Область техники
Изобретение относится к области иммунологии и вирусологии, в частности к препарату для профилактики краснухи, полученного посредством сборки антигенных детерминант вируса краснухи на поверхности структурно модифицированных вирусов растений в качестве универсальной платформы - адъюванта.
Уровень техники
Краснуха - острая, обычно легко протекающая вирусная инфекция, поражающая детей и лиц молодого возраста по всему миру. Ее опасность определяется, в основном, тератогенным потенциалом вируса. Краснуха, возникшая непосредственно перед оплодотворением и в течение раннего периода беременности, приводит в 90% случаев к выкидышу, смерти плода и врожденной патологии, известной как синдром врожденной краснухи (СВК). Для врожденной краснухи характерна хроническая форма инфекции, сопровождающаяся длительной персистенцией вируса. Специфического лечения краснухи не существует. Наиболее эффективным методом борьбы с приобретенной и врожденной краснухой является вакцинация. Живые аттенуированные вакцины против краснухи используются с 1969 года. Однако, несмотря на высокую действенность в предотвращении краснухи и СВК живые аттенуированные вакцины имеют ряд противопоказаний и побочных эффектов. Серьезная проблема, связанная с вакцинацией беременных женщин против краснухи, остается нерешенной из-за возможного тератогенного риска. Вакцинация аттенуированными штаммами также не рекомендована лицам, страдающим выраженным иммунодефицитом, включая ВИЧ-инфекцию. Среди побочных эффектов наиболее распространены хронические артриты и артралгии. Хронический артрит, вызванный вакциной против краснухи, включен в Программу компенсации за осложнения от прививок (Vaccine Injury Compensation Program, VICP). Кроме этого, существуют определенные сложности с хранением и доставкой живых вакцин, связанные с соблюдением условий «холодовой цепи» на всем протяжении логистической цепочки - от производителя до конечного потребителя - все вакцины должны храниться и транспортироваться при температуре (+2) - (+8)°С.
Таким образом, задача создания безопасной стабильной и в тоже время высокоэффективной вакцины против краснухи с использованием современных подходов является актуальной.
Вирус краснухи является единственным представителем рода Rubivirus в семействе Togaviridae. Геном вируса краснухи представляет собой одноцепочечную (+) РНК и содержит 2 открытые рамки считывания: с 5'-конца располагаются гены неструктурных белков, участвующих в репликации вирусного генома, с 3'-конца располагаются гены структурных вирусных белков - капсидный белок С и два поверхностных гликопротеина (Е1, Е2).
Во многих исследованиях показано, что вирусная защита обеспечивается в основном нейтрализующими антителами против гликопротеина Е1, которые сохраняют свою активность в организме человека десятилетиями. Преобладающее значение белка Е1 в защите от реинфекции подтверждается присутствием нейтрализующих эпитопов почти исключительно в белке E1 (Petrova et al. The key role of rubella virus glycoproteins in the formation of immune response, and perspectives on their use in the development of new recombinant vaccines. Vaccine 2016, 34, 1006-1011).
Основные подходы для создания новой вакцины против краснухи, представленные в научной и патентной литературе, сосредоточены на разработке оптимальных систем экспрессии, продукции и очистки белка Е1 или его фрагмента, получении кДНК клонов для ДНК-вакцины и создании синтетической пептидной вакцины.
Описано получение рекомбинантного белка Е1 в бактериальной системе экспрессии на основе E.coli (Londesborough et al. Reactivity of a recombinant rubella El antigen expressed in E.coli. Arch Virol., 1992, 122, 391-397; Terry et al. A bio-engineered rubella El antigen. Arch. Virol. 1989, 104, 63-75; London Biotechnology Limited. Rubella El glycoprotein antigens EP 0299673 A1 18.01.1989), в клеточной линии CHO (Perrenoud et al. A recombinant rubella virus E1 glycoprotein as a rubella vaccine candidate. Vaccine 2004, 23, 480-488), в клетках насекомых, трансфецированных рекомбинантным бакуловирусом ( et al. Diagnostic potential of baculovirus-expressed rubella virus envelope proteins. J. Clin. Microbiol 1991, 29, 1877-1882). В патенте (Котляров и др. Патент РФ №2390563, 27.05.2010) запатентованы синтетические гены, кодирующие модифицированные белки Е1 и Е12 (включающего аминокислоты со 153 по 239 белка Е1 вируса краснухи). Для продукции этих белков в растениях синтетические гены встраивают в вирусный вектор на основе генома X вируса картофеля. Растения инфицируют бактериями Agrobacterium tumefaciens, содержащими полученный рекомбинантный вирус-вектор. Продукция Е1 и Е12 с помощью созданных вирусных систем экспрессии в растениях Nicotiana benthamiana может быть использована для создания кандидатной рекомбинантной вакцины против краснухи. Однако предложенный подход ограничен тем, что полученные в растении белки трудно очистить в достаточной степени от таких растительных компонентов как алкалоиды. Высокая стоимость очистки и низкий выход белка являются недостатками этого метода.
et al. Diagnostic potential of baculovirus-expressed rubella virus envelope proteins. J. Clin. Microbiol 1991, 29, 1877-1882). В патенте (Котляров и др. Патент РФ №2390563, 27.05.2010) запатентованы синтетические гены, кодирующие модифицированные белки Е1 и Е12 (включающего аминокислоты со 153 по 239 белка Е1 вируса краснухи). Для продукции этих белков в растениях синтетические гены встраивают в вирусный вектор на основе генома X вируса картофеля. Растения инфицируют бактериями Agrobacterium tumefaciens, содержащими полученный рекомбинантный вирус-вектор. Продукция Е1 и Е12 с помощью созданных вирусных систем экспрессии в растениях Nicotiana benthamiana может быть использована для создания кандидатной рекомбинантной вакцины против краснухи. Однако предложенный подход ограничен тем, что полученные в растении белки трудно очистить в достаточной степени от таких растительных компонентов как алкалоиды. Высокая стоимость очистки и низкий выход белка являются недостатками этого метода.
Использование кДНК клонов для создания ДНК-вакцин описано в работе (Pougatcheva et al. Development of a rubella virus DNA vaccine. Vaccine. 1999, 9, 2104-2112) и патентах (Frey et al. Highly infectious rubella virus DNA constructs and methods of production US Patent 20030130498 A1, 2003; Frey et al. DNA encoding infectious Rubella virus US 5663065 A, 1997; Прилипов и др. Плазмида, несущая вектор, содержащий кДНК (24S РНК) вируса краснухи. Патент РФ 2167938 27.05.2001) Однако последствия, связанные с введением и длительной экспрессией чужеродного генетического материала в организме недостаточно изучены, и вопросы, связанные с безопасностью ДНК-вакцин, остаются открытыми.
Одно из перспективных направлений в разработке надежных и безопасных вакцин нового поколения основано на идентификации Т- и В-клеточных эпитопов внутри белковых антигенов и создании на их основе эпитопных вакцин. Такие синтетические вакцины должны быть лишены недостатков, которые присущи субъединичным вакцинам, а также вакцинам, создаваемым на основе аттенуированного вируса. При современном уровне техники имеется возможность целенаправленно подбирать иммуногены с прицелом на те иммунные процессы, которые должна запускать будущая вакцина. В патенте (Chong et al. Synthetic peptides for a rubella vaccine US 6037448 A, 14.03.2000), запатентованы синтетические пептиды, которые включают в себя последовательности из структурных вирусных белков E1, Е2 и С, а также синтетические липопептиды, содержащие эпитопы цитотоксических Т-лимфоцитов (CTL) для получения новой кандидатной пептидной вакцины. Основной недостаток таких вакцин - низкая антигенная активность большинства индивидуальных пептидов из-за их малого размера. Кроме того, они часто оказываются нестабильными в сыворотке крови. Часто удается добиться усиления иммуногенности и стабильности таких иммуногенов путем связывания их с высокомолекулярными и высокоиммуногенными носителями, в качестве которых могут выступать вирусные частицы.
В настоящее время для создания новых технологий все чаще используют вирусные частицы, в том числе вирусы растений (Turpen et al. Production of peptides in plants as viral coat protein fusions. US Patent 5977438. Nov. 1999; Werner et al. International Patent Application PCT/EP2006/009029 (WO/2007/031339); Werner et al. Immunoabsorbent nanoparticles based on a tobamovirus displaying protein A. Proc. Natl. Acad. Sci. USA, 2006, 103, 17678-17683; McCormick & Palmer. Genetically engineered Tobacco mosaic virus as nanoparticle vaccines. Expert Rev Vaccines. 2008, 7, 33-41; Steinmetz et al., Structure-based engineering of an icosahedral virus for nanomedicine and nanotechnology. Curr. Top. Microbiol. Immunol. 2009, 327, 23-58; Denis et al., Immunogenicity of papaya mosaic virus-like particles fused to a hepatitis С virus epitope: evidence for the critical function of multimerization. Virology 2007, 363, 59-68; Acosta-Ramirez et al., Translating innate response into long-lasting antibody response by the intrinsic antigen-adjuvant properties of papaya mosaic virus. Immunology, 2007, 124, 186-197; Lacasse et al., Novel plant virus-based vaccine induces protective cytotoxic T-lymphocyte-mediated antiviral immunity through dendritic cell maturation. J. Virology, 2008, 82, 785-794; Leclerc et al., Adjuvant viral particle. US Patent 7641896, Jan. 5 2010).; Атабеков и др. Способ усиления иммунного ответа, патент РФ 2442604, 20.02.2012; Атабеков и др. Новый тип частиц-носителей (платформ) для получения активных комплексов, патент РФ 2441667, 10.02.2012).
Прототипом данного изобретения является иммуногенный комплекс, содержащий целевые антигены вируса краснухи на поверхности сферических частиц-платформ (СЧ), полученных при термической структурной перестройке фитовирусов. Ранее этот комплекс был создан и была охарактеризована только его антигенная специфичность (Атабеков и др. Иммуногенная композиция, содержащая чужеродные антигены на поверхности сферических носителей, полученных при термической денатурации спиральных вирусов, патент РФ 2440140, 20.01.2012). Способность индуцировать образование специфических антител против вируса краснухи не была охарактеризована. Не были представлены данные, характеризующие адъювантные свойства СЧ в составе комплекса с антигенами вируса краснухи по сравнению с другими адъювантами и с коммерческой живой аттенуированной вакциной против краснухи. Кроме того, в патенте не раскрыт способ получения иммунногенной композиции, а именно не приведено оптимальное соотношение антигена вируса краснухи к СЧ и оптимальный размер СЧ, а также не показана стабильность полученного комплекса.
Раскрытие изобретения
Задачей изобретения является создание стабильной рекомбинантной эпитопной вакцины против краснухи посредством внеклеточной блочной сборки антигенов/эпитопов вируса краснухи на поверхности сферических частиц-платформ нового типа.
Поставленная задача решается вакциной против краснухи, включающей сферические частицы вируса табачной мозаики (ВТМ) и антигены вируса краснухи, адсорбированные на поверхности сферических частиц, взятые в массовом соотношении 10:1 с допустимым отклонением от указанного соотношения до 10%, при этом размер полученного комплекса сферическая частица - антиген составляет 300±20 нм.
Предпочтительно, что сферические частицы получены методом термической структурной перестройки вируса табачной мозаики.
Предпочтительно, что вакцина против краснухи включает фосфатно-солевой буфер рН 7,2-7,5.
Также поставленная задача решается способом получения вышеуказанной вакцины, включающего следующие этапы:
- получение сферических частиц вируса табачной мозаики;
- получение белка-антигена вируса краснухи;
- смешение полученных сферических частиц с антигеном в массовом соотношении 10:1 с допустимым отклонением от указанного соотношения до 15%, и формированием комплекса сферическая частица - антиген с размерами 300±20 нм;
при этом сферические частицы получают методом термической структурной перестройки вируса табачной мозаики, используемые в концентрации 2±0.5 мг/мл, которые сначала нагревают до температуры 94±0.5°С и выдерживают в течение 10±0,5 минут, затем охлаждают в ледяной бане с формированием комплекса размером 300±20 нм.
Предпочтительно, что антиген вируса краснухи представляет собой рекомбинантный белок с молекулярной массой 22±0,5 кДа, включающий в себя четыре повтора эпитопа А гликопротеина Е1 вируса краснухи
Техническим результатом предлагаемой группы изобретений является создание вакцины для профилактики краснухи с высокой иммуногенностью и низкой реактогенностью, обладающей стабильностью.
Техническим результатом также является расширение арсенала средств аналогичного назначения.
Краткое описание чертежей
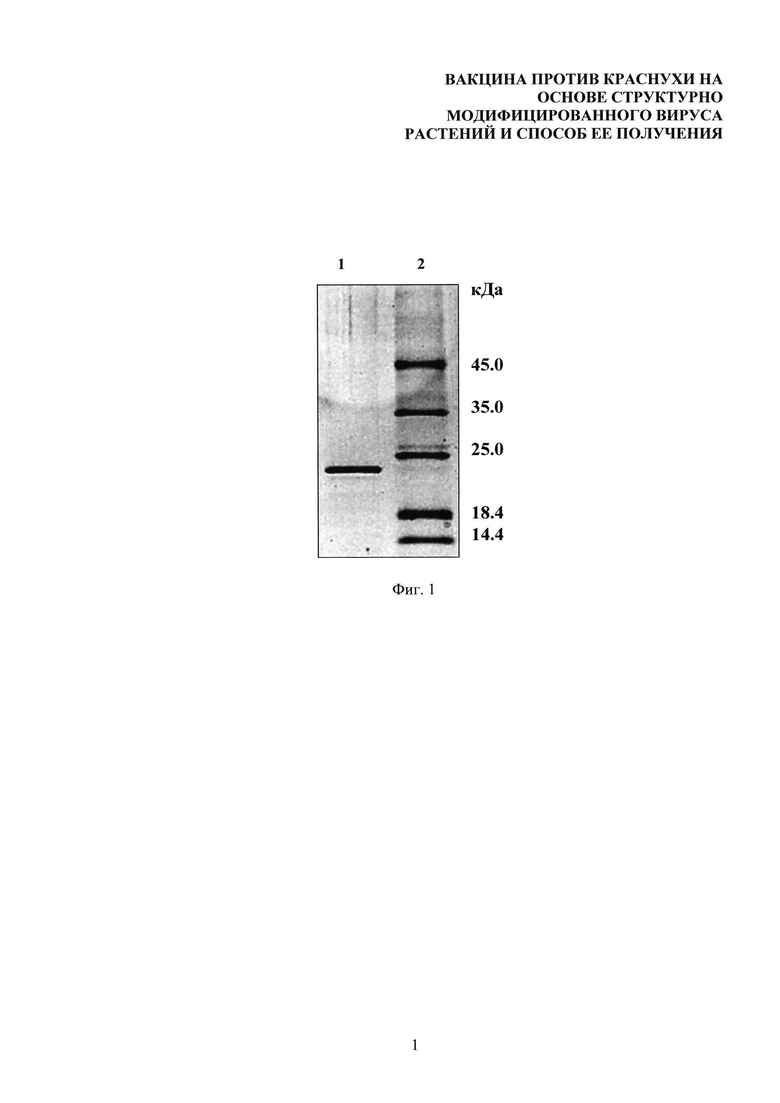
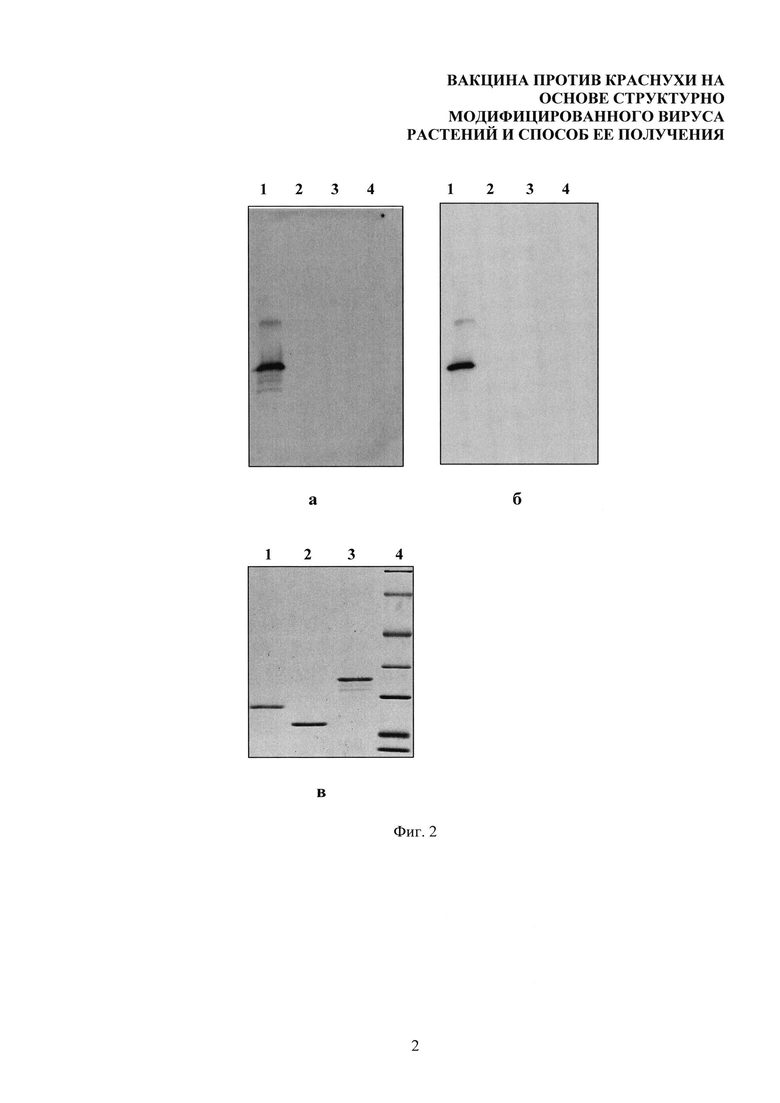
На фиг. 1 представлен электрофоретический анализ тетраэпитопа А после диализа против Milli-Q. 1. Тетраэпитоп А. 2. Маркеры (кДа). Градиент 8-20% ДСН-ПААГ. Гель окрашен кумасси G-250.
На фиг. 2 представлен вестерн-блот анализ эффективности взаимодействия тетраэпитопа А с антисывороткой, которая была к нему получена (а) и с коммерческими антителами к вирусу краснухи (б). 1. Тетраэпитоп А. 2. БО ВТМ (отрицательный контроль). 3. БО ХВК (отрицательный контроль). 4. Белковые маркеры. Белки разделяли методом электрофореза в градиентном ДСН-ПААГ (8-20%), после чего переносили на мембрану Hybond-P (GE Healthcare Life Sciences). Положение на мембране рекомбинантных белков определяли с помощью мышиной антисыворотки к тетраэпитопу А (а) или с коммерческими антителами к вирусу краснухи (б) и вторичных противомышиных антител, коньюгированных с пероксидазой хрена (Promega). Визуализировали с помощью ECL системы (GE Healthcare Life Sciences). Контрольный эксперимент - электрофоретический анализ (гель, окрашенный кумасси G-250) (в).
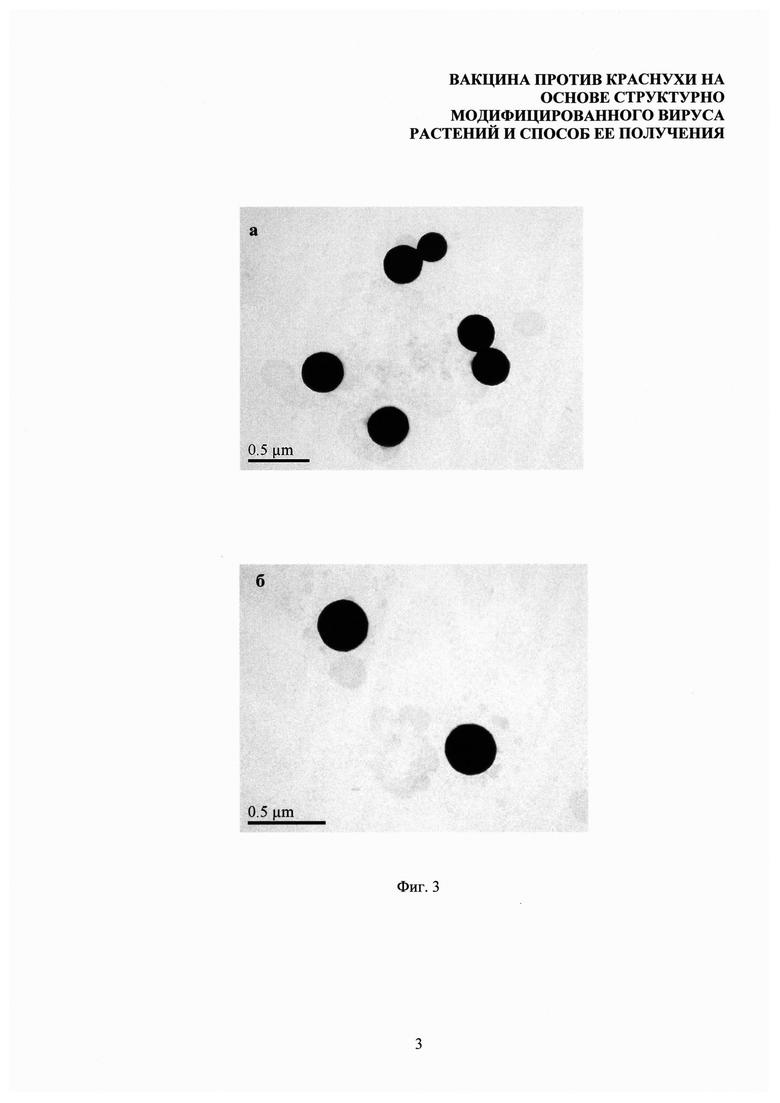
На фиг. 3 представлен сравнительный анализ структурно модифицированных вирусов растений (а) и их композиций с тетраэпитопом А (б). Просвечивающая электронная микроскопия, контрастирование 2% уранилацетатом.
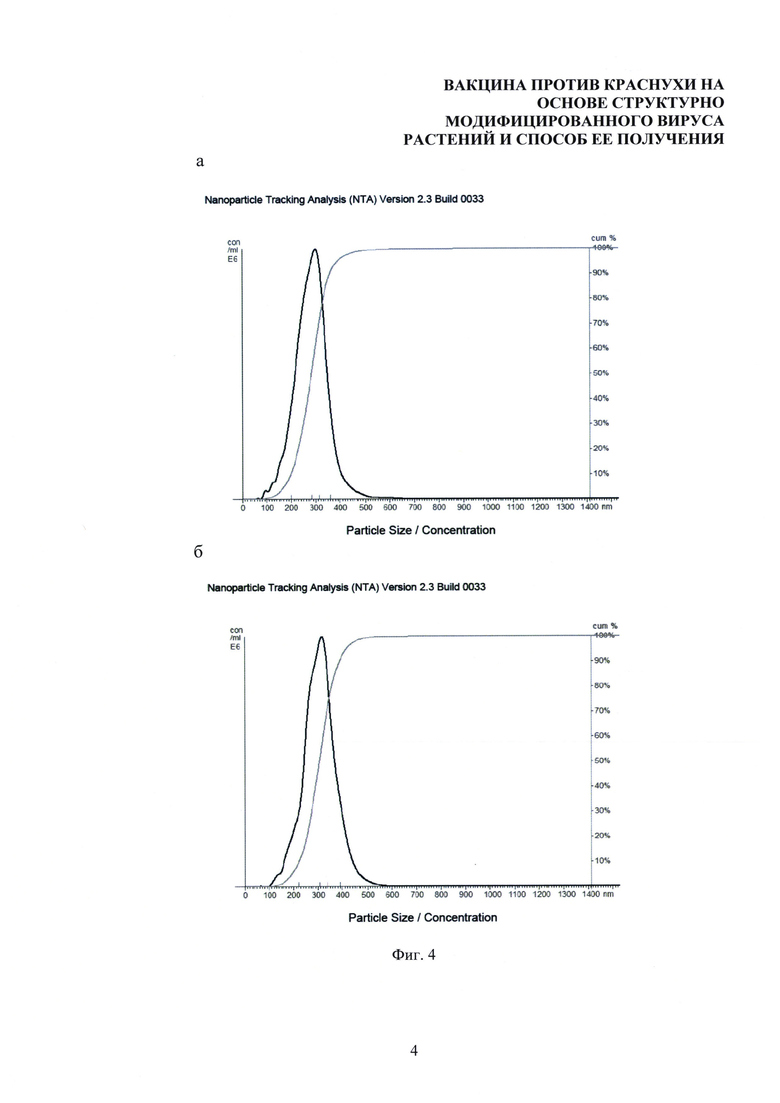
На фиг. 4 представлен сравнительный анализ размеров и агрегационного состояния структурно модифицированных вирусов растений, средний диаметр 299±15 нм (а) и их композиций с тетраэпитопом А, средний диаметр 305±12 нм (б). Метод анализа траекторий наночастиц.
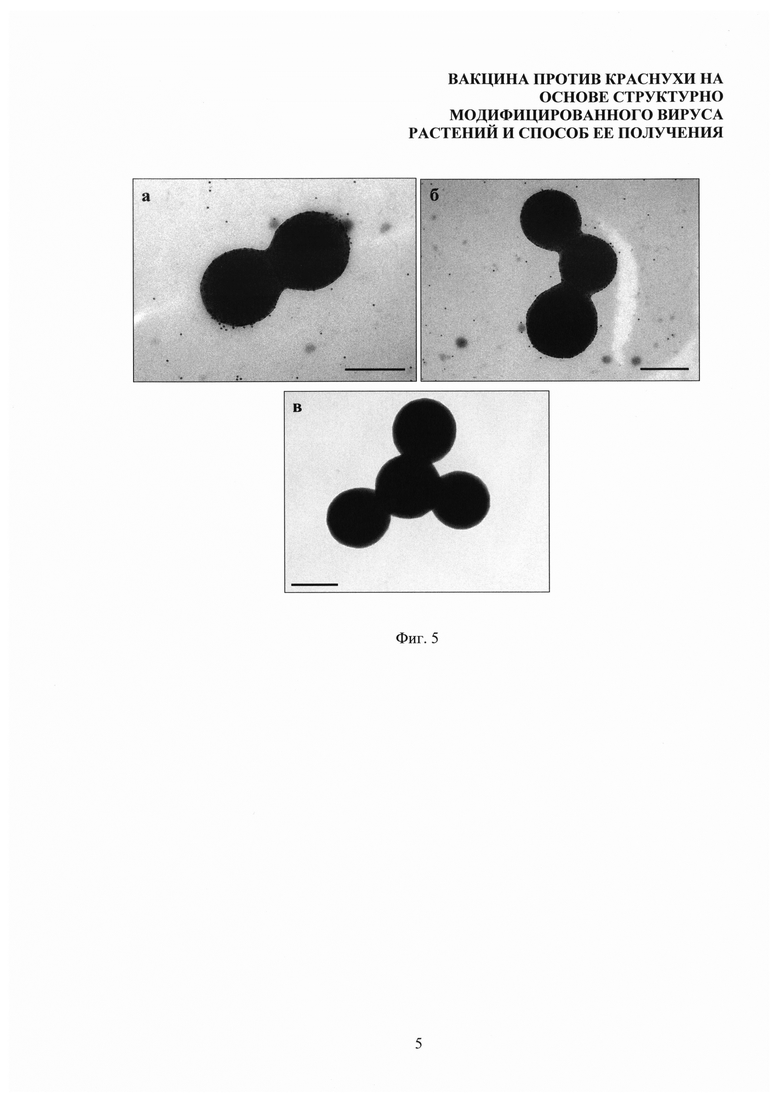
На фиг. 5 представлена иммуноэлектронная микроскопия антигенного комплекса СЧ-тетраэпитоп А вируса краснухи. В качестве первичных антител использовали антисыворотку к тетраэпитопу А гликопротеина Е1 вируса краснухи (а) и поликлональную антисыворотку к вирусу краснухи (б), а в качестве вторичных антител - видоспецифические антитела, конъюгированные с коллоидным золотом, (в) - отрицательный контроль: комплекс, обработанный так же, как в (а) и (б), но без первичных антител, специфичных к белковому антигену. Размер метки 300 нм.
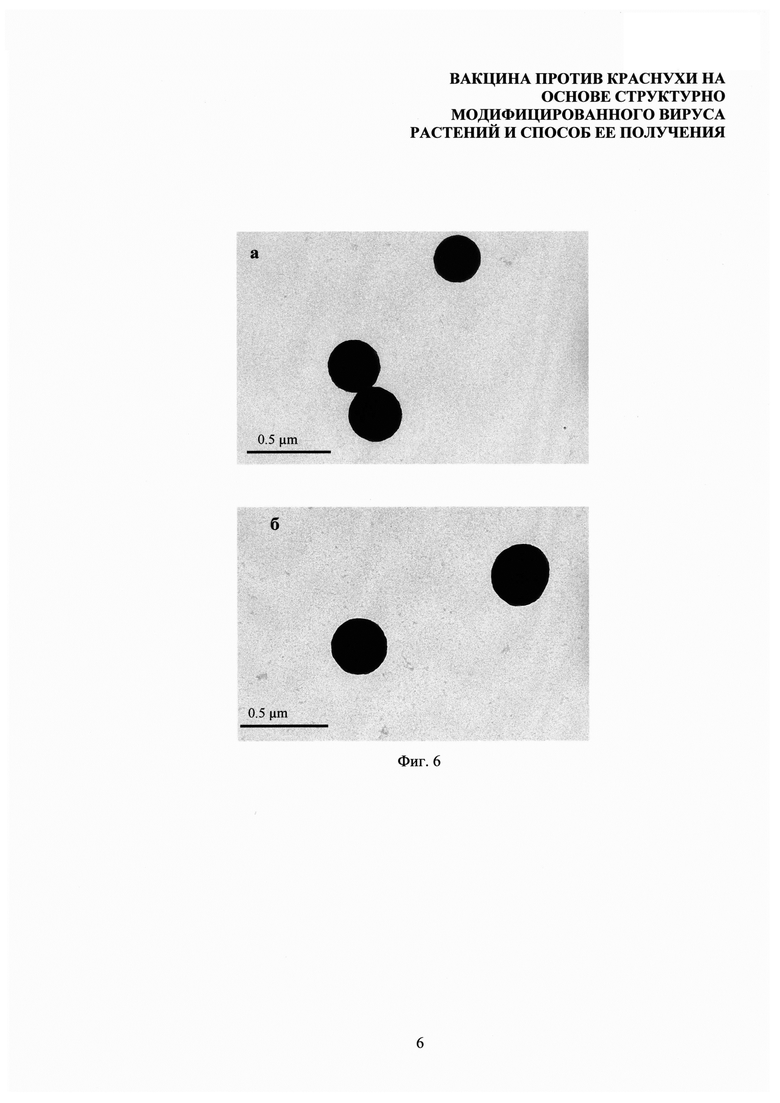
На фиг. 6 представлен анализ стабильности композиций структурно модифицированных вирусов растений с тетраэпитопом А в физиологических условиях. Композиции СЧ-тетраэпитоп в воде (а). Композиции СЧ-тетраэпитоп А в PBS (б). Просвечивающая электронная микроскопия, контрастирование 2% уранилацетатом.
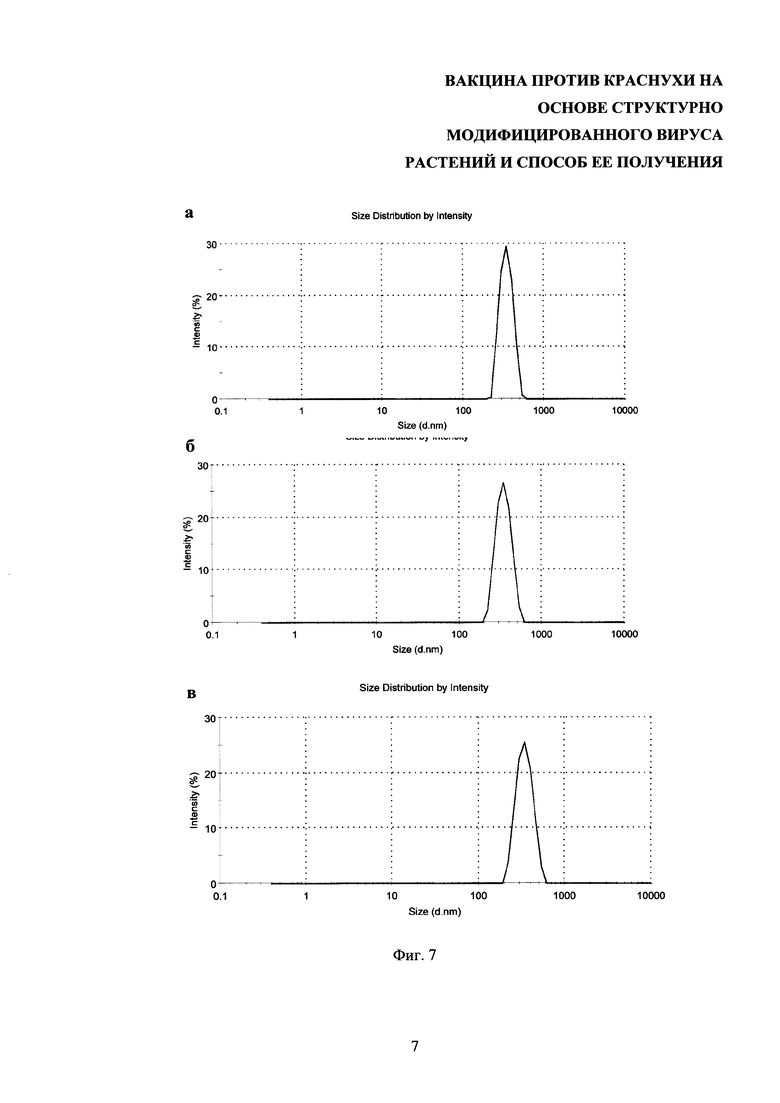
На фиг. 7 представлен анализ стабильности композиций структурно модифицированных вирусов растений с тетраэпитопом А в физиологических условиях. Композиции СЧ-тетраэпитоп А после приготовления в воде (а). Композиции СЧ тетраэпитоп А после приготовления и перевода в PBS буфер (б). Композиции СЧ-тетраэпитоп А после хранения в PBS в течение 3 месяцев (в). Метод динамического рассеяния света
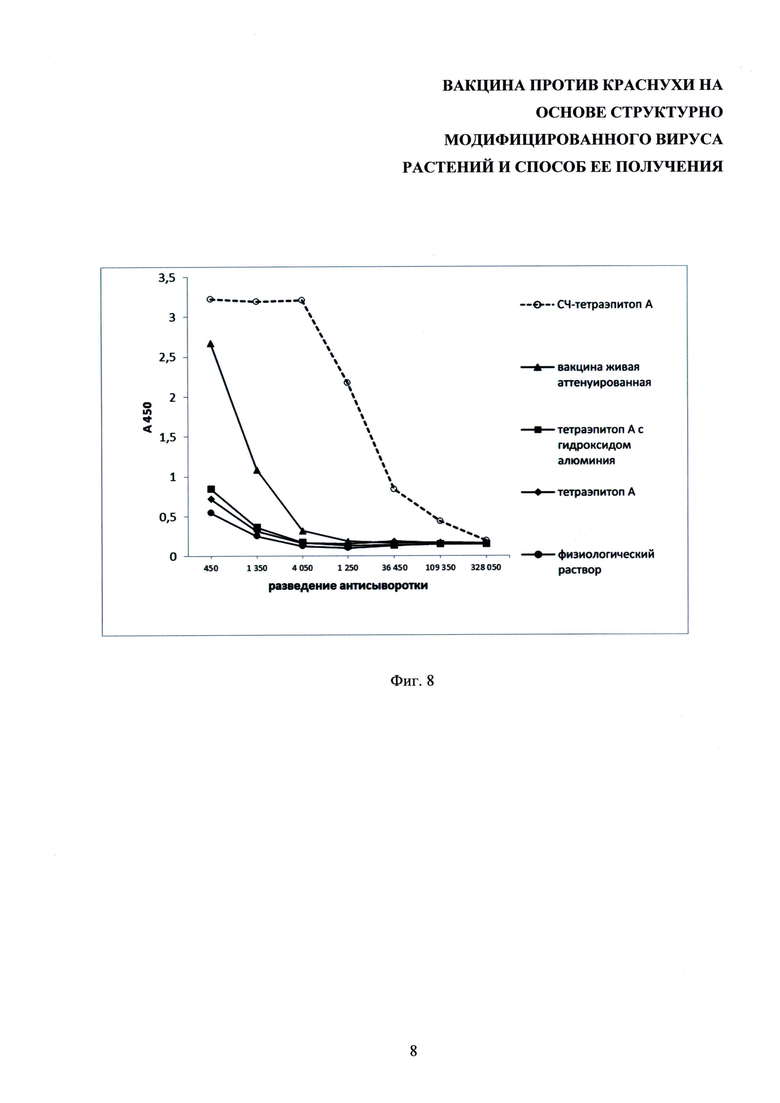
На фиг. 8 представлен график, иллюстрирующий результаты определения титра специфических антител в сыворотках крови мышей, иммунизированных тетраэпитопом А, тетраэпитопом А в смеси с адъювантом, содержащим гидроксид алюминия; кандидатной вакциной (СЧ-тетраэпитоп А) и живой аттенуированной вакциной против краснухи. ИФА.
На фиг. 9 представлена диаграмма, иллюстрирующая результаты определения титра мышиных антисывороток к СЧ (б) и к тетраэпитопу А (а, б, в), полученных при иммунизации животных тетраэпитопом А (а), тетраэпитопом А в смеси с СЧ (б) или с адъювантом, содержащим гидроксид алюминия (в). ИФА.
Осуществление изобретения
В соответствии с настоящим изобретением вакцина для профилактики краснухи представляет собой рекомбинантный белок (пример 1), состоящий из четырех повторов нейтрализующего эпитопа А (тетраэпитоп А) гликопротеина Е1 в комплексе с СЧ. Выбор эпитопа А определялся тем, что он полностью охарактеризован по структуре и антигенным свойствам и является основным антигенным участком гликопротеина Е1 вируса краснухи, на который вырабатываются нейтрализующие антитела (Wolinsky et al. An antibody- and synthetic peptide-defined rubella virus El glycoprotein neutralization domain. J. Virol. 1993, 67, 961-968; Cordoba et al. Evaluation of antibodies against a rubella virus neutralizing domain for determination of immune status. Clia Diaga Lab. Immunol. 2000, 7, 964-966; Petrova et al. The key role of rubella virus glycoproteins in the formation of immune response, and perspectives on their use in the development of new recombinant vaccines. Vaccine 2016, 34, 1006-1011). Показано, что сферические частицы-платформы, полученные путем термической денатурации вируса табачной мозаики (ВТМ), способны эффективно и неспецифически адсорбировать различные белки, в том числе антигенные детерминанты гликопротеида Е1 вируса краснухи, и могут быть использованы в качестве носителя. В то же время сферические частицы являются эффективными иммуностимуляторами. СЧ обладают высокой стабильностью и безопасны для человека, так как растения и животные не имеют общих патогенов. Кандидатная эпитопная вакцина на основе структурно модифицированных вирусов растений может радикально упростить и удешевить процесс производства вакцин против краснухи и устранить причину многих побочных реакций, которые возникают при вакцинации препаратами, изготовленными на основе живых аттенуированных вирусных частиц. Предложенный подход не требует применения ослабленного или убитого патогена, применения дорогих культуральных сред, дорогостоящего оборудования, особых условий стерильности. Стоимость получения СЧ-платформ также невысока, ВТМ накапливается в зараженном растении в больших количествах (10 г на 1 кг листьев), а процесс выделения очищенного вируса очень прост и занимает 1-2 дня.
Изобретение осуществляется следующим способом.
Суспензию частиц ВТМ в воде в концентрации 2±0.5 мг/мл подвергают кратковременной (10±0,5 минут) термической обработке при температуре 94±0.5°С, затем охлаждают в ледяной бане с формированием комплекса размером 300±20 нм. СЧ, образовавшиеся в результате такой обработки, используют в качестве носителя для получения вакцины. Для получения композиции с СЧ используют рекомбинантный белок с молекулярной массой 22±0,5 кДа, включающий в себя четыре повтора эпитопа А гликопротеина Е1 вируса краснухи. СЧ смешивают с антигеном вируса краснухи в массовом соотношении 10:1 с допустимым отклонением от указанного соотношения до 15% с формированием комплекса размером 300±20 нм. При таком соотношении белок полностью связывается с СЧ и не изменяет своих специфических антигенных и иммуногенных свойств. В качестве носителей для инъекционных растворов, предпочтительно используют воду или водные физиологические растворы и водные растворы декстрозы и глицерина. Пригодные фармацевтические носители описаны в «Remington's Pharmaceutical Sciences» под ред. E.W. Martin, 18-е изд. Для применения предпочтительно использовать вакцину стабильную в фосфатно-солевом буфере рН 7,2-7,5. Анализ состава и стабильности полученных соединений, а также их специфических свойств осуществляют с использованием следующих методик: SDS-электрофорез в полиакриламидном геле, иммуноферментный анализ, иммуноблот, метод просвечивающей электронной микроскопии, метод иммунной электронной микроскопии, метод анализа траекторий наночастиц, метод динамического рассеяния света. Действие вакцины исследовано на экспериментальных животных, где показаны иммуногенные свойства препарата.
Приведенные примеры конкретного осуществления изобретения приведены для предоставления специалистам в данной области техники полного описания проведения и применения изобретения, и подразумевают, что приведенные примеры не ограничивают предполагаемый авторами изобретения объем изобретения.
Пример 1
Продукция рекомбинантного белка (тетраэпитоп А), содержащего эпитопы гликопротеина Е1 вируса краснухи в бактериальных клетках
В качестве антигена вируса краснухи выбран рекомбинантный белок (тетраэпитоп А), содержащий эпитопы гликопротеина Е1 вируса краснухи. Эпитоп А известен в литературе как антигенный участок SP15, в состав которого входят аминокислотные остатки белка Е1 с 208 по 239. Это основной антигенный участок гликопротеина вируса краснухи Е1, на который вырабатываются нейтрализующие антитела (Wolinsky et al. An antibody- and synthetic peptide-defined rubella virus El glycoprotein neutralization domain. J. Virol. 1993, 67, 961-968; Cordoba et al. Evaluation of antibodies against a rubella virus neutralizing domain for determination of immune status. Clin. Diagn Lab. Immunol. 2000, 7, 964-966; Petrova et al. The key role of rubella virus glycoproteins in the formation of immune response, and perspectives on their use in the development of new recombinant vaccines. Vaccine 2016, 34, 1006-1011). Тетраэпитоп А несет в своем составе четырехкратный повтор участка SP15. Тетроэпитоп А был получен с использованием генно-инженерной конструкции, клонированной в экспрессионный вектор pQE-30. Данная плазмида позволяет получить рекомбинантные белки, содержащие в своем составе последовательность из шести аминокислотных остатков гистидина, которую можно использовать для дальнейшей хроматографической очистки белка. Для повышения иммуногенности эпитопа А гликопротеина Е1, он был экспрессирован в составе полипептида, содержащего четырехкратный повтор последовательности данного эпитопа. Плазмидой, содержащей последовательность, кодирующую тетраэпитоп А, трансформировали клетки Е.coli штамма SG. Культуру клеток выращивали в течение ночи в небольшом объеме (3 мл) в среде 2×YT (1,6% бактотриптона (Difco, США), 1% дрожжевого экстракта (Difco, США), 0,5% NaCl (Merk, Германия)), содержащей 100 мкг/мл ампициллина (MP Biomedicals, США) и 25 мкг/мл канамицина (MP Biomedicals, США), при +37°С. Ночную культуру переносили в 200 мл свежей 2×YT содержащей 100 мкг/мл ампициллина и 25 мкг/мл канамицина и выращивали при +37С до CD600 0,7-0,9. Экспрессию белка индуцировали добавлением изопропил-β-D-1-тиогалактопиранозоида (ИПТГ) (Anatrace, США) до конечной концентрации 2 мМ. Культуру растили дополнительно 3-5 часов при +37°С. Клетки отделяли от культуральной среды центрифугированием при +4°С (5000 об/мин, 10 минут, центрифуга «Beckman», ротор JA-14).
Для выделения и очистки тетраэпитопа А проводили хромотографию на Ni2+-нитрилотриацетатной агарозе (Ni-HTA) согласно протоколу «The QIAexpressionist» фирмы QIAGEN.
Тетраэпитоп переводили в водный раствор с помощью диализа. Диализ проводили в диализных мешках (Serva, Германия) в течение 4 часов, меняя воду каждый час. Соотношение раствор белка/вода 1:1000. Для диализа использовали воду качества Milli-Q, полученную с помощью системы Simplicity UV (Millipore, США). Тетраэпитоп А после диализа был проанализирован с помощью электрофореза в градиентном 8-20% ДСН-ПААГ (фиг. 1). Представленные на фиг. 1 данные свидетельствуют о том, что в условиях диализа не произошла деградация тетраэпитопа А и что целевой белок был успешно переведен в водный раствор. При анализе электрофореграммы выявляется единичная полоса, соответствующая тетраэпитпу А с молекулярной массой 22 кДа (фиг. 1, дорожка 1), что совпадает с расчетной молекулярной массой этого белка, полученной с использованием программы ProtParam (http://www.expasy.org).
Пример 2
Вестерн-блот анализ антигенной активности тетраэпитопа А вируса краснухи
При помощи метода вестерн-блот анализа была исследована способность тетраэпитопа А взаимодействовать с сывороткой, полученной к тетраэпитопу А, и с коммерческими антителами к вирусу краснухи. Белки фракционировали в градиентном 8-20% ПААГ (фиг. 2а и 2б). В качестве отрицательного контроля был использован белок оболочки вируса табачной мозаики (ВТМ) и белок оболочки X вируса картофеля (ХВК). Контрольный эксперимент (гель, окрашенный кумасси G-250) представлен на фиг. 2в. После электрофореза белки переносили на мембрану Amersham Hybond-P (GE Healthcare Life Sciences, Великобритания) методом электро-переноса (прибор Protean II фирмы Bio-Rad, США) в течение ночи при напряжении 30 В. Состав буфера для переноса: 47,9 мМ триса, 39 мМ глицина, 20% этанола, 0,04% ДСН. После переноса мембрану инкубировали в 5% растворе сухого молока (Sigma-Aldrich, США) и 2% твин-20 (Amresco, США) в буфере TBS (0,01 М трис-HCL, рН 7,4; 0,15 М NaCl) в течение часа. Далее проводили инкубацию с мышиной антисывороткой, полученной к тетраэпитопу А, в разведении 1:10000 или с коммерческими мышиными антителами к вирусу краснухи в разведении 1:10000 в 5% растворе сухого молока и 2% твин-20 в буфере TBS в течение 1 часа. В дальнейшем проводили отмывку (3 раза по 10 минут) в буфере tTBS (TBS, содержащий 0,05% твин-20), каждый раз меняя буфер. После отмывки мембраны инкубировали с вторичными противомышиными антителами, конъюгированными с пероксидазой хрена (Promega, США) в разведении 1:10000 в 5% молоке и 2% твин-20 в буфере TBS (1 час при комнатной температуре). Проводили отмывку 3 раза по 10 минут в буфере tTBS, каждый раз меняя буфер. Последнюю отмывку проводили в течение 10 минут в TBS. Проявляли с помощью системы ECL (GE Healthcare Life Sciences, Великобритания). Хемилюминесцентный сигнал детектировали с помощью гельдокументирующей системы Chemi Doc XRS+ с программным обеспечением Image Lab Software (Bio Rad, США). Из фиг. 2 видно, что тетраэпитоп А (фиг. 2а, дорожка 1) эффективно взаимодействуют как с сывороткой, которая бы к нему получена, так и с коммерческими антителами к вирусу краснухи (фиг. 2б, дорожка 1). Отрицательные контроли (БО ВТМ, БО ХВК) и маркеры молекулярной массы с антисывороткой к тетраэпитопу А и с коммерческими антителами к вирусу краснухи не взаимодействуют. Способность взаимодействия коммерческих антител к вирусу краснухи с тетраэпитопом А свидетельствует о доступности для взаимодействия с антителами эпитопов вируса краснухи в составе полученного рекомбинантного белка.
Пример 3
Получение вакцины
Получение сферических частиц (СЧ)
Для получения СЧ очищенный препарат вируса табачной мозаики (ВТМ) объемом 1 мл в концентрации 2 мг/мл вносят в полипропиленовую пробирку, помещают в термостат, нагретый до 94°С, и выдерживают не более 10 минут. Через 10 минут полипропиленовую пробирку переносят в лед и выдерживают в течение 5 минут до полного охлаждения. Формирование СЧ фиксируется визуально по изменению мутности раствора.
При исходной концентрации ВТМ 2 мг/мл формируются СЧ размером 300±20 нм. Размер и форма СЧ контролируются с помощью метода электронной микроскопии (фиг. 2а) и на анализаторе Zetasizer Nano-ZS (фиг. 3а).
Получение вакцины
Для адсорбции целевого белка (тетраэпитопа А белка Е1 вируса краснухи) на поверхности СЧ в полипропиленовую пробирку с 1 мл суспензии, содержащей 2 мг СЧ, добавляют 0,5 мл водного (Milli-Q) раствора, содержащего 0,2 мг целевого белка. При таком соотношении (10:1) белок полностью связывается с СЧ. Смесь инкубируют в течение 20 минут при температуре 20°С. Выпавший в осадок комплекс СЧ-белок концентрируют центрифугированием в течение 10 минут при 2000 g в центрифуге при температуре 20°С. Затем осадок ресуспендируют в 1 мл воды (Milli-Q).
С помощью метода просвечивающей электронной микроскопии был проведен сравнительный анализ СЧ, свободных от тетраэпитопа А (фиг. 3а), и композиций СЧ-тетраэпитоп А (фиг. 3б). Препараты сорбировали на медных сетках для электронной микроскопии, покрытых формваровой пленкой (при нанесении пленки использовали 0,5% раствор формвара в дихлорэтане) в течение 15-20 секунд, после чего сетки негативно контрастировали 2% раствором уранилацетата. Препараты анализировали с помощью электронного микроскопа JEM-1011 (JEOL, Япония), оснащенного цифровой фотокамерой ES500W Erlangshen (Gatan, Япония).
Данные, представленные на фиг. 3, позволяют сделать вывод о том, что адсорбция белка на поверхности СЧ не влияет на морфологию и размеры частиц.
Микроскопические методы анализа, как правило, связаны с высушиванием образца на подложке, что может привести к искажению получаемых результатов. В связи с этим для изучения размеров и агрегационного состояния композиций структурно модифицированных вирусов растений с тетроэпитопом в жидкости был использован метод анализа траекторий наночастиц (АТН). Анализ препаратов СЧ и композиций СЧ-тетраэпитоп А проводили с использованием прибора NanoSight NS500 и программного обеспечения Nanosight NTA 2.3 (Nanosight, Великобритания). Прибор оснащен лазерным источником с длиной волны 532 нм, мощностью 45 мВт и высокочувствительной черно-белой видеокамерой Luca (640×480, 14 бит на пиксель, переменная частота кадров) (Andor, Великобритания). Измерения проводили в соответствии с процедурой и рекомендациями, описанными в принятом стандарте (ASTM Е2834-12) в 10 повторах. Продемонстрировано, что после адсорбции тетраэпитопа А на поверхности структурно модифицированных вирусов растений не происходит агрегации композиций СЧ-тетраэпитоп А. Размер композиций СЧ-тетраэпитоп А по данным АТН был сопоставим с размерами СЧ (фиг. 4а) и составил 305±12 нм (фиг. 4б). Следовательно, адсорбция тетраэпитопа А на поверхности СЧ не приводила к значительному увеличению диаметра комплексов по сравнению со свободными СЧ.
Пример 4
Определение специфической активности антигена - тетраэпитопа А белка E1 вируса краснухи - в составе комплекса с СЧ методом иммунной электронной микроскопии
Суспензию комплексов СЧ-тетраэпитоп А вируса краснухи сорбировали на никелевых сетках для электронной микроскопии, покрытых формваровой пленкой (при нанесении пленки используют 0,5% раствор формвара в дихлорэтане) в течение 15-20 секунд. Перед инкубацией с первичными антителами неспецифические места связывания были блокированы в течение 20 минут PBS буфером, содержащим 1% бычий сывороточный альбумин (БСА) и 0,05% твин-20. После этого препараты инкубировали с мышиной антисывороткой к тетраэпитопу А гликопротеина Е1 или с поликлональной антисывороткой к вирусу краснухи в разведении 1:100 в блокирующем растворе в течение 30 минут. Контрольные образцы инкубировали в блокирующем растворе без добавления первичных антител. Несвязавшиеся антитела отмывали раствором для отмывки (PBS с 0,25% БСА и 0,05% твин-20) 3 раза по 5 минут. Связывание первичных антител с антигенными комплексами детектировали при помощи вторичных противомышиных антител, конъюгированных с коллоидным золотом 12 нм «Jackson Immunoresearch Inc.» (США) в разведении 1:50. С вторичными антителами инкубацию проводили в течение 30 минут, от несвязавшихся антител отмывали 3 раза по 5 минут в растворе для отмывки. Последнюю отмывку проводили в Milli-Q в течение 5 минут на качалке. После этого сеточки высушивали на воздухе и анализировали с помощью электронного микроскопа «JEM-1400» (JEOL, Япония). Показано, что комплексы СЧ-тетраэпитоп А связаны с множеством частиц коллоидного золота, как в случае использования в качестве первичных антител антисыворотки к тетраэпитопу А гликопротеина Е1 (фиг. 5а), так и в случае использования поликлональной антисыворотки к вирусу краснухи (фиг. 5б). Это свидетельствует о том, что поверхность СЧ равномерно покрыта молекулами белкового антигена и что полученные комплексы СЧ-тетраэпитоп А вируса краснухи обладают антигенными свойствами, сходными с нативным вирусом краснухи. В случае отрицательного контроля на поверхности СЧ частицы коллоидного золота не наблюдаются, что свидетельствует об отсутствии неспецифической реакции вторичных антител с тетраэпитопом А или СЧ (фиг. 5в).
Пример 5
Анализ стабильности вакцинного препарата с помощью метода электронной микроскопии и метода динамического рассеяния света (ДРС)
При попадании в организм композиции структурно модифицированных вирусов растений с антигенами вируса краснухи будут подвергаться воздействию солей. В связи с этим крайне важно было показать стабильность композиций СЧ-тетраэпитоп А в физиологических условиях. Для имитации условий воздействия среды организма комплексы СЧ-тетраэпитоп А переводили из водного раствора в PBS (состав: 7 мМ Na2HPO4, 1,5 мМ KH2PO4, рН 7,4,137 мМ NaCl, 2,7 мМ KCl). Полученные препараты были проанализированы с помощью просвечивающей электронной микроскопии, которая показала, что перевод комплексов СЧ-тетрапитоп А в PBS не приводит к изменению морфологии композиций, а их средний размер сохраняется (фиг. 6).
С помощью метода динамического рассеяния света (фотонная корреляционная спектроскопия) была подтверждена стабильность препаратов композиций структурно модифицированных вирусов растений с антигенами вируса краснухи в жидкости в физиологических условиях.
Метод ДРС позволяет определить коэффициент диффузии дисперсных частиц в жидкости путем анализа характерного времени флуктуаций интенсивности рассеянного света. Далее из коэффициента диффузии математически рассчитывается диаметр наночастиц. Измерения образцов проводили на установке Zetasizer Nano ZS (Malvern Instruments Ltd., Великобритания) с He-Ne лазером (633 нм, 10 мВт) в качестве источника света. Температура образцов поддерживалась в пределах 0,1°С при помощи системы Пельтье термостатирования. Измерения светорассеяния проводили под углом 173°. Детекцию и обработку автокорреляционных функций производили при помощи программного обеспечения Dispersion Technology Software (DTS) version 5.10. Для экспериментов использовали полистироловые кюветы с длиной оптического пути 10 мм. Объем образца в кювете составлял 1 мл. Концентрация образца по белку составляла 50 мкг/мл. Результаты анализа стабильности композиций структурно модифицированных вирусов растений с тетроэпитопом А в физиологических условиях с помощью метода ДРС приведены на фиг. 7. Таким образом, с помощью метода динамического рассеяния света было продемонстрировано, что композиции СЧ-тетраэпитоп А сохраняют свой средний диаметр как при переводе в PBS, так и при хранении в PBS. Важно отметить, что при изменении солевых условий не происходило их агрегации, что свидетельствует о высокой стабильности полученных препаратов в физиологических условиях.
Пример 6
Оценка иммуногенности предлагаемой вакцины в сравнении с живой аттенуированной вакциной против краснухи
Мышей линии BALB/c иммунизировали рекомбинантным белком - тетраэпитопом А гликопротеина Е1 вируса краснухи, тетраэпитопом А вместе с наиболее распространенным в клинической практике адъювантом, содержащим гидроксид алюминия, предлагаемым вакцинным препаратом против краснухи, существующей зарегистрированной вакциной против краснухи и нулевым контролем - стерильным физиологическим раствором. Для всех групп мышей проводили трехкратную иммунизацию с двухнедельным интервалом. Способ получения тетраэпитопа А гликопротеина Е1 вируса краснухи описан в примере 1. Вакцинный препарат получали, как описано в примере 3. Схема иммунизации приведена в таблице 1. В качестве вакцины сравнения использовали живую аттенуированную вакцину против краснухи («ФГУП НПО Микроген»).
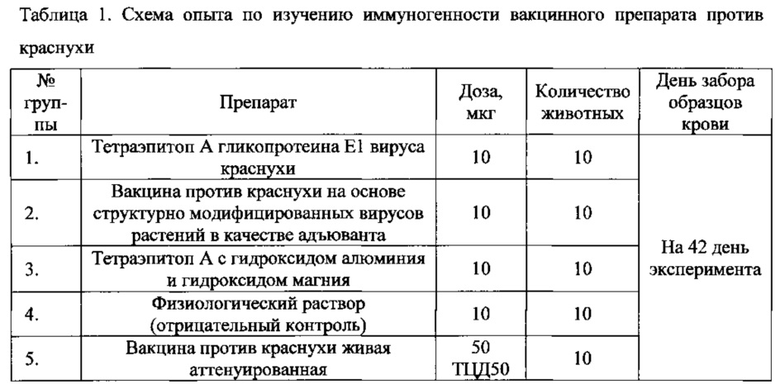
Количество тетраэпитопа А во всех случаях составляло 10 мкг на 1 инъекцию. Доза живой вакцины составляла 50 ТЦД50. Исследуемые растворы вводились по 100 мкл внутримышечно в заднюю мышцу бедра. Забор крови производили после забоя животных. Консервация образцов - отделение сыворотки и заморозка при -70°С. Полученную сыворотку использовали для определения титра антител против тетраэпитопа А гликопротеина Е1 вируса краснухи методом иммуноферментного анализа. С этой целью на планшетах Greiner (Германия) сорбировали антиген - тетраэпитоп А гликопротеина Е1 вируса краснухи в PBS (0,05 М K2HPO4, 0,1 М NaCl, рН 7,4) в концентрации 10 мкг/мл, по 100 мкл в лунку. Планшеты инкубировали 2 часа при комнатной температуре, проводили три отмывки tPBS (PBS, 0,2% твин-20, рН 7,4). Блокировку проводили обезжиренным молоком (Parmalat 0,5% ультрапастеризованное), 2 часа при комнатной температуре. После чего проводили три отмывки tPBS и вносили тестируемые антисыворотки. Сыворотки разводили в буфере для разведения сывороток «Иммуностаб» (ООО "НВО Иммунотех") в тройных серийных разведениях, начиная с разведения 1/450 в двух параллелях. Инкубировали 1 час при 37°С на термостатируемом шейкере, промывали, как описано, и добавляли пероксидазный конъюгат (Promega) в рабочем разведении 1/10000, приготовленном на PBS, содержащим 0,5% бычьего сывороточного альбумина. Инкубировали 1 ч при 37°С на шейкере, промывали, как описано, и добавляли субстрат - ТМВ (ООО "НВО Иммунотех"). Цветную реакцию останавливали, заливая в лунки планшета по 100 мкл 0,5 М серной кислоты. Оптическую плотность в лунках определяли на ридере Biotek Cambrex ELx808 Microplate Reader (США) при длине волны 450 нм. При обсчете за результат измерения оптической плотности принималось среднее арифметическое оптической плотности двух параллелей. Титры рассчитывались программными средствами, доступными на сайте http://www.myassays.com/search.aspx?search=antibody%20titers, методом линейной регрессии. Область учитываемых значений определялась путем визуального анализа графика линейной регрессии и степени дисперсности полученных результатов и включала в себя не менее трех разведений.
Для уточнения границ раститровки индивидуальных образцов были использованы собранные по группам пулы сывороток опытных животных. Был проведен иммуноферментный анализ образцов, полученные результаты определения оптической плотности использовали для построения графика в табличном процессоре Microsoft Excel 2010. Анализ специфической активности антисыворотки, полученной на 42-е сутки иммунизации, показал, что самый высокий уровень антител (1:181400) наблюдали у второй группы животных, иммунизированных кандидатной вакциной против краснухи (фиг. 8).
Для количественного сравнения выработки антител на СЧ и на тетраэпитоп А в составе комплекса СЧ-тетраэпитоп А полученную сыворотку использовали для определения титра антител к СЧ. Для этого на планшетах сорбировали СЧ в концентрации 10 мкг/мл в объеме 100 мкл на лунку и проводили ИФА, как описано выше. Результаты определения титров мышиных антисывороток представлены на фиг. 9. Согласно полученным данным уровень выработки антител на СЧ в составе комплекса СЧ-тетраэпитоп А чрезвычайно низкий. При иммунизации мышей кандидатной вакциной антитела вырабатываются почти исключительно к целевому белку (тетраэпитопу А).
При анализе индивидуальных образцов была проведена статистическая обработка полученных данных с помощью пакета програмного обеспечения STATISTICA (data analysis software system), version 10. StatSoft, Inc. (2011) www.statsoft.com. Программа рекомендована к использованию для статистической обработки клинических и доклинических исследований Управлением по санитарному надзору за качеством пищевых продуктов и медикаментов США и Европейским Агентством лекарственных средств, разработчиком данного программного обеспечения предоставляется услуга валидации установленной версии программного обеспечения на электронно-вычислительной машине заказчика.
Согласно рекомендациям производителя для оценки нескольких независимых групп с неизвестным типом распределения был использован дисперсионный анализ Краскела-Уоллиса и медианный тест, а для попарного сравнения - критерий Данна.
Согласно дисперсионному анализу гипотеза о принадлежности 6 групп данных к одной генеральной совокупности была отвергнута.
При попарном сравнении значимые различия (обозначены «+») были установлены между следующими группами (Таблица 2):
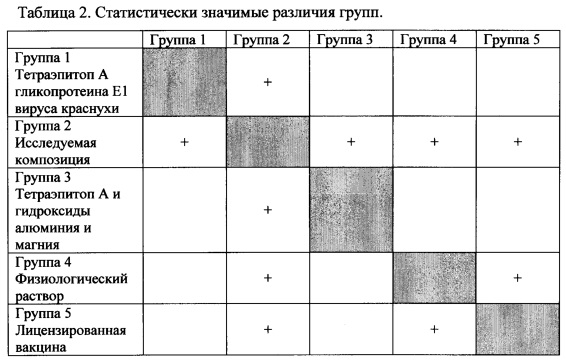
Было обнаружено статистически значимое отличие от нулевого контроля титров антител к белку вируса у животных, получавших кандидатную вакцину, и у животных, получавших зарегистрированную вакцину против краснухи. Так же были выявлены статистически значимо более высокие титры антител у животных, получавших кандидатную вакцину, чем титры антител у животных, получавших зарегистрированную вакцину против краснухи. Отличие групп, получавших тетраэпитоп А гликопротеина Е1 вируса краснухи и тетраэпитоп А с гидроксидами алюминия и магния, от нулевого контроля статистически не значимо.
Таким образом, заявляемая вакцина для профилактики краснухи, включает сферические частицы вируса табачной мозаики и антигены вируса краснухи в массовом соотношении, позволяющем добиться стабильности вакцины, при этом не происходит агрегации комплексов СЧ-тетраэпитоп А.
Кандидатная вакцина показала высокую иммуногенность на животной модели, результаты были статистически обработаны и подтверждены отрицательным и положительным контролем. Полученные данные позволяют экстраполировать достигнутый эффект и на человека, поскольку механизмы функционирования иммунной системы у всех млекопитающих в основном сходны, а лабораторные животные являются стандартной моделью для создания вакцин. Следовательно, полученный препарат может рассматриваться в качестве кандидатной вакцины против краснухи, предназначенной для человека.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИГЕН ДЛЯ РЕКОМБИНАНТНОЙ ВАКЦИНЫ ПРОТИВ ВИРУСА КРАСНУХИ | 2019 |
|
RU2709328C1 |
| ИММУНОГЕННАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ ЧУЖЕРОДНЫЕ АНТИГЕНЫ НА ПОВЕРХНОСТИ СФЕРИЧЕСКИХ НОСИТЕЛЕЙ, ПОЛУЧЕННЫХ ПРИ ТЕРМИЧЕСКОЙ ДЕНАТУРАЦИИ СПИРАЛЬНЫХ ВИРУСОВ | 2010 |
|
RU2440140C1 |
| НОВЫЙ ТИП ЧАСТИЦ-НОСИТЕЛЕЙ (ПЛАТФОРМ) ДЛЯ ПОЛУЧЕНИЯ АКТИВНЫХ КОМПЛЕКСОВ | 2010 |
|
RU2441667C1 |
| СПОСОБ УСИЛЕНИЯ ИММУННОГО ОТВЕТА | 2010 |
|
RU2442604C1 |
| РЕКОМБИНАНТНАЯ ВАКЦИНА ПРОТИВ СИБИРСКОЙ ЯЗВЫ НА ОСНОВЕ ВИРУСОВ РАСТЕНИЙ | 2023 |
|
RU2821917C1 |
| ВИРИОНЫ И ВИРУСОПОДОБНЫЕ ЧАСТИЦЫ ВИРУСА МОЗАИКИ АЛЬТЕРНАНТЕРЫ КАК УСИЛИТЕЛИ ИММУННОГО ОТВЕТА | 2015 |
|
RU2639491C2 |
| Антиген для универсальной рекомбинантной вакцины против вируса гриппа птиц и человека | 2021 |
|
RU2839428C2 |
| РЕКОМБИНАНТНЫЙ ВИРУСНЫЙ ВЕКТОР ДЛЯ ПРОДУКЦИИ В РАСТЕНИЯХ БЕЛКА Е1 ВИРУСА КРАСНУХИ (ВАРИАНТЫ) И СИСТЕМА ЭКСПРЕССИИ БЕЛКА Е1 ВИРУСА КРАСНУХИ В КЛЕТКАХ РАСТЕНИЯ (ВАРИАНТЫ) | 2008 |
|
RU2390563C1 |
| ОЛИГОМЕРНАЯ ЧАСТИЦА, ИНДУЦИРУЮЩАЯ ИММУНИТЕТ ПРОТИВ ВИРУСА ГЕПАТИТА С, СПОСОБ ПОЛУЧЕНИЯ ОЛИГОМЕРНОЙ ЧАСТИЦЫ, КОМПОЗИЦИЯ, СПЕЦИФИЧЕСКОЕ АНТИТЕЛО, НАБОР (ВАРИАНТЫ), ИММУНОЛОГИЧЕСКИЙ АНАЛИЗ И ВАКЦИНА ПРОТИВ ВИРУСА ГЕПАТИТА С | 1999 |
|
RU2247729C2 |
| РЕКОМБИНАНТНЫЙ МОДИФИЦИРОВАННЫЙ ВИРУС ВАКЦИНЫ АНКАРА, СПОСОБНЫЙ ЭКСПРЕССИРОВАТЬ СТРУКТУРНЫЕ АНТИГЕНЫ ВИРУСА ГЕПАТИТА С | 2002 |
|
RU2270860C2 |

Предложены вакцина для профилактики краснухи и способ ее получения. Охарактеризованная вакцина включает сферические частицы вируса табачной мозаики и антигены вируса краснухи, адсорбированные на поверхности сферических частиц, взятые в массовом соотношении 10:1. Размер полученного комплекса сферическая частица - антиген составляет 300±20 нм. Способ получения вакцины включает следующие этапы: получение сферических частиц вируса табачной мозаики посредством метода термической структурной перестройки вируса табачной мозаики, используемого в концентрации 2 мг/мл, который сначала нагревают до температуры 94±0.5°С и выдерживают в течение 10±0,5 минут, затем охлаждают в ледяной бане с формированием комплекса размером 300±20нм. Затем получение белка-антигена вируса краснухи. Смешение полученных сферических частиц с антигеном. Смешение полученного комплекса с фармацевтически приемлемым носителем. Получаемый препарат по своим иммуногенным свойствам не уступает живой аттенуированной вакцине против краснухи и является высоко эффективным. Используемая технология не требует применения ослабленного или убитого патогена, применения дорогих культуральных сред, дорогостоящего оборудования и особых условий стерильности. 2 н. и 1 з.п. ф-лы, 9 ил., 2 табл., 6 пр.
1. Вакцина для профилактики краснухи, характеризующаяся тем, что она включает два компонента: сферические частицы вируса табачной мозаики и антигены вируса краснухи, адсорбированные на поверхности сферических частиц, взятые в массовом соотношении 10:1 с допустимым отклонением от указанного соотношения до 15%, при этом размер полученного комплекса сферическая частица - антиген составляет 300±20 нм, при следующем содержании компонентов в дозе: антиген вируса краснухи - 10 мкг, сферические частицы вируса табачной мозаики - 100 мкг, фосфатно-солевой буфер рН 7,2-7,5 - 100 мкл.
2. Способ получения вакцины по п. 1, характеризующийся тем, что включает следующие этапы:
- получение сферических частиц вируса табачной мозаики,
при этом сферические частицы получают методом термической структурной перестройки вируса табачной мозаики, используемого в концентрации 2 мг/мл, который сначала нагревают до температуры 94±0.5°С и выдерживают в течение 10±0,5 минут, затем охлаждают в ледяной бане с формированием комплекса размером 300±20 нм;
- получение белка-антигена вируса краснухи;
- смешение полученных сферических частиц с антигеном в массовом соотношении 10:1 с допустимым отклонением от указанного соотношения до 15%, и формированием комплекса сферическая частица - антиген с размерами 300±20 нм;
- смешение полученного комплекса с фармацевтически приемлемым носителем для получения инъекционного раствора.
3. Способ получения вакцины по п. 2, характеризующийся тем, что антиген вируса краснухи представляет собой рекомбинантный белок с молекулярной массой 22±0,5 кДа, включающий в себя четыре повтора эпитопа А гликопротеина Е1 вируса краснухи.
| А.А | |||
| ДМИТРИЕВА и др., Разработка кандидатной вакцины против краснухи на основе структурно модифицированных вирусов растений, XXII Международная научная конференция студентов, аспирантов и молодых ученых "Ломоносов-2015", 2015, стр | |||
| Пожарный двухцилиндровый насос | 0 |
|
SU90A1 |
| ИММУНОГЕННАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ ЧУЖЕРОДНЫЕ АНТИГЕНЫ НА ПОВЕРХНОСТИ СФЕРИЧЕСКИХ НОСИТЕЛЕЙ, ПОЛУЧЕННЫХ ПРИ ТЕРМИЧЕСКОЙ ДЕНАТУРАЦИИ СПИРАЛЬНЫХ ВИРУСОВ | 2010 |
|
RU2440140C1 |
| P.K | |||
| COYLE et al., Rubella-Specific Immune Complexes after Congenital Infection and Vaccination, Infection and Immunity, 1982, Vol | |||
| Коридорная многокамерная вагонеточная углевыжигательная печь | 1921 |
|
SU36A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Льномолотилка веялка | 1923 |
|
SU498A1 |

