ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится выделенному полипептиду, обладающему резистентностью к ингибированию метионином по принципу обратной связи и гомосерин-О-сукцинилтрансферазной активностью, микроорганизму, экспрессирующему указанный полипептид, и способу получения О-сукцинилгомосерина с использованием указанного микроорганизма.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Известно, что большинство микроорганизмов, представленных в природе, используют О-сукцинилгомосерин или О-ацетилгомосерин в качестве промежуточного продукта биосинтеза метионина. Обычно О-сукцинилгомосерин образует гомосерин-О-сукцинилтрансфераза (MetA), конъюгирующая сукцинильную группу сукцинил-КоА с гомосерином, и О-ацетилгомосерин образует гомосерин-О-ацетилтрансфераза (MetX), конъюгирующая ацетильную группу ацетил-КоА с гомосерином. То есть, применительно к получению О-сукцинилгомосерина наряду с другими промежуточными продуктами, metA является одним из наиболее важных генов при разработке микроорганизмов, продуцирующих О-сукцинилгомосерин. В то же время известно, что, в отличие от MetA, MetX не подвержен ингибированию по принципу обратной связи и обладает высокой ферментной стабильностью.
О-сукцинилгомосерин может быть получен с использованием штамма с делецией гена metB, кодирующего цистатионин-гамма-синтазу в метаболическом пути биосинтеза метионина. Тем не менее, штамму, продуцирующему О-сукцинилгомосерин, необходим L-метионин. По этой причине происходит ингибирование активности гомосерин-О-сукцинилтрансферазы по принципу обратной связи посредством ингибирования метионином, добавленным в среду, и, в конечном счете, О-сукцинилгомосерин не может быть получен в высокой концентрации.
Соответственно, во многих предшествующих патентах первостепенное внимание было уделено исследованиям по устранению ингибирования metA по принципу обратной связи контрольной системой обратной связи. Тем не менее, для гомосерин-О-сукцинилтрансферазы, кодируемой metA, свойственны проблемы низкой стабильности самого белка дикого типа, и введение мутаций для устранения ингибирования по принципу обратной связи усугубляет нестабильность. Соответственно, для разработки штамма, продуцирующего О-сукцинилгомосерин с высокой продуктивностью, необходимо устранение ингибирования гена metA по принципу обратной связи и сохранение стабильности фермента.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Техническая проблема
Для решения феномена ингибирования metA по принципу обратной связи и проблемы нестабильности фермента, описанной выше, авторы настоящего изобретения предприняли попытку разработать гомосерин-О-сукцинилтрансферазу с сохраненной стабильностью фермента, не подверженную, в то же время, ингибированию метионином по принципу обратной связи, и провели на предмет этого скрининг новых ферментов, обладающих указанной активностью. В результате отбора генов-кандидатов, прошедших такой скрининг, и культивирования в колбах после их введения в Escherichia sp. авторы настоящего изобретения обнаружили, что происходило образование О-сукцинилгомосерина и что отобранные таким образом гены обладали гомосерин-О-сукцинилтрансферазной активностью и резистентностью к ингибированию метионином по принципу обратной связи, завершив посредством этого настоящее изобретение.
Техническое решение
Задачей настоящего изобретения является обеспечение нового выделенного полипептида, обладающего резистентностью к ингибированию метионином по принципу обратной связи и гомосерин-О-сукцинилтрансферазной активностью.
Другой задачей настоящего изобретения является обеспечение полинуклеотида, кодирующего новый выделенный полипептид.
Еще одной задачей настоящего изобретения является обеспечение микроорганизма для получения О-сукцинилгомосерина, экспрессирующего новый выделенный полипептид.
Еще одной задачей настоящего изобретения является обеспечение способа получения О-сукцинилгомосерина с использованием указанного выше микроорганизма.
Полезные эффекты изобретения
Микроорганизм для получения О-сукцинилгомосерина, содержащий новый выделенный полипептид, обладающий резистентностью к ингибированию метионином по принципу обратной связи и гомосерин-О-сукцинилтрансферазной активностью, который может обладать резистентностью к ингибированию метионином по принципу обратной связи и продуцировать О-сукцинилгомосерин с высоким выходом и, таким образом, может быть эффективно использован для получения L-метионина, используемого им в качестве предшественника, с высоким выходом.
Наилучший вариант осуществления изобретения
Для решения указанных выше задач в одном аспекте согласно настоящему изобретению предложен новый выделенный полипептид, обладающий резистентностью к ингибированию метионином по принципу обратной связи и гомосерин-О-сукцинилтрансферазной активностью.
Использованный здесь термин «гомосерин-О-сукцинилтрансферазная активность» относится к активности по превращению гомосерина в О-сукцинилгомосерин в процессе биосинтеза метионина.
Использованный здесь термин «ингибирование по принципу обратной связи» относится к ингибированию активности гомосерин-О-сукцинилтрансферазы метионином в процессе биосинтеза метионина.
Полипептид по настоящему изобретению характеризуется тем, что он имеет аминокислотную последовательность SEQ ID NO: 29, обладающую гомосерин-О-сукцинилтрансферазной активностью и резистентностью к ингибированию метионином по принципу обратной связи. Любой полипептид, последовательность которого на 80% или более, конкретно на 90% или более, конкретнее на 95% или более и еще конкретнее на 97% или более гомологична указанному выше полипептиду, также включен в объем настоящего изобретения, с учетом того, что полипептид обладает гомосерин-О-сукцинилтрансферазной активностью и резистентностью к ингибированию метионином по принципу обратной связи, предложенными в настоящем изобретении. Гомология может быть определена с использованием BLAST 2.0, являющегося эталонным алгоритмом, или FASTA по Pearson [Methods Enzymol., 183, 63(1990), ниже]. На основе алгоритма BLAST разработаны программы, называемые BLASTN и BLASTX [www.ncbi.nlm.nih.gov, ниже].
В другом аспекте согласно настоящему изобретению предложен выделенный полинуклеотид, кодирующий указанный выше полипептид. Конкретно, полипептид может быть кодирован полинуклеотидной последовательностью SEQ ID NO: 36. Ввиду вырожденности кодонов объем настоящего изобретения также включает, без ограничения, полинуклеотиды, последовательность которых по меньшей мере на 80%, конкретно на 90% или более, конкретнее на 95% или более и еще конкретнее на 97% или более гомологична указанной выше последовательности.
В еще одном аспекте согласно настоящему изобретению предложен вектор, содержащий функциональный полинуклеотид.
Использованный здесь термин «вектор» относится к ДНК-конструкции, содержащей нуклеотидную последовательность полинуклеотида, кодирующего интересующий белок, где интересующий белок функционально связан с подходящей регуляторной последовательностью, таким образом, что интересующий белок может быть экспрессирован в подходящем хозяине. Регуляторная последовательность может содержать промотор, способный инициировать транскрипцию, любую операторную последовательность для регуляции транскрипции, последовательность, кодирующую подходящий домен связывания рибосом на мРНК, и последовательность, регулирующую терминацию транскрипции и трансляции. После трансформации подходящего хозяина вектор может быть реплицирован, или может функционировать независимо от генома хозяина, или может быть интегрирован в сам геном хозяина.
Относительно вектора, используемого в настоящем изобретении, может не быть существенных ограничений, при условии, что вектор может быть реплицирован у хозяина, и может быть использован любой вектор, известный в данной области.
В еще одном аспекте согласно настоящему изобретению предложен микроорганизм, продуцирующий О-сукцинилгомосерин, экспрессирующий полипептид, обладающий резистентностью к ингибированию метионином по принципу обратной связи и гомосерин-О-сукцинилтрансферазной активностью.
Использованный здесь термин «микроорганизм, продуцирующий О-сукцинилгомосерин», может относиться к микроорганизму, способному продуцировать О-сукцинилгомосерин и хранить его внутриклеточно и внеклеточно.
Микроорганизм для получения О-сукцинилгомосерина включает штаммы прокариотических и эукариотических микроорганизмов, например, без ограничения, штаммы микроорганизмов, принадлежащие к роду Escherichia, роду Erwinia, роду Serratia, роду Providencia, роду Corynebacterium и роду Brevibacterium. Конкретно, микроорганизм может представлять собой микроорганизм, принадлежащий к роду Escherichia, например, Escherichia coli.
Микроорганизм, продуцирующий О-сукцинилгомосерин, может быть получен с использованием штаммов микроорганизмов, продуцирующих L-лизин, L-треонин или L-изолейцин, и, конкретно, с использованием штамма, продуцирующего L-треонин. Поскольку штамм, продуцирующий L-треонин, представляет собой штамм, способный синтезировать L-треонин и гомосерин в качестве предшественника О-сукцинилгомосерина, с использованием этого штамма может быть синтезировано большое количество предшественников метионина, то есть О-сукцинилгомосерина.
В настоящем изобретении экспрессия полипептида может быть достигнута трансформацией рекомбинантным вектором, содержащим функциональный ген, кодирующий полипептид, или введением полинуклеотида, кодирующего полипептид, в хромосому, однако методы не ограничены указанными выше.
Использованный здесь термин «трансформация» относится к процессу введения вектора, содержащего полинуклеотид, кодирующий целевой белок, в клетку-хозяина, что позволяет экспрессировать полинуклеотид, кодируемый белком, в клетке-хозяине. Неважно, введен ли полинуклеотид, используемый для трансформации, в хромосому клетки-хозяина, будучи расположен в ней, или расположен вне хромосомы, при условии что он может быть экспрессирован в клетке-хозяине. Полинуклеотид может быть введен в любой форме, с учетом того, что он может быть введен в клетку-хозяина и экспрессирован в ней. Например, полинуклеотид может быть введен в клетку-хозяина в форме экспрессионной кассеты, представляющей собой полинуклеотидную конструкцию, содержащую все основные элементы, необходимые для самостоятельной экспрессии. Обычно экспрессионная кассета может содержать промотор, функционально связанный с открытой рамкой считывания (далее - «ORF») гена, сигнал терминации транскрипции, домен связывания рибосом и сигнал терминации трансляции.
Относительно промотора, используемого в настоящем изобретении, может не быть существенных ограничений, при условии, что он способен инициировать транскрипцию полинуклеотида, кодирующего целевой белок, в клетке-хозяине с высокой частотой, и может быть использован любой промотор, известный в данной области. Конкретно, могут быть использованы промотор T7, промотор trc, промотор tac, промотор CJ1 (патент Кореи №0620092) и так далее.
В типичном воплощении настоящего изобретения ген metB, кодирующий цистатионин-гамма-синтазу, у микроорганизма может быть дополнительно удален или ослаблен.
В типичном воплощении настоящего изобретения ген thrB, кодирующий гомосеринкиназу, и ген metA, кодирующий гомосерин-О-сукцинилтрансферазу, у микроорганизма могут быть дополнительно удалены или ослаблены.
Кроме того, в типичном воплощении микроорганизм может представлять собой Escherichia sp. с дополнительно усиленными фосфоенолпируваткарбоксилазой, аспартатаминотрансферазой и аспартатполуальдегиддегидрогеназой.
В настоящем изобретении последовательности генов могут быть получены из баз данных, таких как Национальный центр биотехнологической информации (National Center for Biotechnology Information, NCBI).
Использованный здесь термин «делеция» относится к типу удаления из хромосомы части или всей области нуклеотидной последовательности целевого гена, начиная с нуклеотидной последовательности, соответствующей инициирующему кодону, до нуклеотидной последовательности, соответствующей терминирующему кодону, или части или всей области нуклеотидной последовательности его регуляторной области.
Использованный здесь термин «ослабление» относится к устранению или снижению внутриклеточной активности по меньшей мере одного фермента, кодируемого соответствующим полинуклеотидом, у штамма микроорганизма. Например, экспрессия белка может быть ослаблена модификацией последовательности регуляции экспрессии или нуклеотидной последовательности 5'--UTR (5'--нетранслируемой области) гена, или активность белка может быть ослаблена заменой инициирующего кодона или введением мутации в область ORF соответствующего гена.
Использованный здесь термин «усиление» относится к повышению внутриклеточной активности фермента, кодируемого соответствующим полинуклеотидом. Усиление внутриклеточной активности фермента может быть достигнуто сверхэкспрессией гена или введением модификации в саму полинуклеотидную последовательность.
Сверхэкспрессия полинуклеотида может представлять собой модификацию посредством замены последовательности регуляции экспрессии, или модификацию посредством мутации, замены инициирующего кодона, дополнительного введения полинуклеотида в хромосому или увеличения числа копий посредством введения с использованием вектора, или их комбинацию.
Последовательность регуляции экспрессии представляет собой последовательность, контролирующую экспрессию полинуклеотида, функционально связанного с ней, например, промотор, терминатор, энхансер, сайленсер, последовательность Шайна-Дальгарно и так далее. Инициирующий кодон, состоящий из TTG или GTG, может быть заменен на ATG для повышения ферментативной активности соответствующего гена или снижения ферментативной активности соответствующего гена посредством противоположной замены. Полинуклеотид может представлять собой полинуклеотид, число копий которого увеличено посредством его введения в определенный сайт на хромосоме. Этот определенный сайт может включать, например, транспозон или межгенную область. Кроме того, полинуклеотид может представлять собой полинуклеотид, введенный в вектор экспрессии, который был снова введен в клетку-хозяина, посредством чего число его копий было увеличено.
В другом аспекте согласно настоящему изобретению предложен способ получения О-сукцинилгомосерина, включающий культивирование микроорганизма в среде для получения О-сукцинилгомосерина и получение О-сукцинилгомосерина из микроорганизма или среды.
Культивирование штамма микроорганизма для получения О-сукцинилгомосерина, полученного выше, может быть проведено в соответствии с подходящей средой и условиями культивирования, известными в данной области. Способ культивирования может быть легко адаптирован специалистом в данной области для применения в соответствии с выбранным штаммом. Конкретно, культура может представлять собой, без ограничения, периодическую культуру, непрерывную культуру и адаптированную культуру (fetch culture). Эти различные способы культивирования раскрыты, например, в приведенной ссылке (“Biochemical Engineering” by James M. Lee, Prentice-Hall International Editions, pp 138-176).
Среды, используемые для культивирования, должны подходящим образом соответствовать требованиям для определенных штаммов. Примеры сред для различных микроорганизмов раскрыты, например, в приведенной ссылке (“Manual of Methods for General Bacteriology” by the American Society for Bacteriology, Washington D.C., USA, 1981). Среды могут содержать различные источники углерода, источники азота и микроэлементы. Примеры источников углерода для включения в среды могут включать углеводы, такие как глюкоза, сахароза, лактоза, фруктоза, мальтоза, крахмал и целлюлоза; жиры, такие как соевое масло, подсолнечное масло, касторовое масло и кокосовое масло; жирные кислоты, такие как пальмитиновая кислота, стеариновая кислота и линолевая кислота; спирты, такие как глицерин и этанол; и органические кислоты, такие как уксусная кислота. Эти источники углерода могут быть использованы по отдельности или в комбинации. Примеры источников азота для включения в среды могут включать органические источники азота, такие как пептон, дрожжевой экстракт, мясной бульон, солодовый экстракт, жидкий кукурузный экстракт (CSL) и бобовую муку; и неорганические источники азота, такие как мочевина, сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония. Эти источники азота могут быть использованы по отдельности или в комбинации. В качестве источника фосфора среды могут содержать дигидрофосфат калия, гидроортофосфат калия и соответствующие натрий-содержащие соли. В дополнение, культуральные среды могут содержать металлы, такие как сульфат магния и сульфат железа. Кроме того, могут быть включены аминокислоты, витамины, подходящие предшественники и так далее. Эти культуральные среды или предшественники могут быть добавлены в культуру в форме периодической культуры или непрерывной культуры.
В дополнение, во время культивирования рН культуры можно подходящим образом корректировать добавлением такого соединения, как гидроксид аммония, гидроксид калия, аммиак, фосфорная кислота и серная кислота. Кроме того, образование пузырьков во время культивирования можно предотвращать с использованием пеногасителя, такого как полигликолевый сложный эфир жирной кислоты. В дополнение, в культуру может быть добавлен газообразный кислород или газ, содержащий газообразный кислород (например, воздух), для поддержания в культуре аэробных условий. Температура культуры может входить в диапазон от 20°С до 45°С и, конкретно, от 25°С до 40°С. Культивирование можно продолжать до получения желаемого количества продукта О-сукцинилгомосерина и, конкретно, от 10 часов до 160 часов.
О-сукцинилгомосерин, полученный способом по настоящему изобретению, может быть превращен в метионин цистатионин-гамма-синтазой или О-сукцинилгомосеринсульфгидрилазой. Кроме того, возможно получение янтарной кислоты в качестве побочного продукта, в дополнение к L-метионину, посредством взаимодействия O-сукцинил-L-гомосерина, полученного способом по настоящему изобретению, с CH3SH.
В еще одном аспекте настоящее изобретение относится к применению полипептида, обладающего резистентностью к ингибированию метионином по принципу обратной связи и гомосерин-О-сукцинилтрансферазной активностью, имеющего аминокислотную последовательность SEQ ID NO: 29. Было подтверждено, что новый выделенный полипептид по настоящему изобретению обладает резистентностью к ингибированию метионином по принципу обратной связи и способен продуцировать О-сукцинилгомосерин с высоким выходом, и, таким образом, указанный полипептид может быть использован для получения О-сукцинилгомосерина.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Далее настоящее изобретение будет описано более подробно со ссылкой на следующие Примеры. Тем не менее, эти Примеры приведены лишь в иллюстративных целях, и изобретение не следует ограничивать этими Примерами.
Пример 1: Получение штамма, продуцирующего треонин, на основе исходного штамма
(1) Делеция гена metB
Для описания субстратной специфичности и активности гена metX получали штамм, способный накапливать гомосерин и имеющий делецию по утилизации ацилгомосерина. Штамм конструировали на основе FTR2533 (КССМ 10541), штамма, продуцирующего треонин, раскрытого в международной заявке на патент №WO 05/075625.
Делецию гена metB, кодирующего цистатионинсинтазу, у штамма FTR2533 (КССМ 10541), продуцирующего треонин, проводили методом делеции «FRT-one-step-PCR» {PNAS (2000) vol 97: Р 6640-6645). Делеционную кассету конструировали посредством ПЦР (полимеразной цепной реакции) с использованием праймеров SEQ ID NO: 1 и SEQ ID NO: 2 и вектора pKD3 (PNAS (2000) vol 97: P 6640-6645) в качестве матрицы. ПЦР проводили на протяжении 30 циклов в следующих условиях: денатурация при 94°С в течение 30 с, отжиг при 55°С в течение 30 с и полимеризация при 72°С в течение 1 мин.

Полученный продукт ПЦР подвергали электрофорезу в 1,0%-м агарозном геле и 1,2 т.п.о. полосу ДНК, полученную из него, очищали. Полученный фрагмент ДНК электропорировали в штамм FTR2533, уже трансформированный вектором pKD46 (PNAS (2000) vοl97: Р6640-6645).
Для электропорации штамм FTR2533, трансформированный pKD46, культивировали в среде LB, содержащей 100 мкг/л ампициллина и 5 мМ L-арабинозы, при 30°С до достижения OD600 0,6. Полученный штамм промывали два раза стерильной дистиллированной водой и затем для использования промывали один раз 10%-м глицерином. Электропорацию проводили при 2500 В.
Полученный штамм высевали на чашки со средой LB, содержащей 25 мкг/л хлорамфеникола, культивировали при 37°С в течение ночи и отбирали штамм, резистентный к хлорамфениколу. Отобранный штамм подвергали ПЦР с использованием тех же праймеров и штамма в качестве матрицы и делецию гена metB подтверждали, наблюдая присутствие 1,2 т.п.о. полосы гена в 1,0%-м агарозном геле. Подтвержденный таким образом штамм снова трансформировали вектором рСР20 (PNAS (2000) vol 97: Р6640-6645) и культивировали в среде LB, и снова конечный штамм с делецией гена metB, имеющего уменьшенный размер 150 п. о., подтвержденный в 1,0%-м агарозном геле, конструировали, проводя ПЦР в тех же условиях, и подтверждали удаление хлорамфениколового маркера из штамма. Конструированный таким образом штамм был назван «CJMA1».
(2) Делеция гена thrB
Делецию гена thrB, кодирующего гомосеринкиназу, у конструированного таким образом штамма CJMA1 проводили методом делеции «FRT-one-step-PCR», как в случае с делецией гена metB.
Делеционную кассету thrB конструировали посредством ПЦР с использованием праймеров SEQ ID NO: 3 и SEQ ID NO: 4 и вектора pKD4 (PNAS (2000) vol 97: Ρ 6640-6645) в качестве матрицы. ПЦР проводили на протяжении 30 циклов в следующих условиях: денатурация при 94°С в течение 30 с, отжиг при 55°С в течение 30 с и удлинение при 72°С в течение 1 мин.

Полученный продукт ПЦР подвергали электрофорезу в 1,0%-м агарозном геле и 1,6 т.п.о. полосу ДНК, полученную из него, очищали. Полученный фрагмент ДНК электропорировали в штамм CJMA1, уже трансформированный вектором pKD46. Полученный штамм высевали на чашки со средой LB, содержащей 50 мкг/л канамицина, культивировали при 37°С в течение ночи и отбирали штамм, резистентный к канамицину. Отобранный штамм подвергали ПЦР в тех же условиях с использованием праймеров SEQ ID NO: 3 и 4 и делецию гена thrB подтверждали, наблюдая присутствие 1,6 т.п.о. полосы гена в 1,0%-м агарозном геле. Подтвержденный таким образом штамм снова трансформировали вектором рСР20 и культивировали в среде LB, и конечный штамм с делецией гена thrB, имеющего уменьшенный размер 150 п. о., подтвержденный в 1,0%-м агарозном геле, конструировали, проводя ПЦР в тех же условиях, и подтверждали удаление канамицинового маркера из штамма. Конструированный таким образом штамм был назван «CJMA2».
(3) Делеция гена metA
Для описания субстратной специфичности и активности гена metX, имеющего происхождение от Chromobacterium violaceum, у штамма CJMA2, у штамма FTR2533 (КССМ 10541) проводили делецию исходного гена metA на хромосоме на основе штамма CJMA2 с удаленными генами metB и thrB. Делецию гена metA проводили методом делеции «FRT-one-step-PCR».
Делеционную кассету metA конструировали посредством ПЦР с использованием праймеров SEQ ID NO: 5 и 6 и вектора pKD3 (PNAS (2000) vol 97: Р6640-6645) в качестве матрицы. ПЦР проводили на протяжении 30 циклов в следующих условиях: денатурация при 94°С в течение 30 с, отжиг при 55°С в течение 30 с и удлинение при 72°С в течение 1 мин.
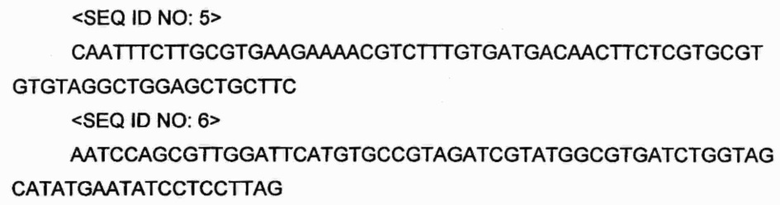
Полученный продукт ПЦР подвергали электрофорезу в 1,0%-ном агарозном геле и 1,2 т.п.о. полосу ДНК, полученную из него, очищали. Полученный фрагмент ДНК электропорировали в штамм CJMA2, уже трансформированный вектором pKD46. Полученный штамм высевали на чашки со средой LB, содержащей хлорамфеникол, культивировали при 37°С в течение ночи и отбирали штамм, резистентный к хлорамфениколу.
Отобранный штамм подвергали ПЦР в тех же условиях с использованием праймеров SEQ ID NO: 5 и 6 и делецию гена metA подтверждали, наблюдая присутствие 1,1 т.п.о. полосы гена в 1,0%-ном агарозном геле. Подтвержденный таким образом штамм снова трансформировали вектором рСР20 и культивировали в среде LB, и конечный штамм с делецией гена thrB, имеющего уменьшенный размер 100 п. о., подтвержденный в 1,0%-ном агарозном геле, конструировали, проводя ПЦР в тех же условиях, и подтверждали удаление хлорамфениколового маркера из штамма. Конструированный таким образом штамм был назван «CJM2».
Штамм CJM2 способен накапливать избыточное количество гомосерина и продуцировать О-ацетилгомосерин или О-сукцинилгомосерин, в зависимости от субстратной специфичности metX во введенной плазмиде.
Пример 2: Отбор полипептидов, обладающих новой О-сукцинилтрансферазной активностью
Для сохранения стабильности и устранения контроля гена metA по принципу обратной связи отбирали 10 типов ортологов, названных metX на интернет-сайте KEGG (//www.genome.jp/kegg/), и клонировали их в вектор pCL1920_PCJ1. Штамм CJM2, полученный в Примере 1, трансформировали векторами 10-ти различных типов.
Полученные таким образом штаммы 10-ти различных типов оценивали методом культивирования в колбах, описанным в Примере 5-(2) ниже. CJM2 представляет собой штамм, способный накапливать гомосерин. При введении гена гомосеринсукцинилтрансферазы в pCL1920 в качестве конечного продукта может быть получен О-сукцинилгомосерин, в то время как при введении гена гомосеринацетилтрансферазы в pCL1920 в качестве конечного продукта может быть получен О-ацетилгомосерин. В этом отношении, ген, представляющий собой ген metX, кодирующий гомосеринсукцинилтрансферазу, получали из уже оцененных 10-ти различных типов. Ген представляет собой ген metX, имеющий происхождение от Chromobacterium violaceum, для которого характерно образование О-сукцинилгомосерина с высоким выходом (аминокислотная последовательность SEQ ID NO: 29, и нуклеотидная последовательность SEQ ID NO: 36), и авторы настоящего изобретения подтвердили, что указанная выше активность является новой активностью, о которой никогда не сообщалось ранее.
Пример 3: Конструирование плазмид
3-1. Конструирование плазмиды, экспрессирующей ген metA, имеющий происхождение от Е. соli дикого типа
Проводили ПЦР с использованием хромосомы E.coliW3110 (регистрационный номер: АТСС 9637), полученной из Американской коллекции типовых культур (American Type Culture Collection, АТСС), в качестве матрицы с праймерами SEQ ID NO: 7 и SEQ ID NO: 8, амплифицируя ген metA, кодирующий гомосерин-О-сукцинилтрансферазу.
Праймеры, использованные при ПЦР, были получены на основе нуклеотидной последовательности хромосомы Е. coli (NC_000913), зарегистрированной в GenBank Национальных институтов здравоохранения (NIH GenBank), и праймеры SEQ ID NO: 7 и SEQ ID NO: 8 имеют сайт рестрикции EcoRV и сайт рестрикции HindIII, соответственно.
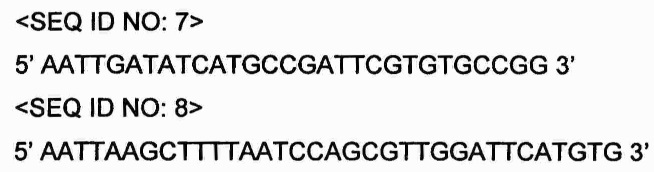
ПЦР проводили посредством денатурации при 94°С в течение 3 мин; 30 циклов денатурации при 94°С в течение 30 с, отжига при 56°С в течение 30 с и полимеризации при 68°С в течение 2 мин; и полимеризации при 68°С в течение 10 мин.
Плазмиду pCL1920, содержащую полученный таким образом ПЦР-продукт и промотор СJ1 (патент Кореи №0620092), клонировали после обработки EcoRV и HindIII, соответственно. Клонированной плазмидой трансформировали Е. coli DH5α и получали плазмиду, отбирая трансформированные Е. coli DH5α из чашек с LB, содержащей спектиномицин (50 мкг/мл). Полученная таким образом плазмида была названа pCL_Pcj1_metA (wt).
3-2. Конструирование плазмиды, экспрессирующей ген metA, резистентный к обратной связи
Ген metA (metA #11), резистентный к действию метионина по принципу обратной связи, конструировали с использованием набора для сайт-направленного мутагенеза (Stratagene, USA) на основе pCL_Pcj1_metA (wt), полученной в Примере 3-1, в качестве матрицы.
Конкретно, в соответствии с описанием международной заявки на патент №WO 2008/127240, 29-ю аминокислоту, серии, заменяли на пролин (S29P) с использованием праймеров SEQ ID NO: 9 и SEQ ID NO: 10; 114-ю аминокислоту, глутаминовую кислоту, заменяли на глицин (E114G) с использованием праймеров SEQ ID NO:11 и SEQ ID NO: 12; 140-ю аминокислоту, фенилаланин, заменяли на серин (F140S) с использованием праймеров SEQ ID NO: 13 и SEQ ID NO: 14. Нуклеотидные последовательности использованных праймеров показаны ниже.


Конструировали плазмиду, содержащую ген metA (#11), в который были последовательно введены все три типа модификаций, и она была названа pCL_Pcj1_metA #11.
3-3. Конструирование плазмиды, экспрессирующей ген metX, имеющий происхождение от Deinococcus radiodurans
Проводили ПЦР с использованием хромосомы Deinococcus radiodurans (регистрационный номер: АТСС BAA-816D), полученной из Американской коллекции типовых культур (АТСС), в качестве матрицы с праймерами SEQ ID NO:15 и SEQ ID NO:16, амплифицируя ген metX, кодирующий гомосерин-О-ацетилтрансферазу.
Праймеры, использованные при ПЦР, были получены на основе нуклеотидной последовательности хромосомы (АЕ000513), зарегистрированной в NIH GenBank, и праймеры SEQ ID NO:15 и SEQ ID NO: 16 имеют сайт рестрикции ЕсоРV и сайт рестрикции HindIII, соответственно.

ПЦР проводили посредством денатурации при 94°С в течение 3 мин; 30 циклов денатурации при 94°С в течение 30 с, отжига при 56°С в течение 30 с и полимеризации при 68°С в течение 5 мин; и полимеризации при 68°С в течение 7 мин.
Плазмиду pCL1920, содержащую полученный таким образом ПЦР-продукт и промотор CJ1 (патент Кореи №0620092), клонировали после обработки EcoRV и HindIII, соответственно. Клонированной плазмидой трансформировали Е. coli DH5α и получали плазмиду, отбирая трансформированные Е. coli DH5α из чашек с LB, содержащей спектиномицин (50 мкг/мл). Полученная таким образом плазмида была названа pCL_Pcj1_dra metX.
3-4. Конструирование плазмиды, экспрессирующей ген metX, имеющий происхождение от Chromobacterium violaceum
Проводили ПЦР с использованием хромосомы Chromobacterium violaceum (регистрационный номер: АТСС 12472), полученной из Американской коллекции типовых культур (АТСС), в качестве матрицы с праймерами SEQ ID NO: 17 и SEQ ID NO: 18, амплифицируя ген metX, имеющий происхождение от Chromobacterium violaceum.
Праймеры, использованные при ПЦР, были получены на основе нуклеотидной последовательности хромосомы Chromobacterium violaceum (NC_005085), зарегистрированной в NIH GenBank, и праймеры SEQ ID NO: 17 и SEQ ID NO: 18 имеют сайт рестрикции EcoRV и сайт рестрикции HindIII, соответственно.
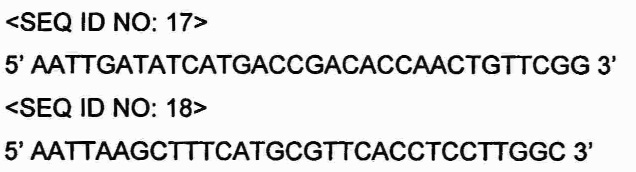
ПЦР проводили посредством денатурации при 94°С в течение 3 мин; 30 циклов денатурации при 94°С в течение 30 с, отжига при 56°С в течение 30 с и полимеризации при 68°С в течение 2 мин; и полимеризации при 68°С в течение 10 мин.
Плазмиду pCL1920, содержащую полученный таким образом ПЦР-продукт и промотор СЛ (патент Кореи №0620092), клонировали после обработки EcoRW и HindIII, соответственно. Клонированной плазмидой трансформировали Е. coli DH5α и получали плазмиду, отбирая трансформированные Е. coli DH5α из чашек с LB, содержащей спектиномицин (50 мкг/мл). Полученная таким образом плазмида была названа pCL_Pcj1_cvi metX.
3-5. Плазмида для конструирования штамма с 2-мя копиями гена для усиления биосинтеза
(1) Конструирование вектора pSG76c для введения гена ррс
В настоящем Примере конструировали pSG76c-2ppc, представляющий собой вектор для введения хромосомной ДНК Е. coli, содержащей ген ррс, кодирующий фосфоенолпируваткарбоксилазу.
Информацию о нуклеотидной последовательности гена ррс получали на основе базы данных NIH GenBank (NCBI Reg. No. gi: 89110074) и на основании этой информации синтезировали праймеры (SEQ ID NO: 19 и SEQ ID NO: 20), содержащие ORF ррс и сайты рестрикции EcoRI и Sad в положении -200 гена ррс, и праймеры (SEQ ID NO: 21 и SEQ ID NO: 22), содержащие сайты рестрикции SacI и КрnI.

Проводили ПЦР с использованием хромосомы Е. coli W3110 в качестве матрицы с праймерами SEQ ID NO: 19 и 20 и SEQ ID NO: 21 и 22. В качестве полимеразы использовали высокоточную ДНК-полимеразу PfuUltra™ (Stratagene) и ПЦР проводили посредством денатурации при 94°С в течение 3 мин; 30 циклов денатурации при 94°С в течение 30 с, отжига при 56°С в течение 30 с и полимеризации при 68°С в течение 5 мин; и полимеризации при 68°С в течение 7 мин. В результате получали амплифицированный ген ррс размером приблизительно 3,1 т.п.о., содержащий сайты рестрикции EcoRI и SacI и сайты рестрикции SacI и КрnI.
После обработки конца гена ррс, полученного посредством ПЦР, рестриктазами EcoRI и SacI, а также SacI и КрnI, полученный ген ррс лигировали с вектором pSG76c (J Bacteriol. 1997 Jul; 179 (13): 4426 - 8), уже обработанным EcoRI и KpnI, и в итоге конструировали рекомбинантный вектор pSG76c-2ppc с двумя клонированными копиями гена ррс.
(2) Конструирование вектора pSG76c для введения aspC
В настоящем Примере конструировали pSG76c-2aspC, представляющий собой вектор для введения хромосомной ДНК Е. coli, содержащей ген aspC, кодирующий аспартатаминотрансферазу.
Информацию о нуклеотидной последовательности гена aspC получали на основе базы данных NIH GenBank (NCBI Reg. No. gi: 85674274) и на основании этой информации синтезировали праймеры (SEQ ID NO: 23 и SEQ ID NO: 24), содержащие ORF aspC и сайт рестрикции SacI в положении -200 гена aspC.
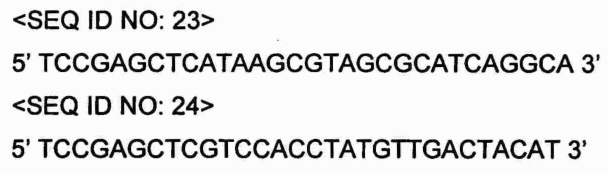
Проводили ПЦР с использованием хромосомы Е. coli W3110 в качестве матрицы с олигонуклеотидными праймерами SEQ ID NO: 23 и 24. В качестве полимеразы использовали высокоточную ДНК-полимеразу PfuUltra™ (Stratagene) и ПЦР проводили посредством денатурации при 94°С в течение 3 мин; 30 циклов денатурации при 94°С в течение 30 с, отжига при 56°С в течение 30 с и полимеризации при 68°С в течение 2 мин; и полимеризации при 68°С в течение 7 мин. В результате получали амплифицированный ген aspC размером приблизительно 1,5 т.п.о., содержащий сайт рестрикции ВаmНI.
После обработки конца гена aspC, полученного посредством ПЦР, рестриктазой ВаmНI, полученный ген aspC лигировали с вектором pSG76c (J Bacterid. 1997 Jul; 179 (13): 4426 - 8), уже обработанным ВаmНI, и в итоге конструировали рекомбинантный вектор pSG76c-2aspC с двумя клонированными копиями гена aspC.
(3) Конструирование вектора pSG76c для введения asd
В настоящем Примере конструировали pSG76c-2asd, представляющий собой вектор для введения хромосомной ДНК Е. coli, содержащей ген asd, кодирующий аспартатполуальдегиддегидрогеназу.
Информацию о нуклеотидной последовательности гена asd получали на основе базы данных NIH GenBank (NCBI Reg. No. gi: 89110578) и на основании этой информации синтезировали праймеры (SEQ ID NO: 25 и SEQ ID NO: 26), содержащие ORF asd и сайты рестрикции ЕсоRI и XbaI в положении -200 гена asd, и праймеры (SEQ ID NO: 27 и SEQ ID NO: 28), содержащие сайты рестрикции XbaI и ЕсоRI.

Проводили ПЦР с использованием хромосомы Е. coli W3110 в качестве матрицы с олигонуклеотидными праймерами SEQ ID NO: 25 и 26 и SEQ ID NO: 27 и 28. В качестве полимеразы использовали высокоточную ДНК-полимеразу PfuUltra™ (Stratagene) и ПЦР проводили на протяжении 30 циклов, состоящих из денатурации при 96°С в течение 30 с, отжига при 50°С в течение 30 с и полимеризации при 68°С в течение 2 мин. В результате получали амплифицированный ген asd размером приблизительно 1,5 т.п.о., содержащий сайты рестрикции ЕсоRI и XbaI и сайты рестрикции XbaI и ЕсоRI.
После обработки конца гена asd, полученного посредством ПЦР, рестриктазами ЕсоRI и XbaI, полученный ген asd лигировали с вектором pSG76c, уже обработанным ЕсоRI, и в итоге конструировали рекомбинантный вектор pSG76c-2asd с двумя клонированными копиями гена asd.
Пример 4: Конструирование исходного штамма на основе штамма дикого типа
(1) Усиление генов ррс, aspC и asd
E.coli W3110 (регистрационный номер: АТСС 9637), полученный из Американской коллекции типовых культур (АТСС), трансформировали векторами pSG76c-2ppc, pSG76c-2aspC и pSG76c-2asd, полученными в Примере 3-5, высевали на чашки со средой LB-Cm (10 г/л дрожжевого экстракта, 5 г/л NaCl, 10 г/л триптона, 25 мкг/л хлорамфеникола и 15 г/л агара) и отбирали колонии, резистентные к хлорамфениколу. Отобранные трансформанты представляли собой штаммы, в ррс-часть генома которых сначала был введен вектор pSG76c-2ppc.
Полученный таким образом штамм с 2-мя введенными копиями гена ррс трансформировали вектором pST76-AsceP, экспрессирующим I-SceI, являющуюся рестриктазой, расщепляющей часть I-SeeI, присутствующую в векторе pSG76c, и высевали на чашки со средой LB-Ap (10 г/л дрожжевого экстракта, 5 г/л NaCl, 10 г/л триптона, 100 мкг/л ампициллина и 15 г/л агара) и отбирали штаммы, которые росли при 30°С.
Выращенные таким образом штаммы могли быть в состоянии, где ген ррс был амплифицирован до 2-х копий или возвращен к одной копии. Штаммы с 2-мя копиями гена ррс с увеличенным размером гена 6,5 т.п.о. отбирали в ходе электрофореза в 1%-м агарозном геле после проведения ПЦР с использованием праймеров SEQ ID NO: 30 и SEQ ID NO: 31. В результате описанного выше процесса происходит дополнительное ведение гена ррс и удаление вектора pSG76c.
Описанным выше способом конструировали штаммы W3110 с амплифицированными копиями генов ррс, asd и aspC, последовательно используя векторы pSG76c-2aspC и pSG76c-2asd. В процессе, конструирование штамма с 2-мя копиями гена aspC подтверждали, идентифицируя ген с увеличенным размером 3,2 т.п.о. электрофорезом в 1%-м агарозном геле после проведения ПЦР с использованием праймеров SEQ ID NO: 32 и SEQ ID NO: 33, в то время как конструирование штамма с 2-мя копиями гена asd подтверждали, идентифицируя ген с увеличенным размером 3,2 т.п.о. в ходе электрофореза в 1%-м агарозном геле после проведения ПЦР с использованием праймеров SEQ ID NO: 34 и SEQ ID NO: 35. Конструированный таким образом штамм был назван CJW2.

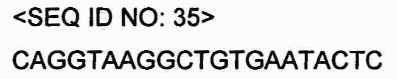
(2) Делеция генов metB, thrB и metA
Штамм с делецией генов metB, thrB и metA конструировали таким же образом, как в Примере 3-1, с использованием штамма CJW2, и полученный штамм был назван CJW2H. Штамм CJW2H представляет собой штамм, способный накапливать избыточное количество гомосерина и продуцировать О-ацетилгомосерин или О-сукцинилгомосерин, в зависимости от субстратной специфичности metX во введенной плазмиде.
Пример 5: Конструирование экспериментальных штаммов
(1) Конструирование штаммов
Из штаммов E. coli CJM2 и CJW2H, конструированных в Примерах 1-(3) и 4-(2), соответственно, получали компетентные клетки и посредством электропорации вводили в них плазмиды четырех различных типов: pCL_Pcj1_metA (wt), pCL_Pcj1_metA#11, pCL_Pcj1_dra metX и pCL_Pcj1_cvi metX, конструированные в Примерах 3-1, 3-2, 3-3 и 3-4, соответственно.
(2) Эксперимент с культивированием в колбах
Затем проводили эксперимент в колбах для сравнения типов предшественников метионина и количество продукта, продуцируемое каждым из штаммов, в которые были введены плазмиды четырех типов, соответственно. Эксперимент в колбах проводили следующим образом: каждый штамм высевали штрихом на чашку со средой LB, культивировали в инкубаторе при 31°С в течение 16 часов и отдельные колонии засевали в 3 мл среды LB и культивировали в инкубаторе при 31°С и 200 об/мин в течение 16 часов.
В колбу объемом 250 мл добавляли 25 мл среды для получения предшественников метионина, показанной в Таблице 1, и затем добавляли по 500 мкл каждого из культуральных бульонов, полученных ранее, соответственно. Затем колбы культивировали в инкубаторе при 31°С и 200 об/мин в течение 40 часов и сравнивали типы и количество предшественников метионина, полученных с использованием каждого из штаммов с каждой из введенных плазмид. Результаты показаны в Таблицах 2 и 3 ниже.
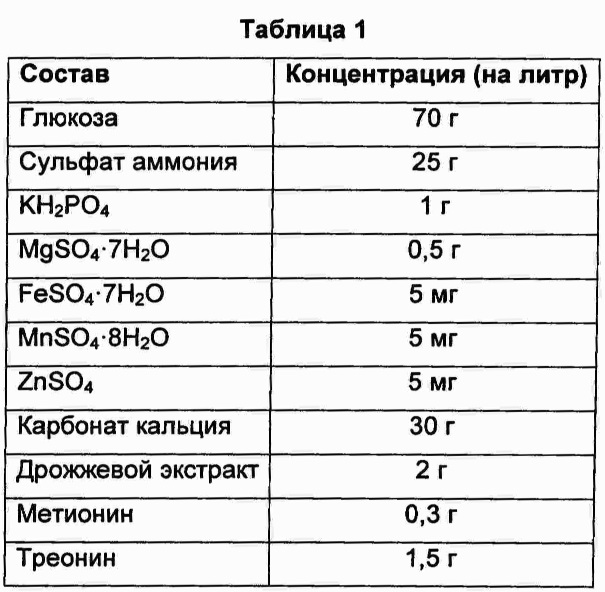
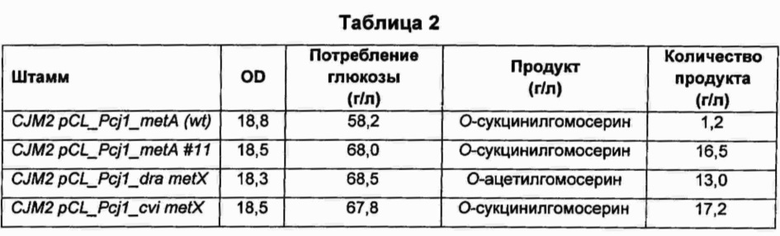
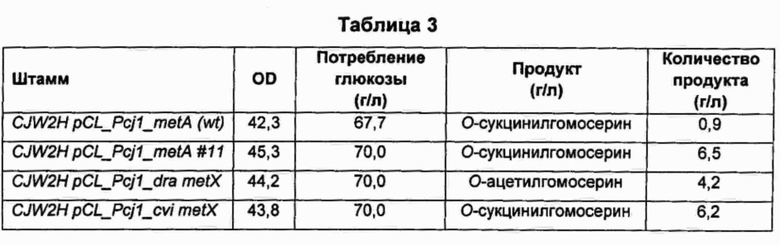
В результате, согласно Таблицам 2 и 3, было подтверждено, что штаммы CJM2 pCL_Pcj1_metA (wt), CJM2 pCL_Pcj1_metA #11, CJW2H pCL_Pcj1_metA (wt) и CJW2H pCL_Pcj1_metA #11, содержащие, соответственно, ген metA Е. coli дикого типа и ген metA#11, резистентный к обратной связи, продуцировали О-сукцинилгомосерин, в то время как штаммы CJM2 pCL_Pcj1_dra metX и CJW2H pCL_Pcj1_dra metX, содержащие, соответственно, ген metX, имеющий происхождение от Deinococcus radiodurans, продуцировали О-ацетилгомосерин.
В случае гена metX, имеющего происхождение от Chrome-bacterium violaceum, ген имеет высокую гомологию с другими гомологичными генами metX (ортологами), по сравнению с ортологами гена metA. Тем не менее, по субстратной специфичности этот ген представляет собой гомосеринсукцинилтрансферазу, образующую сукцинилгомосерин, в отличие от большинства сообщений о гене metX.
Кроме того, в случае введения гена metA Ε. coli дикого типа (metA (wt)) происходило образование О-сукцинилгомосерина в количестве приблизительно 1 г/л из-за феномена ингибирования метионином, добавленным в среду в концентрации 0,3 г/л, по принципу обратной связи, в то время как в случае введения гена metX, имеющего происхождение от Chromobacterium violaceum, образование О-сукцинилгомосерина происходило без феномена ингибирования метионином, добавленным в среду, по принципу обратной связи, даже при использовании дикого типа самого по себе, без введения в ген каких-либо модификаций.
Штамм CJM2 с введенной pCL_Pcj1_cvi metX (CJM2 pCL_Pcj1_cvi metX) был депонирован в Корейском центре культур микроорганизмов (Korean Culture Center of Microorganisms, KCCM), расположенном по адресу 361 - 221, Hongje-1-dong, Seodaemun-gu, Seoul, Korea, являющемся дочерней структурой Корейской федерации коллекций культур (Korean Federation of Culture Collections, KFCC) и признанном международным органом по депонированию согласно Будапештскому соглашению, 20 июня 2013 г. под регистрационным номером КССМ11433Р.
(3) Эксперимент с культивированием в большом ферментере
Для крупномасштабного получения О-сукцинилгомосерина, предшественника метионина, с использованием штаммов CJM2 pCL_Pcj1_cvi metX и CJW2H pCL_Pcj1_cvi metX проводили культивирование в ферментере объемом 5 л.
Штаммы CJM2 pCL_Pcj1_cvi metX и CJW2H pCL_Pcj1_cvi metX высевали на чашки со средой LB, содержащей антибиотик спектиномицин, и культивировали при 31°С в течение ночи. Затем отдельные колонии засевали в 10 мл среды LB, содержащей спектиномицин, культивировали при 31°С в течение 5 часов и 2 мл культуры снова засевали в колбу Эрленмейера объемом 1000 мл, содержащую 200 мл посевной среды. Затем полученную культуру культивировали в инкубаторе при 31°С и 200 об/мин в течение 3-10 часов, 255 мл посевной культуры засевали в 1,7 л основной среды в ферментере объемом 5 л, используя 1,3 л посевной среды, методом подпитываемой культуры, и проводили культивирование в течение 50-100 часов.
Подробный состав среды показан в Таблице 4 ниже. Концентрацию в культивированной таким образом ферментационной жидкости анализировали посредством HPLC (высокоэффективная жидкостная хроматография), и результаты показаны в Таблице 5 ниже.
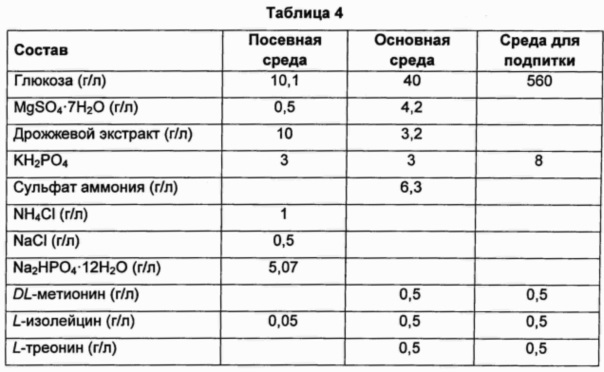
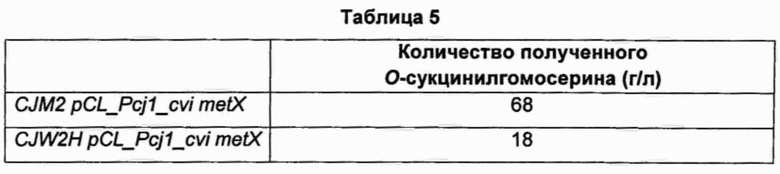
Как показано в Таблице 5 выше, было подтверждено, что штамм CJM2 pCL_Pcj1_cvi metX, в который был введен ген metX, имеющий происхождение от Chrome-bacterium violaceum, на основе штамма, продуцирующего треонин, в качестве исходного штамма, накапливает высокие уровни О-сукцинилгомосерина.
Специалистам в данной области будет ясно, что настоящее изобретение может быть воплощено в других конкретных формах без выхода за рамки его сущности или основных признаков. Описанные воплощения следует рассматривать во всех отношениях только как иллюстративные и не ограничивающие. Таким образом, объем настоящего изобретения определен приложенной формулой изобретения, но не предшествующим описанием. Все изменения в рамках значения и диапазона эквивалентности формулы изобретения следует рассматривать как включенные в объем настоящего изобретения.









Группа изобретений относится к микроорганизму Escherichia sp., продуцирующему О-сукцинилгомосерин, и способу получения О-сукцинилгомосерина. Предложен микроорганизм Escherichia sp., содержащий полипептид, имеющий аминокислотную последовательность, показанную в SEQ ID NO: 29. При этом указанный микроорганизм экспрессирует указанный полипептид, обладающий резистентностью к ингибированию метионином по принципу обратной связи и гомосерин-О-сукцинилтрансферазной активностью. Предложен способ получения О-сукцинилгомосерина, включающий культивирование указанного микроорганизма Escherichia sp., продуцирующего О-сукцинилгомосерин и содержащего полипептид, имеющий аминокислотную последовательность, показанную в SEQ ID NO: 29, в среде с последующим получением О-сукцинилгомосерина из указанного микроорганизма или среды. Группа изобретений обеспечивает получение высокого уровня О-сукцинилгомосерина. 2 н. и 8 з.п. ф-лы, 5 табл., 5 пр.
1. Микроорганизм Escherichia sp., продуцирующий О-сукцинилгомосерин, содержащий полипептид, имеющий аминокислотную последовательность, показанную в SEQ ID NO: 29, где указанный микроорганизм экспрессирует указанный полипептид, обладающий резистентностью к ингибированию метионином по принципу обратной связи и гомосерин-О-сукцинилтрансферазной активностью.
2. Микроорганизм Escherichia sp. по п. 1, представляющий собой Escherichia coli.
3. Микроорганизм Escherichia sp. по п. 1, где ген metB, кодирующий цистатионин-гамма-синтазу, дополнительно удален или ослаблен.
4. Микроорганизм Escherichia sp. по п. 1, где ген thrB, кодирующий гомосеринкиназу, и ген metA, кодирующий гомосерин-О-сукцинилтрансферазу, дополнительно удалены или ослаблены.
5. Микроорганизм Escherichia sp. по п. 1, где фосфоенолпируваткарбоксилаза, аспартатаминотрансфераза и аспартатполуальдегиддегидрогеназа дополнительно усилены.
6. Способ получения О-сукцинилгомосерина, включающий:
а) культивирование микроорганизма Escherichia sp., продуцирующего О-сукцинилгомосерин, содержащего полипептид, имеющий аминокислотную последовательность, показанную в SEQ ID NO: 29, в среде, причем указанный микроорганизм экспрессирует указанный полипептид, обладающий резистентностью к ингибированию метионином по принципу обратной связи и гомосерин-О-сукцинилтрансферазной активностью; и
б) получение О-сукцинилгомосерина из указанного микроорганизма или среды.
7. Способ по п. 6, где микроорганизм Escherichia sp. представляет собой Escherichia coli.
8. Способ по п. 6, где ген metB, кодирующий цистатионин-гамма-синтазу, дополнительно удален или ослаблен в указанном микроорганизме.
9. Способ по п. 6, где ген thrB, кодирующий гомосеринкиназу, и ген metA, кодирующий гомосерин-О-сукцинилтрансферазу, дополнительно удалены или ослаблены в указанном микроорганизме.
10. Способ по п. 6, где фосфоенолпируваткарбоксилаза, аспартатаминотрансфераза и аспартатполуальдегиддегидрогеназа дополнительно усилены в указанном микроорганизме.
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| Механизм подачи к станкам для обработки камней | 1939 |
|
SU58462A1 |
| US 0007851180 B2, 14.12.2010 | |||
| US 20110053253 А1, 03.03.2011 | |||
| КЛЕТКА МИКРООРГАНИЗМА, ПЛАЗМИДНЫЙ ВЕКТОР, СПОСОБ СОЗДАНИЯ КЛЕТКИ МИКРООРГАНИЗМА И СПОСОБ ПОЛУЧЕНИЯ L-МЕТИОНИНА | 2004 |
|
RU2280687C2 |

