ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ПРИЛОЖЕНИЯ
Эта заявка утверждает приоритет заявки США с серийным № 61/563303, поданной 23 ноября 2011 года, раскрытие которой включено посредством ссылки в полном объеме.
ТЕХНИЧЕСКАЯ ОБЛАСТЬ
Данное изобретение относится к способам и материалам для ферментативного синтеза могрозидных соединений и, в частности, к гликозилированию могрола при помощи уридин-5'-дифосфо (УДФ) зависимых гликозилтрансфераз (UGT) для производства различных могрозидных соединений.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
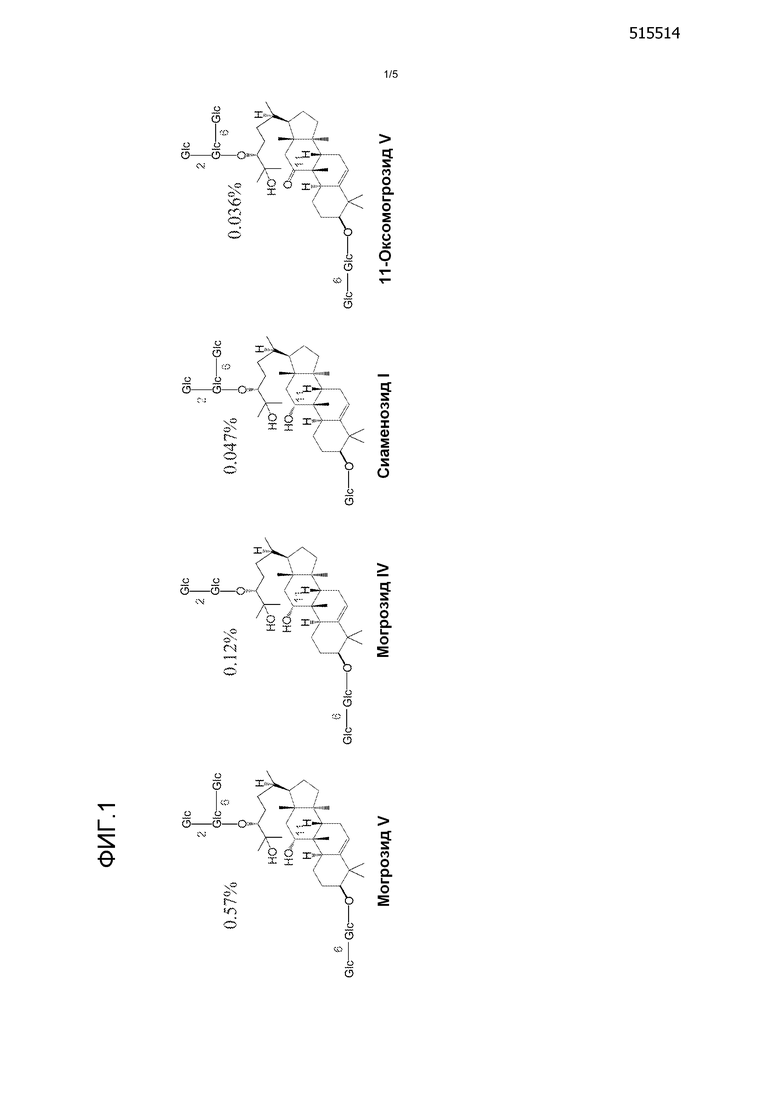
Могрозиды представляют собой семейство тритерпеновых гликозидов, изолированных из плодов Siraitia grosvenorii (Swingle), также известной как Momordica grosvenori (Swingle). Экстракты плодов коммерчески используются как натуральные подсластители. Четыре основных соединения - могрозид V, могрозид IV, сиаменозид I и 11-оксоoмогрозид V - были идентифицированы из плодов Siraitia grosvenorii (Swingle), которые отвечают за сладость плодов. Смотри фиг. 1. Могрозид V является наиболее распространенным из этих четырех соединений, примерно с 0,57% (масс./масс.) сухих фруктов, за которым следуют могрозид IV и сиаменозид I, каждый из которых содержит четыре глюкозные группы. 11-оксoмогрозид V имеет кетоновую группу вместо гидроксила в C-11. Смотри, например, Takemoto, et al., Yakugaku Zasshi, 103, 1151-1154; 1155-1166; 1167-1173, (1983); Kasai, et al., Agric. Biol. Chem. 53, 3347-3349 (1989); Matsumoto, Chem. Pharm. Bull. 38, 2030-2032 (1990); и Prakash, et al., J. Carbohydrate Chem. 30, 16-26 (2011).
Все могрозиды имеют одинаковый тритерпеновый кор, называемый могролом. Агликон могрол гликозилирован с разным количеством глюкозных групп, связанных с формированием различных могрозидных соединений. Считается, что могрозид синтезируется в следующем порядке: синтез кукурбитадиенола из общего тритерпенового предшественника сквалена; Р450-окисления кукурбитадиенола с образованием агликона могрола и гликозилирование могрола с добавлением пяти глюкоз одна за другой с образованием могрозида V. Смотри Tang, et al., BMC Genomics, 12, 343 (2011). Оба интермедиата, как кукурбитадиенол, так и могрол, существуют в плодах, поскольку они были выделены в качестве минорных продуктов. Смотри Ukiya, et al., J. Agric. Food Chem. 50, 6710-6715 (2002). Гликозидные интермедиаты существуют как в серии 11-гидрокси, так и в серии 11-оксо, и постепенно меняются от могрозида I к могрозиду V по мере созревания плодов, что указывает на то, что тритерпеновое ядро полностью окисляется ферментной системой цитохрома Р450 перед последующими гликозилированиями. При этом ферменты, отвечающие за выработку могрозидов, не были идентифицированы.
КРАТКОЕ СОДЕРЖАНИЕ
В одном аспекте в этом документе описывается способ получения могрозидного соединения. Способ включает в себя инкубацию могрола с уридин-5'-дифосфо (УДФ) зависимой гликозилтрансферазой (UGT), чтобы произвести могрозидное соединение (например, могрозид 1а, могрозид 1b или могрозидное соединение, гликозилированное по C25-OH). UGT можно выбрать из группы, состоящей из 73C3, 73C6, 85C2, 73C5 и 73E1. UGT можно продуцировать рекомбинантным способом, или они могут присутствовать в клеточном лизате рекомбинантного хозяина.
В настоящем документе также предлагается способ получения могрозидного соединения. Способ включает в себя контактирование могрола с лизатом клеток, подготовленным из рекомбинантного хозяина, экспрессирующего UGT, для получения могрозидного соединения (например, могрозид 1а, могрозид 1b или могрозидное соединение, гликозилированное по C25-OH). UGT может быть выбран из группы, состоящей из 73C3, 73C6, 85C2, 73C5 и 73E1.
Если не указано иное, все технические и научные термины, использованные в настоящем документе, имеют тот же смысл, как обычно понимается любым специалистом в рассматриваемой области, к которой относится изобретение. Несмотря на то, что способы и материалы, аналогичные или эквивалентные тем, которые описаны в настоящем документе, могут быть использованы в практике изобретения, подходящие способы и материалы описываются ниже. Все публикации, патентные заявки, патенты и другие ссылки, упомянутые в настоящем документе, включены посредством ссылки в полном объеме. В случае конфликта настоящая спецификация, включая определения, будет определяющей. Кроме того, материалы, способы и примеры приведены только для иллюстрации и не предназначены для ограничения. Другие особенности и преимущества изобретения будут очевидны из приведенного ниже подробного описания. Заявители оставляют за собой право альтернативно заявить любое раскрытое изобретение с помощью переходного выражения "включающий", "состоящий в основном из" или "состоящий из" в соответствии со стандартной практикой в области патентного законодательства.
ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг. 1 содержит химическую структуру могрозида V, могрозида IV, сиаменозида I и 11-оксoмогрозида V.
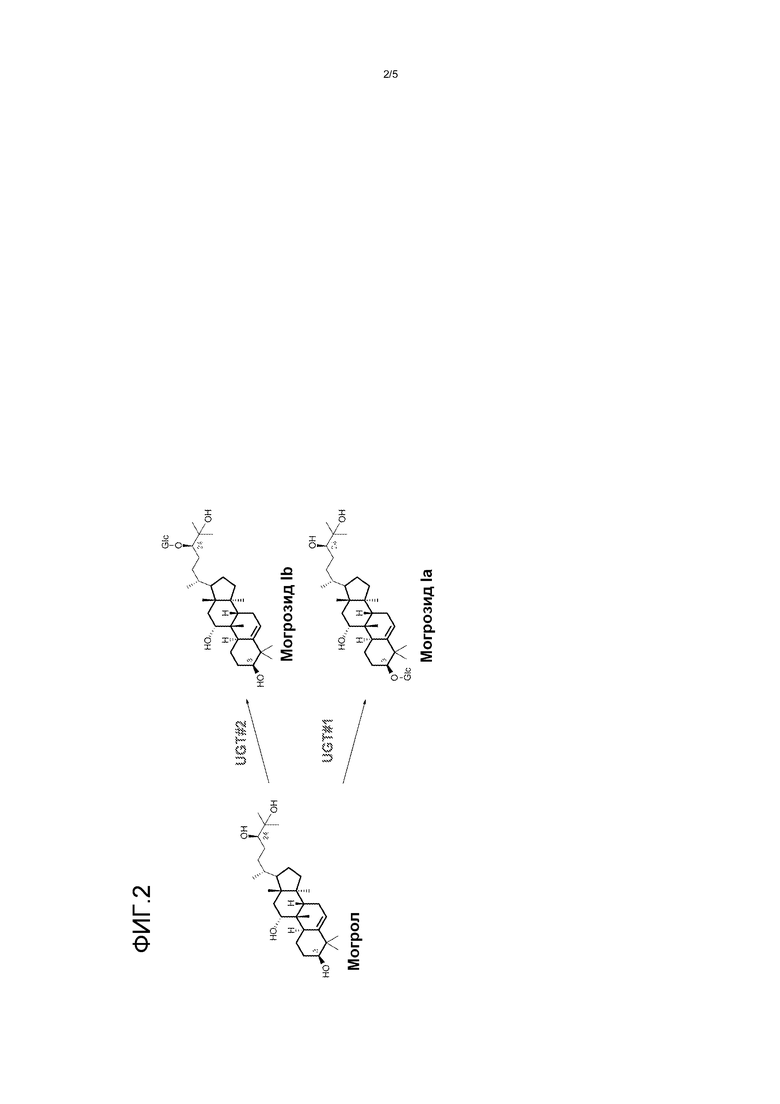
Фиг. 2 представляет собой изображение биосинтеза могрозида Iа и могрозида Ib из могрола с помощью UGT.
Фиг. 3 представляет последовательности аминокислот нижеследующих UGT: UGT73C3, UGT73C5, UGT73C6, UGT73E1 и UGT85C2 (SEQ ID NOs: 1-5 соответственно).
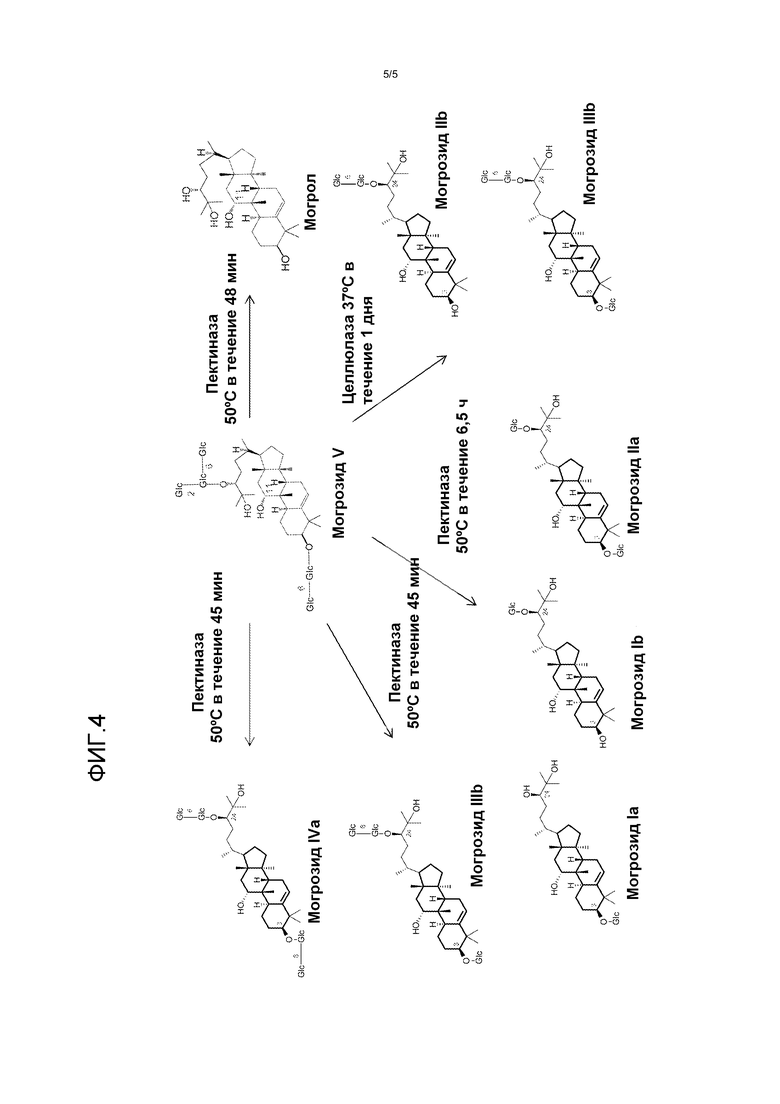
Фиг. 4 представляет собой схематическое изображение продуктов, полученных из могрозида V после инкубации с пектиназой и/или целлюлазой.
ПОДРОБНОЕ ОПИСАНИЕ
Этот документ предоставляет способы и материалы для гликозилирования могрола с использованием одной или более уридин-5'-дифосфо (UDP) зависимых гликозилтрансфераз (UGT). Как указано ниже, было идентифицировано, по меньшей мере, пять UGT, которые гликозилируют агликон могрол. Смотри фиг. 2. Каждая из UGT, идентифицированных в настоящем документе, относится к семейству I гликозилтрансфераз. UGT 73C3, 73C6, 85C2 и 73E1 гликозилируют по положению C24-OH (UGT#2 на фиг. 2), в то время как UGT73C5 гликозилирует по положению как C3-OH (UGT#1 на фиг. 2), так и C24-OH (UGT#2). UGT 73C3, 73C5 и 73C6 происходят из Arabidopsis thaliana. UGT 73E1 и 85C2 происходят из Stevia rebaudiana. Аминокислотные последовательности UGT 73C3, 73C5, 73C6, 73E1 и 85C2 (SEQ ID NOs: 1-5) приведены на фиг. 3.
UGT-полипептиды, описанные в настоящем документе, могут быть произведены с помощью любого подходящего способа. Например, UGT-полипептиды могут быть произведены путем химического синтеза. Кроме того, UGT-полипептид, описанный в настоящем документе, может быть произведен с помощью стандартной рекомбинантной технологии с использованием гетерологичных экспрессионных векторов, кодирующих этот UGT-полипептид. Экспрессионные векторы можно вводить в клетки хозяина (например, путем трансформации или трансфекции) для экспрессии кодируемого полипептида, который затем можно очистить. Системы экспрессии, которые могут быть использованы для малого или крупномасштабного производства UGT-полипептидов, включают, без ограничения, микроорганизмы, такие как бактерии (например, E. coli и B. subtilis), трансформированные рекомбинантной ДНК бактериофага, плазмидной ДНК или векторами экспрессии на основе космидной ДНК, содержащими молекулы нуклеиновых кислот, описанные в настоящем документе. Полезные системы экспрессии также включают системы клеток насекомых, инфицированных рекомбинантными вирусными векторами экспрессии (например, бакуловирус), содержащими молекулы нуклеиновых кислот, описанные в настоящем документе, и клеток растений, инфицированных рекомбинантными вирусными векторами экспрессии (например, вирус табачной мозаики) или трансформированных рекомбинантными плазмидными векторами экспрессии (например, Ti-плазмиды), содержащими молекулы нуклеиновых кислот, описанные в настоящем документе. UGT-полипептиды также могут быть получены с использованием системы экспрессии млекопитающих, несущей рекомбинантные экспрессионные конструкции, содержащие промоторы, полученные из генома клеток млекопитающих (например, металлотионеиновый промотор) или из вирусов млекопитающих (например, поздний промотор аденовируса и промотор цитомегаловируса), наряду с нуклеиновыми кислотами, описанными в настоящем документе. UGT-полипептиды могут иметь N-концевой или С-концевой тэг, как описано ниже.
Этот документ также предоставляет изолированные нуклеиновые кислоты, кодирующие UGT-полипептиды. "Изолированная нуклеиновая кислота" относится к нуклеиновой кислоте, которая отделена от других молекул нуклеиновых кислот, которые присутствуют в геноме, в том числе и нуклеиновых кислот, которые в норме фланкируют нуклеиновую кислоту в геноме с одного фланга или с обеих сторон. Термин "изолированный", как он употребляется в настоящем документе в отношении нуклеиновых кислот, также включает в себя любую последовательность не встречающейся в природе нуклеиновой кислоты, поскольку такие не встречающиеся в природе последовательности не обнаруживаются в природе и не имеют непосредственно прилегающих последовательностей в природном геноме.
Изолированная нуклеиновая кислота может представлять собой, например, молекулу ДНК, предоставленную одной из последовательностей нуклеиновых кислот, в норме обнаруживаемых непосредственно фланкирующими ту молекулу ДНК, удаленную из или отсутствующую в природном геноме. Таким образом, изолированная нуклеиновая кислота включает, без ограничения, молекулу ДНК, которая существует в виде отдельной молекулы (например, химически синтезированная нуклеиновая кислота, или кДНК или фрагмент геномной ДНК, полученный способом ПЦР или обработкой рестрикционной эндонуклеазой) независимо от других последовательностей, а также ДНК, которая встроена в вектор, автономно реплицирующуюся плазмиду, вирус (например, любой парамиксовирус, ретровирус, лентивирус, аденовирус или вирус герпеса) или в геномную ДНК прокариотов или эукариотов. Кроме того, изолированная нуклеиновая кислота может содержать сконструированную нуклеиновую кислоту, такую как молекула ДНК, которая является частью гибридной или слитой нуклеиновой кислоты. Нуклеиновая кислота, существующая среди нуклеиновых кислот в пределах от сотен до миллионов других нуклеиновых кислот, например библиотек кДНК или геномных библиотек, или гелевых фрагментов, содержащих рестрикционный перевар геномной ДНК, не считается изолированной нуклеиновой кислотой.
В некоторых вариантах осуществления последовательность нуклеиновых кислот, кодирующая UGT-полипептид, может включать тег-последовательность, которая кодирует "тег", спроектированный для облегчения последующей манипуляции (например, для облегчения очистки или детекции), секреции или локализации кодируемого полипептида. Тег-последовательности могут быть встроены в последовательность нуклеиновых кислот, кодирующую UGT-полипептид, такой, что кодируемый тег располагается либо в карбоксильном, либо в амино-конце UGT-полипептида. Неограничивающие примеры кодируемых тэгов включают зеленый флуоресцентный белок (GFP), глутатион-S-трансферазу (GST), HIS-тег и FLAG™-тег (Kodak, New Haven, CT). Другие примеры тэгов включают транзитный пептид хлоропластов, транзитный пептид митохондрий, пептид амилопластов, сигнальный пептид или тег секреции.
Функциональные гомологи
Функциональные гомологи полипептидов, описанных выше, пригодны также для использования в способах и рекомбинантных хозяевах, описанных в настоящем документе. Функциональный гомолог представляет собой полипептид, который имеет сходство последовательности с контрольным полипептидом и который осуществляет одну или более биохимических или физиологических функций(ю) контрольного полипептида. Функциональный гомолог и контрольный полипептид могут представлять собой полипептиды природного происхождения, и сходство последовательностей может иметь место из-за конвергентных или дивергентных эволюционных событий. Как таковые, функциональные гомологи иногда обозначаются в литературе как гомологи, или ортологи, или паралоги. Варианты функционального гомолога природного происхождения, такого как полипептиды, кодируемые мутантами кодирующей последовательности дикого типа, сами по себе могут быть функциональными гомологами. Функциональные гомологи также могут быть созданы посредством сайт-направленного мутагенеза последовательности, кодирующей полипептид, либо путем комбинирования доменов кодирующих последовательностей для различных полипептидов природного происхождения ("перестановка доменов"). Способы модификации генов, кодирующих функциональные UGT-полипептиды, описанные в настоящем документе, известны и включают в себя, среди прочего, методики направленной эволюции, методики сайт-направленного мутагенеза и методики случайного мутагенеза и могут быть полезны для повышения специфической активности полипептида, изменения субстратной специфичности, изменения уровней экспрессии, изменения субклеточной локализации или изменения полипептид-полипептидных взаимодействий нужным образом. Такие модифицированные полипептиды считаются функциональными гомологами. Термин "функциональный гомолог" иногда применяется к нуклеиновой кислоте, которая кодирует функционально гомологичный полипептид.
Функциональные гомологи можно определить путем анализа выравниваний нуклеотидных и полипептидных последовательностей. Например, при выполнении запроса к базе данных нуклеотидных или пептидных последовательностей можно определить гомологи UGT-полипептидов. Анализ последовательности может включать анализ BLAST, Reciprocal BLAST или PSI-BLAST резервируемых баз данных с использованием последовательности аминокислот UGT в качестве эталонной последовательности. В некоторых случаях аминокислотная последовательность выводится из нуклеотидной последовательности. Те полипептиды в базе данных, которые имеют больше чем 40% идентичности последовательности, являются кандидатами для дальнейшего оценивания на пригодность в качестве UGT-полипептида. Сходство аминокислотной последовательности предусматривает использование консервативных аминокислотных замещений, как, например, замена одного гидрофобного остатка на другой или замена одного полярного остатка на другой. При желании можно осуществить проверку таких кандидатов вручную для того, чтобы сузить число кандидатов при дальнейшем изучении. Проверку вручную можно производить путем выбора тех кандидатов, которые, как представляется, имеют домены, присутствующие в UGT-полипептидах, например консервативные функциональные домены.
Консервативные области можно определить путем локализации области в рамках первичной последовательности аминокислот полипептида, которая представляет собой повторяющуюся последовательность, образует некоторую вторичную структуру (например, спирали и бета-листы), устанавливает положительно или отрицательно заряженные домены или представляет белковый мотив или домен. Смотри, например, веб-страницу сайта Pfam с описанием консенсусных последовательностей для различных белковых мотивов и доменов на World Wide Web по адресу sanger.ac.uk/Software/Pfam/ и pfam.janelia.org/. Информация, включенная в базу данных Pfam, описана в Sonnhammer et al., Nucl. Acids Res., 26: 320-322 (1998); Sonnhammer et al., Proteins, 28: 405-420 (1997); и Bateman et al., Nucl. Acids Res., 27: 260-262 (1999). Консервативные области можно также определить путем выравнивания последовательностей сходных или родственных полипептидов из близкородственных видов. Близкородственные виды предпочтительно представляют собой виды из того же семейства. В некоторых вариантах осуществления выравнивание последовательностей из двух разных видов является адекватным.
Как правило, полипептиды, которые обладают по меньшей мере около 40% идентичности аминокислотной последовательности, полезны для определения консервативных областей. Консервативные области родственных полипептидов проявляют по меньшей мере 45% идентичности аминокислотной последовательности (например, по меньшей мере 50%, не менее 60%, по меньшей мере 70%, по меньшей мере 80% или по меньшей мере 90% идентичности аминокислотной последовательности). В некоторых вариантах осуществления консервативная область проявляет по меньшей мере 92%, 94%, 96%, 98% или 99% идентичности аминокислотной последовательности. Идентичность последовательностей может быть определена, как указано выше.
Способы получения могрозидных соединений
Могрозидные соединения можно получать путем инкубирования в могроловый субстрат с одним или более UGT-полипептидов, описанных в настоящем документе, в результате чего образуется могрозидный продукт. В некоторых вариантах осуществления реакционная смесь содержит множество UGT-полипептидов, таких, что в реакционном сосуде имеет место множественность гликозилирований. В других вариантах осуществления реакционная смесь содержит один UGT-полипептид, и имеет место одно или более гликозилирований, катализируемых полипептидом. Например, первый реакционный сосуд может содержать субстрат и один или более UGT-полипептидов для получения интермедиата, который может быть внесен в другой реакционный сосуд, содержащий один или более других UGT-полипептидов, для продуцирования следующего интермедиата или могрозидного продукта. Продукт, произведенный во втором реакционном сосуде, затем может быть извлечен.
Каждый из UGT-полипептидов может представлять собой очищенный полипептид, например, может быть добавлен к реакционной смеси в виде раствора, содержащего 80%, 90%, 95% или более 99 мас.% нужного UGT. Кроме того, UGT-полипептид(ы) может присутствовать в клеточном лизате, приготовленном из рекомбинантного хозяина, экспрессирующего UGT(ы), и может быть добавлен в реакционную смесь в качестве клеточного лизата для инкубации с могроловым субстратом.
Уровни продуктов, субстратов и интермедиатов могут быть определены путем экстракции образцов из реакционного сосуда для анализа в соответствии с опубликованными способами. Могрозидные соединения могут быть извлечены из реакционного сосуда, используя различные способы, известные в рассматриваемой области.
Изобретение далее будет охарактеризовано в следующих примерах, не ограничивающих объем изобретения, который описан в пунктах формулы изобретения.
Примеры
Пример 1 - Очистка могрозида V
Могрозид V очищали из коммерчески доступных экстрактов архата (PureLo®, Swanson) следующим образом. Три бутылки PureLo® (240 граммов) растворяли в воде (900 мл), затем нагружали на колонку со смолой НР-20 (400 граммов смолы). Колонку промывали водой (2,5 литра), затем дополнительно промывали 20% метанолом-водой. Продукт элюировали метанолом. После испарения растворителей и сушки в условиях высокого вакуума был получен могрозид V (2,5 грамма, ~80% чистоты; 11-oксoмогрозид V составлял основную примесь).
Пример 2 - Ферментативный синтез могрола из могрозида V
Могрозид V (300 мг) растворяли в 0,1 M натрийацетатном буфере (pH 4,5, 100 мл) и добавляли грубую пектиназу из Aspergillus niger (25 мл, Sigma P2736). Смесь перемешивали при 50°C в течение 48 часов. Реакционную смесь экстрагировали этилацетатом (2×100 мл). Органический экстракт высушивали под вакуумом, затем очищали с помощью препаративной ВЭЖХ. Очищенный могрол (40 мг) был получен, и его структура подтверждена при помощи ЯМР- и масс-спектроскопии. Смотри фиг. 4.
Пример 3 - Ферментативный синтез могрол 3-O-глюкозида (могрозид 1а) и могрол 24-O-глюкозида (могрозид 1b) из могрозида V
Могрозид V (300 мг) растворяли в 0,1 M натрийацетатном буфере (pH 4,5, 100 мл) и добавляли грубую пектиназу из Aspergillus niger (25 мл, Sigma P2736). Смесь перемешивали при 50°C в течение 6,5 часов. Реакционную смесь экстрагировали этилацетатом (2×100 мл). Органический экстракт высушивали под вакуумом, затем очищали с помощью препаративной ВЭЖХ. Очищенный могрозид Ia (11,0 мг) и могрозид Ib (8,0 мг) были получены. Их структуры были подтверждены ЯМР- и масс-спектроскопией. Смотри фиг. 4.
Пример 4 - In vitro UGT-скрининг и реакции
Проводили in vitro реакции могрола с панелью из 230 UGT-ферментов, и продукты анализировали с помощью LC-MS. In vitro UGT реакционные смеси включали 4X Трис-буфер, могрол (250 мкМ), UDP-глюкозу (750 мкМ) и 1% щелочную фосфатазу. Пять мкл каждого частично очищенного UGT-фермента или грубого ферментного экстракта добавляли к реакции, и объем реакции доводили до 50 мкл водой. Реакцию инкубировали в течение ночи при 30°C и проводили в стерилизованных 96-луночных планшетах. После инкубации добавляли 25 мкл ДМСО к каждой реакции, и реакционные планшеты центрифугировали в течение 5 мин. Образцы по сорок мкл отбирали из каждой лунки и фильтровали и использовали для LC-MS-анализа.
Было обнаружено, что UGT 73C3, 73C6 и 85C2 преобразовывали все могроловые субстраты в могрозид Ib. UGT 73C5 дает как могрозид Ia, так и могрозид Ib. В реакции с UGT 73E1, хотя реакция не была полной, могрозид Ib был обнаружен в качестве основного продукта вместе с новым гликозилированным могролом (не могрозид Ia и не могрозид Ib; показанное точное значение массы, как для могрозида I, предположительно вызванного событием гликозилирования по C25-OH).
ДРУГИЕ ВАРИАНТЫ ОСУЩЕСТВЛЕНИЯ
Следует понимать, что хотя изобретение было описано в сочетании с подробным описанием этого, вышеизложенное описание предназначено для иллюстрации и не для ограничения объема изобретения, который определяется объемом прилагаемых пунктов формулы изобретения. Другие аспекты, преимущества и изменения находятся в рамках следующих пунктов формулы изобретения.











| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ И ВЕЩЕСТВА ДЛЯ ОСНОВАННОГО НА РЕКОМБИНАЦИИ ПОЛУЧЕНИЯ СОЕДИНЕНИЙ ШАФРАНА | 2012 |
|
RU2676730C2 |
| БИОСИНТЕТИЧЕСКОЕ ПОЛУЧЕНИЕ СТЕВИОЛОВОГО ГЛИКОЗИДА РЕБАУДИОЗИДА D4 ИЗ РЕБАУДИОЗИДА E | 2017 |
|
RU2764803C2 |
| НОВЫЕ ФУКОЗИЛТРАНСФЕРАЗЫ И ИХ ПРИМЕНЕНИЕ | 2011 |
|
RU2642307C2 |
| РЕКОМБИНАНТНОЕ ПОЛУЧЕНИЕ СТЕВИОЛ-ГЛИКОЗИДОВ | 2014 |
|
RU2706789C2 |
| ИММУНОТЕРАПЕВТИЧЕСКИЙ СПОСОБ ЛЕЧЕНИЯ РАКА ПРОСТАТЫ | 2011 |
|
RU2586774C2 |
| ГИДРОЛИЗ СТЕВИОЛОВЫХ ГЛИКОЗИДОВ С ПОМОЩЬЮ БЕТА-ГЛЮКОЗИДАЗЫ | 2018 |
|
RU2775697C2 |
| ИММУНОГЕННАЯ КОМПОЗИЦИЯ ПРЕЗЕНТАЦИИ МНОЖЕСТВЕННЫХ АНТИГЕНОВ, ОТНОСЯЩИЕСЯ К НЕЙ СПОСОБЫ И ПРИМЕНЕНИЯ | 2012 |
|
RU2619176C2 |
| НОВЫЕ ФУКОЗИЛТРАСФЕРАЗЫ И ИХ ПРИМЕНЕНИЯ | 2011 |
|
RU2628307C2 |
| НОВЫЙ ГЕН ГЛИКОЗИЛТРАНСФЕРАЗЫ И ЕГО ПРИМЕНЕНИЕ | 2013 |
|
RU2636463C2 |
| ФУНКЦИОНАЛЬНЫЕ ЛОКУСЫ FAD2 И СООТВЕТСТВУЮЩИЕ СПЕЦИФИЧНЫЕ ДЛЯ САЙТА-МИШЕНИ СВЯЗЫВАЮЩИЕСЯ БЕЛКИ, СПОСОБНЫЕ ИНДУЦИРОВАТЬ НАПРАВЛЕННЫЕ РАЗРЫВЫ | 2013 |
|
RU2656158C2 |


Группа изобретений относится к биотехнологии, а именно к способам ферментативного синтеза могрозидных соединений. Способы включают добавление одного или нескольких рекомбинантных полипептидов, способных катализировать гликозилирование могрола для получения могрозидного соединения и обладающих аминокислотной последовательностью, представленной в SEQ ID NO: 1-5, или клеточного лизата, полученного из рекомбинантного хозяина, экспрессирующего один или несколько указанных полипептидов, и могрола растительного или синтетического происхождения в реакционную смесь, синтез и выделение могрозидного соединения из реакционной смеси. Изобретения позволяют получать могрозид Iа, могрозид Ib или могрозидное соединение, гликозилированное по C25-OH. 2 н. и 9 з.п. ф-лы, 4 ил., 4 пр.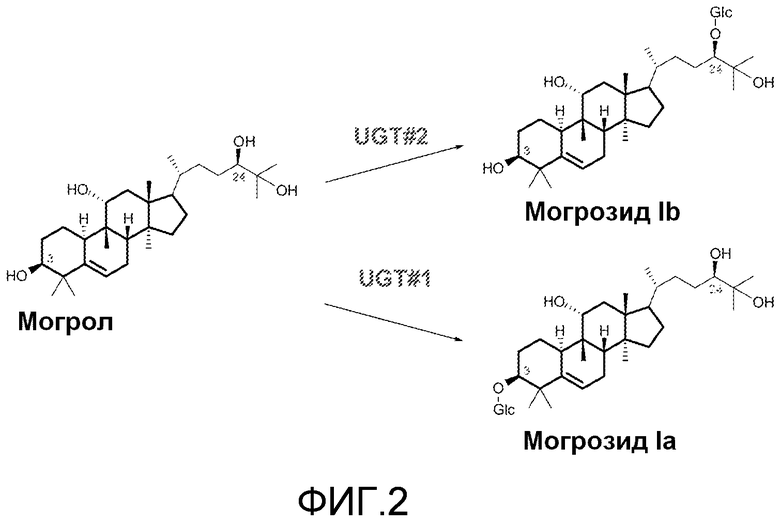
1. Способ получения могрозидного соединения, включающий:
(a) добавление одного или нескольких рекомбинантных полипептидов, способных катализировать гликозилирование могрола для получения могрозидного соединения и обладающих аминокислотной последовательностью, представленной в SEQ ID NO: 1-5, и могрола растительного или синтетического происхождения в реакционную смесь; и, таким образом, синтез могрозидного соединения; и
(b) выделение могрозидного соединения, полученного на стадии (а) из реакционной смеси.
2. Способ получения могрозидного соединения, включающий:
(a) добавление клеточного лизата, полученного из рекомбинантного хозяина, экспрессирующего один или несколько рекомбинантных полипептидов, способных катализировать гликозилирование могрола для получения могрозидного соединения и обладающих аминокислотной последовательностью, представленной в SEQ ID NO: 1-5, и могрола растительного или синтетического происхождения к реакционной смеси; и, таким образом, синтез могрозидного соединения; и
(b) выделение могрозидного соединения, полученного на стадии (а) из реакционной смеси.
3. Способ по п. 1 или 2, в котором:
(a) гликозилирование осуществляется за счет активности полипептида, обладающего аминокислотной последовательностью, представленной в SEQ ID NO: 2, а могрозидным соединением является могрозид Ia;
(b) гликозилирование осуществляется за счет активности одного или нескольких полипептидов, обладающих аминокислотной последовательностью, представленной в SEQ ID NO: 1-5, а могрозидным соединением является могрозид Ib; или
(c) гликозилирование осуществляется за счет активности полипептида, обладающего аминокислотной последовательностью, представленной в SEQ ID NO: 4, а могрозидным соединением является могрозидное соединение, гликозилированное по С25-ОН.
4. Способ по п. 1, где рекомбинантный полипептид получен в рекомбинантном хозяине.
5. Способ по п. 1, где полипептид получен путем химического синтеза.
6. Способ по п. 1 или 2, в котором полипептид, способный катализировать гликозилирование могрола для получения могрозидного соединения, содержит тэг-последовательность, расположенную либо в карбоксильном, либо в амино-конце UGT-полипептида.
7. Способ по п. 6, в котором тэг-последовательность включает зеленый флуоресцентный белок (GFP), глутатион-S-трансферазу (GST), HIS-тег и FLAG-тег, транзитный пептид хлоропластов, транзитный пептид митохондрий, пептид амилопластов, сигнальный пептид или тег-секреции.
8. Способ по п. 2 или 4, где рекомбинантным хозяином является микроорганизм, трансформированный рекомбинантной ДНК бактериофага, плазмидной ДНК или векторами экспрессии на основе космидной ДНК.
9. Способ по п. 2 или 4, где рекомбинантным хозяином является система клеток насекомых, инфицированных рекомбинантными вирусными векторами экспрессии.
10. Способ по п. 2 или 4, где рекомбинантным хозяином является система клеток растений, инфицированных рекомбинантными вирусными векторами экспрессии или трансформированных рекомбинантными вирусными векторами экспрессии.
11. Способ по п. 2 или 4, где рекомбинантным хозяином является система экспрессии млекопитающих, несущая рекомбинантные экспрессионные конструкции, содержащие промоторы, полученные из генома клеток млекопитающих или из вирусов млекопитающих.
| RICHMAN A | |||
| et al | |||
| "Functional genomics uncovers three glucosyltransferases involved in the synthesis of the major sweet glucosides of Stevia rebaudiana", The Plant Journal, 2005, v.41, p.56-67 | |||
| TANG Q | |||
| et al | |||
| "An efficient approach to finding Siraitia grosvenorii triterpene biosynthetic genes by RNA-seq and digital gene expression analysis", BMC GENOMICS, 05.07.2011, v.12, no.343, p.1-13 | |||
| SHAO Hui et al | |||
| "Crystal structures of a multifunctional triterpene/flavonoid glycosyltransferase from Medicago truncatula", The Plant Cell, 2005, v.17, no.11, p.3141-3154 | |||
| Станок для намотки катушек трапецеидальной формы | 1957 |
|
SU112845A1 |
| POPPENBERGER B | |||
| et al | |||
| "Detoxification of the Fusarium mycotoxin deoxynivalenol by a UDP-glucosyltransferase from Arabidopsis thaliana", Journal of Biological Chemistry, 2003, v.278, no.48, p.47905-47914 | |||
| RU 2008123244 A, 27.12.2009. | |||

