ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится выделенному полипептиду, обладающему резистентностью к ингибированию метионином по принципу обратной связи и гомосерин-О-сукцинилтрансферазной активностью, микроорганизму, экспрессирующему указанный полипептид, и способу получения О-сукцинилгомосерина с использованием указанного микроорганизма.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Известно, что большинство микроорганизмов, представленных в природе, используют О-сукцинилгомосерин или О-ацетилгомосерин в качестве промежуточного продукта биосинтеза метионина. Обычно О-сукцинилгомосерин образует гомосерин-О-сукцинилтрансфераза (MetA), конъюгирующая сукцинильную группу сукцинил-КоА с гомосерином, и О-ацетилгомосерин образует гомосерин-О-ацетилтрансфераза (MetX), конъюгирующая ацетильную группу ацетил-КоА с гомосерином. То есть, применительно к получению О-сукцинилгомосерина наряду с другими промежуточными продуктами, metA является одним из наиболее важных генов при разработке микроорганизмов, продуцирующих О-сукцинилгомосерин. В то же время известно, что, в отличие от MetA, MetX не подвержен ингибированию по принципу обратной связи и обладает высокой ферментной стабильностью.
Накопление О-сукцинилгомосерина происходит при блокаде цистатионин-гамма-синтазы в метаболическом пути биосинтеза метионина, и, таким образом, штамму, продуцирующему О-сукцинилгомосерин, необходим L-метионин. Соответственно, в среду добавляют метионин, который ингибирует активность гомосерин-О-сукцинилтрансферазы, и, в конечном счете, О-сукцинилгомосерин не может быть получен в высокой концентрации.
Соответственно, во многих предшествующих патентах первостепенное внимание было уделено исследованиям по устранению ингибирования metA по принципу обратной связи контрольной системой обратной связи. Тем не менее для гомосерин-О-сукцинилтрансферазы, кодируемой metA, свойственны проблемы низкой стабильности самого белка дикого типа, и введение мутаций для устранения ингибирования по принципу обратной связи усугубляет нестабильность. Соответственно, для разработки штамма, продуцирующего О-сукцинилгомосерин с высокой продуктивностью, необходимо устранение ингибирования гена metA по принципу обратной связи и сохранение стабильности фермента.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Техническая проблема
Для решения феномена ингибирования metA по принципу обратной связи и проблемы нестабильности фермента, описанной выше, авторы настоящего изобретения предприняли попытку разработать гомосерин-О-сукцинилтрансферазу с сохраненной стабильностью фермента, не подверженную, в то же время, ингибированию метионином по принципу обратной связи, и провели на предмет этого скрининг новых ферментов, обладающих указанной активностью. В результате отбора генов-кандидатов, прошедших такой скрининг, и культивирования в колбах после их введения в Escherichia sp. авторы настоящего изобретения обнаружили, что происходило образование О-сукцинилгомосерина и что отобранные таким образом гены обладали гомосерин-О-сукцинилтрансферазной активностью и резистентностью к ингибированию метионином по принципу обратной связи, завершив посредством этого настоящее изобретение.
Техническое решение
Задачей настоящего изобретения является обеспечение нового выделенного полипептида, обладающего резистентностью к ингибированию метионином по принципу обратной связи и гомосерин-О-сукцинилтрансферазной активностью.
Другой задачей настоящего изобретения является обеспечение полинуклеотида, кодирующего новый выделенный полипептид.
Еще одной задачей настоящего изобретения является обеспечение микроорганизма для получения О-сукцинилгомосерина, экспрессирующего новый выделенный полипептид.
Еще одной задачей настоящего изобретения является обеспечение способа получения О-сукцинилгомосерина с использованием указанного выше микроорганизма.
Полезные эффекты изобретения
Микроорганизм для получения О-сукцинилгомосерина, содержащий новый выделенный полипептид, обладающий резистентностью к ингибированию метионином по принципу обратной связи и гомосерин-О-сукцинилтрансферазной активностью, который может обладать резистентностью к ингибированию метионином по принципу обратной связи и продуцировать О-сукцинилгомосерин с высоким выходом и, таким образом, может быть эффективно использован для получения L-метионина, используемого им в качестве предшественника, с высоким выходом.
Наилучший вариант осуществления изобретения
Для решения указанных выше задач в одном аспекте согласно настоящему изобретению предложен новый выделенный полипептид, обладающий резистентностью к ингибированию метионином по принципу обратной связи и гомосерин-О-сукцинилтрансферазной активностью.
Использованный здесь термин «гомосерин-О-сукцинилтрансферазная активность» относится к активности по превращению гомосерина в О-сукцинилгомосерин.
Использованный здесь термин «ингибирование по принципу обратной связи» относится к ингибированию активности гомосерин-О-сукцинилтрансферазы метионином.
Полипептид по настоящему изобретению характеризуется тем, что он имеет аминокислотную последовательность SEQ ID NO: 1, обладающую гомосерин-О-сукцинилтрансферазной активностью и резистентностью к ингибированию метионином по принципу обратной связи. Любой полипептид, последовательность которого на 80% или более гомологична указанному выше полипептиду, конкретно на 90% или более, конкретнее на 95% или более и еще конкретнее на 97% или более, также включен в объем настоящего изобретения, с учетом того, что полипептид обладает гомосерин-О-сукцинилтрансферазной активностью и резистентностью к ингибированию метионином по принципу обратной связи, предложенными в настоящем изобретении. Процент гомологии может быть определен с использованием BLAST 2.0, являющегося эталонным алгоритмом, или FASTA от Pearson [Methods Enzymol., 183, 63(1990), ниже]. На основе алгоритма BLAST разработаны программы, называемые BLASTN и BLASTX [www.ncbi.nlm.nih.gov, ниже].
В другом аспекте согласно настоящему изобретению предложен выделенный полинуклеотид, кодирующий указанный выше полипептид. Конкретно, полипептид может быть кодирован полинуклеотидной последовательностью SEQ ID NO: 2 или SEQ ID NO: 3. Ввиду вырожденности кодонов объем настоящего изобретения также включает, без ограничения, полинуклеотиды, последовательность которых по меньшей мере на 80%, конкретно на 90% или более, конкретнее на 95% или более и еще конкретнее на 97% или более гомологична указанной выше последовательности.
В еще одном аспекте согласно настоящему изобретению предложен вектор, содержащий функциональный полинуклеотид.
Использованный здесь термин «вектор» относится к ДНК-конструкции, содержащей нуклеотидную последовательность полинуклеотида, кодирующего интересующий белок, где интересующий белок функционально связан с подходящей регуляторной последовательностью, таким образом, что интересующий белок может быть экспрессирован в подходящем хозяине. Регуляторная последовательность может содержать промотор, способный инициировать транскрипцию, любую операторную последовательность для регуляции транскрипции, последовательность, кодирующую подходящий домен связывания рибосом на мРНК, и последовательность, регулирующую терминацию транскрипции и трансляции. После трансформации в подходящий хозяин вектор может быть реплицирован, или может функционировать независимо от генома хозяина, или может быть интегрирован в сам геном хозяина.
Относительно вектора, используемого в настоящем изобретении, может не быть существенных ограничений, при условии, что вектор может быть реплицирован в хозяине, и может быть использован любой вектор, известный в данной области.
В еще одном аспекте согласно настоящему изобретению предложен микроорганизм, продуцирующий О-сукцинилгомосерин, экспрессирующий полипептид, обладающий резистентностью к ингибированию метионином по принципу обратной связи и гомосерин-О-сукцинилтрансферазной активностью.
Использованный здесь термин «микроорганизм, продуцирующий О-сукцинилгомосерин», может относиться к микроорганизму, способному продуцировать О-сукцинилгомосерин и хранить его внутриклеточно и внеклеточно.
Микроорганизм для получения О-сукцинилгомосерина включает штаммы прокариотических и эукариотических микроорганизмов, например, без ограничения, штаммы микроорганизмов, принадлежащие к роду Escherichia, роду Erwinia, роду Serratia, роду Providencia, роду Corynebacterium и роду Brevibacterium. Конкретно, микроорганизм может представлять собой микроорганизм, принадлежащий к роду Escherichia, например, Escherichia coli.
Микроорганизм, продуцирующий О-сукцинилгомосерин, может быть получен с использованием штаммов микроорганизмов, продуцирующих L-лизин, L-треонин или L-изолейцин, и, конкретно, с использованием штамма, продуцирующего L-треонин. Поскольку штамм, продуцирующий L-треонин, представляет собой штамм, способный синтезировать L-треонин и гомосерин в качестве предшественника О-сукцинилгомосерина, с использованием этого штамма может быть синтезировано большое количество предшественников метионина, то есть О-сукцинилгомосерина.
В настоящем изобретении экспрессия полипептида может быть достигнута трансформацией рекомбинантным вектором, содержащим функциональный ген, кодирующий полипептид, или введением полинуклеотида, кодирующего полипептид, в хромосому, однако методы не ограничены указанными выше.
Использованный здесь термин «трансформация» относится к процессу введения вектора, содержащего полинуклеотид, кодирующий целевой белок, в клетку-хозяина, что позволяет экспрессировать полинуклеотид, кодируемый белком, в клетке-хозяине. Неважно, введен ли полинуклеотид, используемый для трансформации, в хромосому клетки-хозяина, будучи расположен в ней, или расположен вне хромосомы, при условии что он может быть экспрессирован в клетке-хозяине. Полинуклеотид может быть введен в любой форме, с учетом того, что он может быть введен в клетку-хозяина и экспрессирован в ней. Например, полинуклеотид может быть введен в клетку-хозяина в форме экспрессионной кассеты, представляющей собой полинуклеотидную конструкцию, содержащую все основные элементы, необходимые для самостоятельной экспрессии. Обычно экспрессионная кассета может содержать промотор, функционально связанный с открытой рамкой считывания (далее - «ORF») гена, сигнал терминации транскрипции, домен связывания рибосомы и сигнал терминации трансляции.
Относительно промотора, используемого в настоящем изобретении, может не быть существенных ограничений, при условии, что он способен инициировать транскрипцию полинуклеотида, кодирующего целевой белок, в клетке-хозяине с высокой частотой, и может быть использован любой промотор, известный в данной области. Конкретно, но без ограничения, могут быть использованы промотор Т7, промотор trc, промотор tac и промотор cysK (патент Кореи №10-0966324).
В типичном воплощении настоящего изобретения ген metB, кодирующий цистатионин-гамма-синтазу, у микроорганизма может быть дополнительно удален или ослаблен.
В типичном воплощении настоящего изобретения ген thrB, кодирующий гомосеринкиназу, и ген metA, кодирующий гомосерин-О-сукцинилтрансферазу, у микроорганизма могут быть дополнительно удалены или ослаблены.
В настоящем изобретении последовательности генов могут быть получены из баз данных, таких как Национальный центр биотехнологической информации (National Center for Biotechnology Information, NCBI).
Использованный здесь термин «делеция» относится к типу удаления из хромосомы части или всей области нуклеотидной последовательности целевого гена, начиная с нуклеотидной последовательности, соответствующей инициирующему кодону, до нуклеотидной последовательности, соответствующей терминирующему кодону, или части или всей области нуклеотидной последовательности его регуляторной области.
Использованный здесь термин «ослабление» относится к устранению или снижению внутриклеточной активности по меньшей мере одного фермента, кодируемого соответствующей ДНК, у штамма микроорганизма. Например, экспрессия белка может быть ослаблена модификацией промоторной области нуклеотидной последовательности 5'-UTR (5'-нетранслируемой области) гена, или активность белка может быть ослаблена введением мутации в область ORF соответствующего гена.
В другом аспекте согласно настоящему изобретению предложен способ получения О-сукцинилгомосерина, включающий культивирование микроорганизма в среде для получения О-сукцинилгомосерина и получение О-сукцинилгомосерина из микроорганизма или среды.
Культивирование штамма микроорганизма для получения О-сукцинилгомосерина, полученного выше, может быть проведено в соответствии с подходящей средой и условиями культивирования, известными в данной области. Способ культивирования может быть легко адаптирован специалистом в данной области для применения в соответствии с выбранным штаммом. Конкретно, культура может представлять собой, без ограничения, периодическую культуру, непрерывную культуру и адаптированную культуру (fetch culture). Эти различные способы культивирования раскрыты, например, в приведенной ссылке ("Biochemical Engineering" by James M. Lee, Prentice-Hall International Editions, pp. 138-176).
Среды, используемые для культивирования, должны подходящим образом соответствовать требованиям для определенных штаммов. Примеры сред для различных микроорганизмов раскрыты, например, в приведенной ссылке ("Manual of Methods for General Bacteriology by the American Society for Bacteriology, Washington D.C., USA, 1981). Среды могут содержать различные источники углерода, источники азота и микроэлементы. Примеры источников углерода для включения в среды могут включать углеводы, такие как глюкоза, сахароза, лактоза, фруктоза, мальтоза, крахмал и целлюлоза; жиры, такие как соевое масло, подсолнечное масло, касторовое масло и кокосовое масло; жирные кислоты, такие как пальмитиновая кислота, стеариновая кислота и линолевая кислота; спирты, такие как глицерин и этанол; и органические кислоты, такие как уксусная кислота. Эти источники углерода могут быть использованы по отдельности или в комбинации. Примеры источников азота для включения в среды могут включать органические источники азота, такие как пептон, дрожжевой экстракт, мясной бульон, солодовый экстракт, жидкий кукурузный экстракт (CSL) и бобовую муку; и неорганические источники азота, такие как мочевина, сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония. Эти источники азота могут быть использованы по отдельности или в комбинации. В качестве источника фосфора среды могут содержать дигидрофосфат калия, гидроортофосфат калия и соответствующие натрийсодержащие соли. В дополнение, культуральные среды могут содержать металлы, такие как сульфат магния и сульфат железа. Кроме того, могут быть включены аминокислоты, витамины, подходящие предшественники и так далее. Эти культуральные среды или предшественники могут быть добавлены в культуру в форме периодической культуры или непрерывной культуры.
В дополнение, во время культивирования рН культуры можно подходящим образом корректировать добавлением такого соединения, как гидроксид аммония, гидроксид калия, аммиак, фосфорная кислота и серная кислота. Кроме того, образование пузырьков во время культивирования можно предотвращать с использованием пеногасителя, такого как полигликолевый сложный эфир жирной кислоты. В дополнение, в культуру может быть добавлен газообразный кислород или газ, содержащий газообразный кислород (например, воздух), для поддержания в культуре аэробных условий. Температура культуры может входить в диапазон от 20°С до 45°С и, конкретно, от 25°С до 40°С. Культивирование можно продолжать до получения желаемого количества продукта О-сукцинилгомосерина и, конкретно, от 10 часов до 160 часов.
О-сукцинилгомосерин, полученный способом по настоящему изобретению, может быть превращен в метионин цистатионин-гамма-синтазой или О-сукцинилгомосеринсульфгидрилазой. Кроме того, возможно получение янтарной кислоты в качестве побочного продукта, в дополнение к L-метионину, посредством взаимодействия O-сукцинил-L-гомосерина, полученного способом по настоящему изобретению, с CH3SH.
В еще одном аспекте настоящее изобретение относится к применению полипептида, обладающего резистентностью к ингибированию метионином по принципу обратной связи и гомосерин-О-сукцинилтрансферазной активностью, имеющего аминокислотную последовательность SEQ ID NO:1. Было подтверждено, что новый выделенный полипептид по настоящему изобретению обладает резистентностью к ингибированию метионином по принципу обратной связи и способен продуцировать О-сукцинилгомосерин с высоким выходом, и, таким образом, указанный полипептид может быть использован для получения О-сукцинилгомосерина.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Далее настоящее изобретение будет описано более подробно со ссылкой на следующие Примеры. Тем не менее, эти Примеры приведены лишь в иллюстративных целях, и изобретение не следует ограничивать этими Примерами.
Пример 1: Отбор полипептидов, обладающих новой О-сукцинилтрансферазной активностью
В качестве способа устранения контроля гена metA по принципу обратной связи и сохранения его стабильности был разработан metX, имеющий происхождение от Chromobacterium violaceum, на основании того факта, что ген metX (гомосерин-О-ацетилтрансфераза) не подвержен ингибированию L-метионином по принципу обратной связи, несмотря на то, что ген metX имеет структуру, близкую к структуре гена metA.
В этом отношении, для разработки новой гомосерин-О-ацетилтрансферазы авторы настоящего изобретения провели анализ гомологии аминокислотных последовательностей уже разработанных metX, имеющих происхождение от Chromobacterium violaceum, и в итоге отобрали полипептид, имеющий аминокислотную последовательность SEQ ID NO:1, не подверженный ингибированию метионином по принципу обратной связи. Авторы настоящего изобретения недавно подтвердили, что отобранный полипептид, несмотря на то, что он представляет собой metX, имеющий происхождение от Sideroxydans lithotrophicus ES-1, обладает новой активностью, о которой никогда не сообщалось ранее.
Пример 2: Конструирование плазмид
2-1. Синтез гена metX, имеющего происхождение из Pseudogulbenkiania ferrooxidans 2002
Ген metX, имеющий происхождение из Pseudogulbenkiania ferrooxidans 2002 (SEQ ID NO: 3), синтезировали на основе последовательности гена metX (SEQ ID NO: 2) из базы данных NCBI (эталонная последовательность: ZP_03696709.1) с оптимизацией кодонов, чтобы ген можно было экспрессировать в Е. coli.
2-2. Конструирование плазмиды, экспрессирующей ген metX, имеющий происхождение из Pseudogulbenkiania ferrooxidans 2002
Ген metX амплифицировали посредством ПЦР (полимеразная цепная реакция) с использованием праймеров SEQ ID NO: 4 и 5 на основе синтезированной нуклеотидной последовательности SEQ ID NO: 3. Праймер SEQ ID NO: 5 имеет сайт рестрикции HindIII.

ПЦР проводили на протяжении 30 циклов, состоявших из денатурации при 95°С в течение 30 с, отжига при 55°С в течение 30 с и удлинения при 72°С в течение 1 мин. Продукты ПЦР подвергали электрофорезу в 1,0%-м агарозном геле и полосу 1,14 т.п.о. элюировали, очищали и обрабатывали HindIII. Вектор pCL1920, содержащий промотор cysK, обрабатывали EcoRV и HindIII и полученные рестрикционные фрагменты клонировали. Плазмида, экспрессирующая ген metX, имеющий происхождение из Pseudogulbenkiania ferrooxidans, полученная в результате клонирования, была названа «pCL-PcysK-metX (pfe)».
Пример 3: Конструирование экспериментальных штаммов
3-1. Делеция гена metB
Проводили делецию гена metB, кодирующего цистатионин-гамма-синтазу, у штамма Е. coli (К12) W3110 дикого типа. Для делеции гена metB применяли метод делеции «FRT-one-step-PCR» (PNAS (2000) vol 97: Р 6640-6645). Для делеции гена metB конструировали делеционную кассету посредством ПЦР с использованием праймеров SEQ ID NO: 6 и 7 и вектора pKD3 (PNAS (2000) vol 97: P 6640-6645) в качестве матрицы.

ПЦР проводили на протяжении 30 циклов, состоящих из денатурации при 95°С в течение 30 с, отжига при 55°С в течение 30 с и удлинения при 72°С в течение 1 мин. Продукты ПЦР подвергали электрофорезу в 1,0%-м агарозном геле и полосу 1,1 т.п.о. элюировали и очищали. Полученный фрагмент ДНК электропорировали в штамм Е. coli (К12) W3110, уже трансформированный вектором pKD46 (PNAS (2000) vol 97 Р 6640-6645). Для электропорации штамм W3110, трансформированный pKD46, культивировали в среде LB, содержащей 200 мкг/л ампициллина и 5 мМ L-арабинозы, при 30°С до достижения OD600 0,5 и для использования промывали Зраза 10%-м глицерином. Электропорацию проводили при 2500 В. Полученный штамм высевали на чашки со средой LB, содержащей 30 мкг/л хлорамфеникола, культивировали при 37°С в течение 1-2 суток и отбирали штамм, резистентный к хлорамфениколу. Отобранный штамм подвергали ПЦР в тех же условиях, что описаны выше, с использованием праймеров SEQ ID NO: 8 и 9 и делецию гена metB подтверждали, наблюдая присутствие 1,5 т.п.о. полосы гена в 1,0%-м агарозном геле.

Подтвержденный таким образом штамм трансформировали вектором рСР20 (PNAS (2000) vol 97 Р 6640-6645) и культивировали в среде LB, содержащей 100 мкг/л ампициллина. Конечный штамм с делецией гена metB, имеющего уменьшенный размер, подтвержденный в 1,0%-м агарозном геле, конструировали, проводя ПЦР в тех же условиях, и подтверждали удаление хлорамфениколового маркера из штамма. Сконструированный таким образом штамм, которому необходим метионин, был назван «СС03-0131».
3-2. Делеция гена thrB
Была предпринята попытка увеличить количество О-сукцинилгомосерина, синтезируемого из гомосерина, делецией гена thrB, кодирующего гомосеринкиназу. В частности, для использования штамма, продуцирующего треонин, необходима делеция гена thrB, поскольку активность утилизации гомосерина очень высока. Делецию гена thrB в штамме СС03-0131, конструированном выше, проводили методом делеции «FRT-one-step-PCR». Делеционную кассету thrB конструировали посредством ПЦР с использованием праймеров SEQ ID NO: 10 и 11 и вектора pKD3 в качестве матрицы.

ПЦР проводили на протяжении 30 циклов, состоящих из денатурации при 95°С в течение 30 с, отжига при 55°С в течение 30 с и удлинения при 72°С в течение 1 мин. Продукты ПЦР подвергали электрофорезу в 1,0%-м агарозном геле и полосу 1,1 т.п.о. элюировали и очищали. Полученный фрагмент ДНК электропорировали в штамм СС03-0131, уже трансформированный вектором pKD46. Для электропорации штамм СС03-0131, трансформированный pKD46, культивировали в среде LB, содержащей 200 мкг/л ампициллина и 5 мМ арабинозы, при 30°С до достижения OD600 0,5 и для использования промывали 3 раза 10%-м глицерином. Электропорацию проводили при 2500 В. Полученный штамм высевали на чашки со средой LB, содержащей 30 мкг/л хлорамфеникола, культивировали при 37°С в течение 1-2 суток и отбирали штамм, резистентный к хлорамфениколу.
Отобранный штамм подвергали ПЦР в тех же условиях, что описаны выше, с использованием праймеров SEQ ID NO: 12 и 13 и делецию гена thrB подтверждали, наблюдая присутствие 1,5 т.п.о. полосы гена в 1,0%-м агарозном геле.
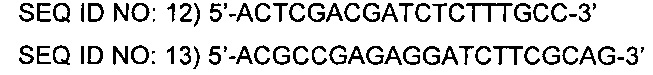
Подтвержденный таким образом штамм трансформировали вектором рСР20 и культивировали в среде LB, содержащей 100 мкг/л ампициллина. Конечный штамм с делецией гена thrB, имеющего уменьшенный размер, подтвержденный в 1,0%-м агарозном геле, конструировали, проводя ПЦР в тех же условиях, и подтверждали удаление хлорамфениколового маркера из штамма. Конструированный таким образом штамм был назван «СС03-0131-2».
3-3. Делеция гена metA
Для описания субстратной специфичности и активности гена metX, имеющего происхождение из Pseudogulbenkiania ferrooxidans 2002, у штамма Е. coli проводили делецию исходного гена metA на хромосоме на основе штамма СС03-0131-2, представляющего собой штамм Е. coli (К12) W3110 с делециями генов metB и thrB. Делецию гена metA проводили методом делеции «FRT-one-step-PCR». Делеционную кассету metA конструировали посредством ПЦР с использованием праймеров SEQ ID NO: 14 и 15 и вектора pKD3 в качестве матрицы.

ПЦР проводили на протяжении 30 циклов, состоящих из денатурации при 95°С в течение 30 с, отжига при 55°С в течение 30 с и удлинения при 72°С в течение 1 мин. Продукты ПЦР подвергали электрофорезу в 1,0%-м агарозном геле и полосу 1,1 т.п.о. элюировали и очищали. Полученный фрагмент ДНК электропорировали в штамм СС03-0131-2, уже трансформированный вектором pKD46. Для электропорации штамм СС03-0131-2, трансформированный pKD46, культивировали в среде LB, содержащей 200 мкг/л ампициллина и 5 мМ арабинозы, при 30°С до достижения OD600 0,5 и для использования промывали 3 раза 10%-м глицерином. Электропорацию проводили при 2500 В. Полученный штамм высевали на чашки со средой LB, содержащей 30 мкг/л хлорамфеникола, культивировали при 37°С в течение 1-2 суток и отбирали штамм, резистентный к хлорамфениколу.
Отобранный штамм подвергали ПЦР в тех же условиях, что описаны выше, с использованием праймеров SEQ ID NO: 16 и 17 и делецию гена metA подтверждали, наблюдая присутствие 1,5 т.п.о. полосы гена в 1,0%-м агарозном геле.

Подтвержденный таким образом штамм трансформировали вектором рСР20 и культивировали в среде LB, содержащей 100 мкг/л ампициллина. Конечный штамм с делецией гена metA, имеющего уменьшенный размер, подтвержденный в 1,0%-м агарозном геле, конструировали, проводя ПЦР в тех же условиях, и подтверждали удаление хлорамфениколового маркера из штамма. Конструированный таким образом штамм был назван «СС03-0132».
3-4. Конструирование штамма с введенной плазмидой, экспрессирующей ген metX, имеющий происхождение из Pseudogulbenkiania ferrooxidans
Для описания субстратной специфичности и активности гена metX, имеющего происхождение из Pseudogulbenkiania ferrooxidans 2002, в штамм СС03-0132, представляющий собой штамм Е. coli (К12) W3110 с делециями генов metB, thrB и metA, вводили плазмиду pCL-PcysK-metX (pfe), конструированную в Примере 2.
Штамм СС03-0132 с введенной pCL-PcysK-metX (pfe) был назван «СС03-0135» и депонирован в Корейском центре культур микроорганизмов (Korean Culture Center of Microorganisms, KCCM), расположенном по адресу 361-221, Hongje-1-dong, Seodaemun-gu, Seoul, Korea, являющемся дочерней структурой Корейской федерации коллекций культур (Korean Federation of Culture Collections, KFCC) и признанном международным органом по депонированию согласно Будапештскому соглашению 10 июня 2013 г., под регистрационным номером КССМ11423Р.
Штамм был конструирован введением плазмиды pCL-PcysK-metA, конструированной таким же образом, как в Примере 2, за исключением того, что в качестве контрольной группы был использован metA дикого типа в штамме СС03-0132. Конструированный таким образом штамм был назван «СС03-0132/pCL-PcysK-metA».
Кроме того, конструировали штамм с использованием штамма CJM002, продуцирующего треонин, не нуждающегося в метионине (регистрационный номер: КССМ-10568), посредством искусственной мутации с использованием NTG на основе штамма TF4076 (регистрационный номер: KFCC-10718), продуцирующего L-треонин, представляющего собой штамм, которому необходим метионин, раскрытый в патенте Кореи №10-0905381, таким же образом, как в Примерах 3-1 - 3-3, и конструированный таким образом штамм был назван «CJM-ВТА».
Плазмиды pCL-PcysK-metX (pfe) и pCL-PcysK-metA, как описано выше, вводили в штамм CJM-ВТА, и конструированные таким образом штаммы были названы «CJM-BTA/pCL-PcysK-metX (pfe)» и «CJM-BTA/pCL-PcysK-metA», соответственно.
Пример 4: Получение О-сукцинилгомосерина с использованием штамма
4-1. Эксперименте культивированием в колбах
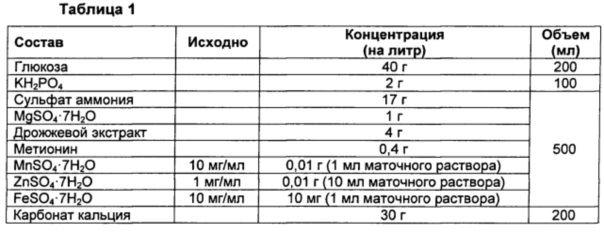
Для описания субстратной специфичности и активности гена metX, имеющего происхождение из Pseudogulbenkiania ferrooxidans 2002, введенного в штамм, конструированный в Примере 3, проводили культивирование в колбах Эрленмейера. Состав культур в колбах показан в Таблице 1 ниже.
Штамм СС03-0132 и штамм CJM-ВТА высевали на чашки со средой LB в качестве контрольных групп. Штамм СС03-0135 (трансформированный вектором экспрессии metX), штамм СС03-0132/pCL-PcysK-metA (трансформированный вектором экспрессии metA, полученный с использованием того же вектора) и два других штамма, CJM-BTAIpCL-PcysK-metX(pfe) и CJM-ВТА/pCL-PcysK-metA (трансформированные вектором экспрессии metX или вектором экспрессии metA на основе штамма CJM-ВТА, соответственно), высевали на чашки со средой LB, содержащей спектиномицин, и культивировали при 33°С в течение ночи. Затем отдельные колонии засевали в 2 мл среды LB, содержащей спектиномицин, культивировали при 33°С в течение 2 часов, засевали снова в колбы Эрленмейера объемом 250 мл, содержащие 25 мл среды, до оптической плотности OD600 0,07, культивировали при 33°С и 200 об/мин в течение 48 часов и количество полученного О-сукцинилгомосерина сравнивали посредством HPLC-анализа (высокоэффективная жидкостная хроматография). Результаты показаны в Таблице 2 ниже.
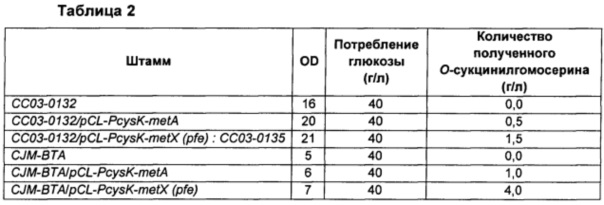
В результате было подтверждено, что ген metX, имеющий происхождение из Pseudogulbenkiania ferrooxidans 2002, как и ген metA Е. coli, продуцирует О-сукцинилгомосерин, используя сукцинил-КоА в качестве субстрата, но не продуцирует О-ацетилгомосерин. При введении гена metX, имеющего происхождение из Pseudogulbenkiania ferrooxidans 2002, не было ингибирования метионином, добавленным в среду, по принципу обратной связи, даже при использовании дикого типа самого по себе, без каких-либо модификаций.
Специалистам в данной области будет ясно, что настоящее изобретение может быть воплощено в других конкретных формах без выхода за рамки его сущности или основных признаков. Описанные воплощения следует рассматривать во всех отношениях только как иллюстративные и не ограничивающие. Таким образом, объем настоящего изобретения определен приложенной формулой изобретения, но не предшествующим описанием. Все изменения в рамках значения и диапазона эквивалентности формулы изобретения следует рассматривать как включенные в объем настоящего изобретения.






Группа изобретений относится к области биотехнологии. Предложено применение полипептида с аминокислотной последовательностью SEQ ID NO: 1 в качестве гомосеринсукцинилтрансферазы или кодирующего его полинуклеотида, имеющего нуклеотидную последовательность SEQ ID NO: 2 или 3, для получения О-сукцинилгомосерина. Предложен микроорганизм Escherichia sp., продуцирующий О-сукцинилгомосерин, экспрессирующий указанный полипептид, обладающий резистентностью к ингибированию метионином по принципу обратной связи и гомосеринсукцинилтрансферазной активностью. Предложен способ получения О-сукцинилгомосерина с использованием указанного микроорганизма. Группа изобретений позволяет получать О-сукцинилгомосерин с высоким выходом. 3 н. и 4 з.п. ф-лы, 2 табл., 4 пр.
1. Применение выделенного полипептида в качестве гомосеринсукцинилтрансферазы для получения О-сукцинилгомосерина, где полипептид имеет аминокислотную последовательность SEQ ID NO: 1.
2. Применение выделенного полинуклеотида, кодирующего полипептид по п.1, для получения О-сукцинилгомосерина, где полинуклеотид имеет нуклеотидную последовательность SEQ ID NO: 2 или 3.
3. Микроорганизм Escherichia sp., продуцирующий О-сукцинилгомосерин, экспрессирующий полипептид, обладающий резистентностью к ингибированию метионином по принципу обратной связи и гомосеринсукцинилтрансферазной активностью, где полипептид имеет аминокислотную последовательность SEQ ID NO: 1.
4. Микроорганизм Escherichia sp., продуцирующий О-сукцинилгомосерин, по п.3, представляющий собой Escherichia coli.
5. Микроорганизм Escherichia sp. по п.3, где ген metB, кодирующий цистатионин-гамма-синтазу, дополнительно удален или ослаблен.
6. Микроорганизм Escherichia sp. по п.3, где ген thrB, кодирующий гомосеринкиназу, или ген metA, кодирующий гомосерин-О-сукцинилтрансферазу, дополнительно удален или ослаблен.
7. Способ получения О-сукцинилгомосерина, включающий:
а) культивирование микроорганизма по любому из пп.3-6 в среде; и
б) получение О-сукцинилгомосерина из микроорганизма или среды.
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| US7851180 B2, 14.12.2010 | |||
| ПРИМЕНЕНИЕ ДИМЕТИЛДИСУЛЬФИДА ДЛЯ ПРОДУКЦИИ МЕТИОНИНА МИКРООРГАНИЗМАМИ | 2006 |
|
RU2413001C2 |

