Область техники, к которой относится изобретение
Настоящее изобретение относится к О-сукцинилгомосерин-продуцирующему микроорганизму и к способу получения О-сукцинилгомосерина с его использованием.
Уровень техники
О-сукцинилгомосерин получают с помощью связывания гомосерина и сукцинила-КоА биосинтетическим путем. Таким образом, при разработке штамма, продуцирующего О-сукцинилгомосерин с высоким выходом, образование гомосерина и сукцинила-КоА имеет важное значение. Из них сукцинил-КоА продуцируется в цикле TCA, и, таким образом, требуется усиление цикла TCA для получения высокой концентрации сукцинила-КоА.
Пентозофосфатный путь (PPP) хорошо известен в качестве основного источника NADPH, и кофактор NADPH требуется в путях биосинтеза аминокислот. Таким образом, для усиления пентозофосфатного пути при разработке штаммов, продуцирующих аминокислоты, ген zwf, кодирующий глюкозо-6-фосфат-1-дегидрогеназу, участвующий на первой стадии пути, обычно усиливают, и эти штаммы описаны в корейских патентных публикациях № 2008-0036608 и 2006-0129352.
Когда экспрессия гена zwf или активность фермента, кодируемого указанным геном, ослаблена, пентозофосфатный путь ослабевает, что приводит к недостаточному поступлению NADPH. В этом случае NADPH может быть частично восполнен сверхэкспрессией изоцитратдегидрогеназы (icd) и малатдегидрогеназы (mae) цикла TCA (Appl Microbiol Biotechnol. 2004 64(1);91-8, Metab Eng. 2004 6(2);164-74, FEBS Letters 581 2007 3771-6).
Авторы настоящего изобретения провели исследование для разработки штамма, способного продуцировать О-сукцинилгомосерин с высоким выходом и высокой эффективностью, и разработали штамм, в котором ген zwf ослаблен и делетирован для получения высокой концентрации сукцинила-КоА как предшественника О-сукцинилгомосерина в E. coli, способном продуцировать О-сукцинилгомосерин. В результате культивирования авторы настоящего изобретения обнаружили, что концентрация О-сукцинилгомосерина была увеличена, в чем и заключается настоящее изобретение.
Подробное описание изобретения
Техническая проблема
Задачей настоящего изобретения является предложить О-сукцинилгомосерин-продуцирующий микроорганизм.
Другой задачей настоящего изобретения является предложить способ получения О-сукцинилгомосерина, включающий стадию культивирования О-сукцинилгомосерин-продуцирующего микроорганизма.
Техническое решение
В одном аспекте настоящее изобретение предлагает О-сукцинилгомосерин-продуцирующий микроорганизм.
В конкретном варианте осуществления настоящего изобретения О-сукцинилгомосерин-продуцирующий микроорганизм может быть микроорганизмом рода Escherichia для продуцирования О-сукцинилгомосерина, в котором активность глюкозо-6-фосфат-1-дегидрогеназы ослаблена или элиминирована по сравнению с ее эндогенной активностью.
Термин «О-сукцинилгомосерин-продуцирующий микроорганизм», употребляемый в настоящем документе, относится к прокариотическому или эукариотическому микроорганизму, способному продуцировать О-сукцинилгомосерин в организме и аккумулирующему О-сукцинилгомосерин. Например, О-сукцинилгомосерин-продуцирующий микроорганизм может быть микроорганизмом, относящимся к роду Escherichia, роду Erwinia, роду Serratia, роду Providencia, роду Corynebacteria, роду Pseudomonas, роду Leptospira, роду Salmonella, роду Brevibacteria, роду Hyphomonas, роду Chromobacterium, роду Norcardia, или к грибам или дрожжам. О-сукцинилгомосерин-продуцирующий микроорганизм может быть, в частности, микроорганизмом, относящимся к роду Escherichia и, более конкретно, Escherichia coli (E.coli).
Глюкозо-6-фосфат-1-дегидрогеназа участвует в пентозофосфатном пути, который представляет собой метаболический путь, обеспечивающий восстановительную способность клеток за счет поддержания концентраций NADPH. Этот фермент катализирует окисление глюкозо-6-фосфата в 6-фосфоглюконолактон путем восстановления NADP в NADPH на первой стадии пентозофосфатного пути. Ген, кодирующий этот фермент, обычно называют zwf. Ослабление или элиминирование этого фермента приводит к потоку через цикл TCA, вызывая усиление цикла TCA.
Глюкозо-6-фосфат-1-дегидрогеназа может иметь аминокислотную последовательность SEQ ID NO: 23. Кроме того, глюкозо-6-фосфат-1-дегидрогеназа может иметь аминокислотную последовательность, имеющую 80% или более высокую, 90%, или более высокую, 95%, или более высокую гомологию с SEQ ID NO: 23.
Термин «гомология», используемый в настоящем описании применительно к последовательности, относится к степени совпадения с определенной аминокислотной последовательностью или последовательностью оснований, и гомология может быть выражена в процентах. В настоящем описании гомологичная последовательность, имеющая активность, которая одинакова или аналогична определенной аминокислотной последовательности или последовательности оснований, выражается в виде «процент гомологии». Например, аминокислотная последовательность, имеющая 80% или более высокую, 90%, или более высокую, 95%, или более высокую гомологию с SEQ ID NO: 23, представляет последовательность, имеющую активность глюкозо-6-фосфат-1-дегидрогеназы.
Термин «эндогенный фермент и активность», употребляемый в настоящем документе, относится к нативному ферменту, естественным образом присутствующему в микроорганизме или клетке, и к его активности, - другими словами, относится к ферменту и к его активности до модификации соответствующего фермента и его активности.
В конкретном варианте осуществления настоящего изобретения микроорганизм может быть микроорганизмом рода Escherichia для продуцирования О-сукцинилгомосерина, в котором активность одной или более из цистатионин-гамма-синтазы и гомосеринкиназы дополнительно ослаблена или элиминирована по сравнению с их эндогенными активностями. В частности, микроорганизм может быть микроорганизмом для продуцирования О-сукцинилгомосерина, в котором активности и цистатионин-гамма-синтазы и гомосеринкиназы ослаблены или элиминированы по сравнению с их эндогенными активностями.
Цистатионин-гамма-синтаза обладает активностью для превращения О-сукцинилгомосерина в цистатионин. Цистатионин-гамма-синтаза кодируется геном metB. Когда активность данного фермента ослаблена или элиминирована, О-сукцинилгомосерин может аккумулироваться без превращения в цистатионин.
Гомосеринкиназа катализирует синтез О-фосфогомосерина из гомосерина, и кодируется геном thrB. Когда активность данного фермента ослаблена или элиминирована, гомосерин может не превращаться в О-фосфогомосерин, и может использоваться для продуцирования О-сукцинилгомосерина.
В конкретном варианте осуществления настоящего изобретения цистатионин-гамма-синтаза может иметь аминокислотную последовательность SEQ ID NO: 24, и гомосеринкиназа может иметь аминокислотную последовательность SEQ ID NO: 25. Кроме того, цистатионин-гамма-синтаза и гомосеринкиназа могут иметь аминокислотные последовательности, имеющие 80% или более высокую, 90% или более высокую, 95% или более высокую гомологию с SEQ ID NO: 24 и SEQ ID NO:25, соответственно.
Термин «ослабление» или «элиминирование ферментативной активности», используемый в настоящем документе, означает, что экспрессия гена, кодирующего соответствующий фермент или активность фермента снижена по сравнению с эндогенной активностью или совсем отсутствует, что может быть обусловлено модификацией всей или части последовательности оснований гена, кодирующего соответствующий фермент, или всей или части последовательности, регулирующей экспрессию гена, путем делеции, замены или инсерции, или путем их сочетания.
Термин «последовательность, регулирующая экспрессию», используемый в настоящем документе, представляет собой последовательность оснований, регулирующую экспрессию гена, и относится к сегменту, способному к увеличению или уменьшению экспрессии конкретного гена у субъекта, и может включать промотор, сайт связывания факторов транскрипции и т.д., но не ограничивается ими.
В конкретном варианте осуществления настоящего изобретения микроорганизм может быть микроорганизмом для продуцирования О-сукцинилгомосерина, в котором активность гомосерин-O-сукцинилтрансферазы дополнительно усилена по сравнению с ее эндогенной активностью.
Гомосерин O-сукцинилтрансфераза представляет собой фермент, который катализирует продуцирование О-сукцинилгомосерина из сукцинила-КоА и гомосерина и участвует в первой стадии пути биосинтеза метионина. Ген, кодирующий данный фермент, обычно называют metA, и его экспрессия подавляется метионином по типу регуляции с обратной связью. Таким образом, может использоваться мутант для экспрессирования гена на высоком уровне за счет устранения регуляции метионином по типу обратной связи.
В конкретном варианте осуществления настоящего изобретения гомосерин-O-сукцинилтрансфераза может иметь аминокислотную последовательность SEQ ID NO: 26. Кроме того, гомосерин-O-сукцинилтрансфераза может иметь аминокислотную последовательность, имеющую 80% или более высокую, 90%, или более высокую, 95%, или более высокую гомологию с SEQ ID NO: 26. В то же время, гомосерин-O-сукцинилтрансфераза, для которой устранена регуляция метионином по типу обратной связи, может иметь аминокислотную последовательность, имеющую 80% или более высокую, 90%, или более высокую, 95%, или более высокую гомологию с SEQ ID NO: 27 (metA11: корейская патентная публикация № 2009-0106365).
Термин «усиление ферментативной активности», используемый в настоящем документе, означает, что активность соответствующего фермента повышается по сравнению с его активностью до модификации. В частности, ферментативная активность возрастает за счет сверхэкспрессии гена, кодирующего соответствующий фермент, по сравнению с ее эндогенной активностью, или активность фермента, кодируемого геном, увеличивается за счет мутации гена, по сравнению с ее эндогенной активностью, и усиление может быть вызвано увеличением числа копий кодирующего гена, заменой промотора гена более сильным промотором, чем эндогенный промотор, или модификацией всей или части последовательности оснований гена на хромосоме или всей или части последовательности, регулирующей его экспрессию, путем делеции, замены или инсерции или путем их сочетания.
В конкретном варианте осуществления настоящего изобретения гомосерин-O-сукцинилтрансфераза может кодироваться геном, в котором промотор гена, кодирующего данный фермент, заменен на более сильный промотор, чем эндогенный промотор. Например, промотор включает известные сильные промоторы Ptac, Ptrc, Ppro, PR, PL, Prmf, PcysK и т.д., но не ограничивается ими.
В конкретном варианте осуществления настоящего изобретения микроорганизмом может быть E. coli. Микроорганизмом может быть E.coli, в котором активность глюкозо-6-фосфат-1-дегидрогеназы ослаблена или элиминирована по сравнению с ее эндогенной активностью, и активности одной или более из цистатионин-гамма-синтазы и гомосеринкиназы ослаблены или элиминированы по сравнению с их эндогенными активностями, соответственно, и активность гомосерин-O-сукцинилтрансферазы усилена по сравнению с ее эндогенной активностью.
В конкретном варианте осуществления настоящего изобретения микроорганизмом может быть E. coli, в котором metA на хромосоме E.coli заменен на metA11 (SEQ ID NO: 27), который является мутантом, полученным с помощью устранения регуляции метионином по типу обратной связи, thrB и metB на хромосоме делетированы, и zwf ослаблен или элиминирован.
В конкретном варианте осуществления настоящего изобретения микроорганизмом может быть штамм E. coli CC03-0156, который депонировали в Корейском центре культур микроорганизмов (KCCM) 22 ноября 2013 года под номером доступа KCCM11487P.
Кроме того, в конкретном варианте осуществления настоящего изобретения микроорганизмом может быть микроорганизм, наделенный способностью ассимилировать сахарозу. Ассимиляция сахарозы означает способность метаболизировать сахарозу в качестве источника углерода или метаболического источника. Способность ассимилировать сахарозу может быть получена с помощью введения метаболического фермента сахарозы, например, фруктокиназы, сахарозной PTS пермеазы, сахарозной гидролазы или инвертазы. Например, способность ассимилировать сахарозу может быть получена с помощью трансформации рекомбинантным вектором (pAscrSM, SEQ ID NO: 28), включающим ген, кодирующий Scr-PTS фермент, полученный из Streptococcus mutans, который описан в корейской патентной публикации № 2010-0099572.
В конкретном варианте осуществления настоящего изобретения микроорганизмом может быть E. coli, в котором metA на хромосоме E.coli заменен на metA11 (SEQ ID NO: 27), который является мутантом, полученным с помощью устранения регуляции метионином по типу обратной связи, thrB и metB на хромосоме делетированы, и zwf элиминирован, и который способен утилизировать сахар-сырец с помощью трансформации рекомбинантным вектором, включающим scrKYABR, кодирующий фруктокиназу, сахарозную PTS пермеазу, сахарозную гидролазу и сахарозный транскрипционный регулятор, полученный из ассимилирующего сахарозу Streptococcus mutans.
В аспекте настоящего изобретения предложен способ продуцирования О-сукцинилгомосерина, включающий стадии культивирования микроорганизма рода Escherichia для продуцирования О-сукцинилгомосерина, в котором активность глюкозо-6-фосфат-1-дегидрогеназы ослаблена или элиминирована по сравнению с ее эндогенной активностью, и извлечения О-сукцинилгомосерина из культуры микроорганизма.
В способе продуцирования О-сукцинилгомосерина в соответствии с конкретным вариантом осуществления настоящего изобретения культивирование О-сукцинилгомосерин- продуцирующего штамма может осуществляться в подходящей среде и условиях, известных в области техники. Процедуры культивирования могут быть легко скорректированы специалистами в данной области техники в соответствии с выбранным штаммом. Примеры процедур культивирования включают способы периодического типа, непрерывного типа и периодического типа с подпиткой, но не ограничиваются ими. Различные процедуры культивирования описаны, например, в литературе («Biochemical Engineering», James M. Lee, Prentice-Hall International Editions, pp.138-176).
Среда, используемая для культивирования, должна соответствовать требованиям культивирования конкретного штамма. Культуральные среды для различных микроорганизмов описаны в литературе («Manual of Methods for General Bacteriology», American Society for Bacteriology, Washington D.C., USA, 1981). Эти среды содержат множество источников углерода, источников азота и следовых элементов. Источники углерода включают углеводы, такие как глюкоза, лактоза, сахароза, фруктоза, мальтоза, крахмал и целлюлоза; жиры, такие как соевое масло, подсолнечное масло, касторовое масло и кокосовое масло; жирные кислоты, такие как пальмитиновая кислота, стеариновая кислота и линолевая кислота; спирты, такие как глицерин и этанол; и органические кислоты, такие как уксусная кислота. Указанные источники углерода могут использоваться по отдельности или в сочетании. Источники азота включают органические источники азота, такие как пептон, дрожжевой экстракт, мясной экстракт, экстракт солода, жидкий кукурузный экстракт (CSL) и соевая мука, и неорганические источники азота, такие как мочевина, сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония. Указанные источники азота могут использоваться по отдельности или в сочетании. Кроме того, среда может содержать первичный кислый фосфат калия, вторичный кислый фосфат калия и соответствующие им натрийсодержащие соли в качестве источника фосфора. Также среда может содержать соль металла, такую как сульфат магния или сульфат железа. Кроме того, также могут быть добавлены аминокислоты, витамины и подходящие предшественники.
Кроме того, для поддержания культуры в аэробных условиях в культуру можно вводить кислород или кислородосодержащий газ (например, воздух). Температура культуры обычно составляет 20-45°С, и в частности, 25-40°С. Культивирование может продолжаться до тех пор, пока образование предшественника L-метионина не достигнет желаемого уровня, и время культивирования может составлять от 10 ч до 160 ч.
Полезные эффекты изобретения
О-сукцинилгомосерин-продуцирующий штамм настоящего изобретения эффективно продуцирует О-сукцинилгомосерин, который может использоваться при получении L-метионина. Полученный таким образом L-метионин может широко применяться в производстве кормов для животных или кормовых добавок, а также пищевых продуктов для человека или пищевых добавок.
Вариант осуществления изобретения
Далее в этом документе настоящее изобретение будет описано более подробно со ссылкой на примеры. Однако данные примеры приводятся только в целях иллюстрации, и объем настоящего изобретения не предполагает ограничения этими примерами.
Ссылочный пример 1. Получение О-сукцинилгомосерин-продуцирующего штамма
1-1 Делеция гена metB
Для повышения аккумуляции О-сукцинилгомосерина штамм получали с помощью делеции гена metB, кодирующего цистатионин-гамма-синтазу, которая участвует в разложении О-сукцинилгомосерина.
В штамме W3110 E. coli (K12) дикого типа ген metB, кодирующий цистатионин-гамма-синтазу, делетировали. Известно, что цистатионин-гамма-синтаза связывается с различными предшественниками метионина в клетках, тем самым образуя различные побочные продукты. Таким образом, сверхэкспрессия цистатионинсинтазы может увеличивать число побочных реакций, снижая эффективность внутриклеточных реакций. Для делеции гена metB осуществляли метод делеции с использованием FRT-одностадийной ПЦР (PNAS (2000), Vol. 97, p. 6640-6645). Сначала ПЦР проводили с использованием праймеров SEQ ID NO: 1 и 2 и вектора pKD3 (PNAS (2000) Vol.l97, pp. 6640-6645) в качестве матрицы для получения делеционной кассеты.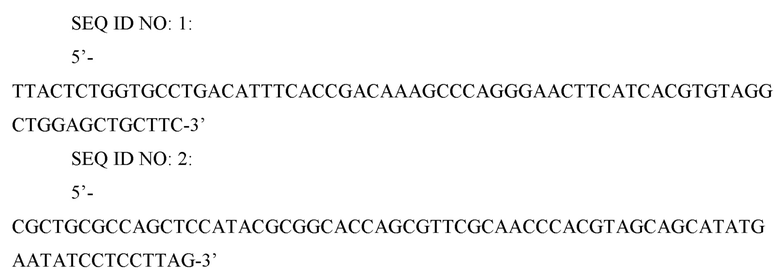
ПЦР проводили в следующих условиях: 30 циклов, состоящих из денатурации при 95°C в течение 30 секунд, отжига при 55°C в течение 30 секунд и удлинения при 72°C в течение 1 минуты. Полученный таким образом продукт ПЦР подвергали электрофорезу на 1,0% агарозном геле, с последующим элюированием и очисткой полосы размером 1,1 т.п.н. Полученный таким образом фрагмент ДНК электропорировали в штамм W3110 E. coli (K12), предварительно трансформированный вектором pKD46 (PNAS (2000) Vol. 97, p. 6640-6645). Перед электропорацией штамм W3110, трансформированный pKD46, культивировали при 30°C в среде LB, содержащей 200 мкг/л ампициллина и 5 мМ L-арабинозы до тех пор, пока OD600 не достигала 0,5. Затем штамм промывали трижды 10% глицерином. Электропорацию проводили при 2500 В. Полученный штамм сеяли штрихом в среду LB для чашек Петри, содержащую 30 мкг/л хлорамфеникола, с последующим культивированием при 37°C в течение 1-2 суток. Затем отбирали штамм, проявляющий устойчивость.
ПЦР проводили с использованием отобранного штамма в качестве матрицы и праймеров SEQ ID NO: 3 и 4 в описанных выше условиях. Делецию гена metB подтверждали идентификацией гена размером 1,5 т.п.н на 1,0% агарозном геле.
SEQ ID NO: 3: 5’-TATTCGCCGCTCCATTCAGC-3’
SEQ ID NO: 4: 5’-TACCCCTTGTTTGCAGCCCG-3’
Штамм, в котором была подтверждена делеция гена metB, далее трансформировали вектором pCP20 (PNAS (2000) vol.97, p. 6640-6645) и культивировали в среде LB, содержащей 100 мкг/л ампицилина. Затем ПЦР проводили в тех же самых условиях, и элиминирование хлорамфеникольного маркера подтверждали наблюдением меньшего по размерам ПЦР-продукта на 1,0% агарозном геле. Наконец, получали штамм с делетированным геном metB. Полученный ауксотрофный по метионину штамм был назван CC03-0132.
1-2. Делеция гена thrB
Для повышения продукции О-сукцинилгомосерина из гомосерина делетировали ген thrB, который является геном, кодирующим гомосеринкиназу. В частности, при использовании треонин-продуцирующего штамма, делеция гена thrB является необходимой, поскольку активность утилизации гомосерина очень высока. Для делеции гена thrB в штамме CC03-0132, полученном в ссылочном примере 1-1, осуществляли метод делеции с использованием FRT-одностадийной ПЦР (PNAS (2000), Vol. 97, p. 6640-6645). Для делеции гена thrB ПЦР проводили с использованием праймеров SEQ ID NO: 5 и 6 и вектора pKD3 (PNAS (2000) Vol. 97, p. 6640-6645) в качестве матрицы для получения делеционной кассеты.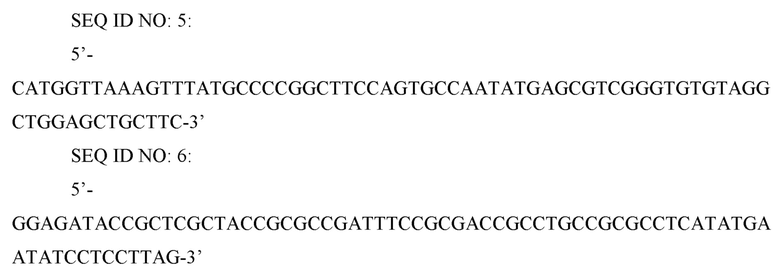
ПЦР проводили в следующих условиях: 30 циклов, состоящих из денатурации при 95°C в течение 30 секунд, отжига при 55°C в течение 30 секунд и удлинения при 72°C в течение 1 минуты. Полученный таким образом продукт ПЦР подвергали электрофорезу на 1,0% агарозном геле, с последующим элюированием и очисткой полосы размером 1,1 т.п.н. Полученный таким образом фрагмент ДНК электропорировали в штамм CC03-0132, предварительно трансформированный вектором pKD46 (PNAS (2000) Vol. 97, p. 6640-6645). Для электропорации штамм CC03-0132, трансформированный pKD46, культивировали при 30°C в среде LB, содержащей 200 мкг/л ампициллина и 5 мМ L-арабинозы до тех пор, пока OD600 не достигала 0,5. Затем штамм промывали трижды 10% глицерином. Электропорацию проводили при 2500 В. Полученный штамм сеяли штрихом в среду LB для чашек Петри, содержащую 30 мкг/л хлорамфеникола, с последующим культивированием при 37°C в течение 1-2 суток. Затем отбирали штамм, проявляющий устойчивость.
ПЦР проводили с использованием отобранного штамма в качестве матрицы и праймеров SEQ ID NO: 7 и 8 в описанных выше условиях. Делецию гена thrB подтверждали идентификацией гена размером 1,5 т.п.н на 1,0% агарозном геле.
SEQ ID NO: 7: 5’-ACTCGACGATCTCTTTGCC-3’
SEQ ID NO: 8: 5’-ACGCCGAGAGGATCTTCGCAG-3’
Подтвержденный таким образом штамм затем трансформировали вектором pCP20 (PNAS (2000) vol.97, p. 6640-6645) и культивировали в среде LB, содержащей 100 мкг/л ампицилина. Затем ПЦР проводили в тех же самых условиях, и элиминирование хлорамфеникольного маркера подтверждали наблюдением меньшего по размерам ПЦР-продукта на 1,0% агарозном геле. Наконец, получали штамм с делетированным геном thrB. Полученный таким образом штамм был назван CC03-0133.
1-3. Делеция гена metA
Для введения устойчивого к регуляции по типу обратной связи гена metA в хромосому, аутентичный хромосомный ген metA на основе штамма CC03-0133, который получали с помощью делеции генов metB и thrB в штамме W3110 E. coli (K12), делетировали. Для делеции гена metA осуществляли метод делеции с использованием FRT-одностадийной ПЦР (PNAS (2000), Vol. 97, p. 6640-6645). Для делеции гена metA ПЦР проводили с использованием праймеров SEQ ID NO: 9 и 10 и вектора pKD3 (PNAS (2000) Vol.97, pp. 6640-6645) в качестве матрицы для получения делеционной кассеты.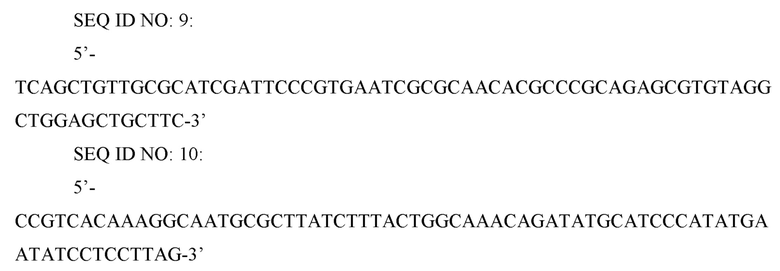
ПЦР проводили в следующих условиях: 30 циклов, состоящих из денатурации при 95°C в течение 30 секунд, отжига при 55°C в течение 30 секунд и удлинения при 72°C в течение 1 минуты. Полученный таким образом продукт ПЦР подвергали электрофорезу на 1,0% агарозном геле, с последующим элюированием и очисткой полосы размером 1,1 т.п.н. Полученный таким образом фрагмент ДНК электропорировали в штамм CC03-0133, предварительно трансформированный вектором pKD46 (PNAS (2000) Vol. 97, p. 6640-6645). Для электропорации штамм CC03-0133, трансформированный pKD46, культивировали при 30°C в среде LB, содержащей 200 мкг/л ампициллина и 5 мМ L-арабинозы до тех пор, пока OD600 не достигала 0,5. Затем штамм промывали трижды 10% глицерином. Электропорацию проводили при 2500 В. Полученный штамм сеяли штрихом в среду LB для чашек Петри, содержащую 30 мкг/л хлорамфеникола, с последующим культивированием при 37°C в течение 1-2 суток. Затем отбирали штамм, проявляющий устойчивость.
ПЦР проводили с использованием отобранного штамма в качестве матрицы и праймеров SEQ ID NO: 11 и 12 в описанных выше условиях. Делецию гена metA подтверждали идентификацией гена размером 1,5 т.п.н. на 1,0% агарозном геле.
SEQ ID NO: 11: 5’-CTCATTAACGTTGGTTGTCA-3’
SEQ ID NO: 12: 5’-TATCTTGCTGCTGCTGAATG-3’
Подтвержденный таким образом штамм затем трансформировали вектором pCP20 (PNAS (2000) vol.97, p. 6640-6645) и культивировали в среде LB, содержащей 100 мкг/л ампицилина. Затем ПЦР проводили в тех же самых условиях, и элиминирование хлорамфеникольного маркера подтверждали наблюдением меньшего по размерам ПЦР-продукта на 1,0% агарозном геле. Наконец, получали штамм с делетированным геном metA. Полученный таким образом штамм был назван CC03-0134.
1.4. Инсерция гена metA11
- Получение вектора pSG для инсерции metA11
Большая часть активности гомосеринсукцинилтрансферазы регулируется ингибированием по типу обратной связи небольшим количеством метионина, добавляемого в среду, и, поэтому, мутант, в котором была устранена регуляция метионином по типу обратной связи, был замещен для повышенного образования предшественника L-метионина, О-сукцинилгомосерина. Для замещения хромосомного гена metA дикого типа, кодирующего гомосеринсукцинилтрансферазу E.coli, на metA11 (SEQ ID NO: 27), кодирующий мутант, в котором устранена регуляция метионином по типу обратной связи, получали pSG-metA11 вектор для инсерции. Согласно корейской патентной публикации № 2009-0106365 была получена информация о последовательности оснований гена metA11, и на основе данной последовательности оснований праймеры (SEQ ID NO: 13 и 14), включающие от ATG до ORF гена metA11 и сайты узнавания для рестрикционных ферментов EcoRI и SacI, были синтезированы. ПЦР проводили с использованием плазмиды pMetA11-CL в качестве матрицы (корейская патентная публикация № 2009-0106365), которая была получена путем лигирования гена metA11 вектором pCL1920, и праймеров следующих SEQ ID NO.
SEQ ID NO: 13: 5’-ggccgaattcatgccgattcgtgtgccgga-3’
SEQ ID NO: 14: 5’-ggccgagctcgttaatccagcgttggattca-3’
ПЦР проводили с использованием pfu-X ДНК-полимеразы (SolGent; SPX16-R250), и условиями ПЦР были: 30 циклов, состоящих из денатурации при 95°C в течение 30 секунд, отжига при 55°C в течение 30 секунд и удлинения при 72°C в течение 2 минут. В результате, был получен продукт ПЦР амплифицированного metA11 ORF, включающий сайты узнавания для рестрикционных ферментов EcoRI и SacI на обоих концах. Полученный в результате ПЦР ген metA11 обрабатывали рестрикционными ферментами EcoRI и SacI и лигировали вектором pSG76-C (Nucleic Acids Res. 1999 Nov 15; 27(22):4409-15), который обрабатывали рестрикционными ферментами EcoRI и SacI для клонирования гена. Наконец, получали рекомбинантный вектор pSG-metA11 клонированного гена metA11.
- Получение штамма с инсертированным геном metA11
Штамм CC03-0134, полученный в ссылочном примере 1-3, трансформировали с получением pSG-metA11, который был вектором для инсерции гена metA11, и культивировали в среде LB Cm (10 г/л дрожжевого экстракта, 5 г/л NaCl, 10 г/л триптона и 30 мкг/л хлорамфеникола). Затем выбирали колонию, проявляющую устойчивость к хлорамфениколу. Выбранный трансформант представлял собой штамм, в котором вектор pSG-metA11 был преимущественно инсертирован в хромосомный metA. Штамм с инсертированным геном metA11 трансформировали вектором pST76-ASceP (Nucleic Acids Res. 1999 Nov 15;27(22):4409-15), экспрессирующим I-SceI, который является рестрикционным ферментом, расщепляющим I-SceI в векторе pSG, и штамм, выращиваемый в LB-Amp (10 г/л дрожжевого экстракта, 5 г/л NaCl, 10 г/л триптона и 100 мкг/л хлорамфеникола), отбирали. Этот штамм, отобранный таким путем, являлся штаммом, в котором metA дикого типа был замещен на metA11 и инсертированный вектор pSG76-C был удален. Полученный штамм был назван E. coli CC03-0038.
Пример 1. Ослабление и делеция гена zwf
1-1. Ослабление гена zwf
- Получение вектора pSG для замещения инициирующего кодона гена zwf
Для ослабления гена zwf в штамме CC03-0038, полученном в ссылочном примере 1-4, применяли способ замещения инициирующего кодона ATG области ORF гена zwf на GTG. ПЦР проводили с использованием праймеров SEQ ID NO: 15 и 16 и SEQ ID NO: 17 и 18, и генома E. coli W3110 в качестве матрицы.
SEQ ID NO: 15: 5’-ggccgaattcctgaaagaaatcgaaatgcag-3’
SEQ ID NO: 16: 5’-cacgtcattctccttaagaattc-3’
SEQ ID NO: 17: 5’-gaattcttaaggagaatgacgtg-3’
SEQ ID NO: 18: 5’-ggccgagctcgggcatggcaaagtagttaatg-3’
ПЦР проводили с использованием pfu-X ДНК-полимеразы (SolGent; SPX16-R250), и ПЦР проводили в следующих условиях: 30 циклов, состоящих из денатурации при 95°C в течение 30 секунд, отжига при 55°C в течение 30 секунд и удлинения при 72°C в течение 1 минуты. ПЦР со сплайсингом перекрывающимися расширениями (SOE, Splicing by Overlap Extension) проводили с использованием двух фрагментов, полученных таким образом в качестве матриц. В результате, были получены область zwf, включающая сайты узнавания для рестрикционных ферментов EcoRI и SacI на обоих концах, и инициирующий кодон GTG. Рестрикционные ферменты EcoRI и SacI были обработаны на концах полученного фрагмента, включая сайты узнавания для ферментов, и клонированы в вектор pSG76-C (Nucleic Acids Res. 1999 Nov 15;27(22):4409-15), обработанный рестрикционными ферментами EcoRI и SacI путем лигирования. Наконец, получали рекомбинантный вектор pSG-zwf(GTG).
- Получение штамма с замещением инициирующего кодона гена zwf
pSG-zwf(GTG), который был получен для замещения инициирующего кодона гена zwf, как описано выше, трансформировали в штамм CC03-0038, полученный в ссылочном примере 1-4, и культивировали в среде LB_Cm (10 г/л дрожжевого экстракта, 5 г/л NaCl, 10 г/л триптона и 30 мкг/л хлорамфеникола). Затем выбирали колонию, проявляющую устойчивость к хлорамфениколу. Выбранный трансформант представлял собой штамм, в котором вектор pSG-zwf(GTG) был преимущественно инсертирован в хромосомный zwf. Выбранный штамм трансформировали pST76-ASceP (Nucleic Acids Res. 1999 Nov 15;27(22):4409-15), экспрессирующим I-SceI, который является рестрикционным ферментом, расщепляющим I-SceI на вектор pSG, и штамм, выращиваемый в LB-Amp (10 г/л дрожжевого экстракта, 5 г/л NaCl, 10 г/л триптона и 100 мкг/л хлорамфеникола), отбирали. Этот штамм, отобранный таким образом, являлся штаммом, в котором ген zwf был ослаблен замещением инициирующего кодона ATG гена zwf на GTG, и затем инсертированный вектор pSG76-C был удален. Полученный штамм был назван CC03-0038zwfGTG.
1-2. Делеция гена zwf
Для делеции гена zwf в штамме CC03-0038, полученном в ссылочном примере 1-4, осуществляли метод делеции с использованием FRT-одностадийной ПЦР (PNAS (2000), Vol. 97, p. 6640-6645). Для делеции гена zwf ПЦР проводили с использованием праймеров SEQ ID NO: 19 и 20 и вектора pKD3 (PNAS (2000) Vol. 97, p. 6640-6645) в качестве матрицы для получения делеционной кассеты.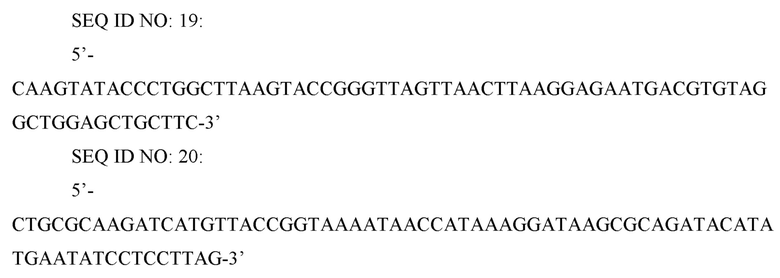
ПЦР проводили в следующих условиях: 30 циклов, состоящих из денатурации при 95°C в течение 30 секунд, отжига при 55°C в течение 30 секунд и удлинения при 72°C в течение 1 минуты. Полученный таким образом продукт ПЦР подвергали электрофорезу на 1,0% агарозном геле, с последующим элюированием и очисткой полосы размером 1,1 т.п.н. Полученный таким образом фрагмент ДНК электропорировали в штамм CC03-0038, предварительно трансформированный вектором pKD46 (PNAS (2000) Vol. 97, p. 6640-6645). Для электропорации штамм CC03-0038, трансформированный pKD46, культивировали при 30°C в среде LB, содержащей 200 мкг/л ампициллина и 5 мМ арабинозы, до тех пор, пока OD600 не достигала 0,5. Затем штамм промывали трижды 10% глицерином. Электропорацию проводили при 2500 В. Полученный штамм сеяли штрихом в среду LB для чашек Петри, содержащую 30 мкг/л хлорамфеникола, с последующим культивированием при 37°C в течение 1-2 суток. Затем отбирали штамм, проявляющий устойчивость.
ПЦР проводили с использованием отобранного штамма в качестве матрицы и праймеров SEQ ID NO: 21 и 22 в описанных выше условиях. Делецию гена zwf подтверждали идентификацией гена размером 2 т.п.н. на 1,0% агарозном геле.
SEQ ID NO: 21: 5’-CATAACATGATCAGTGTCAGAT-3’
SEQ ID NO: 22: 5’-CGCGTAACAATTGTGGATTCAT-3’
Идентифицированный таким образом штамм был назван CC03-0156.
1-3. Получение О-сукцинилгомосерин-продуцирующего штамма на основе треонин-продуцирующего штамма
Треонин-продуцирующий штамм, E.coli KCCM 10541P, описанный в международной патентной публикации WO 2005/075625, использовали для делеции генов metB, thrB и метА таким же образом, как описано в ссылочном примере 1, и затем устойчивый к регуляции по типу обратной связи ген metA11 вводили для получения штамма для продуцирования О-сукцинилгомосерина, который был назван CJM2-A11. Кроме того, штамм с делетированным геном zwf получали таким же образом, как описано в примере 1-2, и назвали CJM2-A11Z.
Пример 2. Ферментация для получения О-сукцинилгомосерина
Для изучения влияния делеции гена zwf в штамме, полученном в примере 1, проводили культивирование в колбе Эрленмейера. Состав среды в колбе был таким, как в приведенной ниже таблице 1.
Далее, плазмиду pAscrSM (SEQ ID NO: 28), имеющую последовательность scrKYABR (ниже называется «scrO»), описанную в корейской патентной публикации № 2009-0018128, вводили для получения штамма, способного утилизировать сахар-сырец, с последующим культивированием в колбе. Состав среды в колбе был таким, как в приведенной ниже таблице 2. Штаммы, полученные указанным выше способом, были названы CC03-0038/pAscrSM, CC03-0038zwfGTG/pAscrSM, CC03-0156/pAscrSM, CJM2-A11/pAscrSM и CJM2-A11Z/pAscrSM, соответственно.
Таблица 1. Состав среды в колбе
Таблица 2. Состав среды в колбе
Каждый из штаммов CC03-0038, CC03-0038zwfGTG, CC03-0156, CJM2-A11, CJM2-A11Z, CC03-0038/pAscrSM, CC03-0038zwfGTG/pAscrSM, CC03-0156/pAscrSM, CJM2-A11/pAscrSM и CJM2-A11Z/pAscrSM инокулировали в среды LB для чашек Петри и культивировали при 33°C в течение ночи. Одиночную колонию инокулировали в 2 мл среды LB, с последующим культивированием при 33°С в течение 2 ч. Культуру инокулировали в 250 мл колбе Эрленмейера, содержащей 25 мл среды в колбе при OD600=0,5, с последующим культивированием при 33°С, 200 об/мин в течение 48 ч. Для сравнения продуцирования О-сукцинилгомосерина проводили ВЭЖХ. Результаты приведены в таблице 3 ниже.
Таблица 3. Продуцирование О-сукцинилгомосерина при культивировании в колбе
Результаты культивирования в колбе показали, что штаммы с ослабленным или делетированным геном zwf CC03-0038zwfGTG, CC03-0156, CJM2-A11Z, CC03-0038zwfGTG/pAscrSM, CC03-0156/pAscrSM, и CJM2-A11Z/pAscrSM демонстрируют продукцию О-сукцинилгомосерина, которая на 8,9-40,0% выше, чем для соответствующих контрольных штаммов. Кроме того, было отмечено уменьшение образования гомосерина, что позволяет предположить, что образование сукцинил-КоА повысилось за счет усиления цикла TCA.
Авторы настоящего изобретения подтвердили, что О-сукцинилгомосерин может быть получен с высоким выходом путем ослабления или делеции гена zwf в штамме CC03-0038 и штамме CJM2-A11. Штамм CC03-0156 депонировали в соответствии с условиями Будапештского договора в Корейском центре культур микроорганизмов (KCCM) 22 ноября 2013 года под номером доступа KCCM11487P.
Что касается применения гена zwf для усиления цикла TCA, следует понимать, что ослабление или делеция гена zwf не ограничивается E. coli, но точно так же применимо для всех микроорганизмов, имеющих цикл TCA, включая род Escherichia, род Corynebacterium, дрожжи и т.д., для получения высокого выхода О-сукцинилгомосерина.
--->
Список последовательностей
<110> CJ Cheiljedang
<120> Microorganisms for production of O-succinyl homoserine and method
for production of O-succinyl homoserine using the same
<130> PX048557
<150> KR 2014/043187
<151> 2014-04-10
<160> 28
<170> KopatentIn 2.0
<210> 1
<211> 70
<212> DNA
<213> Artificial Sequence
<220>
<223> Forward primer of metB deletion cassette
<400> 1
ttactctggt gcctgacatt tcaccgacaa agcccaggga acttcatcac gtgtaggctg 60
gagctgcttc 70
<210> 2
<211> 70
<212> DNA
<213> Artificial Sequence
<220>
<223> Reverse primer of metB deletion cassette
<400> 2
cgctgcgcca gctccatacg cggcaccagc gttcgcaacc cacgtagcag catatgaata 60
tcctccttag 70
<210> 3
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Forward primer for deleted metB
<400> 3
tattcgccgc tccattcagc 20
<210> 4
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Reverse primer for deleted metB
<400> 4
taccccttgt ttgcagcccg 20
<210> 5
<211> 70
<212> DNA
<213> Artificial Sequence
<220>
<223> Forward primer for thrB deletion cassette
<400> 5
catggttaaa gtttatgccc cggcttccag tgccaatatg agcgtcgggt gtgtaggctg 60
gagctgcttc 70
<210> 6
<211> 70
<212> DNA
<213> Artificial Sequence
<220>
<223> Reverse primer for thrB deletion cassette
<400> 6
ggagataccg ctcgctaccg cgccgatttc cgcgaccgcc tgccgcgcct catatgaata 60
tcctccttag 70
<210> 7
<211> 19
<212> DNA
<213> Artificial Sequence
<220>
<223> Forward primer for deleted thrB
<400> 7
actcgacgat ctctttgcc 19
<210> 8
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> Reverse primer for deleted thrB
<400> 8
acgccgagag gatcttcgca g 21
<210> 9
<211> 70
<212> DNA
<213> Artificial Sequence
<220>
<223> Forward primer for metA deletion cassette
<400> 9
tcagctgttg cgcatcgatt cccgtgaatc gcgcaacacg cccgcagagc gtgtaggctg 60
gagctgcttc 70
<210> 10
<211> 70
<212> DNA
<213> Artificial Sequence
<220>
<223> Reverse primer for metA deletion cassette
<400> 10
ccgtcacaaa ggcaatgcgc ttatctttac tggcaaacag atatgcatcc catatgaata 60
tcctccttag 70
<210> 11
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Forwad primer for deleted metA
<400> 11
ctcattaacg ttggttgtca 20
<210> 12
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Reverse primer for deleted metA
<400> 12
tatcttgctg ctgctgaatg 20
<210> 13
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> Forward primer for metA11
<400> 13
ggccgaattc atgccgattc gtgtgccgga 30
<210> 14
<211> 31
<212> DNA
<213> Artificial Sequence
<220>
<223> Reverse primer for metA11
<400> 14
ggccgagctc gttaatccag cgttggattc a 31
<210> 15
<211> 31
<212> DNA
<213> Artificial Sequence
<220>
<223> Forwad primer for start codon substitution of zwf
<400> 15
ggccgaattc ctgaaagaaa tcgaaatgca g 31
<210> 16
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<223> Reverse primer for start codon substitution of zwf
<400> 16
cacgtcattc tccttaagaa ttc 23
<210> 17
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<223> Forward primer 2 for start codon substitution of zwf
<400> 17
gaattcttaa ggagaatgac gtg 23
<210> 18
<211> 32
<212> DNA
<213> Artificial Sequence
<220>
<223> Reverse primer 2 for start codon substitution of zwf
<400> 18
ggccgagctc gggcatggca aagtagttaa tg 32
<210> 19
<211> 70
<212> DNA
<213> Artificial Sequence
<220>
<223> Forward primer for zwf deletion cassette
<400> 19
caagtatacc ctggcttaag taccgggtta gttaacttaa ggagaatgac gtgtaggctg 60
gagctgcttc 70
<210> 20
<211> 72
<212> DNA
<213> Artificial Sequence
<220>
<223> Reverse primer for zwf deletion cassette
<400> 20
ctgcgcaaga tcatgttacc ggtaaaataa ccataaagga taagcgcaga tacatatgaa 60
tatcctcctt ag 72
<210> 21
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Forward primer for deleted zwf
<400> 21
cataacatga tcagtgtcag at 22
<210> 22
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Reverse primer for deleted zwf
<400> 22
cgcgtaacaa ttgtggattc at 22
<210> 23
<211> 491
<212> PRT
<213> Escherichia coli
<220>
<221> PEPTIDE
<222> (1)..(491)
<223> Glucose 6-phosphate-1-dehydrogenase
<400> 23
Met Ala Val Thr Gln Thr Ala Gln Ala Cys Asp Leu Val Ile Phe Gly
1 5 10 15
Ala Lys Gly Asp Leu Ala Arg Arg Lys Leu Leu Pro Ser Leu Tyr Gln
20 25 30
Leu Glu Lys Ala Gly Gln Leu Asn Pro Asp Thr Arg Ile Ile Gly Val
35 40 45
Gly Arg Ala Asp Trp Asp Lys Ala Ala Tyr Thr Lys Val Val Arg Glu
50 55 60
Ala Leu Glu Thr Phe Met Lys Glu Thr Ile Asp Glu Gly Leu Trp Asp
65 70 75 80
Thr Leu Ser Ala Arg Leu Asp Phe Cys Asn Leu Asp Val Asn Asp Thr
85 90 95
Ala Ala Phe Ser Arg Leu Gly Ala Met Leu Asp Gln Lys Asn Arg Ile
100 105 110
Thr Ile Asn Tyr Phe Ala Met Pro Pro Ser Thr Phe Gly Ala Ile Cys
115 120 125
Lys Gly Leu Gly Glu Ala Lys Leu Asn Ala Lys Pro Ala Arg Val Val
130 135 140
Met Glu Lys Pro Leu Gly Thr Ser Leu Ala Thr Ser Gln Glu Ile Asn
145 150 155 160
Asp Gln Val Gly Glu Tyr Phe Glu Glu Cys Gln Val Tyr Arg Ile Asp
165 170 175
His Tyr Leu Gly Lys Glu Thr Val Leu Asn Leu Leu Ala Leu Arg Phe
180 185 190
Ala Asn Ser Leu Phe Val Asn Asn Trp Asp Asn Arg Thr Ile Asp His
195 200 205
Val Glu Ile Thr Val Ala Glu Glu Val Gly Ile Glu Gly Arg Trp Gly
210 215 220
Tyr Phe Asp Lys Ala Gly Gln Met Arg Asp Met Ile Gln Asn His Leu
225 230 235 240
Leu Gln Ile Leu Cys Met Ile Ala Met Ser Pro Pro Ser Asp Leu Ser
245 250 255
Ala Asp Ser Ile Arg Asp Glu Lys Val Lys Val Leu Lys Ser Leu Arg
260 265 270
Arg Ile Asp Arg Ser Asn Val Arg Glu Lys Thr Val Arg Gly Gln Tyr
275 280 285
Thr Ala Gly Phe Ala Gln Gly Lys Lys Val Pro Gly Tyr Leu Glu Glu
290 295 300
Glu Gly Ala Asn Lys Ser Ser Asn Thr Glu Thr Phe Val Ala Ile Arg
305 310 315 320
Val Asp Ile Asp Asn Trp Arg Trp Ala Gly Val Pro Phe Tyr Leu Arg
325 330 335
Thr Gly Lys Arg Leu Pro Thr Lys Cys Ser Glu Val Val Val Tyr Phe
340 345 350
Lys Thr Pro Glu Leu Asn Leu Phe Lys Glu Ser Trp Gln Asp Leu Pro
355 360 365
Gln Asn Lys Leu Thr Ile Arg Leu Gln Pro Asp Glu Gly Val Asp Ile
370 375 380
Gln Val Leu Asn Lys Val Pro Gly Leu Asp His Lys His Asn Leu Gln
385 390 395 400
Ile Thr Lys Leu Asp Leu Ser Tyr Ser Glu Thr Phe Asn Gln Thr His
405 410 415
Leu Ala Asp Ala Tyr Glu Arg Leu Leu Leu Glu Thr Met Arg Gly Ile
420 425 430
Gln Ala Leu Phe Val Arg Arg Asp Glu Val Glu Glu Ala Trp Lys Trp
435 440 445
Val Asp Ser Ile Thr Glu Ala Trp Ala Met Asp Asn Asp Ala Pro Lys
450 455 460
Pro Tyr Gln Ala Gly Thr Trp Gly Pro Val Ala Ser Val Ala Met Ile
465 470 475 480
Thr Arg Asp Gly Arg Ser Trp Asn Glu Phe Glu
485 490
<210> 24
<211> 386
<212> PRT
<213> Escheichia coli
<220>
<221> PEPTIDE
<222> (1)..(386)
<223> Cystathionine gamma synthase
<400> 24
Met Thr Arg Lys Gln Ala Thr Ile Ala Val Arg Ser Gly Leu Asn Asp
1 5 10 15
Asp Glu Gln Tyr Gly Cys Val Val Pro Pro Ile His Leu Ser Ser Thr
20 25 30
Tyr Asn Phe Thr Gly Phe Asn Glu Pro Arg Ala His Asp Tyr Ser Arg
35 40 45
Arg Gly Asn Pro Thr Arg Asp Val Val Gln Arg Ala Leu Ala Glu Leu
50 55 60
Glu Gly Gly Ala Gly Ala Val Leu Thr Asn Thr Gly Met Ser Ala Ile
65 70 75 80
His Leu Val Thr Thr Val Phe Leu Lys Pro Gly Asp Leu Leu Val Ala
85 90 95
Pro His Asp Cys Tyr Gly Gly Ser Tyr Arg Leu Phe Asp Ser Leu Ala
100 105 110
Lys Arg Gly Cys Tyr Arg Val Leu Phe Val Asp Gln Gly Asp Glu Gln
115 120 125
Ala Leu Arg Ala Ala Leu Ala Glu Lys Pro Lys Leu Val Leu Val Glu
130 135 140
Ser Pro Ser Asn Pro Leu Leu Arg Val Val Asp Ile Ala Lys Ile Cys
145 150 155 160
His Leu Ala Arg Glu Val Gly Ala Val Ser Val Val Asp Asn Thr Phe
165 170 175
Leu Ser Pro Ala Leu Gln Asn Pro Leu Ala Leu Gly Ala Asp Leu Val
180 185 190
Leu His Ser Cys Thr Lys Tyr Leu Asn Gly His Ser Asp Val Val Ala
195 200 205
Gly Val Val Ile Ala Lys Asp Pro Asp Val Val Thr Glu Leu Ala Trp
210 215 220
Trp Ala Asn Asn Ile Gly Val Thr Gly Gly Ala Phe Asp Ser Tyr Leu
225 230 235 240
Leu Leu Arg Gly Leu Arg Thr Leu Val Pro Arg Met Glu Leu Ala Gln
245 250 255
Arg Asn Ala Gln Ala Ile Val Lys Tyr Leu Gln Thr Gln Pro Leu Val
260 265 270
Lys Lys Leu Tyr His Pro Ser Leu Pro Glu Asn Gln Gly His Glu Ile
275 280 285
Ala Ala Arg Gln Gln Lys Gly Phe Gly Ala Met Leu Ser Phe Glu Leu
290 295 300
Asp Gly Asp Glu Gln Thr Leu Arg Arg Phe Leu Gly Gly Leu Ser Leu
305 310 315 320
Phe Thr Leu Ala Glu Ser Leu Gly Gly Val Glu Ser Leu Ile Ser His
325 330 335
Ala Ala Thr Met Thr His Ala Gly Met Ala Pro Glu Ala Arg Ala Ala
340 345 350
Ala Gly Ile Ser Glu Thr Leu Leu Arg Ile Ser Thr Gly Ile Glu Asp
355 360 365
Gly Glu Asp Leu Ile Ala Asp Leu Glu Asn Gly Phe Arg Ala Ala Asn
370 375 380
Lys Gly
385
<210> 25
<211> 310
<212> PRT
<213> Escherichia coli
<220>
<221> PEPTIDE
<222> (1)..(310)
<223> Homoserine kinase
<400> 25
Met Val Lys Val Tyr Ala Pro Ala Ser Ser Ala Asn Met Ser Val Gly
1 5 10 15
Phe Asp Val Leu Gly Ala Ala Val Thr Pro Val Asp Gly Ala Leu Leu
20 25 30
Gly Asp Val Val Thr Val Glu Ala Ala Glu Thr Phe Ser Leu Asn Asn
35 40 45
Leu Gly Arg Phe Ala Asp Lys Leu Pro Ser Glu Pro Arg Glu Asn Ile
50 55 60
Val Tyr Gln Cys Trp Glu Arg Phe Cys Gln Glu Leu Gly Lys Gln Ile
65 70 75 80
Pro Val Ala Met Thr Leu Glu Lys Asn Met Pro Ile Gly Ser Gly Leu
85 90 95
Gly Ser Ser Ala Cys Ser Val Val Ala Ala Leu Met Ala Met Asn Glu
100 105 110
His Cys Gly Lys Pro Leu Asn Asp Thr Arg Leu Leu Ala Leu Met Gly
115 120 125
Glu Leu Glu Gly Arg Ile Ser Gly Ser Ile His Tyr Asp Asn Val Ala
130 135 140
Pro Cys Phe Leu Gly Gly Met Gln Leu Met Ile Glu Glu Asn Asp Ile
145 150 155 160
Ile Ser Gln Gln Val Pro Gly Phe Asp Glu Trp Leu Trp Val Leu Ala
165 170 175
Tyr Pro Gly Ile Lys Val Ser Thr Ala Glu Ala Arg Ala Ile Leu Pro
180 185 190
Ala Gln Tyr Arg Arg Gln Asp Cys Ile Ala His Gly Arg His Leu Ala
195 200 205
Gly Phe Ile His Ala Cys Tyr Ser Arg Gln Pro Glu Leu Ala Ala Lys
210 215 220
Leu Met Lys Asp Val Ile Ala Glu Pro Tyr Arg Glu Arg Leu Leu Pro
225 230 235 240
Gly Phe Arg Gln Ala Arg Gln Ala Val Ala Glu Ile Gly Ala Val Ala
245 250 255
Ser Gly Ile Ser Gly Ser Gly Pro Thr Leu Phe Ala Leu Cys Asp Lys
260 265 270
Pro Glu Thr Ala Gln Arg Val Ala Asp Trp Leu Gly Lys Asn Tyr Leu
275 280 285
Gln Asn Gln Glu Gly Phe Val His Ile Cys Arg Leu Asp Thr Ala Gly
290 295 300
Ala Arg Val Leu Glu Asn
305 310
<210> 26
<211> 309
<212> PRT
<213> Escherichia coli
<220>
<221> PEPTIDE
<222> (1)..(309)
<223> Homoserine succinyltransferase
<400> 26
Met Pro Ile Arg Val Pro Asp Glu Leu Pro Ala Val Asn Phe Leu Arg
1 5 10 15
Glu Glu Asn Val Phe Val Met Thr Thr Ser Arg Ala Pro Gly Gln Glu
20 25 30
Ile Arg Pro Leu Lys Val Leu Ile Leu Asn Leu Met Pro Lys Lys Ile
35 40 45
Glu Thr Glu Asn Gln Phe Leu Arg Leu Leu Ser Asn Ser Pro Leu Gln
50 55 60
Val Asp Ile Gln Leu Leu Arg Ile Asp Ser Arg Glu Ser Arg Asn Thr
65 70 75 80
Pro Ala Glu His Leu Asn Asn Phe Tyr Cys Asn Phe Glu Asp Ile Gln
85 90 95
Asp Gln Asn Phe Asp Gly Leu Ile Val Thr Gly Ala Pro Leu Gly Leu
100 105 110
Val Gly Phe Asn Asp Val Ala Tyr Trp Pro Gln Ile Lys Gln Val Leu
115 120 125
Glu Trp Ser Lys Asp His Val Thr Ser Thr Leu Ser Val Cys Trp Ala
130 135 140
Val Gln Ala Ala Leu Asn Ile Leu Tyr Gly Ile Pro Lys Gln Thr Arg
145 150 155 160
Thr Glu Lys Leu Ser Gly Val Tyr Glu His His Ile Leu His Pro His
165 170 175
Ala Leu Leu Thr Arg Gly Phe Asp Asp Ser Phe Leu Ala Pro His Ser
180 185 190
Arg Tyr Ala Asp Phe Pro Ala Ala Leu Ile Arg Asp Tyr Thr Asp Leu
195 200 205
Glu Ile Leu Ala Glu Thr Glu Glu Gly Asp Ala Tyr Leu Phe Ala Ser
210 215 220
Lys Asp Lys Arg Ile Ala Phe Val Thr Gly His Pro Glu Tyr Asp Ala
225 230 235 240
Gln Thr Leu Ala Gln Glu Phe Phe Arg Asp Val Glu Ala Gly Leu Asp
245 250 255
Pro Asp Val Pro Tyr Asn Tyr Phe Pro His Asn Asp Pro Gln Asn Thr
260 265 270
Pro Arg Ala Ser Trp Arg Ser His Gly Asn Leu Leu Phe Thr Asn Trp
275 280 285
Leu Asn Tyr Tyr Val Tyr Gln Ile Thr Pro Tyr Asp Leu Arg His Met
290 295 300
Asn Pro Thr Leu Asp
305
<210> 27
<211> 930
<212> DNA
<213> Artificial Sequence
<220>
<223> metA11
<400> 27
atgccgattc gtgtgccgga cgagctaccc gccgtcaatt tcttgcgtga agaaaacgtc 60
tttgtgatga caacttctcg tgcgcctggt caggaaattc gtccacttaa ggttctgatc 120
cttaacctga tgccgaagaa gattgaaact gaaaatcagt ttctgcgcct gctttcaaac 180
tcacctttgc aggtcgatat tcagctgttg cgcatcgatt cccgtgaatc gcgcaacacg 240
cccgcagagc atctgaacaa cttctactgt aactttgaag atattcagga tcagaacttt 300
gacggtttga ttgtaactgg tgcgccgctg ggcctggtgg ggtttaatga tgtcgcttac 360
tggccgcaga tcaaacaggt gctggagtgg tcgaaagatc acgtcacctc gacgctgtct 420
gtctgctggg cggtacaggc cgcgctcaat atcctctacg gcattcctaa gcaaactcgc 480
accgaaaaac tctctggcgt ttacgagcat catattctcc atcctcatgc gcttctgacg 540
cgtggctttg atgattcatt cctggcaccg cattcgcgct atgctgactt tccggcagcg 600
ttgattcgtg attacaccga tctggaaatt ctggcagaga cggaagaagg ggatgcatat 660
ctgtttgcca gtaaagataa gcgcattgcc tttgtgacgg gccatcccga atatgatgcg 720
caaacgctgg cgcaggaatt tttccgcgat gtggaagccg gactagaccc ggatgtaccg 780
tataactatt tcccgcacaa tgatccgcaa aatacaccgc gagagagctg gcgtagtcac 840
ggtaatttac tgtttaccaa ctggctcaac tattacgtct accagatcgc gccatacgat 900
ctacggcaca tgtatccaac gctggattaa 930
<210> 28
<211> 9960
<212> DNA
<213> Artificial Sequence
<220>
<223> pAscrSM
<400> 28
tgccgaacag ttctctatgc tctgcatata attcatctaa ggggacacct ttataaacac 60
cacttttaac aacagacact ccattaggat gcgcagaaat cgcccaatat tctcccgtcg 120
tttcactagg aatgtcataa ccaaattctt ttctgagccg attgccgccc cagatttttt 180
tatgcatttg tgattgtaaa aacaaaggtt ctgccatagt gttctccttt ctgtttttca 240
atataccact atctagtttc agaggaaatc aagagtaatt atattcactt cattatcctc 300
ataaaatgaa aatttcgttc acgtaaattc taacagttta atattgaata ttagaaggtg 360
ctttaatctt ccaattcatt tttaattaac gcgctgctat ctttttagct aaagcgagat 420
ttcccaatgt tgctgaacca ttttctgcaa cagctggtgt cacaatataa tctttaacat 480
ctggaactgg aagatagtca ttcaaaagtg aagtaaattt ttcacgaacc cgattgagca 540
tatgttcttg tgccataacg cctccgccaa atacaatgac ttgcggacga taaaggacag 600
tcgcttgaat agccgcctga gcaatgtagt atgcctgaat atcccaaact tctgagtttt 660
gctcaattaa ctcaccacga ataccagtac gagcctctaa gctaggaccc gctgcaagtc 720
cttctaaaca gcctttatgg aaaggacagg tgcctacaaa accatgatga acatcattgg 780
gatgcggagc catgtaaacg tgtccagctt ccgtatgtcc cataccgcca atgaattcgc 840
cattttgaat agtccctgct ccaatgcctg ttccaatagt ataataaacc agacttttaa 900
catttgaacg agcaattgtt tccccataag cagaagaatt aacatctgtc gtaaagtaaa 960
atggaatttt aaaatcttta gaaattaagc cgacaaaatc aacgttagcc cagtttggct 1020
ttggtgttga agtaatgtaa ccataagtgt ctgaattttg atcaatatca atagggccaa 1080
aagaaccaat ggcaacactg gctaaatcag cttcaaattt tttaaagaaa gcaactgttt 1140
tttctattgt ttcataaggt gttgttgttg ggaactgaac tttttctaaa atttgaaaat 1200
tttcatcacc tacagcacag acaaattttg ttccgccagc ttcgatgctg ccatataatt 1260
tagacataat aaacctctta tattttactt aacttaccaa atcaattttt ggatttgaat 1320
acccttaatc tacccttaag gactacttaa cattatatca taatttgcta aatttcaagt 1380
attagttagc caaaaggctt taatactatt gttactgaat cttttaagca gtcaaaaaga 1440
ccttacggtc tttctgataa tatttcttat tttttaactt caagtaagct atcacctaca 1500
gcaacagttc ctgaagaagc gagggtttca actgacgcat aatcggctgt attagtaaca 1560
atgaacattg ttgtattatc aagaccagct tctgcaattt tgtcactatc aaatgttccg 1620
agaacgtcac cttttttaat tttttgatct gcttgaactt tttgttcaaa tcctttacct 1680
tccattgata ctgtatcaat accaatatga ataagaatct ccgcaccatt gtctgactta 1740
ataccataag catggccagt atcaaaagca atttgaacag tcccatcaac cggtgcataa 1800
actgtattac cactaggttt gatagcaatc cccttaccca ttgcttcgct agaaaaaaca 1860
gggtcattaa cagaggttaa ttcaacagct tcaccagcaa gaggagccgc aagaacttca 1920
tcagtaactt gtgccttatt tgctgcagaa gcagcttctt ctggaatttc ttgcacttct 1980
tcaatagctt cttcgacagc agcttcagca gcaaagacat caacagcctt cgtcttacca 2040
tagccataag ttactgcaaa agctatagca aaactgatta attcacagac tacataaaat 2100
ggaatagaac ttgccttaat agaaaggaaa ccaagaaaac ctgccgaacc aagggaaacc 2160
gctacaactt gaagtaagcc cgcaattgct gcagcactag ctgagccaat cagagcacaa 2220
aagaatggga aacgatattt caaattcacc ccaaaaagag caggttctgt aataccaaga 2280
agcgctgaca cacctgaaga agatgagaga cctttcatct tcttatcttt tgttaagaag 2340
taaattgcaa aggttgctgc gccttgtgcg acattggcca ttgaagctgt aacaaagata 2400
aagtcaccat gaccagtacc attttggaaa gctgaaatga gttgggtttc aatagctggg 2460
aaactttgat ggagaccagt cataactaca ggtgaataca gagcaccaaa gacacccata 2520
cctaggaatc ccgttgtatc atacagccat acgattccat ttgtaagcca gtcagaaact 2580
tctttcataa caggaccaat aacgataaat gtcaaaaatc cagtaataat aactgacaat 2640
aaaggtgtaa aggtaaagtc aactgctgag ggcaatctct tatggaaaaa tttttctaag 2700
atagataaaa gccatacagc cacaagtact ggaatcactt gataagtata acttgcctga 2760
gtaacatgca acccaaaaat attccaaaat cctgtataag caccaatagt agcagccttt 2820
gaaattggag cattagctgc taaaccaatg atatttgctg cccccggtgc gaccataatc 2880
atgccgattg aagctcctaa gaattggtta gcaccaaaac gtttagctgc tgaaatccca 2940
accaaaatgg gtaagaacca gaaaggcgct gctgacatta attgaatcat gtcagatgaa 3000
cccttaataa tcgggaattg ttgaaccagt gacttagtac caaataaacc ttctgaagtt 3060
aaaaagttat ttaaagccat taagagaccg cctgctacca aggcagggat aattggtaca 3120
aagatatcag acaacagttt gatcaaagcc ataataggat tgaacttttt accactggca 3180
gcaatctttt tcaaatcatc tgttgagact tctgttaaac ctgtttgttt aataatttca 3240
tcataaacaa agttaacatc accagggcca ataataactt gatattgacc atccgtttta 3300
aaagttccct tgacatcagc atttttatcc aacgcttttt gatctacttt actgtcatct 3360
tttaaaacaa gacgaagacg tgtcgcacag tgagcggcag ctacaagatt atctttacca 3420
acagctgtga tgacttcact agctactttg ctataatcca tttgcaaaaa actccttaat 3480
aattattttg ttttgaaagc gtttttttac tttccacgtt attcatttta tcattttttt 3540
ccaatatgtc aatcgtttta cataaaattc ttttttgcaa caaaagattg atttttagaa 3600
agaaaacgtt tactatatta gtatgtagat aaactattat taggagtaga taaacgatga 3660
atttgcctca aaatatcaga tatcgccgct atcaagattg gactgaagaa gaaataaaaa 3720
gtattaaaac caatgtagct ttgtctcctt ggcatacaac gtatcatata gaacctaaaa 3780
caggactcct taatgatcca aacggttttt cctattttaa tggaaaattt aacctttttt 3840
atcaaaattg gccatttgga gcagctcacg gcttaaaatc ttggatccat actgaaagtg 3900
aagacttagt ccatttcaaa gaaacaggta cagtccttta tcccgatact tcccatgaca 3960
gccatggtgc atactcgggc agtgcctatg aaatcggtga tcagcttttt ctcttttata 4020
caggaaatgt ccgagatgaa aattgggttc gtcatccact tcaaatcggc gcttttatgg 4080
ataaaaaagg taatatccaa aaatttactg atgtccttat taaacagcca aatgatgtta 4140
ctgaacactt tcgcgatccc caaattttta attataaagg acaattttac gctattgttg 4200
gagcacaaag tctagataaa aaaggattta tcaaactcta taaagctgtt gataatgata 4260
ttaagaattg gcaagaagtt ggtaatctag actttggcgg aagtaaatct gagtatatga 4320
ttgagtgccc aaatcttgtt tttataaacg aacagcctgt cctgatttat agtcctcagg 4380
gactcagtaa atctgaatta gattatcata atatttatcc taatacttac aaagtatgtc 4440
aatcgtttga cacagaaaag cctgccctag ttgatgcatc ggaaattcaa aatcttgact 4500
tcggatttga atgttatgct acccaagctt tcaatgctcc tgatggccgt gtttatgctg 4560
tctcatggat tgggttacca gatattgatt atccaagtga ttcatacgac tatcaaggag 4620
ctttgagcct cgtcaaagag ctaagcctaa aacacggtaa actctatcaa tatcccgttg 4680
aagctgttcg ttcattacgt tctgaaaaag aagcagtcac ttacaagcca gaaaccaata 4740
atacttatga attagagtta acttttgact cttcatcagt taatgaattg ctcctttttg 4800
ctgataataa aggcaatggt ctagctatta cagttgatac taagatggga accattctga 4860
tcgatcgctc taaagcaggg gagcaatatg ccttagaatt tggaagccaa cgttcttgct 4920
ctatccaagc aaaagagact gttgtcaata tttttgttga caaatcgatt tttgaaattt 4980
ttattaataa gggagaaaaa gtttttactg gacgtgtttt tccaaatgac aaacaaactg 5040
gtattgtgat taaatctgga aagccaagcg gtaattacta cgaattgaaa tattaactat 5100
ggttgcaaaa ttaacagatg tcgcaaaact cgctggcgta agtccaacaa ccgtttcacg 5160
cgtgattaat cgaaaaggct atctctctga aaaaacaatt actaaagtac aggctgccat 5220
gaaaactcta ggatacaagc ccaataatct cgctcgcagc cttcagggga aatctgccaa 5280
gctaattgga cttattttcc ctaatatcag tcacatcttc tattctgaac ttattgaata 5340
tttagaaata gagttgttta aacatggcta caaagccatt atttgtaaca gtcagaataa 5400
tcccgataaa gaacgggatt atctcgaaat gttagaagct aatcaggttg atggcattat 5460
ttccagtagt cacaacctcg gcattgatga ctatgagaaa gtttctgctc ctattattgc 5520
ctttgatcgt aacttagcac ccaatattcc catcgtctct tctgacaatt ttgaaggcgg 5580
acgaatggcc gcaaaacttt taaaaaaaca cggctgccaa catcccatta tgatcgctgg 5640
aaaagataat tctaattctc ggacaggact gcgacaatta ggctttaagt ccgtctttgc 5700
tcaagcacct atttttcatc tatctggaga gctgtccatc attcgtaaag aaatggaaat 5760
aaaagtaatt ctccaaaatg aaaaacctga tggtatcttt ttgtccgatg atatgacagc 5820
tattttaaca atgaaaattg ctaaccagtt aaacattacc attccccatg aacttaaaat 5880
tattgggtac gatggaacac actttgttga gaactactat ccttatctaa caactattag 5940
gcaacctatt aaagatatcg cacatctttt ggtagacgta ttgctaaaga aaattgatca 6000
ccaagatact cctaaagatt atattctccc cgttggtctt ttatcaggag aaagtgttta 6060
gaatcaaaag gtttatttag gtttgtcttt tgtgggttga ttaaaagtcc ttacctaatt 6120
aatgagcatc agttcaatgt caatatttaa tcaactttgc gcaaaagaat agacatacaa 6180
ttcttagttt taaattcaaa aagaagaggt aagatttaag cctaaaatct tacctcttct 6240
tttatcagcc cttattattt tctttcaagc acaaactgac ttaaaacgcc attaataaac 6300
ttactggatt tctcatctga atactttttt gcaatttcaa taatttcatt aagagcaaca 6360
cgttctggtg tttccttatg atatttaatt tcgtataaac ctaaacgcaa caggctctta 6420
tcaatcaaag tcaatcgaga aagcgtccag ccagactagt agggcaaact attaactggc 6480
gaactactta ctctagcttc ccggcaacaa ttaatagact ggatggaggc ggataaagtt 6540
gcaggaccac ttctgcgctc ggcccttccg gctggctggt ttattgctga taaatctgga 6600
gccggtgagc gtgggtctcg cggtatcatt gcagcactgg ggccagatgg taagccctcc 6660
cgtatcgtag ttatctacac gacggggagt caggcaacta tggatgaacg aaatagacag 6720
atcgctgaga taggtgcctc actgattaag cattggtaac tgtcagacca agtttactca 6780
tatatacttt agattgattt aaaacttcat ttttaattta aaaggatcta ggtgaagatc 6840
ctttttgata atctcatgac caaaatccct taacgtgagt tttcgttcca ctgagcgtca 6900
gaccccttaa taagatgatc ttcttgagat cgttttggtc tgcgcgtaat ctcttgctct 6960
gaaaacgaaa aaaccgcctt gcagggcggt ttttcgaagg ttctctgagc taccaactct 7020
ttgaaccgag gtaactggct tggaggagcg cagtcaccaa aacttgtcct ttcagtttag 7080
ccttaaccgg cgcatgactt caagactaac tcctctaaat caattaccag tggctgctgc 7140
cagtggtgct tttgcatgtc tttccgggtt ggactcaaga cgatagttac cggataaggc 7200
gcagcggtcg gactgaacgg ggggttcgtg catacagtcc agcttggagc gaactgccta 7260
cccggaactg agtgtcaggc gtggaatgag acaaacgcgg ccataacagc ggaatgacac 7320
cggtaaaccg aaaggcagga acaggagagc gcacgaggga gccgccaggg ggaaacgcct 7380
ggtatcttta tagtcctgtc gggtttcgcc accactgatt tgagcgtcag atttcgtgat 7440
gcttgtcagg ggggcggagc ctatggaaaa acggctttgc cgcggccctc tcacttccct 7500
gttaagtatc ttcctggcat cttccaggaa atctccgccc cgttcgtaag ccatttccgc 7560
tcgccgcagt cgaacgaccg agcgtagcga gtcagtgagc gaggaagcgg aatatatcct 7620
gtatcacata ttctgctgac gcaccggtgc agcctttttt ctcctgccac atgaagcact 7680
tcactgacac cctcatcagt gccaacatag taagccagta tacactccgc tagcgctgag 7740
gtctgcctcg tgaagaaggt gttgctgact cataccaggc ctgaatcgcc ccatcatcca 7800
gccagaaagt gagggagcca cggttgatga gagctttgtt gtaggtggac cagttggtga 7860
ttttgaactt ttgctttgcc acggaacggt ctgcgttgtc gggaagatgc gtgatctgat 7920
ccttcaactc agcaaaagtt cgatttattc aacaaagcca cgttgtgtct caaaatctct 7980
gatgttacat tgcacaagat aaaaatatat catcatgaac aataaaactg tctgcttaca 8040
taaacagtaa tacaaggggt gttatgagcc atattcaacg ggaaacgtct tgctcgaggc 8100
cgcgattaaa ttccaacatg gatgctgatt tatatgggta taaatgggct cgcgataatg 8160
tcgggcaatc aggtgcgaca atctatcgat tgtatgggaa gcccgatgcg ccagagttgt 8220
ttctgaaaca tggcaaaggt agcgttgcca atgatgttac agatgagatg gtcagactaa 8280
actggctgac ggaatttatg cctcttccga ccatcaagca ttttatccgt actcctgatg 8340
atgcatggtt actcaccact gcgatccccg ggaaaacagc attccaggta ttagaagaat 8400
atcctgattc aggtgaaaat attgttgatg cgctggcagt gttcctgcgc cggttgcatt 8460
cgattcctgt ttgtaattgt ccttttaaca gcgatcgcgt atttcgtctc gctcaggcgc 8520
aatcacgaat gaataacggt ttggttgatg cgagtgattt tgatgacgag cgtaatggct 8580
ggcctgttga acaagtctgg aaagaaatgc ataagctttt gccattctca ccggattcag 8640
tcgtcactca tggtgatttc tcacttgata accttatttt tgacgagggg aaattaatag 8700
gttgtattga tgttggacga gtcggaatcg cagaccgata ccaggatctt gccatcctat 8760
ggaactgcct cggtgagttt tctccttcat tacagaaacg gctttttcaa aaatatggta 8820
ttgataatcc tgatatgaat aaattgcagt ttcatttgat gctcgatgag tttttctaat 8880
cagaattggt taattggttg taacactggc agagcattac gctgacttga cgggacggcg 8940
gctttgttga ataaatcgaa cttttgctga gttgaaggat cagatcacgc atcttcccga 9000
caacgcagac cgttccgtgg caaagcaaaa gttcaaaatc accaactggt ccacctacaa 9060
caaagctctc atcaaccgtg gctccctcac tttctggctg gatgatgggg cgattcaggc 9120
ctggtatgag tcagcaacac cttcttcacg aggcagacct cagcgctcaa agatgcaggg 9180
gtaaaagcta accgcatctt taccgacaag gcatccggca gttcaacaga tcgggaaggg 9240
ctggatttgc tgaggatgaa ggtggaggaa ggtgatgtca ttctggtgaa gaagctcgac 9300
cgtcttggcc gcgacaccgc cgacatgatc caactgataa aagagtttga tgctcagggt 9360
gtagcggttc ggtttattga cgacgggatc agtaccgacg gtgatatggg gcaaatggtg 9420
gtcaccatcc tgtcggctgt ggcacaggct gaacgccgga ggatcctaga gcgcacgaat 9480
gagggccgac aggaagcaaa gctgaaagga atcaaatttg gccgcaggcg taccgtggac 9540
aggaacgtcg tgctgacgct tcatcagaag ggcactggtg caacggaaat tgctcatcag 9600
ctcagtattg cccgctccac ggtttataaa attcttgaag acgaaagggc ctcgtgatac 9660
gcctattttt ataggttaat gtcatgataa taatggtttc ttagacgtca ggtggcactt 9720
ttcggggaaa tgtgcgcgga acccctattt gtttattttt ctaaatacat tcaaatatgt 9780
atccgctcat gagacaataa ccctgataaa tgcttcaata atattgaaaa aggaagagta 9840
tgagtattca acatttccgt gtcgccctta ttcccttttt tgcggcattt tgccttcctg 9900
tttttgctca cccagaaacg ctggtgaaag taaaagatgc tgaagatcag ttgggtgcac 9960
9960
<---
Изобретение относится к способу получения метионина. Предложен способ получения метионина, включающий культивирование микроорганизма рода Escherichia в среде, получение О-сукцинилгомосерина из микроорганизма или среды, превращение О-сукцинилгомосерина в метионин. При этом в указанном микроорганизме активность глюкозо-6-фосфат-1-дегидрогеназы ослаблена или устранена по сравнению с ее эндогенной активностью, активность одной или нескольких цистатион-гамма-синтазы и гомосерин-киназы устранена по сравнению с эндогенными активностями в микроорганизме дикого типа, активность гомосерин-O-сукцинилтрансферазы повышена по сравнению с эндогенной активностью в микроорганизме дикого типа. Изобретение обеспечивает повышение выхода O-сукцинилгомосерина по сравнению с выходом O-сукцинилгомосерина в микроорганизме дикого типа. 7 з.п. ф-лы, 3 табл., 3 пр.
1. Способ получения метионина, включающий:
(а) культивирование микроорганизма в среде;
(b) получение О-сукцинилгомосерина из микроорганизма или среды;
(с) превращение О-сукцинилгомосерина в метионин,
в котором микроорганизм представляет собой микроорганизм рода Escherichia для продуцирования O-сукцинилгомосерина, в котором:
- активность глюкозо-6-фосфат-1-дегидрогеназы ослаблена или устранена по сравнению с ее эндогенной активностью;
- активность одной или нескольких цистатион-гамма-синтазы и гомосерин-киназы устранена по сравнению с эндогенными активностями в микроорганизме дикого типа;
- активность гомосерин-O-сукцинилтрансферазы повышена по сравнению с эндогенной активностью в микроорганизме дикого типа; и
- выход O-сукцинилгомосерина повышен по сравнению с выходом в микроорганизме дикого типа.
2. Способ получения метионина по п. 1, в котором глюкозо-6-фосфат-1-дегидрогеназа имеет аминокислотную последовательность SEQ ID NO: 23.
3. Способ получения метионина по п. 1, в котором цистатион-гамма-синтаза имеет аминокислотную последовательность SEQ ID NO: 24, а гомосеринкиназа имеет аминокислотную последовательность SEQ ID NO: 25.
4. Способ получения метионина по п. 1, в котором гомосерин-О-сукцинилтрансфераза имеет аминокислотную последовательность SEQ ID NO: 26.
5. Способ получения метионина по любому из пп. 1-4, в котором микроорганизм представляет собой Escherichia coli (E.coli).
6. Способ получения метионина по п. 1, в котором ген, кодирующий глюкозо-6-фосфат-1-дегидрогеназу, представляет собой ген zwf.
7. Способ получения метионина по п. 1, в котором ген, кодирующий цистатион-гамма-синтазу, представляет собой ген metB, а ген, кодирующий гомосеринкиназу, представляет собой ген thrB.
8. Способ получения метионина по п. 1, в котором ген, кодирующий гомосерин O-сукцинилтрансферазу, является геном metA и активность гомосерин O-сукцинилтрансферазы усиливается путем замены гена metA на ген metA11.
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| ZHAO J | |||
| ET AL | |||
| Effect of zwf gene knockout on the metabolism of Escherichia coli grown on glucose or acetate // Metabolic Engineering, 2004, 6, pp | |||
| Способ получения суррогата олифы | 1922 |
|
SU164A1 |
| NICOLAS C ET AL | |||
| Response of the central metabolism of Escherichia coli to modified expression of the gene encoding the | |||

