Область техники
Данное изобретение относится к области медицины и вирусологии, в частности, к аттенуированному химерному гриппозному вектору и его применению для профилактики гриппа в качестве мукозальной универсальной гриппозной вакцины.
Уровень техники
Из всех респираторных вирусных заболеваний гриппозная инфекция характеризуется наиболее тяжелой патологией и причиняет наибольший ущерб здоровью населения и экономике. Периодически появляющиеся новые пандемические штаммы, к которым отсутствует популяционный иммунитет, превращают грипп в особо опасную инфекцию. Известно, что испанский грипп 1918 года стал причиной смерти от 30 до 50 млн человек. В настоящее время, по данным Всемирной организации здравоохранения (ВОЗ), во время сезонных эпидемий ежегодно во всем мире заболевает гриппом до 20% населения, в том числе 5-10% взрослых и 20-30% детей (World Health Organization [Электронный ресурс]. URL: http://www.who.int/biologicals/vaccines/influenza/en/ (дата обращения 28.03.2016)). Тяжелые формы отмечаются в 3-5 млн. случаев, летальные исходы составляют от 250 000 до 500 000 случаев. Экономические потери, вызванные гриппом и другими острыми респираторными вирусными инфекциями (ОРВИ), составляют около 77% от ущерба, приходящегося на долю всех инфекционных болезней. Значительные убытки связаны как с прямыми расходами на лечение и реабилитацию, так и с косвенными потерями производственного характера, вызванными снижением производительности труда и сокращением прибыли предприятий. Из общего числа случаев временной нетрудоспособности на грипп и ОРВИ приходится 12-14%.
Вирус гриппа относится к семейству Ortomyxoviridae, в которое входит пять родов: грипп A, B, C, D (Thogotovirus) и Isavirus. Геномы вирусов гриппа А и В структурно схожи, они состоят из 8 геномных сегментов РНК негативной полярности, которые кодируют 12 белков и названы по продукту, транслируемому с основной открытой рамки считывания: PB1, PB2, PA, HA, NP, NA, M и NS (Chou YY, Vafabakhsh R, Doğanay S, Gao Q, Ha T, Palese P. One influenza virus particle packages eight unique viral RNAs as shown by FISH analysis. Proc Natl Acad Sci U S A. 2012 Jun 5;109(23):9101-6. doi: 10.1073/pnas.1206069109. Epub 2012 Apr 30). Полимеразный комплекс PB2, PB1, PA транскрибирует одну мРНК с каждого геномного фрагмента, транслируемую в одноименный белок. Кроме того, мРНК геномных сегментов M и NS подвергается сплайсингу, кодируя белки M2 и NEP в дополнение к белкам М1 и NS1 соответственно. У отдельных штаммов с альтернативной рамки считывания сегмента РВ1 транслируется белок PB1-F2. Все белки, за исключением NS1 и PB1-F2, являются структурными компонентами вирусных частиц. Неструктурный белок NS1 накапливается в цитоплазме зараженных клеток и выполняет функцию ингибитора системы интерферона (Krug RM. Functions of the influenza A virus NS1 protein in antiviral defense. Curr Opin Virol. 2015 Jun;12:1-6. doi: 10.1016/j.coviro.2015.01.007. Epub 2015 Jan 29. Review).
Сегментированность генома вируса гриппа, высокий мутационный фон РНК-ового генома, а также наличие природного резервуара вирусных вариантов у птиц и животных служит неисчерпаемым источником новых штаммов, которые внедряются в человеческую популяцию. Антигенные свойства вируса гриппа определяются поверхностными гликопротеинами, образующими шипы на поверхности вирионов - гемагглютинином (НА) и нейраминидазой (NA). Молекула НА ответственна за механизмы прикрепление вируса к сиаловым рецепторам клетки и слияния вирусной и клеточной мембран для проникновения генома в цитоплазму и ядро клетки. В процессе репродукции вируса происходит расщепление (активация) НА клеточными протеазами на две субъединицы - НА1 и НА2, которые остаются соединенными дисульфидной связью (Bullough PA, Hughson FM, Skehel JJ, Wiley DC. Structure of influenza haemagglutinin at the pH of membrane fusion. Nature 1994; 371, 37-43.). Молекула НА состоит из двух частей: глобулярной, образованной субъединицей НА1, и стволовой, которая состоит в основном из субъединицы НА2 и частично НА1. Глобулярная часть включает рецептор-связывающий сайт и пять антигенных сайтов и является основной мишенью для образования антител. Антитела, блокирующие связывание вируса с рецептором, являются нейтрализующими. Для НА1 субъединицы характерна высокая изменчивость. Стволовая часть НА расположена в непосредственной близости от вирусной мембраны и отличается слабой иммуногенностью. Субъединица НА2 играет основную роль в обеспечении слияния вирусной мембраны с эндосомальной и отличается высокой консервативностью. В соответствии с антигенной специфичностью к настоящему моменту для вируса гриппа А известно 18 подтипов HA и 11 подтипов NA. Подтипы НА H1, H2, H5, H6, H8, H9, H11, H12, H13, H16, H17, H18 относят к первой группе, а H3, H4, H7, H10, H14 и H15 - ко второй группе.
Известно, что специфический иммунитет, вырабатываемый после перенесенного заболевания или вакцинации одним сероподтипом вируса гриппа А, слабо защищает от заражения другим сероподтипом. Иммунитет к любому сероподтипу вируса гриппа А не защищает от вируса гриппа В, и наоборот, иммунизация против гриппа В неэффективна в отношении вируса гриппа А.
Лучшим средством предотвращения распространения гриппозной инфекции считается вакцинация. Существующие вакцины можно разделить на два вида: аттенуированные (живые, содержащие цельные и активные вирусы, проявляющие низкую патогенность при интраназальном введении) и инактивированные (содержащие фрагменты вирусных частиц или цельные неактивные вирусы). Живые аттенуированные вирусы, способные к ограниченной репродукции в клетках респираторного тракта, вызывают как гуморальный, так и клеточный иммунные ответы. Поэтому живые аттенуированные вакцины обладают более широким спектром защиты в отношении антигенных вариантов вируса гриппа по сравнению с инактивированными вакцинами, которые, как правило, стимулируют лишь гуморальную часть иммунной системы.
В основе необычайно быстрой изменчивости вируса гриппа, а значит, и его ускользания от действия нейтрализующих антител лежат два механизма: 1) накопление точечных мутаций, ведущих к изменению антигенной структуры поверхностных гликопротеинов (антигенный дрейф) и 2) реассортация геномных сегментов. Они приводят к появлению новых антигенных вариантов вируса (антигенный шифт), которые могут вызывать пандемию.
Для всех гриппозных вакцин, применяемых в настоящее время, характерен риск низкой эффективности при иммунизации пожилых людей и маленьких детей (Jefferson T, Rivetti A, Di Pietrantonj C, Demicheli V, Ferroni E. Vaccines for preventing influenza in healthy children. Cochrane Database Syst Rev 2012; 8, CD004879.;
Osterholm MT, Kelley NS, Sommer A, Belongia EA. Efficacy and effectiveness of influenza vaccines: a systematic review and meta-analysis. Lancet Infect Dis 2011; 12, 36-44; Pfleiderer M, Trouvin JH, Brasseur D, Granstrom M, Shivji R, Mura M, Cavaleri M. Summary of knowledge gaps related to quality and efficacy of current influenza vaccines. Vaccine 2014; 32, 4586-91). Кроме того, эти вакцины защищают от циркулирующих штаммов только в случае совпадения эпидемического вируса с вакцинным штаммом по антигенным свойствам. Именно высокой изменчивостью поверхностных антигенов вируса гриппа, НА и NA, обусловлена необходимость проведения ежегодной вакцинации и обновления состава вакцин. Следует отметить, что сезонные вакцины, формируемые на основании рекомендаций ВОЗ, неэффективны в случае внезапного появления нового пандемического штамма, кардинально отличающегося от всех циркулирующих вариантов, как это произошло в 2009 г. при появлении пандемического вируса A/California/7/2009 (H1N1pdm09). Еще одним примером может служить низкая эффективность компонента H3N2 сезонной вакцины 2014 г., обусловленная появлением нового антигенного варианта этого сероподтипа в результате антигенного дрейфа (Skowronski DM, Chambers C, Sabaiduc S, De Serres G, Dickinson JA, Winter AL, Drews SJ, Fonseca K, Charest H, Gubbay JB, Petric M, Krajden M, Kwindt TL, Martineau C, Eshaghi A, Bastien N, Li Y. Interim estimates of 2014/15 vaccine effectiveness against influenza A(H3N2) from Canada's Sentinel Physician Surveillance Network, January 2015. Euro Surveill 2015; 20). За последние 60 лет разработано множество вариантов живых и инактивированных гриппозных вакцин, имеющих определенные преимущества и недостатки, однако ни один из известных препаратов не решает проблемы контроля над заболеваемостью гриппом из-за их неспособности вызывать кросс-протективный иммунитет к постоянно эволюционирующим вирусам гриппа. В связи с этим существует настоятельная необходимость в создании универсальной гриппозной вакцины, вызывающей широкий кросс-протективный длительный иммунитет, способный противостоять всем известным известным циркулирующим сероподтипам вирусам гриппа А и В.
Все известные противогриппозные вакцины - инактивированные (цельновирионные, сплит или субъединичные) или живые (аттенуированные холодоадаптированные) - в основном направлены на создание иммунитета к глобулярной части НА. В отличие от изменчивой глобулярной части, стволовая часть НА отличается консервативностью среди вирусов гриппа А (I и II группы) и В. Для антител, индуцированных к этому району НА, известно несколько механизмов как прямой, так и непрямой нейтрализации. Один из механизмов прямой нейтрализации связан с предотвращением конформационного изменения НА, которое необходимо для высвобождения пептида слияния и последующего объединения эндосомальной и вирусных мембран для доставки вирусного генома в клетку. Второй механизм прямой нейтрализации связан с предотвращением активационного расщепления НА на субъединицы НА1 и НА2 с помощью антител, взаимодействующих с участком НА в непосредственной близости от сайта расщепления. В механизмах непрямой нейтрализации задействованы антителозависимая и комплементзависимая цитотоксичность (Terajima M, Cruz J, Co MD, Lee JH, Kaur K, Wrammert J, Wilson PC, Ennis FA. Complement-dependent lysis of influenza a virus-infected cells by broadly cross-reactive human monoclonal antibodies. J Virol 2011; 85, 13463-7.; Jegaskanda S, Weinfurter JT, Friedrich TC, Kent SJ. Antibody-dependent cellular cytotoxicity is associated with control of pandemic H1N1 influenza virus infection of macaques. J Virol 2013; 87, 5512-22).
Антитела к стволовому участку НА при вакцинации практически не образуются и выявляются в небольшом количестве только после естественной инфекции (Moody MA, Zhang R, Walter EB, Woods CW, Ginsburg GS, McClain MT, Denny TN, Chen X, Munshaw S, Marshall DJ, Whitesides JF, Drinker MS, Amos JD, Gurley TC, Eudailey JA, Foulger A, DeRosa KR, Parks R, Meyerhoff RR, Yu JS, Kozink DM, Barefoot BE, Ramsburg EA, Khurana S, Golding H, Vandergrift NA, Alam SM, Tomaras GD, Kepler TB, Kelsoe G, Liao HX, Haynes BF. H3N2 influenza infection elicits more cross-reactive and less clonally expanded anti-hemagglutinin antibodies than influenza vaccination. PLoS ONE 2011; 6, e25797).
Большинство разрабатываемых в настоящее время подходов к получению универсальной вакцины основано на фокусировании иммунного ответа к консервативным участкам белков вируса гриппа. Антитела к консервативным внутренним белкам PB2, PB1, PA, NP и M1 не относятся к нейтрализующим, но могут играть важную роль в обеспечении элиминации вируса за счет антителозависимой цитотоксичности (ADCC).
Известно несколько примеров создания универсальной вакцины на основе субъединицы НА2. Трехкратная иммунизация мышей пептидами, представляющими эктодомен НА2 (23-185 аминокислотные остатки) или пептид слияния (1-38 аминокислотные остатки), в комбинации с адъювантами KLH (keyhole limpet hemocyanin) и Фрейнда индуцировала кросс-реактивный иммунитет, снижающий гибель животных при летальной инфекции гетерологичным штаммом (Stanekova Z, Kiraly J, Stropkovska A, Mikuskova T, Mucha V, Kostolansky F, Vareckova E. Heterosubtypic protective immunity against influenza A virus induced by fusion peptide of the hemagglutinin in comparison to ectodomain of M2 protein. Acta Virol 2011; 55, 61-7). Более эффективная защита развивалась при вакцинации химерными конструкциями НА. Krammer с соавт. показали, что гетеросубтипичный гуморальный иммунитет у мышей индуцируется при иммунизации химерными белками, содержащими глобулярные части НА от штаммов разных сероподтипов и одинаковую стволовую часть НА (Krammer F, Palese P, Steel J. Advances in universal influenza virus vaccine design and antibody mediated therapies based on conserved regions of the hemagglutinin. Curr Top Microbiol Immunol 2014; 386, 301-21.; Krammer F, Hai R, Yondola M, Tan GS, Leyva-Grado VH, Ryder AB, Miller MS, Rose JK, Palese P, Garcia-Sastre A, Albrecht RA. Assessment of influenza virus hemagglutinin stalk-based immunity in ferrets. J Virol 2014; 88, 3432-42). К недостаткам этого подхода следует отнести сложную схему иммунизации, которая включает электропорацию животных с помощью ДНК и двукратную иммунизацию белковыми конструкциями внутримышечно и интраназально с адъювантом poly (I: C).
Другим примером служит использование стабильных конструкций (мини-НА), полученных генно-инженерным способом на основе аминокислотной последовательности стволовой части НА вируса сероподтипа H1N1. Из большой коллекции отбирали только конструкции с самой высокой аффинностью к антителам, обладающим нейтрализующей активностью широкого спектра. Иммунизация мышей такими конструкциями также защищала животных от гибели при инфицировании штаммом I группы - высокопатогенным вирусом гриппа птиц сероподтипа H5N1 (Impagliazzo A, Milder F, Kuipers H, Wagner MV, Zhu X, Hoffman RM, van Meersbergen R, Huizingh J, Wanningen P, Verspuij J, de Man M, Ding Z, Apetri A, Kukrer B, Sneekes-Vriese E, Tomkiewicz D, Laursen NS, Lee PS, Zakrzewska A, Dekking L, Tolboom J, Tettero L, van Meerten S, Yu W, Koudstaal W, Goudsmit J, Ward AB, Meijberg W, Wilson IA, Radosevic K. A stable trimeric influenza hemagglutinin stem as a broadly protective immunogen. Science 2015; 349, 1301-6). Cтопроцентная защита мышей от гибели достигалась в результате двукратной внутримышечной иммунизации очищенным белком мини-НА в дозе 30 мкг с адъювантом Matrix-M производства компании Novavax.
Еще одно из перспективных направлений в разработке универсальной гриппозной вакцины основано на создании самособирающихся наночастиц, которые, как показано ранее, значительно повышают иммуногенные свойства НА (Kanekiyo M, Wei CJ, Yassine HM, McTamney PM, Boyington JC, Whittle JR, Rao SS, Kong WP, Wang L, Nabel GJ. Self-assembling influenza nanoparticle vaccines elicit broadly neutralizing H1N1 antibodies. Nature 2013; 499, 102-6). Животным двукратно или трехкратно внутримышечно вводили наночастицы с добавлением нового адъюванта SAS (Sigma Adjuvant System). Несмотря на отсутствие выработки нейтрализующих антител после иммунизации наночастицами, как мыши, так и хорьки оказались полностью защищены от гибели при заражении высокопатогенным вирусом гриппа птиц сероподтипа H5N1.
Одна из современных технологий получения живой вакцины основана на конструировании вакцинных векторов, в котором один вирус экспрессирует антигены другого вируса. В качестве векторов, экспрессирующих гриппозные антигены, используют различные ДНК-содержащие вирусы, а именно: аденовирус, герпесвирус, бакуловирус или поксвирус (Dudek T, Knipe DM. Replication-defective viruses as vaccines and vaccine vectors. Virology 2006; 344, 230-9.; He F, Madhan S, Kwang J. Baculovirus vector as a delivery vehicle for influenza vaccines. Expert Rev Vaccines 2009; 8, 455-67.; Draper SJ, Cottingham MG, Gilbert SC. Utilizing poxviral vectored vaccines for antibody induction-progress and prospects. Vaccine 2013; 31, 4223-30. Price GE, Soboleski MR, Lo CY, Misplon JA, Pappas C, Houser KV, Tumpey TM, Epstein SL. Vaccination focusing immunity on conserved antigens protects mice and ferrets against virulent H1N1 and H5N1 influenza A viruses. Vaccine 2009; 27, 6512-21). Так, при применении аденовирусного вектора показано, что трехкратная иммунизация плазмидой (50 мкг), содержащей последовательности консервативных белков NP и M2 вируса гриппа A, с последующим интраназальным заражением двумя аденовирусными векторами, экспрессирующими эти же белки, полностью защищала мышей и хорьков от гибели и потери веса при заражении вирулентным вирусом A/FM/1/47 (H1N1) или высокопатогенным штаммом вируса гриппа птиц сероподтипа H5N1.
Таким образом, все перечисленные подходы фокусирования иммунного ответа в отношении консервативных антигенов вируса гриппа подтверждают принципиальную возможность создания гриппозной вакцины, которая способна защищать от заражения различными вариантами вируса гриппа А. Однако, для достижения этой цели использовались сложные схемы многократной вакцинации животных с применением иммунологических адъювантов различной природы. Кроме того, ни один из известных экспериментальных препаратов универсальной гриппозной вакцины не обеспечивал защиту от вируса гриппа В. К этому следует добавить, что перечисленные выше экспериментальные препараты требуют сложных технологических процесов производства многокомпонентных вакцин, сопряженых с неприемлемо высокой стоимостью конечного препарата.
Таким образом, сохраняется потребность в новых эффективных аттенуированных гриппозных векторах, которые могут применяться для профилактики гриппа в виде универсальной гриппозной вакцины, в частности в виде мукозальной гриппозной вакцины.
Раскрытие изобретения
Настоящее изобретение относится к аттенуированному гриппозному вектору, индуцирующему кросс-протективный ответ против вирусов гриппа A и В, содержащему:
нуклеотидную последовательность гена белка PB2, происходящую от вируса гриппа A/PR/8/34 (H1N1), или нуклеотидную последовательность, имеющую по меньшей мере 95% идентичность указанной нуклеотидной последовательности гена белка PB2;
нуклеотидную последовательность гена белка PB1, происходящую от вируса гриппа A/PR/8/34 (H1N1), или нуклеотидную последовательность, имеющую по меньшей мере 95% идентичность указанной нуклеотидной последовательности гена белка PB1;
нуклеотидную последовательность гена белка PA, происходящую от вируса гриппа A/PR/8/34 (H1N1), или нуклеотидную последовательность, имеющую по меньшей мере 95% идентичность указанной последовательности белка РА;
нуклеотидную последовательность гена белка NP, происходящую от вируса гриппа A/PR/8/34 (H1N1), или нуклеотидную последовательность, имеющую по меньшей мере 95% идентичность указанной последовательности белка NP;
нуклеотидную последовательность гена белка M, происходящую от вируса гриппа A/PR/8/34 (H1N1), или нуклеотидную последовательность, имеющую по меньшей мере 95% идентичность указанной последовательности гена белка М;
нуклеотидную последовательность гена белка HA, происходящую от вируса гриппа A/California/7/09-like H1N1pdm), или нуклеотидную последовательность, имеющую по меньшей мере 95% идентичность указанной нуклеотидной последовательности гена белка НА;
нуклеотидную последовательности гена белка NA, происходящую от вируса гриппа A/California/7/09-like H1N1pdm), или нуклеотидную последовательность, имеющую по меньшей мере 95% идентичность указанной нуклеотидной последовательности гена белка NA; и
нуклеотидную последовательность химерного гена белка NS, включающую
рамку считывания белка NS1, происходящую от вируса A/PR/8/34 (H1N1), где указанная рамка считывания является укороченной и кодирует белок NS1 размером 124 аминокислотных остатков,
и последовательность гена белка Nep, происходящую от вируса гриппа А/Singapore/1/57-like (H2N2), или
нуклеотидную последовательность, имеющую по меньшей мере 95% идентичность указанной последовательности химерного гена NS;
где указанная укороченная рамка считывания гена белка NS1 продолжена вставкой нуклеотидной последовательности, кодирующей пептид слияния субъединицы НА2 от вируса гриппа В, и нуклеотидной последовательности, кодирующей консервативный В-клеточный эпитоп нуклеопротеина (NP) вируса гриппа А.
Аттенуированный гриппозный вектор по изобретению индуцирует кросс-протективный ответ против различных антигенных вариантов вируса гриппа A и В после мукозального введения, в частности, после интраназальной иммунизации.
Предпочтительным вариантом является гриппозный вектор, в котором нуклеотидная последовательность химерного гена белка NS представлена в SEQ ID NO: 8.
Еще одним объектом настоящего изобретения является фармацевтическая композиция для профилактики гриппа у субъекта, содержащая в эффективном количестве аттенуированный гриппозный вектор по изобретению и фармацевтически приемлемый носитель.
Одним из вариантов осуществления изобретения является фармацевтическая композиция, содержащая 6 - 8,5 logЭИД 50/мл аттенуированного гриппозного вектора по изобретению и буферный раствор, содержащий 0-1,5 масс.% моновалентной соли, 0-5 масс.% L-карнозина, 0-5 масс.% углеводного компонента, 0-2 масс.% белкового компонента, 0-2 масс.% аминокислотного компонента и 0-10 масс.% гидроксиэтилированного крахмала. Предпочтительным вариантом является фармацевтическая композиция, где буферный раствор содержит 0,5-1,5 масс.% моновалентной соли, 0,01-5 масс.% L-карнозина, 1-5 масс.% углеводного компонента, 0,1-2 масс.% белкового компонента, 0,01-2 масс. % аминокислотного компонента и 1-10 масс.% гидроксиэтилированного крахмала, предпочтительно, где моновалентная соль представляет собой хлорид натрия, углеводный компонент представляет собой сахарозу, трегалозу или лактозу, белковый компонент представляет собой человеческий рекомбинантный альбумин, казитон, гидролизат лактальбумина или желатин, аминокислотный компонент представляет собой аргинин, глицин или глутамат натрия.
В предпочтительном варианте осуществления изобретения субъект представляет собой млекопитающее или птицу, в частности, субъект представляет собой человека.
Изобретение относится также к вакцине против гриппа, содержащей в эффективном количестве аттенуированный гриппозный вектор по изобретению и фармацевтически приемлемый носитель.
Одним из вариантов осуществления изобретения является вакцина, содержащая 6-8,5 logЭИД 50/мл аттенуированного гриппозного вектора по изобретению и буферный раствор, содержащий 0-1,5 масс.% моновалентной соли, 0-5 масс.% L-карнозина, 0-5 масс.% углеводного компонента, 0-2 масс.% белкового компонента, 0-2 масс.% аминокислотного компонента и 0-10 масс.% гидроксиэтилированного крахмала. Предпочтительным вариантом является вакцина, где буферный раствор содержит 0,5-1,5 масс.% моновалентной соли, 0,01-5 масс.% L-карнозина, 1-5 масс.% углеводного компонента, 0,1-2 масс.% белкового компонента, 0,01-2 масс. % аминокислотного компонента и 1-10 масс.% гидроксиэтилированного крахмала, предпочтительно, где моновалентная соль представляет собой хлорид натрия, углеводный компонент представляет собой сахарозу, трегалозу или лактозу, белковый компонент представляет собой человеческий рекомбинантный альбумин, казитон, гидролизат лактальбумина или желатин, аминокислотный компонент представляет собой аргинин, глицин или глутамат натрия.
Изобретение также относится к применению аттенуированного гриппозного вектора по изобретению или фармацевтической композиции по изобретению для профилактики гриппа у субъекта.
В предпочтительном варианте осуществления изобретения субъект представляет собой млекопитающее или птицу, в частности, субъект представляет собой человека.
Техническим результатом настоящего изобретения является получение химерного по NS геномному фрагменту гриппозного вектора, обладающего свойствами универсальной гриппозной вакцины при однократном мукозальном введении в отсутствии адъювантов. Кроме того, техническим результатом является высокий потенциал роста полученного гриппозного вектора в 10-дневных куриных эмбрионах. Еще одним техническим результатом является получение гриппозных векторов, которые обладают свойствами универсальной гриппозной вакцины. Еще одним техническим результатом является снижение стоимости производства вакцины против гриппа ввиду неиспользования адъювантов.
Краткое описание чертежей
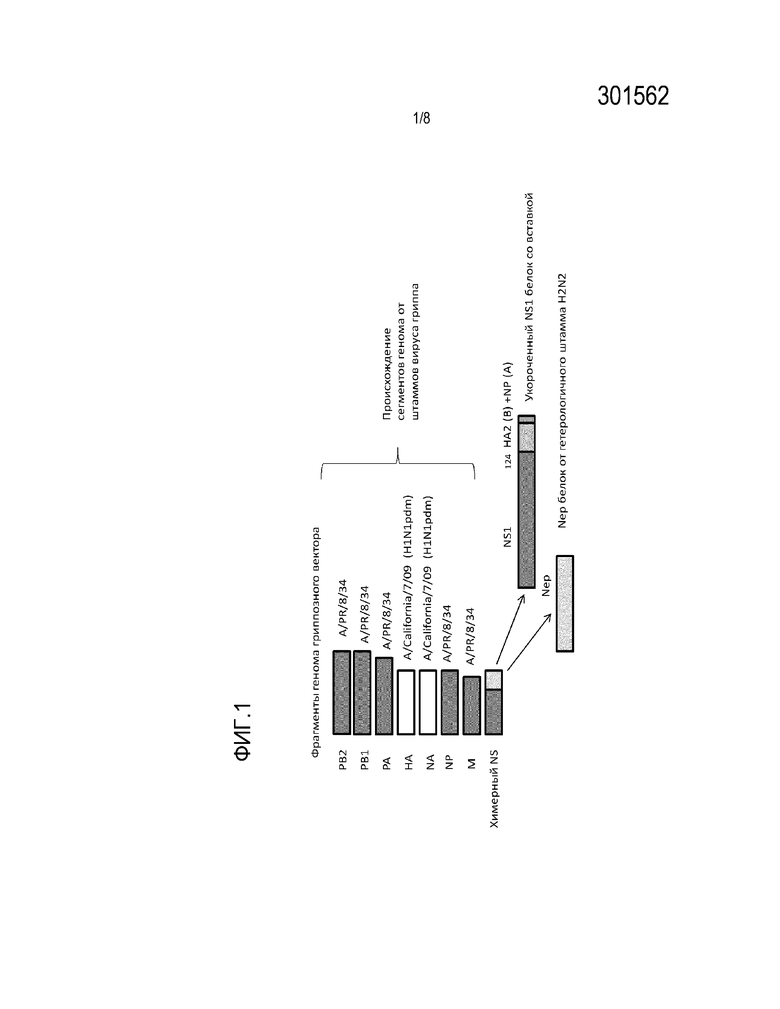
На Фиг.1 показана структура гриппозного аттенуированного вектора. Обозначены 8 фрагментов генома вируса и их особенности.
Показано, что фрагменты генома PB2, PB1, PA, Np и M имеют происхождение от вируса А/PR/8/34 (H1N1); гены поверхностных гликопротеинов HA и NA имеют происхождение от вируса A/California/7/09-like (H1N1pdm); NS геномный фрагмент имеет химерную структуру, кодирующую два белка: 1) укороченный до 124 аминокислотных остатка белок NS1, продолженный вставкой последовательности N- терминальной области НА2 вируса гриппа В и вставкой консервативного В-клеточного эпитопа NP белка вируса гриппа А 2) белок Nep, имеющий последовательность от гетерологичного штамма вируса гриппа А, сероподтипа H2N2 А/Singapore/1/57 -like.
На Фиг.2 представлены нуклеотидные последовательности геномных фрагментов вакцинного вектора: PB2, PB1, PA, NP, M - имеющие происхождение от вируса A/PR/8/34 (H1N1); HA и NA, имеющие происхождение от вируса A/California/7/09-like (H1N1pdm); химерного NS (вставка в рамке считывания NS1 выделена жирным шрифтом).
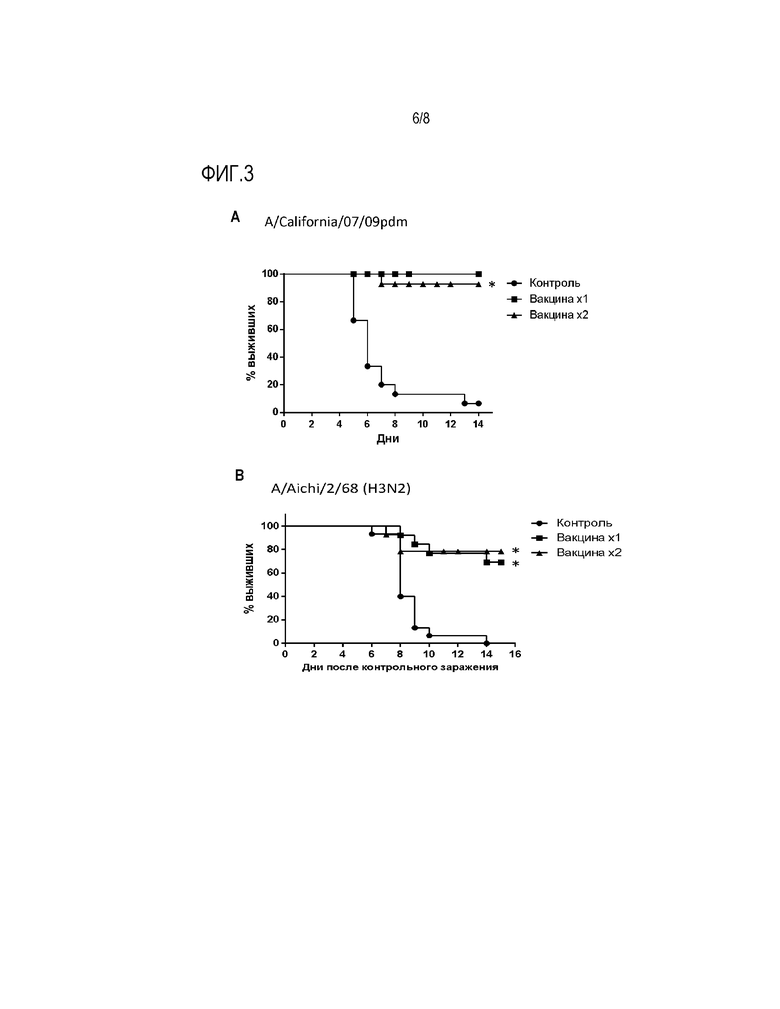
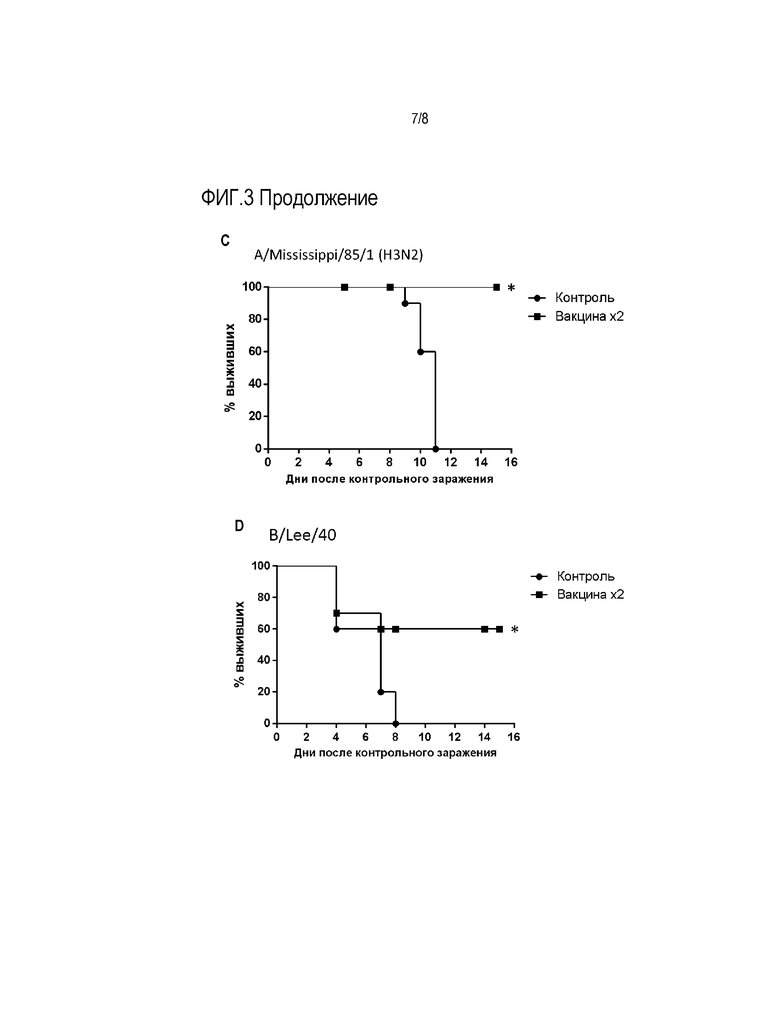
На Фиг.3 представлены результаты, отражающие протективные свойства вакцинного вектора после интраназальной иммунизации мышей в отношении различных вариантов вируса гриппа А и вируса гриппа В. На графиках показан % летальности у вакцинированных мышей после контрольного заражения указаными сероподтипами вируса гриппа А или вирусом гриппа В по сравнению с контрольными животными. Вакцина вводилась однократно - 1х или двукратно - 2х.
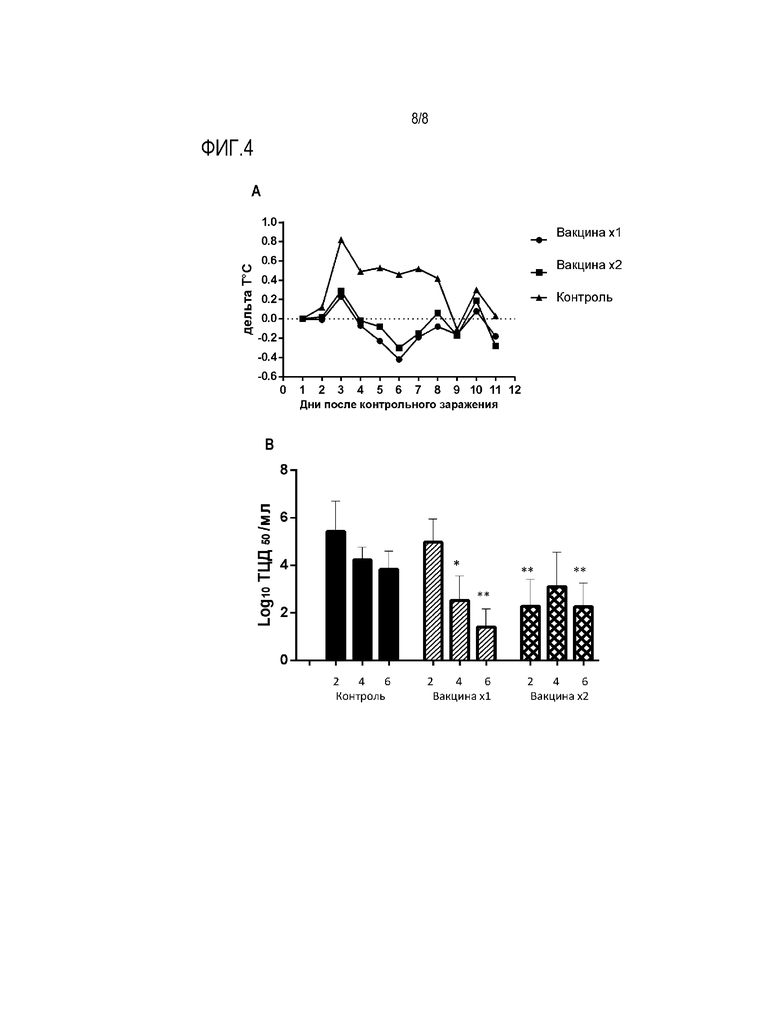
На Фиг.4 представлены данные, демонстрирующие протективные свойства вакцинного вектора после интраназальной иммунизации хорьков. На графике А представлена динамика среднего значения тепературных колебаний после контрольного заражения вирусом A/St.Petersburg/224/2015 (H3N2) у вакцинированных и контрольных животных. На графике В представлены результаты высева контрольного вируса из носовых смывов хорьков на 2,4 и 6 день после заражения. Титры выражены в виде среднего значения концентрации вируса в носовых смывах, выраженных в виде значения 50% цитопатических доз/мл после титрования на клетках MDCK. Вакцина вводилась однократно - 1х или двукратно - 2х.
Подробное описание изобретения
Настоящее изобретение относится к полученному генно-инженерным методом вакцинному вектору на основе вируса гриппа A, который может применяться для профилактики гриппа, вызванного всеми циркулирующими в человеческой популяции штаммами, включая вирусы гриппа А и В. В частности, настоящее изобретение относится к аттенуированному вирусу гриппа А, индуцирующему кросс-протективный ответ против вируса гриппа A и В, содержащему химерный NS фрагмент, включающий укороченную рамку считывания белка NS1 и гетерологичную последовательность гена белка Nep, происходящую от подтипа вируса гриппа А H2N2. Таким образом, подтип вируса А для последовательности, кодирующей укороченный белок NS1, отличается от подтипа вируса А, из которого получена последовательность, кодирующая белок Nep. В частности, в вакцинном векторе укороченная рамка считывания белка NS1 происходит от вируса гриппа подтипа H1N1, а гетeрологичная последовательность гена белка Nep происходит от вируса гриппа подтипа от H2N2.
В частности, настоящее изобретение относится к аттенуированному гриппозному вектору, индуцирующему кросс-протективный ответ против вирусов гриппа A и В, содержащему:
нуклеотидную последовательность гена, кодирующую белок PB2, происходящую от вируса гриппа A/PR/8/34 (H1N1), или нуклеотидную последовательность, имеющую по меньшей мере 95% или больше (например, 96, 97, 98 или 99%) идентичность указанной нуклеотидной последовательности гена белка PB2;
нуклеотидную последовательность гена, кодирующую белок PB1, происходящую от вируса гриппа A/PR/8/34 (H1N1), или нуклеотидную последовательность, имеющую по меньшей мере 95% или больше (например, 96, 97, 98 или 99%) идентичность указанной нуклеотидной последовательности гена белка PB1;
нуклеотидную последовательность гена, кодирующую белок PA, происходящую от вируса гриппа A/PR/8/34 (H1N1), или нуклеотидную последовательность, имеющую по меньшей мере 95% или больше (например, 96, 97, 98 или 99%) идентичность указанной последовательности белка РА;
нуклеотидную последовательность гена, кодирующую белок NP, происходящую от вируса гриппа A/PR/8/34 (H1N1), или нуклеотидную последовательность, имеющую по меньшей мере 95% или больше (например, 96, 97, 98 или 99%) идентичность указанной последовательности белка NP;
нуклеотидную последовательность гена, кодирующую белок M, происходящую от вируса гриппа A/PR/8/34 (H1N1), или нуклеотидную последовательность, имеющую по меньшей мере 95% или больше (например, 96, 97, 98 или 99%) идентичность указанной последовательности гена белка М;
нуклеотидную последовательность гена, кодирующую белок HA, происходящую от вируса гриппа A/California/7/09-like H1N1pdm), или нуклеотидную последовательность, имеющую по меньшей мере 95% или больше (например, 96, 97, 98 или 99%) идентичность указанной нуклеотидной последовательности гена белка НА;
нуклеотидную последовательности гена, кодирующую белок NA, происходящую от вируса гриппа A/California/7/09-like H1N1pdm), или нуклеотидную последовательность, имеющую по меньшей мере 95% или больше (например, 96, 97, 98 или 99%) идентичность указанной нуклеотидной последовательности гена белка NA; и
нуклеотидную последовательность химерного гена белка NS, включающую
рамку считывания белка NS1, происходящую от вируса A/PR/8/34 (H1N1), где указанная рамка считывания является укороченной и кодирует белок NS1 размером 124 аминокислотных остатков,
и последовательность гена белка Nep, происходящую от вируса гриппа А/Singapore/1/57-like (H2N2), или
нуклеотидную последовательность, имеющую по меньшей мере 95% или больше (например, 96, 97, 98 или 99%) идентичность указанной последовательности химерного гена NS;
где указанная укороченная рамка считывания гена белка NS1 продолжена вставкой нуклеотидной последовательности, кодирующей пептид слияния субъединицы НА2 от вируса гриппа В, и нуклеотидной последовательности, кодирующей консервативный В-клеточный эпитоп нуклеопротеина (NP) вируса гриппа А.
Указанная укороченная рамка считывания кодирует белок NS1 размером 124 аминокислотных остатка, который продолжен двумя глицинами, вставкой N-терминальной области второй субъединицы гемагглютинина НА2 вируса гриппа В (23 аминокислотных остатка) и вставкой последовательности консервативного В клеточного эпитопа вируса гриппа А (7 аминокислотных остатков).
Гены поверхностных гликопротеинов указанного вектора происходят от вируса гриппа A/California/7/09 (H1N1pdm). Гены внутренних белков PB2, PB1, РА, NP и М происходят от вируса гриппа A/PR/8/34 (H1N1). Таким образом, гриппозный вектор по изобретению представляет собой сложную генетическую конструкцию, состоящую из геномных последовательностей различных штаммов вируса гриппа, а именно: 1) гены, кодирующие PB2,PB1,PA,NP и M, имеют происхождение от вируса A/PR/8/34 (H1N1) (PB2 (Genbank accession number: AB671295), PB1 (Genbank accession number: CY033583), PA (Genbank accession number: AF389117), NP (Genbank accession number: AF389119), M (Genbank accession number: AF389121)), 2) гены, кодирующие HA и NA, имеют происхождение от вируса A/California/7/09-like H1N1pdm)(HA (GenBank: KM408964.1) и (NA GenBank: KM408965.1), 3) ген NS имеет химерную природу, где рамка считывания белка NS1 от вируса A/PR/8/34 (H1N1) укорочена до 124 аминокислотных остатка и продолжена вставкой последовательности, кодирующей пептид слияния субъединицы НА2 от вируса гриппа В, и последовательности, кодирующей консервативный В-клеточный эпитоп нуклеопротеина (NP) вируса гриппа А, а рамка считывания белка NEP происходит от вируса гриппа подтипа H2N2.
Настоящее изобретение основано в частности на том, что авторы неожиданно обнаружили, что вектор с указанной структурой при интраназальной иммунизации мышей и хорьков без применения адъювантов вызывает защиту животных от контрольного заражения не только вирусами гриппа А (H1N1), но и вирусами гриппа А(H3N2)и гриппа В. Следовательно вакцинный вектор обладает свойствами универсальной гриппозной вакцины.
Термин «универсальная вакцина» в контексте настоящего изобретения означает вакцину, способную защищать от всех известных и неизвестных вариантов вируса гриппа. Обычные «сезонные вакцины» защищают только от вирусов, подобных тем вирусам, что входят в их состав.
Термин «мукозальная вакцина» означает, что вакцину можно вводить в полости респираторного и пищеварительного тракта и наносить на слизистые оболочки рта и носа, т.е. применять интраназально, перорально, подъязычно.
Гриппозный вектор на основе вируса A/PR/8/34, несущий химерный NS геномный фрагмент, оказался неспособным к активной репродукции при 39°C и в легких мышей (фенотип аттенуации), но при этом по-прежнему репродуцировался до высоких титров в 10-дневных куриных эмбрионах.
Фармацевтическая композиция согласно настоящему изобретению может быть составлена в виде вакцины, содержащей в эффективном количестве аттенуированный гриппозный вектор согласно настоящему изобретению и фармацевтически приемлемый носитель.
Термин «субъект» или «животное» в контексте настоящего описания означает позвоночных, которые подвержены инфекции, вызываемой патогенными бактериями, вирусами или простейшими, включая птиц (водоплавающие, куры и т.д.) и представителей различных видов млекопитающих, таких как псовые, кошачьи, волки, хорьки, грызуны (крысы, мыши и т.д.), лошади, коровы, овцы, козы, свиньи и приматы. В одном из вариантов осуществления изобретения субъект представляет собой человека.
Под «эффективным количеством» подразумевается такое количество вектора, которое при введении субъекту, в однократной или двукратной дозе, является эффективным для профилактики гриппа. Это количество может варьироваться в зависимости от состояния здоровья и физического состояния пациента, возраста, таксономической группы индивида, подвергаемого вакцинации, препаративной формы, оценки медицинской ситуации лечащим врачом и других важных факторов. Полагают, что количество может варьировать в относительно широком диапазоне, который специалист в данной области может определить стандартными способами. В частности, фармацевтическая композиция может содержать 6-8,5 logЭИД 50/мл гриппозного вектора согласно изобретению.
Термин «фармацевтически приемлемый носитель» в контексте настоящего изобретения означает любой носитель, используемый в данной области, в частности, воду, физиологический раствор, буферный раствор и т.п. В одном из вариантов осуществления фармацевтически приемлемый носитель представляет собой буферный раствор, содержащий 0-1,5 масс.% моновалентной соли, 0-5 масс.% L-карнозина, 0-5 масс.% углеводного компонента, 0-2 масс.% белкового компонента, 0-2 масс.% аминокислотного компонента и 0-10 масс.% гидроксиэтилированного крахмала, предпочтительно, указанный буферный раствор содержит 0,5-1,5 масс.% моновалентной соли, 0,01-5 масс.% L-карнозина, 1-5 масс.% углеводного компонента, 0,1-2 масс.% белкового компонента, 0,01-2 масс. % аминокислотного компонента и 1-10 масс.% гидроксиэтилированного крахмала, в наиболее предпочтительном варианте, моновалентная соль представляет собой хлорид натрия, углеводный компонент представляет собой сахарозу, трегалозу или лактозу, белковый компонент представляет собой человеческий рекомбинантный альбумин, казитон, гидролизат лактальбумина или желатин, аминокислотный компонент представляет собой аргинин, глицин или глутамат натрия.
Настоящее изобретение относится также к применению аттенуированного аттенуированного гриппозного вектора или фармацевтической композиции согласно настоящему изобретению для профилактики гриппа.
Введение субъекту может осуществляться любыми стандартными методами, в частности, перорально, сублингвально, ингаляционно или интраназально.
Гриппозный вектор или фармацевтическую композицию можно вводить субъекту один, два или более раз, предпочтительным является однократное введение.
Далее изобретение проиллюстрировано примерами осуществления, не ограничивающими объем изобретения.
ПРИМЕРЫ
Пример 1
Получение гриппозного вектора с модифицированным NS геномным фрагментом
Сборка рекомбинантного вируса осуществлялась в несколько этапов. На первом этапе синтетическим путем были получены комплиментарные ДНК (кДНК) копии 5 геномных фрагментов(PB2, PB1, PA, NP, M) вируса гриппа A/PR/8/34 (H1N1) (PB2 (Genbank accession number: AB671295), PB1 (Genbank accession number: CY033583), PA (Genbank accession number: AF389117), NP (Genbank accession number: AF389119), M (Genbank accession number: AF389121)) и 2 геномных фрагмента (HA, NA) вируса A/California/7/09-like(HA (GenBank: KM408964.1) и (NA GenBank: KM408965.1)), а также синтезирован химерный геномный фрагмент NS, составленный из последовательностей, относящихся к вирусу H1N1(ген NS1), вирусу H2N2 (ген Nep) и последовательностей двух пептидов от НА2 вируса гриппа В и NP вируса гриппа А.
На втором этапе синтезированные последовательности были клонированы в двунаправленный вектор на основе плазмиды pHW2000 (Hoffmann E, Neumann G, Kawaoka Y, Hobom G, Webster RG. A DNA transfection system for generation of influenza A virus from eight plasmids. Proc Natl Acad Sci USA. 2000; 97(11):6108-13.). Указанный плазмидный вектор благодаря наличию Pol I и Pol II промоторов обеспечивает одновременную внутриклеточную транскрипцию вирусных и соответствующих матричных РНК при трансфекции клеток млекопитающих. На Фиг.1 представлена генетическая схема гриппозного вектора. На Фиг. 2 представлены нуклеотидные последовательности всех 8 геномных фрагментов гриппозного вектора.
Нуклеотидная последовательность гена PB2
1 agcgaaagca ggtcaattat attcaatatg gaaagaataa aagaactacg aaatctaatg
61 tcgcagtctc gcacccgcga gatactcaca aaaaccaccg tggaccatat ggccataatc
121 aagaagtaca catcaggaag acaggagaag aacccagcac ttaggatgaa atggatgatg
181 gcaatgaaat atccaattac agcagacaag aggataacgg aaatgattcc tgagagaaat
241 gagcaaggac aaactttatg gagtaaaatg aatgatgccg gatcagaccg agtgatggta
301 tcacctctgg ctgtgacatg gtggaatagg aatggaccaa taacaaatac agttcattat
361 ccaaaaatct acaaaactta ttttgaaaga gtcgaaaggc taaagcatgg aacctttggc
421 cctgtccatt ttagaaacca agtcaaaata cgtcggagag ttgacataaa tcctggtcat
481 gcagatctca gtgccaagga ggcacaggat gtaatcatgg aagttgtttt ccctaacgaa
541 gtgggagcca ggatactaac atcggaatcg caactaacga taaccaaaga gaagaaagaa
601 gaactccagg attgcaaaat ttctcctttg atggttgcat acatgttgga gagagaactg
661 gtccgcaaaa cgagattcct cccagtggct ggtggaacaa gcagtgtgta cattgaagtg
721 ttgcatttga ctcaaggaac atgctgggaa cagatgtata ctccaggagg ggaagtgagg
781 aatgatgatg ttgatcaaag cttgattatt gctgctagga acatagtgag aagagctgca
841 gtatcagcag atccactagc atctttattg gagatgtgcc acagcacaca gattggtgga
901 attaggatgg tagacatcct taggcagaac ccaacagaag agcaagccgt ggatatatgc
961 aaggctgcaa tgggactgag aattagctca tccttcagtt ttggtggatt cacatttaag
1021 agaacaagcg gatcatcagt caagagagag gaagaggtgc ttacgggcaa tcttcaaaca
1081 ttgaagataa gagtgcatga gggatatgaa gagttcacaa tggttgggag aagagcaaca
1141 gccatactca gaaaagcaac caggagattg attcagctga tagtgagtgg gagagacgaa
1201 cagtcgattg ccgaagcaat aattgtggcc atggtatttt cacaagagga ttgtatgata
1261 aaagcagtca gaggtgatct gaatttcgtc aatagggcga atcaacgatt gaatcctatg
1321 catcaacttt taagacattt tcagaaggat gcgaaagtgc tttttcaaaa ttggggagtt
1381 gaacctatcg acaatgtgat gggaatgatt gggatattgc ccgacatgac tccaagcatc
1441 gagatgtcaa tgagaggagt gagaatcagc aaaatgggtg tagatgagta ctccagcacg
1501 gagagggtag tggtgagcat tgaccgtttt ttgagaatcc gggaccaacg aggaaatgta
1561 ctactgtctc ccgaggaggt cagtgaaaca cagggaacag agaaactgac aataacttac
1621 tcatcgtcaa tgatgtggga gattaatggt cctgaatcag tgttggtcaa tacctatcaa
1681 tggatcatca gaaactggga aactgttaaa attcagtggt cccagaaccc tacaatgcta
1741 tacaataaaa tggaatttga accatttcag tctttagtac ctaaggccat tagaggccaa
1801 tacagtgggt ttgtaagaac tctgttccaa caaatgaggg atgtgcttgg gacatttgat
1861 accgcacaga taataaaact tcttcccttc gcagccgctc caccaaagca aagtagaatg
1921 cagttctcct catttactgt gaatgtgagg ggatcaggaa tgagaatact tgtaaggggc
1981 aattctcctg tattcaacta taacaaggcc acgaagagac tcacagttct cggaaaggat
2041 gctggcactt taactgaaga cccagatgaa ggcacagctg gagtggagtc cgctgttctg
2101 aggggattcc tcattctggg caaagaagac aagagatatg ggccagcact aagcatcaat
2161 gaactgagca accttgcgaa aggagagaag gctaatgtgc taattgggca aggagacgtg
2221 gtgttggtaa tgaaacggaa acgggactct agcatactta ctgacagcca gacagcgacc
2281 aaaagaattc ggatggccat caattagtgt cgaatagttt aaaaacgacc ttgtttctac
2341 t (SEQ ID NO:1)
Нуклеотидная последовательность гена PB1
1 atggatgtca atccgacctt acttttctta aaagtgccag cacaaaatgc tataagcaca
61 actttccctt atactggaga ccctccttac agccatggga caggaacagg atacaccatg
121 gatactgtca acaggacaca tcagtactca gaaaagggaa gatggacaac aaacaccgaa
181 actggagcac cgcaactcaa cccgattgat gggccactgc cagaagacaa tgaaccaagt
241 ggttatgccc aaacagattg tgtattggag gcgatggctt tccttgagga atcccatcct
301 ggtatttttg aaaactcgtg tattgaaacg atggaggttg ttcagcaaac acgagtagac
361 aagctgacac aaggccgaca gacctatgac tggactctaa atagaaacca acctgctgca
421 acagcattgg ccaacacaat agaagtgttc agatcaaatg gcctcacggc caatgagtct
481 ggaaggctca tagacttcct taaggatgta atggagtcaa tgaacaaaga agaaatgggg
541 atcacaactc attttcagag aaagagacgg gtgagagaca atatgactaa gaaaatgata
601 acacagagaa caatgggtaa aaagaagcag agattgaaca aaaggagtta tctaattaga
661 gcattgaccc tgaacacaat gaccaaagat gctgagagag ggaagctaaa acggagagca
721 attgcaaccc cagggatgca aataaggggg tttgtatact ttgttgagac actggcaagg
781 agtatatgtg agaaacttga acaatcaggg ttgccagttg gaggcaatga gaagaaagca
841 aagttggcaa atgttgtaag gaagatgatg accaattctc aggacaccga actttctttc
901 accatcactg gagataacac caaatggaac gaaaatcaga atcctcggat gtttttggcc
961 atgatcacat atatgaccag aaatcagccc gaatggttca gaaatgttct aagtattgct
1021 ccaataatgt tctcaaacaa aatggcgaga ctgggaaaag ggtatatgtt tgagagcaag
1081 agtatgaaac ttagaactca aatacctgca gaaatgctag caagcatcga tttgaaatat
1141 ttcaatgatt caacaagaaa gaagattgaa aaaatccgac cgctcttaat agaggggact
1201 gcatcattga gccctggaat gatgatgggc atgttcaata tgttaagcac tgtattaggc
1261 gtctccatcc tgaatcttgg acaaaagaga tacaccaaga ctacttactg gtgggatggt
1321 cttcaatcct ctgacgattt tgctctgatt gtgaatgcac ccaatcatga agggattcaa
1381 gccggagtcg acaggtttta tcgaacctgt aagctacttg gaatcaatat gagcaagaaa
1441 aagtcttaca taaacagaac aggtacattt gaattcacaa gttttttcta tcgttatggg
1501 tttgttgcca atttcagcat ggagcttccc agttttgggg tgtctgggat caacgagtca
1561 gcggacatga gtattggagt tactgtcatc aaaaacaata tgataaacaa tgatcttggt
1621 ccagcaacag ctcaaatggc ccttcagttg ttcatcaaag attacaggta cacgtaccga
1681 tgccatagag gtgacacaca aatacaaacc cgaagatcat ttgaaataaa gaaactgtgg
1741 gagcaaaccc gttccaaagc tggactgctg gtctccgacg gaggcccaaa tttatacaac
1801 attagaaatc tccacattcc tgaagtctgc ctaaaatggg aattgatgga tgaggattac
1861 caggggcgtt tatgcaaccc actgaaccca tttgtcagcc ataaagaaat tgaatcaatg
1921 aacaatgcag tgatgatgcc agcacatggt ccagccaaaa acatggagta tgatgctgtt
1981 gcaacaacac actcctggat ccccaaaaga aatcgatcca tcttgaatac aagtcaaaga
2041 ggagtacttg aggatgaaca aatgtaccaa aggtgctgca atttatttga aaaattcttc
2101 cccagcagtt catacagaag accagtcggg atatccagta tggtggaggc tatggtttcc
2161 agagcccgaa ttgatgcacg gattgatttc gaatctggaa ggataaagaa agaagagttc
2221 actgagatca tgaagatctg ttccaccatt gaagagctca gacggcaaaa atagtgaatt
2281 tagcttgt (SEQ ID NO:2)
Нуклеотидная последовательность гена PA
1 agcgaaagca ggtactgatc caaaatggaa gattttgtgc gacaatgctt caatccgatg
61 attgtcgagc ttgcggaaaa aacaatgaaa gagtatgggg aggacctgaa aatcgaaaca
121 aacaaatttg cagcaatatg cactcacttg gaagtatgct tcatgtattc agattttcac
181 ttcatcaatg agcaaggcga gtcaataatc gtagaacttg gtgatccaaa tgcacttttg
241 aagcacagat ttgaaataat cgagggaaga gatcgcacaa tggcctggac agtagtaaac
301 agtatttgca acactacagg ggctgagaaa ccaaagtttc taccagattt gtatgattac
361 aaggagaata gattcatcga aattggagta acaaggagag aagttcacat atactatctg
421 gaaaaggcca ataaaattaa atctgagaaa acacacatcc acattttctc gttcactggg
481 gaagaaatgg ccacaaaggc agactacact ctcgatgaag aaagcagggc taggatcaaa
541 accagactat tcaccataag acaagaaatg gccagcagag gcctctggga ttcctttcgt
601 cagtccgaga gaggagaaga gacaattgaa gaaaggtttg aaatcacagg aacaatgcgc
661 aagcttgccg accaaagtct cccgccgaac ttctccagcc ttgaaaattt tagagcctat
721 gtggatggat tcgaaccgaa cggctacatt gagggcaagc tgtctcaaat gtccaaagaa
781 gtaaatgcta gaattgaacc ttttttgaaa acaacaccac gaccacttag acttccgaat
841 gggcctccct gttctcagcg gtccaaattc ctgctgatgg atgccttaaa attaagcatt
901 gaggacccaa gtcatgaagg agagggaata ccgctatatg atgcaatcaa atgcatgaga
961 acattctttg gatggaagga acccaatgtt gttaaaccac acgaaaaggg aataaatcca
1021 aattatcttc tgtcatggaa gcaagtactg gcagaactgc aggacattga gaatgaggag
1081 aaaattccaa agactaaaaa tatgaagaaa acaagtcagc taaagtgggc acttggtgag
1141 aacatggcac cagaaaaggt agactttgac gactgtaaag atgtaggtga tttgaagcaa
1201 tatgatagtg atgaaccaga attgaggtcg ctagcaagtt ggattcagaa tgagtttaac
1261 aaggcatgcg aactgacaga ttcaagctgg atagagctcg atgagattgg agaagatgtg
1321 gctccaattg aacacattgc aagcatgaga aggaattatt tcacatcaga ggtgtctcac
1381 tgcagagcca cagaatacat aatgaagggg gtgtacatca atactgcctt gcttaatgca
1441 tcttgtgcag caatggatga tttccaatta attccaatga taagcaagtg tagaactaag
1501 gagggaaggc gaaagaccaa cttgtatggt ttcatcataa aaggaagatc ccacttaagg
1561 aatgacaccg acgtggtaaa ctttgtgagc atggagtttt ctctcactga cccaagactt
1621 gaaccacata aatgggagaa gtactgtgtt cttgagatag gagatatgct tataagaagt
1681 gccataggcc aggtttcaag gcccatgttc ttgtatgtga gaacaaatgg aacctcaaaa
1741 attaaaatga aatggggaat ggagatgagg cgttgcctcc tccagtcact tcaacaaatt
1801 gagagtatga ttgaagctga gtcctctgtc aaagagaaag acatgaccaa agagttcttt
1861 gagaacaaat cagaaacatg gcccattgga gagtccccca aaggagtgga ggaaagttcc
1921 attgggaagg tctgcaggac tttattagca aagtcggtat tcaacagctt gtatgcatct
1981 ccacaactag aaggattttc agctgaatca agaaaactgc ttcttatcgt tcaggctctt
2041 agggacaacc ttgaacctgg gacctttgat cttggggggc tatatgaagc aattgaggag
2101 tgcctgatta atgatccctg ggttttgctt aatgcttctt ggttcaactc cttccttaca
2161 catgcattga gttagttgtg gcagtgctac tatttgctat ccatactgtc caaaaaagta
2221 ccttgtttct act (SEQ ID NO:3)
Нуклеотидная последовательность гена NP
1 agcgaaagca ggtagatatt gaaagatgag tcttctaacc gaggtcgaaa cgtacgtact
61 ctctatcatc ccgtcaggcc ccctcaaagc cgagatcgca cagagacttg aagatgtctt
121 tgcagggaag aacactgatc ttgaggttct catggaatgg ctaaagacaa gaccaatcct
181 gtcacctctg actaagggga ttttaggatt tgtgttcacg ctcaccgtgc ccagtgagcg
241 aggactgcag cgtagacgct ttgtccaaaa tgcccttaat gggaacgggg atccaaataa
301 catggacaaa gcagttaaac tgtataggaa gctcaagagg gagataacat tccatggggc
361 caaagaaatc tcactcagtt attctgctgg tgcacttgcc agttgtatgg gcctcatata
421 caacaggatg ggggctgtga ccactgaagt ggcatttggc ctggtatgtg caacctgtga
481 acagattgct gactcccagc atcggtctca taggcaaatg gtgacaacaa ccaatccact
541 aatcagacat gagaacagaa tggttttagc cagcactaca gctaaggcta tggagcaaat
601 ggctggatcg agtgagcaag cagcagaggc catggaggtt gctagtcagg ctagacaaat
661 ggtgcaagcg atgagaacca ttgggactca tcctagctcc agtgctggtc tgaaaaatga
721 tcttcttgaa aatttgcagg cctatcagaa acgaatgggg gtgcagatgc aacggttcaa
781 gtgatcctct cgctattgcc gcaaatatca ttgggatctt gcacttgaca ttgtggattc
841 ttgatcgtct ttttttcaaa tgcatttacc gtcgctttaa atacggactg aaaggagggc
901 cttctacgga aggagtgcca aagtctatga gggaagaata tcgaaaggaa cagcagagtg
961 ctgtggatgc tgacgatggt cattttgtca gcatagagct ggagtaaaaa actaccttgt
1021 ttctact (SEQ ID NO:4)
Нуклеотидная последовательность гена M
1 agcgaaagca ggtagatatt gaaagatgag tcttctaacc gaggtcgaaa cgtacgtact
61 ctctatcatc ccgtcaggcc ccctcaaagc cgagatcgca cagagacttg aagatgtctt
121 tgcagggaag aacactgatc ttgaggttct catggaatgg ctaaagacaa gaccaatcct
181 gtcacctctg actaagggga ttttaggatt tgtgttcacg ctcaccgtgc ccagtgagcg
241 aggactgcag cgtagacgct ttgtccaaaa tgcccttaat gggaacgggg atccaaataa
301 catggacaaa gcagttaaac tgtataggaa gctcaagagg gagataacat tccatggggc
361 caaagaaatc tcactcagtt attctgctgg tgcacttgcc agttgtatgg gcctcatata
421 caacaggatg ggggctgtga ccactgaagt ggcatttggc ctggtatgtg caacctgtga
481 acagattgct gactcccagc atcggtctca taggcaaatg gtgacaacaa ccaatccact
541 aatcagacat gagaacagaa tggttttagc cagcactaca gctaaggcta tggagcaaat
601 ggctggatcg agtgagcaag cagcagaggc catggaggtt gctagtcagg ctagacaaat
661 ggtgcaagcg atgagaacca ttgggactca tcctagctcc agtgctggtc tgaaaaatga
721 tcttcttgaa aatttgcagg cctatcagaa acgaatgggg gtgcagatgc aacggttcaa
781 gtgatcctct cgctattgcc gcaaatatca ttgggatctt gcacttgaca ttgtggattc
841 ttgatcgtct ttttttcaaa tgcatttacc gtcgctttaa atacggactg aaaggagggc
901 cttctacgga aggagtgcca aagtctatga gggaagaata tcgaaaggaa cagcagagtg
961 ctgtggatgc tgacgatggt cattttgtca gcatagagct ggagtaaaaa actaccttgt
1021 ttctact (SEQ ID NO:5)
Нуклеотидная последовательность гена HA
1 atgaaggcaa tactagtagt tctgctatat acatttgcaa ccgcaaatgc agacacatta
61 tgtataggtt atcatgcaaa caattcaaca gacactgtag acacagtact agaaaagaat
121 gtaacagtaa cacactctgt taaccttcta gaagacaagc ataacgggaa actatgcaaa
181 ctaagagggg tagccccatt gcatttgggt aaatgtaaca ttgctggctg gatcctggga
241 aatccagagt gtgaatcact ctccacagca agttcatggt cctacattgt ggaaacatct
301 agttcagaca atggaacgtg ttacccagga gatttcatca attatgagga gctaagagag
361 caattgagct cagtgtcatc atttgaaagg tttgagatat tccccaaaac aagttcatgg
421 cccaatcatg actcgaacaa aggtgtaacg gcagcatgtc ctcacgctgg agcaaaaagc
481 ttctacaaaa atttaatatg gctagttaaa aaaggaaatt catacccaaa gctcagccaa
541 tcctacatta atgataaagg gaaagaagtc ctcgtgctgt ggggcattca ccatccatct
601 actactgctg accaacaaag tctctatcag aatgcagatg catatgtttt tgtggggaca
661 tcaagataca gcaagaagtt caagccggaa atagcaataa gacccaaagt gagggatcaa
721 gaagggagaa tgaactatta ctggacacta gtagagccgg gagacaaaat aacattcgaa
781 gcaactggaa atctagtggt accgagatat gcattcacaa tggaaagaaa tgctggatct
841 ggtattatca tttcagatac accagtccac gattgcaata caacttgtca gacacccgag
901 ggtgctataa acaccagcct cccatttcag aatatacatc cgatcacaat tggaaaatgt
961 ccaaagtatg taaaaagcac aaaattgaga ctggccacag gattgaggaa tgtcccgtct
1021 attcaatcta gaggcctatt cggggccatt gccggcttca ttgaaggggg gtggacaggg
1081 atggtagatg gatggtacgg ttatcaccat caaaatgagc aggggtcagg atatgcagcc
1141 gacctgaaga gcacacaaaa tgccattgac aagattacta acaaagtaaa ctctgttatt
1201 gaaaagatga atacacagtt cacagcagtg ggtaaagagt tcaaccacct ggaaaaaaga
1261 atagagaatt taaataaaaa agttgatgat ggtttcctgg acatttggac ttacaatgcc
1321 gaactgttgg ttctattgga aaatgaaaga actttggact accatgattc aaatgtgaag
1381 aacttgtatg aaaaggtaag aaaccagtta aaaaacaatg ccaaggaaat tggaaacggc
1441 tgctttgaat tttaccacaa atgcgataac acgtgcatgg aaagtgtcaa aaatgggact
1501 tatgactacc caaaatactc agaggaagca aaattaaaca gagaaaaaat agatggggta
1561 aagctggaat caacaaggat ttaccagatt ttggcgatct attcaactgt cgccagttca
1621 ttggtgctgg tagtctccct gggggcaatc agcttctgga tgtgctctaa tgggtctcta
1681 cagtgtagaa tatgtattta a (SEQ ID NO:6)
Нуклеотидная последовательность гена NA
1 atgaatccaa accaaaagat aataaccatt ggttcggtct gtatgacaat tggaatggct
61 aacttaatat tacaaattgg aaacataatc tcaatatgga ttagccactc aattcaagtt
121 gggaatcaaa gtcagatcga aacatgcaat caaagcgtca ttacttatga aaacaacact
181 tgggtaaatc agacatatgt taacatcagc aacaccaact ttgctgctgg gcagccagtg
241 gtttccgtga aattagcggg caattcctct ctctgccctg ttagtggatg ggctatatac
301 agtaaagaca acagtgtaag agtcggttcc aagggggatg tgtttgtcat aagggaacca
361 ttcatatcat gctccccctt ggaatgcaga accttcttct tgactcaagg ggccttgcta
421 aatgacaaac attccaatgg aaccattaaa gacaggagcc catatcgaac cttaatgagc
481 tgtcctattg gtgaagttcc ctctccatac aactcaagat ttgagtcagt cgcttggtca
541 gcaagtgctt gtcatgatgg catcaattgg ctaacaattg gaatttctgg cccagacagt
601 ggggcagtgg ctgtgttaaa gtacaacggc ataataacag acactatcaa gagttggaga
661 aacgatatat tgagaacaca agagtctgaa tgtgcatgtg taaatggttc ttgctttacc
721 ataatgaccg atggaccaag tgatggacag gcctcataca agatcttcag aatagaaaag
781 ggaaagatag tcaaatcagt cgaaatgaat gcccctaatt atcactatga ggaatgctcc
841 tgttatcctg attctagtga aatcacatgt gtgtgcaggg ataactggca tggctcgaat
901 cgaccgtggg tgtctttcaa ccagaatctg gaatatcaga taggatacat atgcagtggg
961 attttcggag acaatccacg ccctaatgat aagacaggca gttgtggtcc agtatcgtct
1021 aatggagcaa atggagtaaa aggattttca ttcaaatacg gcaatggtgt ttggataggg
1081 agaactaaaa gcattagttc aagaaaaggt tttgagatga tttgggatcc aaatggatgg
1141 actgggacag acaataactt ctcaataaag caagatatcg taggaataaa tgagtggtca
1201 ggatatagcg ggagttttgt tcagcatcca gaactaacag ggctggattg tataagacct
1261 tgcttctggg ttgaactaat cagagggcga cccaaagaga acacaatctg gactagtggg
1321 agcagcatat ccttttgtgg tgtaaacagt gacactgtgg gttggtcttg gccagacggt
1381 gctgagttgc catttaccat tgacaagtaa (SEQ ID NO: 7)
Нуклеотидная последовательность гена химерного NS фрагмента (вставка выделена жирным шрифтом)
AGCAAAAGCAGGGTGACAAAGACATAATGGATCCAAACACTGTGTCAAGCTTTCAGGTAGATTGCTTTCTTTGGCATGTCCGCAAACGAGTTGCAGACCAAGAACTAGGTGATGCCCCATTCCTTGATCGGCTTCGCCGAGATCAGAAATCCCTAAGAGGAAGGGGCAGCACTCTTGGTCTGGACATCGAGACAGCCACACGTGCTGGAAAGCAGATAGTGGAGCGGATTCTGAAAGAAGAATCCGATGAGGCACTTAAAATGACCATGGCCTCTGTACCTGCGTCGCGTTACCTAACCGACATGACTCTTGAGGAAATGTCAAGGGAATGGTCCATGCTCATACCCAAGCAGAAAGTGGCAGGCCCTCTTTGTATCAGAATGGACCAGGCGATCATGGGAGGAGGTTTCTTCGGAGCTATTGCTGGTTTCTTGGAAGGAGGATGGGAAGGAATGATTGCAGGTTGGGGAGGAAGAGAGAGCCGGAACCCAGGGAATGCTTGATAATAAGCGGCCGCAGTGTGATTTTTGACCGGCTGGAGACTCTAATATTGCTAAGGGCTTTCACCGAAGAGGGAGCAATTGTTGGCGAAATTTCACCATTGCCTTCTCTTCCAGGACATACTAATGAGGATGTCAAAAATGCAATTGGGGTCCTCATCGGAGGACTTGAATGGAATGATAACACAGTTCGAGTCTCTAAAACTCTACAGAGATTCGCTTGGAGAAGCAGTAATGAGAATGGGAGACCTCCACTCACTCCAAAACAGAAACGGAAAATGGCGAGAACAATTAGGTCAAAAGTTCGAAGAAATAAGATGGCTGATTGAAGAAGTGAGACACAAATTGAAGATAACAGAGAATAGTTTTGAGCAAATAACATTTATACAAGCCTTACAGCTACTATTTGAAGTGGAACAAGAGATAAGAACTTTCTCGTTTCAGCTTATTTAATAATAAAAAACACCCTTGTTTCTACT (SEQ ID NO: 8)
Сборка рекомбинантных вирусов осуществлялась путем трансфекции клеток VERO восемью плазмидами, кодирующими геномные немодифицированные фрагменты вируса гриппа, и химерный NS геномный фрагмент, используя метод электропорации плазмидной ДНК (Cell Line Nucleofector® Kit V (Lonza)) в соответствии с инструкцией к применению. После трансфекции клетки инкубировались в среде Optipro (Invitrogen) в течение 96 часов при 34°C c добавлением трипсина 1 мкг/мл для обеспечения пост-трансляционного расщепления предшественника гемагглютинина на субъединицы НА1 и НА2. Вирусный урожай с клеток Vero использовался для заражения 10-дневных куриных эмбрионов (SPF). Эмбрионы инкубировались в течение 48 часов при 34°C, после чего аллантоисные жидкости, имеющие положительный титр в реакции гемагглютинации, использовались для проведения последующих пассажей на куриных эмбрионах. Аллантоисные жидкости 7 пассажа очищались с помощью тангенциальной фильтрации и лиофилизировались для хранения. Иммунизация животных проводилась после растворения лиофилизата дистилированной водой в эквивалентном объеме.
Пример 2
Протективный ответ в отношении гетерологичных штаммов вируса гриппа А и В при контрольном заражении мышей
С целью определения протективной активности в отношении гетерологичных антигенных вариантов вируса гриппа для иммунизации мышей был использован гриппозный вектор в дозе 6,8log ЭИД50 /мышь, который вводился интраназально в объеме 50 мкл под легкой анастезией, однократно или двукратно с периодом в 3 недели. Через 21 день после последней иммунизации животные были подвергнуты контрольному заражению патогенными для мышей штаммами вируса гриппа: гомологичным А/California/7/09 (H1N1pdm) или гетерологичными A/Aichi/2/68 (H3N2), A/Mississippi/85/1(H3N2) или вирусом гриппа В/Lee/40, взятыми в дозе соответствующей 3-5 ЛД50, соответственно.
Как видно из Фиг.3А, контрольное заражение не иммунных мышей вирусом H1N1pdm приводило к их гибели в 90% случаев. В тоже время, мыши, иммунизированные вирусным препаратом однократно или двукратно, были достоверно защищены от гибели.
Как видно из Фиг.3В, контрольное заражение не иммунных мышей вирусом A/Aichi/2/68 (H3N2) приводило к их гибели в 100% случаев. В тоже время, мыши, иммунизированные вирусным препаратом однократно или двукратно, были в достоверно защищены от гибели.
Как видно из Фиг.3С, контрольное заражение не иммунных мышей вирусом A/Mississippi/85/1(H3N2) приводило к их гибели в 100% случаев. В тоже время, мыши, иммунизированные вирусным препаратом двукратно имели 100% защиту.
Как видно из Фиг.3D, контрольное заражение не иммунных мышей вирусом гриппа В/Lee/40 приводило к гибели животных в 100% случаев. В тоже время, мыши, иммунизированные вирусным препаратом двукратно имели достоверно отличающуюся от контроля 60% защиту.
Таким образом, гриппозный вектор, несущий химерный NS геномный фрагмент, проявлял признаки универсальной гриппозной вакцины, эффективной в отношении как гетерологичных антигенных подтипов вируса гриппа А, так и в отношении вируса гриппа В.
Пример 3
Протективный ответ в отношении гетерологичного штамма вируса гриппа А (H3N2) при контрольном заражении хорьков
Хорьки являются оптимальной, рекомендованной ВОЗ моделью для изучения эффективности противогриппозных вакцин и препаратов. С целью определения протективной активности в отношении гетерологичного антигенного варианта вируса гриппа для иммунизации хорьков (9 на группу) был использован гриппозный вектор в дозе 7,5 log ЭИД50/хорька, который вводился интраназально в объеме 500 мкл под легкой анастезией, однократно или двукратно с периодом в 3 недели. Через 21 день после последней иммунизации животные были подвергнуты контрольному заражению патогенным для хорьков вирусом гриппа A/St.Petersburg/224/2015 (H3N2). Как видно из Фиг.4А, контрольное заражение не иммунных животных приводило к подъему температуры тела на 2 сутки после заражения, в то время как вакцинированные животные не имели температурной реакции.
С целью изучения эффекта вакцинации на репродукцию контрольного вируса в респираторном тракте хорьков у животных были взяты носовые смывы на 2,4 и 6 день с целью определения в них концентрации инфекционного вируса путем титрования 50% цитопатической дозы в культуре клеток MDCK. Как видно из Фиг.4В, контрольное заражение не иммунных хорьков приводило к активной репродукции вируса без существенного снижения титров к 6 дню. При однократной иммунизации хорьков достоверное снижение вирусного титра наблюдалось на 4 и 6 день после контрольного заражения. После двукратной иммунизации достоверное, более чем 100-кратное, снижение вирусного титра регистрировалось уже на 2 день после заражения животных.
Таким образом, даже однократная вакцинация хорьков гриппозным вектором приводила к защите животных от клинических проявлений в виде температурной реакции и способствовала ускоренной элиминации контрольного гетерологичного штамма из респираторного тракта. Повторная иммунизация способствовала ускорению процесса вирусной элиминации.
Пример 4
Состав вакцины на основе гриппозного вектора для интраназальной иммунизации
Вакцина, где гриппозный вектор содержится в количестве от 6,5 до 8,5 log эмбриональных 50% инфекционных доз (ЭИД50)/мл, и буферный стабилизирующий раствор содержит 0,9 масс.% раствора хлорида натрия, 0,5 масс.% L-карнозина, 2,5 масс.% сахарозы, 1 масс.% рекомбинантного альбумина, 1 масс.% L-аргинина и 3 масс.% гидроксиэтилированного крахмала 130/0,4 (молекулярная масса - 130 кДа, степень молярного замещения 0,4).




Изобретения относятся к области медицины и вирусологии. Представлены аттенуированный гриппозный вектор, индуцирующий кросс-протективный ответ против вирусов гриппа А и В, содержащие его фармацевтическая и вакцинная композиции и их применение. Охарактеризованный аттенуированный гриппозный вектор содержит нуклеотидные последовательности генов белков РВ2, РВ1, PA, NP и M, происходящие от вируса гриппа A/PR/8/34 (H1N1), нуклеотидные последовательности генов белков HA и N, происходящие от вируса гриппа A/California/7/09-like (H1N1pdm), нуклеотидную последовательность химерного гена белка NS, включающую укороченную рамку считывания белка NS1 размером 124 аминокислоты, происходящую от вируса A/PR/8/34 (H1N1) и продолженную вставкой нуклеотидной последовательности, кодирующей пептид слияния субъединицы HA2 от вируса гриппа В, и нуклеотидной последовательности, кодирующей консервативный В-клеточный эпитоп нуклеопротеина (ТЗ) вируса гриппа А, и последовательность гена белка Nep, происходящую от вируса А/Singapore/1/57–like (H2N2). Изобретения могут применяться для профилактики гриппа А и В. 5 н. и 11 з.п. ф-лы, 15 ил., 4 пр.
1. Аттенуированный гриппозный вектор, индуцирующий кросс-протективный ответ против вирусов гриппа A и В, содержащий:
нуклеотидную последовательность гена белка PB2, происходящую от вируса гриппа A/PR/8/34 (H1N1), или нуклеотидную последовательность, имеющую по меньшей мере 95% идентичность указанной нуклеотидной последовательности гена белка PB2;
нуклеотидную последовательность гена белка PB1, происходящую от вируса гриппа A/PR/8/34 (H1N1), или нуклеотидную последовательность, имеющую по меньшей мере 95% идентичность указанной нуклеотидной последовательности гена белка PB1;
нуклеотидную последовательность гена белка PA, происходящую от вируса гриппа A/PR/8/34 (H1N1), или нуклеотидную последовательность, имеющую по меньшей мере 95% идентичность указанной последовательности белка РА;
нуклеотидную последовательность гена белка NP, происходящую от вируса гриппа A/PR/8/34 (H1N1), или нуклеотидную последовательность, имеющую по меньшей мере 95% идентичность указанной последовательности белка NP;
нуклеотидную последовательность гена белка M, происходящую от вируса гриппа A/PR/8/34 (H1N1), или нуклеотидную последовательность, имеющую по меньшей мере 95% идентичность указанной последовательности гена белка М;
нуклеотидную последовательность гена белка HA, происходящую от вируса гриппа A/California/7/09-like (H1N1pdm), или нуклеотидную последовательность, имеющую по меньшей мере 95% идентичность указанной нуклеотидной последовательности гена белка НА;
нуклеотидную последовательности гена белка NA, происходящую от вируса гриппа A/California/7/09-like (H1N1pdm), или нуклеотидную последовательность, имеющую по меньшей мере 95% идентичность указанной нуклеотидной последовательности гена белка NA; и
нуклеотидную последовательность химерного гена белка NS, включающую
рамку считывания белка NS1, происходящую от вируса A/PR/8/34 (H1N1), где указанная рамка считывания является укороченной и кодирует белок NS1 размером 124 аминокислотных остатков,
и последовательность гена белка Nep, происходящую от вируса гриппа А/Singapore/1/57–like (H2N2), или
нуклеотидную последовательность, имеющую по меньшей мере 95% идентичность указанной последовательности химерного гена NS;
где указанная укороченная рамка считывания гена белка NS1 продолжена вставкой нуклеотидной последовательности, кодирующей пептид слияния субъединицы НА2 от вируса гриппа В, и нуклеотидной последовательности, кодирующей консервативный В-клеточный эпитоп нуклеопротеина (NP) вируса гриппа А.
2. Вектор по п. 1, где нуклеотидная последовательность химерного гена белка NS представлена в SEQ ID NO: 8.
3. Фармацевтическая композиция для профилактики гриппа А и В, содержащая в эффективном количестве аттенуированный гриппозный вектор по п. 1 или 2 и фармацевтически приемлемый носитель.
4. Фармацевтическая композиция по п. 3, содержащая 6.5–8,5 logЭИД 50/мл аттенуированного гриппозного вектора по п. 1 или 2 и буферный раствор, содержащий 0-1,5 масс.% моновалентной соли, 0-5 масс.% L-карнозина, 0-5 масс.% углеводного компонента, 0-2 масс.% белкового компонента, 0-2 масс.% аминокислотного компонента и 0-10 масс.% гидроксиэтилированного крахмала.
5. Фармацевтическая композиция по п. 4, где буферный раствор содержит 0,5-1,5 масс.% моновалентной соли, 0,01-5 масс.% L-карнозина, 1-5 масс.% углеводного компонента, 0,1-2 масс.% белкового компонента, 0,01-2 масс. % аминокислотного компонента и 1-10 масс.% гидроксиэтилированного крахмала.
6. Фармацевтическая композиция по п. 5, где моновалентная соль представляет собой хлорид натрия, углеводный компонент представляет собой сахарозу, трегалозу или лактозу, белковый компонент представляет собой человеческий рекомбинантный альбумин, казитон, гидролизат лактальбумина или желатин, аминокислотный компонент представляет собой аргинин, глицин или глутамат натрия.
7. Фармацевтическая композиция по любому из пп. 3-6, где субъект представляет собой млекопитающее или птицу.
8. Фармацевтическая композиция по п. 7, где субъект представляет собой человека.
9. Вакцина против гриппа А и В, содержащая в эффективном количестве аттенуированный гриппозный вектор по п. 1 или 2 и фармацевтически приемлемый носитель.
10. Вакцина по п. 9, содержащая 6,5–8,5 logЭИД 50/мл аттенуированного вируса гриппа А по любому из пп. 1-5 или аттенуированного гриппозного вектора по любому из пп. 6-14 и буферный раствор, содержащий 0-1,5 масс.% моновалентной соли, 0-5 масс.% L-карнозина, 0-5 масс.% углеводного компонента, 0-2 масс.% белкового компонента, 0-2 масс.% аминокислотного компонента и 0-10 масс.% гидроксиэтилированного крахмала.
11. Вакцина по п. 10, где буферный раствор содержит 0,5-1,5 масс.% моновалентной соли, 0,01-5 масс.% L-карнозина, 1-5 масс.% углеводного компонента, 0,1-2 масс.% белкового компонента, 0,01-2 масс. % аминокислотного компонента и 1-10 масс.% гидроксиэтилированного крахмала.
12. Вакцина по п. 11, где моновалентная соль представляет собой хлорид натрия, углеводный компонент представляет собой сахарозу, трегалозу или лактозу, белковый компонент представляет собой человеческий рекомбинантный альбумин, казитон, гидролизат лактальбумина или желатин, аминокислотный компонент представляет собой аргинин, глицин или глутамат натрия.
13. Применение аттенуированного гриппозного вектора по п. 1 или 2 для профилактики гриппа А и В у субъекта.
14. Применение фармацевтической композиции по любому из пп. 3-8 для профилактики гриппа А и В у субъекта.
15. Применение по п. 13 или 14, где субъект представляет собой млекопитающее или птицу.
16. Применение по п. 13 или 14, где субъект представляет собой человека.
| Приспособление для уплотнения раструбных стыков нормальных чугунных водопроводных труб | 1929 |
|
SU12965A1 |
| US 20090053264 A1, 26.02.2009 | |||
| US 0006800288 B2, 05.10.2004 | |||
| US 0007037707 B2, 02.05.2006. | |||

