ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к способу исследования множества культивированных клеток на наличие регулярных структур по меньшей мере одного целевого компонента (например, одного или более белка), содержащегося в культивированных клетках.
УРОВЕНЬ ТЕХНИКИ
Некоторые фенотипы серьезных заболеваний миокарда (или скелетных мышц) связаны с повреждением саркомерной структуры в кардиомиоцитах (или миоцитах, соответственно). Для простоты изложения нижеследующее будет относиться только к кардиомиоцитам. В кардиомиоцитах саркомер представляет собой наименьшую единицу, способную сокращаться и расслабляться.
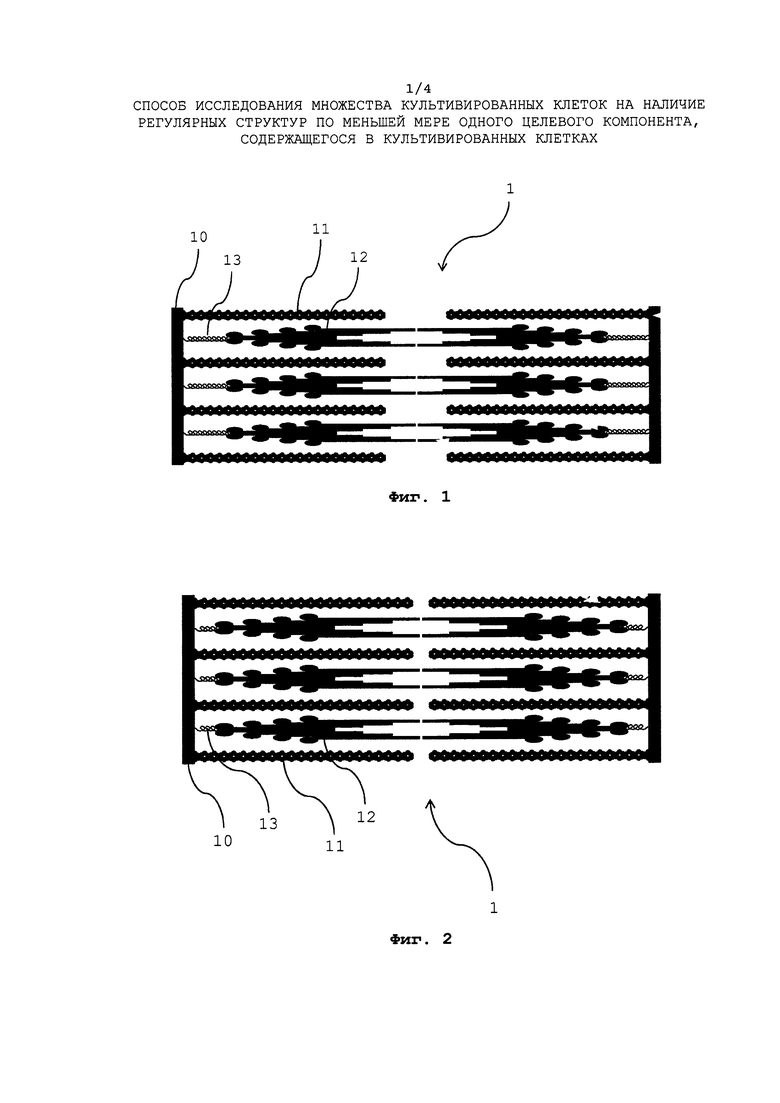
Саркомер схематически показан на Фиг. 1 (в расслабленном состоянии) и на Фиг. 2 (в сокращенном состоянии). Саркомер 1 связан двумя z-дисками 10, а также содержит две чередующихся системы филаментов. Тонкие филаменты первой системы филаментов двух чередующихся систем филаментов состоят из гексамерных актиновых нитей 11, тонкие филаменты второй системы филаментов двух чередующихся систем филаментов состоят из гексамерных миозиновых нитей 12. Миозиновые нити 12 соединены с двумя z-дисками 10 с помощью двух эластичных элементов 13, состоящих из белка титина. При гидролизе АТФ (Аденозинтрифосфат), моторный белок миозин миозиновых нитей 12 претерпевает конформационное изменение, которое преобразуется в силовой удар, который перемещает актиновые нити 11, приводя к сокращению саркомера 1 (см. Фиг. 2). Сокращение, возникающее для большого количества таких саркомеров 1, ведет к сокращению миокарда пока обратный процесс не приведет к расслаблению миокарда.
Такое сокращающее движение может уже наблюдаться в зрелых культивированных живых кардиомиоцитах, которые проявляют спонтанное и синхронное ритмическое сокращение и такое спонтанное и синхронное ритмическое сокращение живых кардиомиоцитов может использоваться для характеристики зрелого состояния развития живых кардиомиоцитов.
В зафиксированных кардиомиоцитах структурная целостность саркомерных структур указывает на зрелое состояние развития живых кардиомиоцитов. Фиксацию кардиомиоцитов можно выполнять, например, путем добавления детергента к живым кардиомиоцитам в результате чего разрушаются клеточные мембраны живых кардиомиоцитов (живые кардиомиоциты умерщвляются) и путем добавления формальдегида, в результате которого белки, содержащиеся в кардиомиоцитах, поперечно сшиваются, однако структуры белков в кардиомиоцитах сохраняются и остаются зафиксированными.
Для идентификации веществ - потенциальных кандидатов в лекарственное средство против заболеваний миокарда, которые связаны с повреждением саркомерных структур, проведены эксперименты in vitro, в которых живые кардиомиоциты подвергались стрессу путем добавления веществ, таких как глюкоза и/или эндотелии до тех пор, пока кардиомиоциты не теряют свою способность пульсировать без умерщвления кардиомиоцитов (имитация кардиомиопатии). После этого к кардиомиоцитам, подвергнутым стрессу добавляют вещество - кандидат в лекарственное средство для того, чтобы определить оказывает ли вещество - кандидат в лекарственное средство восстановительное воздействие на кардиомиоциты, приводящее к тому, что кардиомиоциты снова начинают пульсировать. Для такого исследования небольшой кинофрагмент кардиомиоцитов записывают и анализируют, действительно ли кардиомиоциты снова начали пульсировать (только анализируя кинофрагмент можно оценить циклы пульсирующего сокращения и последующего расслабления). Однако такой анализ кинофрагментов должен выполняться человеком, наблюдающим за кинофрагментами, что является время- и ресурсозатратным. Кроме того, люди, наблюдающие и анализирующие кинофрагменты, должны иметь необходимые образование и навыки.
В другом случае, после того как вещество-кандидат в лекарственное средство было добавлено к живым кардиомиоцитам и после того как они впоследствии были зафиксированы, исследуют зафиксированные кардиомиоциты на наличие регулярных структур саркомеров. Такие регулярные структуры саркомеров являются признаком кардиомиоцитов, восстановившихся до фиксации. Данное может быть реализовано получением изображения зафиксированных кардиомиоцитов, которое должно быть внимательно проанализировано человеком посредством визуального просмотра. Это опять же является время- и ресурсозатратным, а человек, выполняющий визуальный просмотр должен иметь необходимые образование и навыки. Даже в таком случае изображения трудно анализировать.
РАСКРЫТИЕ ИЗОБРЕТЕНИЯ
Таким образом, задача настоящего изобретения в общем состоит в том, чтобы предложить улучшенный способ исследования культивированных клеток на наличие в культивированных клетках регулярных структур определенных компонентов. В частности, задача настоящего изобретения заключается в том, чтобы предложить способ идентификации веществ - потенциальных кандидатов в лекарственное средство для лечения мышечных заболеваний или заболеваний миокарда, связанных с повреждением саркомерных структур.
Согласно одному аспекту, настоящее изобретение предлагает способ исследования множества культивированных клеток на наличие регулярных структур по меньшей мере одного целевого компонента, содержащегося в культивированных клетках. Способ содержит этапы, на которых:
- обеспечивают множество культивированных клеток, подлежащих исследованию,
- фиксируют культивированные клетки, подлежащие исследованию, сохраняя любые структуры по меньшей мере одного целевого компонента, содержащегося в культивированных клетках,
- окрашивают по меньшей мере один целевой компонент, содержащийся в зафиксированных культивированных клетках, используя первый окрашивающий агент, связанный с по меньшей мере одним целевым компонентом и способный при возбуждении испускать свет первой длины волны,
- возбуждают первый окрашивающий агент, что приводит к тому, что он испускает свет первой длины волны,
- получают двухмерное изображение зафиксированных культивированных клеток с первым окрашивающим агентом, испускающим свет первой длины волны,
- из упомянутого двухмерного изображения зафиксированных культивированных клеток извлекают первое отфильтрованное изображение, показывающее яркими только те части, которые содержат по меньшей мере один целевой компонент, с которым связан первый окрашивающий агент,
- осуществляют автокорреляцию первого отфильтрованного изображения, или взаимную корреляцию первого отфильтрованного изображения с двухмерным вторым отфильтрованным изображением, извлеченным из двухмерного изображения зафиксированных культивированных клеток и показывающим ярким только части, отличные от тех, которые содержат по меньшей мере один целевой компонент, с которым связан первый окрашивающий агент, для получения двухмерного корреляционного изображения, и
- устанавливают наличие или отсутствие регулярных структур, по меньшей мере, одного целевого компонента, содержащегося в зафиксированных культивированных клетках, путем определения, присутствуют ли в корреляционном изображении регулярные структуры максимальной и минимальной яркости.
Хотя, вообще способ подходит для in vitro исследования клеток любого вида на наличие в них любых регулярных структур, он в частности подходит для исследования мышечных клеток и, в частности, кардиомиоцитов на наличие регулярных структур саркомеров. Ввиду этого в качестве примера далее приводится исследование кардиомиоцитов на наличие регулярных структур саркомеров.
Обычно фиксируют культивированные кардиомиоциты, подлежащие исследованию, например, живые кардиомиоциты, которые были подвергнуты стрессу добавлением глюкозы и/или эндотелина (или любого другого подходящего вещества) для того, чтобы живые кардиомиоциты потеряли способность пульсировать (имитация кардиомиопатии) и к которым затем было добавлено вещество-кандидат в лекарственное средство. Хотя фиксация кардиомиоцитов повреждает клеточные мембраны и таким образом «умерщвляет» кардиомиоциты, любые регулярные саркомерные структуры, содержащиеся в кардиомиоцитах, сохраняются на стадии фиксации. Хотя кардиомиоциты, подлежащие исследованию, более не являются живыми, их желательно хранить в нейтральной жидкости для предотвращения их высыхания.
Затем окрашивают по меньшей мере один компонент саркомера зафиксированных кардиомиоцитов, например, белок Актинин, содержащийся в Z-дисках саркомеров зафиксированных кардиомиоцитов. Для этого к зафиксированным кардиомиоцитам может быть добавлен подходящий окрашивающий агент. Примером такого подходящего окрашивающего агента может быть Актининовое антитело, снабженное меткой, которая при возбуждении способна испускать свет заданной длины волны. Если Актининовое антитело, снабженное меткой, связано с белком Актинином, содержащимся в кардиомиоцитах, то окрашивающий агент возбуждается, что заставляет метку испускать свет заданной длины волны.
Во время испускания света заданной длины волны получают двухмерное изображение культивированных кардиомиоцитов. Из этого двухмерного изображения зафиксированных культивированных кардиомиоцитов извлекают двухмерное первое отфильтрованное изображение. Данное первое отфильтрованное изображение показывает яркими только те части изображения зафиксированных культивированных кардиомиоцитов, которые содержат белок Актинин с которым связано Актининовое антитело с меткой.
Вообще далее возможны два варианта. Первый вариант заключается в автокорреляции первого отфильтрованного изображения, чтобы сгенерировать двухмерное корреляционное изображение. Второй вариант заключается во взаимной корреляции первого отфильтрованного изображения со вторым отфильтрованным изображением. Второе отфильтрованное изображение также является двухмерным изображением, извлеченным из двухмерного изображения зафиксированных культивированных кардиомиоцитов. Однако в отличие от первого отфильтрованного изображения, второе отфильтрованное изображение показывает яркими только те части зафиксированных культивированных кардиомиоцитов, которые отличны от частей окрашенного белка Актинина. Затем выполняют взаимную корреляцию первого и второго отфильтрованных изображений, чтобы сгенерировать двухмерное корреляционное изображение.
С математической точки зрения автокорреляция первого отфильтрованного изображения соответствует свертке интенсивностей отдельных пикселей первого отфильтрованного изображения с интенсивностями отдельных пикселей первого отфильтрованного изображения, или, говоря иначе, автокорреляция представляет собой свертку первого отфильтрованного изображения с самим собой. Взаимная корреляция первого отфильтрованного изображения и второго отфильтрованного изображения соответствует свертке интенсивностей яркости отдельных пикселей первого и второго отфильтрованных изображений (которые подвергаются антикорреляции определением при нулевом пространственном смещении). Однако имея компьютерное исполнение, математическая свертка интенсивностей яркости отдельных пикселей будет приводить к высоким вычислительным затратам. Несмотря на то, что в принципе такое возможно, существуют способы, более эффективные, чем выполнение математической свертки, и такие более эффективные способы приводят к тому же результату, но требуют гораздо меньших вычислительных затрат. Примеры таких более эффективных способов подробно описаны ниже.
В том случае, когда корреляционное изображение показывает регулярные структуры в максимумах и минимумах яркости, это означает, что присутствуют регулярные структуры белка Актинина, содержащиеся в z-дисках саркомеров зафиксированных кардиомиоцитов. В зависимости от того как выглядят регулярные структуры на корреляционном изображении, из корреляционного изображения регулярных саркомерных структур, содержащихся в кардиомиоцитах, могут быть сделаны выводы, которые будут подробно поясняться ниже на конкретных примерах.
В общем, способ по изобретению предлагает улучшенный способ in vitro (например, может быть осуществлен в лунках стандартных микропланшетов) для исследования культивированных клеток на наличие регулярных структур, содержащихся в культивированных клетках, который более не требует необходимых время и ресурсозатратных стадий способов из уровня техники (запись кинофрагментов, оптический анализ кинофрагментов и определение пульсируют ли кардиомиоциты; сложный оптический анализ зафиксированных культивированных клеток, при наличии возможности). Вместо этого способ по изобретению предлагает получать вышеописанное двухмерное изображение зафиксированных культивированных клеток, извлекать из этого двухмерного изображения зафиксированных культивированных клеток отфильтрованное изображение или отфильтрованные изображения, коррелировать эти отфильтрованные изображения, для того чтобы сгенерировать корреляционное изображение и определять наличие или отсутствие регулярных структур в культивированных клетках путем определения, присутствуют ли в корреляционном изображении регулярные структуры максимальной и минимальной яркости. Указанные этапы могут быть выполнены полностью автоматически.
В некоторых вариантах выполнения способа по изобретению этап, на котором извлекают двухмерное первое и второе отфильтрованные изображения содержит выполнение фильтрации гребней и впадин двухмерного изображения зафиксированных культивированных клеток, чтобы сгенерировать двухмерное изображение гребней и сгенерировать изображение впадин. На изображении гребней полосообразные структуры по меньшей мере одного целевого компонента, с которым связан первый окрашивающий агент, усилены по яркости, а структуры, отличные от полосообразных структур по меньшей мере одного целевого компонента, с которым связан окрашивающий агент, ослаблены по яркости. Изображение гребней образует первое отфильтрованное изображение. На изображении впадин полосообразные структуры, расположенные между полосообразными структурами целевого компонента, с которым связан первый окрашивающий агент, усилены по яркости, а структуры, отличные от полосообразных структур, расположенных между полосообразными структурами, с которыми связан первый окрашивающий агент, ослаблены по яркости. Указанное изображение впадин образует второе отфильтрованное изображение. Затем выполняют взаимную корреляцию изображения гребней и изображения впадин для создания корреляционного изображения.
Указанный вариант осуществления способа по изобретению является предпочтительным для обнаружения полосообразных структур, таких как саркомерные структуры в кардиомиоцитах, поскольку фильтрация гребней и впадин усиливает только полосообразные структуры, ослабляя при этом любые структуры, отличные от полосообразных структур. Таким образом, структуры, отличные от полосообразных структур, находящихся на изображении зафиксированных культивированных кардиомиоцитов, скрываются. Для вышеописанного примера, относящегося к окрашенному белку Актинину, содержащемуся в z-дисках саркомеров кардиомиоцитов (предполагая, что зафиксированные кардиомиоциты содержат саркомерные структуры, которые являются интактными), это означает, что изображение гребней усиливает по яркости только структуры окрашенного белка Актинина (z-диски), в то время как любая другая полосообразная информация, содержащаяся в изображении культивированных клеток, на изображении гребней скрывается (на изображении гребней эти пиксели темные). Однако данная информация содержится на изображении впадин, которое усиливает полосообразные структуры между полосообразными структурами окрашенного белка Актинина. Для того чтобы использовать и эту информацию тоже, при генерировании корреляционного изображения осуществляют взаимную корреляцию изображения гребней с изображением впадин, чтобы сгенерировать корреляционное изображение. Вообще, однако, для того, чтобы сгенерировать коррелированное изображение также возможно осуществить только автокорреляцию изображения гребней. Однако, в этом случае информация, скрытая на изображении гребней, теряется и более не содержится на корреляционном изображении. По этой причине взаимная корреляция изображения гребней и изображения впадин является предпочтительнее, чем автокорреляция изображения гребней.
В некоторых дополнительных варианта способа по изобретению, корреляционное изображение генерируется путем:
- выполнения преобразования Фурье первого отфильтрованного изображения для получения двухмерного Фурье-преобразованного первого отфильтрованного изображения и умножения Фурье-преобразованного изображения само на себя для получения двухмерного Фурье-преобразованного корреляционного изображения, или
- выполнения преобразования Фурье первого отфильтрованного изображения для получения двухмерного Фурье-преобразованного первого отфильтрованного изображения и выполнения преобразования Фурье второго отфильтрованного изображения для получения двухмерного Фурье-преобразованного второго отфильтрованного изображения и умножения Фурье-преобразованного первого отфильтрованного изображения на Фурье-преобразованное второе отфильтрованное изображение для получения двухмерного перемноженного Фурье-преобразованного корреляционного изображения,
- выполнения обратного преобразования Фурье перемноженного Фурье-преобразованного корреляционного изображения для получения двухмерного корреляционного изображения.
Такой вариант выполнения является предпочтительным, поскольку он существенно снижает вычислительные затраты на получение корреляционного изображения или посредством автокорреляции первого отфильтрованного изображения или посредством взаимной корреляции первого отфильтрованного изображения и второго отфильтрованного изображения. Как было рассмотрено выше, с математической точки зрения автокорреляция и взаимная корреляция означает выполнение свертки. Для того чтобы продемонстрировать данное существенное снижение вычислительных затрат на получение корреляционного изображения, далее будет рассмотрен пример.
Полагая, что двухмерное изображение культивированных клеток было получено, и то, что изображение гребней и изображение впадин было из него извлечено, пусть
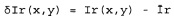 и
и
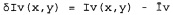
представляют дифференциальные изображения гребней и впадин, где
Ir(х,у) - интенсивность изображения гребней в координатах х, у;
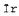 - средняя интенсивность (яркость) изображения гребней;
- средняя интенсивность (яркость) изображения гребней;
Iv(х,у) - интенсивность изображения впадин в координатах х, у;
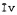 - средняя интенсивность (яркость) изображения впадин.
- средняя интенсивность (яркость) изображения впадин.
Корреляционное изображение затем представляют уравнением
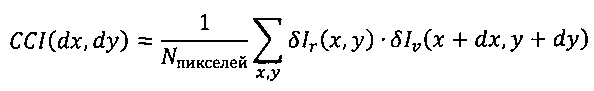
Это означает, что для того чтобы найти величину интенсивности корреляционного изображения для одного конкретного относительного смещения (dx,dy) изображений гребней и впадин, должна быть рассчитана величина интенсивности (яркости) для этого конкретного смещения (dx,dy) в соответствии с уравнением, представленным выше (свертка). Соответственно, для того, чтобы найти на корреляционном изображении величины интенсивности для всех относительных смещений (dx,dy), необходимо выполнить большой объем вычислительной работы с использованием изображений гребней и впадин. Однако такая вычислительная работа значительно сокращается после преобразования Фурье изображений гребней и впадин. Представленная выше свертка изображений гребней и впадин соответствует всего одному умножению Фурье-преобразованных изображений гребней и впадин (без относительного смещения) для получения Фурье-преобразованного корреляционного изображения, а затем должно быть выполнено только обратное преобразование Фурье Фурье-преобразованного корреляционного изображения для получения корреляционного изображения.
В соответствии с предпочтительным вариантом выполнения вышеупомянутого способа, этап, на котором выполняют преобразование Фурье первого и второго отфильтрованных изображений (в вышеописанном примере изображений гребней и впадин), так же как и этап выполнения обратного преобразования Фурье Фурье-преобразованного корреляционного изображения, выполняют путем использования алгоритма быстрого преобразования Фурье. Они представляют собой хорошо известные численные алгоритмы для очень эффективного выполнения преобразования Фурье и обратного преобразования Фурье.
В некоторых вариантах осуществления способа по изобретению яркость двухмерного корреляционного изображения усредняется до определенного радиуса в пределах всей окружности и при этом такое усреднение яркости в пределах всей окружности выполняется при различных радиусах для получения одномерной корреляционной функции. Из этой одномерной корреляционной функции получают степень полосатости в двухмерном изображении зафиксированных культивированных клеток путем определения расстояния между двумя смежными максимумами яркости или путем определения расстояния между максимумом и смежным минимумом яркости одномерной корреляционной функции и путем определения соотношения амплитуд яркости двух смежных максимумов или соотношения амплитуд яркости максимума и смежного минимума одномерной корреляционной функции.
Такой вариант выполнения является предпочтительным, если распределение полосатых структур в двухмерном изображении зафиксированных культивированных клеток является изотропным (случайно ориентированным), поскольку в таком случае при одном конкретном радиусе интенсивность в корреляционном изображении распределена по существу равномерно в пределах окружности с таким конкретным радиусом, так, что никакая информация не уничтожается усреднением. Одномерная корреляционная функция представляет собой свертку числа клеток, содержащих полосатые структуры, амплитуду модуляции их полос и однородность их периодичности. В одномерной корреляционной функции расстояние между двумя смежными максимумами или между максимумом и смежным минимумом яркости, также как и соотношение амплитуд яркости между двумя смежными максимумами или амплитуд яркости максимума и смежного минимума, представляют собой существенные признаки, которые говорят о степени полосатости в двухмерном изображении зафиксированных культивированных клеток.
Некоторые варианты осуществления способа по изобретению дополнительно содержат этап, на котором выполняют преобразование Фурье двухмерного корреляционного изображения для получения двухмерного Фурье-преобразованного корреляционного изображения. Из этого Фурье-преобразованного корреляционного изображения, определяют расстояние между смежными максимумами яркости путем измерения расстояния между полосообразными структурами в двухмерном изображении культивированных клеток. Кроме того, угол, под которым расположены смежные максимумы яркости, определяют как отображение общей ориентации, в соответствии с которой регулярные полосообразные структуры располагаются в двухмерном изображении культивированных клеток.
Данный вариант выполнения является предпочтительным, если распределение полосатых структур является (единообразно) ориентированным в двухмерном изображении зафиксированных культивированных клеток. В случае упорядоченного распределения полосатых структур в двухмерном изображении зафиксированных культивированных клеток, Фурье-преобразованное корреляционное изображение в большей степени указывает на полосатые структуры, чем одномерная корреляционная функция, рассмотренная выше, поскольку информация одномерной корреляционной функции об ориентации уничтожается при усреднении в пределах окружности при конкретном радиусе. Преобразование Фурье корреляционного изображения практически обозначает преобразование х, у - системы координат коррелированного изображения в kx, ky - систему координат (волновое число) Фурье-преобразованного корреляционного изображения. Соответственно, определение волнового числа 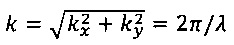 позволяет определить длину волны регулярных полос, из которой можно рассчитать расстояние между регулярными полосами в двухмерном изображении культивированных клеток. Угол, под которым расположен участок регулярных полос, может быть определен из Фурье-преобразованного корреляционного изображения, как
позволяет определить длину волны регулярных полос, из которой можно рассчитать расстояние между регулярными полосами в двухмерном изображении культивированных клеток. Угол, под которым расположен участок регулярных полос, может быть определен из Фурье-преобразованного корреляционного изображения, как 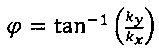 .
.
Кроме того, этап, на котором выполняют преобразование Фурье двухмерного кросскорреляционного изображения, предпочтительно осуществляют с использованием хорошо известного алгоритма быстрого преобразования Фурье, который уже был упомянут выше.
Поскольку невозможно предсказать заранее являются ли любые полосообразные структуры, содержащиеся в двухмерном изображении зафиксированных культивированных клеток, изотропно ориентированными (ориентированными случайным образом) или же (упорядоченно) ориентированными, в предпочтительном варианте выполнения анализируют и одномерную корреляционную функцию, и Фурье-преобразованное изображение корреляционного изображение, и определяют, что содержит более четко выраженную информацию о полосатости, содержащуюся в двухмерном изображении зафиксированных культивированных клеток.
Как уже упоминалось выше, хотя способ по изобретению вообще подходит для исследования клеток любого вида на наличие регулярных структур, содержащихся в них, он в частности подходит для определения содержат ли миоциты или кардиомиоциты регулярные саркомерные структуры.
Соответственно, другой аспект изобретения относится к способу идентификации вещества - потенциального кандидата в лекарственное средство для лечения мышечных заболеваний или заболеваний миокарда, связанных с повреждением саркомерных структур в миоцитах или кардиомиоцитах. Данный способ содержит этапы, на которых:
- обеспечивают множество живых миоцитов или кардиомиоцитов, содержащих саркомерные структуры,
- к множеству живых миоцитов или кардиомиоцитов добавляют вещество, оказывающее стрессовое воздействие, что приводит к разрушению саркомерных структур или по меньшей мере их существенному сокращению,
- после добавления вещества, оказывающего стрессовое воздействие, к множеству живых миоцитов или кардиомиоцитов, содержащих разрушенные или по меньшей мере существенно сокращенные саркомерные структуры, добавляют вещество-кандидат,
- после того как было добавлено вещество-кандидат, культивируют миоциты или кардиомиоциты,
- определяют привело ли добавление вещества-кандидата к образованию саркомерных структур, вновь сформировавшихся в культивированных миоцитах или кардиомиоцитах,
- если добавление вещества-кандидата привело к образованию саркомерных структур, вновь сформировавшихся в культивированных миоцитах или кардиомиоцитах, вещество-кандидат квалифицируют в качестве кандидата в лекарственное средство.
Определение того, привело ли добавление вещества-кандидата к образованию саркомерных структур, вновь сформировавшихся в миоцитах или кардиомиоцитах, содержит использование способа исследования культивированных клеток в соответствии с любым из вариантов выполнения, описанных выше.
Другой аспект изобретения относится к способу идентификации способности вещества - потенциального кандидата лекарственного средства обладать защитным действием против мышечных заболеваний или заболеваний миокарда, связанных с повреждением саркомерных структур в миоцитах или кардиомиоцитах. Способ содержит этапы, на которых:
- обеспечивают множество живых миоцитов или кардиомиоцитов, содержащих саркомерные структуры,
- к множеству живых миоцитов или кардиомиоцитов добавляют вещество-кандидат,
- после добавления вещества-кандидата к множеству живых миоцитов или кардиомиоцитов, добавляют вещество, оказывающее стрессовое воздействие, что приводит к разрушению саркомерных структур или по меньшей мере их существенному сокращению,
- после того как было добавлено вещество, оказывающее стрессовое воздействие, культивируют миоциты или кардиомиоциты,
- определяют, привело ли добавление вещества, оказывающего стрессовое воздействие, к разрушению или, по меньшей мере, существенному сокращению саркомерных структур,
- если добавление вещества, оказывающего стрессовое воздействие, не привело к разрушению или по меньшей мере существенному сокращению саркомерных структур, вещество-кандидат квалифицируют в качестве кандидата в лекарственное средство.
Этап определения того, привело ли добавление вещества, оказывающего стрессовое воздействие, к разрушению или по меньшей мере существенному сокращению саркомерных структур в миоцитах или кардиомиоцитах, содержит использование способа исследования культивированных клеток согласно любому из вариантов выполнения, описанных выше.
Такой способ является полезным для идентификации веществ -потенциальных кандидатов в лекарственное средство, которые могут обладать защитным действием в том смысле, что они или предотвращают или по меньшей мере существенно снижают стресс-реакции миоцитов или кардиомиоцитов, когда миоциты или кардиомиоциты подвергают воздействию веществ (например, глюкозы и/или эндотелина), которые могут приводить к частичному или полному разрушению сарокмерных структур миоцитов или кардиомиоцитов. Затем могут дополнительно оценивать потенциал таких веществ - потенциальных кандидатов в лекарственное средство к разработке лекарственного средства, которое может составлять часть защитной или профилактической терапии.
Дополнительный аспект изобретения относится к способу определения наличия саркомерных структур в живых миоцитах или кардиомиоцитах, полученных из живых индуцированных плюрипотентных стволовых клеток. Способ содержит этапы, на которых:
- обеспечивают множество живых индуцированных плюрипотентных стволовых клеток,
- вызывают дифференцировку живых индуцированных плюрипотентных стволовых клеток в живые миоциты или кардиомиоциты,
- культивируют живые миоциты или кардиомиоциты,
- определяют, содержат ли культивированные миоциты или кардиомиоциты саркомерные структуры,
причем этап, на котором определяют, содержат ли культивированные живые миоциты или кардиомиоциты сакромерные структуры, содержит использование способа исследования культивированных клеток в соответствии с любым из вариантов выполнения, описанных выше.
Данный способ позволяет оценить успешность дифференцировки живых индуцированных плюрипотентных стволовых клеток в живые миоциты или кардиомиоциты. Индуцированные плюрипотентные стволовые клетки могут быть получены из клеток кожи, например, посредством перепрограммирования и дифференцировки. Структурная целостность саркомера, содержащегося в миоцитах или кардиомиоцитах, полученных таким образом, является признаком зрелого состояния развития миоцитов или кардиомиоцитов, и в данном случае, является признаком успешной дифференцировки плюрипотентных стволовых клеток в миоциты или кардиомиоциты.
Дополнительный вариант выполнения данного способа содержит дополнительные этапы, на которых:
- до культивирования живых миоцитов или кардиомиоцитов, к живым миоцитам и кардиомиоцитам добавляют вещество, оказывающее стрессовое воздействие, что приводит к разрушению саркомерных структур или по меньшей мере их существенному сокращению, и
- после осуществления способа исследования культивированных клеток в соответствии с любым из вариантов выполнения, описанных выше, определяют, привело ли добавление вещества, оказывающего стрессовое воздействие к разрушению или по меньшей мере существенному сокращению саркомерных структур в культивированных миоцитах или кардиомиоцитах.
В таком варианте выполнения способа можно получить информацию о стресс-реакции миоцитов или кардиомиоцитов, полученных вышеописанным способом. Если стресс-реакция миоцитов или кардиомиоцитов представляет собой ожидаемую реакции миоцитов или кардиомиоцитов, функционирующих должным образом, тогда такой вариант выполнения способа может составлять часть способа диагностики. В таком способе диагностики, например, клетки кожи могут быть взяты у пациента, перепрограммированы и дифференцированы. Миоциты или кардиомиоциты, полученные таким образом, затем могут быть использованы для определения того, склонен ли данный пациент к заболеваниям мышц или миокарда, связанных с повреждением саркомерных структур при воздействии определенных веществ (например, глюкозы и/или эндотелина). Такой способ является не обременительным для пациента.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Дополнительные предпочтительные аспекты изобретения становятся очевидными из следующего описания вариантов выполнения изобретения с использованием чертежей, на которых:
Фиг. 1 показывает схематичный вид саркомера в расслабленном состоянии;
Фиг. 2 показывает саркомер с Фиг. 1 в сокращенном состоянии;
Фиг. 3 схематично показывает изображение зафиксированного кардиомиоцита с единообразно ориентированными саркомерными структурами;
Фиг. 4 показывает корреляционное изображение, сгенерированное посредством взаимной корреляции изображений гребней и впадин, извлеченных из изображения зафиксированного кардиомиоцита с Фиг. 3;
Фиг. 5 показывает Фурье-преобразованное корреляционное изображение с Фиг. 4;
Фиг. 6 схематично показывает изображение зафиксированного кардиомиоцита с саркомерными структурами, ориентированными случайным образом (изотропно);
Фиг. 7 показывает корреляционное изображение, сгенерированное посредством взаимной корреляции изображений гребней и впадин, извлеченных из изображения зафиксированного кардиомиоцита с Фиг. 6;
Фиг. 8 показывает Фурье-преобразованное корреляционное изображение с Фиг. 7;
Фиг. 9 схематично показывает изображение зафиксированного кардиомиоцита без саркомерных структур;
Фиг. 10 показывает корреляционное изображение, сгенерированное посредством взаимной корреляции изображений гребней и впадин, извлеченных из изображения зафиксированного кардиомиоцита с Фиг. 9;
Фиг. 11 показывает Фурье-преобразованное корреляционное изображение с Фиг. 10;
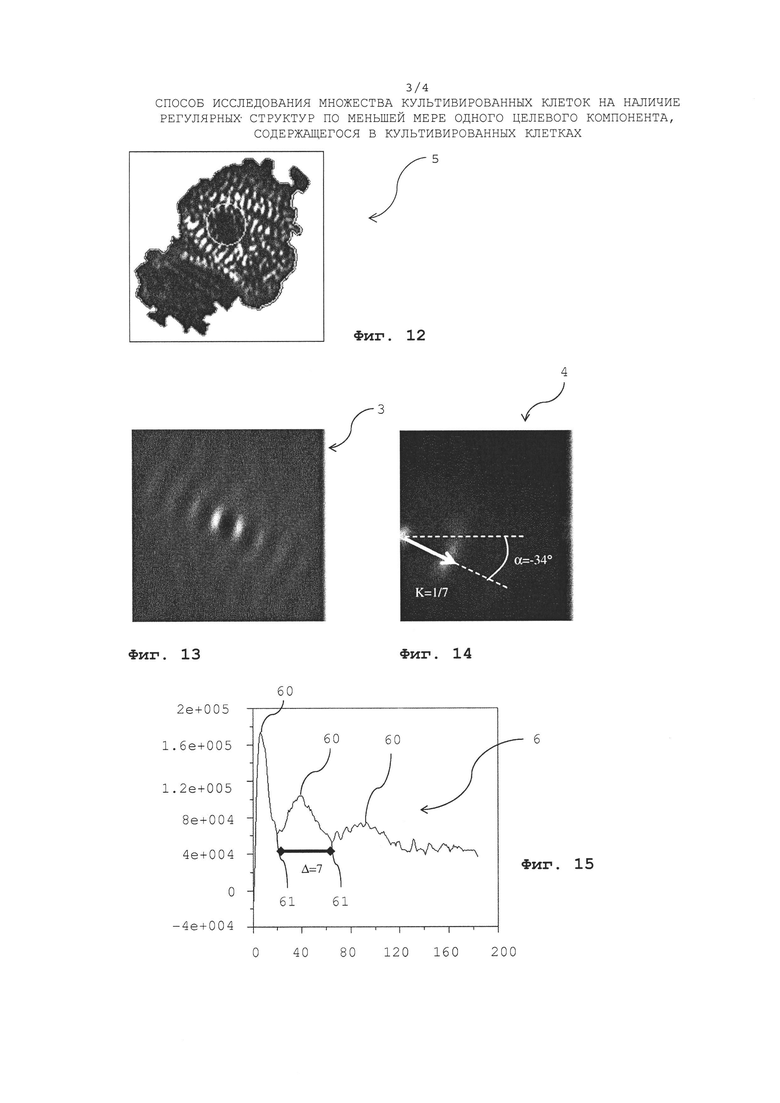
Фиг. 12 показывает изображение окрашивания Актинина реального кардиомиоцита с единообразно ориентированными саркомерными структурами;
Фиг. 13 показывает корреляционное изображение, сгенерированное посредством взаимной корреляции изображений гребней и впадин, извлеченных из изображения реального кардиомиоцита с Фиг. 12;
Фиг. 14 показывает Фурье-преобразованное корреляционное изображение с Фиг. 13;
Фиг. 15 показывает одномерную корреляционную функцию, сгенерированную из корреляционного изображения с Фиг. 13;
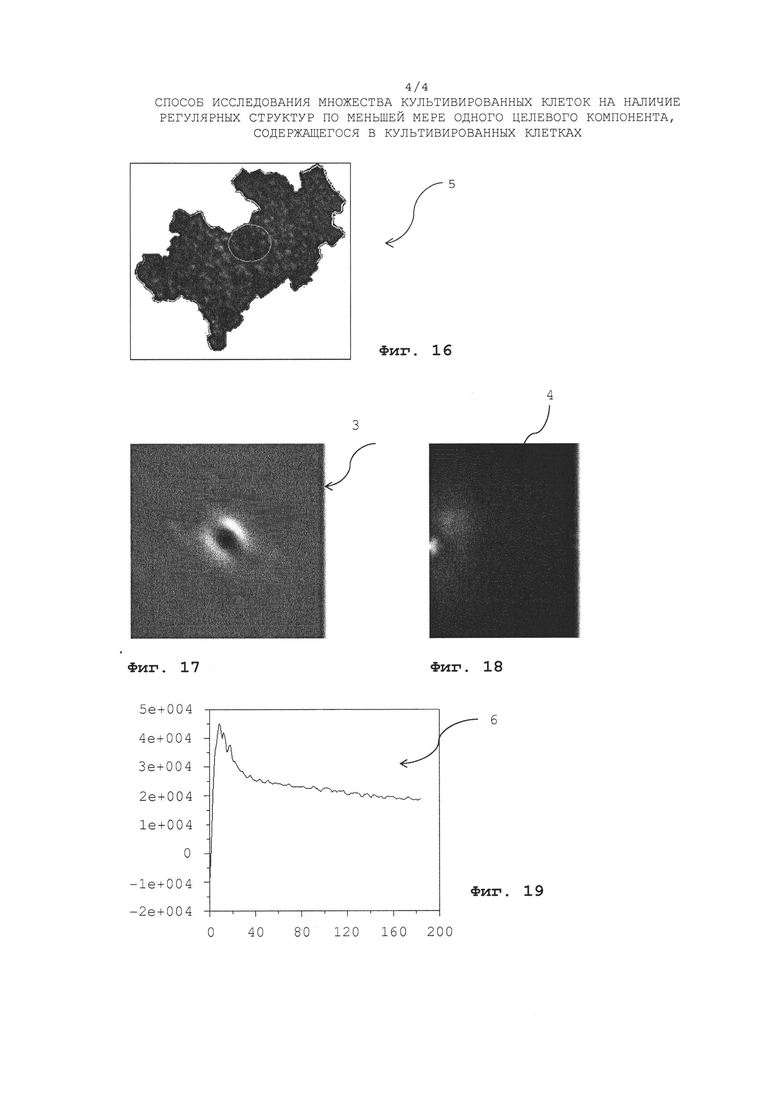
Фиг. 16 показывает изображение окрашивания Актинина реального кардиомиоцита без саркомерных структур;
Фиг. 17 показывает корреляционное изображение, сгенерированное посредством взаимной корреляции изображений гребней и впадин, извлеченных из изображения кардиомиоцита с Фиг. 16;
Фиг. 18 показывает Фурье-преобразованное корреляционное изображение с Фиг. 17;
Фиг. 19 показывает одномерную корреляционную функцию, сгенерированную из корреляционного изображения с Фиг. 17.
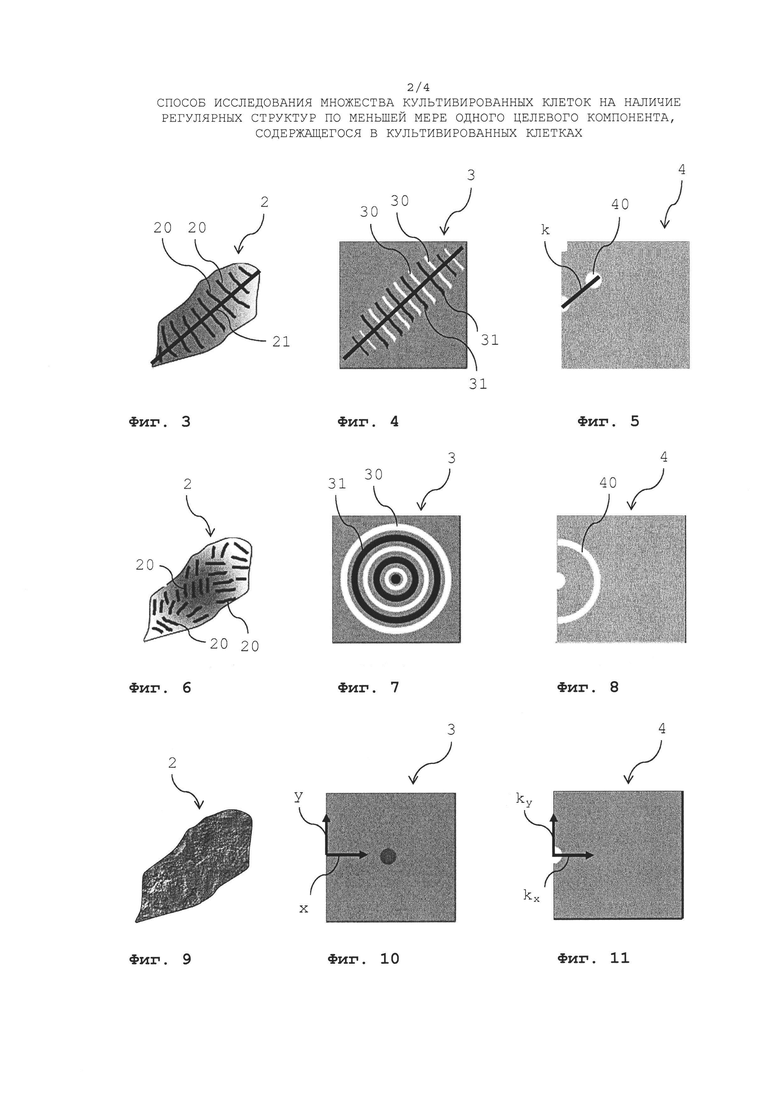
Сарокмер 1 и его общая структура, показанные на Фиг. 1 и Фиг. 2, уже пояснялись выше, потому данное пояснение здесь не повторяется. Изображение 2 зафиксированного миоцита или кардиомиоцита показывает регулярные сарокмерные структуры 20, которые расположены с единой ориентацией, как показано на схематичном двухмерном изображении 2 кардиомиоцита, показанного на Фиг. З. Направление 21 единой ориентации регулярных саркомерных структур 1 на изображении 2 кардиомиоцита показано прямой линией, которая не является частью изображения 2 кардиомиоцита.
Двухмерное изображение 2 кардиомиоцита, показанное на Фиг. З, затем подвергается процессу фильтрации гребней и впадин. При этом генерируется двухмерное изображение гребней, показывающее яркими любые саркомерные полосообразные структуры, находящиеся на изображении 2 кардиомиоцита. Любые структуры, отличные от сарокмерных полосообразных структур, скрываются благодаря процессу фильтрации гребней. Кроме того, генерируется изображение впадин, показывающее яркими любые полосообразные структуры, расположенные между полосообразными структурами изображения гребней, тогда как структуры, отличные от них, скрываются благодаря процессу фильтрации впадин. Изображение гребней и изображение впадин здесь не показаны, поскольку они являются только промежуточными изображениями, которые сами по себе не анализируются (изображение гребней показано, например, на Фиг. 12). По сути, они подвергаются антикорреляции, а их задача заключается в том, чтобы усиливать полосообразные структуры, содержащихся в изображении, тогда как структуры, отличные от полосообразных структур подавляются так, чтобы периодические структуры, содержащиеся в изображении 2 кардиомиоцита, приводили к более четко выраженным признакам, указывающим на присутствие таких регулярных структур в образующемся корреляционном изображении.
Изображения гребней и впадин подвергаются взаимной корреляции. Как уже упоминалось, взаимная корреляция изображений гребней и впадин выполняется для того, чтобы убедиться, что любая информация о полосообразных структурах, содержащаяся в любом из изображений гребней или впадин, учитывается при генерировании корреляционного изображения. В качестве альтернативы, вообще также возможно авто коррелировать, например, изображение гребней. Однако автокорреляция изображения гребней только подразумевает, что любая информация о полосообразных структурах, содержащаяся в изображении впадин, которая не содержится в изображении гребней (в силу того, что она подавлена в нем), не учитывается при генерировании корреляционного изображения.
Как было описано выше, взаимная корреляция изображений гребней и впадин подразумевает, что должна быть выполнена свертка изображений гребней и впадин. Как уже описывалось выше, такая свертка представляет собой математическую операцию, требующая большого объема вычислительной работы. Однако большой объем вычислительной работы может быть значительно сокращен за счет выполнения преобразований Фурье изображения гребней и изображения впадин в Фурье пространстве, и за счет умножения Фурье-преобразованного изображения гребней на Фурье-преобразованное изображение впадин (точнее: величины яркости пикселей Фурье-преобразованного изображения гребней и Фурье-преобразованного изображения впадин), поскольку свертка изображений гребней и впадин соответствует умножению Фурье-преобразованного изображения гребней на Фурье-преобразованное изображение впадин, в Фурье пространстве. Такое умножение Фурье-преобразованного изображения гребней на Фурье-преобразованное изображение впадин, дает в результате Фурье-преобразованное корреляционное изображения, для которого должно быть выполнено обратное преобразование Фурье для получения корреляционного изображения, показанного на Фиг. 4. Или же, как упоминалось, такое корреляционное изображение также может быть получено сверткой изображений гребней и впадин (не преобразованных по Фурье).
Корреляционное изображение 3, показанное на Фиг. 4, (сгенерированное посредством взаимной корреляции изображений гребней и впадин, как указано выше) демонстрирует регулярное расположение максимумов (30) и минимумов (31) яркости. Такое регулярное расположение максимумов (30) и минимумов (31) яркости, как правило, единообразно ориентировано в том же направлении 32 (указано прямой линией на Фиг. 4, которая не является частью корреляционного изображения), что и регулярные саркомерные структуры 2 0 на изображении 2 кардиомиоцита (см. Фиг. 3). Амплитуды яркости зависят от согласованности регулярных саркомерных структур 1 на изображении 2 зафиксированного кардиомиоцита (Фиг. 3).
Фиг. 5 показывает корреляционное Фурье-преобразованное изображение 4, которое представляет собой преобразование Фурье корреляционного изображения 3, показанного на Фиг. 4. Такое преобразование Фурье соответствует преобразованию х, у-пространства в корреляционном изображении 3 в kx, ky-пространство в Фурье-преобразованном корреляционном изображении 4. В таком Фурье-преобразованном корреляционном изображении амплитуда модуляции (соотношение двух смежных максимумов яркости или соотношение максимума яркости и смежного минимума), частота и направление, говорящее о степени полосатости, содержащейся в изображении 2 кардиомиоцита. Как можно видеть, удлиненная область параллельных полос (саркомерных структур 20) на изображении 2 кардиомиоцита (см. Фиг. 3) преобразуется в локальный максимум 40, расположенный на расстоянии 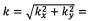
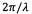 от нулевой точки (показано на Фиг. 5 с помощью черной линии, которая не является частью Фурье-преобразованного изображения 4, с λ, являющееся «длиной волны» регулярных полос, содержащихся в изображении 2) и под углом
от нулевой точки (показано на Фиг. 5 с помощью черной линии, которая не является частью Фурье-преобразованного изображения 4, с λ, являющееся «длиной волны» регулярных полос, содержащихся в изображении 2) и под углом 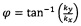 . Фурье-преобразованное изображение 4 содержит четко выраженные признаки и, следовательно, особенно подходит для анализа исследуемых зафиксированных клеток, которые содержат единообразно ориентированные полосатые структуры.
. Фурье-преобразованное изображение 4 содержит четко выраженные признаки и, следовательно, особенно подходит для анализа исследуемых зафиксированных клеток, которые содержат единообразно ориентированные полосатые структуры.
Фиг. 6 показывает изображение 2 зафиксированного миоцита или кардиомиоцита также содержащего регулярные саркомерные структуры 20, однако данные регулярные саркомерные структуры 2 0 не расположены с единой ориентацией, а скорее ориентированы случайным образом (изотропно) в различных направлениях. Как следствие, корреляционное изображение 3, показанное на Фиг. 7, сгенерированное посредством взаимной корреляции изображений гребней и впадин, показывает максимумы 30 и минимумы 31 яркости, однако в силу случайной ориентации (или изотропной ориентации) регулярных саркомерных структур во всех направлениях на изображении 2, максимумы 30 и минимумы 31 образуют в корреляционном изображении кольца с уменьшающейся амплитудой яркости (данные кольца показывают распределение структур во всех направлениях). Амплитуда яркости зависит от числа «участков» регулярных саркомерых структур, а ширина колец зависит от ширины и искажения отдельных регулярных саркомерных структур. Таким образом, Фурье-преобразованное корреляционное изображение 4, показанное на Фиг. 8, показывает не предпочтительное направление, а максимум 40 все еще расположен на вышеописанном расстоянии в kx, ky - пространстве.
Фиг. 9 показывает изображение 2 зафиксированного миоцита или кардиомиоцита, которое вообще не показывает какие-либо регулярные саркомерные структуры. Таким образом, корреляционное изображение 3 не показывает какие-либо структуры максимумов и минимумов (см. Фиг. 10), это также справедливо для Фурье-преобразованного корреляционного изображения 4 (см. Фиг. 11).
Фиг. 12 показывает изображение гребней реального зафиксированного кардиомиоцита, содержащего саркомерные структуры, которые на Фиг. 12 выглядят яркими. На Фиг. 13 показано реальное коррелированное изображение 3, сгненерированное посредством взаимной корреляции изображения гребней и изображения впадин кардиомиоцита (не показан), а Фиг. 14 показывает соответствующее Фурье-преобразованное коррелированное изображение 4, из которого может быть определена величина К как обозначено стрелкой (данная величина К представляет «длину волны» саркомерных структур, содержащихся в изображении кардиомиоцита; в вышеупомянутых примерах данная величина К обозначена как k) и угол α общего направления ориентации саркомерной структуры на изображении кардиомиоцита (угол α в вышеупомянутых примерах обозначен ϕ). Поскольку изображение 5 окрашивания Актинина показывает саркомерную структуру, является очевидным, что направление ориентации на Фиг. 14 является правильным (данное направление ориентации также можно найти, взглянув на корреляционное изображение 3 на Фиг. 13). На Фиг. 15 показана одномерная корреляционная функция 6 и такая одномерная корреляционная функция получена усреднением яркости корреляционного изображения при конкретном радиусе в пределах всей окружности, с последующим повторением этого усреднения для различных радиусов. Поскольку при некоторых конкретных радиусах наблюдается увеличенная яркость, присутствующая в корреляционном изображении 3, в то время как при других конкретных радиусах яркость отсутствует или наблюдается только низкая яркость, одномерная корреляционная функция характеризуется максимумами 60 и минимумами 61 яркости. Однако в силу того, что усреднение выполняется при конкретном радиусе в пределах всей окружности, любая информация об ориентации регулярных саркомерных структур не содержится в корреляционной функции 6. Расстояние между двумя смежными максимумами 60 и минимумами 61 яркости одномерной корреляционной функции 6, соотношение амплитуд яркости двух смежных максимумов 60 или соотношение амплитуд максимума 60 и смежного минимума 61 и однородность периодичности максимумов 60 и минимумов 61, свидетельствуют о степени полосатости, содержащейся в изображении кардиомиоцита.
Как уже упоминалось ранее в случае саркомерных структур, случайным образом распределенных на изображении зафиксированного кардиомиоцита, корреляционная функция 6 является более репрезентативной и содержит больше четко выраженных признаков, поскольку в таком случае процесс усреднения не удаляет много информации, в силу того, что практически отсутствует предпочтительная ориентация регулярных структур. В случае единообразно ориентированных регулярных саркомерных структур на изображении зафиксированного кардиомиоцита Фурье-преобразованное изображение 4 корреляционной функции является более репрезентативным и содержит больше ярко выраженных признаков, поскольку усреднение, выполненное для получения корреляционной функции 6 затем удаляет информацию о предпочтительных направлениях ориентации. Поскольку невозможно предсказать заранее какой анализ является более перспективным, выполняют и анализ корреляционной функции 6, и анализ Фурье-преобразованного корреляционного изображения 4, а из результатов обоих анализов затем можно определить, какой из них является более репрезентативным.
Фиг. 16 показывает изображение 5 окрашивания Актинина реальной зафиксированной клетки, не содержащей регулярные структуры. Следовательно, корреляционное изображение 3, показанное на Фиг. 17, соответствующее Фурье-преобразованное коррелированное изображение 4, показанное на Фиг. 18 и одномерная корреляционная функция 6, показанная на Фиг. 19, не показывают каких-либо конкретных признаков, которые можно было бы использовать для анализа, так что вывод заключается в том, изображение зафиксированной клетки не содержит регулярных структур.
Как уже было изложено, вышеописанные варианты способа исследования культивированных клеток имеют особенно предпочтительную область применения в обнаружении веществ потенциальных кандидатов в лекарственное средство для лечения мышечных заболеваний или заболеваний миокарда, которые связаны с повреждением саркомерных структур в миоцитах или кардиомиоцитах, поскольку живые миоциты и кардиомиоциты, содержащие саркомерные структуры, могут сначала подвергаться стрессу путем добавления к живым миоцитам или кардиомиоцитам одного или более веществ, таких как глюкоза или эндотелии, для разрушения или по меньшей мере значительного сокращения сарокмерных структур, а затем может быть добавлено вещество - потенциальный кандидат в лекарственное средство. После этого миоциты и кардиомиоциты снова культивируют и затем эти культивированные клетки фиксируют и анализируют в соответствии с вышеописанными вариантами выполнения.
Пример
Человеческие кардиомиоциты засевают из расчета 35000/лунку в микропланшетах Becton Dickinson Falcon на 96 лунок с тонким дном, покрытых желатином или фибронектином. Кардиомиоциты могут прикрепляться в течение 2 дней. Удаляют среду и добавляют на 2 дня новую среду, содержащую 10 мМ глюкозы и 10 нМ эндотелина. Затем выполняют фиксацию 4% пара-формальдегидом в течение 15 минут, затем пермеабилизацию 0,1% Tween-20 в течение 15 минут. Затем выполняют окрашивание. Используются следующие антитела: первичное АВ9465 клон ЕА-53-моноклональное к α-Актинину человека, разбавленное 1/120 (доступно у Abeam pic 330, Кембридж, Великобритания), первичное АВ45932 поликлональное кроличье к тропонину Т человека, разбавленное 1/120 (доступно у Abeam plc, Кембридж, Великобритания), вторичное АВ козье антимышинное AlexaFluor488 А-11029, разбавленное 1/200 (доступно у Life Technologies, Карлсбад, Калифорния, США), вторичное АВ ослиное антикроличье AlexaFluor647 А-11029, разбавленное 1/200 (доступно у Life Technologies, Карлсбад, Калифорния, США). Все этапы окрашивания выполняют при комнатной температуре в течение 30 минут с последующей трехкратной промывкой фосфатным забуференным физиологическим раствором (PBS). Окрашивание Актинина антителами дает рисунки вытянутых регулярных полос в зрелых миоцитах, но не дает или дает неструктурированное окрашивание в незрелых или поврежденных клетках.
Изображения получают в системе высокоскоростного скрининга Opera™ QEHS (коммерчески доступной у PerkinElmer Cellular Technologies). Такая система содержит многоцветный автоматически вращающийся диск конфокального микроскопа для многолуночных планшетов. Возбуждение возникает через водно-иммерсионный объектив 20х NA 0,7 с помощью лазеров с 640 нм, 488 нм и 405 нм. Для каждого канала метки записывается одно изображение последовательно с временем выдержки 2 с, 1 с и 0,8 с для метки Тропонина Т (красный канал), метки Актинина (зеленый канал) и метки Хехст ДНК (синий канал), соответственно. Фильтры эмиссии выбирают как компромисс между максимальной эффективностью обнаружения и минимальными помехами.
Затем выполняют анализ изображения с использованием специализированного скриптового языка Acapella™ (PerkinElmer Cellular Technologies), включенного в систему высокоскоростного скрининга Opera™ QEHS. После настройки регулируемых параметров, определяемых пользователем, анализ выполняется без вмешательства человека одинаковым образом для всех изображений.
Два цитоскелетных маркерных канала Актинина и Тропонина С не демонстрируют полосатый рисунок одинаковой величины и контраста. Более четко выраженный сигнал Актинина был выбран для количественной оценки полосатой структуры. Изображение Актинина подвергалось фильтрации с применением специализированных фильтров гребней и впадин из «SER» функции («Пятна, края и гребни») в библиотеке выделения признаков текстуры Acapella™ (PerkinElmer) для извлечения изображения гребней и изображения впадин, которые усиливают светлые и темные линии. Такие изображения гребней и впадин затем коррелируют путем преобразования Фурье изображений гребней и впадин с применением алгоритма быстрого преобразования Фурье, перемножения Фурье-преобразованных изображений гребней и впадин для генерирования Фурье-преобразованного корреляционного изображения, последующего выполнения обратного преобразования Фурье для генерирования корреляционного изображения. Корреляционное изображение, Фурье-преобразованное корреляционное изображение и одномерную корреляционную функцию генерируют и анализируют так, как было описано выше.
Варианты выполнения изобретения были описаны выше с использованием чертежей и примера. Однако изобретение не ограничивается этими вариантами, а скорее, различными модификациями и изменениями возможными без отступления от идеи настоящего изобретения. Таким образом, объем охраны не ограничен вариантами выполнения изобретения, а скорее, определяется прилагаемой формулой изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ КОНСТРУИРОВАНИЯ МАССЫ МИОКАРДИАЛЬНЫХ КЛЕТОК И ПРИМЕНЕНИЕ МАССЫ МИОКАРДИАЛЬНЫХ КЛЕТОК | 2008 |
|
RU2467066C2 |
| СПОСОБ ОТБОРА КАРДИОМИОЦИТОВ С ИСПОЛЬЗОВАНИЕМ ВНУТРИКЛЕТОЧНЫХ МИТОХОНДРИЙ В КАЧЕСТВЕ ИНДИКАТОРА | 2005 |
|
RU2371478C2 |
| СПОСОБ СЕЛЕКЦИИ КАРДИОМИОЦИТОВ (ВАРИАНТЫ) | 2007 |
|
RU2426784C2 |
| СПОСОБ ИНДУКЦИИ ДИФФЕРЕНЦИАЦИИ СТВОЛОВЫХ КЛЕТОК В МИОКАРДИАЛЬНЫЕ | 2004 |
|
RU2392315C2 |
| КОМПОЗИЦИИ КЛЕТОК И СПОСОБЫ ИХ ПРИМЕНЕНИЯ ДЛЯ ЛЕЧЕНИЯ СЕРДЕЧНОЙ ТКАНИ | 2009 |
|
RU2519762C2 |
| СПОСОБ ПОЛУЧЕНИЯ РЕЗИДЕНТНЫХ СТВОЛОВЫХ КЛЕТОК СЕРДЦА МЛЕКОПИТАЮЩЕГО ИЗ ОБРАЗЦОВ МИОКАРДА | 2012 |
|
RU2505602C1 |
| СПОСОБ ПОЛУЧЕНИЯ ПРОГЕНИТОРНЫХ КЛЕТОК МИОКАРДА | 2013 |
|
RU2542964C1 |
| СПОСОБ ПОЛУЧЕНИЯ И ИНДУКЦИИ НАПРАВЛЕННОЙ ДИФФЕРЕНЦИРОВКИ КУЛЬТУРЫ МУЛЬТИПОТЕНТНЫХ КЛЕТОК СЕРДЦА ДЛЯ КЛЕТОЧНОЙ ТЕРАПИИ И/ИЛИ ТКАНЕВОЙ ИНЖЕНЕРИИ В ЗОНЕ ИШЕМИИ МИОКАРДА | 2008 |
|
RU2366706C1 |
| Устройство для тестирования эффективности биологически активных веществ на клетках | 2021 |
|
RU2777971C1 |
| СПОСОБ ОЧИСТКИ КАРДИОМИОЦИТОВ | 2021 |
|
RU2813532C1 |
Использование: для исследования культивированных клеток на наличие регулярных структур. Сущность изобретения заключается в том, что выполняют этапы, на которых: обеспечивают культивированные клетки, фиксируют культивированные клетки, окрашивают зафиксированные культивированные клетки, используя первый окрашивающий агент, возбуждают первый окрашивающий агент, что приводит к тому, что он испускает свет, получают двухмерное изображение зафиксированных культивированных клеток, осуществляют взаимную корреляцию первого и второго отфильтрованных изображений, полученных из изображения зафиксированных культивированных клеток, для получения корреляционного изображения и определяют наличие или отсутствие регулярных структур в зафиксированных культивированных клетках путем определения, присутствуют ли в корреляционном изображении регулярные структуры в максимумах и минимумах яркости. Технический результат: обеспечение возможности более оперативно и менее ресурсозатратно исследовать культивированные клетки на наличие в культивированных клетках регулярных структур определенных компонентов. 4 н. и 11 з.п. ф-лы, 19 ил.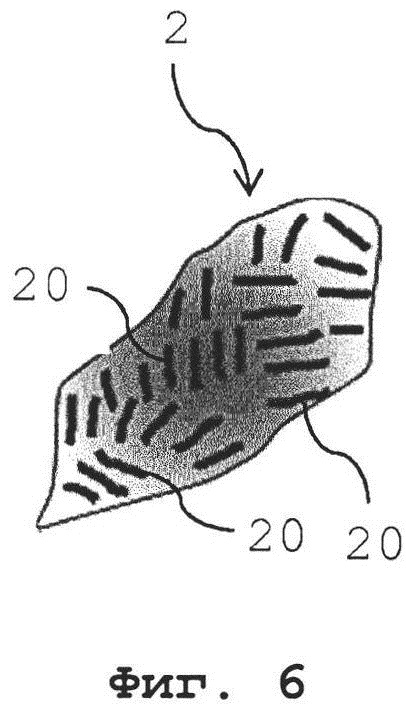
1. Способ исследования множества культивированных клеток на наличие регулярных структур по меньшей мере одного целевого компонента, содержащегося в культивированных клетках, причем способ содержит этапы, на которых:
- обеспечивают множество культивированных клеток, подлежащих исследованию,
- фиксируют культивированные клетки, подлежащие исследованию, сохраняя любые структуры по меньшей мере одного целевого компонента, содержащегося в культивированных клетках,
- окрашивают по меньшей мере один целевой компонент, содержащийся в зафиксированных культивированных клетках, используя первый окрашивающий агент, связанный с по меньшей мере одним целевым компонентом и способный при возбуждении испускать свет первой длины волны,
- возбуждают первый окрашивающий агент, что приводит к тому, что он испускает свет первой длины волны,
- получают двухмерное изображение (2) зафиксированных культивированных клеток с первым окрашивающим агентом, испускающим свет первой длины волны,
- из упомянутого двухмерного изображения (2) зафиксированных культивированных клеток извлекают двухмерное первое отфильтрованное изображение (5), показывающее яркими только те части, которые содержат по меньшей мере один целевой компонент, с которым связан первый окрашивающий агент,
- осуществляют автокорреляцию первого отфильтрованного изображения (5) или взаимную корреляцию первого отфильтрованного изображения со вторым двухмерным отфильтрованным изображением, извлеченным из двухмерного изображения зафиксированных культивированных клеток и показывающим яркими только части, отличные от тех, которые содержат по меньшей мере один целевой
компонент, с которым связан первый окрашивающий агент, для получения двухмерного корреляционного изображения (3), и
- определяют наличие или отсутствие регулярных структур по меньшей мере одного целевого компонента, содержащегося в зафиксированных культивированных клетках, путем определения, присутствуют ли в корреляционном изображении (3) регулярные структуры в максимумах (30) и минимумах (31) яркости.
2. Способ по п. 1, в котором этап, на котором извлекают двухмерные первое и второе отфильтрованные изображения содержит выполнение фильтрации гребней и впадин двухмерного изображения зафиксированных культивированных клеток,
для того чтобы сгенерировать двухмерное изображение (5) гребней, в котором полосообразные структуры по меньшей мере одного целевого компонента, с которым связан окрашивающий агент, усилены по яркости и в котором любые структуры, отличные от полосообразных структур по меньшей мере одного целевого компонента, с которым связан окрашивающий агент, ослаблены по яркости, указанное изображение гребней образует первое отфильтрованное изображение,
и для того чтобы сгенерировать двухмерное изображение впадин, в котором полосообразные структуры, расположенные между полосообразными структурами по меньшей мере одного целевого компонента, с которым связан окрашивающий агент, усилены по яркости и в котором структуры, отличные от полосообразных структур, расположенных между полосообразными структурами по меньшей мере одного целевого компонента, с которым связан окрашивающий агент, ослаблены по яркости, указанное изображение впадин образует второе отфильтрованное изображение,
и в котором осуществляют взаимную корреляцию изображения (5) гребней и изображения впадин, чтобы сгенерировать корреляционное изображение (3).
3. Способ по любому из предыдущих пунктов, в котором корреляционное изображение (3) генерируют путем:
- выполнения преобразования Фурье первого отфильтрованного изображения для получения двухмерного Фурье-преобразованного первого отфильтрованного изображения и умножения Фурье-преобразованного первого отфильтрованного изображения само на себя для получения двухмерного Фурье-преобразованного корреляционного изображения или
- выполнения преобразования Фурье первого отфильтрованного изображения и выполнения преобразования Фурье второго отфильтрованного изображения для получения двухмерного Фурье-преобразованного второго отфильтрованного изображения и умножения Фурье-преобразованного первого отфильтрованного изображения на Фурье-преобразованное второе отфильтрованное изображение для получения двухмерного Фурье-преобразованного корреляционного изображения,
- выполнения обратного преобразования Фурье Фурье-преобразованного корреляционного изображения для получения двухмерного корреляционного изображения (3).
4. Способ по п. 3, в котором этап на котором выполняют преобразование Фурье первого и второго отфильтрованных изображений, так же как и этап выполнения обратного преобразования Фурье Фурье-преобразованного корреляционного изображения, выполняют с использованием алгоритма быстрого преобразования Фурье.
5. Способ по любому из пп. 1, 2 или 4, в котором яркость двухмерного корреляционного изображения (3) усредняют до определенного радиуса в пределах всей окружности, и в котором это усреднение яркости в пределах всей окружности выполняют при различных радиусах для получения одномерной корреляционной функции (6), и в котором из этой одномерной корреляционной функции (6) степень полосатости в двухмерном изображении (2) зафиксированных культивированных клеток получают
путем определения расстояния между двумя смежными максимумами (60) яркости или путем определения расстояния между максимумом (60) и смежным минимумом (61) яркости одномерной корреляционной функции (6) и путем определения соотношения амплитуд яркости двух смежных максимумов (60) или соотношения амплитуд яркости максимума (60) и смежного минимума (61) одномерной корреляционной функции (6).
6. Способ по п. 3, в котором яркость двухмерного корреляционного изображения (3) усредняют до определенного радиуса в пределах всей окружности, и в котором это усреднение яркости в пределах всей окружности выполняют при различных радиусах для получения одномерной корреляционной функции (6), и в котором из этой одномерной корреляционной функции (6) степень полосатости в двухмерном изображении (2) зафиксированных культивированных клеток получают путем определения расстояния между двумя смежными максимумами (60) яркости или путем определения расстояния между максимумом (60) и смежным минимумом (61) яркости одномерной корреляционной функции (6) и путем определения соотношения амплитуд яркости двух смежных максимумов (60) или соотношения амплитуд яркости максимума (60) и смежного минимума (61) одномерной корреляционной функции (6).
7. Способ по любому из пп. 1, 2, 4 или 6, который дополнительно содержит этап, на котором выполняют преобразование Фурье двухмерного корреляционного изображения для получения двухмерного Фурье-преобразованного корреляционного изображения (4), в котором из этого Фурье-преобразованного корреляционного изображения (4) определяют расстояние (k, K) между смежными максимумами (40) яркости измеряя расстояние между полосообразными структурами в двухмерном изображении (2) культивированных клеток и в котором также угол (ϕ, α), под которым расположены смежные максимумы яркости, определяют как отображающий единую ориентацию (21), в соответствии с которой регулярные полосообразные структуры (20) располагаются в двухмерном изображении (2) культивированных клеток.
8. Способ по п. 3, который дополнительно содержит этап, на котором выполняют преобразование Фурье двухмерного корреляционного изображения для получения двухмерного Фурье-преобразованного корреляционного изображения (4), в котором из этого Фурье-преобразованного корреляционного изображения (4) определяют расстояние (k, K) между смежными максимумами (40) яркости, измеряя расстояние между полосообразными структурами в двухмерном изображении (2) культивированных клеток, и в котором также угол (ϕ, α), под которым расположены смежные максимумы яркости, определяют как отображающий единую ориентацию (21), в соответствии с которой регулярные полосообразные структуры (20) располагаются в двухмерном изображении (2) культивированных клеток.
9. Способ по п. 5, который дополнительно содержит этап, на котором выполняют преобразование Фурье двухмерного корреляционного изображения для получения двухмерного Фурье-преобразованного корреляционного изображения (4), в котором из этого Фурье-преобразованного корреляционного изображения (4) определяют расстояние (k, K) между смежными максимумами (40) яркости измеряя, расстояние между полосообразными структурами в двухмерном изображении (2) культивированных клеток, и в котором также угол (ϕ, α), под которым расположены смежные максимумы яркости, определяют как отображающий единую ориентацию (21), в соответствии с которой регулярные полосообразные структуры (20) располагаются в двухмерном изображении (2) культивированных клеток.
10. Способ по п. 7, в котором этап, на котором выполняют преобразование Фурье двухмерного корреляционного изображения (3), выполняют, используя алгоритм быстрого преобразования Фурье.
11. Способ по п. 8 или 9, в котором этап, на котором выполняют преобразование Фурье двухмерного корреляционного изображения (3), выполняют, используя алгоритм быстрого преобразования Фурье.
12. Способ идентификации вещества - потенциального кандидата в лекарственное средство для лечения мышечных заболеваний или заболеваний миокарда, связанных с повреждением саркомерных структур в миоцитах или кардиомиоцитах, причем способ содержит этапы, на которых:
- обеспечивают множество живых миоцитов или кардиомиоцитов, содержащих саркомерные структуры,
- к множеству живых миоцитов или кардиомиоцитов добавляют вещество, оказывающее стрессовое воздействие, что приводит к разрушению саркомерных структур или по меньшей мере их существенному сокращению,
- после добавления вещества, оказывающего стрессовое воздействие, к множеству живых миоцитов или кардиомиоцитов, содержащих разрушенные или по меньшей мере существенно сокращенные саркомерные структуры, добавляют вещество-кандидат,
- после того как было добавлено вещество-кандидат, культивируют миоциты или кардиомиоциты,
- определяют, привело ли добавление вещества-кандидата к образованию саркомерных структур, вновь сформировавшихся в культивированных миоцитах или кардиомиоцитах,
- если добавление вещества-кандидата привело к образованию саркомерных структур, вновь сформировавшихся в культивированных миоцитах или кардиомиоцитах, вещество-кандидат квалифицируют в качестве кандидата в лекарственное средство,
причем этап, на котором определяют, привело ли добавление вещества-кандидата к образованию саркомерных структур, вновь сформировавшихся в миоцитах или кардиомиоцитах, содержит использование способа по любому из пп. 1-11.
13. Способ идентификации способности вещества - потенциального кандидата лекарственного средства обладать защитным действием против мышечных заболеваний или заболеваний миокарда, связанных
с повреждением саркомерных структур в миоцитах или кардиомиоцитах, причем способ содержит этапы, на которых:
- обеспечивают множество живых миоцитов или кардиомиоцитов, содержащих саркомерные структуры,
- к множеству живых миоцитов или кардиомиоцитов добавляют вещество-кандидат,
- после добавления вещества-кандидата к множеству живых миоцитов или кардиомиоцитов добавляют вещество, оказывающее стрессовое воздействие, что приводит к разрушению саркомерных структур или по меньшей мере их существенному сокращению,
- после того как было добавлено вещество, оказывающее стрессовое воздействие, культивируют миоциты или кардиомиоциты,
- определяют, привело ли добавление вещества, оказывающего стрессовое воздействие, к разрушению или по меньшей мере существенному сокращению саркомерных структур,
- если добавление вещества, оказывающего стрессовое воздействие, не привело к разрушению или по меньшей мере существенному сокращению саркомерных структур, вещество-кандидат квалифицируют в качестве кандидата в лекарственное средство,
причем этап, на котором определяют, привело ли добавление указанного вещества к разрушению или по меньшей мере существенному сокращению саркомерных структур в миоцитах или кардиомиоцитах, содержит использование способа по любому из пп. 1-11.
14. Способ определения наличия саркомерных структур в живых миоцитах или кардиомиоцитах, полученных из индуцированных плюрипотентных стволовых клеток, причем способ содержит этапы, на которых:
- обеспечивают множество живых индуцированных плюрипотентных стволовых клеток,
- вызывают дифференцировку живых индуцированных плюрипотентных стволовых клеток в живые миоциты или кардиомиоциты,
- культивируют живые миоциты или кардиомиоциты,
- определяют, содержат ли культивированные миоциты или кардиомиоциты саркомерные структуры,
причем этап, на котором определяют, содержат ли культивированные живые миоциты или кардиомиоциты сакромерные структуры, содержит использование способа по любому из пп. 1-11.
15. Способ по п. 14, дополнительно содержащий этапы, на которых:
- до культивирования живых миоцитов или кардиомиоцитов к живым миоцитам и кардиомиоцитам добавляют вещество, оказывающее стрессовое воздействие, что приводит к разрушению саркомерных структур или по меньшей мере их существенному сокращению, и
- после осуществления способа по любому из пп. 1-11 определяют, привело ли добавление вещества, оказывающего стрессовое воздействие, к разрушению или по меньшей мере существенному сокращению саркомерных структур в культивированных миоцитах или кардиомиоцитах.
| US 20020022716 A1, 21.02.2002 | |||
| Устройство для нагревания жидкостей | 1942 |
|
SU66985A1 |
| JP 2006068002 A, 16.03.2006 | |||
| JP 2013094064 A, 20.05.2013 | |||
| СПОСОБ И УСТРОЙСТВО ДЛЯ АНАЛИЗА ЧАСТИЦ В ЖИДКОМ ОБРАЗЦЕ | 2009 |
|
RU2452957C2 |
| Способ оценки кинетики клеточных популяций IN VIтRо | 1986 |
|
SU1451164A1 |

