Область техники, к которой относится настоящее изобретение
Предоставлены отличные от человека животные с заменой эндогенных генов IL-6 и/или рецептора IL-6 отличного от человека животного. Гены IL-6 и/или рецептора IL-6 отличного от человека животного в эндогенных не принадлежащих человеку локусах заменены генами IL-6 человека и/или генами гуманизированного рецептора IL-6, содержащими последовательность человека. Отличные от человека животные, несущие гены IL-6 человека и/или гуманизированного рецептора IL-6, где у отличных от человека животных не выявляют ни одной или более патологий, характерных для отличных от человека животных, трансгенных по IL-6 человека.
Предшествующий уровень техники настоящего изобретения
В данной области известны мыши, трансгенные по гену IL-6 человека. Однако случайная вставка трансгена IL-6 человека в геном мыши приводит к плохорегулируемой экспрессии белка IL-6 человека, которая проявляет себя во множестве патологий у таких трансгенных мышей, включая в качестве неограничивающих примеров, плазмацитоз и гломерулонефрит. В результате эти мыши имеют ограниченную применимость.
Существует необходимость в экспрессии у отличных от человека животных, например, у мышей и крыс, IL-6 человека или гуманизированного IL-6 и/или рецептора IL-6 человека или гуманизированного рецептора IL-6. Существует необходимость в таких гуманизированных мышах, у которых не выявлено ни одной или более патологий, демонстрируемых трансгенными по hIL-6 мышами.
Краткое раскрытие настоящего изобретения
Согласно одному из аспектов предоставлены генетически модифицированные отличные от человека животные, у которых проведена замена эндогенных локусов генов IL-6 и/или рецептора IL-6, кодирующих эндогенные IL-6 и/или рецептор IL-6, на гены, кодирующие IL-6 человека или гуманизированный IL-6 и/или рецептор IL-6. Предоставлены животные семейства мышиных, у которых проведена замена эндогенного гена IL-6 в эндогенном локусе IL-6 мыши на ген IL-6 человека; и/или у которых проведена замена эндогенного гена рецептора IL-6 (или нуклеотидной последовательности, кодирующей его внеклеточный домен) на ген рецептора IL-6 человека (или нуклеотидную последовательность, кодирующую его внеклеточный домен).
Согласно одному из аспектов предоставлены генетически модифицированные животные семейства мышиных, которые экспрессируют ген IL-6 человека под контролем эндогенного промотора мыши и/или эндогенных регуляторных элементов мыши с эндогенного локуса IL-6 мыши.
Согласно одному из аспектов предоставлены генетически модифицированные животные семейства мышиных, которые экспрессируют ген рецептора IL-6 человека (или ген, кодирующий внеклеточный домен человека и трансмембранный и внутриклеточный домены мыши) под контролем эндогенного промотора мыши и/или эндогенных регуляторных элементов мыши с эндогенного локуса рецептора IL-6 мыши.
Согласно одному из аспектов предоставлены генетически модифицированные животные (например, животные семейства мышиных, например, мышь или крыса), которые экспрессируют белок IL-6 человека, где у отличного от человека животного не выявляют патологии, выбранной из плазмацитоза, гломерулонефрита, гломерулосклероза, мезангиального пролиферативного гломерулонефрита, лимфомы кишечника, лимфомы почка, спленомегалии, увеличения лимфоузлов, увеличения печени, мегакариоцитов в костном мозге, уплотненных аномальных плазматических клеток, инфильтрации плазматических клеток в легкие, или печень, или почки, мезангиальной клеточной пролиферации в почки, церебральной сверхэкспрессии IL-6, ветвящихся микроглиальных клеток в белом веществе мозга, реактивных астроцитов в головном мозге, почечной недостаточности, повышенного количества мегакариоцитов в селезенке, мышечной атрофии (например, атрофии икроножной мышцы), повышенного количества мышечных катепсинов В и B+L (например, приблизительно в 20 раз и в 6 раз) и их сочетания.
Согласно одному из вариантов осуществления отличное от человека животное несет нормальную популяцию В-клеток. Согласно одному из вариантов осуществления нормальная популяция В-клеток по количеству и антигенному фенотипу приблизительно соответствует животному дикого типа, например, мыши дикого типа.
Согласно одному из вариантов осуществления отличное от человека животное является представителем семейства мышиные (например, мышью или крысой) и экспрессирует IL-6 человека (hIL-6) в сыворотке на уровне приблизительно ниже 800 пг/мл, приблизительно ниже 700, 600, 500, 400, 300 или 200 пг/мл. В конкретном варианте осуществления животное семейства мышиных экспрессирует hIL-6 в сыворотке на уровне приблизительно от 50 до приблизительно не более 200 пг/мл, согласно другому варианту осуществления приблизительно 75-125 пг/мл, согласно другому варианту осуществления приблизительно 100 пг/мл.
Согласно одному из аспектов предоставлено отличное от человека животное, которое экспрессирует hIL-6 и/или hIL-6R, где отличное от человека животное экспрессирует hIL-6 и/или hIL-6R с эндогенного не принадлежащего человеку локуса IL-6 и/или эндогенного не принадлежащего человеку локуса hIL-6R. В конкретном варианте осуществления отличное от человека животное является представителем семейства мышиных (например, мышью или крысой).
Согласно одному из аспектов предоставлена генетически модифицированная мышь, которая экспрессирует hIL-6 с эндогенного локуса IL-6 мыши, где эндогенный ген IL-6 мыши заменен геном hIL-6.
Согласно одному из вариантов осуществления мышь несет клетку, которая экспрессируют рецептор IL-6 (IL-6R), который на поверхности клетки содержит внеклеточный домен человека. Согласно одному из вариантов осуществления клетка представляет собой лимфоцит. Согласно одному из вариантов осуществления лимфоцит представляет собой В-клетку.
Согласно одному из вариантов осуществления приблизительно 6,8 т.п.н. в эндогенном локусе IL-6 мыши, включая экзоны с 1 по 5 и а 3'-нетранслируемую последовательность, удалены и замещены последовательностью гена IL-6 человека длиной приблизительно 4,8 т.п.н., содержащей экзоны с 1 по 5 гена IL-6 человека. В конкретном варианте осуществления ген IL-6 человека содержит экзоны с 1 по 5 гена IL-6 человека ВАС человека CTD-2369M23.
Согласно одному из аспектов предоставлена генетически модифицированная мышь, которая экспрессирует IL-6 с гена IL-6 человека, где мышь экспрессирует IL-6 человека в сыворотке.
Согласно одному из вариантов осуществления сыворотка у мышей выявляют сывороточную концентрацию IL-6 человека приблизительно от 25 до приблизительно 300 пг/мл, от 50 до приблизительно 250 пг/мл, от 75 до приблизительно 200 пг/мл или от 100 до приблизительно 150 пг/мл. В конкретном варианте осуществления уровень IL-6 человека в сыворотке мыши составляет приблизительно 100 пг/мл.
Согласно одному из вариантов осуществления уровень общего специфичного для В-клеток маркера в костном мозге мыши является приблизительно таким же, как уровень специфичного для В-клеток маркера в костном мозге мыши дикого типа. Согласно одному из вариантов осуществления уровень общего специфичного для В-клеток маркера в селезенке является приблизительно таким же, как уровень специфичного для В-клеток маркера в селезенке мыши дикого типа. Согласно одному из вариантов осуществления общий специфичный для В-клеток маркер выбран из В220, CD 19, CD20, CD22, CD79a, CD79b, L26 и Pax-5 (BSAP).
Согласно одному из аспектов предоставлена генетически модифицированная мышь, которая экспрессирует hIL6, где у мыши не выявляют свойства, выбранного из плазмацитоза, спленомегалии, увеличения лимфоузлов, уплотненных аномальных плазматических клеток и их сочетания.
Согласно одному из вариантов осуществления мышь обладает селезенкой, масса которой составляет приблизительно такую же массу (от массы тела), что и у мыши дикого типа. Согласно одному из вариантов осуществления масса лимфоузлов мыши составляет приблизительно такую же массу (от массы тела), что и у мыши дикого типа. Согласно одному из вариантов осуществления плазматические клетки мыши не демонстрируют характеристик плазмацитоза, характерных для мышей со сверхэкспрессией IL-6 человека.
Согласно одному из вариантов осуществления у мыши не выявляют гломерулонефрита.
Согласно одному из вариантов осуществления у мыши определяют уровень мезангиальных клеток, сравнимый с мышью дикого типа.
Согласно одному из аспектов предоставлена генетически модифицированная мышь, которая экспрессирует hIL6 с эндогенного локуса IL-6 мыши, где эндогенный ген IL-6 мыши заменен геном hIL-6, где у мыши не выявляют свойства, выбранного из морфологически детектируемой нейропатологии, реактивных астроцитов и их сочетания. Согласно одному из вариантов осуществления мышь обладает головным мозгом, который морфологически неотличим от головного мозга мыши дикого типа. Согласно одному из вариантов осуществления мышь обладает тканью головного мозга, которая демонстрирует уровень реактивных астроцитов, не выше чем уровень реактивных астроцитов мыши дикого типа.
Согласно одному из вариантов осуществления мышь не экспрессирует IL-6 человека в нейронах. Согласно одному из вариантов осуществления у мыши присутствуют уровни активированных астроцитов, сравнимые с уровнями активированных астроцитов у мыши дикого типа.
Согласно одному из вариантов осуществления мышь обладает ветвящимися микроглиальными клетками в белом веществе, где ветвящиеся микроглиальные клетки присутствуют в количестве, эквивалентном количеству ветвящихся микроглиальных клеток у мыши дикого типа.
Согласно одному из вариантов осуществления у мыши не выявляют реактивного астроцитоза. Согласно одному из вариантов осуществления белое вещество мыши морфологически неотличимо от белого вещества мыши дикого типа. Согласно одному из вариантов осуществления белое вещество мыши гистологически неотличима от белого вещества мыши дикого типа в отношении гистохимического окрашивания реактивных астроцитов.
Согласно одному из вариантов осуществления мышь обладает головным мозгом, который морфологически неотличим от головного мозга мыши дикого типа. Согласно одному из вариантов осуществления мышь обладает тканью головного мозга, которая демонстрирует уровень реактивных астроцитов не выше, чем уровень реактивных астроцитов у мыши дикого типа.
Согласно одному из аспектов предоставлена генетически модифицированная мышь, которая экспрессирует hIL6 с эндогенного локуса IL-6 мыши, где эндогенный ген IL-6 мыши заменен геном hIL-6, где у мыши не выявляют свойства, выбранного из продолжительности жизни, укороченной приблизительно на 50% или более, почечной недостаточности, гипергаммаглобулинемии, повышенного количества мегакариоцитов в селезенке, повышенного количества мегакариоцитов в костном мозге, плазмацитоза селезенки, плазмацитоза тимуса, плазмацитоза лимфоузлов, гломерулонефрита, гломерулосклероза и их сочетания.
Согласно одному из вариантов осуществления продолжительность жизни мышей превосходит 20 недель. Согласно одному из вариантов осуществления продолжительность жизни мышей превосходит 30 недель, 40 недель или 50 недель. Согласно одному из вариантов осуществления мыши демонстрируют продолжительность жизни приблизительно равную продолжительности жизни мышей дикого типа той же линии.
Согласно одному из вариантов осуществления у мышей определяют уровень мегакариоцитов в селезенке, который приблизительно не превышает уровень мегакариоцитов в селезенки у мышей дикого типа.
Согласно одному из вариантов осуществления мыши обладают лимфоидными органами, которые по существу не содержат аномальных и плотно расположенных плазмацитоидных клеток.
Согласно одному из вариантов осуществления у мышей определяют уровни гамма-глобулинов в сыворотке, эквивалентные уровням гамма-глобулинов в сыворотке у мышей дикого типа. Согласно одному из вариантов осуществления уровни α1- и β-глобулинов в сыворотке мышей эквивалентны уровням α1- и β-глобулинов в сыворотке мышей дикого типа той же линии.
Согласно одному из аспектов предоставлена генетически модифицированная мышь, которая экспрессирует IL-6 человека с эндогенного локуса IL-6 мыши, где эндогенный ген IL-6 мыши заменен геном hIL-6, где у мыши не выявляют свойства, выбранного из мышечной атрофии, повышенного уровень катепсина В по сравнению с мышью дикого типа той же линии, повышенного уровня катепсинов А+В по сравнению с мышью дикого типа той же линии, увеличенной массы печени по сравнению с мышью дикого типа той же линии и их сочетания.
Согласно одному из вариантов осуществления масса печени мыши на 12 неделе составляет приблизительно 800-900 мг.
Согласно одному из вариантов осуществления у мыши на всем протяжении ее жизни выявляют уровень катепсина В, который приблизительно не превышает уровня, наблюдаемого у мышей дикого типа. Согласно одному из вариантов осуществления у мыши на всем протяжении ее жизни выявляют уровень катепсинов А+В, который приблизительно не превышает уровня, наблюдаемого у мышей дикого типа.
Согласно одному из вариантов осуществления у мыши во взрослом возрасте определяют массу икроножной мышцы, которая находится в пределах приблизительно 10% от массы у мыши дикого типа той же линии. Согласно одному из вариантов осуществления у мыши во взрослом возрасте определяют массу икроножной мышцы, которая приблизительно является такой же, как масса икроножной мышцы мыши дикого типа.
Согласно одному из аспектов предоставлена мышь, которая несет нуклеотидную последовательность, кодирующую белок IL-6 человека, где нуклеотидная последовательность, кодирующая белок IL-6 человека, полностью или частично замещает эндогенную нуклеотидную последовательность, кодирующую эндогенный белок IL-6 мыши.
Согласно одному из аспектов предоставлена мышь, которая несет замену в эндогенном локус рецептора IL-6 мыши внеклеточного домена IL-6Rα мыши на последовательность внеклеточного домена IL-6Rα человека с формированием химерного гена IL-6Rα человека/мыши.
Согласно одному из вариантов осуществления химерный ген IL-6Rα находится под контролем промотора мыши и/или регуляторных элементов мыши в эндогенном локусе IL-6Rα мыши.
Согласно одному из вариантов осуществления последовательность длиной приблизительно 35,4 т.п.н., кодирующая внеклеточный домен IL-6Rα мыши, замещена последовательностью длиной приблизительно 45,5 т.п.н., кодирующей внеклеточный домен IL-6R человека.
Согласно одному из вариантов осуществления последовательность, кодирующая внеклеточный домен IL-6R человека, содержит первый (ATG) кодон от экзона 1 до экзона 8.
Согласно одному из вариантов осуществления замещаемая последовательность IL-6Rα мыши содержит непрерывную последовательность, которая содержит экзоны с 1 по 8. В конкретном варианте осуществления удалены экзоны с 1 по 8 и часть интрона 8.
Согласно одному из аспектов предоставлена генетически модифицированная мышь, несущая замену в эндогенном локусе IL-6 мыши гена мыши, кодирующего IL-6 на ген человека, кодирующий IL-6 человека, где ген человека, кодирующий IL-6 человека, находится под контролем эндогенных регуляторных элементов мыши в эндогенном локусе IL-6 мыши.
Согласно одному из вариантов осуществления ген человека, кодирующий IL-6 человека представляет собой ген IL-6 человека ВАС ID CTD-2369M23.
Согласно одному из вариантов осуществления мышь экспрессирует IL-6Rα мыши. Согласно одному из вариантов осуществления мышь экспрессирует IL-6Rα человека. Согласно одному из вариантов осуществления гуманизированный IL-6Rα содержит внеклеточный домен человека. Согласно одному из вариантов осуществления гуманизированный IL-6Rα содержит трансмембранный домен мыши и цитоплазматический домен мыши. Согласно одному из вариантов осуществления мышь экспрессирует гуманизированный IL-6Rα, который включает гуманизирование внеклеточного домена, но не трансмембранного и/или цитозольного домена.
Согласно одному из вариантов осуществления у мыши не выявляют свойства, выбранного из плазмацитоза, гломерулосклероза, гломерулонефрита, почечной недостаточности, гипергаммаглобулинемии, повышенного количество мегакариоцитов в селезенке, повышенного количества мегакариоцитов в костном мозге, спленомегалии, увеличение лимфоузлов, уплотненных аномальных плазматических клеток и их сочетания.
Согласно одному из аспектов предоставлена генетически модифицированная мышь, несущая гуманизированный эндогенный ген IL-6Rα мыши, где гуманизирование включает замену последовательности, кодирующей внеклеточный домен IL-6Rα мыши, на последовательность, кодирующую внеклеточный домен IL-6Rα человека, в эндогенном локусе IL-6Rα мыши.
Согласно одному из вариантов осуществления непрерывная последовательность мыши, содержащая экзоны с 1 по 8 мыши, замещена непрерывным геномным фрагментом последовательности IL-6Rα человека, кодирующей внеклеточный домен IL-6Rα человека. Согласно одному из вариантов осуществления непрерывный геномный фрагмент последовательности IL-6Rα человека, кодирующей внеклеточный домен, происходит из BACCTD-2192J23.
Согласно одному из вариантов осуществления мышь дополнительно несет гуманизированный ген IL-6. Согласно одному из вариантов осуществления у мыши в эндогенном локусе IL-6 мыши присутствует замена гена IL-6 мыши на ген IL-6 человека. Согласно одному из вариантов осуществления гуманизированный ген IL-6 находится под контролем эндогенных регуляторных элементов мыши.
Согласно одному из аспектов предоставлен способ получения гуманизированной мыши, включающий замену последовательности гена мыши, кодирующей IL-6 мыши на ген человека, кодирующий IL-6 человека.
Согласно одному из вариантов осуществления замену проводят в эндогенном локусе IL-6 мыши, и ген человека, кодирующий IL-6 человека, функционально связан с эндогенными регуляторными последовательностями мыши.
Согласно одному из аспектов предоставлен способ получения гуманизированной мыши, включающий замену экзонов мыши, кодирующих последовательности внеклеточного домена мыши IL-6Rα геномным фрагментом человека, кодирующим последовательности внеклеточного домена IL-6Rα человека с получением гуманизированного гена IL-6Rα.
Согласно одному из вариантов осуществления замену проводят в эндогенном локусе IL-6Rα мыши, и гуманизированный ген IL-6Rα функционально связан с эндогенными регуляторными последовательностями мыши.
Согласно одному из аспектов предоставлена генетически модифицированная мышь, несущая гуманизированный ген IL-6Rα, содержащий замену последовательности, кодирующей внеклеточный домен мыши, последовательностью внеклеточного домена человека, где гуманизированный ген IL-6Rα содержит трансмембранную последовательность мыши и цитоплазматическую последовательность мыши; где мышь дополнительно несет ген, кодирующий IL-6 человека, где ген, кодирующий IL-6 человека, находится под контролем эндогенных регуляторных элементов IL-6 мыши.
Согласно одному из вариантов осуществления мышь не способна к экспрессии полностью принадлежащий мыши IL-6Rα и не способна к экспрессии IL-6 мыши.
В различных аспектах генетически модифицированная мыши, описываемая в настоящем документе, несет генетические модификации в зародышевой линии.
Согласно одному из аспектов предоставлены ткань, клетка или фрагмент мембраны мыши, как описано в настоящем документе.
Согласно одному из вариантов осуществления ткань или клетка получены у мыши, которая экспрессирует белок IL-6 человека, но не экспрессирует белок IL-6 мыши. Согласно одному из вариантов осуществления ткань или клетка получены у мыши, которая экспрессирует гуманизированный белок IL-6Rα, но не белок IL-6Rα мыши. Согласно одному из вариантов осуществления гуманизированный белок IL-6Rα содержит внеклеточный домен человека и трансмембранный домен мыши и цитозольный домен мыши. Согласно одному из вариантов осуществления ткань или клетка получены у мыши, которая экспрессирует IL-6 человека, гуманизированный IL-6Rα и не экспрессирует IL-6 мыши и не экспрессирует IL-6Rα, который содержит внеклеточный домен мыши.
Согласно одному из аспектов предоставлен комплекс клетки мыши ex vivo, несущей гуманизированный IL-6Rα (внеклеточный домен человека и трансмембранный домен мыши и цитоплазматический домен мыши) и IL-6 человека.
Согласно одному из аспектов предоставлен эмбрион мыши, несущий генетическую модификацию, как описано в настоящем документе.
Согласно одному из аспектов предоставлен эмбрион мыши-хозяина, который содержит донорскую клетку, несущую генетическую модификацию, как описано в настоящем документе.
Согласно одному из аспектов предоставлена плюрипотентная или тотипотентная клетка отличного от человека животного, несущая генетическую модификацию, как описано в настоящем документе. Согласно одному из вариантов осуществления клетка представляет собой клетку мыши. Согласно одному из вариантов осуществления клетка представляет собой ES клетку.
Согласно одному из аспектов предоставлена яйцеклетка мыши, где яйцеклетка мыши несет эктопическую хромосому мыши, где эктопическая хромосома мыши содержит генетическую модификацию, как описано в настоящем документе.
Согласно одному из аспектов мышь, эмбрион, яйцеклетка или клетка, которая генетически модифицированы, чтобы нести ген IL-6 человека или ген IL-6α человека или гуманизированный ген IL-6Rα человека принадлежат мышам линии C57BL, выбранной из C57BL/A, C57BL/An, C57BL/GrFa, C57BL/KaLwN, C57BL/6, C57BL/6J, C57BL/6ByJ, C57BL/6NJ, C57BL/10, C57BL/10ScSn, C57BL/10Cr и C57BL/01a. Согласно другому варианту осуществления мышь принадлежит линии 129, выбранной из группы, состоящей из линий, представляющих собой 129Р1, 129Р2, 129РЗ, 129Х1, 129S1 (например, 129S1/SV, 12981/SvIm), 129S2, 129S4, 129S5, 129S9/SvEvH, 129S6 (129/SvEvTac), 129S7, 129S8, 129Т1, 129Т2 (см., например, Festing et al. (1999) Revised nomenclature for strain 129 mice. Mammalian Genome 10:836, также см., Auerbach et al (2000) Establishment and Chimera Analysis of 129/SvEv- and C57BL/6-Derived Mouse Embryonic Stem Cell Lines). В конкретном варианте осуществления генетически модифицированная мышь представляет собой метиса указанной выше линии 129 и указанной выше линии C57BL/6. В другом конкретном варианте осуществления мышь представляет собой метиса указанных выше линий 129 или метиса указанных выше линий BL/6. В конкретном варианте осуществления линия 129 для метиса представляет собой линию 129S6 (129/SvEvTac). Согласно другому варианту осуществления мышь принадлежит линии BALB, например, линии BALB/c. В еще одном варианте осуществления мышь представляет собой метиса линии BALB и другой указанной выше линии. Согласно одному из вариантов осуществления мышь представляет собой мышь Swiss или Swiss Webster.
Каждый из аспектов и вариантов осуществления, описываемых в настоящем документе, можно использовать вместе, если недвусмысленно или явно не исключено из контекста варианта осуществления или аспекта.
Краткое описание чертежей
На фиг. 1 предоставлена иллюстрация без соблюдения масштаба геномных локусов IL-6 человека (вверху) и мыши (внизу). Экзоны I, II, III, IV и V (у человека и мышь) указаны закрытыми рамками в правой части фигуры. Выбранные предполагаемые регуляторные области указаны открытыми рамками в левой части фигуры.
На фиг. 2 представлен ответ острой фазы (уровень mSAA) в присутствии или отсутствие скипидара у мышей дикого типа, мышей с гуманизированным внеклеточным доменом IL-6R и мышей с гуманизированными генами IL-6 и IL-6R.
На фиг. 3 представлен зависимый от скипидара ответ острой фазы (SAA) у мышей дикого типа в отсутствии или присутствии антитела к IL-6R мыши (слева); и зависимый от скипидара ответ острой фазы у мышей с гуманизированными IL-6/IL-6R в отсутствие или присутствии антитела к IL-6R человека (справа).
На фиг. 4 представлен анализ FACS для В-клеток селезенки мышей с IL-6 дикого типа и гуманизированным IL-6; общий маркер В-клеток.
На фиг. 5 представлен анализ FACS для Т-клеток селезенки мышей с IL-6 дикого типа и гуманизированным IL-6; Т-клетки хелперы и цитотоксические Т-клетки.
На фиг. 6 представлен анализ FACS для клеток селезенки мышей с IL-6 дикого типа и гуманизированным IL-6; Ly6G/C(Gr1).
На фиг. 7 представлен анализ FACS для клеток селезенки мышей с IL-6 дикого типа и гуманизированным IL-6; NK клетки и грану лоциты (Ly6Ghi+/CD116hi+).
На фиг. 8 представлен анализ FACS В-клеток крови мышей с IL-6 дикого типа и гуманизированным IL-6; общий маркер В-клеток.
На фиг. 9 представлен анализ FACS Т-клеток крови мышей с IL-6 дикого типа и гуманизированным IL-6; Т-клетки хелперы и цитотоксические Т-клетки.
На фиг. 10 представлен анализ FACS для миелоидных клеток крови мышей с IL-6 дикого типа и гуманизированным IL-6; клетки Gr1+.
На фиг. 11 представлен анализ FACS для миелоидных клеток крови мышей с IL-6 дикого типа и гуманизированным IL-6; CD11b в сравнении с.Ly6G/C(Gr1).
На фиг. 12 представлен анализ FACS для миелоидных клеток крови мышей с IL-6 дикого типа и гуманизированным IL-6; клетки, несущие DX5, в сравнении с клетками, несущими CD11b.
На фиг. 13 представлен анализ FACS костного мозга IgM/CD24/B220 для мышей с IL-6 дикого типа и гуманизированным IL-6. Вверху: нормальное развитие в костном мозге. Внизу: анализ FACS для дикого типа, гетерозигот hIL-6 и гомозигот hIL-6 (окрашивание IgM).
На фиг. 14 представлен анализ FACS костного мозга IgM/CD24/B220 для мышей с IL-6 дикого типа и гуманизированным IL-6. Вверху: нормальное развитие в костном мозге. Внизу: анализ FACS для дикого типа, гетерозигот hIL-6 и гомозигот hIL-6 (окрашивание CD24).
На фиг. 15 представлен анализ FACS костного мозга CD43 и В220 для мышей с IL-6 дикого типа и гуманизированным IL-6. Вверху: нормальное развитие в костном мозге. Внизу: анализ FACS для дикого типа, гетерозигот hIL-6 и гомозигот hIL-6 (окрашивание CD43).
Подробное раскрытие настоящего изобретения
IL-6 и IL-6R
Рецептор IL-6 (IL-6R) давно охарактеризован, как рецептор для стимулирующего В-клетки фактора (BSF-2 или фактор стимуляции В-клеток 2; также, BCDF или фактор дифференцировки В-клеток), ответственного за индукцию синтеза иммуноглобулина В-клетками (Yamasaki et al. (1988) Cloning and Expression of the Human Interleukin-6(BSF-2/IFNβ 2) Receptor, Science 241: 825-828). IL-6 впервые описан как интерферон-β2 как результат его открытия при поиске индуцируемого вирусами белка, названного интерферон-β, при обработке фибробластов человека дцРНК поли(I)поли(С) с индукцией противовирусного ответа (Weissenbach et al. (1980) Two interferon mRNAs in human fibroblasts: In vitro translation and Escherichia coli cloning studies, Proc. Natl Acad. Sci. USA 77(12): 7152-7156; Keller et al. (1996) Molecular and Cellular Biology of Interleukin-6 and Its Receptor, Frontiers in Bioscience 1: d340-357).
кДНК человека кодирует белок из 468 аминокислот, содержащий 19-членную сигнальную последовательность и цитоплазматический домен приблизительно из 82 аминокислот, в котором отсутствует тирозинкиназный домен (см. там же). N-концевой (внеклеточный) домен белка содержит домен Ig суперсемейства приблизительно из 90 аминокислот, домен из 250 аминокислот между доменом Ig суперсемейства и мембраной, трансмембранный участок приблизительно из 28 аминокислот (см. там же). Внеклеточный домен рецептора связывается с его лигандом IL-6, который инициирует ассоциацию с gp130 в мембране, и образуется комплекс, который передает сигнал; цитоплазматический домен по опубликованным данным не передает сигнал (Taga et al. (1989) Interleukin-6 Triggers the Association of Its Receptor with a Possible Signal Transducer, gp130, Cell 58: 573-581)). Фактически, растворимая форма IL-6R с отсутствием цитоплазматического домена может ассоциировать с IL-6 и связывать gp130 на поверхности клетки и эффективно передавать сигнал (там же).
Гомология hIL-6R и mIL-6R на уровне белка составляет приблизительно только 54%; гомология трансмембранных доменов составляет приблизительно 79%, тогда как гомология цитоплазматических доменов составляет приблизительно 54% (Sugito et al. (1990)).
Природный лиганд IL-6R, IL-6, впервые выделен из культур трансформированных HTLV-1 Т-клеток (см., Hirano et al. (1985) Purification to homogeneity and characterization of human В cell differentiation factor (BCDF or BSFβ-2), Proc. Natl. Acad. Sci. USA 82: 5490-5494). кДНК для гена IL-6 человека клонировали по меньшей мере дважды, один раз как BSF-2 (см., Hirano et al. (1086) Complementary DNA fro a novel human interleukin (BSF-2) that induces В lymphocytes to produce immunoglobulin. Nature 324: 73-76) и один раз как IFNβ 2 (см., Zilberstein et al. (1986) Structure and expression of cDNA and genes for human interferon-beta-2, a distinct species inducible by growth-stimulatory cytokines, EMBO 5: 2529-2537), хотя с тех пор показано, что у рекомбинантного IL-6 человека не выявляют детектируемой активности IFN.
IL-6 человека представляет собой белок из 184 аминокислот, у которого выявлено только приблизительно 42% гомологии с IL-6 мыши, хотя геномная организация генов человека и мыши в основном является сходной, и промоторные области генов человека и мыши содержат участок в 400 п.н., который является высококонсервативным (см., Tanabe et al. (1988) Genomic Structure of the Murine IL-6 Gene: High Degree Conservation of Potential Regulatory Sequences between Mouse and Human, J. Immunol. 141(11): 3875-3881).
Ген IL-6 человека приблизительно состоит из 5 т.п.н. (Yasukawa et al. (1987) Structure and expression of human В cell stimulatory factor-2 (BSC-2/IL-6) gene, EMBO J. 6(10): 2939-2945), тогда как ген IL-6 мыши состоит приблизительно из 7 т.п.н. (Tanabe et al. (1988) Genomic Structure of the Murine IL-6 Gene: High Degree Conservation of Potential Regulatory Sequences between Mouse and Human, J. Immunol. 141(11): 3875-3881). Гены IL-6 мыши и человека по опубликованным данным содержат высококонсервативную 5'-фланкирующую последовательность, важную для регуляции. На фиг. 1 представлена схематическая диаграмма геномные локусы IL-6 человека и мыши (без соблюдения масштаба). Экзоны I, II, III, IV и V (у человека и мыши) указаны закрытыми рамками в правой части фигуры. Выбранные предполагаемые регуляторные области указаны открытыми рамки в левой части фигуры. Предполагаемые регуляторные области у людей представляют собой, слева направо, глюкокортикоидный элемент от -557 до -552; короную последовательность IFN-энхансера от -472 до -468; глюкокортикоидный элемент от -466 до -461; АТ-богатую область от -395 до -334, консенсусный участок связывания АР-1 от -383 до -277; коровую последовательность IFN-энхансера от -253 до -248; содержащий GGAAA мотив от -205 до -192; последовательность, гомологичную SRE c-fos от -169 до -82, содержащую коровую последовательность IFN-энхансера, элемент ответа на цАМФ, мотив GGAAA, участок ССААТ и GC-богатую область; и участок связывания АР-1 от -61 до -55; и участок ССААТ от -34 до -30. Предполагаемые регуляторные области у мыши представляют собой, слева направо, GC-богатую область от -553 до -536, глюкокортикоидный элемент от -521 до -516 и от -500 до -495; участок Z-ДНК от -447 до -396; участок связывания АР-1, перекрывающий коровую последовательность IFN-энхансера от -277 до -288, мотив GGAAA, перекрывающий коровую последовательность IFN-энхансера от -210 до -195; область гомологии с SRE c-fos от -171 до -82, содержащий элемент ответа на цАМФ, мотив GGAAA, перекрывающий коровую последовательность IFN-энхансера, и GC-богатую область; и, участок связывания АР-1 от -61 до -55. Длина кодонов мыши I-V составляют 19, 185, 114, 150 и 165, соответственно. Длины интронов мыши составляют: I-II, 162 п.н.; II-III, 1253 п.н.; III-IV, 2981 п.н.; IV-V, 1281 п.н. Длина кодонов человека I-V составляет 19, 191, 114, 147 и 165. Длины интронов человека составляют I-II, 154; II-III, 1047; III-IV, 706; IV-V, 1737. Данные по организации генома взяты из Tanabe et al. (1988) и Yasukawa et al. (1987) Structure and expression of human В cell stimulatory factor-2 (BSF-2/IL-6) gene, EMBO J. 9(10): 2939-2945.
На основе сходства 5'-фланкирующей последовательности генов IL-6 мыши и человека можно обоснованно полагать, что гены IL-6 мыши и человека, по-видимому, регулируются сходным образом. У множества типов клеток в ответ на IL-1, TNF, PDGF, IFNJ3, сыворотку, поли(1)поли(С) и циклогексимид увеличивается экспрессия IL-6 (см. Tanabe et al. (1988). IL-6 у людей опосредует ответ острой фазы, гемопоэз, В-клеточную дифференцировку, активацию Т-клеток, рост, и/или дифференцировку, и/или активацию множества типов клеток (например, гепатоцитов, фибробластов, эндотелиальных клеток, нейронов, клеток гипофиза, лимфом, миелом, карцином молочной железы, NK клеток, макрофагов, остеокластов и т.д.) (рассмотрено, например, в Heinrich et al. (1990), Kishimoto et al. (1989) и Keller et al. (1996); Sugita et al. (1990) Functional Murine Interleukin Receptor with Intracisternal A Particle Gene Product at its Cytoplasmic Domain, J. Exp. Med. 171: 2001-2009).
Однако на практике у мышей, трансгенных по IL-6 человека, выявляют множество существенных и инвалидизирующих патологий, отражающих значительную плейотропию гена IL-6. Трансгенные мыши, несущие фрагмент 6,6 т.п.н., содержащий ген IL-6 человека и энхансер μ (Eμ), продуцируют высокие концентрации hIL-6 и экстремально высокие уровни IgGI (в 120-400 раз выше, чем у мышей дикого типа), отражая нарушение регуляции IL-6, которое сопровождается плазмацитозом, мезангиальным пролиферативным гломерулонефритом и высокими уровнями мегакариоцитов в костном мозге (Suematsu et al. (1989) IgGI plasmacytosis in interleukin 6 transgenic mice, Proc. Nati Acad. Sci. USA 86: 7547-7551). Нарушенная регуляция IL-6 и/или IL-6R ассоциирована с миеломами, плазмацитомами, ревматоидным артритом, болезнью Кастлемана, мезангиальным пролиферативным гломерулонефритом, миксомой сердца, неоплазиями плазматических клеток, псориазом и другими нарушениями (см., Kishimoto, Т. (1989) The Biology of Interleukin-6, Blood 74(1): 1-10; Sugita et al. (1990); also, Hirano et al. (1990) Biological and clinical aspects of interleukin 6, Immunology Today 11(12): 443-449)). IL-6 также участвует в поддержание уровней внутрипростатических андрогенов при лечении пациентов с раком предстательной железы посредством депривации андрогенов по паракринному и/или аутокринному механизмам, потенциально приводя к устойчивому к кастрации росту опухолей предстательной железы (Chun et al. (2009) Interleukin-6 Regulates Androgen Synthesis in Prostate Cancer Cells, Clin. Cancer Res. 15: 4815-4822).
Белок человека закодирован как белок из 212 аминокислот, в зрелой форме, после отщепления сигнальной последовательности из 28 аминокислот, белок из 184 аминокислот. Он содержит два участка N-гликозилирования и два участка O-гликозилирования, и IL-6 человека в некоторых клетках фосфорилирован. Белок мыши закодирован как белок из 211 аминокислот, в зрелой форме, после отщепления сигнальной последовательности из 23 аминокислот, белок из 187 аминокислот. O-участки гликозилирования присутствуют, а участки N-гликозилирования - нет. (См. обзоры об IL-6, например, Heinrich et al. (1990) Interleukin-6 and the acute phase response, Biochem. J. 265: 621-636).
IL-6 обладает плейотропным действием. Рецептор IL-6 выявлен на активированных В-клетках, но по опубликованным данным не на покоящихся В-клетках. В отличие от этого, IL-6R выявлен на покоящихся Т-клетках и по опубликованным данным может стимулировать дифференцировку, активацию и пролиферацию Т-клеток, включая дифференцировку Т-клеток в цитотоксические Т-лимфоциты в присутствии IL-2.
Мыши с гуманизированными IL-6/внеклеточным доменом IL-6R и опосредуемый IL-6 ответ острой фазы
У людей IL-6 индуцирует ответ острой фазы. Ранние исследования с гепатоцитами человека установили, что IL-6 индуцирует белки острой фазы, например, такие как С-реактивный белок (CRP) и сывороточный амилоид A (SAA) зависимым от дозы и времени образом (рассмотрено в Heinrich et al. (1990) Interleukin-6 and the acute phase response, Biochem. J. 265: 621-636). Таким образом, отличные от человека животные, например, мыши или крысы, содержащие гуманизированные гены IL-6 и IL-6R, являются подходящими системами для измерения ответа острой фазы, опосредуемого IL-6 человека. Такие животные также подходят для определения индукции веществом опосредуемого IL-6 ответа острой фазы, посредством воздействия веществом на животное с гуманизированными IL-6/IL-6R, как описано в настоящем документе, и измерения уровня одного или более белков (или РНК) ответа острой фазы. Согласно одному из вариантов осуществления на гуманизированное животное воздействуют веществом в присутствии антагониста IL-6R человека и измеряют уровень одного или более белков (или РНК) ответа острой фазы, где снижение уровня белка (или РНК) ответа острой фазы в присутствии антагониста IL-6R человека указывает на опосредуемый IL-6R человека ответ острой фазы.
IL-6 человека может связывать IL-6R человека и IL-6R мыши; IL-6 мыши связывается с IL-6R мыши, но не с IL-6R человека (отсутствие связывания mIL-6 с hIL-6R можно выявлять, тогда как hIL-6 может конкурировать с mIL-6 за связывание с mIL-6R; Coulie et al. (1989) High- and low-affinity receptors for murine interleukin 6. Distinct distribution on В and Т cells, Eur. J. Immunol. 19: 2107-211); также см., например, Peters et al. (1996) The Function of the Soluble Interleukin 6 (IL-6) Receptor In Vivo: Sensitization of Human Soluble IL-6 Receptor Transgenic Mice Towards IL-6 and Prolongation of the Plasma Half-life of IL-6, J. Exp. Med. 183: 1399-1406). Таким образом, клетки человека, несущие hIL-6R, у мыши (например, в ксеногенном трансплантате), не могут зависеть от эндогенного mIL-6 для осуществления опосредуемых IL-6 функций, включая в качестве неограничивающих примеров роль IL-6 в развитии клеток крови или лимфоцитов (например, гемопоэз, активация В-клеток, активация Т-клеток и т.д.).
В смешанной системе in vivo, содержащей ген IL-6 мыши дикого типа и ген IL-6R человека (но не ген IL-6R мыши), не ожидается, что индуктор ответа острой фазы будет индуцировать детектируемые уровни белков острой фазы, которые будут указывать на ответ острой фазы. Однако гуманизированная мышь, как описано в настоящем документе, несущая гуманизированный ген IL-6 и ген IL-6R, содержащий гуманизированную последовательность внеклеточного домена, будет отвечать на индуктор ответа острой фазы и демонстрировать белки ответа острой фазы в сыворотке. Мыши дикого типа по IL-6/IL-6R, тестируемые на белки острой фазы в присутствии или отсутствие индуктора острой фазы, скипидара, демонстрировали зависимое от скипидара увеличение уровня белков острой фазы. У мышей с гуманизированным геном IL-6, но не с IL-6R, не выявляли ответа острой фазы в присутствии скипидара. Но у мышей, несущих ген IL-6 человека и ген IL-6R с гуманизированным внеклеточным доменом, выявляли сильный ответ острой фазы (фиг. 2). Опосредуемый IL-6 ответ острой фазы зависел от IL-6 у мышей дикого типа (фиг. 3, вверху) и у мышей с гуманизированными IL-6/внеклеточным доменом IL-6R (фиг. 3, внизу), о чем свидетельствует способность соответствующего антитела к IL-6R при достаточно высокой дозе антитела устранять ответ острой фазы. Таким образом, двойное гуманизирование IL-6 и IL-6R воспроизводит опосредуем IL-6 дикого типа ответ острой фазы в отношении сывороточных белков острой фазы.
Генетически модифицированные мыши
Предоставлены генетически модифицированные мыши, которые экспрессируют IL-6 человека и/или гуманизированный рецептор IL-6 с эндогенных локусов мыши, где эндогенный ген IL-6 мыши и/или эндогенный ген рецептора IL-6 мыши заменены на ген IL-6 человека и/или последовательность человека, содержащую последовательность, кодирующую внеклеточный домен рецептора IL-6 человека. Генетически модифицированные мыши экспрессируют IL-6 человека и/или гуманизированный рецептор IL-6 с гуманизированных эндогенных локусов, которые находятся под контролем промоторов мыши и/или регуляторных элементов мыши. Замена(ы) эндогенных локусов мыши позволяет получать отличных от человека животных, которые экспрессируют IL-6 человека и гуманизированный рецептор IL-6 таким образом, что это не приводит к множеству существенных патологий, наблюдаемых у трансгенных по IL-6 мышей, известных в данной области.
Трансгенные мыши, экспрессирующие IL-6 человека, известны в данной области. Однако, как правило, они страдают значительными патологиями, которые сильно ограничивают их пользу. Гуманизированные мыши, как описано в настоящем документе, экспрессируют IL-6 человека и/или гуманизированный рецептор IL-6 под контролем эндогенных регуляторных элементов мыши в эндогенных локусах IL-6 и IL-6Rα мыши. У этих мышей, наоборот, определяют профили экспрессии эти генов, которые отличаются от профилей экспрессии трансгенных мышей, известных в данной области.
Замена не принадлежащих человеку генов у отличного от человека животного гомологичными или ортологичными генами человека или последовательностями человека в эндогенном не принадлежащем человеку локусе и под контролем эндогенных промоторов и/или регуляторных элементов может позволять получать отличного от человека животного с качествами и характеристиками, которые могут значительно отличаться от типичного нокаутного и трансгенного животного. У типичного нокаутного и трансгенного животного эндогенный локус удален или поврежден и в геном животного встроен и предположительно случайно интегрирован полностью человеческий трансген. Как правило, положение интегрированного трансгена неизвестно; экспрессию белка человека определяют посредством транскрипции гена человека, и/или анализа белка, и/или функционального анализа. Очевидно, полагают, что включение в трансген человека последовательностей человека выше и/или ниже по ходу транскрипции является достаточным для обеспечения подходящей поддержки для экспрессии и/или регуляции трансгена в любом месте встраивания трансгена в геном животного. Но во многих случаях трансген с регуляторными элементами человека экспрессируется таким образом, который не является нефизиологическим или иным образом неудовлетворительным и фактически может быть вредным для животного. В отличие от этого, авторы изобретения демонстрируют, что замена последовательностью человека в эндогенном локусе под контролем эндогенных регуляторных элементов обеспечивает физиологически соответствующие профиль экспрессии и уровень, позволяющие получить в результате подходящее гуманизированное животное, физиология которого в отношении замещенного гена является полноценной и допустимой и подходящей для физиологии гуманизированного животного.
Оплодотворенные яйцеклетки мыши с инъецированной конструкцией, содержащей промотор МНС класса I H2 и интрон β-глобина, направляющие экспрессию 695 п.н. гена IL-6 мыши по опубликованным данным приводят к получению мышей, которые конститутивно экспрессируют IL-6 мыши на относительно высоких уровнях (по сравнению с мышами дикого типа) (см. Woodrofe et al. (1992) Long-Term Consequences of Interleukin-6 Overexpression in Transgenic Mice, DNA and Cell Biology 11(8): 587-592). Ho эти мыши подвержены развитию лимфомы, ассоциированной с кишечником, лимфоузлами и почками, а также отложению амилоида в почках. У них также выявляют аномальное созревание В-клеток (см. Woodrofe et al., там же), таким образом, исследования функции В-клеток нарушено. В отличие от этого, мыши, как описано в настоящем документе, несущие замену гена IL-6 мыши на ген IL-6 человека в локусе IL-6 мыши не подвержены развитию этих лимфом, и у мышей определяют очевидные нормальные популяции В-клеток.
В литературе описаны мыши (C57BL/6), трансгенные по hIL-6 после случайной вставки участка ДНК человека длиной 6,6 т.п.н. (фрагмент BamHI-Pvu II), содержащего ген hIL-6 в сочетании с энхансером IgM (см. Suematsu et al. (1989) IgGI plasmocytosis in interleukin 6 transgenic mice, Proc. Natl. Acad. Sci. USA 86: 7547-7551). Мыши экспрессируют hIL-6 в сыворотке на уровне от 800 пг/мл до 20000 пг/мл, тогда как мыши дикого типа, как правило, экспрессируют только приблизительно 100 пг/мл IL-6. У мышей выявляют возрастание Ig в сыворотке (в 120-400 раз по сравнению с мышами дикого типа) и снижение альбумина по мере их взросления. Мыши страдают от тяжелого плазмацитоза, у них выявляют спленомегалию и увеличение лимфоузлов, а также у них выявляют плазматические клетки и увеличенное количество мегакариоцитов в костном мозге. При исследовании того, что выглядит увеличенными лимфоузлами, вместо этого в них сосредоточены уплотненные аномальные плазматические клетки. В селезенке и тимусе выявляют массовую пролиферацию плазматических клеток, которые также инфильтрируют части легких, печени и почек. В почках у этих мышей также выявляют стимулируемую IL-6 пролиферацию мезангиальных клеток, типичную для мезангиального пролиферативного гломерулонефрита. Подобным образом, у мышей (BALB/c), трансгенных по укороченной кДНК hIL-6 под контролем промотора H-2Ld мыши, случайно встроенной в геном, выявляют тяжелый плазмацитоз (см. Suematsu et al. (1992) Generation of plasmacytomas with the chromosomal translocation t(12;15) in interleukin 6 transgenic mice, Proc. Natl. Acad. Sci. USA 89: 232-235). Хотя у мышей C57BL/6 со сверхэкспрессией hIL-6 не развиваются трансплантационные плазмацитомы (у них выявляют плазмацитоз), у трансгенных мышей BL/6, подвергаемых обратному скрещиванию с мышами BALB/c, они, по опубликованным данным, развиваются.
Случайный трансгенез кДНК hIL-6 под контролем промотора гена глиального фибриллярного кислого белка (GFAP) по опубликованным данным приводит к сверхэкспрессии hIL-6 в центральной нервной системе мышей, что также приводит к значительным патологиям (см. Campbell et al. (1993) Neurologic disease induced in transgenic mice by cerebral overexpression of interleukin 6, Proc. Natl. Acad. Sci. USA 90: 10061-10065). У этих мышей выявляют обширную нейропатологию и реактивные астроциты, являющиеся результатом экспрессии IL-6 в ЦНС вследствие потери контроля в результате случайной интеграции трансгена IL-6 в очевидный пермиссивный для ЦНС транскрипционный локус. Хотя экспрессия кДНК hIL-6, сцепленной с 3'-UTR β-глобина и находящейся под контролем промотора нейронспецифической енолазы, микроинъецированной в оплодотворенные яйцеклетки мыши (Fl C57BL/6 х BALB/c), приводила получению мышей с нормальной продолжительностью жизни и без видимых неврологических деффектов, у которых hIL-6 экспрессировался в нейронах, но больше нигде (см. Fattor et al. (1994) IL-6 Expression in Neurons of Transgenic Mice Causes Reactive Astrocytosis and Increase in Ramified Microglial Cells But No Neuronal Damage, Eur. J. Neuroscience 7: 2441-2449), у мышей выявляли высокие уровни (в 20-30 раз выше, чем у дикого типа) активированных и увеличенных астроцитов с увеличенными отростками во всем головном мозге, а также возрастание в 10-15 раз ветвящихся микроглиальных клеток в белом веществе. Таким образом, по опубликованным данным экспрессия IL-6 в головном мозге приводит к состояниям, которые находятся в диапазоне от реактивных астроцитов до явной и глубокой нейропатологии.
Микроинъекция в оплодотворенные яйцеклетки потомства F1 после скрещивания мышей C57BL/6x"DBAII" 639 п.н. кДНК hIL-6, сцепленной с 3'-UTR β-глобина и промотором МТ-1 мыши по опубликованным данным приводила к трансгенной мыши, у которой случайно встроенный ген hIL-6 приводил к получению ослабленной и подверженной заболеваниям мыши, которая погибала в молодом возрасте от почечной недостаточности (см. Fattori et al. (1994) Blood, Development of Progressive Kidney Damage and Myeloma Kidney in Interleukin-6 Transgenic Mice, Blood 63(9): 2570-2579). Трансгенные мыши погибали через 12-20 недель, и у них выявляли повышенные уровни α1- и β-глобулинов в плазме, гипергаммаглобулинемию, повышенное количество мегакариоцитов в селезенке (в 3 раза выше, чем у дикого типа) и костном мозге, плазмацитоз лимфоидных органов (селезенка, тимус и лимфоузлы), характеризуемый аномальными и плотно расположенными плазмацитоидными клетками, и гломерулонефрит, приводящий к гломерулосклерозу, сходному со множественной миеломой.
Микроинъекция в оплодотворенные яйцеклетки мыши C57BL/6J кДНК hIL-6 под контролем H-2L приводила к зависимой от IL-6 мышечной атрофии у мышей, частично характеризуемой значительно меньшей массой икроножной мышцы у трансгенных мышей по сравнению с совпадающим по массе контролем, различие, которое снижалось при обработке антагонистом IL-6 (см., Tsujinaka et al. (1996) Interleukin 6 Receptor Antibody Inhibits Muscle Atrophy and Modulates Proteolytic Systems in Interleukin 6 Transgenic Mice, J. Clin. Invest. 97(1): 244-249). Ha 12 неделе у этих мышей выявляли уровни hIL-6 в сыворотке более 600000 пг/мл. У трансгенных мышей также выявляли печень с массой приблизительно 1,242 мг в сравнении с контрольной печенью с массой приблизительно 862 мг. У трансгенных мышей, обработанных антагонистом IL-6, масса печени составляла приблизительно 888 мг. У трансгенных мышей были значительно повышены уровни мышечных катепсинов В и B+L (в 20 раз и в 6,2 раз) по сравнению с контролем, признак, который исчезал у трансгенных мыши при обработке антагонистом IL-6. Определяли, что мРНК катепсинов по сравнению с мышами дикого типа В и L составляла приблизительно 277% и 257% соответственно; различие значительно уменьшалось при обработке антагонистом IL-6.
У мышей, несущих миниген hIL-6 под контролем промотора H-2Ld MHC класса I мыши и миниген hIL-6R под контролем промотора β-актина курицы, и ген gp130, выявляли патологии, типичные для мышей, трансгенных по hIL-6 (например, гипергаммаглобулинемию, спленомегалию, мезангиальный пролиферативный гломерулонефрит, лимфоидная инфильтрация легких), а также гипертрофию желудочков (Hirota et al. (1995) Continuous activation of gp130, a signal-transducing receptor component for interleukin 6-related cytokines, causes myocardial hypertrophy in mice, Proc. Nati Acad. Sci. USA 92: 4862-4866). Полагают, что гипертрофия желудочка опосредована непрерывной активацией gp130 (там же). Роль IL-6 по опубликованным данным состоит в помощи стабилизации комплекса цитокинового рецептора и индукции димеризации gp130, который представляет собой передающий сигнал компонент, ответственный за передачу сигнала IL-6 (Paonessa et al. (1995) Two distinct and independent sites on IL-6 trigger gp130 dimer formation and signalling, EMBO J. 14(9): 1942-1951). Полагают, что активированный комплекс представляет собой гексамер, состоящий из двух два IL-6, где каждый IL-6 связан с одним IL-6Rα, и двух gp130 (каждый IL-6 содержит два независимых участка связывания gp130) демонстрируя стехиометрию 2:2:2, где димеризация gp130 вызывает активацию тирозинкиназы JAK-Tyk, фосфорилирование gp130 и семейства факторов транскрипции STAT и других внутриклеточных субстратов (там же; Stahl, N. (1994) Association and Activation of Jak-Tyk Kinases by CNTF-LIF-OSM-IL-6 β Receptor Components, Science 263: 92-95), в соответствии с общей моделью формирования комплекса цитокиновых рецепторов (см. Stahl, N. and Yancopoulos, G. (1993) The Alphas, Betas, and Kinases of Cytokine Receptor Complexes, Cell 74:587-590; Davis et al. (1993) LIFRβ and gp130 as Heterodimerizing Signal Transducers of the Tripartite CNTF Receptor, Science 260: 1805-1808; Murakami et al. (1993) IL-6-Induced Homodimerization of gp130 and Associated Activation of a Tyrosine Kinase, Science 2601808-1810).
Мыши, трансгенные по sIL-6R человека под управлением промотора PEP крысы и IL-6 человека под управлением промотора металлотионеина-1 мыши, по опубликованным данным были заметно меньше, чем мыши, трансгенные только по IL-6 человека или только sIL-6R человека (Peters et al. (1997) Extramedullary Expansion of Hematopoietic Progenitor Cells in Interleukin(IL-)-6-sIL-6R Double Transgenic Mice, J. Exp. Med. 185(4):755-766), что отражалось в сниженной жировой ткани и сниженной массе (20-25 г в отличие от 40 г). По опубликованным данным у двойных трансгенных мышей также выявляли увеличение селезенки (в 5 раз) и печени (в 2 раза) по сравнению с по опубликованным данным для нормальных масс органов у мышей с одним трансгеном, по-видимому вследствие экстрамедуллярной пролиферации гемопоэтических клеток в селезенке и печени, но не в костном мозге, а также повышенного количества мегакариоцитов в селезенке и инфильтратов плазматических клеток во всех паренхиматозных органах (там же). У двойных трансгенных животных также выявляли печень с увеличенным в количестве приблизительно от 200 до приблизительно 300 раз уровнем гранулоцитов, макрофагов, клеток-предшественников и В-клеток по сравнению с животными с одним трансгеном; в отличие от этого у мышей с одним трансгеном IL-6 выявляли меньшее количество макрофагов (в 15 раз) и В-клеток (в 45 раз) (там же). Необычные результаты получены предположительно вследствие стимуляции роста и дифференцировки гемопоэтических клеток-предшественников при активации сигнала gp130 (там же).
Кроме того, у двойных трансгенных мышей (hIL-6 под управлением промотора металлотионеина MbiniH/hIL-6R под управлением промотора PEP карбоксикиназы крысы) выявляют гиперплазию клеток печени, которая по опубликованным данным идентична узелковой регенеративной гиперплазии человека с длительной пролиферацией гепатоцитов, что дает веские основания полагать, что IL-6 ответственен за пролиферацию гепатоцитов и патогенную трансформацию клеток печени (Maione et al. (1998) Coexrpession of IL-6 and soluble IL-6R causes nodular regenerative hyperplasia and adenomas of the liver, EMBO J. 17(19): 5588-5597). Так как гиперплазию клеток печени по опубликованным данным не наблюдали у мышей с одним трансгеном hIL-6 и hIL-6 может связывать mIL-6R, результат может выглядеть парадоксальным, пока не предположить, что два трансгена могут приводить к более высоким уровням hIL-6 в комплексе с растворимым IL-6R (в настоящем документе, растворимым hIL-6R), где этот комплекс является более мощным ингибитором, чем IL-6 отдельно (там же).
В отличие от мышей, трансгенных по IL-6 человека, у мышей с гуманизированным IL-6, которые несут замену в эндогенном локусе IL-6 мыши, который сохраняет регуляторные элементы мыши, но в нем производят гуманизирование кодирующей IL-6 последовательности, не выявляют тяжелых патологий мышей, полученных на известном уровне техники. Генетически модифицированные мыши, которые были гетерозиготными или гомозиготными по hIL-6 росли нормально.
Мышей с гуманизированным геном IL-6 (MAID 760), как описано в примерах, подвергали иммунофенотипированию и при анализах FACS (с отбором лимфоцитов) В-клеток селезенки с использованием общего маркера В-клеток (CD445R(B220)) выявили у них нормальные количества В-клеток (фиг. 4). Для селезенки у мышей дикого типа выявляли 63% В-клеток; у гетерозиготных по hIL-6 мышей выявляли 63% В-клеток и у мышей гомозиготных по hIL-6 в эндогенном локусе мыши выявляли 63% В-клеток. Количества В-клеток у гомозиготных по hIL-6 мышей, иммунизированных TNP-KLH также было нормальным (65% для дикого типа и 61% для гомозигот по hIL-6).
Количество Т-клеток селезенки также являлось приблизительно таким же, как у дикого типа (фиг. 5). Процентное содержание Т-клеток селезенки для Т-хелперов/цитотоксических Т-клеток для дикого типа составляло 20%/40% (отношение 1,4:1); для гетерозигот по hIL-6 - 23%/14% (отношение 1,6:1); для гомозигот по hIL-6 - 21%/15% (отношение 1,4:1) (маркерами являлись CD8a-APC; CD4-FITC). У гомозиготных по hIL-6 мышей, иммунизированных TNP-KLH, выявляли сходные с мышами дикого типа количества Т-клеток селезенки, т.е. Т-хелперы/цитотоксические Т-клетки составляли 22%/20% (отношение 1,1:1) по сравнению с 21%/19% для дикого типа (отношение также составляло 1,1:1).
У мышей с гуманизированным IL-6 при анализе FACS (CD11b и DX5) также выявляли приблизительно нормальные уровни NK клеток селезенки (фиг. 7). У гетерозигот по hIL-6 выявляли 2,2% NK клеток, и у гомозигот по hIL-6 выявляли 1,8% NK клеток, тогда как у мышей дикого типа выявляли 2,4% NK клеток. После иммунизации TNP-KLH, у гомозигот выявляли 1,6% NK клеток селезенки, тогда как у мышей дикого типа выявляли 2,1% NK клеток селезенки.
У мышей с гуманизированным IL-6 также выявляли нормальные уровни клеток Ly6G/C(Gr1) селезенки (фиг. 6). У гетерозигот по hIL-6 выявляли 7,0% клеток GR1+ (1,3% Gr1hi); у гомозигот выявляли 6,8% клеток Gr1+ (0,9% Gr1hi), тогда как у мышей дикого типа выявляли 8,0% клеток Gr1+ (1,8%Cr1hi). У иммунизированных гомозигот по IL-6 (иммунизировали TNP-KLH) выявляли 11% клеток Gr1+ (4,0% Gr1hi), тогда как у мышей дикого типа выявляли 10% клеток Gr1+ (3,0% Gr1hi).
У мышей с гуманизированным IL-6 при анализе FACS также выявляли нормальные количества В- и Т-клеток в крови (фиг. 8 и 9). FACS с общим маркером В-клеток (CD445R(B220)) выявил, что у гомозиготных по hIL-6 мыши можно наблюдать 52% В-клеток по сравнению с 53% у дикого типа; у гетерозигот можно наблюдать 38% (среднее двух различных линий с 29% и 47%). Гомозиготные по hIL-6 мыши, иммунизированные TNP-KLH, давали сходные количества В-клеток (43% по сравнению с 45% для мышей дикого типа).
У мышей с гуманизированным IL-6 при анализе FACS выявляли нормальный кровь Т-клеток как измеряли при окрашивании CD8a и CD4. У гетерозиготных hIL-6 мышей выявляли количества Т-хелперов/цитотоксических Т-клеток 39%/26% (отношение 1,5:1); у гомозиготных по hIL-6 мышей выявляли количества Th/Tc 24%/20% (отношение 1,2:1), тогда как у мышей дикого типа выявляли количества Th/Tc 26%/20% (отношение 1,3:1). У гомозиготных по hIL-6 мышей, иммунизированных TNP-KLH, выявляли количества Th/Tc 29%/21% (отношение 1,4:1), тогда как у иммунизированных мышей дикого типа мыши выявляли количества Th/Tc 28%/23% (1,2:1).
У мышей с гуманизированным IL-6 также выявляли количества миелоидных клеток в крови, которые были сходны с количествами у мышей дикого типа как измеряли посредством анализа FACS в крови наивных и иммунизированных мышей с окраской Ly6G/C(Gr1) и CD11b, а также CD11b и DX5 (фиг. 10, 11 и 12). У гетерозиготных по hIL-6 мышей выявляли % Gr+ клеток 10,8%, у гомозигот - 6,9%, тогда как у мышей дикого типа выявляли 9,7%. У иммунизированных гомозигот по hIL-6 выявляли количества M1(Ly6G/C(Gr) 10l-104)/M2(Ly6G/C(Gr) с окраской приблизительно 102-103) 43%/34%, тогда как у мышей дикого типа выявляли количества 45%/38%. Диаграммы FACS зависимости CD11b (вертикальная ось) от Ly6G/C (горизонтальная ось) для иммунизированных гомозиготных по hIL-6 мышей продемонстрировали процентное содержание клетка в квадрантах (в верхнем левом/верхнем правом/нижнем правом) 16%/8%/3%, что было идентично количествам в квадрантах у иммунизированных мышей дикого типа.
У гомозиготных мышей, иммунизированных TNP-KLH, с гуманизированным IL-6 получали диаграммы окрашивания FACS зависимости CD11b от DX5(NK), которые были сходны с диаграммами у иммунизированных мышей дикого типа. Анализ квадрантов диаграмм FACS крови (CD11b - вертикальная ось, DX5(NK) - горизонтальная ось) выявил количества в верхнем левом/верхнем правом/нижнем правом квадрантах 9,5%/17%/10% для гомозигот по hIL-6 и 6,5%/17,3%/14% для мышей дикого типа.
У мышей с гуманизированным IL-6 выявляли изотипический ответ, который по существу был таким же, как наблюдали у мышей дикого типа. Ранние и конечные уровни IgGI, IgG2a, IgG2b, IgG3, IgA, IgE и IgM являлись приблизительно такими же, как наблюдали у мышей дикого типа. В одном из экспериментов конечный уровень IgM был немного выше у гуманизированных мышей; у гуманизированных мышей также был повышен конечный уровень IgG3.
Развитие В-клеток у наивных по hIL-6 мышей на основе анализа FACS костного мозга с окрашиванием IgM/CD24/B220 по существу не отличалось от развития у мышей дикого типа (фиг. 13). Иммунофенотипирование иммунных мышей выявило, что группы маркеров различных типов клеток при прохождении развития В-клеток у мышей с hIL-6 были по существу нормальными. Развитие клеток из гемопоэтических стволовых клеток, общих предшетсвенников лимфоидных клеток, ProB клеток, PreB клеток и незрелых и зрелых В-клеток у мышей с hIL-6 было нормальным (фиг. 14 и 15).
ПРИМЕРЫ
Пример 1: Замена эндогенного гена IL-6 мыши на ген hIL-6
Геном IL-6 человека длиной 4,8 т.п.н., содержащим экзоны с 1 по 4 гена IL-6 человека, заменяли 6,8 т.п.н. локуса гена IL-6 мышей.
Направленную конструкцию для замены гена IL-6 мыши на ген человека за один этап взаимодействия конструировали с использованием технологии генетической инженерии VELOCIGENE® (см., Valenzuela et al. (2003) High-throughput engineering of the mouse genome coupled with high-resolution expression analysis, Nature Biotech, 21(6): 652-659). ДНК IL-6 мыши и человека получали из клона 368С3 бактериальной искусственной хромосомы (ВАС) RPCI-23 и из клона 2369М23 ВАС CTD соответственно. В кратком изложении, линеаризованную NotI направленную конструкцию, полученную посредством клонирования с репарацией пробелов, содержащую участки гомологии выше и ниже IL-6 мыши, фланкирующие последовательность IL-6 человека длиной 4,8 т.п.н., содержащую участки от ATG в экзоне 1 до экзона 5 с 16 нуклеотидами расположенной ниже 3'-конца последовательности (геномные координаты: NCBIh37.1: от ch7:22.766.882 до 22.771.637) и селекционной кассетой neo, фланкированной LoxP, электропорировали в эмбриональные стволовые (ES) клетки мыши F1H4 (гибрид Fl C57BL/6 × 129). ES с корректной вставкой (MAID 790) дополнительно электропорировали транзиторным вектором, экспрессирующим Cre для удаления лекарственной селекционной кассеты. Клоны модифицированных ES клеток без кассеты лекарственного средства (MAID 1428) вводили в эмбрион мыши на стадии 8 клеток способом VELOCIMOUSE® (см., патенты США №№7294754, 7576259, 7659442 и Poueymirou et al. (2007) FO generation mice that are essentially fully derived from the donor gene-targeted ES cells allowing immediate phenotypic analyses Nature Biotech. 25(1): 91-99). VELOCIMICE® (мыши FO полностью происходящие из донорских ES клеток), несущих гуманизированный ген IL-6 идентифицировали посредством генотипирования на потерю аллеля мышей и получение аллеля человека с использованием анализа модификации аллелей (см. Valenzuela et al. (2003)).
Клоны клеток ES с корректной вставкой идентифицировали посредством анализа потери нативного аллеля (LONA) (Valenzuela et al. 2003), в котором количество копий нативного, немодифицированного гена Il6 определяли посредством двух реакций количественной полимеразной цепной реакции (qPCR) TaqMan™, специфичных для последовательностей гена Il6 мыши, которые были предназначены для делеции. Анализы qPCR включали следующие наборы праймеров-зондов (записанных в направлении от 5' к 3'): расположенный выше по ходу транскрипции прямой праймер, TTGCCGGTTTTCCCTTTTCTC (SEQ ID NO: 1); расположенный выше по ходу транскрипции обратный праймер, AGGGAAGGCCGTGGTTGTC (SEQ ID NO: 2); расположенный выше по ходу транскрипции зонд, FAM-CCAGCATCAGTCCCAAGAAGGCAACT-BHQ (SEQ ID NO: 3); расположенный ниже по ходу транскрипции прямой праймер, TCAGAGTGTGGGCGAACAAAG (SEQ ID NO: 4); расположенный ниже по ходу транскрипции обратный праймер, GTGGCAAAAGCAGCCTTAGC (SEQ ID NO: 5); расположенный ниже по ходу транскрипции зонд, FAM-TCATTCCAGGCCCTTCTTATTGCATCTG-BHQ (SEQ ID NO: 6); где FAM относится к флуоресцентному зонду 5-карбоксифлуоресцеину, a BHQ относится к гасителю флуоресценции, принадлежащему к гасителям типа черной дыры (Biosearch Technologies). ДНК, выделенную из клонов ES клеток, захвативших направленный вектор и встроивших его в свой геном, комбинировали с основной смесью TaqMan™ Gene Expression Master Mix (Life Technologies) в соответствии с рекомендациями производителя в 384-луночном планшете для ПЦР (384-луночный планшет для реакций MicroAmp™ Optical, Life Technologies) и проводили циклическую реакцию в Applied Biosystems Prism 7900HT, собирающем данные флуоресценции в течение PCR и определяющем пороговый цикл (Ct), частичный цикл ПЦР, при котором накопленная флуоресценция достигает предустановленного порога. Для каждого образца ДНК проводили qPCR, специфичные для расположенных выше и расположенных ниже по ходу транскрипции Il6 участков, и две qPCR для не являющихся мишенью контрольных генов. Рассчитывали различия в значениях Ct (ΔCt) между каждой qPCR из специфичных для Il6 и каждой qPCR для контрольного гена, а затем для каждого образца рассчитывали различие между каждым ΔCt и средним ΔCt для всех анализируемых образцов с получением значений ΔΔCt для каждого образца. Количество копий гена Il6 в каждом образце рассчитывали по следующей формуле: количество копий = 2⋅2-ΔΔCt. Клон с корректной вставкой с потерей одной из нативных копий содержит количество копий гена Il6, равное единице. Подтверждение того, что последовательность гена IL6 человека замещала удаленную последовательность гена Il6 мыши в гуманизированном аллеле проводили посредством анализа qPCR TaqMan™, включающего следующие наборы праймеров-зондов (записанных в направлении от 5' к 3'): прямой праймер для последовательности человека, CCCCACTCCACTGGAATTTG (SEQ ID NO: 7); обратный праймер для последовательности человека, GTTCAACCACAGCCAGGAAAG (SEQ ID NO: 8) и зонд для последовательности человека, FAM-AGCTACAACTCATTGGCATCCTGGCAA-BHQ (SEQ ID NO: 9).
Тот же анализ LONA использовали для анализа ДНК, выделенной из материала хвостовой биопсии у мышей, происходящих из модифицированных ES клеток для определения их генотипов Il6 и подтверждения того, что гуманизированный аллель Il6 передается через зародышевую линию. Размножали двоих детенышей, гетерозиготных по замене, для получения мышей, гомозиготных по замене эндогенного гена IL-6 мыши на ген IL-6 человека. Для фенотипирования используют детенышей, гомозиготных по замене.
Предусмотрено, что расположенное выше по ходу транскрипции соединения локуса мыши и последовательности, содержащей ген hIL-6, находится в пределах участка 5'-AATTAGAGAG TTGACTCCTA ATAAATATGA GACTGGGGAT GTCTGTAGCT CATTCTGCTC TGGAGCCCAC CAAGAACGAT AGTCAATTCC AGAAACCGCT ATGAACTCCT TCTCCACAAG TAAGTGCAGG AAATCCTTAG CCCTGGAACT GCCAGCGGCG GTCGAGCCCT GTGTGAGGGA GGGGTGTGTG GCCCAGG (SEQ ID NO: 10), где конечный нуклеотид мышь перед первым нуклеотидом гена человека представляет собой "Т" в CCGCT, а первый нуклеотид последовательности человека представляет собой первый "А" в ATGAA. Предусмотрено, что расположенное ниже по ходу транскрипции соединения последовательности, содержащей ген hIL-6 и локуса мыши, находится в пределах участка 5'-TTTTAAAGAA ATATTTATAT TGTATTTATA TAATGTATAA ATGGTTTTTA TACCAATAAA TGGCATTTTA AAAAATTCAG CAACTTTGAG TGTGTCACGC TCCCGGGCTC GATAACTATA ACGGTCCTAA GGTAGCGACT CGAGATAACT Т-3' (SEQ ID NO: 11), где конечный нуклеотид последовательности человека представляет собой конечный "G" в TCACG, а первый нуклеотид последовательности мыши представляет собой первый "С" в СТССС; расположенная ниже по ходу транскрипции область соединения также находится в контакте с участком LoxP на 3'-конце (начало которого показано) для удаления фланкированной LoxP кассеты neo под контролем промотора убиквитина. Предусмотрено, что соединение кассеты neo с локусом IL-6 мыши находится в пределах участка 5'-TATACGAAGT TATCCTAGGT TGGAGCTCCT AAGTTACATC САААСАТССТ СССССАААТС AATAATTAAG CACTTTTTAT GACATGTAAA GTTAAATAAG AAGTGAAAGC TGCAGATGGT GAGTGAGA (SEQ ID NO: 12), где конечный "С" AGCTC представляет собой конечный нуклеотид кассеты neo; первый нуклеотид генома мыши после кассеты представляет собой начальный "С" в CTAAG.
Пример 2: Иммунофенотипирование наивных и иммунизированных мышей с hIL-6: В-клетки
У мышей, гомозиготных по замене на ген hIL-6, анализировали В-клетки (DC445R(B220). Фракции, обогащенные лимфоцитами, из препаратов клеток селезенки наивных и иммунизированных (TNP-KLH) мышей с hIL-6, окрашивали и подвергали иммунофенотипированию с использованием проточной цитометрии. Анализ FACS продемонстрировал, что процентное содержание В-клеток в препаратах клеток селезенки, как измеряли посредством окрашивания CD45R(B220)-FITC, у препаратов, полученных у наивных мышей дикого типа, гетерозигот по hIL-6 и гомозигот по hIL-6 являлось приблизительно одинаковым (63% клеток). У иммунизированных мышей В-клетки составляли приблизительно 65% от всех клеток препарата клеток селезенки у мышей дикого типа, и приблизительно 61% от всех клеток у гомозигот hIL-6. Селезенка мышей с hIL-6 (наивных и иммунизированных) содержала популяцию В-клеток, которая являлась приблизительно такого же размера как популяция В-клеток селезенки у мышей дикого типа.
Костный мозг мышей дикого типа, гетерозигот по hIL-6 и гомозигот по hIL-6 окрашивали на В-клеточные маркеры (CD45R(B220)-APC, CD24(HSA)-PE, или CD43, конъюгированные с красителем и/или IgM (IgM-FITC). По мере развития клеток из стволовых клеток до ранних про-В-клеток до поздних про-В-клеток, до больших пре-В-клетки до малых пре-В-клетки до незрелых В-клетки и наконец, до зрелых В-клеток развитие В-клеток в костном мозге нормальных мышей отражается в поверхностных маркерах. Общие предшественники лимфоцитов про-В-клетки экспрессируют CD45R, а на поздних стадиях экспрессируют IgM, как незрелые и, позднее, как зрелые В-клетки. Таким образом, окрашивание В-клеток на CD45R и на IgM должно выявлять профиль, характерный для развития В-клеток. Костный мозг гетерозигот и гомозигот по hIL-6 демонстрировал профиль окрашивания CD45R(B220)-APC и антителами к IgM-FITC, который по существу был неотличим от костного мозга дикого типа, демонстрируя популяции В-клеток, которые положительно окрашивались на CD45R(B220) и IgM или только на CD45R(B220). Субпопуляции В-клеток в костном мозге мышей с hIL-6, выявленные посредством окрашивания FACS были сходными с субпопуляциями В-клеток у мышей дикого типа (таблица 1; также см. фиг. 13).

Окрашивание на CD24 (см. фиг. 14) выявило (нормальный) профиль, представленный в таблице 2, указывающий на нормальное развитие в костном мозге.
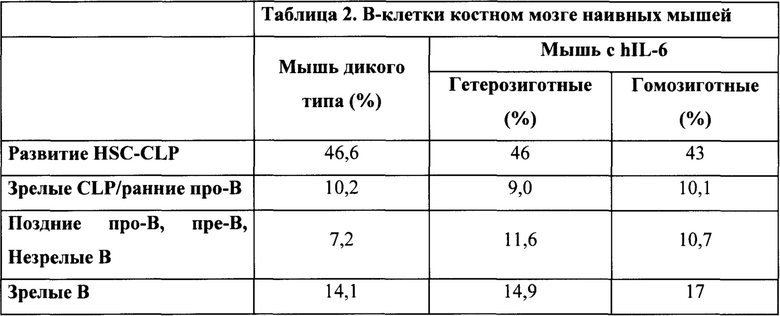
Окрашивание на CD43 (см. фиг. 15) выявило (нормальный) профиль, представленный в таблице 3, указывающий на нормальное развитие в костном мозге.
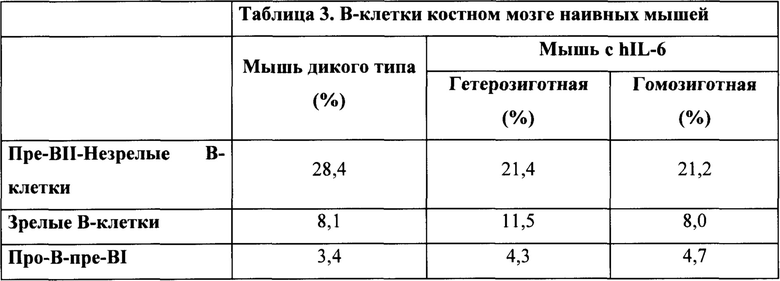
Таким образом, иммунофенотипирование наивных мышей с hIL-6 выявило, что развитие В-клеток у таких мышей по существу является нормальным.
Пример 3: Замена эндогенной генной последовательности внеклеточного домена IL-6Rα мыши на генную последовательность внеклеточного домена hIL-6Rα
Последовательностью гена IL-6Rα человека длиной 45 т.п.н., содержащей экзоны с 1 по 8 гена IL-6Rα человека, замещали локус гена IL-6Rα мыши длиной 35,4 т.п.н. Экзоны мыши 9 и 10 сохраняли; гуманизированными являлись только экзоны 1-8. Всего 35384 п.н. последовательности мыши заменили на 45047 п.н. последовательности человека.
Направленную конструкцию для замены гена IL-6Rα мыши на ген человека за один этап взаимодействия конструировали с использованием технологии генетической инженерии VELOCIGENE® (см. Valenzuela et al. (2003) High-throughput engineering of the mouse genome coupled with high-resolution expression analysis, Nature Biotech, 21(6): 652-659). ДНК IL-6 мыши и человека получали из клона 125J8 бактериальной искусственной хромосомы (ВАС) RPCI-23 и из клона 2192J23 ВАС CTD, соответственно. В кратком изложении, линеаризованную NotI направленную конструкцию, полученную посредством клонирования с репарацией пробелов, содержащую участки гомологии выше и ниже IL-6Rα мыши, фланкирующие последовательность IL-6Rα человека длиной 45 т.п.н., содержащую участки от ATG в экзоне 1 до экзона 8 с 69 нуклеотидами расположенной ниже 3'-конца последовательности и селекционной кассетой neo, фланкированной LoxP, электропорировали в эмбриональные стволовые (ES) клетки мыши F1H4 (гибрид F1 C57BL/6 × 129). ES с корректной вставкой (MAID 794) дополнительно электропорировали транзиторным вектором, экспрессирующим Cre для удаления лекарственной селекционной кассеты. Клоны модифицированных ES клеток без кассеты лекарственного средства (MAID 1442) вводили в эмбрион мыши на стадии 8 клеток способом VELOCIMOUSE® (см. патенты США №№7294754, 7576259, 7659442 и Poueymirou et al. (2007) FO generation mice that are essentially fully derived from the donor gene-targeted ES cells allowing immediate phenotypic analyses Nature Biotech. 25(1): 91-99). VELOCIMICE® (мыши FO полностью происходящие из донорских ES клеток), несущих гуманизированный ген IL-6Rα идентифицировали посредством генотипирования на потерю аллеля мышей и получение аллеля человека с использованием анализа модификации аллелей (см. Valenzuela et al. (2003)).
Клоны клеток ES с корректной вставкой идентифицировали посредством анализа потери нативного аллеля (LONA) (Valenzuela et al. 2003), в котором количество копий нативного, немодифицированного гена IL-6Rα определяли посредством двух реакций количественной полимеразной цепной реакции (qPCR) TaqMan™, специфичных для последовательностей гена IL-6Rα мыши, которые были предназначены для делеции. Анализы qPCR включали следующие наборы праймеров-зондов (записанных в направлении от 5' к 3'): расположенный выше по ходу транскрипции прямой праймер, GCCCTAGCATGCAGAATGC (SEQ ID NO: 13); расположенный выше по ходу транскрипции обратный праймер, AAGAGGTCCCACATCCTTTGC (SEQ ID NO: 14); расположенный выше по ходу транскрипции зонд, СССАСАТССАТСССААТССТ GTGAG (SEQ ID NO: 15); расположенный ниже по ходу транскрипции прямой праймер, GAGCTTGCCCCCAGAAAGG (SEQ ID NO: 16); расположенный ниже по ходу транскрипции обратный праймер, CGGCCACATCTCTGGAAGAC (SEQ ID NO: 17); расположенный ниже по ходу транскрипции зонд, CATGCACTGCCCCAAGTCTGGTTTCAGT (SEQ ID NO: 18). ДНК, выделенную из клонов ES клеток, захвативших направленный вектор и встроивших его в свой геном, комбинировали с основной смесью TaqMan™ Gene Expression Master Mix (Life Technologies) в соответствии с рекомендациями производителя в 384-луночном планшете для ПЦР (384-луночный планшет для реакций MicroAmp™ Optical, Life Technologies) и проводили циклическую реакцию в Applied Biosystems Prism 7900HT, собирающем данные флуоресценции в течение PCR и определяющем пороговый цикл (Ct), частичный цикл ПЦР, при котором накопленная флуоресценция достигает предустановленного порога. Для каждого образца ДНК проводили qPCR, специфичные для расположенных выше и расположенных ниже по ходу транскрипции IL-6Rα участков, и две qPCR для не являющихся мишенью контрольных генов. Рассчитывали различия в значениях Ct (ΔCt) между каждой qPCR из специфичных для IL-6Rα и каждой qPCR для контрольного гена, а затем для каждого образца рассчитывали различие между каждым ΔCt и средним ΔCt для всех анализируемых образцов с получением значений ΔΔCt для каждого образца. Количество копий гена IL-6Rα в каждом образце рассчитывали по следующей формуле: количество копий = 2⋅2-ΔΔCt. Клон с корректной вставкой с потерей одной из нативных копий содержит количество копий гена IL-6Rα, равное единице. Подтверждение того, что последовательность гена IL-6Rα человека замещала удаленную последовательность гена IL-6Rα мыши в гуманизированном аллеле проводили посредством анализа qPCR TaqMan™, включающего следующие наборы праймеров-зондов (записанных в направлении от 5' к 3'): прямой праймер для последовательности человека, GGAGAGGGCAGAGGCACTTAC (SEQ ID NO: 19); обратный праймер для последовательности человека, GGCCAGAGCCCAAGAAAAG (SEQ ID NO: 20) и зонд для последовательности человека, CCCGTTGACTGTAATCTGCCCCTGG (SEQ ID NO: 21).
Тот же анализ LONA использовали для анализа ДНК, выделенной из материала хвостовой биопсии у мышей, происходящих из модифицированных ES клеток для определения их генотипов IL-6Rα и подтверждения того, что гуманизированный аллель IL-6Rα передается через зародышевую линию. Размножали детенышей, гетерозиготных по замене, для получения мышей, гомозиготных по замене эндогенного гена IL-6Rα мыши на ген (внеклеточный домен) IL-6Rα человека. Для фенотипирования используют детенышей, гомозиготных по замене.
Предусмотрено, что расположенное выше по ходу транскрипции соединения локуса мыши и последовательности, содержащей ген hIL-6Ra, находится в пределах участка 5'-CGAGGGCGAC TGCTCTCGCT GCCCCAGTCT GCCGGCCGCC CGGCCCCGGC TGCGGAGCCG CTCTGCCGCC CGCCGTCCCG CGTAGAAGGA AGCATGCTGG CCGTCGGCTG CGCGCTGCTG GCTGCCCTGC TGGCCGCGCC GGGAGCGGCG CTGGCCCCAA GGCGCTGCCC TGCGCAGGGT AAGGGCTTCG G (SEQ ID NO: 22), где конечный нуклеотид мышь перед первым нуклеотидом гена человека представляет собой "С" в GAAGC, а первый нуклеотид последовательности человека представляет собой первый "А" в ATGCT. Предусмотрено, что расположенное ниже по ходу транскрипции соединения последовательности, содержащей ген hIL-6Rα и локуса мыши, находится в пределах участка 5'-CAAGATTATT GGAGTCTGAA ATGGAATACC TGTTGAGGGA AATCTTTATT TTGGGAGCCC TTGATTTCAA TGCTTTTGAT ТСССТАТССС TGCAAGACCC GGGCTCGATA ACTATAACGG TCCTAAGGTA GCGACTCGAG АТААСТТС-3' (SEQ ID NO: 23), где конечный нуклеотид последовательности человека представляет собой конечный "А" в CAAGA, а первый нуклеотид последовательности мыши представляет собой первый "С" в CCCGG; расположенная ниже по ходу транскрипции область соединения также находится в контакте с участком LoxP на 3'-конце для удаления фланкированной LoxP кассеты neo под контролем промотора убиквитина. Первый нуклеотид участка Loxp представляет собой первый "А" в АТААС. Предусмотрено, что соединение кассеты neo с локусом IL-6Rα мыши находится в пределах участка 5'-TATACGAAGT TATCCTAGGT TGGAGCTCTA CTCCATATGC TCACTTGCCG TTGTTTGCTA CGATACGGTG AGGCCCGTGC GAAGAGTGGC ACAGATCAGG AGGCTTATGT GGTCAGTCCA CAGTATGGC (SEQ ID NO: 24), где конечный "С" AGCTC представляет собой конечный нуклеотид кассеты neo; первый нуклеотид генома мыши после кассеты представляет собой начальный "Т" в ТАСТС.
| название | год | авторы | номер документа |
|---|---|---|---|
| ГУМАНИЗИРОВАННЫЕ IL-6 И РЕЦЕПТОР IL-6 | 2012 |
|
RU2751240C2 |
| ЖИВОТНЫЕ С ГУМАНИЗИРОВАННЫМ ГЕНОМ IL-15 | 2014 |
|
RU2674910C2 |
| АНТИТЕЛА ВЫСОКОЙ АФФИННОСТИ К IL-6-РЕЦЕПТОРУ ЧЕЛОВЕКА | 2007 |
|
RU2433138C2 |
| ГРЫЗУНЫ С ГУМАНИЗИРОВАННЫМ IL-7 | 2013 |
|
RU2654565C2 |
| ЖИВОТНЫЕ С ГУМАНИЗИРОВАННЫМИ IL-4 И IL-4Rα | 2015 |
|
RU2788523C2 |
| ЖИВОТНЫЕ С ГУМАНИЗИРОВАННЫМИ IL-4 И IL-4Rα | 2015 |
|
RU2703139C2 |
| ГУМАНИЗИРОВАННЫЕ МЫШИ С НОКИНОМ SIRPA-IL15 И СПОСОБЫ ИХ ИСПОЛЬЗОВАНИЯ | 2016 |
|
RU2730599C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ В ОТНОШЕНИИ ГЛАВНОГО КОМПЛЕКСА ГИСТОСОВМЕСТИМОСТИ МЫШИ | 2012 |
|
RU2783984C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ В ОТНОШЕНИИ ГЛАВНОГО КОМПЛЕКСА ГИСТОСОВМЕСТИМОСТИ МЫШИ | 2012 |
|
RU2653433C2 |
| ЖИВОТНЫЕ, СОДЕРЖАЩИЕ ГУМАНИЗИРОВАННУЮ ДИПЕПТИДИЛПЕПТИДАЗУ IV (DPP4) | 2015 |
|
RU2648166C1 |





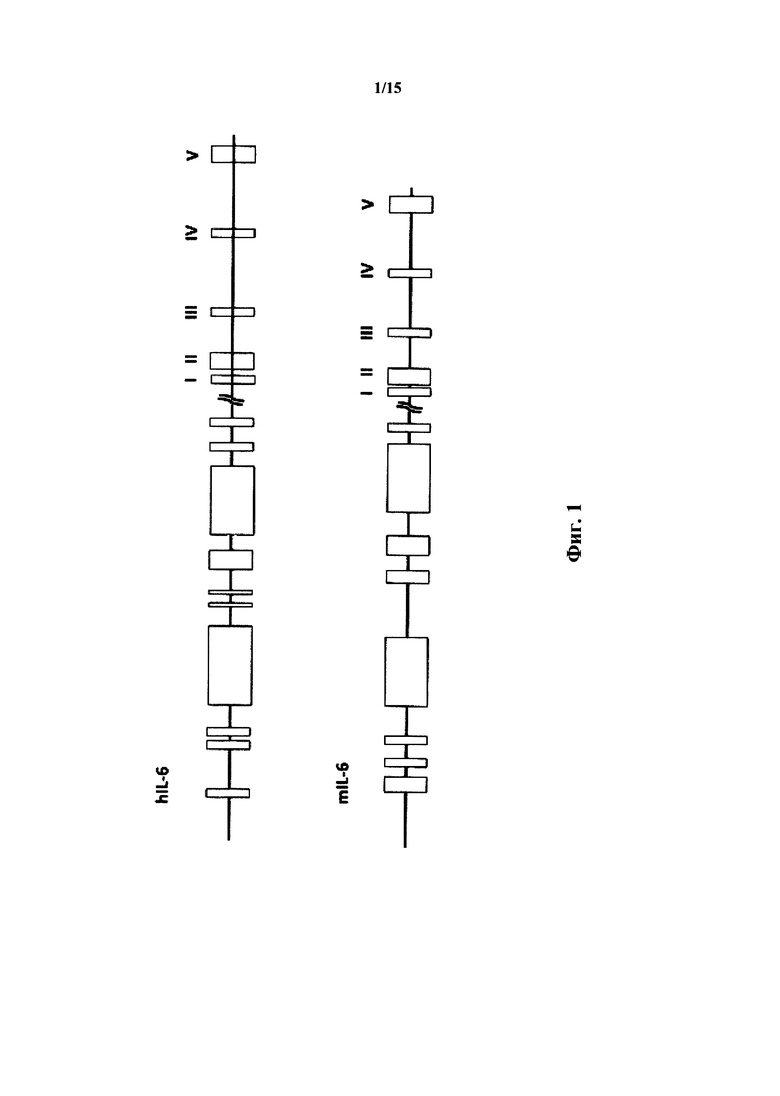

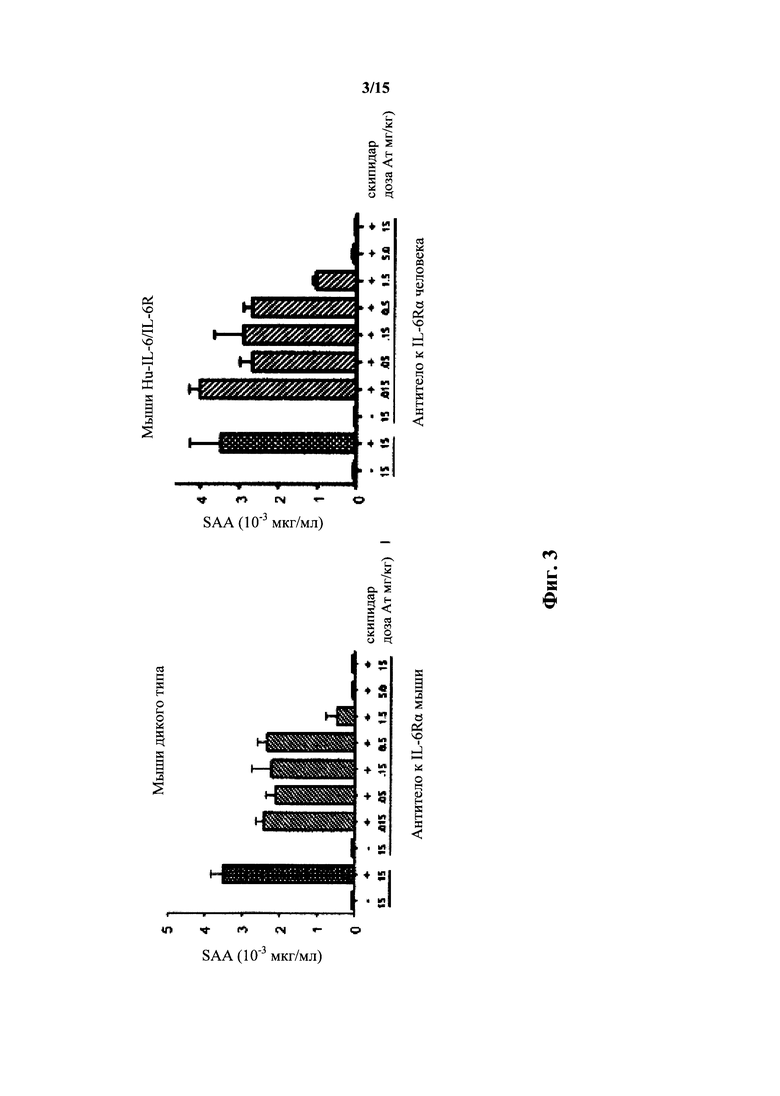
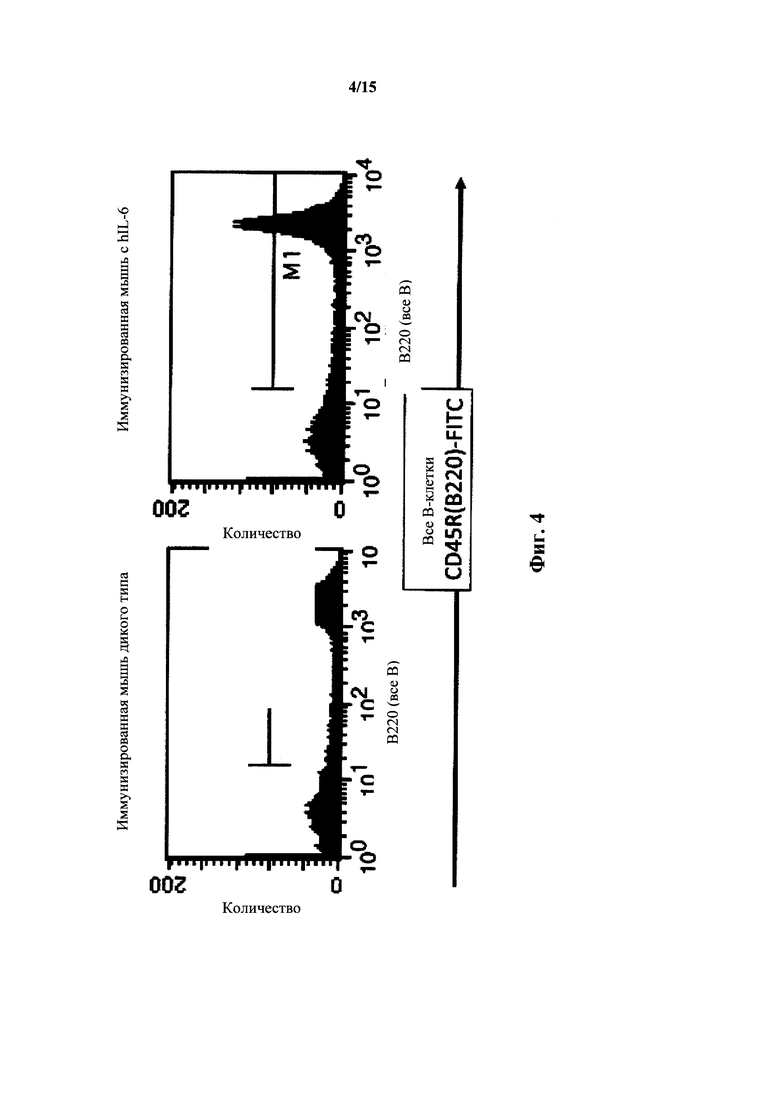
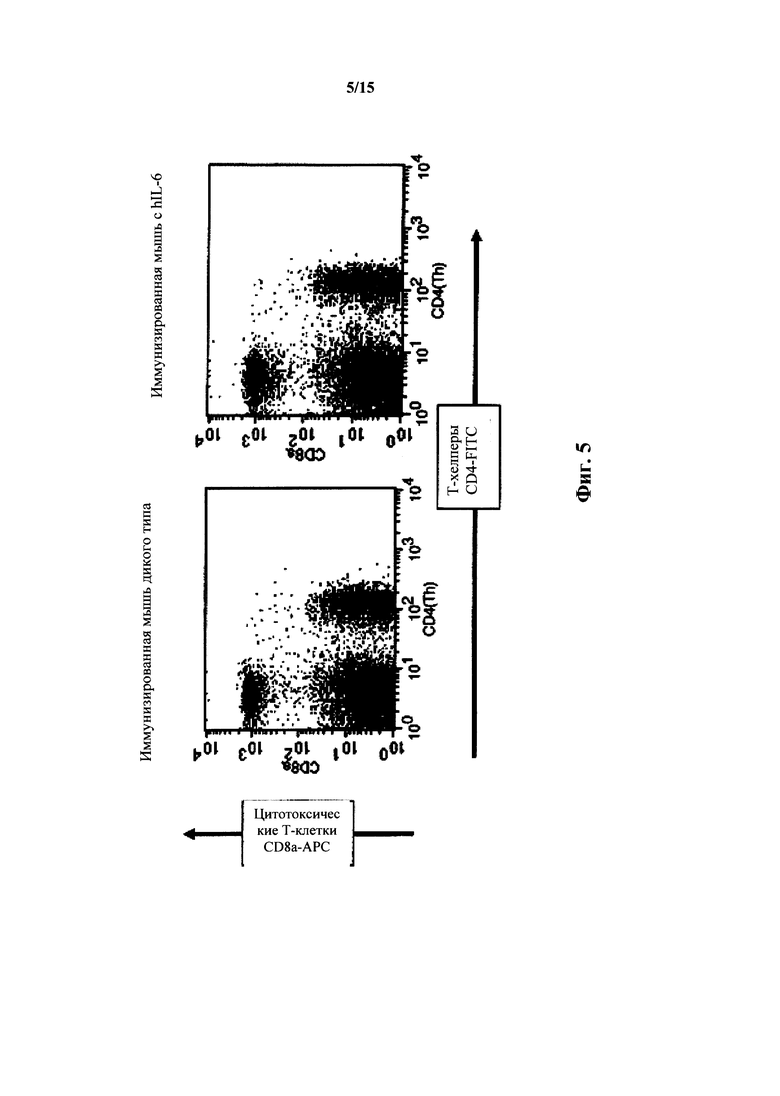
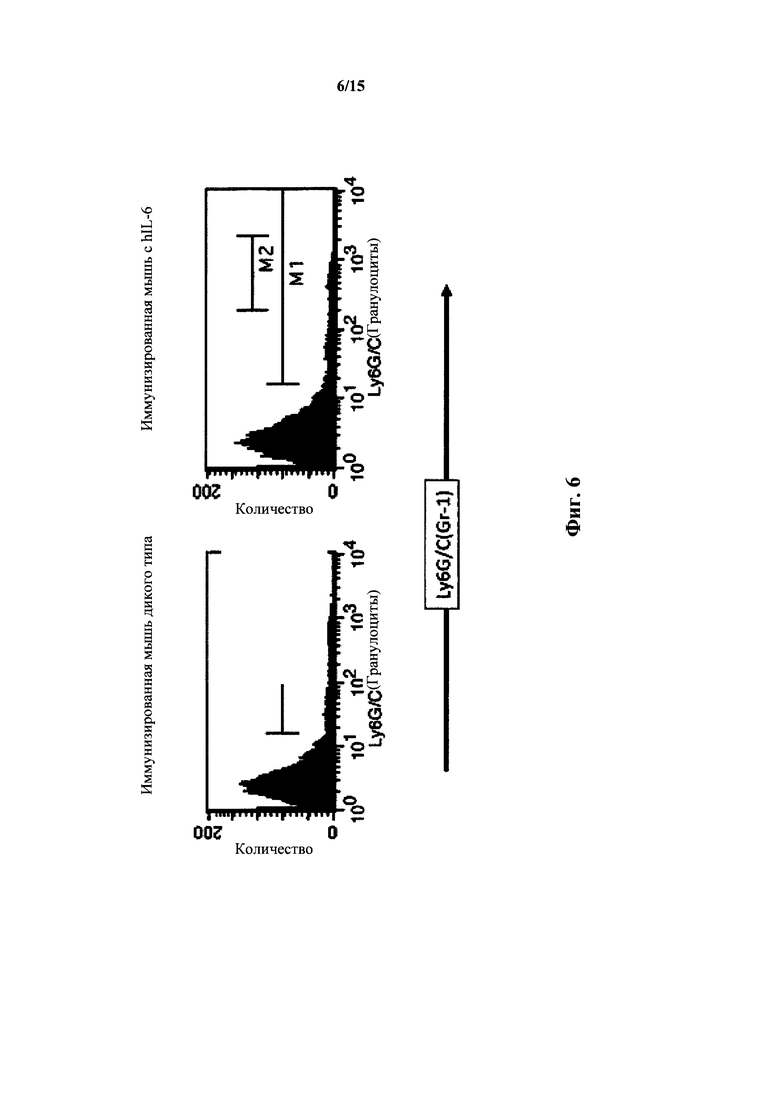
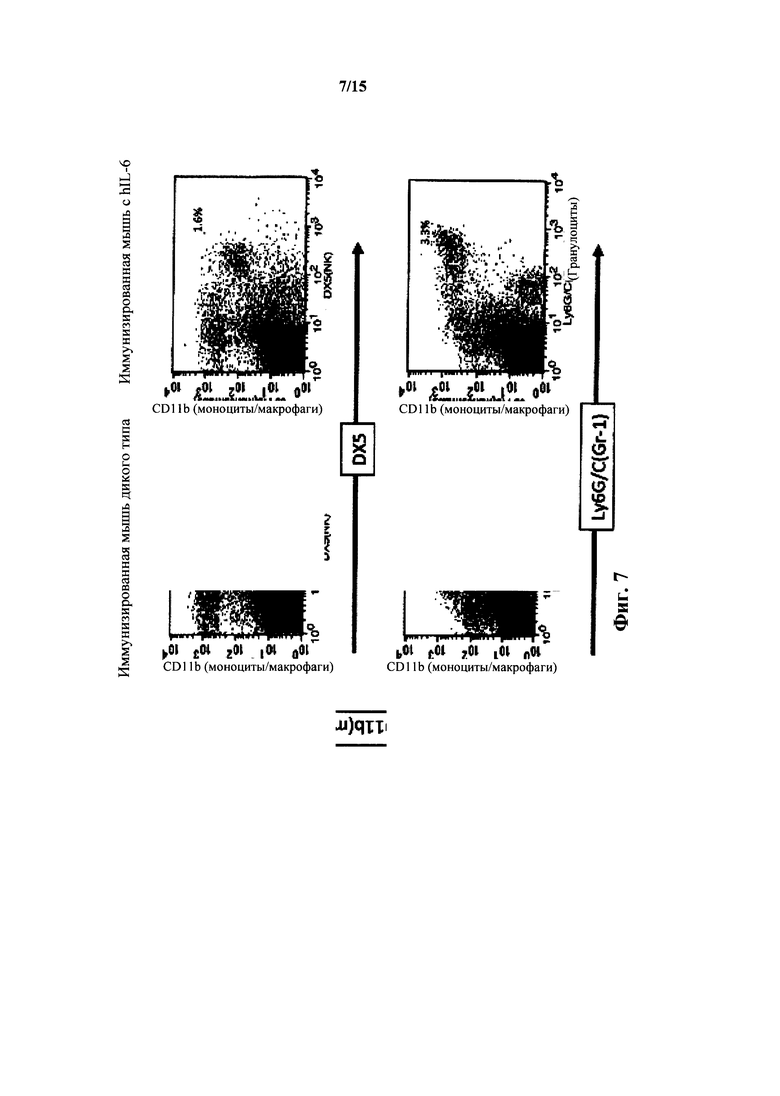
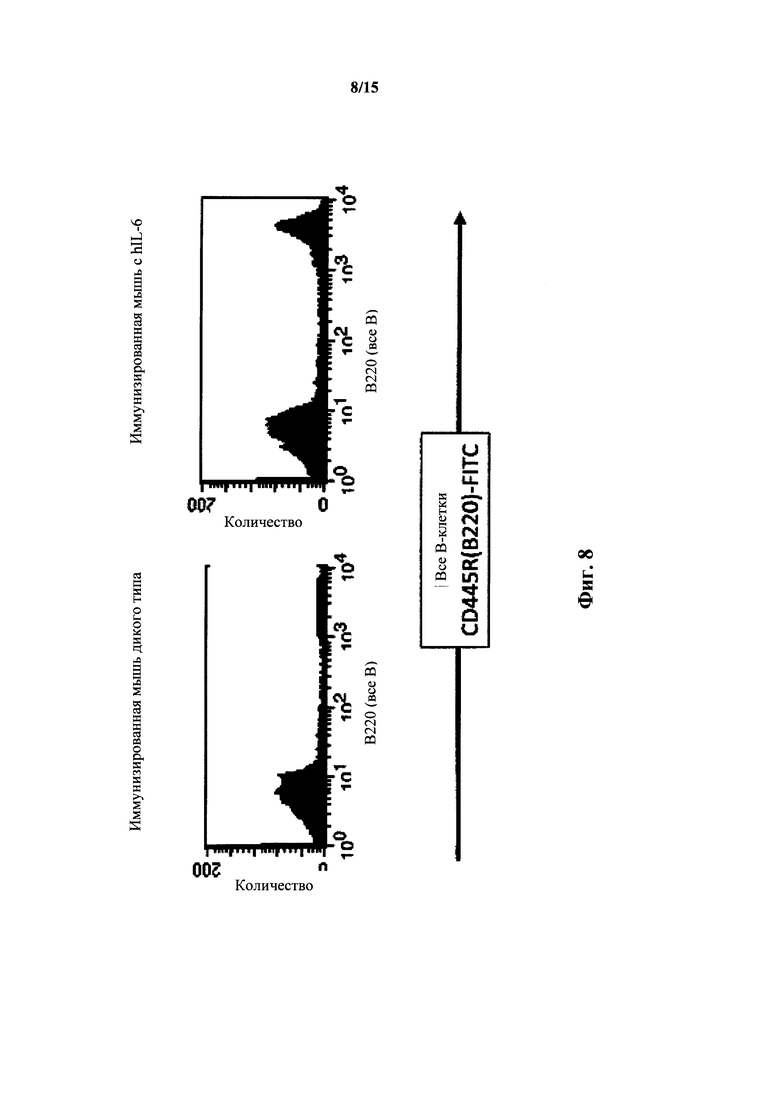
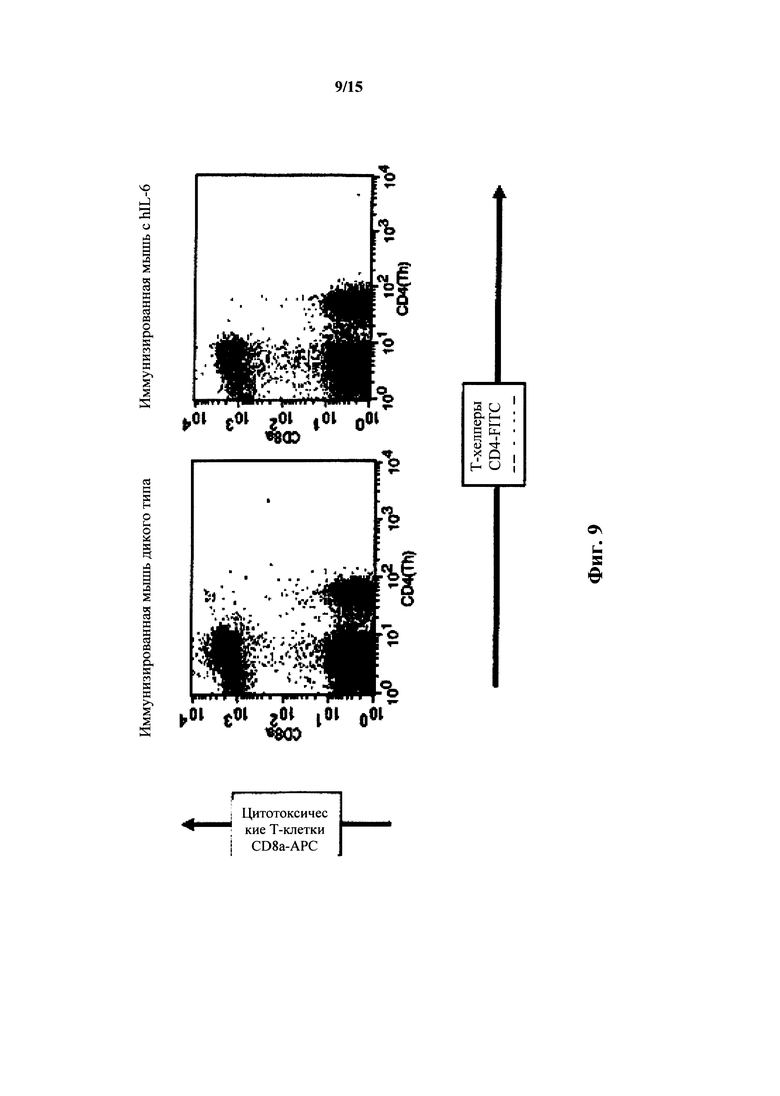
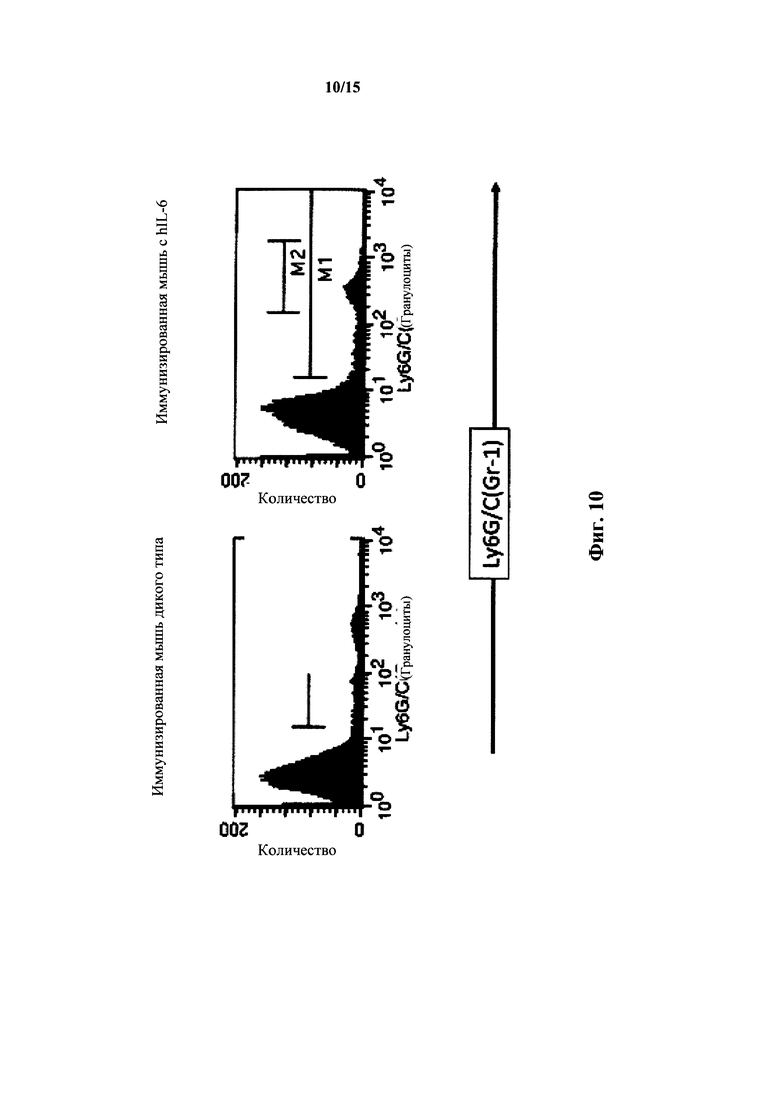
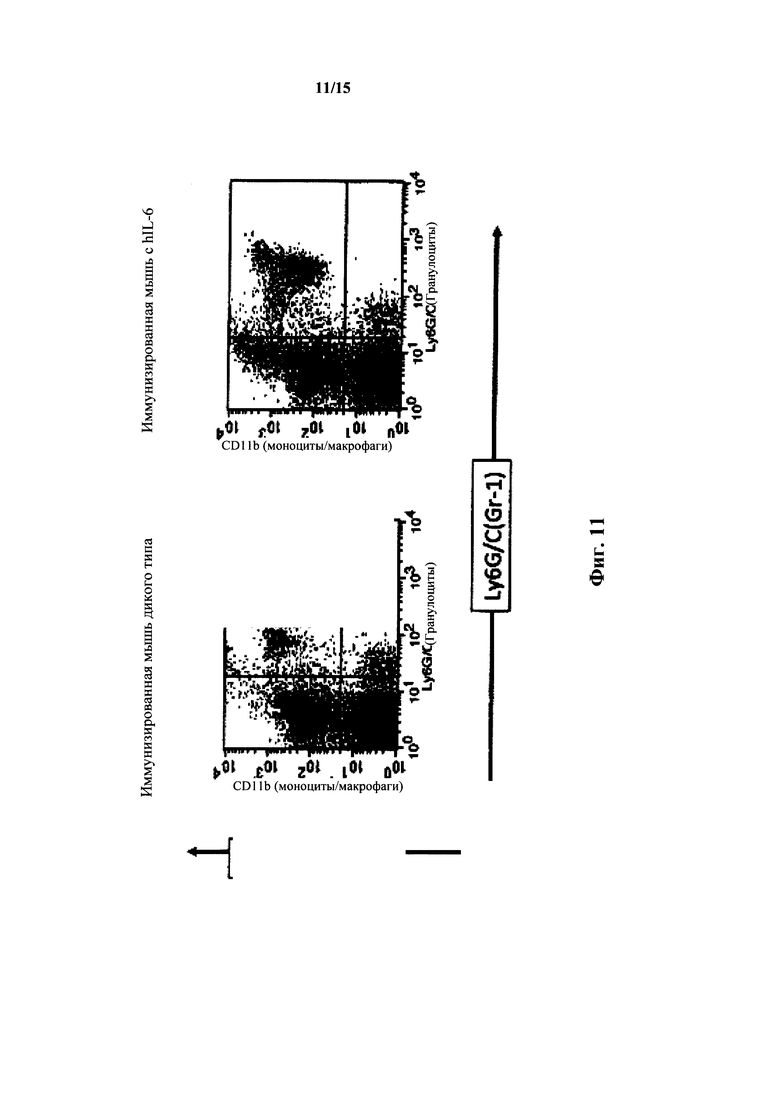
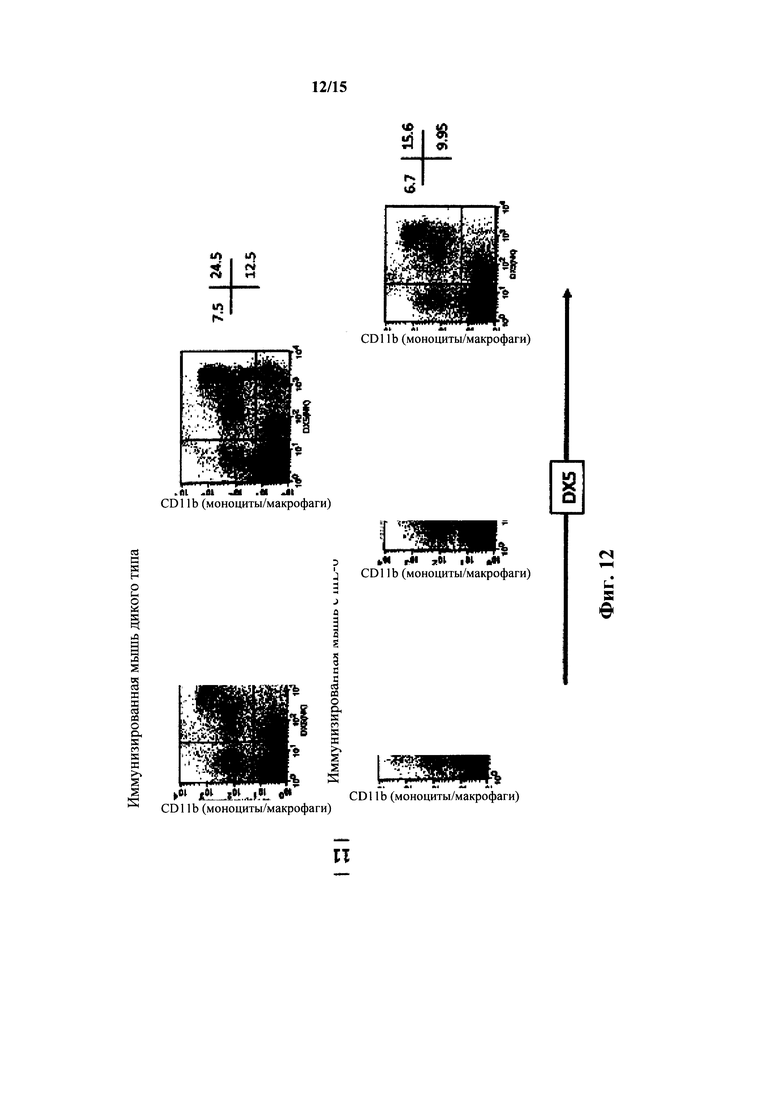
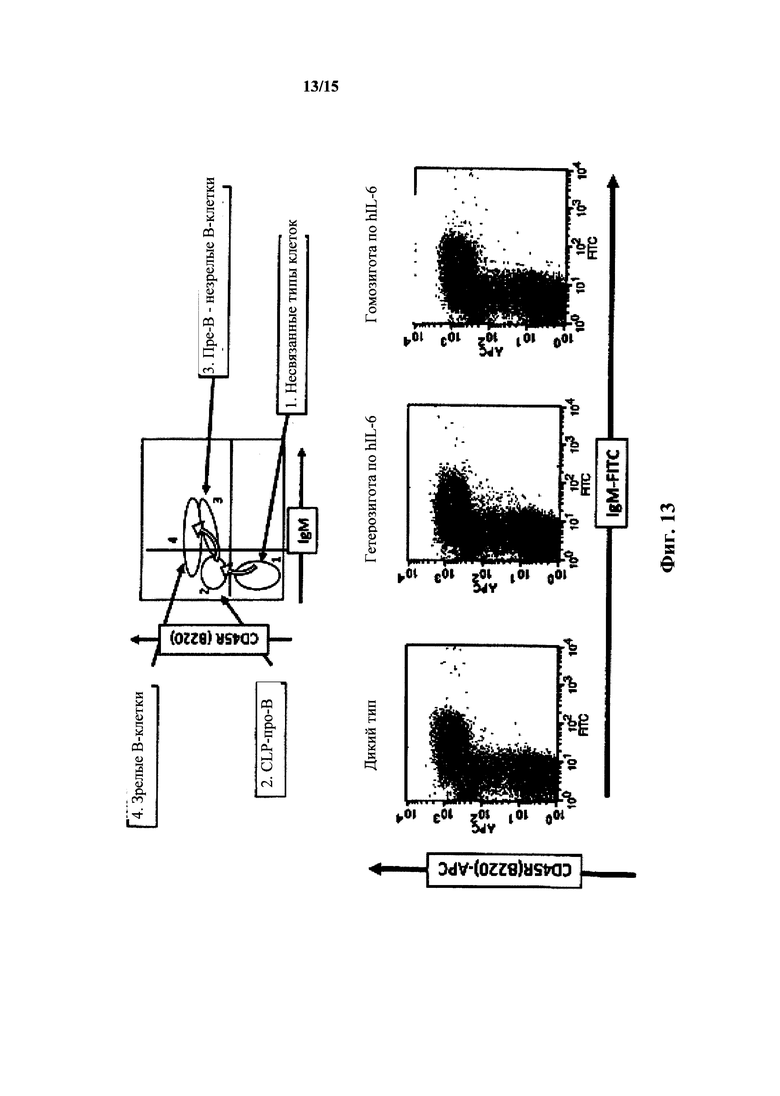
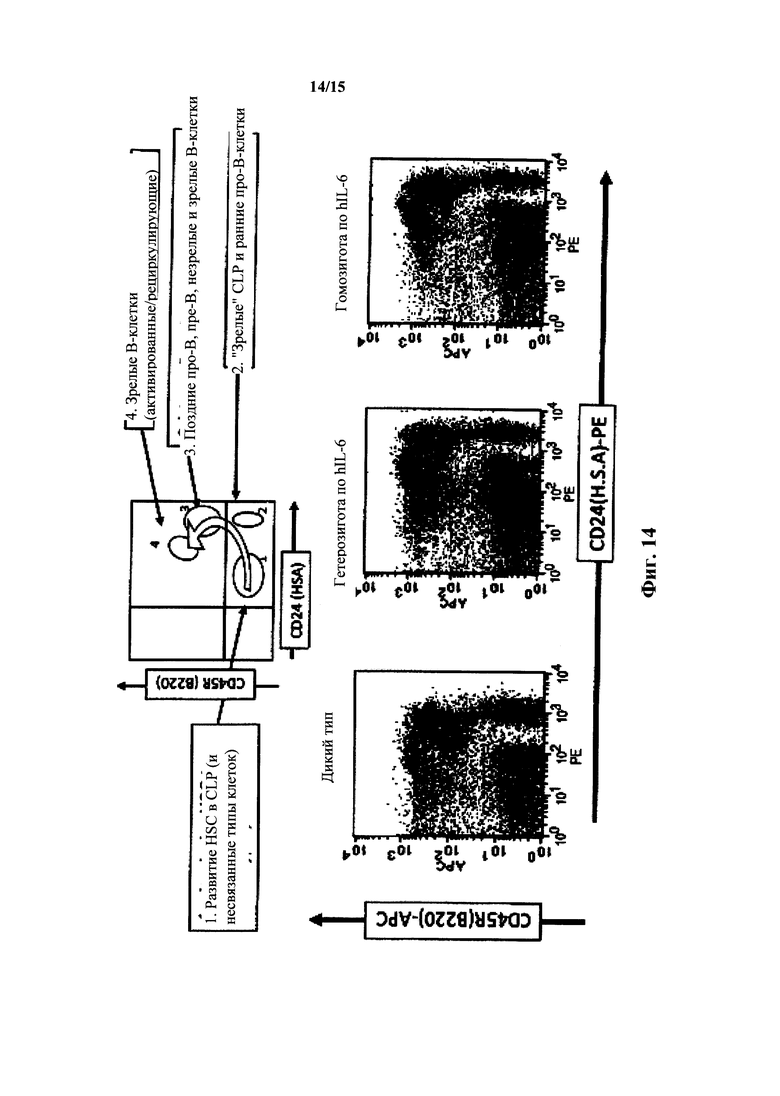
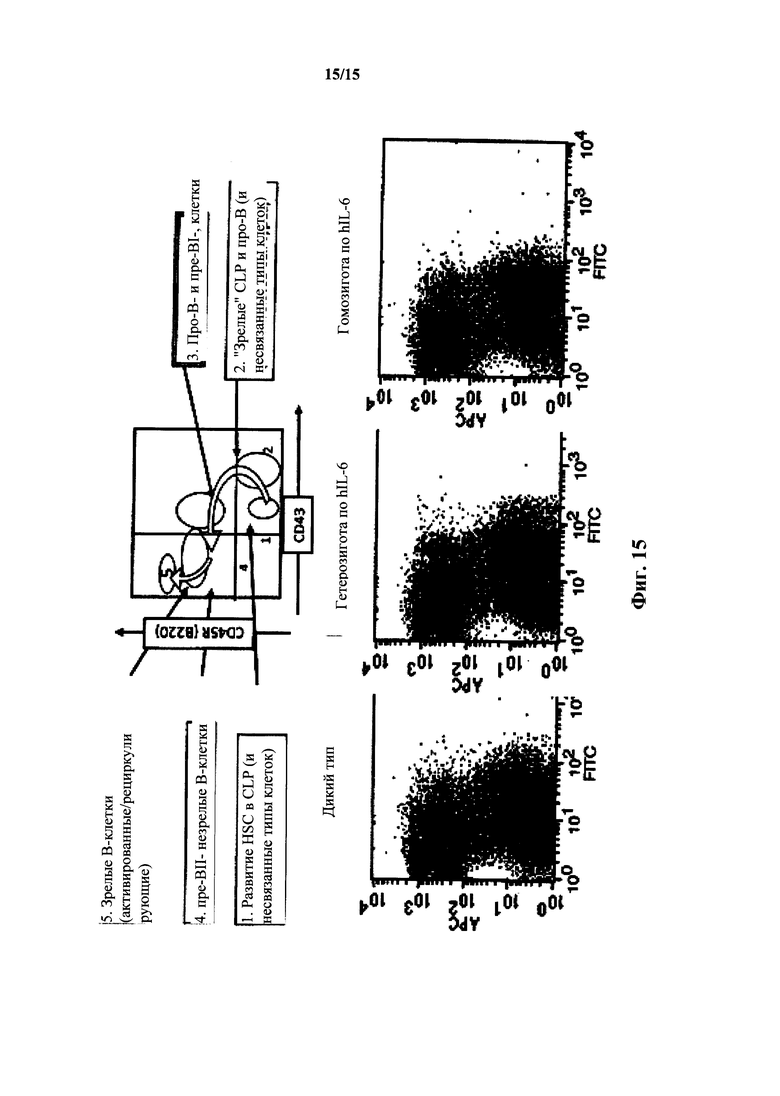
Изобретение относится к биотехнологии, генетической инженерии, в частности к генетически модифицированным животным семейства мышиных, а именно к мышам и крысам, которые экспрессируют IL-6 человека и дополнительно могут экспрессировать гуманизированный IL-6Rα. Указанное животное несет замену своего гена, кодирующего IL-6, в его эндогенном локусе IL-6 на ген человека, кодирующий IL-6 человека, и дополнительно замену последовательности, кодирующей внеклеточный домен IL-6Rα животного, в его эндогенном локусе IL-6Rα, на последовательность, кодирующую внеклеточный домен IL-6Rα человека, с получением гуманизированного гена IL-6Rα. При этом ген человека, кодирующий IL-6 человека, и гуманизированный ген IL-6Rα находятся под контролем эндогенных регуляторных элементов указанного животного в его эндогенном локусе IL-6 и IL-6Rα соответственно. Указанный гуманизированный ген IL-6Rα содержит эндогенные трансмембранную и цитоплазматическую последовательности IL-6Rα указанного животного. Изобретение также раскрывает способ получения указанных животных и выделенные эмбриональные стволовые клетки для получения таких животных. Изобретение позволяет получать трансгенных указазных мышей и крыс без каких-либо патологий. 5 н. и 13 з.п. ф-лы, 15 ил., 3 табл., 3 пр.
1. Генетически модифицированное животное семейства мышиных, которое экспрессирует IL-6 человека, несущее замену гена животного семейства мышиных, кодирующего IL-6, в эндогенном локусе IL-6 животного семейства мышиных на ген человека, кодирующий IL-6 человека, где ген человека, кодирующий IL-6 человека, находится под контролем эндогенных регуляторных элементов животного семейства мышиных в эндогенном локусе IL-6 животного семейства мышиных, при этом животное семейства мышиных представляет собой мышь или крысу.
2. Животное семейства мышиных по п. 1, где животное семейства мышиных представляет собой мышь.
3. Животное семейства мышиных по п. 1, где животное семейства мышиных представляет собой крысу.
4. Животное семейства мышиных по любому из пп.1-3, где ген человека, кодирующий IL-6 человека, содержит экзоны с 1 по 5 гена IL-6 человека, происходящие из бактериальной искусственной хромосомы CTD-2369M23.
5. Животное семейства мышиных по любому из пп.1-3, где животное семейства мышиных дополнительно содержит гуманизированный IL-6Rα, который содержит внеклеточный домен IL-6Rα человека.
6. Животное семейства мышиных по п. 5, где гуманизированный IL-6Rα содержит эндогенный трансмембранный домен IL-6Rα животного семейства мышиных и эндогенный цитоплазматический домен IL-6Rα животного семейства мышиных.
7. Животное семейства мышиных по любому из пп. 1-3, где у животного семейства мышиных не выявляют свойства, выбранного из плазмоцитоза, гломерулосклероза, гломерулонефрита, почечной недостаточности, гипергаммаглобулинемии, повышенного количества мегакариоцитов в селезенке, повышенного количества мегакариоцитов в костном мозге, спленомегалии, увеличения лимфоузлов, уплотненных аномальных плазматических клеток и их сочетания.
8. Генетически модифицированное животное семейства мышиных, которое экспрессирует IL-6 человека и гуманизированный IL-6Rα, несущее:
замену гена животного семейства мышиных, кодирующего IL-6, в эндогенном локусе животного семейства мышиных на ген человека, кодирующий IL-6 человека, где ген человека, кодирующий IL-6 человека, находится под контролем эндогенных регуляторных элементов животного семейства мышиных в эндогенном локусе IL-6 животного семейства мышиных, и
замену последовательности, кодирующей внеклеточный домен IL-6Rα животного семейства мышиных, в эндогенном локусе IL-6Rα животного семейства мышиных, на последовательность, кодирующую внеклеточный домен IL-6Rα человека, с получением гуманизированного гена IL-6Rα, где гуманизированный ген IL-6Rα содержит эндогенную трансмембранную последовательность IL-6Rα животного семейства мышиных и эндогенную цитоплазматическую последовательность IL-6Rα животного семейства мышиных, и гуманизированный ген IL-6Rα находится под контролем эндогенных регуляторных элементов животного семейства мышиных в локусе IL-6Rα животного семейства мышиных,
при этом животное семейства мышиных представляет собой мышь или крысу.
9. Животное семейства мышиных по п.8, где мышь не экспрессирует IL-6Rα животного семейства мышиных и не экспрессирует IL-6 животного семейства мышиных.
10. Животное семейства мышиных по п.8 или 9, где животное семейства мышиных представляет собой мышь.
11. Животное семейства мышиных по п.8 или 9, где животное семейства мышиных представляет собой крысу.
12. Способ получения генетически модифицированного животного семейства мышиных, включающий замену гена животного семейства мышиных, кодирующего IL-6, в эндогенном локусе IL-6 животного семейства мышиных, на ген человека, кодирующий IL-6 человека, где ген IL-6 человека, кодирующий IL-6 человека, находится под контролем эндогенных регуляторных элементов животного семейства мышиных в эндогенном локусе IL-6 животного семейства мышиных, при этом животное семейства мышиных представляет собой мышь или крысу.
13. Способ по п.12, где животное семейства мышиных представляет собой мышь.
14. Способ по п.12, где животное семейства мышиных представляет собой крысу.
15. Выделенная эмбриональная стволовая клетка (ES) животного семейства мышиных для получения животного семейства мышиных, которая содержит замену гена животного семейства мышиных, кодирующего IL-6, в эндогенном локусе IL-6 животного семейства мышиных на ген человека, кодирующий IL-6 человека, где ген человека, кодирующий IL-6 человека, находится под контролем эндогенных регуляторных элементов животного семейства мышиных в эндогенном локусе IL-6 животного семейства мышиных, при этом животное семейства мышиных представляет собой мышь или крысу.
16. Выделенная ES-клетка животного семейства мышиных для получения животного семейства мышиных, которая содержит замену гена животного семейства мышиных, кодирующего IL-6, в эндогенном локусе IL-6 животного семейства мышиных на ген человека, кодирующий IL-6 человека, где ген человека, кодирующий IL-6 человека, находится под контролем эндогенных регуляторных элементов животного семейства мышиных в эндогенном локусе IL-6 животного семейства мышиных, и указанная ES-клетка животного семейства мышиных дополнительно содержит замену последовательности, кодирующей внеклеточный домен IL-6Rα животного семейства мышиных, в эндогенном локусе IL-6Rα животного семейства мышиных, на последовательность, кодирующую внеклеточный домен IL-6Rα человека, с получением гуманизированного гена IL-6Rα, где гуманизированный ген IL-6Rα содержит эндогенную трансмембранную последовательность IL-6Rα животного семейства мышиных и эндогенную цитоплазматическую последовательность IL-6Rα животного семейства мышиных, и гуманизированный ген IL-6Rα находится под контролем эндогенных регуляторных элементов животного семейства мышиных в локусе IL-6Rα животного семейства мышиных, при этом животное семейства мышиных представляет собой мышь или крысу.
17. ES-клетка по п.15 или 16, где животное семейства мышиных представляет собой мышь.
18. ES-клетка по п.15 или 16, где животное семейства мышиных представляет собой крысу.
| GOYA S | |||
| et al | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| HIROTA H | |||
| et al | |||
| Реверсивный дисковый культиватор для тросовой тяги | 1923 |
|
SU130A1 |
| NAKA T | |||
| et al | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| РЫБЧИН В.Н | |||
| Основы генетической инженерии | |||
| Учебник для ВУЗов, Санкт-Петербург | |||
| Издательство СПбГТУ, 2002, 522 с., с.410-417 | |||
| RU 2003123598 A, 10.02.2005. | |||

