Настоящее изобретение относится, в общем, к молекулярной биологии и касается способа повышения урожайности растений. Более конкретно, настоящее изобретение касается способа повышения урожайности растений, особенно урожайности семян, путем введения в растение нуклеиновой кислоты, кодирующей циклин-зависимую киназу D-типа (CDKD - D-type Cyclin-Dependent Kinase). Настоящее изобретение также касается растений, полученных способами по изобретению, при этом растения имеют повышенную урожайность по сравнению с соответствующими растениями дикого типа. Изобретение также касается конструкций, используемых в способах по изобретению.
Постоянно растущее народонаселение в мире и уменьшение пахотных земель, пригодных для сельскохозяйственных культур, стимулирует исследования, направленные на улучшение эффективности сельского хозяйства. Обычными средствами улучшения сельскохозяйственных и садовых культур является использование методик селекционного разведения для идентификации растений, имеющих желаемые свойства. Однако такие методики селекционного разведения имеют различные недостатки, а именно, эти методики обычно трудоемкие, и полученные в результате растения часто содержат гетерогенные генетические компоненты, которые не всегда дают в результате желаемые черты при переходе из родительских растений. Успехи в молекулярной биологии позволили человечеству модифицировать гермаплазму животных и растений. Генная инженерия растений включает выделение и манипуляции с генетическим материалом (обычно в форме ДНК или РНК) и последующее введение этого генетического материала в растение. Такая технология привела к разработке растений, имеющих различные улучшенные экономические, агрономические или садоводческие признаки. Признаком, особенно экономически интересным, является урожайность. Урожайность обычно определяют как измеримое получение экономического объема из культуры. Она может быть определена в терминах количества и/или качества. Урожайность прямо зависит от различных факторов, например числа и размера органов, строения растения (например, числа веток), выработки семян и т.д. Развитие корней, поглощение питательных веществ и устойчивость к стрессам также являются важными факторами, определяющими урожайность. Урожайность сельскохозяйственных культур может быть путем оптимизации одного из вышеперечисленных факторов, что может быть сделано путем модификации врожденных механизмов роста растений.
Врожденные механизмы роста растений неотъемлемы от высокоупорядоченной последовательности событий, вместе известных как «клеточный цикл». Продвижение через клеточный цикл является фундаментальным для роста и развития всех многоклеточных организмов и является ключевым в пролиферации клетки. Главные компоненты клеточного цикла хорошо сохраняются в дрожжах, млекопитающих и растениях. Клеточный цикл обычно делится на следующие последовательные фазы: G0 - G1 - S - G2 - М. Репликация ДНК или синтез обычно протекают во время S фазы («S» - для синтеза ДНК), а митотическая сегрегация хромосом имеет место во время М фазы («М» - для митоза), с промежуточными гэп фазами, G1 (во время которого клетки растут перед репликацией ДНК) и G2 (период после репликации ДНК, во время которого клетка готовится к делению). Деление клетки завершается после цитокинеза, последнего этапа М фазы. Клетки, которые вышли из клеточного цикла и которые становятся покоящимися, находятся в вышеуказанной G0 фазе. Клетки в этой фазе могут быть стимулированы, чтобы начать клеточный цикл на G1 фазе. "G" в G1, G2 и G0 положения для "гэп". Завершение процесса клеточного цикла позволяет каждой дочерней клетке во время деления клетки получить полную копию родительского генома.
Деление клетки контролируют два основных события клеточного цикла, а именно инициация синтеза ДНК и инициация митоза. Каждый переход к каждому из этих ключевых событий контролируется с помощью контрольных точек (реперов), представленных специфическими белковыми комплексами (вовлеченными в репликацию ДНК и деление). Экспрессия генов, необходимая для синтеза ДНК на границе G1/ S, регулируется с помощью E2F семейства транскрипционных факторов в клетках млекопитающих и растительных клетках (La Thangue, 1994; Muller et al., 2001; De Veylder et al., 2002). Вступление в клеточный цикл регулируется/инициируется с помощью E2F/Rb комплекса, который объединяет сигналы и разрешает активацию транскрипции генов клеточного цикла. Переход между различными фазами клеточного цикла и, следовательно, продвижение через клеточный цикл стимулируется путем формирования и активации различных гетеродимерных серин/треонин белковых киназ, главным образом, относящихся к циклин-зависимым киназам (CDKs). Предварительным условием активности этих киназ является физическая ассоциация со специфическим циклином, выбор времени активации сильно зависит от экспрессии циклина. Связывание циклина вызывает конформационные изменения в N-концевой доле привязки CDK и способствует локализации и субстратной специфичности комплекса. Мономерные CDKs активируются, когда они ассоциированы с циклинами, и тогда обладают киназной активностью. Уровни циклинового белка колеблются в клеточном цикле и поэтому являются главным фактором при определении выбора времени активации CDK. Периодическая активация этих комплексов, содержащих циклины и CDK, во время клеточного цикла опосредует временную регуляцию переходов клеточного цикла (контрольные точки). Другие факторы, регулирующие активность CDK, включают CDK ингибиторы (CKIs или ICKs, KIPs, CIPs, INKs), CDK активирующие киназы (CAK), CDK фосфатазу (Cdc25) и CDK субъединицы (CKS) (Mironov et al. 1999; Reed 1996).
До настоящего времени в растениях были изучены два главных класса CDK, известные как А-тип и В-тип CDKs. А-тип CDKs регулирует и G1-to-S и G2-to-M переходы, в то время как В-тип CDKs контролирует только G2-to-M контрольную точку (Hemerly et al., 1995; Magyar et al., 1997; Porceddu et al., 2001). Кроме того, сообщалось о наличии С-типа CDKs и CDK-активирующих киназ (CAKs) (Magyar et al., 1997; Umeda et al., 1998; Joubes et al., 2001). Vandepoele et al., 2002, идентифицировали четыре CAKs с помощью основанного на гомологии анотационного метода. Этими CAKs были три D типа CAKs (Arath; CDKD; 1, Arath; CDKD; 2 и Arath;CDKD;3); и один F-тип CAK (Arath; CDKF; 1).
Yamaguchi et al. (PNAS Vol.100 (13) 8019-8023, 2003) описывает переэкспрессию рисовой R2 кДНК (которая кодирует CAK) в эксплантаты табачных листьев. Они сообщили, что временная экспрессия R2 в течение первых 7 дней выращивания стимулирует формирование каллуса в отсутствии цитокинина. Yamaguchi et al. также провели контроль in vitro органогенеза с помощью CDK.
Fabian-Marwedel et al. (The Plant Cell, Vol.14, 197-210, 2002) сообщают, что рисовая CAK, R2, регулирует S-фазовое продвижение и общий темп роста в суспензионных клетках.
Способность влиять на клеточный цикл растения и тем самым модифицировать различные характеристики роста растений может быть применена в различных областях, таких как повышение урожайности, размножение растений, в производстве декоративных культур, местных культур, садовых культур, лесоводстве, производстве водорослей для использования в биореакторах (для биотехнологического получения таких веществ, как фармацевтические вещества, антитела или вакцины, или для биоконсервирования органических отходов) и в других таких областях.
Было найдено, что введение в растение нуклеиновой кислоты, кодирующей CDKD, дает растения с повышенной урожайностью по сравнению с соответствующими растениями дикого типа. Поэтому согласно одному из вариантов настоящего изобретения предлагается способ повышения урожайности растений, включающий введение в растение нуклеиновой кислоты, кодирующей CDKD.
Термин «повышенная урожайность», как он определен здесь, означает одну или более из следующих характеристик, каждая относительно соответствующего растения дикого типа: (i) повышенная биомасса (масса) одной или более частей растения, в частности, надземных (обеспечивающих урожай) частей, повышенная биомасса корней или повышенная биомасса любой другой обеспечивающей урожай части; (ii) повышенный выход семян, который может быть результатом увеличения биомассы семени (массы семени) и который может быть за счет увеличения массы семян на растении или за счет увеличения отдельных семян, и при этом увеличение массы семян может быть обусловлено изменением размеров семени, таких как длина семени и/или ширина семени и/или площадь семени; (iii) увеличенное число (наполненных) семян; (iv) увеличенный размер семян, который может также влиять на состав семян; (v) увеличенный объем семян, который может также влиять на состав семян; (vi) увеличенный индекс урожайности, который выражается как отношение выхода обеспечивающих урожай частей, таких как семена, к общей биомассе; и (vii) увеличенная масса тысячи зерен (TKW), которая экстраполируется из подсчитанного числа наполненных зерен и их общей массы. Повышенная TKW может быть результатом увеличенного размера семян и/или удельного веса семян.
Согласно предпочтительному варианту осуществления изобретения повышение урожайности охватывает повышение урожайности на уровне семени, как определено в любом одном или более вышеуказанных (ii)-(vii).
Взяв в качестве примера кукурузу, повышенную урожайность можно представить как одну или более из следующих характеристик: увеличение числа растений на гектар или акр, увеличение числа початков на растение, увеличение числа рядов, увеличение числа зерен в ряду, массы зерна, массы тысячи зерен, длина/диаметр початка и др. Взяв рис в качестве примера, увеличение выхода можно выразить путем увеличения одной или более из следующих характеристик: число растений на гектар или акр, число метелок на растение, число колосков в метелке, число цветков на метелку, увеличение скорости наполнения семян, повышение массы тысячи зерен и др. Повышение урожайности также может дать в результате изменение строения или может быть результатом измененного строения.
Согласно предпочтительному варианту осуществление способов по изобретению приводит в результате к растениям с повышенной урожайностью, которая выражается с помощью, по меньшей мере, одного из признаков: увеличенная площадь надземной части, увеличенная TKW, увеличенное число наполненных семян, повышенная масса семени и повышенный индекс урожайности, каждый относительно контроля или соответствующих растений дикого типа.
Осуществление способов по изобретению преимущественно ведет к повышенной урожайности в любых растениях.
Используемый здесь термин «растение» охватывает целые растения, предки и потомство растений и части растений, включая семена, побеги, стебли, листья, корни (включая клубни), и растительные клетки, ткани и органы, где каждый из вышеупомянутых содержит интересующий ген. Термин «растение» также охватывает зародыши, меристематические области, гаметофиты, спорофиты, пыльцу и микроспоры, где снова каждый из вышеупомянутых содержит интересующий ген. Используемый здесь термин «растение» не включает суспензионные культуры и ткани каллуса.
Способы по изобретению могут быть осуществлены на любом растении, предпочтительны все растения, принадлежащие к суперсемейству Viridiplantae, в особенности однодольные и двудольные растения, включая кормовые или фуражные бобы, декоративные растения, продовольственные культуры, деревья или кустарники, выбранные из списка, включающего Acacia spp., Acer spp., Actinidia spp., Aesculus spp., Agathis australis, Albizia amara, Alsophila tricolor, Andropogon spp., Arachis spp., Areca catechu, Astelia fragrans, Astragalus cicer, Baikiaea plurijuga, Betula spp., Brassica spp., Bruguiera gymnorrhiza, Burkea africana, Butea frondosa, Cadaba farinosa, Calliandra spp., Camellia sinensis, Canna indica, Capsicum spp., Cassia spp., Centroema pubescens, Chaenomeles spp., Cinnamomum cassia, Co/Tea arabica, Colophospermum mopane, Cotonillia varia, Cotoneaster serotina, Crataegus spp., Cucumis spp., Cupressus spp., Cyathea dealbata, Cydonia oblonga, Cryptomeria japonica, Cymbopogon spp., Cynthea dealbata, Cydonia oblonga, Dalbergia monetaria, Davallia divaricata, Desmodium spp., Dicksonia squarosa, Diheteropogon amplectens, Dioclea spp., Dolichos spp., Dorycnium rectum, Echinochloa pyramidalis, Ehrartia spp., Eleusine coracana, Eragrestis spp., Erythrina spp., Eucalyptus spp., Euclea schimperi, Eulalia villosa, Fagopyrum spp., Feijoa sellowiana, Fragaria spp., Flemingia spp., Freycinetia banksii, Geranium thunbergii, Ginkgo biloba, Glycine javanica, Gliricid/a spp., Gossypium hirsutum, Grew/tea spp., Guibourtia coleosperma, Hedysarum spp., Hemarthia altissima, Heteropogon contortus, Hordeum vulgare, Hyparrhenia rufa, Hypericum erectum, Hyperthelia dissoluta, Indigo incamata, Iris spp., Leptarrhena pyrolifolia, Lespediza spp., Lettuca spp., Leucaena leucocephala, Loudetia simplex, Lotonus bainesii, Lotus spp., Macrotyloma axillare, Malus spp., Manihot esculenta, Medicago sativa, Metasequoia glyptostroboides, Musa sapientum, Nicotianum spp., Onobrychis spp., Omithopus spp., Oryza spp., Peltophorum africanum, Pennisetum spp., Persea gratissima, Petunia spp., Phaseolus spp., Phoenix canariensis, Phormium cookianum, Photinia spp., Picea glauca, Pinus spp., Pisum sativum, Podocarpus totara, Pogonarthria fleckii, Pogonarthria squarmsa, Populus spp., Prosopis cineraria, Pseudotsuga menziesii, Pterolobium stellatum, Pynis communis, Quercus spp., Rhaphiolepsis umbellata, Rhopalostylis sapida, Rhus natalensis, Ribes grossularia, Ribes spp., Robinia pseudoacacia, Rosa spp., Rubus spp., Salix spp., Schyzachyrium sanguineum, Sciadopitys verticillata, Sequoia sempervirens, Sequoiadendron giganteum, Sorghum bicolor, Spinacia spp., Sporobolus fimbriatus, Stiburus alopecuroides, Stylosanthos humilis, Tadehagi spp., Taxodium distichum, Themeda triandra, Trifolium spp., Triticum spp., Tsuga heterophylla, Vaccinium spp., Vicia spp., Vitis vinifera, Watsonia pyramidata, Zantedeschia aethiopica, Zea mays, амарант, артишок, аспарагус, брокколи, брюссельскую капусту, качанную капусту, канолу, морковь, цветную капусту, сельдерей, листовую капусту, лен, капусту кормовую, чечевицу, семена масличного рапса, окру, лук, картофель, рис, сою, землянику, сахарную свеклу, сахарный тростник, подсолнечник, томат, тыкву, чай, водоросли и др. Согласно предпочтительному варианту настоящего изобретения растение является сельскохозяйственным растением, таким как соя, подсолнечник, канола, люцерна, семена рапса, хлопок, томат, картофель или табак. Более предпочтительно растение является однодольным растением, таким как сахарный тростник. Наиболее предпочтительно растение является зерновой культурой, такой как рис, кукуруза, пшеница, просо, ячмень, рожь, сорго или овес.
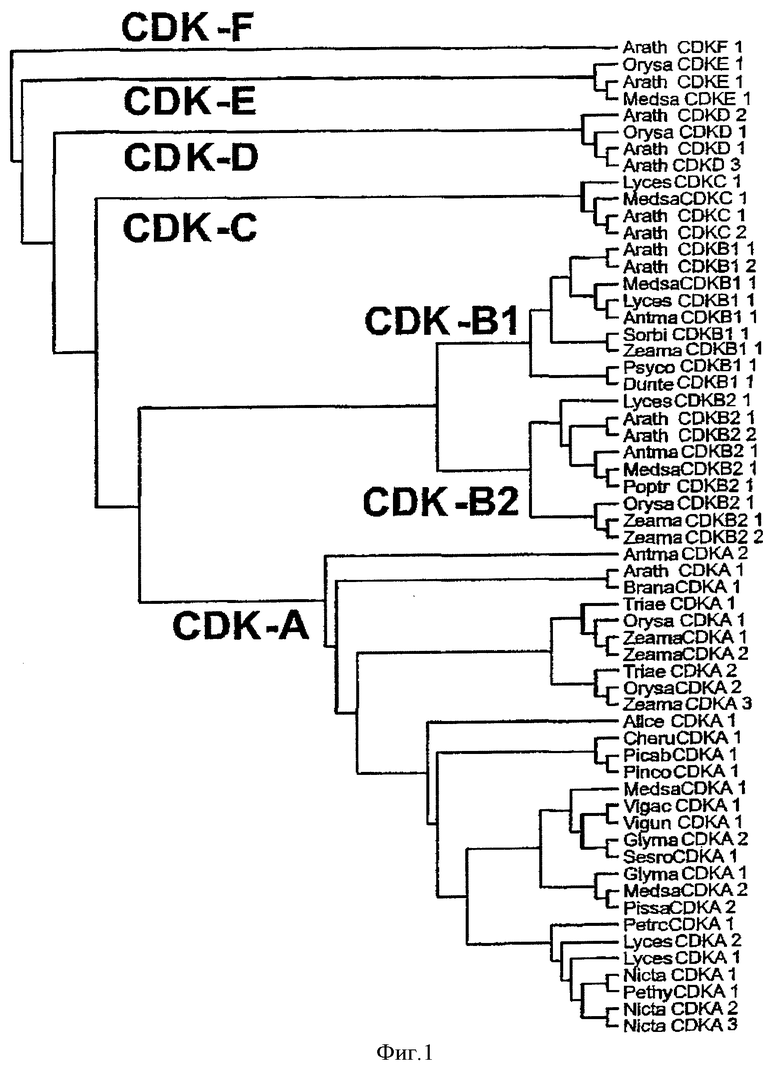
Термины " CDK D-типа" или "CDKD" используются взаимозаменяемо и относятся к любой аминокислотной последовательности, которая, когда используется в конструкции CDK филогенетического дерева, такого как указано на Фиг.1, скапливается вокруг или в группу, которая включает CDKs D-типа, но не другие типы CDK, такие как А-, В-, С-, Е- или F-тип CDKs. Упоминаемая здесь нуклеиновая кислота, кодирующая CDKD, является нуклеиновой кислотой, кодирующей CDKD аминокислоту, как определено выше.
Специалист в данной области может легко определить, любая ли из рассматриваемых последовательностей аминокислот попадает под вышеуказанное определение, используя известные для изготовления такого филогенетического дерева методики и компьютерные программы, такие как GCG, EBI или CLUSTAL пакет, используя параметры по умолчанию. В конструкции такого филогенетического дерева последовательности, скапливающиеся вокруг или в группе CDK D-типа будут рассматриваться, как попадающие под определение «CDK D-типа». Нуклеиновые кислоты, кодирующие такие последовательности, будут использоваться в выполняемых способах по изобретению.
CDKs D-типа обычно обладают способностью фосфорилировать и активировать CDKs, и также могут фосфорилировать и активировать РНК полимеразу II. CDKs D-типа могут также проявлять одну или более, а предпочтительно все, из следующих черт: (i) NXTALRE звено, где Х - любая аминокислота; (ii) каталитический киназный домен; и (iii) способность связывать циклин Н.
CDKD можно легко отличить от любой другой CDK, т.к. звено NXTALRE является специфичным для этого типа CDK (согласно теперешним знаниям). В отличие от него, согласно теперешним знаниям, CDK А-типа будет иметь PSTAIRE звено; CDK В-типа - P(P/S)T(A/T)(L/M)RE звено; а CDK С-типа - PITAIRE звено; CDK Е-типа будет иметь SPTARE звено; и CDK F-типа будет иметь XSAXRE звено.
Специалист в данной области может легко оценить активность киназы, например, на очищенных субстратах, таких как человеческая CAK2 или Arabidopsis thaliana РНК полимераза II карбокси-концевая. Способность CDKD связывать циклин Н может быть легко определена путем соосаждения комплекса CDKD-циклин Н из очищенной CDKD и циклина Н, или путем использования двух гибридных оценок.
В Arabidopsis thaliana CDKDs кодируются 3 различными генами: CDKD; 1, CDKD; 2 и CDKD; 3, каждый ген кодирует белок, который содержит звено NXTALRE, где Х - любая аминокислота.
Преимущественно способы по изобретению могут быть осуществлены с использованием любой нуклеиновой кислоты, кодирующей CDKD, как определено выше. Введение в растение кодирующей CDKD нуклеиновой кислоты дает измененную экспрессию (предпочтительно повышенную экспрессию) в растение такой нуклеиновой кислоты и/или измененные (предпочтительно повышенные) активность и/или уровни в растении CDKD полипептида. Активность CDKD может быть увеличена путем повышения уровней полипептида. Альтернативно, активность может быть повышенной, когда уровни CDKD полипептида не изменяются, или даже когда уровни CDKD полипептида снижены. Это может произойти, когда существенные свойства полипептида изменяются, например, путем изготовления мутантной версии, которая является более активной, чем полипептид дикого типа.
Нуклеиновая кислота, кодирующая CDKD, предпочтительно операбельно связана с конститутивным промотором для сверхэкспрессии в растении. Конститутивный промотор предпочтительно является GOS2 промотором, более предпочтительно GOS2 промотором из риса. Важно, что применимость настоящего изобретения не ограничивается использованием CDKD из Arabidopsis thaliana, ни CDKD, представленной в виде SEQ ID NO: 1, также применимость настоящего изобретения не ограничивается экспрессией CDKD-кодирующей нуклеиновой кислоты, когда побуждается с помощью GOS2 промотора.
Согласно предпочтительному варианту настоящего изобретения, рассматривается усиленная или повышенная экспрессия CDKD нуклеиновой кислоты. Способы достижения усиленной или повышенной экспрессии генов или генных продуктов хорошо описаны в уровне техники и включают, например, сверхэкспрессию, побуждаемую с помощью сильного промотора, использование транскрипционных энхансеров или трансляционных энхансеров.
Нуклеиновая кислота, кодирующая CDKD, может быть произведена из любого источника. Нуклеиновая кислота/ген, кодирующая CDKD, может быть выделена из микробного источника, такого как бактерия, дрожжи или грибы, или из растения, водоросли или животного (включая человека) источника. Эта кислота может быть модифицирована из ее природной формы в композиции или геномной среде путем преднамеренных манипуляций человека. Нуклеиновая кислота является предпочтительно гомологичной нуклеиновой кислотой, т.е. кислота получена либо из растения, либо из растения того же вида, в которое она вводится, либо из различных видов растений. Нуклеиновая кислота может быть выделена из двудольных видов, предпочтительно семейства Brassicaceae, более предпочтительно из Arabidopsis thaliana. Более предпочтительно кодирующая CDKD нуклеиновая кислота, выделенная из Arabidopsis thaliana, представляет собой CDKD; 1, CDKD; 2 или CDKD; 3. Наиболее предпочтительно CDKD является CDKD; 1 из Arabidopsis thaliana, в частности последовательностью нуклеиновой кислоты, как представленная в SEQ ID NO: 1 и соответствующей аминокислотной последовательностью, представленной в виде SEQ ID NO: 2.
Преимущественно осуществление настоящего изобретения не ограничивается использованием CDKD; 1 из Arabidopsis, как представленная в SEQ ID NO: 1. Способы по настоящему изобретению могут быть осуществлены с использованием функциональных вариантов CDKD, как описанные выше, или с использованием функциональных вариантов кодирующих CDKD нуклеиновых кислот. Предпочтительными функциональными вариантами являются варианты последовательности нуклеиновой кислоты, представленной в виде SEQ ID NO: 1 или функциональные варианты аминокислотной последовательности, представленной в виде SEQ ID NO: 2.
Используемый здесь термин «функциональный вариант» означает вариант, который попадает под приведенное ранее определение CDKD. Предпочтительно функциональный вариант также обладает способностью фосфорилировать и активировать CDKs, и фосфорилировать и активировать РНК полимеразу II. Предпочтительно функциональный вариант CDKs D-типа проявляет одну или более, а предпочтительно все, из следующих черт: (i) NXTALRE звено, где Х - любая аминокислота; (ii) каталитический киназный домен; и (iii) способность связывать циклин Н. Специалист в данной области легко определит, является ли частный вариант функциональным (в смысле способен ли он повышать выход растения) путем простой замены последовательности, описанной ниже в разделе Примеры, вариантом, тестируемым на функцию.
Подходящий вариант нуклеиновой кислоты и аминокислотных последовательностей, используемый при осуществлении способа по настоящему изобретению, включает:
(i) функциональную часть кодирующей CDKD нуклеиновой кислоты;
(ii) последовательности, способные к гибридизации кодирующей CDKD нуклеиновой кислоты;
(iii) альтернативные сплайс-варианты кодирующей CDKD нуклеиновой кислоты;
(iv) аллельные варианты кодирующей CDKD нуклеиновой кислоты;
(v) гомологи, производные и активные фрагменты CDKD аминокислоты.
Каждый из вышеупомянутых вариантов является функциональным вариантом, как он определен ранее.
Специалисту в данной области очевидно, что использование кодирующей CDKD последовательности ДНК полной длины не является необходимым условием для выполнения способов по изобретению. Способы по изобретению преимущественно могут быть осуществлены с использованием функциональной части кодирующей CDKD ДНК/нуклеиновой кислоты, предпочтительно функциональной части последовательности нуклеиновой кислоты, как представленная в SEQ ID NO: 1. Функциональная часть является кодирующей CDKD нуклеиновой кислотой, попадающей под определение функциональных вариантов, приведенное выше. Часть может быть получена, например, путем проведения одной или более делеций кодирующей CDKD нуклеиновой кислоты, такой как последовательность нуклеиновой кислоты, представленная в SEQ ID NO: 1, с использованием методик, хорошо известных из уровня техники.
Поэтому согласно изобретению предлагается способ повышения урожайности растения, особенно выхода семян, включающий введение в растение части кодирующей CDKD нуклеиновой кислоты.
Другим вариантом является последовательность, способная к гибридизации кодирующей CDKD нуклеиновой кислоты. Такими гибридизирующими последовательностями являются последовательности, попадающие под определение функциональных вариантов, приведенное выше. Особенно предпочтительными являются последовательности, способные к гибридизации кодирующей CDKD нуклеиновой кислоты при жестких условиях, особенно кодирующая CDKD нуклеиновая кислота, представленная в виде SEQ ID NO: 1.
Термин «гибридизация», как он определен здесь, означает процесс, где по существу гомологичные комплементарные нуклеотидные последовательности аннелируют друг друга. Процесс гибридизации может быть осуществлен полностью в растворе, т.е. обе комплементарные нуклеиновые кислоты находятся в растворе. Методы молекулярной биологии, на которых основан такой процесс, включают полимеразную цепную реакцию (PCR; и все основанные на ней методы), субстрактивную гибридизацию, случайное (рандомное) праймерное расширение, нуклеаза S1 картирование, праймерное расширение, обратную транскрипцию, синтезы кДНК, различные развертки РНК, и определение последовательности ДНК. Процесс гибридизации также может быть осуществлен с одной из комплементарных нуклеиновых кислот, иммобилизованной в матрице, такой как магнитные гранулы, гранулы Сефарозы или любой другой смолы.
Методы молекулярной биологии, на которых основан такой процесс, включают выделение поли (А*) мРНК. Процесс гибридизации, кроме того, может быть осуществлен с одной из комплементарных нуклеиновых кислот, иммобилизованной в твердой подложке, такой как нитроцеллюлоза или нейлоновая мембрана, или иммобилизованной, например, путем фотолитографии, например, на кремнийсодержащей стеклянной подложке (последние известны как нуклеиновокислотные матрицы или микроматрицы или как нуклеиновокислотные чипы). Методы молекулярной биологии, на которых основан такой процесс, включают РНК и ДНК гелевые блот-анализы, гибридизацию колоний микроорганизмов, гибридизацию тромбоцитов, in situ гибридизацию и микроматричную гибридизацию. Для того чтобы осуществить возможную гибридизацию, молекулы нуклеиновой кислоты обычно термически или химически денатурируют, чтобы перевести двойную цепь в две единичные цепи и/или удалить «шпильки» или другие вторичные структуры из единичных цепных нуклеиновых кислот. На строгость гибридизации влияют условия, такие как температура, концентрация соли и состав буфера гибридизации. Гибридизацию предпочтительно осуществляют при жестких условиях. Жесткими условиями являются те, при которых (1) используют низкую ионную концентрацию и высокую температуру промывки, например, 0,5М натрийфосфатный буфер с рН 7.2, 1 мМ ЭДТА рН 8.0 в 7% SDS при температуре или 65°С или 55°С, или (2) используют во время гибридизации денатурирующий агент, такой как формамид, например, 50% (об/об) формамид с 0.1% бычьим сывороточным альбумином, 0.1% Ficoll, 0.1% поливиниполипирролидон, 0.05 М натрийфосфатный буфер с рН 6.5 с 0.75 М NaCl, 0.075 М натрий цитрат при 42°С. Специфические примеры включают использование 50% формамида, 5XSSC (0.75 M NaCl, 0.075 М натрий цитрата), 50 мМ натрий фосфата (рН 6.8), 0.1% натрий пирофосфата, 5Х Denhard's раствора, обработанной ультразвуком ДНК из молок лососевых (50 nm/мл), 0.1% SDS и 10% декстран сульфата при 55°С, с промывками при 55°С в 0.2XSSC и 0.1% SDS. Специалист может легко определить и изменить жесткость условий, подходящих для получения ясного и обнаруживаемого сигнала гибридизации.
Поэтому согласно изобретению предлагается способ повышения урожайности растения, включающий введение в растение последовательности способной к гибридизации, предпочтительно при жестких условиях, кодирующей CDKD нуклеиновой кислоты.
Другим вариантом, используемым в способах по изобретению, является альтернативный сплайс-вариант кодирующей CDKD нуклеиновой кислоты. Подходящими сплайс-вариантами являются варианты, попадающие под определение функциональных вариантов, приведенное выше. Используемый здесь термин «альтернативный сплайс-вариант» охватывает варианты нуклеиновой кислоты, в которой выбранные интроны и/или экзоны исключены, замещены или добавлены. Такими вариантами будут те, в которых биологическая активность белка сохраняется нетронутой, что может быть достигнуто путем селективного сохранения функциональных сегментов белка. Такие сплайс-варианты могут быть найдены в природе или могут быть изготовлены человеком. Способы изготовления таких сплайс-вариантов хорошо известны из уровня техники. Сплайс-варианты SEQ ID NO: 1 являются особенно предпочтительными для использования в способах по изобретению.
Поэтому изобретение также предлагает способ повышения урожайности растения, включающий введение в растение альтернативного сплайс-варианта кодирующей CDKD нуклеиновой кислоты.
Другим вариантом, используемым в способах по изобретению, является аллельный вариант кодирующей CDKD нуклеиновой кислоты. Подходящими аллельными вариантами являются варианты, попадающие под определение функциональных вариантов, приведенное выше. Аллельные варианты существуют в природе, и использование в способах по настоящему изобретению является использованием этих природных аллелей. Аллельные варианты охватывают единичные нуклеотидные полиморфизмы (SNPs), а также малые инсерция/делеция полиморфизмы (INDELs). Размер INDELs обычно меньше чем 100 bp.SNPs и INDELs формируют наибольший набор вариантов последовательностей существующих в природе полиморфных цепей большинства организмов. Аллельные варианты SEQ ID NO: 1 являются особенно предпочтительными для использования в способах по изобретению.
Поэтому изобретение также предлагает способ повышения урожайности растения, включающий введение в растение аллельного варианта кодирующей CDKD нуклеиновой кислоты.
Кроме того, преимущественно способы по настоящему изобретению могут быть осуществлены с использованием гомологов, производных или активных фрагментов CDKD. Нуклеиновые кислоты, кодирующие гомологи, производные или активные фрагменты аминокислоты, такой как представленная в виде SEQ ID NO: 2, могут быть легко определены с использованием обычных методик, хорошо известных специалистам в данной области. Какие нуклеиновые кислоты подходят для использования в способах по изобретению можно легко определить, как описано выше.
«Гомологи» белка охватывают пептиды, олигопептиды, полипептиды, белки и энзимы, имеющие аминокислотные замещения, делеции и/или инсерции относительно данного не модифицированного белка и имеющие схожую биологическую и функциональную активность с не модифицированным белком, из которого они произведены. Для получения таких гомологов аминокислоты белка могут быть замещены другими аминокислотами, имеющими схожие свойства (такими как схожие гидрофобность, гидрофильность, антигенность, склонность к образованию или разрыву α-спиральных структур или β-пластинчатых структур). Таблицы консервационного замещения хорошо известны из уровня техники (см., например, Creighton (1984) Proteins. W.H.Freeman and Company).
Гомологи, используемые в способах по настоящему изобретению, имеют преимущественно, по меньшей мере, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% идентичности последовательности с аминокислотной последовательностью, представленной в виде SEQ ID NO: 2. CDKDs проявляют около 65% идентичности друг другу и проявляют менее 40% идентичности с другими CDKs. Поэтому гомолог, имеющий, по меньшей мере, 50% идентичности с CDK, как представленная SEQ ID NO: 2, не будет охватывать любую другую CDK, кроме CDK D-типа.
Термином «гомологи» также охватываются две специальные формы гомологии, которые включают ортологические последовательности и паралогические последовательности, которые охватывают эволюционные концепции, используемые для описания наследственных соотношений генов. Термин «паралогический» относится к ген-дупликациям внутри генома вида, ведущего к паралогическим генам. Термин «ортологический» относится к гомологичным генам в различных организмах, обусловленным наследственными взаимоотношениями.
Ортологи, например, в видах однодольных растений, могут быть легко найдены с помощью так называемого реципрокного бласт-поиска. Это может быть сделано с помощью первого бласта, включающего бластирование данной последовательности (например, SEQ ID NO: 1 или SEQ ID NO: 2) по отношению к любой последовательности из базы данных, такой как открыто доступной базе данных NCBI, которую можно найти на: http://www.ncbi.nlm.nih.gov. Если бы разыскивались ортологи в рисе, данная последовательность была бы бластирована, например, по отношению к клонам кДНК полной длины 28469 из Oryza sativa Nipponbare, доступных из NCBI. Может быть использован BLASTn, когда исходят из нуклеотидов, или TBLASTX, когда исходят из белка, при стандартных значениях по умолчанию (ожидание 10, выравнивание 50). Бласт-результаты могут быть отфильтрованы. Полные последовательности либо из отфильтрованных результатов, либо из не фильтрованных результатов затем повторно бластируют (второй бласт) по отношению к данной последовательности (SEQ ID NO: 1 или 2). Результаты первого и второго бластов затем сравнивают. В случае больших семейств используют ClustalW с последующей визуализацией в виде дерева ближайших соединений.
Гомолог может быть в форме «замещенного варианта» белка, т.е. когда, по меньшей мере, один остаток в аминокислотной последовательности удален и на его место вставлен другой остаток. Аминокислотные замещения обычно бывают из единичных остатков, но могут быть кластерными, в зависимости от функциональных ограничений, накладываемых на полипептид; инсерции обычно бывают порядка от 1 до 10 аминокислотных остатков, а делеции будут в интервале от 1 до 20 остатков. Предпочтительно аминокислотные замещения содержат консервативные аминокислотные замещения.
Гомолог может быть в форме «инсерционного варианта» белка, т.е. когда один или более аминокислотных остатков введены в предопределенный сайт в белке. Инсерции могут содержать амино-концевые и/или карбокси-концевые слияния, а также инсерции внутрь последовательности единичной или множества аминокислот. В общем, инсерции внутри аминокислотной последовательности должны быть меньшими, чем амино- или карбокси-концевые слияния, порядка от 1 до 10 остатков. Примеры амино- или карбокси-терминально слитых белков или пептидов включают связывающий домен или активирующий домен транскрипционного активатора, как используемые в дрожжевой двухгибридной системе, белки оболочки фагов, (гистидин)6-метка, глутатион-S-трансфераза-метка, белок А, мальтоза-связывающий белок, дигидрофолат редуктаза, эпитоп Tag·100, c-myc эпитоп, FLAG® - эпитоп, lacZ, CMP (калмодулин-связывающий белок), эпитоп НА, эпитоп белка С и эпитоп VSV.
Гомологи в форме «делеционных вариантов» белка характеризуются удалением одной или более аминокислот из белка.
Аминокислотные варианты белка могут быть легко изготовлены с использованием методик синтеза пептидов, хорошо известных из уровня техники, таких как твердофазный пептидный синтез и аналогичные, или путем манипуляций рекомбинантных ДНК. Способы манипулирования ДНК последовательностями для получения замещенных, инсерционных или делеционных вариантов белков хорошо известны из уровня техники. Например, методики изготовления замещенных мутаций при предопределенных сайтах в ДНК хорошо известны специалистам в данной области и включают М13 мутагенез, T7-Gen in vitro мутагенез (USB, Cleveland, ОН), быстро обменный сайт-специфический мутагенез (Stratagene, San Diego, CA), PCR-опосредованный сайт-специфический мутагенез или другие протоколы сайт-специфического мутагенеза.
«Производные» включают пептиды, олигопептиды, полипептиды, белки и энзимы, которые могут содержать замещения, делеции или добавления природных или не имеющихся в природе аминокислотных остатков, по сравнению с аминокислотными последовательностями имеющихся в природе форм белка, например, как представленные в SEQ ID NO: 2. «Производные» белка охватывают пептиды, олигопептиды, полипептиды, белки и энзимы, которые могут содержать природно-существующие измененные, гликозилированные, ацилированные, или не существующие в природе аминокислотные остатки, по сравнению с аминокислотными последовательностями имеющихся в природе форм полипептида. Производное может также содержать один или более не аминокислотный заместитель, по сравнению с аминокислотной последовательностью, из которой оно произведено, например молекула репортер или другой лиганд, ковалентно или не ковалентно связанный с аминокислотной последовательностью, такой как молекула репортер, которая связана для облегчения ее обнаружения, и не существующие в природе аминокислотные остатки, родственные аминокислотным последовательностям природно-существующего белка.
«Активные фрагменты» CDKD белка охватывают аминокислотные остатки, подходящие для группирования вокруг или в группе CDK D-типа на конструкции филогенетического дерева, такого как показано на Фиг.1. Когда используемые фрагменты в филогенетическом дереве похожи с теми, с которыми должны быть сравнены, это значит, что должны быть использованы соответствующие фрагменты других CDK для изготовления дерева.
Способы поиска и идентификации гомологов CDKD должны быть хорошо известны специалистам в данной области техники. Методы выборки последовательностей для сравнения хорошо известны специалистам в данной области, такие методы включают GAP, BESTFIT, BLAST, FASTA и TFASTA. GAP использует алгоритм Needleman and Wunsch (J. Mol. Biol. 48: 443-453, 1970) для выборки из двух полных последовательностей последовательности с максимальным числом пар и минимальным числом пробелов. Алгоритм BLAST рассчитывает процент идентичности последовательности и выполняет статистический анализ схожести между двумя последовательностями.
Программное обеспечение для выполнения BLAST анализов доступно через Национальный центр биотехнологической информации. Гомологи, подходящие для использования в способе по изобретению, т.е. имеющие, по меньшей мере, 50% идентичности последовательности с аминокислотной последовательностью, представленной в виде SEQ ID NO: 2, могут быть идентифицированы с помощью последовательностей белка CDK полной длины путем их сравнения с использованием программного обеспечения ClustalX1.81, используя параметры по умолчанию. Матрица расстояний может быть затем рассчитана из этого сравнения с использованием программного обеспечения BOXSHADE, также используя параметры по умолчанию. Оба программных обеспечения общедоступны.
Таким образом, изобретение также предлагает способ повышения урожайности растения, включающий введение в растение нуклеиновой кислоты, кодирующей гомолог, производное или активный фрагмент CDKD, такой как CDKD, представленная в виде SEQ ID NO: 2, причем гомолог, производное или активный фрагмент имеют преимущественно, по меньшей мере, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% идентичности последовательности с аминокислотной последовательностью, представленной в виде SEQ ID NO: 2.
Изобретение также предлагает генетические конструкции и векторы для облегчения введения и/или экспрессии нуклеотидных последовательностей, используемых в способах по изобретению.
Таким образом, предлагается генетическая конструкция, содержащая:
(i) нуклеиновую кислоту, кодирующую CDKD, предпочтительно, как представленная в виде SEQ ID NO: 1, или ее функциональный вариант (как определенный выше);
(ii) одну или более управляющих последовательностей, способных к регулированию экспрессии последовательности нуклеиновой кислоты из (i); и необязательно
(iii) транскрипционную концевую последовательность.
Конструкции, используемые в способах по настоящему изобретению, могут быть созданы с использованием метода рекомбинантных ДНК, хорошо известного специалистам в данной области. Генные конструкции могут быть вставлены в векторы, которые могут быть коммерчески доступными, подходящие для трансформирования растений и подходящие для экспрессии интересующего гена в трансформируемые клетки.
Растения были трансформированы вектором, содержащим интересующую последовательность (т.е. нуклеиновую кислоту, как представленная в виде SEQ ID NO: 1, или ее функциональный вариант (как определенный выше)). Интересующая последовательность является операбельно связанной с одной или более управляющих последовательностей (по меньшей мере, промотором). Используемые здесь термины «регуляторный элемент», «управляющая последовательность» и «промотор» являются взаимозаменяемыми и взятые в широком контексте относятся к регуляторным последовательностям нуклеиновых кислот, способным к эффективной экспрессии последовательностей, с которыми они лигированы. Вышеуказанными терминами охватываются транскрипционные регуляторные последовательности, произведенные из классического эукариотического геномного гена (включая ТАТА-бокс, который требуется для точной инициации транскрипции, с или без ССААТ-бокс последовательностью) и дополнительные регуляторные элементы (т.е. выше активирующие последовательности, энхансеры и глушители), которые изменяют экспрессию гена в ответ на эволюционные и/или внешние стимулы или ткане-специфическим образом. Термин также включает транскрипционную регуляторную последовательность классического прокариотического гена, который может включать -35 бокс последовательность и/или -10 бокс транскрипционные регуляторные последовательности. Термин «регуляторный элемент» также охватывает синтетическую слитую молекулу или производное, которое предоставляет, активирует или усиливает экспрессию молекулы нуклеиновой кислоты в клетку, ткань или орган. Используемый здесь термин «операбельно связанный» относится к функциональному связыванию между последовательностью промотора и интересующим геном, так что последовательность промотора способна инициировать транскрипцию интересующего гена.
Предпочтительно нуклеиновая кислота, кодирующая CDKD или ее функциональный вариант, является операбельно связанной с конститутивным промотором. Термин «конститутивный промотор», как он здесь определен, относится к промотору, который экспрессируется преимущественно, по меньшей мере, в одну ткань или орган и преимущественно на любой стадии жизненного цикла растения. Предпочтительно конститутивный промотор экспрессируется преимущественно через растение. Предпочтительно конститутивный промотор является GOS2 промотором из риса.
Примеры других конститутивных промоторов, подходящих для использования в способах по изобретению, приведены в таблице А ниже.
Необязательно одна или более терминаторных последовательностей также могут быть использованы в конструкции, вводимой в растение. Термин «терминатор» охватывает управляющую последовательность, которая представляет собой последовательность ДНК на конце транскрипционной единицы, которая подает сигнал 3' процессингу и полиаденилированию первичного транскрипта и окончанию транскрипции. Дополнительные регуляторные элементы могут включать транскрипционные, а также трансляционные энхансеры. Специалисты в данной области осведомлены о терминаторных и энхансерных последовательностях, которые могут быть подходящими для использования в осуществлении изобретения. Такие последовательности известны или могут быть легко получены специалистом в данной области.
Генетическая конструкция по изобретению может также включать источник репликационной последовательности, которая требуется для сохранения и/или репликации в специфическом клеточном типе. Одним из примеров является, когда генетическая конструкция должна быть сохранена в бактериальной клетке в качестве эписомального генетического элемента (например, молекулы плазмиды или космиды). Предпочтительные источники репликации включают, но не ограничиваются ими, fl-ori и colE1.
Генетическая конструкция необязательно может содержать селектируемый маркерный ген. Используемый здесь термин «селектируемый маркерный ген» включает любой ген, который обеспечивает фенотип клетки, в которую он экспрессируется, чтобы облегчить идентификацию и/или отбор клеток, которые трансфектируются или трансформируются конструкцией нуклеиновой кислоты по изобретению. Подходящие маркеры могут быть выбраны из маркеров, которые придают устойчивость к антибиотикам или гербицидам, что вносит новое метаболическое свойство или позволяет визуальный отбор. Клетки, содержащие рекомбинантную ДНК, будут таким образом сохранять жизнеспособность в присутствии таких концентраций антибиотика или гербицида, которые убивают нетрансформированные клетки.
Примеры селектируемых маркерных генов включают bar ген, который придает устойчивость к гербициду Basta; npt ген, придающий устойчивость к антибиотику канамицину; hpt ген, который обеспечивает устойчивость к гигромицину. Визуальные маркеры, такие как Green Fluorescent Protein - зеленый флуоресцентный белок (GFP, Haseloff et al., 1997), β-глюкуронидаза (GUS) или люцифераза, также могут быть использованы в качестве селектируемых маркеров. Другие примеры подходящих селектируемых маркерных генов включают ген устойчивости к ампициллину (Ampr), ген устойчивости к тетрациклину (Tcr), бактериальный ген устойчивости к канамицину (Kanr), ген устойчивости к фосфинотрицину, неомицин фосфотрансферазный ген (nptll), ген устойчивости к гигромицину, ген, и хлорамфеникол ацетилтрансферазный ген (CAT) и др.
Настоящее изобретение также охватывает растения, получаемые с помощью способов по изобретению. Настоящее изобретение поэтому предлагает растения, получаемые способом по изобретению, причем растения имеют повышенную урожайность и измененные активность и/или уровни белка CDKD и/или измененную экспрессию нуклеиновой кислоты, кодирующей белок CDKD, относительно соответствующих растений дикого типа.
Изобретение также предлагает способ получения трансгенных растений, имеющих повышенную урожайность, включающий введение и экспрессию в растение молекулы нуклеиновой кислоты, кодирующей CDKD или ее функционального варианта (как определенный выше).
Более конкретно, настоящее изобретение предлагает способ получения трансгенных растений с повышенной урожайностью, при этом способ включает:
(i) введение в растение или растительную клетку последовательности нуклеиновой кислоты, кодирующей CDKD или ее функционального варианта (как определенный выше);
(ii) культивирование растительной клетки в условиях, промотирующих регенерацию и созревание растения.
Нуклеиновая кислота может быть введена непосредственно в растительную клетку или в само растение (включая введение в ткань, орган или любую другую часть растения). Согласно предпочтительному варианту настоящего изобретения нуклеиновая кислота предпочтительно вводится в растение путем трансформации.
Термин «трансформация», употребляемый здесь, охватывает перенос экзогенного полинуклеотида в клетку-хозяин, независимо от способа, используемого для переноса. Растительная ткань, способная к последующему клональному размножению, путем или органогенеза или эмбриогенеза, может быть трансформирована генетической конструкцией по настоящему изобретению и из нее регенерируется целое растение. Отдельные выбранные ткани будут различаться в зависимости от используемых систем клонального размножения, и наиболее подходящие отдельные виды трансформируют. Образцы тканевых мишеней включают диски листьев, пыльцу, зародыши, семядоли, гипокотили, макрогаметофиты, каллюсовую ткань, существующие меристемные ткани (например, верхушечная меристема, пазушные почки и корневые меристемы) и полученные меристемные ткани (например, семядольная меристема и гипокотильная меристема). Полинуклеотид может быть временно или устойчиво введен в клетку-хозяин и может сохраняться неинтегрированным, например, как плазмида. Альтернативно, он может быть интегрированным в геном хозяина. Полученная в результате трансформированная растительная клетка затем может быть использована для регенерации трансформированного растения способом, известным специалистам в данной области.
Трансформация растительных видов является сейчас достаточно стандартной методикой. Преимущественно любой из различных способов трансформации может быть использован для введения интересующего гена в подходящую клетку-прототип. Способы трансформации включают применение липосом, электропорации, химикатов, которые усиливают свободный ввод ДНК, инжекцию ДНК прямо в растение, бомбардирование частицами, трансформацию с использованием вирусов или пыльцы или микропроекцию. Способы могут быть выбраны из кальций/полиэтиленгликолевого способа для протопластов (Krens, F.A. et al, 1882, Nature 296, 72-74; Negrutiu I., et al., June 1987, Plant Mol. Biol. 8, 363-373); электропорации протопластов (Shillito R.D. et al., 1985 Bio/Technol 3, 1099-1102); микроинъекции в растительный материал (Crossway A. et al., 1986, Mol. Gen Genet 202, 179-185); бомбардирования частицами, покрытыми ДНК или РНК (Klein T.M. et al., 1987, Nature 327, 70), заражения (не интеграционного) вирусами и т.п. Трансгенные рисовые растения, экспрессирующие нуклеиновую кислоту, кодирующую CDKD, предпочтительно получают путем Agrobacterium-опосредованной трансформации с использованием любого известного способа трансформации риса, такого как описанные в любом из следующих источников: опубликованная европейская патентная заявка ЕР 1198985 Al, Aldemita and Hodges (Planta, 199, 612-617, 1996); Chan et al. (Plant Mol. Biol. 22 (3) 491-506, 1993), Hiei et al. (Plant J. 6 (2) 271-282, 1994), раскрытие которых включено здесь с помощью ссылок, как полностью изложенное. В случае трансформации зерновых предпочтительным способом является способ, описанный либо Ishida et al. (Nat. Biotechnol. 1996 Jun; 14(6); 745-50), либо Frame et al. (Plant Physiol. 2002 May; 129(1): 13-22), раскрытие которых включено здесь с помощью ссылок, как полностью изложенное.
В основном после трансформации выбирают растительные клетки или группы клеток по наличию одного или более маркеров, которые кодируются растительно-экспрессируемыми генами, переносимыми совместно с интересующим геном, с последующей регенерацией трансформированного материала в целое растение.
Последующие перенос ДНК и регенерация предположительно трансформированных растений могут быть оценены, например, с использованием саузерн-анализа, на наличие интересующего гена, числа генов и/или геномной организации. Альтернативно или дополнительно уровни экспрессии вновь введенной ДНК могут быть отслежены с использованием нозерн- и/или вестерн-анализа, обе методики хорошо известны среднему специалисту в данной области.
Генерированные трансформированные растения могут быть размножены любыми способами, такими как клональное размножение или классические способы разведения. Например, трансформированное растение первой генерации (или Т1) может давать гомозиготные трансформанты второй генерации (или Т2), и Т2 растения затем размножают классическими способами разведения.
Генерированные трансформированные организмы могут принимать различные формы. Например, они могут быть химерами трансформированных клеток и не трансформированных клеток; клональными трансформантами (например, все клетки трансформировали для включения кассеты экспрессии); трансплантатами трансформированных и не трансформированных тканей (например, в растениях трансформированный корневой побег прививали на нетрансформированный побег растения).
Настоящее изобретение распространяется на любые растительные клетки и растения, полученные любым из описанных здесь способов, и на все части растений и их побеги. Настоящее изобретение распространяется далее так, что охватывает потомство первично трансформированных или трансфектированных клеток, тканей, органов или целого растения, которые были получены любым из вышеуказанных методов, единственное требование, чтобы потомство показывало тот же генотип и/или фенотипические свойства, что и родители, из которых произведено способами по изобретению. Изобретение также включает клетки-хозяева, содержащие выделенную нуклеиновую кислоту, кодирующую CDKD. Предпочтительными клетками-хозяевами согласно изобретению являются растительные клетки. Изобретение также распространяется на обеспечивающие урожай части растений, такие как перечисленные далее, но не ограничивается ими: семена, листья, фрукты, цветы, корневища, клубни и луковицы.
Настоящее изобретение также охватывает применение нуклеиновых кислот, кодирующих CDKDs и применение CDKD полипептидов.
Одно из таких применений, конечно, относится к применению CDKD для повышения урожайности растений, в частности для повышения урожайности семян. Урожайность семян может включать одну или более из следующих характеристик: увеличенное число наполненных семян, увеличенная масса семени, увеличенный индекс урожайности и увеличенная TKW и др. CDKD может быть нуклеиновой кислотой, как представленная в виде SEQ ID NO: 1, или ее функциональным вариантом, как он определен выше; или CDKD может быть нуклеиновой кислотой, как представленная в виде SEQ ID NO: 2, или ее функциональным вариантом, как он определен выше.
Нуклеиновые кислоты, кодирующие CDKDs, и CDKD полипептиды могут также найти применение в программах размножения. CDKD может быть нуклеиновой кислотой, как представленная в виде SEQ ID NO: 1, или ее функциональным вариантом, как он определен выше; или CDKD может быть нуклеиновой кислотой, как представленная в виде SEQ ID NO: 2, или ее функциональным вариантом, как он определен выше. Например, нуклеиновая кислота, кодирующая CDKD, или ее функциональный вариант могут быть на хромосоме (или ее части), предпочтительно с одним или более членов родственного семейства. В примере такой программы размножения идентифицируется маркер ДНК, который может быть генетически связанным с нуклеиновой кислотой, кодирующей CDKD белок, или ее функциональным вариантом. Этот ДНК маркер затем может быть использован в программах размножения для отбора растений с повышенной урожайностью.
Аллельные варианты CDKD также могут быть использованы в определенных программах размножения, таких как программы с использованием маркера. Такие программы размножения иногда требуют введения аллельного варианта путем мутагенной обработки растения. Одним из подходящих мутагенных способов является EMS мутагенез. Затем имеет место идентификация аллельных вариантов, например, с помощью PCR. Это следующий селекционный этап отбора наилучших аллельных вариантов данной последовательности, которые дают растения с повышенной урожайностью. Селекция обычно осуществляется путем мониторинга выхода растений, содержащих различные аллельные варианты данной последовательности, например различные аллельные варианты SEQ ID NO: 1. Мониторинг урожайности может быть сделан в теплице или в поле. Другие необязательные этапы включают скрещивание растений, в которых был идентифицирован наилучший аллельный вариант с другим растением. Это может быть использовано, например, для получения комбинации интересующих фенотипических черт.
Нуклеиновые кислоты, кодирующие CDKDs, и CDKD полипептиды могут также найти применение в качестве регуляторов роста. CDKD может быть нуклеиновой кислотой, как представленная в виде SEQ ID NO: 1, или ее функциональным вариантом, как он определен выше; или CDKD может быть нуклеиновой кислотой, как представленная в виде SEQ ID NO: 2, или ее функциональным вариантом, как он определен выше. Т.к. эти CDKDs используются для повышения урожайности растений, CDKDs будут также использованы регуляторами роста, такими как гербициды или стимуляторы роста. Настоящее изобретение предлагает композицию, содержащую CDKD вместе с подходящим носителем, разбавителем или эксципиентом, для использования в качестве регулятора роста.
Способы по настоящему изобретению также могут быть осуществлены без введения нуклеиновой кислоты, кодирующей CDKD, в растение. Это может быть достигнуто путем введения генетической модификации (предпочтительно в локус гена, кодирующего CDKD). Локус гена, как определено здесь, означает геномную область, которая включает интересующий ген и 10 kb выше- или нижерасположенных от кодирующей области.
Генетическая модификация может быть выполнена, например, одним (или более) из следующих методов: Т-ДНК активация, TILLING, сайт-специфический мутагенез, гомологичная рекомбинация, или, как обсуждалось выше, путем введения и экспрессии в растении (клетке) нуклеиновой кислоты, кодирующей CDKD.
Т-ДНК активационное мечение (Hayashi et al. Science (1992) 1350-1353) включает инсерцию Т-ДНК, обычно содержащую промотор (может также быть трансляционный энхансер или интрон), в геномную область интересующего гена или 10 KB расположенных выше или ниже кодирующей области гена в такой конфигурации, что такой промотор направляет экспрессию нацеливающего гена. Обычно, регуляция экспрессии нацеливающего гена с помощью его природного промотора нарушается, и ген подпадает под контроль вновь введенного промотора. Промотор обычно внедряется в Т-ДНК. Эта Т-ДНК случайным образом вставляется в геном растения, например, через заражение Agrobacterium, и приводит к сверхэкспрессии генов рядом со вставленной Т-ДНК. Полученные в результате трансгенные растения проявляют доминантные фенотипы, обусловленные сверхэкспрессией генов, близких с введенным промотором. Введенным промотором может быть любой промотор, способный направлять экспрессию гена в желаемом организме, в данном случае, в растении. Например, конститутивные, ткане-специфические, клеточно-специфические и индуцибельные промоторы - все подходят для использования в Т-ДНК активации.
Генетические модификации могут быть также введены в локус кодирующей CDKD нуклеиновой кислоты/гена с использованием методики TILLING (Targeted Induced Local Lesions IN Genomes). Это технология мутагенеза, используемая для генерации и/или идентификации, и, в конечном счете, для выделения мутагенных вариантов нуклеиновой кислоты, кодирующей CDKD, способных к проявлению CDKD биологической активности. TILLING также дает возможность селекции растений, несущих такие мутантные варианты. Эти мутантные варианты могут даже проявлять CDKD активность выше, чем проявляет ген в его природной форме. TILLING объединяет высокоплотный мутагенез с высокопроизводительными методами скрининга. Обычно стадиями TILLING являются: (a) EMS мутагенез (Redei and Koncz (1992); Feldmann et al., (1994); Lightner and Caspar (1998; (b) получение ДНК и объединение индивидуальных; (с) PCR амплификация интересующей области; (d) денатурация и аннелирование для образования гетеродуплексов; (е) DHPLC, где наличие гетеродуплекса в пуле определяется как высший пик на хроматограмме; (f) идентификация индивидуального мутанта; и (g) секвенирование мутантного PCR продукта. Способы TILLING хорошо известны из уровня техники (McCallum, Nat Biotechnol. 2000 Apr; 18(4):455-7, описанные Stemple, 2004 (TILLING-a high-throughput harvest for functional genomics. Nat Rev Genet. 2004 Feb; 5(2): 145-50.)).
Сайт-специфический мутагенез может быть использован для генерирования вариантов нуклеиновых кислот, кодирующих CDKD. Пригодны различные способы достижения сайт-специфического мутагенеза, наиболее общими являются методы, основанные на PCR (данные протоколы в молекулярной биологии. Wiley Eds. http://www.4ulr.com/products/currentprotocols/index.html).
Т-ДНК активация, TILLING и сайт-специфический мутагенез являются примерами технологий, которые делают возможной генерацию новых аллелей и вариантов CDKD.
Гомологичная рекомбинация делает возможным введение в геном выбранной нуклеиновой кислоты в определенное выбранное положение. Гомологичная рекомбинация является стандартной технологией, обычно используемой в биологических исследованиях низших организмов, таких как дрожжи или мох Physcomitrella. Способы осуществления гомологичной рекомбинации в растениях описаны не только для модельных растений (Offringa et al. Extrachromosomal homologous recombination and gene targeting in plant cells after Agrobacterium-mediated transformation. 1990 ЕМ ВО J. 1990 Oct; 9(10):3077-84), но также и для сельскохозяйственных культур, например риса (Terada R, Urawa H, Inagaki Y, Tsugane K, lida S. Efficient gene targeting by homologous recombination in rice. Nat Biotechnol. 2002. Iida and Terada: A tale of two integrations, transgene and T-DNA: gene targeting by homologous recombination in rice. Curr Opin Biotechnol. 2004 Apr; 15(2): 132-8). Для нацеливания нуклеиновой кислоты (которой может быть молекула нуклеиновой кислоты, кодирующей CDKD или ее вариант, как он определен ранее) нет необходимости нацеливания в локус гена, кодирующего CDKD, но она может быть введена, например, в область высокой экспрессии. Нуклеиновой кислотой для нацеливания может быть улучшенная аллель, используемая для замены эндогенного гена, или может быть введенный дополнительно эндогенный ген.
Способы по настоящему изобретению дают в результате растения с повышенной урожайностью, как описано выше. Эти преимущества эффектов урожайности могут быть также объединены с другими экономически привлекательными чертами, такими как другие повышающие урожайность черты, устойчивость к различным стрессам, черты, изменяющие различные структурные признаки и/или биохимические и/или физиологические признаки.
Описание чертежей
Настоящее изобретение будет теперь описываться со ссылкой на следующие чертежи, где:
Фиг.1 представляет дерево, показывающее различные растительные CDKs. Последовательности белка CDK полной длины были сравнены с использованием программного обеспечения «ClustalX1.81», используя параметры по умолчанию. Дерево ближайших соединений было рассчитано из этого сравнения с использованием программного обеспечения «ClustalX1.81», используя параметры по умолчанию. Дерево было нарисовано с использованием «рисовальной» программы из пакета «Phylip3.5», используя параметры по умолчанию.
Фиг.2 представляет бинарный вектор для экспрессии в Oryza sativa из Arabidopsis thaliana CDKD; 1 гена под контролем GOS2 промотора.
Фиг.3 представляет примеры последовательностей, используемых при осуществлении способов по настоящему изобретению.
Примеры
Настоящее изобретение будет теперь описываться со ссылкой на следующие примеры, которые представлены единственно для иллюстрации.
ДНК манипуляции: если не указано иначе, методы рекомбинантных ДНК осуществлялись согласно стандартным протоколам, описанным в (Sambrook (2001) Molecular Cloning: a laboratory manual, 3rd Edition Cold Spring Harbor Laboratory Press, CSH, New York) или в томах 1 и 2 Ausubel et al. (Current Protocols in Molecular Biology, Current Protocols). Стандартные материалы и способы для молекулярной работы с растениями описаны в Plant Molecular Biology Labfase (1993) by R.D.D. Croy, опубликованы BIOS Scientific Publications Ltd (UK) и Blackwell Scientific Publications (UK).
Пример 1: Клонирование гена
Arabidopsis CDKD1; 1 был амплифицирован с помощью PCR, с использованием в качестве матрицы Arabidopsis thaliana рассады библиотеки кДНК (Invitrogen, Paisley, UK). После обратной транскрипции РНК, экстрагированной из рассады, кДНК клонировали в pCMV Sport 6.0. Средний размер вставки из банка был 1.5 kb, и первоначальное число клонов было 1.59×107 cfu. Первоначальный tHer был определен как 9.6×105 cfu/мл, после первой амплификации 6×1011 cfu/мл. После экстракции плазмиды 200 нг матрицы использовали в 50 µ1 PCR смеси. Праймеры prm2676 (смысловой, стартовый кодон жирным шрифтом, AttB1 сайт курсивом: 5' GGGGACAAGTTTGTACAAAAAAGCAGGCTTCACA ATGGAACAGCCGAAGAAAG 3')
и prm3677 (реверсивный, комплементарный, терминирующий кодон жирным шрифтом, AttB2 сайт курсивом: 5' GGGGA CCACTTTGTACAAGAAAGCTGGGTCCTATAGGAACTCGAGATCAAGTT
3'), которые включают AttB сайты для Gateway рекомбинации, были использованы для PCR амплификации. PCR осуществляли с использованием Hifi Taq ДНК полимеразы в стандартных условиях. PCR фрагмент из 1256 bp амплифицировали и очищали также с использованием стандартных методов. Затем был выполнен первый этап Gateway процедуры, BP реакция, во время которой PCR фрагмент рекомбинировал in vivo с pDONR201 плазмидой с получением, согласно терминологии Gateway, "входного клона", р2777. Плазмида pDONR201 была приобретена из Invitrogen, как часть Gateway® технологии.
Пример 2: Векторная конструкция
Входной клон р2777 впоследствии использовали в LR реакции с р0640, целевым вектором, используемым для трансформации Oryza sativa. Этот вектор внутри Т-ДНК краев содержал: растительный селектируемый маркер; скринабельный маркер; и Gateway кассету, предназначенную для in vivo LR рекомбинации с интересующей последовательностью, уже клонированной во входной клон. Рисовый GOS2 промотор для конститутивной экспрессии был локализован выше этой Gateway кассеты.
После этапа LR рекомбинации результирующий вектор экспрессии, как показанный на Фиг.2 (CDK D1; 1::GOS2-upregulation), был трансформирован в Agrobacterium штамм LBA4404 и впоследствии в растения Oryza sativa. Трансформированные растения риса были выращены и затем оценены по параметрам, описанным в примере 3.
Пример 3: Оценка и результаты
Были генерированы приблизительно 15-20 самостоятельных Т0 рисовых трансформантов. Первичные трансформанты переносили из тканевых культуральных камер в теплицу для выращивания и получения урожая Т1 семян. Были сохранены 6 случаев, в которых Т1 потомство разделялось 3:1 по присутствию/отсутствию трансгенов. Для каждого из этих случаев отобрали приблизительно 10 Т1 сеянцев, содержащих трансген (гетеро- и гомозиготы), и приблизительно 10 Т1 сеянцев, с недостатком трансгена (нуль-зиготы) путем контрольной экспрессии визуального маркера.
Статистический анализ: t-тест и F-тест
Двухфакторный ANOVA (analysis of variants - анализ вариантов) использовали в качестве статистической модели для полной оценки фенотипических характеристик растений. F-тест выполняли на все измеренные параметры всех растений всех случаев, трансформированных геном по настоящему изобретению. F-тест выполняли, чтобы определить влияние гена во всех случаях трансформации и подтвердить общее влияние гена, называемое также «общий генный эффект». Порог достоверности для верного общего генного эффекта установлен 5% уровня вероятности для F-теста. Значительные данные F-теста для генного эффекта показывают, что не только присутствие или положение гена являются причиной различий в фенотипе.
Для контроля влияния гена в пределах события, т.е. линейно-специфического эффекта, был выполнен t-тест в пределах каждого события с использованием наборов данных из трансгенных растений и соответствующих нулевых растений. «Нулевые растения» или «нулевые сегреганты» - это растения, обработанные также, как трансгенные растения, но из которых трансгены были выделены. Нулевые растения могут быть также описаны как гомозиготные негативные трансформанты. Порог достоверности для t-теста установлен 10% уровня вероятности. Внутри одной совокупности из 5 трансформационных событий, некоторые события могут быть ниже ли выше этого порога t-теста. Это основывается на предположении, что ген может оказывать влияние только в определенном положении в геноме, и что месторасположение этого положения, от которого зависит влияние, не является исключительным. Этот вид генного эффекта также известен как линейное влияние гена. Р-значение было получено путем сравнения t-значения с t-распределением, или альтернативно, путем сравнения F-значения с F-распределением. Р-значение поддерживает вероятность того, что нулевая гипотеза (нулевая гипотеза - «не имеется влияния трансгена») является корректной.
4.1 Измерения вегетативного роста:
Отобранные Т1 растения (приблизительно 10 с трансгеном и приблизительно 10 без трансгена) переносили в теплицу. Каждое растение получало уникальную штриховую метку, чтобы однозначно связать фенотипические данные с соответствующим растением. Отобранные Т1 растения выращивали в грунте в горшках диаметром 10 см при следующих параметрах окружающей среды: фотопериод = 11,5 ч, интенсивность дневного света = 30000 люкс и более, дневная температура = 28°С и выше, ночная температура = 22°С, относительная влажность = 60-70%. Трансгенные растения и соответствующие нуль-зиготы выращивали рядом при беспорядочном расположении. От стадии посева до стадии созревания каждое растение несколько раз проходило через кабинет цифрового формирования изображения и экспонировалось. Каждый раз точка цифрового изображения (2048×1536 пикселей, 16 миллионов цветов) бралась, по меньшей мере, под 6 разными углами. Параметры, описанные ниже, были получены автоматическим путем из всех цифровых изображений всех растений с использованием программы анализа изображения.
4.1.1 Площадь надземной части растения
Площадь надземной части растения была определена путем подсчета общего числа пикселей надземной части растения, выделенных их фона. Это значение было усреднено для картинок, полученных из одной и той же точки под разными углами, и было превращено в физическое значение поверхности, выраженное в квадратных мм с помощью калибровки. Эксперименты показали, что площадь надземной части растения, измеренная этим способом, коррелирует с биомассой надземных частей растения. 3 наилучшие линии по Т1 оценке были затем оценены на Т2 этапе. Результаты Т2 оценки показаны в Таблице 1 ниже. Как видно, одна из линий показывает статистически значительное увеличение площади надземной части (р-значение из t-теста 0,0107) по сравнению с соответствующими нуль-зиготами.
Каждая строка соответствует одному событию, для которого площадь надземной части была определена для трансгенных (TR) и нулевых линий (null), выраженная в единицах. Числовая разница между позитивными растениями и негативными растениями дана в (dif), а процент различия между этими растениями в (%dif). Р-значение установлено для вероятности, полученной с помощью t-теста для каждой линии растений. Последняя строка представляет средние числа для всех событий. Там р-значение установлено для р-значения, полученного из F-теста.
4.2 Измерения параметров, относящихся к семенам
Зрелые первичные метелки собирали, раскладывали по мешкам, метили штрих-кодом и сушили в течение трех дней в сушильном шкафу при 37°С. Метелки затем молотили и все семена собирали и подсчитывали. Наполненная шелуха отделялась от пустой с использованием воздуходувного устройства. Пустая шелуха отбрасывалась и оставшаяся фракция снова подсчитывалась. Наполненная шелуха взвешивалась на аналитических весах. Эта процедура позволила получить ряд относящихся к семени параметров, описанных ниже.
4.2.0 Количество наполненных семян
Количество наполненных семян определяли путем подсчета количества наполненной шелухи, которая осталась после стадии разделения. Снова 3 лучших растения по Т1 оценке были затем оценены на Т2 этапе. Результаты Т2 оценки показаны в Таблице 2 ниже. Как видно, 2 линии показали значительное увеличение числа наполненных семян в трансгенных растениях относительно числа наполненных семян в соответствующих не трансгенных растениях. Имелся также общий генный эффект, как заключили по существенному р-значению из F-теста, равному 0.
Каждая строка соответствует одному событию, для которого количество наполненных семян было определено для трансгенных (TR) и нулевых линий (null), выраженное в единицах. Числовая разница между позитивными растениями и негативными растениями дана в (dif), а процент различия между этими растениями в (%dif). Р-значение установлено для вероятности, полученной с помощью t-теста для каждой линии растений. Последняя строка представляет средние числа для всех событий. Там р-значение установлено для р-значения, полученного из F-теста.
4.2.1 Общий выход семян на растение
Общий выход семян был измерен путем взвешивания всей наполненной шелухи, собранной с растения. Снова 3 лучших растения по Т1 оценке были затем оценены на Т2 этапе. Результаты Т2 оценки показаны в Таблице 3 ниже. Как видно, 2 линии показали значительное увеличение массы семян в трансгенных растениях относительно массы семян в соответствующих не трансгенных растениях. Имелся также общий генный эффект, как заключили по существенному р-значению из F-теста, равному 0.
Каждая строка соответствует одному событию, для которого общий выход семян был определен для трансгенных (TR) и нулевых линий (null), выраженный в единицах. Числовая разница между позитивными растениями и негативными растениями дана в (dif), а процент различия между этими растениями в (%dif). Р-значение установлено для вероятности, полученной с помощью t-теста для каждой линии растений. Последняя строка представляет средние числа для всех событий. Там р-значение установлено для р-значения, полученного из F-теста.
4.2.3 Индекс урожайности растений
Индекс урожайности в настоящем изобретении определяется как отношение общего выхода семян и площади надземной части (мм), умноженное на коэффициент 106. Три лучших растения по Т1 оценке были затем оценены на Т2 этапе, и результаты Т2 оценки показаны в Таблице 4 ниже. Как видно, 1 линия показала увеличенный индекс урожайности в трансгенных растениях относительно индекса урожайности в соответствующих не трансгенных растениях с р-значением из t-теста, равным 0. Общий генный эффект был также очевиден по р-значению из F-теста, равному 0.
Каждая строка соответствует одному событию, для которого индекс урожайности был определен для трансгенных (TR) и нулевых линий (null), выраженный в единицах. Числовая разница между позитивными растениями и негативными растениями дана в (dif), а процент различия между этими растениями в (%dif). Р-значение установлено для вероятности, полученной с помощью t-теста для каждой линии растений. Последняя строка представляет средние числа для всех событий. Там р-значение установлено для р-значения, полученного из F-теста.
4.2.1 Масса тысячи зерен (TKW)
Этот параметр экстраполировали из рассчитанного количества наполненных семян и их общей массы. Три лучших растения по Т1 оценке были затем оценены на Т2 этапе, и результаты Т2 оценки показаны в Таблице 5 ниже. Как видно, 1 линия показала увеличение TKW в трансгенных растениях относительно соответствующих не трансгенных растениях с р-значением из t-теста, равным 0,0455.
Каждая строка соответствует одному событию, для которого TKW был определен для трансгенных (TR) и нулевых линий (null), выраженный в единицах. Числовая разница между позитивными растениями и негативными растениями дана в (dif), а процент различия между этими растениями в (% dif). Р-значение установлено для вероятности, полученной с помощью t-теста для каждой линии растений. Последняя строка представляет средние числа для всех событий. Там р-значение установлено для р-значения, полученного из F-теста.
| название | год | авторы | номер документа |
|---|---|---|---|
| РАСТЕНИЯ, ИМЕЮЩИЕ УСИЛЕННЫЕ ПРИЗНАКИ, СВЯЗАННЫЕ С УРОЖАЙНОСТЬЮ, И СПОСОБ ИХ ПОЛУЧЕНИЯ | 2008 |
|
RU2503721C2 |
| ИДЕНТИФИКАЦИЯ И ПРИМЕНЕНИЕ МУТАНТНЫХ KRP У РАСТЕНИЙ | 2012 |
|
RU2631790C2 |
| РАСТЕНИЯ, ХАРАКТЕРИЗУЮЩИЕСЯ ПОВЫШЕННОЙ УРОЖАЙНОСТЬЮ, И СПОСОБ ИХ ПОЛУЧЕНИЯ | 2006 |
|
RU2463351C2 |
| СПОСОБЫ УВЕЛИЧЕНИЯ УРОЖАЯ И СТРЕССОУСТОЙЧИВОСТИ У РАСТЕНИЯ | 2012 |
|
RU2632569C2 |
| ИНСЕКТИЦИДНЫЕ БЕЛКИ | 2017 |
|
RU2765722C2 |
| СКОНСТРУИРОВАННЫЕ ПЕСТИЦИДНЫЕ БЕЛКИ И СПОСОБЫ КОНТРОЛЯ ВРЕДИТЕЛЕЙ РАСТЕНИЙ | 2017 |
|
RU2772947C2 |
| СПОСОБЫ ТРАНСФОРМАЦИИ РАСТЕНИЙ ДЛЯ ЭКСПРЕССИИ ДЕЛЬТА-ЭНДОТОКСИНОВ BACILLUS THURINGIENSIS | 1999 |
|
RU2234531C2 |
| ПРОМОТОР ЦЕЛОГО СЕМЕНИ | 2010 |
|
RU2561463C2 |
| СПОСОБЫ ГЕНЕТИЧЕСКОГО КОНТРОЛЯ ПОРАЖЕНИЯ РАСТЕНИЙ НАСЕКОМЫМИ И ПРИМЕНЯЕМЫЕ ДЛЯ ЭТОГО КОМПОЗИЦИИ | 2006 |
|
RU2478710C2 |
| НОВЫЙ КЛАСС ГЕНОВ УСТОЙЧИВОСТИ К ГЛИФОСАТУ | 2013 |
|
RU2634411C2 |


Изобретение относится к биотехнологии. Описан способ увеличения урожайности обеспечивающих урожай частей растений относительно соответствующих растений дикого типа, включающий введение в растение нуклеиновой кислоты, кодирующей циклин-зависимую киназу D-типа (CDKD), содержащую NXTALRE звено, где Х означает любую аминокислоту. Раскрыт способ производства трансгенного растения с повышенной урожайностью, включающий введение в растение или растительную клетку кодирующей CDKD нуклеиновой кислоты, содержащую NXTALRE звено, где Х означает любую аминокислоту, и культивирование растительной клетки в условиях, промотирующих регенерацию и рост зрелого растения. Изобретение позволяет повышать урожайность растений. 2 н. и 12 з.п. ф-лы, 3 ил., 5 табл.
1. Способ увеличения урожайности обеспечивающих урожай частей растений относительно соответствующих растений дикого типа, включающий введение в растение нуклеиновой кислоты, кодирующей циклин-зависимую киназу D-типа (CDKD), содержащую NXTALRE звено, где Х означает любую аминокислоту.
2. Способ по п.1, где указанная увеличенная урожайность является увеличенной урожайностью семян.
3. Способ по п.1, где указанная увеличенная урожайность выбрана из группы, состоящей из (i) повышенной биомассы одной или более частей растения; (ii) повышенной биомассы семени; (iii) увеличенного числа (наполненных) семян; (iv) увеличенного размера семени; (v) увеличенного объема семени; (vi) увеличенного индекса урожайности; и (vii) увеличенной массы тысячи зерен (TKW).
4. Способ по п.1, где указанная циклин-зависимая киназа D-типа (CDKD) дополнительно содержит каталитический киназный домен и/или обладает способностью связывать циклин Н.
5. Способ по п.1, где указанная нуклеиновая кислота, кодирующая циклин-зависимую киназу D-типа (CDKD), является способной к гибридизации при жестких условиях кодирующей CDKD нуклеиновой кислоты, представленной в виде SEQ ID NO: 1.
6. Способ по п.1, где указанная циклин-зависимая киназа D-типа (CDKD) имеет по повышающейся степени предпочтительности, по меньшей мере, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90% или 95% идентичности последовательности с аминокислотной последовательностью, представленной в виде SEQ ID NO: 2.
7. Способ по п.1, где указанная нуклеиновая кислота, кодирующая CDKD, получена из растения.
8. Способ по п.7, где указанная нуклеиновая кислота, кодирующая CDKD, представляет собой CDKD; 1, CDKD; 2 или CDKD; 3 из Arabidopsis thaliana.
9. Способ по п.7 или 8, где указанная нуклеиновая кислота, кодирующая CDKD, представлена в виде SEQ ID NO: 1, и где CDKD полипептид представлен в виде SEQ ID NO: 2.
10. Способ по п.1, где указанная последовательность нуклеиновой кислоты, кодирующей CDKD, сверхэкспрессируется в растение.
11. Способ по п.10, где сверхэкспрессия указанной нуклеиновой кислоты, кодирующей CDKD, стимулируется конститутивным промотором.
12. Способ производства трансгенного растения с повышенной урожайностью, включающий
(i) введение в растение или растительную клетку кодирующей CDKD нуклеиновой кислоты, содержащую NXTALRE звено, где Х означает любую аминокислоту, и
(ii) культивирование растительной клетки в условиях, промотирующих регенерацию и рост зрелого растения.
13. Способ по п.12, где указанная повышенная урожайность является повышенной урожайностью семян.
14. Способ по п.12 или 13, где указанная увеличенная урожайность выбрана из группы, состоящей из (i) повышенной биомассы одной или более частей растения; (ii) повышенной биомассы семени; (iii) увеличенного числа (наполненных) семян; (iv) увеличенного размера семени; (v) увеличенного объема семени; (vi) увеличенного индекса урожайности; и (vii) увеличенной массы тысячи зерен (TKW).
| WO 03027299, 03.04.2003 | |||
| DATA BASE, GenBank, AY063843, 18.10.2002 | |||
| DATA BASE, GenPept, AAG52081, 19.01.2001 | |||
| KLAAS VANDEPOELE et al: «Genome-Wide Analysis of Core Cell Cycle Genes in Arabidopsis» Plant Cell, 2002 14(4): 903-916 | |||
| НУКЛЕОТИДНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ (ВАРИАНТЫ), РЕКОМБИНАНТНАЯ МОЛЕКУЛА ДНК, СПОСОБ ПОЛУЧЕНИЯ ОПТИМИЗИРОВАННОЙ ДЛЯ КУКУРУЗЫ КОДИРУЮЩЕЙ ПОСЛЕДОВАТЕЛЬНОСТИ ИНСЕКТИЦИДНОГО ПРОТЕИНА, СПОСОБ ЗАЩИТЫ РАСТЕНИЙ КУКУРУЗЫ ОТ НАСЕКОМЫХ-ВРЕДИТЕЛЕЙ | 1992 |
|
RU2202611C2 |

