Настоящее изобретение, главным образом, относится к области молекулярной биологии и касается способа повышения урожайности растений по сравнению с таковой соответствующих растений дикого типа. Более конкретно, настоящее изобретение относится к способу повышения урожайности растений, предусматривающему модулирование экспрессии растением нуклеиновой кислоты, кодирующей полипептид транслокации синовиальной саркомы (synovial sarcoma translocation (SYT) polypeptide) или его гомолог. Настоящее изобретение также относится к растениям, характеризующимся измененной экспрессией нуклеиновой кислоты, кодирующей полипептид SYT или его гомолог, причем растения характеризуются повышенной урожайностью по сравнению с таковой соответствующих растений дикого типа. Изобретение также относится к конструкциям, применимым в соответствии со способами, согласно изобретению.
Постоянно растущее население мира и сокращающийся фонд сельскохозяйственных пахотных земель стимулируют исследования, направленные на повышение эффективности сельского хозяйства. Стандартные средства повышения урожайности сельскохозяйственных и садовых культур включают в себя селекционные методики, основанные на выявлении растений, характеризующихся желаемыми признаками. Однако, таким селекционным методикам присущи некоторые недостатки, а именно, заключающиеся в том, что такие методики, как правило, трудоемки и позволяют получать растения, зачастую содержащие разнородные генные компоненты, которые не всегда обеспечивают передачу желаемого признака от исходных растений. Успехи молекулярной биологии позволили человеку модифицировать генотип животных и растений. Генная инженерия растений основана на выделении и манипуляции генетическим материалом (как правило, в виде ДНК или РНК) и последующем введении этого генетического материала в растение. Такая технология позволяет создавать культуры или растения, характеризующиеся различными улучшенными экономическими, агротехническими и садовотехническими признаками.
Особенно экономически важным признаком является урожайность, а для многих растений семенная продуктивность. Урожайность обычно определяют как количественное выражение экономической ценности культуры. Она может быть определена в количественном и/или качественном выражении. Семена растений являются важным источником питательных веществ для человека и животных. Более половины общей калорийности рациона человечества потребляется с такими культурами, как кукуруза, рис, пшеница, канола и соя, либо при непосредственном потреблении семян как таковых, либо при потреблении мясных продуктов, произведенных на обработанных семенах. Они также являются источником сахаров, масел и различных видов метаболитов, используемых в промышленных процессах. Семена содержат зародыш, источник новых побегов и корней после прорастания, и эндосперм, источник питательных веществ для роста зародыша, на стадии прорастания и на ранних стадиях роста всходов. В процесс развития семян вовлечены многие гены, и для его протекания требуется перенос метаболитов из корней, листьев и стеблей в растущее семя. В частности, эндосперм поглощает метаболические предшественники полимерных углеводов, масла и белки и синтезирует из них запасаемые макромолекулы, заполняющие зерно. Способность повышать семенную продуктивность растений посредством увеличения количества семян, увеличения биомассы семян, ускорения развития семян, усиления налива семян или усиления любого другого относящегося к семенам признака имело бы множество применений в сельском хозяйстве и даже множество несельскохозяйственных применений, таких как применение для биотехнологического получения таких веществ, как фармацевтические средства, антитела или вакцины.
Урожайность также может зависеть от таких факторов, как количество и размер органов, строение растения (например, количество ветвей), семенная продуктивность и т.д. Важными факторами, определяющими урожайность, также могут быть развитие корневой системы, усвоение питательных веществ и стрессоустойчивость. Таким образом, оптимизация этих факторов может способствовать повышению урожайности культур.
Было выявлено, что модулирование экспрессии нуклеиновой кислоты, кодирующей полипептид SYT или его гомолог, в растении позволяет получать растения, характеризующиеся более высокой урожайностью по сравнению с таковой соответствующих растений дикого типа.
SYT представляет собой коактиватор транскрипции, который в растениях образует функциональный комплекс с активаторами транскрипции семейства белков GRF (фактора регуляции роста, growth-regulating factor) (Kim H.J., Kende H. (2004) Proc. Nat. Acad. Sc. 101:13374-9). SYT также называют GIF, т.е. GRF-взаимодействующий фактор (GRF-interacting factor). Активаторы транскрипции GRF содержат структурные домены (в N-концевой области), сходные с белками SWI/SNF хроматин-ремоделирующих комплексов дрожжей (van der Knaap E. et al. (2000) Plant Phys. 122:695-704). Было высказано предположение о том, что коактиваторы транскрипции из этих комплексов вовлечены в рекрутирование комплексов SWI/SNF в энхансерные и промоторные области для осуществления локального ремоделирования хроматина (см. Näär A.M. et al. (2001) Annu. Rev. Biochem. 70:475-501). Изменение локальной структуры хроматина модулирует активацию транскрипции. Точнее говоря, высказано предположение о том, что SYT взаимодействует с комплексом SWI/SNF растения, оказывая влияние на активацию транскрипции гена-мишени(генов-мишеней) GRF (Kim H.J., Kende H. (2004) Proc. Nat. Acad. Sc. 101:13374-9).
SYT принадлежит к семейству генов, состоящему у Arabidopsis из трех членов. Полипептид SYT характеризуется гомологией с SYT человека. Было показано, что полипептид SYT человека является коактиватором транскрипции (Thaete et al. (1999) Hum. Molec. Genet. 8:585-591). Для структуры полипептида SYT млекопитающих характерно наличие трех доменов:
(i) N-концевого домена SNH (N-концевой гомологии SYT, SYT N-terminal homology), консервативного у млекопитающих, растений, нематод и рыб;
(ii) С-концевого богатого QPGY домена, состоящего преимущественно из глицина, пролина, глутамина и тирозина, встречающихся с переменными интервалами;
(iii) богатого метионином (Met-богатого) домена, расположенного между указанными выше двумя доменами.
В полипептидах SYT растений домен SNH высококонсервативен. С-концевой домен богат глицином и глутамином, но содержит мало пролина и тирозина. Таким образом, его назвали богатым QG доменом в отличие от богатого QPGY домена млекопитающих. Как и в случае SYT млекопитающих, богатый Met домен может быть идентифицирован в направлении N-конца от богатого QG домена. Богатый QG домен можно считать по существу С-концом белка (за исключением домена SHN); богатый Met домен, как правило, содержится в пределах первой половины богатого QG домена (от N-конца к С-концу). В полипептидах SYT растений богатый Met домен может предшествовать домену SNH (см. фиг.1).
Сообщалось, что у мутанта с выключенной функцией SYT и трансгенных растений с ослабленной экспрессией SYT развиваются мелкие и узкие листья и лепестки, содержащие меньшее количество клеток (Kim H.J., Kende H. (2004) Proc. Nat. Acad. Sc. 101:13374-9).
В соответствии с настоящим изобретением раскрыт способ повышения урожайности растения, предусматривающий модулирование экспрессии нуклеиновой кислоты, кодирующей полипептид SYT или его гомолог, в растении.
Подразумевается, что в настоящем описании ссылка на "соответствующие растения дикого типа" означает любое подходящее контрольное растение или растения, выбор которого полностью находится в пределах квалификации специалиста в данной области техники и может включать в себя, например, соответствующие растения дикого типа или соответствующие растения, не несущие интересующий ген. Используемый в настоящем описании термин "контрольное растение" относится не только к целым растениям, но также и к частям растений, включая семена и части семян.
Предпочтительно, осуществление способов согласно настоящему изобретению приводит к получению растений, характеризующихся повышенной урожайностью, особенно семенной продуктивностью, по сравнению с таковой соответствующих растений дикого типа.
Подразумевается, что в настоящем описании термин "повышенная урожайность" означает наличие любого одного или нескольких из следующих признаков по сравнению с соответствующими растениями дикого типа: (i) повышенной биомассы (массы) одной или нескольких частей растения, особенно надземных (пригодных для уборки) частей, повышенной биомассы корня или повышенной биомассы любой другой пригодной для уборки части (такой как плоды, орехи и бобы); (ii) повышенной общей семенной продуктивности, что включает в себя повышенную биомассу семян (массу семян), что может заключаться в увеличении массы семян с отдельного растения или в увеличении массы отдельных семян; (iii) повышенного количества (налитых) семян; (iv) повышенного размера семян, что также может сказываться на составе семян; (v) повышенного объема семян, что также может сказываться на составе семян (включая общее содержание и состав масел, белков и углеводов); (vi) повышенной площади поверхности отдельных семян; (vii) повышенных длины или ширины отдельных семян; (viii) повышенного процентного отношения массы урожая к полной массе растений, выражаемого как отношение урожая пригодных для уборки частей, таких как семена, к общей биомассе; и (ix) повышенной массы тысячи зерен (TKW), которую получают путем экстраполяции количества подсчитанных налитых семян и их общей массы. Повышение значения TKW может иметь место благодаря увеличению размера семян и/или массы семян. Повышение значения TKW может иметь место благодаря увеличению размера зародыша и/или размера эндосперма. Увеличение размера семян, объема семян, площади поверхности семян, периметра семян, ширины семян и длины семян может иметь место благодаря увеличению отдельных частей семян, например, благодаря увеличению размера зародыша, и/или эндосперма, и/или алейронового слоя, и/или щитка зародыша, или других частей семян.
На примере кукурузы повышение урожайности может быть определено, помимо прочего, по одному или нескольким следующим признакам: повышение количества растений на один гектар или акр, повышение количества початков на одно растение, повышение количества рядов, количества зерен в ряду, массы зерна, массы тысячи зерен, длины/диаметра початка, повышение скорости налива зерна (рассчитываемой как количество налитых зерен, деленное на общее количество зерен и умноженное на 100). На примере риса повышение урожайности может быть определено, помимо прочего, по повышению одного или нескольких следующих параметров: количества растений на один гектар или акр, количества метелок на одно растение, количества вторичных колосков на одну метелку, количества цветков на один колосок (выраженного как отношение количества налитых семян к количеству метелок), повышению скорости налива зерна (рассчитываемой как количество налитых зерен, деленное на общее количество зерен и умноженное на 100), повышению массы тысячи зерен.
Повышение урожайности также может привести к измененному строению или может быть результатом измененного строения.
В соответствии с предпочтительным вариантом выполнения осуществление способов согласно изобретению позволяет получить растения, характеризующиеся повышенной семенной продуктивностью. Таким образом, в соответствии с настоящим изобретением раскрыт способ повышения семенной продуктивности растения, причем указанный способ предусматривает модулирование экспрессии нуклеиновой кислоты, кодирующей полипептид SYT или его гомолог, в растении.
Поскольку трансгенные растения согласно настоящему изобретению характеризуются повышенной урожайностью, вероятно, что у этих растений наблюдается повышенная скорость роста (по меньшей мере, в течение части их жизненного цикла) по сравнению со скоростью роста соответствующих растений дикого типа на соответствующей стадии их жизненного цикла. Повышение скорости роста может специфично происходить в отношении одной или нескольких частей растения (включая семена) или может иметь место по существу в отношении всего растения. У растений, характеризующихся повышенной скоростью роста, даже может наблюдаться ранее цветение. Повышение скорости роста может иметь место на одной или нескольких стадиях жизненного цикла растения или по существу в течение всего жизненного цикла растения. Повышенная скорость роста на ранних стадиях жизненного цикла растения может свидетельствовать о повышенной жизненности. Повышение скорости роста может изменить цикл урожая растения, что позволяет сеять растения позже и/или убирать позже, что в других обстоятельствах было бы невозможно. Если скорость роста повышена в достаточной степени, это может позволить провести дополнительный посев семян того же вида растения (например, сев и уборка растений риса с последующими севом и уборкой следующих растений риса в течение одного стандартного периода роста). Аналогично, если скорость роста повышена в достаточной степени, это может позволить провести дополнительный посев семян других видов растений (например, сев и уборка растений риса, например, с последующими севом и необязательной уборкой сои, картофеля или любого другого подходящего растения). Также для некоторых сельскохозяйственных культур может оказаться возможной неоднократная уборка урожая с одного корневища. Изменение цикла урожая растения может приводить к увеличению годичного производства биомассы с каждого акра (благодаря увеличению количества возможных циклов (скажем, в течение года) выращивания и уборки урожая каждого конкретного растения). Повышение скорости роста также может позволить культивировать трансгенные растения в более широкой географической зоне, чем их аналоги дикого типа, поскольку территориальные ограничения выращивания культуры зачастую определяются неблагоприятными условиями окружающей среды либо на момент сева (в начале вегетационного периода), либо на момент уборки (в конце вегетационного периода). Таких неблагоприятных условий можно избежать при сокращении продолжительности цикла урожая. Скорость роста может быть определена по различным параметрам кривых роста, такими параметрами, помимо прочих, могут выступать T-Mid (время, необходимое для достижения растениями 50% от их максимального размера) и Т-90 (время, необходимое для достижения растениями 90% от их максимального размера).
Осуществление способов согласно изобретению позволяет получить растения, характеризующиеся повышенной скоростью роста по сравнению с таковой соответствующих растений дикого типа. Таким образом, в соответствии с настоящим изобретением раскрыт способ повышения скорости роста растений, причем указанный способ предусматривает модулирование экспрессии нуклеиновой кислоты, кодирующей полипептид SYT или его гомолог, в растении.
Повышение (семенной) продуктивности и/или скорости роста возникает либо когда растение находится в бесстрессовых условиях, либо когда растение подвергается различным стрессам по сравнению с подходящими контрольными растениями. Растения, как правило, реагируют на стресс замедлением роста. В условиях сильного стресса растение может полностью прекратить рост. С другой стороны, умеренный стресс определяется в настоящем описании как любой стресс, будучи подвергнутым которому, растение не прекращает рост полностью без возможности возобновления роста. Благодаря развитию агротехнических приемов (орошения, внесения удобрений, обработки пестицидами) при культивировании сельскохозяйственных культур сильные стрессы возникают нечасто. В результате вызванный умеренным стрессом нарушенный рост часто является нежелательным явлением в сельском хозяйстве. Умеренные стрессы представляют собой типичные стрессовые воздействия, которым может подвергаться растение. Такие могут представлять собой каждодневные биотические и/или абиотические (средовые) стрессовые факторы, которым подвергается растение. Типичные абиотические, или средовые, стрессовые факторы включают в себя температурные стрессы, вызываемые необычно высокими или холодными температурами/заморозками; солевой стресс; водный стресс (засуха или избыток воды). Химические вещества также могут вызывать абиотические стрессы. Биотические стрессы, как правило, представляют собой стрессы, вызываемые такими патогенами, как бактерии, вирусы, грибы и насекомые.
Предпочтительно, может быть изменена урожайность любого растения.
Используемый в настоящем описании термин "растение" охватывает целые растения, предшественники и потомство растений и части растений, включая семена, побеги, стебли, листья, корни (включая клубни), цветки и ткани и органы, причем все они содержат интересующий трансген. Термин "растение" также охватывает клетки растений, суспензионные культуры, каллюсную ткань, зародыши, меристемы, гаметофиты, спорофиты, пыльцу и микроспоры, причем все они, опять же, содержат интересующий трансген.
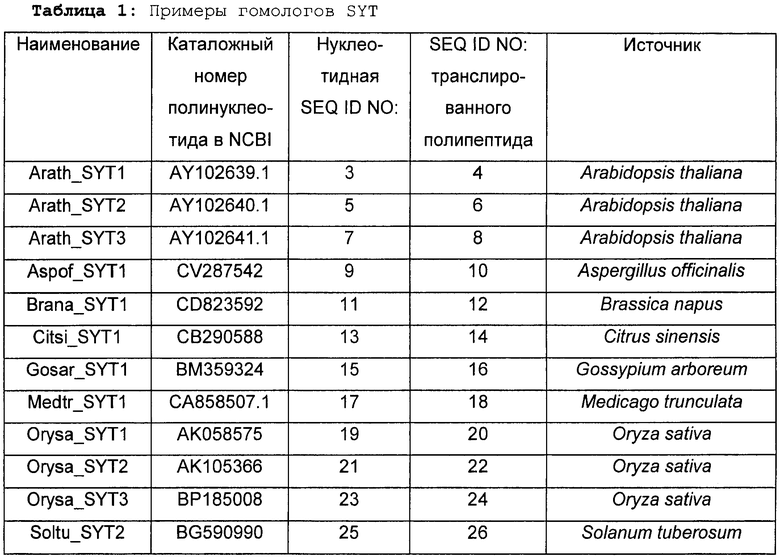
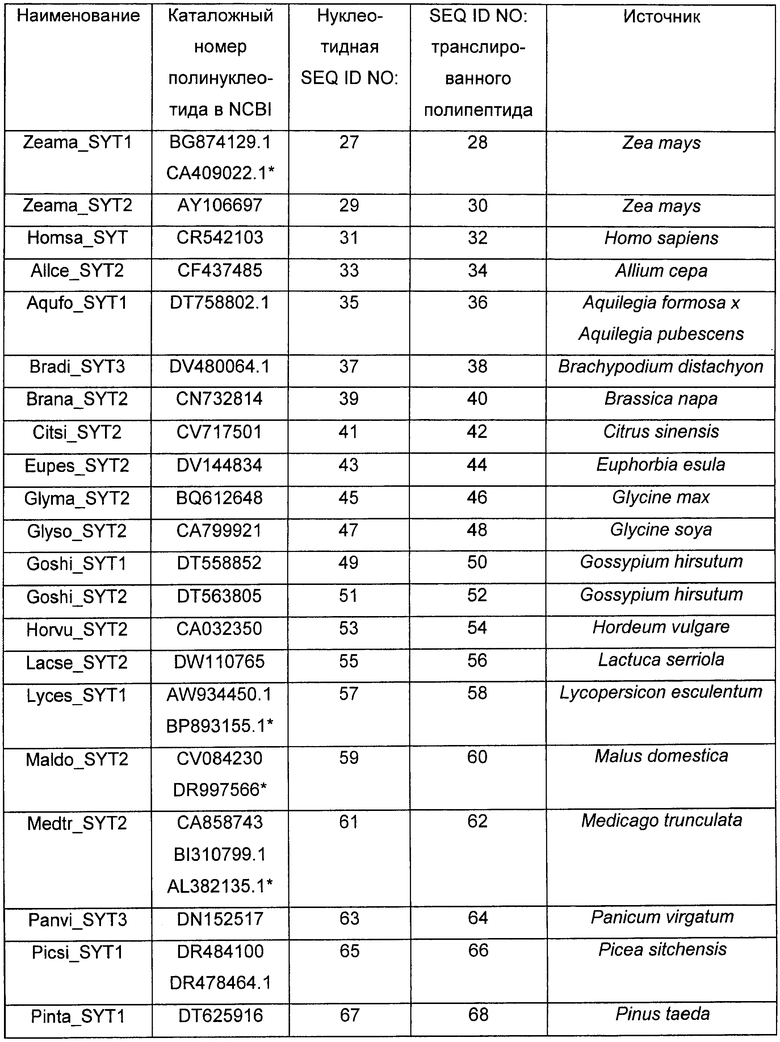
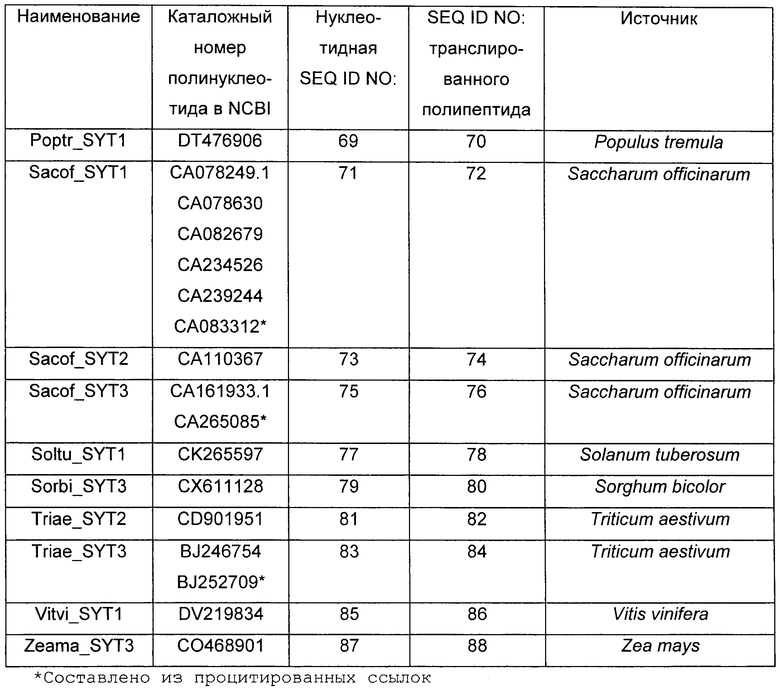
Растения, особенно применимые в соответствии со способами согласно изобретению, включают в себя все растения, принадлежащие к надсемейству Viridiplantae, в частности, однодольные и двудольные растения, включающие в себя кормовые, или фуражные, бобовые, декоративные растения, продовольственные культуры, деревья и кустарники, выбранные из перечня, содержащего, помимо прочего. Acacia spp., Acer spp., Actinidia spp., Aesculus spp., Agathis australis, Albizia amara, Alsophila tricolor, Andropogon spp., Arachis spp., Areca catechu, Astelia fragrans. Astragalus cicer, Baikiaea plurijuga, Betula spp., Brassica spp., Bruguiera gymnorrhiza, Burkea africana, Butea frondosa, Cadaba farinosa, Calliandra spp., Camellia sinensis, Canna indica, Capsicum spp., Cassia spp., Centroema pubescens, Chaenomeles spp., Cinnamomum cassia, Coffea arabica, Colophospermum mopane, Coronillia varia, Cotoneaster serotina, Crataegus spp., Cucumis spp., Cupressus spp., Cyathea dealbata, Cydonia oblonga, Cryptomeria japonica, Cymbopogon spp., Cynthea dealbata, Cydonia oblonga, Dalbergia monetaria, Davallia divaricata, Desmodium spp., Dicksonia squarosa, Diheteropogon amplectens, Dioclea spp., Dolichos spp., Dorycnium rectum, Echinochloa pyramidalis, Ehrartia spp., Eleusine coracana, Eragrestis spp., Erythrina spp., Eucalyptus spp., Euclea schimperi, Eulalia villosa, Fagopyrum spp., Feijoa sellowiana, Fragaria spp., Flemingia spp., Freycinetia banksii, Geranium thunbergii, Ginkgo biloba, Glycine javanica, Gliricidia spp., Gossypium hirsutum, Grevillea spp., Guibourtia coleosperma, Hedysarum spp., Hemarthia altissima, Heteropogon contortus, Hordeum vulgare, Hyparrhenia rufa, Hypericum erectum, Hyperthelia dissoluta, Indigo incarnata. Iris spp., Leptarrhena pyrolifolia, Lespediza spp., Lettuca spp., Leucaena leucocephala, Loudetia simplex, Lotonus bainesii, Lotus spp., Macrotyloma axillare, Malns spp., Manihot esculenta, Medicago saliva, Metasequoia glyptostroboides, Musa sapientum, Nicotianum spp., Onobrychis spp., Ornitthpus spp., Oryza spp., Peltophorum africanum, Pennisetum spp., Persea gratissima, Petunia spp., Phaseolus spp., Phoenix canariensis, Phormium cookianum, Photinia spp., Picea glauca, Pinus spp., Pisum sativum, Podocarpus totara, Pogonarthria fleckii, Pogonarthria squarrosa, Populus spp., Prosopis cineraria, Pseudotsuga menziesii, Pterolobium stellatum, Pyrus communis, Quercus spp., Rhaphiolepsis umbellata, Rhopalostylis sapida, Rhus natalensis, Ribes grossularia, Ribes spp., Robinia pseudoacacia, Rosa spp., Rubus spp., Salix spp., Schyzachyrium sanguineum, Sciadopitys verticillata, Sequoia sempervirens, Sequoiadendron giganteum, Sorghum bicolor, Spinacia spp., Sporobolus fimbriatus, Stiburus alopecuroides, Stylosanthos humilis, Tadehagi spp., Taxodium distichum, Themeda triandra, Trifolium spp., Triticum spp., Tsuga heterophylla, Vaccinium spp., Vicia spp., Vitis vinifera, Watsonia pyramidata, Zantedeschia aethiopica, Zea mays, щирицу, артишок, спаржу, брокколи, брюссельскую капусту, капусту, канолу, морковь, цветную капусту, сельдерей, кормовую капусту, лен, браунколь, чечевицу, масличный рапс, бамию, лук, картофель, рис, сою, землянику, сахарную свеклу, сахарный тростник, подсолнечник, томат, тыкву, чай и водоросли. Согласно предпочтительному варианту выполнения настоящего изобретения растение представляет собой сельскохозяйственную культуру. Примеры сельскохозяйственных культур включают в себя, помимо прочего, сою, подсолнечник, канолу, люцерну, рапс, хлопок, томат, картофель или табак. Arabidopsis thaliana обычно не считают сельскохозяйственной культурой. Еще более предпочтительно, растение представляет собой однодольное растение, такое как сахарный тростник. Более предпочтительно, растение представляет собой зерновую культуру, такую как рис, кукуруза, пшеница, ячмень, просо, рожь, сорго или овес.
Используемый в настоящем описании термин "полипептид SYT или его гомолог" относится к полипептиду, содержащему в направлении от N-конца к С-концу: (i) домен SNH, характеризующийся в возрастающем порядке предпочтительности, по меньшей мере, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% идентичностью последовательности по отношению к домену SNH SEQ ID NO:2; и (ii) богатый Met домен; и (iii) богатый QG домен.
Предпочтительно, домен SNH, характеризующийся, по меньшей мере, 40% идентичностью по отношению к домену SNH SEQ ID NO:2, содержит остатки, показанные черным на фиг.2. Более предпочтительно, домен SNH представлен SEQ ID NO:1.
Кроме того, полипептид SYT или его гомолог могут содержать один или несколько из следующего: (a) SEQ ID NO:90; (b) SEQ ID NO:91; и (с) богатого Met домена на N-конце перед доменом SNH.
Полипептид SYT или его гомолог, как правило, взаимодействуют с полипептидами GRF (факторами регуляции роста, growth-regulating factor) в системах дрожжевых двойных гибридов. Анализы взаимодействия дрожжевых двойных гибридов хорошо известны из предшествующего уровня техники (см. Field et al. (1989) Nature 340(6230):245-246). Например, полипептид SYT, представленный SEQ ID NO:4, способен взаимодействовать с AtGRF5 и с AtGRF9. Авторы настоящего изобретения показали, что полипептид SYT или его гомолог повышают урожайность, особенно семенную продуктивность, растений.
Полипептид SYT или его гомолог кодируются нуклеиновой кислотой/геном SYT. Таким образом, используемый в настоящем описании термин "нуклеиновая кислота/ген SYT" относится к любым нуклеиновой кислоте/гену, кодирующим определенные выше полипептид SYT или его гомолог.
Полипептиды SYT или их гомологи могут быть легко идентифицированы в соответствии со стандартными методиками, хорошо известными из предшествующего уровня техники, например, по методу выравнивания последовательностей. Методы выравнивания последовательностей для целей сравнения хорошо известны из предшествующего уровня техники, такие методы включают в себя GAP, BESTFIT, BLAST, FASTA и TFASTA. В GAP для определения выравнивания двух полных последовательностей, максимизирующего количество совпадений и минимизирующего количество пропусков использован алгоритм Needleman и Wunsch ((1970) J. Mol. Biol. 48:443-453). В соответствии с алгоритмом BLAST (Altschul et al. (1990) J. Mol. Biol. 215:403-10) рассчитывается процентная идентичность последовательностей и проводится статистический анализ сходства двух последовательностей. Программное обеспечение для проведения анализа BLAST общедоступно в National Centre for Biotechnology Information. Гомологи SYT, содержащие домен SNH, характеризующийся, по меньшей мере, 40% идентичностью последовательности по отношению к домену SNH SEQ ID NO:2, и/или содержащие SEQ ID NO:90 и/или SEQ ID NO:91, могут быть легко идентифицированы с помощью, например, алгоритма выравнивания множественных последовательностей ClustalW (версия 1.83), доступного по адресу http://clustalw.genome.jp/sit-bin/nph-clustalw, с использованием параметров попарного выравнивания по умолчанию и метода оценки в процентах. Последовательность, характеризующаяся 40% идентичностью по отношению к домену SNH SEQ ID NO:2, достаточна для идентификации последовательности как SYT.
Кроме того, легко может быть также идентифицировано наличие богатого Met домена или богатого QG домена. Как показано на фиг.3, богатый Met домен и богатый QG домен следует за доменом SNH. Богатый QG домен можно считать по существу С-концом белка (за исключением домена SHN); богатый Met домен, как правило, содержится в пределах первой половины богатого QG домена (от N-конца к С-концу). Первичный аминокислотный состав (в %) для определения того, является ли полипептидный домен богатым конкретными аминокислотами, может быть рассчитан с помощью программного обеспечения с сервера ExPASy (Gasteiger E et al. (2003) ExPASy: the proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 31:3784-3788), в частности, с помощью инструмента ProtParam. Состав интересующего белка может быть сравнен со средним аминокислотным составом (в %) в базе данных белковых последовательностей Swiss-Prot. В этой базе данных среднее содержание Met (M) составляет 2,37%, среднее содержание Gln (Q) составляет 3,93% и среднее содержание Gly (G) составляет 6,93%. Как определено в настоящем описании, богатый Met домен или богатый QG домен характеризуются содержанием Met (в %) или содержанием Gln и Gly (в %), превышающим средние значения аминокислотного состава (в %) базы данных белковых последовательностей Swiss-Prot.


Следует понимать, что последовательности, подпадающие под определение "полипептида SYT или его гомолога", не ограничены последовательностями, представленными SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8, SEQ ID NO:10, SEQ ID NO:12, SEQ ID NO:14, SEQ ID NO:16, SEQ ID NO:18, SEQ ID NO:20, SEQ ID NO:22, SEQ ID NO:24, SEQ ID NO:26, SEQ ID NO:28, SEQ ID NO:30, SEQ ID NO:32, SEQ ID NO:34, SEQ ID NO:36, SEQ ID NO:38, SEQ ID NO:40, SEQ ID NO:42, SEQ ID NO:44, SEQ ID NO:46, SEQ ID NO:48, SEQ ID NO:50, SEQ ID NO:52, SEQ ID NO:54, SEQ ID NO:56, SEQ ID NO:58, SEQ ID NO:60, SEQ ID NO:62, SEQ ID NO:64, SEQ ID NO:66, SEQ ID NO:68, SEQ ID NO:70, SEQ ID NO:72, SEQ ID NO:74, SEQ ID NO:76, SEQ ID NO:78, SEQ ID NO:80, SEQ ID NO:82, SEQ ID NO:84, SEQ ID NO:86, SEQ ID NO:88, и что любой полипептид, содержащий в направлении от N-конца к С-концу: (i) домен SNH, характеризующийся, по меньшей мере, 40% идентичностью последовательности по отношению к домену SNH SEQ ID NO:2; и (ii) богатый Met домен; и (iii) богатый QG домен, может быть признан подходящим для осуществления способов согласно изобретению.
Примеры нуклеиновых кислот SYT включают в себя без ограничения нуклеиновые кислоты, представленные любой из SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:15, SEQ ID NO:17, SEQ ID NO:19, SEQ ID NO:21, SEQ ID NO:23, SEQ ID NO:25, SEQ ID NO:27, SEQ ID NO:29, SEQ ID NO:31, SEQ ID NO:33, SEQ ID NO:35, SEQ ID NO:37, SEQ ID NO:39, SEQ ID NO:41, SEQ ID NO:43, SEQ ID NO:45, SEQ ID NO:47, SEQ ID NO:49, SEQ ID NO:51, SEQ ID NO:53, SEQ ID NO:55, SEQ ID NO:57, SEQ ID NO:59, SEQ ID NO:61, SEQ ID NO:63, SEQ ID NO:65, SEQ ID NO:67, SEQ ID NO:69, SEQ ID NO:71, SEQ ID NO:73, SEQ ID NO:75, SEQ ID NO:77, SEQ ID NO:79, SEQ ID NO:81, SEQ ID NO:83, SEQ ID NO:85, SEQ ID NO:87. Для осуществления способов согласно изобретению могут подходить нуклеиновые кислоты/гены SYT или их варианты. Варианты нуклеиновых кислот/генов SYT, как правило, представляют собой нуклеиновые кислоты/гены, обладающие той же функцией, что и природные нуклеиновые кислоты/гены SYT, которая может представлять собой ту же биологическую функцию или функцию повышения урожайности при модулировании экспрессии нуклеиновых кислот/генов в растении. Такие варианты содержат участки нуклеиновой кислоты/гена SYT и/или нуклеиновых кислот, способные гибридизоваться с определенными ниже нуклеиновой кислотой/геном SYT.
Используемый в настоящем описании термин "часть" относится к отрезку ДНК, кодирующему полипептид, содержащий в направлении от N-конца к С-концу: (i) домен SNH, характеризующийся в возрастающем порядке предпочтительности, по меньшей мере, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% идентичностью последовательности по отношению к домену SNH SEQ ID NO:2; и (ii) богатый Met домен; и (iii) богатый QG домен. Часть может быть получена, например, путем внесения одной или нескольких делеций в нуклеиновую кислоту SYT. Части могут быть использованы в выделенном виде, либо они могут быть слиты с другими кодирующими (или не кодирующими) последовательностями, например, для того, чтобы продуцировать белок, сочетающий несколько активностей. При слиянии с другими кодирующими последовательностями полученный транслированный полипептид может быть иметь более крупный размер, чем таковой, предсказанный для фрагмента SYT. Предпочтительно, частью является часть нуклеиновой кислоты, представленная любой из SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:15, SEQ ID NO:17, SEQ ID NO:19, SEQ ID NO:21, SEQ ID NO:23, SEQ ID NO:25, SEQ ID NO:27, SEQ ID NO:29, SEQ ID NO:31, SEQ ID NO:33, SEQ ID NO:35, SEQ ID NO:37, SEQ ID NO:39, SEQ ID NO:41, SEQ ID NO:43, SEQ ID NO:45, SEQ ID NO:47, SEQ ID NO:49, SEQ ID NO:51, SEQ ID NO:53, SEQ ID NO:55, SEQ ID NO:57, SEQ ID NO:59, SEQ ID NO:61, SEQ ID NO:63, SEQ ID NO:65, SEQ ID NO:67, SEQ ID NO:69, SEQ ID NO:71, SEQ ID NO:73, SEQ ID NO:75, SEQ ID NO:77, SEQ ID NO:79, SEQ ID NO:81, SEQ ID NO:83, SEQ ID NO:85, SEQ ID NO:87. Наиболее предпочтительно, часть нуклеиновой кислоты представлена SEQ ID NO:3, SEQ ID NO:5 или SED IQ NO:7.
Другим вариантом нуклеиновой кислоты/гена SYT является нуклеиновая кислота, способная гибридизоваться в условиях низкой жесткости, предпочтительно, в жестких условиях с определенными выше нуклеиновой кислотой/геном SYT, причем указанная гибридизующаяся последовательность кодирует полипептид, содержащий в направлении от N-конца к С-концу: (i) домен SNH, характеризующийся в возрастающем порядке предпочтительности, по меньшей мере, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% идентичностью последовательности по отношению к домену SNH SEQ ID NO:2; и (ii) богатый Met домен; и (iii) богатый QG домен. Предпочтительно, гибридизующаяся последовательность представляет собой последовательность, способную гибридизоваться с нуклеиновой кислотой, представленной любой из SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:15, SEQ ID NO:17, SEQ ID NO:19, SEQ ID NO:21, SEQ ID NO:23, SEQ ID NO:25, SEQ ID NO:27, SEQ ID NO:29, SEQ ID NO:31, SEQ ID NO:33, SEQ ID NO:35, SEQ ID NO:37, SEQ ID NO:39, SEQ ID NO:41, SEQ ID NO:43, SEQ ID NO:45, SEQ ID NO:47, SEQ ID NO:49, SEQ ID NO:51, SEQ ID NO:53, SEQ ID NO:55, SEQ ID NO:57, SEQ ID NO:59, SEQ ID NO:61, SEQ ID NO:63, SEQ ID NO:65, SEQ ID NO:67, SEQ ID NO:69, SEQ ID NO:71, SEQ ID NO:73, SEQ ID NO:75, SEQ ID NO:77, SEQ ID NO:79, SEQ ID NO:81, SEQ ID NO:83, SEQ ID NO:85, SEQ ID NO:87, или с частью любой из упомянутых выше определенных в настоящем описании последовательностей. Наиболее предпочтительно, гибридизующаяся последовательность нуклеиновой кислоты представлена SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:7.
Используемый здесь термин "гибридизация" относится к процессу, в ходе которого по существу гомологичные комплементарные нуклеотидные последовательности гибридизуются друг с другом. Процесс гибридизации может полностью протекать в растворе, т.е. когда обе комплементарные нуклеиновые кислоты находятся в растворе. Процесс гибридизации также может протекать, когда одна из комплементарных нуклеиновых кислот иммобилизована на матриксе, таком как магнитные бусы, сефарозные бусы или любая другая смола. Кроме того, процесс гибридизации может протекать, когда одна из комплементарных нуклеиновых кислот иммобилизована на твердой подложке, такой как нитроцеллюлозная или найлоновая мембрана, или иммобилизована, например, методом фотолитографии, например, на подложку из кварцевого стекла (последний вариант известен как массивы или микромассивы нуклеиновых кислот или чипы нуклеиновых кислот). Для обеспечения протекания гибридизации молекулы нуклеиновых кислот, как правило, термально или химически денатурируют для плавления двухцепочечной молекулы на две одинарные цепи и/или для удаления шпилек или других вторичных структур из одноцепочечных нуклеиновых кислот. На строгость гибридизации оказывают влияние такие условия, как температура, концентрация солей, ионная сила и состав гибридизационного буфера.
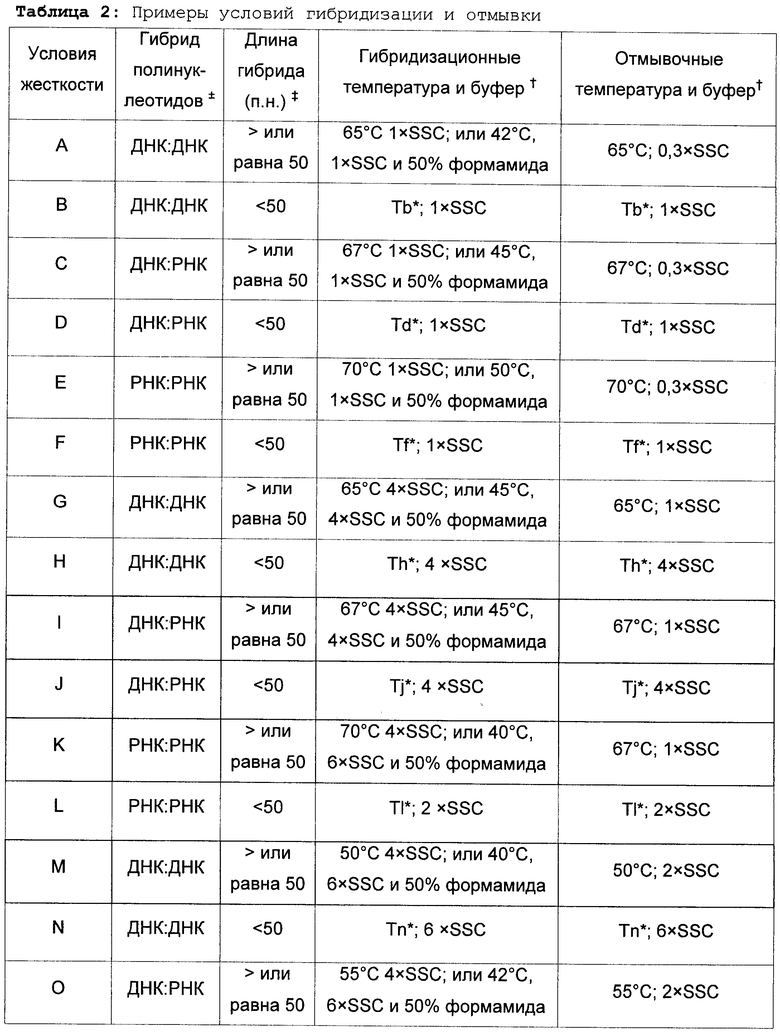
"Жесткие условия гибридизации" и "жесткие условия гибридизационной отмывки" в контексте экспериментов по гибридизации нуклеиновых кислот, таких как саузерн- и нозерн-гибридизации, зависят от последовательностей и различны при разных средовых параметрах. Специалисту в данной области техники известны различные параметры, которые могут быть изменены в ходе гибридизации и отмывки и которые способствуют либо поддержанию, либо изменению условий жесткости.
Тпл представляет собой температуру, при которой при определенных значениях ионной силы и pH 50% последовательности-мишени гибридизуется с полностью соответствующим зондом. Тпл зависит от условий в растворе и нуклеотидного состава и длины зонда. Например, длинные последовательности специфично гибридизуются при более высоких температурах. Максимальный уровень гибридизации достигается приблизительно при температуре от 16°С до 32°С ниже Тпл. Наличие моновалентных катионов в гибридизационном растворе ослабляет электростатическое отталкивание между двумя цепями нуклеиновой кислоты, тем самым способствуя образованию гибрида; этот эффект наблюдается при концентрациях ионов натрия до 0,4 М. Формамид снижает температуру плавления дуплексов ДНК-ДНК и ДНК-РНК на 0,6-0,7°C с каждым процентом концентрации формамида, а добавление 50% формамида позволяет проводить гибридизацию при 30-45°С, хотя уровень гибридизации будет снижен. Несоответствия пар нуклеотидов снижает уровень гибридизации и термальную стабильность дуплексов. В среднем и для крупных зондов Тпл снижается примерно на 1°C с каждым % несоответствия нуклеотидов. Тпл может быть рассчитана в соответствии со следующими уравнениями в зависимости от типов гибридов:
1. Гибриды ДНК-ДНК (Meinkoth and Wahl, Anal. Biochem., 138:267-284, 1984):
Тпл=81,5°С+16,6×log[Na+]a+0,41×%[G/Cb]-500×[Lc]-1-0,61×% формамида
2. Гибриды ДНК-РНК или РНК-РНК:
Тпл=79,8+18,5(log10[Na+]a)+0,58(%G/Cb)+11,8(%G/Cb)2-820/Lc
3. Гибриды олиго-ДНК или олиго-РНКd:
Для <20 нуклеотидов: Тпл=2 (ln)
Для 20-35 нуклеотидов: Тпл=22+1,46 (ln)
а или другого одновалентного катиона, но только строго в диапазоне 0,01-0,4 М.
b %GC только строго в диапазоне от 30% до 75%.
c L = длина дуплекса в парах нуклеотидов.
d Олиго, олигонуклеотид; ln, расчетная длина праймера = 2×(число G/C)+(число А/Т).
Примечание: с каждым 1% формамида Тпл снижается приблизительно на 0,6-0,7°С, тогда как присутствие 6 М мочевины снижает Тпл приблизительно на 30°С.
Специфичность гибридизации, как правило, зависит от постгибридизационных отмывок. Для устранения фона, связанного с неспецифичной гибридизацией, образцы отмывали разбавленными солевыми растворами. Ключевые факторы таких отмывок включают в себя ионную силу и температуру конечного отмывочного раствора: чем ниже концентрация солей и чем выше температура отмывки, тем выше жесткость отмывки. Обычно создают условия отмывки на уровне жесткой гибридизации или ниже. Как правило, условия подходящей жесткости для гибридизационных анализов нуклеиновых кислот или методик детекции амплификации генов соответствуют изложенным выше. Также могут быть выбраны условия большей или меньшей жесткости. Как правило, выбираемые условия низкой жесткости предусматривают температуру приблизительно на 50°С ниже, чем температура плавления (Тпл) для конкретной последовательности при определенных значениях ионной силы и pH. Условия средней жесткости предусматривают температуру на 20°С ниже Тпл, а условия высокой жесткости предусматривают температуру на 10°С ниже Тпл. Например, жесткими условиями являются условия, по меньшей мере, уровня жесткости A-L; а менее жесткими условиями являются условия, по меньшей мере, уровня жесткости M-R. Неспецифическое связывание может быть подавлено в соответствии с одной из ряда известных методик таких как, например, блокирование мембраны белоксодержащими растворами, добавления в гибридизационный буфер гетерологичных РНК, ДНК и SDS и обработка РНКазой. Примеры условий гибридизации и отмывки приведены ниже в таблице 2.
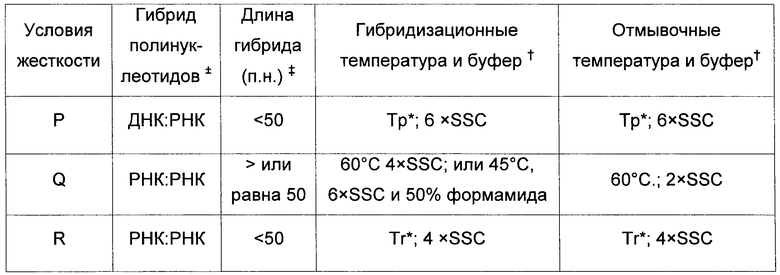

Для отсылки к сведениям, касающимся определения уровня жесткости, можно сослаться на Sambrook et al. (2001) Molecular Cloning: a Laboratory Manual, 3rd Edition, Cold Spring Harbor Laboratory Press, CSH, New York или на Current Protocols in Molecular Biology, John Wiley & Sons, N.Y. (1989).
Нуклеиновая кислота SYT или ее вариант могут быть получены из любого искусственного источника или природного источника, такого как растение, водоросли или животное. Такая нуклеиновая кислота может быть модифицирована из нативной формы в композиции и/или геномном окружении путем произвольных манипуляций исследователя. Нуклеиновая кислота предпочтительно имеет растительное происхождение либо из растения того же вида (например, из вида растения, в которое она должна быть введена), либо из растения другого вида. Предпочтительно, нуклеиновая кислота растительного происхождения кодирует SYT1. Альтернативно, нуклеиновая кислота может кодировать SYT2 или SYT3, которые близкородственны друг другу на уровне полипептидов. Нуклеиновая кислота может быть выделена из двудольных растений, предпочтительно из семейства Brassicaceae, более предпочтительно, из Arabidopsis thaliana. Более предпочтительно, три нуклеиновые кислоты SYT, выделенные из Arabidopsis thaliana, представлены SEQ ID NO:3, SEQ ID NO:5 и SEQ ID NO:7, а три аминокислотные последовательности SYT представлены SEQ ID NO:4, SEQ ID NO:6 и SEQ ID NO:8.
Экспрессия нуклеиновой кислоты, кодирующей полипептид SYT или его гомолог, может быть модулирована путем введения генной модификации (предпочтительно, в локус гена SYT). Используемый в настоящем описании термин "локус гена" означает область генома, содержащую интересующий ген и участок размером 10 т.п.н. левее или правее кодирующей области.
Генная модификация может быть введена, например, в соответствии с одним (или несколькими) из следующих методов: активация Т-ДНК, TILLING, сайт-направленный мутагенез, направленная эволюция и гомологичная рекомбинация, либо путем введения и экспрессии в растении нуклеиновой кислоты, кодирующей полипептид SYT или его гомолог. После внесения генной модификации следует стадия отбора по модулированной экспрессии нуклеиновой кислоты, кодирующей полипептид SYT или его гомолог, причем указанная модулированная экспрессия позволяет получить растения, характеризующиеся повышенной урожайностью, особенно, повышенной семенной продуктивностью.
Активирующее мечение Т-ДНК (Hayashi et al. Science (1992) 1350-1353) предусматривает встраивание Т-ДНК, обычно содержащей промотор (также может содержаться энхансер трансляции или интрон), в геномную область интересующего гена или в 10 т.п.н. левее или правее от кодирующей области гена в такой конфигурации, чтобы промотор управлял экспрессией гена-мишени. Как правило, регуляция экспрессии гена-мишени его естественным промотором прекращается, и ген оказывается под контролем вновь встроенного промотора. Промотор, как правило, встроен в Т-ДНК. Эта Т-ДНК случайным образом встраивается в геном растения, например, с помощью инфекции Agrobacterium, и вызывает усиленную экспрессию генов, расположенных поблизости от встроенной Т-ДНК. Полученные трансгенные растения обладают доминантным фенотипом благодаря усиленной экспрессии генов, расположенных поблизости от встроенного промотора. Подлежащим встраиванию промотором может являться любой промотор, способный управлять экспрессией гена в желаемом организме, в данном случае, в растении. Например, для использования в активации Т-ДНК подходит любой из конститутивных, тканепредпочтительных, специфичных в отношении определенного типа клеток и индуцибельных промоторов.
Генная модификация также может быть проведена в локусе гена SYT в соответствии с методом TILLING (направленных индуцированных локальных повреждений геномов). Это основанная на мутагенезе технология, применимая для создания и/или идентификации и в итоге выделения претерпевших мутагенез вариантов нуклеиновой кислоты SYT, кодирующих белок с повышенной активностью SYT. TILLING также позволяет осуществлять отбор растений, несущих такие мутантные варианты. Эти мутантные варианты могут даже характеризоваться повышенной активностью SYT по сравнению с таковой, характерной для природной формы гена. TILLNG объединяет высокоплотный мутагенез с высокопродуктивными скрининговыми методами. Как правило, стадии TILLING представляют собой следующее: (a) EMS мутагенез (Redei GP and Koncz С (1992) In Methods in Arabidopsis Research, Koncz C, Chua NH, Schell J, eds. Singapore, World Scientific Publishing Co, pp.16-82; Feldmann et al., (1994) In Meyerowitz EM, Somerville CR, eds., Arabidopsis. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, pp.137-172; Lightner J and Caspar Т (1998) In J Martinez-Zapater, J Salinas, eds., Methods on Molecular Biology, Vol.82. Humana Press, Totowa, NJ, pp.91-104); (b) получение ДНК и группировка субъектов; (с) PCR-амплификация интересующей области; (d) денатурация и гибридизация с образованием гетеродуплексов; (е) DHPLC, в ходе которой присутствие гетеродуплекса в пуле определяется по дополнительному пику на хроматограмме; (f) идентификация мутантного субъекта; и (g) секвенирование мутантного продукта PCR. Методики TILLING хорошо известны из предшествующего уровня техники (McCallum et al., (2000) Nat. Biotechnol. 18:455-457; обзор Stemple (2004) Nat. Rev. Genet. 5(2):145-50).
Для создания вариантов нуклеиновых кислот SYT может быть использован сайт-направленный мутагенез. Известно несколько методик проведения сайт-направленного мутагенеза, самыми распространенными из которых являются методики, основанные на PCR (Current Protocols in Molecular Biology. Wiley Eds. http://www.4ulr.corn/products/currentprotocols/index.html).
Для создания вариантов нуклеиновых кислот SYT также может быть использована направленная эволюция. Она предусматривает многократные перестановки ДНК с последующим скринингом и/или отбором для получения вариантов нуклеиновых кислот SYT или их частей, кодирующих полипептиды SYT или их гомологи или части, характеризующиеся модифицированной биологической активностью (Castle et al., (2004) Science 304(5674):1151-4; патенты США №№5811238 и 6395547).
Активация Т-ДНК, TILLING, сайт-направленный мутагенез и направленная эволюция представляют собой примеры технологий, позволяющих создавать новые аллели и варианты SYT.
Гомологичная рекомбинация позволяет вводить в определенное выбранное положение в геноме выбранную нуклеиновую кислоту. Гомологичная рекомбинация представляет собой стандартный метод, рутинно используемый в биологических экспериментах на низших организмах, таких как дрожжи или мох Physcomitrella. Методики проведения гомологичной рекомбинации у растений описаны не только для модельных растений (Offringa et al. (1990) ЕМВО J. 9 (10):3077-84), но и для сельскохозяйственных культур, например, риса (Terada et al. (2002) Nat. Biotech. 20(10):1030-4; Iida and Terada (2004) Curr. Opin. Biotech. 15 (2):132-8). Направляемую нуклеиновую кислоту (которой могут являться определенные выше нуклеиновая кислота SYT или ее вариант) направляют в локус гена SYT. Направляемая нуклеиновая кислота может представлять собой улучшенный аллель, используемый для замены эндогенного гена, или может быть введена в дополнение к эндогенному гену.
Предпочтительный способ введения генной модификации (которая, в таком случае, не обязана находиться в локусе гена SYT) состоит во введении и экспрессии в растении нуклеиновой кислоты, кодирующей полипептид SYT или его гомолог. Полипептид SYT или его гомолог определены как полипептид, содержащий в направлении от N-конца к С-концу: (i) домен SNH, характеризующийся в возрастающем порядке предпочтительности, по меньшей мере, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% идентичностью последовательности по отношению к домену SNH SEQ ID NO:2; и (ii) богатый Met домен; и (iii) богатый QG домен.
Предпочтительно, домен SNH, характеризующийся, по меньшей мере, 40% идентичностью по отношению к домену SNH SEQ ID NO:2, содержит остатки, показанные на фиг.2 черным. Более предпочтительно, домен SNH представлен SEQ ID NO:1.
Вводимая в растение нуклеиновая кислота может представлять собой полноразмерную нуклеиновую кислоту или может представлять собой определенные выше часть или гибридизующуюся последовательность.
"Гомологами" белка являются пептиды, олигопептиды, полипептиды, белки и ферменты, содержащие аминокислотные замены, делеции и/или инсерции по сравнению с интересующим немодифицированным белком и характеризующиеся биологической и функциональной активностью, сходной с таковой немодифицированного белка, из которого они получены. Для получения таких гомологов аминокислоты белка могут быть заменены другими аминокислотами, имеющими сходные свойства (такие как сходную гидрофобность, гидрофильность, антигенность, склонность к образованию или разрушению α-спиралей или β-складок). Таблицы консервативных замен хорошо известны из предшествующего уровня техники (см., например, Creighton (1984) Proteins. W.H. Freeman and Company и таблицу 3 ниже).
Гомологи включают в себя ортологи и паралоги, на которых основаны эволюционные концепции, используемые для описания родственных связей генов. Паралоги представляют собой гены в пределах одного вида, возникшие в результате удвоения гена-предшественника, а ортологи представляют собой гены различных организмов, возникшие в процессе видообразования.
Ортологи, например, в видах однодольных растений могут быть легко выявлены путем проведения так называемого двустороннего BLAST-поиска. Это может быть сделано с помощью первого BLAST-поиска, предусматривающего скрининг по заданной последовательности (например, SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:5, SEQ ID NO:6, SEQ ID NO:7 или SEQ ID NO:8) любой базы данных, такой как общедоступная база данных NCBI, в которую можно зайти по адресу http://www.ncbi.nlm.nih.gov. При исследовании нуклеотидной последовательности могут быть использованы BLASTN или TBLASTX (с использованием стандартных параметров по умолчанию), а при исследовании аминокислотной последовательности могут быть использованы BLASTP или TBLASTN (с использованием стандартных параметров по умолчанию). Результаты BLAST необязательно могут быть отфильтрованы. По полноразмерным последовательностям либо из отфильтрованных результатов, либо из неотфильтрованных результатов затем проводят обратный BLAST-поиск (второй BLAST) среди последовательностей организма, из которого происходит задаваемая последовательность (если задаваемой последовательностью является SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:5, SEQ ID NO:6, SEQ ID NO:7 или SEQ ID NO:8, второй BLAST-поиск, следовательно, будет проводиться среди последовательностей Arabidopsis). Затем результаты первого и второго BLAST-поисков сравнивают. Паралог считают идентифицированным, если наилучший хит второго BLAST-поиска относится к тому же виду, из которого происходит заданная последовательность; ортолог считают идентифицированным, если наилучший хит не относится к тому же виду, из которого происходит заданная последовательность. Наилучшими хитами являются хиты, характеризующиеся низким Е-значением. Чем ниже Е-значение, тем более значима оценка (или, другими словами, тем ниже вероятность того, что хит был найден случайно). Процедура расчета Е-значения хорошо известна из предшествующего уровня техники. В случае крупных семейств может быть использован ClustalW с последующим построением дерева методом присоединения соседа для облегчения визуализации кластеризации близких генов и идентификации ортологов и паралогов.
Гомолог может представлять собой "вариант замены" белка, т.е. вариант, в котором, по меньшей мере, один остаток аминокислотной последовательности удален, а отличный остаток помещен на его место. Аминокислотные замены, как правило, представляют собой замены отдельных остатков, но могут и образовывать кластеры в зависимости от функциональных ограничений, налагаемых на полипептид; инсерции обычно затрагивают порядка 1-10 аминокислотных остатков. Предпочтительно, аминокислотные замены представляют собой консервативные аминокислотные замены. Таблицы консервативных замен легко доступны из предшествующего уровня техники. В приведенной ниже таблицы показаны примеры консервативных аминокислотных замен.
Гомолог также может представлять собой "инсерционный вариант" белка, т.е. вариант, в котором в предварительно определенный участок белка введены один или несколько аминокислотных остатков. Инсерции могут включать в себя N-концевые и/или С-концевые слияния, равно как и инсерции отдельных или множественных аминокислот внутри последовательности. Как правило, инсерции внутри аминокислотной последовательности меньше, чем N- или С-концевые слияния, приблизительно порядка 1-10 остатков. Примеры гибридных по N- или С-концу белков или пептидов включают в себя связывающий домен или активирующий домен активатора транскрипции, используемого в системе дрожжевых двойных гибридов, белки оболочки фагов, метку (гистидин)6, глутатион-S-трансферазную метку, белок А, связывающий мальтозу белок, дигидрофолатредуктазу, эпитоп Tag·100, эпитоп c-myc, эпитоп FLAG®, lacZ, CMP (кальмодулинсвязывающий белок), эпитоп НА, эпитоп белка С и эпитоп VSV.
Гомологи в виде "делеционных вариантов" белка характеризуются отсутствием в белке одной или нескольких аминокислот.
Аминокислотные варианты белка могут быть легко получены в соответствии с методами пептидного синтеза, хорошо известными из предшествующего уровня техники, такими как твердофазный пептидный синтез и т.п., или с помощью манипуляций рекомбинантными ДНК. Методы манипулирования последовательностями ДНК для осуществления вариантов замен, инсерций и делеций белка хорошо известны из предшествующего уровня техники. Например, специалистам в данной области техники хорошо известны методики введения мутаций в виде замены в предварительно определенные участки ДНК, и такие методики включают в себя М13-мутагенез, мутагенез T7-Gen in vitro (USB, Cleveland, ОН), сайт-направленный мутагенез QuickChange (Stratagene, San Diego, CA), PCR-опосредованный сайт-направленный мутагенез или другие протоколы сайт-направленного мутагенеза.
Полипептид SYT или его гомолог могут быть производным. "Производные" включают в себя пептиды, олигопептиды, полипептиды, белки и ферменты, которые могут содержать замены, делении или добавления неприродных аминокислотных остатков по сравнению с аминокислотной последовательностью природной формы белка, например, представленного SEQ ID NO:4, SEQ ID NO:6, SEQ ID NO:8, SEQ ID NO:10, SEQ ID NO:12, SEQ ID NO:14, SEQ ID NO:16, SEQ ID NO:18, SEQ ID NO:20, SEQ ID NO:22, SEQ ID NO:24, SEQ ID NO:26, SEQ ID NO:28, SEQ ID NO:30, SEQ ID NO:32, SEQ ID NO:34, SEQ ID NO:36, SEQ ID NO:38, SEQ ID NO:40, SEQ ID NO:42, SEQ ID NO:44, SEQ ID NO:46, SEQ ID NO:48, SEQ ID NO:50, SEQ ID NO:52, SEQ ID NO:54, SEQ ID NO:56, SEQ ID NO:58, SEQ ID NO:60, SEQ ID NO:62, SEQ ID NO:64, SEQ ID NO:66, SEQ ID NO:68, SEQ ID NO:70, SEQ ID NO:72, SEQ ID NO:74, SEQ ID NO:76, SEQ ID NO:78, SEQ ID NO:80, SEQ ID NO:82, SEQ ID NO:84, SEQ ID NO:86 и SEQ ID NO:88.
"Производные" белка включают в себя пептиды, олигопептиды, полипептиды, белки и ферменты, которые могут содержать природные измененные, гликозилированные, ацилированные, пренилированные или неприродные аминокислотные остатки по сравнению с аминокислотной последовательностью природной формы полипептида. Производное также может содержать один или несколько неаминокислотных заместителей по сравнению с аминокислотной последовательностью, из которой оно получено, например, репортерную молекулу или другой лиганд, ковалентно или нековалентно связанные с аминокислотной последовательностью, такие как репортерная молекула, присоединенная для облегчения обнаружения, или неприродные аминокислотные остатки по сравнению с аминокислотной последовательностью природного белка.
Полипептид SYT или его гомолог могут кодироваться альтернативным сплайс-вариантом нуклеиновой кислоты/гена SYT. Используемый в настоящем описании термин "альтернативный сплайс-вариант" охватывает варианты последовательности нуклеиновой кислоты, в которой избранные интроны и/или экзоны могут быть вырезаны, заменены или добавлены или в которой интроны укорочены или удлинены. Такие варианты будут сохранять биологическую активность белка, чего можно достичь, избирательно сохранив функциональные сегменты белка. Такие сплайс-варианты могут быть природными или искусственными. Способы получения таких сплайс-вариантов хорошо известны из предшествующего уровня техники. Предпочтительными сплайс-вариантами являются сплайс-варианты нуклеиновой кислоты, кодирующей полипептид, содержащий в направлении от N-конца к С-концу: (i) домен SNH, характеризующийся в возрастающем порядке предпочтительности, по меньшей мере, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% идентичностью последовательности по отношению к домену SNH SEQ ID NO:2; и (ii) богатый Met домен; и (iii) богатый QG домен. Предпочтительно, домен SNH, характеризующийся, по меньшей мере, 40% идентичностью по отношению к домену SNH SEQ ID NO:2, содержит остатки, показанные черным на фиг.2. Более предпочтительно, домен SNH представлен SEQ ID NO:1.
Кроме того, полипептид SYT или его гомолог могут содержать один или несколько из следующего: (a) SEQ ID NO:90; и/или (b) SEQ ID NO:91; и/или (с) богатого Met домена на N-конце перед доменом SNH.
Другие предпочтительные сплайс-варианты нуклеиновых кислот представлены SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:15, SEQ ID NO:17, SEQ ID NO:19, SEQ ID NO:21, SEQ ID NO:23, SEQ ID NO:25, SEQ ID NO:27, SEQ ID NO:29, SEQ ID NO:31, SEQ ID NO:33, SEQ ID NO:35, SEQ ID NO:37, SEQ ID NO:39, SEQ ID NO:41, SEQ ID NO:43, SEQ ID NO:45, SEQ ID NO:47, SEQ ID NO:49, SEQ ID NO:51, SEQ ID NO:53, SEQ ID NO:55, SEQ ID NO:57, SEQ ID NO:59, SEQ ID NO:61, SEQ ID NO:63, SEQ ID NO:65, SEQ ID NO:67, SEQ ID NO:69, SEQ ID NO:71, SEQ ID NO:73, SEQ ID NO:75, SEQ ID NO:77, SEQ ID NO:79, SEQ ID NO:81, SEQ ID NO:83, SEQ ID NO:85 и SEQ ID NO:87. Наиболее предпочтительными являются сплайс-варианты нуклеиновой кислоты/гена SYT, представленные SEQ ID NO:3, SEQ ID NO:5 и SEQ ID NO:7.
Гомолог также может кодироваться аллельным вариантом нуклеиновой кислоты, кодирующей полипептид SYT или его гомолог, предпочтительно, аллельный вариант нуклеиновой кислоты, кодирующей полипептид, содержащий в направлении от N-конца к С-концу: (i) домен SNH, характеризующийся в возрастающем порядке предпочтительности, по меньшей мере, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% идентичностью последовательности по отношению к домену SNH SEQ ID NO:2; и (ii) богатый Met домен; и (iii) богатый QG домен. Предпочтительно, домен SNH, характеризующийся, по меньшей мере, 40% идентичностью по отношению к домену SNH SEQ ID NO:2, содержит остатки, показанные черным на фиг.2. Более предпочтительно, домен SNH представлен SEQ ID NO:1. Кроме того, полипептид SYT или его гомолог могут содержать один или несколько из следующего: (a) SEQ ID NO:90; и/или (b) SEQ ID NO:91; и/или (с) богатого Met домена на N-конце перед доменом SNH.
Также предпочтительно, аллельный вариант представляет собой аллельный вариант нуклеиновой кислоты, представленный SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:15, SEQ ID NO:17, SEQ ID NO:19, SEQ ID NO:21, SEQ ID NO:23, SEQ ID NO:25, SEQ ID NO:27, SEQ ID NO:29, SEQ ID NO:31, SEQ ID NO:33, SEQ ID NO:35, SEQ ID NO:37, SEQ ID NO:39, SEQ ID NO:41, SEQ ID NO:43, SEQ ID NO:45, SEQ ID NO:47, SEQ ID NO:49, SEQ ID NO:51, SEQ ID NO:53, SEQ ID NO:55, SEQ ID NO:57, SEQ ID NO:59, SEQ ID NO:61, SEQ ID NO:63, SEQ ID NO:65, SEQ ID NO:67, SEQ ID NO:69, SEQ ID NO:71, SEQ ID NO:73, SEQ ID NO:75, SEQ ID NO:77, SEQ ID NO:79, SEQ ID NO:81, SEQ ID NO:83, SEQ ID NO:85 и SEQ ID NO:87. Более предпочтительно, аллельный вариант представляет собой аллельный вариант нуклеиновой кислоты, представленный любой из SEQ ID NO:3, SEQ ID NO:5 и SEQ ID NO:7.
Аллельные варианты существуют в природе, и способы согласно настоящему изобретению предусматривают применение этих природных аллелей. Аллельные варианты включают в себя однонуклеотидные полиморфизмы (SNP), равно как и малые инсерционные/делеционные полиморфизмы (INDEL). Размер INDEL обычно составляет менее 100 п.н. SNP и INDEL составляют наибольшую группу вариантов последовательностей в природных полиморфных штаммах большинства организмов.
В соответствии с предпочтительным аспектом настоящего изобретения модулированная экспрессия нуклеиновой кислоты SYT или ее варианта представляет собой усиленную экспрессию. Усиление экспрессии может приводить к повышению содержания мРНК или полипептида SYT, что может быть приравнено к повышенной активности полипептида SYT; либо активность может быть также повышена при неизменном содержании полипептида или даже при пониженном содержании полипептида. Такой эффект может иметь место, если изменены внутренние свойства полипептида, например, в результате создания мутантных вариантов, более активных, нежели полипептид дикого типа. Методы усиления экспрессии генов или генных продуктов подробно описаны в предшествующем уровне техники и включают в себя, например, усиленную экспрессию, управляемую подходящими промоторами, использование энхансеров транскрипции или энхансеров трансляции. Выделенные нуклеиновые кислоты, которые служат промоторными или энхансерными элементами, могут быть введены в подходящее положение (как правило, левее) негетерологичной формы полинуклеотида с тем, чтобы положительно регулировать экспрессию нуклеиновой кислоты SYT или ее варианта. Например, эндогенные промоторы могут быть изменены in vivo путем мутации, делении и/или замены (см., Kmiec, патент США №5565350; Zarling et al., PCT/US93/03868), либо выделенные промоторы могут быть введены в растительную клетку в правильной ориентации и на надлежащем расстоянии от гена согласно настоящему изобретению с тем, чтобы контролировать экспрессию гена. Методы ослабления экспрессии генов или генных продуктов подробно описаны в предшествующем уровне техники.
Если желаема экспрессия полипептида, как правило, желательно поместить на 3'-конец кодирующей области полинуклеотида сайт полиаденилирования. Сайт полиаденилирования может быть получен из нативного гена, из множества других генов растения или из Т-ДНК. Добавляемая 3'-концевая последовательность может быть получена, например, из генов нопалинсинтазы или октопинсинтазы или, альтернативно, из другого гена растения или, менее предпочтительно, из любого другого эукариотического гена.
Интронная последовательность также может быть добавлена в 5'-нетранслируемую область или кодирующую последовательность частичной кодирующей последовательности для увеличения количества зрелого мессенджера, накапливающегося в цитозоле. Было показано, что включение сплайсируемого интрона в транскриптон экспрессирующих конструкций как растительного, так и животного происхождения вызывает до 1000-кратного усиления экспрессию гена как на уровне мРНК, так и на белковом уровне, Buchman and Berg, Mol. Cell. Biol. 8:4395-4405 (1988); Callis et al., Genes Dev. 1:1183-1200 (1987). Такое интронное усиление генной экспрессии наиболее существенно, когда интрон помещают вблизи 5'-конца транскриптона. Из предшествующего уровня техники известно использование интронов кукурузы Adh1-S 1, 2 и 6, интрона Bronze-1. С общими сведениями можно ознакомиться в The Maize Handbook, Chapter 116, Freeling and Walbot, Eds., Springer, N.Y. (1994).
Изобретение также относится к генным конструкциям и векторам, облегчающим введение и/или экспрессию нуклеотидных последовательностей, применимых в соответствии со способами согласно изобретению.
Таким образом, раскрыта генная конструкция, содержащая
(i) любые определенные выше нуклеиновую кислоту SYT или ее вариант;
(ii) одну или несколько контрольных последовательностей, способных управлять экспрессией последовательности нуклеиновой кислоты (i); и необязательно
(iii) последовательность терминации транскрипции.
Предпочтительной конструкцией является конструкция, в которой контрольная последовательность представляет собой промотор, имеющий растительное происхождение, предпочтительно, из однодольного растения.
Конструкции, применимые в соответствии со способами согласно настоящему изобретению, могут быть сконструированы в соответствии с технологией рекомбинантных ДНК, хорошо известной специалистам в данной области техники. Генные конструкции могут быть встроены в векторы, которые могут быть коммерчески доступны, подходящие для трансформации растений и подходящие для экспрессии интересующего гена в трансформированных клетках.
Растения трансформируют вектором, содержащим интересующую последовательность (т.е. нуклеиновую кислоту, кодирующую полипептид SYT или его гомолог). Интересующая последовательность функционально связана с одной или несколькими контрольными последовательностями (по меньшей мере, с промотором). Термины "регуляторный элемент", "контрольная последовательность" и "промотор" используются в настоящем описании взаимозаменяемо и в широком контексте должны пониматься как относящиеся к регуляторным последовательностям нуклеиновых кислот, способным эффективно экспрессировать последовательности, с которыми они лигированы. Указанные выше термины охватывают последовательности регуляции транскрипции, полученные из классического эукариотического геномного гена (включая ТАТА-бокс, служащий для точной инициации транскрипции при участии последовательности ССААТ-бокса или без него), и дополнительные регуляторные элементы (т.е. расположенные левее активирующие последовательности, энхансеры и сайленсеры), которые изменяют экспрессию гена в ответ на дифференцировочные и/или внешние стимулы или тканеспецифично. Также под определение термина подпадает последовательность регуляции транскрипции классического прокариотического гена, и в этом случае она может содержать последовательности регуляции транскрипции бокс -35 и/или бокс -10. Термин "регуляторный элемент" также относится к синтетическим гибридной молекуле или производному, обеспечивающим, активирующим или усиливающим экспрессию молекулы нуклеиновой кислоты в клетке, ткани или органе. Используемый в настоящем описании термин "функционально связанный" относится к функциональной связи между промоторной последовательностью и интересующим геном, так что промоторная последовательность способна инициировать транскрипцию интересующего гена.
Предпочтительно, для управления экспрессией последовательности нуклеиновой кислоты может быть использован любой тип промотора. Промотор может представлять собой индуцибельный промотор, т.е. индуцировать или усиливать инициацию транскрипции в ответ на дифференцировочный, химический, средовой или физический стимул. Примером индуцибельного промотора является индуцируемый стрессом промотор, т.е. промотор, активируемый при воздействии на растение различных стрессовых факторов. Дополнительно или альтернативно, промотор может представлять собой тканепредпочтительный промотор, т.е. промотор, способный инициировать транскрипцию преимущественно в определенных тканях, таких как листья, корни, ткани семян и т.д. Промоторы, способные инициировать транскрипцию только в определенных тканях, называются в настоящем описании "тканеспецифичными".
Предпочтительно, нуклеиновая кислота SYT или ее вариант функционально связаны с конститутивным промотором. Конститутивный промотор транскрипционно активен в течение большей части, но необязательно всех, фаз роста и развития растения и экспрессируется по существу повсеместно. Предпочтительно, промотор имеет растительное происхождение, также предпочтительно, из однодольного растения. Наиболее предпочтительным является использование промотора GOS2 (из риса) (SEQ ID NO:89). Следует понимать, что применимость настоящего изобретения не ограничивается нуклеиновой кислотой SYT, представленной SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:7, равно как применимость изобретения не ограничивается экспрессией нуклеиновой кислоты SYT, управляемой промотором GOS2. Примеры других конститутивных промоторов, которые также могут быть использованы для управления экспрессией нуклеиновой кислоты SYT, приведены в таблице 4 ниже.
Необязательно, в конструкции, вводимой в растение, также могут быть использованы одна или несколько терминирующих последовательностей. Термин "терминатор" относится к контрольной последовательности, представляющей собой последовательность ДНК на конце транскриптона, которая дает сигнал на 3'-процессинг и полиаденилирование первичного транскрипта и терминацию транскрипции. Дополнительные регуляторные элементы могут включать в себя энхансеры транскрипции и трансляции. Специалистам в данной области техники известны последовательности терминаторов и энхансеров, которые могут подходить для использования в выполнении изобретения. Такие последовательности известны или могут быть легко получены специалистом в данной области техники.
Генные конструкции согласно изобретению могут дополнительно содержать последовательность инициации репликации, которая необходима для поддержания функции и/или репликации в конкретном типе клеток. Одним примером является ситуация, когда необходимо присутствие генной конструкции в бактериальной стенке в виде эписомного генного элемента (например, плазмидной или космидной молекулы). Предпочтительные последовательности инициации репликации включают в себя без ограничения f1-ori и colE1.
Генная конструкция может необязательно содержать селектируемый маркерный ген. Используемый в настоящем описании термин "селектируемый маркерный ген" означает любой ген, который придает клетке фенотипический признак, который при экспрессии облегчает идентификацию и/или отбор клеток, трансфицированных или трансформированных нуклеотидной конструкцией согласно изобретению. Подходящие маркеры могут быть выбраны из маркеров, которые придают устойчивость к антибиотикам или гербицидам, детерминируют новый метаболический признак или обеспечивают визуальный отбор. Примеры селектируемых маркерных генов включают в себя гены, придающие устойчивость к антибиотикам (такие как nptII, обеспечивающий фосфорилирование неомицина и канамицина, или hpt, обеспечивающий фосфорилирование гигромицина), к гербицидам (например, bar, обеспечивающий устойчивость к Basta; aroA или дох, обеспечивающие устойчивость к глифозату), или гены, детерминирующие новый метаболический признак (такой как manA, который позволяет растениям использовать маннозу в качестве единственного источника углерода). Визуальные маркерные гены приводит к появлению окраски (например, ген β-глюкуронидазы, GUS), люминесценции (такой как ген люциферазы) или флуоресценции (гены зеленого флуоресцентного белка, GFP, и его производных).
Настоящее изобретение также относится к растениям, получаемым в соответствии со способами согласно настоящему изобретению. Таким образом, настоящее изобретение относится к растениям, частям растений и клеткам растений, получаемым в соответствии со способами согласно настоящему изобретению, причем указанные растения содержат введенные в них нуклеиновую кислоту SYT или ее вариант и указанные растения, части растений и клетки растений предпочтительно относятся к сельскохозяйственной культуре, также предпочтительно, к однодольному растению.
Изобретение также относится к способу получения трансгенных растений, характеризующихся повышенной урожайностью, предусматривающему введение и экспрессию растением нуклеиновой кислоты SYT или ее варианта.
Более конкретно, настоящее изобретение также относится к способу получения трансгенных растений, предпочтительно, однодольных растений, характеризующихся повышенной урожайностью, причем указанный способ предусматривает
(i) введение и экспрессию растением или клеткой растения нуклеиновой кислоты SYT или ее варианта;
(ii) культивирование клетки растения в условиях, способствующих росту и развитию растения.
Последующие поколения растений, полученных на стадии культивирования (ii), могут быть размножены многими способами, такими как клональное размножение или методики классического скрещивания. Например, трансгенное растение первого поколения (или Т1) может быть подвергнуто самоопылению с получением гомозиготных трансформантов второго поколения (или Т2), а растения Т2 размножают далее в соответствии с методиками классического скрещивания.
Нуклеиновая кислота может быть введена непосредственно в клетку растения или в само растение (включая введение в ткань, орган или любую другую часть растения). В соответствии с предпочтительным вариантом выполнения настоящего изобретения нуклеиновую кислоту вводят в растение методом трансформации.
Используемый в настоящем описании термин "трансформация" относится к переносу экзогенного полинуклеотида в клетку-хозяина безотносительно к методу переноса. Ткань растения, способная к дальнейшему клональному. размножению путем либо органогенеза, либо эмбриогенеза, может быть трансформирована генной конструкцией согласно настоящему изобретению, а из нее может быть регенерировано целое растение. Выбор конкретной ткани будет зависеть от систем клонального размножения, доступных для конкретного вида трансформируемого растения или наиболее подходящих для него. Примеры тканей-мишеней включают в себя листовые диски, пыльцу, зародыши, семядоли, гипокотили, мегагаметофиты, ткань каллюса, первичная меристематическая ткань (например, верхушечная меристема, пазушные почки и корневые меристемы) и индуцированная меристематическая ткань (например, меристема семядоли и меристема гипокотиля). Полинуклеотид может быть транзиторно или стабильно введен в клетку-хозяина и может находиться в ней без интеграции, например, в виде плазмиды. Альтернативно, он может быть интегрирован в геном хозяина. Полученная трансформированная клетка растения может затем быть использована для регенерации трансформированного растения в соответствии с методикой, известной специалистам в данной области техники.
На сегодняшний день трансформация растения стала абсолютно стандартным методом. Преимущественно, для введения интересующего гена в подходящую клетку-предшественника может быть использована любая из нескольких методик трансформации. Методики трансформации предусматривают использование липосом, электропорации, химических агентов, усиливающих поглощение свободной ДНК, введения ДНК непосредственно в растение, бомбардировки частицами, трансформации с помощью вирусов или пыльцы и микропроекции. Методики могут быть выбраны из метода на основе кальция/полиэтиленгликоля для протопластов (Krens F.A. et al., (1982) Nature 296:72-74; Negrutiu I et al. (1987) Plant Mol. Biol. 8:363-373); элетропорации протопластов (Shillito R.D. et al., 1985 Bio/Technol. 3:1099-1102); микроинъекции в растительный материал (Crossway A. et al., (1986) Mol. Gen. Genet. 202:179-185); бомбардировки покрытыми ДНК или РНК частицами (Klein TM et al., (1987) Nature 327:70), инфекции (неинтегрирующимися) вирусами и т.п. Трансгенные растения риса, экспрессирующие нуклеиновую кислоту/ген SYT, предпочтительно, получают путем опосредованной Agrobacterium трансформации в соответствии с любой из хорошо известных методик трансформации риса, таких как описаны в следующих публикациях: опубликованной заявке на выдачу Европейского патента №ЕР 1198985 A1, Aldemita and Hodges (Planta 199:612-617, 1996); Chan et al. (Plant Mol. Biol. 22 (3):491-506, 1993), Hiei et al. (Plant J. 6(2):271-282, 1994), содержание которых включено в настоящее описание во всей полноте посредством ссылки. Предпочтительной методикой трансформации кукурузы является методика, описанная либо в Ishida et al. (Nat. Biotechnol. 14(6):745-50, 1996), либо Frame et al. (Plant Physiol. 129 (1):13-22, 2002), содержание которых включено в настоящее описание во всей полноте посредством ссылки.
Как правило, после трансформации клетки или группы клеток растения отбирают по наличию одного или нескольких маркеров, кодируемых экспрессируемыми в растениях генами, переносимыми совместно с интересующим геном, с последующей регенерацией трансформированного материала в целое растение.
После переноса ДНК и регенерации предположительно трансформированные растения могут быть проверены, например, методом саузерн-блоттинга, на предмет наличия интересующего гена, числа копий и/или организации генома. Альтернативно или дополнительно, уровни экспрессии вновь введенной ДНК могут отслеживаться с помощью нозерн- и/или вестерн-блоттинга, количественной PCR, причем эти методы хорошо известны средним специалистам в данной области техники.
Полученные трансформированные растения могут быть размножены многими способами, такими как клональное размножение или методики классического скрещивания. Например, транформированное растение первого поколения (или Т1) может быть подвергнуто самоопылению с получением гомозиготных трансформантов второго поколения (или Т2), а растения Т2 размножают далее в соответствии с методиками классического скрещивания.
Полученные трансформированные организмы могут принимать различные формы. Например, они могут представлять собой химеры трансформированных клеток и нетрансформированных клеток; клональные трансформанты (например, все трансформированные клетки, содержащие экспрессирующую кассету); трансплантаты трансформированных или нетрансформированных тканей (например, у растений трансформированный привой, привитый на нетрансформированный подвой).
Настоящее изобретение, очевидно, относится к любым клетке растения или растению, полученным в соответствии с любым из описанных здесь методов, и ко всем частям растения и их частям для вегетативного размножения. Настоящее изобретение дополнительно относится к потомству первичных трансформированных или трансфицированных клетки, ткани, органа или целого растения, полученных в соответствии с любым из указанных выше методов, причем единственным требованием является наличие у потомства таких же генотипических и/или фенотипических характеристик, что и характеристики, проявленные родительским организмом при осуществлении способов согласно изобретению. Изобретение также относится к клеткам-хозяевам, содержащим выделенные нуклеиновую кислоту SYT или ее вариант. Предпочтительными клетками-хозяевами согласно изобретению являются клетки растения. Изобретение также относится к убираемым частям растения, таким как без ограничения семена, листья, плоды, цветки, стебли, корневища, клубни и луковицы. Кроме того, изобретение относится к продуктам, полученным, предпочтительно, непосредственно полученным, из убираемой части такого растения, таким как сухие брикеты или порошки, мука, масло, жир и жирные кислоты, крахмал или белки.
Настоящее изобретение также относится к нуклеиновым кислотам SYT или их вариантам и применению полипептидов SYT или их гомологов или к применению определенной выше конструкции для повышения урожайности растения, особенно семенной продуктивности. Семенная продуктивность определена выше и предпочтительно включает в себя повышенную общую семенную продуктивность или повышенную TKW.
Нуклеиновые кислоты SYT или их варианты или полипептиды SYT или их гомологи могут находить применение в программах скрещивания, в соответствии с которыми идентифицируют ДНК-маркер, который может быть генетически связан с геном SYT или его вариантом. Нуклеиновые кислоты/гены SYT или их варианты или полипептиды SYT или их гомологи могут быть использованы для определения молекулярного маркера. Этот ДНК- или белковый маркер может быть затем использован в программах скрещивания для отбора растений, характеризующихся повышенной урожайностью. Ген SYT или его вариант могут, например, представлять собой нуклеиновую кислоту, представленную любой из SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:15, SEQ ID NO:17, SEQ ID NO:19, SEQ ID NO:21, SEQ ID NO:23, SEQ ID NO:25, SEQ ID NO:27, SEQ ID NO:29, SEQ ID NO:31, SEQ ID NO:33, SEQ ID NO:35, SEQ ID NO:37, SEQ ID NO:39, SEQ ID NO:41, SEQ ID NO:43, SEQ ID NO:45, SEQ ID NO:47, SEQ ID NO:49, SEQ ID NO:51, SEQ ID NO:53, SEQ ID NO:55, SEQ ID NO:57, SEQ ID NO:59, SEQ ID NO:61, SEQ ID NO:63, SEQ ID NO:65, SEQ ID NO:67, SEQ ID NO:69, SEQ ID NO:71, SEQ ID NO:73, SEQ ID NO:75, SEQ ID NO:77, SEQ ID NO:79, SEQ ID NO:81, SEQ-ID NO:83, SEQ ID NO:85 и SEQ ID NO:87.
Аллельные варианты нуклеиновой кислоты/гена SYT также могут находить применение в основанных на использовании маркеров программах скрещивания. Для проведения таких программ скрещивания иногда требуется введение аллельной вариации путем мутагенной обработки растений с использованием, например, EMS-мутагенеза; альтернативно, проведение программы может начинаться со сбора непроизвольно возникших аллельных вариантов так называемого "природного" происхождения. Затем проводят идентификацию аллельных вариантов, например, с использованием PCR. Далее следует стадия отбора лучших аллельных вариантов интересующей последовательности, которые позволяют достигать повышенной урожайности. Отбор, как правило, проводят путем мониторинга эффективности роста растений, содержащих различные аллельные варианты интересующей последовательности, например, различных аллельных вариантов любой из SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:15, SEQ ID NO:17, SEQ ID NO:19, SEQ ID NO:21, SEQ ID NO:23, SEQ ID NO:25, SEQ ID NO:27, SEQ ID NO:29, SEQ ID NO:31, SEQ ID NO:33, SEQ ID NO:35, SEQ ID NO:37, SEQ ID NO:39, SEQ ID NO:41, SEQ ID NO:43, SEQ ID NO:45, SEQ ID NO:47, SEQ ID NO:49, SEQ ID NO:51, SEQ ID NO:53, SEQ ID NO:55, SEQ ID NO:57, SEQ ID NO:59, SEQ ID NO:61, SEQ ID NO:63, SEQ ID NO:65, SEQ ID NO:67, SEQ ID NO:69, SEQ ID NO:71, SEQ ID NO:73, SEQ ID NO:75, SEQ ID NO:77, SEQ ID NO:79, SEQ ID NO:81, SEQ ID NO:83, SEQ ID NO:85 и SEQ ID NO:87. Эффективность роста можно отслеживать в теплице или в поле. Дополнительные необязательные стадии включают в себя скрещивание растения, в котором был выявлен лучший аллельный вариант, с другим растением. Это может быть использовано, например, для получения сочетания интересующих фенотипических признаков.
Нуклеиновая кислота SYT или ее вариант также могут быть использованы в качестве зондов для генетического и физического картирования генов, в состав которых они входят, и в качестве маркеров признаков, сцепленных с этими генами. Такая информация может быть полезна в селекции растений для получения линий с желаемыми фенотипами. Для такого использования нуклеиновых кислот SYT или их вариантов требуется лишь последовательность нуклеиновой кислоты, состоящая, по меньшей мере, из 15 нуклеотидов. Нуклеиновые кислоты SYT или их варианты также могут быть использованы в качестве маркеров полиморфизма длин фрагментов рестрикции (RFLP). Нуклеиновые кислоты SYT или их варианты также могут выступать в качестве зондов для саузерн-блотов (Sambrook J, Fritsch EF and Maniatis Т (1989) Molecular Cloning, A Laboratory Manual) рестрицированной геномной ДНК растений. Полученные профили формирования полос могут затем быть подвергнуты генетическим анализам с использованием компьютерных программ, таких как MapMaker (Lander et al. (1987) Genomics 1:174-181), для конструирования генетической карты. Кроме того, нуклеиновые кислоты могут быть использованы в качестве зондов для саузерн-блотов, содержащих обработанные рестриктазами геномные ДНК группы организмов, представляющих родителей и потомков относительно определенного генетического гибрида. Расщепление полиморфизмов ДНК регистрируют и используют для расчета положения нуклеиновой кислоты SYT или ее варианта на генетической карте, предварительно определенной для этой популяции (Botstein et al. (1980) Am. J. Hum. Genet. 32:314-331).
Получение и применение происходящих из генов растений зондов для использования в генетическом картировании описано в Bematzky and Tanksley (1986) Plant Mol. Biol. Reporter 4:37-41. Во множестве публикаций описано генетическое картирование клонов конкретных кДНК в соответствии с упомянутой выше методикой и ее вариантами. Например, для картирования могут быть использованы F2 популяции перекрестного скрещивания, популяции межгибридного скрещивания, популяции обратного скрещивания, популяции случайного скрещивания, околоизогенные линии и другие группы организмов. Такие методики хорошо известны специалистам в данной области техники.
Нуклеотидные зонды также могут быть использованы для физического картирования (т.е. размещения последовательностей на физических картах; см. Hoheisel et al. In: Non-mammalian Genomic Analysis: A Practical Guide, Academic press 1996, pp.319-346 и процитированные там ссылки).
В другом варианте выполнения нуклеотидные зонды могут быть использованы в картировании методом прямой флуоресцентной гибридизации in situ (FISH) (Trask (1991) Trends Genet. 7:149-154). Хотя современные методики FISH-картирования предусматривают использование больших клонов (от нескольких т.п.н. до нескольких сотен т.п.н.; см. Laan et al. (1995) Genome Res. 5:13-20), повышение чувствительности может обеспечить эффективность FISH-картирования при использовании более коротких зондов.
С использованием нуклеиновых кислот может быть осуществлено множество методов генетического или физического картирования на основе амплификации нуклеиновых кислот. Примеры включают в себя аллель специфичную амплификацию (Kazazian (1989) J. Lab. Clin. Med. 11:95-96), полиморфизм PCR-амплифицированных фрагментов (CAPS; Sheffield et al. (1993) Genomics 16:325-332), аллельспецифичное лигирование (Landegren et al. (1988) Science 241:1077-1080), реакции удлинения полинуклеотидов (Sokolov (1990) Nucleic Acid Res. 18:3671), картирование с помощью облученных гибридов (Walter et al. (1997) Nat. Genet. 7:22-28) и Happy Mapping (Dear and Cook (1989) Nucleic Acid Res. 17:6795-6807). В этих методах последовательность нуклеиновой кислоты используют для конструирования и получения пар праймеров для использования в реакции амплификации или реакциях удлинения праймера. Конструирование таких праймеров хорошо известно специалистам в данной области техники. Для осуществления методов, в которых используется генетическое картирование на основе PCR, может потребоваться идентифицировать различия последовательностей ДНК между родителями гибрида в области, соответствующей настоящей последовательности нуклеиновой кислоты. Однако, как правило, для осуществления методов картирования этого не требуется.
Способы согласно настоящему изобретению позволяют получить растения, характеризующиеся повышенной урожайностью, как описано выше. Эти признаки повышения урожайности также могут быть объединены с другими экономически выгодными признаками, такими как дополнительные признаки повышения урожайности, переносимость различных стрессов, признаки изменения различных свойств строения и/или биохимических и/или физиологических свойств.
Описание чертежей
Настоящее изобретение теперь будет описано со ссылками на следующие фигуры.
На фиг.1 показана типичная доменная структура полипептидов SYT растений и млекопитающих. Консервативный домен SNH расположен на N-конце белка. Оставшаяся С-концевая часть домена белка состоит из богатого QG домена в полипептидах SYT растений и из богатого QPGY домена в полипептидах SYT млекопитающих. Богатый Met домен, как правило, располагается в первой половине богатого QG домена (в направлении от N-конца к С-концу) в растениях или богатого QPGY домена у млекопитающих. Второй богатый Met домен может находиться перед доменом SNH в полипептидах SYT растений.
На фиг.2 проиллюстрировано множественное выравнивание N-конца нескольких полипептидов SYT с помощью программы множественного выравнивания VNTI AlignX на основе модифицированного алгоритма ClustalW (InforMax, Bethesda, MD, http://www.informaxinc.com) с использованием настроек по умолчанию, представляющих собой штраф за открытие пропуска 10 и продление пропуска 0,05). Домен SNH выделен рамкой среди полипептидов SYT растений и человека. В последней строке выравнивания приведена консенсусная последовательность, полученная из выравниваемых последовательностей.
На фиг.3 проиллюстрировано множественное выравнивание нескольких полипептидов SYT растений с помощью программы множественного выравнивания VNTI AlignX на основе модифицированного алгоритма ClustalW (InforMax, Bethesda, MD, http://www.informaxinc.com) с использованием настроек по умолчанию, представляющих собой штраф за открытие пропуска 10 и продление пропуска 0,05). Два основных домена, в направлении от N-конца к С-концу, выделены рамкой и идентифицированы как домен SNH и богатый Met/богатый QG домен. Кроме того, также выделен рамкой N-концевой богатый Met домен, а положения SEQ ID NO:90 и SEQ ID NO:91 подчеркнуты толстой линией.
На фиг.4 показано дерево, построенное методом присоединения соседа на основании выравнивания множества полипептидов SYT с помощью CLUSTALW 1.83 (http://align.genome.jp/sit-bin/clustalw). Ветви SYT1 и SYT2/SYT3 выделены скобками.
На фиг.5 показан бинарный вектор р0523 для экспрессии AtSYT1 Arabidopsis thaliana в Oryza sativa под контролем промотора GOS2 (внутренняя ссылка PR00129).
На фиг.6 показан бинарный вектор р0524 для экспрессии AtSYT2 Arabidopsis thaliana в Oryza sativa под контролем промотора GOS2 (внутренняя ссылка PRO0129).
На фиг.7 показан бинарный вектор р0767 для экспрессии AtSYT3 Arabidopsis thaliana в Oryza sativa под контролем промотора GOS2 (внутренняя ссылка PRO0129).
На фиг.8 приведены примеры последовательностей, применимых для осуществления способов согласно настоящему изобретению. Последовательности нуклеиновых кислот SYT приведены от точки инициации транскрипции до точки терминации транскрипции. Большинство этих последовательностей получено путем секвенирования EST более низкого качества. Таким образом, в нуклеиновых кислотах могут встречаться замены.
Примеры
Настоящее изобретение теперь будет описано со ссылкой на следующие примеры, которые приведены исключительно в иллюстративных целях.
Манипуляции ДНК: если иное не указано особо, методики рекомбинантных ДНК осуществляют в соответствии со стандартными протоколами, описанными в (Sambrook (2001) Molecular Cloning: A Laboratory Manual, 3rd Edition Cold Spring Harbor Laboratory Press, CSH, New York) или в томах 1 и 2 Ausubel et al. (1994), Current Protocols in Molecular Biology, Current Protocols. Стандартные материалы и методы для работы с молекулами растительного происхождения описаны в Plant Molecular Biology Labfase (1993) R.D.D. Croy, опубликованной BIOS Scientific Publications Ltd. (UK) и Blackwell Scientific Publications (UK).
Пример 1: Клонирование генов AtSYT1, AtSYT2 и AtSYT3
Ген AtSYT1 Arabidopsis thaliana амплифицировали по методу PCR с использованием в качестве матрицы библиотеку кДНК всходов Arabidopsis thaliana (Invitrogen, Paisley, UK). После обратной транскрипции РНК, выделенной из всходов, кДНК клонировали в pCMV Sport 6.0. Средний размер вставки из банка составлял 1,5 т.п.н., а исходное количество клонов составляло порядка 1,59×107 к.о.е. Исходный титр определяли на уровне 9,6×105 к.о.е./мл после первой амплификации 6×1011 к.о.е./мл. После выделения плазмиды 200 нг матрицы использовали в 50 мкл PCR-смеси. Для PCR-амплификации использовали праймеры prm06681 (SEQ ID NO:92; смысловой, стартовый кодон указанным жирным шрифтом, сайт AttB1 указан курсивом: 5'-

которые содержали сайты AttB для рекомбинации Gateway. PCR проводили с использованием ДНК-полимеразы Hifi Taq в стандартных условиях. PCR-фрагмент длиной 727 п.н. (включая сайты attB) амплифицировали и очищали также в соответствии со стандартными методами. Затем проводили первую стадию методики Gateway, реакцию ВР, в ходе которой PCR-фрагмент рекомбинировал in vivo с плазмидой pDONR201 с получением в соответствии с терминологией Gateway, "вводного клона", р07466. Плазмиду pDONR201 приобретали у Invitrogen в виде части набора для проведения методики Gateway®.
Ген AtSYT2 Arabidopsis thaliana амплифицировали путем PCR в соответствии с той же методикой, по которой амплифицировали ген AtSYT1 Arabidopsis thaliana. Для PCR-амплификации использовали праймеры prm06685 (SEQ ID NO:94; смысловой,

содержали сайты AttB для рекомбинации Gateway. PCR проводили с использованием ДНК-полимеразы Hifi Taq в стандартных условиях. PCR-фрагмент длиной 66,6 п.н. (включая сайты attB) амплифицировали и очищали, как описано выше. Вводному клону присваивали номер р07467.
Ген AtSYT3 Arabldopsis thaliana амплифицировали путем PCR в соответствии с той же методикой, по которой амплифицировали гены AtSYT1 и AtSYT2 Arabidopsis thaliana. Для PCR-амплификации использовали праймеры prm06683 (SEQ ID NO:96; смысловой, стартовый кодон указанным жирным шрифтом, сайт AttB1 указан
сайты AttB для рекомбинации Gateway. PCR проводили с использованием ДНК-полимеразы Hifi Taq в стандартных условиях. PCR-фрагмент длиной 745 п.н. (включая сайты attB) амплифицировали и очищали, как описано выше. Вводному клону присваивали номер р07604.
Пример 2: Конструирование векторов
Вводные клоны р07466, р07467 и р07604 затем использовали в LR-реакции с р00640, вектором назначения, используемым для трансформации Oryza sativa. Этот вектор содержал в качестве функциональных элементов в пределах Т-ДНК растительный селектируемый маркер; экспрессирующую кассету скринируемого маркера; и кассету Gateway, предназначенную для in vivo LR-рекомбинации с интересующей последовательностью, уже клонированной в вводный клон. Промотор GOS2 риса (SEQ ID NO:89) для конститутивной экспрессии (PRO0129) располагался левее этой кассеты Gateway.
После стадии LR-рекомбинации полученными экспрессирующими векторами, соответственно р0523 для AtSYT1, p0524 для AtSYT2 и р0767 для AtSYT3 (фиг.5-7), трансформировали штамм Agrobacterium LBA4044 и далее растения Oryza saliva. Трансформированным растениям риса давали вырасти и затем измеряли параметры, описанные в примере 3.
Пример 3: Оценка, и результаты функция AtSYT1, AtSYT2 и AtSYT3 под контролем промотора GOS2 ряса.
Получали приблизительно от 15 до 20 независимых трансформантов риса Т0. Первичные трансформанты переносили из камеры для культивирования тканей в теплицу для роста и сбора семян Т1. По шести событиям относительно присутствия/отсутствия трансгена потомство Т1 расщеплялось 3:1. Для каждого из этих событий путем мониторинга экспрессии визуального маркера отбирали примерно 10 ростков Т1, содержащих трансген (гетеро- и гомозиготы) и примерно 10 ростков Т1, не содержащих трансген (нуль-зиготы).
Статистический анализ: F-тест
В качестве статистической модели для общей оценки фенотипических характеристик растений использовали двухфакторный ANOVA (анализ вариантов). F-тест проводили по всем параметрам всех событий, измеренным у всех растений, трансформированных геном согласно настоящему изобретению. F-тест проводили для проверки эффекта гена при каждом событии трансформации и общего эффекта гена, также известного как глобальный эффект гена. Порог значимости для истинного глобального эффекта гена для F-теста устанавливали на уровне 5% вероятности. Значимое значение F-теста указывает на эффект гена, что означает, что фенотипические различия вызывают не только наличие или положение гена.
Измерение относящихся к семенам параметров
Зрелые первичные метелки убирали, фасовали в мешки, метили штрихкодом и затем сушили в течение трех суток в печи при 37°С. Затем метелки обмолачивали и все семена собирали и подсчитывали. Налитые семена отделяли от пустой шелухи с помощью устройства для продувки воздухом. Пустую шелуху отбрасывали, а оставшуюся фракцию пересчитывали. Налитые семена взвешивали на аналитических весах. Количество налитых семян определяли путем подсчета количества налитых семян, оставшихся после стадии разделения. Общую семенную продуктивность измеряли путем взвешивания всех налитых семян, собранных с растения. Общую семенную продуктивность на каждое растение измеряли путем подсчета количества семян, собранных с растения. Массу тысячи зерен (TKW) экстраполировали из подсчитанного количества налитых семян и их общей массы.
Параметры отдельных семян (включая ширину, длину, площадь поверхности, массу) измеряли с помощью изготовленного по заказу устройства, состоящего из двух основных компонентов, взвешивающего и визуализирующего устройств, сопряженного с программным обеспечением для анализа изображения.
3.1 Результаты измерения общей семенной продуктивности и TKW для выращенных в теплице трансгенных растений
Результаты измерения общей семенной продуктивности и TKW для несущих трансгены AtSYT1, AtSYT2 и AtSYT3 растений в поколении Т1 приведены соответственно в таблицах 5-7. Указано количество линий, характеризующихся увеличением какого-либо параметра. Также показана процентная разница между трансгенными растениями и соответствующими нуль-зиготами, равно как и значения Р F-теста.
Как общая семенная продуктивность, так и TKW значительно возросли в поколении Т1 несущих трансгены AtSYT1, AtSYT2 и ATSYT3 растений (таблицы 5-7, соответственно).
3.2 Результаты измерении размеров семян поколения Т2 несущих трансген AtSYT1 растении
Параметры отдельных семян (ширину, длину и площадь поверхности) измеряли у семян поколения Т2 с помощью изготовленного по заказу устройства, состоящего из двух основных компонентов, взвешивающего и визуализирующего устройств, сопряженного с программным обеспечением для анализа изображения. Проводили измерения как необрушенных, так и обрушенных семян.
Средние результаты измерения площади поверхности, длины и ширины отдельных семян Т3 (собранных с растений Т2) несущих трансген AtSYT1 растений Oryza sativa приведены в таблице 8. Показана процентная разница между трансгенными растениями и соответствующими нуль-зиготами, равно как и количество событий увеличения данного параметра и значения Р F-теста.
Средние площадь поверхности, длина и ширина отдельных как необрушенных, так и обрушенных семян Т3 (собранных с несущих трансген AtSYT1 растений Oryza sativa Т2) были значительно увеличены по сравнению с соответствующими нуль-зиготами (таблица 8).
3.3 Результата намерения размера зародыша, и эндосперма семян поколения Т2 несущих трансген AtSYT1 растений
Размер зародыша и эндосперма также измеряли путем продольного рассечения обрушенных семян напополам и окрашивания половин семян в течение 2-3 часов при 35°С красителем, хлоридом 2,3,5-трифенилтетразолия. После окрашивания две половины помещали на агарозный гель в чашке Петри, готовой к визуализации. Принимались во внимание три независимые события, и для каждого события анализировали 120 семян, гомозиготных по трансгену, и 120 семян без трансгена. Делали цифровые фотографии семян и изображения анализировали с помощью программного обеспечения ImagePro. Результаты исследования трех событий приведены ниже.
Для всех трех событий зародыши семян, гомозиготных по трансгену, были крупнее, чем зародыши семян без трансгена. Отмечалось значительное увеличение средней площади поверхности зародыша семян для всех трех событий при значениях p t-теста, составляющих 0,0325, <0,0001 и <0,0001. Аналогично, наблюдалось значительное увеличение среднего периметра зародыша семян для всех трех событий при значениях p t-теста, составляющих 0,0176, <0,0001 и <0,0001. Кроме того, отмечалось значительное увеличение средней площади поверхности и периметра эндосперма семян для всех трех событий при значении p во всех трех случаях, составляющем <0,0001.
3.4 Результаты измерения TKW для несущих трансген AtSYT1 растений, выращенных в поле
Гомозиготные по AtSYT1 трансгенные растения и соответствующие контрольные растения пересаживали в поле в сентябре и убирали в декабре. По каждому вводу высаживали четыре повторения (четыре события) при высаживании 104 растений в каждом повторении. Расстояние между растениями составляло 20 на 20 см. Поле затапливали и орошали. После уборки семян их TKW измеряли, как описано выше. Результаты этих измерений представлены в таблице 9.
TKW увеличивалось при всех трансгенных событиях, оцененных в поле.
| название | год | авторы | номер документа |
|---|---|---|---|
| РАСТЕНИЯ С ПОВЫШЕННОЙ УРОЖАЙНОСТЬЮ И СПОСОБ ИХ ПОЛУЧЕНИЯ | 2005 |
|
RU2384621C2 |
| РАСТЕНИЯ, ИМЕЮЩИЕ УСИЛЕННЫЕ ПРИЗНАКИ, СВЯЗАННЫЕ С УРОЖАЙНОСТЬЮ, И СПОСОБ ИХ ПОЛУЧЕНИЯ | 2008 |
|
RU2503721C2 |
| СПОСОБЫ УВЕЛИЧЕНИЯ УРОЖАЯ И СТРЕССОУСТОЙЧИВОСТИ У РАСТЕНИЯ | 2012 |
|
RU2632569C2 |
| ИНСЕКТИЦИДНЫЕ БЕЛКИ | 2017 |
|
RU2765722C2 |
| ПРОМОТОР ЦЕЛОГО СЕМЕНИ | 2010 |
|
RU2561463C2 |
| НОВЫЕ БЕЛКИ, ПРОЯВЛЯЮЩИЕ ИНГИБИРУЮЩУЮ АКТИВНОСТЬ ПО ОТНОШЕНИЮ К НАСЕКОМЫМ | 2016 |
|
RU2765310C2 |
| СКОНСТРУИРОВАННЫЕ ПЕСТИЦИДНЫЕ БЕЛКИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2017 |
|
RU2772947C2 |
| РАСТЕНИЯ С ИЗМЕНЕННЫМ РОСТОМ И/ИЛИ РАЗВИТИЕМ И СПОСОБ ИХ СОЗДАНИЯ | 2009 |
|
RU2522480C2 |
| ГЕН АХМ1554, КОДИРУЮЩИЙ ДЕЛЬТА-ЭНДОТОКСИН, И СПОСОБЫ ЕГО ПРИМЕНЕНИЯ | 2016 |
|
RU2755282C2 |
| ИНСЕКТИЦИДНЫЕ БЕЛКИ | 2017 |
|
RU2765304C2 |






























































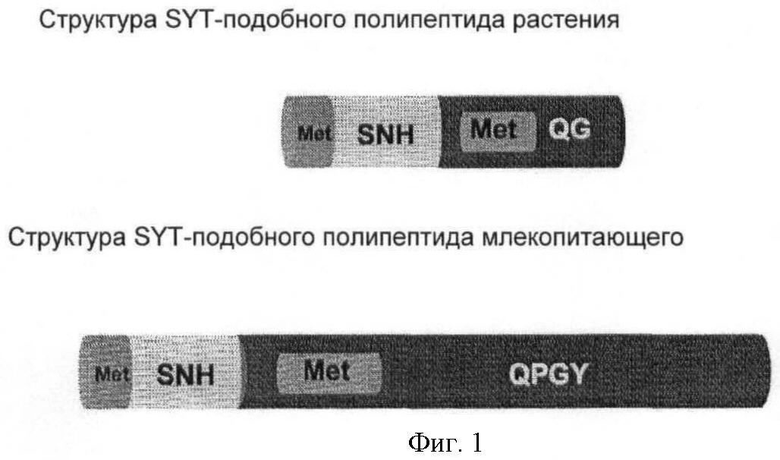

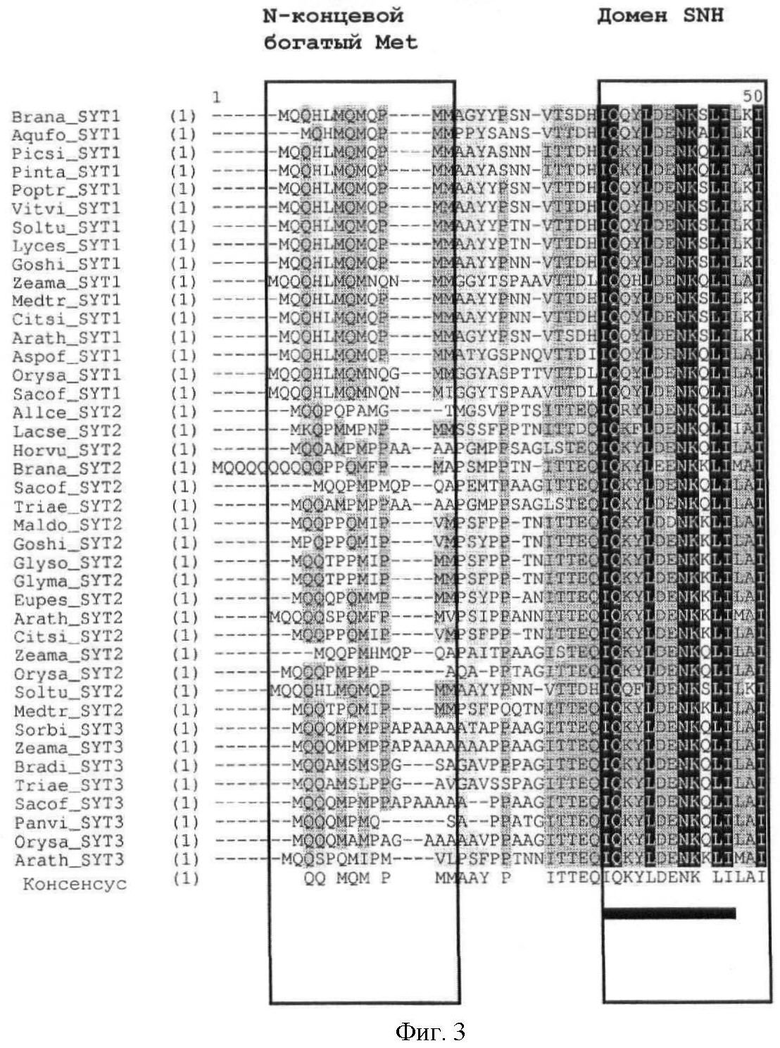
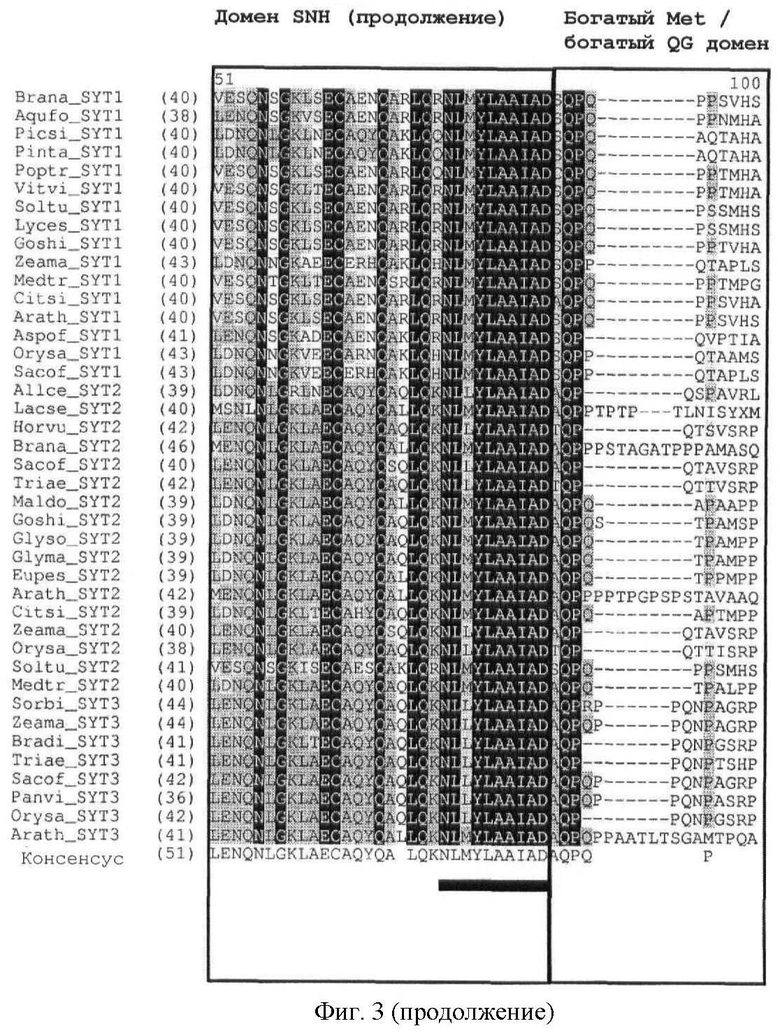
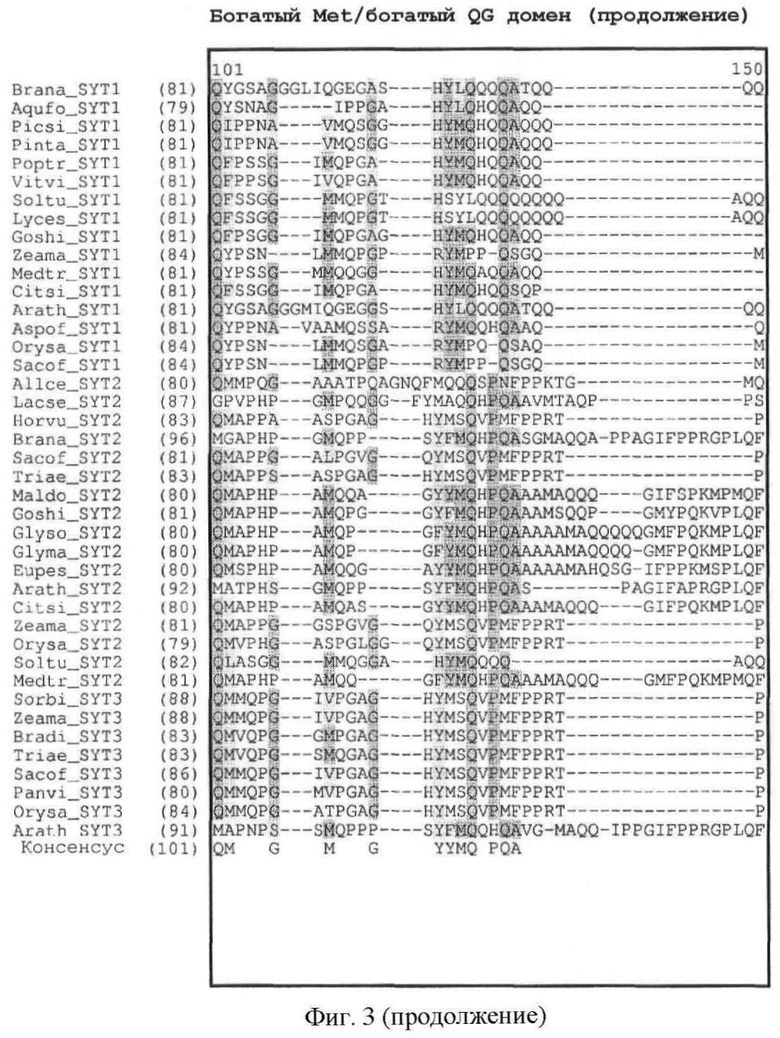
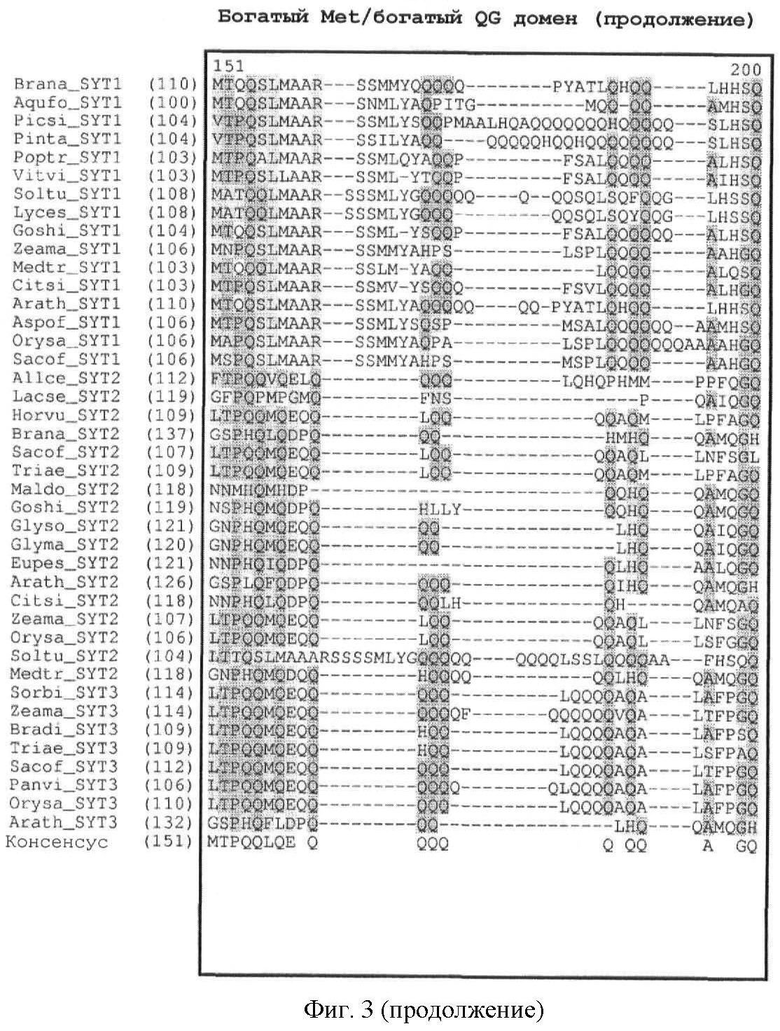

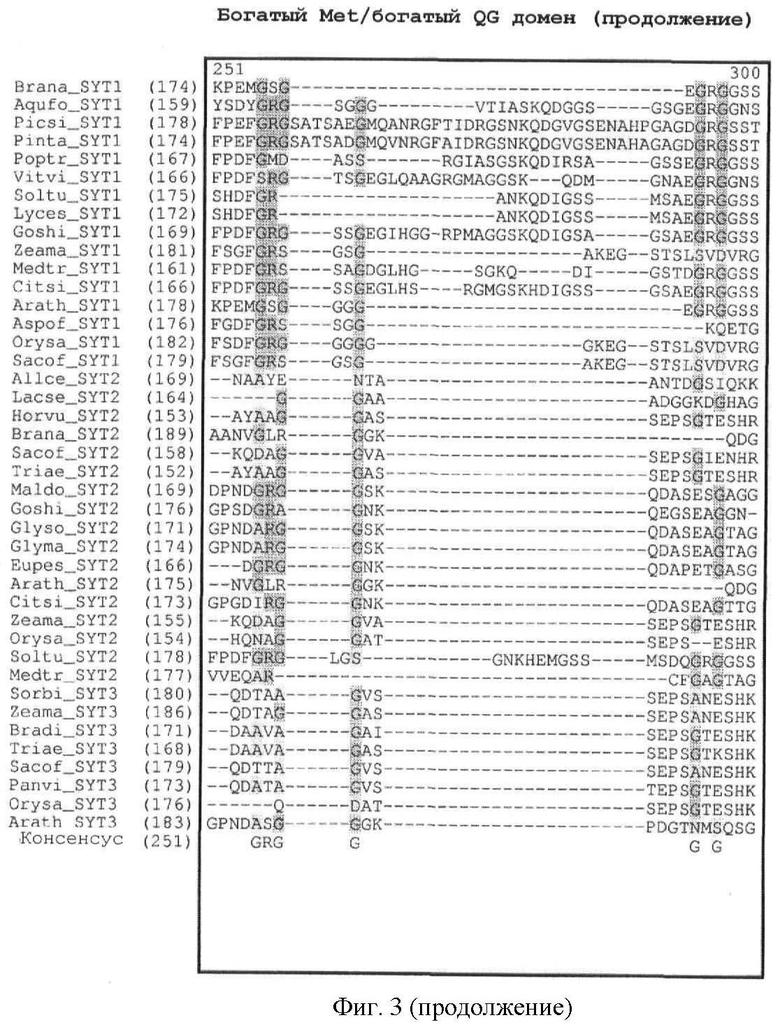
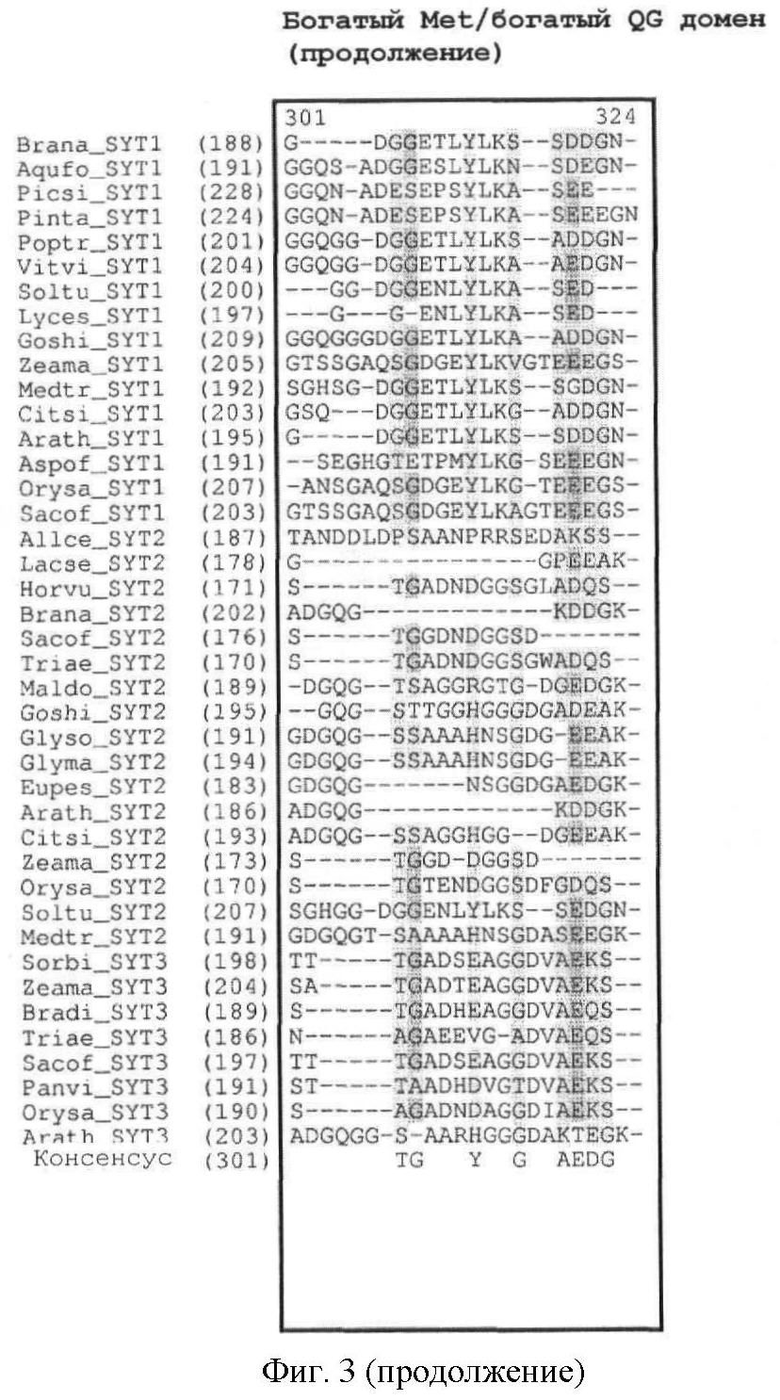
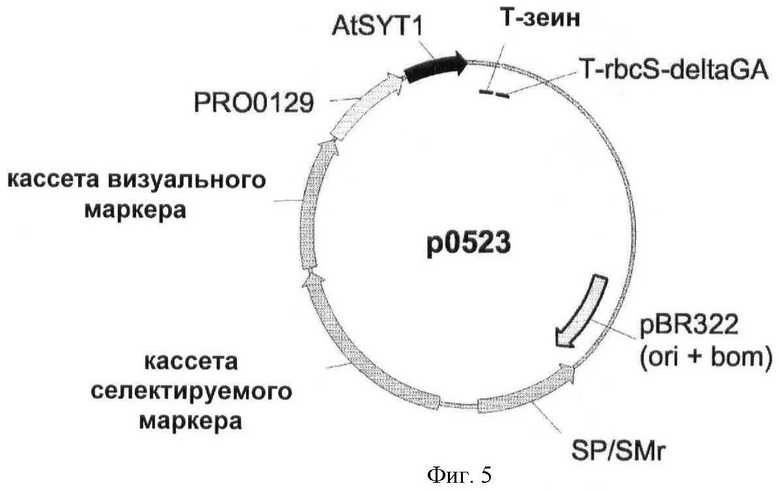

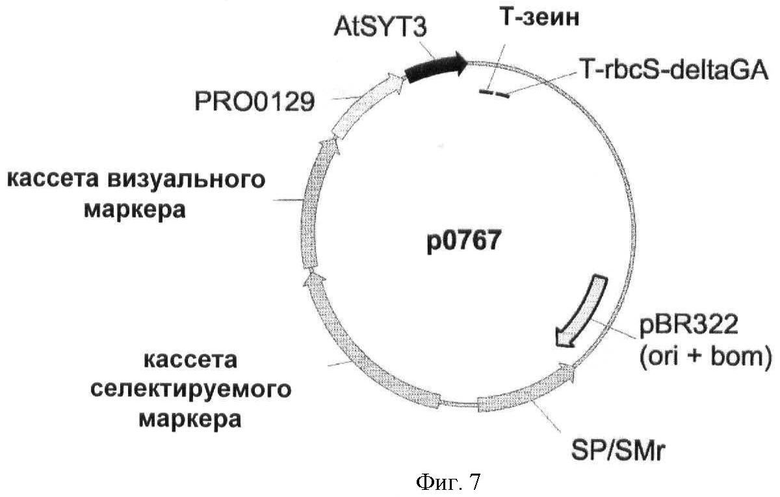























Настоящее изобретение относится к способу повышения семенной продуктивности и/или биомассы корня в растениях по сравнению с таковой в растениях дикого типа, путем трансформирования растения конструкцией на основе нуклеиновой кислоты, кодирующей полипептид транслокации синовиальной саркомы (SYT) или ее гомолога с соответствующей структурой. Описанная конструкция содержит SYT, который содержит от N-конца к С-концу домен SNH, богатый Met домен, богатый QG домен. Раскрыто трансгенное растение, несущее такую конструкцию и его соответствующая часть. Использование изобретения обеспечивает растения, характеризующиеся повышенной урожайностью по сравнению с таковой соответствующих растений дикого типа и может найти применение в сельском хозяйстве. 4 н. и 8 з.п. ф-лы, 8 ил., 9 табл., 3 пр.
1. Способ повышения одного или нескольких признаков, таких как семенная продуктивность и биомасса корня, в растениях по сравнению с таковой соответствующих растений дикого типа, предусматривающий трансформирование растения конструкцией, содержащей
(а) нуклеиновую кислоту SYT или ее вариант, кодирующую полипептид транслокации синовиальной саркомы (SYT) или его гомолог, причем указанные полипептид SYT или гомолог содержат в направлении от N-конца к С-концу: (i) домен SNH, характеризующийся, по меньшей мере, 40% идентичностью последовательности по отношению к домену SNH SEQ ID NO:2, содержащий остатки, показанные черным на фиг.2; и (ii) богатый Met домен; и (iii) богатый QG домен;
(б) одну или несколько контрольных последовательностей, способных управлять экспрессией последовательности нуклеиновой кислоты (а) и представляющих собой конститутивные промоторы; и необязательно
(с) последовательность терминации транскрипции.
2. Способ по п.1, предусматривающий отбор растений, характеризующихся повышенной урожайностью.
3. Способ по п.1, в котором указанный домен SNH представлен SEQ ID NO:1.
4. Способ по п.1, в котором указанный полипептид SYT или его гомолог дополнительно содержат богатый Met домен на N-конце перед доменом SNH.
5. Конструкция для трансформирования растения, содержащая
(а) нуклеиновую кислоту SYT или ее вариант, кодирующую полипептид транслокации синовиальной саркомы (SYT) или его гомолог, причем указанные полипептид SYT или гомолог содержат в направлении от N-конца к С-концу: (i) домен SNH, характеризующийся, по меньшей мере, 40% идентичностью последовательности по отношению к домену SNH SEQ ID NO:2, содержащий остатки, показанные черным на фиг.2; и (ii) богатый Met домен; и (iii) богатый QG домен;
(б) одну или несколько контрольных последовательностей, способных управлять экспрессией последовательности нуклеиновой кислоты (а) и представляющих собой конститутивные промоторы; и необязательно
(с) последовательность терминации транскрипции.
6. Конструкция по п.5, в которой указанная контрольная последовательность, представляющая собой конститутивный промотор, представляет собой конститутивный промотор, полученный из однодольного растения.
7. Конструкция по п.6, в которой указанный конститутивный промотор представляет собой промотор GOS2.
8. Конструкция по п.7, в которой указанный промотор GOS2 представлен SEQ ID NO:89.
9. Трансгенное растение, характеризующееся повышением одного или нескольких признаков, таких как семенная продуктивность и биомасса корня, в результате введения в указанное растение конструкта по п.5, причем указанная урожайность повышена по сравнению с таковой соответствующих растений дикого типа.
10. Трансгенное растение по п.9, причем указанное растение представляет собой однодольное растение, такое как сахарный тростник, или растение представляет собой зерновую культуру, такую как рис, кукуруза, пшеница, ячмень, просо, рожь, овес или сорго.
11. Часть трансгенного растения, характеризующаяся повышением одного или нескольких признаков, таких как семенная продуктивность и биомасса корня, в результате введения в указанную часть растения конструкта по п.5, причем указанная урожайность повышена по сравнению с таковой соответствующих растений дикого типа.
12. Часть трансгенного растения по п.11, представляющая собой семя.
| KIM et al., "The AtGRF family of putative transcription factors is involved in leaf and cotyledon growth in Arabidopsis", The Plant Journal, 2003, 36, стр.94-104 | |||
| KIM et al., "A transcriptional coactivator, AtGIF1, is involved in regulating leaf growth and morphology in Arabidopsis", Proc | |||
| Nat | |||
| Acad | |||
| Sc., 2004, vol.101, no.36, с.13374-13379 | |||
| СПОСОБ УВЕЛИЧЕНИЯ УРОЖАЙНОСТИ РАСТЕНИЯ, СПОСОБ КОНТРОЛИРОВАНИЯ РЕЖИМА ЦВЕТЕНИЯ РАСТЕНИЯ И КОНСТРУКЦИЯ ДНК ДЛЯ ИХ ОСУЩЕСТВЛЕНИЯ | 1998 |
|
RU2241753C2 |
| JP 2004350553 A, 16.12.2004. | |||

