Настоящая заявка выделена из заявки №2016130707 на выдачу патента РФ на изобретение, поданной 29.12.2014 г., с испрашиванием приоритета по дате подачи заявки PCT/CN2014/070183, поданной 06.01.2014 г.
Область изобретения
Данное изобретение относится к модулю-переносчику через гематоэнцефалический барьер, который обладает одной специфичностью связывания, которая специфически связывается с рецептором гематоэнцефалического барьера (blood brain barrier receptor, BBBR) и которая является одновалентной относительно этой специфичности связывания, а также к способам применения этой конструкции в качестве переносчика через гематоэнцефалический барьер и в лечении неврологических нарушений.
Уровень техники
Проникновение в мозг лекарственных препаратов для лечения неврологических нарушений, таких как, например, крупные биотерапевтические препараты или низкомолекулярные препараты с низким проникновением в мозг, строго ограничено обширным и непроницаемым гематоэнцефалическим барьером (blood brain barrier, ВВВ) вместе с другим клеточным компонентом в нейрососудистом блоке (neurovascular unit, NVU). Для преодоления этого препятствия были испытаны многие стратегии, и одна из них заключается в использовании путей трансцитоза, опосредованных эндогенными рецепторами, экспрессированными на эндотелии капилляров мозга (рецепторами гематоэнцефалического барьера). Против этих рецепторов были разработаны рекомбинантные белки, такие как моноклональные антитела или пептиды, для обеспечения рецептор-опосредованной доставки биотерапевтических препаратов в мозг. Тем не менее, неисследованными остаются стратегии максимизации поглощения мозгом при минимизации неправильной сортировки в эндотелиальных клетках головного мозга (brain endothelial cells, ВЕС), а также степень накопления в определенных органеллах (особенно в органеллах, которые приводят к деградации биотерапевтических препаратов) в ВЕС.
Моноклональные антитела и другие биотерапевтические препараты имеют огромный терапевтический потенциал для лечения патологии в центральной нервной системе (ЦНС, central nervous system, CNS). Тем не менее, их путь в мозг предотвращается гематоэнцефалическим барьером. Предыдущие исследования продемонстрировали, что очень небольшой процент (примерно 0,1%) IgG, введенных в кровоток, способен проникать в компартмент ЦНС (FeIgenhauer, Klin. Wschr. 52 (1974) 1158-1164). Это, безусловно, ограничивает любой фармакологический эффект из-за низкой концентрации антитела в ЦНС.
Таким образом, существует потребность в системах доставки через ВВВ лекарственных средств для лечения неврологических нарушений, эффективно переносящих лекарственные препараты в мозг.
В WO 2014/033074 сообщается о переносчике через гематоэнцефалический барьер.
Мышиное антитрансферриновое антитело 8D3 и вариант его вариабельного домена легкой цепи (VL) (L596V и L598I) описаны в Boado, R.J., et al. (Biotechnol. Bioeng. 102 (2009) 1251-1258).
Краткое описание
Одним из аспектов данного изобретения является модуль-переносчик через гематоэнцефалический барьер, содержащий группировку, действующую в мозге (эффекторную группировку), линкер и одну одновалентную связывающую группировку, которая связывается с рецептором гематоэнцефалического барьера, где линкер соединяет эффекторную группировку с одновалентной связывающей группировкой, которая связывается с рецептором гематоэнцефалического барьера, где одновалентная связывающая группировка не содержит вариабельные домены антитела против рецептора трансферрина 8D3 (SEQ ID NO 01 и SEQ ID NO 02) или вариантного антитела против рецептора трансферрина 8D3v (SEQ ID NO 01 и SEQ ID NO 03).
Антитело против рецептора трансферрина 8D3 имеет вариабельный домен тяжелой цепи со следующей аминокислотной последовательностью:
EVQLVESGGG LVQPGNSLTL SCVASGFTFS NYGMHWIRQA PKKGLEWIAM IYYDSSKMNY ADTVKGRFTI SRDNSKNTLY LEMNSLRSED TAMYYCAVPT SHYWDVWGQ GVSVTVSS
(SEQ ID NO 01).
Антитело против рецептора трансферрина 8D3 имеет вариабельный домен легкой цепи со следующей аминокислотной последовательностью:
DIQMTQSPAS LSASLEEIVT ITCQASQDIG NWLAWYQQKP GKSPQLLIYG ATSLADGVPS RFSGSRSGTQ FSLKISRVQV EDIGIYYCLQ AYNTPWTFGG GTKLELK
(SEQ ID NO 02).
Вариантное антитело против рецептора трансферрина 8D3v имеет тот же вариабельный домен тяжелой цепи, что и антитело 8D3, и вариабельный домен легкой цепи с мутантами L104V и L106I, который имеет следующую аминокислотную последовательность:
DIQMTQSPAS LSASLEEIVT ITCQASQDIG NWLAWYQQ KP GKSPQLLIYG ATSLADGVPS RFSGSRSGTQ FSLKISRVQV EDIGIYYCLQ AYNTPWTFGG GTKVEIK (SEQ ID NO 03).
В одном из воплощений модуля-переносчика через гематоэнцефалический барьер одновалентная связывающая группировка, которая связывается с рецептором гематоэнцефалического барьера, представляет собой полипептид.
В одном из воплощений модуля-переносчика через гематоэнцефалический барьер одновалентная связывающая группировка, которая связывается с рецептором гематоэнцефалического барьера, содержит молекулу, выбранную из группы, состоящей из лиганда рецептора гематоэнцефалического барьера, полноразмерного антитела, scFv, Fv, scFab и VHH.
В одном из воплощений модуля-переносчика через гематоэнцефалический барьер рецептор гематоэнцефалического барьера выбран из группы, состоящей из: рецептора трансферрина, рецептора инсулина; рецептора инсулиноподобного фактора роста; белка 8, связанного с рецептором липопротеинов низкой плотности; белка 1, связанного с рецептором липопротеинов низкой плотности и гепаринсвязывающего фактор роста, подобного эпидермальному фактору роста. В одном воплощении рецептор гематоэнцефалического барьера представляет собой рецептор трансферрина.
В одном из воплощений одновалентная связывающая группировка специфически связывается с рецептором трансферрина человека и рецептором трансферрина яванского макака.
В одном из воплощений модуля-переносчика через гематоэнцефалический барьер одновалентная связывающая группировка, которая связывается с рецептором гематоэнцефалического барьера, содержит один scFab, направленный против рецептора трансферрина, более конкретно, scFab, который специфически связывается с эпитопом на рецепторе трансферрина, содержащимся в аминокислотной последовательности SEQ ID NO 04, 05 или 06.
В одном из воплощений модуля-переносчика через гематоэнцефалический барьер одновалентная связывающая группировка, которая связывается с рецептором гематоэнцефалического барьера, включает один scFv, направленный против рецептора трансферрина, более конкретно, scFv, распознающий эпитоп на рецепторе трансферрина, содержащийся в аминокислотной последовательности SEQ ID NO 04, 05 или 06.
В одном из воплощений модуля-переносчика через гематоэнцефалический барьер действующая в мозге группировка выбрана из группы, состоящей из лекарственных препаратов для лечения неврологических нарушений, нейротрофических факторов, факторов роста, ферментов, цитотоксических агентов, антител, направленных против мишеней в мозге, моноклональных антител, направленных против мишеней в мозге, пептидов, направленных против мишеней в мозге.
В одном из воплощений модуля-переносчика через гематоэнцефалический барьер мишень в мозге выбрана из группы, состоящей из бета-секретазы 1, Аβ (Abeta), эпидермального фактора роста, рецептора эпидермального фактора роста 2, тау-белка, фосфорилированного тау-белка, фосфорилированного тау(р8422)-белка, аполипопротеина Е4, альфа-синуклеина, олигомерных фрагментов альфа-синуклеина, CD20, хантингтина, прионного белка, киназы 2 с богатым лейцином повтором, паркина, пресенилина 2, гамма-секретазы, рецептора смерти 6, белка-предшественника амилоида, рецептора нейротрофинов р75 и каспазы 6.
В конкретном воплощении модуля-переносчика через гематоэнцефалический барьер действующая в мозге группировка представляет собой полипептид.
В одном из воплощений модуля-переносчика через гематоэнцефалический барьер одновалентная связывающая группировка, которая связывается с рецептором гематоэнцефалического барьера, представляет собой полипептид, и одновалентная связывающая группировка конъюгирована с С-концом действующей в мозге группировки либо напрямую, либо через линкер.
В одном из воплощений модуля-переносчика через гематоэнцефалический барьер действующая в мозге группировка содержит полноразмерное антитело, направленное против мишени в мозге. В одном воплощении полноразмерное антитело представляет собой IgG.
В одном предпочтительном воплощении модуля-переносчика через гематоэнцефалический барьер переносчик через гематоэнцефалический барьер содержит полноразмерное антитело IgG в качестве действующей в мозге группировки, линкер и один scFab в качестве одновалентной связывающей группировки, которая связывается с рецептором гематоэнцефалического барьера, где scFab конъюгирован с С-концом Fc-области одной из тяжелых цепей антитела IgG через линкер.
В одном из воплощений модуля-переносчика через гематоэнцефалический барьер первая тяжелая цепь антитела в переносчике через гематоэнцефалический барьер, направленного на мишень в мозге, включает первый модуль димеризации, а вторая тяжелая цепь антитела в переносчике гематоэнцефалического барьера, направленного на мишень в мозге, содержит второй модуль димеризации, которые делают возможной гетеродимеризацию двух тяжелых цепей.
В одном из воплощений модуля-переносчика через гематоэнцефалический барьер первый модуль димеризации первой тяжелой цепи антитела в переносчике через гематоэнцефалический барьер, направленного против мишени в мозге, включает "ключ", а модуль димеризации второй тяжелой цепи антитела в переносчике через гематоэнцефалический барьер, направленного против мишени в мозге, включает "замок" в соответствии со стратегией "ключ в замке".
В одном из воплощений модуля-переносчика через гематоэнцефалический барьер линкер представляет собой пептидный линкер. В одном из воплощений пептидный линкер имеет аминокислотную последовательность длиной по меньшей мере 25 аминокислот.В одном из воплощений пептидный линкер имеет аминокислотную последовательность длиной от 30 до 50 аминокислот.В одном из воплощений пептидный линкер представляет собой (G4S)6G2 (SEQ ID NO 07) или (G4S)4 (SEQ ID NO 08).
Следующие три воплощения направлены на модуль-переносчик через гематоэнцефалический барьер, где действующая в мозге группировка представляет собой полипептид, при условии, что действующая в мозге группировка не является полноразмерным антителом, в частности, не является полноразмерным IgG.
В одном из воплощений модуля-переносчика через гематоэнцефалический барьер одновалентная связывающая группировка, которая связывается с рецептором гематоэнцефалического барьера, содержит СН2-СН3-Ig-группировку и один scFab (содержащий первый линкер), который связывается с рецептором гематоэнцефалического барьера, где scFab соединен с С-концом CH2-CH3-Ig-группировки вторым линкером.
В одном из воплощений модуля-переносчика через гематоэнцефалический барьер переносчик через гематоэнцефалический барьер содержит действующую в мозге группировку, линкер, СН2-СН3-Ig-домен, второй линкер и один scFab, который связывается с рецептором гематоэнцефалического барьера, где действующая в мозге группировка конъюгирована с помощью первого линкера с N-концом CH2-CH3-Ig-домена, a scFab конъюгирован с С-концом СН2-СН3-Ig-домена с помощью второго линкера.
В одном из воплощений модуля-переносчика через гематоэнцефалический барьер СН2-СН3-Ig-группировка представляет собой СН2-СН3-группировку IgG.
Другими аспектами данного изобретения являются (изолированная) нуклеиновая кислота, кодирующая модуль-переносчик через гематоэнцефалический барьер, описанный в данном документе, клетка-хозяин, содержащая (изолированную) нуклеиновую кислоту, кодирующую модуль-переносчик через гематоэнцефалический барьер, а также фармацевтическая композиция, содержащая модуль-переносчик через гематоэнцефалический барьер.
Модуль-переносчик через гематоэнцефалический барьер, описанный в данном документе, может быть использован в качестве лекарственного средства, в частности, он может быть использован для лечения неврологических нарушений, таких как, например, болезнь Альцгеймера.
Модуль-переносчик через гематоэнцефалический барьер, описанный в данном документе, может быть использован для транспортировки действующей в мозге группировки через гематоэнцефалический барьер.
В конкретном воплощении изобретения тяжелая цепь IgG-антитела модуля-переносчика через гематоэнцефалический барьер, описанного в данном документе, конъюгированная на своем С-конце Fc-области с scFab в качестве одновалентной связывающей группировки, которая связывается с рецептором гематоэнцефалического барьера, имеет следующую структуру:
- тяжелая цепь IgG,
- линкер, конъюгирующий С-конец Fc-области тяжелой цепи IgG с N-концом VL-домена scFab,
- вариабельный домен легкой цепи (VL) и домен С-каппа легкой цепи scFab,
- линкер, конъюгирующий С-конец домена С-каппа легкой цепи scFab с N-концом VH-домена scFab,
- вариабельный домен тяжелой цепи (VH) scFab антитела и СН1-домена тяжелой цепи IgG.
Одним из аспектов, описанных в данном документе, является гибридный полипептид для транспортировки действующей в мозге группировки через гематоэнцефалический барьер, включающий СН2-СН3-Ig-группировку, линкер и один scFab, который специфически связывается с рецептором гематоэнцефалического барьера, где scFab конъюгирован с С-концом СН2-СН3-Ig-группировки через линкер, где scFab не содержит вариабельные домены антитела 8D3 против рецептора трансферрина (SEQ ID NO 01 и SEQ ID NO 02) или вариантного антитела 8D3v против рецептора трансферрина (SEQ ID NO 01 и SEQ ID NO 03).
Одним из аспектов, описанных в данном документе, является гибридный полипептид для транспортировки через гематоэнцефалический барьер действующей в мозге группировки, включающий СН2-СН3-Ig-группировку, линкер и один scFv, который специфически связывается с рецептором гематоэнцефалического барьера, где scFv конъюгирован с С-концом СН2-СН3-Ig-группировки через линкер, где scFv не содержит вариабельные домены антитела 8D3 против рецептора трансферрина (SEQ ID NO 01 и SEQ ID NO 02) или вариантного антитела 8D3v против рецептора трансферрина (SEQ ID NO 01 и SEQ ID NO03).
В одном из воплощений гибридный полипептид также содержит линкер на N-конце СН2-СН3-Ig-группировки для конъюгации действующей в мозге группировки с N-концом СН2-СН3-Ig-группировки.
В одном из воплощений гибридного полипептида действующая в мозге группировка выбрана из группы, состоящей из лекарственных препаратов для лечения неврологических нарушений, нейротрофических факторов, факторов роста, ферментов, цитотоксических агентов, фрагментов антител или пептидов, направленных против мишени в мозге, выбранных из группы, состоящей из scFv, Fv, scFab, Fab, VHH, F(ab')2.
В одном из воплощений гибридного полипептида scFab или scFv, которые специфически связываются с рецептором гематоэнцефалического барьера, специфически связываются с рецептором трансферрина. В одном воплощении scFab или scFv специфически связываются с эпитопом на рецепторе трансферрина, содержащимся в аминокислотной последовательности SEQ ID NO 04, 05 или 06.
В воплощении гибридного полипептида линкер представляет собой пептидный линкер. В одном из воплощений пептидный линкер имеет аминокислотную последовательность длиной по меньшей мере 15 аминокислот.В одном из воплощений пептидный линкер имеет длину от 20 до 50 аминокислот.В одном из воплощений пептидный линкер имеет аминокислотную последовательность (G4S)6G2 (SEQ ID NO 07) или (G4S) 4 (SEQ ID NO 08).
В одном из воплощений гибридного полипептида СН2-СН3-Ig-группировка представляет собой СН2-СН3-группировку IgG.
Другими аспектами данного изобретения являются изолированная нуклеиновая кислота, кодирующая гибридный полипептид, описанный в данном документе, и клетка-хозяин, содержащая нуклеиновую кислоту, кодирующую гибридный полипептид, описанный в данном документе.
Одним из аспектов, описанных в данном документе, является конъюгат, содержащий гибридный полипептид, описанный в данном документе, и действующую в мозге группировку, конъюгированную через линкер с N-концом CH2-CH3-Ig-группировки гибридного полипептида, описанного в данном документе.
В одном из воплощений конъюгата действующая в мозге группировка является нейротрофическим фактором, а линкер, конъюгирующий нейротрофический фактор с N-концом СН2-СН3-Ig-группировки, является пептидным линкером.
Другими аспектами, описанными в данном документе, являются фармацевтический состав, содержащий конъюгат, описанный в данном документе, и фармацевтический носитель, а также применение конъюгата, описанного в данном документе, в частности, применение конъюгата для лечения нейродегенеративного нарушения, в частности, болезни Альцгеймера.
Одновалентная связывающая группировка, которая специфически связывается с рецептором гематоэнцефалического барьера, может быть конъюгирована с любым концом легкой или тяжелой цепи антитела либо напрямую, либо через пептидный линкер. В одном воплощении одновалентная связывающая группировка конъюгирована с С-концом тяжелой цепи.
С-конец тяжелой цепи антитела может быть полным С-концом, заканчивающимся аминокислотными остатками PGK. С-конец тяжелой цепи может быть укороченным С-концом, в котором один или два С-концевых аминокислотных остатка удалены. В одном воплощении С-конец тяжелой цепи является укороченным С-концом, который заканчивается аминокислотными остатками PG.
Одновалентная связывающая группировка может быть конъюгирована с соответствующей цепью антитела либо напрямую, либо через пептидный линкер. В одном из воплощений пептидный линкер имеет аминокислотную последовательность GGSGGGGSGGGGSGGGGS (SEQ ID NO 09).
Одновалентная связывающая группировка может представлять собой scFv-фрагмент антитела. В одном воплощении одновалентная связывающая группировка представляет собой scFv, содержащий в направлении от N-конца к С-концу: вариабельный домен легкой цепи - константный домен легкой цепи - пептидный линкер - вариабельный домен тяжелой цепи - константный домен 1 тяжелой цепи.
В одном воплощении одновалентная связывающая группировка представляет собой scFv-фрагмент антитела против рецептора трансферрина с (C4S)6-пептидным линкером (SEQ ID NO 10).
В одном воплощении рецептор гематоэнцефалического барьера выбран из группы, состоящей из рецептора трансферрина; рецептора инсулина; рецептора инсулиноподобного фактора роста; белка 8, связанного с рецептором липопротеинов низкой плотности; белка 1, связанного с рецептором липопротеинов низкой плотности и гепаринсвязывающего фактора роста, подобного эпидермальному фактору роста. В одном воплощении рецептор гематоэнцефалического барьера является человеческим рецептором гематоэнцефалического барьера. В одном воплощении рецептор гематоэнцефалического барьера является рецептором трансферрина, и антитело не ингибирует связывание рецептора трансферрина с трансферрином. В одном воплощении рецептор гематоэнцефалического барьера является рецептором трансферрина человека.
В одном из воплощений пептидный линкер, конъюгирующий одновалентную связывающую группировку с действующей в мозге группировкой, представляет собой аминокислотную последовательность длиной по меньшей мере 15 аминокислот.В одном из воплощений пептидный линкер имеет длину от 18 до 25 аминокислот.
В одном из воплощений действующая в мозге группировка представляет собой полноразмерное антитело. В одном из воплощений действующая в мозге группировка представляет собой полноразмерное антитело подкласса IgG1 или IgG4.
В одном из воплощений одновалентная связывающая группировка представляет собой антитело против рецептора гематоэнцефалического барьера или его фрагмент, связывающий рецептор гематоэнцефалического барьера. В одном из воплощений антитело против рецептора гематоэнцефалического барьера или его фрагмент не нарушают связывание рецептора гематоэнцефалического барьера с одним или более чем одним из его нативных лигандов. В другом воплощении антитело против рецептора гематоэнцефалического барьера специфически связывается с рецептором трансферрина человека таким образом, что оно не ингибирует связывание рецептора трансферрина человека с трансферрином человека.
В одном из воплощений модуль-переносчик через гематоэнцефалический барьер является молчащим эффектором.
В одном из воплощений действующая в мозге группировка представляет собой полноразмерное антитело, содержащее Fc-область, где в случае, когда Fc-область принадлежит человеческому подклассу IgG1, Fc-область содержит мутации L234A, L235A и P329G (нумерация в соответствии с ЕС-индексом Kabat), или в случае, когда Fc-область принадлежит человеческому подклассу IgG4, Fc-область содержит мутации S228P, L235E и P329G (нумерация в соответствии с ЕС-индексом Kabat).
Подробное описание воплощений изобретения
I. Определения
"Акцепторная человеческая каркасная область" в контексте данного изобретения является каркасной областью, включающей аминокислотную последовательность каркасной области вариабельного домена легкой цепи (VL) или каркасной области вариабельного домена тяжелой цепи (VH), полученной из каркасной области человеческого иммуноглобулина или человеческой консенсусной каркасной области, как определено ниже. Акцепторная человеческая каркасная область, "полученная из" каркасной области человеческого иммуноглобулина или консенсусной человеческой каркасной области, может содержать такую же аминокислотную последовательность, или она может содержать аминокислотные замены. В некоторых воплощениях число аминокислотных замен составляет 10 или менее, 9 или менее, 8 или менее, 7 или менее, 6 или менее, 5 или менее, 4 или менее, 3 или менее или 2 или менее. В некоторых воплощениях человеческая акцепторная каркасная область VL по своей последовательности идентична последовательности каркасной области VL человеческого иммуноглобулина или последовательности консенсусной человеческой каркасной области.
Понятие "аффинность" относится к силе суммарных общих нековалентных взаимодействий между одним сайтом связывания молекулы (например, антитела) и его партнером по связыванию (например, антигеном). Если не указано иное, то термин "аффинность связывания", используемый в данном документе, относится к внутренней аффинности связывания, которая отражает взаимодействие 1:1 между элементами пары связывания (например, между антителом и антигеном). Аффинность молекулы X в отношении ее партнера Y может в целом быть выражена константой диссоциации (Kd). Аффинность можно измерить обычными способами, известными в данной области, включая описанные в данном документе. Далее будут описаны конкретные иллюстративные и типичные воплощения измерения аффинности связывания.
Понятие "антитела с созревшей аффинностью" относится к антителу с одним или более чем одним изменением в одной или более чем одной гипервариабельной области (HVR) по сравнению с родительским антителом, которое не содержит таких изменений, причем такие изменения приводят к повышению аффинности антитела к антигену.
Термин "антитело" в данном документе используется в самом широком смысле и охватывает различные структуры антител, в том числе, но не ограничиваясь ими, моноклональные антитела, поликлональные антитела, мультиспецифические антитела (например, биспецифические антитела) и фрагменты антител, до тех пор, пока они проявляют нужную антигенсвязывающую активность.
Понятие "фрагмент антитела" относится к молекуле, отличающейся от интактного антитела, которая содержит часть интактного антитела, связывающуюся с антигеном, с которым связывается интактное антитело. Примеры фрагментов антител включают, но не ограничиваясь ими, Fv, Fab, Fab', Fab'-SH, F(ab')2; димеры; линейные антитела; одноцепочечные молекулы антител (например, scFv); и мультиспецифические антитела, образованные из фрагментов антител.
Термин "химерное" антитело относится к антителу, в котором часть тяжелой и/или легкой цепи получена из конкретного источника или вида, в то время как остальная часть тяжелой и/или легкой цепи получена из другого источника или вида.
Термин "класс" антитела относится к типу константного домена или константной области, которой обладает его тяжелая цепь. Есть пять основных классов антител: IgA, IgD, IgE, IgG и IgM, и некоторые из них могут быть разделены на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелой цепи, которые соответствуют различным классам иммуноглобулинов, называются α, δ, ε, γ и μ, соответственно.
Понятие "эффекторные функции" относится к биологическим активностям, приписываемым Fc-области антитела, которая варьирует в зависимости от класса антитела. Примеры эффекторных функций антител включают: C1q-связывание и комплемент-зависимую цитотоксичность (CDC); Fc-рецепторное связывание; антитело-зависимую клеточную цитотоксичность (ADCC); фагоцитоз; понижающую регуляцию рецепторов клеточной поверхности (например, В-кпеточного рецептора); и В-клеточную активацию.
Понятие "эффективного количества" агента, например, фармацевтического состава, относится к количеству, эффективному в дозах и в течение периода времени, необходимого для достижения желаемого терапевтического или профилактического результата.
Термин "Fc-область" в данном документе используется для определения С-концевой области тяжелой цепи иммуноглобулина, которая содержит по меньшей мере часть константной области. Термин включает Fc-области с нативной последовательностью или вариантные Fc-области. В одном воплощении Fc-область тяжелой цепи человеческого IgG продолжается от Cys226 или от Pro230 до карбоксильного конца тяжелой цепи. Тем не менее, С-концевой лизин (Lys447) Fc-области может присутствовать или отсутствовать. Если в данном документе не указано иное, то нумерация аминокислотных остатков в Fc-области или константной области осуществляется в соответствии с системой нумерации ЕС, также называемой индексом ЕС, как описано в Kabat, Е.А. et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991), NIH Publication 91-3242.
Термин "каркасная область", сокращенно "FR", относится к остаткам вариабельного домена за исключением остатков гипервариабельной области (HVR). FR вариабельного домена обычно состоит из четырех FR-доменов: FR1, FR2, FR3 и FR4. Соответственно, последовательности HVR и FR обычно идут в следующей последовательности в VH (или VL): FR1-H1(L1)-FR2-H2(L2)-FR3-H3(L3)-FR4.
Термины "полноразмерное антитело", "интактное антитело" и "целое антитело" используются в данном документе взаимозаменяемо и относятся к антителу, имеющему структуру, которая по существу подобна структуре нативного антитела, или имеющему тяжелые цепи, которые содержат Fc-область, определенную в данном описании.
Термины "клетка-хозяин", "клеточная линия-хозяин" и "клеточная культура-хозяин" используются взаимозаменяемо и относятся к клеткам, в которые была введена экзогенная нуклеиновая кислота, включая потомство таких клеток. Клетки-хозяева включают "трансформанты" и "трансформированные клетки", которые включают первичные трансформированные клетки и потомство, полученное из них, независимо от числа пассажей. Потомство может не быть полностью идентичным родительской клетке по содержанию нуклеиновых кислот, а может содержать мутации. Мутантное потомство, которое имеет такую же функцию или биологическую активность при скрининге или селекции, как у исходно трансформированной клетки, включено в данный документ.
"Человеческая консенсусная каркасная область" является каркасной областью, которая представляет наиболее часто встречающиеся аминокислотные остатки в выборе каркасных последовательностей VL или VH иммуноглобулина человека. Как правило, выбор последовательностей VL или VH человеческого иммуноглобулина проводится из подгруппы последовательностей вариабельных доменов. Как правило, подгруппа последовательностей является подгруппой, описанной в Kabat, Е.А. et al., Sequences of Proteins of Immunological Interest, 5th ed., Bethesda MD (1991), NIH Publication 91-3242, Vols. 1-3. В одном воплощении для VL подгруппа является подгруппой каппа I, как описано в Kabat et al., см, выше, В одном воплощении для VH подгруппа является подгруппой III, как описано в Kabat et al., см, выше.
Понятие "гуманизированного" антитела относится к химерному антителу, содержащему аминокислотные остатки из нечеловеческих HVR и аминокислотные остатки из человеческих FR. В некоторых воплощениях гуманизированное антитело будет содержать по существу все из по меньшей мере одного, а обычно двух вариабельных доменов, в которых все или по существу все HVR (например, CDR) соответствуют таковым в нечеловеческом антителе, и все или по существу все FR соответствуют таковым в человеческом антителе. Гуманизированное антитело, возможно, может содержать по меньшей мере часть константной области антитела, полученной из человеческого антитела. "Гуманизированная форма" антитела, например, нечеловеческого антитела, относится к антителу, которое подверглось гуманизации.
Термины "гипервариабельная область" или "HVR", используемые в данном документе, относятся к каждой из областей вариабельного домена антитела, которые являются гипервариабельными в последовательности ("области, определяющие комплементарность", или "CDR") и/или формируют структурно определенные петли ("гипервариабельные петли") и/или содержат контактирующие с антигеном остатки ("антигенные контакты"). Как правило, антитела содержат шесть HVR, три в VH (Н1, Н2, H3)htphbVL (L1, L2, L3).
HVR в данном документе включают
(a) гипервариабельные петли, возникающие на аминокислотных остатках 26-32 (L1), 50-52 (L2), 91-96 (L3), 26-32 (Н1), 53-55 (Н2) и 96-101 (Н3). (Chothia, С.and Lesk, A.M., J. Mol. Biol. 196 (1987) 901-917);
(b) CDR, возникающие на аминокислотных остатках 24-34 (L1), 50-56 (L2), 89-97 (L3), 31-35b (Н1), 50-65 (Н2) и 95-102 (Н3) (Kabat, Е.А. et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991), NIH Publication 91-3242);
(c) антигенные контакты, возникающие на аминокислотных остатках 27 с-36 (L1), 46-55 (L2), 89-96 (L3), 30-35b (Н1), 47-58 (Н2) и 93-101 (Н3) (MacCallum et al. J. Mol. Biol. 262: 732-745 (1996)); и
(d) комбинации (a), (b) и/или (с), включая аминокислотные остатки HVR 46-56 (L2), 47-56 (L2), 48-56 (L2), 49-56 (L2), 26-35 (Н1), 26-35b (Н1), 49-65 (Н2), 93-102 (Н3) и 94-102 (Н3).
Если не указано иное, то HVR-остатки и другие остатки в вариабельном домене (например, FR-остатки) пронумерованы в данном документе в соответствии с Kabat et aI., см. выше.
"Индивидуум" или "субъект" является млекопитающим. Млекопитающие включают, но не ограничиваясь ими, домашних животных (например, коров, овец, кошек, собак и лошадей), приматов (например, людей и приматов, таких как обезьяны), кроликов и грызунов (например, мышей и крыс). В некоторых воплощениях индивидуум или субъект является человеком.
"Изолированное" антитело является таким антителом, которое было отделено от компонента его природной среды. В некоторых воплощениях антитело очищают до чистоты более 95% или 99%, что определено, например, с помощью электрофореза (например, SDS-PAGE, изоэлектрофокусировки (IEF), капиллярного электрофореза) или хроматографии (например, ионнообменной или обращено-фазовой HPLC). Обзор методик оценки чистоты антител см., например, в Flatman, S. et al., J. Chromatogr. В 848 (2007) 79-87.
Понятие "изолированной" нуклеиновой кислоты относится к нуклеиновокислотной молекуле, которая была отделена от компонента ее природной среды. Изолированная нуклеиновая кислота включает нуклеиновокислотную молекулу, содержащуюся в клетках, которые обычно содержат данную нуклеиновокислотную молекулу, но нуклеиновокислотная молекула находится вне хромосомы или в том месте хромосомы, которое отличается от ее природной хромосомной локализации.
Термин "моноклональное антитело", используемый в данном документе, относится к антителу, полученному из популяции по существу гомогенных антител, т.е. популяции антител, состоящей из отдельных антител, которые являются идентичными и/или связывают один и тот же эпитоп, за исключением возможных вариантных антител, например, содержащих встречающиеся в природе мутации или образованных во время производства препарата моноклонального антитела, которые, как правило, могут присутствовать в незначительных количествах. В отличие от препаратов поликлональных антител, которые обычно включают различные антитела, направленные против различных детерминант (эпитопов), каждое моноклональное антитело в препарате моноклонального антитела направлено против одной единственной детерминанты на антигене. Таким образом, указатель "моноклональное" указывает на характер антитела, которое было получено из популяции по существу гомогенных антител, и не должен быть истолкован как требующий получения антитела каким-либо конкретным способом. Например, моноклональные антитела, используемые в соответствии с данным изобретением, могут быть получены с помощью различных методик, включая, но не ограничиваясь ими, гибридомную методику, рекомбинантные ДНК-методики, методики фагового дисплея, а также методики с использованием трансгенных животных, содержащих все или часть человеческих иммуноглобулиновых локусов, и такие методики и другие иллюстративные методики получения моноклональных антител описаны в данном документе.
Понятие "нативных антител" относится к природным молекулам иммуноглобулинов с различными структурами. Например, нативные IgG-антитела являются гетеротетрамерными гл и ко протеина ми весом примерно 150000 дальтон, состоящими из двух идентичных легких цепей и двух идентичных тяжелых цепей, которые связаны дисульфидной связью. От N-конца к С-концу каждая тяжелая цепь содержит вариабельную область (VH), также называемую вариабельным тяжелым доменом или вариабельным доменом тяжелой цепи, а затем три константных домена (СН1, СН2 и СН3). Аналогичным образом от N-конца к С-концу каждая легкая цепь имеет вариабельную область (VL), также называемую вариабельным легким доменом или вариабельным доменом легкой цепи, с последующим константным легким (CL) доменом. Легкая цепь антитела может быть отнесена к одному из двух типов, называемых каппа (κ) и лямбда (λ), на основании аминокислотной последовательности ее константного домена.
Понятие "вкладыш в упаковку" используется для обозначения инструкций, обычно включаемых в коммерческие упаковки терапевтических продуктов, которые содержат информацию о показаниях, применении, дозировке, введении, комбинированной терапии, противопоказаниях и/или предупреждениях, касающихся применения таких терапевтических продуктов.
"Процент (%) идентичности аминокислотной последовательности" по отношению к референсной полипептидной последовательности определяется как процент аминокислотных остатков в кандидатной последовательности, которые идентичны аминокислотным остаткам в референсной полипептидной последовательности после выравнивания последовательностей и введения при необходимости пробелов для достижения максимального процента идентичности последовательностей, и без учета любых консервативных замен как части идентичности последовательности. Выравнивание с целью определения процента идентичности аминокислотных последовательностей может быть достигнуто различными способами, которые доступны специалистам в данной области, например, с использованием общедоступного программного обеспечения, такого как программное обеспечение BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Специалисты в данной области могут определить соответствующие параметры для выравнивания последовательностей, в том числе любые алгоритмы, необходимые для достижения максимального выравнивания по всей длине сравниваемых последовательностей. Тем не менее, для целей данного изобретения значения % идентичности аминокислотных последовательностей получают с использованием компьютерной программы для сравнивания последовательностей ALIGN-2. Компьютерная программа для сравнивания последовательностей ALIGN-2 была разработана Genentech, Inc., и исходный код был подан с пользовательской документацией в бюро авторских прав США, Вашингтон, округ Колумбия, 20559, где он был зарегистрирован под номером TXU510087. Программа ALIGN-2 находится в открытом доступе от Genentech, Inc., Саут-Сан-Франциско, Калифорния, или может быть скомпилирована из исходного кода. Программа ALIGN-2 должна быть скомпилирована для применения на операционной системе UNIX, включая цифровую UNIX V4.0D. Все параметры сравнивания последовательностей установлены программой ALIGN-2 и не меняются.
В ситуациях, когда для сравнивания аминокислотных последовательностей используется ALIGN-2, % идентичности аминокислотной последовательности данной аминокислотной последовательности А с данной аминокислотной последовательностью В (альтернативно можно сказать, что данная аминокислотная последовательность А имеет или содержит определенный % идентичности аминокислотной последовательности с данной аминокислотной последовательностью В) вычисляется следующим образом:
100 × доля X/Y,
где X обозначает число аминокислотных остатков, оцененных как идентичные совпадения с помощью программы для выравнивания последовательностей ALIGN-2 при выравнивании в этой программе А и В, и где Y обозначает общее число аминокислотных остатков в В. Следует иметь в виду, что если длина аминокислотной последовательности А не равна длине аминокислотной последовательности В, то % идентичности аминокислотной последовательности А с В не будет равен % идентичности аминокислотной последовательности В с А. Если специально не указано иное, то все значения % идентичности аминокислотной последовательности, используемые в данном описании, получены так, как описано в предыдущем абзаце, с использованием компьютерной программы ALIGN-2.
Термин "фармацевтический состав" относится к препарату, который находится в такой форме, которая разрешает биологическую активность активного ингредиента, содержащегося в нем, и который не содержит дополнительных компонентов, которые являются неприемлемо токсичными для субъекта, которому будет осуществляться введение состава.
Понятие "фармацевтически приемлемого носителя" относится к ингредиенту в фармацевтическом составе, помимо активного ингредиента, который является нетоксичным для субъекта. Фармацевтически приемлемые носители включают, но не ограничиваясь ими, буфер, эксципиент, стабилизатор или консервант.
Используемое в данном документе понятие "лечение" (и его грамматические варианты, такие как "лечить") относится к клиническому вмешательству в попытке изменить естественное развитие заболевания у индивидуума, которого лечат, которое может быть выполнено либо для профилактики, либо в ходе клинической патологии. Желательные эффекты лечения включают, но не ограничиваясь ими, предотвращение возникновения или рецидива заболевания, ослабление симптомов, уменьшение любого прямого или косвенного патологического последствия заболевания, предотвращение метастазирования, уменьшение скорости прогрессирования заболевания, улучшение или временное облегчение болезненного состояния, достижение ремиссии или улучшение прогноза. В некоторых воплощениях антитела согласно данному изобретению используются для задержки развития заболевания или для замедления прогрессии заболевания.
Термин "вариабельная область" или "вариабельный домен" обозначает домен тяжелой или легкой цепи антитела, который участвует в связывании антитела с антигеном. Вариабельные домены тяжелой цепи и легкой цепи (VH и VL, соответственно) нативного антитела, как правило, имеют аналогичную структуру, при этом каждый домен содержит четыре консервативные каркасные области (FR) и три гипервариабельные области (HVR). (См., например, Kindt, T.J. et al., Kuby Immunology, 6th ed., W.H. Freeman and Co., N.Y. (2007), page 91). Один VH- или VL-домен может быть достаточным для придания антигенсвязывающей специфичности. Кроме того, антитела, которые связывают конкретный антиген, могут быть выделены с помощью VH- или VL-домена из антитела, которое связывается с антигеном, для скрининга библиотеки комплементарных VL- или VH-доменов, соответственно. См., например, Portolano, S. et al., J. Immunol. 150 (1993) 880-887; Clackson, T. et al., Nature 352 (1991) 624-628).
Термин "вектор", используемый в данном документе, обозначает нуклеиновокислотную молекулу, способную размножать другую нуклеиновую кислоту, с которой она связана. Этот термин включает вектор как самореплицирующуюся нуклеиновокислотную структуру, а также как вектор, включенный в геном клетки-хозяина, в которую он был введен. Некоторые векторы способны направлять экспрессию нуклеиновых кислот, с которыми они функционально связаны. Такие векторы называют в данном документе "экспрессионными векторами".
Термин "гематоэнцефалический барьер" (ВВВ) обозначает физиологический барьер между периферическим кровообращением и головным мозгом и спинным мозгом, который образован плотными соединениями плазматических мембран эндотелиальных клеток капилляров мозга, создающими плотный барьер, который ограничивает перенос молекул в мозг, даже очень малых молекул, таких как мочевина (60 Да). ВВВ в головном мозге, гемато-спинномозговой барьер в спинном мозге и гемато-ретинальный барьер в сетчатке являются смежными капиллярными барьерами в ЦНС, и в настоящем документе совместно называются гематоэнцефалический барьером, или ВВВ. ВВВ также охватывает барьер кровь-CSF (сосудистое сплетение), где барьер состоит из эпендимальных клеток, а не эндотелиальных клеток капилляров.
Термин "центральная нервная система" (ЦНС, central nervous system, CNS) обозначает комплекс нервных тканей, которые контролируют физическую функцию, и включает головной и спинной мозг.
Термин "рецептор гематоэнцефалического барьера" (blood-brain barrier receptor, BBBR) обозначает внеклеточный связанный с мембранной белок рецептора, экспрессированный на эндотелиальных клетках головного мозга, который способен транспортировать молекулы через ВВВ или использоваться для транспортировки экзогенно вводимых молекул. Примеры BBBR включают, но не ограничиваясь ими, рецептор трансферрина (TfR), рецептор инсулина, рецептор инсулиноподобного фактора роста (IGF-R), рецепторы липопротеинов низкой плотности, включая, но не ограничиваясь ими, белок 1, связанный с рецептором липопротеинов низкой плотности (LRP1), и белок 8, связанный с рецептором липопротеинов низкой плотности (LRP8), и гепаринсвязывающий фактор роста, подобный эпидермальному фактору роста (НВ-EGF). Иллюстративным BBBR является рецептор трансферрина (TfR).
Термин "действующая в мозге группировка" ("эффекторная группировка") означает молекулу, которая должна быть транспортирована в мозг через ВВВ. Эффекторная группировка, как правило, имеет характерную терапевтическую активность, которую нужно доставить в мозг.Эффекторные группировки включают лекарственные препараты для лечения неврологических нарушений и цитотоксические агенты, такие как, например, полипептиды и антитела, в частности, моноклональные антитела или их фрагменты, направленные на мишень в мозге.
Термин "одновалентная связывающая группировка" означает молекулу, способную связываться с BBBR специфически и в режиме одновалентного связывания. Модуль-переносчик через гематоэнцефалический барьер и/или конъюгат, описанный в данном документе, характеризуются наличием одной единицы одновалентной связывающей группировки, т.е. модуль-переносчик через гематоэнцефалический барьер и/или конъюгат согласно данному изобретению содержат ровно одну единицу одновалентной связывающей группировки. Одновалентная связывающая группировка включает, но не ограничиваясь ими, полипептиды, полноразмерные антитела, фрагменты антител, включая Fab, Fab', Fv-фрагменты, молекулы одноцепочечных антител, такие как, например, одноцепочечный Fab, scFv. Одновалентная связывающая группировка может быть, например, каркасным белком, разработанным с помощью стандартных в данной области методик, таких как фаговый дисплей или иммунизация. Одновалентная связывающая группировка также может представлять собой полипептид. В некоторых воплощениях одновалентная связывающая группировка содержит СН2-СН3-Ig-домен и одноцепочечный Fab (scFab), направленный против рецептора гематоэнцефалического барьера. scFab соединен с С-концом СН2-СН3-Ig-домена с помощью линкера. В некоторых воплощениях scFab направлен против рецептора трансферрина.
Термин "режим одновалентного связывания" обозначает специфическое связывание с BBBR, где взаимодействие между одновалентной связывающей группировкой и BBBR происходит через один единственный эпитоп.Режим одновалентного связывания предотвращает димеризацию/мультимеризацию BBBR благодаря одной точке взаимодействия с эпитопом. Режим одновалентного связывания предотвращает изменения внутриклеточной сортировки BBBR.
Термин "эпитоп" означает любую полипептидную детерминанту, способную специфически связываться с антителом. В некоторых воплощениях антигенные детерминанты включают химически активные поверхностные группировки молекул, такие как аминокислоты, сахарные боковые цепи, фосфорильные или сульфонильные группировки, а в некоторых воплощениях они могут иметь специфические трехмерные структурные характеристики и/или специфические характеристики заряда. Эпитоп представляет собой область антигена, которая связывается антителом.
"Рецептор трансферрина" (TfR) представляет собой трансмембранный гликопротеин (с молекулярной массой примерно 180000 Да), который состоит из двух связанных дисульфидным мостиком субъединиц (каждая имеет кажущуюся молекулярную массу примерно 90000 Да) и участвует в поглощении железа у позвоночных. В одном из воплощений TfR в данном документе является человеческим TfR, содержащим аминокислотную последовательность, указанную в Schneider et al. (Nature 311 (1984) 675 - 678).
Термин "неврологическое нарушение" означает заболевание или нарушение, которое влияет на ЦНС, и/или которое имеет этиологию в ЦНС.Иллюстративные заболевания или нарушения ЦНС включают, но не ограничиваясь ими, нейропатию, амилоидоз, рак, глазное заболевание или нарушение, вирусную или бактериальную инфекцию, воспаление, ишемию, нейродегенеративное заболевание, апоплексический удар, поведенческие нарушения и лизосомальную болезнь накопления. В контексте данной заявки следует понимать, что ЦНС включает глаз, который, как правило, отделен от остальной части организма гемато-ретинальным барьером. Конкретные примеры неврологических нарушений включают, но не ограничиваясь ими, нейродегенеративные заболевания (включая, но не ограничиваясь ими, болезнь с тельцами Леви, постполиомиелитный синдром, синдром Шая - Дрейджера, оливопонтоцеребральную атрофию, болезнь Паркинсона, множественную системную атрофию, стриатонигральную дегенерацию, тауопатии (включая, но не ограничиваясь ими, болезнь Апьцгеймера и супрануклеарный паралич), прионные болезни (включая, но не ограничиваясь ими, губчатую энцефалопатию крупного рогатого скота, скрепи, синдром Крейтцфельда - Якоба, куру, болезнь Герстмана - Штраусслера - Шейнкера, хроническую изнуряющую болезнь и фатальную семейную бессонницу), бульбарный паралич, болезнь двигательного нейрона, а также гетеродегенеративные нарушения нервной системы (включая, но не ограничиваясь ими, болезнь Канавана, болезнь Хантингтона, нейронный восковидный липофусциноз, болезнь Александера, синдром Туретта, синдром курчавых волос Менкеса, синдром Коккейна, синдром Галлервордена - Шпатца, болезнь Лафоры, синдром Ретта, гепатолентикулярную дегенерацию, синдром Леша - Нихена и синдром Унферрихта - Лундборга), деменцию (включая, но не ограничиваясь ими, болезнь Пика и спиноцеребеллярную атаксию), рак (например, ЦНС и/или головного мозга, включая метастазы в головном мозге в результате рака в других частях организма).
Термин "лекарственный препарат для лечения неврологического нарушения" обозначает лекарственное средство или терапевтический агент, который лечит одно или более чем одно неврологическое нарушение. Лекарственные препараты для лечения неврологических нарушений включают, но не ограничиваясь ими, низкомолекулярные соединения, антитела, пептиды, белки, природные лиганды одной или более чем одной мишени ЦНС, модифицированные варианты природных лигандов одной или более чем одной мишени ЦНС, аптамеры, ингибирующие нуклеиновые кислоты (т.е. малые ингибирующие РНК (siRNA) и короткие шпилечные РНК (shRNA)), рибозимы и малые молекулы или активные фрагменты любого из вышеперечисленных соединений. Примеры лекарственных препаратов для лечения неврологического нарушения описаны в данном документе и включают, но не ограничиваясь ими: антитела, аптамеры, белки, пептиды, ингибирующие нуклеиновые кислоты и малые молекулы и активные фрагменты любого из вышеперечисленных соединений, которые либо выступают сами по себе, либо специфически распознают и/или воздействуют (т.е. ингибируют, активируют или обнаруживают) на антиген ЦНС или молекулы-мишени, такие как, но не ограничиваясь ими, белок-предшественник амилоида или его части, бета-амилоид, бета-секретаза, гамма-секретаза, тау-белок, альфа-синуклеин, паркин, хантингтин, DR6, пресенилин, АпоЕ, маркеры глиомы или другого рака ЦНС и нейротрофины. Неограничивающие примеры лекарственных препаратов для лечения неврологических нарушений и соответствующие нарушения, для лечения которых они могут быть использованы: нейротрофический фактор головного мозга (BDNF, от англ. -Brain-derived neurotrophic factor), хроническое повреждение головного мозга (нейрогенез), фактор роста фибробластов 2 (FGF-2, от англ. - Fibroblast growth factor 2), рак мозга и антитело против рецептора эпидермального фактора роста (EGFR), нейронный фактор из линии глиальных клеток (GDNF, от англ. - glial cell-line derived neural factor) при болезни Паркинсона, нейротрофический фактор из головного мозга (BDNF) при боковом амиотрофическом склерозе, депрессия, лизосомальный фермент и лизосомные болезни накопления мозга, цилиарный нейротрофический фактор (CNTF, от англ. - Ciliary neurotrophic factor) и боковой амиотрофический склероз, нейрегулин-1 и шизофрения, анти-HER2-антитело (например, трастузумаб) и метастазы HER2-положительного рака в головном мозге.
Термин "агент для визуализации" обозначает соединение, которое имеет одно или чем одно более свойство, позволяющее обнаруживать его присутствие и/или местонахождение прямо или косвенно. Примеры таких агентов для визуализации включают белки и низкомолекулярные соединения, включающие меченую группировку, которую можно обнаружить.
Термины "антиген ЦНС" и "мишень в головном мозге" обозначают антиген и/или молекулу, экспрессированную в ЦНС, включая головной мозг, на которую может быть нацелено антитело или малая молекула. Примеры таких антигенов и/или молекул включают, но не ограничиваясь ими: бета-секретазу 1 (ВАСЕ1), бета-амилоид (Abeta), рецептор эпидермального фактора роста (EGFR), рецептор человеческого эпидермального фактора роста 2 (HER2, от англ. - human epidermal growth factor receptor 2), тау-белок, аполипопротеин E4 (ApoE4), альфа-синуклеин, CD20, хантингтин, прионный белок (РгР, от англ. - prion protein), киназу 2 с богатым лейцином повтором (LRRK2, от англ. - leucine rich repeat kinase 2), паркин, пресенилин 1, пресенилин 2, гамма-секретазу, рецептор смерти 6 (DR6, от англ. - death receptor 6), белок-предшественник амилоида (АРР, от англ. - amyloid precursor protein), рецептор нейротрофина р75 (p75NTR, от англ. - р75 neurotrophin receptor) и каспазу 6. В одном из воплощений антиген представляет собой ВАСЕ1.
Термин "которое специфически связывает" означает антитело, селективно или предпочтительно связывающееся с антигеном. Аффинность связывания обычно определяют с помощью стандартного анализа, например, анализа Скэтчарда или поверхностного плазмонного резонанса (например, с использованием Biacore®).
Термин "СН2-СН3-Ig-группировка", используемый в данном документе, относится к белковой группировке, полученной из СН2- или СН3-домена иммуноглобулина. "СН2-СН3-Ig-группировка" включает два полипептида "СН2-СН3", формирующих димер. Иммуноглобулин может представлять собой IgG, IgA, IgD, IgE или IgM. В одном из воплощений СН2-СН3-Ig-группировка получена из иммуноглобулина IgG и упоминается в данном описании как "CH2-CH3-IgG-группировка". Термин включает нативную последовательность доменов СН2-СН3 и вариантные домены СН2-СН3. В одном воплощении "СН2-СН3-Ig-группировка" получена из человеческого СН2-СН3-домена тяжелой цепи IgG, который тянется от Cys226 или от Pro230 до карбоксильного конца тяжелой цепи. Тем не менее, С-концевой лизин (Lys447) Fc-области может присутствовать или может отсутствовать. Если не указано иное, то нумерация аминокислотных остатков в области СН2-СН3-доменов или константной области осуществляется в соответствии с системой нумерации ЕС, также называемой индексом ЕС, как описано в Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD, 1991.
"Конъюгат" представляет собой гибридный белок согласно данному изобретению, конъюгированный с одной или более чем одной гетерологичной молекулой, включая, но не ограничиваясь ими, метку, лекарственный препарат для лечения неврологического нарушения или цитотоксический агент.
Термин "линкер" означает химический линкер или одноцепочечный пептидный линкер, который ковалентно связывает различные группировки модуля-переносчика через гематоэнцефалический барьер и/или гибридный полипептид и/или конъюгат, описанный в данном документе. Линкер связывает, например, действующую в мозге группировку с одновалентной связывающей группировкой. Например, если одновалентная связывающая группировка содержит СН2-СН3-Ig-группировку и scFab, направленный против рецептора гематоэнцефалического барьера, то линкер конъюгирует scFab с С-концом СН3-СН2-Ig-группировки. Линкер, конъюгирующий действующую в мозге группировку с одновалентной связывающей группировкой (первый линкер), и линкер, соединяющий scFab с С-концом СН2-СН3-Ig-домена (второй линкер), могут быть одинаковыми или разными.
Могут быть использованы одноцепочечные пептидные линкеры, включающие от одного до двадцати аминокислотных остатков, соединенных пептидными связями. В некоторых воплощениях аминокислоты выбраны среди двадцати природных аминокислот.В некоторых других воплощениях одна или более чем одна из аминокислот выбрана среди глицина, аланина, пролина, аспарагина, глутамина и лизина. В других воплощениях линкер представляет собой химический линкер. В некоторых воплощениях линкер представляет собой одноцепочечный пептидный линкер с аминокислотной последовательностью длиной по меньшей мере 25 аминокислотных остатков, в одном предпочтительном воплощении длиной от 32 до 50 аминокислотных остатков. В одном из воплощений пептидный линкер представляет собой (G×S)n-линкер, где G - глицин, S - серии, (× равен 3, n равен 8, 9 или 10) или (х равен 4 и n равен 6, 7 или 8), в одном из воплощений × равен 4, n равен 6 или 7, в одном предпочтительном воплощении × равен 4, n равен 7. В одном из воплощений линкер представляет собой (G4S)4 (SED ID № 08). В одном из воплощений линкер представляет собой (G4S)6G2 (SEQ ID NO 07).
Конъюгация может быть выполнена с использованием различных химических линкеров. Например, одновалентная связывающая группировка или гибридный полипептид и действующая в мозге группировка могут быть конъюгированы с помощью множества бифункциональных агентов, связывающих белки, таких как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные имидоэфиров (такие как диметиладипимидат HCl), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаральдегид), бис-азидо-соединения (такие как бис(п-азидобензоил)гександиамин), производные бис-диазония (такие как бис(п-диазониябензоил)этилендиамин), диизоцианаты (такие как толуол-2,6-диизоцианат) и бис-активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). Линкер может представлять собой "расщепляемый линкер", облегчающий высвобождение эффекторной группировки после доставки в мозг.Например, может быть использован кислотно-лабильный линкер, чувствительный к пептидазе линкер, фотолабильный линкер, диметиловый линкер или линкер, содержащий дисульфид (Chari et al, Cancer Res. 52 (1992) 127-131; US 5208020).
Ковалентная конъюгация может быть осуществлена напрямую или через линкер. В некоторых воплощениях прямая конъюгации осуществляется путем конструирования гибридного полипептида (т.е. путем генетического слияния двух генов, кодирующих одновалентную связывающую группировку против BBBR и эффекторную группировку, и экспрессии в виде единого полипептида (цепи)). В некоторых воплощениях прямая конъюгация осуществляется путем формирования ковалентной связи между реакционноспособной группой на одной из двух частей одновалентной связывающей группировки против BBBR и соответствующей группой или акцептором на действующей в мозге группировке. В некоторых воплощениях прямая конъюгация осуществляется путем модификации (т.е. генетической модификации) одной из двух молекул, которые должны быть конъюгированы, в ходе которой включают реакционноспособную группу (в качестве неограничивающих примеров, сульфгидрильную группу или карбоксильную группу), которая формирует ковалентную связь с другой молекулой, которая должна быть конъюгирована в соответствующих условиях. В качестве неограничивающего примера, молекула (т.е. аминокислота) с нужной реакционноспособной группой (т.е. остатком цистеина) может быть введена, например, в одновалентную связывающую группировку против антитела BBBR, и дисульфидная связь формируется с препаратом для лечения неврологического нарушения. Способы ковалентной конъюгации нуклеиновых кислот с белками также известны в данной области (т.е. фотохимическое сшивание, см., например, Zatsepin et al. Russ. Chem. Rev. 74 (2005) 77-95). Конъюгация также может быть осуществляется с использованием различных линкеров. Например, одновалентная связывающая группировка и эффекторная группировка могут быть конъюгированы с помощью множества бифункциональных агентов, связывающих белки, таких как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные имидоэфиров (такие как диметиладипимидат HCl), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаральдегид), бис-азидо-соединения (такие как бис(п-азидобензоил)гександиамин), производные бис-диазония (такие как бис(п-диазониябензоил)этилендиамин), диизоцианаты (такие как толуол-2,6-диизоцианат) и бис-активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). Также могут быть использованы пептидные линкеры, содержащие от одного до двадцати аминокислотных остатков, соединенных пептидными связями. В некоторых таких воплощениях аминокислотные остатки выбраны среди двадцати природных аминокислот. В некоторых других таких воплощениях одна или более чем одна из аминокислот выбрана среди глицина, аланина, пролина, аспарагина, глутамина и лизина. Линкер может представлять собой "расщепляемый линкер", облегчающий высвобождение эффекторной группировки после доставки в мозг.Например, может быть использован кислотно-лабильный линкер, чувствительный к пептидазе линкер, фотолабильный линкер, диметиловый линкер или линкер, содержащий дисульфид (Chari et al, Cancer Res. 52 (1992) 127-131; US 5208020).
II. Композиции и способы
В одном из аспектов данное изобретение частично основано на обнаружении того факта, что модули-переносчики через гематоэнцефалический барьер, описанные в данном документе, могут быть использованы для доставки действующей в мозге группировки через гематоэнцефалический барьер в головной мозг.В некоторых воплощениях модуль-переносчик через гематоэнцефалический барьер включает одновалентную связывающую группировку, которая специфически связывается с рецептором гематоэнцефалического барьера, таким как рецептор трансферрина. Модули-переносчики через гематоэнцефалический барьер, описанные в данном документе, могут быть использованы, например, для диагностики или лечения неврологических нарушений, таких как болезнь Альцгеймера, болезнь Паркинсона и болезнь Альцгеймера с сопутствующей болезнью Паркинсона.
А. Иллюстративные антитела против рецептора гематоэнцефалического барьера
Одновалентные связывающие группировки, которые специфически связываются с рецептором гематоэнцефалического барьера, можно охарактеризовать в отношении свойств их связывания и трансцитоза:
- эффективное связывание клеток, экспрессирующих BBBR, в качестве одновалентной связывающей группировки,
- эффективный трансцитоз in vitro в качестве одновалентной связывающей группировки,
- перекрестная реактивность человек - яванский макак (например, в экспериментах BIAcore и FACS).
Скрининг трансцитоза проводили в анализе, основанном на hCMEC/D3. Анализ проводили в режиме вытеснения метки. Эндотелиальные клетки мозга hCMEC/D3 инкубировали с одновалентной связывающей группировкой в течение 1 часа, промывали, а затем определяли следующие параметры через 0 часов и 4 часа после промывания:
i) количество одновалентной связывающей группировки, захваченное клетками во время фазы загрузки,
ii) базолатеральное количество одновалентной связывающей группировки через 4 часа после загрузки и промывания;
iii) апикальное количество одновалентной связывающей группировки через 4 часа после загрузки и промывания;
iv) количество одновалентной связывающей группировки в клетках (по лизису клеток) через 0 ч и 4 ч после загрузки и промывания;
v) общее количество одновалентной связывающей группировки через 0 часов и 4 часа после загрузки и промывания.
Для того чтобы выступать в качестве одновалентной связывающей группировки в модуле-переносчике через гематоэнцефалический барьер, описанном в данном документе, одновалентная связывающая группировка должна быть i) захвачена клетками hCMEC/D3 (эндоцитоз), ii) транспортирована вне клеток hCMEC/D3 (экзоцитоз), и iii) стабильна внутри клеток hCMEC/D3 (отсутствие или низкий уровень транспорта в эндосомы для деградации).
Таким образом, в одном воплощении одновалентная связывающая группировка характеризуется в анализе на основании hCMEC/D3 i) (существенным) поглощением hCMEC/D3-клетками в течение периода загрузки длительностью один час, ii) высвобождением в апикальном и/или базолатеральном компартменте после этапа загрузки и этапа промывания через 4 часа после промывания, и iii) низкой (внутриклеточной) скоростью деградации.
В одном воплощении загрузка осуществляется в концентрации одновалентной связывающей группировки примерно 2,67 мкг/мл в течение одного часа.
Было обнаружено, что одновалентная связывающая группировка для того, чтобы выступать в качестве одновалентной связывающей группировки в модуле-переносчике через гематоэнцефалический барьер, описанном в данном документе, должна показать в описанном выше анализе на основе hCMEC/D3 следующие пороговые значения:
i) количество одновалентной связывающей группировки, захваченной клетками во время фазы загрузки, 400 пг или более,
ii) базолатеральное количество одновалентной связывающей группировки через 4 ч после загрузки и промывания - 100 пг или более, и
iii) апикальное количество одновалентной связывающей группировки через 4 часа после загрузки и промывания - 150 пг или более.
Мышиное антитело против человеческого рецептора трансферрина 128.1 (последовательности вариабельных областей см. в WO 93/10819 и SEQ ID NO 11 и 12) может быть взято в качестве референсного. В этом случае одновалентная связывающая группировка для того, чтобы выступать в качестве одновалентной связывающей группировки в модуле-переносчике через гематоэнцефалический барьер, описанном в данном документе, должна показать в описанном выше анализе на основе hCMEC/D3 следующие пороговые значения:
i) количество одновалентной связывающей группировки, захваченной клетками во время фазы загрузки, 20% или более от загрузки антитела 128.1,
ii) базолатеральное количество одновалентной связывающей группировки через 4 ч после загрузки и промывания - 15% или более от базолатерального количества антитела 128.1; и
iii) апикальное количество одновалентной связывающей группировки через 4 часа после загрузки и промывания - 15% или более от апикального количества антитела 128.1.
Анализ на основе hCMEC/D3 проводили следующим образом (это одно из воплощений всех аспектов, описанных в данном документе):
Среда и добавки для hCMEC/D3 (см. WO 2006/056879 и Weksler, В.В., et al., FAS ЕВ J. 19 (2005) 1872-1874) могут быть получены из Lonza. Клетки hCMEC/D3 (пассажи 26-29) культивируют/могут культивировать до конфлюэнтности на покрытых коллагеном покровных стеклах (для микроскопии) или в колбах в среде ЕВМ2, содержащей 2,5% FBS и четверть от поставляемых факторов роста и полностью дополненной поставляемых гидрокортизоном, гентамицином и аскорбиновой кислотой.
Для всех анализов трансцитоза в 12-луночных планшетах для культивирования клеток используются/могут использоваться вставные фильтры (размер пор 0,4 мкм, диаметр 12 мм) с РЕТ-мембраной с высокой плотностью пор (1×108 пор/см2). Объемы среды рассчитывают как 400 мкл и 1600 мкл для апикальной и базолатеральной камер, соответственно. Апикальные камеры вставных фильтров покрывают/могут покрывать коллагеном I крысиного хвоста (7,5 мкг/см2), а затем фибронектином (5 мкг/мл), при этом каждую инкубацию осуществляют в течение 1 ч при комнатной температуре. Клетки hCMEC/D3 выращивают/могут выращивать до конфлюэнтных монослоев (примерно 2×105 клеток/см2) в течение 10-12 дней в среде ЕВМ2. Пустые фильтры блокируют/могут блокировать с помощью PBS, содержащего 1% BSA, в течение 1 ч или в течение ночи (о/n) перед анализом, а затем калибруют в ЕВМ2 в течение по меньшей мере 1 часа перед анализом.
Анализ (схему анализа см. на фиг.1) проводили в бессывороточной среде ЕВМ2, которую в противном случае восстанавливали, как описано в данном документе. В день анализа клетки инкубировали в бессывороточных условиях в течение 60 минут для истощения природного лиганда рассматриваемого рецептора гематоэнцефалического барьера. Вставные фильтры с клетками или без них (но блокированные в течение ночи в полной среде) инкубировали апикально с рассматриваемыми моноклональными антителами (одновалентная связывающая группировка) в течение 1 ч при 37°С. Монослои промывали при комнатной температуре (RT) в бессывороточной среде апикально (400 мкп) и базолатерально (1600 мкп) три раза в течение 3-5 минут каждый. Предварительно согретую среду добавляли в апикальную камеру и фильтры переносили в свежий 12-луночный планшет (заблокированный в течение ночи с помощью PBS, содержащего 1% BSA), содержащий 1600 мкл предварительно согретой среды. В этот момент фильтры с клетками или без клеток лизировали в 500 мкл буфера RIPA, чтобы определить поглощение специфического антитела (одновалентной связывающей группировки). Оставшиеся фильтры инкубировали при температуре 37°С или 4°С и собирали образцы в различные моменты времени, чтобы определить апикальное и/или базолатеральное высвобождение антитела (одновалентной связывающей группировки). Содержание антител в образцах можно количественно оценить с использованием высокочувствительного ELISA на IgG (см. пример 10). Для каждой временной точки данные должны быть получены от двух пустых фильтров и трех фильтров с клеточными культурами.
Результаты для 69 антител против рецептора трансферрина показаны в приведенной ниже таблице. Антитело 128.1 был использовано в качестве референсного.

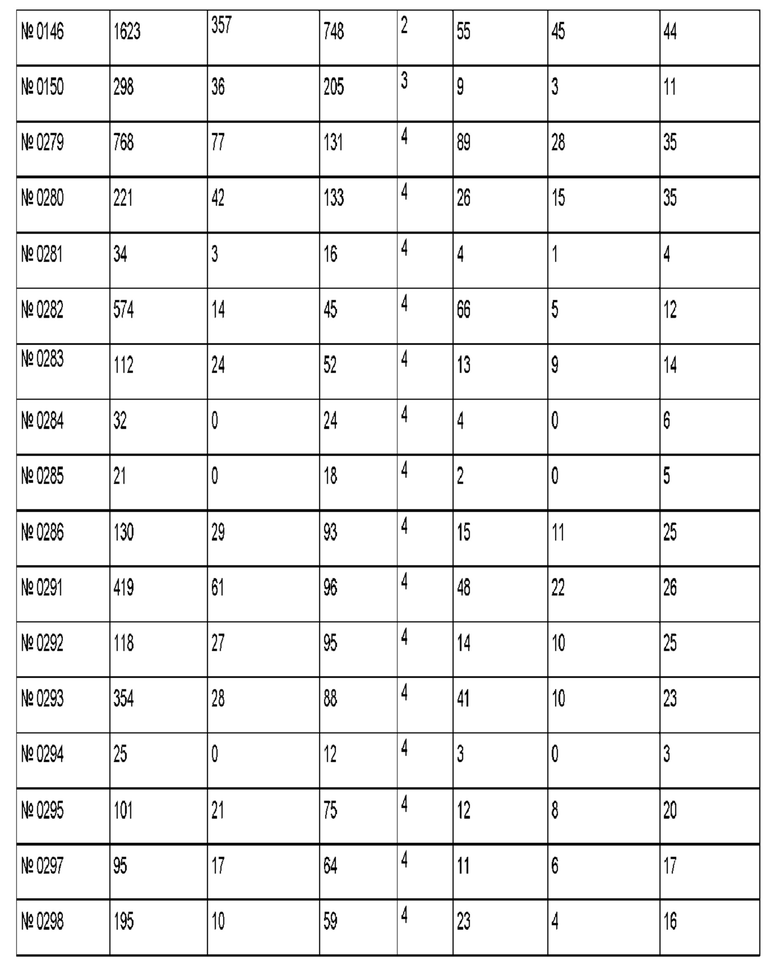
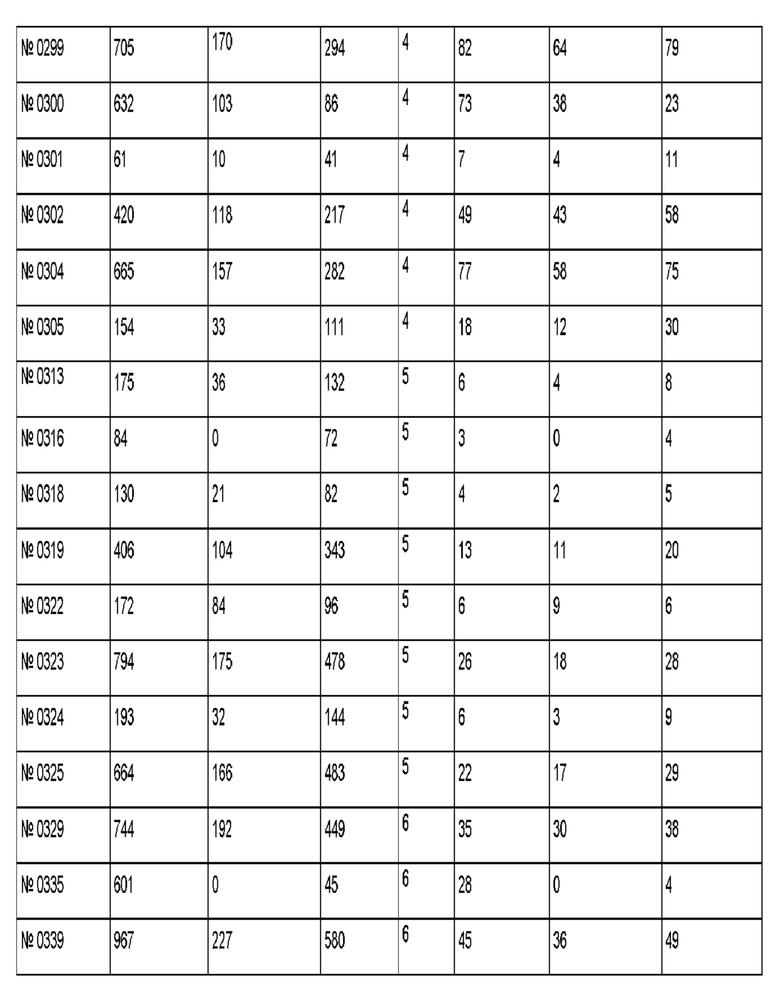
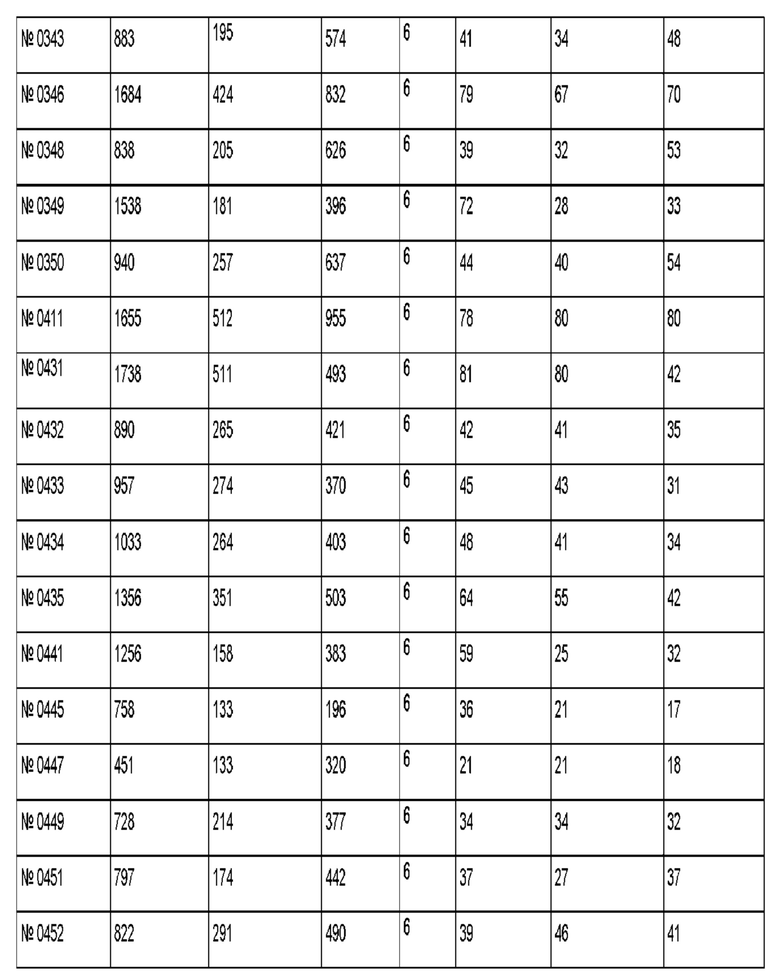
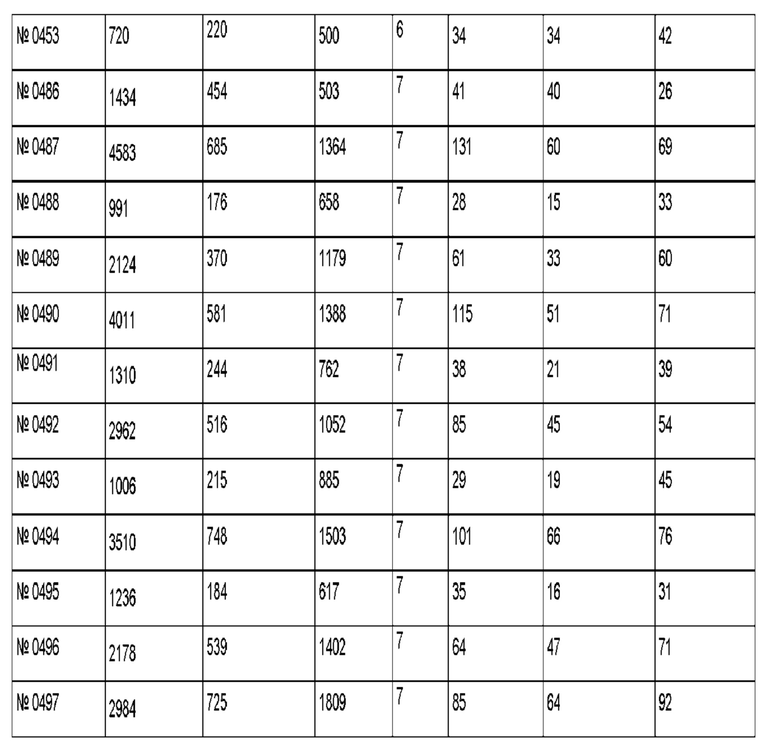
Антитело 299 демонстрирует трансцитозную загрузку 705 пг, где через 4 ч 170 пг (соответствует 24% загрузки) можно обнаружить в базолатеральном компартменте, и 294 пг (соответствует 42% загрузки) можно обнаружить в апикальном компартменте.
Антитело 494 демонстрирует трансцитозную загрузку 3510 пг, где через 4 ч 748 пг (соответствует 21% загрузки) можно обнаружить в базолатеральном компартменте, и 1503 пг (соответствует 43% загрузки) можно обнаружить в апикальном компартменте.
Антитела, которые отвечают критериям, указанным выше, являются воплощениями данного изобретения.
Таким образом, один из аспектов, описанных в данном документе, представляет собой антитело против рецептора трансферрина или его фрагмент, связывающий рецептор трансферрина, содержащий
(1) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 13, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 14, или
(2) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 15, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 16, или
(3) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 17, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 18, или
(4) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 19, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 20, или
(5) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 21, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 22, или
(6) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 23, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 24, или
(7) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 25, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 26, или
(8) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 27, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 28, или
(9) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 29, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 30, или
(10) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 31, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 32, или
(11) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 33, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 34, или
(12) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 35, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 36, или
(13) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 37, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 38, или
(14) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 39, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 40, или
(15) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 41, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 42, или
(16) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 43, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 44, или
(17) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 45, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 46, или
(18) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 47, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 48, или
(19) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 49, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 50, или
(20) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 51, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 52, или
(21) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 53, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 54, или
(22) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 55, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 56, или
(23) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 57, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 58, или
(24) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 59, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 60, или
(25) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 61, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 62, или
(26) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 63, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 64, или
(27) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 65, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 66, или
(28) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 67, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 68, или
(29) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 69, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 70, или
(30) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 71, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 72, или
(31) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 73, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 74, или
(32) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 75, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 76, или
(33) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 77, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 78, или
(34) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 79, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 80, или
(35) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 81, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 82, или
(36) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 83, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 84, или
(37) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 85, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 86, или
(38) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 87, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 88, или
(39) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 89, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 90, или
(40) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 91, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 92.
(41) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 93, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 94, or
(42) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 95, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 96, or
(43) вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 97, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 98.
Одним из предпочтительных аспектов, описанных в данном документе, является антитело против рецептора трансферрина или его фрагмент, связывающий рецептор трансферрина, содержащий вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 23, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 24.
Одним из предпочтительных аспектов, описанных в данном документе, является антитело против рецептора трансферрина или его фрагмент, связывающий рецептор трансферрина, содержащий вариабельный домен тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 91, и вариабельный домен легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 92.
Соответствующие аминокислотные последовательности показаны в следующей таблице.




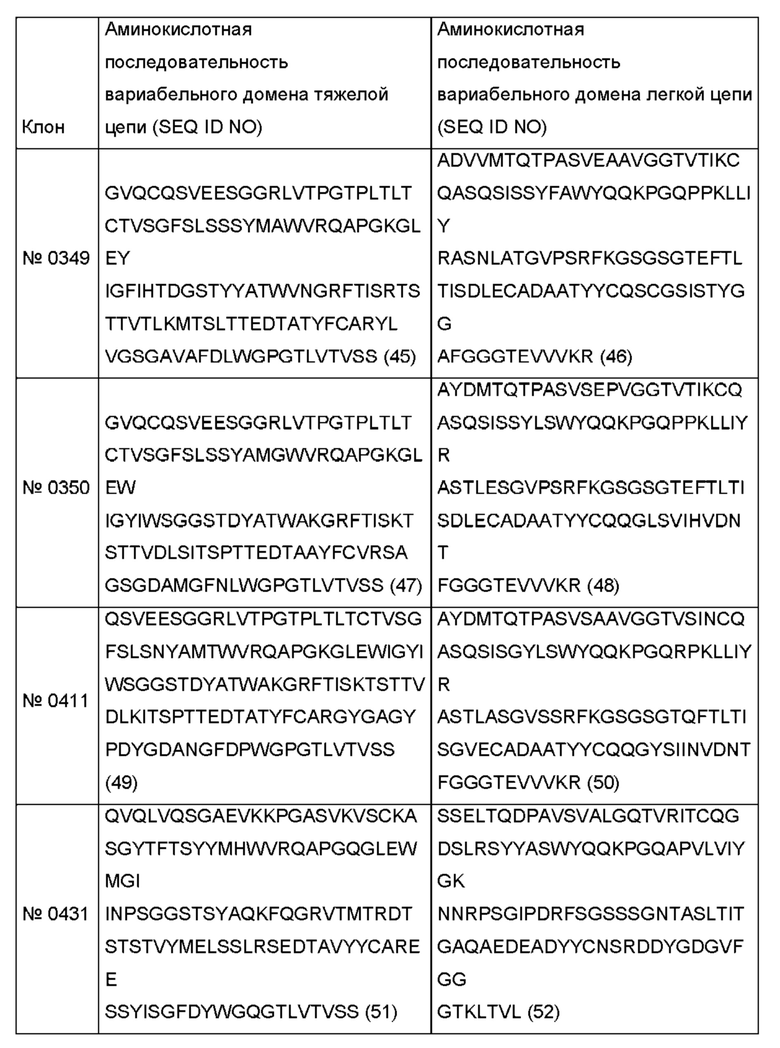


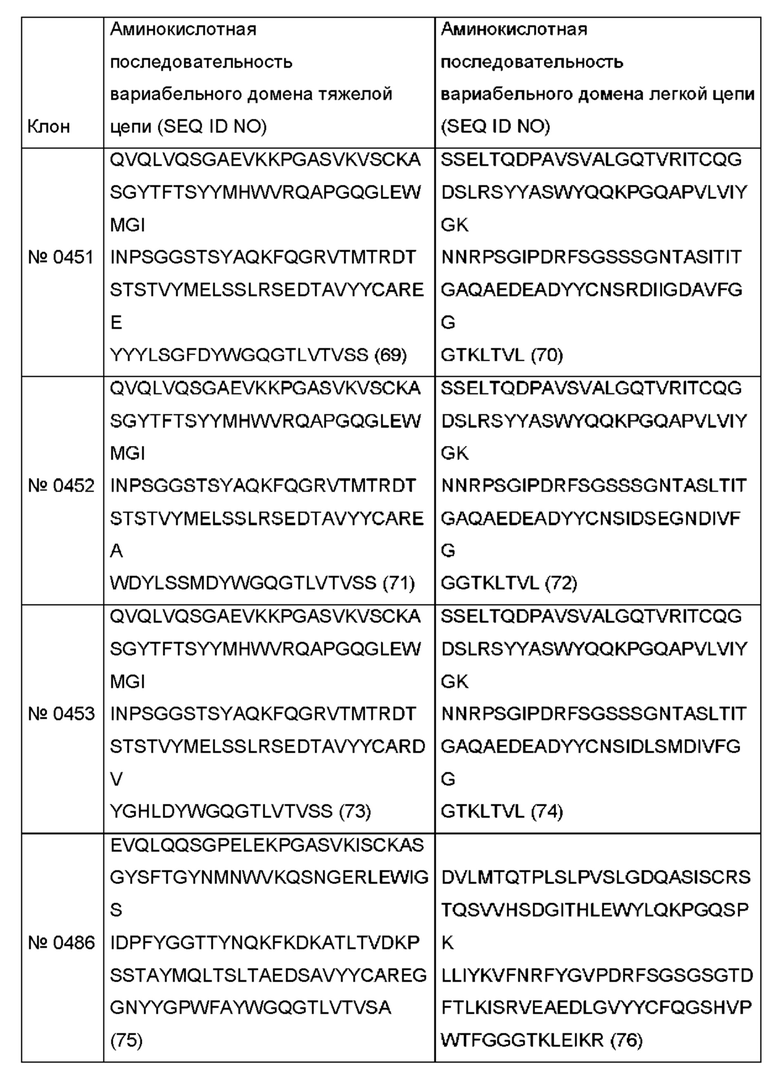



Одним из аспектов, описанных в данном документе, является гуманизированное антитело против рецептора трансферрина или его фрагмент, связывающий рецептор трансферрина, содержащий
(1) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 13, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 14, или
(2) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 15, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 16, или
(3) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 17, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 18, или
(4) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 19, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 20, или
(5) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 21, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 22, или
(6) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 23, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 24, или
(7) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 25, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 26, или
(8) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 27, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 28, или
(9) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 29, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 30, или
(10) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 31, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 32, или
(11) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 33, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 34, или
(12) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 35, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 36, или
(13) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 37, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 38, или
(14) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 39, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 40, или
(15) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 41, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 42, или
(16) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 43, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 44, или
(17) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 45, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 46, или
(18) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 47, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 48, или
(19) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 49, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 50, или
(20) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 51, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 52, или
(21) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 53, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 54, или
(22) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 55, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 56, или
(23) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 57, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 58, или
(24) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 59, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 60, или
(25) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 61, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 62, или
(26) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 63, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 64, или
(27) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 65, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 66, или
(28) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 67, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 68, или
(29) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 69, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 70, или
(30) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 71, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 72, или
(31) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 73, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 74, или
(32) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 75, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 76, или
(33) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 77, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 78, или
(34) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 79, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 80, или
(35) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 81, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 82, или
(36) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 83, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 84, или
(37) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 85, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 86, или
(38) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 87, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 88, или
(39) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 89, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 90, или
(40) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 91, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 92.
(41) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 93, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 94, or
(42) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 95, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 96, or
(43) гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 97, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 98.
Одним из предпочтительных аспектов, описанных в данном документе, является гуманизированное антитело против рецептора трансферрина или его фрагмент, связывающий рецептор трансферрина, содержащий гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 23, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 24.
Одним из аспектов, описанных в данном документе, является гуманизированное антитело против человеческого рецептора трансферрина, содержащее (a) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO 109; (b) HVR-H2, содержащий аминокислотную последовательность SEQ ID NO 110; и (с) HVR-H3, содержащий аминокислотную последовательность SEQ ID NO 112. В одном воплощении антитело также содержит (d) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO 113; (e) HVR-L2, содержащий аминокислотную последовательность SEQ ID NO 114; и (f) HVR-L3, содержащий аминокислотную последовательность SEQ ID NO 115.
Одним из аспектов, описанных в данном документе, является гуманизированное антитело против человеческого рецептора трансферрина, содержащее (a) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO 109; (b) HVR-H2, содержащий аминокислотную последовательность SEQ ID NO 111; и (с) HVR-H3, содержащий аминокислотную последовательность SEQ ID NO 112. В одном воплощении антитело также содержит (d) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO 113; (е) HVR-L2, содержащий аминокислотную последовательность SEQ ID NO 114; и (f) HVR-L3, содержащий аминокислотную последовательность SEQ ID NO 115.
В одном аспекте изобретение предусматривает антитело против рецептора трансферрина, содержащее по меньшей мере, один, два, три, четыре, пять или шесть HVR, выбранных среди следующих: (a) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO 109; (b) HVR-H2, содержащий аминокислотную последовательность SEQ ID NO 110; (с) HVR-H3, содержащий аминокислотную последовательность SEQ ID NO 112; (d) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO 113; (е) HVR-L2, содержащий аминокислотную последовательность SEQ ID NO 114; и (f) HVR-L3, содержащий аминокислотную последовательность SEQ ID NO 115.
В одном аспекте изобретение предусматривает антитело, содержащее по меньшей мере одну, две или все три HVR-последовательности VH, выбранные среди следующих: (a) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO 109; (b) HVR-H2, содержащий аминокислотную последовательность SEQ ID NO 110; и (с) HVR-H3, содержащий аминокислотную последовательность SEQ ID NO 112. В одном воплощении антитело также содержит HVR-H3, содержащий аминокислотную последовательность SEQ ID NO 112. В другом воплощении антитело также содержит HVR-H3, содержащий аминокислотную последовательность SEQ ID NO 112, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO 115. В другом воплощении антитело также содержит HVR-H3, содержащий аминокислотную последовательность SEQ ID NO 112, HVR-L3, содержащий аминокислотную последовательность SEQ ID NO 115, и HVR-H2, содержащий аминокислотную последовательность SEQ ID NO 111. В другом воплощении антитело также содержит (a) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO 109; (b) HVR-H2, содержащий аминокислотную последовательность SEQ ID NO 111; и (с) HVR-Н3, содержащий аминокислотную последовательность SEQ ID NO 112.
В другом аспекте изобретение предусматривает антитело, содержащее по меньшей мере одну, две или все три VL HVR-последовательности, выбранные среди следующих: (a) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO 113; (b) HVR-L2, содержащий аминокислотную последовательность SEQ ID NO 114; и (с) HVR-L3, содержащий аминокислотную последовательность SEQ ID NO 115. В одном воплощении, антитело также содержит (a) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO 113; (b) HVR-L2, содержащий аминокислотную последовательность SEQ ID NO 114; и (с) HVR-L3, содержащий аминокислотную последовательность SEQ ID NO 115.
В другом аспекте антитело согласно изобретению также содержит (а) VH-домен, содержащий по меньшей мере одну, две или все три HVR-последовательности VH, выбранные среди следующих: (i) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO 109, (ii) HVR-H2, содержащий аминокислотную последовательность SEQ ID NO 110, и (iii) HVR-H3, содержащий аминокислотную последовательность, выбранную среди SEQ ID NO 112; и (b) VL-домен, содержащий по меньшей мере одну, две или все три HVR-последовательности VL, выбранные среди следующих: (i) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO 113, (ii) HVR-L2, содержащий аминокислотную последовательность SEQ ID NO 114, и (с) HVR-L3, содержащий аминокислотную последовательность SEQ ID NO 115.
В другом аспекте изобретение предусматривает антитело, содержащее (a) HVR-Н1, содержащий аминокислотную последовательность SEQ ID NO 109; (b) HVR-H2, содержащий аминокислотную последовательность SEQ ID NO 110; (с) HVR-H3, содержащий аминокислотную последовательность SEQ ID NO 112; (d) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO 113; (е) HVR-L2, содержащий аминокислотную последовательность SEQ ID NO 114; и (f) HVR-L3, содержащий аминокислотную последовательность, выбранную из SEQ ID NO 115.
Одним из предпочтительных аспектов, описанных в данном документе, является гуманизированное антитело против рецептора трансферрина или его фрагмент, связывающий рецептор трансферрина, содержащий гуманизированный вариабельный домен тяжелой цепи, полученный из вариабельного домена тяжелой цепи, который имеет аминокислотную последовательность SEQ ID NO 91, и гуманизированный вариабельный домен легкой цепи, полученный из вариабельного домена легкой цепи, который имеет аминокислотную последовательность SEQ ID NO 92.
Одним из аспектов, описанных в данном документе, является гуманизированное антитело против человеческого рецептора трансферрина, содержащее (a) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO 116; (b) HVR-H2, содержащий аминокислотную последовательность SEQ ID NO 117; и (с) HVR-H3, содержащий аминокислотную последовательность SEQ ID NO 119. В одном воплощении антитело также содержит (d) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO 120; (е) HVR-L2, содержащий аминокислотную последовательность SEQ ID NO 121; и (f) HVR-L3, содержащий аминокислотную последовательность SEQ ID NO 122.
Одним из аспектов, описанных в данном документе, является гуманизированное антитело против человеческого рецептора трансферрина, содержащее (a) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO 116; (b) HVR-H2, содержащий аминокислотную последовательность SEQ ID NO 118; и (с) HVR-H3, содержащий аминокислотную последовательность SEQ ID NO 119. В одном воплощении антитело также содержит (d) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO 120; (е) HVR-L2, содержащий аминокислотную последовательность SEQ ID NO 121; и (f) HVR-L3, содержащий аминокислотную последовательность SEQ ID NO 122.
В одном аспекте изобретение предусматривает антитело против рецептора трансферрина, содержащее по меньшей мере, один, два, три, четыре, пять или шесть HVR, выбранных среди следующих: (a) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO 116; (b) HVR-H2, содержащий аминокислотную последовательность SEQ ID NO 117; (с) HVR-H3, содержащий аминокислотную последовательность SEQ ID NO 119; (d) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO 120; (е) HVR-L2, содержащий аминокислотную последовательность SEQ ID NO 121; и (f) HVR-L3, содержащий аминокислотную последовательность SEQ ID NO 122.
В одном аспекте изобретение предусматривает антитело, содержащее по меньшей мере одну, две или все три HVR-последовательности VH, выбранные среди следующих: (a) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO 116; (b) HVR-H2, содержащий аминокислотную последовательность SEQ ID NO 117; и (с) HVR-H3, содержащий аминокислотную последовательность SEQ ID NO 119. В одном воплощении, антитело также содержит HVR-H3, содержащий аминокислотную последовательность SEQ ID NO 119. В другом воплощении антитело также содержит HVR-H3, содержащий аминокислотную последовательность SEQ ID NO 119, и HVR-L3, содержащий аминокислотную последовательность SEQ ID NO 122. В другом воплощении антитело также содержит HVR-H3, содержащий аминокислотную последовательность SEQ ID NO 119, HVR-L3, содержащий аминокислотную последовательность SEQ ID NO 122, и HVR-H2, содержащий аминокислотную последовательность SEQ ID NO 118. В другом воплощении антитело также содержит (a) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO 116; (b) HVR-H2, содержащий аминокислотную последовательность SEQ ID NO 118; и (с) HVR-Н3, содержащий аминокислотную последовательность SEQ ID NO 119.
В другом аспекте изобретение предусматривает антитело, содержащее по меньшей мере одну, две или все три HVR-последовательности VL, выбранные среди следующих: (a) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO 120; (b) HVR-L2, содержащий аминокислотную последовательность SEQ ID NO 121; и (с) HVR-L3, содержащий аминокислотную последовательность SEQ ID NO 122. В одном воплощении, антитело также содержит (a) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO 120; (b) HVR-L2, содержащий аминокислотную последовательность SEQ ID NO 121; и (с) HVR-L3, содержащий аминокислотную последовательность SEQ ID NO 122.
В другом аспекте антитело согласно изобретению также содержит (а) VH-домен, содержащий по меньшей мере одну, две или все три HVR-последовательности VH, выбранные среди следующих: (i) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO 116, (ii) HVR-H2, содержащий аминокислотную последовательность SEQ ID NO 117, и (iii) HVR-H3, содержащий аминокислотную последовательность, выбранную среди SEQ ID NO 119; и (b) VL-домен, содержащий по меньшей мере одну, две или все три VL HVR-последовательности, выбранные среди следующих: (i) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO 120, (ii) HVR-L2, содержащий аминокислотную последовательность SEQ ID NO 121, и (с) HVR-L3, содержащий аминокислотную последовательность SEQ ID NO 122.
В другом аспекте изобретение предусматривает антитело, содержащее (a) HVR-Н1, содержащий аминокислотную последовательность SEQ ID NO 116; (b) HVR-H2, содержащий аминокислотную последовательность SEQ ID NO 117; (с) HVR-H3, содержащий аминокислотную последовательность SEQ ID NO 119; (d) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO 120; (е) HVR-L2, содержащий аминокислотную последовательность SEQ ID NO 121; и (f) HVR-L3, содержащий аминокислотную последовательность, выбранную среди SEQ ID NO 122.
Одним из аспектов, описанных в данном документе, является модуль-переносчик трансферрина, содержащий полноразмерное антитело IgG в качестве действующей в мозге группировки, линкер и один scFab в качестве одновалентной связывающей группировки, которая связывается с рецептором трансферрина, где scFab конъюгирован с С-концом Fc-области одной из тяжелых цепей антитела IgG через линкер.
Одним из аспектов, описанных в данном документе, является модуль-переносчик трансферрина, содержащий полноразмерное антитело IgG в качестве действующей в мозге группировки, линкер и один scFv в качестве одновалентной связывающей группировки, которая связывается с рецептором трансферрина, где scFv конъюгирован с С-концом Fc-области одной из тяжелых цепей антитела IgG через линкер.
В одном предпочтительном воплощении одновалентная связывающая группировка содержит HVR с SEQ ID NO 109, 110, 112, 113, 114 и 115 или с SEQ ID NO 116, 117, 119, 120, 121 и 122.
В другом аспекте антитело против рецептора трансферрина согласно любому из описанных выше воплощений может включать любой из признаков, по отдельности или в комбинации, как описано в разделах 1-6 ниже:
1. Аффинность антитела
В одном воплощении Kd определяется в анализе связывания антигена, меченного радиоактивной меткой (radiolabeled antigen binding assay, RIA). В одном воплощении RIA выполняется с Fab-версией антитела, представляющего интерес, и его антигеном. Например, аффинность связывания Fab с антигеном в растворе измеряется путем уравновешивания Fab с минимальной концентрацией Обмеченного антигена в присутствии серии титров немеченого антигена, затем захвата связанного антигена на покрытом aHTH-Fab-антителом планшете (см., например, Chen, Y. et al., J. Mol. Biol. 293 (1999) 865-881). Чтобы создать условия для анализа, многолуночные планшеты MICROTITER® (Thermo Scientific) покрывали в течение ночи анти-Fab-антителом захвата (Cappel Labs) в концентрации 5 мкг/мл в 50 мМ растворе карбоната натрия (рН 9,6), а затем блокировали 2% (вес/объем) бычьим сывороточным альбумином в PBS в течение 2-5 часов при комнатной температуре (примерно 23°С). В неадсорбирующем планшете (Nunc №269620) смешивали 100 пМ или 26 пМ [125I]-антигена с серийными разведениями Fab, представляющего интерес (например, в соответствии с оценкой анти-VEGF-антитела, Fab-12, в Presta, L.G. et al., Cancer Res. 57 (1997) 4593-4599). Затем Fab, представляющий интерес, инкубировали в течение ночи; тем не менее, инкубацию можно продолжать в течение более долгого периода (например, примерно 65 часов), чтобы гарантировать достижение равновесия. После этого смеси переносили в планшет для захвата и инкубировали при комнатной температуре (например, в течение одного часа). Затем раствор удаляли и планшет промывали восемь раз с помощью 0,1% полисорбата 20 (Tween-20®) в PBS. Когда планшеты высыхали, в них добавляли 150 мкл/лунка сцинтиллянта (MICROSCINT-20™; Packard), и проводили анализ на гамма-счетчике TOPCOUNT™ (Packard) в течение 10 минут. Концентрации каждого Fab, которые давали 20% от максимального связывания или меньше, выбирали для применения в анализах конкурентного связывания.
В соответствии с другим воплощением Kd измеряли в анализе поверхностного плазмонного резонанса с использованием BIACORE®-2000 или BIACORE®-3000 (BIAcore, Inc., Пискатавэй, Нью-Джерси) при 25°С на СМ5-чипах с иммобилизованным антигеном в количестве примерно 10 единиц ответа (RU). В одном воплощении биосенсорные чипы из карбоксиметилированного декстрана (СМ5, BIACORE, Inc) активировали N-этил-N'-(3-диметиламинопропил)карбодиимида гидрохлоридом (EDC) и N-гидроксисукцинимидом (NHS) в соответствии с инструкциями производителя. Антиген разводили в 10 мМ растворе ацетата натрия, рН 4,8, до концентрации 5 мкг/мл (примерно 0,2 мкМ), а затем вводили со скоростью потока 5 мкл/мин для достижения примерно 10 единиц ответа (RU) связанного белка. После введения антигена вводили 1 М этаноламин для блокировки непрореагировавших групп.Для кинетических измерений вводили двукратные серийные разведения Fab (от 0,78 нМ до 500 нМ) в PBS с 0,05% поверхностно-активного вещества полисорбата 20 (TWEEN-20™) (PBST) при 25°С со скоростью потока примерно 25 мкл/мин. Скорости ассоциации (kon) и скорости диссоциации (koff) рассчитывали с помощью простой модели связывания Ленгмюра 1:1 (программное обеспечение BIACORE® Evaluation Software версии 3.2) путем одновременной подгонки сенсограмм ассоциации и диссоциации. Константу равновесия диссоциации (Kd) рассчитывали как отношение koff/kon (см., например, Chen, Y. et al., J. Mol. Biol. 293 (1999) 865-881). Если on-скорость превышала 106 M-1 с-1 в анализе поверхностного плазмонного резонанса, описанном выше, то оп-скорость могла быть определена с помощью методики тушения флуоресценции, которая измеряет увеличение или уменьшение интенсивности флуоресцентного излучения (возбуждение равно 295 нм; эмиссия равна 340 нм; 16 нм полосовой фильтр) при 25°С с 20 нМ антитела против антигена (Fab-форма) в PBS, рН 7,2, в присутствии возрастающих концентраций антигена, измеряемых спектрометром, таком как спектрофотометр, оборудованный системой быстрого смешивания, с последующей остановкой (stop-flow) (Aviv Instruments) или спектрофотометр SLM-AMINCO™ серии 8000 (ThermoSpectronic) со встряхивающейся кюветой.
2. Фрагменты антител
В некоторых воплощениях антитело, описанное в данном документе, представляет собой фрагмент антитела. Фрагменты антител включают, но не ограничиваясь ими, Fab-, Fab'-, Fab'-SH-, F(ab')2-, Fv- и scFv-фрагменты, а также другие фрагменты, описанные ниже. Обзор определенных фрагментов антител см. в Hudson, P.J. et al., Nat. Med. 9 (2003) 129-134. Обзор scFv-фрагментов см., например, в Plueckthun, A., In; The Pharmacology of Monoclonal Antibodies, Vol.113, Rosenburg and Moore (eds.), Springer-Verlag, New York (1994), pp.269-315; см. также WO 93/16185; US 5571894 и US 5587458. Для обсуждения Fab- и Р(ab')2-фрагментов, содержащих рецепторсвязывающие антигенные детерминанты реутилизации и имеющих повышенный период полужизни in vivo, см. US 5869046.
Димерные антитела представляют собой фрагменты антител с двумя антигенсвязывающими сайтами, которые могут быть двухвалентными или биспецифическими. См., например, ЕР 0404097; WO 1993/01161; Hudson, P.J. et al., Nat. Med. 9 (2003) 129-134; и Holliger, P. et al., Proc. Natl. Acad. Sci. USA 90 (1993) 6444-6448. В Hudson, P.J. et al., Nat. Med. 9 (2003) 129-134) также описаны тримерные и тетрамерные антитела.
Однодоменные антитела представляют собой фрагменты антител, содержащие все или часть вариабельных доменов тяжелой цепи или все или часть вариабельных доменов легкой цепи антитела. В некоторых воплощениях однодоменное антитело является человеческим однодоменным антителом (Domantis, Inc., Уолтем, Массачусетс; см., например, US 6248516).
Фрагменты антител могут быть получены с помощью различных методик, включая, но не ограничиваясь ими, протеолитическое расщепление интактного антитела, а также продукцию рекомбинантными клетками-хозяевами (например, Е, coli или фагом), как описано в данном документе.
3. Химерные и гуманизированные антитела
В некоторых воплощениях антитело, представленное в данном документе, является химерным антителом. Некоторые химерные антитела описаны, например, в US 4816567; и в Morrison, S.L. et al., Proc. Natl. Acad. Sci. USA 81 (1984) 6851-6855. В одном примере химерное антитело содержит нечеловеческую вариабельную область (например, вариабельную область, полученную от мыши, крысы, хомяка, кролика или примата, например, обезьяны) и человеческую константную область. В другом примере химерное антитело представляет собой антитело "с переключением класса", в котором класс или подкласс был изменен в сравнении с родительским антителом. Химерные антитела включают их антигенсвязывающие фрагменты.
В некоторых воплощениях химерное антитело представляет собой гуманизированное антитело. Как правило, нечеловеческое антитело гуманизируют, чтобы уменьшить иммуногенность для людей, сохраняя при этом специфичность и аффинность родительского нечеловеческого антитела. Как правило, гуманизированное антитело содержит один или более чем один вариабельный домен, в котором HVR, например, CDR (или их части), получены из нечеловеческого антитела, a FR (или их части) получены из последовательностей человеческого антитела. Гуманизированное антитело, возможно, также будет содержать по меньшей мере часть человеческой константной области. В некоторых воплощениях некоторые FR-остатки в гуманизированном антителе замещают соответствующими остатками из нечеловеческого антитела (например, антитела, из которого получены HVR-остатки), например, для восстановления или повышения специфичности или аффинности антитела.
Гуманизированные антитела и способы их получения приведены, например, в Almagro, J.С.and Fransson, J., Front. Biosci. 13 (2008) 1619-1633, а также описаны, например, в Riechmann, I. et al., Nature 332 (1988) 323-329; Queen, C. et al., Proc. Natl. Acad. Sci. USA 86 (1989) 10029-10033; US 5821337, US 7527791, US 6982321 и US 7087409; Kashmiri, S.V. et al., Methods 36 (2005) 25-34 (описывает прививание области, определяющей специфичность (specificity determining region, SDR)); Padlan, E.A., Mol. Immunol. 28 (1991) 489-498 (описывает "перекладку"); Dall'Acqua, W.F. et al., Methods 36 (2005) 43-60 (описывает "перетасовку FR"); и Osbourn, J. et al., Methods 36 (2005) 61-68 и Klimka, A. et al., Br. J. Cancer 83 (2000) 252-260 (описывают подход "направляемой селекции" к перетасовке FR).
Человеческие каркасные области, которые могут быть использованы для гуманизации, включают, но не ограничиваясь ими: каркасные области, выбранные с помощью методики "наилучшего соответствия" (см., например, Sims, M.J. et al., J. Immunol. 151 (1993) 2296-2308); каркасные области, полученные из консенсусной последовательности человеческих антител конкретной подгруппы вариабельных областей легкой или тяжелой цепей (см., например, Carter, P. et al., Proc. Natl. Acad. Sci. USA 89 (1992) 4285-4289; и Presta, L.G. et al., J. Immunol. 151 (1993) 2623-2632); человеческие зрелые (соматически мутантные) каркасные области или человеческие зародышевые каркасные области (см., например, Almagro, J.C. and Fransson, J., Front. Biosci. 13 (2008) 1619-1633); и каркасные области, полученные при скрининге FR-библиотеки (см., например, Baca, М. et al., J. Biol. Chem. 272 (1997) 10678-10684, и Rosok, M.J. etal., J. Biol. Chem. 271 (19969 22611-22618).
4. Антитела, полученные из библиотек
Антитела согласно данному изобретению могут быть выделены путем скрининга комбинаторных библиотек на антитела с нужной активностью или активностями. Например, в данной области известны различные способы создания библиотек фагового дисплея и скрининга таких библиотек антител, обладающих желаемыми характеристиками связывания. Такие методики описаны, например, в Hoogenboom, H.R. et al., Methods in Molecular Biology 178 (2001) 1-37, а также, например, в the McCafferty, J. et al., Nature 348 (1990) 552-554; Clackson, T. et al., Nature 352 (1991) 624-628; Marks, J.D. et al., J. Mol. Biol. 222 (1992) 581-597; Marks, J.D. and Bradbury, A., Methods in Molecular Biology 248 (2003) 161-175; Sidhu, S.S. et al., J. Mol. Biol. 338 (2004) 299-310; Lee, C.V. et al., J. Mol. Biol. 340 (2004) 1073-1093; Fellouse, F.A., Proc. Natl. Acad. Sci. USA 101 (2004) 12467-12472; и Lee, C.V. et al., J. Immunol. Methods 284 (2004) 119-132.
В некоторых методиках фагового дисплея репертуар VH- и VL-генов отдельно клонировали с помощью полимеразной цепной реакции (PCR) и рекомбинировали в случайном порядке в фаговых библиотеках, которые затем могут быть подвергнуты скринингу в отношении антигенсвязывающего фага, как описано в Winter, G. et al., Ann. Rev. Immunol. 12 (1994) 433-455. Фаг, как правило, представляет фрагменты антитела либо как одноцепочечные Fv-фрагменты (scFv), либо как Fab-фрагменты. Библиотеки из иммунизированных источников обеспечивают высокоаффинные антитела к иммуногену без необходимости конструирования гибридом. Альтернативно, можно клонировать наивный репертуар (например, от человека), чтобы получить единый источник антител к широкому спектру чужих, а также собственных антигенов без какой-либо иммунизации, как описано в Griffiths, A.D. et al., EMBO J. 12 (1993) 725-734. Наконец, наивные библиотеки также могут быть получены синтетически путем клонирования неперестроенных сегментов V-гена из стволовых клеток и с использованием ПЦР-праймеров, содержащих случайную последовательность для кодирования высоко вариабельных CDR3-областей и для выполнения перестройки in vitro, как описано в Hoogenboom, H.R. and Winter, G., J. Mol. Biol. 227 (1992) 381-388. Патентные публикации, описывающие фаговые библиотеки человеческих антител, включают, например: US 5,750,373 и US 2005/0079574, US 2005/0119455, US 2005/0266000, US 2007/0117126, US 2007/0160598, US 2007/0237764, US 2007/0292936 и US 2009/0002360.
5. Мультиспецифические антитела
В некоторых воплощениях антитело, представленное в данном документе, является мультиспецифическим антителом, например, биспецифическим антителом. Мультиспецифические антитела представляют собой моноклональные антитела, которые обладают специфичностью связывания по меньшей мере с двумя различными сайтами. В некоторых воплощениях одна из специфичностей связывания является специфичностью в отношении рецептора трансферрина, а другая в отношении любого другого антигена. Биспецифические антитела также могут быть использованы для локализации цитотоксических агентов на клетках, которые экспрессируют рецептор трансферрина. Биспецифические антитела могут быть получены в виде полноразмерных антител или фрагментов антител.
Методики изготовления мультиспецифических антител включают, но не ограничиваясь ими, рекомбинантную коэкспрессию двух пар тяжелых цепей и легких цепей иммуноглобулина, имеющих различные специфичности (см. Milstein, С.and Cuello, А.С., Nature 305 (1983) 537-540, WO 93/08829, и Traunecker, A. et al., EMBO J. 10 (1991) 3655-3659), и инженерию типа "ключ в замке" (см., например, US 5731168). Мультиспецифические антитела также могут быть получены за счет использования инженерных электростатических перемешивающих влияний для изготовления Fc-гетеродимерных молекул (WO 2009/089004); поперечного сшивания двух или более антител или фрагментов (см., например, US 4676980, и Brennan, М. et al., Science 229 (1985) 81-83); использования лейциновых застежек для получения биспецифических антител (см., например, Kostelny, S.A. et al., J. Immunol. 148 (1992) 1547-1553); использования методики "димерного антитела" для изготовления фрагментов биспецифических антител (см., например, Holliger, P. et al., Proc. Natl. Acad. Sci. USA 90 (1993) 6444-6448); и использования одноцепочечных Fv(sFv)-flHMepoB (см., например, Gruber, М et al., J. Immunol. 152 (1994) 5368-5374); и получения триспецифических антител, как описано, например, в Tutt, A. et al., J. Immunol. 147 (1991) 60-69.
Разработанные антитела с тремя и более чем тремя функциональными антигенсвязывающими сайтами, в том числе "антитела Octopus", также включены в данный документ (см., например, US 2006/0025576).
Антитело или фрагмент в данном документе также включает "Fab двойного действия", или "DAF" (dual acting Fab), содержащий антигенсвязывающий сайт, который связывается с рецептором трансферрина, а также с другим, отличным от него, антигеном (см. US 2008/0069820, например).
Антитело или фрагмент в данном документе также включает мультиспецифические антитела, описанные в WO 2009/080251, WO 2009/080252, WO 2009/080253, WO 2009/080254, WO 2010/112193, WO 2010/115589, WO 2010/136172, WO 2010/145792 и WO 2010/145793.
В одном воплощении всех аспектов, описанных в данном документе, антитело против рецептора трансферрина является биспецифическим антителом.
Одним из аспектов, описанных в данном документе, является двухвалентное биспецифическое антитело, содержащее
a) первую легкую цепь и первую тяжелую цепь антитела, специфически связывающегося с первым антителом, и
b) вторую легкую цепь и вторую тяжелую цепь антитела, специфически связывающегося со вторым антителом, где вариабельные домены VL и VH второй легкой цепи и второй тяжелой цепи заменяют друг друга,
где первый антиген или второй антиген является человеческим рецептором трансферрина.
Антитело согласно а) не содержит модификации согласно b), и тяжелая цепь и легкая цепь согласно а) являются изолированными цепями.
В антителе согласно b)
в легкой цепи
вариабельный домен легкой цепи VL заменен вариабельным доменом тяжелой цепи VH указанного антитела, и
в тяжелой цепи
вариабельный домен тяжелой цепи VH заменен вариабельным доменом легкой цепи VL указанного антитела. В одном воплощении
i) в константном домене CL первой легкой цепи согласно а) аминокислота в позиции 124 (нумерация согласно Kabat) замещена положительно заряженной аминокислотой, а в константном домене СН1 первой тяжелой цепи согласно а) аминокислота в позиции 147 или аминокислота в позиции 213 (нумерация согласно ЕС-индексу Kabat) замещена отрицательно заряженной аминокислотой,
или
ii) в константном домене CL второй легкой цепи согласно b) аминокислота в позиции 124 (нумерация согласно Kabat) замещена положительно заряженной аминокислотой, а в константном домене СН1 второй тяжелой цепи согласно b) аминокислота в позиции 147 или аминокислота в позиции 213 (нумерация согласно ЕС-индексу Kabat) замещена отрицательно заряженной аминокислотой.
В одном предпочтительном воплощении
i) в константном домене CL первой легкой цепи согласно а) аминокислота в позиции 124 замещена независимо лизином (K), аргинином (R) или гистидином (Н) (нумерация согласно Kabat) (в одном предпочтительном воплощении независимо лизином (K) или аргинином (R)), а в константном домене СН1 первой тяжелой цепи согласно а) аминокислота в позиции 147 или аминокислота в позиции 213 замещена независимо глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация согласно ЕС-индексу Kabat),
или
ii) в константном домене CL второй легкой цепи согласно b) аминокислота в позиции 124 замещена независимо лизином (K), аргинином (R) или гистидином (Н) (нумерация согласно Kabat) (в одном предпочтительном воплощении независимо лизином (K) или аргинином (R)), а в константном домене СН1 второй тяжелой цепи согласно b) аминокислота в позиции 147 или аминокислота в позиции 213 замещена независимо глутаминовой кислотой (Е) или аспарагиновой кислотой (D) (нумерация согласно ЕС-индексу Kabat).
В одном воплощении в константном домене CL второй тяжелой цепи аминокислоты в позициях 124 и 123 замещены K (нумерация согласно ЕС-индексу Kabat).
В одном воплощении в константном домене СН1 второй легкой цепи аминокислоты в позициях 147 и 213 замещены Е (нумерация согласно ЕС-индексу Kabat).
В одном предпочтительном воплощении в константном домене CL первой легкой цепи аминокислоты в позициях 124 и 123 замещены K, а в константном домене СН1 первой тяжелой цепи аминокислоты в позициях 147 и 213 замещены Е (нумерация согласно ЕС-индексу Kabat).
В одном воплощении в константном домене CL второй тяжелой цепи аминокислоты в позициях 124 и 123 замещены K, в константном домене СН1 второй легкой цепи аминокислоты в позициях 147 и 213 замещены Е, в вариабельном домене VL первой легкой цепи аминокислота в позиции 38 замещена K, в вариабельном домене VH первой тяжелой цепи аминокислота в позиции 39 замещена Е, в вариабельном домене VL второй тяжелой цепи аминокислота в позиции 38 замещена K, а в вариабельном домене VH второй легкой цепи аминокислота в позиции 39 замещена Е (нумерация согласно ЕС-индексу Kabat).
Один из аспектов, описанных в данном документе, представляет собой двухвалентное биспецифическое антитело, содержащее
a) первую легкую цепь и первую тяжелую цепь антитела, специфически связывающегося с первым антителом, и
b) вторую легкую цепь и вторую тяжелую цепь антитела, специфически связывающегося со вторым антителом, где вариабельные домены VL и VH второй легкой цепи и второй тяжелой цепи заменяют друг друга, и где константные домены CL и СН1 второй легкой цепи и второй тяжелой цепи заменяют друг друга,
где первый антиген или второй антиген является человеческим рецептором трансферрина.
Антитело согласно а) не содержит модификации согласно b), а тяжелая цепь и легкая цепь согласно а) являются изолированными цепями. В антителе согласно b) в легкой цепи
вариабельный домен легкой цепи VL заменен вариабельным доменом тяжелой цепи VH указанного антитела, а константный домен легкой цепи CL заменен константным доменом тяжелой цепи СН1 указанного антитела;
а в тяжелой цепи
вариабельный домен тяжелой цепи VH заменен вариабельным доменом легкой цепи VL указанного антитела, а константный домен тяжелой цепи СН1 заменен константным доменом легкой цепи CL указанного антитела.
Одним из аспектов, описанных в данном документе, является двухвалентное биспецифическое антитело, содержащее
a) первую легкую цепь и первую тяжелую цепь антитела, специфически связывающегося с первым антителом, и
b) вторую легкую цепь и вторую тяжелую цепь антитела, специфически связывающегося со вторым антителом, где константные домены CL и СН1 второй легкой цепи и второй тяжелой цепи заменяют друг друга,
где первый антиген или второй антиген является человеческим рецептором трансферрина.
Антитело согласно а) не содержит модификации согласно b), а тяжелая цепь и легкая цепь согласно а) являются изолированными цепями. В антителе согласно b) в легкой цепи
константный домен легкой цепи CL заменен константным доменом тяжелой цепи СН1 указанного антитела; а в тяжелой цепи
константный домен тяжелой цепи СН1 заменен константным доменом легкой цепи CL указанного антитела.
Одним из аспектов, описанных в данном документе, является мутиспецифическое антитело, содержащее
a) полноразмерное антитело, специфически связывающееся с первым антигеном и состоящее из двух тяжелых цепей антитела и двух легких цепей антитела, и
b) один, два, три или четыре одноцепочечных Fab-фрагмента, специфически связывающихся с 1-4 другими антигенами (т.е. вторым и/или третьим, и/или четвертым и/или пятым антигеном, предпочтительно, специфически связывающихся с одним дополнительным антигеном, т.е. со вторым антигеном),
где указанные одноцепочечные Fab-фрагменты согласно b) слиты с указанным полноразмерным антителом согласно а) через пептидный линкер на С- или N-конце тяжелой или легкой цепи указанного полноразмерного антитела,
где первый антиген или один из дополнительных антигенов является человеческим рецептором трансферрина.
В одном из воплощений один или два идентичных одноцепочечных Fab-фрагмента, связывающиеся со вторым антигеном, слиты с указанным полноразмерным антителом через пептидный линкер на С-конце тяжелых или легких цепей указанного полноразмерного антитела.
В одном из воплощений один или два идентичных одноцепочечных Fab-фрагмента, связывающиеся со вторым антигеном, слиты с указанным полноразмерным антителом через пептидный линкер на С-конце тяжелых цепей указанного полноразмерного антитела.
В одном из воплощений один или два идентичных одноцепочечных Fab-фрагмента, связывающиеся со вторым антигеном, слиты с указанным полноразмерным антителом через пептидный линкер на С-конце легких цепей указанного полноразмерного антитела.
В одном из воплощений два идентичных одноцепочечных Fab-фрагмента, связывающиеся со вторым антигеном, слиты с указанным полноразмерным антителом через пептидный линкер на С-конце каждой тяжелой или легкой цепи указанного полноразмерного антитела.
В одном из воплощений два идентичных одноцепочечных Fab-фрагмента, связывающиеся со вторым антигеном, слиты с указанным полноразмерным антителом через пептидный линкер на С-конце каждой тяжелой цепи указанного полноразмерного антитела.
В одном из воплощений два идентичных одноцепочечных Fab-фрагмента, связывающиеся со вторым антигеном, слиты с указанным полноразмерным антителом через пептидный линкер на С-конце каждой легкой цепи указанного полноразмерного антитела.
Одним из аспектов, описанных в данном документе, является трехвалентное биспецифическое антитело, содержащее
a) полноразмерное антитело, специфически связывающееся с первым антигеном и состоящее из двух тяжелых цепей антитела и двух легких цепей антитела,
b) первый полипептид, состоящий из
ba) вариабельного домена тяжелой цепи антитела (VH), или
bb) вариабельного домена тяжелой цепи антитела (VH) и константного домена антитела 1 (СН1),
где указанный первый полипептид слит N-концом своего VH-домена через пептидный линкер с С-концом одной из двух тяжелых цепей указанного полноразмерного антитела,
c) второй полипептид, состоящий из
са) вариабельного домена легкой цепи антитела (VL),
или
cb) вариабельного домена легкой цепи антитела (VL) и константного домена легкой цепи антитела (CL),
где указанный второй полипептид слит N-концом своего VL-домена через пептидный линкер с С-концом другой из двух тяжелых цепей указанного полноразмерного антитела,
и
где вариабельный домен тяжелой цепи антитела (VH) первого полипептида и вариабельный домен легкой цепи антитела (VL) второго полипептида вместе формируют антигенсвязывающий сайт, специфически связывающийся со вторым антителом,
и
где первый антиген или второй антиген является человеческим рецептором трансферрина.
В одном из воплощений вариабельный домен тяжелой цепи антитела (VH) полипептида согласно b) и вариабельный домен легкой цепи антитела (VL) полипептида согласно с) связаны и стабилизированы с помощью межцепочечного дисульфидного мостика путем введения дисульфидной связи между следующими позициями:
i) позиция 44 вариабельного домена тяжелой цепи и позиция 100 вариабельного домена легкой цепи, или
ii) позиция 105 вариабельного домена тяжелой цепи и позиция 43 вариабельного домена легкой цепи, или
iii) позиция 101 вариабельного домена тяжелой цепи и позиция 100 вариабельного домена легкой цепи (нумерация всегда согласно ЕС-индексу Kabat).
Методики введения неприродных дисульфидных мостиков для стабилизации описаны, например, в WO 94/029350, Rajagopal, V., et al., Prot. Eng. (1997) 1453-59; Kobayashi, H., et al., Nuclear Medicine & Biology, Vol.25, (1998) 387-393; или Schmidt, M., et al., Oncogene (1999) 18 1711-1721. В одном воплощении возможная дисульфидная связь между вариабельными доменами полипептидов согласно b) и с) находится между позицией 44 вариабельного домена тяжелой цепи и позицией 100 вариабельного домена легкой цепи. В одном воплощении возможная дисульфидная связь между вариабельными доменами полипептидов согласно b) и с) находится между позицией 105 вариабельного домена тяжелой цепи и позицией 43 вариабельного домена легкой цепи (нумерация всегда согласно ЕС-индексу Kabat). В одном воплощении предпочтительным является трехвалентное биспецифическое антитело без указанной возможной стабилизации дисульфидными мостиками между вариабельными доменами VH и VL одноцепочечных Fab-фрагментов.
Одним из аспектов, описанных в данном документе, является триспецифическое или тетраспецифическое антитело, содержащее
a) первую легкую цепь и первую тяжелую цепь полноразмерного антитела, которое специфически связывается с первым антигеном, и
b) вторую (модифицированную) легкую цепь и вторую (модифицированную) тяжелую цепь полноразмерного антитела, которое специфически связывается со вторым антигеном, где вариабельные домены VL и VH заменяют друг друга, и/или где константные домены CL и СН1 заменяют друг друга,
c) где 1-4 антигенсвязывающих пептида, которые специфически связываются с 1-4 антигенами (т.е с третьим и/или четвертым антигеном), слиты через пептидный линкер с С- или N-концом легких цепей или тяжелых цепей а) и/или b),
где первый антиген или второй антиген или один из дополнительных антигенов является человеческим рецептором трансферрина.
Антитело согласно а) не содержит модификации согласно b), а тяжелая цепь и легкая цепь согласно а) являются изолированными цепями.
В одном воплощении триспецифическое или тетраспецифическое антитело также содержит согласно с) один или два антигенсвязывающих пептида, которые специфически связываются с одним или двумя дополнительными антигенами.
В одном воплощении антигенсвязывающие пептиды выбраны из группы scFv-фрагмента и scFab-фрагмента.
В одном воплощении антигенсвязывающие пептиды являются scFv-фрагментами.
В одном воплощении антигенсвязывающие пептиды являются scFab-фрагментами.
В одном воплощении антигенсвязывающие пептиды слиты с С-концом тяжелых цепей а) и/или b).
В одном воплощении триспецифическое или тетраспецифическое антитело содержит согласно с) один или два антигенсвязывающих пептида, которые специфически связываются с 1-4 антигенами.
В одном воплощении триспецифическое или тетраспецифическое антитело содержит согласно с) два идентичных антигенсвязывающих пептида, которые специфически связываются с третьим антигеном. В одном предпочтительном воплощении такие два идентичных антигенсвязывающих пептида слиты через один и тот же пептидный линкер с С-концом тяжелых цепей а) и b). В одном предпочтительном воплощении два идентичных антигенсвязывающих пептида являются либо scFv-фрагментами, либо scFab-фрагментами.
В одном воплощении триспецифическое или тетраспецифическое антитело содержит согласно с) два антигенсвязывающих пептида, которые специфически связываются с третьим и четвертым антигеном. В одном воплощении указанные два антигенсвязывающих пептида слиты через один и тот же пептидый линкер с С-концом тяжелых цепей а) и b). В одном предпочтительном воплощении указанные два антигенсвязывающих пептида являются либо scFv-фрагментами, либо scFab-фрагментами.
Одним из аспектов, описанных в данном документе, является биспецифическое тетравалентное антитело, содержащее
a) две легкие цепи и две тяжелые цепи антитела, которые специфически связываются с первым антигеном (и содержат два Fab-фрагмента),
b) два дополнительных Fab-фрагмента антитела, которые специфически связываются со вторым антигеном, где указанные дополнительные Fab-фрагменты слиты оба через пептидный линкер либо с С-, либо с N-концами тяжелых цепей а),
и
где в Fab-фрагментах были сделаны следующие модификации
i) в обоих Fab-фрагментах из а) или в обоих Fab-фрагментах из b) вариабельные домены VL и VH заменяют друг друга, и/или константные домены CL и СН1 заменяют друг друга,
или
ii) в обоих Fab-фрагментах из а) вариабельные домены VL и VH заменяют друг друга, и константные домены CL и СН1 заменяют друг друга,
и
в обоих Fab-фрагментах из b) вариабельные домены VL и VH заменяют друг друга, или константные домены CL и СН1 заменяют друг друга, или
iii) в обоих Fab-фрагментах из а) вариабельные домены VL и VH заменяют друг друга или константные домены CL и СН1 заменяют друг друга,
и
в обоих Fab-фрагментах из b) вариабельные домены VL и VH заменяют друг друга, и константные домены CL и СН1 заменяют друг друга, или
iv) в обоих Fab-фрагментах из а) вариабельные домены VL и VH заменяют друг друга, и в обоих Fab-фрагментах из b) константные домены CL и СН1 заменяют друг друга,
или
v) в обоих Fab-фрагментах из а) константные домены CL и СН1 заменяют друг друга, и в обоих Fab-фрагментах из b) вариабельные домены VL и VH заменяют друг ДРУга,
где первый антиген или второй антиген является человеческим рецептором трансферрина.
В одном из воплощений указанные дополнительные Fab-фрагменты слиты оба через пептидный линкер либо с С-концами тяжелых цепей из а), либо с N-концами тяжелых цепей из а).
В одном из воплощений указанные дополнительные Fab-фрагменты слиты оба через пептидный линкер с С-концами тяжелых цепей из а).
В одном из воплощений указанные дополнительные Fab-фрагменты слиты оба через пептидный линкер с N-концами тяжелых цепей из а).
В одном из воплощений в Fab-фрагментах проведены следующие модификации:
i) в обоих Fab-фрагментах из а) или в обоих Fab-фрагментах из b) вариабельные домены VL и VH заменяют друг друга, и/или
константные домены CL и СН1 заменяют друг друга.
В одном из воплощений в Fab-фрагментах проведены следующие модификации:
i) в обоих Fab-фрагментах из а) вариабельные домены VL и VH заменяют друг друга,
и/или
константные домены CL и СН1 заменяют друг друга.
В одном из воплощений в Fab-фрагментах проведены следующие модификации:
i) в обоих Fab-фрагментах из а) константные домены CL и СН1 заменяют друг друга.
В одном из воплощений в Fab-фрагментах проведены следующие модификации:
i) в обоих Fab-фрагментах из b) вариабельные домены VL и VH заменяют друг друга,
и/или
константные домены CL и СН1 заменяют друг друга.
В одном из воплощений в Fab-фрагментах проведены следующие модификации:
i) в обоих Fab-фрагментах из b) константные домены CL и СН1 заменяют друг друга.
Одним из аспектов, описанных в данном документе, является биспецифическое тетравалентное антитело, содержащее:
a) (модифицированную) тяжелую цепь первого антитела, которое специфически связывается с первым антигеном и содержит первую пару доменов VH-СН1, где с С-концом указанной тяжелой цепи N-конец второй пары доменов VH-CH1 указанного первого антитела слит через пептидный линкер,
b) две легкие цепи указанного первого антитела из а),
c) (модифицированную) тяжелую цепь второго антитела, которое специфически связывается со вторым антигеном и содержит первую пару доменов VH-CL, где с С-концом указанной тяжелой цепи N-конец второй пары доменов VH-CL указанного второго антитела слит через пептидный линкер, и
d) две (модифицированные) легкие цепи указанного второго антитела из с), каждая из которых содержит пару доменов CL-CH1,
где первый антиген или второй антиген является человеческим рецептором трансферрина.
Одним из аспектов, описанных в данном документе, является биспецифическое антитело, содержащее
a) тяжелую цепь и легкую цепь первого полноразмерного антитела, которое специфически связывается с первым антигеном, и
b) тяжелую цепь и легкую цепь второго полноразмерного антитела, которое специфически связывается со вторым антигеном, где N-конец тяжелой цепи соединен с С-концом легкой цепи через пептидный линкер,
где первый антиген или второй антиген является человеческим рецептором трансферрина.
Антитело согласно а) не содержит модификации согласно b), а тяжелая цепь и легкая цепь являются изолированными цепями.
Одним из аспектов, описанных в данном документе, является биспецифическое антитело, содержащее
a) полноразмерное антитело, специфически связывающееся с первым антигеном и состоящее из двух тяжелых цепей антитела и двух легких цепей антитела, и
b) Fv-фрагмент, специфически связывающийся со вторым антителом и содержащий VH2-домен и VL2-домен, где оба домена соединены друг с другом через дисульфидный мостик,
где только либо VH2-домен, либо VL2-домен слит через пептидный линкер с тяжелой или легкой цепью полноразмерного антитела, специфически связывающегося с первым антигеном,
где первый антиген или второй антиген является человеческим рецептором трансферрина.
В биспецифическом антителе тяжелые цепи и легкие цепи согласно а) являются изолированными цепями.
В одном воплощении другой VH2-домен или VL2-домен не слит через пептидный линкер с тяжелой или легкой цепью полноразмерного антитела, специфически связывающегося с первым антигеном.
Во всех аспектах, приведенных в данном документе, первая легкая цепь содержит домен VL и домен CL, а первая тяжелая цепь содержит домен VH, домен СН1, шарнирную область, домен СН2 и домен СН3.
В одном из воплощений всех аспектов изобретения антитело, описанное в данном документе, представляет собой мультиспецифическое антитело, которое требует гетеродимеризации по меньшей мере двух тяжелых полипептидных цепей, где антитело специфически связывается с рецептором человеческого трансферрина и вторым антигеном, не являющимся человеческим рецептором трансферрина.
Для поддержания гетеродимеризации было описано несколько подходов к СН3-модификации, например, в WO 96/27011, WO 98/050431, ЕР 1870459, WO 2007/110205, WO 2007/147901, WO 2009/089004, WO 2010/129304, WO 2011/90754, WO 2011/143545, WO 2012/058768, WO 2013/157954, WO 2013/096291, которые включены в данный документ посредством ссылки. Как правило, в подходах, известных в данной области, СН3-домен первой тяжелой цепи и СН3-домен второй тяжелой цепи сконструированы комплементарным образом так, что тяжелая цепь, содержащая один сконструированный СН3-домен, не гомодимеризуется с другой тяжелой цепью с такой же структурой (например, СН3-сконструированная первая тяжелая цепь больше не может гомодимеризоваться с другой СН3-сконструированной первой тяжелой цепью; и СН3-сконструированная вторая тяжелая цепь больше не может гомодимеризоваться с другой СН3-сконструированной второй тяжелой цепью). При этом тяжелая цепь, содержащая один сконструированный СН3-домен, вынуждена гетеродимеризоваться с другой тяжелой цепью, содержащей СН3-домен, который сконструирован комплементарным образом. Для этого воплощения изобретения СН3-домен первой тяжелой цепи и СН3-домен второй тяжелой цепи сконструированы комплементарным образом с помощью аминокислотных замен таким образом, что первая тяжелая цепь и вторая тяжелая цепь вынуждены гетеродимеризоваться, в то время как первая тяжелая цепь и вторая тяжелая цепь больше не гомодимеризуются (например, по стерическим причинам).
Различные подходы к поддержанию гетеродимеризации тяжелой цепи, известные в данной области, которые были процитированы и включены выше, рассматриваются в качестве других альтернатив, используемых в мультиспецифическом антителе в соответствии с изобретением, которое содержит "не пере крестную Fab-область", полученную из первого антитела, которая специфически связывается с первым антигеном, и "перекрестную Fab-область", полученную из второго антитела, которое специфически связывается со вторым антигеном, в сочетании с конкретными аминокислотными заменами, описанными выше в данном изобретении.
СН3-домены мультиспецифического антитела, описанного в данном документе, могут быть изменены по методике "ключ в замке", которая подробно описана в нескольких примерах, например, в WO 96/027011, Ridgway, J.В., et al., Protein Eng. 9 (1996) 617-621; и Merchant, A.M., etal., Nat Biotechnol 16 (1998) 677-681. В этом способе взаимодействующие поверхности двух СН3-доменов изменены таким образом, чтобы увеличить гетеродимеризацию обеих тяжелых цепей, содержащих эти два СН3-домена. Каждый из двух СН3-доменов (двух тяжелых цепей) может быть "ключом", а другой "замком". Введение дисульфидного мостика также стабилизирует гетеродимеры (Merchant, A.M., et al., Nature Biotech 16 (1998) 677-681; Atwell, S., et al., J. Mol. Biol. 270 (1997) 26-35) и увеличивает выход.
В одном предпочтительном воплощении мультиспецифическое антитело, описанное в данном документе, содержит мутацию T366W в СН3-домене "цепи-ключа" и мутации T366S, L368A, Y407V в СН3-домене "цепи-замка" (нумерация в соответствии с ЕС-индексом согласно Kabat). Также между СН3-доменами может быть введен дополнительный межцепочечный дисульфидный мостик (Merchant, А.М, et al., Nature Biotech 16 (1998) 677-681), например, путем введения мутации Y349C в СН3-домен "цепи-ключа" и мутации Е356С или мутации S354C в СН3-домен "цепи-замка". Так, в другом предпочтительном воплощении мультиспецифическое антитело, описанное в данном документе, содержит мутации Y349C и T366W в одном из двух СН3-доменов и мутации Е356С, T366S, L368A и Y407V в другом из двух СН3-доменов, либо мультиспецифическое антитело, описанное в данном документе, содержит мутации Y349C и T366W в одном из двух СН3-доменов и мутации S354C, T366S, L368A и Y407V в другом из двух СН3-доменов (дополнительная мутация Y349C в одном СН3-домене и дополнительная мутация Е356С или S354C в другом СН3-домене для формирования межцепочечного дисульфидного мостика) (нумерация в соответствии с ЕС-индексом Kabat).
Альтернативно или дополнительно также могут быть использованы и другие методики типа "ключ в замке", такие как описанные в ЕР 1870459А1. В одном воплощении мультиспецифическое антитело, описанное в данном документе, содержит мутации R409D и К370Е в СН3-домене "цепи-ключа" и мутации D399K и Е357К в СН3-домене "цепи-замка" (нумерация в соответствии с ЕС-индексом Kabat).
В одном воплощении мультиспецифическое антитело, описанное в данном документе, содержит мутацию T366W в СН3-домене "цепи-ключа" и мутации T366S, L368A и Y407V в СН3-домене "цепи-замка", и дополнительно мутации R409D и К370Е в СН3-домене "цепи-ключа" и мутации D399K и Е357К в СН3-домене "цепи-замка" (нумерация в соответствии с ЕС-индексом Kabat).
В одном воплощении мультиспецифическое антитело, описанное в данном документе, содержит мутации Y349C и T366WB одном из двух СН3-доменов и мутации S354C, T366S, L368A и Y407V в другом из двух СН3-доменов, или мультиспецифическое антитело, описанное в данном документе, содержит мутации Y349C и T366W в одном из двух СН3-доменов и мутации S354C, T366S, L368A, Y407V в другом из двух СН3-доменов, и дополнительные мутации R409D и К370Е в СН3-домене "цепи-ключа" и мутации D399K и Е357К в СН3-домене "цепи-замка" (нумерация в соответствии с ЕС-индексом Kabat).
Помимо методики "ключ в замке" в данной области известны и другие методики модификации СН3-доменов тяжелых цепей мультиспецифического антитела для обеспечения гетеродимеризации. Эти методики, особенно те, которые описаны в WO 96/27011, WO 98/050431, ЕР 1870459, WO 2007/110205, WO 2007/147901, WO 2009/089004, WO 2010/129304, WO 2011/90754, WO 2011/143545, WO 2012/058768, WO 2013/157954 и WO 2013/096291, рассматриваются в данном документе в качестве альтернатив методике "ключ в замке" в сочетании с мультиспецифическим антителом, описанным в данном документе.
В одном из воплощений мультиспецифического антитела, описанного в данном документе, подход, описанный в ЕР 1870459, используется для поддержания гетеродимеризации первой тяжелой цепи и второй тяжелой цепи мультиспецифического антитела. Этот подход основан на введении заряженных аминокислот с противоположными зарядами в определенных аминокислотных позициях в поверхности взаимодействия СН3/СН3-доменов между обеими, первой и второй, тяжелой цепями.
Соответственно, это воплощение относится к мультиспецифическому антителу, описанному в данном документе, где в третичной структуре антитела СН3-домен первой тяжелой цепи и СН3-домен второй тяжелой цепи образуют поверхность взаимодействия, которая расположена между соответствующими СН3-доменами антитела, где каждая из соответствующих аминокислотных последовательностей СН3-домена первой тяжелой цепи и СН3-домена второй тяжелой цепи содержит ряд аминокислот, который расположен внутри указанной поверхности взаимодействия в третичной структуре антитела, где из ряда аминокислот, которые расположены в поверхности взаимодействия в СН3-домене одной тяжелой цепи, первая аминокислота замещается положительно заряженной аминокислотой, а из ряда аминокислот, которые расположены в поверхности взаимодействия в СН3-домене другой тяжелой цепи, вторая аминокислота замещается отрицательно заряженной аминокислотой. Мультиспецифическое антитело согласно данному воплощению также упоминается в данном документе как "СН3(+/-)-инженерное мультиспецифическое антитело" (где аббревиатура "+/-" обозначает противоположно заряженные аминокислоты, которые были введены в соответствующие СН3-домены).
В одном воплощении указанного СН3(+/-)-инженерного мультиспецифического антитела, описанного в данном документе, положительно заряженная аминокислота выбрана среди K, R и Н, а отрицательно заряженная аминокислота выбрана среди Е или D.
В одном воплощении указанного СН3(+/-)-инженерного мультиспецифического антитела, описанного в данном документе, положительно заряженная аминокислота выбрана среди К и R, а отрицательно заряженная аминокислота выбрана среди Е или D.
В одном воплощении указанного СН3(+/-)-инженерного мультиспецифического антитела, описанного в данном документе, положительно заряженная аминокислота представляет собой K, а отрицательно заряженная аминокислота представляет собой Е.
В одном воплощении указанного СН3(+/-)-инженерного мультиспецифического антитела, описанного в данном документе, в СН3-домене одной тяжелой цепи аминокислота R в позиции 409 замещена D, и аминокислота К в позиции замещена Е, а в СН3-домене другой тяжелой цепи аминокислота D в позиции 399 замещена K, и аминокислота Е в позиции 357 замещена К (нумерация согласно ЕС-индексу Kabat).
В одном воплощении мультиспецифического антитела, описанного в данном документе, подход, описанный в WO 2013/157953, используется для поддержания гетеродимеризации первой тяжелой цепи и второй тяжелой цепи мультиспецифического антитела. В одном воплощении указанного мультиспецифического антитела, описанного в данном документе, в СН3-домене одной тяжелой цепи аминокислота Т в позиции 366 замещена K, а в СН3-домене другой тяжелой цепи аминокислота L в позиции 351 замещена D (нумерация согласно ЕС-индексу Kabat). В другом воплощении указанного мультиспецифического антитела, описанного в данном документе, в СН3-домене одной тяжелой цепи аминокислота Т в позиции 366 замещена K, и аминокислота L в позиции 351 замещена K, а в СН3-домене другой тяжелой цепи аминокислота L в позиции 351 замещена D (нумерация согласно ЕС-индексу Kabat).
В другом воплощении указанного мультиспецифического антитела, описанного в данном документе, в СН3-домене одной тяжелой цепи аминокислота Т в позиции 366 замещена K, и аминокислота L в позиции 351 замещена K, а в СН3-домене другой тяжелой цепи аминокислота L в позиции 351 замещена D (нумерация согласно ЕС-индексу Kabat). Кроме того, по меньшей мере одна из следующих замен содержится в СН3-домене другой тяжелой цепи: аминокислота Y в позиции 349 замещена Е, аминокислота Y в позиции 349 замещена D, и аминокислота L в позиции 368 замещена Е (нумерация согласно ЕС-индексу Kabat). В одном воплощении аминокислота L в позиции 368 замещена Е (нумерация согласно ЕС-индексу Kabat).
В одном воплощении мультиспецифического антитела, описанного в данном документе, подход, описанный в WO 2012/058768, используется для поддержания гетеродимеризации первой тяжелой цепи и второй тяжелой цепи мультиспецифического антитела. В одном воплощении указанного мультиспецифического антитела, описанного в данном документе, в СН3-домене одной тяжелой цепи аминокислота L в позиции 351 замещена Y, и аминокислота Y в позиции 407 замещена А, а в СН3-домене другой тяжелой цепи аминокислота Т в позиции 366 замещена А, и аминокислота К в позиции 409 замещена F (нумерация согласно ЕС-индексу Kabat). В другом воплощении помимо описанных выше замен в СН3-домене другой тяжелой цепи замещена по меньшей мере одна из аминокислот в позициях 411 (исходно Т), 399 (исходно D), 400 (исходно S), 405 (исходно F), 390 (исходно N) и 392 (исходно K) (нумерация согласно ЕС-индексу Kabat). Предпочтительными заменами являются:
замещение аминокислоты Т в позиции 411 на аминокислоту, выбранную среди N, R, Q, K, D, Е и W (нумерация согласно ЕС-индексу Kabat),
замещение аминокислоты D в позиции 399 на аминокислоту, выбранную среди R, W, Y и K (нумерация согласно ЕС-индексу Kabat),
замещение аминокислоты S в позиции 400 на аминокислоту, выбранную среди Е, D, R и K (нумерация согласно ЕС-индексу Kabat),
замещение аминокислоты F в позиции 405 на аминокислоту, выбранную среди I, М, Т, S, V и W (нумерация согласно ЕС-индексу Kabat);
замещение аминокислоты N в позиции 390 на аминокислоту, выбранную среди R, K и D (нумерация согласно ЕС-индексу Kabat);
- замещение аминокислоты K в позиции 392 на аминокислоту, выбранную среди V, М, R, L, F и Е (нумерация согласно ЕС-индексу Kabat).
В другом воплощении указанного мультиспецифического антитела, описанного в данном документе (сконструированного согласно WO 2012/058768), в СН3-домене одной тяжелой цепи аминокислота L в позиции 351 замещена Y, и аминокислота Y в позиции 407 замещена А, а в СН3-домене другой тяжелой цепи аминокислота Т в позиции 366 замещена V, и аминокислота К в позиции 409 замещена F (нумерация согласно ЕС-индексу Kabat). В другом воплощении указанного мультиспецифического антитела, описанного в данном документе, в СН3-домене одной тяжелой цепи аминокислота Y в позиции 407 замещена А, а в СН3-домене другой тяжелой цепи аминокислота Т в позиции 366 замещена А, и аминокислота К в позиции 409 замещена F (нумерация согласно ЕС-индексу Kabat). В последнем из указанных выше воплощений в СН3-домене указанной другой тяжелой цепи аминокислота К в позиции 392 замещена Е, аминокислота Т в позиции 411 замещена Е, аминокислота D в позиции 399 замещена R, и аминокислота S в позиции 400 замещена R (нумерация согласно ЕС-индексу Kabat).
В одном из воплощений мультиспецифического антитела, описанного в данном документе, подход, описанный в публикации WO 2011/143545, используется для поддержания гетеродимеризации первой тяжелой цепи и второй тяжелой цепи мультиспецифического антитела. В одном из воплощений указанного мутиспецифического антитела, описанного в данном документе, аминокислотные модификации в СН3-доменах обеих тяжелых цепей введены в позициях 368 и/или 409 (нумерация в соответствии с ЕС-индексом Kabat).
В одном из воплощений мультиспецифического антитела, описанного в данном документе, подход, описанный в публикации WO 2011/090762, используется для поддержания гетеродимеризации первой тяжелой цепи и второй тяжелой цепи мультиспецифического антитела. WO 2011/090762 относится к аминокислотным модификациям в соответствии с методикой "ключ в замке". В одном воплощении указанного СНЗ(К1Н)-инженерного мультиспецифического антитела, описанного в данном документе, в СН3-домене одной тяжелой цепи аминокислота Т в позиции 366 замещена W, а в СН3-домене другой тяжелой цепи аминокислота Y в позиции 407 замещена А (нумерация в соответствии с ЕС-индексом Kabat). В другом воплощении указанного СН3(KiH)-инженерного мультиспецифического антитела, описанного в данном документе, в СН3-домене одной тяжелой цепи аминокислота Т в позиции 366 замещена Y, а в СН3-домене другой тяжелой цепи аминокислота Y в позиции 407 замещена Т (нумерация в соответствии с ЕС-индексом Kabat).
В одном из воплощений мультиспецифического антитела, описанного в данном документе, которое имеет изотип IgG2, для поддержания гетеродимеризации первой тяжелой цепи и второй тяжелой цепи мультиспецифического антитела используется подход, описанный в WO 2011/090762.
В одном из воплощений мультиспецифического антитела, описанного в данном документе, для поддержания гетеродимеризации первой тяжелой цепи и второй тяжелой цепи мультиспецифического антитела используется подход, описанный в WO 2009/089004. В одном воплощении указанного мультиспецифического антитела, описанного в данном документе, в СН3-домене одной тяжелой цепи аминокислота К или N в позиции 392 замещена отрицательно заряженной аминокислотой (в одном предпочтительном воплощении замещена Е или D, в одном предпочтительном воплощении D), а в СН3-домене другой тяжелой цепи аминокислота D в позиции 399, аминокислота Е или D в позиции 356 или аминокислота Е в позиции 357 замещена положительно заряженной аминокислотой (в одном предпочтительном воплощении замещент К или R, в одном предпочтительном воплощении K, в одном предпочтительном воплощении аминокислоты в позициях 399 или 356 замещены K) (нумерация согласно ЕС-индексу Kabat). В одном дополнительном воплощении помимо указанных выше замен в СН3-домене одной тяжелой цепи аминокислота К или R в позиции 409 замещена отрицательно заряженной аминокислотой (в одном предпочтительном воплощении Е или D, в одном предпочтительном воплощении D) (нумерация согласно ЕС-индексу Kabat). В еще одном дополнительном воплощении помимо или альтернативно указанным выше заменам в СН3-домене одной тяжелой цепи аминокислота К в позиции 439 и/или аминокислота К в позиции 370 независимо замещена отрицательно заряженной аминокислотой (в одном предпочтительном воплощении Е или D, в одном предпочтительном воплощении D) (нумерация согласно ЕС-индексу Kabat).
В одном из воплощений мультиспецифического антитела, описанного в данном документе, подход, описанный в WO 2007/147901, используется для поддержания гетеродимеризации первой тяжелой цепи и второй тяжелой цепи мультиспецифического антитела. В одном из воплощений указанного мультиспецифического антитела, описанного в данном документе, в СН3-домене одной тяжелой цепи аминокислота К в позиции 253 замещена Е, аминокислота D в позиции 282 замещена K, и аминокислота К в позиции 322 замещена D, а в СН3-домене другой тяжелой цепи аминокислота D в позиции 239 замещена K, аминокислота Е в позиции 240 замещена K, и аминокислота К в позиции 292 замещена D (нумерация согласно ЕС-индексу Kabat).
В одном из воплощений мультиспецифического антитела, описанного в данном документе, для поддержания гетеродимеризации первой тяжелой цепи и второй тяжелой цепи мультиспецифического антитела используется подход, описанный в WO 2007/110205.
В одном из воплощений всех аспектов и воплощений, описанных в данном документе, мультиспецифическое антитело представляет собой биспецифическое антитело или триспецифическое антитело. В одном предпочтительном воплощении данного изобретения мультиспецифическое антитело представляет собой биспецифическое антитело.
В одном из воплощений всех аспектов, описанных в данном документе, антитело представляет собой двухвалентное или трехвалентное антитело. В одном из воплощений антитело представляет собой двухвалентное антитело.
В одном из воплощений всех аспектов, описанных в данном документе, мультиспецифическое антитело имеет структуру константного домена из антитела типа IgG. В еще одном воплощении всех аспектов, описанных в данном документе, мультиспецифическое антитело характеризуется тем, что указанное мультиспецифическое антитело относится к человеческому подклассу IgG1 или человеческому подклассу IgG1 с мутациями L234A и L235A. В еще одном воплощении всех аспектов, описанных в данном документе, мультиспецифическое антитело характеризуется тем, что указанное мультиспецифическое антитело относится к человеческому подклассу IgG2. В еще одном воплощении всех аспектов, описанных в данном документе, мультиспецифическое антитело характеризуется тем, что указанное мультиспецифическое антитело относится к человеческому подклассу IgG3. В еще одном воплощении всех аспектов, описанных в данном документе, мультиспецифическое антитело характеризуется тем, что указанное мультиспецифическое антитело относится к человеческому подклассу IgG4 или человеческому подклассу IgG4 с дополнительной мутацией S228P. В еще одном воплощении всех аспектов, описанных в данном документе, мультиспецифическое антитело характеризуется тем, что указанное мультиспецифическое антитело относится к человеческому подклассу IgG1 или человеческому подклассу IgG4. В еще одном воплощении всех аспектов, описанных в данном документе, мультиспецифическое антитело характеризуется тем, что указанное мультиспецифическое антитело относится к человеческому подклассу IgG1 с мутациями L234A и L235A (нумерация согласно ЕС-индексу Kabat). В еще одном воплощении всех аспектов, описанных в данном документе, мультиспецифическое антитело характеризуется тем, что указанное мультиспецифическое антитело относится к человеческому подклассу IgG1 с мутациями L234A, L235A и P329G (нумерация согласно ЕС-индексу Kabat). В еще одном воплощении всех аспектов, описанных в данном документе, мультиспецифическое антитело характеризуется тем, что указанное мультиспецифическое антитело относится к человеческому подклассу IgG4 с мутациями S228P и L235E (нумерация согласно ЕС-индексу Kabat). В еще одном воплощении всех аспектов, описанных в данном документе, мультиспецифическое антитело характеризуется тем, что указанное мультиспецифическое антитело относится к человеческому подклассу IgG4 с мутациями S228P, L235E и P329G (нумерация согласно ЕС-индексу Kabat).
В одном из воплощений всех аспектов, описанных в данном документе, антитело, содержащее тяжелую цепь, включающую СН3-домен, как он определен в данном документе, содержит дополнительный С-концевой дипептид глицин-лизин (G446 и К447, нумерация согласно ЕС-индексу Kabat). В одном из воплощений всех аспектов, описанных в данном документе, антитело, содержащее тяжелую цепь, включающую СН3-домен, как он определен в данном документе, содержит дополнительный С-концевой остаток глицина (G446, нумерация согласно ЕС-индексу Kabat).
6. Варианты антител
В некоторых воплощениях рассматриваются варианты аминокислотных последовательностей антител, представленных в данном документе. Например, может быть желательным улучшение аффинности связывания и/или других биологических свойств антитела. Варианты аминокислотной последовательности антитела могут быть получены путем введения соответствующих модификаций в нуклеотидную последовательность, кодирующую антитело, или путем пептидного синтеза. Такие модификации включают, например, делеции и/или вставки и/или замены остатков в аминокислотных последовательностях антитела. Можно использовать любую комбинацию делеций, вставок и замен, чтобы получить конечную конструкцию, при условии, что конечная конструкция обладает нужными характеристиками, например, связыванием антигена.
а) Варианты с заменой, вставкой и делецией
В некоторых воплощениях предложены варианты антител, имеющие одну или более чем одну аминокислотную замену. Сайты для мутагенеза путем замен, представляющие интерес, включают HVR и FR. Консервативные замены представлены в таблице 1 под заголовком "консервативные замены". Более существенные изменения представлены в таблице 1 под заголовком "иллюстративные замены" и дополнительно описаны ниже со ссылкой на классы боковых цепей аминокислот.Аминокислотные замены могут быть введены в антитело, представляющее интерес, и продукты могут быть проверены на желаемую активность, например, на сохранившееся/улучшенное связывание антигена, снижение иммуногенности или улучшение ADCC или CDC.
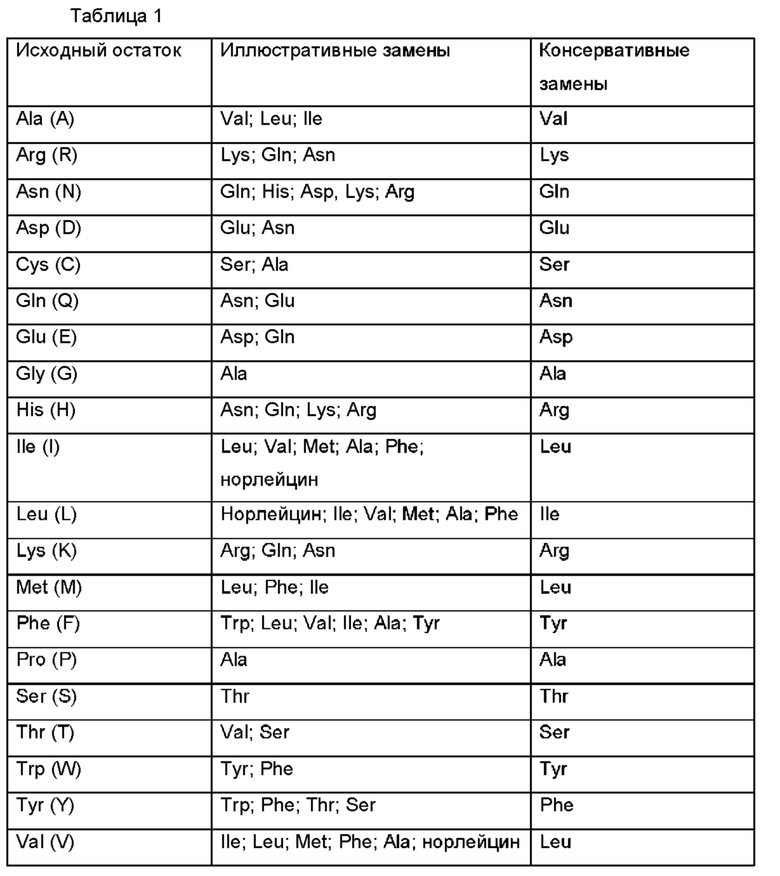
Аминокислоты могут быть сгруппированы в соответствии с общими свойствами боковой цепи:
(1) гидрофобные: норлейцин, Met, Ala, Val, Leu, Ile;
(2) нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gin;
(3) кислые: Asp, Glu;
(4) основные: His, Lys, Arg;
(5) остатки, которые влияют на ориентацию цепи: Gly, Pro;
(6) ароматические: Trp, Tyr, Phe.
Неконсервативные замены влекут за собой замену члена одного из этих классов на член другого класса.
Один тип варианта с заменой предполагает замену одного или более чем одного остатка гипервариабельной области родительского антитела (например, гуманизированного или человеческого антитела). Как правило, полученный вариант(ы), выбранный для дальнейшего изучения, будет иметь модификации (например, улучшения) в определенных биологических свойствах (например, повышенную аффинность, сниженную иммуногенность) относительно родительского антитела и/или будет иметь по существу сохранившиеся определенные биологические свойства родительского антитела. Примером варианта с заменой является антитело с созревшей аффинностью, которое можно легко получить, например, с помощью методик созревания аффинности на основе фагового дисплея, таких как описанные в данном документе. Вкратце, один или более чем один остаток HVR подвергается мутированию, и вариантные антитела выставляют на фаге и проверяют на конкретную биологическую активность (например, аффинность связывания).
Изменения (например, замены) могут быть сделаны в HVR, например, для улучшения аффинности антитела. Такие изменения могут быть сделаны в "горячих точках" HVR, т.е. в остатках, кодируемых кодонами, которые подвергаются мутации с высокой частотой в ходе процесса соматического созревания (см., например, Chowdhury, P.S., Methods Mol. Biol. 207 (2008) 179-196), и/или в остатках, которые контактируют с антигеном, при этом получают вариантные VH или VL, которые исследуют на аффинность связывания. Созревание аффинности путем конструирования и повторного выбора из вторичных библиотек было описано, например, в Hoogenboom, H.R. et al. in Methods in Molecular Biology 178 (2002) 1-37. В некоторых воплощениях созревания аффинности разнообразие вводится в вариабельные гены, выбранные для созревания, с помощью любой из разнообразных методик (например, ПЦР с внесением ошибок, перестановки цепей или олигонуклеотид-направленного мутагенеза). Затем создается вторичная библиотека. Затем эту библиотеку подвергают скринингу для выявления любых вариантов антител с нужной аффинностью. Другой способ внесения разнообразия включает HVR-направленные подходы, в которых рандомизируют несколько остатков HVR (например, 4-6 остатков за один раз). HVR-остатки, участвующие в связывании антигена, могут быть конкретно выявлены, например, с помощью сканирующего аланином мутагенеза или моделирования. CDR-H3 и CDR-L3, в частности, часто становятся мишенями.
В некоторых воплощениях замены, вставки или делеции могут возникать в одном или более чем одном HVR до тех пор, пока такие изменения не вызывают существенного снижения способности антитела связывать антиген. Например, в HVR могут быть сделаны консервативные изменения (например, консервативные замены, предложенные в данном документе), которые существенно не снижают аффинность связывания. Такие изменения, например, могут быть в HVR вне остатков, контактирующих с антигеном. В некоторых воплощениях вариантных VH- и VL-последовательностей, приведенных выше, каждый HVR либо не меняется или содержит не более чем одну, две или три аминокислотных замены.
Способ, который может быть использован для выявления остатков или областей антитела, которые могут быть мишенями мутагенеза, называется "сканирующим аланином мутагенезом" и описан в Cunningham, B.C. and Wells, J.A., Science 244 (1989) 1081-1085. В этом способе остаток или группа остатков-мишеней (например, заряженные остатки, такие как Arg, Asp, His, Lys и Glu) определяют и заменяют нейтральной или отрицательно заряженной аминокислотой (например, аланином или полиаланином), чтобы определить, влияют ли они на взаимодействие антитела с антигеном. Дальнейшие замены могут быть введены в местах аминокислот, демонстрирующих функциональную чувствительность к первоначальным заменам. Альтернативно или дополнительно определяют кристаллическую структуру комплекса "антиген-антитело" для выявления точек контакта между антителом и антигеном. Такие контактные остатки и соседние остатки могут стать мишенями или могут быть устранены в качестве кандидатов на замену. Варианты можно подвергнуть скринингу, чтобы определить, проявляют ли они нужные свойства.
Вставки в аминокислотную последовательность включают амино- и/или карбокси-концевые слияния длиной от одного остатка до полипептидов, содержащих сто или более ста остатков, а также вставки внутри последовательности длиной от одного или множества аминокислотных остатков. Примеры концевых вставок включают антитела с N-концевым остатком метионина. Другие варианты молекулы антитела со вставками включают слияние N- или С-конца антитела с ферментом (например, ADEPT) или полипептидом, который увеличивает время полужизни антитела в сыворотке.
b) Варианты гликозилирования
В некоторых воплощениях антитело, представленное в данном документе, изменяют для увеличения или уменьшения степени, в которой антитело гликозилировано. Добавление к антителу или удаление сайтов гликозилирования можно удобно провести путем изменения аминокислотной последовательности таким образом, чтобы создавался или удалялся один или более чем один сайт гликозилирования.
Если антитело содержит Fc-область, то углевод, прикрепленный к ней, может быть изменен. Нативные антитела, продуцируемые клетками млекопитающих, обычно содержат разветвленный двухантенный олигосахарид, как правило, прикрепленный с помощью N-связи к Asn297 СН2-домена Fc-области (см., например, Wright, A. and Morrison, S.L., TIBTECH 15 (1997) 26-32). Олигосахарид может включать различные углеводы, например, маннозу, N-ацетилглюкозамин (GlcNAc), галактозу и сиаловую кислоту, а также фукозу, прикрепленную к GlcNAc в "стволе" двухантенной олигосахаридной структуры. В некоторых воплощениях модификации олигосахаридов в антителе изобретения могут быть сделаны для того, чтобы получить варианты антитела с определенными улучшенными свойствами.
В одном воплощении предложены варианты антитела, имеющие углеводную структуру, в которой отсутствует фукоза, прикрепленная (напрямую или опосредованно) к Fc-области. Например, количество фукозы в таком антителе может составлять от 1% до 80%, от 1% до 65%, от 5% до 65% или от 20% до 40%. Количество фукозы определяется путем вычисления среднего количества фукозы в сахарной цепи на Asn297 по отношению к сумме всех гл и ко структур, прикрепленных к Asn 297 (например, сложных, гибридных и высокоманнозных структур), измеренного путем масс-спектрометрии MALDI-TOF, как описано, например, в WO 2008/077546. Asn297 относится к остатку аспарагина, расположенному примерно в позиции 297 в Fc-области (ЕС-нумерация остатков Fc-области); тем не менее, Asn297 также может быть расположен в позиции примерно плюс/минус 3 аминокислоты до или после позиции 297, т.е. между позициями 294 и 300, в связи с минорными изменениями последовательности в антителах. Такие варианты фукозилирования могут иметь улучшенную ADCC-функцию. См., например, US 2003/0157108; US 2004/0093621. Примеры публикаций, касающихся "дефукозилированных" или "фукозодефицитных" вариантов антител, включают: US 2003/0157108; WO 2000/61739; WO 2001/29246; US 2003/0115614; US 2002/0164328; US 2004/0093621; US 2004/0132140; US 2004/0110704; US 2004/0110282; US 2004/0109865; WO 2003/085119; WO 2003/084570; WO 2005/035586; WO 2005/035778; WO 2005/053742; WO 2002/031140; Okazaki, A. et al., J. Mol. Biol. 336 (2004) 1239-1249; Yamane-Ohnuki, N. et al., Biotech. Bioeng. 87 (2004) 614-622. Примеры клеточных линий, способных продуцировать дефукозилированные антитела, включают СНО-клетки Lec13, дефицитные по белковому фукозилированию (Ripka, J. et al., Arch. Biochem. Biophys. 249 (1986) 533-545; US 2003/0157108; и WO 2004/056312, в частности, пример 11), и нокаутные клеточные линии, такие как СНО-клетки с нокаутом гена альфа-1,6-фукозилтрансферазы, FUT8 (см., например, Yamane-Ohnuki, N. et al., Biotech. Bioeng. 87 (2004) 614-622; Kanda, Y. et al., Biotechnol. Bioeng. 94 (2006) 680-688; и WO 2003/085107).
Также предложены варианты антител с олигосахаридами с симметричным разветвлением, например, в которых двухантенный олигосахарид, присоединенный к Fc-области антитела, разделен пополам посредством GlcNAc. Такие варианты антител могут обладать уменьшенным фукозилированием и/или улучшенной ADCC-функцией. Примеры таких вариантов антител описаны, например, в WO 2003/011878; US 6602684; и US 2005/0123546. Также предусмотрены варианты антител по меньшей мере с одним остатком галактозы в олигосахариде, прикрепленном к Fc-области. Такие варианты антител могут обладать улучшенной CDC-функцией. Такие варианты антител описаны, например, в WO 1997/30087; WO 1998/58964; и WO 1999/22764.
с) варианты Fc-области
В некоторых воплощениях в Fc-область антитела, предложенного в данном документе, может быть введена одна или более чем одна аминокислотная модификация с получением, тем самым, варианта Fc-области. Вариант Fc-области может содержать последовательность человеческой Fc-области (например, человеческой Fc-области IgG1, IgG2, IgG3 или IgG4), содержащую аминокислотную модификацию (например, замену) в одной или более чем одной аминокислотной позиции.
В некоторых воплощениях изобретение предусматривает вариант антитела, который обладает некоторыми, но не всеми эффекторными функциями, что делает его желательным кандидатом для применений, в которых время полужизни антитела in vivo является важным, а еще некоторые эффекторные функции (такие как комплемент и ADCC) являются не необходимыми или вредными. Можно провести анализ цитотоксичности in vitro и/или in vivo, чтобы подтвердить уменьшение/истощение CDC-и/или ADCC-активности. Например, можно провести анализы Fc-рецепторного (FcR) связывания для того, чтобы убедиться, что антитело не связывает FcyR (и, следовательно, скорее всего не обладает ADCC-активностью), но сохраняет FcRn-связывающую способность. Первичные клетки для опосредования ADCC, NK-клетки, экспрессируют только FcyRIII, тогда как моноциты экспрессируют FcyRI, FcyRII и FcyRIII. Экспрессия FcR на гемопоэтических клетках суммирована в таблице 3 на странице 464 в Ravetch, J.V. and Kinet, J.P., Annu. Rev. Immunol. 9 (1991) 457-492. Неограничивающие примеры анализов in vitro для оценки ADCC-активности молекулы, представляющей интерес, описаны в US 5500362 (см., например, Hellstrom, I. et al., Proc. Natl. Acad. Sci. USA 83 (1986) 7059-7063; и Hellstrom, I. et al., Proc. Natl. Acad. Sci. USA 82 (1985) 1499-1502); US 5821337 (см. Bruggemann, M. et al., J. Exp.Med. 166 (1987) 1351-1361). Альтернативно, могут быть использованы нерадиоактивные способы анализа (см., например, нерадиоактивный анализ цитотоксичности ACTI™ для проточной цитометрии (CellTechnology Inc., Маунтин-Вью, Калифорния); и нерадиоактивный анализ цитотоксичности CytoTox 96® (Promega, Мэдисон, Висконсин)). Используемые эффекторные клетки для таких анализов включают мононуклеарные клетки периферической крови (РВМС) и натуральные киллеры (NK-клетки). Альтернативно или дополнительно, ADCC-активность молекулы, представляющей интерес, можно оценить in vivo, например, на животной модели, такой как описана в Clynes, R. et al., Proc. Natl. Acad. Sci. USA 95 (1998) 652-656. Также можно выполнить анализы C1q-связывания, чтобы подтвердить, что антитело не может связывать C1q и, следовательно, не обладает CDC-активностью. См., например, C1q- и СЗс-связывающий ELISA в WO 2006/029879 и WO 2005/100402. Для оценки активации комплемента можно выполнить анализ CDC (см., например, Gazzano-Santoro, Н. et al., J. Immunol. Methods 202 (1996) 163-171; Cragg, M.S. et al., Blood 101 (2003) 1045-1052; и Cragg, M.S. and M.J. Glennie, Blood 103 (2004) 2738-2743). Определение FcRn-связывания и клиренса/периода полужизни in vivo также можно проводить с использованием способов, известных в данной области (см., например, Petkova, S.B. et al., Int. Immunol. 18 (2006) 1759-1769).
Антитела со сниженной эффекторной функцией включают таковые с заменой одного или более чем одного из остатков Fc-области среди 238, 265, 269, 270, 297, 327 и 329 (US 6737056). Такие Fc-мутанты включают Fc-мутанты с заменами в двух или более аминокислотных позициях, выбранных среди 265, 269, 270, 297 и 327, включая так называемый "DANA" Fc-мутант с заменой остатков 265 и 297 на аланин (US 7332581).
Описаны некоторые варианты антител с улучшенным или уменьшенным FcR-связыванием. (См., например, US 6737056; WO 2004/056312, и Shields, R.L. et al., J. Biol. Chem. 276 (2001) 6591-6604).
В некоторых воплощениях вариант антитела содержит Fc-область с одной или более чем одной аминокислотной заменой, которая улучшает ADCC, например, с заменами в позициях 298, 333 и/или 334 в Fc-области (ЕС-нумерация остатков).
В некоторых воплощениях в Fc-области сделаны изменения, которые приводят к измененному (т.е. повышенному или пониженному) Clq-связыванию и/или комплементзависимой цитотоксичности (CDC), например, как описано в US 6194551, WO 99/51642 и в Idusogie, Е.Е. et al., J. Immunol. 164 (2000) 4178-4184.
Антитела с повышенным периодом полужизни и повышенным связыванием с неонатальным Fc-рецептором (FcRn), который отвечает за перенос материнских IgG к плоду (Guyer, R.L. et al., J. Immunol. 117 (1976) 587-593; и Kim, J.K. et al., J. Immunol. 24 (1994) 2429-2434), описаны в US 2005/0014934. Эти антитела содержат Fc-область с одной или более чем одной заменой, которая улучшает связывание Fc-области с FcRn. Такие Fc-варианты включают варианты с заменами в одном или более чем одном остатке Fc-области: 238, 256, 265, 272, 286, 303, 305, 307, 311, 312, 317, 340, 356, 360, 362, 376, 378, 380, 382, 413, 424 или 434, например, с заменой остатка 434 в Fc-области (US 7371826).
См. также Duncan, A.R. and Winter, G., Nature 322 (1988) 738-740; US 5648260; US 5624821 и WO 94/29351, касающиеся других примеров вариантов Fc-области.
d) Цистеин-инженерные варианты антител
В некоторых воплощениях может быть желательным создание цистеин-инженерных антител, например, "тиоМКА" ("thioMAb"), в которых один или более чем один из остатков антитела замещен остатком цистеина. В конкретных воплощениях замещенные остатки находятся в доступных сайтах антитела. При замещении этих остатков цистеином реактивные тиольные группы, таким образом, располагаются в доступных участках антитела и могут быть использованы для конъюгирования антитела с другими группировками, такими как лекарственные группировки или линкер-лекарственные группировки, для образования иммуноконъюгата, как описано далее. В некоторых воплощениях один или более чем один из следующих остатков может быть замещен цистеином: V205 (нумерация по Kabat) легкой цепи; А118 (ЕС-нумерация) тяжелой цепи; и S400 (ЕС-нумерация) Fc-области тяжелой цепи. Цистеин-инженерные антитела могут быть получены, как описано, например, в US 7521541.
e) Производные антител
В некоторых воплощениях антитело, представленное в данном документе, может быть дополнительно модифицировано так, чтобы содержать дополнительные небелковые группировки, которые известны в данной области и легко доступны. Группировки, пригодные для получения производных антитела, включают, но не ограничиваясь ими, водорастворимые полимеры. Неограничивающие примеры водорастворимых полимеров включают полиэтиленгликоль (ПЭГ), сополимеры этиленгликоля/пропиленгликоля, карбоксиметилцеллюлозу, декстран, поливиниловый спирт, поливинилпирролидон, поли-1,3-диоксолан, поли-1,3,6-триоксан, сополимеры этилена/малеинового ангидрида, полиаминокислоты (гомополимеры или статистические сополимеры) и декстран или поли(н-винилпирролидон)полиэтиленгликоль, гомополимеры полипропиленгликоля, сополимеры полипропиленоксида/этиленоксида, полиоксиэтилированные полиолы (например, глицерин), поливиниловый спирт и их смеси. Полиэтиленгликоля пропиональдегид может давать преимущества в производстве благодаря его стабильности в воде. Полимер может иметь любую молекулярную массу и может быть разветвленным или неразветвленным. Число полимеров, присоединенных к антителу, может варьировать, и если присоединено больше одного полимера, то они могут быть одинаковыми или различными молекулами. Как правило, число и/или тип полимеров, используемых для получения производных, может быть определен на основании ряда соображений, включая, но не ограничиваясь ими, конкретные свойства или функции антитела, которые будут улучшены, будет ли использоваться производное антитела в терапии при определенных состояниях и т.д.
В другом воплощении предложены конъюгаты антитела и небелковой группировки, которая может селективно нагреваться при воздействии радиации. В одном из воплощений небелковая группировка представляет собой углеродную нанотрубку (Kam, N.W. et al., Proc. Natl. Acad. Sci. USA 102 (2005) 11600-11605). Излучение может быть любой длины волны, включая, но не ограничиваясь ими, длины волн, которые не повреждают обычные клетки, но нагревают небелковую группировку до температуры, при которой клетки, ближайшие к небелковой группировке антитела, погибают.
В. Рекомбинантные способы и композиции
Антитела могут быть получены с использованием рекомбинантных способов и композиций, например, описанных в US 4816567. В одном воплощении предложена изолированная нуклеиновая кислота, кодирующая антитело против рецептора трансферрина, описанное в данном документе. Такая нуклеиновая кислота может кодировать аминокислотную последовательность, содержащую VL, и/или аминокислотную последовательность, содержащую VH антитела (например, легкую и/или тяжелую цепи антитела). В другом воплощении предложен один или более чем один вектор (например, экспрессионный вектор), содержащий такую нуклеиновую кислоту. В другом воплощении предложена клетка-хозяин, содержащая такую нуклеиновую кислоту. В одном таком воплощении клетка-хозяин содержит (например, была трансформирована ими): (1) вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VL антитела, и аминокислотную последовательность, содержащую VH антитела, или (2) первый вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VL антитела, и второй вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VH антитела. В одном воплощении клетка-хозяин является эукариотической, например, клеткой яичника китайского хомячка (СНО) или лимфоидной клеткой (например, Y0, NS0, Sp2/0). В одном воплощении предложен способ получения антитела против рецептора трансферрина, который включает культивирование клетки-хозяина, содержащей нуклеиновую кислоту, кодирующую антитело, как указано выше, в условиях, подходящих для экспрессии антитела, и, возможно, выделение антитела из клетки-хозяина (или культуральной среды клетки-хозяина).
Для рекомбинантной продукции антитела против рецептора трансферрина нуклеиновую кислоту, кодирующую антитело, например, описанное выше, выделяют и встраивают в один или более чем один вектор для дальнейшего клонирования и/или экспрессии в клетке-хозяине. Такая нуклеиновая кислота может быть легко выделена и секвенирована с использованием обычных процедур (например, с использованием олигонуклеотидных зондов, которые способны специфически связываться с генами, кодирующими тяжелые и легкие цепи антитела).
Подходящие клетки-хозяева для клонирования или экспрессии векторов, кодирующих антитела, включают прокариотические или эукариотические клетки, описанные в данном документе. Например, антитела могут быть получены в бактериях, в частности, когда не нужны гликозилирование и Fc-эффекторные функции. Для экспрессии фрагментов антитела и полипептидов в бактериях см., например, US 5648237, US 5789199 и US 5840523. (См. также Charlton, К.А., In: Methods in Molecular Biology, Vol. 248, Lo, B.K.C. (ed.), Humana Press, Totowa, NJ (2003), pp. 245-254, где описывается экспрессия фрагментов антител в Е. coli). После экспрессии антитело может быть выделено из массы бактериальных клеток в растворимой фракции и может быть далее очищено.
Помимо прокариотов подходящими клонирующими или экспрессирующими хозяевами для векторов, кодирующих антитела, являются эукариотические микроорганизмы, такие как нитчатые грибы или дрожжи, включая штаммы грибов и дрожжей, у которых пути гликозилирования являются "гуманизированными", что приводит к продукции антитела с частично или полностью человеческим характером гликозилирования (см. Gerngross, T.U., Nat. Biotech. 22 (2004) 1409-1414; и Li, Н. et al., Nat. Biotech. 24 (2006) 210-215).
Подходящие клетки-хозяева для экспрессии гликозилированного антитела также получают из многоклеточных организмов (беспозвоночных и позвоночных). Примеры клеток беспозвоночных включают клетки растений и насекомых. Были идентифицированы многочисленные бакуловирусные штаммы, которые могут быть использованы в сочетании с клетками насекомых, в частности, для трансфекции клеток Spodoptera frugiperda.
Культуры растительных клеток также могут быть использованы в качестве хозяев. См., например, US 5959177, US 6040498, US 6420548, US 7125978 и US 6417429 (описывающие технологию PLANTIBODIES™ для получения антител в трансгенных растениях).
Клетки позвоночных также могут быть использованы в качестве хозяев. Например, могут быть использованы клеточные линии млекопитающих, приспособленные к росту в суспензии. Другими примерами используемых клеточных линий-хозяев от млекопитающих являются линия почки обезьяны CV1, трансформированная SV40 (COS-7); человеческая эмбриональная почечная линия (293 или клетки 293, описанные, например, в Graham, F.L. et al., J. Gen Virol. 36 (1977) 59-74); клетки почек детенышей хомяка (ВНК); мышиные клетки Сертоли (ТМ4-клетки, описанные, например, в Mather, J.P., Biol. Reprod. 23 (1980) 243-252); клетки почки обезьяны (CV1); клетки почки африканской зеленой обезьяны (VERO-76); клетки шейки матки человеческой карциномы (HELA); клетки почки собаки (MDCK); клетки печени крысы buffalo (BRL 3А); клетки легких человека (W138); клетки печени человека (Hep G2); клетки опухоли молочной железы мыши (ММТ 060562); TRI-кпетки, описанные, например, в Mather, J.P. et al., Annals N.Y. Acad. Sci. 383 (1982) 44-68; клетки MRC5 и клетки FS4. Другие используемые линии клеток-хозяев от млекопитающих включают клетки яичника китайского хомячка (СНО), в том числе DHFR-CHO-клетки (Urlaub, G. et al., Proc. Natl. Acad. Sci. USA 77 (1980) 4216-4220); и клеточные линии миеломы, такие как Y0, NS0 и Sp2/0. Обзор некоторых линий клеток-хозяев от млекопитающих, подходящих для продукции антител, см., например, в Yazaki, P. and Wu, A.M., Methods in Molecular Biology, Vol.248, Lo, B.K.C. (ed.), Humana Press, Totowa, NJ (2004), pp.255-268.
С. Анализы
Антитела против рецептора трансферрина, представленные в данном документе, могут быть идентифицированы, подвергнуты скринингу или охарактеризованы на предмет их физических/химических свойств и/или биологической активности с помощью различных анализов, известных в данной области.
1. Анализ связывания
В одном из аспектов антитело согласно изобретению анализируют на активность связывания с антигеном, например, с помощью известных способов, таких как ELISA, alphaLISA, вестерн-блот, микрочипы для прямого (с антителами) и обратно-фазового анализа белка и т.д.
В иллюстративном анализе ELISA или alphaLISA рецептор трансферрина в растворе (клеточный супернатант, лизаты клеток или тканей, жидкости организма и т.д.) связывается антителом захвата, которое специфически связывается с первым эпитопом на рецепторе трансферрина или с рецептором трансферрина в определенной конформации, и с антителом обнаружения, связанным с группировкой для обнаружения, которое специфически связывается со вторым эпитопом или конформацией рецептора трансферрина. Регистрация основана на обнаружении группировки (хемилюминисценция, флуоресценция, люминесценция, индуцированная переносом энергии и т.п.).
В случае микрочипа с антителами эти антитела наносят точками на стеклянные или нитроцеллюлозные чипы. Стекла блокируют и инкубируют с раствором, содержащим рецептор трансферрина, промывают для удаления несвязанных антител, и связанные антитела обнаруживают с помощью соответствующего вторичного антитела, меченного флуоресцентной меткой. Флуоресцентный сигнал измеряют с помощью сканера для флуоресцентных стекол. Аналогично, для обратно-фазовых микрочипов на стеклянные или нитроцеллюлозные чипы наносят точками рекомбинантный рецептор трансферрина, клеточный супернатант, лизаты клеток или тканей, жидкости организма и т.д. Стекла блокируют и отдельные микрочипы инкубируют с антителами против конкретного эпитопа на рецепторе трансферрина. Несвязанные антитела отмывают, а связанные антитела обнаруживают с помощью соответствующего вторичного антитела, меченного флуоресцентной меткой. Флуоресцентный сигнал измеряют с помощью сканера для флуоресцентных стекол (Dernick, G., et al., J. Lipid Res. 52 (2011) 2323-2331).
D. Способы и композиции для диагностики и обнаружения
В некоторых воплощениях любые антитела против рецептора трансферрина, представленные в данном документе, используются для обнаружения присутствия рецептора человеческого трансферрина в биологическом образце. Термин "обнаружение", используемый в данном документе, включает количественное или качественное обнаружение. В некоторых воплощениях биологический образец содержит клетку или ткань, например, ткань головного мозга.
В одном воплощении предложено антитело против рецептора трансферрина для применения в способе диагностики или обнаружения. В другом аспекте предложен способ обнаружения присутствия рецептора трансферрина в биологическом образце. В некоторых воплощениях способ включает контактирование биологического образца с антителом против рецептора трансферрина, описанным в данном документе, в условиях, разрешающих связывание антитела против рецептора трансферрина с рецептором трансферрина, и обнаружение того, образуется ли комплекс между антителом против рецептора трансферрина и рецептором трансферрина. Такой способ может быть способом in vitro или in vivo. В одном воплощении антитело против рецептора трансферрина используется для выбора субъектов, имеющих право на терапию антителом против рецептора трансферрина, например, когда рецептор трансферрина является биомаркером для выбора пациентов.
Примеры нарушений, которые могут быть диагностированы с использованием антитела изобретения, включают нейродегенерацию с накоплением железа в головном мозге типа 1 (NBIA1), идиопатическую вегетативную недостаточность, синдром Дауна, комплекс Гуама, а также несколько нарушений с тельцами Лев и, таких как болезнь диффузных телец Леви (DLBD), вариант болезни Альцгеймера с тельцами Леви (LBVAD), некоторые формы болезни Гоше и болезнь Паркинсона сдеменцией (PDD).
В некоторых воплощениях предложены меченые антитела против рецептора трансферрина. Метки включают, но не ограничиваясь ими, метки или группировки, которые обнаруживаются напрямую (такие как флуоресцентные, хромофорные, электронно-плотные, хемилюминесцентные и радиоактивные метки), а также группировки, такие как ферменты или лиганды, которые обнаруживаются косвенно, например, через ферментативную реакцию или молекулярное взаимодействие. Примеры меток включают, но не ограничиваясь ими, радиоизотопы 32Р, 14С, 125I, 3Н и 131I, флуорофоры, такие какхелаты редкоземельных элементов или флуоресцеин и его производные, родамин и его производные, дансил, умбеллиферон, люциферазы, например, люциферазу светлячка и бактериальную люциферазу (US 4737456), люциферин, 2,3-дигидрофталазиндионы, пероксидазу хрена (HRP), щелочную фосфатазу, (3-галактозидазу, глюкоамилазу, лизоцим, оксидазы сахаридов, например, глюкозооксидазу, галактозооксидазу и глюкозо-6-фосфатдегидрогеназу, гетероциклические оксидазы, например, уриказу и ксантиноксидазу, в сочетании с ферментом, который использует пероксид водорода для окисления предшественника метки, таким как HRP, лактопероксидазу или микропероксидазу, биотин/авидин, спиновые метки, бактериофаговые метки, стабильные свободные радикалы и т.п.
Е. Фармацевтические составы
Фармацевтические составы антитела против рецептора трансферрина, описанного в данном документе, получают путем смешивания такого антитела, имеющего требуемую степень чистоты, с одним или более чем одним возможным фармацевтически приемлемым носителем (Remington's Pharmaceutical Sciences, 16th edition, Osol, A. (ed.) (1980)), в виде лиофилизированных составов или водных растворов. Фармацевтически приемлемые носители, как правило, нетоксичны для реципиентов в используемых дозах и концентрациях и включают, но не ограничиваясь ими: буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как октадецилдиметилбензиламмония хлорид; гексаметония хлорид; бензалкония хлорид; бензетония хлорид; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцин; циклогексанол, 3-пентанол и м-крезол); низкомолекулярные (менее чем примерно 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как EDTA; сахара, такие как сахарозу, маннит, трегалозу или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например, Zn-белковые комплексы); и/или неионные поверхностно-активные вещества, такие как полиэтиленгликоль (ПЭГ). Иллюстративные фармацевтически приемлемые носители в данном документе также включают интерстициальные агенты, диспергирующие лекарственные средства, такие как растворимые нейтрально-активные гиалуронидазные гликопротеины (sHASEGP), например, человеческие растворимые гиалуронидазные гликопротеины РН-20, такие как rhuPH20 (HYLENEX®, Baxter International, Inc.). Некоторые примеры sHASEGP и способы их применения, в том числе rhuPH20, описаны в US 2005/0260186 и US 2006/0104968. В одном аспекте sHASEGP комбинирован с одной или более чем одной дополнительной гликозамингликаназой, такой какхондроитиназа.
Иллюстративные лиофилизированные составы антител описаны в US 6267958. Водные составы антител включают те, которые описаны в US 6171586 и WO 2006/044908, последние составы включают гистидин-ацетатный буфер.
Состав в данном документе может также содержать более чем один активный ингредиент, необходимый при лечении по конкретным показаниям, особенно с дополняющими активностями, которые не влияют друг на друга. Такие активные ингредиенты соответствующим образом присутствуют в комбинации в количествах, которые эффективны для данной цели.
Активные ингредиенты могут быть заключены в микрокапсулы, полученные, например, с помощью методик коацервации или путем межфазной полимеризации, например, в гид роксиметил целлюлозные или желатиновые микрокапсулы и полиметилметакрилатные микрокапсулы, соответственно, в коллоидных системах доставки лекарственных средств (например, липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсиях. Такие методики раскрыты в Remington's Pharmaceutical Sciences 16th edition, Osol, A. (ed.), (1980).
Могут быть изготовлены препараты с замедленным высвобождением. Подходящие примеры препаратов с замедленным высвобождением включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащие антитело или конъюгат, при этом матрицы находятся в виде формованных изделий, например пленок или микрокапсул.
Составы, используемые для введения in vivo, обычно являются стерильными. Стерильность может быть легко достигнута, например, путем фильтрации через стерилизующие фильтрационные мембраны.
III. Продукты производства
В другом аспекте изобретения предусмотрен продукт производства, содержащий материалы, используемые для лечения, профилактики и/или диагностики нарушений, описанных выше. Продукт производства содержит контейнер и этикетку или вкладыш в упаковку, прикрепленный или связанный с контейнером. Подходящие контейнеры включают, например, бутылки, флаконы, шприцы, пакеты для внутривенно вводимых растворов и т.д. Контейнеры могут быть изготовлены из различных материалов, таких как стекло или пластик. Контейнер содержит композицию саму по себе или в сочетании с другой композицией, эффективной для лечения, профилактики и/или диагностики данного состояния, и может иметь стерильное входное отверстие (например, контейнер может представлять собой пакет для внутривенно вводимого раствора или флакон с пробкой, прокалываемой иглой для подкожной инъекции). По меньшей мере один активный агент в композиции представляет собой антитело согласно данному изобретению. На этикетке или вкладыше в упаковку указывается, что композиция используется для лечения выбранного состояния. Кроме того, продукт производства может включать: (а) первый контейнер с композицией, содержащейся в нем, где композиция содержит антитело согласно данному изобретению; и (b) второй контейнер с композицией, содержащейся в нем, где композиция также содержит цитотоксический или другой терапевтический агент.Продукт производства в этом воплощении изобретения может также содержать вкладыш в упаковку, указывающий, что композиции могут быть использованы для лечения конкретного состояния. Альтернативно или дополнительно продукт производства может также включать второй (или третий) контейнер, содержащий фармацевтически приемлемый буфер, например, бактериостатическую воду для инъекций (BWFI), забуференный фосфатом физиологический раствор, раствор Рингера и раствор декстрозы. Он также может включать другие материалы, желательные с коммерческой и потребительской точки зрения, в том числе другие буферы, разбавители, фильтры, иглы и шприцы.
Понятно, что любой из указанных выше продуктов производства может включать иммуноконъюгат согласно данному изобретению вместо антитела против рецептора трансферрина или в дополнение к нему.
VI. Примеры
Ниже приведены примеры способов и композиций согласно изобретению. Понятно, что на практике могут применяться различные другие воплощения, учитывая общее описание, данное выше.
Материалы и методы
Методики рекомбинантной ДНК
Для манипуляций с ДНК были использованы стандартные способы, описанные в Sambrook, J., et al., Molecular cloning: A laboratory manual; Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1989. Молекулярно-биологические реагенты использовали в соответствии с инструкциями производителя.
Генный и олигонуклеотидный синтез
Нужные генные сегменты получали путем химического синтеза на Geneart GmbH (Регенсбург, Германия). Синтезированные фрагменты генов клонировали в плазмиду для размножения/амплификации в Е. coli. Последовательности ДНК субклонированных фрагментов генов подтверждали секвенированием ДНК. Альтернативно, короткие синтетические фрагменты ДНК собирали путем отжига химически синтезированных олигонуклеотидов или с помощью ПЦР. Соответствующие олигонуклеотиды получали с помощью Metabion GmbH (Planegg-Martinsried, Германия)
Реагенты
Все коммерческие химические реагенты, антитела и наборы использовали в соответствии с протоколами производителей, если не указано иное.
Пример 1
Иммунизация кроликов и мышей
Иммунизация мышей
Мышей NMRI иммунизировали генетически, используя экспрессионный плазмидный вектор, кодирующий полноразмерный TfR человека или яванского макака, путем внутрикожного введения векторной ДНК в дозе 100 мкг с последующей электропорацией (2 импульса прямоугольной формы, 1000 В/см, длительность 0,1 мс, интервал 0,125 с; затем 4 импульса прямоугольной формы, 287,5 В/см, длительность 10 мс, интервал 0,125 сек). Мышей подвергали 6 или 7 последовательным иммунизациям в дни 0, 14, 28, 42, 56, 70 и 84. Четвертую и шестую иммунизации выполняли вектором, кодирующим TfR яванского макака; вектор, кодирующий человеческий TfR, использовали для всех других иммунизации. Кровь брали в дни 36, 78 и 92, и из нее получали сыворотки, которые использовали для определения титра путем ELISA (см. ниже). Животных с самыми высокими титрами отбирали для бустирования в день 96 путем внутривенного введения либо 106 человеческих клеток TF-1, либо 50 мкг рекомбинантного человеческого растворимого TfR, лишенного спирального домена (внеклеточный домен человеческого TfR, начинающийся с Leu122, заканчивающийся на Asn608, экспрессированный в клетках HEK293F в виде N-концевого гибрида с человеческой Fc-областью и очищенный путем аффинной хроматографии на белке А и эксклюзионной хроматографии); моноклональные антитела выделяли путем гибридомной технологии, основанной на их способности связывать рецептор трансферрина человека и яванского макака, экспрессированный на поверхности стабильно трансфицированных клеток СНО-К1 (см. пример 4).
Иммунизация кроликов
Новозеландских белых кроликов или трансгенных кроликов, экспрессирующих репертуар гуманизированных антител, иммунизировали генетически, используя экспрессионный плазмидный вектор, кодирующий полноразмерный TfR человека или яванского макака, путем внутрикожного введения векторной ДНК в дозе 400 мкг с последующей электропорацией (5 импульсов прямоугольной формы, 750 В/см, длительность 10 мс, интервал 1 с). Кроликов подвергали 6 последовательным иммунизациям в дни 0, 14, 28, 56, 84 и 112. Четвертую и шестую иммунизации выполняли вектором, кодирующим TfR яванского макака; для всех остальных иммунизации использовали вектор, кодирующий человеческий TfR. Кровь (10% от предполагаемого общего объема крови) брали в дни 35, 63, 91 и 119. Получали сыворотки, которые использовали для определения титра путем ELISA (см. ниже), а также выделяли мононуклеарные клетки периферической, которые использовали в качестве источника антиген-специфических В-клеток в процессе клонирования В-клеток (см. пример 2).
Определение сывороточных титров (ELISA)
Человеческий рекомбинантный растворимый TfR (R&D Systems, кат. №. 2474-ТР) иммобилизовали на 96-луночном планшете NUNC Maxisorb в дозе 3 мкг/мл, 100 мкл/лунка, в PBS, а затем планшет блокировали с помощью 2% CroteinC в PBS, 200 мкл/лунка; вносили серийные разведения антисывороток в дубликатах, в 0,5% CroteinC в PBS, 100 мкл/лунка; обнаружение проводили с помощью (1) HRP-конъюгированного козлиного противомышиного антитела (Jackson Immunoresearch/Dianova 115-036-071; 1/16000) для всех мышиных сывороток, (2) HRP-конъюгированного ослиного противокроличьего IgG-антитела (Jackson Immunoresearch/Dianova 711-036-152; 1/16000) для всех кроличьих сывороток, (3) кроличьего противочеловеческого IgG-антитела (Pierce/Thermo Scientific 31423; 1/5000) только для сывороток от трансгенных кроликов, (4) биотинилированного козлиного антитела против человеческой каппа-области (Southern Biotech/Biozol 2063-08, 1/5000) и стрептавидин-HRP только для сывороток от трансгенных кроликов; разведенного в 0,5% CroteinC в PBS, 100 мкл/лунка. На всех этапах планшеты инкубировали в течение 1 ч при 37°С. Между всеми этапами планшеты промывали 3 раза с помощью 0,05% Tween 20 в PBS. Сигнал получали путем добавления растворимого субстрата ВМ Blue POD Substrate (Roche), 100 мкл/лунка; и останавливали добавлением 1 М HCl, 100 мкл/лунка. Абсорбцию считывали при 450 нм против 690 нм в качестве референса. Титр определяли как разведение антисыворотки, дающее половину максимального сигнала.
Пример 2
Клонирование В-клеток от кроликов
Выделение кроличьих мононуклеарных клеток периферической крови (РВМС)
Образцы крови брали в сумме от 6 животных (2 кролика дикого типа (wt) и 4 трансгенных (tg) кролика). Этих кроликов получали из двух различных программ иммунизации: первая программа с двумя кроликами wt и двумя кроликами tg и вторая программа с двумя кроликами tg (см. также пример "Иммунизация кроликов"). Цельную кровь, содержащую EDTA, разводили в два раза в 1× PBS (РАА, Пашинг, Австрия) и центрифугировали в градиенте плотности с использованием лимфоцитов млекопитающих (Cedarlane Laboratories, Берлингтон, Онтарио, Канада) в соответствии с инструкциями производителя. РВМС дважды промывали 1× PBS.
Среда EL-4 В5
Среда RPMI 1640 (Pan Biotech, Айденбах, Германия), дополненная 10% FCS (Hyclone, Логан, Юта, США), 2 мМ глутамином, 1% раствором пенициллина/стрептомицина (РАА, Пашинг, Австрия), 2 мМ пируватом натрия, 10 мМ HEPES (PAN Biotech, Айденбах, Германия) и 0,05 мМ (3-меркаптоэтанолом (Gibco, Пейсли, Шотландия)
Истощение клеток
Первая программа иммунизации: стерильные 6-луночные планшеты (для культивирования клеток), покрытые конфлюэнтным монослоем СНО-клеток, использовали для истощения макрофагов/моноцитов за счет неспецифической адгезии, а также неспецифического связывания лимфоцитов.
Вторая программа иммунизации: пропускали этап истощения с помощью лунок, покрытых СНО-клетками, так как мы не могли исключить, что В-клетки, продуцирующие антитела, которые перекрестно реагируют с антителами против рецептора трансферрина хомяка, не будут удалены. Таким образом, использовали пустые стерильные 6-луночные планшеты (для культивирования клеток) для истощения макрофагов и моноцитов за счет неспецифической адгезии, позволяя потенциальным В-лимфоцитам, продуцирующим антитела с перекрестной реактивностью к хомяку (и, возможно, с перекрестной реактивностью к мыши), достигать следующего этапа в рабочем процессе.
Для каждой программы иммунизации: в каждую лунку вносили максимум 4 мл среды и РВМС от иммунизированного кролика в количестве до 6×106, и оставляли для связывания в течение 1 ч при 37°С в инкубаторе. Клетки в супернатанте (лимфоциты периферической крови (PBL)) использовали на этапе пэннинга антигена.
Обогащение В-клеток на рецепторе трансферрина человека
6-луночные планшеты для тканевых культур, покрытые монослоем клеток СНО, положительных на рецептор трансферрина человека, засевали до плотности 6×106 PBL на 4 мл среды и давали связываться в течение 1 ч при 37°С в инкубаторе. Неприкрепленные клетки удаляли путем тщательного промывания лунок 1-2 раза с помощью 1× PBS. Оставшиеся прикрепленные клетки отделяли путем обработки трипсином в течение 10 минут при 37°С в инкубаторе. Трипсинизацию останавливали средой EL-4 В5. Клетки хранили на льду до иммунофлуоресцентного окрашивания. Иммунофлуоресцентное окрашивание и проточная цитометрия Анти-IgG-FITC (ABD Serotec, Дюссельдорф, Германия) использовали для сортировки единичных клеток. Для поверхностного окрашивания клетки из этапа истощения и обогащения инкубировали с анти-IgG-FITC-антителом в PBS и инкубировали в течение 45 минут в темноте при температуре 4°С. После окрашивания РВМС промывали два раза ледяным PBS. Наконец, РВМС ресуспендировали в ледяном PBS и немедленно подвергали FACS-анализу. Перед проведением FACS-анализа добавляли пропидия йодид в концентрации 5 мкг/мл (BD Pharmingen, Сан-Диего, Калифорния, США) для различения мертвых и живых клеток.
Для сортировки единичных клеток использовали Becton Dickinson FACSAria, оснащенный компьютером и программным обеспечением FACSDiva (BD Biosciences, США).
Культивирование В-клеток
Культивирование кроличьих В-клеток проводили аналогично тому, как описано в Zubler et al. (1985). Вкратце, одиночные отсортированные кроличьи В-клетки инкубировали в 96-луночных планшетах с 200 мкл/лунка среды EL-4 В5, содержащей клетки Pansorbin (1:100000) (Calbiochem (Merck), Дармштадт, Германия), 5% супернатант кроличьих тимоцитов (TSN-M13 (10242), MicroCoat, Бернрид, Германия) и гамма-облученные мышиные клетки тимомы EL-4-B5 (2,5 × 104/лунка) в течение 7 дней при 37°С в атмосфере 5% СО2 в инкубаторе. Супернатанты культивируемых В-клеток удаляли для скрининга, а оставшиеся клетки сразу собирали и замораживали при -80°С в 100 мкл буфера RLT (Qiagen, Хильден, Германия).
Пример 3
Фаговый дисплей для отбора и продукции одновалентных анти-TfR-IgG
Отбор с помощью фагового дисплея
Получение антител, связывающихся с TfR человека и яванского макака, осуществляли путем фагового дисплея с использованием стандартных протоколов (Silacci et al, Proteomics, 5 (2005) 2340-2350). Синтезированный ген для антигена hTfR-Fc(KiH)-Avi (KiH - "ключ в замке", Avi - Avi-метка) клонировали путем соединения hTfR ECD с N-концевым шарниром человеческой Fc-области типа "замок", который несет С-концевую Avi-метку (SEQ ID NO 99), и лигировали в вектор для экспрессии в клетках млекопитающих. Все наши векторы для экспрессии в клетках млекопитающих несут промотор MPSV для инициации транскрипции и трансляции, где транскрипция заканчивается синтетической сигнальной полиА-последовательностью, расположенной после ORF. Кроме того, вектор содержит последовательность EBV оп'Р для автономной репликации в клеточных линиях, экспрессирующих EBV-EBNA. Правильную ORF подтверждали секвенированием. Этот вектор использовали для экспрессии в суспензионных клетках HEK293 EBNA путем коэкспрессии с пустой Fc-областью типа "ключа" и белком BirA и добавления 1 мМ биотина в культуральную среду. Полученный биотинилированный белок очищали путем аффинной хроматографии на белке А с последующей эксклюзионной хроматографией. Антиген cyTfR-Fc(KiH)-Avi получали соответственно (SEQ ID NO 100).
Используемая библиотека была полностью синтетической библиотекой, использующей человеческие каркасные области семейств VH1, 3, 4, и 5, а также V-каппа-1, -2 и -3 и V-лямбда-3. Рандомизированные аминокислоты находились в CDR-Н3 и CDR-L3. Пулы библиотек создавали, используя каждую тяжелую цепь VH вместе со всеми различными библиотеками легких цепей. Отбор проводили в растворе в соответствии со следующей процедурой: 1. связывание примерно 1012 фагмидных частиц из каждой библиотеки с 100 нМ биотинилированным TfR-avi-his в течение 0,5 ч в общем объеме 1 мл; 2. захват биотинилированного TfR-avi-his и специфически связанных фаговых частиц путем добавления 5,4 × 107 магнитных гранул, покрытых стрептавидином, в течение 10 минут; 3. промывание гранул 5-10 раз с помощью 1 мл PBS/Tween 20 и 5-10 раз с помощью 1 мл PBS; 4. элюирование фаговых частиц путем добавления 1 мл 100 мМ TEA (триэтиламина) в течение 10 минут и нейтрализация путем добавления 500 мкл 1 М Tris/HCl, рН 7,4; и 5. повторное инфицирование экспоненциально растущих бактерий Е, coli TG1, инфицирование фагом-хелпером VCSM13 и последующая преципитация фагмидных частиц с помощью ПЭГ/NaCl для использования в последующих раундах отбора. Отбор проводили в течение 3-5 раундов с использованием постоянной или уменьшающейся (от 10-7 М до 2×10-9 М) концентрации антигена. Во втором раунде захват комплексов антигена/фага проводили с использованием планшетов с нейтравидином вместо гранул со стрептавидином. Так как связывающие вещества, полученные против рекомбинантного растворимого антигена, часто показывали только слабое связывание на клетках, мы ввели два дополнительных этапа отбора с использованием клеток, демонстрирующих нативный антиген, после второго или третьего раунда обогащения на рекомбинантном антигене.
Были использованы две клеточные линии, TF-1 и NCl-H460 (обе доступны в АТСС). Вкратце, 1×106 клеток инкубировали примерно с 1012 фаговых частиц в течение 1 ч на льду, чтобы избежать опосредованной рецептором интернализации. Промывание проводили путем 5-10 этапов центрифугирования с использованием PBST-буфера (PBS, содержащего 1% Tween-20). Целые клетки использовали для инфицирования бактерий TG1 для высвобождения фага.
Конкретные связывающие агенты идентифицировали с помощью ELISA следующим образом: 100 мкл 10 нМ биотинилированного TfR-avi-his на лунку наносили на планшеты с нейтравидином. Добавляли Fab-содержащие бактериальные супернатанты и регистрировали связывание Fab за счет их FLAG-меток с помощью анти-FLAG/HRP-BTopn4Horo антитела. ELISA-положительные клоны экспрессировали в бактериях в виде растворимых Fab-фрагментов в 96-луночном формате, и супернатанты подвергали эксперименту кинетического скрининга с помощью SPR-анализа с использованием ProteOn XPR36 (BioRad). Выявляли клоны, экспрессирующие Fab с самыми высокими константами аффинности, и секвенировали соответствующие фагмиды.
Очистка Fab-антител
Клетки Тор10 индивидуально трансфицировали фагмидными плазмидами из всех клонов, положительных в клеточном ELISA. Клетки выращивали в среде и индуцировали продукцию Fab-антител. Fab-антитела выделяли путем получения периплазмы и очищали с помощью IMAC.
FACS-анализ
Очищенные Fab-антитела добавляли к клеткам TF-1 в различных концентрациях. Связанные с клетками Fab-антитела обнаруживали с помощью флуоресцентного меченного анти-Fab-антитела и измеряли с помощью FACS. Рассчитывали значения ЕС50.
Продукция и характеризация отобранных клонов
Преобразование в формат IgG
Для преобразования в формат IgG были выбраны отобранные клоны с периодом полужизни комплекса (t1/2) от 6 до 20 минут или ЕС50 на клетках от 10 нМ до 500 нМ, с отчетливым сигналом связывания клеток либо на FACS, либо в клеточном ELISA, и с перекрестно-реактивным связыванием в отношении обоих антигенов, hTfR-Fc(KiH)-Avi и cyTfR-Fc(KiH)-Avi.
Для этого домен VL амплифицировали с помощью ПЦР и клонировали в вектор для экспрессии в клетках млекопитающих, как описано выше, непосредственно перед доменом CL. Кроме того, домен VH амплифицировали с помощью ПЦР и клонировали непосредственно перед доменом CH-Fc. Определяли последовательности обоих экспрессионных векторов.
Продукция антител IgG
Суспензионные клетки НЕК293 EBNA трансфицировали обеими плазмидами, кодирующими LC и НС, и культивировали в течение 7 дней. Супернатант осветляли путем стерильной фильтрации. Концентрацию IgG определяли с помощью хроматографии на белке А.
Определение ЕС50 с использованием методики TagLite
Ген ECD hTfR или cyTfR, соответственно, включая трансмембранный домен, лигировали в вектор для экспрессии в клетках млекопитающих после SNAP-метки. Этот вектор использовали для трансфекции суспензионных клеток НЕК293 EBNA, получая дисплей гибридного TfR с С-концевой SNAP-меткой (SEQ ID NO 101, SEQ ID NO 102). SNAP-метку специфически метили с помощью SNAP-Lumi4Tb (Cisbio). Эффективность мечения определяли путем измерения эмиссии тербия при 615 нм. Меченые клетки хранили при -80°С.
В присутствии IgG-антитела против человеческого Fc-d2 меченые клетки инкубировали с IgG-супернатантом в различных разведениях и через 4 часа измеряли FRET-сигналы (эмиссия метки-донора Lumi4Tb: 615 нм и метки-акцептора d2: 665 нм). Рассчитывали значения ЕС50, получая 25 IgG, которые связывались как с hTfR, так и с cyTfR.
Определение связывания клеток с помощью FACS
Ген полноразмерного hTfR или cyTfR, соответственно, клонировали в вектор для экспрессии в клетках млекопитающих. Этот вектор использовали для трансфекции суспензионных клеток СНО EBNA. IgG-супернатант непосредственно наносили на клетки, несущие TfR. После промывания PBST комплекс "антиген-антитело" обнаруживали с использованием конъюгата анти-huFc-IgG-HRP-антитело, с последующим развитием окраски с помощью 3,3'-5,5'-тетраметилбензидина. Функциональный дисплей проверяли с использованием коммерчески доступного анти-TfR-антитела. Все 25 Tag Lite-положительных клона демонстрировали сильный связывающий сигнал.
Продукция и очистка одновалентных IgG-подобных молекул Домен VH клонировали в вектор для экспрессии в клетках млекопитающих, кодирующий человеческую НС IgG1 типа "ключ", которая несет мутации L234A, L235A и P329G. Этот вектор использовали для экспрессии в суспензионных клетках НЕК293 EBNA, коэкспрессирующих LC (вектор, описанный выше) и пустой Fc-домен типа "замок", несущий мутации L234A, L235A, P329G, H435R и Y436F. Полученный белок очищали путем аффинной хроматографии на белке А. В случаях, когда мономерный белок составлял более 95%, что определяли путем аналитической эксклюзионной хроматографии, белок далее очищали путем эксклюзионной хроматографии.
Пример 4
Идентификация антител, связывающих TfR человека и яванского макака, с помощью клеточного ELISA
Для скрининга кроличьих В-клеток или супернатантов мышиных гибридом на наличие антител, распознающих TfR человека и яванского макака, использовали клеточный ELISA со стабильно трансфицированными клетками СНО-К1. Стабильные трансфектанты получали путем трансфекции клеток СНО-К1 экспрессионными плазмидами, содержащими экспрессионные кассеты для TfR человека и яванского макака, а также для неомицин-фосфотрансферазы. После трансфекции клетки разводили в ростовой среде, содержащей 500 мкг/мл G418 (Life Technologies). После появления растущих клонов клетки отделяли, окрашивали МЕМ-75 (Abeam) или 13Е4 (Life Technologies) и РЕ-мечеными вторичными антителами против TfR человека и яванского макака, и сильно флуоресцирующие клетки сортировали в виде единичных клеток в лунки 96-луночного планшета (FACS Aria). После 7 дней роста клоны снова проверяли на экспрессию TfR, и клоны с наилучшей экспрессией отбирали для экспериментов в клеточном ELISA.
Вкратце, 15000 клеток на лунку высевали в 384-луночный планшет и инкубировали в течение 18 ч при температуре 37°С, 5% СО2. Супернатант удаляли с использованием автоматизированного промывателя (BIOTEK), и в каждую лунку добавляли 30 мкл супернатанта, содержащего антитело, а затем 24 мкл ростовой среды. После 2 часов инкубации лунки опорожняли и добавляли 30 мкл 0,05% глутаральдегида в PBS на 45 минут при комнатной температуре. После 3 промываний с помощью PBS/0,025% Tween 20 (PBST) вносили 30 мкл противокроличьей HRP или противомышиной HRP (Southern Biotech), разведенной 1: 5000 в блокирующем буфере, и планшеты инкубировали в течение 1 часа при комнатной температуре. Лунки промывали 6 раз с помощью PBST, получали сигнал, добавляя 30 мкл ТМВ на лунку, и измеряли абсорбцию при 450 нм.
Пример 5
Клонирование и экспрессия aHTH-TfR-антител
Методики рекомбинантной ДНК
Для манипуляций с ДНК были использованы стандартные способы, описанные в Sambrook, J., et al., Molecular cloning: A laboratory manual; Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1989. Молекулярно-биологические реагенты использовали в соответствии с инструкциями изготовителя.
Генный и олигонуклеотидный синтез
Нужные генные сегменты получали путем химического синтеза на Geneart GmbH (Регенсбург, Германия). Синтезированные фрагменты генов клонировали в плазмиду для размножения/амплификации в Е. coli. Последовательности ДНК субклонированных фрагментов генов подтверждали секвенированием ДНК. Альтернативно, короткие синтетические фрагменты ДНК собирали путем отжига химически синтезированных олигонуклеотидов или с помощью ПЦР. Соответствующие олигонуклеотиды получали с помощью Metabion GmbH (Planegg-Martinsried, Германия)
ПЦР-амплификация V-доменов
Тотальную РНК получали из лизатов В-клеток (ресуспендировали в буфере RLT, Qiagen, кат. №79216) с помощью набора NucleoSpin 8/96 RNA (Macherey&Nagel; 740709.4, 740698) в соответствии с протоколом производителя. РНК элюировали в 60 мкл без РНКазной воды. 6 мкл РНК использовали для получения кДНК с помощью реакции обратной транскрипции с использованием реагентов Superscript III First-Strand Synthesis SuperMix (Invitrogen 18080-400) и олиго-dT-праймера в соответствии с инструкциями производителя. Все этапы выполняли на приборе Hamilton ML Star System. 4 мкл кДНК использовали для амплификации вариабельных областей тяжелой и легкой цепей иммуноглобулина (VH и VL) с помощью реагентов AccuPrime SuperMix (Invitrogen 12344-040) в конечном объеме 50 мкл с использованием праймеров rbHC.up и rbHC.do для тяжелой цепи, rbLC.up и rbLC.do для легкой цепи кроличьих В-клеток дикого типа, и BcPCR_FHLC_leader.fw и BcPCR_huCkappa.rev для легкой цепи трансгенных кроличьих В-клеток (см. таблицу ниже). Все прямые праймеры были специфичными для сигнального пептида (соответственно, VH и VL), тогда как обратные праймеры были специфичными для константных областей (соответственно, VH и VL). Условия ПЦР для RbVH+RbVL были следующими: горячий старт при 94°С в течение 5 минут; 35 циклов: 20 секунд при 94°С, 20 секунд при 70°С, 45 секунд при 68°С; финальная элонгация при 68°С в течение 7 минут. Условия ПЦР для HuVL были следующими: горячий старт при 94°С в течение 5 минут; 40 циклов: 20 секунд при 94°С, 20 секунд при 52°С, 45 секунд при 68°С; финальная элонгация при 68°С в течение 7 минут.


8 мкл из 50 мкл ПЦР-смеси загружали в гель 48 E-Gel 2% (Invitrogen G8008-02). Положительные ПЦР-реакции очищали с помощью набора NucleoSpin Extract II (Macherey&Nagel; 740609250) в соответствии с протоколом производителя и элюировали в 50 мкп буфера для элюции. Все этапы очистки выполняли на приборе Hamilton ML Starlet System.
Рекомбинантная экспрессия кроличьих моноклональных двухвалентных антител
Для рекомбинантной экспрессии кроличьих моноклональных двухвалентных антител ПЦР-продукты, кодирующие VH или VL, клонировали в виде кДНК в экспрессионные векторы с помощью методики клонирования с выступами ("липкими концами") (RS Haun et al., BioTechniques (1992) 13, 515-518; MZ Li et al., Nature Methods (2007) 4, 251-256). Экспрессионные векторы содержали экспрессионную кассету, состоящую из 5'-СМ\/-промотора, включая интрон А, и 3'-последовательности полиаденилирования из BGH. Помимо экспрессионной кассеты плазмиды содержали сайт инициации репликации, полученный из pUC18, и ген бета-лактамазы, придающий устойчивость к ампициллину, для амплификации плазмиды в Е. coli. Были использованы три варианта основной плазмиды: одна плазмида, содержащая кроличью константную область IgG, предназначенная для введения VH-областей, и две дополнительные плазмиды, содержащие кроличью или человеческую константную область LC каппа для введения VL-областей.
Линеаризованные экспрессионные плазмиды, кодирующие константную область каппа или гамма и VL/VH-вставки, амплифицировали с помощью ПЦР с использованием перекрывающихся праймеров.
Очищенные ПЦР-продукты инкубировали с Т4 ДНК-полимеразой, которая создавала однонитевые выступы. Реакцию останавливали путем добавления дЦТФ.
На следующем этапе плазмиду и вставку объединяли и инкубировали с гесА, который индуцировал сайт-специфическую рекомбинацию. Рекомбинированными плазмидами трансформировали Е. coli. На следующий день выросшие колонии собирали и анализировали на правильность рекомбинированной плазмиды путем выделения плазмиды, рестрикционного анализа и ДНК-секвенирования.
Для экспрессии антитела выделенными НС- и LC-плазмидами временно котрансфицировали клетки НЕК293, и супернатанты собирали через 1 неделю.
Создание векторов для экспрессии кроличьих монокпональных одновалентных антител
Для рекомбинантной экспрессии отобранных кандидатов в виде монокпональных одновалентных антител кроличьи константные области всех VH-цепей преобразовывали в человеческие константные области, окружающие мутацию типа "ключ" в СН3-сегменте. Для VL-цепей, полученных из кроличьих В-клеток дикого типа, кроличьи константные области С-каппа преобразовывали в человеческие. 4 мкл кДНК отобранных кандидатов использовали для амплификации вариабельных областей тяжелой и легкой цепи иммуноглобулина с помощью реагентов AccuPrime SuperMix (Invitrogen 12344-040) в конечном объеме 50 мкл с прямыми праймерами, специфичными для сигнального пептида, и обратными праймерами, специфичными для CDR3-J-o6nacrn с (3'-концевой) перекрывающейся последовательностью (20 п. н.), гомологичной человеческим константным областям (соответственно, VH и VL). Условия ПЦР для амплификации цепей VH и VL были следующими: горячий старт при 94°С в течение 5 минут; 35 циклов: 20 секунд при 94°С, 20 секунд при 68°С, 45 секунд при 68°С; финальная элонгация при 68°С в течение 7 минут.
ПЦР-продукты, кодирующие VH или VL, клонировали в виде кДНК в экспрессионные векторы с помощью методики клонирования с выступами ("липкими концами") (RS Haun et al., BioTechniques (1992) 13, 515-518; MZ Li et al., Nature Methods (2007) 4, 251-256). Экспрессионные векторы содержали экспрессионную кассету, состоящую из 5'-CMV-промотора, включая интрон А, и 3'-последовательности полиаденилирования из BGH. Помимо экспрессионной кассеты плазмиды содержали сайт инициации репликации, полученный из pUC18, и ген бета-лактамазы, придающий устойчивость к ампициллину, для амплификации плазмиды в Е. coli. Были использованы два варианта основной плазмиды: одна плазмида, содержащая человеческую константную область IgG, созданная для введения новой амплифицированной VH-цепи, и вторая плазмида, содержащая человеческую константную область LC-каппа для введения VL-цепи.
Линеаризованные экспрессионные плазмиды, кодирующие константную область каппа или гамма и VL/VH-вставки, амплифицировали с помощью ПЦР с использованием перекрывающихся праймеров.
Очищенные ПЦР-продукты инкубировали с Т4 ДНК-полимеразой, которая создавала однонитевые выступы. Реакцию останавливали путем добавления дЦТФ.
На следующем этапе плазмиду и вставку объединяли и инкубировали с recA, который индуцировал сайт-специфическую рекомбинацию. Рекомбинированными плазмидами трансформировали Е. coli. На следующий день выросшие колонии собирали и анализировали на правильность рекомбинированной плазмиды путем выделения плазмиды, рестрикционного анализа и ДНК-секвенирования.
Пример 6
Временная экспрессия одновалентных анти-TfR-антител
Антитела получали in vivo во временно трансфицированных клетках НЕК293 (полученных из клеточной линии эмбриональной почки человека 293), культивируемых в среде F17 (Invitrogen Corp.). Для трансфекции использовали реагент "293-Free" Transfection Reagent (Novagen). Антитела и модифицированные молекулы на основе антител, описанные выше, экспрессировали из отдельных экспрессионных плазмид. Трансфекцию выполняли, как указано в инструкции производителя. Клеточные культуральные супенатанты, содержащие рекомбинантный белок, собирали через 3-7 дней после трансфекции. Супернатанты хранили при пониженной температуре (например, -80°С) до очистки.
Общая информация, касающаяся рекомбинантной экспрессии человеческих иммуноглобулинов, например, в клетках НЕК293, дана в Meissner, P. et al., Biotechnol. Bioeng. 75 (2001) 197-203.
Пример 7
Высокопроизводительная очистка неполных антител (с одной ветвью) против рецептора трансферрина
50 мл осветленных супернатантов, содержащих антитела с одной ветвью, загружали в 96-луночные глубокие планшеты на 200 мкл колонки MabSelectSuRe. После этапа промывания с помощью PBS при рН 7,4 белки элюировали с помощью 2,5 мМ HCl с использованием системы Tecan/Atoll, получая 0,5 мл элюата. Элюат нейтрализовали с помощью 2 М Tris с рН 8. Очищенные белки количественно оценивали, используя спектрофотометр Nanodrop, и анализировали путем CE-SDS в денатурирующих и восстанавливающих условиях и путем аналитической SEC. Для получения белка с высокой степенью чистоты (более 95%) значительную долю антител нужно было очищать далее путем эксклюзионной хроматографии, чтобы отделить от полуантител, антител "ключ-ключ" и высокомолекулярных агрегатов. Затем 500 мкл образцов вводили на колонку Superdex200 10/300GL в 20 мМ гистидине, содержащем 140 мМ NaCl, рН 6,0, с использованием Dionex UltiMate 3000. Этот способ позволяет фракционировать 25-30 образцов/день и, следовательно, позволяет очистить большое число подвергающихся скринингу попаданий в формате с одной ветвью. Фракции собирали и анализировали снова, как описано выше.
Пример 8
Культура клеток hCMEC/D3 для анализов трансцитоза
Среду и добавки для hCMEC/D3 (Weksler, В. В. et al., FAS ЕВ J. 19 (2005), 1872-1874) получали от фирмы Lonza. Клетки hCMEC/D3 (пассажи 26-29) культивировали до конфлюэнтности на покрытых коллагеном покровных стеклах (для микроскопии) или в колбах в среде ЕВМ2, содержащей 2,5% FBS, на четверть дополненной поставляемыми факторами роста и полностью дополненной поставляемым гидрокортизоном, гентамицином и аскорбиновой кислотой.
Для всех анализов трансцитоза в 12-луночных планшетах для культивирования клеток использовали вставные фильтры (размер пор 0,4 мкм, диаметр 12 мм) с РЕТ-мембраной с высокой плотностью пор (1×108 пор/см2). Оптимальные объемы среды рассчитывали как 400 мкл и 1600 мкл для апикальной и базолатеральной камер, соответственно. Апикальные камеры вставных фильтров покрывали коллагеном I крысиного хвоста (7,5 мкг/см2), а затем фибронектином (5 мкг/мл), при этом каждую инкубацию осуществляли в течение 1 ч при комнатной температуре. Клетки hCMEC/D3 выращивали до конфлюэнтных монослоев (примерно 2×105 клеток/см2) в течение 10-12 дней в среде ЕВМ2.
Пример 9
Анализ трансцитоза одновалентных антител
Весь анализ проводили в бессывороточной среде ЕВМ2, которую в противном случае восстанавливали, как описано в примере 1. Вставные фильтры с клетками инкубировали апикально с одновалентными антителами (концентрация: 2,67 мкг/мл) в течение 1 ч при 37°С, после чего собирали все апикальные и базолатеральные среды. Из этих значений рассчитывали парацеллюлярный ток. Монослои промывали при комнатной температуре в бессывороточной среде апикально (400 мкл) и базолатерально (1600 мкл) 3 раза по 3-5 минут каждый. Все смывки собирали для контроля эффективности удаления несвязанного антитела. В апикальную камеру добавляли предварительно согретую среду и переносили фильтры в свежий 12-луночный планшет (заблокированный в течение ночи с помощью PBS, содержащего 1% BSA), содержащий 1600 мкл предварительно согретой среды. В этот момент клетки на фильтрах лизировали в 500 мкл буфера RIPA для того, чтобы определить специфическое поглощение антитела. Остальные фильтры инкубировали при 37°С и образцы собирали в различные моменты времени, чтобы определить апикальное и/или базолатеральное высвобождение антитела. Содержание антитела в образцах оценивали количественно с использованием высокочувствительного ELISA на IgG (см. пример 3). Для каждой временной точки получали данные от клеточных культур с трех фильтров.
Пример 10
Чувствительный ELISA на IgG после анализов трансцитоза
Всю процедуру проводили при комнатной температуре с использованием автоматизированного промывателя на этапах промывания. 384-луночный планшет покрывали 30 мкл/лунка противочеловеческого/противомышиного IgG, Fc-специфического, с концентрацией 1 мкг/мл в PBS в течение 2 ч, а затем в течение 1 ч инкубировали в блокирующем буфере PBS, содержащем 1% BSA или 1% CroteinC в анализах человеческого и мышиного IgG, соответственно). Серийно разведенные образцы из анализа трансцитоза и стандартные концентрации антитела, используемые в анализе трансцитоза, вносили в планшет и инкубировали в течение 2 ч. После четырех промываний вносили 30 мкл/лунка противочеловеческого/противомышиного F(ab)2-биотина с концентрацией 50 нг/мл в блокирующем буфере и инкубировали в течение еще 2 ч. После 6 промываний добавляли 30 мкл/лунка 50 нг/мл (анализ huIgG) или 100 нг/мл (анализ mIgG) поли-НРР40-стрептавидина (Fitzgerald, в PBS, содержащем 1% BSA и 0,05% Tween-20) и инкубировали в течение 30 минут. После 4 промываний обнаруживали иммунные комплексы путем добавления 30 мкл/лунка хемилюминесцентного субстрата ВМ Chemiluminescence Substrate (Roche). Сигнал люминесценции измеряли с помощью люминесцентного планшетного ридера и рассчитывали концентрации с использованием подогнанной стандартной кривой. Чувствительность анализа варьировала от 10 пг/мл до 10 нг/мл.
Пример 11
Картирование эпитопов с помощью клеточного ELISA на клетках СНО, трансфицированных мутантными hTfR
Чтобы иметь возможность определить эпитопные области на человеческом рецепторе трансферрина (hTfR), в последовательность hTfR были введены мутации в позициях, где кластер экспонированных на поверхности аминокислот имел отличающиеся аминокислоты по сравнению с выровненной последовательностью мышиного TfR (см. таблицу ниже), следуя тому обоснованию, что, несмотря на значительную гомологию между человеческим и мышиным TfR (77% идентичности), не известны такие антитела, направленные против внеклеточной части, которые демонстрировали бы высокую степень перекрестной реактивности между обоими ортологами. Клонирование плазмид с соответствующими мутациями описано выше. Чтобы картировать связывание молекул, связывающих человеческий TfR, с этими эпитопами, клетки СНО-К1 временно трансфицировали описанными плазмидами и измеряли связывание антитела в клеточном ELISA. Вкратце, за день до эксперимента в 96-луночный планшет высевали 104 клеток на лунку в нормальной ростовой среде (RPMI/10% FCS). На другой день среду заменяли на среду OPTI-MEM со сниженным содержанием сыворотки (Gibco), и после предварительной инкубации в течение 30 минут в лунки вносили 10 мкл смеси, состоящей из 1200 мкл OPTI-MEM, 12 мкг плазмидной ДНК и 12 мкл реагента для трансфекции XtremeGENE (Roche). Клетки инкубировали в течение 2 дней при 37°С/7,5% СО2, а затем среду удаляли и добавляли TfR-антитела в концентрациях от 1 нМ до 100 нМ в ростовой среде, после чего инкубировали в течение 2 ч при 4°С. Затем растворы антител заменяли на 0,05% глутаральдегид в PBS и клетки фиксировали в течение 15 минут при комнатной температуре, а затем дважды промывали PBS и инкубировали с HRP-конъюгированным вторичным антителом против человеческого Fc (BioRad; 1:2000 в блокирующем реагенте для ELISA (Roche)) в течение 1,5 ч при комнатной температуре. После трех промываний в PBS получали сигнал, внося 50 мкл ТМВ на лунку, и измеряли поглощение при 450 нм.

Пример 12
Анализ связывания на основе поверхностного плазмонного резонанса в отношении взаимодействия антитела против TfR человека
Эксперимент по связыванию проводили на приборе BIAcore В 4000 (GE Healthcare), оборудованном сенсорным чипом С1 (GE Healthcare, кат. №BR1005-35), предварительно обработанным антителом против человеческого Fab (GE Healthcare, кат. №28-9583-25), с использованием стандартной процедуры аминного связывания согласно руководству поставщика.
Для кинетических измерений антитело иммобилизовали путем нанесения со временем контактирования 60 секунд и скоростью потока 10 мкл/мин в фосфатно-солевом буфере, рН 7,4, 0,05% Tween 20, при 25°С. Рекомбинантный Н1э6-меченый рецептор человеческого трансферрина (R&D Systems, кат. №2474-TR-050) наносили в возрастающих концентрациях и сигнал контролировали в течение некоторого времени. Было зарегистрировано среднее время ассоциации 150 секунд и время диссоциации 600 секунд при скорости потока 30 мкл/мин. Данные подгоняли с использованием модели связывания 1:1 (изотерма Ленгмюра).
| название | год | авторы | номер документа |
|---|---|---|---|
| ОДНОВАЛЕНТНЫЕ МОДУЛИ-ПЕРЕНОСЧИКИ ЧЕРЕЗ ГЕМАТОЭНЦЕФАЛИЧЕСКИЙ БАРЬЕР | 2014 |
|
RU2694659C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА К CD19 ЧЕЛОВЕКА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2731156C1 |
| БИСПЕЦИФИЧНЫЕ АНТИТЕЛА ПРОТИВ ГАПТЕНА/ПРОТИВ РЕЦЕПТОРА ГЕМАТОЭНЦЕФАЛИЧЕСКОГО БАРЬЕРА, ИХ КОМПЛЕКСЫ И ИХ ПРИМЕНЕНИЕ В КАЧЕСТВЕ ЧЕЛНОКОВ ГЕМАТОЭНЦЕФАЛИЧЕСКОГО БАРЬЕРА | 2014 |
|
RU2693438C2 |
| АНТИТЕЛА ПРОТИВ 5-БРОМ-2'-ДЕЗОКСИУРИДИНА И СПОСОБЫ ПРИМЕНЕНИЯ | 2015 |
|
RU2705299C2 |
| АНТИТЕЛА К АЛЬФА-СИНУКЛЕИНУ И СПОСОБЫ ПРИМЕНЕНИЯ | 2014 |
|
RU2697098C1 |
| АНТИТЕЛА К АЛЬФА-СИНУКЛЕИНУ И СПОСОБЫ ПРИМЕНЕНИЯ | 2014 |
|
RU2718990C1 |
| АНТИТЕЛА ПРОТИВ ТРАНСФЕРРИНОВОГО РЕЦЕПТОРА, ОБЛАДАЮЩИЕ ИНДИВИДУАЛИЗИРОВАННОЙ АФФИННОСТЬЮ | 2016 |
|
RU2831724C2 |
| АНТИТЕЛА ПРОТИВ ТРАНСФЕРРИНОВОГО РЕЦЕПТОРА, ОБЛАДАЮЩИЕ ИНДИВИДУАЛИЗИРОВАННОЙ АФФИННОСТЬЮ | 2016 |
|
RU2729416C2 |
| ШАТТЛ ДЛЯ ГЕМАТОЭНЦЕФАЛИЧЕСКОГО БАРЬЕРА | 2013 |
|
RU2663120C2 |
| КОВАЛЕНТНО СВЯЗАННЫЕ КОНЪЮГАТЫ ХЕЛИКАР-АНТИТЕЛО ПРОТИВ ХЕЛИКАРА И ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2694981C2 |
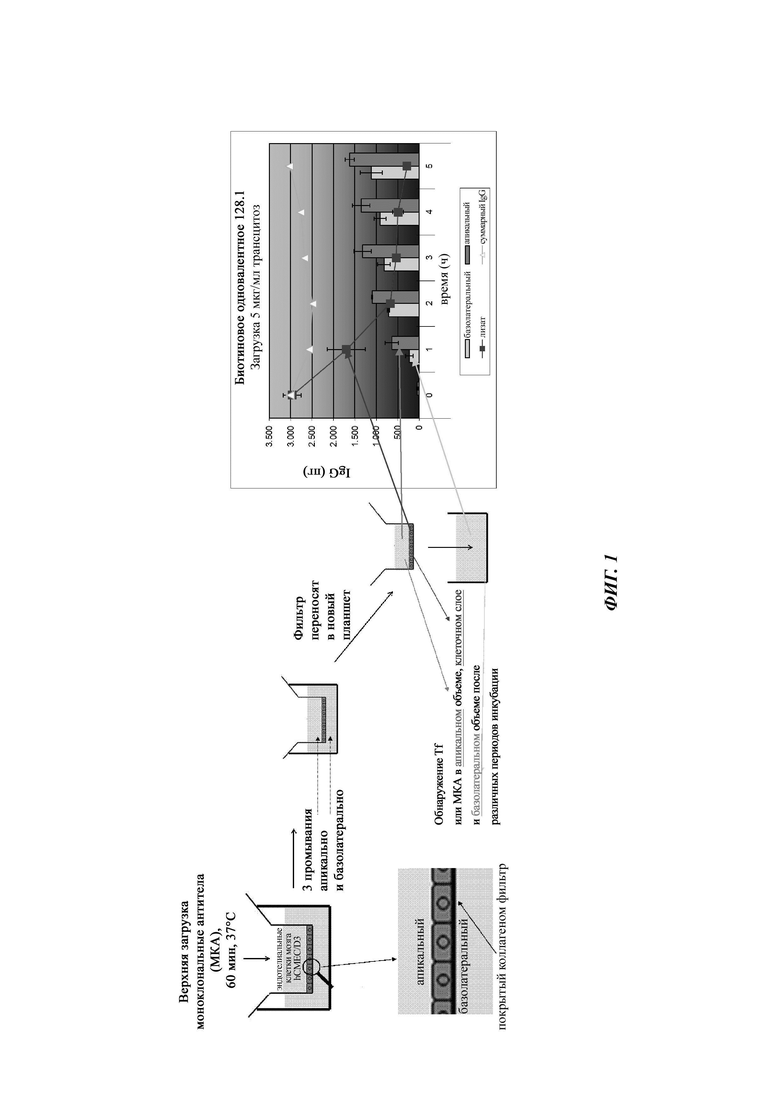
Изобретение относится к области биотехнологии. Предложен модуль-переносчик через гематоэнцефалический барьер, содержащий молекулу, подлежащую транспортировке в мозг через гематоэнцефалический барьер, и одно одновалентное антитело против рецептора трансферрина или его фрагмент, связывающий рецептор трансферрина. Также изобретение относится к фармацевтической композиции, включающей указанный модуль-переносчик, и к применению модуля-переносчика для транспортировки молекулы, подлежащей транспортировке в мозг. Изобретение может быть использовано в медицине для эффективного транспорта эффекторной молекулы через гематоэнцефалический барьер. 3 н. и 9 з.п. ф-лы, 1 ил., 1 табл., 12 пр.
1. Модуль-переносчик через гематоэнцефалический барьер, содержащий молекулу, подлежащую транспортировке в мозг через гематоэнцефалический барьер, и ровно одно (одновалентное) антитело против рецептора трансферрина или его фрагмент, связывающий рецептор трансферрина, где антитело против рецептора трансферрина или его фрагмент, связывающий рецептор трансферрина, содержит (а) HVR-H1, содержащий аминокислотную последовательность SEQ ID NO: 109; (b) HVR-H2, содержащий аминокислотную последовательность SEQ ID NO: 110; (с) HVR-H3, содержащий аминокислотную последовательность SEQ ID NO: 112; (d) HVR-L1, содержащий аминокислотную последовательность SEQ ID NO: 113; (е) HVR-L2, содержащий аминокислотную последовательность SEQ ID NO: 114; и (f) HVR-L3, содержащий аминокислотную последовательность SEQ ID NO: 115,
где указанная молекула, подлежащая транспортировке в мозг через гематоэнцефалический барьер, выбрана из группы, состоящей из лекарственных препаратов для лечения неврологических нарушений, нейротрофических факторов, факторов роста, ферментов, цитотоксических агентов, антител, направленных против мишеней в мозге, моноклональных антител, направленных против мишеней в мозге, пептидов, направленных против мишеней в мозге.
2. Модуль-переносчик через гематоэнцефалический барьер по п. 1, где модуль-переносчик через гематоэнцефалический барьер дополнительно содержит линкер и где линкер соединяет молекулу, подлежащую транспортировке в мозг через гематоэнцефалический барьер, с одновалентным антителом против рецептора трансферрина или его фрагментом, связывающим рецептор трансферрина.
3. Модуль-переносчик через гематоэнцефалический барьер по п. 1, где одновалентное антитело против рецептора трансферрина или его фрагмент, связывающий рецептор трансферрина, содержит молекулу, выбранную из группы, состоящей из полноразмерного антитела, scFv, Fv, scFab и VHH.
4. Модуль-переносчик через гематоэнцефалический барьер по п. 1, где одновалентное антитело против рецептора трансферрина или его фрагмент, связывающий рецептор трансферрина, специфически связывается с рецептором трансферрина человека и с рецептором трансферрина яванского макака.
5. Модуль-переносчик через гематоэнцефалический барьер по п. 1, где одновалентное антитело против рецептора трансферрина или его фрагмент, связывающий рецептор трансферрина, содержит один scFab, направленный против рецептора трансферрина.
6. Модуль-переносчик через гематоэнцефалический барьер по п. 1, где одновалентное антитело против рецептора трансферрина или его фрагмент, связывающий рецептор трансферрина, содержит один scFv, направленный против рецептора трансферрина.
7. Модуль-переносчик через гематоэнцефалический барьер по п. 1, где мишень в мозге выбрана из группы, состоящей из: бета-секретазы 1, Аβ (Abeta), эпидермального фактора роста, рецептора эпидермального фактора роста 2, тау-белка, фосфорилированного тау-белка, фосфорилированного Tay(pS422)-белка, аполипопротеина Е4, альфа-синуклеина, олигомерных фрагментов альфа-синуклеина, CD20, хантингтина, прионного белка, киназы 2 с богатым лейцином повтором, паркина, пресенилина 2, гамма-секретазы, рецептора смерти 6, белка-предшественника амилоида, рецептора нейротрофинов р75 и каспазы 6.
8. Модуль-переносчик через гематоэнцефалический барьер по п. 1, в котором одновалентное антитело против рецептора трансферрина или его фрагмент, связывающий рецептор трансферрина, конъюгировано с С-концом действующей в мозге группировки либо непосредственно, либо через линкер.
9. Модуль-переносчик через гематоэнцефалический барьер по п. 1, в котором молекула, подлежащая транспортировке в мозг через гематоэнцефалический барьер, содержит полноразмерное антитело, направленное против мишени в мозге.
10. Фармацевтическая композиция для транспортировки молекулы, подлежащей транспортировке в мозг через гематоэнцефалический барьер, выбранной из группы, состоящей из лекарственных препаратов для лечения неврологических нарушений, нейротрофических факторов, факторов роста, ферментов, цитотоксических агентов, антител, направленных против мишеней в мозге, моноклональных антител, направленных против мишеней в мозге, пептидов, направленных против мишеней в мозге, где композиция содержит эффективное количество модуля-переносчика через гематоэнцефалический барьер по любому из пп. 1-9.
11. Применение модуля-переносчика через гематоэнцефалический барьер по любому из пп. 1-9 в качестве лекарственного средства для транспортировки молекулы, подлежащей транспортировке в мозг через гематоэнцефалический барьер, выбранной из группы, состоящей из лекарственных препаратов для лечения неврологических нарушений, нейротрофических факторов, факторов роста, ферментов, цитотоксических агентов, антител, направленных против мишеней в мозге, моноклональных антител, направленных против мишеней в мозге, пептидов, направленных против мишеней в мозге.
12. Применение по п. 11, где лекарственное средство предназначено для лечения неврологического нарушения.
| US 2010290985 A1, 18.11.2010 | |||
| WO 2007044323 A2, 19.04.2007 | |||
| RUBEN J BOADO et al., Engineering and Expression of a Chimeric Transferrin Receptor Monoclonal Antibody for Blood-Brain Barrier Delivery in the Mouse, BIOTECHNOLOGY AND BIOENGINEERING, 2009, vol | |||
| Транспортер для перевозки товарных вагонов по трамвайным путям | 1919 |
|
SU102A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| ШАМЕНКОВ Д.А., СВИСТУНОВ А.А., Перспективы использования | |||
