Введение
Настоящее изобретение относится к модификации клеток-естественных киллеров (NK) и линий NK-клеток для получения их производных с фенотипом повышенной цитотоксичности. Кроме того, настоящее изобретение относится к способам получения модифицированных NK-клеток и линий NK-клеток, композициям, содержащим эти клетки и их линии, а также использованию этих композиций для лечения рака.
Уровень техники изобретения
Как правило, иммунным клеткам требуется клетка-мишень для внедрения антигена через главный комплекс гистосовместимости (ГКГС) до провоцирования иммунной реакции, приводящей к гибели клетки-мишени. Это позволяет раковым клеткам, не представляющим собой ГКГС класса I, избежать большинства иммунных реакций.
При этом NK-клетки могут распознавать раковые клетки при отсутствии выраженности ГКГС класса I. Вследствие этого они играют критически важную роль в защите организма от рака.
С другой стороны, в некоторых обстоятельствах раковые клетки демонстрируют способность снижать цитотоксичную активность NK-клеток посредством экспрессии лиганд, связывающих ингибирующие рецепторы на оболочке NK-клеток. Устойчивость к раку может включать в себя наличие равновесия между этими и другими факторами.
В этом случае цитотоксичность относится к способности иммунных эффекторных клеток, например, NK-клеток, вызывать смерть раковых клеток, например, посредством выработки цитолитических соединений или связывания рецепторов оболочек раковых клеток, а также посредством апоптоза этих раковых клеток. На цитотоксичность влияют не только сигналы, провоцирующие выработку цитолитических соединений, но также сигналы, ингибирующие их выработку. Следовательно, повышение цитотоксичности приведет к более эффективному уничтожению раковых клеток с меньшим шансом снижения цитотоксичной активности NK-клетки раковой клеткой, как приведено выше.
В качестве способа повышения цитотоксичности NK-клеток в отношении раковых клеток, в которых отсутствует экспрессия ГКГС класса I, но которые могут снижать цитотоксичность NK-клеток, был предложен способ генетических модификаций для устранения функции ингибирующего рецептора NK-клеток (Боддулуру и соавт. 2012). В качестве ингибирующего рецептора, функцию которого стоило бы устранить в данных обстоятельствах, был выбран NKG2A, так как известно, что определенные раковые клетки вырабатывают MICA, связывающий NKG2A, и ингибирующий цитотоксичность NK-клеток при отсутствии экспрессии ГКГС класса I (Шук и соавт. 2011; WO 2006/023148).
В другом способе снижения экспрессии NKG2A была показана связь трансфекции гена, кодирующего IL-15, в клетках NK-92, со снижением экспрессии NKG2A (Чжан и соавт. 2004). При этом, несмотря на наблюдаемое увеличение цитотоксичности NK-клеток, оно, вероятнее всего, стало результатом сопутствующего увеличения экспрессии активирующего рецептора NKG2D. Это подтверждается наблюдениями того, что блокировка рецепторов NKG2A в клетках NK-92 не была связана с повышением цитотоксичности в отношении клеток множественной миеломы (Хайденрайх и соавт. 2012). Тем не менее, необходимо отметить, что линия клеток NK-92 обладает высокой цитотоксичностью с очень низкой экспрессией ингибирующих рецепторов. Следовательно, любое увеличение цитотоксичности, связанное с пониженной экспрессией NKG2A, могло оказаться слишком незначительным для того, чтобы его можно было обнаружить.
Аналогичные исследования проводились на мышах. Например, мыши выделяют рецептор, именуемый Ly49, в NK-клетках, являющийся аналогом человеческих ингибирующих KIR-рецепторов. Было продемонстрировано, что при блокировке рецептора Ly49 фрагментами антител NK-клетки обладают повышенной цитотоксичностью и могут уничтожать клетки лейкемии мышей in vitro и in vivo (Кох и соавт. 2001).
При этом именно вследствие снижения функции ингибирующего рецептора «нормальные» клетки в организме также становятся более предрасположены к воздействию модифицированных NK-клеток, так как способность этих NK-клеток отличать «нормальные» клетки от раковых снижается. Это значительный недостаток снижения «классической» функции ингибирующего рецептора.
Другой известный способ, при котором NK-клетки уничтожают раковые клетки, заключается в экспрессии TRAIL на их поверхности. Лиганд TRAIL может связывать рецепторы TRAIL на раковых клетках и вызывать их апоптоз. В одном предполагаемом подходе описывается сверхэкспрессирование TRAIL в NK-клетках для получения преимуществ этого противоракового механизма (ЕР 1621550). Кроме того, имеются свидетельства того, что IL-12 вызывает повышение экспрессии TRAIL в NK-клетках (Смит и соавт 2001).
Тем не менее, в раковых клетках выработались механизмы уклонения и защиты от NK-клеток с экспрессирующим TRAIL. Рецепторы-приманки TRAIL часто экспрессируются на оболочках раковых клеток, и привязка TRAIL к этим рецепторам-приманкам не может вызвать апоптоз. Способы обхода таких механизмов до настоящего момента не разрабатывались.
Острый миелоидный лейкоз (AML) - это гемопоэтическое злокачественное новообразование, включающее развитие в костном мозгу прекурсорных клеток, и составляющее значительную часть случаев заболевания острой лейкемией как у взрослых (90%), так и у детей (15-20%) (Хурвиц, Маунс и соавт. 1995; Ловенберг, Даунинг и соавт. 1999). Несмотря на то, что у 80% пациентов наблюдается ремиссия при стандартной химиотерапии (Хурвиц, Маунс и соавт. 1995; Рибейро, Раззук и соавт. 2005), выживаемость остается неудовлетворительной вследствие высокой частоты рецидивов при минимальном остаточном заболевании (MRD). Пятилетняя выживаемость зависит от возраста. Она составляет 60% для детей (Рубниц 2012), 40% для взрослых возрастом до 65 лет (Ловенберг, Даунинг и соавт. 1999) и 10% для взрослых возрастом старше 65 лет (Феррара и Шиффер 2013). Эти результаты могут быть улучшены, если у пациентов имеется соответствующий донор гемопоэтических клеток, но у многих его нет, что подчеркивает необходимость в альтернативном подходе к лечению.
Клетки - естественные киллеры (NK) - цитотоксические лимфоциты с выраженными фенотипами и эффекторными функциями, отличающимися, например, от Т-клеток-естественных киллеров (NK-T). Например, в то время как клетками NK-T экспрессируются антигенные рецепторы как CD3, так и Т-клеток (TCR), NK-клетки этого не делают. В целом, установлено, что NK-клетки экспрессируют маркеры CD16 и CD56, причем CD16 действует как рецептор Fc и медиирует антителозависимую клеточноопосредованную цитотоксичность (ADCC), о которой пойдет речь ниже. В данном случае KHYG-1 - существенное исключение. Несмотря на то, что NK-клетки естественно цитотоксичны, были выделены их линии с повышенной цитотоксичностью. NK-92 и KHYG-1 представляют собой две линии NK-клеток, подвергавшихся подробному изучению, и представляющие перспективу при лечении рака (Свифт и соавт. 2011; Свифт и соавт. 2012).
Адоптивная клеточная иммунотерапия при лечении рака зачастую включает в себя введение естественных и модифицированных Т-клеток пациенту. Т-клетки могут быть модифицированы несколькими способами, например, генетически так, чтобы экспрессировались рецепторы и/или лиганды, связываемые с конкретными раковыми клетками-мишенями. Трансфекция Т-клеток с высокоафинными рецепторами этих клеток (TCR) и химерными антигенными рецепторами (CAR), свойственными антигенам раковых клеток, может вызвать высокореакционноспособный отклик Т-клеток на рак. Главным ограничением настоящего иммунотерапевтического подхода является то, что Т-клетки должны браться либо у пациента для автологического размножения способом ex vivo, либо должны быть использованы Т-клетки, совпадающие с ГКГС, для предотвращения иммунологической ликвидации незамедлительно после переноса клеток пациенту или, в некоторых случаях, до развития реакции отторжения трансплантата (GVHD). В дополнение к этому, успешно перенесенные Т-клетки зачастую выживают в кровотоке в течение продолжительных периодов времени, усложняя контроль долгосрочных побочных эффектов вследствие лечения.
При гаплотипной трансплантации считается, что эффект «трансплантат против лейкемии» опосредуется NK-клетками при несовпадении ингибирующих KIR-рецепторов и лиганда, что может привести к повышению выживаемости при лечении AML (Руггери, Капании и соавт. 2002; Руггери, Манкуси и соавт. 2005).
Более того, быстрое восстановление NK связано с лучшим результатом и эффектом «трансплантат против лейкемии» (GVL) у пациентов, проходящих трансплантацию гаплотипных гемопоэтических клеток без Т-элемента (НСТ) при AML (Савани, Мельке и соавт. 2007). При других испытаниях использовались гаплоидентичные NK-клетки, размноженные ex vivo, для лечения AML у взрослых (Миллер, Суанье и соавт. 2005) и детей (Рубниц, Инаба и соавт. 2010).
Были установлены несколько постоянных линий NK-клеток, и самые примечательные - NK-92, взятые у пациента с неходжкинской лимфомой, в которых экспрессируются маркеры NK-клеток, за исключением CD16 (гамма-рецептор Fc III). NK-92 прошел широкие преклинические испытания и демонстрирует повышенный лизис в отношении широкого спектра опухолей по сравнению с активированными NK-клетками и лимфокин-активированными клетками-киллерами (LAK) (Гун, Маки и соавт. 1994). Была установлена цитотоксичность клеток NK-92 в отношении, в основном, AML (Ян, Штайнхерц и соавт. 1998).
Другая линия NK-клеток, KHYG-1, была определена как потенциальный кандидат на клиническое применение (Сак и соавт. 2005), но она обладает сниженной цитотоксичностью, и поэтому ей было уделено меньше внимания, чем NK-92. Известно, что клетки KHYG-1 преактивированы. В отличие от эндогенных NK-клеток клетки KHYG-1 время от времени поляризуются, из-за чего возрастает их цитотоксичность, и они быстрее реагируют на внешние раздражители. Клетки NK-92 обладают большей базовой цитотоксичностью, чем клетки KHYG-1.
Следовательно, ясно, что на текущие протоколы адоптивной иммунотерапии отрицательно влияет смена доноров в плане количества и качества эффекторных клеток - переменных, которые могли бы быть устранены, если бы действующие линии клеток могли обеспечивать более стандартизированную терапию.
Значительный объем исследований цитотоксичности NK-клетки был проведен с использованием мышиных моделей. Один из примеров - установление того, что мРНК перфорина и гранзима В конституитивно транскрибируются в NK-клетки мышей, но до стимуляции или активации NK-клеток наблюдаются минимальные уровни белка (Фенигер и соавт. 2007). Несмотря на то, что данная работа, а также другая работа с использованием NK-клеток мышей, представляют интерес, на них нельзя положиться как на неоспоримое доказательство цитотоксичности NK-клеток у людей. В отличие от примера, приведенного выше, человеческие NK-клетки экспрессируют высокие уровни перфорина и белка гранзима В до начала стимулирования (Леонг и соавт. 2011). Результат: когда либо мышиные, либо человеческие NK-клетки недавно изолированы в культуре, мышиные NK-клетки обладают слабой цитолитической активностью, в то время как человеческие NK-клетки демонстрируют сильные цитолитические способности.
Мышиные и человеческие NK-клетки также сильно отличаются по экспрессионным маркерам, сигнальным каскадам и распределению в тканях. Например, CD56 используется в качестве маркера NK-клеток человека, в то время как NK-клетки мышей вовсе не экспрессируют этот маркер. Кроме того, устоявшийся механизм регулирования цитотоксичности NK-клеток осуществляется через лиганд, связывающий активацию NK-клетки и ингибирующих рецепторов. Два из наиболее выдающихся рецепторов, активирующих NK-клетки человека - NKp30 и NKp44, ни один из которых не экспрессируется в мышиных NK-клетках. Что касается ингибирующих рецепторов NK, в то время как NK-клетки человека экспрессируют KIR, распознающие ГКГС класса I, и снижают цитотоксичную активность, KIR в мышиных NK-клетках не экспрессируется вовсе, но вместо этого экспрессируется Ly49s (Траусдейл и соавт. 2001). В результате, несмотря на то, что мышиные NK-клетки стали выполнять ту же функцию, что и NK-клетки человека в естественной физиологической среде, механизмы, обеспечивающие такое действие, значительно отличаются между видами.
Таким образом, существует необходимость в альтернативном варианте и, предпочтительно, в улучшенных NK-клетках человека и их линиях, например, с более цитотоксичными свойствами.
Цель изобретения - предоставление NK-клеток и их линий с фенотипом с повышенной цитотоксичностью. Дополнительной целью является предоставление способов получения модифицированных NK-клеток и линий NK-клеток, композиций, содержащих эти клетки или их линии, а также вариантов применения этих композиций для лечения рака. Конкретные варианты осуществления направлены на предложение способов лечения определенных раковых заболеваний, например, рака крови, такого как лейкемия. Конкретные варианты осуществления изобретения направлены на сочетание двух или более модификаций NK-клеток и их линий для дополнительного улучшения цитотоксичности модифицированных клеток.
Сущность изобретения
В настоящем документе представлены модифицированные NK-клетки и их линии с фенотипом с повышенной цитотоксичностью, а также способы получения этих клеток и их линий. Также предоставляются композиции модифицированных NK-клеток и их линий, а также варианты применения этих композиций для лечения рака.
В настоящем изобретении предлагаются способы модификации NK-клеток и их линий с помощью, например, генной инженерии, для нокаута генов, кодирующих ингибирующие рецепторы, экспрессии генов, кодирующих лиганды и варианты TRAIL, а также для экспрессии генов, кодирующих химерные антигенные рецепторы (CAR) и/или рецепторы Fc.
Кроме того, композиции по настоящему изобретению включают NK-клетки и их линии, в которых имеются две или несколько модификаций, причем многочисленные модификации дополнительно повышают цитотоксическую активность композиции.
В соответствии с настоящим изобретением имеются дополнительные способы лечения рака, например, рака крови с помощью модифицированных линий NK-клеток, например, производных клеток KHYG-1. При этом модифицированные линии NK-клеток получают с отсутствием ингибирующих рецепторов контрольной точки для экспрессии вариантов лигандов TRAIL и/или экспрессии CAR и/или рецепторов Fc.
В частности, к заболеваниям, поддающимся лечению в соответствии с настоящим изобретением, относятся раковые заболевания, рак крови, лейкемия и, в особенности, острый миелоидный лейкоз. При этом могут поддаваться лечению опухоли и раковые заболевания человека. Ссылки на опухоли в настоящем документе включают ссылки на новообразования.
Подробное описание изобретения
Соответственно, в настоящем изобретении предлагается клетка-естественный киллер (NK) или линия NK-клетки, которая была генетически модифицирована с целью повышения цитотоксичности.
Согласно подробному описанию в примерах ниже, NK-клетки и линии NK-клеток были подвержены генетической модификации для увеличения их цитотоксической активности против рака.
Совместно NK-клетки и линии NK-клеток в настоящем изобретении будут именоваться NK-клетками (если контекстом не подразумевается иное).
В определенных вариантах осуществления настоящего изобретения предусматриваются NK-клетки с пониженной функцией ингибирующих рецепторов контрольной точки или без нее. Таким образом, в нижеуказанных примерах NK-клетки образуются при условии нокаута одного или более генов ингибирующих рецепторов контрольной точки. Предпочтительно, чтобы эти рецепторы являлись определенными ингибирующими рецепторами контрольной точки. Также предпочтительно, чтобы один, несколько или все из этих ингибирующих рецепторов контрольной точки принадлежали к CD96 (TACTILE), CD152 (CTLA4), CD223 (LAG-3), CD279 (PD-1), CD328 (SIGLEC7), SIGLEC9, TIGHT и/или TIM-3.
В других вариантах осуществления предусматриваются NK-клетки, в которых один или несколько сигнальных путей ингибирующих рецепторов находятся в нокауте или проявляют пониженную функцию - в результате функция ингибирующего рецептора также понижается или отсутствует. Например, сигнальные пути, опосредованные SHP-1, SHP-2 и/или SHIP, нокаутируются посредством генетической модификации клеток.
Полученные NK-клетки проявляют улучшенную цитотоксичность и, таким образом, повышенную применимость в терапии рака, особенно при злокачественном заболевании крови, в особых способах лечения лейкемии и множественной миеломы.
В одном из вариантов осуществления изобретения генетическая модификация происходит до видоизменения клетки в NK-клетку. Например, плюрипотенциальные стволовые клетки (например, iPSC) могут генетически модифицироваться для потери способности экспрессии ингибирующих рецепторов контрольной точки. Затем модифицированные iPSC видоизменяются для получения генетически модифицированных NK-клеток с повышенной цитотоксичностью.
Предпочтительно снижать функцию ингибирующих рецепторов контрольной точки в отношении других ингибирующих рецепторов, в связи с экспрессией первых из упомянутых, которая происходит после активации NK-клеток. Нормальные или «классические» ингибирующие рецепторы, например, большинство KIR-семейства, NKG2A и LIR-2, связывают ГКГС класса I и, следовательно, в первую очередь задействованы в понижении проблемы самонаведения. Таким образом, предпочтительно нокаутировать ингибирующие рецепторы контрольной точки. Пониженная функция этих рецепторов или ее отсутствие по настоящему изобретению не позволяет раковым клеткам супрессировать функцию иммунных эффекторов (которая в ином случае может возникнуть в случае полной функциональности рецепторов). Таким образом, ключевое преимущество этих вариантов осуществления настоящего изобретения заключается в NK-клетках, которые являются менее восприимчивыми к супрессии их цитотоксичной активности раковыми клетками, что приводит к практическим результатам в лечении рака.
В контексте настоящего документа ссылки на ингибирующие рецепторы, как правило, относятся к рецепторам, экспрессированным на цитоплазматической оболочке иммунной эффекторной клетки, например, NK-клетки; после привязки ее дополнительного лиганда внутриклеточные сигналы отвечают за снижения цитотоксичности иммунной эффекторной клетки. Эти ингибирующие рецепторы экспрессируются как в состоянии «покоя», так и в «активном» состоянии иммунных эффекторных клеток, и часто связаны с обеспечением механизма аутотолерантности иммунной системы, который ингибирует цитотоксичную ответную реакцию на клетки и ткани организма. Примером является ингибирующий рецептор KIR-семейства, который экспрессируется на NK-клетках, и распознает ГКГС класса I, экспрессированный на здоровых клетках организма.
Также в контексте настоящего документа ингибирующие рецепторы контрольной точки обычно считаются разновидностью ингибирующих рецепторов, указанных выше. При этом в отличие от других ингибирующих рецепторов ингибирующие рецепторы контрольной точки экспрессируются на более высоких уровнях при продолжительном периоде активации и цитотоксичности иммунной эффекторной клетки, например NK-клетки. Данный феномен является полезным для ослабления хронической цитотоксичности, например, в очаге воспаления. Примеры включают ингибирующие рецепторы контрольной точки PD-1, CTLA-4 и CD96, каждый из которых экспрессируется на NK-клетках.
Настоящее изобретение также предусматривает NK-клетку без гена, отвечающего за ингибирующий рецептор контрольной точки, выбранный из CD96 (TACTILE), CD152 (CTLA4), CD223 (LAG-3), CD279 (PD-1), CD328 (SIGLEC7), SIGLEC9, TIGIT и TIM-3.
NK-клетка без гена может относиться к полной или частичной делеции, мутации или, в противном случае, может привести к отсутствию экспрессии продукта функционального гена. В некоторых вариантах осуществления изобретения NK-клетки не содержат гены, ответственные за два или более ингибирующих рецепторов.
Более конкретные варианты осуществления изобретения включают NK-клетку без гена, ответственного за ингибирующие рецепторы контрольной точки, выбранные из CD96 (TACTILE), CD152 (CTLA4) и CD279 (PD-1). Предпочтительные варианты осуществления включают NK-клетку, полученную из KHYG-1.
В примерах, описанных ниже, авторы настоящего изобретения в достаточной степени показали цитотоксический эффект при использовании малой интерферирующей РНК для нокдауна экспрессии ингибирующего рецептора контрольной точки CD96 в клетках KHYG-1. Нокдаун клеток (KD) KHYG-1 посредством CD96 продемонстрировал повышенную цитотоксичность в отношении клеток лейкемии при различных соотношениях эффектор : мишень (Э : М).
В других вариантах осуществления настоящего изобретения предусматриваются NK-клетки, экспрессирующие лиганд TRAIL или, предпочтительно, мутировавший (вариантный) лиганд TRAIL. Согласно описанию в примерах ниже, модификации, повышающие цитотоксичность NK-клеток, также включают повышенную экспрессию лиганда TRAIL и/или вариантов мутировавшего лиганда TRAIL.
Полученные NK-клетки проявляют повышенную привязку к рецепторам TRAIL и благодаря этому повышенную цитотоксичность в отношении раковых заболеваний, особенно злокачественных заболеваниях крови, определенных видах лейкемии.
Предпочтительно, чтобы мутации/варианты имели низкую (или фактически не имели) аффинность для рецепторов-приманок в сравнении с привязкой TRAIL дикого типа к рецепторам-приманкам. Такие рецепторы-приманки представляют класс рецепторов TRAIL, которые связывают лиганд TRAIL, но не могут инициировать гибель клеток, а также, в некоторых случаях, противодействовать сигнальному пути апоптоза. Мутировавшие/вариантные лиганды TRAIL могут подготавливаться в соответствии с WO 2009/077857.
Мутации/варианты могут отдельно повышать аффинность для рецепторов TRAIL, например, DR4 и DR5. TRAIL дикого типа обычно содержит KD>2 нМ для DR4,>5 нМ для DR5 и >20 нМ для рецептора-приманки DcR1 (WO 2009/077857; измерено с помощью поверхностного плазмонного резонанса), или около 50-100 нМ для DR4, 1-10 нМ для DR5 и 175-225 нМ для DcR1 (Трунех, А. и соавт. 2000; измерено с помощью изотермической титрационной калориметрии и ELISA). Следовательно, повышенная аффинность для DR4 надлежащим образом определяется как KD<2 нМ или <50 нМ, соответственно, в то время как повышенная аффинность для DR5 надлежащим образом определяется как KD<5 нМ или <1 нМ, соответственно. Пониженная аффинность для рецептора-приманки DcR1 надлежащим образом определяется как KD>50 нМ или >225 нМ, соответственно. В любом случае повышение или понижение аффинности, которая проявляется посредством варианта/мутации TRAIL, относится к базовой аффинности, которая проявляется посредством TRAIL дикого типа. Предпочтительно, чтобы аффинность увеличивалась минимум на 10%, более предпочтительно - минимум на 25%, в сравнении с проявлением посредством TRAIL дикого типа.
Предпочтительно, чтобы вариант TRAIL имел повышенную аффинность для DR5 в сравнении с ее аффинностью для DR4, DcR1 и DcR2. Предпочтительно, чтобы аффинность для DR5 превышала аффинность для одного или нескольких DR4, DcR1 и DcR2 как минимум в 1,5, 2, 5, 10, 100 или даже в 1000 раз или более. Более предпочтительно, чтобы аффинность для DR5 превышала аффинность для минимум двух и, желательно, всех DR4, DcR1 и DcR2 как минимум в 1,5, 2, 5, 10, 100 или даже в 1000 раз или более.
Ключевое преимущество этих вариантов осуществления настоящего изобретения заключается в том, что NK-клетки являются более эффективными в уничтожении раковых клеток.
Далее конкретные варианты осуществления включают NK-клетку, экспрессирующую мутировавший лиганд TRAIL, аффинность которого для рецепторов-приманок TRAIL понижена или отсутствует. Предпочтительно, чтобы NK-клетка была получена из KHYG-1. Далее конкретные варианты осуществления включают NK-клетку, экспрессирующую мутировавший лиганд TRAIL, аффинность которого для рецепторов-приманок TRAIL понижена или отсутствует, а для DR4 и/или DR5 - повышена.
В примерах настоящего изобретения, более подробно описанных ниже, NK-клетки были генетически модифицированы для экспрессии мутировавшего TRAIL. Модифицированные клетки KHYG-1 экспрессировали мутировавший TRAIL и NK-92 экспрессировали мутировавший TRAIL. Модифицированные клетки KHYG-1 проявляли улучшенную цитотоксичность в отношении линий раковых клеток по технологии in vitro. Клетки KHYG-1 экспрессируют рецепторы TRAIL (например, DR4 и DR5), но на более низких уровнях. В других предпочтительных вариантах осуществления модифицированных NK-клеток рецепторы TRAIL не экспрессируются вообще или экспрессируются в недостаточной степени, или только на низком уровне - достаточно низком, чтобы на жизнеспособность модифицированных NK-клеток не оказывала негативное влияние экспрессия мутировавшего TRAIL.
В дополнительном варианте осуществления лечение рака с помощью модифицированных NK-клеток, экспрессирующих TRAIL или вариант TRAIL, усиливается посредством ввода пациенту агента, способного повысить экспрессию рецепторов смерти TRAIL на раковых клетках. Данный агент может вводиться перед, в сочетании или после ввода модифицированных NK-клеток. При этом предпочтительно, чтобы агент вводился перед вводом модифицированных NK-клеток.
В предпочтительном варианте осуществления агент вызывает повышение экспрессии DR5 на раковых клетках. Дополнительно в качестве агента может использоваться химиотерапевтический препарат, например, Бортезомиб, который вводится небольшой дозой, способной вызывать повышение экспрессии DR5 на раковых клетках.
Данное изобретение не ограничивается какими-либо конкретными агентами, способными вызывать повышение эксперессии DR5; следующие агенты, содержащие DR5, приведены в качестве примеров: Бортезомиб, гефитиниб, пайперлонгумин, доксорубицин, альфа-токоферол сукцинат и ингибиторы HDAC.
В соответствии с предпочтительным вариантом осуществления настоящего изобретения мутировавший/вариативный лиганд TRAIL связывается с одним или несколькими костимулирующими доменами NK-клетки, например, 41BB/CD137, CD3zeta/CD247, DAP12 или DAP10. Привязка варианта к рецептору на клетке-мишени способствует апоптическим сигналам в пределах клетки-мишени, а также стимулирует цитотоксичные сигналы в NK-клетке.
В дополнительных предпочтительных вариантах осуществления настоящего изобретения предусматриваются NK-клетки, которые имеют пониженную функцию ингибирующих рецепторов контрольной точки, а также экспрессируют мутировавший лиганд TRAIL, согласно более подробному описанию ниже, в отношении этих соответствующих модификаций NK-клеток. В более предпочтительных вариантах осуществления NK-клетки экспрессируют мутировавший лиганд TRAIL с пониженным содержанием или отсутствием аффинности для рецепторов-приманок TRAIL и могут быть получены из KHYG-1, также без гена, кодирующего ингибирующий рецептор контрольной точки, выбранный из CD96 (TACTILE), CD152 (CTLA4), CD223 (LAG-3), CD279 (PD-1), CD328 (SIGLEC7), SIGLEC9, TIGIT и TIM-3.
В настоящем изобретении также предусматриваются NK-клетки и линии NK-клеток, предпочтительно клеток KHYG-1, а также их производные, модифицированные для экспрессии одного или нескольких CAR.
С расчетом на применение в терапии рака, CAR специфически связываются с одним или несколькими лигандами на раковых клетках, например, CS1 (SLAMF7) на клетках миеломной болезни. Для использования в лечении конкретных видов раковых заболеваний, например, множественной миеломы, CAR может связываться с CD38. Например, CAR может включать свойства связывания различных областей, полученных от, схожих или идентичных производным от известного моноклонального антитела (даратумумаб). Такие NK-клетки могут использоваться в терапии рака в сочетании с агентом, ингибирующим ангиогенез, например, леналидомидом. Для использования в терапии раковых заболеваний, особенно лейкемии и AML, CAR могут связываться CLL-1.
CAR-NK могут быть биспецифичными, причем, их аффинность применяется для двух отдельных лигандов/антигенов. Биспецифичные CAR-NK могут использоваться либо для увеличения количества потенциальных участков связывания на раковых клетках, либо, в качестве альтернативного варианта, для локализации раковых клеток для других иммунных эффекторных клеток, которые экспрессируют лиганды специально для NK-CAR. Для использования в терапии рака биспецифичный CAR может связываться с опухолевой клеткой-мишенью и эффекторной клеткой, например, Т-клеткой, NK-клеткой или макрофагоцитом. Так, например, в случае множественной миеломы биспецифичный CAR может сзываться с антигеном Т-клетки (например, CD3 и т.д.) и маркером опухолевой клетки (например, CD38 и т.д.). В качестве альтернативного варианта биспецифичный CAR может связываться с двумя отдельными маркерами опухолевых клеток, повышая аффинность связывания NK-клетки для опухолевых клеток мишеней. Это может снизить риск раковых клеток, развивая сопротивление с помощью одного или нескольких антигенов-мишеней. В качестве примера в данном случае (случае множественной меиломы) можно привести привязку CAR к CD38 и CS-1/SLAMF7. Другой маркер опухолевой клетки, на который нацелен CAR, является маркером типа «не ешь меня» на опухолях, примером которого является CD47.
Дополнительные признаки изобретения включают обеспечение дальнейших модификаций NK-клеток и их линий, описанных выше, причем, например, рецептор Fc (который может представлять собой CD16, CD32 или CD64, включая подтипы и производные), экспрессируется на поверхности клетки. При использовании эти клетки могут характеризоваться повышенным распознаванием раковых клеток, покрытых антителами, и улучшать активацию цитотоксичной ответной реакции.
Дополнительные признаки изобретения включают адаптацию модифицированных NK-клеток и их линий для улучшения наведения в конкретные целевые части тела. NK-клетки в настоящем изобретении могут быть нацелены на конкретные участки раковых клеток. В предпочтительных вариантах осуществления для лечения рака крови, NK-эффекторы в настоящем изобретении адаптированы для наведения в костный мозг. Конкретные NK-клетки модифицированы путем фукозилирования и/или сиалирования для наведения в костный мозг. Это может быть достигнуто путем генетической модификации NK-клеток для экспрессии фукозилтрансферазы и/или сиалилтрансферазы, соответственно. Улучшенное наведение NK-эффекторных клеток в опухолевые участки также может быть обеспечено путем нарушения сосудистой сети опухоли, например, методом метрономической химиотерапии или с использованием препаратов, нацеленных на ангиогенез (Мелеро и соавт. 2014) для нормализации инфильтрации NK-клеток через раковые кровеносные сосуды.
Еще одним дополнительным признаком изобретения является обеспечение модифицированных NK-клеток и их линий с повышенной эндогенной способностью быстрого роста и разрастания культуры. Например, это может быть достигнуто путем трансфицирования клеток для сверхэкспрессирования стимулирующих рост цитокинов IL-2 и IL-15. Кроме того, это дополнительное изменение обеспечивает экономически эффективную альтернативу пополнению среды для роста с цитокинами на постоянной основе.
Настоящее изобретение также предусматривает способ получения модифицированной NK-клетки или ее линии, включая генетическую модификацию клетки или ее линии, как описано в настоящем документе, с целью повышения ее цитотоксичности. Данная генетическая модификация может быть стабильным нокаутом гена, например, CRISPR, или временным нокдауном гена, например, малая интерферирующая РНК.
В предпочтительном варианте осуществления используется способ стабильной генетической модификации, например, CRISPR, для получения новой линии NK-клетки с повышенной цитотоксичностью, например, производная клеток KHYG-1.
В некоторых вариантах осуществления изобретения способ предусмотрен для получения NK-клетки или ее линии, которая была модифицирована для снижения функции ингибирующего рецептора. Предпочтительно, чтобы эти ингибирующие рецепторы являлись ингибирующими рецепторами контрольной точки.
Более конкретные варианты осуществления изобретения включают способ получения NK-клетки или ее линии с пониженной функцией ингибирующего рецептора, причем, ингибирующие рецепторы контрольной точки выбираются из CD96 (TACTILE), CD152 (CTLA4), CD223 (LAG-3), CD279 (PD-1), CD328 (SIGLEC7), SIGLEC9, TIGIT и TIM-3.
В предпочтительных вариантах осуществления способ включает модификацию NK-клеток для снижения функции двух или более ингибирующих рецепторов.
Настоящее изобретение также предусматривает способ получения модифицированной NK-клетки или ее линии, включая генетическую модификацию клетки или ее линии для экспрессии лиганда TRAIL или мутировавшего (вариантного) лиганда TRAIL.
В некоторых вариантах осуществления изобретения способ включает модификацию NK-клетки или ее линии для экспрессии мутировавшего лиганда TRAIL с повышенной аффинностью для рецепторов TRAIL. Предпочтительно, рецепторами TRAIL являются DR4 и/или DR5. Предпочтительные варианты осуществления изобретения предусматривают способ модификации NK-клеток или их линий для экспрессии мутировавшего лиганда TRAIL с пониженной аффинностью для рецепторов-приманок TRAIL.
В дополнительных предпочтительных вариантах осуществления способ включает модификацию NK-клетки или ее линии для исключения функции ингибирующего рецептора контрольной точки, а также экспрессии мутировавшего лиганда TRAIL с пониженной аффинностью или без аффинности связывания для рецепторов-приманок TRAIL.
Дополнительные стандартные варианты осуществления предусматривают способ получения NK-клетки или ее линии, при котором функция одного или более ингибирующих рецепторов контрольной точки исключена и/или экспрессируется мутировавший лиганд TRAIL с пониженной аффинностью или без аффинности связывания для рецепторов-приманок TRAIL, а клетка дополнительно модифицируется для экспрессии CAR или биспецифичного CAR. Свойства CAR являются дополнительными, как представлено выше.
В некоторых вариантах осуществления изобретения способ включает получение NK-клетки или ее линии, при котором функция одного или более ингибирующих рецепторов контрольной точки исключена и/или экспрессируется мутировавший лиганд TRAIL с пониженной аффинностью или без аффинности связывания для рецепторов-приманок TRAIL, а клетка дополнительно модифицируется для экспрессии CAR или биспецифичного CAR, а также клетка дополнительно модифицируется для экспрессии одного или более рецепторов Fc. Соответствующие рецепторы Fc выбираются из CD16 (FcRIII), CD32 (FcRII) и CD64 (FcRI).
Предпочтительные варианты осуществления изобретения из вышеуказанного включают способ получения NK-клеток и их линий, полученных из KHYG-1.
Согласно целям настоящего изобретения модифицированная NK-клетка, ее линия или состав с повышенной цитотоксичностью предусмотрены для использования при лечении рака у пациента, в частности рака крови.
В предпочтительных вариантах осуществления модифицированная NK-клетка, ее линия или состав предусматривается для использования при лечении рака крови, включая острый лимфоцитарный лейкоз (ALL), острый миелоидный лейкоз (AML), хронический лимфоцитарный лейкоз (CLL), хронический миелоидный лейкоз (CML), ходжкинскую лимфому, неходжкинскую лимфому, включая Т-клеточные и В-клеточные лимфомы, асимптомную миелому, вялотекущую множественную миелому (SMM), активную миелому или миелому легкой цепи.
В более предпочтительных вариантах осуществления изобретение предусматривает линию NK-клетки, полученную из KYHG-1 путем снижения функции ингибирующих рецепторов контрольной точки в клетке KHYG-1 или экспрессии мутировавшего лиганда TRAIL в клетке KHYG-1, или обоих, для использования при лечении рака крови.
Модифицированные NK-клетки, их линии и составы, описанные в настоящем документе, выше и ниже, подходят для лечения рака, в частности, раковых заболеваний человека, например, для лечения рака крови или солидного рака. Предпочтительно, NK-клетками и производными являются NK-клетки человека. Для лечения пациента предпочтительно используются NK-клетки человека.
Различные способы ввода хорошо известны специалисту для введения активных веществ и их комбинаций в организм пациента. Варианты осуществления настоящего изобретения предусмотрены для лечения рака крови. Введение модифицированных NK-клеток и/или их линий может быть системным или локальным, например, интраперитонеальное введение.
В других вариантах осуществления настоящего изобретения активное вещество вводится напрямую. Таким образом, введение может быть внутриопухолевым, в частности, подходит для солидных опухолей.
Как правило, NK-клетки считаются пригодными для способов, типов применения и композиции по настоящему изобретению. Сообразно клеткам, используемым в определенных примерах, NK-клеткой может быть клетка, полученная из линии раковой клетки. Предпочтительно, NK-клетка, прошедшая обработку для снижения онкогенности, например, путем получения мортальной клетки и/или клетки, которая не может делиться, может быть получена из линии кровяной раковой клетки и может использоваться в способах по изобретению для лечения рака крови.
Для получения раковой клетки, которая является более приемлемой для использования в терапевтических целях, ее обычно обрабатывают или предварительно обрабатывают определенным образом для снижения или исключения возможности образования опухолей у пациента. Конкретные линии модифицированных NK-клеток, используемые в примерах, являются безопасными, поскольку они не могут делиться; они облучены и сохраняют свою цитотоксическую активность, но погибают примерно в течение 3-4 дней. Таким образом, конкретные клетки и их линии не могут разрастаться, например, в результате облучения. Способы обработки потенциальных NK-клеток для использования в способах, описанных в настоящем документе, включают облучение во избежание их деления и образования опухоли in vivo и генетическую модификацию для снижения онкогенности, например, для последовательности, кодирующей «суицидальный» ген, который может быть активирован во избежание деления клеток и образования опухоли in vivo. «Суицидальные» гены могут быть активированы экзогенными, например, циркулирующими веществами, которые вызывают гибель клеток в клетках, экспрессирующих ген. Еще одной альтернативой является использование моноклональных антител, нацеленных на конкретные NK-клетки для терапии. Например, CD52 экспрессируется на клетках KHYG-1, а привязка моноклональных антител к этому маркеру может вызвать антителозависимую клеточноопосредованную цитотоксичность (ADCC) и гибель клеток KHYG-1.
Как указано в статье, которую опубликовали Сак и соавт. 2006, раковые NK-клетки и их линии легко облучаются с использованием облучателей, например, Gammacell 3000 Elan. Источник цезия-137 используется для контроля дозировки облучения, а кривая зависимости «доза-эффект», например, между 1 Gy и 50 Gy может использоваться для определения оптимальной дозы для исключения пролиферативной способности клеток, сохраняя при этом преимущества повышенной цитотоксичности. Это достигается с помощью анализа клеток на цитотоксичность после введения каждой дозы облучения.
Наблюдаются существенные преимущества использования линии облученной NK-клетки для адоптивной клеточной иммунотерапии в сравнении с устоявшимся подходом аутологичных клеток или Т-клеток, совпадающих с ГКГС. Во-первых, использование линии NK-клетки с высокой пролиферативной способностью подразумевает, что рост линий модифицированных NK-клеток может быть обеспечен проще на коммерческом уровне. Облучение линии модифицированной NK-клетки может быть выполнено до введения клеток в организм пациента. Эти облученные клетки, сохраняющие свою полезную цитотоксичность, имеют ограниченный жизненный цикл и, в отличие от модифицированных Т-клеток, не циркулируют в течение длительного периода времени, вызывая при этом долгосрочные побочные эффекты.
Кроме того, использование аллогенных модифицированных NK-клеток и их линий подразумевает, что клетки экспрессии ГКГС класса I в организме пациента не могут ингибировать цитотоксичную ответную реакцию NK-клеток таким же образом, как для цитотоксичной ответной реакции аутологичных NK-клеток. Преимуществом использования аллогенных NK-клеток и их линий для уничтожения раковых клеток является ранее указанный эффект GVL и, в отличие от Т-клеток, аллогенные NK-клетки и их линии не стимулируют развитие реакции GVHD, что делает их предпочтительным вариантом лечения рака с использованием адоптивной клеточной иммунотерапии.
Как указано в формуле изобретения и в другом месте данного документа, в настоящем изобретении предусмотрены следующие варианты осуществления:
1. Клетка-естественный киллер (NK) или линия NK-клетки, которая была генетически модифицирована с целью повышения ее цитотоксичности.
2. NK-клетка или ее линия по варианту осуществления 1, модифицированная для обеспечения пониженной функции одного или более ингибирующих рецепторов.
3. NK-клетка или ее линия по варианту осуществления 2, отличающаяся тем, что ингибирующими рецепторами являются ингибирующие рецепторы контрольной точки.
4. NK-клетка или ее линия по варианту осуществления 3, отличающаяся тем, что ингибирующие рецепторы контрольной точки выбираются из CD96 (TACTILE), CD152 (CTLA4), CD223 (LAG-3), CD279 (PD-1), CD328 (SIGLEC7), SIGLEC9, TIGIT и TIM-3.
5. NK-клетка или ее линия по любому из вариантов осуществления 2-4, модифицированная для обеспечения пониженной функции двух или более ингибирующих рецепторов.
6. NK-клетка или ее линия по любому из вариантов осуществления 1 - 5, модифицированная для экспрессии лиганда TRAIL.
7. NK-клетка или ее линия по варианту осуществления 6, отличающаяся тем, что лигандом TRAIL является мутировавший лиганд TRAIL.
8. NK-клетка или ее линия по варианту осуществления 7, отличающаяся тем, что мутировавший лиганд TRAIL имеет повышенную аффинность для рецепторов TRAIL, например, DR4 и/или DR5.
9. NK-клетка или ее линия по любому из вариантов осуществления 7-8, отличающаяся тем, что мутировавший лиганд TRAIL имеет пониженную аффинность для рецепторов-приманок TRAIL.
10. NK-клетка или ее линия по любому из предыдущих вариантов осуществления, модифицированная для исключения функции ингибирующего рецептора контрольной точки, а также экспрессии мутировавшего лиганда TRAIL с пониженной аффинностью или без аффинности связывания для рецепторов-приманок TRAIL.
11. NK-клетка или ее линия по любому из предыдущих вариантов осуществления, экспрессирующая химерный антигенный рецептор (CAR).
12. NK-клетка или ее линия по варианту осуществления 11, отличающаяся тем, что CAR является биспецифичный CAR.
13. NK-клетка или ее линия по варианту осуществления 12, отличающаяся тем, что биспецифичный CAR связывает два лиганда по одному типу клетки.
14. NK-клетка или ее линия по варианту осуществления 12, отличающаяся тем, что биспецифичный CAR связывает один лиганд по любому из двух отдельных типов клеток.
15. NK-клетка или ее линия по вариантам осуществления 11 и 12, отличающаяся тем, что лиганд(-ы) для CAR или биспецифичного CAR экспрессируются на раковой клетке.
16. NK-клетка или ее линия по варианту осуществления 13, отличающаяся тем, что лиганды для биспецифичного CAR экспрессируются на раковой клетке.
17. NK-клетка или ее линия по варианту осуществления 14, отличающаяся тем, что лиганды для биспецифичного CAR экспрессируются на раковой и иммунной эффекторной клетке.
18. NK-клетка или ее линия по любому из предыдущих вариантов осуществления, модифицированная для экспрессии одного или более рецепторов Fc.
19. NK-клетка или ее линия по варианту осуществления 18, отличающаяся тем, что рецепторы Fc выбираются из CD16 (FcRIII), CD32 (FcRII) и CD64 (FcRI).
20. NK-клетка или ее линия по любому из предыдущих вариантов осуществления, отличающаяся тем, что линия клетки получена из линии клетки KHYG-1.
21. NK-клетка без гена, отвечающего за ингибирующий рецептор контрольной точки, выбранный из CD96 (TACTILE), CD152 (CTLA4), CD223 (LAG-3), CD279 (PD-1), CD328 (SIGLEC7), SIGLEC9, TIGIT и TIM-3.
22. NK-клетка по варианту осуществления 21 без генов, отвечающих за два или более ингибирующих рецептора контрольной точки, выбранных из CD96 (TACTILE), CD152 (CTLA4), CD223 (LAG-3), CD279 (PD-1), CD328 (SIGLEC7), SIGLEC9, TIGIT и TIM-3.
23. NK-клетка по варианту осуществления 21 или 22, отличающаяся тем, что ингибирующий рецептор контрольной точки выбирается из CD96 (TACTILE), CD152 (CTLA4) и CD279 (PD-1).
24. NK-клетка по любому из вариантов осуществления 21-23, полученная из KHYG-1.
25. NK-клетка, экспрессирующая мутировавший лиганд TRAIL, аффинность которого для рецепторов-приманок TRAIL понижена или отсутствует.
26. NK-клетка по варианту осуществления 25, полученная из KHYG-1.
27. NK-клетка по варианту осуществления 25 или 26 без гена, отвечающего за ингибирующий рецептор контрольной точки, выбранный из CD96 (TACTILE), CD152 (CTLA4), CD223 (LAG-3), CD279 (PD-1), CD328 (SIGLEC7), SIGLEC9, TIGIT и TIM-3.
28. NK-клетка или ее линия по любому из предыдущих вариантов осуществления, которая не может разрастаться, например, в результате облучения.
29. Способ получения модифицированной NK-клетки или ее линии, включая генетическую модификацию клетки или ее линии с целью повышения ее цитотоксичности.
30. Способ по варианту осуществления 29, отличающийся тем, что NK-клетка или ее линия модифицируется для снижения функции ингибирующего рецептора.
31. Способ по варианту осуществления 30, отличающийся тем, что ингибирующими рецепторами являются ингибирующие рецепторы контрольной точки.
32. Способ по варианту осуществления 31, отличающийся тем, что ингибирующие рецепторы контрольной точки выбираются из CD96 (TACTILE), CD152 (CTLA4), CD223 (LAG-3), CD279 (PD-1), CD328 (SIGLEC7), SIGLEC9, TIGIT и TIM-3.
33. Способ по любому из вариантов осуществления 29-32, включающий модификацию NK-клеток для снижения функции двух или более ингибирующих рецепторов.
34. Способ по любому из вариантов осуществления 29-33, включающий модификацию NK-клетки или ее линии для экспрессии лиганда TRAIL или мутировавшего лиганда TRAIL.
35. Способ по варианту осуществления 34, отличающийся тем, что мутировавший лиганд TRAIL имеет повышенную аффинность для рецепторов TRAIL.
36. Способ по варианту осуществления 35, отличающийся тем, что рецепторами TRAIL являются DR4 и/или DR5.
37. Способ по любому из вариантов осуществления 34-36, отличающийся тем, что мутировавший лиганд TRAIL имеет пониженную аффинность для рецепторов-приманок TRAIL.
38. Способ по любому из вариантов осуществления 29-37, отличающийся тем, что NK-клетка или ее линия модифицируется для исключения функции ингибирующего рецептора контрольной точки, а также экспрессии мутировавшего лиганда TRAIL с пониженной аффинностью или без аффинности связывания для рецепторов-приманок TRAIL.
39. Способ по варианту осуществления 38, отличающийся тем, что NK-клетка или ее линия модифицируется для экспрессии CAR или биспецифичного CAR.
40. Способ по варианту осуществления 39, отличающийся тем, что биспецифичный CAR связывает два лиганда по одному типу клетки.
41. Способ по варианту осуществления 39, отличающийся тем, что биспецифичный CAR связывает один лиганд по любому из двух отдельных типов клеток.
42. Способ по варианту осуществления 39, отличающийся тем, что лиганд(-ы) для CAR или биспецифичного CAR экспрессируются на раковой клетке.
43. Способ по варианту осуществления 40, отличающийся тем, что лиганды для биспецифичного CAR экспрессируются на раковой клетке.
44. Способ по варианту осуществления 41, отличающийся тем, что лиганды для биспецифичного CAR экспрессируются на раковой и иммунной эффекторной клетке.
45. Способ по любому из вариантов осуществления 29-44, отличающийся тем, что NK-клетка или ее линия модифицируется для экспрессии одного или более рецепторов Fc.
46. Способ по варианту осуществления 45, отличающийся тем, что рецепторы Fc выбираются из CD16 (FcRIII), CD32 (FcRII) и CD64 (FcRI).
47. Способ по любому из вариантов осуществления 29-46, отличающийся тем, что линия клетки получена из линии клетки KHYG-1.
48. NK-клетка или ее линия, полученная способом по любому из вариантов осуществления 29 - 47.
49. KHYG-1, полученная способом по любому из вариантов осуществления 29-48.
50. Модифицированная NK-клетка, ее линия или состав с повышенной цитотоксичностью для использования при лечении рака у пациента.
51. NK-клетка или ее линия по любому из вариантов осуществления 1-28, либо полученная по любому из вариантов осуществления 29-49, для использования по варианту осуществления 50.
52. Модифицированная NK-клетка, ее линия или состав для использования по варианту осуществления 50 или 51, отличающаяся тем, что типом рака является рак крови.
53. Модифицированная NK-клетка, ее линия или состав для использования по варианту осуществления 52, отличающаяся тем, что типом рака крови является острый лимфоцитарный лейкоз (ALL), острый миелоидный лейкоз (AML), хронический лимфоцитарный лейкоз (CLL), хронический миелоидный лейкоз (CML), ходжкинская лимфома, неходжкинская лимфома, включая Т-клеточные и В-клеточные лимфомы, асимптомную миелому, вялотекущую множественную миелому (SMM), активную миелому или миелому легкой цепи.
54. Линия NK-клетки, полученная из KYHG-1 путем снижения функции ингибирующих рецепторов контрольной точки в клетке KHYG-1 или экспрессии мутировавшего лиганда TRAIL в клетке KHYG-1, или обоих, для использования при лечении рака крови.
Примеры
Настоящее изобретение имеет более подробное и конкретное описание получения производных NK-клеток KHYG-1, измененных для проявления большей цитотоксической активности и, таким образом, возможности приводить к гибели клетки лейкемии в клинической практике.
Данное изобретение иллюстрируется с помощью конкретных вариантов осуществления со ссылкой на прилагаемые чертежи, где:
На фиг. 1 показана последовательность ДНК целевой области гена LIR2 и отмечены фланкирующие области «руководящей» РНК;
На фиг. 2 показана последовательность ДНК целевой области гена CTLA4 и отмечены фланкирующие области «руководящей» РНК;
На фиг. 3 показана структура РНК (экспрессирующий вектор), используемая для трансфекции;
На фиг. 4 показаны полосы после электрофореза в геле для первичной и мутировавшей ДНК LIR2 перед и после трансфекции;
На фиг. 5 показаны полосы после электрофореза в геле для первичной и мутировавшей ДНК CTLA4 перед и после трансфекции;
Фиг. 6А представляет собой график FACS, на котором показан успешный нокдаун CD96 посредством электроимпульсного открытия клеточных пор;
Фиг. 6В представляет собой график FACS, на котором показан успешный нокдаун CD96 посредством электроимпульсного открытия клеточных пор;
Фиг. 7 представляет собой гистограмму, на которой показана повышенная цитотоксичность нокдауна клеток KHYG-1 посредством CD96, по сравнению с клетками K562 при различных соотношениях Э : М;
На фиг. 8 показан нокдаун CD328 (Siglec-7) в клетках NK-92;
На фиг. 9 показана цитотоксичность NK-клеток, повышенная посредством нокдауна CD328 (Siglec-7);
На фиг. 10 показан график FACS базовой экспрессии TRAIL на клетках KHYG-1;
На фиг. 11 показан график FACS экспрессии TRAIL и варианта TRAIL после трансфекции клеток KHYG-1;
На фиг. 12 показан график FACS экспрессии CD107a после трансфекции клеток KHYG-1;
На фиг. 13 показано воздействие трансфицирования клеток KHYG-1 с TRAIL и вариантом TRAIL на жизнеспособность клетки;
На фиг. 14 показан график FACS базовой экспрессии DR4, DR5, DcR1 и DcR2 на обеих клетках KHYG-1 и клетках NK-92;
На фиг. 15, 16 и 17 показано воздействие экспрессии TRAIL или варианта TRAIL в клетках KHYG-1 на апоптоз популяций трех клеток-мишеней: K562, RPMI8226 и MM1.S, соответственно;
На фиг. 18 показано два графика FACS экспрессии DR5 в клетках RPMI8226 и клетках MM1.S, соответственно; кроме того, показано воздействие обработки препаратом Бортезомиб на экспрессию DR5;
На фиг. 19 показаны графики FACS апоптоза в клетках MM1.S, прошедших предварительную обработку/непрошедших обработку препаратом Бортезомиб, культивированных с клетками KHYG-1, с вариантом/без варианта TRAIL.
На фиг. 20 показан график FACS экспрессии уровней перфорина в клетках KHYG-1, которые проходили обработку 100 нМ СМА на протяжении 2 часов;
На фиг. 21 показаны графики FACS жизнеспособности клеток KHYG-1 после обработки 100 нМ СМА или наполнителем;
На фиг. 22 показаны графики FACS апоптоза в клетках MM1.S, культивированных с клетками KHYG-1, с вариантом/без варианта TRAIL, прошедших предварительную обработку/непрошедших обработку СМА;
На фиг. 23 показаны графики FACS апоптоза в клетках K562, культивированных с клетками KHYG-1, с малой интерферирующей РНК CD96 и/или экспрессией варианта TRAIL; а также
На фиг. 24 показаны графики FACS апоптоза в клетках MM1.S, культивированных с клетками KHYG-1, с малой интерферирующей РНК CD96 и/или экспрессией варианта TRAIL;
ДНК, РНК и аминокислотные последовательности приведены ниже, где:
SEQ ID №: 1 - это полная последовательность ДНК LIR2;
SEQ ID №: 2 - это аминокислотная последовательность LIR2;
SEQ ID №: 3 - это последовательность «руководящей» РНК g9 LIR2;
SEQ ID №: 4 - это последовательность «руководящей» РНК g18 LIR2;
SEQ ID №: 5 - это последовательность прямого праймера LIR2;
SEQ ID №: 6 - это последовательность обратного праймера LIR2;
SEQ ID №: 7 - это полная последовательность ДНК CTLA4;
SEQ ID №: 8 - это аминокислотная последовательность CTLA4;
SEQ ID №: 9 - это последовательность «руководящей» РНК g7 CTLA4;
SEQ ID №: 10 - это последовательность «руководящей» РНК g15 CTLA4;
SEQ ID №: 11 - это последовательность прямого праймера CTLA4; а также
SEQ ID №: 12 - это последовательность обратного праймера CTLA4.
Пример 1 - Нокаут функции ингибирующих рецепторов
CRISPR/Cas9
Клетки подготавливались следующим образом, после удаления функции ингибирующих рецепторов. Структуры «руководящей» РНК были сконструированы и подготовлены для воздействия с генами, кодирующими «классический» ингибирующий рецептор LIR2 и ингибирующий рецептор «контрольной точки» CTLA4 в человеческом геноме NK-клеток. После этого для нокаута генов-мишеней LIR2 и CTLA4 использовалось редактирование генома CRISPR/Cas9.
Для каждого гена-мишени было выбрано два «руководящих» РНК-кандидата и определена их расщепляющая активность в K562. Последовательности «руководящих» РНК-кандидатов показаны в таблице 1, а мотив, прилегающий к протоспейсеру (РАМ), относится к последним 3 основам последовательности. Фланкирующие области последовательностей «руководящей» РНК на гене LIR2 (SEQ ID №: 1) и гене CTLA4 (SEQ ID №: 7) показаны на фигурах 1 и 2, соответственно.

Таблица 1. Последовательности и «руководящие» РНК-кандидаты
Клетки K562 были трансфицированы с помощью подготовленных структур «руководящей» РНК (фигура 3) и последовательно отобраны для амплификации PCR. Присутствие экспрессии GFP использовалось для сообщения об успешном вводе структуры «руководящей» РНК в клетки K562. Таким образом, была подтверждена экспрессия гена Cas9 и, следовательно, возможность нокаута экспрессии генов LIR2 и CTLA4.
Расщепляющая активность структур «руководящей» РНК была определена in vitro с помощью анализа на обнаружение несовместимости. Т7Е1 эндонуклеаза I распознает и расщепляет неидеально совместимые ДНК, тем самым обеспечивая возможность сравнивания исходных генов LIR2 и CTLA4 с мутировавшими генами после трансфекции CRISPR/Cas9 и негомологичного соединения концов (NHEJ).
На фигуре 4 показаны полосы, полученные в результате электрофореза в агарозном геле после нокаута гена LIR2 с помощью последовательностей «руководящей» РНК g9 и g18. Три полосы, соответствующие каждой мутации, относятся к каждому исходному гену и двум полученным структурам после обнаружения несовместимости в последовательности ДНК по завершению трансфекции. В результате последовательности «руководящей» РНК g9 коэффициент успешности выполнения трансфекции составил 11%, в то время как в результате последовательности «руководящей» РНК g18 коэффициент успешности составил 10%.
На фигуре 5 показаны полосы, полученные в результате электрофореза в агарозном геле после нокаута гена CTLA4 с помощью последовательностей «руководящей» РНК g7 и g15. В результате последовательности «руководящей» РНК g7 коэффициент успешности выполнения трансфекции составил 32%, в то время как в результате последовательности «руководящей» РНК g15 коэффициент успешности составил 26%.
После успешного нокаута LIR2 и CTLA4 в клетках K562, клетки KHYG-1 трансфицировались с помощью структур «руководящей» РНК.
Были определены клоны, полученные из KHYG-1, имеющие гомозиотные делеции. С этой целью использовалась экспрессия Cas9 / пуромицин ацетилтрансфераза (РАС). Успешно трансфицированные клетки определялись, основываясь на их сопротивлении антибиотическому пуромицину.
Cas9 RNP
Другим протоколом, используемым для нокаута ингибирующих рецепторов контрольной точки NK-клеток, является трансфекция Cas9 RNP. Преимущество использования данного протокола заключалось в том, что достигалась аналогичная эффективность трансфекции, но со значительно меньшим уровнем токсичности, по сравнению с использованием плазмид ДНК протокола CRISPR/Cas9.
Клетки KHYG1 1×106 отбирались для каждого эксперимента трансфекции. Клетки промывались с помощью PBS и раскручивались в центрифуге. После чего супернатант удалялся. Затем материалы CRISPR RNP (связывающий белок РНК) подготавливались следующим образом:
(1) готовился раствор 20 мкМ необходимых синтезированных крРНК и тРНК (приобретались в Dharmacon).
(2) вместе смешивались 4 мкл крРНК (20 мкМ) и 4 мкл РНК (20 мкМ).
(3) Затем смесь добавлялась в 2 мкл белка Cas9 (5 мкг/мкл).
(4) Все компоненты смешивались и выдерживались при комнатной температуре в течение 10 минут.
После обработки в системе трансфекции Neon®, клетки смешивались с Cas9 RNP, и производилось электроимпульсное открытие клеточных пор с помощью следующих параметров:
Напряжение: 1450 В
Длительность импульса: 30 мс
Количество импульсов: 1
Затем клетки переносились в одну лунку 12-луночного планшета, содержащую среду для роста (включая IL-2 и IL-15).
Клетки отбирались после 48-72 часов для подтверждения эффективности редактирования гена посредством анализа Т7 эндонуклеазы и/или метода Сэнгера. Было подтверждено наличие вставок, что указывает на успешный нокаут CTLA4, PD1 и CD96 в клетках KHYG1.
Сайт-специфичные нуклеазы
Другим протоколом, используемым для нокаута ингибирующих рецепторов контрольной точки NK-клеток, является трансфекция XTN TALEN. Преимущество использования данного протокола заключалось в том, что имелась возможность достижения особенно высокого уровня специфичности по сравнению с CRISPR дикого типа.
Этап 1: Подготовка реагентов
Клетки KHYG-1 анализировались на наличие определенных характерных особенностей, включая эффективность трансфекции, эффективность клонирования одной клетки и хромосомный набор/число копий. После этого клетки культивировались в соответствии с рекомендациями поставщика.
В зависимости от ингибирующего рецептора контрольной точки, нокаут которого выполнялся, нуклеазы подготавливались с учетом индивидуальных требований, по меньшей мере, из двух пар XTN TALEN. Этап определения индивидуальных требований включает в себя оценку генного локуса, числа копий и функциональную оценку (т.е. оценка гомологов, нецелевая оценка).
Этап 2: Получение линии клетки
Клетки трансфицировались с помощью нуклеаз этапа 1; этот этап повторялся 3 раза, чтобы достичь высоких уровней резания, культуры разделялись, а промежуточные культуры поддерживались в определенном состоянии до начала трансфекции.
Предварительный отбор происходил через несколько дней после каждой трансфекции; популяции клеток испытывались на эффективность резания посредством анализа Cel-1. По достижению приемлемых уровней или пологого участка характеристики резания после повторных трансфекций, клетки считались готовыми к клонированию одной клетки.
Объединенные клетки сортировались по одной клетке на лунку 96-луночного планшета; число планшетов для каждой популяции зависело от эффективности клонирования одной клетки, которая определяется на этапе 1. Планшеты выдерживались в течение 3-4 недель.
Этап 3 - Отбор и расширение
По завершению слияния клеток на 96-луночном планшете, культуры объединялись и разделялись на три 96-луночные планшета; один планшет замораживался в качестве резервного, на другом планшете была выполнена повторная посадка, чтобы продолжить наращивание клонов, а последний планшет использовался для подтверждения генотипа.
Каждый клон в планшете для подтверждения генотипа анализировался на потерю сигнала количественной PCR, таким образом подтверждая то, что все аллеломорфы были изменены. Все клоны с отрицательным результатом проходили амплификацию PCR и клонировались, чтобы определить характер вставок и отсутствие любого дикого типа или вставок внутри рамки.
Клоны с подтвержденным нокаутом объединялись на планшетах с количеством лунок не больше 24, а затем дополнительно наращивались; обычно, в результате одного нокаута получается от 5 до 10 криопробирок, содержащих 1×106 клеток на одну пробирку для 5 отдельных клонов.
Этап 4 - Утверждение
Клетки хранились в асептических условиях.
Основной критерий отбора для всех клеток, которые хранились, включал в себя число жизнеспособных клеток (перед заморозкой и после оттаивания), подтверждение идентификации посредством STR, обеспечение общей стерильности и испытание на присутствие микроплазмы; другие критерии выпуска применялись в случае необходимости (хромосомный набор, экспрессивность поверхностного маркера, стерильность высокого уровня, оценка матрицы или белка посредством нокаута, и т.д.).
Пример 2 - Нокдаун функции ингибирующего рецептора контрольной точки CD96 посредством РНК-интерференции
Нокдаун CD96 в клетках KHYG-1 с использованием малой интерферирующей РНК производился посредством электроимпульсного открытия клеточных пор. Использовался набор Т Nucleofection, в сочетании с Amaxa Nucleofector II от компании Lonza, так как данное устройство подходит для использования с линиями клеток и может выполнять трансфекцию как реплицирующихся, так и нереплицирующихся клеток, а также достигает эффективности трансфекции до 90%.
Контрольная малая интерферирующая РНК (номер по каталогу: sc-37007) и малая интерферирующая РНК CD96 (номер по каталогу: sc-45460) были получены от компании Santa Cruz Biotechnology. Безантибиотическая среда RPMI-1640, содержащая 10% FBS и 2 мМ L-глютамина использовались для культивирования после нуклеофекции. Мышиное антитело CD96-APC (номер по каталогу: 338409) было получено от компании Biolegend для окрашивания.
Было подготовлено 20 мкМ основного раствора малой интерферирующей РНК. Лиофилизированный дуплекс малой интерферирующей РНК перерастворялся в 33 мкл воды без РНКазы (буфер для разведения малой интерферирующей РНК: sc-29527) для контроля FITC/контрольной малой интерферирующей РНК, в 165 мкл воды без РНКазы для малой интерферирующей РНК гена-мишени (малая интерферирующая РНК CD96). Трубка нагревалась до 90°С в течение 1 минуты, а затем выдерживалась при 37°С в течение 60 минут. После этого основной раствор малой интерферирующей РНК хранился при -20°С до появления необходимости в нем.
Клетки KHYG-1 пассировались один или два дня перед нуклеофекцией, так как клетки должны находиться в фазе логарифмического роста.
Раствор Nucleofector подогревался до комнатной температуры (100 мкл на один образец).
Аликвота культурной среды, содержащей сыворотку и добавки, также предварительно нагревалась при 37°С в трубке 50 мл. В 6-луночные планшеты добавлялось 1,5 мл культурной среды, содержащей сыворотку и добавки. Планшеты предварительно выдерживались в инкубаторе СО2, с влажностью 5%, при 37°С.
100 мкл раствора для нуклеофекции с клетками 2×106 было смешано с 4 мкл раствора, содержащего 20 мкМ малой интерферирующей РНК (1,5 мкг малой интерферирующей РНК). Образование пузырей воздуха при смешивании избегалось. Смесь переносилась в кюветы, сертифицированные для работы с Amaxa, которые помещались в держатель кювет Nucleofector, и выбиралась программа U-001.
Подтверждалось завершение программы, после чего образцы сразу изымались из кювет. Затем в каждую кювету добавлялось 500 мкл предварительно взвешенной культурной среды. После этого образцы в каждой кювете аккуратно переносились в соответствующую лунку подготовленного 6-луночного планшета, чтобы в каждой лунке получить 2 мл конечного объема.
Потом клетки выдерживались в инкубаторе СО2, с влажностью 5%, при 37°С, до начала выполнения анализа трансфекции. Анализ проточной цитометрии производился через 16-24 часа после электроимпульсного открытия клеточных пор для того, чтобы измерить уровни экспрессии CD96. Данный протокол электроимпульсного открытия клеточных пор выполнялся несколько раз, и было обнаружено, что в результате всегда происходит нокдаун CD96 в клетках KHYG-1 (см., например, фигуры 6А и 6В).
Пример 3 - Цитотоксичность NK-клеток, повышенная посредством нокдауна CD328
Клетки KHYG-1, как с, так и без нокдауна при помощи CD96, совместно культивировались с клетками K562 при разных эффекторах : мишень (Э : М).
Цитотоксичность измерялась через 4 часа после совместной культивации, с использованием набора для измерения цитотоксичности DELFIA EuTDA от компании PerkinElmer (номер по каталогу: AD0116).
Клетки-мишени K562 культивировались в среде RPMI-1640, содержащей 10% FBS, 2 мкл L-глютамина и антибиотики. 96-луночные планшеты с V-образным днищем (номер по каталогу: 83.3926) покупались у компании SARSTEDT. Для осаждения планшета использовалась центрифуга 581 OR Eppendorf (с ротором для планшетов). Для измерения сигнала флуоресценции, который образовывается растворенными клетками K562, использовался VARIOSKAN FLASH (с программным обеспечением Scanlt версии 2.4.3).
Клетки K562 промывались культурной средой, и с ее помощью число клеток доводилось до 1×106 клеток/мл. От 2 до 4 мл клеток добавлялось в 5 мкл реагента BATDA и выдерживались в течение 10 минут при 37°С. Сложноэфирные связи в клетках подвергаются гидролизу и формируют лиганд, легко поглощающий воду и растворяющийся в воде, который больше не выходит за пределы мембраны. Клетки центрифугировались на скорости 1 500 об/мин в течение 5 минут для промывки нагруженных клеток K562. Процедура повторялась от 3 до 5 раз с использованием среды, содержащей 1 мМ пробенецида (Sigma Р8761). После окончательной промывки, клеточный осадок перерастворялся в культурной среде и доводился приблизительно до 5×104 клеток/мл.
Лунки подготавливались для обнаружения фона, спонтанных высвобождений и максимального высвобождения. 100 мкл нагруженных клеток-мишеней (5000 клеток) переносилось в лунки планшета с V-образным днищем и 100 мкл эффекторных клеток (клетки KHYG-1) добавлялось при различных концентрациях клеток, чтобы получить эффектор для мишеней от 1:1 до 20:1. Планшет центрифугировался при 100 х г в течение 1 минуты, и выдерживались в течение 4 часов в атмосфере CO2 с влажностью 5%, при 37°С. За 15 минут до сбора среды, в каждую лунку максимального высвобождения добавлялось 10 мкл лизирующего буфера. Планшет центрифугировался при 500 х г, в течение 5 минут.
20 мкл супернатанта переносилось на 96-луночный планшет с плоским днищем, после чего добавлялось 200 мкл предварительно нагретого раствора европия. Планшет выдерживался при комнатной температуре, в течение 15 минут, с использованием планшетного шейкера. После растворения клеток K562 с помощью клеток KHYG-1, они выпускают в среду лиганд. Затем этот лиганд вступает в реакцию с раствором европия для того, чтобы сформировать флуоресцентный хелат, который напрямую связан с числом растворенных клеток.
Флуоресценция измерялась в флуорометре с временным разрешением, с использованием VARIOSKAN FLASH. Специфическое высвобождение рассчитывалось с помощью следующих формул:
% специфическое высвобождение = Экспериментальное высвобождение - Спонтанное высвобождение/Максимальное высвобождение - Спонтанное высвобождение
Статистический анализ производился с использованием программного обеспечения Graphpad Prism 6.04. Парный t-критерий использовался для сравнения разницы между нокдауном CD96 в клетках KHYG-1 с использованием малой интерферирующей РНК и контрольных групп (n=3).
Значительный рост специфического высвобождения был обнаружен в совместных культурах, содержащих нокдаун CD96 в клетках KHYG-1. Это происходило при всех мишенях Э : М (см. фигуру 7).
Так как флуоресценция непосредственно связана с растворением клеток, было подтверждено, что нокдаун экспрессии CD96 в клетках KHYG-1 приводит к увеличению их способности к уничтожению раковых клеток-мишеней K562.
Пример 4 - Цитотоксичность NK-клеток, повышенная посредством нокдауна CD328 (Siglec-7)
Нокдаун CD328 в клетках NK-92 с использованием среды малой интерферирующей РНК
Материалы, реагенты и приборы
Контрольная малая интерферирующая РНК (номер по каталогу: sc-37007) и малая интерферирующая РНК CD328 (номер по каталогу: sc-106757) покупались у компании Santa Cruz Biotechnology. Для достижения эффективности трансфекции до 90% с высокой жизнеспособностью клетки (>75%) в клетках NK-92, использовались устройство Nucleofector™ (Nucleofector II, Lonza) и набор Т Nucleofector™ от компании Lonza. RPMI-1640, содержащая 10% FBS и 2 мМ L-глютамина, без антибиотиков, использовались для культивирования после нуклеофекции. Мышиное антитело CD328-APC (номер по каталогу: 339206) покупалось у компании Biolegend.
Протокол
Для получения 10 мкМ основного раствора малой интерферирующей РНК
 Перерастворить лиофилизированный дуплекс малой интерферирующей РНК в 66 мкл воды без РНКазы (буфер для разведения малой интерферирующей РНК: sc-29527) для контроля FITC/контрольной малой интерферирующей РНК, в 330 мкл воды без РНКазы для малой интерферирующей РНК гена-мишени (малая интерферирующая РНК CD328).
Перерастворить лиофилизированный дуплекс малой интерферирующей РНК в 66 мкл воды без РНКазы (буфер для разведения малой интерферирующей РНК: sc-29527) для контроля FITC/контрольной малой интерферирующей РНК, в 330 мкл воды без РНКазы для малой интерферирующей РНК гена-мишени (малая интерферирующая РНК CD328).
 Нагреть трубку до 90°С за 1 минуту.
Нагреть трубку до 90°С за 1 минуту.
 Выдерживать при 37°С в течение 60 минут.
Выдерживать при 37°С в течение 60 минут.
 Хранить основной раствор малой интерферирующей РНК при -20°С, если не используется сразу после получения.
Хранить основной раствор малой интерферирующей РНК при -20°С, если не используется сразу после получения.
 Один образец Nucleofection содержит (на 100 мкл стандартных кювет)
Один образец Nucleofection содержит (на 100 мкл стандартных кювет)
 Число клеток: 2×106 клеток
Число клеток: 2×106 клеток
 Малая интерферирующая РНК: 4 мкл от 10 мкМ основного раствора
Малая интерферирующая РНК: 4 мкл от 10 мкМ основного раствора
 Раствор Nucleofector: 100 мкл
Раствор Nucleofector: 100 мкл
Нуклеофекция
 Культивировать требуемое число клеток. (Пассировать один или два дня перед нуклеофекцией, так как клетки должны находиться в фазе логарифмического роста).
Культивировать требуемое число клеток. (Пассировать один или два дня перед нуклеофекцией, так как клетки должны находиться в фазе логарифмического роста).
 Подготовить малую интерферирующую РНК для каждого образца.
Подготовить малую интерферирующую РНК для каждого образца.
 Предварительно нагреть раствор Nucleofector до комнатной температуры (100 мкл на образец).
Предварительно нагреть раствор Nucleofector до комнатной температуры (100 мкл на образец).
 Предварительно нагреть аликвоту культурной среды, содержащей сыворотку и добавки, при 37°С в 50 мл трубке. Подготовить 6-луночные планшеты, наполнив их 1,5 мл культурной среды, содержащей сыворотку и добавки, а также предварительно выдерживать в инкубаторе СО2 с уровнем влажности 5%, при 37°.
Предварительно нагреть аликвоту культурной среды, содержащей сыворотку и добавки, при 37°С в 50 мл трубке. Подготовить 6-луночные планшеты, наполнив их 1,5 мл культурной среды, содержащей сыворотку и добавки, а также предварительно выдерживать в инкубаторе СО2 с уровнем влажности 5%, при 37°.
 Взять аликвоту культуры клеток и подсчитать клетки, чтобы определить их плотность.
Взять аликвоту культуры клеток и подсчитать клетки, чтобы определить их плотность.
 Центрифугировать необходимое число клеток при 1500 об/мин в течение 5 мин. Полностью удалить супернатант, чтобы никакая остаточная среда не покрывала клеточный осадок.
Центрифугировать необходимое число клеток при 1500 об/мин в течение 5 мин. Полностью удалить супернатант, чтобы никакая остаточная среда не покрывала клеточный осадок.
 Повторно растворить клеточный осадок в растворе Nucleofector при комнатной температуре, чтобы получить окончательную концентрацию 2×106 клеток/100 мкл. Избегать выдерживания суспензии клеток в растворе Nucleofector больше 15-20 мин, так как это снижает жизнеспособность клеток и эффективность переноса генов.
Повторно растворить клеточный осадок в растворе Nucleofector при комнатной температуре, чтобы получить окончательную концентрацию 2×106 клеток/100 мкл. Избегать выдерживания суспензии клеток в растворе Nucleofector больше 15-20 мин, так как это снижает жизнеспособность клеток и эффективность переноса генов.
 Смешать 10 мкл суспензии клеток с малой интерферирующей РНК.
Смешать 10 мкл суспензии клеток с малой интерферирующей РНК.
 Переместить образец в кювету, сертифицированную для работы с Amaxa. Убедиться, что образец покрывает днище кюветы, избегать образования пузырей воздуха при отмеривании пипеткой. Закрыть кювету синим колпачком.
Переместить образец в кювету, сертифицированную для работы с Amaxa. Убедиться, что образец покрывает днище кюветы, избегать образования пузырей воздуха при отмеривании пипеткой. Закрыть кювету синим колпачком.
 Выбрать соответствующую программу в устройстве Nucleofector (А-024 для клеток NK-92). Вставить кювету в держатель кювет (Nucleofector II: необходимо повернуть карусель по часовой стрелке в крайнее положение) и нажать кнопку «х» для запуска программы.
Выбрать соответствующую программу в устройстве Nucleofector (А-024 для клеток NK-92). Вставить кювету в держатель кювет (Nucleofector II: необходимо повернуть карусель по часовой стрелке в крайнее положение) и нажать кнопку «х» для запуска программы.
 Во избежание повреждения клеток, необходимо изъять образцы из кюветы сразу по завершению работы программы (на дисплее будет отображаться «ОК»). Добавить 500 мкл предварительно нагретой культурной среды в кювету и переместить образец на подготовленный 6-луночный планшет.
Во избежание повреждения клеток, необходимо изъять образцы из кюветы сразу по завершению работы программы (на дисплее будет отображаться «ОК»). Добавить 500 мкл предварительно нагретой культурной среды в кювету и переместить образец на подготовленный 6-луночный планшет.
 Выдерживать клетки в инкубаторе СО2 с влажностью 5%, при 37°С. Через 16-24 часа провести проточный цитометрический анализ и взять пробу на цитотоксичность.
Выдерживать клетки в инкубаторе СО2 с влажностью 5%, при 37°С. Через 16-24 часа провести проточный цитометрический анализ и взять пробу на цитотоксичность.
Результаты: был выполнен вышеизложенный протокол и произведен проточный цитометрический анализ уровня экспрессии CD328 в клетках NK-92. Результаты одного из наглядных экспериментов показаны на фиг. 8, что подтверждает успешный нокдаун.
Нокдаун CD328 увеличивает цитотоксичность
Материалы, реагенты и приборы
Набор для измерения цитотоксичности DELFIA EuTDA на основе лиганда, усиливающего флуоресценцию (номер по каталогу: AD0116), покупался у компании PerkinElmer. Клетки-мишени K562 культивировались в среде RPMI-1640, содержащей 10% FBS, 2 мкл L-глютамина и антибиотики. 96-луночные планшеты с V-образным днищем (номер по каталогу: 83.3926) покупались у компании SARSTEDT. Для осаждения планшета использовалась центрифуга 581 OR Eppendorf (с ротором для планшетов). Для измерения сигнала флуоресценции, который образовывается растворенными клетками K562, использовался VARIOSKAN FLASH (с программным обеспечением Scanlt версии 2.4.3).
Протокол
 Нагрузить клетки K562 реагентом BATDA DELFIA лиганда, увеличивающего флуоресценцию.
Нагрузить клетки K562 реагентом BATDA DELFIA лиганда, увеличивающего флуоресценцию.
 Промыть клетки K562 культурной средой и с ее помощью довести число клеток до 1×106 клеток/мл. Добавить от 2 до 4 мл клеток в 5 мкл реагента BATDA, выдерживать в течение 10 минут при 37°С.
Промыть клетки K562 культурной средой и с ее помощью довести число клеток до 1×106 клеток/мл. Добавить от 2 до 4 мл клеток в 5 мкл реагента BATDA, выдерживать в течение 10 минут при 37°С.
 Производить осаждение при скорости 1500 об/мин в течение 5 минут для промывки нагруженных клеток K562 от 3 до 5 раз с использованием среды, содержащей 1 мМ пробенецида (Sigma Р8761).
Производить осаждение при скорости 1500 об/мин в течение 5 минут для промывки нагруженных клеток K562 от 3 до 5 раз с использованием среды, содержащей 1 мМ пробенецида (Sigma Р8761).
 После окончательной промывки, повторной растворить клеточный осадок в культурной среде и довести приблизительно до 5×104 клеток/мл.
После окончательной промывки, повторной растворить клеточный осадок в культурной среде и довести приблизительно до 5×104 клеток/мл.
Взятие проб на цитотоксичность
 Подготовить лунки для обнаружения фона, спонтанных высвобождений и максимального высвобождения.
Подготовить лунки для обнаружения фона, спонтанных высвобождений и максимального высвобождения.
 Отобрать пипеткой 100 мкл нагруженных клеток (5000 клеток) в планшет с V-образным днищем.
Отобрать пипеткой 100 мкл нагруженных клеток (5000 клеток) в планшет с V-образным днищем.
 Добавить 100 мкл эффекторных клеток (NK-92) различных концентраций клеток. Получить эффектор для мишеней от 1:1 до 20:1.
Добавить 100 мкл эффекторных клеток (NK-92) различных концентраций клеток. Получить эффектор для мишеней от 1:1 до 20:1.
 Производить осаждение планшета при 100 х г RCF в течение 1 минуты.
Производить осаждение планшета при 100 х г RCF в течение 1 минуты.
 Выдерживать в течение 2 часов в атмосфере СО2 с влажностью 5%, при 37°С. За 15 минут до сбора среды, в каждую лунку максимального высвобождения добавить 10 мкл лизирующего буфера.
Выдерживать в течение 2 часов в атмосфере СО2 с влажностью 5%, при 37°С. За 15 минут до сбора среды, в каждую лунку максимального высвобождения добавить 10 мкл лизирующего буфера.
 Производить осаждение планшета при 500 х г в течение 5 минут.
Производить осаждение планшета при 500 х г в течение 5 минут.
 Переместить 20 мкл супернатанта на 96-луночный планшет с плоским днищем, добавить 200 мкл предварительно нагретого раствора европия, выдерживать при комнатной температуре, в течение 15 минут, с использованием планшетного шейкера.
Переместить 20 мкл супернатанта на 96-луночный планшет с плоским днищем, добавить 200 мкл предварительно нагретого раствора европия, выдерживать при комнатной температуре, в течение 15 минут, с использованием планшетного шейкера.
 Измерить флуоресценцию в флуорометре с временным разрешением, с использованием VARIOSKAN FLASH. Специфическое высвобождение рассчитывалось с помощью следующих формул:
Измерить флуоресценцию в флуорометре с временным разрешением, с использованием VARIOSKAN FLASH. Специфическое высвобождение рассчитывалось с помощью следующих формул:
 % специфическое высвобождение = Экспериментальное высвобождение - Спонтанное высвобождение/Максимальное высвобождение - Спонтанное высвобождение
% специфическое высвобождение = Экспериментальное высвобождение - Спонтанное высвобождение/Максимальное высвобождение - Спонтанное высвобождение
Результаты: был выполнен вышеизложенный протокол для определения воздействия нокдауна CD328 на цитотоксичность. Результаты одного из наглядных экспериментов показаны на фигуре 9. Как видно, цитотоксичность в отношении клеток-мишеней повысилась в клетках с нокдауном CD328.
Пример 5 - Протокол терапии рака крови нокдауном / нокаутом ингибирующих рецепторов контрольной точки
Как показано в вышеприведенных примерах, нокдаун или нокаут функции ингибирующего рецептора контрольной точки может быть предусмотрен различными способами. Следующий протокол был разработан для использования при лечении пациентов, больных раком крови:
После того как у пациента был диагностирован рак, подлежащий лечению по настоящему изобретению, перед введением в организм пациента аликвоту модифицированных NK-клеток можно подвергать оттаиванию и культивации.
Как вариант, временная мутация может быть предусмотрена, например, с использованием малой интерферирующей РНК в течение одного или двух дней, как описано выше. Платформа MaxCyte Flow Electroporation предусматривает соответствующее решение для обеспечения в клинике быстрых крупномасштабных трансфекций.
Исключение определенных ингибирующих рецепторов контрольной точки может быть более эффективным в сравнении с другими способами. Вероятно, это зависит от пациента и формы рака. По этой причине, метод диагностики рака не всегда относится к биопсийному, а раковые клетки выращиваются в культуре ex vivo. Таким образом, ряд NK-клеток с различными модификациями ингибирующих рецепторов контрольной точки может быть проверен на цитотоксичность в отношении конкретного вида раковых заболеваний. Эта стадия может быть использована для выбора наиболее подходящей NK-клетки или ее производной для терапии.
После успешной модификации перерастворение клеток выполняется в соответствующем носителе (например, физиологический раствор) для внутривенного и/или внутриопухолевого введения в организм пациента.
Пример 6 - Нокин KHYG-1 TRAIL / варианта TRAIL
Клетки KHYG-1 были трансфицированы с помощью TRAIL и варианта TRAIL для оценки жизнеспособности и способности уничтожения раковых клеток после трансфекции.
Используемый вариант TRAIL представляет собой вариант, описанный в WO 2009/077857. Он кодируется по гену TRAIL дикого типа, который включает мутацию D269H/E195R. Данная мутация существенно повышает аффинность варианта TRAIL для DR5, снижая при этом аффинность для обоих рецепторов-приманок (DcR1 и DcR2).
Базовая экспрессия TRAIL
Базовая экспрессия TRAIL (CD253) в клетках KHYG-1 анализировалась с использованием проточной цитометрии.
Мышиное антитело CD253-APC (номер по каталогу Biolegend: 308210) и изотипическое контрольное антитело (номер по каталогу Biolegend: 400122) использовались для окрашивания образцов клеток и анализировались посредством проточного цитометра BD FACS Canto II.
Клетки KHYG-1 культивировались в среде RPMI 1640, содержащей 10% FBS, 2 мМ L-глютамина, пенициллин (100 ед/мл)/стрептомицин (100 мг/мл) и IL-2 (10 нг/мл). 0,5-1,0×106 клетки/пробы были собраны путем центрифугирования (1500 об./мин × 5 минут), а супернатант был аспирирован. Клетки (суспензия отдельных клеток) промывались с помощью 4 мл охлажденного буфера FACS (PBS, 0,5-1% BSA, 0,1% NaN3 азида натрия). Клетки повторно суспендировались в 100 мкл охлажденного буфера FACS, в каждую трубку было добавлено 5 мкл антитела с выдержкой в заморозке в течение 30 минут. Клетки промывались 3 раза путем центрифугирования при 1500 об./мин в течение 5 минут. Затем клетки повторно суспендировались в 500 мкл охлажденного буфера FACS и временно выдерживались в заморозке в темноте.
Впоследствии клетки анализировались посредством проточного цитометра (BD FACS Canto II), а обработка полученных данных выполнялась с помощью ПО FlowJo 7.6.2.
На фиг. 10 видно, что анализ FACS выявил слабую базовую экспрессию TRAIL на поверхности клетки KHYG-1.
Нокин TRAIL / варианта TRAIL путем электроимпульсного открытия клеточных пор
мРНК TRAIL дикого типа и мРНК варианта TRAIL (D269H/195R) синтезировались TriLink Bio Technologies, разделялись на аликвоты и хранились при температуре -80°С. Мышиное антитело CD253-APC (номер по каталогу Biolegend: 308210) и изотипическое контрольное антитело (номер по каталогу Biolegend: 400122), а также мышиное антитело CD107a-PE (номер по каталогу eBioscience: 12-1079-42) и изотипические контрольные антитела (номер по каталогу eBioscience: 12-4714) использовались для окрашивания образцов клеток и анализировались посредством проточного цитометра BD FACS Canto II. Использовался краситель ДНК SYTOX-Green (номер по каталогу Life Technologies: S7020; 5 мМ раствора в DMSO). Для достижения эффективности трансфекции до 90% с высокой жизнеспособностью клетки в клетках KHYG-1, использовались устройство Nucleofector™ (Nucleofector II, Lonza) и набор Т Nucleofector™ от компании Lonza. Безантибиотическая среда RPMI-1640, содержащая 10% FBS, L-глютамин (2 мМ) и IL-2 (10 нг/мл) использовалась для культивирования после нуклеофекции.
Клетки KHYG-1 и NK-92 пассировались один или два дня перед нуклеофекцией, так как клетки должны находиться в фазе логарифмического роста. Раствор Nucleofector был предварительно нагрет до комнатной температуры (100 мкл на образец) с аликвотой культурной среды, содержащей сыворотку и добавки при температуре 37°С в трубке 50 мл. В 6-луночные планшеты добавлялось 1,5 мл культурной среды, содержащей сыворотку и добавки и выполнялась предварительная выдержка в инкубаторе СО2 с влажностью 5%, при 37°С. Была подготовлена аликвота культуры клеток и проведен подсчет клеток для определения их плотности. Перед полным удалением супернатанта необходимое количество клеток центрифугировалось при 1500 об./мин в течение 5 минут. Клеточный осадок повторно растворялся в растворе Nucleofector комнатной температуры до получения окончательной концентрации 2×106 клеток/100 мкл (максимальный период нахождения в суспензии = 20 минут). 100 мкл суспензии клеток смешали с 10 мкг мРНК (объем РНК<10 мкл). Образец был перенесен в кюветы, сертифицированные для работы с Amaxa (убедившись, что образец покрыл днище кюветы и избегая образования пузырей воздуха). Была выбрана соответствующая программа в устройстве Nucleofector (т.е. U-001 для клеток KHYG-1). Затем кюветы были вставлены в держатель кювет. В кювету было добавлено 500 мкл предварительно нагретой культурной среды, а в подготовленный 6-луночный планшет сразу по завершению работы программы был перемещен образец во избежание повреждения клеток. Клетки выдерживались в инкубаторе СО2 с влажностью 5%, при 37°С. Проточный цитометрический анализ и взятие проб на цитотоксичность были выполнены через 12-16 часов после электроимпульсного открытия клеточных пор. Проточное цитометрическое окрашивание выполнено, как указано выше.
На фиг. 11 и 12 видно, что экспрессия TRAIL / варианта TRAIL и CD107a (маркер активации NK-клетки) повысила посттрансфекцию, подтверждая успешный нокин генов TRAIL в клетках KHYG-1.
На фиг. 13 представлено доказательство жизнеспособности клеток KHYG-1 до и после трансфекции посредством электроимпульсного открытия клеточных пор. Видно, что статистически значимые расхождения в отношении жизнеспособности клеток после трансфекции клеток с использованием TRAIL / варианта TRAIL не наблюдаются, подтверждая нетоксичность экспрессии TRAIL дикого типа или варианта TRAIL в отношении клеток. Данное наблюдение противоречит соответствующим выводам относительно клеток NK-92, согласно которым нокин генов варианта TRAIL является токсичным для клеток (данные не показаны). Тем не менее, вероятно, это объясняется относительно высокими уровнями экспрессии рецепторов TRAIL DR4 и DR5 на поверхности клеток NK-92 (см. фиг. 14).
Воздействие TRAIL / варианта TRAIL на цитотоксичность клеток KHYG-1 Использовалось мышиное антитело CD2-APC (номер по каталогу BD Pharmingen: 560642). Использовалось антитело Annexin V-FITC (номер по каталогу Immuno Tools: 31490013). Использовался краситель ДНК SYTOX-Green (номер по каталогу Life Technologies: S7020). Использовался 24-луночный планшет для культуры клеток (номер по каталогу SARSTEDT AG: 83.3922). Линия клеток миелоидного лейкоза K562, линия клеток множественной миеломы RPMI8226 и MM1.S использовались в качестве клеток-мишеней. K562, RPMI8226, MM1.S культивировались в среде RPMI 1640, содержащей 10% FBS, 2 мМ L-глютамина и пенициллин (100 ед/мл)/стрептомицин (100 мг/мл).
Как описано выше, клетки KHYG-1 были трансфицированы с помощью TRAIL / варианта TRAIL.
Клетки-мишени были промыты и осаждены центрифугированием при 1 500 об./мин в течение 5 минут. Трансфицированные клетки KHYG-1 были разбавлены до 0,5×106/мл. Затем плотность клеток-мишеней была отрегулирована в предварительно нагретой среде RPMI 1640, чтобы получить соотношение эффектор : мишень (Э : М) 1:1.
0,5 мл клеток KHYG-1 и 0,5 мл клеток-мишеней смешали в 24-луночном планшете для культур и поместили в инкубатор СО2 с влажностью 5%, при 37°С на 12 часов. Затем проточный цитометрический анализ был предусмотрен для взятия проб на цитотоксичность клеток KHYG-1; клетки, культивированные вместе (в различные периоды времени) были промыты, а затем окрашены антителом CD2-APC (5 мкл/проба), Annexin V-FITC (5 мкл/проба) и SYTOX-Green (5 мкл/проба) с использованием буферного раствора Annexin V.
Далее данные были проанализированы с использованием ПО FlowJo 7.6.2. Были предусмотрены положительные и отрицательные «ворота» CD2, которые представляют популяции клеток KHYG-1 и клеток-мишеней, соответственно. Затем для апоптоза, вызванного TRAIL, были проанализированы клетки с положительной реакцией на Annexin V-FITC и SYTOX-Green в популяции с отрицательным показателем CD2.
На фиг. 15, 16 и 17 показано воздействие обоих клеток KHYG-1, экспрессирующих TRAIL или вариант TRAIL на апоптоз линий трех клеток-мишеней: K562, RPMI8226 и MM1.S, соответственно. Очевидно, что для всех популяций клеток-мишеней экспрессия TRAIL на клетках KHYG-1 повысила уровень апоптоза при сравнении с нормальными клетками KHYG-1 (не трансфицированы с помощью TRAIL). Кроме того, экспрессия варианта TRAIL на клетках KHYG-1 еще больше повысила уровень апоптоза во всех линиях клеток-мишеней при сравнении с клетками KHYG-1, трансфицированными с помощью TRAIL дикого типа.
Клетки в настоящем изобретении, экспрессирующие вариант TRAIL, характеризуются существенным преимуществом в терапии рака благодаря более высокой аффинности для рецептора смерти DR5. При задействовании клеток по настоящему изобретению исключается развитие защитных стратегий раковых клеток для предотвращения их гибели определенным способом. Таким образом, формы рака не могут обеспечить эффективное предотвращение гибели клеток, вызванной TRAIL, за счет увеличения количества рецепторов-приманок TRAIL, поскольку клетки по настоящему изобретению модифицированы таким образом, при котором в данных обстоятельствах они остаются цитотоксичными.
Пример 7 - Протокол терапии рака крови с использованием NK-клеток с помощью вариантов TRAIL, подверженных нокину
Клетки KHYG-1 были трансфицированы с помощью варианта TRAIL, как приводится в примере 6 выше. Следующий протокол был разработан для использования при лечении пациентов, больных раком крови:
После того как у пациента был диагностирован рак, подлежащий лечению по настоящему изобретению, перед введением модифицированных NK-клеток вводится агент, содержащий DR5, например, Бортезомиб, и, следовательно, используется в небольших дозах для повышения экспрессии DR5 на раковых клетках, делая терапию с использованием модифицированных NK-клеток более эффективной.
Затем аликвота модифицированных NK-клеток подвергается оттаиванию, культивации и введению в организм пациента.
Поскольку вариант TRAIL, который экспрессируется NK-клетками, используемыми в терапии, имеет низкую аффинность для рецепторов-приманок в сравнении с TRAIL дикого типа, на поверхности раковых клеток наблюдается повышенная привязка рецепторов смерти, и, следовательно, больший апоптоз раковых клеток.
Перед внедрением вышеизложенного протокола, еще одним вариантом является биопсийный метод диагностики рака и культивация раковых клеток ех vivo. Эта стадия может быть использована для идентификации форм рака, экспрессирующих особо высокие уровни рецепторов-приманок и/или низкие уровни рецепторов смерти, чтобы определить, подходит ли агент, содержащий DR5, данному пациенту. Эта стадия также может быть предусмотрена во время терапии с использованием вышеизложенного протокола, поскольку данная форма рака может адаптироваться, например, для снижения экспрессии DR5, и, следовательно, может быть подходящей для лечения с помощью агента, содержащего DR5, в ходе терапии.
Пример 8 - Небольшая доза Бортезомиба сенсибилизирует раковые клетки к NK-клеткам, экспрессирующим вариант TRAIL
Бортезомиб (Bt) - ингибитор протеасом (препарат с химиотерапевтическим эффектом), целесообразный для лечения множественной миеломы (ММ). Известно, что Бортезомиб может повышать экспрессию DR5 у некоторых типов раковых клеток, включая клетки ММ.
Перед использованием для целевого воздействия на клетки ММ при воздействии Бортезомиба и без него клетки KHYG-1 были трансфицированы с помощью варианта TRAIL, как описывалось выше в примере 6.
Экспрессия DR5 под действием Бортезомиба
Бортезомиб был приобретен у компании Millennium Pharmaceuticals. Мышиное антитело DR5-AF647 (номер по каталогу: 565498) было приобретено у BD Pharmingen. Окрашенные образцы клеток были проанализированы на BD FACS Canto II.
(1) Линии клеток MM RPMI8226 и MM1.S были выращены в среде RPMI1640 (Sigma, Сент-Луис, шт. Миссури, США) с добавлением 2 мМ L-глютамина, 10 мМ ГЭПЭС, 24 мМ бикарбоната натрия, 0,01% антибиотиков и 10% эмбриональной телячьей сыворотки (Sigma, Сент-Луис, шт. Миссури, США) в атмосфере с 5% CO2 при 37°С.
(2) Клетки ММ были отсеяны на 6-луночные планшеты 1×106/мл, 2 мл/лунку.
(3) Затем клетки ММ обрабатывались разными дозами Бортезомиба в течение 24 часов.
(4) После этого экспрессия DR5 в клетках ММ, обработанных/необработанных Бортезомибом, была проанализирована посредством проточной цитометрии (фиг. 18).
Было установлено, что небольшая доза при лечении Бортезомибом повышает экспрессию DR5 в обеих линиях клеток ММ (фиг. 18). Повышение DR5 было связано со слабой индукцией апоптоза (данные не показаны). При этом было установлено, что экспрессию DR5 было невозможно повысить высокими дозами Бортезомиба из-за высокой токсичности, приводящей к гибели большинства клеток ММ.
Сенсибилизация раковых клеток под действием Бортезомиба
Клетки KHYG-1 были трансфицированы с помощью варианта TRAIL (D269H/E195R TRAIL), как приводится в примере 6 выше.
(1) В качестве клеток-мишеней были использованы клетки MM1.S, обработанные/необработанные Бортезомибом. Клетки MM1.S были обработаны 2,5 нМ Бортезомиба или наполнителем (контрольным) в течение 24 часов.
(2) Через 6 часов после электроимпульсного открытия клеточных пор варианта TRAIL мРНК клетки KHYG-1 были культивированы клетками ММ в 12-луночных планшетах. После промывки концентрации клеток были отрегулированы до 1×106/мл перед смешиванием клеток KHYG-1 и MM1.S в соотношении 1:1 для взращивания в течение 12 часов.
(3) Был проведен проточный цитометрический анализ циотоксичности клеток KHYG-1. Клетки, культивированные вместе, были промыты, а затем окрашены антителом CD2-APC (5 мкл/проба), AnnexinV-FITC (5 мкл/проба) и SYTOX-Green (5 мкл/проба) с использованием буферного раствора AnnexinV.
(4) Далее данные были проанализированы с использованием ПО FlowJo 7.6.2. Клетки MM1.S представлены популяцией с отрицательным показателем CD2.
Клетки KHYG-1 демонстрируют выраженно положительную реакцию на CD2. В заключение были проанализированы клетки с положительной реакцией на AnnexinV-FITC и SYTOX-Green в популяции с отрицательным показателем CD2.
В отношении клеток MM1.S с предварительной обработкой/без обработки Бортезомибом, культивированных вместе с клетками KHYG-1, прошедших электроимпульсное открытие клеточных пор с помощью/без варианта TRAIL (фиг. 19), был проведен проточный цитометрический анализ апоптоза.
Было установлено, что Бортезомиб вызвал чувствительность клеток ММ к клеткам KHYG-1, экспрессирующим вариант TRAIL. Следовательно, данные указали на то, что агент, вызвавший экспрессию DR5, был эффективен в модели увеличения цитотоксичности в отношении раковых клеток и, тем самым, может быть полезен при расширении терапии рака по настоящему изобретению.
Пример 9 - Подтверждение апоптоза, вызванного вариантом TRAIL
Несмотря на неоспоримые доказательства повышения цитотоксичности NK-клеток в результате экспрессии варианта TRAIL в предыдущих примерах, также требовалось проверить, стала ли повышенная цитотоксичность результатом индукции апоптоза раковых клеток (наиболее вероятно) или непроизвольной активации NK-клеток, которые проявили более цитотоксичный фенотип и, тем самым, уничтожали раковые клетки посредством секреции перфорина.
Было продемонстрировано, что конканамицин А (СМА) может вызывать цитотоксичную активность NK-клеток, опосредованную перфорином, в основном ввиду ускоренной деградации перфорина посредством повышения уровня рН литических гранул. Было проведено исследование относительно того, может ли быть выделена цитотоксичность клеток KHYG-1, экспрессирующих вариант TRAIL, если цитотоксичность, опосредованная перфорином, была частично ослаблена СМА.
Снижение экспрессии перфорина. вызванное СМА
Мышиное антитело к перфорину AF647 (номер по каталогу: 563576) было приобретено у компании BD Pharmingen. Конканамицин А (номер по каталогу:
SC-202111) был приобретен у компании Santa Cruz Biotechnology. Окрашенные образцы клеток были проанализированы с помощью BD FACS Canto II.
Клетки KHYG-1 культивировались в среде RPMI 1640, содержащей 10% FBS (эмбриональной телячьей сыворотки), 2 мМ L-глютамина, пенициллин (100 ед/мл)/стрептомицин (100 мг/мл) и IL-2 (10 нг/мл).
(2) Далее клетки KHYG-1 были обработаны 100 нМ СМА или наполнителем (DMSO) аналогичного объема в течение 2 часов (культивированы в среде RPMI1640 без пенициллина/стрептомицина через 6 часов после электроимпульсного открытия клеточных пор).
(3) Клетки были собраны (1,0×106 клеток/проба) путем центрифугирования (1500 об./мин × 5 минут), а супернатант был аспирирован.
(4) Клетки фиксировались в параформальдегиде 4% в растворе PBS при комнатной температуре в течение 15 минут.
(5) Клетки были дважды промыты в 4 мл буферного раствора FACS (фосфатно-солевой буферный раствор, 0,5-1% BSA, 0,1% азида натрия).
(6) Клетки пермеабилизировались в 1 мл PBS/буферного раствора сапонина 0,1% в течение 30 минут при комнатной температуре.
(7) Клетки были промыты в 4 мл PBS/буферного раствора сапонина 0,1%.
(8) Перед добавлением в каждую трубку 5 мкл антител и выдержки в заморозке в течение 30 минут клетки повторно суспендировались в 100 мкл PBS/буферном растворе сапонина 0,1%.
(9) Клетки промывались 3 раза PBS/буферным раствором сапонина 0,1% путем центрифугирования при 1500 об./мин в течение 5 минут.
(10) До проведения анализа клетки повторно суспендировались в 500 мкл охлажденного буфера FACS и кратковременно выдерживались в темноте на льду или при 4°С в холодильнике.
(11) Клетки были проанализированы на проточном цитометре (BD FACS Canto II). Данные были обработаны с использованием ПО Flow J о 7.6.2.
Обработка СМА значительно снизила уровень экспрессии перфорина в клетках KHYG-1 (фиг. 20) и не оказала отрицательного воздействия на жизнеспособность клеток KHYG-1 (фиг. 21).
Цитотоксичность вариантов TRAIL NK-клеток при наличии СМА
Клетки KHYG-1 были трансфицированы с помощью варианта TRAIL (D269H/E195R TRAIL), как приводится в примере 6 выше.
(1) В качестве клеток-мишеней были использованы клетки MM1.S.
(2) Через 6 часов после электроимпульсного открытия клеточных пор мРНК TRAIL клетки KHYG-1 были обработаны 100 мМ СМА или наполнителем аналогичного объема в течение 2 часов.
(3) Клетки KHYG-1 были промыты в среде RPMI1640 путем центрифугирования и повторно суспендированы в среде RPM11640 с содержанием IL-2 с регулировкой концентрации клеток до 1×106/мл.
(4) Клетки MM1.S были повторно суспендированы в среде RPMI1640 с содержанием IL-2 с регулировкой концентрации клеток до 1×106/мл.
(5) Клетки KHYG-1 и MM1.S были смешаны в соотношении 1:1 и подвержены совместной культивации в течение 12 часов.
(6) Был проведен проточный цитометрический анализ циотоксичности клеток KHYG-1. Клетки, культивированные вместе, были промыты и окрашены антителом CD2-APC (5 мкл/проба).
(7) После промывки было проведено дополнительное окрашивание с использованием AnnexinV-FITC (5 мкл/проба) и SYTOX-Green (5 мкл/проба) с использованием буферного раствора AnnexinV.
(8) Далее данные были проанализированы с использованием ПО FlowJo 7.6.2. Клетки MM1.S представлены популяцией с отрицательным показателем CD2. Клетки KHYG-1 демонстрируют выраженно положительный показатель CD2. Затем были проанализированы клетки с положительной реакцией на AnnexinV-FITC и SYTOX-Green в популяции с отрицательным показателем CD2.
Снова было выявлено, что NK-клетки, экспрессирующие вариант TRAIL, демонстрируют более высокую цитотоксичность, чем контрольные клетки, в которых отсутствует экспрессия варианта TRAIL (фиг. 22). При этом в настоящем примере было продемонстрировано, что СМА не смог значительно снизить цитотоксичную активность NK-клеток, экспрессирующих вариант TRAIL, в отличие от результата, установленного в отношении контрольных NK-клеток, обработанных СМА.
Было установлено, что NK-клетки без варианта TRAIL (контрольные или ложные NK-клетки) вызывают гибель 48% раковых клеток при отсутствии СМА, и 35,9% - с СМА (фиг. 22). NK-клетки, экспрессирующие вариант TRAIL, смогли вызвать гибель большего количества раковых клеток, чем контрольные NK-клетки, как при наличии, так и при отсутствии СМА. Фактически даже при наличии СМА NK-клетки, экспрессирующие вариант TRAIL, вызвали гибель большего количества раковых клеток, чем контрольные NK-клетки при отсутствии СМА.
Таким образом, настоящие данные демонстрируют важность варианта TRAIL в повышении цитотоксичности NK-клетки в отношении раковых клеток посредством механизма, менее подверженного снижению численности клеточных компонентов, связанному с перфорином. Так как перфорин, в основном, используется NK-клетками для уничтожения клеток-мишеней, и многие раковые клетки выработали механизмы снижения экспрессии перфорина NK-клеток, для предотвращения цитотоксичного воздействия NK-клетки по настоящему изобретению представляют собой хорошую альтернативу, менее подверженную аттенюации со стороны раковых клеток.
Пример 10 - Комбинированная экспрессия мутировавшего варианта TRAIL и нокдаун ингибирующего рецептора контрольной точки CD96 в клетках KHYG-1
При нокдауне экспрессии ингибирующего рецептора контрольной точки CD96, а также при экспрессировании варианта TRAIL наблюдалось повышение цитотоксичности в NK-клетках. Также было проведено исследование с комбинированием генетических модификаций для провоцирования синергического воздействия на цитотоксичность NK-клеток.
Был выполнен нокдаун экспрессии CD96 в клетках KHYG-1, как представлено в примере 2.
Клетки KHYG-1 были трансфицированы с помощью варианта TRAIL (D269H/E195R TRAIL), как приводится в примере 6 выше.
(1) Через 12 часов после электроимпульсного открытия клеточных пор клетки KHYG-1 были совместно культивированы с клетками-мишенями (K562 или MM1.S) при концентрации 1×106/мл в 12-луночных планшетах (2 мл/лунка) в течение 12 часов. Соотношение Э : М составляло 1:1.
(2) Через 12 часов после совместной культивации клетки были собраны, промыты, окрашены антителом CD2-APC, снова промыты и дополнительно окрашены AnnexinV-FITC (5 мкл/проба) и SYTOX-Green (5 мкл/проба) с использованием буферного раствора AnnexinV.
(3) Образцы клеток были проанализированы с помощью проточного цитометра BD FACS Canto II. Далее данные были проанализированы с использованием ПО FlowJo 7.6.2. Клетки MM1.S представлены популяцией с отрицательным показателем CD2. Клетки KHYG-1 демонстрируют выраженно положительную реакцию на CD2. Затем были проанализированы клетки с положительной реакцией на AnnexinV-FITC и SYTOX-Green в популяции с отрицательным показателем CD2.
Было установлено, что одновременный нокдаун экспрессии CD96 и экспрессия варианта TRAIL в клетках KHYG-1 синергетически повышают цитотоксичность клеток в отношении как клеток-мишеней K562 (фиг. 23), так и клеток-мишеней MM1.S (фиг. 24). На это указывал факт того, что в обеих группах клеток-мишеней большая часть случаев гибели произошла в результате одновременной генной модификации, а не ввиду отдельных модификаций в изоляции.
В то же время, были получены дополнительные доказательства того, что нокдаун CD96 повышает цитотоксичность NK-клеток (фиг. 23 и 24), в дополнение к доказательствам того, что экспрессия мутировавшего/вариативного TRAIL также повышает цитотоксичность NK-клеток (фиг. 23 и 24).
Таким образом, в настоящем изобретении предлагаются NK-клетки и линии клеток, а также их продукты для использования в терапии рака крови.
--->
SEQUENCE LISTING
<110> Onkimmune Limited
<120> MODIFIED NATURAL KILLER CELLS AND NATURAL KILLER CELL LINES
HAVING INCREASED CYTOTOXICITY
<130> P40153EP
<160> 12
<170> PatentIn version 3.5
<210> 1
<211> 9540
<212> DNA
<213> Homo sapiens
<400> 1
ctctggcctc tgttctttct tgtgagtccg tctacacttg gggtttccac atgtcttttt 60
ctgctcatga ccttgatact ctgggtattt cagaaatgct acacatacgt ttctccatta 120
cggtcagatg tgacatcttg agtggactca tcaatcacct acagaatgtg gagtccaaca 180
gcaagatcct ctcacgtccc aaagcctcag gtcttaccct ggtctggaaa tcaagcacaa 240
atgagcccct cccaatgtcc caggcaccac tgaccccaca accactgtga cgagtgggat 300
tcatgacaac aatctgcaaa ggaagaaact gaggctcagt gatgggacat tacaaaccaa 360
ggtcacgtag gcagcggatg ataaccagtc atcaaataaa tatcaactcc ctcccccact 420
ccccaaatca aagctcaaac ataagtcatt gttcccaaaa tgttgaccag gaattgaggt 480
gcagagggac ggctaaggac gcaatgggca ccgaggaggc aggaaagact cagaggtttc 540
ttcccggggg ggagggagtg gacgctggag caaaaacatt taaaaagggg aagttaagag 600
gggactattt ggttgaaaga aaacccacaa tccagtgtca agaaagaagt caacttttct 660
tcccctactt ccctgcattt ctcctctgtg ctcactgcca cacacagctc aacctggaca 720
gcacagccag aggcgagatg cttctctgct gatctgagtc tgcctgcagc atggacctgg 780
gtcttccctg aagcatctcc agggctggag ggacgactgc catggtaagg accccacaac 840
gctgtgctga tggatgggct gaaggaggga gggtgaccat gtgggaagct gtgagaagga 900
aggggaagcc actgctaccc tcatcaggaa gggcagacac aagaagcacc agttctattt 960
gctgctacat cccggctctc ggtgagacga ggagaaacca gacagacagt ggctgggggt 1020
caggaaagac cccattacag tctgaaatgt ctgcagaggg cccagtgcct gcccccacct 1080
cagctctaaa agaatgagag tcaggctcct gggagggcag ttccgcttct tgtgtggctg 1140
cagatgacaa caccccatga gaaggaccca gcctctgagt gtccacacag ggtgggaagg 1200
aggggaggct atttctctct gtgtgtctct gtcccgccag caccgagggc tcatccatcc 1260
gcagagcagg gcagtgggag gagacgccat gacccccatc gtcacagtcc tgatctgtct 1320
cggtgagatt tgaagagaga ggggagcttc taacctagga gggacctcac cccacagcca 1380
aactctggtc cctaaggaga ccccaggggc tcacaaagat cccagggagg ggaggacctg 1440
ctcaggcttc agggggcaaa tccctcacag ggaactctct tccagggctg agtctgggcc 1500
ccaggacccg cgtgcagaca ggtgagtctg tccccagctc tcccaggtcc ctcctcctca 1560
ctggggacaa ggggccacct ccgtgcagct ggggatgggg attagaagtt ctggactgac 1620
tgatgggggc atctggaggg tcctgggctg agagctgaga tctgttgggt gggaaatgac 1680
ttcgaatctg acctttgatt tccttccagg gaccatcccc aagcccaccc tgtgggctga 1740
gccagactct gtgatcaccc aggggagtcc cgtcaccctc agttgtcagg ggagccttga 1800
agcccaggag taccgtctat atagggagaa aaaatcagca tcttggatta cacggatacg 1860
accagagctt gtgaagaacg gccagttcca catcccatcc atcacctggg aacacacagg 1920
gcgatatggc tgtcagtatt acagccgcgc tcggtggtct gagctcagtg accccctggt 1980
gctggtgatg acaggtgaga ggacactcag ggatcccagc cccaggctct gccctcagga 2040
aggaggctct caggggtgtc tccctctcac agcccagccc tggggatgat gtgggaggtg 2100
ggagccccat ttaacacgat gcctccttct ctcctaggag cctacccaaa acccaccctc 2160
tcagcccagc ccagccctgt ggtgacctca ggaggaaggg tgaccctcca gtgtgagtca 2220
caggtggcat ttggcggctt cattctgtgt aaggaaggag aagatgaaca cccacaatgc 2280
ctgaactccc agccccatgc ccgtgggtcg tcccgcgcca tcttctccgt gggccccgtg 2340
agcccgaatc gcaggtggtc gcacaggtgc tatggttatg acttgaactc tccctatgtg 2400
tggtcttcac ccagtgatct cctggagctc ctggtcccag gtgagaaatt cacagcattg 2460
tctggagttc cctgagtctc cctgagtctc caggcaggtg gggagcagcc gtgtctcagg 2520
gcagttccag gtgggatgat gttggggcga gagggctcag gggtcctggg gccagagaca 2580
caggaagatc agcagtggtg aggcaccggg ggagagggag ggtttgtggg gaagcctgag 2640
ggtcggctcc tggaaaccat gagcaccttt tcccaggtgt ttctaagaag ccatcactct 2700
cagtgcagcc gggtcctgtc atggcccctg gggaaagcct gaccctccag tgtgtctctg 2760
atgtcggcta tgacagattt gttctgtaca aggaggggga acgtgacctt cgccagctcc 2820
ctggccggca gccccaggct gggctctccc aggccaactt caccctgggc cctgtgagcc 2880
gctcctacgg gggccagtac agatgctacg gtgcacacaa cctctcctct gagtgctcgg 2940
cccccagcga ccccctggac atcctgatca caggtgagga gcccagcggg ttcagtcagg 3000
gacccagact ctgcacaggc cctgccgggg gaatccaatt agtgatggcc aggatgaggc 3060
ggggggtggt cccaagggag ggagagacag agagagagac aggggatggg tggggagggg 3120
aagactcaga gaaaacagag acagaggctc ctagagaggc ctggggaggt ctcagctcag 3180
agcaaggtgg ggcagcccct cacccatcct tcttctctcc aggacagatc cgtggcacac 3240
ccttcatctc agtgcagcca ggccccacag tggcctcagg agagaacgtg accctgctgt 3300
gtcagtcatg gcggcagttc cacactttcc ttctgaccaa ggcgggagca gctgatgccc 3360
cactccgtct aagatcaata cacgaatatc ctaagtacca ggctgaattc cccatgagtc 3420
ctgtgacctc agcccacgcg gggacctaca ggtgctacgg ctcactcaac tccgacccct 3480
acctgctgtc tcaccccagt gagcccctgg agctcgtggt ctcaggtggg ggccttgacc 3540
ctgtcctctc tgagctcaaa ggctcagctc aggccctgcc ccccaggaga gctctgggct 3600
gggatggagt gagcgggggt ctgagcgggg ctcagccagt gggagactca ccctcagagg 3660
gaaggaggac aacaggccct cccaggcctg cgcacactca gcggcatcgc cagcatcatg 3720
gacaggagag gcgggtggag ggaggggcct ggggaggcca cagggcccat gtagagaaat 3780
ttggtttgag gtggagactt caggaaagcc ccagctcctc accctcctct cattctttca 3840
cccaggaccc tccatgggtt ccagcccccc acccaccggt cccatctcca cacctggtga 3900
gtccctgagg cctctggctc gaagggagcg cagcgacccc cagggcagct ttgagtgtcc 3960
aggaggatcc cattcccttc agggactcaa tcaagggctt ctgtccaggg agctgggcag 4020
agccagagga ggggccacag ggtccccagg gctctgaggc tgggctggtg aggggtgggg 4080
ggtcaaggca gagagaaatg ttggggccca gcctggggga ggagcagccg ggctgatgtg 4140
gggagcaggg cagccccagc cctcacctcc ccgtcctgac ccagcaggcc ctgaggacca 4200
gcccctcacc cccactgggt cggatcccca aagtggtgag tgaggggctc tgagtgggag 4260
gtgggcgggg tcccggggag gcaggggtgg gttctgtcct aggttcaggc tcctctggag 4320
gtggtgatgt agacaggctc ctcccctgcc tgggcctcag tttctccaag tgtaaaggag 4380
agaggcctgc aggtgggaaa gttcctttca gctctcactc ccagctgtga cctcctggga 4440
gaggaggccc ctcagggaag actccaagac tcgattccgc gggggcctgt cccgtcccac 4500
ctgcagcaga gacggtgacc tggggcaggg gaggggagca gagtcgtggt tcaggacggt 4560
aaggctcttt ccctgcagct ccggggctcg gctctggtgc aggaacaagg gctgcaggtc 4620
agactcccag gctcccttcc cagctctgcc gcttcctggc tgggggcccg gggcaggcga 4680
ttcccctctc tgagcgtcag tttttcatct gtagagtggg tggggtggat gtttgtgtgc 4740
tgcacgactg ttgtgggggt tggaggtggt gaacagaagg tccagcagtc acctgcacac 4800
agtaggcgct catttcaatg acatcacccc catccctgac atcatcgtgc tcaaggtctg 4860
ggaaggcacc tgggggttgt gatcggcatc ttggtggccg tcgtcctact gctcctcctc 4920
ctcctcctcc tcttcctcat cctccgacat cgacgtcagg gcaaacactg gacatcgagt 4980
gagtagggaa ggggaaaccc tgtgggccga ccgagggtgg gctcagggca cagccaaaga 5040
gaatccaaac cactgggcaa atgcagcttt gagaaactgt tccagcattt ctcaccaggt 5100
gaatggagaa agcacttaac gtcagtccca tctacaaata taaagtgtcc tccgggctca 5160
gtcccatcta caaatgtaaa gtgtccttcg gactctgtcc atctcatgag gcatttggaa 5220
catggaggca ggagtgtttt taggtttcct tccttacctt cgagctgtgt gtgcagggca 5280
gggggctcca atgttcccag ggctgaggct ctgtccttct tcccccagcc cagagaaagg 5340
ctgatttcca acatcctgca ggggctgtgg ggccagagcc cacagacaga ggcctgcagt 5400
ggaggtaatt ctgcccgaag accccagact cccacctgct cgtggcccat acactgcccc 5460
taaagctccc attcctcccc caggtccagc ccagctgccg acgcccagga agaaaacctc 5520
tgtgagtgag aggaagaggt gaccagccag gagggagata ggggccccga agtttccgta 5580
gcaatgggga aaggggcacc ggctggaaag ggtctggggc tcagggtgag atcatctcac 5640
cccacactgt gggacctcag ggacattgca gcccctccct gcatctcagt agccccatct 5700
gggagcaggg caggggctgg caggactcag aggtcccagg gaaccttccc aagagacgaa 5760
ccccttgctc tgccccagca gatgctgccg tgaaggacac acagcctgaa gatggggtgg 5820
agatggacac tcgggtgaga ccccgcccct gtcccaggca ccaaaggcct cctggtgcca 5880
gatctaatcc agcaggactt ctctgtcctc cttcccccgg ctctcagcat cgtcacggtg 5940
gacccctcct tgtccagcac gctgcctccc gcctgctgtg acctcactct ctcctgctgt 6000
cctgggacct cgtgggcctc ctcccgggtc cccttcctgc tcctcatcct ctgtttggcc 6060
gtctggttgt tagagcgctc cccaggcctc tggaggatga ggaataaatg aaccaccccg 6120
gtcccctggg ctccccttca ttcattcaac cagtgagtgt tcccagggag ctcactgtgg 6180
atcaggctcc ccatgggagc tgcagacaca gcagggagca aagccgcccc cgcctcctga 6240
gctcacctca tggtgggaga caaaatgcaa ataaatgcgc catgtccagg agtgcaacgt 6300
gcttaaagga acatacacca gggaaagggc agagagtgtg gggcagtggg gccagtctga 6360
atggaagggg agggctgtct gctcagctgt catctgagaa gcctggacag agtggggcac 6420
acgatcctct aatggacgag cccctgcagg cagaggaaac agccgtgcaa aggccccgag 6480
gcagcagcga gctcttgcgg gaaggcccat gaggctgcag ccaaatgggc aaggtcaaag 6540
tgaggagcag aggccagaac cacaggaagg gagcggccag accctccacg gccttagggc 6600
gtccctgaga ttccatcggg aaagggatgt aatcggatca ccccgggaac agtgaggaaa 6660
attgactcca ggaggtcagg gggactcaag gacacccccc accactgtct ctctccagca 6720
gagcccacat gatgaagacc cccaggcagt gacatatgcc ccggtgaaac actccagacc 6780
taggagagaa atggcctctc ctccctcccc actgtccggg gaattcctgg acacaaagga 6840
cagacaggca gaagaggaca gacagatgga cactgagaga gtcctttcct ctccaggccc 6900
ccaggcctcc cccaccccca ccacgttcct tacctctcac tctcccccgc tgcaggctgc 6960
tgcatctgaa gccccccagg atgtgaccta cgcccagctg cacagcttga ccctcagacg 7020
gaaggcaact gagcctcctc catcccagga aagggaacct ccagctgagc ccagcatcta 7080
cgccaccctg gccatccact agcccggagg gtacgcagac tccacactca gtagaaggag 7140
actcaggact gctgaaggca cgggagctgc ccccagtgga caccaatgaa ccccagtcag 7200
cctggacccc taacaaagac catgaggaga tgctgggaac tttgggactc acttgattct 7260
gcagtcgaaa taactaatat ccctacattt tttaattaaa gcaacagact tctcaataat 7320
caatgagtta accgagaaaa ctaaaatcag aagtaagaat gtgctttaaa ctgaatcaca 7380
atataaatat tacacatcac acaatgaaat tgaaaaagta caaaccacaa atgaaaaaag 7440
tagaaacgaa aaaaaaaaac taggaaatga atgacgttgg ctttcgtata aggaatttag 7500
aaaaagaata accaattatt ccaaatgaag gtgtaagaaa gggaataaga agaagaagag 7560
ttgctcatga ggaaaaacca aaacttgaaa attcaacaaa gccaatgaag ctcattcttg 7620
aaaatattaa ttacagtcat aaatcctaac tacattgagc aagagaaaga aagagcaggc 7680
acgcatttcc atatgggagt gagccagcag acagcccagc agatcctaca cacattttca 7740
caaactaacc ccagaacagg ctgcaaacct ataccaatat actagaaaat gcagattaaa 7800
tggatgaaat attcaaaact ggagtttaca taatgaacgt aagagtaatc agagaatctg 7860
actcatttta aatgtgtgtg tatgtgtgtg tatatatatg tgtgtgtgtg tgtgtgtgtg 7920
tgtgtgtgaa aaacattgac tgtaataaaa atgttcccat cgtatcaact ccagttcagg 7980
aagtttcact ggtgatttct tacaaatatt gacgcactaa tgaaacacac aaacacaccc 8040
agagcatcac aaatgtttct tgagaataga aaaagaggca atgtgcccgg gtgcggtggc 8100
tcacgcctgt aatctcaaca cctagggagg cagaggccac agattacttg aggccgggag 8160
ttcaagacca gcatggccaa caaggcaaaa ccccatctct actaaaaata caaaaattag 8220
ctggacatgg tggcgcacgc tgcaatccca gctacttggg aggcagaggc aggaggatca 8280
cttgaatgaa cccgggaggt ggaggttgaa gtgagcaaaa acaaaccccc tacaattcag 8340
cctaggatat gtttattaaa tttacatttg tctttttgct taagattgct ttggtattca 8400
tcctcttttt ggttccatat gaattttagg atttttttct aattctgtga aaaaaatgat 8460
gttgatattt tgatgggaat tgcattgaac ctaaatattg ctttgggaag tgtgatcatt 8520
ttcacaatat tgattctgcc aatccatgag catgggatat atttctattt tgctgtgtca 8580
tctacgattt ctttctgcag cattttgttg ttcttcttgt agagatcttt cacctcctca 8640
gttaggtata ttcttagata tttttaattt tttgcaactg atgtacaagg gattgagttt 8700
tgcagcaacc tggatgagct ggaggccatt attcatgaca ccacatccag ctaatttttg 8760
tatttcttgt agagatgagg ttttgccatg ttgcccaggc tggtcttgaa ctcctgggcc 8820
caagtgaccc gcccgccttg acctcccaaa gtgctgggac tgcaggcatg agccacggtg 8880
cctggcccat catagcactt ttgatcatta ggataattcc ttctccttgt catttttgga 8940
cacatgcttc ccacatgcct catcttccag agagggtttc caccagggct gtgctgggag 9000
ttaaggctgg aaaaggggag atggttccac ctgccagtgc cacatgagtc tactcagggc 9060
tgtaaccagc agggagggtc cagtgtgagc ctcagactcg catgtgggac agacgcccat 9120
gtgtgacaac gctgcagtga atctgtttca cacacatgga ggaggcggct cagggctgac 9180
catggacctg agtcaatgag cagagatatc ccagtgccat ccacaaacac aggggagaag 9240
gagccacaac ttcccacttt catccaaaac cccgacccct ccctgtctgt gagggccctg 9300
gggttctcct ctgtctcata cagaggcaga aacctccccc ttagtgaccc ccagctttgc 9360
aagtcaccag cagcccctcg gcgctggcat cttctgcttc ttaaggtttc ctgcctatga 9420
caggaagtct catttctcat tttcttcatt ggaccatggc tacatatttc agacacatta 9480
taagtaggtt ttcccagtgt taggagcaga tgtgggctgt tgagcacata agtcactcac 9540
<210> 2
<211> 101
<212> PRT
<213> Homo sapiens
<400> 2
Gly Ala Tyr Pro Lys Pro Thr Leu Ser Ala Gln Pro Ser Pro Val Val
1 5 10 15
Thr Ser Gly Gly Arg Val Thr Leu Gln Cys Glu Ser Gln Val Ala Phe
20 25 30
Gly Gly Phe Ile Leu Cys Lys Glu Gly Glu Asp Glu His Pro Gln Cys
35 40 45
Leu Asn Ser Gln Pro His Ala Arg Gly Ser Ser Arg Ala Ile Phe Ser
50 55 60
Val Gly Pro Val Ser Pro Asn Arg Arg Trp Ser His Arg Cys Tyr Gly
65 70 75 80
Tyr Asp Leu Asn Ser Pro Tyr Val Trp Ser Ser Pro Ser Asp Leu Leu
85 90 95
Glu Leu Leu Val Pro
100
<210> 3
<211> 23
<212> DNA
<213> Homo sapiens
<400> 3
gagtcacagg tggcatttgg cgg 23
<210> 4
<211> 23
<212> DNA
<213> Homo sapiens
<400> 4
cgaatcgcag gtggtcgcac agg 23
<210> 5
<211> 20
<212> DNA
<213> Homo sapiens
<400> 5
gggagcccca tttaacacga 20
<210> 6
<211> 20
<212> DNA
<213> Homo sapiens
<400> 6
gggagactca gggaactcca 20
<210> 7
<211> 7375
<212> DNA
<213> Homo sapiens
<400> 7
ctttggacct tcttcaactc tgttttgtct ctgttgagtt aaggctttta agaacacctg 60
aattctttcc ttctgcaaaa ccagaggcag cttcttttcc gcctattttc agtttatttc 120
ttgtgatttt agtttttttc tcttaaccaa atgctaaatg gatttaggag aaataaactt 180
atttgtaaag ctgtcaaggg accattagaa ggatggtgct tcacagatag aatacagttt 240
ttattaatga tgcctagaca aatcctgcca ttagcccaag ggctcagaaa gttagcagcc 300
tagtagtttt ggagttgtca atgaaatgaa ttggactgga tggttaagga tgcccagaag 360
attgaataaa attgggattt aggaggaccc ttgtactcca ggaaattctc caagtctcca 420
cttagttatc cagatcctca aagtgaacat gaagcttcag tttcaaattg aatacatttt 480
ccatccatgg attggcttgt tttgttcagt tgagtgcttg aggttgtctt ttcgacgtaa 540
cagctaaacc cacggcttcc tttctcgtaa aaccaaaaca aaaaggcttt ctattcaagt 600
gccttctgtg tgtgcacatg tgtaatacat atctgggatc aaagctatct atataaagtc 660
cttgattctg tgtgggttca aacacatttc aaagcttcag gatcctgaaa ggttttgctc 720
tacttcctga agacctgaac accgctccca taaagccatg gcttgccttg gatttcagcg 780
gcacaaggct cagctgaacc tggctaccag gacctggccc tgcactctcc tgttttttct 840
tctcttcatc cctgtcttct gcaaaggtga gtgagacttt tggagcatga agatggagga 900
ggtgtttctc ctacctgggt ttcatttgtt tcagcagtca aaggcagtga tttatagcaa 960
agccagaagt taaaggtaaa actccaatct ggcttggctg gctctgtatt ccagggccag 1020
cagggagcag ttgggcggca gcaaataagg caaagagata gctcagaaca gagcgccagg 1080
tatttagtag gggcttcatg aatgcatgtg agttggttta gtagagagac acaggcaatt 1140
tcagaccctt ctatgagact ggaagtgatt taagagggaa aggatagcca tagtcctgaa 1200
tacatttgag ctgggtttca ggatgagctc acaagttcct ttaaaaaaaa ttgacttaag 1260
caaatcctgg gaagagtttt tttgctatac aattcaaggt tttaaggtcc tcggattcat 1320
atactttata aatgaattag ccagcttgtt taaaatgtag ggaaattgtg ggaagaatgc 1380
cttctttact taattcaagg ttttaaggtt ctcttaatca attctactag ctaattagcc 1440
aattatttaa aaataaaagt ttgaaattgc caaaaaaaaa agacaaggaa aaggaaagaa 1500
agaaagccac cagtctgttt ggcatacaat acttaattgt tgcctgacct acgtgtgggt 1560
ttcagatgca gatcctcagt tttcagctct tcagagactg acaccaggtt tgttacacgg 1620
cttaaaatga tgagtatatc cattgaatct caaccttatc tctctctaga ccttcttggt 1680
taagaaacca tgtagtttgt atgaagtagg tactcaaaag atatttgatg atttaatttt 1740
tactggagaa gaaatattca tatatgtttt cttattttta catgttttaa atatgtaaag 1800
attaaataaa cactcttaga agtatttaaa tttcctaaag taaatttatc tcaaccagta 1860
acaggaccct cccaatactg gaaagttgag tgtgaccgca tttagtggtg atgagtgtga 1920
gcttgcttgg ggagagggca ggacatttag gatttcttaa gcttagagtc aatacaataa 1980
agattattga gtgctcactt gggtgggcta taatcactgc tcacaggagt tcatgaacca 2040
caagtaaaag agtgaggaga tatgattagc tcacaaataa ctttaataca gagcagaaag 2100
taatgaacta ctgcaatgga gttatcacag tgctaaggat gctcagaggg catctctgat 2160
aggcagaggt gagggttagg gaaggaagct gtagtctagc tagctagagc tgctggaata 2220
gacatgacaa tggctgctgc caaactgttt tctcttctga ggacagatgt cccgtgcaag 2280
tggcttggtg gaagggacta gtgtctctaa tatagggtga tttataagca ggaaagtgtg 2340
tcctagaaat tcagaccaga gtgatagatt ggaattggat catgggggac tcattgaatg 2400
ttatttattg tatttgtttt tgcgatcagt gttagtaaag tgtcaaaggg attgagcaga 2460
tgagtgacat catgcaacac aagttttgag tttcacttgt cagactgact ggagaggggc 2520
ctggttagtt acaggaaggt aatttggcat gcagccacta tttttgagtt gatgcaagcc 2580
tctctgtatg gagagctggt ctcctttatc ctgtgggaaa agagaacaaa ggagcatggg 2640
agtgttcaag ggaaggagaa ataaagggca gagaggcagc ggtggtgtca ggggaagccc 2700
acaggagtta acagcagggt tgcctcaacc tagagaggaa gcgacctggt gccctcggct 2760
ctgtggcttc cttcatctaa caacatcttc cactctacaa caatgccagg gaaggcggag 2820
gctggtacag tgcatcaaga cacagctact cctgggtgac agaggttcag ggccagctca 2880
ctaagtaggc agaagttttt gacatatact ttgagagata aagcaagatt ctgtacctca 2940
accttcagaa tttcccctac cactcattat agttccggag ctatatagct cctatcattc 3000
tatcataacc ttagaatacc agagaacata tcatctcatc taattatctc ttactatatg 3060
tgaaaaaaat gaaggacatg ggggaagtgt gacttgcccc aaatcacata tttcatggta 3120
gagccaggtc ttctgtttgt catatcagtg ttcttcctgc cacaaccatc ttgaagaatc 3180
tatttctcag taagaaaata tctttatgga gagtagctgg aaaacagttg agagatggag 3240
gggaggctgg gggtgtggag aggggaaggg gtaagtgata gattcgttga aggggggaga 3300
aaaggccgtg gggatgaagc tagaaggcag aagggcttgc ctgggcttgg ccatgaagga 3360
gcatgagttc actgagttcc ctttggcttt tccatgctag caatgcacgt ggcccagcct 3420
gctgtggtac tggccagcag ccgaggcatc gccagctttg tgtgtgagta tgcatctcca 3480
ggcaaagcca ctgaggtccg ggtgacagtg cttcggcagg ctgacagcca ggtgactgaa 3540
gtctgtgcgg caacctacat gatggggaat gagttgacct tcctagatga ttccatctgc 3600
acgggcacct ccagtggaaa tcaagtgaac ctcactatcc aaggactgag ggccatggac 3660
acgggactct acatctgcaa ggtggagctc atgtacccac cgccatacta cctgggcata 3720
ggcaacggaa cccagattta tgtaattggt gagcaaagcc atttcactga gttgacacct 3780
gttgcattgc agtcttctat gcacaaaaac agttttgttc cttaatttca ggaggtttac 3840
ttttaggact gtggacattc tctttaagag ttctgtacca catggtagcc ttgcttattg 3900
tgggtggcaa ccttaatagc attctgactg taaaataaaa tgatttgggg aagttggggc 3960
tctcgctctg gagtgctaac catcatgacg tttgatctgt acttttgata tgatatgatg 4020
ctcctgggga agtagtccca aatagccaaa cctattggtg ggctacccat gcaatttagg 4080
ggtggacctc aaggcctgga agctctaatg tccttttttc accaatgttg gggagtagag 4140
ccctagagtt taaaactgtc tcagggaggc tctgctttgt tttctgttgc agatccagaa 4200
ccgtgcccag attctgactt cctcctctgg atccttgcag cagttagttc ggggttgttt 4260
ttttatagct ttctcctcac agctgtttct ttgagcaaaa tggtgagtgt ggtgctgatg 4320
gtgcaccatg tctgatgggg atacctttag tggtatcaac tggccaaaag atgatgttga 4380
gtttagtgtt cttgagatga gatgaggcaa taaatgaaga ggaaggacag tggtaaagaa 4440
cgcactagaa ccgtaggcat tggcatttga ggtttcagaa tgactaatat tttagatgaa 4500
tttgtttgac attgaatgtt catgtgcttc tgagcagggt ttcaatttga gtaaccgttg 4560
caataacatg gggcagctgt tttgctcttt gtcttcatga caactgtact taagctaaca 4620
gccctgaaac atgagattag gctgggcaga atgctgctag agaggaccac ttggatggtc 4680
tttattctcc ttctccatgt ccctctccat cacctggaag tcacctctgg gtgccactct 4740
ggtgccttcc ttgtcgaagc tgtagctgct cacatgacac ctatccctgt tatccagttt 4800
gcttgactgg gacgttttgc cttccccttc agccaggaag tgaaagtccc agtttttatt 4860
tatcacaggt gttggtattg gtggtagaag aggtagaatt atggaatcag gcctcctgtc 4920
aggatttctt tttgacagtc cctctcagac acctctgcct aaggccagct ttgccattac 4980
aaactctccc ttctccctct ctcccttctt ctcttcctct tccttcttct cgctctttct 5040
ctctctctct ttctccctct ctgtctctta tacacataca caaagatata ctctattcca 5100
acatcctcta cccaacctga cagagatgtc ctttgctgta ggttcagcag tggggatgag 5160
aaatacagct ctcaaacagg ataactaaag cttattatct tatcaagctt gttcccttgc 5220
agacaagatt gatcaattat cataggcttt ctgggtgttc tttctgaagc tttctcaaag 5280
tctctttctc ctatcttcca ttcaaggcaa atgattgcca tttaacatca aaatcacagt 5340
tatttatcta aaataaattt taatagctga atcaagaaaa tctcctgagg tttataattc 5400
tgtatgctgt gaacattcat ttttaaccag ctagggaccc aatatgtgtt gagttctatt 5460
atggttagaa gtggcttccg tattcctcag tagtaattac tgtttctttt tgtgtttgac 5520
agctaaagaa aagaagccct cttacaacag gggtctatgt gaaaatgccc ccaacagagc 5580
cagaatgtga aaagcaattt cagccttatt ttattcccat caattgagaa accattatga 5640
agaagagagt ccatatttca atttccaaga gctgaggcaa ttctaacttt tttgctatcc 5700
agctattttt atttgtttgt gcatttgggg ggaattcatc tctctttaat ataaagttgg 5760
atgcggaacc caaattacgt gtactacaat ttaaagcaaa ggagtagaaa gacagagctg 5820
ggatgtttct gtcacatcag ctccactttc agtgaaagca tcacttggga ttaatatggg 5880
gatgcagcat tatgatgtgg gtcaaggaat taagttaggg aatggcacag cccaaagaag 5940
gaaaaggcag ggagcgaggg agaagactat attgtacaca ccttatattt acgtatgaga 6000
cgtttatagc cgaaatgatc ttttcaagtt aaattttatg ccttttattt cttaaacaaa 6060
tgtatgatta catcaaggct tcaaaaatac tcacatggct atgttttagc cagtgatgct 6120
aaaggttgta ttgcatatat acatatatat atatatatat atatatatat atatatatat 6180
atatatatat atatatattt taatttgata gtattgtgca tagagccacg tatgtttttg 6240
tgtatttgtt aatggtttga atataaacac tatatggcag tgtctttcca ccttgggtcc 6300
cagggaagtt ttgtggagga gctcaggaca ctaatacacc aggtagaaca caaggtcatt 6360
tgctaactag cttggaaact ggatgaggtc atagcagtgc ttgattgcgt ggaattgtgc 6420
tgagttggtg ttgacatgtg ctttggggct tttacaccag ttcctttcaa tggtttgcaa 6480
ggaagccaca gctggtggta tctgagttga cttgacagaa cactgtcttg aagacaatgg 6540
cttactccag gagacccaca ggtatgacct tctaggaagc tccagttcga tgggcccaat 6600
tcttacaaac atgtggttaa tgccatggac agaagaaggc agcaggtggc agaatggggt 6660
gcatgaaggt ttctgaaaat taacactgct tgtgttttta actcaatatt ttccatgaaa 6720
atgcaacaac atgtataata tttttaatta aataaaaatc tgtggtggtc gttttccgga 6780
gttgtcttta tcatccttgc atttgaatat tgtgttcaaa tttttgattg attcattcag 6840
tatctggtgg agtctccaat attagaaata ctggaaacaa actgaaaaac cacaaaagga 6900
caaataatgc ttcatgagtc agctttgcac cagccattac ctgcaagtca ttcttggaag 6960
gtatccatcc tctttccttt tgatttcttc accactattt gggatataac gtgggttaac 7020
acagacatag cagtccttta taaatcaatt ggcatgctgt ttaacacagg ttcttcacct 7080
cccctttctt accgcctgct ttctcagctc aactatcaca ggcattacag ttgtcatggc 7140
aaccccaatg ttggcaacca cgtcccttgc agccattttg atctgccttc ctgaaatata 7200
gagcttttcc ctgtggcttc caaatgaact attttgcaaa tgtggggaaa acacacacct 7260
gtggtcctat gttgctatca gctggcacac ctaggcctgg cacactaagc cctctgtgat 7320
tcttgcttaa ccaatgtata gtctcagcac atttggtttc cacttaaggt ttcct 7375
<210> 8
<211> 36
<212> PRT
<213> Homo sapiens
<400> 8
Met Ala Cys Leu Gly Phe Gln Arg His Lys Ala Gln Leu Asn Leu Ala
1 5 10 15
Thr Arg Thr Trp Pro Cys Thr Leu Leu Phe Phe Leu Leu Phe Ile Pro
20 25 30
Val Phe Cys Lys
35
<210> 9
<211> 23
<212> DNA
<213> Homo sapiens
<400> 9
cactcacctt tgcagaagac agg 23
<210> 10
<211> 23
<212> DNA
<213> Homo sapiens
<400> 10
ccttgtgccg ctgaaatcca agg 23
<210> 11
<211> 29
<212> DNA
<213> Homo sapiens
<400> 11
aggacccttg tactccagga aattctcca 29
<210> 12
<211> 26
<212> DNA
<213> Homo sapiens
<400> 12
agcccctact aaatacctgg cgctct 26
<---
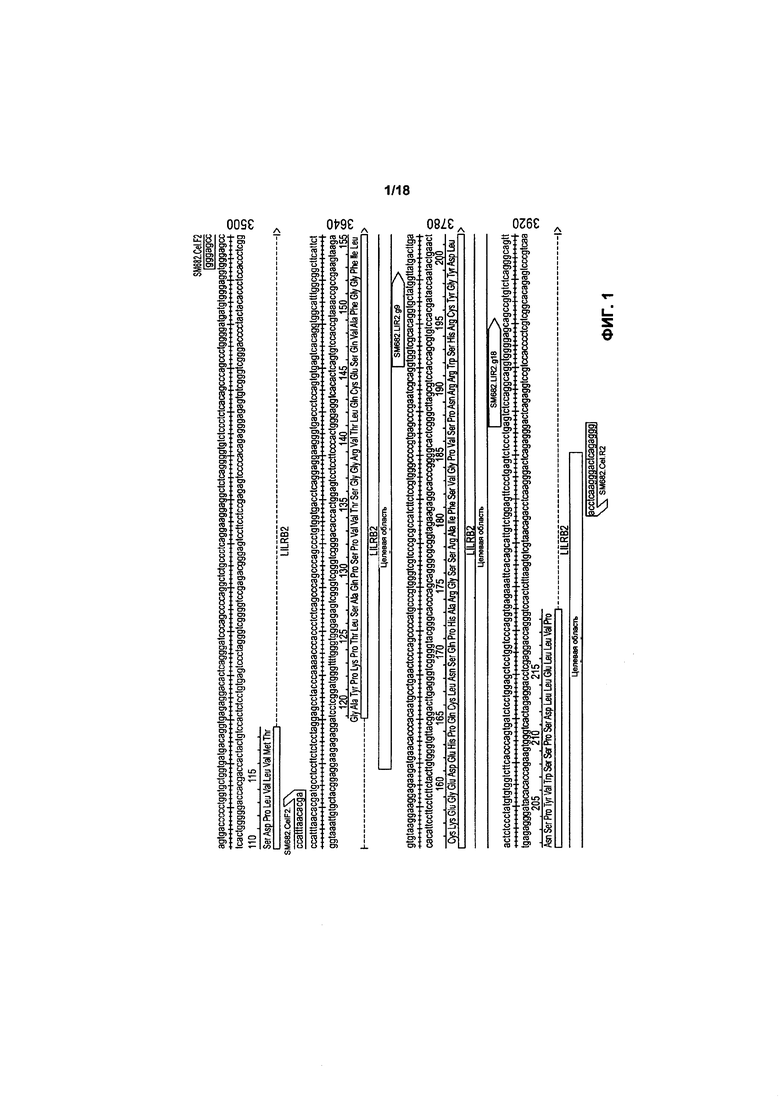

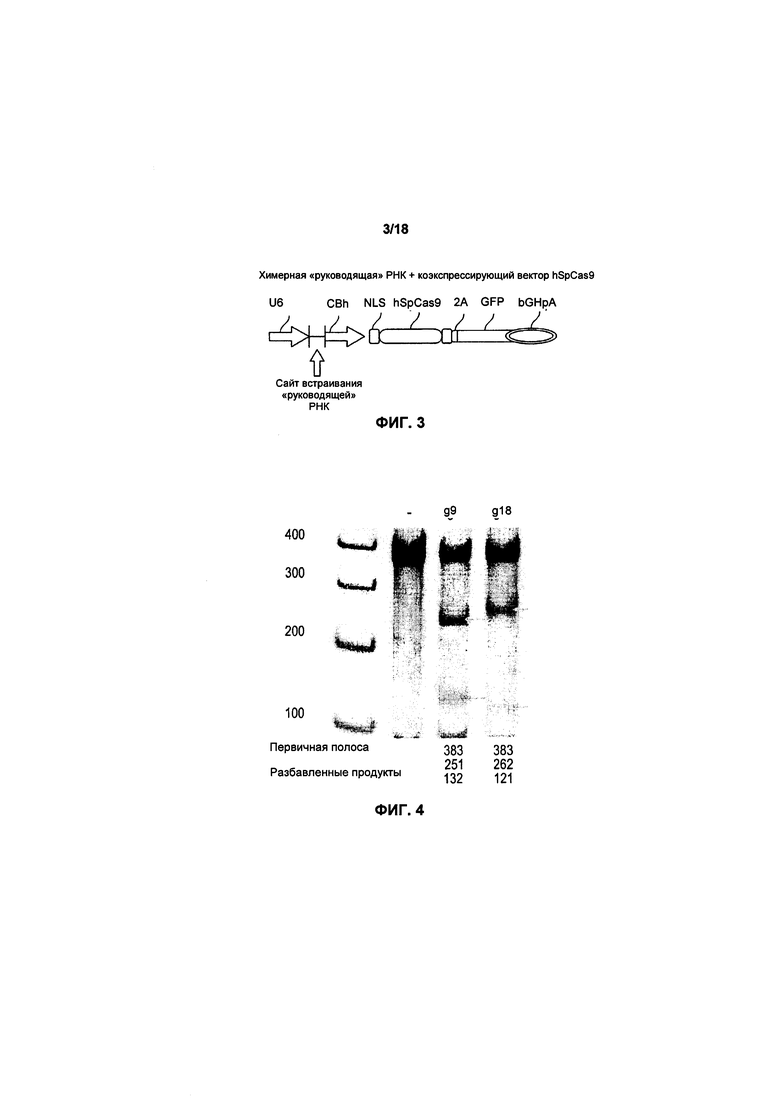
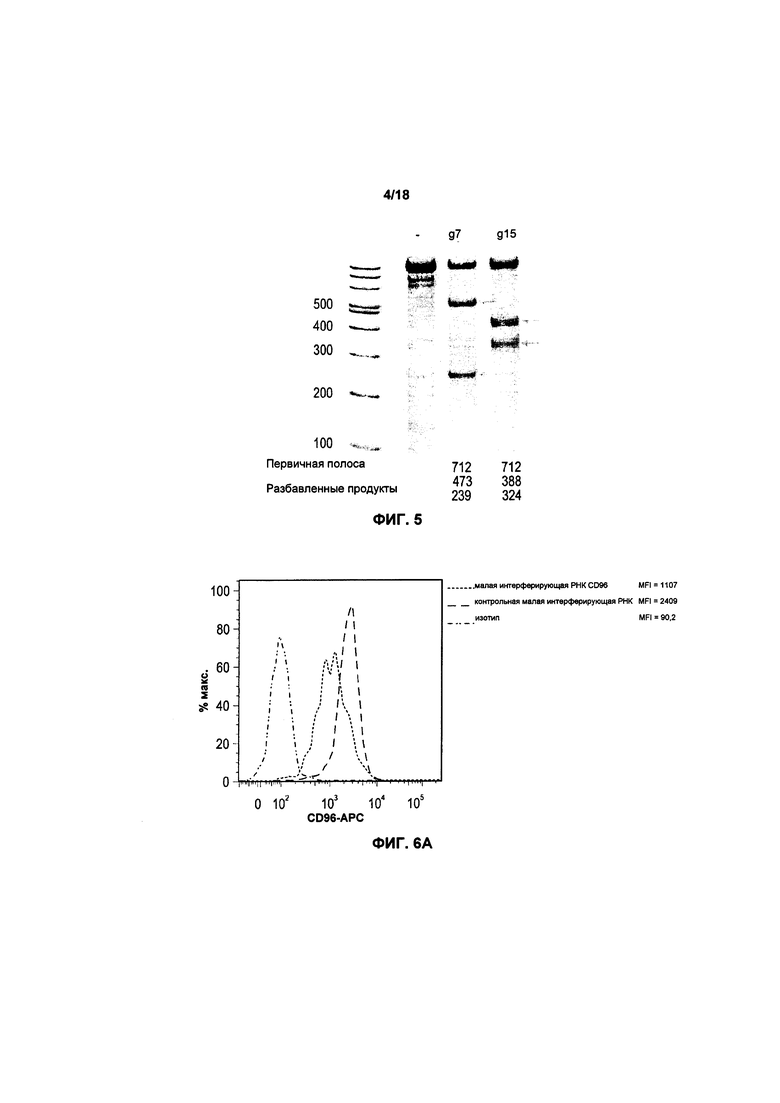
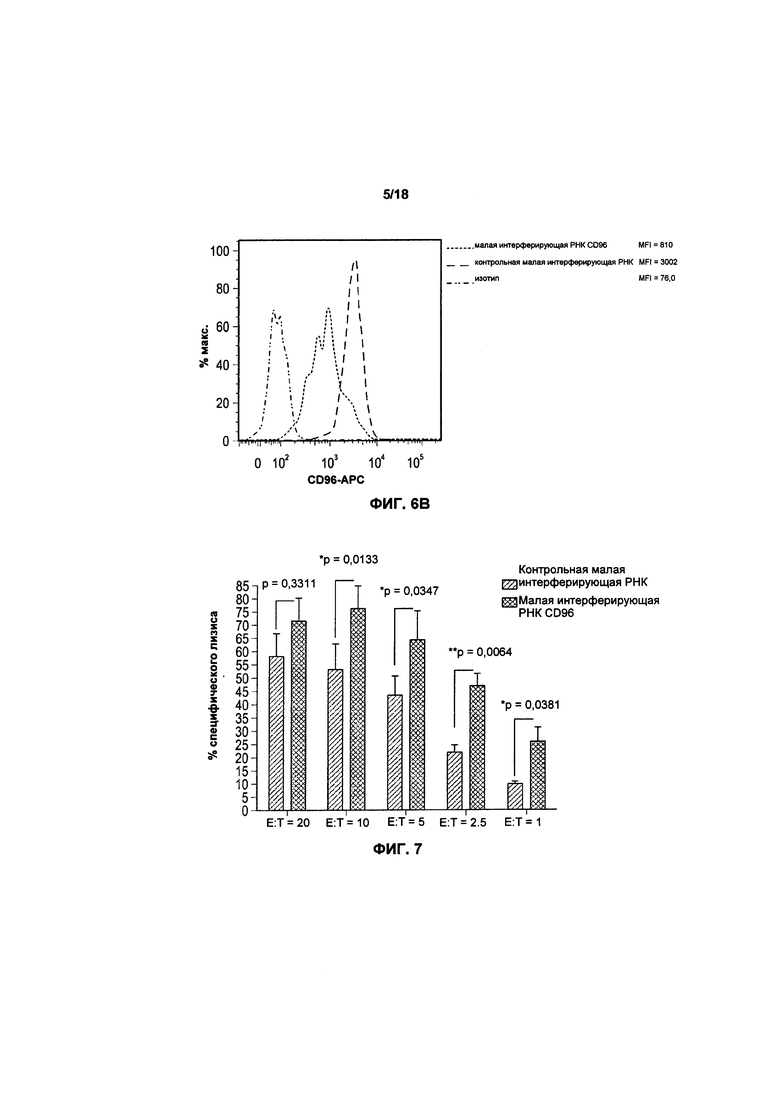
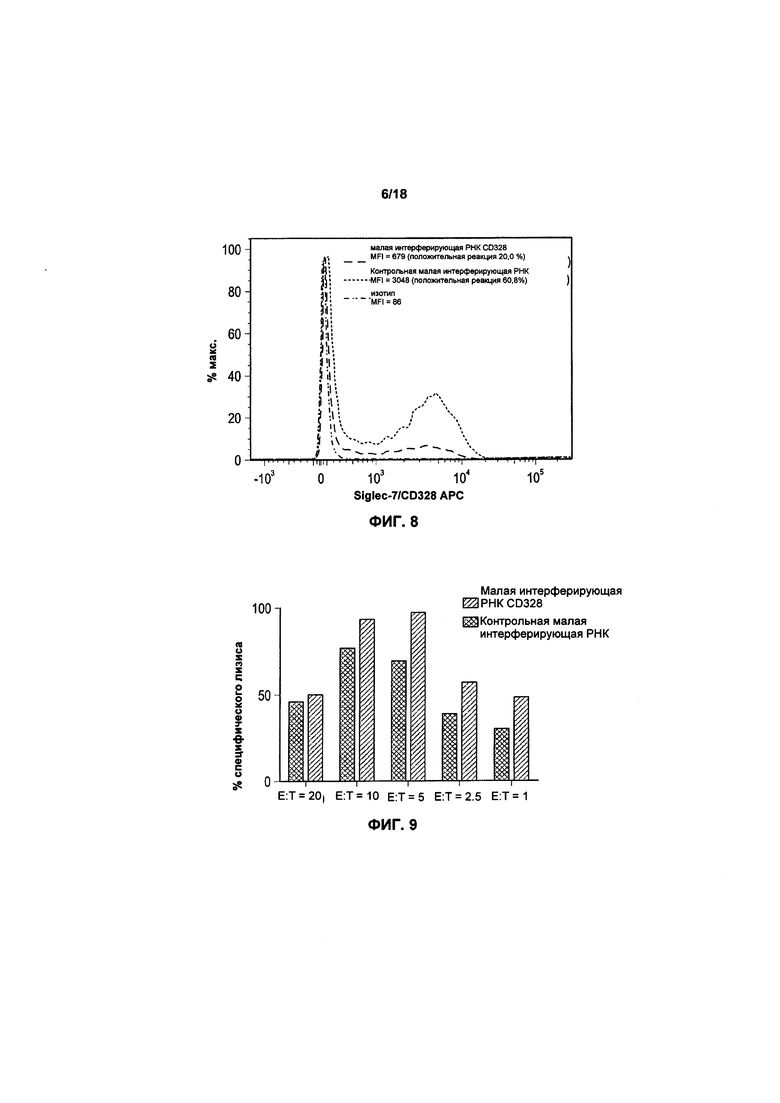
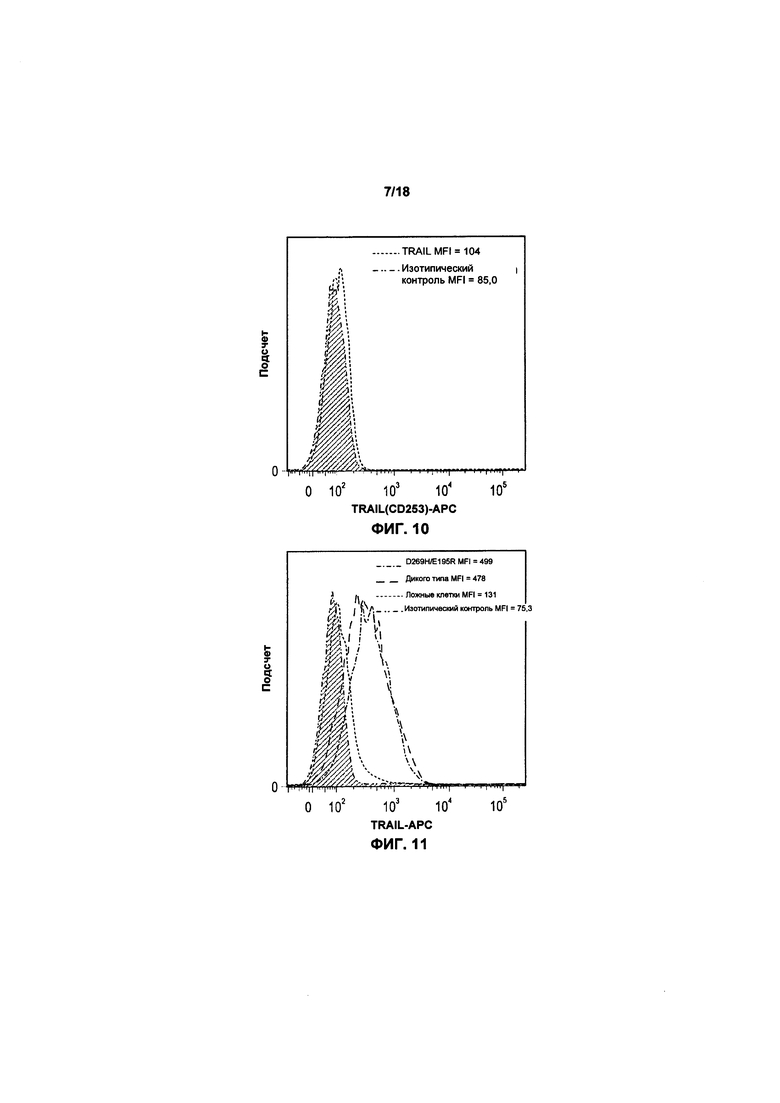
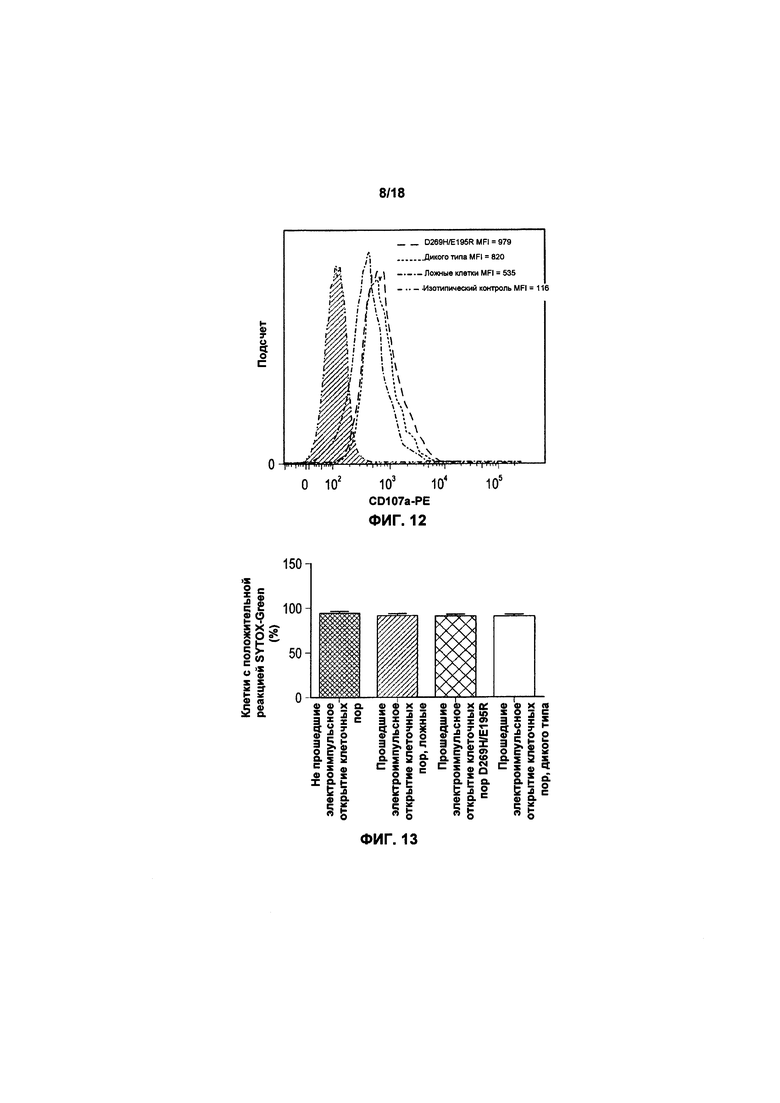
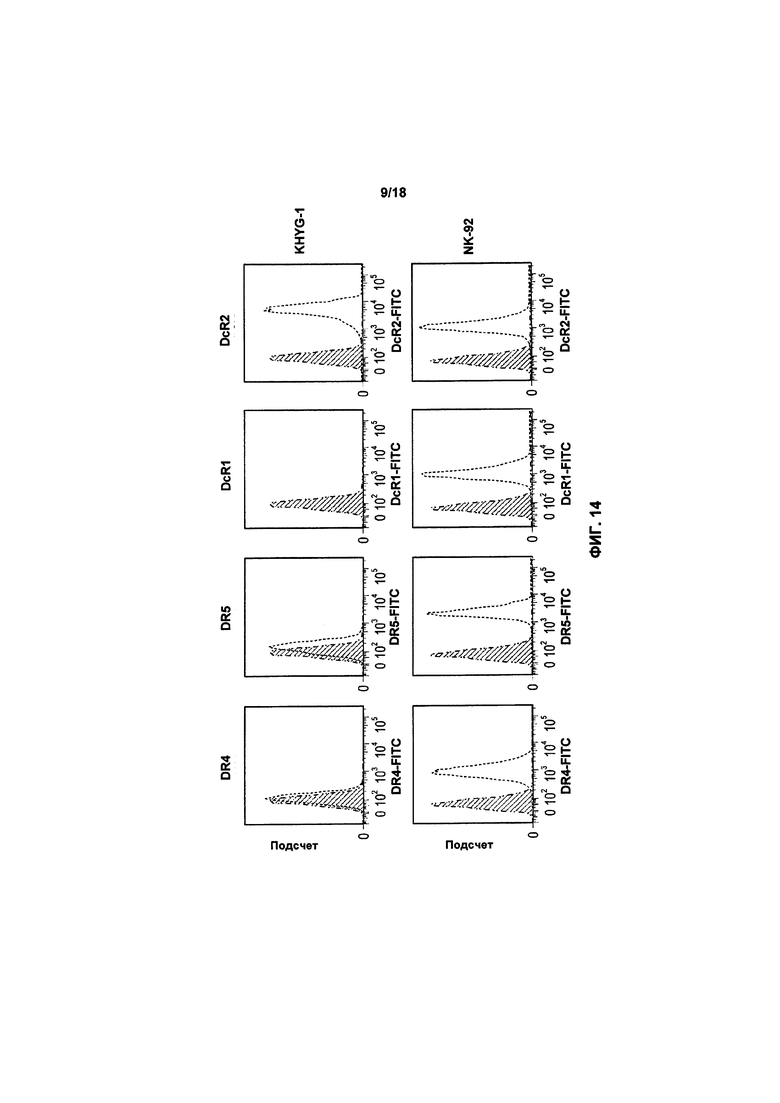
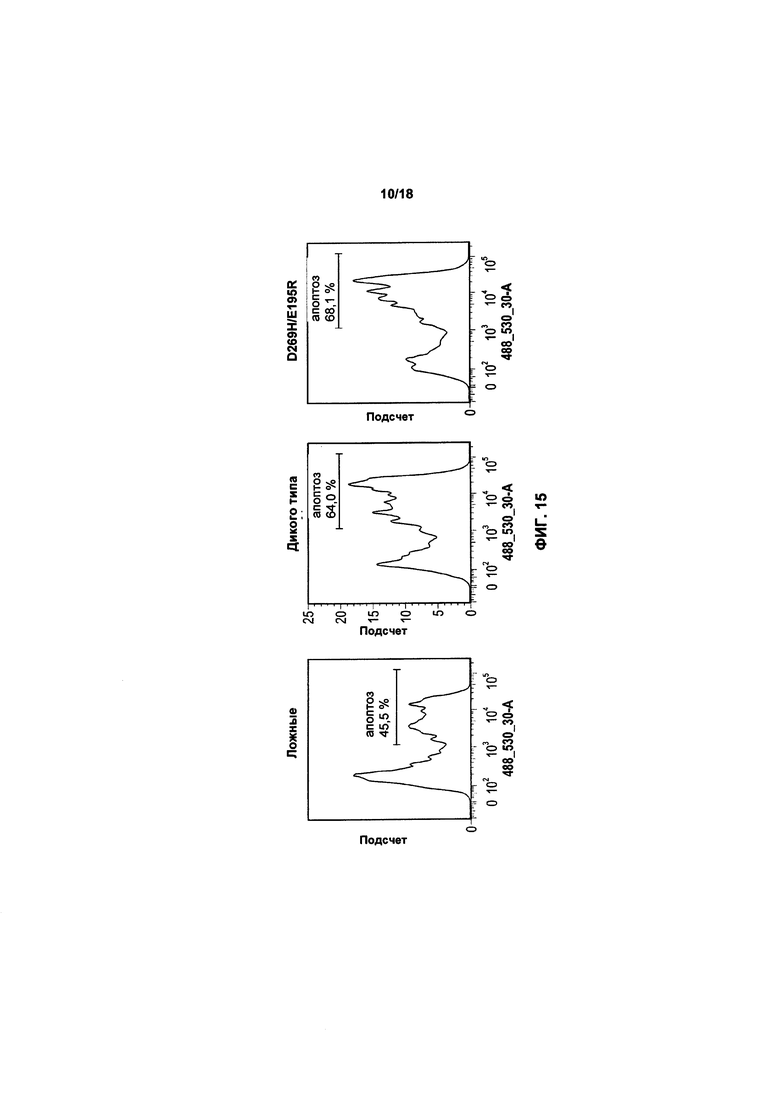
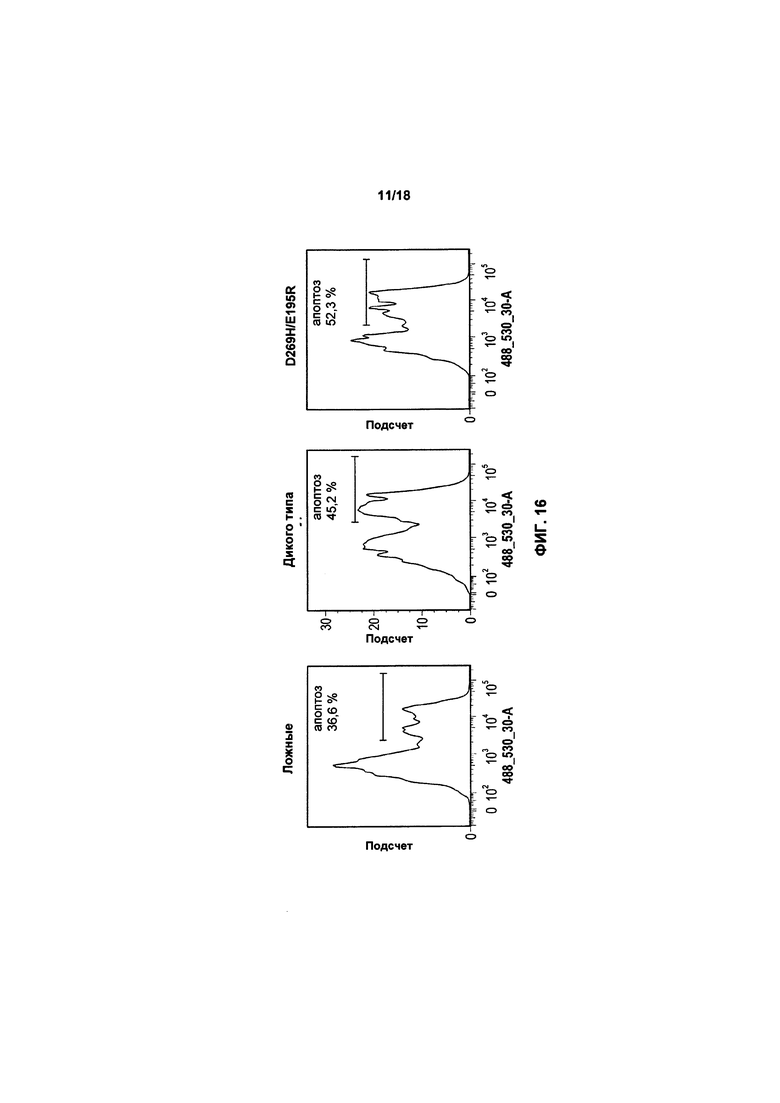
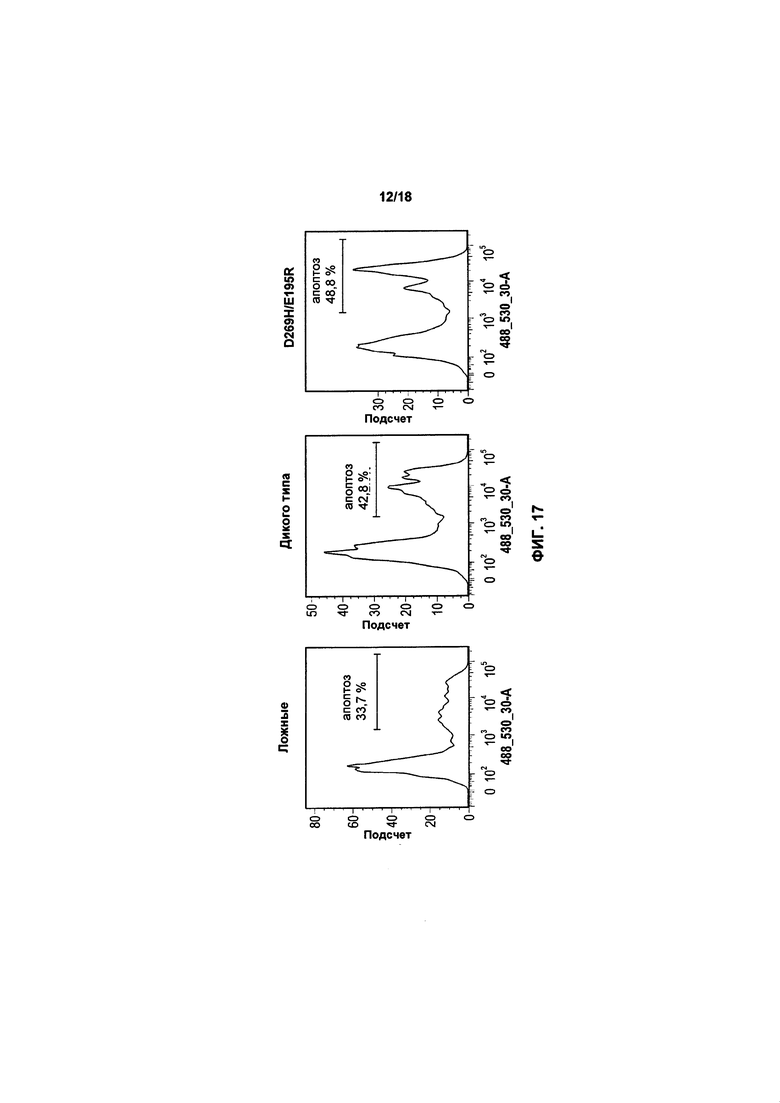
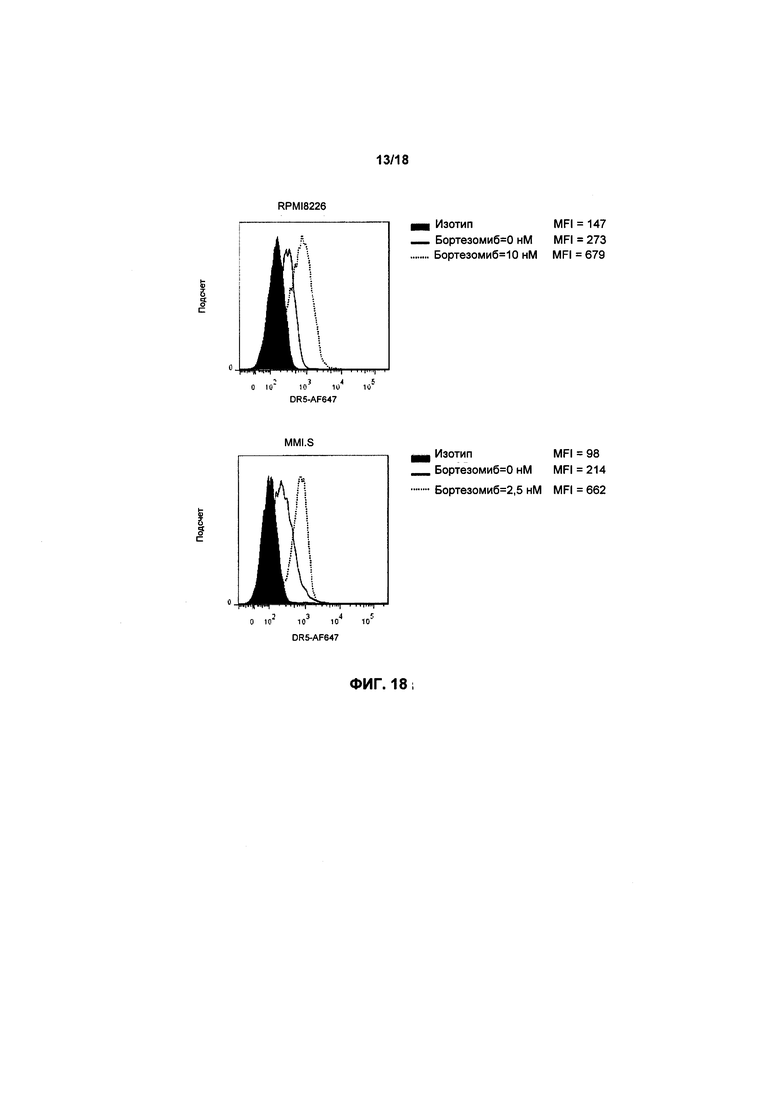
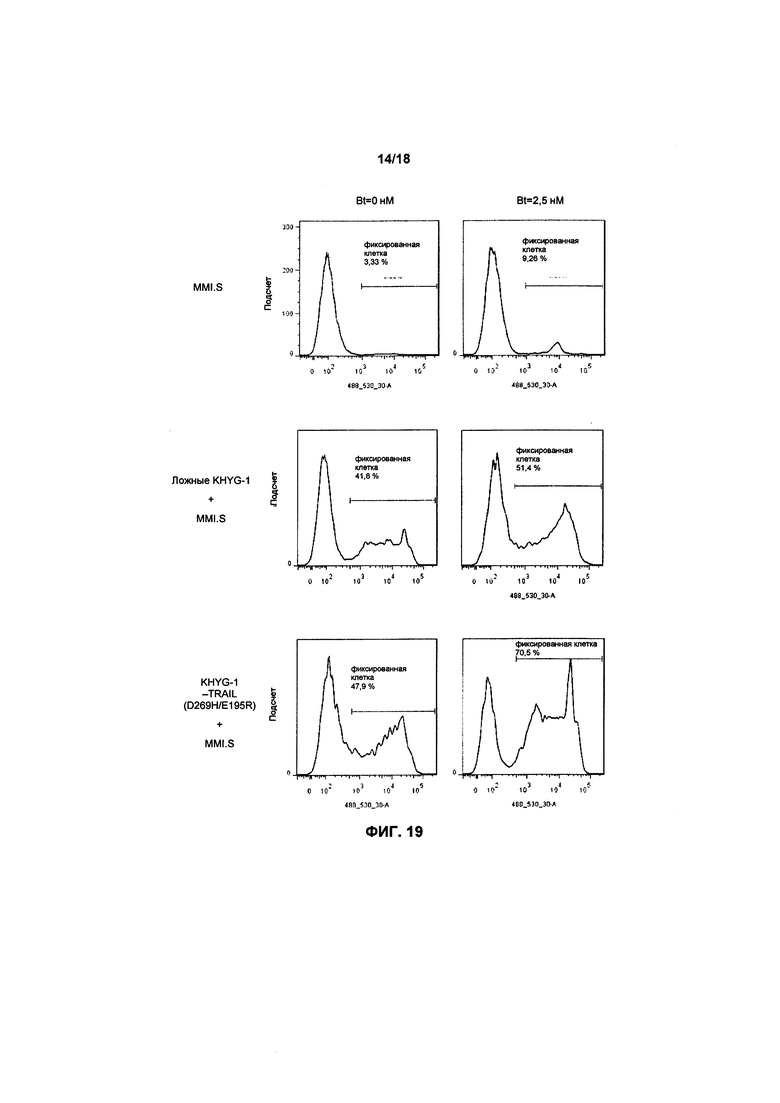
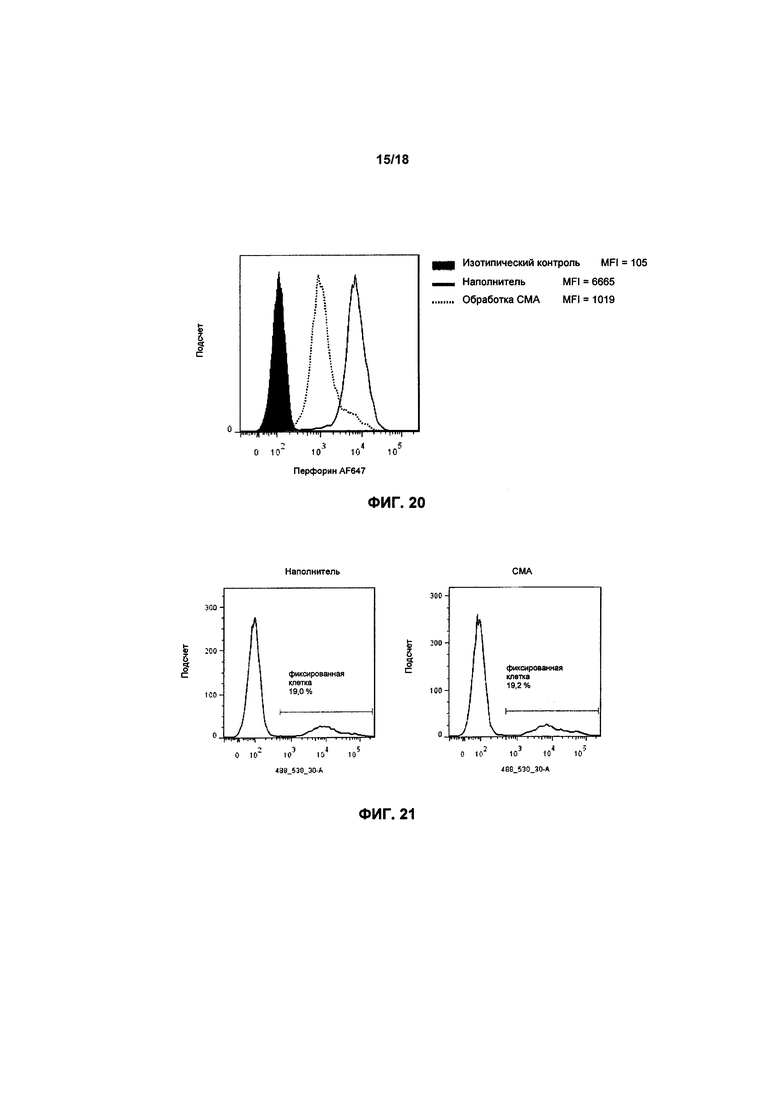
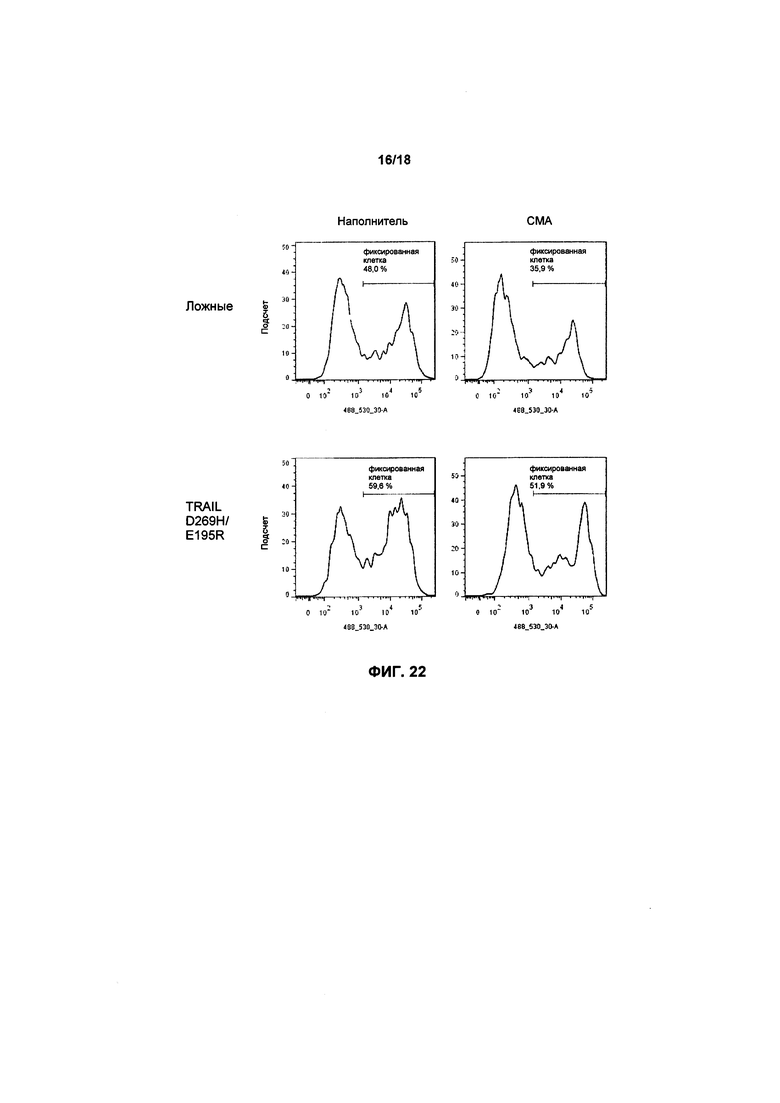
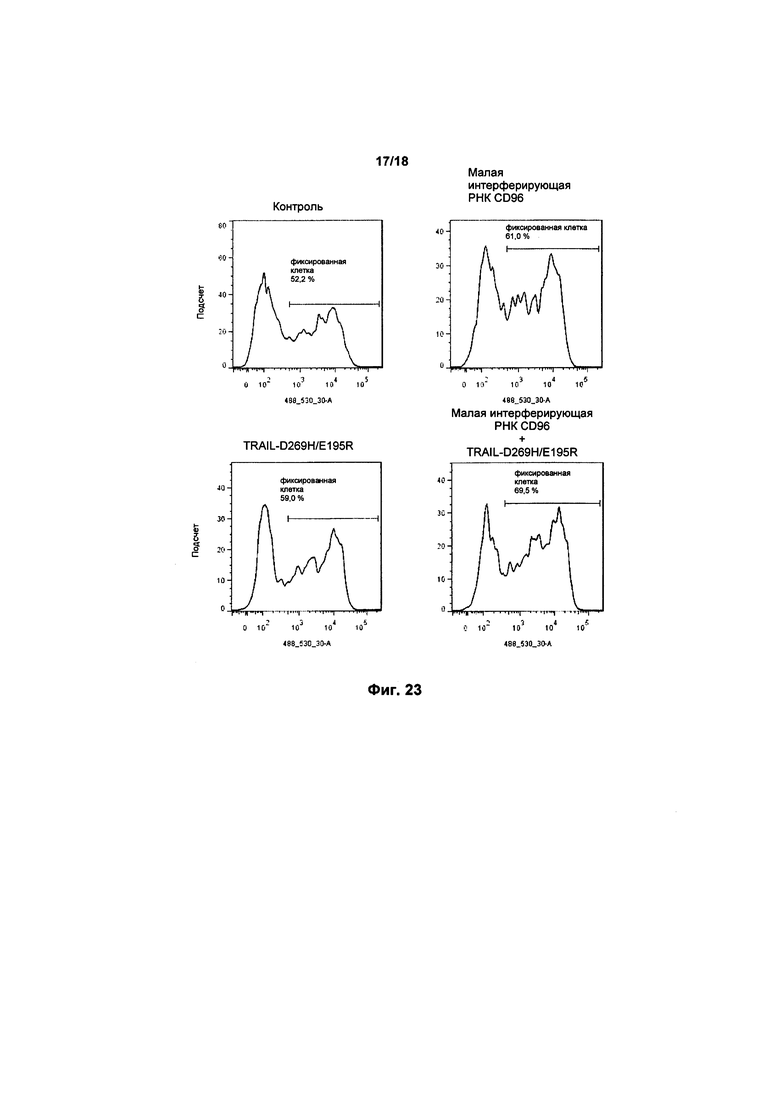
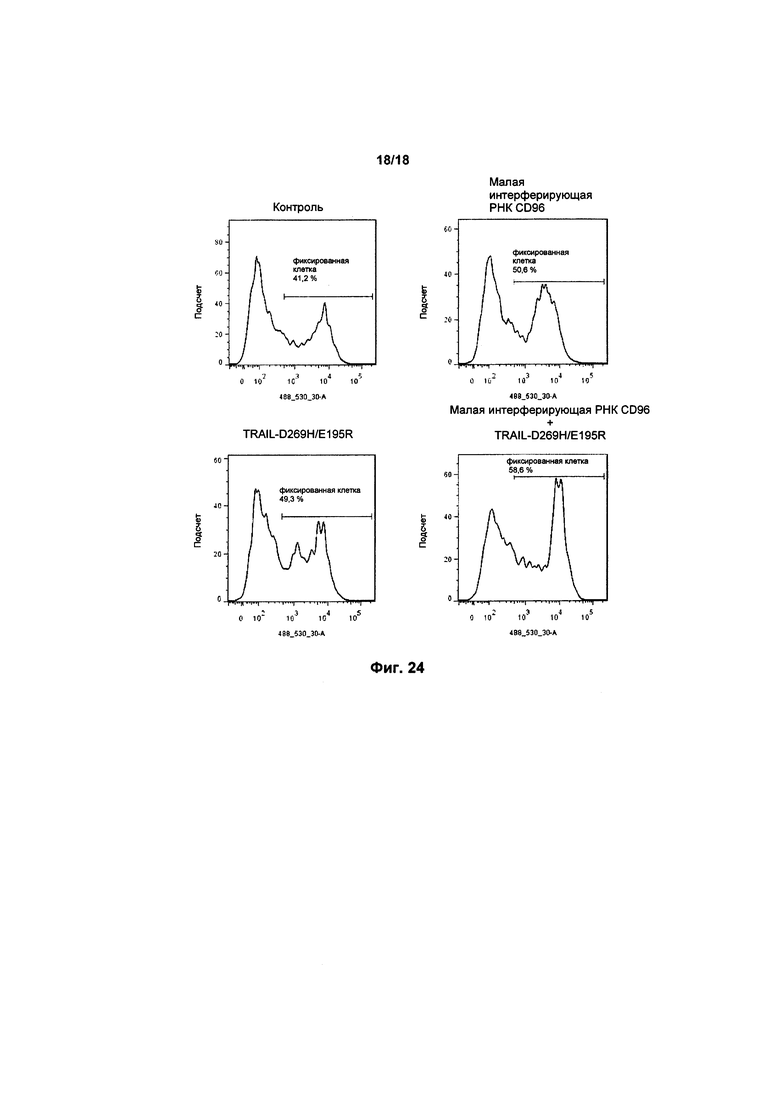
Изобретение относится к области биотехнологии, в частности к человеческой клетке - естественному киллеру (NK), генетически модифицированной для экспрессии варианта TRAIL, включающего мутацию D269H/E195R. Изобретение эффективно для лечения рака. 13 з.п. ф-лы, 24 ил., 1 табл., 10 пр.
1. Человеческая клетка - естественный киллер (NK), предназначенная для лечения рака, генетически модифицированная для экспрессии варианта TRAIL, включающего мутацию D269H/E195R.
2. Человеческая NK-клетка по п. 1, предназначенная для лечения рака, генетически модифицированная путем трансфекции или трансдукции вектором, включающим вариант гена TRAIL D269H/E195R.
3. Человеческая NK-клетка по любому из пп. 1, 2, предназначенная для лечения рака, дополнительно генетически модифицированная в отношении NK-клетки дикого типа для снижения экспрессии одного или более ингибирующих рецепторов контрольной точки, выбранных из CD96 (TACTILE), CD152 (CTLA4), CD223 (LAG-3), CD279 (PD-1), CD328 (SIGLEC7), SIGLEC9, TIGIT или TIM-3.
4. Человеческая NK-клетка по любому из пп. 1-3, предназначенная для лечения рака, отличающаяся тем, что NK-клетка нацелена на костный мозг.
5. Человеческая NK-клетка по любому из пп. 1-4, предназначенная для лечения рака, отличающаяся тем, что NK-клетка происходит из клеточной линии KHYG-1.
6. Человеческая NK-клетка по любому из пп. 1-5, предназначенная для лечения рака, отличающаяся тем, что вариант TRAIL связан с одним или более костимулирующими доменами NK-клетки.
7. Человеческая NK-клетка по п. 6, предназначенная для лечения рака, отличающаяся тем, что костимулирующим доменом NK-клетки является 41BB/CD137, CD3zeta/CD247, DAP12 или DAP10.
8. Человеческая NK-клетка по любому из пп. 1-7, предназначенная для лечения рака, которая дополнительно генетически модифицирована для сверхэкспрессирования IL-2 или IL-15, например, с помощью трансфекции или трансдукции.
9. Человеческая NK-клетка по любому из пп. 1-8, предназначенная для лечения рака, отличающаяся тем, что типом рака является солидный рак.
10. Человеческая NK-клетка по любому из пп. 1-9, отличающаяся тем, что типом рака является рак крови.
11. Человеческая NK-клетка, предназначенная для лечения рака крови по п. 10, отличающаяся тем, что типом рака крови является острый лимфоцитарный лейкоз (ALL), острый миелоидный лейкоз (AML), хронический лимфоцитарный лейкоз (CLL), хронический миелоидный лейкоз (CML), ходжкинская лимфома, неходжкинская лимфома, включая Т-клеточные и В-клеточные лимфомы, асимптомная миелома, вялотекущая множественная миелома (SMM), активная миелома или миелома легкой цепи.
12. Человеческая NK-клетка по любому из пп. 1-11, предназначенная для лечения рака, отличающаяся тем, что лечение включает введение агента, способного повысить экспрессию рецепторов смерти TRAIL на раковых клетках.
13. Человеческая NK-клетка по п. 12, предназначенная для лечения рака, отличающаяся тем, что агент способен повышать DR5.
14. Человеческая NK-клетка по п. 13, предназначенная для лечения рака, отличающаяся тем, что в качестве агента используется Бортезомиб.
| WO 2009077857 A2, 25.06.2009 | |||
| WO 2006023148 A2, 02.03.2006 | |||
| EP 1621550 A1, 01.02.2006 | |||
| LAKSHMI NARENDRA BODDULURU et al., Natural killer cells: the journey from puzzles in biology to treatment of cancer, Cancer Letters, 2014 | |||
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ПОВЫШЕНИЯ ЭФФЕКТИВНОСТИ АНТИТЕЛ ДЛЯ ЛЕЧЕБНЫХ ЦЕЛЕЙ С ИСПОЛЬЗОВАНИЕМ СОЕДИНЕНИЙ, ПОТЕНЦИИРУЮЩИХ NK-КЛЕТКИ | 2004 |
|
RU2396981C2 |

