Заявление о государственной поддержке
Данное изобретение создано при государственной поддержке по гранту №W81XWH-19-1-0371, предоставленном министерством обороны. Правительство имеет определенные права на изобретение.
Перекрестная ссылка на связанные заявки
Эта заявка заявляет приоритет в соответствии с 35 U.S.C. § 119 (е) по Предварительной заявке США №62/808 264, поданной 20 февраля 2019 г., содержание которой полностью включено в настоящее описание ссылкой.
Предшествующий уровень техники
Перенос генов в качестве лечения заболеваний (то есть генная терапия) теперь стал реальностью. В 2017 году FDA одобрило Luxturna, аденоассоциированный вирус (AAV), разработанный для стабильной экспрессии нормальной копии гена RPE65 в клетках сетчатки для лечения врожденной дистрофии сетчатки. В 2019 году FDA одобрило Zolgensma, AAV, предназначенный для экспрессии SMN1 в мотонейронах для лечения спинальной мышечной атрофии. Имеются весьма обнадеживающие результаты текущих клинических испытаний использования векторов AAV для замены функции генов при других заболеваниях, включая мини-версию гена дистрофина для мышечной дистрофии Дюшенна и гены факторов VIII и IX для гемофилии A и B, соответственно.
Во всех приведенных выше примерах целью является долговременная экспрессия гена для замены отсутствующего или дефектного гена. Тем не менее, могут быть применения генной терапии, в которых только кратковременная экспрессия трансгена (ген экспрессируется как терапевтическое средство в генной терапии), например, при лечении ограниченного по времени заболевания, такого как инфекция или онкологическое заболевание. Другим иллюстративным примером является экспрессия бактериального белка Cas9 для индукции редактирования гена CRISPR/Cas9, при котором только временная экспрессия Cas9 является идеальной для минимизации потенциальной иммунной реакции на Cas9 и для минимизации мутаций нецелевого гена. Другим применением может быть ситуация, при которой генная терапия может вызвать нежелательные побочные эффекты, и возникнет потребность инактивировать ген. В этих случаях необходим способ активации и/или репрессии экспрессии трансгена.
Таким образом, необходим новый способ, чтобы иметь возможность активировать экспрессию трансгена, чтобы использовать платформу генной терапии для краткосрочной экспрессии генов (от недель до месяцев). В идеальной системе экспрессия трансгена должна находиться под контролем лекарственного средства, так что введение лекарственного средства активирует экспрессию гена, а при отмене лекарственного средства происходит возврат к неактивной экспрессии гена. В таком сценарии, если есть побочные эффекты или если экспрессия трансгена больше не требуется, введение лекарства больше не требуется. Это раскрытие удовлетворяет эту потребность, а также обеспечивает связанные преимущества.
Сущность изобретения
В этом раскрытии предложен способ или вектор для экспрессии терапевтического гена для заболевания или состояния, необязательно под внешним контролем. В некоторых случаях заболевание или состояние представляет собой онкологическое заболевание. В некоторых случаях вектор экспрессирует полипептид (называемый в данном документе «Dimert»), который служит для соединения иммунной клетки-исполнителя с раковой клеткой, чтобы вызвать уничтожение раковой клетки.
В настоящем документе предложен рекомбинантный полинуклеотид или вектор, содержащий, или альтернативно состоящий в основном из, или дополнительно состоящий из: (а) первой полинуклеотидной последовательности, содержащей первую часть открытой рамки считывания, кодирующей первое антитело или его антигенсвязывающий фрагмент; (b) второй полинуклеотидной последовательности, содержащей вторую часть открытой рамки считывания, кодирующую первое антитело или его антигенсвязывающий фрагмент; (c) третьей полинуклеотидной последовательности, кодирующей второе антитело или его антигенсвязывающий фрагмент; и (d) полинуклеотидной последовательности генной регуляции, расположенной между первым полинуклеотидом и вторым полинуклеотидом. В дополнительном аспекте полинуклеотидная последовательность регуляции гена включает, или состоит в основном из, или дополнительно состоит из донорного сайта сплайсинга, вышележащего интрона, экзона, содержащего более одной последовательности стоп-кодона в их соответствующих рамках считывания, нижележащего интрона и акцепторного сайта сплайсинга. В дополнительном аспекте полинуклеотидная последовательность, регулирующая ген, содержит одну или несколько связывающих последовательностей для антисмыслового олигонуклеотида. В дополнительном аспекте антисмысловой олигонуклеотид представляет собой морфолиноолигонуклеотид. В дополнительном аспекте связывающая последовательность для морфолиноолигонуклеотида включает полинуклеотидную последовательность, по меньшей мере на 95% идентичную SEQ ID NO. 24 (AATATGATCCAACAATAGAGGTAAATCTTG) или SEQ ID NO. 25 (GATCCAACAATAGAGGTAAATCTTGTTTTA). В другом воплощении стоп-кодон включает олигонуклеотид из группы: TAA, TAG или TGA. В дополнительном аспекте последовательность стоп-кодона включает полинуклеотидную последовательность TAAxTAGxTGAxTAGxTAAxTGAx (SEQ ID NO. 1), где x представляет собой любой нуклеотид или, альтернативно, последовательность стоп-кодона включает полинуклеотидную последовательность TAATTAGTTGATTAGTTAATTGAT (SEQ ID NO. 2) или ее эквивалент.В другом воплощении рекомбинантный полинуклеотид или вектор кодирует биспецифический или триспецифический рекрутер. В другом воплощении биспецифический клеточный рекрутер представляет собой биспецифический рекрутер. В другом воплощении биспецифический клеточный рекрутер представляет собой триспецифический рекрутер.
В дополнительных воплощениях первое антитело или его антигенсвязывающий фрагмент специфически связывается с активирующим антигеном на иммунной эффекторной клетке, а второе антитело или его антигенсвязывающий фрагмент связывается с опухолевым антигеном. В другом аспекте первое антитело или его антигенсвязывающий фрагмент специфически связывается с опухолевым антигеном, а второе антитело или его антигенсвязывающий фрагмент связывается с активирующим антигеном на иммунной эффекторной клетке. В дополнительном аспекте рекомбинантный полинуклеотид или вектор также содержит четвертую полинуклеотидную последовательность, кодирующую третье антитело или его антигенсвязывающий фрагмент, где третье антитело или его антигенсвязывающий фрагмент связывается с активирующим антигеном на иммунной эффекторной клетке или опухолевым антигеном. В одном аспекте иммунная эффекторная клетка включает дендритную клетку, клетку-натуральный киллер («NK»), макрофаг, Т-клетку, В-клетку, нейтрофил, эозинофил, базофил, тучную клетку или их комбинацию. В одном аспекте эффекторная иммунная клетка представляет собой Т-клетку. В другом аспекте иммунная эффекторная клетка представляет собой NK-клетку. В другом воплощении третье антитело или его антигенсвязывающий фрагмент связывается с активирующим антигеном на NK-клетке и индуцирует иммунный ответ, опосредованный NK-клеткой.
В одном воплощении активирующий антиген представляет собой молекулу поверхности Т-клетки. В другом воплощении активирующий антиген представляет собой молекулу поверхности NK-клетки. Неограничивающие примеры активирующих антигенов на иммунной эффекторной клетке включают CD3, CD2, CD4, CD8, LFA1, CD45, NKG2D, NKp44, NKp46, NKp30, EphA2, DNAM, BT-H3, CD20, CD22 или их комбинации.
В одном из аспектов dimert (например, биспецифический или триспецифический клеточный рекрутер) включает полипептидную последовательность, по меньшей мере на 95% идентичную любой из SEQ ID NO. 7-10. В другом воплощении полипептидная последовательность кодирует антигенсвязывающий фрагмент для CD3, CD2, CD4, CD8, LFA1, CD45, IL21R, NKG2D, NKp44, NKp46, NKp30 или DNAM. В другом воплощении полипептидная последовательность кодирует антигенсвязывающий фрагмент для CD3, CD19, GD2 или NKG2D. В другом воплощении полипептид кодирует первый антигенсвязывающий фрагмент и второй антигенсвязывающий фрагмент. В другом воплощении первый антигенсвязывающий фрагмент связывается с CD3, а второй антигенсвязывающий фрагмент связывается с CD19. В другом воплощении первый антигенсвязывающий фрагмент связывается с CD3, а второй антигенсвязывающий фрагмент связывается с GD2. В другом воплощении первый антигенсвязывающий фрагмент связывается с NKG2D, а второй антигенсвязывающий фрагмент связывается с GD2.
В одном воплощении dimert (например, триспецифический рекрутер или антитело) включает первый антигенсвязывающий фрагмент, второй антигенсвязывающий фрагмент и третий антигенсвязывающий фрагмент. В другом воплощении триспецифический рекрутер или антитело содержит три антигенсвязывающих фрагмента, которые индивидуально связываются с NKG2D, IL21R и GD2. В дополнительном аспекте триспецифический рекрутер или антитело содержит полипептидную последовательность, по меньшей мере, на 95% идентичную SEQ ID NO. 11.
В одном воплощении антигенсвязывающий фрагмент, который связывается с IL-21R, представляет собой IL-21. Последовательности аминокислот и кДНК IL-12 показаны в SEQ ID NO. 3 и SEQ ID NO. 4, соответственно.
SEQ ID NO. 3: аминокислотная последовательность IL-21
MRSSPGNMERIVICLMVIFLGTLVHKSSSQGQDRHMIRMRQLIDIVDQLKNYVNDLVPEFLPAPEDVETNCEWSAFSCFQKAQLKSANTGNNERIINVSIKKLKRKPPSTNAGRRQKHRLTCPSCDSYEKKPPKEFLERFKSLLQKMIHQHLSSRTHGSEDS
SEQ ID NO. 4: последовательность кДНК IL-21.
ATGAGAAGCAGCCCCGGCAACATGGAGAGAATCGTGATCTGCCTGATGGTGATCTTCCTGGGCACCCTGGTGCACAAGAGCAGCAGCCAGGGCCAGGACAGACACATGATCAGAATGAGACAGCTGATCGACATCGTGGACCAGCTGAAGAACTACGTGAACGACCTGGTGCCCGAGTTCCTGCCCGCCCCCGAGGACGTGGAGACCAACTGCGAGTGGAGCGCCTTCAGCTGCTTCCAGAAGGCCCAGCTGAAGAGCGCCAACACCGGCAACAACGAGAGAATCATCAACGTGAGCATCAAGAAGCTGAAGAGAAAGCCCCCCAGCACCAACGCCGGCAGAAGACAGAAGCACAGACTGACCTGCCCCAGCTGCGACAGCTACGAGAAGAAGCCCCCCAAGGAGTTCCTGGAGAGATTCAAGAGCCTGCTGCAGAAGATGATCCACCAGCACCTGAGCAGCAGAACCCACGGCAGCGAGGACAGC
В одном воплощении антигенсвязывающий фрагмент, который связывается с NKG2D, включает MICA, MICB, ULBP1, ULBP2, ULBP3, ULBP4, ULBP5, ULBP6, Rae-1α, Rae-1β, Rae-1γ, Rae-1δ, Rae-1ε, H60a, H60b, H60c, MULT1 или их фрагмент. В одном воплощении антигенсвязывающий фрагмент, который связывается с NKG2D, представляет собой MICA или его фрагмент. Последовательности аминокислот и кДНК MICA показаны в SEQ ID NO. 5 и SEQ ID NO. 6, соответственно.
SEQ ID NO. 5 аминокислотная последовательность MICA
EPHSLRYNLTVLSWDGSVQSGFLAEVHLDGQPFLRYDRQKCRAKPQGQWAEDVLGNKTWDRETRDLTGNGKDLRMTLAHIKDQKEGLHSLQEIRVCEIHEDNSTRSSQHFYYDGELFLSQNLETEEWTVPQSSRAQTLAMNVRNFLKEDAMKTKTHYHAMHADCLQELRRYLESSVVLRRTVPPMVNVTRSEASEGNITVTCRASSFYPRNIILTWRQDGVSLSHDTQQWGDVLPDGNGTYQTWVATRICRGEEQRFTCYMEHSGNHSTHPVPS
SEQ ID NO. 6 последовательность кДНК MICA
gagccccaca gtcttcgtta taacctcacg gtgctgtcct gggatggatc tgtgcagtca gggtttcttg ctgaggtaca tctggatggt cagcccttcc tgcgctatga caggcagaaa tgcagggcaa agccccaggg acagtgggca gaagatgtcc tgggaaataa gacatgggac agagagacca gggacttgac agggaacgga aaggacctca ggatgaccct ggctcatatc aaggaccaga aagaaggctt gcattccctc caggagatta gggtctgtga gatccatgaa
gacaacagca ccaggagctc ccagcatttc tactacgatg gggagctctt cctctcccaa aacctggaga ctgaggaatg gacagtgccc cagtcctcca gagctcagac cttggccatg aacgtcagga atttcttgaa ggaagatgcc atgaagacca agacacacta tcacgctatg catgcagact gcctgcagga actacggcga tatctagaat ccagcgtagt cctgaggaga acagtgcccc ccatggtgaa tgtcacccgc agcgaggcct cagagggcaa catcaccgtg acatgcaggg cttccagctt ctatccccgg aatatcatac tgacctggcg tcaggatggg gtatctttga gccacgacac ccagcagtgg ggggatgtcc tgcctgatgg gaatggaacc taccagacct gggtggccac caggatttgc cgaggagagg agcagaggtt cacctgctac atggaacaca gcgggaatca cagcactcac cctgtgccct ct
Последовательность MICA в одном воплощении включает мутант дикого типа. В одном воплощении мутант MICA представляет собой вариант последовательности MUC-30, который содержит мутацию метионина вместо аланина в положении 129 последовательности MICA дикого типа. Последовательности аминокислот и кДНК варианта MUC-30 показаны в SEQ ID NO. 7 и SEQ ID NO. 8, соответственно.
SEQ ID NO. 7 аминокислотная последовательность MUC-30
EPHSLRYNLTVLSWDGSVQSGFLAEVHLDGQPFLRCDRQKCRAKPQGQWAEDVLGNKTWDRETRDLTGNGKDLRMTLAHIKDQKEGLHSLQEIRVCEIHEDNSTRSSQHFYYDGELFLSQNLETEEWT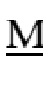 PQSSRAQTLAMNIRNFLKEDAMKTKTHYHAMHADCLQELRRYLKSGVVLRRTVPPMVNVTRSEASEGNITVTCRASGFYPWNITLSWRQDGVSLSHDTQQWGDVLPDGNGTYQTWVATRICQGEEQRFTCYMEHSGNHSTHPVPS
PQSSRAQTLAMNIRNFLKEDAMKTKTHYHAMHADCLQELRRYLKSGVVLRRTVPPMVNVTRSEASEGNITVTCRASGFYPWNITLSWRQDGVSLSHDTQQWGDVLPDGNGTYQTWVATRICQGEEQRFTCYMEHSGNHSTHPVPS
SEQ ID NO. 8 последовательность кДНК MUC-30
gagccccaca gtcttcgtta taacctcacg gtgctgtcct gggatggatc tgtgcagtca
gggtttctcg ctgaggtaca tctggatggt cagcccttcc tgcgctgtga caggcagaaa
tgcagggcaa agccccaggg acagtgggca gaagatgtcc tgggaaataa gacatgggac
agagagacca gggacttgac agggaacgga aaggacctca ggatgaccct ggctcatatc
aaggaccaga aagaaggctt gcattccctc caggagatta gggtctgtga gatccatgaa
gacaacagca ccaggagctc ccagcatttc tactacgatg gggagctctt cctctcccaa
aacctggaga ctgaggaatg gacaatgccc cagtcctcca gagctcagac cttggccatg
aacatcagga atttcttgaa ggaagatgcc atgaagacca agacacacta tcacgctatg
catgcagact gcctgcagga actacggcga tatctaaaat ccggcgtagt cctgaggaga
acagtgcccc ccatggtgaa tgtcacccgc agcgaggcct cagagggcaa cattaccgtg
acatgcaggg cttctggctt ctatccctgg aatatcacac tgagctggcg tcaggatggg
gtatctttga gccacgacac ccagcagtgg ggggatgtcc tgcctgatgg gaatggaacc
taccagacct gggtggccac caggatttgc caaggagagg agcagaggtt cacctgctac
atggaacaca gcgggaatca cagcactcac cctgtgccct ct
В дополнительном аспекте экспрессирует пре-мРНК, которая кодирует триспецифическое антитело, когда пре-мРНК находится в контакте с морфолиноолигонуклеотидом.
В одном аспекте антигенсвязывающий домен представляет собой одноцепочечный вариабельный фрагмент или антитело.
В дополнительном аспекте рекомбинантный полинуклеотид или вектор содержит полинуклеотидную последовательность, кодирующую секреторный пептид. В другом аспекте рекомбинантный полинуклеотид или вектор дополнительно содержит полинуклеотидную последовательность, кодирующую домен димеризации. В другом аспекте рекомбинантный полинуклеотид или вектор содержит 5’-концевой инвертированный повтор (ITR) и 3’-ITR. В другом аспекте вектор содержит последовательность SEQ ID NO. 4, 6, 8, 12, 14, 16-23, 30-33 или 40-46. Неограничивающие примеры такого вектора включают: рекомбинантный вирусный вектор, который включает основной вектор, выбранный из группы ретровирусного вектора, лентивирусного вектора, вектора вируса лейкоза мышей («MLV»), вектора вируса Эпштейна-Барр («EBV»), аденовирусного вектора, вектора вируса герпеса («HSV»), вектора аденоассоциированного вируса («AAV»), вектора AAV или, необязательно, самокомплементарного вектора AAV.
Рекомбинантный полинуклеотид или вектор может содержаться в клетках-хозяевах, например, в прокариотических или эукариотических клетках.
Рекомбинантный полинуклеотид, векторы и клетки могут содержаться в композиции, которая содержит вектор и/или клетку-хозяин и носитель, например, фармацевтически приемлемый носитель. Они могут быть составлены для различных способов введения и содержать эффективное количество полинуклеотида, вектора и/или клетки-хозяина, которое эффективно для пациента, расстройства или заболевания, вектора и способа введения. В одном аспекте способ введения является системным или внутривенным. В другом аспекте введение является местным путем прямой инъекции. В одном аспекте морфолиноолигонуклеотид контактирует одновременно или после полинуклеотида или вектора, например, в виде системной или местной инъекции. Альтернативно, контакт происходит до полинуклеотида или вектора.
Рекомбинантные полинуклеотиды или векторы полезны для лечения множества заболеваний или расстройств. В одном аспекте предлагается способ доставки трансгена. Способ включает введение эффективного количества рекомбинантного полинуклеотида или вектора, содержащего трансген, в клетку, ткань или пациента, подлежащего лечению. В одном аспекте эффективное количество морфолиноолигонуклеотида вводится в клетку, ткань или пациента, подлежащего лечению. Предлагаются неограничивающие примеры трансгенов, выбранные в зависимости от цели способа. Клетки или ткань могут принадлежать млекопитающим, например человеку. В одном из аспектов морфолиноолигонуклеотид контактирует одновременно с вектором или после него. В качестве альтернативы морфолиноолигонуклеотид предшествует вектору. В другом аспекте вектор вводят в клетку путем трансфекции, инфицирования, трансформации, электропорации, инъекции, микроинъекции или их комбинации.
Предлагается способ лечения рака у объекта, который в этом нуждается. Способ включает или, альтернативно, состоит по существу или, кроме того, состоит из введения объекту эффективного количества рекомбинантного полинуклеотида, вектора или клетки, как описано в данном документе. В дополнительном способе дополнительно вводят объекту эффективное количество морфолиноолигонуклеотида. В одном аспекте объекту вводят эффективное количество противоопухолевого агента. Неограничивающие примеры противоопухолевых агентов включают противоопухолевые пептиды, полипептиды, молекулы нуклеиновых кислот, низкомолекулярные вещества, вирусные частицы или их комбинации. В другом аспекте вектор вводят в клетку путем трансфекции, инфицирования, трансформации, электропорации, инъекции, микроинъекции или их комбинации.
В одном из аспектов раскрытых способов вирусная частица представляет собой онколитическую частицу HSV.
Дополнительный способ, обеспечиваемый этим раскрытием, представляет собой способ получения биспецифического антитела или триспецифического антитела в клетке, включающий, или состоящий по существу из, или кроме того состоящий из контакта клетки, содержащей полинуклеотид или вектор, как описано в данном документе, с эффективное количество морфолиноолигонуклеотида. В одном из аспектов морфолиноолигонуклеотид контактирует одновременно с вектором или после него. Альтернативно, он предшествует полинуклеотиду или вектору. В одном аспекте морфолиноолигонуклеотид содержит последовательность по меньшей мере на 95% идентичную SEQ ID NO. 27 или 28. Неограничивающий пример биспецифического антитела включает полипептидную последовательность, по меньшей мере на 95% идентичную SEQ ID NO. 13 или 15. В одном воплощении биспецифическое антитело кодируется полинуклеотидной последовательностью, по меньшей мере на 95% идентичной SEQ ID NO. 14, 16, 22, 23, 30-33 или 40-46. В другом аспекте триспецифическое антитело содержит полипептидную последовательность, по меньшей мере на 95% идентичную SEQ ID NO. 11. В одном воплощении триспецифическое антитело кодируется полинуклеотидной последовательностью, по меньшей мере на 95% идентичной SEQ ID NO. 12.
В другом аспекте полинуклеотид или вектор вводят в клетку путем трансфекции, инфицирования, трансформации, электропорации, инъекции, микроинъекции или их комбинации. Неограничивающие примеры клеток включают фибробласт, скелетную клетку, эпителиальную клетку, мышечную клетку, нервную клетку, эндокринную клетку, меланоцит, клетку крови или их комбинацию.
Кроме того, предлагаются наборы, содержащие один или несколько из числа полинуклеотида, вектора, клетки или композиции, как описано в данном документе, необязательно с инструктивным материалом.
Краткое описание чертежей
Фиг.1 демонстрирует обзор стратегии общей концепции сплайсинга с использованием генной структуры с 3 экзонами. Экзоны обозначены цифрами 1, 2 или 3, а интроны представлены прямой линией. Различные паттерны сплайсинга показаны строчными буквами (a, b, c), а структуры полученных РНК показаны в зависимости от того, какие из них используются. Олигонуклеотиды, которые могут мешать работе донорных (1D, 2D) или акцепторных (2A, 3A) сайтов сплайсинга, показаны короткими синими линиями, а ожидаемая экспрессия различных возможных изоформ РНК показана как «+», в зависимости от того, какие сайты блокируются олигонуклеотидом(ами).
Фиг.2 демонстрирует обзор стратегии фиг.1, с использованием сплайсинга типа 1, в котором экзон 2 обычно включен в транскрипт гена. Ожидается, что транскрипт, который включает все 3 сплайсированных вместе экзона, будет преобладающим, но в присутствии олигонуклеотидов, которые блокируют акцепторные или донорные сайты сплайсинга экзона 2, будет преобладать транскрипт с экзоном 1, слитым с экзоном 3.
Фиг.3 иллюстрирует обзор стратегии №1 для создания сконструированного интрона-экзонаСТОП-интрона в пределах трансгена. Те транскрипты, которые сохраняют интроны, будут нефункциональными, потому что интроны будут иметь преждевременные стоп-кодоны и/или будут вне рамки считывания. Транскрипт «a+b» будет нефункциональным из-за стоп-кодонов в экзоне 2. Функционален только транскрипт, в котором экзон 2 пропущен (с использованием сплайсинга c), который будет низким на исходном уровне и активируется олигонуклеотидами, которые блокируют сайты сплайсинга на экзоне 2.
Фиг.4 демонстрирует карту регуляции трансгена в стратегии, показанной на фиг.1.
Фиг.5 иллюстрирует вторую стратегию создания сконструированного интрона-экзонСТОП-интрона в трансгене с использованием «сплайсинга типа 2», при котором экзон 2 обычно исключен в раковых клетках, но включен в нормальные клетки. В этом сценарии олигонуклеотиды, пропускающие экзон, активируют экспрессию трансгена (экзон 1+3) в нормальных клетках, оставляя ее неизменной (или, возможно, даже выше) при определенном пропуске клеточного гена вне мишени в качестве потенциально «дополнительного» терапевтического эффекта.
Фиг.6 иллюстрирует третью стратегию создания сконструированного интрона-экзонаСТОП-интрона в трансгене с использованием «сплайсинга типа 3», в котором экзон 2 обычно включен в определенные раковые клетки, но исключен в нормальных клетках. В этом сценарии в нормальных клетках наблюдается высокий исходный уровень активного трансгена (экзон 1+3), а олигонуклеотиды, пропускающие экзон, активируют экспрессию в опухолевых клетках.
Фиг.7 иллюстрирует примерные конструкции AAV CD3xGD2-HDD.
Фиг.8 иллюстрирует схематическое изображение анализов для определения структуры и функции Dimert, раскрытых в данном документе.
Фиг.9 иллюстрирует SDS-PAGE лизатов целых клеток из трансфицированных клеток 293T. Три конструкции на фиг.7, №1101, №1040 и №1124, которые содержат пептид секреции, показали меньшее количество CD3xGD2-HDD Dimert, удерживаемого в трансфицированных клетках 293T, по сравнению с конструкцией №1104 и контрольной конструкцией pcDNA3-GFP.
Фиг.10 иллюстрирует CD3xGD2-HDD Dimert, секретируемый вектором AAV, который связывает и активирует Т-клетки человека.
Фиг.11 иллюстрирует секретируемый CD3xGD2-HDD Dimert, который активирует Т-клетки человека.
Фиг.12A иллюстрирует связывание CD3xGD2-HDD с Т-клетками в зависимости от дозы.
Фиг.12B иллюстрирует гистограмму медианных уровней окрашивания, нормализованных к неокрашенным контролям.
Фиг.13 иллюстрирует схематическое изображение анализа связывания для анти-GD2-плеча CD3xGD2-HDD Dimert. Этот анализ подтвердил, что остатки связывания как с GD2, так и с CD3 находятся на одной молекуле.
На фиг.14 показан пример CD3xGD2-HDD Dimert, который связывается как с CD3, так и с GD2.
Фиг.15 иллюстрирует блок-схему анализа для определения, индуцирует ли CD3xGD2-HDD уничтожение GD2+клеток-мишеней Т-клетками.
Фиг.16 иллюстрирует секретируемый CD3xGD2-HDD, индуцирующий Т-клеточное уничтожение клеток нейробластомы.
Фиг.17 иллюстрирует опосредованную Т-клетками цитотоксичность CD3xGD2-HDD Dimert, которая связана с экспрессией GD2.
Фиг.18А показаны карты типичных конструкций AAV для тестирования экспрессии CD19xCD3 Dimert.
Фиг.18B иллюстрирует типичную геномную структуру CD19 TransJoin. На фигуре показаны элементы трансгена, кодируемого AAV.
Фиг.18C иллюстрирует схематическое изображение CD19 dimert, взаимодействующего с раковой клеткой и Т-клеткой. CD19 dimert продуцируется клеткой, содержащей CD19 TransJoin. Показанное на фигуре имеет следующие значения: «Са» представляет раковую клетку, а «Т» представляет собой Т-клетку.
Фиг.19 иллюстрирует надосадочные жидкости клеток, трансфицированных конструкцией AAV CD19xCD3, содержащей последовательность секреции, связывающую и активирующую Т-клетки.
Фиг.20 иллюстрирует надосадочные жидкости клеток, трансфицированных векторами AAV, содержащими пептид секреции, активирующий Т-клетки человека.
Фиг.21 иллюстрирует CD19xCD3, секретируемый AAV, специфически связывает CD19, но не CD45.
Фиг.22A иллюстрирует связывание CD19xCD3 с Т-клетками в зависимости от дозы.
Фиг.22B иллюстрирует гистограмму средних уровней окрашивания, нормализованных к неокрашенным контролям.
Фиг.23A иллюстрирует связывание CD19xCD3 с B-клетками в зависимости от дозы.
Фиг.23B иллюстрирует гистограмму медианных уровней окрашивания, нормализованных к неокрашенным контролям.
Фиг.24 иллюстрирует, что CD3 Dimert активируют Т-клетки с помощью анти-CD28 костимуляции лучше, чем анти-CD3 антитела.
Фиг.25 иллюстрирует клетки 293T, обеспечивающие наивысшую эффективность трансдукции для вектора AAV8.
Фиг.26 иллюстрирует, что концентрация Dimert в надосадочных жидкостях клеток, инфицированных AAV8, зависит от дозы AAV и эффективности трансдукции.
Фиг.27 иллюстрирует однократную внутривенную инъекцию CD19xCD3 TransJoin, которая избирательно удаляет В-клетки у гуманизированных мышей.
Фиг.28 иллюстрирует однократную внутривенную инъекцию CD19xCD3 TransJoin, которая приводит к пролонгированному истощению В-клеток у гуманизированных мышей.
Фиг.29 иллюстрирует однократную внутривенную инъекцию CD19xCD3 TransJoin, которая устраняет лимфому CD19+у гуманизированных мышей.
Фиг.30 иллюстрирует обзор OncoSkip и TransSkip, описанных в данном документе.
Фиг.31 иллюстрирует антисмысловые морфолиноолигонуклеотиды KRAS OncoSkip, индуцирующие пропуск экзонов эндогенного KRAS в клетках рака легкого.
Фиг.32 иллюстрирует примерные векторные карты AAV CD3xGD2-HDD TransSkip, показывающие реконструированные интроны, фланкирующие экзон, вставленный в кодирующую последовательность CD3xGD2-HDD Dimert.
Фиг.33 иллюстрирует примерную стратегию тестирования активности OncoSkip и TransSkip, описанных в данном документе.
Фиг.34 иллюстрирует антисмысловые морфолиноолигонуклеотиды, которые вызывают пропуск экзонов трансгена CD3xGD2-HDD TransSkip.
Фиг.35 иллюстрирует KRAS OncoSkip, который индуцирует секретируемую экспрессию CD3xGD2-HDD Dimert в клетках, трансфицированных вектором CD3xGD2-HDD TransSkip AAV.
Фиг.36 иллюстрирует пропуск экзона CD3xGD2-HDD TransSkip с помощью KRAS. OncoSkip является целевым эффектом.
Фиг.37 иллюстрирует индукцию экспрессии CD3xGD2-HDD Dimert, что, как определено связыванием Т-клеток, является целевым эффектом.
Фиг.38 иллюстрирует карты генома AAV типичных вариантов сплайсинга TransSkip для уменьшения базовой «утечки» TransSkip, но для поддержания индуцируемого пропуска экзонов.
Фиг.39 иллюстрирует вариант сплайсинга CD3xGD2-HDD TransSkip K3, который исключает базовый, но поддерживает индуцируемый пропуск экзонов.
Фиг.40 иллюстрирует вариант K3 CD3xGD2-HDD TransSkip, который не показывает исходного продуцирования Dimert, но более заметен с помощью KRAS OncoSkip по сравнению с другими протестированными вариантами TransSkip.
Фиг.41 иллюстрирует, что OncoSkip-опосредованная индукция экспрессии Dimert из вариантов K3 и K5 CD3xGD2-HDD TransSkip, является целевым эффектом.
Фиг.42 иллюстрирует секретируемый Dimert, индуцированный OncSkip из векторов AAV CD3xGD2-HDD TransSkip, является функциональным в опосредовании уничтожения Т-клетками клеток нейробластомы.
Фиг.43 иллюстрирует, что пропуск экзона трансгена в клетках, инфицированных вариантом K3 AV CD3xGD2-HDD TransSkip индуцируется на мишени с помощью KRAS OncoSkip.
Фиг.44A иллюстрирует карты генома AAV типичных вариантов сплайсинга CD19xCD3 TransSkip.
Фиг.44B иллюстрирует типичную геномную структуру CD19 TransSkip.На фигуре показаны элементы трансгена, кодируемого AAV.
Фиг.44C иллюстрирует схематическое изображение CD19 dimert, взаимодействующего с раковой клеткой и Т-клеткой. CD19 dimert продуцируется клеткой, содержащей CD19 TransSkip.Показанное на фигуре имеет следующие значения: «Са» представляет раковую клетку, а «Т» представляет собой Т-клетку.
Фиг.45 иллюстрирует OncoSkip-морфолиноолигонуклеотиды KTS1 и KTS2, индуцирующие пропуск экзона-мишени CD19xCD3 TransSkip K1.
Фиг.46 иллюстрирует, что KRAS OncoSkip индуцирует секретируемую экспрессию CD19xCD3 Dimert в клетках, трансфицированных вектором CD19xCD3 TransSkip K1 AAV.
Фиг.47 иллюстрирует вариант сплайсинга CD19xCD3 TransSkip K3, исключающий базовый, но сохраняющий индуцируемый пропуск экзонов.
Фиг.48 иллюстрирует индукцию экспрессии CD19xCD3 Dimert из CD19xCD3 TransSkip K3, что, как определено связыванием Т-клеток, является целевым эффектом.
Фиг.49 иллюстрирует, что индукция экспрессии CD19xCD3 Dimert из CD19xCD3 TransSkip K3, является воспроизводимой.
Подробное описание
Воплощения согласно настоящему раскрытию будут описаны более полно ниже. Однако аспекты раскрытия могут быть воплощены в различных формах и не должны рассматриваться как ограниченные воплощениями, изложенными в данном документе. Скорее, эти воплощения предоставлены таким образом, чтобы это раскрытие было доскональным и полным и полностью передавало объем изобретения специалистам в данной области техники. Терминология, используемая в данном описании, предназначена только для описания конкретных воплощений и не предназначена для ограничения.
Если не указано иное, все термины (включая технические и научные термины), используемые в данном документе, имеют то же значение, которое обычно понимают специалисты в области техники, к которой относится данное изобретение. Далее будет понятно, что термины, такие как те, которые определены в обычно используемых словарях, должны интерпретироваться как имеющие значение, которое соответствует их значению в контексте настоящей заявки и соответствующего уровня техники, и не должны интерпретироваться как идеализированные или имеющие чрезмерно формальный смысл, если это прямо не определено в данном документе. Хотя эти термины не определены явно ниже, их следует толковать в соответствии с их общим значением.
Терминология, используемая в описании в данном документе, предназначена только для описания конкретных воплощений и не предназначена для ограничения изобретения. Все публикации, заявки на патенты, патенты и другие источники, упомянутые в данном документе, полностью включены ссылкой.
Практическое осуществление настоящей технологии будет использовать, если не указано иное, общепринятые способы культивирования тканей, иммунологии, молекулярной биологии, микробиологии, клеточной биологии и рекомбинантной ДНК, которые известны специалистам в данной области.
Если в контексте не указано иное, специально подразумевается, что различные признаки изобретения, описанные в данном документе, могут использоваться в любой комбинации. Кроме того, раскрытие также предполагает, что в некоторых воплощениях любой признак или комбинация признаков, изложенных в данном документе, могут быть исключены или опущены. Чтобы проиллюстрировать, если в описании указано, что комплекс содержит компоненты A, B и C, специально подразумевается, что любой из A, B или C или их комбинация может быть опущен и отвергнут по отдельности или в любой комбинации.
Если явно не указано иное, все указанные воплощения, признаки и термины предназначены для включения как изложенного воплощения, признака или термина, так и их биологических эквивалентов.
Все числовые обозначения, например pH, температура, время, концентрация и молекулярная масса, включая диапазоны, являются приблизительными, которые изменяются (+) или (-) с шагом 1,0 или 0,1, в зависимости от ситуации, или, альтернативно, изменением+/- 15%, или альтернативно 10%, или альтернативно 5%, или альтернативно 2%. Следует понимать, хотя и не всегда явно указано, что всем числовым обозначениям предшествует термин «около». Также следует понимать, хотя это и не всегда явно указано, что реагенты, описанные в настоящем документе, являются просто примерными и что эквиваленты такие известны в данной области.
В этом раскрытии различные публикации, патенты и опубликованные описания патентов обозначаются ссылками на них или арабскими цифрами. Полные ссылки на публикации, обозначенные арабскими цифрами, находятся непосредственно перед формулой изобретения. Описание данных публикаций, патентов и опубликованных патентных описаний включено в данную заявку ссылкой с целью более полного описания области техники, к которой принадлежит данной изобретение.
Определения
Практическое осуществление настоящей технологии будет использовать, если не указано иное, обычные методы органической химии, фармакологии, иммунологии, молекулярной биологии, микробиологии, клеточной биологии и рекомбинантной ДНК, которые находятся в пределах квалификации в данной области. См., например, Sambrook, Fritsch and Maniatis, Molecular Cloning: A Laboratory Manual, 2nd edition (1989); Current Protocols In Molecular Biology (F. M. Ausubel, et al. eds., (1987)); серия Methods in Enzymology (Academic Press, Inc.): PCR 2: A Practical Approach (M.J. MacPherson, B.D. Hames and G.R. Taylor eds. (1995)), Harlow and Lane, eds. (1988) Antibodies, a Laboratory Manual, and Animal Cell Culture (R.I. Freshney, ed. (1987)).
Используемые в описании изобретения и прилагаемой формуле изобретения формы единственного числа предназначены для включения и форм множественного числа, если контекст явно не указывает иное.
Используемый в данном документе термин «содержащий» предназначен для обозначения того, что композиции и способы включают перечисленные элементы, но не исключают другие. В данном контексте переходная фраза, состоящая в основном из (и грамматических вариантов), должна интерпретироваться как охватывающая перечисленные материалы или стадии, а также те, которые существенно не влияют на основную и новую характеристику(и) указанного воплощения. Таким образом, используемый в данном документе термин «состоящий по существу из» не следует интерпретировать как эквивалент «содержащий». «Состоящий из» означает исключение более чем следовых элементов других ингредиентов и существенных стадий способа введения композиций, раскрытых в данном документе. Аспекты, определенные каждым из этих переходных терминов, находятся в пределах объема настоящего раскрытия.
Термин «около», используемый в данном документе по отношению к измеряемой величине, такой как количество или концентрация и т.п., предназначен для охвата вариаций в 20%, 10%, 5%, 1%, 0,5% или даже 0,1%. от указанного количества.
Термины или «приемлемый», «эффективный» или «достаточный», когда они используются для описания выбора любых компонентов, диапазонов, дозировочных форм и т.д., раскрытых в данном документе, подразумевают, что указанный компонент, диапазон, дозировочная форма и т.д. подходят для раскрытой цели.
Также, при использовании в описании изобретения, «и/или» относится и охватывает любые возможные комбинации одного или нескольких связанных перечисленных элементов, а также отсутствие комбинаций при интерпретации в альтернативе («или»).
Используемый в данном документе термин «аденоассоциированный вирус» или «AAV» относится к представителю класса вирусов, ассоциированных с этим названием и принадлежащих к роду Dependoparvovirus, семейству Parvoviridae. Известно, что для доставки генов подходят несколько серотипов этого вируса; все известные серотипы могут инфицировать клетки из различных типов тканей. В данной области известно по меньшей мере 11 последовательно пронумерованных серотипов AAV. Неограничивающие иллюстративные серотипы, используемые в раскрытых в данном документе способах, включают любой из 11 серотипов, например, AAV2, AAV8, AAV9, или вариантные серотипы, например, AAV-DJ и AAV PHP.B. Частица AAV состоит из трех основных вирусных белков: VP1, VP2 и VP3. В одном воплощении AAV относится к серотипу AAV1, AAV2, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAV12, AAV13, AAV PHP.B, AAV rh74 или AAV-DJ (химера, полученная перетасовкой восьми различных AAV дикого типа).
В некоторых случаях серотип AAV предпочтительно нацелен на определенный тип ткани. В некоторых случаях следующая диаграмма иллюстрирует приводимые в качестве примера типы тканей и серотипы AAV, которые нацелены на каждый тип ткани.
Используемый в данном документе термин «клетка» может относиться либо к прокариотической, либо к эукариотической клетке, необязательно полученной от объекта или из коммерчески доступного источника.
«Эукариотические клетки» включают все царства жизни, кроме дробянок. Их легко отличить по мембраносвязанному ядру. Животные, растения, грибы и простейшие - это эукариоты или организмы, клетки которых организованы в сложные структуры с помощью внутренних мембран и цитоскелета. Наиболее характерная мембраносвязанная структура - ядро. Если не указано иное, термин «хозяин» включает эукариотического хозяина, включая, например, дрожжи, клетки высших растений, насекомых и млекопитающих. Неограничивающие примеры эукариотических клеток или хозяев включают обезьян, быков, свиней, мышей, крыс, птиц, рептилий и человека, например, клетки HEK293 и клетки 293T.
«Прокариотические клетки», которые обычно лишены ядра или каких-либо других мембраносвязанных органелл и делятся на два домена: бактерии и археи. Помимо хромосомной ДНК, эти клетки могут также содержать генетическую информацию в кольцевой петле, называемой эписомой. Бактериальные клетки очень маленькие, размером примерно с митохондрию животного (около 1-2 мкм в диаметре и 10 мкм в длину). Прокариотические клетки имеют три основные формы: стержневидную, сферическую и спиральную. Вместо того, чтобы проходить сложные процессы репликации, как у эукариот, бактериальные клетки делятся путем бинарного деления. Примеры включают, без ограничения указанным, бактерии Bacillus, E.coli и бактерии Salmonella.
Термин «кодировать» в применении к последовательностям нуклеиновых кислот относится к полинуклеотиду, который, как говорят, «кодирует» полипептид, если в его нативном состоянии или при манипулировании методами, хорошо известными специалистам в данной области, его можно транскрибировать и/или транслируется для получения мРНК полипептида и/или его фрагмента. Антисмысловая цепь представляет собой комплемент такой нуклеиновой кислоты, и кодирующая последовательность может быть выведена из нее.
Термины «эквивалент» или «биологический эквивалент» используются взаимозаменяемо при обращении к конкретной молекуле, биологическому или клеточному материалу и предназначены для тех, которые имеют минимальную гомологию, при этом сохраняя желаемую структуру или функциональность. Неограничивающие примеры эквивалентных полипептидов включают полипептид, содержащий по меньшей мере 60%, или, альтернативно, по меньшей мере 65%, или, альтернативно, по меньшей мере 70%, или, альтернативно, по меньшей мере 75%, или альтернативно 80%, или, альтернативно, по меньшей мере 85%, или, альтернативно, по меньшей мере 90%, или альтернативно, по меньшей мере, 95% идентичности, или альтернативно, по меньшей мере, 96% идентичности, или альтернативно, по меньшей мере, 97% идентичности, или альтернативно, по меньшей мере, 98% идентичности, или альтернативно, по меньшей мере, 99% идентичности для полипептидных последовательностей, или полипептид, который кодируется полинуклеотидом или его комплементом, который гибридизуется в условиях высокой жесткости с полинуклеотидом, кодирующим такие полипептидные последовательности, которые имеют по существу идентичную или идентичную функцию, как референсный полипептид, и в одном аспекте кодирует референсный полипептид. Условия высокой жесткости описаны в данном документе и включены в данный документ ссылкой. Альтернативно, его эквивалентом является полипептид, кодируемый полинуклеотидом или его комплементом, имеющий по меньшей мере 70% или альтернативно по меньшей мере 75%, или альтернативно 80%, или альтернативно по меньшей мере 85%, или альтернативно по меньшей мере 90%, или альтернативно, по меньшей мере, 95% идентичности, или по меньшей мере 96% идентичности, или по меньшей мере 97% идентичности последовательностей, или, альтернативно, по меньшей мере, 98% идентичности, или альтернативно, по меньшей мере, 99% идентичности с референсным полинуклеотидом, например полинуклеотидом дикого типа или упомянутым полинуклеотидом.
Неограничивающие примеры эквивалентных полинуклеотидов включают полинуклеотид, имеющий по меньшей мере 60% или, альтернативно, по меньшей мере 65%, или, альтернативно, по меньшей мере 70%, или, альтернативно, по меньшей мере 75%, или альтернативно 80%, или, альтернативно, по меньшей мере 85%, или, альтернативно, по меньшей мере 90%, или альтернативно, по меньшей мере, 95%, или по меньшей мере 96% идентичности, или по меньшей мере 97% идентичности, или, альтернативно, по меньшей мере, 98% идентичности, или альтернативно, по меньшей мере, 99% идентичности с референсным полинуклеотидом. Эквивалент также подразумевает полинуклеотид или его комплемент, который гибридизуется в условиях высокой жесткости с референсным полинуклеотидом.
Полинуклеотидная или полинуклеотидная область (или полипептид или полипептидная область), имеющая определенный процент (например, 80%, 85%, 90% или 95%) «идентичности последовательности» с другой последовательностью, означает, что при выравнивании этот процент оснований (или аминокислот) одинаковы при сравнении двух последовательностей. Выравнивание и процент гомологии или идентичности последовательностей можно определить с помощью программ, известных в данной области, например, программ, описанных в Current Protocols in Molecular Biology (Ausubel et al., Eds. 1987) Supplement 30, section 7.7.18, Table 7.7.1. В некоторых воплощениях для выравнивания используются параметры по умолчанию. Неограничивающей примерной программой выравнивания является BLAST с использованием параметров по умолчанию. В частности, примерные программы включают BLASTN и BLASTP с использованием следующих параметров по умолчанию: генетический код=стандартный; фильтр=нет; цепь=обе; отсечка=60; ожидание=10; Матрица=BLOSUM62; Описания=50 последовательностей; сортировать по=НАИБОЛЬШЕМУ БАЛЛУ; Базы данных=неизбыточные, GenBank+EMBL+DDBJ+PDB+GenBank CDS translations+SwissProtein+SPupdate+PIR. Подробную информацию об этих программах можно найти по следующему интернет-адресу: ncbi.nlm.nih.gov/cgi-bin/BLAST. Идентичность последовательностей и процент идентичности можно определить, включив их в clustalW (доступно по веб-адресу: genome.jp/tools/clustalw/, last accessed on Jan. 13, 2017).
«Гомология», или «идентичность», или «сходство» относится к сходству последовательностей между двумя пептидами или между двумя молекулами нуклеиновой кислоты. Гомология может быть определена путем сравнения положения в каждой последовательности, которая может быть выровнена для целей сравнения. Когда положение в сравниваемой последовательности занято одним и тем же основанием или аминокислотой, молекулы гомологичны в этом положении. Степень гомологии между последовательностями является функцией количества совпадающих или гомологичных положений, общих для последовательностей. «Неродственная» или «негомологичная» последовательность имеет менее 40% идентичности или, альтернативно, менее 25% идентичности с одной из последовательностей настоящего раскрытия.
«Гомология», «идентичность» или «сходство» также может относиться к двум молекулам нуклеиновой кислоты, которые гибридизуются в жестких условиях.
«Гибридизация» относится к реакции, в которой один или несколько полинуклеотидов реагируют с образованием комплекса, который стабилизируется за счет водородной связи между основаниями нуклеотидных остатков. Водородная связь может происходить из-за спаривания оснований Уотсона-Крика, связывания по Хугстину или любым другим специфичным для последовательности способом. Комплекс может включать две цепи, образующие дуплексную структуру, три или более цепей, образующих многонитевой комплекс, одну самогибридизирующуюся цепь или любую их комбинацию. Реакция гибридизации может представлять собой стадию более обширного процесса, такого как инициирование реакции ПЦР или ферментативное расщепление полинуклеотида рибозимом.
Примеры жестких условий гибридизации включают: температуры инкубации от около 25°C до около 37°C; концентрации гибридизационного буфера от около 6× солевой раствор цитрата натрия (SSC) до около 10× SSC; концентрации формамида от около 0% до около 25%; и промывочные растворы от около 4× SSC до около 8× SSC. Примеры умеренных условий гибридизации включают: температуры инкубации от около 40°C до около 50°C; концентрации буфера от около 9× SSC до около 2× SSC; концентрации формамида от около 30% до около 50%; и промывочные растворы от около 5× SSC до около 2× SSC. Гибридизация высокой жесткости относится к состоянию, при котором гибридизация олигонуклеотида с целевой последовательностью не содержит несовпадений (или полной комплементарности). Примеры условий высокой жесткости включают: температуры инкубации от около 55°C до около 68°C; концентрации буфера от около 1× SSC до около 0,1× SSC; концентрации формамида от около 55% до около 75%; и промывочные растворы около 1× SSC, 0,1× SSC или деионизированная вода. Как правило, время инкубации при гибридизации составляет от 5 минут до 24 часов с 1, 2 или более стадиями промывки, а время инкубации при промывании составляет около 1, 2 или 15 минут.SSC представляет собой 0,15 М NaCl и 15 мМ цитратный буфер. Понятно, что можно использовать эквиваленты SSC с использованием других буферных систем.
Используемый в данном документе термин «экспрессия» относится к процессу, с помощью которого полинуклеотиды транскрибируются в мРНК, и/или процессу, посредством которого транскрибируемая мРНК впоследствии транслируется в пептиды, полипептиды или белки. Если полинуклеотид получен из геномной ДНК, экспрессия может включать сплайсинг мРНК в эукариотической клетке.
«Ген» относится к полинуклеотиду, содержащему по меньшей мере одну открытую рамку считывания (ORF), которая способна кодировать конкретный полипептид или белок после транскрибирования и трансляции. «Продукт гена» или, альтернативно, «продукт экспрессии гена» относится к аминокислоте (например, пептиду или полипептиду), образуемой при транскрипции и трансляции гена.
«Под транскрипционным контролем» - это термин, хорошо понятный в данной области, и указывает, что транскрипция полинуклеотидной последовательности, обычно последовательности ДНК, зависит от ее функциональной связи с элементом, который способствует инициации или способствует транскрипции. «Функционально связанные» означает, что полинуклеотиды расположены таким образом, чтобы они могли функционировать в клетке. В одном аспекте данное изобретение обеспечивает промоторы, функционально связанные с последовательностями, расположенными ниже по ходу цепи.
Термин «экзон» относится к последовательности нуклеиновой кислоты, которая содержит последовательность, кодирующую белок. Ген обычно включает более одного экзона, разделенных интроном между ними.
Используемый в данном документе термин «интрон» относится к последовательности нуклеиновой кислоты, фланкированной донорным сайтом сплайсинга на 5’ конце и акцепторным сайтом сплайсинга на 3’ конце. В некоторых воплощениях интрон сплайсируется или удаляется из последовательности РНК или мРНК, экспрессируемой из вектора, в котором интрон присутствует.
Термин «донорный сайт сплайсинга» означает последовательность или домен нуклеиновой кислоты на 5’ конце интрона. Донорный сайт сплайсинга в одном воплощении отмечает начало интрона и/или границы интрона с непосредственно предшествующей кодирующей последовательностью (или экзоном).
Используемый в данном документе термин «акцепторный сайт сплайсинга» относится к последовательности или домену нуклеиновой кислоты на 3’ конце интрона. В одном воплощении акцепторный сайт сплайсинга отмечает начало интрона и его границу со следующей кодирующей последовательностью-экзоном. В другом воплощении акцепторный сайт сплайсинга содержит точку ветвления интрона, т.е. точку, с которой 5’ конец интрона присоединяется в процессе сплайсинга. В некоторых воплощениях акцепторная последовательность сплайсинга и сайт разветвления интрона расположены рядом друг с другом как единое целое. В некоторых воплощениях акцепторная последовательность сплайсинга и сайт разветвления интрона могут быть дополнительно разделены путем перемещения сайта разветвления дальше 5’ от акцепторной последовательности сплайсинга.
Используемый в данном документе термин «сайт сплайсинга» относится к последовательности или домену нуклеиновой кислоты, присутствующей либо на 5’-конце, либо на 3’-конце интрона, как определено выше.
Используемый в данном документе термин «пропуск экзона» относится к модификации сплайсинга пре-мРНК путем нацеливания на донорные и/или акцепторные сайты сплайсинга в пре-мРНК с одним или несколькими комплементарными антисмысловыми олигонуклеотидами. Блокируя доступ сплайсосомы к одному или нескольким донорным или акцепторным сайтам сплайсинга, один или несколько комплементарных антисмысловых олигонуклеотидов могут предотвратить реакцию сплайсинга, тем самым вызывая делецию одного или нескольких экзонов из полностью процессированной мРНК. В одном воплощении пропуск экзона достигается в ядре во время процесса созревания пре-мРНК. Он включает маскирование ключевых последовательностей, участвующих в сплайсинге экзонов-мишеней, с использованием антисмысловых олигонуклеотидов, комплементарных последовательностям донора сплайсинга в пре-мРНК.
Используемый в данном документе термин «последовательность регуляции гена» относится к последовательности нуклеиновой кислоты, способной контролировать транскрипцию, сплайсинг или модификацию гена, открытой рамки считывания или экзона или интрона. Последовательность регуляции гена по изобретению может включать промотор, сайт связывания антисмыслового олигонуклеотида и/или энхансер. Следовательно, помещение гена под регуляторный контроль промотора или регуляторного элемента означает размещение гена таким образом, чтобы экспрессия гена контролировалась регуляторной последовательностью(ями). Таким образом, при конструировании комбинаций промотор-ген промотор предпочтительно располагается выше гена и на расстоянии от сайта начала транскрипции, которое приблизительно равно расстоянию между промотором и геном, который он контролирует в естественных условиях. Изменение этого расстояния можно допустить без потери функции промотора. Точно так же предпочтительное расположение регуляторного элемента, такого как энхансер, по отношению к гетерологичному гену, находящемуся под его контролем, отражает его естественное положение по отношению к структурному гену, который он естественным образом регулирует.Энхансеры, как полагают, могут находится независимо относительно положения и ориентации в отличие от элементов промотора. В некоторых воплощениях последовательность регуляции генов включает в себя один или более из последовательности связывания для антисмыслового олигонуклеотида, последовательности связывания для доксициклина, или полинуклеотидной последовательности, кодирующей рибосвитч. В некоторых воплощениях антисмысловой олигонуклеотид (ASO) содержит один или несколько модифицированных нуклеотидов. В одном воплощении антисмысловой олигонуклеотид представляет собой морфолиноолигонуклеотид.
В некоторых воплощениях антисмысловой олигонуклеотид (ASO), описанный в настоящем документе, содержит от около 8 до около 50 нуклеотидов в длину. В некоторых случаях ASO включает от около 8 до около 30, от около 8 до около 25, от около 8 до около 20, от около 8 до около 18, от около 8 до около 15, от около 10 до около 50, от около 10 до около 30, от около 10 до около 25, от около 10 до около 20, от около 10 до около 18, от около 10 до около 15, от около 12 до около 50, от около 12 до около 30, от около от 12 до около 25, от около 12 до около 20, от около 12 до около 18 или от около 12 до около 15 нуклеотидов в длину. В некоторых воплощениях ASO включает 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 30, 40 или 50 нуклеотидов в длину.
В некоторых случаях ASO включает один или несколько модифицированных нуклеотидов. В некоторых случаях ASO включает около 5, 10, 20, 30, 40, 50, 60, 70, 80, 90 или 100% модифицированных нуклеотидов. В других случаях ASO включает около 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 20, 25, 30, 35, 40 или более модифицированных нуклеотидов. В некоторых случаях модификация относится к 2’-гидроксильной группе рибозного фрагмента. В некоторых случаях модификация включает H, OR, R, галоген, SH, SR, NH2, NHR, NR2 или CN, где R представляет собой алкильный фрагмент. Примерный алкильный фрагмент включает, без ограничения указанным, галогены, серу, тиолы, тиоэфиры, тиоэфиры, амины (первичные, вторичные или третичные), амиды, простые эфиры, сложные эфиры, спирты и кислород. В некоторых случаях алкильный фрагмент дополнительно включает модификацию. В некоторых случаях модификация включает азогруппу, кетогруппу, альдегидную группу, карбоксильную группу, нитрогруппу, нитрозо, группу, нитрильную группу, гетероциклическую (например, имидазол, гидразино или гидроксиламино) группу, изоцианатную или цианатную группу или серосодержащую группу (например, сульфоксид, сульфон, сульфид и дисульфид). В некоторых случаях алкильный фрагмент дополнительно включает гетерозамещение. В некоторых случаях углерод гетероциклической группы замещен азотом, кислородом или серой. В некоторых случаях гетероциклическое замещение включает, без ограничения указанным, морфолино, имидазол и пирролидино. В некоторых случаях модификация 2’-гидроксильной группы представляет собой 2’-O-метильную модификацию или 2’-O-метоксиэтил (2’-O-MOE) модификацию. В некоторых случаях модифицированный нуклеотид представляет собой заблокированную или мостиковую модификацию рибозы (например, заблокированную нуклеиновую кислоту или LNA), этиленовую нуклеиновую кислоту (ENA) (например, нуклеиновую кислоту с 2’-4’-этиленовым мостиком), пептид-нуклеиновую кислоту или морфолино. В некоторых случаях модифицированный нуклеотид дополнительно содержит одну или несколько модифицированных межнуклеотидных связей. Примерная модифицированная межнуклеотидная связь включает, без ограничения указанным, фосфоротиоаты, фосфородитиоаты, метилфосфонаты, 5’-алкиленфосфонаты, 5’-метилфосфонат, 3’-алкиленфосфонаты, боронтрифторидаты, боранофосфатные эфиры и селенофосфаты 3′-5′ связи или 2′-5′ связи, фосфотриэфиры, тионоалкилфосфотриэфиры, гидрофосфонатные связи, алкилфосфонаты, алкилфосфонотиоаты, арилфосфонотиоаты, фосфороселеноаты, фосфородиселеноаты, фосфинаты, фосфорамидаты, 3’-алкилфосфорамидаты, аминоалкилфософорамидаты, тионофсфорамидаты, фосфоропиперазидаты, фсфороанилотиоаты, фосфороанилидаты, кетоны, сульфоны, сульфонамиды, арбонаты, карбаматы, метиленгидразо, метилендиметилгидразо, формацетали, тиоформацетали, оксимы, метиленимино, метиленметилимино, тиоамидаты, связи с рибоацетильными группами, аминоэтилглицин, силильные или силоксановые связи, алкильные или циклоалкильные связи с или без гетероатомов, например, с 1-10 атомами углерода, которые являются насыщенными или ненасыщенными и/или замещенными, и/или содержат гетероатомы, связи со структурами морфолиноолигонуклеотида, амидами, полиамидами, в которых основания прямо или косвенно присоединены к аза нитрогенам основной цепи, и их комбинации.
Используемый в данном документе термин «морфолиноолигонуклеотид» относится к полимерной молекуле, имеющей основу, которая поддерживает основания, способные образовывать водородную связь с полинуклеотидом. В некоторых воплощениях полимер на морфолиноолигонуклеотиде лишен фрагмента основной цепи пентозного сахара и, более конкретно, основной цепи рибозы, связанной фосфодиэфирными связями, что типично для нуклеотидов и нуклеозидов. В одном воплощении морфолиноолигонуклеотид содержит азотистое кольцо. В другом воплощении морфолиноолигонуклеотид представляет собой стереочистый олигонуклеотид (например, см. Wavelifesciences.com, последний доступ 25 января 2019 г.) или его производные. В другом воплощении морфолиноолигонуклеотид содержит последовательность, по меньшей мере на 95% идентичную стереочистому полинуклеотиду.
В другом воплощении морфолиноолигонуклеотид содержит структуру от около 8 до около 50, от около 8 до около 30, от около 10 до около 50, от около 10 до около 30 или от около 12 до около 30 нуклеотидов, включая целевую последовательность оснований, которая комплементарна целевой области выбранной предварительно обработанной мРНК или пре-мРНК, такой как интронная область пре-мРНК. В другом воплощении антисмысловой морфолиноолигонуклеотид способствует сплайсингу экзона-мишени, что приводит к транскрипту, в котором отсутствует экзон-мишень.
В некоторых случаях антисмысловой олигонуклеотид (ASO) упоминается в данном документе как OncoSkip.Используемый в данном документе термин «OncoSkip» относится к ASO, предназначенному для индукции пропуска экзона-мишени во время сплайсинга трансгена-мишени, тем самым вызывая экспрессию трансгена-мишени. В некоторых случаях трансген-мишень кодирует полипептид, который связывается с поверхностным полипептидом (например, поверхностным рецептором) клетки-мишени. В некоторых случаях клетка-мишень представляет собой опухолевую клетку или иммунную клетку. В некоторых случаях трансген-мишень является онкогеном. В таких случаях использование OncoSkip вызывает пропуск экзона-мишени во время сплайсинга, чтобы индуцировать экспрессию онкогена.
В некоторых воплощениях OncoSkip, описанный в настоящем документе, содержит от около 8 до около 50 нуклеотидов в длину. В некоторых случаях OncoSkip включает от около 8 до около 30, от около 8 до около 25, от около 8 до около 20, от около 8 до около 18, от около 8 до около 15, от около 10 до около 50, от около 10 до около 30, от около 10 до около 25, от около 10 до около 20, от около 10 до около 18, от около 10 до около 15, от около 12 до около 50, от около 12 до около 30, от около от 12 до около 25, от около 12 до около 20, от около 12 до около 18 или от около 12 до около 15 нуклеотидов в длину. В некоторых воплощениях OncoSkip содержит 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 30, 40 или 50 нуклеотидов в длину.
В некоторых воплощениях OncoSkip содержит один или несколько модифицированных нуклеотидов, например, содержит около 5, 10, 20, 30, 40, 50, 60, 70, 80, 90 или 100% модифицированных нуклеотидов. В некоторых случаях OncoSkip включает около 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 20, 25, 30, 35, 40 или более модифицированных нуклеотидов. В некоторых случаях OncoSkip содержит один или несколько морфолино-модифицированных нуклеотидов. В некоторых случаях OncoSkip содержит около 5, 10, 20, 30, 40, 50, 60, 70, 80, 90 или 100% морфолино-модифицированных нуклеотидов. В некоторых случаях OncoSkip включает около 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 20, 25, 30, 35, 40 или более морфолино-модифицированных нуклеотидов.
В некоторых случаях OncoSkip работает совместно с TransSkip.В контексте настоящего описания термин «TransSkip» относится к рекомбинантному вектору (например, рекомбинантному вирусному вектору, такому как вектор AAV), который содержит трансген, прерываемый областью интрон-экзон-интрон, в которой экзон содержит стоп-кодон, который предотвращает нормальную экспрессию полипептида, кодируемого трансгеном. В некоторых случаях трансген дополнительно охвачен конструкцией, которая содержит полинуклеотид, кодирующий dimert. В некоторых случаях, в сочетании с OncoSkip, OncoSkip пропускает экзон из области интрон-экзон-интрон во время сплайсинга, чтобы генерировать мРНК, которая делает возможной экспрессию полипептида, кодируемого трансгеном. В отсутствие OncoSkip экспрессия трансгена из TransSkip подавляется из-за присутствия стоп-кодона в области интрон-экзон-интрон.
Используемый в данном документе термин «dimert» относится к сконструированной белковой молекуле, содержащей два или более одноцепочечных вариабельных фрагмента (scFv), в которых каждый scFv распознает поверхностный полипептид (например, поверхностный рецептор), экспрессируемый на клетке, а вторая клетка является другой. В некоторых случаях dimert нацелен на два разных типа клеток, например, на раковую клетку и иммунную клетку, или на два разных типа иммунных клеток. Типичные типы иммунных клеток включают дендритные клетки, клетки естественные киллеры (NK), макрофаги, Т-клетки, В-клетки, моноциты или нейтрофилы. В некоторых случаях иммунная клетка является эффекторной иммунной клеткой. В некоторых случаях эффекторная иммунная клетка включает эффекторную Т-клетку (Teff) (также называемую в данном документе Т-клеткой, инфильтрирующей опухоль). Примеры клеток Teff включают CD8+Т-клетки и нерегуляторные CD4+хелперные Т-клетки. В некоторых случаях dimert нацелен на раковые клетки и эффекторные иммунные клетки. В некоторых случаях dimert нацелен на раковые клетки и клетки Teff. В некоторых случаях dimert нацелен на две разные раковые клетки, например, две разные раковые клетки одного и того же рака.
В некоторых случаях два или более scFv из dimert связаны линкером. В некоторых случаях линкер представляет собой пептидный линкер, который облегчает связывание каждого scFv с его соответствующим полипептидом-мишенью. В некоторых случаях линкер включает серию поли-Ala, поли-Gly или их комбинацию. В некоторых случаях линкер поли-Ala, линкер поли-Gly или пептидный линкер, содержащий комбинацию Ala и Gly, каждый независимо имеет длину от около 2 остатков до около 50 остатков. В некоторых случаях линкер поли-Ala, линкер поли-Gly или пептидный линкер, содержащий комбинацию Ala и Gly, каждый независимо составляет от около 2 остатков до около 45 остатков, от около 4 остатков до около 45 остатков, от около 5 до около 45 остатков, от около 8 до около 45 остатков, от около 10 до около 45 остатков, от около 15 до около 45 остатков, от около 20 до около 45 остатков, от около 30 до около 45 остатков остатки, от около 2 остатков до около 40 остатков, от около 4 остатков до около 40 остатков, от около 5 остатков до около 40 остатков, от около 8 остатков до около 40 остатков, от около 10 остатков до около 40 остатков, от около 15 остатков около до 40 остатков, от около 20 остатков до около 40 остатков, от около 30 остатков до около 40 остатков, от около 2 остатков до около 30 остатков, от около 4 остатков до около 30 остатков, от около 5 остатков до около 30 остатков, от около 8 остатков т o около 30 остатков, от около 10 остатков до около 30 остатков, от около 15 остатков до около 30 остатков, от около 20 остатков до около 30 остатков, от около 2 остатков до около 20 остатков, от около 4 остатков до около 20 остатков, от около 5 остатков до около 20 остатков, от около 8 остатков до около 20 остатков, от около 10 остатков до около 20 остатков или от около 15 остатков до около 20 остатков в длину. В некоторых случаях линкер поли-Ala, линкер поли-Gly или пептидный линкер, содержащий комбинацию Ala и Gly, каждый независимо составляет около 2, 4, 5, 6, 8, 10, 12, 14, 15, 16, 18, 20, 25, 30, 35, 40, 45 или 50 остатков в длину.
В некоторых случаях линкер представляет собой линкер (Gly4Ser)n, где n представляет собой целое число от 1 до 10. В некоторых случаях n представляет собой целое число от 1 до 6, от 1 до 4 или от 1 до 3. В некоторых случаях n равно 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10. В некоторых случаях линкер имеет длину от около 10 до около 50 аминокислотных остатков, необязательно от около 10 до около 30, от около 10 до около 25 или от около 10 до около 20 аминокислотных остатков в длину. В некоторых случаях линкер содержит одну или несколько неприродных аминокислот.
В некоторых случаях dimert дополнительно включает дополнительный полипептид. В некоторых случаях дополнительный полипептид увеличивает авидность dimert по отношению к клетке-мишени. В некоторых случаях дополнительный полипептид представляет собой домен димеризации ядерного фактора 1α гепатоцитов человека (HNF1α). В некоторых случаях HNF1α содержит полипептидную последовательность, имеющую по меньшей мере 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности с MVSKLSQLQTELLAALLESGLSKEALIQALGE (SEQ ID NO: 47). В некоторых случаях HNF1α кодируется полинуклеотидом, имеющим по меньшей мере 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности с ATGGTGAGCAAGCTGAGCCAGCTGCAGACCGAGCTGCTGGCCGCCCTGCTGGAGAGCGGCCTGGCCGCCGCTGCTGGAGAGCGGCCTGAGCAGCTAG. В некоторых случаях дополнительный полипептид не вызывает дополнительной иммуногенности или токсичности.
В некоторых воплощениях дополнительный полипептид связан с остальной частью dimert линкером. В некоторых случаях линкер включает GSGGAP. В контексте настоящего описания пептид GSGGAP также называется спейсером. В некоторых случаях линкер содержит TPLGDTTHTSG. В некоторых случаях пептид TPLGDTTHTSG происходит из шарнирной области IgG3.
Используемый в данном документе термин «TransJoin» относится к рекомбинантному вектору (например, рекомбинантному вирусному вектору, такому как вектор AAV), который содержит полипептид, кодирующий dimert, описанный в данном документе, но без присутствия области интрон-экзон-интрон, в которой экзон содержит стоп-кодон. Таким образом, TransJoin отличается от TransSkip тем, что TransJoin может экспрессировать dimert без необходимости в OncoSkip.
В некоторых воплощениях TransJoin, описанный в настоящем документе, оптимизирован для доставки dimert в интересующую клетку-мишень или ткань-мишень. В некоторых случаях TransJoin содержит пару промотор и энхансер или пару промотор и регуляторный элемент, которые оптимизированы для доставки и/или экспрессии в клетку-мишень или ткань-мишень. В некоторых случаях TransJoin оптимизирован (например, с парой промотор и энхансер или парой промотор и регуляторный элемент) для конститутивной экспрессии dimert в течение определенного периода времени. В некоторых случаях TransJoin оптимизирован (например, с парой промотора и энхансера или парой промотора и регуляторного элемента) для конститутивной и стабильной экспрессии dimert в течение определенного периода времени. В некоторых случаях TransJoin содержит промотор и регуляторный элемент, такой как посттранскрипционный регуляторный элемент (WPRE) вируса гепатита сурков (WHP) для усиленной экспрессии, конститутивной экспрессии, стабильной экспрессии или их комбинации.
В некоторых воплощениях TransJoin, описанный в данном документе, содержит пару промотора и энхансера или пару промотор и регуляторный элемент, оптимизированную для сбалансированной экспрессии dimert в клетке-мишени или ткани-мишени. В некоторых случаях сбалансированная экспрессия относится к диапазону экспрессии, в котором экспрессия, превышающая этот диапазон, будет вызывать токсичность, а ниже этого диапазона не будет оказывать терапевтического эффекта. В некоторых случаях промотор и энхансер или промотор и регуляторный элемент (например, WPRE) действуют совместно, чтобы сбалансировать экспрессию dimert для достижения целевого диапазона. В некоторых случаях сбалансированная экспрессия включает широкий диапазон, например, чтобы обеспечить широкое терапевтическое окно для dimert.
В некоторых воплощениях TransJoin, описанный в настоящем документе, дополнительно содержит секреторную консенсусную последовательность, описанную ниже, которая оптимизирована для сбалансированной экспрессии dimert в клетке-мишени или ткани-мишени. В некоторых случаях сбалансированная экспрессия относится к диапазону экспрессии, в котором экспрессия, превышающая этот диапазон, будет вызывать токсичность, а ниже этого диапазона не будет оказывать терапевтического эффекта. В некоторых случаях секреторная консенсусная последовательность действует совместно с промотором, энхансером, регуляторным элементом (например, WPRE) или их комбинацией, чтобы сбалансировать экспрессию dimert для достижения целевого диапазона. В некоторых случаях сбалансированная экспрессия включает широкий диапазон, например, чтобы обеспечить широкое терапевтическое окно для dimert.
При использовании выше, период времени включает один день, два дня, три дня, четыре дня, пять дней, 7 дней, 21 день, 28 дней, одну неделю, две недели, три недели, четыре недели, один месяц, два месяца, три месяца, четыре месяца, пять месяцев, шесть месяцев, 8 месяцев, 10 месяцев, 1 год, 2 года или более.
Используемый в данном документе термин «выделенный» относится к молекулам, биологическим или клеточным материалам, которые по существу не содержат других материалов.
Используемый в данном документе термин «функциональный» может использоваться для модификации любой молекулы, биологического или клеточного материала, чтобы подразумевать, что он достигает определенного, определенного эффекта.
В данном контексте термины «последовательность нуклеиновой кислоты» и «полинуклеотид» используются взаимозаменяемо для обозначения полимерной формы нуклеотидов любой длины, будь то рибонуклеотиды или дезоксирибонуклеотиды. Таким образом, этот термин включает, без ограничения указанным, одно-, двух- или многонитевую ДНК или РНК, геномную ДНК, кДНК, гибриды ДНК-РНК или полимер, содержащий пуриновые и пиримидиновые основания или другие природные химически или биохимически модифицированные, неприродные или дериватизированные нуклеотидные основания.
Используемый в данном документе термин «промотор» относится к любой последовательности, которая регулирует экспрессию кодирующей последовательности, такой как ген. Промоторы могут быть, например, конститутивными, индуцибельными, репрессируемыми или тканеспецифичными. «Промотор» представляет собой контрольную последовательность, которая представляет собой область полинуклеотидной последовательности, в которой контролируются инициация и скорость транскрипции. Он может содержать генетические элементы, с которыми могут связываться регуляторные белки и молекулы, такие как РНК-полимераза и другие факторы транскрипции. Неограничивающие примеры промоторов включают промотор LTR вируса саркомы Рауса (RSV) (необязательно с энхансером RSV), промотор цитомегаловируса (CMV), промотор SV40, промотор дигидрофолатредуктазы, промотор β-актина, промотор фосфоглицеринкиназы (PGK), промотор U6 или промотор короткой формы EF1 альфа (EFS).
В некоторых воплощениях промотор представляет собой промотор короткой формы альфа-альфа (EFS) EF1. В некоторых случаях, промотор короткой формы EF1-альфа (EFS) включает
GTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGCTTTTTTGAATTCGCTAGCTAGGTCTTGAAAGGAGTGGGAATTGGCTCCGGTGCCCGTCAGTGGGCAGAGCGCACATCGCCCACAGTCCCCGAGAAGTTGGGGGGAGGGGTCGGCAATTGATCCGGTGCCTAGAGAAGGTGGCGCGGGGTAAACTGGGAAAGTGATGTCGTGTACTGGCTCCGCCTTTTTCCCGAGGGTGGGGGAGAACCGTATATAAGTGCAGTAGTCGCCGTGAACGTTCTTTTTCGCAACGGGTTTGCCGCCAGAACACAGG (SEQ ID NO: 49), или ее эквивалент.
Дополнительные неограничивающие иллюстративные промоторы с определенной целевой специфичностью представлены в данном документе ниже, включая, без ограничения указанным, цитомегаловирус (CMV), фактор элонгации полипептидной цепи (EF1a) человека, SV40, фосфоглицераткиназу (PGK), такую как PGK1 (человек или мышь), P5, Ubc, бета-актин человека, CAG, TRE, UAS, Ac5, полиэдрин, CaMKIIa, Gal1, TEF1, GDS, ADH1, CaMV35S, убиквитин (Ubi), такой как убиквитин C (UbiC), H1, U6, альфа-1-антитрипсин, вирус некроза селезенки (SFFV), и куриный бета-актин (CBA). Промоторы синтетического происхождения можно использовать для повсеместной или тканеспецифической экспрессии. Кроме того, промоторы, полученные из вирусов, некоторые из которых указаны выше, могут быть полезны в способах, раскрытых в данном документе, например промоторы CMV, HIV, аденовируса и AAV.
В некоторых воплощениях промотор представляет собой тканеспецифичный промотор. В некоторых случаях тканеспецифический промотор является эндогенным промотором или промотором, который происходит от генов, экспрессируемых исключительно в клетках-мишенях. Примеры тканеспецифических промоторов включают, без ограничения указанным, печеночно-специфические промоторы, такие как ApoE/hAAT, LP1, SV40/hAlb (InvivoGen); специфичные для фоторецепторов промоторы, такие как родопсинкиназа человека (GRK1) и аррестин колбочек (CAR); промотор, специфичный для В-клеток, такой как B29 (InvivoGen); промотор, специфичный для гематопоэтических клеток, такой как промотор CD45 и SV40/CD45 от InvivoGen; промотор, специфичный для мышечных клеток, такой как промотор десмина (InvivoGen); промотор, специфичный для ацинарных клеток поджелудочной железы, такой как промотор эластазы-1 (InvivoGen); промотор, специфичный для эндотелиальных клеток, такой как промотор Flt-1 (InvivoGen); и нейрон-специфический промотор, такой как промотор SYN1 (InvivoGen).
В некоторых воплощениях промотор связан с энхансером для повышения эффективности транскрипции. Неограничивающие примеры энхансеров включают энхансер RSV, энхансер CMV и энхансер α-фетопротеина MERII.
Энхансер - это регуляторный элемент, который увеличивает экспрессию целевой последовательности. «Промотор/энхансер» представляет собой полинуклеотид, который содержит последовательности, способные обеспечивать функции как промотора, так и энхансера. Например, длинные концевые повторы ретровирусов содержат функции как промотора, так и энхансера. Энхансер/промотор может быть «эндогенным», «экзогенным» или «гетерологичным». «Эндогенный» энхансер/промотор - это энхансер/промотор, который естественным образом связан с данным геном в геноме. «Экзогенный» или «гетерологичный» энхансер/промотор - это энхансер/промотор, который помещают рядом с геном посредством генетической манипуляции (то есть молекулярно-биологическими методами), так что транскрипция этого гена направляется связанным энхансером/промотором.
В некоторых воплощениях вектор, используемый в данном документе (например, вирусный вектор, такой как вектор AAV), дополнительно содержит один или несколько дополнительных регуляторных элементов. Примеры регуляторных элементов включают, без ограничения указанным, терминаторы транскрипции, сайты полиаденилирования и инвертированные концевые повторы (ITR), такие как 5’ ITR и 3’ ITR. В некоторых случаях регуляторный элемент включает посттранскрипционный регуляторный элемент (WPRE) вируса гепатита сурков (WHP). В некоторых случаях примерный WPRE содержит последовательность нуклеиновой кислоты SEQ ID NO: 50 или ее эквивалент. Последовательность SEQ ID NO: 50 представлена ниже:
TCGACAATCAACCTCTGGATTACAAAATTTGTGAAAGATTGACTGGTATTCTTAACTATGTTGCTCCTTTTACGCTATGTGGATACGCTGCTTTAATGCCTTTGTATCATGCTATTGCTTCCCGTATGGCTTTCATTTTCTCCTCCTTGTATAAATCCTGGTTGCTGTCTCTTTATGAGGAGTTGTGGCCCGTTGTCAGGCAACGTGGCGTGGTGTGCACTGTGTTTGCTGACGCAACCCCCACTGGTTGGGGCATTGCCACCACCTGTCAGCTCCTTTCCGGGACTTTCGCTTTCCCCCTCCCTATTGCCACGGCGGAACTCATCGCCGCCTGCCTTGCCCGCTGCTGGACAGGGGCTCGGCTGTTGGGCACTGACAATTCCGTGGTGTTGTCGGGGAAGCTGACGTCCTTTCCATGGCTGCTCGCCTGTGTTGCCACCTGGATTCTGCGCGGGACGTCCTTCTGCTACGTCCCTTCGGCCCTCAATCCAGCGGACCTTCCTTCCCGCGGCCTGCTGCCGGCTCTGCGGCCTCTTCCGCGTCTTCGCCTTCGCCCTCAGACGAGTCGGATCTCCCTTTGGGCCGCCTCCCCGCCTG (SEQ ID NO: 50)
Термины «белок», «пептид» и «полипептид» используются взаимозаменяемо и в самом широком смысле для обозначения соединения из двух или более субъединиц аминокислот, аналогов аминокислот или пептидомиметиков. Субъединицы могут быть связаны пептидными связями. В другом аспекте субъединица может быть связана другими связями, например, сложным эфиром, простым эфиром и т.д. Белок или пептид должен содержать по меньшей мере две аминокислоты, и никакие ограничения не налагаются на максимальное количество аминокислот, которое может включать последовательность белка или пептида. Используемый в данном документе термин «аминокислота» относится либо к природным, либо к неприродным, либо к синтетическим аминокислотам, включая глицин и оптические изомеры D и L, аналоги аминокислот и пептидомиметики.
Используемый в данном документе термин «вектор» относится к нехромосомной нуклеиновой кислоте, содержащей интактный репликон, так что вектор может реплицироваться при помещении в клетку, например, в процессе трансформации. Векторы могут быть вирусными или невирусными. Вирусные векторы включают ретровирусы, аденовирусы, герпесвирус, бакуловирусы, модифицированные бакуловирусы, паповавирусы или иным образом модифицированные природные вирусы. Примерные невирусные векторы для доставки нуклеиновой кислоты включают «оголенную» ДНК; ДНК в комплексе с катионными липидами, отдельно или в комбинации с катионными полимерами; анионные и катионные липосомы; ДНК-белковые комплексы и частицы, содержащие ДНК, конденсированную с катионными полимерами, такими как гетерогенный полилизин, олигопептиды определенной длины и полиэтиленимин, в некоторых случаях содержащиеся в липосомах; и использование тройных комплексов, содержащих вирус и полилизин-ДНК. В другом воплощении вектор представляет собой рекомбинантный вирусный вектор, содержащий основной вектор, выбранный из группы ретровирусного вектора, лентивирусного вектора, вектора вируса лейкоза мышей («MLV»), вектора вируса Эпштейна-Барр («EBV»), аденовирусный вектор, вектор вируса герпеса («HSV») или вектор аденоассоциированного вируса («AAV»). В другом воплощении вектор представляет собой вектор AAV или, необязательно, самокомплементарный вектор AAV.
«Вирусный вектор» определяется как рекомбинантно полученный вирус или вирусная частица, которая содержит полинуклеотид, который должен быть доставлен в клетку-хозяин in vivo, ex vivo или in vitro. Примеры вирусных векторов включают ретровирусные векторы, векторы AAV, лентивирусные векторы, аденовирусные векторы, альфавирусные векторы и т.п.Альфавирусные векторы, такие как векторы на основе вируса леса Семлики и векторы на основе вируса Синдбис, также были разработаны для использования в генной терапии и иммунотерапии. См. Schlesinger and Dubensky (1999) Curr. Opin. Biotechnol. 5: 434-439 и Ying, et al. (1999) Nat. Med., 5(7): 823-827. В некоторых случаях вирусный вектор представляет собой вектор AAV, например, AAV1, AAV2, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAV12, AAV13, AAV PHP.B, AAV rh74 или AAV-DJ. В некоторых случаях вирусным вектором является AAV rh74. В некоторых случаях AAV rh74 содержит последовательность вектора с регистрационным номером GenBank LP899424.1 (по состоянию на 7 февраля 2020 г.).
В некоторых воплощениях у вирусного вектора (например, вектора AAV) бывает ограниченная несущая способность. Например, вектор AAV имеет предел несущей способности 4,7 тыс.н. Таким образом, комбинация dimert вместе с промотором, энхансером и другими регуляторными элементами должна быть в пределах 4,7 т.п.н. В таких случаях используемый в данном документе промотор выбирается на основе его длины нуклеиновой кислоты, чтобы обеспечить упаковку dimert и других регуляторных элементов в вирусный вектор, например, вектор AAV. В некоторых случаях промотором для такого применения является SFFV, EF1α, PGK, UbiC, CMV, CBA или EFS. В некоторых случаях промотором является EFS.
В другом воплощении промотор представляет собой индуцибельный промотор. В конкретном родственном воплощении промотор представляет собой индуцибельный тетрациклиновый промотор. Системы экспрессии генов Tet-Off и Tet-On предоставляют исследователям легкий доступ к регулируемым системам экспрессии генов высокого уровня, описанным Gossen & Bujard (1992; Tet-Off) и Gossen et al. (1995; Tet-On). В системе Tet-Off экспрессия генов включается, когда тетрациклин (Tc) или доксициклин (Dox; производное Tc) удаляется из культуральной среды. Напротив, экспрессия в системе Tet-On включается добавлением Dox. Обе системы позволяют жестко регулировать экспрессию генов в ответ на меняющиеся концентрации Tc или Dox. Максимальные уровни экспрессии в системах Tet очень высоки и выгодно отличаются от максимальных уровней, получаемых от сильных конститутивных промоторов млекопитающих, таких как CMV (Yin et al., 1996). В отличие от других индуцибельных экспрессирующих систем млекопитающих, регуляция генов в Tet-системах очень специфична, поэтому интерпретация результатов не усложняется плейотропными эффектами или неспецифической индукцией. В E.coli белок-репрессор Tet (TetR) отрицательно регулирует гены оперона устойчивости к тетрациклину на транспозоне Tn10. TetR блокирует транскрипцию этих генов путем связывания с последовательностями оператора tet (tetO) в отсутствие Tc. TetR и tetO обеспечивают основу регуляции и индукции для использования в экспериментальных системах на млекопитающих. В системе Tet-On регуляторный белок основан на «обратном» репрессоре Tet (rTetR), который был создан заменой четырех аминокислот в TetR (Hillen & Berens, 1994; Gossen et al., 1995). Полученный белок, rtTA (обратный tTA, также называемый белком-активатором тетрациклина), кодируется плазмидой-регулятором pTet-On.
В связанном воплощении вектор дополнительно содержит или, альтернативно, состоит по существу из, или кроме того состоит из нуклеиновой кислоты, кодирующей белок-активатор тетрациклина; и промотор, регулирующий экспрессию белка-активатора тетрациклина.
Другие индуцибельные системы, применимые в векторах, выделенных клетках, вирусных упаковочных системах и способах, описанных в данном документе, включают регуляцию экдизоном, эстрогеном, прогестероном, химическими индукторами димеризации и изопропил-бета-D1-тиогалактопиранозидом (EPTG).
Используемый в данном документе термин «рекомбинантная система экспрессии» или «рекомбинантный вектор» относится к генетической конструкции или конструкциям для экспрессии определенного генетического материала, образованного путем рекомбинации.
«Носитель для доставки генов» определяется как любая молекула, которая может нести встроенные полинуклеотиды в клетку-хозяин. Примерами носителей для доставки генов являются липосомы, мицеллы, биосовместимые полимеры, включая природные полимеры и синтетические полимеры; липопротеины; полипептиды; полисахариды; липополисахариды; искусственные вирусные оболочки; металлические частицы; и бактерии или вирусы, такие как бакуловирус, аденовирус и ретровирус, бактериофаг, космида, плазмида, грибковые векторы и другие рекомбинантные носители, обычно используемые в данной области, которые были описаны для экспрессии в различных эукариотических и прокариотических хозяевах, и могут быть используется для генной терапии, а также для простой экспрессии белков.
Раскрытый в данном документе полинуклеотид может быть доставлен в клетку или ткань с использованием носителя для доставки генов. «Доставка гена», «перенос гена», «трансдукция» и тому подобное в контексте настоящего описания - это термины, относящиеся к введению экзогенного полинуклеотида (иногда называемого «трансгеном») в клетку-хозяин, независимо от способа, используемого для введения. Такие способы включают множество хорошо известных методов, таких как вектор-опосредованный перенос генов (например, посредством вирусной инфекции/трансфекции или различных других комплексов доставки генов на основе белков или липидов), а также методы, облегчающие доставку «голых» полинуклеотидов (такие как электропорация, доставка «генной пушкой» и различные другие методы, используемые для введения полинуклеотидов). Введенный полинуклеотид может стабильно или временно сохраняться в клетке-хозяине. Стабильное поддержание обычно требует, чтобы введенный полинуклеотид либо содержал точку начала репликации, совместимую с клеткой-хозяином, либо интегрировался в репликон клетки-хозяина, такой как внехромосомный репликон (например, плазмида), ядерную или митохондриальную хромосому. Известно, что ряд векторов способен опосредовать перенос генов в клетки млекопитающих, как это известно в данной области и описано в данном документе.
«Плазмида» представляет собой молекулу внехромосомной ДНК, отдельную от хромосомной ДНК, которая способна реплицироваться независимо от хромосомной ДНК. Во многих случаях она бывает круглой и двухцепочечной. Плазмиды обеспечивают механизм для горизонтального переноса гена внутри популяции микробов и, как правило, обеспечивают селективное преимущество в данном состоянии окружающей среды. Плазмиды могут нести гены, которые обеспечивают устойчивость к природным антибиотикам в конкурентной экологической нише, или, альтернативно, продуцируемые белки могут действовать как токсины при аналогичных обстоятельствах.
«Плазмиды», используемые в генной инженерии, называются «плазмидными векторами». Многие плазмиды коммерчески доступны для таких применений. Реплицируемый ген вставляется в копии плазмиды, содержащей гены, которые делают клетки устойчивыми к определенным антибиотикам, и сайт множественного клонирования (MCS, или полилинкер), который представляет собой короткую область, содержащую несколько часто используемых сайтов рестрикции, позволяющих легко встраивать ДНК-фрагменты в этом месте. Еще одно важное применение плазмид - продуцирование большого количества белков. В этом случае исследователи выращивают бактерии, содержащие плазмиду, несущую интересующий ген. Так же, как бактерия вырабатывает белки, придающие ей устойчивость к антибиотикам, ее также можно заставить производить большое количество белков из встроенного гена.
В аспектах, где перенос гена опосредуется ДНК-вирусным вектором, таким как аденовирус (Ad) или аденоассоциированный вирус (AAV), векторная конструкция относится к полинуклеотиду, содержащему вирусный геном или его часть, и трансгену. Аденовирусы (Ads) - это относительно хорошо охарактеризованная однородная группа вирусов, включающая более 50 серотипов. Аденовирусы не требует интеграции в геном клетки-хозяина. Также были сконструированы векторы, полученные из рекомбинантных Ad, особенно те, которые снижают потенциал рекомбинации и генерацию вируса дикого типа. Такие векторы коммерчески доступны из таких источников, как Takara Bio USA (Маунтин-Вью, Калифорния), Vector Biolabs (Филадельфия, Пенсильвания) и Creative Biogen (Ширли, Нью-Йорк). AAV дикого типа обладает высокой инфекционностью и специфичностью, интегрируясь в геном клетки-хозяина. См. Wold and Toth (2013) Curr. Ген. Ther. 13(6): 421-433, Hermonat & Muzyczka (1984) Proc. Natl. Acad. Sci. USA 81: 6466-6470 и Lebkowski et al. (1988) Mol. Cell. Biol. 8: 3988-3996.
Векторы, которые содержат как промотор, так и сайт клонирования, с которым может быть функционально связан полинуклеотид, хорошо известны в данной области. Такие векторы способны транскрибировать РНК in vitro или in vivo и коммерчески доступны из таких источников, как Agilent Technologies (Санта-Клара, Калифорния) и Promega Biotech (Мэдисон, Висконсин). Для оптимизации экспрессии и/или транскрипции in vitro может потребоваться удаление, добавление или изменение 5 ‘и/или 3’ нетранслируемых частей клонов для устранения дополнительных, потенциально неподходящих альтернативных кодонов инициации трансляции или других последовательностей, которые могут мешать экспрессии или уменьшать экспрессию на уровне транскрипции или трансляции. Альтернативно, консенсусные сайты связывания рибосом могут быть вставлены непосредственно 5’ от стартового кодона для усиления экспрессии.
Используемый в данном документе термин «рекрутер», «рекрутирующая молекула» или «активирующий антиген» относится к молекуле, которая секретируется клеткой, которая способна активировать иммунные клетки. В конкретных воплощениях, рекрутер специфически активирует иммунную эффекторную клетку в соответствии с доменами, присутствующими в рекрутере. Иллюстративные примеры клеток, которые секретируют рекрутеры, без ограничения указанным, включают Т-клетку, NK-клетку, NKT-клетки, CAR T-клетку, мезенхимальную стволовую клетку (MSC), нейрональную стволовую клетку, гемопоэтическую стволовую клетку или их смесь. В другом воплощении иммунные эффекторные клетки включают дендритную клетку, клетку-натуральный киллер («NK»), макрофаг, Т-клетку, В-клетку или их комбинацию.
Термин «домен распознавания антигена» или «антигенсвязывающий домен» относится к части молекулы рекрутера, которая распознает антиген. В конкретных воплощениях антигены могут иметь любую природу, включая, без ограничения указанным, белки, углеводы и/или синтетические молекулы.
Используемый в данном документе термин «активирующий антиген» или «домен активации» относится к части молекул рекрутеров, которая взаимодействует с иммунными клетками и индуцирует положительный или отрицательный иммуномодулирующий сигнал. Иллюстративные примеры положительных иммуномодулирующих сигналов включают сигналы, которые индуцируют пролиферацию клеток, секрецию цитокинов или цитолитическую активность. Иллюстративные примеры отрицательных иммуномодулирующих сигналов включают сигналы, которые ингибируют пролиферацию клеток, ингибируют секрецию иммуносупрессивных факторов или вызывают гибель клеток.
Используемый в данном документе термин «нативная иммунная клетка» относится к иммунной клетке, которая естественным образом встречается в иммунной системе. Иллюстративные примеры включают, без ограничения указанным, Т-клетки, NK-клетки, NKT-клетки, B-клетки и дендритные клетки.
Используемый в данном документе термин «сконструированная иммунная клетка» относится к генетически модифицированной иммунной клетке.
Используемый в данном документе термин «Т-клетка» включает наивные Т-клетки, CD4+Т-клетки, CD8+Т-клетки, Т-клетки памяти, активированные Т-клетки, анергические Т-клетки, толерантные Т-клетки, химерные В-клетки и антиген-специфические Т-клетки.
Используемый в данном документе термин «антитело» означает не только интактные молекулы антител, но также фрагменты молекул антител, которые сохраняют иммуноген-связывающую способность. Такие фрагменты также хорошо известны в данной области и регулярно используются как in vitro, так и in vivo. Соответственно, используемый в данном документе термин «антитело» означает не только интактные молекулы иммуноглобулина, но также хорошо известные активные фрагменты F(ab’)2 и Fab. F(ab’)2 и Fab-фрагменты, в которых отсутствует Fc-фрагмент интактного антитела, быстрее выводятся из кровотока и могут иметь меньшее неспецифическое тканевое связывание интактного антитела (Wahl et al., J. Nucl. Med., 24: 316-325 (1983)). Антитела по изобретению включают цельные нативные антитела, моноклональные антитела, человеческие антитела, гуманизированные антитела, верблюжьи антитела, мультиспецифические антитела, биспецифические антитела, химерные антитела, Fab, Fab’, одноцепочечные фрагменты V-области (scFv), однодоменные антитела (например, нанотела и однодоменные антитела верблюжьих), фрагменты VNAR, антитела-биспецифические рекрутеры Т-клеток (BiTE), минитела, дисульфидно-связанные Fv (sdFv) и антиидиотипические (анти-Id) антитела, интратела, слитые полипептиды, неклассические антитела и антигенсвязывающие фрагменты любого из вышеперечисленных. В частности, антитела включают иммуноглобулиновые молекулы и иммунологически активные фрагменты иммуноглобулиновых молекул, то есть молекулы, которые содержат антигенсвязывающий участок. Молекулы иммуноглобулинов могут быть любого типа (например, IgG, IgE, IgM, IgD, IgA и IgY), класса (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2) или подкласса.
В некоторых воплощениях антитело представляет собой гликопротеин, содержащий по меньшей мере две тяжелые (H) цепи и две легкие (L) цепи, соединенные дисульфидными связями. Каждая тяжелая цепь состоит из вариабельной области тяжелой цепи (сокращенно VH) и константной области тяжелой цепи (CH). Константная область тяжелой цепи состоит из трех доменов: CH1, CH2 и CH3. Каждая легкая цепь состоит из вариабельной области легкой цепи (сокращенно VL) и константной области CL легкой цепи. Константная область легкой цепи содержит один домен, CL. Области VH и VL могут быть далее разделены на области гипервариабельности, называемыми областями, определяющими комплементарность (CDR), которые перемежаются областями, которые являются более консервативными, называемыми каркасными областями (FR). Каждая VH и VL состоит из трех CDR и четырех FR, расположенных от аминоконца до карбокси-конца в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Вариабельные области тяжелой и легкой цепей содержат связывающий домен, который взаимодействует с антигеном. Константные области антител могут опосредовать связывание иммуноглобулина с тканями или факторами хозяина, включая различные клетки иммунной системы (например, эффекторные клетки) и первый компонент (Cl q) классической системы комплемента. Используемые в данном документе взаимозаменяемо, термины «антигенсвязывающая часть», «антигенсвязывающий фрагмент» или «антигенсвязывающая область» антитела относятся к области или части антитела, которая связывается с антигеном и придает антигенную специфичность к антителу; фрагменты антигенсвязывающих белков, например антитела, включают один или несколько фрагментов антитела, которые сохраняют способность специфически связываться с антигеном (например, комплекс пептид/HLA). Было показано, что антигенсвязывающая функция антитела может быть осуществлена с помощью фрагментов полноразмерного антитела. Примеры антигенсвязывающих частей, охватываемых термином «фрагменты антитела» антитела, включают фрагмент Fab, моновалентный фрагмент, состоящий из доменов VL, VH, CL и CHI; фрагмент F(ab)2, двухвалентный фрагмент, содержащий два фрагмента Fab, связанных дисульфидным мостиком в шарнирной области; фрагмент Fd, состоящий из доменов VH и CHI; фрагмент Fv, состоящий из доменов VL и VH одного плеча антитела; фрагмент dAb (Ward et al., Nature 341: 544-546 (1989)), который состоит из домена VH; и выделенную область, определяющую комплементарность (CDR).
Антитела и фрагменты антител могут быть полностью или частично получены от млекопитающих (например, людей, не являющихся человеком приматов, коз, морских свинок, хомяков, лошадей, мышей, крыс, кроликов и овец) или животных, продуцирующих антитела, не относящихся к млекопитающим (например, кур, уток, гусей, змей, хвостатых амфибий). Антитела и фрагменты антител можно продуцировать у животных или продуцировать вне животных, например, из дрожжей или фагов (например, в виде одного антитела или фрагмента антитела или как часть библиотеки антител).
Кроме того, хотя два домена Fv-фрагмента, VL и VH, кодируются отдельными генами, они могут быть соединены с помощью рекомбинантных способов с помощью синтетического линкера, который позволяет им быть в виде единой белковой цепи, в которой VL и пары областей VH образуют одновалентные молекулы. Они известны как одноцепочечные Fv (scFv); см., например, Bird et al., Science 242: 423-426 (1988); и Huston et al., Proc. Natl. Acad. Sci. 85: 5879-5883 (1988). Эти фрагменты антител получают с использованием обычных методов, известных специалистам в данной области, и фрагменты подвергаются скринингу на пригодность таким же образом, как и интактные антитела.
«Выделенное антитело» или «выделенный антигенсвязывающий белок» - это антитело, которое было идентифицировано и отделено и/или извлечено из компонента его естественного окружения. «Синтетические антитела» или «рекомбинантные антитела» обычно получают с использованием рекомбинантной технологии или методов пептидного синтеза, известных специалистам в данной области.
Используемый в данном документе термин «одноцепочечный вариабельный фрагмент» или «scFv» представляет собой гибридный белок вариабельных областей тяжелой (VH) и легкой цепей (VL) иммуноглобулина (например, мыши или человека), ковалентно связанных с образуют гетеродимер VH:VL. Тяжелая (VH) и легкая цепи (VL) либо соединены напрямую, либо соединены линкером, кодирующим пептид (например, около 10, 15, 20, 25 аминокислот), который соединяет N-конец VH с C -концом VL или C-конец VH с N-концом VL. Линкер обычно обогащен глицином для гибкости, а также серином или треонином для растворимости. Линкер может связывать вариабельную область тяжелой цепи и вариабельную область легкой цепи внеклеточного антигенсвязывающего домена.
Несмотря на удаление константных областей и введение линкера, белки scFv сохраняют специфичность исходного иммуноглобулина. Антитела одноцепочечного Fv полипептида могут быть экспрессированы из нуклеиновой кислоты, содержащей последовательности, кодирующие VH и VL, как описано Huston, et al. (Proc. Nat. Acad. Sci. USA, 85: 5879-5883 (1988)). См. также пат.США №5 091 513, 5 132 405 и 4 956 778; и пат.публ. США №20050196754 и 20050196754. Описаны антагонистические scFv, обладающие ингибирующей активностью (см., например, Zhao et al., Hybridoma (Larchmt) 27(6): 455-51 (2008); Peter et al., J Cachexia Sarcopenia Muscle (2012); Shieh et al., J. Imunol 183(4): 2277-85 (2009); Giomarelli et al., Thromb Haemost 97(6): 955-63 (2007); Fife eta., J Clin Invst 116(8): 2252-61 (2006); Brocks et al., Immunotechnology 3(3): 173-84 (1997); Moosmayer et al., Ther Immunol 2(10): 31- 40 (1995). Были описаны агонистические scFv, обладающие стимулирующей активностью (см., например, Peter et al., J Biol Chem 25278(38): 36740-7 (2003); Xie et al., Nat Biotech 15(8): 768-71 (1997); Ledbetter et al., Crit Rev Immunol 17(5-6): 427-55 (1997); Ho et al., Bio Chim Biophys Acta 1638(3): 257-66 (2003)).
В данном контексте «F(ab)» относится к фрагменту структуры антитела, который связывается с антигеном, но является моновалентным и не имеет части Fc, например, антитело, расщепленное ферментом папаином, дает два F(ab)-фрагмента и Fc-фрагмент (например, константная область тяжелой (H) цепи; Fc-область, которая не связывается с антигеном).
Используемый в данном документе термин «F(ab’)2» относится к фрагменту антитела, полученному перевариванием пепсином целых антител IgG, где этот фрагмент имеет две антигенсвязывающие (ab’) (двухвалентные) области, где каждая (ab1) область включает две отдельные аминокислотные цепи, часть H-цепи и легкая (L) цепь, связанные SS-связью для связывания антигена, и где оставшиеся части H-цепи связаны вместе. Фрагмент «F(ab’)2» можно разделить на два отдельных фрагмента Fab’.
Используемый в данном документе термин «CDR» определяется как аминокислотные последовательности определяющей комплементарность области антитела, которые представляют собой гипервариабельные области тяжелой и легкой цепей иммуноглобулина. См., например, Kabat et al., Sequences of Proteins of Immunological Interest, 4th U.S. Department of Health and Human Services, National Institutes of Health (1987). Обычно антитела содержат три CDR тяжелой цепи и три CDR легкой цепи или области CDR в вариабельной области. CDR обеспечивают большинство контактных остатков для связывания антитела с антигеном или эпитопом. В некоторых воплощениях области CDR описаны с помощью системы Kabat (Kabat, E. A., et al. Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242(1991)).
Используемый в данном документе термин «аффинность» означает меру силы связывания. Без привязки к теории полагают, что аффинность зависит от близости стереохимического соответствия между комбинированными сайтами антител и антигенной детерминантой, от размера площади контакта между ними и от распределения заряженных и гидрофобных групп.Аффинность также включает термин «авидность», который относится к силе связи антиген-антитело после образования обратимых комплексов (например, одновалентных или поливалентных). Способы расчета аффинности антитела к антигену известны в данной области, включая использование экспериментов по связыванию для расчета аффинности. Активность антител в функциональных анализах (например, в анализе проточной цитометрии) также отражает аффинность антител. Антитела и аффинности можно фенотипически охарактеризовать и сравнить с помощью функциональных анализов (например, анализа проточной цитометрии). Молекулы нуклеиновой кислоты, используемые в настоящем изобретении, включают любую молекулу нуклеиновой кислоты, которая кодирует полипептид или его фрагмент.В некоторых воплощениях молекулы нуклеиновой кислоты, применимые в настоящем изобретении, включают молекулы нуклеиновой кислоты, которые кодируют антитело или его антигенсвязывающую часть. Такие молекулы нуклеиновой кислоты не обязательно должны быть на 100% идентичными с последовательностью эндогенной нуклеиновой кислоты, но обычно будут демонстрировать существенную идентичность. Полинуклеотиды, имеющие «существенную гомологию» или «существенную идентичность» с эндогенной последовательностью, обычно способны гибридизоваться по меньшей мере с одной цепью двухцепочечной молекулы нуклеиновой кислоты. Под «гибридизацией» подразумевается пара для образования двухцепочечной молекулы между комплементарными полинуклеотидными последовательностями (например, геном, описанным в данном документе) или их частями в различных условиях жесткости. (См., например, Wahl, G. M. and S. L. Berger, Methods Enzymol. 152: 399 (1987); Kimmel, A. R. Methods Enzymol. 152: 507 (1987)).
Термины «по существу гомологичный» или «практически идентичный» означают молекулу полипептида или нуклеиновой кислоты, которая проявляет по меньшей мере 50% или более гомологию или идентичность с эталонной аминокислотной последовательностью (например, любой одной из аминокислотных последовательностей, описанных в данном документе) или последовательность нуклеиновой кислоты (например, любая из описанных в данном документе последовательностей нуклеиновой кислоты). Например, такая последовательность составляет по меньшей мере около 60%, около 65%, около 70%, около 75%, около 80%, около 85%, около 90%, около 95%, или около 96%, или около 97%, или около на 98%, или около на 99% гомологичен или идентичен на уровне аминокислот или нуклеиновой кислоты последовательности, используемой для сравнения (например, последовательности дикого типа или нативной последовательности). В некоторых воплощениях практически гомологичный или практически идентичный полипептид содержит одну или несколько аминокислотных замен, вставок или делеций относительно последовательности, используемой для сравнения. В некоторых воплощениях по существу гомологичный или по существу идентичный полипептид содержит одну или несколько неприродных аминокислот или аналогов аминокислот, включая D-аминокислоты и ретроинверсо амино, для замены гомологичных последовательностей.
Гомологию последовательностей или идентичность последовательностей обычно измеряют с помощью программного обеспечения для анализа последовательностей (например, пакета программного обеспечения для анализа последовательностей Genetics Computer Group, University of Wisconsin Biotechnology Center, 1710 University Avenue, Madison, Wis. 53705, программы BLAST, BESTFIT, GAP или PILEUP/PRETTYBOX). Такое программное обеспечение сопоставляет идентичные или похожие последовательности, присваивая степени гомологии различным заменам, делециям и/или другим модификациям. В иллюстративном подходе к определению степени идентичности может использоваться программа BLAST с вероятностной оценкой между e-3 и e-100, указывающей на близкородственную последовательность.
Используемый в данном документе термин «аналог» относится к структурно родственному полипептиду или молекуле нуклеиновой кислоты, выполняющей функцию эталонного полипептида или молекулы нуклеиновой кислоты.
Используемый в данном документе термин «консервативная модификация последовательности» относится к модификации аминокислоты, которая не оказывает значительного влияния или не изменяет характеристики связывания описанного в данном документе сконструированного рецептора (например, внеклеточного антигенсвязывающего домена сконструированного рецептора), содержащего аминокислотную последовательность. Консервативные модификации могут включать аминокислотные замены, добавления и делеции. В scFv человека, описанного в данном документе сконструированного рецептора можно ввести модификации стандартными методами, известными в данной области, такими как сайт-направленный мутагенез и ПЦР-опосредованный мутагенез. Аминокислоты можно разделить на группы в соответствии с их физико-химическими свойствами, такими как заряд и полярность. Консервативные аминокислотные замены - это замены, при которых аминокислотный остаток заменяется аминокислотой в той же группе. Например, аминокислоты можно классифицировать по заряду: положительно заряженные аминокислоты включают лизин, аргинин, гистидин, отрицательно заряженные аминокислоты включают аспарагиновую кислоту, глутаминовую кислоту, аминокислоты с нейтральным зарядом включают аланин, аспарагин, цистеин, глутамин, глицин, изолейцин, лейцин, метионин, фенилаланин, пролин, серин, треонин, триптофан, тирозин и валин. Кроме того, аминокислоты можно классифицировать по полярности: полярные аминокислоты включают аргинин (основной полярный), аспарагин, аспарагиновую кислоту (кислый полярный), глутаминовую кислоту (кислый полярный), глутамин, гистидин (основной полярный), лизин (основной полярный)., серин, треонин и тирозин; неполярные аминокислоты включают аланин, цистеин, глицин, изолейцин, лейцин, метионин, фенилаланин, пролин, триптофан и валин. Таким образом, один или несколько аминокислотных остатков в области CDR могут быть заменены другими аминокислотными остатками из той же группы, и измененное антитело может быть протестировано на сохранность функции (т.е. функции, указанные в пунктах (c) - (1) выше) с использованием функциональных анализов, описанных в данном документе. В некоторых воплощениях не более одного, не более двух, не более трех, не более четырех, не более пяти остатков в указанной последовательности или области CDR.
Используемый в данном документе термин «лиганд» относится к молекуле, которая связывается с рецептором. В частности, лиганд связывает рецептор на другой клетке, обеспечивая распознавание и/или взаимодействие между клетками.
Используемый в данном документе термин «костимулирующий сигнальный домен» или «костимулирующий домен» относится к части сконструированного рецептора, включающей внутриклеточный домен костимулирующей молекулы. Костимулирующие молекулы представляют собой молекулы клеточной поверхности, отличные от рецепторов антигена или рецепторов Fc, которые обеспечивают второй сигнал, необходимый для эффективной активации и функционирования Т-лимфоцитов при связывании с антигеном. Примеры таких костимулирующих молекул включают CD27, CD28, 4-1BB (CD137), OX40 (CD134), CD30, CD40, PD-1, ICOS (CD278), LFA-1, CD2, CD7, LIGHT, NKD2C, B7-H2 и лиганд, который специфически связывает CD83. Соответственно, хотя в настоящем описании представлены иллюстративные костимулирующие домены, происходящие из CD28 и 4-1BB, другие костимулирующие домены рассматриваются для применения с описанными в данном документе сконструированными рецепторами. Включение одного или нескольких костимулирующих сигнальных доменов может повысить эффективность и рост Т-клеток, экспрессирующих сконструированные рецепторы. Внутриклеточные сигнальные и костимулирующие сигнальные домены могут быть связаны в любом порядке тандемно с карбоксильным концом трансмембранного домена.
Используемый в данном документе термин «химерный костимулирующий рецептор» или «CCR» относится к химерному рецептору, который связывается с антигеном и обеспечивает костимулирующие сигналы, но не обеспечивает сигнал активации Т-клеток.
Термин «биспецифическое антитело» или «триспецифическое антитело» относится к антителу, имеющему две разные антигенсвязывающие области (биспецифическое антитело) или три разных антигенсвязывающих области (триспецифическое антитело). В некоторых воплощениях биспецифическое антитело включает формат антитела (или структуру антитела), как показано на фиг.2 из Brinkmann et al., “The making of bispecific antibodies,» MABS 9(2): 182-212 (2017); или фиг.2 из Labrijn, et al., “Bispecific antibodies: a mechanistic review of the pipeline,» Nature Reviews 18: 585-608 (2019).
В некоторых воплощениях по меньшей мере одна из антигенсвязывающих областей биспецифического антитела или триспецифического антитела связывается с активирующим антигеном на иммунной эффекторной клетке. Это можно понимать как связывания разных мишеней, но также включает связывание с разными эпитопами на одной мишени. Неограничивающие примеры активирующих антигенов включают, без ограничения указанным, CD3, CD2, CD4, CD8, CD19, LFA1, CD45, NKG2D, NKp44, NKp46, NKp30, DNAM, B7-H3 (CD276), CD20, CD22, или их комбинацию. Неограничивающие примеры биспецифических антител включают, без ограничения указанным, антитела к CD3xCD19, CD3xGD2, CD3xEphA2 и NKG2DxGD2.
В одном воплощении описанный выше dimert включает биспецифическое антитело. В некоторых случаях dimert содержит антитело-биспецифический рекрутер Т-клеток («BiTE»), антитело-биспецифический рекрутер клеток-киллеров («BiKE»), или другие биспецифические антитела, описанные в данном документе, такие как антитела, ассоциированные с другой иммунной клеткой. В другом воплощении dimert включает триспецифическое антитело. В некоторых случаях dimert включает антитело-триспецифический рекрутер Т-клеток («TriTE»), или антитело-триспецифический рекрутер клеток-киллеров («TriKE»), или другие триспецифические антитела, описанные в данном документе, например, связанные с другой иммунной клеткой.
Термин «биспецифический рекрутер Т-клеток» или «BiTE» в контексте настоящего описания относится к биспецифическому моноклональному антителу, которое включает первый антигенсвязывающий фрагмент, который связывается с рецептором Т-клетки, и второй антигенсвязывающий фрагмент, который связывается с опухолевой клеткой через опухоль-специфичную молекулу. Термин «триспецифический рекрутер Т-клеток» («TiTE» или «TriTE») в контексте настоящего описания относится к триспецифическому моноклональному антителу, которое содержит первый антигенсвязывающий фрагмент, который связывается с рекрутером Т-клеток, вторым антигенсвязывающим фрагментом, который связывается с опухолевой клеткой через опухолеспецифическую молекулу и третий антигенсвязывающий фрагмент, который связывается с Т-клеткой рекрутером или цитокиновым доменом, активирующим Т-клетки. В одном воплощении опухолеспецифическая молекула содержит опухолевый антиген, выбранный из числа рецептора 2 эфрина типа A (EphA2), интерлейкина (IL)-13r альфа 2, EGFR VIII, PSMA, EpCAM, GD2 или GD3, фукозил GM1, PSCA, PLAC1, точечного разрыва саркомы, опухоли Вильмса 1, альфафетопротеина (AFP), карциноэмбрионального антигена (CEA), CA-125, MUC-1, эпителиального опухолевого антигена (ETA), тирозиназы, меланома- ассоциированного антигена (MAGE), антигена гематологической дифференцировки, поверхностного гликопротеина, ганглиозидов (GM2), рецептора фактора роста, стромального антигена, сосудистого антигена, орфанного рецептора 1, подобного рецепторной тирозинкиназе (ROR1), мезотелина, CD38, CD123, рецептора 2 эпидермального фактора роста человека (HER2), антигена созревания В-клеток (BCMA), белка активации фибробластов (FAP) альфа или их комбинации. Иллюстративные примеры BiTE включают, без ограничения указанным, блинатумомаб (МТ103) и солитомаб (МТ110).
Термин «биспецифический рекрутер клеток-киллеров» или «BiKE», используемый в данном документе, относится к биспецифическому моноклональному антителу, которое содержит первый антигенсвязывающий фрагмент, который связывается с доменом-рекрутером натуральных киллеров (NK), и второй антигенсвязывающий фрагмент, который связывается с опухолевой клеткой через опухолеспецифическую молекулу. Термин «триспецифический рекрутер NK-клеток» («TiKE» или «TriKE») в контексте настоящего описания относится к триспецифическому моноклональному антителу, которое содержит первый антигенсвязывающий фрагмент, который связывается с рекрутером NK-клеток, вторым антигенсвязывающим фрагментом, который связывается с опухолевой клеткой через опухолеспецифическую молекулу и третий антигенсвязывающий фрагмент, который связывается с рекрутером NK-клеток или цитокиновым доменом, активирующим NK-клетки. Антитело NKG2DxIL21RxGD2 представляет собой пример антитела TriKE. Примеры доменов, рекрутирующих NK-клетки, включают, без ограничения указанным, лиганды или молекулы, которые связывают CD16, CD16+CD2, CD16+DNAM и CD16+NKp46. Примеры NK-активирующих доменов цитокинов включают, без ограничения указанным, IL-15, IL-12, IL-18, IL-21 или другие цитокины, хемокины и/или активирующие молекулы, усиливающие NK-клетки. Взаимодействующий с NK домен может включать любой фрагмент, который связывается с NK-клеткой и/или активирует ее, и/или любой фрагмент, который блокирует ингибирование NK-клетки. В некоторых воплощениях домен, рекрутирующий NK, может включать антитело, которое избирательно связывается с компонентом поверхности NK-клетки. В других воплощениях домен, рекрутирующий NK, может включать лиганд или небольшую молекулу, которая избирательно связывается с компонентом поверхности NK-клетки.
В некоторых воплощениях домен, рекрутирующий NK, может избирательно связываться с рецептором, по меньшей мере, частично расположенным на поверхности NK-клетки. В некоторых воплощениях домен, рекрутирующий NK, может выполнять функцию связывания NK-клетки и тем самым приближать NK в пространственную близость с мишенью, с которой целевые домены выборочно связываются. Однако в некоторых воплощениях домен, рекрутирующий NK может избирательно связываться с рецептором, который активирует NK-клетку и, следовательно, также обладает активирующей функцией. Как описано выше, активация рецептора CD16 может вызывать антителозависимую клеточную цитотоксичность. Таким образом, в некоторых воплощениях домен, рекрутирующий NK, может включать по меньшей мере часть антитела против рецептора CD16, эффективного для селективного связывания с рецептором CD16. В других воплощениях домен, рекрутирующий NK-клетки, может прерывать механизмы, которые ингибируют NK-клетки. В таких воплощениях домен, рекрутирующий NK, может включать, например, анти-PD1/PDL1, анти-NKG2A, анти-TIGIT, анти-иммуноглобулиноподобный рецептор клеток-киллеров (KIR) и/или любой другой домен, блокирующий ингибирование.
В конкретных воплощениях клетки генетически модифицированы (включая иммунные клетки) с помощью молекул-рекрутеров, содержащих по меньшей мере домен распознавания антигена и домен активации и, необязательно, цитокин, костимулирующий домен и/или домен для ингибирования молекул, отрицательно регулирующих активацию Т-клеток. Домен распознавания антигена молекулы-рекрутера связывается с одной или несколькими молекулами, присутствующими в и/или на клетках-мишенях, или которые секретируются клетками-мишенями. В конкретных аспектах клетки-мишени представляют собой раковые клетки, включая по меньшей мере клетки солидных опухолей. После связывания молекул-рекрутеров с молекулой-мишенью они могут активировать клетки, экспрессирующие молекулу, распознаваемую доменом активации. Молекулы-рекрутеры могут активировать клетки, которые генетически модифицированы молекулами-рекрутерами, или могут активировать немодифицированные клетки.
В зависимости от желаемого эффекта активация может дать положительный или отрицательный сигнал. Примеры положительных сигналов включают сигналы, которые вызывают пролиферацию клеток, секрецию цитокинов или цитолитическую активность. Примеры отрицательных сигналов включают сигналы, которые ингибируют пролиферацию Т-клеток, ингибируют секрецию иммуносупрессивных факторов или вызывают гибель клеток.
В частности, иммунные клетки, которые секретируют молекулы-рекрутеры, способны перенаправлять резидентные (естественно эндогенные для конкретного индивидуума) иммунные клетки к раковым клеткам.
Воплощения настоящего изобретения обеспечивают доставку модифицированных иммунных клеток, которые секретируют молекулу-рекрутера, индивидууму, нуждающемуся в этом (заведомо имеющему онкологическое заболевание или подозреваемому в наличии онкологического заболевания, включая конкретный рак), в отличие от доставки молекулы-рекрутера только самому индивидууму (в отсутствие продуцирования модифицированными иммунными клетками). В настоящем раскрытии индивидуум получает модифицированные иммунные клетки, которые позволяют продуцировать молекулу(ы)-рекрутер(ы). В конкретных воплощениях клетки продуцируют иммуностимулирующие цитокины; пролиферируют антиген-специфическим образом; убивают соответствующие клетки-мишени; перенаправляют сторонние иммунные клетки (включая, по меньшей мере, Т-клетки или NK-клетки) на раковые клетки; при активации секретируют молекулы-рекрутеры; и/или эффективны против рака локально-региональным или системным образом. Фиг.3 иллюстрирует примеры модифицированных Т-клеток или NK-клеток, которые секретируют молекулы-рекрутеры. Хотя конкретная Т-клетка или NK-клетка может продуцировать рекрутер, который может нацеливаться на один и тот же специфический для раковых клеток антиген, домены активации для T-клетки или NK-клетки должны быть разными, поскольку NK-клетки не экспрессируют CD3. Примеры доменов активации для NK-клетки включают, например, по меньшей мере CD16, NKG2D или NKp30.
Носители для доставки генов также включают комплексы ДНК/липосомы, мицеллы и целевые комплексы вирусный белок-ДНК. Липосомы, которые также содержат нацеленное антитело или его фрагмент, можно использовать в способах, раскрытых в данном документе. В дополнение к доставке полинуклеотидов в клетку или популяцию клеток, прямое введение белков, описанных в данном документе, в клетку или популяцию клеток может быть выполнено с помощью неограничивающего метода трансфекции белков, альтернативно в условиях культивирования, которые могут усилить экспрессию и/или способствовать активности раскрытых в данном документе белков являются другими неограничивающими способами.
Используемый в данном документе термин «сигнальный пептид» или «сигнальный полипептид» означает аминокислотную последовательность, обычно присутствующую на N-конце вновь синтезированных секреторных или мембранных полипептидов или белков. Он действует, направляя полипептид в конкретное клеточное место, например, через клеточную мембрану, в клеточную мембрану или в ядро. В некоторых воплощениях сигнальный пептид удаляется после локализации. Примеры сигнальных пептидов хорошо известны в данной области. Неограничивающие примеры описаны в Пат.США №8 853 381, 5 958 736 и 8 795 965.
Используемый в данном документе термин «вирусный капсид» или «капсид» относится к белковой оболочке или оболочке вирусной частицы. Функции капсидов заключаются в инкапсидировании, защите, транспортировке и высвобождении вирусного генома в клетку-хозяин. Капсиды обычно состоят из олигомерных структурных субъединиц белка («белки капсида»). Используемый в данном документе термин «инкапсидированный» означает заключенный в вирусный капсид.
Используемый в данном документе термин «хелпер» в отношении вируса или плазмиды относится к вирусу или плазмиде, используемым для обеспечения дополнительных компонентов, необходимых для репликации и упаковки вирусной частицы или рекомбинантной вирусной частицы, такой как модифицированный AAV, описанный в данном документе. Компоненты, кодируемые вирусом-помощником, могут включать любые гены, необходимые для сборки, инкапсидации, репликации генома и/или упаковки вириона. Например, вирус-помощник может кодировать необходимые ферменты для репликации вирусного генома. Неограничивающие примеры вспомогательных вирусов и плазмид, подходящих для использования с конструкциями AAV, включают pHELP (плазмида), аденовирус (вирус) или герпесвирус (вирус).
Используемый в данном документе термин «AAV» является стандартной аббревиатурой для аденоассоциированного вируса. Аденоассоциированный вирус - это парвовирус с одноцепочечной ДНК, который растет только в клетках, в которых определенные функции выполняются сопутствующим вирусом-помощником. Общую информацию и обзоры AAV можно найти, например, в Carter, 1989, Handbook of Parvoviruses, Vol.1, pp.169-228, and Berns, 1990, Virology, pp.1743-1764, Raven Press, (New York). Полностью ожидается, что те же принципы, описанные в этих обзорах, будут применимы к дополнительным серотипам AAV, охарактеризованным после дат публикации обзоров, поскольку хорошо известно, что различные серотипы довольно тесно связаны, как структурно, так и функционально, даже по генетическому уровню. (См., например, Blacklowe, 1988, pp.165-174 of Parvoviruses and Human Disease, J. R. Pattison, ed.; and Rose, Comprehensive Virology 3: 1-61 (1974)). Например, все серотипы AAV, по-видимому, проявляют очень похожие свойства репликации, опосредованные гомологичными генами rep; и все они несут три родственных капсидных белка, таких как экспрессируемые в AAV2. Степень родства дополнительно подтверждается гетеродуплексным анализом, который показывает обширную перекрестную гибридизацию между серотипами по длине генома; и наличие аналогичных сегментов самоотжига на концах, которые соответствуют «последовательностям инвертированных концевых повторов» (ITR). Сходные паттерны инфекционности также предполагают, что функции репликации в каждом серотипе находятся под аналогичным регуляторным контролем.
Используемый в данном документе термин «вектор AAV» относится к вектору, содержащему один или несколько представляющих интерес полинуклеотидов (или трансгенов), которые фланкированы последовательностями концевых повторов AAV (ITR). Такие векторы AAV можно реплицировать и упаковывать в инфекционные вирусные частицы, если они присутствуют в клетке-хозяине, трансфицированной вектором, кодирующим и экспрессирующим продукты генов rep и cap.
«Вирион AAV», или «вирусная частица AAV», или «частица вектора AAV» относится к вирусной частице, состоящей по меньшей мере из одного капсидного белка AAV и инкапсидированного полинуклеотидного вектора AAV. Если частица содержит гетерологичный полинуклеотид (т.е. полинуклеотид, отличный от генома AAV дикого типа, такого как трансген, который должен быть доставлен в клетку млекопитающего), ее обычно называют «частицей вектора AAV» или просто «вектор AAV». Таким образом, получение частицы вектора AAV обязательно включает продуцирование вектора AAV, поскольку такой вектор содержится в частице вектора AAV.
В некоторых воплощениях AAV представляет собой парвовирус с дефицитом репликации, геном одноцепочечной ДНК которого имеет длину около 4,7 т.п.о., включая два инвертированных концевых повтора (ITR) по 145 нуклеотидов. Существует несколько серотипов AAV. Нуклеотидные последовательности геномов серотипов AAV известны. Например, полный геном AAV-1 представлен под учетным номером GenBank NC_002077; полный геном AAV-2 представлен под учетным номером GenBank NC_001401 и Srivastava et al., J. Virol., 45: 555-564 (1983); полный геном AAV-3 представлен под учетным номером GenBank NC_1829; полный геном AAV-4 представлен под учетным номером GenBank NC_001829; геном AAV-5 представлен под учетным номером GenBank AF085716; полный геном AAV-6 представлен под учетным номером GenBank NC_00 1862; по меньшей мере части геномов AAV-7 и AAV-8 представлен под учетным номером GenBank AX753246 и AX753249, соответственно; геном AAV-9 представлен в Gao et al., J. Virol., 78: 6381-6388 (2004); геном AAV-10 представлен в Mol. Ther., 13 (1): 67-76 (2006); а геном AAV-11 представлен в Virology, 330 (2): 375-383 (2004). Последовательность генома AAV rh.74 предоставлена в пат.США 9434928, включенный в настоящий документ ссылкой. Патент США №9434928 также раскрывает последовательности белков капсида и самокомплементарный геном. В одном аспекте геном представляет собой самокомплементарный геном. Цис-действующие последовательности, управляющие репликацией вирусной ДНК, инкапсидацией/упаковкой и интеграцией хромосомы клетки-хозяина, содержатся в ITR AAV. Три промотора AAV (названные p5, pl9 и p40 из-за их относительного расположения на карте) управляют экспрессией двух внутренних открытых рамок считывания AAV, кодирующих гены rep и cap.Два промотора rep (p5 и pi 9) в сочетании с дифференциальным сплайсингом одного интрона AAV (в нуклеотидах 2107 и 2227) приводят к продуцированию четырех белков rep (rep 78, rep 68, rep 52 и rep 40) из гена rep.Rep-белки обладают множеством ферментативных свойств, которые в конечном итоге отвечают за репликацию вирусного генома. Ген cap экспрессируется с промотора p40 и кодирует три капсидных белка VP1, VP2 и VP3. Альтернативный сплайсинг и неконсенсусные стартовые сайты трансляции ответственны за продуцирование трех родственных белков капсида. Единый консенсусный сайт полиаденилирования расположен в положении 95 карты генома AAV. Жизненный цикл и генетика AAV рассмотрены в Muzyczka, Current Topics in Microbiology and Immunology, 158: 97-129 (1992).
AAV обладает уникальными свойствами, которые делают его привлекательным в качестве вектора для доставки чужеродной ДНК в клетки, например, в генной терапии. Инфекция клеток в культуре AAV не является цитопатической, а естественное инфицирование людей и других животных протекает скрыто и бессимптомно. Более того, AAV инфицирует многие клетки млекопитающих, что дает возможность воздействовать на многие различные ткани in vivo. Более того, AAV трансдуцирует медленно делящиеся и неделящиеся клетки и может сохраняться, по существу, в течение всей жизни этих клеток как транскрипционно активная ядерная эписома (внехромосомный элемент). Провирусный геном AAV встраивается в плазмиды в виде клонированной ДНК, что делает возможным создание рекомбинантных геномов. Кроме того, поскольку сигналы, управляющие репликацией AAV и инкапсидацией генома, содержатся в ITR генома AAV, часть или весь внутренний геном длиной приблизительно 4,3 тыс.н. (кодирующие белки репликации и структурного капсида, rep-cap) могут быть заменены чужеродными ДНК. Для создания векторов AAV белки rep и cap могут быть предоставлены в транс.Еще одна важная особенность AAV заключается в том, что это чрезвычайно стабильный и обильный вирус.Он легко выдерживает условия, используемые для инактивации аденовируса (от 56° до 65°C в течение нескольких часов), что делает холодную консервацию AAV менее критичной. AAV можно даже лиофилизировать. Наконец, инфицированные AAV клетки не устойчивы к суперинфекции.
Многочисленные исследования продемонстрировали долгосрочную (>1,5 года) экспрессию рекомбинантного AAV-белка в мышцах. См. Clark et al., Hum Gene Ther, 8: 659-669 (1997); Kessler et al., Proc Nat. Acad Sc. USA, 93: 14082-14087 (1996); и Xiao et al., J Virol, 70: 8098-8108 (1996). См. также Chao et al., Mol Ther, 2: 619-623 (2000) и Chao et al., Mol Ther, 4: 217-222 (2001). Более того, поскольку мышцы сильно васкуляризованы, рекомбинантная трансдукция AAV привела к появлению продуктов трансгена в системном кровотоке после внутримышечной инъекции, как описано в Herzog et al., Proc Natl Acad Sci USA, 94: 5804-5809 (1997) и Murphy et al., Proc Natl Acad Sci USA, 94: 13921- 13926 (1997). Более того, Lewis et al., J Virol, 76: 8769-8775 (2002) продемонстрировали, что скелетные миофибриллы обладают необходимыми клеточными факторами для правильного гликозилирования, фолдинга и секреции антител, что указывает на то, что мышцы способны к стабильной экспрессии терапевтических секретируемых белков. ДНК AAV в геномах rAAV может происходить из любого серотипа AAV, для которого может быть получен рекомбинантный вирус, включая, без ограничения указанным, серотипы AAV-1, AAV-2, AAV-3, AAV-4, AAV-5, AAV-6, AAV-7, AAV-8, AAV-9, AAV- 10, AAV-11, AAV- 12, AAV-13, AAV PHP.B, AAV rh74 и AAV-DJ. Производство псевдотипированного rAAV раскрыто, например, в WO 01/83692. Также рассматриваются другие варианты rAAV, например rAAV с мутациями капсида. См., например, Marsic et al., Molecular Therapy, 22 (11): 1900-1909 (2014). Нуклеотидные последовательности геномов различных серотипов AAV известны в данной области.
Используемый в данном документе термин «внешний» по отношению к белку вирусного капсида относится к поверхности, домену, области или концевому концу белка капсида, который обращен наружу в собранном вирусном капсиде. Термин «внутренний» по отношению к белку вирусного капсида относится к поверхности, домену, области или концевому концу (аминоконцевой конец или карбокси-конец) белка капсида, который обращен внутрь собранного вирусного капсида. При использовании в отношении собранного вирусного капсида термин «внутренний» относится к инкапсидированному пространству внутри вирусного капсида и обращенной внутрь поверхности капсида, которая открыта для замкнутого пространства. Внутреннее пространство инкапсидировано белками вирусного капсида и может содержать нуклеиновые кислоты, такие как вирусный геном, вирусные белки, белки клетки-хозяина или упаковочной клетки, а также любые другие компоненты или факторы, упакованные или инкапсидированные во время репликации, сборки вириона, инкапсидации и/или упаковки.
Используемый в данном документе термин «метка» означает определяемое прямо или косвенно соединение или композицию, которые прямо или косвенно конъюгированы с детектируемой композицией, например, полинуклеотид или белок, такой как антитело, для создания «меченой» композиции. Термин также включает последовательности, конъюгированные с полинуклеотидом, которые будут обеспечивать сигнал при экспрессии вставленных последовательностей, таких как зеленый флуоресцентный белок (GFP) и т.п.Метка может быть обнаружена сама по себе (например, радиоизотопные метки или флуоресцентные метки) или, в случае ферментативной метки, может катализировать химическое изменение субстратного соединения или композиции, которое поддается обнаружению. Метки могут быть подходящими для мелкомасштабного обнаружения или более подходящими для высокопроизводительного скрининга. Как таковые подходящие метки включают, без ограничения указанным, радиоизотопы, флуорохромы, хемилюминесцентные соединения, красители и белки, включая ферменты. Метка может быть просто обнаружена или может быть определена количественно. Отклик, который просто обнаруживается, обычно включает отклик, существование которого просто подтверждается, тогда как количественно определяемый отклик обычно включает отклик, имеющий поддающееся количественной оценке (например, числовое значение), такое как интенсивность, поляризация и/или другие свойства. В анализах люминесценции или флуоресценции выявляемый ответ может быть получен непосредственно с использованием люминофора или флуорофора, связанного с компонентом анализа, фактически участвующего в связывании, или косвенно с использованием люминофора или флуорофора, связанного с другим (например, репортером или индикатором) компонентом.
Примеры люминесцентных меток, которые производят сигналы, включают, без ограничения указанным, биолюминесценцию и хемилюминесценцию. Обнаруживаемый отклик люминесценции обычно включает изменение или возникновение сигнала люминесценции. Подходящие способы и люминофоры для люминесцентного мечения компонентов анализа известны в данной области и описаны, например, в Haugland, Richard P. (1996) Handbook of Fluorescent Probes and Research Chemicals (6th ed.). Примеры люминесцентных зондов включают, без ограничения указанным, экуорин и люциферазы.
Примеры подходящих флуоресцентных меток включают, без ограничения указанным, флуоресцеин, родамин, тетраметилродамин, эозин, эритрозин, кумарин, метил-кумарины, пирен, малацитовый зеленый, стильбен, люцифер желтый, Каскад Синий TM и техасский красный. Другие подходящие оптические красители описаны в Haugland, Richard P. (1996) Handbook of Fluorescent Probes and Research Chemicals (6th ed.).
В другом аспекте флуоресцентная метка функционализирована для облегчения ковалентного прикрепления к клеточному компоненту, присутствующему в или на поверхности клетки или ткани, такому как маркер клеточной поверхности. Подходящие функциональные группы, включая, без ограничения указанным, изотиоцианатные группы, аминогруппы, галогенацетильные группы, малеимиды, сукцинимидиловые эфиры и сульфонилгалогениды, все из которых могут быть использованы для присоединения флуоресцентной метки ко второй молекуле. Выбор функциональной группы флуоресцентной метки будет зависеть от сайта присоединения к линкеру, агенту, маркеру или второму метящему агенту.
Присоединение флуоресцентной метки может происходить либо непосредственно к клеточному компоненту или соединению, либо, альтернативно, через линкер. Подходящие пары связывания для применения при косвенном связывании флуоресцентной метки с промежуточным продуктом включают, без ограничения указанным, антигены/антитела, например, родамин/анти-родамин, биотин/авидин и биотин/стрепавидин.
Фраза «твердая подложка» относится к неводным поверхностям, таким как «планшеты для культивирования», «генные чипы» или «микроматрицы». Такие генные чипы или микрочипы можно использовать для диагностических и терапевтических целей рядом методов, известных специалисту в данной области. В одном методе олигонуклеотиды присоединяют и размещают на генном чипе для определения последовательности ДНК с помощью подхода гибридизации, такого как описанный в Пат. США №6 025 136 и 6 018 041. Полинуклеотиды по настоящему изобретению можно модифицировать в зонды, которые, в свою очередь, можно использовать для обнаружения генетической последовательности. Такие методы описаны, например, в Пат. США №5 968 740 и 5 858 659. Зонд также может быть прикреплен или закреплен к поверхности электрода для электрохимического обнаружения последовательностей нуклеиновых кислот, как описано Kayem et al. Патент США №952 172 и Kelley et al. (1999) Nucleic Acids Res. 27: 4830-4837.
«Композиция» предназначена для обозначения комбинации активного полипептида, полинуклеотида или антитела и другого соединения или композиции, инертного (например, детектируемая метка) или активного (например, носителя для доставки гена).
Подразумевается, что «фармацевтическая композиция» включает комбинацию активного полипептида, полинуклеотида или антитела с носителем, инертным или активным, таким как твердая подложка, что делает композицию подходящей для диагностического или терапевтического использования in vitro, in vivo или ex vivo.
Используемый в данном документе термин «фармацевтически приемлемый носитель» охватывает любой из стандартных фармацевтических носителей, такой как забуференный фосфатом физиологический раствор, вода и эмульсии, такие как эмульсия масло/вода или вода/масло, а также различные типы смачивающих агентов. Композиции также могут включать стабилизаторы и консерванты. Примеры носителей, стабилизаторов и адъювантов см. в Martin (1975) Remington’s Pharm. Sci., 15th Ed. (Mack Publ. Co., Easton).
Используемый в данном документе термин «рак» включает солидные опухоли и гематологические злокачественные новообразования. Примеры солидных опухолей включают, без ограничения указанным, рак мочевого пузыря, рак костей, рак мозга (например, глиобластому), рак молочной железы, колоректальный рак, рак пищевода, рак глаза, рак головы и шеи, рак почки, рак легкого, меланому, мезотелиому, рак яичников, рак поджелудочной железы, рак предстательной железы или рак желудка. Примеры гематологических злокачественных новообразований включают, без ограничения указанным, острый миелоидный лейкоз (AML), хронический лимфоцитарный лейкоз (CLL), малую лимфоцитарную лимфому (SLL), фолликулярную лимфому (FL), диффузную крупноклеточную B-клеточную лимфому (DLBCL), лимфому мантийных клеток (MCL), макроглобулинемию Вальденстрема, множественную миелому, В-клеточную лимфому экстранодальной маргинальной зоны, узловую B-клеточную лимфому из клеток краевой зоны, лимфому Беркитта, В-клеточную лимфому не-Беркитта высокой степени злокачественности, первичную медиастинальную В-клеточную лимфому (PMBL), иммунобластную крупноклеточную лимфому, B-лимфобластную лимфому из клеток-предшественников, В-клеточный пролимфоцитарный лейкоз, лимфоплазмоцитарную лимфому, лимфому маргинальной зоны селезенки, плазматическую миелому, плазмоцитому, медиастинальную (тимическую) В-крупноклеточную лимфому, внутрисосудистую В-крупноклеточную лимфому, первичную выпотную лимфому или лимфогранулематоз. В некоторых случаях рак представляет собой метастатический рак (например, метастатическую солидную опухоль или метастатическое гематологическое злокачественное новообразование). В некоторых случаях рак представляет собой рецидивирующий или резистентный рак (например, рецидивирующую или резистентную солидную опухоль или рецидивирующую или рефрактерную гематологическую злокачественную опухоль).
В некоторых воплощениях рак характеризуется повышенной экспрессией белка активации фибробластов (FAP) (в котором аминокислотная последовательность человеческого FAP раскрыта в учетном номере GenPept I38593, по состоянию на 10 февраля 2020 г.). FAP, также известный как FAP-альфа и пролилэндопептидаза FAP, представляет собой мембранно-связанный гликопротеин и часть семейства дипептидилпептидаз (DPP). FAP обладает как постпролинэкзопептидазной активностью, так и активностью желатиназы. В некоторых случаях рак, который характеризуется повышенной экспрессией FAP (FAP-положительные раки), включает, без ограничения указанным, рак костей, рак мозга, рак молочной железы, колоректальный рак, рак пищевода, рак желудка, рак печени, рак легкого, рак полости рта, рак яичников, рак поджелудочной железы, рак паращитовидной железы и рак почек. В некоторых случаях FAP-положительные раки имеют высокий уровень фиброза. В некоторых случаях этот уровень сравнивают с эквивалентным раком, при котором не происходит активации FAP. В дополнительных случаях этот уровень сравнивают с уровнем фиброза нормального объекта.
В некоторых случаях описанный в данном документе dimert связывается с внеклеточной частью FAP-альфа. В некоторых случаях dimert связывается с человеческим FAP-альфа (например, с внеклеточной частью FAP-альфа с учетным номером GenPept I38593 или его эквивалентом). В некоторых случаях dimert связывается с FAP-альфа и иммунной клеткой-мишенью, например, полипептидом клеточной поверхности, экспрессируемым на Т-клетке или NK-клетке. В некоторых случаях dimert связывается с FAP-альфа и, необязательно, с мишенью для иммунных клеток (например, полипептид клеточной поверхности, экспрессируемый на Т-клетке или NK-клетке) для лечения FAP-положительного рака, необязательно рака поджелудочной железы, дополнительно, необязательно, FAP-положительного рака, характеризующегося высоким уровнем фиброза.
В некоторых воплощениях рак характеризуется экспрессией и/или активацией антигена созревания B-клеток (BCMA) (также известного как 17 представитель суперсемейства рецепторов фактора некроза опухоли, TNFRSF17 и BCM), рецептора клеточной поверхности суперсемейства рецепторов TNF. В некоторых случаях аминокислотная последовательность человеческого ВСМА раскрыта под учетным номером GenPept BAB60895.1 (по состоянию на 10 февраля 2020 г.). В некоторых случаях онкологическое заболевание, характеризующийся экспрессией и/или активацией BCMA, представляет собой миелому (или множественную миелому).
В некоторых случаях описанный в данном документе dimert связывается с внеклеточной частью BCMA. В некоторых случаях dimert связывается с человеческим BCMA (например, с внеклеточной частью BCMA с учетным номером GenPept BAB60895.1 или его эквивалентом). В некоторых случаях dimert связывается с ВСМА и иммунной клеткой-мишенью, например, полипептидом клеточной поверхности, экспрессируемым на Т-клетке или NK-клетке. В некоторых случаях dimert связывается с ВСМА и, необязательно, с мишенью для иммунных клеток (например, полипептид клеточной поверхности, экспрессируемый на Т-клетке или NK-клетке) для лечения миеломы.
В некоторых воплощениях онкологическое заболевание характеризуется экспрессией или активацией мутанта рецептора эпидермального фактора роста (EGFR), например, варианта III EGFR (EGFRvIII). EGFRvIII относится к мутации EGFR, которая включает делецию экзонов 2-7 гена EGFR. В некоторых случаях раки, которые характеризуются экспрессией или активацией EGFRvIII (EGFRvIII-положительный рак), включают, без ограничения указанным, глиобластому, рак мочевого пузыря, рак молочной железы, колоректальный рак, рак пищевода, плоскоклеточную карциному головы и шеи (HNSCC), рак легкого, меланому, рак яичников, опухоль оболочки периферических нервов (PNST), рак предстательной железы, саркому и рак щитовидной железы.
В некоторых случаях описанный в данном документе dimert связывается с внеклеточной частью EGFRvIII. В некоторых случаях dimert связывается с внеклеточной частью человеческого EGFRvIII. В некоторых случаях dimert связывается с EGFRvIII и иммунной клеткой-мишенью, например, полипептидом клеточной поверхности, экспрессируемым на Т-клетке или NK-клетке. В некоторых случаях dimert связывается с EGFRvIII и, необязательно, с мишенью для иммунных клеток (например, полипептидом клеточной поверхности, экспрессируемым на Т-клетке или NK-клетке) для лечения EGFRvIII-положительного рака, необязательно глиобластомы.
В некоторых воплощениях рак характеризуется повышенной регуляцией рецептора 2 эпидермального фактора роста человека (HER2), также известного как HER2/neu, рецепторной тирозин-протеинкиназы erbB-2, CD340 и ERBB2. В некоторых случаях аминокислотная последовательность HER2 человека раскрыта под учетным номером GenPept NP-004439.2 (по состоянию на 10 февраля 2020 г.). В некоторых случаях HER2-положительные раки включают, без ограничения указанным, рак молочной железы, рак яичников, рак желудка, колоректальный рак, рак поджелудочной железы и рак эндометрия.
В некоторых случаях описанный в данном документе dimert связывается с внеклеточной частью HER2. В некоторых случаях dimert связывается с внеклеточной частью HER2 человека (например, содержащей аминокислотную последовательность, как указано в учетном номере GenPept NP-004439.2, или его эквиваленте). В некоторых случаях dimert связывается с HER2 и иммунной клеткой-мишенью, например, полипептидом клеточной поверхности, экспрессируемым на Т-клетке или NK-клетке. В некоторых случаях dimert связывается с HER2 и, необязательно, с мишенью-иммунной клеткой (например, полипептидом клеточной поверхности, экспрессируемым на Т-клетке или NK-клетке) для лечения HER2-положительного рака, необязательно рака молочной железы.
В некоторых воплощениях рак характеризуется повышенной регуляцией CD123, также известного как рецептор интерлейкина-3 или IL-3RA. В некоторых случаях аминокислотная последовательность человеческого CD123 раскрыта под учетным номером GenPept NP_002174.1 (по состоянию на 10 февраля 2020 г.). В некоторых случаях CD123-положительные раки включают, без ограничения указанным, острый миелоидный лейкоз (AML), острый лимфоцитарный лейкоз (ALL), новообразование бластных плазмоцитоидных дендритных клеток и волосатоклеточный лейкоз.
В некоторых случаях описанный в данном документе dimert связывается с внеклеточной частью CD123. В некоторых случаях dimert связывается с внеклеточной частью человеческого CD123 (например, содержащим аминокислотную последовательность, указанную в учетном номере GenPept NP_002174.1, или его эквиваленте). В некоторых случаях dimert связывается с CD123 и иммунной клеткой-мишенью, например, полипептидом клеточной поверхности, экспрессируемым на Т-клетке или NK-клетке. В некоторых случаях dimert связывается с CD123 и, необязательно, с иммунной клеткой-мишенью (например, полипептид клеточной поверхности, экспрессируемый на Т-клетке или NK-клетке) для лечения CD123-положительного рака, необязательно AML.
В некоторых воплощениях рак характеризуется повышенной регуляцией CD38, также известной как АДФ-рибозилциклаза 1 или ADPRC 1. В некоторых случаях аминокислотная последовательность человеческого CD38 раскрыта под учетным номером GenPept BAA18966.1 (по состоянию на 10 февраля 2020 г.). В некоторых случаях CD38-положительные раки включают, без ограничения указанным, множественную миелому, острый миелоидный лейкоз, рак предстательной железы и рак легкого.
В некоторых случаях описанный в данном документе dimert связывается с внеклеточной частью CD38. В некоторых случаях dimert связывается с внеклеточной частью человеческого CD38 (например, содержит аминокислотную последовательность, указанную в учетном номере GenPept BAA18966.1, или его эквивалент). В некоторых случаях dimert связывается с CD38 и мишенью иммунной клетки, например полипептидом клеточной поверхности, экспрессируемым на Т-клетке или NK-клетке. В некоторых случаях dimert связывается с CD38 и, необязательно, с мишенью иммунной клетки (например, полипептидом клеточной поверхности, экспрессируемым на Т-клетке или NK-клетке) для лечения CD38-положительного рака, необязательно AML.
В некоторых воплощениях рак характеризуется повышенной регуляцией мезотелина (также известного как MSLN). В некоторых случаях аминокислотная последовательность человеческого мезотелина раскрыта под учетным номером GenPept AAV87530.1 (по состоянию на 10 февраля 2020 г.). В некоторых случаях мезотелин-положительные виды рака включают, без ограничения указанным, мезотелиому, рак поджелудочной железы, рак яичников, рак эндометрия, рак желчных путей, рак желудка, аденокарциному легких и острый миелоидный лейкоз у детей.
В некоторых случаях описанный в данном документе dimert связывается с внеклеточной частью мезотелина. В некоторых случаях dimert связывается с внеклеточной частью мезотелина человека (например, содержащей аминокислотную последовательность, как указано в учетном номере GenPept AAV87530.1, или его эквиваленте). В некоторых случаях dimert связывается с мезотелином и мишенью иммунной клетки, например полипептидом клеточной поверхности, экспрессируемым на Т-клетке или NK-клетке. В некоторых случаях dimert связывается с мезотелином и, необязательно, с мишенью иммунной клетки (например, полипептидом клеточной поверхности, экспрессируемым на Т-клетке или NK-клетке) для лечения мезотелин-положительного рака, необязательно мезотелиомы.
В некоторых воплощениях рак характеризуется повышенной регуляцией рецептора интерлейкина-13 альфа (IL13R альфа). В некоторых случаях IL13R альфа включает IL13R альфа 1 (IL13R α1) и IL13R альфа 2 (IL13R α2). В некоторых случаях аминокислотная последовательность человеческого IL13R α1 раскрыта под учетным номером GenPept P78552.1 (по состоянию на 10 февраля 2020 г.). В некоторых случаях аминокислотная последовательность человеческого IL13R α2 раскрыта под учетным номером GenPept Q14627.1 (по состоянию на 10 февраля 2020 г.). В некоторых случаях IL13R альфа-положительные раки включают, без ограничения указанным, рак мозга (например, глиобластому) и почечно-клеточную карциному (ПКР).
В некоторых случаях описанный в данном документе dimert связывается с внеклеточной частью IL13R альфа. В некоторых случаях dimert связывается с внеклеточной частью человеческого IL13R альфа (например, содержащей аминокислотную последовательность, указанную в учетном номере GenPept P78552.1, Q14627.1 или их эквиваленте). В некоторых случаях dimert связывается с IL13R альфа и иммунной клеткой-мишенью, например, полипептидом клеточной поверхности, экспрессируемым на Т-клетке или NK-клетке. В некоторых случаях dimert связывается с IL13R альфа и, необязательно, с мишенью для иммунных клеток (например, полипептидом клеточной поверхности, экспрессируемым на Т-клетке или NK-клетке) для лечения IL13R альфа-положительного рака, необязательно рака мозга.
В некоторых воплощениях рак характеризуется положительной регуляцией B7-H3, также известного как CD276, представителя иммунных контрольных точек. В некоторых случаях аминокислотная последовательность человеческого B7-H3 раскрыта под учетным номером GenPept CAE47548.1 (по состоянию на 10 февраля 2020 г.). В некоторых случаях B7-H3-положительные раки включают, без ограничения указанным, рак легкого (например, немелкоклеточный рак легкого), рак молочной железы, рак предстательной железы, почечно-клеточную карциному, рак мозга, рак поджелудочной железы, почки. рак, рак желудка, рак яичников, меланома и рак щитовидной железы.
В некоторых случаях описанный в данном документе dimert связывается с внеклеточной частью B7-H3. В некоторых случаях dimert связывается с внеклеточной частью человеческого B7-H3 (например, содержащей аминокислотную последовательность, указанную в учетном номере GenPept CAE47548.1, или ее эквиваленте). В некоторых случаях dimert связывается с B7-H3 и мишенью-Т-клеткой или NK-клеткой, например полипептидом клеточной поверхности, экспрессируемым на T-клетке или NK-клетке. В некоторых случаях dimert связывается с B7-H3 и необязательно с T-клеткой или NK-клеткой-мишенью (например, полипептид клеточной поверхности, экспрессируемый на T-клетке или NK-клетке) для лечения B7-H3-положительного рака, необязательно, рака легкого (например, немелкоклеточного рака легкого), рака молочной железы, рака предстательной железы, почечно-клеточной карциномы, рака мозга, рака поджелудочной железы, рака почки, рака желудка, рака яичников, меланомы или рака щитовидной железы.
В некоторых воплощениях рак характеризуется повышенной регуляцией рецепторной тирозинкиназы, подобной орфанному рецептору 1 (ROR1), также известной как нейротрофическая тирозинкиназа, рецептор-родственная 1 или NTRKR1. В некоторых случаях аминокислотная последовательность человеческого ROR1 раскрыта под учетным номером GenPept NP_005003 (по состоянию на 10 февраля 2020 г.). В некоторых случаях ROR1-положительные раки включают, без ограничения указанным, рак молочной железы, рак легкого, рак желудка, рак яичников, хронический лимфоцитарный лейкоз (CLL) и острый лимфолейкоз (ALL).
В некоторых случаях описанный в данном документе dimert связывается с внеклеточной частью ROR1. В некоторых случаях dimert связывается с внеклеточной частью человеческого ROR1 (например, содержит аминокислотную последовательность, указанную в учетном номере GenPept NP_005003, или его эквивалент). В некоторых случаях dimert связывается с ROR1 и мишенью иммунной клетки, например, полипептидом клеточной поверхности, экспрессируемым на Т-клетке или NK-клетке. В некоторых случаях dimert связывается с ROR1 и, необязательно, с мишенью иммунной клетки (например, полипептидом клеточной поверхности, экспрессируемым на Т-клетке или NK-клетке) для лечения ROR1-положительного рака, необязательно рака молочной железы, рака легкого, рака желудка, рака яичников, CLL или ALL.
В некоторых воплощениях рак характеризуется повышенной регуляцией рецептора 2 эфрина типа A (EphA2), также известного как рецептор А2 EPH, рецепторная тирозин-протеинкиназа ECK или рецепторная протеинтирозинкиназа эпителиальных клеток. В некоторых случаях аминокислотная последовательность человеческого EphA2 раскрыта под учетным номером GenPept NP_004422.2 (по состоянию на 10 февраля 2020 г.). В некоторых случаях EphA2-положительные раки включают, без ограничения указанным, рак молочной железы, рак мочевого пузыря, рак предстательной железы, рак кожи, рак легкого, рак яичников, рак мозга, мезотелиому, рак щитовидной железы, колоректальный рак, рак желудка, рак пищевода, рак эндометрия, рак шейки матки, рак поджелудочной железы, меланомы, почечно-клеточный рак и рак печени.
В некоторых случаях описанный в данном документе dimert связывается с внеклеточной частью EphA2. В некоторых случаях dimert связывается с внеклеточной частью EphA2 человека (например, содержащей аминокислотную последовательность, указанную в учетном номере GenPept NP_004422.2, или его эквиваленте). В некоторых случаях dimert связывается с EphA2 и иммунной клеткой-мишенью, например, полипептидом клеточной поверхности, экспрессируемым на Т-клетке или NK-клетке. В некоторых случаях dimert связывается с EphA2 и, необязательно, с мишенью иммунной клетки (например, полипептидом клеточной поверхности, экспрессируемым на Т-клетке или NK-клетке) для лечения EphA2-положительного рака, необязательно рака молочной железы, рака мочевого пузыря, рака предстательной железы, рака кожи, рака легкого, рака яичников, рака мозга, мезотелиомы, рака щитовидной железы, колоректального рака, рака желудка, рака пищевода, рака эндометрия, рака шейки матки, рака поджелудочной железы, меланомы, почечно-клеточного рака или рака печени.
В некоторых воплощениях рак представляет собой В-клеточный лейкоз или В-клеточную лимфому. Примеры В-клеточного лейкоза включают В-клеточный хронический лимфоцитарный лейкоз (или В-клеточную малую лимфоцитарную лимфому); острый лимфобластный лейкоз из зрелых B-клеток; В-клеточный пролимфоцитарный лейкоз; лимфобластный лейкоз из предшественников В-лимфоцитов; и волосатоклеточный лейкоз. В некоторых случаях В-клеточный лейкоз, В-клеточная лимфома или их комбинация характеризуются повышенной экспрессией CD20 и/или CD22 на В-клетках. В некоторых случаях описанный в данном документе dimert связывается с CD20 или CD22. В некоторых случаях dimert связывается с CD20, экспрессируемым на В-клетке. В некоторых случаях dimert связывается с CD22, экспрессируемым на В-клетке. В некоторых случаях dimert дополнительно связывается с другой клеткой-мишенью, например, полипептидом клеточной поверхности, экспрессируемым на раковойклетке, или полипептидом клеточной поверхности, экспрессируемым на Т-клетке или NK-клетке. В некоторых случаях dimert связывается с CD20 или CD22 и, необязательно, с другой клеткой-мишенью (например, полипептид клеточной поверхности, экспрессируемый на раковой клетке, или полипептид клеточной поверхности, экспрессируемый на T-клетке или NK-клетке) для лечения B-клеточного лейкоза или В-клеточной лимфомы.
Используемый в данном документе термин «терапия первой линии» включает первичное лечение объекта, необязательно больного раком. В некоторых случаях в условиях онкологического заболевания, рак - это первичный рак. В других случаях онкологическое заболевание представляет собой метастатический или рецидивирующий рак. В некоторых случаях терапия первой линии включает химиотерапию. В других случаях лечение первой линии включает лучевую терапию. Квалифицированный специалист легко поймет, что разные виды лечения первой линии могут быть применимы к разным типам рака.
Используемый в данном документе термин «терапия второй линии» включает лечение, которое применяется после прекращения лечения первичной или первой линии. Терапия третьей линии, терапия четвертой линии или терапия пятой линии включают последующие виды лечения. Как указано в соглашении о наименовании, терапия третьей линии включает курс лечения, на основании которого прекращена терапия первой и второй линий.
«Объектом» диагностики или лечения является клетка или животное, такое как млекопитающее или человек. Объект не ограничен конкретным видом и включает животных, не относящихся к человеку, подлежащих диагностике или лечению, а также животных, подверженных инфекциям, или животных моделей, например, обезьяньих, мышевидных, таких как крысы, мыши, шиншиллы, псовых, такие как собаки, зайцевых, таких как кролики, домашний скот, спортивных животных и животных-компаньонов. В этот термин также входят пациенты-люди.
Термин «ткань» используется в настоящем документе для обозначения ткани живого или умершего организма или любой ткани, полученной или предназначенной для имитации живого или умершего организма. Ткань может быть здоровой, больной и/или иметь генетические мутации. Биологическая ткань может включать любую отдельную ткань (например, совокупность клеток, которые могут быть взаимосвязаны) или группу тканей, составляющих орган или часть или область тела организма. Ткань может содержать гомогенный клеточный материал или может представлять собой сложную структуру, такую как структура, которая находится в областях организма, включая грудную клетку, которая, например, может включать ткань легкого, скелетную ткань и/или мышечную ткань. Примеры тканей включают, без ограничения указанным, ткани, полученные из печени, легких, щитовидной железы, кожи, поджелудочной железы, кровеносных сосудов, мочевого пузыря, почек, мозга, системы желчных протоков, двенадцатиперстной кишки, брюшной аорты, подвздошной вены, сердца и кишечника, включая любую их комбинацию.
Используемый в данном документе термин «специфически связывается», или «специфически связывается с», или «специфически поражает» означает полипептид или его фрагмент, который распознает и связывает интересующую биологическую молекулу (например, полипептид), но который по существу не распознает и не связывает другие молекулы в образце, например, биологическом образце, который включает или экспрессирует опухолевый антиген.
Используемый в данном документе термин «лечить» или «лечение» заболевания у объекта относится к (1) предотвращению появления симптомов или заболевания у объекта, который предрасположен или еще не проявляет симптомов заболевания; (2) подавление заболевания или прекращение его развития; или (3) улучшение или вызывание регрессии заболевания или симптомов заболевания. Как понятно из уровня техники, «лечение» представляет собой подход для получения полезных или желаемых результатов, включая клинические результаты. Для целей настоящей технологии полезные или желаемые результаты могут включать одно или несколько, без ограничения указанным, из числа ослабления или улучшения одного или нескольких симптомов, уменьшения степени состояния (включая заболевание), стабилизированного (т.е. не ухудшенное) состояния (включая заболевание), задержки или замедления состояния (включая заболевание), прогрессирования, улучшения или смягчения состояния (включая заболевание), состояния и ремиссию (частичную или полную), детектируемые или не детектируемые. В одном аспекте термин «лечение» исключает профилактику.
Используемый в данном документе термин «эффективное количество» означает количество, достаточное для достижения желаемого эффекта. В контексте терапевтических или профилактических применений эффективное количество будет зависеть от типа и тяжести рассматриваемого состояния и характеристик отдельного объекта, таких как общее состояние здоровья, возраст, пол, масса тела и переносимость фармацевтических композиций. В контексте генной терапии в некоторых воплощениях эффективное количество представляет собой количество, достаточное для восстановления части или полной функции гена, дефицитного у объекта. В других воплощениях эффективное количество рекомбинантного полинуклеотида, вектора или вирусной частицы AAV является количеством, достаточным для того, чтобы вызвать экспрессию гена у объекта. В некоторых воплощениях эффективное количество представляет собой количество, необходимое для увеличения метаболизма галактозы у нуждающегося в этом объекта. Квалифицированный специалист сможет определить подходящие количества в зависимости от этих и других факторов.
В некоторых воплощениях эффективное количество будет зависеть от размера и природы рассматриваемого применения. Это также будет зависеть от характера и чувствительности целевого объекта и используемых способов. Квалифицированный специалист сможет определить эффективное количество на основе этих и других соображений. Эффективное количество может включать одно или несколько введений композиции в зависимости от воплощения.
Используемый в данном документе термин «вводить» или «введение» предназначен для обозначения доставки вещества объекту, такому как животное или человек. Введение может быть осуществлено в одной дозе, непрерывно или периодически в течение всего курса лечения. Способы определения наиболее эффективных средств и дозировки известны специалистам в данной области техники и будут варьировать в зависимости от композиции, используемой для терапии, цели терапии, а также возраста, состояния здоровья или пола объекта, которого лечат.Однократное или многократное введение можно проводить, причем уровень дозы и паттерн выбирает лечащий врач или, в случае животных-компаньонов и животных, которых лечит ветеринар. Подходящие лекарственные формы и способы введения агентов известны в данной области. Путь введения также может быть определен, и способ определения наиболее эффективного пути введения известен специалистам в данной области и будет варьировать в зависимости от композиции, используемой для лечения, цели лечения, состояния здоровья или стадии заболевания подвергаемого лечению объекта и клетки-мишени или ткани-мишени. Неограничивающие примеры пути введения включают внутривенный, внутриартериальный, внутримышечный, внутрисердечный, интратекальный, субвентрикулярный, эпидуральный, внутримозговой, интрацеребровентрикулярный, субретинальный, интравитреальный, внутрисуставной, внутриглазный, внутрибрюшинный, внутриматочный, внутрикожный, подкожный, чрескожный, чрезслизистый и ингаляционный.
Способы осуществления изобретения
Раковые заболевания являются второй ведущей причиной смерти во всем мире: в 2018 году, по оценкам, умерло 9,6 миллиона человек. В клинической разработке и на рынке имеется множество различных типов лечения раковых заболеваний, например, иммунотерапия, гормональная терапия, таргетная лекарственная терапия, адоптивная клеточная терапия и химиотерапия. Хотя во многих терапевтических областях достигнуты успехи, все еще существуют проблемы, связанные с такими факторами, как период полувыведения и токсичность, которые влияют на терапевтическую эффективность. Например, терапия на основе онколитических вирусов обеспечивает адресную доставку полезных нагрузок в интересующее микросреду опухоли. Однако эти онколитические вирусы экспрессируют свои полезные нагрузки в опухолевых клетках и вызывают эффект уничтожения клеток, тем самым ограничивая экспрессию полезных нагрузок максимум несколькими днями, часто ограничиваясь несколькими часами. Таким образом, для получения устойчивого и пролонгированного терапевтического эффекта необходимо многократное введение, которое увеличивает токсичность и приводит к неблагоприятным иммунным ответам. Адоптивная клеточная терапия, такая как терапия химерными антигенными рецепторами (CAR)-T-клетками, обеспечивает индивидуализированный вариант лечения для объекта, страдающего от рака. Однако некоторые из наиболее распространенных побочных эффектов терапии CAR Т-клетками включают синдром высвобождения цитокинов; неврологические явления, такие как энцефалопатия, афазия, судороги и потеря равновесия; нейтропения; и анемия. Кроме того, получение CAR-T-клеток включает несколько недель культивирования и размножения перед введением, время, которое может иметь решающее значение, в частности, для пациента на поздней стадии рака.
В некоторых воплощениях в данном документе раскрыты способы доставки терапевтического трансгена (например, dimert, раскрытого в данном документе) с использованием вектора для генной терапии. В некоторых воплощениях вектор для генной терапии обеспечивает стабильную устойчивую экспрессию терапевтического трансгена (например, dimert). В дополнительных воплощениях вектор для генной терапии обеспечивает конститутивную экспрессию. В дополнительных воплощениях вектор для генной терапии обеспечивает регулируемую экспрессию. В некоторых случаях вектор генной терапии трансдуцируется в нормальных клетках (например, в клетках органов, таких как клетки печени или мышечные клетки). В некоторых случаях однократного введения вектора для генной терапии достаточно для индукции стабильной устойчивой экспрессии терапевтического трансгена (например, dimert). В дополнительных случаях вектор для генной терапии обеспечивает непрерывную долгосрочную экспрессию терапевтического трансгена (например, dimert), который оказывает долгосрочное давление на раковые клетки.
В некоторых воплощениях в настоящем документе раскрывается способ доставки терапевтического трансгена (например, dimert, раскрытого в настоящем документе) с использованием TransJoin. В некоторых случаях TransJoin обеспечивает конститутивную экспрессию терапевтического трансгена (например, dimert). В некоторых случаях TransJoin обеспечивает устойчивую, стабильную, долгосрочную экспрессию терапевтического трансгена (например, dimert). В некоторых случаях долговременная экспрессия включает около одну неделю, две недели, три недели, четыре недели, один месяц, 2 месяца, 3 месяца, 4 месяца, 5 месяцев, 6 месяцев, 8 месяцев, 10 месяцев, один год, или более. В некоторых случаях TransJoin трансдуцируется в нормальных клетках (например, в клетках органов, таких как клетки печени или мышечные клетки). В некоторых случаях однократного введения TransJoin достаточно для индукции стабильной устойчивой экспрессии терапевтического трансгена (например, dimert). В дополнительных случаях TransJoin обеспечивает непрерывную долговременную экспрессию терапевтического трансгена (например, dimert), который оказывает долговременное давление на е раковые клетки.
В некоторых воплощениях в настоящем документе раскрывается новый способ, который активирует экспрессию трансгена (например, dimert, раскрытый в настоящем документе), который можно использовать в качестве платформы генной терапии для регулируемой экспрессии, например, краткосрочной экспрессии гена (например, от недель до месяцев, дополнительно, необязательно, одна неделя, две недели, три недели, четыре недели, один месяц, 2 месяца, 3 месяца, 4 месяца, 5 месяцев, 6 месяцев, 8 месяцев или более). В некоторых случаях в способе используется TransSkip.В одном аспекте экспрессия трансгена находится под контролем лекарственного средства или внешнего агента (например, OncoSkip), так что введение лекарственного средства или агента модулирует (активирует или инактивирует) экспрессию гена. С другой стороны, отсутствие введения или отмена лекарственного средства или агента возвращается к исходному состоянию экспрессии гена до введения. Например, когда лекарство или агент могут активировать экспрессию гена, если есть побочные эффекты или если экспрессия трансгена больше не требуется, отмена введения может инактивировать экспрессию гена, тем самым минимизируя побочные эффекты, если таковые имеются. В некоторых случаях TransSkip трансдуцируется в нормальных клетках (например, в клетках органов, таких как клетки печени или мышечные клетки). В некоторых случаях однократного введения TransSkip достаточно для индукции стабильной устойчивой экспрессии терапевтического трансгена (например, dimert). В дополнительных случаях TransSkip обеспечивает регулируемую, но непрерывную экспрессию терапевтического трансгена (например, dimert), который оказывает долгосрочное давление на раковые клетки.
Пропуск экзона - это технология, используемая для лечения определенных генетических заболеваний, при которых короткая область гена является дефектной. Мутации ДНК приводят к повреждению белка либо потому, что в нем появляются неправильные аминокислоты, либо они вызывают стоп-мутацию, приводящую к укороченному белку, либо они изменяют рамку считывания, генерируя и то, и другое. Поскольку гены млекопитающих обычно кодируются в экзонах, то есть они разделены на несколько генных сегментов (экзонов), которые сращиваются во время процессирования мРНК, мутации ДНК (изменения или делеции пар оснований), лежащие в основе многих заболеваний, содержатся в пределах одиночного экзона. Если этот экзон можно пропустить и не включить в конечную сплайсинговую мРНК, то мутированная область не будет включена в конечный белок. Хотя белок будет короче и может не иметь некоторых частей, он все равно будет «в рамке» и может сохранить некоторые из своих функций.
Например, мышечная дистрофия Дюшенна (DMD) и мышечная дистрофия Беккера (BMD) являются наиболее распространенными формами мышечной дистрофии у детей, которые вызваны генетическими дефектами гена DMD, кодирующего дистрофин, мышечный белок, необходимый для взаимодействия между цитоскелетом и внеклеточным матриксом для поддержания стабильности мышечных волокон во время сокращения. Мутации DMD в гене дистрофина характеризуются вставками или делециями со сдвигом рамки считывания или точечными нонсенс-мутациями, что приводит к отсутствию функционального дистрофина. Мутации BMD в целом сохраняют рамку считывания нетронутой, позволяя синтез частично функционального дистрофина. Терапия на основе пропуска экзонов привела к трансформации мутаций вне рамки считывания, присутствующих у пациентов с DMD, в мутации внутри рамки считывания, которые кодируют частично функциональный дистрофин. В 2016 году FDA одобрило первое лекарственное средство для пропуска экзонов, Eteplirsen™ (Sarepta Therapeutics) для лечения мышечной дистрофии Дюшенна с мутациями в 51 экзоне гена дистрофина. Когда вводится лекарственное средство (которое представляет собой модифицированный короткий участок ДНК, также называемый олиго), 51 экзон «пропускается», чтобы восстановить почти полноразмерный и более функциональный белок. Аналогичная технология разрабатывается, чтобы пропустить экзоны других дистрофинов, а также экзоны других генов, вызывающих заболевания.
В отличие от пропуска экзонов в настоящем описании используется активация трансгена в генно-терапевтических применениях. Большинство, если не все подходы к генной терапии в настоящее время используют комплементарную (кДНК) последовательность гена; то есть генетическая последовательность трансгена представляет собой только кодирующую последовательность (только экзоны) и не включает никаких промежуточных (интронных) последовательностей. Таким образом, ген не подвергается сплайсингу РНК.
В этом раскрытии и технологии используются промежуточные последовательности, которые содержат донорные и акцепторные сайты сплайсинга и сайты связывания сплайсосомы РНК, чтобы заставить трансген подвергаться нормальному сплайсингу экзонов. Таким образом, обеспечивается «реконструированная» (искусственная) структура гена экзон-интрон-экзон в трансгене, который подвергается сплайсингу, когда он экспрессируется в клетке-мишени. Однако сплайсинг регулируется на основе принципов пропуска экзонов. В одном аспекте намеренно вставленный экзон, который содержит стоп-кодон в середине гена, регулирует экспрессию гена, т.е. только когда этот экзон пропущен, нормальный функциональный трансген будет экспрессироваться искусственной структурой. В одном воплощении функциональный трансген кодирует антитело. В другом воплощении антитело представляет собой биспецифическое или триспецифическое антитело (например, dimert). В другом воплощении антитело (например, dimert) представляет собой биспецифический рекрутер Т-клеток (BiTE), биспецифический рекрутер NK-клеток (BiKE), триспецифический рекрутер Т-клеток (TriTE) или триспецифический рекрутер NK-клеток (TriKE).
В дополнительном аспекте степень и тип сплайсинга изменяются в зависимости от типа клетки, поскольку многие гены обычно подвергаются альтернативному сплайсингу, который иногда зависит от типа клетки. Кроме того, сплайсинг иногда изменяется в определенных раковых клетках (некоторые экзоны определенных генов могут быть включены или исключены в нормальных клетках по сравнению с раковыми клетками). Эта технология может использовать эти функции для достижения контроля над трансгенами, который отличается в нормальных и раковых клетках. Также антитела, например биспецифические или триспецифические антитела, кодируемые функциональными трансгенами, можно использовать для лечения рака. Таким образом, в одном аспекте модуляция сплайсинга трансгена в данном настоящем раскрытии обеспечивает способ лечения рака.
Структурные воплощения
В определенных воплощениях в настоящем документе предлагается полинуклеотид или вектор, содержащий или альтернативно состоящий в основном из или дополнительно состоящий из: (а) первой полинуклеотидной последовательности, содержащей первую часть открытой рамки считывания, кодирующей первый полипептид; (b) второй полинуклеотидной последовательности, содержащей вторую часть открытой рамки считывания, кодирующую первый полипептид; (c) третьей полинуклеотидной последовательности, кодирующей второй полипептид; и (d) полинуклеотидной последовательности, регулирующей ген, расположенный между первым полинуклеотидом и вторым полинуклеотидом. В некоторых случаях первый полипептид связывается с поверхностным полипептидом (например, поверхностным рецептором) первой клетки-мишени, а второй полипептид связывается с поверхностным полипептидом (например, поверхностным рецептором) второй клетки-мишени. В некоторых случаях первая клетка-мишень и вторая клетка-мишень отличаются. Например, первая клетка-мишень может быть раковой клеткой, а вторая клетка-мишень может быть иммунной клеткой. Во втором примере первая клетка-мишень может быть иммунной клеткой, а вторая клетка-мишень может быть раковой клеткой. В третьем примере первая клетка-мишень может быть первой иммунной клеткой, вторая клетка-мишень может быть второй иммунной клеткой, и первая иммунная клетка представляет собой тип клетки, отличный от второй иммунной клетки. В четвертом примере первая клетка-мишень представляет собой первую раковую клетку, а вторая клетка-мишень представляет собой раковую клетку, а первая раковая клетка и раковая опухолевая клетка относятся к одному и тому же типу ракового заболевания, например, относятся к одному и тому же генетическому дефекту или, альтернативно, одному и тому же типу ткани.
В некоторых воплощениях первый полипептид представляет собой первое антитело или его связывающий фрагмент, а второй полипептид представляет собой второе антитело или его связывающий фрагмент.В некоторых случаях в настоящем документе предлагается полинуклеотид или вектор, содержащий или альтернативно состоящий в основном из, или дополнительно состоящий из: (а) первой полинуклеотидной последовательности, содержащей первую часть открытой рамки считывания, кодирующей первое антитело или антиген- его связывающий фрагмент; (b) второй полинуклеотидной последовательности, содержащей вторую часть открытой рамки считывания, кодирующую первое антитело или его антигенсвязывающий фрагмент; (c) третьей полинуклеотидной последовательности, кодирующей второе антитело или его антигенсвязывающий фрагмент; и (d) полинуклеотидной последовательности, регулирующей ген, расположенный между первым полинуклеотидом и вторым полинуклеотидом. Также предлагается комплиментарный полинуклеотид. В одном аспекте полинуклеотид, его комплементарный нуклеотид и/или вектор помечены детектируемой меткой. В некоторых случаях первое антитело связывается с первой мишенью, а второе антитело связывается со второй мишенью. В некоторых случаях первая мишень представляет собой поверхностный полипептид (например, поверхностный рецептор) на первой клетке, а вторая мишень представляет собой поверхностный полипептид (например, поверхностный рецептор) второй клетки. В некоторых случаях первая клетка-мишень и вторая клетка-мишень отличаются. Например, первая клетка-мишень может быть раковой клеткой, а вторая клетка-мишень может быть иммунной клеткой. Во втором примере первая клетка-мишень может быть иммунной клеткой, а вторая клетка-мишень может быть раковой клеткой. В третьем примере первая клетка-мишень может быть первой иммунной клеткой, вторая клетка-мишень может быть второй иммунной клеткой, и первая иммунная клетка представляет собой тип клетки, отличный от второй иммунной клетки. В четвертом примере первая клетка-мишень представляет собой первую раковую клетку, а вторая клетка-мишень представляет собой вторую раковую клетку, а первая раковая клетка и вторая раковая клетка относятся к одному и тому же типу онкологического заболевания, например, относятся к одному и тому же генетическому дефекту или, альтернативно, одному и тому же типу ткани. В некоторых случаях первая мишень является первым эпитопом, а вторая мишень - вторым эпитопом, и оба эпитопа присутствуют на одном и том же антигене. В некоторых случаях первое и второе антитела имеют разные аминокислотные последовательности. В одном аспекте полинуклеотид содержится в векторе экспрессии гена, неограничивающие примеры таких включают плазмиды, ДНК-вирусные векторы или носители для доставки генов.
В некоторых воплощениях в настоящем документе также раскрывается вектор для применения в генной терапии, содержащий: первую полинуклеотидную последовательность, кодирующую первое антитело или его антигенсвязывающий фрагмент; и вторую полинуклеотидную последовательность, кодирующую второе антитело или его антигенсвязывающий фрагмент.Также предлагается комплиментарный полинуклеотид. В одном аспекте полинуклеотид, его комплементарный нуклеотид и/или вектор помечены детектируемой меткой. В некоторых случаях первое антитело связывается с первой мишенью, а второе антитело связывается со второй мишенью. В некоторых случаях первая мишень представляет собой поверхностный полипептид (например, поверхностный рецептор) на первой клетке, а вторая мишень представляет собой поверхностный полипептид (например, поверхностный рецептор) второй клетки. В некоторых случаях первая клетка-мишень и вторая клетка-мишень отличаются. Например, первая клетка-мишень может быть раковой клеткой, а вторая клетка-мишень может быть иммунной клеткой. Во втором примере первая клетка-мишень может быть иммунной клеткой, а вторая клетка-мишень может быть раковой клеткой. В третьем примере первая клетка-мишень может быть первой иммунной клеткой, вторая клетка-мишень может быть второй иммунной клеткой, и первая иммунная клетка представляет собой тип клетки, отличный от второй иммунной клетки. В четвертом примере первая клетка-мишень представляет собой первую раковую клетку, а вторая клетка-мишень представляет собой вторую раковую клетку, а первая раковая клетка и вторая раковая клетка относятся к одному и тому же типу ракового заболевания, например, относятся к одному и тому же генетическому дефекту или, альтернативно, одному и тому же типу ткани. В некоторых случаях первая мишень является первым эпитопом, а вторая мишень - вторым эпитопом, и оба эпитопа присутствуют на одном и том же антигене. В некоторых случаях первое и второе антитела имеют разные аминокислотные последовательности.
В дополнительном аспекте полинуклеотидная последовательность регуляции гена содержит донорный сайт сплайсинга, вышележащий интрон, экзон, содержащий последовательности стоп-кодона во всех трех рамках считывания, нижележащий интрон и акцепторный сайт сплайсинга. В дополнительном аспекте полинуклеотидная последовательность, регулирующая ген, содержит одну или несколько связывающих последовательностей для антисмыслового олигонуклеотида. В дополнительном аспекте антисмысловым олигонуклеотидом является морфолино. В дополнительном аспекте связывающая последовательность для морфолиноолигонуклеотида включает полинуклеотидную последовательность, по меньшей мере на 95% идентичную SEQ ID NO: 24 (AATATGATCCAACAATAGAGGTAAATCTTG) или SEQ ID NO. 25 (GATCCAACAATAGAGGTAAATCTTGTTTTA), или, альтернативно, идентичную по меньшей мере на 96% или альтернативно по меньшей мере 97%, или альтернативно 98%, или альтернативно, по меньшей мере, 99%. В одном воплощении морфолиноолигонуклеотид содержит полинуклеотидную последовательность, идентичную по меньшей мере на 95% SEQ ID NO. 27 (CAAGATTTACCTCTATTGTTGGATCATATT) или SEQ ID NO. 28 (TAAAACAAGATTTACCTCTATTGTTGGATC), или, альтернативно, идентичности по меньшей мере на 96%, или альтернативно, по меньшей мере, 97%, или альтернативно 98%, или альтернативно, по меньшей мере, 99% с каждой из них. Донорный сайт сплайсинга и акцепторный сайт сплайсинга хорошо известны в данной области. Специалисту в данной области техники должны быть известны последовательности, например консенсусные последовательности, для донорного сайта сплайсинга и акцепторного сайта сплайсинга. Примерная консенсусная последовательность сайта сплайсинга для интронов U2 может включать 5’-сайт сплайсинга MAG-GTRAGT, в котором M представляет собой A или C; R представляет собой A или G, подчеркнутые нуклеотиды означают, что «GT» инвариантен; а тире «-» обозначает место стыковки. 3’-сайтом сплайсинга для интрона U2 может быть CAG-G, в котором подчеркнутые нуклеотиды означают, что «AG» инвариантен; а тире «-» обозначает место стыковки. Дополнительные примеры последовательностей консенсусных сайтов сплайсинга включают, без ограничения указанным, следующие:
5’-сайт сплайсинга экзона 10 p53 (донор): CAG-gtgagt, в котором тире «-» обозначает сайт сплайсинга;
Brd2 экзон 3 5’ss: AAG-gtgagt, в котором тире «-» обозначает сайт сплайсинга;
5’-сайт сплайсинга экзона 22 BRCA1: CAG-gtaagt, в котором тире «-» обозначает сайт сплайсинга;
SMN1 экзон 1 5’ss: CAG-gtgagg, в котором тире «-» обозначает сайт сплайсинга;
BRD2 3’ акцепторный сайт сплайсинга интрона 1 (нижний регистр)/экзон 2 (верхний регистр): cccatctttacag-GCTCCC, в котором тире «-» обозначает сайт сплайсинга;
BCL-X 3’ акцепторный сайт сплайсинга интрона 2 (нижний регистр)/экзон 3 (верхний регистр): tctctccctgcag-GATACT, в котором тире «-» обозначает сайт сплайсинга;
3’ акцепторный сайт сплайсинга интрона 28 фибронектина (нижний регистр)/экзон29 (верхний регистр): ctttttcatacag-GAGGAA, в котором тире «-» обозначает сайт сплайсинга; и
3’ акцепторный сайт сплайсинга интрона 2 сурвивина (нижний регистр)/экзон3 (верхний регистр): tctttatttccagGCAAAG, в котором тире» - «обозначает сайт сплайсинга.
В некоторых воплощениях консенсусная последовательность сайта сплайсинга получена из //science.umd.edu/labs/mount/RNAinfo/matrices.html.
В некоторых воплощениях стоп-кодон включает олигонуклеотид из группы: TAA, TAG или TGA. В дополнительном аспекте последовательность стоп-кодона включает полинуклеотидную последовательность TAAxTAGxTGAxTAGxTAAxTGAx (SEQ ID NO. 1), где x представляет собой любой нуклеотид или, альтернативно, последовательность стоп-кодона включает полинуклеотидную последовательность TAATTAGTTGATTAGTTAATTGAT (SEQ ID NO. 2). В еще одном дополнительном аспекте полинуклеотид, регулирующий ген, содержит полинуклеотидную последовательность, по меньшей мере на 95% идентичную SEQ ID NO. 2, или, альтернативно, по меньшей мере на 96%, или альтернативно, по меньшей мере, 97%, или альтернативно 98%, или альтернативно, по меньшей мере, 99% идентичную SEQ ID NO. 2.
В дополнительных воплощениях первое антитело или его антигенсвязывающий фрагмент специфически связывается с активирующим антигеном на иммунной эффекторной клетке, а второе антитело или его антигенсвязывающий фрагмент связывается с опухолевым антигеном. В другом аспекте первое антитело или его антигенсвязывающий фрагмент специфически связывается с опухолевым антигеном, а второе антитело или его антигенсвязывающий фрагмент связывается с активирующим антигеном на иммунной эффекторной клетке. В дополнительном аспекте вектор также содержит четвертую полинуклеотидную последовательность, кодирующую третье антитело или его антигенсвязывающий фрагмент, где третье антитело или его антигенсвязывающий фрагмент связывается с активирующим антигеном на эффекторной иммунной клетке или опухолевом антигене. В одном аспекте иммунная эффекторная клетка включает дендритную клетку, клетку-натуральный киллер («NK»), макрофаг, Т-клетку, В-клетку или их комбинацию. Неограничивающие примеры иммунных эффекторных клеток включают Т-клетку или NK-клетку.
Неограничивающие примеры активирующего антигена на иммунной эффекторной клетке включают CD3, CD2, CD4, CD8, CD19, LFA1, CD45, NKG2D, NKp44, NKp46, NKp30, DNAM или их комбинацию.
Неограничивающие примеры антигена-мишени на антигенпрезентирующей клетке включают, без ограничения указанным, B7-H3 (CD276).
Неограничивающие примеры антигена-мишени на B-клетке включают, без ограничения указанным, CD20 и CD22.
Неограничивающие примеры опухолевых антигенов включают один или несколько из числа рецептора 2 эфрина типа A (EphA2), интерлейкина (IL)-13r альфа 2, EGFR VIII, PSMA, EpCAM, GD2 или GD3, фукозил GM1, PSCA, PLAC1, точечного разрыва саркомы, опухоли Вильмса 1, альфафетопротеина (AFP), карциноэмбрионального антигена (CEA), CA-125, MUC-1, эпителиального опухолевого антигена (ETA), тирозиназы, меланома- ассоциированного антигена (MAGE), антигена гематологической дифференцировки, поверхностного гликопротеина, ганглиозидов (GM2), рецептора фактора роста, стромального антигена, сосудистого антигена, орфанного рецептора 1, подобного рецепторной тирозинкиназе (ROR1), мезотелина, CD38, CD123, рецептора 2 эпидермального фактора роста человека (HER2), антигена созревания В-клеток (BCMA), белка активации фибробластов (FAP) альфа или их комбинации. Дополнительные примеры можно найти в данной области, см., например, включенные в данный документ ссылкой. В другом аспекте рекомбинантный вектор экспрессирует пре-мРНК, которая кодирует dimert, описанный в данном документе. В некоторых случаях dimert представляет собой биспецифическое антитело или триспецифическое антитело, когда пре-мРНК находится в контакте с морфолиноолигонуклеотидом. Неограничивающими примерами dimert являются группа: биспецифического рекрутера Т-клеток (BiTE), биспецифического рекрутера NK-клеток (BiKE), триспецифического рекрутера Т-клеток (TriTE) или триспецифического рекрутера NK-клеток (TriKE). В одном аспекте триспецифическое антитело включает первое антитело или его антигенсвязывающий фрагмент и второе антитело или его антигенсвязывающий фрагмент.
В одном аспекте dimert (например, биспецифический или триспецифический клеточный рекрутер) содержит полипептидную последовательность, идентичную последовательности с SEQ ID NO: 11, по меньшей мере на 95%, необязательно, идентичную последовательности с SEQ ID NO: 11 по меньшей мере на 96%, по меньшей мере 97%, по меньшей мере 98%, или по меньшей мере 99%. В другом воплощении полипептидная последовательность кодирует антигенсвязывающий фрагмент CD3; CD2; CD4; CD8; CD19; антиген 1, связанный с функцией лимфоцитов (LFA1); CD45; рецептор интерлейкина 21 (IL21R); группа 2 натуральных киллеров, представитель D (NKG2D); рецептор естественной цитотоксичности (NCR), такой как NKp44, NKp46 или NKp30; или вспомогательная молекула DNAX -1(DNAM или DNAM-1; также обозначаемая как CD226, или антиген 1 активации тромбоцитов и Т-клеток (PTA1)). В другом воплощении полипептидная последовательность кодирует антигенсвязывающий фрагмент для CD3, CD19, GD2 или NKG2D. В другом воплощении полипептид кодирует первый антигенсвязывающий фрагмент и второй антигенсвязывающий фрагмент.В другом воплощении первый антигенсвязывающий фрагмент связывается с CD3, а второй антигенсвязывающий фрагмент связывается с CD19. В другом воплощении первый антигенсвязывающий фрагмент связывается с CD3, а второй антигенсвязывающий фрагмент связывается с GD2. В другом воплощении первый антигенсвязывающий фрагмент связывается с NKG2D, а второй антигенсвязывающий фрагмент связывается с GD2.
В одном воплощении триспецифический агент или антитело содержит первый антигенсвязывающий фрагмент, второй антигенсвязывающий фрагмент и третий антигенсвязывающий фрагмент.В другом воплощении триспецифический рекрутер или антитело содержит три антигенсвязывающих фрагмента, которые индивидуально связываются с NKG2D, IL21R и GD2. В дополнительном аспекте триспецифический агент или антитело содержит полипептидную последовательность, по меньшей мере, на 95% идентичность последовательности с SEQ ID NO. 11, или, альтернативно, по меньшей мере 96%, или альтернативно, по меньшей мере, 97%, или альтернативно 98%, или альтернативно, по меньшей мере, 99% идентичности последовательности с SEQ ID NO: 11.
В одном воплощении антигенсвязывающий фрагмент, который связывается с IL-21R, представляет собой IL-21. Последовательности аминокислот и кДНК IL-12 показаны в SEQ ID NO. 3 и SEQ ID NO. 4, соответственно. В одном воплощении антигенсвязывающий фрагмент, который связывается с NKG2D, включает MICA, MICB, ULBP1, ULBP2, ULBP3, ULBP4, ULBP5, ULBP6, Rae-1α, Rae-1β, Rae-1γ, Rae-1δ, Rae-1ε, H60a, H60b, H60c, MULT1 или их фрагмент.В одном воплощении антигенсвязывающий фрагмент, который связывается с NKG2D, представляет собой MICA (SEQ ID NO. 5) или его фрагмент или его эквивалент.
Последовательность MICA в одном воплощении включает мутант дикого типа. В одном воплощении мутант MICA представляет собой вариант последовательности MICA дикого типа (например, последовательность MICA дикого типа, указанную в SEQ ID NO: 5). В другом воплощении мутант MICA представляет собой вариант последовательности MUC-30 (SEQ ID NO. 7), который содержит метиониновую мутацию вместо аланина в положении 129 последовательности MICA дикого типа (MICA-129Met). Эквивалент мутанта MICA (MICA-129Met) сохраняет мутацию метионина в положении 129 MICA дикого типа. В другом воплощении антигенсвязывающие фрагменты биспецифического или триспецифического рекрутера или антитела разделены линкерной последовательностью. Одно воплощение линкерной последовательности содержит, по существу или состоит из GGGGSGGGGSGGGGS (SEQ ID NO. 9) или ее эквивалента. Линкерная последовательность кодируется полинуклеотидной последовательностью GGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGC (SEQ ID NO. 10) или ее эквивалентом. В одном воплощении два антигенсвязывающих фрагмента, разделенных линкерной последовательностью, представляют собой IL21 и MICA (например, MICA дикого типа или мутант MICA, такой как MUC-30 или MICA-129Met), или их фрагменты или эквиваленты каждого из них. В одном воплощении два антигенсвязывающих фрагмента, разделенных линкерной последовательностью, представляют собой GD2 и MICA (например, MICA дикого типа или мутант MICA, такой как MUC-30 или MICA-129Met) или их фрагменты или эквиваленты каждого из них. В одном воплощении два антигенсвязывающих фрагмента, разделенных линкерной последовательностью, представляют собой IL21 и GD2 или их фрагменты или эквиваленты каждого из них. В другом воплощении линкер вставлен между антигенсвязывающими фрагментами любого из следующих триспецифических рекрутеров:
IL21-MICA129-GD2
MICA129-IL21-GD2
GD2-IL21-MICA129
GD2-MICA129-IL21
IL21-MICA/V129M-GD2-HDD
MICA/V12M-IL21-GD2-HDD
GD2-IL21-MICA129-HDD
GD2-MICA129-IL21-HDD
В другом воплощении dimert (например, биспецифический или триспецифический рекрутер) содержит консенсусную последовательность секреции (также называемую в данном документе sec0A). В некоторых случаях консенсусная последовательность секреции (sec0A) имеет по меньшей мере 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательностей с MWWRLWLLLLLLLLWPMVWA (SEQ ID NO: 51) или состоит из SEQ ID NO: 51. В некоторых случаях sec0A кодируется полинуклеотидом, содержащим ATGTGGTGGAGACTGTGGTGGCTGCTGCTGCTGCTGCTGCTGCTGTGGCCCATGGTGTGGGCC (SEQ ID NO: 52) или его эквивалентом.
В некоторых случаях консенсусная последовательность секреции дополнительно содержит один, два, три, четыре или более остатков на С-конце последовательности. В некоторых случаях один, два, три, четыре или несколько остатков представляют собой остатки с алифатическими боковыми цепями (например, Ala, Met, Ile, Val или Leu). В некоторых случаях один, два, три, четыре или более остатков представляют собой остатки Ala, остатки Gly, остатки Val, остатки Ile или их комбинацию. В некоторых случаях один, два, три, четыре или более остатков представляют собой остатки Ala, остатки Gly, остатки Val или их комбинацию. В некоторых случаях один, два, три, четыре или несколько остатков представляют собой остатки Ala, остатки Gly или их комбинацию. В некоторых случаях секреторная консенсусная последовательность дополнительно содержит один, два, три, четыре или более остатков Ala на С-конце последовательности. В некоторых случаях секреторная консенсусная последовательность дополнительно содержит один, два, три, четыре или более остатков Gly на С-конце последовательности. В некоторых случаях секреторная консенсусная последовательность дополнительно содержит один, два, три, четыре или более остатков Val на С-конце последовательности. В некоторых случаях секреторная консенсусная последовательность дополнительно содержит один, два, три, четыре или более остатков Ile на С-конце последовательности.
В некоторых случаях консенсусная последовательность секреции дополнительно содержит один, два или три остатка с алифатическими боковыми цепями (например, Ala, Met, Ile, Val или Leu). В некоторых случаях один, два или три остатка представляют собой остатки Ala, остатки Gly, остатки Val, остатки Ile или их комбинацию. В некоторых случаях один, два или три остатка представляют собой остатки Ala, остатки Gly, остатки Val или их комбинацию. В некоторых случаях один, два или три остатка представляют собой остатки Ala, остатки Gly или их комбинацию. В некоторых случаях секреторная консенсусная последовательность дополнительно содержит один, два или три остатка Ala на С-конце последовательности. В некоторых случаях секреторная консенсусная последовательность дополнительно содержит один, два или три остатка Gly на С-конце последовательности. В некоторых случаях секреторная консенсусная последовательность дополнительно содержит один, два или три остатка Val на С-конце последовательности. В некоторых случаях секреторная консенсусная последовательность дополнительно содержит один, два или три остатка Ile на С-конце последовательности.
В некоторых случаях секреторная консенсусная последовательность дополнительно содержит один или два остатка Gly на С-конце последовательности. В некоторых случаях один или два остатка представляют собой остатки Ala, остатки Gly, остатки Val, остатки Ile или их комбинацию. В некоторых случаях секреторная консенсусная последовательность дополнительно содержит один или два остатка Gly на С-конце последовательности. В некоторых случаях один или два остатка представляют собой остатки Ala, остатки Gly или их комбинацию. В некоторых случаях секреторная консенсусная последовательность дополнительно содержит один или два остатка Ala на С-конце последовательности. В некоторых случаях секреторная консенсусная последовательность дополнительно содержит один или два остатка Gly на С-конце последовательности. В некоторых случаях секреторная консенсусная последовательность дополнительно содержит один или два остатка Val на С-конце последовательности. В некоторых случаях секреторная консенсусная последовательность дополнительно содержит один или два остатка Ile на С-конце последовательности.
В некоторых случаях консенсусная последовательность секреции дополнительно содержит один остаток с алифатической боковой цепью (например, Ala, Met, Ile, Val или Leu). В некоторых случаях один остаток представляет собой Ala, Gly, Val или Ile. В некоторых случаях секреторная консенсусная последовательность дополнительно содержит один Ala на С-конце последовательности. В некоторых случаях секреторная консенсусная последовательность дополнительно содержит один Gly на С-конце последовательности. В некоторых случаях секреторная консенсусная последовательность дополнительно содержит один Val на С-конце последовательности. В некоторых случаях секреторная консенсусная последовательность дополнительно содержит один Ile на С-конце последовательности.
В некоторых случаях консенсусная последовательность секреции дополнительно содержит один или два остатка Ala на С-конце консенсусной последовательности, и такие последовательности называются секрекон1A (или sec1A) с одним Ala на C-конце и секрекон2A (или sec2A) с два остатка Ala на С-конце. В некоторых случаях sec1A имеет по меньшей мере 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательностей с MWWRLWWLLLLLLLLWPMVWAA (SEQ ID NO: 53). В некоторых случаях sec1A кодируется полинуклеотидом, содержащим ATGTGGTGGAGACTGTGGTGGCTGCTGCTGCTGCTGCTGCTGCTGTGGCCCATGGTGTGGGCCGCC (SEQ ID NO: 54) или его эквивалентом. В некоторых случаях sec2A включает по меньшей мере 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательности с MWWRLWWLLLLLLLLWPMVWAAA (SEQ ID NO: 55). В некоторых случаях sec2A кодируется полинуклеотидом, содержащим ATGTGGTGGAGACTGTGGTGGCTGCTGCTGCTGCTGCTGCTGCTGTGGCCCATGGTGTGGGCCGCCGCC (SEQ ID NO: 56) или его эквивалентом.
В некоторых воплощениях секреторная консенсусная последовательность модулирует экспрессию и/или секрецию dimert. В некоторых случаях секреторная консенсусная последовательность (например, sec1A или sec2A) усиливает экспрессию и/или секрецию dimert. В некоторых случаях секреторная консенсусная последовательность (например, sec1A или sec2A) модулирует (например, усиливает) экспрессию dimert в около 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 50 раз, или более. В некоторых случаях секреторная консенсусная последовательность (например, sec1A или sec2A) модулирует (например, усиливает) секрецию dimert в около 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 20, 50 раз, или более.
В некоторых воплощениях dimert, описанный в данном документе (например, триспецифический рекрутер), содержит консенсусную последовательность секреции (например, sec0A, sec1A или sec2A), последовательность IL21 (например, SEQ ID NO: 3), последовательность MICA (например, последовательность дикого типа, MUC-30 или MICA-129Met), последовательность пептида против GD2 или эквивалент одного или нескольких из них. В некоторых случаях dimert (например, триспецифический рекрутер) включает последовательность секрекон1A, последовательность IL21, последовательность MICA129, последовательность пептида антиGD2 или эквивалент одного или нескольких из них. В одном воплощении dimert (например, триспецифический рекрутер) включает пептид секрекон1A-линкер-IL21-линкер-MICA129-линкер-антиGD2 или эквивалент одного или нескольких из них. В одном воплощении dimert (например, триспецифический рекрутер) содержит полипептидную последовательность SEQ ID NO: 11.
SEQ ID NO. 11
MWWRLWWLLLLLLLLWPMVWAARSSPGNMERIVICLMVIFLGTLVHKSSSQGQDRHMIRMRQLIDIVDQLKNYVNDLVPEFLPAPEDVETNCEWSAFSCFQKAQLKSANTGNNERIINVSIKKLKRKPPSTNAGRRQKHRLTCPSCDSYEKKPPKEFLERFKSLLQKMIHQHLSSRTHGSEDSGGGGSGGGGSGGGGSEPHSLRYNLTVLSWDGSVQSGFLAEVHLDGQPFLRCDRQKCRAKPQGQWAEDVLGNKTWDRETRDLTGNGKDLRMTLAHIKDQKEGLHSLQEIRVCEIHEDNSTRSSQHFYYDGELFLSQNLETEEWTMPQSSRAQTLAMNIRNFLKEDAMKTKTHYHAMHADCLQELRRYLKSGVVLRRTVPPMVNVTRSEASEGNITVTCRASGFYPWNITLSWRQDGVSLSHDTQQWGDVLPDGNGTYQTWVATRICQGEEQRFTCYMEHSGNHSTHPVPSGGGGSGGGGSGGGGSQVQLQQSGPELVKPGASVKISCKTSGYKFTEYTMHWVKQSHGKCLEWIGGINPNNGGTNYNQKFKGKATLTVDKSSSTAYMELRSLTSEDSAVYYCARDTTVPYAYWGQGTTVTVSSGGGGSGGGGSGGGGSDIELTQSPAIMSASPGEKVTMTCSASSSISYMHWYQQKPGTSPKRWIYDTSKLASSVPARFSGSGSGTSYSLTISSMEAEDAATYYCHQRSSYPLTFGCGTKLEIKRASTKGP, или ее эквивалент.
В одном воплощении dimert (например, триспецифический рекрутер) кодируется последовательностью SEQ ID NO. 12.
SEQ ID NO. 12
ATGTGGTGGAGACTGTGGTGGCTGCTGCTGCTGCTGCTGCTGCTGTGGCCCATGGTGTGGGCCGCCAGAAGCAGCCCCGGCAACATGGAGAGAATCGTGATCTGCCTGATGGTGATCTTCCTGGGCACCCTGGTGCACAAGAGCAGCAGCCAGGGCCAGGACAGACACATGATCAGAATGAGACAGCTGATCGACATCGTGGACCAGCTGAAGAACTACGTGAACGACCTGGTGCCCGAGTTCCTGCCCGCCCCCGAGGACGTGGAGACCAACTGCGAGTGGAGCGCCTTCAGCTGCTTCCAGAAGGCCCAGCTGAAGAGCGCCAACACCGGCAACAACGAGAGAATCATCAACGTGAGCATCAAGAAGCTGAAGAGAAAGCCCCCCAGCACCAACGCCGGCAGAAGACAGAAGCACAGACTGACCTGCCCCAGCTGCGACAGCTACGAGAAGAAGCCCCCCAAGGAGTTCCTGGAGAGATTCAAGAGCCTGCTGCAGAAGATGATCCACCAGCACCTGAGCAGCAGAACCCACGGCAGCGAGGACAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGAGCCCCACAGTCTTCGTTATAACCTCACGGTGCTGTCCTGGGATGGATCTGTGCAGTCAGGGTTTCTCGCTGAGGTACATCTGGATGGTCAGCCCTTCCTGCGCTGTGACAGGCAGAAATGCAGGGCAAAGCCCCAGGGACAGTGGGCAGAAGATGTCCTGGGAAATAAGACATGGGACAGAGAGACCAGGGACTTGACAGGGAACGGAAAGGACCTCAGGATGACCCTGGCTCATATCAAGGACCAGAAAGAAGGCTTGCATTCCCTCCAGGAGATTAGGGTCTGTGAGATCCATGAAGACAACAGCACCAGGAGCTCCCAGCATTTCTACTACGATGGGGAGCTCTTCCTCTCCCAAAACCTGGAGACTGAGGAATGGACAATGCCCCAGTCCTCCAGAGCTCAGACCTTGGCCATGAACATCAGGAATTTCTTGAAGGAAGATGCCATGAAGACCAAGACACACTATCACGCTATGCATGCAGACTGCCTGCAGGAACTACGGCGATATCTAAAATCCGGCGTAGTCCTGAGGAGAACAGTGCCCCCCATGGTGAATGTCACCCGCAGCGAGGCCTCAGAGGGCAACATTACCGTGACATGCAGGGCTTCTGGCTTCTATCCCTGGAATATCACACTGAGCTGGCGTCAGGATGGGGTATCTTTGAGCCACGACACCCAGCAGTGGGGGGATGTCCTGCCTGATGGGAATGGAACCTACCAGACCTGGGTGGCCACCAGGATTTGCCAAGGAGAGGAGCAGAGGTTCACCTGCTACATGGAACACAGCGGGAATCACAGCACTCACCCTGTGCCCTCTGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCCAGGTGCAGCTGCAGCAGAGCGGCCCCGAGCTGGTGAAGCCCGGCGCCAGCGTGAAGATCAGCTGCAAGACCAGCGGCTACAAGTTCACCGAGTACACCATGCACTGGGTGAAGCAGAGCCACGGCAAGTGCCTGGAGTGGATCGGCGGCATCAACCCCAACAACGGCGGCACCAACTACAACCAGAAGTTCAAGGGCAAGGCCACCCTGACCGTGGACAAGAGCAGCAGCACCGCCTACATGGAGCTGAGAAGCCTGACCAGCGAGGACAGCGCCGTGTACTACTGCGCCAGAGACACCACCGTGCCCTACGCCTACTGGGGCCAGGGCACCACCGTGACCGTGAGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGACATCGAGCTGACCCAGAGCCCCGCCATCATGAGCGCCAGCCCCGGCGAGAAGGTGACCATGACCTGCAGCGCCAGCAGCAGCATCAGCTACATGCACTGGTACCAGCAGAAGCCCGGCACCAGCCCCAAGAGATGGATCTACGACACCAGCAAGCTGGCCAGCAGCGTGCCCGCCAGATTCAGCGGCAGCGGCAGCGGCACCAGCTACAGCCTGACCATCAGCAGCATGGAGGCCGAGGACGCCGCCACCTACTACTGCCACCAGAGAAGCAGCTACCCCCTGACCTTCGGCTGCGGCACCAAGCTGGAGATCAAGAGAGCCAGCACCAAGGGCCCCTAG, или ее эквивалент.
В одном воплощении dimert (например, биспецифический рекрутер) включает последовательность секрекон1A (sec1A), последовательность CD3 и последовательность пептида анти-GD2 или эквивалент одного или нескольких из них. В одном воплощении биспецифический рекрутер содержит полипептидную последовательность SEQ ID NO. 13, организованный как sec1A-анти-CD3-линкер-анти-GD2-HDD. Часть sec1A подчеркнута. Анти-CD3 часть выделена жирным шрифтом. Анти-GD2 часть подчеркнута и выделена курсивом. Заштрихованная область серым цветом обозначает часть HDD, которая содержит пептид HDD жирным шрифтом, шарнирную область маленькой буквой перед пептидом HDD и спейсерную область маленькой буквой ниже пептида HDD.
SEQ ID NO. 13

В другом воплощении dimert (например, биспецифический рекрутер), содержащий последовательность sec1A, последовательность анти-CD3 и последовательность пептида анти-GD2, или эквивалент одного или нескольких из них, и в одном аспекте кодируется последовательностью SEQ ID NO. 14.
SEQ ID NO. 14
ATGTGGTGGAGACTGTGGTGGCTGCTGCTGCTGCTGCTGCTGCTGTGGCCCATGGTGTGGGCCGCCCAGGTGCAGCTGCAGCAGAGCGGCCCCGAGCTGGTGAAGCCCGGCGCCAGCGTGAAGATCAGCTGCAAGACCAGCGGCTACAAGTTCACCGAGTACACCATGCACTGGGTGAAGCAGAGCCACGGCAAGTGCCTGGAGTGGATCGGCGGCATCAACCCCAACAACGGCGGCACCAACTACAACCAGAAGTTCAAGGGCAAGGCCACCCTGACCGTGGACAAGAGCAGCAGCACCGCCTACATGGAGCTGAGAAGCCTGACCAGCGAGGACAGCGCCGTGTACTACTGCGCCAGAGACACCACCGTGCCCTACGCCTACTGGGGCCAGGGCACCACCGTGACCGTGAGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGACATCGAGCTGACCCAGAGCCCCGCCATCATGAGCGCCAGCCCCGGCGAGAAGGTGACCATGACCTGCAGCGCCAGCAGCAGCATCAGCTACATGCACTGGTACCAGCAGAAGCCCGGCACCAGCCCCAAGAGATGGATCTACGACACCAGCAAGCTGGCCAGCAGCGTGCCCGCCAGATTCAGCGGCAGCGGCAGCGGCACCAGCTACAGCCTGACCATCAGCAGCATGGAGGCCGAGGACGCCGCCACCTACTACTGCCACCAGAGAAGCAGCTACCCCCTGACCTTCGGCTGCGGCACCAAGCTGGAGATCAAGAGAGCCAGCACCAAGGGCCCCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCCAGGTGCAGCTGGTGCAGAGCGGCGGCGGCGTGGTGCAGCCCGGCAGAAGCCTGAGACTGAGCTGCAAGGCCAGCGGCTACACCTTCACCAGATACACCATGCACTGGGTGAGACAGGCCCCCGGCAAGGGCCTGGAGTGGATCGGCTACATCAACCCCAGCAGAGGCTACACCAACTACAACCAGAAGTTCAAGGACAGATTCACCATCAGCAGAGACAACAGCAAGAACACCGCCTTCCTGCAGATGGACAGCCTGAGACCCGAGGACACCGGCGTGTACTTCTGCGCCAGATACTACGACGACCACTACTGCCTGGACTACTGGGGCCAGGGCACCCCCGTGACCGTGAGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGACATCCAGATGACCCAGAGCCCCAGCAGCCTGAGCGCCAGCGTGGGCGACAGAGTGACCATCACCTGCAGCGCCAGCAGCAGCGTGAGCTACATGAACTGGTACCAGCAGACCCCCGGCAAGGCCCCCAAGAGATGGATCTACGACACCAGCAAGCTGGCCAGCGGCGTGCCCAGCAGATTCAGCGGCAGCGGCAGCGGCACCGACTACACCTTCACCATCAGCAGCCTGCAGCCCGAGGACATCGCCACCTACTACTGCCAGCAGTGGAGCAGCAACCCCTTCACCTTCGGCCAGGGCACCAAGCTGCAGATCACCAGAACCCCCCTGGGCGACACCACCCACACCAGCGGCATGGTGAGCAAGCTGAGCCAGCTGCAGACCGAGCTGCTGGCCGCCCTGCTGGAGAGCGGCCTGAGCAAGGAGGCCCTGATCCAGGCCCTGGGCGAGGGCAGCGGCGGCGCCCCCTAG, или ее эквивалент.
В другом воплощении dimert (например, биспецифический рекрутер) содержит последовательность секрекон1A, анти-CD19 последовательность и анти-CD3 последовательность, организованную как sec1A-анти-CD19-линкер-анти-CD3. Биспецифический рекрутер содержит полипептидную последовательность SEQ ID NO. 15. Часть sec1A подчеркнута.
SEQ ID NO. 15
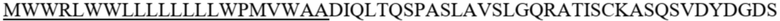
YDGDSYLNWYQQIPGQPPKLLIYDASNLVSGIPPRFSGSGSGTDFTLNIHPVEKVDAATYHCQQSTEDPWTFGGGTKLEIKGGGGSGGGGSGGGGSQVQLQQSGAELVRPGSSVKISCKASGYAFSSYWMNWVKQRPGQGLEWIGQIWPGDGDTNYNGKFKGKATLTADESSSTAYMQLSSLASEDSAVYFCARRETTTVGRYYYAMDYWGQGTTVTVSSGGGGSDIKLQQSGAELARPGASVKMSCKTSGYTFTRYTMHWVKQRPGQGLEWIGYINPSRGYTNYNQKFKDKATLTTDKSSSTAYMQLSSLTSEDSAVYYCARYYDDHYCLDYWGQGTTLTVSSVEGGSGGSGGSGGSGGVDDIQLTQSPAIMSASPGEKVTMTCRASSSVSYMNWYQQKSGTSPKRWIYDTSKVASGVPYRFSGSGSGTSYSLTISSMEAEDAATYYCQQWSSNPLTFGAGTKLELKHHHHHHTPLGDTTHTSGMVSKLSQLQTELLAALLESGLSKEALIQALGEGSGGAP, или ее эквивалент.
В другом воплощении dimert (например, биспецифический рекрутер), содержащий последовательность секрекон1A, последовательность анти-CD19 и последовательность анти-CD3, организованную как sec1A-анти-CD19-линкер-анти-CD3, или эквивалент одного или нескольких из них. В одном аспекте он кодируется полинуклеотидной последовательностью SEQ ID NO. 16.
SEQ ID NO. 16
ATGTGGTGGAGACTGTGGTGGCTGCTGCTGCTGCTGCTGCTGCTGTGGCCCATGGTGTGGGCCGCCGACATCCAGCTGACCCAGAGCCCCGCCAGCCTGGCCGTGAGCCTGGGCCAGAGAGCCACCATCAGCTGCAAGGCCAGCCAGAGCGTGGACTACGACGGCGACAGCTACCTGAACTGGTACCAGCAGATCCCCGGCCAGCCCCCCAAGCTGCTGATCTACGACGCCAGCAACCTGGTGAGCGGCATCCCCCCCAGATTCAGCGGCAGCGGCAGCGGCACCGACTTCACCCTGAACATCCACCCCGTGGAGAAGGTGGACGCCGCCACCTACCACTGCCAGCAGAGCACCGAGGACCCCTGGACCTTCGGCGGCGGCACCAAGCTGGAGATCAAGGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCCAGGTGCAGCTGCAGCAGAGCGGCGCCGAGCTGGTGAGACCCGGCAGCAGCGTGAAGATCAGCTGCAAGGCCAGCGGCTACGCCTTCAGCAGCTACTGGATGAACTGGGTGAAGCAGAGACCCGGCCAGGGCCTGGAGTGGATCGGCCAGATCTGGCCCGGCGACGGCGACACCAACTACAACGGCAAGTTCAAGGGCAAGGCCACCCTGACCGCCGACGAGAGCAGCAGCACCGCCTACATGCAGCTGAGCAGCCTGGCCAGCGAGGACAGCGCCGTGTACTTCTGCGCCAGAAGAGAGACCACCACCGTGGGCAGATACTACTACGCCATGGACTACTGGGGCCAGGGCACCACCGTGACCGTGAGCAGCGGCGGCGGCGGCAGCGACATCAAGCTGCAGCAGAGCGGCGCCGAGCTGGCCAGACCCGGCGCCAGCGTGAAGATGAGCTGCAAGACCAGCGGCTACACCTTCACCAGATACACCATGCACTGGGTGAAGCAGAGACCCGGCCAGGGCCTGGAGTGGATCGGCTACATCAACCCCAGCAGAGGCTACACCAACTACAACCAGAAGTTCAAGGACAAGGCCACCCTGACCACCGACAAGAGCAGCAGCACCGCCTACATGCAGCTGAGCAGCCTGACCAGCGAGGACAGCGCCGTGTACTACTGCGCCAGATACTACGACGACCACTACTGCCTGGACTACTGGGGCCAGGGCACCACCCTGACCGTGAGCAGCGTGGAGGGCGGCAGCGGCGGCAGCGGCGGCAGCGGCGGCAGCGGCGGCGTGGACGACATCCAGCTGACCCAGAGCCCCGCCATCATGAGCGCCAGCCCCGGCGAGAAGGTGACCATGACCTGCAGAGCCAGCAGCAGCGTGAGCTACATGAACTGGTACCAGCAGAAGAGCGGCACCAGCCCCAAGAGATGGATCTACGACACCAGCAAGGTGGCCAGCGGCGTGCCCTACAGATTCAGCGGCAGCGGCAGCGGCACCAGCTACAGCCTGACCATCAGCAGCATGGAGGCCGAGGACGCCGCCACCTACTACTGCCAGCAGTGGAGCAGCAACCCCCTGACCTTCGGCGCCGGCACCAAGCTGGAGCTGAAGCACCACCACCACCACCACACCCCCCTGGGCGACACCACCCACACCAGCGGCATGGTGAGCAAGCTGAGCCAGCTGCAGACCGAGCTGCTGGCCGCCCTGCTGGAGAGCGGCCTGAGCAAGGAGGCCCTGATCCAGGCCCTGGGCGAGGGCAGCGGCGGCGCCCCCTA, или ее эквивалент.
В некоторых воплощениях dimert содержит сигнал секреции, называемый секреконAA (sec2A). В некоторых случаях dimert включает секреконAA, связанный в тандеме с аминокислотной последовательностью из Блинатумомаба (который нацелен на CD3 и CD19). В некоторых случаях секреконAA-Блинатумомаб (или sec2A-CD19xCD3) содержит полипептидную последовательность SEQ ID NO: 29, в которой часть секреконAA подчеркнута.
SEQ ID NO: 29

VDYDGDSYLNWYQQIPGQPPKLLIYDASNLVSGIPPRFSGSGSGTDFTLNIHPVEKVDAATYHCQQSTEDPWTFGGGTKLEIKGGGGSGGGGSGGGGSQVQLQQSGAELVRPGSSVKISCKASGYAFSSYWMNWVKQRPGQGLEWIGQIWPGDGDTNYNGKFKGKATLTADESSSTAYMQLSSLASEDSAVYFCARRETTTVGRYYYAMDYWGQGTTVTVSSGGGGSDIKLQQSGAELARPGASVKMSCKTSGYTFTRYTMHWVKQRPGQGLEWIGYINPSRGYTNYNQKFKDKATLTTDKSSSTAYMQLSSLTSEDSAVYYCARYYDDHYCLDYWGQGTTLTVSSVEGGSGGSGGSGGSGGVDDIQLTQSPAIMSASPGEKVTMTCRASSSVSYMNWYQQKSGTSPKRWIYDTSKVASGVPYRFSGSGSGTSYSLTISSMEAEDAATYYCQQWSSNPLTFGAGTKLELKHHHHHH, или ее эквивалент.
В некоторых воплощениях dimert, содержащий sec2A-CD19xCD3, кодируется полинуклеотидной последовательностью SEQ ID NO: 30.
SEQ ID NO: 30
ATGTGGTGGAGACTGTGGTGGCTGCTGCTGCTGCTGCTGCTGCTGTGGCCCATGGTGTGGGCCGCCGCCGACATCCAGCTGACCCAGAGCCCCGCCAGCCTGGCCGTGAGCCTGGGCCAGAGAGCCACCATCAGCTGCAAGGCCAGCCAGAGCGTGGACTACGACGGCGACAGCTACCTGAACTGGTACCAGCAGATCCCCGGCCAGCCCCCCAAGCTGCTGATCTACGACGCCAGCAACCTGGTGAGCGGCATCCCCCCCAGATTCAGCGGCAGCGGCAGCGGCACCGACTTCACCCTGAACATCCACCCCGTGGAGAAGGTGGACGCCGCCACCTACCACTGCCAGCAGAGCACCGAGGACCCCTGGACCTTCGGCGGCGGCACCAAGCTGGAGATCAAGGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCCAGGTGCAGCTGCAGCAGAGCGGCGCCGAGCTGGTGAGACCCGGCAGCAGCGTGAAGATCAGCTGCAAGGCCAGCGGCTACGCCTTCAGCAGCTACTGGATGAACTGGGTGAAGCAGAGACCCGGCCAGGGCCTGGAGTGGATCGGCCAGATCTGGCCCGGCGACGGCGACACCAACTACAACGGCAAGTTCAAGGGCAAGGCCACCCTGACCGCCGACGAGAGCAGCAGCACCGCCTACATGCAGCTGAGCAGCCTGGCCAGCGAGGACAGCGCCGTGTACTTCTGCGCCAGAAGAGAGACCACCACCGTGGGCAGATACTACTACGCCATGGACTACTGGGGCCAGGGCACCACCGTGACCGTGAGCAGCGGCGGCGGCGGCAGCGACATCAAGCTGCAGCAGAGCGGCGCCGAGCTGGCCAGACCCGGCGCCAGCGTGAAGATGAGCTGCAAGACCAGCGGCTACACCTTCACCAGATACACCATGCACTGGGTGAAGCAGAGACCCGGCCAGGGCCTGGAGTGGATCGGCTACATCAACCCCAGCAGAGGCTACACCAACTACAACCAGAAGTTCAAGGACAAGGCCACCCTGACCACCGACAAGAGCAGCAGCACCGCCTACATGCAGCTGAGCAGCCTGACCAGCGAGGACAGCGCCGTGTACTACTGCGCCAGATACTACGACGACCACTACTGCCTGGACTACTGGGGCCAGGGCACCACCCTGACCGTGAGCAGCGTGGAGGGCGGCAGCGGCGGCAGCGGCGGCAGCGGCGGCAGCGGCGGCGTGGACGACATCCAGCTGACCCAGAGCCCCGCCATCATGAGCGCCAGCCCCGGCGAGAAGGTGACCATGACCTGCAGAGCCAGCAGCAGCGTGAGCTACATGAACTGGTACCAGCAGAAGAGCGGCACCAGCCCCAAGAGATGGATCTACGACACCAGCAAGGTGGCCAGCGGCGTGCCCTACAGATTCAGCGGCAGCGGCAGCGGCACCAGCTACAGCCTGACCATCAGCAGCATGGAGGCCGAGGACGCCGCCACCTACTACTGCCAGCAGTGGAGCAGCAACCCCCTGACCTTCGGCGCCGGCACCAAGCTGGAGCTGAAGCACCACCACCACCACCACTAG, или ее эквивалент.
В некоторых воплощениях dimert содержит последовательность секреконA (sec1A), CD19 и CD3, расположенные в sec1A-CD19xCD3, или эквивалент одного или нескольких из них. В одном аспекте он кодируется полинуклеотидной последовательностью SEQ ID NO: 31.
SEQ ID NO: 31
ATGTGGTGGAGACTGTGGTGGCTGCTGCTGCTGCTGCTGCTGCTGTGGCCCATGGTGTGGGCCGCCGACATCCAGCTGACCCAGAGCCCCGCCAGCCTGGCCGTGAGCCTGGGCCAGAGAGCCACCATCAGCTGCAAGGCCAGCCAGAGCGTGGACTACGACGGCGACAGCTACCTGAACTGGTACCAGCAGATCCCCGGCCAGCCCCCCAAGCTGCTGATCTACGACGCCAGCAACCTGGTGAGCGGCATCCCCCCCAGATTCAGCGGCAGCGGCAGCGGCACCGACTTCACCCTGAACATCCACCCCGTGGAGAAGGTGGACGCCGCCACCTACCACTGCCAGCAGAGCACCGAGGACCCCTGGACCTTCGGCGGCGGCACCAAGCTGGAGATCAAGGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCCAGGTGCAGCTGCAGCAGAGCGGCGCCGAGCTGGTGAGACCCGGCAGCAGCGTGAAGATCAGCTGCAAGGCCAGCGGCTACGCCTTCAGCAGCTACTGGATGAACTGGGTGAAGCAGAGACCCGGCCAGGGCCTGGAGTGGATCGGCCAGATCTGGCCCGGCGACGGCGACACCAACTACAACGGCAAGTTCAAGGGCAAGGCCACCCTGACCGCCGACGAGAGCAGCAGCACCGCCTACATGCAGCTGAGCAGCCTGGCCAGCGAGGACAGCGCCGTGTACTTCTGCGCCAGAAGAGAGACCACCACCGTGGGCAGATACTACTACGCCATGGACTACTGGGGCCAGGGCACCACCGTGACCGTGAGCAGCGGCGGCGGCGGCAGCGACATCAAGCTGCAGCAGAGCGGCGCCGAGCTGGCCAGACCCGGCGCCAGCGTGAAGATGAGCTGCAAGACCAGCGGCTACACCTTCACCAGATACACCATGCACTGGGTGAAGCAGAGACCCGGCCAGGGCCTGGAGTGGATCGGCTACATCAACCCCAGCAGAGGCTACACCAACTACAACCAGAAGTTCAAGGACAAGGCCACCCTGACCACCGACAAGAGCAGCAGCACCGCCTACATGCAGCTGAGCAGCCTGACCAGCGAGGACAGCGCCGTGTACTACTGCGCCAGATACTACGACGACCACTACTGCCTGGACTACTGGGGCCAGGGCACCACCCTGACCGTGAGCAGCGTGGAGGGCGGCAGCGGCGGCAGCGGCGGCAGCGGCGGCAGCGGCGGCGTGGACGACATCCAGCTGACCCAGAGCCCCGCCATCATGAGCGCCAGCCCCGGCGAGAAGGTGACCATGACCTGCAGAGCCAGCAGCAGCGTGAGCTACATGAACTGGTACCAGCAGAAGAGCGGCACCAGCCCCAAGAGATGGATCTACGACACCAGCAAGGTGGCCAGCGGCGTGCCCTACAGATTCAGCGGCAGCGGCAGCGGCACCAGCTACAGCCTGACCATCAGCAGCATGGAGGCCGAGGACGCCGCCACCTACTACTGCCAGCAGTGGAGCAGCAACCCCCTGACCTTCGGCGCCGGCACCAAGCTGGAGCTGAAGCACCACCACCACCACCACTAG, или ее эквивалент.
В некоторых воплощениях dimert, содержащий последовательность secrecon (sec0A), CD19 и CD3, или эквивалент одного или нескольких из них, что является одним аспектом, организованным в виде sec0A-CD19xCD3, кодируется полинуклеотидной последовательностью SEQ ID NO: 32.
SEQ ID NO: 32
ATGTGGTGGAGACTGTGGTGGCTGCTGCTGCTGCTGCTGCTGCTGTGGCCCATGGTGTGGGCCGACATCCAGCTGACCCAGAGCCCCGCCAGCCTGGCCGTGAGCCTGGGCCAGAGAGCCACCATCAGCTGCAAGGCCAGCCAGAGCGTGGACTACGACGGCGACAGCTACCTGAACTGGTACCAGCAGATCCCCGGCCAGCCCCCCAAGCTGCTGATCTACGACGCCAGCAACCTGGTGAGCGGCATCCCCCCCAGATTCAGCGGCAGCGGCAGCGGCACCGACTTCACCCTGAACATCCACCCCGTGGAGAAGGTGGACGCCGCCACCTACCACTGCCAGCAGAGCACCGAGGACCCCTGGACCTTCGGCGGCGGCACCAAGCTGGAGATCAAGGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCCAGGTGCAGCTGCAGCAGAGCGGCGCCGAGCTGGTGAGACCCGGCAGCAGCGTGAAGATCAGCTGCAAGGCCAGCGGCTACGCCTTCAGCAGCTACTGGATGAACTGGGTGAAGCAGAGACCCGGCCAGGGCCTGGAGTGGATCGGCCAGATCTGGCCCGGCGACGGCGACACCAACTACAACGGCAAGTTCAAGGGCAAGGCCACCCTGACCGCCGACGAGAGCAGCAGCACCGCCTACATGCAGCTGAGCAGCCTGGCCAGCGAGGACAGCGCCGTGTACTTCTGCGCCAGAAGAGAGACCACCACCGTGGGCAGATACTACTACGCCATGGACTACTGGGGCCAGGGCACCACCGTGACCGTGAGCAGCGGCGGCGGCGGCAGCGACATCAAGCTGCAGCAGAGCGGCGCCGAGCTGGCCAGACCCGGCGCCAGCGTGAAGATGAGCTGCAAGACCAGCGGCTACACCTTCACCAGATACACCATGCACTGGGTGAAGCAGAGACCCGGCCAGGGCCTGGAGTGGATCGGCTACATCAACCCCAGCAGAGGCTACACCAACTACAACCAGAAGTTCAAGGACAAGGCCACCCTGACCACCGACAAGAGCAGCAGCACCGCCTACATGCAGCTGAGCAGCCTGACCAGCGAGGACAGCGCCGTGTACTACTGCGCCAGATACTACGACGACCACTACTGCCTGGACTACTGGGGCCAGGGCACCACCCTGACCGTGAGCAGCGTGGAGGGCGGCAGCGGCGGCAGCGGCGGCAGCGGCGGCAGCGGCGGCGTGGACGACATCCAGCTGACCCAGAGCCCCGCCATCATGAGCGCCAGCCCCGGCGAGAAGGTGACCATGACCTGCAGAGCCAGCAGCAGCGTGAGCTACATGAACTGGTACCAGCAGAAGAGCGGCACCAGCCCCAAGAGATGGATCTACGACACCAGCAAGGTGGCCAGCGGCGTGCCCTACAGATTCAGCGGCAGCGGCAGCGGCACCAGCTACAGCCTGACCATCAGCAGCATGGAGGCCGAGGACGCCGCCACCTACTACTGCCAGCAGTGGAGCAGCAACCCCCTGACCTTCGGCGCCGGCACCAAGCTGGAGCTGAAGCACCACCACCACCACCACTAG, или ее эквивалент.
В некоторых воплощениях dimert, содержащий последовательность CD19 и CD3 без последовательности секрекона, организованный как CD19xCD3, или эквивалент одного или обоих из них, который в одном аспекте кодируется полинуклеотидной последовательностью SEQ ID NO: 33.
SEQ ID NO: 33
ATGGACATCCAGCTGACCCAGAGCCCCGCCAGCCTGGCCGTGAGCCTGGGCCAGAGAGCCACCATCAGCTGCAAGGCCAGCCAGAGCGTGGACTACGACGGCGACAGCTACCTGAACTGGTACCAGCAGATCCCCGGCCAGCCCCCCAAGCTGCTGATCTACGACGCCAGCAACCTGGTGAGCGGCATCCCCCCCAGATTCAGCGGCAGCGGCAGCGGCACCGACTTCACCCTGAACATCCACCCCGTGGAGAAGGTGGACGCCGCCACCTACCACTGCCAGCAGAGCACCGAGGACCCCTGGACCTTCGGCGGCGGCACCAAGCTGGAGATCAAGGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCCAGGTGCAGCTGCAGCAGAGCGGCGCCGAGCTGGTGAGACCCGGCAGCAGCGTGAAGATCAGCTGCAAGGCCAGCGGCTACGCCTTCAGCAGCTACTGGATGAACTGGGTGAAGCAGAGACCCGGCCAGGGCCTGGAGTGGATCGGCCAGATCTGGCCCGGCGACGGCGACACCAACTACAACGGCAAGTTCAAGGGCAAGGCCACCCTGACCGCCGACGAGAGCAGCAGCACCGCCTACATGCAGCTGAGCAGCCTGGCCAGCGAGGACAGCGCCGTGTACTTCTGCGCCAGAAGAGAGACCACCACCGTGGGCAGATACTACTACGCCATGGACTACTGGGGCCAGGGCACCACCGTGACCGTGAGCAGCGGCGGCGGCGGCAGCGACATCAAGCTGCAGCAGAGCGGCGCCGAGCTGGCCAGACCCGGCGCCAGCGTGAAGATGAGCTGCAAGACCAGCGGCTACACCTTCACCAGATACACCATGCACTGGGTGAAGCAGAGACCCGGCCAGGGCCTGGAGTGGATCGGCTACATCAACCCCAGCAGAGGCTACACCAACTACAACCAGAAGTTCAAGGACAAGGCCACCCTGACCACCGACAAGAGCAGCAGCACCGCCTACATGCAGCTGAGCAGCCTGACCAGCGAGGACAGCGCCGTGTACTACTGCGCCAGATACTACGACGACCACTACTGCCTGGACTACTGGGGCCAGGGCACCACCCTGACCGTGAGCAGCGTGGAGGGCGGCAGCGGCGGCAGCGGCGGCAGCGGCGGCAGCGGCGGCGTGGACGACATCCAGCTGACCCAGAGCCCCGCCATCATGAGCGCCAGCCCCGGCGAGAAGGTGACCATGACCTGCAGAGCCAGCAGCAGCGTGAGCTACATGAACTGGTACCAGCAGAAGAGCGGCACCAGCCCCAAGAGATGGATCTACGACACCAGCAAGGTGGCCAGCGGCGTGCCCTACAGATTCAGCGGCAGCGGCAGCGGCACCAGCTACAGCCTGACCATCAGCAGCATGGAGGCCGAGGACGCCGCCACCTACTACTGCCAGCAGTGGAGCAGCAACCCCCTGACCTTCGGCGCCGGCACCAAGCTGGAGCTGAAGCACCACCACCACCACCACTAG, или ее эквивалент.
В одном аспекте биспецифический рекрутер Т-клеток, содержит полипептидную последовательность, по меньшей мере на 95% идентичную любой из SEQ ID NO. 12 и 13, или, альтернативно, идентичную по меньшей мере на 96%, или альтернативно, по меньшей мере, 97%, или, альтернативно, 98%, или альтернативно, по меньшей мере, 99%. В одном воплощении триспецифическое антитело включает первое антитело или его антигенсвязывающий фрагмент, второе антитело или его антигенсвязывающий фрагмент и третье антитело или его антигенсвязывающий фрагмент.В дополнительном аспекте триспецифическое антитело содержит полипептидную последовательность, имеющую по меньшей мере 95% идентичности последовательностей с SEQ ID NO. 11, или, альтернативно, имеющую по меньшей мере 96%, или альтернативно, по меньшей мере, 97%, или альтернативно 98%, или альтернативно, по меньшей мере, 99% идентичности последовательностей с SEQ ID NO: 11.
В дополнительном аспекте рекомбинантный полинуклеотид экспрессирует пре-мРНК, которая кодирует триспецифическое антитело, когда пре-мРНК находится в контакте с морфолиноолигонуклеотидом.
В одном аспекте антигенсвязывающий домен представляет собой одноцепочечный вариабельный фрагмент антитела.
В дополнительном аспекте рекомбинантный полинуклеотид или вектор дополнительно содержит полинуклеотидную последовательность, кодирующую секреторный пептид. В другом аспекте вектор дополнительно содержит полинуклеотидную последовательность, кодирующую домен димеризации. В другом аспекте вектор содержит 5’-концевой инвертированный повтор (ITR) и 3’-ITR. В другом аспекте вектор содержит последовательность SEQ ID NO. 4, 6, 8, 12, 15, 16, 30-33 или их эквивалент, или полинуклеотид, или альтернативно идентичен по меньшей мере на 95%, или, альтернативно по меньшей мере 96%, или альтернативно по меньшей мере, 97% или альтернативно 98%, или альтернативно по меньшей мере 99%. Неограничивающие примеры вектора включают: рекомбинантный вирусный вектор, который включает основной вектор, выбранный из группы ретровирусного вектора, лентивирусного вектора, вектора вируса лейкоза мышей («MLV»), вектора вируса Эпштейна-Барр («EBV»), аденовирусного вектора, вектора вируса герпеса («HSV»), вектора аденоассоциированного вируса («AAV»), вектора AAV или, необязательно, самокомплементарного вектора AAV. Они необязательно помечены детектируемой меткой. Также предлагаются наборы полинуклеотидов, которые необязательно помечены детектируемой меткой.
Рекомбинантные полинуклеотидные векторы могут содержаться в клетках-хозяевах, например, в прокариотических или эукариотических клетках. Клетки можно использовать для рекомбинантной экспрессии или репликации полинуклеотидов путем культивирования клеток, содержащих полинуклеотиды, в условиях, которые позволяют репликацию полинуклеотидов и, необязательно, экспрессию полинуклеотидов. Полинуклеотиды и/или продукты экспрессии необязательно выделяют из культуры клеток.
Изобретение также предлагает вирусную упаковочную систему, содержащую: вектор, как описано выше, в котором каркас происходит из плазмиды, вируса; упаковочную плазмиду; и плазмиду оболочки. Упаковочная плазмида содержит нуклеозидный, капсидный и матричный белки. Примеры упаковочных плазмид также описаны в патентной литературе, например, Пат.США №7 262 049; 6 995 258; 7 252 991 и 5 710 037. Система также содержит плазмиду, кодирующую белок оболочки, обеспечиваемый плазмидой оболочки.
Это раскрытие также обеспечивает подходящую упаковочную клеточную линию. В одном аспекте линия упаковочных клеток представляет собой линию клеток HEK-293. Другие подходящие клеточные линии известны в данной области, например, они описаны в патентной литературе в Пат.США №7 070 994; 6 995 919; 6 475 786; 6 372 502; 6 365 150; и 5 591 624.
В этом изобретении дополнительно предложен способ получения частицы AAV, включающей, или альтернативно состоящей в основном из, или дополнительно состоящей из трансдукции линии упаковочных клеток вирусной системой, как описано выше, в условиях, подходящих для упаковки вирусного вектора. Такие условия известны в данной области и кратко описаны в данном документе. Вирусная частица может быть выделена из клеточной надосадочной жидкости с использованием способов, известных специалистам в данной области, например центрифугирования. Такие выделенные частицы дополнительно предлагаются данным изобретением.
В данном изобретении также предлагаются выделенные вирусные частицы AAV, полученным этим способом. Вирусная частица содержит полинуклеотид, как описано в данном документе, или, альтернативно, состоит из него, или дополнительно состоит из него.
Клетки-хозяева
Дополнительно предлагается выделенная клетка или популяция клеток, содержащая или альтернативно состоящая в основном из или дополнительно состоящая из выделенных полинуклеотидов, вирусных частиц, векторов и упаковочных систем, как описано выше и включено в настоящий документ и ссылкой. В одном аспекте выделенная клетка представляет собой упаковочную клеточную линию.
Также предлагается выделенная клетка или популяция клеток, включающая или, альтернативно, состоящая в основном из или дополнительно состоящая из полинуклеотидной последовательности, как описано в данном документе.
Выделенные клетки, описанные в настоящем документе, могут быть любой клеткой вида из группы: мыши, крысы, кролики, обезьяны, коровы, овцы, свиньи, собаки, кошки, сельскохозяйственные животные, спортивные животные, животные-компаньоны, лошади и приматы, и, в частности, человеческой клеткой.
Векторы и клетки могут содержаться в композиции, которая содержит вектор и/или клетку-хозяин и носитель, например, фармацевтически приемлемый носитель. Они могут быть составлены для различных способов введения и содержать эффективное количество вектора и/или клетки-хозяина, которое эффективно для пациента, расстройства или заболевания, вектора и способа введения. В одном аспекте способ введения является системным или внутривенным. В другом аспекте введение является местным путем прямой инъекции. В одном из аспектов морфолиноолигонуклеотид контактирует одновременно с вектором или после него. Альтернативно, морфолиноолигонуклеотид контактирует до вектора.
Способы применения
Полинуклеотиды и векторы полезны для лечения множества заболеваний или расстройств. В одном аспекте предлагается способ доставки трансгена. Способ включает введение эффективного количества полинуклеотида или вектора, содержащего трансген, в клетку, ткань или пациента, подлежащего лечению. В одном аспекте эффективное количество антисмыслового олигонуклеотида (например, морфолиноолигонуклеотида) вводят в клетку, ткань или пациента, подлежащего лечению. Предлагаются неограничивающие примеры трансгенов, выбранные в зависимости от цели способа. Клетки или ткань могут быть из млекопитающих, например человека. В одном аспекте антисмысловой олигонуклеотид (например, морфолиноолигонуклеотид) контактирует одновременно с вектором или после него. В качестве альтернативы морфолиноолигонуклеотид предшествует вектору. В другом аспекте вектор вводят в клетку путем трансфекции, инфицирования, трансформации, электропорации, инъекции, микроинъекции или их комбинации.
В настоящем документе также предлагается способ лечения рака у нуждающегося в этом объекта. Способ включает, или, альтернативно, состоит в основном, или, кроме того, состоит из введения объекту эффективного количества рекомбинантного вирусного вектора или клетки, как описано в данном документе. В дополнительном аспекте способ дополнительно включает введение объекту эффективного количества антисмыслового олигонуклеотида (например, морфолиноолигонуклеотида). В одном аспекте объекту вводят эффективное количество противоракового агента. Неограничивающие примеры противораковых агентов включают противораковые пептиды, полипептиды, молекулы нуклеиновых кислот, низкомолекулярные вещества, вирусные частицы или их комбинации. В другом аспекте вектор вводят в клетку путем трансфекции, инфицирования, трансформации, электропорации, инъекции, микроинъекции или их комбинации. Терапия может проводиться как терапия первой линии, второй линии, третьей линии, четвертой линии или пятой линии. Терапия может быть адъювантной или комбинированной с другими видами лечения раковых заболеваний.
В одном из аспектов раскрытых способов вирусная частица представляет собой онколитическую частицу HSV.
Введение
Введение рекомбинантного полинуклеотида и/или вектора (например, AAV), вирусной частицы или композиций по настоящему раскрытию можно осуществлять одной дозой, непрерывно или периодически на протяжении всего курса лечения. Введение может осуществляться любым подходящим способом введения, включая, без ограничения указанным: внутривенный, внутриартериальный, внутримышечный, внутрисердечный, интратекальный, субвентрикулярный, эпидуральный, внутримозговой, интрацеребровентрикулярный, субретинальный, интравитреальный, внутрисуставной, внутриглазный, внутрибрюшинный, внутриматочный, внутрикожный, подкожный, чрескожный, чрезслизистый и ингаляционный. В некоторых случаях способ введения включает парентеральное введение. В одном воплощении рекомбинантный полинуклеотид, или вектор, или композиция вводят внутримышечной инъекцией или внутривенной инъекцией. В другом воплощении рекомбинантный полинуклеотид, или вектор, или композицию вводят системно. В другом воплощении рекомбинантный полинуклеотид, или вектор, или композицию вводят парентерально путем инъекции, инфузии или имплантации.
Способы определения наиболее эффективных способов и дозировки известны специалистам в данной области и будут варьировать в зависимости от композиции, используемой для терапии, цели терапии и объекта, подвергаемого лечению. Можно проводить однократное или многократное введение, при этом уровень дозы и характер дозы выбирает лечащий врач. Следует отметить, что на дозировку может влиять способ введения. Подходящие лекарственные формы и способы введения агентов известны в данной области. Неограничивающие примеры таких подходящих доз могут составлять от 1E+9 векторных геномов до 1E+17 векторных геномов на одно введение.
В некоторых воплощениях способов, описанных в данном документе, количество вирусных частиц (например, AAV), вводимых объекту, варьирует от около 109 до около 1017. В конкретных воплощениях от около 1010 до около 1016, от около 1010 до около 1015, от около 1010 до около 1012, от около 1011 до около 1013, от около 1011 до около 1012, от около 1011 до около 1014, от около 1011 до около 1015, от около 1011 до около 1016, от около 5x1011 до около 5x1012 или от около 1012 до около 1013 вирусных частиц вводят объекту. В некоторых случаях объекту вводят от около 1011 до около 1012 вирусных частиц. В некоторых случаях объекту вводят от около 1013 до около 1015 вирусных частиц. В некоторых случаях объекту вводят от около 109 до около 1012 вирусных частиц. В некоторых случаях объекту вводят от около 109 до около 1011 вирусных частиц. В некоторых случаях количество вводимой вирусной частицы зависит от массы пациента. Специалист в данной области поймет, как регулировать количество доставляемых вирусных частиц так, чтобы общее количество, доставляемое объекту, составляло от около 109 до около 1017, необязательно от около 1010 до около 1016, от около 1010 до около 1015, от около 1010 до около 1012, от около 1011 до около 1013, от около 1011 до около 1012, от около 1011 до около 1014, от около 1011 до около 1015, от около 1011 до около 1016, от около 5× 1011 до около 5× 1012 или от около 1012 до около 1013 вирусных частиц. В некоторых случаях объектом является объект детского возраста (например, объект младше 18 лет). В некоторых случаях педиатру вводят от около 109 до около 1012, от около 1010 до около 1012, от около 1011 до около 1012 или от около 109 до около 1010 вирусных частиц.
В дополнительном аспекте вирусные частицы и композиции по настоящему описанию можно вводить в сочетании с другими видами лечения, например, теми одобренными методами лечения, которые подходят для рака и связанных с ним расстройств или состояний.
Успешное лечение и/или восстановление определяется, когда обнаруживается одно или несколько из следующего: облегчение или улучшение одного или нескольких симптомов заболевания, расстройства или состояния у подвергающегося лечению объекта, уменьшение степени заболевания, расстройства или состояния объекта, стабилизированное (т.е. не ухудшающееся) состояние заболевания, расстройства или состояния, задержка или замедление прогрессирования заболевания, расстройства или состояния, а также улучшение или временное облегчение заболевания, расстройства или состояния. В некоторых воплощениях успех лечения определяется путем обнаружения присутствия исправленного полинуклеотида-мишени в одной или нескольких клетках, тканях или органах, выделенных от объекта. В некоторых воплощениях успех лечения определяется путем обнаружения полипептида, кодируемого исправленным полинуклеотидом-мишенью, в одной или нескольких клетках, тканях или органах, выделенных из объекта.
Наборы
Агенты, векторы или композиции, описанные в данном документе, в некоторых воплощениях могут быть собраны в фармацевтические, диагностические или исследовательские наборы для облегчения их использования в терапевтических, диагностических или исследовательских целях. В некоторых воплощениях наборы по настоящему раскрытию включают один или несколько из следующих: модифицированные белки вирусного капсида, выделенные полинуклеотиды, векторы, клетки-хозяева, рекомбинантные вирусные частицы, рекомбинантные системы экспрессии, модифицированные AAV, модифицированные клетки, выделенные ткани, композиции или фармацевтические композиции, как описано в данном документе.
В некоторых воплощениях набор дополнительно включает инструкции по применению. В частности, такие наборы могут включать один или несколько агентов, описанных в данном документе, вместе с инструкциями, описывающими предполагаемое применение и правильное применение этих агентов. В качестве примера, в одном воплощении набор может включать инструкции по смешиванию одного или нескольких компонентов набора и/или по выделению и смешиванию образца и применению к объекту. В некоторых воплощениях агенты в наборе находятся в фармацевтическом составе и дозировке, подходящей для конкретного применения и для способа введения агентов. Наборы для исследовательских целей могут содержать компоненты в соответствующих концентрациях или количествах для проведения различных экспериментов.
Набор может быть разработан для облегчения применения описанных в данном документе способов и может принимать различные формы. Каждая из композиций набора, где это применимо, может быть предоставлена в жидкой форме (например, в растворе) или в твердой форме (например, в сухом порошке). В некоторых случаях некоторые из композиций могут быть составными или обработанными иным образом (например, до активной формы), например, путем добавления подходящего растворителя или других веществ (например, воды или среды для культивирования клеток), которые могут входит или не входить в набор. В некоторых воплощениях композиции могут быть представлены в растворе для консервации (например, в растворе для криоконсервации). Неограничивающие примеры консервирующих растворов включают ДМСО, параформальдегид и CryoStor® (Stem Cell Technologies, Ванкувер, Канада). В некоторых воплощениях консервирующий раствор содержит некоторое количество ингибиторов металлопротеиназы.
Используемый в данном документе термин «инструкции» может определять компонент инструкции и/или продвижения и обычно включает письменные инструкции на упаковке или связанные с упаковкой заявленных способов, рекомбинантных векторов или композиций. Инструкции также могут включать любые устные или электронные инструкции, предоставленные любым способом, чтобы пользователь мог четко понять, что инструкции должны быть связаны с набором, например, аудиовизуальные (например, видеокассета, DVD и т.д.), интернет и/или общение через интернет и т.д. В некоторых воплощениях письменные инструкции представлены в форме, предписанной правительственным агентством, регулирующим производство, использование или продажу фармацевтических или биологических продуктов, причем эти инструкции также могут отражать одобрение агентства по производству, применению или продаже для введения животным.
В некоторых воплощениях набор содержит один или несколько компонентов, описанных в данном документе, в одном или нескольких контейнерах. Таким образом, в некоторых воплощениях набор может включать в себя контейнер, содержащий агенты, описанные в данном документе. Агенты могут быть в форме жидкости, геля или твердого вещества (порошка). Агенты могут быть приготовлены стерильно, упакованы в шприцы и отправлены охлажденными. В качестве альтернативы он может быть помещен во флакон или другой контейнер для хранения. Во втором контейнере могут быть стерильно приготовлены другие агенты. В качестве альтернативы набор может включать активные агенты, предварительно смешанные и отправленные в шприце, флаконе, пробирке или другом контейнере. Набор может содержать один или несколько или все компоненты, необходимые для введения агентов объекту, такие как шприц, устройства для местного нанесения или трубка для внутривенной иглы и мешок.
Терапия, описанная в данном документе, может быть объединена с соответствующими диагностическими методами для выявления и отбора пациентов для лечения.
Способ получения
Дополнительный способ, предлагаемый данным раскрытием, представляет собой способ продуцирования биспецифического антитела или триспецифического антитела в клетке, включающий контакт клетки, содержащей вектор, как описано в данном документе, с эффективным количеством морфолиноолигонуклеотида. В одном воплощении морфолиноолигонуклеотид содержит полинуклеотидную последовательность, идентичную по меньшей мере на 95% SEQ ID NO. 27 (CAAGATTTACCTCTATTGTTGGATCATATT) или SEQ ID NO. 28 (TAAAACAAGATTTACCTCTATTGTTGGATC) или эквиваленту каждой из них. В одном из аспектов морфолиноолигонуклеотид контактирует одновременно с вектором или после него. В качестве альтернативы морфолиноолигонуклеотид предшествует вектору. Неограничивающий пример биспецифического антитела включает полипептидную последовательность, идентичную по меньшей мере на 95% любой из SEQ ID NO: 13 и 15, или, альтернативно, идентичную по меньшей мере на 96%, или по меньшей мере 97%, или по меньшей мере 98%, или по меньшей мере 99% последовательности любой из SEQ ID NO: 13 и 15. В другом аспекте триспецифическое антитело содержит полипептидную последовательность, по меньшей мере на 95% SEQ ID NO: 11, или, альтернативно, по меньшей мере на 96%, или по меньшей мере 97%, или по меньшей мере на 98%, или по меньшей мере на 99% идентичную последовательности с SEQ ID NO: 11.
В другом аспекте вектор вводят в клетку путем трансфекции, инфицирования, трансформации, электропорации, инъекции, микроинъекции или их комбинации. Неограничивающие примеры клеток включают фибробласт, скелетную клетку, эпителиальную клетку, мышечную клетку, нервную клетку, эндокринную клетку, меланоцит, клетку крови или их комбинацию.
Кроме того, предусмотрены наборы, содержащие один или несколько векторов, клеток или композиций, как описано в данном документе, необязательно с инструктивными документами или материалами.
Указанные воплощения
Есть несколько способов модулировать экспрессию гена с помощью принципов пропуска экзонов.
В одном аспекте необходимы по меньшей мере три экзона, чтобы иметь возможность пропустить экзон в этой стратегии. В трехэкзонной структуре имеется два донорных сайта сплайсинга и два акцепторных сайта сплайсинга, как показано на фиг.1. Первый донорный сайт сплайсирован с первым акцепторным сайтом («а» на фиг.1) или, альтернативно, со вторым акцепторным сайтом («с» на фиг.1). Второй донорный сайт может сплайсироваться только со вторым акцепторным сайтом («b» на фиг.1). Таким образом, существует 5 возможных структур РНК, которые могут возникнуть в результате такой структуры из трех экзонов/двух интронов, в зависимости от того, происходит ли сплайсинг, происходит только одно событие сплайсинга (a, b или c) или происходит два сплайсинга (a+b).
Конструируются специфичные для последовательности олигонуклеотиды («олигонуклеотиды»), которые специфически мешают каждому из событий сплайсинга, обычно в сайте донорных или акцепторных сайтов сплайсинга или в сайте связывания для одного из компонентов сплайсосомы (иногда в середине интрон). Для целей этого воплощения и как показано на фиг.1 каждый олиго классифицируется в зависимости от того, мешает ли он донору или акцептору: 1-донор (1D), 2-акцептор (1A), 2-донор (2D) или 3-акцептор (3D).
Например, из-за альтернативного сплайсинга или вмешательства олигонуклеотидов экзон 2 гена пропускается в раковой клетке, но остается в нормальной клетке. В этом воплощении нормальный ген, содержащий пропущенный экзон, не экспрессируется в раковых клетках, но экспрессирует белок в нормальных клетках. Последовательности интронов, фланкирующих экзон 2 в этой конструкции, можно использовать для повторения того же паттерна сплайсинга в трансгене. Отсюда следует, что конструкция, содержащая трансген в типичной структуре интрон-экзон-интрон, которая сплайсируется как в нормальных, так и в опухолевых клетках, поддерживает относительно эффективный сплайсинг и приводит к высоким уровням полностью сплайсированного транскрипта (экзоны 1+2+3, используя сайты сплайсинга a+b). Кроме того, когда олигонуклеотиды блокируют сплайсинг интрона a или интрона b (или обоих), экзон 2 пропускается, с получением транскрипта, в котором слиты экзон 1 с экзоном 3. Относительное ожидаемое количество «теоретически возможных» транскриптов, показанное на фиг.1, таким образом модифицированы на фиг.2, чтобы показать транскрипты, которых, вероятно, будет много.
В одном воплощении, основанном на вышеупомянутых концепциях сплайсинга, олигонуклеотиды и технология «пропуска экзона» используются для переключения трансгена с транскрипта, содержащего экзон 1, экзон 2 и экзон 6, на транскрипт, содержащий экзон 1 и экзон 3. Таким образом, конструкция или вектор создают функциональный полипептид, который кодируется транскриптом с экзоном 1, слитым с экзоном 3. В одном воплощении стоп-кодон сконструирован так, чтобы он был функционально связан с экзоном 2, так что без вмешательства олигонуклеотида в экзон 2 транскрипция останавливается на стоп-кодоне и до трансляции экзона 3. Преждевременный стоп-кодон приводит к нефункциональному транскрипту, который содержит экзон 1, экзон 2 и экзон 3. В одном воплощении стоп-кодон помещен во все три рамки считывания для обеспечения полной остановки. В этом случае базовый уровень нефункционального транскрипта (экзоны 1+2+3) переключается на функциональный транскрипт (экзон 1+2), как показано на фиг.3, потому что рибосома останавливает трансляцию транскрипта после стоп-кодона, который включает экзон 3.
Конструирование такого трансгена довольно просто, но выбирается структура интрон-экзон-СТОП-интрон, которая при нормальных обстоятельствах соответствующим образом «сплайсируется». Интрон-экзонСТОП-интрон может быть сконструирован с использованием последовательностей, фланкирующих нормально сплайсированный экзон, и клонирован в конкретный сайт трансгена, что приводит к сплайсингу донорных и акцепторных консенсусных последовательностей во фланкирующих основаниях. Поскольку экзон в нормальном гене, из которого была взята последовательность, также будет пропущен, экзон следует выбирать осторожно. Например, интрон-экзон-интрон может происходить из гена сплайсинга ДНК вируса, и в этом случае не ожидается, что это повлияет на нормальные клеточные гены. В качестве альтернативы, если приложение генной терапии предназначено для лечения рака, можно рассмотреть возможность применения границы интрон-экзон-интрон из онкогена, что может привести к дефектному онкогену. Такая стратегия будет использовать нецелевой пропуск клеточного гена как потенциально «бонусный» терапевтический эффект.
Некоторые экзоны в раковых клетках регулируются по-разному по сравнению с нормальными клетками. Например, экзон 2 типа сплайсинга «сплайсирован» в нормальных клетках, но исключен из того же гена в некоторых раковых клетках. В этом случае, если в конструкции используется граница интрон-экзон-интрон экзона 2 типа сплайсинга (см. фиг.4), исходная и пропущенная экспрессия трансгена будет отличаться в нормальных и раковых клетках, как показано на фиг.5. Поскольку сконструированный экзонСТОП обычно отсутствует в раковых клетках, трансген будет активирован в этих клетках, но не в нормальных клетках (фиг.5). Такой экзон можно использовать для селективной экспрессии трансгена в раковых клетках, но не в нормальных клетках, даже в отсутствие какого-либо пропуска экзона. Такой сценарий может быть желательным, например, для гена токсина или пролекарственного фермента. Пропуск экзона 2 с использованием олигонуклеотида активирует трансген в нормальных клетках и может увеличить экспрессию в раковых клетках, если пропуск базового уровня в этих клетках составляет менее 100%.
И наоборот, экзон 3 типа сплайсинга исключен в нормальных клетках, но «сплайсирован» в некоторых раковых клетках. Таким образом, использование регулируемого сплайсинга такого экзона даст результат, противоположный сплайсингу типа 2, как показано на фиг.6.
Таким образом, была раскрыта стратегия достижения следующего в отношении контроля экспрессии трансгена в контексте генной терапии:
Стратегия №1: Активация экспрессии трансгена зависит от введения олигонуклеотида, пропускающего экзон. Эффект наблюдается как в нормальных, так и в раковых клетках.
Стратегия №2: Активация экспрессии трансгена обычно высока в некоторых раковых клетках (только при определенных формах рака) и низка в нормальных клетках в отсутствие какого-либо лекарственного средства или внешнего добавленного агента. Это применение можно использовать для раково-специфичной экспрессии для тех видов раковых заболеваний, которые имеют надлежащим образом регулируемый сплайсинг.Введение олигонуклеотида с пропуском экзона активирует экспрессию в нормальных клетках в дополнение к специфическим раковым клеткам.
Стратегия №3: Активация экспрессии трансгена высокая в нормальных клетках, но низкая в некоторых раковых клетках. Введение олигонуклеотида с пропуском экзона активирует экспрессию в этих раковых клетках.
Хотя принципы контролируемой экспрессии генов могут быть применены к любому применению генной терапии, генная терапия для лечения рака является специфическим применением. В этом воплощении терапевтическое лекарственное средство, которое экспрессируется и секретируется нормальными клетками, может быть активировано при необходимости путем введения соответствующего олигонуклеотида с пропуском экзона.
Для этих целей заявитель сконструировал кассету интрон-экзонСТОП-интрон из экзона 1 гена KRAS в качестве примера конструкции Стратегии №1 для вставки в интересующий трансген для достижения контролируемой экспрессии гена. Олигонуклеотиды, которые вызывают пропуск экзонов, выбирают для терапевтического применения.
В одном аспекте заявитель использует структуру гена KRAS, который является одним из наиболее часто мутировавших генов при раке. KRAS не является мишенью для лекарственных средств, потому что он не является ферментом и не имеет явного связывающего лекарство кармана на своей поверхности. Таким образом, насколько известно заявителю, эта технология пропуска является первым способом нацеливания на KRAS при раке. Для этих целей конструкция заявителя использует первый экзон KRAS в 5’-нетранслируемой области, а стартовый кодон ATG для трансляции белка находится внутри 2-го экзона. (Таким образом, 2-й экзон часто называют экзоном 1, а первый экзон - экзоном 0). Олигонуклеотид, которое вызывает пропуск экзона экзона, содержащего ATG, приводит к транскрипту, который не транслируется нормально, потому что отсутствует стартовый кодон ATG.
Интрон-экзонСТОП-интрон на основе KRAS
Из последовательности Genbank (референсная последовательность NCBI: NG_007524.1, последний доступ 20 февраля 2019 г.) экзоны KRAS расположены в положениях: 4990..5170, 10526..10647, 28509..28687, 30148..30307, 46010..51132, с учетом того, что кДНК соединяется в положениях: 10537..10647, 28509..28687, 30148..30307, 46010..46126.
Первый ATG нормального гена KRAS находится в положении 10537.
Пропускаемый экзон находится между 10526-10647 (122 п.н.).
Конструкция содержит поддельную версию со СТОП-кодонами в каждой рамке считывания.
В одном из воплощений для экзона KRAS 1 заштрихованные и подчеркнутые нуклеотиды (включая ATG из KRAS) заменены стоп-кодонами:

AGGCAAGAGTGCCTTGACGATACAGCTAATTCAGAATCATTTTGTGGACGAATATGATCCAACAATAGAG (SEQ ID NO: 17).
Примеры стоп-кодонов: TAA, TAG и TGA. В одном аспекте все три рамки считывания представляют собой: TAAxTAGxTGA, где x представляет собой любой нуклеотид.
Чтобы сконструировать все 3 стоп-кодона дважды подряд, можно вставить стоп-кодон. Кроме того, можно избежать повторения 11 пар оснований, используя последовательность: TAAxTAGxTGAxTAGxTAAxTGAx (24 п.н.) (SEQ ID No: 1), где x представляет собой любой нуклеотид. Конкретное воплощение представляет собой: TAATTAGCTGAGTAGATAAGTGAT (SEQ ID No: 2). В результате, одно воплощение, содержащее экзон Kras1СТОП (стоп-кодоны подчеркнуты), представляет собой:
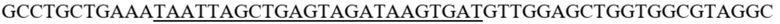
AGGCAAGAGTGCCTTGACGATACAGCTAATTCAGAATCATTTTGTGGACGAATATGATCCAACAATAGAG (SEQ ID NO. 18).
Нормальный передний интрон - это 5171-10525 (5355 п.н.) референсной последовательности NCBI NG_007524.1: «gtacg… ataag», где остальная часть вышележащего интрона обозначена как «…».
Нормальным нижележащим интроном является 10648-28508 (17 861 п.н.) референсной последовательности NCBI NG_007524.1: «gtaaa… ctcag», где остальная часть нижележащего интрона обозначена как «…».
В одном аспекте включено около 75 пар оснований («п.о.») на верхнем и нижнем концах обоих интронов. В одном из воплощений последовательность интрона выше по течению имеет следующий вид:
GTACGGAGCGGACCACCCCTCCTGGGCCCCTGCCCGGGTCCCGACCCTCTTTGCCGGCGCCGGGCGGGGCCGGCGGAGTATTTGATAGTGTATTAACCTTATGTGTGACATGTTCTAATATAGTCACATTTTCATTATTTTTATTATAAG (SEQ ID NO. 19).
В одном из воплощений нижележащая последовательность интрона имеет следующий вид:
GTAAATCTTGTTTTAATATGCATATTACTGGTGCAGGACCATTCTTTGATACAGATAAAGGTTTCTCTGACCATTTTGAGTTGTATATAACACCTTTTTTGAAGTAAAAGGTGCACTGTAATAATCCAGACTGTGTTTCTCCCTTCTCAG (SEQ ID NO. 20).
Таким образом, в одном воплощении, следующая последовательность представляет собой последовательность интрон-экзонKras1СТОП-интрон (последовательность, без выделения находится выше интрона, последовательность серого цвета представляет собой интрон, содержащий стоп-последовательности, а подчеркнутая последовательность представляет собой нижележащий интрон):

В одном аспекте может быть добавлена серия олигонуклеотидов, которые охватывают соединение границы экзона-нижележащего интрона из-за их способности вызывать пропуск экзона. Олигонуклеотиды, пропускающие экзоны, обычно состоят из 20-30 пар оснований, антисмысловых по отношению к ДНК.
В одном воплощении сайт связывания морфолиноолигонуклеотида представляет собой сайт морфолиноолигонуклеотида интрон-экзонСТОП-интрон, полученный из Kras1 (SEQ ID NO. 26). Последовательность 3’-соединения K1ЭкзонСтопИнтрон указана ниже (экзон помечен серым цветом, а интрон подчеркнут):
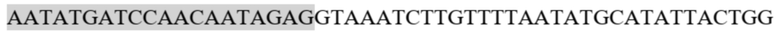
(SEQ ID NO. 26).
Заявитель создал и протестировал два морфолиноолигонуклеотидных сайта связывания, названные KTS1 (SEQ ID NO. 24) и KTS2 (SEQ ID NO. 25). Последовательность морфолиноолигонуклеотида KTS1 включает последовательность SEQ ID NO. 27. Последовательность морфолиноолигонуклеотида KTS2 включает последовательность SEQ ID NO. 28. Заявитель использовал инвертированную морфолиноолигонуклеотидную последовательность KTS2 (SEQ ID NO. 29) в качестве отрицательного контроля.
SEQ ID NO. 24 - KTS1 (KRAS TransSkip 1) (экзон отмечен серым цветом, а интрон подчеркнут)

SEQ ID NO. 29 - морфолиноолигонуклеотидная инвертированная последовательность KTS2.
CTAGGTTGTTATCTCCATTTAGAACAAAAT
В некоторых случаях донорный сайт сплайсинга включает A/C AG ***GT A/G AGT (SEQ ID NO: 34), в котором «***» обозначает границу экзон-интрон, а также сайт встраивания последовательности интрон-экзонСТОП-интрон.
В некоторых случаях акцептор сплайсинга включает (Py)XCAG ***G G/T (SEQ ID NO: 35), в котором «***» обозначает границу интрон-экзон.
В некоторых случаях иллюстративные донорные сайты сплайсинга для вставки последовательности интрон-экзонСТОП-интрон включают AAG-GG (SEQ ID NO: 36), CAG-GG (SEQ ID NO: 37), AAG-GT (SEQ ID NO: 38) или CAG-GT (SEQ ID NO: 39), в котором «-» указывает сайт вставки.
Конструкция CATAAVERT
Для создания вирус-кодируемого регулируемого агента, связывающего T-клетки (CATVERT), нацеленного на рак (также называемого «TransSkip»), последовательность интрон-экзонKras1СТОП-интрон вставляют в сайты кодирующей области, которая генерирует консенсусные донорные и акцепторные сайты сплайсинга. В одном воплощении вектор, содержащий регулируемый CATAAVERT, экспрессирует CD3 и GD2.
В другом воплощении вектор содержит последовательность, кодирующую dimert секрекон-AA-CD3xGD2-HDD. Для этой конструкции предлагается последовательность секрекон-AA-CD3xGD2-HDD dimert включает все потенциальные сайты (выделены серым цветом) для вставки интрон-экзон Kras1 стоп-интрон для создания консенсусного донора/акцептора сплайсинга (вставочная последовательность после 3 из 5 п.н. одного или нескольких из них):

AGCAGAGACAACAGCAAGAACACCGCCTTCCTGCAGATGGACAGCCTGAGACCCGAGGACACCGGCGTGTACTTCTGCGCCAGATACTACGACGACCACTACTGCCTGGACTA
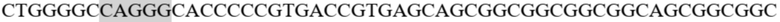
GGCGGCAGCGGCGGCGGCGGCAGCGACATCCAGATGACCCAGAGCCCCAGCAGCCTGAGCGCCAGCGTGGGCGACAGAGTGACCATCACCTGCAGCGCCAGCAGCAGCGTGAGCTACATGAACTGGTACCAGCAGACCCCCGGCAAGGCCCCCAAGAGATGGATCTACGACACCAGCAAGCTGGCCAGCGGCGTGCCCAGCAGATTCAGCGGCAGCGGCAGCGGCACCGACTACACCTTCACCATCAGCAGCCTGCAGCCCGAGGACATCGCCACCTACTACTGCCAGCAGTGGAGCAGCAACCCCTTCACCTTCGGCCAGGGCACCAAGCTGCAGATCACCAGAACCCCCCTGGGCGACACCACCCACACCAGCGGCATGGTGAGCAAGCTGAGCCAGCTGCAGACCGAGCTGCTGGCCGCCCTGCTGGAGAGCGGCCTGAGCAAGGAGGCCCTGATCCAGGCCCTGGGCGAGGGCAGCGGCGGCGCCCCCTAG (SEQ ID NO. 22)
В одном из аспектов вставка производится в самый вышележащий сайт для минимизации длины CD3xGD2 dimert, который транслируется до стоп-кодона (подчеркнутая последовательность представляет собой границу сплайсинга, которая сохраняется в мРНК при сплайсинге интрона, последовательность с двойным подчеркиванием представляет собой донорный сайт сплайсинга в начале последовательности вышележащего интрона, последовательность, выделенная серым цветом, представляет собой экзон, содержащий последовательности СТОП, а последовательность, выделенная жирным шрифтом, представляет собой нижележащий интрон). В некоторых случаях эта последовательность называется CD3xGD2 K1 dimert:

CGCCTACATGGAGCTGAGAAGCCTGACCAGCGAGGACAGCGCCGTGTACTACTGCGCCAGAGACACCACCGTGCCCTACGCCTACTGGGGCCAGGGCACCACCGTGACCGTGAGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGACATCGAGCTGACCCAGAGCCCCGCCATCATGAGCGCCAGCCCCGGCGAGAAGGTGACCATGACCTGCAGCGCCAGCAGCAGCATCAGCTACATGCACTGGTACCAGCAGAAGCCCGGCACCAGCCCCAAGAGATGGATCTACGACACCAGCAAGCTGGCCAGCAGCGTGCCCGCCAGATTCAGCGGCAGCGGCAGCGGCACCAGCTACAGCCTGACCATCAGCAGCATGGAGGCCGAGGACGCCGCCACCTACTACTGCCACCAGAGAAGCAGCTACCCCCTGACCTTCGGCTGCGGCACCAAGCTGGAGATCAAGAGAGCCAGCACCAAGGGCCCCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCCAGGTGCAGCTGGTGCAGAGCGGCGGCGGCGTGGTGCAGCCCGGCAGAAGCCTGAGACTGAGCTGCAAGGCCAGCGGCTACACCTTCACCAGATACACCATGCACTGGGTGAGACAGGCCCCCGGCAAGGGCCTGGAGTGGATCGGCTACATCAACCCCAGCAGAGGCTACACCAACTACAACCAGAAGTTCAAGGACAGATTCACCATCAGCAGAGACAACAGCAAGAACACCGCCTTCCTGCAGATGGACAGCCTGAGACCCGAGGACACCGGCGTGTACTTCTGCGCCAGATACTACGACGACCACTACTGCCTGGACTACTGGGGCCAGGGCACCCCCGTGACCGTGAGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGACATCCAGATGACCCAGAGCCCCAGCAGCCTGAGCGCCAGCGTGGGCGACAGAGTGACCATCACCTGCAGCGCCAGCAGCAGCGTGAGCTACATGAACTGGTACCAGCAGACCCCCGGCAAGGCCCCCAAGAGATGGATCTACGACACCAGCAAGCTGGCCAGCGGCGTGCCCAGCAGATTCAGCGGCAGCGGCAGCGGCACCGACTACACCTTCACCATCAGCAGCCTGCAGCCCGAGGACATCGCCACCTACTACTGCCAGCAGTGGAGCAGCAACCCCTTCACCTTCGGCCAGGGCACCAAGCTGCAGATCACCAGAACCCCCCTGGGCGACACCACCCACACCAGCGGCATGGTGAGCAAGCTGAGCCAGCTGCAGACCGAGCTGCTGGCCGCCCTGCTGGAGAGCGGCCTGAGCAAGGAGGCCCTGATCCAGGCCCTGGGCGAGGGCAGCGGCGGCGCCCCCTAG (SEQ ID NO. 23).
В некоторых случаях модифицируется вышележащий сайт связывания фактора сплайсинга (измененная область выделена курсивом и жирным шрифтом). Подчеркнутая последовательность представляет собой границу сплайсинга, которая сохраняется в мРНК при сплайсинге интрона, двойная подчеркнутая последовательность представляет собой донорный сайт сплайсинга в начале последовательности вышележащего интрона, последовательность, выделенная серым цветом, представляет собой экзон, содержащий последовательности СТОП, а последовательность, выделенная жирным шрифтом, представляет собой нижележащий интрон. В некоторых случаях эту последовательность называют CD3xGD2 K2 dimert:
ATGTGGTGGAGACTGTGGTGGCTGCTGCTGCTGCTGCTGCTGCTGTGGCCCATGGTGTGGGCCGCCGCCCAGGTGCAGCTGCAGCAGAGCGGCCCCGAGCTGGTGAAGCCCGGCGCCAGCGTGAAGATCAGCTGCAAGACCAGCGGCTACAAGTTCACCGAGTACACCATGCACTGGGTGAAGCAGAGCCACGGCAAGTGCCTGGAGTGGATCGGCGGCAT

CGCCTACATGGAGCTGAGAAGCCTGACCAGCGAGGACAGCGCCGTGTACTACTGCGCCAGAGACACCACCGTGCCCTACGCCTACTGGGGCCAGGGCACCACCGTGACCGTGAGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGACATCGAGCTGACCCAGAGCCCCGCCATCATGAGCGCCAGCCCCGGCGAGAAGGTGACCATGACCTGCAGCGCCAGCAGCAGCATCAGCTACATGCACTGGTACCAGCAGAAGCCCGGCACCAGCCCCAAGAGATGGATCTACGACACCAGCAAGCTGGCCAGCAGCGTGCCCGCCAGATTCAGCGGCAGCGGCAGCGGCACCAGCTACAGCCTGACCATCAGCAGCATGGAGGCCGAGGACGCCGCCACCTACTACTGCCACCAGAGAAGCAGCTACCCCCTGACCTTCGGCTGCGGCACCAAGCTGGAGATCAAGAGAGCCAGCACCAAGGGCCCCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCCAGGTGCAGCTGGTGCAGAGCGGCGGCGGCGTGGTGCAGCCCGGCAGAAGCCTGAGACTGAGCTGCAAGGCCAGCGGCTACACCTTCACCAGATACACCATGCACTGGGTGAGACAGGCCCCCGGCAAGGGCCTGGAGTGGATCGGCTACATCAACCCCAGCAGAGGCTACACCAACTACAACCAGAAGTTCAAGGACAGATTCACCATCAGCAGAGACAACAGCAAGAACACCGCCTTCCTGCAGATGGACAGCCTGAGACCCGAGGACACCGGCGTGTACTTCTGCGCCAGATACTACGACGACCACTACTGCCTGGACTACTGGGGCCAGGGCACCCCCGTGACCGTGAGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGACATCCAGATGACCCAGAGCCCCAGCAGCCTGAGCGCCAGCGTGGGCGACAGAGTGACCATCACCTGCAGCGCCAGCAGCAGCGTGAGCTACATGAACTGGTACCAGCAGACCCCCGGCAAGGCCCCCAAGAGATGGATCTACGACACCAGCAAGCTGGCCAGCGGCGTGCCCAGCAGATTCAGCGGCAGCGGCAGCGGCACCGACTACACCTTCACCATCAGCAGCCTGCAGCCCGAGGACATCGCCACCTACTACTGCCAGCAGTGGAGCAGCAACCCCTTCACCTTCGGCCAGGGCACCAAGCTGCAGATCACCAGAACCCCCCTGGGCGACACCACCCACACCAGCGGCATGGTGAGCAAGCTGAGCCAGCTGCAGACCGAGCTGCTGGCCGCCCTGCTGGAGAGCGGCCTGAGCAAGGAGGCCCTGATCCAGGCCCTGGGCGAGGGCAGCGGCGGCGCCCCCTAG (SEQ ID NO: 40).
В некоторых случаях конструкция CD3xGD2 dimert содержит одну или несколько дополнительных модификаций полинуклеотидной последовательности. В некоторых случаях конструкция CD3xGD2 dimert представляет собой sec2A-CD3xGD2-HDD-K3, а последовательность sec2A-CD3xGD2-HDD-K3 представлена как SEQ ID NO: 41. Подчеркнутая последовательность представляет собой границу сплайсинга, которая сохраняется в мРНК при сплайсинге интрона, двойная подчеркнутая последовательность представляет собой донорный сайт сплайсинга в начале последовательности вышележащего интрона, последовательность, выделенная серым цветом, представляет собой экзон, содержащий последовательности СТОП, а последовательность, выделенная жирным шрифтом, представляет собой нижележащий интрон.
SEQ ID NO: 41
ATGTGGTGGAGACTGTGGTGGCTGCTGCTGCTGCTGCTGCTGCTGTGGCCCATGGTGTGGGCCGCCGCCCAGGTGCAGCTGCAGCAGAGCGGCCCCGAGCTGGTGAAGCCCGGCGCCAGCGTGAAGATCAGCTGCAAGACCAGCGGCTACAAGTTCACCGAGTACACCATGCACTGGGTGAAGCAGAGCCACGGCAAGTGCCTGGAGTGGATCGGCGGCAT

CGCCTACATGGAGCTGAGAAGCCTGACCAGCGAGGACAGCGCCGTGTACTACTGCGCCAGAGACACCACCGTGCCCTACGCCTACTGGGGCCAGGGCACCACCGTGACCGTGAGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGACATCGAGCTGACCCAGAGCCCCGCCATCATGAGCGCCAGCCCCGGCGAGAAGGTGACCATGACCTGCAGCGCCAGCAGCAGCATCAGCTACATGCACTGGTACCAGCAGAAGCCCGGCACCAGCCCCAAGAGATGGATCTACGACACCAGCAAGCTGGCCAGCAGCGTGCCCGCCAGATTCAGCGGCAGCGGCAGCGGCACCAGCTACAGCCTGACCATCAGCAGCATGGAGGCCGAGGACGCCGCCACCTACTACTGCCACCAGAGAAGCAGCTACCCCCTGACCTTCGGCTGCGGCACCAAGCTGGAGATCAAGAGAGCCAGCACCAAGGGCCCCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCCAGGTGCAGCTGGTGCAGAGCGGCGGCGGCGTGGTGCAGCCCGGCAGAAGCCTGAGACTGAGCTGCAAGGCCAGCGGCTACACCTTCACCAGATACACCATGCACTGGGTGAGACAGGCCCCCGGCAAGGGCCTGGAGTGGATCGGCTACATCAACCCCAGCAGAGGCTACACCAACTACAACCAGAAGTTCAAGGACAGATTCACCATCAGCAGAGACAACAGCAAGAACACCGCCTTCCTGCAGATGGACAGCCTGAGACCCGAGGACACCGGCGTGTACTTCTGCGCCAGATACTACGACGACCACTACTGCCTGGACTACTGGGGCCAGGGCACCCCCGTGACCGTGAGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGACATCCAGATGACCCAGAGCCCCAGCAGCCTGAGCGCCAGCGTGGGCGACAGAGTGACCATCACCTGCAGCGCCAGCAGCAGCGTGAGCTACATGAACTGGTACCAGCAGACCCCCGGCAAGGCCCCCAAGAGATGGATCTACGACACCAGCAAGCTGGCCAGCGGCGTGCCCAGCAGATTCAGCGGCAGCGGCAGCGGCACCGACTACACCTTCACCATCAGCAGCCTGCAGCCCGAGGACATCGCCACCTACTACTGCCAGCAGTGGAGCAGCAACCCCTTCACCTTCGGCCAGGGCACCAAGCTGCAGATCACCAGAACCCCCCTGGGCGACACCACCCACACCAGCGGCATGGTGAGCAAGCTGAGCCAGCTGCAGACCGAGCTGCTGGCCGCCCTGCTGGAGAGCGGCCTGAGCAAGGAGGCCCTGATCCAGGCCCTGGGCGAGGGCAGCGGCGGCGCCCCCTAG
В некоторых случаях димерная конструкция CD3xGD2 представляет собой sec2A-CD3xGD2-HDD-K4, а последовательность sec2A-CD3xGD2-HDD-K4 представлена как SEQ ID NO: 42. Подчеркнутая последовательность представляет собой границу сплайсинга, которая сохраняется в мРНК при сплайсинге интрона, двойная подчеркнутая последовательность представляет собой донорный сайт сплайсинга в начале последовательности вышележащего интрона, последовательность, выделенная серым цветом, представляет собой экзон, содержащий последовательности СТОП, а последовательность, выделенная жирным шрифтом, представляет собой нижележащий интрон.
SEQ ID NO: 42
ATGTGGTGGAGACTGTGGTGGCTGCTGCTGCTGCTGCTGCTGCTGTGGCCCATGGTGTGGGCCGCCGCCCAGGTGCAGCTGCAGCAGAGCGGCCCCGAGCTGGTGAAGCCCGGCGCCAGCGTGAAGATCAGCTGCAAGACCAGCGGCTACAAGTTCACCGAGTACACCATGCACTGGGTGAAGCAGAGCCACGGCAAGTGCCTGGAGTGGATCGGCGGCATCAACCCCAACAACGGCGGCACCAACTACAACCAGAAGTTC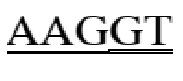 ACGGAGCGGACCACCCCTCCTGGGCCCCTGCCCGGGTCCCGACCCTCTTTGCCGGCGCCGGGCGGGGC
ACGGAGCGGACCACCCCTCCTGGGCCCCTGCCCGGGTCCCGACCCTCTTTGCCGGCGCCGGGCGGGGC

CCTACATGGAGCTGAGAAGCCTGACCAGCGAGGACAGCGCCGTGTACTACTGCGCCAGAGACACCACCGTGCCCTACGCCTACTGGGGCCAGGGCACCACCGTGACCGTGAGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGACATCGAGCTGACCCAGAGCCCCGCCATCATGAGCGCCAGCCCCGGCGAGAAGGTGACCATGACCTGCAGCGCCAGCAGCAGCATCAGCTACATGCACTGGTACCAGCAGAAGCCCGGCACCAGCCCCAAGAGATGGATCTACGACACCAGCAAGCTGGCCAGCAGCGTGCCCGCCAGATTCAGCGGCAGCGGCAGCGGCACCAGCTACAGCCTGACCATCAGCAGCATGGAGGCCGAGGACGCCGCCACCTACTACTGCCACCAGAGAAGCAGCTACCCCCTGACCTTCGGCTGCGGCACCAAGCTGGAGATCAAGAGAGCCAGCACCAAGGGCCCCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCCAGGTGCAGCTGGTGCAGAGCGGCGGCGGCGTGGTGCAGCCCGGCAGAAGCCTGAGACTGAGCTGCAAGGCCAGCGGCTACACCTTCACCAGATACACCATGCACTGGGTGAGACAGGCCCCCGGCAAGGGCCTGGAGTGGATCGGCTACATCAACCCCAGCAGAGGCTACACCAACTACAACCAGAAGTTCAAGGACAGATTCACCATCAGCAGAGACAACAGCAAGAACACCGCCTTCCTGCAGATGGACAGCCTGAGACCCGAGGACACCGGCGTGTACTTCTGCGCCAGATACTACGACGACCACTACTGCCTGGACTACTGGGGCCAGGGCACCCCCGTGACCGTGAGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGACATCCAGATGACCCAGAGCCCCAGCAGCCTGAGCGCCAGCGTGGGCGACAGAGTGACCATCACCTGCAGCGCCAGCAGCAGCGTGAGCTACATGAACTGGTACCAGCAGACCCCCGGCAAGGCCCCCAAGAGATGGATCTACGACACCAGCAAGCTGGCCAGCGGCGTGCCCAGCAGATTCAGCGGCAGCGGCAGCGGCACCGACTACACCTTCACCATCAGCAGCCTGCAGCCCGAGGACATCGCCACCTACTACTGCCAGCAGTGGAGCAGCAACCCCTTCACCTTCGGCCAGGGCACCAAGCTGCAGATCACCAGAACCCCCCTGGGCGACACCACCCACACCAGCGGCATGGTGAGCAAGCTGAGCCAGCTGCAGACCGAGCTGCTGGCCGCCCTGCTGGAGAGCGGCCTGAGCAAGGAGGCCCTGATCCAGGCCCTGGGCGAGGGCAGCGGCGGCGCCCCCTAG
В некоторых случаях конструкция CD3xGD2 dimert представляет собой sec2A-CD3xGD2-HDD-K5, а последовательность sec2A-CD3xGD2-HDD-K5 представлена как SEQ ID NO: 43. Подчеркнутая последовательность представляет собой границу сплайсинга, которая сохраняется в мРНК при сплайсинге интрона, двойная подчеркнутая последовательность представляет собой донорный сайт сплайсинга в начале последовательности вышележащего интрона, последовательность, выделенная серым цветом, представляет собой экзон, содержащий последовательности СТОП, а последовательность, выделенная жирным шрифтом, представляет собой нижележащий интрон.
SEQ ID NO: 43
ATGTGGTGGAGACTGTGGTGGCTGCTGCTGCTGCTGCTGCTGCTGTGGCCCATGGTGTGGGCCGCCGCCCAGGTGCAGCTGCAGCAGAGCGGCCCCGAGCTGGTGAAGCCCGGCGCCAGCGTGAAGATCAGCTGCAAGACCAGCGGCTACAAGTTCACCGAGTACACCATGCACTGGGTGAAGCAGAGCCACGGCAAGTGCCTGGAGTGGATCGGCGGCATCAACCCCAACAACGGCGGCACCAACTACAACCAGAAGTTC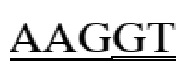 ACGGAGCGGACCACCCCTCCTGGGCCCCTGCCCGGGTCCCGACCCTCTTTGCCGGCGCCGGGCGGGGCCGGCGGAGTATTTGATAGTGTATTAACCTTATGTGTGACATGTTCTAATATAGTCAC
ACGGAGCGGACCACCCCTCCTGGGCCCCTGCCCGGGTCCCGACCCTCTTTGCCGGCGCCGGGCGGGGCCGGCGGAGTATTTGATAGTGTATTAACCTTATGTGTGACATGTTCTAATATAGTCAC

CGCCTACATGGAGCTGAGAAGCCTGACCAGCGAGGACAGCGCCGTGTACTACTGCGCCAGAGACACCACCGTGCCCTACGCCTACTGGGGCCAGGGCACCACCGTGACCGTGAGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGACATCGAGCTGACCCAGAGCCCCGCCATCATGAGCGCCAGCCCCGGCGAGAAGGTGACCATGACCTGCAGCGCCAGCAGCAGCATCAGCTACATGCACTGGTACCAGCAGAAGCCCGGCACCAGCCCCAAGAGATGGATCTACGACACCAGCAAGCTGGCCAGCAGCGTGCCCGCCAGATTCAGCGGCAGCGGCAGCGGCACCAGCTACAGCCTGACCATCAGCAGCATGGAGGCCGAGGACGCCGCCACCTACTACTGCCACCAGAGAAGCAGCTACCCCCTGACCTTCGGCTGCGGCACCAAGCTGGAGATCAAGAGAGCCAGCACCAAGGGCCCCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCCAGGTGCAGCTGGTGCAGAGCGGCGGCGGCGTGGTGCAGCCCGGCAGAAGCCTGAGACTGAGCTGCAAGGCCAGCGGCTACACCTTCACCAGATACACCATGCACTGGGTGAGACAGGCCCCCGGCAAGGGCCTGGAGTGGATCGGCTACATCAACCCCAGCAGAGGCTACACCAACTACAACCAGAAGTTCAAGGACAGATTCACCATCAGCAGAGACAACAGCAAGAACACCGCCTTCCTGCAGATGGACAGCCTGAGACCCGAGGACACCGGCGTGTACTTCTGCGCCAGATACTACGACGACCACTACTGCCTGGACTACTGGGGCCAGGGCACCCCCGTGACCGTGAGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGACATCCAGATGACCCAGAGCCCCAGCAGCCTGAGCGCCAGCGTGGGCGACAGAGTGACCATCACCTGCAGCGCCAGCAGCAGCGTGAGCTACATGAACTGGTACCAGCAGACCCCCGGCAAGGCCCCCAAGAGATGGATCTACGACACCAGCAAGCTGGCCAGCGGCGTGCCCAGCAGATTCAGCGGCAGCGGCAGCGGCACCGACTACACCTTCACCATCAGCAGCCTGCAGCCCGAGGACATCGCCACCTACTACTGCCAGCAGTGGAGCAGCAACCCCTTCACCTTCGGCCAGGGCACCAAGCTGCAGATCACCAGAACCCCCCTGGGCGACACCACCCACACCAGCGGCATGGTGAGCAAGCTGAGCCAGCTGCAGACCGAGCTGCTGGCCGCCCTGCTGGAGAGCGGCCTGAGCAAGGAGGCCCTGATCCAGGCCCTGGGCGAGGGCAGCGGCGGCGCCCCCTAG
В некоторых воплощениях dimert, описанный в данном документе, содержит последовательность CD3, последовательность CD19 и, необязательно, последовательность секрекона. В некоторых случаях dimert включает конструкцию CD3xCD19, изложенную в SEQ ID NO: 44, с выделением всех потенциальных сайтов для встраивания последовательности интрон-экзон-Kras1 стоп-интрон для создания консенсусного донорского/акцепторного сайта сплайсинга. Каждый потенциальный сайт для вставки выделен серым цветом.
SEQ ID NO: 44
ATGTGGTGGAGACTGTGGTGGCTGCTGCTGCTGCTGCTGCTGCTGTGGCCCATGGTGTGGGCCGCCGACATCCAGCTGACCCAGAGCCCCGCCAGCCTGGCCGTGAGCCTGGGCCAGAGAGCCACCATCAGCTGCAAGGCCAGCCAGAGCGTGGACTACGACGGCGACAGCTACCTGAACTGGTACCAGCAGATCCCCGGCCAGCCCCCCAAGCTGCTGATCTACGACGCCAGCAACCTGGTGAGCGGCATCCCCCCCAGATTCAGCGGCAGCGGCA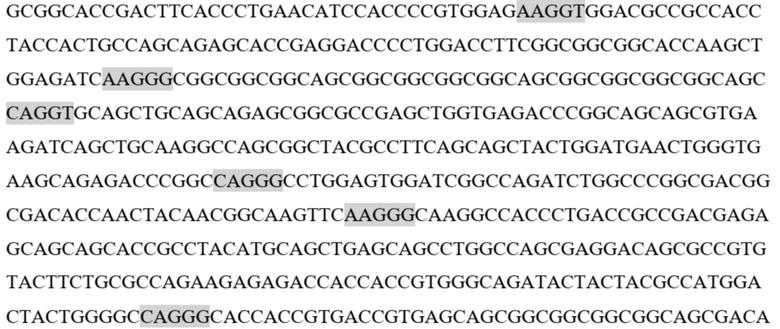

В некоторых случаях вставка выполняется в самый вышележащий сайт.В некоторых случаях dimert дополнительно содержит последовательность секрекона. В некоторых случаях конструкция CD3xCD19 представляет собой sec1A-CD3xCD19-K1, и ее последовательность представлена как SEQ ID NO: 45. Как показано ниже, подчеркнутая последовательность представляет собой границу сплайсинга, которое сохраняется в мРНК при сплайсинге интрона, выделенная курсивом и жирным шрифтом последовательность представляет собой последовательность вышележащего интрона, последовательность, выделенная серым цветом, представляет собой экзон, содержащий последовательности СТОП, а последовательность, выделенная жирным шрифтом, является последовательностью нижележащего интрона.
SEQ ID NO: 45
ATGTGGTGGAGACTGTGGTGGCTGCTGCTGCTGCTGCTGCTGCTGTGGCCCATGGTGTGGGCCGCCGACATCCAGCTGACCCAGAGCCCCGCCAGCCTGGCCGTGAGCCTGGGCCAGAGAGCCACCATCAGCTGCAAGGCCAGCCAGAGCGTGGACTACGACGGCGACAGCTACCTGAACTGGTACCAGCAGATCCCCGGCCAGCCCCCCAAGCTGCTGATCTACGACGCCAGCAACCTGGTGAGCGGCATCCCCCCCAGATTCAGCGGCAGCGGCA


CCTGGACCTTCGGCGGCGGCACCAAGCTGGAGATCAAGGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCCAGGTGCAGCTGCAGCAGAGCGGCGCCGAGCTGGTGAGACCCGGCAGCAGCGTGAAGATCAGCTGCAAGGCCAGCGGCTACGCCTTCAGCAGCTACTGGATGAACTGGGTGAAGCAGAGACCCGGCCAGGGCCTGGAGTGGATCGGCCAGATCTGGCCCGGCGACGGCGACACCAACTACAACGGCAAGTTCAAGGGCAAGGCCACCCTGACCGCCGACGAGAGCAGCAGCACCGCCTACATGCAGCTGAGCAGCCTGGCCAGCGAGGACAGCGCCGTGTACTTCTGCGCCAGAAGAGAGACCACCACCGTGGGCAGATACTACTACGCCATGGACTACTGGGGCCAGGGCACCACCGTGACCGTGAGCAGCGGCGGCGGCGGCAGCGACATCAAGCTGCAGCAGAGCGGCGCCGAGCTGGCCAGACCCGGCGCCAGCGTGAAGATGAGCTGCAAGACCAGCGGCTACACCTTCACCAGATACACCATGCACTGGGTGAAGCAGAGACCCGGCCAGGGCCTGGAGTGGATCGGCTACATCAACCCCAGCAGAGGCTACACCAACTACAACCAGAAGTTCAAGGACAAGGCCACCCTGACCACCGACAAGAGCAGCAGCACCGCCTACATGCAGCTGAGCAGCCTGACCAGCGAGGACAGCGCCGTGTACTACTGCGCCAGATACTACGACGACCACTACTGCCTGGACTACTGGGGCCAGGGCACCACCCTGACCGTGAGCAGCGTGGAGGGCGGCAGCGGCGGCAGCGGCGGCAGCGGCGGCAGCGGCGGCGTGGACGACATCCAGCTGACCCAGAGCCCCGCCATCATGAGCGCCAGCCCCGGCGAGAAGGTGACCATGACCTGCAGAGCCAGCAGCAGCGTGAGCTACATGAACTGGTACCAGCAGAAGAGCGGCACCAGCCCCAAGAGATGGATCTACGACACCAGCAAGGTGGCCAGCGGCGTGCCCTACAGATTCAGCGGCAGCGGCAGCGGCACCAGCTACAGCCTGACCATCAGCAGCATGGAGGCCGAGGACGCCGCCACCTACTACTGCCAGCAGTGGAGCAGCAACCCCCTGACCTTCGGCGCCGGCACCAAGCTGGAGCTGAAGCACCACCACCACCACCACTAG
В некоторых случаях конструкция CD3xCD19 представляет собой sec1A-CD3xCD19-K3, и ее последовательность представлена как SEQ ID NO: 46. Как показано ниже, подчеркнутая последовательность представляет собой границу сплайсинга, которая сохраняется в мРНК при сплайсинге интрона, выделенная курсивом и жирным шрифтом область представляет собой модифицированную область вышележащего сайта связывания фактора сплайсинга, последовательность, выделенная серым цветом, представляет собой экзон, содержащий последовательности СТОП, и последовательность, выделенная жирным шрифтом, представляет собой нижележащий интрон.
SEQ ID NO: 46
ATGTGGTGGAGACTGTGGTGGCTGCTGCTGCTGCTGCTGCTGCTGTGGCCCATGGTGTGGGCCGCCGACATCCAGCTGACCCAGAGCCCCGCCAGCCTGGCCGTGAGCCTGGGCCAGAGAGCCACCATCAGCTGCAAGGCCAGCCAGAGCGTGGACTACGACGGCGACAGCTACCTGAACTGGTACCAGCAGATCCCCGGCCAGCCCCCCAAGCTGCTGATCTACGACGCCAGCAACCTGGTGAGCGGCATCCCCCCCAGATTCAGCGGCAGCGGCA

CCCTGGACCTTCGGCGGCGGCACCAAGCTGGAGATCAAGGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCGGCGGCGGCGGCAGCCAGGTGCAGCTGCAGCAGAGCGGCGCCGAGCTGGTGAGACCCGGCAGCAGCGTGAAGATCAGCTGCAAGGCCAGCGGCTACGCCTTCAGCAGCTACTGGATGAACTGGGTGAAGCAGAGACCCGGCCAGGGCCTGGAGTGGATCGGCCAGATCTGGCCCGGCGACGGCGACACCAACTACAACGGCAAGTTCAAGGGCAAGGCCACCCTGACCGCCGACGAGAGCAGCAGCACCGCCTACATGCAGCTGAGCAGCCTGGCCAGCGAGGACAGCGCCGTGTACTTCTGCGCCAGAAGAGAGACCACCACCGTGGGCAGATACTACTACGCCATGGACTACTGGGGCCAGGGCACCACCGTGACCGTGAGCAGCGGCGGCGGCGGCAGCGACATCAAGCTGCAGCAGAGCGGCGCCGAGCTGGCCAGACCCGGCGCCAGCGTGAAGATGAGCTGCAAGACCAGCGGCTACACCTTCACCAGATACACCATGCACTGGGTGAAGCAGAGACCCGGCCAGGGCCTGGAGTGGATCGGCTACATCAACCCCAGCAGAGGCTACACCAACTACAACCAGAAGTTCAAGGACAAGGCCACCCTGACCACCGACAAGAGCAGCAGCACCGCCTACATGCAGCTGAGCAGCCTGACCAGCGAGGACAGCGCCGTGTACTACTGCGCCAGATACTACGACGACCACTACTGCCTGGACTACTGGGGCCAGGGCACCACCCTGACCGTGAGCAGCGTGGAGGGCGGCAGCGGCGGCAGCGGCGGCAGCGGCGGCAGCGGCGGCGTGGACGACATCCAGCTGACCCAGAGCCCCGCCATCATGAGCGCCAGCCCCGGCGAGAAGGTGACCATGACCTGCAGAGCCAGCAGCAGCGTGAGCTACATGAACTGGTACCAGCAGAAGAGCGGCACCAGCCCCAAGAGATGGATCTACGACACCAGCAAGGTGGCCAGCGGCGTGCCCTACAGATTCAGCGGCAGCGGCAGCGGCACCAGCTACAGCCTGACCATCAGCAGCATGGAGGCCGAGGACGCCGCCACCTACTACTGCCAGCAGTGGAGCAGCAACCCCCTGACCTTCGGCGCCGGCACCAAGCTGGAGCTGAAGCACCACCACCACCACCACTAG
Примеры
Эти примеры предоставлены только для иллюстративных целей, а не для ограничения объема формулы изобретения, представленной в данном документе.
Пример 1. Создание CD3xGD2-HDD Dimert с использованием примерных CD3xGD2-HDD TransJoin
Карты типичных конструкций AAV для тестирования экспрессии CD3xGD2-HDD.
В качестве доказательства принципа была экспрессирована ранее описанная биспецифическая молекула, которая нацелена на CD3 человека на Т-клетках и дизиалоганглиозид GD2 на нейробластоме и других типах раковых клеток. Этот биспецифический белок имеет аминокислотную последовательность для вариабельной области тяжелой и легкой цепи против CD3 человека (с использованием формата для одноцепочечного вариабельного фрагмента, scFv), полученную из клона OKT3, слитого с помощью короткого линкера (L), с конструкцией scFv против GD2, полученной из клона 5F11, связанного как димер доменом димеризации (HDD) HNF1a, как описано в Ahmed et al., OncoImmunology, 4: 4, e989776, DOI: 10.4161/2162402X.2014.989776. Инструмент оптимизации кодонов vectorbuilder.com (//en.vectorbuilder.com/tool/codon-optimization.html) был использован для обратного проектирования оптимальной кодирующей последовательности ДНК человека для CD3xGD2-HDD. Полученная последовательность ДНК, начинающаяся со стартового кодона ATG, была синтезирована и клонирована ниже промотора куриного актина-β-глобина (CAGp) в экспрессирующей кассете адено-ассоциированного вируса с инвертированными концевыми повторами, полученными из AAV2. Три другие версии, которые содержат консенсусный секреторный сигнальный домен («секрекон», на основе Barash et al., Biochem Biophys Res Commun. 2002; 294: 835-842) ниже начального сайта ATG и заканчивая 0, 1 или 2 аланинами, были получены на основании открытия того, что аланины усиливают секрецию в зависимости от белка (Güler-Gane et al., PLoS ONE 11(5): e0155340. doi:10.1371/journal.pone.0155340). Белки, полученные из этих конструкций, были названы гетеродимерными scFv или Dimert. ФИГ. 7 иллюстрирует 4 примерных конструкций CD3xGD2-HDD dimert.
Анализы для определения структуры и функции Dimert.
Клетки 293T трансдуцировали векторами экспрессии AAV (или контролями), собирали и хранили надосадочные жидкости. Надосадочные жидкости тестировали на наличие белка правильного размера посредством электрофореза (SDS-PAGE), на связывание с CD3 с помощью анализа конкуренции связывания с использованием проточной цитометрии, на активацию Т-клеток с помощью проточной цитометрии и на уничтожение опухолевых клеток при совместной инкубации с Т-клетками. См. фиг.8, которая иллюстрирует схематическое изображение анализов для определения структуры и функции dimert, раскрытых в данном документе.
Три конструкции, содержащие пептид секреции, показывают меньшее количество CD3xGD2-HDD Dimert, удерживаемого в клетках.
Цельноклеточные лизаты клеток 293T, трансфицированных различными экспрессирующими плазмидами AAV CD3xGD2-HDD, собирали через 48 часов после трансфекции. Полиакриламидный электрофорез (PAGE) выполняли с использованием 50 мкг общего белка на дорожку и окрашивали гель Ponceau S. Результаты показали больше белка с использованием конструкции №1104, в которой отсутствует последовательность секреции, тогда как все 3 конструкции с последовательностью секреции показали меньше белка. Контроли включали только клетки или клетки, трансфицированные контрольной плазмидой (pcDNA3-GFP). КДа, килодальтон; Sec, секреторный домен; GFP, зеленый флуоресцентный белок; MW, молекулярная масса. См. фиг.9.
Только надосадочные жидкости от клеток, трансфицированных конструкциями AAV CD3xGD2-HDD, содержащими секреторную последовательность, связываются и активируют Т-клетки.
Надосадочные жидкости тестировали на связывание с человеческими Т-клетками и их активацию. Связывание определяли по конкуренции с флуоресцентно меченным антителом против CD3, предварительно связанным с Т-клетками (Jurkat). Окрашенные клетки показали 77,6%, 79,24% и 78,7% (Q2+Q3) CD3-положительных в контрольных группах (DMEM, Ctrl, GFP), соответственно, и 77,7% для вектора AAV №1104, в котором отсутствует пептид секреции. Напротив, связывание флуоресцентно меченного антитела против CD3 снижалось до 2,15%, 3,67% и 3,99% (Q2+Q3) надосадочными жидкостями из клеток, трансфицированных каждым из векторов, содержащих пептид секреции. Кроме того, клетки были совместно окрашены анти-CD69 в качестве маркера активации Т-клеток. Все контрольные группы и вектор AAV №1104 показали менее 13,4% положительных по CD69 (Q1+Q2), тогда как три разных вектора, содержащих AAV, показали 58,87-68,5% положительных по CD69. См. фиг.10.
Подробности способа: клетки Jurkat 2e5/лунку в 400 мкл RPMI+10% FBS высевали в 24-луночные планшеты, затем в каждую лунку добавляли 100 мкл надосадочных жидкостей от трансфицированных клеток 293Т (20% от общего количества культуральной среды). После 48 часов инкубации клетки Jurkat центрифугировали, промывали 1X PBS, затем окрашивали PE-Anti-hCD69 (1: 100) и PerCP-anti-hCD3e (1: 300 #OKT3) в течение 30 минут на льду. После промывки буфером FACS каждый образец фиксировали в 1% PFA и анализировали проточной цитометрией. DMEM - это среда, добавленная к Jurkat, Ctrl - надосадочную жидкость, добавленная из нетрансфицированных клеток 293T, и GFP - это надосадочная жидкость, добавленная из клеток 293T, трансфицированных плазмидой AAV, экспрессирующей GFP.
Только надосадочные жидкости из клеток, трансфицированных векторами AAV, содержащими пептид секреции, активируют Т-клетки человека.
Человеческие Т-клетки (Jurkat) инкубировали с надосадочными жидкостями из клеток 293Т, трансфицированных различными векторами AAV, окрашивали по CD69 с помощью PE-меченого антитела и исследовали с помощью флуоресцентной микроскопии. См. фиг.11, на котором левые панели представляют собой фазовый контраст, а правые панели - флуоресценцию. Ни контрольный вектор EGFP, ни вектор без пептида секреции №1104 не показали положительного окрашивания, тогда как другие три вектора показали высокие уровни окрашивания (красный). Подробности способа: клетки Jurkat 2e5/лунку в 400 мкл RPMI+10% FBS высевали в 24-луночные планшеты, затем в каждую лунку добавляли 100 мкл надосадочных жидкостей от трансфицированных клеток 293Т (20% от общего количества культуральной среды). После 48 часов инкубации клетки Jurkat центрифугировали, промывали 1X PBS, затем окрашивали фикоэритрин (PE)-анти-hCD69 (1:100) в течение 30 минут на льду. После промывки буфером FACS каждый образец фиксировали в 1% PFA в течение 5 минут.Масштабная метка: 100 мкм.
Связывание CD3xGD2-HDD с Т-клетками зависит от дозы.
Различные количества надосадочной жидкости из клеток 293T, трансфицированных плазмидами AAV, содержащими различные пептиды секреции, инкубировали и тестировали на конкуренцию с анти-hCD3, конъюгированным с перидинин-хлорофилл-протеиновым комплексом (PerCP), на Т-клетках Jurkat. См. ФИГ. 12A, на котором на левой панели показаны графики FACS, а на правой панели показаны числовые показатели медианного уровня окрашивания. Серая штриховка на левых панелях - это неокрашенные, а темно-серая линия на каждой панели указывает максимальное количество окрашенных клеток без добавления надосадочных жидкостей. Фиг.12B показывает гистограмму медианных уровней окрашивания, нормализованных к неокрашенным контролям.
Подробности способа: клетки Jurkat 2e5/лунку в 400 мкл RPMI+10% FBS высевали в 24-луночные планшеты, затем в каждую лунку добавляли 10 мкл, 25 мкл, 50 мкл или 100 мкл надосадочных жидкостей из трансфицированных клеток 293Т (2 ~ 20% от общего количества культуральной среды). После 24 часов инкубации клетки Jurkat центрифугировали, промывали 1X PBS, затем окрашивали PerCP-anti-hCD3e (1: 300 #OKT3) в течение 30 минут на льду. После промывки буфером FACS каждый образец фиксировали в 1% PFA в течение 5 минут перед анализом проточной цитометрией. Sec=секреторный пептид, A=аланин.
Анализ связывания для анти-GD2 плеча CD3xGD2-HDD Dimert и подтверждение того, что связывание GD2 и CD3 происходит на одной молекуле.
Связывание CD3xGD2-HDD с GD2 измеряли косвенно, сначала инкубируя надосадочные жидкости от трансфицированных клеток 293T либо с GD2-положительными, либо с GD2-отрицательными клетками, затем повторяя анализ конкуренции связывания CD3 T-клеток. Белки, связывающие GD2, должны абсорбироваться GD2-положительными клетками, но не GD2-отрицательными клетками, что приводит к потере конкуренции за связывание с Т-клетками. Этот анализ был разработан как таковой для подтверждения того, что связывание GD2 связано со связыванием CD3, другими словами, что отдельная молекула является биспецифичной. См. фиг.13 для графического изображения анализа связывания.
CD3xGD2-HDD связывает как CD3, так и GD2.
Как показано на фиг.13, надосадочные жидкости, полученные из клеток 293T, трансфицированных вектором #1101 CD3xGD2-HDD AAV, собирали и предварительно инкубировали с GD2-положительными SK-N-Be (2) или GD2-отрицательными клетками Raji. После центрифугирования и промывки был проведен анализ конкуренции связывания для клеток CD3 T (Jurkat).
На фиг.14 показан пример CD3xGD2-HDD Dimert, который связывается как с CD3, так и с GD2. Серая штриховка на верхней панели с черным контуром - неокрашенные Т-клетки Jurkat, темно-серая линия - Т-клетки Jurkat, полностью окрашенные на CD3 без добавления надосадочной жидкости. Образец Jurkat_cd3e показал полностью завершенное связывание CD3 с использованием надосадочной жидкости без предварительной инкубации, почти перекрываясь с надосадочной жидкостью, предварительно инкубированной с GD2-отрицательными клетками Raji. Напротив, образец Jurkat_cd3e sknbe2 показал потерю конкуренции, когда надосадочная жидкость была предварительно инкубирована с GD2-положительными клетками нейробластомы SK-N-Be(2), подтверждая, что одни и те же молекулы связывают как с CD3, так и с GD2.
Анализ для того, чтобы определить, индуцирует ли CD3xGD2-HDD уничтожение GD2+клеток-мишеней Т-клетками.
Фиг.15 иллюстрирует блок-схему анализа для определения, индуцирует ли CD3xGD2-HDD уничтожение GD2+клеток-мишеней Т-клетками. Как показано на фиг.15, GD2+клетки нейробластомы (SK-N-Be(2)) высевают в лунки, за которыми следуют первичные человеческие Т-клетки (приобретенные у StemExpress) и надосадочную жидкость от клеток 293Т, трансфицированных вектором AAV. Жизнеспособность клеток оценивали с помощью CellTiter-Glo от Promega (Мэдисон, Висконсин).
Секретируемый CD3xGD2-HDD индуцирует опосредованное Т-клетками уничтожение клеток нейробластомы.
Используя анализ, показанный на фиг.15, цитотоксичность человеческих Т-клеток плюс надосадочные жидкости (от клеток 293Т, трансфицированных различными векторными плазмидами AAV) тестировали на GD2+клетках нейробластомы SK-N-Be (2). Использовали соотношение Т-клеток к клеткам-мишеням 10:1. Клетки проверяли на жизнеспособность через 48 часов совместного культивирования. По сравнению с надосадочными жидкостями от нетрансфицированных клеток («без Dimert») не было заметной цитотоксичности при использовании надосадочных жидкостей от клеток, трансфицированных конструкцией, экспрессирующей неродственный dimert (CD19xCD3) или вектор, лишенный секретирующего пептида (№1104) (фиг.16). Напротив, надосадочные жидкости клеток, трансфицированных каждым из векторов, содержащим пептид секреции, вызывали статистически значимую цитотоксичность (p<0,001), убивая 25-30% клеток (фиг.16).
Опосредованная Т-клетками цитотоксичность CD3xGD2-HDD Dimert связана с экспрессией GD2.
Панель клеточных линий нейробластомы тестировали на их чувствительность к уничтожению человеческими Т-клетками в сочетании с надосадочными жидкостями от вектора AAV #1101 (верхние графики на фиг.17) и измеряли их экспрессию GD2 с помощью проточной цитометрии (нижние графики на фиг.17, заштрихованные кривые - изотипические контроли). Клетки CHP-134 показали наибольшую цитотоксичность и самую высокую экспрессию GD2.
Пример 2. Получение CD19xCD3 Dimert с использованием примерных CD19xCD3 TransJoin
Карты типичных конструкций AAV для тестирования экспрессии CD19xCD3 Dimert.
Одобренный FDA терапевтический белок, известный как блинатумомаб, так называемый биспецифический рекрутер Т-клеток (BiTE), был создан для нацеливания на CD19 человека на В-клетках/В-клеточных злокачественных новообразованиях и CD3 человека на Т-клетках. Была использована общедоступная аминокислотная последовательность блинатумомаба (//www.drugbank.ca/drugs/DB09052) и дополнительно был использован инструмент оптимизации кодонов vectorbuilder.com (//en.vectorbuilder.com/tool/codon-optimization.html). используется для обратного проектирования оптимальной кодирующей последовательности ДНК человека для CD19xCD3. Полученная последовательность ДНК, начинающаяся со стартового кодона ATG, была синтезирована и клонирована ниже промотора куриного актина-b-глобина (CAGp) в экспрессирующей кассете адено-ассоциированного вируса с инвертированными концевыми повторами, полученными из AAV2. Три другие версии, которые содержат консенсусный секреторный сигнальный домен («секрекон», на основе Barash et al., Biochem Biophys Res Commun. 2002; 294: 835-842) ниже начального сайта ATG и заканчивая 0, 1 или 2 аланинами, были получены на основании открытия того, что аланины усиливают секрецию в зависимости от белка (Güler-Gane et al., PLoS ONE 11(5): e0155340. doi:10.1371/journal.pone.0155340). Белки, полученные из этих конструкций, были названы гетеродимерными scFv или Dimert. Фиг.18A и фиг.18B иллюстрируют 5 примерных конструкций CD19xCD3.
Фиг.18C иллюстрирует схематическое изображение CD19 dimert, взаимодействующего с раковой клеткой и Т-клеткой. CD19 dimert продуцируется клеткой, содержащей CD19 TransJoin. При введении объекту, например, внутривенно, AAV TransJoin (например, AAV CD19 TransJoin, как показано на этой фигуре) проникает в нормальные клетки, такие как печень или мышцы, и экспрессирует полипептид, который он кодирует.Сигнальный пептид секреции расщепляется во время секреции, оставляя активный dimert, который связывает раковую клетку (Са) на одном конце и Т-иммунную клетку на другом конце.
Только надосадочные жидкости из клеток, трансфицированных конструкциями AAV CD19xCD2, содержащими секреторную последовательность, связываются и активируют Т-клетки.
Надосадочные жидкости тестировали на связывание с человеческими Т-клетками и их активацию. Связывание определяли по конкуренции с флуоресцентно меченным антителом против CD3, предварительно связанным с человеческими Т-клетками (Jurkat). Окрашенные клетки показали 77,6%, 79,24% и 78,7% (Q2+Q3) CD3-положительных в контрольных группах (DMEM, Ctrl, GFP), соответственно, и 79,4% из вектора #1323, в котором отсутствует пептид секреции. Напротив, связывание флуоресцентно меченного анти-CD3 антитела снижалось до 23,88%, 18,59% и 30,76% (Q2+Q3) надосадочными жидкостями из клеток, трансфицированных каждым из векторов, содержащих пептид секреции. Кроме того, клетки были совместно окрашены анти-CD69 в качестве маркера активации Т-клеток. Все контрольные группы и #1323 показали менее 13,38% положительных по CD69 (Q1+Q2), тогда как три разных вектора, содержащих секрецию, показали 63,9%, 67,3% и 66,6% положительных по CD69 (фиг.19).
Подробности способа: клетки Jurkat 2e5/лунку в 400 мкл RPMI+10% FBS высевали в 24-луночные планшеты, затем в каждую лунку добавляли 100 мкл надосадочных жидкостей от трансфицированных клеток 293Т (20% от общего количества культуральной среды). После 48 часов инкубации клетки Jurkat центрифугировали, промывали 1X PBS, затем окрашивали PE-Anti-hCD69 (1: 100) и PerCP-anti-hCD3e (1: 300 #OKT3) в течение 30 минут на льду. После промывки буфером FACS каждый образец фиксировали в 1% PFA и анализировали проточной цитометрией. DMEM - это среда, добавленная к Jurkat, Ctrl - надосадочную жидкость, добавленная из нетрансфицированных клеток 293T, и GFP - это надосадочная жидкость, добавленная из клеток 293T, трансфицированных плазмидой AAV, экспрессирующей GFP.
Только надосадочные жидкости из клеток, трансфицированных векторами AAV, содержащими пептид секреции, активируют Т-клетки человека.
Человеческие Т-клетки (Jurkat) инкубировали с надосадочными жидкостями из клеток 293Т, трансфицированных различными векторами AAV, окрашивали по CD69 с помощью PE-меченого антитела и исследовали с помощью флуоресцентной микроскопии. См. фиг.20, левые панели - фазовый контраст, а правые панели - флуоресценция. Ни контрольный вектор EGFP, ни вектор AAV, не содержащий пептида секреции #1323, не показали положительного окрашивания, тогда как три других вектора показали высокие уровни окрашивания (красный).
Подробности способа: клетки Jurkat 2e5/лунку в 400 мкл RPMI+10% FBS высевали в 24-луночные планшеты, затем в каждую лунку добавляли 100 мкл надосадочных жидкостей от трансфицированных клеток 293Т (20% от общего количества культуральной среды). После 48 часов инкубации клетки Jurkat центрифугировали, промывали 1X PBS, затем окрашивали фикоэритрин (PE)-анти-hCD69 (1:100) в течение 30 минут на льду. После промывки буфером FACS каждый образец фиксировали в 1% PFA в течение 5 минут.Масштабная метка: 100 мкм.
CD19xCD3, секретируемый AAV, специфически связывает CD19, но не CD45.
Анализ конкуренции связывания использовали для определения того, мешают ли надосадочные жидкости надосадочных жидкостей клеток 293T, трансфицированных вектором AAV, окрашиванию В-клеток человека, трансформированных вирусом Эпштейна-Барр (EBV), двумя разными антителами, одно из которых окрашивает маркер В-клеток CD19, а другое окрашивает панлейкоцитарный маркер CD45.
См. фиг.21, в котором клетки в 3 контрольных группах (DMEM, Ctrl, GFP) показали 93%, 93,1% и 93,2% положительного окрашивания по CD19 (Q1+Q2), соответственно, и надосадочная жидкость от клеток, трансфицированных вектором AAV #1323, который не содержит пептид секреции, не конкурировал с этим окрашиванием, показывая положительную реакцию на CD19 на 93,2%. Напротив, каждый из векторов AAV, содержащих пептид секреции, подавлял сигнал CD19 до 33,33%, 31,07% и 35,88%. Обратите внимание, что порог был установлен таким образом, что неокрашенные клетки показали 14,58% положительную реакцию по CD19, что позволяет предположить, что надосадочная жидкость из клеток, трансфицированных этими тремя векторами AAV, конкурировала в 2 раза выше фона. Напротив, ни один из векторов не конкурировал за окрашивание CD45, которое было на 78,89%, 79,14% и 78,32% положительным (Q2+Q3) в 3 контролях, на 82,9% положительным в векторе, не содержащем пептида секреции, и 80,5%, 78,5%. % и на 79,4% положительным при использовании надосадочных жидкостей клеток, трансфицированных тремя другими векторами.
Подробности способа: трансформированные EBV В-клетки под названием NB122R (Gene Ther. 2013 Jul;20(7):761-9. doi: 10.1038/gt.2012.93) при 2e5 клетки/лунку в 400 мкл RPMI+10% FBS высевали в 24-луночный планшет, затем в каждую лунку добавляли 100 мкл надосадочных жидкостей из трансфицированных клеток 293Т (2 ~ 20% всей культуры). После 24 часов инкубации клетки центрифугировали, промывали 1 раз PBS, затем окрашивали PEcy7-анти-hCD45 (1: 100) и APC-анти-hCD19 (1: 300) в течение 30 минут на льду. После промывки буфером FACS каждый образец фиксировали в 1% PFA в течение 5 минут перед анализом проточной цитометрии. DMEM - это среда, добавленная к Jurkat, Ctrl - надосадочную жидкость, добавленная из нетрансфицированных клеток 293T, и GFP - это надосадочная жидкость, добавленная из клеток 293T, трансфицированных плазмидой AAV, экспрессирующей GFP.
Связывание CD19xCD3 с Т-клетками зависит от дозы, и вектор, содержащий единственный аланин, расположенный ниже консенсусной последовательности секретирующего пептида, превосходит другие тестируемые векторы.
Различные количества надосадочной жидкости из клеток 293T, трансфицированных плазмидами AAV, содержащими различные пептиды секреции, инкубировали и тестировали на конкуренцию с анти-hCD3, конъюгированным с перидинин-хлорофилл-протеиновым комплексом (PerCP), на Т-клетках Jurkat. См. ФИГ. 22A, на которой на левой панели показаны графики FACS, а на правой панели показаны значения среднего уровня окрашивания. Серым цветом на каждой из левых панелей представлены неокрашенные клетки, а темно-серая линия на каждой левой панели представляет максимально окрашенные клетки без добавления надосадочных жидкостей. Фиг.22B показывает гистограмму медианных уровней окрашивания, нормализованных к неокрашенным контролям. Вектор с одним аланином ниже консенсусной последовательности секретирующего пептида, #1325, показал наибольшую конкуренцию по сравнению с другими тестируемыми векторами и был выбран для дальнейших экспериментов.
Подробности способа: клетки Jurkat 2e5/лунку в 400 мкл RPMI+10% FBS высевали в 24-луночные планшеты, затем в каждую лунку добавляли 10 мкл, 25 мкл, 50 мкл или 100 мкл надосадочных жидкостей из трансфицированных клеток 293Т (2 ~ 20% от общего количества культуральной среды). После 24 часов инкубации клетки Jurkat центрифугировали, промывали 1 раз PBS, затем окрашивали PerCP-anti-hCD3e (1: 300 #OKT3) в течение 30 минут на льду. После промывки буфером FACS каждый образец фиксировали в 1% PFA в течение 5 минут перед анализом проточной цитометрией. MFI=средняя интенсивность флуоресценции, Sec=секреторный пептид, A=аланин.
Связывание CD19xCD3 с В-клетками зависит от дозы, и вектор, содержащий единственный аланин, расположенный ниже консенсусной последовательности секретирующего пептида, превосходит другие тестируемые векторы.
Различные количества надосадочной жидкости из клеток 293T, трансфицированных плазмидами AAV, содержащими различные пептиды секреции, инкубировали и тестировали на конкуренцию с анти-hCD19, конъюгированным с аллофикоцианином (APC), на В-клетках человека. См. фиг.23A, на которой на левой панели показаны графики FACS, а на правой панели показаны значения среднего уровня окрашивания. Серая заливка на каждой из левых панелей была неокрашенной, а темно-серая линия на каждой из левых панелей была максимально окрашенными клетками без добавления надосадочных жидкостей. Фиг.23B показывает гистограмму средних уровней окрашивания, нормализованных к неокрашенным контролям. Вектор с одним аланином на конце пептида секреции, #1325, показал наибольшую конкуренцию, согласующуюся с результатами по связыванию с Т-клетками, показанными на фиг.22A и фиг.22B и был выбран для дальнейших экспериментов.
Подробности способа: В-клетки человека NB122R в количестве 2e5 клеток/лунку в 400 мкл RPMI+10% FBS высевали в 24-луночные планшеты, затем в каждую лунку добавляли 10 мкл, 25 мкл, 50 мкл или 100 мкл надосадочных жидкостей из трансфицированных клеток 293Т (2~20% от общего количества культуральной среды). После 24 часов инкубации клетки центрифугировали, промывали 1X PBS, затем окрашивали APC-анти-hCD19 (1: 300) в течение 30 минут на льду. После промывки буфером FACS каждый образец фиксировали в 1% PFA в течение 5 минут перед анализом проточной цитометрией. MFI=средняя интенсивность флуоресценции, Sec=секреторный пептид, A=аланин.
CD3 Dimert активируют Т-клетки с помощью костимуляции против CD28 лучше, чем антитела против CD3.
Человеческие Т-клетки (Jurkat) инкубировали совместно с антителами к CD28 и надосадочными жидкостями, полученными из клеток 293Т, трансфицированных вектором AAV (левые панели на фиг.24), или с возрастающими концентрациями анти-CD3-антител (правые панели на фиг.24). Собирали клеточную мРНК и проводили количественную полимеразную цепную реакцию с обратной транскриптазой (RT-PCR) для мРНК IL-2 (верхние панели фиг.24) и IL-8 (нижние панели фиг.24) и рассчитывали экспрессию относительно мРНК гена домашнего хозяйства, GAPDH. Только надосадочные жидкости из клеток, трансфицированных векторными конструкциями AAV, содержащими секреторный домен, показали стимуляцию любого гена выше контрольных (gfp, 293t) и выше векторных конструкций без секреторного домена (#1104 для CD3xGD2-HDD, #1323 для CD19xCD3).
Клетки 293T обеспечивают самую высокую эффективность трансдукции для AAV8.
Четыре разные клеточные линии инфицировали 2 различными концентрациями AAV8, экспрессирующего зеленый флуоресцентный белок (GFP), при множественности инфицирования (MOI)=1e4 или 1e5 копий генома (gc) с культуральной средой, содержащей 2% фетальной бычьей сыворотки. Сигнал GFP оценивали через 48 часов после заражения вирусом с помощью флуоресцентной микроскопии. См. фиг.25. AML-12: нормальные клетки печени мыши; 293T: эмбриональные клетки почек человека, трансформированные антигеном SV40-T; H441-CRM: клетки рака легкого человека с мутацией KRAS; SK-N-Be (2): клетки нейробластомы человека с амплифицированным NMYC.
Концентрация dimert в надосадочных жидкостях клеток, инфицированных AAV8, зависит от дозы AAV и эффективности трансдукции.
Анализ связывания T (Jurkat) выполняли на надосадочных жидкостях клеток 293T и H441, трансфицированных векторами TransJoin или контролем AAV-GFP. См. фиг.26, на которой серая штриховка на панелях представляет собой неокрашенные контрольные клетки, а темно-серые линии на каждой панели - Т-клетки, полностью окрашенные анти-CD3-антителом. Хотя CD19xCD3 Dimert оказался намного более эффективным, чем CD3xGD2-HDD Dimert в конкуренции за связывание, эффект был дозозависимым (кривые смещены дальше влево с более высокой MOI) и не наблюдались в линии клеток с плохой эффективностью трансдукции AAV8 (клетки H441). MOI, множественность заражения.
Однократная внутривенная инъекция CD19xCD3 TransJoin избирательно устраняет В-клетки у гуманизированных мышей.
Были приобретены иммунодефицитные мыши (NSG-SGM3, Jackson Labs), которых облучали и внутривенно вводили человеческие CD34+гемопоэтические стволовые клетки. К 12 неделям мыши показали приживление клеток крови человека с помощью проточной цитометрии периферической крови, включая Т- и В-лимфоциты. Однократная инъекция CD19xCD3 TransJoin (AAV8-Sec1A-CAG-193, вектор №1325, упакованный в капсиды AAV8) вводили в различных дозах мышам и анализировали кровь с помощью проточной цитометрии на лимфоциты человека, собранные в разные моменты времени, как показано на фиг.27. Хотя не было никакого эффекта при низких дозах (5e9 и 5e10 векторных геномов vg(vg) на килограмм (кг) массы тела), более высоких доз (5e11 и 5e12 vg/кг) было достаточно для устранения циркулирующих B-клеток без воздействия на CD4+или CD8+T-клетки.
Однократная внутривенная инъекция CD19xCD3 TransJoin приводит к длительному истощению В-клеток у гуманизированных мышей.
Гуманизированных мышей обрабатывали однократной дозой CD19xCD3 AAV8 TransJoin, и впоследствии измеряли субпопуляции лимфоцитов в крови. Как показано на фиг.28 для одной мыши, длительное и избирательное истощение В-клеток наблюдалось у всех проанализированных мышей до тех пор, пока эти мыши оставались живыми.
Однократная внутривенная инъекция CD19xCD3 TransJoin устраняет лимфому CD19+у гуманизированных мышей.
Человеческие CD19+клетки Raji (полученные от пациента с лимфомой Беркитта) имплантировали в бока двух гуманизированных мышей и ждали, пока они не достигнут размера более 250 мм3. Затем в хвостовую вену одного животного инъецировали контрольный вирус AAV, AAVGFP, а в хвостовую вену второго животного инъецировали CD19xCD3 AAV8 TransJoin. Опухоль у контрольной мыши в конечном итоге быстро росла, и животное нужно было усыпить. Опухоль у мышей, которым вводили TransJoin, росла медленно, а затем в конечном итоге почти полностью сократилась. См. фиг.29. Животное нужно было усыпить из-за симптомов болезни трансплантат против хозяина, которая часто встречается у гуманизированных мышей, но эксперимент демонстрирует доказательство принципа того, что одна инъекция TransJoin может лечить онкологическое заболевание.
Пример 3. Использование OncoSkip и TransSkip для одновременного нацеливания на экспрессию онкогена и активации терапевтической экспрессии трансгена
Обзор OncoSkip и TransSkip описан в данном документе.
OncoSkip - это антисмысловой морфолиноолигонуклеотид, который вызывает пропуск экзона онкогена. В некоторых воплощениях OncoSkip сконструирован как антисмысловой морфолиноолигонуклеотид, который индуцирует пропуск критического экзона в онкогене (левая сторона фиг.30). Для доказательства принципа использовали KRAS (экзоны 1, 2, 3, представленные черно-золотисто-серым цветом на левой стороне фиг.30) и тестировали морфолины (показанные на фиг.30 в виде небольшой светло-серой полосы на вершине онкогенов) разработанные для того, чтобы пропускать экзон 2 KRAS, которые содержат стартовый сайт ATG, так что нормальная экспрессия онкогена будет снижена. Затем создавали производное того же экзона, который нужно пропустить, но с мутацией, содержащей множественные стоп-кодоны в каждой рамке считывания вместе с фланкирующими последовательностями интронов. Новый интрон-экзон (СТОП)-интрон затем вставляли в кодирующую последовательность трансгена в сайтах, которые воссоздают сайты сплайсинга донора и акцептора, так что он встраивается в мРНК трансгена и прерывает нормальную кодирующую последовательность. В присутствии антисмыслового морфолиноолигонуклеотида (небольшая светло-серая полоса на вершине онкогенов) вновь вставленный экзон пропускается, повторно соединяя кодирующую последовательность для генерации функционального продукта. Класс векторов AAV, содержащих гены, прерванные последовательностью интрон-экзон-интрон, называют вирусами TransSkip, а класс морфолино, сконструированным для подавления онкогена, называют OncoSkip.
Антисмысловые морфолины KRAS OncoSkip вызывают пропуск экзонов эндогенного KRAS в клетках рака легкого.
Клеточные линии рака легкого A549 и H441 (не показаны) инкубировали с антисмысловыми морфолиноолигонуклеотидами KTS1 и KTS2 OncoSkip, а также с инвертированным контрольным морфолиноолигонуклеотидом для KTS2. KTS1 и KTS2 оба связываются на 3’-конце соединения экзона-интрона для экзона 2 KRAS и были разработаны для индукции пропуска экзона 2, поскольку он содержит стартовый сайт ATG. Затем эндогенную мРНК KRAS анализировали с помощью обратной транскриптазы RT-PCR на наличие экзона 2 с помощью PCR с праймерами, присутствующими в экзоне 1 (прямой) и экзоне 2 (обратный). Праймеры в экзоне 4 использовали в качестве контроля для общей мРНК KRAS. Дозозависимое снижение количества транскриптов, содержащих экзон 2 («экзон 1+2»), наблюдали для обоих морфолиноолигонуклеотидов OncoSkip.См. фиг.31.
Векторные карты AAV для CD3xGD2-HDD TransSkip, показывающие реконструированные интроны, фланкирующие экзон, вставленный в кодирующую последовательность CD3xGD2-HDD Dimert.
3’ последовательность человеческого интрона перед экзоном 2 KRAS (Ki1), производное экзона 2 KRAS, содержащее мутированные множественные стоп-кодоны во всех трех рамках считывания (СТОП), и 5’ последовательность человеческого интрона лежащего ниже экзона 2 (Ki) KRAS, и кассету клонировали в последовательность гена, кодирующую GD2 Dimert, в конкретной последовательности, которая повторяет консенсусные донорные и акцепторные сайты сплайсинга. См. фиг.32. Когда новый СТОП экзон встраивается в транскрипт, функциональный dimert не образуется. Когда клетки подвергаются воздействию антисмыслового морфолиноолигонуклеотида, который связывает 3’ соединение экзон-интрон, СТОП экзон пропускается и экспрессируется мРНК Dimert полной длины. Последовательности KRAS были выбраны таким образом, чтобы антисмысловой морфолиноолигонуклеотид, предназначенный для пропуска экзона 2, одновременно изменял бы сплайсинг мРНК трансгена TransSkip, что приводило к экспрессии полноразмерного Dimert в трансдуцированных клетках и уменьшало или устраняло экспрессию нативного KRAS в раковых клетках, поскольку экзон 2 содержит стартовый кодон KRAS ATG.
Примерная стратегия тестирования активности OncoSkip и TransSkip.
Как показано на фиг.33, осадок клеток и надосадочные жидкости собирали из клеток 293Т, трансдуцированных векторами AAV Dimert. мРНК выделяли из осадка клеток и подвергали RT-PCR с обратной транскриптазой для определения степени, в которой искусственный экзон в трансгене был сплайсирован в мРНК. Надосадочную жидкость для экспрессии Dimert анализировали с помощью анализа связывания Т-клетками и анализа уничтожения.
Антисмысловые морфолиновые олигомеры вызывают пропуск экзона трансгена CD3xGD2-HDD TransSkip.
Клетки 293T трансфицировали CD3xGD2-HDD Dimert-плазмидой #1042. Клетки собирали через 48 часов после трансфекции для выделения общей РНК. Для RT-PCR использовали около 1 мкг РНК. См. фиг.34, в которой дорожка №9 показывает полноразмерную трансгенную РНК между праймерами без сплайсинга с использованием ДНК-плазмиды в качестве матрицы. Дорожки 7 и 8 и контрольные дорожки не содержат антисмыслового морфолиноолигонуклеотида и показывают, что большинство транскриптов включает внутренний экзон (прямоугольник с полосами). Есть некоторые транскрипты с исключенным экзоном (наименьшая полоса), что позволяет предположить, что эта конструкция имеет некоторую «утечку» активированных транскриптов. Включение любого из морфолиноолигонуклеотидов (KTS1, KTS2) снижает долю инактивированного транскрипта с экзоном (219 п.н.) дозозависимым образом и увеличивает активированный транскрипт (97 п.н.), в котором исключен экзон.
KRAS OncoSkip индуцирует секретируемую экспрессию CD3xGD2-HDD Dimert в клетках, трансфицированных вектором CD3xGD2-HDD TransSkip AAV.
Надосадоные жидкости собирали из трансфицированных клеток 293Т и тестировали на связывание с Т-клетками (интерференция с флуоресцентно меченными анти-CD3 антителами). Как показано на фиг.35, серая линия на верхней панели это неокрашенные Т-клетки (Jurkat), темно-серая линия на верхней панели - полностью окрашенные Т-клетки с анти-CD3 антителом. В качестве положительного контроля надосадочная жидкость клеток, трансфицированных конститутивно экспрессируемым CD3xGD2-HDD TransJoin #1011, почти полностью подавляла окрашивание анти-CD3. Надосадочная жидкость из клеток, трансфицированных CD3xGD2-HDD TransSkip #1042, показала промежуточную конкуренцию, соответствующую некоторой «утечке» мРНК Dimert с пропущенным экзоном, и была дополнительно индуцирована морфолиноолигонуклеотидом OncoSkip (KTS1, KTS2).
Пропуск экзонов CD3xGD2-HDD TransSkip с помощью KRAS OncoSkip - это целевой эффект.
Пропуск экзона, индуцированный направленным антисмысловым морфолиноолигонуклеотидом KTS2, сравнивали с морфолиноолигонуклеотидом, содержащим те же основания, но с обратной последовательностью (фиг.36). KTS2 индуцировал пропуск экзонов почти до 100% транскриптов (97 п.н.), тогда как обращенный контрольный морфолиноолигонуклеотид не имел никакого эффекта по сравнению с контролем только Endoporter (носитель для морфолиноолигонуклеотидов).
Индукция экспрессии CD3xGD2-HDD Dimert, определяемая связыванием Т-клеток, является целевым эффектом.
Были собраны надосадочные жидкости из клеток 293T, трансфицированных CD3xGD2-HDD TransJoin (положительный контроль) и CD3xGD2-HDD TransSkip, инкубированных без или с различными антисмысловыми морфолиновыми олигомерами, и был проведен анализ связывания человеческих T-клеток (Jurkat) с помощью проточной цитометрии (конкуренция за связывание с флуоресцентно мечеными анти-CD3 антителами). См. фиг.37, на которой серая линия на верхней панели представляет собой неокрашенные Т-клетки, а темно-серая линия на верхней панели представляет собой полностью окрашенные Т-клетки. Линия, указывающая #1101 CD3xGD2-HDD TransJoin, является почти полностью завершенным сигналом от конститутивно экспрессируемого GD3 TransJoin. Как и ранее, надосадочная жидкость из клеток, трансфицированных CD3xGD2-HDD TransSkip, показала небольшую конкуренцию на исходном уровне (#1042 CD3xGD2-HDD TransSkip, без морфолиноолигонуклеотида), которая не была изменена контрольным морфолиноолигонуклеотидом (CD3xGD2-HDD TransSkip+«KTS2-Invert ctl»), но была индуцирована до конкуренции с большим сигналом с помощью целевого морфолиноолигонуклеотида «OncoSkip», KTS2 (CD3xGD2-HDD TransSkip+KTS2).
Карты генома AAV образцовых вариантов сплайсинга TransSkip созданы и протестированы для уменьшения исходной «утечки» TransSkip, но при сохранении индуцируемого пропуска экзонов.
Сплайсинговые варианты TransSkip были разработаны для уменьшения базового пропускания экзонов, которое наблюдалось с CD3xGD2-HDD TransSkip.Посттранскрипционный регуляторный элемент вируса гепатита сурка (WPRE), расположенный ниже кодирующей последовательности, использовали для увеличения экспрессии трансгена. Из-за ограничений размера упаковки AAV включение последовательностей WPRE потребовало использования более короткого промотора, в силу этого такая новая серия векторов была создана с промотором, полученным из короткой формы промотора α1 фактора элонгации трансляции 1 эукариот человека (EFSp). Варианты были созданы путем изменения конкретных пар оснований в сайте связывания вспомогательного фактора U2 (U2AF), расположенном в полипиримидиновой дорожке на 3’-конце первого интрона (Ki1-5). См. фиг.38.
Вариант сплайсинга K3 CD3xGD2-HDD TransSkip исключает пропуск экзонов на исходном уровне, но поддерживает индуцируемый пропуск экзонов.
Клетки 293T трансфицировали новой панелью из 5 различных конструкций CD3xGD2-HDD TransSkip без и с антисмысловым морфолиноолигонуклеотидом «OncoSkip» KTS2 и сравнивали их с контролем «только эндо», который включает носитель морфолиноолигонуклеотидов, но не морфолиноолигонуклеотид. Транскрипты мРНК анализировали с помощью RT-PCR с обратной транскриптазой и обнаружили различные фенотипы. Как показано на фиг.39, вариант K4 показал аналогичный (или даже немного больший) базовый уровень пропуска экзона относительно K1 и был индуцирован к пропуску даже больше, чем K1. Варианты K2, K3 и K5 практически не показали пропусков базовой линии. Из этих трех вариант K3 оказался способен индуцировать наибольшее количество пропусков экзона.
Вариант K3 CD3xGD2-HDD TransSkip не показывает исходного продуцирования Dimert, но является наиболее индуцируемым с помощью KRAS OncoSkip.
Панель вариантов сплайсинга CD3xGD2-HDD TransJoin тестировали на продуцирование экспрессии секретируемого димера CD3xGD2-HDD без или с антисмысловым морфолиноолигонуклеотидом KRAS KTS2 OncoSkip с использованием анализа связывания T-клеток (Jurkat) (крайний справа). Как показано на фиг.40, серая линия на каждой из правых панелей - это неокрашенные Т-клетки, темно-серая - полностью окрашенные Т-клетки, синяя - это конститутивно экспрессируемый CD3xGD2-HDD TransJoin, зеленый - соответствующий CD3xGD2-HDD TransSkip без OncoSkip («только Endo»). Последовательность показывает серым цветом (подчеркнутым и курсивом) сайт связывания полипиримидина U2AF со встроенными вариантами пары оснований относительно последовательности интрона KRAS дикого типа (K1). Верхняя полоса в геле RT-PCR - это транскрипт, содержащий экзон, нижняя полоса - транскрипт с пропущенным экзоном. Варианты K1 и K4 показывают некоторый детектируемый пропуск базовых экзонов с помощью RT-PCR и заметную базовую экспрессию Dimert (зеленый, крайний правый), тогда как варианты K2, K3 и K5 все не показывают пропуска базовых экзонов с помощью RT-PCR и экспрессию Dimert за счет конкуренции за связывание (зеленый, крайний справа). Все три из этих вариантов демонстрируют небольшое количество индуцируемого пропуска экзонов при экспрессии RT-PCR в присутствии KRAS-антисмыслового морфолиноолигонуклеотида OncoSkip KTS2 и различной степени дозозависимой экспрессии Dimert в присутствии KRAS OncoSkip (светлый и темно-фиолетовый представляют, соответственно более низкие и более высокие концентрации). Вариант K3 оказался наиболее индуцибельным, поэтому K3 был выбран в качестве ведущей конструкции для дальнейшего изучения.
Опосредованная OncoSkip индукция экспрессии Dimert из вариантов K3 и K5 CD3xGD2-HDD TransSkip является целевым эффектом.
Анализы RT-PCR и связыванияT-клетками (Jurkat) повторяли с вариантами K2, K3 и K5 TransSkip CD3xGD2-HDD и включали антисмысловой морфолиноолигонуклеотид «инвертированный KTS2» в качестве контроля. См. фиг.41. Инвертированный KTS2 содержит те же нуклеотидные основания, что и KTS2, но в противоположном порядке. Инвертированный KTS2 не смог вызвать пропуск экзона (нижняя полоса на геле) как в K3, так и в K5, тогда как правильная последовательность KTS2 индуцировала пропуск экзона как в K3, так и в K5 (ни один из них не индуцировал его в варианте K2). В анализе связывания Т-клеток для экспрессии Dimert (крайние правые панели) серый цвет - это неокрашенные Т-клетки, темно-серый - полностью окрашенные Т-клетки, синий - конститутивно экспрессируемый CD3xGD2-HDD TransJoin, фиолетовый - с KRAS OncoSkip KTS2 (светлый - более низкая доза, темный - более высокая доза), а желтый - инвертированный контроль KTS2. Анализ белка Dimert согласуется с RT-PCR; ни один из морфолиноолигонуклеотидов не индуцировал экспрессию Dimert из варианта K2, и только KTS2 OncoSkip, но не инвертированный контроль, индуцировал экспрессию Dimert из вариантов K3 и K5.
Секретируемый Dimert, индуцированный OncoSkip из векторов TransSkip AAV CD3xGD2-HDD, является функциональным в опосредовании уничтожения Т-клетками клеток нейробластомы.
Была протестирована цитотоксичность человеческих Т-клеток плюс надосадочные жидкости от клеток 293Т, трансфицированных различными вариантами AAV CD3xGD2-HDD TransSkip без и с KRAS OncoSkip KTS2 на клетках нейробластомы GDS2+SK-N-Be(2) (фиг.42). Использовали соотношение Т-клеток к клеткам-мишеням 10:1. Клетки проверяли на жизнеспособность через 48 часов совместного культивирования. Результаты были нормализованы к надосадочным жидкостям от нетрансфицированных клеток. Варианты K1 и K4 CD3xGD2-HDD TransSkip показали цитотоксичность в отсутствие KRAS OncoSkip KTS2, что согласуется с исходным пропуском экзонов и дополнительной индуцируемой цитотоксичностью с использованием K4. Вариант K2 CD3xGD2-HDD TransSkip не показал ни исходной, ни индуцибельной цитотоксичности, что согласуется с его известным эффективным сплайсингом и включением экзона. Напротив, варианты K3 и K5 TransSkip CD3xGD2-HDD не показали исходного уровня, но действительно показали индуцибельную цитотоксичность, будучи дозозависимыми с вариантом K3.
Пропуск экзона трансгена в клетках, инфицированных вариантом K3 AAV CD3xGD2-HDD TransSkip, индуцируется целевым образом с помощью KRAS OncoSkip.
Пропуск экзона в контексте инфицирования вирусом AAV был исследован для того, чтобы определить, совпадает ли он с тем, что наблюдалось при трансфекции плазмид AAV в других исследованиях. Варианты K1 и K3 CD3xGD2-HDD TransSkip упаковывали в AAV8 и анализировали включение («инактивированный») и исключение («активированный») экзона с помощью RT-PCR через 48 часов после инфицирования AAV клеток 293T. Клетки инкубировали без («только Endo») или с антисмысловыми морфолиноновыми олигомерами KRAS OncoSkip KTS2 или инвертированным контролем («InvtKTS2 Ct1»). Как показано на фиг.43 и с векторными плазмидами вариант AAV GD2 K1 показал пропуск экзона на исходном уровне, который дополнительно индуцировался KRAS OncoSkip KTS2, но не контролем. Напротив, пропуск экзона на исходном уровне не наблюдался с вариантом K3, но наблюдалось, что он индуцировался с помощью OncoSkip KTS2 (как отдельно, так и в комплексе с носителем, «Vivo-KTS2»). Напротив, пропуск не индуцировался контрольным морфолиноолигонуклеотидом.
Карты генома AAV для вариантов сплайсинга CD19xCD3 TransSkip.
На основании результатов исследований CD3xGD2-HDD TransSkip, в которых вариант K3 показал более низкий исходный уровень по сравнению с K1, но все еще индуцируемую экспрессию Dimert, были созданы варианты интронов K1 и K3, вставленные в трансген CD19xCD3 Dimert. Как и в случае с серией GD2, для увеличения экспрессии трансгена использовали WPRE ниже кодирующей последовательности, что потребовало применения более короткого промотора EFSp.Ki, последовательность, полученная из части интрона KRAS. В СТОП, экзон 2 из KRAS вносили мутации так, чтобы включить стоп-кодоны во всех трех рамках считывания. См. фиг.44A и фиг.44B для иллюстративных примеров конструкций CD19xCD3 TransSkip.
Фиг.44C иллюстрирует схематическое изображение CD19 dimert, взаимодействующего с раковой клеткой и Т-клеткой. CD19 dimert продуцируется клеткой, содержащей CD19 TransSkip.При введении объекту, например, внутривенно, AAV TransSkip (например, AAV CD19 TransSkip, как показано на этой фигуре) проникает в нормальные клетки, такие как печень или мышцы, но он НЕ экспрессирует полипептид из-за вставки интронов и экзона, содержащего стоп-кодоны, в нормальную кодирующую последовательность трансгена. В присутствии антисмыслового полинуклеотида (например, морфолинового антисмыслового олигонуклеотида), который вызывает пропуск экзона, транскрипт мРНК, процессируемый антисмысловым олигонуклеотидом, содержит интактный трансген, который не прерывается СТОП-экзоном. При таких обстоятельствах полипептид синтезируется и секретируется, сигнальный пептид секреции расщепляется во время секреции, оставляя активный dimert, который связывает раковые клетки (Са) на одном конце и Т-иммунную клетку на другом конце.
Оба морфолиноолигонуклеотида OncoSkip KTS1 и KTS2 индуцируют целевой пропуск экзона CD19xCD3 TransSkip K1.
Как показано на фиг.45, клетки 293T трансфицировали векторами AAV, включая CD19xCD3 TransJoin (положительный контроль, #1325) и CD19xCD3 TransSkip K1 (#1098) без (дорожка #4) или с совместной инкубацией с контрольным морфолиноолигонуклеотидом (дорожка №3) или двумя разными морфолиновыми олигомерами KRAS OncoSkip (дорожки 1 и 2). CD19xCD3 TransSkip K1 демонстрирует некоторый исходный пропуск экзонов (наличие нижней «активированной» полосы на дорожках 3 и 4), что согласуется с нашими результатами в параллельных конструкциях для CD3xGD2-HDD TransSkip.Эти результаты показывают, что созданный дизайн TransSkip одинаково работает со множеством различных трансгенов.
KRAS OncoSkip индуцирует секретируемую экспрессию CD19xCD3 Dimert в клетках, трансфицированных вектором AAV CD19xCD3 TransSkip K1.
Надосадоные жидкости собирали из трансфицированных клеток 293Т и тестировали на связывание с Т-клетками (интерференция с флуоресцентно меченными анти-CD3 антителами). Как показано на фиг.46, серая линия на верхней панели - это неокрашенные Т-клетки (Jurkat), темно-серая линия на верхней панели - полностью окрашенные Т-клетки с антителом к CD3. В качестве положительного контроля надосадочная жидкость клеток, трансфицированных конститутивно экспрессируемым CD19xCD3 TransJoin #1325, конкурировала за окрашивание с анти-CD3 (#1325). Надосадочная жидкость из клеток, трансфицированных CD19xCD3 TransSkip #1098 в присутствии контрольного морфолиноолигонуклеотида, демонстрировала некоторую исходную конкуренцию (KTS2-Inverted Ct1), соответствующую «утечке» мРНК Dimert с пропущенным экзоном, и была дополнительно индуцирована морфолиноолигонуклеотидом OncoSkip (KTS1; KTS2) до уровней, достигаемых с помощью конститутивного CD19xCD3 TransJoin.
Вариант сплайсинга CD19xCD3 TransSkip K3 исключает пропуск экзонов на исходном уровне, но поддерживает индуцируемый пропуск экзонов.
Как показано на фиг.47, клетки 293T трансфицировали конструкциями K1 и K3 CD19xCD3 TransSkip без и с антисмысловым морфолиноновым олигомером «OncoSkip» KTS2 или контрольным морфолиноолигонуклеотидом («InvtKTS2 Ct1») и сравнивали их с контролем «только эндо», который включает носитель для морфолиноолигонуклеотидов, но не содержит морфолиноолигонуклеотида. Транскрипты мРНК анализировали с помощью RT-PCR с обратной транскриптазой. В соответствии с параллельными конструкциями CD3xGD2-HDD TransJoin вариант K1 демонстрировал исходный пропуск экзонов. Напротив, варианты K3 не показали пропуска на исходном уровне («только эндо») или пропуска с контрольным морфолиноолигонуклеотидом. Пропуск наблюдался с KTS2 OncoSkip с вариантом K3 (#1168), что было подтверждено анализом экспрессии белка с помощью анализа связывания Т-клеток.
Индукция экспрессии CD19xCD3 Dimert из CD19xCD3 TransSkip K3, определенная по связыванию с Т-клетками, является целевым эффектом.
Надосадочные жидкости собирали из клеток 293T, трансфицированных CD3xGD2-HDD TransJoin (положительный контроль), и CD19xCD3 TransSkip, инкубированных без или с различными антисмысловыми морфолиновыми олигомерами, и проводили анализ связывания T-клеток человека (Jurkat) с помощью проточной цитометрии (конкуренция за связывание с флуоресцентно мечеными анти-CD3 антителами). Как показано на фиг.48, серый цвет на каждой из верхних панелей - неокрашенные Т-клетки, темно-серый - полностью окрашенные Т-клетки, розовый - полностью окрашенные Т-клетки в присутствии надосадочной жидкости от нетрансфицированных клеток 293Т, синий - конкурирующий сигнал надосадочной жидкости из трансфицированных клеток, конститутивно экспрессирующих CD19xCD3 TransJoin #1073. Все надосадочные жидкости из CD19xCD3 TransSkip K1 #11166 демонстрировали высокую экспрессию, эквивалентную положительному контролю CD19xCD3 TransJoin на исходном уровне и с контрольными морфолиновыми олигомерами, демонстрируя, что эта конструкция не подходит для контроля экспрессии гена. В отличие от опыта с CD3xGD2-HDD TransSkip и в соответствии с ним, вариант K3 #1168 CD19xCD3 TransSkip не показал экспрессии на исходном уровне (зеленый) или экспрессии, индуцированной контрольным морфолиноолигонуклеотидом (светло-темно-оранжевый, «IntCtl»), но показал дозозависимую экспрессию от морфолиноолигонуклеотида KTS2 KRAS OncoSkip (светлый и темно-фиолетовый).
Индукция экспрессии CD19xCD3 Dimert из CD19xCD3 TransSkip K3 является воспроизводимой.
Анализ пропуска экзонов и анализ связывания Т-клеток повторяли для подтверждения отсутствия утечки и наличия индуцибельности с помощью K3 CD19xCD3 TransSkip по сравнению с K1 CD19xCD3 TransSkip (фиг.49). Также см. фиг.48.
Воплощения
Воплощение 1: вектор для применения в генной терапии, включающий: первую полинуклеотидную последовательность, кодирующую первое антитело или его антигенсвязывающий фрагмент; и вторую полинуклеотидную последовательность, кодирующую второе антитело или его антигенсвязывающий фрагмент.
Воплощение 2: вектор по воплощению 1, где вектор представляет собой рекомбинантный вектор.
Воплощение 3: вектор по воплощению 1 или 2, где вектор представляет собой вирусный вектор.
Воплощение 4: вектор по любому из воплощений 1-3, где вирусный вектор представляет собой ретровирусный вектор.
Воплощение 5: вектор по любому из воплощений 1-4, где вирусный вектор представляет собой аденовирусный вектор, вектор аденоассоциированного вируса (AAV), лентивирусный вектор, вектор вируса лейкоза мышей («MLV»), Вектор вируса Эпштейна-Барр («EBV») или вектор вируса герпеса («HSV»).
Воплощение 6: вектор по любому из воплощений 1-5, в котором вектор AAV представляет собой AAV1, AAV2, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAV12, AAV13, AAV PHP.B, AAV rh74, или вектор AAV-DJ.
Вариант 7: вектор по варианту 6, где вектор AAV представляет собой AAV rh74 (номер доступа GenBank LP899424.1).
Воплощение 8: вектор по любому из воплощений 1-7, где первое антитело или его антигенсвязывающий фрагмент специфически связывается с активирующим антигеном на иммунной эффекторной клетке, а второе антитело или его антигенсвязывающий фрагмент связывается с опухолевым антигеном.
Воплощение 9: вектор по любому из воплощений 1-7, где первое антитело или его антигенсвязывающий фрагмент специфически связывается с опухолевым антигеном, а второе антитело или его антигенсвязывающий фрагмент связывается с активирующим антигеном на иммунной эффекторной клетке.
Воплощение 10: вектор по любому из воплощений 1-9, дополнительно содержащий третью полинуклеотидную последовательность, кодирующую третье антитело или его антигенсвязывающий фрагмент, где третье антитело или его антигенсвязывающий фрагмент связывается с активирующим антигеном на иммунной эффекторной клетке или опухолевым антигеном.
Воплощение 11: вектор по любому из воплощений 8-10, где иммунная эффекторная клетка включает дендритную клетку, клетку-натуральный киллер («NK»), макрофаг, Т-клетку или В-клетку.
Воплощение 12: вектор по любому из воплощений 8-11, где иммунная эффекторная клетка представляет собой Т-клетку или NK-клетку.
Воплощение 13: вектор по любому из воплощений 8-12, где активирующий антиген на иммунной эффекторной клетке включает CD3, CD2, CD4, CD8, CD19, LFA1, CD45, NKG2D, NKp44, NKp46, NKp30, DNAM, B7. -H3, CD20, CD22 или их комбинацию.
Воплощение 14: вектор по любому из воплощений 8-13, где опухолевый антиген содержит один или несколько из числа рецептора 2 эфрина типа A (EphA2), интерлейкина (IL)-13r альфа 2, EGFR VIII, PSMA, EpCAM, GD2 или GD3, фукозил GM1, PSCA, PLAC1, точечного разрыва саркомы, опухоли Вильмса 1, альфафетопротеина (AFP), карциноэмбрионального антигена (CEA), CA-125, MUC-1, эпителиального опухолевого антигена (ETA), тирозиназы, меланома- ассоциированного антигена (MAGE), антигена гематологической дифференцировки, поверхностного гликопротеина, ганглиозидов (GM2), рецептора фактора роста, стромального антигена, сосудистого антигена, орфанного рецептора 1, подобного рецепторной тирозинкиназе (ROR1), мезотелина, CD38, CD123, рецептора 2 эпидермального фактора роста человека (HER2), антигена созревания В-клеток (BCMA), белка активации фибробластов (FAP) альфа или их комбинации.
Воплощение 15: вектор по любому из воплощений 1-14, где первое антитело или его антигенсвязывающий фрагмент и второе антитело или его антигенсвязывающий фрагмент образуют dimert.
Воплощение 16: вектор по воплощению 15, где dimert представляет собой биспецифическое антитело.
Воплощение 17: вектор по воплощению 15, где dimert представляет собой триспецифическое антитело.
Воплощение 18: вектор согласно воплощению 15 или 16, где биспецифическое антитело содержит полипептидную последовательность, по меньшей мере на 95% идентичную любой из SEQ ID NO: 13 или 15.
Воплощение 19: вектор согласно воплощению 15 или 17, где триспецифическое антитело содержит полипептидную последовательность, по меньшей мере на 95% идентичную SEQ ID NO: 11.
Воплощение 20: вектор по любому из воплощений 1-19, где вектор дополнительно содержит полинуклеотидную последовательность, кодирующую секреторный пептид.
Воплощение 21: вектор согласно воплощению 20, где секреторный пептид содержит секреторную консенсусную последовательность.
Воплощение 22: вектор по воплощению 20 или 21, где секреторная консенсусная последовательность имеет по меньшей мере 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательностей с SEQ ID NO: 51.
Воплощение 23: вектор по воплощению 20 или 21, где секреторная консенсусная последовательность состоит из SEQ ID NO: 51.
Воплощение 24: вектор по любому из воплощений 20-23, где секреторная консенсусная последовательность кодируется полинуклеотидом, содержащим SEQ ID NO: 52, или его эквивалентом.
Воплощение 25: вектор по любому из воплощений 20-24, где секреторная консенсусная последовательность дополнительно содержит один, два, три, четыре или более остатков на С-конце последовательности.
Воплощение 26: вектор по любому из воплощений 20-25, где секреторная консенсусная последовательность дополнительно содержит один, два, три, четыре или более остатков Ala на С-конце последовательности.
Воплощение 27: вектор по любому из воплощений 20-26, где секреторная консенсусная последовательность дополнительно содержит один, два или три остатка Ala на С-конце последовательности.
Воплощение 28: вектор по любому из воплощений 20-27, где секреторная консенсусная последовательность дополнительно содержит два остатка Ala на С-конце последовательности.
Воплощение 29: вектор по воплощению 28, где секреторная консенсусная последовательность имеет по меньшей мере 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательностей с SEQ ID NO: 55 или состоит из SEQ ID NO: 55.
Воплощение 30: вектор по воплощению 28 или 29, где секреторная консенсусная последовательность кодируется полинуклеотидом, содержащим SEQ ID NO: 56, или его эквивалентом.
Воплощение 31: вектор по любому из воплощений 20-30, где секреторная консенсусная последовательность дополнительно содержит один остаток Ala на С-конце последовательности.
Воплощение 32: вектор по воплощению 31, где секреторная консенсусная последовательность имеет по меньшей мере 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательностей с SEQ ID NO: 53 или состоит из SEQ ID NO: 53.
Воплощение 33: вектор по воплощению 31 или 32, где секреторная консенсусная последовательность кодируется полинуклеотидом, содержащим SEQ ID NO: 54, или его эквивалентом.
Воплощение 34: вектор по любому из воплощений 1-33, где секреторная консенсусная последовательность модулирует экспрессию и/или секрецию dimert.
Воплощение 35: вектор по любому из воплощений 1-34, где секреторная консенсусная последовательность усиливает экспрессию и/или секрецию dimert.
Воплощение 36: вектор по любому из воплощений 1-35, где вектор дополнительно содержит полинуклеотидную последовательность, кодирующую домен димеризации.
Воплощение 37: вектор согласно воплощению 36, где домен димеризации включает домен димеризации ядерного фактора 1α гепатоцитов человека (HNF1α).
Воплощение 38: вектор согласно воплощению 37, где домен димеризации HNF1α содержит полипептидную последовательность, имеющую по меньшей мере 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичности последовательностей с SEQ ID NO: 47.
Воплощение 39: вектор по любому из воплощений 1-38, где вектор дополнительно содержит промотор.
Воплощение 40: вектор по воплощению 39, где промотор представляет собой конститутивный промотор.
Воплощение 41: вектор по воплощению 39 или 40, где промотор представляет собой тканеспецифический промотор.
Воплощение 42: вектор по любому из воплощений 39-41, где промотор включает промотор LTR вируса саркомы Рауса (RSV), промотор цитомегаловируса (CMV), промотор SV40, промотор дигидрофолатредуктазы, промотор β-актина, промотор фосфоглицеринкиназы (PGK), промотор U6, промотор короткой формы EF1 альфа (EFS), промотор фосфоглицераткиназы (PGK), промотор убиквитина C (UbiC), альфа-1-антитрипсин, промотор вируса некроза селезенки (SFFV) или промотор куриного бета-актина (CBA).
Воплощение 43: вектор по любому из воплощений 39-42, где промотор представляет собой EFS, необязательно содержащий SEQ ID NO: 49 или ее эквивалент.
Воплощение 44: вектор по любому из воплощений 1-43, где вектор дополнительно содержит энхансер.
Воплощение 45: вектор согласно воплощению 44, где энхансер представляет собой энхансер RSV, энхансер CMV и энхансер MERII α-фетопротеина.
Воплощение 46: вектор по любому из воплощений 1-45, где вектор также содержит один или несколько дополнительных регуляторных элементов.
Воплощение 47: вектор по любому из воплощений 1-46, где вектор включает регуляторный элемент, содержащий посттранскрипционный регуляторный элемент (WPRE) вируса гепатита сурка (WHP), необязательно SEQ ID NO: 50 или ее эквивалент.
Воплощение 48: вектор по любому из воплощений 1-47, где вектор содержит 5’ инвертированный концевой повтор (ITR) и 3’ ITR.
Воплощение 49: вектор по любому из воплощений 1-48, где вектор содержит последовательность, представленную в SEQ ID NO: 4, 6, 8, 12, 14, 16-23, 30-33 или 40-46.
Воплощение 50: композиция содержит вектор по любому из воплощений 1-49 и носитель, необязательно фармацевтически приемлемый носитель.
Воплощение 51: композиция по воплощению 50, где композиция составлена для системного введения.
Воплощение 52: композиция по воплощению 50, где композиция составлена для местного введения.
Воплощение 53: композиция по любому из воплощений 50-52, где композиция составлена для парентерального введения.
Воплощение 54: способ лечения онкологического заболевания у объекта, нуждающегося в этом, включающий введение объекту эффективного количества вектора по любому из воплощений 1-49 или фармацевтической композиции по любому из воплощений 50-53, где вектор экспрессирует терапевтическое противоопухолевое антитело или его антигенсвязывающий фрагмент.
Воплощение 55: способ воплощения 54 дополнительно включает введение объекту противоопухолевого агента.
Воплощение 56: способ по воплощению 55, где противоопухолевый агент включает агент, выбранный из пептида, полипептида, молекулы нуклеиновой кислоты, низкомолекулярного вещества, вирусной частицы или их комбинаций.
Воплощение 57: способ по воплощению 56, где вирусная частица представляет собой онколитическую частицу HSV.
Воплощение 58: способ по любому из воплощений 54-57, где объектом является млекопитающее.
Воплощение 59: способ по любому из воплощений 54-58, где объектом является человек.
Воплощение 60: способ получения биспецифического антитела или триспецифического антитела в клетке, включающий контакт клетки с вектором по любому из воплощений 1-49.
Воплощение 61: способ по воплощению 60, где приведение в контакт включает трансфекцию, инфекцию, трансформацию, электропорацию, инъекцию, микроинъекцию или их комбинацию.
Воплощение 62: способ воплощения 60 или 61, где клетка включает фибробласт, скелетную клетку, эпителиальную клетку, мышечную клетку, нервную клетку, эндокринную клетку, меланоцит, клетку крови или их комбинацию.
Воплощение 63: способ по любому из воплощений 60-62, где биспецифическое антитело содержит полипептидную последовательность, по меньшей мере на 95% идентичную SEQ ID NO: 13 или 15.
Воплощение 64: способ по любому из воплощений 60-62, где триспецифическое антитело содержит полипептидную последовательность, по меньшей мере на 95% идентичную SEQ ID NO: 11.
Воплощение 65: способ по любому из воплощений 60-62, где биспецифическое антитело кодируется полинуклеотидной последовательностью, по меньшей мере на 95% идентичную SEQ ID NO: 14, 16, 22, 23, 30-33 или 40- 46.
Воплощение 66: способ по любому из воплощений 60-62, где триспецифическое антитело кодируется полинуклеотидной последовательностью, по меньшей мере на 95% идентичную SEQ ID NO: 12.
Воплощение 67: набор, содержащий вектор по любому из воплощений 1-49 или фармацевтическую композицию по любому из воплощений 50-53.
Воплощение 68: набор по воплощению 67 дополнительно включает инструктивный материал.
Эквиваленты
Следует понимать, что хотя раскрытие было описано в связи с вышеупомянутыми воплощениями, вышеприведенное описание и примеры предназначены для иллюстрации, а не для ограничения объема раскрытия. Другие аспекты, преимущества и модификации в пределах объема раскрытия будут очевидны специалистам в данной области техники, к которой относится раскрытие.
Если не указано иное, все технические и научные термины, используемые в данном документе, имеют то же значение, которое обычно понимается специалистом в области, к которой относится это раскрытие. Все нуклеотидные последовательности, представленные в настоящем документе, представлены в направлении от 5 ‘до 3’.
Воплощения, иллюстративно описанные в данном документе, могут подходящим образом применяться на практике при отсутствии какого-либо элемента или элементов, ограничения или ограничений, конкретно не раскрытых в данном документе. Таким образом, например, термины «содержащий», «включающий», «охватывающий» и т.д. следует читать широко и без ограничения. Кроме того, используемые в данном документе термины и выражения использовались как термины описания, а не как ограничения, и нет намерения в применении таких терминов и выражений для исключения каких-либо эквивалентов, показанных и описанных функций или их частей, но при этом признается, что в объеме раскрытия возможны различные модификации.
Таким образом, следует понимать, что, хотя настоящее раскрытие было конкретно раскрыто посредством конкретных воплощений и дополнительных функций, к модификациям, усовершенствованию и вариациям раскрытых в нем воплощений могут прибегать специалисты в данной области техники, и что такие модификации, улучшения и вариации считаются находящимися в рамках этого раскрытия. Материалы, способы и примеры, представленные в данном документе, являются репрезентативными для конкретных воплощений, являются иллюстративными и не предназначены для ограничения объема раскрытия.
Объем раскрытия был описан в данном документе широко и в общих чертах. Каждый из более узких видов и подродовых групп, подпадающих под общее раскрытие, также составляет часть раскрытия. Это включает общее описание с условием или отрицательным ограничением, исключающим любой объект из рода, независимо от того, указан ли специально или не указан в данном документе исключенный материал.
Кроме того, если признаки или аспекты раскрытия описаны в терминах групп Маркуша, специалисты в данной области техники поймут, что воплощения раскрытия также могут быть описаны в терминах любого отдельного члена или подгруппы членов группы Маркуша.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110>RESEARCH INSTITUTE AT NATIONWIDE CHILDREN'S HOSPITAL
<120>ВИРУС-КОДИРУЕМЫЕ РЕГУЛИРУЕМЫЕ АГЕНТЫ, СВЯЗЫВАЮЩИЕ T-КЛЕТКИ (CATVERT) ИЛИ
NK-КЛЕТКИ (CATVERN), НАЦЕЛЕННЫЕ НА ЗЛОКАЧЕСТВЕННЫЕ ОПУХОЛИ
<130>106887-7610
<140>PCT/US2020/018806
<141>2020-02-19
<150>62/808,264
<151>2019-02-20
<160>71
<170>Версия PatentIn 3.5
<210>1
<211>24
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
олигонуклеотид
<220>
<221>modified_base
<222>(4)..(4)
<223>a, c, t, g, неизвестное или другое
<220>
<221>modified_base
<222>(8)..(8)
<223>a, c, t, g, неизвестное или другое
<220>
<221>modified_base
<222>(12)..(12)
<223>a, c, t, g, неизвестное или другое
<220>
<221>modified_base
<222>(16)..(16)
<223>a, c, t, g, неизвестное или другое
<220>
<221>modified_base
<222>(20)..(20)
<223>a, c, t, g, неизвестное или другое
<220>
<221>modified_base
<222>(24)..(24)
<223>a, c, t, g, неизвестное или другое
<400>1
taantagntg antagntaan tgan 24
<210>2
<211>24
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
олигонуклеотид
<400>2
taattagttg attagttaat tgat 24
<210>3
<211>162
<212>PRT
<213>Неизвестный
<220>
<223>Описание неизвестного:
последовательность IL-21
<400>3
Met Arg Ser Ser Pro Gly Asn Met Glu Arg Ile Val Ile Cys Leu Met
1 5 10 15
Val Ile Phe Leu Gly Thr Leu Val His Lys Ser Ser Ser Gln Gly Gln
20 25 30
Asp Arg His Met Ile Arg Met Arg Gln Leu Ile Asp Ile Val Asp Gln
35 40 45
Leu Lys Asn Tyr Val Asn Asp Leu Val Pro Glu Phe Leu Pro Ala Pro
50 55 60
Glu Asp Val Glu Thr Asn Cys Glu Trp Ser Ala Phe Ser Cys Phe Gln
65 70 75 80
Lys Ala Gln Leu Lys Ser Ala Asn Thr Gly Asn Asn Glu Arg Ile Ile
85 90 95
Asn Val Ser Ile Lys Lys Leu Lys Arg Lys Pro Pro Ser Thr Asn Ala
100 105 110
Gly Arg Arg Gln Lys His Arg Leu Thr Cys Pro Ser Cys Asp Ser Tyr
115 120 125
Glu Lys Lys Pro Pro Lys Glu Phe Leu Glu Arg Phe Lys Ser Leu Leu
130 135 140
Gln Lys Met Ile His Gln His Leu Ser Ser Arg Thr His Gly Ser Glu
145 150 155 160
Asp Ser
<210>4
<211>486
<212>DNA
<213>Неизвестный
<220>
<223>Описание неизвестного:
последовательность IL-21
<400>4
atgagaagca gccccggcaa catggagaga atcgtgatct gcctgatggt gatcttcctg 60
ggcaccctgg tgcacaagag cagcagccag ggccaggaca gacacatgat cagaatgaga 120
cagctgatcg acatcgtgga ccagctgaag aactacgtga acgacctggt gcccgagttc 180
ctgcccgccc ccgaggacgt ggagaccaac tgcgagtgga gcgccttcag ctgcttccag 240
aaggcccagc tgaagagcgc caacaccggc aacaacgaga gaatcatcaa cgtgagcatc 300
aagaagctga agagaaagcc ccccagcacc aacgccggca gaagacagaa gcacagactg 360
acctgcccca gctgcgacag ctacgagaag aagcccccca aggagttcct ggagagattc 420
aagagcctgc tgcagaagat gatccaccag cacctgagca gcagaaccca cggcagcgag 480
gacagc 486
<210>5
<211>274
<212>PRT
<213>Неизвестный
<220>
<223>Описание неизвестного:
последовательность MICA
<400>5
Glu Pro His Ser Leu Arg Tyr Asn Leu Thr Val Leu Ser Trp Asp Gly
1 5 10 15
Ser Val Gln Ser Gly Phe Leu Ala Glu Val His Leu Asp Gly Gln Pro
20 25 30
Phe Leu Arg Tyr Asp Arg Gln Lys Cys Arg Ala Lys Pro Gln Gly Gln
35 40 45
Trp Ala Glu Asp Val Leu Gly Asn Lys Thr Trp Asp Arg Glu Thr Arg
50 55 60
Asp Leu Thr Gly Asn Gly Lys Asp Leu Arg Met Thr Leu Ala His Ile
65 70 75 80
Lys Asp Gln Lys Glu Gly Leu His Ser Leu Gln Glu Ile Arg Val Cys
85 90 95
Glu Ile His Glu Asp Asn Ser Thr Arg Ser Ser Gln His Phe Tyr Tyr
100 105 110
Asp Gly Glu Leu Phe Leu Ser Gln Asn Leu Glu Thr Glu Glu Trp Thr
115 120 125
Val Pro Gln Ser Ser Arg Ala Gln Thr Leu Ala Met Asn Val Arg Asn
130 135 140
Phe Leu Lys Glu Asp Ala Met Lys Thr Lys Thr His Tyr His Ala Met
145 150 155 160
His Ala Asp Cys Leu Gln Glu Leu Arg Arg Tyr Leu Glu Ser Ser Val
165 170 175
Val Leu Arg Arg Thr Val Pro Pro Met Val Asn Val Thr Arg Ser Glu
180 185 190
Ala Ser Glu Gly Asn Ile Thr Val Thr Cys Arg Ala Ser Ser Phe Tyr
195 200 205
Pro Arg Asn Ile Ile Leu Thr Trp Arg Gln Asp Gly Val Ser Leu Ser
210 215 220
His Asp Thr Gln Gln Trp Gly Asp Val Leu Pro Asp Gly Asn Gly Thr
225 230 235 240
Tyr Gln Thr Trp Val Ala Thr Arg Ile Cys Arg Gly Glu Glu Gln Arg
245 250 255
Phe Thr Cys Tyr Met Glu His Ser Gly Asn His Ser Thr His Pro Val
260 265 270
Pro Ser
<210>6
<211>822
<212>DNA
<213>Неизвестный
<220>
<223>Описание неизвестного:
последовательность MICA
<400>6
gagccccaca gtcttcgtta taacctcacg gtgctgtcct gggatggatc tgtgcagtca 60
gggtttcttg ctgaggtaca tctggatggt cagcccttcc tgcgctatga caggcagaaa 120
tgcagggcaa agccccaggg acagtgggca gaagatgtcc tgggaaataa gacatgggac 180
agagagacca gggacttgac agggaacgga aaggacctca ggatgaccct ggctcatatc 240
aaggaccaga aagaaggctt gcattccctc caggagatta gggtctgtga gatccatgaa 300
gacaacagca ccaggagctc ccagcatttc tactacgatg gggagctctt cctctcccaa 360
aacctggaga ctgaggaatg gacagtgccc cagtcctcca gagctcagac cttggccatg 420
aacgtcagga atttcttgaa ggaagatgcc atgaagacca agacacacta tcacgctatg 480
catgcagact gcctgcagga actacggcga tatctagaat ccagcgtagt cctgaggaga 540
acagtgcccc ccatggtgaa tgtcacccgc agcgaggcct cagagggcaa catcaccgtg 600
acatgcaggg cttccagctt ctatccccgg aatatcatac tgacctggcg tcaggatggg 660
gtatctttga gccacgacac ccagcagtgg ggggatgtcc tgcctgatgg gaatggaacc 720
taccagacct gggtggccac caggatttgc cgaggagagg agcagaggtt cacctgctac 780
atggaacaca gcgggaatca cagcactcac cctgtgccct ct 822
<210>7
<211>274
<212>PRT
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
полипептид
<400>7
Glu Pro His Ser Leu Arg Tyr Asn Leu Thr Val Leu Ser Trp Asp Gly
1 5 10 15
Ser Val Gln Ser Gly Phe Leu Ala Glu Val His Leu Asp Gly Gln Pro
20 25 30
Phe Leu Arg Cys Asp Arg Gln Lys Cys Arg Ala Lys Pro Gln Gly Gln
35 40 45
Trp Ala Glu Asp Val Leu Gly Asn Lys Thr Trp Asp Arg Glu Thr Arg
50 55 60
Asp Leu Thr Gly Asn Gly Lys Asp Leu Arg Met Thr Leu Ala His Ile
65 70 75 80
Lys Asp Gln Lys Glu Gly Leu His Ser Leu Gln Glu Ile Arg Val Cys
85 90 95
Glu Ile His Glu Asp Asn Ser Thr Arg Ser Ser Gln His Phe Tyr Tyr
100 105 110
Asp Gly Glu Leu Phe Leu Ser Gln Asn Leu Glu Thr Glu Glu Trp Thr
115 120 125
Met Pro Gln Ser Ser Arg Ala Gln Thr Leu Ala Met Asn Ile Arg Asn
130 135 140
Phe Leu Lys Glu Asp Ala Met Lys Thr Lys Thr His Tyr His Ala Met
145 150 155 160
His Ala Asp Cys Leu Gln Glu Leu Arg Arg Tyr Leu Lys Ser Gly Val
165 170 175
Val Leu Arg Arg Thr Val Pro Pro Met Val Asn Val Thr Arg Ser Glu
180 185 190
Ala Ser Glu Gly Asn Ile Thr Val Thr Cys Arg Ala Ser Gly Phe Tyr
195 200 205
Pro Trp Asn Ile Thr Leu Ser Trp Arg Gln Asp Gly Val Ser Leu Ser
210 215 220
His Asp Thr Gln Gln Trp Gly Asp Val Leu Pro Asp Gly Asn Gly Thr
225 230 235 240
Tyr Gln Thr Trp Val Ala Thr Arg Ile Cys Gln Gly Glu Glu Gln Arg
245 250 255
Phe Thr Cys Tyr Met Glu His Ser Gly Asn His Ser Thr His Pro Val
260 265 270
Pro Ser
<210>8
<211>822
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
полинуклеотид
<400>8
gagccccaca gtcttcgtta taacctcacg gtgctgtcct gggatggatc tgtgcagtca 60
gggtttctcg ctgaggtaca tctggatggt cagcccttcc tgcgctgtga caggcagaaa 120
tgcagggcaa agccccaggg acagtgggca gaagatgtcc tgggaaataa gacatgggac 180
agagagacca gggacttgac agggaacgga aaggacctca ggatgaccct ggctcatatc 240
aaggaccaga aagaaggctt gcattccctc caggagatta gggtctgtga gatccatgaa 300
gacaacagca ccaggagctc ccagcatttc tactacgatg gggagctctt cctctcccaa 360
aacctggaga ctgaggaatg gacaatgccc cagtcctcca gagctcagac cttggccatg 420
aacatcagga atttcttgaa ggaagatgcc atgaagacca agacacacta tcacgctatg 480
catgcagact gcctgcagga actacggcga tatctaaaat ccggcgtagt cctgaggaga 540
acagtgcccc ccatggtgaa tgtcacccgc agcgaggcct cagagggcaa cattaccgtg 600
acatgcaggg cttctggctt ctatccctgg aatatcacac tgagctggcg tcaggatggg 660
gtatctttga gccacgacac ccagcagtgg ggggatgtcc tgcctgatgg gaatggaacc 720
taccagacct gggtggccac caggatttgc caaggagagg agcagaggtt cacctgctac 780
atggaacaca gcgggaatca cagcactcac cctgtgccct ct 822
<210>9
<211>15
<212>PRT
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
пептид
<400>9
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210>10
<211>45
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
олигонуклеотид
<400>10
ggcggcggcg gcagcggcgg cggcggcagc ggcggcggcg gcagc 45
<210>11
<211>732
<212>PRT
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
полипептид
<400>11
Met Trp Trp Arg Leu Trp Trp Leu Leu Leu Leu Leu Leu Leu Leu Trp
1 5 10 15
Pro Met Val Trp Ala Ala Arg Ser Ser Pro Gly Asn Met Glu Arg Ile
20 25 30
Val Ile Cys Leu Met Val Ile Phe Leu Gly Thr Leu Val His Lys Ser
35 40 45
Ser Ser Gln Gly Gln Asp Arg His Met Ile Arg Met Arg Gln Leu Ile
50 55 60
Asp Ile Val Asp Gln Leu Lys Asn Tyr Val Asn Asp Leu Val Pro Glu
65 70 75 80
Phe Leu Pro Ala Pro Glu Asp Val Glu Thr Asn Cys Glu Trp Ser Ala
85 90 95
Phe Ser Cys Phe Gln Lys Ala Gln Leu Lys Ser Ala Asn Thr Gly Asn
100 105 110
Asn Glu Arg Ile Ile Asn Val Ser Ile Lys Lys Leu Lys Arg Lys Pro
115 120 125
Pro Ser Thr Asn Ala Gly Arg Arg Gln Lys His Arg Leu Thr Cys Pro
130 135 140
Ser Cys Asp Ser Tyr Glu Lys Lys Pro Pro Lys Glu Phe Leu Glu Arg
145 150 155 160
Phe Lys Ser Leu Leu Gln Lys Met Ile His Gln His Leu Ser Ser Arg
165 170 175
Thr His Gly Ser Glu Asp Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
180 185 190
Ser Gly Gly Gly Gly Ser Glu Pro His Ser Leu Arg Tyr Asn Leu Thr
195 200 205
Val Leu Ser Trp Asp Gly Ser Val Gln Ser Gly Phe Leu Ala Glu Val
210 215 220
His Leu Asp Gly Gln Pro Phe Leu Arg Cys Asp Arg Gln Lys Cys Arg
225 230 235 240
Ala Lys Pro Gln Gly Gln Trp Ala Glu Asp Val Leu Gly Asn Lys Thr
245 250 255
Trp Asp Arg Glu Thr Arg Asp Leu Thr Gly Asn Gly Lys Asp Leu Arg
260 265 270
Met Thr Leu Ala His Ile Lys Asp Gln Lys Glu Gly Leu His Ser Leu
275 280 285
Gln Glu Ile Arg Val Cys Glu Ile His Glu Asp Asn Ser Thr Arg Ser
290 295 300
Ser Gln His Phe Tyr Tyr Asp Gly Glu Leu Phe Leu Ser Gln Asn Leu
305 310 315 320
Glu Thr Glu Glu Trp Thr Met Pro Gln Ser Ser Arg Ala Gln Thr Leu
325 330 335
Ala Met Asn Ile Arg Asn Phe Leu Lys Glu Asp Ala Met Lys Thr Lys
340 345 350
Thr His Tyr His Ala Met His Ala Asp Cys Leu Gln Glu Leu Arg Arg
355 360 365
Tyr Leu Lys Ser Gly Val Val Leu Arg Arg Thr Val Pro Pro Met Val
370 375 380
Asn Val Thr Arg Ser Glu Ala Ser Glu Gly Asn Ile Thr Val Thr Cys
385 390 395 400
Arg Ala Ser Gly Phe Tyr Pro Trp Asn Ile Thr Leu Ser Trp Arg Gln
405 410 415
Asp Gly Val Ser Leu Ser His Asp Thr Gln Gln Trp Gly Asp Val Leu
420 425 430
Pro Asp Gly Asn Gly Thr Tyr Gln Thr Trp Val Ala Thr Arg Ile Cys
435 440 445
Gln Gly Glu Glu Gln Arg Phe Thr Cys Tyr Met Glu His Ser Gly Asn
450 455 460
His Ser Thr His Pro Val Pro Ser Gly Gly Gly Gly Ser Gly Gly Gly
465 470 475 480
Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser Gly Pro
485 490 495
Glu Leu Val Lys Pro Gly Ala Ser Val Lys Ile Ser Cys Lys Thr Ser
500 505 510
Gly Tyr Lys Phe Thr Glu Tyr Thr Met His Trp Val Lys Gln Ser His
515 520 525
Gly Lys Cys Leu Glu Trp Ile Gly Gly Ile Asn Pro Asn Asn Gly Gly
530 535 540
Thr Asn Tyr Asn Gln Lys Phe Lys Gly Lys Ala Thr Leu Thr Val Asp
545 550 555 560
Lys Ser Ser Ser Thr Ala Tyr Met Glu Leu Arg Ser Leu Thr Ser Glu
565 570 575
Asp Ser Ala Val Tyr Tyr Cys Ala Arg Asp Thr Thr Val Pro Tyr Ala
580 585 590
Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly Gly Gly Gly
595 600 605
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Glu Leu Thr
610 615 620
Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu Lys Val Thr Met
625 630 635 640
Thr Cys Ser Ala Ser Ser Ser Ile Ser Tyr Met His Trp Tyr Gln Gln
645 650 655
Lys Pro Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp Thr Ser Lys Leu
660 665 670
Ala Ser Ser Val Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Ser
675 680 685
Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu Asp Ala Ala Thr Tyr
690 695 700
Tyr Cys His Gln Arg Ser Ser Tyr Pro Leu Thr Phe Gly Cys Gly Thr
705 710 715 720
Lys Leu Glu Ile Lys Arg Ala Ser Thr Lys Gly Pro
725 730
<210>12
<211>2199
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
полинуклеотид
<400>12
atgtggtgga gactgtggtg gctgctgctg ctgctgctgc tgctgtggcc catggtgtgg 60
gccgccagaa gcagccccgg caacatggag agaatcgtga tctgcctgat ggtgatcttc 120
ctgggcaccc tggtgcacaa gagcagcagc cagggccagg acagacacat gatcagaatg 180
agacagctga tcgacatcgt ggaccagctg aagaactacg tgaacgacct ggtgcccgag 240
ttcctgcccg cccccgagga cgtggagacc aactgcgagt ggagcgcctt cagctgcttc 300
cagaaggccc agctgaagag cgccaacacc ggcaacaacg agagaatcat caacgtgagc 360
atcaagaagc tgaagagaaa gccccccagc accaacgccg gcagaagaca gaagcacaga 420
ctgacctgcc ccagctgcga cagctacgag aagaagcccc ccaaggagtt cctggagaga 480
ttcaagagcc tgctgcagaa gatgatccac cagcacctga gcagcagaac ccacggcagc 540
gaggacagcg gcggcggcgg cagcggcggc ggcggcagcg gcggcggcgg cagcgagccc 600
cacagtcttc gttataacct cacggtgctg tcctgggatg gatctgtgca gtcagggttt 660
ctcgctgagg tacatctgga tggtcagccc ttcctgcgct gtgacaggca gaaatgcagg 720
gcaaagcccc agggacagtg ggcagaagat gtcctgggaa ataagacatg ggacagagag 780
accagggact tgacagggaa cggaaaggac ctcaggatga ccctggctca tatcaaggac 840
cagaaagaag gcttgcattc cctccaggag attagggtct gtgagatcca tgaagacaac 900
agcaccagga gctcccagca tttctactac gatggggagc tcttcctctc ccaaaacctg 960
gagactgagg aatggacaat gccccagtcc tccagagctc agaccttggc catgaacatc 1020
aggaatttct tgaaggaaga tgccatgaag accaagacac actatcacgc tatgcatgca 1080
gactgcctgc aggaactacg gcgatatcta aaatccggcg tagtcctgag gagaacagtg 1140
ccccccatgg tgaatgtcac ccgcagcgag gcctcagagg gcaacattac cgtgacatgc 1200
agggcttctg gcttctatcc ctggaatatc acactgagct ggcgtcagga tggggtatct 1260
ttgagccacg acacccagca gtggggggat gtcctgcctg atgggaatgg aacctaccag 1320
acctgggtgg ccaccaggat ttgccaagga gaggagcaga ggttcacctg ctacatggaa 1380
cacagcggga atcacagcac tcaccctgtg ccctctggcg gcggcggcag cggcggcggc 1440
ggcagcggcg gcggcggcag ccaggtgcag ctgcagcaga gcggccccga gctggtgaag 1500
cccggcgcca gcgtgaagat cagctgcaag accagcggct acaagttcac cgagtacacc 1560
atgcactggg tgaagcagag ccacggcaag tgcctggagt ggatcggcgg catcaacccc 1620
aacaacggcg gcaccaacta caaccagaag ttcaagggca aggccaccct gaccgtggac 1680
aagagcagca gcaccgccta catggagctg agaagcctga ccagcgagga cagcgccgtg 1740
tactactgcg ccagagacac caccgtgccc tacgcctact ggggccaggg caccaccgtg 1800
accgtgagca gcggcggcgg cggcagcggc ggcggcggca gcggcggcgg cggcagcgac 1860
atcgagctga cccagagccc cgccatcatg agcgccagcc ccggcgagaa ggtgaccatg 1920
acctgcagcg ccagcagcag catcagctac atgcactggt accagcagaa gcccggcacc 1980
agccccaaga gatggatcta cgacaccagc aagctggcca gcagcgtgcc cgccagattc 2040
agcggcagcg gcagcggcac cagctacagc ctgaccatca gcagcatgga ggccgaggac 2100
gccgccacct actactgcca ccagagaagc agctaccccc tgaccttcgg ctgcggcacc 2160
aagctggaga tcaagagagc cagcaccaag ggcccctag 2199
<210>13
<211>572
<212>PRT
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
полипептид
<400>13
Met Trp Trp Arg Leu Trp Trp Leu Leu Leu Leu Leu Leu Leu Leu Trp
1 5 10 15
Pro Met Val Trp Ala Ala Gln Val Gln Leu Gln Gln Ser Gly Pro Glu
20 25 30
Leu Val Lys Pro Gly Ala Ser Val Lys Ile Ser Cys Lys Thr Ser Gly
35 40 45
Tyr Lys Phe Thr Glu Tyr Thr Met His Trp Val Lys Gln Ser His Gly
50 55 60
Lys Cys Leu Glu Trp Ile Gly Gly Ile Asn Pro Asn Asn Gly Gly Thr
65 70 75 80
Asn Tyr Asn Gln Lys Phe Lys Gly Lys Ala Thr Leu Thr Val Asp Lys
85 90 95
Ser Ser Ser Thr Ala Tyr Met Glu Leu Arg Ser Leu Thr Ser Glu Asp
100 105 110
Ser Ala Val Tyr Tyr Cys Ala Arg Asp Thr Thr Val Pro Tyr Ala Tyr
115 120 125
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Gly Gly Gly Gly Ser
130 135 140
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Glu Leu Thr Gln
145 150 155 160
Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu Lys Val Thr Met Thr
165 170 175
Cys Ser Ala Ser Ser Ser Ile Ser Tyr Met His Trp Tyr Gln Gln Lys
180 185 190
Pro Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp Thr Ser Lys Leu Ala
195 200 205
Ser Ser Val Pro Ala Arg Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr
210 215 220
Ser Leu Thr Ile Ser Ser Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr
225 230 235 240
Cys His Gln Arg Ser Ser Tyr Pro Leu Thr Phe Gly Cys Gly Thr Lys
245 250 255
Leu Glu Ile Lys Arg Ala Ser Thr Lys Gly Pro Gly Gly Gly Gly Ser
260 265 270
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Val Gln
275 280 285
Ser Gly Gly Gly Val Val Gln Pro Gly Arg Ser Leu Arg Leu Ser Cys
290 295 300
Lys Ala Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Arg
305 310 315 320
Gln Ala Pro Gly Lys Gly Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser
325 330 335
Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Arg Phe Thr Ile
340 345 350
Ser Arg Asp Asn Ser Lys Asn Thr Ala Phe Leu Gln Met Asp Ser Leu
355 360 365
Arg Pro Glu Asp Thr Gly Val Tyr Phe Cys Ala Arg Tyr Tyr Asp Asp
370 375 380
His Tyr Cys Leu Asp Tyr Trp Gly Gln Gly Thr Pro Val Thr Val Ser
385 390 395 400
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
405 410 415
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
420 425 430
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
435 440 445
Asn Trp Tyr Gln Gln Thr Pro Gly Lys Ala Pro Lys Arg Trp Ile Tyr
450 455 460
Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
465 470 475 480
Gly Ser Gly Thr Asp Tyr Thr Phe Thr Ile Ser Ser Leu Gln Pro Glu
485 490 495
Asp Ile Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Phe Thr
500 505 510
Phe Gly Gln Gly Thr Lys Leu Gln Ile Thr Arg Thr Pro Leu Gly Asp
515 520 525
Thr Thr His Thr Ser Gly Met Val Ser Lys Leu Ser Gln Leu Gln Thr
530 535 540
Glu Leu Leu Ala Ala Leu Leu Glu Ser Gly Leu Ser Lys Glu Ala Leu
545 550 555 560
Ile Gln Ala Leu Gly Glu Gly Ser Gly Gly Ala Pro
565 570
<210>14
<211>1719
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
полинуклеотид
<400>14
atgtggtgga gactgtggtg gctgctgctg ctgctgctgc tgctgtggcc catggtgtgg 60
gccgcccagg tgcagctgca gcagagcggc cccgagctgg tgaagcccgg cgccagcgtg 120
aagatcagct gcaagaccag cggctacaag ttcaccgagt acaccatgca ctgggtgaag 180
cagagccacg gcaagtgcct ggagtggatc ggcggcatca accccaacaa cggcggcacc 240
aactacaacc agaagttcaa gggcaaggcc accctgaccg tggacaagag cagcagcacc 300
gcctacatgg agctgagaag cctgaccagc gaggacagcg ccgtgtacta ctgcgccaga 360
gacaccaccg tgccctacgc ctactggggc cagggcacca ccgtgaccgt gagcagcggc 420
ggcggcggca gcggcggcgg cggcagcggc ggcggcggca gcgacatcga gctgacccag 480
agccccgcca tcatgagcgc cagccccggc gagaaggtga ccatgacctg cagcgccagc 540
agcagcatca gctacatgca ctggtaccag cagaagcccg gcaccagccc caagagatgg 600
atctacgaca ccagcaagct ggccagcagc gtgcccgcca gattcagcgg cagcggcagc 660
ggcaccagct acagcctgac catcagcagc atggaggccg aggacgccgc cacctactac 720
tgccaccaga gaagcagcta ccccctgacc ttcggctgcg gcaccaagct ggagatcaag 780
agagccagca ccaagggccc cggcggcggc ggcagcggcg gcggcggcag cggcggcggc 840
ggcagccagg tgcagctggt gcagagcggc ggcggcgtgg tgcagcccgg cagaagcctg 900
agactgagct gcaaggccag cggctacacc ttcaccagat acaccatgca ctgggtgaga 960
caggcccccg gcaagggcct ggagtggatc ggctacatca accccagcag aggctacacc 1020
aactacaacc agaagttcaa ggacagattc accatcagca gagacaacag caagaacacc 1080
gccttcctgc agatggacag cctgagaccc gaggacaccg gcgtgtactt ctgcgccaga 1140
tactacgacg accactactg cctggactac tggggccagg gcacccccgt gaccgtgagc 1200
agcggcggcg gcggcagcgg cggcggcggc agcggcggcg gcggcagcga catccagatg 1260
acccagagcc ccagcagcct gagcgccagc gtgggcgaca gagtgaccat cacctgcagc 1320
gccagcagca gcgtgagcta catgaactgg taccagcaga cccccggcaa ggcccccaag 1380
agatggatct acgacaccag caagctggcc agcggcgtgc ccagcagatt cagcggcagc 1440
ggcagcggca ccgactacac cttcaccatc agcagcctgc agcccgagga catcgccacc 1500
tactactgcc agcagtggag cagcaacccc ttcaccttcg gccagggcac caagctgcag 1560
atcaccagaa cccccctggg cgacaccacc cacaccagcg gcatggtgag caagctgagc 1620
cagctgcaga ccgagctgct ggccgccctg ctggagagcg gcctgagcaa ggaggccctg 1680
atccaggccc tgggcgaggg cagcggcggc gccccctag 1719
<210>15
<211>575
<212>PRT
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
полипептид
<400>15
Met Trp Trp Arg Leu Trp Trp Leu Leu Leu Leu Leu Leu Leu Leu Trp
1 5 10 15
Pro Met Val Trp Ala Ala Asp Ile Gln Leu Thr Gln Ser Pro Ala Ser
20 25 30
Leu Ala Val Ser Leu Gly Gln Arg Ala Thr Ile Ser Cys Lys Ala Ser
35 40 45
Gln Ser Val Asp Tyr Asp Gly Asp Ser Tyr Leu Asn Trp Tyr Gln Gln
50 55 60
Ile Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr Asp Ala Ser Asn Leu
65 70 75 80
Val Ser Gly Ile Pro Pro Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp
85 90 95
Phe Thr Leu Asn Ile His Pro Val Glu Lys Val Asp Ala Ala Thr Tyr
100 105 110
His Cys Gln Gln Ser Thr Glu Asp Pro Trp Thr Phe Gly Gly Gly Thr
115 120 125
Lys Leu Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
130 135 140
Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Val
145 150 155 160
Arg Pro Gly Ser Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ala
165 170 175
Phe Ser Ser Tyr Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln Gly
180 185 190
Leu Glu Trp Ile Gly Gln Ile Trp Pro Gly Asp Gly Asp Thr Asn Tyr
195 200 205
Asn Gly Lys Phe Lys Gly Lys Ala Thr Leu Thr Ala Asp Glu Ser Ser
210 215 220
Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Ala Ser Glu Asp Ser Ala
225 230 235 240
Val Tyr Phe Cys Ala Arg Arg Glu Thr Thr Thr Val Gly Arg Tyr Tyr
245 250 255
Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
260 265 270
Gly Gly Gly Gly Ser Asp Ile Lys Leu Gln Gln Ser Gly Ala Glu Leu
275 280 285
Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Thr Ser Gly Tyr
290 295 300
Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg Pro Gly Gln
305 310 315 320
Gly Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn
325 330 335
Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser
340 345 350
Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser
355 360 365
Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr Cys Leu Asp
370 375 380
Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Val Glu Gly Gly
385 390 395 400
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Val Asp Asp Ile
405 410 415
Gln Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu Lys
420 425 430
Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met Asn Trp
435 440 445
Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp Thr
450 455 460
Ser Lys Val Ala Ser Gly Val Pro Tyr Arg Phe Ser Gly Ser Gly Ser
465 470 475 480
Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu Asp Ala
485 490 495
Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr Phe Gly
500 505 510
Ala Gly Thr Lys Leu Glu Leu Lys His His His His His His Thr Pro
515 520 525
Leu Gly Asp Thr Thr His Thr Ser Gly Met Val Ser Lys Leu Ser Gln
530 535 540
Leu Gln Thr Glu Leu Leu Ala Ala Leu Leu Glu Ser Gly Leu Ser Lys
545 550 555 560
Glu Ala Leu Ile Gln Ala Leu Gly Glu Gly Ser Gly Gly Ala Pro
565 570 575
<210>16
<211>1727
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
полинуклеотид
<400>16
atgtggtgga gactgtggtg gctgctgctg ctgctgctgc tgctgtggcc catggtgtgg 60
gccgccgaca tccagctgac ccagagcccc gccagcctgg ccgtgagcct gggccagaga 120
gccaccatca gctgcaaggc cagccagagc gtggactacg acggcgacag ctacctgaac 180
tggtaccagc agatccccgg ccagcccccc aagctgctga tctacgacgc cagcaacctg 240
gtgagcggca tcccccccag attcagcggc agcggcagcg gcaccgactt caccctgaac 300
atccaccccg tggagaaggt ggacgccgcc acctaccact gccagcagag caccgaggac 360
ccctggacct tcggcggcgg caccaagctg gagatcaagg gcggcggcgg cagcggcggc 420
ggcggcagcg gcggcggcgg cagccaggtg cagctgcagc agagcggcgc cgagctggtg 480
agacccggca gcagcgtgaa gatcagctgc aaggccagcg gctacgcctt cagcagctac 540
tggatgaact gggtgaagca gagacccggc cagggcctgg agtggatcgg ccagatctgg 600
cccggcgacg gcgacaccaa ctacaacggc aagttcaagg gcaaggccac cctgaccgcc 660
gacgagagca gcagcaccgc ctacatgcag ctgagcagcc tggccagcga ggacagcgcc 720
gtgtacttct gcgccagaag agagaccacc accgtgggca gatactacta cgccatggac 780
tactggggcc agggcaccac cgtgaccgtg agcagcggcg gcggcggcag cgacatcaag 840
ctgcagcaga gcggcgccga gctggccaga cccggcgcca gcgtgaagat gagctgcaag 900
accagcggct acaccttcac cagatacacc atgcactggg tgaagcagag acccggccag 960
ggcctggagt ggatcggcta catcaacccc agcagaggct acaccaacta caaccagaag 1020
ttcaaggaca aggccaccct gaccaccgac aagagcagca gcaccgccta catgcagctg 1080
agcagcctga ccagcgagga cagcgccgtg tactactgcg ccagatacta cgacgaccac 1140
tactgcctgg actactgggg ccagggcacc accctgaccg tgagcagcgt ggagggcggc 1200
agcggcggca gcggcggcag cggcggcagc ggcggcgtgg acgacatcca gctgacccag 1260
agccccgcca tcatgagcgc cagccccggc gagaaggtga ccatgacctg cagagccagc 1320
agcagcgtga gctacatgaa ctggtaccag cagaagagcg gcaccagccc caagagatgg 1380
atctacgaca ccagcaaggt ggccagcggc gtgccctaca gattcagcgg cagcggcagc 1440
ggcaccagct acagcctgac catcagcagc atggaggccg aggacgccgc cacctactac 1500
tgccagcagt ggagcagcaa ccccctgacc ttcggcgccg gcaccaagct ggagctgaag 1560
caccaccacc accaccacac ccccctgggc gacaccaccc acaccagcgg catggtgagc 1620
aagctgagcc agctgcagac cgagctgctg gccgccctgc tggagagcgg cctgagcaag 1680
gaggccctga tccaggccct gggcgagggc agcggcggcg cccccta 1727
<210>17
<211>122
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
полинуклеотид
<400>17
gcctgctgaa aatgactgaa tataaacttg tggtagttgg agctggtggc gtaggcaaga 60
gtgccttgac gatacagcta attcagaatc attttgtgga cgaatatgat ccaacaatag 120
ag 122
<210>18
<211>122
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
полинуклеотид
<400>18
gcctgctgaa ataattagct gagtagataa gtgatgttgg agctggtggc gtaggcaaga 60
gtgccttgac gatacagcta attcagaatc attttgtgga cgaatatgat ccaacaatag 120
ag 122
<210>19
<211>150
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
полинуклеотид
<400>19
gtacggagcg gaccacccct cctgggcccc tgcccgggtc ccgaccctct ttgccggcgc 60
cgggcggggc cggcggagta tttgatagtg tattaacctt atgtgtgaca tgttctaata 120
tagtcacatt ttcattattt ttattataag 150
<210>20
<211>150
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
полинуклеотид
<400>20
gtaaatcttg ttttaatatg catattactg gtgcaggacc attctttgat acagataaag 60
gtttctctga ccattttgag ttgtatataa cacctttttt gaagtaaaag gtgcactgta 120
ataatccaga ctgtgtttct cccttctcag 150
<210>21
<211>422
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
полинуклеотид
<400>21
gtacggagcg gaccacccct cctgggcccc tgcccgggtc ccgaccctct ttgccggcgc 60
cgggcggggc cggcggagta tttgatagtg tattaacctt atgtgtgaca tgttctaata 120
tagtcacatt ttcattattt ttattataag gcctgctgaa ataattagct gagtagataa 180
gtgatgttgg agctggtggc gtaggcaaga gtgccttgac gatacagcta attcagaatc 240
attttgtgga cgaatatgat ccaacaatag aggtaaatct tgttttaata tgcatattac 300
tggtgcagga ccattctttg atacagataa aggtttctct gaccattttg agttgtatat 360
aacacctttt ttgaagtaaa aggtgcactg taataatcca gactgtgttt ctcccttctc 420
ag 422
<210>22
<211>1722
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
полинуклеотид
<400>22
atgtggtgga gactgtggtg gctgctgctg ctgctgctgc tgctgtggcc catggtgtgg 60
gccgccgccc aggtgcagct gcagcagagc ggccccgagc tggtgaagcc cggcgccagc 120
gtgaagatca gctgcaagac cagcggctac aagttcaccg agtacaccat gcactgggtg 180
aagcagagcc acggcaagtg cctggagtgg atcggcggca tcaaccccaa caacggcggc 240
accaactaca accagaagtt caagggcaag gccaccctga ccgtggacaa gagcagcagc 300
accgcctaca tggagctgag aagcctgacc agcgaggaca gcgccgtgta ctactgcgcc 360
agagacacca ccgtgcccta cgcctactgg ggccagggca ccaccgtgac cgtgagcagc 420
ggcggcggcg gcagcggcgg cggcggcagc ggcggcggcg gcagcgacat cgagctgacc 480
cagagccccg ccatcatgag cgccagcccc ggcgagaagg tgaccatgac ctgcagcgcc 540
agcagcagca tcagctacat gcactggtac cagcagaagc ccggcaccag ccccaagaga 600
tggatctacg acaccagcaa gctggccagc agcgtgcccg ccagattcag cggcagcggc 660
agcggcacca gctacagcct gaccatcagc agcatggagg ccgaggacgc cgccacctac 720
tactgccacc agagaagcag ctaccccctg accttcggct gcggcaccaa gctggagatc 780
aagagagcca gcaccaaggg ccccggcggc ggcggcagcg gcggcggcgg cagcggcggc 840
ggcggcagcc aggtgcagct ggtgcagagc ggcggcggcg tggtgcagcc cggcagaagc 900
ctgagactga gctgcaaggc cagcggctac accttcacca gatacaccat gcactgggtg 960
agacaggccc ccggcaaggg cctggagtgg atcggctaca tcaaccccag cagaggctac 1020
accaactaca accagaagtt caaggacaga ttcaccatca gcagagacaa cagcaagaac 1080
accgccttcc tgcagatgga cagcctgaga cccgaggaca ccggcgtgta cttctgcgcc 1140
agatactacg acgaccacta ctgcctggac tactggggcc agggcacccc cgtgaccgtg 1200
agcagcggcg gcggcggcag cggcggcggc ggcagcggcg gcggcggcag cgacatccag 1260
atgacccaga gccccagcag cctgagcgcc agcgtgggcg acagagtgac catcacctgc 1320
agcgccagca gcagcgtgag ctacatgaac tggtaccagc agacccccgg caaggccccc 1380
aagagatgga tctacgacac cagcaagctg gccagcggcg tgcccagcag attcagcggc 1440
agcggcagcg gcaccgacta caccttcacc atcagcagcc tgcagcccga ggacatcgcc 1500
acctactact gccagcagtg gagcagcaac cccttcacct tcggccaggg caccaagctg 1560
cagatcacca gaacccccct gggcgacacc acccacacca gcggcatggt gagcaagctg 1620
agccagctgc agaccgagct gctggccgcc ctgctggaga gcggcctgag caaggaggcc 1680
ctgatccagg ccctgggcga gggcagcggc ggcgccccct ag 1722
<210>23
<211>2144
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
полинуклеотид
<400>23
atgtggtgga gactgtggtg gctgctgctg ctgctgctgc tgctgtggcc catggtgtgg 60
gccgccgccc aggtgcagct gcagcagagc ggccccgagc tggtgaagcc cggcgccagc 120
gtgaagatca gctgcaagac cagcggctac aagttcaccg agtacaccat gcactgggtg 180
aagcagagcc acggcaagtg cctggagtgg atcggcggca tcaaccccaa caacggcggc 240
accaactaca accagaagtt caaggtacgg agcggaccac ccctcctggg cccctgcccg 300
ggtcccgacc ctctttgccg gcgccgggcg gggccggcgg agtatttgat agtgtattaa 360
ccttatgtgt gacatgttct aatatagtca cattttcatt atttttatta taaggcctgc 420
tgaaataatt agctgagtag ataagtgatg ttggagctgg tggcgtaggc aagagtgcct 480
tgacgataca gctaattcag aatcattttg tggacgaata tgatccaaca atagaggtaa 540
atcttgtttt aatatgcata ttactggtgc aggaccattc tttgatacag ataaaggttt 600
ctctgaccat tttgagttgt atataacacc ttttttgaag taaaaggtgc actgtaataa 660
tccagactgt gtttctccct tctcagggca aggccaccct gaccgtggac aagagcagca 720
gcaccgccta catggagctg agaagcctga ccagcgagga cagcgccgtg tactactgcg 780
ccagagacac caccgtgccc tacgcctact ggggccaggg caccaccgtg accgtgagca 840
gcggcggcgg cggcagcggc ggcggcggca gcggcggcgg cggcagcgac atcgagctga 900
cccagagccc cgccatcatg agcgccagcc ccggcgagaa ggtgaccatg acctgcagcg 960
ccagcagcag catcagctac atgcactggt accagcagaa gcccggcacc agccccaaga 1020
gatggatcta cgacaccagc aagctggcca gcagcgtgcc cgccagattc agcggcagcg 1080
gcagcggcac cagctacagc ctgaccatca gcagcatgga ggccgaggac gccgccacct 1140
actactgcca ccagagaagc agctaccccc tgaccttcgg ctgcggcacc aagctggaga 1200
tcaagagagc cagcaccaag ggccccggcg gcggcggcag cggcggcggc ggcagcggcg 1260
gcggcggcag ccaggtgcag ctggtgcaga gcggcggcgg cgtggtgcag cccggcagaa 1320
gcctgagact gagctgcaag gccagcggct acaccttcac cagatacacc atgcactggg 1380
tgagacaggc ccccggcaag ggcctggagt ggatcggcta catcaacccc agcagaggct 1440
acaccaacta caaccagaag ttcaaggaca gattcaccat cagcagagac aacagcaaga 1500
acaccgcctt cctgcagatg gacagcctga gacccgagga caccggcgtg tacttctgcg 1560
ccagatacta cgacgaccac tactgcctgg actactgggg ccagggcacc cccgtgaccg 1620
tgagcagcgg cggcggcggc agcggcggcg gcggcagcgg cggcggcggc agcgacatcc 1680
agatgaccca gagccccagc agcctgagcg ccagcgtggg cgacagagtg accatcacct 1740
gcagcgccag cagcagcgtg agctacatga actggtacca gcagaccccc ggcaaggccc 1800
ccaagagatg gatctacgac accagcaagc tggccagcgg cgtgcccagc agattcagcg 1860
gcagcggcag cggcaccgac tacaccttca ccatcagcag cctgcagccc gaggacatcg 1920
ccacctacta ctgccagcag tggagcagca accccttcac cttcggccag ggcaccaagc 1980
tgcagatcac cagaaccccc ctgggcgaca ccacccacac cagcggcatg gtgagcaagc 2040
tgagccagct gcagaccgag ctgctggccg ccctgctgga gagcggcctg agcaaggagg 2100
ccctgatcca ggccctgggc gagggcagcg gcggcgcccc ctag 2144
<210>24
<211>30
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
олигонуклеотид
<400>24
aatatgatcc aacaatagag gtaaatcttg 30
<210>25
<211>30
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
олигонуклеотид
<400>25
gatccaacaa tagaggtaaa tcttgtttta 30
<210>26
<211>51
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
олигонуклеотид
<400>26
aatatgatcc aacaatagag gtaaatcttg ttttaatatg catattactg g 51
<210>27
<211>30
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
олигонуклеотид
<400>27
caagatttac ctctattgtt ggatcatatt 30
<210>28
<211>30
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
олигонуклеотид
<400>28
taaaacaaga tttacctcta ttgttggatc 30
<210>29
<211>527
<212>PRT
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
полипептид
<400>29
Met Trp Trp Arg Leu Trp Trp Leu Leu Leu Leu Leu Leu Leu Leu Trp
1 5 10 15
Pro Met Val Trp Ala Ala Ala Asp Ile Gln Leu Thr Gln Ser Pro Ala
20 25 30
Ser Leu Ala Val Ser Leu Gly Gln Arg Ala Thr Ile Ser Cys Lys Ala
35 40 45
Ser Gln Ser Val Asp Tyr Asp Gly Asp Ser Tyr Leu Asn Trp Tyr Gln
50 55 60
Gln Ile Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr Asp Ala Ser Asn
65 70 75 80
Leu Val Ser Gly Ile Pro Pro Arg Phe Ser Gly Ser Gly Ser Gly Thr
85 90 95
Asp Phe Thr Leu Asn Ile His Pro Val Glu Lys Val Asp Ala Ala Thr
100 105 110
Tyr His Cys Gln Gln Ser Thr Glu Asp Pro Trp Thr Phe Gly Gly Gly
115 120 125
Thr Lys Leu Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
130 135 140
Gly Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu
145 150 155 160
Val Arg Pro Gly Ser Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr
165 170 175
Ala Phe Ser Ser Tyr Trp Met Asn Trp Val Lys Gln Arg Pro Gly Gln
180 185 190
Gly Leu Glu Trp Ile Gly Gln Ile Trp Pro Gly Asp Gly Asp Thr Asn
195 200 205
Tyr Asn Gly Lys Phe Lys Gly Lys Ala Thr Leu Thr Ala Asp Glu Ser
210 215 220
Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Ala Ser Glu Asp Ser
225 230 235 240
Ala Val Tyr Phe Cys Ala Arg Arg Glu Thr Thr Thr Val Gly Arg Tyr
245 250 255
Tyr Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser
260 265 270
Ser Gly Gly Gly Gly Ser Asp Ile Lys Leu Gln Gln Ser Gly Ala Glu
275 280 285
Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Thr Ser Gly
290 295 300
Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg Pro Gly
305 310 315 320
Gln Gly Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr
325 330 335
Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys
340 345 350
Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp
355 360 365
Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr Cys Leu
370 375 380
Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Val Glu Gly
385 390 395 400
Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Val Asp Asp
405 410 415
Ile Gln Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu
420 425 430
Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met Asn
435 440 445
Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp
450 455 460
Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg Phe Ser Gly Ser Gly
465 470 475 480
Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu Asp
485 490 495
Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr Phe
500 505 510
Gly Ala Gly Thr Lys Leu Glu Leu Lys His His His His His His
515 520 525
<210>30
<211>1584
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
полинуклеотид
<400>30
atgtggtgga gactgtggtg gctgctgctg ctgctgctgc tgctgtggcc catggtgtgg 60
gccgccgccg acatccagct gacccagagc cccgccagcc tggccgtgag cctgggccag 120
agagccacca tcagctgcaa ggccagccag agcgtggact acgacggcga cagctacctg 180
aactggtacc agcagatccc cggccagccc cccaagctgc tgatctacga cgccagcaac 240
ctggtgagcg gcatcccccc cagattcagc ggcagcggca gcggcaccga cttcaccctg 300
aacatccacc ccgtggagaa ggtggacgcc gccacctacc actgccagca gagcaccgag 360
gacccctgga ccttcggcgg cggcaccaag ctggagatca agggcggcgg cggcagcggc 420
ggcggcggca gcggcggcgg cggcagccag gtgcagctgc agcagagcgg cgccgagctg 480
gtgagacccg gcagcagcgt gaagatcagc tgcaaggcca gcggctacgc cttcagcagc 540
tactggatga actgggtgaa gcagagaccc ggccagggcc tggagtggat cggccagatc 600
tggcccggcg acggcgacac caactacaac ggcaagttca agggcaaggc caccctgacc 660
gccgacgaga gcagcagcac cgcctacatg cagctgagca gcctggccag cgaggacagc 720
gccgtgtact tctgcgccag aagagagacc accaccgtgg gcagatacta ctacgccatg 780
gactactggg gccagggcac caccgtgacc gtgagcagcg gcggcggcgg cagcgacatc 840
aagctgcagc agagcggcgc cgagctggcc agacccggcg ccagcgtgaa gatgagctgc 900
aagaccagcg gctacacctt caccagatac accatgcact gggtgaagca gagacccggc 960
cagggcctgg agtggatcgg ctacatcaac cccagcagag gctacaccaa ctacaaccag 1020
aagttcaagg acaaggccac cctgaccacc gacaagagca gcagcaccgc ctacatgcag 1080
ctgagcagcc tgaccagcga ggacagcgcc gtgtactact gcgccagata ctacgacgac 1140
cactactgcc tggactactg gggccagggc accaccctga ccgtgagcag cgtggagggc 1200
ggcagcggcg gcagcggcgg cagcggcggc agcggcggcg tggacgacat ccagctgacc 1260
cagagccccg ccatcatgag cgccagcccc ggcgagaagg tgaccatgac ctgcagagcc 1320
agcagcagcg tgagctacat gaactggtac cagcagaaga gcggcaccag ccccaagaga 1380
tggatctacg acaccagcaa ggtggccagc ggcgtgccct acagattcag cggcagcggc 1440
agcggcacca gctacagcct gaccatcagc agcatggagg ccgaggacgc cgccacctac 1500
tactgccagc agtggagcag caaccccctg accttcggcg ccggcaccaa gctggagctg 1560
aagcaccacc accaccacca ctag 1584
<210>31
<211>1581
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
полинуклеотид
<400>31
atgtggtgga gactgtggtg gctgctgctg ctgctgctgc tgctgtggcc catggtgtgg 60
gccgccgaca tccagctgac ccagagcccc gccagcctgg ccgtgagcct gggccagaga 120
gccaccatca gctgcaaggc cagccagagc gtggactacg acggcgacag ctacctgaac 180
tggtaccagc agatccccgg ccagcccccc aagctgctga tctacgacgc cagcaacctg 240
gtgagcggca tcccccccag attcagcggc agcggcagcg gcaccgactt caccctgaac 300
atccaccccg tggagaaggt ggacgccgcc acctaccact gccagcagag caccgaggac 360
ccctggacct tcggcggcgg caccaagctg gagatcaagg gcggcggcgg cagcggcggc 420
ggcggcagcg gcggcggcgg cagccaggtg cagctgcagc agagcggcgc cgagctggtg 480
agacccggca gcagcgtgaa gatcagctgc aaggccagcg gctacgcctt cagcagctac 540
tggatgaact gggtgaagca gagacccggc cagggcctgg agtggatcgg ccagatctgg 600
cccggcgacg gcgacaccaa ctacaacggc aagttcaagg gcaaggccac cctgaccgcc 660
gacgagagca gcagcaccgc ctacatgcag ctgagcagcc tggccagcga ggacagcgcc 720
gtgtacttct gcgccagaag agagaccacc accgtgggca gatactacta cgccatggac 780
tactggggcc agggcaccac cgtgaccgtg agcagcggcg gcggcggcag cgacatcaag 840
ctgcagcaga gcggcgccga gctggccaga cccggcgcca gcgtgaagat gagctgcaag 900
accagcggct acaccttcac cagatacacc atgcactggg tgaagcagag acccggccag 960
ggcctggagt ggatcggcta catcaacccc agcagaggct acaccaacta caaccagaag 1020
ttcaaggaca aggccaccct gaccaccgac aagagcagca gcaccgccta catgcagctg 1080
agcagcctga ccagcgagga cagcgccgtg tactactgcg ccagatacta cgacgaccac 1140
tactgcctgg actactgggg ccagggcacc accctgaccg tgagcagcgt ggagggcggc 1200
agcggcggca gcggcggcag cggcggcagc ggcggcgtgg acgacatcca gctgacccag 1260
agccccgcca tcatgagcgc cagccccggc gagaaggtga ccatgacctg cagagccagc 1320
agcagcgtga gctacatgaa ctggtaccag cagaagagcg gcaccagccc caagagatgg 1380
atctacgaca ccagcaaggt ggccagcggc gtgccctaca gattcagcgg cagcggcagc 1440
ggcaccagct acagcctgac catcagcagc atggaggccg aggacgccgc cacctactac 1500
tgccagcagt ggagcagcaa ccccctgacc ttcggcgccg gcaccaagct ggagctgaag 1560
caccaccacc accaccacta g 1581
<210>32
<211>1578
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
полинуклеотид
<400>32
atgtggtgga gactgtggtg gctgctgctg ctgctgctgc tgctgtggcc catggtgtgg 60
gccgacatcc agctgaccca gagccccgcc agcctggccg tgagcctggg ccagagagcc 120
accatcagct gcaaggccag ccagagcgtg gactacgacg gcgacagcta cctgaactgg 180
taccagcaga tccccggcca gccccccaag ctgctgatct acgacgccag caacctggtg 240
agcggcatcc cccccagatt cagcggcagc ggcagcggca ccgacttcac cctgaacatc 300
caccccgtgg agaaggtgga cgccgccacc taccactgcc agcagagcac cgaggacccc 360
tggaccttcg gcggcggcac caagctggag atcaagggcg gcggcggcag cggcggcggc 420
ggcagcggcg gcggcggcag ccaggtgcag ctgcagcaga gcggcgccga gctggtgaga 480
cccggcagca gcgtgaagat cagctgcaag gccagcggct acgccttcag cagctactgg 540
atgaactggg tgaagcagag acccggccag ggcctggagt ggatcggcca gatctggccc 600
ggcgacggcg acaccaacta caacggcaag ttcaagggca aggccaccct gaccgccgac 660
gagagcagca gcaccgccta catgcagctg agcagcctgg ccagcgagga cagcgccgtg 720
tacttctgcg ccagaagaga gaccaccacc gtgggcagat actactacgc catggactac 780
tggggccagg gcaccaccgt gaccgtgagc agcggcggcg gcggcagcga catcaagctg 840
cagcagagcg gcgccgagct ggccagaccc ggcgccagcg tgaagatgag ctgcaagacc 900
agcggctaca ccttcaccag atacaccatg cactgggtga agcagagacc cggccagggc 960
ctggagtgga tcggctacat caaccccagc agaggctaca ccaactacaa ccagaagttc 1020
aaggacaagg ccaccctgac caccgacaag agcagcagca ccgcctacat gcagctgagc 1080
agcctgacca gcgaggacag cgccgtgtac tactgcgcca gatactacga cgaccactac 1140
tgcctggact actggggcca gggcaccacc ctgaccgtga gcagcgtgga gggcggcagc 1200
ggcggcagcg gcggcagcgg cggcagcggc ggcgtggacg acatccagct gacccagagc 1260
cccgccatca tgagcgccag ccccggcgag aaggtgacca tgacctgcag agccagcagc 1320
agcgtgagct acatgaactg gtaccagcag aagagcggca ccagccccaa gagatggatc 1380
tacgacacca gcaaggtggc cagcggcgtg ccctacagat tcagcggcag cggcagcggc 1440
accagctaca gcctgaccat cagcagcatg gaggccgagg acgccgccac ctactactgc 1500
cagcagtgga gcagcaaccc cctgaccttc ggcgccggca ccaagctgga gctgaagcac 1560
caccaccacc accactag 1578
<210>33
<211>1518
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
полинуклеотид
<400>33
atggacatcc agctgaccca gagccccgcc agcctggccg tgagcctggg ccagagagcc 60
accatcagct gcaaggccag ccagagcgtg gactacgacg gcgacagcta cctgaactgg 120
taccagcaga tccccggcca gccccccaag ctgctgatct acgacgccag caacctggtg 180
agcggcatcc cccccagatt cagcggcagc ggcagcggca ccgacttcac cctgaacatc 240
caccccgtgg agaaggtgga cgccgccacc taccactgcc agcagagcac cgaggacccc 300
tggaccttcg gcggcggcac caagctggag atcaagggcg gcggcggcag cggcggcggc 360
ggcagcggcg gcggcggcag ccaggtgcag ctgcagcaga gcggcgccga gctggtgaga 420
cccggcagca gcgtgaagat cagctgcaag gccagcggct acgccttcag cagctactgg 480
atgaactggg tgaagcagag acccggccag ggcctggagt ggatcggcca gatctggccc 540
ggcgacggcg acaccaacta caacggcaag ttcaagggca aggccaccct gaccgccgac 600
gagagcagca gcaccgccta catgcagctg agcagcctgg ccagcgagga cagcgccgtg 660
tacttctgcg ccagaagaga gaccaccacc gtgggcagat actactacgc catggactac 720
tggggccagg gcaccaccgt gaccgtgagc agcggcggcg gcggcagcga catcaagctg 780
cagcagagcg gcgccgagct ggccagaccc ggcgccagcg tgaagatgag ctgcaagacc 840
agcggctaca ccttcaccag atacaccatg cactgggtga agcagagacc cggccagggc 900
ctggagtgga tcggctacat caaccccagc agaggctaca ccaactacaa ccagaagttc 960
aaggacaagg ccaccctgac caccgacaag agcagcagca ccgcctacat gcagctgagc 1020
agcctgacca gcgaggacag cgccgtgtac tactgcgcca gatactacga cgaccactac 1080
tgcctggact actggggcca gggcaccacc ctgaccgtga gcagcgtgga gggcggcagc 1140
ggcggcagcg gcggcagcgg cggcagcggc ggcgtggacg acatccagct gacccagagc 1200
cccgccatca tgagcgccag ccccggcgag aaggtgacca tgacctgcag agccagcagc 1260
agcgtgagct acatgaactg gtaccagcag aagagcggca ccagccccaa gagatggatc 1320
tacgacacca gcaaggtggc cagcggcgtg ccctacagat tcagcggcag cggcagcggc 1380
accagctaca gcctgaccat cagcagcatg gaggccgagg acgccgccac ctactactgc 1440
cagcagtgga gcagcaaccc cctgaccttc ggcgccggca ccaagctgga gctgaagcac 1500
caccaccacc accactag 1518
<210>34
<211>9
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
олигонуклеотид
<400>34
maggtragt 9
<210>35
<211>7
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
олигонуклеотид
<220>
<221>misc_feature
<222>(1)..(1)
<223>Пиримидиновый тракт, охватывающий по меньшей мере один нуклеотид
<220>
<221>modified_base
<222>(2)..(2)
<223>a, c, t, g, неизвестное или другое
<220>
<223>См. поданное описание для конкретизации
замен и предпочтительных воплощений
<400>35
yncaggk 7
<210>36
<211>5
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
олигонуклеотид
<400>36
aaggg 5
<210>37
<211>5
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
олигонуклеотид
<400>37
caggg 5
<210>38
<211>5
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
олигонуклеотид
<400>38
aaggt 5
<210>39
<211>5
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
олигонуклеотид
<400>39
caggt 5
<210>40
<211>2144
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
полинуклеотид
<400>40
atgtggtgga gactgtggtg gctgctgctg ctgctgctgc tgctgtggcc catggtgtgg 60
gccgccgccc aggtgcagct gcagcagagc ggccccgagc tggtgaagcc cggcgccagc 120
gtgaagatca gctgcaagac cagcggctac aagttcaccg agtacaccat gcactgggtg 180
aagcagagcc acggcaagtg cctggagtgg atcggcggca tcaaccccaa caacggcggc 240
accaactaca accagaagtt caaggtacgg agcggaccac ccctcctggg cccctgcccg 300
ggtcccgacc ctctttgccg gcgccgggcg gggccggcgg agtatttgat agtgtattaa 360
ccttatgtgt gacatgttct aatatagtca cattttcctt cttttttttt tcaggcctgc 420
tgaaataatt agctgagtag ataagtgatg ttggagctgg tggcgtaggc aagagtgcct 480
tgacgataca gctaattcag aatcattttg tggacgaata tgatccaaca atagaggtaa 540
atcttgtttt aatatgcata ttactggtgc aggaccattc tttgatacag ataaaggttt 600
ctctgaccat tttgagttgt atataacacc ttttttgaag taaaaggtgc actgtaataa 660
tccagactgt gtttctccct tctcagggca aggccaccct gaccgtggac aagagcagca 720
gcaccgccta catggagctg agaagcctga ccagcgagga cagcgccgtg tactactgcg 780
ccagagacac caccgtgccc tacgcctact ggggccaggg caccaccgtg accgtgagca 840
gcggcggcgg cggcagcggc ggcggcggca gcggcggcgg cggcagcgac atcgagctga 900
cccagagccc cgccatcatg agcgccagcc ccggcgagaa ggtgaccatg acctgcagcg 960
ccagcagcag catcagctac atgcactggt accagcagaa gcccggcacc agccccaaga 1020
gatggatcta cgacaccagc aagctggcca gcagcgtgcc cgccagattc agcggcagcg 1080
gcagcggcac cagctacagc ctgaccatca gcagcatgga ggccgaggac gccgccacct 1140
actactgcca ccagagaagc agctaccccc tgaccttcgg ctgcggcacc aagctggaga 1200
tcaagagagc cagcaccaag ggccccggcg gcggcggcag cggcggcggc ggcagcggcg 1260
gcggcggcag ccaggtgcag ctggtgcaga gcggcggcgg cgtggtgcag cccggcagaa 1320
gcctgagact gagctgcaag gccagcggct acaccttcac cagatacacc atgcactggg 1380
tgagacaggc ccccggcaag ggcctggagt ggatcggcta catcaacccc agcagaggct 1440
acaccaacta caaccagaag ttcaaggaca gattcaccat cagcagagac aacagcaaga 1500
acaccgcctt cctgcagatg gacagcctga gacccgagga caccggcgtg tacttctgcg 1560
ccagatacta cgacgaccac tactgcctgg actactgggg ccagggcacc cccgtgaccg 1620
tgagcagcgg cggcggcggc agcggcggcg gcggcagcgg cggcggcggc agcgacatcc 1680
agatgaccca gagccccagc agcctgagcg ccagcgtggg cgacagagtg accatcacct 1740
gcagcgccag cagcagcgtg agctacatga actggtacca gcagaccccc ggcaaggccc 1800
ccaagagatg gatctacgac accagcaagc tggccagcgg cgtgcccagc agattcagcg 1860
gcagcggcag cggcaccgac tacaccttca ccatcagcag cctgcagccc gaggacatcg 1920
ccacctacta ctgccagcag tggagcagca accccttcac cttcggccag ggcaccaagc 1980
tgcagatcac cagaaccccc ctgggcgaca ccacccacac cagcggcatg gtgagcaagc 2040
tgagccagct gcagaccgag ctgctggccg ccctgctgga gagcggcctg agcaaggagg 2100
ccctgatcca ggccctgggc gagggcagcg gcggcgcccc ctag 2144
<210>41
<211>2144
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
полинуклеотид
<400>41
atgtggtgga gactgtggtg gctgctgctg ctgctgctgc tgctgtggcc catggtgtgg 60
gccgccgccc aggtgcagct gcagcagagc ggccccgagc tggtgaagcc cggcgccagc 120
gtgaagatca gctgcaagac cagcggctac aagttcaccg agtacaccat gcactgggtg 180
aagcagagcc acggcaagtg cctggagtgg atcggcggca tcaaccccaa caacggcggc 240
accaactaca accagaagtt caaggtacgg agcggaccac ccctcctggg cccctgcccg 300
ggtcccgacc ctctttgccg gcgccgggcg gggccggcgg agtatttgat agtgtattaa 360
ccttatgtgt gacatgttct aatatagtca cattttcctt cttttttttt taaggcctgc 420
tgaaataatt agctgagtag ataagtgatg ttggagctgg tggcgtaggc aagagtgcct 480
tgacgataca gctaattcag aatcattttg tggacgaata tgatccaaca atagaggtaa 540
atcttgtttt aatatgcata ttactggtgc aggaccattc tttgatacag ataaaggttt 600
ctctgaccat tttgagttgt atataacacc ttttttgaag taaaaggtgc actgtaataa 660
tccagactgt gtttctccct tctcagggca aggccaccct gaccgtggac aagagcagca 720
gcaccgccta catggagctg agaagcctga ccagcgagga cagcgccgtg tactactgcg 780
ccagagacac caccgtgccc tacgcctact ggggccaggg caccaccgtg accgtgagca 840
gcggcggcgg cggcagcggc ggcggcggca gcggcggcgg cggcagcgac atcgagctga 900
cccagagccc cgccatcatg agcgccagcc ccggcgagaa ggtgaccatg acctgcagcg 960
ccagcagcag catcagctac atgcactggt accagcagaa gcccggcacc agccccaaga 1020
gatggatcta cgacaccagc aagctggcca gcagcgtgcc cgccagattc agcggcagcg 1080
gcagcggcac cagctacagc ctgaccatca gcagcatgga ggccgaggac gccgccacct 1140
actactgcca ccagagaagc agctaccccc tgaccttcgg ctgcggcacc aagctggaga 1200
tcaagagagc cagcaccaag ggccccggcg gcggcggcag cggcggcggc ggcagcggcg 1260
gcggcggcag ccaggtgcag ctggtgcaga gcggcggcgg cgtggtgcag cccggcagaa 1320
gcctgagact gagctgcaag gccagcggct acaccttcac cagatacacc atgcactggg 1380
tgagacaggc ccccggcaag ggcctggagt ggatcggcta catcaacccc agcagaggct 1440
acaccaacta caaccagaag ttcaaggaca gattcaccat cagcagagac aacagcaaga 1500
acaccgcctt cctgcagatg gacagcctga gacccgagga caccggcgtg tacttctgcg 1560
ccagatacta cgacgaccac tactgcctgg actactgggg ccagggcacc cccgtgaccg 1620
tgagcagcgg cggcggcggc agcggcggcg gcggcagcgg cggcggcggc agcgacatcc 1680
agatgaccca gagccccagc agcctgagcg ccagcgtggg cgacagagtg accatcacct 1740
gcagcgccag cagcagcgtg agctacatga actggtacca gcagaccccc ggcaaggccc 1800
ccaagagatg gatctacgac accagcaagc tggccagcgg cgtgcccagc agattcagcg 1860
gcagcggcag cggcaccgac tacaccttca ccatcagcag cctgcagccc gaggacatcg 1920
ccacctacta ctgccagcag tggagcagca accccttcac cttcggccag ggcaccaagc 1980
tgcagatcac cagaaccccc ctgggcgaca ccacccacac cagcggcatg gtgagcaagc 2040
tgagccagct gcagaccgag ctgctggccg ccctgctgga gagcggcctg agcaaggagg 2100
ccctgatcca ggccctgggc gagggcagcg gcggcgcccc ctag 2144
<210>42
<211>2143
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
полинуклеотид
<400>42
atgtggtgga gactgtggtg gctgctgctg ctgctgctgc tgctgtggcc catggtgtgg 60
gccgccgccc aggtgcagct gcagcagagc ggccccgagc tggtgaagcc cggcgccagc 120
gtgaagatca gctgcaagac cagcggctac aagttcaccg agtacaccat gcactgggtg 180
aagcagagcc acggcaagtg cctggagtgg atcggcggca tcaaccccaa caacggcggc 240
accaactaca accagaagtt caaggtacgg agcggaccac ccctcctggg cccctgcccg 300
ggtcccgacc ctctttgccg gcgccgggcg gggccggcgg agtatttgat agtgtattaa 360
ccttatgtgt gacatgttct aatatagtca cattttcctt ctttttatta taagcctgct 420
gaaataatta gctgagtaga taagtgatgt tggagctggt ggcgtaggca agagtgcctt 480
gacgatacag ctaattcaga atcattttgt ggacgaatat gatccaacaa tagaggtaaa 540
tcttgtttta atatgcatat tactggtgca ggaccattct ttgatacaga taaaggtttc 600
tctgaccatt ttgagttgta tataacacct tttttgaagt aaaaggtgca ctgtaataat 660
ccagactgtg tttctccctt ctcagggcaa ggccaccctg accgtggaca agagcagcag 720
caccgcctac atggagctga gaagcctgac cagcgaggac agcgccgtgt actactgcgc 780
cagagacacc accgtgccct acgcctactg gggccagggc accaccgtga ccgtgagcag 840
cggcggcggc ggcagcggcg gcggcggcag cggcggcggc ggcagcgaca tcgagctgac 900
ccagagcccc gccatcatga gcgccagccc cggcgagaag gtgaccatga cctgcagcgc 960
cagcagcagc atcagctaca tgcactggta ccagcagaag cccggcacca gccccaagag 1020
atggatctac gacaccagca agctggccag cagcgtgccc gccagattca gcggcagcgg 1080
cagcggcacc agctacagcc tgaccatcag cagcatggag gccgaggacg ccgccaccta 1140
ctactgccac cagagaagca gctaccccct gaccttcggc tgcggcacca agctggagat 1200
caagagagcc agcaccaagg gccccggcgg cggcggcagc ggcggcggcg gcagcggcgg 1260
cggcggcagc caggtgcagc tggtgcagag cggcggcggc gtggtgcagc ccggcagaag 1320
cctgagactg agctgcaagg ccagcggcta caccttcacc agatacacca tgcactgggt 1380
gagacaggcc cccggcaagg gcctggagtg gatcggctac atcaacccca gcagaggcta 1440
caccaactac aaccagaagt tcaaggacag attcaccatc agcagagaca acagcaagaa 1500
caccgccttc ctgcagatgg acagcctgag acccgaggac accggcgtgt acttctgcgc 1560
cagatactac gacgaccact actgcctgga ctactggggc cagggcaccc ccgtgaccgt 1620
gagcagcggc ggcggcggca gcggcggcgg cggcagcggc ggcggcggca gcgacatcca 1680
gatgacccag agccccagca gcctgagcgc cagcgtgggc gacagagtga ccatcacctg 1740
cagcgccagc agcagcgtga gctacatgaa ctggtaccag cagacccccg gcaaggcccc 1800
caagagatgg atctacgaca ccagcaagct ggccagcggc gtgcccagca gattcagcgg 1860
cagcggcagc ggcaccgact acaccttcac catcagcagc ctgcagcccg aggacatcgc 1920
cacctactac tgccagcagt ggagcagcaa ccccttcacc ttcggccagg gcaccaagct 1980
gcagatcacc agaacccccc tgggcgacac cacccacacc agcggcatgg tgagcaagct 2040
gagccagctg cagaccgagc tgctggccgc cctgctggag agcggcctga gcaaggaggc 2100
cctgatccag gccctgggcg agggcagcgg cggcgccccc tag 2143
<210>43
<211>2144
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
полинуклеотид
<400>43
atgtggtgga gactgtggtg gctgctgctg ctgctgctgc tgctgtggcc catggtgtgg 60
gccgccgccc aggtgcagct gcagcagagc ggccccgagc tggtgaagcc cggcgccagc 120
gtgaagatca gctgcaagac cagcggctac aagttcaccg agtacaccat gcactgggtg 180
aagcagagcc acggcaagtg cctggagtgg atcggcggca tcaaccccaa caacggcggc 240
accaactaca accagaagtt caaggtacgg agcggaccac ccctcctggg cccctgcccg 300
ggtcccgacc ctctttgccg gcgccgggcg gggccggcgg agtatttgat agtgtattaa 360
ccttatgtgt gacatgttct aatatagtca cattttcatt attttttttt tcaggcctgc 420
tgaaataatt agctgagtag ataagtgatg ttggagctgg tggcgtaggc aagagtgcct 480
tgacgataca gctaattcag aatcattttg tggacgaata tgatccaaca atagaggtaa 540
atcttgtttt aatatgcata ttactggtgc aggaccattc tttgatacag ataaaggttt 600
ctctgaccat tttgagttgt atataacacc ttttttgaag taaaaggtgc actgtaataa 660
tccagactgt gtttctccct tctcagggca aggccaccct gaccgtggac aagagcagca 720
gcaccgccta catggagctg agaagcctga ccagcgagga cagcgccgtg tactactgcg 780
ccagagacac caccgtgccc tacgcctact ggggccaggg caccaccgtg accgtgagca 840
gcggcggcgg cggcagcggc ggcggcggca gcggcggcgg cggcagcgac atcgagctga 900
cccagagccc cgccatcatg agcgccagcc ccggcgagaa ggtgaccatg acctgcagcg 960
ccagcagcag catcagctac atgcactggt accagcagaa gcccggcacc agccccaaga 1020
gatggatcta cgacaccagc aagctggcca gcagcgtgcc cgccagattc agcggcagcg 1080
gcagcggcac cagctacagc ctgaccatca gcagcatgga ggccgaggac gccgccacct 1140
actactgcca ccagagaagc agctaccccc tgaccttcgg ctgcggcacc aagctggaga 1200
tcaagagagc cagcaccaag ggccccggcg gcggcggcag cggcggcggc ggcagcggcg 1260
gcggcggcag ccaggtgcag ctggtgcaga gcggcggcgg cgtggtgcag cccggcagaa 1320
gcctgagact gagctgcaag gccagcggct acaccttcac cagatacacc atgcactggg 1380
tgagacaggc ccccggcaag ggcctggagt ggatcggcta catcaacccc agcagaggct 1440
acaccaacta caaccagaag ttcaaggaca gattcaccat cagcagagac aacagcaaga 1500
acaccgcctt cctgcagatg gacagcctga gacccgagga caccggcgtg tacttctgcg 1560
ccagatacta cgacgaccac tactgcctgg actactgggg ccagggcacc cccgtgaccg 1620
tgagcagcgg cggcggcggc agcggcggcg gcggcagcgg cggcggcggc agcgacatcc 1680
agatgaccca gagccccagc agcctgagcg ccagcgtggg cgacagagtg accatcacct 1740
gcagcgccag cagcagcgtg agctacatga actggtacca gcagaccccc ggcaaggccc 1800
ccaagagatg gatctacgac accagcaagc tggccagcgg cgtgcccagc agattcagcg 1860
gcagcggcag cggcaccgac tacaccttca ccatcagcag cctgcagccc gaggacatcg 1920
ccacctacta ctgccagcag tggagcagca accccttcac cttcggccag ggcaccaagc 1980
tgcagatcac cagaaccccc ctgggcgaca ccacccacac cagcggcatg gtgagcaagc 2040
tgagccagct gcagaccgag ctgctggccg ccctgctgga gagcggcctg agcaaggagg 2100
ccctgatcca ggccctgggc gagggcagcg gcggcgcccc ctag 2144
<210>44
<211>1581
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
полинуклеотид
<400>44
atgtggtgga gactgtggtg gctgctgctg ctgctgctgc tgctgtggcc catggtgtgg 60
gccgccgaca tccagctgac ccagagcccc gccagcctgg ccgtgagcct gggccagaga 120
gccaccatca gctgcaaggc cagccagagc gtggactacg acggcgacag ctacctgaac 180
tggtaccagc agatccccgg ccagcccccc aagctgctga tctacgacgc cagcaacctg 240
gtgagcggca tcccccccag attcagcggc agcggcagcg gcaccgactt caccctgaac 300
atccaccccg tggagaaggt ggacgccgcc acctaccact gccagcagag caccgaggac 360
ccctggacct tcggcggcgg caccaagctg gagatcaagg gcggcggcgg cagcggcggc 420
ggcggcagcg gcggcggcgg cagccaggtg cagctgcagc agagcggcgc cgagctggtg 480
agacccggca gcagcgtgaa gatcagctgc aaggccagcg gctacgcctt cagcagctac 540
tggatgaact gggtgaagca gagacccggc cagggcctgg agtggatcgg ccagatctgg 600
cccggcgacg gcgacaccaa ctacaacggc aagttcaagg gcaaggccac cctgaccgcc 660
gacgagagca gcagcaccgc ctacatgcag ctgagcagcc tggccagcga ggacagcgcc 720
gtgtacttct gcgccagaag agagaccacc accgtgggca gatactacta cgccatggac 780
tactggggcc agggcaccac cgtgaccgtg agcagcggcg gcggcggcag cgacatcaag 840
ctgcagcaga gcggcgccga gctggccaga cccggcgcca gcgtgaagat gagctgcaag 900
accagcggct acaccttcac cagatacacc atgcactggg tgaagcagag acccggccag 960
ggcctggagt ggatcggcta catcaacccc agcagaggct acaccaacta caaccagaag 1020
ttcaaggaca aggccaccct gaccaccgac aagagcagca gcaccgccta catgcagctg 1080
agcagcctga ccagcgagga cagcgccgtg tactactgcg ccagatacta cgacgaccac 1140
tactgcctgg actactgggg ccagggcacc accctgaccg tgagcagcgt ggagggcggc 1200
agcggcggca gcggcggcag cggcggcagc ggcggcgtgg acgacatcca gctgacccag 1260
agccccgcca tcatgagcgc cagccccggc gagaaggtga ccatgacctg cagagccagc 1320
agcagcgtga gctacatgaa ctggtaccag cagaagagcg gcaccagccc caagagatgg 1380
atctacgaca ccagcaaggt ggccagcggc gtgccctaca gattcagcgg cagcggcagc 1440
ggcaccagct acagcctgac catcagcagc atggaggccg aggacgccgc cacctactac 1500
tgccagcagt ggagcagcaa ccccctgacc ttcggcgccg gcaccaagct ggagctgaag 1560
caccaccacc accaccacta g 1581
<210>45
<211>2003
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
полинуклеотид
<400>45
atgtggtgga gactgtggtg gctgctgctg ctgctgctgc tgctgtggcc catggtgtgg 60
gccgccgaca tccagctgac ccagagcccc gccagcctgg ccgtgagcct gggccagaga 120
gccaccatca gctgcaaggc cagccagagc gtggactacg acggcgacag ctacctgaac 180
tggtaccagc agatccccgg ccagcccccc aagctgctga tctacgacgc cagcaacctg 240
gtgagcggca tcccccccag attcagcggc agcggcagcg gcaccgactt caccctgaac 300
atccaccccg tggagaaggt acggagcgga ccacccctcc tgggcccctg cccgggtccc 360
gaccctcttt gccggcgccg ggcggggccg gcggagtatt tgatagtgta ttaaccttat 420
gtgtgacatg ttctaatata gtcacatttt cattattttt attataaggc ctgctgaaat 480
aattagctga gtagataagt gatgttggag ctggtggcgt aggcaagagt gccttgacga 540
tacagctaat tcagaatcat tttgtggacg aatatgatcc aacaatagag gtaaatcttg 600
ttttaatatg catattactg gtgcaggacc attctttgat acagataaag gtttctctga 660
ccattttgag ttgtatataa cacctttttt gaagtaaaag gtgcactgta ataatccaga 720
ctgtgtttct cccttctcag gtggacgccg ccacctacca ctgccagcag agcaccgagg 780
acccctggac cttcggcggc ggcaccaagc tggagatcaa gggcggcggc ggcagcggcg 840
gcggcggcag cggcggcggc ggcagccagg tgcagctgca gcagagcggc gccgagctgg 900
tgagacccgg cagcagcgtg aagatcagct gcaaggccag cggctacgcc ttcagcagct 960
actggatgaa ctgggtgaag cagagacccg gccagggcct ggagtggatc ggccagatct 1020
ggcccggcga cggcgacacc aactacaacg gcaagttcaa gggcaaggcc accctgaccg 1080
ccgacgagag cagcagcacc gcctacatgc agctgagcag cctggccagc gaggacagcg 1140
ccgtgtactt ctgcgccaga agagagacca ccaccgtggg cagatactac tacgccatgg 1200
actactgggg ccagggcacc accgtgaccg tgagcagcgg cggcggcggc agcgacatca 1260
agctgcagca gagcggcgcc gagctggcca gacccggcgc cagcgtgaag atgagctgca 1320
agaccagcgg ctacaccttc accagataca ccatgcactg ggtgaagcag agacccggcc 1380
agggcctgga gtggatcggc tacatcaacc ccagcagagg ctacaccaac tacaaccaga 1440
agttcaagga caaggccacc ctgaccaccg acaagagcag cagcaccgcc tacatgcagc 1500
tgagcagcct gaccagcgag gacagcgccg tgtactactg cgccagatac tacgacgacc 1560
actactgcct ggactactgg ggccagggca ccaccctgac cgtgagcagc gtggagggcg 1620
gcagcggcgg cagcggcggc agcggcggca gcggcggcgt ggacgacatc cagctgaccc 1680
agagccccgc catcatgagc gccagccccg gcgagaaggt gaccatgacc tgcagagcca 1740
gcagcagcgt gagctacatg aactggtacc agcagaagag cggcaccagc cccaagagat 1800
ggatctacga caccagcaag gtggccagcg gcgtgcccta cagattcagc ggcagcggca 1860
gcggcaccag ctacagcctg accatcagca gcatggaggc cgaggacgcc gccacctact 1920
actgccagca gtggagcagc aaccccctga ccttcggcgc cggcaccaag ctggagctga 1980
agcaccacca ccaccaccac tag 2003
<210>46
<211>2003
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
полинуклеотид
<400>46
atgtggtgga gactgtggtg gctgctgctg ctgctgctgc tgctgtggcc catggtgtgg 60
gccgccgaca tccagctgac ccagagcccc gccagcctgg ccgtgagcct gggccagaga 120
gccaccatca gctgcaaggc cagccagagc gtggactacg acggcgacag ctacctgaac 180
tggtaccagc agatccccgg ccagcccccc aagctgctga tctacgacgc cagcaacctg 240
gtgagcggca tcccccccag attcagcggc agcggcagcg gcaccgactt caccctgaac 300
atccaccccg tggagaaggt acggagcgga ccacccctcc tgggcccctg cccgggtccc 360
gaccctcttt gccggcgccg ggcggggccg gcggagtatt tgatagtgta ttaaccttat 420
gtgtgacatg ttctaatata gtcacatttt ccttcttttt tttttaaggc ctgctgaaat 480
aattagctga gtagataagt gatgttggag ctggtggcgt aggcaagagt gccttgacga 540
tacagctaat tcagaatcat tttgtggacg aatatgatcc aacaatagag gtaaatcttg 600
ttttaatatg catattactg gtgcaggacc attctttgat acagataaag gtttctctga 660
ccattttgag ttgtatataa cacctttttt gaagtaaaag gtgcactgta ataatccaga 720
ctgtgtttct cccttctcag gtggacgccg ccacctacca ctgccagcag agcaccgagg 780
acccctggac cttcggcggc ggcaccaagc tggagatcaa gggcggcggc ggcagcggcg 840
gcggcggcag cggcggcggc ggcagccagg tgcagctgca gcagagcggc gccgagctgg 900
tgagacccgg cagcagcgtg aagatcagct gcaaggccag cggctacgcc ttcagcagct 960
actggatgaa ctgggtgaag cagagacccg gccagggcct ggagtggatc ggccagatct 1020
ggcccggcga cggcgacacc aactacaacg gcaagttcaa gggcaaggcc accctgaccg 1080
ccgacgagag cagcagcacc gcctacatgc agctgagcag cctggccagc gaggacagcg 1140
ccgtgtactt ctgcgccaga agagagacca ccaccgtggg cagatactac tacgccatgg 1200
actactgggg ccagggcacc accgtgaccg tgagcagcgg cggcggcggc agcgacatca 1260
agctgcagca gagcggcgcc gagctggcca gacccggcgc cagcgtgaag atgagctgca 1320
agaccagcgg ctacaccttc accagataca ccatgcactg ggtgaagcag agacccggcc 1380
agggcctgga gtggatcggc tacatcaacc ccagcagagg ctacaccaac tacaaccaga 1440
agttcaagga caaggccacc ctgaccaccg acaagagcag cagcaccgcc tacatgcagc 1500
tgagcagcct gaccagcgag gacagcgccg tgtactactg cgccagatac tacgacgacc 1560
actactgcct ggactactgg ggccagggca ccaccctgac cgtgagcagc gtggagggcg 1620
gcagcggcgg cagcggcggc agcggcggca gcggcggcgt ggacgacatc cagctgaccc 1680
agagccccgc catcatgagc gccagccccg gcgagaaggt gaccatgacc tgcagagcca 1740
gcagcagcgt gagctacatg aactggtacc agcagaagag cggcaccagc cccaagagat 1800
ggatctacga caccagcaag gtggccagcg gcgtgcccta cagattcagc ggcagcggca 1860
gcggcaccag ctacagcctg accatcagca gcatggaggc cgaggacgcc gccacctact 1920
actgccagca gtggagcagc aaccccctga ccttcggcgc cggcaccaag ctggagctga 1980
agcaccacca ccaccaccac tag 2003
<210>47
<211>32
<212>PRT
<213>Homo sapiens
<400>47
Met Val Ser Lys Leu Ser Gln Leu Gln Thr Glu Leu Leu Ala Ala Leu
1 5 10 15
Leu Glu Ser Gly Leu Ser Lys Glu Ala Leu Ile Gln Ala Leu Gly Glu
20 25 30
<210>48
<211>96
<212>DNA
<213>Homo sapiens
<400>48
atggtgagca agctgagcca gctgcagacc gagctgctgg ccgccctgct ggagagcggc 60
ctgagcaagg aggccctgat ccaggccctg ggcgag 96
<210>49
<211>313
<212>DNA
<213>Неизвестный
<220>
<223>Описание неизвестного:
последовательность промотора EFS
<400>49
gtccgttatc aacttgaaaa agtggcaccg agtcggtgct tttttgaatt cgctagctag 60
gtcttgaaag gagtgggaat tggctccggt gcccgtcagt gggcagagcg cacatcgccc 120
acagtccccg agaagttggg gggaggggtc ggcaattgat ccggtgccta gagaaggtgg 180
cgcggggtaa actgggaaag tgatgtcgtg tactggctcc gcctttttcc cgagggtggg 240
ggagaaccgt atataagtgc agtagtcgcc gtgaacgttc tttttcgcaa cgggtttgcc 300
gccagaacac agg 313
<210>50
<211>597
<212>DNA
<213>Неизвестный
<220>
<223>Описание неизвестного:
последовательность WPRE
<400>50
tcgacaatca acctctggat tacaaaattt gtgaaagatt gactggtatt cttaactatg 60
ttgctccttt tacgctatgt ggatacgctg ctttaatgcc tttgtatcat gctattgctt 120
cccgtatggc tttcattttc tcctccttgt ataaatcctg gttgctgtct ctttatgagg 180
agttgtggcc cgttgtcagg caacgtggcg tggtgtgcac tgtgtttgct gacgcaaccc 240
ccactggttg gggcattgcc accacctgtc agctcctttc cgggactttc gctttccccc 300
tccctattgc cacggcggaa ctcatcgccg cctgccttgc ccgctgctgg acaggggctc 360
ggctgttggg cactgacaat tccgtggtgt tgtcggggaa gctgacgtcc tttccatggc 420
tgctcgcctg tgttgccacc tggattctgc gcgggacgtc cttctgctac gtcccttcgg 480
ccctcaatcc agcggacctt ccttcccgcg gcctgctgcc ggctctgcgg cctcttccgc 540
gtcttcgcct tcgccctcag acgagtcgga tctccctttg ggccgcctcc ccgcctg 597
<210>51
<211>21
<212>PRT
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
пептид
<400>51
Met Trp Trp Arg Leu Trp Trp Leu Leu Leu Leu Leu Leu Leu Leu Trp
1 5 10 15
Pro Met Val Trp Ala
20
<210>52
<211>63
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
олигонуклеотид
<400>52
atgtggtgga gactgtggtg gctgctgctg ctgctgctgc tgctgtggcc catggtgtgg 60
gcc 63
<210>53
<211>22
<212>PRT
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
пептид
<400>53
Met Trp Trp Arg Leu Trp Trp Leu Leu Leu Leu Leu Leu Leu Leu Trp
1 5 10 15
Pro Met Val Trp Ala Ala
20
<210>54
<211>66
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
олигонуклеотид
<400>54
atgtggtgga gactgtggtg gctgctgctg ctgctgctgc tgctgtggcc catggtgtgg 60
gccgcc 66
<210>55
<211>23
<212>PRT
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
пептид
<400>55
Met Trp Trp Arg Leu Trp Trp Leu Leu Leu Leu Leu Leu Leu Leu Trp
1 5 10 15
Pro Met Val Trp Ala Ala Ala
20
<210>56
<211>69
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
олигонуклеотид
<400>56
atgtggtgga gactgtggtg gctgctgctg ctgctgctgc tgctgtggcc catggtgtgg 60
gccgccgcc 69
<210>57
<211>24
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
олигонуклеотид
<400>57
taattagctg agtagataag tgat 24
<210>58
<211>30
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
олигонуклеотид
<400>58
ctaggttgtt atctccattt agaacaaaat 30
<210>59
<211>50
<212>PRT
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
полипептид
<220>
<221>MISC_FEATURE
<222>(1)..(50)
<223>This sequence may encompass 1-10 "Gly Gly Gly Gly Ser"
repeating units
<400>59
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
20 25 30
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
35 40 45
Gly Ser
50
<210>60
<211>6
<212>PRT
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
пептид
<400>60
Gly Ser Gly Gly Ala Pro
1 5
<210>61
<211>11
<212>PRT
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
пептид
<400>61
Thr Pro Leu Gly Asp Thr Thr His Thr Ser Gly
1 5 10
<210>62
<211>19
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
олигонуклеотид
<400>62
cccatcttta caggctccc 19
<210>63
<211>19
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
олигонуклеотид
<400>63
tctctccctg caggatact 19
<210>64
<211>19
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
олигонуклеотид
<400>64
ctttttcata caggaggaa 19
<210>65
<211>19
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
олигонуклеотид
<400>65
tctttatttc caggcaaag 19
<210>66
<211>11
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
олигонуклеотид
<220>
<221>modified_base
<222>(4)..(4)
<223>a, c, t, g, неизвестное или другое
<220>
<221>modified_base
<222>(8)..(8)
<223>a, c, t, g, неизвестное или другое
<400>66
taantagntg a 11
<210>67
<211>21
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
олигонуклеотид
<400>67
tttcattatt tttattataa g 21
<210>68
<211>21
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
олигонуклеотид
<400>68
tttccttctt ttttttttca g 21
<210>69
<211>21
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
олигонуклеотид
<400>69
tttccttctt ttttttttaa g 21
<210>70
<211>21
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
олигонуклеотид
<400>70
tttccttctt tttattataa g 21
<210>71
<211>21
<212>DNA
<213>Искусственная последовательность
<220>
<223>Описание искусственной последовательности: Синтетический
олигонуклеотид
<400>71
tttcattatt ttttttttca g 21
<---
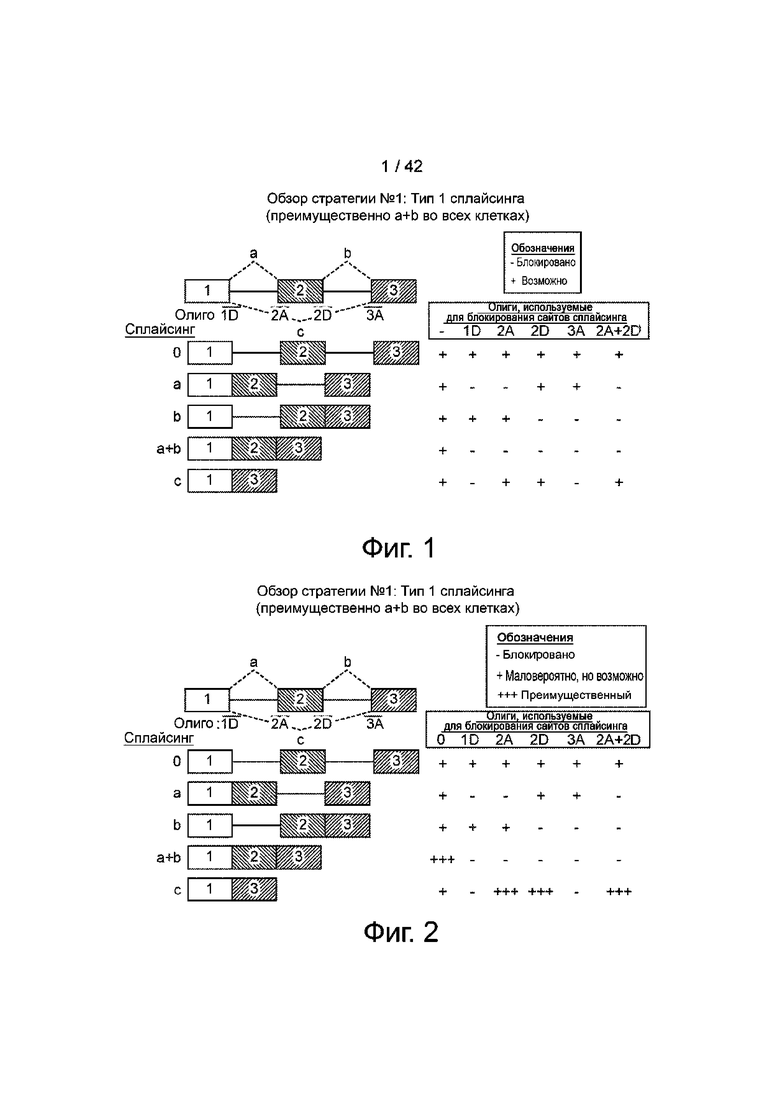
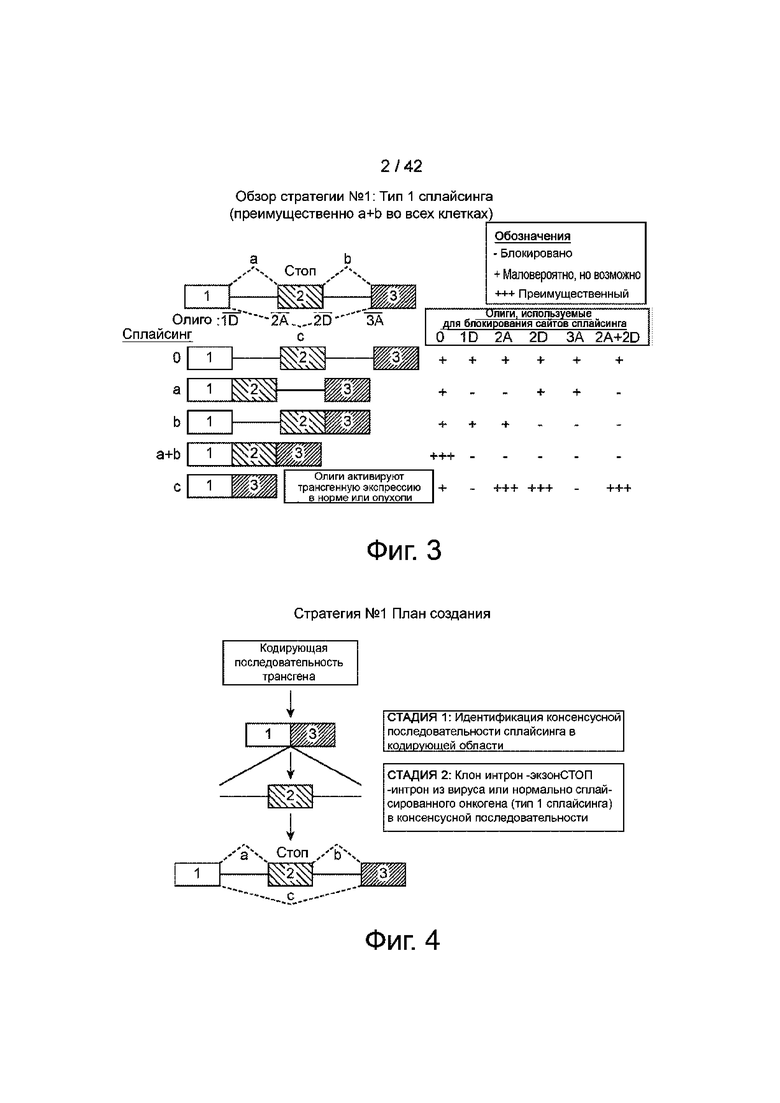
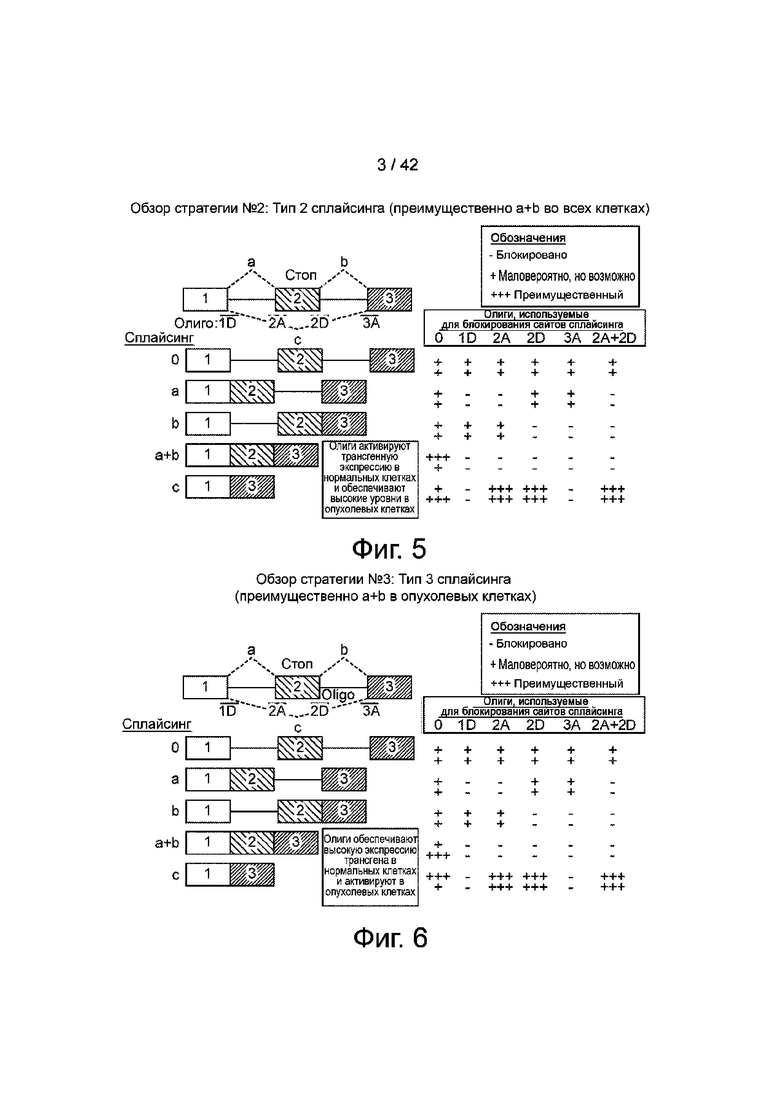
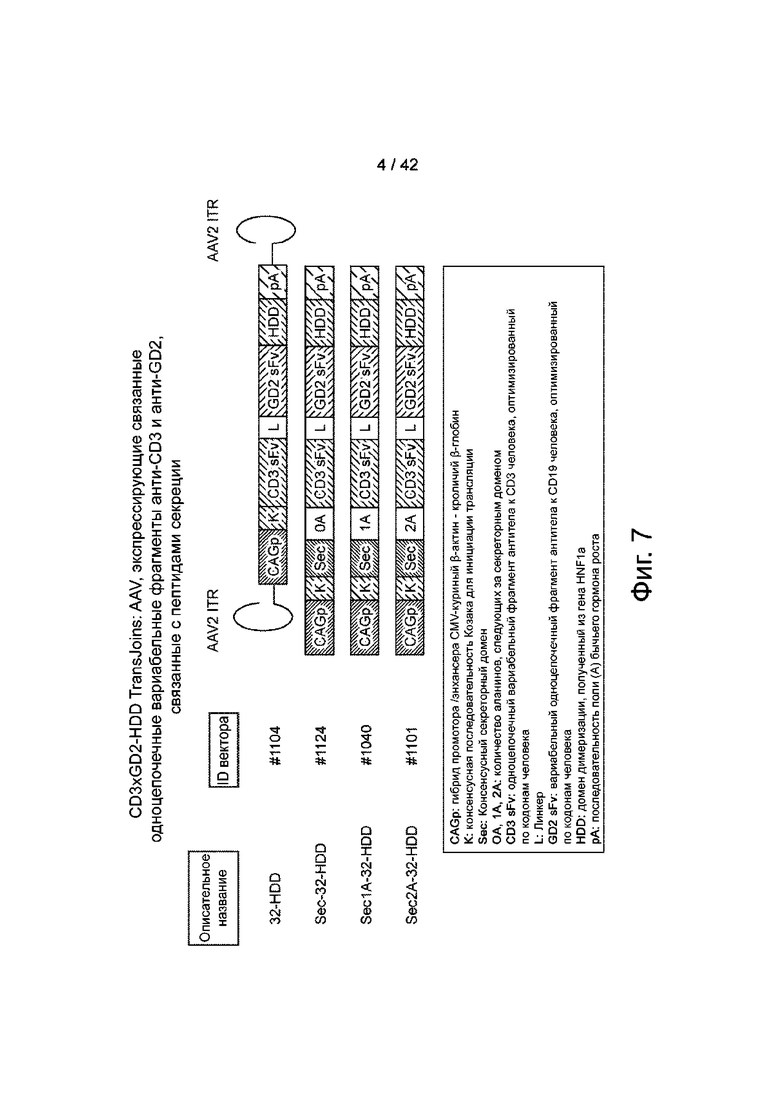
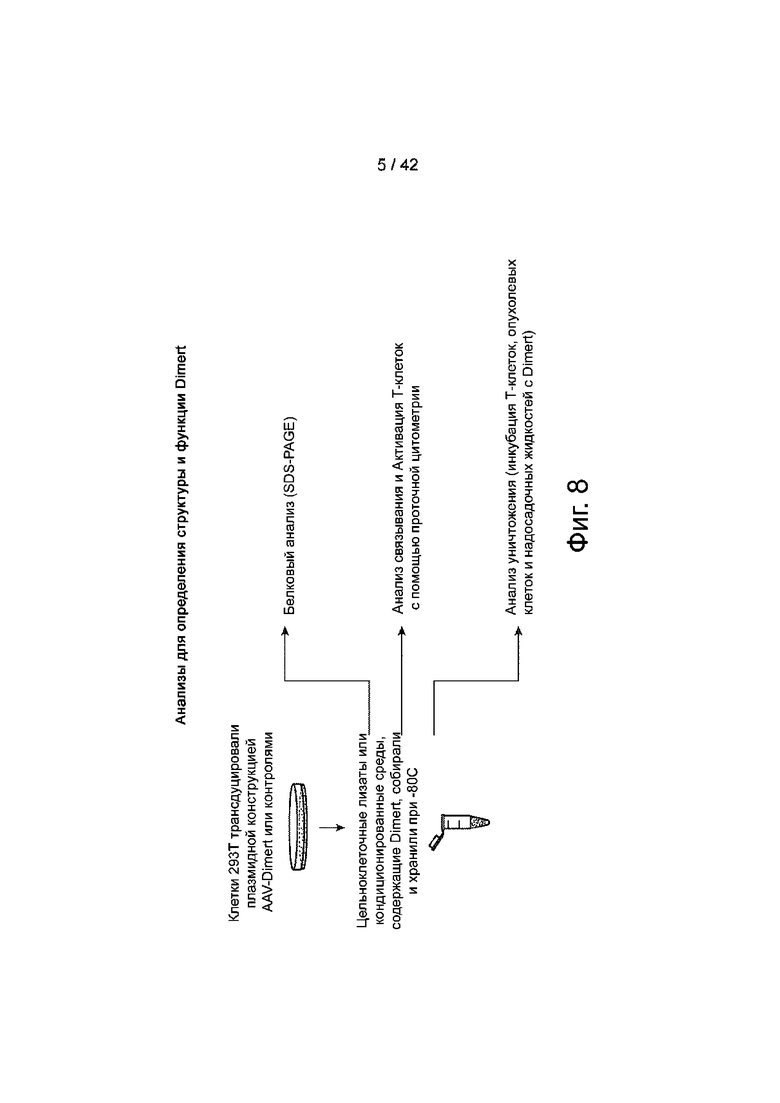

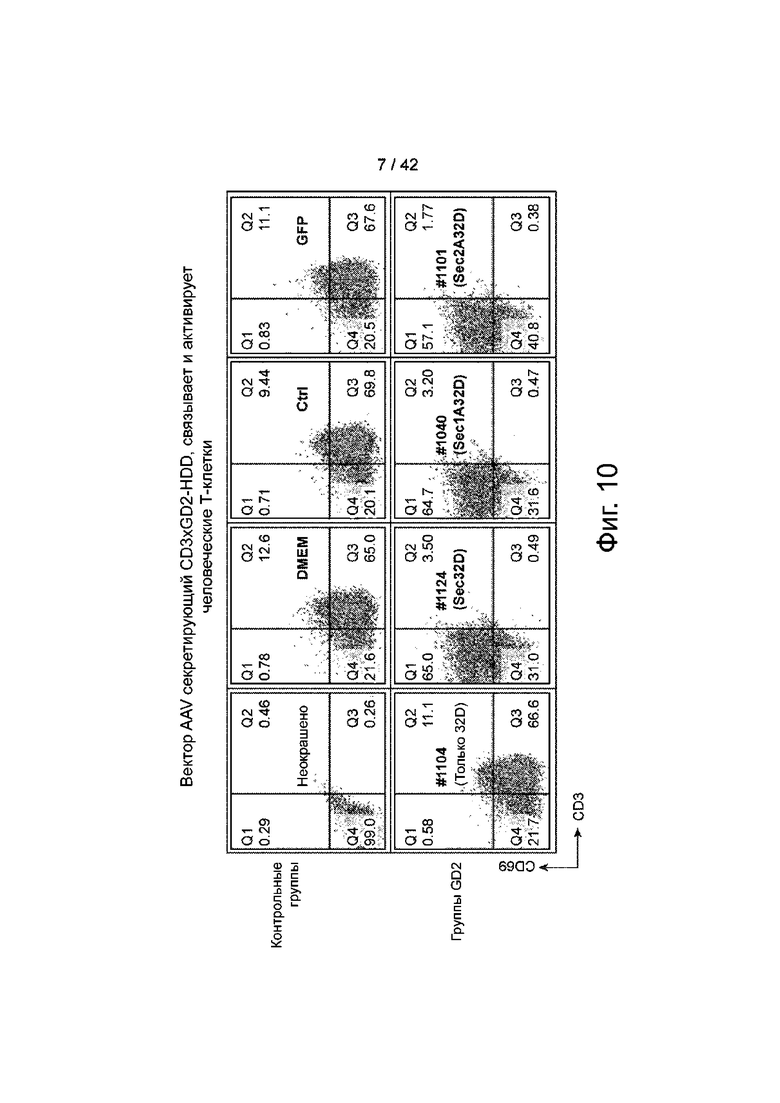
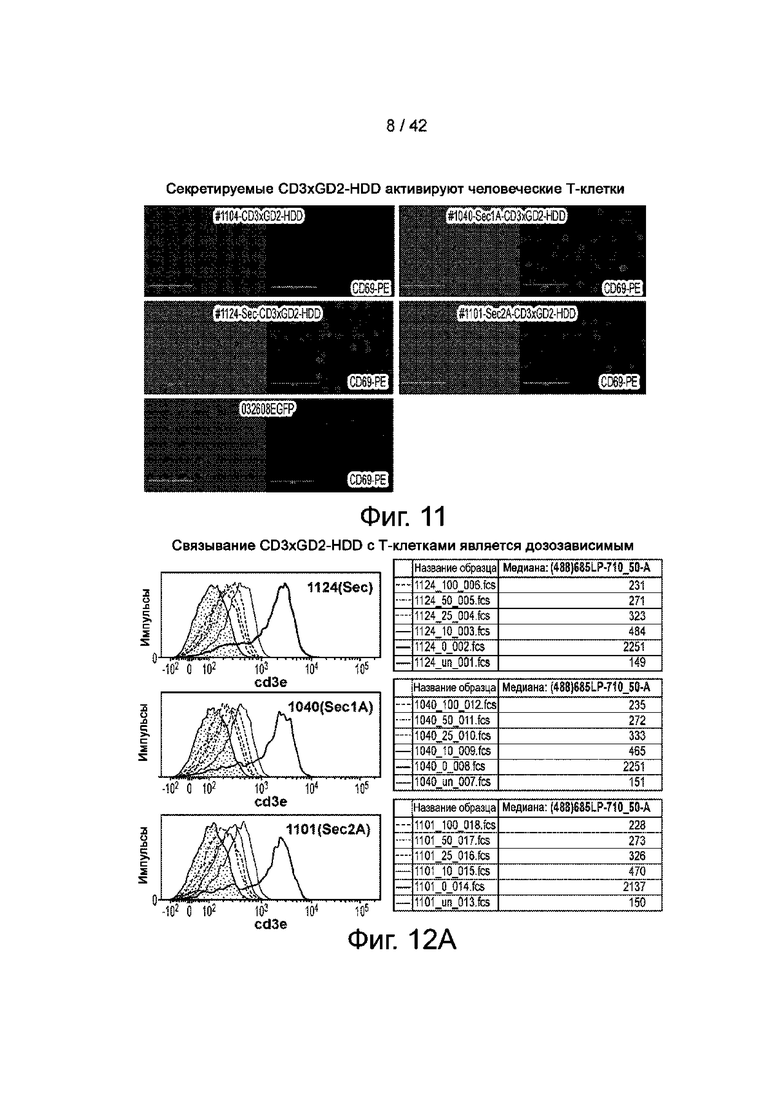
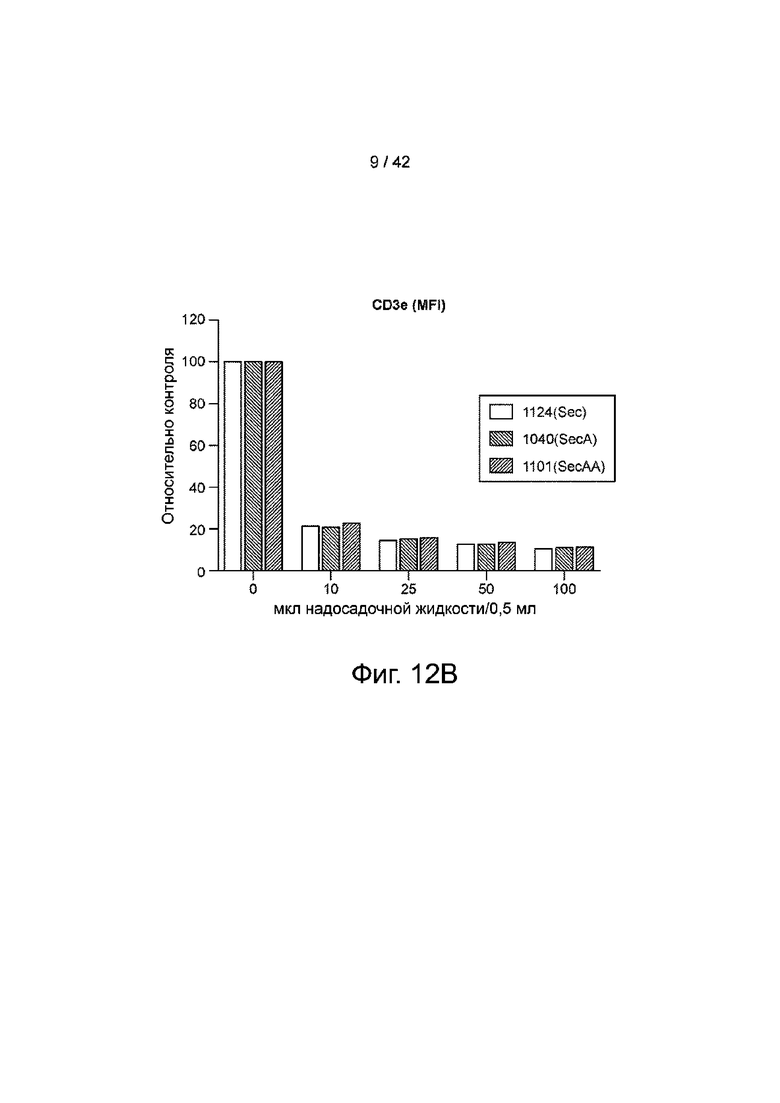
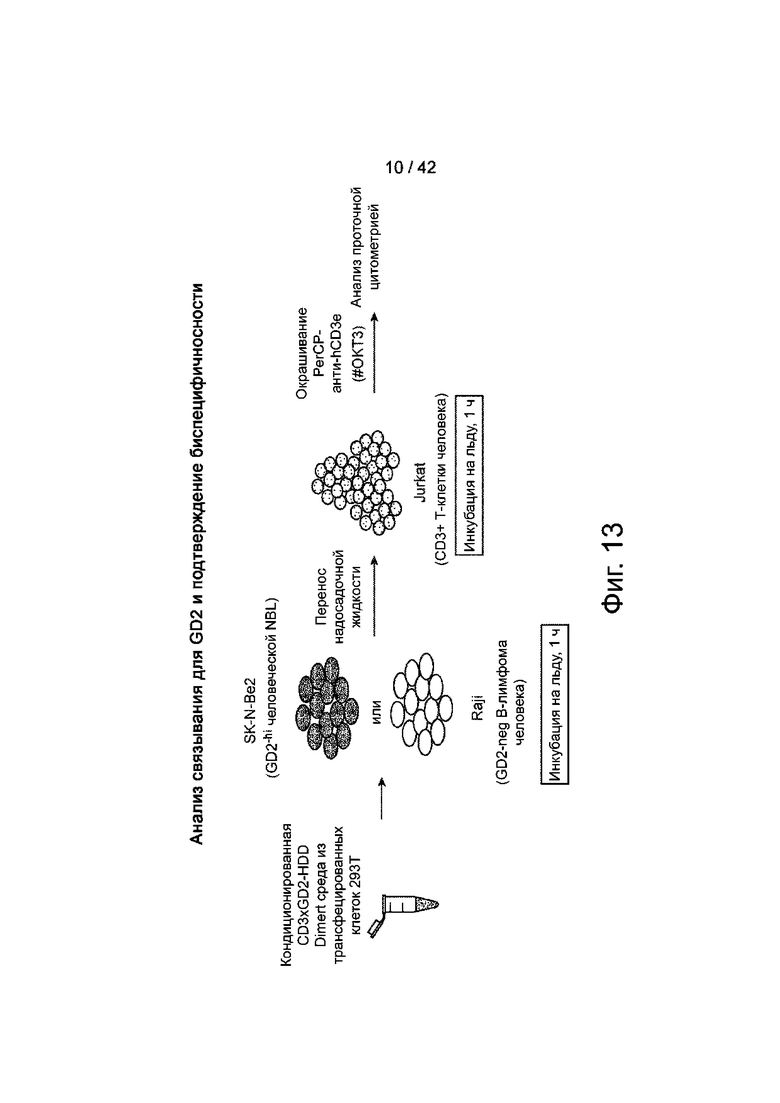

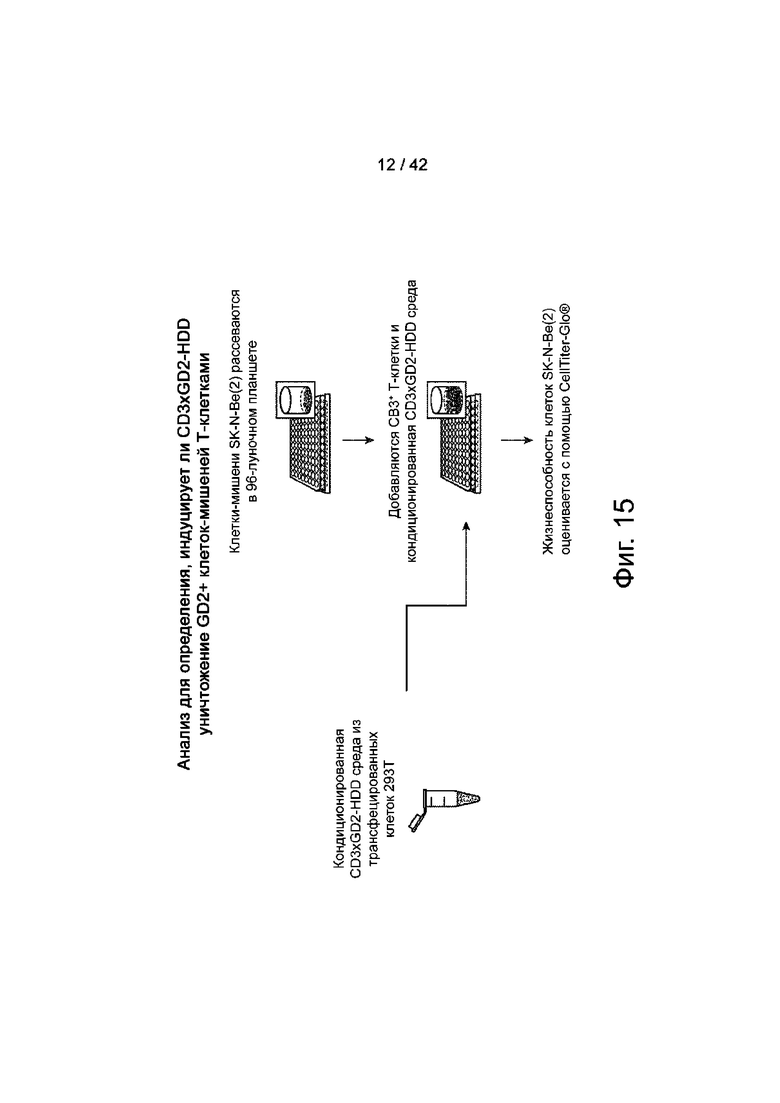
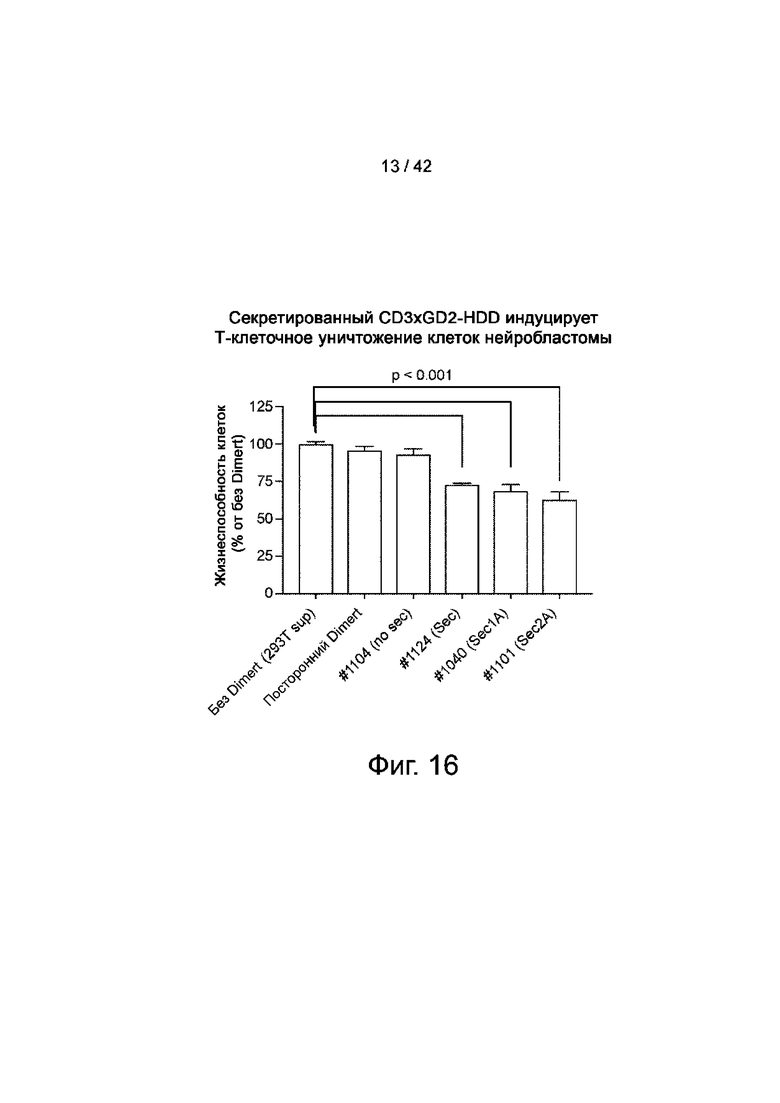
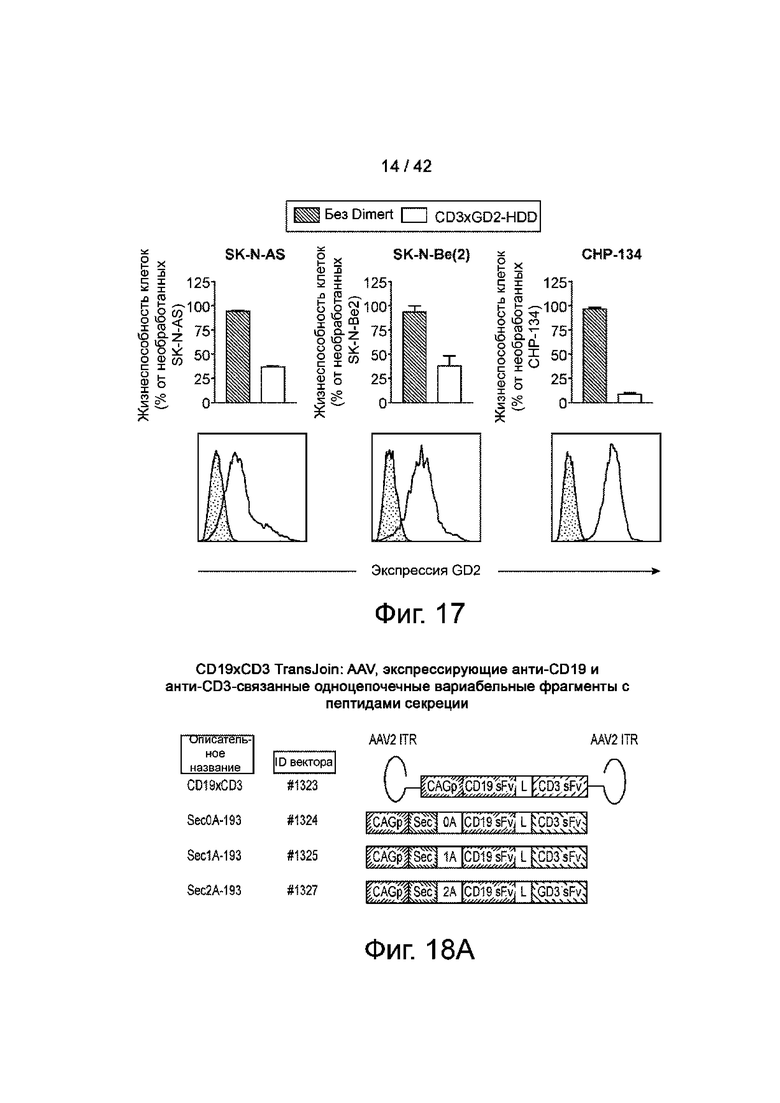
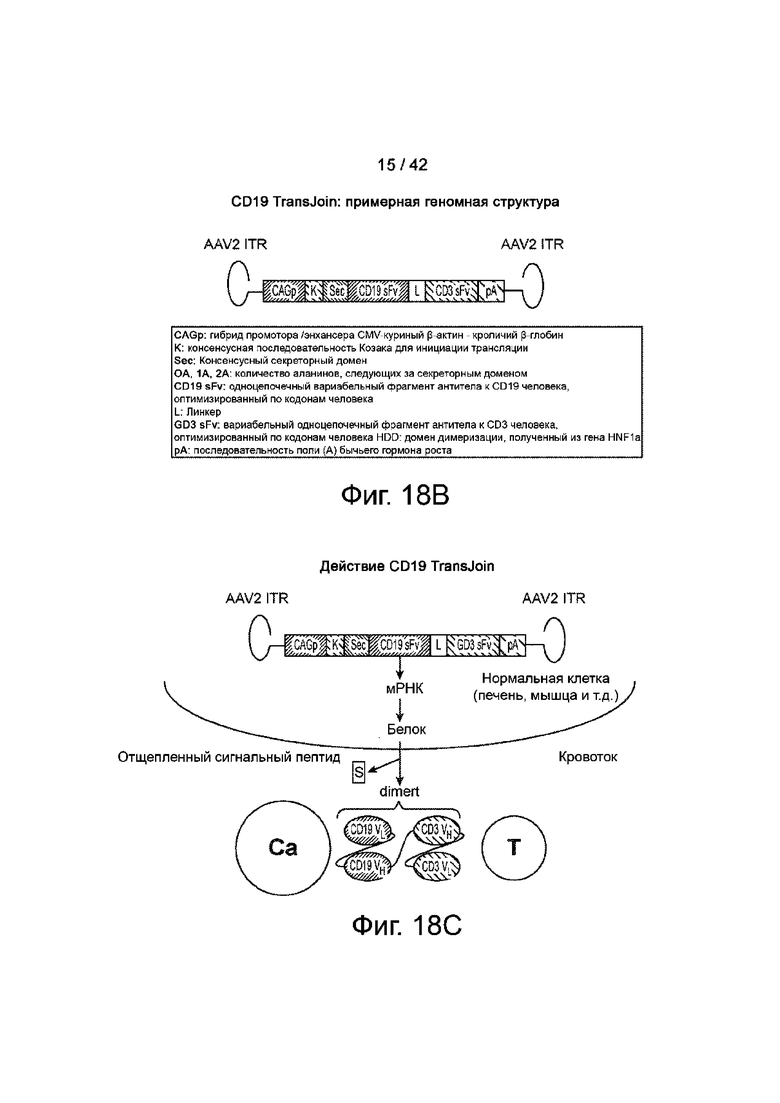
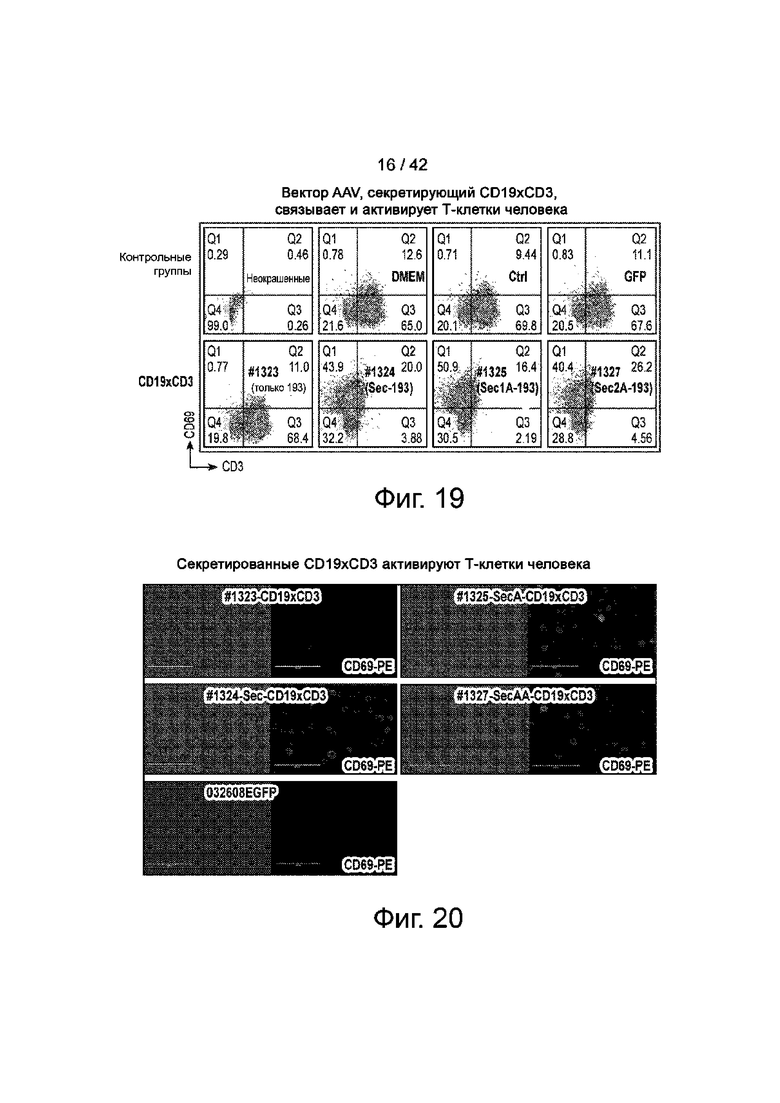
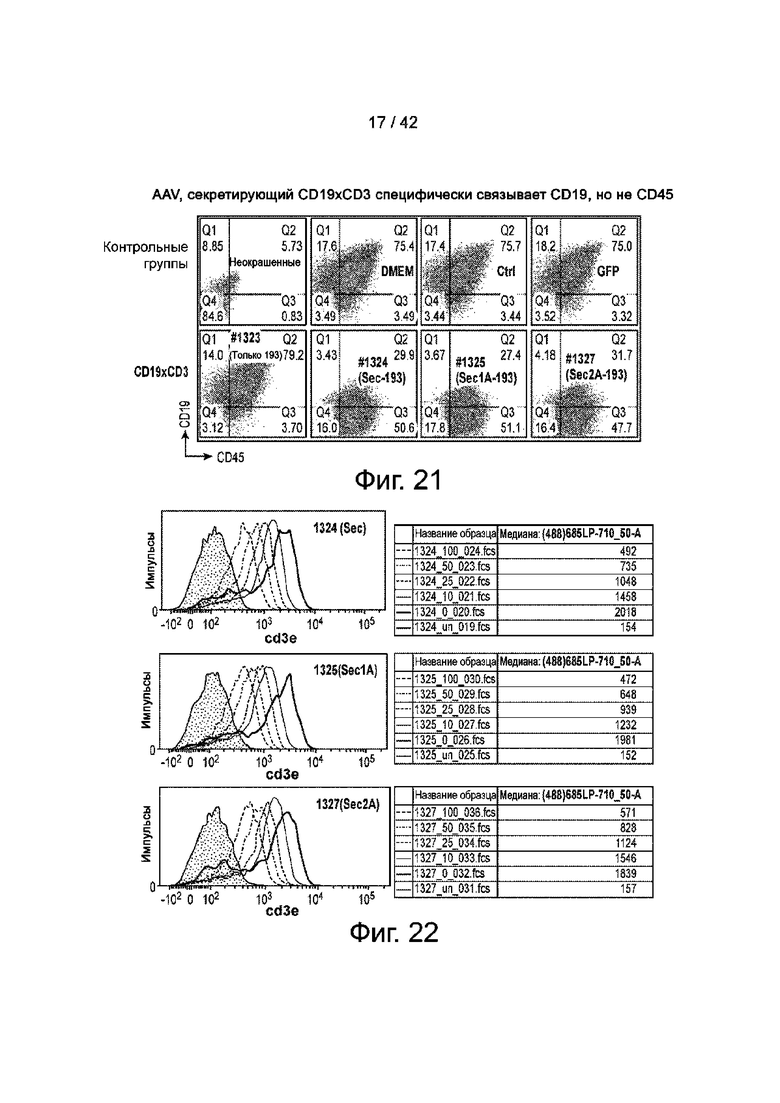
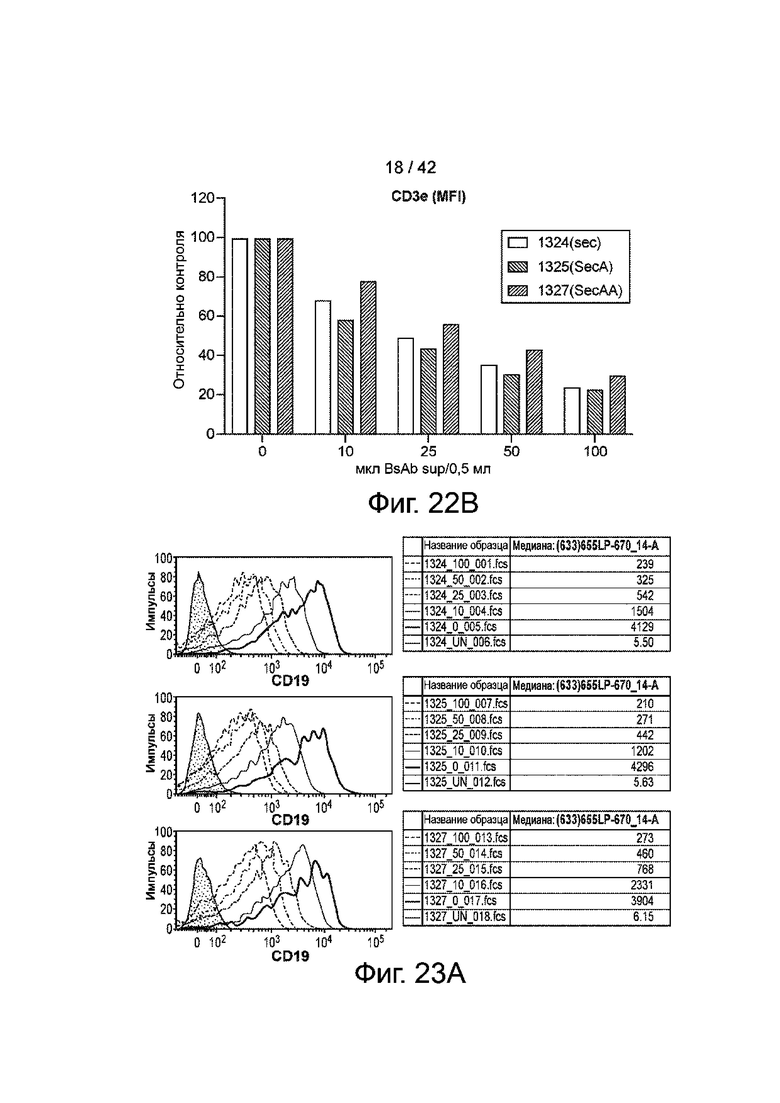
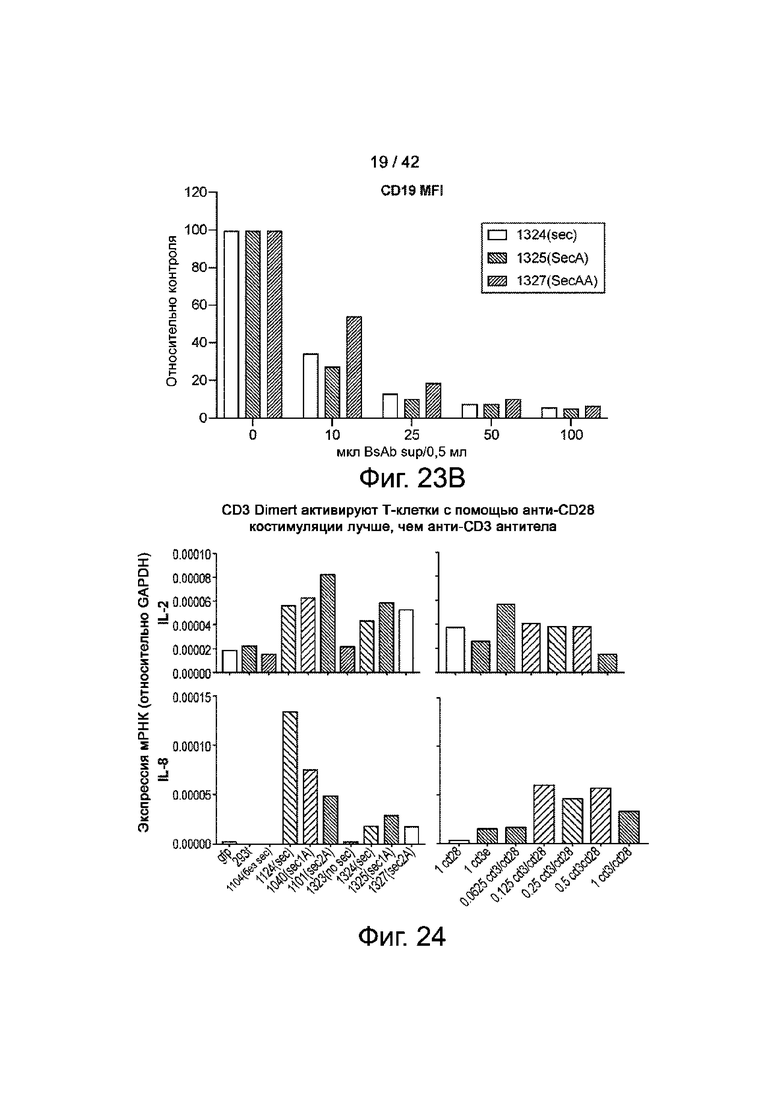
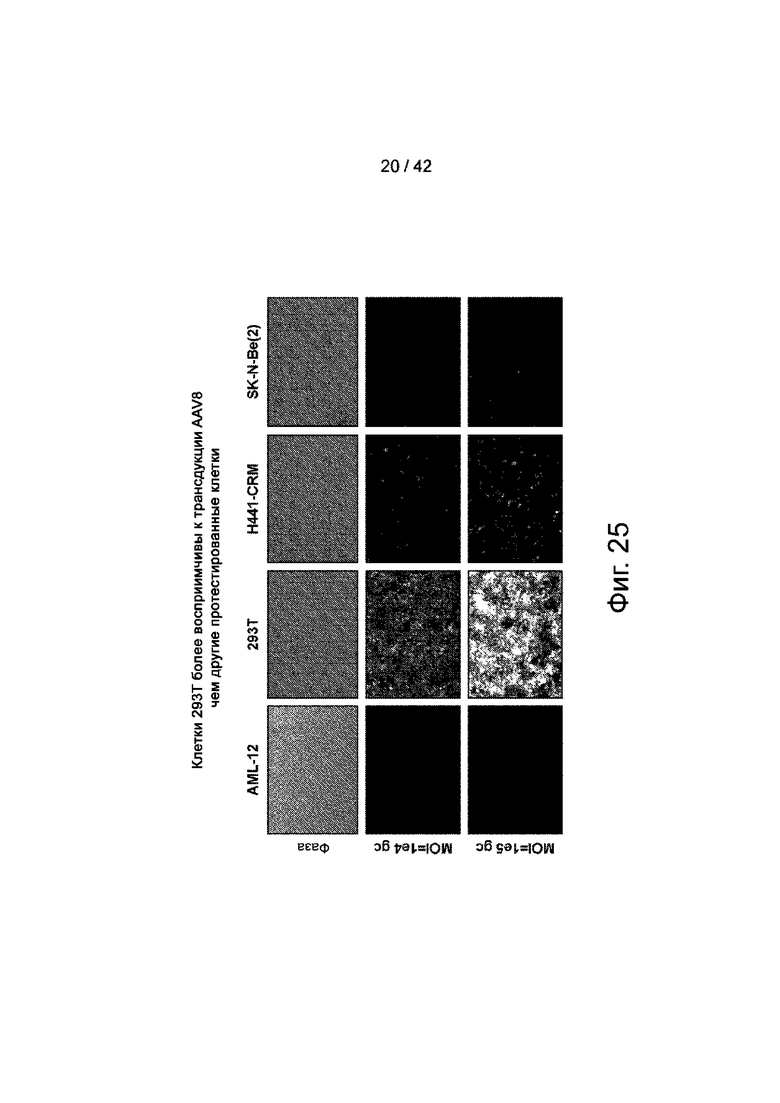
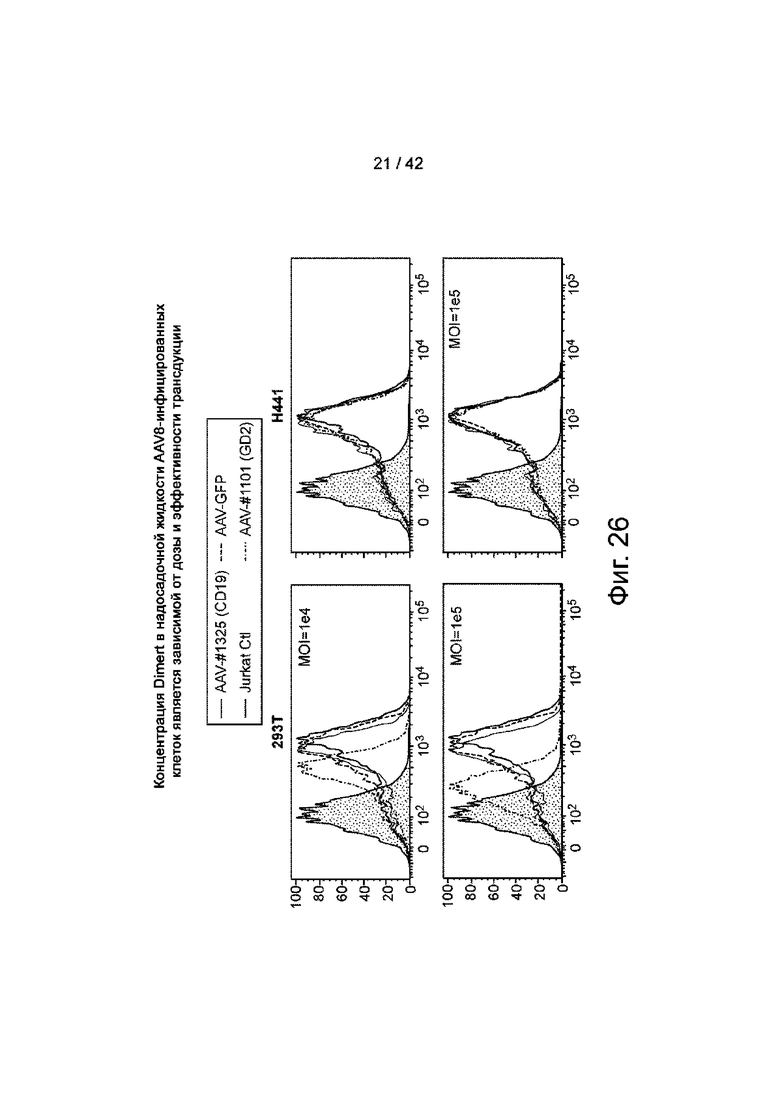
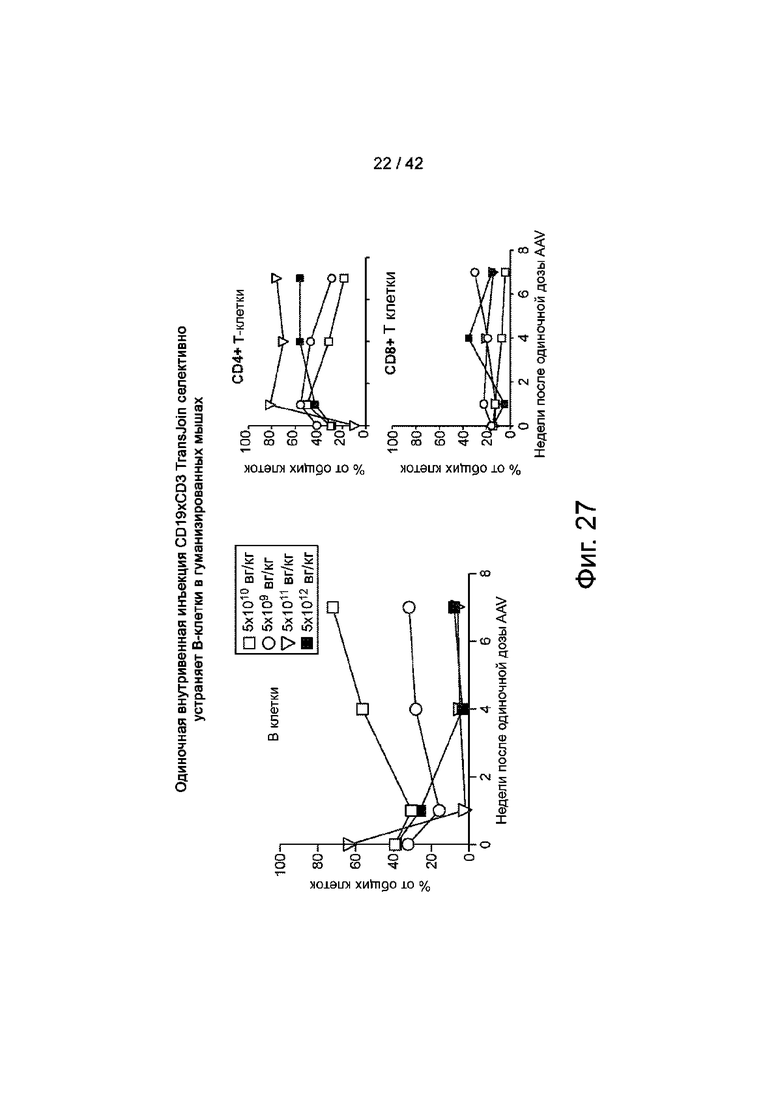
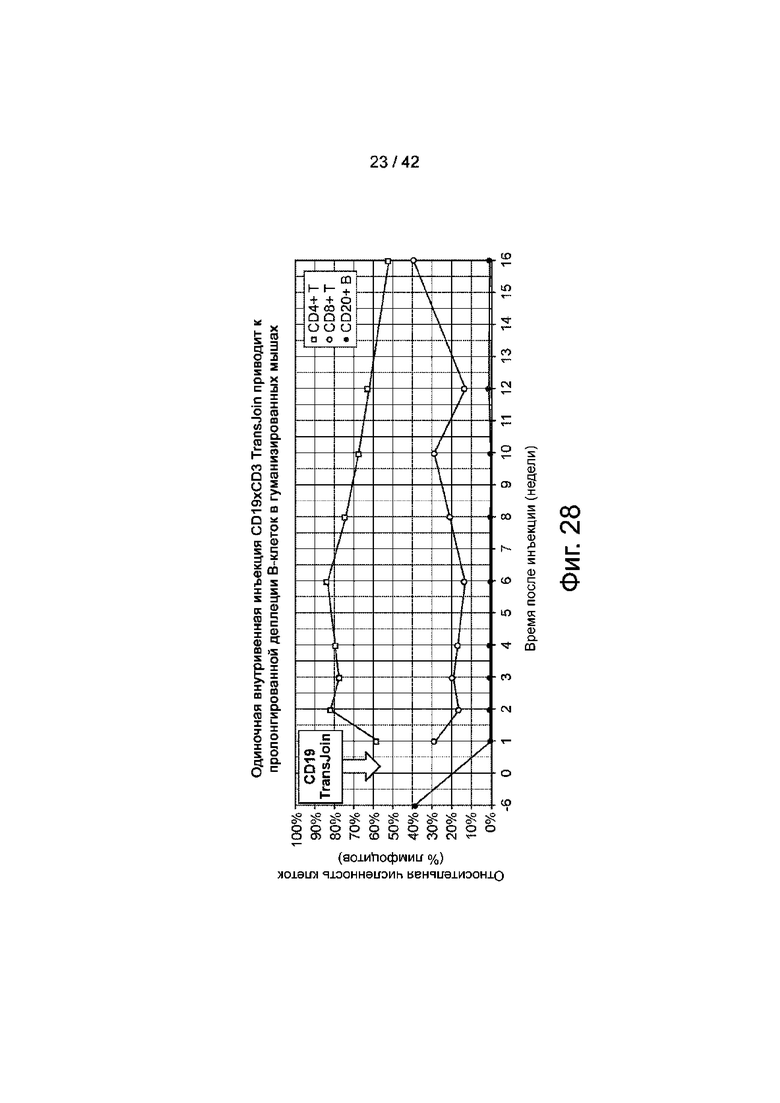

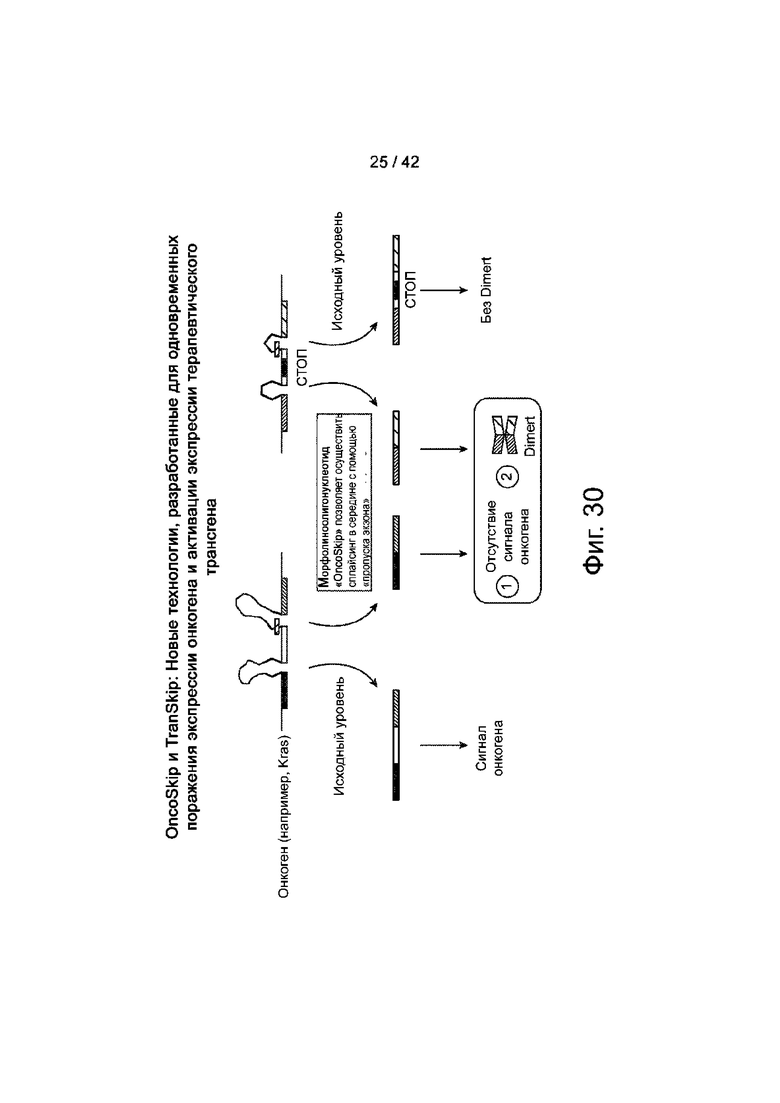

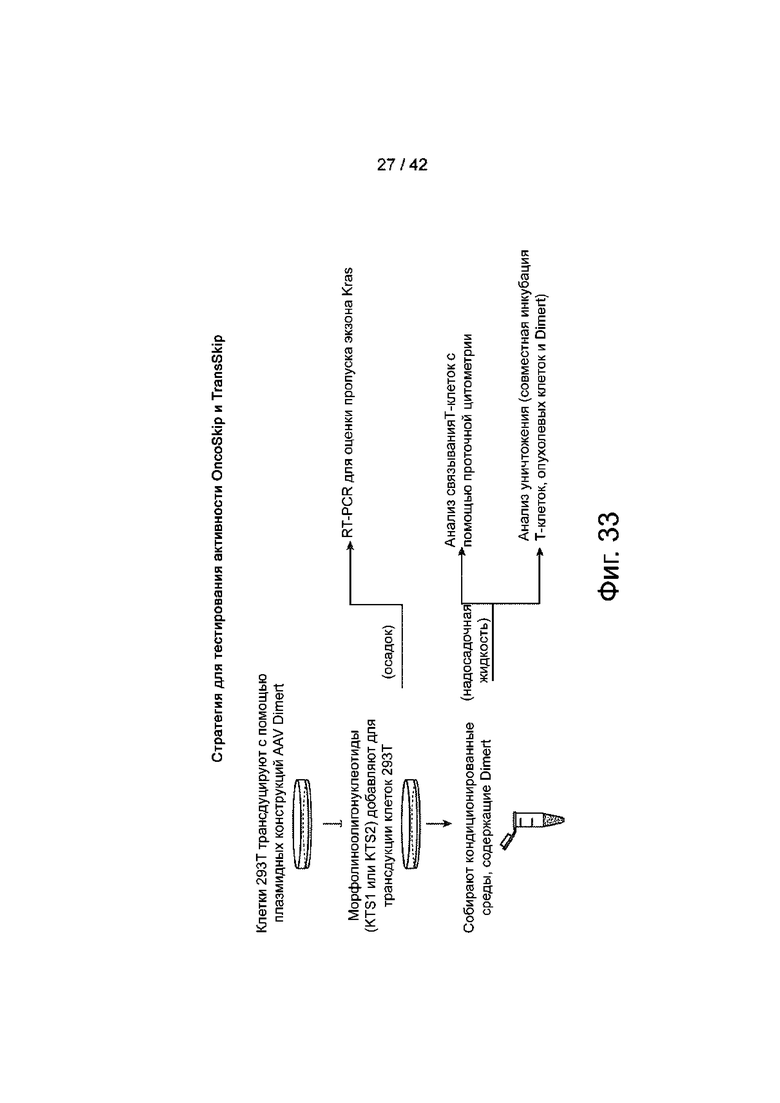
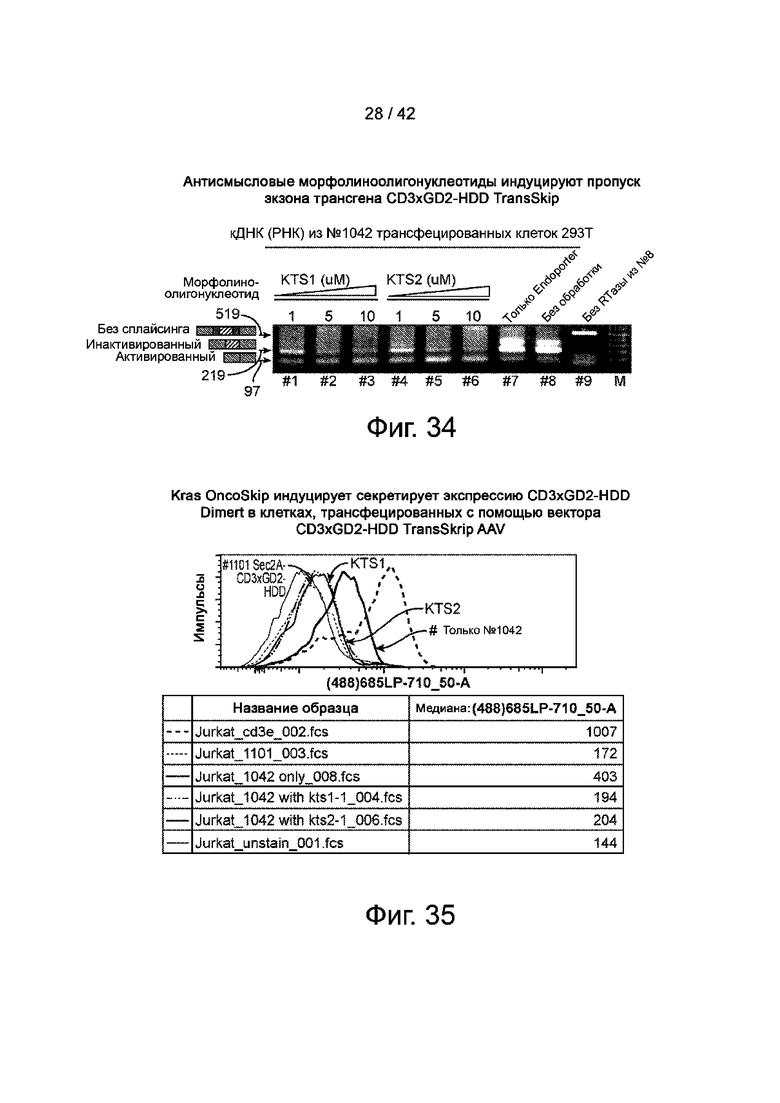
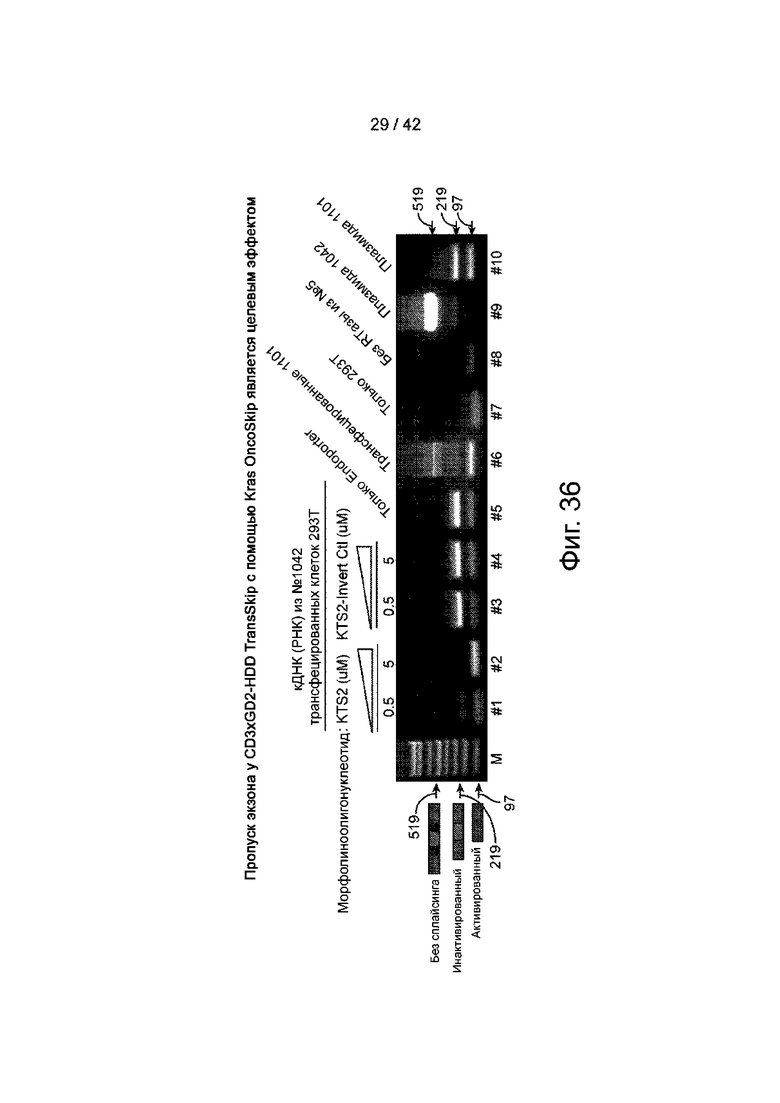
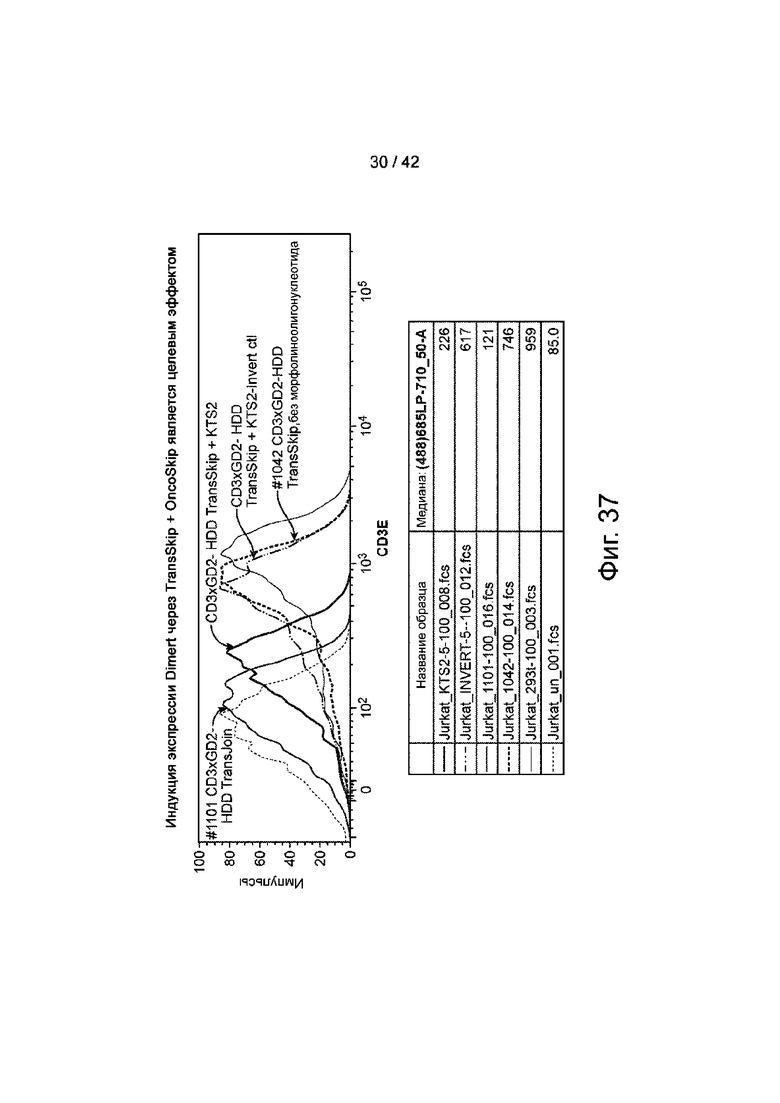
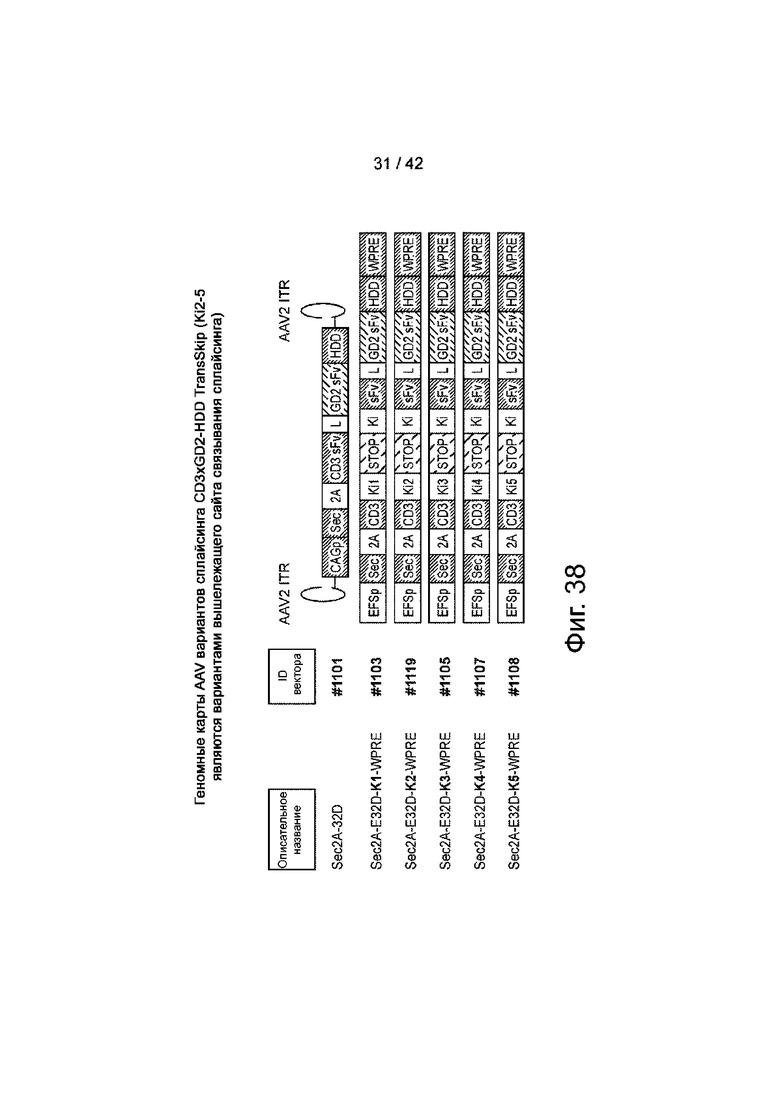
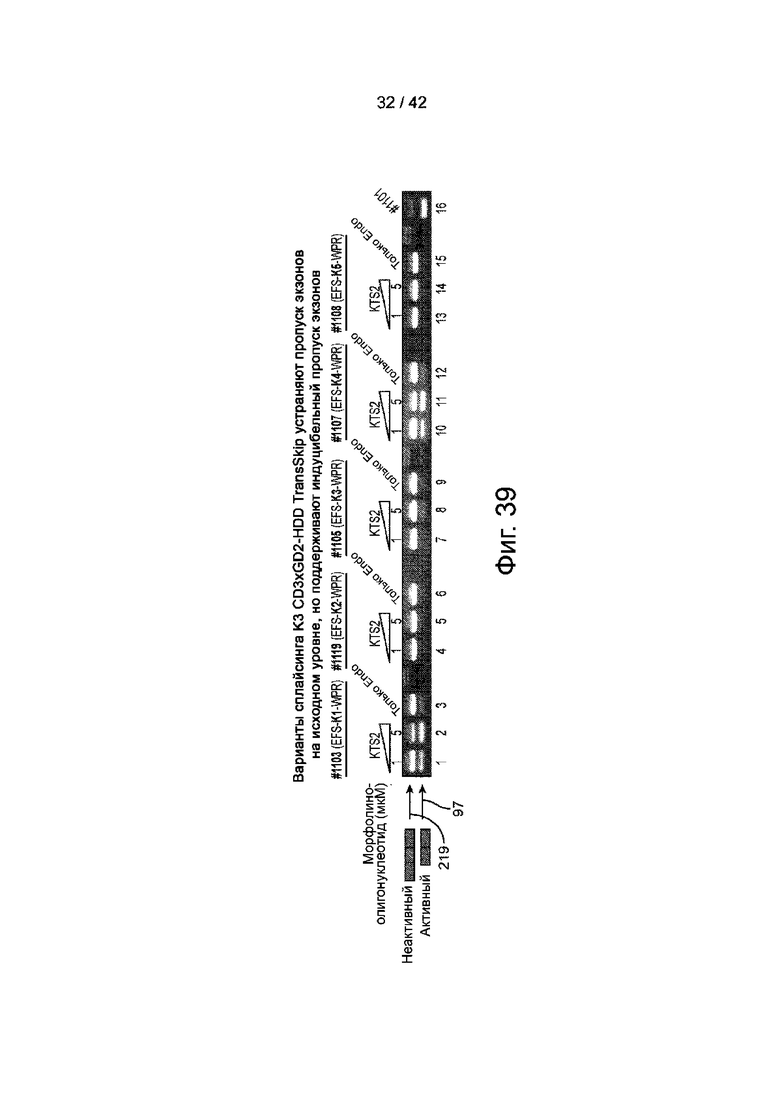
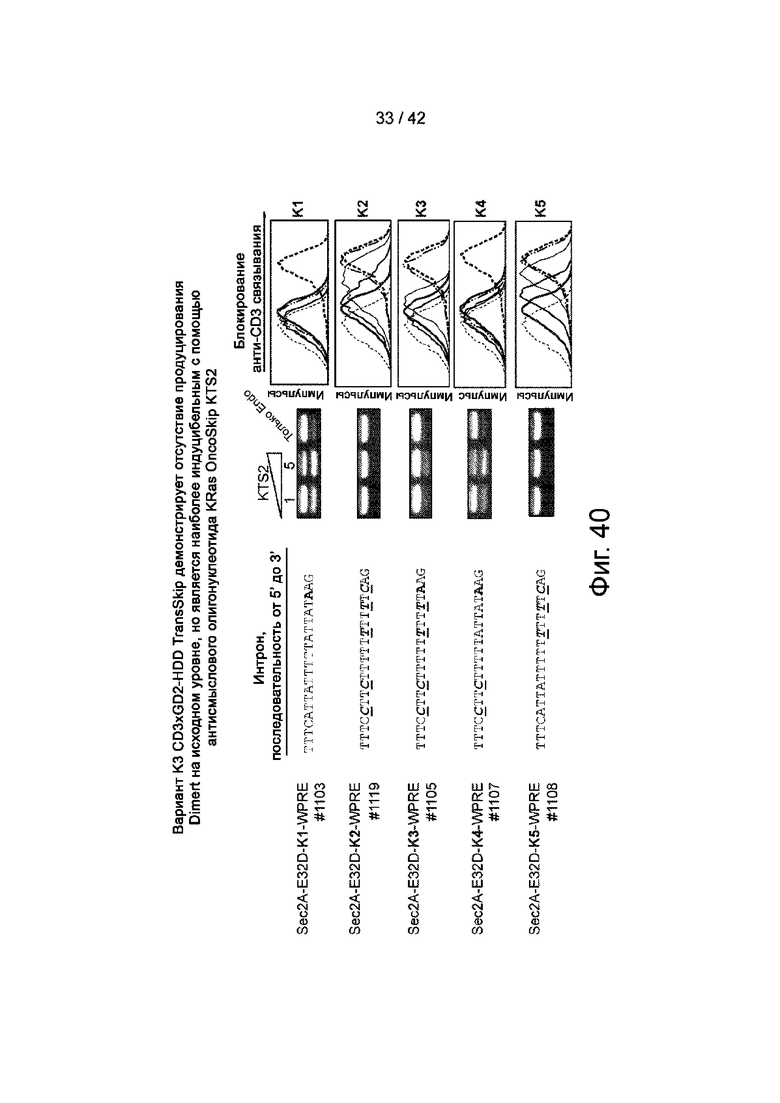
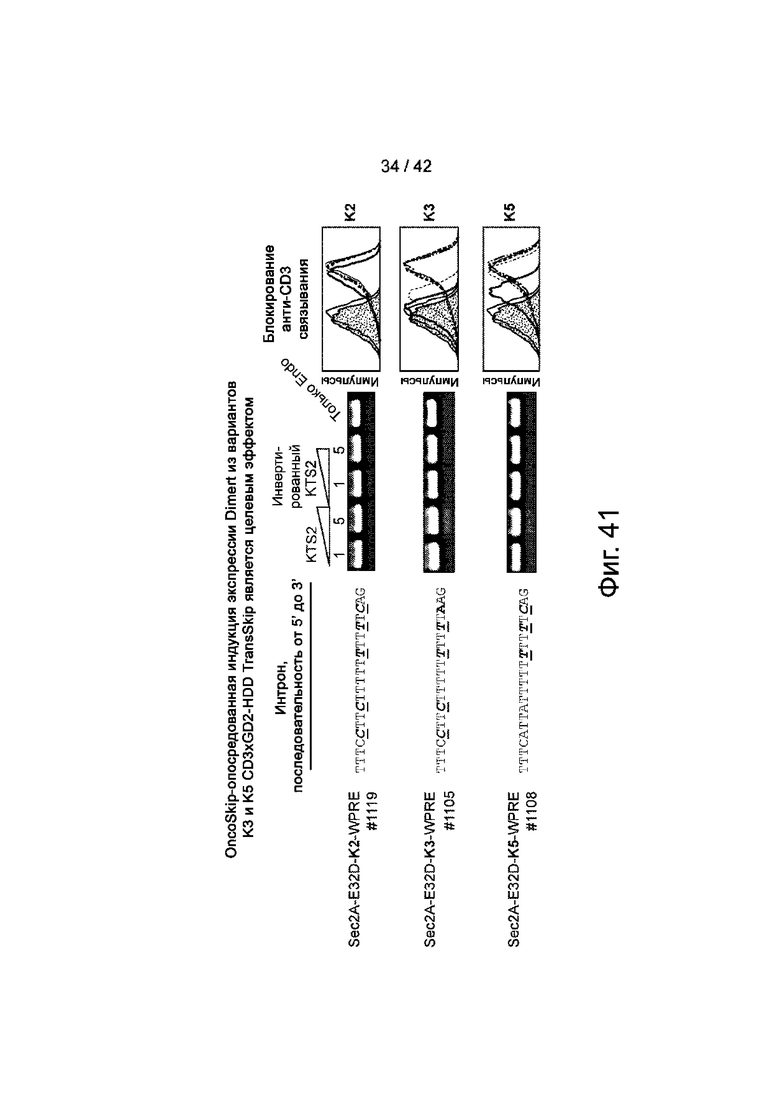
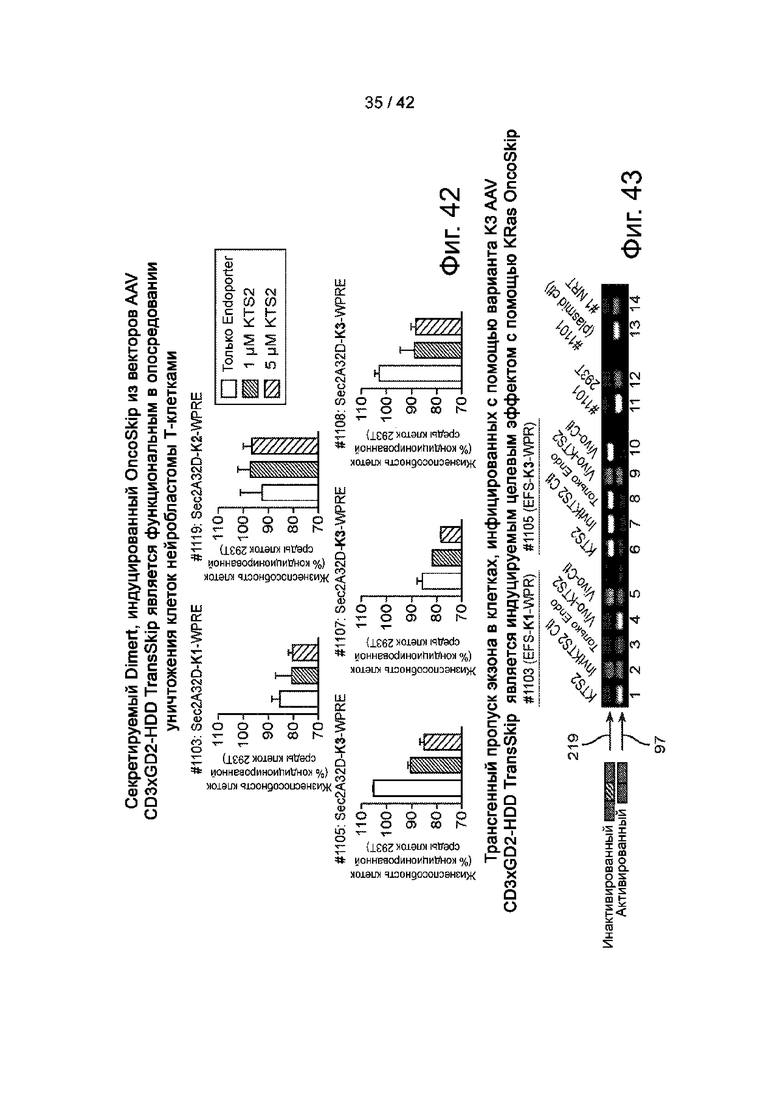
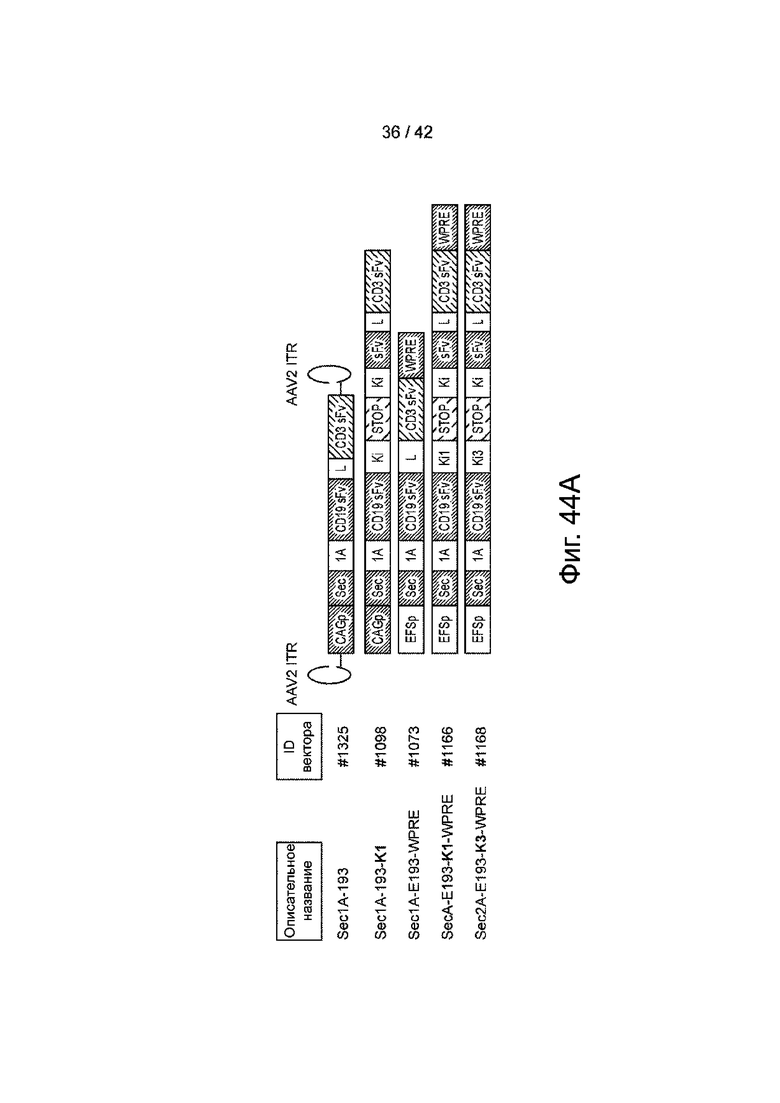

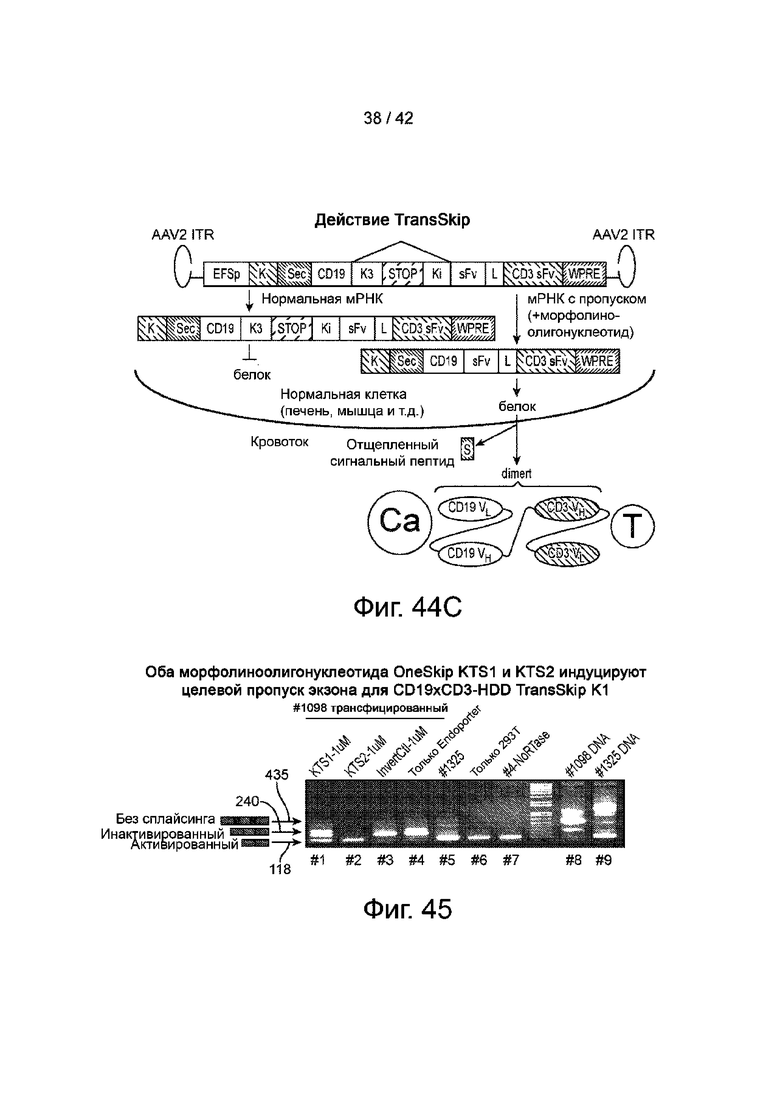
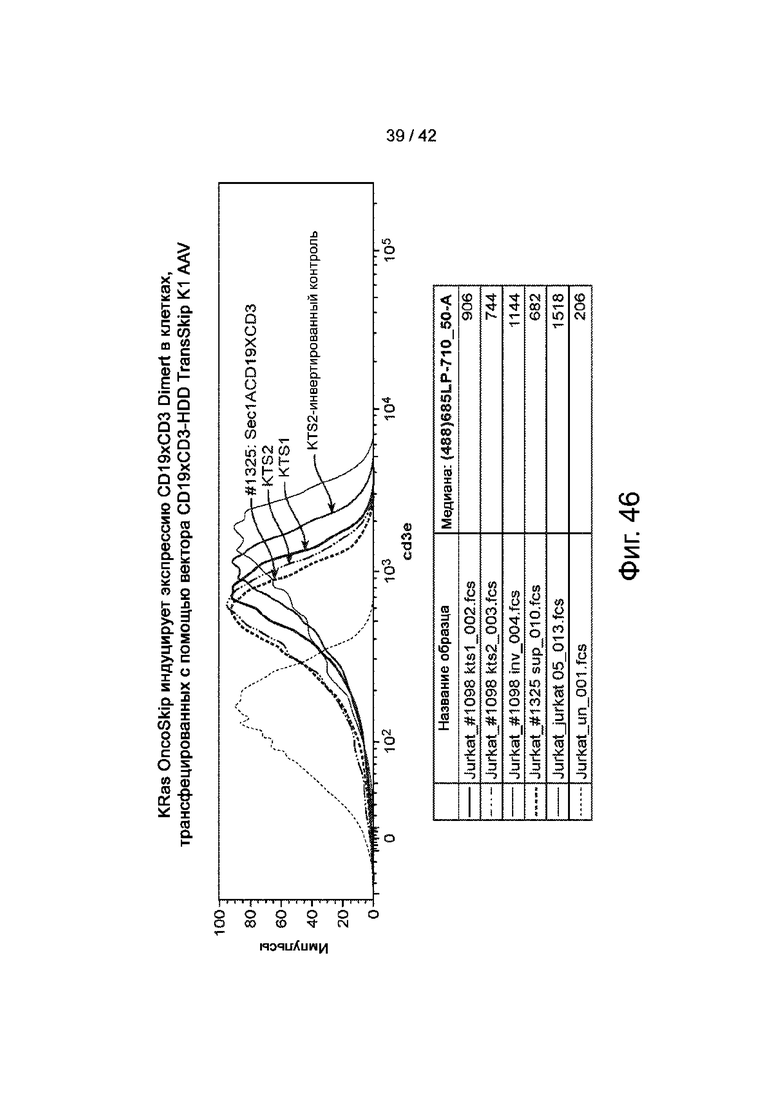

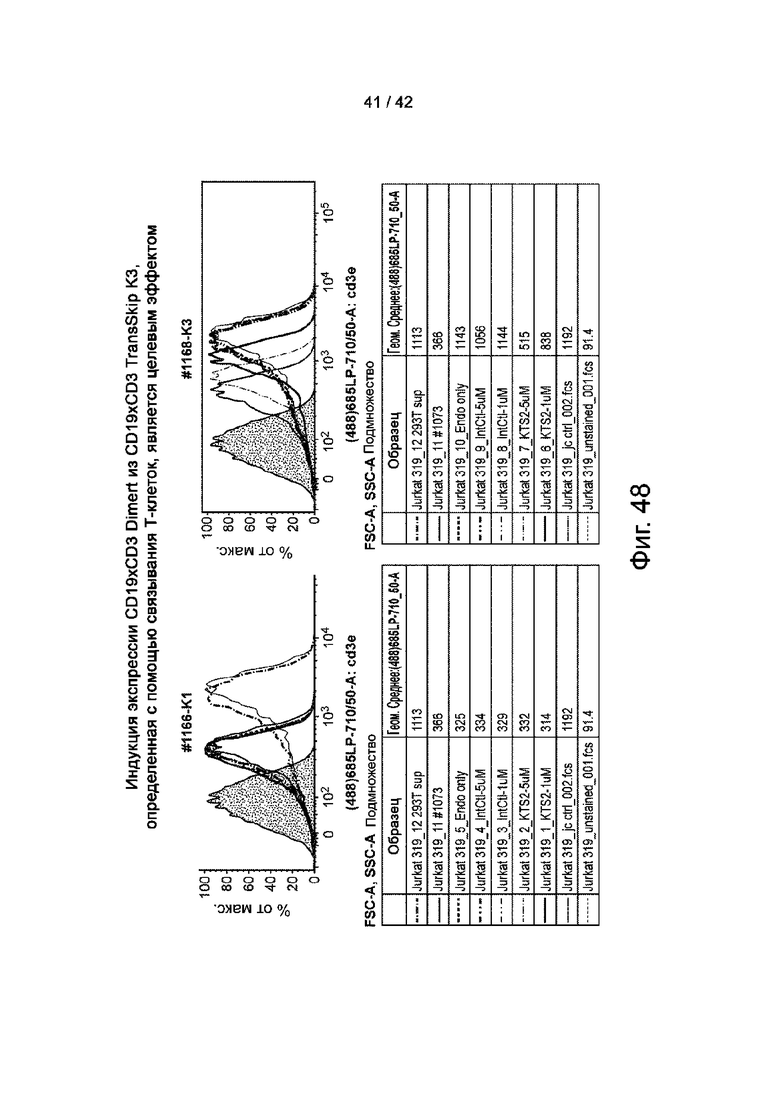
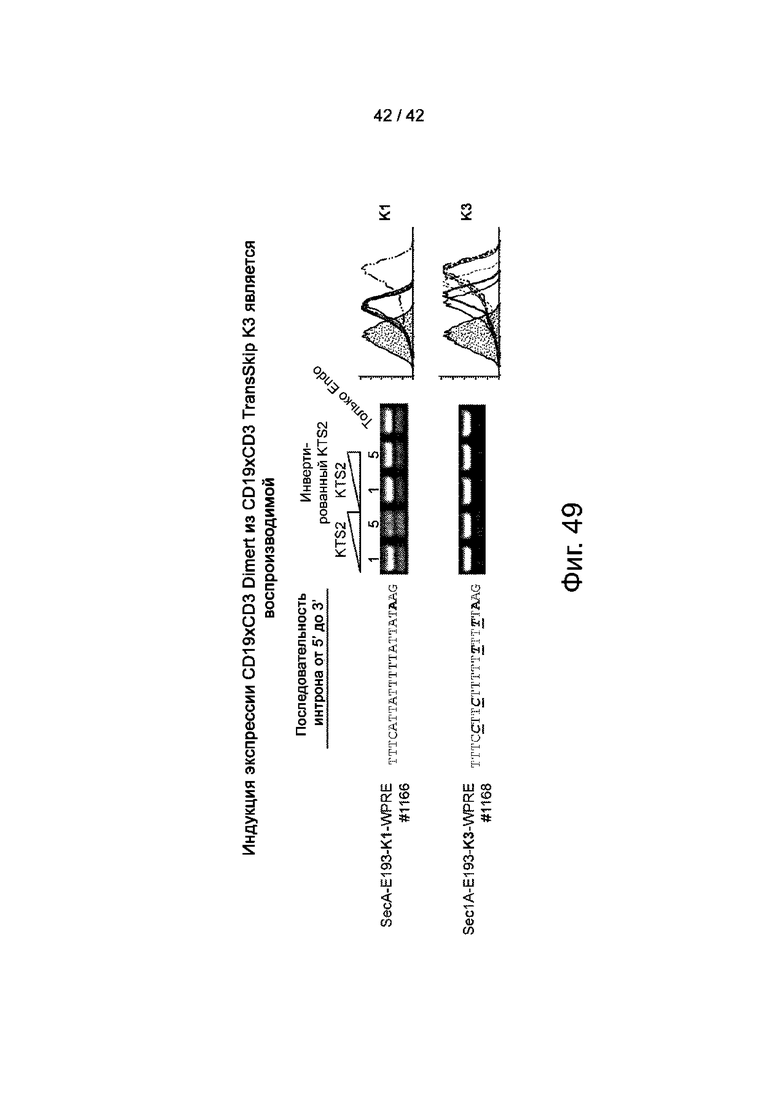
Изобретение относится к биотехнологии. Раскрыт полинуклеотид для регуляции генов, включающий: (а) первую полинуклеотидную последовательность, содержащую первую часть открытой рамки считывания, кодирующую первый полипептид; (b) вторую полинуклеотидную последовательность, содержащую вторую часть открытой рамки считывания, кодирующую первый полипептид; (c) третью полинуклеотидную последовательность, кодирующую второй полипептид; и (d) полинуклеотидную последовательность регуляции гена, расположенную между первым полинуклеотидом и вторым полинуклеотидом. Также описан экспрессирующий вектор, содержащий указанный полинуклеотид. Описана фармацевтическая композиция, содержащая полинуклеотид или вектор и носитель, необязательно фармацевтически приемлемый носитель. Описан способ лечения рака у объекта, который в этом нуждается, включающий введение объекту эффективного количества рекомбинантного полинуклеотида или вектора или фармацевтической композиции, где полинуклеотид или вектор экспрессирует терапевтическое противоопухолевое антитело или его антигенсвязывающий фрагмент. Раскрыт способ получения биспецифического антитела или триспецифического антитела в клетке, включающий контакт клетки, содержащей полинуклеотид или вектор, с эффективным количеством антисмыслового олигонуклеотида. Раскрыт набор для регуляции генов, содержащий полинуклеотид, или вектор, или фармацевтическую композицию. Изобретение расширяет арсенал средств для экспрессии терапевтического гена для заболевания или состояния. 6 н. и 49 з.п. ф-лы, 49 ил., 3 пр.
1. Полинуклеотид для регуляции генов, включающий:
(а) первую полинуклеотидную последовательность, содержащую первую часть открытой рамки считывания, кодирующую первый полипептид;
(b) вторую полинуклеотидную последовательность, содержащую вторую часть открытой рамки считывания, кодирующую первый полипептид;
(c) третью полинуклеотидную последовательность, кодирующую второй полипептид; и
(d) полинуклеотидную последовательность регуляции гена, расположенную между первым полинуклеотидом и вторым полинуклеотидом.
2. Полинуклеотид по п. 1, отличающийся тем, что полинуклеотидная последовательность регуляции гена включает донорный сайт сплайсинга, вышележащий интрон, экзон, содержащий более одной последовательности стоп-кодона в их соответствующих рамках считывания, нижележащий интрон и акцепторный сайт сплайсинга.
3. Полинуклеотид по п. 1 или 2, отличающийся тем, что полинуклеотидная последовательность, регулирующая ген, содержит одну или несколько последовательностей связывания для антисмыслового олигонуклеотида, последовательности связывания для доксициклина или полинуклеотидной последовательности, кодирующей рибосвитч.
4. Полинуклеотид по любому из пп. 1-3, отличающийся тем, что антисмысловой олигонуклеотид представляет собой морфолиноолигонуклеотид.
5. Полинуклеотид по п. 4, отличающийся тем, что связывающая последовательность для морфолиноолигонуклеотида включает полинуклеотидную последовательность, по меньшей мере на 95% идентичную SEQ ID NO. 24 или 25.
6. Полинуклеотид по п. 4, отличающийся тем, что морфолиноолигонуклеотид содержит полинуклеотидную последовательность, по меньшей мере на 95% идентичную SEQ ID NO: 27 или 28.
7. Полинуклеотид по п. 2, отличающийся тем, что стоп-кодон содержит:
олигонуклеотид из группы:TAA, TAG или TGA;
полинуклеотидную последовательность TAAxTAGxTGAxTAGxTAAxTGAx (SEQ ID NO. 1), где x представляет собой любой нуклеотид; или
полинуклеотидную последовательность TAATTAGTTGATTAGTTAATTGAT (SEQ ID NO. 2) или их эквивалент.
8. Полинуклеотид по любому из пп. 1-7, отличающийся тем, что полинуклеотид, регулирующий ген, содержит полинуклеотидную последовательность, по меньшей мере на 95% идентичную SEQ ID NO. 21.
9. Полинуклеотид по любому из пп. 1-8, отличающийся тем, что первый полипептид представляет собой первое антитело или его антигенсвязывающий фрагмент, а второй полипептид представляет собой второе антитело или его антигенсвязывающий фрагмент.
10. Полинуклеотид по любому из пп. 1-9, отличающийся тем, что первое антитело или его антигенсвязывающий фрагмент специфически связывается с активирующим антигеном на иммунной эффекторной клетке, а второе антитело или его антигенсвязывающий фрагмент связывается с опухолевым антигеном.
11. Полинуклеотид по любому из пп. 1-9, отличающийся тем, что первое антитело или его антигенсвязывающий фрагмент специфически связывается с опухолевым антигеном, а второе антитело или его антигенсвязывающий фрагмент связывается с активирующим антигеном на иммунной эффекторной клетке.
12. Полинуклеотид по любому из пп. 1-11, дополнительно содержащий четвертую полинуклеотидную последовательность, кодирующую третье антитело или его антигенсвязывающий фрагмент, при этом третье антитело или его антигенсвязывающий фрагмент связывается с активирующим антигеном на иммунной эффекторной клетке или опухолевым антигеном.
13. Полинуклеотид по любому из пп. 10-12, отличающийся тем, что иммунная эффекторная клетка включает дендритную клетку, клетку-натуральный киллер («NK»), макрофаг, Т-клетку, В-клетку или их комбинацию.
14. Полинуклеотид по любому из пп. 10-12, отличающийся тем, что эффекторная иммунная клетка представляет собой Т-клетку или NK-клетку.
15. Полинуклеотид по любому из пп. 10-12, отличающийся тем, что активирующий антиген на иммунной эффекторной клетке включает CD3, CD2, CD4, CD8, CD19, LFA1, CD45, NKG2D, NKp44, NKp46, NKp30, DNAM, B7-H3, CD20, CD22 или их комбинацию.
16. Полинуклеотид по любому из пп. 10-12, отличающийся тем, что опухолевый антиген включает один или несколько из числа рецептора 2 эфрина типа A (EphA2), интерлейкина (IL)-13r альфа 2, EGFR VIII, PSMA, EpCAM, GD2 или GD3, фукозил GM1, PSCA, PLAC1, точечного разрыва саркомы, опухоли Вильмса 1, альфафетопротеина (AFP), карциноэмбрионального антигена (CEA), CA-125, MUC-1, эпителиального опухолевого антигена (ETA), тирозиназы, меланома-ассоциированного антигена (MAGE), антигена гематологической дифференцировки, поверхностного гликопротеина, ганглиозидов (GM2), рецептора фактора роста, стромального антигена, сосудистого антигена, орфанного рецептора 1, подобного рецепторной тирозинкиназе (ROR1), мезотелина, CD38, CD123, рецептора 2 эпидермального фактора роста человека (HER2), антигена созревания В-клеток (BCMA), белка активации фибробластов (FAP) альфа или их комбинации.
17. Полинуклеотид по любому из пп. 9-16, отличающийся тем, что первое антитело и второе антитело, каждое независимо, представляют собой одноцепочечный вариабельный фрагмент.
18. Полинуклеотид по любому из пп. 1-17, дополнительно содержащий полинуклеотидную последовательность, кодирующую секреторный пептид, необязательно секреторную консенсусную последовательность.
19. Полинуклеотид по любому из пп. 1-18, дополнительно содержащий полинуклеотидную последовательность, кодирующую домен димеризации.
20. Полинуклеотид по любому из пп. 1-19, дополнительно содержащий 5’-концевой инвертированный повтор (ITR) и 3’-ITR.
21. Экспрессирующий вектор, содержащий полинуклеотид по любому из пп. 1-20.
22. Вектор по п. 21, отличающийся тем, что вектор представляет собой рекомбинантный вектор, необязательно вирусный вектор, и способен экспрессировать пре-мРНК, кодирующую dimert, когда пре-мРНК находится в контакте с антисмысловым олигонуклеотидом, необязательно морфолиноолигонуклеотидом.
23. Вектор по п. 22, отличающийся тем, что dimert представляет собой биспецифическое антитело.
24. Вектор по п. 22, отличающийся тем, что dimert представляет собой триспецифическое антитело.
25. Вектор по п. 23 или 24, отличающийся тем, что биспецифическое антитело или триспецифическое антитело включает первое антитело или его антигенсвязывающий фрагмент и второе антитело или его антигенсвязывающий фрагмент.
26. Вектор по п. 23, отличающийся тем, что биспецифическое антитело содержит полипептидную последовательность, по меньшей мере на 95% идентичную любой из SEQ ID NO. 13 или 15.
27. Вектор по любому из пп. 21-26, отличающийся тем, что вектор экспрессирует пре-мРНК, кодирующую триспецифическое антитело, когда пре-мРНК находится в контакте с антисмысловым олигонуклеотидом, необязательно с морфолиноолигонуклеотидом.
28. Вектор по п. 27, отличающийся тем, что триспецифическое антитело включает первое антитело или его антигенсвязывающий фрагмент, второе антитело или его антигенсвязывающий фрагмент и третье антитело или его антигенсвязывающий фрагмент.
29. Вектор по п. 27, отличающийся тем, что триспецифическое антитело содержит полипептидную последовательность, по меньшей мере на 95% идентичную SEQ ID NO. 11.
30. Вектор по любому из пп. 21-29, отличающийся тем, что первое антитело и второе антитело, каждое независимо, представляют собой одноцепочечный вариабельный фрагмент.
31. Вектор по любому из пп. 21-30, дополнительно содержащий полинуклеотидную последовательность, кодирующую секреторный пептид, необязательно секреторную консенсусную последовательность.
32. Вектор по любому из пп. 21-31, дополнительно содержащий полинуклеотидную последовательность, кодирующую домен димеризации.
33. Вектор по любому из пп. 21-32, дополнительно содержащий 5’-концевой инвертированный повтор (ITR) и 3’-ITR.
34. Вектор по п. 21, где вектор содержит последовательность, изложенную в SEQ ID NO: 4, 6, 8, 12, 14, 16-23, 30-33 или 40-46.
35. Вектор по любому из пп. 21-34, отличающийся тем, что вектор представляет собой рекомбинантный вирусный вектор, который содержит вектор основной цепи, выбранный из группы, состоящей из ретровирусного вектора, лентивирусного вектора, вектора вируса мышиного лейкоза («MLV»), вектор вируса Эпштейна-Барр («EBV»), вектор аденовируса, вектор вируса герпеса («HSV») или вектор аденоассоциированного вируса («AAV»).
36. Вектор по любому из пп. 21-35, отличающийся тем, что вектор представляет собой вектор AAV, необязательно самокомплементарный вектор AAV, дополнительно необязательно вектор AAV rh74.
37. Фармацевтическая композиция, содержащая полинуклеотид по любому из пп. 1-20 или вектор по любому из пп. 21-36 и носитель, необязательно фармацевтически приемлемый носитель.
38. Способ лечения рака у объекта, который в этом нуждается, включающий введение объекту эффективного количества рекомбинантного полинуклеотида по любому из пп. 1-20 или вектора по любому из пп. 21-36 или фармацевтической композиции по п. 37, где полинуклеотид или вектор экспрессирует терапевтическое противоопухолевое антитело или его антигенсвязывающий фрагмент.
39. Способ по п. 38, дополнительно включающий введение объекту эффективного количества антисмыслового олигонуклеотида, необязательно морфолиноолигонуклеотида.
40. Способ по п. 38 или 39, дополнительно включающий введение объекту противоракового агента.
41. Способ по п. 49, отличающийся тем, что противораковый агент включает агент, выбранный из пептида, полипептида, молекулы нуклеиновой кислоты, небольшой молекулы, вирусной частицы или их комбинаций.
42. Способ по п. 41, отличающийся тем, что вирусная частица представляет собой онколитическую частицу HSV.
43. Способ по любому из пп. 37-42, отличающийся тем, что объектом является млекопитающее.
44. Способ по любому из пп. 37-43, отличающийся тем, что объектом является человек.
45. Способ получения биспецифического антитела или триспецифического антитела в клетке, включающий контакт клетки, содержащей полинуклеотид по любому из пп. 1-20 или вектор по любому из пп. 21-36, с эффективным количеством антисмыслового олигонуклеотида.
46. Способ по п. 45, где антисмысловой олигонуклеотид представляет собой морфолиноолигонуклеотид.
47. Способ по п. 46, отличающийся тем, что морфолиноолигонуклеотид содержит последовательность, по меньшей мере на 95% идентичную стереочистому полинуклеотиду.
48. Способ по п. 45, отличающийся тем, что вектор вводят в клетку путем трансфекции, инфицирования, трансформации, электропорации, инъекции, микроинъекции или их комбинации.
49. Способ по любому из пп. 45-48, отличающийся тем, что клетка включает фибробласт, скелетную клетку, эпителиальную клетку, мышечную клетку, нервную клетку, эндокринную клетку, меланоцит, клетку крови или их комбинацию.
50. Способ по любому из пп. 45-49, отличающийся тем, что биспецифическое антитело содержит полипептидную последовательность, по меньшей мере на 95% идентичную SEQ ID NO. 13 или 15.
51. Способ по любому из пп. 45-50, отличающийся тем, что триспецифическое антитело содержит полипептидную последовательность, по меньшей мере на 95% идентичную SEQ ID NO. 11.
52. Способ по любому из пп. 45-49, отличающийся тем, что биспецифическое антитело кодируется полинуклеотидной последовательностью, по меньшей мере на 95% идентичной SEQ ID NO. 14, 16, 22, 23, 30-33 или 40-46.
53. Способ по любому из пп. 45-50, отличающийся тем, что триспецифическое антитело кодируется полинуклеотидной последовательностью, по меньшей мере на 95% идентичной SEQ ID NO. 12.
54. Набор для регуляции генов, содержащий полинуклеотид по любому из пп. 1-20, или вектор по любому из пп. 21-36, или фармацевтическую композицию по п. 37.
55. Набор по п. 54 дополнительно включает инструктивный материал.
| US 20170266243 A1, 21.09.2017 | |||
| WO 2018041827 A1, 08.03.2018 | |||
| US 20110092573 A1, 21.04.2011 | |||
| WO 2017177337 A1, 19.10.2017 | |||
| WO 2018120842 A1, 05.07.2018 | |||
| ИНДУЦИРУЮЩИЙ ЦИТОТОКСИЧНОСТЬ ТЕРАПЕВТИЧЕСКИЙ АГЕНТ | 2011 |
|
RU2663123C2 |

