Описание
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится, главным образом, к способу получения клеточных масс путем агрегации очищенных кардиомиоцитов, полученных из плюрипотентной стволовой клетки, полученной путем диспергирования до отдельных клеток, а также настоящее изобретение относится к способу лечения болезней сердца путем приживления полученных клеточных масс кардиомиоцитов в сердечной ткани и к способу получения пластов клеточных масс с использованием клеточных масс кардиомиоцитов.
УРОВЕНЬ ТЕХНИКИ
Кардиомиоциты взрослых утрачивают пролиферативную активность, и трансплантация сердечной ткани является единственным путем для лечения серьезных болезней сердца, таких как инфаркт миокарда и кардиомиопатия. На самом деле при этом из-за проблемы отсутствия доноров сердечной ткани имеется срочная необходимость в разработке способа лечения, кроме трансплантации сердечной ткани.
Напротив, ожидается, что использование кардиомиоцитов, полученных вне живого организма, для замены ими пораженной части кардиомиоцитов станет наиболее многообещающим способом спасения пациентов, ожидающих трансплантацию сердечной ткани. Этот подход к лечению называют клеточной терапией сердца. Для того чтобы эта терапия стала реальностью, были проведены различные способы проб и ошибок. Эти способы включают: использование кардиомиоцитов или скелетных миобластов и клеток костного мозга или им подобных клеток, которые были экстрагированы из тканей эмбриона, новорожденного или взрослого; использование дифференцированных эмбриональных стволовых клеток; и получение стволовых клеток (таких как соматические стволовые клетки), которые, как полагают, существуют в живом организме, и индуцирование их дифференцировки (непатентный документ 1: Zhonghua Yi Xue Za Zhi 2003, 83, 1818-22).
Такие способы могут быть разделены на два типа подходов. Один из подходов включает трансплантацию кардиомиоцитов в виде клеток, и в этом способе кардиомиоциты, диспергированные до отдельных клеток, непосредственно вводят в ткань посредством иглы для инъекции (этот способ в настоящем документе обозначен как "инъекционный способ"). Другой подход включает конструирование ткани или органа вне живого организма (способ, который в настоящем документе обозначен как "способ тканевой инженерии"), и эту искусственную ткань или орган переносят в организм.
Для проведения способа тканевой инженерии были предприняты различные попытки, и они включают: 1) способ, в котором кардиомиоциты образуют подобную пласту структуру, которая затем прикрепляется на ткани (непатентный документ 2: Circulation Research 2002, 90(3):e-40); 2) способ, в котором кардиомиоциты и другие типы клеток, смешивают в такой же пропорции, в какой они находятся в сердечной ткани, и трехмерная структура, образованная из смеси, используется для замены ткани; 3) способ, в котором трехмерная структура образуется из кардиомиоцитов, диспергированных до отдельных клеток, с последующим конструированием сосудистой структуры, и трехмерной структурой заменяют ткань; и 4) способ, в котором вместо замены сердечной ткани новый вспомогательный орган, который принимает участие в осуществлении присущей органу функции, эктопически трансплантируют в мишеневый сайт (непатентный документ 3: Circulation Research 2007 2, 100: 263-272).
Однако в настоящее время, когда разнообразные способы проб и ошибок используются на пути разработки клинического терапевтического применения, ни один из способов не применяется на практике. Это связано с тем, что с трансплантацией кардиомиоцитов в сердце связано несколько проблем, таких как включение клеток, не являющихся кардиомиоцитами, низкая степень приживления трансплантированных кардиомиоцитов и неспособность удаления компонентов, полученных из других видов.
Для использования кардиомиоцитов в качестве клеточных масс при трансплантации известны способы, с помощью которых возможно конструирование клеточных масс, включающих эмбриональные и неонатальные кардиомиоциты грызунов, и согласно недавней публикации клеточные массы были сконструированы с использованием всех клеток (включая не только кардиомиоциты), которые были получены из сердца эмбриона (непатентный документ 4: Developmental Dynamics 235; 2200-2209, 2006). Что касается трансплантации кардиомиоцитов, то сообщался случай, когда было подтверждено приживление эмбриональных кардиомиоцитов мыши, трансплантированных в сердце взрослых мышей (непатентный документ Science 1994, 264(5155): 98-101). Однако этот способ трансплантации кардиомиоцитов включал использование всех клеток, до которых были диспергированы эмбриональные сердца под действием коллагеназы, поэтому трансплантированные клетки состояли из популяции клеток, включающей смесь кардиомиоцитов и других типов клеток. Также известно, что неочищенные кардиомиоциты, полученные из живого организма, могут быть трансплантированы в сердце (непатентный документ 5: Science 1994, 264(5155): 98-101; и непатентный документ 1: Zhonghua Yi Xue Za Zhi 2003, 83, 1818-22).
Также известен способ, в котором в процессе дифференцировки эмбриоидных тел из клеток ES эмбриоидные тела не полностью обрабатывали протеолитическим ферментом, посредством чего получали популяцию, содержащую клеточные массы, насыщенные и не насыщенные кардиомиоцитами, и затем подвергали их центрифугированию в градиенте плотности, получая, таким образом, клеточные массы, которые содержали кардиомиоциты в количестве, составляющем примерно до 70% (патентный документ 1: US 2005-0214938 A).
Однако каждый из этих способов включает использование популяции клеток, которая также содержит клетки, не являющиеся кардиомиоцитами, и загрязнение такими клетками, не являющимися кардиомиоцитами, может вызывать серьезные непредсказуемые побочные эффекты, угрожающие жизни пациента после трансплантации. При данных обстоятельствах существует необходимость использовать в трансплантационной терапии кардиомиоциты, подвергшиеся очистке.
В нескольких публикациях описаны результаты трансплантации в сердце неочищенных, полученных из клеток ES кардиомиоцитов и возможность их последующей трансплантации (непатентный документ 6: Cardiovasc Res. 2007 May 17; непатентный документ 7: Stem Cells. 2007 May 31; и непатентный документ 8: FASEB J. 2007 Apr 13). Однако согласно недавней публикации, в которой описана очистка полученных из клеток ES кардиомиоцитов и их инъекция в сердце, степень приживления трансплантированных кардиомиоцитов была крайне низкой, и не было обнаружено прижившихся кардиомиоцитов (т.e. кардиомиоцитов, выживших в органе-хозяине и оставшихся прикрепленными на нем на протяжении продолжительного периода времени); как оказалось, очищенные полученные из клеток ES кардиомиоциты не способны к приживлению после трансплантации индивидууму (в живой организм) (непатентный документ 9: J Exp Med. 2006; 203:2315-27.)
Эта публикация пролила свет на трудности приживления очищенных кардиомиоцитов после трансплантации. Для решения этой проблемы в той же публикации был описан способ, который включает трансплантацию полученных из клеток ES кардиомиоцитов вместе с эмбриональными фибробластами мышей, для усиления степени их приживления после трансплантации (непатентный документ 9: J Exp Med. 2006 Oct 2; 203(10): 2315-27). Это подтверждает, что не существует способов, способных обеспечить трансплантацию очищенных, полученных из клеток ES кардиомиоцитов, и их приживление, при этом сохраняя чистоту клеток.
Кроме того, для получения клеточных трансплантатов, предназначенных для использования в терапии человека, должны быть удалены сывороточные и другие факторы, полученные из других животных. В способе получения кардиомиоцитов, используемых для трансплантации, культивирование обычно осуществляют в присутствии сыворотки; но известно также, что в условиях культивирования в среде, не содержащей сыворотку, клетки ES человека могут образовывать эмбриоидные тела, которые содержат кардиомиоциты в количествах, сравнимых с кардиомиоцитами, полученными обычным культивированием в присутствии сыворотки (непатентный документ 10: Stem cells and development 15:931-941, 2006). Однако нет публикаций, включая и эту публикацию, описывающих случай трансплантации кардиомиоцитов, полученных без использования таких факторов, как сыворотка, полученных из других животных.
Таким образом, для успешной трансплантации кардиомиоцитов в сердце должны быть решены некоторые проблемы, такие как включение клеток, не являющихся кардиомиоцитами, низкая степень приживления трансплантированных кардиомиоцитов и невозможность удаления компонентов, полученных из других видов.
Более того, в связи с трансплантацией кардиомиоцитов в сердечную ткань предполагается трансплантировать их в форме так называемых "клеточных пластов". Что касается получения клеточных пластов, то известно, что неонатальные кардиомиоциты используют для образования однослойного пласта, и in vitro могут наслаиваться до трех таких пластов (непатентный документ 11: FASEB J. 2006 Apr; 20(6): 708-10). Однако в указанном документе также утверждается, что вследствие ограниченной проницаемости кислорода клеточные пласты не могут быть сделаны любой толщины без неоваскуляризации клеточного пласта, и до сих пор невозможно получить желаемый клеточный пласт, который бы соответствовал размеру пораженной сердечной ткани.
Как описано выше, данные, известные из уровня техники, указывают на то, что получение кардиомиоцитов, используемых при трансплантации, и собственно трансплантация кардиомиоцитов нуждается в дополнительном улучшении с точки зрения практической реализации.
Патентный документ 1: US 2005-0214938 A
Непатентный документ 1: Zhonghua Yi Xue Za Zhi 2003, 83, 1818-22
Непатентный документ 2: Circulation Research 2002, 90(3):e-40
Непатентный документ 3: Circulation Research 2007 2, 100: 263-272
Непатентный документ 4: Developmental Dynamics 235; 2200-2209, 2006
Непатентный документ 5: Science 1994, 264(5155): 98-101
Непатентный документ 6: Cardiovasc Res. 2007 May 17 (Flk1(+) cardiac stem/progenitor cells derived from embryonic stem cells improve cardiac function in a dilated cardiomyopathy mouse model)
Непатентный документ 7: Stem Cells. 2007 May 31 (Differentiation in vivo of Cardiac Committed Human Embryonic Stem Cells in Post-Myocardial Infarcted Rats)
Непатентный документ 8: FASEB J. 2007 Apr 13 (Identification and selection of cardiomyocytes during human embryonic stem cell differentiation)
Непатентный документ 9: J Exp Med. 2006 Oct 2; 203(10): 2315-27
Непатентный документ 10: Stem Cell and Development 15: 931-941, 2006
Непатентный документ 11: FASEB J. 2006 Apr; 20(6): 708-10
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
ПРОБЛЕМЫ, РЕШАЕМЫЕ ИЗОБРЕТЕНИЕМ
Следовательно, авторы настоящего изобретения исследовали необходимые условия, которые допустимы, по меньшей мере в настоящее время, для клинического использования культивированных кардиомиоцитов, и обнаружили в результате следующие трудности.
(1) Очистка кардиомиоцитов: при получении кардиомиоцитов из живого организма или из плюрипотентных стволовых клеток их безопасность не может быть гарантирована, если они загрязнены неизвестными клетками, поэтому в любом случае кардиомиоциты обязательно должны иметь высокую степень чистоты. Для поддержания постоянного уровня очистки таких имеющих высокую степень чистоты кардиомиоцитов необходимо, чтобы кардиомиоциты полученные из живого организма или из плюрипотентных стволовых, клеток, были диспергированы до дискретных клеток (таких как отдельные клетки), причем отдельные клетки должны отличаться друг от друга так, что можно было выбрать только кардиомиоциты.
(2) Природа кардиомиоцитов: Различия в свойствах кардиомиоцитов, которые существуют у разных видов, не только вызывают проблему иммунного отторжения и этические проблемы, но также серьезно влияют на клиническую безопасность и эффективность, поэтому важно использовать клетки доноров того же вида, что и реципиент.
(3) Удаление факторов, полученных из животных других видов: для того чтобы исключить иммуногенность и загрязнение неизвестными патогенами, должны быть удалены примеси, такие как сыворотка, полученные из животного другого вида.
(4) Приживление трансплантированных кардиомиоцитов: для функционирования трансплантированных кардиомиоцитов таким же образом, как кардиомиоцитов в организме-хозяине, в первую очередь они должны приживиться в кардиомиоцитах хозяина (поддерживаться приживленными в течение продолжительного периода времени).
(5) На следующей стадии клетки должны созреть (вырасти до большего размера).
Коротко, целью настоящего изобретения являются средства, с помощью которых кардиомиоциты, которые были очищены так, что являются свободными от клеток, не являющихся кардиомиоцитами, и от любых компонентов, полученных из другого вида, могут быть трансплантированы и иметь высокую степень приживления, что стимулирует их созревание.
СРЕДСТВА, РЕШАЮЩИЕ УКАЗАННЫЕ ЗАДАЧИ
Для повышения степени приживления кардиомиоцитов, которые были выделены из живого организма или плюрипотентных стволовых клеток и которые очищены до той степени, когда они свободны от клеток, не являющихся кардиомиоцитами, и от любых компонентов, полученных из другого вида, авторы настоящего изобретения исследовали возможность конструирования клеточных масс из очищенных кардиомиоцитов. В результате было обнаружено, что проблема, указанная выше, может быть решена благодаря способу получения клеточных масс кардиомиоцитов, полученных из плюрипотентных эмбриональных стволовых клеток (клеток ES) или из индуцированных плюрипотентных стволовых клеток (клеток iPS), отличающихся тем, что клеточные массы, содержащие кардиомиоциты, которые были дифференцированы и индуцированы из клеток ES или из клеток iPS, были диспергированы до отдельных клеток, с получением таким образом очищенных кардиомиоцитов, которые затем культивируют в культуральной среде без сыворотки, таким образом, чтобы они повторно агрегировали.
Авторы настоящего изобретения сначала использовали кардиомиоциты, полученные из живого организма, и исследовали возможность решения проблемы, указанной выше. Более конкретно, используя кардиомиоциты, полученные из живого организма, которые были очищены до такой степени, что они свободны от клеток, не являющихся кардиомиоцитами, и от любых компонентов, полученных из другого вида, авторы настоящего изобретения исследовали возможность повышения степени приживления таких кардиомиоцитов после трансплантации.
Как оказалось, очищенные кардиомиоциты, полученные из живого организма, не способны к образованию клеточных масс даже после культивирования в течение 24 часов в культуральной среде, содержащей 10%-сыворотку. Этот результат подтвердил, что известный способ формирования клеточных масс кардиомиоцитов, при получении их из живого организма, которые эффективны в неочищенном состоянии, жестко зависит от вспомогательного действия клеток, не являющихся кардиомиоцитами. В другом экспериментальном исследовании, проведенном для исследования, каким образом очищенные кардиомиоциты, полученные из живого организма, будут себя вести в условиях без сыворотки, такие клетки были не способны к образованию клеточных масс; напротив, они сами подвергались клеточной смерти. Эти результаты ведут к выводу, что "конструирование клеточных масс неочищенных кардиомиоцитов, полученных из сердца живого организма", является неосуществимым способом и что невозможно предсказать поведение в условиях без сыворотки очищенных кардиомиоцитов, полученных из сердца живого организма.
Кроме того, в известных способах получения клеточных масс, агрегированных кардиомиоцитов, использовали клетки, полученные из сердца новорожденных животных или из сердца эмбриона, которые культивировали в культуральной среде, содержащей сыворотку. Этот способ основан на известном факте, что сыворотка, по существу, обладает сильным протективным эффектом, независимо от типа клеток. Однако, как указано ниже, в случае терапии человека необходимо избегать использования факторов (таких как сыворотка), полученных из других животных. Следовательно, используя вместо сыворотки бессывороточную среду, которая содержит различные добавки, содержащие заместители сыворотки, авторы настоящего изобретения исследовали способность агрегации кардиомиоцитов, полученных из живого организма. Однако при любых тестированных условиях в отсутствие сыворотки кардиомиоциты, полученные из живого организма, были не способны формировать желаемые клеточные массы, и степень их выживаемости также была низкой.
Учитывая данные об известном способе, заключающиеся в том, что при культивировании кардиомиоцитов, полученных из живого организма, в виде клеточных масс, выживаемость кардиомиоцитов сохраняется в течение длительного периода времени, авторы настоящего изобретения сделали попытку получить клеточные массы очищенных кардиомиоцитов, полученных из живого организма, в условиях отсутствия сыворотки. Однако, как оказалось, никаких клеточных масс не образовалось даже после пяти дней культивирования. В результате проведения этого эксперимента были получены новые данные о том, что в условиях отсутствия сыворотки очищенные кардиомиоциты, полученные из живого организма, не способны формировать клеточные массы.
Следовательно, авторы настоящего изобретения предположили, что решением этого технологического затруднения может стать изменение источника получения кардиомиоцитов. В результате проведения подобных исследований с различными источниками получения клеток авторы настоящего изобретения обнаружили, что кардиомиоциты, полученные из очищенных эмбриональных стволовых клеток, присоединяются друг к другу даже в условиях отсутствия сыворотки достаточно быстро в течение 12 часов, в результате чего могут быть получены трехмерные клеточные массы, и что кардиомиоциты в это время начинали агрегировать синхронно.
Это демонстрирует, что кардиомиоциты, полученные из эмбриональных стволовых клеток, способны к эффективному конструированию клеточной адгезии между очищенными кардиомиоцитами, и это также демонстрирует, что очищенные кардиомиоциты, полученные из эмбриональных стволовых клеток, обладают высокой способностью агрегировать в условиях отсутствия сыворотки. Поскольку эти результаты воспроизводились с клетками более чем одного вида, используя эмбриональные стволовые клетки, полученные из соответствующих видов, свойство, описанное выше, по-видимому, является естественным, то есть обычным для кардиомиоцитов, полученных у человека, и, в другом случае, полученных из эмбриональных стволовых клеток. На основании этого естественного свойства, характерного для этой эмбриональной стволовой клетки в условиях отсутствия сыворотки, авторы настоящего изобретения впервые в данной области успешно сформировали клеточные массы очищенных кардиомиоцитов в условиях отсутствия сыворотки.
Более того, на основании известного сообщения ожидалось, что кардиомиоциты, полученные из эмбриональных стволовых клеток, обладающие низкой степенью приживления, также обладают низкой способностью к повторной агрегации (Transplantation 70:1310-1317, 2000); с другой стороны, было сделано предположение, что попытки агрегации очищенных кардиомиоцитов, полученных из эмбриональных стволовых клеток, являются едва ли осуществимыми и сделан вывод о том, что они не могут агрегировать. Тем не менее, несмотря на сделанное предположение, авторы настоящего изобретения впервые продемонстрировали в данной области, что очищенные кардиомиоциты, полученные из эмбриональных стволовых клеток, могут формировать клеточные массы без помощи других клеток и что благодаря сконструированным клеточным массам может быть достигнуто значительное улучшение степени приживления клеток при трансплантации. Коротко, авторы настоящего изобретения обнаружили, что кардиомиоциты, полученные из эмбриональных стволовых клеток, обладают абсолютно другими характеристиками по сравнению с кардиомиоцитами, полученными из сердца живого организма, а именно они могут быть очищены до отдельных клеток и могут формировать клеточные массы даже в условиях отсутствия сыворотки.
Более того, для создания способа, с помощью которого из кардиомиоцитов, полученных из эмбриоанальных стволовых клеток, могут быть получены клеточные массы, более подходящие для трансплантации, авторы настоящего изобретения исследовали примеси, которые могут быть добавлены в культуральную среду в качестве средства, с помощью которого кардиомиоциты, полученные из эмбриональных стволовых клеток, могут более эффективно создавать клеточные массы в условиях отсутствия сыворотки. В результате при добавлении в качестве примеси инсулина, трансферрина и селена (ITS) было показано, что клеточные массы сильнее спонтанно пульсируют, чем клеточные массы, к которым не добавляли ITS, и это явление имело достаточную воспроизводимость. Коротко, было показано, что добавление ITS (конкретно, инсулина) является желательным для получения клеточных масс с использованием очищенных кардиомиоцитов, полученных из эмбриональных стволовых клеток. Следует отметить, что инсулин представляет собой наиболее важный фактор среди ITS, при этом трансферрин и селен играют вспомогательную роль.
Затем авторы настоящего изобретения добавляли ITS к базальной культуральной среде, не содержащей сыворотки (в настоящем документе обозначен как базальная культуральная среда), и дополнительно добавляли основной фактор роста фибробластов (bFGF) и/или инсулиноподобный фактор роста 1 (IGF1) к базальной культуральной среде; как оказалось, при добавлении только bFGF клеточные массы существенно увеличивались в диаметре на 5 день после их образования, что указывает на то, что клетки, представляющие интерес, были защищены. Это явление не было обнаружено при культивировании в среде, содержащей сыворотку, и оно представляет собой эффект защиты и пролиферации клеток, который являлся характерным для условий культивирования в отсутствие сыворотки при добавлении только bFGF. Известно, что bFGF обладает функцией защиты клеток, функцией стимулирования роста клеток и аналогичными функциями в отношении кардиомиоцитов, культивированных в таких экспериментальных условиях, при которых формирование монослойной культуры осуществляли при добавлении сыворотки (J Mol Cell Cardiol. 2007 Jan; 42(1): 222-33; и Cardiovasc Res. 2004; 64:516-25), но было абсолютно неизвестно, что продолжительный рост и защита кардиомиоцитов могут происходить и в условиях отсутствия сыворотки.
Этот эффект был сильнее, чем эффект, который наблюдался при формировании клеточных масс с использованием культуральной среды с добавлением 10%-сыворотки; с другой стороны, в случае добавления bFGF, но отсутствия образования клеточных масс до монослойной культуры, эффект, представляющий интерес, был слабее, чем эффект, наблюдаемый, когда клеточные массы формировались с использованием культуральной среды с добавлением 10% сыворотки. На основании результатов этих двух экспериментов было сделано предположение, что для обеспечения продолжительного перспективного эффекта bFGF на клетки и стимулирования роста свыше эффекта, полученного для формирующихся клеточных масс с использованием культуральной среды с добавлением 10% сыворотки, необходимо получить клеточные массы.
Следовательно, для подтверждения того, что конструирование клеточных масс является обязательным условием для осуществления указанных эффектов, авторы настоящего изобретения провели культивирование монослойной адгезионной культуры кардиомиоцитов, полученных из очищенных эмбриональных стволовых клеток мыши, с добавлением bFGF к культуральной среде и проанализировали эффекты добавления bFGF.
В монослойной культуре, в которой не образовывались клеточные массы, очищенные кардиомиоциты едва ли могли выжить, поэтому к базальной культуральной среде добавляли ITS. Однако при добавлении bFGF было обнаружено, что выживает и прикрепляется к культуральному планшету большее количество клеток. Этот эффект был слабее эффекта, который наблюдали, когда клеточные массы образовывались с использованием культуральной среды с добавлением 10% сыворотки.
При конструировании клеточных масс очищенных кардиомиоцитов, полученных из эмбриональных стволовых клеток, сравнивали влияние культивирования при отсутствии сыворотки+ITS+bFGF с влиянием продолжительного культивирования в присутствии 10% сыворотки; в культуральной среде, содержащей 10% сыворотку, размер клеточных масс существенно уменьшался, при этом он значительно увеличивался в группе с условиями культивирования в отсутствие сыворотки+ITS+bFGF. Это указывает на то, что влияние bFGF, как обнаружено авторами настоящего изобретения, является синергическим эффектом всех трех элементов: очистки, отсутствия сыворотки и наличия клеточных масс.
На основании этого результата авторы настоящего изобретения обнаружили, что влияние ITS и/или bFGF на культуральную среду, не содержащую сыворотки, обеспечивает клеточным массам кардиомиоцитов возможность поддерживать их состояние в течение продолжительного периода времени. На основании этого обнаружено, что степень выживаемости кардиомиоцитов может быть увеличена до 90% или более, исключительно положительное улучшение до значения, составляющего более чем 60-70% для стандартного способа культивирования монослойной культуры.
Перед осуществлением настоящего изобретения клеточные массы кардиомиоцитов не могли быть получены в условиях высокой чистоты и отсутствия сыворотки; однако на основании вышеизложенного авторы настоящего изобретения решили эту проблему благодаря использованию кардиомиоцитов, полученных из эмбриональных стволовых клеток; также было обнаружено, что клеточные массы кардиомиоцитов могли быть получены наиболее эффективно в присутствии добавленных ITS и bFGF.
Таким образом, в одном из вариантов осуществления изобретения авторами настоящего изобретения также предлагается способ получения клеточных масс кардиомиоцитов, полученных из эмбриональных стволовых клеток, отличающийся тем, что очищенные кардиомиоциты, полученные из эмбриональных стволовых клеток, полученных путем диспергирования агрегированных клеточных масс, содержащих кардиомиоциты, дифференцированные и индуцированные из эмбриональных стволовых клеток до отдельных клеток, культивируют в культуральной среде в условиях отсутствия сыворотки так, что они повторно агрегируют. Культуральная среда, используемая для культуры, описанной выше, предпочтительно также содержит по меньшей мере инсулин из ITS и в более предпочтительном варианте осуществления изобретения также может содержать bFGF. В способе, описанном выше, клеточные массы, диспергированные до отдельных клеток, необязательно культивировать в отдельной зоне, как это делают в известных способах, но они могут быть разделены на группы, максимально состоящие из 10000 клеток, и культивировать в соответствующем количестве независимых зон.
При загрязнении пролиферирующими клетками, не являющимися кардиомиоцитами, клеточные массы агрегированных клеток будут расти до исключительно большого размера, а также изменят морфологию. Кроме того, может быть использовано окрашивание флуоресцентным красителем, который накапливается в митохондриях, для идентификации пролиферирующих клеток, которые идентифицируют как клетки, в которых затруднено накопление красителя. Клеточные массы, загрязненные клетками, не являющимися кардиомиоцитами, могут быть удалены при применении в трансплантационной терапии или в подобной терапии, для гарантии, что малейшее загрязнение пролиферирующими клетками, не являющимися кардиомиоцитами, удалено в имплантированных клетках.
Кроме того, ранее было опубликовано, что очищенные кардиомиоциты, полученные из эмбриональных стволовых клеток, диспергированных до отдельных клеток (индивидуальных клеток), не приживлялись при трансплантации сами по себе в сердечную ткань индивидуума (живого организма), и авторами настоящего изобретения получен такой же результат. Из уровня техники известно, что эта проблема решалась путем смешивания очищенных кардиомиоцитов, полученных из эмбриональных стволовых клеток, вместе с вспомогательными клетками, что указывает, что в живом организме защитная функция клеток, не являющихся кардиомиоцитами, является необходимой для выживания кардиомиоцитов. Однако загрязнение клетками, не являющимися кардиомиоцитами, может вызывать тяжелые непредсказуемые побочные эффекты, которые могут подвергать угрозе жизнь пациента после трансплантации, поэтому авторы настоящего изобретения предполагают, что будет обеспечена большая безопасность и более сильный терапевтический эффект, если очищенные кардиомиоциты будут трансплантировать индивидууму (живому организму) без смешивания с вспомогательными клетками, но с поддерживанием высокой степени чистоты кардиомиоцитов.
Следовательно, авторам настоящего изобретения пришла идея создания клеточных масс очищенных кардиомиоцитов, полученных из эмбриональной стволовой клетки, полученной способом, описанным выше, и авторы настоящего изобретения обнаружили, что при трансплантации таких клеточных масс в сердечную ткань индивидуума (живого организма) степень приживления после трансплантации может быть существенно увеличена. Другими словами, авторы настоящего изобретения обнаружили, что при культивировании очищенных кардиомиоцитов, полученных из эмбриональной стволовой клетки, полученной путем диспергирования клеточных масс до отдельных клеток, в культуральной среде в условиях отсутствия сыворотки так, чтобы они повторно агрегировали с формированием клеточных масс, степень приживления после трансплантации очищенных кардиомиоцитов, полученных из эмбриональной стволовой клетки, может быть существенно увеличена.
Следовательно, другой вариант осуществления настоящего изобретения относится к способу лечения болезней сердца, отличающемуся тем, что клеточные массы, полученные с помощью повторно агрегированных кардиомиоцитов, полученных из эмбриональных стволовых клеток, и которые были очищены путем диспергирования до отдельных клеток, трансплантируют в сердечную ткань (в частности, в пораженную часть сердечной ткани) индивидуума (живого организма) так, чтобы они приживились. При использовании в настоящей заявке термин "приживление" обозначает выживаемость в органе-хозяине и сохранение в нем в прикрепленном состоянии в течение продолжительного периода времени.
Кроме того, как описано выше, существует известный способ, который обеспечивает возможность наслаивания до трех монослойных пластов неонатальных кардиомиоцитов, но более толстые пласты кардиомиоцитов не могут быть получены.
Для решения этой проблемы авторам настоящего изобретения пришла идея создания клеточных масс очищенных кардиомиоцитов, полученных из эмбриональной стволовой клетки, полученных способом, описанным выше. Клеточные массы, представляющие интерес, конструировали способом, описанным выше, и полученные клеточные массы извлекали и высевали на разбитую на секции поверхность культуральной посуды, не обладающей способностью к адгезии клеток, в отсутствие пространства между клеточными массами, так чтобы соседние клеточные массы находились в постоянном контакте друг с другом, с последующим поддержанием суспензионной культуры. В результате клеточные массы конъюгируют вместе с течением времени с образованием пласта клеточных масс кардиомиоцитов толщиной 50-300 мкм; таким образом, было обнаружено, что вне живого организма может быть получен так называемый "пласт клеток", имеющий большую толщину, чем максимальная толщина, известная из уровня техники. Таким образом, понятно, что в реальных способах применения желательное количество клеточных масс очищенных кардиомиоцитов желательного размера, полученных из эмбриональных стволовых клеток, может использоваться для получения пласта клеток желаемого размера.
Следовательно, следующим вариантом осуществления настоящего изобретения является способ получения пласта клеточных масс кардиомиоцитов (пласта клеток), причем способ отличается тем, что клеточные массы очищенных кардиомиоцитов, полученных из эмбриональных стволовых клеток, подвергаются суспензионному культивированию, поскольку они помещаются с небольшими интервалами в один и тот же планшет, и отличается тем, что суспензионную культуру поддерживают до тех пор, пока клеточные массы не конъюгируют вместе с получением желаемой толщины, которая находится в интервале 50-300 мкм.
Авторы настоящего изобретения обнаружили, что клеточные массы очищенных кардиомиоцитов, полученных из эмбриональной стволовой клетки, имеющей свойства, описанные выше, могут быть трансплантированы в сердечную ткань так, чтобы они приживлялись. Эти клеточные массы могут использоваться в качестве медицинского устройства для трансплантации, которой может быть трансплантация в организм животных, включая организм человека.
Следовательно, в другом варианте осуществления настоящее изобретение относится к медицинскому устройству, содержащему клеточные массы кардиомиоцитов, полученных из эмбриональной стволовой клетки, которая была получена способом, который включает получение клеточных масс агрегированных клеток, которые содержат кардиомиоциты, дифференцированные и индуцированные из эмбриональных стволовых клеток, диспергирование клеточных масс до отдельных клеток с получением, таким образом, очищенных кардиомиоцитов, полученных из эмбриональной стволовой клетки, и культивирование кардиомиоцитов в культуральной среде в условиях отсутствия сыворотки так, чтобы они повторно агрегировали. Такое медицинское устройство предназначено для применения в трансплантации, когда это средство трансплантируется в сердечную ткань индивидуума, и при этом приживляется; демонстрирует значительный эффект в приживлении и может использоваться у пациента, нуждающегося в трансплантации сердечной ткани.
Таким образом, авторы настоящего изобретения провели интенсивные исследования условий культивирования, которые могут значительно повысить степени выживаемости кардиомиоцитов, полученных из эмбриональных стволовых клеток, которые были диспергированы до отдельных клеток (отдельных клеток) для их полной очистки; в результате было обнаружено, что кардиомиоциты обладают новыми характеристиками, заключающимися в агрегации с образованием клеточных масс при культивировании в культуральной среде при условии, когда среда не содержит сыворотки животного происхождения (т.e. в условиях отсутствия сыворотки), предпочтительно в культуральной среде, содержащей инсулин, более предпочтительно в культуральной среде, содержащей трансферрин, селен и/или основной фактор роста фибробластов, кроме инсулина. Авторы настоящего изобретения трансплантировали эти клетки в сердечную ткань индивидуума (живого организма) и получили новые данные о том, что степень приживления этих клеток в ткани была значительно повышена. Авторы настоящего изобретения получили другие данные о том, что, используя эти клетки, можно получить пласт клеточных масс кардиомиоцитов большей толщины, чем толщина, ожидаемая в результате использования известного способа, а также можно получить медицинское устройство, содержащее эти клеточные массы; эти данные привели в результате к созданию настоящего изобретения.
Эти данные были получены путем культивирования эмбриональных стволовых клеток, и аналогичные данные также могут быть получены при использовании других плюрипотентных стволовых клеток вместо эмбриональных стволовых клеток. Более подробно, когда плюрипотентные стволовые клетки, которые для их полной очистки были диспергированы до состояния суспензии отдельных клеток (индивидуальных клеток), культивировали в культуральной среде при условии отсутствия в среде сыворотки животного происхождения (т.e. в условиях отсутствия сыворотки), предпочтительно в культуральной среде, содержащей инсулин, более предпочтительно в культуральной среде, содержащей трансферрин, селен, и/или основной фактор роста фибробластов дополнительно к инсулину, эти клетки были способны агрегировать с образованием клеточных масс, и степень выживаемости кардиомиоцитов могла значительно повышаться. Плюрипотентные стволовые клетки, которые могут быть использованы, включают не только эмбриональные стволовые клетки, но также все другие плюрипотентные стволовые клетки, обладающие свойствами, подобными свойствам эмбриональных стволовых клеток, как полученных из клеток органов и тканей взрослых млекопитающих, а также из их клеток костного мозга, клеток крови и даже из эмбриональных и фетальных клеток; примеры представляют собой эмбриональные зародышевые клетки (EG-клетки), стволовые зародышевые клетки (GS-клетки) и индуцированные плюрипотентные стволовые клетки (клетки iPS).
Таким образом, ниже перечислены объекты настоящего изобретения.
(1) Способ получения клеточных масс кардиомиоцитов, полученных из плюрипотентных стволовых клеток, отличающийся тем, что очищенные кардиомиоциты, полученные из плюрипотентной стволовой клетки, полученной путем диспергирования агрегированных клеточных масс, которые содержат кардиомиоциты, дифференцированные и индуцированные из плюрипотентных стволовых клеток (таких как эмбриональные стволовые клетки, эмбриональные зародышевые клетки, зародышевые стволовые клетки или индуцированные плюрипотентные стволовые клетки) до отдельных клеток, культивируют в культуральной среде в условиях отсутствия сыворотки так, что они повторно агрегируют.
(2) Способ, описанный в пункте (1) выше, в котором культуральная среда содержит инсулин.
(3) Способ, описанный в пункте (1) или (2) выше, в котором культуральная среда содержит по меньшей мере одно вещество, выбранное из группы, состоящей из трансферрина, селена, основного фактора роста фибробластов (bFGF), фактора роста эпителиальных клеток (EGF), фактора-BB роста тромбоцитов (PDGF-BB) и эндотелина-1 (ET-1).
(4) Способ, описанный в любом из пунктов (1)-(3) выше, в котором в культуральной среде содержится 0,1-10 мг/л инсулина, 0,1-10 мкг/л трансферрина, 0,1-10 мкг/л селена, 1 нг/мл-100 нг/мл основного фактора роста фибробластов, 1 нг/мл-1000 нг/мл фактора роста эпителиальных клеток, 1 нг/мл-1000 нг/мл фактора роста тромбоцитов и 1×10-8-1×10-6 M эндотелина-1 (ET-1).
(5) Способ лечения болезни сердца, отличающийся тем, что клеточные массы, полученные с помощью повторной агрегации кардиомиоцитов, полученных из очищенных плюрипотентных стволовых клеток, диспергированных до отдельных клеток, трансплантируют в сердечную ткань индивидуума так, чтобы они приживались.
(6) Способ, описанный в пункте (5) выше, в котором клеточные массы кардиомиоцитов представляют собой клеточные массы, полученные способом, описанным в любом из пунктов (1)-(4) выше.
(7) Способ, описанный в пунктах (5) или (6) выше, в котором трансплантация включает инъекцию клеточных масс кардиомиоцитов в сердечную ткань.
(8) Способ, описанный в пунктах (5) или (6) выше, в котором трансплантация включает трансплантацию пласта клеточных масс кардиомиоцитов на сердечную ткань.
(9) Способ получения пласта клеточных масс кардиомиоцитов, отличающийся тем, что клеточные массы очищенных кардиомиоцитов, полученных из плюрипотентных стволовых клеток, высевают на разделенную стенками культуральную посуду, не обладающую способностью к адгезии клеток, в отсутствие пространства между клеточными массами, так чтобы соседние клеточные массы находились в постоянном контакте друг с другом, с последующим поддержанием суспензионной культуры, которую поддерживают до тех пор, пока клеточные массы не конъюгируют вместе и не образуют желаемую толщину, составляющую 50-300 мкм.
(10) Способ, описанный в пункте (9) выше, в котором клеточные массы кардиомиоцитов представляют собой клеточные массы, полученные способом, описанным в любом из пунктов (1)-(4) выше.
(11) Медицинское устройство, содержащее клеточные массы кардиомиоцитов, полученные из плюрипотентной стволовой клетки, предназначенное для использования для трансплантации в сердечную ткань индивидуума, так чтобы клеточные массы кардиомиоцитов приживились, причем медицинское устройство получают способом, включающим стадии получения клеточных масс агрегированных клеток, которые содержат кардиомиоциты, дифференцированные и индуцированные из плюрипотентных стволовых клеток, диспергирования клеточных масс до отдельных клеток с получением в результате очищенных кардиомиоцитов, полученных из плюрипотентной стволовой клетки, и культивирования кардиомиоцитов в культуральной среде в условиях отсутствия сыворотки так, что они повторно агрегируют.
(12) Медицинское устройство, описанное в пункте (11) выше, в котором трансплантация включает инъекцию клеточных масс кардиомиоцитов в сердечную ткань.
(13) Медицинское устройство, описанное в пункте (11) выше, где трансплантация включает трансплантацию пласта клеточных масс кардиомиоцитов на сердечную ткань.
ПРЕИМУЩЕСТВА ИЗОБРЕТЕНИЯ
Авторами настоящего изобретения было обнаружено, что кардиомиоциты, полученные из плюрипотентных стволовых клеток, которые были очищены диспергированием до отдельных клеток, способны повторно агрегировать при культивировании в условиях отсутствия сыворотки. Благодаря созданию клеточных масс способом по настоящему изобретению кардиомиоциты, представляющие интерес, могут культивироваться в течение продолжительного периода времени с поддержанием на высоком уровне степени их выживаемости и способности к росту. Кроме того, если эти кардиомиоциты трансплантируют в сердечную ткань индивидуума (живого организма), было обнаружено, что степень их приживления в сердечной ткани значительно увеличивается, и они остаются приживленными в течение продолжительного периода времени, не смешиваясь с другими клетками. Этот способ делает возможным реализацию способа лечения, который является многообещающим способом клеточной терапии, и делает медицинское устройство, которое содержит клеточные массы кардиомиоцитов.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг. 1 продемонстрирован способ получения клеточных масс кардиомиоцитов с использованием очищенных кардиомиоцитов, полученных из эмбриональных стволовых клеток мыши, экспрессирующих усиленный зеленый флуоресцентный белок (EGFP), и способ получения клеточных масс, полученных с использованием 313-10000 очищенных кардиомиоцитов, полученных из эмбриональных стволовых клеток мышей.
На фиг. 2 продемонстрировано создание клеточных масс кардиомиоцитов с использованием очищенных кардиомиоцитов, полученных из эмбриональных стволовых клеток мармозетки через 24 часа (фиг. 2A) или через 48 часов (фиг. 2B) после выделения стволовых клеток.
На фиг. 3 продемонстрированы результаты ведения монослойной культуры очищенных кардиомиоцитов, полученных из эмбриональных стволовых клеток мышей, конкретно продемонстрирована идентификация оптимального адгезионного субстрата, предназначенного для использования в монослойной культуре, на основе сравнения степени выживаемости клеток с помощью адгезионных субстратов (фиг. 3A), и продемонстрировано сравнение клеточных масс и степени выживаемости кардиомиоцитов монослойной культуры в зависимости от присутствия сыворотки в культуральной среде (фиг. 3B и 3C).
На фиг. 4 продемонстрирован способ (1) обнаружения эмбриональных стволовых клеток, которыми загрязнены клеточные массы, образованные из очищенных кардиомиоцитов, полученных из эмбриональных стволовых клеток мыши; было обнаружено, что клеточные массы, отмеченные красной рамкой, содержат гигантские клеточные массы, полученные в результате клеточной пролиферации.
На фиг. 5 продемонстрирован способ (2) обнаружения эмбриональных стволовых клеток, которыми загрязнены клеточные массы, образованные очищенными кардиомиоцитами, полученными из эмбриональных стволовых клеток мыши; загрязняющие и пролифирирующие клетки, не являющиеся кардиомиоцитами, могут обнаруживаться на основании слабого флуоресцентного сигнала, полученного благодаря TMRM (реагента, который специфически окрашивает митохондрии), становится понятно, что в отличие от клеточных масс нормальных кардиомиоцитов (фиг. 5A) клеточные массы аномальных кардиомиоцитов загрязняются пролифирирующими клетками, не являющимися кардиомиоцитами.
На фиг. 6A продемонстрировано, что при культивировании на круглодонных 96-луночных плашках, не обладающих адгезионной способностью, очищенные кардиомиоциты, полученные из сердца новорожденной крысы, не способны к конструированию клеточных масс даже через 24 часа культивирования, и на фиг. 6B продемонстрировано, что очищенные кардиомиоциты, полученные из сердца новорожденной крысы, при культивировании в условиях отсутствия сыворотки не образуют клеточных масс, а погибают после пассирования в течение 5 дней.
На фиг. 7 продемонстрированы результаты культивирования на круглодонных 96-луночных плашках, не обладающих адгезионной способностью, с использованием среды, не содержащей сыворотку, к которой были добавлены ITS, bFGF и другие разнообразные добавки; обнаружили протективный эффект клеток и активность стимулирования роста клеток в результате этих добавок путем обнаружения эффектов, который они оказывали на увеличение диаметра клеточной массы.
На фиг. 8 продемонстрированы результаты ведения монослойной адгезионной культуры очищенных кардиомиоцитов, полученных из эмбриональных стволовых клеток мыши, на культуральных планшетах, покрытых фибронектином; выживаемость клеток была существенно низкой в группе, соответствующей условиям отсутствия сыворотки+ITS, и в группе, соответствующей условиям ITS+bFGF.
На фиг. 9 продемонстрированы результаты измерения выживаемости в ткани клеток очищенных отдельных кардиомиоцитов, полученных из эмбриональных стволовых клеток мыши, после того как клетки были трансплантированы в сердце без применения способа повторной агрегации, но они сохранялись в диспергированном состоянии.
На фиг. 10 продемонстрировано, что когда очищенные кардиомиоциты, полученные из эмбриональных стволовых клеток мыши, в виде образующихся в клеточные массы способом повторной агрегации метили с помощью красного красителя (Mitotracker Red) и трансплантировали в сердце, кардиомиоциты эффективно выживали.
На фиг. 11 продемонстрированы результаты трансплантации в сердце клеточных масс очищенных кардиомиоцитов, полученных из эмбриональных стволовых клеток мыши, экспрессирующих EGFP, после повторной агрегации. На фиг. 11A и 11B продемонстрирован результат анализа степени выживаемости клеток в сердечной ткани с помощью измерения количества клеток в сердечной ткани, и на фиг. 11C и 11D продемонстрировано, что клеточные массы кардиомиоцитов мыши сохраняются приживленными в сердце-хозяине в течение продолжительного периода времени и могут созревать с течением времени.
На фиг. 12 продемонстрирован результат получения клеточных масс очищенных кардиомиоцитов, полученных из эмбриональных стволовых клеток мармозетки, в условиях отсутствия сыворотки (фиг. 12A), условие отсутствия сыворотки и с добавлением KSR (фиг. 12B), и условие отсутствия сыворотки и с добавлением ITS (фиг. 12C).
На фиг. 13 продемонстрировано получение пластов кардиомиоцитов желаемых размеров, имеющих желаемую толщину, с использованием клеточных масс очищенных кардиомиоцитов, полученных из эмбриональных стволовых клеток мармозетки.
На фиг. 14 продемонстрировано, что клеточные массы кардиомиоцитов, полученных из очищенных эмбриональных стволовых клеток человека, могут быть получены в условиях отсутствия сыворотки.
На фиг. 15 продемонстрировано, что кардиомиоциты, полученные из стволовых клеток человека, которые были трансплантированы в сердце иммунодефицитных мышей, могут выживать в сердечной ткани в течение 2 недель.
На фиг. 16 продемонстрировано, что кардиомиоциты, полученные из стволовых клеток человека, которые были трансплантированы в сердце иммунодефицитных мышей, могут выживать в сердечной ткани в течение 5 недель, что продемонстрировано с помощью красного красителя, использованного для слежения за трансплантированными клетками.
На фиг. 17 продемонстрировано, что кардиомиоциты, полученные из стволовых клеток человека, которые были трансплантированы в сердце иммунодефицитных мышей, могут выживать в сердечной ткани в течение 5 недель, что продемонстрировано с помощью красителя (Microtracker Red: красный цвет), Nkx2.5 (водянисто-голубой цвет), с помощью антитела к саркомерному актинину (зеленый цвет), которые использовали для слежения за трансплантированными клетками.
На фиг. 18 продемонстрировано, что кардиомиоциты, полученные из стволовых клеток человека, которые были трансплантированы в сердце иммунодефицитных мышей, могут выживать в сердечной ткани в течение 5 недель, что продемонстрировано с помощью красителя Nkx2.5 (водянисто-голубой цвет), и с помощью антитела против антитела человека (зеленый цвет).
На фиг. 19 продемонстрированы результаты получения клеточных масс очищенных кардиомиоцитов, полученных из клеток iPS мыши при культивировании в условиях отсутствия сыворотки, в условиях отсутствия сыворотки и с добавлением ITS и в условиях отсутствия сыворотки и с добавлением KSR.
На фиг. 20 продемонстрированы результаты культивирования очищенных кардиомиоцитов, полученных из клеток ES человека, в условиях отсутствия сыворотки в присутствии добавленного bFGF или других факторов роста; bFGF является предпочтительным для эффектов защиты клеток и активации роста.
На фиг. 21 продемонстрирована экспрессия генов bFGF, EGF, PDGE-BB, и ET-1 в сердце-хозяине, поскольку она связана с механизмом созревания для случая, где клеточные массы кардиомиоцитов мыши сохраняются приживленными в сердце-хозяине в течение продолжительного периода времени.
НАИЛУЧШИЙ СПОСОБ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Специалистам в данной области для осуществления настоящего изобретения необходимо знать способы молекулярной биологии, способы генетической инженерии, такие как технология рекомбинантной ДНК, общие способы клеточной биологии, а также известный уровень техники, могут, если не указано иначе, ссылаться на стандартную литературу в этих областях. Примеры такой литературы включают: "Molecular Cloning: A Laboratory Manual, 3rd Edition" (Sambrook & Russell, Cold Spring Harbor Laboratory Press, 2001); "Current Protocols in Molecular biology" (Ed. by Ausubel et al., John Wiley & Sons, 1987); "Methods in Enzymology in series" (Academic Press); "PCR Protocols: Methods in Molecular Biology" (Ed. by Bartlett & Striling, Humana Press, 2003); "Animal Cell Culture: A Practical Approach, 3rd Edition" (Ed. by Masters, Oxford University Press, 2000); и "Antibodies: A Laboratory Manual" (Ed. by Harlow et al. & Lane, Cold Spring Harbor Laboratory Press, 1987). Реагенты и наборы реагентов для применения в культивировании клеток и в экспериментах клеточной биологии, на которые в настоящей заявке сделаны ссылки, доступны из коммерческих источников, таких как Sigma, Aldrich, Invitrogen/GIBCO, Clontech и Stratagene.
(1) Плюрипотентные стволовые клетки
Специалистам в данной области для осуществления настоящего изобретения необходимо знать способы культивирования клеток с использованием плюрипотентных стволовых клеток и общие способы для экспериментов в биологии развития и в клеточной биологии, могут, если не указано иначе, ссылаться на стандартную литературу в этих областях. Примеры такой литературы включают: "Guide to Techniques in Mouse Development" (Ed. by Wasserman et al., Academic Press, 1993); "Embryonic Stem Cell Differentiation in vitro" (M.V. Wiles, Meth. Enzymol. 225:900, 1993); "Manipulating the Mouse Embryo: A laboratory manual" (Ed. by Hogan et al., Cold Spring Harbor Laboratory Press, 1994); "Embryonic Stem Cells" (Ed. by Turksen, Humana Press, 2002). Реагенты и наборы реагентов для применения в культивировании клеток и в экспериментах по биологии развития и клеточной биологии, на которые в настоящей заявке сделаны ссылки, доступны из коммерческих источников, таких как Invitrogen/GIBCO и Sigma.
Для способов получения, последовательного культивирования и консервации плюрипотентных стволовых клеток мыши или человека уже созданы стандартные протоколы, и специалисты в данной области, желающие осуществить настоящее изобретение, могут использовать плюрипотентные стволовые клетки с помощью ссылки на множество справочных документов и им подобных дополнительно к справочной литературе, перечисленной в предыдущих разделах. Такие документы включают следующие: Matsui et al., Cell 70:841, 1992; Thomson et al., United States Patent 5843780; Thomson et al., Science 282: 114, 1998; Shamblott et al., Proc. Natl. Acad. Sci. USA 95:13726, 1998; Shamblott et al., Патент США 6090622; Reubinoff et al., Nat. Biotech. 18:399, 2000; и Международную Публикацию WO 00/27995 A1. Для других видов животных, таких как обезьяна (Thomson et al., Патент США 5843780; и Proc. Natl. Acad. Sci. USA, 92, 7844, 1996), крыса (Iannaccone et al., Dev. Biol. 163:288, 1994; и Loring et al., Международная Публикация WO 99/27076 A1), курица (Pain et al., Development 122:2339, 1996; Патент США 5340740 и Патент США 5656479), и свинья (Wheeler et al., Reprod. Fertil. Dev. 6:563, 1994; и Shim et al., Biol. Reprod. 57:1089, 1997), известны способы, с помощью которых можно создавать плюрипотентные клетки, такие как эмбриональные стволовые клетки и клетки, подобные эмбриональным стволовым клеткам, и эти плюрипотентные стволовые клетки, которые могут быть использованы в настоящем изобретении, могут быть получены или использованы согласно способам, описанным в этих документах.
Способ по настоящему изобретению может использоваться в отношении плюрипотентных стволовых клеток, полученных из любых млекопитающих. Например, он может использоваться в отношении плюрипотентных стволовых клеток, полученных из мыши, коровы, козы, собаки, кошки, мармозетки, макаки-резус и человека; однако он не ограничен плюрипотентными стволовыми клетками, полученными из этих видов животных. Примерами плюрипотентных стволовых клеток, используемых по настоящему изобретению, могут являться эмбриональные стволовые клетки (клетки ES), полученные из млекопитающих, таких как мышь, обезьяна и человек, которые уже широко используются в качестве культивируемых клеток.
Конкретные примеры эмбриональных стволовых клеток, полученных из мыши, включают клетки EB3, E14, D3, CCE, R1, 129SV и клетки J1. Эмбриональные стволовые клетки, полученные из мыши, по настоящему изобретению доступны из американской коллекции типовых культур (ATCC), Chemicon, Cell & Molecular Technologies и т.д.
Что касается эмбриональных стволовых клеток, полученных из обезьяны, то клеточные линии, созданные из макаки-резус (Macaca mulatta) (Thomson et al., Proc. Natl. Acad. Sci. USA 1995; 92:7844), Яванская макака (Macaca fascicularis) (Suemori et al., Dev. Dyn. 2001; 222: 273-279) и обычной мармозетки (Callithrix jacchus) (Sasaki et al., Stem Cells. 2005; 23: 1304-1313), опубликованы и являются доступными. Например, эмбриональные стволовые клетки мармозетки также доступны из Центрального института экспериментальных животных (на правовой основе).
На настоящий момент в мире создано более чем несколько десятков линий эмбриональных стволовых клеток человека; например, в списке Национальных Институтов Здоровья США (US National Institutes of Health) (http://stemcells.nih.gov/registry/index.asp) зарегистрированы для общественного использования многочисленные клеточные линии, и другие клеточные линии доступны из коммерческих источников, включающих Cellartis, ES Cell International, Wisconsin Alumni Research Foundation и т.д. В Японии эмбриональные стволовые клетки человека также доступны из Исследовательского центра стволовых клеток, принадлежащего учреждению Института передовых медицинских наук, Киотского университета (национальной корпорации университета) (Suemori et al., Biochem. Biophys. Res. Commun., 2006; 345: 926-932).
Также было опубликовано, что линии эмбриональных стволовых клеток созданы для коровы (Mitalipova et al., Cloning 2001; 3: 59-67), птицы (Petitte et al., Mech. Dev. 2004; 121: 1159-1168), ии Данио рерио (Fishman, M. C., Science 2001; 294: 1290-1291).
В то время как линии эмбриональных стволовых клеток, как правило, создаются с помощью культивирования ранних эмбрионов, они также могут быть получены из ранних эмбрионов, в которые были перенесены ядра соматических клеток (Munsie et al., Curr. Biol. 10:989, 2000; Wakayama et al., Science 292:740, 2001 и Hwang et al., Science 303: 1669, 2004). Также были опубликованы данные о попытке развития партеногенетических эмбрионов до стадии, сравнимой со стадией бластоциста, и получения эмбриональных стволовых клеток на этой стадии (Патентная публикация США 02/168763 A1 и Vrana K et al., Proc. Natl. Acad. Sci. USA 100:11911-6), и был опубликован способ, в котором эмбриональная стволовая клетка сливается с соматической клеткой с получением эмбриональной стволовой клетки, несущей генетическую информацию ядра соматической клетки (Международная публикация WO 00/49137 A1 и Tada et al., Curr. Biol. 11:1553, 2001). Эмбриональные стволовые клетки, которые могут использоваться по настоящему изобретению, также включают клетки, которые были получены способами, описанными выше, а также клетки, в которых гены, локализованные на соответствующих хромосомах, были модифицированы способами генетической инженерии.
Плюрипотентные стволовые клетки, которые могут использоваться в способе по настоящему изобретению, не ограничены эмбриональными стволовыми клетками, но включают все другие плюрипотентные стволовые клетки, обладающие свойствами, подобными свойствам эмбриональных стволовых клеток, как клетки, полученные из клеток органов и тканей взрослых млекопитающих, так и клетки костного мозга, клетки крови и даже эмбриональные и фетальные клетки. В этом случае термин "свойства, подобные свойствам эмбриональных стволовых клеток" может определяться с помощью клеточных биологических свойств, которые специфичны для эмбриональных стволовых клеток и которые иллюстрируются присутствием поверхностного (антигена) маркера, специфичного для эмбриональных стволовых клеток, экспрессией гена, специфичного для эмбриональных стволовых клеток, а также способностью образования тератомы и способностью образования химерной мыши. Конкретные примеры других применяемых плюрипотентных стволовых клеток включают эмбриональные зародышевые клетки (EG-клетки), полученные из первичных зародышевых клеток, стволовые зародышевые клетки (GS-клетки), полученные из зародышевых клеток яичек, и индуцированные плюрипотентные стволовые клетки (клетки iPS), полученные из соматических клеток, таких как фибробласты, с помощью специальной генной манипуляции. Примеры индуцированных плюрипотентных стволовых клеток включают клетки, которые могут быть получены путем введения специфических факторов в соматические клетки, и они могут быть получены способами, описанными в статье, написанной исследовательской группой Профессора Shinya Yamanaka Киотского университета (K. Takahashi, et al., "Induction of Pluripotent Stem Cells from Adult Human Fibroblasts by Defined Factors" Cell 2007 131: 861-872), и в статье, написанной исследовательской группой Thomson Университета Висконсина (J. Yu, et al., "Induced Pluripotent Stem Cell Lines Derived from Human Somatic Cells" Science 2007 318:1917-1920). Конкретно, по меньшей мере, один ген, выбранный из генов Oct3/4, Sox2, c-Myc, Klf4, Nanog и LIN28, переносят в данную соматическую клетку, детектируют экспрессию гена или белка, специфичного для плюрипотентных стволовых клеток, и те клетки, которые экспрессируют такой ген, выбирают в качестве плюрипотентных стволовых клеток. Подобно эмбриональным стволовым клеткам индуцированные плюрипотентные стволовые клетки, полученные таким образом, могут культивироваться вместе с основным фактором роста фибробластов в присутствии фибробластов мыши, дезактивированных по росту, или в присутствии клеток, которые могут их заменить, и культивируемые клетки могут использоваться в качестве плюрипотентных стволовых клеток подобно эмбриональным стволовым клеткам.
Ранее было обнаружено, что индуцированные плюрипотентные стволовые клетки, описанные выше, обладают теми же свойствами, что и эмбриональные стволовые клетки по части характеристик дифференцировки в разнообразные ткани и по части экспрессии генов в клетках (Park I.H. et al., Nature, 2008, 451, 141-147), и условия индукции дифференцировки эмбриональных стволовых клеток в разнообразные ткани могут непосредственно применяться к индуцированным плюрипотентным стволовым клеткам (Takahashi and Yamanaka, Saibou Kogaku (Cell Engineering), Vol. 27, No. 3, 252-253, 2008).
(2) Способы индукции дифференцировки плюрипотентных стволовых клеток в кардиомиоциты
Следующее описание относится к эмбриональным стволовым клеткам (клеткам ES) как к примеру плюрипотентных стволовых клеток. Когда эмбриональные стволовые клетки, способные дифференцироваться до кардиомиоцитов, подвергают соответствующей обработке для индукции дифференцировки до кардиомиоцитов, они начинают дифференцироваться до кардиомиоцитов. Например, дифференцировка эмбриональных стволовых клеток мыши в кардиомиоциты может быть индуцирована способом висячей капли, в котором эмбриональные стволовые клетки подвергаются культивированию в условиях суспензионной культуры в культуральной среде, не содержащей фактор ингибирования лейкоза (LIF) до тех пор, пока не образуются клеточные массы (эмбриоидные тела). Альтернативно, эмбриональные стволовые клетки мармозетки или эмбриональные стволовые клетки человека также могут быть подвергнуты обработке для индукции дифференцировки до кардиомиоцитов. Для индукции дифференцировки эмбриональных стволовых клеток до кардиомиоцитов могут использоваться любые известные способы. Например, способ индукции дифференцировки в присутствии вещества, которое подавляет сигнальный путь BMP (WO 2005/033298), и способ индукции дифференцировки в присутствии вещества, которое стимулирует активацию канонического сигнального пути Wnt (PCT/JP 2007/59242, опубликовано как WO 2007/126077).
(3) Очистка кардиомиоцитов
После индукции дифференцировки эмбриональных стволовых клеток до кардиомиоцитов способом, описанным в пункте (2) выше, кардиомиоциты могут быть очищены (селектированы) любым способом, способным диспергировать кардиомиоциты на отдельные клетки (отдельные клетки) и очищать их до состояния индивидуальных кардиомиоцитов. Например, для очистки (селекции) только кардиомиоцитов может использоваться способ селекции с использованием митохондрий в кардиомиоцитах в качестве показателя (WO 2006/022377) и способ селекции клеток, которые выживают в условиях дефицита питательных веществ (PCT/JP 2007/051563, опубликовано как WO 2007/088874).
(4) Получение клеточных масс кардиомиоцитов
Очищенные кардиомиоциты, полученные из эмбриональных стволовых клеток, которые были получены диспергированием до отдельных клеток согласно способу, описанному в пункте (3) выше, могут культивироваться в условиях отсутствия сыворотки так, чтобы они могли агрегировать с получением клеточных масс кардиомиоцитов, полученных из эмбриональных стволовых клеток. Предпочтительно культуральная среда, используемая для данных условий культивирования, содержит по меньшей мере одно вещество, выбранное из группы, состоящей из инсулина (0,1-10 мг/л), трансферрина (0,1-10 мкг), селена (0,1-10 мкг/л), основного фактора роста фибробластов (bFGF; 1 нг/мл-100 нг/мл), фактора роста эпителиальных клеток (1 нг/мл-1000 нг/мл), фактора роста тромбоцитов (1 нг/мл-1000 нг/мл) и эндотелина-1 (ET-1) (1×10-8-1×10-6 M).
Клеточные массы очищенных кардиомиоцитов, полученных из эмбриональных стволовых клеток, которые были получены способом, описанным выше, содержат пролиферирующие клетки в виде небольшого количества примеси; если такие пролиферирующие клетки исключают из клеток для трансплантации, то в дальнейшем может быть гарантирована безопасность трансплантации. Известные в настоящее время способы очистки кардиомиоцитов включают предварительное введение определенных маркерных генов в геном стволовых клеток (FASEB J. 2000; 14: 2540-2548). Каждый из этих способов может обеспечить 99% чистоты, но они не способны гарантировать 100±0% чистоты. Например, если для лечения инфаркта миокарда человека требуется 1011 кардиомиоцитов, то 99% чистоты обозначает загрязнение клетками, не являющимися кардиомиоцитами, в количестве 109. Таким образом, даже способ, который может быть описан как почти совершенное средство очистки в свете известного состояния уровня техники, не дает возможности 100% очистки кардиомиоцитов и должен быть объединен с другими способами очистки или должен применяться с другими способами, которые гарантируют безопасность.
Следовательно, авторы настоящего изобретения повторили вышеописанный способ после специального смешивания клеточных масс недифференцированных кардиомиоцитов с эмбриональными стволовыми клетками. Как оказалось, недифференцированные эмбриональные стволовые клетки, которые более способны к росту, чем кардиомиоциты, создают отдельные клеточные массы большего размера вне клеточных масс кардиомиоцитов. Клеточные массы кардиомиоцитов, загрязненные недифференцированными эмбриональными стволовыми клетками, могут точно обнаруживаться с помощью контроля полного размера клеточных масс. Авторы настоящего изобретения также добавляли митохондриальный индикатор (например, TMRM) к клеточным массам, представляющим интерес, после чего кардиомиоциты, которые богаты митохондриями, обнаруживали по яркости, тогда как эмбриональные стволовые клетки и другие пролиферирующие клетки, которые не богаты митохондриями, обнаруживали по их темноте. Удаление клеточных масс, имеющих большее различие флуоресценции, может проводиться автоматизировано с помощью приборов Arrayscan (Cellomics), Incell 1000 (GE/Amersham Biosciences, Cardiff, UK), Scanalyzer (Scanalyzer LemnaTec, Aachen Germany), "ImageXpress MICRO" (Molecular Devices, Union City, USA), "Pathway HT" (Becton Dickinson Biosciences), "Scan^R" (Olympus Soft Imaging Solutions, Germany) и т.д. Таким образом, способ, описанный выше, обеспечивает простой и автоматизированный путь идентификации загрязнения недифференцированными эмбриональными стволовыми клетками. Коротко, пролифирирующие клетки, которые в небольшой степени смешаны с очищенными кардиомиоцитами, полученными из эмбриональных стволовых клеток, которые образуются в виде агрегатов в клеточные массы в условиях культивирования при отсутствии сыворотки, могут быть идентифицированы с использованием размера и формы таких клеточных масс в качестве показателей, которые необязательно объединяют с митохондриальным показателем и последующей идентификацией с использованием интенсивности флуоресценции и ее распределения в клеточных массах в качестве показателей. Таким образом, клеточные массы, загрязненные клетками, не являющимися кардиомиоцитами, могут быть удалены из клеток для трансплантации с достижением, таким образом, большей безопасности.
(5) Трансплантация клеточных масс кардиомиоцитов в сердечную ткань и их приживление
Используя клеточные массы, полученные путем агрегации способом, описанным выше, а именно клеточные массы очищенных кардиомиоцитов, полученных из эмбриональных стволовых клеток, специалист может трансплантировать только кардиомиоциты в сердечную ткань индивидуума (живого организма). Например, кардиомиоциты могут быть непосредственно инъецированы в сердечную ткань шприцем; в этом случае инъекция выполняется с использованием тонкой (с толщиной 29- или 30-G), следовательно, менее инвазивной иглы. Степень приживления кардиомиоцитов, трансплантированных способом, описанным выше, значительно повышается по сравнению с известными способами. Термин "приживление" обозначает, что трансплантированные клетки выживают внутри органа-хозяина и сохраняются прикрепленными внутри органа в течение продолжительного периода времени.
(6) Пласты для трансплантации, полученные из клеточных масс кардиомиоцитов
Посредством известных способов за один раз нельзя получить пласт кардиомиоцитов, толщиной более чем в три клетки, даже если используются неонатальные кардиомиоциты. Однако в настоящем изобретении после конструирования клеточных масс очищенных кардиомиоцитов, полученных из эмбриональных стволовых клеток, полученные клеточные массы извлекают, высевают на разделенную стенками культуральную посуду, не обладающую способностью к адгезии клеток, в отсутствие пространства между клеточными массами, так чтобы соседние клеточные массы находились в постоянном контакте друг с другом, с последующим поддержанием суспензионной культуры, которая поддерживается до тех пор, пока клеточные массы кардиомиоцитов не конъюгируют вместе, с образованием с течением времени пласта клеточных масс кардиомиоцитов (пласта клеток), имеющего толщину, составляющую 50-300 мкм. Следовательно, культивирование осуществляют до тех пор, пока не образуется желаемая толщина. В результате, в реальных способах применения, могут использоваться в желаемом количестве желаемого размера клеточные массы очищенных кардиомиоцитов, полученных из эмбриональных стволовых клеток, с получением пласта клеток желаемого размера.
ПРИМЕРЫ
Настоящее изобретение подробно иллюстрируется с помощью ссылки на следующие примеры.
Пример 1: Получение Кардиомиоцитов, полученных из Эмбриональных Стволовых Клеток Мыши, и Очистка Кардиомиоцитов с Использованием Митохондриального Способа
Целью данного примера было получение кардиомиоцитов из эмбриональных стволовых клеток мыши и исследование того, возможно ли очищать полученные кардиомиоциты с использованием митохондриального индикатора.
В качестве эмбриональных стволовых клеток использовали клеточную линию EB3 (Niwa H, et al., Nat Genet 2000; 24: 372-376). В клеточную линию EB3 вводили элемент, экспрессирующий EGFP, посредством плазмиды, получали и создавали клеточную линию из клеток, экспрессирующих EGFP. Полученные таким образом эмбриональные стволовые клетки, экспрессирующие EGFP (EB3-клетки), суспендировали в культуральной среде α-MEM (Sigma), так чтобы концентрация эмбриональных стволовых клеток достигала 75 клеток/35 мкл; в культуральную среду α-MEM добавляли инактивированную нагреванием фетальную бычью сыворотку (55°C × 30 мин) с конечной концентрацией, составляющей 10%. Затем полученную таким образом суспензию эмбриональных стволовых клеток мыши распределяли в коммерческом культуральном 384-луночном планшете (продукт компании Greiner, модель 788161; в.д. устья каждой лунки, 3,0 мм) и получали эмбриоидные тела согласно следующему способу.
384-луночный планшет имеет номинальный допустимый объем жидкости 25 мкл на лунку, но с целью подъема уровня жидкости выше устья лунки с помощью эффекта поверхностного натяжения, суспензию распределяли в объеме, составляющем 35 мкл на лунку. В результате распределяли по 75 эмбриональных стволовых клеток на лунку. В этом случае суспензию представляли в объеме 28 мкл с целью достижения горизонтального уровня в каждом устье и в дополнительном объеме 7 мкл для подъема выше, чем горизонтальный уровень. Для распределения суспензии использовали многоканальную пипетку Theremo Labsystems (Лот No. 4610070) или распределяющий прибор компании BioTech Co., Ltd. (Модель LD-01).
Планшет, в котором распределяли культуральную среду, содержащую эмбриональные стволовые клетки, до тех пор, пока она не поднималась выше устьев лунок, переворачивали вверх дном так, чтобы культуральная среда выступала вниз от нижних кромок устьев лунок. Как только планшет оставляли в таком состоянии, планшет накрывали крышкой и осуществляли культивирование в инкубаторе при 37°C
в атмосфере 5% CO2 до тех пор, пока эмбриональные стволовые клетки не вырастали в выступах от нижних кромок устьев лунок. Через день после начала культивирования планшет с выступающим уровнем жидкости культуральной среды, перевернутый вниз, поддерживали чистыми пинцетами или чем-то подобным и выступающую культуральную среду приводили в контакт с поверхностью культуральной среды α-MEM (Sigma), заполняющей отдельный флакон большего размера, в которую добавляли инактивированную нагреванием фетальную бычью сыворотку (55°C×30 мин) до конечной концентрации 10%; клеточным массам давали возможность оседания под их собственным весом в культуральной среде во флаконе большего размера, посредством чего извлекали эмбриоидные тела или клеточные массы, полученные из эмбриональных стволовых клеток.
Извлеченные эмбриоидные тела культивировали в чашках, не обладающих адгезионной способностью (Asahi Techno Glass, стерильная чашка Петри #SH90-15; или Eiken Chemical Co., Ltd., стерильная прямоугольная чашка Петри типа 2) в течение дополнительных 2 или 3 дней. Культивированные эмбриоидные тела извлекали в центрифужную пробирку и после замены суспензии на культуральную среду, не содержащую сыворотку (культуральная среда α-MEM (#MO644 SIGMA) с добавлением раствора ITS (GIBCO #41400-045) после разведения 1/100 (раствор ITS, использованный в настоящем изобретении, содержал 1 г/л инсулина, 0,55 г/л трансферрина и 0,67 мг/л хлорида селена)), эмбриоидные тела культивировали в стерильной культуральной чашке, обладающей адгезионной способностью (FALCON #353003).
Культуральную среду меняли каждый день до 15-го дня культивирования для дифференцировки. На 15 день к образцу добавляли митохондриальный индикатор TMRM (Invitrogen #T668) с конечной концентрацией 10 мМ, который инкубировали в течение 2 часов. Далее с использованием физиологического буфера (116 мМ NaCl, 20 мМ Hepes, 12,5 мМ NaH2PO4, 5,6 мМ глюкозы, 5,4 мМ KCl, 0,8 мМ MgSO4, pH 7,35), содержащего коллагеназу (Wortington Типа 3) и трипсин (DIFCO #215240), каждый добавляли с конечной концентрацией 0,1%, культивируемые клетки диспергировали до отдельных клеток в культуральной среде с помощью встряхивания. Образец или суспензию отдельных клеток загружали в клеточный сортер с возбуждением флуоресценции (FACS) с тем, чтобы извлечь группы клеток с сильной флуоресценцией (WO 2006/022377). Очищенные клетки подсчитывали на предмет количества живых и мертвых клеток гематоцитометром. Как оказалось, пропорция живых клеток составила примерно 75%.
Пример 2: Получение Клеточных Масс с Использованием Кардиомиоцитов, полученных из Эмбриональных Стволовых Клеток Мыши
Целью данного примера было узнать, возможно ли получить клеточные массы с использованием кардиомиоцитов, полученных из эмбриональных стволовых клеток мыши, которые были получены в примере 1.
Очищенные кардиомиоциты, полученные из эмбриональных стволовых клеток мыши, которые были получены в примере 1, распределяли в круглодонных 96-луночных планшетах, не обладающих адгезионной способностью (SUMITOMO BAKELIKE CO.,LTD.; CELLFECTIGHT SPHEROID), так чтобы в каждой лунке было представлено 10000, 5000, 2500, 1250, 625 или 313 клеток. Культуральная среда представляла собой α-MEM с добавлением 10% фетальной бычьей сыворотки. За распределенными клетками наблюдали в динамике по времени; через 10 часов образовались клеточные массы и начали спонтанно разбиваться синхронным образом. Через двадцать четыре часа все клеточные массы приняли почти совершенную сферическую форму и через 10 дней проявилось цикличное, синхронное и спонтанное разбиение (фиг. 1).
Эти результаты продемонстрировали, что после того как кардиомиоциты, полученные из эмбриональных стволовых клеток мыши, диспергировались до отдельных клеток, они могли повторно агрегировать с образованием клеточных масс.
Пример 3: Получение Клеточных Масс с Использованием Кардиомиоцитов, полученных из Эмбриональных Стволовых Клеток Мармозетки
Целью данного примера было узнать, возможно ли получить клеточные массы с использованием кардиомиоцитов, полученных из эмбриональных стволовых клеток мармозетки, которые были получены в примере 1.
Эмбриональные стволовые клетки мармозетки получали из Центрального института экспериментальных животных (Sasaki E, et al., Stem Cells. 2005; 23(9): 1304-13). С использованием эмбриональных фибробластов мыши (MEF), рост которых инактивировали обработкой митамицином C, эти эмбриональные стволовые клетки мармозетки культивировали так, чтобы они оставались недифференцированными. Культуральная среда состояла из KO-DMEM (GIBCO), 20% KO-СЫВОРОТКИ (GIBCO), 1,6 мМ L-глутамина, 0,1 мМ заменимых аминокислот (MEM), 0,2 мМ β-меркаптоэтанола (2-ME; Sigma), 100 МЕ/мл пенициллина, 100 мкг/мл стрептомицин сульфата, рекомбинантного человеческого фактора ингибирования лейкоза (LIF; Chemicon) и рекомбинантного человеческого основного фактора роста фибробластов (bFGF; Peprotech) по 8 нг/мл каждого. Для последовательного пассажа колонии эмбриональных стволовых клеток разделяли путем обработки 0,1% коллагеназой типа III (Wortington) при 37°C в течение 10 минут.
Затем с целью отделения эмбриональных стволовых клеток от MEF культуральную среду, содержащую клеточные массы, пропускали сквозь фильтр с размером пор, составляющим 100 мкм, затем пропускали через фильтр с размером пор 40 мкм для удаления ненужной фракции; извлекали клеточные массы из фракции большего размера. Извлеченные клеточные массы представляли собой клеточные массы чистых эмбриональных стволовых клеток. Для дифференцировки культивировали как эмбриоидные тела по 50-1000 эмбриональных стволовых клеток на ЭТ на бактериальных чашках, не обладающих адгезионной способностью (Asahi Techno Glass; стерильная чашка Петри) общим сроком в течение 15-30 дней так, чтобы они дифференцировались в эмбриоидные тела, включающие кардиомиоциты. Культуральная среда, использованная для этой дифференцировки, представляла собой ту же среду, что идентифицирована выше, за исключением того, что она не содержала bFGF, т.e. среда состояла из KO-DMEM (GIBCO), 20% KO-СЫВОРОТКИ (GIBCO), 1,6 мМ L-глутамина, 0,1 мМ заменимых аминокислот (MEM), 0,2 мМ β-меркаптоэтанола (2-ME; Sigma), 100 МЕ/мл пенициллина, 100 мкг/мл стрептомицин сульфата и рекомбинантного человеческого фактора ингибирования лейкоза (LIF; Chemicon) в количестве 8 нг/мл.
Через один или два месяца после их получения эмбриоидные тела были взяты и обработаны согласно способу, описанному в WO 2006/022377, для очистки кардиомиоцитов. Более конкретно, эмбриоидные тела обрабатывали коллагеназой и трипсином с получением отдельных клеток. К культуральной среде в виде клеточной суспензии добавляли митохондриальный индикатор TMRM (Invitrogen #T66) с конечной концентрацией 10 мМ и смесь оставляли стоять при 37°C в течение 15 минут, промывали три раза и немедленно подвергали FACS-анализу. Клетки (кардиомиоциты), демонстрирующие более высокую интенсивность флуоресценции, чем основная популяция клеток, отделяли и извлекали.
Отделенные кардиомиоциты обрабатывали согласно тому же способу, как описано в примере 2, с получением клеточных масс кардиомиоцитов. Более конкретно, очищенные кардиомиоциты, полученные из эмбриональных стволовых клеток мармозетки, распределяли в круглодонном 96-луночном планшете, не обладающем адгезионной способностью (SUMITOMO BAKELIKE CO.,LTD.; CELLFECTIGHT SPHEROID) так, чтобы на одну лунку было представлено 2000 клеток. За распределенными клетками наблюдали в динамике по времени; через 24 часа образовались клеточные массы (фиг. 2A) и начали спонтанно разбиваться синхронным образом. Через сорок восемь часов все клеточные массы приняли почти совершенную сферическую форму (фиг. 2B), и через 10 дней проявилось цикличное, синхронное и спонтанное разбиение (фиг. 2).
Эти результаты продемонстрировали, что после того как кардиомиоциты, полученные из эмбриональных стволовых клеток мармозетки, диспергировались до отдельных клеток, они могли повторно агрегировать с образованием клеточных масс.
Пример 4: Измерение Степени Выживаемости Клеточных Масс, Образованных с Использованием Кардиомиоцитов, полученных из Эмбриональных Стволовых Клеток Мыши, и Сравнение с Результатом Адгезионного Культивирования
Целью данного примера было исследование адгезионного субстрата с использованием в качестве показателя силы защитной функции в условиях монослойной культуры и сравнение степени выживаемости очищенных кардиомиоцитов, полученных из эмбриональных стволовых клеток, между монослойной адгезионной культурой и культурой клеточных масс; монослойное адгезионное культивирование осуществляли с использованием сыворотки, обладающей сильной функцией защиты клеток, и культивирование клеточных масс осуществляли в условии присутствия и в отсутствие сыворотки; было обнаружено, что функция защиты клеток была выражена сильнее, когда культивирование клеточных масс осуществляли в условиях отсутствия сыворотки.
В примере 4 кардиомиоциты, полученные из эмбриональных стволовых клеток мыши, очищали согласно примеру 1.
Очищенные кардиомиоциты высевали на пластиковые культуральные чашки (продукт BD), покрытые или (1) желатином, или (2) фибронектином, в присутствии сыворотки (фиг. 3A). Кроме того, с целью образования клеточных масс, (3) очищенные кардиомиоциты распределяли на круглодонных 96-луночных планшетах, не обладающих адгезионной способностью, согласно примеру 2 так, чтобы на одну лунку было представлено 1000 клеток, и суспензии центрифугировали при 120g в течение 5 минут. Кроме того, затем (4) очищенные кардиомиоциты распределяли на круглодонных 96-луночных планшетах, не обладающих адгезионной способностью, согласно примеру 2, за исключением условий культивирования в отсутствие сыворотки для конструирования клеточных масс, распределяли так, чтобы на одну лунку было представлено 1000 клеток, и суспензии центрифугировали при 120g в течение 5 минут. Образцы (1)-(4) культивировали в том же инкубаторе в течение 12 часов и 4 дней. Через четыре дня измеряли количество клеток, прикрепленных к каждой чашке (количество живых клеток), и не прикрепленные клетки в суспензионном состоянии (количество мертвых клеток). Результаты изображены на фиг. 3 (см. фиг. 3A для условий (1) и (2), а также см. фиг. 3B и 3C для условий (3) и (4) соответственно).
Как оказалось, выживаемость клеток в культуре клеточных масс в условиях отсутствия сыворотки была очевидно выше, чем максимальное значение для монослойной адгезионной культуры в присутствии сыворотки (приблизительно 60% в случае (2)), т.e. 99,2% выживаемости в случае (3) и 90,4% выживаемости в случае (4).
Пример 5: Получение Клеточных Масс с Использованием Кардиомиоцитов, полученных из Очищенных Эмбриональных Стволовых Клеток Мыши, и обнаружение Загрязнения Эмбриональными Стволовыми Клетками
Целью данного примера было обнаружение клеток, не являющихся кардиомиоцитами, которые загрязняли клеточные массы, образованные из очищенных кардиомиоцитов, полученных из эмбриональных стволовых клеток мыши.
Клеточные массы очищенных кардиомиоцитов получали способом согласно примерам 1 и 2, при условии, что перед последним высеванием на 96-луночный планшет к суспензии кардиомиоцитов добавляли 2% недифференцированных эмбриональных стволовых клеток. Клеточные массы культивировали в растворе среды α-MEM в отсутствие сыворотки, который содержал 1 мг/мл инсулина и 10 нМ TMRM; через 14 дней получали флуоресцентные изображения и фазово-контрастные изображения для всех лунок.
В результате в двух лунках, которые составляли примерно 2% всех лунок, наблюдали большее количество клеточных масс (более чем в два раза размера нормальных клеточных масс) (фиг. 4), и было обнаружено, что часть этих несферических клеточных масс образовала клеточную популяцию, которая испускала слабый выделяемый TMRM флуоресцентный сигнал (т.e. клетки, не являющиеся кардиомиоцитами) и которая состояла из аномальных кардиомиоцитов (фиг. 5B). Все клеточные массы нормальных кардиомиоцитов испускали выделяемый TMRM флуоресцентный сигнал (фиг. 5A).
Таким образом, способ, представленный в примере 5, дает возможность идентификации с высокой чувствительностью загрязнения клетками, не являющимися кардиомиоцитами.
Пример 6: Получение Клеточных Масс с Использованием Очищенных Кардиомиоцитов, полученных из Сердца Новорожденных Крыс
Целью данного примера было узнать, возможно ли получение клеточных масс с использованием очищенных кардиомиоцитов, полученных из сердца новорожденных крыс.
Новорожденных крыс через 0-2 дня после рождения анестизировали эфиром. Сердце удаляли и сердечную ткань диспергировали до отдельных клеток 0,1% коллагеназой (Wortington). Клетки окрашивали 10 нМ TMRM и затем обрабатывали FACS для очистки кардиомиоцитов.
Количество очищенных кардиомиоцитов подсчитывали и культивировали в 96-луночном планшете, не обладающем адгезионной способностью (SUMITOMO BAKELITE), высевая по 3000 клеток на лунку. Культуральная среда состояла из DMEM
с высоким содержанием глюкозы (Invitrogen) с добавлением 10% FBS (JRH).
Вид клеток через 24 часа культивирования изображено на фиг. 6. Очищенные кардиомиоциты, полученные из сердца новорожденной крысы, не образовывали клеточных масс, но выстраивались вдоль круглого дна каждой лунки (фиг. 6A). Очищенные кардиомиоциты, полученные из сердца новорожденной крысы, на 5 день культивирования фактически погибли (фиг. 6B) в условиях отсутствия сыворотки только в присутствии базальной культуральной среды α-MEM (левая панель фиг. 6B), или в присутствии α-MEM+ITS (правая панель фиг. 6B)).
Пример 7: Состав Культуральной Среды, оптимальный для Формирования Клеточных Масс с Использованием Кардиомиоцитов, полученных из Эмбриональных Стволовых Клеток Мыши
Целью данного примера было проанализировать разнообразные свойства первичных кардиомиоцитов новорожденных крыс и кардиомиоцитов, полученных из эмбриональных стволовых клеток мыши с тем, чтобы выявить культуральную среду, наиболее подходящую для кардиомиоцитов, полученных из эмбриональных стволовых клеток.
Очищенные кардиомиоциты, полученные из эмбриональных стволовых клеток мыши, и кардиомиоциты новорожденных крыс получали, как описано выше; затем их культивировали при каждом из 10 различных условий: одно соответствовало только α-MEM, и другие девять состояли из α-MEM+ITS, α-MEM+ITS+50 нг/мл bFGF (peprotech), α-MEM+ITS+50 нг/мл IGF-1 (Wako), α-MEM+ITS+50 нг/мл bFGF+50 нг/мл IGF-1, α-MEM+5% KSR (замещение сыворотки при нокауте: Invitrogen), α-MEM+10% KSR (замещение сыворотки при нокауте: Invitrogen), α-MEM+1% FBS (Equitech Bio),
α-MEM+5% FBS и α-MEM+10% FBS соответственно.
Когда очищенные кардиомиоциты, полученные из эмбриональных стволовых клеток мыши, культивировали в круглодонном 96-луночном планшете, не обладающем адгезионной способностью, клеточные массы образовывались во всех культуральных средах точно через 12 часов (фиг. 7A). Однако кардиомиоциты из сердца новорожденных крыс были не способны образовывать клеточные массы даже через 24 часа при всех условиях культивирования. Культуральная среда с добавлением 10% сыворотки приведена в качестве примера. Через 4 дня кардиомиоциты из сердца новорожденных крыс образовывали клеточные массы только в среде с добавлением 5% FBS и 10% FBS соответственно.
Наблюдали клеточные массы очищенных кардиомиоцитов, полученных из эмбриональных стволовых клеток мыши, которые образовывались через 6 дней культивирования в круглодонных 96-луночных планшетах, не обладающих адгезионной способностью (фиг. 7B); значительное увеличение диаметра клеточных масс по сравнению с диаметром только что образовавшихся клеточных масс было обнаружено только в культуральной среде α-MEM+ITS+50 нг/мл bFGF (см. фиг. 7C, конкретно, пожалуйста, обращайтесь к колонке, отмеченной звездочкой). Даже в среде, содержащей сыворотку, диаметр клеточных масс составлял примерно половину значения для ранних стадий (фиг. 7C).
На основе вышесказанного предполагается, что культуральная среда, не содержащая сыворотки, с добавлением ITS и bFGF обладает очень сильной функцией защиты клеток и демонстрирует уникальное свойство индукции пролиферации кардиомиоцитов.
Пример 8: Состав Культуральной Среды, Оптимальный для Образования Клеточных Масс с Использованием Кардиомиоцитов, полученных из Эмбриональных Стволовых Клеток Мыши
В примере 7 было обнаружено, что культуральная среда, не содержащая сыворотки, с добавлением ITS и bFGF обладала очень сильной функцией защиты клеток и демонстрировала уникальное свойство индукции пролиферации кардиомиоцитов. Следовательно, эксперимент примера 8 осуществляли с целью продемонстрировать, какое влияние оказывают bFGF и инсулин, компонент раствора ITS, на увеличение диаметра клеточных масс.
По существу, эксперименты проводили, как в примере 7, за исключением того, что использовали следующие культуральные среды: индивидуальная α-MEM; α-MEM+50 нг/мл bFGF; α-MEM+10 мкг/мл инсулина +5 нг/мл bFGF; и α-MEM+1 мкг/мл инсулина+1 нг/мл bFGF.
Через шесть дней наблюдали клеточные массы кардиомиоцитов, полученных из эмбриональных стволовых клеток мыши; в результате увидели значительное увеличение диаметра клеточных масс в каждой из сред α-MEM+50 нг/мл bFGF, α-MEM+10 мкг/мл инсулина+5 нг/мл bFGF и α-MEM+1 мкг/мл инсулина+1 нг/мл bFGF по сравнению с клеточными массами в культуральной среде, состоящей из индивидуальной α-MEM (фиг. 7B). Кроме того, было обнаружено сильное влияние на сократительную активность кардиомиоцитов, и было обнаружено такое же влияние, какое достигалось при добавлении ITS в примере 7.
На основе вышесказанного предположили, что культуральная среда, не содержащая сыворотки, с добавлением bFGF или инсулин+bFGF обладала очень сильной функцией защиты клетки и демонстрировала уникальное свойство индукции пролиферации кардиомиоцитов.
Пример 9: Влияние "Отсутствия Сыворотки" и bFGF в Монослойной Адгезионной Системе Культивирования Очищенных Кардиомиоцитов, полученных из Эмбриональных Стволовых Клеток Мыши
Кардиомиоциты, полученные из эмбриональных стволовых клеток мыши, очищали согласно примеру 1. Очищенные кардиомиоциты высевали на такое же количество покрытых фибронектином культуральных чашек и подвергали их культивированию в условиях монослойной адгезионной культуры в разнообразных культуральных средах. Все разнообразные культуральные среды включали α-MEM в качестве базальной культуральной среды, но они содержали соответственно следующие добавленные в нее компоненты: только 10% FBS, 10% FBS+50 нг/мл bFGF, 10% KSR (замещение сыворотки при нокауте: Invitrogen), ITS, и ITS+50 нг/мл bFGF. Клетки, посеянные в этих условиях, культивировали общим сроком в течение 5 дней и затем фотографировали (фиг. 8A). Как оказалось, группы, соответствующие условию отсутствия сыворотки + ITS, и группы, соответствующие условию ITS+bFGF, обладали значительно меньшей выживаемостью клеток, чем клетки в группах, соответствующих условиям добавления 10% FBS или 10% KSR.
Пример 10: Трансплантация Очищенных Кардиомиоцитов, полученных из Эмбриональных Стволовых Клеток Мыши, в Миокардиальную Ткань Иммунодефицитной Крысы и Измерение Степени Их Приживления
Во-первых, проводили следующий эксперимент с целью измерения степени выживаемости очищенных кардиомиоцитов, полученных из эмбриональных стволовых клеток мыши, в случае, где не применяли способ повторной агрегации.
Всего трансплантировали 2×105 клеток в левую вентрикулярную свободную стенку иммунодефицитной мыши (NOD-SCID). Индуцировали анестезию у мыши и поддерживали с использованием воздуха, содержащего 2% изофлурана, подаваемого через искусственный респиратор. Мышь подвергали торакотомии (в третьем межреберном пространстве) под глубокой анестезией, и для воздействия на сердце разрывали сердечную сумку пинцетом. Инъецировали физиологический солевой раствор (30 мкл), содержащий клеточные массы кардиомиоцитов, посредством шприца с иглой 30G. Для инъекции иглу вставляли в верхнюю часть сердца, из которой иглу продвигали сквозь свободную сердечную стенку приблизительно на 3 мм к основанию сердца. После трансплантации грудную клетку закрывали быстро и после возвращения самопроизвольного биения сердца мышь возвращали в клетку.
Через три недели после трансплантации сердце фиксировали в процессе перфузии и получали замороженные секции. Секции подвергали иммуноокрашиванию антителом к саркомерному актинину и получали изображения с помощью флуоресцентного микроскопа (фиг. 9A). Как оказалось, живые клеточные трансплантаты были едва видны и можно было обнаружить только реакцию с маркерным красным красителем (фиг. 9A и 9D).
Затем каждую из клеточных масс, состоящих из 2000 кардиомиоцитов, как они были созданы в условиях отсутствия сыворотки, описанных в примере 7 (общим количеством клеток 2×105), трансплантировали в левую вентрикулярную свободную стенку иммунодефицитной мыши (NOD-SCID). Трансплантацию проводили, как в эксперименте, описанном выше, и через 3 недели сердце фиксировали в процессе перфузии и получали замороженные секции. Секции подвергали иммуноокрашиванию антителом к саркомерному актинину и получали изображения с помощью флуоресцентного микроскопа (фиг. 10). На основе изображений, полученных флуоресцентным микроскопом, подсчитывали количество клеток, приживленных на ткани сердца-хозяина, содержащихся в одной клеточной массе.
Как оказалось, допуская, что каждая из трансплантированных клеточных масс точно состояла из 2000 кардиомиоцитов, было обнаружено приживленными 92,05±11,1% кардиомиоцитов (n=4) (фиг. 11A и 11B). Напротив, когда инъецировали очищенные отдельные клетки как таковые (как диспергированные), не могли обнаружить приживленных клеток. Этот результат обозначает, что степень пост-трансплантационного приживления кардиомиоцитов кардинально повышается от 0% до 92%.
Далее, с целью подтверждения изменения в кардиомиоцитах во время продолжительной трансплантации осуществляли исследование на основе иммуноокрашивания сердца через 3 и 8 недель после трансплантации. Как оказалось, объем цитоплазмы кардиомиоцитов заметно увеличивался через 3 и 8 недель после трансплантации по сравнению с кардиомиоцитами перед трансплантацией ("Перед трансплантацией" на фиг. 11C) и к тому же трансплантированные кардиомиоциты выстраивались в том же направлении, что и кардиомиоциты хозяина (фиг. 11C и 11D). Это демонстрирует, что трансплантированные клеточные массы кардиомиоцитов созревают в ткани сердца-хозяина.
Пример 11: Получение Клеточных Масс с использованием Очищенных кардиомиоцитов, полученных из Эмбриональных Стволовых Клеток Мармозетки
Целью данного примера было получение клеточных масс очищенных кардиомиоцитов, полученных из эмбриональных стволовых клеток мармозетки, в условиях отсутствия сыворотки или в условиях добавления ITS или KSR.
Коротко, клеточные массы очищенных кардиомиоцитов, полученных из эмбриональных стволовых клеток мармозетки, получали согласно примеру 3, при условии, что клеточные массы культивировали в культуральной среде, не содержащей сыворотку (фиг. 12A), или в культуральной среде, не содержащей сыворотку, с добавлением 10% KSR (фиг. 12B) или ITS (фиг. 12C). Клеточные массы, созданные через 12 часов или через 3 дня после момента трансплантации, продемонстрированы на фиг. 12.
Пример 12: Создание "Толстых" Пластов Клеток с Использованием Клеточных Масс Очищенных Кардиомиоцитов, полученных из Эмбриональных Стволовых Клеток Мармозетки
Клеточные массы очищенных кардиомиоцитов, полученных из эмбриональных стволовых клеток мармозетки, которые были получены в примере 11, культивировали в условиях суспензионной культуры в том же планшете. По прошествии периода времени, составляющего от 0 до 12 часов, соседние клеточные массы конъюгировали вместе с образованием "толстого" пласта клеток кардиомиоцитов. Пример 12 описывает модельный эксперимент, предназначенный для демонстрации применимости способа по настоящему изобретению. В реальном применении вариантов осуществления изобретения имеющие желаемый размер клеточные массы очищенных кардиомиоцитов, полученных из эмбриональных стволовых клеток, могут использоваться в желаемом количестве для создания имеющего желаемый размер пласта клеток (фиг. 13). Кроме того, имеющий желаемую толщину пласт клеток может быть образован в зависимости от размера используемых клеточных масс.
Пример 13: Трансплантация Кардиомиоцитов, полученных из Эмбриональных Стволовых Клеток Человека, в Сердце Иммунодефицитной Мыши
В этом примере эксперименты проводили для определения, имеют ли клеточные массы кардиомиоцитов, которые были получены дифференцировкой эмбриональных стволовых клеток человека в кардиомиоциты, способность приживляться в сердечной ткани.
Эмбриональные стволовые клетки человека получали из Центра Исследований Стволовых Клеток, принадлежащего учреждению Института Передовых Медицинских Наук, Киотского Университета (Центр Эмбриональных Стволовых Клеток финансировался согласно Проекту Национальных Биоресурсов).
С использованием эмбриональных фибробластов мыши (MEF), рост которых инактивировали обработкой митамицином C, эти эмбриональные стволовые клетки человека культивировали так, чтобы они оставались недифференцированными. Культуральная среда состояла из F12/DMEM (1:1) (SIGMA, Лот No. D6421), 20% KO-СЫВОРОТКИ (GIBCO), 1,6 мМ L-глутамина, 0,1 мМ заменимых аминокислот (MEM), 0,1 мМ β-меркаптоэтанола (2-ME; Sigma), 100 МЕ/мл пенициллина, 100 мкг/мл стрептомицин сульфата и рекомбинантного человеческого основного фактора роста фибробластов (bFGF; Peprotech). Для последовательного пассажа колонии эмбриональных стволовых клеток разделяли путем обработки 0,1% коллагеназой типа III (Wortington) при 37°C в течение 10 минут.
Затем с целью отделения эмбриональных стволовых клеток от MEF культуральную среду, содержащую клеточные массы, пропускали сквозь фильтр с размером пор, составляющим 40 мкм, и извлекали клеточные массы из фракции большего размера. Извлеченные клеточные массы представляли собой клеточные массы чистых эмбриональных стволовых клеток. Для дифференцировки культивировали как эмбриоидные тела по 50-1000 эмбриональных стволовых клеток на ЭТ на бактериальных чашках, не обладающих адгезионной способностью (Asahi Techno Glass; стерильная чашка Петри) общим сроком в течение 15-30 дней так, чтобы они дифференцировались в эмбриоидные тела, включающие кардиомиоциты. Культуральная среда, использованная для этой дифференцировки, представляла собой ту же среду, что идентифицирована выше, за исключением того, что она не содержала bFGF, т.e. среда состояла из F12/DMEM (1:1) (SIGMA, Лот No. D6421), 20% KO-СЫВОРОТКИ (GIBCO), 1,6 мМ L-глутамина, 0,1 мМ заменимых аминокислот (MEM), 0,1 мМ β-меркаптоэтанола (2-ME; Sigma), 100 МЕ/мл пенициллина, 100 мкг/мл стрептомицин сульфата.
Кардиомиоциты, полученные из эмбриональных стволовых клеток человека, очищали согласно примеру 1. Затем, согласно результатам примера 8, клеточные массы, каждая из которых содержала 1000 очищенных кардиомиоцитов, получали с использованием раствора среды α-MEM, не содержащей сыворотки, который содержал 1 мкг/мл инсулина и 1 нг/мл bFGF (см. фиг. 14).
Затем клеточные массы трансплантировали в сердечную ткань иммунодефицитной мыши согласно примеру 10. Через две недели после трансплантации получали замороженные секции согласно примеру 10. Полученные таким образом секции подвергали иммуноокрашиванию с помощью Nkx2.5 и антитела к саркомерному актинину и получали изображения с помощью флуоресцентного микроскопа.
Было обнаружено, что некоторые клеточные массы окрашиваются красным красителем, использованным в качестве маркера трансплантированных клеток. Секции детектировали на наличие Nkx2.5 и антитела к саркомерному актинину иммунологическим способом. Результат продемонстрирован на фиг. 15. Было продемонстрировано, что трансплантированные клетки, при окрашивании красным маркером красителя, являются преимущественно красного цвета. Иммуноокрашивание актинином также продемонстрировало окрашивание в виде полос. Также продемонстрировано, что Nkx2.5, маркер кардиомиоцитов, был обнаружен преимущественно в ядрах трансплантированных кардиомиоцитов. Это явление представляется характерным для незрелых кардиомиоцитов. Кроме того, ядра кардиомиоцитов, полученных из эмбриональных стволовых клеток человека, крупнее, чем ядра окружающих кардиомиоцитов мыши, и, следовательно, могут быть различимы от последних (фиг. 15).
Затем через 5 недель после трансплантации замороженные секции получали согласно примеру 10. Было обнаружено, что некоторые клеточные массы окрашивались красным красителем, использованным в качестве маркера трансплантированных клеток (фиг. 16). Полученные таким образом секции подвергали иммуноокрашиванию Nkx2.5 (водянисто-голубой цвет) и антителом к саркомерному актинину (зеленый цвет) или Nkx2.5 (водянисто-голубой цвет) и ядерным антигеном к антителу человека (зеленый: связывание с антигеном, который не присутствует в мышином ядре, но присутствует только у приматов) и получали изображения с помощью флуоресцентного микроскопа. Результаты продемонстрированы на фиг. 17 и 18. Митохондрии трансплантированных клеток окрашивались маркерным красителем, давая возможность детектирования красного красителя в виде пятен. Иммуноокрашивание актинином также продемонстрировало окрашивание в виде полос (фиг. 17). Чтобы продемонстрировать, что эти популяции клеток, составляющие одну и ту же область, выделены из клеток человека, осуществляли детектирование с использованием антитела человека, и снова было продемонстрировано, что трансплантированные клетки представляют собой клетки человека, конкретно кардиомиоциты, которые окрашивали вместе с Nkx2.5 (фиг. 18).
Пример 14: Получение Клеточных Масс с Использованием Очищенных Кардиомиоцитов, полученных из Индуцированных Плюрипотентных Стволовых (iPS) Клеток Мыши
Целью данного примера было получение клеточных масс очищенных кардиомиоцитов, полученных из клеток iPS мыши в условиях отсутствия сыворотки с добавлением или без добавления ITS или KSR.
Клетки iPS мыши получали из Института Передовых Медицинских Наук Киотского Университета. Дифференцировку клеток iPS мыши проводили, как в примере 1. В том примере было обнаружено, что оптимальное количество исходных клеток для образования одного эмбриоидного тела составляет 1000 клеток.
На фиг. 19A продемонстрирован результат FACS-анализа, который проводили с митохондриальным индикатором TMRM с целью очистки кардиомиоцитов. Прямоугольник в диаграмме представляет область кардиомиоцитов. Полученные таким образом кардиомиоциты подвергали культивированию в условиях адгезионной культуры и подвергали иммуноокрашиванию (с актинином и Nkx2.5); результат продемонстрирован на фиг. 19B. Так как было продемонстрировано, что кардиомиоциты, индуцированные клетками iPS мыши, образуют агрегированные клеточные массы, то было обнаружено, что клеточные фракции, извлеченные с помощью FACS-анализа, состояли почти из 100% кардиомиоцитов. Затем на фиг. 19C продемонстрирован вид очищенных кардиомиоцитов через 24 часа после того, как их высевали на 96-луночный культуральный планшет, не обладающий адгезионной способностью. В этом случае использовали культуральную среду, не содержащую сыворотки. Как оказалось, очищенные кардиомиоциты, полученные из клеток iPS мыши, также могут использоваться для конструирования клеточных масс способом по настоящему изобретению.
Пример 15: Состав Культуральной Среды, Оптимальный для Образования Клеточных Масс с Использованием Кардиомиоцитов, полученных из Эмбриональных Стволовых Клеток Человека
В примере 7 было обнаружено, что культуральная среда, не содержащая сыворотки, с добавлением ITS и bFGF обладает очень сильной защитной функцией на клетки, полученные у мышей и демонстрирует уникальное свойство индукции пролиферации кардиомиоцитов. Следовательно, пример 15 проводили с целью продемонстрировать эффективность bFGF в кардиомиоцитах, полученных из клеток ES человека, и для более тщательной проверки его эффективности путем сравнения с другими факторами роста.
По существу, среду α-MEM+ITS использовали в качестве культуральной среды. В эту базальную культуральную среду добавляли 25 нг/мл bFGF (Peprotech, Inc., Rocky Hill, NJ, USA), 25 нг/мл кислого FGF (aFGF), 25 нг/мл FGF-4, 20 нг/мл фактора роста кератиноцитов (KGF), 100 нг/мл фактора стволовых клеток (SCF), 100 нг/мл васкулярного эндотелиального фактора роста (VEGF), 10 нг/мл фактора ингибирования лейкоза (LIF) (Millipore Corporation, Billerica, MA, USA), 100 нг/мл нейротрофического фактора, полученного из линии глиальных клеток (GDNF), 20 нг/мл фактора роста гепатоцитов (HGF), 10 нг/мл инсулиноподобного фактора роста (IGF)-1, 100 нг/мл эпидермального фактора роста (EGF), 1×10-7 M эндотелина-1 (ET-1), 10 нг/мл фактора роста тромбоцитов (PDGF)-AA или 100 нг/мл PDGF-BB (все реагенты, для которых не указано, где они были получены, были приобретены у R&D systems). Клетки ES человека культивировали с использованием каждой из культуральных сред с получением клеточных агрегатов.
Диаметр клеточных масс измеряли через 3, 8, 25 и 40 дней после их получения. Как оказалось, клеточные массы, полученные в присутствии bFGF, имели наибольший диаметр в каждый из дней (фиг. 20A). Также было обнаружено, что эта защитная функция продолжалась до 40 дней. Вместо bFGF в культуральную среду добавляли каждое из вышеуказанных разнообразных веществ и контролировали их эффект на образование клеточных масс; было обнаружено, что EGF, PDFG-BB и ET-1 эффективны, но не настолько эффективны, как bFGF (фиг. 20B).
В результате, как оказалось, даже в случае дифференцировки кардиомиоцитов, полученных из клеток ES человека, bFGF обладал активностями защиты клеток и стимулирования роста в условиях отсутствия сыворотки и что эти активности более сильные, чем соответствующие активности других факторов роста.
Затем с целью выяснения механизма, с помощью которого кардиомиоциты, трансплантированные в сердце-хозяин, могут созревать в сердечной ткани после трансплантации, bFGF, EGF, PDGF-BB и ET-1 тестировали с помощью ПЦР в реальном времени (Aplied Biosystems) на предмет возможности экспрессии генов в сердце-хозяине. Праймеры и пробы для соответствующих генов были приобретены у Applied Biosystems (TaqMan-анализ экспрессии генов); более конкретно, использовали bFGF (Mm0128715_m1), EGF (Mm01316967_m1), PDGF-BB (Mm01298577_m1) и ET-1 (Mm01351840_g1). Реагенты, использованные для анализа и рабочей процедуры, соответствовали руководству по применению, предоставленному Applied Biosystems. В результате, как оказалось, вышеуказанные гены экспрессировались в сердце-хозяине (фиг. 21).
На основе результатов примера 15 предположили, что группа факторов роста, требующихся для выживаемости и созревания клеточных масс кардиомиоцитов, трансплантированных в сердце, обеспечивается за счет сердца-хозяина.
ПРОМЫШЛЕННАЯ ПРИМЕНИМОСТЬ
По настоящему изобретению было обнаружено, что кардиомиоциты, полученные из эмбриональных стволовых клеток, которые были очищены путем диспергирования до отдельных клеток, обладают такими новыми характеристиками, что они способны агрегировать при культивировании в условиях отсутствия сыворотки. С помощью создания клеточных масс с использованием способа по настоящему изобретению может осуществляться продолжительное культивирование, причем степень выживаемости или способность пролиферации этих кардиомиоцитов поддерживается на высоком уровне. Кроме того, было обнаружено, что, когда эти клетки трансплантируют в сердечную ткань индивидуума (живого организма), степень их приживления в сердечной ткани значительно повышается, в результате чего кардиомиоциты не смешиваются с клетками, не являющимися кардиомиоцитами, но смогут оставаться приживленными внутри сердечной ткани в течение продолжительного периода времени. Таким образом, с помощью настоящего изобретения повысилась возможность реализации предоставления кардиомиоцитов для трансплантации, а также возможность реализации способа клеточной терапии на сердце, который является альтернативой трансплантации сердца, в качестве лечения болезней сердца путем трансплантации кардиомиоцитов, которые были получены вне живого организма, и возможность реализации медицинского устройства, включающего клеточные массы кардиомиоцитов.
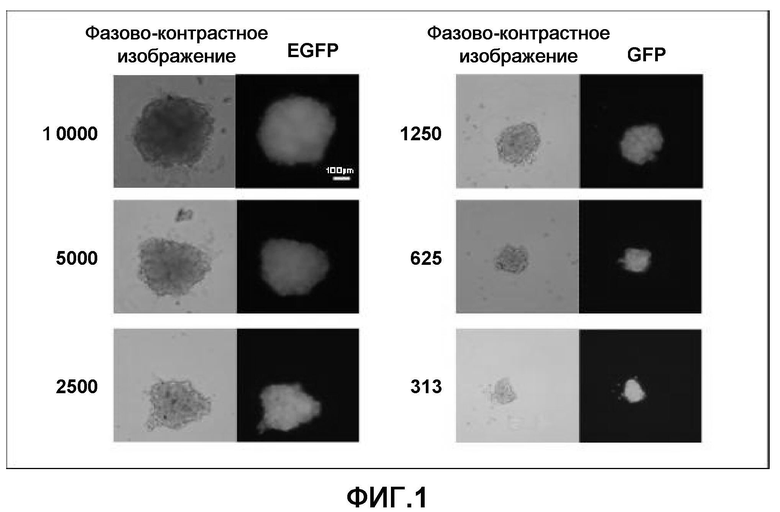
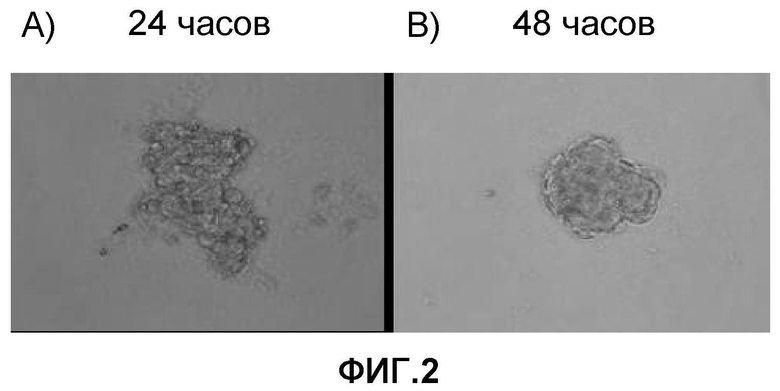
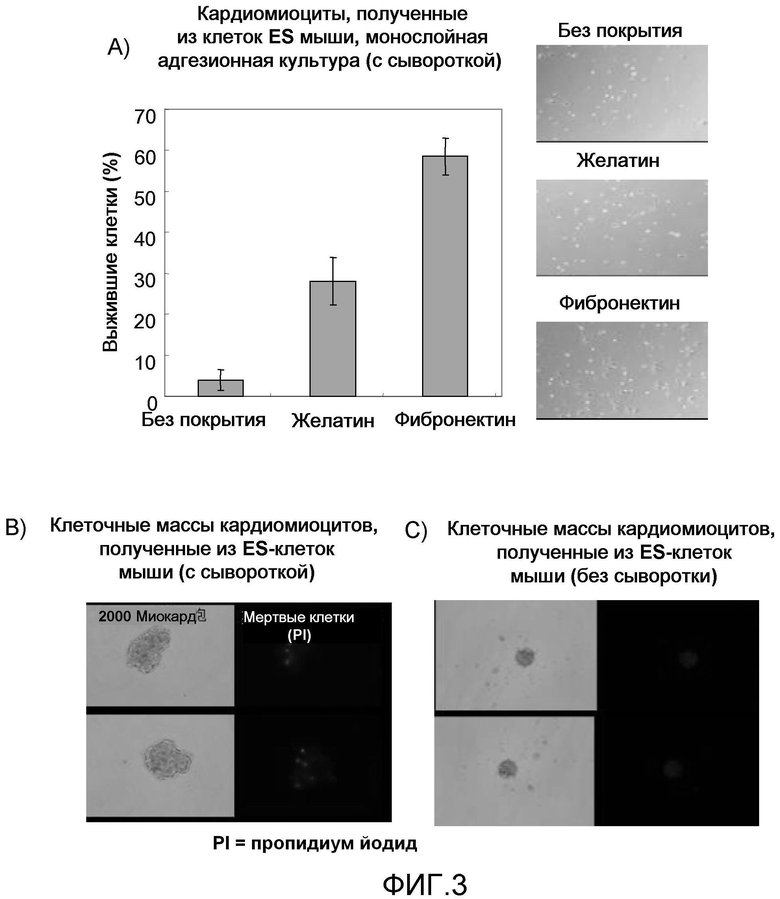


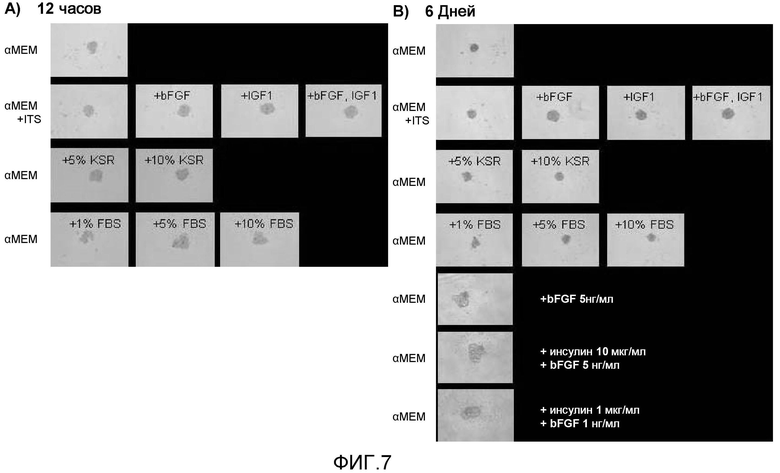
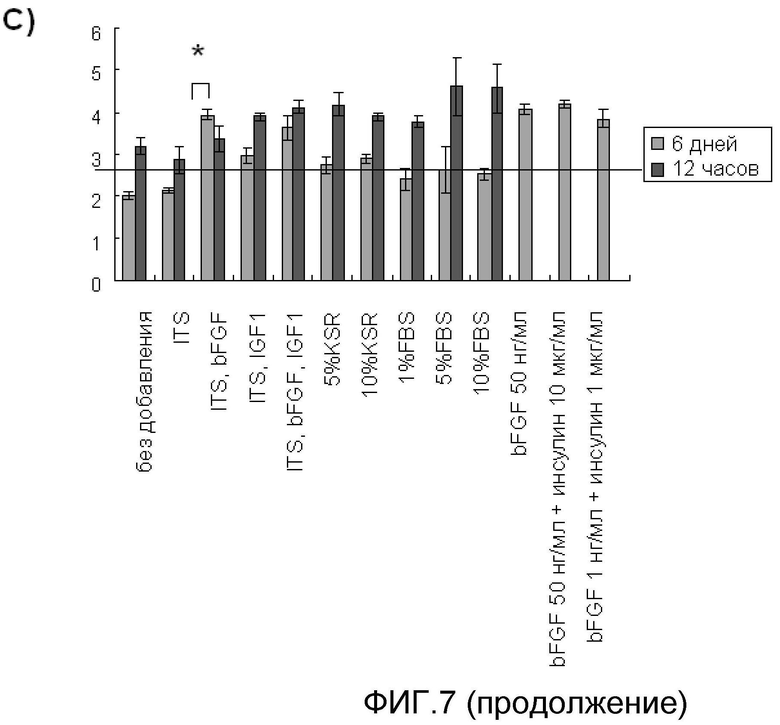
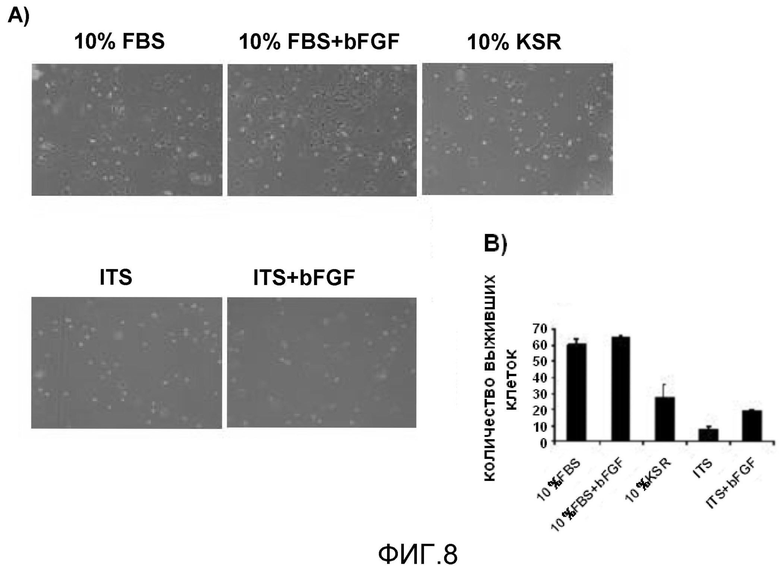
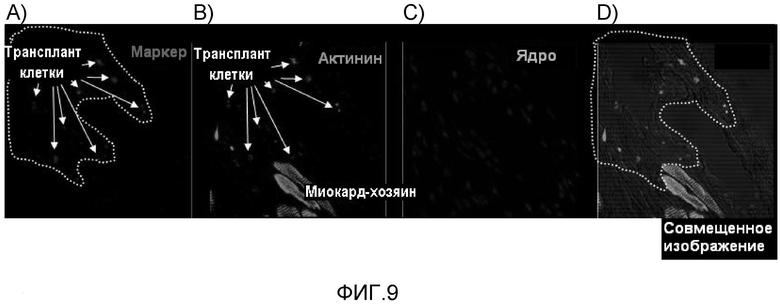
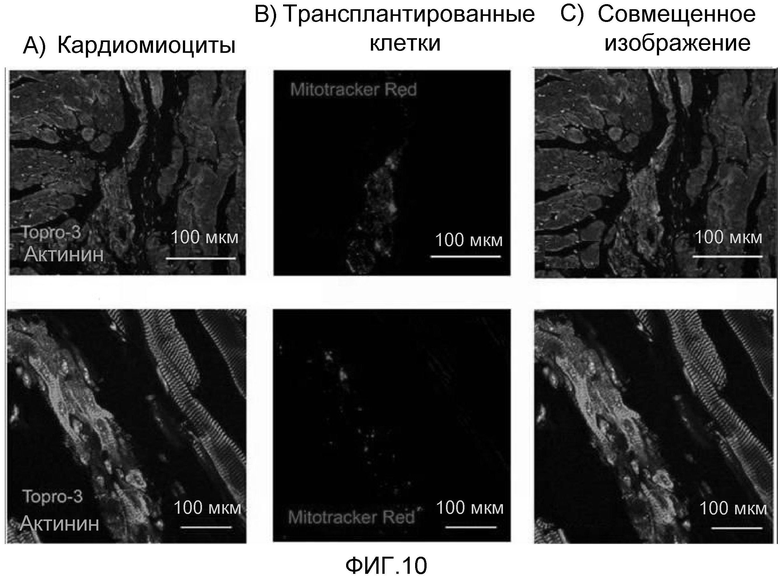
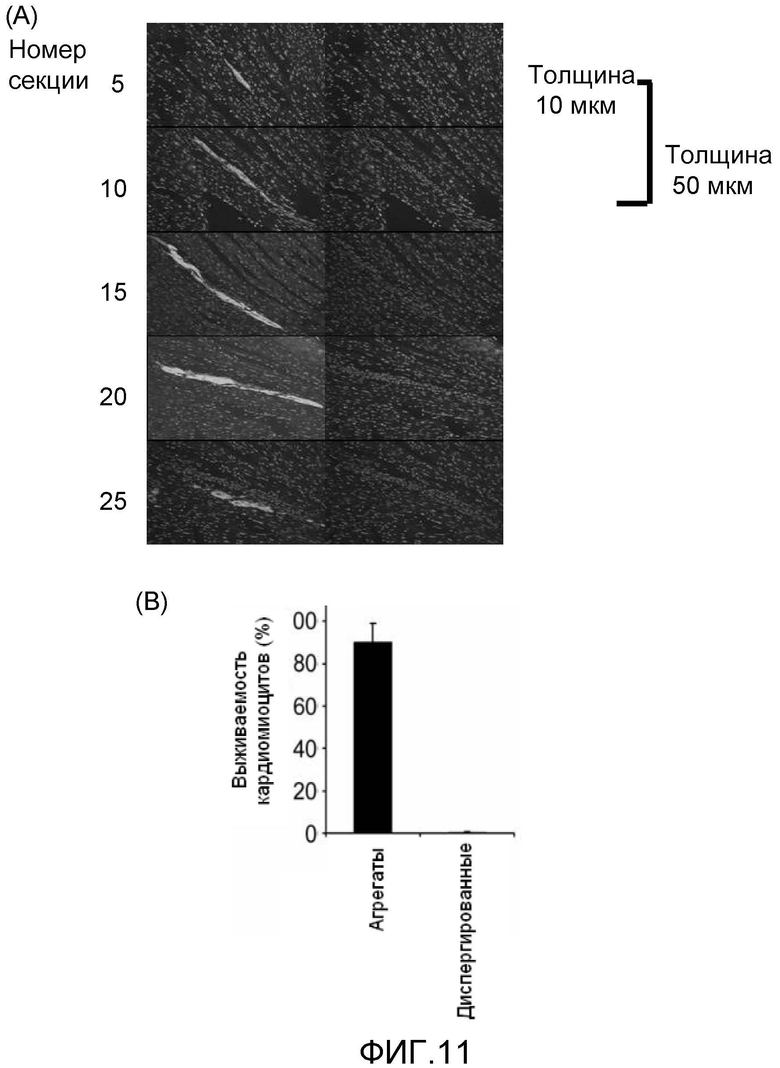

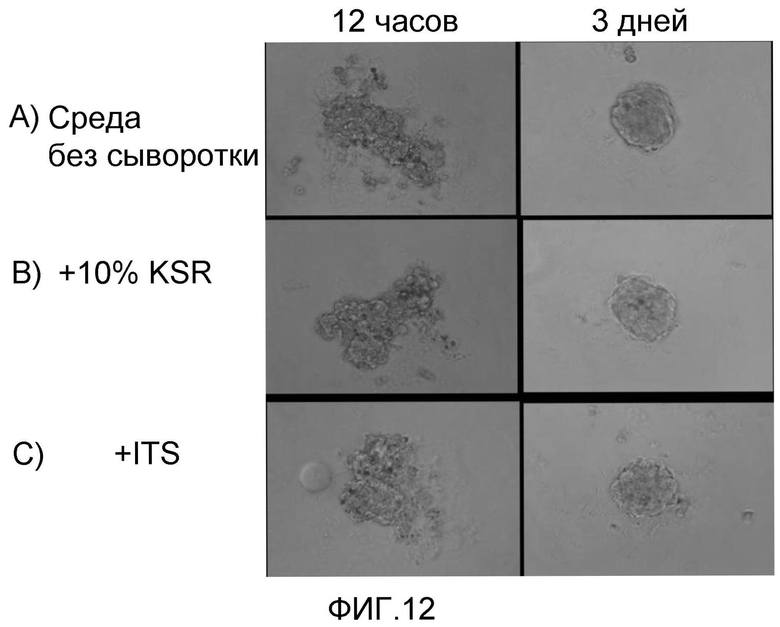
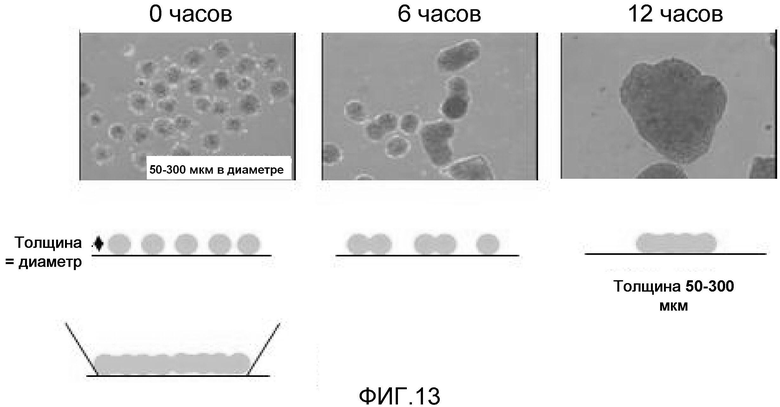
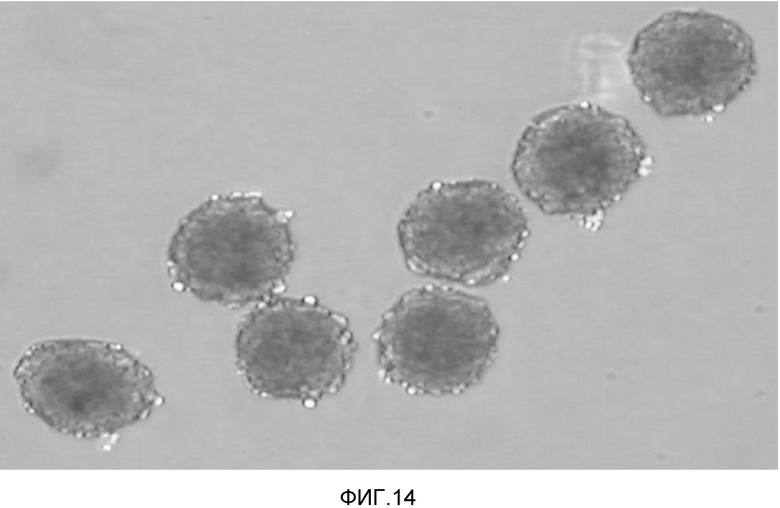
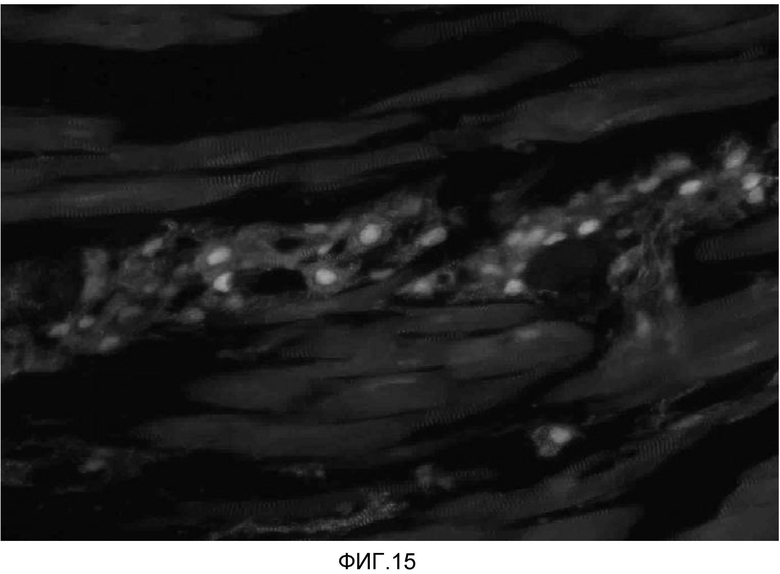
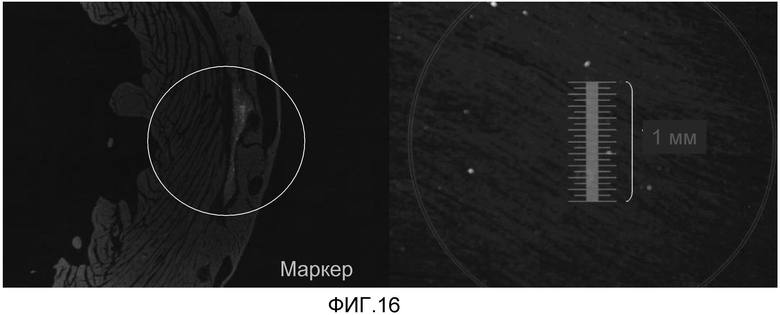
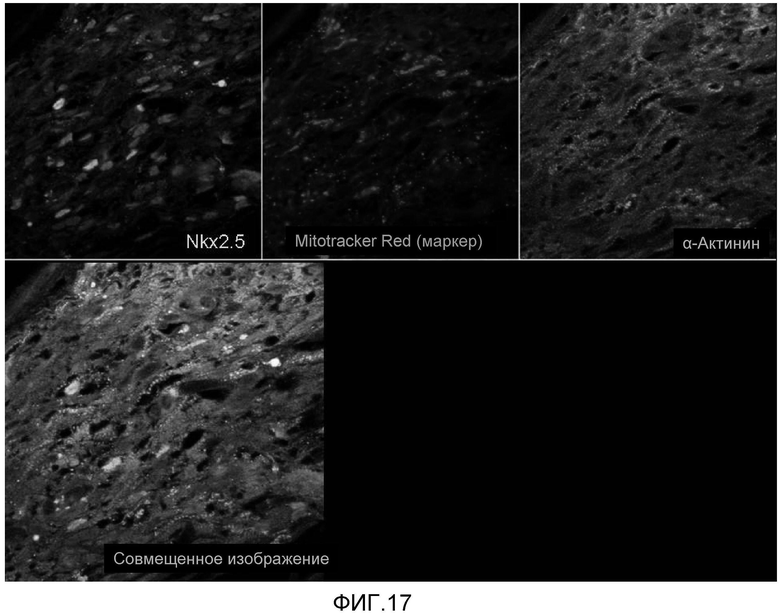
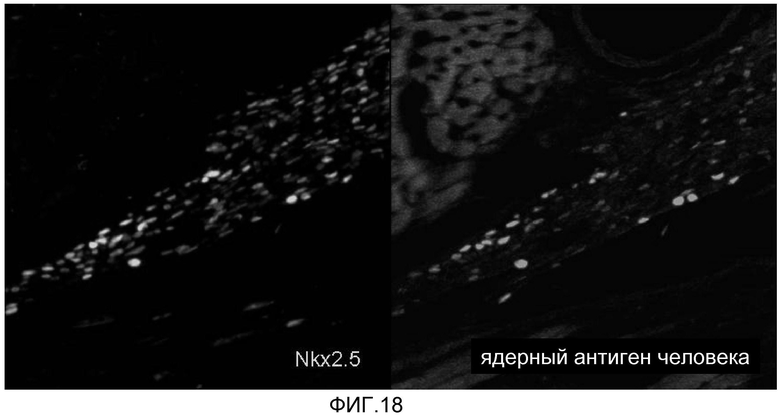
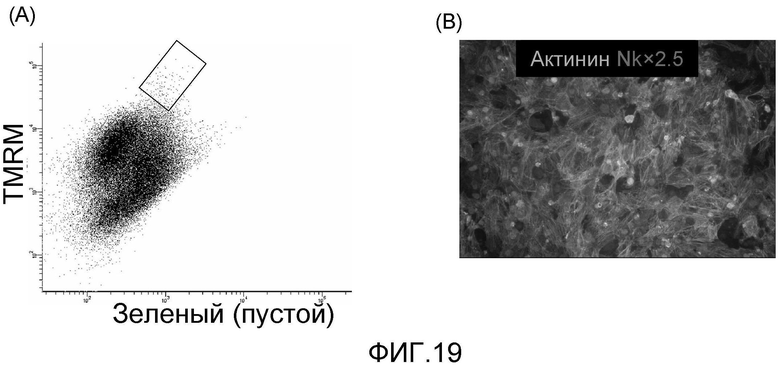
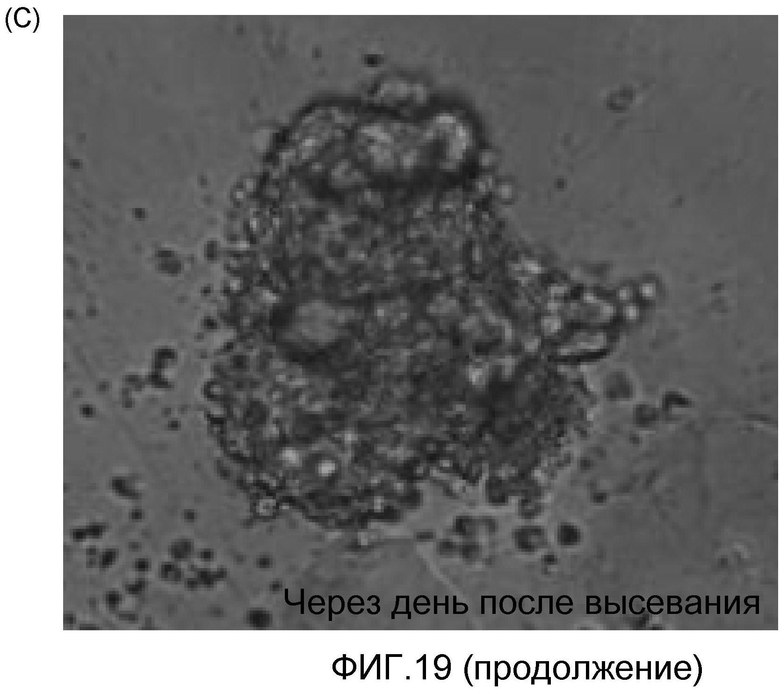
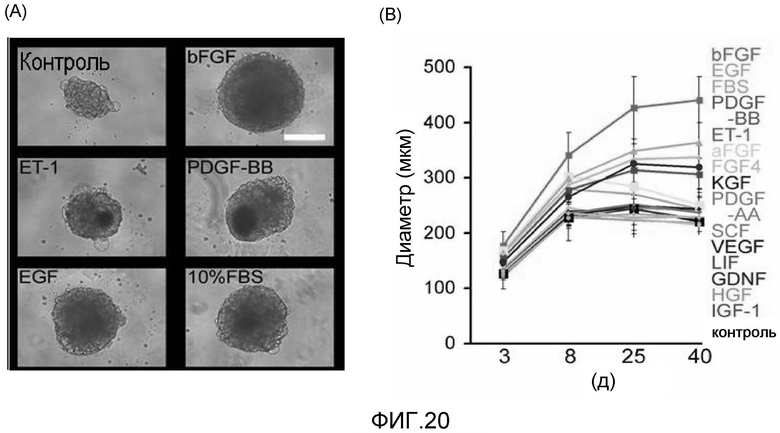

Изобретение относится к области биотехнологии, клеточной технологии, медицине и трансплантологии. Предложен способ получения клеточных масс кардиомиоцитов, получаемых из плюрипотентных стволовых клеток. Способ предусматривает дифференцировку и индукцию плюрипотетных стволовых клеток с получением клеточных масс кардиомиоцитов. Далее проводят диспергирование клеточных масс до отдельных клеток и их культивирование в культуральной среде в условиях отсутствия сыворотки для их повторной агрегации. Способ позволяет получать кардиомиоциты высокой степени очистки, свободных от клеток, не являющихся кардиомиоцитами, и от любых компонентов, полученных из другого вида, которые обладают повышенной пост-трансплантационной приживляемостью. 2 н. и 9 з.п. ф-лы, 34 ил., 15 пр.
1. Способ получения клеточных масс кардиомиоцитов, полученных из плюрипотентных стволовых клеток, где эмбриональные стволовые клетки человека получены из зарегистрированных линий эмбриональных стволовых клеток человека, отличающийся тем, что очищенные кардиомиоциты, полученные путем диспергирования до отдельных клеток агрегированных клеточных масс, которые получены в результате дифференцировки и индукции из плюрипотентных стволовых клеток, культивируют в культуральной среде в условиях отсутствия сыворотки так, что они повторно агрегируют способом висячей капли или с использованием неадгезивного для клеток круглодонного луночного планшета.
2. Способ по п.1, где плюрипотентные стволовые клетки выбраны из группы, состоящей из эмбриональных стволовых клеток, эмбриональных зародышевых клеток, зародышевых стволовых клеток и индуцированных плюрипотентных стволовых клеток.
3. Способ по п.1, где культуральная среда содержит инсулин.
4. Способ по любому из пп.1-3, где культуральная среда содержит по меньшей мере одно вещество, выбранное из группы, состоящей из трансферрина, селена, основного фактора роста фибробластов (bFGF), фактора роста эпителиальных клеток (EGF), фактора роста тромбоцитов- ВВ (PDGF-BB) и эндотелина-1 (ЕТ-1).
5. Способ по любому из пп.1-3, где культуральная среда содержит 0,1-10 мг/л инсулина, 0,1-10 мкг/л трансферрина, 0,1-10 мкг/л селена, 1 нг/мл - 100 нг/мл основного фактора роста фибробластов, 1 нг/мл - 1000 нг/мл фактора роста эпителиальных клеток, 1 нг/мл - 1000 нг/мл фактора роста тромбоцитов, и 1·10-8-1·10-6 М эндотелина-1 (ЕТ-1).
6. Способ лечения болезней сердца, при которых необходима трансплантация кардиомиоцитов, отличающийся тем, что клеточные массы, полученные способом по любому из пп.1-5, трансплантируют в сердечную ткань индивидуума, так чтобы они приживились.
7. Способ по п.6, где плюрипотентные стволовые клетки выбраны из группы, состоящей из эмбриональных стволовых клеток, эмбриональных зародышевых клеток, зародышевых стволовых клеток и индуцированных плюрипотентных стволовых клеток.
8. Способ по любому из пп.6 или 7, где трансплантация включает инъекцию клеточных масс кардиомиоцитов в сердечную ткань.
9. Способ по любому из пп.6 или 7, где трансплантация включает трансплантацию пласта клеточных масс кардиомиоцитов на сердечную ткань.
10. Способ получения пласта клеточных масс кардиомиоцитов, отличающийся тем, что клеточные массы очищенных кардиомиоцитов, полученных способом по любому из пп.1-5, высевают на разделенную стенками культуральную посуду, не обладающую способностью к адгезии клеток, в отсутствие пространства между клеточными массами, так чтобы соседние клеточные массы находились в постоянном контакте друг с другом, с последующим поддержанием суспензионной культуры, которая поддерживается до конъюгирования клеточных масс, с получением желаемой толщины, составляющей 50-300 мкм.
11. Способ по п.10, где плюрипотентные стволовые клетки выбраны из группы, состоящей из эмбриональных стволовых клеток, эмбриональных зародышевых клеток, зародышевых стволовых клеток и индуцированных плюрипотентных стволовых клеток.
| WO 2005090558 А1, 29.09.2005 | |||
| SHIMIZU Т | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| RES | |||
| Пожарный двухцилиндровый насос | 0 |
|
SU90A1 |
| GUO X.M | |||
| et al | |||
| "Creation of engineered cardiac tissue in vitro from mouse embryonic stem cells", Circulation | |||
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |

