Изобретение относится к области пассивной иммунотерапии онкологических заболеваний, в частности к методу адоптивного переноса клеток, в котором используют генетически модифицированные аутологичные или аллогенные цитотоксические Т-лимфоциты, обладающие приобретенной иммуногенностью против клеток опухоли. И может быть использовано в Т-клеточной терапии рака.
В настоящее время определено большое количество опухоль-асоциированных антигенов, которые по отдельности или в сочетании могут быть использованы для индукции иммунного ответа, направленного против опухолевой ткани. Разрабатываемые подходы иммунотерапии, такие как вакцинация и адоптивный перенос иммунных клеток продемонстрировали эффективность в ряде доклинических и клинических исследований [1, 2]. Однако, несмотря на огромный интерес исследователей к иммунотерапевтическим подходам и большой потенциал иммунотерапии, случаи полного излечения от раковых заболеваний с помощью таких подходов остаются редкими событиями.
Особое место в иммунотерапии занимает метод адоптивного переноса клеток, в котором используют генетически модифицированные аутологичные или аллогенные цитотоксические Т-лимфоциты, обладающие рецепторами, специфичными к антигенам клеток опухоли [2]. Исследования показали, что наиболее перспективным является получение лимфоцитов, экспрессирующих химерные иммуноглобулиновые мономолекулярные Т-клеточные рецепторы, в которых антиген-распознающая часть, представляющая собой вариабельный фрагмент антитела, объединена с эффекторным фрагментом Т-клеточного рецептора (обычно с ζ-цепью) [3]. Распознавание антигена таким химерным рецептором за счет иммуноглобулиновой части является независимым от представления антигена клетками главного комплекса гистосовместимости (ГКГ), связывание с антигеном приводит к секреции модифицированными Т-лимфоцитами интерлейкина-2 (ИЛ-2) и ряда других цитокинов и проявлению цитотоксического действия, а объединение в химерном рецепторном комплексе различных ко-стимулирующих доменов позволяет повысить выживаемость лимфоцитов и усилить цитотоксический эффект [4, 5, 6].
На сегодняшний день сконструировано множество химерных иммунорецепторов к различным опухолевым антигенам, таким как антиген Gd2 клеток нейробластомы [7], простатспецифический мембранный антиген (ПСМА) [8, 9], раково-эмбриональный антиген (РЭА) [10] и др.
РЭА - гликопротеин, молекулярной массой 180 кДа, экспрессирующийся во всех аденокарциномах желудочно-кишечного тракта, в 50% случаев рака молочной железы и в 70% случаев немелкоклеточного рака легких и в клетках некоторых других видов рака [11]. В норме РЭА экспрессируется в тканях эмбриона в ходе внутриутробного развития, а после рождения его продукция дектируется в клетках желудочных ямок, эпителия толстой кишки и в других отделах желудочно-кишечного тракта, в основном на поверхности микроворсинок. Однако уровень экспрессии РЭА в опухолевых клетках значительно превышает его уровень экспрессии в нормальной ткани (в 35 раз и выше) [12]. Несмотря на успех доклинических экспериментальных исследований, описанные в литературе химерные Т-клеточные рецепторы к РЭА имеют свои недостатки, не позволяющие перейти к стадии клинического применения [13, 14]. В связи с этим остается актуальным создание новых конструкций химерных иммуноглобулиновых Т-клеточных рецепторов к другим эпитопам РЭА. Кроме того, в проявлении цитотоксического действия модифицированных клеток, содержащих химерные рецепторы, остается высокой гибель лимфоцитов после связывания антигена, обусловленная активацией лимфоцитов, точно не установлен набор сигнальных последовательностей, усиливающих цитотоксическое действие, весьма актуально повышение аффинности химерных рецепторов к антигену.
Задачей данного изобретения является получение лимфоцитов, экспрессирующих химерные иммуноглобулиновые мономолекулярные Т-клеточные рецепторы к РЭА, с усиленным цитотоксическим действием.
Техническим результатом изобретения является увеличение более, чем на 45%, цитотоксического эффекта по отношению к опухолевым клеткам при использовании генетически модифицированных созданной конструкцией Т-лимфоцитов.
Технический результат достигается благодаря получению на основе уникальных участков, кодирующих аминокислотную последовательность, проявляющую высокий уровень связывания РЭА в том числе и в растворе, генетических конструкций, кодирующих мономолекулярные химерные Т-клеточные рецепторы, распознающие различные эпитопы РЭА, трансфекция которыми активированных лимфоцитов человека приводит к экспрессии этих рецепторов на поверхности лимфоцитов.
В одном из вариантов осуществления настоящее изобретение относится к одноцепочечному антителу к раково-эмбриональному антигену, которое кодируется нуклеотидной последовательностью SEQ ID NO: 1.
В одном из вариантов осуществления настоящее изобретение относится к химерному мономолекулярному Т-клеточному рецептору, специфичному к раково-эмбриональному антигену, включающему одноцепочечное антитело, кодируемое нуклеотидной последовательностью SEQ ID NO: 1.
В предпочтительном варианте химерный мономолекулярный Т-клеточный рецептор кодируется нуклеотидной последовательностью SEQ ID NO: 3.
В одном из вариантов осуществления изобретение относится к вектору для обеспечения экспрессии химерного мономолекулярного Т-клеточного рецептора, специфичного к раково-эмбриональному антигену, включающему нуклеотидную последовательность, представленную на SEQ ID NO: 1.
В одном из вариантов осуществления изобретение относится к клетке-хозяину, продуцирующей химерный мономолекулярный Т-клеточный рецептор, специфичный к раково-эмбриональному антигену.
В одном из вариантов осуществления изобретение относится к способу диагностики или лечения заболеваний, характеризующихся наличием антигенов, способных связываться с химерным мономолекулярным Т-клеточным рецептором.
Используемый в настоящем документе термин «одноцепочечное антитело» обозначает рекомбинантное антитело, т.н. одноцепочечный вариабельный фрагмент scFv (singl chain variable fragments), кодируется одним геном и содержит только один антиген-связывающий участок, состоящий из вариабельных доменов легкой (VL) и тяжелой (VH) цепей, соединенных гибким пептидным линкером.


scFv
3с8
Nucleotide Seq SEQ ID NO 1

Am Ac Seq SEQ ID NO 2

Используемый в настоящем документе термин «химерный мономолекулярный Т-клеточный рецептор» включает в себя антиген-распознающую часть, представляющую собой вариабельный фрагмент антитела, объединенную с эффекторным фрагментом Т-клеточного рецептора (обычно с ζ-цепью) [3]. Распознавание антигена таким химерным рецептором за счет иммуноглобулиновой части является независимым от представления антигена клетками главного комплекса гистосовместимости (ГКГ), связывание с антигеном приводит к секреции модифицированными Т-лимфоцитами интерлейкина-2 (ИЛ-2) и ряда других цитокинов и проявлению цитотоксического действия, а объединение в химерном рецепторном комплексе различных ко-стимулирующих доменов позволяет повысить выживаемость лимфоцитов и усилить цитотоксический эффект [4, 5, 6].


SEQ ID NO 3
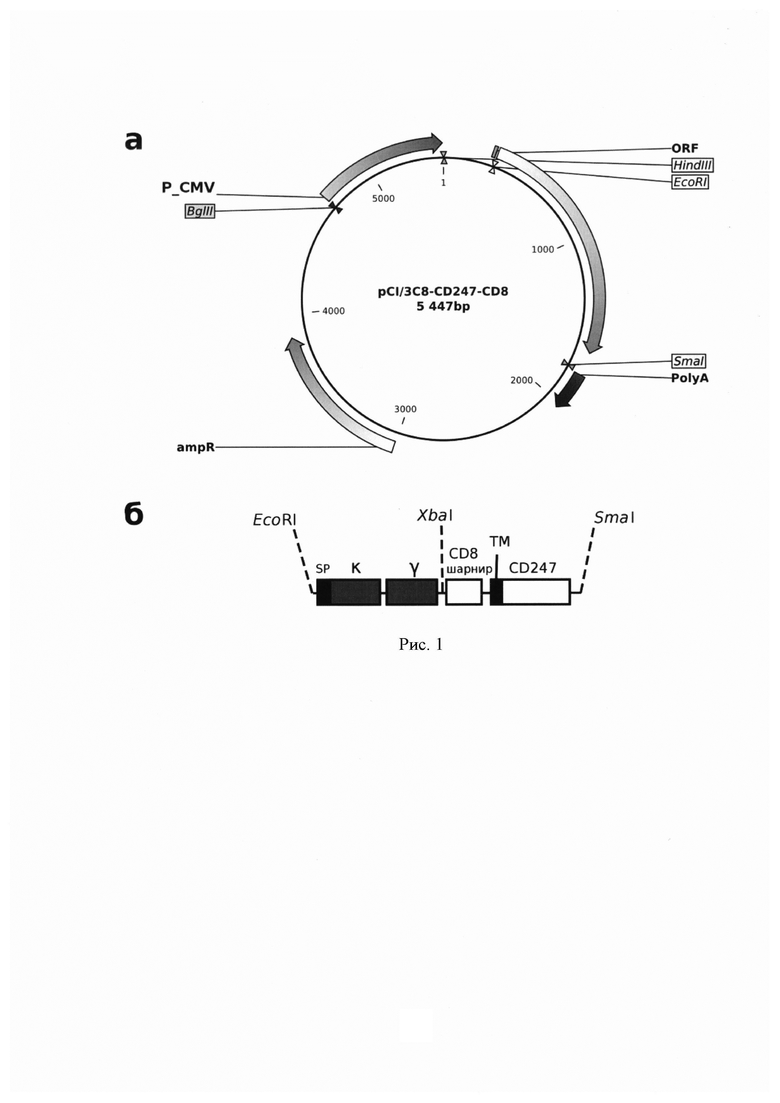
Используемый в настоящем документе термин «экспрессионный вектор» включает в себя конструкцию рСЕА-3, созданую на основе распознающих участков антитела 3С8, путем клонирования в плазмиду рСI участков, кодирующих i) вариабельные фрагменты κ- (легких) и γ- (тяжелых) цепей выбранных моноклональных антител к РЭА, разделенных линкерным пептидом (G4S)3 (одноцепочечные миниантитела в scFv-формате), ii) «шарнирный» участок белка CD8 и iii) участок белка CD247 (ζ-цепь Т-клеточного рецептора), включающий трансмембранный и цитоплазматический домены (рис. 1).
Используемый в настоящем документе термин «клетка-хозяин» включает в себя трансфицированные указанными конструкциями лимфоцитов периферической крови человека, характеризующихся высокой цитотоксической активностью в отношении РЭА-экспрессирующих клеток.
Используемый в настоящем документе термин «заболеваний, характеризующихся наличием антигенов, способных связываться с химерным мономолекулярным Т-клеточным рецептором» включает в себя заболевания, при которых уровень экспрессии РЭА значительно превышает его уровень экспрессии в нормальной ткани (в 35 раз и выше) (12).
Сущностью предлагаемого изобретения является создание оригинальной генетической конструкции, кодирующей химерный мономолекулярный Т-клеточный рецептор, в котором эффекторный фрагмент Т-клеточного рецептора объединен с антиген-распознающей частью, представляющей собой вариабельный фрагмент антитела к раково-эмбриональному антигену (РЭА). Было показано, что после трансфекции данные рецепторы экспрессировались на поверхности клеток и связывались с РЭА. Также продемонстрирована высокая цитотоксическая активность трансфицированных описанными конструкциями лимфоцитов периферической крови человека в отношении РЭА-экспрессирующих клеток.
Серия мышиных моноклональных биотинилированных антител (изотип IgG1), способных связываться с РЭА в растворе, была предоставлена ООО «Хема-Медика» (Россия, Москва). В серию вошли следующие антитела: 2С5, 1С10, 1С11, 1С13, 1С3, 1С6, 3С1, 3С13, 3С2, 3С4, 3С5, 3С6, 3С8 [17]. Для анализа способности антител имеющейся серии связывать РЭА на поверхности экспрессирующих его клеток были выбраны три клеточные линии человека: НТ29, А549 и НСТ116. Антитело 3С8 было отобрано как проявляющее высокий уровень связывания РЭА со всеми исследуемыми клеточными линиями. Фрагменты кДНК иммуноглобулиновых генов, кодирующие вариабельные участки антител (домены VL и VH) из гибридомы 3С8, были ПЦР-амплифицированы и клонированы для определения полной нуклеотидной последовательности.
Конструкция рСЕА-3 была создана на основе распознающих участков антитела 3С8 путем клонирования в плазмиду рСI участков, кодирующих i) вариабельные фрагменты κ- (легких) и γ- (тяжелых) цепей выбранных моноклональных антител к РЭА, разделенных линкерным пептидом (G4S)3 (одноцепочечные миниантитела в scFv-формате), ii) «шарнирный» участок белка CD8 и iii) участок белка CD247 (ζ-цепь Т-клеточного рецептора), включающий трансмембранный и цитоплазматический домены (рис. 1).
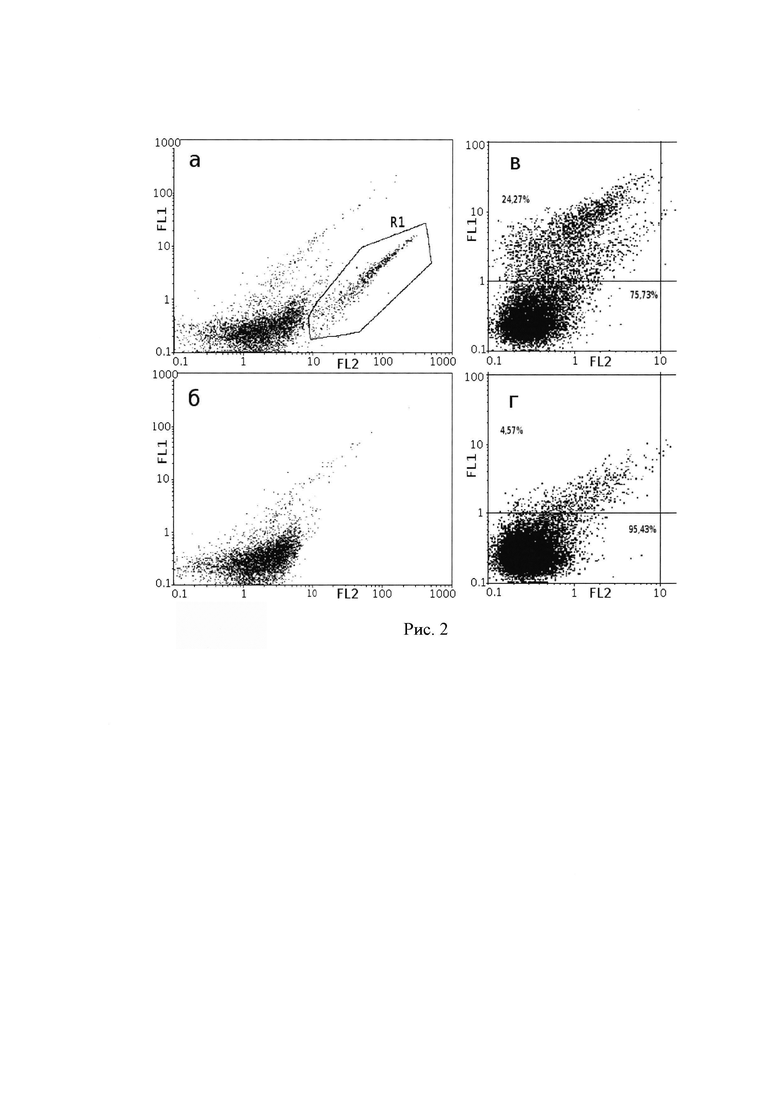
Для подтверждения правильного функционирования химерных иммунорецепторов, кодируемых созданными конструкциями, была выполнена трансфекция клеток НЕК293. Через 24 ч после трансфекции методом проточной цитометрии было исследовано связывание с клетками антител к ζ-цепи Т-клеточного рецептора. Было показано, что положительный сигнал обнаруживался в трансфицированных клетках, тогда как сигнал в контрольной группе клеток, окрашенных изотипическими контрольными антителами, отсутствовал (рис. 2 а, б). Кроме этого, трансфицированные клетки НЕК293, связывали РЭА в растворе, что было показано после инкубации в течение 30 мин клеток с раствором РЭА, меченного ФИТЦ (рис. 2 в, г).
Далее была проведена трансфекция созданными генетическими конструкциями лимфоцитов человека. В работе использовали мононуклеарные клетки крови, стимулированные к пролиферации с помощью ФГА и активированные рекомбинантным ИЛ-2 человека (50 Ед/мл) в течение 3 суток. По истечении этого времени количество CD3+ Т-лимфоцитов среди всей популяции составляло около 60%. Спустя 24 ч после трансфекции, в ходе проведения ОТ-ПЦР, в лимфоцитах была выявлена экспрессия генов, кодирующих химерные иммунорецепторы.
Способность трансфицированных лимфоцитов связывать РЭА, спустя 24 ч после электропорации, была подтверждена методом непрямого иммуноцитохимического окрашивания (рис. 3). При этом, количество модифицированных лимфоцитов, связавшихся с РЭА, было примерно в 4 раза выше для предварительно активированных ИЛ-2 лимфоцитов по сравнению с нестимулированными клетками.
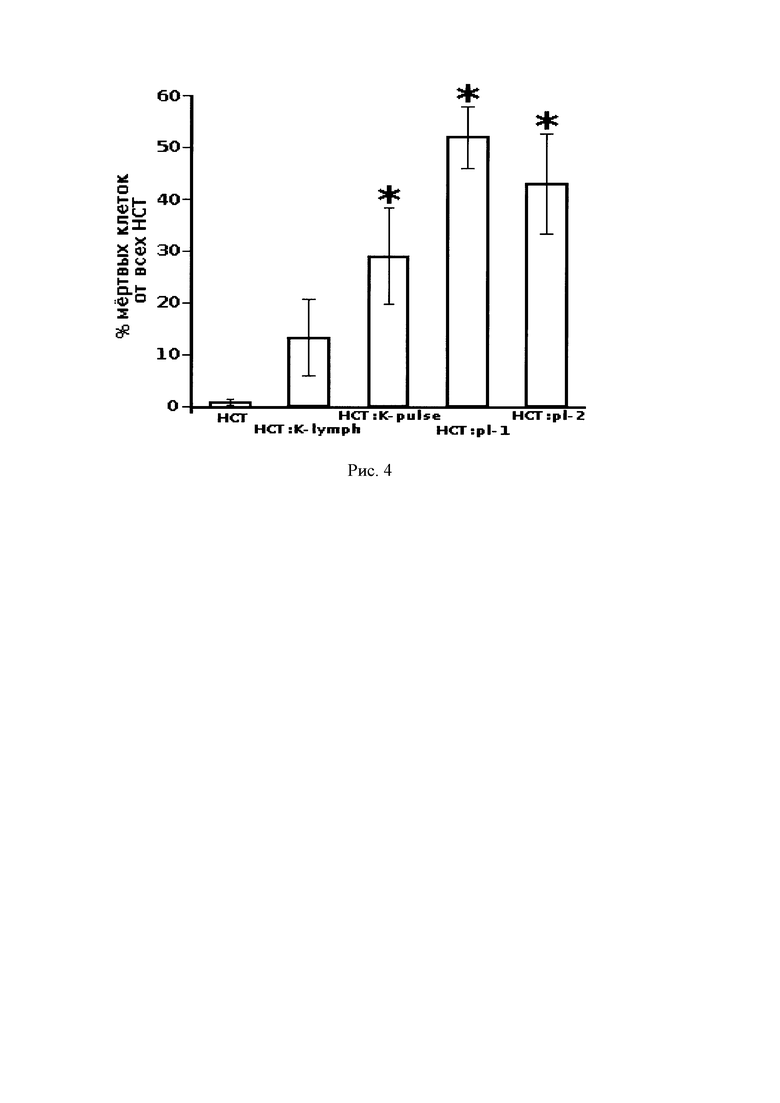
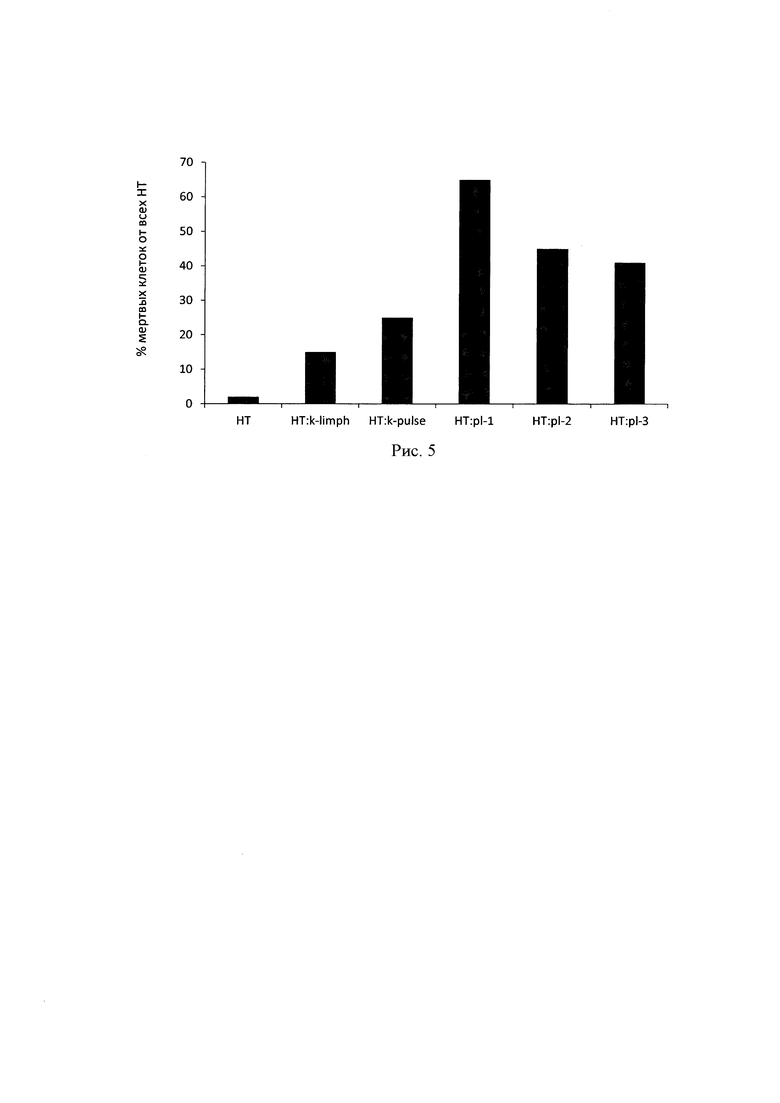
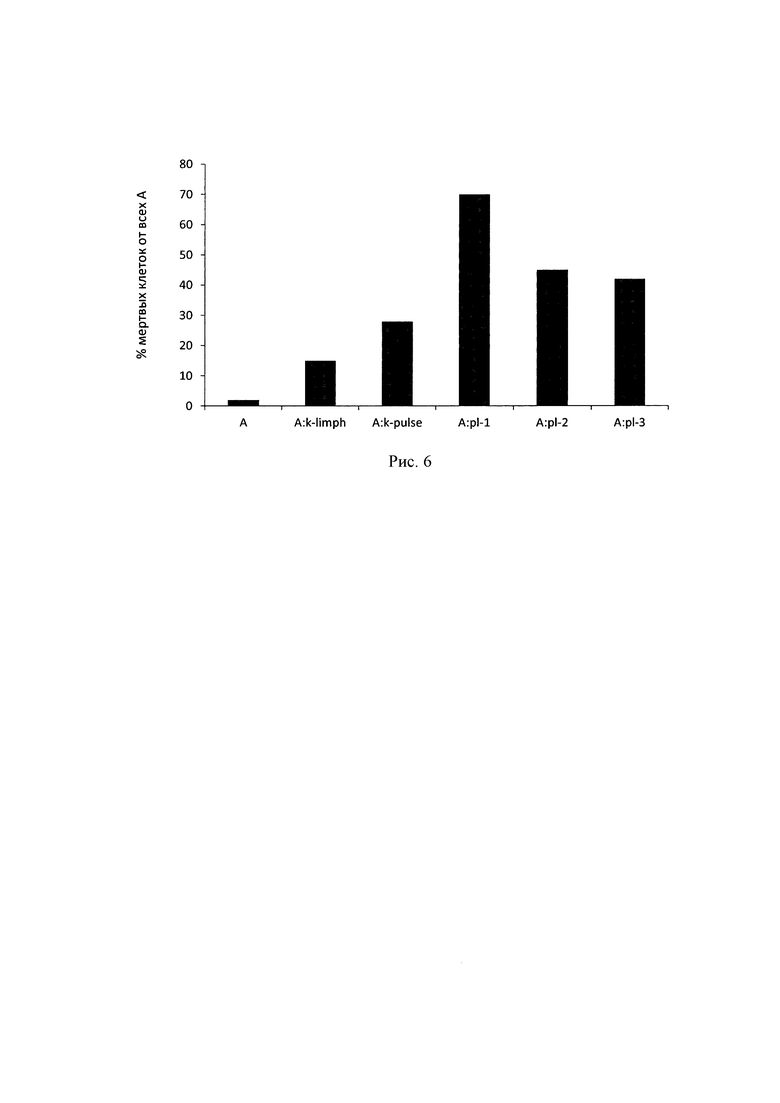
Исследование цитотоксичности лимфоцитов, трансфицированных исследуемыми плазмидами, проводили с использованием метода двойной окраски CFDA-SE/PI клеток НСТ116, экспрессирующих поверхностный РЭА. Было показано, что при добавлении к клеткам НСТ 116, окрашенных с помощью CFDA-SE, трансфицированных лимфоцитов в соотношении 1:10 максимальный цитотоксический эффект достигался по истечении 5 ч совместной инкубации. При этом процент живых клеток НСТ116 после воздействия лимфоцитов, трансфицированных плазмидой рСЕА-3, составил 47.9%, а в случае с лимфоцитами, трансфицированными рСЕА-х, - 56.9% (рис. 4). Средний процент живых клеток НСТ116, инкубируемых в присутствии контрольных лимфоцитов, составил 86.7%, при инкубации с лимфоцитами, подверженными электрическому импульсу, но без добавления ДНК, процент живых НСТ116 равнялся 70.9%.
Краткое описание чертежей
Рис. 1. Схема плазмиды, кодирующей химерные иммунорецепторы (а) и детализи рованная структура гена химерного иммунорецептора (б), sp - сигнальный пептид, κ, γ - вариабельные фрагменты кДНК легких и тяжелых цепей антител соответственно; CD8 - «шарнирная» область антигена CD8; CD247 - ζ-цепь Т-клеточного рецептора; ТМ - трансмембранный домен; PCMV - промотор CMV; SV40 polyА - сигнал полиаденилирования SV40; Ampr - ген устойчивости к ампициллину; BglII, HinDIII EcoRl, Xbal, Smal - сайты соответствующих эндонуклеаз рестрикции.
Рис. 2. Данные цитофлуорометрического анализа клеток НЕК293, спустя 24 часа после трансфекции мономолекулярными химерными Т-клеточными рецепторами, а, б - связывание с клетками антител к ζ-цепи Т-клеточного рецептора на примере плазмиды рСЕА-3: а - клетки при добавлении антител к ζ-цепи; б - клетки при добавлении изотипических контрольных антител. R1 - область клеток, связавшихся с антителами к ζ-цепи; в, г - связывание клеток, модифицированных плазмидой рСЕА-3 с РЭА в растворе: в - трансфицированные клетки; г - нетрансфицированные клетки (отрицательный контроль). По осям отложены значения флуоресценции в каналах: по оси абсцисс FL2, по оси ординат FL1.
Рис. 3. Данные цитофлуорометрического анализа лимфоцитов человека, активированных ИЛ-2 и трансфицированных мономолекулярными химерными Т-клеточными рецепторами, по связыванию с РЭА в растворе, спустя 24 часа после трансфекции. а, б - лимфоциты, трансфицированные плзмидами рСЕА-3 и рСЕА-х соответственно; в - нетрансфицированные лимфоциты (отрицательный контроль). RN1 - области клеток с флуоресценцией выше пороговой. По оси абсцисс отложены значения флуоресценции в канале FL2, по оси ординат число флуоресцирующих частиц.
Рис. 4. Цитотоксичность лимфоцитов, трансфицированных плазмидами рСЕА-3 (НСТ:рl-1) и рСЕА-х (НСТ:рl-2), в отношении экспрессирующих РЭА клеток НСТ116 (через 5 ч после добавления, соотношение эффекторных клеток и клеток-мишеней 10:1). Приведены средние проценты лизируемых клеток от общего количества клеток-мишеней и стандартные отклонения. HCT:K-pulse - цитотоксичность лимфоцитов, подверженных электрическому пульсу без добавления ДНК. НСТ - клетки НСТ116 без добавления лимфоцитов. * - р<0.05 по сравнению с контрольными нетрансфицированными лимфоцитами (HCT:K-lymph).
Пример 1. Клеточные линии
В работе использовались клеточные линии НЕК293 (клетки почки эмбриона человека), Jurkat (CD4(+) клеточная линия Т-клеточного лейкоза), любезно предоставленные ГУ НИИ эпидемиологии и микробиологии имени Н.Ф. Гамалеи.
Вся работа с культурами клеток выполнялась в стерильных условиях в ламинарном боксе в специально оборудованной для работы с клетками комнате.
Культивирование НЕК293
Среда для культивирования НЕК293 имела следующий состав: DMEM (ПанЭко, Россия), 10% эмбриональная бычья сыворотка (ПанЭко) и антибиотики пенициллин, и стрептомицин в стандартной концентрации. Клетки культивировали в условиях высокой влажности при 37°С и 5% СО2. При достижении 80-90% конфлюэнтности клетки пассировали. Для этого клетки 2 раза промывали раствором стерильного фосфатно-солевого буфера (FBS, Invitrogen), обрабатывали 0,25% раствором трипсина-ЭДТА. Для повышения эффективности работы ферментов на этой стадии флаконы с клетками помещали на 37°С на 4-5 минут. После открепления всех клеток от подложки трипсин инактивировали добавлением среды с сывороткой, затем клетки разбивали до получения однородной суспензии, отбирали требуемое количество клеток, а к оставшимся клеткам добавляли соответствующее количество полной культуральной среды и продолжали культивирование.
Заморозка и оттаивание НЕК293
Среда для заморозки имела следующий состав: 90% полной среды для культивирования и 10% ДМСО (ПанЭко).
От культивируемых клеток отбирали среду, промывали 2 раза PBS, добавляли 0,25% раствора трипсина-ЭДТА. Инкубировали при 37°С в течение 5 минут, пипетировали для получения одноклеточной суспензии, нейтрализовали трипсин полной культуральной средой, содержащей сыворотку. Полученную суспензию центрифугировали 5 минут при 1000 об./мин. Надосадочную жидкость удаляли, осадок суспендировали в соответствующем количестве среды для заморозки, переносили в криопробирки в концентрации 106 клеток в 1 мл среды и сразу помещали на -70°С на 24 часа. Через 24 часа криопробирки переносили в жидкий азот для продолжительного хранения. В одной криопробирке замораживали 1.0-2.0 млн клеток.
Криопробирки оттаивали на водяной бане при 37С, стерилизовали 70% этанолом, затем их содержимое переносили в пробирку, содержащую среду для культивирования прогретую до 37С, центрифугировали 5 минут при 1000 об./мин, надосадочную жидкость удаляли, осадок суспендировали в соответствующем количестве среды и переносили в флакон со средой. Клетки равномерно распределяли по культуральной поверхности и помещали в инкубатор на 37°С, 5% СО2. Через 24 ч, когда клетки прикреплялись к подложке и начинали пролиферировать, заменяли среду для устранения погибших в процессе заморозки клеток.
Культивирование лимфоцитов
Среда для культивирования лимфоцитов имела состав идентичный составу среды для культивирования Jurkat.
Выделение лимфоцитов из периферической крови человека
Лимфоциты человека получали следующим образом. Свежую кровь человека отбирали в 5 мл. стерильную пробирку, обрабатывали антикоагулянтом (цитрат натрия). Цельную кровь либо отстаивали при комнатной температуре в течение часа для осаждения эритроцитов, а затем отбирали плазму, содержащую лейкоциты, либо брали кровь непосредственно после выделения и разбавляли в PBS в соотношении 1:2, полученный раствор наслаивали на раствор фиколла-урографина (ПанЭко) в соотношении 2:1. Разделение клеток проводили центрифугированием в течение 30 мин. при 1500 об./мин., после чего отбирали кольцо лимфоцитов. Отмывали лимфоциты от фиколла в 5 мл PBS 2 раза, клетки осаждали центрифугированием в течение 10 мин. при 1000 об./мин. Осадок суспендировали в соответствующем количестве среды для культивирования лимфоцитов. Чистоту выделения популяции лимфоцитов и подсчет концентрации проводили на приборе Micros. Обычно среди выделенных клеток процент лимфоцитов составлял около 80-90%. Лимфоциты либо непосредственно использовали для опыта, либо помещали в культуральные чашки и инкубировали в стандартных условиях (37°С, 5% СО2).
Пример 2. Получение генетических конструкций
При осуществлении изобретения клеточные линии человека НСТ116 (карцинома толстой кишки), НТ29 (аденокарцинома толстой кишки), А549 (карцинома легкого) и НЕК293 (почка эмбриона) культивировали в среде DMEM. Мононуклеары периферической крови выделяли из гепаринизированной периферической крови пациентов ФГУ "Российский научный центр рентгенорадиологии" центрифугированием в градиенте фиколла. Лимфоциты культивировали в среде RPMI. Лимфоциты стимулировали к пролиферации добавлением 1 мкг/мл фитогемагглютинина (ФГА) и активировали 50 Ед/мл ИЛ-2 (БИОТЕХ) в течение 72-120 ч.
Тотальную РНК из лимфоцитов и культур гибридом выделяли при помощи набора RNeasy Mini Kit. Синтез первой цепи кДНК проводили с использованием праймера олиго-dT и ревертазы RevertAid Н- (Fermentas). ПЦР проводили с помощью полимеразы TaqSE (СибЭнзим) и олигонуклеотидных праймеров, перечисленных в Табл. 1.
Таблица 1. Олигонуклеотидные праймеры. Подчеркнуты сайты распознавания рестриктаз XbaI, SmaI, ЕсоRI. Курсивом выделены последовательности, кодирующие линкерный пептид (G4S)3.

Фрагменты кДНК легких и тяжелых цепей, кодирующие вариабельные домены VL и VH иммуноглобулинов IgG1, были амплифицированы из препаратов РНК гибридомы 3С8 с использованием набора вырожденных праймеров и методов, описанных ранее [15]. Полученные ПЦР-продукты клонировали в плазмидный вектор pAL-TA (Евроген) при помощи стандартных методов [16] и секвенировали.
Для получения конструкций, кодирующих химерные иммунорецепторы, фрагменты кДНК генов CD8 и CD247 амплифицировали на матрице кДНК из лимфоцитов человека с использованием праймеров CD8-F, CD8-R, CD247-F, CD247-R. Полученные продукты лигировали между собой, реамплифицировали с использованием праймеров CD8-F, CD247-R, и клонировали в вектор pCI (GE-Amersham) по сайтам рестрикции XbaI и SmaI. Вариабельные фрагменты кДНК легких (κ) и тяжелых (γ) цепей антитела 3С8 амплифицировали с использованием праймеров из Табл. 1. ПЦР-продукты смешивали попарно для отжига перекрывающихся терминальных участков и реамплифицировали при помощи пар праймеров 3C8G-F/3C8K-R. Полученные фрагменты клонировали в плазмиду pCI/CD8-CD247 по сайтам ЕсоRI. и XbaI. Сконструированную в итоге плазмиду pCI/3C8-CD8-CD247 сокращенно назвали рСЕА-3. Секвенирование рекомбинантной плазмиды проводили с использованием автоматического секвенатора CEQ8000 (Beckman-Coulter).
Пример 3. Введение экзогенной ДНК в клетки
Трансфекцию клеток НЕК293 проводили с помощью реагента PolyFect (QIAGEN) согласно протоколу производителя. После обработки клетки инкубировали в стандартных условиях в течение 24-48 ч. В качестве положительного контроля использовали плазмиду, кодирующую зеленый флуоресцентный белок (GFP).
Трансфекцию лимфоцитов периферической крови человека проводили методом электропорации на приборе Amaxa Nucleofector с использованием набора Human Т Cell Nucleofector Kit (Lonza) согласно инструкциям производителя. Для трансфекции одного образца использовали 4-5 млн клеток и 2 мкг плазмидной ДНК. Непосредственно после импульса клетки культивировали в полной среде RPMI в стандартных условиях в течение 4-5 ч, после чего к клеткам добавляли ИЛ-2 до концентрации 100 Ед/мл. Через 24 ч после электропорации культуральную среду, содержащую ИЛ-2, заменяли новой без добавления интерлейкина. Клетки культивировали еще сутки, после чего подвергали анализу. Экспрессию конструкций, кодирующих химерные Т-клеточные рецепторы, подтверждали методом полимеразной цепной реакции, сопряженной с обратной транскрипцией (ОТ-ПЦР) через 48 ч после трансфекции.
Трансфекция НЕК293
Липосомальная трансфекция
PolyFect (Quagene). Трансфекция PolyFect (Quagene) проводилась согласно прилагаемому протоколу производителя. Клетки пассировали за 1 день до трансфекции, рассаживая около 1,0-1,2-105 клеток на лунку 24-луночного планшета в 500 мкл культуральной среды и инкубировали при 37°С, 5% СО2. Процент конфлюэнтности на день трансфекции составлял около 60-80%. Всегда использовали 2 контрольных образца: первый - клетки, трансфецированные плазмидной ДНК, кодирующией зеленый флуоресцентный белок (pmaxGFP (Amaxa)), второй - нетрансфецированнные клетки. Трансфекция одного образца, согласно протоколу, проводилась следующим образом. В бессывороточной среде, не содержащей атнтибиотиков (DMEM), растворяли 0,4 мкг плазмиды до суммарного объема 15 мкл. К раствору ДНК добавляли 4 мкл агента PolyFect, тщательно суспендировали, инкубировали 5-10 мин. при комнатной температуре. Клеткам заменяли среду свежей полной культуральной средой до объема 400 мкл. По истечении 5-10 минут к комплексам ДНК-PolyFect добавляли 100 мкл полной среды, пипетировали и немедленно добавляли к клеткам. Помешивали, покачивая планшет, и помещали в инкубатор на 24-48 часов.
Lipofectin (Invitrogene). Трансфекция с использованием lipofectin проводилась согласно прилагаемому протоколу производителя. Клетки рассаживали за 24 часа до трансфекции в количестве около 1,0-1,2-105 клеток на лунку 24-луночного планшета в 500 мкл культуральной среды и инкубировали стандартных условиях. Процент конфлюэнтности на день трансфекции составлял около 60-80%. Всегда использовали 2 контрольных образца: первый - клетки, трансфецированные плазмидной ДНК, кодирующией зеленый флуоресцентный белок (pmaxGFP), второй - нетрансфецированнные клетки. Для одной трансфекции 0.4 мкг плазмидной ДНК растворяли в 20 мкл безсывороточной среды в отсутствие антибиотиков. 4 мкл Lipofectin растворяли в 20 мкл среды без сыворотки и антибиотиков, оставляли раствор при комнатной температуре на 30-45 мин. После этого соединяли раствор ДНК и Lipofectin, осторожно их перемешивали и инкубировали при комнатной температуре в течение 10-15 мин. От клеток отбирали среду, промывали 400 мкл среды без сыворотки и антибиотиков. По истечении времени инкубации к комплексам ДНК-lipofectin добавляли 160 мкл (на одну лунку) безсывороточной среды, осторожно перемешивали и добавляли к клеткам. Клетки инкубировали в стандартных условиях 24 часа, затем заменяли среду полной средой для культивирования и инкубировали еще 24 часа. Затем клетки снимали с субстрата стандартным образом и анализировали экспрессию плазмидной ДНК.
Lipofectamine (Invitrogene) Трансфекция с использованием lipofectamine проводили согласно протоколу производителя. Клетки рассаживали за 24 часа до трансфекции в количестве около 1,0-1,2-105 клеток на лунку 24-луночного планшета в 500 мкл культуральной среды и инкубировали при 37°С, 5% СО2. Процент конфлюэнтности на день трансфекции составлял около 60-80%. Всегда использовали 2 контрольных образца: первый - клетки, трансфецированные плазмидной ДНК, кодирующией зеленый флуоресцентный белок (pmaxGFP (Amaxa), второй - нетрансфецированнные клетки. Для одной трансфекции 0.4 мкг плазмидной ДНК растворяли в 25 мкл безсывороточной среды в отсутствие антибиотиков. Lipofectamine осторожно перемешивали, на одну трансфекцию брали 4 мкл Lipofectamine, растворяли в 25 мкл среды без сыворотки и антибиотиков, осторожно перемешивали. Затем соединяли раствор ДНК и Lipofectamine, осторожно перемешивали и инкубировали при комнатной температуре в течение 15-45 мин. После этого к комплексам ДНК-Lipofectamine добавляли 150 мкл безсывороточной среды. Среду у клеток заменяли 200 мкл среды без сыворотки и антибиотиков, добавляли комплексы ДНК-Lipofectamine и помешивали, покачивая планшет. Клетки инкубировали в стандартных условиях около 5 часов, затем добавляли среды и соответствующее количество сыворотки до 10% и суммарного объема среды 500-600 мкл. Анализ экспрессии плазмидной ДНК проводили далее спустя 24 часа.
Электропорация
Электропорация НЕК293 проводилась на приборе Amaxa Nucleofector (Lonzabio), с использованием набора «Human Т Cell Nucleofector Kit» (Amaxa), согласно прилагаемому протоколу. Клетки пассировали за 24 часа до трансфекции стандартным образом в концентрации достаточной для того, чтобы на день трансфекции достигалось 80-90% конфлюэнтности. Всегда использовали 2 контрольных образца: первый - клетки, трансфецированные плазмидной ДНК, кодирующией зеленый флуоресцентный белок (pmaxGFP), второй - нетрасфецированнные клетки. Предварительно готовили и разогревали до комнатной температуры Nucleofection Solution (для трасфекции одного образца добавляли 18 мкл. Supplement к 82 мкл Human Т Cell Nucleofector Solution из набора Human Т Cell Nucleofector Kit). Клетки снимали с субстрата стандартным образом, подсчитывали их количество, помещали нужное количество клеток в отдельную пробирку и осаждали центрифугированием при 1000 об/мин в течение 5 мин. На одну трансфекцию брали около 1-2 млн клеток. После центрифугирования от осадка клеток отбирали досуха среду, добавляли 100 мкл раствора Nucleofection Solution и суспендировали в нем клетки для получения однородной суспензии. Затем в клеточную суспензию добавляли 2 мкг плазмидной ДНК и переносили в кювету (Human Т Cell Nucleofector Kit), избегая появления пузырей. Кювету помещали в держатель прибора Amaxa Nucleofector. Для трансфекции использовали программу Q-001. Сразу же после пульса в кювету добавляли 500 мкл предварительно прогретой до комнатной температуры культуральной среды, переносили в чашки Петри диаметром 3 см, содержащие 1,5 мл. культуральной среды, предварительно согретой до 37°С. Затем клетки помещали в инкубатор на 37°С, 5% СО2 на 24-48 часов.
Трансфекция лимфоцитов человека
Трансфекцию проводили с использованием программы V-024, обеспечивающей высокую эффективность трансфекции Т-лимфоцитов на приборе Amaxa Nucleofector. Для трансфекции одного образца использовали 2-3 млн клеток. Остальные условия и процедура трансфекции лимфоцитов человека были идентичны условиям для НЕК293.
Цитофлуорометрический анализ
Цитофлуорометрический анализ проводился на проточном цитофлуориметре DAKO Galaxy (Дания), в котором в качестве источника излучения используется аргоновый лазер (λ=488 нм). Полученные данные обрабатывали, используя программу FloMax, версия 3.0. Клетки осаждали центрифугированием при 1000 об/мин в течение 5-10 мин. Осадок промывали в 1 мл раствора PBS дважды. К полученному осадку добавляли 2 мл PBS и тщательно пипетировали. Полученный препарат клеток либо сразу анализировали на проточном цитофлуориметре, либо клетки затем подвергали фиксации и пермеабилизации и окраске с помощью аннтител (см. ниже).
Мечение антителами ζ-цепи Т-клеточного рецептора
Фиксацию и пермеабилизацию клеток для мечения с помощью антител проводили с использованием набора «Fixation and permeabilization kit for flow cytometry» (DakoCytomation) согласно прилагаемой инструкции. Сухой остаток анализируемых клеток, содержащий не более 2 млн клеток, суспендировали в 100 мкл раствора PBS, разделяли на 2 равные части (опытный образец и изотипический контроль) и к каждой части добавляли 100 мкл фиксирующего реагента А. Клетки тщательно суспендировали и инкубировали при комнатной температуре в течение 15 мин. Образцы промывали в 2 мл PBS, клетки осаждали центрифугированием при 1000 об/мин в течение 5 минут, удаляли супернатант до жидкого остатка в 50 мкл. Затем клетки суспендировали для получения однородной суспензии, добавляли по 100 мкл пермобилизирующего раствора В к каждому образцу и тут же к образцам добавляли антитела на ζ-цепь (Dako), мышиные антитела на IgG (Dako) в качестве изотопического контроля соответственно, тщательно суспендировали. Образцы инкубировали в течение 15 мин в темноте, затем промывали в 2 мл PBS, осаждали клетки центрифугированием при 1000 об/мин в течение 5 минут. К полученному осадку добавляли 2 мл PBS и тщательно пипетировали. После этого препараты сразу же анализировали на проточном цитофлуориметре способом, описанным выше.
Пример 4. Связывание экспрессируемых рецепторов с РЭА-ФИТЦ
Для оценки способности экспрессируемых химерных Т-клеточных рецепторов связывать РЭА клетки через 24 и 48 часов после трансфекции, инкубировали с раствором раково-эмбрионального антигена, меченного флуоресцентным красителем ФИТЦ (РЭА-ФИТЦ), в течение 30 минут. После чего клетки анализировали на проточном цитофлуориметре.
Цитофлуорометрические исследования проводили на проточном цитофлуориметре DAKO Galaxy (Dako). Фиксацию и пермеабилизацию клеток для мечения с помощью антител проводили с использованием соответствующего набора фирмы DakoCytomation. В работе использовали антитела к ζ-цепи Т-клеточного рецептора человека, мышиные антитела к IgG (Dako), биотинилированные антитела 1С11 и 3С5 (Beckman Coulter) поликлональные антитела к РЭА AS229 (Beckman Coulter).
Для оценки способности изучаемых мономолекулярных химерных Т-клеточных рецепторов, экспрессируемых клетками НЕК293, связывать РЭА, клетки спустя 24 после трансфекции инкубировали 30 мин с раствором РЭА, меченного флуоресцентным красителем ФИТЦ (РЭА-ФИТЦ, в конечной концентрации 0.06 мкг/мкл) и анализировали на проточном цитофлуориметре.
Оценку способности связывать поверхностно экспрессируемый РЭА биотинилированными моноклональными антителами на различные эпитопы РЭА, а также оценку способности сконструированных мономолекулярных химерных Т-клеточных рецепторов связывать РЭА после трансфекции лимфоцитов проводили методом непрямого иммуноцитохимического окрашивания. Для этого экспрессирующие РЭА клетки или лимфоциты, экспрессирующие мономолекулярные химерные рецепторы, через 48 ч после электропорации промывали раствором ФСБ и окрашивали биотнилированными антителами (около 3 мкл на 106 клеток) в течение 20 мин при комнатной температуре. Для выявления связавшихся антител отмытые клетки окрашивали стрептавидином, меченным фикоэритрином (РЕ), в соответствии с рекомендациями производителя (Beckman Coulter). Окрашенные клетки анализировали на проточном цитофлуориметре. В качестве отрицательного контроля использовали клетки, к которым добавляли только стрептавидин, конъюгированный с РЕ.
Анализ цитотоксического действия лимфоцитов с помощью двойной окраски CFDA-SE:PI. За 48 ч до постановки эксперимента клетки НСТ116 снимали с подложки стандартным методом, подсчитывали количество и окрашивали сукцинимидным эфиром карбоксифлуоресцеин-диацетата (CFDA-SE) в ФСБ в концентрации 0.625 мкг/мл в течение 7-8 мин в инкубаторе. Реакцию останавливали добавлением равного объема сыворотки. Клетки отмывали от невключившегося красителя, и рассаживали в чашки Петри в концентрации 2-3×105 кл/мл. Часть неокрашенных клеток рассаживали в такой же концентрации в качестве контроля. В день постановки эксперимента клетки НСТ116 неокрашенные и окрашенные (HCT116/CFDA-SE) снимали с субстрата стандартным способом и смешивали с лимфоцитам в требуемых соотношениях. Эксперименты по цитотоксичности проводили в среде RPMI с добавлением 10% ЭТС. Смешанные культуры инкубировали в стандартных условиях необходимое время. По истечении времени инкубации среду с лимфоцитами отбирали и сохраняли, а прикрепленные клетки НСТ116 промывали и снимали с субстрата минимальным количеством раствора трипсина: ЭДТА стандартным образом. Затем отделенные клетки объединяли с ранее отобранной суспензией лимфоцитов, окрашивали йодидом пропидия (PI, 2.5 мкг/мл) и анализировали на проточном цитофлуориметре.
Литература
1. Murphy A., Westwood J.A., Teng M.W., Moeller М., Darcy Р.К., Kershaw M.H. Gene modification strategies to induce tumor immunity // Immunity. - 2005. - V. 22. - N. 4. - P. 403-414.
2. Yee C. Adoptive therapy using antigen-specific T-cell clones // Cancer J. - 2010. - V. 16. -N. 4. - P. 367-373.
3. Chhabra A. TCR-engineered, customized, antitumor T cells for cancer immunotherapy: advantages and limitations // ScientificWorldJournal. - 2011. - V. 11. - P. 121-129.
4. Altenschmidt U., Moritz D., Groner B. Specific cytotoxic T lymphocytes in gene therapy // J. Mol. Med. - 1997. - V. 75. - N. 4. - P. 259-266.
5. Chen J., Dave S.K., Simmons A. Prevention of genital herpes in a guinea pig model using a glycoprotein D-specific single chain antibody as a microbicide // Virol J. - 2004. - V. 1. - P. 11.
6. Ma Q.Z., Gonzalo-Daganzo R., Junghans R.P. Genetically engineered T cells as adoptive immunotherapy of cancer // In: Giaccone, R.; Schlinsky, R.; Sondel, P., editors. Cancer Chemotherapy & Biological Response Modifiers - Annual 20. Oxford: Elsevier Science. - 2002. - P. 319-345.
7. Rossig C, Bollard С.М., Nuchtern J.D., Merchant D.A., Brenner M.K. Targeting of GD2-positive tumor cells by human T lymphocytes engineered to express chimeric T-cell receptor genes // Int J Cancer. - 2001. - V. 94. - N. 2. - P. 228-236.
8. Gade T.P., Hassen W., Santos E., Gunset G., Saudemont A., Gong M.C., Brentjens R., Zhong X.S., Stephan M., Stefanski J., Lyddane C, Osborne J.R., Buchanan I.M., Hall S.J., Heston W.D., Rivi re I., Larson S.M., Koutcher J.A., Sadelain M. Targeted elimination of prostate cancer by genetically directed human T lymphocytes // Cancer Res. - 2005. - V. 65. - N. 19. - P. 9080-9088.
re I., Larson S.M., Koutcher J.A., Sadelain M. Targeted elimination of prostate cancer by genetically directed human T lymphocytes // Cancer Res. - 2005. - V. 65. - N. 19. - P. 9080-9088.
9. Ma Q., Safar M., Holmes E., Wang Y., Boynton A.L., Junghans R.P. Anti-prostate specific membrane antigen designer T cells for prostate cancer therapy // Prostate. - 2004. - V. 61. - N. 1. - P. 12-25.
10. Ma Q., DeMarte L., Wang Y., Stanners C.P., Junghans R.P. Carcinoembryonic antigen-immunoglobulin Fc fusion protein (CEA-Fc) for identification and activation of anti-CEA immunoglobulin-T-cell receptor-modified T cells, representative of a new class of Ig fusion proteins // Cancer Gene Ther. - 2004. - V. 11. - N. 4. - P. 297-306.
11. Schwartz M.K. Cancer markers // In: DeVita, VT.; Hellman, S.; Rosenberg, SA., editors. Cancer: Principles and Practice of Oncology. Lippincott. - 1993. - P. 531-542.
12. Hammarstrom S. The carcinoembryonic antigen (CEA) family: structures, suggested functions and expression in normal and malignant tissues // Seminars in Cancer Biology. - 1999. -V. 9.-N. 2. - P. 67-81.
13. Emtage P.C., Lo A.S., Gomes E.M., Liu D.L., Gonzalo-Daganzo R.M., Junghans R.P. Second-generation anti-carcinoembryonic antigen designer T cells resist activation-induced cell death, proliferate on tumor contact, secrete cytokines, and exhibit superior antitumor activity in vivo: a preclinical evaluation // Clin Cancer Res. - 2008. - V. 14. - N. 24. - P. 8112-8122.
14. Ma Q., DeMarte L., Wang Y., Stanners C.P., Junghans R.P. Carcinoembryonic antigen-immunoglobulin Fc fusion protein (CEA-Fc) for identification and activation of anti-CEA immunoglobulin-T-cell receptor-modified T cells, representative of a new class of Ig fusion proteins // Cancer Gene Ther. - 2004. - V. 11. - N. 4. - P. 297-306.
15. Chen J., Dave S.K., Simmons A. Prevention of genital herpes in a guinea pig model using a glycoprotein D-specific single chain antibody as a microbicide // Virol J. - 2004. - V. 1. -P. 11.
16. Sambrook J., Russell D.W. Molecular cloning: a laboratory manual // Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York. - 2001.
17. Bjerner J., Lebedin Y., Bellanger L., Kuroki M., Shively J.E., Varaas Т., Nustad K., Hammarstr m S., Bormer O.P. Protein epitopes in carcinoembryonic antigen. Report of the ISOBM TD8 workshop // Tumour Biol. - 2002. - V. 23. - N. 4. - P. 249-262.
m S., Bormer O.P. Protein epitopes in carcinoembryonic antigen. Report of the ISOBM TD8 workshop // Tumour Biol. - 2002. - V. 23. - N. 4. - P. 249-262.
Список последовательностей
SEQ ID NO1
atgggatggagctgtatcatcctcttcttggtatcaacagctacagtccactccGACATCAAGATGACCCAGTCTCCATCCTCCATGTATGCATCGCTGGGAGAGAGAGTCACTATCACTTGCAAGAAGAGTCAGGACATTAAAACATATTTAAGCTGGTACCAGCAGAAACCATGGAAATCTCCTAAGACCCTGATTTATTATGCAACAACCCAGGCAGAAGGGGTCCCGTCAAGATTCAGTGGCAGTGGATCTGGGCAAGATTTTTCTCTAACCATCAGCAGCCTGGAGTctGAAGATACAGCAACCAACTACTGTCTACAACATggtgagagccctggtggaggcggttcaggcggaggtggctctggcggtggcggatcgCAGGTCCAACTGCAGCAGCCTGGGGCTGAGCTGGTGAGGCCTGGGGCTTCAGTGAAACTGTCCTGCAAGGCTTCTGGCTACACCTTCACCAGCTACTGGATAAACTGGGTGAAGCAGAGGCCTGGACAAGGCCTTGAGTGGATCGGAAATATTTTTCCTTCTGATGGTTATACTAACTACAATCAAAAGTTCAAGGACAAGGCCACATTGACTTTAGACAAATCCTCCAGCACAGCCTACATGCAGCTCAGCAGCCCGACGTCTGAGGACTCTGCGGTCTATTACTGTACAAGAGGGGGATTACTTCAGGCTATGGACTACTGGGGTCAAGGAACCTCAGTCACCGTCTCCTCA
SEQ ID NO 2
MGWSCIILFLVSTATVHSQVQLQQPGAELVRPGASVKLSCKASGYTFTSYWINWVKQRPGQGLEWIGNIFPSDGYTNYNQKFKDKATLTLDKSSSTAYMQLSSPTSEDSAVYYCTRGGLLQAMDYWGQGTSVTVSSGGGGSGGGGSGGGGSDIKMTQSPSSMYASLGERVTITCKKSQDIKTYLSWYQQKPWKSPKTLIYYATTQAEGVPSRFSGSGSGQDFSLTISSLESEDTATNYCLQHGESP
SEQ ID NO 3
atgggatggagctgtatcatcctcttcttggtatcaacagctacagtccactccGACATCAAGATGACCCAGTCTCCATCCTCCATGTATGCATCGCTGGGAGAGAGAGTCACTATCACTTGCAAGAAGAGTCAGGACATTAAAACATATTTAAGCTGGTACCAGCAGAAACCATGGAAATCTCCTAAGACCCTGATTTATTATGCAACAACCCAGGCAGAAGGGGTCCCGTCAAGATTCAGTGGCAGTGGATCTGGGCAAGATTTTTCTCTAACCATCAGCAGCCTGGAGTctGAAGATACAGCAACCAACTACTGTCTACAACATggtgagagccctggtggaggcggttcaggcggaggtggctctggcggtggcggatcgCAGGTCCAACTGCAGCAGCCTGGGGCTGAGCTGGTGAGGCCTGGGGCTTCAGTGAAACTGTCCTGCAAGGCTTCTGGCTACACCTTCACCAGCTACTGGATAAACTGGGTGAAGCAGAGGCCTGGACAAGGCCTTGAGTGGATCGGAAATATTTTTCCTTCTGATGGTTATACTAACTACAATCAAAAGTTCAAGGACAAGGCCACATTGACTTTAGACAAATCCTCCAGCACAGCCTACATGCAGCTCAGCAGCCCGACGTCTGAGGACTCTGCGGTCTATTACTGTACAAGAGGGGGATTACTTCAGGCTATGGACTACTGGGGTCAAGGAACCTCAGTCACCGTCTCCTCATCTAGAGCGAAGCCCACCACGACGCCAGCGCCGCGACCACCAACACCGGCGCCCACCATCGCGTCGCAGCCCCTGTCCCTGCGCCCAGAGGCGTGCCGGCCAGCGGCGGGGGGCGCAGTGCACACGAGGGGGCTGGACTTCGCCTGTGATGATCCCAAACTCTGCTACCTGCTGGATGGAATCCTCTTCATCTATGGTGTCATTCTCACTGCCTTGTTCCTGAGAGTGAAGTTCAGCAGGAGCGCAGACGCCCCCGCGTACCAGCAGGGCCAGAACCAGCTCTATAACGAGCTCAATCTAGGACGAAGAGAGGAGTACGATGTTTTGGACAAGAAACGTGGCCGGGACCCTGAGATGGGGGGAAAGCCGAGAAGGAAGAACCCTCAGGAAGGCCTGTACAATGAACTGCAGAAAGATAAGATGGCGGAGGCCTACAGTGAGATTGGGATGAAAGGCGAGCGCCGGAGGGGCAAGGGGCACGATGGCCTTTACCAGGGTCTCAGTACAGCCACCAAGGACACCTACGACGCCCTTCACATGCAGGCCCTGCCCCCTCGCTAACAGCCAGGGGATTTCACCACTCAAAGGCCAGACCTGCAGACGCCCAGATTATGAGACACAGGATGAAGCATTTACAACCCGGTTCACTCTTCTCAGCCACTGAAGTATCCCGGG
| название | год | авторы | номер документа |
|---|---|---|---|
| ОДНОЦЕПОЧЕЧНОЕ АНТИТЕЛО К РАКОВО-ЭМБРИОНАЛЬНОМУ АНТИГЕНУ, ХИМЕРНЫЙ МОНОМОЛЕКУЛЯРНЫЙ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР, ВЕКТОР И КЛЕТКА-ХОЗЯИН ДЛЯ ОБЕСПЕЧЕНИЯ ЭКСПРЕССИИ ТАКОГО РЕЦЕПТОРА И СПОСОБ ДИАГНОСТИКИ ИЛИ ЛЕЧЕНИЯ. | 2012 |
|
RU2522004C2 |
| МОНОМОЛЕКУЛЯРНЫЙ ХИМЕРНЫЙ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР К РАКОВО-ЭМБРИОНАЛЬНОМУ АНТИГЕНУ | 2015 |
|
RU2652955C1 |
| МОНОМОЛЕКУЛЯРНЫЙ ХИМЕРНЫЙ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР К РАКОВОМУ АНТИГЕНУ СА125 | 2018 |
|
RU2747095C2 |
| СПОСОБ НАПРАВЛЕНИЯ КЛЕТОЧНОГО ИММУННОГО ОТВЕТА ПРОТИВ ВИЧ-ИНФИЦИРОВАННОЙ КЛЕТКИ МЛЕКОПИТАЮЩЕГО, БЕЛКОВЫЙ МЕМБРАННОСВЯЗАННЫЙ ХИМЕРНЫЙ РЕЦЕПТОР, ДНК | 1995 |
|
RU2173167C2 |
| СПОСОБ ПОДАВЛЕНИЯ ВИЧ-ИНФИЦИРОВАННЫХ КЛЕТОК МЛЕКОПИТАЮЩЕГО И БЕЛКОВЫЙ РЕКОМБИНАНТНЫЙ РЕЦЕПТОР ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 1995 |
|
RU2165703C2 |
| ПЕРЕНАПРАВЛЕНИЕ КЛЕТОЧНОГО ИММУНИТЕТА ПОСРЕДСТВОМ ХИМЕРНЫХ РЕЦЕПТОРОВ | 1996 |
|
RU2167676C2 |
| ЛИНИЯ КЛЕТОК МЕЛАНОМЫ ЧЕЛОВЕКА KG, СЕКРЕТИРУЮЩИХ РЕКОМБИНАНТНЫЙ ГРАНУЛОЦИТАРНО-МАКРОФАГАЛЬНЫЙ КОЛОНИЕСТИМУЛИРУЮЩИЙ ФАКТОР | 2008 |
|
RU2362805C1 |
| Модифицированные клетки-естественные киллеры и линии клеток-естественных киллеров с повышенной цитотоксичностью | 2016 |
|
RU2772348C2 |
| NK-клеточная линия, экспрессирующая PSMA-специфический химерный антигенный рецептор и секретирующая блокатор взаимодействия CD47/SIRPa | 2020 |
|
RU2757353C1 |
| КЛЕТОЧНАЯ ТЕРАПИЯ, ОСНОВАННАЯ НА УЛУЧШЕННЫХ КЛЕТКАХ-ЕСТЕСТВЕННЫХ КИЛЛЕРАХ | 2017 |
|
RU2776890C2 |

Изобретение относится к области иммунологии. Предложено одноцепочечное антитело к раково-эмбриональному антигену, а также включающий его химерный мономолекулярный Т-клеточный рецептор, вектор и клетка-хозяин для получения химерного мономолекулярного Т-клеточного рецептора. Данное изобретение может найти применение в пассивной иммунотерапии онкологических заболеваний. 4 н. и 1 з.п. ф-лы, 6 ил., 1 табл., 4 пр.
1. Одноцепочечное антитело к раково-эмбриональному антигену, которое кодируется нуклеотидной последовательностью SEQ ID NO: 1.
2. Химерный мономолекулярный Т-клеточный рецептор, специфичный к раково-эмбриональному антигену, включающий одноцепочечное антитело, кодируемое нуклеотидной последовательностью SEQ ID NO: 1, по п. 1.
3. Химерный мономолекулярный Т-клеточный рецептор по п. 2, кодируемый нуклеотидной последовательностью SEQ ID NO: 3.
4. Вектор для обеспечения экспрессии химерного мономолекулярного Т-клеточного рецептора, специфичного к раково-эмбриональному антигену, включающий нуклеотидную последовательность, представленную на SEQ ID NO: 1.
5. Клетка-хозяин, продуцирующая химерный мономолекулярный Т-клеточный рецептор, специфичный к раково-эмбриональному антигену, содержащая вектор по п. 4.
| ОДНОЦЕПОЧЕЧНОЕ АНТИТЕЛО К РАКОВО-ЭМБРИОНАЛЬНОМУ АНТИГЕНУ, ХИМЕРНЫЙ МОНОМОЛЕКУЛЯРНЫЙ Т-КЛЕТОЧНЫЙ РЕЦЕПТОР, ВЕКТОР И КЛЕТКА-ХОЗЯИН ДЛЯ ОБЕСПЕЧЕНИЯ ЭКСПРЕССИИ ТАКОГО РЕЦЕПТОРА И СПОСОБ ДИАГНОСТИКИ ИЛИ ЛЕЧЕНИЯ. | 2012 |
|
RU2522004C2 |
| БОЖЕНКО В.К | |||
| и др | |||
| "Исследование свойств новых мономолекулярных химерных Т-клеточных рецепторов к раково-эмбриональному антигену" | |||
| Клеточные технологии в биологии и медицине, 2013; 3: 174-180 | |||
| WOERN A., PLUECKTHUN A | |||
| "Stability engineering of antibody single-chain Fv fragments." Journal of molecular biology, 2001; 305(5): 989-1010 | |||
| BJERNER J | |||
| et al | |||
| "Protein epitopes in carcinoembryonic antigen." Tumor biology, 2002; 23(4): 249-262 | |||
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |

