Изобретение относится к медицине и может быть использовано для получения рекомбинантных вакцинных штаммов для профилактики и лечения туберкулеза.
Настоящее изобретение направлено на решение одной из важнейших задач современной медицины - создание новой, более эффективной, чем БЦЖ, вакцины против туберкулеза. Необходимость создания такой вакцины продиктована несколькими причинами. Во-первых, единственная применяемая в настоящее время вакцина БЦЖ эффективна для профилактики туберкулеза у детей, но, по многочисленным данным, не способна предотвращать реактивацию заболевания у взрослых людей, которая является причиной большинства случаев легочного туберкулеза (Trends Immunol, 2001; 22:160-8). Во-вторых, введение живой вакцины БЦЖ может представлять серьезную угрозу для привитых с нарушениями иммунитета, например, у ВИЧ-инфицированных больных в стадии СПИДа. В этих случаях аттенуированный штамм микобактерии способен вызывать истинный инфекционный процесс с тяжелыми осложнениями (Clin. Infect. Dis., 1997; 24:1139-46).
В настоящее время известны несколько новых подходов к усовершенствованию существующей БЦЖ-вакцины:
1) рекомбинантные аттенуированные микобактерии, представляющие из себя сконструированные генно-инженерными методами делеционные мутанты вирулентных штаммов микобактерии туберкулеза (Science, 1998; 282:759-762). В отличие от БЦЖ, полученной эмпирическим путем, данные штаммы несут меньшие по протяженности делеции участков генома, ответственных за факторы вирулентности, но экспрессируют важные протективные антигены М. tuberculosis и М. bovis, утраченные БЦЖ. В то же время такие вакцины потенциально небезопасны для людей с иммуносупрессией;
2) рекомбинантные БЦЖ-вакцины, экспрессирующие дополнительные протективные белки, утраченные в процессе длительного культивирования данного штамма (Nature medicine, 2003; V.9, N5: 533-539). Данному типу вакцин присущи недостатки, свойственные вакцине БЦЖ, в плане ее небезопасности;
3) субъединичные белковые препараты на основе протективных белков, вводимые парентерально. Следует отметить, что имеющийся опыт по созданию субъединичных и пептидных вакцин, указывает на необходимость введения в состав вакцин мощных адъювантов, поскольку сами по себе белковые или пептидные антигены недостаточно иммуногенны и часто создают лишь кратковременный иммунитет даже при многократных иммунизациях (Infect. Immun., 2000; 68:791-795). Другим недостатком является неспособность подобного типа вакцин стимулировать выработку цитотоксического клеточного иммунитета;
4) ДНК-вакцины. Предназначены для внутриклеточной экспрессии протекивных антигенов и поэтому способны стимулировать Т-клеточное звено иммунитета (Infect. Immun., 1999; 67: 4780-4786). Однако ДНК-вакцины низкоиммуногенны, требуют многократного введения и до сих пор не были лицензированы для применения у людей.
Альтернативой этим подходам, позволяющей повысить иммуногенность протективных микобактериальных антигенов, может служить использование векторной системы на основе рекомбинантного аттенуированного гриппозного вируса, экспрессирующего ранний секреторный микобактериальный антиген ESAT-6, в качестве интраназальной вакцины. С данным антигеном связывают активацию лимфоцитов и начало продукции интерферона, которые играют важную роль в активации макрофагов, тем самым оказывая патогенетическое влияние на формирование иммунитета при инфицировании микобактериями туберкулеза (Infect. Immun., 62:2536-2544).
В отличие от вакцины БЦЖ, предназначенной, прежде всего, для парентерального введения, гриппозная векторная вакцина может применяться интраназально в виде капель или аэрозоля. В результате достигается выработка иммунитета слизистых оболочек (мукозный иммунитет) во входных воротах туберкулезной инфекции. Существование множества антигенных подтипов вируса гриппа позволяет рассчитывать на возможность проведения нескольких иммунизации для достижения максимального профилактического или лечебного эффекта.
Цель изобретения - создание рекомбинантных вакцинных штаммов, обеспечивающих развитие протективного иммунитета против туберкулеза при использовании их в качестве профилактической вакцины или служащих цели лечения туберкулеза, то есть использование их в качестве терапевтической вакцины.
Задача решена с помощью методов обратной генетики, а именно созданием живых рекомбинантных штаммов вируса гриппа А (гриппозных векторов), содержащих вставку чужеродной генетической последовательности в гене NS1 вируса гриппа и экспрессирующих протективные антигены Mycobacterium tuberculosis.
Сущностью данного изобретения является получение химерного продукта на основе штаммов вируса гриппа А различных антигенных подтипов и посторонней генетической последовательности, иммунодоминантного микобактериального белка ESAT-6, применимого в медицинской практике.
Достижение основной цели изобретения иллюстрируется следующими примерами.
ПОЛУЧЕНИЕ И ХАРАКТЕРИСТИКА РЕКОМБИНАНТНЫХ ШТАММОВ ВИРУСА ГРИППА.
Рекомбинантные штаммы вирусы гриппа, экспрессирующие микобактериальный антиген ESAT-6, получены с помощью методов обратной генетики (J. Virol., 1996; 70: 4188-4192; Adv. Vims Res., 1999; 53: 265-300; J. Virol., 1999; 73: 9679-82).
Конструирование плазмид. Геномная РНК вируса гриппа A/PR8/34 была экстрагирована с помощью реагента Ultraspec (Biotecx Laboratories). кДНК копия NS гена была получена с помощью обратной транскрипции с использованием праймера (5'-ACTACTTCTAGAGAAGACAAAGCAAAAGCAGGGTGACA-3') и M-MuLV обратной транскриптазы (Fermentas). Полученная кДНК использовалась для амплификации фрагмента NS1 гена в полимеразной цепной реакции (ПЦР) с помощью следующих праймеров:
прямой: (5'-ACTACTTCTAGAGAAGACAAAGCAAAAGCAGGGTGACA-3') и
обратный: (5'-ACTACTCTGCAGATTAACCCTCACTAAAAGTAGAAACAAG-3').
Амплифицированный фрагмент был клонирован в плазмидный вектор pQBI 25/50-fC1(Qubiogene) по тупым концам между Poll промотором и последовательностью рибозима гепатита дельта (HDV), играющего роль терминатора транскрипции (J. Virol., 1999; 73:9679-9682; Adv. Virus Res., 1999; 53:265-300; J.Virol, 1996; 70(6): 4188-4192). Полученная плазмида получила название pPolI-NS-HDV.
Синтез нуклеотидной последовательности ESAT-6 был осуществлен путем гибридизации эквимолярного количества шести синтетических перекрывающихся олигонуклеотидов (по 10 пкмоль/мл каждого) (CODON Genetic Systems) при нагревании до температуры 100°С в течение 4 минут с последующим медленным охлаждением смеси до комнатной температуры.
Полученный ДНК-фрагмент, 288 нуклеотидов в длину, амплифицировали с помощью концевых праймеров в ПЦР (прямой праймер: 5'ATGACAGAGCAGCAGTG 3' и обратный праймер: 3'CTATGCGAACATCCCAG 5'), и клонировали в плазмиду pPolI-NS-HDV между нуклеотидами 400-401 в открытой рамке считывания NS1 гена вируса гриппа без удаления участков гена. Данная конструкция получила название pPol-NS-ESAT-6-HDV.
С целью посттрансляционного разделения ESAT-6 белка с N-терминальной областью NS1 белка, между ними была вставлена синтезированная последовательность (51 нуклеотидов), кодирующая участок сайта узнавания клеточными протеазами белка 2А (NFDLLKLAGDVESNLG/P) пикорновируса, который обладает свойством саморазрезания перед последним пролином (J. ViroL, 1994; 68:4486-4492). Данная конструкция получила название pPol-NS125-2A-ESAT-6-HDV.
Структура рекомбинантного NS1 гена представлена на схеме (Фиг.1). После нуклеотидной последовательности NS1 гена, кодирующей 125 аминокислот белка NS1 вируса гриппа, следует последовательность, кодирующая участок белка 2А (NFDLLKLAGDVESNLG/P) пикорновируса вместе с последовательностью ESAT-6 (95 аминокислот). Стрелкой отмечен сайт саморазрезания перед последним пролином, обеспечивающий посттрансляционный выход ESAT-6 белка.
Экспрессионные плазмиды для РВ1, РВ2, РА и NP белков вируса гриппа были получены путем клонирования соответствующих PR8 генов в плазмиду pTriEx-1 (Novagene) под активный промотор цыпленка и названы pTriEx-PB1, pTriEx-PB2, pTriEx-PA и pTriEx-NP соответственно.
Получение рекомбинантных штаммов вирусов гриппа. Система получения вирусов гриппа (H1N1, H2N2), содержащих рекомбинантный NS1 ген представлена на схеме (Фиг.2). Клетки Vero (American Type Culture Collection (ATCC) были трансфецированы плазмидой pPol-NS125-2A-ESAT-6-HDV (1), экспрессирующей химерную РНК NS гена вируса гриппа (2), совместно с набором плазмид pTriEx-PBl, pTriEx-PB2, pTriEx-PA, и pTriEx-NP, экспрессирующих белки полимеразного комплекса вируса гриппа (по 1 мкг каждой плазмиды). Трансфекцию проводили путем электропарации (Nucleofection technique, Amaxa) в соответствии с инструкцией по использованию. 24 часа спустя трансфецированные клетки Vero были заражены хелперными вирусами гриппа A/PR/8-delNS (H1N1) или A/Sing/delNS87 (H2N2) (3), которые содержали частично или полностью удаленный NS1 ген и потому были чувствительны к действию интерферона (Virology; 2004, 324(1):67-73). Через 8 часов после заражения в качестве селективного препарата добавляли человеческий лейкоцитарный интерферон-альфа (NIBSC 1st International Standard 1999; 3 ед./мл культуральной среды) и инкубировали клетки в течение 48 часов при 37°С. Полученный урожай вирусов был дважды пропассирован на клетках Vero в присутствии интерферона-альфа в концентрации 3 ед./мл. Конечная популяция вирусов (4) содержала рекомбинантный NS1 ген со вставкой ESAT-6, что подтверждалось секвенированием продукта ОТ-ПЦР. После проведения двукратной очистки вирусы изолировали.
Рекомбинантный вирус гриппа, относящийся к подтипу H1N1, получил название FLU/H1-ESAT-6, рекомбинантный вирус гриппа, относящийся к подтипу H2N2 получил название FLU/H2-ESAT-6.
Рекомбинантный вирус гриппа подтипа H3N2 был получен путем генетической реассортации вируса FLU/H1-ESAT-6 с вирусом A/Aichi/1/68 (H3N2) на клетках Vero в присутствии кроличьей антисыворотки к вирусу A/PR/8/34 (H1N1). Полученный вирусный клон содержал 6 фрагментов генома (РВ2, РВ1, PA, NP, M, recNS) от вектора A/PR8-2A-ESAT-6, а гены гликопротеинов (НА, NA) от штамма A/Aichi/1/68 (H3N2). Генотипирование реассортантов проводилось методом ОТ-ПЦР с последующим сравнительным рестрикционным анализом кДНК копий каждого сегмента. Полученный вирус получил название FLU/H3-ESAT-6.
Пример 1. Рекомбинантный вирус A/PR8-2A-ESAT-6 (H1N1) получен с помощью методов обратной генетики на клеточной культуре Vero и характеризуется наличием рекомбинантного NS1 гена, содержащим вставку нуклеотидной последовательности ESAT-6 M. tuberculosis между позициями 400-401 открытой рамки считывания NS гена, что подтверждено секвенированием продукта ОТ-ПЦР.
Вследствие наличия вставки, вирус A/PR8-2A-ESAT-6 отличается от вируса гриппа дикого типа наличием увеличенного размера геномного фрагмента NS 1, что было определено с помощью электрофореза в агарозном геле после амплификации в ПЦР со специфическими праймерами:
5'-ACTACTTCTAGAGAAGACAAAGCAAAAGCAGGGTGACA-3' и 5'-ACTACTCTGCAGATTAACCCTCACTAAAAGTAGAAACAAG-3' (Фиг.3). На чертеже видно, что размер амплифицированного рекомбинантного фрагмента NS 1 превышает размер амплифицированного фрагмента NS1 гена вируса гриппа дикого типа на расчетную величину.
Другим генетическим маркером, определяющим рекомбинантную природу данного вируса, является экспрессия ESAT-6 белка, определяемого при иммунофлюоресцентном анализе зараженных клеток с помощью моноклональных антител (Фиг.4А). На чертеже отчетливо видно свечение ESAT-6 белка, накопившегося в ядре зараженных рекомбинантным вирусом клеток Vero.
Экспрессия вставки ESAT-6 при заражении клеток Vero подтверждена также анализом лизатов зараженных вирусом клеток методом Вестернблот с использованием моноклональных антител против белка ESAT-6 (Фиг.4В). На чертеже видны окрашенные зоны, соответствующие продуктам с молекулярным весом в 26 kDa и 24 kDa. Наличие нескольких зон определяется посттрансляционным отщеплением карбоксильной части ESAT-6, ранее показанным для этого белка.
Полученный вирус способен размножаться в культуре клеток Vero и MDCK до титров 7.5 log БОЕ/мл и в 10 дневных куриных эмбрионах до титра 9.2 log ЭИД 50/мл при оптимальной температуре 37°С. Вирус не является температурочувствительным и репродуцируются до аналогичных титров при температуре 39°С.
При интраназальном заражении мышей в дозе 5.5 log БОЕ/мышь полученный вирус обладает способностью размножения в легких животных до титров 1,5 БОЕ/мл 10% суспензии легких, не вызывая при этом гибели животных т.е. является аттенуированным по сравнению с исходным родительским штаммом. Отсутствие патологии в легких иммунизированных животных и отсутствие снижения веса мышей подтверждают аттенуацию. Вирус обладает генетической стабильностью в течение, как минимум, 5 последовательных пассажей в культуре клеток Vero.
Характеристика полученного авторами рекомбинантного штамма вируса гриппа A/PR8-2A-ESAT-6 (H1N1) приведена в прилагаемом удостоверении о депонировании штамма (№147) в коллекцию Музея вирусов ГУ НИИ гриппа РАМН.
Пример 2. Рекомбинантный вирус A/Singapore/-2A-ESAT-6 (H2N2) получен с помощью методов обратной генетики на клеточной культуре Vero и характеризуется наличием рекомбинантного NS1 гена, содержащим вставку нуклеотидной последовательности ESAT-6 M.tuberculosis между позициями 400-401 открытой рамки считывания NS гена, что подтверждено секвенированием продукта ОТ-ПЦР.
При заражении клеток Vero данным вирусным штаммом наблюдается экспрессия ESAT-6 белка, что подтверждено иммунофлюоресцентным анализом и иммуноблотингом лизатов зараженных клеток с использованием моноклональных антител к ESAT-6 белку (Фиг.4).
Полученный вирус способен размножаться в культуре клеток Vero и MDCK до титров 7.5 log БОЕ/мл и в 10 дневных куриных эмбрионах до титра 8 log ЭИД 50/мл при оптимальной температуре 37°С. Вирус не является температурочувствительным и репродуцируются до аналогичных титров при температуре 39°С.
При интраназальном заражении мышей в дозе 5.5 log БОЕ/мышь полученный вирус обладает способностью размножения в легких животных до титров 3,5 БОЕ/мл 10% суспензии легких, не вызывая при этом гибели животных т.е. является аттенуированным по сравнению с исходным родительским штаммом.
Вирус обладает генетической стабильностью в течение, как минимум, 5 последовательных пассажей в культуре клеток Vero.
Характеристика полученного авторами рекомбинантного штамма вируса гриппа A/Singapore/-2A-ESAT-6 (H2N2) приведена в прилагаемом удостоверении о депонировании штамма (№149) в коллекцию Музея вирусов ГУ НИИ гриппа РАМН.
Пример 3. Рекомбинантный вирус гриппа A/Aichi-2A-ESAT-6 (H3N2) был получен путем генетической реассортации вируса A/PR8-2A-ESAT-6 с вирусом A/Aichi/1/68 (H3N2) на клетках Vero в присутствии кроличьей антисыворотки к вирусу A/PR/8/34 (H1N1). Полученный вирусный клон содержал 6 фрагментов генома (РВ2, РВ1, PA, NP, M, recNS) от вектора A/PR8-2A-ESAT-6, а гены гликопротеинов (НА, NA) от штамма A/Aichi/1/68 (H3N2).
Вследствие наличия вставки последовательности ESAT-6 полученный вирус отличался от вируса гриппа дикого типа наличием увеличенного размера геномного фрагмента NS1, что было определено с помощью электрофореза в агарозном геле после его амплификации в ПНР со специфическими праймерами:
5'-ACTACTTCTAGAGAAGACAAAGCAAAAGCAGGGTGACA-3' и
5'-ACTACTCTGCAGATTAACCCTCACTAAAAGTAGAAACAAG-3' (Фиг.3). На чертеже видно, что амплифицированный рекомбинантный фрагмент NS1 имеет вставку последовательности ESAT-6, превышая размер NS1 гена дикого типа вируса на величину размера вставки.
Данный вирус является температурочувствительным и адаптирован к росту в культуре клеток Vero.
При заражении клеток Vero вирусным штаммом A/Aichi-2A-ESAT-6 (H3N2) наблюдается экспрессия ESAT-6 белка, что подтверждено иммунофлюоресцентным анализом и иммуноблотингом лизатов зараженных клеток с использованием моноклональных антител к ESAT-6 белку (Фиг.4).
Полученный вирус способен размножаться в культуре клеток Vero и MDCK до титров 7.5 log БОЕ/мл при оптимальной температуре 34°С, репродуцируется до титров 3 log БОЕ/мл при температуре 39°С.
При интраназальном заражении мышей в дозе 5.5 log БОЕ/мышь полученный вирус обладает способностью размножения в легких животных до титров 1,5 БОЕ/мл 10% суспензии легких, не вызывая при этом гибели животных т.е. является аттенуированным по сравнению с исходным родительским штаммом. Отсутствие патологии в легких иммунизированных животных и отсутствие снижения веса мышей подтверждают аттенуацию.
Вирус обладает генетической стабильностью в течение, как минимум, 5 последовательных пассажей в культуре клеток Vero.
Характеристика полученного авторами рекомбинантного штамма вируса гриппа A/Aichi-2A-ESAT-6 (H3N2) приведена в прилагаемом удостоверении о депонировании штамма (№148) в коллекцию Музея вирусов ГУ НИИ гриппа РАМН.
СПОСОБ ПРОФИЛАКТИКИ ТУБЕРКУЛЕЗА ЛЕГКИХ.
В настоящее время отмечается значительное ухудшение эпидемиологической обстановки по туберкулезу в России и во всем мире в связи с неблагоприятной экологической обстановкой, негативными социальными факторами, ростом иммунодефицитных заболеваний различного генеза и отсутствием надежной специфической профилактики заболевания.
Классическая вакцина БЦЖ, представляющая собой живой аттенуированный штамм M.bovis, эффективна для профилактики туберкулеза у детей, но не способна предотвращать реактивацию заболевания у взрослых людей, которая является причиной большинства случаев легочного туберкулеза (Trends Immunol. 2001; 22:160-8).
Известно, что протективный эффект при развитии туберкулезной инфекции обеспечивают цитокины, вырабатываемые Т-хелперами 1 типа (ТХ-1) - гамма-интерферон (ИФН-γ) интерлейкин-2 (ИЛ-2), которые являются медиаторами клеточно-опосредованного иммунитета. При активной прогрессирующей инфекции обнаруживается стимуляция Т-2 хелперной субпопуляции лимфоцитов (ТХ-2), с высоким уровнем продукции ИЛ-4, расцениваемой в последнее время как показатель снижения сопротивляемости к туберкулезу и о снижении активности CД4+ ТХ-1.
Среди многообразия предположений, объясняющих неспособность вакцины БЦЖ защитить популяцию взрослых от туберкулеза, можно выделить отсутствие важных (протективных) антигенов в геноме БЦЖ (в том числе ESAT-6), неспособность стимулировать оптимальное сочетание субпопуляций Т-лимфоцитов для создания протекции, нежелательную стимуляцию выработки ИЛ-4.
К тому же при введении вакцины БЦЖ иммунная система организма человека сталкивается с исключительно сложным набором антигенов, конкурирующих за презентирующие клетки, а иммунодоминирующие антигены не всегда индуцируют максимальную протекцию (J.Immunol, 1996; 157:3039-3045) или их экспрессия транзиторна (Parasite Immunol., 1993; 15:187-193).
Обоснованным и перспективным направлением при разработке новых вакцин против туберкулеза является формирование Th1 иммунного ответа, характерного для благоприятного течения и обратного развития инфекции, обеспечение его высокой активности и целенаправленного переключения на Тh1 в случае доминирования Th2 цитокинов.
Пример 4. Влияние профилактической иммунизации рекомбинантными гриппозными векторами на развитие и течение острой туберкулезной инфекции у мышей.
Исследование проведено на 240 беспородных белых мышах-самцах с исходной массой тела 18-20 г, полученных из питомника «Рапполово», Всеволожского района. Ленинградской области. Для иммунизации использовались вируссодержащие суспензии рекомбинантных гриппозных векторов FLU/ESAT-6, относящихся к различным антигенным подтипам вируса гриппа A (H1N1, H3N2 и H2N2).
Животные были распределены по 5-и группам. В первой группе мышей двукратно иммунизировали смесью трех рекомбинантных гриппозных векторов различных антигенных подтипов (FLU/ESAT-6(Tri+Tri)). Во второй группе первая иммунизация проводилась рекомбинантным гриппозным вектором подтипа H1N1, вторая - гриппозным вектором разновидности H3N2 (FLU/ESAT-6(H1+H3)). В третьей опытной группе первая иммунизация проводилась рекомбинантным гриппозным вектором подтипа H1N1, вторая - гриппозным вектором разновидности H2N2 (FLU/ESAT-6(H1+H2)). Иммунизацию проводили интраназально, без наркоза, в дозе 106 БОЕ/мышь, с интервалом в 3 недели. Контролем служили интактные животные и невакцинированные зараженные МБТ мыши (контроль заражения).
Генерализованный туберкулез вызывали введением в латеральную хвостовую вену мышей 0,2 мл взвеси трехнедельной культуры М. bovis bovinus 8 в дозе 0,1 мг/мышь через 6 недель после первой иммунизации. Использовался вирулентный штамм М. bovis bovinus 8, полученный в виде лиофилизированной культуры из ГосНИИ стандартизации и контроля медицинских биологических препаратов им. Тарасевича (лаборатория препаратов для профилактики и диагностики туберкулеза и микобактериозов). При заражении белых беспородных мышей этим штаммом очаги специфического воспаления в легких образуются на 14-20 день, а гибель животных отмечается на 30-45 день с момента инокуляции инфекта, что близко к соответствующим показателям у мышей, зараженных М. tuberculosis H37Rv (Першин Г.Н., 1971).
Эффект протективного действия вакцины оценивали по следующим показателям тяжести течения экспериментального туберкулезного процесса: динамика массы мышей (взвешивание 1 раз в неделю), макроскопическая оценка легких и селезенки (индексы поражения легких, коэффициенты масс легких и селезенки), гистологическое исследование срезов легких. Сравнение с контрольными показателями (интактные животные и контроль заражения) проводили в динамике через 30 и 40 дней после заражения МБТ.
Индекс поражения легких высчитывали в соответствии с количеством и выраженностью очагов специфического воспаления, выявляемых при визуальном осмотре. При этом единичные субмилиарные очаги оценивались в 0,5 балла; многочисленные субмилиарные очаги (не более 20) - в 1 балл; многочисленные субмилиарные (более 20) - в 1,5 балла; единичные милиарные - в 1,75 балла; многочисленные сливающиеся субмилиарные и единичные милиарные - в 2 балла; милиарные (не более 10) - в 2,25 балла; многочисленные милиарные, сливающиеся - 2,75 балла, появление мелких казеозных некротических фокусов - в 3 балла; обширный казеоз - 4 балла; сплошное поражение легких - 5 баллов. В случаях серозного пропитывания ткани легких к вычисленному индексу поражения прибавлялись от 0,25 до 1 баллов в зависимости от площади поражения.
Коэффициенты масс органов вычисляли исходя из соотношения массы органа и массы тела животного (КМ=масса органа (г)×100/масса животного (г)) и выражали в условных единицах (усл. ед.).
Для гистологического изучения легких орган фиксировали в 10% формалине. После фиксации и проводки материал заливали в парафин и из парафиновых блоков изготовляли ступенчатые гистологические срезы. Использовались окраски гематоксилином и эозином.
Полученные результаты подвергали статистической обработке, достоверность различий оценивали по непарному t-критерию Стьюдента.
Анализ полученных результатов показал, что у мышей, двукратно иммунизированных рекомбинантными гриппозными векторами, развитие экспериментальной туберкулезной инфекции замедлено, а течение - более благоприятно, чем у зараженных невакцинированных животных.
На 30-й день после заражения у вакцинированных FLU/ESAT-6 мышей отмечалась тенденция к снижению коэффициентов массы органов (легких и селезенки) и статистически значимое (р<0,001, р<0,02 и р<0,05) уменьшение индексов поражения легких по сравнению с зараженными невакцинированными животными, что отражено в таблице 1.
Показатели тяжести течения экспериментального туберкулеза на 30-й день после заражения МБТ
На 40-й день после заражения протективный эффект отмечен при использовании трех вариантов исследуемой вакцины. Коэффициенты массы легких, как и на предыдущем сроке наблюдения, имели лишь тенденцию к снижению, а индекс поражения легких в двух опытных группах был достоверно ниже, чем в контрольной группе (р<0,05). К этому времени у мышей, иммунизированных трехвалентной гриппозной вакциной и вакциной FLU/ESAT-6 (H1+H2), отмечено и снижение коэффициента массы селезенки (р<0,02), что отражено в таблице 2.
Показатели тяжести течения экспериментального туберкулеза на 40-й день после заражения МБТ
При этом в легких мышей контрольной группы определялись множественные, хорошо выраженные туберкулезные очаги, большое количество крупных сливных некротических очагов, что отражено на макрофотографиях легких мышей сравниваемых групп (Фиг.5А). Туберкулезные изменения в легких вакцинированных животных были выражены слабее и были представлены единичными некрупными туберкулезными очагами серого цвета (Фиг.5 В). Необходимо отметить, что по величине индекса поражения легких ко второму контрольному сроку (40-й день после заражения) во всех группах животных отмечалось нарастание распространенности специфического поражения легких, но у вакцинированных мышей оно было менее выражено и отсрочено по времени (табл.1, 2).
Исследование динамики массы мышей выявило, что к окончанию срока наблюдения масса тела вакцинированных FLU/ESAT-6 (Tri+Tri) животных соответствовала данным мышей интактной группы (27,9 г против 28,4 г) (Фиг.6). При этом процент прибавки массы тела у вакцинированных мышей ко дню забоя (12,9-35,4%) был в 1,9 (FLU/ESAT-6 (H1+H3), в 2,8 (FLU/ESAT-6 (H1+H2) и в 5,2 (FLU/ESAT-6 (Tri+Tri) раз соответственно выше, чем в контрольной группе (7,8%), что свидетельствует о меньшей выраженности симптомов интоксикации в обеих группах вакцинированных мышей.
Гистологическое исследование срезов легких на 40-й день после заражения показало, что у всех зараженных животных в ткани легкого имелись специфические инфильтративные изменения различной степени выраженности с накоплением в альвеолах серозного, а чаще фибринозного экссудата, крупных вакуолизированных макрофагов и лимфоидных элементов (альвеолярно-макрофагальный грануломатоз). У зараженных невакцинированных мышей они были представлены полями и крупными сливными очагами инфильтрации (табл.3). При этом преобладали свежие инфильтративные изменения, распространяющиеся как на воздушную паренхиму, так и на перибронхиальные и периваскулярные пространства. Альвеолы, межальвеолярные перегородки и просветы расширенных капилляров были инфильтрированы скоплениями макрофагов, эпителиоидных клеток, лимфоидными элементами.
Распространенность специфического поражения легочной ткани у мышей с экспериментальным туберкулезом
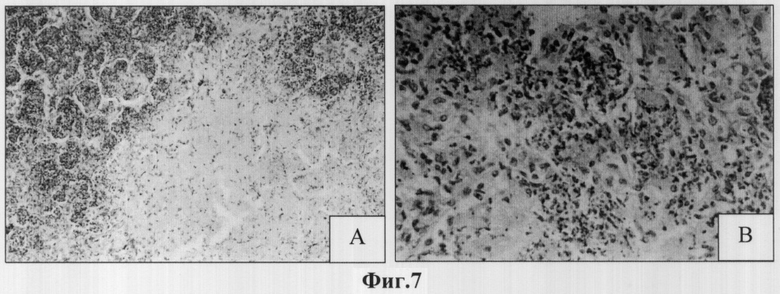
При этом у всех животных контрольной группы в паренхиме легких обнаруживались скопления нейтрофильных гранулоцитов, которые в 6 из 10 случаев частично распадались и образовывали ядерный детрит, а в 2-х случаях - фокусы деструкции в центре гранулемы, что отражено на представленных микрофотографиях гистологического среза легкого мыши контрольной группы (Фиг.7А).
Особенности клеточного состава участков альвеолярно-макрофагального гранулематоза у мышей с экспериментальным туберкулезом
Клеточный состав гранулемы был представлен участками казеозного некроза, ядерным детритом, скоплениями распадающихся нейтрофильных гранулоцитов, макрофагами, эпителиоидными клетками, единичными лимфоцитами (Фиг.7В).
В просвете бронхов у животных с выраженными некротическими изменениями обнаруживались ядерный детрит и слущенные эпителиальные клетки. Лимфогистиоцитарная инфильтрация, свидетельствующая о напряженности местного иммунитета легочной ткани, была выражена очень слабо, значительные периваскулярные инфильтраты зарегистрированы только в 4 из 10 (40%) случаев, крупные перибронхиальные инфильтраты не отмечены (табл.5).
Выраженность признаков напряженности местного иммунитета легочной ткани у мышей с экспериментальным туберкулезом
У вакцинированных мышей опытных групп по сравнению с контрольной группой животных наблюдалось повышение воздушности легочной ткани, снижение распространенности специфического воспаления, которое в большинстве случаев (59%-86%) было представлено небольшими отдельными очагами инфильтрации (табл.3). В очагах инфильтрации практически не обнаруживалось скоплений нейтрофильных гранулоцитов, не выявлялись ядерный детрит и участки некроза, значительно уменьшилась встречаемость единичных не распадающихся нейтрофилов (табл.4). Только в одном случае (14%) у мышей, вакцинированных FLU/ESAT-6 (H1+H3), отмечены мелкие скопления нейтрофильных гранулоцитов.
В качестве иллюстрации (Фиг.8) на микрофотографии препарата ткани легкого (в двух увеличениях) одной из вакцинированных FLU/ESAT-6 (Н1+Н3) мышей представлен небольшой очаг альвеолярно-макрофагального гранулематоза с сохранившимися альвеолами, имеющими несколько утолщенные перегородки и инфильтрированные лимфоцитами (Фиг.8А). Клеточный состав инфильтрата представлен лимфоидными элементами, макрофагами, единичными эпителиоидными клетками и единичными не распадающимися нейтрофилами (Фиг.8В).
У всех опытных животных более часто, чем в контрольной группе обнаруживались крупные периваскулярные и перибронхиальные лимфогистиоцитарные инфильтраты, что свидетельствует об активации напряженности местного иммунитета ткани легкого (табл.5).
Таким образом, двукратная интраназальная вакцинация белых беспородных мышей гриппозными векторами, экспрессирующими протективный микобактериальный антиген ESAT-6, проведенная за 3 недели до заражения мышей вирулентным штаммом М. bovis bovinus 8, привела к задержке развития экспериментальной туберкулезной инфекции и более благоприятному ее течению по сравнению с контрольной группой зараженных невакцинированных животных.
Значительный протективный эффект отмечен как при двукратном интраназальном введении смеси трех рекомбинантных гриппозных векторов (FLU/ESAT-6 (Tri+Tri), так и при последовательной интраназальной иммунизации животных рекомбинантными гриппозными векторами, относящимися к H1N1 и H3N2, а также H1N1 и H2N2 антигенным подтипам вируса гриппа.
Под воздействием вакцинации наблюдалось выраженное снижение тяжести течения инфекции: благоприятная динамика массы тела, более низкие коэффициенты массы легких (р<0,05) и селезенки (р<0,02), уменьшение индексов поражения легких (р<0,001).
Оценка морфодинамики острой туберкулезной инфекции белых беспородных мышей выявила в легких вакцинированных животных регрессию воспалительных изменений, что проявилось в уменьшении распространенности специфического воспаления и снижении его альтеративного компонента, а также активации местного иммунитета ткани легкого.
Пример 5. Иммуногенность гриппозных векторов и эффективность профилактической иммунизации рекомбинантными гриппозными векторами при вялотекущей туберкулезной инфекции у С57 black/6 мышей.
Сравнительная оценка эффективности вакцинации рекомбинантными гриппозными штаммами FLU/ESAT-6 и стандартной вакциной БЦЖ на развитие и течение вялотекущей туберкулезной инфекции проведена на 165 мышах-самцах линии С57 black/6 с исходной массой тела 18-20 г, полученных из питомника «Рапполово», Всеволожского района, Ленинградской области.
С целью индукции системного иммунного ответа и иммунитета слизистых оболочек мышей одной из опытных групп последовательно иммунизировали гриппозными векторами, относящимися к двум антигенным подтипам вируса гриппа: (H1N1) и (H3N2). Контролями служили зараженные невакцинированные мыши (контроль заражения); мыши, иммунизированные диким вирусом гриппа A/PR/8/34 (контроль вакцины), БЦЖ-вакцинированные мыши и интактные животные.
Двукратную иммунизацию FLU/ESAT-6 (H1+H3) и диким вирусом гриппа проводили интраназально, без наркоза, в дозе 106 БОЕ/мышь с интервалом в 3 недели. Вакцина БЦЖ вводилась однократно подкожно в дозе 105 БОЕ/мышь в день первой иммунизации.
Генерализованный туберкулез вызывали введением в латеральную хвостовую вену мышей взвеси трехнедельной культуры М. bovis bovinus 8 (106 КОЕ/мышь) через 6 недель после первой иммунизации.
Эффект протективного действия исследуемой вакцины оценивали на 22-й и 35-й дни после заражения МБТ по следующим показателям тяжести течения экспериментального туберкулезного процесса: высеваемость МБТ из легких и селезенки, макроскопическая оценка легких и селезенки (индексы поражения легких, коэффициенты масс легких и селезенки), летальность. Полученные данные сравнивали с аналогичными показателями в контрольных группах животных.
При бактериологическом исследовании легких и селезенки осуществляли дозированный посев гомогената ткани органа на плотную яичную среду Левенштейна-Йенсена методом 10-кратных серийных разведений. Массивность роста микобактерий туберкулеза (МБТ) выражали в десятичных логарифмах (Log10) от числа колониеобразующих единиц (КОЕ).
Иммунологическое исследование проводили через 3 недели после второй иммунизации (до заражения животных M. bovis) и в динамике инфекционного процесса (на 15-й и 35-й дни после заражения M. bovis).
Пролиферация лимфоцитов селезенки и трахеобронхиальных лимфоузлов мышей оценивалась на 96-луночных планшетах в реакции бласттрансформации при индукции клеток КонА (1 мкг/мл) и рекомбинантного белка ESAT-6 (Fusion Antibodies Ltd, Northern Ireland; 5 мкг/мл) на жидкостном сцинтилляционном счетчике по количеству 3H-тимидина, включившегося в ДНК пролиферирующих клеток. Результаты по каждой пробе выражали в виде абсолютных величин пролиферативной активности клеток (число импульсов за 1 минуту).
Определение продукции ИЛ-4 и ИФН-γ в супернатантах культур клеток селезенки и трахеобронхиальных лимфоузлов проводили с использованием иммуноферментных тест систем Quantikine™ (R&D Systems, Minneapolis) в соответствии с инструкцией по использованию. Уровень ИФН-γ и ИЛ-4 (пкг/мл) в исследуемых образцах рассчитывали по калибровочной кривой зависимости оптической плотности от концентрации цитокина.
Определение уровня ИЛ-2 (спонтанной и индуцированной) определяли в первичных культурах спленоцитов после 24 часовой культивации клеток при 37°С в атмосфере, содержащей 5% СО2 по уровню пролиферации ИЛ-2 - зависимой клеточной линии CTLL-2 (J.Immunol., 120(6): 2027-2032; 1978).
Статистическая обработка данных проводилась с использованием параметрического теста Стьюдента.
Иммуногенность рекомбинантных гриппозных векторов.
С целью индукции системного иммунного ответа мышей линии С57black/6 двукратно иммунизировали рекомбинантными штаммами вируса гриппа А, экспрессирующими ранний секреторный микобактериальный антиген (ESAT-6). Для усиления иммунного ответа повторную иммунизацию проводили гриппозным вектором другого подтипа.
Т-клеточный иммунный ответ определялся путем измерения пролиферативной активности спленоцитов и их способности к выработке ИФН-γ в ответ на стимуляцию рекомбинантным ESAT-6 белком (rESAT-6). Через 3 недели после второй иммунизации в суспензии клеток селезенки иммунизированных FLU/ESAT-6 (H1+H3) мышей отмечалась существенная выработка ИФН-γ в количестве 13 нг/мл супернатанта, что представлено на Фиг.9. При этом продукции цитокина в ответ на ESAT-6 в супернатантах культур клеток селезенки мышей, иммунизированных вирусами дикого типа (контроль вакцины) или вакциной БЦЖ, отмечено не было. Таким образом, двукратная иммунизация рекомбинантными гриппозными векторами, относящимися к различным антигенным подтипам (H1N1, H3N2) и экспрессирующими микобактериальный белок ESAT-6, позволила получить выраженный специфический Т-клеточный ответ к вставке (ESAT-6 антигену) на системном уровне при интраназальном введении вакцины.
Эффективность профилактической иммунизации рекомбинантными гриппозными векторами при вялотекущей туберкулезной инфекции у C57 black/6 мышей.
Влияние иммунизации рекомбинантными гриппозными векторами на развитие и течение экспериментальной туберкулезной инфекции исследовали на мышах C57BL/6, которые являются низкоотвечающей линией мышей к заражению микобактериями туберкулеза. Это определяло развитие вялотекущей туберкулезной инфекции в данном опыте.
Результаты исследования показали, что иммунизация FLU/ESAT-6 (Н1+Н3) ощутимо повлияла на летальность и обсемененность микобактериями органов мышей - интегральных показателей тяжести течения инфекции.
Оценка высеваемости МБТ из легких и селезенки на 22-й день после заражения выявила существенное снижение роста МБТ в группе мышей, иммунизированных FLU/ESAT-6, по сравнению с контролем заражения (на 1,37 log10 КОЕ в легких; на 0,6 log10 КОЕ в селезенке), что отражено в таблице 6. Снижения роста МБТ из паренхиматозных органов при использовании дикого вируса гриппа (контроль вакцины) не отмечено.
Примечательно, что защитный эффект FLU/ESAT-6 по данным высеваемости (индекс защиты) превышал действие стандартной вакцины БЦЖ (в легких 1.37 log10 против 1,17 log10; в селезенке 0.6 log10 против 0.52 log10), что подтвердилось данными морфометрической оценки органов на данном сроке наблюдения.
Рост M. bovis bovinus из паренхиматозных органов (легкие, селезенка) мышей на 22-й день после заражения
Достоверное снижение коэффициента массы легких (0.98±0.06 усл.ед. против 0.83±0.05 усл.ед., р<0,05) и тенденция к снижению индекса поражения легких (1.75±0.11 усл.ед. против 1.66±0.09 усл.ед.) по сравнению с контролем заражения были зарегистрированы только в группе иммунизированных гриппозными векторами мышей.
Снижение показателей тяжести течения туберкулезного процесса в группе животных, иммунизированных FLU/ESAT-6, наблюдалось и на 35-й день после заражения. К этому времени у вакцинированных мышей по сравнению с зараженными невакцинированными животными отмечалась тенденция к снижению коэффициентов массы органов (легких и селезенки) и статистически значимое уменьшение индекса поражения легких (2.4±0.11 усл.ед. против 2.1±0.05 усл.ед., р<0,05).
Анализ данных выживаемости животных показал, что в группе FLU/ESAT-6 начало гибели от туберкулеза отмечено на 59 день после заражения, тогда как у зараженных невакцинированных животных и вакцинированных БЦЖ вакциной оно было зафиксировано на 69 и 64 дни соответственно (Фиг.10). Это позволяет говорить об отсрочке гибели мышей исследуемой группы. При этом к 137 дню после заражения, когда в группе контроля заражения летальность достигла 100%, в группе иммунизированных FLU/ESAT-6 оставалось 45%, а в группе БЦЖ-вакцинированных - 25% выживших животных. К концу периода наблюдения (>250 дней после заражения), в группе привитых исследуемой вакциной оставалось 10% выживших мышей. К этому времени летальность в группе мышей, получивших вакцину БЦЖ, достигла 100%. Увеличение продолжительности жизни иммунизированных FLU/ESAT-6 мышей по сравнению с группой контроля заражения было статистически значимым по Log Rank test (р<0.0008).
Таким образом, показан значительный протективный эффект иммунизации рекомбинантными гриппозными векторами при вялотекущем туберкулезе у C57BL/6 мышей. Примечательно, что по данным высеваемости МБТ из паренхиматозных органов и выживаемости животных защитный эффект FLU/ESAT-6 превышал действие стандартной вакцины БЦЖ.
Кинетика иммунного ответа в ходе развития туберкулезной инфекции.
Иммунологическое обследование мышей выявило выраженные сдвиги показателей иммунитета в ходе развития туберкулезного процесса.
Уже на 15 день после заражения, когда при визуальном осмотре легких зараженных МБТ мышей еще не выявлялись выраженные очаги туберкулезного поражения, в культурах их спленоцитов регистрировалось достоверное угнетение как спонтанной (1327±132 cpm против 6034±733 cpm, р<0,001), так и индуцированной (Con А) пролиферативной активности лимфоидных клеток по сравнению с интактными животными (13406±1835 cpm против 52399±3087 cpm, p<0,01).
При этом у вакцинированных мышей уровень спонтанной пролиферативной активности лимфоидных клеток селезенки был достоверно выше, чем у животных из группы контроля заражения (4106±778 cpm против 1327±132 cpm соответственно, р<0,02).
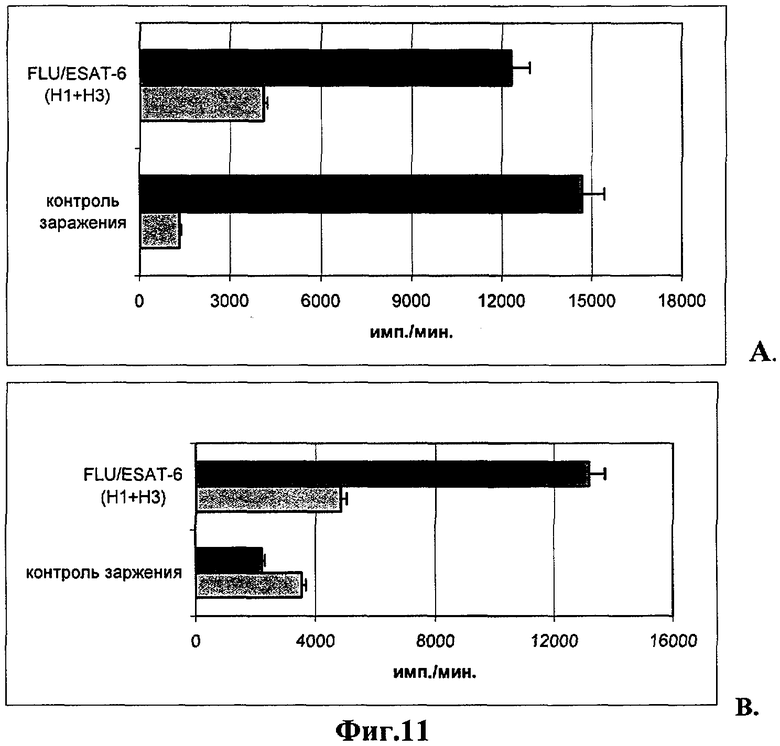
В пролиферативном ответе клеток на специфический антиген ESAT-6 существенных различий между вакцинированными животными и контролем заражения не отмечено, что указывает на формирование пула лимфоцитов, распознающих данный антиген и отвечающих in vitro на стимуляцию ESAT-6 белком усилением митотической активности клеток на раннем сроке развития туберкулезной инфекции (Фиг.11А).
К 35 дню после заражения степень угнетения пролиферации (как спонтанной, так и ConA-индуцированной) у зараженных невакцинированных мышей еще больше увеличилась и достоверность различий с интактной группой достигла<0,001 по непарному t-критерию Стьюдента.
В пролиферативном ответе клеток на специфический антиген ESAT-6 отмечена достоверная стимуляция (р<0,002) митотической активности спленоцитов вакцинированных животных по сравнению с контролем заражения (13185±1720 cpm против 2223±1080 cpm) (Фиг.11В).
Таким образом, в динамике инфекционного процесса наличие антиген-специфического пролиферативного ответа спленоцитов была характерна для относительно ранних сроков инфекции (15 день от заражения); на более позднем сроке (35 день от заражения) на фоне развития иммуносупрессии, обусловленной нарастанием туберкулезного процесса, способность спленоцитов зараженных невакцинированных животных к ответу на ESAT-6 уменьшалась.
При оценке продукции IFN-γ спленоцитами на 15 день развития инфекции у всех зараженных животных отмечено достоверное снижение спонтанного уровня продукции цитокина и повышение Con А- и ESAT-6-индуцированного (р<0,001). Данные по повышению уровня индуцированной продукции IFN-γ на первом сроке обследования согласуются с литературными, где отмечается высокий уровень продукции IFN-γ Т-лимфоцитами на ранних сроках развития туберкулезной. При этом различия между группами были недостоверными.
На 35 день после заражения существенная стимуляция Con A-индуцированной продукции IFN-γ отмечалась в супернатантах культур клеток селезенки группе мышей, вакцинированных FLU/ESAT-6, при сравнении с зараженными невакцинированными животными, у которых продукция цитокина была достоверно угнетена при сравнении с интактной группой (р<0,002).
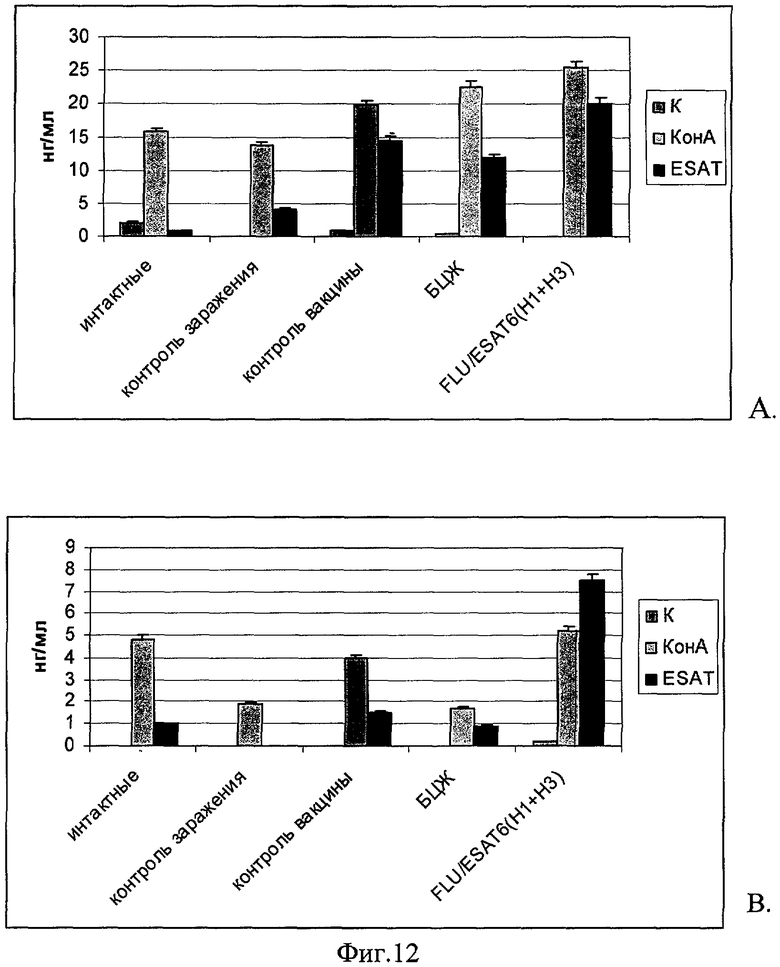
На 35 день после заражения при индукции спленоцитов recESAT-6 (5μg/ml) уровень секреции IFN-γ у всех вакцинированных животных был достоверно выше, чем у зараженных невакцинированных мышей (Фиг.12А), при этом максимальная выработка цитокина (20,1 нг/мл супернатанта) определялась в группе FLU/ESAT-6 иммунизированных мышей при уровне продукции цитокина 4,1 нг/мл в группе контроле заражения.
Наибольшие различия в продукции IFN-γ в ответ на rESAT на данном сроке отмечены в супернатантах культур лимфоцитов трахеобронхиальных лимфатических узлов. Существенная выработка ИФН-γ в количестве 7,5 нг/мл супернатанта отмечалась у иммунизированных FLU/ESAT-6 (H1+H3) мышей, что представлено на Фиг.12В. При этом продукция ИФН-γ в супернатантах культур клеток лимфатических узлов мышей, иммунизированных вирусами дикого типа (контроль вакцины) или вакциной БЦЖ в ответ на ESAT-6 была достоверно ниже (р<0,001), а в группе контроля выработки цитокина отмечено не было.
Снижение в результате вакцинации FLU/ESAT-6 (H1+H3) выраженности ингибиции Th-1 ответа лимфоцитов, сопровождающей развитие туберкулезной инфекции, подтверждается и результатами, полученными при оценке продукции IL-2 в супернатантах культур спленоцитов. При этом на 15 день после заражения, когда угнетение продукции цитокина в ответ на Con А у зараженных невакцинированных мышей по отношению к интактным было еще недостоверным, повышение данного показателя у мышей, вакцинированных FLU/ESAT-6 (H1+H3), было незначительным. На 35 день, когда ингибиция секреции IL-2 в контрольной группе стала значимой (р<0,01), продукция IL-2 в ответ на Con А в группе мышей, вакцинированных FLU/ESAT-6, была на уровне интактных животных. При этом, подсчет продукции IL-2 в % от уровня интактной группы показал, что в то время, как у мышей контроля заражения с развитием инфекции на 22-й и 35-й дни после заражения наблюдалось снижение уровня секреции цитокина (72,5% и 57,8%), то у вакцинированных этого уменьшения не зарегистрировано (77,5% и 82,1% соответственно).
Иммунизация FLU/ESAT-6 предотвратила и угнетение продукции IL-2 в ответ на специфический антиген recESAT 6, отмеченную при естественном течении инфекции к 35 дню после заражения. У вакцинированных мышей в ответ на recESAT-6 продукция IL-2 была на достоверно более высоком, чем у контроля заражения уровне (р<0,001).
На продукцию спленоцитами IL-4 влияние иммунизации FLU/ESAT-6 обнаружено на 15 день после заражения на фоне повышенной продукции цитокина у всех зараженных животных. Оно проявилось в достоверном снижении уровня секреции IL-4 в ответ на Con А у иммунизированных мышей по сравнению с контрольной группой животных (88,9±1,9 пг/мл против 141,8±10,9 пг/мл супернатанта).
Отмеченные сдвиги продукции IFN-γ и IL-4 подтверждаются изменениями индекса IFN-γ/IL-4 на 15-й день туберкулезной инфекции. В культуре спленоцитов при индукции Con А у зараженных невакцинированных животных индекс был достоверно снижен (р<0,02), в то время, как при вакцинации FLU/ESAT-6 (H1+H3) - на уровне интактной группы. Самый высокий индекс IFN-γ/IL-4 у данной группы мышей отмечен и в ответ на recESAT-6.
Таким образом, иммунологическое исследование в ходе развития туберкулезного процесса показало, что иммунизация гриппозными векторами, экспрессирующими ESAT-6 антиген, вызвала антигенспецифичный клеточный ответ, модифицировавший Th-1 иммунный ответ на инфицирование мышей МБТ.
Под влиянием вакцинации отмечены: на ранних сроках развития инфекции - достоверное снижение повышенного уровня секреции IL-4 и восстановление индекса IFN-γ/IL-4 на Con А в культуре спленоцитов; на более поздних, на фоне выраженного угнетения Th-1 лимфоцитов у невакцинированных зараженных мышей - стимуляция спонтанной и индуцированной (Con А и recESAT-6) продукции IFN-γ спленоцитами, повышение уровня их пролиферации (спонтанной и в ответ на rESAT 6), активация recESAT-6-индуцируемой продукции спленоцитами IL-2. Наиболее существенная стимуляция секреции IFN-γ в ответ на recESAT-6 отмечалась в трахеобронхиальных лимфатических узлах иммунизированных FLU/ESAT-6 животных.
Заключение. Иммунизация гриппозными векторами, экспрессирующими ESAT-6 белок, проведенная за 3 недели до заражения C57BL/6 мышей М. bovis, привела к развитию выраженного антиген специфичного клеточного ответа и обеспечила значительный протективный эффект при экспериментальной вялотекущей туберкулезной инфекции. Примечательно, что по данным высеваемости МБТ из паренхиматозных органов и выживаемости животных защитный эффект FLU/ESAT-6 превышал действие стандартной вакцины БЦЖ.
Реализация профилактического эффекта, по-видимому, осуществляется за счет направленной коррекции установленной нами при данном течении туберкулезного процесса выраженной ингибиции Тх-1 со значительным снижением продукции цитокинов, вырабатываемых Т-хелперами 1 типа (ТХ-1) - гамма-интерферона (ИФН-γ), интерлейкин-2 (ИЛ-2), которые являются медиаторами клеточно-опосредованного иммунитета и определяют благоприятное течение и обратное развитие инфекции.
Пример 6. Профилактическое действие рекомбинантных гриппозных векторов при туберкулезной инфекции у морских свинок.
Сравнительное исследование защитного эффекта рекомбинантных гриппозных векторов (FLU/ESAT-6) и стандартной вакцины БЦЖ при туберкулезе проведено в опыте на морских свинках, которые ввиду высокой восприимчивости к инфекции считаются наиболее подходящими животными для опытов по лечению и профилактике туберкулеза. В данном опыте проведена оценка эффективности профилактической иммунизации при интраназальном и подкожном путях введения гриппозной вакцины.
Исследование проведено на 16 морских свинках-самцах с исходной массой тела 250-300 г, полученных из питомника «Рапполово», Всеволожского района, Ленинградской области. Животные были распределены по 4-м группам: иммунизированные гриппозной вакциной, вводимой интраназально; иммунизированные гриппозной вакциной, вводимой подкожно; иммунизированные вакциной БЦЖ; невакцинированные зараженные МБТ животные (контроль заражения). Животные содержались в отдельных клетках (по 4 морские свинки в клетке) и имели свободный доступ к воде и пище.
Двукратную иммунизацию FLU/ESAT-6 проводили без наркоза, в дозе 106 БОЕ/морскую свинку с интервалом в 3 недели. Для иммунизации использовалась вируссодержащая суспензия смеси рекомбинантных гриппозных векторов, относящихся к различным антигенным подтипам вируса гриппа A (H1N1, H3N2 и H2N2). Вакцина БЦЖ вводилась однократно подкожно в левую паховую область в дозе 104 КОЕ/морскую свинку в день первой иммунизации.
Генерализованный туберкулез вызывали подкожным введением 10 КОЕ М. Tuberculosis H37Rv в правую паховую область животных через 6 недель после первой иммунизации.
Оценку влияния иммунизации на развитие и течение туберкулезной инфекции проводили через 9 недель после заражения М. tuberculosis H37Rv, когда одна из морских свинок из группы контроля заражения погибла от генерализованной туберкулезной инфекции.
Эффект защитного действия вакцины оценивали по следующим показателям тяжести течения экспериментального туберкулеза: степени поражения внутренних органов и лимфоузлов (индексы поражения органов); изменения весовых коэффициентов органов; высеваемости МБТ из паренхиматозных органов (легкие, селезенка).
При бактериологическом исследовании легких и селезенки осуществляли дозированный посев гомогената ткани органа на плотную яичную среду Левенштейна-Йенсена методом 10-кратных серийных разведений. Массивность роста МБТ выражали в десятичных логарифмах (Log10) от числа колониеобразующих единиц (КОЕ).
Величина суммарного индекса поражения рассчитывалась по результатам визуальной оценки поражения органов морских свинок туберкулезным процессом (Першин Г.Н., "Методы экспериментальной химиотерапии (практическое руководство)" - М., 1971, стр.182). Он является суммой индексов поражения легких, селезенки, печени и паховых лимфатических узлов (правого и левого). В каждом органе единичные субмилиарные очаги оценивались в 0,5 балла; единичные милиарные - в 1 балл; многочисленные сливающиеся субмилиарные либо до 20 милиарных - в 2 балла; многочисленные сливающиеся милиарные либо до 20 казеозных некротических фокусов - в 3 балла; более 20 казеозных некротических фокусов либо крупные участки некроза - в 4 балла. Максимальное значение индекса поражения для каждого органа составляло 4 балла, суммарного индекса - 20 баллов. Полученные результаты обрабатывались с использованием непараметрического критерия Вилкоксона-Манна-Уитни.
Коэффициенты массы легких и селезенки высчитывали исходя из соотношения общего веса животного и массы органа и выражали в условных единицах (усл.ед.). Достоверность различий оценивали по непарному t-критерию Стьюдента.
Для гистологического изучения легких орган фиксировали в 10% формалине. После фиксации и проводки материал заливали в парафин и из парафиновых блоков изготовляли ступенчатые гистологические срезы. Использовали окраски гематоксилином и эозином.
Как показали результаты исследования, через 9 недель после заражения М. Tuberculosis H37Rv, одна морская свинка из группы контроля заражения погибла от генерализованной туберкулезной инфекции. К этому сроку все животные в опытных группах были живы. Визуальная оценка распространенности туберкулезного поражения органов морских свинок (легких, печени, селезенки и паховых лимфоузлов) к этому сроку показала снижение суммарного индекса поражения животных во всех вакцинированных группах (табл.7).
Показатели тяжести течения экспериментального туберкулеза у морских свинок через 9 недель после заражения М. tuberculosis H37 Rv
Примечательно, что наименьший суммарный индекс поражения органов наблюдался в группе животных, иммунизированных подкожно FLU/ESAT-6 (6,53 балла при 8,5 баллов у БЦЖ-вакцинированных и интраназально иммунизированных гриппозной вакциной), что было связано с меньшей степенью поражения селезенки и печени, специфическое поражение которых было представлено единичными субмилиарными очагами, и отсутствием очагов специфического воспаления в коллатеральных паховых лимфоузлах у животных данной группы.
Параллельно с этим, при биометрической оценке органов было выявлено достоверное снижение коэффициентов массы легких у всех вакцинированных животных по сравнению с контролем заражения, тогда как коэффициент массы селезенки был достоверно меньше только в группе морских свинок, подкожно иммунизированных FLU/ESAT-6 вакциной. В то же время в группах, вакцинированных БЦЖ и иммунизированных интраназально FLU/ESAT-6, коэффициенты массы селезенки имели лишь тенденцию к снижению (табл.8).
Коэффициенты масс органов (легкие, селезенка) морских свинок через 9 недель после заражения M. tuberculosis H37Rv
Оценка высеваемости МБТ из легких и селезенки через 9 недель после заражения выявила существенное снижение роста МБТ в группах вакцинированных животных по сравнению с контролем заражения (табл.9). При этом индекс защиты был выше в группах, иммунизированных вакциной БЦЖ и гриппозной вакциной при ее подкожном введении по сравнению с группой животных, иммунизированных интраназально. Эти результаты подтвердились данными анализа гистологических срезов легких морских свинок (представлены ниже).
Высеваемость МБТ из паренхиматозных органов (легкие, селезенка) морских свинок через 9 недель после заражения М. tuberculosis H37Rv
В ткани легких зараженных невакцинированных морских свинок специфические изменения были представлены в виде сливающихся бугорков с очагами казеозного некроза в центре и зонами макрофагально-альвеолярного грануломатоза перифокально (Фиг.13А). Инфильтративные изменения очень значительно снижали воздушность легочной ткани и распространялись как на паренхиму легких, так и на перибронхиальные и периваскулярные пространства. В просвете бронхов найдены скопления слущенных эпителиальных клеток, некротические массы и отдельные лимфоидные элементы. Лимфогистиоцитарная инфильтрация не отмечена.
Интраназальное введение FLU/ESAT-6 слабо отразилось на распространенности специфического воспаления в легких. Только в двух из четырех случаев отмечались периваскулярно и перибронхиально расположенные некрупные очаги макрофагально-альвеолярной инфильтрации, не имевшие скоплений ядерного детрита и участков некротизированной легочной ткани. В двух других случаях сливающиеся бугорки и инфильтративные изменения имели такой же характер, как и у контроля заражения. Однако в препаратах этой группы были обнаружены крупные периваскулярные лимфогистиоцитарные инфильтраты, что является одним из признаков активации местного иммунитета легочной ткани.
У морских свинок, вакцинированных БЦЖ и FLU/ESAT-6 (при подкожном введении), как и в опытах с мышами, отмечалось снижение распространенности поражения легочной ткани и повышение ее воздушности. Фокусы специфического воспаления были представлены очагами макрофагально-альвеолярного грануломатоза, имели незначительные размеры, рассеянный характер, без ярко выраженного локального строения (Фиг.13В). Альтеративный компонент воспаления в них встречался гораздо реже, чем у невакцинированных животных. У двух из четырех морских свинок в обеих группах в просвете альвеол и межальвеолярных перегородках в очагах поражения отмечались скопления альвеолярных макрофагов, гистиоцитов и эпителиоидных клеток, в одном случае в фокусах инфильтрации были обнаружены отдельные нейтрофильные гранулоциты и только у одной из четырех морских свинок в гистологическом срезе ткани легкого были найдены скопления ядерного детрита. Крупные периваскулярные и перибронхиальные лимфогистиоцитарные инфильтраты отмечены у всех вакцинированных животных, что свидетельствует об активации местного иммунитета ткани легкого.
Таким образом, двукратная иммунизация морских свинок смесью трех антигенно различных рекомбинантных гриппозных векторов, проведенная за 6 недель до заражения М. Tuberculosis H37Rv, привела к более благоприятному течению экспериментальной туберкулезной инфекции по сравнению с контрольной группой зараженных невакцинированных животных. Под воздействием вакцинации отмечено снижение тяжести течения инфекции, что отразилось в достоверном снижении индексов поражения органов и лимфоузлов у вакцинированных животных по сравнению с контролем заражения (р<0.05), отсутствием смертности в опытных группах, более низких весовых коэффициентах. Оценка морфодинамики туберкулезной инфекции выявила в легких вакцинированных морских свинок регрессию воспалительных изменений, что проявилось в уменьшении распространенности специфического воспаления и снижении его альтеративного компонента, а также активации местного иммунитета ткани легкого.
Неожиданным фактом явилось преимущество подкожного пути введения гриппозной вакцины над интраназальным, что может быть связано с пониженной восприимчивостью морских свинок к заражению гриппозной инфекцией (Med. Microbiol. Immunol. (Berl.) 1979; 165: 241-54).
Примечательно, что по ряду показателей защитный эффект исследуемой вакцины при ее подкожном введении превышал протективное действие стандартной вакцины БЦЖ, что отразилось в более низких индексах поражения органов и лимфоузлов и лучших весовых коэффициентах в группе подкожно иммунизированных FLU/ESAT-6 животных.
СПОСОБ ЛЕЧЕНИЯ ТУБЕРКУЛЕЗА ЛЕГКИХ.
В настоящее время ведутся поиски новых подходов к лечению туберкулеза с использованием терапевтических вакцин. Ожидается, что применение терапевтической вакцины даст возможность сократить длительность применения туберкулостатических препаратов, что позволит отсрочить появление мутантных форм микобактерий туберкулеза, резистентных к комплексной терапии антибиотиками, и добиться перерыва в многомесячных курсах лечения. Целью терапевтической вакцинации при туберкулезе является, прежде всего, повышение надежности и эффективности применяемой этиотропной лекарственной терапии.
Проведено сравнительное исследование результатов использования рекомбинантных гриппозных векторов и вакцины БЦЖ в терапии туберкулезной инфекции.
Оценка терапевтического эффекта FLU/ESAT-6 на течение экспериментальной туберкулезной инфекции.
Исследование лечебного эффекта монотерапии FLU/ESAT-6 на течение туберкулезного процесса проведено на 50 мышах-самцах линии C57black/6, весом 16-18 г, полученных из питомника «Рапполово», Всеволожского района. Ленинградской области.
Использовалась модель вялотекущего генерализованного туберкулеза, вызванного инокуляцией в хвостовую латеральную вену культуры M.Erdman в дозе 104 КОЕ/мышь. Последовательная иммунизация FLU/ESAT-6 гриппозными векторами, относящимися к антигенным подтипам вируса гриппа (H1N1) и (H3N2), проводилась двукратно (подкожно либо интраназально), без наркоза, с трехнедельным перерывом с 15-ого дня после заражения, на фоне обнаружения на пробном вскрытии зараженных мышей множественных субмилиарных очагов специфического воспаления. Терапевтический эффект FLU/ESAT-6 сравнивали с действием вакцины БЦЖ (106 КОЕ/мышь), которая вводилась однократно подкожно в день первой иммунизации FLU/ESAT-6.
Терапевтическое действие FLU/ESAT-6 на фоне этиотропной терапии оценивалось на 100 мышах-самцах линии C57BL/6 при моделировании классического экспериментального туберкулеза, вызванного введением в хвостовую латеральную вену культуры M.Erdman в дозе 106 КОЕ/мышь. Иммунизация FLU/ESAT-6 с контролем вакцины проводилась двукратно (подкожно либо интраназально), с трехнедельным перерывом с 11-го дня после заражения, на фоне обнаружения на пробном вскрытии зараженных мышей множественных субмилиарных очагов и бактериологического контроля обсемененности МБТ. Иммунизация вакциной БЦЖ (106 КОЕ/мышь) проводилась однократно подкожно в день первого введения FLU/ESAT-6. Этиотропное лечение (изониазид 10 мг/кг, ежедневно, п/к) начато со дня первой иммунизации FLU/ESAT-6.
Эффект лечебного действия оценивали по следующим показателям тяжести течения экспериментального туберкулезного процесса: летальность, динамика массы мышей, макроскопическая оценка легких и селезенки (индекс поражения легких, коэффициенты масс легких и селезенки), гистологическое исследование срезов легких, высеваемость МБТ из легких и селезенки.
Для гистологического изучения легких орган фиксировали в 10% формалине. После фиксации и проводки материал заливали в парафин и из парафиновых блоков изготовляли ступенчатые гистологические срезы. Использовали окраски гематоксилином и эозином.
При бактериологическом исследовании селезенки осуществляли дозированный посев гомогената ткани органа на плотную яичную среду Левенштейна-Йенсена методом серийных разведений. Массивность роста МБТ выражали в десятичных Log10 от числа КОЕ.
Для оценки фагоцитарной активности клеточную взвесь перитонеальных макрофагов (пМф) получали при промывании брюшной полости средой 199 («Биолот») с 10% сывороткой крупного рогатого скота и 5 ед/мл гепарина. Клетки (1·106 клеток/ мл) инкубировали при 37°С в одноразовых чашках Петри (1 час в атмосфере, содержащей 5% CO2). После удаления неприкрепившихся клеток к монослою пМф добавляли взвесь дрожжей рода Saccharomyces cerevisiae (1·107 дрожжей на чашку), предварительно опсонизированных сывороткой мышей, инкубировали в течение 1 и 2,5 часов, двукратно отмывали, фиксировали 70° этанолом, окрашивали гематоксилином и эозином. Подсчет проводился при 80-кратном увеличении. По полученным данным вычисляли следующие показатели: фагоцитарную активность Мф (ФА) - процент Мф, вовлеченных в фагоцитоз; фагоцитарное число (ФЧ) - среднее количество дрожжей, поглощенных одной фагоцитирующей клеткой; показатель завершенности фагоцитоза (ПЗФ) - количество дрожжей, переваренных макрофагами за 1,5 часа культивирования; индекс завершенности фагоцитоза (ИЗФ) - отношение фагоцитарного числа за 1 час культивирования к фагоцитарному числу за 2,5 часа культивирования.
Пример 7. Влияние терапевтической вакцинации FLU/ESAT-6 на течение экспериментальной туберкулезной инфекции.
Оценка лечебного эффекта монотерапии FLU/ESAT-6 проведена при использовании низкой инфицирующей дозы M.Erdman, что вызвало развитие вялотекущей туберкулезной инфекции. Появление очагов специфического воспаления в данном опыте зарегистрировано на 15 день после заражения, начало гибели в группе контроля заражения - на 19-й день после инокуляции инфекта.
Как показали результаты исследования, введение FLU/ESAT-6 в режиме монотерапии положительно отразилось на показателе летальности мышей. К концу периода наблюдения (71 день после заражения) в группе нелеченных животных (гр.10) и в группе 5 (БЦЖ) от генерализованного туберкулезного процесса погибло по 3 из 8 мышей. При введении FLU/ESAT-6 в подкожном режиме (8 мышей) гибели не наблюдалось, при интраназальном введении погибла одна мышь из 8.
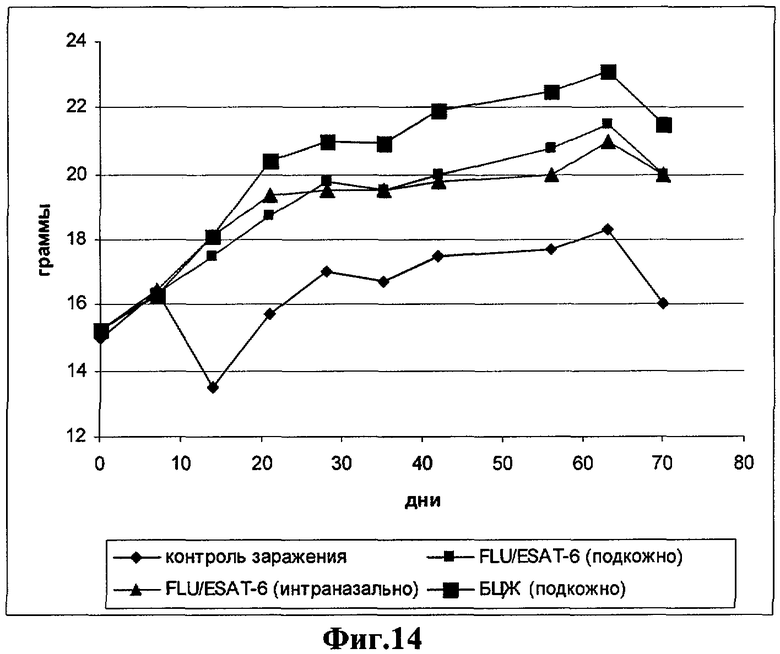
Оценка динамики массы мышей (Фиг.14) выявила тенденцию к снижению тяжести течения инфекции при использовании FLU/ESAT-6. Так, процент прибавки массы тела к концу периода наблюдения у иммунизированных FLU/ESAT-6 мышей составил 24,5% при подкожном введении вакцины, 18,3% при интраназальном и 35% при использовании вакцины БЦЖ. В группе зараженных нелеченных животных прибавки массы тела к этому сроку практически не наблюдалось (2%), что свидетельствует о выраженности симптомов интоксикации в данной группе животных.
При гистологическом исследовании срезов легких, проведенных через 10 недель после заражения МБТ (8 недель от начала терапии FLU/ESAT-6), у трех выживших нелеченных мышей в легочной паренхиме, периваскулярно и перибронхиально выявлено большое количество свежих инфильтративных изменений. Очаги инфильтрации в основном были некрупные, местами сливающиеся, с размытыми границами без четкой пространственной ориентации клеток. Альвеолы и межальвеолярные перегородки в них заполнены серозным и фибринозным экссудатом и рыхло инфильтрированы лимфоцитами, крупными вакуолизированными макрофагами, эпителиоидными клетками и нейтрофильными гранулоцитами. В то же время в легочной ткани обнаружены и гранулемы - плотные очаги инфильтрации с четкими границами, где лимфоциты были концентрически расположены вокруг очень крупных скоплений эпителиоидных клеток. В инфильтратах отмечено большое количество скоплений нейтрофильных лейкоцитов, в одном из них клетки частично распадались и образовывали ядерный детрит. Периваскулярные и перибронхиальные инфильтраты были выражены очень слабо.
Лечение мышей БЦЖ вакциной увеличило распространенность поражения ткани легких. У всех животных очаги инфильтрации были очень распространенными, носили сливной характер и включали наряду с клетками лимфоцитарно-макрофагального ряда нейтрофильные лейкоциты и их скопления, в 3 из 5 случаев распадающиеся до ядерного детрита. Гранулемы, как и у нелеченных мышей, состояли из большого количества эпителиоидных клеток и содержали нейтрофильные лейкоциты и их скопления.
Монотерапия FLU/ESAT-6 снизила пораженность легочной ткани как при подкожном (гр. 3А), так и при интраназальном введении (гр. 3В). Легкие мышей в этих группах были более воздушными, в 1 из 4 случаев в гр. 3А и в 2 из 4 случаев в гр. 3В встречались только небольшие по площади очаги инфильтрации, расположенные вокруг бронхов и сосудов. Гранулемы состояли преимущественно из эпителиоидных клеток. Под влиянием монотерапии FLU/ESAT-6 в очагах инфильтрации наблюдалось снижение выраженности альтеративного компонента воспаления. У мышей гр. 3А (подкожное введение вакцины) нейтрофильные гранулоциты в очагах инфильтрации обнаружены в 2 из 4 случаев, их скопления - только в одном случае. В гр. 3В (интраназальное введение вакцины) признаки альтерации были более существенными, но частота их встречаемости - ниже, чем в контроле заражения: единичные нейтрофильные лейкоциты зарегистрированы у 3 из 4 мышей, скопления нейтрофилов - у 2 животных, образование ядерного детрита и очагов казеоза - у одной мыши. Лимфогистиоцитарная инфильтрация отмечалась, однако крупных периваскулярных и перибронхиальных инфильтратов не зарегистрировано.
Таким образом, введение FLU/ESAT-6 в режиме монотерапии как подкожно, так и интраназально выявило терапевтический эффект иммунизации животных на течение туберкулезного процесса по данным уменьшения летальности и привела к снижению выраженности альтеративного компонента воспаления при гистологическом исследовании легких. Использование вакцины БЦЖ в терапевтическом режиме по сравнению с контролем заражения привело к увеличению распространенности специфического поражения ткани легких.
Пример 8. Эффективность терапевтической вакцинации FLU/ESAT-6, проводимой на фоне этиотропной терапии, на течение экспериментального туберкулеза.
Эффект включения FLU/ESAT-6 в комплексную терапию экспериментального туберкулеза оценивался на модели классического течения туберкулезной инфекции у мышей. На пробных вскрытиях нелеченных мышей, которые проводились каждые два-три дня (с 7-го дня после инокуляции инфекта), макроскопически заметные единичные субмилиарные очаги специфического воспаления обнаружены с 9-ого дня, а множественные субмилиарные - с 11-ого дня после заражения. Гибель нелеченных мышей зарегистрирована с 17-ого дня после заражения.
Результаты иммунизации FLU/ESAT-6 в этих условиях при подкожном (гр. 4А) и интраназальном введении (гр. 4В) сравнивались с данными группы контроля терапии (гр. 1), групп контроля вакцины (подкожно - гр. 7А, интраназально - гр. 7 В), контроля заражения (гр. 10) и группы мышей, иммунизированных вакциной БЦЖ (гр. 6). Все мыши опытных групп, за исключением контроля заражения (гр. 10), получали изониазид в средней терапевтической дозе (10 мг/кг, ежедневно, подкожно).
Анализ показателей тяжести течения инфекции выявил отчетливое действие FLU/ESAT-6 на величину индекса поражения легких и высеваемости из них МБТ.
Показатели тяжести течения экспериментального туберкулеза через 12 недель от начала комплексной терапии с использованием FLU/ESAT-6
P1-4a<0,01
Р1-7в<0,01
На рост М. Erdman в селезенке мышей иммунизация FLU/ESAT-6 оказала существенный эффект уже через 6 недель от начала лечения. При этом наиболее высоким был индекс защиты в гр. 4А при подкожном использовании вакцины (2,32 Log10). Некоторое повышение показателя наблюдалось и в соответствующей группе контроля вакцины (0,47 Log10), что отражено в таблице 11.
Рост М. Erdman в легких мышей, получавших FLU/ESAT-6 в составе комплексной терапии
На втором контрольном сроке (12 недель от начала лечения) роста М. Erdman в легких не отмечено в обеих основных опытных группах (индекс защиты составил 2,851 Log10). В контроле вакцины при подкожном введении (гр. 7А) защитный эффект также отмечался, хотя индекс защиты был несколько ниже и составил 0,531 Log10.
Гистологическое исследование срезов легких, проведенных через 10 недель после заражения (8 недель от начала терапии FLU/ESAT-6), показало, что этиотропная терапия выраженно отразилась на распространенности специфического воспаления, потерявшего сливной характер и представленного у всех мышей, получавших противотуберкулезные препараты, отдельными некрупными очагами инфильтрации.
При использовании только специфической терапии (контроль лечения, гр. 1) в инфильтратах, состоящих из лимфоцитов, макрофагов, эпителиальных клеток и их скоплений, регистрировали отчетливые признаки альтеративного характера воспалительного процесса: в 3 из 6 случаях - скопления нейтрофильных гранулоцитов, у двух мышей - скопления ядерного детрита, у одной - очаги казеозного некроза. Многочисленные гранулемы в легких всех мышей этой группы содержали крупные скопления эпителиоидных клеток, то есть имели преимущественно эпителиоидно-клеточный характер.
Введение мышам вакцины БЦЖ на фоне этиотропной терапии (гр. 6) практически не уменьшило специфического воспаления в легочной ткани. Как и в группе контроля лечения в очагах инфильтрации отмечались нейтрофильные гранулоциты (в 4 из 6 случаях), их скопления (в 3 из 6), участки ядерного детрита (в 1 случае). Гранулемы также носили преимущественно эпителиоидно-клеточный характер.
У мышей, которым в составе терапии вводился FLU/ESAT-6 (подкожно - гр. 4А; интраназально - гр. 4В), в клеточном составе инфильтратов нейтрофильные гранулоциты, ядерный детрит и некротические очаги не обнаружены. Гранулемы в этих группах мышей также имели свои особенности: они состояли преимущественно из лимфоцитов, у 4 из 6 животных концентрически расположенных вокруг единичных (двух трех) эпителиоидных клеток, и только в двух случаях - вокруг их некрупных скоплений. Следовательно, можно говорить о преимущественно лимфоидном характере гранулем.
В группах, получавших на фоне противотуберкулезной терапии контроль вакцины (гр. 7А и гр. 7В), направленность изменений была сходной с основной опытной группой, но выраженность сдвигов - меньшей: показатели альтерации - нейтрофильные гранулоциты, их скопления и участки ядерного детрита были зарегистрированы у одной из мышей; частота встречаемости преимущественно лимфоидных гранулем - более низкой (гр. 7А - в 2-х случаях из 5, гр. 7В - в 2-х из 4-х случаев).
Влияние иммунизации FLU/ESAT-6 на фагоцитарную активность перитонеальных макрофагов.
Анализ особенностей фагоцитоза пМф показал, что у зараженных нелеченных мышей (гр.10) на фоне распространенного туберкулезного поражения легочной ткани (53-й день после заражения) развивается достоверное угнетение как поглотительной, так и переваривающей способности Мф по всем изученным показателям. Фагоцитарная активность снизилась при этом в среднем до 32,3% против 54,3% у интактных мышей (р<0,05), фагоцитарное число - до 4,14 дрожжевых клеток против 6,39 дрожжевых клеток (р<0,05), ПЗФ - до 30,0 дрожжевых клеток против 217,67 дрожжевых клеток (р<0,05), индекс завершенности фагоцитоза - до 1,03 усл.ед. против 1,65 усл.ед. (р<0,05).
Терапия мышей противотуберкулезными препаратами (контроль лечения, гр. 1) через 6 недель от начала лечения привела к достоверному повышению 3-х из 4-х изученных показателей: фагоцитарного числа, ПЗФ и ИЗФ, не восстановив до уровня интактной группы только величину ПЗФ (157,0 дрожжевых клеток против 217,67 дрожжевых клеток).
К полной нормализации активности фагоцитоза привело использование в составе комплексной терапии FLU/ESAT-6 при интраназальном введении препарата (гр. 4В) и вакцины БЦЖ (гр. 6). Подкожное введение FLU/ESAT-6 (гр. 4А) на этом сроке оказало менее выраженный эффект, несколько повысив по сравнению с контролем заражения лишь индекс завершенности фагоцитоза (до 1,94 усл.ед. против 1,59 усл.ед.).
Некоторое подавляющее действие на поглотительную функцию фагоцитов, наблюдалось в группе 7В (интраназальное введение контроля вакцины), где по сравнению с гр. 1 были достоверно снижены ПЗФ (до 83,4 дрожжевых клеток против 157,0 дрожжевых клеток, р<0,05) и ИЗФ (до 1,12 усл.ед. против 1,59 усл.ед., р<0,05).
Через 12 недель от начала лечения активность фагоцитоза перитонеальных Мф в группе контроля лечения полностью восстановилась до уровня интактных животных. В этих условиях использованные вакцины во всех вариантах их применения вызвали стимуляцию как поглотительной, так и переваривающей способности Мф. Так, у мышей, леченных FLU/ESAT-6, наблюдалось значимое повышение выше данных интактной группы фагоцитарной активности, фагоцитарного числа и показателя завершенности фагоцитоза. По сравнению с гр. контроля лечения (гр. 1) введение FLU/ESAT-6 интраназально (гр. 4В) достоверно усилило фагоцитарное число, а при подкожном введении вакцины наблюдалось достоверное усиление как фагоцитарного числа, так и показателя завершенности фагоцитоза.
Таким образом, использование на фоне противотуберкулезных препаратов FLU/ESAT-6 привело к снижению распространенности туберкулезного воспаления в легких по индексу поражения, к санации легочной ткани от М. Erdman, а также к полному исчезновению альтеративного компонента воспаления и изменению характера клеточного состава гранулем с эпителиоидного на преимущественно лимфоидный.
Применение FLU/ESAT-6 в составе комплексной терапии привело к стимуляции активности фагоцитоза клеточной взвеси дрожжей перитонеальными макрофагами. В период подавления реакций фагоцитоза FLU/ESAT-6 ускорил их восстановление, в период нормализации этой функции - усилил ее до уровня, достоверно более высокого, чем у интактных мышей.
Активирующий эффект FLU/ESAT-6 на функциональное состояние Мф, вероятно, является следствием его стимулирующего действия на дифференцировку и функциональную активность Th-1, которые, продуцируя IFN-γ, реализуют один из важнейших путей активации Мф в ходе развития туберкулезной инфекции.
| название | год | авторы | номер документа |
|---|---|---|---|
| Рекомбинантный штамм вируса гриппа A/PR8-NS124-TB10.4-2A-HspX и способ специфической профилактики туберкулеза легких с использованием вакцины мукозального применения на его основе | 2019 |
|
RU2726106C1 |
| Рекомбинантный штамм вируса гриппа A/PR8/HK-NS80E85A, экспрессирующий фрагменты антигенов ESAT-6 и Ag85A M.tuberculosis, для получения векторной вакцины против туберкулеза | 2018 |
|
RU2678175C1 |
| РЕКОМБИНАНТНЫЙ ВИРУС ГРИППА, ПРЕДНАЗНАЧЕННЫЙ ДЛЯ ПРОФИЛАКТИКИ COVID-19 И ГРИППА, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2022 |
|
RU2802058C1 |
| ВАКЦИНА ПРОТИВ ТУБЕРКУЛЕЗА | 2017 |
|
RU2647831C1 |
| УНИВЕРСАЛЬНАЯ ВАКЦИНА ПРОТИВ ВИРУСА ГРИППА ПТИЦ | 2007 |
|
RU2358981C2 |
| АТТЕНУИРОВАННЫЕ ГРИППОЗНЫЕ ВЕКТОРЫ ДЛЯ ПРОФИЛАКТИКИ И/ИЛИ ЛЕЧЕНИЯ ИНФЕКЦИОННЫХ ЗАБОЛЕВАНИЙ, А ТАКЖЕ ДЛЯ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2015 |
|
RU2628690C2 |
| РЕКОМБИНАНТНАЯ ПРОТИВОГРИППОЗНАЯ ВАКЦИНА С ШИРОКИМ СПЕКТРОМ ЗАЩИТЫ И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2017 |
|
RU2757013C2 |
| ПРОТИВОТУБЕРКУЛЕЗНАЯ ВАКЦИНА | 2007 |
|
RU2443773C2 |
| ВАКЦИННАЯ КОМПОЗИЦИЯ ДЛЯ ПРИМЕНЕНИЯ В ПОПУЛЯЦИЯХ СУБЪЕКТОВ С ОСЛАБЛЕННЫМ ИММУНИТЕТОМ | 2013 |
|
RU2661408C2 |
| НАНОАНТИТЕЛО "ANTI-FLU", РЕКОМБИНАНТНЫЕ ВИРУСНЫЕ ВЕКТОРЫ И ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ ДЛЯ ПРОФИЛАКТИКИ И ТЕРАПИИ ГРИППА ТИПА А | 2011 |
|
RU2502745C2 |
Изобретение относится к области медицины. Получены живые аттенуированные рекомбинантные штаммы вируса гриппа, экспрессирующие ранний секреторный антиген Mycobacterium tuberculosis ESAT-6. Изобретение может быть использовано для профилактики и лечения туберкулеза. По ряду показателей протективный эффект рекомбинантных гриппозных векторов превышает защитное действие стандартной вакцины БЦЖ. 5 н. и 2 з.п. ф-лы, 14 ил., 11 табл.
| SHI C.H | |||
| et al | |||
| Устройство для выпрямления опрокинувшихся на бок и затонувших у берега судов | 1922 |
|
SU85A1 |
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| WARDS B.J | |||
| et al | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| Tuber Lung Dis | |||
| ЩИТОВОЙ ДЛЯ ВОДОЕМОВ ЗАТВОР | 1922 |
|
SU2000A1 |

