ПЕРЕКРЕСТНАЯ ССЫЛКА НА СМЕЖНЫЕ ЗАЯВКИ
Настоящая заявка испрашивает приоритет по предварительной заявке на патент США №62/005,476, поданной 30 мая 2014 г., предварительной заявке на патент США №62/051,626, поданной 17 сентября 2014 г., и предварительной заявке на патент США №62/072,692, поданной 30 октября 2014 г., причем описания каждой из заявок полностью включены в настоящий документ путем ссылки.
ОБЛАСТЬ ПРИМЕНЕНИЯ ИЗОБРЕТЕНИЯ
Не относящиеся к человеку животные, содержащие нуклеотидные последовательности, кодирующие белок дипептидилпептидазу IV (DPP4), который содержит человеческую последовательность. Трансгенные не относящиеся к человеку животные, содержащие ген DPP4, который полностью или частично является человеческим. Не относящиеся к человеку животные, которые экспрессируют человеческие или гуманизированные белки DPP4. Способы получения и использования не относящихся к человеку животных, содержащих человеческие или гуманизированные нуклеотидные последовательности DPP4.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Дипептидилпептидаза IV (DPP4) представляет собой мишень для терапевтического воздействия при лечении различных заболеваний, расстройств или состояний у человека, включая, например, гипергликемию (см., например, Gerich (2013) Pathogenesis and Management of Postpandrial Hyperglycemia: Role of Incretin-Based Therapies, Intl. J. Gen. Med. 6:877-895) и инфекцию коронавируса ближневосточного респираторного синдрома (MERS-CoV) (см., например, Raj et al. (2013) Dipeptidyl Peptidase 4 is a Functional Receptor for the Emerging Human Coronovirus-EMC, Nature 495(7440):251-254).
По отношению к не относящимся к человеку животным, например грызунам, таким как мыши или крысы, на регулярной основе выполняется оценка фармакокинетики (ФК) и фармакодинамики (ФД) терапевтических молекул, которые специфически воздействуют на человеческий белок DPP4. Однако ФД таких молекул невозможно правильно определить у некоторых не относящихся к человеку животных, если эти терапевтические молекулы также не воздействуют на эндогенный белок Dpp4.
Более того, оценку in vivo терапевтической эффективности специфичных к человеческому DPP4 низкомолекулярных, пептидных или белковых (т.е. биологических) антагонистов в моделях заболеваний с использованием не относящихся к человеку животных сложно выполнить у некоторых не относящихся к человеку животных, у которых видоспецифичный антагонист не взаимодействует с эндогенным белком Dpp4. Кроме того, оценку in vivo терапевтической эффективности низкомолекулярных, пептидных или белковых (т.е. биологических) антагонистов, воздействующих на молекулы-мишени, которые специфически взаимодействуют с человеческим белком DPP4, также сложно выполнить у некоторых не относящихся к человеку животных, у которых сама терапевтическая молекула-мишень не взаимодействует с эндогенным белком Dpp4.
Таким образом, существует потребность в не относящихся к человеку животных, например грызунах, таких как мыши или крысы, которые содержат человеческий или гуманизированный ген DPP4. Например, существует потребность в не относящихся к человеку животных, например грызунах, таких как мыши или крысы, у которых ген Dpp4 не относящегося к человеку животного является полностью или частично гуманизированным или заменен (например в эндогенных нечеловеческих локусах) на человеческий ген DPP4, содержащий последовательности, кодирующие человеческий или гуманизированный белок DPP4.
Также существует потребность в не относящихся к человеку животных, содержащих ген DPP4 (например человеческий или гуманизированный), у которых ген DPP4 контролируется нечеловеческими регуляторными элементами (например эндогенными регуляторными элементами), например, во 5'-фланкирующей области, например промоторе или энхансере (-ах), или в 3'-нетранслируемой области гена DPP4.
Также существует потребность в не относящихся к человеку животных, содержащих ген DPP4 (например человеческий или гуманизированный), у которых ген DPP4 контролируется человеческими регуляторными элементами, например, во 5'-фланкирующей области, например промоторе или энхансере (-ах), или в 3'-нетранслируемой области человеческого гена DPP4.
Также существует потребность в не относящихся к человеку животных, которые экспрессируют человеческий или гуманизированный белок DPP4 на поверхности иммунных клеток, например Т-клеток, и/или на поверхности клеток в одной или более тканях, например плацентарной, почечной, легочной, печеночной, скелетно-мышечной, сердечной, мозговой и/или панкреатической ткани, в концентрации, аналогичной концентрации белка Dpp4 на поверхности иммунных клеток, например Т-клеток, и/или на поверхности клеток в одной или более тканях, например плацентарной, почечной, легочной, печеночной, скелетно-мышечной, сердечной, мозговой и/или панкреатической ткани, у не относящегося к человеку животного такого же возраста, которое экспрессирует функциональный белок Dpp4, но не содержит человеческий или гуманизированный ген DPP4.
Кроме того, существует потребность в не относящихся к человеку животных, которые экспрессируют человеческий или гуманизированный белок DPP4 на поверхности иммунных клеток, например Т-клеток, и/или на поверхности клеток в одной или более тканях, например плацентарной, почечной, легочной, печеночной, скелетно-мышечной, сердечной, мозговой и/или панкреатической ткани, в концентрации, которая больше или меньше концентрации белка Dpp4 на поверхности иммунных клеток, например Т-клеток, и/или на поверхности клеток в одной или более тканях, например плацентарной, почечной, легочной, печеночной, скелетно-мышечной, сердечной, мозговой и/или панкреатической ткани, у не относящегося к человеку животного такого же возраста, которое экспрессирует функциональный белок Dpp4, но не содержит человеческий или гуманизированный ген DPP4.
В настоящем описании приводятся ссылки на различные патенты, заявки на патент и другие типы публикаций (например статьи в журналах, записи в электронных базах данных и т.п.). Описания всех патентов, заявок на патент и других публикаций, упомянутых в настоящем документе, полностью включены в настоящий документ путем ссылки для любых целей.
ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Предложены не относящиеся к человеку животные, содержащие нуклеотидные последовательности, кодирующие белок DPP4, который содержит человеческую последовательность.
Предложены трансгенные не относящиеся к человеку животные, содержащие ген DPP4, который полностью или частично является человеческим.
Предложены не относящиеся к человеку животные, которые экспрессируют человеческий или гуманизированный белок DPP4.
Предложены не относящиеся к человеку животные, у которых полностью или частично заменен эндогенный ген Dpp4 не относящегося к человеку животного.
Предложены не относящиеся к человеку животные, содержащие полностью или частично гуманизированный DPP4 в локусе эндогенного нечеловеческого Dpp4.
Предложены не относящиеся к человеку животные, которые имеют человеческий или гуманизированный ген DPP4, причем не относящиеся к человеку животные не экспрессируют эндогенный белок Dpp4, и при этом они экспрессируют человеческий или гуманизированный белок DPP4 на поверхности иммунных клеток, например Т-клеток, и/или на поверхности клеток в одной или более тканях, включая плацентарную, почечную, легочную, печеночную, скелетно-мышечную, сердечную, мозговую и/или панкреатическую ткань, в концентрации, аналогичной концентрации белка Dpp4 на поверхности иммунных клеток, например Т-клеток, и/или на поверхности клеток в одной или более тканях, включая плацентарную, почечную, легочную, печеночную, скелетно-мышечную, сердечную, мозговую и/или панкреатическую ткань, у не относящегося к человеку животного такого же возраста, которое экспрессирует функциональный эндогенный белок Dpp4, но не содержит замены.
В одном аспекте предложены не относящиеся к человеку животные, содержащие человеческую или гуманизированную нуклеотидную последовательность DPP4.
В одном аспекте предложены генетически модифицированные не относящиеся к человеку животные, у которых в эндогенном локусе Dpp4 ген, кодирующий эндогенный белок Dpp4, заменен на ген, кодирующий человеческий или гуманизированный белок DPP4. Предложены грызуны, например мыши или крысы, у которых эндогенный ген Dpp4 в эндогенном локусе Dpp4 заменен на человеческий ген DPP4. В одном варианте осуществления грызун является гетерозиготным по замененному участку в эндогенном локусе Dpp4 эндогенного гена Dpp4, кодирующего кодирующий человеческий или гуманизированный белок DPP4. В одном варианте осуществления грызун является гомозиготным по замененному участку в эндогенном локусе Dpp4 эндогенного гена Dpp4, кодирующего кодирующий человеческий или гуманизированный белок DPP4. В одном варианте осуществления грызун представляет собой мышь. В одном варианте осуществления грызун представляет собой крысу.
В одном аспекте предложены генетически модифицированные грызуны, например мыши или крысы, у которых выполнена гуманизация эндогенного гена Dpp4 грызуна, причем гуманизация включает замену в эндогенном локусе Dpp4 грызуна гена грызуна, кодирующего экзон гена Dpp4, на нуклеотидную последовательность, кодирующую по меньшей мере один экзон человеческого гена DPP4, с получением модифицированного гена DPP4, при этом экспрессия модифицированного гена DPP4 находится под контролем регуляторных элементов грызуна в эндогенном локусе Dpp4 грызуна.
В одном варианте осуществления грызун является гетерозиготным по нуклеотидной последовательности, кодирующей по меньшей мере один экзон человеческого гена DPP4, с образованием модифицированного гена DPP4. В одном варианте осуществления грызун является гомозиготным по нуклеотидной последовательности, кодирующей по меньшей мере один экзон человеческого гена DPP4, с образованием модифицированного гена DPP4.
В одном варианте осуществления грызун представляет собой мышь или крысу. В одном варианте осуществления грызун представляет собой мышь. В одном варианте осуществления грызун представляет собой крысу.
В одном варианте осуществления человеческий ген DPP4, кодирующий человеческий или гуманизированный белок DPP4, содержит экзоны 2-26 человеческого гена DPP4.
В одном варианте осуществления гуманизированный белок DPP4 содержит внеклеточный домен человеческого белка DPP4.
В одном варианте осуществления гуманизированный белок DPP4 содержит трансмембранный домен и цитоплазматический домен мышиного белка DPP4.
В одном варианте осуществления грызун представляет собой мышь, которая неспособна экспрессировать мышиный белок Dpp4.
В одном варианте осуществления грызун представляет собой мышь, причем непрерывный геномный фрагмент мышиной последовательности Dpp4, кодирующей экзоны 2-26 мышиного Dpp4, заменен на непрерывный геномный фрагмент человеческой последовательности DPP4, кодирующей экзоны 2-26 человеческого DPP4.
В одном аспекте предложены генетически модифицированные грызуны, например мышь или крыса, которые экспрессируют человеческий или гуманизированный белок DPP4, причем грызун, который экспрессирует человеческий или гуманизированный белок DPP4, имеет нормальную иммунную систему, т.е. число иммунных клеток, например Т-клеток, в крови, плазме или сыворотке грызуна, экспрессирующего человеческий или гуманизированный белок DPP4, аналогично числу иммунных клеток, например Т-клеток, в крови, плазме или сыворотке грызуна, экспрессирующего функциональный эндогенный белок Dpp4. В одном варианте осуществления грызун представляет собой мышь. В одном варианте осуществления грызун представляет собой крысу.
В одном варианте осуществления кровь грызуна, который экспрессирует человеческий или гуманизированный белок DPP4, имеет аналогичное число иммунных клеток, например Т-клеток, что и грызун, который экспрессирует функциональный эндогенный белок Dpp4, например мыши или крысы дикого типа. В одном варианте осуществления грызун представляет собой мышь. В одном варианте осуществления грызун представляет собой крысу.
В одном варианте осуществления у мыши, экспрессирующей человеческий или гуманизированный DPP4 на поверхности Т-клеток, количество Т-клеток, присутствующих в крови, составляет по меньшей мере приблизительно 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100%, 110%, 120%, 130%, 140%, 150%, 160%, 170%, 180%, 190% или 200% от количества Т-клеток, присутствующих в крови мыши такого же возраста, которая экспрессирует функциональный эндогенный белок Dpp4, но не содержит человеческого гена DPP4 вместо эндогенного гена Dpp4 в локусе эндогенного мышиного Dpp4.
В одном варианте осуществления у мыши, экспрессирующей человеческий или гуманизированный белок DPP4 на поверхности Т-клеток, количество Т-клеток в крови находится в диапазоне от приблизительно 20% до приблизительно 200%, от приблизительно 40% до приблизительно 160% или от приблизительно 80% до приблизительно 120% от количества Т-клеток, присутствующих в крови мыши такого же возраста, которая экспрессирует функциональный эндогенный белок Dpp4, но не содержит человеческого гена DPP4 вместо эндогенного гена Dpp4 в локусе эндогенного мышиного Dpp4.
В одном аспекте предложены генетически модифицированные грызуны, например мышь или крыса, которые экспрессируют человеческий или гуманизированный белок DPP4, причем грызун экспрессирует человеческий или гуманизированный белок DPP4 на поверхности иммунных клеток, например Т-клеток, и/или на поверхности клеток в одной или более тканях, например плацентарной, почечной, легочной, печеночной, скелетно-мышечной, сердечной, мозговой и/или панкреатической ткани, грызуна такого же возраста, который экспрессирует функциональный эндогенный белок Dpp4. В одном варианте осуществления грызун представляет собой мышь. В одном варианте осуществления грызун представляет собой крысу.
В одном варианте осуществления на поверхности иммунных клеток, например Т-клеток, грызуна, который экспрессирует человеческий или гуманизированный белок DPP4, находится приблизительно такая же концентрация белка DPP4, что и у иммунных клеток, например Т-клеток, грызуна, который экспрессирует функциональный эндогенный белок Dpp4, например мыши или крысы дикого типа. В одном варианте осуществления грызун представляет собой мышь. В одном варианте осуществления грызун представляет собой крысу.
В одном варианте осуществления мышь экспрессирует человеческий или гуманизированный белок DPP4 на поверхности иммунных клеток, например Т-клеток, в концентрации, составляющей по меньшей мере приблизительно 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100%, 110%, 120%, 130%, 140%, 150%, 160%, 170%, 180%, 190% или 200% от концентрации белка Dpp4 на поверхности иммунных клеток, например Т-клеток, у мыши такого же возраста, которая экспрессирует функциональный эндогенный белок Dpp4, но не содержит человеческого гена DPP4 вместо эндогенного гена Dpp4 в локусе эндогенного мышиного Dpp4.
В одном варианте осуществления мышь экспрессирует человеческий или гуманизированный белок DPP4 на поверхности иммунных клеток, например Т-клеток, в концентрации, находящейся в диапазоне от приблизительно 20% до приблизительно 200%, от приблизительно 40% до приблизительно 160% или от приблизительно 80% до приблизительно 120% от концентрации мышиного белка Dpp4 на поверхности иммунных клеток, например Т-клеток, у мыши такого же возраста, которая экспрессирует функциональный эндогенный белок Dpp4, но не содержит человеческого гена DPP4 вместо эндогенного гена Dpp4 в локусе эндогенного мышиного Dpp4.
В одном варианте осуществления на поверхности клеток в одной или более тканях, например плацентарной, почечной, легочной, печеночной, скелетно-мышечной, сердечной, мозговой и/или панкреатической ткани, грызуна, который экспрессирует человеческий или гуманизированный белок DPP4, находится приблизительно такая же концентрация белка DPP4, что и у клеток в одной или более тканях, например плацентарной, почечной, легочной, печеночной, скелетно-мышечной, сердечной, мозговой и/или панкреатической ткани, грызуна, который экспрессирует функциональный эндогенный белок Dpp4, например мыши или крысы дикого типа. В одном варианте осуществления грызун представляет собой мышь. В одном варианте осуществления грызун представляет собой крысу.
В одном варианте осуществления мышь экспрессирует человеческий или гуманизированный белок DPP4 на поверхности клеток в одной или более тканях, например плацентарной, почечной, легочной, печеночной, скелетно-мышечной, сердечной, мозговой и/или панкреатической ткани, в концентрации, составляющей по меньшей мере приблизительно 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100%, 110%, 120%, 130%, 140%, 150%, 160%, 170%, 180%, 190% или 200% от концентрации белка Dpp4 на поверхности клеток в одной или более тканях, например плацентарной, почечной, легочной, печеночной, скелетно-мышечной, сердечной, мозговой и/или панкреатической ткани, у мыши такого же возраста, которая экспрессирует функциональный эндогенный белок Dpp4, но не содержит человеческого гена DPP4 вместо эндогенного гена Dpp4 в локусе эндогенного мышиного Dpp4.
В одном варианте осуществления мышь экспрессирует человеческий или гуманизированный белок DPP4 на поверхности клеток в одной или более тканях, например плацентарной, почечной, легочной, печеночной, скелетно-мышечной, сердечной, мозговой и/или панкреатической ткани, в концентрации, находящейся в диапазоне от приблизительно 20% до приблизительно 200%, от приблизительно 40% до приблизительно 160% или от приблизительно 80% до приблизительно 120% от концентрации мышиного белка Dpp4 на поверхности клеток в одной или более тканях, например плацентарной, почечной, легочной, печеночной, скелетно-мышечной, сердечной, мозговой и/или панкреатической ткани, у мыши такого же возраста, которая экспрессирует функциональный эндогенный белок Dpp4, но не содержит человеческого гена DPP4 вместо эндогенного гена Dpp4 в локусе эндогенного мышиного Dpp4.
В одном аспекте предложен генетически модифицированный грызун, содержащий гуманизированный ген DPP4, который содержит человеческую последовательность, кодирующую внеклеточный домен DPP4, вместо последовательности грызуна, кодирующей внеклеточный домен Dpp4, причем гуманизированный ген DPP4 содержит трансмембранную последовательность Dpp4 грызуна и цитоплазматическую последовательность Dpp4 грызуна, при этом гуманизированный ген DPP4 контролируется регуляторными элементами эндогенного Dpp4 грызуна в эндогенном локусе Dpp4.
В одном варианте осуществления грызун является гетерозиготным по гуманизированному гену DPP4. В одном варианте осуществления грызун является гомозиготным по гуманизированному гену DPP4.
В одном варианте осуществления грызун представляет собой мышь или крысу. В одном варианте осуществления грызун представляет собой мышь. В одном варианте осуществления грызун представляет собой крысу.
В одном варианте осуществления мышь неспособна экспрессировать мышиный белок Dpp4.
В одном варианте осуществления регуляторные элементы или последовательности грызуна в локусе эндогенного Dpp4 грызуна взяты у мыши или крысы.
В одном варианте осуществления регуляторные элементы или последовательности грызуна представляют собой эндогенные регуляторные элементы или последовательности грызуна в локусе Dpp4 грызуна и взяты у мыши или крысы.
В одном аспекте предложено не относящееся к человеку животное, например грызун, такой как мышь или крыса, которое экспрессирует человеческий или гуманизированный белок DPP4, причем не относящееся к человеку животное экспрессирует человеческий или гуманизированный белок DPP4 из локуса эндогенного нечеловеческого Dpp4. В варианте осуществления не относящееся к человеку животное относится к семейству грызунов. В варианте осуществления грызун представляет собой мышь. В варианте осуществления грызун представляет собой крысу. В одном варианте осуществления грызун является гетерозиготным по локусу эндогенного нечеловеческого Dpp4, который экспрессирует человеческий или гуманизированный белок DPP4. В одном варианте осуществления грызун является гомозиготным по локусу эндогенного нечеловеческого Dpp4, который экспрессирует человеческий или гуманизированный белок DPP4.
В одном аспекте предложена генетически модифицированная мышь, которая экспрессирует человеческий или гуманизированный белок DPP4 из локуса эндогенного мышиного Dpp4, причем эндогенный мышиный ген Dpp4 полностью или частично заменен на человеческий ген DPP4.
В одном варианте осуществления приблизительно 78,8 тыс.п.н. в локусе эндогенного мышиного Dpp4, включая участок от экзона 2 до стоп-кодона в экзоне 26, удалены и заменены на человеческую последовательность гена DPP4 размером приблизительно 81,8 тыс.п.н., содержащей участок от экзона 2 до экзона 26 и участок нетранслируемой последовательности со стороны 3'-конца человеческого гена DPP4. В конкретном варианте осуществления человеческий ген DPP4 содержит участок от экзона 2 до экзона 26 и участок нетранслируемой последовательности со стороны 3'-конца человеческого гена DPP4 человеческой ВАС RP11-68L22. В конкретном варианте осуществления ген DPP4 содержит регуляторные элементы со стороны 5'-конца мышиного гена Dpp4, экзон 1 мышиного Dpp4, включая первые две аминокислоты метионин и лизин мышиного белка Dpp4, и регуляторные элементы со стороны 3'-конца Dpp4, экзон 1 мышиного Dpp4, включая первые две аминокислоты метионин и лизин мышиного белка Dpp4, и регуляторные элементы со стороны 3'-конца мышиного гена Dpp4 (например нетранслируемую последовательность со стороны 3'-конца) и участок человеческого гена DPP4 от экзона 2 до экзона 26, т.е. последовательности, кодирующие человеческий белок DPP4, за исключением первых двух аминокислот, которые взяты от экзона 1 мышиного Dpp4.
В одном аспекте предложена генетически модифицированная мышь, которая содержит нуклеотидную последовательность, кодирующую человеческий или гуманизированный белок DPP4, причем нуклеотидная последовательность, кодирующая человеческий или гуманизированный белок DPP4, полностью или частично заменяет эндогенную нуклеотидную последовательность, кодирующую эндогенный мышиный белок Dpp4.
В одном варианте осуществления мышь является гетерозиготной по нуклеотидной последовательности, кодирующей человеческий или гуманизированный белок DPP4. В одном варианте осуществления мышь является гомозиготной по нуклеотидной последовательности, кодирующей человеческий или гуманизированный белок DPP4.
В одном аспекте предложен способ получения грызуна с гуманизированным DPP4, включающий замену последовательности гена Dpp4 грызуна, кодирующей белок Dpp4 грызуна, человеческой последовательностью гена DPP4, содержащей один или более экзонов человеческой последовательности гена DPP4, кодирующей человеческий или гуманизированный белок DPP4, причем замена выполнена в локусе эндогенного Dpp4 грызуна, и человеческая последовательность гена DPP4, содержащая один или более экзонов человеческой последовательности гена DPP4, кодирующей человеческий или гуманизированный белок DPP4, функционально связана с регуляторными элементами или последовательностями грызуна в локусе эндогенного Dpp4 грызуна.
В одном варианте осуществления грызун является гетерозиготным по нуклеотидной последовательности, кодирующей человеческий или гуманизированный белок DPP4. В одном варианте осуществления грызун является гомозиготным по нуклеотидной последовательности, кодирующей человеческий или гуманизированный белок DPP4.
В одном варианте осуществления грызун представляет собой мышь или крысу. В одном варианте осуществления грызун представляет собой мышь. В одном варианте осуществления грызун представляет собой крысу.
В одном варианте осуществления регуляторные элементы или последовательности грызуна взяты от мыши. В одном варианте осуществления регуляторные элементы или последовательности грызуна взяты от крысы.
В одном варианте осуществления регуляторные элементы или последовательности грызуна представляют собой эндогенные регуляторные элементы или последовательности грызуна в локусе Dpp4 грызуна. В одном варианте осуществления грызун представляет собой мышь. В одном варианте осуществления грызун представляет собой крысу.
В одном варианте осуществления человеческая последовательность гена DPP4, заменяющая последовательность гена Dpp4 грызуна, содержит по меньшей мере один экзон человеческой последовательности гена DPP4. В других вариантах осуществления человеческая последовательность гена DPP4, заменяющая последовательность гена Dpp4 грызуна, содержит по меньшей мере 2, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5, по меньшей мере 6, по меньшей мере 7, по меньшей мере 8, по меньшей мере 9, по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24 или по меньшей мере 25 экзонов человеческой последовательности гена DPP4. В одном варианте осуществления человеческая последовательность гена DPP4, заменяющая последовательность гена Dpp4 грызуна, содержит все 26 экзонов человеческой последовательности гена DPP4. В одном варианте осуществления грызун представляет собой мышь. В одном варианте осуществления грызун представляет собой крысу.
В одном варианте осуществления человеческая или гуманизированная последовательность гена DPP4, заменяющая последовательность гена Dpp4 грызуна, кодирует белок, который приблизительно на 85%, 90%, 95%, 96%, 97%, 98% или приблизительно 99% идентичен человеческому DPP4.
В одном варианте осуществления человеческая или гуманизированная последовательность гена DPP4, заменяющая последовательность гена Dpp4 грызуна, содержит по меньшей мере один экзон человеческой последовательности гена DPP4, кодирующей внеклеточный домен человеческого белка DPP4. В других вариантах осуществления человеческая последовательность гена DPP4, заменяющая последовательность гена Dpp4 грызуна, содержит по меньшей мере 2, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5, по меньшей мере 6, по меньшей мере 7, по меньшей мере 8, по меньшей мере 9, по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22 или по меньшей мере 23 экзона человеческой последовательности гена DPP4, кодирующей внеклеточный домен человеческого белка DPP4. В одном варианте осуществления человеческая последовательность гена DPP4, заменяющая последовательность гена Dpp4 грызуна, содержит все 24 экзона человеческой последовательности гена DPP4, кодирующей внеклеточный домен человеческого белка DPP4. В одном варианте осуществления грызун представляет собой мышь. В одном варианте осуществления грызун представляет собой крысу.
В одном варианте осуществления человеческая или гуманизированная последовательность гена DPP4, заменяющая последовательность гена Dpp4 грызуна, кодирует внеклеточный домен белка DPP4, который приблизительно на 85%, 90%, 95%, 96%, 97%, 98% или приблизительно 99% идентичен внеклеточному домену человеческого белка DPP4.
В одном варианте осуществления замена выполнена в локусе эндогенного Dpp4 грызуна, и человеческая последовательность гена DPP4, содержащая один или более экзонов человеческой последовательности гена DPP4, кодирующей человеческий или гуманизированный белок DPP4, функционально связана с эндогенными регуляторными элементами или последовательностями грызуна в локусе эндогенного Dpp4 грызуна.
В одном аспекте предложен способ получения мыши с гуманизированным DPP4, включающий замену последовательности мышиного гена Dpp4, кодирующей мышиный белок Dpp4, человеческой последовательностью гена DPP4, кодирующей человеческий или гуманизированный белок DPP4.
В одном варианте осуществления замена выполнена в локусе эндогенного Dpp4 мыши, и человеческий ген DPP4, кодирующий человеческий или гуманизированный белок DPP4, функционально связан с регуляторными элементами или последовательностями мыши в локусе эндогенного Dpp4 мыши.
В одном варианте осуществления замена выполнена в локусе эндогенного Dpp4 мыши, и человеческий ген DPP4, кодирующий человеческий или гуманизированный белок DPP4, функционально связан с эндогенными регуляторными элементами или последовательностями мыши в локусе эндогенного Dpp4 мыши.
В различных аспектах генетически модифицированные не относящиеся к человеку животные, например грызуны, такие как мыши или крысы, описанные в настоящем документе, содержат генетические модификации в их зародышевой линии.
В одном аспекте предложен эмбрион не относящегося к человеку животного, например грызуна, такого как мышь или крыса, содержащий генетическую модификацию, описанную в настоящем документе.
В одном аспекте предложен эмбрион-носитель не относящегося к человеку животного, например грызуна, такого как мышь или крыса, который содержит донорскую клетку, которая содержит генетическую модификацию, описанную в настоящем документе.
В одном аспекте предложена плюрипотентная или тотипотентная клетка не относящегося к человеку животного, например грызуна, такого как мышь или крыса, содержащая генетическую модификацию, описанную в настоящем документе. В одном варианте осуществления клетка представляет собой клетку грызуна. В одном варианте осуществления клетка представляет собой клетку мыши. В одном варианте осуществления клетка представляет собой эмбриональную стволовую (ЭС) клетку грызуна. В одном варианте осуществления клетка представляет собой ЭС-клетку.
В одном аспекте предложена яйцеклетка не относящегося к человеку животного, например грызуна, такого как мышь или крыса, причем яйцеклетка не относящегося к человеку животного содержит эктопическую хромосому не относящегося к человеку животного, при этом эктопическая хромосома не относящегося к человеку животного содержит генетическую модификацию, описанную в настоящем документе. В одном варианте осуществления не относящееся к человеку животное относится к семейству грызунов. В одном варианте осуществления грызун представляет собой мышь. В одном варианте осуществления грызун представляет собой крысу.
В одном аспекте мышиный эмбрион, яйцеклетка или клетка, генетически модифицированная с внедрением человеческого гена DPP4, представляет собой эмбрион, яйцеклетку или клетку мыши линии C57BL, которую выбирают из C57BL/A, C57BL/An, C57BL/GrFa, C57BL/KaLwN, C57BL/6, C57BL/6J, C57BL/6ByJ, C57BL/6NJ, C57BL/10, C57BL/10ScSn, C57BL/10Cr и C57BL/01a. В другом варианте осуществления мышь представляет собой мышь линии 129, которую выбирают из группы, состоящей из 129Р1, 129Р2, 129Р3, 129X1, 129S1 (например, 129S1/SV, 129Sl/Svlm), 129S2, 129S4, 129S5, 129S9/SvEvH, 129S6 (129/SvEvTac), 129S7, 129S8, 129Т1, 129Т2 (см., например, Festing et al. (1999) Revised nomenclature for strain 129 mice, Mammalian Genome 10:836, см. также Auerbach et al (2000) Establishment and Chimera Analysis of 129/SvEv- and C57BL/6-Derived Mouse Embryonic Stem Cell Lines). В конкретном варианте осуществления генетически модифицированная мышь представляет собой смесь вышеупомянутой линии 129 и вышеупомянутой линии C57BL/6. В другом конкретном варианте осуществления мышь представляет собой смесь вышеупомянутых линий 129 или смесь вышеупомянутых линий BL/6. В конкретном варианте осуществления линия 129 смеси представляет собой линию 129S6 (129/SvEvTac). В другом варианте осуществления мышь представляет собой мышь линии BALB, например линии BALB/c. В другом варианте осуществления мышь представляет собой смесь линии BALB с другой вышеупомянутой линией. В одном варианте осуществления мышь представляет собой швейцарскую мышь или швейцарскую мышь Вебстер.
В различных аспектах не относящихся к человеку животных, содержащих человеческую или гуманизированную нуклеотидную последовательность DPP4, выбирают из млекопитающих или птиц. В одном варианте осуществления не относящиеся к человеку животные представляют собой млекопитающих. В одном варианте осуществления млекопитающие представляют собой грызунов. В одном варианте осуществления грызуны представляют собой мышей или крыс.
В одном аспекте предложен грызун, который содержит нуклеотидную последовательность, содержащую человеческий ген DPP4 или его фрагмент, причем человеческий ген DPP4 или его фрагмент содержит по меньшей мере один экзон человеческого гена DPP4, и при этом человеческий ген DPP4 или его фрагмент кодирует человеческий или гуманизированный белок DPP4.
В одном варианте осуществления человеческий ген DPP4 или его фрагмент содержит по меньшей мере 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 экзонов человеческого гена DPP4.
В одном варианте осуществления человеческий ген DPP4 или его фрагмент содержит все 26 экзонов человеческого гена DPP4.
В одном варианте осуществления нуклеотидная последовательность дополнительно содержит 5'-фланкирующую область человеческого гена DPP4. В одном варианте осуществления человеческий ген DPP4 или его фрагмент функционально связан с 5'-фланкирующей областью человеческого гена DPP4. В одном варианте осуществления длина 5'-фланкирующей области человеческого гена DPP4 составляет по меньшей мере 1 тыс.п.н. (например, длина составляет по меньшей мере 1, 2, 3, 4, 5, 10, 20, 30, 40, 50 тыс.п.н. или более). В одном варианте осуществления длина 5'-фланкирующей области человеческого гена DPP4 составляет по меньшей мере 10 тыс.п.н. В одном варианте осуществления длина 5'-фланкирующей области человеческого гена DPP4 составляет по меньшей мере 40 тыс.п.н.
В одном варианте осуществления экспрессия человеческого гена DPP4 или его фрагмента контролируется 5'-фланкирующей областью человеческого гена DPP4.
В одном варианте осуществления человеческий или гуманизированный белок DPP4 содержит аминокислотную последовательность SEQ ID NO: 24 или ее фрагмент.
В одном варианте осуществления грызун экспрессирует человеческий или гуманизированный белок DPP4 на поверхности Т-клеток в концентрации, составляющей по меньшей мере приблизительно 20% (например, по меньшей мере приблизительно 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 98%, 99% или более) от концентрации белка Dpp4 грызуна, присутствующего на поверхности Т-клеток грызуна такого же возраста, который экспрессирует функциональный эндогенный белок Dpp4 грызуна, но не содержит человеческого гена DPP4 или его фрагмента.
В одном варианте осуществления грызун экспрессирует человеческий или гуманизированный белок DPP4 на поверхности клеток в одной или более тканях, выбранных из группы, состоящей из плацентарной, почечной, легочной, печеночной, скелетно-мышечной, сердечной, мозговой и панкреатической ткани, в концентрации, составляющей по меньшей мере приблизительно 20% (например, по меньшей мере приблизительно 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 98%, 99% или более) от концентрации белка Dpp4 грызуна, присутствующего на поверхности одной или более тканей грызуна такого же возраста, который экспрессирует функциональный эндогенный белок Dpp4 грызуна, но не содержит человеческого гена DPP4 или его фрагмента.
В одном варианте осуществления грызун экспрессирует функциональный эндогенный белок Dpp4 грызуна.
В одном варианте осуществления грызун представляет собой мышь или крысу.
В одном аспекте предложен способ получения гуманизированного трансгенного грызуна, включающий встраивание нуклеотидной последовательности, содержащей один или более экзонов человеческой последовательности гена DPP4, в хромосому грызуна, причем один или более экзонов человеческой последовательности гена DPP4 кодируют человеческий или гуманизированный белок DPP4.
В одном варианте осуществления человеческий ген DPP4 или его фрагмент содержит по меньшей мере 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 экзонов человеческого гена DPP4.
В одном варианте осуществления человеческий ген DPP4 или его фрагмент содержит все 26 экзонов человеческого гена DPP4.
В одном варианте осуществления нуклеотидная последовательность дополнительно содержит 5'-фланкирующую область человеческого гена DPP4.
В одном варианте осуществления человеческая последовательность гена DPP4 функционально связана с 5'-фланкирующей областью человеческого гена DPP4.
В одном варианте осуществления человеческий или гуманизированный белок DPP4 содержит аминокислотную последовательность SEQ ID NO: 24 или ее фрагмент.
В одном варианте осуществления грызун представляет собой мышь или крысу. В дополнительных аспектах предложены способы определения in vivo терапевтической эффективности специфичного для человека антагониста DPP4 у любого из грызунов с гуманизированным DPP4, описанных в настоящем документе, включающие введение грызуну антагониста DPP4, причем грызун инфицирован коронавирусом ближневосточного респираторного синдрома (MERS-CoV); и определение того, обеспечивает ли антагонист DPP4 лечение или предотвращение одного или более симптомов инфекции MERS-CoV по сравнению с контрольными грызунами, инфицированными MERS-CoV, которым не вводили антагонист DPP4.
В одном варианте осуществления антагонист DPP4 выбран из группы, состоящей из малых молекул, пептидов и антител.
В одном варианте осуществления антагонист DPP4 представляет собой антитело к белку MERS-CoV.
В одном варианте осуществления белок MERS-CoV представляет собой шипиковый белок MERS-CoV.
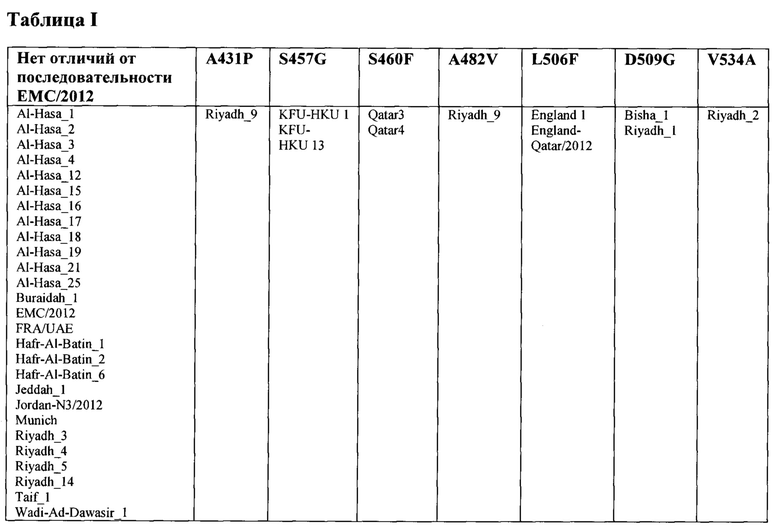
В одном варианте осуществления грызун инфицирован одним или более штаммами MERS-CoV, выбранных из группы, состоящей из Al-Hasa_1, Al-Hasa_2, Al-Hasa_3, Al-Hasa_4, Al-Hasa_12, Al-Hasa_15, Al-Hasa_16, Al-Hasa_17, Al-Hasa_18, Al-Hasa_19, Al-Hasa_21, Al-Hasa_25, Buraidah_1, EMC/2012, FRA/UAE, Hafr-Al-Batin_1, Hafr-Al-Batin_2, Hafr-Al-Batin_6, Jeddah_1, Jordan-N3/2012, Munich, Riyadh_3, Riyadh_4, Riyadh_5, Riyadh_14, Taif_1, Wadi-Ad-Dawasir_1, Riyadh_9, KFU-HKU 1, KFU-HKU 13, Qatar3, Qatar4, England 1, England-Qatar/2012, Bisha_1, Riyadh_1 и Riyadh_2.
В одном варианте осуществления антагонист вводят до инфицирования MERS-CoV. В одном варианте осуществления антагонист вводят после инфицирования MERS-CoV.
В одном варианте осуществления антагонист вводят одновременно с инфицированием MERS-CoV.
В одном варианте осуществления симптомом инфекции MERS-CoV является титр вируса или концентрация РНК.
В одном варианте осуществления титр вируса или концентрацию РНК оценивают с помощью одного или более способов, выбранных из группы, состоящей из количественной полимеразной цепной реакции (кПЦР), Нозерн-блоттинга, анализа бляшкообразования и гибридизации in situ.
В одном варианте осуществления симптомом инфекции MERS-CoV является воспаление легких.
В одном варианте осуществления воспаление легких оценивается гистохимическими методами.
В одном варианте осуществления симптомом инфекции MERS-CoV является снижение веса.
В одном варианте осуществления грызун представляет собой мышь или крысу. В одном варианте осуществления грызун представляет собой мышь. В одном варианте осуществления грызун представляет собой крысу.
Все аспекты и варианты осуществления, описанные в настоящем документе, могут использоваться совместно, кроме случаев, когда это в прямой форме исключено из контекста варианта осуществления или аспекта.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
На Фиг. 1А-В приведены иллюстрации (без соблюдения масштаба) принципа гуманизации локуса Dpp4. На Фиг. 1А схематически показано, что 78,8 тыс.п.н. мышиного гена Dpp4 (сверху), включая участок от экзона 2 до стоп-кодона в экзоне 26, удалены и заменены на 81,7 тыс.п.н. человеческого гена DPP4 (снизу), включая участок от экзона 2 до экзона 26 и участок 3'-нетранслируемой области. На Фиг. 1В схематически показано, что мышь с гуманизированным DPP4 содержит (i) 5'-фланкирующую область мышиного гена Dpp4, включая регуляторные последовательности, например промотор и сайт инициации транскрипции, и экзон 1, включая инициаторный кодон ATG, (ii) человеческий ген DPP4, включающий участок от экзона 2 до экзона 26 вместе со стоп-кодоном и участок 3'-нетранслируемой области вместе с сайтом loxP, и (iii) нетранслируемую область стороны 3'-конца мышиного гена Dpp4, начиная от непосредственно 3'-конца до стоп-кодона.
На Фиг. 2 показана аминокислотная последовательность (SEQ ID NO: 17) гуманизированного белка DPP4, экспрессированная у мышей с гуманизированным DPP4.
На Фиг. 3 показаны результаты ПЦР в реальном времени, выполненной на РНК, полученной из легочной ткани ложноинфицированных (фосфатно-солевой буфер (PBS)) или MERS-CoV-инфицированных (штамм Jordan) мышей F0 с гуманизированным DPP4 через 4 дня после инфицирования.
На Фиг. 4 показано окрашивание гематоксилином и эозином дыхательных путей (увеличение 10Х и 40Х) и альвеол (увеличение 40Х) из легких ложноинфицированных (PBS) или MERS-CoV-инфицированных (штамм Jordan) мышей F0 с гуманизированным DPP4 через 4 дня после инфицирования.
На Фиг. 5 показано выравнивание аминокислотной последовательности (SEQ ID NO: 25) мышиного Dpp4 (mDpp4) с человеческим белком DPP4 (hDPP4), кодируемым трансгенными мышами MAID 7326/7327 (SEQ ID NO: 26). Негомологичные остатки, которые отличаются между последовательностями, подчеркнуты, гомологичные остатки, которые отличаются между последовательностями, выделены жирным шрифтом и курсивом, а гэпы обозначены дефисом. Остатки, идентичные в двух последовательностях, показаны в виде неформатированного текста.
Фиг. 6 представляет собой таблицу, в которой показаны ген, последовательность и хромосомная информация для человеческого DPP4.
На Фиг. 7 схематически показаны границы человеческого гена DPP4 и фланкирующие геномные последовательности для каждой из искусственных бактериальных хромосом (ВАС): ВАС RP11-345J9 и ВАС RP11-68L22. Также показаны местоположения в пределах человеческого гена DPP4 и областей промотора, в которых выполняется отжиг с использованием наборов праймеров и зондов TaqMan™ для человека (7333 hTU и 7333 hTD для наборов, выполняющих отжиг со стороны 5'-конца и 3'-конца).
Фиг. 8 представляет собой таблицу, в которой показаны наборы праймеров и зондов, используемые для анализов приобретения аллеля TaqMan™ у человека, причем 7333 hTU относится к набору со стороны 5'-конца, а 7333 hTD относится к набору со стороны 3'-конца.
На Фиг. 9 показана аминокислотная последовательность гуманизированного белка DPP4, кодируемого трансгенными мышами MAID 7333 и 7334 (SEQ ID NO: 24).
Фиг. 10А представляет собой столбиковую диаграмму, на которой показаны измерения с помощью количественной ПЦР генома MERS-CoV (РНК UpE) у инфицированных мышей через 2 и 4 дня после инфицирования (dpi). Фиг. 10В представляет собой столбиковую диаграмму, на которой показаны измерения с помощью количественной ПЦР транскрипта мРНК (лидерная РНК) у инфицированных мышей через 2 dpi и 4 dpi. Фиг. 10С представляет собой столбиковую диаграмму, на которой показаны результаты количественной оценки титра вируса MERS-CoV в легком инфицированной мыши через 2 dpi и 4 dpi. Концентрации MERS-CoV в гомогенизированном легком мыши количественно определяли с помощью анализа дозы, инфицирующей 50% культуры ткани (TCID50), и выражали в виде бляшкообразующих единиц (pfu) на мл. Фиг. 10D представляет собой группу гистологических изображений легких инфицированной MERS-CoV мыши, окрашенных гематоксилином и эозином. Показаны дыхательные пути (10Х), сосуды (10Х) и интерстиций (40Х) мышей, которым вводили фосфатно-солевой буфер (PBS), и мышей через 2 dpi и 4 dpi.
Фиг. НА представляет собой столбиковую диаграмму, на которой показаны измерения с помощью количественной ПЦР генома MERS-CoV (РНК UpE) из легких мышей, которым перед инфицированием вирусом предварительно вводили антитела против шипикового белка MERS-CoV (Ab 2 или Ab 4). Фиг. 11В представляет собой столбиковую диаграмму, на которой показаны измерения с помощью количественной ПЦР транскрипта мРНК (лидерная РНК) из легких мышей, которым перед инфицированием вирусом предварительно вводили антитела против шипикового белка MERS-CoV. РНК количественно определяли с использованием праймеров, специфичных к геному MERS-CoV, и сравнивали с мышами изотипического контроля hIgG1. Все образцы сравнивали с контрольной группой hIgG1, принятой за 100%.
Фиг.11С представляет собой столбиковую диаграмму, на которой показан титр вируса в легких мышей, которым перед инфицированием вирусом предварительно вводили антитела против шипикового белка MERS-CoV. Титр вируса количественно определяли с помощью анализа бляшкообразования и выражали в виде pfu/мл.
Фиг. 12А представляет собой группу гистологических изображений легких мышей B6/hDPP4, инфицированных MERS-CoV, которым предварительно вводили антитела против шипикового белка MERS-CoV (Ab 2 или Ab 4). При этом на участках легкого мыши, окрашенных гематоксилином и эозином, видны дыхательные пути, сосуды и интерстиций репрезентативной мыши из каждой группы. Фиг. 12В представляет собой столбиковую диаграмму, на которой показано количество баллов при гистологической оценке легких мыши, показанных на Фиг. 12А. Количество баллов является средним значением для всех мышей в каждой экспериментальной группе в определенной временной отметке.
Фиг. 13А представляет собой столбиковую диаграмму, на которой показаны измерения с помощью количественной ПЦР генома MERS-CoV (РНК UpE) из инфицированных легких. Сравнивали эффекты антител против шипикового белка MERS-CoV (Ab 2 или Ab 4), которые вводили за один день до или через один день после инфицирования вирусом. Фиг. 13 В представляет собой столбиковую диаграмму, на которой показаны измерения с помощью количественной ПЦР транскрипта мРНК (лидерная РНК) MERS-CoV из инфицированных легких. Сравнивали эффекты антител против шипикового белка MERS-CoV (Ab 2 или Ab 4), которые вводили за один день до или через один день после инфицирования вирусом. На Фиг. 13А-В количественное определение РНК выполняли с использованием праймеров, специфичных к геному MERS-CoV, а результаты сравнивали с измерениями у мышей изотипического контроля hIgG1. Все образцы сравнивали с контрольной группой hIgG1, принятой за 100%. Фиг. 13С представляет собой столбиковую диаграмму, на которой показан титр вируса в легких мышей, которым после инфицирования вирусом вводили антитела против шипикового белка MERS-CoV (Ab 2 или Ab 4). Титр вируса количественно определяли с помощью анализа бляшкообразования и выражали в виде pfu/мл. Сравнивали эффекты антител, которые вводили за один день до или через один день после инфицирования вирусом.
Фиг. 14А представляет собой группу гистологических изображений легких мышей B6/hDPP4, инфицированных MERS-CoV, которым вводили антитела против шипикового белка MERS-CoV, Ab 2, через 1 день после инфицирования. При этом на участках легкого мыши, окрашенных гематоксилином и эозином, видны дыхательные пути, сосуды и интерстиций репрезентативной мыши из каждой группы. Фиг. 14В представляет собой столбиковую диаграмму, на которой показано количество баллов при гистологической оценке легких мыши на Фиг. 14А.
На Фиг. 15 показан график зависимости массы тела от времени после инфицирования MERS-CoV для мышей с гуманизированным DPP4. Одна группа содержит 4-5 мышей.
На Фиг. 16 показан график зависимости массы тела от времени после инфицирования MERS-CoV (1×106 pfu/мышь; 4-5 мышей на группу) для мышей, гетерозиготных и гомозиготных по гуманизированному DPP4.
На Фиг. 17 показаны патологические изменения (воспаление), наблюдаемые при гистологическом исследовании на 7 день мыши с гуманизированным DPP4, инфицированной большой дозой вируса (1×106 pfu/мышь), по сравнению с контролями, которым вводили PBS.
ПОДРОБНОЕ ОПИСАНИЕ
Ген и белок DPP4
Ген DPP4 кодирует трансмембранный белок типа II, трипептидилпептидазу IV (DPP4) (также известен под названием CD26, вспомогательный белок-2 аденозиндезаминазы (ADCP2), белок, связывающий аденозиндеаминазу (ADABP) и TP103), который обладает сериновой экзопептидазной активностью и играет важную роль в активации Т-клеток и внутриклеточных каскадах передачи сигнала в нескольких других типах клеток.
Человеческий DPP4. № гена в NCBI: 1803; основной источник: HGNC:3009; транскрипт RefSeq: NM_001935.3; № UniProt: Р27487; сборка генома: GRCh38; местоположение: chr2:162,848,755-162,931,052 - нить. (См. Фиг. 6).
Человеческий ген DPP4 расположен на хромосоме 2 в 2q24.3. Человеческий ген DPP4 имеет 26 экзонов и кодирует трансмембранный полипептид типа II длиной 766 аминокислот, включая цитоплазматический домен из 6 аминокислот на N-конце, трансмембранный домен из 22 аминокислот и внеклеточный домен из 738 аминокислот на С-конце. Внеклеточный домен (т.е. эктодомен) человеческого белка DPP4 кодируется кодирующими экзонами 3-26 человеческого гена DPP4.
Мышиный Dpp44. №гена в NCBI: 13482; основной источник: HGNC:94919; транскрипт RefSeq: NM_010074.3; № UniProt: Р28843; сборка генома: GRCm38; местоположение: chr2:62,330,073-62,412,591 - нить.
Мышиный ген DPP4 расположен на хромосоме 2 в 2 35.85 сМ. Мышиный ген Dpp4 имеет 26 экзонов и кодирует трансмембранный полипептид типа II длиной 760 аминокислот, включая цитоплазматический домен из 6 аминокислот на N-конце, трансмембранный домен из 22 аминокислот и внеклеточный домен из 732 аминокислот на С-конце. Внеклеточный домен (т.е. эктодомен) мышиного белка Dpp4 кодируется кодирующими экзонами 3-26 мышиного гена Dpp4.
Видовая специфичность белка DPP4
Как описано ниже в примере 2, человеческий (но не мышиный) белок DPP4 представляет собой функциональный рецептор для инфекции коронавируса ближневосточного респираторного синдрома (MERS-CoV).
Фармакокинетику (ФК) и фармакодинамику (ФД) обладающих видоспецифичностью потенциальных терапевтических молекул, которые воздействуют на человеческий белок DPP4, или обладающих видоспецифичностью молекул-мишеней, таких как MERS-CoV, которые взаимодействуют с человеческим белком DPP4, обычно оценивают у не относящихся к человеку животных, например грызунов, таких как мыши или крысы. Такие терапевтические молекулы также тестируют на наличие терапевтической эффективности in vivo у не относящихся к человеку животных, например грызунов, таких как мыши или крысы, в моделях человеческих заболеваний, расстройств или состояний, в которых DPP4 играет определенную роль.
Однако ФД и/или терапевтическую эффективность in vivo терапевтических молекул, специфичных к человеческому белку DPP4, например специфичных для человека ингибиторов DPP4, невозможно правильно оценить у грызунов, в частности у мышей, в связи с отсутствием мишеней для этих терапевтических молекул.
Более того, терапевтическую эффективность in vivo терапевтических молекул, специфичных к мишеням, которые взаимодействуют исключительно с человеческим белком DPP4, например, к MERS-CoV, специфичному к человеческому DPP4, невозможно правильно оценить у грызунов, в частности у мышей, в связи с отсутствием мишеней (например, рецептора, пары для взаимодействия) для этих терапевтических молекул-мишеней.
Соответственно, в различных вариантах осуществления для оценки ФД и/или терапевтической эффективности in vivo специфичного для человека антагониста или ингибитора белка DPP4 у не относящихся к человеку животных, например грызунов, таких как мыши или крысы, желательно заменить эндогенный белок Dpp4 человеческим или гуманизированным белком DPP4. В различных вариантах осуществления для оценки терапевтической эффективности in vivo низкомолекулярных, пептидных или биологических антагонистов или ингибиторов молекулы-мишени, которые взаимодействуют исключительно с человеческим белком DPP4, у не относящихся к человеку животных, например грызунов, таких как мыши или крысы, желательно заменить эндогенный белок Dpp4 человеческим или гуманизированным белком Dpp4.
Дополнительно в различных вариантах осуществления для предотвращения потенциальных проблем, связанных с избыточной или недостаточной экспрессией человеческого или гуманизированного белка DPP4 и/или неправильной экспрессией человеческого или гуманизированного белка DPP4 в клетках или тканях, в которых эндогенный белок Dpp4 обычно не экспрессируется, желательно полностью или частично вставить человеческий ген DPP4 в геном не относящихся к человеку животных, например грызунов, таких как мыши или крысы, в локусы эндогенного гена Dpp4 и экспрессировать человеческий или гуманизированный белок DPP4 в не относящихся к человеку животных, например грызунах, таких как мыши или крысы, под по меньшей мере частичным контролем эндогенных регуляторных элементов Dpp4.
В некоторых вариантах осуществления желательной является направленная замена эндогенного, например, мышиного или крысиного, гена Dpp4 человеческим геном DPP4 или его фрагментом.
В других вариантах осуществления вместо замены эндогенного гена Dpp4 человеческим геном DPP4 или его фрагментом человеческий ген DPP4 или его фрагмент произвольно вставляют в геном грызуна, например мыши или крысы. В некоторых вариантах осуществления у грызунов, например мышей или крыс, у которых человеческий ген DPP4 или его фрагмент произвольно вставлен в геном, сохранена экспрессия эндогенного Dpp4 грызуна.
В настоящем изобретении предложены не относящиеся к человеку животные, например грызуны, такие как мыши или крысы, которые содержат человеческий ген DPP4 или его фрагмент либо в локусе эндогенного Dpp4 (т.е. выполнена замена), либо в одном или более других локусах. Также в настоящем изобретении предложены не относящиеся к человеку животные, например грызуны, такие как мыши или крысы, которые содержат человеческий ген DPP4 или его фрагмент как в локусе эндогенного Dpp4 (т.е. выполнена замена), так и в дополнительном локусе/локусах.
В некоторых вариантах осуществления фрагмент человеческого гена DPP4 содержит 200 тысяч пар нуклеотидов (тыс.п.н.) или менее, например 180, 160, 140, 120, 100, 80, 70, 60, 50, 40, 30, 20, 10, 5, 2.5, 1 тыс.п.н. или менее, например 1000, 800, 600, 400, 200 или менее пар нуклеотидов.
Создание клеток и не относящихся к человеку животных с человеческим DPP4
Для направленной замены эндогенного нечеловеческого гена Dpp4 или его фрагмента человеческим геном DPP4 или его фрагментом создана таргетирующая конструкция. См., например, Valenzuela et al. Nature Biotech, 21.6(2003):652-659; US 6,586,251 и US 8,759,105. Например, таргетирующая конструкция содержит гомологичные участки, фланкирующие заменяющий человеческий ген DPP4 или его фрагмент.
В некоторых вариантах осуществления заменяющий человеческий ген DPP4 или его фрагмент содержит весь человеческий ген DPP4. В других вариантах осуществления заменяющий человеческий ген DPP4 или его фрагмент содержит часть человеческого гена DPP4. Например, заменяющий человеческий ген DPP4 или его фрагмент содержит один или более экзонов человеческого гена DPP4, например 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или 26 экзонов человеческого гена DPP4. Например, заменяющий человеческий ген DPP4 или его фрагмент содержит экзоны 1-26 человеческого гена DPP4. В других вариантах осуществления заменяющий человеческий ген DPP4 или его фрагмент содержит экзоны 2-26 человеческого гена DPP4. Например, заменяющий человеческий ген DPP4 или его фрагмент содержит интрон 1, расположенный в направлении 5'-конца от экзонов 2-26 человеческого гена DPP4. В некоторых вариантах осуществления заменяющий человеческий ген DPP4 или его фрагмент дополнительно содержит человеческий (-ие) регуляторный (-ые) элемент (элементы), например, часть человеческой нетранслируемой области (UTR) в направлении 3'-конца от человеческого гена DPP4, например, по меньшей мере 1 тыс.п.н. расположенной справа области (например, по меньшей мере 1, 2, 3, 4, 5, 10, 20, 30, 40 тыс.п.н. или более), и/или область человеческого промотора или энхансера в направлении 5'-конца от человеческого гена DPP4, например, по меньшей мере 10 тыс.п.н. расположенной слева области (например, по меньшей мере 10, 20, 30, 40, 50 тыс.п.н. или более).
Гомологичные участки представляют собой последовательности, которые являются гомологичными по отношению к хромосомным нуклеотидным последовательностям, фланкирующим желательную генетическую модификацию/замену, например, фланкирующие эндогенный ген Dpp4 или его фрагмент, который нужно заменить. Гомологичные нуклеотидные последовательности могут представлять собой две или более нуклеотидных последовательностей, которые являются либо идентичными, либо аналогичными настолько, чтобы можно было выполнять их гибридизацию друг с другом или межмолекулярный обмен. Из-за гомологии между гомологичными участками и соответствующей эндогенной последовательностью гомологичные участки направляют таргетирующую конструкцию к специфическому местоположению на хромосоме в пределах генома, например, к локусу эндогенного гена Dpp4. См., например, Valenzuela et al. Nature Biotech, 21.6(2003):652-659; US 6,586,251 и US 8,759,105.
В некоторых случаях таргетирующая конструкция дополнительно содержит маркер отбора, например, между двумя гомологичными участками. Примеры маркеров отбора включают маркеры устойчивости к антибиотикам (например неомицину или канамицину) и флуоресцентные белки. В некоторых вариантах осуществления маркер отбора фланкирован, например, двумя сайтами loxP. Маркер отбора, фланкированный сайтами loxP, можно удалить посредством добавления рекомбиназы Cre, которая катализирует вырезание фланкированного сайтами loxP сегмента, например, содержащего маркер отбора.
Вектор/конструкции
Трансгенные не относящиеся к человеку животные (например грызуны, такие как мыши или крысы) изобретения можно получить с использованием различных векторов и/или конструкций. В некоторых вариантах осуществления таргетирующая конструкция представляет собой круглый фрагмент двухцепочечной ДНК, например искусственную бактериальную хромосому (ВАС), плазмиду или искусственную хромосому, полученную из ДНК бактериофага Р1 (РАС).
Для создания нечеловеческой клетки, в которой направленно заменен локус эндогенного Dpp4, таргетирующую конструкцию, содержащую человеческий ген DPP4 или его фрагмент, описанный в настоящем документе, встраивают в нечеловеческую клетку (например, клетку грызуна, такого как мышь или крыса), например эмбриональную стволовую (ЭС) клетку.
Для создания нечеловеческой клетки, содержащей человеческий ген DPP4 или его фрагмент, произвольно вставленный в геном, круглую конструкцию ДНК, например ВАС, содержащую человеческий ген DPP4 или его фрагмент, встраивают в нечеловеческую клетку (например, клетку грызуна, такого как мышь или крыса), например ЭС-клетку. В некоторых случаях круглая конструкция ДНК, например ВАС, дополнительно содержит человеческий регуляторный элемент DPP4, например область человеческого промотора или энхансера, расположенную в направлении 5'-конца и/или 3'-конца от человеческого гена DPP4. Например, круглая конструкция ДНК содержит по меньшей мере 10 тыс.п.н. (например, по меньшей мере 10, 20, 30, 40, 50 тыс.п.н. или более) области промотора/энхансера в направлении 5'-конца от инициаторного кодона ATG человеческого гена DPP4. Кроме того или альтернативно, круглая конструкция ДНК содержит по меньшей мере 1 тыс.п.н. (например, по меньшей мере 1, 2, 3, 4, 5, 10, 20, 30, 40 тыс.п.н. или более) нетранслируемой области в направлении 3'-конца от человеческого гена DPP4. Например, человеческий ген DPP4 или его фрагмент функционально связан с человеческим регуляторным элементом DPP4.
В некоторых вариантах осуществления человеческий ген DPP4 или его фрагмент в круглой конструкции ДНК (например ВАС) содержит весь человеческий ген DPP4. В других вариантах осуществления человеческий ген DPP4 или его фрагмент содержит часть человеческого гена DPP4. Например, человеческий ген DPP4 или его фрагмент содержит один или более экзонов человеческого гена DPP4, например 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или 26 экзонов человеческого гена DPP4. Например, человеческий ген DPP4 или его фрагмент содержит экзоны 1-26 человеческого гена DPP4. В других вариантах осуществления человеческий ген DPP4 или его фрагмент содержит экзоны 2-26 человеческого гена DPP4. Например, человеческий ген DPP4 или его фрагмент содержит интрон 1, расположенный в направлении 5'-конца от экзонов 2-26 человеческого гена DPP4.
Например, этап введения в клетку выполнен посредством электропорации или опосредованной липидами трансфекции.
В некоторых случаях перед введением в клетку выполняется линеаризация круглой конструкции ДНК, например ВАС. Например, линеаризация выполняется с помощью редкощепящих рестриктаз, например SgrDI, SfiI, NotI, PacI или SwaI.
В случаях, когда таргетирующая конструкция содержит маркер отбора по устойчивости к антибиотикам (например, неомицину), клетки, которые приняли таргетирующую конструкцию, необязательно отбирают в среде, содержащей неомицин/G418. Отбирают клетки, которые выживают и/или размножаются в среде, содержащей неомицин/G418. Эти клетки содержат таргетирующую конструкцию.
В некоторых вариантах осуществления клеточную популяцию подвергают скринингу для выявления клеток, в геном которых встроен человеческий ген DPP4 или его фрагмент, например, произвольно вставлен в геном или направленно внедрен в локус эндогенного Dpp4 (например, с помощью таргетирующей конструкции).
Способы скрининга включают количественную ПЦР и флуоресцентную гибридизацию in situ. См., например, US 6,586,251 В2 и US 8,759,105 В2. Например, способы скрининга включают обнаружение наличия человеческого гена DPP4 или его фрагмента. В некоторых вариантах осуществления способы скрининга включают обнаружение уменьшения числа копий эндогенного гена Dpp4 или его фрагмента и/или увеличение числа копий человеческого гена DPP4 или его фрагмента. Примеры способов скрининга описаны в примерах 1 и 3.
В некоторых вариантах осуществления, в которых таргетирующая конструкция содержит маркер отбора, фланкированный сайтами loxP, клетки с правильно выполненным встраиванием, необязательно дополнительно подвергаются электропорации с использованием вектора, экспрессирующего Cre, например, временно экспрессирующего рекомбиназу Cre, для удаления маркера отбора, фланкированного сайтами loxP.
Для создания трансгенных животных положительные ЭС-клеточные клоны, например, без маркера отбора, фланкированного сайтами loxP, содержащие человеческий ген DPP4 или его фрагмент, вводят в эмбрион грызуна, например эмбрион мыши или крысы, такой как мышиный эмбрион на 8-клеточной стадии. Например, этап введения выполняют с использованием технологии инъекции бластоцист, методик агрегации, ядерной передачи и клонирования и/или способа VelociMouse®. См., например, US 8,759,105 В2, US 7,294,754, US 7,576,259 и US 7,659,442. Например, ЭС-клеточный клон представляет собой субпопуляцию клеток, полученную из одной клетки ЭС-клеточной популяции после введения ДНК и последующего отбора.
В некоторых случаях ДНК от трансгенных не относящихся к человеку животных подвергают скринингу способами, аналогичными описанным выше способам, для подтверждения прохождения человеческого гена DPP4 или его фрагмента через зародышевую линию.
В некоторых вариантах осуществления грызуны с гуманизированным DPP4, описанные в настоящем документе, являются гетерозиготными по человеческому аллелю DPP4. Таким образом, у этих грызунов имеется один человеческий аллель DPP4 и один аллель DPP4 дикого типа грызуна. В других вариантах осуществления грызуны с гуманизированным DPP4 являются гомозиготными по человеческому аллелю DPP4.
Применение грызунов с гуманизированным DPP4
Грызуны с гуманизированным DPP4, например мыши или крысы, могут использоваться для оценки фармакодинамики (ФД) специфичных для человека антагонистов DPP4, например низкомолекулярных, пептидных или биологических ингибиторов, используемых для лечения гипергликемии.
Анализы фармакокинетики (ФК) и ФД у грызунов с гуманизированным DPP4, например мышей или крыс, выполняются в соответствии со стандартными процедурами, известными в данной области.
Грызуны с гуманизированным DPP4, например мыши или крысы, могут использоваться для оценки терапевтической эффективности in vivo специфичных для человека антагонистов DPP4, например низкомолекулярных, пептидных или биологических ингибиторов, при лечении гипергликемии.
Грызуны с гуманизированным DPP4, например мыши или крысы, могут использоваться для проверки терапевтической эффективности in vivo антагонистов, например низкомолекулярных, пептидных или биологических ингибиторов, например нейтрализующих антител, специфичных к молекулам-мишеням, например MERS-CoV (например шипиковый белок (S) MERS-CoV, например рецептор-связывающий домен шипикового белка MERS-CoV), которые взаимодействуют исключительно с человеческим DPP4, при лечении или предотвращении (или профилактике) инфекции MERS-CoV. В некоторых вариантах осуществления грызуны, гетерозиготные по человеческому аллелю DPP4, используются для проверки терапевтической эффективности in vivo одного или более антагонистов при лечении или предотвращении (или профилактике) инфекции MERS-CoV. В других вариантах осуществления грызуны с DPP4, гомозиготные по человеческому аллелю DPP4, используются для проверки терапевтической эффективности in vivo одного или более антагонистов при лечении или предотвращении (или профилактике) инфекции MERS-CoV.
Примерами штаммов MERS-CoV являются штамм MERS-CoV Jordan (№ последовательности в GenBank: КС776174.1, MERS-CoV- Hu/Jordan-N3/2012) и штамм MERS-CoV EMC/2012 (№ последовательности в GenBank: JX869059.2). В некоторых вариантах осуществления вирус MERS-CoV, описанный в настоящем документе, содержит клинический изолят MERS-CoV. В других вариантах осуществления вирус MERS-CoV, описанный в настоящем документе, содержит штамм, содержащий ту же последовательность рецептор-связывающего домена (RBD) шипикового белка, что и клинический изолят, описанный в настоящем документе. Примеры клинических изолятов приведены в таблице ниже. В таблице показаны вариации аминокислотной последовательности в пределах рецептор-связывающего домена (RBD) шипикового белка у нескольких клинических изолятов MERS-CoV. Последовательности клинических изолятов MERS-CoV, хранящиеся в Национальном центре биотехнологической информации (NCBI), выравнивали по аминокислотам 367-606 и сравнивали с последовательностями штамма ЕМС/2012. В таблице показаны клинические изоляты, содержащие замещения А431Р, S457G, S460F, A482V, L506F, D509G и V534A (причем аминокислота (однобуквенное обозначение) перед числом относится к штамму ЕМС/2012, а аминокислота (однобуквенное обозначение) после числа относится к клиническому изоляту).
В некоторых вариантах осуществления антагонист вводят до (например, по меньшей мере за 1, 2, 4, 6, 12, 24, 48 часов, 2, 3, 4, 5, 6 или 7 дней или более до) инфицирования грызуна MERS-CoV. В других вариантах осуществления антагонист вводят после (например, по меньшей мере через 1, 2, 4, 6, 12, 24, 48 часов, 2, 3, 4, 5, 6 или 7 дней или более после) инфицирования грызуна MERS-CoV.
В некоторых вариантах осуществления, в которых антагонист вводят грызуну после инфицирования MERS-CoV, уменьшение титра вируса или концентрации РНК (например, концентрации вирусной UpE или лидерной последовательности РНК) у грызуна после введения антагониста, например снижение по меньшей мере в 5 раз (например, по меньшей мере в 5, 10, 50, 100, 500, 1000, 104, 105, 106, 107 раз или более) по сравнению с контрольной концентрацией, указывает, что антагонист является эффективным средством лечения инфекции MERS-CoV. Например, контрольная концентрация представляет собой титр вируса или концентрацию РНК у грызуна до введения антагониста. В других примерах контрольная концентрация представляет собой титр вируса или концентрацию РНК у инфицированного вирусом грызуна, которому не вводили антагонист.
В некоторых вариантах осуществления, в которых антагонист вводят грызуну до инфицирования MERS-CoV, уменьшение титра вируса или концентрации РНК (например, концентрации вирусной UpE или лидерной последовательности РНК) у грызуна после введения антагониста, например снижение по меньшей мере в 5 раз (например, по меньшей мере в 5, 10, 50, 100, 500, 1000, 104, 105, 106, 107 раз или более) по сравнению с контрольной концентрацией, указывает, что антагонист является эффективным средством предотвращения инфекции MERS-CoV. Например, контрольная концентрация представляет собой титр вируса или концентрацию РНК у грызуна, инфицированного MERS-CoV, которому не вводили антагонист.
В некоторых вариантах осуществления концентрации вирусной РНК в легких грызуна можно определить с помощью экстракции РНК из легких грызуна посредством гомогенизации в растворе, содержащем фенол, например в растворе, содержащем фенол и гуанидинизотиоцианат (например, Trizol® (Life Technologies, Inc)). Например, легкие можно гомогенизировать с помощью прибора Magnalyzer (Roche) в соответствии с инструкциями производителя. В некоторых вариантах осуществления концентрации РНК MERS-CoV можно оценить посредством количественной ПЦР (кПЦР) с использованием праймеров, которые нацелены на геном MERS-CoV, и/или праймеров, которые нацелены на транскрипт мРНК MERS-CoV. Например, при кПЦР можно применять вирусную одноэтапную реакционную смесь Taqman® Fast (Applied Biosystems) в соответствии с инструкциями производителя с использованием пары праймеров от компании Life Technologies, нацеленных на область генома, расположенную по направлению к 5'-концу от гена оболочечного белка (UpE) или лидерной последовательности информационной РНК нуклеокапсида (лидерный праймер), и по сравнению с эндогенным контролем, таким как 18S рРНК грызуна (например, мыши). Например, реакции кПЦР в быстрых оптических реакционных планшетах Microamp® (Applied Biosystems) можно регистрировать с помощью прибора для ПЦР в реальном времени 7500 fast DX real-time PCR (Applied Biosystems). В некоторых примерах данные кПЦР можно анализировать с помощью способа дельта Ct, приняв результаты для неинфицированных контролей за 1. Например, обнаруженное количество РНК MERS-CoV можно выразить в процентах по отношению к концентрациям РНК, обнаруженным у инфицированных мышей, по отношению к которым использовали контрольный антагонист, например контрольные антитела такого же изотипа, в случаях, когда антагонистом является антитело против MERS-CoV.
В некоторых вариантах осуществления титр вируса в легких грызуна можно определить с помощью гомогенизации легких грызуна в буфере (например, фосфатном-солевом буфере (PBS)) после центрифугирования (например, при 10000 об./мин). Надосадочную жидкость можно оценить посредством анализа бляшкообразования на клетках млекопитающих, таких как клетки Vero (например, клетки Vero Е6), с целью количественного определения концентраций вируса, оставшихся после использования антагониста. Можно использовать стандартный анализ бляшкообразования, такой как анализ бляшкообразования, описанный в Page et al. (2012) Induction of alternatively activated macrophages enhances pathogenesis during severe acute respiratory syndrome coronavirus infection, J Virol 86:13334-13349. В некоторых примерах анализ бляшкообразования Page et al. можно модифицировать, оставив планшеты на 3 дня для образования бляшек.
В некоторых вариантах осуществления ослабление воспаления (например интерстициального или периваскулярного воспаления), снижение числа воспалительных клеток и/или уменьшение бронхиолярной инфильтрации, например, определенной посредством гистологического анализа, в образце легких грызуна, по отношению к которому использовали антагонист до или после инфицирования MERS-CoV, по сравнению с контрольным образцом легких может указывать на то, что антагонист является эффективным средством лечения или предотвращения инфекции MERS-CoV. Например, контрольный образец легких может быть получен от грызуна, инфицированного MERS-CoV, которому не вводили антагонист, например, которому не вводили антагонист до и/или после инфицирования.
Грызуны, описанные в настоящем документе, могут использоваться для эффективного тестирования лекарственных средств и вакцин против MERS-CoV, например для демонстрации безопасности и эффективности перед клиническими испытаниями у людей. Они позволяют быстро выявлять и/или выполнять валидацию терапевтических/профилактических средств, например, в течение нескольких недель тестирования.
Определения
Гомологичные аминокислотные остатки представляют собой остатки, которые обладают аналогичными характеристиками или свойствами. Характеристики или свойства аминокислотных остатков зависят, например, от структуры полипептидного остова (например, листовая или спиральная конформация), заряда или гидрофобности остатка и/или размера боковой (-ых) цепи (-ей). Например, гомологичные остатки по свойствам аналогичны боковой цепи, например по полярности, заряду, размеру, ароматичности и/или гидрофобности.
Термин «приблизительно» обозначает указанное значение плюс или минус другое количество, таким образом определяя диапазон значений. В определенных вариантах осуществления термин «приблизительно» обозначает диапазон по отношению к базовой (или основной, или контрольной) величине или количеству плюс или минус до 15%, 14%, 13%, 12%, 11%, 10%, 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2%, 1%, 0,75%, 0,5%, 0,25% или 0,1%. Например, приблизительно обозначает диапазон +/- 5% ниже и выше указанных чисел, например чисел пар нуклеотидов.
Термин «функционально связанный» в контексте настоящего документа обозначает, что положения описываемых компонентов, например нуклеотидных последовательностей, находятся в такой взаимосвязи, которая позволяет им функционировать предполагаемым образом.
В контексте настоящего документа термин «белок» включает полипептиды, пептиды, фрагменты полипептидов и гибридные полипептиды.
В контексте настоящего документа термин «нуклеиновая кислота» обозначает два или более дезоксирибонуклеотида и/или рибонуклеотида, ковалентно связанных друг с другом в одноцепочечной или двухцепочечной форме.
Термин «замена» в отношении замены гена обозначает размещение экзогенного генетического материала в эндогенном генетическом локусе, что приводит к замене всего или части эндогенного гена ортологичной или гомологичной нуклеотидной последовательностью. В одном случае эндогенный нечеловеческий ген или его фрагмент заменяется соответствующим человеческим геном или его фрагментом. Соответствующий человеческий ген или его фрагмент представляет собой человеческий ген или фрагмент, который является ортологом или гомологом или по существу идентичен или аналогичен по структуре и/или функции заменяемому эндогенному нечеловеческому гену или его фрагменту.
В контексте настоящего документа термин «грызун» обозначает любого представителя отряда грызунов (Rodentia). Примеры грызунов включают, без ограничений, мышей, крыс, белок, луговых собачек, дикобразов, бобров, морских свинок и хомяков. В одном варианте осуществления грызун представляет собой крысу. В другом варианте осуществления грызун представляет собой мышь.
Если в настоящем документе не оговорено иное, все используемые в настоящем документе технические и научные термины имеют общепринятое значение, понятное любому специалисту в области, к которой относится настоящее изобретение.
В рамках настоящего документа применение терминов в единственном числе включает ссылку на множественное число, если из контекста четко не следует иное.
Подразумевается, что каждое максимальное числовое ограничение, встречающееся в данном описании, включает также каждое меньшее числовое ограничение, как если бы такие меньшие числовые ограничения были в прямой форме указаны в настоящем документе. Каждое минимальное числовое ограничение, встречающееся в данном описании, будет включать каждое большее числовое ограничение, как если бы такие большие числовые ограничения были в прямой форме указаны в настоящем документе. Каждый числовой диапазон, встречающийся в настоящем описании, будет включать каждый более узкий числовой диапазон, который находится в пределах такого более широкого числового диапазона, как если бы все такие более узкие числовые диапазоны были в прямой форме указаны в настоящем документе.
Приведенные ниже примеры предложены для целей иллюстрации и не должны каким-либо образом ограничивать изобретение.
ПРИМЕРЫ
Пример 1
Замена эндогенного мышиного гена Dpp4 человеческим геном DPP4
Человеческий ген DPP4 размером 81,8 тыс.п.н., содержащий участок с экзона 2 до экзона 26 и участок 3'-нетранслируемой области человеческого гена DPP4, заменил мышиный локус гена Dpp4 размером 78,8 тыс.п.н. от экзона 2 до стоп-кодона в экзоне 26. См. Фиг. 1А и 1В.
Таргетирующая конструкция для замены мышиного гена Dpp4 человеческим за один этап была получена с использованием технологии генной инженерии VelociGene® (см. Valenzuela et al. (2003) High-throughput engineering of the mouse genome coupled with high-resolution expression analysis, Nature Biotech, 21(6):652-659). Мышиная и человеческая ДНК DPP4 были получены из клонов искусственной бактериальной хромосомы (ВАС) RP23-362N15 и RP11-68L22 соответственно. Вкратце, линеаризованную с помощью SgrDI таргетирующую конструкцию, созданную посредством клонирования с репарацией гэпа и содержащую гомологичные участки мышиного Dpp4 со стороны 5'-конца и 3'-конца, фланкирующие человеческую последовательность DPP4 размером 82 тыс.п.н. от интрона 1 слева от экзонов 2-26, включая стоп-кодон и участок нетранслируемой области на стороне 3'-конца (геномные координаты полного человеческого гена DPP4: GRCh38: chr2:162,848,755-162,931,052 (-нить)), и фланкированную сайтами loxP кассету отбора по устойчивости к неомицину подвергали электропорации с внедрением в мышиные эмбриональные стволовые (ЭС) клетки VGB6 (полученные от мышей C57BL/6N). ЭС-клетки (MAID 7326) с правильно выполненным внедрением подвергли дальнейшей электропорации с вектором, временно экспрессирующим Cre, для удаления кассеты отбора по устойчивости к лекарственному средству. ЭС-клеточные клоны с внедренным человеческим геном и без кассеты отбора по устойчивости к лекарственному средству (MAID 7327) внедрили в эмбрион мыши SW на 8-клеточной стадии с помощью способа VelociMouse® (см. патенты США №№7,294,754, 7,576,259, 7,659,442 и Poueymirou et al. (2007) F0 generation mice that are essentially fully derived from the donor gene-targeted ES cells allowing immediate phenotypic analyses Nature Biotech. 25(1):91-99). Мыши VelociMice® (мыши F0, полученные полностью из донорской ЭС-клетки), имеющие гуманизированный ген DPP4, были выявлены посредством генотипирования для определения утраты мышиного аллеля и приобретения человеческого аллеля с использованием модифицированного анализа аллелей (см. Valenzuela et al. (2003)).
ЭС-клеточные клоны с правильно выполненным внедрением были выявлены с помощью анализа на утрату нативного аллеля (LONA) (Valenzuela et al. 2003), в котором число копий нативного немодифицированного гена Dpp4 определили с помощью количественных полимеразных цепных реакций (кПЦР) TaqMan™, специфичных для последовательностей в мышином гене Dpp4, которые были удалены. Анализы кПЦР включали следующие наборы праймеров и зондов (указаны от 5'-конца к 3'-концу): прямой праймер со стороны 5'-конца TCGCCACTGT GCCTAACATA G (SEQ ID NO: 1); обратный праймер со стороны 5'-конца CCGGGACTAA ACTGGAACAT ТС (SEQ ID NO: 2); зонд со стороны 5'-конца FAM-TCAGTCAACT TCTTCTGGGT TGTTTCC-BHQ (SEQ ID NO: 3); прямой праймер со стороны 3'-конца CAGCTCTGGT GGAGAACTAG AC (SEQ ID NO: 4); обратный праймер со стороны 3'-конца GGAGGTCCTC GGTCTTTAGA AG (SEQ ID NO: 5); зонд со стороны 3'-конца FAM-TCACACTTAG GCTTATAAAC CATTCCCGT-BHQ (SEQ ID NO: 6); в котором FAM обозначает 5-карбоксифлуоресцеиновый флуоресцентный зонд, a BHQ обозначает гаситель флуоресценции типа Black Hole (Biosearch Technologies). ДНК, очищенную из ЭС-клеточных клонов, которые захватили таргетирующий вектор и встроили его в свои геномы, добавляли в реакционную смесь TaqMan™ Gene Expression Master Mix (Life Technologies) в соответствии с рекомендациями производителя в 384-луночном планшете для ПЦР (оптический 384-луночный реакционный планшет MicroAmp™ Optical 384-Well Reaction Plate, Life Technologies) и выполняли ряд циклов в приборе Applied Biosystems Prism 7900НТ, который собирает данные флуоресценции в процессе ПЦР и определяет пороговый цикл (Ct), фракционный цикл ПЦР, в котором накопленная флуоресценция достигает предварительно установленного порогового значения. Для каждого образца ДНК провели прямые и обратные DPP4-специфичные кПЦР и две кПЦР для неизмененных контрольных генов. Рассчитывали различия в значениях Ct (ΔCt) между каждой DPP4-специфичной кПЦР и каждой кПЦР контрольного гена, а затем рассчитывали различия между каждым ΔCt и медианным ΔCt для всех проанализированных образцов для получения значений ΔΔCt для каждого образца. Рассчитывали число копий гена DPP4 в каждом образце по следующей формуле: число копий = 2×2-ΔΔCt. Клон с правильно выполненным внедрением, потерявший одну из своих нативных копий, будет иметь число копий гена Dpp4, равное единице. Подтверждение того, что человеческая последовательность гена DPP4 заменила удаленную мышиную последовательность гена Dpp4 в гуманизированном аллеле, было выполнено с помощью анализа кПЦР TaqMan™, который содержит следующие наборы праймеров и зондов (от 5'-конца к 3'-концу): человеческий прямой праймер со стороны 5'-конца GCGGTCTCCC TCTTCTAACG (SEQ ID NO: 7); человеческий обратный праймер со стороны 5'-конца GCAAGCCGAG CAGATCAAG (SEQ ID NO: 8); человеческий зонд со стороны 5'-конца FAM-ACTCCCACCT GCAAATCCTG CTGC-BHQ (SEQ ID NO: 9); человеческий прямой праймер со стороны 3'-конца AACCGCACTG GCATATGGA (SEQ ID NO: 10); человеческий обратный праймер со стороны 3'-конца TACAAGGTAG TCTGGGATTA CTAACAAAA (SEQ ID NO: 11); человеческий зонд со стороны 3'-конца FAM-ACATTTATCT AGAAAGGCTC-BHQ (SEQ ID NO: 12).
Аналогичный анализ LONA используется для оценки ДНК, очищенной из биопсий хвоста, у мышей, полученных из ЭС-клеток с внедренным человеческим геном, для определения генотипов DPP4 и подтверждения проникновения аллеля гуманизированного DPP4 через зародышевую линию. Двух детенышей, гетерозиготных по замене, скрестили для получения мыши, гомозиготной по замене эндогенного мышиного гена Dpp4 человеческим геном DPP4. Детеныши, гомозиготные по замене, используются для фенотипирования.
Расположенное со стороны 5'-конца место соединения мышиного локуса Dpp4 с последовательностью, содержащей человеческий ген DPP4, выполнено с возможностью нахождения в пределах 5'-AGGAGAGAAG CCAACAAGAT CATAAGATCA TGCTCAGGGC СААААТТСАА GGGCTTCTGC (CGTCGACG) GCCTTAGAGA ACTCCAACTG GCGCACTCCA GACGCCACCC CCACCCCCAG CCCGCGGTCT CCCTCTTCTA ACGCACTCCC АС С TGCA ААТ (SEQ ID NO: 13), причем человеческие последовательности DPP4 выделены курсивом, а сайт рестрикции для SgrDI взят в скобки. Расположенное со стороны 3'-конца место соединения последовательности, содержащей человеческий ген DPP4, с фланкированной сайтами loxP кассетой отбора по чувствительности к неомицину выполнено с возможностью нахождения в пределах 5'-TTATTCCAGG GAACTATGATGAGGCTTATA TAAGAACGAA TAAGATCAGA ААТАТСАТТС TGGCAGTTCT TATGGCTCAG ctcgag(ataa cttcgtataa tgtatgctat acgaagttat) atgcatggcc tccgcgccgg gttttggcgc ctcccgcggg (SEQ ID NO: 14), причем человеческие последовательности DPP4 выделены курсивом, последовательности кассеты резистентности к неомицину записаны строчными буквами, а сайт loxP взят в скобки. Расположенное со стороны 3'-конца место соединения последовательности фланкированной сайтами loxP кассеты отбора по чувствительности к неомицину с мышиным локусом Dpp4 выполнено с возможностью нахождения в пределах 5'-atgtctgga(a taacttcgta taatgtatgc tatacgaagt tatjgctagta actataacgg tcctaaggta gcgagctagc CAGCATAGCT CTCCATAGCT TATTTAAGAC CACATTTGTT CTCATTATCT CAAAAGTGCA CTGTTAAGAT GAAGATCTTA (SEQ ID NO: 15), причем последовательности кассеты резистентности к неомицину записаны строчными буквами, а сайт loxP взят в скобки. После удаления кассеты отбора по чувствительности к неомицину место соединения последовательности, содержащей человеческий ген DPP4, сайта loxP, оставшегося после удаления кассеты отбора по чувствительности к неомицину, и мышиного локуса Dpp4 выполнено с возможностью нахождения в пределах 5'- TATTCCAGGG AACTATGATG AGGCTTATATAAGAACGAAT AAGATCAGAA АТАТСАТТСТ GGCAGTTCTTATGGCTCAG ctcgag(ataa cttcgtataa tgtatgctat acgaagttat) gctagtaact ataacggtcc taaggtagcg agctagcCA GCATAGCTCT CCATAGCTTA TTTAAGACCA CATTTGTTCT CATTATCTCA AAAGTGCACT GTTAAGATGA AGATCTTAAT AATGTTGCAT TGAGACATTT CAGGCTGCTT TCTCCAGTTT TACACCTGCA ATCCTAACTA AGGATGCCTG TCCCCAGAAC (SEQ ID NO: 16), причем человеческие последовательности DPP4 выделены курсивом, последовательности кассеты резистентности к неомицину записаны строчными буквами, а сайт loxP взят в скобки.
На Фиг. 2 показана аминокислотная последовательность DPP4, кодируемая гуманизированной нуклеотидной последовательностью DPP4 в MAID 7326/7327 (SEQ ID NO: 17), которая идентична человеческой последовательности DPP4, так как кодон 1 мышиного Dpp4 кодирует только первые две аминокислоты DPP4 метионин и лизин, которые аналогичны аминокислотам, кодируемым кодоном 1 человеческого DPP4.
Пример 2
Инфицирование MERS-CoV мышей с гуманизированным DPP4
Коронавирус ближневосточного респираторного синдрома (MERS-CoV) представляет собой новый распространяющийся вирус, который вызывает тяжелое острое респираторное заболевание. Рецептором для MERS-CoV является дипептидилпептидаза IV (DPP4) (см. Raj et al. (2013) Dipeptidyl Peptidase 4 is a Functional Receptor for the Emerging Human Coronavirus-EMC, Natuere 495(7440):251-254). Для тестирования in vivo противовирусных молекул необходима животная модель, например небольшая животная модель, такая как грызун (например мышь или крыса), восприимчивый к инфекции MERS-CoV. Однако последние исследования показали, что мышиный Dpp4 неспособен реагировать на инфицирование MERS-CoV (см., например, Cockrell et al. (2014) Mouse Dipeptidyl Peptidase is Not a Functional Receptor for Middle East Respiratory Syndrome Coronavirus (MERS-CoV) Infection, J. Virol. 88(9):5195-5199; и Coleman et al. (2014) Wild-Type and Innate Immune-Deficient Mice are Not Susceptible to the Middle East Respiratory Syndrome Coronavirus, J. Gen. Virol. 95(2):408-412), по меньшей мере частично из-за того, что шипиковый белок MERS-CoV взаимодействует с человеческим (но не мышиным) DPP4 (см., например, Coleman et al. (2013); и Raj et al. (2013)). При сравнении последовательностей мышиного и человеческого DPP4 было выявлено, что аминокислоты, которые ранее были определены как участки контакта между шипиковым белком MERS-CoV и рецептором, различаются у двух видов. Кроме того, экспрессия человеческой DPP4 в мышиных клетках обеспечивает проникновение и распространение вируса MERS-CoV. Это указывает на то, что проникновение вируса является ограничивающим этапом в инфицировании мышиных клеток и что отсутствие взаимодействия между мышиной DPP4 и гликопротеином MERS-CoV определяет видовой тропизм in vitro. См., например, Lu et al. (2013) Molecular basis of binding between novel human coronavirus MERS-CoV and its receptor CD26, Nature. 500(7461):227-31; и Cockrell et al. (2014) Mouse Dipeptidyl Peptidase is Not a Functional Receptor for Middle East Respiratory Syndrome Coronavirus (MERS-CoV) Infection, J. Virol. 88(9):5195-5199.
В результате, нормальные линии мышей нельзя использовать для оценки эффективности терапевтических средств, воздействующих на MERS-CoV. В Zhao et al. (2014) Rapid Generation of a Mouse Model for Middle East Respiratory Syndrome, Proc. Natl. Acad. Sci. USA 111(13):4970-4975 экспрессировали человеческую DPP4 у мышей посредством аденовирусной трансдукции, обеспечив тем самым развитие инфекции MERS-CoV. Однако такая аденовирусная модель имеет несколько ограничений, включая следующие ограничения: (а) вирус быстро удаляется из инфицированной мыши; (b) со временем наблюдается утрата экспрессии человеческой DPP4; (с) распределение в тканях трансдуцированной вирусом DPP4 не отражает экспрессии, наблюдаемой у мышей или людей; и (d) аденовирусная инфекция индуцирует интерфероновую реакцию.
Для создания мышиной модели инфекции MERS-CoV получали мышей с гуманизированным DPP4, как описано выше в примере 1. Как показано на Фиг. 1В, экзоны 2-26 мышиного Dpp4 были заменены на соответствующие последовательности человеческого DPP4. Поскольку оставшийся кодирующий экзон 1 мышиного Dpp4 кодирует только первые две аминокислоты Dpp4 метионин и лизин, аналогичные аминокислотам в соответствующем экзоне 1 человеческого DPP4, белок DPP4, экспрессируемый у мышей с гуманизированным DPP4, является полностью человеческим (см. Фиг. 2, SEQ ID NO: 17). Таким образом, мыши с гуманизированным DPP4 экспрессируют полностью человеческую DPP4 под контролем эндогенных мышиных регуляторных последовательностей, а именно последовательностей 5'-фланкирующей области (промотор и энхансер (-ы)) и 3'-нетранслируемой области. Предполагается, что гуманизированная DPP4 экспрессируется в тех же клетках и тканях и в тех же или аналогичных концентрациях, что и мышиная Dpp4, экспрессируемая у мышей дикого типа, не содержащих человеческих нуклеиновых последовательностей DPP4.
Двух мышей F0 с гуманизированным DPP4 инфицировали интраназально MERS-CoV (штамм Jordan) или вводили PBS для имитации инфицирования. Через четыре дня после инфицирования выполнили количественное определение РНК MERS-CoV в легких посредством ПЦР в реальном времени с использованием праймеров, специфичных для репликативной формы генома MERS-CoV. Данные нормализовали по количеству продукта ПЦР, полученного из легких ложноинфицированных мышей (произвольно принятому за 1). На Фиг. 3 показано, что РНК MERS-CoV можно амплифицировать из легких мышей с гуманизированным DPP4, инфицированных MERS-CoV. Также выполнили окрашивание гематоксилином и эозином с использованием легочной ткани от ложноинфицированных и инфицированных MERS-CoV мышей. На Фиг. 4 показано, что инфекция MERS-CoV у мышей с гуманизированным DPP4 не поражает дыхательные пути, но приводит к утолщению стенок альвеол и уменьшению пространства между альвеолярными клетками, что указывает на связь воспаления в легких с инфекцией MERS-CoV.
Кроме того, по отношению к мышам в возрасте 6-8 недель выполнили интраназальную инокуляцию MERS-CoV, например, 2×105 pfu MERS-CoV (Jordan). Вплоть до 4 дня после инокуляции отсутствовали смертельные случаи или клинические признаки заболевания. На 2 и 4 день после инокуляции мышей умертвили и извлекли их легкие. Для измерения концентраций РНК вируса легкие гомогенизировали в Trizol®, РНК экстрагировали и выполнили анализ посредством ПЦР в реальном времени с использованием праймеров, специфичных к MERS-CoV (Фиг. 10А и 10В). Набор праймеров был специфичным к области вирусного генома со стороны 5'-конца от гена оболочечного белка (UpE), а другой набор праймеров был специфичным к лидерной последовательности мРНК нуклеокапсида (лидерный праймер). Мышиную 18S рРНК использовали в качестве эндогенного контроля.
Для измерения титров вируса легкие гомогенизировали в фосфатно-солевом буфере (PBS), осветленном посредством центрифугирования, и титровали на клетках Vero Е6 (Фиг. 10С). Например, надосадочную жидкость проанализировали с помощью анализа бляшкообразования на клетках Vero Е6 для количественного определения концентраций вируса в легких. Например, анализ бляшкообразования выполнили, как описано в Page et al. (2012) Induction of alternatively activated macrophages enhances pathogenesis during severe acute respiratory syndrome coronavirus infection, J Virol 86:13334-13349, причем планшеты оставляли на 3 дня для образования бляшек.
Устойчивую репликацию MERS-CoV в легких наблюдали на 2 и 4 дни после инфицирования. Количественный анализ РНК с использованием набора праймеров, специфичных к лидерной последовательности MERS-CoV, который был разработан только для амплификации реплицирующегося MERS-CoV, продемонстрировал высокие концентрации реплицирующейся РНК MERS-CoV в извлеченных на 2 день легких, и эти концентрации сохранялись до 4 дня после инфицирования (Фиг. 10А-В). В количественном анализе бляшкообразования гомогената легких на клетках Vero Е6 были получены концентрации MERS-CoV (Jordan) ~7,27×104 pfu/мл легкого на 2 день и ~3,75×105 pfu/мл легкого на 4 день после инфицирования (Фиг. 10С), что указывает на активную репликацию MERS-CoV в легких инфицированных мышей с гуманизированным DPP4.
Также легкие мышей с гуманизированным DPP4, по отношению к которым выполняли интраназальную инокуляцию MERS-CoV (штамм Jordan) или PBS (ложноинфицированные), проанализировали на наличие патологических изменений (Фиг. 10D). На 2 день после инфицирования выявляли перибронхиолярное воспаление с изменениями структуры бронхиолярных клеток во всех областях легких. В данную временную отметку наблюдали минимальные периваскулярные воспалительные реакции и изменения структуры альвеол. На 4 день после инфицирования наблюдали интерстициальную инфильтрацию, а также периваскулярную инфильтрацию и обширное утолщение альвеол. Также присутствовали бронхиолярные изменения. См. Фиг. 10D. Данные патологические изменения согласуются с рентгенографической картиной развития интерстициальной пневмонии и выраженного легочного заболевания, наблюдаемого у людей с MERS-CoV.
Приведенные выше данные показывают, что мыши с гуманизированным DPP4, такие как мыши, описанные в настоящем документе, восприимчивы к инфекции MERS-CoV. Данные также демонстрируют, что мыши с гуманизированным DPP4, описанные в настоящем документе, представляют собой модель in vivo инфекции MERS-CoV, которая повторяет патологические процессы, например патологические последствия, наблюдаемые при инфекции MERS-CoV у людей.
Таким образом, мыши с гуманизированным DPP4 представляют собой надежную модель MERS-CoV, которую можно использовать для оценки лечения MERS-CoV in vivo. Например, мыши с гуманизированным DPP4 являются подходящими животными-носителями для измерения фармакокинетики, фармакодинамики и терапевтической эффективности терапевтических молекул, которые воздействуют на MERS-CoV.
На Фиг. 5 представлено выравнивание белковой последовательности Dpp4 мыши (SEQ ID NO: 25) с человеческой DPP4 (кодируется трансгенными мышами 7326/7327) (SEQ ID NO: 26).
Затем для мышей с гуманизированной DPP4 построили график зависимости массы тела от времени после инфицирования MERS-CoV. Мышей инфицировали MERS-CoV (штамм Jordan) или PBS (ложноинфицированные), как описано выше, и анализировали на потерю массы тела, которая является признаком продуктивной инфекции, в течение семи дней. Как показано на Фиг. 14, у мышей с гуманизированной DPP4 развивалась продуктивная инфекция (т.е. наблюдались патологические изменения, характерные для проявившегося заболевания), причем уменьшение массы тела началось на 4 день после инфицирования. В одну группу включали от четырех до пяти мышей. На Фиг. 15 показано, что мыши, гетерозиготные по аллелю гуманизированного DPP4, имели одинаковую восприимчивость к инфекции MERS-CoV, так как у них была аналогичная степень потери массы тела по сравнению с гомозиготами. Этот результат является важным, так как указывает на то, что исследования можно выполнять с использованием гетерозиготных мышей. Кроме того, использование гетерозиготных мышей позволяет избежать любых потенциальных проблем, связанных с инактивацией функциональной мышиной Dpp4, которая потенциально может происходить у гомозиготных мышей с гуманизированной DPP4.
Также проводили гистологическое исследование легких этих мышей на наличие воспаления в соответствии с описанными ниже способами. Как показано на Фиг. 16, у мышей с hDPP4, которым вводили высокую дозу вируса (1×106 pfu/мышь), наблюдали более выраженные патологические изменения по сравнению с контролями (PBS).
Пример 3
Создание трансгенных мышей, содержащих человеческий ген DPP4, путем произвольного встраивания искусственных бактериальных хромосом (ВАС)
Создавали трансгенных мышей, содержащих человеческий ген DPP4, для которого информация о нуклеотидной последовательности и геноме показана на Фиг. 6. Использовали две различные перекрывающиеся ВАС, содержащие человеческий ген DPP4: ВАС RP11-68L22 и ВАС RP11-345J9 (Фиг. 7). Обе ВАС содержали кодирующую область человеческого гена DPP4, а также более 40 тыс.п.н. области промотора со стороны 5'-конца от инициаторного ко дона ATG гена DPP4 и несколько тысяч пар нуклеотидов со стороны 3'-конца от стоп-ко дона гена DPP4 (Фиг. 7).
Для создания трансгенных мышей с использованием ВАС ДНК каждой ВАС подвергали электропорации с внедрением в мышиные эмбриональные стволовые (ЭС) клетки VGB6 (полученные от мышей C57BL/6N). ЭС-клетки, содержащие кодирующую область человеческого гена DPP4, а также области промотора гена, внедряли в эмбрион мыши SW на 8-клеточной стадии с помощью способа VelociMouse® (см., например, патенты США Ms 7,294,754, 7,576,259, 7,659,442 и Poueymirou et al. (2007) F0 generation mice that are essentially fully derived from the donor gene-targeted ES cells allowing immediate phenotypic analyses Nature Biotech. 25(1):91-99).
Анализы приобретения аллеля, применяемые у человека, использовали для скрининга ЭС-клеточных клонов для выявления клеток, содержащих копии человеческого гена DPP4 вместе с областями промотора гена. Анализы приобретения аллеля, применяемые у человека, использовали для выявления мышей VelociMice® (мыши F0, полученные полностью из донорской ЭС-клетки), имеющих гуманизированный ген DPP4 вместе с областями промотора гена.
Вкратце, геномную ДНК экстрагировали из ЭС-клеточных клонов с использованием стандартных способов и протестировали в анализе количественной ПЦР (кПЦР) TaqMan™ с использованием двух наборов праймеров и зондов для обнаружения человеческой последовательности ДНК со стороны 5'-конца (7333 hTU) и 3'-конца (7333 hTD) от человеческой кодирующей последовательности DPP4 (Фиг. 8). Местоположения в пределах человеческого гена DPP4 и фланкирующих областей (например областей промотора), в которых с помощью набора праймеров и зондов выполняется отжиг, показаны на Фиг. 7. Результаты измерения флуоресценции напротив фона в анализе кПЦР TaqMan™ указывают на наличие человеческого гена DPP4 и по меньшей мере 40 тыс.п.н. 5'-фланкирующей области от человеческого гена DPP4, которые были встроены в геном трансгенной мыши.
Набор праймеров и зондов 7333 hTU состоял из (от 5'-конца к 3'-концу): человеческого прямого праймера со стороны 5'-конца TGGCTTATTCTCTATTCCTCACCTA (SEQ ID NO: 18); человеческого зонда со стороны 5'-конца FAM-TGCTTTCCCTCCTCCCTTCTGA-BHQ (SEQ ID NO: 19); человеческого обратного праймера со стороны 5'-конца GGCCTTAGCCCAGAAACTG (SEQ ID NO: 20). Набор праймеров и зондов 7333 hTD состоял из (от 5'-конца к 3'-концу): человеческого прямого праймера со стороны 3'-конца TGCAGACTTGTCTTGACATTCATA (SEQ ID NO: 21); человеческого зонда со стороны 3'-конца CAL-AGCCTCTGCAGACACAGGAATGGC-BHQ (SEQ ID NO: 22) и человеческого обратного праймера со стороны 3'-конца TCTGGGCACTGGTGTACTC (SEQ ID NO: 23); в котором FAM и CAL обозначают 5-карбоксифлуоресцеиновый флуоресцентный зонд и флуоресцентный зонд CAL Orange соответственно, a BHQ обозначает гаситель флуоресценции типа Black Hole (Biosearch Technologies).
Например, геномную ДНК из Эс-клеточных клонов добавляли в реакционную смесь TaqMan™ Gene Expression Master Mix (Life Technologies) в соответствии с рекомендациями производителя в 384-луночном планшете для ПЦР (оптический 384-луночный реакционный планшет Micro Amp™ Optical 384-Well Reaction Plate, Life Technologies) и выполняли ряд циклов в приборе Applied Biosystems Prism 7900НТ, который собирает данные флуоресценции в процессе ПЦР и определяет пороговый цикл (Ct), фракционный цикл ПЦР, в котором накопленная флуоресценция достигает предварительно установленного порогового значения. Для каждого образца ДНК проводили прямые и обратные DPP4-специфичные кПЦР и две кПЦР для контрольных генов, не содержащих DPP4. Рассчитывали различия в значениях Ct (ΔCt) между каждой DPP4-специфичной кПЦР и каждой кПЦР контрольного гена, а затем рассчитывали различия между каждым ΔCt и медианным ΔCt для всех проанализированных образцов для получения значений ΔΔCt для каждого образца. Рассчитывали число копий гена DPP4 в каждом образце по следующей формуле: число копий = 2×2-Δ ΔCt. У клона, содержащего по меньшей мере одну копию человеческого DPP 4 плюс области промотора, встроенные в хромосому, число копий гена DPP4 было больше или равно единице.
Аналогичный анализ приобретения аллеля, применяемый у человека, использовали для анализа ДНК, очищенной из биопсий хвоста, у мышей, полученных из ЭС-клеток, для подтверждения проникновения аллеля гуманизированного DPP4 вместе с человеческими 5'-фланкирующими областями через зародышевую линию.
С использованием способов встраивания ВАС и скрининга, описанных в настоящем документе, получили две трансгенных мыши с ДНК, кодирующей человеческую DPP4. ВАС RP11-68L22 использовали для создания ЭС-клеточных клонов и трансгенных мышей под названием MAID 7333, а ВАС RP11-345J9 использовали для создания ЭС-клеточных клонов и трансгенных мышей под названием MAID 7334.
Аминокислотная последовательность белка, кодируемого гуманизированной нуклеотидной последовательностью DPP4, у мышей MAID 7333 и 7334 показана на Фиг. 9 (SEQ ID NO: 24) и аналогична аминокислотной последовательности человеческой DPP4 (кодируемой транскриптом NM_001935.3).
Пример 4
Лечение мышей с гуманизированным DPP4, которые были инфицированы вирусом MERS-CoV
Трансгенных мышей с гуманизированным геном DPP4 и фланкирующими областями промотора протестировали на их способность инфицироваться MERS-CoV и выступать в качестве модели для оценки терапевтических молекул, предназначенных для лечения или предотвращения MERS-CoV.
Трансгенных мышей MAID 7333 (например, созданных с помощью способов, описанных в примере 3) лечили антителами против шипикового белка MERS-CoV или изотипическими контролями в дозе 200 мкг путем интраперитонеальной инъекции. Через один день после инъекции антител по отношению к мышам выполняли интраназальное инфицирование MERS-CoV. Через четыре дня после инфицирования извлекали легкие мышей и измеряли концентрации вирусной РНК с помощью ПЦР в реальном времени. В частности, измеряли концентрации геномной РНК (UpE) или реплицирующейся РНК (лидерной) (специфичной для репликативной формы генома MERS-CoV) MERS-CoV.
Данные ПЦР в реальном времени приведены в таблице ниже.
Лечение трансгенных мышей антителами уменьшило концентрации вирусной РНК (как UpE, так и лидерной) от приблизительно 50 раз до приблизительно 500 раз.
Данные, описанные в настоящем документе, показывают, что трансгенные мыши, созданные с помощью способов нацеленного встраивания человеческого DPP4 (пример 1) и способов произвольного встраивания ВАС (пример 3) с человеческим DPP4, были восприимчивы к инфекции MERS-CoV. Кроме того, антитела против MERS-CoV блокировали инфекцию у трансгенных мышей in vivo. Таким образом, трансгенных мышей с человеческой DPP4 (например, созданных с помощью способов, описанных в настоящем документе) можно использовать для оценки эффективности терапевтических средств (например, антител), которые воздействуют на вирус MERS-CoV.
Пример 5
Профилактическое действие антител против MERS-CoV на инфекцию MERS-CoV у мышей с гуманизированным DPP4
Мышей с гуманизированным DPP4, описанных в настоящем документе, использовали для оценки профилактической эффективности двух моноклональных антител in vivo. Мышам интраперитонеально вводили антитела против MERS-CoV (в дозе 200 мкг, 20 мкг или 2 мкг антител 2 против шипикового белка MERS-CoV (Ab 2), антител 4 против шипикового белка MERS-CoV (Ab 4) или 200 мкг человеческих антител IgG1 (hIgG1) изотипического контроля) за 24 часа до интраназального инфицирования MERS-CoV (штамм Jordan) в дозе 1×105 pfu. Ab 2 и Ab 4 были полностью человеческими антителами против шипикового белка MERS-CoV. РНК экстрагировали из легких мышей и проанализировали с помощью количественной ПЦР, как описано выше. Например, данные кПЦР проанализировали с помощью способа дельта Ct, приняв результаты для неинфицированных контролей за 1. Обнаруженное количество РНК MERS-CoV можно выразить в процентах по отношению к концентрациям РНК, обнаруженным у инфицированных мышей, по отношению к которым использовали контрольные антитела такого же изотипа (Фиг. 11А-В). Кроме того, титры вируса из легких мышей определили, как описано выше.
Оба типа антител значительно уменьшали концентрации специфической РНК MERS-CoV в легких более чем на 2 log при дозе 200 мкг на мышь по сравнению с контрольными антителами такого же изотипа (Фиг. 11А-В). Антитела Ab 2 оказались более эффективны в отношении уменьшения концентраций РНК MERS-CoV при дозе 20 мкг по сравнению с антителами Ab 4 при аналогичной дозе. Введение 2 мкг антител любого типа было неэффективным в отношении уменьшения концентраций вирусной РНК по сравнению с мышами изотипического контроля. При анализе титра MERS-CoV в легких (Фиг. 11С) как доза 200 мкг, так и 20 мкг Ab 2 уменьшала концентрации вируса практически до предела обнаружения данного анализа (2×103 pfu/мл). Антитела Ab 4 были такими же эффективными в дозе 200 мкг, как антитела Ab 2, причем дозы 20 мкг и 2 мкг продемонстрировали дозозависимое ингибирование вируса. Эти данные показывают, что антитела против MERS-CoV, например Ab 2 и Ab 4, эффективно блокировали инфекцию MERS-CoV in vivo.
На 4 день после инфицирования также выполняли гистологический анализ на легких мышей, по отношению к которым за 24 часа до инфицирования использовали антитела Ab 2, Ab 4 или антитела изотипического контроля hIgG1 (Фиг. 12А). Например, степени выраженности интерстициального, перибронхиолярного и периваскулярного воспаления оценивали по шкале от 0 до 5 баллов. Также проанализировали другие гистологические характеристики, такие как наличие бронхиолярного эпителия и повреждения альвеол, изменения в плевре и выраженность перибронховаскулярного воспаления. Для каждой экспериментальной группы рассчитывали среднее значение общего количества баллов по шкале оценки воспаления для каждой мыши, и эти баллы были представлены в виде среднего количества баллов в каждой группе и в каждой временной отметке (Фиг. 12В).
В легких от мышей, которым предварительно вводили изотипический контроль hIgG1, обнаружили выраженные патологические изменения с усилением интерстициального воспаления, периваскулярной инфильтрацией и утолщением стенок альвеол. У мышей, по отношению к которым использовали антитела Ab 2 или Ab 4 в дозе 200 мкг, наблюдали уменьшение воспаления с минимальными скоплениями воспалительных клеток в интерстиций, незначительной бронхиолярной инфильтрацией и менее выраженным утолщением стенок альвеол. У мышей, которым предварительно вводили Ab 2 или Ab 4 в дозе 20 мкг, выявили средние уровни периваскулярной инфильтрации и интерстициального воспаления по сравнению с группой, которым вводили антитела в более высокой дозе. В группе, по отношению к которой предварительно использовали антитела в дозе 2 мкг, наблюдали аналогичные патологические изменения по сравнению с изотипическим контролем hIgG1, характеризующимся выраженным интерстициальным воспалением и преобладающим периваскулярным воспалением. В гистологическом исследовании в слепом режиме было продемонстрировано уменьшение количества баллов по шкале оценки воспаления у получавших лечение мышей (Фиг. 12В). Такие результаты показывают, что антитела против MERS-CoV, такие как Ab 2 или Ab 4, обеспечивают дозозависимое уменьшение патологических изменений в легких при инфекции MERS-CoV, подтверждая данные о концентрациях вирусной РНК и титрах вируса у мышей.
Таким образом, антитела против MERS-CoV, такие как Ab 2 или Ab 4, оказались эффективны в модели in vivo инфекции MERS-CoV. Антитела блокировали инфекцию MERS-CoV и вызываемое заболевание in vivo при введении до инфицирования, например за 1 день до инфицирования.
Пример 6
Лечение антителами мышей с гуманизированным DPP4, которые были инфицированы MERS-CoV
Для определения терапевтического эффекта (например, способности подавлять репликацию MERS-CoV и патологические изменения в легких после инфицирования) антител против MERS-CoV (например, Ab 2 или Ab 4) мышей с гуманизированным DPP4 инфицировали MERS-CoV. Через 24 часа после инфицирования мышам интраперитонеально вводили 500 мкг изотипического контроля hIgG1 или Ab 2 в дозе 500 мкг или 200 мкг. На 4 день после инфицирования мышей умертвили и в мышиных легких оценили концентрацию вирусной РНК, титр вируса и патологические изменения легких. Антитела Ab 2 как в дозе 500 мкг, так и 200 мкг уменьшали концентрации вирусной РНК приблизительно в 10 раз в легких мышей по сравнению с мышами, по отношению к которым использовали контрольные антитела (Фиг. 13А-В). Титры вируса в легких у одних и тех же мышей продемонстрировали выраженное уменьшение, причем на 4 день после инфицирования уменьшение было более чем на 2 log (Фиг. 13С). Эти данные демонстрируют, что после инфицирования, например через 24 часа после инфицирования, антитела против MERS-CoV (например Ab 2) существенно подавляли репликацию вируса.
Гистологический анализ также выполняли у мышей, которым через 24 часа после инфицирования вводили контрольные антитела hIgG1, антитела Ab 2 в дозе 500 мкг или антитела Ab 2 в дозе 200 мкг (Фиг. 14А-В). У мышей, по отношению к которым использовали контрольные антитела, обнаружили аналогичные патологические изменения по сравнению с контрольными животными в примерах 2 и 5, характеризующиеся выраженным интерстициальным воспалением, периваскулярной инфильтрацией и утолщением стенок альвеол. У мышей, по отношению к которым использовали антитела Ab 2 в дозе 200 мкг или 500 мкг, наблюдали минимальное интерстициальное воспаление и менее выраженное и только фокальное периваскулярное воспаление во всех областях легких. В гистологическом исследовании в слепом режиме было продемонстрировано уменьшение количества баллов по шкале оценки воспаления у получавших лечение мышей (Фиг. 14В). Полученные данные показывают, что терапевтические дозы антител против MERS-CoV (например, Ab 2) уменьшали индуцированные MERS-CoV патологические изменения даже при введении после инфицирования, например через 24 часа после инфицирования.
| название | год | авторы | номер документа |
|---|---|---|---|
| НЕ ОТНОСЯЩИЕСЯ К ЧЕЛОВЕКУ ЖИВОТНЫЕ, ЭКСПРЕССИРУЮЩИЕ ГУМАНИЗИРОВАННЫЙ КОМПЛЕКС CD3 | 2015 |
|
RU2726446C2 |
| ГУМАНИЗИРОВАННЫЕ ЖИВОТНЫЕ ПО С5 И С3 | 2015 |
|
RU2713327C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ ЖИВОТНЫЕ, ОТЛИЧНЫЕ ОТ ЧЕЛОВЕКА, ЭКСПРЕССИРУЮЩИЕ EPO ЧЕЛОВЕКА | 2015 |
|
RU2711744C1 |
| ЖИВОТНЫЕ С ГУМАНИЗИРОВАННЫМИ IL-4 И IL-4Rα | 2015 |
|
RU2788523C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ В ОТНОШЕНИИ ГЛАВНОГО КОМПЛЕКСА ГИСТОСОВМЕСТИМОСТИ МЫШИ | 2012 |
|
RU2783984C2 |
| ЖИВОТНЫЕ С ГУМАНИЗИРОВАННЫМ ГЕНОМ IL-15 | 2014 |
|
RU2674910C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ МЫШИ, ЭКСПРЕССИРУЮЩИЕ ХИМЕРНЫЕ МОЛЕКУЛЫ ГЛАВНОГО КОМПЛЕКСА ГИСТОСОВМЕСТИМОСТИ | 2012 |
|
RU2660564C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ В ОТНОШЕНИИ ГЛАВНОГО КОМПЛЕКСА ГИСТОСОВМЕСТИМОСТИ МЫШИ | 2012 |
|
RU2653433C2 |
| ЖИВОТНЫЕ С ГУМАНИЗИРОВАННЫМИ IL-4 И IL-4Rα | 2015 |
|
RU2703139C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЕ МЫШИ, ЭКСПРЕССИРУЮЩИЕ ХИМЕРНЫЕ МОЛЕКУЛЫ ГЛАВНОГО КОМПЛЕКСА ГИСТОСОВМЕСТИМОСТИ | 2012 |
|
RU2797549C2 |

















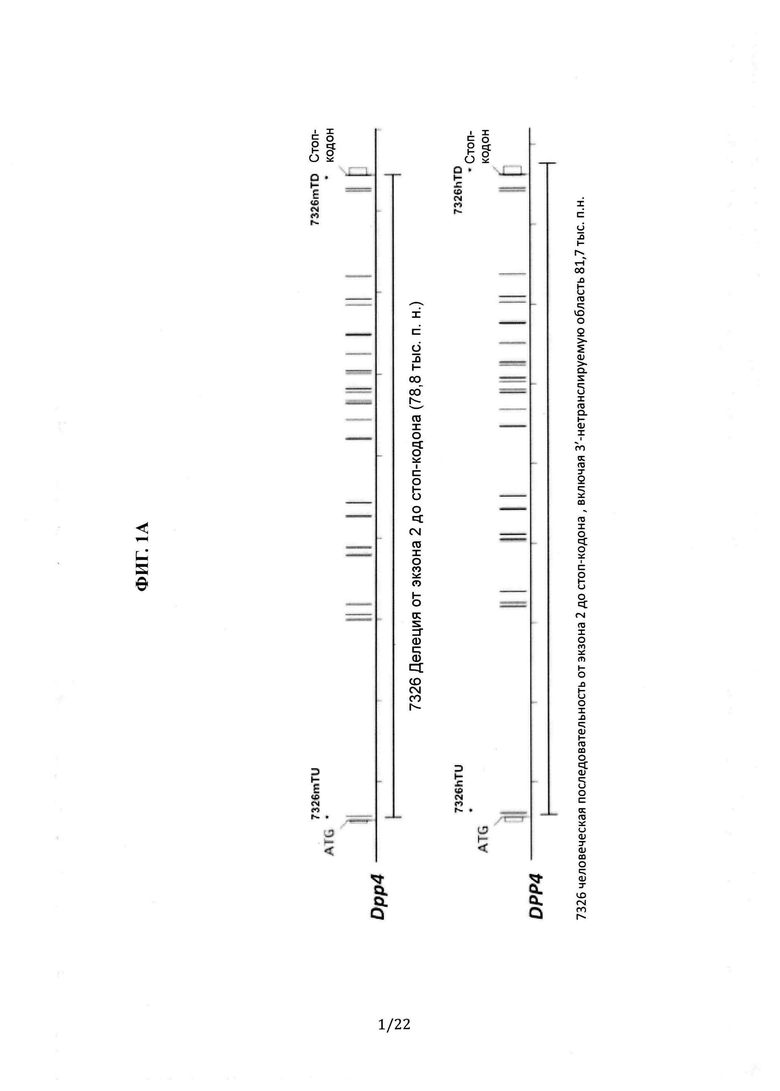
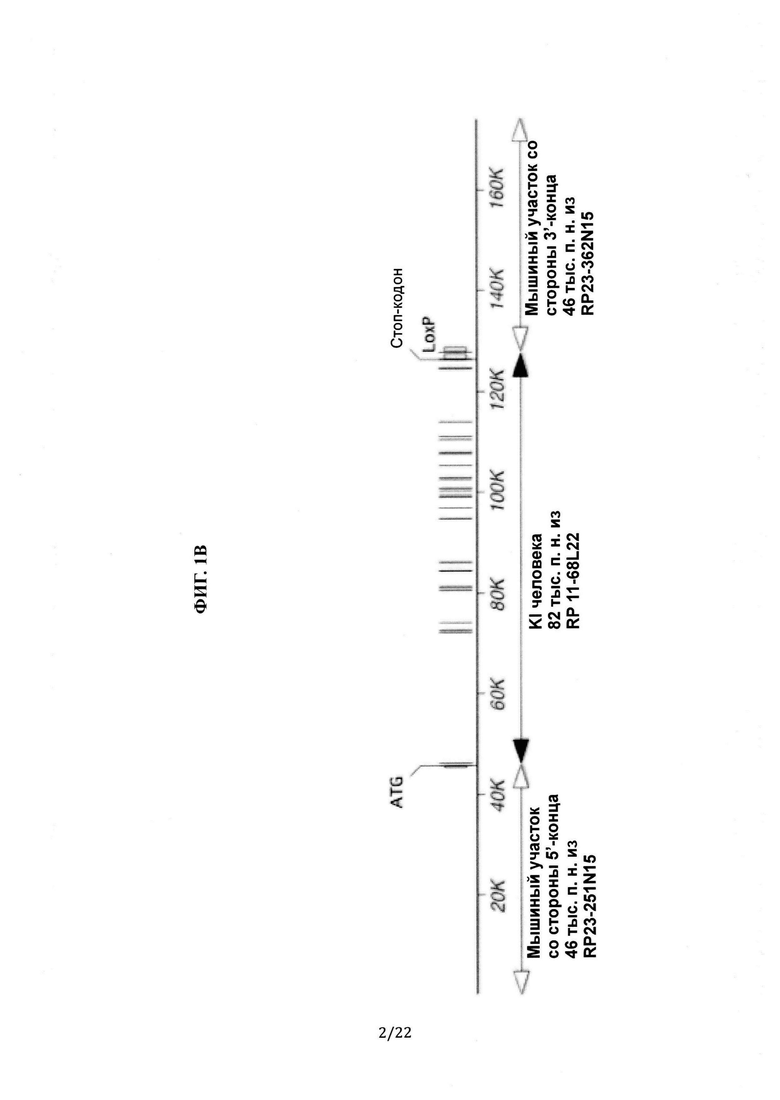

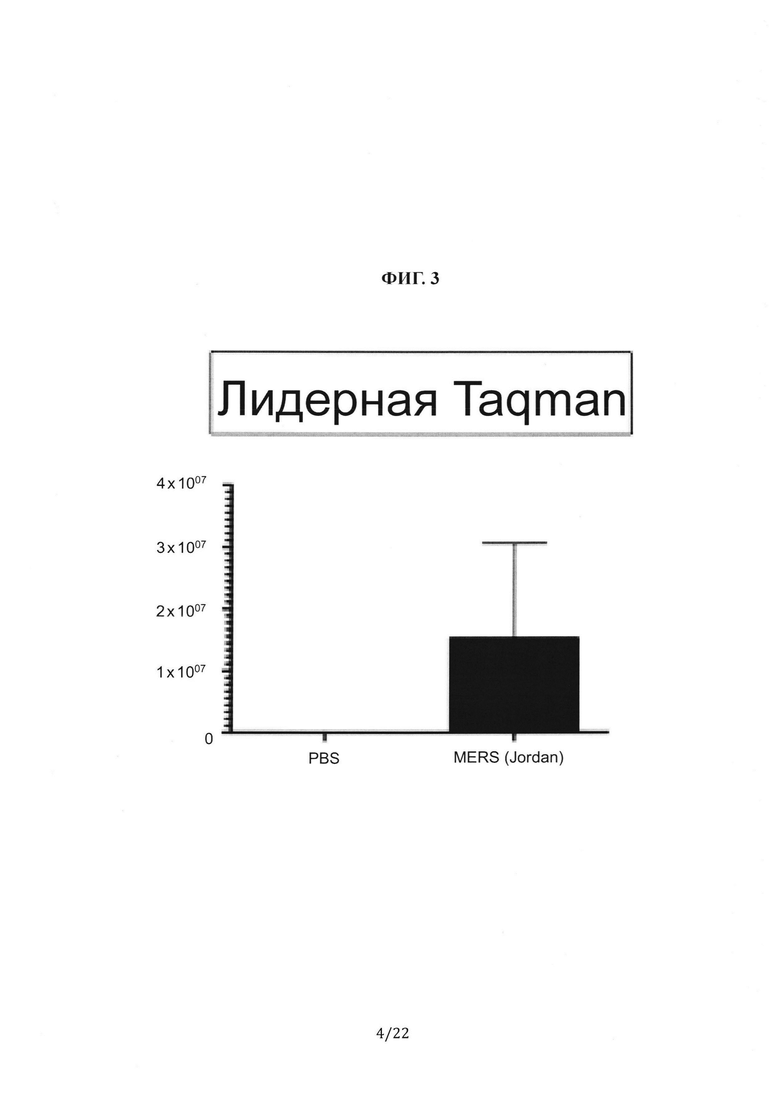
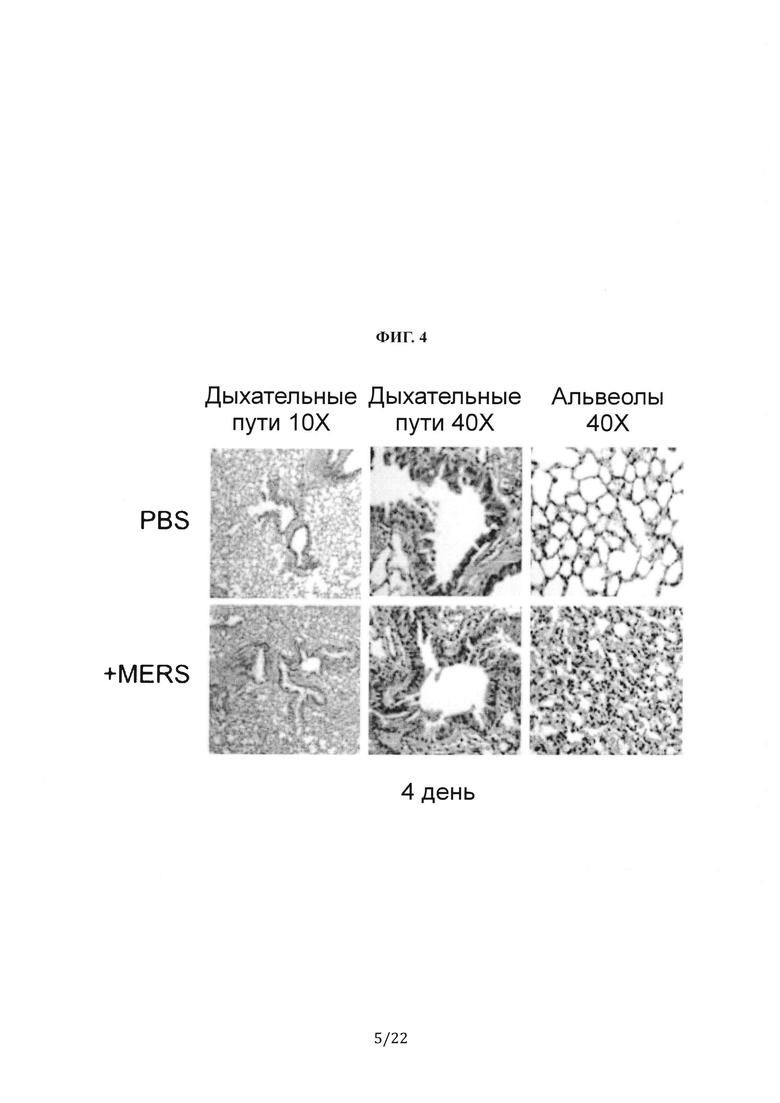

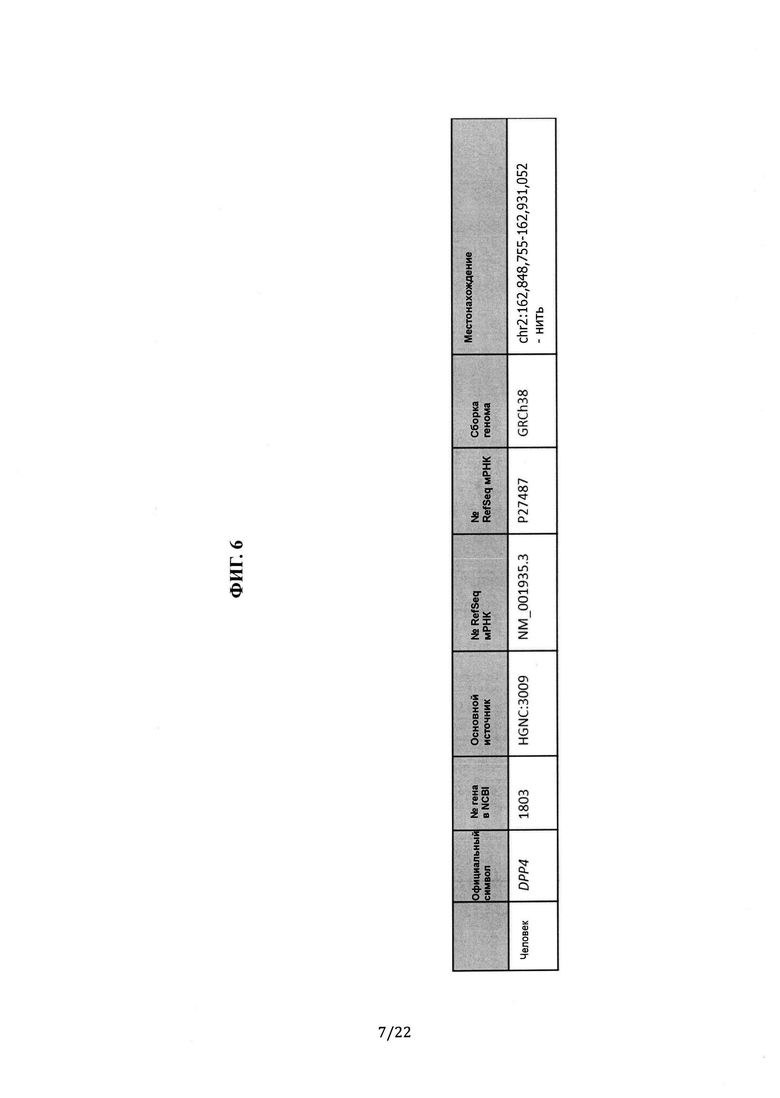
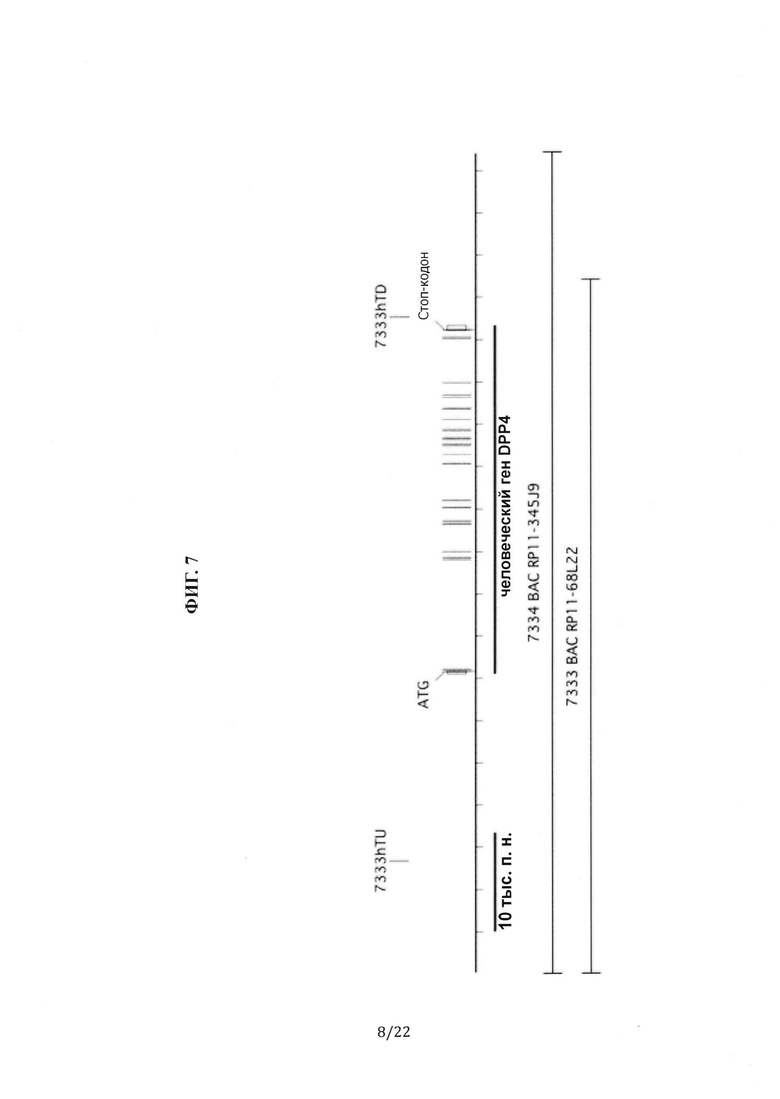


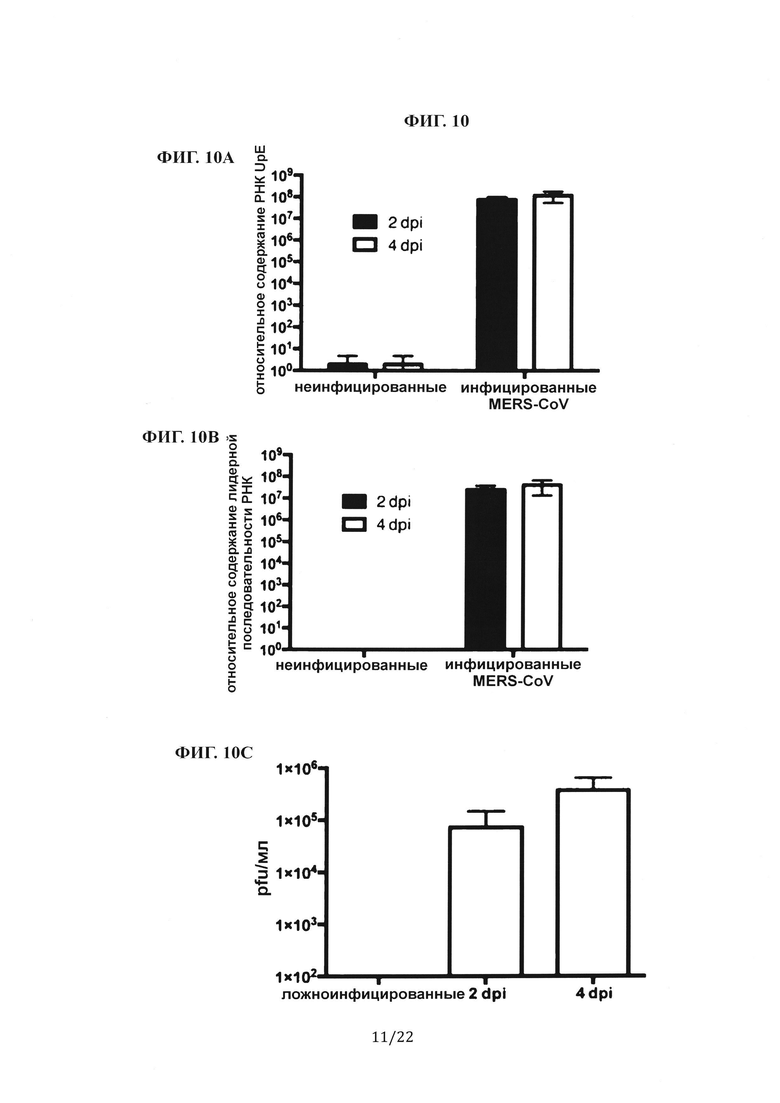
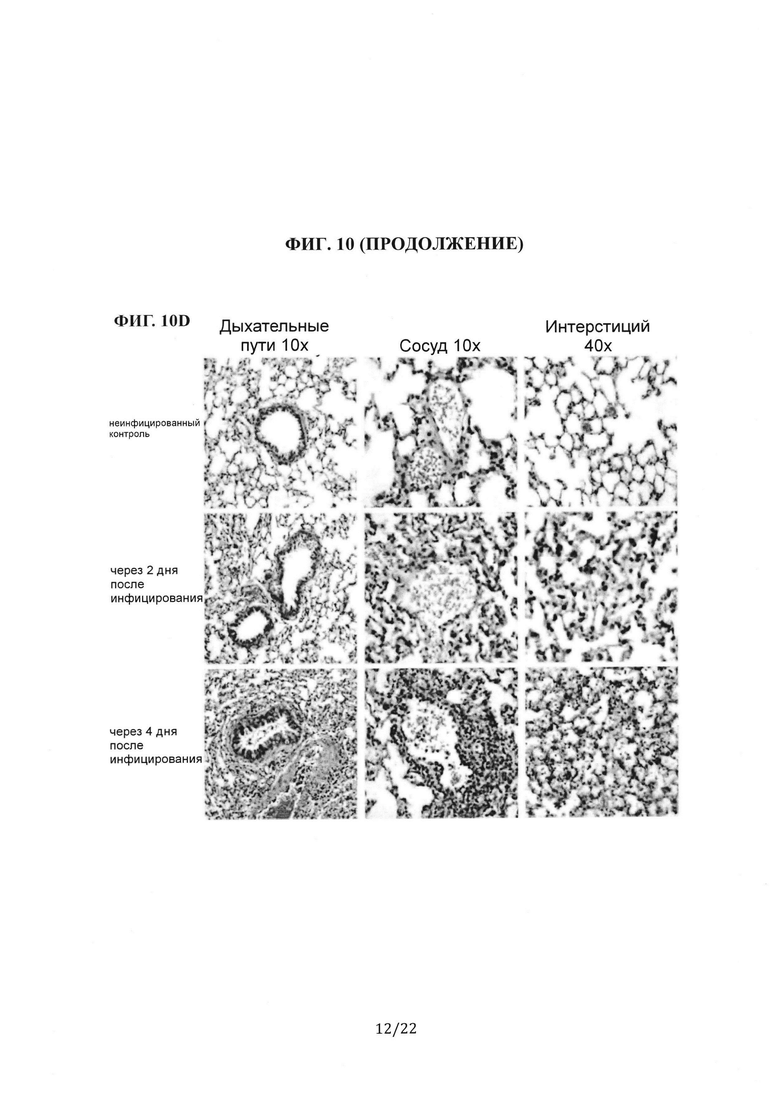
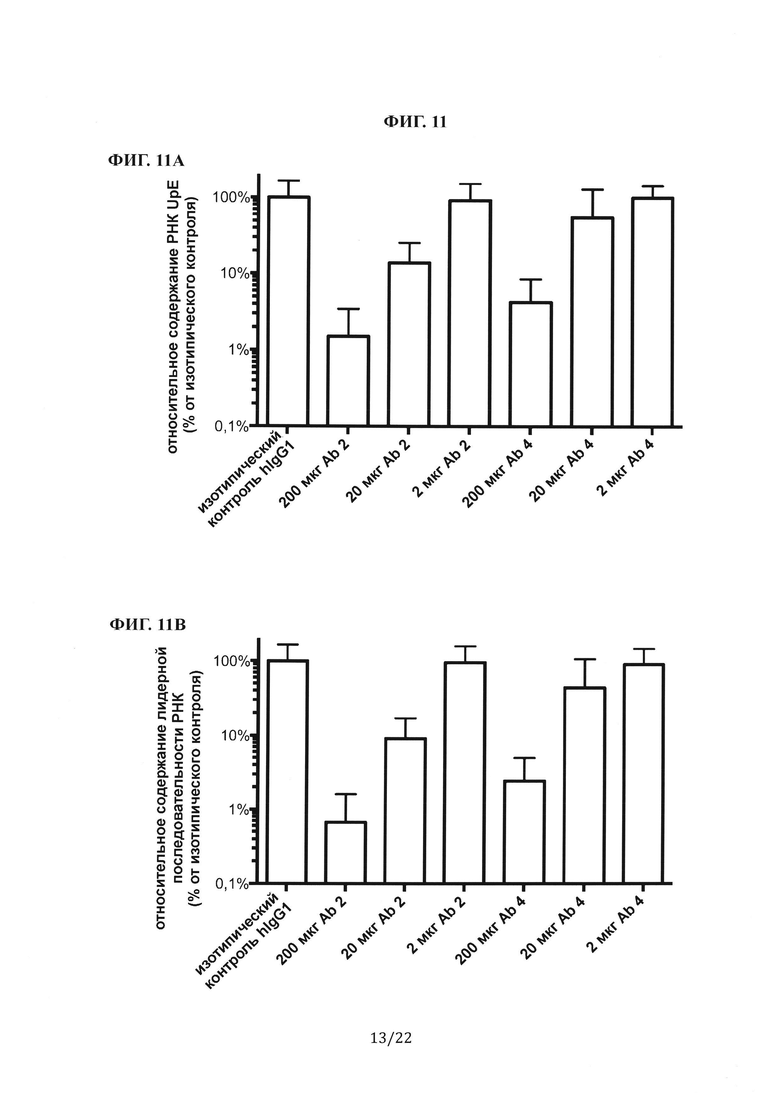
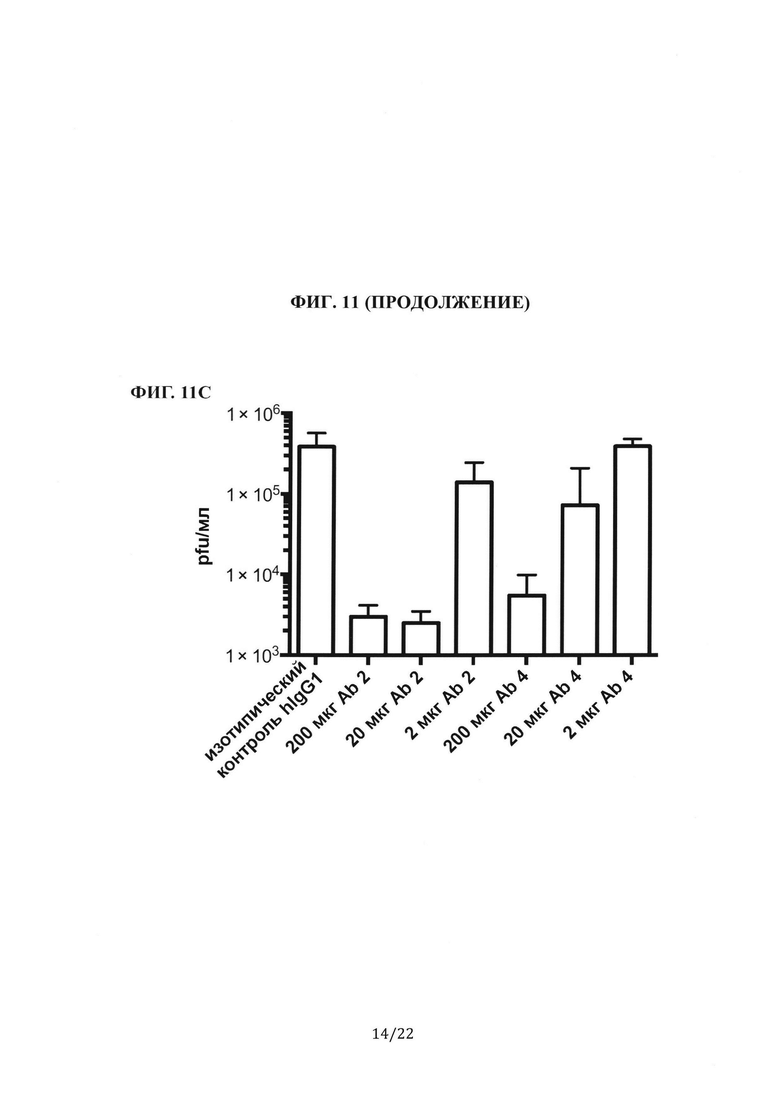

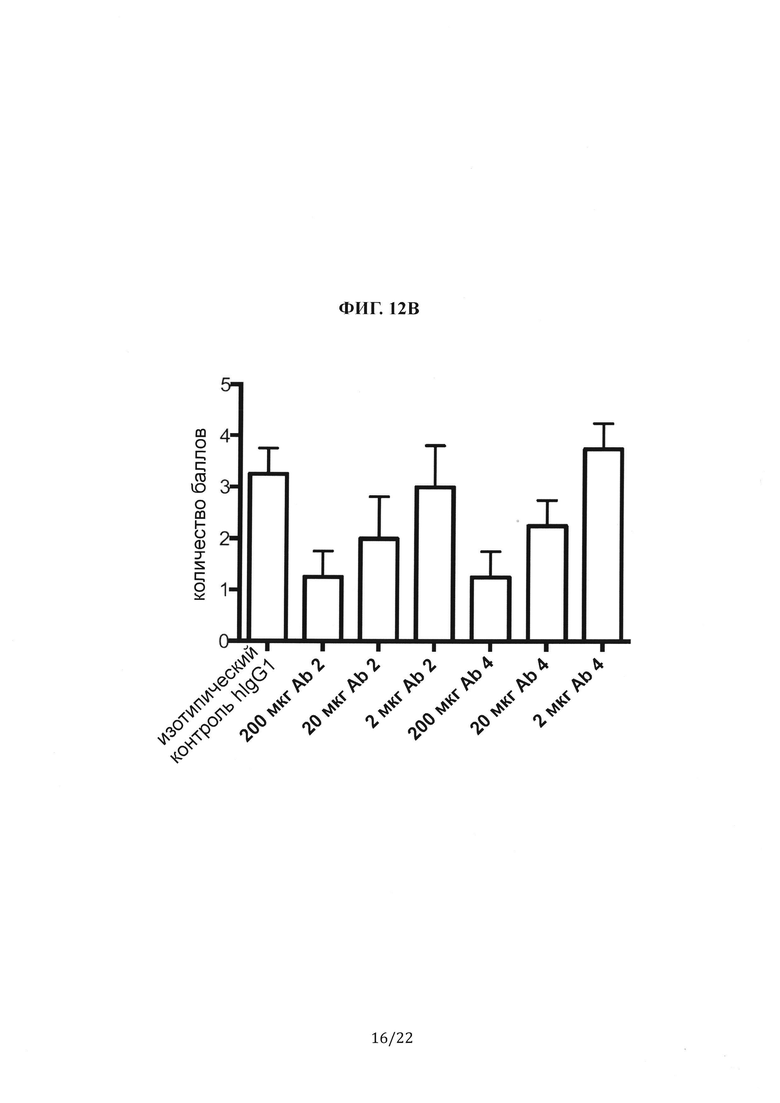
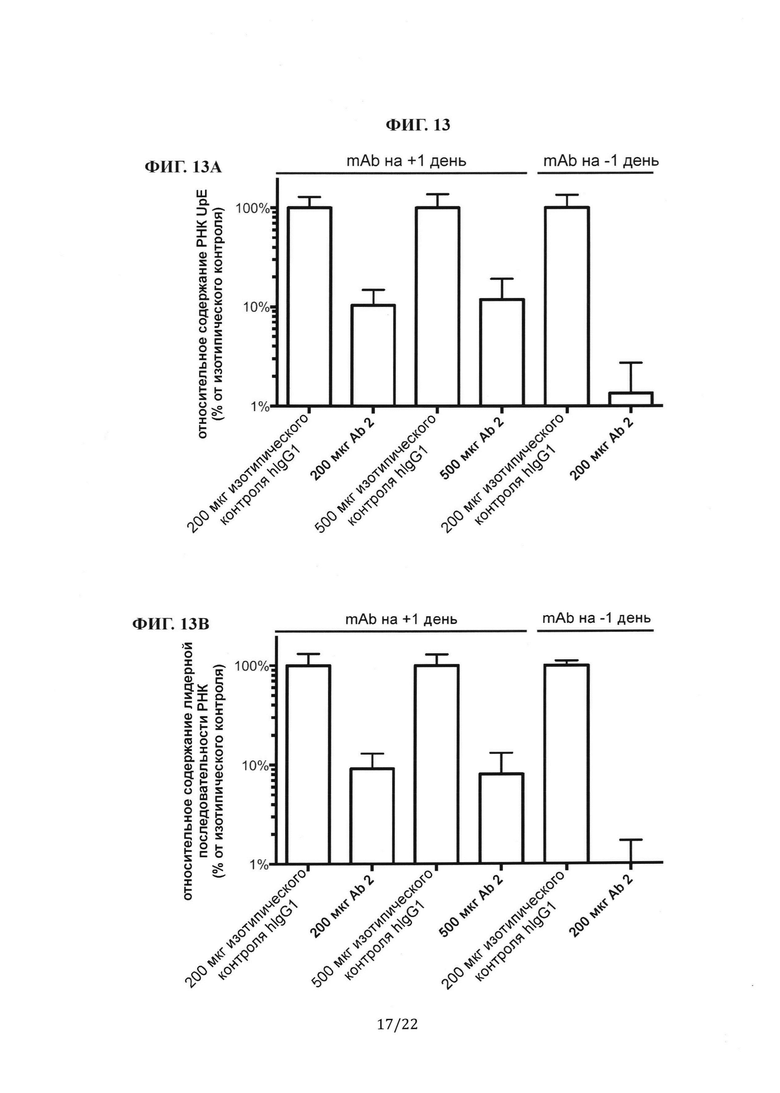
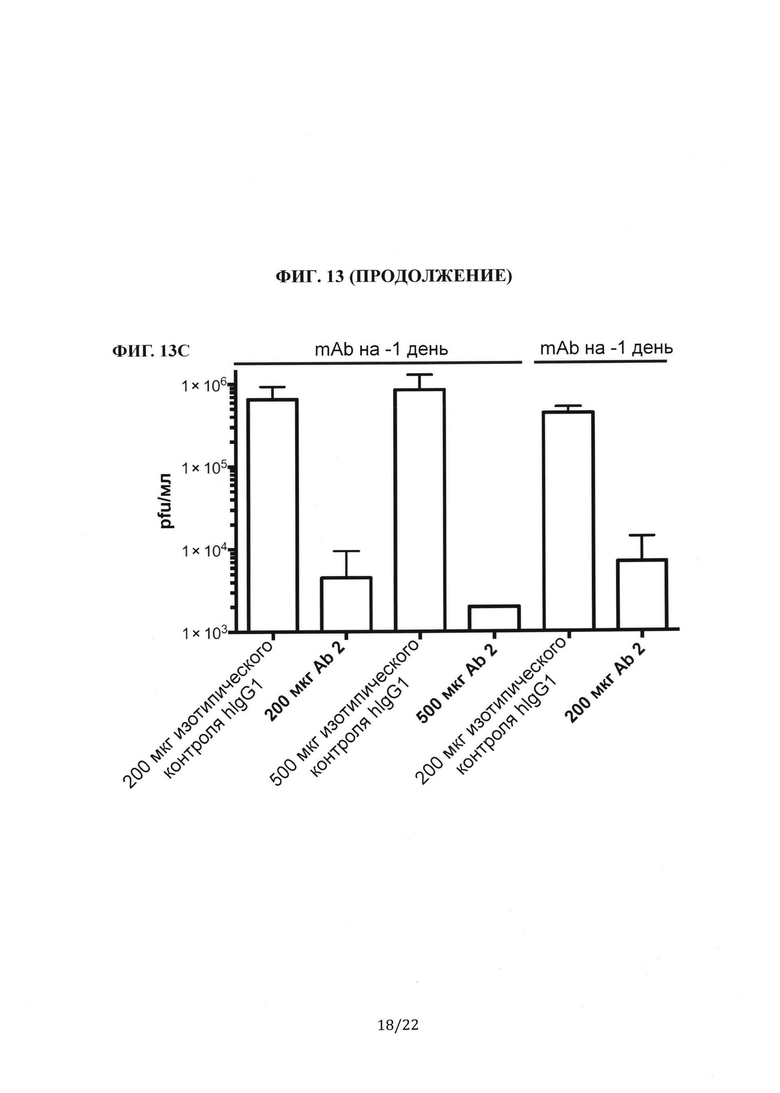
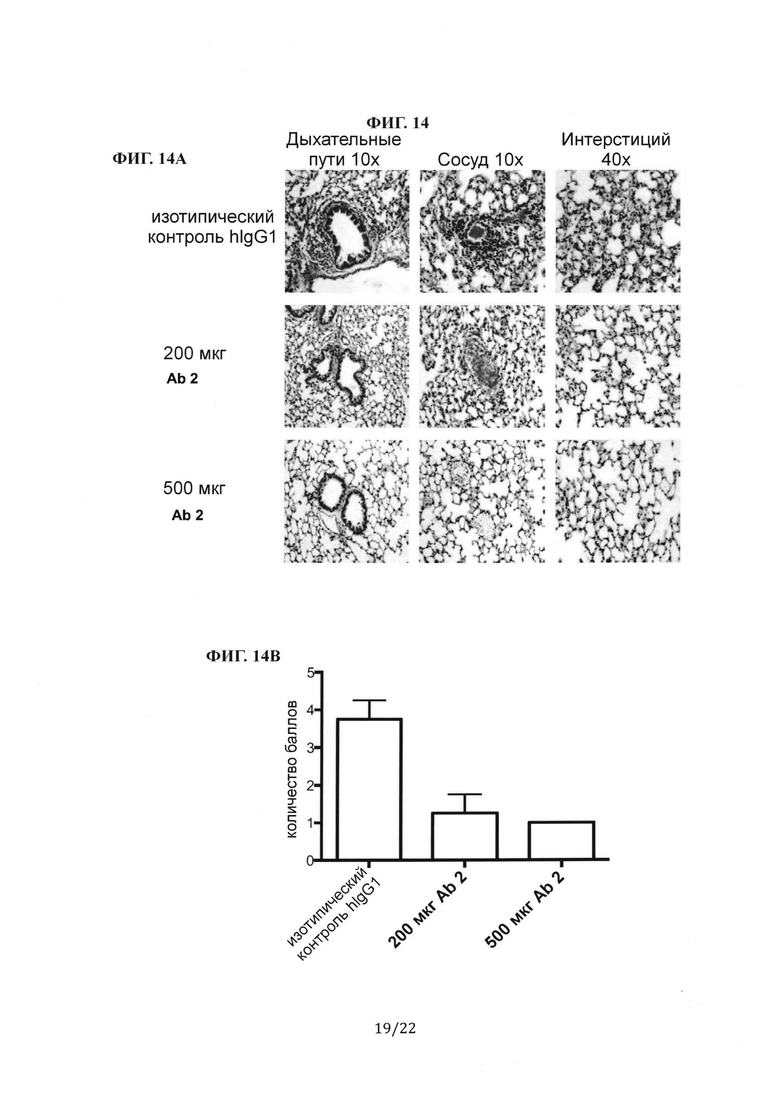
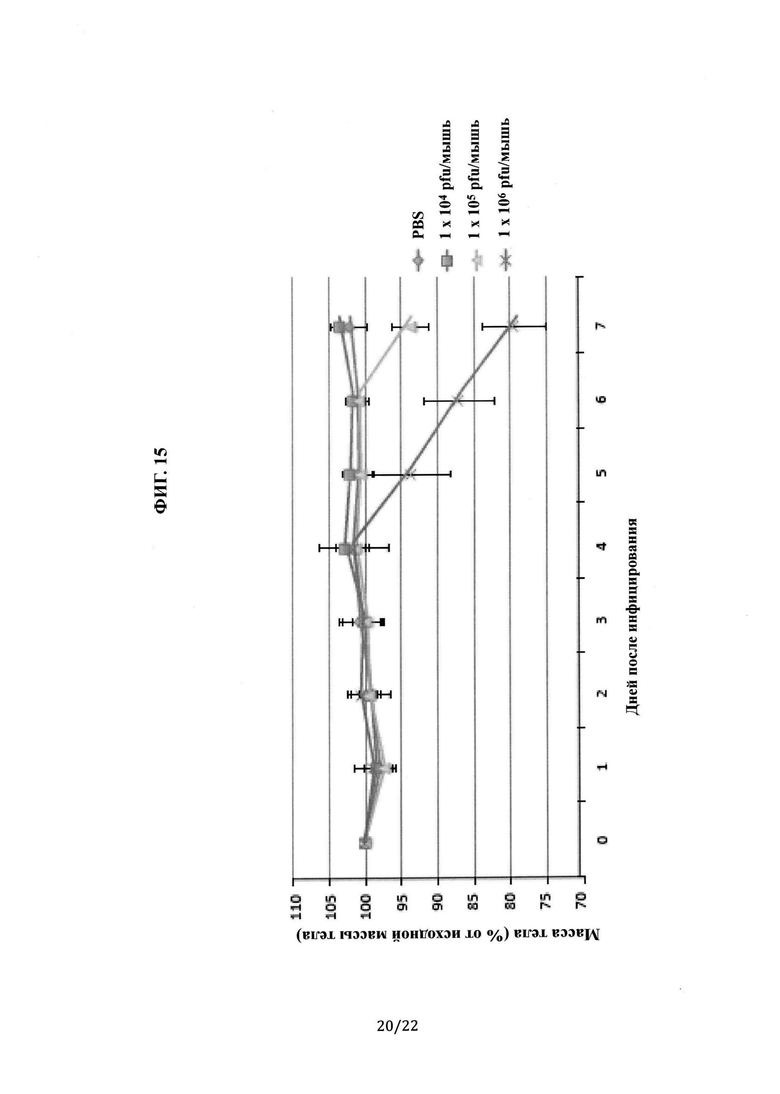
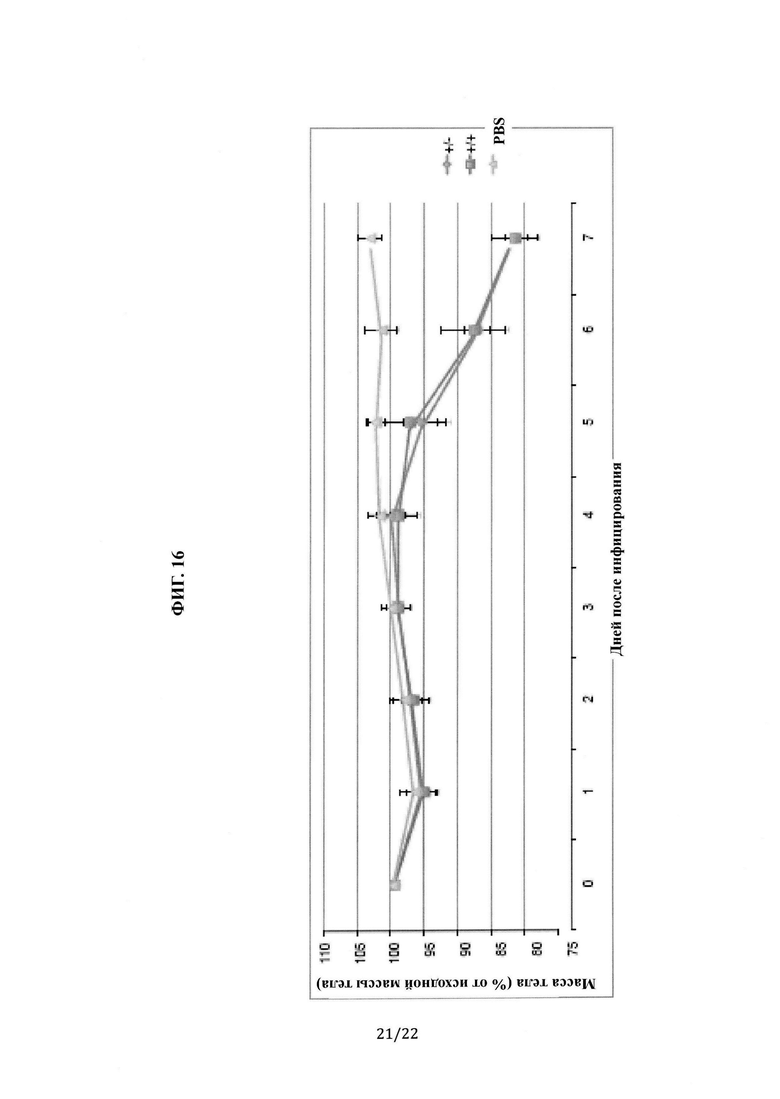
Настоящее изобретение относится к генетической инженерии, в частности к трансгенному грызуну, мыши или крысе, которого используют для экспрессии человеческого белка DPP4. Геном указанного грызуна содержит замену геномного фрагмента гена Dpp4 грызуна в эндогенном локусе Dpp4 грызуна на геномный фрагмент человеческого гена DPP4 с получением модифицированного гена DPP4. Указанный модифицированный ген DPP4 кодирует человеческий белок DPP4, а его экспрессия находится под контролем регуляторных элементов грызуна в эндогенном локусе Dpp4 грызуна. При этом у грызуна проявляется воспаление легких при инфицировании коронавирусом ближневосточного респираторного синдрома (MERS-CoV). Настоящее изобретение также раскрывает способ получения указанного грызуна и способ определения терапевтической эффективности in vivo специфичного для человека антагониста DPP4 с использованием указанного грызуна. Настоящее изобретение позволяет улучшить качество оценки in vivo эффективности специфичных к человеческому DPP4 антагонистов, воздействующих на молекулы-мишени, в моделях заболеваний человека. 4 н. и 18 з.п. ф-лы, 17 ил., 1 табл., 6 пр.
1. Грызун для экспрессии человеческого белка DPP4, геном которого содержит замену геномного фрагмента гена Dpp4 грызуна в эндогенном локусе Dpp4 грызуна на геномный фрагмент человеческого гена DPP4 с получением модифицированного гена DPP4, при этом модифицированный ген DPP4 кодирует человеческий белок DPP4, при этом экспрессия модифицированного гена DPP4 находится под контролем регуляторных элементов грызуна в эндогенном локусе Dpp4 грызуна, при этом грызун представляет собой мышь или крысу, и при этом у грызуна проявляется воспаление легких при инфицировании коронавирусом ближневосточного респираторного синдрома (MERS-CoV).
2. Грызун по п. 1, у которого геномный фрагмент человеческого гена DPP4 содержит экзоны 2-26 человеческого гена DPP4.
3. Грызун по п. 2, у которого геномный фрагмент заменяемого гена Dpp4 грызуна содержит экзоны 2-26 гена Dpp4 грызуна.
4. Грызун по п. 2, который представляет собой мышь, и при этом модифицированный ген DPP4 содержит экзон 1 эндогенного мышиного Dpp4, функционально связанный с геномным фрагментом человеческого гена DPP4.
5. Грызун по п. 1, который представляет собой мышь, которая не способна экспрессировать мышиный белок Dpp4.
6. Грызун по п. 1, причем грызун является гетерозиготным по модифицированному гену DPP4.
7. Грызун по п. 1, причем грызун является гомозиготным по модифицированному гену DPP4.
8. Способ получения гуманизированного грызуна, включающий замену геномного фрагмента гена Dpp4 грызуна в эндогенном локусе Dpp4 грызуна на геномный фрагмент человеческого гена DPP4 с получением модифицированного гена DPP4, при этом модифицированный ген DPP4 кодирует человеческий белок DPP4, при этом экспрессия модифицированного гена DPP4 находится под контролем регуляторных элементов грызуна в эндогенном локусе Dpp4 грызуна, при этом грызун представляет собой мышь или крысу, и при этом у грызуна проявляется воспаление легких при инфицировании MERS-CoV.
9. Способ получения гуманизированного грызуна, включающий:
(i) замену геномного фрагмента гена Dpp4 грызуна в эндогенном локусе Dpp4 грызуна в эмбриональной стволовой клетке грызуна на геномный фрагмент человеческого гена DPP4 с получением модифицированного гена DPP4, при этом модифицированный ген DPP4 кодирует человеческий белок DPP4, при этом экспрессия модифицированного гена DPP4 находится под контролем регуляторных элементов грызуна в эндогенном локусе Dpp4 грызуна, и при этом грызун представляет собой мышь или крысу; и
(ii) получение грызуна из эмбриональной стволовой клетки из этапа (i), при этом у грызуна проявляется воспаление легких при инфицировании MERS-CoV.
10. Способ по п. 9, в котором геномный фрагмент человеческого гена DPP4 содержит экзоны 2-26 человеческого гена DPP4.
11. Способ по п. 10, в котором геномный фрагмент заменяемого гена Dpp4 грызуна содержит экзоны 2-26 гена Dpp4 грызуна.
12. Способ по п. 10, в котором грызун представляет собой мышь, и при этом модифицированный ген DPP4 содержит экзон 1 эндогенного мышиного Dpp4, функционально связанный с геномным фрагментом человеческого гена DPP4.
13. Способ по п. 9, в котором грызун представляет собой мышь, которая неспособна экспрессировать мышиный белок Dpp4.
14. Способ по п. 9, в котором грызун является гетерозиготным по модифицированному гену DPP4.
15. Способ по п. 9, в котором грызун является гомозиготным по модифицированному гену DPP4.
16. Способ определения терапевтической эффективности in vivo специфичного для человека антагониста DPP4 у грызуна, включающий следующие этапы:
(a) введение антагониста DPP4 грызуну по п. 1 и
(b) определение того, способен ли антагонист DPP4 лечить или предотвращать один или более симптомов инфекции MERS-CoV при инфицировании MERS-CoV по сравнению с контрольными грызунами, инфицированными MERS-CoV, но без введения антагониста DPP4.
17. Способ по п. 16, в котором антагонист DPP4 выбран из группы, состоящей из малых молекул, пептидов и антител.
18. Способ по п. 17, в котором антагонист DPP4 представляет собой антитело к белку MERS-CoV.
19. Способ по п. 18, в котором белок MERS-CoV представляет собой шипиковый белок MERS-CoV.
20. Способ по п. 16, дополнительно включающий инфицирование грызуна MERS-CoV, при этом антагонист вводят до, одновременно с или после инфицирования MERS-CoV.
21. Способ по п. 16, в котором определение включает измерение титра вируса или концентрации РНК.
22. Способ по п. 16, в котором один или более симптомов инфекции MERS-CoV включают воспаление легких или снижение массы тела.
| COLEMAN CHRISTOPHER M., et al., Wild-type and innate immune-deficient mice are not susceptible to the Middle East respiratory syndrome coronavirus, J Gen Virol., 2014 Feb, 95(Pt 2), pp.408-412 | |||
| SIMEONI LUCA, et al., Human CD26 expression in transgenic mice affects murine T-cell populations and modifies their subset distribution, Hum Immunol., 2002, 63(9), pp.719-730 | |||
| WO 2004104216 A2, 02.12.2004 | |||
| РЫБЧИН В.Н | |||
| Основы генетической инженерии, Учебник дляВУЗов, Санкт-Петербург, Издательство СПбГТУ, 2002, 522с., с.410-417. |

