ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
[001] Настоящее изобретение относится, в общем, к области молекулярной биологии и рассматривает различные полинуклеотиды, полипептиды и способы применения, которые могут использоваться для повышения урожая трансгенных растений. Трансгенные растения, содержащие какой-либо из полинуклеотидов или полипептидов, описанных в данном документе, могут проявлять какой-либо из признаков, состоящих из увеличенного урожая, увеличенной устойчивости к абиотическому стрессу, увеличенного клеточного роста, увеличенного коэффициента использования воды и увеличенного коэффициента использования питательных веществ.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
[002] Увеличение населения мира и истощающийся запас пахотных земель, доступных для сельского хозяйства, активизирует необходимость исследования в области увеличения эффективности сельского хозяйства. Традиционные способы повышения урожая посевных и садовых культур применяют техники селекционного разведения для выявления растений, имеющих необходимые характеристики. Тем не менее, такие техники селекционного разведения имеют определенные недостатки, а именно то, что эти техники зачастую являются трудоемкими и дают в результате растения, которые часто содержат гетерогенные генетические компоненты, которые не всегда могут приводить к передаче необходимого признака от родительских растений. Успехи в молекулярной биологии позволили людям модифицировать идиоплазму животных и растений. Генная инженерия растений предусматривает выделение и манипуляцию с генетическим материалом (типично в форме ДНК или РНК) и последующее введение этого генетического материала в геном растения. Такая технология обладает способностью предоставлять сельскохозяйственные культуры или растения, имеющие различные улучшенные экономические, агрономические или садоводческие признаки.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[003] Изложенное ниже краткое описание изобретения перечисляет некоторые варианты осуществления рассматриваемого изобретения и во многих случаях перечисляет изменения и перестановки этих вариантов осуществления. Данное краткое описание изобретения является исключительно иллюстративным для многочисленных и разнообразных вариантов осуществления. Упоминание одного или нескольких репрезентативных признаков данного варианта осуществления также является иллюстративным. Такой вариант осуществления может типично иметь место с упомянутым(и) признаком(ами) или без него(них); аналогично, эти признаки могут применяться к другим вариантам осуществления настоящего изобретения, независимо от того, перечислены ли они в кратком описании изобретения или нет. Чтобы избежать чрезмерного повторения, настоящее краткое описание изобретения не перечисляет или предполагает все возможные комбинации таких признаков.
[004] Настоящее изобретение предлагает нуклеотидные последовательности, кодирующие полипептиды трегалозного пути, которые будучи трансгенно экспрессированными в растении, увеличивают урожай. Полипептиды Т6РР, описанные в данном документе, содержат модификации, изменяющие активность белков трегалозо-6-фосфатфосфатазы (Т6РР), причем активность снижается по сравнению с немодифицированной Т6РР. Модифицированные Т6РР могут содержать консенсусную последовательность, проиллюстрированную в SEQ ID NO: 9, и могут иметь по меньшей мере одну модифицированную аминокислоту. Модифицированная Т6РР может иметь модификацию, произведенную в консервативной аминокислоте, включая консервативные аминокислоты, описанные в консенсусной последовательности SEQ ID NO: 9. Модифицированные Т6РР могут вызывать in vitro активацию Т6РР дикого типа. Модификация в Т6РР может находиться в пределах по меньшей мере одного из доменов CAP, фосфатазы, А-фосфатазы или В-фосфатазы. Т6РР, имеющие сниженное связывание с субстратом трегалозо-6-фосфатом (Т6Р), демонстрируют увеличенную урожайность и увеличенную стрессоустойчивость, будучи трансгенно экспрессированными в растениях, а также наличие повышенной устойчивости к стрессу, включая засуху. Растения, экспрессирующие модифицированную Т6РР, включают однодольные или двудольные растения, включая растения, выбранные из группы, состоящей из следующего: маис, сахарный тростник, соя, рис, сорго или пшеница. Модификации полипептидов Т6РР могут включать одну или комбинацию аминокислотных замен, аминокислотных делеций или аминокислотных вставок. Модифицированная Т6РР может быть представителем надсемейства галогенокислой дегалогеназы (HAD) фосфатаз. Ферментативная активность Т6РР может быть снижена по меньшей мере на 50%, 60%, 70%, 80%, 90%, 95%, 99% или 100%.
[005] Любой из описанных полипептидов Т6РР может применяться для получения полинуклеотидов. Полинуклеотиды, кодирующие модифицированные полипептиды Т6РР, могут быть введены в клетку хозяина. Клетка хозяина может включать клетку растения. Полинуклеотиды могут быть включены в кассеты экспрессии, которые обеспечивают транскрипцию полинуклеотида в растении. Кассеты экспрессии могут включать промотор, который экспрессирует полинуклеотид в репродуктивной ткани растения. Кроме того, репродуктивная ткань может быть выбрана из группы, состоящей из следующего: ткань колоска, узел початка, ткань прицветника, ткань меристемы колоска, ткань цветоножки соцветия и незрелая ткань цветка. Промотор, экспрессирующий полинуклеотиды Т6РР, может включать промотор OsMADS или промотор OsMADS6. Экстракт, содержащий полинуклеотиды или полипептиды, может быть получен из чего-либо из клеток хозяина, растений, частей растения или тканей растения.
[006] Способы, раскрытые в данном документе, дополнительно включают введение растениям полинуклеотидов, например, как раскрыто в данном документе. Трансгенные растения, содержащие полинуклеотиды, раскрытые в данном документе, могут проявлять увеличенный клеточный рост, увеличенную мощность растений и/или всходов, увеличенную урожайность, увеличенный вес семян, увеличенный коэффициент использования воды и/или увеличенную биомассу. Предусматривается, что растения, полученные с помощью способов, раскрытых в данном документе, имеют увеличенную устойчивость к абиотическому стрессу. Трансгенные растения, описанные в данном документе, могут производить более высокий урожай, как в отношении урожайности биомассы, так и урожайности зерна растения. Один аспект настоящего изобретения предлагает различные модификации, которые могут быть проведены на любой данной генной последовательности Т6РР, которая может использоваться в трансгенных растениях для обеспечения увеличенного урожая. Альтернативно, другие многочисленные способы могут применяться для увеличения уровней Т6Р для обеспечения увеличенного урожая и стрессоустойчивости у растения.
[007] Способы увеличения урожая, увеличения устойчивости растения к абиотическому стрессу или снижения бесплодия у растения, увеличения количества початков на растение и/или зерен на растение в условиях дефицита воды могут включать введение кассеты экспрессии, содержащей модифицированную Т6РР, в растительную клетку и затем получение трансгенного растения. Растение может представлять собой однодольное растение, например, может представлять собой растение маиса, риса, пшеницы, сорго, сахарного тростника или газонной травы. Это может быть двудольное растение, такое как соя. Для способов увеличения устойчивости растения к абиотическому стрессу стресс может быть выбран из группы, состоящей из стресса, вызванного недостатком воды, теплового стресса или стресса, вызванного низкими температурами. Стресс, вызванный недостатком воды, может быть обусловлен засухой.
[008] Кроме того, включены способы модификации уровня трегалозо-6-фосфата в растении путем выявления полипептида в трегалозном пути; модификации полипептида так, чтобы он имел измененную ферментативную активность; введения в растение кассеты экспрессии, содержащей полинуклеотид, кодирующий модифицированный полипептид; и получения растения с модифицированными уровнями трегалозо-6-фосфата. Полипептид может иметь активность, выбранную из группы, состоящей из трегалозо-6-фосфат-синтазы; трегалозо-6-фосфатфосфатазы и трегалазы. Модифицированные уровни трегалозо-6-фосфата могут приводить к увеличению экспрессии генов в пентозофосфатном шунтовом пути в течение стресса, вызванного засухой, в ткани узла початка растения. Дополнительно или альтернативно, модифицированные уровни трегалозо-6-фосфата могут вызывать увеличение уровня карбоангидразы в ткани узла початка растения. Модифицированный полипептид может быть экспрессирован в репродуктивных тканях растения, включая, ткань колоска, узел початка, ткань прицветника, ткань меристемы колоска, ткань цветоножки соцветия и незрелую ткань цветка. Модифицированный полипептид модифицированного фермента трегалозного пути может иметь сниженную ферментативную активность и/или вызывать in vitro активацию трегалозо-6-фосфатфосфатазы дикого типа.
[009] Эти и другие признаки, цели и преимущества настоящего изобретения станут более понятными из последующего описания. В описании приведена ссылка на прилагаемые последовательности, являющися его частью и в которых показаны с целью иллюстрации, но не ограничения, варианты осуществления настоящего изобретения. Описание предпочтительных вариантов осуществления не предусмотрено для ограничения настоящего изобретения, чтобы охватить все модификации, эквиваленты и альтернативы. Следовательно, необходимо привести ссылку на варианты осуществления, перечисленные в данном документе для интерпретации объема настоящего изобретения.
КРАТКОЕ ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ В СПИСКЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
nt = нуклеотидная последовательность
pt = белковая последовательность
SEQ ID NO: 1 трегалоза-6-фосфатфосфатаза (nt), OsT6PP-WT
SEQ ID NO: 2 трегалоза-6-фосфатфосфатаза (pt), OsT6PP-WT
SEQ ID NO: 3 трегалозо-6-фосфатфосфатаза-одинарная модификация (nt), OsT6PP-H244D
SEQ ID NO: 4 трегалоза-6-фосфатфосфатаза-одинарная модификация (pt), OsT6PP-H244D
SEQ ID NO: 5 трегалозо-6-фосфатфосфатаза-двойная модификация a (nt), OsT6PP-I129F
SEQ ID NO: 6 трегалоза-6-фосфатфосфатаза-двойная модификация a (pt), OsT6PP-I129F
SEQ ID NO: 7 трегалозо-6-фосфатфосфатаза-тройная модификация b (nt)
SEQ ID NO: 8 трегалоза-6-фосфатфосфатаза-тройная модификация b (pt)
SEQ ID NO: 9 консенсусная последовательность трегалозо-6-фосфатфосфатазы
SEQ ID NO: 10 фосфатазный домен трегалозо-6-фосфатфосфатазы
SEQ ID NO: 11 бокс А-фосфатазы трегалозо-6-фосфатфосфатазы
SEQ ID NO: 12 бокс В-фосфатазы трегалозо-6-фосфатфосфатазы
SEQ ID NO: 13 промотор OsMADS6
SEQ ID NO: 14 трегалоза-6-фосфатфосфатаза Arabidopsis thaliana 19925
SEQ ID NO: 15 трегалоза-6-фосфатфосфатаза Arabidopsis thaliana 19926
SEQ ID NO: 16 трегалоза-6-фосфатфосфатаза Oryza sativa 19924
SEQ ID NO: 17 подобная трегалозо-6-фосфатфосфатазе Thermoplasma acidophilum (Q9HIW7)
ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[010] На фиг. 1А-1С показано исходное выравнивание последовательностей Т6РР из Arabidopsis thaliana (19925; SEQ ID NO: 14), Arabidopsis thaliana (19926; SEQ ID NO: 15), Oryza sativa (19924; SEQ ID NO: 16), Oryza sativa (15777; OsT6PP-WT дикого типа; SEQ ID NO: 2) и Thermoplasma acidophilum (Q9HIW7; SEQ ID NO: 17). Последовательности выравнивали с помощью Vector NTI с применением способа ClustalW.
[011] На фиг. 2 показано филогенетическое дерево, полученное от выравнивания множественных белков Т6РР.
[012] На фиг. 3 показано описание трегалозного пути. Трегалозный путь состоит из двух ферментов биосинтеза, трегалозо-6-фосфатсинтазы (TPS) и трегалозо-6-фосфатфосфатазы (Т6РР), и одного гидролизирующего фермента, трегалазы, вовлеченного в распад.
[013] На фиг. 4 показан трегалозный путь с дополнительными последующими продуктами. UDPG получают напрямую посредством сахарозосинтазы; G6P получают из глюкозы посредством инвертазы и гексокиназы или посредством фруктозы и фруктозо-6-фосфата (F6P) через сахарозосинтазу, фруктокиназу и фосфоглюкозоизомеразу. UDPG и G6P также представляют собой два центральных активированных предшественника, из которых в конечном счете могут быть получены многие клеточные функциональные соединения. UDPG представляет собой предшественник для основных полисахаридов клеточной стенки и для гликолипидов. G6P представляет собой предшественник для синтеза крахмала, NADPH путем окислительной части пентозофосфатного пути и ATP путем гликолиза, цикла Кребса и окислительного фосфорилирования. Кроме того, G6P может быть превращен во фруктозо-6-фосфат и вместе с UDPG может применяться для синтеза сахарозо-6-фосфата и затем сахарозы. Будучи произведенным из UDPG и G6P, Т6Р находится на пересечениях основных углеводных потоках в растениях.
[014] На фиг. 5 описано влияние связывания Т6Р с SnRK1. SnRK1 представляет собой гетеротримерный белок и представляет собой растительный гомолог AMP-активированной протеинкиназы животных и дрожжевой неферментирующей сахарозу протеинкиназы (SnF1).
[015] На фиг. 6 показано вовлечение SnRK1 в многочисленные метаболические пути в растениях.
[016] На фиг. 7 показано ленточное графическое изображение гомологичной модели Т6РР риса. Фермент содержит два домена: домен α/β-гидролазы и меньший "lid-домен". Активный сайт лежит ан верхней части домена α/β-гидролазы (dot); углубление для связывания с субстратом находится на границе между двумя доменами. Два домена соединены гибким линкером, позволяющим lid-домену открываться и закрываться в ходе катализа. Поскольку матрица для гомологичного моделирования, бактериальный T6PP-связанный белок (код PDB 1U02), является апоферментом с пустым активным сайтом, вероятно, что относительная ориентация lid-домена и гидролазного домена слегка отличается в настоящем фермент-субстратном комплексе.
[017] На фиг. 8 показано расположение 6-фосфатной группы Т6Р. Один из атомов кислорода 6-фосфатной группы расположен в экваториальном положении в координационной сфере Mg2+. Asp121, принимающий фосфатную группу в ходе катализа для образования ковалентного промежуточного соединения, принимает аксиальное положение. Lys289 напрямую связан водородной связью с фосфатной группой и His181 находится в непосредственной близости. Оба остатка оптимально расположены для стабилизации образующегося отрицательного заряда на фосфатном фрагменте в ходе катализа.
[018] На фиг. 9 показаны способные к вращению связи в субстрате Т6Р. В то время как положение и ориентация пиранозного кольца, ближайшего к 6-фосфатной группе, точно определены с помощью зафиксированной фосфатной группы, существует значительная гибкость между первым и вторым кольцами вследствие присутствия двух способных к вращению гликозидных связей (стрелки). Диапазон возможных положений для второго кольца был получен путем вращения двух связей. Конформации, в которых второе кольцо нежелательным образом накладывается на белок, выбраковывали.
[019] На фиг. 10 показана консенсусная последовательность для Т6РР (SEQ ID NO: 9). Остатки, выделенные жирным шрифтом и подчеркнутые, определяют высоко консервативные участки. Отмеченные "X" положения в консенсусной последовательности указывают на участки изменчивости, где любая другая отдельная буква относится к общепринятому однобуквенному обозначению аминокислот, применяемому в настоящей области техники. Подчеркнутые аминокислотные остатки указывают на участки, которые могут быть модифицированы для конструирования полипептидов Т6РР, обеспечивающих увеличенную урожайность и/или увеличенную стрессоустойчивость у трансгенного растения. Остатки, обозначенные DYDGTLSPIV, кодируют бокс В-фосфатазы.
[020] На фиг. 11A-11C показано окончательное выравнивание последовательностей Т6РР для Arabidopsis thaliana (19925; SEQ ID NO: 14), Arabidopsis thaliana (19926; SEQ ID NO: 15), Oryza sativa (19924; SEQ ID NO: 16), и Oryza sativa (15777; OsT6PP-WT дикого типа; SEQ ID NO: 2) и родственного T6PP белка из Thermoplasma acidophilum (Q9HIW7; SEQ ID NO: 17).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[021] Практическое осуществление настоящего изобретения будет использовать, если не указано иное, общепринятые техники ботаники, микробиологии, тканевой культуры, молекулярной биологии, химии, биохимии, генетики количественных признаков растений, статистики и рекомбинантной ДНК-технологии, находящиеся в пределах навыков специалистов в настоящей области техники. Такие техники объясняются в полном объеме в литературе. Смотрите, например, Langenheim and Thimann, (1982) Botany: Plant Biology and Its Relation to Human Affairs, John Wiley; Cell Culture and Somatic Cell Genetics of Plants, vol. 1, Vasil, ed. (1984); Stanier, et al., (1986) The Microbial World, 5th ed., Prentice-Hall; Dhringra and Sinclair, (1985) Basic Plant Pathology Methods, CRC Press; Maniatis, et al., (1982) Molecular Cloning: A Laboratory Manual; DNA Cloning, vols. I and II, Glover, ed. (1985); Oligonucleotide Synthesis, Gait, ed. (1984); Nucleic Acid Hybridization, Hames and Higgins, eds. (1984); и серию Methods in Enzymology, Colowick and Kaplan, eds, Academic Press, Inc., San Diego, Calif.
[022] Единицы, приставки и символы могут быть указаны в их приемлемой для системы SI форме. Если не указано иное, нуклеиновые кислоты написаны слева направо в ориентации 5ʹ-3ʹ; аминокислотные последовательности написаны слева направо в ориентации от амино- к карбокси-концу, соответственно. Числовые диапазоны включают в себя числа, определенные в диапазоне. Аминокислоты в данном документе могут быть обозначены либо по их общеизвестным трехбуквенным символам, либо по однобуквенным символам, рекомендованным Комиссией по биохимической номенклатуре IUPAC-IUB. Аналогичным образом, нуклеотиды могут быть обозначены по их общепринятым однобуквенным кодам. Выражения, определенные ниже, более полно определены посредством ссылки на описание изобретения в целом.
[023] Если не определено иное, все технические и научные термины, применяемые в данном документе, имеют такое же значение, которое обычно понимается специалистом в настоящей области техники, к которой принадлежит настоящее изобретение.
[024] Следует понимать, что настоящее изобретение не ограничено конкретной методологией, протоколами, клеточными линиями, видами или родами растений, конструктами и реагентами, описанными по существу. Следует также понимать, что терминология, применяемая в данном документе, представлена исключительно с целью описания конкретных вариантов осуществления, и не предусмотрена для ограничения объема настоящего изобретения.
[025] Применяемые в данном документе формы единственного числа включают ссылку на формы множественного числа, если иное ясно не продиктовано контекстом. Таким образом, например, ссылка на "вектор" представляет собой ссылку на один или несколько векторов и включает их эквиваленты, известные специалистам в настоящей области техники.
[026] Выражение "около" используется в данном документе для обозначения приблизительно, грубо, примерно или в районе. Когда выражение "около" используется в связи с числовым диапазоном, оно модифицирует этот диапазон путем расширения границ выше и ниже указанных числовых значений. В общем, выражение "около" используется в данном документе для модификации числового значения выше и ниже указанного значения с помощью отклонения на 20 процентов.
[027] Применяемое в данном документе слово "или" означает любого одного представителя конкретного перечня, а также включает любую комбинацию представителей из данного перечня.
[028] Выражения "содержит", "содержащий", "включает", "включающий", "имеющий" и родственные им по значению слова означают "включая, но не ограничиваясь следующим". Выражение "состоящий из" означает "включая и ограничиваясь следующим".
[029] Выражение "состоящий, по сути, из" означает, что композиция, способ или структура могут включать дополнительные ингредиенты, этапы и/или части, но только если дополнительные ингредиенты, этапы и/или части не изменяют существенно основные и новые характеристики заявленной композиции, способа или структуры.
[030] Во всех случаях, когда указывается числовой диапазон в данном документе, это означает, что он включает любое перечисленное число (дробное или целое) в пределах указанного диапазона. Фразы "в диапазоне/диапазоны между" первым указанным числом и вторым указанным числом и "в диапазоне/диапазонах от" первого указанного числа "до" второго указанного числа применяются в данном документе взаимозаменяемо и означают включение первого и второго указанных чисел и всех дробных и целых чисел между ними. Применяемое в данном документе выражение "способ" относится к образам действия, средствам, техникам и процедурам для осуществления данной задачи, включая, но без ограничения, такие образы действия, средства, техники и процедуры, или известные, или легко разрабатываемые на основе известных образов действия, средств, техник и процедур специалистами в химической, фармакологической, биологической, биохимической и медицинской областях техники. Следует понимать, что определенные признаки настоящего изобретения, описанные для ясности в контексте отдельных вариантов осуществления, могут также быть представлены в комбинации в одном варианте осуществления. Напротив, различные признаки настоящего изобретения, описанные для краткости в контексте одного варианта осуществления, могут также быть представлены отдельно или в любой подходящей подкомбинации или в форме, подходящей в любом другом описанном варианте осуществления настоящего изобретения. Определенные признаки, описанные в контексте различных вариантов осуществления, не должны рассматриваться как существенные признаки этих вариантов осуществления, если только вариант осуществления не является неработоспособным без этих элементов.
[031] Под выражением "микроб" понимают любой микроорганизм (включая как эукариотические, так и прокариотические микроорганизмы), такие как грибы, дрожжи, бактерии, актиномицеты, водоросли и простейшие, а также другие одноклеточные структуры.
[032] Под выражением "амплифицированный" понимают конструкцию множественных копий последовательности нуклеиновой кислоты или множественных копий, комплементарных последовательности нуклеиновой кислоты, применяя по меньшей мере одну из последовательностей нуклеиновых кислот в качестве матрицы. Системы амплификации включают систему полимеразной цепной реакции (ПЦР), систему лигазной цепной реакции (ЛЦР), основанную на последовательности нуклеиновых кислот амплификацию (NASBA, Cangene, Миссиссога, Онтарио), системы О-бета-репликазы, основанную на транскрипции систему амплификации (TAS) и амплификацию с перемещением цепи (SDA). Смотрите, например, Diagnostic Molecular Microbiology: Principles and Applications, Persing, et al., eds., American Society for Microbiology, Washington, D.C. (1993). Продукт амплификации имеет название ампликон.
[033] Выражение "консервативно модифицированные варианты" применяется как в отношении аминокислотных последовательностей, так и в отношении последовательностей нуклеиновых кислот. В отношении конкретных последовательностей нуклеиновых кислот, консервативно модифицированные варианты относятся к тем нуклеиновым кислотам, которые кодируют идентичные или консервативно модифицированные варианты аминокислотных последовательностей. Вследствие вырожденности генетического кода большое число функционально идентичных нуклеиновых кислот кодируют любой данный белок. Например, все кодоны GCA, GCC, GCG и GCU кодируют аминокислоту аланин. Таким образом, в любом положении, где аланин определяется кодоном, кодон может быть изменен на любой из соответствующих описанных кодонов без изменения кодируемого полипептида. Такие вариации нуклеиновых кислот представляют собой "молчащие вариации" и представляют один вид консервативно модифицированной вариации. Каждая последовательность нуклеиновой кислоты в данном документе, кодирующая полипептид, также описывает каждую возможную молчащую вариацию нуклеиновой кислоты. Специалисту в настоящей области техники будет понятно, что каждый кодон в нуклеиновой кислоте (за исключением AUG, который, как правило, является единственным кодоном для метионина; одно исключение представляет собой Micrococcus rubens, для которого GTG представляет собой метиониновый кодон (Ishizuka, et al., (1993) J. Gen. Microbiol. 139: 425-32) может быть модифицирован для получения функционально идентичной молекулы. Соответственно, каждая молчащая вариация нуклеиновой кислоты, кодирующая полипептид согласно настоящему изобретению, подразумевается в каждой описанной полипептидной последовательности и включена в данный документ посредством ссылки.
[034] Выражение "контрольное растение" или "контроль", применяемое в данном документе, может представлять собой нетрансгенное растение родительской линии, применяемой для создания трансгенного растения в данном документе. Контрольное растение может в некоторых случаях быть трансгенной растительной линией, включающей пустой вектор или маркерный ген, но не содержащей рекомбинантный полинуклеотид согласно настоящему изобретению, который экспрессируется в трансгенном растении, подлежащем оцениванию. Контрольное растение в других случаях представляет собой трансгенное растение, экспрессирующее ген с конститутивным промотором. Как правило, контрольное растение представляет собой растение той же линии или сорта, что и трансгенное растение, подлежащее исследованию, не имеющее специфической обеспечивающей признак рекомбинантной ДНК, характеризующей трансгенное растение. Такое растение-предшественник, не содержащее такой специфической обеспечивающей признак рекомбинантной ДНК, может представлять собой естественное растение, растение дикого типа, элитное, нетрансгенное растение или трансгенное растение без специфической обеспечивающей признак рекомбинантной ДНК, характеризующей трансгенное растение. Растение-предшественник, не содержащее специфическую, обеспечивающую признак рекомбинантную ДНК, может представлять собой сибс трансгенного растения, имеющий специфическую, обеспечивающую признак рекомбинантную ДНК. Такое растение-сибс, являющееся предшественником, может включать другую рекомбинантную ДНК.
[035] Как и в отношении аминокислотных последовательностей, специалисту в настоящей области техники понятно, что отдельные замены, делеции или вставки в последовательность нуклеиновой кислоты, пептид, полипептид или белок, которые изменяют, добавляют или удаляют одну аминокислоту или небольшой процент аминокислот в кодируемой последовательности, представляет собой "консервативно модифицированный вариант", когда изменение приводит к замещению аминокислоты химически сходной аминокислотой. Таким образом, любое число аминокислотных остатков, выбранное из группы целых чисел, состоящей из 1-15, может быть изменено таким образом. Таким образом, может быть произведено, например, 1, 2, 3, 4, 5, 7 или 10 изменений. Консервативно модифицированные варианты типично обеспечивают биологическую активность, аналогичную таковой у немодифицированной полипептидной последовательности, из которой они произошли. Например, специфичность к субстрату, активность фермента или связывание лиганда с рецептором, как правило, составляет по меньшей мере 30%, 40%, 50%, 60%, 70%, 80% или 90%, предпочтительно 60-90% нативного белка для его нативного субстрата. Таблицы консервативных замен, обеспечивающих функционально сходные аминокислоты, являются хорошо известными в настоящей области техники.
[036] Каждая из следующих шести групп содержит аминокислоты, представляющие собой консервативные замены друг для друга:
1) Аланин (А), Серин (S), Треонин (Т);
2) Аспарагиновая кислота (D), Глутаминовая кислота (Е);
3) Аспарагин (N), Глутамин (Q);
4) Аргинин (R), Лизин (K);
5) Изолейцин (I), Лейцин (L), Метионин (М), Валин (V) и
6) Фенилаланин (F), Тирозин (Y), Триптофан (W).
7) См. также Creighton, Proteins, W.H. Freeman and Co. (1984).
[037] Применяемые в данном документе выражения "модифицированный" или "модификация" взаимозаменяемо относятся к преднамеренным или случайным заменам, делениям или вставкам в последовательность нуклеиновой кислоты, пептида, полипептида или белка, изменяющую, добавляющую или удаляющую по меньшей мере один аминокислотный остаток в пределах данного полипептида. Выражение "модифицированная Т6РР", применяемое в данном документе, относится к любой нуклеиновой кислоте, кодирующей Т6РР или пептиды, полипептиды или белок, имеющий активность Т6РР, любой из которых модифицируют так, чтобы полученная Т6РР обеспечивала модифицированную активность Т6РР и/или модифицированное связывание с Т6Р, приводя к улучшенному урожаю и/или устойчивости к абиотическому стрессу у растения по сравнению с немодифицированной Т6РР.
[038] Применяемое в данном документе выражение "гомологичное положение" относится к положению одной или нескольких аминокислот в полипептиде или одной или нескольких пар оснований в полинуклеотидной последовательности, которые находятся в аналогичном или эквивалентном положении во втором полипептиде или полинуклеотиде, который является ортологом, паралогом или гомологом исходной последовательности. Положение аминокислот может находиться в одинаковом функциональном участке двух белков, но может не являться точным численным положением аминокислоты между двумя полипептидными последовательностями. Гомологичное положение аминокислот на двух белках может быть определено с помощью нескольких способов, хорошо известных в настоящей области техники, включая, например, выравнивание последовательностей (например, BLAST), трехмерное моделирование белков (смотрите, например, Sander С. and Scheider R., (1991) PROTEINS: Structure, Function and Genetics 9: 56-68) и подобное.
[039] Под выражением "кодирующий" или "кодируемый" в отношении определенной нуклеиновой кислоты, понимают содержащий информацию для трансляции в определенный белок. Нуклеиновая кислота, кодирующая белок, может содержать нетранслируемые последовательности (например, интроны) в пределах транслируемых участков нуклеиновой кислоты или может не содержать такие промежуточные нетранслируемые последовательности (например, как в кДНК). Информацию, с помощью которой кодируется белок, определяют путем использования кодонов. Типично, аминокислотная последовательность кодируется нуклеиновой кислотой, применяя "универсальный" генетический код. Тем не менее, варианты универсального кода, такие как присутствуют в некоторых растительных, животных и грибковых митохондриях, бактерии Mycoplasma capricolumn (Yamao, et al., (1985) Proc. Natl. Acad. Sci. USA 82: 2306-9) или инфузории Macronucleus, могут применяться, когда нуклеиновая кислота экспрессируется с применением этих организмов.
[040] Когда нуклеиновую кислоту получают или изменяют синтетически, можно использовать преимущество от использования известных предпочтительных кодонов предусмотренного хозяина, в котором будет экспрессироваться нуклеиновая кислота. Например, хотя последовательности нуклеиновых кислот согласно настоящему изобретению могут экспрессироваться как в однодольных, так и двудольных видах растений, последовательности могут быть модифицированы так, чтобы учитывать предпочтения в отношении специфических кодонов и предпочтения в отношении содержание GC для однодольных растений или двудольных растений, поскольку было показано, что эти предпочтения, как было показано, отличаются (Murray, et al., (1989) Nucleic Acids Res. 17: 477-98 и в данный документ включена посредством ссылки). Таким образом, предпочтительный кодон маиса для конкретной аминокислоты может быть получен от известных генных последовательностей из маиса. Частота использования кодона маиса для 28 генов из растений маиса перечислена в Таблице 4 Murray, et al., выше.
[041] Применяемое в данном документе выражение "гетерологичный" в отношении нуклеиновой кислоты представляет собой нуклеиновую кислоту, которая происходит из чужеродного вида, или, если она происходит из того же вида, то существенно модифицирована по сравнению с ее нативной формой в композиции и/или геномном локусе путем преднамеренного вмешательства человека. Например, промотор, функционально связанный с гетерологичным структурным геном происходит из вида, отличного от того, из которого был получен структурный ген или, если он происходит из того же вида, то один или оба являются существенно модифицированными по сравнению с их исходной формой. Гетерологичный белок может происходить их чужеродного вида или, если он происходит из того же вида, то он существенно модифицирован по сравнению с его исходной формой путем преднамеренного вмешательства человека.
[042] Под "клеткой хозяина" понимают клетку, которая содержит гетерологичную последовательность нуклеиновой кислоты настоящего изобретения, которая содержит вектор и поддерживает репликацию и/или экспрессию вектора экспрессии. Клетки хозяина могут представлять собой прокариотические клетки, такие как Е.coli, или эукариотические клетки, такие как клетки дрожжей, насекомых, растений, амфибий или млекопитающих. Предпочтительно, клетки хозяина представляют собой клетки однодольных или двудольных растений, включая, но без ограничения, маис, сорго, подсолнечник, соя, пшеницу, люцерну, рис, хлопок, канолу, ячмень, просо и помидор. Особенно предпочтительная однодольная клетка хозяина представляет собой клетку хозяина маиса.
[043] Выражение "комплекс гибридизации" включает ссылку на структуру двойной спирали нуклеиновых кислот, образованной двумя одноцепочечными последовательностями нуклеиновых кислот, селективно гибридизированных друг с другом.
[044] Выражение "введенный" в контексте введения нуклеиновой кислоты в клетку, с помощью любого средства, такого как "трансфекция", "трансформация" или "трансдукция" и включает ссылку на встраивание нуклеиновой кислоты в эукариотическую или прокариотическую клетку, где нуклеиновая кислота может быть встроена в геном клетки (например, хромосомную, плазмидную, пластидную или митохондриальную ДНК), превращена в автономный репликон, как часть минихромосомы или временно экспрессирована (например, трансфицированная мРНК).
[045] Применяемое в данном документе выражение "пакет генов" относится к введению двух или более генов в геном организма. В определенных аспектах настоящего изобретения может быть желательным подвергнуть стэкингу любой ген абиотического стресса (например, белки холодового шока, гены, связанные с реакцией ABA) с Т6РР, описанной в данном документе. Аналогично, также может быть желательным подвергнуть стэкингу гены трегалозного пути, описанные в данном документе, с генами, обеспечивающими устойчивость к воздействию насекомых, устойчивость к болезням, увеличенную урожайность или любой другой благоприятный признак (например, увеличенная высота растения и т.д.), известный в настоящей области техники. Альтернативно, трансгенные растения, содержащие модифицированный ген трегалозного пути, могут быть подвергнуты стэкингу с аллелями нативного признака, которые обеспечивают дополнительные признаки, такие как, улучшенное использование воды, увеличенную устойчивость к болезням и подобное. В одном варианте осуществления растения, экспрессирующие модифицированные гены трегалозного пути, подвергаются стэкингу с аллелями, описанными в WO 2011/079277. Признаки могут быть подвергнуты стэкингу путем введения кассет экспрессии с множественными генами или выведения/скрещивания растений с одним или несколькими признаками с другими растениями, содержащими один или несколько дополнительных признаков.
[046] Выражения "выделенный" относится к материалу, такому как нуклеиновая кислота или белок, который существенно или по сути не содержит компоненты, обычно сопровождающих его или взаимодействующих с ним, как это встречается в его естественном окружении. Выделенный материал необязательно содержит материал, не встречающийся вместе с материалом в его естественном окружении. Нуклеиновые кислоты, являющиеся "выделенными", как определено в данном документе, также имеют название "гетерологичные" нуклеиновые кислоты. Если иное не заявлено, то выражение "NUE нуклеиновая кислота" означает нуклеиновую кислоту, содержащую полинуклеотид ("NUE полинуклеотид"), кодирующий NUE полипептид полной или частичной длины.
[047] Применяемая в данном документе "нуклеиновая кислота" включает ссылку на дезоксирибонуклеотидный или рибонуклеотидный полимер в форме одинарной или двойной цепи и, если нет иных ограничений, включает известные аналоги, имеющие основную природу естественных нуклеотидов в том отношении, что они гибридизируются с одноцепочечными нуклеиновыми кислотами способом, аналогичным таковому у встречающихся в природе нуклеотидов (например, пептидные нуклеиновые кислоты).
[048] Под выражением "библиотека нуклеиновых кислот" понимают коллекцию выделенных молекул ДНК или РНК, которые содержат в одном случае основное представление всей транскрибируемой фракции генома определенного организма. Конструкция иллюстративных библиотек нуклеиновых кислот, таких как геномные и кДНК библиотеки, представлена в стандартных ссылках по молекулярной биологии, таких как Berger and Kimmel, (1987) Guide To Molecular Cloning Techniques, из серии Methods in Enzymology, vol. 152, Academic Press, Inc., San Diego, Calif.; Sambrook, et al., (1989) Molecular Cloning: A Laboratory Manual, 2nd ed., vols. 1-3; и Current Protocols in Molecular Biology, Ausubel, et al., eds, Current Protocols, a joint venture between Greene Publishing Associates, Inc. and John Wiley & Sons, Inc. (1994 Supplement). В другом случае выражение "библиотека нуклеиновых кислот", как определено в данном документе, может также подразумевать представление библиотек, содержащих установленную фракцию или скорее не существенно представляющие весь геном определенного организма. Например, малые РНК, мРНК и метилированная ДНК. Библиотека нуклеиновых кислот, определенная в данном документе, может также включать варианты конкретной молекулы (например, коллекцию вариантов для конкретного белка).
[049] Применяемое в данном документе выражение "функционально связанный" включает ссылку на функциональную связь между первой последовательностью, такой как промотор, и второй последовательностью, где промоторная последовательность инициирует и опосредует транскрипцию ДНК, соответствующей второй последовательности. Обычно "функционально связанный" означает, что последовательности связанных нуклеиновых кислот являются смежными и, если есть необходимость соединить две области, кодирующие белок, они являются смежными и находятся в одной и той же рамке считывания.
[050] Применяемое в данном документе выражение "растение" включает ссылку на целые растения, органы растения (например, листья, стебли, корни и т.д.), семена и клетки растения и их потомство. Растительная клетка, применяемая в данном документе включает без ограничения семена, суспензионные культуры, зародыши, меристематические участки, каллюсную ткань, листья, корни, побеги, гаметофиты, спорофиты, пыльцу и микроспоры. Класс растений, которые могут применяться в способах настоящего изобретения, является, как правило, настолько широким, как класс высших растений, поддающихся трансформационным техникам, включая как однодольные и двудольные растения, включая виды из родов: Cucurbita, Rosa, Vitis, Juglans, Fragaria, Lotus, Medicago, Onobrychis, Trifolium, Trigonella, Vigna, Citrus, Linum, Geranium, Manihot, Daucus, Arabidopsis, Brassica, Raphanus, Sinapis, Atropa, Capsicum, Datura, Hyoscyamus, Lycopersicon, Nicotiana, Solanum, Petunia, Digitalis, Majorana, Ciahorium, Helianthus, Lactuca, Bromus, Asparagus, Antirrhinum, Heterocallis, Nemesis, Pelargonium, Panieum, Pennisetum, Ranunculus, Senecio, Salpiglossis, Cucumis, Browaalia, Glycine, Pisum, Phaseolus, Lolium, Oryza, Avena, Hordeum, Secale, Allium и Triticum. Особенно предпочтительным растением является Zea mays.
[051] Применяемое в данном документе выражение "урожайность" может включать ссылку на бушели на акр зерновой культуры при уборке, с поправкой на влажность зерна (15% типично для маиса, например), и объем образованной биомассы (для кормовых сельскохозяйственных культур, таких как люцерна и размер корня растения для сельскохозяйственных культур, дающих несколько урожаев в год). Влажность зерна измеряют в зерне при уборке. Скорректированный натурный вес зерна определяют как вес в фунтах на бушель, с поправкой на уровень влажности зерна при уборке. Биомассу измеряют как вес образованного пригодного для уборки растительного материала. На урожайность могут влиять многие свойства, включая без ограничения высоту растения, число стручков, положение стручка на растении, число междоузлий, частота растрескивания стручков, размер зерна, эффективность образования клубеньков и фиксации азота, эффективность ассимиляции питательных веществ, ассимиляции углерода, архитектуру растения, процент прорастание семян, мощность всходов и ювенильные признаки. На урожайность также может влиять эффективность прорастания (включая прорастание в неблагоприятных внешних условиях), скорость роста (включая скорость роста в неблагоприятных внешних условиях), число початков, число семян на початок, размер семени, состав семени (крахмал, масло, белок) и характеристики налива зерна. Урожайность растения может быть измерена различными образами, включая натурный вес, количество семян на растение, вес семян, количество семян на единицу площади (т.е. семена, или вес семян, на акр), бушели на акр, тонны на акр или килограммы на гектар. Например, урожайность кукурузы может быть измерена в виде продукции обмолоченных кукурузных зерен на единицу площади продукции, например, в бушелях на акр или метрических тоннах на гектар, часто сообщается на основании коррекции по влажности, например, при 15,5 процентах влажности. Более того, бушель кукурузы определяется по закону штата Айова как 56 фунтов по весу, полезный коэффициент перевода для урожая кукурузы представляет собой: 100 бушелей на акр эквивалентны 6,272 метрическим тоннам на гектар. Другие измерения для урожая являются общепринятой практикой в настоящей области техники. В определенных вариантах осуществления настоящего изобретения урожайность может быть увеличена в неблагоприятных и/или не являющихся неблагоприятными внешних условиях.
[052] Применяемое в данном документе выражение "полинуклеотид" включает ссылку на дезоксирибополинуклеотид, рибополинуклеотид или их аналоги, которые имеют основную природу естественного рибонуклеотида, заключающуюся в том, что они гибридизируются, при жестких условиях гибридизации, с, по существу, такой же нуклеотидной последовательностью, что и встречающиеся в природе нуклеотиды и/или обеспечивают трансляцию в такую(ие) же аминокислоту(ы), что и встречающийся(еся) в природе нуклеотид(ы). Полинуклеотид может быть полноразмерным или подпоследовательностью нативного или гетерологичного структурного или регуляторного гена. Если не указано иное, выражение включает ссылку на определенную последовательность, а также ее комплементарную последовательность. Таким образом, ДНК или РНК с каркасами, модифицированными для стабильности или по другим причинам, представляют собой "полинуклеотиды", как это выражение предусмотрено в данном документе. Более того, ДНК или РНК, содержащие необычные основания, такие как инозин или модифицированные основания, такие как тритилированные основания, чтобы назвать просто два примера, являются полинуклеотидами, как данное выражение используют в данном документе. Следует понимать, что большое разнообразие модификаций было произведено в отношении ДНК и РНК, которые случат для многих полезных целей, известных специалистам в настоящей области техники. Выражение "полинуклеотид", как оно применяется в данном документе, охватывает такие химически, ферментативно или метаболически модифицированные формы полинуклеотидов, а также химические формы ДНК и РНК, характерные для вирусов и клеток, включая среди прочего, простые и сложные клетки.
[053] Выражения "полипептид", "пептид" и "белок" используются взаимозаменяемо в данном документе для обозначения полимера аминокислотных остатков. Выражения применяются к аминокислотным полимерам, в которых один или несколько аминокислотных остатков представляет собой искусственный химический аналог соответствующей встречающейся в природе аминокислоты, а также встречающихся в природе аминокислота полимеров.
[054] Применяемый в данном документе "промотор" включает ссылку на участок ДНК выше начала транскрипции и вовлеченный в распознавание и связывание РНК-полимеразы и других белков для инициации транскрипции. "Растительный промотор" представляет собой промотор, способный инициировать транскрипцию в растительных клетках. Иллюстративные растительные промоторы включают без ограничения такие, которые получают из растений, растительных вирусов и бактерий, которые содержат гены, экспрессируемые в растительных клетках, такие как Agrobacterium или Rhizobium. Примеры представляют собой промоторы, которые предпочтительно инициируют транскрипцию в определенных тканях, таких как листья, корни, семена, волокна, сосуды ксилемы, трахеиды или склеренхима. Такие промоторы имеют название "тканепредпочтительные". Специфический для "типа клеток" промотор в первую очередь управляет экспрессией в определенных клеточных типах в одном или нескольких органах, например, сосудистых клетках в корнях или листьях. "Индуцируемый" или "регулируемый" промотор представляет собой промотор, которые находится под контролем окружающей среды. Примеры условий окружающей среды, которые могут воздействовать на транскрипцию с помощью индуцируемых промоторов, включают анаэробные условия или присутствие света. Другой тип промотора представляет собой регулируемый стадией роста промотор, например, промотор, который управляет экспрессией в ходе развития пыльцы. Тканепредпочтительные, специфические для типа клеток, регулируемые стадией роста и индуцируемые промоторы составляют класс "неконститутивных" промоторов. "Конститутивный" промотор представляет собой промотор, которые является активным при большинстве условий окружающей среды в большинстве клеток.
[055] Любая подходящая промоторная последовательность может использоваться конструктом нуклеиновых кислот согласно настоящему изобретению. Согласно некоторым вариантам осуществления настоящего изобретения промотор представляет собой конститутивный промотор, тканеспецифический или индуцируемый абиотическим стрессом промотор.
[056] Подходящие конститутивные промоторы включают, например, промотор 35S CaMV (SEQ ID NO: 1546; Odell et al., Nature 313: 810-812, 1985); Arabidopsis At6669 промотор (SEQ ID NO: 1652; смотрите PCT публикацию № WO 04081173 A2); Ubi 1 маиса (Christensen et al., Plant Mol. Biol. 18: 675-689, 1992); актин риса (McElroy et al., Plant Cell 2: 163-171, 1990); pEMU (Last et al., Theor. Appl. Genet. 81: 581-588, 1991); CaMV 19S (Nilsson et al., Physiol. Plant 100: 456-462, 1997); GOS2 (de Pater et al., Plant J November; 2 (6): 837-44, 1992); убиквитин (Christensen et al., Plant Mol. Biol. 18: 675-689, 1992); циклофилин риса (Bucholz et al., Plant Mol Biol. 25 (5): 837-43, 1994); Н3 гистон маиса (Lepetit et al., Mol. Gen. Genet. 231: 276-285, 1992); актин 2 (An et al., Plant J. 10 (1); 107-121, 1996), конститутивный промотор корневого кончика CT2 (SEQ ID NO: 1535; см. также PCT заявку № IL/2005/000627) и синтетический Super MAS (Ni et al., The Plant Journal 7: 661-76, 1995). Другие конститутивные промоторы включают описанные в Патентах США №№5659026, 5608149; 5608144; 5604121; 5569597: 5466785; 5399680; 5268463 и 5608142.
[057] Подходящие тканеспецифические промоторы включают, но без ограничения, специфические для листьев промоторы [такие как описанные, например, Yamamoto et al., Plant J. 12: 255-265, 1997; Kwon et al., Plant Physiol. 105: 357-67,1994; Yamamoto et al., Plant Cell Physiol. 35: 773-778, 1994; Gotor et al., Plant J. 3: 509-18, 1993; Orozco et al., Plant Mol. Biol. 23: 1129-1138, 1993; и Matsuoka et al., Proc. Natl. Acad. Sci. USA 90: 9586-9590, 1993], семяпредпочтительные промоторы [например, из семяспецифических генов (Simon, et al., Plant Mol. Biol. 5. 191, 1985; Scofield, et al., J. Biol. Chem. 262: 12202, 1987; Baszczynski, et al., Plant Mol. Biol. 14: 633, 1990), альбумин из бразильского ореха (Pearsonʹ et al., Plant Mol. Biol. 18: 235-245, 1992), легумин (Ellis, et al. Plant Mol. Biol. 10: 203-214, 1988), глютелин (рис) (Takaiwa, et al., Mol. Gen. Genet. 208: 15-22, 1986; Takaiwa, et al., FEBS Letts. 221: 43-47, 1987), зеин (Matzke et al., Plant Mol Biol, 143). 323-32 1990), napA (Stalberg, et al., Planta 199: 515-519, 1996), SPA пшеницы (Albanietal, Plant Cell, 9: 171-184, 1997), олеозин подсолнечника (Cummins, etal., Plant Mol. Biol. 19: 873-876, 1992)], специфические для эндосперма промоторы [например, LMW и HMW пшеницы, глютенин-1 (Mol Gen Genet 216: 81-90, 1989; NAR 17: 461-2), глиадины a, b и g пшеницы (EMBO3: 1409-15, 1984), промотор ltrl ячменя, гордеин B1, С, D ячменя (Theor Appl Gen 98: 1253-62, 1999; Plant J 4: 343-55, 1993; Mol Gen Genet 250: 750-60, 1996), DOF ячменя (Mena et al., The Plant Journal, 116 (1): 53-62, 1998), Biz2 (EP 99106056.7), синтетический промотор (Vicente-Carbajosa et al., Plant J. 13: 629-640, 1998), проламин NRP33 риса, глобулин Glb-1 риса (Wu et al., Plant Cell Physiology 39 (8) 885-889, 1998), альфа-глобулин REB/OHP-1 риса (Nakase et al. Plant Mol. Biol. 33: 513-S22, 1997), ADP-глюкоза PP риса (Trans Res 6: 157-68, 1997), семейство генов ESR маиса (Plant J 12: 235-46, 1997), гамма-кафирин сорго (Plant Mol. Biol 32: 1029-35, 1996)], специфические для зародышей промоторы [например, OSH1 риса (Sato et al., Proc. Nati. Acad. Sci. USA, 93: 8117-8122), KNOX (Postma-Haarsma of al, Plant Mol. Biol. 39: 257-71, 1999), олеозин риса (Wu et at, J. Biochem., 123: 386, 1998)], и специфические для цветов промоторы [например, AtPRP4, chalene-синтаза (chsA) (Van der Meer, et al., Plant Mol. Biol. 15, 95-109, 1990), LAT52 (Twell et al., Mol. Gen Genet. 217: 240-245; 1989), апетала-3; репродуктивные ткани растений [например, промоторы OsMADS (Заявка на патент США №2007/0006344)].
[058] Подходящие индуцируемые абиотическим стрессом промоторы включают, но без ограничения, индуцируемые солью промоторы, такие как RD29A (Yamaguchi-Shinozalei et al., Mol. Gen. Genet. 236: 331-340, 1993); индуцируемые засухой промоторы, такие как промотор rab17 гена маиса (Pla et. al., Plant Mol. Biol. 21: 259-266, 1993), промотор rab28 гена маиса (Busk et. al., Plant J. 11: 1285-1295, 1997) и промотор Ivr2 гена маиса (Pelleschi et. al., Plant Mol. Biol. 39: 373-380, 1999); индуцируемые тепловым воздействием промоторы, такие как тепловой hsp80-промотор помидора из помидора (Патент США №5187267).
[059] Выражение "ферментативная активность" подразумевает включение деметилирования, гидроксилирования, эпоксидирования, N-окисления, сульфоксидации, N-, S-, и О-деалкилирований, десульфатации, деаминирования и восстановление азо, нитро и N-оксидных групп. Выражение "нуклеиновая кислота" относится к дезоксирибонуклеотидному или рибонуклеотидному полимеру в одноцепочечной либо в двухцепочечной форме, или сенсовому либо антисенсовому, и если нет иных ограничений, охватывает известные аналоги естественных нуклеотидов, которые гибридизируются с нуклеиновыми кислотами способом, аналогичным таковому у встречающихся в природе нуклеотидов. Если не указано иное, конкретная последовательность нуклеиновой кислоты включает ее комплементарную последовательность.
[060] "Структурный ген" представляет собой такую часть гена, содержащую сегмент ДНК, кодирующий белок, полипептид или его часть, и исключающую 5ʹ последовательность, управляющую инициацией транскрипции. Структурный ген может альтернативно кодировать нетранслируемый продукт. Структурный ген может представлять собой ген, который в норме встречается в клетке, или ген, который в норме не встречается в клетке или месте в клетке, где он введен, в этом случае он имеет название "гетерологичный ген". Гетерологичный ген может быть получен в целом или частично из любого источника, известного в данной области техники, включая бактериальный геном или эписому, эукариотическую, ядерную или плазмидную ДНК, кДНК, вирусную ДНК или химически синтезированную ДНК. Структурный ген может содержать одну или несколько модификаций, которые могут воздействовать на биологическую активность или ее характеристики, биологическую активность или химическую структуру продукта экспрессии, скорость экспрессии или способ контроля экспрессии. Такие модификации включают без ограничения мутации, вставки, делеции и замены одного или нескольких нуклеотидов. Структурный ген может составлять непрерывную кодирующую последовательность или он может включать один или несколько интронов, ограниченных соответствующими границами сплайсинга. Структурный ген может являться транслируемым или нетранслируемым, включая ген в антисенсовой ориентации. Структурный ген может быть составной частью сегментов, полученных из множества источников и из множества генных последовательностей (встречающихся в природе или синтетических, где синтетический относится к ДНК, которая является химически синтезированной).
[061] Выражение "полученный из" используют в значении взятый, произведенный, полученный, восстановленный, реплицированный или происходящий из источника (химического и/или биологического). Производное может быть получено с помощью химической или биологической манипуляции (включая, но без ограничения, замену, добавление, вставку, делецию, экстракцию, выделение, мутацию и репликацию) исходного источника.
[062] Выражение "химически синтезированная" в отношении последовательности ДНК означает, что части составных нуклеотидов собраны in vitro. Неавтоматизированный химический синтез ДНК может быть осуществлен с применением общепринятых процедур (Caruthers, Methodology of DNA and RNA Sequencing, (1983), Weissman (ed.), Praeger Publishers, New York, Chapter 1); автоматизированный химический синтез может быть проведен с применением ряда коммерчески доступных инструментов.
[063] Применяемое в данном документе выражение "рекомбинантный" включает ссылку на клетку или вектор, который был модифицирован путем введения гетерологичной нуклеиновой кислоты или ссылку на то, что клетка получена из клетки, модифицированной таким образом. Таким образом, например, рекомбинантные клетки экспрессируют гены, которые не встречаются в идентичной форме в нативной (нерекомбинантной) форме клетки или экспрессируют нативные гены, которые иным образом аномально экспрессируются, сверхэкспрессируются или не экспрессируются вообще в результате преднамеренного вмешательства человека или могут иметь сниженную или устраненную экспрессию нативного гена. Выражение "рекомбинантный", применяемое в данном документе, не охватывает изменение клетки или вектора с помощью встречающихся в природе событий (например, спонтанной мутации, естественной трансформации/трансдукции/транспозиции), таких как события, происходящие без преднамеренного вмешательства человека.
[064] Применяемое в данном документе выражение "кассета экспрессии" представляет собой конструкт нуклеиновой кислоты, образованный рекомбинантно или синтетически, с серией определенных элементов нуклеиновых кислот, которые позволяют произойти транскрипции конкретной нуклеиновой кислоты в целевой клетке. Кассета экспрессии может быть встроена в плазмидную, хромосомную, митохондриальную ДНК, пластидную ДНК, фрагмент вируса или нуклеиновой кислоты. Типично, часть кассеты экспрессии вектора экспрессии включает среди прочего другие последовательности, нуклеиновую кислоту, подлежащую транскрипции, и промотор.
[065] Выражения "остаток" или "аминокислотный остаток" или "аминокислота" используются взаимозаменяемо в данном документе для обозначения аминокислоты, которую встраивают в белок, полипептид или пептид (обобщенно "белок"). Аминокислота может являться встречающейся в природе аминокислотой и, если нет иных ограничений, может включать известные аналоги естественных аминокислот, которые могут функционировать способом, аналогичным таковому у встречающихся в природе аминокислот.
[066] Выражение "селективно гибридизируется" включает ссылку на гибридизацию, при жестких условиях гибридизации, последовательности нуклеиновой кислоты с определенной целевой последовательностью нуклеиновой кислоты в заметно большей степени (например, по меньшей мере в 2 раза по сравнению с исходным уровнем), чем ее гибридизация с нецелевыми последовательностями нуклеиновых кислот и с существенным исключением нецелевых нуклеиновых кислот. Селективно гибридизирующиеся последовательности типично имеют около по меньшей мере 40% идентичности последовательностей, предпочтительно 60-90% идентичности последовательностей и наиболее предпочтительно 100% идентичности последовательностей (т.е. комплементарная) друг другу.
[067] Выражения "жесткие условия" или "жесткие условия гибридизации" включают ссылку на условия, при которых зонд будет гибридизироваться с его целевой последовательностью, в заметно большей степени, чем другие последовательности (например, по меньшей мере в 2 раза по сравнению с исходным уровнем). Жесткие условия зависят от последовательности и будут различаться в различных обстоятельствах. С помощью контроля жесткости условий гибридизации и/или отмывки могут быть выявлены целевые последовательности, которые могут быть до 100% комплементарными зонду (гомологичное зондирование). Альтернативно, условия жесткости могут быть отрегулированы, чтобы обеспечить некоторое несовпадение в последовательностях так, чтобы более низкие степени сходства были обнаружены (гетерологичное зондирование). Оптимально, чтобы зонд составлял приблизительно 500 нуклеотидов в длину, но его длина может значительно варьироваться от менее чем 500 нуклеотидов до равного полной длине целевой последовательности.
[068] Типично, жесткие условия будут такими, при которых концентрация соли составляет менее чем около 1,5 М концентрация ионов Na, типично около 0,01-1,0 М концентрация ионов Na (или других солей) при pH 7,0-8,3 и температура составляет по меньшей мере около 30°C для коротких зондов (например, 10-50 нуклеотидов) и по меньшей мере около 60°C для длинных зондов (например, более чем 50 нуклеотидов). Жесткие условия могут также быть достигнуты путем добавления дестабилизирующих средств, таких как формамид или раствор Денхардта. Иллюстративные условия низкой жесткости включают гибридизацию с буферным раствором 30-35% формамида, 1 М NaCl, 1% SDS (додецилсульфат натрия) при 37°C и отмывку в 1×-2×SSC (20×SSC=3,0 М NaCl/0,3 М тринатрийцитрата) при 50-55°C. Иллюстративные условия умеренной жесткости включают гибридизацию в 40-45% формамида, 1 М NaCl, 1% SDS при 37°C и отмывку в 0,5×-1×SSC при 55-60°C. Иллюстративные условия высокой жесткости включают гибридизацию в 50% формамида, 1 М NaCl, 1% SDS при 37°C и отмывку в 0,1×SSC при 60-65°C. Специфичность типично зависит от постгибридизационных отмывок, причем критическими факторами является ионная сила и температура финального раствора отмывки. Для гибридов ДНК-ДНК Tm может быть приближенно выражена из уравнения Meinkoth and Wahl, (1984) Anal. Biochem., 138: 267-84: Tm=81,5°C+16,6 (log M)+0,41 (% GC)-0,61 (% форм.)-500/L; где M представляет собой молярность моновалентных катионов, % GC представляет собой процентное отношение гуанозиновых и цитозиновых нуклеотидов в ДНК, % форм, представляет собой процентное отношение формамида в растворе гибридизации, и L представляет собой длину гибрида в парах оснований. Tm представляет собой температуру (при определенной ионной силе и pH), при которой 50% комплементарной целевой последовательности гибридизируется с абсолютно совпадающим зондом. Tm снижается на около 1°C для каждого 1% несовпадения; таким образом, Tm, условия гибридизации и/или отмывки могут быть отрегулированы для гибридизации с последовательностями необходимой идентичности. Например, если искомыми являются последовательности с >90% идентичностью, то Tm может быть снижена на 10°C. В общем, жесткие условия выбирают так, чтобы они были на около 5°C ниже, чем температурная точка плавления (Tm) для специфической последовательности и комплементарной ей последовательности при заданной ионной силе и pH. Тем не менее, условия сильной жесткости могут использовать гибридизацию и/или отмывку при температуре на 1, 2, 3 или 4°C ниже, чем температурная точка плавления (Tm); условия умеренной жесткости могут использовать гибридизацию и/или отмывку при температуре на 6, 7, 8, 9 или 10°C ниже, чем температурная точка плавления (Tm); условия низкой жесткости могут использовать гибридизацию и/или отмывку при температуре на 11, 12, 13, 14, 15 или 20°C ниже, чем температурная точка плавления (Tm). Применяя уравнение, составы для гибридизации и отмывки и необходимую Tm, специалистам будет очевидно, что вариации в жесткости гибридизации и/или растворах отмывки по существу описаны. Если необходимая степень несовпадения приводит к Tm менее чем 45°C (водный раствор) или 32°C (раствор формамида), предпочтительно увеличить концентрация SSC так, чтобы могла применяться более высокая температура. Подробное руководство по гибридизации нуклеиновых кислот можно найти в Tijssen, Laboratory Techniques in Biochemistry and Molecular Biology-Hybridization with Nucleic Acid Probes, part I, chapter 2, "Overview of principles of hybridization and the strategy of nucleic acid probe assays," Elsevier, New York (1993); и Current Protocols in Molecular Biology, chapter 2, Ausubel, et al., eds, Greene Publishing and Wiley-Interscience, New York (1995). Если не указано иное, в настоящей заявке высокую жесткость определяют как гибридизацию в 4×SSC, 5×раствора Денхардта (5 г Фиколла, 5 г поливинилпирролидона, 5 г альбумина бычьей сыворотки в 500 мл воды), 0,1 мг/мл кипяченой ДНК из молок лососевых и 25 мМ фосфата натрия при 65°C и отмывку в 0,1×SSC, 0,1% SDS при 65°C.
[069] Применяемое в данном документе выражение "трансгенное растение" включает ссылку на растение, которое содержит в его геноме гетерологичный полинуклеотид. Как правило, гетерологичный полинуклеотид стабильно интегрирован в геном так, чтобы полинуклеотид передавался последующим поколениям. Гетерологичный полинуклеотид может быть интегрирован в геном отдельно или как часть рекомбинантной кассеты экспрессии. Выражение "трансгенный" используют в данном документе для включения любой клетки, клеточной линии, каллюса, ткани, части растения или растения, генотип которого был изменен путем присутствия гетерологичной нуклеиновой кислоты, включая трансгены, изначально измененные таким образом, а также трансгены, созданные путем половых скрещиваний или бесполого размножения из исходного трансгена. Выражение "трансгенный", применяемое в данном документе, не включает изменение генома (хромосомного или внехромосомного) с помощью традиционных способов разведения растений или с помощью встречающихся в природе событий, таких как случайное перекрестное опыление, нерекомбинантная вирусная инфекция, нерекомбинантная бактериальная трансформация, нерекомбинантная транспозиция или спонтанная мутация.
[070] Применяемое в данном документе выражение "вектор" включает ссылку на нуклеиновую кислоту, применяемую в трансфекции клетки хозяина и в которую может быть введен полинуклеотид. Векторы зачастую являются репликонами. Векторы экспрессии обеспечивают транскрипцию нуклеиновой кислоты, введенной в них.
[071] Выражение "сверхэкспрессия" относится к уровню экспрессии в трансгенных организмах, который превышает уровни экспрессии в нормальных или нетрансформированных организмах.
[072] Выражение "ткань растения" включает дифференцированные и недифференцированные ткани или растения, включая, но без ограничения, корни, стебли, побеги, листья, пыльцу, семена, опухолевую ткань и различные формы клеток и культуры, такие как отдельные клетки, протопласт, зародыши и каллюсная ткань. Ткань растения может находиться в растениях или в органе, тканевой или клеточной культуре.
[073] Выражения "предпочтительная экспрессия", "преимущественная транскрипция" или "предпочтительная транскрипция" взаимозаменяемо относится к экспрессии генных продуктов, которые предпочтительно экспрессируются на более высоком уровне в одной или нескольких тканях растения (пространственное ограничение) и/или на одной или нескольких стадиях развития растения (временное ограничение), тогда как в других тканях/стадиях развития имеет место относительно низкий уровень экспрессии.
[074] Выражения "первичный трансформант" и "поколение ТО" относятся к трансгенным растениям, которые происходят из одного генетического поколения в виде ткани, которая была изначально трансформирована (т.е., не проходя через мейоз и опыление после трансформации). Выражения "вторичные трансформанты" и "поколения T1, Т2, T3 и т.д." относятся к трансгенным растениям, полученным их первичных трансформантов посредством одного или нескольких мейотических циклов и циклов опыления. Они могут быть получены путем самоопыления первичных или вторичных трансформантов или скрещиваний первичных или вторичных трансформантов с другими трансформированными или нетрансформированными растениями.
[075] Выражение "селектируемый маркерный ген" относится к гену, чья экспрессия в растительной клетке дает клетке селективное преимущество. Селективное преимущество, которым обладают клетки, трансформированные с селектируемым маркерным геном, может быть обусловлено их способностью к росту в присутствии отрицательного селективного средства, такого как антибиотик или гербицид, по сравнению со способностью к росту нетрансформированных клеток. Селективное преимущество, которым обладают трансформированные клетки, может также быть обусловлено их усиленной способность, относительно нетрансформированных клеток, использовать добавленные соединения в качестве питательного вещества, фактора роста или источника энергии. Селективное преимущество, которым обладает трансформированная клетка, может также быть обусловлено потерей гена, который имелся ранее, в так называемой "отрицательной селекции". При этом добавляют соединение, токсичное только для клеток, которые не потеряли специфический ген (отрицательный селектируемый маркерный ген), присутствующий в родительской клетке (типично, в трансгене).
[076] Выражение "трансформация" относится к переносу фрагмента нуклеиновой кислоты в геном клетки хозяина, приводя к генетически стабильному наследованию. Выражение "временно трансформированные" относится к клеткам, в которых были введены трансгены и чужеродная ДНК (например, с помощью таких способов, как опосредованная Agrobacterium трансформация или биолистическая бомбардировка), но не отобранные в отношении стабильного поддержания. Выражение "стабильно трансформированные" относится к клеткам, которые были отобраны и регенерированы на селекционных средах после трансформации.
[077] Выражения "трансформированный /трансгенный/ рекомбинантный" относятся к организму хозяина, такому как бактерия или растение, в который была введена гетерологичная молекула нуклеиновой кислоты. Молекула нуклеиновой кислоты может быть стабильно интегрирована в геном хозяина или молекула нуклеиновой кислоты может также присутствовать как внехромосомная молекула. Такая внехромосомная молекула может являться самореплицирующейся. Предусматривается, что трансформированные клетки, ткани или растения включают не только конечный продукт процесса трансформации, но также его трансгенное потомство, "Нетрансформированный", "нетрансгенный" или "нерекомбинантный" хозяин относится к организму дикого типа, например, бактерии или растению, которое не содержит гетерологичную молекулу нуклеиновой кислоты.
[078] Выражение "трансляционная энхансерная последовательность" относится к такой части последовательности ДНК гена между промотором и кодирующей последовательностью, которая транскрибируется в РНК и присутствует в полностью процессированной мРНК против хода транскрипции (5ʹ) трансляционного старт-кодона. Трансляционная энхансерная последовательность может воздействовать на процессинг первичного транскрипта в мРНК, стабильность мРНК или эффективность трансляции. "Видимый маркер" относится к гену, чья экспрессия не обеспечивает преимущество трансформированной клетке, но может сделать ее обнаруживаемой или видимой. Примеры видимых маркеров включают без ограничения β-глюкуронидазу (GUS), люциферазу (LUC) и зеленый флюоресцентный белок (GFP).
[079] "Дикий тип" относится к нормальному гену, вирусу или организму, встречающемуся в природе без какой-либо мутации или модификации.
[080] Применяемые в данном документе, "растительный материал," "часть растения" или "ткань растения" означает растительные клетки, протопласты растений, тканевые культуры клеток растений, из которых могут быть регенерированы растения, каллюсы растений, скопления растений и растительные клетки, которые являются интактными в растениях или частях растений, таких как зародыши, пыльца, семязачатки, семена, листья, цветы, ветви, плоды, зерна, початки, стержни кукурузных початков, оболочки, стебли, корни, корневые кончики, пыльники, клубни, ризомы и подобное.
[081] Применяемое в данном документе выражение "белковый экстракт" относится к частичному или общему белку, экстрагированному из части растения. Способы экстракции растительного белка хорошо известны в настоящей области техники.
[082] Применяемое в данном документе выражение "образец растения" относится или к интактной, или не интактной (например, молотая ткань семени или растения, рубленая ткань растения, лиофилизированная ткань) ткани растения. Это также может быть экстракт, содержащий интактную или не интактную ткань семени или растения.
[083] Следующие выражения используют для описания взаимоотношений последовательностей между двумя или более нуклеиновыми кислотами или полинуклеотидами или полипептидами: (а) "эталонная последовательность", (b) "окно сравнения", (с) "идентичность последовательностей", (d) "процентное отношение идентичности последовательностей" и (е) "существенная идентичность."
[084] Применяемое в данном документе выражение "эталонная последовательность" представляет собой заданную последовательность, применяемую в качестве основы для сравнения последовательностей. Эталонная последовательность может представлять собой разновидность или целостность определенной последовательности; например, в виде сегмента полноразмерной кДНК или генной последовательности или полной кДНК или генной последовательности.
[085] Применяемое в данном документе выражение "окно сравнения" подразумевает включение ссылки на смежный и определенный сегмент полинуклеотидной последовательности, где полинуклеотидная последовательность может сравниваться с эталонной последовательностью и где часть полинуклеотидной последовательности в окне сравнения может содержать вставки или делеции (т.е. гэпы) по сравнению с эталонной последовательностью (которая не содержит вставки или делеции) для оптимального выравнивания двух последовательностей. Как правило, окно сравнения составляет по меньшей мере 20 смежных нуклеотидов в длину и необязательно может составлять 30, 40, 50 и 100 или более. Специалистам в настоящей области техники понятно, что во избежание высокого сходства с эталонной последовательностью вследствие включения гэпов в полинуклеотидную последовательность, типично вводят штраф на введение гэпа и вычитают его из числа совпадений.
[086] Способы выравнивания нуклеотидных и аминокислотных последовательностей для сравнения являются хорошо известными в настоящей области техники. Алгоритм локальной гомологии (BESTFIT) Smith and Waterman, (1981) Adv. Appl. Math 2: 482, может проводить оптимальное выравнивание последовательностей для сравнения; с помощью алгоритма гомологичного выравнивание (GAP) Needleman and Wunsch, (1970) J. Mol. Biol. 48: 443-53; с помощью способа поиска сходства (Tfasta and Fasta) Pearson and Lipman, (1988) Proc. Natl. Acad. Sci. USA 85: 2444; с помощью компьютеризированных реализаций этих алгоритмов, включая, но без ограничения: CLUSTAL в программе PC/Gene от Intelligenetics, Mountain View, Calif., GAP, BESTFIT, BLAST, FASTA и TFASTA в пакете программного обеспечения Wisconsin Genetics, версия 8 (доступна от Genetics Computer Group (программы GCG® (Accelrys, Inc., San Diego, Calif.).). Программа CLUSTAL хорошо описана Higgins and Sharp, (1988) Gene 73: 237-44; Higgins and Sharp, (1989) CABIOS 5: 151-3; Corpet, et al., (1988) Nucleic Acids Res. 16: 10881-90; Huang, et al., (1992) Computer Applications in the Biosciences 8: 155-65 и Pearson, et al., (1994) Meth. Mol Biol. 24: 307-31. Предпочтительной программой для применения для оптимального глобального выравнивания множественных последовательностей является PileUp (Feng and Doolittle, (1987) J. Mol. Evol., 25: 351-60, которая аналогична способу, описанному Higgins and Sharp, (1989) CABIOS 5: 151-53 и включена в настоящий документ посредством ссылки). Семейство программ BLAST, которое может применяться для поисков сходства в базах данных, включает: BLASTN для нуклеотидных последовательностей запроса среди нуклеотидных последовательностей базы данных; BLASTX для нуклеотидных последовательностей запроса среди белковых последовательностей базы данных; BLASTP для белковых последовательностей запроса среди белковых последовательностей базы данных; TBLASTN для белковых последовательностей запроса среди нуклеотидных последовательностей базы данных; и TBLASTX для нуклеотидных последовательностей запроса среди нуклеотидных последовательностей базы данных. См. Current Protocols in Molecular Biology, глава 19, Ausubel et al., eds., Greene Publishing and Wiley-Interscience, New York (1995).
[087] GAP применяет алгоритм Needleman and Wunsch, ранее, для обнаружения выравнивания двух полных последовательностей, который максимизирует число совпадений и минимизирует число гэпов. GAP рассматривает все возможные выравнивания и положения гэпов и создает выравнивание с наибольшим числом совпавших оснований и наименьшими гэпами. Это предусматривает обеспечение штрафа за введение гэпа и штраф за продолжение гэпа в единицах совпавших оснований. GAP должен извлекать выгоду из числа штрафов за введение гэпа совпадений для каждого гэпа, который он вводит. Если выбирают штраф за продолжение гэпа больше чем ноль, то GAP должен, кроме того, извлекать выгоду для каждого введенного гэпа, равную длине гэпа, умноженной на штраф за продолжение гэпа. Значения по умолчанию штрафа за введение гэпа и значения штрафа за продолжение гэпа в версии 10 пакета программного обеспечения Wisconsin Genetics составляет 8 и 2, соответственно. Штрафы за введение гэпа и продолжения гэпа могут быть выражены как целое, выбранное из группы целых чисел, состоящих из 0-100. Таким образом, например, штрафы за введение гэпа и продолжения гэпа могут представлять собой 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 30, 40 и 50 или более.
[088] GAP представляет одного представителя семейства лучших выравниваний. Может существовать многие представители этого семейства, но ни один другой представитель не имеет лучшего качества. GAP представляет четыре графика критерия для выравниваний: качество, соотношение, идентичность и сходство. Качество представляет собой показатель, максимизированный для того чтобы выровнять последовательности. Соотношение представляет собой качество, деленное на число оснований в более коротком сегменте. Процент идентичности представляет собой процент символов, которые фактически совпадают. Процент сходства представляет собой процент символов, которые являются сходными. Символы, которые находятся напротив гэпов, не учитывают. Сходство подсчитывают, когда значение матрицы замен для пары символов составляет более чем или равно 0,50, пороговое значение сходства. Матрица замен, применяемая в версии 10 пакета программного обеспечения Wisconsin Genetics, представляет собой BLOSUM62 (смотрите, Henikoff and Henikoff, (1989) Proc. Natl. Acad. Sci. USA 89: 10915).
[089] Если не указано иное, значения идентичности/сходства последовательностей, представленные в данном документе, относятся к значению, полученному с применением пакета программ BLAST 2.0 с применением параметров по умолчанию (Altschul, et al., (1997) Nucleic Acids Res. 25: 3389-402).
[090] Как будет понятно специалистам в настоящей области техники, поиски с помощью BLAST предполагают, что белки могут быть смоделированы как случайные последовательности. Тем не менее, многие существующие белки содержат участки неслучайных последовательностей, которые могут быть гомополимерными трактами, кратковременными повторами или участками, обогащенными одной или несколькими аминокислотами. Такие участки низкой сложности могут выравниваться между несвязанными белками, даже если другие участки белка являются полностью несходными. Ряд программ-фильтров для участков низкой сложности может применяться для снижения таких выравниваний низкой сложности. Например, SEG (Wooten and Federhen, (1993) Comput. Chem. 17: 149-63) и XNU (Claverie and States, (1993) Comput. Chem. 17: 191-201) фильтры низкой сложности могут применяться отдельно или в комбинации.
[091] Применяемое в данном документе выражение "идентичность последовательностей" или "идентичность" в контексте двух последовательностей нуклеиновых кислот или полипептидных последовательностей включает ссылку на остатки в двух последовательностях, которые являются одинаковыми при выравнивании для максимального соответствия на определенном окне сравнения. Когда процентное отношение идентичности последовательностей применяют со ссылкой на белки, это означает, что положения остатков, которые не являются идентичными, часто отличаются на консервативные аминокислотные замены, где аминокислотные остатки замещены другими аминокислотными остатками со сходными химическими свойствами (например, зарядом или гидрофобностью) и, следовательно, не изменяют функциональные свойства молекулы. Если последовательности отличаются по консервативным заменам, то процент идентичности последовательностей может быть отрегулирован в сторону повышения, чтобы скорректировать консервативную природу замены. Последовательности, отличающиеся по таким консервативным заменам, как говорят, обладают "сходством последовательностей" или "сходством". Средства для осуществления такой регуляции хорошо известны специалистам в настоящей области техники. Типично это включает оценивание консервативной замены в качестве частичного, а не полного несовпадения, тем самым увеличивая процентное отношение идентичности последовательностей. Таким образом, например, если идентичной аминокислоте присваивают балл 1 и неконсервативной замене присваивают балл ноль, то консервативной замене присваивают балл от ноля до 1. Присвоение баллов консервативным заменам рассчитывают, например, согласно алгоритму Meyers and Miller, (1988) Computer Applic. Biol. Sci. 4: 11-17, например, как реализовано в программе PC/GENE (Intelligenetics, Mountain View, Calif., USA).
[092] Применяемое в данном документе выражение "процентное отношение идентичности последовательностей" означает значение, определенной с помощью сравнения двух оптимально выровненных последовательностей в окне сравнения, где часть полинуклеотидной последовательности в окне сравнения может содержать вставки или делеции (т.е. гэпы) по сравнению с эталонной последовательностью (которая не содержит вставок или делеций) для оптимального выравнивания двух последовательностей. Процентное отношение рассчитывают путем определения числа положений, в которых идентичное основание нуклеиновой кислоты или аминокислотный остаток встречается в обеих последовательностях для получения числа совпавших положений, разделяя число совпавших положений на общее число положений в окне сравнения и умножая результат на 100 для получения процентного отношения идентичности последовательностей.
[093] Выражение "существенная идентичность" полинуклеотидных последовательностей означает, что полинуклеотид содержит последовательность, которая имеет 50-100% идентичность последовательностей, предпочтительно по меньшей мере 50% идентичность последовательностей, предпочтительно по меньшей мере 60% идентичность последовательностей, предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95%, по сравнению с эталонной последовательностью с применением одной из программ выравнивания, описанных с применением стандартных параметров. Специалисту в настоящей области техники будет понятно, что эти значения могут быть соответствующим образом скорректированы для определения соответствующей идентичности белков, кодируемых двумя нуклеотидными последовательностями, учитывая вырожденность кодонов, аминокислотное сходство, расположение рамки считывания и подобного. Существенная идентичность аминокислотных последовательностей для этих целей в норме означает идентичность последовательностей, равную 55-100%, предпочтительно по меньшей мере 55%, предпочтительно по меньшей мере 60%, более предпочтительно по меньшей мере 70%, 80%, 90% и наиболее предпочтительно по меньшей мере 95%.
[094] Другим признаком того, что нуклеотидные последовательности являются существенно идентичными, является гибридизация двух молекул друг с другом при жестких условиях. Вырожденность генетического кода предусматривает многие аминокислотные замены, приводящие к разнообразию в нуклеотидной последовательности, кодирующей одну и ту же аминокислоту, поэтому возможно, что последовательность ДНК может кодировать такой же полипептид, не гибридизироваться друг с другом при жестких условиях. Это может происходить, например, когда копия нуклеиновой кислоты создается с применением максимальной вырожденности кодона, разрешенной генетическим кодом. Один признак того, что две последовательности нуклеиновых кислот являются существенно идентичными, заключается в том, что полипептид, кодирующий первую нуклеиновую кислоту, является иммунологически перекрестно реагирует с полипептидом, кодируемым второй нуклеиновой кислотой.
[095] Фраза "абиотический стресс", применяемая в данном документе, относится к любому неблагоприятному эффекту на метаболизм, рост, размножение и/или жизнеспособность растения со стороны абиотических факторов (т.е. водообеспеченность, тепловое воздействие, холод и т.д.). Соответственно, абиотический стресс может быть вызван субоптимальными окружающими условиями роста, такими как, например, засоленность, водное голодание, водный дефицит, засуха, наводнение, замораживание, низкая или высокая температура (например, охлаждение или чрезмерное нагревание), загрязнение токсичными химическими веществами, токсичность тяжелых металлов, анаэробиоз, недостаточность питательных веществ, избыток питательных веществ, атмосферное загрязнение или УФ-облучение.
[096] Фраза "устойчивость к абиотическому стрессу", применяемая в данном документе, относится к способности растения выдерживать абиотический стресс, не испытывая существенного изменения в метаболизме, росте, продуктивности и/или жизнеспособности.
[097] Применяемое в данном документе выражение "водный дефицит" означает период, когда вода, доступная растению, не восполняется со скоростью, с которой она потребляется растением. Длительный период водного дефицита в разговорной речи называется засухой. Отсутствие дождя или орошения может не приводить немедленно к стрессу, вызванному недостатком воды, если имеется доступный резервуар грунтовых вод для поддержания скорости роста растений. Растения, выращенные в почве с достаточным запасом грунтовых вод, могут выживать в течение дней без дождя или орошения без неблагоприятных воздействий на урожайность. Растения, выращенные в сухой почве, вероятно, перенесут неблагоприятные воздействия с минимальными периодами водного дефицита. Стресс, вызванный тяжелым водным дефицитом, может вызвать увядание и гибель растений; умеренная засуха может снизить урожайность, сдерживать рост или сдерживать развитие. Растения могут восстановиться после нескольких периодов стресса, вызванного водным дефицитом, без существенного воздействия на урожайность. Тем не менее, водный дефицит во время опыления может уменьшить или снизить урожайность. Таким образом, применимый период в жизненном цикле кукурузы, например, для наблюдения за реакцией или устойчивостью к водному дефициту, представляет собой позднюю вегетативную стадию роста перед появлением метелки или переходом к репродуктивному развитию. Устойчивость к водному дефициту определяют путем сравнения с контрольными растениями. Например, растения согласно настоящему изобретению могут производить более высокий урожай, чем контрольные растения при воздействии водного дефицита. В лабораторных и в полевых испытаниях засуху можно имитировать путем подачи растениям согласно настоящему изобретению и контрольным растениям меньшего количества воды, чем подают контрольным растениям с достаточным уровнем полива и измерения различий в признаках. Например, урожайность можно разделить на такие компоненты, как растения на акр, количество рядов зерен на початок, количество початков на растение, количество зерен на растение и вес на зерно. Бесплодие или сниженное образование семян может также повышаться при стрессе, вызванном недостатком воды. Один аспект настоящего изобретения предлагает растения, сверхэкспрессирующие гены, раскрытые в данном документе, которые обеспечивают более высокую устойчивость к водному дефициту.
[098] Применяемая в данном документе фраза "оптимизация воды" относится к любому параметру растения, его частей или его структуры, который может быть измерен и/или количественно определен для того чтобы оценить степень или скорость роста и развития растений при различных условиях водообеспеченности. В связи с этим, "признак оптимизации воды" представляет собой любой признак, который, как может быть показано, влияет на урожайность растения при различных наборах условий роста, относящихся к водообеспеченности. Иллюстративные параметры оптимизации воды представляют собой урожайность зерна при стандартном процентном отношении влажности (YGSMN), влажность зерна при уборке (GMSTP), вес зерна на делянку (GWTPN), и процентное восстановление урожая (PYREC).
[099] Применяемые в данном документе фразы "засухоустойчивость" и "засухоустойчивый" относятся к способности растений выдерживать и/или успешно произрастать при условиях, когда водообеспеченность является субоптимальной. Как правило, растение называются "засухоустойчивым", если оно проявляет "усиленную засухоустойчивость". Применяемая в данном документе фраза "усиленная засухоустойчивость" относится к измеряемому улучшению, усилению или увеличению одного или нескольких фенотипов оптимизации воды по сравнению с одним или несколькими контрольными растениями.
[0100] Коэффициент использования воды (WUE) представляет собой параметр, часто применяемый для оценки отношения между потреблением воды и поглощением CO2/ростом (Kramer, 1983, Water Relations of Plants, Academic Press p. 405). WUE был определен и измерен многочисленными способами. Одним подходом является расчет отношения сухого веса целого растения к воде, потребляемой растением в течение его жизни (Chu et al., 1992, Oecologia 89: 580). Другой вариант - это применение более короткого временного интервала, когда измеряют накопление биомассы и использование воды (Mian et al., 1998, Crop Sci. 38: 390). Другой подход представляет собой использование измерений от ограниченных частей растения, например, измерение только роста надземных частей и использования воды (Nienhuis et al 1994 Amer J Bot 81: 943). WUE также был определен как отношение поглощения CO2 к потере с испарением воды из листа или части листа, часто измеряемых в течение очень короткого периода времени (например, секунд/минут) (Kramer, 1983, р. 406). Соотношение 13С/12С, зафиксированного в ткани растения и измеренного с помощью масс-спектрометра для измерения изотопного соотношения, также применялось для оценки WUE у растений с применением С-3 фотосинтеза (Martin et al., 1999, Crop Sci. 1775). Применяемое в данном документе выражение "коэффициент использования воды" относится к количеству органических веществ, произведенных растением, разделенному на количество воды, использованной растением в их производстве, т.е. к сухому весу растения по отношению к использованию воды растением. Применяемое в данном документе выражение "сухой вес" относится к чему-либо в растении, отличному от воды, и включает, например, углеводы, белки, масла и минеральные питательные вещества. Предусматривается, что трансгенные растения, полученные с помощью способов, описанных в данном документе, будут обеспечивать увеличение коэффициента использования воды.
[0101] Фраза "биотический стресс", применяемая в данном документе, относится к любому неблагоприятному эффекту на метаболизм, рост, репродукцию и/или жизнеспособность растения со стороны биотических факторов (т.е. давление со стороны насекомых, заболевание и т.д.).
[0102] Фраза "устойчивость к биотическому стрессу", применяемая в данном документе, относится к способности растения выдерживать биотический стресс, не испытывая существенного изменения метаболизма, роста, репродукции и/или жизнеспособности.
[0103] Применяемая в данном документе фраза "растительная биомасса" относится к количеству (измеренное в граммах воздушно-сухой или сухой ткани) ткани, полученной из растения в вегетационный период, которое может также определить или воздействовать на урожайность растений или урожайность на площадь выращивания.
[0104] Применяемая в данном документе фраза "мощность растений" относится к количеству (измеренному на вес) ткани, произведенной растением в заданное время. Следовательно, увеличенная мощность может определять или воздействовать на урожайность растений или урожайность на время выращивания или площадь выращивания.
[0105] Выражение "ранняя мощность" относится к активному здоровому устойчивому росту особенно в течение ранних стадий роста растения, и может быть результатом увеличенной приспособляемости растения вследствие того, например, что растения лучше адаптированы к их окружающей среде (оптимизируя применение источников энергии и распределяя между побегом и корнем). Растения, имеющую раннюю мощность также демонстрируют увеличенное выживание всходов и лучшую приживаемость сельскохозяйственной культуры, что часто приводит к полям с высокой однородностью (например, сельскохозяйственные культуры, растущие равномерным образом, например, сельскохозяйственные культуры, достигающие различных стадий развития в одно время), и зачастую к более высоким урожаям. Следовательно, ранняя мощность может быть определена с помощью измерения различных факторов, таких как масса тысячи зерен, процентное отношение прорастания, процентное отношение всхожести, рост всходов, высота всходов, длина корня, биомасса корня и побега и многие другие.
[0106] Применяемое в данном документе выражение "мощность всходов" относится к характеристике растений, при которой растение появляется из почвы быстрее, имеет увеличенную скорость прорастания (т.е., прорастает быстрее), имеет более быстрый и больший рост всходов и/или прорастает быстрее в условиях холода по сравнению с диким типом или контролем при аналогичных условиях. Мощность всходов часто определяли как включающую свойства семян, которые определяют "потенциал быстрого, однородного появления и развития нормальных всходов в широком диапазоне полевых условий".
[0107] Жизненный цикл цветущих растений, как правило, может быть разделен на три фазы роста: вегетативную фазу, фазу бутонизации и фаза цветения (поздняя фаза бутонизации). В вегетативной фазе апикальная меристема побега (SAM) образует листья, которые позже будут обеспечивать источники, необходимые для создания фертильного потомства. При получении соответствующих сигналов от окружающей среды и стадии развития растение переходит к росту цветка, или репродуктивному росту, и SAM вступает в фазу бутонизации (I) и дает начало соцветию с помощью примордия цветка. В течение этой фазы направление развития SAM и вторичных побегов, которые возникают в пазухах листьев, определяется набором геном меристемной идентичности, некоторые из которых предотвращают, а некоторые из которых стимулируют развитие меристем цветка. Как только это появилось, растение входит в позднюю стадию бутонизации, когда формируются цветковые органы. Если присутствуют соответствующие сигналы окружающей среды и стадии развития, растение переходит к росту цветка, или репродуктивному росту. Если такие сигналы нарушаются, растение не будет способным входить в репродуктивный рост, следовательно, поддерживать вегетативный рост.
[0108] Термин "идиоплазма" относится к генетическому материалу особи или от нее (например, растения), группы особей (например, линия, сорт или семейство растений) или клона, полученного из линии, сорта, вида или культуры. Идиоплазма может представлять собой часть организма или клетки, или может быть выделена из организма или клетки. Как правило, идиоплазма обеспечивает генетический материал со специфическим молекулярным составом, который обеспечивает физическую основу для некоторых или всех наследственных качеств организма или культуры клеток. Применяемое в данном документе выражение "идиоплазма" включает клетки, семя или ткани, из которых могут быть выращены новые растения или части растений, такие как листья, стебли, пыльца или клетки, которые могут быть культивированы в целое растение.
[0109] Антитела согласно настоящему изобретению включают поликлональные и моноклональные антитела и их смеси, которые могут представлять собой любое из IgG, IgA, IgM, IgE, IgD и любой их изотип, например, IgG1, IgG2, IgG3 или IgG4. В случае моноклонального антитела иллюстративный класс антитела представляет собой IgG. Подклассы IgG включают, например, IgG1, IgG2, IgG3 и IgG4. Антитела включают интактные и химерные иммуноглобулиновые молекулы с двумя полноразмерными тяжелыми цепями и двумя полноразмерными легкими цепями (например, зрелая часть последовательностей вариабельной области тяжелой и легкой цепей), а также подпоследовательности/фрагменты тяжелой или легкой цепи, которые сохраняют по меньшей мере часть функции (например, специфичность связывания Т6РР или аффинность связывания Т6РР) родительского интактного антитела, которые специфически связывает Т6РР и, в частности, модифицированную Т6РР. Подпоследовательности могут иметь такую же или по существу такую же специфичность связывания, аффинность связывания, что и родительские интактные и химерные антитела.
[0110] Продукция моноклональных или поликлональных антител может быть осуществлена с помощью техник, хорошо известных в настоящей области техники. "Поликлональные" антитела представляют собой антитела, полученные из различных источников В-клеток. Они представляют собой комбинацию молекул иммуноглобулина, которые секретируют специфический антиген, каждый из которых определяет различный эпитоп. Поликлональные антитела типично получают путем инокуляции подходящему млекопитающему, такому как мышь, кролик или коза. Например, композиция, содержащая белок Т6РР, способный производить антиген, вводят инъекционно млекопитающему. Присутствие нового белка индуцирует В-лимфоциты производить иммуноглобулины IgG, специфические для антигена Т6РР. Поликлональные IgG иммуноглобулины затем очищают от сыворотки млекопитающего.
[0111] Выражение "моноклональное антитело", применяемое в данном документе, относится к препарату молекул антител одного молекулярного состава. Моноклональное антитело проявляет одну специфичность и аффинность связывания к конкретному эпитопу. Способ получения моноклональных антител включает получение иммунных соматических клеток с потенциалом к продукции антитела, в частности, В-лимфоцитов, которые были предварительно иммунизированы с помощью антигена, представляющего интерес, in vivo или in vitro и которые являются подходящими для слияния с линий В-клеточной миеломы.
[0112] Лимфоциты млекопитающих типично иммунизируют с помощью in vivo иммунизации животного (например, мыши) с помощью необходимого белка или полипептида, например, с помощью Т6РР согласно настоящему изобретению. Такие иммунизации повторяют при необходимости с интервалами до нескольких недель для получения достаточного титра антител. Будучи иммунизированными, животные могут применяться в качестве источника производящих антитела лимфоцитов. После последнего вторичного введения антигена животных умерщвляют и удаляют клетки селезенки. Мышиные лимфоциты дают более высокое процентное отношение стабильных слияний с мышиными миеломными линиями, описанными в данном документе. Из них BALB/c мышь является предпочтительной. Тем не менее, другие штаммы мышей, кролик, хомяк, овца и лягушка могут также применяться в качестве хозяев для получения производящих антитела клеток. Смотрите; Goding (в Monoclonal Antibodies: Principles and Practice, 2d ed., pp. 60-61, Orlando, Fla., Academic Press, 1986).
[0113] Те производящие антитела клетки, которые находятся в стадии делящегося плазмобласта, сливаются предпочтительно. Соматические клетки могут быть получены из лимфатических узлов, селезенок и периферической крови примированных антигеном животных, и лимфатические клетки выбора зависят в большой степени от их эмпирической полезности в конкретной системе слияния. Секретирующие антитела лимфоциты затем сливают с (мышиными) клетками В-клеточной миеломы или трансформированными клетками, которые способны к неограниченной репликации в культуре клеток, тем самым производя бессмертную, секретирующую иммуноглобулин клеточную линию. Полученные слитые клетки, или гибридомы, культивируют и полученные колонии подвергают скринингу в отношении продукции необходимых моноклональных антител. Колонии, производящие такие антитела, клонируют и выращивают in vivo или in vitro для получения больших количеств антитела. Описание теоретического основания и практической методологии слияния таких клеток изложено в Kohler and Milstein, Nature 256: 495 (1975), которая включена в настоящий документ посредством ссылки.
[0114] Увеличенная урожайность сельскохозяйственной культуры представляет собой признак, представляющий значительный экономический интерес во всем мире. Урожайность в норме определяют как измеряемое производство экономической ценности из сельскохозяйственной культуры. Он может быть определен в контексте количества и/или количества. Урожайность напрямую зависит от нескольких факторов, например, числа и размера органов, структуры растения (например, числа ветвей), продукции семян, старения листьев и других. Развитие корня, поглощение питательных веществ, стрессоустойчивость и ранняя мощность могут также представлять собой важные факторы в определении урожая. Кроме того, в сельском хозяйстве очень желательным является разработать сельскохозяйственные культуры, которые могут демонстрировать увеличенную урожайность в оптимальных условиях роста, а также в неоптимальных условиях роста (например, засухе, в условиях абиотического стресса). Оптимизация вышеупомянутых факторов, следовательно, может вносить вклад в увеличение урожая сельскохозяйственных культур.
[0115] Урожайность семян представляет собой особенно важный признак, поскольку семена многих растений являются важными для питания людей и животных. Сельскохозяйственные культуры, такие как, кукуруза, рис, пшеница, канола и соя, отвечают более чем за половину общей калорийности, потребляемой человеком, независимо от того, осуществляется ли это посредством прямого потребления самих семян или посредством потребления сельскохозяйственных животных, выращенных на обработанных семенах. Семена растений также являются источником сахаров, масел и многих видов метаболитов, применяемых в различных промышленных процессах. Семена состоят из зародыша (источника новых побегов и корней) и эндосперма (источника питательных веществ для роста зародыша в ходе прорастания и в ходе раннего роста всходов). В развитие семени вовлечены многие гены и оно требует переноса метаболитов из корней, листьев и стеблей в развивающееся семя. Эндосперм ассимилирует метаболические предшественники углеводов, масел и белков и синтезирует их в виде запасных макромолекул для наполнения зерна.
[0116] Ранняя мощность представляет собой другой важный аспект урожая. Улучшение ранней мощности представляет собой важную цель современных программ разведения риса у культиваров риса как умеренного, так и тропического пояса. Длинные корни важны для водного риса для его правильного закрепления в почве. Водный рис высевают непосредственно в затопленные поля, где растения должны быстро появиться из воды, таким образом, более длинные побеги обеспечивают увеличенное выживание молодых всходов. Если практикуют рядовой посев, более длинные мезокотили и колеоптили важны для хорошего появления всходов. Способность внедрить в растения раннюю мощность будет иметь большое значение в сельском хозяйстве. Например, слабая ранняя мощность оказалась ограничением для введения гибридов маиса (Zea mays L.) на основе идиоплазмы Кукурузного пояса в Атлантической части Европы. В другом аспекте ранняя мощность может также создавать возможность для более ранних периодов уборки у некоторых сельскохозяйственных культур, что может служить преимуществом при многих сельскохозяйственных применениях.
[0117] Растения, разработанные для улучшенного урожая при различных биотических и абиотических стрессах, представляют особый интерес в области сельского хозяйства. Например, абиотический стресс представляет собой основную причину потери урожая во всем мире, снижая средние уровни урожая для наиболее важных культурных растений более чем на 50% (Wang et al., Planta (2003) 218: 1-14). Абиотические стрессы могут быть вызваны засухой, наводнениями, засоленностью, экстремальными показателями температуры, химической токсичностью и окислительным стрессом. Способность улучшать устойчивость растений к абиотическому стрессу будет представлять собой большое экономическое преимущество для фермеров во всем мире и будет обеспечивать культивирование сельскохозяйственных культур в течение неблагоприятных условий и на территориях, где культивирование сельскохозяйственных культур иным образом может быть невозможным.
[0118] В некоторых случаях урожайность растений соотносится с количеством растительной биомассы, которую может произвести конкретное растение. Большее растение с большей площадью листа может типично поглотить больше света, питательных веществ и углекислого газа, чем растение меньшего размера и, следовательно, будет, вероятно, давать больший вес в течение одинакового периода (Fasoula & Tollenaar 2005 Maydica 50: 39). Увеличенная растительная биомасса может также быть очень желательна в таких процессах, как превращение биомассы (например, кукурузы, трав, сорго, тростника) в топлива, такие как, например, этанол или бутанол.
[0119] Способность увеличивать урожайность растений будет иметь множество применений в таких областях, как сельское хозяйство, продукция декоративных растений, древоводстве, садоводстве, получении биотоплива, фармацевтике, производствах ферментов, которые используют растения как фабрики для этих молекул, и лесоводстве. Увеличение урожая может также находить применение в получении микробов или водорослей для применения в биореакторах (для биотехнологического получения веществ, таких как фармацевтические средства, антитела, вакцины, топливо или для биологического преобразования органических отходов) и других таких областях.
[0120] Селекционеры растений часто интересуются улучшением специфических аспектов урожая в зависимости от изучаемой культуры или растения, и части этого растения или культуры, которая имеет относительную экономическую ценность. Например, селекционер растений может специфически искать улучшений в растительной биомассе (весе) одной или нескольких частей растения, которые могут включать надземные (пригодные для уборки) части и/или пригодные для уборки части под землей. Это особенно уместно там, где надземные части или подземные части растения имеют потребительское значение. Для многих сельскохозяйственных культур, особенно злаков, улучшение урожая семян является очень желательным. Увеличенная урожайность семян может проявлять себя многими путями, причем каждый отдельный аспект урожая семян имеет переменную важность для селекционера растений в зависимости от изучаемой культуры или растения и его конечного применения.
[0121] Для селекционера растений будет большим преимуществом быть способным отсортировать и выбрать аспекты урожая, подлежащие изменению. Также может быть очень желательной способность отобрать ген, подходящий для изменения конкретного аспекта урожайности (например, урожайность семян, вес биомассы, коэффициент использования воды, урожайность при стрессовых условиях). Например, увеличение скорости налива, комбинированное с увеличенным весом тысячи зерен, будет очень желательным для культуры, такой как кукуруза. Для риса и пшеницы комбинация увеличенной скорости налива, процентного отношения массы урожая к общей массе растений и увеличенного веса тысячи зерен будет очень желательным.
[0122] Трегалоза представляет собой очень стабильный дисахарид, состоящий из двух молекул глюкозы. Она не распадается при нагревании до 100°C в течение 24 часов при широком диапазоне pH. Ее низкая реакционная способность, физиохимические свойства и стабильность делают его исключительным осмопротектором. При отсутствии воды она стабилизирует белки, мембраны и другие клеточные структуры путем замещения воды посредством водородных связей на полярные остатки. При обезвоживании трегалоза образует сетчатую стеклоподобную структуру, которая ограничивает движение молекул, что предотвращает агрегацию белков и диффузию свободных радикалов (Brumfield G (2004) Nature 428: 14-15, Paul MJ (2004) Plant Biotechnol J, 2 pp. 71-82).
[0123] Ранее считалось, что только определенные растения, такие как устойчивые к обезвоживанию виды, например, плауны скальные, накапливают трегалозу, поскольку они производят ее в количествах, достаточных для простого измерения. Недавно полученные генетические и геномные доказательства, тем не менее, показывают, что все растения, исследованные к настоящему времени, обладают способностью синтезировать трегалозу, но типично в очень малых количествах (Paul MJ, et al. (2008) Annu Rev Plant Biol 59: 417-441). Более того, оказалось, что метаболизм трегалозы является существенным, так как устранение способности растения производить трегалозу, например, посредством мутации, является летальным (Eastmond PJ, et al. (2002) Plant J 29: 225-235). Большинство культурных видов содержат следовые количества трегалозы (измеренные на низком микромолярном или даже наномолярном уровне), хотя типичное накопление является неизвестным для маиса.
[0124] Выражение "трегалозный путь", применяемое в данном документе, состоит из двух биосинтетических ферментов, трегалозо-6-фосфатсинтазы (TPS) и трегалозо-6-фосфатфосфатазы (Т6РР), и одного гидролизующего фермента, трегалазы, вовлеченного в распад, пожалуйста, см. фиг. 3. В настоящее время неизвестно, сколько существует изоформ (TPS) или (Т6РР) у маиса. В модельном растении Arabidopsis в общем существует 11 генов TPS (Paul et al. (2008) Annu Rev Plant Biol 59: 417-441), тем не менее, каталитическая активность была показана только для AtTPS1 (Vandesteene L, et al. (2010) Mol Plant 3: 406-419). Функция других 10 гомологов TPS Arabidopsis остается неясной. Существуют 10 генов Т6РР у Arabidopsis, и хотя большинство не были хорошо охарактеризованы, все они, вероятно, имеют активность Т6РР (Schluepmann Н, et al. (2012) J Exp Bot: 1-12; Paul et al., (2008) Annu Rev Plant Biol 59: 417-441). Наоборот, Arabidopsis содержит только один ген, кодирующий трегалазу (Müller J, et al. (2001) Plant Physiol 125: 1086-1093; Frison M, et al. (2007) FEBS Letters 581: 4010-4016; Paul et al. 2008). Аналогичная ситуацию наблюдается у риса (Shima S, et al. (2007) FEBS J 274: 1192-1201; Zang et al., (2011) Plant Mol Biol 76: 507-522). Данные профилей экспрессии показывают, что представители семейства генов избирательно экспрессируются в ходе развития растения (Scheulpmann et al., (2004) Plant Physiol 135: 879-890; Zang et al., (2011) Plant Mol Biol 76: 507-522). Разнообразие в тканеспецифической экспрессии и трансляции среди некоторых представителей в семействе генов Т6РР и TPS представляет ясное доказательство того, что эти ферменты осуществляют важные и, возможно, различные функции в растении (Satoh-Nagasawa et al., (2006) Nature 441: 227-230; Mustroph et al., (2009) Proc Natl Acad Sci USA 106: 18843-18848; Ramon et al., (2009) Plant Cell Environ 32: 1015-1032).
[0125] Ферменты трегалозного пути также регулируются на посттрансляционном уровне. Белки TPS представляют собой субстраты для протеинкиназ, родственных неферментирующим сахарозу (SNF1) (SnRK1) (Zhang et al., (2009) Plant Physiol 149: 1860-1871). Некоторые TPS (AtTPS5, 6 & 7) фосфопротеины связывают 14-3-3 белки (Harthill et al., (2006) Plant J 47: 211-223). Координационное фосфорилирование и кэппирование сайта фосфорилирования с помощью 14-3-3 белков могут быть возможным механизмом, с помощью которого регулируется активность TPS. Нитратредуктаза регулируется с помощью аналогичного механизма, основанного на светотемновом переходе (Huber et al., (2002) Plant Mol Biol 50: 1053-1063). Harthill et al. предполагают, что все из нитратредуктазы, 6-фосфофрукто-2-киназы/фруктоза-2,6-бисфосфатазы (F2KP) и TPS модифицируются с помощью одинаковой киназы (вероятно, SnRK1), связывают 14-3-3 белки и в некоторой мере могут регулироваться светотемновым циклом (Harthill et al. (2006) Plant J 47: 211-223).
[0126] Сложность в экспрессии и регуляции дает веские основания предполагать, что белки в пути метаболизма трегалозы должны быть сгруппированы с ферментами, важными для метаболизма растения, которые выполняют как каталитические, так и регуляторные роли. Их точная роль полностью не ясна, но перспективная модель может ассоциироваться с тремя центральными функциями: передача сигнала с участием сахаров, развитие растений и защита от стресса.
[0127] Основываясь на опубликованных сообщениях о микробиологических системах (Wiemken, А. (1990) J Gen Microbiol 58: 209-217), где стабилизация мембран и другие преимущества коррелируют с высокими уровнями трегалозы (1-10% сухого веса, г/г), ранняя трансгенная работа фокусировалась на применении трегалозы в качестве осмопротектора. Некоторые опубликованные исследования предполагали, что трегалоза защищает растения от засухи и других осмотических стрессов (Garg et al., (2002) PNAS 99: 15898-15903; Paul et al., (2008) Annu Rev Plant Biol 59: 417-441; Ge et al., 2008; Li et al., 2011). Тем не менее, когда уровни трегалозы измеряют в трансгенных растениях, разработанных для чрезмерного накопления трегалозы, было обнаружено, что они являлись очень низкими, порядка микрограммов на грамм сырого веса. Филогенетическая информация также показывает, что несмотря на пролиферацию генов трегалозного пути в растениях только небольшое число растений накапливает метаболиты трегалозы на уровнях, необходимых для непосредственного действия в качестве осмопротектора или стабилизатора клеточных структур.
[0128] Деятельность в середине 1990-ых по получению трегалозы в растениях (для пищевой промышленности) отличалась от деятельности по получению молекул других 'чужеродных' сахаров в растениях (Eastmand and Graham, (2003) Current Opin Plant Biol 6: 231-235; Pellny et al. (2004) Plant Biotechnol J, 2 pp. 71-82; Paul M.J. (2007) Current Opinion in Plant Biology Volume 10, Issue 3, June, Pages 303-309). Изменение метаболизма трегалозы в трансгенных растениях часто приводило к неожиданным плейотропным дефектам роста (Lordachescu and Imai, (2008) J Integrative Plant Biol 50: 1223-1229).
[0129] P.J. Eastmond et al. первыми показали неопровержимую необходимость в активном гене TPS растения, TPS1 Arabidopsis для развития зародыша (Eastmond P.J. et al., (2002) Plant J 29: 225-235). Гомозиготные tps1 мутанты не развивали зрелые семена. Schluepmann et al. показали, что tps1 мутант может быть спасен с помощью экспрессии гена Е.coli otsA, кодирующего трегалозо-6-фосфатсинтазу, позволяя предположить, что активный компонент трегалозного пути, который регулирует метаболизм растения, представляет собой Т6Р. Концентрация Т6Р в растительных клетках является очень низкой, указывая на то, что она функционирует в качестве сигнальной молекулы. (Scheulpmann et al., (2004) Plant Physiol 135: 879-890). Небольшое изменение в ее концентрации может иметь значительный "гормоноподобный" эффект на регуляцию метаболических процессов в растении (Rolland et al. (2002) Plant Cell: S185-S205).
[0130] В настоящее время имеется растущее количество доказательств для подтверждения вовлечения трегалозного пути в передачу сигналов с участием сахаров, которая является необходимой для всех организмов. Он обеспечивает регуляцию уровней сахаров между крайними значениями насыщения и голода (Eveland and Jackson, (2012) J Exp Bot: 1-11). Например, у млекопитающих регуляция инсулином уровней сахара в крови является необходимой для поддержания оптимального состояния здоровья.
[0131] Принцип, согласно которому трегалозные сахара вовлечены в получение сигнала от сахара и энергетического статуса, установлен у микроорганизмов, в частности, у дрожжей, и в настоящее время подтверждается у растений (Schluepmann et al., (2012) J Exp Bot: 1-12). У растений T6P синтезируется из G6P и UDPG, последующих продуктах распада сахарозы. UDPG производится напрямую посредством сахарозосинтазы; G6P производится из глюкозы посредством инвертазы и гексокиназы или посредством фруктозы и фруктоза-6-фосфата (F6P) посредством сахарозосинтазы, фруктокиназы и фосфоглюкозоизомеразы. UDPG и G6P также представляют собой центральные активированные предшественники, из которых к итоге могут быть получены многие клеточные функциональные соединения (Фигура 4). UDPG представляет собой предшественник для большинства полисахаридов клеточной стенки и для гликолипидов. G6P представляет собой предшественник для синтеза крахмала, NADPH путем окислительной части пентозофосфатного пути (Masakapalli S.K. et al., (2010) Plant Physiol 152: 602-619), и ATP путем гликолиза, цикла Кребса и окислительного фосфорилирования. Кроме того, G6P может быть превращен в фруктозо-6-фосфат и, вместе с UDPG, может применяться для синтеза сахарозо-6-фосфата и затем сахарозы. Будучи произведенным из UDPG и G6P, Т6Р находится на пересечениях основных углеводных потоков в растениях. Таким образом, уровни Т6Р у растительных клетках могут отражать доступность гексозофосфатов, UDPG и сахарозы.
[0132] Важное открытие показывает, что Т6Р связывается с SnRK1 для ингибирования ее активности (Zhang et al., 2009) Plant Physiol 149: 1860-1871), пожалуйста, см. фиг. 5. SnRK1 представляет собой гетеротримерный белок и является растительным гомологом AMP-активированной протеинкиназы животных и дрожжевой неферментирующей сахарозу протеинкиназы (SnF1) (Paul et al. (2008) Annu Rev Plant Biol 59: 417-441; Schluepmann et al., (2012) J Exp Bot: 1-12; Polge and Thomas (2007) Trends in Plant Science Volume 12, Issue 1, January 2007, Pages 20-28). Глобальная генная экспрессия в растениях, сверхэкспрессирующих ген субъединицы SnRK1 имеет сходство с реакцией растения на вызванный недостатком углеродного питательного вещества или энергетический стресс, вызванный длительным темновым периодом, ингибированием фотосинтаза или погружением в воду (Baena-González et al., (2007) Nature 448: 938-942). Увеличенный уровень SnRK1, следовательно, опосредует реакции на низкоэнергетический стресс. В то же время инактивация SnRK1 стимулирует процессы, необходимые для утилизации и анаболизма углерода (Фигура 6).
[0133] У Arabidopsis, Т6Р связывался опосредованно с зависимыми от NADPH процессами (Kolbe et al., (2005) PNAS 102: 11118-11123). Уровни T6P у Arabidopsis имеют тенденцию к обратно-пропорциональной зависимости от уровней G6P (Schleupmann et al., (2003) Proc Natl Acad Sci USA 100: 6849-6854), и прямо-пропорциональной зависимости от уровней сахарозы (Martinez-Barajas et al., 2011). У Arabidopsis, получающих питание в виде сахарозы или трегалозы, увеличиваются уровни Т6Р и активируется включение углерода в крахмал, что представляет собой утилизацию углерода для запасания. Т6Р вовлечен в окислительно-восстановительную активацию AGPазы в гетеротрофных тканях (Schleupmann et al., (2012) J Exp Bot: 1-12). Кроме того, AGPa3a существенно активируется путем опосредованного тиоредоксином восстановления, когда уровни Т6Р увеличиваются с помощью экспрессии трансгена. Фермент NAPDH-тиоредоксинредуктаза С (NTRC) использует NADPH для окислительно-восстановительной активации AGPазы. Мутанты в NTRC не реагируют на подкормку трегалозой, указывая на то, что Т6Р действует на фермент NTRC или предшествующий элемент в пути для окислительно-восстановительной активации AGPазы. Рассматривая вместе, функция Т6Р в растениях воздействует на передачу энергетического сигнала, а также окислительно-восстановительную передачу сигнала с помощью NADPH; Т6Р необходим, поскольку он контролирует доступность NADPH-субстрата для восстановительного биосинтеза. Тем не менее механизмы, с помощью которых Т6Р проявляет свой эффект на зависимую от NADPH окислительно-восстановительную передачу сигнала, неизвестны у растений. Природа вовлечения SnRK1 неизвестна в этом случае (Schleupmann et al., (2012) J Exp Bot: 1-12). SnRK1 необходима для активации AGPазы (Kolbe et al., (2005) PNAS 102: 11118-11123) и по-видимому, активна как выше, так и ниже Т6Р в окислительно-восстановительной активации AGPазы в гетеротрофных тканях.
[0134] Т6Р представляет собой необходимый метаболит, так как он контролирует доступность субстрата для роста. Рост всходов Arabidopsis ингибируется, когда они растут на средах с трегалозой и эффект подавляется при подкормке сахарозой с трегалозой. Прекращение роста на трегалоза, как было показано, вызывается накоплением Т6Р; ингибирование трегалозы коррелирует с 10-кратным увеличением Т6Р (Schluepmann et al., (2004) Plant Physiol 135: 879-890). Ингибирование роста всходов Arabidopsis связано с существенным накоплением крахмала в семядолях. Напротив, корневые кончики не демонстрируют типичного накопления крахмала. Подкормка трегалозой всходов нарушает распределение углерода, где весь углерод остается в семядолях - его источнике и наблюдается видимое углеродное голодание на верхушках. Активация Т6Р AGPазы и включение углерода в крахмал, тем не менее, не является причиной ингибирования роста. Вместо этого, ингибирование Т6Р активности SnRK1, как было показано, вызывает ингибирование роста на трегалозе, поскольку всходы, сверхэкспрессирующие каталитическую субъединицу (KIN10) SnRK1, выращивали на трегалозе (Delatte et al., (2011) Plant Physiol 157: 160-174). Таким образом, по-видимому, всходы перестают расти на трегалозе, поскольку накопление Т6Р ингибирует активность SnRK1.
[0135] Это может показаться парадоксальным, что как слишком мало Т6Р, как в tps1 мутанте, так и слишком много Т6Р при отсутствии экзогенно поставляемого углерода останавливает рост растения. Механизм, с помощью которого T6P/SnRK1 проявляет контроль над метаболизмом в растении далек от понимания. Тем не менее, не ограничиваясь теорией, вгетеротрофных тканях Т6Р увеличивается, когда доступна сахароза. Увеличение Т6Р ингибирует активность SnRK1 так, чтобы клетки переключились на анаболические процессы, необходимые для роста. Когда снижаются уровни сахарозы или доступность углерода, то уровень Т6Р падает, снимая контроль с активности SnRK1; анаболические процессы ингибируются. Также известно, что активность SnRK1 активируется в ходе реакций на углеродный и энергетический стресс (Фигура 6). SnRK1 принимает сигнал о низком уровне энергии или углерода и опосредует процессы, чтобы сделать углерод доступным в клетках-реципиентах для роста. Активность SnRK1 необходима, пока углерод не станет снова доступным в гетеротрофных тканях. Затем синтезируется сахароза и уровень Т6Р увеличивается достаточно, чтобы ингибировать активность SnRK1.
[0136] Данный путь передачи сигнала T6P/SnRK1 никоим образом не является универсальным. У Arabidopsis связывание Т6Р с SnRK1 нуждается во все еще неясной растворимой белковой молекуле, факторе 1 (Zhang et al., (2009) Plant Physiol 149: 1860-1871), которая не может быть обнаружена в зрелом листе. Кроме того, субъединицы комплекса SnRK1 гетерогенно распределены в тканей растения (Bitrián et al., (2011) Plant J 65: 829-842). Гетерогенность, следовательно, предполагается в Т6Р ингибировании SnRK1 в гетеротрофных тканях и фотоавтотрофных тканях (Bitrian et al., (2011) Plant J 65: 829-842). Также T6P представляет собой заряженный метаболит и, следовательно, вряд ли свободно экспортируется в другие клетки (Schleupmann et al., (2012) J Exp Bot: 1-12).
[0137] Генетические и биохимические данные показывают, что трегалозный путь является необходимым для развития растений и контролирует клеточный морфогенез. Потеря ферментативной активности TPS1 у Arabidopsis является летальной для зародышей (Eastmond et al., (2002) Plant J 29: 225-235; Paul et al., (2008) Annu Rev Plant Biol 59: 417-441). Экспрессия AtTPS1 была связана со специфическими характерными для стадии развития фенотипами, такими как замедленное развитие зародыша, измененный рост корней и побегов и измененный переход к цветению. Данные показывают, что отсутствие Т6Р ослабляет способность зародыша к переходу в фазу развития - клеточный рост (Paul et al., (2008) Annu Rev Plant Biol 59: 417-441).
[0138] У маиса RAMOSA3 (RA3), который является важным для ветвления соцветия, кодирует состоящий из одного домена белок Т6РР (Satoh-Nagasawa et al. (2006) Nature 441: 227-230). Считается, что экспрессия Т6РР в областях, окружающих пазушные меристемы, может способствовать передаче подвижного, имеющего короткий диапазон сигнала для регуляции меристемной идентичности и детерминированности. Потеря активности RA3 вызывает тяжелые аномалии початков, которые включают длинные пазушные побеги в основании початка в раннем развитии (Eveland & Jackson (2012) J Exp Bot: 1-11).
[0139] Chary et al. описывает выявление мутанта фенотипа формы клетки-1 (csp-1) Arabidopsis thaliana. CSP-1 показал большое количество дефектов в процессах развития растения, таких как стебель растения, морфология листа, развитие корня, сниженное ветвление побегов и замедленное цветение (Chary et al. (2008) Plant Physiol146: 97-107). Мутация была обнаружена в гене AtTPS6. Это обеспечивает существенное доказательство, показывающее, что ген TPS является важным модификатором клеточной морфологии развития всего растения. Неясно, какой механизм действия лежит в основе этих серьезных эффектов на развитие. Возможно, что они, по меньшей мере частично, опосредованы путем передачи сигнала T6P/SnRK1.
[0140] Т6Р маиса присутствует в концентрации <10 мкг/GFW в большинстве образцов, что представляет сложность для точного измерения Т6Р. Кроме того, семейство генов Т6РР маиса сходно по размеру с таковым у риса и Arabidopsis, и естественное накопление Т6РР является высоким в большинстве тканей. Это указывает на то, что трансгенная экспрессия Т6РР риса в маисе, вероятно, оказывает свой эффект пространственным и временным образом.
[0141] Сейчас было обнаружено, что экспрессия некоторых форм трегалозо-6-фосфатфосфатазы (Т6РР) в растениях обеспечивает существенное увеличение урожая, а также придает устойчивость к различным типам стресса (т.е. абиотическому стрессу). В ходе курса анализа различных вариантов Т6РР, трансгенно экспрессирующихся в растениях, было обнаружено, что трансгенные растения, показывающие самую высокую урожайность семян, также содержали Т6РР с модификациями в аминокислотных остатках, связанных со связыванием субстрата. Не ограничиваясь теорией, эти белки могут иметь сниженную ферментативную активность при прямом сравнении с Т6РР, не содержащей эти модификации. Более того, не ограничиваясь теорией, вероятно, что экспрессия Т6РР в растении с модифицированной ферментативной активностью приводит к предпочтительному фенотипу, имеющему существенно увеличенную урожайность как при неблагоприятных (например, засухе), так и не являющихся неблагоприятными полевых условиях. Снова, не ограничиваясь теорией, это может указывать на то, что менее активная форма Т6РР служит в качестве молекулярного сигнала, следовательно, обеспечивая в растении увеличение крахмала, сахара и увеличенного урожая семян. В теории этот молекулярный сигнал может представлять собой белок-белковое взаимодействие с трегалозо-6-фосфатсинтазой (TPS), образуя комплекс, который затем может передавать сигнал растению. В теории этот сигнал может представлять собой измененные уровни трегалозо-6-фосфата (Т6Р). Не ограничиваясь теорией, одна возможность заключается в том, что модифицированная Т6РР при экспрессии в растении образует белок-белковый комплекс с эндогенными формами TPS и/или Т6РР. В теории это связывание изменяет способность Т6РР гидролизовать его субстрат Т6Р и активирует или модулирует TPS для получения Т6Р. Это взаимодействие затем обеспечивает измененные уровни Т6Р, которые затем передаются в качестве сигнала растению для запасания большего количества сахаров в семени и/или биомассе. Альтернативно, но не ограничиваясь теорией, модифицированная Т6РР, имеющая сниженную активность, взаимодействует с некоторым другим компонентом, еще не выясненным, что приводит к инициации каскада для передачи сигнала растению для запасания большего количества сахаров в урожае и/или биомассе. Основываясь на изложенных выше теориях, многие способы могут применяться для получения растения с увеличенным урожаем при любом условии. Также предусматривается, что можно применять способы, описанные в данном документе, для модулирования продукции сахаров в растении и дополнительно применять техники молекулярной инженерии для транспорта сахаров к различным частям растений для увеличения, например, таких показателей, как растительная биомасса, урожайность семян, содержание сахара в надземном растительном материале (т.е. таком как сахарный тростник), и пригодных для потребления частях растений. Дополнительно эти гены трегалозо-6-фосфатфосфатазы могут найти применение в обеспечении многих более желательных признаков, таких как ранняя мощность, стрессоустойчивость, засухоустойчивость, увеличенный коэффициент использования питательных веществ, увеличенная масса корня и увеличенный коэффициент использования воды.
[0142] Трегалоза, нередуцирующий дисахарид, состоящий из двух молекул глюкозы, соединенных посредством альфа-1,1 связей. Сахар трегалоза может быть обнаружен во многих различных организмах во многих царствах (например, растения, бактерии, насекомые и т.д.). Было показано, что трегалоза вовлечена в функцию запасания углеводов и дополнительно была связана с тем, что выполняет роль в стрессоустойчивости у бактерий, грибов и насекомых. У растений, как считалось вначале, трегалоза содержится только в экстремофилах, таких как плаун скальный Selaginella lepidophylla, тем не менее в настоящее время общепризнанным является то, что метаболизм трегалозы является повсеместно распространенным в царстве растений.
[0143] Трегалоза синтезируется из UDP-глюкозы и глюкоза-6-фосфата в двух ферментативных реакциях. Вначале UDP-глюкоза и глюкоза-6-фосфат превращаются в UDP (уридиндифосфат) и альфа, альфа-трегалозо-6-фосфат (Т6Р) с помощью фермента TPS (трегалозафосфатсинтаза). Во втором этапе, который катализируется ферментом Т6РР (трегалозо-фосфатфосфатаза), Т6Р дефосфорилируется для получения трегалозы и ортофосфата. (См. фиг. 3)
[0144] У дрожжей две ферментативных активности (TPS и Т6РР активность) присущи большому белковому комплексу, содержащему активные субъединицы, TPS1 и TPS2, и регуляторные субъединицы, с TPS1, имеющей активность TPS и TPS2, имеющей активность Т6РР. У Е.coli две ферментативных активности обнаруживаются в отдельных белковых комплексах. У растений белковый комплекс к настоящему времени не был охарактеризован.
[0145] У Arabidopsis thaliana, биосинтетические ферменты трегалозы были классифицированы на три класса:
[0146] Класс I: содержащий четыре гена, AtTPS1-AtTPS4, имеющие высокое сходство с ScTPS1;
[0147] Класс II: имеющий семерых представителей, AtTPS5-AtTPS11, с высоким сходством последовательностей с ScTPS2; и
[0148] Класс III: содержащий 10 представителей, AtT6PPA-AtT6PPJ, кодирующих белки со сходством с Е.coli TPS2 и С-концом ScTPS2 белков.
[0149] Гены, кодирующие белки в этих классах, также присутствуют в других видах растений.
[0150] В пределах Класса I и Класса II ферментативная активность была однозначно определена только для AtTPS1, проявляющего активность TPS (Blazquez et al. Plant J. March 1998; 13 (5): 685-9.). Неожиданно, к настоящему времени не сообщалось о какой-либо активности Т6РР для любого из других белков TPS Класса II. Напротив, активность Т6РР ранее была описана для AtT6PPA и AtT6PPB, двух из представителей Класса III (Vogel et al. Plant J. March 1998; 13 (5): 673-83). Растительные T6PP Класса III содержат два мотива консенсусной последовательности фосфатазы, обнаруженные во всех ферментах Т6РР, описанных к настоящему времени (Thaller et al. Protein Sci. July 1998; 7 (7): 1647-52).
[0151] Сообщалось, что генетическая манипуляция с генами биосинтеза трегалозы приводила к улучшенной стрессоустойчивости у растений, а также, вызывала четко выраженные изменения в развитии. Сверхэкспрессия генов OtsA и OtsB Е.coli (эквивалентов Т6РР и TPS) в трансгенных растениях табака и картофеля, как сообщалось, вызывала отклонения в развитии корнях и листьях, а также угнетала рост растений. Меньшее количество семян производилось в OtsA трансгенных растениях табака и OtsB трансгенные растения картофеля не производили клубни (Goddijn et al. Plant Physiol. January 1997; 113 (1): 181-90). Сходные результаты были описаны другими авторами (Holmstrom et al. Nature, 379, 683-684; Romero et al. Planta, 201, 293-297; Pilont-Smits et al. 1998; J Plant Physiol. 152: 525-532; Schluepmann et al. Proc Natl. Acad. Sci. USA. 2003; 100 (11): 6849-54). Также сообщалось, что мутанты с дефектом в генах TPS и Т6РР показывают дефекты развития. Мутанты с нокаутированным TPS1 у Arabidopsis показали замедленное развитие зародышей (Eastmond et al. Plant J. January 2002; 29 (2): 225-35). McSteen et al. (Plant Cell 2006; 18; 518-522) упоминают о том, что выделение и получение характеристик гена RAMOSA3 (RA3) маиса, как сообщается, отвечает за развитие меристемы и развитие соцветий, включая ветвление. Это указывает на то, что ген, продукт гена и регуляторные участки могут применяться для управления ветвлением, ростом меристемы, развитием и расположением соцветий. Отрицательные фенотипы, связанные с экспрессией трансгена, могут иметь неблагоприятные воздействия на относительную урожайность растения. Например, без образования семян, налива семян, фертильности растения и т.д. не будет увеличения урожая семян. Заявка на патент США 2007/0006344 является первой записью, касающейся сведений в отношении применения способа, что описывает экспрессию Т6РР в растении с обеспечением увеличения урожайности растений без каких-либо отрицательных фенотипов и/или неблагоприятных эффектов на биологическое функционирование растений. В заявке на патент США 2007/0006344 описывается применение трегалоза-6-фосфатфосфатазы, функционально связанной с промотором OsMADS, который управляет предпочтительной экспрессией Т6РР в отношении материнской репродуктивной ткани растения, что приводит в результате к существенному увеличению урожайности маиса в условиях дефицита воды и условиях хорошего орошения. Далее изобретение в целом включает идентификацию Т6РР, имеющей модификации, которые обеспечивают улучшенную урожайность и полевую производительность сельскохозяйственных культур, и дополнительно описываются способы, которые можно применять для увеличения урожайности растения путем использование растением Т6РР.
[0152] Настоящее изобретение предлагает нуклеотидные последовательности, которые при трансгенной экспрессии в растении увеличивают мощность растений, урожайность, использование азота и/или биомассу при стрессовых и безстрессовых условиях. Было обнаружено, что белки Т6РР, содержащие модификации, которые изменяют активность белка Т6РР, где активность снижена при прямом сравнении с родственной Т6РР, не имеющей модификаций, обеспечивают существенное повышение как урожайности, так и/или повышенную устойчивость к стрессу, когда эти модифицированные Т6РР трансгенно экспрессируются в растении. Не ограничиваясь теорией, эти модификации, вероятно, выполнены в ключевых положениях белка Т6РР, которые оказывают влияние на связывание Т6РР с ее субстратом Т6Р. Теоретически, Т6РР, раскрытые в данном документе, обладающие сниженной активностью, служат в качестве сигнала для растения или инициируют в растении определенные метаболические каскады или метаболические пути, которые обеспечивают в растении увеличенный транспорт сахаров к плодам, зернам, листьям, корням или другим частям, тем самым приводя к повышенной урожайности семян, биомассы и/или росту корня.
[0153] Не ограничиваясь теорией, экспрессия Т6РР, модифицированной для снижения активности и/или модифицированной так, чтобы она имела увеличенный потенциал связывания с эндогенным белком TPS, может применяться для инициации комплекса T6PP/TPS, причем затем присутствие указанного комплекса T6PP/TPS обеспечивает в растении повышенную урожайность и/или повышенную устойчивость к стрессу. В другом варианте осуществления, не ограничиваясь теорией, комплекс T6PP/TPS обеспечивает в растении измененные уровни Т6Р, которые затем обеспечивают у растения повышенную урожайность и/или повышенную устойчивость к стрессу. В одном аспекте настоящего изобретения трансгенный комплекс T6PP/TSPS может трансгенно экспрессироваться в растении с обеспечением повышенной урожайности и/или повышенной стрессоустойчивости у растения. В другом варианте осуществления предполагается, что посредством способов, хорошо известных в соответствующей области техники, могут быть созданы различные химические структуры и/или молекулярные структуры для имитации функции комплекса T6PP/TSPS, такие как описанные в данном документе. В одном варианте осуществления, не ограничиваясь теорией, любую Т6РР можно модифицировать так, чтобы она имела уменьшенное связывание с субстратом Т6Р, что затем будет обеспечивать повышенную урожайность и полевую производительность, когда модифицированные Т6РР трансгенно экспрессируются в растениях. В некоторых случаях нуклеотидные последовательности кодируют белок, вовлеченный в путь биосинтеза трегалозы, при этом в одном варианте осуществления белок кодирует Т6РР. В другом варианте осуществления настоящего изобретения Т6РР выделяют из генома покрытосеменного растения, а затем модифицируют, как описано в данном документе. В дополнительном варианте осуществления настоящего изобретения ген Т6РР выделяют из однодольного растения. В одном аспекте гены Т6РР, раскрытые в данном документе, выделяют из генома растения риса.
[0154] Гены Т6РР, описанные в данном документе, являются, главным образом, представителями надсемейства фосфатаз HAD или содержат белковую консенсусную последовательность, отображенную в SEQ ID NO: 9. Надсемейство фосфатаз HAD, которое также можно называть надсемейством фосфатаз DDDD, включает консервативный альфа/бета-коровый домен, а также кэп-домен, чья функция может изменяться (т.е. связывание с субстратом и т.д.). В одном варианте осуществления настоящего изобретения модификация локализована в любом из кэп-домена, домена фосфатазы, домена А-фосфатазного бокса или домена В-фосфатазного бокса Т6РР, где указанная модификация изменяет связывание с ее субстратом (например, Т6Р). В другом варианте осуществления модификации могут включать одно или несколько из замены, делеции или вставки одного или нескольких аминокислотных остатков для родственной консенсусной последовательности белка Т6РР, представленной в SEQ ID NO: 9, где замена, делеция или вставка одного или нескольких аминокислотных остатков обеспечивает изменение активности указанной Т6РР и/или увеличение уровней Т6Р, когда Т6РР трансгенно экспрессируется в растении. Не ограничиваясь теорией, считают, что создание вариантов белка Т6РР, обладающих сниженной каталитической активностью (т.е. уменьшенным гидролизом Т6Р) создаст сигнал в растении, обеспечивающий повышенную урожайность и/или повышенную устойчивость к стрессу (например, засухе, теплу, холоду). Теоретически, Т6РР, обладающая сниженной активностью, может взаимодействовать с TPS, образуя белок-белковый комплекс.
[0155] Теоретически, Т6РР, модифицированная так, чтобы она обладала сниженной ферментативной активностью, может служить в качестве метаболического сигнала растению направлять сахара на образование семени и/или сахаров. Боле того, эти модифицированные Т6РР могут также служить в качестве метаболических сигналов для инициации процессов, которые обеспечивают в растении повышенную устойчивость к абиотическим и/или биотическим стрессам. Опять-таки, не ограничиваясь теорией, полагают, что растения, экспрессирующие модифицированные Т6РР, обладающие сниженной активностью, будут иметь модифицированный уровень трегалозы по сравнению с контрольным растением или по сравнению с трансгенным растением, сверхэкспрессирующим активную форму Т6РР.
[0156] Предусматривается, что любая Т6РР или фермент, обладающий активностью Т6РР, могут быть разработаны, чтобы обладать сниженной активностью и, таким образом, применяться для создания трансгенных растений или организмов (например, водорослей), имеющих повышенную урожайность. В одном варианте осуществления настоящего изобретения белки, содержащие консенсусную последовательность, такую как представлена в SEQ ID NO: 9, могут применяться в способах, описанных в данном документе. Применяемое в данном документе выражение "сниженная активность" относится к любому уменьшению активности Т6РР. Например, фермент, модифицированный, чтобы иметь 1%, 5%, 10%, 20%, 30%, 40%, 50%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% уменьшение активности Т6РР относительно контрольного или нативного гена Т6РР, может найти применение в получении трансгенных растений с повышенной урожайностью. В одном варианте осуществления настоящего изобретения полипептид Т6РР содержит аминокислотную последовательность, имеющую 50%, 60%, 70%, 80%, 90%, 95% или 99% идентичность последовательности с консенсусной последовательностью, отображенной в SEQ ID NO: 9. В дополнительном варианте осуществления Т6РР содержит консенсусную последовательность, отображенную в SEQ ID NO: 9, где проводится по меньшей мере одна модификация (включая аминокислотную замену, аминокислотную делецию, аминокислотную вставку) и указанная модификация снижает активность указанной Т6РР и/или увеличивает вероятность указанной Т6РР образовывать белок-белковый комплекс с TPS. В другом варианте осуществления растения, экспрессирующие вариант Т6РР, обладающий сниженной активностью, и/или Т6РР, характеризующуюся увеличенной вероятностью образования белок-белкового комплекса с TPS, будут иметь модифицированные уровни Т6Р по сравнению с контролем, не экспрессирующим вариант Т6РР, что не обладает сниженной активностью, где модифицированный уровень Т6Р обеспечивает у растения повышенную урожайность (например, урожайность семян, биомассу, мощность), клеточный рост, содержание сахара, содержание крахмала и повышенную устойчивость к стрессу (засухе, теплу, холоду, засоленности, затоплению, абиотическому и/или биотическому стрессу). В другом варианте осуществления настоящего изобретения экспрессия варианта Т6РР, обладающего сниженной активностью в растении, направлена на репродуктивную ткань растения. В другом варианте осуществления настоящего изобретения паттерны генной экспрессии, вероятно, являются важными в увеличении длительности засухоустойчивости у растения. Например, можно сконструировать кассеты экспрессии для экспрессии модифицированной Т6РР во время цветения растения, тем самым обеспечивая засухоустойчивость на этой стадии развития растения. Аналогично, можно также сконструировать кассеты экспрессии, которые конститутивно экспрессируют модифицированную Т6РР в растении, тем самым обеспечивая засухоустойчивость в течение всего жизненного цикла растения. Предполагается, что Т6РР могут быть смоделированы, чтобы они обладали сниженной активностью, посредством модификаций вне консервативных аминокислотных остатков консенсусной последовательности, представленной в SEQ ID NO: 9, где модификация уменьшает связывание Т6РР с Т6Р и/или указанные модификации увеличивают вероятность того, что Т6РР будет образовывать белок-белковое взаимодействие с TPS.
[0157] В одном варианте осуществления можно снизить активность Т6РР путем облучения ДНК и, таким образом, получая варианты, обладающие сниженной активностью при экспрессии в растении. Аналогично, предполагается, что для получения вариантов Т6РР, обладающих сниженной активностью, можно применять общепринятые методики, такие как перестановка генов. Предусматривается, что можно создать химические ортогональные структуры, которые могут до некоторой степени имитировать структуру неактивного белка Т6РР и в одном варианте осуществления образовывать комплекс с TPS растения, тем самым обеспечивая повышенную урожайность растения. В другом варианте осуществления можно создать химическую ортогональную структуру, что может имитировать структуру комплекса T6PP/TPS, которую затем можно применять по отношению к растениям (например, применение химического продукта) для обеспечения повышенной урожайности и стрессоустойчивости (т.е. засухоустойчивости). Предусматривается, что химические ортогональные структуры можно применять по отношению к растению для регуляции транспорта сахаров на данной репродуктивной стадии или стадии развития растения.
[0158] Предполагается, что гены Т6РР из любого класса или семейства можно применять в любом из способов и/или аспектов настоящего изобретения, раскрытых в данном документе. Также предполагается, что гены Т6РР со сходной гомологией последовательностей (например, большей или равной 50% 75%, 80% или 85% идентичности последовательностей с SEQ ID NO: 1 и/или молекулярной структурой, выделенной или синтезированной, можно применять в вариантах осуществления, описанных в данном документе. Было показано, что Т6РР повсеместно распространена среди представителей царства растений и во многих случаях, как было показано, имеет консервативную функцию у многочисленных родов и видов растений. Следовательно, предполагается, что синтетическая нуклеиновая кислота, кодирующая белок Т6РР, обеспечивающий сниженную активность, и/или уменьшенное связывание с Т6Р, и/или увеличенное связывание с TPS, может применяться в способах, раскрытых в данном документе, для обеспечения растений с подходящими характеристиками (например, повышенной урожайностью, увеличенной биомассой, увеличенной мощностью растений), описанными в данном документе.
[0159] Способы, раскрытые в данном документе, дополнительно описывают трансформацию растений с полинуклеотидами, такими как раскрытые в данном документе. Трансгенные растения, содержащие полинуклеотиды, раскрытые в данном документе, могут проявлять увеличенный клеточный рост, увеличенную мощность растений и/или всходов, повышенную урожайность, увеличенный вес семян и увеличенную биомассу. Предполагается, что растения, полученные с помощью способов по данному документу, имеют повышенную устойчивость к абиотическому стрессу. Предусматривается, что трансгенные растения по настоящему изобретению будут обеспечивать увеличенный рост корня, что может иметь много положительных последствий касательно использования воды и коэффициента использования питательных веществ растения, а также снижения частоты возникновения эрозии почвы на коммерческих сельскохозяйственных угодьях. Также предполагается, что растения, содержащие полинуклеотиды, описанные в данном документе, могут также проявлять более высокую устойчивость к поеданию насекомыми вследствие увеличенного роста. В некоторых случаях растения, описанные в настоящем изобретении, будут иметь повышенную стойкость к полеганию и/или сниженный риск полегания культуры, тем самым обеспечивая трансгенные растения, которые могут переносить погодные условия, такие как сильные ветра или град. Трансгенные растения, описанные в данном документе, могут давать большой урожай как в отношении урожайности биомассы, так и урожайности зерна растения. Один аспект настоящего изобретения предлагает различные модификации, которые можно выполнить на данной последовательности гена Т6РР, которые могут применяться в трансгенных растениях для обеспечения повышенной урожайности. Трансгенные растения по данному документу могут обеспечивать повышение урожайности как при оптимальных, так и/или при неоптимальных условиях произростания.
[0160] Любая ссылка в данном документе ниже на "белок, применимый в способах настоящего изобретения" используется для обозначения полипептида трегалозного пути, определенного в данном документе, включая без ограничения полипептид Т6РР. Любая ссылка в данном документе ниже на "нуклеиновую кислоту, применимую в способах настоящего изобретения" используется для обозначения нуклеиновой кислоты, способной кодировать полипептид трегалозного пути, такой как полипептид Т6РР.
[0161] Предпочтительный способ регулирования экспрессии нуклеиновой кислоты, кодирующей белок Т6РР, применимый в способах настоящего изобретения, заключается во введении в растение и экспрессии нуклеиновой кислоты, кодирующей белок, применимый в способах настоящего изобретения, определенных ниже.
[0162] Нуклеиновая кислота, подлежащая введению в растение (и, следовательно, применимая в проведении способов настоящего изобретения), представляет собой любую нуклеиновую кислоту, кодирующую фермент трегалозного пути. Выражение "полипептид Т6РР", определенное в данном документе, относится к любому полипептиду с трегалоза-6-фосфатфосфатазной активностью, содержащему по меньшей мере один домен трегалоза-фосфатазы. Домены трегалоза-фосфатазы обычно составляют от 200 до 250 аминокислот в длину и обычно содержат мотив консенсусной последовательности фосфатазы, который обнаруживается во всех ферментах Т6РР, описанных к настоящему времени (Thaller et al. 1998). Аминокислотная последовательность домена трегалоза-фосфатазы представлена в SEQ ID NO: 10. Специалист в данной области без труда сможет выявить присутствие домена трегалоза-фосфатазы с применением инструментов и методик, известных в данной области. Этот мотив консенсусной последовательности фосфатазы обычно содержит два фосфатазных бокса, названных А и В-фосфатазным боксом. SEQ ID NO: 11 представляет консенсусную последовательность для фосфатазного бокса A, a SEQ ID NO: 12 представляет консенсусную последовательность для фосфатазного бокса В.
[0163] Предпочтительно, нуклеиновая кислота, подлежащая введению в растение, кодирует белок Т6РР, имеющий по меньшей мере 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательности с последовательностью, выбранной из SEQ ID NO 1, 3, 5 и 7. Наиболее предпочтительно, нуклеиновая кислота класса Т6РР является такой, как представлено любой из SEQ ID NO 1, 3, 5 и 7.
[0164] Примеры белков, применимых в способах настоящего изобретения, и нуклеиновых кислот, кодирующих их, представлены в данном документе в таблице примера 5.
[0165] Также применимыми в способах настоящего изобретения являются гомологи любой из аминокислотных последовательностей, представленных в таблице примера 5. "Гомологи" белка охватывают пептиды, олигопептиды, полипептиды, белки и ферменты, имеющие аминокислотные замены, делеции и/или вставки по сравнению с рассматриваемым немодифицированным белком и обладающие биологической и функциональной активностью, как у немодифицированного белка, из которого они получены.
[0166] Делеция относится к удалению одной или нескольких аминокислот из белка.
[0167] Вставка относится к одному или нескольким аминокислотным остаткам, вводимым в заданный сайт белка. Вставки могут содержать N-концевые и/или С-концевые слияния, а также вставки одной или множества аминокислот внутри последовательности.
[0168] Как правило, вставки в пределах аминокислотной последовательности будут меньше, чем N- или С-концевые слияния, порядка примерно 1-10 остатков. Примеры N- или С-концевых частей слияния белков или пептидов включают домен связывания или домен активации активатора транскрипции, применяемого в дрожжевой двухгибридной системе, белки оболочки фага, метку (гистидин)-6, метку глутатитон-S-трансферазы, белок А, белок, связывающий мальтозу, дигидрофолатредуктазу, эпитоп Tag⋅100, эпитоп с-myc, эпитоп FLAG®, lacZ, CMP (пептид, связывающий кальмодулин), эпитоп HA, эпитоп белка С и эпитоп VSV.
[0169] Замена относится к замещению аминокислот белка другими аминокислотами, имеющими сходные свойства (такие как сходную гидрофобность, гидрофильность, антигенность, предрасположенность к образованию или разрушению α-спиральных структур или β-складчатых структур). Аминокислотные замены обычно являются отдельными остатками, но могут быть объединены в кластеры в зависимости от функциональных ограничений, наложенных на полипептид; вставки будут составлять, как правило, порядка примерно 1-10 аминокислотных остатков. Аминокислотные замены предпочтительно представляют собой консервативные аминокислотные замены. Таблицы консервативных замен хорошо известны в данной области техники (см., например, Creighton (1984) Proteins. W.H. Freeman and Company, и таблицу ниже).
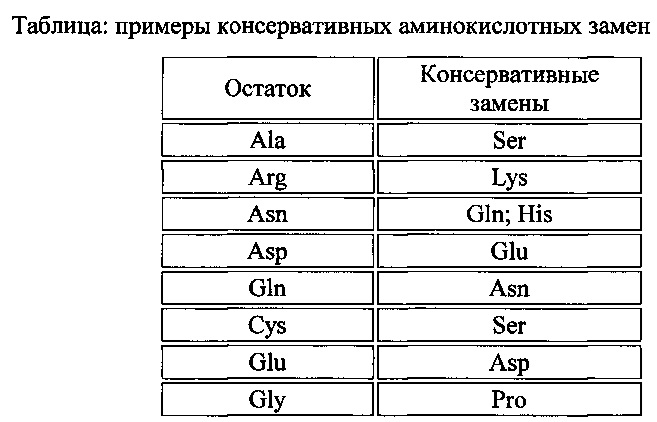
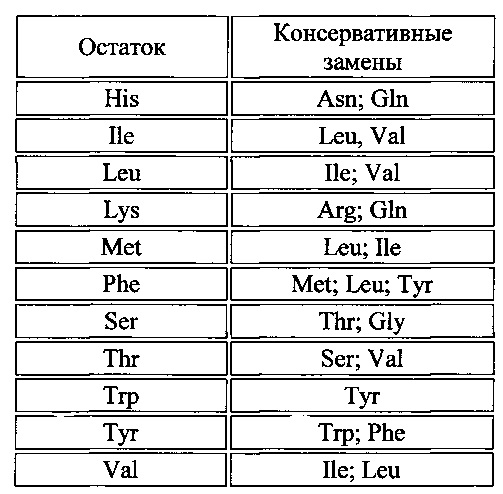
[0170] Аминокислотные замены, делеции и/или вставки могут без труда осуществляться с применением методик пептидного синтеза, хорошо известных в данной области техники, таких как твердофазный пептидный синтез и подобное, или путем манипуляции с рекомбинантной ДНК. Способы манипуляции с последовательностями ДНК для получения вариантов замены, вставки или делеции у белка, хорошо известны в данной области техники. Например, методики создания модификаций замены на заданных сайтах в ДНК хорошо известны специалистам в данной области и включают мутагенез М13, in vitro мутагенез T7-Gen (USB, Кливлэнд, Огайо), сайт-направленный мутагенез QuickChange (Stratagene, Сан-Диего, Калифорния), опосредованный PCR сайт-направленный мутагенез или другие протоколы сайт-направленного мутагенеза.
[0171] Также применимыми в способах настоящего изобретения являются производные любого из полипептидов, представленных в таблице примера 5, или ортологи или паралоги любого из полипептидов, представленных в таблице примера 5, или производные любых ортологов или паралогов любого из полипептидов, представленных в таблице примера 5. "Производные" включают пептиды, олигопептиды, полипептиды, которые могут, по сравнению с аминокислотной последовательностью встречающейся в природе формы белка, содержать замены аминокислот на не встречающиеся в природе аминокислотные остатки или вставки не встречающихся в природе аминокислотных остатков. Производные полипептидов, представленных в таблице примера 5, представляют собой дополнительные примеры, которые могут быть подходящими для применения в способах настоящего изобретения. Некоторые аспекты настоящего изобретения включают модификацию остатков, что влияет на активность молекулы Т6РР. В одном варианте осуществления модификация уменьшает активность молекулы Т6РР при анализе in vitro. В другом аспекте модификация снижает аффинность связывания Т6РР с ее субстратом. В другом аспекте модификации могут быть выполнены в активном сайте белка Т6РР таким образом, что активность белка снижается по сравнению с белком Т6РР без модификации.
[0172] "Производные" полипептида включают пептиды, олигопептиды, полипептиды, которые могут, по сравнению с аминокислотной последовательностью встречающейся в природе формы белка, такой как белок, представляющий интерес, содержать замещения аминокислот на не встречающиеся в природе аминокислотные остатки или вставки не встречающихся в природе аминокислотных остатков. "Производные" белка также охватывают пептиды, олигопептиды, полипептиды, которые содержат встречающиеся в природе измененные (гликозилированные, ацилированные, пренилированные, фосфорилированные, миристоилированные, сульфатированные и т.д.) или не встречающиеся в природе измененные аминокислотные остатки по сравнению с аминокислотной последовательностью встречающейся в природе формы полипептида. Производное может также содержать один или несколько не аминокислотных заместителей или вставок по сравнению с аминокислотной последовательностью, из которой оно получено, например, репортерную группу или другой лиганд, ковалентно или нековалентно связанный с аминокислотной последовательностью, такую как репортерная группа, которая связывается для облегчения его обнаружения, и не встречающиеся в природе аминокислотные остатки по сравнению с аминокислотной последовательностью встречающегося в природе белка. Кроме того, "производные" также включают слияния встречающейся в природе формы белка с пептидами для мечения, такими как FLAG, HIS6 или тиоредоксин (для обзора пептидов для мечения см. Terpe, Appl. Microbiol. Biotechnol. 60, 523-533, 2003).
[0173] Изобретение показано путем экспрессирования в растении различных белков Т6РР, имеющих модификации, которые уменьшают активность молекулы Т6РР. На удивление, эти модификации и/или уменьшенная активность молекул Т6РР, как описано в данном документе, показывают повышенную урожайность, а также полевую производительность, состоящую в том, что растения являются здоровыми и не показывают неблагоприятных фенотипических эффектов. Выполнение изобретения не ограничено этими последовательностями. Способы по настоящему изобретению можно преимущественно выполнять при помощи любой нуклеиновой кислоты, кодирующей белок, применимый в способах по настоящему изобретению, определенных в данном документе, включая нуклеиновые кислоты, кодирующие ортологи, паралоги и гомологи, такие как (без ограничения) любые последовательности нуклеиновых кислот, данные в таблице примера 5.
[0174] Ортологи и паралоги охватывают эволюционные концепции, используемые для описания родственных отношений генов. Паралоги представляют собой гены в пределах таких же видов, которые получены вследствие дупликации предкового гена, а ортологи представляют собой гены различных организмов, которые получены вследствие видообразования и также происходят от общего предкового гена. [0175] Ортологи и паралоги можно легко найти путем выполнения так называемого взаимно-обратного поиска blast. Как правило, он включает первый BLAST, включающий проведение BLAST интересующей последовательности (например, используя любую из последовательностей, перечисленных в таблице примера 5) по отношению к любой последовательности из базы данных, такой как общедоступная база данных NCBI. BLASTN или TBLASTX (используя стандартные величины по умолчанию) обычно применяют, когда начинают с нуклеотидной последовательности, a BLASTP или TBLASTN (используя стандартные величины по умолчанию), когда начинают с белковой последовательности. Результаты BLAST можно необязательно отфильтровать. Затем проводят обратно BLAST полноразмерных последовательностей либо отфильтрованных результатов, либо неотфильтрованных результатов (второй BLAST) по отношению к последовательностям из организма, из которого получена интересующая последовательность. Результаты первого и второго BLAST затем сравнивают. Паралог идентифицируют тогда, когда высокоранговое совпадение из первого blast состоит не из тех же частиц, из которых получена интересующая последовательность, обратный BLAST в идеальном случае затем приводит к интересующей последовательности с наибольшим совпадением; ортолог идентифицируют тогда, когда высокоранговое совпадение в первом BLAST состоит не из тех же частиц, из которых получена интересующая последовательность, и предпочтительно приводит при обратном BLAST к интересующей последовательности, находящейся среди наибольших совпадений.
[0176] Высокоранговыми совпадениями являются такие, которые имеют низкое Е-значение. Чем ниже Е-значение, тем более достоверный балл (или другими словами, ниже вероятность, что совпадение было обнаружено случайно). С вычислением Е-значения хорошо знакомы в данной области техники. В дополнение к Е-значениям сходство также оценивают по процентной идентичности. Процентная идентичность относится к числу идентичных нуклеотидов (или аминокислот) в двух сравниваемых последовательностях нуклеиновой кислоты (или полипептида) на конкретной длине. В случае больших семейств для облегчения визуализации образования кластеров родственных генов и идентификации ортологов и паралогов можно использовать ClustalW с последующим построением дерева методом ближайшего связывания.
[0177] В таблице примера 5 приведены примеры ортологов и паралогов белков Т6РР, представленных SEQ ID NO 2, 4, 6 и 8. Дополнительные ортологи и паралоги можно легко идентифицировать при помощи процедуры BLAST, описанной выше.
[0178] Белки по настоящему изобретению являются идентифицируемыми из-за наличия консервативного трегалоза-фасфатазного домена(ов). Выражение "домен" относится к набору консервативных аминокислот в конкретных положениях вдоль выравнивания последовательностей эволюционно родственных белков. Несмотря на то, что аминокислоты в других положениях могут отличаться от гомологов, аминокислоты, которые являются высококонсервативными в конкретных положениях, указывают на аминокислоты, которые являются важными в отношении структуры, стабильности или активности белка. Будучи идентифицированными по их высокой степени консервативности в выравненных последовательностях семейства гомологов белков, они могут применяться в качестве идентификаторов для определения того, принадлежит ли любой интересующий полипептид ранее идентифицированному семейству полипептидов (в этом случае белки, применимые в способах по настоящему изобретению, и нуклеиновые кислоты, кодирующие их, определенные в данном документе). В предпочтительном варианте осуществления настоящего изобретения в эти высоко консервативные домены могут быть введены одна или несколько модификаций для уменьшения активности конкретного белка Т6РР, что тем самым, при экспрессии в растении, обеспечивает повышение урожайности.
[0179] Выражение "мотив", или "консенсусная последовательность", или "сигнатура" относится к короткому консервативному участку в последовательности эволюционно родственных белков. Мотивы зачастую являются высококонсервативными частями доменов, но также могут включать только часть домена или быть расположенными за пределами консервативного домена (если все аминокислоты мотива оказываются за пределами определенного домена).
[0180] Специализированные базы данных также существуют для идентификации доменов, например, SMART (Schultz et al. (1998) Proc. Natl. Acad. Sci. USA 95, 5857-5864; Letunic et al. (2002) Nucleic Acids Res 30, 242-244, InterPro (Mulder et al., (2003) Nucl. Acids. Res. 31, 315-318, Prosite (Bucher and Bairoch (1994), A generalized profile syntax for biomolecular sequences motifs and its function in automatic sequence interpretation. (In) ISMB-94; Proceedings 2nd International Conference on Intelligent Systems for Molecular Biology. Airman R., Brutlag D., Karp P., Lathrop R., Searls D., Eds., pp 53-61, AAAIPress, Menlo Park; Hulo et al., Nucl. Acids. Res. 32: D134-D137, (2004), или Pfam (Bateman et al., Nucleic Acids Research 30 (1): 276-280 (2002). Набор инструментов для in silico анализа белковых последовательностей доступен на сервере протеомики ExPASY (размещенном Швейцарским институтом биоинформатики (Gasteiger et al., ExPASy: the proteomics server for in-depth protein knowledge and analysis, Nucleic Acids Res. 31: 3784-3788 (2003)).
[0181] Домены можно также идентифицировать при помощи обычных методик, таких как выравнивание последовательностей. В данной области техники хорошо известны способы выравнивания последовательностей для сравнения, при этом такие способы включают GAP, BESTFIT, BLAST, FASTA и TFASTA. В GAP используется алгоритм Needleman и Wunsch ((1970) J Mol Biol 48: 443-453), для обнаружения глобального (т.е. перекрытия полных последовательностей) выравнивания двух последовательностей, что максимизирует число совпадений и минимизирует число пробелов. Алгоритм BLAST (Altschul et al. (1990) J Mol Biol 215: 403-10) рассчитывает процентную идентичность последовательности и осуществляет статистический анализ сходства между двумя последовательностями. Программное обеспечение для проведения анализа BLAST общедоступно благодаря Национальному центру биотехнологической информации (NCBI). Гомологи можно легко идентифицировать при помощи, например, алгоритма множественного выравнивания последовательностей ClustalW (версия 1.83) при отсутствии параметров попарного выравнивания и способа оценки в процентах. Общие проценты сходства и идентичности можно также определить при помощи одного из способов, доступных в пакете программного обеспечения MatGAT (Campanella et al., BMC Bioinformatics. July 2003 10: 4-29. MatGAT: приложение, которое генерирует матрицы сходства/идентичности при помощи белковых последовательностей или последовательностей ДНК). Для оптимизации выравнивания между консервативными мотивами можно проводить незначительное редактирование вручную, что будет понятно специалисту в данной области. Кроме того, для идентификации гомологов вместо использования полноразмерных последовательностей также можно использовать конкретные домены (такие как трегалоза-фосфатазный домен или один из мотивов, определенных выше).
[0182] Кроме того, белки Т6РР (по меньшей мере в их нативной форме) обычно обладают трегалоза-6-фосфатфосфатазной активностью. Полипептиды с трегалоза-6-фосфатфосфатазной активностью согласно классификации Комиссии по ферментам Комитета по номенклатуре Международного союза по биохимии и молекулярной биологии (NC-IUBMB) принадлежат классу ферментов EC:3.1.3.12. Ферменты класса EC:3.1.3.12 катализируют реакцию: трегалоза-6-фосфат+H2O=трегалоза+фосфат. Предполагается, что любую Т6РР или белок, обладающий активностью Т6РР, посредством способов, таких как точечная мутация(и), облучение и пр., можно модифицировать для уменьшения активности с целью обеспечения положительных эффектов, описанных в данном документе, при трансгенной экспрессии в растении.
[0183] Активность трегалоза-6-фосфатфосфатазного белка может быть измерена путем определения уровней подвергнутого обработке субстрата и уровней накопленного в in vitro реакции продукта, а именно, путем определения уровня использования трегалоза-6-фосфата и/или накопления трегалозы в результате реакции. Ферментные способы измерения трегалозы могут основываться на гидролизе трегалозы до глюкозы, такие как описанные Van Dijck et al. Biochem J. August 2002 15; 366 (Pt 1): 63-71, и Zentella et al. Plant Physiol. April 1999; 119 (4): 1473-82.
[0184] Уровни трегалоза-6-фосфата также можно измерить при помощи способов HPLC (высокоэффективная жидкостная хроматография), описанных Avonce et al. Plant Physiol. November 2004; 136 (3): 3649-59; Schluepmann et al. 2003. Также описаны альтернативные способы, основанные на определении высвобождения неорганического фосфата из трегалоза-6-фосфата, Klutts et al. J Biol Chem. (2003) 278 (4): 2093-100. Альтернативный способ определения уровней трегалоза-6-фосфата при помощи жидкостной хроматографии совместно с MS-Q3 (тройная квадрупольная MS) был описан Lunn et al. Biochem J. (2006) 397 (1): 139-48.
[0185] Примеры нуклеиновых кислот, пригодных для использования в выполнении способов по настоящему изобретению, включают последовательности нуклеиновых кислот, приведенных в таблице примера 5, но не ограничены этими последовательностями. В осуществлении способов по настоящему изобретению также могут быть пригодными варианты нуклеиновых кислот. Примеры таких вариантов нуклеиновых кислот включают части нуклеиновых кислот, кодирующих белок, применимый в способах по настоящему изобретению, нуклеиновые кислоты, гибридизующиеся с нуклеиновыми кислотами, кодирующими белок, применимый в способах по настоящему изобретению, сплайс-варианты нуклеиновых кислот, кодирующих белок, применимый в способах по настоящему изобретению, аллельные варианты нуклеиновых кислот, кодирующих белок, применимый в способах по настоящему изобретению, и варианты нуклеиновых кислот, кодирующих белок, применимый в способах по настоящему изобретению, которые получены перетасовкой генов. Выражения часть, гибридизующаяся последовательность, сплайс-вариант, аллельный вариант и перетасовка генов не будут описываться.
[0186] Нуклеиновые кислоты, кодирующие белки, применимые в способах по настоящему изобретению, не должны быть полноразмерными нуклеиновыми кислотами, поскольку выполнение способов по настоящему изобретению не рассчитано на использование последовательностей полноразмерных нуклеиновых кислот. Участки, применимые в способах по настоящему изобретению, кодируют полипептид, подпадающий под определение нуклеиновой кислоты, кодирующей белок, применимый в способах по настоящему изобретению, как определено в данном документе. Предпочтительно участок представляет собой участок любой из нуклеиновых кислот, приведенных в таблице примера 5. Участок обычно составляет по меньшей мере 625 последовательных нуклеотидов в длину, предпочтительно по меньшей мере 825 последовательных нуклеотидов в длину, или по меньшей мере 1025 последовательных нуклеотидов в длину, или по меньшей мере 1125 последовательных нуклеотидов в длину, причем последовательные нуклеотиды, принадлежащие любой из последовательностей нуклеиновых кислот, приведенных в таблице примера 5, в комбинации с предложенной мутацией(ями), раскрытой в данном документе, будут уменьшать активность родственного белка Т6РР по сравнению с такой же Т6РР, не имеющей одной или нескольких модификаций. В одном аспекте изобретения участок представляет собой участок нуклеиновой кислоты SEQ ID NO: 1. Предпочтительно, участок кодирует аминокислотную последовательность, которая при использовании в построении филогенетического дерева T6PP/TPS, такого как представлено на фиг. 2, имеет тенденцию образовывать кластер с группой белков Т6РР, содержащих аминокислотную последовательность, представленную SEQ ID NO: 2, а не с любой другой группой.
[0187] Участок нуклеиновой кислоты, кодирующей белок Т6РР, определенный в данном документе, можно получить, например, путем осуществления одной или нескольких делеций нуклеиновой кислоты. Участки могут использоваться в выделенной форме или они могут быть слиты с другими кодирующими (или некодирующими) последовательностями для, например, получения белка, который совмещает в себе несколько активностей. При слиянии с другими кодирующими последовательностями образующийся полипептид, получаемый при трансляции, может быть больше полипептида, прогнозируемого для части белка Т6РР. Также предполагается, что молекулы Т6РР, не обладающие активностью Т6РР, могут обеспечивать повышение урожайности при трансгенной экспрессии в растении.
[0188] Согласно настоящему изобретению предусматривается способ улучшения связанных с урожайностью свойств растений, в частности повышения урожайности семян, включающий введение и экспрессию в растении части любой из последовательностей нуклеиновых кислот, приведенных в таблице примера 5, или части нуклеиновой кислоты, кодирующей ортолог, паралог или гомолог любой из аминокислотных последовательностей, приведенных в таблице примера 5, где белки были модифицированы таким образом, чтобы снизить активности белка Т6РР.
[0189] Другой вариант нуклеиновой кислоты, пригодный в способах по настоящему изобретению, представляет собой нуклеиновую кислоту, способную к гибридизации при условиях уменьшенной жесткости, предпочтительно при жестких условиях, с нуклеиновой кислотой, кодирующей белок Т6РР, определенный в данном документе, или с частью, определенной в данном документе. В предпочтительном аспекте белок Т6РР будет модифицироваться для обладания уменьшенной активностью по сравнению с немодифицированной версией такого же белка Т6РР, тем самым обеспечивая у трансгенного растения любое из следующего: повышенную урожайность, увеличенную биомассу, повышенную устойчивость к абиотическому и биотическому стрессу, повышенную мощность растения, повышенное содержание сахара, повышенное содержания масла, увеличенный вес семян, увеличенный рост корня, повышенную эффективность использования воды, повышенную засухоустойчивость, повышенную стойкость к полеганию, повышенную эффективность использования питательных веществ (например, азота) и более быстрое развитие растений.
[0190] Гибридизируемые последовательности, пригодные в способах по настоящему изобретению, кодируют полипептид, имеющий трегалоза-фосфатазный домен и обладающий практически такой же биологической активностью, что и белки Т6РР, представленные любой из аминокислотных последовательностей, приведенных в таблице примера 5. Гибридизируемая последовательность обычно представляет собой по меньшей мере 625 последовательных нуклеотидов в длину, предпочтительно по меньшей мере 825 последовательных нуклеотидов в длину, более предпочтительно по меньшей мере 1025 последовательных нуклеотидов в длину и наиболее предпочтительно по меньшей мере 1125 последовательных нуклеотидов в длину, причем последовательные нуклеотиды принадлежат любой из последовательностей нуклеиновых кислот, приведенных в таблице примера 5. Предпочтительно, гибридизируемая последовательность является такой, которая способна к гибридизации с любой из нуклеиновых кислот, приведенных в таблице примера 5, или с частью любой из этих последовательностей, причем часть определена выше. Наиболее предпочтительно, гибридизируемая последовательность способна к гибридизации с нуклеиновой кислотой, представленной SEQ ID NO: 1, или с ее частью. В другом предпочтительном аспекте последовательность содержит по меньшей мере одну модификацию, которая снижает активность родственного белка Т6РР.
[0191] Предпочтительно, гибридизируемая последовательность кодирует аминокислотную последовательность, которая при использовании в построении филогенетического дерева T6PP/TPS, такого как изображено на фиг. 2А, имеет тенденцию образовывать кластер с группой белков Т6РР, содержащих аминокислотную последовательность, представленную SEQ ID NO: 2, а не с любой другой группой.
[0192] Наиболее предпочтительно, выделенная молекула полинуклеотида способна к гибридизации при жестких условиях с последовательностью, представленной одной из SEQ ID NO 1, 3, 5 и 7.
[0193] Согласно настоящему изобретению предусматривается способ улучшения связанных с урожайностью свойств растений, в частности повышения урожайности семян, включающий введение и экспрессию в растении нуклеиновой кислоты, способной к гибридизации с любой из нуклеиновых кислот, приведенных в таблице примера 5, или включающий введение и экспрессию в растении нуклеиновой кислоты, способной к гибридизации с нуклеиновой кислотой, кодирующей ортолог, паралог или гомолог любой из последовательностей нуклеиновых кислот, приведенных в таблице примера 5, где последовательность была модифицирована таким образом, чтобы снизить активность Т6РР закодированного белка.
[0194] Другой вариант нуклеиновой кислоты, пригодный в способах по настоящему изобретению, представляет собой сплайс-вариант, кодирующий белок Т6РР, определенный в данном документе выше. Выражение "сплайс-вариант", как используется в данном документе, охватывает варианты последовательности нуклеиновой кислоты, в которой выбранные интроны и/или экзоны были отсечены, заменены, замещены или добавлены, или в которой интроны были укорочены или удлинены. Эти варианты будут такими, при которых биологическая активность белка практически сохраняется или снижается во многих аспектах настоящего изобретения; это может быть достигнуто путем селективного удержания или делеции функциональных сегментов белка. Такие сплайс-варианты могут встречаться в природе или могут быть получены человеком. Способы прогнозирования и выделения таких сплайс-вариантов хорошо известны в данной области техники (смотри, например, Foissac and Schiex (2005) BMC Bioinformatics 6: 25).
[0195] Согласно настоящему изобретению предусматривается способ улучшения связанных с урожайностью свойств растений, в частности повышения урожайности семян, включающий введение и экспрессию в растении сплайс-варианта любой из последовательностей нуклеиновых кислот, приведенных в таблице примера 5, или сплайс-варианта нуклеиновой кислоты, кодирующей ортолог, паралог или гомолог любой из аминокислотных последовательностей, приведенных в таблице примера 5, где эти молекулы дополнительно модифицируются, чтобы они обладали сниженной активностью.
[0196] Предпочтительными сплайс-вариантами являются сплайс-варианты нуклеиновой кислоты, представленной SEQ ID NO: 1, или сплайс-вариант нуклеиновой кислоты, кодирующей ортолог или паралог SEQ ID NO: 2. Предпочтительно аминокислотная последовательность, закодированная сплайс-вариантом, содержит любой один или несколько мотивов или доменов, определенных в данном документе. Предпочтительно, аминокислотная последовательность, закодированная сплайс-вариантом, при использовании в построении филогенетического дерева T6PP/TPS, такого как изображено на фиг. 2А, имеет тенденцию образовывать кластер с группой белков класса Т6РР, содержащих аминокислотную последовательность, представленную SEQ ID NO: 2, а не с любой другой группой.
[0197] Другой вариант нуклеиновой кислоты, пригодный для выполнения способов по настоящему изобретению, представляет собой аллельный вариант нуклеиновой кислоты, кодирующей белок Т6РР, определенный в данном документе выше. Аллели или аллельные варианты являются альтернативными формами данного гена, расположенными в таком же хромосомном положении. Аллельные варианты существуют в природе, и охваченным в способах по настоящему изобретению является использование этих природных аллелей. Аллельные варианты охватывают полиморфизмы отдельных нуклеотидов (SNP), а также полиморфизмы небольшой вставки/делеции (INDEL). Размер INDEL обычно составляет меньше 100 п.о. SNP и INDEL образуют наибольший набор вариантов последовательностей во встречающихся в природе полиморфных штаммах большинства организмов. Аллельные варианты, пригодные в способах по настоящему изобретению, обладают практически такой же биологической активностью, что и белок Т6РР SEQ ID NO: 2, и могут быть мутированными в положениях в пределах белка, которые соответствуют любому из связывания с субстратом, ферментативной активности, вторичной или третичной структуры.
[0198] Согласно настоящему изобретению предусматривается способ улучшения связанных с урожайностью свойств растений, в частности повышения урожайности семян, включающий введение и экспрессию в растении аллельного варианта любой из нуклеиновых кислот, приведенных в таблице примера 5, или включающий введение и экспрессию в растении аллельного варианта нуклеиновой кислоты, кодирующей ортолог, паралог или гомолог любой из аминокислотных последовательностей, приведенных в таблице примера 5.
[0199] Предпочтительно, аллельный вариант представляет собой аллельный вариант SEQ ID NO: 1, 3, 4, 5 или аллельный вариант нуклеиновой кислоты, кодирующей ортолог или паралог SEQ ID NO: 2. Предпочтительно, аминокислотная последовательность, закодированная аллельным вариантом, содержит любой один или несколько мотивов или доменов, определенных в данном документе. Предпочтительно, аминокислотная последовательность, закодированная аллельным вариантом, при использовании в построении филогенетического дерева TPS/T6PP, такого как изображено на фиг. 2, имеет тенденцию образовывать кластер с группой белков класса Т6РР, содержащих аминокислотную последовательность, представленную SEQ ID NO: 2, а не с любой другой группой.
[0200] Дополнительный вариант нуклеиновой кислоты, пригодный в способах по настоящему изобретению, представляет собой вариант нуклеиновой кислоты, полученный перетасовкой генов. Перетасовка генов или направленная эволюция может также применяться для образования вариантов нуклеиновых кислот, кодирующих белки Т6РР со сниженной активностью, как определено выше. Это состоит из итераций перетасовки ДНК с последующим подходящим скринингом и/или отбором с образованием вариантов нуклеиновых кислот или их частей, кодирующих белки Т6РР, определенные выше, обладающие модифицированной (сниженной) биологической активностью (Castle et al., (2004) Science 304 (5674): 1151-4; патенты США №№5811238 и 6395547).
[0201] Согласно настоящему изобретению, предусматривается способ улучшения связанных с урожайностью свойств растений, включающий введение и экспрессию в растении варианта любой из последовательностей нуклеиновых кислот, приведенных в таблице примера 5, или включающий введение и экспрессию в растении варианта нуклеиновой кислоты, кодирующей ортолог, паралог или гомолог любой из аминокислотных последовательностей, приведенных в таблице примера 5, причем вариантную нуклеиновую кислоту получают перетасовкой генов. В предпочтительном варианте осуществления будет проводиться касательно Т6РР, имеющих более низкую степень связывания с субстратом и/или активность.
[0202] Предпочтительно, вариантная нуклеиновая кислота, полученная перетасовкой генов, кодирует аминокислотную последовательность, содержащую любой один или несколько мотивов или доменов, определенных в данном документе. Предпочтительно, аминокислотная последовательность, кодируемая вариантной нуклеиновой кислотой, полученной перетасовкой генов, при использовании в построении филогенетического дерева TPS/T6PP, такого как изображено на фиг. 2А, имеет тенденцию образовывать кластер с группой белков Т6РР, содержащих аминокислотную последовательность, представленную SEQ ID NO: 2, а не с любой другой группой.
[0203] Кроме того, варианты нуклеиновых кислот можно также получить сайт-направленным мутагенезом. Для достижения сайт-направленного мутагенеза доступны несколько способов, при этом наиболее общими являются способы, основанные на PCR (Current Protocols in Molecular Biology. Wiley Eds.). Один аспект настоящего изобретения заключается в использовании сайт-направленного мутагенеза таким образом, чтобы снизить способность белка Т6РР связываться с его субстратом.
[0204] Нуклеиновые кислоты, кодирующие белки Т6РР, могут быть получены из любого природного или искусственного источника. Нуклеиновая кислота может быть модифицирована из ее нативной формы в композицию и/или геномную среду посредством намеренной обработки человеком. Предпочтительно нуклеиновая кислота, кодирующая Т6РР, получена из растения, более предпочтительно из однодольного растения, еще более предпочтительно из семейства Oryza; наиболее предпочтительно нуклеиновая кислота получена из риса.
[0205] Настоящее изобретение также охватывает растения или их части (включая семена), получаемые способами согласно настоящему изобретению. Растения или их части содержат трансген нуклеиновой кислоты, кодирующий белок Т6РР, который был специально или неспециально модифицирован так, чтобы он обладал сниженной активностью.
[0206] Изобретение также предусматривает неизвестные до сих пор последовательности нуклеиновых кислот Т6РР и белковые последовательности Т6РР, которые можно модифицировать, как описано в данном документе, для способов обеспечения у растений повышенной урожайности. Эти последовательности также могут быть пригодны при выполнении способов по настоящему изобретению.
[0207] Согласно следующему варианту осуществления по настоящему изобретению, в данном документе в силу вышеизложенного предусматривается выделенная молекула нуклеиновой кислоты, содержащая:
(i) нуклеиновую кислоту, представленную SEQ ID NO: 1, SEQ ID NO: 2 или SEQ ID NO: 7;
(ii) комплемент любой из SEQ ID NO, приведенных в (i);
(iii) нуклеиновую кислоту, кодирующую белок Т6РР, имеющий, в порядке возрастания предпочтительности, по меньшей мере 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательности с любой из аминокислотных последовательностей, приведенных в SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 6 или SEQ ID NO: 8.
(iv) нуклеиновую кислоту, способную к гибридизации при жестких условиях с любой из нуклеиновых кислот, приведенных в (i), (ii) или (iii) выше.
[0208] Согласно следующему варианту осуществления настоящего изобретения также предусматривается выделенный полипептид, содержащий:
(i) аминокислотную последовательность, представленную любой из SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 6 или SEQ ID NO: 8;
(ii) аминокислотную последовательность, имеющую, в порядке возрастания предпочтительности, по меньшей мере 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичность последовательности с любой из аминокислотных последовательностей, приведенных в SEQ ID NO: 2, SEQ ID NO: 4, SEQ ID NO: 6 или SEQ ID NO: 8;
(iii) производные любой из аминокислотных последовательностей, приведенных в (i) или (ii) выше.
[0209] Настоящее изобретение также предусматривает генные конструкты, кассеты экспрессии и векторы для облегчения введения и/или экспрессии в растении последовательностей нуклеиновых кислот, пригодных в способах согласно настоящему изобретению. Генные конструкты можно внедрить в векторы, которые могут быть коммерчески доступными, подходящими для трансформации в растения и подходящими для экспрессии гена, представляющего интерес, в трансформированных клетках. Настоящее изобретение также предусматривает использование генного конструкта, определенного в данном документе, в способах по настоящему изобретению.
Настоящее изобретение также предусматривает конструкт, содержащий:
(i) нуклеиновую кислоту, кодирующую белок Т6РР, определенный выше;
(ii) одну или несколько контрольных последовательностей, способных управлять экспрессией последовательности нуклеиновой кислоты(а); и необязательно
(iii) транскрипцию конечной последовательности.
[0210] Предпочтительно нуклеиновая кислота в конструкте согласно настоящему изобретению представляет собой молекулу полинуклеотида, кодирующую белок Т6РР, с аминокислотной последовательностью, в порядке возрастания предпочтительности, по меньшей мере 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности с последовательностью, представленной одной из SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5 и SEQ ID NO: 7. Наиболее предпочтительно, молекула полинуклеотида Т6РР является любой из нуклеотидных последовательностей SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5 и SEQ ID NO: 7.
[0211] Растения трансформируют с вектором, содержащим последовательность, представляющую интерес (т.е., нуклеиновая кислота, кодирующая полипептид Т6РР, который был специально или неспециально модифицирован так, чтобы он обладал сниженной активностью). Специалист в данной области хорошо осведомлен о генетических элементах, которые должны присутствовать в векторе для успешной трансформации, выбора и размножения клеток-хозяев, содержащих последовательность, представляющую интерес. Последовательность, представляющая интерес, функционально связана с одной или несколькими контрольными последовательностями (по меньшей мере с промотором). Выражения "регуляторный элемент", "контрольная последовательность" и "промотор" используются в данном документе взаимозаменяемо, и их следует рассматривать в широком контексте при упоминании регуляторных последовательностей нуклеиновых кислот, способных влиять на экспрессию последовательностей, с которыми они лигированы. Выражение "промотор" обычно относится к контрольной последовательности нуклеиновой кислоты, расположенной выше относительно начала транскрипции гена, и которая вовлечена в распознавание и связывание РНК-полимеразы и других белков, тем самым управляя транскрипцией функционально связанной нуклеиновой кислоты. Вышеуказанные выражения охватывают последовательности регуляции транскрипции, происходящие от классического эукариотического геномного гена (включая TATA-бокс, который требуется для правильной инициации транскрипции, с последовательностью CCAAT-бокса или без нее), и дополнительные регуляторные элементы (т.е. выше расположенные активирующие последовательности, энхансеры и сайленсеры), которые изменяют генную экспрессию в ответ на связанные с развитием и/или внешние стимулы или тканеспецифическим образом. Также выражением учитывается транскрипционная регуляторная последовательность классического прокариотического гена, в случае чего она может включать -35-бокс-последовательность и/или -10-бокс-транскрипционные регуляторные последовательности. Выражение "регуляторный элемент" также охватывает синтетическую слитую молекулу или производное, что обеспечивает, активирует или усиливает экспрессию молекулы нуклеиновой кислоты в клетке, ткани или органе. Выражение "функционально связанный", как используется в данном документе, относится к функциональной связи между промоторной последовательностью и геном, представляющим интерес, так что промоторная последовательность способна инициировать транскрипцию гена, представляющего интерес.
[0212] Преимущественно, любой тип промотора можно использовать для управления экспрессией последовательности нуклеиновой кислоты. Выражение "промотор" относится к контрольной последовательности нуклеиновой кислоты, расположенной выше относительно начала транскрипции гена, и которая вовлечена в распознавание и связывание РНК-полимеразы и других белков, тем самым управляя транскрипцией функционально связанной нуклеиновой кислоты. "Промотор растения" содержит регуляторные элементы, которые опосредуют экспрессию сегмента кодирующей последовательности в клетках растения. Соответственно, промотор растения может не быть растительного происхождения, но может происходить от вирусов или микроорганизмов, например, от вирусов, которые поражают клетки растений. "Промотор растения" может также происходить от растительной клетки, например от растения, которое трансформируется с помощью последовательности нуклеиновой кислоты, что подлежит экспрессии. Это также распространяется на другие регуляторные сигналы "растений", такие как терминаторы "растений". Промоторы, расположенные выше нуклеотидных последовательностей, пригодных в способах по настоящему изобретению, можно модифицировать одним или несколькими нуклеотидными замещениями, вставками и/или делециями без препятствования функциональности или активности каждого из промоторов, открытой рамки считывания (ORF) или 3ʹ-регуляторного участка, такого как терминаторы, или других 3ʹ-регуляторных участков, которые расположены вдали от ORF. Также возможно, что активность промоторов увеличивается путем модификации их последовательности или всецелого их замещения на более активные промоторы, даже промоторы гетерологических организмов. Касательно экспрессии в растениях, молекула нуклеиновой кислоты является функционально связанной с подходящим промотором.
[0213] Промотор может быть конститутивным промотором, который относится к промотору, что является транскрипционно активным во время большинства, но не обязательно во время всех, фаз его роста и развития и при большинстве условий окружающей среды, по меньшей мере в одной клетке, ткани или одном органе. Альтернативно, промотор может быть индуцируемым промотором, т.е. характеризующимся индуцируемой или повышенной инициацией транскрипции в ответ на химический (для обзора смотри Gatz 1997, Annu. Rev. Plant Physiol. Plant Mol. Biol., 48: 89-108), относящийся к окружающей среде или физический стимулы. Другим примером индуцируемого промотора является индуцируемый стрессом промотор, т.е. промотор, активируемый, когда растение подвергается различным стрессовым условиям, или индуцируемый патогеном промотор.
[0214] Дополнительно или альтернативно, промотор может быть органоспецифическим или тканеспецифическим промотором, т.е. таким, который способен инициировать транскрипцию преимущественно в конкретных органах или тканях, таких как листья, корни, ткань семян и пр.; или же промотор может быть повсеместным промотором, который активен практически во всех тканях или клетках организма, или промотор может быть регулируемым при развитии, вследствие чего являясь активным во время конкретных стадий развития или в частях растения, которые подвергаются связанным с развитием изменениям. Промоторы, способные инициировать транскрипцию только в конкретных органах или тканях, называются в данном документе "органоспецифическими" или "тканеспецифическими", соответственно; подобным образом промоторы, способные инициировать транскрипцию только в конкретных клетках, называются в данном документе "клеткоспецифическими".
[0215] В одном аспекте настоящего изобретения нуклеиновая кислота Т6РР или ее вариант функционально связаны с промотором, который управляет экспрессией Т6РР в материнской репродуктивной ткани растения, такой как ткань стебля растения, узел початка, ткань колоска, неопыленную ткань цветка и ткань стебля цветка. Промоторы, такие как промоторы OsMADS, описанные в публикации заявки на патент США 2007/0006344, включенной в данный документ посредством ссылки, можно использовать в конкретных аспектах настоящего изобретения.
[0216] Для идентификации функционально эквивалентных промоторов силу промотора и/или паттерн экспрессии кандидатного промотора можно анализировать, например, путем функционального связывания промотора с репортерным геном и анализа уровня экспрессии и паттерна репортерного гена в различных тканях растения. Подходящие хорошо известные репортерные гены включают, например, β-глюкуронидазу или β-галактозидазу. Активность промотора анализируют путем измерения ферментативной активности β-глюкуронидазы или β-галактозидазы. Силу промотора и/или паттерн экспрессии можно затем сравнить с таковым промотора сравнения (такого как используемый в способах по настоящему изобретению). Альтернативно, силу промотора можно анализировать путем количественного определения уровней мРНК или путем сравнения уровней мРНК нуклеиновой кислоты, используемой в способах по настоящему изобретению, с уровнями мРНК гена "домашнего хозяйства", например 18S рРНК, при помощи способов, известных в данной области техники, таких как нозерн-блоттинг с денситометрическим анализом авторадиограмм, количественная PCR в режиме реального времени или RT-PCR (Heid et al., 1996 Genome Methods 6: 986-994). Обычно выражение "слабый промотор" относится к промотору, который управляет экспрессией кодирующей последовательности на низком уровне, уровнях от приблизительно 1/10000 транскриптов до приблизительно 1/100000 транскриптов, до приблизительно 1/5000000 транскриптов на клетку. Наоборот, "сильный промотор" управляет экспрессией кодирующей последовательности на высоком уровне или от приблизительно 1/10 транскриптов до приблизительно 1/100 транскриптов, до приблизительно 1/1000 транскриптов на клетку.
[0217] Предполагается, что регуляторные элементы можно использовать для модулирования длительности, в течение которой заданное свойство присутствует. Например, специалист может сконструировать кассеты экспрессии для экспрессии модифицированной Т6РР во время цветения растения, таким образом обеспечивая засухоустойчивость на этой стадии развития растения. Подобным образом специалист также может сконструировать кассеты экспрессии, которые конститутивно экспрессируют модифицированную Т6РР в растении, таким образом, не ограничиваясь теорией, придавая засухоустойчивость на протяжении жизненного цикла растения.
[0218] Необязательно, в конструкте, вводимом в растение, можно использовать одну или несколько терминаторных последовательностей. Выражение "терминатор" охватывает контрольную последовательность, которая представляет собой последовательность ДНК на конце единицы транскрипции, после стоп-кодона, что сигнализирует 3ʹ процессинг и полиаденилирование первичного транскрипта и окончание транскрипции. Терминатор можно получить из природного гена, из множества других генов растений или из Т-ДНК. Терминатор, который необходимо добавить, можно получить из, например, генов нопалинсинтазы или октопинсинтазы или, альтернативно, из другого гена растений, или менее предпочтительно из любого другого эукариотического гена. Дополнительные регуляторные элементы могут включать энхансеры транскрипции, а также трансляции. Специалисты в данной области техники будут осведомлены о последовательностях терминатора и энхансера, которые могут быть подходящими для использования при осуществлении настоящего изобретения. Такие последовательности будут известны или могут быть легко получены специалистом в данной области.
[0219] Интронную последовательность можно также добавить к 5ʹ-нетранслируемой области (UTR) или в кодирующую последовательность для увеличения количества зрелой мРНК, которая накапливается в цитозоле. Вставка сплайсируемого интрона в единицу транскрипции в конструктах экспрессии как для растений, так и для животных была показана как увеличивающая генную экспрессию как на уровне мРНК, так и на уровне белка до 1000-кратной (Buchman and Berg, Mol. Cell Biol. 8: 4395-4405 (1988); Callis et al., Genes Dev. 1: 1183-1200 (1987)). Такое усиление интронов генной экспрессии обычно является наибольшим при размещении около 5ʹ-конца единицы транскрипции. Использование интронов маиса, интронов 1, 2 и 6 Adh1-S, интрона Bronze-1 известно в уровне техники. Для получения общих сведений смотри The Maize Handbook, Chapter 116, Freeling and Walbot, Eds., Springer, N.Y. (1994). В некоторых аспектах настоящего изобретения специалист может пожелать увеличить экспрессию Т6РР, модифицированной так, чтобы она обладала пониженными способностями к связыванию с субстратом в растении для придания повышенной урожайности.
[0220] Другие контрольные последовательности (кроме промоторных, энхансерных, сайленсерных, интронных последовательностей, 3ʹUTR и/или 5ʹUTR областей) могут быть белковыми и/или стабилизирующими РНК элементами. Такие последовательности будут известны или могут быть легко получены специалистом в данной области.
[0221] Генетические конструкты настоящего изобретения могут дополнительно включать источник последовательности репликации, что требуется для поддержания и/или репликации в конкретном типе клеток. Одним из примеров является ситуация, когда генетический конструкт необходимо поддерживать в бактериальной клетке в качестве эписомального генетического элемента (например, молекула плазмиды или космиды). Предпочтительные источники репликации включают без ограничения f1-ori и colE1.
[0222] Для обнаружения успешного переноса последовательностей нуклеиновых кислот, как используется в способах по настоящему изобретению, и/или отбора трансгенных растений, содержащих эти нуклеиновые кислоты, предпочтительно использовать маркерные гены (или репортерные гены). Таким образом, генетический конструкт может необязательно содержать селектируемый маркерный ген. Как используется в данном документе, выражение "селектируемый маркер", "селектируемый маркерный ген" или "репортерный ген" включает любой ген, который придает фенотип клетке, в которой он экспрессируется, для облегчения идентификации и/или отбора клеток, что являются трансфицируемыми или трансформируемыми с помощью конструкта нуклеиновой кислоты по настоящему изобретению. Эти маркерные гены позволяют идентифицировать успешный перенос молекул нуклеиновых кислот посредством ряда различных принципов. Подходящие маркеры можно выбрать из маркеров, которые придают стойкость к антибиотикам или гербицидам, которые вводят новое метаболическое свойство или которые позволяют проводить визуальный отбор. Примеры селектируемых маркерных генов включают гены, придающие стойкость к антибиотикам (такие как nptII, который фосфорилирует неомицин и канамицин, или hpt, фосфорилирующий гигромицин, или гены, придающие стойкость к, например, блеомицину, стрептомицину, тетрациклину, хлорамфениколу, ампициллину, гентамицину, генетицину (G418), спектиномицину или бластицидину), к гербицидам (например, полоска, которая обеспечивает стойкость к Basta®; aroA или gox, обеспечивающий стойкость против глифосата, или гены, придающие стойкость к, например, имидазолинону, фосфинотрицину или сульфонилмочевине), или гены, которые обеспечивают метаболическое свойство (такие как manA, который позволяет растениям использовать маннозу в качестве единственного источника углерода или ксилозаизомеразу для использования ксилозы, или непитательные маркеры, например, стойкости к 2-дезоксиглюкозе). Экспрессия визуальных маркерных генов приводит в результате к образованию цвета (например, β-глюкуронидаза, GUS или β-галактозидаза с их окрашенными субстратами, например, X-Gal), люминесценции (такой как система люциферин/люцифераза) или флуоресценции (зеленый флуоресцентный белок, GFP и их производные). Этот список представляет только небольшое число возможных маркеров. Квалифицированный специалист знаком с такими маркерами. В зависимости от организма и способа отбора предпочтительны различные маркеры.
[0223] Известно, что при стабильном или временном встраивании нуклеиновых кислот в клетки растений только незначительное количество клеток поглощает чужеродную ДНК и, при необходимости, встраивает ее в свой геном, в зависимости от используемого вектора экспрессии и используемой техники трансфекции. Для идентификации и отбора этих интегрантов, ген, кодирующий селектируемый маркер (такой как описанный выше), обычно вводят в клетки-хозяева вместе с геном, представляющим интерес. Эти маркеры можно, например, использовать в мутантах, где эти гены являются нефункциональными из-за, например, делеции обычными Способами. Кроме того, молекулы нуклеиновых кислот, кодирующие селектируемый маркер, можно вводить в клетку-хозяин в том же векторе, что содержит последовательность, кодирующую полипептиды настоящего изобретения, или использовать в способах по настоящему изобретению, или иначе в отдельном векторе. Клетки, которые были стабильно трансфицированы введением нуклеиновой кислоты, можно идентифицировать, например, при помощи отбора (например, клетки, в которые был встроен селектируемый маркер, выживали, тогда как другие клетки погибали).
[0224] Поскольку маркерные гены, в частности гены для стойкости к антибиотикам и гербицидам, сразу после того, как нуклеиновые кислоты были успешно введены, больше не требуются или нежелательны в трансгенной клетке-хозяине, в способе согласно настоящему изобретению для введения нуклеиновых кислот предпочтительно используются методики, которые позволяют удалить или вырезать эти маркерные гены. Один такой способ известен как котрансформация. В способе котрансформации для трансформации используют два вектора одновременно, один вектор несет нуклеиновую кислоту согласно настоящему изобретению, а второй несет маркерный ген(ы). Большая часть трансформантов принимает или, в случае растений, содержит (до 40% или больше трансформантов) оба вектора. В случае трансформации с агробактериями, трансформанты обычно получают только часть вектора, т.е. последовательность, фланкированную Т-ДНК, которая обычно представляет кассету экспрессии. Маркерные гены можно затем удалить из трансформированного растения путем проведения скрещивания. В другом способе маркерные гены, встроенные в транспозон, пригодны для трансформации вместе с желаемой нуклеиновой кислотой (известен как технология Ac/Ds). Трансформанты можно скрещивать с источником транспозазы, или же трансформанты трансформируют с конструктом нуклеиновой кислоты, обеспечивая экспрессию транспозазы, временно или постоянно. В некоторых случаях (приблиз. 10%) сразу после того, как успешно произошла трансформация, транспозон выскакивает из генома клетки-хозяина и теряется. В следующем ряде случаев транспозон перескакивает на другое положение. В этих случаях маркерный ген необходимо удалить проведением скрещивания. В микробиологии разработаны методики, которые делают возможным, или облегчают, делецию таких объектов. Дополнительный предпочтительный способ основан на так называемых системах рекомбинации, преимущество которого заключается в том, что можно обойтись без удаления скрещиванием. Наиболее известная система этого типа известна как система Cre/lox. Cre1 является рекомбиназой, которая удаляет последовательности, расположенные между последовательностями 1охР. Если маркерный ген встроен между последовательностями 1охР, сразу после того, как успешно произошла трансформация, он удаляется экспрессией рекомбиназы. Дополнительные системы рекомбинации представляют собой HIN/HIX, FLP/FRT и систему REP/STB (Tribble et al., J. Biol. Chem., 275, 2000: 22255-22267; Velmurugan et al., J. Cell Biol., 149, 2000: 553-566). Возможна сайт-специфическое встраивание в геном растения последовательностей нуклеиновых кислот согласно настоящему изобретению. Конечно же, эти способы можно также применять к микроорганизмам, таким как дрожжи, грибы или бактерии.
[0225] Изобретение также предусматривает способ получения трансгенных растений с улучшенными связанными с урожайностью свойствами, в частности повышенной урожайностью семян, относительно контрольных растений, включающий введение и экспрессию в растении любой нуклеиновой кислоты, кодирующей белок Т6РР, определенный в данном документе выше.
[0226] Один аспект настоящего изобретения предусматривает кассеты экспрессии, содержащие:
(a) последовательности нуклеиновых кислот, кодирующие белки, пригодные в способах по настоящему изобретению, или
(b) генетическую контрольную последовательность(и), которая функционально связана с последовательностью нуклеиновой кислоты согласно настоящему изобретению, например, промотором, или
(c) a) и b).
[0227] Более конкретно, настоящее изобретение предусматривает способ получения трансгенных растений с повышенной урожайностью, причем способ включает:
(i) введение и экспрессию в растении, части растения или клетке растения нуклеиновой кислоты Т6РР или ее варианта, где Т6РР обладает уменьшенной активностью связывания с субстратом; и
(ii) культивирование клетки растения при условиях, ускоряющих рост и развитие растения.
[0228] Нуклеиновую кислоту можно вводить непосредственно в клетку растения или в само растение (включая введение в ткань, орган или любую другую часть растения). Согласно предпочтительной характеристике настоящего изобретения нуклеиновую кислоту предпочтительно вводить в растение трансформацией.
[0229] Перенос чужеродных генов в геном растения называется трансформацией. Трансформация видов растений является в данное время довольно обычной методикой. Преимущественно, для введения гена, представляющего интерес, в подходящую клетку-предшественник можно применять любой из нескольких способов трансформации. Способы, описанные касательно трансформации и регенерации растений из тканей растений или клеток растений, можно применять для временной или постоянной трансформации. Способы трансформации включают применение липосом, электропорации, химических веществ, которые усиливают свободный ввод ДНК, введение ДНК непосредственно в растение, бомбардировку генной пушкой, трансформацию с применением вирусов или пыльцы и микропроекцию. Способы могут быть выбраны из кальций/полиэтиленгликолевого способа для протопластов (Krens, F.A. et al., (1982) Nature 296, 72-74; Negrutiu I et al. (1987) Plant Mol Biol 8: 363-373); электропорации протопластов (Shillito R.D. et al. (1985) Bio/Technol 3, 1099-1102); микроинъекции в растительный материал (Crossway A et al., (1986) Mol. Gen Genet 202: 179-185); бомбардировки частицами, покрытыми ДНК или РНК (Klein Т М et al., (1987) Nature 327: 70); заражения (неинтегрированными) вирусами и подобного. Трансгенные растения, включая трансгенные сельскохозяйственные культуры, предпочтительно получают посредством опосредованной Agrobacterium трансформации. Преимущественным способом трансформации является трансформация in planta. С этой стороны, можно, например, позволить агробактериям действовать на семена растений или инокулировать меристему растения агробактериями. Особенно целесообразным оказалось в соответствии с настоящим изобретением обеспечить действие суспензии трансформированных агробактерий на интактное растение или по меньшей мере на завязь цветков. Растение затем растет до тех пор, пока не будут получены семена обработанного растения (Clough and Bent, Plant J. (1998) 16, 735-743). Способы для опосредованной Agrobacterium трансформации риса включают хорошо известные способы для трансформации риса, такие как описанные в любом из следующего: Европейская патентная заявка EP 1198985 A1, Aldemita and Hodges (Planta 199: 612-617, 1996); Chan et al. (Plant Mol Biol 22 (3): 491-506, 1993), Hiei et al. (Plant J 6 (2): 271-282, 1994), раскрытия которых включены в данный документ посредством ссылки, словно полностью изложены. В случае трансформации кукурузы предпочтительным способом является описанный либо в Ishida et al. (Nat. Biotechnol 14 (6): 745-50, 1996), либо Frame et al. (Plant Physiol 129 (1): 13-22, 2002), раскрытия которых включены в данный документ посредством ссылки, словно полностью "изложены. Указанные способы также описаны в качестве примера в В. Jenes et al., Techniques for Gene Transfer, в Transgenic Plants, Vol. 1, Engineering and Utilization, eds. S.D. Kung and R. Wu, Academic Press (1993) 128-143, и в Potrykus Annu. Rev. Plant Physiol. Plant Molec. Biol. 42 (1991) 205-225). Нуклеиновые кислоты или конструкт, подлежащие экспрессии, предпочтительно клонируют в вектор, что подходит для трансформации Agrobacterium tumefaciens, например, pBin19 (Bevan et al., Nucl. Acids Res. 12 (1984) 8711). Агробактерии, трансформированные таким вектором, затем можно применять известным способом для трансформации растений, таких как растения, применяемые в качестве модели, подобно Arabidopsis, или сельскохозяйственные культуры, такие как, например, растения табака, например, посредством погружения раздробленных листьев или нарубленных листьев в раствор агробактерий и затем культивирования их в подходящей среде. Трансформация растений посредством Agrobacterium tumefaciens описана, например, Höfgen and Willmitzer in Nucl. Acid Res. (1988) 16, 9877, или частично известна из F.F. White, Vectors for Gene Transfer in Higher Plants; в Transgenic Plants, Vol. 1, Engineering and Utilization, eds. S.D. Kung and R. Wu, Academic Press, 1993, pp. 15-38.
[0230] В дополнение к трансформации соматических клеток, которые затем необходимо регенерировать в интактные растения, также можно трансформировать клетки меристем растения и, в частности, такие клетки, которые развиваются в гаметы. В этом случае трансформированные гаметы следуют природному циклу развития растения, что приводит к получению трансгенных растений. Таким образом, например, семена Arabidopsis обрабатывают агробактериями и получают семена от развивающихся растений, конкретная часть которых трансформируется и поэтому является трансгенной [Feldman, К A and Marks М D (1987). Mol Gen Genet 208: 274-289; Feldmann К (1992). In: С Koncz, N-H Chua and J Shell, eds, Methods in Arabidopsis Research. Word Scientific, Singapore, pp. 274-289]. Альтернативные способы основаны на повторном удалении соцветий и инкубации вырезанного участка в центре розетки с трансформированными агробактериями, при этом трансформированные семена могут таким же образом быть получены в более поздний момент времени (Chang (1994). Plant J. 5: 551-558; Katavic (1994). Mol Gen Genet, 245: 363-370). Однако особенно эффективным способом является способ вакуумной инфильтрации с его модификациями, такими как способ "погружения цветка". В случае вакуумной инфильтрации Arabidopsis интактные растения под пониженным давлением обрабатывают суспензией агробактерий (Bechthold, N (1993). С R Acad. Sci. Paris Life Sci., 316: 1194-1199), тогда как в случае способа "погружения цветка" развивающуюся ткань цветка инкубируют в течение короткого периода времени обработанной поверхностно-активным веществом суспензией агробактерий [Clough, S J und Bent, A F (1998). The Plant J. 16, 735-743]. Определенную долю трансгенных семян собирают в обоих случаях, и эти семена могут отличаться от нетрансгенных семян по росту при вышеуказанных условиях отбора. Кроме того, стабильная трансформация пластид является преимуществом, поскольку пластиды у большинства сельскохозяйственных культур наследуются по материнской линии, снижая или устраняя риск утечки трансгена через пыльцу. Трансформацию генома хлоропластов обычно осуществляют посредством процесса, который был схематически представлен в Klaus et al., 2004 [Nature Biotechnology 22 (2), 225-229]. Кратко говоря, последовательности, подлежащие трансформации, клонируют вместе с селектируемым маркерным геном между фланкирующими последовательностями, гомологичными геному хлоропласта. Эти гомологичные фланкирующие последовательности управляют сайт-специфическим встраиванием в пластом. Трансформация пластид была описана для множества различных видов растений, и обзор приведен в Воск (2001) Transgenic plastids in basic research and plant biotechnology. J Mol Biol. September 2001 21; 312 (3): 425-38, или Maliga, P (2003) Progress towards commercialization of plastid transformation technology. Trends Biotechnol. 21, 20-28. Как о биотехнологическом прогрессе недавно сообщили о трансформантах пластид без маркера, которые можно получить при помощи переходного совместно встраиваемого маркерного гена (Klaus et al., 2004, Nature Biotechnology 22 (2), 225-229).
[0231] Генетически модифицированные клетки растений можно регенерировать посредством всех способов, с которыми ознакомлен квалифицированный специалист. Подходящие способы можно найти в вышеуказанных публикациях S.D. Kung and R. Wu, Potrykus или Höfgen and Willmitzer.
[0232] Обычно после трансформации клетки растений или группировки клеток отбирают на наличие одного или нескольких маркеров, которые закодированы экспрессируемыми растениями генами, котрансформированными с геном, представляющий интерес, после чего трансформированный материал регенерируется в целое растение. Для выбора трансформированных растений растительный материал, полученный при трансформации, как правило, подвергают условиям отбора, чтобы трансформированные растения можно было отличить от нетрансформированных растений. Например, семена, полученные вышеописанным способом, можно высеять и, после начального периода вегетации, подвергнуть подходящему отбору путем обрызгивания. Дополнительная перспектива заключается в выращивании семян, если это уместно после стерилизации, на агаровых пластинах с применением подходящего средства для отбора, чтобы только трансформированные семена могли дорасти до растений. Альтернативно, трансформированные растения проверяют на наличие селектируемого маркера, такого как описанные выше.
[0233] После переноса ДНК и регенерации предполагаемо трансформированные растения также можно охарактеризовать, например, с применением саузерн-анализа, на наличие гена, представляющего интерес, число копий и/или геномную организацию. Альтернативно или дополнительно, уровни экспрессии только что введенной ДНК можно отследить с применением нозерн- и/или вестерн-анализа, при этом обе методики хорошо известны специалистам, квалифицированным в данной области.
[0234] Полученные трансформированные растения можно размножать различными способами, такими как клональная пролиферация или классические методики селекции. Например, первое поколение (или Т1) трансформированного растения может самоопыляться, а гомозиготные трансформанты второго поколения (или Т2) выбирают, после чего растения Т2 можно в дальнейшем размножить классическими методиками селекции.
[0235] Полученные трасформированные организмы могут принимать различные формы. Например, они могут быть химерными организмами трансформированных клеток и нетрансформированных клеток; клональными трансформантами (например, все клетки трансформировались для содержания кассеты экспрессии); привитой частью из трансформированных и нетрансформированных тканей (например, в растениях трансформированное корневище, привитое на нетрансформированный привой).
[0236] Настоящее изобретение несомненно распространяется на любую клетку растения или растение, полученные любым из способов, описанных в данном документе, и на все части растений и их побеги. Настоящее изобретение распространяется также для охвата потомства первично трансформированной или трансфицированной клетки, ткани, органа или всего растения, которое было получено любым из вышеуказанных способов, причем единственным требованием является то, чтобы потомство проявляло такую же генотипическую и/или фенотипическую характеристику(и), как проявляется у родителя в способах согласно настоящему изобретению.
[0237] Настоящее изобретение также включает клетки-хозяева, содержащие выделенную нуклеиновую кислоту, кодирующую белок Т6РР, определенный в данном документе выше. Предпочтительными клетками-хозяевами согласно настоящему изобретению являются клетки растений.
[0238] Растения-хозяева для нуклеиновых кислот или вектора, применяемого в способах согласно настоящему изобретению, кассета экспрессии, или конструкт, или вектор являются, в принципе, предпочтительными во всех растениях, которые способны синтезировать полипептиды, применяемые в способах настоящего изобретения.
[0239] Таким образом, под трансгенным растением в целях настоящего изобретения подразумевается, согласно вышеуказанного, что нуклеиновые кислоты, применяемые в способе настоящего изобретения, находятся не на их естественном локусе в геноме указанного растения, причем в отношении нуклеиновых кислот вероятно, что они экспрессируются гомологично или гетерологично. Однако, как указано, трансгенный также означает, что хотя нуклеиновые кислоты согласно настоящему изобретению или применяемые в способе настоящего изобретения находятся в их естественных положениях в геноме растения, последовательность была модифицирована относительно естественной последовательности, и/или что регуляторные последовательности естественных последовательностей были модифицированы. Подразумевается, что трансгенный предпочтительно означает экспрессию нуклеиновых кислот согласно настоящему изобретению в неестественном локусе в геноме, т.е. имеет место гомологичная или, предпочтительно, гетерологичная экспрессия нуклеиновых кислот. Предпочтительные трансгенные растения указаны в данном документе.
[0240] Изобретение также распространяется на пригодные для заготовки части растений, такие как, без ограничения, семена, листья, плоды, цветы, стебли, ризомы, клубни и луковицы. Настоящее изобретение также относится к продуктам, полученным, предпочтительно непосредственно полученным, из пригодных для заготовки частей такого растения, таким как сухие брикеты или порошки, масло, жир и жирные кислоты, крахмал или белки.
[0241] Согласно одному признаку настоящего изобретения регулируемая экспрессия представляет собой увеличенную экспрессию. Способы увеличения экспрессии нуклеиновых кислот или генов, или генных продуктов, как следует описаны в документах, касающихся данной области техники, и включают, например, сверхэкспрессию, управляемую подходящими промоторами, использование энхансеров транскрипции или энхансеров трансляции. Выделенные нуклеиновые кислоты, которые служат промоторными или энхансерными элементами, можно вводить в подходящее положение (обычно выше) негетерологичной формы полинуклеотида для активации экспрессии. Например, эндогенные промоторы можно изменять in vivo мутацией, делецией и/или замещением (смотри Kmiec, патент США №5565350; Zarling et al., PCT/US93/03868) или выделенные промоторы можно вводить в клетку растения при подходящей ориентации и расстоянии от гена настоящего изобретения, чтобы они управляли экспрессией гена.
[0242] Если необходима экспрессия полипептида, обычно желательно вводить область полиаденилирования на 3ʹ-конце кодирующей области полинуктеотида. Область полиаденилирования можно получить из природного гена, из множества генов других растений или из Т-ДНК. Последовательность 3ʹ-конца, которую необходимо добавить, можно получить из, например, генов нопалинсинтазы или октопинсинтазы, или альтернативно из другого гена растения, или менее предпочтительно из любого другого эукариотического гена.
[0243] Также можно добавить интронную последовательность, как описано выше.
[0244] Другие контрольные последовательности (кроме промоторных, энхансерных, сайленсерных, интронных последовательностей, 3ʹUTR и/или 5ʹUTR областей, целевых сайтов микро-РНК) могут быть белковыми и/или стабилизирующими РНК элементами.
[0245] Как указано выше, способ регулирования экспрессии нуклеиновой кислоты, кодирующей белок Т6РР, модифицированный, чтобы он имел сниженную активность, заключается во введении и экспрессии в растении нуклеиновой кислоты, кодирующей белок Т6РР; однако эффектов от выполнения способа, т.е. улучшения связанных с урожайностью свойств, можно также достичь при помощи других хорошо известных методик. Описание некоторых из этих методик следует далее.
[0246] Одной такой методикой является активационное мечение Т-ДНК (Hayashi et al. Science (1992) 1350-1353), которое включает вставку Т-ДНК, что обычно содержит промотор (также может быть энхансер трансляции или интрон), в геномную область гена, представляющего интерес, или 10 т.н. выше или ниже кодирующей области гена в такой конфигурации, чтобы промотор управлял экспрессией целевого гена. Обычно регулирование экспрессии целевого гена его природным промотором нарушено, и ген попадает под контроль только что введенного промотора. Промотор обычно находится в Т-ДНК. Эта Т-ДНК произвольно встраивается в геном растения, например, посредством заражения Agrobacterium, и приводит к модифицированной экспрессии генов около вставленной Т-ДНК. Полученные в результате трансгенные растения проявляют доминантные фенотипы вследствие модифицированной экспрессии генов вблизи введенного промотора.
[0247] Эффекты настоящего изобретения можно также воспроизвести при помощи методики TILLING (нацеленные индуцированные местные поражения в геноме (Targeted Induced Local Lesions In Genomes)). Это является техникой мутагенеза, пригодной для получения и/или идентификации нуклеиновой кислоты, кодирующей белок Т6РР III класса с модифицированной экспрессией и/или активностью. TILLING также позволяет проводить отбор растений, несущих такие мутантные варианты. Эти мутантные варианты могут характеризоваться модифицированной экспрессией, либо по силе, либо по локализации, либо по времени (если, например, мутации затрагивают промотор). Эти мутантные варианты могут проявлять более высокую активность белка Т6РР III класса, чем проявляемая геном в его естественной форме. TILLING объединяет высокую частоту мутагенеза со способами высокопроизводительного скрининга. Как правило, этапами TILLING являются следующие: (а) мутагенез EMS (Redei G Р and Koncz С (1992) In Methods in Arabidopsis Research, Koncz C, Chua NH, Schell J, eds. Singapore, World Scientific Publishing Co, pp. 16-82; Feldmann et al., (1994) In Meyerowitz E M, Somerville С R, eds, Arabidopsis. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., pp 137-172; Lightner J and Caspar T (1998) In J Martinez-Zapater, J Salinas, eds, Methods on Molecular Biology, Vol. 82. Humana Press, Totowa, N.J., pp 91-104); (b) получение ДНК и объединение отдельных объектов; (с) PCR-амплификация области, представляющей интерес; (d) денатурация и отжиг с образованием гетеродуплексов; (е) DHPLC, где присутствие гетеродуплекса в пуле определяется по дополнительному пику на хроматограмме; (f) идентификация мутантного объекта и (g) секвенирование мутантного продукта PCR. В данной области техники хорошо известны способы TILLING (McCallum et al., (2000) Nat Biotechnol 18: 455-457; обзор Stemple (2004) Nat Rev Genet 5 (2): 145-50).
[0248] Эффекты настоящего изобретения можно также воспроизвести при помощи гомологичной рекомбинации, которая позволяет вводить в геном выбранную нуклеиновую кислоту в определенное выбранное положение. Гомологичная рекомбинация является стандартной техникой, обычно применяемой в биологических экспериментах на низших организмах, таких как дрожжи или мох Physcomitrella. Способы проведения гомологичной рекомбинации у растений описаны не только для модельных растений (Offringa et al. (1990) EMBO J 9 (10): 3077-84), но и для растений сельскохозяйственных культур, например, риса (Terada et al. (2002) Nat Biotech 20 (10): 1030-4; Iida and Terada (2004) Curr. Opin. Biotech 15 (2): 132-8).
[0249] Упоминание в данном документе улучшенных связанных с урожайностью свойств используется для обозначения увеличения биомассы (веса) одной или нескольких частей растения, которые могут включать надземные (пригодные для заготовки) части и/или подземные (пригодные для заготовки) части. В частности, такие пригодные для заготовки части представляют собой семена, и осуществление способов настоящего изобретения приводит в результате к тому, что растения характеризуются повышенной урожайностью семян по сравнению с урожайностью семян соответствующих контрольных растений.
[0250] Выражения "повышение", "улучшение" или "улучшать" являются взаимозаменяемыми и будут означать в случае использования по меньшей мере на 5%, 6%, 7%, 8%, 9% или 10%, предпочтительно по меньшей мере на 15% или 20%, более предпочтительно на 25%, 30%, 35% или 40% большую урожайность и/или прирост по сравнению с растением дикого типа, определенным в данном документе.
[0251] Повышенная урожайность семян может проявляться как таковая одним или несколькими из следующего: a) увеличением биомассы семян (общий вес семян), что может выражаться на основании отдельного семени, и/или на растение, и/или на гектар или акр; b) увеличенным числом цветков на растение; с) увеличенным числом (налитых) семян; d) увеличенной нормой налитых семян (что выражается как отношение числа налитых семян, деленного на общее число семян); е) увеличенным индексом урожайности, который выражается как отношение урожайности пригодных для заготовки частей, таких как семена, поделенной на общее количество биомассы; и f) увеличенной абсолютной массой зерен (TKW), которую экстраполируют на основании числа подсчитанных налитых семян и их общего веса. Увеличенная TKW может являться следствием увеличенного размера семян и/или веса семян, а также может являться следствием увеличения размера зародышей и/или эндосперма.
[0252] Увеличение урожайности семян может также проявляться увеличением размера семян и/или объема семян. Кроме того, увеличение урожайности семян может также проявляться как таковое увеличением площади семян, и/или длины семян, и/или ширины семян, и/или окружности семян.
[0253] Принимая кукурузу в качестве примера, повышение урожайности может проявляться одним или нескольким из следующего: увеличением числа растений, которое приходится на гектар или акр, увеличением числа початков на растении, увеличением числа рядков, числа зерен на рядок, веса зерен, абсолютной массы зерен, длины/диаметра початка, увеличением нормы налитых семян (что является числом налитых семян, деленным на общее число семян и умноженным на 100), среди прочего. Принимая рис в качестве примера, повышение урожайности может проявляться увеличением одного или нескольких из следующего: числа растений на гектар или акр, числа метелок на растение, числа колосков на метелку, числа цветков (отдельных цветков) на метелку (что выражается как отношение числа налитых семян относительно числа основных метелок), увеличением нормы налитых семян (что является числом налитых семян, деленным на общее число семян и умноженным на 100), увеличением абсолютной массы зерен, среди прочего.
[0254] Поскольку трансгенные растения согласно настоящему изобретению обладают повышенной урожайностью, вполне вероятно, что эти растения проявляют повышенную скорость роста (во время по меньшей мере части их жизненного цикла) по сравнению со скоростью роста контрольных растений на соответствующей стадии их жизненного цикла. Повышенная скорость роста может быть специфичной для одной или нескольких частей растения (включая семена), или может касаться практически всего растения. Растения, обладающие повышенной скоростью роста, могут иметь более короткий жизненный цикл. Жизненный цикл растения можно принять для обозначения времени, необходимого для роста от сухого зрелого семени до стадии, когда растение образует сухие зрелые семена, подобное исходному материалу. На этот жизненный цикл могут влиять факторы, такие как раннее прорастание, скорость роста, индекс незрелости, период цветения и скорость созревания семян. Повышение скорости роста может иметь место на одной или нескольких стадиях жизненного цикла растения или на протяжении практически всего жизненного цикла растения. Повышенная скорость роста во время ранних стадий жизненного цикла растения может свидетельствовать о увеличенной мощности. Повышение скорости роста может изменить цикл урожаев растения, позволяя сеять растения позже и/или убирать раньше, чем это было бы возможно при других обстоятельствах (подобный эффект можно получить в отношении времени начала цветения). Если скорость роста повышена в достаточной степени, это может позволить провести дополнительный посев семян того же вида растения (например, посев и уборка растений риса с последующим посевом и уборкой следующих растений риса в течение одного стандартного периода вегетации). Аналогично, если скорость роста повышена в достаточной степени, это может позволить провести дополнительный посев семян других видов растений (например, посев и уборка зерновых растений с последующим, например, посевом и необязательной уборкой сои, картофеля или любого другого подходящего растения). В случае некоторых сельскохозяйственных культур возможна неоднократная уборка урожая с одного корневища. Изменение цикла урожаев растения может привести к увеличению годового производства биомассы с акра (благодаря увеличению количества раз (скажем, в течение года) выращивания и уборки любого конкретного растения). Повышение скорости роста также может позволить культивировать трансгенные растения в более широкой географической зоне, чем их аналоги дикого типа, поскольку территориальные ограничения выращивания культуры зачастую определяются неблагоприятными условиями окружающей среды либо на момент посева (в начале вегетационного периода), либо на момент уборки (в конце вегетационного периода). Таких неблагоприятных условий можно избежать при сокращении продолжительности цикла урожаев. Скорость роста может быть определена путем получения различных параметров из кривых роста, при этом такими параметрами могут быть: T-Mid (время, необходимое для достижения растениями 50% от их максимального размера) и Т-90 (время, необходимое для достижения растениями 90% от их максимального размера), среди прочего.
[0255] Выполнение способов настоящего изобретения дает растения с повышенной скоростью роста относительно контрольных растений. Таким образом, согласно настоящему изобретению предусматривается способ повышения скорости роста растений, причем способ включает регулирование экспрессии, предпочтительно увеличение экспрессии, в растении нуклеиновой кислоты, кодирующей белок Т6РР, определенный в данном документе.
[0256] Увеличение урожайности и/или скорости роста происходит либо когда растение находится в бесстрессовых условиях, либо когда растение подвергается различным стрессам по сравнению с контрольными растениями. Растения, как правило, реагируют на воздействие стресса замедлением роста. В условиях сильного стресса растение может полностью прекратить рост. С другой стороны, умеренный стресс определяется в данном документе как любой стресс, будучи подвергнутым которому, растение в результате не прекращает рост полностью без возможности возобновления роста. Умеренный стресс в контексте настоящего изобретения приводит к снижению роста подвергнутых стрессу растений менее чем на 40%, 35% или 30%, предпочтительно менее чем на 25%, 20% или 15%, более предпочтительно менее чем на 14%, 13%, 12%, 11% или 10% или менее по сравнению с контрольным растением, не подвергнутым стрессовым условиям. Благодаря развитию агротехнических приемов (орошения, внесения удобрений, обработки пестицидами) при культивировании сельскохозяйственных культур сильные стрессы возникают нечасто. Поэтому вызванный умеренным стрессом нарушенный рост часто является нежелательным явлением в сельском хозяйстве. Умеренные стрессы представляют собой типичные биотические/абиотические (относящиеся к окружающей среде) стрессовые воздействия, которым может подвергаться растение. Абиотический стресс может возникать из-за засухи или избытка воды, анаэробного стресса, стресса, вызванного засолением, химической токсичности, окислительного стресса и высоких, низких температур или температур замерзания. Абиотический стресс может быть осмотическим стрессом, вызванным стрессом из-за недостатка воды (в частности из-за засухи), стрессом, вызванным засолением, окислительным стрессом или ионным стрессом. Биотические стрессы, как правило, представляют собой стрессы, вызванные такими патогенами, как бактерии, вирусы, грибы и насекомые. Другой абиотический стресс может произойти в результате дефицита питательных веществ, такого как нехватка азота, фосфора и калия.
[0257] Способы настоящего изобретения можно выполнять при безстрессовых условиях или в условиях умеренной засухи для придания растениям свойств повышенной урожайности относительно контрольных растений. Как сообщалось у Wang et al. (Planta (2003) 218: 1-14), абиотический стресс приводит к ряду морфологических, физиологических, биохимических и молекулярных изменений, которые неблагоприятно влияют на рост растений и продуктивность. Засуха, засоленность, экстремальные температуры и окислительный стресс, как известно, взаимосвязаны, и могут вызвать нарушение роста и повреждение клеток путем подобных механизмов. Rabbani et al. (Plant Physiol (2003) 133: 1755-1767), описывает очень высокую степень "взаимного влияния" между стрессом, вызванным засухой, и стрессом, вызванным высокой засоленностью. Например, засуха и/или засоление показаны в первую очередь как осмотический стресс, приводящий к нарушению гомеостаза и ионного распределения в клетке. Окислительный стресс, который часто сопровождает высокую или низкую температуру, засоленность или стресс, вызванный засухой, может вызвать денатурацию функциональных и структурных белков. Как следствие, эти многообразные относящиеся к окружающей среде стрессы часто активируют подобные клеточные сигнальные пути и клеточные ответы, такие как выработку белков стресса, активацию антиоксидантов, накопление совместимых осмолитов и задержку роста. Выражение "безстрессовые" условия, как используется в данном документе, являются такими условиями окружающей среды, которые обеспечивают оптимальный рост растений. Специалисты в данной области осведомлены о нормальных почвенные условия и климатических условиях для данного местоположения.
[0258] Проведение способов настоящего изобретения придает растениям, растущим при безстрессовых условиях или в условиях засухи, увеличенную урожайность относительно соответствующих контрольных растений, растущих при сопоставимых условиях. Таким образом, согласно настоящему изобретению предусматривается способ улучшения связанных с урожайностью свойств растений, растущих при безстрессовых условиях или в условиях засухи, причем способ включает увеличение экспрессии в растении нуклеиновой кислоты, кодирующей модифицированный полипептид Т6РР, характеризующийся уменьшенным связыванием с субстратом и/или активностью.
[0259] В одном варианте осуществления настоящего изобретения улучшенное связанное с урожайностью свойство проявляется как увеличение одного или нескольких из следующего: общего числа семян на растение, числа налитых семян на растение и веса семян на растение. Предпочтительно, эти улучшения обнаруживают у растений, растущих при безстрессовых условиях.
[0260] Способы изобретения предпочтительно применимы к любому растению.
[0261] Растения, особенно применимые в способах настоящего изобретения, включают все растения, принадлежащие к надсемейству Viridiplantae, в частности, однодольные и двудольные растения, включающие в себя кормовые растения или кормовые бобовые культуры, декоративные растения, продовольственные культуры, деревья или кустарники, выбранные из списка, включающего: Acer spp., Actinidia spp., Abelmoschus spp., Agropyron spp., Allium spp., Amaranthus spp., Ananas comosus, Annona spp., Apium graveolens, Arachis spp, Artocarpus spp., Asparagus officinalis, Avena spp. (например, Avena sativa, Avena fatua, Avena byzantina, Avena fatua var. sativa, Avena hybrida), Averrhoa carambola, Benincasa hispida, Bertholletia excelsea, Beta vulgaris, Brassica spp. (например, Brassica napus, Brassica rapa ssp. [канола, масличный рапс, турнепс]), Cadaba farinosa, Camellia sinensis, Canna indica, Capsicum spp., Carex elata, Carica papaya, Carissa macrocarpa, Carya spp., Carthamus tinctorius, Castanea spp., Cichorium endivia, Cinnamomum spp., Citrullus lanatus, Citrus spp., Cocos spp., Coffea spp., Colocasia esculenta, Cola spp., Coriandrum sativum, Corylus spp., Crataegus spp., Crocus sativus, Cucurbita spp., Cucumis spp., Cynara spp., Daucus carota, Desmodium spp., Dimocarpus longan, Dioscorea spp., Diospyros spp., Echinochloa spp., Elaeis (например, Elaeis guineensis, Elaeis oleifera), Eleusine coracana, Eriobotrya japonica, Eugenia uniflora, Fagopyrum spp., Fagus spp., Ficus carica, Fortunella spp., Fragaria spp., Ginkgo biloba, Glycine spp. (например, Glycine max, Soja hispida или Soja max), Gossypium hirsutum, Helianthus spp. (например, Helianthus annum), Hemerocallis fulva, Hibiscus spp., Hordeum spp. (например, Hordeum vulgare), Ipomoea batatas, Juglans spp., Lactuca sativa, Lathyrus spp., Lens culinaris, Linum usitatissimum, Litchi chinensis, Lotus spp., Luffa acutangula, Lupinus spp., Luzula sylvatica, Lycopersicon spp. (например, Lycopersicon esculentum, Lycopersicon lycopersicum, Lycopersicon pyriforme), Macrotyloma spp., Malus spp., Malpighia emarginata, Mammea americana, Mangifera indica, Manihot spp., Manilkara zapota, Medicago sativa, Melilotus spp., Mentha spp., Miscanthus spp., Momordica spp., Morus nigra, Musa spp., Nicotiana spp., Olea spp., Opuntia spp., Ornithopus spp., Oryza spp. (например, Oryza sativa, Oryza latifolia), Panicum miliaceum, Passiflora edulis, Pastinaca sativa, Persea spp., Petroselinum crispum, Phaseolus spp., Phoenix spp., Physalis spp., Pinus spp., Pistacia vera, Pisum spp., Poa spp., Populus spp., Prosopis spp., Prunus spp., Psidium spp., Punica granatum, Pyrus communis, Quercus spp., Raphanus sativus, Rheum rhabarbarum, Ribes spp., Ricinus communis, Rubus spp., Saccharum spp., Sambucus spp., Secale cereale, Sesamum spp., Sinapis sp., Solanum spp. (например, Solanum tuberosum, Solanum integrifolium или Solanum lycopersicum), Sorghum bicolor, Spinacia spp., Syzygium spp., Tagetes spp., Tamarindus indica, Theobroma cacao, Trifolium spp., Triticosecale rimpaui, Triticum spp. (например, Triticum aestivum, Triticum durum, Triticum turgidum, Triticum hybernum, Triticum macha, Triticum sativum или Triticum vulgare), Tropaeolum minus, Tropaeolum majus, Vaccinium spp., Vicia spp., Vigna spp., Viola odorata, Vitis spp., Zea mays, Zizania palustris, Ziziphus spp., среди других.
[0262] Согласно предпочтительному варианту осуществления настоящего изобретения растение представляет собой сельскохозяйственную культуру. Примеры растений сельскохозяйственных культур включают сою, подсолнечник, канолу, люцерну, рапс, хлопок, томат, картофель и табак. Также предпочтительно, растение представляет собой однодольное растение. Примеры однодольных растений включают сахарный тростник. Более предпочтительно растение является злаком. Примеры злаков включают рис, маис, пшеницу, ячмень, просо, рожь, сорго и овес.
[0263] Настоящее изобретение также охватывает использование нуклеиновых кислот, кодирующих белок Т6РР, описанный в данном документе, и использование этих белков Т6РР в улучшении связанных с урожайностью свойств у растений.
[0264] Нуклеиновые кислоты, кодирующие белок Т6РР, описанный в данном документе, или белки Т6РР как таковые могут найти применение в программах селекции, в которых определяют маркер ДНК, который может быть генетически связан с кодирующим геном Т6РР, где белок Т6РР уменьшает активность или связывание с субстратом. Для определения молекулярного маркера можно использовать нуклеиновые кислоты/гены или белки Т6РР как таковые. Этот ДНК или белковый маркер затем можно использовать в программах селекции для отбора растений, характеризующихся улучшенными связанными с урожайностью свойствами, как определено в данном документе выше, в способах по настоящему изобретению.
[0265] Нуклеиновые кислоты, кодирующие белки Т6РР, можно также использовать в качестве проб для генетического и физического картирования генов, частями которых они являются, и в качестве маркеров для свойств, связанных с этими генами. Такая информация может быть полезна в селекции растений для разработки линий с желаемыми фенотипами. Такое использование кодирующих белки Т6РР нуклеиновых кислот требует только последовательности нуклеиновой кислоты из по меньшей мере 15 нуклеотидов в длину. Кодирующие белки Т6РР III класса нуклеиновые кислоты можно использовать как маркеры полиморфизма длин рестрикционных фрагментов (RFLP). Саузерн-блоты (Sambrook J, Fritsch Е F and Maniatis T (1989) Molecular Cloning, A Laboratory Manual) геномной ДНК растений, расщепленной при помощи рестрикционных ферментов, можно изучать при помощи кодирующих белок Т6РР нуклеиновых кислот. Полученный в результате характер исчерченности можно затем подвергать генетическому анализу при помощи компьютерных программ, таких как MapMaker (Lander et al. (1987) Genomics 1: 174-181), для построения генетической карты. Кроме того, нуклеиновые кислоты можно использовать для исследования саузерн-блотов, содержащих обработанные эндонуклеазой рестрикции геномные ДНК из набора отдельных объектов, представляющих родителей и потомство определенного генетического скрещивания. Отмечается сегрегация полиморфизмов ДНК и используется для расчета положения кодирующей белок Т6РР нуклеиновой кислоты на генетической карте, ранее полученной при помощи этой популяции (Botstein et al. (1980) Am. J. Hum. Genet. 32: 314-331).
[0266] Получение и использование проб, полученных из генов растений, для применения в генетическом картировании описано в Bernatzky and Tanksley (1986) Plant Mol. Biol. Reporter 4: 37-41. Множество публикаций описывает генетическое картирование конкретных клонов кДНК при помощи методологии, отмеченной выше, или ее вариантов. Например, для картирования можно использовать популяции перекрестного опыления F2, популяции бэккросса, произвольно скрещенные популяции, близкоизогенные линии и другие наборы отдельных субъектов. Такие методологии хорошо известны специалистам в данной области.
[0267] Пробы нуклеиновых кислот можно также использовать для физического картирования (т.е. размещения последовательностей на физических картах; смотри Hoheisel et al. In: Non-mammalian Genomic Analysis: A Practical Guide, Academic press 1996, pp. 319-346, и ссылки, цитируемые там).
[0268] В другом варианте осуществления пробы нуклеиновых кислот можно использовать в прямом картировании флюоресцентной гибридизации in situ (FISH) (Trask (1991) Trends Genet. 7: 149-154). Хотя современные способы картирования FISH помогают использовать большие клоны (от нескольких т.н. до нескольких сотен т.н.; смотри Laan et al. (1995) Genome Res. 5: 13-20), улучшения в чувствительности могут позволить проводить картирование FISH при помощи более коротких проб.
[0269] Множество основанных на амплификации нуклеиновых кислот способов для генетического и физического картирования можно проводить при помощи нуклеиновых кислот. Примеры включают аллельспецифическую амплификацию (Kazazian (1989) J. Lab. Clin. Med 11: 95-96), полиморфизм PCR-амплифицированных фрагментов (CAPS; Sheffield et al. (1993) Genomics 16:325-332), аллельспецифическое сшивание (Landegren et al. (1988) Science 241: 1077-1080), реакции достройки нуклеотидов (Sokolov (1990) Nucleic Acid Res. 18: 3671), радиационное гибридное картирование (Walter et al. (1997) Nat. Genet. 7: 22-28) и Happy Mapping (Dear and Cook (1989) Nucleic Acid Res. 17: 6795-6807). В отношении этих способов последовательность нуклеиновой кислоты используют для разработки и получения пар праймеров для применения в реакции амплификации или в реакциях достройки праймера. Разработка таких праймеров хорошо известна специалистам в данной области. В способах, в которых используют основанное на PCR генетическое картирование, может быть необходимым идентифицирование различий в ДНК последовательностях между родительским скрещиванием на карте в области, соответствующей текущей последовательности нуклеиновой кислоты. Тем не менее, это в целом не является необходимым для способов картирования.
[0270] В одном аспекте настоящего изобретения, не ограничиваясь теорией, содержащие Т6РР мутации, описанные в данном документе, характеризуются сниженной активностью, а также образуют комплекс белок-белок с TPS. Не ограничиваясь теорией, одним возможным аспектом настоящего изобретения является комплекс белок-белок T6PP/TPS, который можно использовать для подтверждения, что у растения увеличенная урожайность и повышенная устойчивость к стрессу. В другом аспекте специалист сможет модифицировать любой полипептид Т6РР, имеющий консенсусную последовательность SEQ ID NO: 9, чтобы он имел более низкую аффинность связывания с Т6Р, тем самым увеличивая частоту возникновения белкового комплекса T6PP/TPS. Предполагается, что в одном аспекте может быть возможным создание или использование существующего химического вещества, которое может имитировать комплекс или создавать ортогональные структуры, которые можно использовать для имитации комплекса T6PP/TPS. В одном аспекте, не ограничиваясь теорией, одну или несколько мутаций можно проводить в В-фосфатазном боксе любого заданного белка Т6РР, чтобы убедиться в увеличенной урожайности растения.
[0271] Результатом способов согласно настоящему изобретению являются растения с улучшенными связанными с урожайностью свойствами, описанными в данном документе выше. Эти свойства можно также объединять с другими экономически выгодными свойствами, такими как свойства, дополнительно увеличивающие урожайность, устойчивость к другим абиотическим и биотическим стрессам, свойства, модифицирующие различные структурные особенности и/или биохимические и/или физиологические признаки.
Настоящее изобретение далее будет описано со ссылкой на нижеследующие фигуры.
Примеры
Пример 1: Идентификация и клонирование последовательности кДНК Т6РР риса в бинарный вектор
[0272] Вначале гены трегалоза-6-фосфатфосфатазы сосудистого растения клонировали из Arabidopsis thaliana путем комплементации делеционного мутанта TPS2 дрожжей (Vogel et al. 1998). В то же время было показано, что гены, обозначенные AtT6PPA и AtT6PPB (номера доступа GenBank AF007778 и AF007779), обладали трегалоза-6-фосфатфосфатазной активностью. Белковые последовательности AtT6PPA и AtTTPB использовали в запросах TBLASTN в базах данных последовательностей маиса и риса. Путем выравнивания последовательностей устанавливали совпадения в отдельных генах. Идентифицировали три гомолога Т6РР у маиса и три гомолога у риса. Последовательность кДНК Т6РР риса (OsT6PP), указанную в SEQ ID NO. 1, амплифицировали с применением высокоточной ПЦР. 50 мкл реакционной смеси, состоящей из 1 мкл библиотеки кДНК риса (полученной из мРНК каллюса в Lambda Unizap Vector от Stratagene, размер первичной библиотеки >1×106 БОЕ, титр амплифицированной библиотеки >1×1012 БОЕ/мл), 200 мкМ dNTP, 1 мкл 20 мкМ олигонуклеотидного праймера Т6РР-EC-5 (5ʹ-catggaccatggatttgagcaatagctcac-3ʹ) и 1 мкл 20 мкМ олигонуклеотидного праймера Т6РР-EC-3 (5ʹ-atcgcagagctcacactgagtgcttcttcc-3ʹ), 5 мкл 10Х буфера Cloned PFU и 2,5 единиц Pfuturbo ДНК-полимеразы. Программа термоциклирования составляла 95°C в течение 2 минут, а затем 40 циклов (94°C в течение 15 секунд, 50°C в течение 1 минуты, 72°C в течение 1 минуты), а затем 72°C в течение 10 минут. T6PP-продукт риса клонировали при помощи набора для ПЦР-клонирования Zero Blunt TOPO. pCR-Blunt-II-TOPO-OsT6PP идентифицировали путем расщепления 5 мкл минипрепа ДНК pCR-Blunt-II-TOPO-OsT6PP с помощью EcoRI в 20 мкл реакционной смеси, содержащей 2 мкг BSA и 2 мкл 10Х буфера рестрикционной эндонуклеазы EcoRI. Реакционную смесь инкубировали при 37°C в течение 2 часов и продукты pCR-Blunt-II-TOPO-OsT6PP (EcoRI) разделяли на 1% TAE-агарозе. Клон pCR-Blunt-II-TOPO-OsT6PP затем секвенировали. кДНК OsT6PP фланкировали с помощью сайтов рестрикционных эндонуклеаз NcoI/SacI. Затем OsT6PP дополнительно клонировали в бинарный вектор, как описано в примере 8 публикации заявки на патент США 2007/0006344 (называемой в ней OsT6PP-3 и обозначенной нуклеотидной SEQ ID NO: 531 и белковой SEQ ID NO: 532).
Пример 2. Начальная оценка объектов маиса с Т6РР риса в теплице
[0273] Объекты маиса с Т6РР риса, содержащие SEQ ID NO: 1, функционально связанную с промотором, характеризующимся предпочтительной экспрессией в материнской репродуктивной ткани (т.е. промотором OsMADS), получали и дополнительно оценивали как в теплице, так и в поле, как описано в примерах 8-13 в публикации заявки на патент США 2007/0006344. С помощью начальной оценки объектов маиса в теплице и в поле обнаружили несколько трансформантов, обладающих повышенной урожайностью как в незасушливых, так и в засушливых условиях (см. публикацию заявки на патент США 2007/0006344, включенную в данный документ посредством ссылки).
Пример 3: Оценка и идентификация высокоурожайных объектов маиса с Т6РР
[0274] Объекты маиса, для которых было показано, что они передают увеличение урожайности в испытаниях, описанных в примере 2 и более конкретно в публикации заявки на патент США 2007/0006344, были дополнительно описаны в отношении урожайности и полевой производительности. Эти объекты содержали один из бинарных конструктов 15777 или 15769, как это описано в публикации заявки на патент США 2007/0006344. Фактически, бинарный конструкт 15769 содержит кассету экспрессии с OsT6PP (указанной в SEQ ID NO: 1 данной заявки), функционально связанную с промотором OsMADS6 (SEQ ID NO: 13). Бинарный конструкт 15777 содержит такую же кассету экспрессии (промотор OsMADS6 и кодирующую последовательность OsT6PP) с добавлением усилителей транскрипции выше промотора OsMADS6. Подробное описание и специфические особенности обоих этих конструктов опять-таки можно найти в публикации заявки на патент США 2007/0006344. В целом из бинарного конструкта 15769 получили 645 объектов Т0 маиса, а из бинарного конструкта 15777 получили 587 объектов. Отбор стабильных объектов, показывающих хороший рост в теплице и полевых условиях, приводил к относительно небольшому количеству объектов, которые переводили в полевые испытания. Высокий уровень отсева приводил только к 17 объектам, показывающим полевую производительность. Объекты, полученные из растений маиса, трансформированных бинарным конструктом 15777, оказались наиболее производительными при полевом тестировании. Два объекта (в данном документе называемые "объект 1" и "объект 2") отбирали на основании выживаемости и продуктивности в управляемых условиях стресса (сохранение урожайности при засухе во время цветения) и при агрономических испытаниях, при которых измеряли урожайность. В целом, наиболее продуктивные объекты, которые были трансформированы конструктом 15777, показали значительное преимущество урожайности в бушелях на акр относительно контрольных проверочных образцов.
Пример 4: Анализ последовательностей объектов 1 и 2
[0275] Объект 1 и объект 2 дополнительно анализировали с помощью секвенирования CDS Т6РР. ПЦР использовали для амплификации интегрированной кодирующей последовательности OsT6PP при помощи праймеров, которые отжигались по 5ʹ- и 3ʹ-области кодирующей последовательности. Соответствующие ПЦР-ампликоны имели размер бэнда приблизительно 1,1 т.п.о., как и следовало ожидать исходя из кодирующей последовательности OsT6PP, показанной в SEQ ID NO: 1, в данном документе известной как OsT6PP-WT, которая представляла собой последовательность, которая содержалась в соответствующей кассете экспрессии. Ампликоны дополнительно секвенировали, общепринятым в данной области техники способом. Данные секвенирования показали, что оба объекта (1 и 2) содержали модификации. Объект 1, содержащий точечную мутацию в нуклеотиде 730 относительно SEQ ID NO: 1 (Т*CATTA, где * указывает на точку мутации), в данном документе называется OsT6PP-H244D. Эта точечная мутация приводила к мутации аминокислоты в остатке 244 относительно SEQ ID NO: 2, изменяя остаток His на остаток Asp, как представлено в SEQ ID NO: 3, в данном документе эквивалентно называемой "точечная модификация Т6РР". Объект 2, как было обнаружено, содержал две модификации в нуклеотидах 305 (TG*CTTCC, где * указывает на точку мутации) и 388 (CGCC*ATT, где * указывает на точку мутации) относительно SEQ ID NO: 1, в данном документе называется OsT6PP-I129F. Двухточечные мутации нуклеотидов, идентифицированные у объекта 2, приводили к изменению Ala на Val в положении 102, и Ile менялся на Phe в положении 129, или к полипептиду Т6РР, представленному SEQ ID NO 5, в данном документе эквивалентно называемому "двойная модификация Т6РР". Интересно, как показано в последующих примерах, две из этих мутаций расположены в высококонсервативных доменах белка Т6РР. Замена Ala на Val в положении 102 находится вне высококонсервативных областей. Высококонсервативные домены вовлечены в связывание субстрата и/или взаимодействия белок-белок при образовании комплекса.
Пример 5: Идентификация Т6РР, имеющих подобную гомологию и функцию, которые можно улучшать путем введения мутаций
[0276] Последовательности (кДНК полной длины, EST или геномная), родственные SEQ ID NO: 1, и/или белковые последовательности, родственные SEQ ID NO: 2, можно идентифицировать среди сохраненных в базе данных Entrez Nucleotides в Национальном центре биотехнологической информации (NCBI) при помощи средств для поиска последовательностей в базах данных, таких как Basic Local Alignment Tool (BLAST) (Altschul et al. (1990) J. Mol. Biol. 215: 403-410; и Altschul et al. (1997) Nucleic Acids Res. 25: 3389-3402). Программу можно использовать для поиска областей локального подобия между последовательностями путем сравнения последовательностей нуклеиновой кислоты или полипептидных последовательностей с базами данных последовательностей и путем расчета статистической значимости совпадений. Полипептид, кодируемый SEQ ID NO: 1, можно использовать с алгоритмом TBLASTN со значениями по умолчанию и отмеченным фильтром для игнорирования последовательностей с низкой сложностью. Вывод данных анализа можно наблюдать в виде попарного сравнения и ранжированного согласно оценке вероятности (Е-значению), где оценка отражает вероятность того, что конкретное выравнивание происходит случайно (чем ниже Е-значение, тем более значимое совпадение). Кроме Е-значений сравнения можно также оценивать по проценту идентичности. Процент идентичности отражает число идентичных нуклеотидов (или аминокислот) между двумя сравниваемыми последовательностями нуклеиновой кислоты (или полипептидными) последовательностями в пределах конкретной длины. В некоторых случаях параметры по умолчанию можно регулировать для модификации точности поиска.
[0277] Таблица ниже представляет перечень последовательностей нуклеиновой кислоты и белковых последовательностей, родственных последовательности нуклеиновой кислоты, представленной SEQ ID NO: 1, и белковой последовательности, представленной SEQ ID NO: 2, которые могут быть применимы в различных аспектах настоящего изобретения. Последовательности идентифицировали посредством поиска по гомологии последовательностей, а также с помощью молекул Т6РР, приведенных в уровне техники.
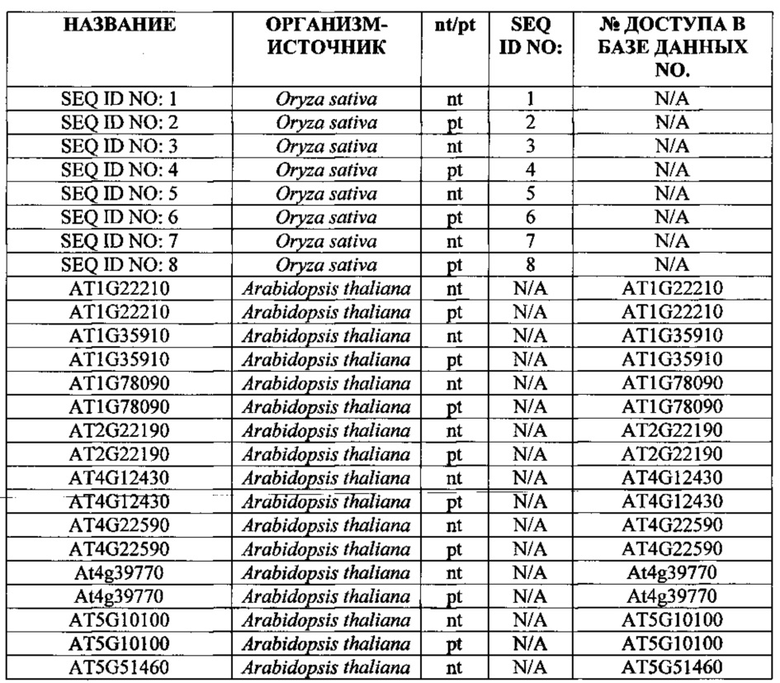
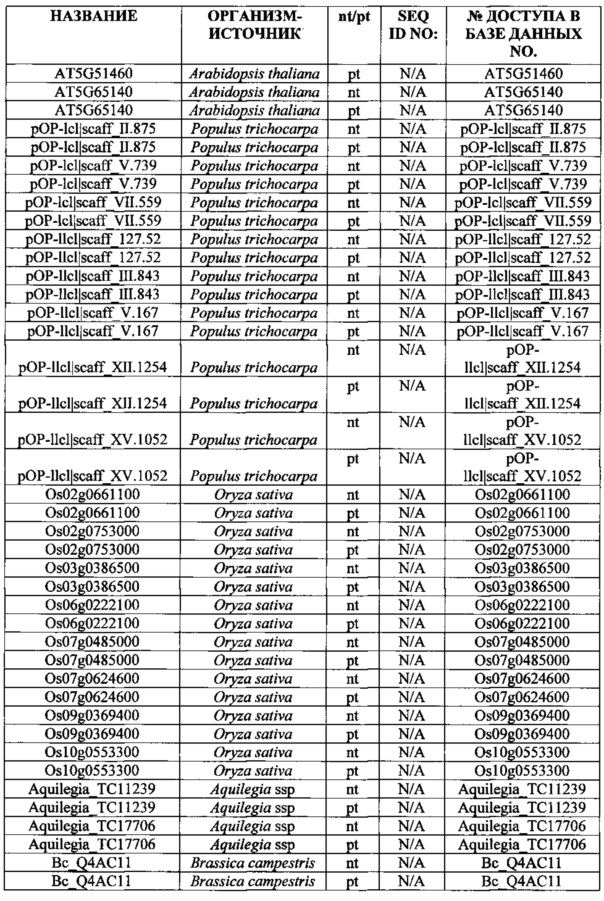
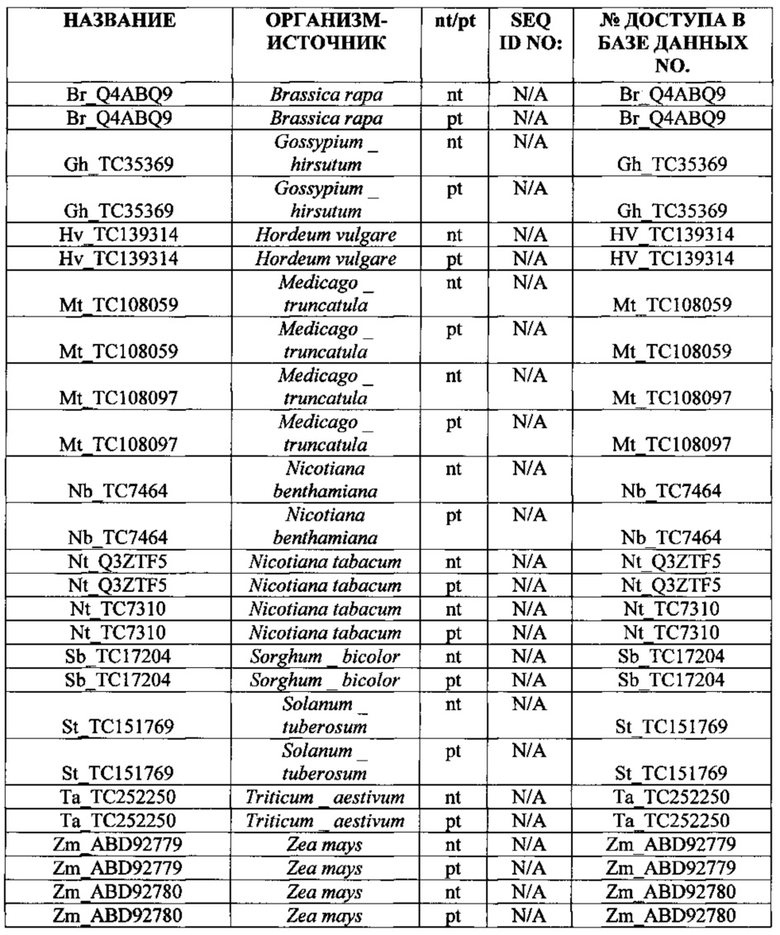
nt = нуклеотидная
pt = белковая
N/A = нет данных
[0278] После идентификации Т6РР, перечисленных в таблице выше, эти Т6РР затем выравнивали с SEQ ID NO: 1, 3 и 5 (при помощи средств выравнивания Vector NTI от INVITROGEN Inc) для определения того, имеются ли мутации в консервативных областях белка. При выравнивании идентифицировали большое количество консервативных последовательностей среди различных белков Т6РР, и их применяли для разработки консенсусной последовательности, изображенной в SEQ ID NO: 9. На фигуре 2 изображена взаимосвязь идентифицированных Т6РР посредством филогенетического древа. Консенсусную последовательность можно применять для модификации любой из нуклеотидных последовательностей и/или белков Т6РР, перечисленных в таблице выше, таким образом, что при использовании в трансгенном растении они будут приводить к повышенной урожайности и/или повышенной устойчивости к стрессу, а также демонстрировать продуктивность в поле. Также подразумевается, что любой белок, обладающий консенсусной последовательностью, характеризующейся 50%, 60%, 70%, 80%, 90%, 95%, 99% или 100% идентичностью последовательности с SEQ ID NO: 9, также показанной на фигуре 10, можно модифицировать, как описано в данном документе, и экспрессировать в растениях, как описано в данном документе.
[0279] Консенсусная последовательность обнаруживает высококонсервативные остатки, присутствующие у множества полипептидов Т6РР, перечисленных в таблице выше. Также подразумевается, что эта консенсусная последовательность будет также соответствовать другим Т6РР, не перечисленным в таблице выше. Положения, обозначенные "X" в консенсусной последовательности выше, указывают на области изменчивости, где может находиться любая отдельная буква, относящаяся к общепринятому однобуквенному обозначению аминокислоты, общеупотребительному в данной области техники. Подчеркнутые аминокислотные остатки указывают на области, которые можно модифицировать для конструирования полипептидов Т6РР, которые придают трансгенному растению повышенную урожайность и/или улучшенную устойчивость к стрессу. В одном аспекте любые из остатков DYDGTLSPIV. кодирующие В-фосфатазный бокс, можно модифицировать для получения белков, которые при экспрессии в растении могут придавать повышенную урожайность и/или улучшенную устойчивость к стрессу. В варианте осуществления настоящего изобретения I (изолейцин) в аминокислотной последовательности, кодирующей В-фосфатазный бокс "DYDGTLSPIV" из консенсусной последовательности выше, меняется на F (фенилаланин), и кодирует при этом (DYDGTLSPFV). В другом варианте осуществления "X" в консенсусной последовательности выше представляет собой V (валин), который является заменой аланина в исходной последовательности. Это замена находится вне консервативного домена и может не влиять на ферментативную активность. Другой вариант осуществления будет содержать такое изменение, "Н" (гистидин) в консенсусной последовательности "CVSVHFRCV" выше, заменяется на D (аспарагиновую кислоту), меняя консенсусную последовательность на последовательность "CVSVDFRCV". Подразумевается, что любую модификацию по меньшей мере одного консервативного аминокислотного остатка SEQ ID NO: 9, где модификация приводит в результате к снижению активности Т6РР, можно экспрессировать в растении для придания повышенной урожайности в условиях стресса и условиях без стресса. Альтернативно, все три модификации можно вводить в ген Т6РР, описываемый SEQ ID NO: 7 (ДНК) и 8 (белок).
[0280] Пример 6: Способы для разработки и тестирования модифицированных белков Т6РР
[0281] Литература, использованная для разработки вариантов T6PP: The α/β hydrolase fold. David L. Ollis et al. (1992) Protein Engineering. vol. 5 197-211. Of barn owls and bankers: a lush variety of α/β hydrolases. Heikinheimo P., Goldman, A., Jeffries, C. and Ollis, D.L. (1999) Structure. Vol 6 141-146. Crystal structure of trehalose-6-phosphate phosphatase-related protein: biochemical and biological implications. Rao, K.N., Kumaran, D., Seetharaman, J., Bonanno, J.B., Burley, S.K., Swaminathan, S. (2006) Protein Sci. 15: 1735-1744. Insights on the evolution of trehalose biosynthesis. Avonce, N. Mendoza-Vargas, A., Morett, E. and Iturriaga, G. (2006) BMC Evolutionary Biology. 6, 109. A single Active Trehalose-6P Synthase (TPS) and a Family of Putative Regulatory TPS-Like Proteins in Arabidopsis. Vandesteene, L., Ramon, M., Le Roy, K., Van Dijck, P. and Rolland, F. (2010) Molecular Plant. Vol 3, Number 2, 406-419. The X-ray crystallographic structure and specificity profile of HAD superfamily phosphohydrolase BT1666: Comparison of paralogous functions in B.thetaiotaomicron. Lu, Z., Dunaway-Mariano, D. and Allen, K.N. (2011) Proteins: Structure, Function and Bioinformatics. 79 (11) 3099-3107.
[0282] Способ
[0283] На основании литературных данных по кристаллической структуре белок, родственный трегалоза-6-фосфатфосфатазе, из Thermoplasma acidophilum идентифицировали как имеющий самую близкую гомологию структуры белка с модельной трегалоза-6-фосфатфосфатазой (Т6РР) риса. Код доступа этого белка в базе данных по структуре белка (PDB) составляет 1U02. См., Crystal structure of trehalose-6-phosphate phosphatase-related protein: biochemical and biological implications Rao, K.N., et al. (2006) Protein Sci. 15: 1735-1744 а код Uniprot: Q9HIW7.
[0284] Начальное выравнивание последовательностей
[0285] Последовательности Т6РР Arabidopsis thaliana (19925; SEQ ID NO: 14), Arabidopsis thaliana (19926; SEQ ID NO: 15), Oryza sativa (19924; SEQ ID NO: 16) и Oryza sativa (15777; OsT6PP-WT; SEQ ID NO: 2) использовали для выравнивания, a также SEQ ID NO: 4. Эти белковые последовательности вместе с белком, родственным Т6РР, из Thermoplasma acidophilum (Q9HIW7) сравнивали с помощью Vector NTI при помощи способа ClustalW для получения начального выравнивания, см. фигуры 1А-1С. Начальное выравнивание белковых последовательностей белка, родственного Т6РР (Q9HIW7), и белковых последовательностей Arabidopsis thaliana (19925, SEQ ID NO: 14), Arabidopsis thaliana (19926; SEQ ID NO: 15), Oryza sativa (19924, SEQ ID NO: 16) и Oryza sativa (15777, SEQ ID NO: 2). Остатки, выделенные жирным шрифтом и курсивом в выравнивании белковых последовательностей ниже, представляют собой значимые остатки для более крупного суперсемейства галогендегидрогеназ, которые помогают подтвердить качество выравнивания.
[0286] Подчеркнутые остатки на фигурах 1А-1С представляют собой значимые остатки, которые помогают подтвердить качество выравнивания. Большинство этих остатков представляют собой каталитические остатки (номера всех остатков ссылаются на номера остатков в белковой последовательности 15777 OsT6PP-WT Oryza sativa): остаток аспарагиновой кислоты 121 образует фосфоаспартатное промежуточное соединение с субстратом. Треонин 318) (в более крупном суперсемействе этот остаток может быть серином) связывается с помощью водородной связи с фосфорилкислородом в субстрате. Аспарагиновые кислоты 315 и 123 образуют координационную связь с ионом Mg2+ за счет своих боковых цепей и карбонила скелета, соответственно. Лизин 289 стабилизирует переходное состояние реакции.
[0287] Получение структуры белка 1U02 для гомологичного моделирования
[0288] Кристаллическую структуру белка, родственного Т6РР (код по PDB 1U02), загружали в Maestro (Schrödinger, Inc.) и анализировали, а также готовили для использования (добавляя протоны, выстраивая дисульфидные мостики, устанавливая Наряды металлов, оптимизируя водородные связи).
[0289] В то же самое время получили и проанализировали экспериментальную карту электронной плотности для элемента 1U02 из PDB. Хотя большинство признаков в карте были смоделированы правильно для осажденной структуры, электронная плотность в положении молекулы глицерина вблизи активного сайта не соответствовала ожидаемым признакам плотности глицерина. Вместо этого, карта ясно указывала на в этом положении двух молекул воды. Таким образом, молекулу глицерина удалили из модели, а две молекулы воды добавили в их правильные положения. Перед началом процесса гомологичного моделирования из структуры удалили все молекулы воды, кроме двух молекул воды, непосредственно связанных координационной связью с ионом Mg2+ в активном сайте.
[0290] Такой результат моделирования в работе Rao et al. имел важные следствия. Поскольку белок, родственный Т6РР, кристаллизовался лишь в виде апофермента и не содержал связанный субстрат, авторы использовали положение молекулы глицерина для определения ими процедуры докинга, осуществляемой вручную. Действительно, авторы делали выводы, что субстрат будет связываться в сайте связывания глицерина, поскольку глицерин отчасти подобен по своим свойствам пиранозам. Однако, поскольку глицерин не обнаружили в активном сайте в структуре, объясняемой Rao et al., их докинг субстрата Т6Р с апоферментом была изменена в следующей модели.
[0291] гомологичное моделирование
[0292] Начальное выравнивание последовательностей считывали с помощью PRIME (Schrödinger) вместе со структурой белка-шаблона. При более подробном анализе выравнивание белковой последовательности модифицировали с целью оптимизации местоположения вставки петель (вставок в направлении внешней поверхности фермента) и значимых остатков из литературы. См. выравнивание (выше) касательно начального и выравнивание (ниже) касательно конечного выравнивания белковой последовательности.
[0293] Конечное выравнивание использовали в PRIME для получения гомологичных моделей Oryza sativa 15777 и Oryza sativa 19924. Эти модели ослабляли с применением минимизации энергии в MacroModel (Schrödinger), сначала для атомов водорода, затем включая боковые цепи белков, наконец включая скелет белков. Конечная модель показана на фигуре 7. Фермент содержит два домена: α/β-гидролазный домен и более мелкий "домен lid". Активный сайт лежит на верхушке α/β-гидролазного домена (точечная отметка); щель связывания субстрата находится на границе между двумя доменами. Два домена соединены гибким линкером, что дает возможность домену lid открываться и закрываться во время катализа. Поскольку шаблон для гомологичного моделирования, бактериальный белок, родственный Т6РР (PDB код 1U02), представляет собой апофермент с пустым активным сайтом, вероятно, относительная ориентация lid и гидролазного доменов в действительном комплексе фермент-субстрат несколько отличается.
[0294] Конечное выравнивание белковой последовательности белка, родственного Т6РР белка (Q9HTW7), и белковых последовательностей Arabidopsis thaliana (19925), Arabidopsis thaliana (19926), Oryza sativa (19924) и Oryza sativa (15777) можно увидеть на фигурах 11A-11С. Выравнивание оптимизировали с применением данных из литературы и анализа каркаса белковой структуры. Остатки, выделенные жирным шрифтом и курсивом в выравнивании белковых последовательностей ниже, представляют собой значимые остатки в более крупном семействе галогендегидрогеназ, которые помогают подтвердить качество выравнивания.
[0295] Как описано выше, положение Т6Р, сообщаемое Rao et al., было основано на неправильной предпосылке и не может использоваться в качестве отправной точки для рациональной разработки мутантов Т6РР. Вместо этого мы решили состыковывать Т6Р в гомологичную модель вручную с помощью поэтапного подхода, следуя принципам механизма реакции и принимая во внимание биохимические данные.
[0296] Начальная отправная точка основывалась на том факте, что один из отрицательно заряженных атомов кислорода группы 6-фосфата должен связываться с помощью координационной связи с каталитическим ионом Mg2+ в активном сайте. Это является необходимым условием механизма катализа и общим признаком семейства фосфатаз HAD (см., "The X-ray crystallographic structure and specificity profile of HAD superfamily phosphohydrolase BT1666: Comparison of paralogous functions in B.thetaiotaomicron." Lu, Z., Dunaway-Mariano, D. and Allen, K.N. (2011) Proteins: Structure, Function and Bioinformatics. 79 (11): 3099-3107). В то же самое время, кислород фосфата должен находиться в непосредственной близости с Asp121, который акцептирует фосфатную группу и образует переходное фосфор-аспартатное промежуточное соединение во время катализа. Помещение кислорода фосфата в экваториальное положение в октаэдрической координационной сфере Mg2+ удовлетворяет оба требования. Это взаимодействие прикрепляет молекулу Т6Р к белку, и его использовали для определения процесса докинга.
[0297] Исходя из начального расположения кислорода фосфата в координационной сфере Mg, становится ясным, что имелась только одна ориентация группы 6-фосфата, которая позволила бы кислороду сложного фосфоэфира направляться в сторону щели активного сайта, а не сталкиваться с остатками, окружающими активный сайт. В этой ориентации один из остальных кислородов фосфата образует водородную связь с Lys289, которая будет позволять лизину стабилизировать возникновение отрицательного заряда на фосфатной группе во время катализа (фигура 8). Один из атомов кислорода группы 6-фосфата находится в экваториальном положении в координационной сфере Mg2+. Asp121, который акцептирует фосфатную группу во время катализа с образованием ковалентного промежуточного соединения, предполагается в осевом положении. Lys289 непосредственно связан водородной связью с фосфатной группой, и His181 находится в непосредственной близости. Оба остатка расположены идеально для стабилизации возникновения отрицательного заряда на фосфатном фрагменте во время катализа. Гомологичная модель Т6РР риса изображена зелеными палочками, а субстрат Т6Р желтыми.
[0298] Когда ясно определили положение и ориентацию группы 6-фосфата, затем минимизировали субстрат в активном сайте при помощи MacroModel (Schrödinger) с жестким удержанием белка. Наконец, две способные к вращению связи между пиранозными кольцами сканировали при помощи Maestro (Schrödinger) и полученные конформеры оценивали визуально, чтобы идентифицировать возможных взаимодействий с белком (фигура 9). Идентифицировали несколько конформаций, в которых второе пиранозное кольцо находилось в достаточно хорошем положении для образования контактов с белком. Наконец, конформер, в котором второе кольцо было приблизительно равноудалено от остатков домена lid, выбрали в качестве наиболее вероятного и использовали в последующих анализах.
[0299] Хотя положение и ориентацию пиранозного кольца, ближайшего к группе 6-фосфата, явно определили с помощью закрепленной фосфатной группы, существует значительная гибкость между первым и вторым кольцами из-за присутствия двух способных к вращению гликозидных связей (синие стрелки). Ряд возможных положений второго кольца получили вращением двух связей. Конформации, в которых второе кольцо сталкивается с белком, отбросили.
[0300] Разработка вариантов OsT6PP
[0301] Разрабатывали точечные мутации, которые снижали бы или полностью устраняли каталитическую активность Т6РР риса (OsT6PP-WT). В общих чертах, спроектированные мутанты относятся к трем классам, перекрывая диапазон от активности, близкой к дикому типу, до полностью неактивных:
[0302] Мутация ключевых каталитических остатков, основанная на ферментативном механизме. Мутации этого класса, как ожидается, обладают сильным воздействием на каталитическую активность ("тяжелые мутанты").
[0303] Мутация остатков, вовлеченных в связывание и распознавание субстрата. Мутации этого класса, как ожидается, будут снижать оборот и/или изменять субстратную специфичность, но не полностью прекращать каталитическую активность. Для идентификации этих остатков требуется хорошая оценка того, где субстрат располагается в щели активного сайта ("мягкие мутанты").
[0304] Контрольные мутации. Мутации этого класса, как прогнозируется, обладают очень умеренным воздействием на каталитическую активность, поскольку они располагаются далеко от сайта связывания субстрата и не будут влиять на стабильность белка ("очень мягкие мутанты").
[0305] Для всех трех классов использовали следующие правила разработки, чтобы убедиться, что наблюдаемая у мутантов активность была результатом определенных замен, произведенных в белке, а не являлась лишь следствием общей дестабилизации:
[0306] Если интересующий остаток является гидрофобным и, как предполагается, образует неполярное взаимодействие с субстратом, сохранить гидрофобный характер остатка и увеличить объем. Это будет приводить к пространственному сталкиванию с субстратом и снижать аффинность связывания.
[0307] Если остаток обладает способностью к образованию водородных связей (либо в качестве донора, либо в качестве акцептора), что будет позволять ему взаимодействовать с одной из гидроксильных групп пиранозы, следует либо укоротить остаток, в то же самое время сохраняя способность к образованию водородных связей, или сохранить длину приблизительно такой же, но убрать способность к образованию водородных связей.
[0308] Ни одна модификация остатков не предполагает образования междоменных солевых мостиков. В белке, родственном Т6РР, существует ряд солевых мостиков, образованных положительно заряженным остатком в домене lid и отрицательно заряженным остатком в α/β-гидролазном домене, и vice versa. Мутирование этих остатков вероятно будет дестабилизировать белок без специфичного изменения субстратной аффинности или каталитической активности.
[0309] Никакого обращения заряда: положительно заряженные остатки не следует мутировать в отрицательно заряженные (и vice versa), а скорее в нейтральный остаток. Мутанты с обращением заряда, как известно, вызывают локальную дестабилизацию структуры белка, и, следовательно, их следует избегать.
[0310] Из-за неопределенности положения второго пиранозного кольца, и того факта, что относительная ориентация домена lid и α/β-гидролазного домена может меняться при связывании субстрата (см. фиг. 4), мы считали, что остатки находятся на 10 Å от смоделированного субстрата Т6Р. Работая в концентрических оболочках снаружи от субстрата, мы анализировали остатки, окружающие субстрат и рассматривали их в отношении мутации, следуя правилам, описанным выше.
[0311] Список точечных мутаций Т6РР, разработанных для снижения ее трегалоза-6-фосфатфосфатазной активности
[0312] Тяжелые мутанты
[0313] D315N: Asp315 является одним из двух остатков Asp в активном сайте, которые образуют хелат с каталитическим ионом Mg2+ (фиг. 5). Замена его на изостерический аспарагин будет удалять его способность к связыванию Mg2+ и приводить к полной потере активности.
[0314] K289A: Lys289, вероятно, образует водородную связь с группой 6-фосфата Т6Р и служит для стабилизации возникающего отрицательного заряда при катализе (фиг. 5). Замена его на Ala будет значительно замедлять реакцию, приводя практически к отсутствию активности.
[0315] H181A: His181 находится в непосредственной близости от группы 6-фосфата Т6Р и, вероятно, служит в качестве неспецифической кислоты при катализе. Мутация на Ala будет удалять неспецифическую кислоту и замедлять катализ практически до отсутствия активности.
[0316] T318V: Thr318 является консервативным остатком в суперсемействе HAD (Lu et al. 2011) и образует водородную связь с группой 6-фосфата. Замена его на меньший неполярный остаток валин удаляет способность к образованию водородных связей и делает связывание с сильно заряженной группой нежелательным. Эта мутация, как ожидается, приводит к практически полной потере активности.
[0317] Мягкие мутанты
[0318] S159A: Ser159 находится в 5,8 Å от субстрата и может быть вовлечен во взаимодействия с образованием водородных связей с субстратом, либо в качестве донора, либо в качестве акцептора. Замена его на аланин будет прекращать образование водородных связей и снижать субстратную аффинность.
[0319] E235D: Glu235 находится в домене lid и в 10,6 Å от субстрата. Он вероятно функционирует в качестве донора при образовании водородной связи; замена его на аспартат будет сохранять его свойства образования водородных связей, но увеличивать расстояние до субстрата, приводя к неидеальной геометрии образования водородной связи и сниженной субстратной аффинности.
[0320] S242D, S242E: Ser242 находится в домене lid и в 8,2 Å от субстрата. Он, как предполагается, образует взаимодействия в форме водородных связей с субстратом, и замещение его на аспартат или глутамат будет сохранять полярный характер и способность к образованию водородных связей, но приводить к пространственному столкновению с субстратом, вследствие того, что аспартат и глутамат имеют более длинные боковые цепи.
[0321] Т273Е, T273V: Thr273 находиться лишь в 4 Å от субстрата и может быть вовлечен в полярное взаимодействие посредством гидроксильной группы или в неполярное взаимодействие с боковой поверхностью пиранозного остатка. Если взаимодействие полярное, тогда как мутация с глутаматом (пространственное препятствование), так и мутация с валином (удаление полярной группы и потеря способности к образованию водородных связей) будет приводить к сниженной активности. Если взаимодействие неполярное, тогда только первая мутация будет приводить к потере активности, в то время как вторая будет иметь нейтральный эффект.
[0322] V278F, V278M: Val278 находится в 3,4 Å от субстрата и, вероятно, образует неполярное взаимодействие с плоскостью пиранозного кольца. Мутация на фенилаланин или метионин будет сохранять гидрофобный характер боковой цепи, но приводить к сильному пространственному столкновению с субстратом и сниженной активности.
[0323] Е280А: Glu280 находится в домене lid и в 6,7 Å от субстрата. Он, вероятно, служит в качестве акцептора при образовании водородной связи, и его мутирование в аланин и будет прекращать его способность к образованию водородных связей и снижать связывание субстрата.
[0324] D316H: Asp316 находится в гидролазном домене и в непосредственной близости к субстрату, но может быть вовлечен в солевой мостик с R276, а не в связывание субстрата. Замещение на гистидин будет удалять его отрицательный заряд и разрушить солевой мостик, что будет иметь лишь умеренные эффекты. Однако, если Asp316 вовлечен в связывание субстрата путем акцептирования водородной связи, тогда воздействие на каталитическую активность будет сильным.
[0325] I129A, I129N: Ile129 находится в непосредственной близости от первого пиранозного кольца и, возможно, образует неполярное взаимодействие с субстратом. Обе мутации, как предполагается, будут приводить к сниженной активности, первая из-за удаления гидрофобных взаимодействий, вторая из-за введения полярной функциональной группы.
[0326] H244S: His244 находится в домене lid в приблизительно 10 Å от субстрата. Мутация H244D уже известна, как обладающая сильно сниженной каталитической активностью, которая может быть обусловлена потерей связывания субстрата через гистидин или обусловлена непрямым эффектом обращения заряда на смежных остатках, которые могут быть вовлечены в связывание субстрата сами по себе. Мутацию с серином проводили для тестирования, действительно ли His244 непосредственно вовлечен в связывание субстрата. Если это так, тогда укорачивание боковой цепи, в то же самое время при сохранении ее полярного характера, должно приводить к снижению в связывании субстрата.
[0327] Очень мягкие/ контрольные мутанты
[0328] R276A: Arg276 расположен на домене lid и может быть вовлечен в связывание субстрата, но он, возможно, направлен от субстрата и также может быть вовлечен в солевой мостик с Asp316 на гидролазном домене. Замещение на аланин, как ожидается, приводит лишь к умеренным эффектам.
[0329] V293A: Val293 находится в 6 Å от субстрата, но развернут наружу в направлении основной массы растворителя и, маловероятно, обладает какой-либо ролью в связывании субстрата. Мутация с аланином является очень консервативной и, как ожидается, не обладает каким-либо эффектом.
[0330] Исходя из вышеописанной стратегии разработали 22 варианта OsT6PP, и они описаны в таблице ниже.
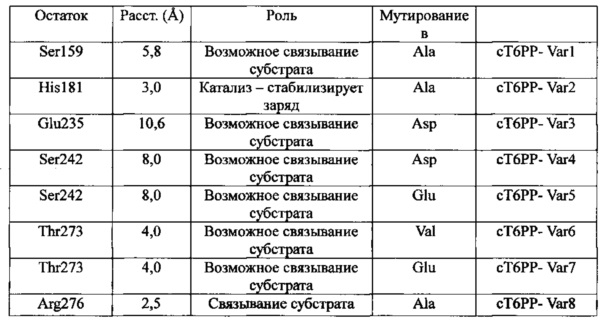
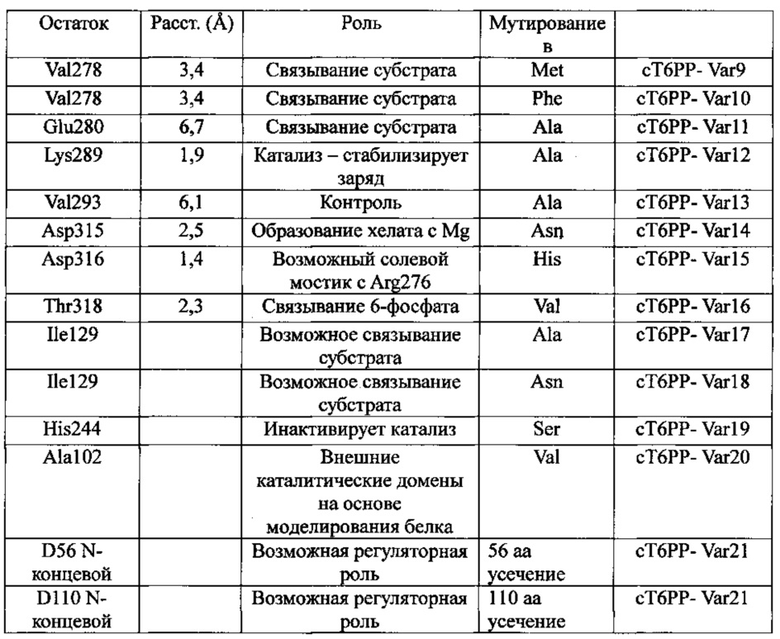
[0331] Что касается нумерации остатков в этой таблице, обратитесь к номерам остатков в белковой последовательности 15777 OsT6PP-WT Oryza sativa. Варианты Δ56 N-концевой и Δ110 N-концевой с N-концевым усечением из 56 и 110 аминокислот, соответственно, были разработаны для тестирования возможной регуляторной роли этой части молекулы белка. Как очевидно из вышеприведенных выравниваний, эта 110 N-концевая часть Т6РР присуща только Т6РР растений и может не являться необходимой для фосфатазного катализа, исходя из моделирования остатков активного сайта.
[0332] Конструирование вектора и трансформация Е.coli
[0333] Кодирующую последовательность вариантов Т6РР синтезировали и клонировали в вектор экспрессии Е.coli pGEX-6P_3 для экспрессии в качестве GST-слитого белка: GST находится на N-конце белка Т6РР. Нуклеотидные последовательности вставленных в вектор ДНК секвенировали и подтверждали.
[0334] Плазмиды для экспрессии вариантов OsT6PP трансформировали в клетки BL21 (DE3) pLysS Е.coli на основе протокола Invitrogen® для экспрессии рекомбинантных белков. Кратко, размораживали клетки Е.coli на льду и по 10 нг (в 10 мкл) каждой плазмидной ДНК добавляли к клеткам и аккуратно перемешивали с помощью постукивания. Пробирки держали на льду в течение 15 мин и осуществляли тепловой шок в течение 30 с при 42°C в водяной бане. Пробирки держали на льду после теплового шока. В каждую пробирку добавляли по 250 мк SOC и инкубировали, со встряхиванием, в течение 1 часа при 37°C. По 50 мкл продуктов реакции трансформации наносили на чашки с LB, содержащей ампициллин (50 мкг/мл) и хлорамфеникол (34 мкг/мл). Чашки переворачивали и инкубировали в течение ночи при 37°C.
[0335] Экспрессия и частичная очистка рекомбинантных OsT6PP и его вариантов
[0336] Клетки Е.coli [BL21(DE3)pLysS], несущие вектор для экспрессии рекомбинантного варианта Т6РР, выращивали до A600нм 0,6 в среде Luria Broth, содержащей 50 мкг/мл ампициллина и 25 мкг/мл хлорамфеникола. Экспрессию рекомбинантного белка индуцировали добавлением 0,5 мМ IPTG в течение 2 ч при 37°C. Клетки собирали при 6000 Xg (ротор JA) или 3000 об/мин в центрифуге Allegra 6R. Дебрис повторно суспендировали в буфере (20 мМ Tris-HCl, pH 7,5, 150 мМ NaCl, ингибитор протеазы (GE Healthcare®) 1 таблетка на 50 мл буфера, 2 мкл ДНКазы I/мл буфера). Клетки разрушали обработкой ультразвуком (10 циклов из 10 с включения - 30 с выключения, установленная мощность на выходе 3,5 (18 Вт). Пробирки держали на льду во время обработки ультразвуком. Лизаты центрифугировали при 15000 Xg в течение 15 мин. Супернатанты переносили в 15 мл пробирки falcon и выдерживали на льду перед добавлением гранул Glutathione Sepharose® 4В (Thermo Scientific®).
[0337] Гранулы Glutathione Sepharose 4B предварительно уравновешивали 3 раза промывочным буфером (20 мМ Tris-HCl, pH 7,5, 150 мМ NaCl, 1 мМ DDT). 250 мкл предварительно уравновешенных гранул добавили в каждую пробирку с лизатом (7 мл) и инкубировали в течение 1:30 ч в 4 при медленном покачивании. Гранулы Glutathione Sepharose 4В, содержащие лизаты, перенесли в хроматографические колонки poly-prep (10 мл BioRad®) и бульону позволяли полностью вытечь. Смолу, связавшую белок, промывали промывочным буфером (3 раза по 5 мл) с последующей промывкой (5 мл, 3X) буфером для отщепления PreScission® (20 мМ Tris-HCl, pH 7,0, 150 мМ NaCl и 1 мМ DTT). Фермент-протеазу PreScission (10 мкл в 300 мкл буфера) добавляли к гранулам, связавшим белок, и инкубировали в течение 16 ч при 4°C с медленным покачиванием. Супернатант собирали и рециркулировали 3 раза через колонку и последний сбор держали на льду для вытекания геля и измерения концентрации.
[0338] Концентрацию белка измеряли при помощи наборов BCA [Thermo Scientific®]. 20 мкл каждого исходного раствора белка смешивали с 4Х красителем SDS и проводили SDS-PAGE (4-12% градиентный гель) с применением буфера MOPS при 200 В для достижения обогащения и обеспечения однородности очищенного белка. Конечную процентную чистоту частично очищенной Т6РР и ее вариантов определяли при помощи автоматизированного электрофореза Bio-Rad Experion®.
[0339] Активность OsT6PP и ее мутантов
[0340] Активность OsT6PP и ее вариантов измеряли на основе стандартного протокола анализа. Кратко, подходящее количество белка получали из исходного раствора и помещали в 96-луночные планшеты. Реакционные смеси (буфер с 2 мМ конечной концентрацией Т6Р в реакционной смеси; состав буфера, 10 мМ Tris, pH 6,7, 2 мМ MgCl2 и 0,5 мг/мл BSA) получали и держали на льду перед смешиванием с препаратом фермента. Подходящее количество разбавленного ферментного белка добавляли к реакционным смесям (применяли концентрацию белка 0,05, 0,1 или 1 мкг в общем реакционном объеме 100 мкл) в 96-луночном планшете. Реакционные смеси инкубировали при 37°C в течение различных временных интервалов (5, 10 и 20 минут) и фермент в реакционной смеси деактивировали путем нагревания при 95°C в течение 5 мин. Инактивированные реакционные смеси в 96-луночном планшете центрифугировали в течение 1 минуты при 700 об/мин, затем 80 мкл каждого реакционного раствора переносили в лунки, содержащие 80 мкл воды. Смеси переносили в лунки планшета для фильтрации MicroScreen-HV и 96-луночный планшет Dionex использовали для сбора фильтрата для анализа HPLC. Специфичную активность рассчитывали на основе стандартной процедуры.
[0341] Dionex HPLC
[0342] Систему для ионной хроматографии Dionex ICS-3000 использовали для обнаружения трегалозного продукта, образованного дефосфорилированием Т6Р-субстрата в присутствии фермента Т6РР. Трегалозу отделяли при помощи аналитической колонки CarboPac® PA1 с градиентом элюирования 15 минут при скорости потока 1 мл/мин. Образцы вводили в колонку CarboPac PA1, уравновешенную 100 мМ NaOH, при скорости потока 1 мл/мин, с последующим 0-200 мМ градиентом ацетата натрия в 100 мМ NaOH от 3 до 8 мин, затем 200 мМ-750 мМ градиентом ацетата натрия в 100 мМ NaOH от 8 до 10 мин и вновь 100 мМ NaOH от 10,5 до 15 мин. Трегалозу, отделенную при помощи хроматографии, количественно определяли путем интегрирования области пика с применением программного обеспечения Chromeleon®.
[0343] График данных и оценка специфических активностей вариантов Т6РР
[0344] Аналитические реакционные смеси инкубировали в течение трех различных интервалов времени, и каждая точка на графике представляет собой среднее от трех независимых аналитических реакционных смесей. Фоновое количество (образование трегалозы в отсутствие фермента, отрицательный контроль) отслеживали и вычитали из трегалозы, полученной в аналитической смеси с добавленным ферментом. К нанесенным на график данным подбирали полиномиальное уравнение (Y=AX2+bX). Скорость реакции оценивали на основании угла наклона (2AX+b) вышеуказанного уравнения в 1 минуту.
[0345] Скорость реакции = полученная трегалоза (мкМ)/мин и преобразованная в мкмоль/мин.
[0346] Специфическая активность = скорость образования трегалозы (мкмоль/мин)/мг Т6РР.
[0347] Сравнение трегалоза-6-фосфатфосфатазных активностей вариантов Т6РР, определенных с помощью аналитической процедуры, описанной выше.
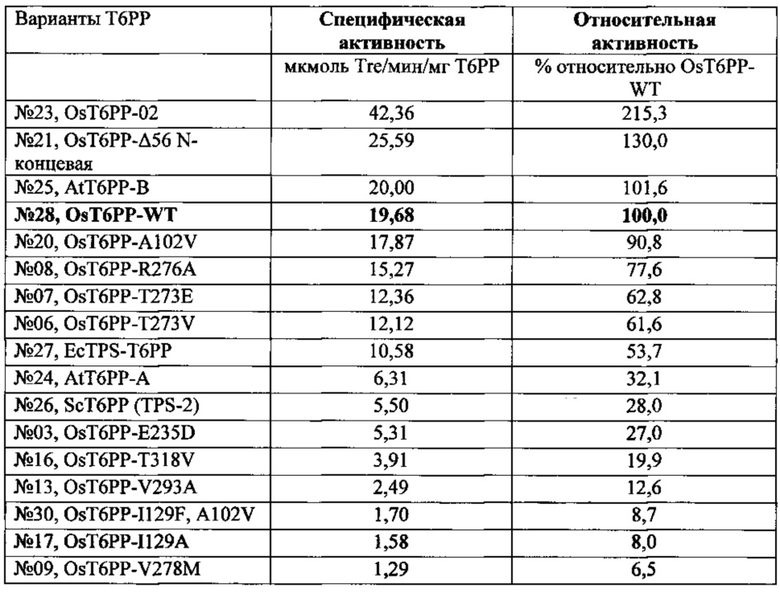
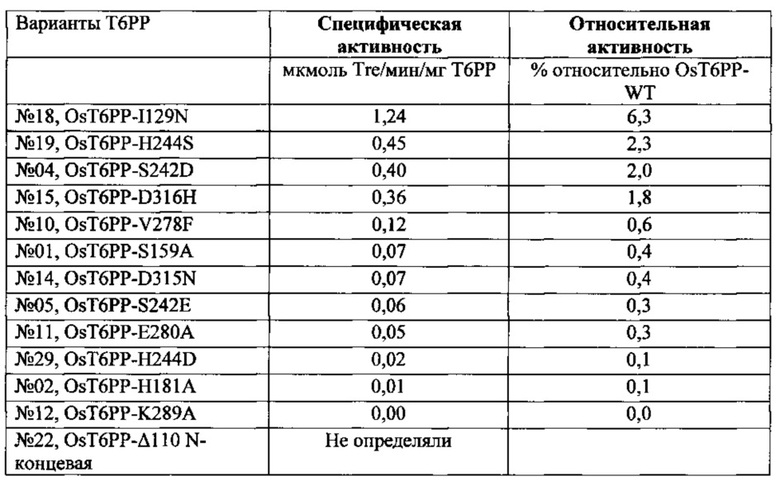
[OsT6PP-02, Oryza sativa T6PP-02; AtT6PP-A, Arabidopsis thaliana T6PP-A; AtT6PP-B, Arabidopsis thaliana T6PP-A; EcTPS-T6PP, слитый белок трегалоза-6-фосфатсинтазы и фосфатазы Escherichia coli и ScT6PP[TPS-2], Saccharomyces cerevisiae, трегалоза-6-фосфатфосфатаза]
Пример 7: In vitro оценка возможных взаимодействий белок-белок между Т6РР и TPS и дополнительная оценка дальнейших эффектов.
[0348] Для оценки того, может ли Т6РР из риса взаимодействовать с различными компонентами TPS-комплекса дрожжей, будет разработана дрожжевая модельная система. Vandercammen et. al. (Eur. J. Biochem. 182, 613-620 (1989)) вследствие совместной очистки TPS и T6PP из Saccharomyces cerevisiae установили, что эти белки являются частью одного бифункционального белка. Bell et. al. (The Journal of Biological Chem. 273; No. 50, pp 33311-33319 (1998)) дополнительно описали состав и провели функциональный анализ комплекса T6PP/TPS в Saccharomyces cerevisiae. Также было показано, что TPS дрожжей может образовывать комплекс с Т6РР, полученным из Arabidopsis (см. Vogel et. al. The Plant Journal 13; No. 5, pp. 673-683). TPS-комплекс дрожжей содержит четыре элемента TPS1, TPS2, TPS3 и TSL1. TPS1 содержит белковый домен с TPS-активностью, a TPS2 содержит белковый домен с Т6РР-активностью. Функцией как TPS3, так и TSL1 является регуляция функции TPS комплекса. Не ограничиваясь теорией, подразумевается, что с помощью разработки дрожжевой модели можно оценивать дополнительные модификации in vitro, что может применяться для разработки белков, пептидов, полипептидов или химических веществ, которые могут повышать образование Т6Р, таким образом, регулируя поток сахаров в репродуктивные ткани и/или запуская различные другие каскады, которые придают растению повышенную урожайность и/или стрессоустойчивость при экспрессии или применении в растении. Целью способа является: (А) тестирование того, может ли Т6РР риса взаимодействовать с различными компонентами TPS-комплекса дрожжей, и дополнительное измерение как эти взаимодействия влияют на каталитические активности каждого компонента путем измерения продуктов ниже по каскаду (т.е. трегалозы, Т6Р); (В) разработка in vitro анализа для тестирования того, взаимодействует ли Т6РР риса с TPS риса; С) экспрессия модифицированной Т6РР риса в маисе для выделения и описания характеристик полученного Т6РР/TPS-комплекса, (D) использование данных, полученных из (А), (В) и/или (С) для проведения биоинформатического анализа для идентификации дополнительных модификаций, которые можно производить в молекуле Т6РР для дальнейшего облегчения получения Т6Р. Также предполагается, что нуклеотиды, кодирующие сам комплекс, можно разработать из данных, собранных в ходе указанных экспериментов.
[0349] Оценка взаимодействия Т6РР риса с компонентами TPS-комплекса дрожжей, меченные TAP
[0350] Векторы экспрессии будут отдельно сконструированы для экспрессии либо в дрожжах, либо в Е.coli, которые содержат полинуклеотидную последовательность, кодирующую немодифицированный белок Т6РР, как представлено в SEQ ID NO: 2, и модифицированный белок Т6РР, как представлено в SEQ ID NO: 6. Конструкт также можно разработать для других модифицированных Т6РР для оценки различных эффектов мутаций, например, нуклеотидной последовательности, кодирующей Т6РР, такой как показанный SEQ ID NO: 4, или любой другой Т6РР, который был модифицирован, чтобы иметь сниженную активность. Эти конструкты можно затем трансформировать как в бактерии, так и в дрожжи для дальнейшей оценки.
[0351] В таблице ниже идентифицированы различные общедоступные штаммы дрожжей, имеющие белки, меченные TAP, которые можно использовать для оценки взаимодействий между экспрессированными Т6РР риса и различными компонентами TPS-комплекса дрожжей. Штаммы дрожжей с меткой TAP можно получить от поставщика, такого как Thermo Scientific. Белки, меченные TAP, можно легко обнаружить с помощью вестерн-блоттинга, что хорошо известно в данной области техники. Для определения того, взаимодействуют ли варианты Т6РР риса (дикого типа и модифицированные) с каким-либо компонентом TPS-комплекса дрожжей, можно получать экстракты, содержащие как белок Т6РР риса, так и родственные белковые компоненты TPS-комплекса дрожжей, меченные TAP. Затем белки можно извлекать из этих экстрактов и проводить вестерн-блоттинг для определения того, происходит ли взаимодействие белок-белок между какими-либо модифицированными Т6РР риса и компонентами TPS-комплекса дрожжей (т.е. на взаимодействие белок-белок будет указывать больший, чем ожидалось, размер бэнда) посредством обнаружения с помощью антитела к TAP. Следующая таблица описывает штаммы дрожжей, содержащих белки, меченные TAP, которые можно обнаруживать с помощью анализа вестерн-блоттинг при помощи антитела к TAP.
[0352] После идентификации взаимодействий белок-белок между Т6РР риса и компонентами TPS-комплекса дрожжей будут проводиться эксперименты для определения того, как различные модифицированные и немодифицированные Т6РР влияют на функцию TPS-комплекса дрожжей. TPS2-компонент дрожжей (т.е. часть, обладающая T6PP-активностью) TPS-комплекса будут замещать либо на немодифицированный белок Т6РР риса (SEQ ID NO: 1), либо модифицированные белки Т6РР риса (SEQ ID NO: 3, 5, 7). Затем эти модифицированные комплексы можно экспрессировать в дрожжах для дальнейшей оценки того, как эти замещения влияют на общее функционирование TPS-комплекса с помощью измерения уровней Т6Р и трегалозы. Уровни Т6Р и уровни трегалозы можно измерять в экстрактах из дрожжей при помощи стандартных способов HPLC, известных в данной области техники, для определения различных влияний каждого замещения. Теоретически полагают, что комплексы, содержащие модифицированную Т6РР риса, будут показывать увеличение в уровнях Т6Р по сравнению с комплексом, содержащим немодифицированную Т6РР риса.
[0353] Пример 8: Оценка модифицированных белков Т6РР в растительной ткани с помощью временной экспрессии в растении.
[0354] Для дополнительной оценки влияния модифицированных Т6РР (т.е. модифицированных Т6РР со сниженной активностью) на образование Т6Р и трегалозы в растительной ткани будут применяться анализы временной экспрессии в растении. Способ анализа временной экспрессии в растении, описанный в заявке на патент США 2010/0319089, включенной в данный документ посредством ссылки, можно применять для оценки различных Т6РР и их влияния на образование Т6Р и трегалозы. В общем, бинарные конструкты, содержащие модифицированные Т6РР, такие как описанные в SEQ ID NO 2, 4 или 6, будут инфильтровываться и временно экспрессироваться, фактически как описано в заявке на патент США 2010/0319089. После временной экспрессии модифицированных Т6РР уровни Т6Р и трегалозы будут отслеживаться в течение диапазона интервалов времени в несколько дней (т.е. 1-10 дней) при помощи HPLC. Кроме того, различные TPS/T6PP-комплексы можно конструировать и оценивать с помощью временной экспрессии каждого родственного комплекса в растительной ткани.
[0355] Скрининг двойных гибридов дрожжей: Кроме того, для идентификации полипептидов или других молекул, которые взаимодействуют или связываются при совместной экспрессии в клетке можно применять подходы с двойными гибридными клетками млекопитающих или дрожжей (см., например, Bartel, P.L. et. al. Methods Enzymol, 254: 241 (1995)). Отбор двойных гибридов дрожжей можно проводить для дополнительной идентификации и определения характеристик каких-либо взаимодействий белок-белок, возникающих между модифицированными белками Т6РР и эндогенными растительными белками.
Пример 9: Факторный анализ урожайности растений, экспрессирующих модифицированные белки Т6РР
[0356] Стресс, вызванный засухой при цветении, снижает урожайность в результате увеличения бесплодия, уменьшения числа початков на растение и зерен на растение. Таким образом, трансгенные растения маиса, экспрессирующие модифицированный белок, показанный в SEQ ID NO: 4, под контролем промотора OsMADS6, тестировали на двух полевых участках в условиях обработки с четырьмя различными вариантами стресса 1) хорошее увлажнение без потери урожайности вследствие стресса, вызванного засухой, 2) умеренный стресс, 30-45% потеря запланированной урожайности контрольного растения вследствие стресса, вызванного засухой при цветении, 3) средний стресс, 60% потеря запланированной урожайности контрольного растения вследствие стресса, вызванного засухой при цветении, и 3) сильный стресс, 80% потеря запланированной урожайности контрольного растения вследствие стресса, вызванного засухой при цветении. Данные собирали относительно числа растений на акр, числа рядов зерен на початок, числа початков на растение, числа зерен на растение и веса на зерно. Тестировали восемь различных гибридов.
[0357] Все трансгенные и контрольные растения тестировали в смежных попарных опытных участков в поле. Пары опытных участков с густотой стояния менее 50 растений исключали. Различия вычисляли для каждого признака. Вероятности рассчитывали при помощи парного t-теста для отдельных гибридов в пределах каждого испытания в каждом участке, а также для групп потери урожайности для каждого сорта. Четыре из восьми протестированных гибридов показали положительный ответ в отношении урожайности при условиях сильного стресса, вызванного засухой при цветении. Два гибрида показали отрицательный ответ в отношении урожайности. Рост урожайности у четырех гибридов был, соответственно, обусловлен увеличением числа початков на растение и числа зерен на растение. Два из четырех гибридов также показали снижение бесплодия. Присутствие модифицированного белка Т6РР помогло смягчить влияние стресса, вызванного засухой при цветении, путем снижения бесплодия, увеличения числа початков на растение и увеличения числа зерен на растение.
Пример 10: In Vitro активация OsT6PP-WT в присутствии OsT6PP-H244D
Повышенная трегалоза-6-фосфатфосфатазная активность (in vitro) OsT6PP-WT в присутствии ее каталитически неэффективного варианта, OsT6PP-H244D (SEQ ID NO: 4)
[0358] Влияние добавленного OsT6PP-H244D на каталитическое свойство OsT6PP-WT измеряли при помощи анализа трегалоза-6-фосфатфосфатазной активности, описанного выше в примере 6. Кратко, подходящее количество (как указано в надписях таблицы) белков (OsT6PP-WT и OsT6PP-H244D), частично очищенных из Е.coli, экспрессирующей рекомбинантные варианты OsT6PP, инкубировали на льду в течение 10 минут в 96-луночных планшетах. Субстрат трегалоза-6-фосфат (конечная концентрация 2 мМ) смешивали в реакционном буфере (состав буфера 10 мМ Tris, рН 7,0, и 2 мМ MgCbJ и выдерживали на льду перед смешиванием с ферментом. 85 мкл реакционных смесей добавляли к ферментативным смесям (общий объем реакции 100 мкл) в 96-луночном планшете. Реакционные смеси инкубировали при 37°C в течение различных временных интервалов (как указано в надписях таблицы). Каждую реакцию проводили в трех повторностях. Фермент в реакционной смеси деактивировали нагреванием при 95°C в течение 5 минут. Неактивные реакционные смеси в 96-луночном планшете центрифугировали в течение 2 минут при 700 об/мин, затем по 80 мкл каждого реакционного раствора переносили в лунки, содержащие по 80 мкл воды. Смеси переносили в лунки планшета для фильтрации MicroScreen-HV и фильтрат собирали для анализа HPLC и оценки трегалозной формы путем реакции с фосфатазой.
[0359] Таблица: Повышенную трегалоза-6-фосфатфосфатазную активность OsT6PP-WT наблюдали в присутствии варианта OsT6PP, OsT6PP-H244D.
[0360] Количество трегалозы (мМ), образовавшейся в реакционной смеси вследствие дефосфорилирования трегалоза-6-фосфата при инкубации (время инкубации - колонка 1) с OsT6PP-WT ((1 мкг/мл; колонка 2), OsT6PP-H244D (10 мкг/мл; колонка 3) или смесью (колонка 4) OsT6PP-WT (1 мкг/мл) и OsT6PP-H244D (10 мкг/мл).
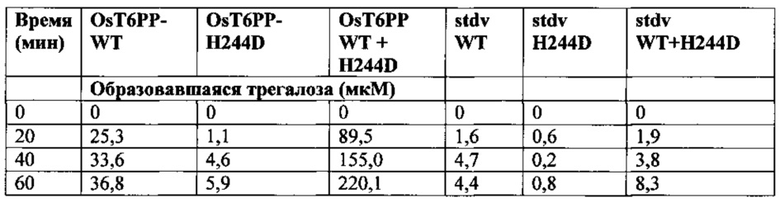
stdv - стандартное отклонение, рассчитанное на основании набора данных трех повторностей для каждой реакции.
Таблица: Влияние концентрации OsT6PP-H244D на активацию трегалоза-6-фосфатфосфатазной активности OsT6PP-WT.
[0361] Количество трегалозы (мкМ, колонка 2), образовавшейся в реакционной смеси вследствие дефосфорилирования трегалоза 6-фосфата при инкубации OsT6PP-WT (1 мкг/мл) с указанной концентрацией OsT6PP-H244D (колонка 1). Количество трегалозы (мкМ, колонка 3), образовавшейся при инкубации OsT6PP-H244D в реакционной смеси отдельно, в отсутствие OsT6PP-WT, при концентрации, указанной в колонке 1. Реакции осуществляли в течение 20 минут.

Т6РР активность усредняли на основании набора данных трех повторностей для каждой реакции, nd - не определено
[0362] Повышенную трегалоза-6-фосфатфосфатазную активность OsT6PP-WT (in vitro) наблюдали в присутствии ее каталитически неэффективных вариантов, включая OsT6PP-H244D, OsT6PP-H244S и OsT6PP-S242E. Наблюдаемая активация Т6РР в присутствии OsT6PP-H244D зависит от концентрации OsT6PP-H244D в реакционной смеси. Степень активации Т6РР варьировала среди протестированных вариантов Т6РР. Заряд на аминокислотных остатках вблизи 244-242 может вовлекаться в это аллостерическое взаимодействие. Приблизительно 3-кратное увеличение Т6РР активности наблюдали, когда OsT6PP-WT анализировали в присутствии OsT6PP-H244D. При сравнении 1,7-кратное увеличение Т6РР активности наблюдали, когда N-концевой усеченный OsT6PP-D56NT смешивали с OsT6PP-H244D. Это может указывать на возможную роль N-концевой части OsT6PP в аллостерическом взаимодействии.
Пример 11: Экспрессия модифицированного Т6РР в пшенице
[0363] Пять конструктов (см. таблицу ниже) вариантов OsT6PP трансформировали в пшеницу посредством протокола трансформации с помощью Agrobacterium (WO 11013764) при помощи системы селектируемых маркеров PMI (патент США №5767378; патент США №5994629) для получения стабильных трансгенных объектов для оценки признака. Три варианта OsT6PP: OsT6PP, последовательность дикого типа; OsT6PP-H244D (SEQ ID NO: 3) с одной модификацией H244D и OsT6PP-I129F (SEQ ID NO: 5) с двумя модификациями A102V и I129F. Среди этих пяти конструктов, 20569 и 20833 разрабатывали с применением + энхансеров, чтобы попытаться изменить уровень экспрессии OsT6PP и нацелить на различные ткани в растениях по сравнению с 20571 и 20832 без энхансеров. 12194 служил в качестве контроля дикого типа для сравнения с вариантами Т6РР.

eFMV представляет собой энхансер из промотора гена 35S вируса мозаики норичника e35S представляет собой энхансер из промотора гена 35S вируса мозаики цветной капусты
[0364] Трансформанты анализировали посредством первичных анализов Taqman для идентификации объектов с низким числом копий и не содержащих векторные остовы. Идентифицированные объекты переносили в теплицу для высаживания. Все другие положительные объекты с большим числом копий и наличием векторного остова исключали. Объекты с наличием признака дополнительно анализировали с помощью вторичного Taqman для подтверждения одной копии внедренного гена и с помощью ELISA и qRTPCR в отношении экспрессии PMI и Т6РР для отбора объектов. ELISA и qRTPCR в отношении PMI и/или Т6РР применяли для отбора трех наборов объектов, экспрессировавших белок или транскрипт (с низким уровнем, средним и высоким), для получения поколения Т1. Всего 154 объектов получили с конструктом 20571. Всего 129 объектов получили с конструктом 20832. Всего 84 объекта получили с конструктом 12194. Всего 161 объект получили с конструктом 20569. Всего 89 объектов получили с конструктом 20833.
[0365] Стресс, вызванный условиями окружающей среды, может произойти в любое время жизненного цикла растения в поле. Засуха во время ранней части цикла роста и на стадии цветения может вызывать значительные проблемы с урожайностью во многих регионах выращивания пшеницы. Молодые растения пшеницы не могут противостоять ограниченному увлажнению, и слабое завязывание семян может быть обусловлено стрессом, вызванным недостатком воды, на стадии цветения. Устойчивость к засухе во время раннего роста растения и при цветении очень желательна. Таким образом, трансгенные по Т6РР растения будут подвергать стрессу на этих двух стадиях развития.
[0366] Ответ растения на недостаток воды можно оценивать при помощи измерения водного режима в листьях (относительного влагосодержания), температуры растительного полога, фотосинтеза, транспирации, проводимости листьев и использования почвенной влаги. После стрессовых обработок у растений измеряли биомассу (высоту растений) и количество побегов, или при созревании определяли урожайность надземного сухого вещества, урожайность зерен и массу зерна для определения компонентов урожайности, а также индекса урожайности в сравнении с нетрансгенными контрольными или нулевыми растениями для оценки повышения урожайности и компонента урожайности с помощью генов признака. При помощи системы получения изображений путем серийной съемки с временным интервалом ответ на стресс, вызванный недостатком воды, можно фиксировать, чтобы показать ответ популяции растений на стресс, вызванный недостатком воды, в расщепляющейся популяции Т1.
[0367] Оценку устойчивости к стрессу, вызванному недостатком воды на стадиях прорастания и сеянцев в лаборатории и вегетационной камере можно использовать для скрининга культиваров пшеницы и тритикале с применением PEG (Sapra, et al., 1991, Crop Science 167, 23-28, Blum et al., 1980, Euphytica 29: 727-736). Прорастающие семена и гидропонно-выращиваемые проростки можно подвергать воздействию раствора PEG с повышающимся градиентом концентраций, что обеспечивает осмотический стресс -0,3--0,6 МПа, или не обеспечивать водой для проверки трансгенных организмов на устойчивость к засухе.
[0368] Следующие фенотипические данные (1, 2 и 3) собирали при помощи анализа отсутствия обеспечения проростков водой. Генотипические данные (зиготность) для 30 растений из каждого объекта записывали и отслеживали. Фенотипические данные выбранных растений записывали и устанавливали их корреляцию с зиготностью. Любую очевидную фенотипическую сегрегацию в условиях стресса тестировали с помощью критерия хи-квадрат в отношении одного вставки с предполагаемым соотношением 3:1. Можно предполагать, что объекты с ожидаемым сегрегационным соотношением, обладают улучшенной устойчивостью к стрессу.
Высота растения и подсчет количества побегов:
Высота растения: От кончика пучка стеблей до основания стебля.
Количество побегов: Подсчет числа новых побегов >2 см.
Оценка влияния засухи на фазе прорастания: (IRRI, 1996)
Измерение относительного влагосодержания: (RWC)
RWC (%)=[(FW-DW)/(TW-DW)]×100
FW (сырой вес): 4-6 полностью распустившихся флаговых листьев/листовых дисков - 10-15°C и взвесить.
TW (вес при туторе): Гидратировать лист в течение 4 ч в чашке Петри с помощью dd-воды при комнатной температуре и освещении и взвесить
DW (сухой вес): Высушить в печи при 80°C в течение 72 ч и взвесить после охлаждения в эксикаторе
Schonfeld et al. (1988) Crop Science 28: 526-531.
[0369] Двадцать один объект 20571 Т0 отбирали для скрининга проростков в отсутствие воды и для получения семян Т2. Семь объектов отбирали как объекты с низкой экспрессией трансгена, семь объектов отбирали как объекты со средней экспрессией трансгена, и семь объектов отбирали как объекты с сильной экспрессией трансгена. Аналогичное число трансформантов Т0 будут отбирать для всех остальных четырех конструктов (20832, 12194, 20833 и 20569) для скрининга в отсутствие воды на стадии сеянца Т1 и стадии цветения Т2.
[0370] Осуществляли экспрессию транскрипта при помощи qRTPCR в отношении OsT6PP-I129F и PMI в пяти объектах 20832, и они показали, что T6PP-I129F экспрессировался только в репродуктивных тканях 3 из 5 объектов, тогда как PMI экспрессировался во всех растениях.
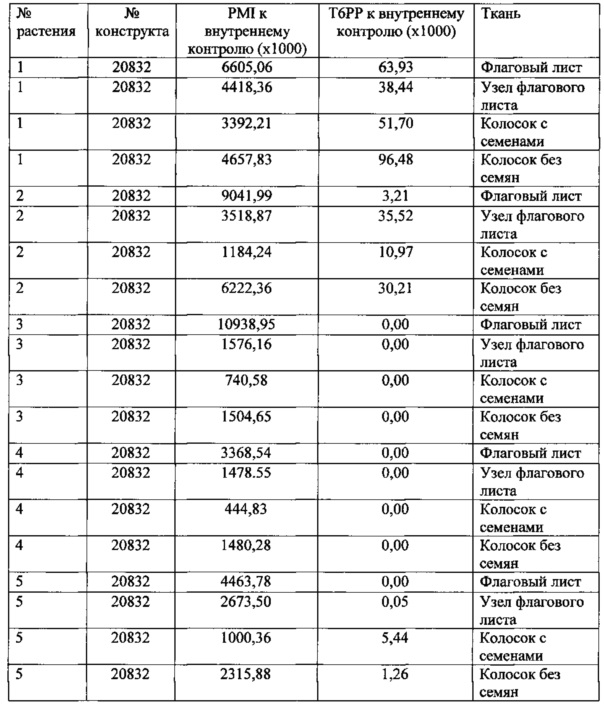
Растение номер 4 является отрицательным контролем, содержащим PMI, а не ген Т6РР.
Пример 12: Экспрессия модифицированной Т6РР в сое
[0371] Трегалозный путь можно использовать для улучшения урожайности двудольных растений. Например, соя (Glycine max) реагирует на засуху сбрасыванием цветков, сбрасыванием бобов, уменьшением количества семян на боб, недоразвитием семян внутри боба и уменьшением размера семени (Liu et al., 2003). Получение устойчивого к засухе растения сои будет повышать урожайность вследствие предотвращения утраты цветков, бобов, семян и/или увеличения размера семян. Эту технологию можно применять на сельскохозяйственной культуре, такой как соя, путем идентификации регуляторной последовательности гена признака, которая специфично функционирует в развивающихся цветках, разработке кассеты экспрессии на основе одного или нескольких кандидатных генов и связывании кассеты экспрессии с трегалоза-6-фосфатфосфатазой дикого типа или модифицированной. Семейство генов MADS является превосходным источником регуляторной последовательности гена признака, которая активна в развивающихся цветках (Jack Т (2001) Plant Mol Biol 46: 515-520). Семейство генов MADS является высококонсервативным среди растений (Munster et al., (2002) Maydica 47: 287-301; De Bolt et al., (2003) Trends Plant Science 8: 475-483). Первичную генетическую информацию из модельных растений можно использовать для идентификации ортологичных генов сои (Hecht et al. (2005) Plant Physiol 137: 1420-1434). Сочетание информации о белковой последовательности и последовательности ДНК с генетическими данными или данными экспрессии позволяет специалистам в данной области разрабатывать прочнорегулируемые кассеты экспрессии, которые активны в цветках сои, такие как промоторы GmSEP1 (Huang et al. (2009) Gene 438: 40-48); APETALA1-подобные промоторы (Chi et al., (2011) J Plant Physiol 168: 2251-2259); промотор OsMADS (патент США 8129588). Например, кассету экспрессии OsMADS6 или ее варианта можно применять непосредственно в трансгенной сое для обеспечения возможности проявления признака.
[0372] После разработки подходящего вектора экспрессии можно получать трансгенное растение сои, содержащее кассету экспрессии, при помощи хорошо известных методик, таких как способ трансформации, опосредованный бомбардировкой частиц (Hadi et al. (1996) Plant Cell Reports 15: 500-505) или опосредованная Agrobacterium трансформация с применением зрелых или незрелых целевых семян (заявка на патент США 20040034889; WO 08112044). Например, кассеты экспрессии, содержащие OsT6PP и вариантную кодирующую последовательность под контролем промотора AGAMOUS Arabidopsis, связывают с кассетой селектируемого маркерного гена EPSPS с образованием бинарного вектора для опосредованной Agrobacterium трансформации. Регенерированные саженцы тестируют на присутствие как маркерного гена EPSPS, так и трансгенных последовательностей Т6РР путем количественного анализа TaqMan PCR (Ingham et al., (2001) Biotechniques 31 (1) 132-4, 136-140). Трансгенные растения выращивают в теплице для завязывания семян.
[0373] Трансгенные растения сои затем можно подвергать скринингу в отношении реакции на дефицит воды при помощи следующего анализа: Семена Т1, отобранные как содержащие одну копию трансгена, выращивают до начала развития репродуктивных органов. В этой момент растения разделяют на две группы, первую группу не подвергают стрессу, а вторая группа получает половину воды, требуемой для поддержания роста в отсутствие стресса. Каждая группа состоит из нулевых, гемизиготных и гомозиготных сиблингов. Растения поддерживают при этих условиях в течение трех-четырех недель. Во время этого периода ежедневно отслеживают развитие цветков и завязывание бобов. В конце периода исследования данные, полученные для каждого генотипа, сравнивают. Вероятно между генотипами в группе, не подвергаемой стрессу, будут небольшие отличия или отличия будут отсутствовать. Группа дефицита воды будет демонстрировать, что положительные в отношении проявления признака растения имеют больше цветков и больше бобов на растение по сравнению с нулевыми растениями. Альтернативно, гомозиготные положительные в отношении проявления признака и нулевые сиблинги можно выращивать в тестовых опытных участках с применением стандартных процедур культивирования. Опытные участки разделяют на блоки, не подвергаемые стрессу, и дефицитные по воде блоки. Дефицитные по воде блоки будут получать половину требуемой воды во время периода цветения. Данные по урожайности для каждого опытного участка собирают в конце периода роста и сравнивают. Опытные участки положительные в отношении проявления признака будут проявлять улучшенную урожайность по сравнению с опытными участками с нулевыми растениями в группе дефицита воды.
Пример 13: Определение профиля экспрессии трансгенных растений OsT6PP с высокой урожайностью
Исследованные типы и образцы тканей
[0374] Собирали семь типов тканей, а именно; лист, лист ниже початка, узел, основной стебель, основной початок, вторичный початок и стебель. Исследовали три педигри-линии: контроль, содержащий нетрансформированный сорт, OsT6PP-I129f (SEQ ID NO: 5) и OsT6PP-H244D (SEQ ID NO: 3). Три педигри-линии либо выращивали в условиях хорошего полива, либо подвергали стрессу, вызванному засухой. Для каждой ткани/состояния отбирали по десять биологических повторностей, что привело к общему количеству в 420 образцов. Однако, по различным техническим причинам, отбирали не все образцы. Всего 407 образцов стали доступны для очистки и мечения РНК для гибридизации на микрочипах.
[0375] Для очистки, оценки качества, мечения и гибридизации РНК применяли стандартные протоколы. Не все образцы характеризовались достаточным качеством или количеством РНК для гибридизации на микрочипах. Всего обрабатывали и сканировали 392 чипа.
[0376] Стандартизация и предварительная обработка данных:
[0377] Для этого исследования применяли Maize Affymetrix GeneChip, sySYNG007a. Необработанные файлы формата CEL для всех экспериментов анализировали с применением специального протокола, который анализирует качество микрочипов на основе ряда критериев, исполнений и поправок на фон, и сжимает данные при помощи алгоритма Bioconductor RMA. В результате этого анализа 6 предметных стекол оказались потенциально низкого качества. Однако общие изображения чипов были достаточными для анализа, и контрольные соотношения зондов 5ʹ/3ʹ были пригодными, так что данные эксперименты не исключали. Для этого исследования применяли специальный CDF (Chip Description File), разработанный для картирования зондов относительно генома маиса. CDF также картирует относительно Unigenes от NCBI и кДНК полной длины от MaizeGDB. Для определения фонового уровня шума анализировали ряд контрольных наборов зондов. Импульсы контролей из генов Bacillus не использовали во время гибридизации, поэтому эти наборы зондов можно рассматривать в качестве фонового шума. Фон определяли как два стандартных отклонения выше среднего значения всех наборов зондов Bacillus. Уровень фонового шума, который определили, использовали как пороговое значение для данных. Пороговые значения устанавливали для исключения любых наборов зондов, которые характеризовались интенсивностью сигналов ниже, чем фон во всех экспериментах. Наборы зондов оставляли для анализа, если сигнал превышал фоновый по меньшей мере в 3 экспериментах. Для оценки качества экспериментов и определения любых возможных отклонений применяли анализ главных компонент и иерархическую кластеризацию.
[0378] Анализ путей
[0379] В попытке придать биологический смысл результатам одномерных статистических тестов, проводили анализы насыщения, применяя подобные эксперименты. Существует множество способов проведения анализа пути, однако для этих экспериментов наиболее подходящим способом был Анализ Насыщения Генов (GSEA), использующий программное обеспечение, доступное от Broad Institute, называемое GSEA. Алгоритм GSEA требует, чтобы гены были объединены в наборы генов согласно некоторым биологическим критериям. Для этого испытания использовали два таких набора генов. Во-первых, наборы генных моделей из генома маиса распределили на два набора генов согласно группировке в пути, доступной от MaizeCyc (на следующем сайте: pathway.gramene.org/MAIZE). Второй список наборов генов был составлен из наборов зондов на специальном разработанном GeneChip маиса, применяя специального файла CDF, используемого для сжатия информации о зондах для этого испытания. Наборы зондов, представляющие транскрипты генных моделей генома маиса, сгруппировали в наборы генов согласно условиям Gene Ontology, приписанных генным моделям от Maize GDB (на следующем сайте: maizegdb.org/). GSEA алгоритм упорядочивает гены из 2 различных образцов для создания ранжированного списка генов и эту информацию используют для определения того, находятся ли гены в пределах ранжированных списках в доступных наборах генов. Р-значения рассчитывают на основании вероятности того, могут ли гены быть найдены в ранжированных списках наборов генов посредством вследствие случайности. Затем Р-значения корректируют путем множественного тестирования для получения уровня ложноположительных результатов для всех наборов генов. Для этого анализа все парные сравнения подвергали GSEA с применением путей из MaizeCyc и Gene Ontology (GO) от MaizeGDB. Кроме того, анализировали только наборы зондов, представляющие отдельные гены. А именно, наборы зондов, вероятно картирующие множественные генов, отбрасывали для GSEA.
[0380] Многомерная статистика
[0381] Ряд многомерных анализов проводили для определения наибольших источников дисперсии данных. Анализ главных компонент (PCA) всех данных показал, что наибольшим источником дисперсии был тип ткани. Лист и ткань листа под початком (ткани-источники) явно отделены от других акцептирующих тканей.
[0382] Для дополнительного определения источников дисперсии отдельные типы тканей анализировали при помощи PCA. Результаты показали, что засуха имела сильное влияние на изменчивость генной экспрессии во всех протестированных тканях, с возможным исключением для тканей вторичного початка. Следует отметить, что меньшее количество образцов тканей вторичного початка было доступно. Вследствие неравномерного взятия образцов для этого типа ткани генная экспрессия могла быть измерена не точно. Систематическая дисперсия, не вызванная педигри-линией или стрессом, а например, обусловленная взятием образцов или расположением опытного участка, могла иметь значительное влияние на генную экспрессию в ткани вторичного початка. Хотя педигри-линия не была основным источником дисперсии в пределах отдельных типов ткани, она имела явный влияние и была очевидной во второй главной компоненте в большинстве тканей. Примечательно, что большую дисперсию вследствие педигри-линии обнаружили в образцах, подвергнутых стрессу, вызванному засухой. Хотя не всегда присутствовала явная компонента, объясняемая только педигри-линией, из РСА было ясно, что имелось большая дисперсия между педигри-линиями при стрессе, вызванном засухой, по сравнению с условиями хорошего полива. Например, из PCA ясно, что первая PC разделяет образцы, выращенные в условиях хорошего полива от таковых в условиях стресса, вызванного засухой. Вторая главная компонента, объясняющая наибольший источник дисперсии после удаления PC 1, показывает разделение подвергаемых засухе образцов согласно педигри-линии. Это не так явно как разделение образцов на подвергаемые стрессу, вызванному засухой, по сравнению с хорошо поливаемыми растениями. Однако существует намного более плотное группировка (т.е. с меньшей дисперсией) контрольной и OsT6PP-H244D педигри-линий, протестированных из хорошо политых образцов по сравнению с образцами, подвергаемыми засухе. Тенденцию компоненты 2 можно главным образом объяснить разделением образцов из контрольных и OsT6PP-H244D педигри-линий при стрессе, вызванном засухой. Это можно интерпретировать как то, что контрольные и OsT6PP-H244D педигри-линии имели аналогичные паттерны генной экспрессии в условиях хорошего полива, но отличные профили генной экспрессии при стрессе, вызванном засухой. Это согласуется с обеспечением пользы в отношении урожайности при стрессе, вызванном засухой. Аналогичное увеличение дисперсии при стрессе, вызванном засухой, обнаруживали в других протестированных тканях и большую часть дисперсии можно объяснить педигри-линией.
[0383] В общем, многомерные анализы идентифицировали основные источники отличий, такие как тип ткани и стресс, вызванный засухой, с меньшим эффектом на отличия в генной экспрессии, вызванные педигри-линией (т.е. присутствием трансгена). Влияние педигри-линии протестировали сравнением OsT6PP-H244D с контрольной гибридной линией. В большинстве протестированных тканей, как акцептирующих, так и тканях-источниках, имелась большая дисперсия, чем та, которую можно было объяснить педигри-линией в образцах, собранных из растений, подвергаемых стрессу, вызванному засухой, чем в образцах хорошо политых растений. Таким образом, различия в генной экспрессии между основным объектом и контрольной линией были наиболее очевидными в растениях, подвергаемых стрессу, вызванному засухой.
[0384] Экспрессия генов Т6РР и TPS
[0385] Поскольку охарактеризованный объект включал ген Т6РР, экспрессированный с помощью промотора OsMADS6, интересно было определить уровень экспрессии трансгена и/или экспрессии эндогенных генов Т6РР. Также на экспрессию Т6РР может влиять экспрессия трегалоза-6-фосфатсинтазы. Оба белка, как известно, кодируются несколькими членами семейства генов в маисе. Делались попытки проанализировать только наборы зондов, которые картируют либо отдельные изоформы транскриптов, либо множественные изоформ в одном локусе, для определения экспрессии отдельных членов семейства генов. Всего восемь наборов зондов, представляющих Т6РР, и четырнадцать наборов зондов, представляющих TPS, анализировали в отношении уровня экспрессии во всех тканях как подвергаемых, так и не подвергаемых стрессу, как из контроля, так и из OsT6PP-H244D. Обнаружили, что ни Т6РР, ни TPS наборы зондов не имели особенно сильной экспрессии в каких-либо тканях в случае OsT6PP-H244D, что позволяет предполагать, что трансген в трансформанте не имеет сильного влияния на гены трегалозного пути, и не имеется заметно более высоких уровней экспрессии какого-либо эндогенного гена Т6РР. Представляется неясным, достаточно ли гибридизуется какой-либо набор зондов с трансгенной Т6РР в объекте для обнаружения надежного сигнала. Можно ожидать более высокий уровень экспрессии Т6РР в линии трансгенного объекта. Эти результаты подтверждали прямыми TTEST в каждой ткани, тестируемой в отношении дифференциальной экспрессии генов Т6РР и TPS между контролем и OsT6PP-H244D. Ни в одном из тестов не было ни одной изоформы, характеризующейся более чем 2-кратнымо отличием в экспрессии между 2 педигри-линиями в любой ткани с любой степенью статистической значимости (значение Р<0,1). Однако имело место явное доказательство того, что изоформы обоих генов в некоторых тканях были чувствительны к стрессу, вызванному засухой. Примечательно, что экспрессия одной из изоформ Т6РР, GRMZM2G347280_T01, была подавлена в ответ на стресс, вызванный засухой, в ткани листьев (источник), однако, экспрессия увеличивалась в ответ на стресс, вызванный засухой, в ткани основного початка и тканях основного стебля. Также, имела место определенная тканевая специфичность характерных наборов зондов Т6РР. Например, ген GRMZM2G080354, кодирующий Т6РР, наиболее сильно экспрессировался в ткани вторичного початка; GRMZM2G112830 едва обнаруживался во всех тканях за исключением стебля, с одним альтернативным транскриптом, GRMZM2G112830_Т07 вероятно также экспрессировался в узле; GRMZM2G174396_T01 наиболее сильно экспрессировался в ткани узла, а GRMZM2G178546_T01 наиболее сильно экспрессировался в основном початке. Были подобные тканеспецифичные и чувствительные к засухе члены семейства TPS, которые были представлены наборами зондов. Интересно, что ген GRMZM2G019183, который характеризовался низким уровнем экспрессии во всех тканях, все же включался в ответ на стресс, вызванный засухой, в тканях узла, основного початка и основного стебля. Относительно высокие уровни экспрессии нескольких генов TPS обнаружили в ткани вторичного початка. Примечательно, что значительного отличия в экспрессии этих генов в ответ на стресс, вызванный засухой, во вторичном початке не было, даже для генов, которые были чувствительными к засухе в других тканях. Поскольку для конструкта, использованного для этого объекта, использовали ген Т6РР из риса, вероятно, что ни один из наборов зондов не соответствовал последовательности достаточно для гибридизации с наборами зондов маиса на чипе. Таким образом, представленные результаты наглядно представляют уровень экспрессии только эндогенных генов.
[0386] Одномерный анализ и попарные сравнения
[0387] Ряд попарных сравнений проводили при помощи Т-теста Уэлча для определения гена, дифференциально экспрессировавшегося у 2 образцов (контроль и OsT6PP-H244D). Произведенные сравнения были сосредоточены на исходной экспериментальной разработке для тестирования отличия в экспрессии между контрольной и OsT6PP-H244D линией в условиях хорошего полива или засухи во всех тканях (7 тестов для каждого состояния) и также тестировали отличия в генной экспрессии в растениях, выращенных в условиях хорошего полива или засухи в каждой педигри-линии во всех тканях (14 сравнений). Результаты тестов приведены в таблицах ниже. Как сообщалось, наборы зондов были дифференциально экспрессированы при Т-тесте Уэлча, Р-значение было менее 0,01, и кратность изменения (соотношение средних значений биологических повторностей) была больше 2. Кроме того, для определения существования каких-либо генов, которые имели значительно отличающийся ответ на засуху, как результат влияния педигри-линии, запускали линейную модель для тестирования эффектов взаимодействия. Результаты в таблице 4 указывают на номера наборов зондов, которые имели значительное влияние взаимодействия обеспечение водой X педигри-линия с FDR менее 0,05. В общем, результаты полностью подтверждают результат многомерных анализов для всех тканей, кроме вторичного початка, показывая большое влияние засухи на экспрессию генов. Примечательно, что во всех тканях было больше генов, чувствительных к засухе в объекте OsT6PP-H244D по сравнению с контрольной линией (таблица 1). Эти результаты могут указывать, что объект OsT6PP-H244D является более чувствительным к стрессу, вызванному засухой, на уровне изменений генной экспрессии.
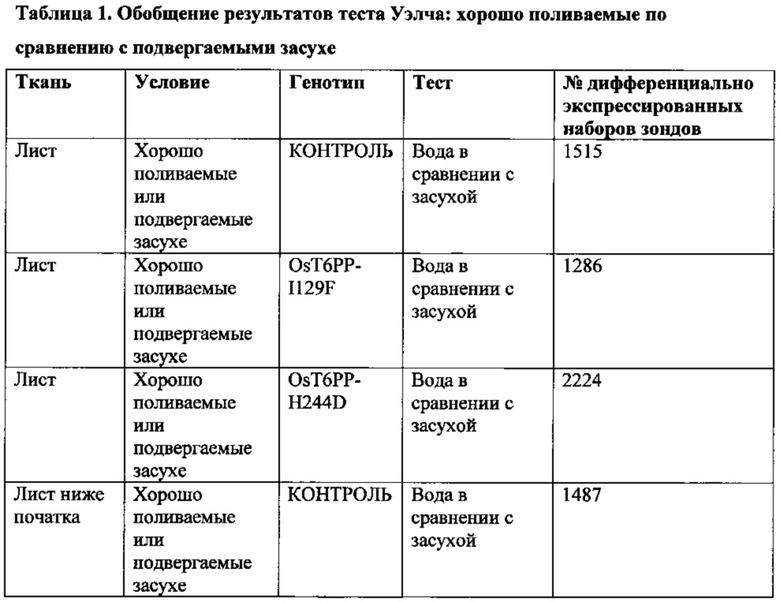
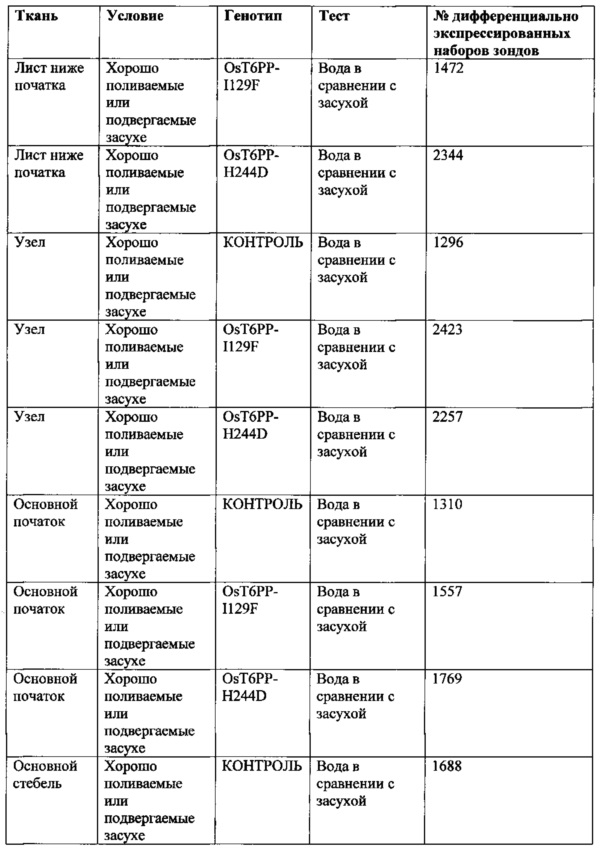
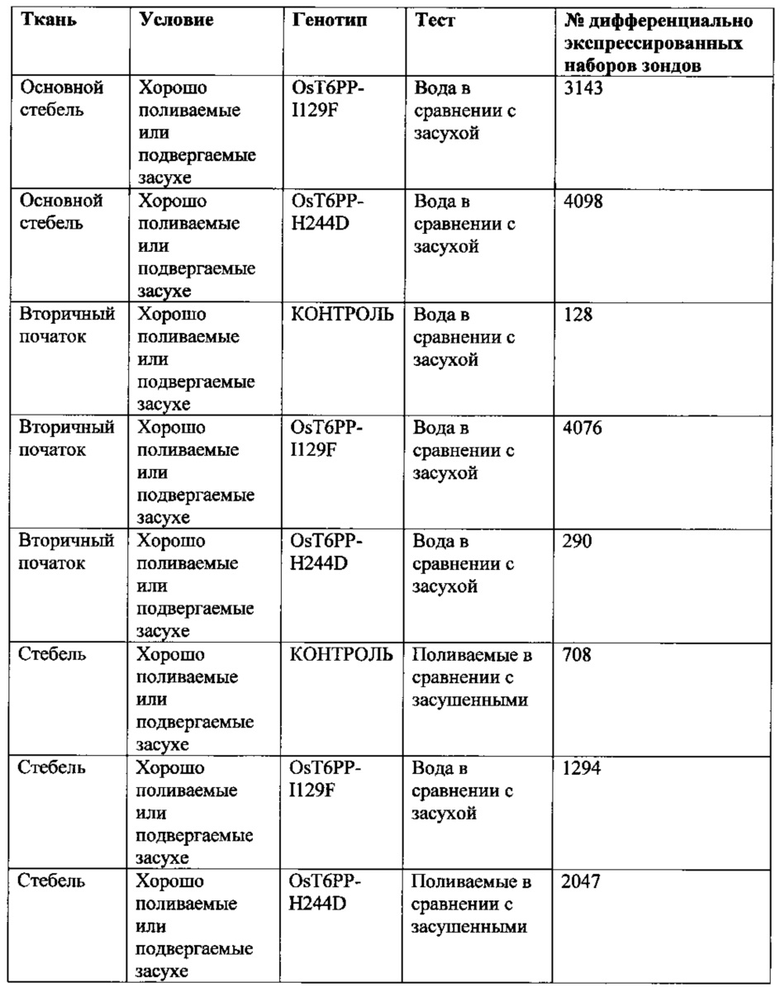
Дифференциально экспрессированные наборы зондов = наборы зондов со значением Р теста Уэлча<0,01 и изменение кратности (соотношение средних значений образцов из 2 протестированных групп)>2.
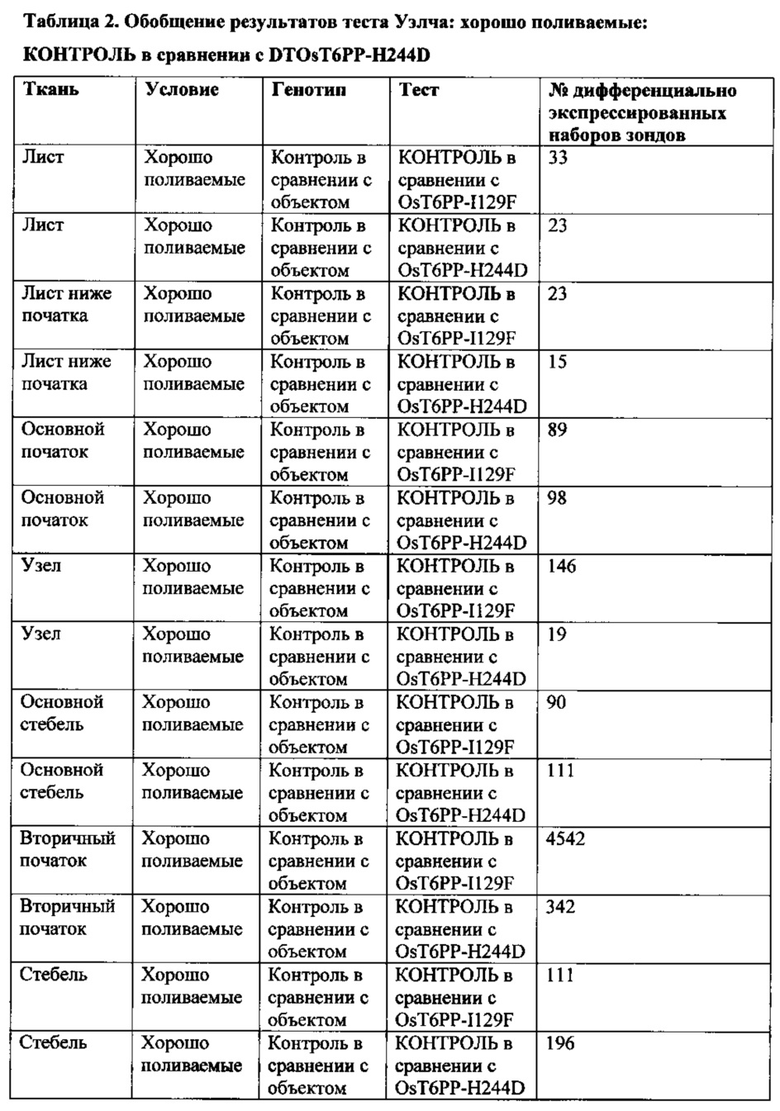
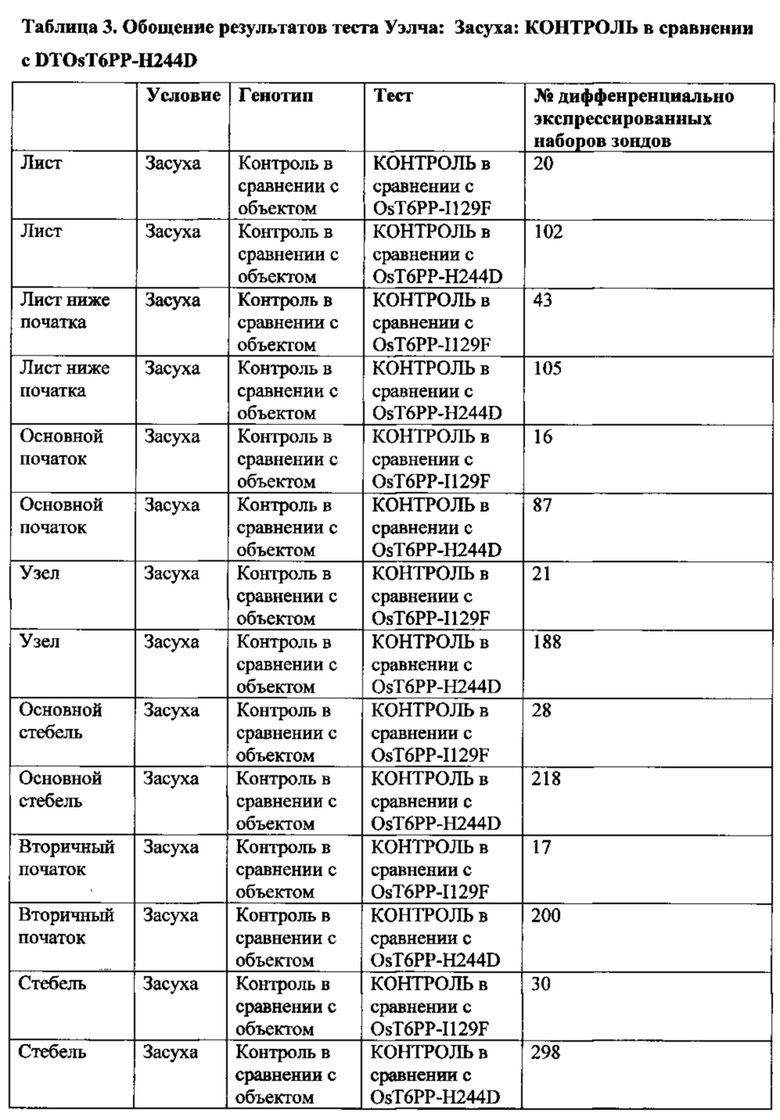
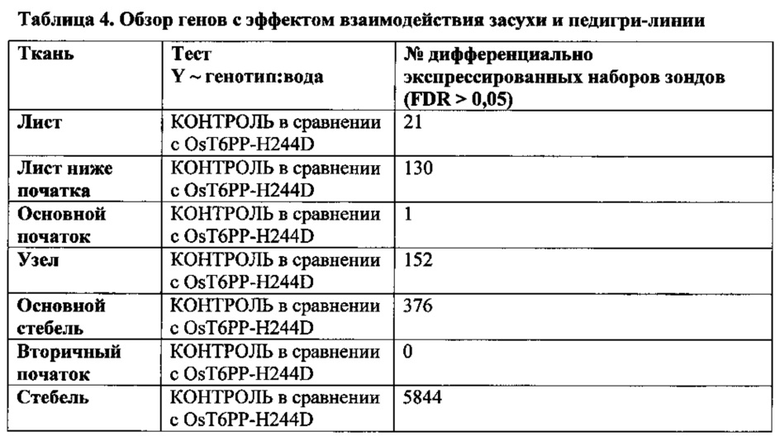
[0388] Хотя многие гены, чувствительные к засухе, были общими для обеих педигри-линий, было много генов, которые обнаруживали только в качестве дифференциально экспрессировавшихся под воздействием стресса, вызванного засухой, в OsT6PP-H244D. Многие гены оказались специфическими для OsT6PP-H244D в ответ на стресс, вызванный засухой. Результаты подобны результатам линейной модели для обнаружения генов, которые имели эффект взаимодействия между двумя экспериментальными факторами засухи и педигри-линии. Причиной отличия в числе генов из 2 анализов является, вероятно, то, что отсечение, использованное в TTEST, было со значением Р менее 0,01, а статистическое отсечение для данных в таблице 4 составляло уровень ложноположительных результатов менее 0,05. Уровень ложноположительных результатов, рассчитанный из поправки для множественных сравнений, является более точным и более надежным способом определения генов, которые дифференциально экспрессируются. Линейная модель является более хорошим тестом в отношении эффектов взаимодействия, чем перекрывающиеся результаты генов, обнаруженных как дифференциально экспрессированные из Т-тестов, поскольку модель включает все данные в факторной схеме эксперимента. Однако объединение результатов обоих тестов может представлять собой мощный способ идентификации генов с различными ответами на 2 фактора, как будет описано более подробно.
[0389] Основные различия между педигри-линиями обнаруживали во время стресса, вызванного засухой, а также в акцептирующей ткани, включая узловую ткань. Линейные модели показывали значительные эффекты взаимодействия в узле, основном стебле, а также стебле. Объединенные результаты позволили предположить, что большинство эффектов взаимодействия происходили вследствие различий в том, какие гены изменялись при стрессе, вызванном засухой, в двух протестированных педигри-линиях. В условиях хорошего полива было меньше различий в генной экспрессии между контролем и OsT6PP-H244D.
[0390] Анализ пути
[0391] В попытке придать биологический смысл результатам одномерных статистических тестов проводили анализы насыщения при помощи аналогичных парных тестов. GSEA проводили с данными для тестирования на насыщения путей MaizeCyc и и условиям GO от MaizeGDB. Поскольку отмечалось, что узловая ткань представляет особый интерес, результаты GSEA можно легче интерпретировать на основании этой ткани. Следует отметить, что анализы путей, выделенных здесь, сосредоточены на попарных сравнениях при помощи всех данных в модели GSEA для увеличения мощности теста на насыщение по сравнению с применением заранее вычисленных списков генов. Проводимые попарные сравнения сосредоточены на различиях контроля и OsT6PP-H244D, в которых предварительные указанные тесты Уэлча имели относительно немного различий в генной экспрессии. Таким образом, уровень ложноположительных результатов (FDR), рассчитанный для большинства сравнений, был в общем высоким, хотя некоторые рассчитанные Р-значения показали статистически значимые результаты. Следует отметить, что хотя Р-значения существенны, значения q FDR могут быть более лучшей индикацией статистического подобия путей или насыщенных условий GO в заданном наборе генов. Таблицы 5 и 6 указывают наборы генов, насыщенных в OsT6PP-H244D, по сравнению с контролем в узловой ткани при стрессе, вызванном засухой.


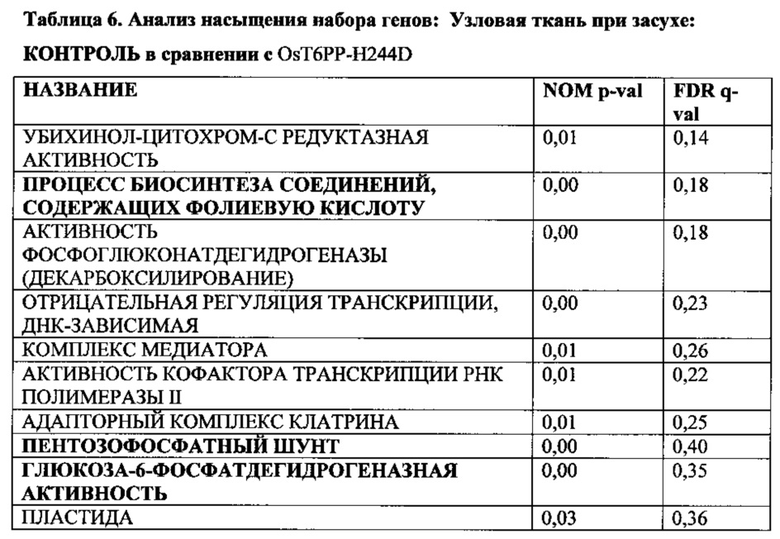
[0392] Другие ткани тестировали, однако в узловой ткани в условиях стресса обнаружили путь, представляющий особый интерес, а именно, пентозофосфатный шунтовый путь. Примечательно, что он представляет собой энергопроизводящий путь, который может быть затронут только в растениях с OsT6PP-H244D при стрессе, вызванном засухой. Хотя, не стремясь ограничиваться теорией, он может представлять собой одним из потенциальных механизмов, которые позволяют объекту OsT6PP-H244D поддерживать энергообразование с получением более высокой урожайности в условиях засухи. Он явно показывает альтернативный путь энергообразования, подвергнутый стрессу, вызванному засухой, только в объекте и необязательно у контрольной гибридной линии. Ферменты в пентозофосфатном пути кодируются множеством генов, и их транскрипция была показана как чувствительная ко множеству различных воздействий. Путь, как известно, вовлечен в производство вторичных метаболитов. Другой путь, идентифицированный в различных акцептирующих тканях как дифференциально регулируемый в OsT6PP-H244D, представлял собой поглощение нитратов или метаболизм азота. Было интересно узнать, из-за результатов предыдущих испытаний, включая испытания измененного метаболизма линий маиса при стрессе, вызванном засухой, что определило аминокислотный биосинтез, в частности накапливающие N аминокислоты, при высокой концентрации в подвергнутых засухе растениях. Кроме того, в недавнем испытании обнаружили, что растения, подвергаемые постоянному N-стрессу, были менее чувствительны к дефициту воды, чем растения, выращиваемые при оптимальных N-условиях. Хотя, не стремясь ограничиваться теорией, растения, подвергаемые стрессу, вызванному засухой, могут использовать нормальные метаболические пути для N, в частности продукцию накапливающих N аминокислот для ассимилирования возможно токсичных концентраций аммиака, который может накапливаться в тканях растений, подвергаемых стрессу, вызванному засухой. Таким образом, эти данные дают подтверждение гипотезе детоксикации аммиака и позволяют предполагать, что возможно экспрессирующий Т6РР объект OsT6PP-H244D может быть в лучшем состоянии для использования N механизма обмена веществ адаптации к дефициту воды.
[0393] Идентифицированный транскрипт карбоангидразы с повышенной экспрессией в OsT6PP-H244D.
[0394] Были исследованы две гипотезы о том, как анализ профиля генной экспрессии может отражать различия между объектом OsT6PP-H244D и контролем. Первая гипотеза состоит в том, что отдельный набор генов меняется специфично в OsT6PP-H244D при стрессе, вызванном засухой, а не в контроле. Такой набор генов может играть роль в более высокой урожайности линии OST6PP-H244D по сравнению с контролем при стрессе, вызванном засухой. Эту гипотезу исследовали при помощи результатов от парных сравнений и из линейной модели. Результаты Т-тестов из таблицы 1 сравнивали с генами, определенными в линейной модели (таблица 4). Два образца тканей тестировали, поскольку они имели значительное число генов, которые имели эффект взаимодействия 2 факторов стресса, вызванного засухой, и педигри-линии. В узловой ткани всего 1127 генов имели измененную экспрессию при стрессе, вызванном засухой, в OST6PP-H244D, что не изменялось значительно при стрессе, вызванном засухой, в контроле. Пересечение этого списка генов и 152 генов с эффектом взаимодействия засуха X педигри-линия из линейной модели узловых тканей (таблица 4) определило 22 набора зондов, специально подвергаемых стрессу, вызванному засухой, только в трансформанте OsT6PP-H244D. Одним из идентифицированных генов был ген карбоангидразы. Также включенными в список были несколько наборов зондов, обозначенных как серинпептидазы, а также ген аквапорин TIP4. Отдельный анализ такой же линейной модели определил карбоангидразу как имеющую большую экспрессию в OsT6PP-H244D независимо от типа ткани или состояний обеспечения водой. В действительности, карбоангидраза была представлена 2 наборами зондов в коротком списке из наборов зондов с более чем 2-кратной или более экспрессией в OsT6PP-H244D среди всех типов тканей. Многие другие гены в этом списке были неизвестными генами. Эти данные показывают, что по меньшей мере один член семейства генов карбоангидраз экспрессируется на более высоком уровне в OsT6PP-H244D, чем в контроле. Экспрессию этой же карбоангидразы анализировали даже повторно, OsT6PP-I129F обнаружили как специфичную к OsT6PP-H244D.
[0395] Результаты анализов по всем тканям и условиям дали относительно короткий список наборов зондов (28), которые присутствовали в значительной степени в линии OsT6PP-H244D по сравнению с контрольной линией. По меньшей мере два набора зондов в этом списке были помечены как карбоангидразы. Карбоангидраза активна при С4-фотосинтезе, помогая в фиксации диоксида углерода и ассимиляции углерода растением. Очень значительное увеличение экспрессии обнаружили в OsT6PP-H244D по меньшей мере одной изоформы одного члена семействат CA.
[0396] Хотя изложенное выше изобретение было описано довольно подробно в качестве иллюстрации и примера с целями ясности и понимания, для специалистов в данной области техники будет понятно, что конкретные изменения и модификации могут быть осуществлены в пределах объема приложенной формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| РАСТЕНИЯ, ИМЕЮЩИЕ УСИЛЕННЫЕ ПРИЗНАКИ, СВЯЗАННЫЕ С УРОЖАЙНОСТЬЮ, И СПОСОБ ИХ ПОЛУЧЕНИЯ | 2008 |
|
RU2503721C2 |
| СПОСОБЫ МОДУЛЯЦИИ ПРОВОДИМОСТИ УСТЬИЦА И РАСТИТЕЛЬНЫЕ ЭКСПРЕССИОННЫЕ КОНСТРУКЦИИ ДЛЯ ИХ ОСУЩЕСТВЛЕНИЯ | 2012 |
|
RU2664461C2 |
| РАСТЕНИЯ, ХАРАКТЕРИЗУЮЩИЕСЯ ПОВЫШЕННОЙ УРОЖАЙНОСТЬЮ, И СПОСОБ ИХ ПОЛУЧЕНИЯ | 2006 |
|
RU2463351C2 |
| РАСТЕНИЯ С ПОВЫШЕННОЙ УРОЖАЙНОСТЬЮ И СПОСОБ ИХ ПОЛУЧЕНИЯ | 2005 |
|
RU2384621C2 |
| ПРОМОТОР ЦЕЛОГО СЕМЕНИ | 2010 |
|
RU2561463C2 |
| СКОНСТРУИРОВАННЫЕ ПЕСТИЦИДНЫЕ БЕЛКИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2017 |
|
RU2772947C2 |
| УСТОЙЧИВЫЕ К БАКТЕРИЯМ ТРАНСГЕННЫЕ РАСТЕНИЯ | 2012 |
|
RU2640246C2 |
| РЕГУЛЯТОРНЫЕ ЭЛЕМЕНТЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2639275C2 |
| ИНСЕКТИЦИДНЫЕ БЕЛКИ | 2017 |
|
RU2765722C2 |
| КОНСТИТУТИВНЫЕ ПРОМОТОРЫ СОИ | 2014 |
|
RU2714724C2 |























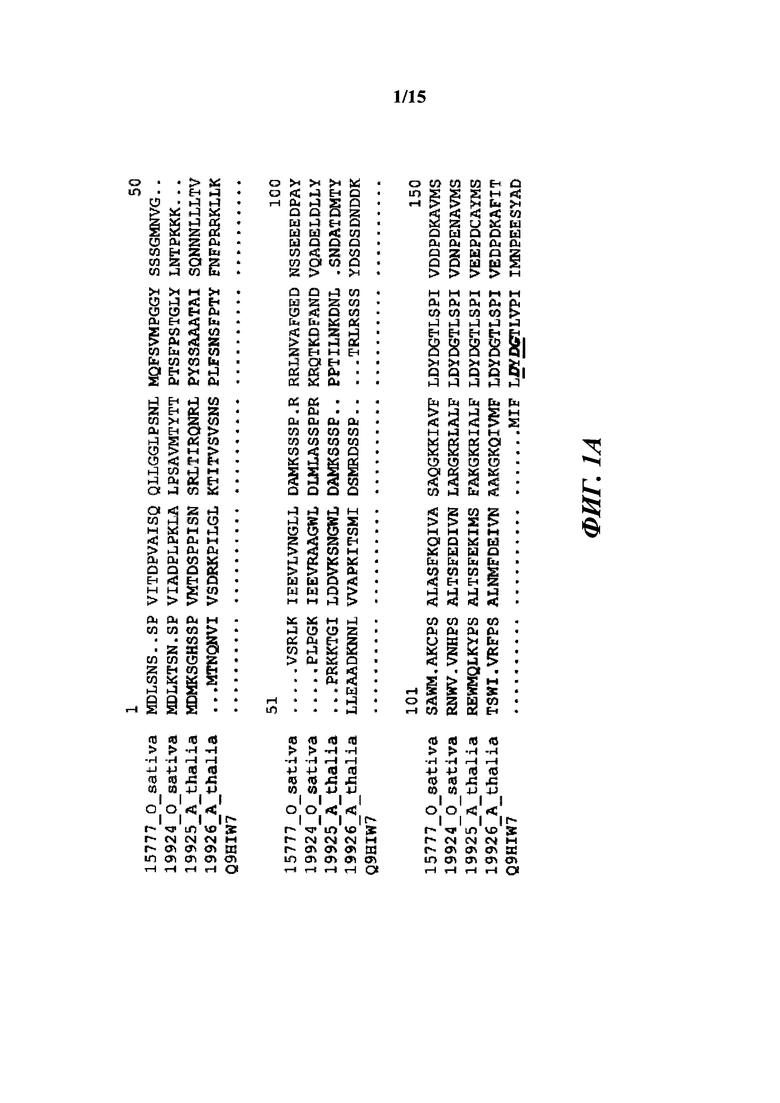


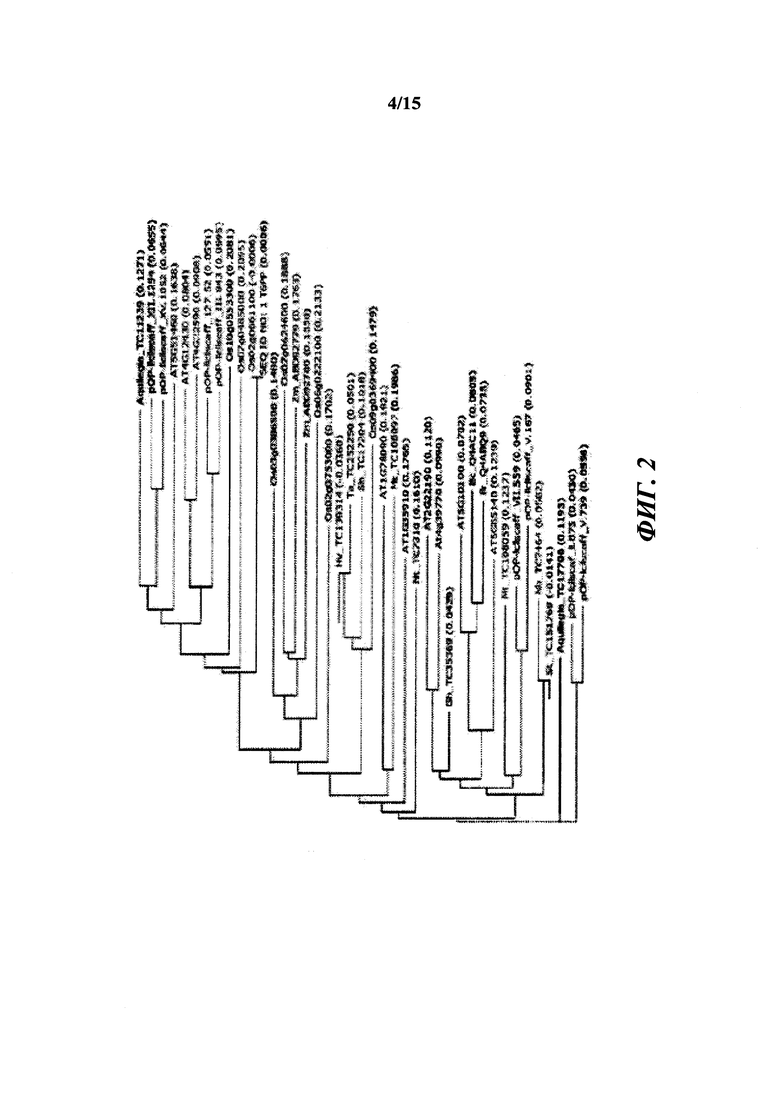
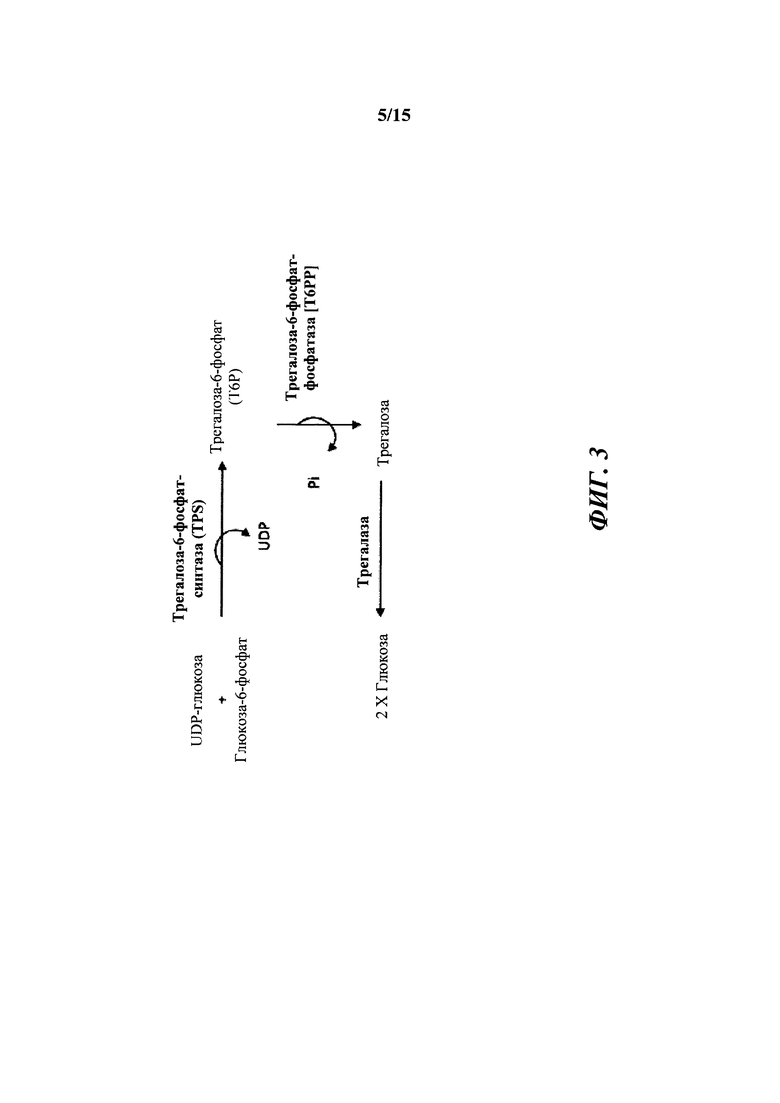
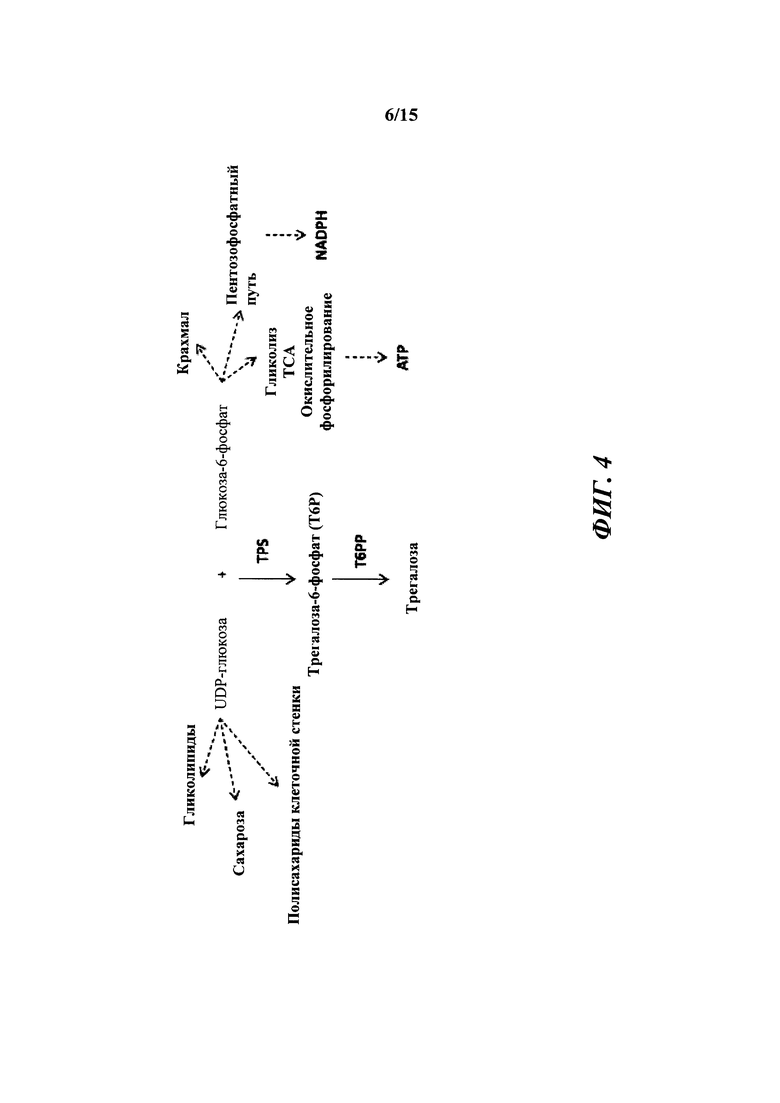
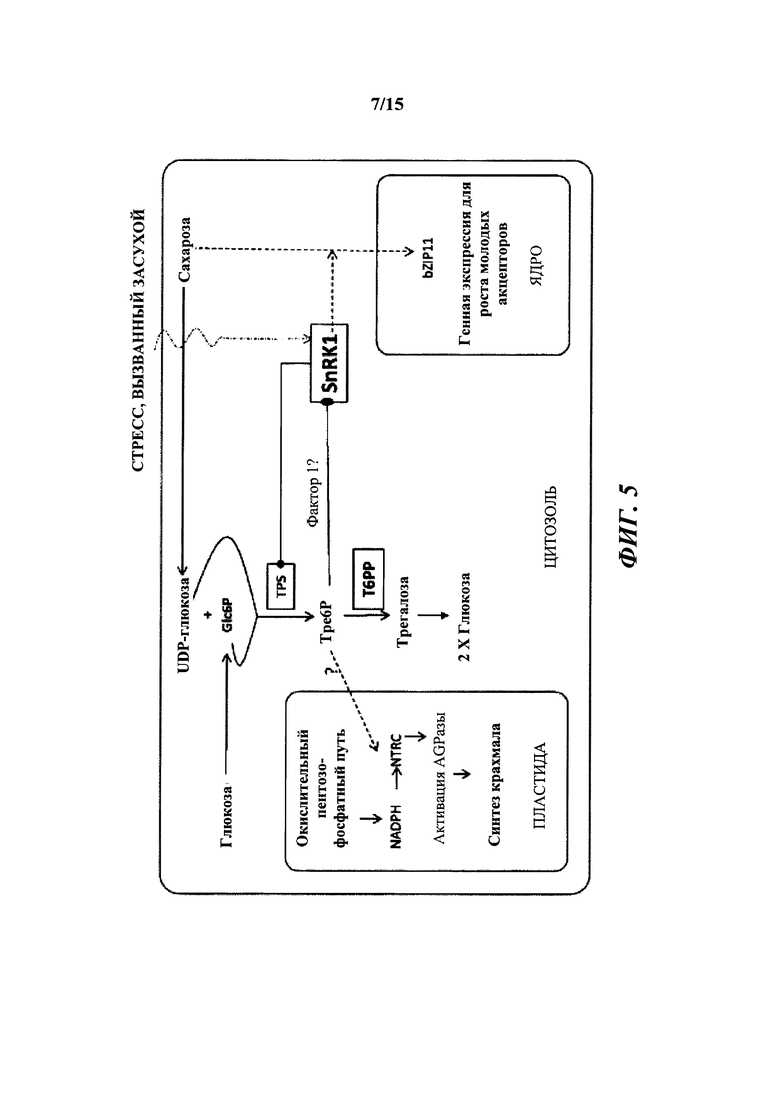
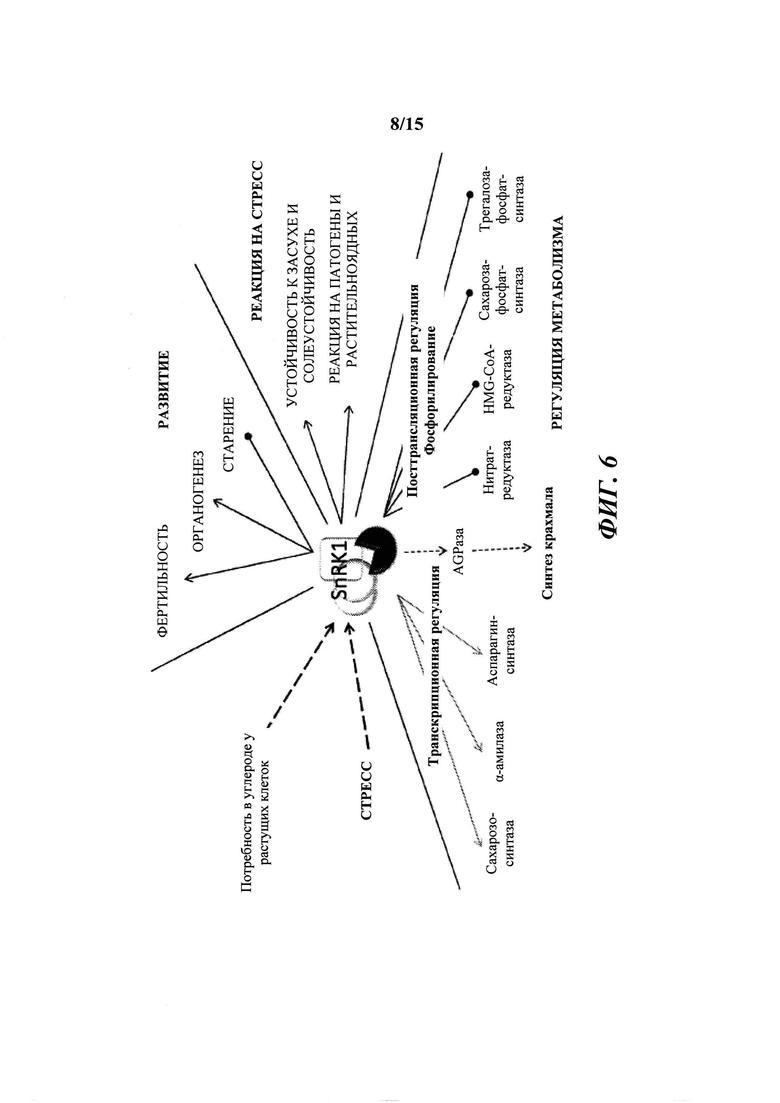
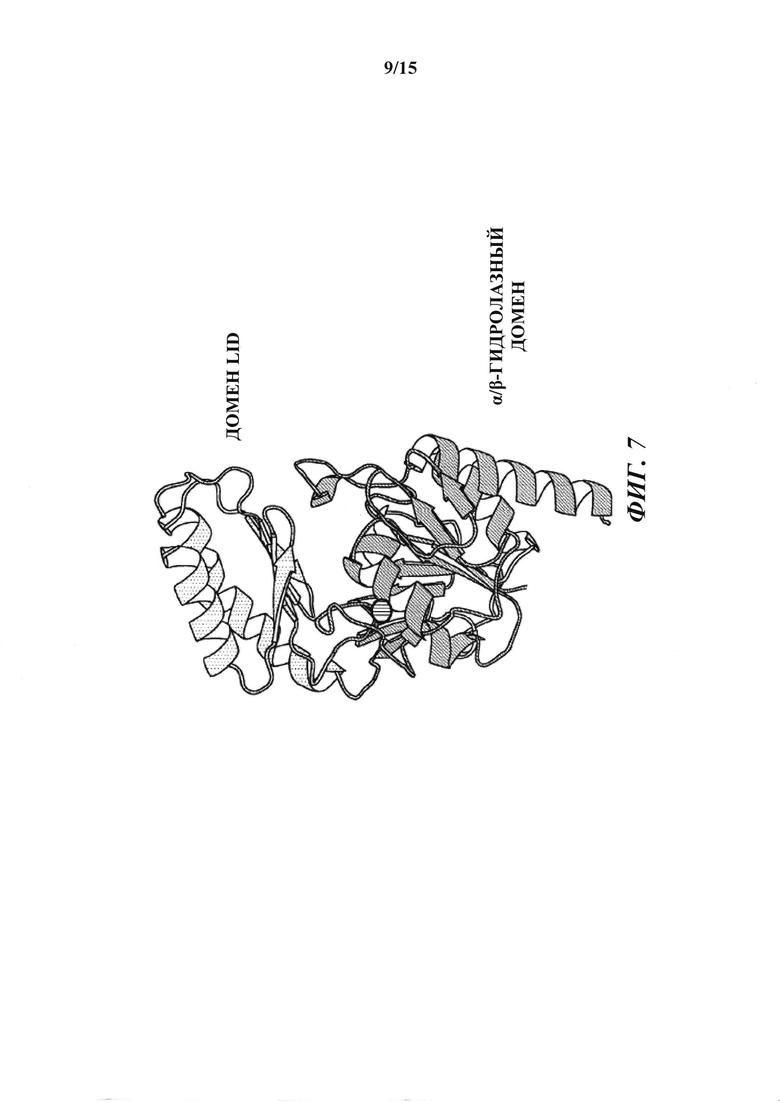
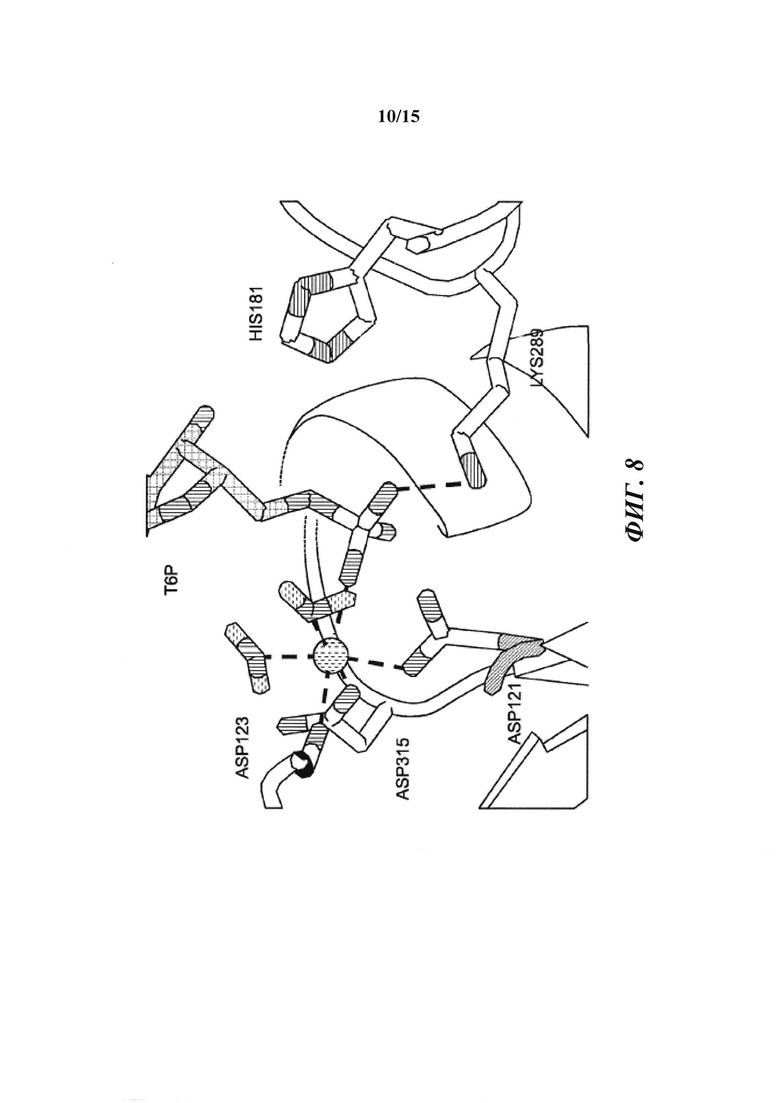
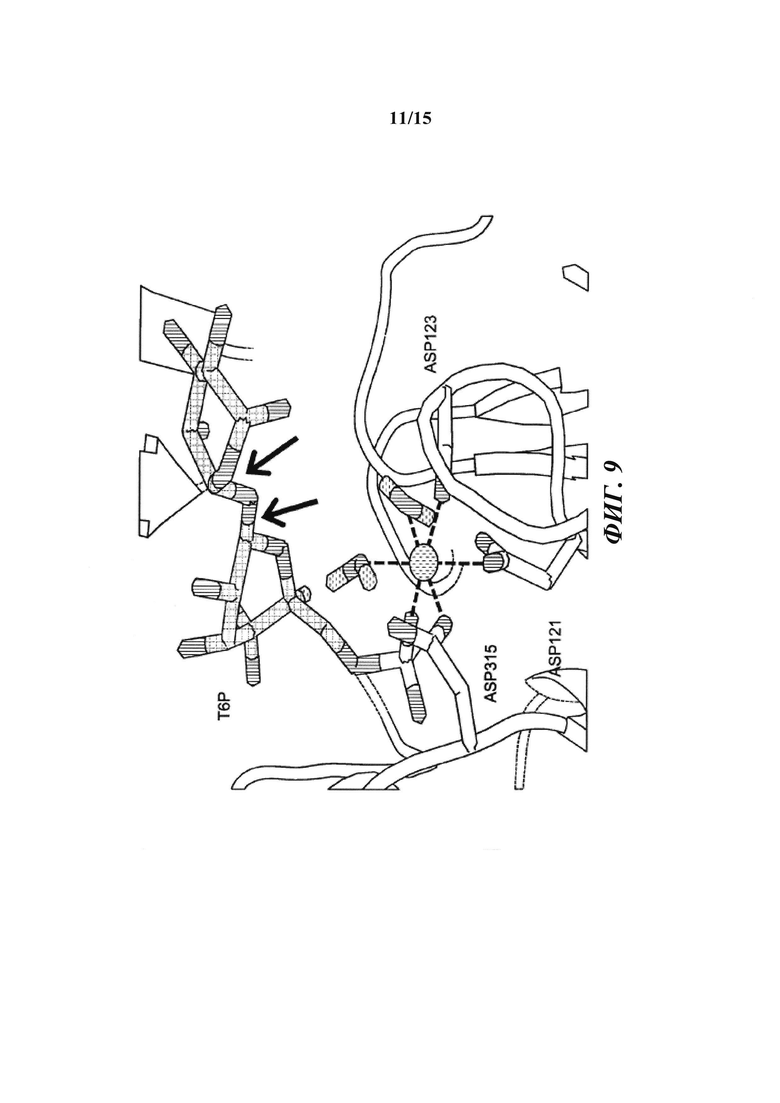

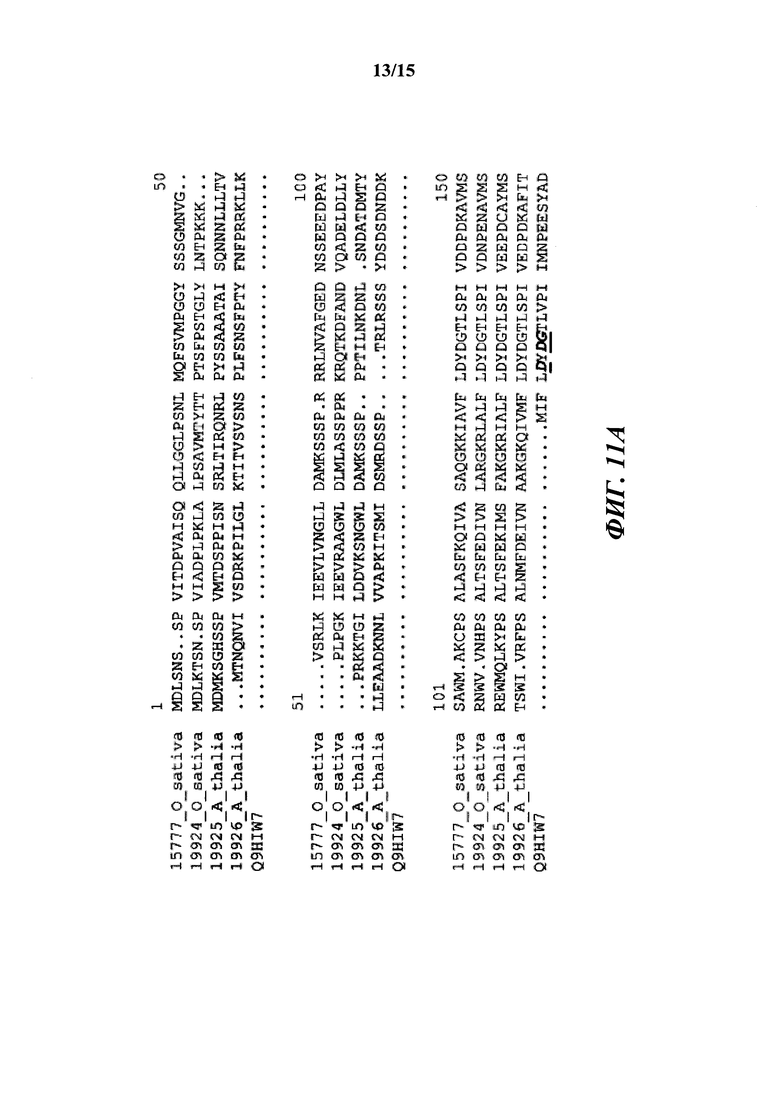


Группа изобретений относится к области биотехнологии. Предложен полинуклеотид, кодирующий трегалоза-6-фосфатфосфатазный полипептид. Предложены растение кукурузы, обладающее повышенной урожайностью, кукурузная клетка-хозяин, ткань растения кукурузы, бинарный вектор экспрессии, включающие указанный полинуклеотид. Предложена кассета экспрессии для трансформации кукурузных клеток, включающая указанный полинуклеотид, функционально связанный с промотором, контролирующим его транскрипцию. Предложен способ повышения урожайности у растений кукурузы с использованием вышеуказанной кассеты экспрессии. Группа изобретений позволяет повысить урожайность растения кукурузы и смягчить влияние стресса, вызванного засухой при цветении, путем снижения бесплодия, увеличения числа початков на растение и увеличения числа зерен на растение. 7 н. и 4 з.п. ф-лы, 15 ил., 15 табл., 13 пр.
1. Полинуклеотид, кодирующий трегалоза-6-фосфатфосфатазный полипептид, где полипептид включает последовательности SEQ ID NO: 4 или SEQ ID NO: 6.
2. Растение кукурузы, обладающее повышенной урожайностью, включающее полинуклеотид по п. 1.
3. Кукурузная клетка-хозяин для получения растения кукурузы по п. 2, включающая полинуклеотид по п. 1.
4. Ткань растения кукурузы для получения растения кукурузы по п. 2, включающая полинуклеотид по п. 1.
5. Бинарный вектор экспрессии, включающий полинуклеотид по п. 1.
6. Кассета экспрессии для трансформации кукурузных клеток, включающая полинуклеотид, функционально связанный с промотором, который контролирует транскрипцию указанного полинуклеотида, где указанный полинуклеотид кодирует трегалоза-6-фосфатфосфатазный полипептид, где полипептид включает последовательности SEQ ID NO: 4 или SEQ ID NO: 6.
7. Кассета экспрессии по п. 6, где промотор экспрессирует указанный полинуклеотид в материнской репродуктивной ткани.
8. Кассета экспрессии по п. 7, где материнскую репродуктивную ткань выбирают из группы, состоящей из ткани колоска, ткани прицветника, меристемной ткани колоска, ткани цветоножки соцветия, узла початка и незрелой ткани цветка.
9. Кассета экспрессии по п. 6, где промотор является промотором OsMADS.
10. Кассета экспрессии по п. 9, где OsMADS промотор является промотором OsMADS 6.
11. Способ повышения урожайности у растения кукурузы, где способ включает:
а) введение в клетку растения кукурузы кассеты экспрессии по п. 6;
б) регенерацию растения кукурузы из клетки растения кукурузы согласно а);
в) получение растения кукурузы с повышенной урожайностью.
| WO 2008071767 A1, 19.06.2008 | |||
| WO 2005102034 A2, 03.11.2005 | |||
| GARG A.K | |||
| et al., Trehalose accumulation in rice plants confers high tolerance levels to different abiotic stresses, Proceedings of the National Academy of Sciences of the United States of America (PNAS) Vol | |||
| Прибор, замыкающий сигнальную цепь при повышении температуры | 1918 |
|
SU99A1 |
| US 7560613 B2, 14.07.2009 | |||
| RU 98121810 A, 20.12.2000. | |||

