Изобретение было сделано при поддержке Правительства по Контракту N A1 27849, присужденному Национальными Институтами Здоровья. Правительство имеет определенные права в этом изобретении.
Данное изобретение касается функциональных химер между фрагментами CD4 и рецепторами иммунокомпетентных клеток (иммуноцитов), которые способны направлять иммунокомпетентные клетки на лизис инфицированных ВИЧ клеток, но не делают эти иммунокомпетентные клетки чувствительными к инфекции ВИЧ. Таким образом, это изобретение обеспечивает новое и эффективное терапевтическое средство против ВИЧ.
Предпосылки изобретения
Узнавание T-клетками антигена через T-клеточный рецептор является основой ряда иммунологических феноменов. T-клетки управляют так называемым клеточно-опосредованным иммунитетом. Он включает в себя разрушение клетками иммунной системы чужеродных тканей или инфицированных клеток. Существуют различные T-клетки, в том числе "хелперные" и "супрессорные" клетки, которые модулируют иммунный ответ, и цитотоксические (или "убийцы") клетки, которые могут непосредственно убивать патологические клетки.
T-клетка, которая узнает и связывает уникальный антиген, обнаруживаемый на поверхности другой клетки, становится активированной; она может затем размножаться и, если она является цитотоксической клеткой, она может убить связанную ею клетку.
ВИЧ и иммунопатогенез
В 1984 году было показано, что ВИЧ является этиологическим агентом СПИДа. С этого времени определение СПИДа пересматривали несколько раз в отношении того, какие критерии должны быть включены в диагноз. Однако, несмотря на неустойчивость в диагностических параметрах, простое общее название СПИДа - это инфекция ВИЧ и последующее развитие стойких конституциональных симптомов и определяющих СПИД заболеваний, таких как вторичные инфекции, неоплазмы и неврологическое заболевание. Harrison's Principles of Internal Medicine, 12th ed., McGraw Hill (1991).
ВИЧ является человеческим ретровирусом группы лентивирусов. Четыре признанных человеческих ретровирусов принадлежат к двум различным группам: T-лимфотропные (или лейкозные) ретровирусы человека, HTLV-1 и HTLV-2, и вирусы иммунодефицита человека, HIV-1 и HIV-2 (ВИЧ-1 и ВИЧ-2). Первые являются трансформирующими вирусами, тогда как последние являются цитопатическими вирусами.
ВИЧ-1 был идентифицирован как наиболее общая причина СПИДа по всему миру. Гомология последовательности между ВИЧ-1 и ВИЧ-2 составляет приблизительно 40%, причем ВИЧ-1 более близок к некоторым членам группы вирусов иммунодефицита обезьян (SIV). CM. Curran, J. et al. Science, 329:1357- 1359 (1985); Weiss, R. et al. Nature, 324:572-575 (1986).
ВИЧ имеет обычные ретровирусные гены  , а также шесть экстрагенов, участвующих в репликации и других биологических активностях этого вируса. Как утверждалось ранее, общим определением СПИДа является глубокая иммуносупрессия преимущественно клеточно-опосредованного иммунитета. Эта иммуносупрессия (супрессия иммунного ответа) приводит к различным заболеваниям, вызываемым условно-патогенными микроорганизмами, в частности некоторым инфекциям и неоплазмам.
, а также шесть экстрагенов, участвующих в репликации и других биологических активностях этого вируса. Как утверждалось ранее, общим определением СПИДа является глубокая иммуносупрессия преимущественно клеточно-опосредованного иммунитета. Эта иммуносупрессия (супрессия иммунного ответа) приводит к различным заболеваниям, вызываемым условно-патогенными микроорганизмами, в частности некоторым инфекциям и неоплазмам.
В качестве основной причины иммунной недостаточности в СПИДе была идентифицирована количественная и качественная недостаточность в субпопуляции образуемых тимусом (Т) лимфоцитов, популяции T4. Эта субпопуляция клеток определяется фенотипически присутствием поверхностной молекулы CD4, которая, как было показано, является клеточным рецептором ВИЧ. Dalgleish et al. Nature 312:763 (1984). Хотя T4-клетка является основным клеточным типом, инфицируемым ВИЧ, по существу любая клетка человека, экспрессирующая молекулу CD4 на ее поверхности, способна связываться с ВИЧ и инфицироваться ВИЧ.
Традиционно, CD4+Т-клеткам приписывают роль хелпера/индуктора, что указывает на их функцию в обеспечении активирующего сигнала для B-клеток или в индуцировании T-лимфоцитов, несущих соответствующий CD8-маркер, с превращением их в цитотоксичные/супрессорные клетки. Reinherz and Schlossman, Cell 19:821-827 (1980); Goldstein et al., Immunol. Rev. 68: 5-42 (1982).
ВИЧ связывается специфически и с высокой аффинностью, через участок аминокислот в оболочке вируса (gp120), с частью района VI молекулы CD4, расположенной вблизи ее N-конца. После связывания вирус сливается с клеточной мембраной-мишенью- и интернализуется. После интернализации он использует фермент обратную транскриптазу для транскрибирования его геномной РНК в ДНК, которая интегрируется в клеточную ДНК, где и существует в течение жизни этой клетки в виде "провируса".
Провирус может оставаться латентным или активироваться для транскрибирования мРНК и геномной РНК, что ведет к синтезу, сборке белка, образованию новых вирионов и активной репликации вируса (бадинга) в результате проникновения из клеточной поверхности в клетку. Хотя точный механизм, при помощи которого вирус индуцирует смерть клетки, не был установлен, считают, что основным механизмом является массированный вирусный бадинг из клеточной поверхности, приводящий к разрушению плазменной мембраны и к нарушению осмотического баланса.
Во время процесса инфицирования организм хозяина вырабатывает антитела против вирусных белков, в том числе против основных гликопротеинов gp120 и gp41 оболочки. Несмотря на этот гуморальный иммунитет, болезнь прогрессирует, приводя к летальной иммуносупрессии, характеризующейся множественными инфекциями, вызываемыми условно-патогенными микроорганизмами, паразитемией, деменцией и смертью. Неспособность антивирусных антител хозяина остановить прогрессирование заболевания является одним из наиболее неприятных и тревожных аспектов этой инфекции и дает плохие прогнозы для усилий по вакцинации, основанной на общепринятых подходах.
Два фактора могут играть роль в эффективности гуморального ответа на вирусы иммунодефицита. Во-первых, подобно другим РНК-вирусам (и, в частности, подобно ретровирусам) вирусы иммунодефицита обнаруживают высокую скорость мутации в ответ на иммунологический контроль хозяина. Во-вторых, сами гликопротеины оболочки являются сильно гликозилированными молекулами, предоставляющими немного эпитопов, пригодных для высокоаффинного связывания антител. Бедная антигенами мишень, которую представляет вирусная оболочка, дает малую возможность хозяину для ограничения вирусной инфекции посредством продуцирования специфических антител.
Клетки, инфицированные вирусом ВИЧ, экспрессируют на их поверхности гликопротеин gp120. gp120 медиирует события слияния среди CD4+-клеток посредством реакции, сходной с реакцией, при помощи которой вирус входит в неинфицированные клетки, что приводит к образованию короткоживущих многоядерных гигантских клеток. Образование синцития зависит от прямого взаимодействия гликопротеина оболочки gp120 с белком CD4. Dalgleish et al., supra; Klatzman, D. et al., Nature; 312:763 (1984); McDougal, J.S. et al., Science 231: 382 (1986); Sodroski, J. et al. Nature 322:470 (1986); Lifson J.D. et al. Nature 323:725 (1986); Sodroski, J. et al., Nature 321:412 (1986).
Доказательство, что связывание CD4-gp120 ответственно за вирусную инфекцию клеток, несущих антиген CD4, включает в себя открытие, что между gp120 и CD4 образуется специфический комплекс. McDougal et al., supra. Другие исследователи показали, что клеточные линии, которые были неинфицируемыми для ВИЧ, превращались в инфицируемые клеточные линии после трансфекции и экспрессии кДНК-гена CD4 человека. Maddon et al. Cell 46:333-348 (1986).
Были предложены и успешно продемонстрированы in vitro рядом групп терапевтические программы, основанные на растворимом CD4 в качестве пассивного агента для противодействия вирусного поглощения и опосредованной синцитием клеточной передачи вируса (Deen et al. Nature 331:82-84 (1988); Fisher et al. Nature 331:76- 78 (1988); Hussey et al. Nature 331:78-81 (1988); Smith et al. Science 238: 1704-1707 (1987); Traunecker et al. Nature 331:84-86 (1988)); и впоследствии были обеспечены слитые белки CD4-иммуноглобулина с увеличенными полупериодами жизни и умеренной биологической активностью (Capon et al. Nature 337:525-531 (1989); Traunecker et al. Nature 339:68-70 (1989); Byrn et al., Nature 344:667-670 (1990); Zettlmeissl et al., DNA Cell Biol. 9: 347-353 (1990)). Хотя конъюгаты CD4-иммунотоксин или слитые белки обнаруживают сильную цитотоксичность для инфицированных клеток in vitro (Chaudary et al. Nature 335: 369-372 (1988); Till et al. Science 242:1166-1168 (1988)), латентное состояние синдрома иммунодефицита делает маловероятным, что какая-либо одного типа терапия будет эффективной в устранении вирусной нагрузки, и антигенность чужеродных слитых белков, по-видимому, ограничивает их приемлемость в лечении, требующем повторяющегося дозирования. Испытания на мартышках, пораженных вирусом иммунодефицита обезьян (SIV) показали, что растворимый CD4 при введении без заметной CD4 цитопении может снижать титр SIV и улучшать измерения in vitro миелоидного потенциала (Watanabe et al. Nature 337:267-270 (1989)). Однако после прерывания обработки наблюдали незамедлительное повторное появление вируса, что заставляет предполагать, что для предотвращения прогрессивного истощения иммунной системы может быть необходимым введение в течение всей жизни.
T-клеточные и Fc-рецепторы
Экспрессия клеточной поверхностью наиболее обильной формы T-клеточного рецептора антигенов (TCR) требует одновременной экспрессии по меньшей мере 6 отличающихся полипептидных цепей (Weiss et al., J. Exp. Med. 160:1284-1299 (1984); Orloffhashi et al. Nature 316:606-609 (1985); Berkhout et al., J. Biol. Chem. 263:8528-8536 (1988); Sussman et al. Cell 52:85-95 (1988)), α/β антиген связывающих цепей, трех полипептидов комплекса CD3 и ζ. Если любая из этих цепей отсутствует, то стабильная экспрессия остальных членов комплекса не происходит. ζ является лимитирующим полипептидом поверхностной экспрессии полного комплекса (Sussman et al. Cell 52:85-95 (1988)), и считают, что эта цепь медиирует по меньшей мере часть клеточной программы активации, запускаемой узнаванием лиганда рецептором (Weissman et al., EМBO J. 8: 3651-3656 (1989); Frank et al. Science 249:174-177 (1990)). Интегральный мембранный гомодимер 32 кДа типа 1, ζ (дзета), имеет внеклеточный домен из 9 остатков без сайтов для добавления (присоединения) N-связанного гликана и внутриклеточный домен из 112 остатков (мышь) или 113 остатков (человек) (Weissman et al. Science 238:1018-1020 (1988); Weissman et al., Proc. Natl. Acad. Sci. USA 85:9709-9713 (1988)). Изоформа ζ, названная η (эта) (Baniyash et al. , J. Biol. Chem. 263:9874-9878 (1988); Orloff et al., J. Biol. Chem. 264: 14812-14817 (1989)), которая возникает из чередующегося пути сплайсинга мРНК (Jin et al., Proc. Natl. Acad. Sci. USA 87:3319-3233 (1990)), присутствует в уменьшенных количествах в клетках, экспрессирующих рецептор антигена. Гетеродимеры ζ-η, как считают, опосредуют образование инозитфосфатов, а также инициируемую рецептором программированную смерть клеток, называемую апоптозом (Mercep et al. Science 242:571-574 (1988); Mercep et al., Science 246:1162-1165 (1989)).
Подобно ζ и η, ассоциированная с рецептором Fc γ-цепь, экспрессируется в комплексах клеточной поверхности с дополнительными полипептидами, некоторые из которых медиируют узнавание лиганда, а другие имеют неопределенную функцию. γ (гамма) имеет гомодимерную структуру и общую организацию, очень похожую на структуру и организацию ζ, и является компонентом высокоаффинного рецептора IgE мастоцитов/базофилов, FceRI, который состоит по меньшей мере из трех отличающихся полипептидных цепей (Blank et al. Nature 337:187-189 (1989); Ra et al. Nature 241:752-754 (1989)), а также компонентом рецепторов низкой аффинности для IgG, представленных в мышах Fc γ RII α (Ra et al., J. Biol. Chem. 264: 15323-15327 (1989)), в человеке экспрессией подтипа CD16 макрофагами и природными клетками-убийцами, CD16TM (CD16 трансмембранный) (Lanier et al. Nature 342:803-805 (1989); Anderson et al., Proc. Natl. Acad. Sci. USA 87: 2274-2278 (1990)) и полипептидом неидентифицированной функции (Anderson et al., Proc. Natl. Acad. Sci. USA 87:2274-2278 (1990)). Недавно сообщалось, что γ экспрессируется мышиной T-клеточной линией, CTL, в которой он образует гомодимеры, а также гетepoдимepы γ-ζ и γ-η (Orloff et al. Nature 347:189-191 (1990)).
Fc-рецепторы медиируют фагоцитоз иммунных комплексов, трансцитоз и зависимую от антител клеточную цитотоксичность (ADCC) (Ravetch and Kinet, Annu. Rev. Immunol. 9:457-492 (1991); Unkeles et al., Annu. Rev. Immunol. 6: 251-281 (1988); и Mellman, Curr. Opin. Immunol. 1:16-25 (1988)). Недавно было показано, что одна из мышиных изоформ рецептоpa Fc низкой аффинности, FcR γ IIIBl, медиирует интернализацию покрытых Ig мишеней в покрытые клатрином углубления и что другой рецептор низкой аффинности, Fci γ lllA, медиирует ADCC посредством его связывания с одним или более членами небольшого семейства "триггерных молекул" (Miettinen et al., Cell 58:317-327 (1989); и Huziker and Mellman, J. Cell Biol. 109:3291-3302 (1989)). Эти триггерные (запускающие) молекулы, T-клеточный рецептор (TCR), ζ -цепь, η -цепь TCR и γ -цепь рецептора Fc, взаимодействуют с доменами узнавания лигандов различных рецепторов иммунной системы и могут автономно инициировать клеточные эффекторные программы, в том числе цитолиз, после агрегации (Samelson et al. Cell 43: 223-231 (1985); Weissman et al. Science 239:1018-1020 (1988); Jin et al., Proc. Natl. Acad. Sci. USA 87:3319-3323 (1990); Blank et al. Nature 337: 187-189 (1989); Lanier et al. Nature 342:803-805 (1989); Kurosaki and Ravetch, Nature 342: 805-807 (1989); Hibbs et al. Science 246:1608-1611 (1989); Anderson et al., Proc. Natl. Acad. Sci. USA 87:2274-2278 (1990); и Irwing and Weiss, Cell 64:891-901 (1991)).
Однако при проведении параллелей между семействами рецепторов Fc низкой аффинности мышей и человека становится ясно, что FcR γ IIA и C-изоформы человека не имеют копии в мышах. Отчасти вследствие этого их функция еще должна быть определена.
Вследствие того, что гуморальные агенты на основе одного CD4 имеют ограниченную применимость in vivo, предшествующие исследования выясняли возможность увеличения клеточного иммунитета к ВИЧ. Были идентифицированы препараты белковых химер, в которых внеклеточный домен CD4 слит с трансмембранным и/или внутриклеточным доменами T-клеточного рецептора, рецептора Fc lgG или элементами трансдукции сигнала B-клеточного рецептора (U.S.S.N. 07/847566 и 07/665961). Цитолитические T-клетки, экспрессирующие химеры, которые включают в себя внеклеточный домен CD4, обнаруживают сильную MHC-независимую деструкцию клеточных мишений, экспрессирующих белки оболочки ВИЧ. Чрезвычайно важным и новым компонентом этого подхода была идентификация отдельных цепей T-клеточного рецептора, рецептора Fc и B-клеточного рецептора, агрегация которых достаточна для инициации клеточного ответа. Одним из особенно ценных приложений этого подхода было изобретение химер между CD4 и ζ,η, или γ, которые нацеливают цитолитические T-лимфоциты на узнавание и убивание клеток, экспрессирующих gp120 ВИЧ (U.S.S.N. 07/847 566 и 07/665 961).
Краткое изложение существа изобретения
В общем, данное изобретение описывает способ направления клеточного иммунного ответа против инфицированной ВИЧ клетки в млекопитающем. Способ предусматривает введение млекопитающему эффективного количества терапевтических клеток, экспрессирующих мембрано-связанный белковый химерный рецептор, содержащий (а) внеклеточную часть, которая включает в себя фрагмент CD4, способный специфически узнавать и связывать инфицированную ВИЧ клетку, но не медиирующий инфекцию ВИЧ, и (b) внутриклеточную часть, которая способна давать сигнал терапевтической клетке о разрушении связанной рецептором инфицированной ВИЧ клетки.
В родственном способе изобретение описывает использование терапевтических клеток, экспрессирующих мембрано-связанный белковый химерный рецептор, включающий в себя (а) внеклеточную часть, содержащую фрагмент CD4, который способен специфически узнавать и связывать инфицированную ВИЧ клетку, но не медиирует инфекцию ВИЧ, и (b) внутриклеточную часть, которая способна давать сигнал терапевтической клетке для разрушения связанной рецептором инфицированной ВИЧ клетки, в приготовлении лекарственного средства для лечения родственных ВИЧ заболеваний.
Во втором аспекте, изобретение описывает клетку, которая экспрессирует белковый мембрано-связанный химерный рецептор, который содержит (а) внеклеточную часть, которая включает в себя фрагмент CD4, который способен специфически узнавать и связывать инфицированную ВИЧ клетку, но не медиирует инфекцию ВИЧ, и (b) внутриклеточную часть, которая способна давать сигнал терапевтической клетке для разрушения связанной рецептором инфицированной ВИЧ клетки.
В предпочтительных вариантах обоих аспектов фрагмент CD4 представляет собой аминокислоты 1-394 CD4 или аминокислоты 1-200 CD4; этот фрагмент CD4 отделен от внутриклеточной части трансмембранным доменом CD7, показанным на фиг. 26, или шарнирной областью, CH2, и доменами CH3 молекулы lgGl человека, показанными на фиг. 25; рецептор включает в себя трансмембранную часть CD7; рецептор включает в себя трансмембранную часть CD5; рецептор включает в себя трансмембранную часть CD34; фрагмент CD4 отделен от мембраны терапевтической клетки одной или более белковыми α - спиралями; фрагмент CD4 отделен от мембраны терапевтической клетки по меньшей мере 48 ангстремами (10-10 м) или по меньшей мере 72 ангстремами; внутриклеточная часть является трансдуцирующей сигнал частью белка T-клеточного рецептора (например, ζ ) белка B-клеточного рецептора или белка рецептора Fc; и терапевтические клетки выбраны из группы, состоящей из: (а) T- лимфоцитов; (b) цитотоксических T-лимфоцитов; (c) природных клеток-убийц; (d) нейтрофилов; (e) гранулоцитов; (f) макрофагов; (g) мастоцитов; (h) HeLa клеток; и (i) эмбриональных стволовых клеток (ES).
В других аспектах, изобретение описывает ДНК, кодирующую химерный рецептор данного изобретения; и вектор, включающий в себя ДНК химерного рецептора.
Хотя характерным вариантом данного изобретения является химера между CD4 и дзета, любая цепь рецептора, имеющая функцию, подобную функции этих молекул, например, в гранулоцитах или B- лимфоцитах, могла бы быть использована для описанных здесь целей. Отличительные признаки желаемой триггерной молекулы иммунокомпетентной клетки включают в себя способность автономной экспрессии (т. е. в виде отдельной цепи), способность сливаться с внеклеточным доменом CD4, так что полученная химера присутствует на поверхности терапевтической клетки, и способность инициировать клеточные эффекторные программы при вторичной агрегации для столкновения с лигандом-мишенью.
В настоящее время наиболее удобным способом доставки химерных белков к клеткам иммунной системы является использование формы генной терапии. Однако реконституирование клеток иммунной системы с химерными рецепторами смешиванием этих клеток с солюбилизированным подходящим образом очищенным химерным белком привело бы также к образованию популяции сконструированных клеток, способных отвечать на ВИЧ-инфицированные мишени. Подобные подходы использовали, например, для введения молекулы CD4 в эритроциты для терапевтических целей. В этом случае популяция сконструированных клеток не была бы способна к самообновлению.
Данное изобретение касается функциональных и упрощенных химер между фрагментами CD4 и T-клеточным рецептором, B-клеточным рецептором и субъединицами рецептора Fc, которые способны направлять иммунокомпетентные клетки для узнавания и лизиса ВИЧ- инфицированных клеток. Способ направления клеточного ответа в млекопитающих предусматривает введение эффективного количества терапевтических клеток (например, цитотоксических T-лимфоцитов) млекопитающему, причем эти клетки способны узнавать и разрушать ВИЧ-инфицированную клетку.
Изобретение также включает в себя химерные рецепторные белки, которые направляют цитотоксические T-лимфоциты для узнавания и лизиса ВИЧ-инфицированных клеток, клетки-хозяева, трансформированные вектором, содержащим эти химерные рецепторы, и антитела, направленные против химерных рецепторов.
Эти и другие нелимитирующие варианты данного изобретения будут очевидны специалистам в этой области из следующего далее подробного описания изобретения.
В следующем далее подробном описании будут сделаны ссылки на различные методологии, известные специалистам в области молекулярной биологии и иммунологии. Публикации и другие материалы, представляющие такие известные методологии, на которые делается ссылка, включены здесь в виде полных ссылок.
Стандартные справочные работы, излагающие основные принципы технологии рекомбинантных ДНК, включают в себя Watson et al. Molecular Biology of the Gene, Volumes I and II, the Benjamin/Cummings Publishing Company, Inc., publisher, Menio Park, CA (1987); Darnell et al. Molecular Cell Biology, Scientific American Books, Inc., Publisher, New York, N.Y. (1986); Lewin, Genesll, John Wiley & Sons, publishers, New York, N.Y. (1985); Old et al. Principles of Gene Manipulation: An Introduction to Genetic Ergineering, 2d edition. University of California Press, publisher, Berkeley, CA (1981); Maniatis et al. Molecular Cloning: A Laboratory Manual, 2nd Ed. Cold Spring Harbor Laboratory, publisher, Cold Spring Harbor, N.Y. (1989); и Ausubel et al. , Current Protocols in Molecular Biology, Wiley Press, New York, N.Y. (1989).
ОПРЕДЕЛЕНИЯ
Под "клонированием" имеют в виду использование способов рекомбинации in vitro для встраивания определенного гена или другой последовательности ДНК в векторную молекулу.
Под "кДНК" подразумевают комплементарную или копию-ДНК, продуцируемую из РНК-матрицы под действием РНК-зависимой ДНК- полимеразы (обратной транскриптазы). Таким образом, "клон кДНК" обозначает дуплексную последовательность ДНК, комплементарную представляющей интерес молекуле РНК, переносимую в клонирующем векторе.
Под "библиотекой кДНК" подразумевают коллекцию рекомбинантных молекул ДНК, содержащих встроенные кДНК, которые содержат ДНК, являющиеся копиями мРНК, экспрессируемыми клеткой при создании этой библиотеки кДНК. Такая библиотека кДНК может быть получена способами, известными специалистам в этой области и описанными, например, в Ausubel et al., supra и Maniatis et al. , supra. В общем виде, РНК сначала выделяют из клеток организма, из генома которого желательно клонировать конкретный ген. Для целей данного изобретения предпочтительны клеточные линии лимфоцитов млекопитающего и, в частности, человека. В настоящее время предпочтительным вектором для этой цели является штамм WR вируса коровьей оспы.
"Вектором" называют молекулу ДНК, полученную, например, из плазмиды, бактериофага или вируса млекопитающего или насекомого, в которую могут быть встроены или клонированы фрагменты ДНК. Вектор должен содержать один или более уникальных сайтов рестрикции и может быть способным автономно реплицироваться в определенном организме хозяина или переносчика, так что клонированная последовательность может размножаться. Таким образом, под "экспрессирующим ДНК-вектором" подразумевают любой автономный элемент, способный управлять синтезом рекомбинантного пептида. Такие экспрессирующие ДНК- векторы включают в себя бактериальные плазмиды и фаги и плазмины и вирусы млекопитающих и насекомых.
"Фактически чистым" называют соединение, например белок, полипептид или антитело, которые фактически не содержат компонентов, сопровождающих их в природе. В общем виде, соединение является фактически чистым, если по меньшей мере 60%, более предпочтительно по меньшей мере 75% и наиболее предпочтительно по меньшей мере 90% всего материала в пробе представляют собой целевое соединение. Чистота может быть измерена любым подходящим способом, например колоночной хроматографией, электрофорезом в полиакриламидном геле или анализом ЖХВР (HPLC). В связи с последовательностью нуклеиновой кислоты "фактически чистая" означает последовательность нуклеиновой кислоты, сегмент или фрагмент которой не является непосредственно смежным (т.е. ковалентно связанным) с обеими кодирующими последовательностями, с которыми он непосредственно связан (т.е. с одной при 5'-конце и с другой при 3'-конце) в природно существующем геноме организма, из которого получена ДНК изобретения.
Под "фрагментом" молекулы, такой как любая из последовательностей кДНК данного изобретения, подразумевают любую часть смежных нуклеотидов этой молекулы. "Аналог" молекулы обозначает неприродную молекулу, существенно сходную либо с целой молекулой, либо с ее фрагментом. О молекуле говорят, что она "существенно сходна" с другой молекулой, если последовательность аминокислот в обеих молекулах по существу одинакова. Существенно сходные аминокислотные молекулы будут обладать сходной биологической активностью. В применении здесь, молекулу называют "химическим производным" другой молекулы, когда она содержит химические части молекулы, не являющиеся обычно частью этой молекулы. Такие части молекулы могут улучшать растворимость молекулы, абсорбцию, биологический полупериод жизни и т.п. Эти части молекулы могут, альтернативно, уменьшать токсичность молекулы, исключать или аттенюировать нежелательное побочное действие молекулы и т.д. Молекулярные части, способные медиировать такие эффекты, описаны, например, в Remington's Phanmaceutical Sciences, 16th ed. Mack Publishing Co., Easton, Penn. (1980).
"Функциональным производным" химерного гена рецептора данного изобретения называют "фрагменты" или "аналоги" этого гена, которые "существенно сходны" по нуклеотидной последовательности и кодируют молекулу, обладающую сходной активностью, например, с химерой рецептора T-клетки, B-клетки или Fc. Наиболее предпочтительно, чтобы это производное имело 90%, более предпочтительно 70%, и предпочтительно 40% активности химеры рецептора дикого типа. Активность функционального производного химерного рецептора включает в себя специфическое связывание (с его внеклеточной частью CD4) с ВИЧ-инфицированной клеткой и в конечном счете разрушение этой клетки; кроме того, химерный рецептор не делает несущую рецептор клетку восприимчивой к инфекции ВИЧ. Активность химерного рецептора может определяться, например, любым из описанных здесь способов.
Последовательность ДНК, кодирующая рецепторную химеру CD4 данного изобретения или ее функциональное производное, может быть рекомбинирована с векторной ДНК в соответствии с общепринятыми способами, включающими в себя затупленные или ступенчатые концы для лигирования, расщепление рестриктазами для обеспечения подходящих концов, заполнение в "липкие" концы, обработку щелочной фосфатазой во избежание нежелательного соединения и лигирование подходящими лигазами. Способы для таких манипуляций описаны Maniatis, Т., et al., supra и хорошо известны в этой области.
О молекуле нуклеиновой кислоты, такой как ДНК, говорят, что она "способна экспрессировать" полипептид, если она содержит нуклеотидные последовательности, которые включают в себя регуляторную информацию для транскрипции и трансляции, и такие регуляторные последовательности "оперативно соединены" с нуклеотидными последовательностями, кодирующими данный полипептид. "Оперативным соединением" является такое соединение, в котором регуляторные последовательности ДНК и последовательности ДНК, которые должны быть экспрессированы, соединены таким образом, чтобы сделать возможной экспрессию гена. Точная природа регуляторных районов, необходимых для генной экспрессии, может варьировать от организма к организму, но в общем виде будет включать в себя промотор (который управляет инициацией транскрипции РНК), а также последовательности ДНК, которые при транскрибировании в РНК будут сигнализировать об инициации синтеза белка. Такие районы обычно включают в себя 5'-некодирующие последовательности, участвующие в инициации транскрипции и трансляции, такие как TATA-блок, кэпирующие последовательность, последовательность CAAT и т.п.
Если желательно, некодирующий район 3 (справа) от генной последовательности, кодирующей белок, может быть получен описанными выше способами. Этот район может быть сохранен для его регуляторных последовательностей терминации транскрипции. Так, путем сохранения этого 3'-района, в природе смежного с последовательностью ДНК, кодирующей белок, могут быть обеспечены сигналы терминации транскрипции. В случае, если эти сигналы терминации транскрипции не функционируют удовлетворительно в экспрессирующей клетке-хозяине, вместо них может быть введен 3'- район, функциональный в клетке-хозяине.
Две последовательности ДНК (такие, как последовательность промоторного района и последовательность, кодирующая химерный рецептор CD4) называют оперативно соединенными, если природа соединения между этими двумя последовательностями ДНК (1) не приводит к введению мутации сдвига рамки, (2) не мешает способности последовательности промоторного района направлять транскрипцию последовательности гена рецепторной химеры или (3) не мешает способности последовательности гена химеры рецептора транскрибироваться последовательностью промоторного района. Промоторный район будет оперативно соединен с последовательностью ДНК, если промотор способен инициировать транскрипцию этой ДНК. Таким образом, для экспрессии белка необходимы сигналы транскрипции и трансляции, узнаваемые соответствующим хозяином.
Данное изобретение включает в себя экспрессию химерного белка рецептора CD4 (или его функционального производного) в прокариотических или эукариотических клетках, хотя предпочтительна эукариотическая экспрессия (и, в частности, в лимфоцитах человека).
Антитела в соответствии с данным изобретением могут быть получены любым из множества способов. Например, клетки, экспрессирующие химерный белок рецептора CD4 или его функциональное производное, могут быть введены животному для индуцирования образования сыворотки, содержащей поликлональные антитела, способные связывать эти химерные белки.
В предпочтительном способе антитела в соответствии с данным изобретением являются моноклональными антителами. Такие моноклональные антитела могут быть получены с применением технологии гибридом (Kohler et al. Nature 256:495 (1975); Kohler et al., Eur. J. Immunol. 6:511 (1976); Kohler et al., Eur. J. Immunol. 6:292 (1976); Haimnerling et al. In: Monoclonal Antibodies and T-cell Hybridomas, Else-vier, N.Y., pp. 563-684 (1981)). В общем виде, такие процедуры включают в себя иммунизацию животного антигеном химеры рецептора CD4. Спленоциты таких животных экстрагируют и сливают с подходящей линией миеломных клеток. Любая подходящая линия миеломных клеток может быть использована в соответствии с данным изобретением. После слияния полученные гибридомные клетки селективно поддерживают в HAT-среде и затем клонируют серийным разведением, как описано Wands et al. (Gastroenterology 80:225-232 (1981)). Гибридомные клетки, полученные таким отбором, затем анализируют для идентификации клонов, которые секретируют антитела, способные связывать химеры.
Антитела согласно данному изобретению также могут быть поликлональными или предпочтительно район-специфическими поликлональными антителами.
Антитела против химеры рецептора CD4 в соответствии с данным изобретением могут быть использованы для мониторинга количества химерного рецептора (или клеток, несущих химерный рецептор) в больном. Такие антитела очень удобны для использования в стандартном иммунодиагностическом тесте, известном в этой области, включающем в себя такие иммунометрические или "сэндвич" анализы, как прямой сэндвич, обратный сэндвич и одновременные сэндвич- тесты. Эти антитела могут быть применены в любом количестве комбинаций, что может быть определено специалистами без чрезмерного экспериментирования для проведения иммунотестов приемлемой специфичности, чувствительности и точности.
Стандартные работы, излагающие общие принципы иммунологии, включают в себя Roitt, Essential Immunology, 6th ed., Blackwell Scientific Publications, Publisher, Oxford (1988); Kimball, Introduction to Immunology, 2d ed., Macrnillan Publishing Co., Publisher, New York (1986); Roitt et al. Immunology, GoWer Medical Publishing Ltd. , Publisher, London (1985); Campbell, "Monoclonal Antibody Technology", in Burdon et al., eds. Laboratory Techniques in Biochemistry and Molecular Biology, Volume 13, Elsevier, Publisher, Amsterdam (1984); Klein, Immunology: The Science of Self-Nonself Discrimination, John Wiley & Sons, Publisher, New York (1982); и Kennet et al., eds. , Monoclonal Antibodies, Nyb-ridoma: A New Dimension In Biological Analyses, Plenum Press, Publisher, New York (1980).
Под "обнаружением" ("детектированием") подразумевают определение присутствия или отсутствия вещества или определение количества вещества. Термин, следовательно, относится к применению материалов, композиций и способов данного изобретения для качественных и количественных определений.
Антитела и фактически очищенный антиген данного изобретения идеально пригодны для изготовления кита (набора). Такой кит может содержать носитель (держатель), в котором размещены один или несколько контейнеров, таких как флаконы, пробирки и т.п., причем каждый из контейнеров содержит отдельные элементы анализа, который должен быть выполнен.
Многие типы тестов могут быть включены в киты, например конкурентный и неконкурентный анализы. Типичными примерами анализов, которые могут использовать антитела этого изобретения, являются радиоиммуноанализ РИА (RIA), ферментные иммуноанализы (EIA), твердофазные иммуноферментные анализы (ELISA) и иммунометрические, или сэндвич, иммуноанализы.
Под термином "иммунометрический анализ" (количественный иммуноанализ), или "сэндвич-иммуноанализ", подразумевают одновременный сэндвич-, прямой сэндвич-, обратный сэндвич-иммуноанализ. Эти термины хорошо понятны специалистам в данной области. Им также понятно, что антитела в соответствии с данным изобретением будут применимы в других вариациях и формах анализов, которые известны в настоящее время и которые могут быть разработаны в будущем. Подразумевается, что они входят в объем данного изобретения.
Фраза "специфически узнает и связывает" означает, что антитело узнает и связывает химерный рецепторный полипептид, но фактически не узнает и не связывает другие, не относящиеся к этому полипептиду молекулы в пробе, например в биологической пробе.
"Терапевтической клеткой" называют клетку, которая была трансформирована химерой рецептора CD4 этого изобретения таким образом, что она способна узнавать и разрушать ВИЧ-инфицированную клетку; предпочтительно такие терапевтические клетки являются клетками гемопоэтической системы.
Термин "внеклеточный" означает, что по меньшей мере часть молекулы экспонирована на поверхности клетки. "Внутриклеточный" означает имеющий по меньшей мере часть молекулы, находящуюся в цитоплазме терапевтической клетки. "Трансмембранный" означает, что по меньшей мере часть молекулы находится в мембране плазмы. "Внеклеточная часть", "внутриклеточная часть" и "трансмембранная часть" могут включать в себя фланкирующие аминокислотные последовательности, которые простираются в прилегающие клеточные компартменты.
"Олигомеризоваться" означает образовывать комплекс с другими белками с образованием димеров, тримеров, тетрамеров или олигомеров более высокого порядка. Такие олигомеры могут быть гомоолигомерами или гетероолигомерами. "Олигомеризующая часть" представляет собой район молекулы, который управляет образованием комплекса (т.е. олигомера).
"Цитолитический" означает способный разрушать клетку (например, ВИЧ-инфицированную клетку) или способный разрушать инфекционный агент (например, вирус ВИЧ).
Под "вирусом иммунодефицита" подразумевают ретровирус, который в форме дикого типа способен инфицировать T4-клетки хозяина-примата и обладает вирусным морфогенезом и морфологией, характерными для подсемейства лентивирусов. Этот термин включает в себя, без ограничений, все варианты HIV (ВИЧ) и SIV, в том числе HIV-1, HIV-2, SIVmac, SIVagm, SIVmnd, SIVsmm, SIVman, SIVmand и SIVcpz.
"MHC-независимый" означает, что клеточный цитолитический ответ не требует присутствия антигена главного комплекса гистосовместимости (MHC) класса II на поверхности клетки-мишени.
"Функциональное передающее (трансдуцирующее) цитолитический сигнал производное" обозначает функциональное производное (определенное выше), которое способно направлять по меньшей мере 40%, более предпочтительно 70% или наиболее предпочтительно 90% биологической активности молекулы дикого типа. В применении здесь, "функциональное передающее цитолитический сигнал производное" может действовать путем прямой подачи сигнала терапевтической клетке разрушить связанный рецептором агент или клетку (например, в случае внутриклеточной части химерного рецептора) или может действовать опосредованно путем усиления олигомеризации с передающими цитолитический сигнал белками терапевтической клетки (например, в случае трансмембранного домена). Такие производные могут быть испытаны на их эффективность, например, с применением описанных здесь тестов in vitro.
"Функциональным связывающим оболочку ВИЧ производным" называют функциональное производное (определенное выше), которое способно связывать любой белок оболочки ВИЧ.
Функциональные производные могут быть идентифицированы с применением, например, описанных здесь тестов in vitro.
ТЕРАПЕВТИЧЕСКОЕ ВВЕДЕНИЕ
Трансформированные клетки данного изобретения применяют для терапии вируса иммунодефицита. Текущие способы введения таких трансформированных клеток включают в себя адоптивную иммунотерапию или терапию переноса клеток. Эти способы позволяют возврат трансформированных клеток иммунной системы в кровоток. Rosenberg, Scientific American 62 (May 1009); Rosenberg et al. The New England Journal of Medicine 323(9):570 (1990).
Фармацевтические композиции этого изобретения могут быть введены любому животному, которое может испытывать благоприятное действие соединений этого изобретения. Основными среди таких животных являются люди, хотя изобретение не лимитировано таким образом.
Подробное описание
Сначала будут описаны чертежи
Краткое описание чертежей
Фиг. 1A - аминокислотная последовательность около сайта слияния между CD4 (остатки 1-369) и различными цепями рецептора. Подчеркнутая последовательность показывает положение аминокислот, кодируемых внутри сайта BamHl, используемого для конструирования слияния. Начало трансмембранного домена отмечено вертикальной чертой. Последовательность η идентична последовательности ζ при амино-конце, но отличается при карбоксильном конце (Jin et al., Proc. Natl. Acad. Sci. USA 87:3319-3323 (1990)).
Фиг. 1B - проточный цитометрический анализ поверхностной экспрессии CD4, CD4: ζ CD4: γ и CD4: η в клетках CVI. Клетки инфицировали вирусом, экспрессирующим химеры CD4 или CD16PI, инкубировали в течение 9 часов при 37oC и окрашивали конъюгированными с фикоэритрином анти-CD4 MAb Leu3A.
Фиг. 2 показывает поверхностную экспрессию CD16TM после ко-инфицирования одним CD16TM (плотные точки) или ко- инфицирования вирусом, экспрессирующим CD4: γ (штрихи) или CD4: ζ (сплошная линия). Редкие точки - клетки, инфицированные одним CD4: ζ окрашенные 3G8 (Fleit et al., Proc. Natl. Acad. Sci. USA 79:3275-3279 (1982)) (анти-CD16 MAb).
Фиг. 3 показывает поверхностную экспрессию CD16TM после ко-инфицирования вирусами, экспрессирующими CD16TM и следующими ζ химерами: CD4: ζ (жирная линия), CD4: ζ CllG (сплошная линия); CD4: ζ (штриховая линия); CD4: ζ C11G/D15G (плотные точки); без ко-инфицирования (CD16TM только, редкие точки). Клетки инкубировали с анти-CD16 MAb 3G8 и конъюгированными с фикоэритрином Fab'2 козьими антителами к мышиному IgG. Уровень экспрессии ζ химер был по существу идентичен для различных анализированных мутантов и ко-инфицирование клеток вирусами, экспрессирующими CD16TM и ζ химерами не изменяло ощутимо поверхностную экспрессию химер.
Фиг. 4A-D показывают увеличение внутриклеточного иона свободного кальция после сшивания мутантных ζ химер в T-клеточной линии. Клетки Jurkat E6 (Weiss et al., J. Immunology 133:123-128 (1984)) инфицировали рекомбинантными вирусами коровьей оспы и анализировали проточной цитометрией. Представлены результаты для популяции CD4+, так что анализировали только клетки, экспрессирующие соответствующий химерный белок (установкой дискриминационного окна в сортере). Среднее отношение фиолетовой к синей флуоресценции Indo-I отражает концентрацию внутриклеточного свободного кальция в популяции в целом, а процент отвечающих клеток отражает фракцию клеток, которая превышает заданное пороговое отношение (установленное таким образом, что 10% необработанных клеток являются положительными).
Фиг. 4A и 4B показывают клетки Jurkat, экспрессирующие CD4: ζ (сплошная линия) или CD16: ζ (штриховая линия), которые выдерживали с анти-CD4 MAb Leu3a (конъюгированными с фикоэритрином) с последующим сшиванием с козьими антителами к мышиному IgG. Линия в виде точек показывает ответ неинфицированных клеток на анти-CD3 MAb ОКТ3. Фиг. 4C и 4D показывают клетки Jurkat, экспрессирующие CD4: ζ D15G (сплошная линия); CD4: ζ C11G/D15G (штрихи) или CD4: ζ C11G (точки), которые были обработаны и анализировались, как на фиг. 4A и 4B.
Фиг. 5A-C показывают, что рецепторы CD4: ζ, CD4: η и CD4: γ позволяют цитолитическим T-лимфоцитам (CTL) убивать
мишени, экспрессирующие gp120/41 ВИЧ-1. Фиг. 5A: сплошные кружки - CTL, экспрессирующие CD4: ζ , инкубировали с клетками HeLa, экспрессирующими gp120/41; открытые кружки - CTL, экспрессирующие CD4: ζ , инкубировали с неинфицированными клетками HeLa; сплошные квадраты - неинфицированные CTL, инкубированные с клетками HeLa, экспрессирующими gp120/41; открытые квадраты - неинфицированные CTL, инкубированные с неинфицированными клетками HeLa. Фиг. 5B: сплошные кружки - CTL, экспрессирующие CD4: η , инкубированные с клетками HeLa, экспрессирующими gp120/41; открытые кружки - CTL, экспрессирующие CD4: γ , инкубированные с клетками HeLa, экспрессирующими gp120/41; открытые квадраты - CTL, экспрессирующие C11G/D15G двойную мутантную химеру CD4: ζ , инкубированные с клетками HeLa, экспрессирующими gp120/41. Фиг. 5C: проточный цитометрический анализ экспрессии CD4 CTL, используемыми в фиг. 5B. Для корректировки отношений мишени к эффектору процент клеток, экспрессирующих химерный CD4, определяли вычитанием масштабированной негативной (неинфицированной) популяции наложением гистограмм; для целей сравнения на этом рисунке неинфицированные клетки были определены как произвольный порог, что дает грубо ту же самую фракцию, позитивную, для других клеточных популяций, какую дало бы вычитание гистограмм.
Фиг. 6A-B показывают специфичность CD4-направляемого цитолиза. Фиг. 6A: сплошные кружки - CTL, экспрессирующие CD4: ζ, инкубированные с клетками HeLa, экспрессирующими CD16PI; открытые кружки - CTL, экспрессирующие CD4, инкубированные с клетками HeLa, экспрессирующими gp120/41. Фиг. 6B: сплошные кружки - CTL, экспрессирующие CD4: ζ , инкубированные с клетками Raji (MHC класса II+); открытые кружки - неинфицированные CTL клетки, инкубированные с RJ2.2.5 (мутант MHC класса II- Raji) клетками; сплошные квадраты - неинфицированные CTL, инкубированные с клетками Raji (MHC класса 11+); открытые квадраты - CTL, экспрессирующие CD4: ζ , инкубированные с клетками RJ2.2.5 (MHC класса II-). Масштаб ординаты увеличен.
Фиг. 7A-B дают характеристику химерного рецептора CD16: ζ. Фиг. 7A - схематическая диаграмма слитого белка CD16: ζ. Внеклеточная часть соединенной с фосфатидилинозитом формы мономерного CD16 была соединена с димерным ζ непосредственно снаружи по отношению к трансмембранному домену. Последовательность белка при соединении показана в нижней части. Фиг. 7B показывает проточный цитометрический анализ мобилизации кальция после сшивания химеры CD16: ζ либо в TCR-положительной, либо в TCR-отрицательной клеточной линии. Среднее отношение фиолетовой к голубой флуоресценции (мера относительной концентрации иона кальция) показано среди клеточных популяций, обработанных антителами, при времени 0. Сплошные квадраты - ответ клеток Jurkat на анти-CD3 MAb ОКТ3; сплошные треугольники - ответ CD16: ζ на анти- CD16 MAb 3G8, сшитых в мутанте REX33A TCR-; открытые квадраты - ответ на сшивание CD16: ζ в TCR- мутантной линии клеток Jurkat JRT3.T3.5; открытые треугольники - ответ на сшивание CD16: ζ в клетках Jurkat; крестики - ответ на нехимерный CD16 в клетках Jurkat; точки - ответ на нехимерный CD16 в REX33A TCR клеточной линии.
Фиг. 8A-B показывает делеционный анализ цитолитического потенциала. Фиг. 8A показывает местоположения концевых точек делеций ζ. Здесь, как и в других местах, мутации в ζ представлены так: первоначальный остаток - местоположение - остаток мутанта, так что D66*, например, обозначает замену Asp-66 кодоном терминации. Фиг. 8B показывает результаты анализа цитолиза неделетированного CD16: ζ и выделяющихся делеций ζ. Гибридомные клетки, экспрессирующие поверхностные антитела к CD16, нагружали 51Cr и инкубировали с увеличивающимися количествами цитолитических лимфоцитов (CTL) человека, инфицированных рекомбинантами вируса коровьей оспы, экспрессирующими химеры CD16: ζ Процент выделенного 51Cr нанесен на график как функция отношения эффекторных клеток (CTL) к клеткам-мишеням (гибридомным клеткам) (e/t). Сплошные кружки - цитолиз, медиируемый клетками, экспрессирующими CD16: ζ (mfi 18.7); сплошные квадраты - цитолиз, медиируемый клетками, экспрессирующими CD16: ζ Asp66* (mfi 840.2); открытые квадраты - цитолиз, медиируемый клетками, экспрессирующими CD16: ζ Glu60+ (mfi 16.0); открытые кружки - цитолиз, медиируемый клетками, экспрессирующими CD16: ζ Tyr51* (mfi 17.4); сплошные треугольники - цитолиз, медиируемый клетками, экспрессирующими CD16: ζ Phe34* (mfi 17.8); и открытые треугольники - цитолиз, медиируемый клетками, экспрессирующими нехимерный CD16 (mfi 591). Хотя в этом эксперименте экспрессия CD16: ζ Asp66* не была сравнима с экспрессией других слитых белков, цитолиз клетками, экспрессирующими CD16: ζ при эквивалентных уровнях в том же самом эксперименте дал результаты, по существу одинаковые с результатами, обнаруживаемыми клетками, экспрессирующими CD16: ζ Asp66.
Фиг. 9A-D показывают, что элиминация потенциала для трансмембранных взаимодействий обнаруживает короткий сегмент ζ, способный медиировать цитолиз. Фиг. 9A - схематическая диаграмма мономерных состоящих из двух и трех частей химер. В верхней части находится конструкция CD16: ζ, укороченная при остатке 65 и не имеющая трансмембранных остатков Cys и Asp. Ниже находятся конструкции CD16:CD5: ζ и CD16: CD7: ζ и соответствующие контроли. Пептидные последовательности внутриклеточных доменов показаны ниже. Фиг. 9B показывает цитолитическую активность мономерных химерных делеционных мутантов. Цитолитическую активность клеток, экспрессирующих CD16: ζ (сплошные кружки; mfi 495), сравнивали с цитолитической активностью клеток, экспрессирующих CD16: ζ Asp66* (сплошные квадраты; mfi 527) или мутантов CD16: ζ Cys11Gly/Asp15Gly/Asp66* (открытые квадраты; mfi 338) и CD16: ζ Cys11Gly/Asp15Gly/Glu60* (заполненные треугольники; mfi 259). Фиг. 9C показывает цитолитическую активность, медиируемую слитыми белками, состоящими из трех частей. Сплошные треугольники - CD16: ζ Asp66*; открытые квадраты - CD16: 5: ζ (48-65); сплошные квадраты - CD16:7: ζ (48-65); открытые треугольники - CD16: 7: ζ (48-59); открытые кружки - CD16:5; сплошные кружки - CD16: 5. Фиг. 9D показывает мобилизацию кальция мутантами и состоящими из трех частей химерами в TCR- линии мутантных клеток Jurkat JRT3.T3.5. Открытые кружки - ответ клеток, экспрессирующих димерный CD16: ζ Asp66*; сплошные квадраты - ответ клеток, экспрессирующих CD16: ζ Cys11Gly/Asp15Gly/Asp66*; открытые квадраты - ответ клеток, экспрессирующих CD16: ζ Cys11Gly/Asp15Gly/Glu60*; сплошные треугольники - ответ клеток, экспрессирующих CD16: 7: ζ (48-65); и открытые треугольники - ответ клеток, экспрессирующих CD16: ζ (48-59).
Фиг. 10A-F показывают вклад отдельных аминокислот в активность передающего цитолитический сигнал мотива из 18 остатков. Фиг. 10A и 10B показывают цитолитическую активность, а фиг. 10C показывает мобилизацию иона кальция, медиируемую химерами, несущими точковые мутации вблизи карбоксил-концевого тирозина (Y62). Фиг. 10A и 10B представляют данные, полученные на клетках, экспрессирующих низкие и высокие количества, соответственно, слитых белков CD16: ζ. Идентичные символы использованы для мобилизации кальция и анализов цитолиза и они показаны в однобуквенном коде справа. Сплошные кружки - клетки, экспрессирующие CD16: ζ (mfi в A, 21; В,376); сплошные квадраты - клетки, экспрессирующие CD16:7: ζ (48-65) (mfi А, 31; В, 82); открытые квадраты - CD16: 7: ζ (48-65)Glu60Gln (mfi A, 33; В, 92); крестики - CD16:7: ζ (48-65)Asp63Asn (mfi A, 30; В, 74); сплошные треугольники - CD16:7: ζ (48-65)Tyr62Phe (mfi A, 24; В, 88); открытые кружки - CD16:7: ζ (48-65)Glu6lGln (mfi A, 20; В, 62); и открытые треугольники - CD16:7: ζ (48-65)Tyr62Ser (mfi В, 64). Фиг. 10D и 10E показывают цитолитическую активность, а фиг. 10F показывает мобилизацию иона кальция химерами, несущими точковые мутации вблизи амино-концевого тирозина (Y51). Идентичные символы использованы для мобилизации кальция и анализов цитолиза и они показаны справа. Сплошные кружки-клетки, экспрессирующие CD16: ζ (mfi в D, 21.2; в E, 672); сплошные квадраты-клетки, экспрессирующие CD16: 7: ζ (48-65) (mfi D, 31.3; E, 179); сплошные треугольники - CD16:7: ζ (48-65)-Asn48Ser (mfi D, 22.4; E, 209); открытые квадраты - CD16: 7: ζ (48-65)Leu50Ser (mfi D, 25.0; E, 142); и открытые треугольники - CD16:7: ζ (48-65)Tyr51Phe (mfi D, 32.3; E, 294).
Фиг. 11A-B показывают сравнительный анализ внутренних повторов ζ и сравнение их способности поддерживать цитолиз. Фиг. 11A - схематическая диаграмма химер, образованных разделением внутриклеточного домена ζ на трети и присоединением их к трансмембранному домену химеры CD16:7. Последовательности этих внутриклеточных доменов показаны ниже, с общими остатками, заключенными в блоки, и близкими остатками, отмеченными звездочками. Фиг. 11B показывает цитолитическую активность трех субдоменов ζ. Сплошные кружки - клетки, экспрессирующие CD16: ζ (mfi 476); сплошные квадраты - CD16:7: ζ (71-104) (mfi 114); и сплошные треугольники - CD16:7: ζ (104-138) (mfi 104).
Фиг. 12 - схема химер CD16:FcRγII.
Фиг. 13A-B показывает мобилизацию кальция после сшивания химер CD4:FcR γ II и CD16:FcRγII. Фиг. 13A показывает отношение фиолетовой к синей флуоресценции, испускаемой клетками, загруженными чувствительным к кальцию флуорофором Indo-1, представленное как функция времени после сшивания внеклеточного домена CD16 с антителами. Фиг. 13B показывает подобный анализ увеличения в отношении фиолетовой к синей флуоресценции клеток, несущих химеры CD4: FcRγII, после сшивания с антителами.
Фиг. 14A-B показывает анализ цитолиза химер CD4:FcR γ II и CD16:FcR γ II. Фиг. 14A показывает процент 51Cr, выделенного из анти-CD16 гибридомных клеток при выдерживании клеток с увеличивающимися количествами цитотоксических T-лимфоцитов, экспрессирующих химеры CD16:FcR γ II (эффекторных клеток). Фиг. 14B показывает подобный анализ цитотоксичности, медиируемой химерами CD4: FcR γ II, против клеток-мишеней, экспрессирующих гликопротеины оболочки ВИЧ.
Фиг. 15A-E показывает идентификацию остатков в хвосте FcR γ II А, который важен для цитолиза. Фиг. 15A - схема делеционных конструкций. Фиг. 15B и 15C показывают мобилизацию кальция и цитолиз карбоксил-концевыми делеционными вариантами CD16:FcR γ II А. Фиг. 15B и 15E показывают мобилизацию кальция и цитолиз химерами из трех частей, несущими прогрессивно уменьшающийся амино-конец внутриклеточного хвоста, CD16:FcR γ II А.
Фиг. 16 (SEQ ID N 24) показывает аминокислотную последовательность рецепторного белка CD3 дельта; заключенная в блок последовательность представляет собой предпочтительную передающую сигнал часть.
Фиг. 17 (SEQ ID N 25) показывает аминокислотную последовательность рецепторного белка T3 гамма; заключенная в блок последовательность представляет собой предпочтительную передающую цитолитический сигнал часть.
Фиг. 18 (SEQ ID N 26) показывает аминокислотную последовательность рецепторного белка mbi; заключенная в блок последовательность представляет собой предпочтительную передающую цитолитический сигнал часть.
Фиг. 19 (SEQ ID 27) показывает аминокислотную последовательность рецепторного белка B29; заключенная в блок последовательность представляет собой предпочтительную передающую цитолитический сигнал часть.
Фиг. 20A-E показывает схему химер CD4. Молекула "А" -CD4(D1- D4):Ig:CD7; молекула "В" - CD4(D1,D2):Ig:CD7; молекула "С" - CD4(D1-D4):Ig:CD7: ζ ; молекула "D" - CD4(D1,D2): Ig:CD7: ζ ; и молекула "E" - CD4: ζ . Внеклеточный домен молекулы CD4 человека, соответствующий аминокислотам 1-394 предшественника, соединяли при помощи сайта BamHI с шарнирной
областью, CH1 и CH2 доменами человеческого IgG1, как описано ранее (Zettlmeissl et al., DNA Cell Biol. 9:347 (1990)), за исключением того, что использовали кДНК-версию последовательностей человеческого Ig для того, чтобы происходила экспрессия в рекомбинантных вирусах коровьей оспы. Двухдоменную версию химер CD4 создавали встраиванием BamHI-адаптора в уникальном сайте NheI (соответствующем аминокислоте 200) в кДНК предшественника CD4. Последовательности присоединения к мембране состояли из 22 остатков из первого экзона человеческого мембрано-связанного IgG1, за которыми следовали остатки CD7 146-203. Аминокислоты 55- 163 ζ служили в качестве триггерного мотива конструкций из четырех частей (C и D). В состоящих из четырех частей конструкциях, содержащих цепь ζ, была документирована внутриклеточная экспрессия ζ c коммерчески доступными антителами против внутриклеточного домена (Coulter).
Фиг. 21 показывает цитолиз клеток-мишеней, экспрессирующих гликопротеин оболочки ВИЧ, медиируемый клоном цитотоксических T- клеток, WH3, экспрессирующих различные произведенные из CD4 химеры в качестве эффекторных молекул. Для анализов на цитотоксичность CD8+ CD4-HLA B44 рестриктированную T-клеточную линию человека, WH3, поддерживали в среде IMDM, дополненную 10% человеческой сывороткой, как описано здесь ранее. Клетки стимулировали облученными гамма-лучами (3000 рад), несущими B44 мононуклеарными клетками и фитогемагглютинином (РНА) при концентрации 1 мкг/мл. После одного дня стимулирования PHA разводили до 0.5 мкг/мл добавлением свежей среды; после 3 дней среду заменяли полностью. Клетки выращивали по меньшей мере в течение 10 дней перед использованием в анализах цитотоксичности. Клетки инфицировали подходящими рекомбинантными вирусами коровьей оспы, как описано здесь для VPE16. Инфекции происходили в течение дополнительных 3-4 часов в полной среде, после чего клетки собирали центрифугированием и ресуспендировали при плотности 1 • 107 клеток/мл. По 100 мкл добавляли в каждую лунку микротитрационного планшета с U-образной нижней частью, содержащую 100 мкл на лунку полной среды и разводили в 2-кратных серийных стадиях. Две лунки для каждой пробы не содержали лимфоцитов для измерения спонтанного выделения хрома и общего поглощения хрома. Клетки-мишени, HeLa сублинию S3 (HeLa-S3, ATCC), инфицировали, как описано выше, в чашках 10 см с vPE16 • 106 инфицированных клеток отделяли при помощи PBS и 1 мМ ЭДТА, центрифугировали и ресуспендировали в 100 мкл 51Cr-хромата натрия (1 мКи/мл в PBS) и выдерживали в течение 1 часа при 37oC и затем промывали 3 раза PBS. 100 мкл меченых клеток-мишеней добавляли к каждой лунке. Микротитрационный планшет откручивали при 750 х g в течение 1 минуты и инкубировали в течение 4 часов при 37oC. В конце инкубационного периода клетки в каждой лунке ресуспендировали осторожным пипетированием, брали пробу для определения общего числа включенных импульсов и микротитрационный планшет откручивали при 750 х g в течение 1 минуты. Брали аликвоты (100 мкл) супернатанта и считали в гамма-лучевом сцинтилляционном счетчике. Отношение эффектор: мишень корректировали на процент инфицированных клеток, измеренных при помощи проточной цитометрии.
Фиг. 22 показывает репликацию ВИЧ-1 в трансфицированных клеточных линиях. Клеточные линии, стабильно экспрессирующие CD4 дикого типа и различные рекомбинантные химеры, были созданы в сублинии линии 293 эмбриональных почечных клеток человека. Исходный раствор вируса изолята ВИЧ-1IIIB готовили с титром приблизительно 106 инфекционных частиц/мл, как измерено анализом конечных результатов разведения с применением линии C8166 T-клеток человека в качестве индикатора. Инфицирования проводили при приблизительной множественности заражения 1 в течение периода 8-12 часов при 37oC. На следующий день клетки промывали PBS три раза, трипсинизировали, пересевали в новые чашки и культуральный супернатант брали для определения титра (обозначенного как день 0). С интервалами 3-4 дня после этого супернатанты клеточных культур собирали и сохраняли для анализа p24. Клетки снабжали свежей средой, содержащей гигромицин B в концентрации 100 мкг/мл. Анализ культуральных супернатантов проводили с применением коммерческого кита для анализа на основе ELISA антигена p24 ВИЧ-1 (Coulter) в соответствии с инструкциями, поставляемыми изготовителем. Результаты являются репрезентативными данными двух независимых экспериментов сходной продолжительности.
Фиг. 23 показывает последовательность нуклеиновой кислоты и аминокислотную последовательность доменов D1-D4 CD4 (CD4 Bam).
Фиг. 24 показывает последовательность нуклеиновой кислоты и аминокислотную последовательность доменов D1-D2 CD4 (CD4 Nhe).
Фиг. 25 показывает последовательность нуклеиновой кислоты и аминокислотную последовательность шарнирной области, CH2 и CH3 доменов человеческого IgG1 (Igh23 Bam).
Фиг. 26 показывает последовательность нуклеиновой кислоты и аминокислотную последовательность трансмембранного домена CD7 (ТМ7 Barn Mlu).
Фиг. 27 показывает последовательность нуклеиновой кислоты и аминокислотную последовательность внутриклеточного домена дзета (Zeta Mlu Not).
Фиг. 28 показывает последовательность ДНК и первичную аминокислотную последовательность синтетической альфа-спирали.
ПРИМЕР 1
Конструирование химер человеческий IgGl: рецептор
Последовательности тяжелых цепей IgGl человека получали соединением последовательностей в CH3 домене с фрагментом кДНК, произведенным из 3'-конца трансмембранной формы мРНК антитела. 3'-концевой фрагмент получали при помощи полимеразной цепной реакции с применением библиотеки кДНК миндалин в качестве субстрата и олигонуклеотидов, имеющих последовательности:
CGC GGG GTG АСС GTG CCC ТСС AGC AGC TTG GGC (SEQ ID N 7)
CGC GGG GAT CCG TCG ТСС AGA GCC CGT CCA GCT CCC CGT CCT CCT CA (SEQ ID N 8),
соответствующих 5'- и 3'-концам желаемых фрагментов ДНК соответственно/ 5'-олиго комплементарен сайту в домене CH1 человеческого IgGl, а 3'-олиго комплементарен сайту, расположенному непосредственно 5' от последовательностей, кодирующих пересекающий мембрану домен. Продукт ПЦР расщепляли при помощи BstXI и BamHI и лигировали между сайтами BstXI и BamHI полусинтетического гена IgGl антитела, несущего вариабельную и константную области. После встраивания фрагмента BstXI-BamHI амплифицированные части этой конструкции вводили в сайт Smal в CH3 обменом рестрикционных фрагментов, так что только часть между сайтом SmaI и 3'-олиго была произведена из реакции ПЦР.
Для создания химерного рецептора человеческий IgGl: ζ ген тяжелой цепи, заканчивающийся сайтом BamHI, соединяли с сайтом BamHI описанной ниже ζ -химеры, так что последовательности антитела образовали внеклеточную часть. Проточная цитометрия клеток COS, трансфицированных плазмидой, кодирующей эту химеру, показала высокие уровни экспрессии детерминат антитела при ко-трансфекции экспрессирующей плазмиды, кодирующей кДНК легкой цепи, и умеренную экспрессию детерминант антитела при отсутствии экспрессирующей плазмиды легкой цепи.
Подобные химеры, включающие в себя IgG1 человека, слитый с η или γ (см. ниже) или любой трансдуцирующей сигнал частью белка T-клеточного рецептора или Fc-рецептора, могут быть сконструированы в общем, как описано выше, с использованием стандартных способов молекулярной биологии.
Для создания отдельной транскрипционной единицы, которая позволила бы как тяжелым, так и легким цепям экспрессироваться от одного промотора, была создана плазмида, кодирующая бицистронную мРНК из последовательностей, кодирующих тяжелую цепь и легкую цепь, и 5'-нетранслируемой части этой мРНК, кодирующей регулируемый глюкозой белок 78 кДа, известный иначе как grp78 или BiP. Последовательности grp78 получали при помощи ПЦР из геномной ДНК человека с применением праймеров, имеющих последовательности:
CGC GGG CGG CCG CGA CGC CGG CCA AGA CAG CAC (SEQ ID N 9) и
CGC GTT GAC GAG CAG CCA GTT GGG CAG CAG CAG (SEQ ID IP 10)
при 5'- и 3'- концах соответственно. Полимеразные цепные реакции выполняли с этими олигонуклеотидами в присутствии 10% диметилсульфоксида. Полученный при помощи ПЦР фрагмент расщепляли NotI и HincII и встраивали между NotI и HpaI по ходу транскрипции от последовательностей, кодирующих IgGl человека. Затем последовательности, кодирующие кДНК легкой каппа-цепи IgG человека, встраивали по ходу транскрипции от лидера grp78 с использованием сайта HincII и другого сайта в векторе. Полученная в результате этих манипуляций экспрессионная плазмида состояла из полусинтетического гена тяжелой цепи, за которым следовали лидерные последовательности grp78, а затем последовательности кДНК легкой цепи каппа и сигналы полиаденилирования, полученные из фрагмента ДНК SV40. Трансфекция клеток COS этой экспрессирующей плазмидой дала заметно улучшенную экспрессию детерминант тяжелой цепи по сравнению с трансфекцией плазмиды, кодирующей только детерминанты тяжелой цепи.
Для создания бицистронного гена, содержащего химеру тяжелая цепь/рецептор и легкую цепь, последовательности тяжелой цепи против хода транскрипции могут быть заменены любым описанным здесь химерным геном тяжелой цепи/рецептора.
ПРИМЕР II
Конструирование химер CD4 рецептора
кДНК ζ (Weissman et al. , Proc. Natl. Acad. Sci. USA 85:9709- 9713 (1988)) и γ (Kuster et al., J. Biol. Chem. 265: 6448-6452 (1990)) человека выделяли с применением полимеразной цепной реакции из библиотек, полученных из линии опухолевых клеток HPB- ALL (Aruffo et al., Proc. Natl. Acad. Sci. USA 84: 8573-8577 (1987b)) и из природных клеток-убийц человека, тогда как кДНК η (Jin et al., Proc. Natl. Acad. Sci. USA 87:3319-3323 (1990)) выделяли из библиотеки мышиных тимоцитов, кДНК ζ,η и γ соединяли с внеклеточным доменом сконструированной формы CD4, обладающим сайтом BamHI сразу против хода транскрипции от пересекающего мембрану домена (Aruffo et al., Proc. Natl. Acad. Sci. USA 84:8573-8577 (1987b); Zettlmeissl et al., DNA Cell Biol. 9: 347-353 (1990)), который соединяли с сайтом BamHI, природно присутствующим в кДНК ζ и η в сходном положении на расстоянии нескольких остатков против хода транскрипции от пересекающего мембрану домена (SEQ ID N 1, N 3, N 4 и N 6). Для образования слитого белка с γ сайт BamHI был сконструирован в последовательности при том же самом приблизительном положении (фиг. 1; SEQ ID N 2 и 5). Эти генные слияния вводили в экспрессирующую плазмиду вируса коровьей оспы, несущую ген  Е. coli в качестве селектируемого маркера, и встраивали в геном штамма WR вируса коровьей оспы посредством гомологичной рекомбинации и отбора на рост в микофенольной кислоте (Falkner et al., J. Virol. 62:1849-1854 (1988); Boyle et al. Gene 65:123-128 (1988)). Проточная цитометрия показала, что рекомбинанты вируса коровьей оспы направляют обильное образование слитых белков CD4: ζ и CD4: γ при клеточной поверхности, тогда как экспрессия CD4: η была значительно более слабой (фиг. 1B). Последнее открытие согласуется с недавним сообщением, что трансфекция экспрессионной плазмиды кДНК η в мышиную гибридомную клеточную линию дала значительно меньшую экспрессию, чем трансфекция сравнимой экспрессионной плазмиды ζ (Clayton et al., J. Exp. Med. 172:1243- 1253 (1990)). Иммунопреципитация клеток, инфицированных рекомбинантами вируса коровьей оспы, выявила, что эти слитые белки образуют ковалентные димеры в отличие от природно встречающегося антигена CD4. Было найдено, что мол. массы мономерных слитых белков CD4: ζ и CD4: γ и нативного CD4 равны 63, 55 и 53 кД соответственно. Большие массы слитых белков приблизительно согласуются с большей длиной внутриклеточной части, которая превышает длину внутриклеточной части нативного CD4 на 75 (CD4: ζ) или 5 (CD4: γ ) остатков.
Е. coli в качестве селектируемого маркера, и встраивали в геном штамма WR вируса коровьей оспы посредством гомологичной рекомбинации и отбора на рост в микофенольной кислоте (Falkner et al., J. Virol. 62:1849-1854 (1988); Boyle et al. Gene 65:123-128 (1988)). Проточная цитометрия показала, что рекомбинанты вируса коровьей оспы направляют обильное образование слитых белков CD4: ζ и CD4: γ при клеточной поверхности, тогда как экспрессия CD4: η была значительно более слабой (фиг. 1B). Последнее открытие согласуется с недавним сообщением, что трансфекция экспрессионной плазмиды кДНК η в мышиную гибридомную клеточную линию дала значительно меньшую экспрессию, чем трансфекция сравнимой экспрессионной плазмиды ζ (Clayton et al., J. Exp. Med. 172:1243- 1253 (1990)). Иммунопреципитация клеток, инфицированных рекомбинантами вируса коровьей оспы, выявила, что эти слитые белки образуют ковалентные димеры в отличие от природно встречающегося антигена CD4. Было найдено, что мол. массы мономерных слитых белков CD4: ζ и CD4: γ и нативного CD4 равны 63, 55 и 53 кД соответственно. Большие массы слитых белков приблизительно согласуются с большей длиной внутриклеточной части, которая превышает длину внутриклеточной части нативного CD4 на 75 (CD4: ζ) или 5 (CD4: γ ) остатков.
ПРИМЕР III
Химеры CD4 могут связываться с другими рецепторными цепями
Экспрессия на поверхности клеток типа макрофагов/природных клеток убийц человеческого Fc γ RIII (CD16TM) на трансфектантах облегчается ко-трансфекцией с мышиным (Kurosaki et al. Nature 342:805-807 (1989)) или человеческим (Hibbs el al. Science 246:1608-1611 (1989)) γ, а также человеческим ζ (Lanier et al. Nature 342:803-805 (1989)).
В соответствии с этими сообщениями экспрессия химер также делала возможной поверхностную экспрессию CD16TM при доставке к клетке-мишени либо посредством ко-трансфекции, либо посредством ко-инфицирования с рекомбинантными вирусами коровьей оспы (фиг. 2). Усиление поверхностной экспрессии CD16TM при помощи ζ было более выраженным, чем усиление при помощи γ (фиг. 2) в испытанных клеточных линиях, в то время как нативный CD4 не усиливал поверхностную экспрессию CD16TM.
ПРИМЕР IV
Asp ζ мутанты не связываются с Fc-рецептором
Для создания химер, которые не связывались бы с существующим антигеном или Fc-рецепторами, получали мутантные ζ слитые белки, которые не содержали либо внутримембранного Asp, либо внутримембранного Cys или обоих. Проточная цитометрия показала, что интенсивность поверхностной экспрессии различными мутантными химерами не была ощутимо отличающейся от немутированного предшественника, и эксперименты с иммунопреципитацией показали, что общая экспрессия химерами была одинаковой. Как и ожидалось, было обнаружено, что мутантные химеры, не имеющие трансмембранных остатков цистеина, не образуют связанных дисульфидными связями димеров. Две мутантные химеры, не имеющие Asp, были неспособны поддерживать поверхностную экспрессию CD16TM, тогда как мономерные химеры, не имеющие Cys, но несущие Asp, делали возможной ко-экспрессию CD16TM, но при более низкой эффективности, чем в случае родительского димера (фиг. 3).
ПРИМЕР V
Мутантные рецепторы сохраняют способность инициировать ответную реакцию кальция
Для определения, позволит ли сшивание слитых белков накопление свободного внутриклеточного кальция, подобное накоплению, имеющему место в случае T-клеточного рецептора антигенов, клетки T-клеточной лейкозной линии человека, Jurkat E6 (АТСС Accession N TIB 152, American Type Culture Collection, Rockville, MD) инфицировали рекомбинантами вируса коровьей оспы и измеряли относительную концентрацию кальция в цитоплазме после сшивания внеклеточного домена с антителами. Измерения проточной цитометрии проводили с клетками, загруженными чувствительным к кальцию красителем Indo-1 (Grynkiewicz et al., J. Biol. Chem. 260:3340-3450 (1985); Rabinovitch et al., J. limnunol. 137: 952-961 (1986)). Фиг. 4A-D показывает результаты экспериментов с выделением кальция клетками, инфицированными CD4: ζ и Asp- и Cys- мутантами ζ. Сшивание химер воспроизводимо увеличивало внутриклеточные количества кальция. CD4: η и CD4: γ также делали возможным накопление внутриклеточного кальция в инфицированных клетках. Клетки Jurkat экспрессируют низкие уровни CD4 на клеточной поверхности, однако сшивание нативного CD4 в присутствии или в отсутствие CD16: ζ не изменяет уровней внутриклеточного кальция (фиг. 4A-B).
ПРИМЕР VI
Химеры CD4: ζ,η и γ медиируют цитолиз клеток-мишеней, экспрессирующих gp 120/41 ВИЧ
Для определения, запускают ли химерные рецепторы цитолитические эффекторные программы, была создана система, основанная на узнавании CD4 комплекса gp120/gp41 оболочки ВИЧ. Клетки HeLa инфицировали рекомбинантными вирусами коровьей оспы, экспрессирующими gp120/gp41 (Chakrabarti et al., Nature 320: 535- 537 (1986); Earl et al., J. Virol. 64:2448-2451 (1990)), и метили 51Cr. Меченые клетки инкубировали с клетками из линии аллоспецифеческих (CD8+, CD4-) цитотоксических T-лимфоцитов, которые были инфицированы рекомбинантами вируса коровьей оспы, экспрессирующими химеры CD4: ζ CD4: η или CD4: γ или двойные мутантные химеры CD4: ζ CysllGly: Asp15Gly. Фиг. 5A-C показывает, что клетки HeLa, экспрессирующие gp120/41, специфически лизировались цитотоксическими T-лимфоцитами (CTL), экспрессирующими химеры CD4. Неинфицированные клетки HeLa не были мишенями CTL, несущими химеры CD4: ζ, а клетки HeLa, экспрессирующие gp120/41 не узнавались неинфицированными CTL. Для сравнения эффективности разных химер отношения эффектор: мишень корректировали на фракцию CTL, экспрессирующих химеры CD4, и на фракцию клеток NeLa, экспрессирующих gp120/41, измеренных проточной цитометрией. Фиг. 5C показывает цитометрический анализ экспрессии CD4 CTL, используемыми в эксперименте по цитолизу, показанному в фиг. 5A и 5B. Хотя средняя плотность поверхностного CD4: ζ значительно превышала среднюю плотность CD4: η, цитолитические эффективности клеток, экспрессирующих любую форму, были сходными. При корректировании на фракцию клеток-мишеней, экспрессирующих gp20, эффективность цитолиза, медиируемого белками CD4; ζ и CD4: η, была сравнима с наивысшими эффективностями, сообщенными для пар мишень: эффектор специфического T-клеточного рецептора (среднее отношение эффектор: мишень для 50% выделения T-клетками, экспрессирующими CD4: ζ, было 1.9 ± 0.99, n = 10. Слитый белок CD4: γ был менее активным, как и слитый белок CD4: ζ, не имеющий трансмембранных остатков Asp и Cys. Однако в обоих случаях наблюдали значительный цитолиз (фиг. 5B-C).
Для контролирования того, что инфекция вирусом коровьей оспы может усиливать артефактное узнавание CTL, сходные эксперименты по цитолизу выполняли с клетками-мишенями, инфицированными рекомбинантами вируса коровьей оспы, экспрессирующими связанную с фосфатидилинозитом форму CD16 (CD16PI) и меченными 51Cr, и с CTL, инфицированными контрольными рекомбинантами, экспрессирующими либо CD16PI, либо CD16: ζ Фиг. 6A показывает, что T-клетки, экспрессирующие не-CD4-химеры, не узнают клетки HeLa, экспрессирующие другие поверхностные белки, кодируемые вирусом коровьей оспы. Кроме того, CTL, экспрессирующие нехимерный CD4, не лизируют значительно клетки HeLa, экспрессирующие gp120/41 (фиг. 6A).
ПРИМЕР VII
Клетки, несущие MHC класса II, не являются мишенями химер
Считают, что CD4 взаимодействует с неполиморфной последовательностью, экспрессируемой антигеном MHC класса II (Gay et al. Nature 328:626-629 (1987); SlecKman et al. , Nature 328:351-353 (1987)). Хотя специфическое взаимодействие между CD4 и антигеном класса II никогда не было документировано с очищенными белками, при определенных условиях можно продемонстрировать адгезию между клетками, экспрессирующими CD4, и клетками, экспрессирующими молекулы класса II (Doyle et al. Nature 330:256-259 (1987); Clayton et al. , J. Exp. Med. 172:1243-1253 (1990); Lamarre et al., Science 245: 743-746 (1989)). Затем нужно было испытать, можно ли обнаружить убивание клеток, несущих антиген класса II. Фиг. 6B показывает, что нет специфического цитолиза, направленного химерой CD4: ζ против B-клеточной линии Raji, которая экспрессирует большие количества антигена класса II. Хотя и наблюдали умеренный цитолиз (приблизительно 5%), не содержащий антигена класса II мутант Raji, RJ2.2.5 (Accolla, J. Exp. Med. 157:1053-1058 (1983)) обнаруживает сходную чувствительность, что и клетки Raji, инкубированные с неинфицированными T-клетками.
ПРИМЕР VIII
Требования к последовательностям для индукции цитолиза химерной T-клеточный антиген/дзета цепь Fc-рецептора
Хотя химеры между CD4 и ζ могут вооружить цитотоксические T-лимфоциты (CTL) для убивания клеток-мишеней, экспрессирующих gp120 ВИЧ, были предприняты поиски альтернативы для CD4, чтобы однозначно сравнить свойства дзета-химер, вводимых в линии T-клеток человека. Такие линии могут экспрессировать CD4, затрудняя специфическое определение взаимоотношения между типом или степенью мобилизации кальция и цитотоксическим потенциалом различных химер. Для преодоления этого были созданы химеры между ζ и CD16, в которых внеклеточный домен CD16 присоединен к трансмембранной и внутриклеточной последовательностям ζ (фиг. 7A). Эти генные слияния вводили в экспрессионную плазмиду вирус коровьей оспы, несущую ген  Е. coli в качестве селектирующего маркера, и встраивали в геном штамма WR вируса коровьей оспы гомологичной рекомбинацией и отбором на рост в микофенольной кислоте (Falkner and Moss, J. Virol. 62:1849 (1988); Boyle and Coupar, Gene 65:123 (1988)).
Е. coli в качестве селектирующего маркера, и встраивали в геном штамма WR вируса коровьей оспы гомологичной рекомбинацией и отбором на рост в микофенольной кислоте (Falkner and Moss, J. Virol. 62:1849 (1988); Boyle and Coupar, Gene 65:123 (1988)).
T-клеточные линии инфицировали рекомбинантами вируса коровьей оспы и измеряли относительные концентрации свободного иона кальция в цитоплазме после сшивания внеклеточных доменов с антителами. Как спектрофлуориметрические (общая популяция), так и проточно-цитометрические (отдельные клетки) измерения проводили с клетками, загруженными красителем Indo-I (Glynkiewicz et al. , J. Biol. Chem. 260:3440 (1985); Rabinovitch et al., J.Immunol. 137:952 (1986)). Фиг. 7B показывает анализ данных, собранных от клеток T-клеточной лейкозной линии человека, Jurkat, инфицированных рекомбинантами вируса коровьей оспы, экспрессирующих слитый белок CD16: ζ. Сшивание этих химер воспроизводимо увеличивало внутриклеточный кальций, тогда как подобная обработка клеток, экспрессирующих нехимерный CD16, дала небольшой эффект или отсутствие эффекта. При экспрессии этой химеры в мутантных клеточных линиях, не имеющих рецептора антигена, либо REX33A (Breitmeyer et al., J. Immunol. 138: 726 (1987); Sancho et al., J. Biol. Chem. 264:20760 (1989)), либо Jurkat мутанте JRT3. T3.5 (Weiss et al. , J. Immunol. 135:123 (1984)), наблюдали сильный ответ на сшитый с антителом CD16. Подобные данные были получены на мутантной клеточной линии REX20A (Breitmeyer et al., supra, 1987; Blumberg et al., J. Biol. Chem. 265:14036 (1990)) и CD3/Ti негативном мутанте клеток Jurkat, полученном в этой лаборатории. Инфицирование рекомбинантами, экспрессирующими CD16: ζ, не восстанавливало ответной реакции на aнти-CD3 антитела, показывая, что этот слитый белок не действовал путем спасения внутриклеточных цепей комплекса CD3.
Для оценки способности этих химер изменять направленность клеточно-опосредованного иммунитета CTL инфицировали рекомбинантами вируса коровьей оспы, экспрессирующими химеры CD16, и использовали для специфического лизиса гибридомных клеток, экспрессирующих мембрано-связанные анти-CD16 антитела. Этот анализ является продолжением анализа гибридомной цитотоксичности, разработанного первоначально для анализа эффекторных механизмов клеток, несущих Fc-рецепторы (Graziano and Fanger, J. Immunol. 138:945, 1987; Graziano and Fanger, J. Immunol. 129: 35-36, 1987; Shen et al., Mol. Immunol, 26:959, 1989; Fander et al., Immunol. Today 10:92, 1989). Фиг. 8B показывает, что экспрессия CD16: ζ цитотоксических T-лимфоцитах позволяет CTL с рецептором убивать 3G8 (анти-CD16; Fleit et al., Proc. Nail. Acad. Sci. USA 79:3275, 1982) гибридомные клетки, тогда как CTL, экспрессирующие соединенную с фосфатидилинозитом форму CD16, являются неактивными. CTL, соединенные с CD16: ζ, также не убивают гибридомные клетки, экспрессирующие не относящиеся к рассматриваемым антитела.
Для идентификации минимальных последовательностей ζ, необходимых для цитолиза, получали ряд делеционных мутантов, в которых последовательно все большую часть внутриклеточного домена ζ удаляли из карбоксил-конца (фиг. 8A). Большую часть внутриклеточного домена дзета можно было удалить с малыми последствиями для цитолитического потенциала; полноразмерная химера CD16: ζ была по существу равна по эффективности химере, делетированной до остатка 65, CD16: ζ Asp66* (фиг. 8B). Существенное уменьшение цитотоксичности наблюдали при делеции до остатка 59 ζ (химера CD16: ζ Glu60*) и дальнейшая делеция до остатка 50 приводила к слегка более низкой активности. Однако полной потери активности не наблюдали, даже при уменьшении внутриклеточного домена до трех остатков трансмембранного якоря (фиг. 8B).
Поскольку ζ является связанным дисульфидной связью димером, одним из объяснений сохранения цитолитической активности было то, что эндогенный ζ образовывал гетеродимеры с химерной ζ делецией, тем самым восстанавливая активность. Для проверки этой идеи остатки 11 и 15 ζ были заменены (вместо Asp и Cys был введен Gly) (CysllGly/Asp15Gly) и иммунопреципитации проводили следующим образом. Приблизительно 2 • 106 клеток CVI инфицировали в течение 1 часа в бессывороточной среде DME рекомбинантным вирусом коровьей оспы при множественности заражения (moi) по меньшей мере 10. Спустя 6-8 часов после инфицирования клетки отделяли от чашек при помощи PBS/1 мМ ЭДТА и поверхностно метили 0.2 мКи 125I на 2 • 106 клеток с применением лактопероксидазы и H2O2 по методу Кларка и Айнфельда (Laucocyte Typing II, рр 155-167, Springer-Verlag, NY, 1986). Меченые клетки собирали центрифугированием и лизировали в 1% NP-40, 0.1% ДСН, 0.15 М NaCl, 0.05 М Трис, pH 8.0, 5 мМ MgCl2, 5 мМ KCl, 0.2 М иодацетамид и 1 мМ PMSF. Ядра удаляли центрифугированием и белки CD16 иммунопреципитировали агарозой с антителом 3G8 (Fleit et al., supra, 1982; Medarex) и с антимышиными lgG (Cappel, Durham, NC). Пробы подвергали электрофорезу на 8% полиакриламидном геле с ДСН при невосстанавливающих условиях или на 10% геле при восстанавливающих условиях. Эти иммунопреципитации подтвердили, что химера CD16: ζ CysllGly/Asp15Gly не ассоциирована в связанные дисульфидной связью димерные структуры.
Также тестировали цитолитическую активность мутантных рецепторов. Мутированная химера, делетированная до остатка 65 (CD16: ζ CysllGly/Asp15Gly/Asp66*), была, в зависимости от условий тестирования, в 2-8 раз менее активна в анализе цитолиза, чем сравнимая немутированная химера (CD16: ζ Asp66*), которая была обычно примерно в 2 раза активнее или не отличима по активности от CD16: ζ (фиг. 9B). Снижение активности мутантных химер сравнимо со снижением, наблюдаемым с химерами CD4 сходной структуры (см. выше) и, наиболее вероятно, связано с более низкой эффективностью мономеров ζ по сравнению с димерами. В противоположность этому Asp-, Cys- мутированная химера, делетированная до остатка 59, не имела цитолитической активности, подтверждая гипотезу о том, что ассоциация с другими цепями, медиируемая трансмембранными остатками Cys и/или Asp, была ответственна за слабую стойкость цитолитической активности при делеции амино-конца далее, чем до остатка 65.
Исследования с применением проточной цитометрии показали, что делеционные мутанты, не имеющие трансмембранных остатков Asp и Cys, вес еще могли ускорять увеличение в количестве свободного иона кальция внутри клетки в ответ на сшивание с антителами в TCR мутантной Jurkat клеточной линии (фиг. 9D). Подобные результаты были получены для химер, экспрессируемых в родительской линии Jurkat. В случае CD16: ζ CysllGly/Asp15Gly/Glu60* эти данные показывают, что способность медиировать "кальциевый" ответ может быть мутационно отделена от способности поддерживать цитолиз.
Для определенного исключения возможного вклада трансмембранных остатков ζ трансмембранные и первые 17 цитоплазматических остатков ζ были заменены последовательностями, кодирующими проходящие через мембрану и первые 14 или первые 17 цитоплазматических остатков CD5- или CD-антигенов, соответственно (фиг. 9А). Полученные слитые из трех частей белки CD16:5: ζ (48-65) и CD16: 7: ζ (48-65) не образовали связанных дисульфидом димеров, которые образуют более простые химеры CD16: ζ , поскольку они не содержат остатка цистеина в трансмембранном домене ζ . Обе состоящие из трех частей химеры были способны мобилизовать кальций в линиях клеток Jurkat и TCR- -клеток (фиг. 9D) и увеличивать цитолитический ответ в CTL (фиг. 9C и не представленные данные). Однако укорочение ζ части до остатка 59 в химере CD16: ζ (48-59) отменяет способность состоящего из трех частей слитого белка вызывать реакцию мобилизации кальция в TCR позитивных или негативных клетках Jurkat или цитолиз в зрелых CTL (фиг. 9C и 9D и не представленные данные).
Для исследования вкладов отдельных остатков внутри мотива из 18 остатков мы получили ряд мутантных вариантов путем сайтспецифического мутагенеза и оценили их способность медиировать направленное рецептором убивание в условиях низкой (фиг. 10A и 10D) или высокой (фиг. 10B и 10E) экспрессии химерного рецептора. Фиг. 10A-F показывает, что хотя ряд относительно консервативных замещений (т.е. замещений кислых остатков их родственными амидами или замены тирозина фенилаланином) в остатках 59-63 дал умеренное снижение цитолитической эффективности, в целом эти варианты сохраняли способность мобилизовать кальций. Однако вместе взятые эти остатки содержат важный субмотив ввиду того, что их делеция исключает цитолитическую активность. Превращение Tyr 62 либо в Phe, либо в Ser исключало как цитолитическую ответную реакцию, так и реакцию мобилизации кальция. При амино-конце сегмента из 18 остатков замена Tyr 51 на Phe отменяла как мобилизацию кальция, так и цитолитическую активность, тогда как замена Leu на Ser при положении 50 исключала ответную реакцию кальция, тогда как лишь частично ослабляла цитолиз. Без привязывания к конкретной гипотезе, предполагается, что неспособность мутанта Leu50Ser мобилизовать кальций в кратковременных проточноцитометрических анализах неполно отражает его способность медиировать значительное увеличение в свободном внутриклеточном кальции в более длинном промежутке времени анализа цитолиза. Однако нечувствительная к кальцию цитолитическая активность сообщалась для некоторых цитолитических T-клеточных линий, и не исключена возможность, что подобное явление лежит в основе описанных здесь результатов. Замена Asn 48 на Ser ослабляла цитотоксичность в некоторых экспериментах, в то время как в других экспериментах оказывала слабое действие.
Для исследования потенциальной роли избыточных элементов последовательности внутриклеточный домен ζ делили на три сегмента, охватывающие остатки 33-65, 71-104 и 104-138. Каждый из этих сегментов присоединяли к химере CD16: CD7 посредством сайта MluI, введенным дистально по отношению к основным мембранным якорным последовательностям внутриклеточного домена CD7 (см. ниже; Фиг. 11A). Сравнение цитолитической эффективности этих трех элементов показало, что они по существу эквипотентны (фиг. 11B). Сравнение последовательностей (фиг. 11A) показывает, что второй мотив несет 11 остатков между тирозинами, тогда как первый и второй мотивы несут 10.
Хотя точная оценка процесса T-клеточной активации не была сделана, ясно, что агрегация рецептора антигена или рецепторных химер, несущих внутриклеточные последовательности ζ, запускает мобилизацию кальция, выделение цитокина и гранул и появление поверхностных маркеров активации. Активный сайт ζ, короткая линейная пептидная последовательность, возможно, слишком маленькая, чтобы иметь собственную ферментативную активность, по-видимому, взаимодействует с одним или, самое большее, с несколькими белками для медиирования клеточной активации. Ясно также, что мобилизация свободного кальция сама по себе недостаточна для клеточной активации, т.к. способность медиировать цитолиз можно при помощи мутаций отделить от способности медиировать накопление кальция.
Как показано здесь, добавление 18 остатков из внутриклеточного домена ζ к трансмембранному и внутриклеточному домену двух неродственных белков позволяет полученным химерам заново направлять цитолитическую активность против клеток-мишеней, которые связываются с внеклеточной частью слитых белков. Хотя химеры, несущие мотив из 18 остатков, приблизительно в 8 раз менее активны, чем химеры на основе полноразмерного ζ, пониженная активность может быть отнесена за счет потери трансмембранных взаимодействий, которые обычно позволяют ζ дикого типа образовать связанные дисульфидными связями димеры. Т.е. делеционные конструкции ζ , которые имеют такой же карбоксил-конец, что и этот мотив, и не имеют трансмембранных остатков Cys и Asp, обычно обнаруживают несколько меньшую активность, чем химеры, несущие только мотив из 18 остатков.
Элемент цитолитической компетентности, на котором мы сконцентрировали внимание, имеет два тирозина и не имеет остатков серина или треонина, исключающих возможные вклады фосфорилирования в активность. Мутация любого тирозина разрушает активность, и хотя предварительные эксперименты не показывают значительного фосфорилирования тирозина после сшивания с химерными поверхностными антигенами, несущими мотив из 18 остатков, возможное участие такого фосфорилирования при низком уровне не может быть исключено. Кроме отмеченных эффектов в отношении двух остатков тирозина, ряд замен аминокислот при амино- и карбокси- конце этого мотива ослабляет активность в условиях низкой плотности рецептора.
Последовательности, сходные с активным мотивом ζ, могут быть обнаружены в цитоплазматических доменах некоторых других трансмембранных белков, в том числе CD3 δ- и γ-молекулах, белков mb1 и B29, находящихся на поверхности клеток и связанных с IgM, и β- и γ-цепей высокоаффинного рецептора IgE, Fc ∈ RI (Reth, Nature 338:383, 1989). Хотя функция этих последовательностей неясна, при их эффективной экспрессии каждый из них может быть способным к автономной активации T-клеток, и такая активность может объяснять остаточную ответную реакцию TCR, наблюдаемую в дзета-негативной мутантной клеточной линии (Sussman et al. Cell 52:85, 1988).
Сам дзета несет три таких последовательности, приблизительно на одинаковом расстоянии одна от другой, и грубое рассечение на три части внутриклеточного домена показывает, что каждый из них способен инициировать цитолитический ответ. η образованная сплайсингом изоформа ζ (Jin et. al., supra, 1990; Clayton et al., Proc. Natl. Acad. Sci. USA 88:5202, 1991), не имеет карбоксил-половины третьего мотива. Поскольку удаление карбоксил-половины первого мотива уничтожает активность, кажется вероятным, что основная часть биологической эффективности η может быть связана с первыми двумя мотивами. Хотя по различным измерениям η так же активен, как ζ , в усилении медиируемого антигеном выделения цитокинов (Bauer et al., Proc. Natl. Acad. Sci. USA 88:3842, 1991) или изменении направления цитолиза (см. выше), η не фосфорилируется в ответ на стимуляцию рецептора (Bauer et al., supra, 1991). Таким образом, либо для фосфорилирования требуются все три мотива, либо третий мотив является предпочтительным субстратом для неидентифицированной пока тирозинкиназы.
ПРИМЕР IX
Трансдукция (передача) цитолитического сигнала Fc- рецептором человека
Для оценки действий различных подтипов Fc-рецептора человека были созданы химерные молекулы, в которых внеклеточный домен антигенов CD4, CD5 или CD16 человека были соединены с трансмембранным и внутриклеточным доменами подтипов B1, B2 и С FcR11 γ A (номенклатура Ravetch and Kinet, Ann. Rev. Immunol. 9:457, 1991). В частности, последовательности кДНК, соответствующие трансмембранному и цитоплазматическому доменам описанных ранее изоформ B1 и B2 FcR11A, амплифицировали из предсуществующего клона PC23 или из библиотеки кДНК минадин человека (сконструированной стандартными способами) с применением следующих синтетических олигонуклеотидных праймеров:
CCC GGA ТСС CAG CAT GGG CAG CTC TT (SEQ ID N 18; FcR11 А прямой);
CGC GGG GCG GCC GCT TTA GTT ATT ACT GTT GAC ATG GTC GTT (SEQ ID N 19; FcR11 А обратный);
GCG GGG GGA ТСС CAC TGT CCA AGC TCC CAG CTC TTC ACC G (SEQ ID N 20; FcR11 B1 и FcR11 B2 обратный);
GCG GGG GCG GCC GCC TAA ATA CGG TTC TGG TC (SEQ ID N 21; FcR11 B1 и FcR11 B2 обратный).
Эти праймеры содержали сайты расщепления для ферментов BamHI и NotI, соответственно, выступающие 6 остатков от 5'-конца. За сайтом NotI непосредственно следовал антисмысловой стоп-кодон, либо CTA, либо TTA. Все праймеры содержали 18 или более остатков, комплементарных 5'- и 3'-концам желаемых фрагментов. Фрагмент кДНК, соответствующий цитоплазматическому домену FcR11 γ C, который отличается от изоформы 11A только по одной аминокислоте (L вместо P при остатке 268), был получен сайт-специфическим мутагенезом при помощи перекрывающейся ПЦР с использованием праймеров, имеющих последовательности:
TCA GAA AGA GAC AAC CTG AAG AAA CCA ACA А (SEQ ID N 22) и
TTG TTG GTT TCT TCA GGT TGT GTC TTT CTG А (SEQ ID N 23).
Эти фрагменты PCR (ПЦР) встраивали в экспрессирующие векторы (вирусы коровьей оспы), которые содержали внеклеточные домены CD16 или CD4 соответственно, и затем встраивали в вирус коровьей оспы дикого типа путем рекомбинации в локусе тимидинкиназы, с использованием отбора на ко-интеграцию  Е. coli для облегчения идентификации желаемых рекомбинантов. Идентичность всех изоформ (показанных в фиг. 12) подтверждали дидезокси-секвенированием.
Е. coli для облегчения идентификации желаемых рекомбинантов. Идентичность всех изоформ (показанных в фиг. 12) подтверждали дидезокси-секвенированием.
Получение химерных рецепторных белков далее подтверждалось исследованиями иммунопреципитации. Приблизительно 107 клеток JRT3.T3.5 инфицировали в течение 1 часа в бессывороточной среде IMDM рекомбинантным вирусом коровьей оспы при множественности заражения (moi) по меньшей мере 10. Спустя 12 часов после инфекции клетки собирали и поверхностно метили 0.5 мКи 125I на 107 клеток с применением лактопероксидазно/глюкозооксидазного метода (Clark and Einfeld, supra). Меченые клетки собирали центрифугированием и лизировали 1% NP-40, 0.1 мМ MgCl2, 5 мМ KCl, 0.2 М иодацетамид и 1 мМ PMSF. Ядра удаляли центрифугированием и слитые белки CD16 иммунопреципитировали агарозой с антителами 4G8 и антимышиным IgG. Пробы подвергали электрофорезу при восстанавливающих условиях. Все иммунопреципитированные химерные рецепторные молекулы имели ожидаемые молекулярные массы.
Для испытания способности химерных рецепторов опосредовать увеличение в цитоплазматическом свободном ионе кальция эти рекомбинантные вирусы использовали для инфицирования TCR- мутантной клеточной линии Jurkat JRT3.T3.5 (описанной выше) и цитоплазматический свободный кальций измеряли в этих клетках (как описано здесь) после сшивания внеклеточных доменов рецептора с моноклональным телом 3G8 или Leu-3A (как описано здесь). Эти эксперименты обнаружили, что внутриклеточные домены FcR γ 11 А и С были способны опосредовать увеличение в цитоплазматическом свободном ионе кальция после сшивания внеклеточных доменов, тогда как внутриклеточные домены FcR γ 11 B1 и B2 были неактивными при сравнимых условиях (фиг. 13A и 13B). Гибридомы CD4, CD5 и CD16 FcR γ 11 А разделяли фактически равную способность усиливать кальциевый ответ (фиг. 13A-B). Другие клеточные линии, как моноцитарной, так и лимфоцитарной линий дифференцировки, были способны отвечать на сигнал, инициируемый сшиванием внеклеточных доменов.
Для выяснения участия различных внутриклеточных доменов FcR γ 11 в цитолизе цитотоксические T-лимфоциты человека (CTR) инфицировали рекомбинантами вируса коровьей оспы, экспрессирующими химеры CD16:FcR γ 11 А, B1, B2 и С. Затем инфицированные клетки ко-культивировали с нагруженными 51Cr гибридомными клетками (т.е. клетками 3G8 10-2), которые экспрессировали на клеточной поверхности антитело к CD16. В этом анализе CTL, несущие химеру CD16, убивали гибридомные клетки-мишени (позволяя выделяться свободному 51Cr), если внеклеточный домен химеры был соединен с внутриклеточным сегментом, способным активировать эффекторную программу лимфоцитов; этот анализ цитолиза описан подробно ниже. Фиг. 14A показывает, что CTL, соединенные с CD16: FcR γ 11A и С, но не FcR γ 11 B1 или B2, способны лизировать клетки-мишени, экспрессирующие поверхностные анти-CD16 антитела.
Для исключения возможности, что этот специфический цитолиз был связан с взаимодействием с CD16-частью, проводили эксперименты по цитолизу, в которых внутриклеточные домены FcR11 были присоединены к внеклеточному домену CD4. В этом случае клетками-мишенями были клетки HeLa, экспрессирующие белки gp120/41 оболочки ВИЧ (в частности, клетки HeLa, инфицированные вектором вируса коровьей оспы vPE16 (доступным из National Institute of Allergy and Infections Disease AIDS Depository, Bethesda, MD). Как и в системе CD16, клетки-мишени, экспрессирующие антигены оболочки ВИЧ, были чувствительны к лизису T-клетками, экспрессирующими химеру CD4:FcR γ 11 А, но не FcR γ 11 B1 или B2 (фиг. 14B).
Внутриклеточные домены FcR γ 11 А и С не имеют поддающейся оценке гомологии последовательности с каким-либо другим белком, в том числе с членами расширенного семейства FcR γ/TCR ζ. Для определения элементов последовательности, ответственных за индукцию цитолиза, получали 5'- и 3'-делеции кодирующих внутриклеточный домен последовательностей (описанных ниже и показанных на фиг. 15A) и их оценивали на эффективность в мобилизации кальция и цитолизе (как описано здесь). В экспериментах, в которых удаляли амино-концевую часть внутриклеточного домена, трансмембранный домен FcR γ 11 заменяли трансмембранным доменом неродственного антигена CD7 для исключения возможного вклада взаимодействий, опосредованных пересекающим мембрану доменом.
Фиг. 15B и 15C показывают, что удаление 14 карбоксил-концевых остатков, в том числе тирозина 298, приводило к полной потере цитолитической способности и существенному уменьшению потенциала мобилизации кальция. Далее делеция до положения перед тирозином 282 дала идентичный фенотип (фиг. 15B и 15C). Делеция из N-конца внутриклеточного домена до остатка 268 не оказывала значительного действия ни на профиль кальция, ни на цитолитическую активность, тогда как делеция до остатка 275 заметно ослабляла выделение свободного кальция, но мало влияла на цитолиз (фиг. 15D и 15E). Дальнейшая делеция, до остатка 282, дала хвосты FcR γ 11, которые не обладали способностью ни мобилизовать кальций, ни запускать цитолиз (фиг. 15D и 15E). "Активный элемент", определяемый этими грубыми измерениями, является относительно большим (36 аминокислот) и содержит два тирозина, разделенных 16 остатками.
ПРИМЕР X
Направленный цитолиз лимфоцитами, несущими химерные CD4-рецепторы, которые не поддерживают инфекцию
Как обсуждалось выше, могут быть сконструированы эффекторные молекулы, которые направляют цитолитическую активность CTL MHC-независимым образом. Например, химера, составленная из внеклеточного домена CD4, слитого с ζ -цепью в клоне CTL человека, WH3, специфически убивает клетки-мишени, обнаруживающие поверхностный гликопротеин оболочки ВИЧ-1, gp120. Однако, поскольку внеклеточный домен молекулы CD4 придает чувствительность к инфекции ВИЧ, такие CTL с перенесенными на них иммуноглобулиновыми рецепторами могут стать мишенями для вируса, что приводит к уменьшению их активности (Dalg-leish et al. Nature 312:767 (1984); Klatzman et al. Nature 312:767 (1984)). Для предотвращения такого результата были сконструированы эффекторные молекулы на основе CD4, которые эффективны в специфическом нацеливании на инфицированные ВИЧ клетки для клеточно- опосредованного убивания, но которые не придают чувствительность к инфекции ВИЧ.
Слитый белок из трех частей был создан генетической аппозицией внеклеточного домена CD4 (фиг. 23) к шарнирной области, второму и третьему константным доменам тяжелой цепи lgG1 человека (Zettlmeissl et al., DNA Cell Biol. 9: 347 (1990)) (фиг. 25), которые в этом случае были присоединены к части первого трансмембранного экзона мембрано-связанного lgGI человека, за которым следовала часть антигена CD7 человека, состоящая из последовательностей между единственным Ig-подобным доменом и стоп- последовательностью передачи после трансмембранного домена (Aruffo and Seed, EMBO J. 6:3313 (1987)) (фиг. 26). Первичная аминокислотная последовательность внеклеточной части сегмента CD7 состояла из богатого пролином района, предполагающего стеблеподобную структуру, которая выдвигает Ig-подобный домен прочь от поверхности клетки (Aruffo and Seed EMBO J. 6:3313 (1987)) (фиг. 26). Рекомбинантные вирусы коровьей оспы готовили для экспрессии этого слитого белка и родственных химер, как описано здесь. В частности, рекомбинантные вирусы коровьей оспы получали гомологичной рекомбинацией в клетках CV-1. По меньшей мере два цикла визуализации бляшек выполняли для каждого штамма перед получением штаммов с высоким титром в клетках CV-1.
Химера из трех частей (CD4(D1-D4):lg:CD7) (фиг. 20, молекула "А") обнаружила эффективную поверхностную экспрессию и была испытана на способность действовать как рецептор ВИЧ в тесте образования синцитиев на основе вируса коровьей оспы (Lifson et al. Nature 323:725 (1986)); Ashorn et al., J. Virol. 64:2149 (1990)). Клетки HeLa, инфицированные peкомбинантным вирусом коровьей оспы (VPE16), кодирующим гликопротеин оболочки ВИЧ-1 (Earl et al., J. Virol. 64: 2448 (1990)), ко-культивировали с клетками HeLa, инфицированными либо CD4, CD4: ζ, либо CD4(D1-D4):Iq:CD7. Чашки 6 см с клетками HeLa (ATCC, Rockville, MD) при 50% конфлюэнтности инфицировали в бессывороточной среде в течение 1 часа при приблизительной множественности заражения (moi) 10. Клетки инкубировали еще в течение 5-6 часов в полной среде и затем отделяли содержащим фосфат солевым раствором (PBS) с 1 мМ ЭДТА. Клетки, экспрессирующие оболочку и химеру CD4, смешивали в отношении 1:1 и пересевали в чашки 6 см с полной средой. Синцитии оценивали в баллах при 6-8 часа совместного культивирования и фотографировали.
Ко-культуры CD4 и vPE16 привели к образованию легко детектируемых многоядерных гигантских клеток. Также, химера, состоящая из внеклеточного домена CD4, слитого с ζ-цепью TCR (фиг. 27), (CD4: ζ) была способна опосредовать образование синцитиев, тогда как клетки, экспрессирующие CD4(D1-D4): Ig:CD7, не показали признаков клеточного слияния. Мы также испытывали конструкцию, экспрессирующую только первый и второй домены CD4 (фиг. 24), CD4(D1,D2):Ig: CD7 (фиг. 20, молекула "В"), так как в другой связи было показано, что амино-концевые два домена CD4 являются необходимыми для инфективности ВИЧ (Landau et al. Nature 334:159 (1988)). Эта молекула также показала невосприимчивость к индуцируемому ВИЧ образованию синцитиев. Исследованию связывания с растворимым 125I-меченым gpl20 показали, что как CD4(D1-D4):Ig:CD7, так и CD4(D1,D2):Ig:CD7 имели ненарушенную аффинность в отношении gp120.
Затем мы определили, будут ли способны химерные молекулы, которые противостояли образованию синцитиев, к нацеливанию убивания клеток при обеспечении их триггерной частью, как описано здесь. Мы слили внутриклеточный домен ζ (фиг. 27) с 3'-концом CD4(D1-D4):Ig:CD7 и CD4(D1,D2):Ig:CD7 и приготовили соответствующие вирусы коровьей оспы. Эти конструкции, CD4(D1-D4):Ig:CD7: ζ CD4(D1, D2): Ig:CD7: ζ (фиг. 20, молекулы "С" и "D") экспрессировали в клоне WH3 CTL человека и тестировали на их способность нацеливаться и убивать клетки HeLa, экспрессирующие поверхностный гликопротеин оболочки ВИЧ (при помощи описанных здесь способов). Фиг. 21 показывает, что внутриклеточный домен ζ, слитый либо с CD4-(D1-D4):Ig:CD7, либо с CD4(D1,D2):Ig:CD7, может сообщать способность убивания; конструкции, не имеющие ζ - цепи, были неспособны опосредовать эту активность. CD4: ζ, положительный контроль, опосредовал слегка более эффективную цитотоксичность, а CD4(D1,D2):Ig:CD7: ζ - несколько менее эффективную цитотоксичность, чем CD4 (D1-D4):Ig:CD7: ζ (фиг. 21). Однако ясно, что как CD4(D1-D4):Ig:CD7: ζ, так и CD4(D1,D2):Ig:CD7:ζ имеют способность медиировать специфическое убивание клеток, экспрессирующих белки оболочки ВИЧ на их поверхности. Эти химеры, состоящие из четырех частей, стойко неспособны медиировать образование синцитиев в тесте на основе вируса коровьей оспы. Мы также продемонстрировали, что единственного ζ -мотива, показанного в фиг. 11А, достаточно для придания цитолитической активности химере CD4(D1-D4).
Эксперименты по радиоиммунопреципитации показали, что слитые молекулы были преимущественно (если не всегда) димерами. В этих экспериментах использовали гранулы протеин-А-агарозы для иммунопреципитации солюбилизированного экстракта метаболически меченных клеток HeLa, инфицированных рекомбинантным вирусом коровьей оспы, экспрессирующим химеры CD4-(D1-D4):Ig:CD7:ζ и CD4(D1, D2): Ig: CD7: ζ. Иммунопреципитированный материал фракционировали электрофорезом в полиакриламидном геле при восстанавливающих и невосстанавливающих условиях. В частности, приблизительно 5 • 106 клеток HeLa-S3 инфицировали, как описано выше для vPE16, подходящим штаммом вируса коровьей оспы. Клетки были метаболически меченными 200 мкКи/мл Tran35S- Меткой (ICN Radiochemicals, Irvine, CA) в течение 6-8 часов в не содержащей цистеина и метионина среде. Их отделяли при помощи PBS, содержащего 1 мМ ЭДТА. Затем клетки осаждали и лизировали в 150 мМ NaCl, 50 мМ Трисе pH 7.5, 5 мМ ЭДТА, 0.5% NP-40, 0.1% ДСН, 5 мМ ЭДТА, 1 мМ PMSF.
После удаления ядер центрифугированием одну пятую каждого клеточного экстракта адсорбировали на промытых гранулах конъюгированной с протеином-А агарозы в течение 2 часов при 4oC. Затем гранулы промывали PBS, содержащим 1% NP-40 и элюировали в буфере для проб, содержащем ДСН, в присутствии или в отсутствии меркаптоэтанола. Результаты этих экспериментов показали, что большая часть иммунопреципитированных химер CD4(D1,D2):Ig:CD7: ζ мигрировала в виде димеров ожидаемой молекулярной массы при невосстанавливающих условиях.
Для прямой оценки способности клеток, экспрессирующих слитые молекулы CD4, поддерживать инфекцию ВИЧ, мы проводили долгосрочные исследования инфективности на трансфектантах, экспрессирующих CD4(D1-D4):Ig:CD7 и CD4:(D1, D2):Ig:CD7. Стабильные трансфектанты CD4(D1-D4):lg:CD7 и CD4(D1,D2):lg:CD7 и CD4 готовили в сублинии клеток 293, легко трансфицируемой клеточной линии эмбриональных почечных клеток человека. Химерные молекулы субклонировали в двунаправленных векторах, в которых ген гигромицина В был под контролем промотора тимидинкиназы вируса простого герпеса. Чашки 10 см с 60-70% конфлюэнтности трансфицировали 10 мкг этой плазмидной ДНК ко-преципитацией фосфатом кальция. Перед трансфекцией плазмиды линеаризовали при уникальном сайте Sfi 1 и концы дополняли при помощи ДНК-полимеразы T4. При 24 часах после трансфекции клетки разводили в 4 раза и при 48 часах после трансфекции проводили отбор с гигромицином В (Sigma, St. Louis, МО) при 400 мкг/мл. Каждые 3-4 дня клетки снабжали свежей средой, содержащей гигромицин.
Устойчивые колонии выскребали, размножали и их экспрессию оценивали непрямой иммунофлуоресценцией с применением конъюгированного с флуоресцеином антител против Fc IgG человека (Organon Teknika, West Chester, PA) или Q4120, антитела, реактивного с CD4 человека (Sigma), с последующей проточной цитометрией (Coulter, Hialeah, FH). Для анализа были отобраны два независимых клона из каждой конструкции с уровнями поверхностного CD4, сравнимыми с уровнем, обнаруживаемым другими клеточными линиями. Фиг. 22 показывает, что после выдерживания с ВИЧ р24 обнаруживали в стабильных CD4 трансфицированных культурах уже при 3 днях после инфицирования. Присутствие многоядерных гигантских клеток и характерного раздувания наблюдали так рано, как 5 дней после инфицирования в этих культурах. Не обнаруживали значительных уровней p24 или многоядерных гигантских клеток в нетрансфицированных родительских клетках или в любом из двух независимо полученных изолятов трансфектантов CD4(D1-D4):Ig:CD7 и CD4(D1,D2):Ig:CD7 после 32 дней культивирования (фиг. 22).
После завершения исследований инфективности клетки анализировали на экспрессию CD4 на клеточной поверхности. Плотность поверхностного эпитопа CD4 была значительно снижена в инфицированных культурах, экспрессирующих CD4, что согласуется с вирусной отрицательной модуляцией, но была ненарушенной в культурах, экспрессирующих CD4(D1-D4):Ig:CD7 и CD4(D1,D2):Ig:CD7. Эти эксперименты показывают, что можно создать химерные молекулы, несущие два апикальных домена CD4, которые при слиянии с ζ -цепью T-клеточного рецептора способны нацеливаться на ВИЧ-инфицированные клетки и убивать их, не поддерживая в то же время CD4-мeдииpуeмoй инфекции ВИЧ.
Дополнительные эксперименты позволяют предположить, что именно физическое расстояние между внеклеточным доменом молекулы CD4 и липидным бислоем придает способность противостоять инфекции ВИЧ. В первом эксперименте мы сконструировали химерную молекулу, имеющую делецию стебля и трансмембранного домена CD7; эта деления удаляла богатый пролином район трансмембранной части CD7. При слиянии этого домена с внеклеточным доменом CD4 он сохранял свою способность эффективно прикреплять внеклеточный домен молекулы CD4, как было измерено по поверхностной экспрессии молекулы CD4 (как описано здесь). Однако способность противостоять образованию синцитиев, индуцируемому гликопротеином оболочки ВИЧ, была утрачена. Таким образом, делеция богатой пролином части молекулы CD7, района, способного, по-видимому, образовывать α -спиральную закрученную структуру, эффективно уменьшала расстояние между внеклеточным доменом CD4 и липидным бислоем и уничтожала способность химеры противостоять образованию синцитиев.
Во втором эксперименте мы показали, что способность противостоять индуцируемому ВИЧ образованию синцитиев может быть обеспечена для химеры CD4/CD5, которая, как было показано ранее, служит трансмембранным якорем для внеклеточного домена CD4, но не способна противостоять индуцируемому ВИЧ образованию синцития. В этом эксперименте шарнирная область, CH2- и CH3-домены тяжелой цепи человеческого IgG1 были встроены в молекулу CD4/CD5; полученная химера противостояла образованию синцития, что опять-таки предполагает, что расстояние, предоставляемое доменами иммуноглобулина, является достаточным для придания устойчивости к индуцируемому ВИЧ образованию синцития.
В третьем эксперименте домен CD4 был удален на варьирующие расстояния от клеточной мембраны с применением синтетических α - спиралей различной длины. В частности, были сконструированы синтетические олигонуклеотиды, несущие повторяющиеся α -спиральные мотивы остатков лизина и глутаминовой кислоты, фланкированные двумя остатками аланина (см. фиг. 28 в отношении первичных последовательностей нуклеиновой кислоты и аминокислот). В более ранних исследованиях было найдено, что такие аминокислотные последовательности встречаются с высокой частотой в альфа-спиралях, что позволяет предположить, что такие повторяющиеся мотивы могли бы принимать альфа-спиральную конформацию и что помещение таких альфа-спиралей между трансмембранным доменом и внеклеточными доменами CD4 выдвигало бы CD4 прочь от клеточной мембраны. Путем варьирования длины альфа- спирального сегмента было рассчитано расстояние выдвигания, необходимое для противостояния вхождению ВИЧ, на основании известных величин высоты альфа-спирали и витка. Эти результаты представлены в Таблице 1.
В этой таблице "CD4" обозначает CD4(D1-D4), если нет других указаний; "H", "CH2" и "CH3" обозначают шарнирную область, районы CH2 и CH3 тяжелой цепи IgG1 человека, соответственно; "CD7-tm" и "stk" обозначают трансмембранный район и район стебля CD7; "CD7tm (long version)" и "CD7tm (short version)" обозначают, соответственно, трансмембранный район CD7 и трансмембранный район CD7, имеющий делецию богатого пролином домена, как обсуждалось выше; "CD5tm" обозначает трансмембранный район CD5; и "CD34tm" обозначает трансмембранный район CD34. В строках J-L длина альфа-спирального района дана в ангстремах; эти величины основаны на факте, что 3.6 остатка приходятся на виток альфа-спирали, что соответствует 5.4 ангстрема (или 1.5 А на остаток).
В соответствии с этим альфа-спираль из 16 остатков должна выдвигать внеклеточный домен CD4 приблизительно на 24 ангстрема. Были сконструированы альфа-спирали 48 и 72 ангстрем путем последовательной конкатемеризации фрагмента BstYl в уникальный сайт BamH1 фрагмента (см.фиг. 28) с последующим отбором клонов с правильной ориентацией.
Образование синцитиев оценивали в тестах совместного культивирования с клетками HeLa, экспрессирующими гликопротеин оболочки ВИЧ-1, из конструкции вируса коровьей оспы vPE-16 (см. выше).
Экспрессию Thy-1 измеряли следующим образом. Конструировали живой ретровирусный вектор на основе клона hxb.2 ВИЧ-1. В этом векторе несущественный ген nef заменяли кодирующей последовательностью крысиного thy-1, эффективно экспрессируемой клеточной поверхностью молекулы, которая прикрепляется к мембране фосфатидилинозитной связью. Вирус, полученный из этого молекулярного клона, обозначенный hxb/thy-1, был инфекционным, как было показано цитопатологическими эффектами и продуцированием p24 в культуральных супернатантах инфицированных клеток C8166 (CD4+ лейкозная T-клеточная линия человека). Кроме того, при выдерживании с hxb/thy-1 клетки HeLa, временно трансфицированные CD4, обнаружили признаки экспрессии thy-1 уже при 18 часах после инфицирования, как это и ожидалось от сигнала, регулируемого по способу регуляции nef гена. Сигналы, кодируемые геном nef, обычно относятся к классу вирусных регуляторных белков, которые подвергаются множественному сплайсингу и не содержат отвечающего на rev элемента. Такие сигналы могут накапливаться конститутивно в цитоплазме в виде продуктов ранних вирусных генов. Ожидалось, что thy-1-сигналы регулируются подобным образом, т.е. появляются рано в жизненном цикле вируса. Короче говоря, эта система облегчала анализ вхождения ВИЧ, поскольку экспрессия thy-1 была применена как заменитель вхождения вируса. Различные основанные на CD4 химеры нестабильно трансфицировали в клетки HeLa с применением стандартных ДЭАЭ-декстрановых методов. Трансфицированные клетки подвергали действию вируса hxb/thy-1 при 48 часах после трансфекции и оценивали на экспрессию thy-1 при 24-48 часах после инфицирования. В результатах, показанных в таблице 1, экспрессию thy-1 измеряли при 24 часах после инфекции с применением коммерчески доступных моноклональных антител против Thy-1.
На основании данных, представленных в Таблице 1, мы сделали вывод, что внеклеточные домены CD4 оптимально должны быть отодвинуты от клеточной мембраны по меньшей мере на 48 ангстремов и предпочтительно по меньшей мере на 72 ангстрема для противостояния инфицированию ВИЧ-1.
С применением стратегии, подобной описанной здесь общей стратегии, могут быть сконструированы химеры на основе антител против оболочки ВИЧ, которые нацелены на ВИЧ-инфицированные клетки. Примеры таких антител описаны в Gorny et al., Proc. Natl. Acad. Sci. USA 86:1624 (1989) и Marasco et al., J. Clin. Invest. 90:1467 (1992).
ПРИМЕР XI
Дополнительные триггерные белки T-клеточного рецептора и B-клеточного рецептора
Другие внутриклеточные и трансмембранные передающие сигнал домены, в соответствии с этим изобретением, могут быть произведены из белков T-клеточного рецептора, CD3 дельта и T3 гамма, и белков B-клеточного рецептора, mbi и B29. Аминокислотные последовательности этих белков показаны на фиг. 16 (CD3 дельта; SEQ ID N 24), фиг. 17 (T3 гамма; SEQ ID N 25), фиг. 18 (mD1; SEQ ID N 26) и фиг. 19 (B29; SEQ ID N 27). Части этих последовательностей, достаточные для трансдукции цитолитического сигнала (и, следовательно, предпочтительно включаемые в химерный рецептор этого изобретения) показаны в скобках (блоках). Химерные рецепторы, включающие в себя эти белковые домены, конструируются и используются в терапевтических способах этого изобретения, как описано в общем виде выше.
ПРИМЕР XII
ЭКСПЕРИМЕНТАЛЬНЫЕ МЕТОДЫ
Инфицирование и радиоиммунопреципитация для вируса коровьей оспы
Приблизительно 5 • 106 клеток CVI инфицировали в течение 1 часа в бессывороточной среде DME рекомбинантным вирусом коровьей оспы при множественности заражения (moi) не менее 10. Клетки помещали в свежую среду после инфицирования и метили метаболически 200 мкКи/мл 35S-метионином плюс цистеином (Tran-36S-метка, ICN; Costo Mesa, CA) в не содержащей метионина и цистеина DMEM (Gibco; Grand Island, NY) в течение 6 часов. Меченые клетки отделяли PBS, содержащим 1 мМ ЭДТА, собирали центрифугированием и лизировали в 1% NP-40, 1% ДСН, 0.15 М NaCl, 0.05 М Трис pH 8.0, 5 мМ ЭДТА и 1 мМ PMSF. Ядра удаляли центрифугированием и белки CD4 иммунопреципитировали с агарозой с антителами ОКТ4 и антимышиным IqG (Cappel, Durham, NC). Пробы подвергали электрофорезу на 8% полиакриламидных с ДСН гелях при невосстанавливающих (NR) и восстанавливающих (R) условиях. Гели, содержащие 35S-меченые пробы, пропитывали En3Hance (New England Nuclear, Boston, MA) перед радиоавтографией. Облегченную экспрессию трансмембранной формы CD16, CD16TM, измеряли сравнением ее экспрессии в клетках CVI, отдельно инфицированных CD16TM, с экспрессией в клетках, совместно инфицированных вирусами, кодирующими CD16TM и химеры ζ или γ. После инфекции и инкубирования в течение 6 часов или более клетки отделяли из чашек при помощи PBS, 1 мМ ЭДТА и экспрессию CD16TM и химер измеряли непрямой флуоресценцией и проточной цитометрией.
Тест выхода кальция
Клетки сублинии Jurkat E6 (Weiss et al., J. Immunol. 133:123-128 (1984)) инфицировали рекомбинантными вирусами коровьей оспы в течение 1 часа в бессывороточной среде IMDM при множественности заражения (moi) 10 и инкубировали в течение 3-9 часов в IMDM, 10% плодной телячьей сыворотке (FBS). Клетки собирали центрифугированием и ресуспендировали при 3 • 106 клеток/мл в полной среде, содержащей 1 мМ ацетометоксиэфир Indo-1 (Grynkiewicz et al., J. Biol. Chem. 260: 3340-3450 (1985)) (Молекулярные зонды) и инкубировали при 37oC в течение 45 минут. Нагруженные Indo-I клетки осаждали и ресуспендировали при 1 • 106 клеток/мл в бессывороточной среде IMDM и хранили при комнатной температуре в темноте. Клетки анализировали на свободный ион кальция путем одновременного измерения фиолетовой и синей флуоресценции при помощи проточной цитометрии (Rabinovitch et al., J. Immunol. 137:952-961 (1986)). Для инициации выхода кальция к клеточной суспензии добавляли конъюгированные с фикоэритрином (РЕ) антитела Leu-3A (антитела против CD4) (Becton Dickinson, Lincoln Park, NJ) при 1 мкг/мл с последующим добавлением 10 мкг-мл неконъюгированного козьего антимышиного IgG при времени 0 или неконъюгированного моноклонального антитела 3G8 (анти-CD16) при 1 мкг/мл, с последующим добавлением 10 мкг/мл конъюгированного с фикоэритрином (РЕ) Fab2, козьего антимышиного IgG при времени 0. Гистограммы отношения фиолетового к синему излучения получали из РЕ позитивной (инфицированной) клеточной популяции, которая в типичном случае составляла 40-80% всех клеток. Ответная реакция T-клеточного рецептора антигенов в неинфицированных клетках запускалась антителом ОКТ3, без сшивания. Для экспериментов с участием CD16 химерных рецепторов пробы, показывающие смещение базовой линии в направлении более низкого внутриклеточного кальция (без антител), исключали из анализа. Данные гистограмм затем анализировали преобразованием бинарных данных в ASC11 с применением программного обеспечения Write Hand Man (Cooper City, FL), с последующим анализом с коллекцией программ FORTRAN. Отношение фиолетовой к синей эмиссии перед добавлением второго антитела использовали для установления нормализованного первоначального отношения, принимаемого за единицу, и порогового отношения покоя, устанавливаемого таким образом, чтобы 10% покоящейся популяции превышали пороговое значение.
Анализ цитолиза
Человеческую T-клеточную линию WH3, CD8+ CD4- HLA B44 рестриктированную цитолитическую линию, поддерживали в IMDM с 10% человеческой сывороткой с 100 E/мл интерлейкина-2 (IL-2) и периодически стимулировали либо неспецифически облученными (3000 рад) не содержащими HLA лимфоцитами периферической крови и 1 мкг/мл фитогемагглютинина либо специфически облученными несущими B44 мононуклеарными клетками. После одного дня неспецифической стимуляции PHA (ФГА) (фитогемагглютинин) разводили до 0.5 мкг/мл добавлением свежей среды и через 3 дня среду заменяли. Клетки выращивали по меньшей мере в течение 10 дней после стимуляции перед использованием в анализах цитотоксичности. Эти клетки инфицировали рекомбинантным вирусом коровьей оспы при множественности заражения не менее 10 в течение 1 часа в бессывороточной среде с последующим инкубированием в полной среде в течение 3 часов. Клетки собирали центрифугированием и ресуспендировали при плотности 1 • 107 клеток/мл. 200 мкл добавляли к каждой лунке микротитрационного планшета с U-образным дном, содержащей 100 мкл/лунку полной среды. Клетки разводили в серийных 2-кратных разведениях. Две лунки для каждой пробы не содержали лимфоцитов, чтобы получить самопроизвольное выделение хрома и измерить общее поглощение хрома. Клетки-мишени, из сублинии S3HeLa, инфицировали в чашках 6.0 или 10.0 см при множественности заражения 10 в течение 1 часа в бессывороточной среде и инкубировали затем в полной среде в течение 3 часов. Затем их отделяли из клеток при помощи PBS, 1 мМ ЭДТА и считали. Аликвоту 106 клеток-мишеней (HeLa, Raji или RJ2.2.5 для экспериментов с CD4-химерным рецептором и клетки 3G8 10-2 (Shel et al., Mol. Immunol. 26:959 (1989) для экспериментов с CD16-химерным рецептором) центрифугировали и ресуспендировали в 50 мкл стерильного 51Cr-хромата натрия (1 мКи/мл, Dupont Wilmington, DE) в течение 1 часа при 37oC с периодическим перемешиванием, затем промывали три раза PBS. 100 мкл меченых клеток, ресуспендированных в среде при 105 клеток/мл, добавляли к каждой лунке. Клетки-мишени Raji и RJ2.2.5 метили так же, как и клетки HeLa. Микротитрационный планшет откручивали при 750 х g в течение 1 минуты и инкубировали в течение 4 часов при 37oC. В конце инкубационного периода клетки в каждой лунке ресуспендировали осторожным пипетированием, брали пробу для определения общего включения и микротитрационный планшет откручивали при 750 х g в течение 1 минуты. Аликвоты 100 мкл супернатанта удаляли и считали в гамма-лучевом сцинтилляционном счетчике. Процент убивания корректировали на фракцию инфицированных клеток-мишеней (обычно 50-90%), измеренную проточной цитометрией. Для инфицированных эффекторных клеток отношение эффектор/мишень корректировали на процент инфицированных клеток (обычно 20-50% для экспериментов с CD4-химерным рецептором и более 70% для экспериментов с CD16- химерным рецептором).
Мутагенез in vitro последовательности ζ (дзета)
Для создания точковых мутаций в аминокислотных остатках 11 и/или 15 последовательности ζ получали синтетические олигонуклеотидные праймеры, простирающиеся от сайта BamHI
против хода транскрипции от трансмембранного домена ζ и превращающие остаток 11 нативного ζ из Cys в Gly (CllG) или остаток 15 из Asp в Gly (D15G) или оба (CllG/D15G). Их использовали в ПЦР-реакциях для генерирования мутированных фрагментов, которые встраивали в конструкции CD4: ζ дикого типа.
Для создания делеций ζ последовательности кДНК ζ амплифицировали при помощи ПЦР с применением синтетических олигонуклеотидных праймеров, сконструированных для создания стоп-кодона (UAG) после остатков 50, 59 или 65. Эти праймеры содержали сайт расщепления для фермента NotI в виде выступающих 5 или 6 остатков от 5'-конца, обычно в последовательности вида CGC GGG CGG CCG CTA (SEQ ID N 11), где последние три остатка соответствуют стоп-антикодону. За последовательностями NotI и стоп-антикодона следовали 18 или более остатков, комплементарных целевому 3'-концу этого фрагмента. Полученные химеры были названы CD16: ζ Y51*, CD16: ζ E60* и CD16: ζ D66* соответственно. Сайт BamHI против хода транскрипции от трансмембранного домена и сайт NotI использовали для генерирования фрагментов, которые повторно вводили в конструкцию CD16: ζ дикого типа. Мономерные ζ -химеры создавали высвобождением транс-мембранных и мембрано-проксимальных внутриклеточных последовательностей ζ расщеплением при помощи BamHI и SacI конструкции Asp- и Cys- CD4: ζ, описанной выше, и встраиванием этого фрагмента в конструкции CD16: ζ E60* и CD16: ζ D66* соответственно.
Состоящие из трех частей химерные конструкции CD16:7: ζ (48-65) и CD16: 7: ζ (48-59)
Для получения конструкции CD16: ζ D66* последовательность кДНК ζ, соответствующую трансмембранному домену и 17 следующим остаткам цитоплазматического домена, заменяли соответствующим трансмембранным и цитоплазматическим доменом, полученным из кДНК CD5 и CD7. Фрагменты CD5 и CD7 получали при помощи реакции ПЦР с применением прямых олигонуклеотидов, включающих в себя сайт расщепления BamHI и соответствующих району против хода транскрипции от трансмембранного домена CD5 и CD7 соответственно и следующих обратных олигонуклеотидов, перекрывающих последовательности CD5 и CD7 соответственно и последовательность ζ, содержащую сайт расщепления SacI.
SD5: ζ : CGC GGG CTC CTT ATA GAG CTG GTT CTG GCG CTG CTT CTT CTG (SEQ ID N 12)
CD7: ζ : CGC GGG GAG CTC GTT ATA GAC CTG GTT TGC CGC CGA ATT CTT ATC CCG (SEQ ID N 13).
Продукты ПЦР CD5 и CD7 расщепляли BamHI и SacI и лигировали с расщепленным BamHI и SacI CD16: ζ E60* и заменяли последовательность ζ от BamHI до SacI фрагментом CD7. Для получения конструкций CD16:CD5 и CD16:CD7 фрагменты CD5 и CD7 получали при помощи ПЦР с применением олигонуклеотида, содержащего сайт расщепления NotI и кодирующего стоп-кодон (UAA) после остатка Gln416 и Ala193 CD5 и CD7 соответственно. ПЦР фрагмент CD5 и CD7 расщепляли BamHI и NotI и встраивали в конструкцию CD16: ζ Asp66*.
Мутагенез in vitro N-концевых остатков внутри трансдуцирующего цитолитический сигнал мотива ζ (дзета)
Синтетически олигонуклеотидные праймеры, удлиняющиеся от сайта SacI внутри мотива ζ и превращающие нативный остаток 48 из Asn в Ser (N48S), остаток 50 из Leu в Ser (L50S) и остаток 51 из Tyr в Phe (Y51F), были синтезированы и использованы в реакции ПЦР для генерирования фрагментов, которые вновь вводили в конструкцию CD16:7: ζ (48-65) дикого типа.
Мутагенез in vitro C-концевых остатков внутри трансдуцирующего цитолитический сигнал мотива ζ (дзета)
Синтетические олигонуклеотидные праймеры, удлиняющиеся от сайта NotI 3' по отношению к стоп-кодону и превращающие нативный остаток 60 из Glu в Gln (E60Q), остаток 61 из Glu в Gin (E61Q), остаток 62 из Tyr в Phe или Ser (Y62F или Y62S) и остаток 63 из Asp в Asn (D63N), были синтезированы и использованы в ПЦР для генерирования фрагментов, которые субклонировали в конструкцию дикого типа CD16: ζ D66* от сайта BamHI до сайта NotI.
Химерные конструкции CD16: 7: ζ (33-65), CD16:7: ζ (71-104), DC16:7: ζ (104-137)
Трансмембранный фрагмент CD7, несущий сайты MluI и NotI в месте соединения между трансмембранным и внутриклеточным доменами, получали при помощи ПЦР с использованием олигонуклеотида следующей последовательности: CGC GGG GCG GCC ACG CGT CCT CGC CAG CAC ACA (SEQ ID N 14). Полученный фрагмент ПЦР расщепляли BamHI и NotI и повторно встраивали в конструкцию CD16:7: ζ (48-65). Фрагменты ζ,, кодирующие остатки 33-65, 71-104 и 104-137, получали реакцией ПЦР с применением пары праймеров, содержащих сайты MluI при 5'- конце прямых праймеров и стоп-кодоны, за которыми следуют сайты NotI при 5'-конце обратных праймеров. В каждом случае сайты рестрикции имели 6 остатков от 5'-конца праймера для гарантии расщепления рестриктазой.
ζ 33: CGC GGG ACG CGT TTC AGC CGT CCT CGC CAG CAC ACA (SEQ ID N 15)
ζ 71: CGC GGG ACG CGT GAC CCT GAG ATG GGG GGA AAG (SEQ ID N 16) и
ζ 104: CGC GGG ACG CGT ATT GGG ATG AAA GGC GAG CGC (SEQ ID N 17).
Конструирование делеционных мутантов FcR γ 11A
Карбоксил-концевые делеционные мутанты FcR γ 11A конструировали при помощи ПЦР так же, как и полноразмерные конструкции, превращая последовательности, кодирующие тирозин в положениях 282 и 298, в стоп-кодоны (ТАА). N-концевые делеции генерировали амплификацией фрагментов, кодирующих все меньшую часть внутриклеточного домена, при помощи ПЦР с применением олигонуклеотидов, позволяющих встраивать полученные фрагменты между сайтами рестрикции MluI и NotI в предварительно сконструированную экспрессионную плазмиду, кодирующую внеклеточный домен CD16, слитый с трансмембранным доменом CD7, причем последний заканчивается в сайте MluI и месте соединения между трансмембранным и внутриклеточным доменами.
ДРУГИЕ ВАРИАНТЫ
Описанные выше примеры показывают, что агрегация химер ζ,η или γ является достаточной для инициации цитолитического эффекторного клеточного ответа в T-клетках. Известный диапазон экспрессии ζ,η и γ, который включает в себя T-лимфоциты, природные клетки-убийцы, базофильные гранулоциты, макрофаги и мастоциты, предполагает, что консервативные мотивы последовательности могут взаимодействовать с сенсорным аппаратом, общим для клеток гемопоэтического происхождения, и что важный компонент иммунной защиты организма в иммунной системе может быть медиирован событиями агрегации рецептора.
Активность цитолитического ответа и отсутствие ответа на клетки-мишени, несущие рецепторы MHC класса II, демонстрируют, что химеры на основе ζ,η или γ образуют основу для генетического воздействия на СПИД через адоптивную иммунотерапию. Широкое распространение эндогенных ζ и γ и доказательство того, что Fc-рецепторы, связанные с γ, медиируют цитотоксичность в различных типах клеток (Fanger et al., Immunol. Today 10:92-99 (1989)), позволяют рассматривать многие клетки, как пригодные, возможно, для этой цели. Например, нейтрофильные гранулоциты, которые имеют очень короткую продолжительность жизни (приблизительно 4 часа) в циркуляционном кровотоке и представляют собой интенсивно цитолитические клетки, являются заманчивыми клетками-мишенями для экспрессии химер. Инфицирование нейтрофилов ВИЧ, по-видимому, не приводит к выделению вируса, и обилие этих клеток (наиболее предпочтительно, лейкоцитов) должно облегчать иммунную защиту организма. Другой заманчивой возможностью в отношении клеток-хозяев являются зрелые T-клетки, популяция, доступная в настоящее время для ретровирусной генной инженерии (Rosenberg, S.A. Sci. Am. 262:62-69 (1990)). При помощи рекомбинантного IL-2 T-клеточные популяции могут быть размножены в культуре с относительной легкостью, и размноженные популяции имеют ограниченную продолжительность жизни при реинфузии (Rosenberg et al., N. Engl. J. Med. 323:570-578 (1990)).
При подходящих условиях узнавание ВИЧ клетками, экспрессирующими химеры CD4, должно также обеспечивать митогенные стимулы, предоставляющие возможность клеточной популяции с иммуноглобулиновыми рецепторами динамично отвечать на вирусную нагрузку. Хотя мы сконцентрировали здесь внимание на поведении слитых белков в цитолитических T-лимфоцитах, экспрессия этих химер в хелперных лимфоцитах может обеспечить мобилизуемый ВИЧ источник цитокинов, которые могли бы противодействовать коллапсу субпопуляции хелперных клеток при СПИДе. Недавнее описание нескольких схем для конструирования устойчивости к инфекции на стадиях, иных, чем проникновение вируса (Friedman et al. Nature 335:452-454 (1988); Green et al. Cell 58:215-223 (1989); Malim et al. , Cell 58:205-214 (1989); Trono et al. Cell 59:113-120 (1989); Buonocore et al. Nature 345: 625-628 (1990)), предполагает, что клетки, несущие химеры CD4, могли бы быть сконструированы таким образом, чтобы снижать образование вируса экспрессией соответствующих агентов, имеющих внутриклеточный сайт действия.
Возможность передачи сигналов к T-лимфоцитам через автономные химеры также обеспечивает возможность регуляции сконструированных при помощи ретровирусов лимфоцитов in vivo. Сшитые стимулы, медиируемые, например, специфическими IgM антителами, сконструированными для удаления комплемент-связывающих доменов, могут позволить таким лимфоцитам увеличиться в числе in situ, тогда как обработка подобными специфическими IgM антителами (например, узнающими аминокислотную вариацию, сконструированную в химерной цепи) могла бы селективно истощать сконструированную популяцию. Кроме того, IgM антитела против CD4 не требуют дополнительного сшивания для мобилизации кальция в клетках Jurkat, экспрессирующих химеры CD4: ζ. Возможность регуляции клеточных популяций без обращения к повторяющимся экстракорпореальным амплификациям может значительно расширить диапазон и эффективность текущих применений, предложенных для генетически сконструированных T-клеток.
Хотя это изобретение было описано в связи с его характерными вариантами, должно быть понятно, что оно может быть модифицировано далее, и эта заявка предназначена для охвата вариаций, применений или адаптаций этого изобретения, и включает в себя такие отклонения от данного описания, которые входят в область, к которой относится это изобретение, и могут быть отнесены к основным изложенным здесь признакам, как это следует из объема прилагаемой формулы изобретения.
















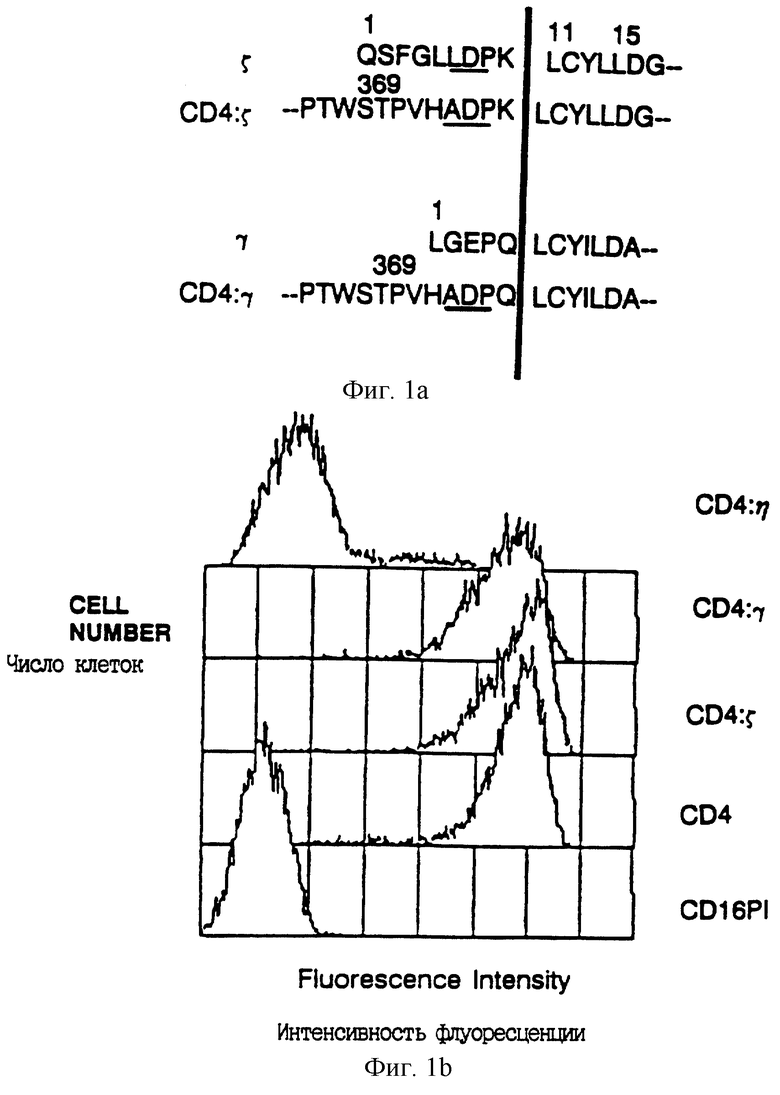
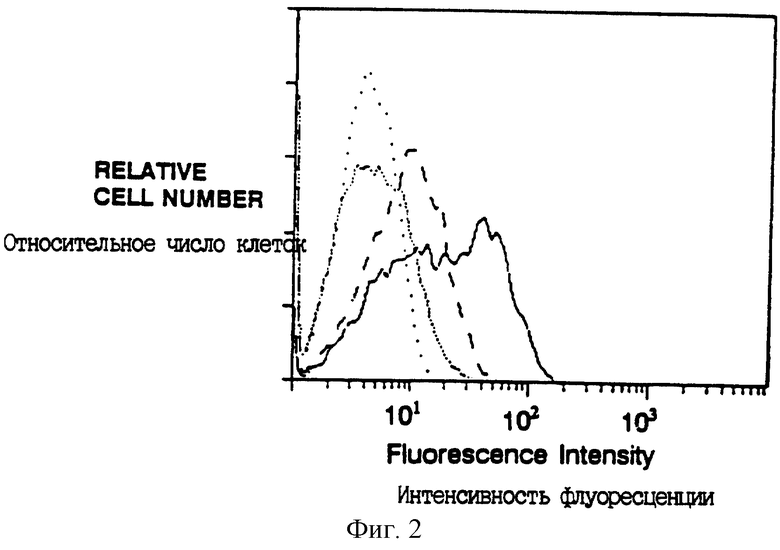
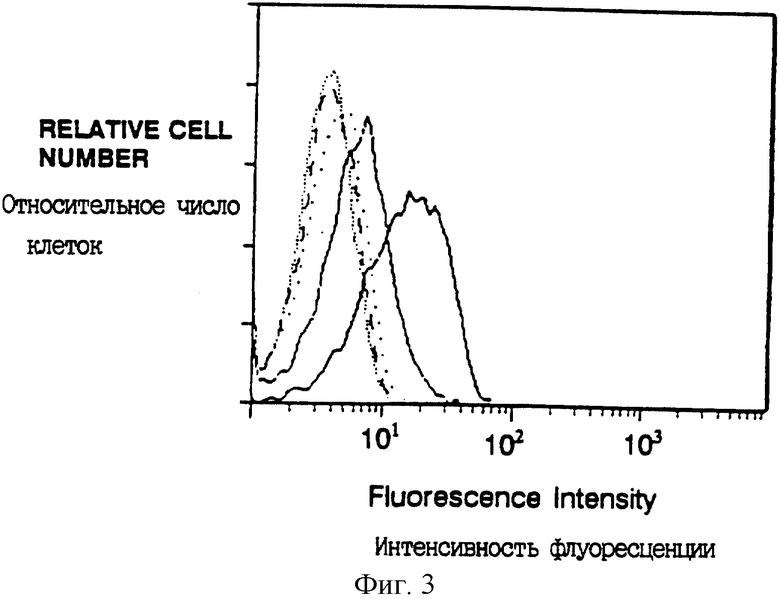
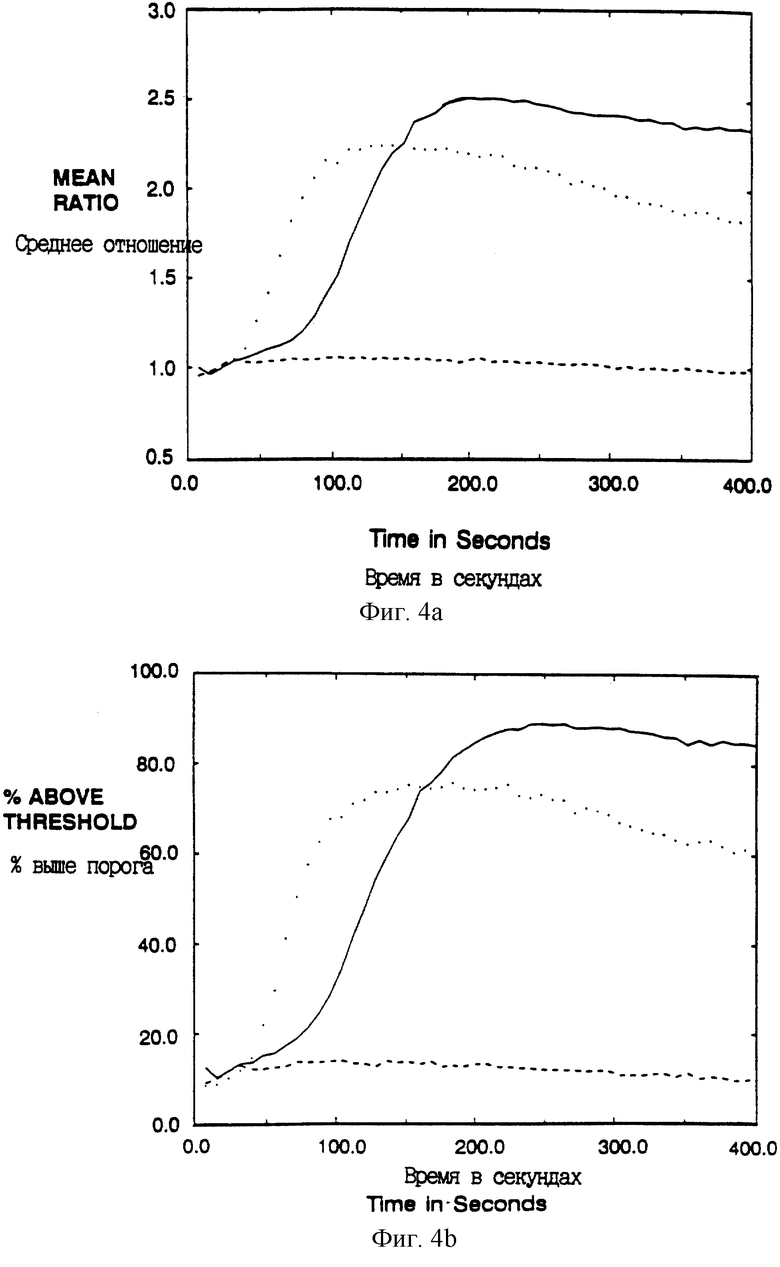
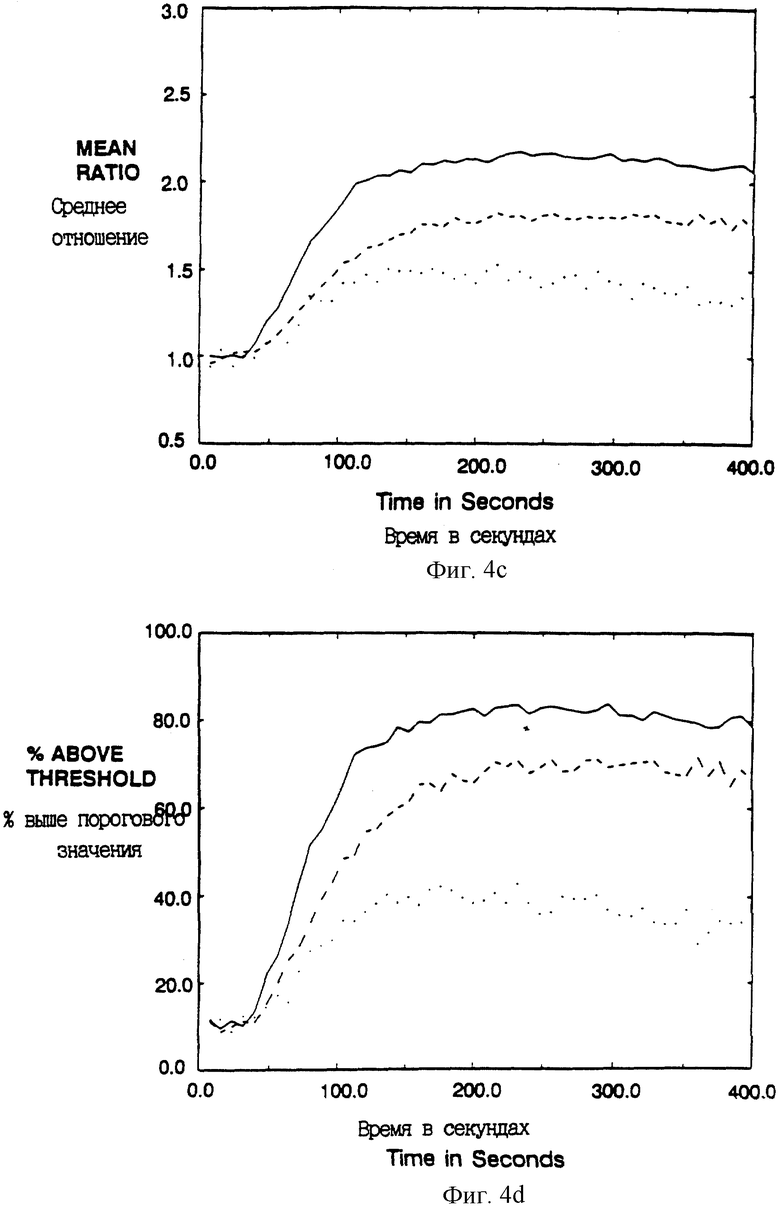
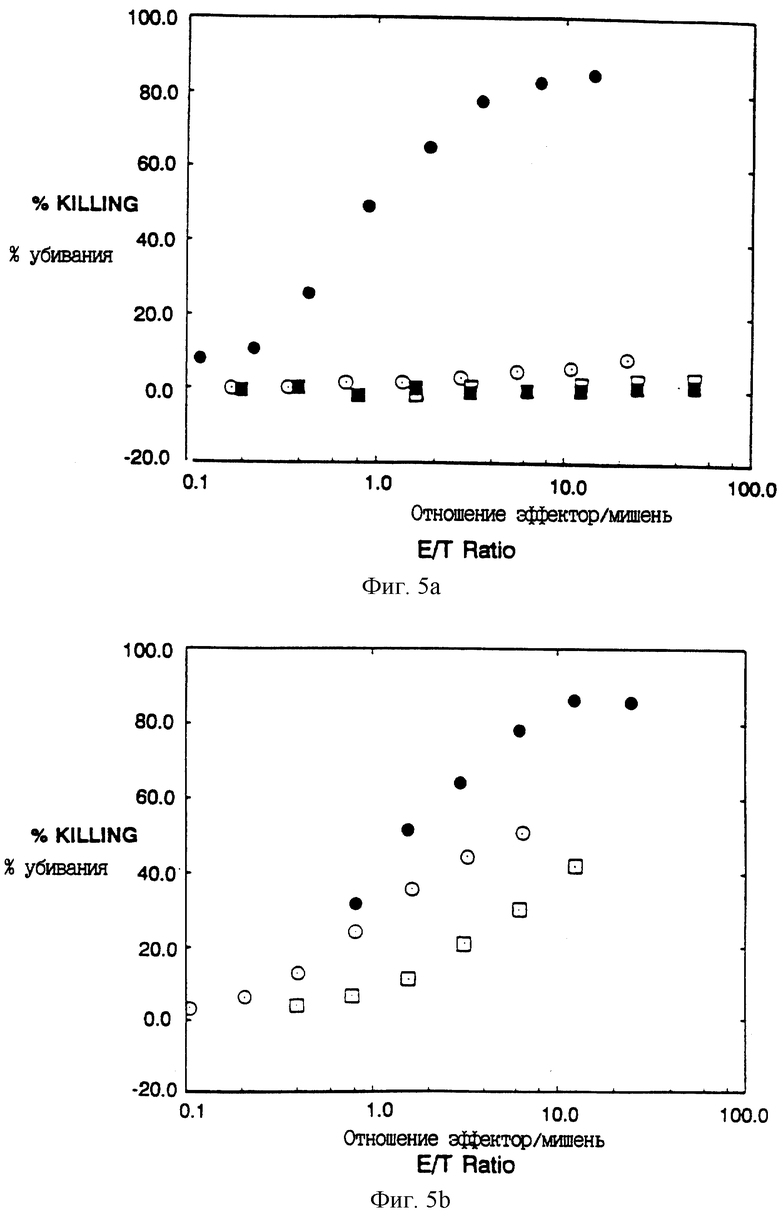
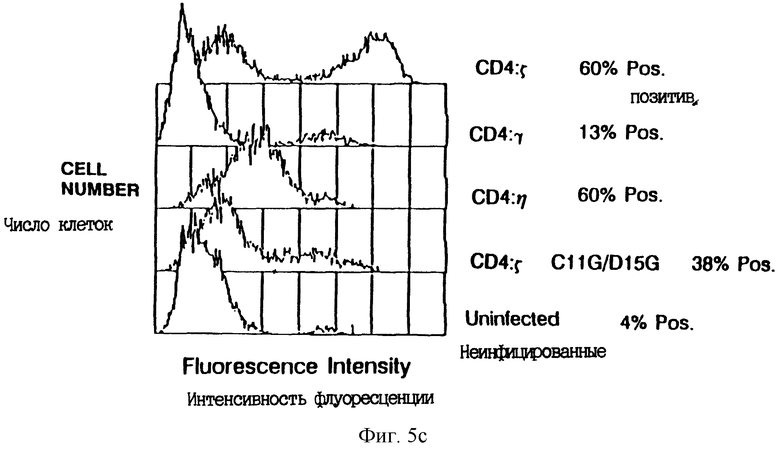
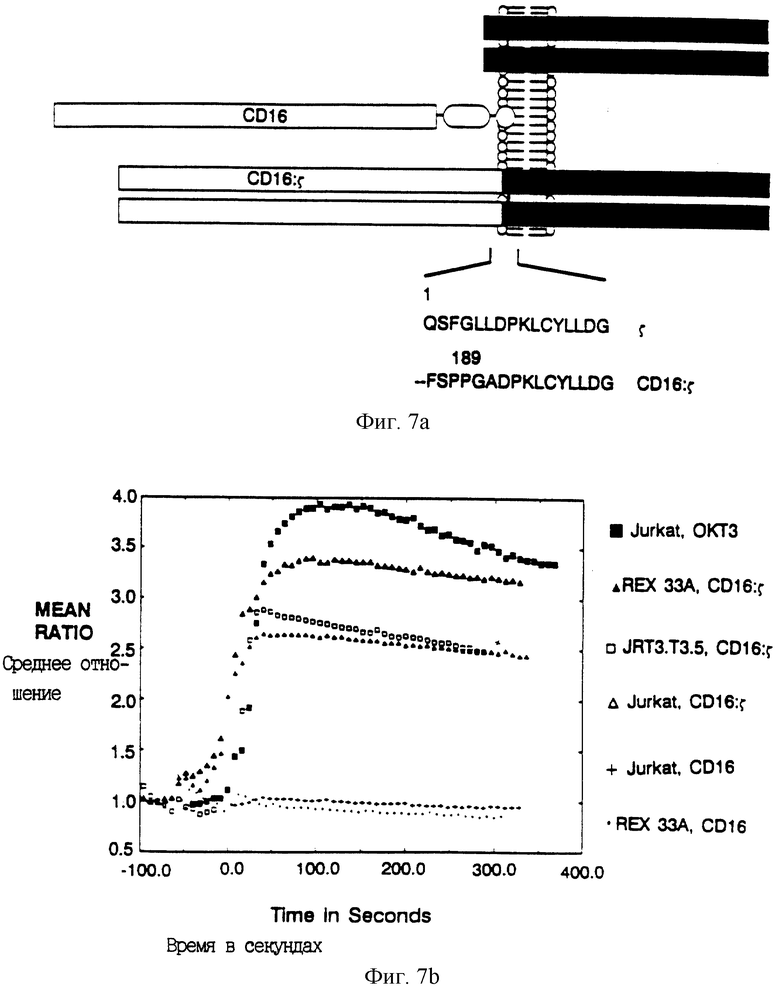
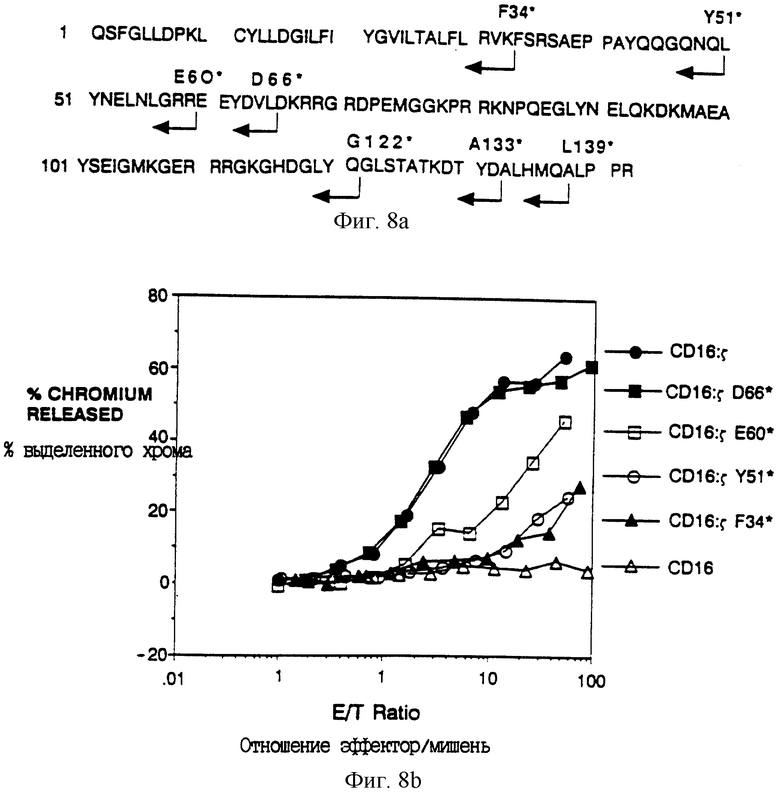
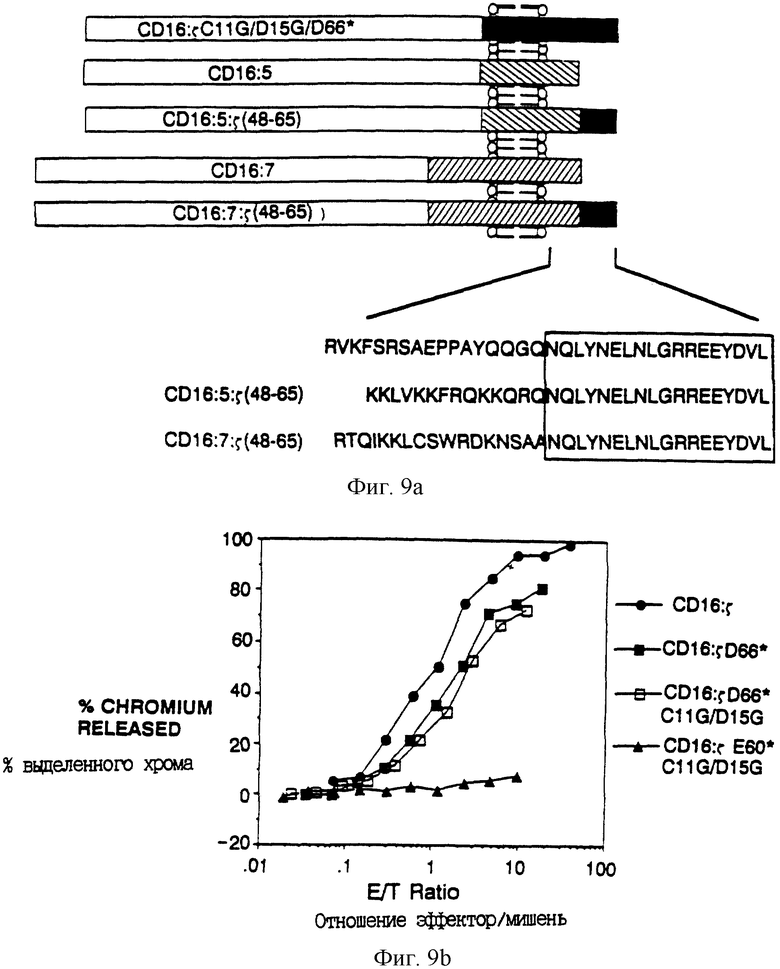
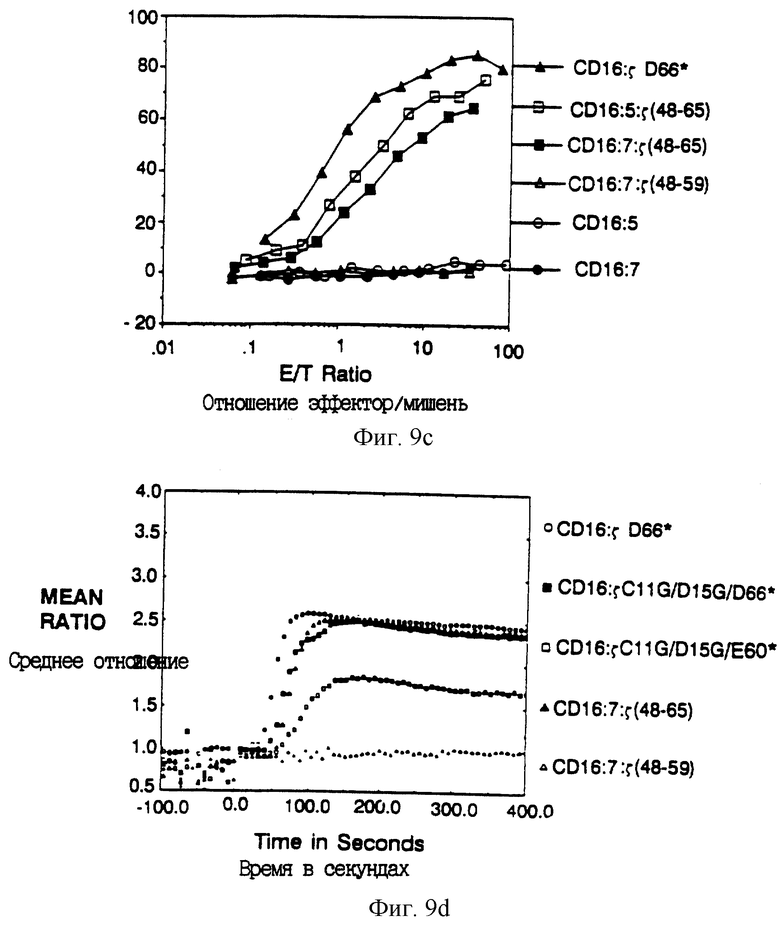
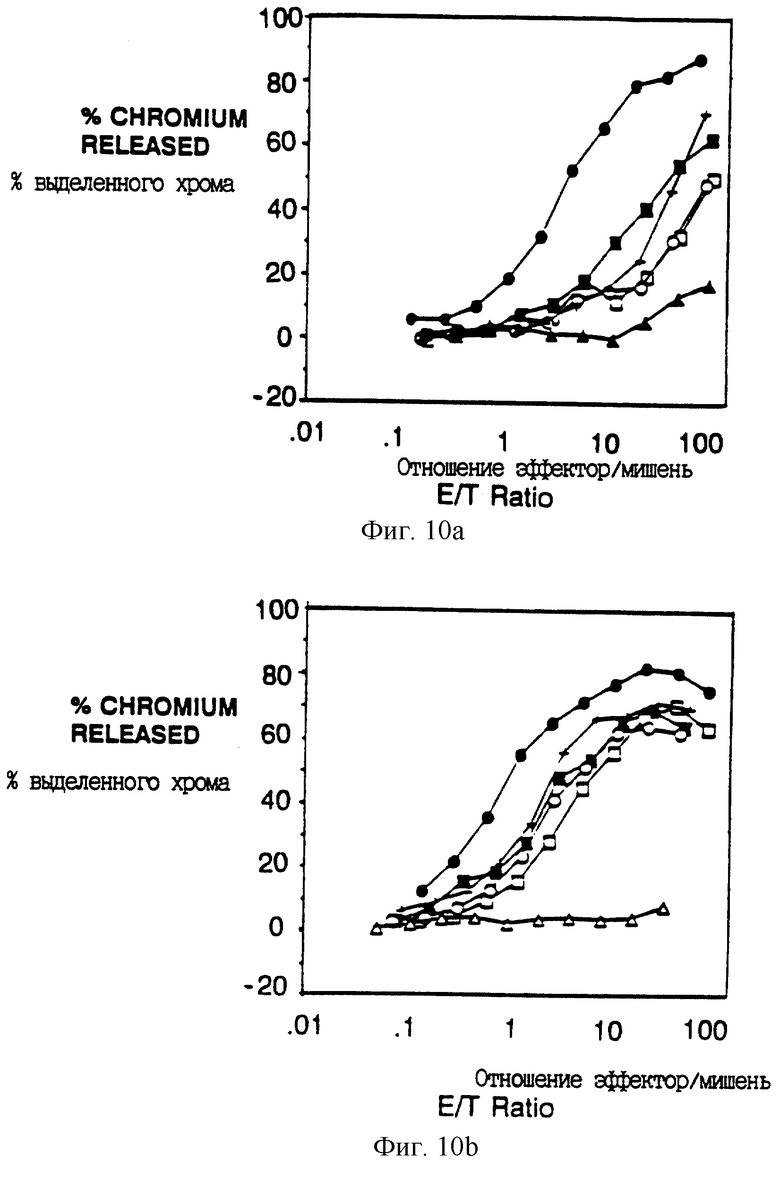

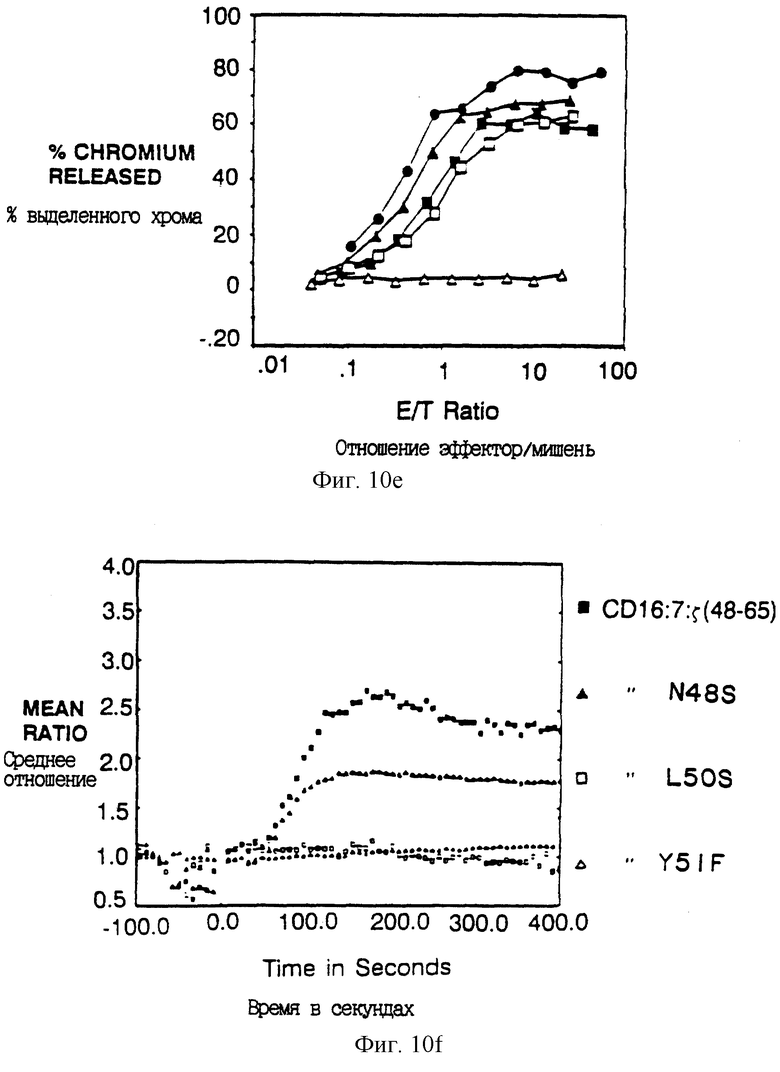
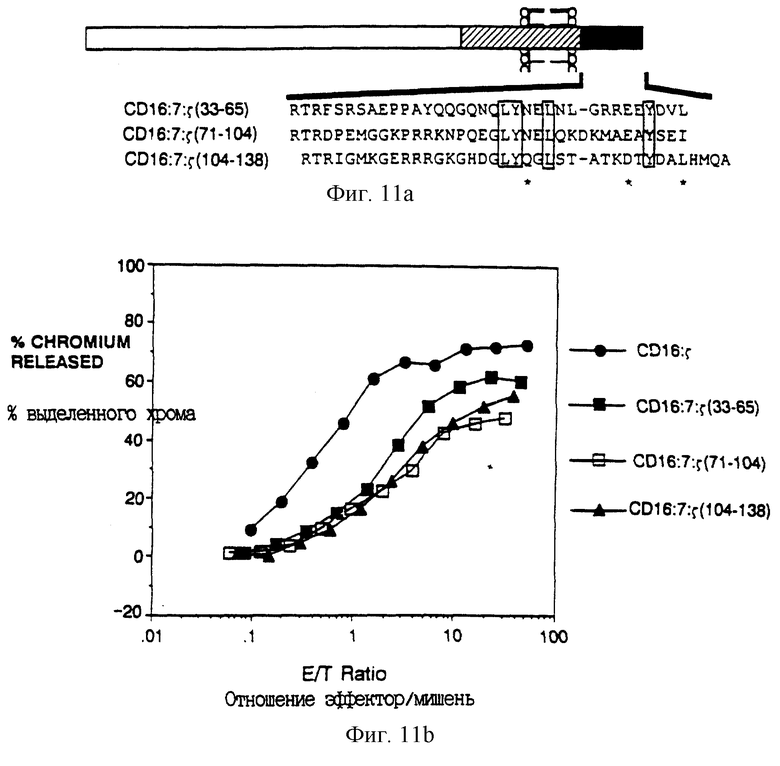
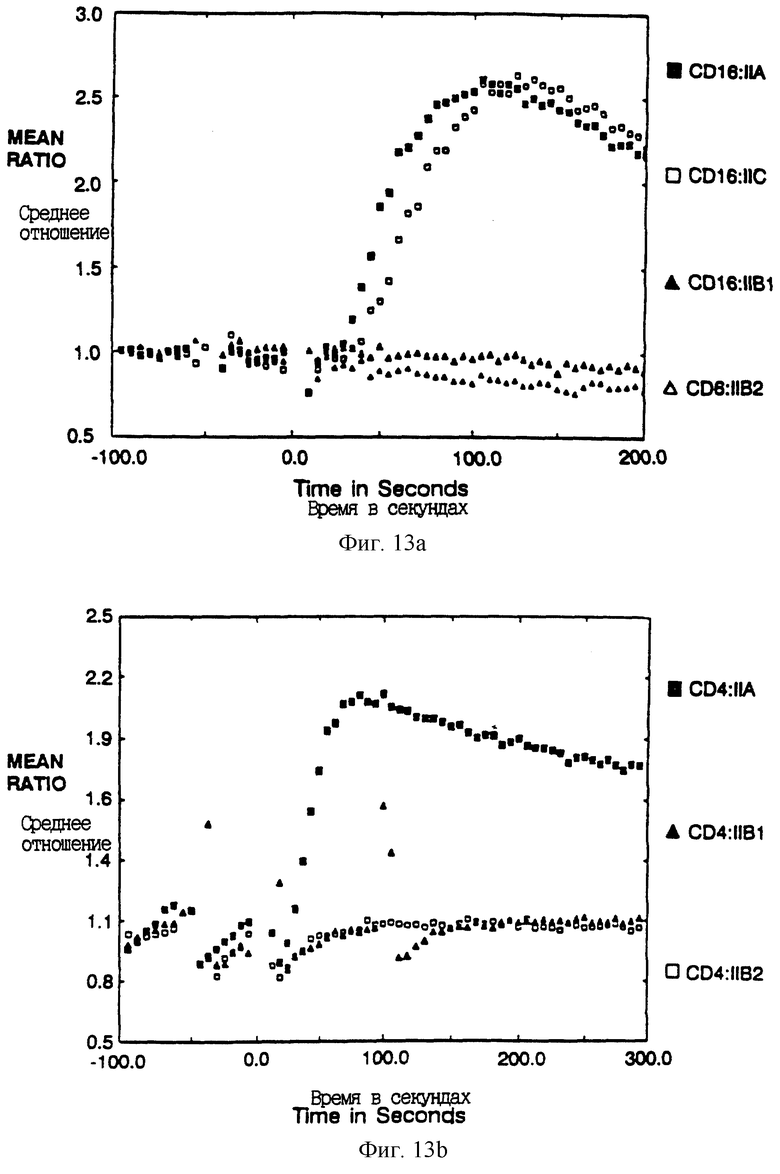
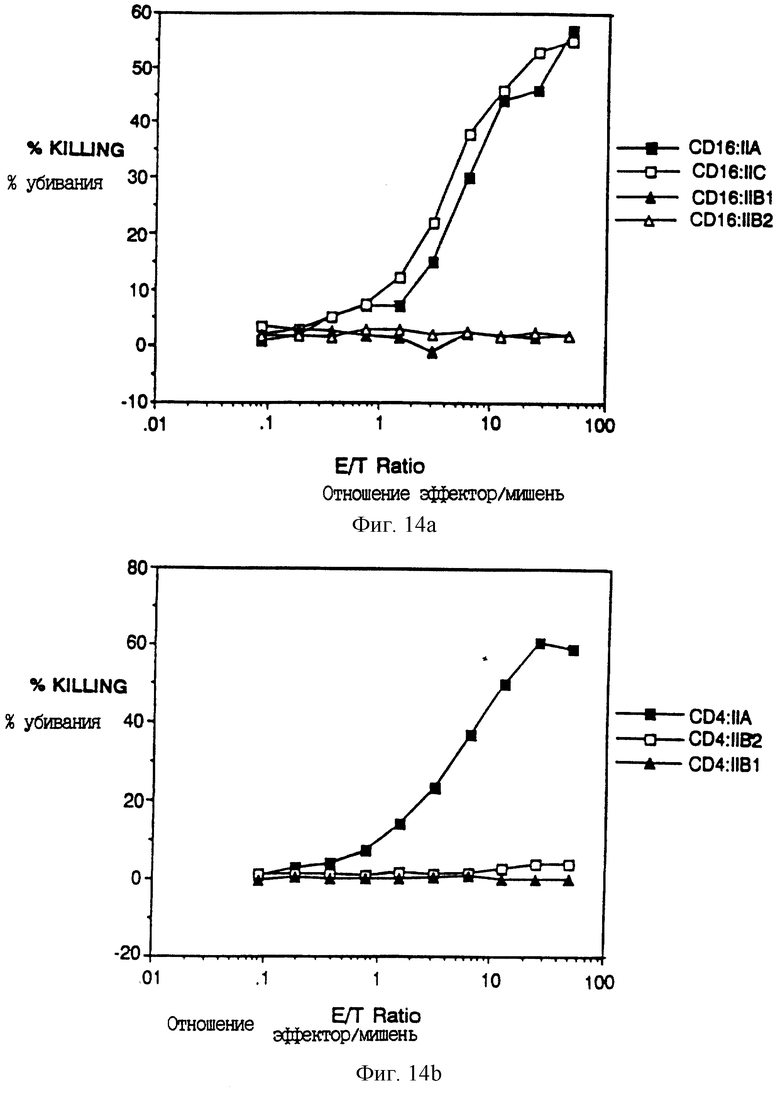
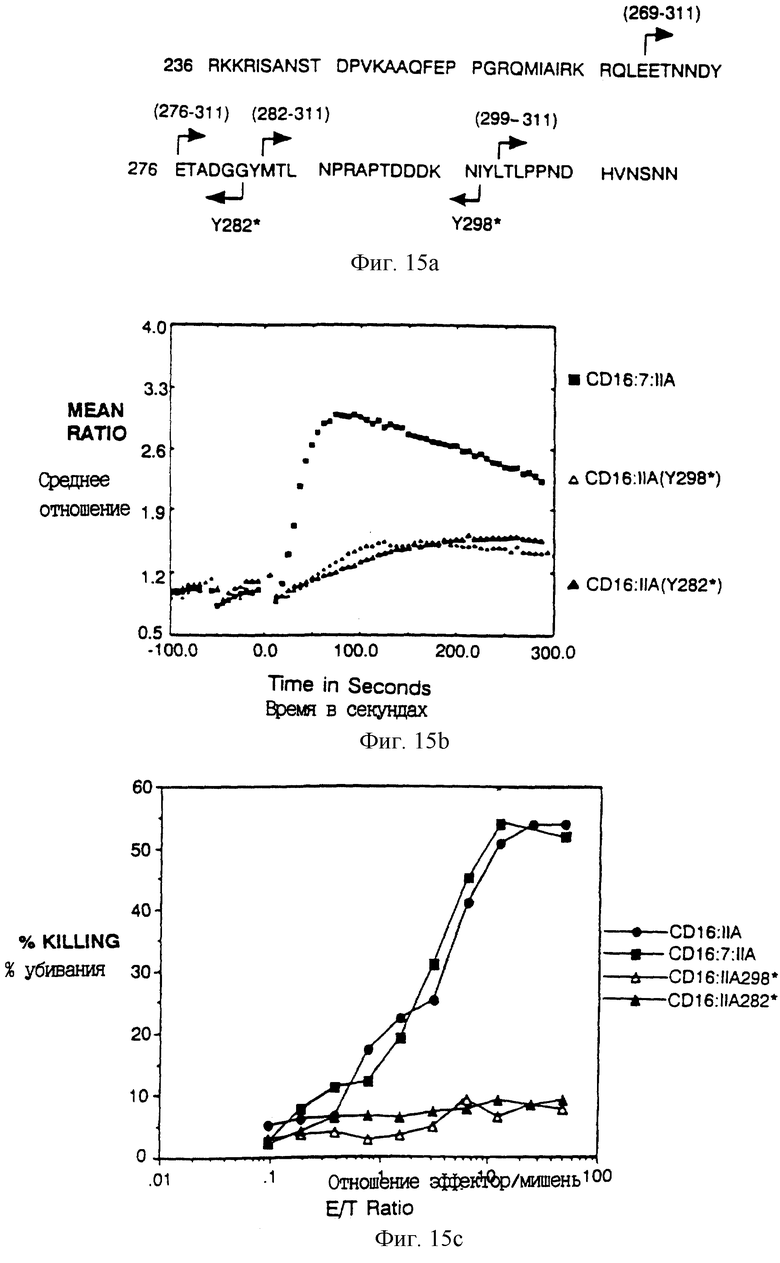
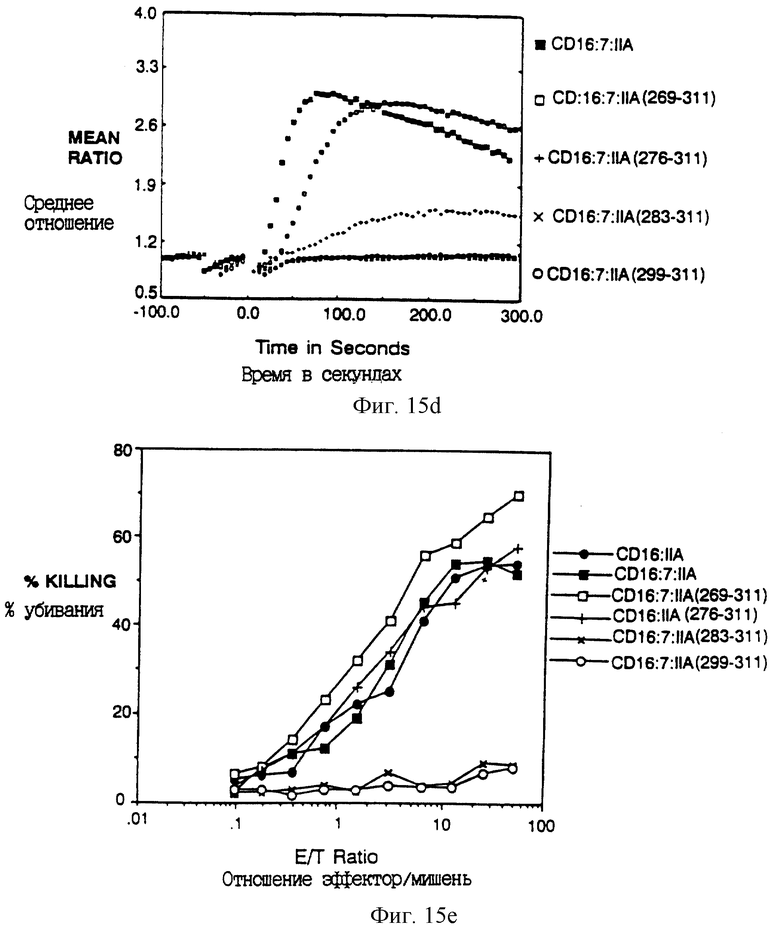
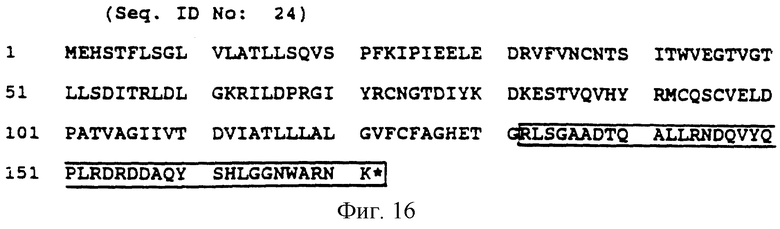

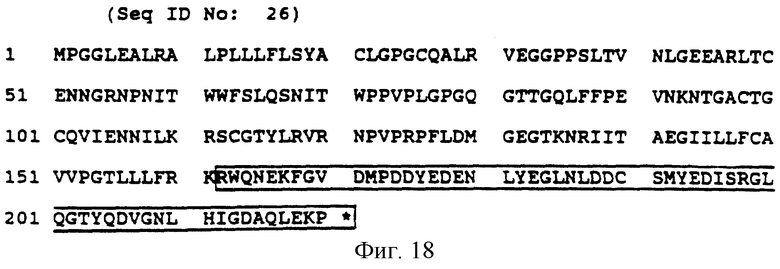

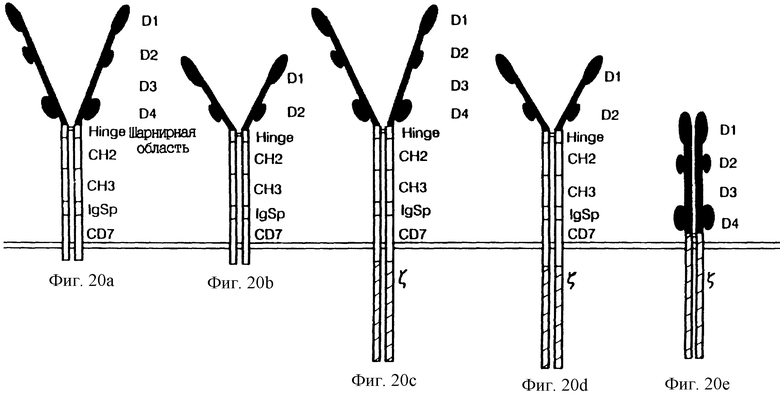
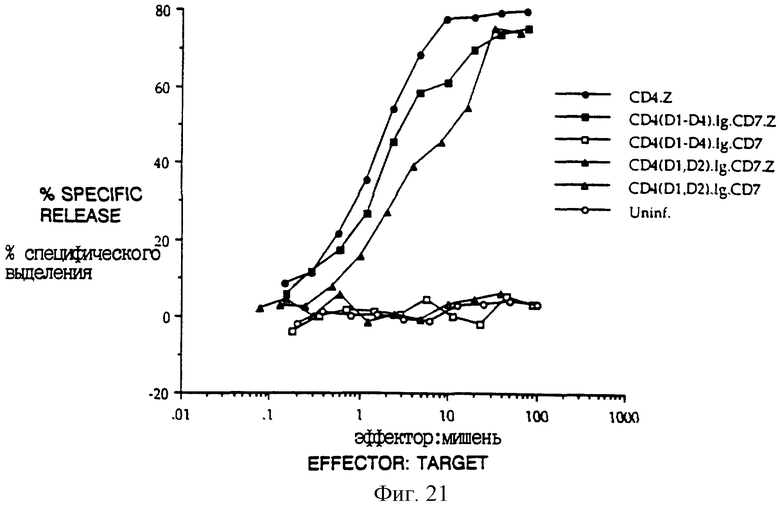
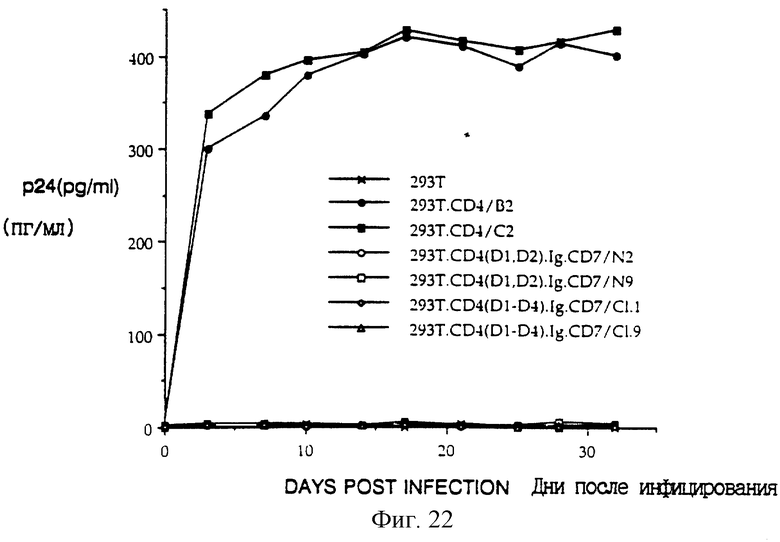






Изобретение относится к медицине, генной инженерии и генной терапии. Способ заключается в направлении клеточного иммунного ответа против ВИЧ-инфицированной клетки млекопитающего. На ВИЧ-инфицированную клетку воздействуют эффективным количеством клеток, которые экспрессируют мембранносвязанный белковый химерный рецептор. Последний содержит (а) внеклеточную часть, которая включает в себя фрагмент CD4, способный специфически узнавать и связывать ВИЧ-инфицированную клетку, но не опосредует инфекцию ВИЧ, и (b) внутриклеточную часть, способную подавать сигнал указанной клетке для разрушения связанной с рецептором ВИЧ-инфицированной клетки. Белковый мембранносвязанный химерный рецептор содержит внеклеточную и внутриклеточную часть. Внеклеточная часть представляет фрагмент CD4. Аминокислотная последовательность приведена в описании. Внутриклеточная часть способна подавать сигнал клеткам, несущим указанный рецептор. Этот сигнал вызывает разрушение связанных рецептором ВИЧ или ВИЧ-инфицированных клеток. Внутриклеточный рецептор является передающей сигнал частью Т-клеточного рецептора, В-клеточного рецептора или Fc-рецептора. Фрагмент CD4 может быть отделен от внутриклеточной части трансмембранным доменом CD7, или CD5, или CD34. Структура трансмембранного домена приведена в описании. Представлена ДНК, кодирующая белковый мембранносвязанный химерный рецептор. Изобретение позволяет создать функциональные химеры, способные направлять иммунокомпетентные клетки на лизис инфицированных ВИЧ клеток. 3 с. и 12 з.п.ф-лы, 28 ил., 1 табл.
13. Химерный рецептор по п.8, отличающийся тем, что указанный рецептор включает в себя трансмембранную часть CD7, трансмембранную часть CD5 или трансмембранную часть CD34.
Приоритет по пунктам:
14.02.1994 по пп.1-3, 5, 6, 8-10, 12, 13, 15;
02.08.1994 по пп.4, 7, 11, 14.
| Домовый номерной фонарь, служащий одновременно для указания названия улицы и номера дома и для освещения прилежащего участка улицы | 1917 |
|
SU93A1 |
| Автоматический огнетушитель | 0 |
|
SU92A1 |
| Автоматический огнетушитель | 0 |
|
SU92A1 |
