Область техники
Настоящее изобретение относится к области биотехнологии и может быть использовано в сельском хозяйстве, в частности - картофелеводстве, и пищевой промышленности в качестве эффективного инструмента лабораторной диагностики бактериальных фитопатогенов, вызывающих болезни сельскохозяйственных культур. Более точно - изобретение позволяет селективно и с высокой чувствительностью определять наличие и проводить количественную оценку в биологических образцах фитопатогенной бактерии вида Dickeya solani в условиях одновременного присутствия в образце других бактерий, например, близкородственных бактерий, таких как штаммы пектолитических бактерий, принадлежащих к роду Pectobacterium - Pectobacterium atrosepticum и Pectobacterium carotovorum subsp. carotovorum. Способ согласно настоящему изобретению основан на методе петлевой изотермической амплификации (LAMP) участка последовательности целевого гена с использованием набора подобранных олигонуклеотидных праймеров.
Описание предшествующего уровня техники
Пектолитические бактерии представляют собой группу близкородственных микроорганизмов, вызывающих у картофеля такие заболевания, как «черная ножка», поражающая вегетирующие растения в полевых и тепличных условиях, и мокрая гниль клубней, которая распространяется при хранении и транспортировке картофеля (Czajkowski R. et al., Detection, identification and differentiation of Pectobacterium and Dickeya species causing potato blackleg and tuber soft rot: a review, Ann Appl Biol., 2015, 166: 18-38). Возбудители этих опасных болезней принадлежат к трем таксономическим рангам: i) третий биовар бактерии вида Dickeya dianthicola, рассматриваемый в настоящее время как новый вид - Dickeya solani (van der Wolf J.M. et al., Dickeya solani sp. nov., a pectinolytic plant-pathogenic bacterium isolated from potato (Solanum tuberosum), Int J Syst Evol Microbiol., 2014, 64: 768-774), поражающий картофель в регионах с влажным и теплым климатом; ii) Pectobacterium atrosepticum - возбудитель «черной ножки» картофеля в полях умеренных климатических поясов; и iii) Pectobacterium carotovorum subsp. carotovorum - подвид, вызывающий мокрую гниль клубней и представляющий особую опасность для картофеля в хранилищах и при транспортировке (Toth I.K. et al., Dickeya species: an emerging problem for potato production in Europe, Plant Pathol, 2011, 60: 385-399). В настоящее время эпифитотическая обстановка свидетельствует, что все три вида имеют общие пути распространения, часто одновременно присутствуют в инфицированном картофеле (Czajkowski R. et al., Detection, identification and differentiation of Pectobacterium and Dickeya species causing potato blackleg and tuber soft rot: a review, Ann Appl Biol., 2015, 166: 18-38) и обладают весьма схожими симптомами проявления инфекции (Toth I.К. et al., Dickeya species: an emerging problem for potato production in Europe, Plant Pathol, 2011, 60: 385-399). Обусловленная адаптацией к различным климатическим условиям агрессивность бактерий Dickeya solani, Pectobacterium atrosepticum и Pectobacterium carotovorum subsp. carotovorum, их широкое распространение и близкие экологические ниши - все эти факторы становятся причиной значительных экономических потерь в сфере картофелеводства и других отраслях растениеводства, в которых указанные выше фитопатогены находят растений-хозяев.
Для молекулярной диагностики бактерий, включающей определение таксономической принадлежности бактерий и их количественную оценку, используют известные специалисту в данной области техники способы увеличения количества копий (амплификации) фрагментов целевой ДНК, в частности - способ, основанный на методе петлевой изотермической амплификации (LAMP, loop-mediated isothermal amplification) участка последовательности целевого гена с использованием набора олигонуклеотидных праймеров. Исторически метод LAMP был разработан для преодоления недостатков полимеразной цепной реакции (ПЦР, Saiki R.K. et al., Primer-directed enzymatic amplification of DNA with a thermostable DNA polymerase, Science, 1988, 239: 487-491) при определении однонуклеотидных полиморфизмов в аллелях целевого гена. Метод LAMP и все его преимущества по сравнению с методом ПЦР подробно описаны в патенте Соединенных Штатов Америки US7494790 В2 (см. также патент-аналог Российской Федерации RU 2252964 С2), а также научных публикациях Notomi Т. et al., Loop-mediated isothermal amplification of DNA, Nucleic Acids Res., 2000, 28(12), е63 и Tomita N. et al., Loop-mediated isothermal amplification (LAMP) of gene sequences and simple visual detection of products, Nat Protoc, 2008, 3(5): 877-782.
Метод LAMP представляет собой один из способов амплификации определенных участков целевой ДНК при постоянной температуре с помощью ДНК-полимеразы и набора, содержащего четыре подобранных олигонуклеотидных праймера, которые комплементарны шести различным участкам на целевой (матричной) ДНК (Notomi Т. et al., Loop-mediated isothermal amplification of DNA, Nucleic Acids Res., 2000, 28(12), е63). Также, возможно использование набора, содержащего шесть подобранных олигонуклеотидных праймеров, которые комплементарны восьми различным участкам на целевой ДНК. Для диагностических целей в методе LAMP используются внутренние праймеры, которые на своих 5'-конце и 3'-конце содержат значительное количество нуклеотидов, комплементарных видоспецифичным позициям нуклеотидов в конкретном молекулярно-генетическом маркере исследуемого генома. В качестве молекулярно-генетического маркера может быть использована любая нуклеотидная последовательность, несущая видоспецифичные признаки в виде различных мутаций. Из механизма реакции амплификации целевой ДНК методом LAMP следует, что если концы внутренних праймеров не будут полностью гибридизоваться с комплементарной областью в выбранной маркерной последовательности, что может иметь место в случае присутствия в реакционной смеси неспецифичной гомологичной нуклеотидной последовательности, то эффективность амплификации будет значительно снижена или будет невозможна. На этом основан механизм избирательного действия набора праймеров по отношению к целевой нуклеотидной последовательности генома. Более того, учитывая большее количество (от четырех до шести) используемых праймеров по сравнению с классической ПЦР, где используется два праймера, метод LAMP обладает более высокой селективностью по отношению к целевой ДНК.
Современные методы молекулярной диагностики фитопатогенных бактерий основаны на различных подходах к выбору молекулярно-генетического маркера, характерного для бактерий, наличие, количество и распространение которых необходимо контролировать. В качестве мишеней для определения бактериальных фитопатогенов могут быть использованы уникальные геномные последовательности, нуклеотидные последовательности генов факторов вирулентности и нуклеотидные последовательности генов основных метаболических путей, обладающих высокой консервативностью и называемые также генами «домашнего хозяйства». Набор олигонуклеотидных праймеров подбирается на основании данных о нуклеотидной последовательности выбранной ДНК-мишени. Уникальные нуклеотидные последовательности и гены факторов вирулентности представляют собой довольно протяженные участки генома, в то время как видоспецифичные участки генов «домашнего хозяйства» образованы за счет различных типов мутаций. Высокая селективность наборов праймеров для определения уникальных нуклеотидных последовательностей обусловлена отсутствием в геномах других организмов сайтов для отжига таких праймеров.
С точки зрения разработки высокоэффективных способов диагностики с использованием метода LAMP наиболее удобны уникальные последовательности, особенно те их них, длина которых составляет более 500 пар оснований. Гены факторов вирулентности лишь зачастую являются уникальными мишенями, поскольку многие патогенные и условно патогенные бактерии обладают способностью к горизонтальному транспорту генов, благодаря которому выбранный для диагностики ген-мишень может оказаться у любой другой бактерии из этой же экологической ниши, давая ложноположительные результаты лабораторной диагностики. Существенным недостатком использования уникальных нуклеотидных последовательностей и генов факторов вирулентности для целей селективного определения целевой фитопатогенной бактерии является отсутствие консервативности - такие участки генома не всегда жизненно необходимы для бактериальной клетки и их нуклеотидная последовательность подвержена мутационной изменчивости под действием различных факторов. Среди таких факторов необходимо учитывать также степень приспособленности бактерии к своему растению-хозяину. Более того, уникальные последовательности могут быть элиминированы из генома патогенного микроорганизма без потери самой бактерией вирулентности (степени патогенности) - в таком случае разработанные наборы праймеров на целевой участок генома потеряют свою применимость в лабораторной диагностике. Отслеживание таких генетических событий представляет собой весьма трудную задачу. Недостатком существующих способов диагностики фитопатогенных бактерий, основанных на применении генов «домашнего хозяйства», является низкая селективность таких способов и, как следствие, наличие высокой вероятности получения ложноположительных результатов лабораторной диагностики, что обусловлено высокой консервативностью и, как следствие, высокой идентичностью нуклеотидных последовательностей генов «домашнего хозяйства» даже у неблизкородственных бактериальных видов.
Раскрыты способы диагностики бактериальных фитопатогенов, таких как, например Clavibacter michiganensis subsp. nebraskensis (патент Китайской народной республики CN 102382882 В) и Ralstonia solanacearum (заявка CN 105238876 А), основанные на применении метода LAMP. Известны другие способы определения фитопатогенных бактерий с помощью метода LAMP, основанные на применении наборов праймеров, комплементарных нуклеотидным последовательностям гена фактора вирулентности Pectobacterium atrosepticum (Li X. et al., Development and evaluation of a loop-mediated isothermal amplification assay for rapid detection and identification of Pectobacterium atrosepticum, Can J Plant Pathol, 2011, 33(4): 447-457), гена β (бета)-субъединицы ДНК-гиразы Pectobacterium atrosepticum (патент Китайской Народной Республики CN 104263841 В; Hu L.X. et al., Sensitive and rapid detection of Pectobacterium atrosepticum by targeting the gyrB gene using a real-time loop-mediated isothermal amplification assay, Lett Appl Microbiol, 2016, 63(4): 289-296) и различных генов биосинтеза бактериальной клеточной стенки Pectobacterium carotovorum (патент Республики Корея KR 101288419 B1; Yasuhara-Bell J et al., Specific detection of Pectobacterium carotovorum by loop-mediated isothermal amplification, Mol Plant Pathol, 2016, 17: 1499-1505).
Однако, отсутствуют ранее представленные данные, которые бы описывали способ селективного определения фитопатогенной бактерии вида Dickeya solani с помощью метода LAMP с использованием в качестве молекулярно-генетического маркера высококонсервативного гена «домашнего хозяйства» infB, кодирующего фактор инициации трансляции IF-2, и набора подобранных олигонуклеотидных праймеров.
Раскрытие сущности изобретения
Задачей настоящего изобретения является разработка селективного и высокочувствительного способа определения в биологических образцах (например, картофеле, почве, воде) фитопатогенной бактерии вида Dickeya solani, основанного на методе петлевой изотермической амплификации (LAMP) участка последовательности целевого гена с использованием набора подобранных олигонуклеотидных праймеров, в присутствии бактерий других видов. Более точно - задачей настоящего изобретения является разработка способа определения бактерии вида Dickeya solani, основанного на методе LAMP участка последовательности целевого гена с использованием набора подобранных олигонуклеотидных праймеров, в присутствии близкородственных пектолитических бактерий рода Pectobacterium, таких как Pectobacterium atrosepticum и Pectobacterium carotovorum subsp. carotovorum.
Таким образом, одним из аспектов настоящего изобретения является предоставление способа определения бактерии вида Dickeya solani методом петлевой изотермической амплификации, отличающегося тем, что в реакции амплификации участка последовательности целевого гена используют набор олигонуклеотидных праймеров, включающий:
(A) праймер FIP, имеющий нуклеотидную последовательность SEQ ID NO: 7, или его гомолог,
(B) праймер BIP, имеющий нуклеотидную последовательность SEQ ID NO: 8, или его гомолог,
(C) праймер F3, имеющий нуклеотидную последовательность SEQ ID NO: 9, или его гомолог,
(D) праймер В3, имеющий нуклеотидную последовательность SEQ ID NO: 10, или его гомолог.
Также аспект настоящего изобретения - предоставление вышеописанного способа, отличающегося тем, что целевой ген есть ген infB, кодирующий фактор инициации трансляции IF-2.
Также аспект настоящего изобретения - предоставление вышеописанного способа, отличающегося тем, что ген infB кодируется нуклеотидной последовательностью SEQ ID NO: 1 или гомологичной ей нуклеотидной последовательностью при условии, что такая гомологичная нуклеотидная последовательность может гибридизоваться с указанными выше праймерами.
Также аспект настоящего изобретения - предоставление вышеописанного способа, отличающегося тем, что гомологи указанных праймеров FIP, BIP, F3 и В3 имеют гомологию не менее чем 95% по сравнению с соответствующими нуклеотидными последовательностями SEQ ID NOs: 7, 8, 9 и 10 при условии, что такие гомологичные праймеры могут гибридизоваться с целевым геном infB.
Также аспект настоящего изобретения - предоставление вышеописанного способа, отличающегося тем, что указанным способом определяют бактерию вида Dickeya solani в образцах картофеля, почвы или воды, или их комбинации.
Следующей задачей настоящего изобретения является предоставление набора олигонуклеотидных праймеров, при использовании которых возможно селективное и высокочувствительное определение фитопатогенной бактерии вида Dickeya solani методом LAMP в биологических образцах (например, клубнях картофеля, почве, воде), которые могут одновременно содержать бактерии других таксономических рангов, например, близкородственные пектолитические бактерии рода Pectobacterium, такие как Pectobacterium atrosepticum и Pectobacterium carotovorum subsp. carotovorum.
Таким образом, другим аспектом настоящего изобретения является предоставление набора праймеров для определения бактерии вида Dickeya solani методом петлевой изотермической амплификации, содержащего:
(E) праймер FIP, имеющий нуклеотидную последовательность SEQ ID NO: 7, или его гомолог,
(F) праймер BIP, имеющий нуклеотидную последовательность SEQ ID NO: 8, или его гомолог,
(G) праймер F3, имеющий нуклеотидную последовательность SEQ ID NO: 9, или его гомолог,
(H) праймер В3, имеющий нуклеотидную последовательность SEQ ID NO: 10, или его гомолог.
Также аспект настоящего изобретения - предоставление вышеописанного набора праймеров, отличающегося тем, что гомологи указанных праймеров FIP, BIP, F3 и В3 имеют гомологию не менее чем 95% по сравнению с соответствующими нуклеотидными последовательностями SEQ ID NOs: 7, 8, 9 и 10 при условии, что такие гомологичные праймеры могут гибридизоваться с целевым геном infB.
Также аспект настоящего изобретения - предоставление вышеописанного набора праймеров, отличающегося тем, что указанный набор дополнительно содержит целевой ген infB, кодирующий фактор инициации трансляции IF-2.
Также аспект настоящего изобретения - предоставление вышеописанного набора праймеров, отличающегося тем, что ген infB кодируется нуклеотидной последовательностью SEQ ID NO: 1 или гомологичной ей нуклеотидной последовательностью при условии, что такая гомологичная нуклеотидная последовательность может гибридизоваться с указанными выше праймерами.
Также аспект настоящего изобретения - предоставление вышеописанного набора праймеров, отличающегося тем, что указанный набор праймеров есть диагностический набор реагентов для анализа биологических образцов картофеля, почвы или воды, или их комбинации.
Технический результат, достигаемый при использовании изобретений, заключается в селективном и высокочувствительном определении фитопатогенной бактерии вида Dickeya solani в биологических образцах (например, картофеле, почве, воде), которые могут одновременно содержать бактерии других таксономических рангов, например, близкородственные пектолитические бактерии рода Pectobacterium, такие как Pectobacterium atrosepticum и Pectobacterium carotovorum subsp.carotovorum. Настоящие изобретения позволяют определить наличие и количество фитопатогенной бактерии вида Dickeya solani в биологических образцах и контролировать ее распространение. При использовании настоящих изобретений возможно повысить контроль за распространением заболеваний растений в открытом и закрытом грунте, а также снизить потери при хранении и транспортировке собранного урожая или товарной продукции. Также, настоящие изобретения позволяют разработать принципиально новые и высокоэффективные методические подходы, направленные на совершенствование системы контроля качества и сертификации семенного материала в некоторых отраслях растениеводства, например, картофелеводстве.
Указанные задачи и технические результаты были достигнуты благодаря обнаружению того факта, что применение в качестве молекулярно-генетического маркера высококонсервативного гена «домашнего хозяйства» infB, кодирующего фактор инициации трансляции IF-2, и набора из четырех подобранных олигонуклеотидных праймеров FIP, BIP, F3 и В3 в методе LAMP возможно селективное определение фитопатогенной бактерии вида Dickeya solani с высокой чувствительностью в условиях одновременного присутствия в биологическом образце близкородственных пектолитических бактерий рода Pectobacterium, таких как Pectobacterium atrosepticum и Pectobacterium carotovorum subsp. carotovorum.
Краткое описание Фигур
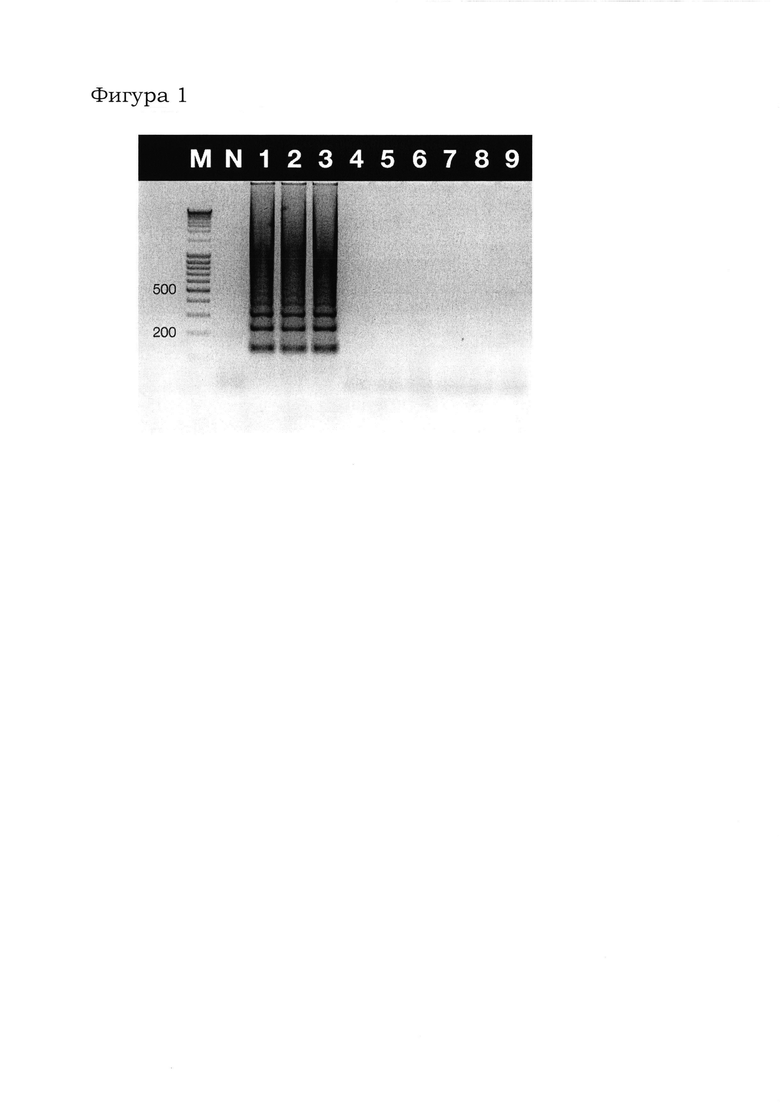
Фигура 1 показывает представленные на электрофореграмме результаты петлевой изотермической амплификации по определению селективности диагностической реакции амплификации участка нуклеотидной последовательности гена infB. Обозначения: М - маркер длин ДНК (подписаны фрагменты длиной 200 и 500 пар оснований); N - отрицательный контроль; 1-3 - результат амплификации с геномной ДНК бактерий Dickeya solani штаммов DSO1001, DSO1002 и DSO1003, соответственно; 4-6 - результат амплификации с геномной ДНК бактерий Pectobacterium atrosepticum штаммов SCRI1043, 21А и PAT 1001, соответственно; 7-9 - результат амплификации с геномной ДНК бактерий Pectobacterium carotovorum subsp.carotovorum штаммов PCC1001, PCC1002 и NCPPB312, соответственно.
Фигура 2 показывает представленные на электрофореграмме результаты петлевой изотермической амплификации по определению чувствительности диагностической реакции амплификации участка нуклеотидной последовательности гена infB. Обозначения: М - маркер длин ДНК (подписаны фрагменты длиной 200 и 500 пар оснований); N - отрицательный контроль; 1-9 - серия последовательных десятикратных разведений плазмиды на основе вектора pAL2-T со вставкой участка гена infB (штамм Dickeya solani DSO1001), начиная с концентрации 1×10-9 г/мкл (лунка №1) до 1×10-17 г/мкл (лунка №9).
Фигура 3 показывает нуклеотидную последовательность на кодирующей цепи гена infB (SEQ ID NO: 1) из генома штамма Dickeya solani IPO2222. Черными прямоугольниками на кодирующей цепи гена выделены участки отжига праймеров и комплементарные им последовательности. Буква «-с» в названии сайтов означает комплементарный участок (например, сайту отжига F1 комплементарна последовательность F1c в составе олигонуклеотидного праймера FIP). Нуклеотидные последовательности сайтов отжига разработанного набора праймеров в гене infB штаммов Dickeya solani IPO2222 и DSO1001 по данным секвенирования идентичны на 100%.
Описание способов осуществления изобретения
Настоящее изобретение более подробно будет описано ниже с приведением не ограничивающих настоящее изобретение примеров.
Ген infB кодирует белковый фактор инициации трансляции IF-2 (идентификаторы в базе данных GenBank; полный геном типового штамма Dickeya solani IPO2222: NZ_CP015137; локализация гена infB в геноме: 2,845,505-2,848,222; аминокислотная последовательность белка IF-2: WP_022632128). Нуклеотидная последовательность гена infB и аминокислотная последовательность белка IF-2, кодируемого геном infB, из штамма Dickeya solani IPO2222 приведены в SEQ ID NO: 1 и SEQ ID NO: 2, соответственно. Другие штаммы бактерии вида Dickeya solani также содержат ген infB и кодируемый им белок IF-2, например, штамм Dickeya solani MK10 (идентификатор в базе данных GenBank; полный геном штамма: NZ_CM001839).
Известны гомологи гена infB и белка IF-2 из штамма Dickeya solani IPO2222, присутствующие в бактерии вида Pectobacterium atrosepticum (идентификаторы в базе данных GenBank; полный геном типового штамма Pectobacterium atrosepticum SCRI1043: NC_004547; локализация гена в геноме: 779,306-782,008; аминокислотная последовательность белка IF-2: WP_011092323). Нуклеотидная последовательность гена infB и аминокислотная последовательность белка IF-2, кодируемого геном infB, из штамма Pectobacterium atrosepticum SCRI1043 приведены в SEQ ID NO: 3 и SEQ ID NO: 4, соответственно. Нуклеотидные последовательности гена infB из бактерии Dickeya solani (SEQ ID NO: 1) и бактерии Pectobacterium atrosepticum (SEQ ID NO: 3) имеют идентичность 82%. Другие штаммы бактерии вида Pectobacterium atrosepticum также содержат ген infB и кодируемый им белок IF-2, например, штамм Pectobacterium atrosepticum 21А (идентификатор в базе данных GenBank; полный геном штамма: NZ_CP009125).
Известны гомологи гена infB и белка IF-2 из штамма Dickeya solani IPO2222, присутствующие в бактерии подвида Pectobacterium carotovorum subsp.carotovorum (идентификаторы в базе данных GenBank; полный геном типового штамма Pectobacterium carotovorum subsp.carotovorum PCI: NC_012917; локализация гена в геноме: 681,463-684,162; аминокислотная последовательность белка IF-2: WP_012773288). Нуклеотидная последовательность гена infB и аминокислотная последовательность белка IF-2, кодируемого геном infB, из штамма Pectobacterium carotovorum subsp.carotovorum PCI приведены в SEQ ID NO: 5 и SEQ ID NO: 6, соответственно. Нуклеотидные последовательности гена infB из бактерии Dickeya solani (SEQ ID NO: 1) и бактерии Pectobacterium carotovorum subsp.carotovorum (SEQ ID NO: 5) имеют идентичность 82%. Другие штаммы бактерии подвида Pectobacterium carotovorum subsp. carotovorum также содержат ген infB и кодируемый им белок IF-2, например, штамм Pectobacterium carotovorum subsp. carotovorum PCC21 (идентификатор в базе данных GenBank; полный геном штамма: NC_018525).
Для специалиста в данной области известно, что могут быть различные штаммы бактерий, принадлежащих к видам Dickeya solani, Pectobacterium atrosepticum и Pectobacterium carotovorum subsp. carotovorum, помимо тех штаммов, что приведены в описании. Такие виды бактерий и их штаммы могут быть использованы согласно настоящему изобретению. Штаммы бактерии вида Dickeya solani, такие как, например, типовой штамм IPO2222, могут быть получены из немецкой коллекции микроорганизмов и клеточных культур, Брауншвейг, Германия (Leibniz-Institut DSMZ-Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH;  7B, 38124 Braunschweig, Germany; веб-сайт: www.dsmz.de; идентификатор: DSM 28711). Штаммы бактерии вида Pectobacterium atrosepticum, такие как, например, типовой штамм SCRI1043 (идентификатор: ВАА-672) и штаммы бактерии подвида Pectobacterium carotovorum subsp.carotovorum, такие как, например, штамм NCPPB312 (идентификатор: 15713) могут быть получены из Американской коллекции типовых культур, Манассас, штат Виргиния, Соединенные Штаты Америки (АТСС, American Туре Culture Collection, Manassas, Virginia, U.S.A.; веб-сайт: www.lgcstandards-atcc.org). Штаммы бактерии вида Pectobacterium atrosepticum, такие как, например, штамм 21 А, могут быть получены из коллекции культур микроорганизмов лаборатории молекулярной биологии Казанского института биохимии и биофизики Казанского научного центра Российской академии наук (Казань, Республика Татарстан, Российская Федерация; веб-сайт: www.kibb.knc.ru/index.php/ru/researchunits/166-labmolbiol). Штаммы бактерии вида Dickeya solani, такие как, например, штаммы DSO1001, DSO1002, DSO1003, штаммы бактерии вида Pectobacterium atrosepticum, такие как, например, штамм PAT 1001, штаммы бактерии подвида Pectobacterium carotovorum subsp. carotovorum, такие как, например, штаммы РСС1001, РСС1002, могут быть получены из коллекции культур микроорганизмов лаборатории молекулярной биоинженерии Института биоорганической химии имени академиков М.М. Шемякина и Ю.А. Овчинникова Российской академии наук (Москва, Российская Федерация; веб-сайт: www.ibch.ru/en/structure/groups/molbioeng).
7B, 38124 Braunschweig, Germany; веб-сайт: www.dsmz.de; идентификатор: DSM 28711). Штаммы бактерии вида Pectobacterium atrosepticum, такие как, например, типовой штамм SCRI1043 (идентификатор: ВАА-672) и штаммы бактерии подвида Pectobacterium carotovorum subsp.carotovorum, такие как, например, штамм NCPPB312 (идентификатор: 15713) могут быть получены из Американской коллекции типовых культур, Манассас, штат Виргиния, Соединенные Штаты Америки (АТСС, American Туре Culture Collection, Manassas, Virginia, U.S.A.; веб-сайт: www.lgcstandards-atcc.org). Штаммы бактерии вида Pectobacterium atrosepticum, такие как, например, штамм 21 А, могут быть получены из коллекции культур микроорганизмов лаборатории молекулярной биологии Казанского института биохимии и биофизики Казанского научного центра Российской академии наук (Казань, Республика Татарстан, Российская Федерация; веб-сайт: www.kibb.knc.ru/index.php/ru/researchunits/166-labmolbiol). Штаммы бактерии вида Dickeya solani, такие как, например, штаммы DSO1001, DSO1002, DSO1003, штаммы бактерии вида Pectobacterium atrosepticum, такие как, например, штамм PAT 1001, штаммы бактерии подвида Pectobacterium carotovorum subsp. carotovorum, такие как, например, штаммы РСС1001, РСС1002, могут быть получены из коллекции культур микроорганизмов лаборатории молекулярной биоинженерии Института биоорганической химии имени академиков М.М. Шемякина и Ю.А. Овчинникова Российской академии наук (Москва, Российская Федерация; веб-сайт: www.ibch.ru/en/structure/groups/molbioeng).
Ген infB и кодируемый им белок IF-2 не ограничены, соответственно, нуклеотидными последовательностями (SEQ ID NOs: 1, 3 и 5) и аминокислотными последовательностями (SEQ ID NOs: 2, 4 и 6), но могут быть использованы также их гомологи ввиду того, что возможно существование некоторых отличий в последовательностях ДНК среди штаммов бактерий, принадлежащих к семейству Enterobacteriaceae, более точно - к родам Dickeya и Pectobacterium, еще более точно - к видам Dickeya solani и Pectobacterium atrosepticum, а также подвиду Pectobacterium carotovorum subsp.carotovorum. Также возможно существование гомологов нуклеотидных последовательностей гена infB ввиду того, что идентичные аминокислотные последовательности могут кодироваться различными нуклеотидными последовательностями ввиду вырожденности генетического кода в соответствии с таблицей стандартного генетического кода (см., например, Griffiths A.J.F. et al., An Introduction to Genetic Analysis. New York: W.H. Freeman; 2000). Также возможно существование гомологов нуклеотидных последовательностей гена infB ввиду возможности направленного изменения нуклеотидной последовательности нативного гена infB (SEQ ID NOs: 1, 3 и 5) с использованием методов генетической инженерии, например, сайт-специфического мутагенеза (см., например, Kunkel Т. A., Rapid and efficient site-specific mutagenesis without phenotypic selection, Proc Natl Acad Sci USA, 1985, 82(2): 488-492).
Такие «гомологи нуклеотидной последовательности гена infB» и «гомологи аминокислотной последовательности IF-2», кодируемой геном infB, могут иметь одно или несколько изменений в нуклеотидной и/или аминокислотной последовательности, соответственно, по сравнению с исходной нуклеотидной последовательностью SEQ ID NOs: 1, 3 и 5, или нативной аминокислотной последовательностью SEQ ID NOs: 2, 4 и 6, причем такими изменениями могут быть замена (также называется как замещение), инсерция (также называется как вставка) или делеция (также называется как удаление) одного или нескольких нуклеотидов и/или аминокислотных остатков при условии, что такая гомологичная нуклеотидная последовательность может быть использована в способе определения бактерии вида Dickeya solani согласно настоящему изобретению также, как и исходный ген infB, или такая гомологичная аминокислотная последовательность имеет такие же функцию и активность, как и нативный белок IF-2, или же трехмерная структура гомологичного белка IF-2 изменена незначительно по сравнению с нативным белком IF-2.
Термин «гомолог нуклеотидной последовательности гена infB» также может означать, но не ограничиваться данными примерами, нуклеотидную последовательность, которая гибридизуется в условиях проведения реакции амплификации с помощью метода LAMP последовательности с олигонуклеотидными праймерами согласно изобретению и может быть использована в способе определения бактерии вида Dickeya solani согласно изобретению. Более точно, термин «гомолог нуклеотидной последовательности гена infB» может означать нуклеотидную последовательность, которая гибридизуется в условиях проведения LAMP с олигонуклеотидными праймерами SEQ ID NOs: 7, 8, 9, 10 и может быть использована в способе определения бактерии вида Dickeya solani согласно изобретению. Гомолог нуклеотидной последовательности гена infB может иметь гомологию, определенную через параметр «идентичность» при использовании компьютерного алгоритма BLAST (Basic Local Alignment Search Tool; веб-сайт: www.ncbi.nlm.nih.gov/BLAST), не менее чем 80%, не менее чем 85%, не менее чем 90%, не менее чем 95%, не менее чем 96%, не менее чем 97%, не менее чем 98% или не менее чем 99% по сравнению с исходной нуклеотидной последовательностью SEQ ID NO: 1, 3 или 5.
Набор олигонуклеотидных праймеров согласно настоящему изобретению содержит, по меньшей мере, четыре праймера, которые способны гибридизоваться в условиях проведения LAMP с нуклеотидной последовательностью гена infB или его гомолога, также обозначаемых как молекулярно-генетических маркер, ген-мишень, ДНК-мишень, целевой ген и тому подобное, которые могут быть использованы взаимозаменяемо, в способе определения бактерии вида Dickeya solani согласно изобретению. Более точно - в частном варианте осуществления настоящего изобретения набор олигонуклеотидных праймеров содержит четыре праймера, условно обозначаемые как FIP (SEQ ID NO: 7), BIP (SEQ ID NO: 8), F3 (SEQ ID NO: 9) и В3 (SEQ ID NO: 10), которые способны гибридизоваться в условиях проведения LAMP с нуклеотидной последовательностью гена infB или его гомолога и могут быть использованы в способе определения бактерии вида Dickeya solani согласно изобретению.
Набор праймеров FIP, BIP, F3 и В3 не ограничен нуклеотидными последовательностями SEQ ID NOs: 7, 8, 9 и 10, но могут быть использованы также их гомологи ввиду того, что возможно существование не только исходного гена infB из бактерии вида Dickeya solani, имеющего нуклеотидную последовательность SEQ ID NO: 1, но и гомологов указанного гена infB, как разъяснено выше. Следовательно, набор праймеров также может включать, но не ограничиваться данными примерами, гомологичные праймеры, имеющие нуклеотидную последовательность, которая гибридизуется в условиях проведения LAMP с геном-мишенью infB или его гомологом в способе определения бактерии вида Dickeya solani согласно изобретению. Более точно - набор праймеров может включать праймеры, имеющие нуклеотидную последовательность, которая гибридизуется в условиях проведения LAMP с нуклеотидной последовательностью SEQ ID NO: 1 или гомологичной ей нуклеотидной последовательностью в способе определения бактерии вида Dickeya solani согласно изобретению. Гомологи праймеров FIP, BIP, F3 и В3 могут иметь гомологию, определенную через параметр «идентичность» при использовании компьютерного алгоритма BLAST, не менее чем 95%, не менее чем 96%, не менее чем 97%, не менее чем 98% или не менее чем 99% по сравнению с исходными нуклеотидными последовательностями SEQ ID NOs: 7, 8, 9 и 10 при условии, что такие гомологичные праймеры способны гибридизоваться в условиях проведения LAMP с геном-мишенью infB или его гомологом согласно изобретению и могут быть использованы в способе определения бактерии вида Dickeya solani согласно изобретению.
Термин «способность гибридизоваться», употребляемый здесь в отношении любой нуклеотидной последовательности, например, целевого гена или праймера, означает, что в условиях проведения LAMP согласно изобретению возможно связывание нуклеотидной последовательности одного или нескольких олигонуклеотидных праймеров (или целевого гена) с нуклеотидной последовательностью целевого гена (или одного, либо нескольких олигонуклеотидных праймеров) с образованием комплекса(-ов), такого(-их) как дуплекс(-ы) ДНК таким образом, что возможно осуществление реакции амплификации целевого гена с помощью метода LAMP.
Термин «условия проведения LAMP согласно изобретению» означает такие условия, при которых возможна гибридизация олигонуклеотидных праймеров FIP, BIP, F3 и В3 с геном-мишенью infB и осуществление реакции амплификации последовательности гена infB с помощью метода LAMP с использованием указанных праймеров. В качестве условий проведения LAMP согласно изобретению могут быть выбраны такие условия, при которых гибридизацию осуществляют на стадии предварительного прогрева реакционной смеси без добавления ДНК-полимеразы при 95°C в течение 10 минут, затем при 60°C в течение 5 минут и последующим охлаждением до 12°C; реакцию амплификации проводят в изотермических условиях в интервале от 60 до 65°C, например, при 65°C, в течение 40 минут; реакцию останавливают при 95°C в течение 2 минут. В качестве ДНК-полимеразы может быть использована Bst-полимераза или ДНК-полимераза, обладающая подобной активностью. Чувствительность способа определения согласно изобретению изменяют, варьируя продолжительность реакции амплификации.
Определение бактерии вида Dickeya solani осуществляют, выявляя образование продукта реакции амплификации участка гена-мишени или его отсутствие. В случае наличия бактерии вида Dickeya solani в анализируемой пробе происходит накопление продукта реакции амплификации. Методы приготовления растворов, праймеров, выделения ДНК, иные условия проведения реакции амплификации, визуализации продукта реакции амплификации и подобные им известны для специалиста в данной области техники, и такие методы описаны, например, в научных публикациях (см., например, Tomita N. et al., Loop-mediated isothermal amplification (LAMP) of gene sequences and simple visual detection of products, Nat Protoc, 2008, 3(5): 877-882; Goto M. et al., Colorimetric detection of loop-mediated isothermal amplification reaction by using hydroxy naphthol blue, Biotechniques, 2009, 46(3): 167-172; Zhang X. et al., Brief review of monitoring methods for loop-mediated isothermal amplification (LAMP), Biosens Bioelectron., 2014, 61: 491-499) или нижеследующих Примерах.
Набор олигонуклеотидных праймеров согласно изобретению содержит указанные выше праймеры FIP, BIP, F3 и В3. С целью повышения селективности анализа набор праймеров может дополнительно содержать другие олигонуклеотидные праймеры, например, другие два праймера, отличные от FIP, BIP, F3 и В3, которые могут быть использованы в способе определения бактерии вида Dickeya solani. Дополнительно набор праймеров может содержать ДНК, имеющую нуклеотидную последовательность гена infB, причем указанная ДНК может быть, например, в виде вставки полной или частичной нуклеотидной последовательности гена infB в плазмидный вектор, в виде очищенной геномной ДНК или в виде одной или нескольких клеток бактерий вида Dickeya solani, содержащей нуклеотидную последовательность гена infB. Таким образом, возможно, что набор праймеров согласно изобретению содержит, не ограничиваясь этим, клетки бактерии вида Dickeya solani или ее геномную ДНК, содержащую нуклеотидную последовательность гена infB, и олигонуклеотидные праймеры FIP, BIP, F3 и В3. В частном и не ограничивающем настоящее изобретение примере набор олигонуклеотидных праймеров согласно изобретению есть диагностический набор реагентов для анализа биологических образцов (например, картофеля, почвы или воды, или их комбинации). Такой диагностический набор реагентов содержит указанные выше праймеры FIP, BIP, F3 и В3, а также в качестве средства положительного контроля реакции амплификации ДНК, имеющую нуклеотидную последовательность гена infB, причем указанная ДНК может быть, например, в виде изолированной полной или частичной нуклеотидной последовательности гена infB, в виде вставки полной или частичной нуклеотидной последовательности гена infB в плазмидный вектор, в виде очищенной геномной ДНК или в виде клеток бактерий вида Dickeya solani. Таким образом, возможно, что диагностический набор содержит, не ограничиваясь этим, клетки бактерии вида Dickeya solani или. ее геномную ДНК, содержащую нуклеотидную последовательность гена infB и олигонуклеотидные праймеры FIP, BIP, F3 и В3.
Также, набор праймеров, включая диагностический набор реагентов, может дополнительно содержать компоненты, необходимые для осуществления способа определения бактерии вида Dickeya solani согласно настоящему изобретению, более точно - для проведения реакции амплификации с помощью метода LAMP участка последовательности целевого гена infB с использованием указанных выше праймеров FIP, BIP, F3 и В3 и гена-мишени infB. В качестве дополнительных компонентов могут быть использованы различные органические и неорганические вещества, необходимые для поддержания требуемого значения кислотности реакционной смеси и ее солевого состава, ферменты (например, ДНК-полимеразу, пирофосфатазу и подобные им ферменты), дезоксирибонуклеозидтрифосфаты (dNTPs), минеральное масло, красители, интеркаляторы и другие компоненты, применяемые в молекулярной диагностике с помощью методов реакции амплификации с целью повышения ее эффективности.
Примеры
Реагенты, расходные материалы и оборудование
Компоненты реакционной смеси: буфер Трис-HCl (Tris Buffer, pH 8,8; Sigma, каталожный номер Т9443), хлорид калия (KCl; Wako Pure Chemicals, каталожный номер 163-03545), гептагидрат сульфата магния (MgSO4×7H2O; Wako Pure Chemicals, каталожный номер 137-00402), сульфат аммония ((NH4)2SO4; Kanto Chemical, каталожный номер 01322-00), Твин-20 (Tween 20; Tokyo Chemical Industry, каталожный номер Т0543), моногидрат бетаина (Betaine, 5 М; Sigma, каталожный номер В0300), дезоксирибонуклеозидтрифосфаты (dNTP Set; Thermo Scientific, каталожный номер R0181), олигонуклеотидные праймеры FIP (SEQ ID NO: 7), BIP (SEQ ID NO: 8), F3 (SEQ ID NO: 9) и В3 (SEQ ID NO: 10), синтез которых был заказан в компании «Евроген», и Bst ДНК-полимераза (Bst DNA Polymerase, 8000 U ml-1 (8000 Е/мл); New England Biolabs, каталожный номер M0275L). Для определения продукта реакции амплификации использовали агарозу (Top Vision Agarose; Thermo Scientific, каталожный номер R0491), раствор бромистого этидия (Ethidium Bromide, 10 mg ml-1 (10 мг/мл); Nippon Gene, каталожный номер 315-9005), маркер длин ДНК (MassRuler DNA Ladder; Thermo Scientific, каталожный номер SM0403). Все реагенты для приготовления компонентов реакционной смеси хранили при -20°C.
Расходные материалы: пробирки объемом 0,5 или 1,5 мл для приготовления компонентов реакционной смеси (Eppendorf), тонкостенные пробирки объемом 0,2 мл для проведения реакции амплификации, стерильные наконечники с фильтром для автоматических пипеток (SSI).
Оборудование: набор автоматических пипеток переменного объема (серия Research Plus; Eppendorf), твердотельный термостат (модель TDB-120; BioSan) или ДНК-амплификатор (модель Т100; BioRad), вортекс (модель MixMate; Eppendorf), микроцентрифуга (модель MiniSpin; Eppendorf), источник питания (модель PowerPac Basic; Bio-Rad), камера для горизонтального электрофореза в агарозном геле (модель Mini-Sub Cell GT cell; Bio-Rad), трансиллюминатор (модель ECX-F20; Vilber Lourmat) или гель-документирующая система (модель ChemiDoc MP; Bio-Rad).
Состав реакционной смеси
Диагностическую реакцию по определению требуемого молекулярно-генетического маркера в заданной бактерии проводили, используя реакционную смесь общим объемом 25 мкл, которая содержала нижеследующие компоненты в конечной концентрации:
Реакционную смесь из перечисленных выше компонентов готовили в день эксперимента.
Реакция амплификации
В тонкостенные пробирки вносили по 24 мкл реакционной смеси, содержащей Bsf-полимеразу. В пробирку с отрицательным контролем вносили 1 мкл стерильной денонсированной воды. В пробирку с положительным контролем добавляли 1 мкл водного раствора очищенной геномной ДНК бактерии Dickeya solani или плазмиды со вставкой сайта отжига диагностических праймеров FIP, BIP, F3, В3 в концентрации 1 нг/мкл. Плазмиду со вставкой участка гена infB изготавливали с помощью набора для быстрого клонирования ПЦР-продуктов в вектор pAL2-T («Евроген»; каталожный номер TAK02) и компетентных клеток штамма Escherichia coli XL 1-Blue для химической трансформации («Евроген»; каталожный номер СС001) с соблюдением идущих в комплекте протоколов. Далее в пробирки вносили по 1 мкл гомогената клубней картофеля, поливной воды или суспензии почвы с полей для выращивания картофеля. Содержимое пробирок перемешивали на вортексе. Реакционные смеси инкубировали при 65°C в течение 40 минут в ячейках твердотельного термостата или ДНК-амплификатора. Реакцию амплификации останавливали, нагревая пробирки до 95°C в течение 2 минут.
Предварительный прогрев реакционный смесей позволял добиться большей чувствительности, если необходимо было детектировать количество пектолитических бактерий, значительно меньшее уровня латентной инфекции. Для проведения такой модификации лабораторной диагностики реакционную смесь готовили без добавления Bst-полимеразы и разливали по пробиркам в объеме 23 мкл. В пробирку с отрицательным контролем вносили 1 мкл стерильной деионизированной воды. В пробирку с положительным контролем добавляли 1 мкл водного раствора очищенной геномной ДНК бактерии Dickeya solani или плазмиды со вставкой сайта отжига диагностических праймеров FIP, BIP, F3, В3 в концентрации 1 нг/мкл. Далее в пробирки вносили по 1 мкл гомогената клубней картофеля, поливной воды или суспензии почвы с полей для выращивания картофеля. Содержимое пробирок перемешивали на вортексе и прогревали при 95°C в течение 10 минут, затем при 60°C в течение 5 минут и охлаждали до 12°C. Испарившуюся жидкость сбрасывали со стенок пробирок, используя микроцентрифугу. В каждую пробирку вносили по 1 мкл Bst-полимеразы и осторожно перемешивали содержимое пробирок. Реакционные смеси инкубировали при 65°C в течение 40 минут в ячейках твердотельного термостата или ДНК-амплификатора. Реакцию амплификации останавливали, нагревая пробирки до 95°C в течение 2 минут.
Визуализация продукта реакции амплификации по пирофосфату магния
Визуальную детекцию положительной реакции амплификации методом LAMP проводили, наблюдая образование взвеси осадка пирофосфата магния белого цвета. В пробирках с отрицательным контролем и отрицательными реакциями содержимое пробирок оставалось прозрачным. Согласно литературным данным, наличие взвеси осадка пирофосфата магния белого цвета находится в соответствии с данными турбидиметрии и означает положительную реакцию (Mori Y. et al., Detection of loop-mediated isothermal amplification reaction by turbidity derived from magnesium pyrophosphate formation, Biochem Biophys Res Commun., 2001, 289(1): 150-154). Данный способ является самым простым из всех существующих на сегодня в приложении к методу LAMP и позволяет провести качественный анализ реакции амплификации.
Визуализация продукта реакции амплификации с помощью электрофореза в агарозном геле
Альтернативным и более чувствительным методом анализа продуктов реакции амплификации является горизонтальный электрофорез в агарозном геле (Zhang X. et al., Brief review of monitoring methods for loop-mediated isothermal amplification (LAMP), Biosens Bioelectron., 2014, 61: 491-499). Содержимое пробирок с положительными реакциями имеет на агарозном геле характерное распределение продуктов амплификации в виде регулярной лесенки. Учитывая образование в ходе реакции амплификации продуктов различной молекулярной массы, для проведения электрофореза в агарозном геле использовали 2% (по массе) агарозу. Электрофорез проводили в течение 30 минут в присутствии 1-кратного Трис-ацетатного буфера (см. методику приготовления, например, в Green М.R. and Sambrook J., Molecular Cloning - A Laboratory Manual, Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press; 2012) при комнатной температуре и напряженности электрического поля 100 В/см. Визуализацию продуктов амплификации проводили на трансиллюминаторе или с помощью гель-документирующей системы за счет разгорания флюоресценции молекул бромистого этидия при интеркаляции в ДНК.
Пример 1. Селективность способа
Селективность способа определения бактерии вида Dickeya solani по отношению к штаммам близкородственных фитопатогенных бактерий оценивали, проводя реакцию амплификации в разных пробирках с добавлением очищенной геномной ДНК штаммов Dickeya solani, Pectobacterium atrosepticum и Pectobacterium carotovorum subsp.carotovorum.
Результаты электрофореза показывают (Фигура 1), что предложенный способ, в котором используются набор олигонуклеотидных праймеров FIP (SEQ ID NO: 7), BIP (SEQ ID NO: 8), F3 (SEQ ID NO: 9) и В3 (SEQ ID NO: 10) и ген-мишень infB, позволяет определить наличие в биологических образцах только требуемой бактерии вида Dickeya solani, при этом близкородственные бактерии рода Pectobacterium указанным способом не определяются. Для специалиста в данной области очевидно, что иные бактерии, не являющиеся близкородственными к бактерии вида Dickeya solani, например, принадлежащие к другим бактериям семейства Enterobacteriaceae или иным семействам, предложенным способом также не будут определяться.
Пример 2. Чувствительность способа
Чувствительность способа определения бактерии вида Dickeya solani оценивали, используя набор праймеров FIP, BIP, F3, В3 и плазмиду pAL2-T, содержащую участок нуклеотидной последовательности гена infB из Dickeya solani, в котором локализованы комплементарные праймерам FIP, BIP, F3 и В3 сайты отжига. Для праймера FIP сайтами отжига являются F1 и F2c (Фигура 3), для праймера BIP-В1 и В2с, для праймера F3-F3c, а для праймера В3-В3с. Нуклеотидную последовательность (SEQ ID NO: 11), представляющую собой участок гена infB (SEQ ID NO: 1) с сайтами отжига праймеров FIP, BIP, F3 и В3, клонировали в плазмиду pAL2-T («Евроген»; каталожный номер ТА002) из генома штамма Dickeya solani DSO1001 (штамм хранится в коллекции культур микроорганизмов лаборатории молекулярной биоинженерии Института биоорганической химии имени академиков М.М. Шемякина и Ю.А. Овчинникова Российской академии наук, Москва, Российская Федерация). Идентичность клонированного фрагмента, соответствующему участку в гене infB, подтверждали с помощью секвенирования и сравнением с нуклеотидной последовательностью гена infB (SEQ ID NO: 1) из полного генома штамма Dickeya solani IPO2222, депонированного в базе данных GenBank (идентификатор: NZ_CP015137).
Результаты электрофореза показывают (Фигура 2), что предложенный способ, в котором используются набор олигонуклеотидных праймеров FIP (SEQ ID NO: 7), BIP (SEQ ID NO: 8), F3 (SEQ ID NO: 9) и В3 (SEQ ID NO: 10) и ген-мишень infB, позволяет определить наличие в биологических образцах требуемой бактерии вида Dickeya solani фемтограммовые количества ДНК (1×10-15 г/мкл), что в пересчете на количество клеток составляет около 250 клеток на одну реакцию амплификации. Количественные данные получены с помощью пересчета данных о концентрации вносимой в реакцию плазмидной ДНК со вставкой сайтов отжига разработанных праймеров и данных о размере генома Dickeya solani IPO2222. При этом увеличение продолжительности реакции амплификации может повысить чувствительность предложенного способа.
Хотя указанное изобретение описано в деталях со ссылкой на наилучшие способы осуществления изобретения, для специалиста в данной области техники очевидно, что могут быть совершены различные изменения и произведены эквивалентные замены, и такие изменения и замены не выходят за рамки настоящего изобретения.





















Изобретение относится к области биохимии. Описана группа объектов, включающая способ определения бактерии вида Dickeya solani методом петлевой изотермической амплификации (LAMP) и набор праймеров для определения бактерии вида Dickeya solani методом петлевой изотермической амплификации (LAMP). В одном из вариантов изобретения в реакции амплификации участка последовательности целевого гена используют набор олигонуклеотидных праймеров, включающий: праймер FIP, имеющий нуклеотидную последовательность SEQ ID NO: 7, или его гомолог, праймер BIP, имеющий нуклеотидную последовательность SEQ ID NO: 8, или его гомолог, праймер F3, имеющий нуклеотидную последовательность SEQ ID NO: 9, или его гомолог, праймер В3, имеющий нуклеотидную последовательность SEQ ID NO: 10, или его гомолог; причем указанные гомологи праймеров FIP, BIP, F3 и В3 имеют гомологию не менее чем 95% по сравнению с соответствующими нуклеотидными последовательностями SEQ ID NOs: 7, 8, 9 и 10 при условии, что такие гомологичные праймеры могут гибридизоваться с целевым геном infB, кодирующим фактор инициации трансляции IF-2. Изобретение расширяет арсенал средств для определения бактерии вида Dickeya solani. 2 н. и 6 з.п. ф-лы, 3 ил., 2 пр.
1. Способ определения бактерии вида Dickeya solani методом петлевой изотермической амплификации (LAMP), отличающийся тем, что в реакции амплификации участка последовательности целевого гена используют набор олигонуклеотидных праймеров, включающий:
(A) праймер FIP, имеющий нуклеотидную последовательность SEQ ID NO: 7, или его гомолог,
(B) праймер BIP, имеющий нуклеотидную последовательность SEQ ID NO: 8, или его гомолог,
(C) праймер F3, имеющий нуклеотидную последовательность SEQ ID NO: 9, или его гомолог,
(D) праймер В3, имеющий нуклеотидную последовательность SEQ ID NO: 10, или его гомолог;
причем указанные гомологи праймеров FIP, BIP, F3 и В3 имеют гомологию не менее чем 95% по сравнению с соответствующими нуклеотидными последовательностями SEQ ID NOs: 7, 8, 9 и 10 при условии, что такие гомологичные праймеры могут гибридизоваться с целевым геном infB, кодирующим фактор инициации трансляции IF-2.
2. Способ по п. 1, отличающийся тем, что ген infB кодируется нуклеотидной последовательностью SEQ ID NO: 1 или гомологичной ей нуклеотидной последовательностью при условии, что такая гомологичная нуклеотидная последовательность может гибридизоваться с указанными праймерами по п. 1.
3. Способ по п. 1, отличающийся тем, что указанным способом определяют бактерию вида Dickeya solani в образцах картофеля, почвы или воды или их комбинации.
4. Набор праймеров для определения бактерии вида Dickeya solani методом петлевой изотермической амплификации (LAMP), содержащий:
(E) праймер FIP, имеющий нуклеотидную последовательность SEQ ID NO: 7, или его гомолог,
(F) праймер BIP, имеющий нуклеотидную последовательность SEQ ID NO: 8, или его гомолог,
(G) праймер F3, имеющий нуклеотидную последовательность SEQ ID NO: 9, или его гомолог,
(H) праймер В3, имеющий нуклеотидную последовательность SEQ ID NO: 10, или его гомолог;
причем указанные гомологи праймеров FIP, BIP, F3 и В3 имеют гомологию не менее чем 95% по сравнению с соответствующими нуклеотидными последовательностями SEQ ID NOs: 7, 8, 9 и 10 при условии, что такие гомологичные праймеры могут гибридизоваться с целевым геном infB, кодирующим фактор инициации трансляции IF-2.
5. Набор праймеров по п. 4, отличающийся тем, что указанные гомологичные праймеры могут гибридизоваться с целевым геном infB, кодируемым нуклеотидной последовательностью SEQ ID NO: 1.
6. Набор праймеров по п. 4, отличающийся тем, что указанный набор дополнительно содержит целевой ген infB, кодирующий фактор инициации трансляции IF-2.
7. Набор праймеров по п. 6, отличающийся тем, что ген infB кодируется нуклеотидной последовательностью SEQ ID NO: 1 или гомологичной ей нуклеотидной последовательностью при условии, что такая гомологичная нуклеотидная последовательность может гибридизоваться с указанными праймерами по п. 4.
8. Набор праймеров по любому из пп. 4-7, отличающийся тем, что указанный набор праймеров - есть диагностический набор реагентов для анализа биологических образцов картофеля, почвы или воды или их комбинации.
| Notomi T., Okayama H., Masubuchi H., Yonekawa T., Watanabe K., Amino N., Hase T | |||
| Loop-Mediated Isothermal Amplification Of DNA // Nucleic Acids Re | |||
| ЩИТОВОЙ ДЛЯ ВОДОЕМОВ ЗАТВОР | 1922 |
|
SU2000A1 |
| - Vol | |||
| Видоизменение прибора с двумя приемами для рассматривания проекционные увеличенных и удаленных от зрителя стереограмм | 1919 |
|
SU28A1 |
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| - P | |||
| Способ приготовления сернистого красителя защитного цвета | 1915 |
|
SU63A1 |
| Saharan P., Dhingolia S., Khatri P., Duhan J.S., Gahlavat S.K | |||
| Loop-mediated isothermal amplification (LAMP) based detection of bacteria: Review // African Journal of Biotechnology | |||
| Способ защиты переносных электрических установок от опасностей, связанных с заземлением одной из фаз | 1924 |
|
SU2014A1 |
| Насос | 1917 |
|
SU13A1 |
| - P | |||
| Контактный детектор | 1924 |
|
SU1920A1 |
| А.Г.Жумина | |||
| Петлевая изотермическая амплификация нуклеиновых кислот: принцип и применение | |||
| Вестник КарГУ Серия Биология | |||
| Медицина | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |

