей мере один элемент мтс-миРНК, выбранный из группы, состоящей из SEQ ID NO: 57-94 и 96-104. В другом варианте воплощения изобретения элемент мтс-миРНК является специфичным для метелки.
[0009] В дополнительном аспекте изобретение предоставляет трансгенное растение, включая конструкцию рекомбинантной ДНК по изобретению, а также семена, клетку или часть трансгенного растения. В одном варианте воплощения изобретения растение является однодольным растением. В другом варианте воплощения изобретения растение является растением кукурузы (Zea mays).
[0010] В дополнительном аспекте изобретение также предоставляет способ избирательной супрессии экспрессии рекомбинантного белка в мужской репродуктивной ткани трансгенного растения путем экспрессии в трансгенном растении конструкции рекомбинантной ДНК, которая включает кодирующую белок последовательность, функционально связанную с последовательностью ДНК, включая элемент мтс-миРНК. В одном варианте воплощения изобретения элемент мтс-миРНК включает по меньшей мере одну последовательность мтс-миРНК. В другом варианте воплощения изобретения мужская репродуктивная ткань является метелкой растения кукурузы. В другом варианте воплощения изобретения элемент мтс-миРНК включает по меньшей мере одну последовательность мтс-миРНК, выбранную из группы, состоящей из SEQ ID NO: 1-56 и 105-149. В другом варианте воплощения изобретения элемент мтс-миРНК является по меньшей мере одним элементом, выбранным из группы, состоящей из SEQ ID NO: 57-94 и 96-104. В другом варианте воплощения изобретения экспрессия рекомбинантного белка в трансгенном растении придает растению по меньшей мере вегетативную толерантность к гербициду. В другом варианте воплощения изобретения рекомбинантный белок является толерантным к глифосату EPSPS.
[0011] Изобретение также предоставляет способ индукции мужской стерильности у трансгенного растения, включая этап применения гербицида по отношению к трансгенному растению, имеющему в своем геноме конструкцию рекомбинантной ДНК, включающую кодирующую белок последовательность, функционально связанную с последовательностью ДНК, включая элемент мтс-миРНК, который придает трансгенному растению по меньшей мере вегетативную толерантность к гербициду, при этом гербицид применяют во время развития мужской репродуктивной ткани трансгенного растения, тем самым индуцируя мужскую стерильность в трансгенном растении. В одном варианте воплощения изобретения трансгенное растение является растением кукурузы. В другом варианте воплощения изобретения применение гербицида предотвращает по меньшей мере опадение пыльцы или выпячивание пыльников в обработанном трансгенном растении. В другом варианте воплощения изобретения стадию развития мужской репродуктивной ткани, во время которой используют гербицид, выбранный из группы, состоящей из стадии развития растения кукурузы V4, V5, V6, V7, V8, V9, V10, V11, V12, V13 и V14. В другом варианте воплощения изобретения гербицид выбран из группы, состоящей из ингибиторов ацетил коэнзим A карбоксилазы (ACCase), ингибиторов ацетолактатсинтазы (ALS), ингибиторов фотосистемы II (PSII), ингибиторов протопорфириногеноксидазы (PPO), ингибиторов 4-гидроксифенилдиоксигеназы (HPPD), ингибиторов 5-энолипирувилшикимат 3-фосфатсинтазы (EPSPS), ингибиторов глутаминсинтетазы (GS) и синтетических ауксинов. В другом варианте воплощения изобретения гербицид является глифосатом, а рекомбинантный белок является толерантным к глифосату EPSPS.
[0012] Изобретение также предоставляет способ получения гибридных семян, включая применение эффективного количества гербицида по отношению к трансгенному растению, включающему в своем геноме конструкцию рекомбинантной ДНК, включающую кодирующую белок последовательность, функционально связанную с последовательностью ДНК, включая элемент мтс-миРНК, при этом гербицид применяют во время развития мужской репродуктивной ткани трансгенного растения, тем самым индуцируя мужскую стерильность у трансгенного растения; опыление трансгенного растения пыльцой от второго растения; и сбор гибридных семян от трансгенного растения. В одном варианте воплощения изобретения трансгенное растение является кукурузой. Эффективное количество гербицида является дозой гербицида, достаточной для придания трансгенному растению, включающему конструкцию рекомбинантной ДНК по изобретению, мужской стерильности (эффективная доза). В другом варианте воплощения изобретения гербицид является глифосатом, а рекомбинантный белок является толерантным к глифосату EPSPS. В другом варианте воплощения изобретения глифосат применяют во время развития в эффективной дозе от примерно 0,125 фунтов кислоты в эквиваленте на акр до примерно 8 фунтов кислоты в эквиваленте на акр. Другие специфические варианты воплощения изобретения описаны в представленном ниже подробном описании. В тексте данной заявки и формуле изобретения, если в контексте не указано иначе, слово «включает» и его вариации, такие как «содержит» и «содержащий», будет пониматься как подразумевающее включение указанного целого числа, элемента или этапа, или группы целых чисел, элементов или этапов, но не исключение любого другого целого числа, элемента или этапа, или группы целых чисел, элементов или этапов.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0013] Фиг.1 отображает картирование последовательностей мтс-миРНК на элементе мтс-миРНК (SEQ ID NO: 85), как это описано в примере 1. Ось Х слева направо представляет ориентацию элемента мтс-миРНК, при этом верхняя спираль представлена в верхней половине диаграммы, а нижняя спираль представлена в нижней половине диаграммы; нуклеотидная позиция в направлении от 5’ до 3’ показана слева направо вверху и справа налево внизу. Последовательности мтс-миРНК показаны в соответствующих позициях выравнивания. Ось Y представляет относительное обилие мтс-миРНК, экспрессируемой в ткани метелки в виде транскриптов на четверть миллиона последовательностей (tpq). Мтс-миРНК, представленные в библиотеке более часто, обведены кружком.
[0014] Фиг.2 отображает анализ нозерн-блоттинг для измерения экспрессии мРНК, специфической для метелки, как это описано в примере 2.
[0015] Фиг.3 отображает картирование последовательностей мтс-миРНК на элементе мтс-миРНК (SEQ ID NO: 87), как это описано в примерах 2 и 8. Ось Х слева направо представляет ориентацию элемента мтс-миРНК, при этом верхняя спираль представлена в верхней половине диаграммы, и нижняя спираль представлена в нижней половине диаграммы; нуклеотидная позиция в направлении от 5’ до 3’ показана слева направо вверху и справа налево внизу. Последовательности мтс-миРНК показаны в их соответствующих позициях выравнивания. Указаны три последовательности мтс-миРНК, используемые для получения трех специфических зондов (sR648011 (SEQ ID NO: 8), sR1372590 (SEQ ID NO: 26) и sR410590 (SEQ ID NO: 33)).
[0016] Фиг.4 отображает анализ нозерн-блоттинг временной экспрессии элемента мтс-миРНК (SEQ ID NO: 87) в зрелой метелке с использованием РНК от различной инбредной зародышевой плазмы, как это описано в примере 2.
[0017] Фиг.5 отображает локализацию экспрессии миРНК in situ в зрелом пыльнике с использованием антисмысловых (слева) или смысловых (справа) зондов для последовательности мтс-миРНК (sR648011, SEQ ID NO: 8), как это описано в примере 2.
[0018] Фиг.6 отображает локализацию белка CP4-EPSPS в пыльниках от необработанных распылением растений, трансгенных на предмет конструкции 3 (фиг.6А) или конструкции 4 (фиг.6В), как это описано в примере 4. Растения кукурузы, трансгенные на конструкцию 3, содержат кассету экспрессии CP4-EPSPS/элемент мтс-миРНК. Растения с конструкцией 4 являются контролем.
[0019] Фиг.7 отображает трансгенные растения кукурузы, полученные из конструкций, содержащих кассету экспрессии CP4-EPSPS/элемент мтс-миРНК, которые были вегетативно толерантны к глифосату и имели индуцированную мужскую стерильность с поздним применением глифосата, как это описано в примере 7. Фиг.7А отображает трансгенные растения кукурузы с и без обработки глифосатом. Фиг.7В отображает метелки от необработанных трансгенных растений, и фиг.7С отображает пыльцевые зерна необработанных трансгенных растений. Фиг.7D отображает метелки от обработанных трансгенных растений, и фиг.7Е отображает пыльцевые зерна обработанных трансгенных растений.
[0020] Фиг.8 отображает данные для одного года полевых исследований, измеряющих показатель мужской фертильности (ПМФ) после поздней обработки глифосатом. Фиг.8А отображает среднее значение ПМФ, полученное при трех различных режимах обработки глифосатом (Trt 1, Trt 2 и Trt 3) для NK603 (CP4-EPSPS трансгенная кукуруза), MON 87427 (CP4-EPSPS трансгенная кукуруза с индуцированной глифосатом мужской стерильностью) и двух явлений, связанных с конструкцией 3, как это описано в примере 5; пунктирная линия указывает на промышленный стандарт для мужской стерильности, ПМФ 2. Фиг.8В отображает метелку растения, у которого обработаны распылением только семена. Фиг.8С отображает метелку растения, обработанного распылением глифосатом на поздней стадии для индукции мужской стерильности.
[0021] Фиг.9 отображает результаты полевого исследования, измеряющие количество растений на делянке, при этом мужская стерильность измерена по выпячиванию пыльника через S90, при S90+3 и S90+6, при двух различных режимах обработки глифосатом способом распыления (Trt 2 и Trt 3) для NK603 (CP4-EPSPS трансгенная кукуруза), MON 87427 (CP4-EPSPS трансгенная кукуруза с индуцированной глифосатом мужской стерильностью) и четырех явлений, связанных с конструкцией 3, как это описано в примере 5.
[0022] Фиг.10 отображает результаты исследований жизнеспособности пыльцы, как это описано в примере 5. Фиг.10А и 10В отображают пример позднего выпячивания пыльников в метелке при явлении распыления при наличии стерильности и конструкции 3. Квадрат на фиг.10А является участком, увеличенным на фиг.10В. Пример позднего выпячивания пыльника обведен кружком на Фигуре 10В. Окрашивание пыльцы из стерильного пыльника с поздним выпячиванием при явлениях распыления конструкции 3 по Alexander выявило только нежизнеспособную пыльцу (полупрозрачный голубой цвет, неправильная форма пыльцевых зерен) (фиг.10С). Пыльца от пыльников, необработанных конструкцией 3, была полностью жизнеспособной и при окрашивании по Alexander была непрозрачной, темно-фиолетовой и сферической (фиг.10D).
[0023] Фиг.11 отображает результаты полевого исследования, изучавшего растения NK603 и явления конструкции 3 для получения инбредных зерен и мужской фертильности, как это описано в примере 6. Урожай инбредных зерен был измерен в значениях бушели/акр, а индуцированная мужская стерильность была измерена как показатель мужской фертильности (ПМФ). Горизонтальные усы указывают промышленный стандарт для мужской стерильности, ПМФ 2. Trt 1, Trt 2 и Trt 3 обозначают режимы обработки 1, 2 и 3.
[0024] Фиг.12 отображает результаты полевого исследования, изучавшего нетрансгенные женские инбредные растения (Null), линия MON87427, и три явления конструкции 3, все с одинаковой генетической основой, которые были перекрестно опылены мужским тестером MON810/MON88017 для получения гибридных семян F1. Урожай гибридных зерен был измерен в значениях бушели/акр. Trt 1, Trt 2 и Trt 3 обозначают режимы обработки 1, 2 и 3.
[0025] Фиг.13 отображает анализ пыльцевых зерен из гибридных растений F1, как это описано в примере 7. Панели отображают результаты окрашивания пыльцы по Alexander, полученной в результате трех различных гибридных скрещиваний F1: нетрансгенная женская особь x MON88017 мужская особь; MON87427 женская особь x MON88017 мужская особь; и явление конструкции 3 женская особь x MON88017 мужская особь. Фертильность метелки была функционально восстановлена в гибридах F1, полученных от растений с явлением конструкции 3 с использованием пыльцы MON88017.
[0026] Фиг.14 отображает схематические чертежи вариантов воплощения конструкций рекомбинантной ДНК (представлено в направлении от 5’ к 3’ слева направо), включая (вверху) кодирующую белок последовательность (например, ДНК, кодирующую устойчивый к глифосату EPSPS), функционально связанную с последовательностью ДНК, включающей элемент мтс-миРНК (например, одну или более, выбранных из группы, состоящей из SEQ ID NO: 57-94 и 96-104) (вверху). В неограничивающем специфическом варианте воплощения изобретения (внизу) конструкция рекомбинантной ДНК включает промотор, функционально связанный, по порядку, с интроном, транзитным пептидом, CP4-EPSPS, кодируемым SEQ ID NO: 95, элементом мтс-миРНК (SEQ ID NO: 81) и 3’UTR.
ДЕТАЛЬНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Конструкции рекомбинантной ДНК
[0027] Изобретение предоставляет композиции и способы для избирательной супрессии экспрессии рекомбинантного белка в репродуктивной ткани мужских особей трансгенных растений и их применение. В одном аспекте изобретение предоставляет конструкцию рекомбинантной ДНК, включающую кодирующую белок последовательность, функционально связанную с последовательностью ДНК, включая элемент мтс-миРНК, т.е. химерный трансген, включающий кодирующую белок последовательность, кодирующую рекомбинантный белок, и по меньшей мере один элемент мтс-миРНК, функционально связанный с кодирующей белок последовательностью. В одном варианте воплощения изобретения такие конструкции рекомбинантной ДНК являются пригодными для избирательной супрессии экспрессии рекомбинантного белка в мужской репродуктивной ткани трансгенного растения. В одном аспекте изобретение предоставляет рекомбинантную молекулу ДНК, включающую конструкцию рекомбинантной ДНК, и способы ее применения. Последовательности нуклеиновых кислот могут быть представлены в качестве ДНК или РНК, в зависимости от того, что указано; описание одной обязательно определяет другую, что известно специалисту в данной области техники. Кроме того, описание данной последовательности нуклеиновой кислоты обязательно определяет точный комплемент такой последовательности, что известно специалисту в данной области техники.
[0028] «Мужская тканеспецифичная миРНК» или «мтс-миРНК» является малой РНК (мРНК) из от примерно 18 до примерно 26 нуклеотидов (например, 18, 19, 20, 21, 22, 23, 24, 25 или 26 нуклеотидов), обогащенной или специфически экспрессируемой в мужской репродуктивной ткани(ях) (например, мужском соцветии) растения, т.е. имеющей мужскую тканеспецифичную схему экспрессии. Мужская тканеспецифичная миРНК встречается естественным образом у растений и может быть определена с использованием известных в данной области техники способов, таких как низкомолекулярный нозерн-анализ. Последовательность ДНК, являющаяся комплементарной мтс-миРНК, обозначается в настоящем описании как «последовательность мтс-миРНК». Примеры последовательностей мтс-миРНК для мтс-миРНК эндогенного растения представлены как SEQ ID NO: 1-56 и 105-149. В варианте воплощения изобретения последовательность мтс-миРНК является точным комплементом ДНК (без несоответствий) для заданной мтс-миРНК. В других вариантах воплощения изобретения последовательность мтс-миРНК варьирует в пределах 1-3 нуклеотидных несоответствий в сравнении с заданной мтс-миРНК, но при этом обладает достаточной комплементарностью для связывания или гибридизации, например, при обычных физиологических условиях, с такой мтс-миРНК. «Комплементарность» обозначает способность нуклеотидов в одной полинуклеотидной цепи образовывать пары с нуклеотидами другой полинуклеотидной цепи в соответствии со стандартными правилами комплементарности Уотсона-Крика (т.е. пары гуанина с цитозином (Г:Ц) и пары аденина с тимином (А:Т) или урацилом (А:У); возможна внутрицепочечная гибридизация между двумя или более комплементарных участков отдельного полинуклеотида. При включении в конструкцию рекомбинантной ДНК, как это описано в настоящем описании, мтс-миРНК способна осуществлять миРНК-опосредованную супрессию или нарушение экспрессии гена и/или белка.
[0029] По меньшей мере один, по меньшей мере два, по меньшей мере три или более трех последовательностей мтс-миРНК могут быть соединены в кластер или даже перекрываться в одной молекуле ДНК. Такая молекула ДНК обозначается в настоящем описании как «мужской тканеспецифичный элемент миРНК» или «элемент мтс-миРНК» и определяется как включающая по меньшей мере одну, по меньшей мере две, по меньшей мере три или более трех последовательностей мтс-миРНК в окне последовательности из примерно 500 нуклеотидов. Элемент мтс-миРНК может иметь любую длину, такую как примерно 20 нуклеотидов (нт), примерно 25 нт, примерно 30 нт, примерно 40 нт, примерно 50 нт, примерно 60 нт, примерно 70 нт, примерно 80 нт, примерно 100 нт, примерно 150 нт, примерно 200 нт, примерно 250 нт, примерно 300 нт, примерно 350 нт, примерно 400 нт, примерно 450 нт, примерно 500 нт, примерно 550 нт или примерно 600 нт.
[0030] Конструкция рекомбинантной ДНК по изобретению является молекулой ДНК, включающей по меньшей мере кодирующую белок последовательность, функционально связанную с последовательностью ДНК, включая элемент мтс-миРНК. Термин «рекомбинантный» обозначает молекулу или клетку, или организм, который был получен искусственно посредством генной инженерии и, таким образом, является продуктом деятельности человека и обычно не встречается иным образом в природе. Как используется в настоящем описании, конструкция рекомбинантной ДНК является молекулой рекомбинантной ДНК, включающей две или более гетерологичных последовательностей ДНК. Термин «гетерологичный» обозначает взаимосвязь между двумя или более последовательностей нуклеиновых кислот или белка, которые были получены из различных источников (например, из разных мест в геноме или от различных видов). В одном примере промотор и кодирующая белок последовательность ДНК являются гетерологичными относительно друг друга, если промотор не является нативным промотором кодирующей белок последовательности ДНК. В другом примере кодирующая белок последовательность является гетерологичной относительно элемента мтс-миРНК, если такая комбинация не встречается естественным образом в природе, например, элемент мтс-миРНК растения, функционально связанный с геном толерантности к гербициду, такому как CP4-EPSPS. Кроме того, отдельная последовательность может быть «гетерологичной» относительно клетки или организма, в которую ее вводят (т.е. последовательность, не встречающаяся естественным образом в такой отдельной клетке или организме).
[0031] Термин «функционально связанный» обозначает две молекулы полинуклеотидов, связанных таким образом, что одна молекула может влиять на экспрессию другой. Например, первая молекула полинуклеотида функционально связана со второй молекулой полинуклеотида, при этом молекулы полинуклеотидов располагаются таким образом, что первая молекула полинуклеотида может влиять на экспрессию другой молекулы полинуклеотида. Две молекулы полинуклеотидов могут быть частью отдельной неразрывной молекулы полинуклеотида и могут быть смежными или разделенными. Например, элемент мтс-миРНК функционально связан с кодирующей белок последовательностью, если, после транскрипции в клетке мужской репродуктивной ткани, присутствие элемента мтс-миРНК приводит к супрессии экспрессии рекомбинантного белка в клетке. Функциональная связь кодирующей белок последовательности и элемента мтс-миРНК может быть достигнута, например, посредством введения элемента мтс-миРНК, прилегающего к кодирующей белок последовательности (такой как расположенная на 5’ или 3’ конце кодирующей белок последовательности, но не обязательно смежным образом), в или прилегающего к нетранслируемому участку (UTR) конструкции рекомбинантной ДНК (такой как расположенная в или после 5’ UTR или 3’ UTR) и/или после кодирующей белок последовательности и перед сигналом полиаденилирования. В одном варианте воплощения изобретения один или более элементов мтс-миРНК располагаются между кодирующей белок последовательностью и последовательностью полиаденилирования, т.е. в направлении 3’ и прилегая к кодирующей белок последовательности. В другом варианте воплощения изобретения один или более элементов мтс-миРНК расположены между стоп-кодоном кодирующей белок последовательности и последовательностью полиаденилирования. В другом варианте воплощения изобретения один или более элементов мтс-миРНК расположены в пределах 3’ UTR последовательности, прилегающей к кодирующей белок последовательности.
[0032] Последовательность ДНК элемента мтс-миРНК может варьировать при использовании различных комбинаций и расположений отдельных последовательностей мтс-миРНК и/или при введении 1-3 нуклеотидных несоответствий в элементе мтс-миРНК (относительно заданной последовательности мтс-миРНК). Примеры элементов мтс-миРНК представлены в настоящем описании как SEQ ID NO: 57-94 и 96-104 и в рабочих Примерах. Элемент мтс-миРНК может функционировать в любом направлении, т.е. он ненаправленный и, таким образом, он может использоваться в направлении от 5’ к 3’ или в направлении от 3’ к 5’ в конструкции рекомбинантной ДНК.
[0033] Элементы мтс-миРНК, последовательности мтс-миРНК и мтс-миРНК могут быть идентифицированы способами, известными специалистам в данной области, например, посредством биоинформационного анализа мРНК растений и библиотек кДНК. Пример такого способа идентификации представлен в примерах ниже. В частности, мтс-миРНК и последовательности мтс-миРНК могут быть идентифицированы из библиотек мРНК. Идентифицированные последовательности мтс-миРНК могут быть сопоставлены с кДНК и/или геномными коллекциями последовательностей для идентификации элементов мтс-миРНК (т.е. участков ДНК, включающих по меньшей мере одну, по меньшей мере две, по меньшей мере три или более трех последовательностей мтс-миРНК в окне из 500 нуклеотидов в последовательности), которые пригодны для разработки конструкций рекомбинантной ДНК, как это описано в настоящем описании.
[0034] В некоторых вариантах воплощения изобретения такие элементы мтс-миРНК синтезируют или модифицируют in vitro для содержания большего, меньшего или различного количества последовательностей мтс-миРНК и/или для изменения относительного положения одной или более последовательностей мтс-миРНК, при этом такая модификация является преимущественной в увеличении или снижении эффекта элемента мтс-миРНК. Способы синтеза или in vitro модификации элемента мтс-миРНК и определения оптимальной вариации требуемого уровня супрессии известны специалистам в данной области. Химерные элементы мтс-миРНК также могут быть получены с использованием способов, известных специалистам в данной области, таких как вставка дополнительных требуемых последовательностей мтс-миРНК внутри элемента мтс-миРНК или присоединение дополнительных последовательностей мтс-миРНК на 5’ или 3’ конце относительно элемента мтс-миРНК. Неограничивающие варианты воплощения химерного элемента мтс-миРНК включают элементы мтс-миРНК, имеющие примерно 80 нт, примерно 100 нт, примерно 150 нт, примерно 200 нт, примерно 250 нт или примерно 300 нт последовательности SEQ ID NO: 86; примерно 80 нт, примерно 100 нт, примерно 150 нт, примерно 200 нт, примерно 250 нт или примерно 300 нт последовательности SEQ ID NO: 87; и/или примерно 80 нт, примерно 100 нт, примерно 150 нт, примерно 200 нт, примерно 250 нт, примерно 300 нт, примерно 350 нт, примерно 400 нт, примерно 450 нт, примерно 500 нт или примерно 550 нт последовательности SEQ ID NO: 85. Дополнительные варианты воплощения изобретения представлены в рабочих Примерах.
[0035] Конструкция рекомбинантной ДНК может использоваться для избирательной супрессии экспрессии рекомбинантного белка в мужских репродуктивных тканях трансгенного растения, экспрессирующих конструкцию, т.е. для получения экспрессии по меньшей мере в вегетативных тканях, но не в мужских репродуктивных тканях. Как используется в настоящем описании, «экспрессия рекомбинантного белка» обозначает выработку рекомбинантного белка из кодирующей белок последовательности и полученного транскрипта (иРНК) в клетке. Как используется в настоящем описании, термин «супрессия» обозначает снижение; например, супрессия экспрессии рекомбинантного белка обозначает снижение уровня рекомбинантного белка, вырабатываемого в клетке, например, посредством посттранскрипционной супрессии гена, опосредованной интерференцией РНК.
[0036] Избирательная супрессия рекомбинантного белка, как используется в настоящем описании, обозначает снижение выработки рекомбинантного белка в клетке или ткани в сравнении с эталонной клеткой или тканью по меньшей мере примерно на 75%, по меньшей мере примерно на 80%, по меньшей мере примерно на 85%, по меньшей мере примерно на 90%, по меньшей мере примерно на 95% или по меньшей мере примерно на 99%. Эталонная клетка или ткань может быть, например, вегетативной клеткой или тканью от одного и того же или подобного трансгенного растения, экспрессирующего рекомбинантный белок, или, например, вегетативной клеткой или тканью трансгенного растения, имеющего подобный трансген для экспрессии рекомбинантного белка, но с отсутствием элемента мтс-миРНК. Супрессия экспрессии белка может быть определена с использованием любого способа, известного специалисту в данной области, такого как непосредственное измерение накопления белка в образце клетки или ткани с использованием способа, такого как ELISA или вестерн-блоттинг, путем измерения ферментативной активности белка или путем фенотипичного определения экспрессии белка. В одном варианте воплощения изобретения избирательная супрессия рекомбинантного белка обозначает достаточное снижение экспрессии рекомбинантного белка, способного придавать толерантность к гербициду в мужской ткани трансгенного растения, что приводит к определяемому фенотипу измененной мужской фертильности у трансгенного растения, по отношению к которому был применен гербицид в качестве воздействия на стерильность. Определение измененной мужской фертильности у такого трансгенного растения, таким образом, будет указывать на избирательную супрессию рекомбинантного белка.
[0037] Как используется в настоящем описании, термин «кодирующая белок последовательность» обозначает молекулу полинуклеотида, имеющую нуклеотидную последовательность, кодирующую полипептидную или белковую последовательность, т.е. полипептидную последовательность, кодирующую рекомбинантный белок. В зависимости от условий нуклеотидная последовательность может фактически транслироваться или не транслироваться в полипептидную молекулу в клетке. Границы кодирующей белок последовательности обычно определяются старт-кодоном трансляции на 5'-конце и стоп-кодоном трансляции на 3'-конце. Кодирующая белок последовательность по изобретению включает, но не ограничивается этим, кодирующую белок последовательность, которая обеспечивает требуемые характеристики, связанные с морфологией, физиологией, ростом и развитием растения, урожайностью, повышением питательности, устойчивостью к болезням или вредителям, толерантностью к гербицидам или толерантностью к окружающей среде или химическим препаратам. В одном варианте воплощения изобретения кодирующая белок последовательность по изобретению кодирует рекомбинантный белок, который, при экспрессии в трансгенном растении, придает толерантность к гербицидам по меньшей мере клетке и/или ткани, в которой возникает экспрессируемый белок; избирательная супрессия белка толерантности к гербициду в мужской репродуктивной ткани трансгенного растения в сочетании со своевременным применением гербицида приводит по меньшей мере к пониженной мужской фертильности или к мужской стерильности. Такая индуцируемая мужская стерильность в комбинации с вегетативной толерантностью к гербицидам может использоваться для повышения эффективности, с которой получают гибридные семена, например, способом элиминации или снижения необходимости физического удаления пестиков у растения кукурузы, используемого в качестве женского в заданном скрещивании во время получения гибридных семян. Системы индуцируемой гербицидами мужской стерильности были описаны, например, в патенте США № 6762344 и публикации патента США 2011/0126310. Примеры гербицидов, пригодных при практическом применении изобретения, включают, но не ограничиваются этим, ингибиторы ацетил коэнзим A карбоксилазы (ACCase) (например, т.н. fops и dims), ингибиторы ацетолактатсинтазы (ALS) (например, сульфонилмочевины (SU) и имидазолиноны (IMI)), ингибиторы фотосистемы II (PSII) (например, триазины и фениловые эфиры), ингибиторы протопорфириногеноксидазы (PPO) (например, флумиоксазин и фомесафен), ингибиторы 4-гидроксифенилпируватдиоксигеназы (HPPD) (например, изоксафлутол и трикетоны, такие как мезотрион), ингибиторы 5-энолипирувилшикимат 3-фосфатсинтазы (EPSPS) (например, глифосат), ингибиторы глутаминсинтетазы (GS) (например, глуфосинат и фосфинотрицин), синтетические ауксины (например, 2,4-D и дикамба). Примеры кодирующих белок последовательностей и/или рекомбинантных белков для использования в практическом применении изобретения включают, но не ограничиваются этим, гены, кодирующие рекомбинантные белки, придающие толерантность к ингибиторам HPPD (таким как нечувствительный к гербицидам HPPD), гены, кодирующие рекомбинантные белки, придающие толерантность к глюфосинату (такому как pat и bar), гены, кодирующие рекомбинантные белки, придающие толерантность к глифосату (такому как толерантный к глифосату EPSPS, известному как CP4-EPSPS, представленному в настоящем описании как SEQ ID NO: 95), и гены, кодирующие рекомбинантные белки, придающие толерантность к дикамбе (такой как дикамба монооксигеназа (DMO)).
[0038] Конструкции рекомбинантной ДНК по изобретению получают способами, известными в данной области техники, и в различных вариантах воплощения изобретения они включены в векторы трансформации растения, плазмиды или ДНК пластид. Такие конструкции рекомбинантной ДНК пригодны для получения трансгенных растений и/или клеток и, таким образом, они также могут содержаться в геномной ДНК трансгенного растения, семени, клетки или части растения. Следовательно, изобретение включает варианты воплощения, в которых конструкция рекомбинантной ДНК расположена в векторе трансформации растения или на биолистической частице для трансформации клетки растения, или в хромосоме, или пластиде клетки трансгенного растения, или в трансгенной клетке, ткани трансгенного растения, семени трансгенного растения, пыльцевом зерне трансгенного растения или в трансгенном или частично трансгенном (например, привитом) растении. Вектором является любая молекула ДНК, которая может использоваться в целях трансформации растения, т.е. введения ДНК в клетку. Конструкции рекомбинантной ДНК по изобретению могут быть, например, вставлены в вектор трансформации растения и использоваться для трансформации растения с целью получения трансгенных растений, семян и клеток. Способы построения векторов трансформации растения хорошо известны в данной области техники. Векторы трансформации растения по изобретению обычно включают, но не ограничиваются этим: подходящий промотор для экспрессии функционально связанной ДНК, функционально связанную конструкцию рекомбинантной ДНК и сигнал полиаденилирования (который может быть включен в последовательность 3’UTR). Промоторы, пригодные для практического применения изобретения, включают промоторы, функционирующие в растении для экспрессии функционально связанного полинуклеотида. Такие промоторы различны и хорошо известны в данной области техники, и включают промоторы, являющиеся индуцируемыми, вирусными, синтетическими, конститутивными, временно регулируемыми, пространственно регулируемыми и/или пространственно-временно регулируемыми. Дополнительные необязательные компоненты включают, но не ограничиваются этим, один или более следующих элементов: 5’ UTR, энхансер, цис-действующий элемент, интрон, сигнальная последовательность, транзитная пептидная последовательность и один или более избирательных маркерных генов. В одном варианте воплощения изобретения вектор трансформации растения включает конструкцию рекомбинантной ДНК.
[0039] Конструкции рекомбинантной ДНК и векторы трансформации растения по данному изобретению получают любым способом, подходящим для намеченного применения, принимая во внимание, например, тип требуемой экспрессии, требуемую кодирующую белок последовательность (и, таким образом, толерантность к гербицидам), а также удобство использования в растении, в котором конструкция рекомбинантной ДНК должна экспрессироваться. Общие способы, пригодные для работы с молекулами ДНК для получения и применения конструкций рекомбинантной ДНК и векторов трансформации растения, хорошо известны в данной области техники и описаны подробно, например, в справочниках и лабораторных указателях, включая Sambrook and Russell, “Molecular Cloning: A Laboratory Manual” (third edition), Cold Spring Harbor Laboratory Press, NY, 2001. Конструкции рекомбинантной ДНК по изобретению могут быть модифицированы способами, известными в данной области техники, полностью или частично, например, для повышенного удобства работы с ДНК (такие как сайты распознавания рестриктазы или сайты клонирования на основе рекомбинации) или для включения преимущественных для растения последовательностей (таких как использование растительного кодона или консенсусных последовательностей Kozak), или для включения последовательностей, пригодных для разработки конструкции рекомбинантной ДНК (таких как последовательности спейсера или линкера). В определенных вариантах воплощения изобретения последовательность ДНК конструкции рекомбинантной ДНК включает последовательность ДНК, которая была оптимизирована кодоном для растения, в котором необходима экспрессия конструкции рекомбинантной ДНК. Например, конструкция рекомбинантной ДНК для экспрессии в растении может иметь все или части оптимизированной кодоном последовательности для экспрессии в растении известными в данной области техники способами. Конструкции рекомбинантной ДНК по изобретению могут находиться совместно с другими рекомбинантными ДНК для придания дополнительных признаков (например, в случае трансформированных растений, признаком, включающим резистентность к гербицидам, резистентность к вредителям, толерантность к прорастанию в холодных условиях, толерантность к дефициту воды), например, способом экспрессии или супрессии других генов.
Клетки трансгенного растения и трансгенные растения
[0040] Аспект изобретения включает клетки трансгенного растения, ткани трансгенного растения и трансгенные растения или семена, которые включают конструкцию рекомбинантной ДНК по изобретению. Дополнительный аспект по изобретению включает искусственные или рекомбинантные хромосомы растения, которые включают конструкцию рекомбинантной ДНК по изобретению. Подходящие способы трансформации клеток растения-хозяина для использования в данном изобретении включают фактически любой способ введения ДНК в клетку (например, если конструкция рекомбинантной ДНК стабильно интегрирована в хромосому растения) и хорошо известны данной области техники. Типичный и широко используемый способ введения конструкции рекомбинантной ДНК в растения представлен системой трансформации Agrobacterium, которая хорошо известна специалистам в данной области. Трансгенные растения могут быть регенерированы из клетки трансформированного растения способами культивации растительной клетки. Трансгенное растение, гомозиготное относительно трансгена, может быть получено половым скрещиванием (самоопылением) независимого сегреганта трансгенного растения, которое содержит одну экзогенную генную последовательность для самого себя, например, растение F0, для получения семян F1. Одна четвертая часть полученных семян F1 будет гомозиготна относительно трансгена. Растения, выращенные из пророщенного семени F1, могут быть тестированы на предмет гетерозиготности обычно с использованием анализа ОНП или анализа термальной амплификации, который позволяет различить гетерозиготы и гомозиготы (т.е. анализ зиготности).
[0041] Изобретение предоставляет трансгенное растение, имеющее в своем геноме конструкцию рекомбинантной ДНК по изобретению, включая, без ограничения, помимо всего прочего, люцерну, хлопок, кукурузу, рапс, рис, сою и пшеницу. Изобретение также предоставляет клетки трансгенного растения, части растения и потомство такого трансгенного растения. В контексте данного изобретения «потомство» включает любое растение, семя, клетку растения и/или часть растения, полученные или регенерированные из растения, семени, клетки растения и/или части растения, которые включают конструкцию рекомбинантной ДНК по изобретению. Трансгенные растения, клетки, части и семена, полученные из таких растений, могут быть гомозиготными или гетерозиготными на предмет конструкции рекомбинантной ДНК по изобретению.
[0042] Кроме того, данное изобретение включает варианты воплощения, в которых конструкция рекомбинантной ДНК находится в готовом продукте, полученном из трансгенного растения, семени или части растения по данному изобретению; такие готовые продукты включают, но не ограничиваются этим, собранные части растения, дробленные или цельные зерна или семена растения, или любой пищевой или непищевой продукт, включающий конструкцию рекомбинантной ДНК по изобретению.
Способы индуцирования мужской стерильности у трансгенных растений и получения гибридных семян
[0043] Другой аспект изобретения включает способ индуцирования мужской стерильности у трансгенного растения, включающий применение эффективного количества гербицида относительно трансгенного растения, включающего конструкцию рекомбинантной ДНК, которая включает кодирующую белок последовательность, кодирующую рекомбинантный белок, который придает трансгенному растению толерантность к гербициду, и функционально связанную с последовательностью ДНК, включающей элемент мтс-миРНК, который придает трансгенному растению по меньшей мере вегетативную толерантность к гербициду, при этом применение гербицида проводится во время развития мужской репродуктивной ткани трансгенного растения, что индуцирует мужскую стерильность у трансгенного растения.
[0044] В одном варианте воплощения изобретения трансгенное растение является растением кукурузы. В одном варианте воплощения изобретения применение гербицида предотвращает по меньшей мере осыпание пыльцы или выпячивание пыльников. В одном варианте воплощения изобретения развитие мужской репродуктивной ткани является стадией, выбранной из группы, состоящей из стадии развития растения кукурузы V4, V5, V6, V7, V8, V9, V10, V11, V12, V13 и V14.
[0045] В одном варианте воплощения изобретения гербицид выбран из группы, состоящей из ингибиторов ацетил коэнзим A карбоксилазы (ACCase), ингибиторов ацетолактатсинтазы (ALS), ингибиторов фотосистемы II (PSII), ингибиторов протопорфириногеноксидазы (PPO), ингибиторов 4-гидроксифенил пируват диоксигеназы (HPPD), ингибиторов 5-энолипирувилшикимат 3-фосфатсинтазы (EPSPS), ингибиторов глутаминсинтетазы (GS) и синтетических ауксинов. В одном варианте воплощения изобретения гербицид является глифосатом, а рекомбинантный белок является толерантным к глифосату EPSPS.
[0046] Дополнительный аспект изобретения включает способ получения гибридных семян, включающий: (а) применение гербицида к трансгенному растению, включающему конструкцию рекомбинантной ДНК, которая включает кодирующую белок последовательность, кодирующую рекомбинантный белок, который придает трансгенному растению толерантность к гербициду, и функционально связанную с последовательностью ДНК, включающей элемент мтс-миРНК, при этом применение гербицида проводится во время развития мужской репродуктивной ткани трансгенного растения, что приводит к индуцированию мужской стерильности в трансгенном растении; (б) опыление трансгенного растения пыльцой второго растения и (в) сбор гибридных семян трансгенного растения. В одном варианте воплощения изобретения трансгенное растение является кукурузой. В одном варианте воплощения изобретения гербицид является глифосатом, а рекомбинантный белок является толерантным к глифосату EPSPS. В одном варианте воплощения изобретения глифосат применяют во время развития в эффективной дозе от примерно 0,125 фунтов кислоты в эквиваленте на акр до примерно 8 фунтов кислоты в эквиваленте на акр.
[0047] В другом аспекте изобретение включает гибридное семя, собранное у трансгенного растения с мужской стерилизацией, которое было опылено пыльцой второго растения, при этом мужское стерильное трансгенное растение включает конструкцию рекомбинантной ДНК, которая включает кодирующую белок последовательность, кодирующую рекомбинантный белок, который придает трансгенному растению толерантность к гербицидам, функционально связанную с последовательностью ДНК, включающей элемент мтс-миРНК, и при этом у трансгенного растения была индуцирована мужская стерильность применением эффективного количества гербицида в течение развития мужской репродуктивной ткани трансгенного растения. В одном варианте воплощения изобретения гибридное семя является гибридным семенем трансгенной кукурузы. В одном варианте воплощения изобретения гербицид является глифосатом, а рекомбинантный белок является толерантным к глифосату EPSPS. В одном варианте воплощения изобретения глифосат применяют во время развития в эффективной дозе от примерно 0,125 фунтов кислоты в эквиваленте на акр до примерно 8 фунтов кислоты в эквиваленте на акр. В одном варианте воплощения изобретения применение гербицида предотвращает по меньшей мере осыпание пыльцы или выпячивание пыльников. В одном варианте воплощения изобретения развитие мужской репродуктивной ткани является стадией, выбранной из группы, состоящей из стадии развития растения кукурузы V4, V5, V6, V7, V8, V9, V10, V11, V12, V13 и V14.
ПРИМЕРЫ
Пример 1
[0048] Данный пример описывает идентификацию мтс-миРНК и элементов мтс-миРНК. Биоинформационный анализ данных секвенирования из нескольких библиотек малых РНК кукурузы идентифицировал группу малых РНК (мРНК), которые находились в избыточном количестве или специфически экспрессировались в метелках кукурузы. Относительное распространение таких мтс-миРНК в метелках кукурузы варьировало от примерно 50 до 631 транскрипта на четверть миллиона последовательностей, что является нормализованным количеством. Такие мРНК идентифицированы в качестве миРНК по причине их длины (18-26 нуклеотидов) и их вероятного происхождения от предшественника дсРНК. По причине своей схемы экспрессии миРНК, специфическую для мужской ткани, обозначают как «мтс-миРНК». В контексте данного изобретения «схема экспрессии» является любой схемой дифференциальной экспрессии ДНК, РНК или белка. Например, специфическая для метелки схема экспрессии обозначает специфическую или повышенную экспрессию ДНК, РНК или белка в ткани метелки и/или клетки. Примеры соответствующей последовательности ДНК для мтс-миРНК, обозначенные в настоящем описании как «последовательности мтс-миРНК», представлены в виде SEQ ID NO: 1-56 и 105-149.
[0049] Такие последовательности мтс-миРНК затем были сопоставлены с коллекциями последовательностей кДНК. Сравнение последовательности мтс-миРНК относительно коллекции генов кукурузы unigene (сборные последовательности кДНК) с использованием BLAST привело к удивительному результату о том, что большое количество мтс-миРНК было собрано в виде кластеров вместе и отмечались даже перекрывания в участке ДНК, находящемся в нескольких близкородственных, но при этом уникальных, последовательностях кДНК. В группе последовательностей кДНК все последовательности содержали такой участок, однако последовательность ДНК участка варьировала по причине различных комбинаций и расположений отдельных последовательностей мтс-миРНК и/или 1-3 нуклеотидных несоответствий отдельным последовательностям мтс-миРНК. Такой участок, определенный как имеющий по меньшей мере одну последовательность мтс-миРНК в окне нуклеотидной последовательности, обозначается в настоящем описании как «элемент мтс-миРНК». В различных вариантах воплощения изобретения окно нуклеотидной последовательности включает по меньшей мере примерно 20 смежных нуклеотидов (нт) (например, по меньшей мере 18, 19, 20, 21, 22, 23 или 24 нт), по меньшей мере примерно 25 нт, по меньшей мере примерно 30 нт, по меньшей мере примерно 40 нт, по меньшей мере примерно 50 нт, по меньшей мере примерно 100 нт или по меньшей мере примерно 150 нт. Примеры последовательности ДНК для элементов мтс-миРНК представлены в настоящем описании как SEQ ID NO: 57-94 и 96-104. Элемент мтс-миРНК может иметь более чем одну последовательность мтс-миРНК, например, по меньшей мере две, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять или более пяти последовательностей мтс-миРНК в заданном окне нуклеотидной последовательности. Две или более последовательностей мтс-миРНК в заданном элементе мтс-миРНК могут перекрываться по причине того, что по меньшей мере часть их нуклеотидных последовательностей является идентичной (см. Таблицу 5, где представлены примеры мтс-миРНК с перекрывающимися нуклеотидными последовательностями).
[0050] Биоинформационный анализ показал, что множественные мтс-миРНК могут быть получены из одного и того же транскрипта РНК, например, транскрипта, полученного из одной из последовательностей кДНК, описанных выше как включающих элемент мтс-миРНК. Также было выявлено, что множество мтс-миРНК имеют 1-3 несоответствия при сравнении с элементами мтс-миРНК в группе близкородственных последовательностей кДНК. Считается, что это указывает на то, что такие мтс-миРНК получают из множественных близкородственных транскриптов, что приводит к получению большой близкородственной группы мтс-миРНК. Таким образом, транскрипт РНК, полученный из кДНК, включающей элемент мтс-миРНК (содержащий множество последовательностей мтс-миРНК), будет комплементарен и, следовательно, способен гибридизировать во множественные мтс-миРНК и/или их комплементы. Таким образом, встречающаяся в природе мтс-миРНК имеет последовательность РНК, которая является идеальным или почти идеальным комплементом последовательности мтс-миРНК (например, когда мтс-миРНК имеет последовательность РНК не более чем примерно с 1-3 несоответствиями относительно последовательности мтс-миРНК); в целом такая мтс-миРНК имеет последовательность РНК, являющуюся идеальным или почти идеальным комплементом к сегменту элемента мтс-миРНК.
[0051] Поиск подобия последовательностей мтс-миРНК относительно базы данных геномной ДНК кукурузы с использованием BLAST выявил множество локусов с подобием по существу относительно элемента мтс-миРНК. Такие локусы затем были проанализированы с использованием открытых рамок считывания (ORF), однако идентифицированные гипотетические полипептиды, как было определено, не имели гомологии по существу с любым известным белком. Биоинформационный анализ мтс-миРНК, приводящих к получению последовательностей кДНК, показал, что значительная гомологичность последовательностей на нуклеотидном уровне относительно любого известного гена растения отсутствует. Такие данные указывают на то, что мтс-миРНК может быть получена из любого такого локуса способом процессинга дсРНК, образованной между транскриптами с противоположной полярностью, или способом процессинга дсРНК от аберрантных транскриптов по причине РНК-зависимой активности РНК-полимеразы. Также возможно, что мтс-миРНК подвергается процессингу из внутренних вторичных структур дсРНК, которые могут быть образованы в некоторых транскриптах, приводящих к образованию мтс-миРНК.
[0052] Обратная транскрипция мтс-миРНК приводила к образованию последовательностей мтс-миРНК, которые были картированы в один из элементов мтс-миРНК (SEQ ID NO: 87). Это представлено на Фигуре 1, где ось Х представляет расположение нуклеотидов в направлении от 5’ к 3’ слева направо вверху и справа налево внизу. Относительный избыток мтс-миРНК представлен в виде транскриптов на четверть миллиона последовательностей (tpq), отложенных на оси Y. Как это можно видеть из Фигуры 1, несколько мтс-миРНК (обведены кружком) широко представлены в специфичной для метелки библиотеке мРНК (ось Y). Прогнозируемые последовательности мтс-миРНК также неоднородно распределены среди элемента мтс-миРНК (ось X).
Пример 2
[0053] Данный пример показывает анализ мтс-миРНК эндогенной экспрессии в метелках. Нативные схемы экспрессии мтс-миРНК in planta были проанализированы с использованием нескольких различных способов. Такие анализы подтвердили, что мРНК, гибридизирующие в элементы мтс-миРНК, находятся в преобладающем количестве и/или специфически экспрессируются в метелках в зародышевой плазме кукурузы (т.е. мтс-миРНК находятся в избыточном количестве и/или специфически экспрессируются в метелках), и что в варианте воплощения изобретения мтс-миРНК находится в преобладающем количестве в и/или экспрессируется специфически в пыльцевом зерне на стадии развития пыльцы в виде одноядерной микроспоры.
[0054] Для доказательства накопления мтс-миРНК специфически в метелках in planta использовали три репрезентативные последовательности мтс-миРНК (SEQ ID NO: 26 (1372590), SEQ ID NO: 8 (648011), SEQ ID NO: 33 (410590)) для разработки зондов для низкомолекулярного (НМ) анализа нозерн-блоттингом с использованием мРНК, полученных из кукурузы или риса. Для таких экспериментов вся РНК была экстрагирована из ткани растения с использованием реагента TRIzol® (Invitrogen, Carlsbad, CA). РНК (7,5 мкг) из каждого образца была денатурирована при 95°C в течение 5 минут перед разделением на геле 17% PAGE, содержащем 7 M мочевины в 0,5X TBE буфере (Allen et al. (2004) Nature Genetics 36:1282-1290). После электрофореза гель был подвергнут блоттингу на мембране Nytran SuPerCharge® (Whatman-Schleicher & Schuell, Florham Park, NJ) с использованием системы для полусухого электрофоретического переноса клеток Trans-Blot® SD Semi-Dry Electrophoretic Transfer Cell (Bio-Rad, Hercules, CA) в соответствии с протоколом производителя. Полученный блот был перекрестным способом соединен при 1200 микроджоулей/см2 × 100 в Stratalinker® 1800 (Stratagene, Cedar Creek, TX). Для приготовления зондов матрица с зондом РНК была получена способом ПЦР и содержала промотор Т7 на одном конце и одну из последовательностей малых РНК на противоположном конце. Последовательности мРНК, включенные в матрицу зонда РНК, включали: [1] Gma-miR159a (miRBase.org номер доступа MI0001773), которую использовали в качестве контроля для загрузки, [2] sR1372590 (SEQ ID NO: 26), [3] sR648011 (SEQ ID NO: 8) и [4] sR410590 (SEQ ID NO: 33). Зонды РНК были транскрибированы с использованием РНК полимеразы Т7 и мечены дигоксигенином (DIG) с использованием набора DIG Northern Starter (Roche, Indianapolis, IN) в соответствии с протоколом производителя. Гибридизацию проводили со 100 нг меченого DIG зонда в буфере гибридизации PerfectHyb™ (Sigma, St. Louis, MO) при 38°C в течение 16 часов. Определение проводили с использованием набора DIG Northern Starter, в соответствии с протоколом производителя, перед воздействием на пленку Kodak ® Biomax™ XAR (Sigma, St. Louis, MO). Протестированные образцы включали все или подгруппу из следующего: лист кукурузы от растений, выращенных при стрессовом воздействии азотом; росток, корень или эндосперм кукурузы от растений, выращенных при стрессовом воздействии холодом; лист и корень кукурузы от растений, выращенных при стрессовом воздействии сухости; кукурузные рыльца; молодые метелки кукурузы; зрелые метелки кукурузы; неопыленные ядра кукурузы; эмбрион кукурузы - 24 дня после опыления (ДПО); ядра кукурузы - 22 ДПО; зрелые ядра кукурузы; эмбрион кукурузы - зрелые (сухие) ядра; эндосперм кукурузы - сухой; рисовое зерно и рисовые проростки. Результаты, полученные с помощью НМ нозерн-блоттинга с использованием по меньшей мере трех разных зондов мтс-миРНК (sR1372590, sR648011 и sR410590), выявили сигнал только в рядах, соответствующих молодой метелке и зрелой метелке, что подтверждает биоинформационный анализ и заключение о том, что экспрессия мтс-миРНК сильно повышена или специфична для ткани метелки.
[0055] Тканевая специфичность и накопление мРНК, которые могут распознавать элемент мтс-миРНК, были оценены с использованием широкого спектра зародышевой плазмы и НМ нозерн-блоттинга. Для этого анализа был отобран элемент мтс-миРНК (SEQ ID NO: 87, содержащий множественные последовательности мтс-миРНК). Такой элемент мтс-миРНК включает три последовательности мтс-миРНК, используемые для разработки зондов миРНК sR1372590, sR648011 и sR410590, что позволяет использовать эти зонды для НМ нозерн-блоттинга образцов зародышевой плазмы кукурузы. Для этих экспериментов была получена РНК от двадцати различных инбредных линий кукурузы с разным генетическим фоном, например, с относительной зрелостью, варьирующей от 83 до 120 (таблица 1). Для трех таких инбредных линий (91DUA6, 01DKD2 и LH244) была собрана ткань из молодой метелки, старой метелки, листка, початка и корня. В таблице 1 представлены соответствующие стадии V и размер метелки при сборе молодой метелки и старой метелки. Вся РНК была экстрагирована с использованием раствора TRIzol®. НМ РНК была выделена с использованием набора для выделения миРНК mirVana™ (№ по каталогу AM1560, Ambion, Austin, TX). НМ нозерн-блоттинг был проведен с использованием акриламидного геля Bio-Rad Criterion™ Precast 15% TBE-мочевина (№ по каталогу 345-0092, BioRad, Hercules, CA). Гель был подвергнут блоттингу на положительно заряженной мембране (№ по каталогу 11209272, Roche Applied Systems, Mannheim, Germany). Зонды были помечены (1) 32-P для случайного примирования или (2) DIG ДНК с использованием набора для мечения Roche PCR, или (3) зондом DIG РНК, как это описано выше. Все зонды, используемые для зондирования нозерн-блотов, были обратно комплементарны эндогенному транскрипту или последовательности кДНК элемента мтс-миРНК. Присутствие мРНК, которая гибридизировала в трансгенный элемент мтс-миРНК, было специфическим для метелки; сигнала для листа, початка или корня для любого из трех инбредных генотипов кукурузы 91DUA6, 01DKD2 и LH244 определено не было (фиг.2).
[0056] Для определения временной схемы экспрессии во время развития мРНК метелки, которая смогла бы распознавать элемент мтс-миРНК (SEQ ID NO: 87), был проведен НМ нозерн-блоттинг. РНК была получена из молодых и старых метелок от различных инбредных линий кукурузы, см. Таблицу 1. Получение РНК и способ НМ нозерн-блоттинга проводили по существу, как это описано выше.
(дюймы)
(дюймы)
[0057] Как это можно видеть на фиг.4, меченый DIG зонд РНК, соответствующий обратному комплементу элемента мтс-миРНК (SEQ ID NO: 87), гибридизировал в мРНК как в молодой, так и в старой метелке, за исключением того, что длина молодой метелки была от 2,5 дюймов до 3 дюймов: ряды 5 (инбред DIDA404), 9 (инбред JEDO115), 10 (инбред FIDA240), 16 (инбред DIDA403), 17 (инбред 64DJD1), 18 (инбред DIQ423) и 20 (инбред LH244). Кроме того, этот эксперимент подтвердил отсутствие определения гибридизации мРНК в элемент мтс-миРНК из образцов листа (ряды 21 и 22) или початка (ряды 23 и 24) от инбредов BIQA208 и LH244. В целом эти данные указывают на то, что мРНК, гибридизирующие в элемент мтс-миРНК, специфически экспрессируются в метелке каждого тестированного инбредного генотипа, когда метелка превышает в длину примерно 3,5 дюймов.
[0058] Анализ гибридизации in situ проводился для изучения клеточно-специфической экспрессии последовательности мтс-миРНК (sR648011, SEQ ID NO: 8). В пыльниках кукурузы микроспоры вырабатываются посредством мейоза, и развиваются в зрелую пыльцу. Микроспорогенез кукурузы может быть четко разделен на следующие стадии: мейоз спорогенных клеток, высвобождение тетрад в виде свободных микроспор, митоз одноядерных микроспор с получением трехклеточной пыльцы и зрелые пыльцевые зерна. Для этих экспериментов использовали метелки кукурузы, собранные до периода цветения у растений кукурузы, выращенных в стандартных условиях в теплице. Зонды запертой нуклеиновой кислоты (LNA) (Integrated DNA Technologies, Coralville, IA) использовали, как это указано ниже, при этом положение LNA указано символом «+». Антисмысловой зонд был разработан для определения мтс-миРНК для sR648011 (SEQ ID NO: 8) (5’-биотин-CAT+GCA+CTG+GTG+AGT+CAC+TGT-3’), в то время как смысловым зондом был обратный комплемент антисмыслового зонда (5’-биотин-ACA+GTG+ACT+CAC+CAG+TGC+ATG-3’) для использования в качестве отрицательного контроля. Зонды LNA позволяют проводить промывание в очень строгих условиях и, следовательно, обеспечивают высокоспецифичную гибридизацию (Válóczi et al., 2006; Nuovo et al., 2009). Все зонды были мечены биотином. Образцы метелки кукурузы были фиксированы в 4% параформальдегиде в 1×ФСБ при 4°C в течение 36 часов и затем дегидратированы при 4°C с использованием установленных серий этанол:H2O. Затем метелки были помещены в 75% EtOH и 25% Histoclear (National Diagnostics, Atlanta, GA) на время 1,5 ч, 50% EtOH и 50% Histoclear на время 1,5 ч, 25% EtOH и 75% Histoclear на время 1,5 ч и 100% Histoclear на время 3×1,5 ч, все при 25°C. Затем Histoclear был постепенно замещен расплавленным парапластом при 50°C, а метелки были перенесены в формы, и их хранили при 4°C перед разделением. Погруженные в парафин метелки были разделены на срезы микротомом толщиной 8 мкм. Серия срезов была получена из одних и тех же пыльников, и затем прилегающие срезы использовали для зондирования смысловым или антисмысловым зондом, соответственно. Прегибридизацию и гибридизацию проводили при 42°C с промыванием при 55°C. Определение меченых биотином зондов LNA, ренатурированных с транскриптом, проводилось с разведением от 1 до 400 антибиотин щелочной фосфатазы (ЩФ) и субстрата BM Purple AP (Roche Applied Science, Indianapolis, IN). Изображения были получены с помощью камеры на микроскопе Olympus (Center Valley, PA). Срезы от одних и тех же пыльников были поделены на две группы - одну использовали для антисмыслового зонда (фиг.5, слева), а вторую - для смыслового зонда (фиг.5, справа). Сигнал гибридизации (темно-фиолетовый) был определен только на срезах, которые были гибридизированы антисмысловым зондом, но не на срезах, которые были инкубированы со смысловым зондом (фиг.5). Сильный сигнал, полученный для антисмыслового зонда, указывает, что такая мтс-миРНК находится в чрезмерном количестве (активно экспрессируется) в пыльцевом зерне на стадии одноядерной микроспоры в ходе развития пыльцы.
Пример 3
[0059] Данный пример иллюстрирует конструкции для трансформации растения и получение трансгенного растения. Элемент мтс-миРНК был введен в 3’UTR кассеты трансгенной экспрессии и использовался для получения трансгенных растений кукурузы для исследования воздействия элемента на трансгенную экспрессию в трансгенных растениях. Элемент мтс-миРНК (SEQ ID NO: 87) был вставлен в 3’UTR кассеты трансгенной экспрессии CP4-EPSPS для трансформации кукурузы. Такой элемент мтс-миРНК был выбран по причине обилия в нем последовательностей мтс-миРНК (фиг.3), включая последовательности трех зондов миРНК (sR1372590, sR648011 и sR410590), используемых для НМ нозерн-блоттинга в примере 2. Такой элемент мтс-миРНК также позволял протестировать эффект несоответствий мтс-миРНК. Протестированный в настоящем описании элемент мтс-миРНК (SEQ ID NO: 87) имеет одно нуклеотидное изменение (C∆T: AAGCTATTGATTCCCTAAGTGCCA) в сравнении с одной из исходных последовательностей мтс-миРНК (SEQ ID NO: 33, использовали для разработки зонда sR410590). Элемент мтс-миРНК был вставлен в трансгенную кассету в обратной комплементарной ориентации относительно своего положения в эндогенной кДНК, однако считается, что элемент функционирует подобным образом в обоих направлениях, поскольку специфичные для метелки молекулы миРНК, комплементарные любой цепи элемента мтс-миРНК, могут быть обнаружены в метелке кукурузы (фиг.1 и 3).
[0060] Было сконструировано несколько кассет экспрессии CP4-EPSPS/элемент мтс-миРНК (таблица 2), которые использовали для трансформации растения кукурузы. В кассетах экспрессии CP4-EPSPS/элемент мтс-миРНК тестировали различные комбинации элементов экспрессии. Элементы экспрессии, такие как промоторы, лидеры, интроны, транзитные пептиды хлоропластов и 3’UTR, необходимые для эффективной и стабильной экспрессии трансгена, хорошо известны в данной области техники. Кассеты экспрессии CP4-EPSPS/элемент мтс-миРНК были разработаны для включения одного из двух отдельных промоторов; функциональной связи с ДНК одного из двух лидеров; функциональной связи с ДНК одного из двух интронов; функциональной связи с одной из двух молекул ДНК, кодирующих один и тот же транзитный пептид хлоропластов (CTP); функциональной связи с молекулой ДНК, полученной из гена aroA от Agrobacterium sp. штамма CP4 и кодирующей белок CP4-EPSPS; функциональной связи с ДНК, кодирующей элемент мтс-миРНК; функциональной связи с 3’UTR одной из двух молекул ДНК. Конструкция 4 содержала ген CP4-EPSPS дикого типа, а все другие векторы содержали версию гена CP4-EPSPS, оптимизированного растительным кодоном. Конструкции 3, 5 и 6 (таблица 2) были разработаны для определения того, будет ли элемент мтс-миРНК, включенный в 3’-UTR, приводить к получению растений со специфичной чувствительностью метелок к глифосату и вегетативной толерантностью к глифосату. Конструкции 4 и 7 являются контрольными конструкциями без элемента мтс-миРНК.
[0061] Трансгенные растения кукурузы, трансформированные каждой из пяти кассет экспрессии, были получены с использованием хорошо известных способов. Вкратце, клетки кукурузы были трансформированы Agrobacterium-опосредованной трансформацией каждой из конструкций, перечисленных в таблице 2 (индивидуально), и регенерированы в интактные растения кукурузы. Отдельные растения, характеризовавшиеся целостностью кассеты трансгенной экспрессии и резистентностью к глифосату, были отобраны из популяции растений. Укоренившиеся растения с нормальными фенотипическими характеристиками были отобраны и перенесены на почву для роста и дальнейшей оценки. Растения R0 были перенесены на почву для роста, орошены 0,75 фунта/акр глифосатом на стадии V3-V4, затем 0,75 фунта/акр глифосатом на стадии V7-V9 и затем опылены перекрестным образом пыльцой нетрансгенных растений кукурузы из одинаковой зародышевой плазмы (для конструкций 3, 5 и 6 явлений) или самоопылены (для конструкций 4 и 7 явлений) с получением семян R1. Затем растения были отобраны комбинацией аналитических способов, включая TaqMan, анализ ПЦР и вегетативную толерантность к распылению гербицидов и пониженному (требуемому) показателю мужской фертильности после распыления гербицида (глифосата).
Пример 4
[0062] Этот пример демонстрирует способы анализа трансгенных растений в теплице. Трансгенные растения, трансформированные кассетой экспрессии CP4-EPSPS/элемент мтс-миРНК, были проанализированы на предмет вегетативной толерантности к глифосату и мужской фертильности. Было выявлено, что трансгенные растения, полученные из конструкций, содержащих кассеты экспрессии CP4-EPSPS/элемент мтс-миРНК, обладают вегетативной толерантностью к глифосату и индуцированной мужской стерильностью при позднем применении глифосата.
[0063] Растения R0 были выращены в двух повторностях в теплице, и не были обработаны или были обработаны глифосатом 0,75 фунта/акр на стадии V6 (ранней), а затем глифосатом 0,75 фунта/акр на стадии V9 (поздней) (Фигура 7 и Таблица 3). Исследуемые явления R0 были явлениями с множественными копиями. Все растения R0, которые не были обработаны, имели нормальное выпячивание пыльников и полностью фертильную пыльцу, как это определено окрашиванием по Alexander. Все растения R0, которые были обработаны, имели вегетативную толерантность к глифосату. Растения R0, полученные из конструкций 4 и 7, которые не содержали элемента мтс-миРНК, не обладали чувствительностью метелки к глифосату или индуцированной мужской стерильностью. Растения R0, полученные из конструкций 3, 5 и 6, которые содержали элемент мтс-миРНК, обладали чувствительностью метелок к глифосату и индуцированной мужской стерильностью; такие растения не имели или имели всего лишь несколько выпячиваний пыльников, и >99% пыльцы было нежизнеспособной, как это определено окрашиванием по Alexander.
Вегетативная толерантность
Индуцированная мужская стерильность
[0064] Такие наблюдения показали, что присутствие элемента мтс-миРНК в 3’UTR трансгенной кассеты приводило к сайленсингу трансгена, специфичному для метелки. Специфичная для метелки потеря транскрипта иРНК, выработанного кассетой экспрессии CP4-EPSPS/элемент мтс-миРНК, привела к получению метелок, которые были чувствительны к глифосату, к получению растения с индуцированной мужской стерильностью, в то время как другие ткани растения были толерантны к глифосату, что привело к развитию вегетативной толерантности к глифосату и хорошей женской фертильности.
[0065] Затем использовали иммунолокализацию для измерения белка CP4-EPSPS в тканях трансгенных растений. От растений, трансформированных конструкцией 3 или конструкцией 4, а также от нетрансгенной кукурузы (LH198) были получены метелки. Растения были выращены в теплице с 14 часами света при 26,67°С(80°F) и 8 часами темноты при 21,11°С(70°F). В каждый горшок было посажено по одному семени. Горшки были случайным образом расположены на полу теплицы. Растения поливали при необходимости и удобряли 20-20-20 смесью азота, калия и фосфора, соответственно. Растения с конструкцией 3 или конструкцией 4 были обработаны распылением глифосатом в количестве 0,75 фунта/акр на стадии V2 для подтверждения вегетативной толерантности к глифосату. Молодые метелки были собраны на стадиях V10-V11 для получения тканей пыльников на стадиях микроспоры материнской клетки и свободной микроспоры; зрелые метелки были собраны на стадии Т7, за 1-2 дня перед опадением пыльцы, для получения тканей пыльников с полностью развившейся пыльцой. Пыльники были извлечены из вторичного колоска метелки с использованием препаровального пинцета, и непосредственно зафиксированы в 3,7% формальдегиде в фосфатно-солевом буфере (ФСБ) при мягком вакууме. После промывания в ФСБ ткани были помещены в заделывающую среду и немедленно заморожены. Замороженные блоки тканей хранили при минус 80°C до момента получения срезов при минус 20°C с использованием микротома, после чего срезы были собраны на заряженных покровных стеклах.
[0066] Срезы тканей были блокированы блокирующим агентом (10% нормальная сыворотка козы, 5% бычий сывороточный альбумин, 0,1% Тритон Х-100 в ФСБ) в течение 2 часов. Срезы были инкубированы с антителом к CP4-EPSPS (1/500 в ФСБ). После трехкратного промывания срезов в ФСБ срезы тканей были инкубированы с вторичным антителом козы к IgG мыши, конъюгированным с флуорофором Alexa 488 (Invitrogen, Eugene, Oregon). Для отрицательного контроля инкубацию антитела CP4-EPSPS не проводили. В качестве положительного контроля антитело к α-тубулину (Sigma, St. Louis, MO), белку цитоскелета, экспрессируемому в большинстве типов клеток, было замещено антителом CP4-EPSPS на отдельных срезах. Первичные и вторичные антитела были инкубированы при комнатной температуре в течение 2-4 часов и затем дополнительно инкубированы в течение ночи при 4°C. После промывания ткани были визуализированы лазерным сканирующим конфокальным микроскопом Zeiss (ЛСМ) 510 META с использованием лазера 488 нм для возбуждения и 500-550 нм для набора эмиссионных фильтров. Такой же самый параметр визуализации применяли для образцов, включая контроль. Флуоресцентные изображения и изображения в ярком поле были сканированы с каждого среза, и объединены с использованием программного обеспечения ЛСМ, впоследствии для получения структурной информации. Сильный сигнал был получен для антитела к CP4-EPSPS в ткани филаментов (фиг.6А, короткая стрелка) и пыльцы (фиг.6А, длинная стрелка) в зрелой метелке от растений, полученных с конструкцией 4 (фиг.6А) с отсутствием элемента мтс-миРНК. Растение с фиг.6А является гемизиготным на предмет кассеты трансгена, таким образом, только примерно 50% пыльцы обладало положительным сигналом CP4-EPSPS. В отличие от этого, сильный сигнал был получен для антитела к CP4-EPSPS только в ткани филаментов (фиг.6В, короткая стрелка), а для пыльцы сигнал отсутствовал (фиг.6В, длинная стрелка) в зрелой метелке от растений, полученных с конструкцией 3, содержащей элемент мтс-миРНК (фиг.6В). Антитело положительного контроля (антитело к альфа-тубулину) характеризовалось сигналом пыльцы в зрелой метелке от растений, полученных с конструкцией 4 или конструкцией 3. Данные для отрицательных контролей характеризовались ожидаемым отсутствием сигнала. Данные для стандартного нетрансгенного контроля характеризовались ожидаемым отсутствием сигнала от окрашивания антителом к CP4-EPSPS и положительным сигналом при окрашивании с антителом к альфа-тубулину. Эти данные указывают на то, что в пыльце или не транслируются, или транслируются всего несколько транскриптов из кассеты трансформации, содержащей элемент мтс-миРНК, однако транскрипт был транслирован в вегетативную ткань филамента. Потеря экспрессии белка CP4-EPSPS в пыльце коррелирует с наблюдаемой и специфичной для метелки чувствительностью к глифосату у растений, полученных из конструкции 3.
Пример 5
[0067] Этот пример иллюстрирует полевое исследование трансгенного растения на предмет мужской фертильности или стерильности. В полевых условиях было исследовано тринадцать подтвержденных уникальных явлений трансгенных растений R1-R3, полученных способом трансформации кассетой экспрессии CP4-EPSPS/элемент мтс-миРНК (конструкция 3), на предмет эффективности кассеты экспрессии. В первый год тринадцать явлений были тестированы при расположении на одном поле. Во второй год восемь явлений были тестированы при расположении в четырех местах на поле. В третий год четыре явления были тестированы при расположении в четырех местах на поле. В течение трех лет полевых исследований средний показатель мужской фертильности (ПМФ) для явлений, полученных из конструкции 3, был приблизительно равен или ниже ПМФ 2, что считается промышленным стандартом мужской стерильности.
[0068] Данные для одногодичных полевых исследований эффективности представлены на фиг.8 со средним показателем ПМФ, полученным при трех различных режимах обработки глифосатом распылением, представленных на графике (фиг.8А) для NK603 (CP4-EPSPS трансгенная кукуруза), MON87427 (CP4-EPSPS трансгенная кукуруза с индуцированной глифосатом мужской стерильностью) и двух явлений из конструкции 3. Фотографии метелок растений, выращенных во время данного отдельного полевого исследования эффективности, иллюстрируют фертильную метелку во время обработки растений распылением глифосатом 0,75 фунта/акр только на стадии V3 (фиг.8В); и стерильную метелку (отсутствие или минимальное выпячивание пыльников) на растениях, обработанных глифосатом распылением 0,75 фунта/акр на стадии V3, затем 0,75 фунта/акр на стадии V8, после чего 0,75 фунта/акр на стадии V10 (фиг.8С). Для такого полевого исследования режимы распыления были следующими: обработка 1 состояла из 0,75 фунта/акр глифосата на стадии V3 (борьба с сорняками); обработка 2 состояла из 0,75 фунта/акр глифосата на стадии V3 (борьба с сорняками), затем 0,75 фунта/акр на стадии V8, затем 0,75 фунта/акр на стадии V10; обработка 3 состояла из 0,75 фунта/акр глифосата на стадии V3 (борьба с сорняками), затем 1,25 фунта/акр на стадии V8, затем 1,25 фунта/акр на стадии V10. Последние две обработки (т.е. на стадиях V8 и V10) обозначаются как обработки для получения стерильности. Эти результаты указывают, что только с обработкой глифосатом распылением 1 для борьбы с сорняками все растения (NK603, MON87427 и явления конструкции 3) имели мужскую фертильность. При обработке глифосатом распылением 2 для получения стерильности растения NK603 имели ПМФ=5, MON87427 были стерильными с ПМФ=2, и явления 2 и 3 конструкции 3 имели частичную мужскую фертильность с ПМФ<3. При обработке глифосатом распылением 3 для получения стерильности растения NK603 имели ПМФ=5, MON87427 имели мужскую стерильность с ПМФ<2, и явления 2 и 3 конструкции 3 имели мужскую стерильность с ПМФ около или ниже значения 2.
[0069] Несмотря на то, что среднее значение ПМФ было примерно или на уровне 2, выпячивание пыльников отмечалось в явлениях конструкции 3 при обработке глифосатом при S90+3 и S90+6 (фиг.9). Для таких данных четыре отдельных явления конструкции 3 были сопоставлены с растениями MON87427 и NK603 при двух режимах обработки распылением глифосатом: обработка 2 состояла из 1,5 фунта/акр глифосата на стадиях V2/V3 (борьба с сорняками), затем 0,75 фунта/акр глифосата на стадии единиц степени роста (ЕСР) 875 (~V8), затем 0,75 фунта/акр глифосата на стадии ЕСР 1025 (~V10), и обработка 3 состояла из 1,5 фунта/акр глифосата на стадиях V2/V3 (борьба с сорняками), затем 1,25 фунта/акр глифосата на стадии ЕСР 875 (~V8), затем 1,25 фунта/акр глифосата на стадии ЕСР 1025 (~V10). Количество растений на делянку (68-74 растения/делянку), демонстрирующих выпячивание пыльников, было подсчитано при S90, S90+3 и S90+6, при этом S90 является днем, когда 90% растений в поле выбросили рыльца; S90+3 является 3 днем после S90; и S90+6 является 6 днем после S90. Как это видно на Фигуре 9, при S90 отмечалось 70(±15) растений NK603 на делянку с выпячиванием пыльников при обоих режимах обработки глифосатом. В отличие от этого, для MON87427 и четырех явлений конструкции 3 отмечалось 1(±12) растение на делянку с выпячиванием пыльников при S90 для обоих режимов обработки глифосатом. При S90+3 и S90+6 отмечалось от 30(±12) до 70(±12) растений на делянку для четырех явлений конструкции 3 с выпячиванием пыльников при любом режиме обработки глифосатом, что примерно то же самое, что и для NK603. Выпячивание пыльников для явления MON87427 оставалось на уровне S90 для временных точек S90+3 и S90+6 и для каждого режима обработки глифосатом. Любое растение с ≥1 выпятившимся пыльником было рассчитано в качестве положительного на предмет выпячивания пыльников. Такое позднее выпячивание пыльников, т.е. S90+3 и S90+6, возникает во время развития кукурузы, когда отмечается максимальная высота роста метелки, и существует значительное расстояние для обеспечения машинного среза метелки с минимальным повреждением двух верхних листьев растения кукурузы, обеспечивая, таким образом, минимальное влияние на выход инбреда. Кроме того, выпячивание пыльников на стадии S90+3 или позднее считается как имеющее незначительное влияние на чистоту семян.
[0070] Анализ жизнеспособности пыльцы проводили для определения того, является ли низкий уровень (при этом стабильное выпячивание пыльников, наблюдаемое от S90+3 до S90+6) индикатором потенциальной поздней мужской фертильности. Фиг.10А и 10В отображают пример позднего выпячивания пыльников в метелке при явлении распыления при наличии стерильности и конструкции 3. Квадрат на фиг.10А является участком, увеличенным на фиг.10В. Пример позднего выпячивания пыльника обведен кружком на Фигуре 10В. Для определения жизнеспособности пыльцы пыльца была собрана от поздних выпятившихся пыльников, и окрашена способом Alexander, Фигура 10С. Пыльца также была собрана от не подвергшихся обработке распылением явлений конструкции 3 в тот же самый день, и окрашена в качестве сравнения способом Alexander, фиг.10D. Результаты такого окрашивания по Alexander показывают только нежизнеспособную пыльцу (полупрозрачный голубой цвет, неправильная форма пыльцевых зерен) от поздно выламывающихся пыльников подвергнутых обработке распылением явлений конструкции 3 (фиг.10С). Полностью жизнеспособная пыльца оказалась непрозрачной, темно-фиолетовой и сферической при окрашивании по Alexander (фиг.10D). Кроме окрашивания пыльцы, собранной от индивидуально выделенных поздно выламывающихся пыльников, мешки для опыления были помещены на некоторые подвергнутые обработке распылением явления конструкции 3 для определения опадения пыльцы. Какого-либо заметного опадения пыльцы в такие мешки для опыления не было определено, что не позволило получить какого-либо семени при использовании для перекрестного опыления початков. Этот результат указывает на отсутствие опадения пыльцы от поздних выпятившихся пыльников или на то, что любая опавшая пыльца является нежизнеспособной.
[0071] В целом эти данные указывают на то, что, несмотря на низкий уровень выпячивания пыльников при обработке распылением для получения стерильности для явлений конструкции 3, такие выпятившиеся пыльники не сбрасывали жизнеспособную пыльцу.
Пример 6
[0072] Этот пример отображает полевые исследования трансгенных растений, определявшие урожайность. Растения конструкции 3 R2 были тестированы на предмет инбредного и гибридного урожая. Для инбредного урожая растения R2 конструкции 3 были тестированы в четырех разных полевых местах на предмет урожайности, вегетативной толерантности к обработке глифосатом и мужской стерильности при обработке глифосатом. Для таких полевых исследований четыре явления конструкции 3 были высажены на делянки в количестве 68-74 растения/делянку. Обработка распылением была следующей: обработка 1 состояла из 1,5 фунта/акр глифосата на стадии V3 (борьба с сорняками); обработка 2 состояла из 1,5 фунта/акр глифосата на стадии V3, затем 0,75 фунта/акр на стадии V8, затем 0,75 фунта/акр на стадии V11; обработка 3 состояла из 1,5 фунта/акр глифосата на стадии V3, затем 1,25 фунта/акр на стадии V8, затем 1,25 фунта/акр на стадии V11. Как это можно видеть на фиг.11, явления конструкции 3 при всех трех режимах обработки глифосатом обладали хорошей вегетативной толерантностью (белые полосы), как это измерено по высоте растения, и хорошим инбредным урожаем (черные полосы), как это измерено в бушелях/акр. Эти же самые явления обладали полной мужской фертильностью при режиме обработки глифосатом для контроля сорняков (обработка 1), но имели мужскую стерильность с показателем ПМФ, равным или менее 2 (серые полосы) при режимах обработки глифосатом 2 или 3. Горизонтальные полосы на фиг.11 указывают промышленный стандарт стерильности, ПМФ 2. NK603 представлено для сравнения. Измерения урожайности инбредных семян для явлений конструкции 3 и NK603 с обработкой глифосатом распылением представлены в таблице 4, где MST = % влажности семян, TWT = натурная масса (показатель плотности, обычно в фунтах на бушель) и S50D является количеством дней до 50% выброса рыльцев из початков на делянке. Значительная разница (нр), измеренная для урожайности при любых из четырех явлений конструкции 3, исследуемых при обработке глифосатом 2 или 3 в сравнении с контролем NK603, отсутствовала.
[0073] Для урожайности гибридного зерна F1 явления R3 конструкции 3 были тестированы в четырех полевых местах. Для таких полевых исследований гибридного урожая нетрансгенные женские инбредные растения (Null), линия MON87427, и три явления конструкции 3, все с одинаковой генетической основой, были перекрестно опылены мужским тестером MON810/MON88017 для получения гибридных семян F1. Гибридные семена F1, полученные от каждого из таких скрещиваний, были высеяны на стандартных делянках в количестве 68-74 растений/делянку. Обработка распылением состояла из обработки 1 без распыления глифосатом; обработки 2 - 2,25 фунта/акр глифосата на стадии V4, затем 2,25 фунта/акр на стадии V7; обработки 3 - 2,25 фунта/акр глифосата на стадии V4, затем 2,25 фунта/акр на стадии V7, затем 2,25 фунта/акр на стадии V10. Растения F1 были открыто опылены для получения зерна F2, что является урожаем, измеренным в бушелях/акр. Все три явления конструкции 3 имели эквивалентную урожайность гибридных зерен F1 при всех режимах обработки глифосатом при сравнении с контрольными скрещиваниями NullxMON810/MON88017 и MON87427xMON810/MON88017 (фиг.12).
Пример 7
[0074] Данный пример иллюстрирует восстановление мужской фертильности в гибридных растениях F1. Гибридные растения F1, полученные от скрещивания явлений конструкции 3, в качестве женского партнера были тестированы на предмет мужской фертильности. Было проведено три различных скрещивания гибридов F1: нетрансгенное женское растение x MON88017 мужское растение; MON87427 женское растение x MON88017 мужское растение; и явление конструкции 3 женское растение x MON88017 мужское растение. Гибридные семена F1 были собраны от каждого из таких трех скрещиваний, высеяны в поле, и обработаны распылением глифосатом 1,125 фунта/акр на стадии V4, затем 1,125 фунта/акр на стадии V10. Мужскую фертильность в F1 оценивали по показателю мужской фертильности (ПМФ) и по определению жизнеспособности пыльцы при окрашивании по Alexander. Для каждого из таких скрещиваний ПМФ гибридных растений F1 составил 5 или отмечалась полная фертильность. Окрашивание жизнеспособной пыльцы по Alexander выявило 50% жизнеспособной пыльцы, произведенной гибридом F1 при каждом скрещивании, как и ожидалось (Фигура 13). Эти данные указывают, что мужская фертильность может быть функционально восстановлена в гибридных растениях F1, полученных из трансгенных растений с индуцированной глифосатом мужской стерильностью, трансформированных кассетой экспрессии CP4-EPSPS/элемент мтс-миРНК.
Пример 8
[0075] Данный пример иллюстрирует вариантную и химерную конструкцию элемента мтс-миРНК. Отдельные мтс-миРНК были картированы на элемент мтс-миРНК, как это представлено на Фигуре 3; ось Х указывает на расположение нуклеотидов в направлении от 5’ к 3’ слева направо вверху и справа налево внизу, и ось Y указывает на относительное обилие мтс-миРНК, указанное в виде транскриптов на четверть миллиона последовательностей (tpq). Мтс-миРНК также были неоднородно распределены среди элемента мтс-миРНК (ось X).
[0076] Используя данную информацию, были получены варианты элемента мтс-миРНК и/или химерные варианты, полученные с использованием одного или более элементов мтс-миРНК, для содержания большего количества (или меньшего) всех последовательностей мтс-миРНК (необязательно или альтернативно одну или более последовательностей мтс-миРНК добавляют или удаляют), что приводит к большему (или меньшему) сайленсингу функционально связанной кодирующей белок последовательности. Такие варианты или химерные элементы мтс-миРНК пригодны для повышения или снижения избирательной супрессии экспрессии рекомбинантного белка в мужской репродуктивной ткани трансгенного растения.
[0077] Примеры вариантов и химер элементов мтс-миРНК были построены с использованием фрагментов SEQ ID NO: 87. Первый вариант (SEQ ID NO: 88) был построен с использованием фрагмента из 104 нуклеотидов от 5’-конца SEQ ID NO: 87. Второй вариант (SEQ ID NO: 89) был построен с использованием фрагмента из 80 нуклеотидов от 3’-половины SEQ ID NO: 87. Химерные элементы мтс-миРНК были построены путем соединения одного фрагмента (SEQ ID NO: 88) с другим фрагментом (SEQ ID NO: 89) с образованием новых химерных элементов мтс-миРНК (SEQ ID NO: 90 и SEQ ID NO: 91). Дополнительные химерные элементы мтс-миРНК были построены путем соединения трех отдельных мтс-миРНК, содержащихся в SEQ ID NO: 87: первая химера (SEQ ID NO: 92) была построена соединением последовательностей мтс-миРНК SEQ ID NO: 26, 27 и 8; вторая химера (SEQ ID NO: 93) была построена соединением последовательностей мтс-миРНК SEQ ID NO: 10, 33 и 5; третья химера (SEQ ID NO: 94) была построена соединением последовательностей мтс-миРНК SEQ ID NO: 26, 10 и 33. Такие варианты и химеры могут быть функционально связаны с кодирующими белок последовательностями для получения конструкций рекомбинантной ДНК (см. фиг.14), которые могут быть тестированы в растениях и растительных клетках на предмет избирательной супрессии рекомбинантного белка, кодируемого кодирующей белок последовательностью в мужской репродуктивной ткани трансгенного растения.
Пример 9
[0078] Данный пример иллюстрирует разработку вариантных и химерных элементов мтс-миРНК. Вариантные и химерные элементы мтс-миРНК были разработаны на основе 300-нуклеотидного (нт) элемента мтс-миРНК, имеющего SEQ ID NO: 81, который подобен 300-нуклеотидным элементам мтс-миРНК, имеющим SEQ ID NO: 82 и 87. Высококонсервативной консенсусной последовательностью для элементов мтс-миРНК SEQ ID NO: 81, 82 и 87 является SEQ ID NO: 96. В отдельности каждый из них также может быть пригодным в качестве элемента мтс-миРНК или в качестве основы для разработки вариантных или химерных элементов мтс-миРНК, например, путем отбора фрагментов элемента мтс-миРНК, идентифицированных из геномной последовательности или кДНК, таких как фрагментов, включающих по меньшей мере одну последовательность мтс-миРНК, и комбинированием или соединением таких фрагментов.
[0079] Было отобрано два фрагмента в SEQ ID NO: 81; фрагмент A (SEQ ID NO: 97) содержал 104 смежных нуклеотида от 5’ участка (положения 1-104) SEQ ID NO: 81, а фрагмент B (SEQ ID NO: 98) содержал 80 смежных нуклеотидов от 3’ участка (положения 215-294) SEQ ID NO: 81; очевидно, что фрагмент A (SEQ ID NO: 97) или фрагмент B (SEQ ID NO: 98) по-отдельности являются элементами мтс-миРНК, содержащими по меньшей мере одну последовательность мтс-миРНК. Расположение фрагментов А и В (указано подчеркнутым текстом) представлено в следующей полной последовательности SEQ ID NO: 81, которая также указывает расположение последовательностей мтс-миРНК (указано курсивом; элементы, указанные курсивом, имеют более 18 смежных нуклеотидов и могут включать более чем одну перекрывающуюся последовательность мтс-миРНК), картированных для данного элемента мтс-миРНК:
GGACAACAAGCACCTTCTTGCCTTGCAAGGCCTCCCTTCCCTATGGTAGCCACTTGAGTGGATGACTTCACCTTAAAGCTATCGATTCCCTAAGTGCCAGACATAATAGGCTATACATTCTCTCTGGTGGCAACAATGAGTCATTTTGGTTGGTGTGGTAGTCTATTATTGAGTTTGTTTTGGCACCGTACTCCCATGGAGAGTACAAGACAAACTCTTCACCGTTGTAGTCGTTGATGGTATTGGTGGTGACGACATCCTTGGTGTGCATGCACTGGTGAGTCACTGTTGTACTCGGCG (SEQ ID NO: 81). Вариантные элементы мтс-миРНК были разработаны с использованием фрагментов «А» и «В», включая элемент мтс-миРНК «А+В» (SEQ ID NO: 99) и элемент мтс-миРНК «В+А» (SEQ ID NO: 100). Химерный элемент (SEQ ID NO: 101) был разработан для включения последовательностей мтс-миРНК (представлено выше курсивом в SEQ ID NO: 81), которые были картированы для элемента мтс-миРНК (SEQ ID NO: 81).
[0080] Подобным образом элемент мтс-миРНК из 251 нт в длину (SEQ ID NO: 102) и элемент мтс-миРНК из 121 нт в длину (SEQ ID NO: 103, фрагмент SEQ ID NO: 102, т.е. непрерывный элемент, расположенный в положениях нуклеотидов 47-167 последовательности SEQ ID NO: 102) были идентифицированы из геномной последовательности кукурузы (Zm_B73_CR10::Segment{75361491..75361742}) в качестве специфичных для метелки и соответствующих мтс-миРНК из молодой метелки (кукуруза LH244, библиотека 347; отдельно идентифицированные мтс-миРНК в некоторых случаях перекрывают большую часть своей последовательности и варьируют только в нескольких нуклеотидах; см. таблицу 5). Основываясь на SEQ ID NO: 102 и 103, был разработан химерный элемент мтс-миРНК (SEQ ID NO: 104).
Пример 10
[0081] Данный пример иллюстрирует векторы и клетки трансгенного растения, ткани и растения, содержащие конструкции рекомбинантной ДНК, включающие кодирующую белок последовательность, кодирующую рекомбинантный белок, и элемент мтс-миРНК, функционально связанный с кодирующей белок последовательностью.
[0082] Вектор трансформации растения, включающий конструкцию рекомбинантной ДНК, используют для опосредованной Agrobacterium трансформации клеток кукурузы. Такой вектор трансформации включает ДНК для опосредованного Agrobacterium трансфера Т-ДНК, кассеты экспрессии (промотора, функционально связанного с интересующей последовательностью ДНК), кассеты избирательного маркера экспрессии (для удобного отбора трансформированных клеток кукурузы или растений) и ДНК для поддержания вектора в E. coli (например, происхождение последовательности репликации от E. coli). В одном варианте воплощения изобретения вектор трансформации включает кассету экспрессии, включающую конструкцию рекомбинантной ДНК, фланкированную правой и левой граничными последовательностями от Agrobacterium, при этом конструкция рекомбинантной ДНК включает трансген толерантности к гербицидам CR-AGRtu.aroA-CP4.nat (представлено как SEQ ID NO: 95) в качестве последовательности ДНК, кодирующей рекомбинантный белок. Трансген толерантности к гербицидам CR-AGRtu.aroA-CP4.nat функционально связан с мтс-миРНК, представленной как SEQ ID NO: 81 в качестве последовательности ДНК, кодирующей элемент мтс-миРНК.
[0083] Векторы трансформации для экспрессии различных конструкций рекомбинантной ДНК построены путем вставки полинуклеотида, включающего элемент мтс-миРНК (например, SEQ ID NO: 57-94 или 97-104), в вектор трансформации растения. Элемент мтс-миРНК вставлен в прилегающем положении к последовательности ДНК, кодирующей рекомбинантный белок, или в 3’-нетранслируемый участок последовательности ДНК, кодирующей рекомбинантный белок. Такие векторы трансформации растения пригодны для получения трансгенных растений, у которых может быть индуцирована мужская стерильность применением гербицида.
[0084] Способы трансформации растений хорошо известны в данной области техники. Например, растения кукурузы трансформируемой линии выращивают в теплице и початки собирают, когда эмбрион достигает в длину от 1,5 до 2,0 мм. Поверхность початков стерилизуют 80% этанолом, затем высушивают на воздухе. Незрелые эмбрионы удаляют из отдельных ядер стерилизованных початков. Перед инокуляцией клеток кукурузы отдельные культуры Agrobacterium, каждая из которых содержит вектор трансформации для экспрессии по меньшей мере одной конструкции рекомбинантной ДНК по данному изобретению, выращивают в течение ночи при комнатной температуре. Клеточные культуры незрелых эмбрионов кукурузы инокулируют Agrobacterium, инкубируют при комнатной температуре с Agrobacterium в течение 5-20 минут, сокультивируют с Agrobacterium в течение 1-3 дней при 23 градусах по Цельсию в темноте, переносят в избирательную среду, и культивируют в течение примерно 2 недель для развития эмбрионального каллюса. Эмбриональный каллюс переносят в питательную среду, содержащую 100 мг/л паромомицина, и субкультивируют при интервале примерно две недели. Множественные явления трансформированных клеток растений восстанавливаются спустя 6-8 недель после начала отбора.
[0085] Трансгенные растения кукурузы регенерируют из каллюса клетки трансгенного растения для каждого из множественных трансгенных явлений, возникших в результате трансформации и отбора, путем помещения трансгенного каллюса каждого явления в среду для инициации развития отростков и корней в проростках, которые переносят в горшковую почву для начального роста в камере роста при 26 градусах по Цельсию, затем роста на скамейке с водяной пылью перед пересадкой в горшки, где растения вырастают до зрелости. Регенерированные растения самоопыляются. Первое поколение семян («R1») собирают. Растения от семян R1 (растения «R2») используют для получения потомства.
Пример 11
[0086] Данный пример иллюстрирует способы выбора последовательностей мтс-миРНК и элементов мтс-миРНК для использования в конструкциях рекомбинантной ДНК, включающих кодирующую белок последовательность, кодирующую рекомбинантный белок, и элемент мтс-миРНК, функционально связанный с кодирующей белок последовательностью.
[0087] Один способ верификации эффективности элемента мтс-миРНК для избирательной супрессии экспрессии рекомбинантного белка в мужской репродуктивной ткани трансгенного растения включает использование анализа протопласта, при этом протопласты растительных клеток сотрансформируют: (а) вектором, содержащим конструкцию рекомбинантной ДНК, включающую кодирующую белок последовательность, и элемент мтс-миРНК, функционально связанный с кодирующей белок последовательностью; и (б) РНК, имеющими последовательности миРНК, соответствующие элементу(ам) мтс-миРНК (или, с другой стороны, последовательности(ям) мтс-миРНК)), при этом уровень экспрессии рекомбинантного белка ожидается обратно пропорциональным степени, до которой элемент мтс-миРНК расщепляется РНК.
[0088] Это иллюстрируется следующим неограничивающим примером. Анализ проводили на двух последовательностях мтс-миРНК (соответствующих двум миРНК, которые сильно экспрессируются в метелке кукурузы). Вкратце, протопласт листа кукурузы сотрансформируют с помощью: (а) плазмиды (3 микрограмма/320000 клеток), содержащей конструкцию рекомбинантной ДНК, включающую кодирующую белок последовательность, кодирующую рекомбинантный белок (CP4-EPSPS, SEQ ID NO: 95), и элемент мтс-миРНК (SEQ ID NO: 81), и (б) первой дсРНК, в которой первая цепь имеет последовательность SEQ ID NO: 150 в направлении от 5’ к 3’, а вторая цепь является комплементарной первой цепи, и второй дсРНК, в которой первая цепь имеет последовательность SEQ ID NO: 151 в направлении от 5’ к 3’, а вторая цепь является комплементарной первой цепи. ДсРНК (от Integrated DNA Technologies, Inc., Coralville, Iowa) были протестированы в количестве 0, 5, 25 или 50 нанограмм/320000 клеток, при этом общее количество РНК, используемое в каждом анализе сотрансформации, было скорректировано с «наполняющей» РНК, состоящей любой из микроРНК395 (в качестве зрелого 21-mer, представленного как дсРНК) или тРНК дрожжей в количестве до 50 нанограмм/320000 клеток. Уровень белка CP4-EPSPS определяли ИФА и использовали для оценки способности тестированных дсРНК супрессировать экспрессию рекомбинантного белка. Результаты приводятся в таблице 6.
[0089] Каждая из дсРНК (SEQ ID NO: 150 и 151) сильно супрессировала экспрессию CP4-EPSPS (указано по сниженному накоплению белка CP4-EPSPS) при сотрансформации с плазмидой, содержащей конструкцию рекомбинантной ДНК, включающую кодирующую белок CP4-EPSPS последовательность, и элемент мтс-миРНК. Наблюдаемая супрессия CP4-EPSPS была дозозависимой от количества дсРНК и независимой от типа наполняющей РНК. Супрессия CP4-EPSPS не отмечалась в контрольных образцах, сотрансформированных наполняющей РНК вместо исследуемых дсРНК.
Пример 12
[0090] Данный пример иллюстрирует конструкции рекомбинантной ДНК, векторы и трансформированные растения по изобретению. Векторы и способы трансформации, подобные описанным в примере 10, использовались для получения стабильно трансформированных растений кукурузы, содержащих в своем геноме конструкцию рекомбинантной ДНК, включающую кодирующую белок последовательность, функционально связанную с последовательностью ДНК, включающей элемент мтс-миРНК. Исследовали шесть комбинаций схема конструкции/элемент мтс-миРНК (см. Таблицу 7). Растения были дважды обработаны распылением (на стадиях V5 и V8) 0,75 фунта/акр/A Roundup WeatherMAX®. Результаты приводятся в таблице 7. Для каждой комбинации схема конструкции/элемент мтс-миРНК примерно 20 растений было не обработано для сравнения с обработанными глифосатом растениями. У необработанных растений была собрана пыльца, и определена хорошая мужская фертильность (данные не указаны). Растения кукурузы, трансформированные конструкцией по схеме В, обладали более выраженной мужской стерильностью, чем растения кукурузы, трансформированные конструкцией по схеме А. Схемы конструкций (от 5’ к 3’, слева направо) были представлены: конструкция A является промотор A/интрон A/транзитный пептид A/CP4-EPSPS (SEQ ID NO: 95)/элемент мтс-миРНК/3’UTR, и конструкция B является промотор B/интрон B/транзитный пептид B/CP4-EPSPS/элемент мтс-миРНК/3’UTR. Как указано ниже, «ни» обозначает «не измерено». Шкала показателя мужской фертильности (ПМФ) следующая: 5 = образование пыльника нормальное, объем пыльцы такой же, как и на необработанных делянках, пыльца может осыпаться или не осыпаться; 4 = образование пыльника нормальное на 50%, но осыпание пыльцы слабое или не осыпается достаточное количество пыльцы; 3 = метелки выглядят нормальными, но отмечается спорадическое выпячивание пыльников (>10 пыльников на метелку), осыпается незначительно пыльцы или пыльца не осыпается; 2,5 = пыльца не осыпается, количество пыльников значительно уменьшено (<10 пыльников на метелку) или появляются слишком поздно (1 неделя) относительно окончания образования рылец; 2 = пыльца не осыпается, пыльников нет или они появляются слишком поздно (1 неделя) относительно окончания образования рылец; и 1 = пыльца не осыпается, метелки имеют аномально утонченный фенотип или образование пыльников замедлено на две или более недель после образования рылец. S90 представлена тогда, когда 90% растений имеют рыльца, готовые для опыления, и S90+3 представлена 3 днями спустя после S90.
[0091] Все описанные в настоящем описании материалы и способы, и представленная формула изобретения могут быть получены и использоваться без излишних экспериментов в соответствии с представленными выше инструкциями. Представленные выше примеры включены для демонстрации вариантов воплощения изобретения. Специалистам в данной области станет очевидно, что описанные в примерах способы, представляющие способы изобретателей, широко используются на практике и, следовательно, могут рассматриваться в качестве неотъемлемых преимущественных вариантов воплощения изобретения на практике. Однако специалистам в данной области, в свете данного изобретения, должна быть очевидна возможность внесения множественных изменений в описанные специфические варианты воплощения изобретения, а также получение подобных или похожих результатов без отклонения от сущности и объема изобретения. Все такие подобные замены и модификации, очевидные для специалистов в данной области, считаются включенными в сущность, объем и концепцию изобретения, как это определено прилагаемой формулой изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ТРАНСГЕННОЕ СОБЫТИЕ MON 87427 МАИСА И ОТНОСИТЕЛЬНАЯ ШКАЛА РАЗВИТИЯ | 2010 |
|
RU2623176C2 |
| ТРАНСГЕННОЕ СОБЫТИЕ MON 87427 МАИСА И ОТНОСИТЕЛЬНАЯ ШКАЛА РАЗВИТИЯ | 2010 |
|
RU2764586C2 |
| ОБЪЕКТ КУКУРУЗЫ MON 87411 | 2013 |
|
RU2648155C2 |
| ИЗОЛИРОВАННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК (ВАРИАНТЫ), РЕКОМБИНАНТНАЯ ДВУНИТЕВАЯ МОЛЕКУЛА ДНК И СПОСОБ ПОЛУЧЕНИЯ ГЕНЕТИЧЕСКИ ТРАНСФОРМИРОВАННЫХ РАСТЕНИЙ | 1991 |
|
RU2146290C1 |
| РАСТЕНИЕ КУКУРУЗЫ DBN9858, УСТОЙЧИВОЕ К ГЕРБИЦИДУ, И НУКЛЕОТИДНАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ И СПОСОБ ДЛЯ ЕЕ ВЫЯВЛЕНИЯ | 2016 |
|
RU2712730C2 |
| РАСТЕНИЕ МАИСА DBN9936 И СПОСОБ ПРИМЕНЕНИЯ В ДЕТЕКТИРОВАНИИ ЕГО ПОСЛЕДОВАТЕЛЬНОСТИ НУКЛЕИНОВОЙ КИСЛОТЫ | 2016 |
|
RU2707527C2 |
| Нуклеиновокислотная последовательность для обнаружения наличия трансгенного трансформанта сои DBN9004 в биологическом образце, набор, содержащий такую последовательность, и способ такого обнаружения | 2017 |
|
RU2743397C2 |
| ГЕНЫ GRG23 И GRG51, ПРИДАЮЩИЕ УСТОЙЧИВОСТЬ К ГЕРБИЦИДАМ | 2006 |
|
RU2393225C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2639275C2 |
| СИНТЕТИЧЕСКИЕ ТРАНЗИТНЫЕ ПЕПТИДЫ ХЛОРОПЛАСТА ИЗ BRASSICA | 2013 |
|
RU2636035C2 |
































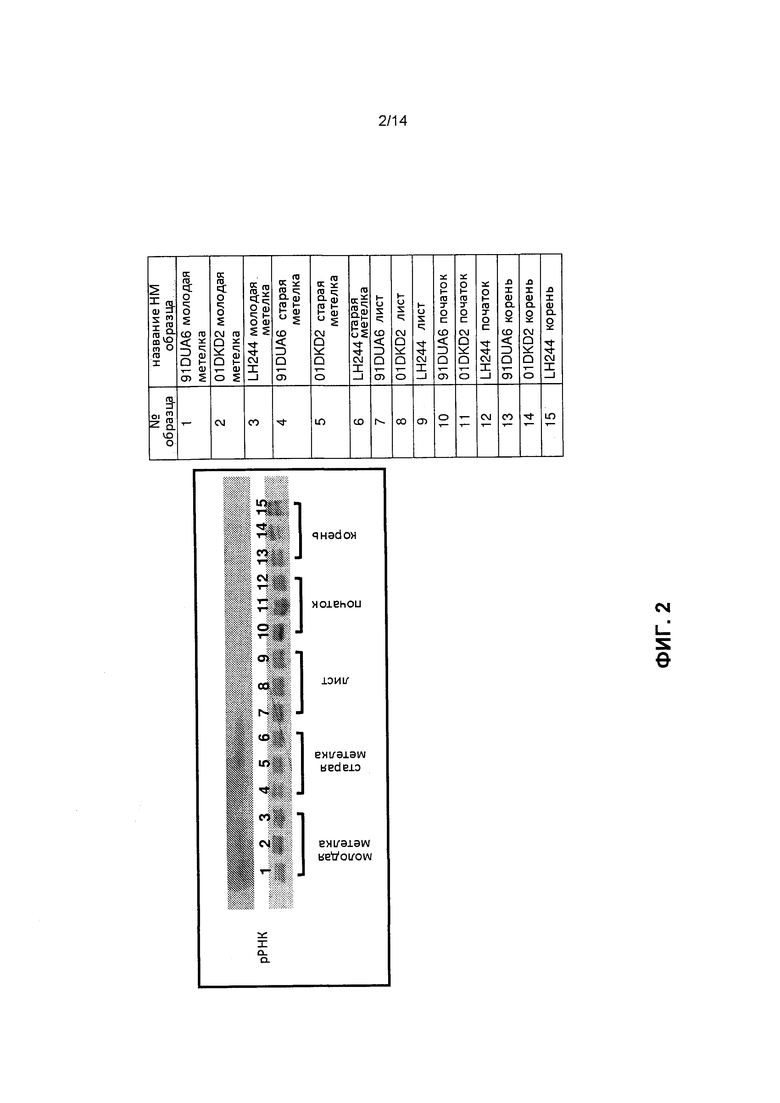

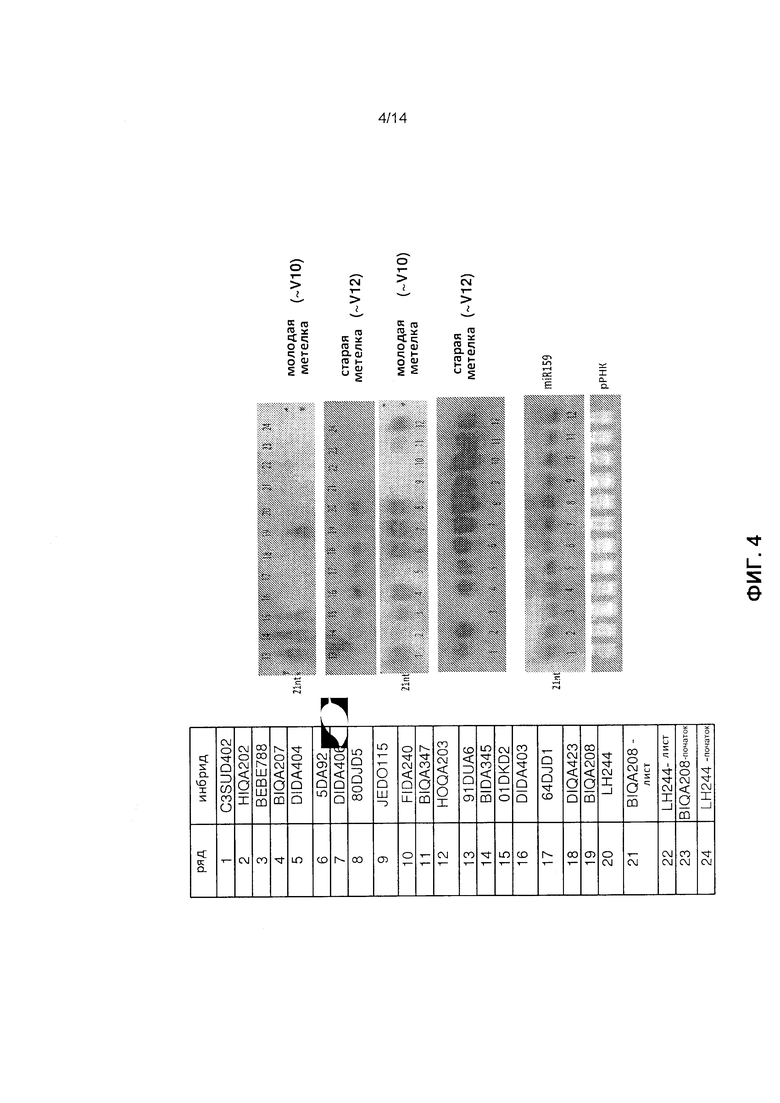
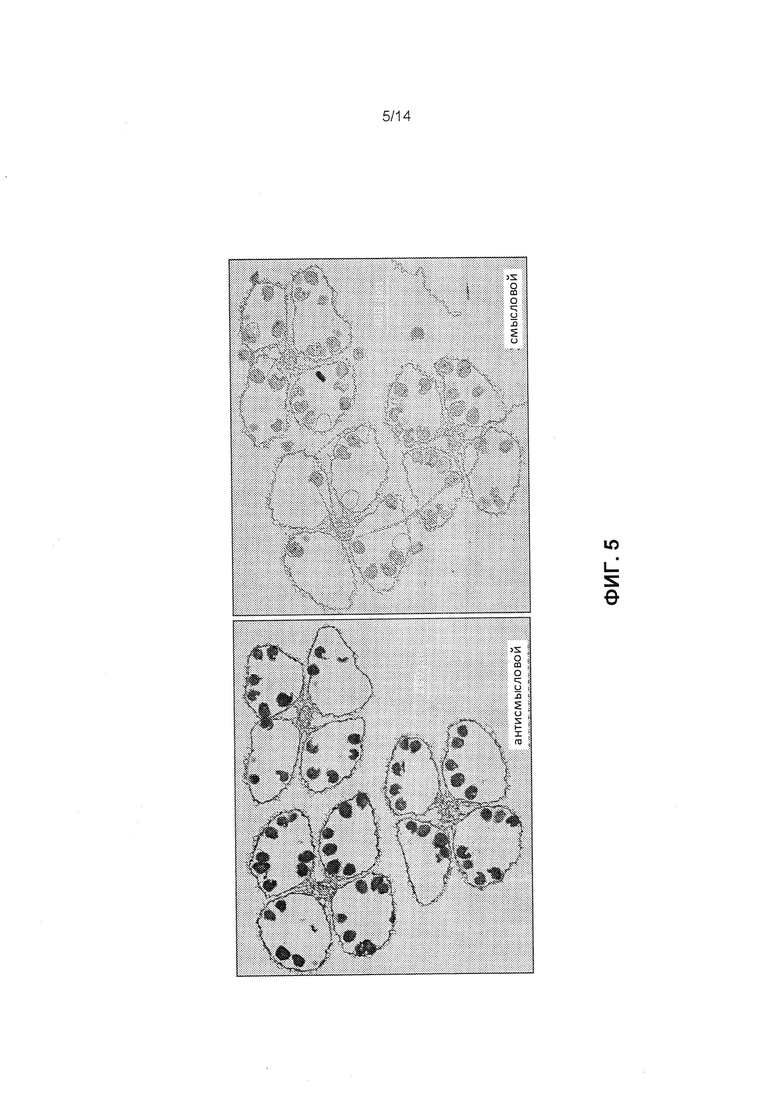
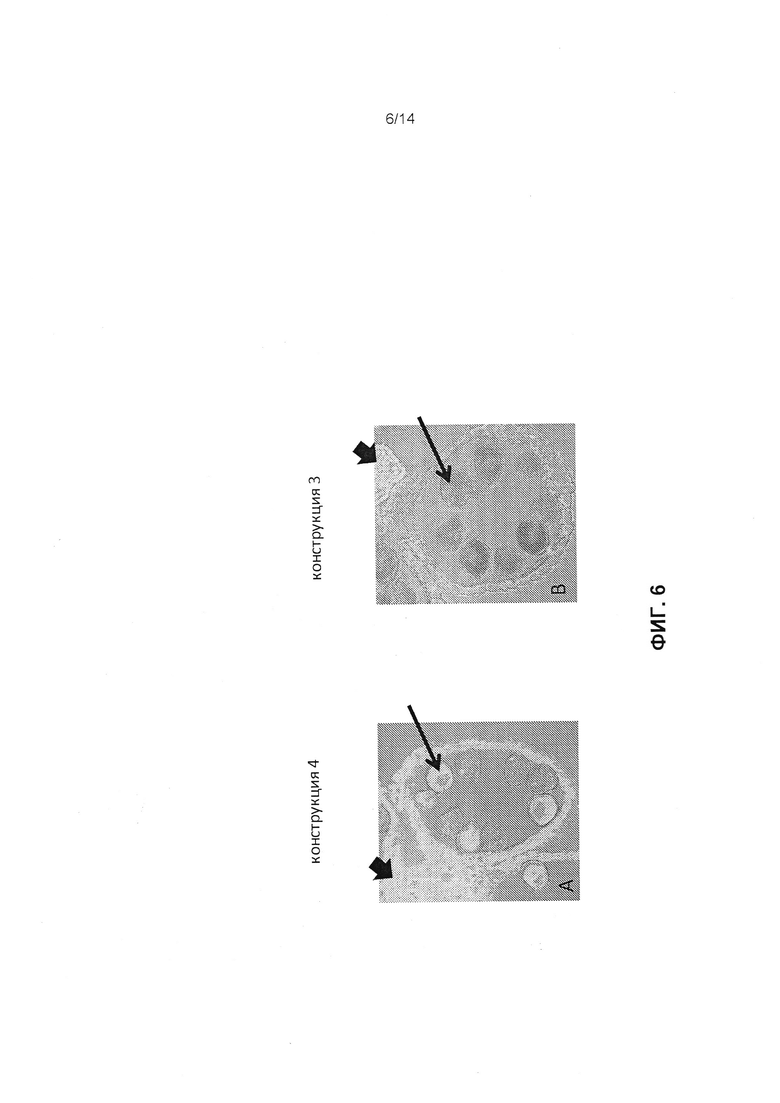
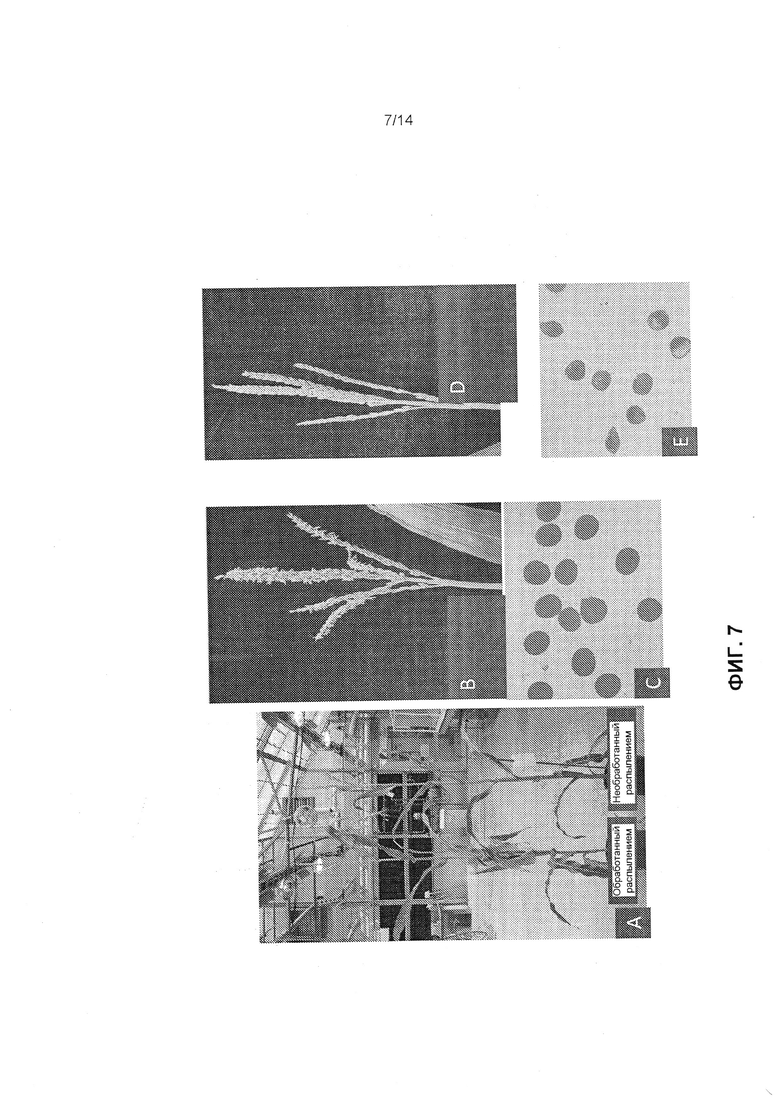
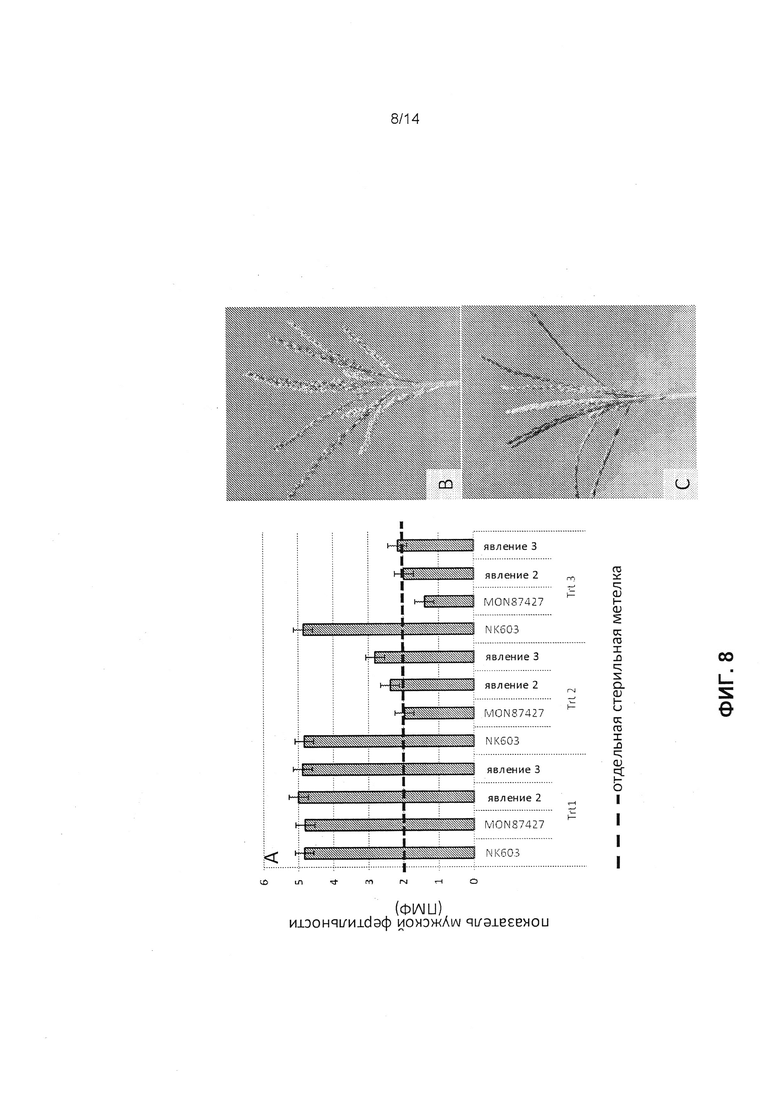

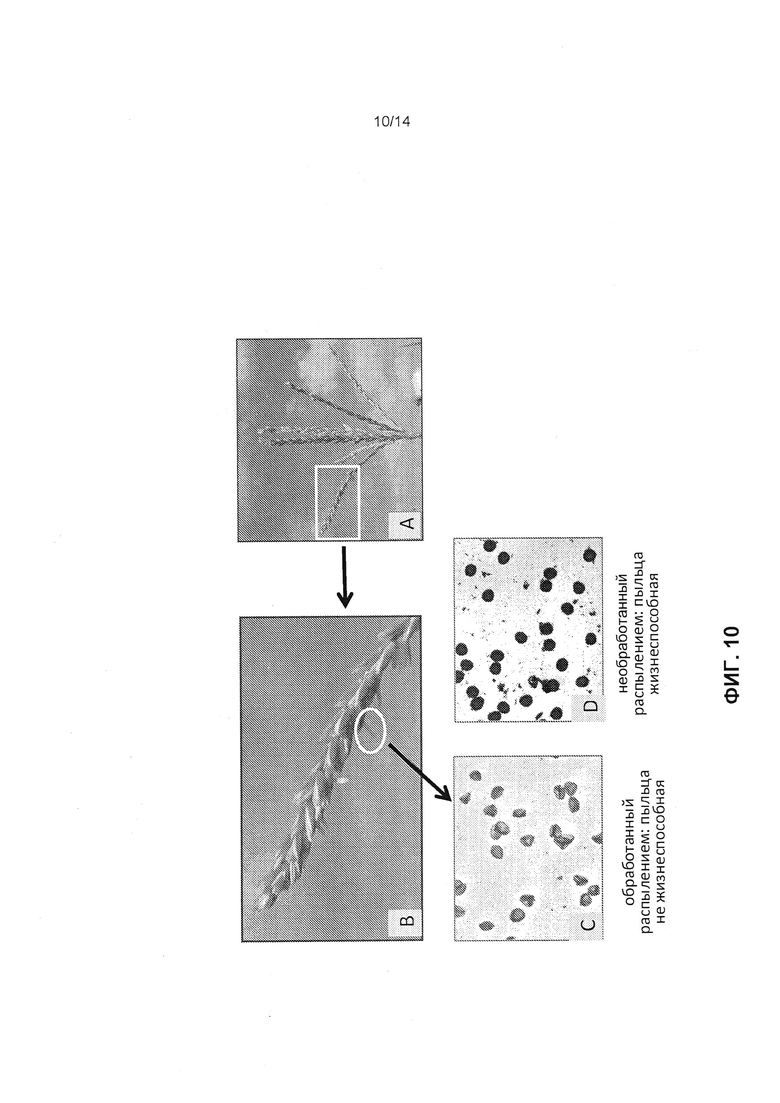
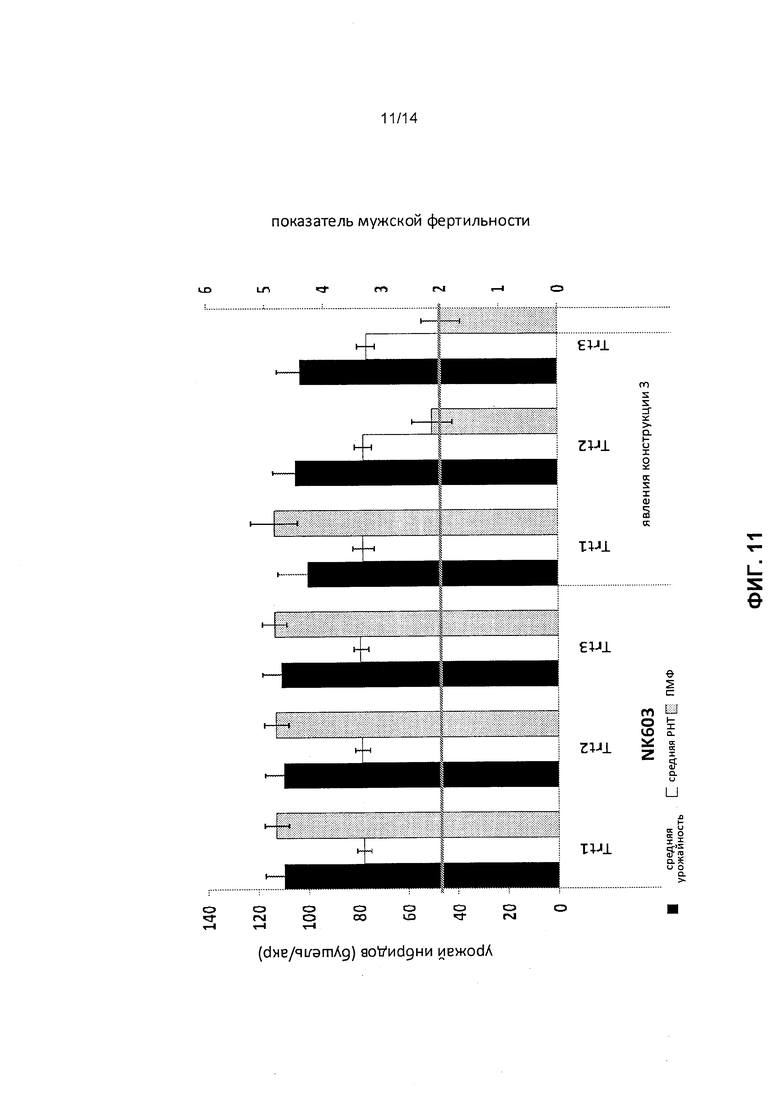
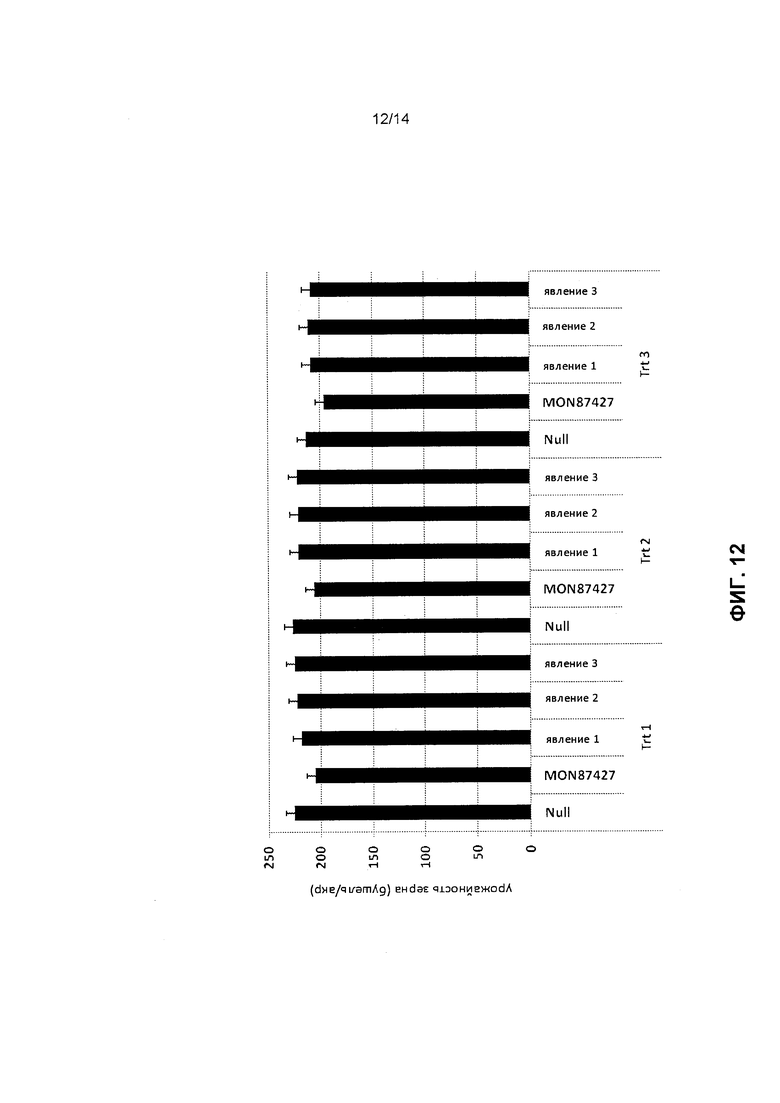


Изобретение относится к области биохимии, в частности к конструкции рекомбинантной ДНК для супрессии экспрессии кодирующей белок последовательности. Также раскрыты трансгенное растение, содержащие указанную конструкцию, семя указанного трансгенного растения, часть указанного трансгенного растения. Раскрыты способ получения указанной конструкции рекомбинантной РНК, способ супрессии экспрессии кодирующей белок последовательности в мужской репродуктивной ткани трансгенного растения, способ индуцирования мужской стерильности в трансгенном растении, способ получения семян, содержащих указанную конструкцию ДНК, способ получения товарного продукта из указанных растения или семени. Изобретение позволяет супрессировать экспрессию кодирующей белок последовательности, придающей растению толерантность к гербициду. 10 н. и 23 з.п. ф-лы, 14 ил., 7 табл., 12 пр.
1. Конструкция рекомбинантной ДНК для супрессии экспрессии кодирующей белок последовательности, придающей растению толерантность к гербициду, в мужской репродуктивной ткани трансгенного растения, экспрессирующего указанную кодирующую белок последовательность, придающую растению толерантность к гербициду, где указанная конструкция включает кодирующую белок последовательность, придающую растению толерантность к гербициду, функционально связанную с последовательностью ДНК, включающей специфический для мужской ткани элемент миРНК (мтс-миРНК), имеющий последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-94 и 96-149, где указанный элемент мтс-миРНК является гетерологичным по отношению к указанной кодирующей белок последовательности, придающей растению толерантность к гербициду.
2. Конструкция рекомбинантной ДНК по п.1, отличающаяся тем, что указанный элемент мтс-миРНК содержит одну последовательность мтс-миРНК.
3. Конструкция рекомбинантной ДНК по п.2, отличающаяся тем, что указанная последовательность мтс-миРНК выбрана из группы, состоящей из SEQ ID NO: 1-56 и 105-149.
4. Конструкция рекомбинантной ДНК по п.1, отличающаяся тем, что указанный элемент мтс-миРНК выбран из группы, состоящей из SEQ ID NO: 57-94 и 96-104.
5. Конструкция рекомбинантной ДНК по п.1, отличающаяся тем, что экспрессия указанной кодирующей белок последовательности в трансгенном растении придает указанному растению по меньшей мере вегетативную толерантность к гербициду.
6. Конструкция рекомбинантной ДНК по п.5, отличающаяся тем, что указанная кодирующая белок последовательность кодирует толерантный к глифосату EPSPS.
7. Способ получения конструкции рекомбинантной ДНК по п.1, включающий идентификацию элемента мтс-миРНК, содержащего последовательность мтс-миРНК, выбранную из группы, состоящей из SEQ ID NO: 1-94 и 96-149, и функциональное связывание такого указанного элемента мтс-миРНК с кодирующей белок последовательностью, где указанный элемент мтс-миРНК является гетерологичным по отношению к указанной кодирующей белок последовательности.
8. Способ по п.7, отличающийся тем, что указанная по меньшей мере одна последовательность мтс-миРНК является по меньшей мере последовательностью, выбранной из группы, состоящей из SEQ ID NO: 1-56 и 105-149, или указанный элемент мтс-миРНК выбирают из группы, состоящей из SEQ ID NO: 57-94 и 96-104.
9. Способ по п.7, отличающийся тем, что указанный элемент мтс-миРНК является специфичным для метелки.
10. Трансгенное растение, проявляющее супрессию экспрессии белка, имеющее в своем геноме конструкцию рекомбинантной ДНК по п.1.
11. Семя трансгенного растения по п.10, проявляющее супрессию экспрессии белка, содержащее конструкцию рекомбинантной ДНК по п.1.
12. Часть трансгенного растения по п.10, проявляющая супрессию экспрессии белка, содержащая конструкцию рекомбинантной ДНК по п.1, где частью растения является пыльца, корни, листья или метелки.
13. Трансгенное растение по п.10, отличающееся тем, что указанное трансгенное растение является однодольным растением.
14. Трансгенное растение по п.10, отличающееся тем, что указанное трансгенное растение является растением кукурузы.
15. Способ супрессии экспрессии кодирующей белок последовательности в мужской репродуктивной ткани трансгенного растения, включающий экспрессию в указанном трансгенном растении конструкции рекомбинантной ДНК по п.1.
16. Способ по п.15, отличающийся тем, что указанная мужская репродуктивная ткань является метелкой растения кукурузы.
17. Способ по п.15, отличающийся тем, что указанный элемент мтс-миРНК содержит три последовательности мтс-миРНК.
18. Способ по п.15, отличающийся тем, что указанный элемент мтс-миРНК включает по меньшей мере одну последовательность мтс-миРНК, выбранную из группы, состоящей из SEQ ID NO: 1-56 и 105-149.
19. Способ по п.15, отличающийся тем, что указанный элемент мтс-миРНК выбирают из группы, состоящей из SEQ ID NO: 57-94 и 96-104.
20. Способ по п.15, отличающийся тем, что экспрессия указанной кодирующей белок последовательности в трансгенном растении придает указанному растению по меньшей мере вегетативную толерантность к гербициду.
21. Способ по п.20, отличающийся тем, что белок, кодируемый указанной последовательностью, является толерантным к глифосату EPSPS.
22. Способ индуцирования мужской стерильности в трансгенном растении, включающий применение эффективного количества гербицида к трансгенному растению, включающему конструкцию рекомбинантной ДНК по п.1, при этом указанное применение гербицида проводится во время развития мужской репродуктивной ткани указанного трансгенного растения, индуцируя тем самым, мужскую стерильность в указанном трансгенном растении.
23. Способ по п.22, отличающийся тем, что указанное трансгенное растение является растением кукурузы.
24. Способ по п.22, отличающийся тем, что указанное применение гербицида предотвращает по меньшей мере осыпание пыльцы или выпячивание пыльника.
25. Способ по п.22, отличающийся тем, что указанное развитие мужской репродуктивной ткани является стадией, выбранной из группы, состоящей из стадии развития растения кукурузы V4, V5, V6, V7, V8, V9, V10, V11, V12, V13 и V14.
26. Способ по п.22, отличающийся тем, что указанный гербицид выбран из группы, состоящей из ингибиторов ацетил коэнзим A карбоксилазы (ACCase), ингибиторов ацетолактатсинтазы (ALS), ингибиторов фотосистемы II (PSII), ингибиторов протопорфириногеноксидазы (PPO), ингибиторов 4-гидроксифенил пируват диоксигеназы (HPPD), ингибиторов 5-энолипирувилшикимат 3-фосфатсинтазы (EPSPS), ингибиторов глутаминсинтетазы (GS) и синтетических ауксинов.
27. Способ по п.22, отличающийся тем, что указанный гербицид является глифосатом, а белок, кодируемый указанной последовательностью, является толерантным к глифосату EPSPS.
28. Способ получения семян, содержащих конструкцию ДНК по п.1, включающий:
a. применение эффективного количества гербицида к трансгенному растению, включающему конструкцию рекомбинантной ДНК по п.1, при этом указанное применение гербицида проводится во время развития мужской репродуктивной ткани указанного трансгенного растения, индуцируя тем самым мужскую стерильность в указанном трансгенном растении;
б. оплодотворение указанного трансгенного растения пыльцой второго растения;
в. сбор семян от указанного трансгенного растения; и
г. обнаружение присутствия конструкции ДНК по п.1 в собранных семенах.
29. Способ по п.28, отличающийся тем, что указанное трансгенное растение является кукурузой.
30. Способ по п.28, отличающийся тем, что указанный гербицид является глифосатом, а белок, кодируемый указанной последовательностью, является толерантным к глифосату EPSPS.
31. Способ по п.30, отличающийся тем, что указанный глифосат применяют во время указанного развития в эффективной дозе от примерно 0,125 фунтов кислоты в эквиваленте на акр до примерно 8 фунтов кислоты в эквиваленте на акр.
32. Семя, полученное способом по п.28, способное к воспроизведению растения, проявляющего подавленную экспрессию кодирующей белок последовательности в мужской репродуктивной ткани, и содержащее конструкцию рекомбинантной ДНК по п.1.
33. Способ получения товарного продукта, включающий получение трансгенного растения, семени или части растения по пп.10, 11 или 12 или семени по п.32 и обработку указанных растений или семян, таким образом получая товарный продукт.
| Кипятильник для воды | 1921 |
|
SU5A1 |
| US 20110035838 A1, 10.02.2011 | |||
| Насос для подачи горючей жидкости в двигателях внутреннего горения | 1926 |
|
SU6761A1 |
| СПОСОБ ИНГИБИРОВАНИЯ ПРОЛИФЕРАЦИИ ГЕПАТОЦИТОВ, СПОСОБ ИНГИБИРОВАНИЯ КЛЕТОЧНОЙ АДГЕЗИИ ГЕПАТОЦИТОВ И СПОСОБ ИНГИБИРОВАНИЯ БИОЛОГИЧЕСКОЙ АКТИВНОСТИ ANGPTL4 В ГЕПАТОЦИТАХ ИЛИ ПРЕДШЕСТВЕННИКАХ ГЕПАТОЦИТОВ | 2005 |
|
RU2380411C2 |
| US 0007355096 B2, 08.04.2008. | |||

