ПЕРЕКРЕСТНЫЕ ССЫЛКИ НА РОДСТВЕННЫЕ ЗАЯВКИ
Настоящая заявка притязает на приоритет предварительной патентной заявки серийный номер 61/378472, поданной 31 августа 2010 г, которая включена в настоящий документ посредством ссылки.
Область применения изобретения
В данном изобретении описываются способы содействия дифференциации плюрипотентных стволовых клеток в клетки, вырабатывающие инсулин. В частности, настоящее изобретение относится к способу получения популяции клеток, в которой более 85% клеток экспрессирует маркеры, характерные для линии сформированной эндодермы.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Последние достижения в области заместительной клеточной терапии для лечения сахарного диабета 1 типа и нехватка островков Лангерганса для трансплантации заставили обратить внимание на разработку источников инсулин-продуцирующих клеток, или β-клеток, подходящих для трансплантации. Одним из подходов является формирование функциональных β-клеток из плюрипотентных стволовых клеток, таких как, например, эмбриональные стволовые клетки.
При эмбриональном развитии позвоночных плюрипотентные клетки дают начало группе клеток, формирующих три зародышевых листка (эктодерму, мезодерму и эндодерму) в ходе процесса, именуемого гаструляцией. Такие ткани, как, например, щитовидная железа, тимус, поджелудочная железа, кишечник и печень, будут развиваться из эндодермы через промежуточную стадию. Промежуточной стадией данного процесса является образование сформированной эндодермы. Клетки сформированной эндодермы экспрессируют ряд маркеров, как например, HNF3 beta, GATA4, MIXL1, CXCR4 и SOX17.
Формирование поджелудочной железы происходит при дифференцировании сформированной эндодермы в панкреатическую эндодерму. Клетки панкреатической эндодермы экспрессируют ген панкреатическо-дуоденального гомеобокса, Pdx1. При отсутствии Pdx1 развитие поджелудочной железы не идет дальше формирования вентрального и дорзального зачатков. Таким образом, экспрессия PDX1 характеризует критическую стадию органогенеза поджелудочной железы. Зрелая поджелудочная железа содержит, помимо других типов клеток, экзокринную ткань и эндокринную ткань. Экзокринная и эндокринная ткани образуются при дифференцировании панкреатической эндодермы.
По имеющимся данным, клетки, обладающие свойствами островковых клеток, были получены из эмбриональных клеток мыши. Например, Lumelsky et al. описывают дифференцирование мышиных эмбриональных стволовых клеток в инсулин-секретирующие структуры, аналогичные островкам поджелудочной железы. Soria et al. (Diabetes 49:157, 2000) описывают инсулин-секретирующие клетки, производные мышиных эмбриональных стволовых клеток, которые нормализуют гликемию у мышей с диабетом, индуцированным стрептозотоцином.
В одном примере, Hori et al. (PNAS 99: 16105, 2002) описывается, что обработка мышиных эмбриональных стволовых клеток ингибиторами фосфоинозитид-3-киназы (LY294002) приводила к получению клеток, подобных β-клеткам.
В другом примере, Blyszczuk et al. (PNAS 100:998, 2003) сообщают о получении инсулин-продуцирующих клеток из мышиных эмбриональных стволовых клеток с конститутивной экспрессией Pax4.
В публикации Micallef et al. сообщается, что ретиноевая кислота может регулировать способность эмбриональных стволовых клеток формировать Pdx1-положительную панкреатическую эндодерму. Ретиноевая кислота с наибольшей эффективностью индуцирует экспрессию Pdx1 при добавлении в культуру на 4 день дифференцирования эмбриональных стволовых клеток в течение периода, соответствующего концу гаструляции эмбриона (Diabetes 54:301, 2005).
В публикации Miyazaki et al. сообщается о линии мышиных эмбриональных стволовых клеток со сверхэкспрессией Pdx1. Результаты показывают, что экспрессия экзогенного Pdx1 очевидно повышает экспрессию генов инсулина, соматостатина, глюкокиназы, нейрогенина 3, p48, Pax6 и HNF6 в образующихся дифференцированных клетках (Diabetes 53: 1030, 2004).
В публикации Skoudy et al. сообщается, что активин A (входящий в суперсемейство TGF-β) повышает экспрессию экзокринных панкреатических генов (p48 и амилаза) и эндокринных генов (Pdx1, инсулин и глюкагон) в эмбриональных стволовых клетках мыши. Максимальный эффект наблюдался при использовании 1 нМ активина A. Кроме того, авторы отметили, что экспрессия инсулина и Pdx1 мРНК не изменялась под действием ретиноевой кислоты; Однако, лечение с использованием 3nM FGF7 привело к повышению уровня транскрипта для Pdx1 (Biochem. J. 379: 749, 2004).
В работе Shiraki et al. изучались эффекты факторов роста, специфически ускоряющих дифференцирование эмбриональных стволовых клеток в Pdx1-положительные клетки. Эти авторы наблюдали, что TGF-β2 приводил к воспроизводимому увеличению доли Pdx1-положительных клеток (Genes Cells. 2005 Jun; 10(6): 503-16.).
В работе Gordon et al. показана индукция образования брахиурических [положительных]/HNF3 бета [положительных] эндодермальных клеток из эмбриональных стволовых клеток мыши в отсутствие сыворотки и в присутствии активина в сочетании с ингибитором сигнального каскада Wnt (патент США № 2006/0003446A1).
Gordon et al. (PNAS, т. 103, с. 16806, 2006) утверждают: «Для образования примитивной первичной полоски одновременно требовались сигнальные пути Wnt и TGF-бета/nodal/активин».
Однако модель развития эмбриональных стволовых клеток на мышах может не имитировать в точности программу развития у высших млекопитающих, например у человека.
В работе Thomson et al. эмбриональные стволовые клетки выделяли из человеческих бластоцист (Science 282:114, 1998). Параллельно, Gearhart и соавторы получили клеточные линии эмбриональных зародышевых клеток человека (hEG) из ткани половых желез эмбриона (Shamblott et al., Proc. Natl. Acad. Sci. USA 95:13726, 1998). В отличие от эмбриональных стволовых клеток мыши, воспрепятствовать дифференцированию которых можно путем простого культивирования с фактором, ингибирующим лейкемию (LIF), эмбриональные стволовые клетки человека необходимо культивировать в крайне специфических условиях (патент США № 6200806; WO 99/20741, WO 01/51616).
D’Amour et al. описывают производство обогащенных культур сформированной эндодермы, производной от человеческих эмбриональных стволовых клеток, в присутствии высокой концентрации активина и низкой концентрации сыворотки (Nature Biotechnology 2005). Трансплантация этих клеток под почечную капсулу мышей привела к их дифференцированию в более зрелые клетки, обладающие характерными особенностями некоторых эндодермальных органов. Клетки сформированной эндодермы, производные от эмбриональных стволовых клеток человека, могут подвергаться дальнейшему дифференцированию в Pdx1-положительные клетки после добавления FGF-10 (US 2005/0266554A1).
D’Amour et al. (Nature Biotechnology -24, 1392-1401 (2006)) утверждают: “Мы разработали процесс дифференцировки, преобразующий эмбриональные клетки человека (hES) в эндокринные клетки, способные синтезировать гормоны поджелудочной железы: инсулин, глюкагон, соматостатин, панкреатический полипептид и грелин. Данный процесс имитирует органогенез поджелудочной железы in vivo, проводя клетки через фазы, напоминающие образование сформированной эндодермы, эндодермы кишечной трубки, панкреатической эндодермы и превращение предшественников эндокринных клеток в клетки, экспрессирующие эндокринные гормоны”.
В другом примере, в публикации Fisk et al., сообщается о системе для производства островковых клеток поджелудочной железы из эмбриональных стволовых клеток человека (US2006/0040387A1). В данном случае процесс дифференцирования был разделен на три стадии. Человеческие эмбриональные стволовые клетки были впервые дифференцированы до эндодермы с помощью сочетания бутирата натрия и активина A. Затем клетки культивировали с антагонистами ФНО-β, например, Noggin, в сочетании с EGF или бетацеллюлином для получения PDX1-положительных клеток. Окончательное дифференцирование запускалось никотинамидом.
Таким образом, сохраняется значительная потребность в разработке лабораторных способов создания функциональной экспрессирующей инсулин клетки более близкой к β-клетке, in vitro. Настоящее изобретение представляет альтернативный подход к повышению эффективности дифференцирования эмбриональных стволовых клеток человека в экспрессирующие инсулин клетки путем получения популяции, в которой более 85% клеток экспрессирует маркеры, характерные для линии сформированной эндодермы
КРАТКОЕ ОПИСАНИЕ
В одном варианте осуществления настоящее изобретение относится к способу получения популяции клеток, в которой более 85% клеток экспрессирует маркеры, характерные для линии сформированной эндодермы.
В одном варианте осуществления изобретения популяции плюрипотентных клеток дифференцировали в популяции клеток, экспрессирующие маркеры, характерные для линии сформированной эндодермы, путем культивирования плюрипотентных стволовых клеток на среде с добавлением БСА и фактора, выбранного из следующей группы: инсулин и IGF-1. В одном варианте осуществления изобретения дифференциация популяции плюрипотентных стволовых клеток в направлении популяции клеток, экспрессирующих маркеры, характерные для линии клеток сформированной эндодермы, достигается обработкой плюрипотентных стволовых клеток активином А и лигандом Wnt.
В одном варианте осуществления изобретения дифференциация популяции плюрипотентных стволовых клеток в направлении популяции клеток, экспрессирующих маркеры, характерные для линии сформированной эндодермы, достигается обработкой плюрипотентных стволовых клеток GDF-8 и, по меньшей мере, одним из факторов, выбранных из следующей группы: анилин-пиридинотриазин, циклический анилин-пиридинотриазин, N-{[1-(фенилметил)азепан-4-ил]метил}-2-пиридин-3-илацетамид, 4-{[4-(4-{[2-(пиридин-2-иламино)этил]амино}-1,3,5-триазин-2-ил)пиридин-2-ил]окси}бутан-1-ол, 3-({3-[4-({2-[метил(пиридин-2-ил)амино]этил}амино)-1,3,5-триазин-2-ил]пиридин-2-ил}амино)пропан-1-ол, N~4~-[2-(3-фторфенил)этил]-N~2~-[3-(4-метилпиперазин-1-ил)пропил]пиридо[2,3-d]пиримидин-2,4-диамин, 1-метил-N-[(4-пиридин-3-ил-2-{[3-(трифторметил)фенил]амино}-1,3-тиазол-5-ил)метил]пиперидин-4-карбоксамид, 1,1-диметилэтил {2-[4-({5-[3-(3-гидроксипропил)фенил]-4H-1,2,4-триазол-3-ил}амино)фенил]этил}карбамат, 1,1-диметилэтил {[3-({5-[5-(3-гидроксипропил)-2-(метилокси)фенил]-1,3-оксазол-2-ил}амино)фенил]метил}карбамат, 1-({5-[6-({4-[(4-метилпиперазин-1-ил)сульфонил]фенил}амино)пиразин-2-ил]тиофен-2-ил}метил)пиперидин-4-ол, 1-({4-[6-({4-[(4-метилпиперазин-1-ил)сульфонил]фенил}амино)пиразин-2-ил]тиофен-2-ил}метил)пиперидин-4-карбоксамид и 2-{[4-(1-метилэтил)фенил]амино}-N-(2-тиофен-2-илэтил)-7,8-дигидропиридо[4,3-d]пиримидин-6(5H)-карбоксамид.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг. 1 показаны результаты анализа экспрессии генов в клетках человеческой эмбриональной линии H1, дифференцированной по способу, описанному в Примере 1, методом ПЦР в реальном времени.
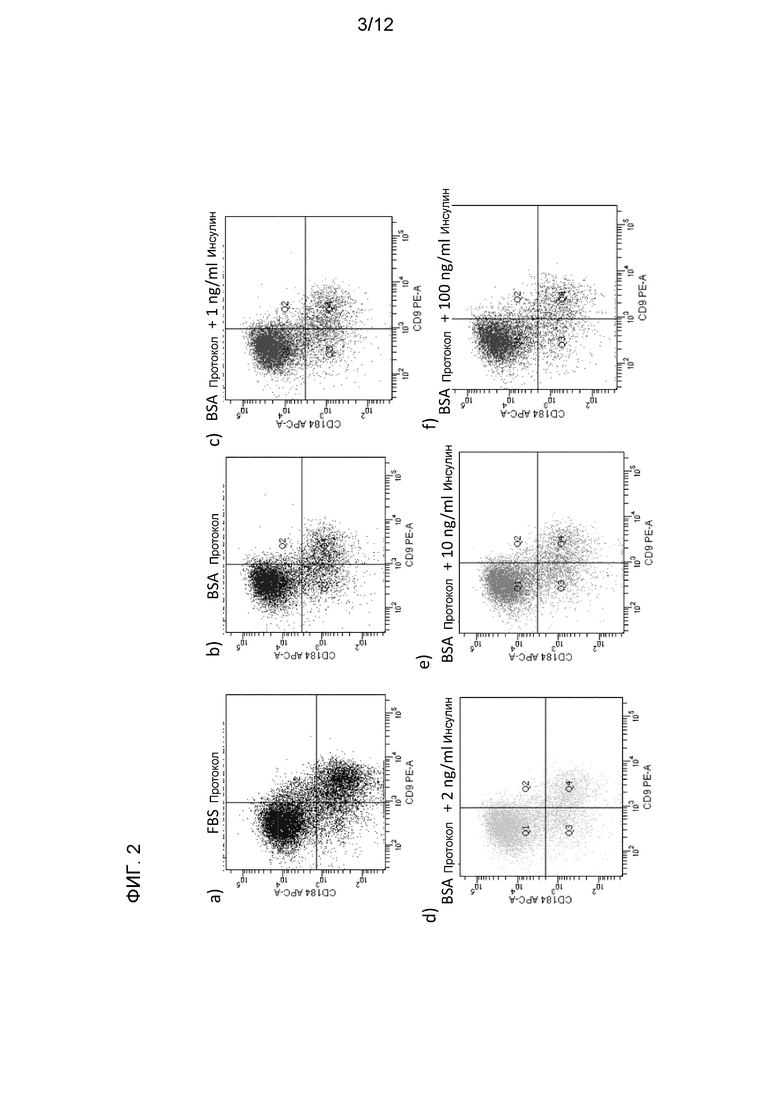
На фиг. 2 показаны результаты анализа экспрессии белков в клетках человеческой эмбриональной линии H1, дифференцированной по способам, описанным в Примере 1, методом флуоресцентной проточной цитометрии.
На фиг. 3 показаны результаты анализа экспрессии генов в клетках человеческой эмбриональной линии H1, дифференцированной по способу, описанному в Примере 2, методом ПЦР в реальном времени.
На фиг. 4 показаны результаты анализа экспрессии белков в SOX17 методом иммунофлуоресценции в клетках человеческой эмбриональной линии H1, дифференцированной по способам, описанным в Примере 2.
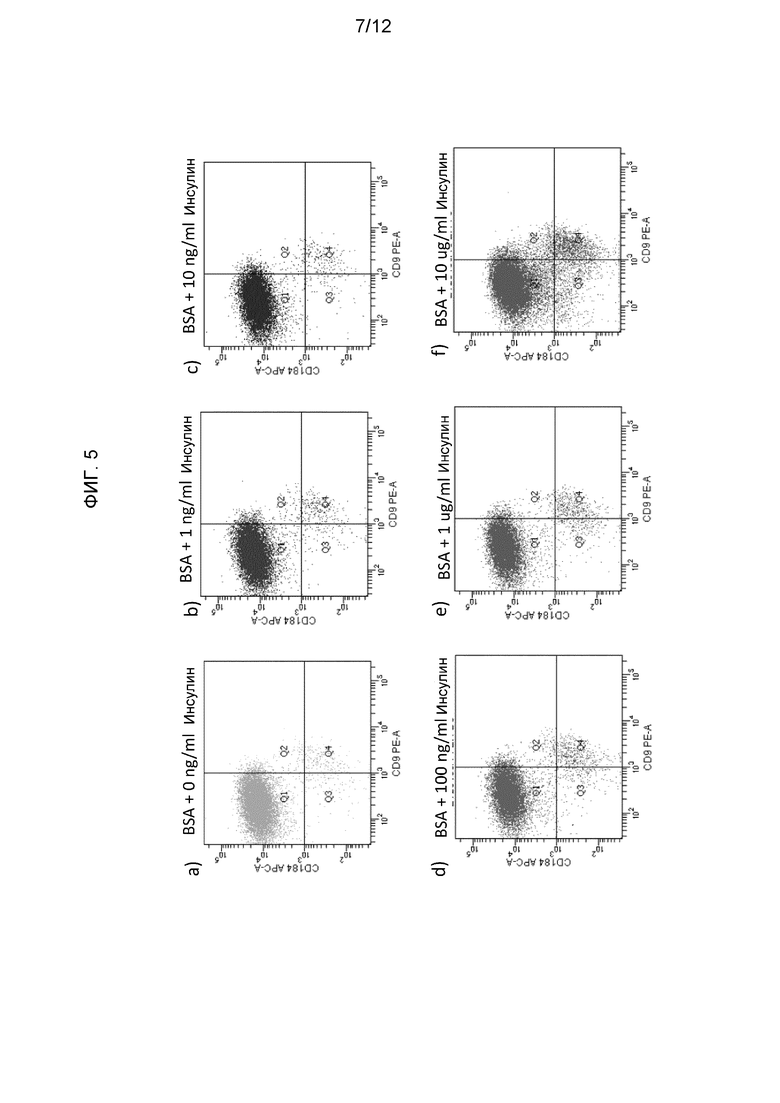
На фиг. 5 показаны результаты анализа экспрессии белков методом флуоресцентной проточной цитометрии в клетках человеческой эмбриональной линии H1, дифференцированной по способам, описанным в Примере 2.
На фиг. 6 показаны результаты анализа экспрессии генов в клетках человеческой эмбриональной линии H1, дифференцированной по способу, описанному в Примере 3, методом ПЦР в реальном времени.
На фиг. 7 показаны результаты анализа экспрессии SOX17 методом иммунофлуоресценции в клетках человеческой эмбриональной линии H1, дифференцированной по способам, описанным в Примере 3.
На фиг. 8 показаны результаты анализа экспрессии SOX17 методом иммунофлуоресценции в клетках человеческой эмбриональной линии H1, дифференцированной по способам, описанным в Примере 3.
На фиг. 9 показаны результаты анализа экспрессии генов в клетках человеческой эмбриональной линии H1, дифференцированной по способам, описанным в Примере 5, методом ПЦР в реальном времени.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Для ясности описания, а не для ограничения изобретения, подробное описание изобретения разделено на следующие подразделы, описывающие или иллюстрирующие определенные особенности, варианты осуществления или области применения настоящего изобретения.
Определения
Стволовые клетки представляют собой недифференцированные клетки, определяемые по их способности на уровне единичной клетки как самообновляться, так и дифференцироваться с образованием клеток-потомков, таких как самообновляющиеся клетки-предшественники, необновляющиеся клетки-предшественники и окончательно дифференцированные клетки. Стволовые клетки также характеризуются способностью дифференцироваться in vitro в функциональные клетки различных клеточных линий дифференцирования из нескольких зародышевых листков (эндодермы, мезодермы и эктодермы), а также после трансплантации давать начало тканям, происходящим от нескольких зародышевых листков, и вносить существенный вклад в формирование большинства, если не всех, тканей после инъекции в бластоцисты.
Стволовые клетки классифицируют по потенциалу развития: (1) тотипотентные, то есть, способные преобразоваться в любой из эмбриональных и внеэмбриональных типов клеток; (2) плюрипотентные, то есть, способные преобразоваться во все типы эмбриональных клеток; (3) мультипотентные, то есть, способные преобразоваться во множество клеточных линий, но в рамках одной ткани, органа или физиологической системы (например, гемопоэтические стволовые клетки (ГСК) могут порождать ГСК (самообновление), олигопотентные ограниченные клетки-предшественники крови и все типы клеток и элементов (например, тромбоциты), являющиеся стандартными составляющими крови); (4) олигопотентные, то есть, способные преобразоваться в более ограниченное подмножество клеточных линий, чем мультипотентные стволовые клетки; и (5) унипотентные, то есть, способные преобразоваться в единственную клеточную линию (например, сперматогенные стволовые клетки).
Дифференцирование представляет собой процесс, при помощи которого неспециализированная («некоммитированная») или менее специализированная клетка приобретает свойства специализированной клетки, например нервной или мышечной клетки. Дифференцированная клетка или клетка с индуцированным дифференцированием представляет собой клетку, занявшую более специализированное («коммитированное») положение в линии дифференцирования клетки. Термин «коммитированная» применительно к процессу дифференцирования обозначает клетку, дошедшую в ходе процесса дифференцирования до стадии, от которой в нормальных условиях она продолжит дифференцироваться до определенного типа клеток или набора типов клеток и не сможет в нормальных условиях дифференцироваться в иной тип клеток или вернуться обратно к менее дифференцированному типу. Дедифференцированием называется процесс, в ходе которого клетка возвращается к менее специализированному (или коммитированному) положению в линии дифференцирования. Используемый в настоящей заявке термин «линия дифференцирования клетки» определяет наследственность клетки, то есть определяет, из какой клетки произошла данная клетка и каким клеткам она может дать начало. В линии дифференцирования клетка помещается в наследственную схему развития и дифференцирования. Маркером, специфичным для линии дифференцирования, называется характерная особенность, специфически ассоциированная с фенотипом клеток конкретной линии дифференцирования, которая может использоваться для оценки дифференцирования некоммитированных клеток в клетки данной линии дифференцирования.
Используемые в настоящей заявке термины «клетки, экспрессирующие маркеры, характерные для линии сформированной эндодермы», «клетки стадии 1» или «стадия 1», относятся к клеткам, экспрессирующим, по меньшей мере, один из следующих маркеров: SOX17, GATA4, HNF3 beta, GSC, CER1, Nodal, FGF8, Brachyury, Mix-подобный гомеобоксовый белок, FGF4 CD48, эомезодермин (EOMES), DKK4, FGF17, GATA6, CXCR4, C-Kit, CD99 или OTX2. К клеткам, экспрессирующим маркеры, характерные для линии сформированной эндодермы, относятся клетки-предшественники первичной полоски, клетки первичной полоски, клетки мезэндодермы и клетки сформированной эндодермы.
Используемый в настоящей заявке термин «клетки с экспрессией маркеров, характерных для линии панкреатической эндодермы» относится к клеткам с экспрессией, по меньшей мере, одного из следующих маркеров: PDX1, NKX6.1, HNF1 beta, PTF1 alpha, HNF6, HNF4 alpha, SOX9, HB9 или PROX1. К клеткам, экспрессирующим маркеры, характерные для линии панкреатической эндодермы, относятся клетки панкреатической эндодермы, клетки первичной кишечной трубки и клетки задней части передней кишки.
Используемый в настоящей заявке термин «сформированная эндодерма» относится к клеткам, обладающим характерными особенностями клеток, происходящих в ходе гаструляции от эпибласта, и формирующим желудочно-кишечный тракт и его производные. Клетки сформированной эндодермы экспрессируют следующие маркеры: HNF3 beta, GATA4, SOX17, Cerberus, OTX2, goosecoid, C-Kit, CD99 и MIXL1.
Используемый в настоящей заявке термин «маркеры» означает молекулы нуклеиновых кислот или полипептидов с дифференциальной экспрессией в интересующих клетках. В данном контексте под дифференциальной экспрессией подразумевается повышение уровня экспрессии для положительного маркера и понижение уровня экспрессии для отрицательного маркера. Поддающийся обнаружению уровень маркерной нуклеиновой кислоты или полипептида в интересующих клетках оказывается значительно выше или ниже по сравнению с другими клетками, что позволяет идентифицировать интересующую клетку и отличить ее от других клеток с помощью любого из множества известных в данной области способов.
«Панкреатической эндокринной клеткой» или «клеткой, экспрессирующей гормон поджелудочной железы» или «клеткой, экспрессирующей характеристики эндокринной линии поджелудочной железы» в настоящем документе называется клетка способная экспрессировать, по меньшей мере, один из следующих гормонов: инсулин, глюкагон, соматостатин и панкреатический полипептид.
Выделение, размножение и культивирование полипотентных стволовых клеток
Характеристика плюрипотентных стволовых клеток
Плюрипотентные стволовые клетки могут экспрессировать один или более стадийно-специфичных эмбриональных антигенов (SSEA) 3 и 4, а также маркеры, определяемые антителами, обозначенными как Tra-1-60 и Tra-1-81 (Thomson et al., Science 282:1145, 1998). Дифференцирование плюрипотентных стволовых клеток in vitro приводит к потере экспрессии SSEA-4, Tra 1-60 и Tra 1-81 (при наличии) и к повышению экспрессии SSEA-1. В недифференцированных полипотентных стволовых клетках, как правило, активна щелочная фосфатаза, которая может быть обнаружена путем фиксации клеток с помощью 4% параформальдегида, с последующим обнаружением с помощью Vector Red, применяемого в качестве субстрата, в соответствии с инструкциями производителя (Vector Laboratories, Burlingame Calif.). Недифференцированные плюрипотентные стволовые клетки также, как правило, экспрессируют OCT4 и TERT, определяемые с помощью ПЦР в реальном времени.
Другим желательным фенотипическим свойством выращенных плюрипотентных стволовых клеток является потенциал дифференцирования в клетки всех трех зародышевых листков: в эндодермальные, мезодермальные и эктодермальные ткани. Полипотентность полипотентных стволовых клеток может быть подтверждена, например, путем инъекции клеток мышам с тяжелым комбинированным иммунодефицитом (SCID), фиксирования образующихся тератом с помощью 4% параформальдегида, и их гистологического исследования для получения доказательств наличия клеточных типов, происходящих от трех зародышевых листков. В качестве альтернативы плюрипотентность можно определить по созданию эмбриоидных телец и анализа их на предмет присутствия маркеров, ассоциирующихся с тремя зародышевыми листками.
Выращенные линии плюрипотентных стволовых клеток могут быть кариотипированы с применением стандартного способа окрашивания с использованием красителя Гимза (G-banding) и сравнения с опубликованными кариотипами соответствующих видов приматов. Желательно получить клетки, имеющие «нормальный кариотип», т.е. эуплоидные клетки, в которых все человеческие хромосомы присутствуют и не имеют видимых изменений.
Источники плюрипотентных стволовых клеток
К типам плюрипотентных стволовых клеток, которые можно использовать, относятся устойчивые линии плюрипотентных клеток, получаемые из формируемой после вынашивания плода ткани, в том числе из преэмбриональной ткани (такой как бластоциста), эмбриональной ткани или ткани плода, взятой в любой момент в ходе вынашивания, как правило, но не обязательно, до срока приблизительно 10-12 недель беременности. Примерами, не ограничивающими настоящее изобретение, являются стабильные линии человеческих эмбриональных стволовых клеток или человеческих эмбриональных зародышевых клеток, например, клеточные линии человеческих эмбриональных стволовых клеток H1, H7 и H9 (WiCell). Также возможно использование описываемых в настоящей заявке составов в ходе первоначального установления или стабилизации таких клеток, в этом случае исходными клетками являются первичные плюрипотентные клетки, взятые напрямую из тканей-источников. Также соответствуют целям настоящего изобретения клетки, взятые из популяции плюрипотентных стволовых клеток, уже культивированных в отсутствие питающих клеток. Также соответствуют целям настоящего изобретения клетки мутантных линий эмбриональных стволовых клеток человека, таких как, например, BG01v (BresaGen, Атенс, Джорджия, США).
В одном варианте осуществления человеческие эмбриональные стволовые клетки готовятся, как описано Thomson et al. патент США № 5843780; Science 282:1145, 1998; Curr. Top. Dev. Biol. 38:133 ff., 1998; Proc. Natl. Acad. Sci. U.S.A. 92:7844, 1995).
Культивирование плюрипотентных стволовых клеток
В одном варианте осуществления плюрипотентные стволовые клетки культивируют на слое питающих клеток, которые поддерживают плюрипотентные стволовые клетки в различных отношениях. Как вариант, полипотентные стволовые клетки культивируются в культуральной системе, по существу не содержащей питающих клеток, но, тем не менее, поддерживающей пролиферацию полипотентных стволовых клеток и не допускающей существенной дифференцировки. Рост плюрипотентных стволовых клеток в свободной от питающих клеток культуральной системе без дифференцирования поддерживается путем использования среды, кондиционированной посредством предварительного культивирования клеток иного типа. В качестве альтернативы рост плюрипотентных стволовых клеток в свободной от питающих клеток культуральной системе без дифференцирования поддерживается путем использования среды с химически определенным составом.
В одном варианте осуществления изобретения плюрипотентные стволовые клетки можно культивировать на питающем слое фибробластов эмбриона мыши в соответствии со способами, изложенными в работе Reubinoff et al (Nature Biotechnology 18: 399-404 (2000)). В альтернативном варианте осуществления изобретения плюрипотентные стволовые клетки можно культивировать на питающем слое фибробластов эмбриона мыши в соответствии со способами, изложенными в работе Thompson et al (Science 6, ноябрь 1998 г.: Vol. 282. no. 5391, pp. 1145-1147). В альтернативном варианте осуществления изобретения плюрипотентные стволовые клетки можно культивировать на любом из слоев питающих клеток, раскрытых в работе Richards et al, (Stem Cells 21: 546-556, 2003).
В одном варианте осуществления изобретения плюрипотентные стволовые клетки можно культивировать на питающем слое клеток человека в соответствии со способами, изложенными в работе Wang et al (Stem Cells 23: 1221-1227, 2005). В альтернативном варианте осуществления изобретения плюрипотентные стволовые клетки можно культивировать на питающем клеток человека, описанном в работе Stojkovic et al (Stem Cells 2005 23: 306-314, 2005). В альтернативном варианте осуществления изобретения плюрипотентные стволовые клетки можно культивировать на питающем слое клеток человека, описанном в работе Miyamoto et al (Stem Cells 22: 433-440, 2004). В альтернативном варианте осуществления изобретения плюрипотентные стволовые клетки можно культивировать на питающем слое клеток человека, описанном в работе Amit et al (Biol. Reprod 68: 2150-2156, 2003). В альтернативном варианте осуществления изобретения плюрипотентные стволовые клетки можно культивировать на питающем слое клеток человека, описанном в работе Inzunza et al (Stem Cells 23: 544-549, 2005).
В одном варианте осуществления плюрипотентные стволовые клетки можно культивировать в культуральной среде, полученной в соответствии со способами, представленными в патенте США № 20020072117. В альтернативном варианте осуществления плюрипотентные стволовые клетки можно культивировать в культуральной среде, полученной в соответствии со способами, представленными в патенте США № 6642048. В альтернативном варианте осуществления изобретения плюрипотентные стволовые клетки можно культивировать в культуральной среде, полученной в соответствии со способами, представленными в WO2005014799. В альтернативном варианте осуществления изобретения плюрипотентные стволовые клетки можно культивировать в культуральной среде, полученной в соответствии со способами, представленными в работе Xu et al (Stem Cells 22: 972-980, 2004). В альтернативном варианте осуществления плюрипотентные стволовые клетки можно культивировать в культуральной среде, полученной в соответствии со способами, представленными в патенте США № 20070010011. В альтернативном варианте осуществления плюрипотентные стволовые клетки можно культивировать в культуральной среде, полученной в соответствии со способами, представленными в патенте США № 20050233446. В альтернативном варианте осуществления плюрипотентные стволовые клетки можно культивировать в культуральной среде, полученной в соответствии со способами, представленными в патенте США № 6800480. В альтернативном варианте осуществления изобретения плюрипотентные стволовые клетки можно культивировать в культуральной среде, полученной в соответствии со способами, описанными в WO2005065354.
В одном варианте осуществления изобретения плюрипотентные стволовые клетки можно культивировать по способам, описанным Cheon et al (BioReprod DOI:10.1095/biolreprod.105.046870, October 19, 2005). В альтернативном варианте осуществления изобретения плюрипотентные стволовые клетки можно культивировать в соответствии со способами, представленными в работе Levenstein et al (Stem Cells 24: 568-574, 2006). В альтернативном варианте осуществления плюрипотентные стволовые клетки можно культивировать в соответствии со способами, представленными в патенте США № 20050148070. В альтернативном варианте осуществления плюрипотентные стволовые клетки можно культивировать в соответствии со способами, представленными в патенте США № 20050244962. В альтернативном варианте осуществления изобретения плюрипотентные стволовые клетки можно культивировать в соответствии со способами, описанными в WO2005086845.
Плюрипотентные стволовые клетки могут быть высеяны на соответствующий культуральный субстрат. В одном из вариантов осуществления соответствующим культуральным субстратом является компонент внеклеточного матрикса, такой как, например, полученный из базальной мембраны или тот, который может участвовать в лиганд-рецепторном взаимодействии с участием молекулы адгезивного слоя. В одном из вариантов осуществления подходящим культуральным субстратом является MATRIGEL® (Becton Dickenson). MATRIGEL® представляет собой растворимый препарат из клеток опухоли Энгельбрета-Холма-Суорма, который при комнатной температуре превращается в гель и образует восстановленную базальную мембрану.
В качестве альтернативы можно использовать другие компоненты внеклеточного матрикса и смеси компонентов. В зависимости от типа пролиферирующих клеток, это может быть ламинин, фибронектин, протеогликан, энтактин, гепарансульфат и т.п., по отдельности или в различных сочетаниях.
Плюрипотентные стволовые клетки могут высеиваться на субстрат с соответствующим распределением по поверхности и в присутствии среды, поддерживающей выживание, размножение и сохранение требуемых характеристик клеток. Все эти характеристики улучшаются при тщательном подходе к распределению клеток при посеве и могут быть определены специалистом в данной области.
Подходящая культуральная среда может быть изготовлена, например, из следующих компонентов модифицированная по способу Дульбекко среда Игла (DMEM), Gibco № 11965-092, нокаутная модифицированная по способу Дульбекко среда Игла (KO DMEM), Gibco №10829-018, базовая среда Хэма F12/50% DMEM, 200 мм L-глутамина, Gibco № 15039-027; раствор неосновных аминокислот, Gibco 11140-050; β-меркаптоэтанол, Sigma № M7522; человеческий рекомбинантный основной фактор роста фибробластов (bFGF), Gibco № 13256-029.
Формирование клеток, экспрессирующих маркеры, характерные для линии сформированной панкреатической эндодермы, из плюрипотентных стволовых клеток
Настоящее изобретение представляет способы получения популяций клеток, экспресирующих маркеры, характерные для сформированной клеточной линии эндодермы, из популяций плюрипотентных стволовых клеток. В одном варианте осуществления настоящее изобретение представляет способы дальнейшей дифференциации клеток, экспрессирующих маркеры линии эндокринных клеток поджелудочной железы. В одном варианте осуществления это достигается пошаговым протоколом дифференцирования, в котором популяции плюрипотентных стволовых клеток сначала дифференцируются в популяции клеток, экспрессирующих маркеры, характерные для сформированного ростка эндодермы. Далее, популяции клеток, экспрессирующих маркеры, характерные для линии сформированной эндодермы, дополнительно дифференцируются в популяции клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы. Далее, популяции клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, дополнительно дифференцируются в популяции клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы.
В настоящем изобретении представлена популяция клеток, в которой более 85% клеток экспрессирует маркеры, характерные для линии сформированной эндодермы. Популяция клеток может подвергаться дальнейшей обработке для получения популяции клеток, экспрессирующих маркеры, характерные для панкреатической эндодермы. Популяция клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, можно дополнительно обрабатывать для получения популяции клеток, экспрессирующих маркеры, характерные для панкреатических эндокринных клеток.
Эффективность дифференцирования может быть определена путем обработки популяции клеток агентом (например, антителом), специфически распознающим белковый маркер, экспрессированный клетками, экспрессирующими маркеры, характерные для желательного вида клеток.
Способы оценки экспрессии маркеров белков и нуклеиновых кислот в культивированных или выделенных клетках являются стандартными для данной области. Сюда относятся количественная ревертазная полимеразная цепная реакция (ОТ-ПЦР), Нозерн-блот, гибридизация in situ (см., например, Current Protocols in Molecular Biology (Ausubel et al., eds. 2001, доп.)), а также способы иммунологического анализа, такие как иммуногистохимический анализ среза материала, Вестерн-блоттинг, а для маркеров, доступных в интактных клетках, - способ проточной цитометрии (FACS) (см., например, Harlow and Lane, Using Antibodies: A Laboratory Manual, New York: Cold Spring Harbor Laboratory Press (1998)).
Характеристики плюрипотентных стволовых клеток хорошо известны специалистам в данной области, и продолжается выявление дополнительных характеристик плюрипотентных стволовых клеток. К маркерам плюрипотентных стволовых клеток относится, например, экспрессия одного или нескольких следующих маркеров: ABCG2, CRIPTO, FOXD3, Connexin43, Connexin45, OCT4, SOX2, Nanog, hTERT, UTF1, ZFP42, SSEA-3, SSEA-4, Tra 1-60, Tra 1-81.
После обработки плюрипотентных стволовых клеток с применением способов, составляющих предмет настоящего изобретения, дифференцированные клетки могут быть выделены путем воздействия на популяцию клеток агентом (например, антителом), специфически распознающим белковый маркер, например CXCR4, экспрессируемый клетками, экспрессирующими маркеры, характерные для линии сформированной эндодермы.
К плюрипотентным стволовым клеткам, которые могут использоваться в настоящем изобретении, относятся, например, человеческие эмбриональные стволовые клетки линии H9 (код NIH: WA09), человеческие эмбриональные стволовые клетки линии H1 (код NIH: WA01), человеческие эмбриональные стволовые клетки линии H7 (код NIH: WA07) и человеческие эмбриональные стволовые клетки линии SA002 (Cellartis, Швеция). Также для использования в рамках настоящего изобретения подходят клетки, экспрессирующие, по меньшей мере, один из следующих маркеров, характерных для плюрипотентных клеток: ABCG2, cripto, CD9, FOXD3, CONNEXIN43, CONNEXIN45, OCT4, SOX2, Nanog, hTERT, UTF1, ZFP42, SSEA-3, SSEA-4, Tra 1-60, и Tra 1-81.
Маркеры характерные для сформированной линии эндодермы выбираются из группы, содержащей SOX17, GATA4, HNF3 beta, GSC, CER1, Nodal, FGF8, Brachyury, Mix-подобный гомеобоксовый белок, FGF4, CD48, эомезодермин (EOMES), DKK4, FGF17, GATA6, CXCR4, C-Kit, CD99 и Otx2. Подходит для использования в настоящем изобретении клетка, экспрессирующая, как минимум, один из маркеров, характерных для линии сформированной эндодермы. В одном из аспектов настоящего изобретения клетка, экспрессирующая маркеры, характерные для линии сформированной эндодермы, представляет собой клетку-предшественник первичной полоски. В другом аспекте настоящего изобретения клетка, экспрессирующая маркеры, характерные для линии сформированной эндодермы, представляет собой мезэндодермальную клетку. В другом аспекте настоящего изобретения клетка, экспрессирующая маркеры, характерные для линии сформированной эндодермы, представляет собой клетку сформированной эндодермы.
Маркеры характерные для линии эндодермы поджелудочной железы, выбираются из группы, содержащей PDX1, NKX6.1, HNF1 beta, PTF1 alpha, HNF6, HNF4 alpha, SOX9, HB9 и PROX1. Подходит для использования в настоящем изобретении клетка, экспрессирующая, как минимум, один из маркеров, характерных для линии панкреатической эндодермы. В одном из аспектов настоящего изобретения клетка, экспрессирующая маркеры, характерные для линии панкреатической эндодермы, представляет собой клетку панкреатической эндодермы.
Маркеры, характерные для линии панкреатических эндокринных клеток, выбирают из группы, состоящей из NGN3, NEUROD, ISL1, PDX1, NKX6.1, PAX4, NGN3 и PTF1 альфа. В одном варианте осуществления панкреатическая эндокринная клетка способна к экспрессии, по меньшей мере, одного из следующих гормонов: инсулин, глюкагон, соматостатин и панкреатический полипептид. Соответствующей целям настоящего изобретения является клетка, экспрессирующая, по меньшей мере, один из маркеров, характерных для линии панкреатических эндокринных клеток. В одном из аспектов настоящего изобретения клетка, экспрессирующая маркеры, характерные для линии панкреатических эндокринных клеток, представляет собой панкреатическую эндокринную клетку. Панкреатическая эндокринная клетка может представлять собой панкреатическую клетку, экспрессирующую гормоны. В альтернативном варианте осуществления панкреатическая эндокринная клетка может представлять собой панкреатическую клетку, секретирующую гормоны.
В одном аспекте настоящего изобретения панкреатическая эндокринная клетка представляет собой клетку с экспрессией маркеров, характерных для линии дифференцирования β-клеток. Клетка с экспрессией маркеров, характерных для линии β-клеток, экспрессирует PDX1 и, по меньшей мере, один из следующих транскрипционных факторов: NGN3, NKX2.2, NKX6.1, NEUROD, ISL1, HNF3 бета, MAFA, PAX4 и PAX6. В одном аспекте настоящего изобретения клетка с экспрессией маркеров, характерных для линии дифференцирования β-клеток, представляет собой β-клетку.
Формирование клеток, экспрессирующих маркеры, характерные для линии сформированной панкреатической эндодермы, из плюрипотентных стволовых клеток
В одном аспекте настоящего изобретения популяции плюрипотентных стволовых клеток могут быть дифференцированы в популяции клеток, экспрессирующих маркеры, характерные для линии сформированной эндодермы, путем культивирования плюрипотентных стволовых клеток на среде без сыворотки с добавлением БСА и фактора, выбранного из следующей группы: инсулин или IGF-1. В одном варианте осуществления дифференциация популяции плюрипотентных стволовых клеток в направлении популяции клеток, экспрессирующих маркеры, характерные для линии клеток сформированной эндодермы, достигается обработкой плюрипотентных стволовых клеток активином А и лигандом Wnt.
В альтернативном варианте осуществления дифференциация популяции плюрипотентных стволовых клеток в направлении популяции клеток, экспрессирующих маркеры, характерные для линии сформированной эндодермы, достигается обработкой плюрипотентных стволовых клеток GDF-8 и, по меньшей мере, одним из факторов, выбранных из следующей группы: анилин-пиридинотриазин, циклический анилин-пиридинотриазин, N-{[1-(фенилметил)азепан-4-ил]метил}-2-пиридин-3-илацетамид, 4-{[4-(4-{[2-(пиридин-2-иламино)этил]амино}-1,3,5-триазин-2-ил)пиридин-2-ил]окси}бутан-1-ол, 3-({3-[4-({2-[метил(пиридин-2-ил)амино]этил}амино)-1,3,5-триазин-2-ил]пиридин-2-ил}амино)пропан-1-ол, N~4~-[2-(3-фторфенил)этил]-N~2~-[3-(4-метилпиперазин-1-ил)пропил]пиридо[2,3-d]пиримидин-2,4-диамин, 1-метил-N-[(4-пиридин-3-ил-2-{[3-(трифторметил)фенил]амино}-1,3-тиазол-5-ил)метил]пиперидин-4-карбоксамид, 1,1-диметилэтил {2-[4-({5-[3-(3-гидроксипропил)фенил]-4H-1,2,4-триазол-3-ил}амино)фенил]этил}карбамат, 1,1-диметилэтил {[3-({5-[5-(3-гидроксипропил)-2-(метилокси)фенил]-1,3-оксазол-2-ил}амино)фенил]метил}карбамат, 1-({5-[6-({4-[(4-метилпиперазин-1-ил)сульфонил]фенил}амино)пиразин-2-ил]тиофен-2-ил}метил)пиперидин-4-ол, 1-({4-[6-({4-[(4-метилпиперазин-1-ил)сульфонил]фенил}амино)пиразин-2-ил]тиофен-2-ил}метил)пиперидин-4-карбоксамид и 2-{[4-(1-метилэтил)фенил]амино}-N-(2-тиофен-2-илэтил)-7,8-дигидропиридо[4,3-d]пиримидин-6(5H)-карбоксамид. Примеры факторов, подходящих для использования, можно найти в патентной заявке США сер. № 12/494789. В одном варианте осуществления, по меньшей мере, один из факторов представляет собой 14-проп-2-ен-1-ил-3,5,7,14,17,23,27-гептаазатетрацикло[19.3.1.1~2,6~.1~8,12~]гептакоза-1(25),2(27),3,5,8(26),9,11,21,23-нонаен-16-он.
Популяция плюрипотентных клеток может культивироваться в бессывороточной среде с добавлением БСА и фактора, выбранного из группы, включающей: IGF-1 от приблизительно одних суток до приблизительно семи суток. Альтернативно, популяция плюрипотентных клеток может культивироваться в бессывороточной среде с добавлением БСА и фактора, выбранного из группы, включающей: IGF-1, от приблизительно одних суток до приблизительно шести суток. Альтернативно, популяция плюрипотентных клеток может культивироваться в бессывороточной среде с добавлением БСА и фактора, выбранного из группы, включающей: IGF-1, от приблизительно одних суток до приблизительно пяти суток. Альтернативно, популяция плюрипотентных клеток может культивироваться в бессывороточной среде с добавлением БСА и фактора, выбранного из группы, включающей: IGF-1, от приблизительно одних суток до приблизительно четырех суток. Альтернативно, популяция плюрипотентных клеток может культивироваться в бессывороточной среде с добавлением БСА и фактора, выбранного из группы, включающей: инсулин и IGF-1, от приблизительно одних суток до приблизительно четырех суток.
В одном из вариантов осуществления GDF-8 используется в концентрации от приблизительно 5 нг/мл до приблизительно 500 нг/мл. В альтернативном варианте осуществления GDF-8 используется в концентрации от приблизительно 5 нг/мл до приблизительно 50 нг/мл. В альтернативном варианте осуществления GDF-8 используется в концентрации от приблизительно 5 нг/мл до приблизительно 25 нг/мл. В альтернативном варианте осуществления GDF-8 используется в концентрации приблизительно 25 нг/мл.
Активин-А может использоваться в концентрациях от приблизительно 1 пг/мл до приблизительно 100 мкг/мл. В альтернативном варианте осуществления концентрация может составлять от примерно 1 пг/мл до примерно 1 мкг/мл. В другом альтернативном варианте осуществления концентрация может составлять от примерно 1 пг/мл до примерно 100 нг/мл. В другом альтернативном варианте осуществления концентрация может составлять от приблизительно 1 пг/мл до приблизительно 100 нг/мл. В другом альтернативном варианте осуществления концентрация может составлять примерно 100 нг/мл.
Лиганд Wnt может выбираться из группы, состоящей из Wnt-1, Wnt-3a, Wnt-5a и Wnt-7a. В одном варианте осуществления лиганд Wnt представляет собой Wnt-1. В альтернативном варианте осуществления лиганд Wnt представляет собой Wnt-3a.
Лиганд Wnt может использоваться в концентрации от приблизительно 1 нг/мл до приблизительно 1000 нг/мл. В альтернативном варианте осуществления лиганд Wnt может использоваться в концентрации от приблизительно 10 нг/мл до приблизительно 100 нг/мл. В одном варианте осуществления лиганд Wnt используется в концентрации приблизительно 20 нг/мл.
В одном из вариантов осуществления инсулин используется в концентрации от приблизительно 1 нг/мл до приблизительно 100 нг/мл.
В одном из вариантов осуществления IGF-1 используется в концентрации от приблизительно 1 нг/мл до приблизительно 200 нг/мл.
Образование клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы
В одном варианте осуществления изобретения популяция клеток, экспрессирующих маркеры, характерные для линии сформированной эндодермы, полученные методами настоящего изобретения, далее дифференцируют в популяции клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы любым известным способом.
Например, популяции клеток, экспрессирующих маркеры, характерные для линии сформированной эндодермы, полученные по способам настоящего изобретения, могут быть дополнительно дифференцированы в популяции клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, путем обработки популяции клеток, экспрессирующих маркеры, характерные для линии сформированной эндодермы, в соответствии со способами, представленными в работе D’ Amour et al., Nature Biotechnology, 24, 1392-1401 (2006)
Например, популяции клеток экспрессирующих маркеры, характерные для линии сформированной эндодермы, полученные по способам настоящего изобретения, могут быть дополнительно дифференцированы в популяции клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, путем обработки популяции клеток, экспрессирующих маркеры, характерные для линии сформированной эндодермы, в соответствии со способами, представленными в патентной заявке США сер. № 11/736908.
Создание клеток, экспрессирующих маркеры, характерные для линии панкреатических эндокринных клеток
В одном варианте осуществления популяции клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, дополнительно дифференцируются в популяции клеток, экспрессирующих маркеры, характерные для линии панкреатических эндокринных клеток, с использования любого способа, известного специалистам в данной области.
Например, популяции клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, могут быть дополнительно дифференцированы в популяции клеток, экспрессирующих маркеры, характерные для линии панкреатических эндокринных клеток, путем обработки популяции клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, в соответствии со способами, представленными в работе D’Amour et al., Nature Biotechnology, 2006.
Например, популяции клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, могут быть дополнительно дифференцированы в популяции клеток, экспрессирующих маркеры, характерные для линии панкреатических эндокринных клеток, путем обработки популяции клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, в соответствии со способами, представленными в работе D’Amour et al., Nature Biotechnology, 2006.
Например, популяции клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, могут далее дифференцироваться в популяции клеток, экспрессирующих маркеры, характерные для линии эндокринных панкреатических клеток, путем обработки популяции клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, в соответствии с методами, описанными в патентной заявке США сер. № 11/736908.
Например, популяции клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, могут далее дифференцироваться в популяции клеток, экспрессирующих маркеры, характерные для линии эндокринных панкреатических клеток, путем обработки популяции клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, в соответствии со способами, описанными в патентной заявке США сер. № 11/779311.
Например, популяции клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, могут далее дифференцироваться в популяции клеток, экспрессирующих маркеры, характерные для линии эндокринных панкреатических клеток, путем обработки популяции клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, в соответствии со способами, описанными в патентной заявке США сер. № 60/953178.
Например, популяции клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, могут далее дифференцироваться в популяции клеток, экспрессирующих маркеры, характерные для линии эндокринных панкреатических клеток, путем обработки популяции клеток, экспрессирующих маркеры, характерные для линии панкреатической эндодермы, в соответствии со способами, описанными в патентной заявке США сер. № 60/990529.
Настоящее изобретение далее иллюстрируется, помимо прочих, следующими примерами.
ПРИМЕРЫ
Пример 1
Роль инсулина в дифференциации человеческих плюрипотентных клеток в клетки, экспрессирующие маркеры, характерные для линии сформированной эндодермы: Посев клеточных скоплений
Предыдущие исследования показали, что высокие концентрации фетальной бычьей сыворотки (ФБС) пагубны для формирования сформированной эндодермы (СЭ) из эмбриональных стволовых клеток. См., например, D’Amour et al., Nature Biotechnology, 2005, где индукция образования сформированной эндодермы из человеческих эмбриональных клеток значительно усиливалась при снижении концентрации ФБС с 10% до 0,5-2%. В других работах описаны сходные наблюдения, когда добавление 25 нг/мл IGF или 200 нг/мл инсулина к 2% ФБС при культуре эмбриональных стволовых клеток на MEF-CM (среда, кондиционированная мышиными фибробластами) уменьшало экспрессию SOX17 после обработки активином А примерно на 70%. См. McLean et al, Stem Cells 25:29-38, 2007.
Наблюдаемое ингибирующее действие, вероятно, было обусловлено присутствием инсулина или IGF в ФБС, что способствовало пути фосфатидилинозит-3-киназы. См. McLean et al, Stem Cells 25:29-38, 2007. Блокада сигнального пути PI-3 киназы увеличивает процент Sox17-положительных клеток среди человеческих эмбриональных стволовых клеток, культивируемых в среде MEF-CM (среда, кондиционированная эмбриональными фибробластами мыши). См. McLean et al, Stem Cells 25:29-38, 2007.
Эти данные позволяют предположить, что добавление даже 25 нг/мл IGF или 200 нг/мл инсулина к среде, содержащей активин A и низкую концентрацию ФБС (0,5-2%), будет блокировать образование сформированной эндодермы. Типичные концентрации IGF и инсулина в ФБС - приблизительно 70 нг/мл (J. Clin. Invest. 76:4, 1985) и приблизительно 60 нг/мл (In Vitro Cell Dev Biol. 32:8-12, 1996), соответственно. Это переводится в примерно 1,4 нг/мл IGF и приблизительно 1,2 нг/мл инсулина в 2% ФБС.
Стволовые клетки человеческой эмбриональной линии H1 (p40-p52) культивировали на чашках, покрытых субстратом MATRIGEL®(разведение 1:30) (BD Biosciences; Кат. № 356231) в среде MEF-CM (седа, кондиционированная мышиными эмбриональными фибробластами) с добавлением 16 нг/мл FGF2 (кат. № 100-18B, PeproTech, NJ) и дифференцировали в клетки, экспрессирующие маркеры, характерные для линии сформированной эндодермы, следующим образом:
a. Среда RPMI с добавлением 2% БСА без жирных кислот (Кат. № 68700, Proliant, Айова) и 100 нг/мл активина A (R&D Systems, Миннесота)+20 нг/мл WNT-3a (Кат. № 1324-WN-002, R&D Systems, Миннесота) одни сутки, затем
b. Среда RPMI с 2% БСА и 100 нг/мл активина А еще трое суток.
В некоторых культурах клетки обрабатывали следующими разведениями ITS-X (кат № 51500-056, Invitrogen, Калифорния): 0, 1:106, 1:5×105, 1:105, 1:104. ITS-X представляет собой заменитель сыворотки, содержащий 1 мг/мл инсулина, 0,55 мг/мл трансферрина, 0,00067 мг/мл натрия селенита и 0,2 мг/мл этаноламина. Диапазон разведений ITS-X соответствовал 0, 2, 10 и 100 нг/мл инсулина. В качестве контроля использовали 0,2% ФБС (Кат. № SH30070.03, Hyclone, Юта) в первый день дифференциации, 0,5% ФБС на 2 день и 2% ФБС на 3-4 дни. В культуры с добавлением ФБС не добавляли ITS-X.
На 4 день отбирали пробы для флуоресцентной проточной цитометрии и анализа генной экспрессии методом ПЦР в реальном времени. Удивительно, см. фиг. 1, что добавление 1-100 нг/мл инсулина к среде, использовавшейся для дифференциации клеток, значительно не влияло на экспрессию маркеров сформированной эндодермы (FOXA2, SOX17 и CXCR4), маркеров мезенхимы (T, также известный как Brach) или экстраэмбриональных маркеров (SOX7, AFP). Кроме того, культуры на средах с добавлением 2% БСА значительно лучше экспрессировали маркеры, связанные со сформированной эндодермой, чем культуры на среде с добавлением 0,5-2% ФБС.
Эти наблюдения были далее подтверждены экспрессией CXCR4 и CD9 по результатам флуоресцентной проточной цитометрии на средах разного состава. См. фиг. 2. Ранее было показано, что рецептор клеточной поверхности CXCR4 является маркером сформированной эндодермы. CD9 представляет собой маркер недифференцированных эмбриональных стволовых клеток. Следовательно, усиление экспрессии CXCR4 и ослабление экспрессии CD9 в популяции клеток показательно для формирования сформированной эндодермы. Как показано в таблице I, в клетках на среде с добавлением БСА не было отмечено изменений экспрессии CXCR4 или CD9 ни при одной из исследованных концентраций инсулина. Эти данные позволяют предположить, что инсулин не обладает ингибирующим действием в исследованных концентрациях в питательных средах, использовавшихся в данном исследовании.
Пример 2
Роль инсулина в дифференциации человеческих плюрипотентных стволовых клеток в клетки, экспрессирующие маркеры, характерные для линии сформированной эндодермы: Посев одиночных клеток
Стволовые клетки человеческой эмбриональной линии H1 (p40-p52) высевали поодиночке из суспензии с концентрацией 100000 кл./см2 на чашки, покрытые субстратом MATRIGEL® (разведение 1:30) (BD Biosciences; Кат. № 356231) в среду MEF-CM (среда, кондиционированная мышиными эмбриональными фибробластами) с добавлением 16 нг/мл FGF2 (Кат. № 100-18B, PeproTech, NJ) и 10 мкМ Y27632 (ингибитор Rock, Кат. № Y0503, Sigma, Миссури). Через 72 ч после посева культуры дифференцировали в сформированную эндодерму (СЭ) следующим образом:
a. Среда MCDB-131 (Кат. № 10372-019, Invitrogen, Калифорния) с 2% БСА без жирных кислот (Кат. № 68700, Proliant, Айова), 0,0025 г/мл натрия бикарбоната (Кат. № S3187, Sigma, Миссури), 1X ГлутаМакса™ (Кат. № 35050-079, Invitrogen, Калифорния), 100 нг/мл активина A (RD plusSystems, Миннесота), 20 нг/мл WNT-3a (Кат. № 1324-WN-002, RD Systems, Миннесота) в течение суток, затем
b. Среда MCDB-131 с 2% БСА, бикарбонатом натрия, Глутамаксом и 100 нг/мл активина A в течение следующих 3 суток.
К некоторым культурам добавляли следующие концентрации инсулина (Кат. № I9278, Sigma, Миссури): 0, 1, 10, 100, 1000 или 10000 нг/мл На 4 день отбирали пробы для флуоресцентной проточной цитометрии и анализа генной экспрессии методом ПЦР в реальном времени.
Добавление 1-100 нг/мл инсулина к среде для дифференциации клеток значительно не влияло на экспрессию маркеров, характерных для сформированной эндодермы (FOXA2, SOX17, CER1 и CXCR4). Сходным образом, экспрессия эмбриональных маркеров (NANOG) или экстраэмбриональных маркеров (SOX7, AFP) также не изменялась. См. фиг. 3. Однако добавление 1-10 мкг/мл инсулина усиливало экспрессию NANOG. Эти данные дополнительно подтверждены иммунофлуоресцентным окрашиванием на маркер сформированной эндодермы SOX17 (кат. № AF1924, R&D systems, Миннесота) (фиг. 4).
На фиг. 5 показаны графики экспрессии CXCR4 и CD9 в средах разного состава по результатам флуоресцентной проточной цитометрии. Как показано в таблице II, только при наивысшей концентрации инсулина(1-10 мкг/мл) наблюдалось снижение процентного содержания клеток CXCR4+CD9 и усиление экспрессии фракции CXCR4-CD9. Эти данные позволяют предположить, что в условиях настоящего исследования только концентрации инсулина больше физиологических подавляют образование клеток сформированной эндодермы.
Пример 3
Роль IGF в дифференциации человеческих плюрипотентных клеток в клетки, экспрессирующие маркеры, характерные для линии сформированной эндодермы: Посев одиночных клеток
Стволовые клетки человеческой эмбриональной линии H1 (p40-p52) высевали на среду одиночно из суспензии с концентрацией 100000 кл./см2 на чашки, покрытые субстратом MATRIGEL® (разведение 1:30) (BD Biosciences; Кат. № 356231) в среду MEF-CM (среда, кондиционированная мышиными эмбриональными фибробластами) с добавлением 16 нг/мл FGF2 (Кат. № 100-18B, PeproTech, NJ) и 10 мкМ Y27632 (ингибитор Rock, Кат. № Y0503, Sigma, Миссури). Через 72 ч после посева культуры дифференцировали в сформированную эндодерму (СЭ) следующим образом:
a. Среда MCDB-131 (Кат. № 10372-019, Invitrogen, Калифорния) с добавлением 2% БСА без жирных кислот (Кат. № 68700, Proliant, Айова), 0,0025 г/мл бикарбоната (кат. № S3187, Sigma, Миссури), 1X ГлутаМакса™ (кат. № 35050-079, Invitrogen, Калифорния) и 100 нг/мл GDF8 (Кат. № 120-00, PeproTech, NJ)+2,5 мкМ ингибитора GSK3B 14-проп-2-ен-1-ил-3,5,7,14,17,23,27-гептаазатетрацикло[19.3.1.1~2,6~.1~8,12~]гептакоза-1(25),2(27),3,5,8(26),9,11,21,23-нонаен-16-он в течение суток, затем
b. Среда MCDB-131 с добавлением 2% БСА, натрия бикарбоната, Глутамакса и 10 нг/мл GDF-8 еще трое суток.
К некоторым культурам добавляли следующие концентрации IGF (Кат. № AF100, PeproTech Нью Джерси): 0, 1, 10, 50 или 200 нг/мл. В качестве контроля вместо БСА использовали 0,2 ФБС (Кат. № SH30070.03, Hyclone, Юта) в первые сутки дифференциации и 2% ФБС на 2-4 сутки. К некоторым культурам с ФБС также добавляли разные концентрации IGF.
На 4 день отбирали пробы для флуоресцентной проточной цитометрии и анализа генной экспрессии методом ПЦР в реальном времени.
Удивительно, что добавление 1-200 нг/мл IGF к культурам с БСА незначительно не влияло на экспрессию маркеров сформированной эндодермы (FOXA2, SOX17, CER1 и CXCR4) при сравнении с контрольными образцами без добавления IGF (Фиг. 6). Сходные результаты наблюдались с экстраэмбриональными маркерами (SOX7, AFP). Добавление 50-200 нг/мл IGF не усиливало экспрессию эмбрионального маркера NANOG.
Культуры на средах с добавлением ФБС были намного чувствительнее к ингибирующему действию IGF. В этих культурах экспрессия SOX17, HNF3B и CXCR4 уменьшалась с повышением концентрации IGF. Эти наблюдения дополнительно подтверждены иммунофлуоресцентным окрашиванием на маркер сформированной эндодермы SOX17 (кат № AF1924, R&D systems, Миннесота) (Фиг. 8-9).
Как кратко представлено в таблице III, только концентрации IGF, превышающие физиологические (50-200 нг/мл) в культурах с добавлением БСА, вызывали снижение экспрессии CXCR4+CD9- и повышение экспрессии фракции CXCR4-CD9+. Однако с повышением доз IGF в культурах с добавлением ФБС показано более значительное падение экспрессии CXCr4+CD9- по сравнению с культурами с добавлением БСА. Все вышеописанные примеры показывают, что в отсутствии ФБС физиологические концентрации IGF или инсулина не оказывают ингибирующего действия на индукцию маркеров СЭ.
Пример 4
Концентрации IGF в разных партиях ФБС
Исследование проводили с помощью набора для определения IGF-1 производства Diagnostic Systems Laboratories (DSL) (кат. № DSL-10-2800). Для исследования брали 20 мкл предварительно обработанной сыворотки (в двух повторностях). Для анализа среды использовали 20 μл пробы непосредственно. Исследование проводили по инструкции к набору. Набор позволяет обнаруживать и человеческий, и бычий IGF-1, так как содержит 2 моноклональных антитела к гомологичным пептидным последовательностям.
Опытные образцы. Использовали следующие образцы:
4А/5А; Сыворотка новорожденных телят Hyclone; Партия AKM12868;
4B/5B:ФБС NIH (из аликвот, хранившихся при -20°С)
4/5C: Сыворотка новорожденных телят Hyclone, партия:
ATK33398: 4D/5D: Сыворотка новорожденных телят Hyclone, партия:
AUK 54924
4Е/5Е: Человеческая сыворотка, лот:.
A70184, производства Valley Biomedical Inc.
4F/5F: Нокаутная сыворотка: Invitrogen, партия:
557914 4F/5F: F12 DMEM, инвиторген, партия:
692281: 4Н/5Н: Кондиционирующая среда MEF, партия:
011410 (день 5)
Контрольные образцы: Использовались следующие контрольные образцы: Основные: 0” IGF-1 (отрицательный контроль, из набора).
Образец F12, указанный выше, также служил отрицательным контролем. 2 положительных контроля (127 нг/мл и 241 нг/мл; из набора). Результаты: Чувствительность метода анализа сыворотки была более 10 нг/мл, а для среды - более 0,1 нг/мл.
Пример 5
Роль инсулина/IGF и ФБС в дифференциации человеческих плюрипотентных клеток в клетки, экспрессирующие маркеры, характерные для линии сформированной эндодермы: Посев одиночных клеток
Стволовые клетки человеческой эмбриональной линии H1 (p40-p52) высевали одиночно из суспензии с концентрацией 100000 кл./см2 на чашки, покрытые субстратом MATRIGEL® (разведение 1:30) (BD Biosciences; Кат. № 356231) в среду MEF-CM (среда, кондиционированная мышиными эмбриональными фибробластами) с добавлением 16 нг/мл FGF2 (Кат. № 100-18B, PeproTech, NJ) и 10 μM Y27632 (ингибитор Rock, Кат. № Y0503, Sigma, Миссури). Через 72 ч после посева культуры дифференцировали в сформированную эндодерму (СЭ) следующим образом:
а. Среда MCDB-131 (кат. № 10372-019, Invitrogen, Калифорния) с добавлением 0,2% ФБС (кат. № SH30070.03, Hyclone, Юта), 0,0025 г/мл натрия бикарбоната (кат. № S3187, Sigma, Миссури), 1X ГлутаМакса™ (кат. № 35050-079, Invitrogen, Калифорния) и 100 нг/мл GDF8 (кат. № 120-00, PeproTech, Нью Джерси)+2,5 мкМ ингибитора GSK3B 14-проп-2-ен-1-ил-3,5,7,14,17,23,27-гептаазотетрацикло[19.3.1.1~2,6~.1~8,12~]гептакоза-1(25),2(27),3,5,8(26),9,11,21,23-нонаен-16-она в течение суток, затем b.среда MCDB-131 с добавлением ФБС, натрия бикарбоната, Глутамакс и 10 нг/мл GDF8 еще трое суток.
На вторые сутки использовали 0,5% ФБС, а на 3-4 сутки 2% ФБС. Кроме обычной ФБС, также исследовали термообработанную ФБС (кат. № F4135, Sigma, Миссури) и ФБС, очищенную активированным углем (кат. № F6765, Sigma, Миссури).
К некоторым из культур с добавлением ФБС также добавляли различные концентрации IGF (10-100 нг/мл или инсулина (10-100 нг/мл). На 4 день отбирали пробы для анализа методом флуоресцентной проточной цитометрии и ПЦР в реальном времени. В отличие от культур с добавлением БСА (см. предыдущие примеры) в присутствии ФБС, добавление инсулина или IGF снижало экспрессию SOX17 и CCXR4 в зависимости от дозы. См. фиг. 9. Экспрессия маркера плюрипотентности NANOG также ингибировалась. Хотя различные аспекты изобретения иллюстрируются выше ссылками на примеры и предпочтительные варианты осуществления, подразумевается, что область изобретения ограничивается не упомянутым выше описанием, а следующими пунктами формулы изобретения, составленными в соответствии с принципами патентного законодательства.










Изобретение относится к области биохимии, в частности к способам получения популяции клеток, в которой более 85% клеток экспрессируют маркеры, характерные для линии сформированной эндодермы. Указанную популяцию клеток получают путем дифференцирования популяции человеческих эмбриональных стволовых клеток линии H1. Дифференцирование происходит в бессывороточной среде с добавлением: (а) БСА, (b) либо (i) GDF-8 и 14-проп-2-ен-1-ил-3,5,7,14,17,23,27-гептаазотетрацикло[19.3.1.1~2,6~.1~8,12~]гептакоза-1(25),2(27),3,5,8(26),9,11,21,23-нонаен-16-она, либо (ii) активина А и лиганда Wnt; и (с) фактора, выбранного из инсулина и IGF-1. Изобретение обеспечивает высокую эффективность дифференцирования эмбриональных стволовых клеток. 3 н. и 17 з.п. ф-лы, 9 ил., 5 табл., 5 пр.
1. Способ получения популяции клеток, в которой более 85% клеток экспрессируют маркеры, характерные для линии сформированной эндодермы, включающий следующие стадии:
a) культивирование популяции человеческих эмбриональных стволовых клеток линии H1;
b) дифференцирование популяции человеческих эмбриональных стволовых клеток линии H1 в популяцию клеток, в которой более 85% экспрессирует маркеры, характерные для линии сформированной эндодермы в бессывороточной среде с добавлением: (а) БСА, (b) либо (i) GDF-8 и 14-проп-2-ен-1-ил-3,5,7,14,17,23,27-гептаазотетрацикло[19.3.1.1~2,6~.1~8,12~]гептакоза-1(25),2(27),3,5,8(26),9,11,21,23-нонаен-16-она, либо (ii) активина А и лиганда Wnt; и (с) фактора, выбранного из группы, состоящей из инсулина и IGF-1, где инсулин представлен в концентрации от 1 нг/мл до 100 нг/мл и IGF-1 представлен в концентрации от 1 нг/мл до 200 нг/мл.
2. Способ по п.1, в котором популяция человеческих эмбриональных стволовых клеток линии H1 дифференцируется в среде без сыворотки с добавлением БСА и фактора, выбранного из группы, состоящей из инсулина и IGF-1, не менее 6 суток.
3. Способ по п.1, в котором популяция человеческих эмбриональных стволовых клеток линии H1 дифференцируется в среде без сыворотки с добавлением БСА и фактора, выбранного из группы, содержащей инсулин и IGF-1, не менее 7 суток.
4. Способ по п.1, в котором фактором является инсулин.
5. Способ по п.1, в котором фактором является IGF-1.
6. Способ по п.1, в котором популяция человеческих эмбриональных стволовых клеток линии H1 дифференцируется в среде без сыворотки с добавлением БСА, GDF-8, 14-проп-2-ен-1-ил-3,5,7,14,17,23,27-гептаазотетрацикло[19.3.1.1~2,6~.1~8,12~]гептакоза-1(25),2(27),3,5,8(26),9,11,21,23-нонаен-16-она и фактора, выбранного из группы, состоящей из инсулина и IGF-1.
7. Способ по п.6, в котором фактором является инсулин.
8. Способ по п.6, в котором фактором является IGF-1.
9. Способ получения популяции клеток, в которой более 85% клеток экспрессируют маркеры, характерные для линии сформированной эндодермы, включающий дифференцирование популяции человеческих эмбриональных стволовых клеток линии H1 в популяцию клеток, в которой более 85% экспрессирует маркеры, характерные для линии сформированной эндодермы, в бессывороточной среде с добавлением: (а) БСА, (b) либо (i) GDF-8 и 14-проп-2-ен-1-ил-3,5,7,14,17,23,27-гептаазотетрацикло[19.3.1.1~2,6~.1~8,12~]гептакоза-1(25),2(27),3,5,8(26),9,11,21,23-нонаен-16-она, либо (ii) активина А и лиганда Wnt; и (с) фактора, выбранного из группы, состоящей из инсулина и IGF-1, где инсулин представлен в концентрации от 1 нг/мл до 100 нг/мл и IGF-1 представлен в концентрации от 1 нг/мл до 200 нг/мл.
10. Способ по п.9, в котором фактором является инсулин.
11. Способ по п.9, в котором фактором является IGF-1.
12. Способ по п.9, в котором популяция человеческих эмбриональных стволовых клеток линии H1 дифференцируется в бессывороточной среде с добавлением БСА, GDF-8, 14-проп-2-ен-1-ил-3,5,7,14,17,23,27-гептаазотетрацикло[19.3.1.1~2,6~.1~8,12~]гептакоза-1(25),2(27),3,5,8(26),9,11,21,23-нонаен-16-она, и фактора, выбранного из группы, состоящей из инсулина и IGF-1.
13. Способ по п.12, в котором фактором является инсулин.
14. Способ по п.12, в котором фактором является IGF-1.
15. Способ получения популяции клеток линии сформированной эндодермы, включающий культивирование популяции человеческих эмбриональных стволовых клеток линии H1 в бессывороточной среде с дополнением: (а) БСА; b) либо (i) GDF-8 и 14-проп-2-ен-1-ил-3,5,7,14,17,23,27-гептаазотетрацикло[19.3.1.1~2,6~.1~8,12~]гептакоза-1(25),2(27),3,5,8(26),9,11,21,23-нонаен-16-она, либо (ii) активина А и лиганда Wnt; и (с) фактора, выбранного из группы, состоящей из инсулина и IGF-1.
16. Способ по п.15, в котором фактором является инсулин.
17. Способ по п.15, в котором фактором является IGF-1.
18. Способ по п.15, отличающийся тем, что способ включает культивирование популяции человеческих эмбриональных стволовых клеток линии H1 в бессывороточной среде, с дополнением БСА, GDF-8, 14-проп-2-ен-1-ил-3,5,7,14,17,23,27-гептаазотетрацикло[19.3.1.1~2,6~.1~8,12~]гептакоза-1(25),2(27),3,5,8(26),9,11,21,23-нонаен-16-она и фактора, выбранного из группы, состоящей из инсулина и IGF-1.
19. Способ по п.18, в котором фактором является инсулин.
20. Способ по п.18, в котором фактором является IGF-1.
| US 20090298178 A1, 03.12.2009 | |||
| US 0007510876 B2, 31.03.2009 | |||
| US 20090304642 A1, 10.12.2009 | |||
| US 20100124781 A1, 20.05.2010 | |||
| WO 2009012428 A2, 22.01.2009 | |||
| WO 2010096496 A2, 26.08.2010 | |||
| СПОСОБ ИНДУКЦИИ ДИФФЕРЕНЦИАЦИИ СТВОЛОВЫХ КЛЕТОК В МИОКАРДИАЛЬНЫЕ | 2004 |
|
RU2392315C2 |
| СПОСОБ ПОЛУЧЕНИЯ ЭНДОТЕЛИАЛЬНЫХ КЛЕТОК ИЗ ЭМБРИОНАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК ЧЕЛОВЕКА (ВАРИАНТЫ) | 2008 |
|
RU2359030C1 |

