Область техники, к которой относится изобретение
Настоящее изобретение относится к O-сукцинилгомосерин-продуцирующему микроорганизму и к способу получения O-сукцинилгомосерина с его использованием.
Уровень техники
O-сукцинилгомосерин получают с помощью связывания гомосерина и сукцинила-КоА биосинтетическим путем. Таким образом, при разработке штамма, продуцирующего O-сукцинилгомосерин с высоким выходом, образование гомосерина и сукцинила-КоА имеет важное значение. Из них сукцинил-КоА продуцируется в цикле TCA, и, таким образом, требуется усиление цикла TCA для получения высокой концентрации сукцинила-КоА.
Пентозофосфатный путь (PPP) хорошо известен в качестве основного источника NADPH, и кофактор NADPH требуется в путях биосинтеза аминокислот. Таким образом, для усиления пентозофосфатного пути при разработке штаммов, продуцирующих аминокислоты, ген zwf, кодирующий глюкозо-6-фосфат-1-дегидрогеназу, участвующий на первой стадии пути, обычно усиливают, и эти штаммы описаны в корейских патентных публикациях №№2008-0036608 и 2006-0129352.
Когда экспрессия гена zwf или активность фермента, кодируемого указанным геном, ослаблена, пентозофосфатный путь ослабевает, что приводит к недостаточному поступлению NADPH. В этом случае NADPH может быть частично восполнен сверхэкспрессией изоцитратдегидрогеназы (icd) и малатдегидрогеназы (mae) цикла TCA (Appl Microbiol Biotechnol. 2004 64(1); 91-8, Metab Eng. 2004 6(2); 164-74, FEBS Letters 581 2007 3771-6).
Авторы настоящего изобретения провели исследование для разработки штамма, способного продуцировать O-сукцинилгомосерин с высоким выходом и высокой эффективностью, и разработали штамм, в котором ген zwf ослаблен и делегирован для получения высокой концентрации сукцинила-КоА как предшественника O-сукцинилгомосерина в Е. coli, способном продуцировать O-сукцинилгомосерин. В результате культивирования авторы настоящего изобретения обнаружили, что концентрация O-сукцинилгомосерина была увеличена, в чем и заключается настоящее изобретение.
Подробное описание изобретения
Техническая проблема
Задачей настоящего изобретения является предложить O-сукцинилгомосерин-продуцирующий микроорганизм.
Другой задачей настоящего изобретения является предложить способ получения O-сукцинилгомосерина, включающий стадию культивирования O-сукцинилгомосерин-продуцирующего микроорганизма.
Техническое решение
В одном аспекте настоящее изобретение предлагает O-сукцинилгомосерин-продуцирующий микроорганизм.
В конкретном варианте осуществления настоящего изобретения O-сукцинилгомосерин-продуцирующий микроорганизм может быть микроорганизмом рода Escherichia для продуцирования O-сукцинилгомосерина, в котором активность глюкозо-6-фосфат-1-дегидрогеназы ослаблена или элиминирована по сравнению с ее эндогенной активностью.
Термин «O-сукцинилгомосерин-продуцирующий микроорганизм», употребляемый в настоящем документе, относится к прокариотическому или эукариотическому микроорганизму, способному продуцировать O-сукцинилгомосерин в организме и аккумулирующему O-сукцинилгомосерин. Например, O-сукцинилгомосерин-продуцирующий микроорганизм может быть микроорганизмом, относящимся к роду Escherichia, роду Erwinia, роду Serratia, роду Providencia, роду Corynebacteria, роду Pseudomonas, роду Leptospira, роду Salmonella, роду Brevibacteria, роду Hyphomonas, роду Chromobacterium, роду Norcardia, или к грибам или дрожжам. O-сукцинилгомосерин-продуцирующий микроорганизм может быть, в частности, микроорганизмом, относящимся к роду Escherichia и, более конкретно, Escherichia coli (E. coli).
Глюкозо-6-фосфат-1-дегидрогеназа участвует в пентозофосфатном пути, который представляет собой метаболический путь, обеспечивающий восстановительную способность клеток за счет поддержания концентраций NADPH. Этот фермент катализирует окисление глюкозо-6-фосфата в 6-фосфоглюконолактон путем восстановления NADP в NADPH на первой стадии пентозофосфатного пути. Ген, кодирующий этот фермент, обычно называют zwf. Ослабление или элиминирование этого фермента приводит к потоку через цикл TCA, вызывая усиление цикла TCA.
Глюкозо-6-фосфат-1-дегидрогеназа может иметь аминокислотную последовательность SEQ ID NO: 23. Кроме того, глюкозо-6-фосфат-1-дегидрогеназа может иметь аминокислотную последовательность, имеющую 80% или более высокую, 90% или более высокую, 95% или более высокую гомологию с SEQ ID NO: 23.
Термин «гомология», используемый в настоящем описании применительно к последовательности, относится к степени совпадения с определенной аминокислотной последовательностью или последовательностью оснований, и гомология может быть выражена в процентах. В настоящем описании гомологичная последовательность, имеющая активность, которая одинакова или аналогична определенной аминокислотной последовательности или последовательности оснований, выражается в виде «% гомологии». Например, аминокислотная последовательность, имеющая 80% или более высокую, 90% или более высокую, 95% или более высокую гомологию с SEQ ID NO: 23, представляет последовательность, имеющую активность глюкозо-6-фосфат-1-дегидрогеназы.
Термин «эндогенный» фермент и активность, употребляемый в настоящем документе, относится к нативному ферменту, естественным образом присутствующему в микроорганизме или клетке, и к его активности, - другими словами, относится к ферменту и к его активности до модификации соответствующего фермента и его активности.
В конкретном варианте осуществления настоящего изобретения микроорганизм может быть микроорганизмом рода Escherichia для продуцирования O-сукцинилгомосерина, в котором активность одной или более из цистатионин-гамма-синтазы и гомосеринкиназы дополнительно ослаблена или элиминирована по сравнению с их эндогенными активностями. В частности, микроорганизм может быть микроорганизмом для продуцирования O-сукцинилгомосерина, в котором активности и цистатионин-гамма-синтазы и гомосеринкиназы ослаблены или элиминированы по сравнению с их эндогенными активностями.
Цистатионин-гамма-синтаза обладает активностью для превращения O-сукцинилгомосерина в цистатионин. Цистатионин-гамма-синтаза кодируется геном metB. Когда активность данного фермента ослаблена или элиминирована, O-сукцинилгомосерин может аккумулироваться без превращения в цистатионин.
Гомосеринкиназа катализирует синтез O-фосфогомосерина из гомосерина, и кодируется геном thrB. Когда активность данного фермента ослаблена или элиминирована, гомосерин может не превращаться в O-фосфогомосерин, и может использоваться для продуцирования O-сукцинилгомосерина.
В конкретном варианте осуществления настоящего изобретения цистатионин-гамма-синтаза может иметь аминокислотную последовательность SEQ ID NO: 24, и гомосеринкиназа может иметь аминокислотную последовательность SEQ ID NO: 25. Кроме того, цистатионин-гамма-синтаза и гомосеринкиназа могут иметь аминокислотные последовательности, имеющие 80% или более высокую, 90% или более высокую, 95% или более высокую гомологию с SEQ ID NO: 24 и SEQ ID NO: 25, соответственно.
Термин «ослабление» или «элиминирование» ферментативной активности, используемый в настоящем документе, означает, что экспрессия гена, кодирующего соответствующий фермент или активность фермента снижена по сравнению с эндогенной активностью или совсем отсутствует, что может быть обусловлено модификацией всей или части последовательности оснований гена, кодирующего соответствующий фермент, или всей или части последовательности, регулирующей экспрессию гена, путем делеции, замены или инсерции, или путем их сочетания.
Термин «последовательность, регулирующая экспрессию», используемый в настоящем документе, представляет собой последовательность оснований, регулирующую экспрессию гена, и относится к сегменту, способному к увеличению или уменьшению экспрессии конкретного гена у субъекта, и может включать промотор, сайт связывания факторов транскрипции и т.д., но не ограничивается ими.
В конкретном варианте осуществления настоящего изобретения микроорганизм может быть микроорганизмом для продуцирования O-сукцинилгомосерина, в котором активность гомосерин-O-сукцинилтрансферазы дополнительно усилена по сравнению с ее эндогенной активностью.
Гомосерин O-сукцинилтрансфераза представляет собой фермент, который катализирует продуцирование O-сукцинилгомосерина из сукцинила-КоА и гомосерина и участвует в первой стадии пути биосинтеза метионина. Ген, кодирующий данный фермент, обычно называют metA, и его экспрессия подавляется метионином по типу регуляции с обратной связью. Таким образом, может использоваться мутант для экспрессирования гена на высоком уровне за счет устранения регуляции метионином по типу обратной связи.
В конкретном варианте осуществления настоящего изобретения гомосерин-O-сукцинилтрансфераза может иметь аминокислотную последовательность SEQ ID NO: 26. Кроме того, гомосерин-O-сукцинилтрансфераза может иметь аминокислотную последовательность, имеющую 80% или более высокую, 90% или более высокую, 95% или более высокую гомологию с SEQ ID NO: 26. В то же время, гомосерин-O-сукцинилтрансфераза, для которой устранена регуляция метионином по типу обратной связи, может иметь аминокислотную последовательность, имеющую 80% или более высокую, 90% или более высокую, 95% или более высокую гомологию с SEQ ID NO: 27 (metA11: корейская патентная публикация №2009-0106365).
Термин «усиление» ферментативной активности, используемый в настоящем документе, означает, что активность соответствующего фермента повышается по сравнению с его активностью до модификации. В частности, ферментативная активность возрастает за счет сверхэкспрессии гена, кодирующего соответствующий фермент, по сравнению с ее эндогенной активностью, или активность фермента, кодируемого геном, увеличивается за счет мутации гена, по сравнению с ее эндогенной активностью, и усиление может быть вызвано увеличением числа копий кодирующего гена, заменой промотора гена более сильным промотором, чем эндогенный промотор, или модификацией всей или части последовательности оснований гена на хромосоме или всей или части последовательности, регулирующей его экспрессию, путем делеции, замены или инсерции, или путем их сочетания.
В конкретном варианте осуществления настоящего изобретения гомосерин-O-сукцинилтрансфераза может кодироваться геном, в котором промотор гена, кодирующего данный фермент, заменен на более сильный промотор, чем эндогенный промотор. Например, промотор включает известные сильные промоторы Ptac, Ptrc, Ppro, PR, PL, Prmf, PcysK, и т.д., но не ограничивается ими.
В конкретном варианте осуществления настоящего изобретения микроорганизмом может быть Е. coli. Микроорганизмом может быть E. coli, в котором активность глюкозо-6-фосфат-1-дегидрогеназы ослаблена или элиминирована по сравнению с ее эндогенной активностью, и активности одной или более из цистатионин-гамма-синтазы и гомосеринкиназы ослаблены или элиминированы по сравнению с их эндогенными активностями, соответственно, и активность гомосерин-O-сукцинилтрансферазы усилена по сравнению с ее эндогенной активностью.
В конкретном варианте осуществления настоящего изобретения микроорганизмом может быть Е. coli, в котором metA на хромосоме E. coli заменен на metA11 (SEQ ID NO: 27), который является мутантом, полученным с помощью устранения регуляции метионином по типу обратной связи, thrB и metB на хромосоме делегированы, и zwf ослаблен или элиминирован.
В конкретном варианте осуществления настоящего изобретения микроорганизмом может быть штамм Е. coli CC03-0156, который депонировали в Корейском центре культур микроорганизмов (KCCM) 22 ноября 2013 года под номером доступа KCCM11487P.
Кроме того, в конкретном варианте осуществления настоящего изобретения микроорганизмом может быть микроорганизм, наделенный способностью ассимилировать сахарозу. Ассимиляция сахарозы означает способность метаболизировать сахарозу в качестве источника углерода или метаболического источника. Способность ассимилировать сахарозу может быть получена с помощью введения метаболического фермента сахарозы, например, фруктокиназы, сахарозной PTS пермеазы, сахарозной гидролазы или инвертазы. Например, способность ассимилировать сахарозу может быть получена с помощью трансформации рекомбинантным вектором (pAscrSM, SEQ ID NO: 28), включающим ген, кодирующий Scr-PTS фермент, полученный из Streptococcus mutans, который описан в корейской патентной публикации №2010-0099572.
В конкретном варианте осуществления настоящего изобретения микроорганизмом может быть Е. coli, в котором metA на хромосоме E. coli заменен на metA11 (SEQ ID NO: 27), который является мутантом, полученным с помощью устранения регуляции метионином по типу обратной связи, thrB и metB на хромосоме делегированы, и zwf элиминирован, и который способен утилизировать сахар-сырец с помощью трансформации рекомбинантным вектором, включающим scrKYABR, кодирующий фруктокиназу, сахарозную PTS пермеазу, сахарозную гидролазу и сахарозный транскрипционный регулятор, полученный из ассимилирующего сахарозу Streptococcus mutans.
В аспекте настоящего изобретения предложен способ продуцирования O-сукцинилгомосерина, включающий стадии культивирования микроорганизма рода Escherichia для продуцирования O-сукцинилгомосерина, в котором активность глюкозо-6-фосфат-1-дегидрогеназы ослаблена или элиминирована по сравнению с ее эндогенной активностью, в среде, и извлечения O-сукцинилгомосерина из культуральной среды или культивируемого микроорганизма.
В способе продуцирования O-сукцинилгомосерина в соответствии с конкретным вариантом осуществления настоящего изобретения культивирование O-сукцинилгомосерин- продуцирующего штамма может осуществляться в подходящей среде и условиях, известных в области техники. Процедуры культивирования могут быть легко скорректированы специалистами в данной области техники в соответствии с выбранным штаммом. Примеры процедур культивирования включают способы периодического типа, непрерывного типа и периодического типа с подпиткой, но не ограничиваются ими. Различные процедуры культивирования описаны, например, в литературе («Biochemical Engineering», James М. Lee, Prentice-Hall International Editions, pp. 138-176).
Среда, используемая для культивирования, должна соответствовать требованиям культивирования конкретного штамма. Культуральные среды для различных микроорганизмов описаны в литературе («Manual of Methods for General Bacteriology», American Society for Bacteriology, Washington D.C., USA, 1981). Эти среды содержат множество источников углерода, источников азота и следовых элементов. Источники углерода включают углеводы, такие как глюкоза, лактоза, сахароза, фруктоза, мальтоза, крахмал и целлюлоза; жиры, такие как соевое масло, подсолнечное масло, касторовое масло и кокосовое масло; жирные кислоты, такие как пальмитиновая кислота, стеариновая кислота и линолевая кислота; спирты, такие как глицерин и этанол; и органические кислоты, такие как уксусная кислота. Указанные источники углерода могут использоваться по отдельности или в сочетании. Источники азота включают органические источники азота, такие как пептон, дрожжевой экстракт, мясной экстракт, экстракт солода, жидкий кукурузный экстракт (CSL) и соевая мука, и неорганические источники азота, такие как мочевина, сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония. Указанные источники азота могут использоваться по отдельности или в сочетании. Кроме того, среда может содержать первичный кислый фосфат калия, вторичный кислый фосфат калия и соответствующие им натрийсодержащие соли в качестве источника фосфора. Также среда может содержать соль металла, такую как сульфат магния или сульфат железа. Кроме того, также могут быть добавлены аминокислоты, витамины и подходящие предшественники.
Кроме того, для поддержания культуры в аэробных условиях в культуру можно вводить кислород или кислородосодержащий газ (например, воздух). Температура культуры обычно составляет 20-45°C, и в частности, 25-40°C. Культивирование может продолжаться до тех пор, пока образование предшественника L-метионина не достигнет желаемого уровня, и время культивирования может составлять от 10 ч до 160 ч.
Полезные эффекты изобретения
O-сукцинилгомосерин-продуцирующий штамм настоящего изобретения эффективно продуцирует O-сукцинилгомосерин, который может использоваться при получении L-метионина. Полученный таким образом L-метионин может широко применяться в производстве кормов для животных или кормовых добавок, а также пищевых продуктов для человека или пищевых добавок.
Вариант осуществления изобретения
Далее в этом документе настоящее изобретение будет описано более подробно со ссылкой на примеры. Однако данные примеры приводятся только в целях иллюстрации, и объем настоящего изобретения не предполагает ограничения этими примерами.
Ссылочный пример 1. Получение O-сукцинилгомосерин-продуцирующего штамма
1-1 Делеция гена metB
Для повышения аккумуляции O-сукцинилгомосерина штамм получали с помощью делеции гена metB, кодирующего цистатионин-гамма-синтазу, которая участвует в разложении O-сукцинилгомосерина.
В штамме W3110 Е. coli (K12) дикого типа ген metB, кодирующий цистатионин-гамма-синтазу, делетировали. Известно, что цистатионин-гамма-синтаза связывается с различными предшественниками метионина в клетках, тем самым образуя различные побочные продукты. Таким образом, сверхэкспрессия цистатионинсинтазы может увеличивать число побочных реакций, снижая эффективность внутриклеточных реакций. Для делеции гена metB осуществляли метод делеции с использованием FRT-одностадийной ПЦР (PNAS (2000), Vol. 97, p. 6640-6645). Сначала ПЦР проводили с использованием праймеров SEQ ID NO: 1 и 2 и вектора pKD3 (PNAS (2000) Vol. 197, pp. 6640-6645) в качестве матрицы для получения делеционной кассеты.
SEQIDNO: 1:
5'-TTACTCTGGTGCCTGACATTTCACCGACAAAGCCCAGGGAACTTCATCACGTGTAGGCTGGAGCTGCTTC-3'
SEQ ID NO: 2:
5'-CGCTGCGCCAGCTCCATACGCGGCACCAGCGTTCGCAACCCACGTAGCAGCATATGAATATCCTCCTTAG-3'
ПЦР проводили в следующих условиях: 30 циклов, состоящих из денатурации при 95°C в течение 30 секунд, отжига при 55°C в течение 30 секунд и удлинения при 72°C в течение 1 минуты. Полученный таким образом продукт ПЦР подвергали электрофорезу на 1,0% агарозном геле, с последующим элюированием и очисткой полосы размером 1,1 т.п.н. Полученный таким образом фрагмент ДНК электропорировали в штамм W3110 Е. coli (K12), предварительно трансформированный вектором pKD46 (PNAS (2000) Vol. 97, p. 6640-6645). Перед электропорацией штамм W3110, трансформированный pKD46, культивировали при 30°C в среде LB, содержащей 200 мкг/л ампициллина и 5 мМ L-арабинозы до тех пор, пока OD600 не достигала 0,5. Затем штамм промывали трижды 10% глицерином. Электропорацию проводили при 2500 В. Полученный штамм сеяли штрихом в среду LB для чашек Петри, содержащую 30 мкг/л хлорамфеникола, с последующим культивированием при 37°C в течение 1-2 суток. Затем отбирали штамм, проявляющий устойчивость.
ПЦР проводили с использованием отобранного штамма в качестве матрицы и праймеров SEQ ID NO: 3 и 4 в описанных выше условиях. Делецию гена metB подтверждали идентификацией гена размером 1,5 т.п.н на 1,0% агарозном геле.
SEQ ID NO: 3: 5'-TATTCGCCGCTCCATTCAGC-3'
SEQ ID NO: 4: 5'-TACCCCTTGTTTGCAGCCCG-3'
Штамм, в котором была подтверждена делеция гена metB, далее трансформировали вектором рСР20 (PNAS (2000) vol. 97, p. 6640-6645) и культивировали в среде LB, содержащей 100 мкг/л ампицилина. Затем ПЦР проводили в тех же самых условиях, и элиминирование хлорамфеникольного маркера подтверждали наблюдением меньшего по размерам ПЦР-продукта на 1,0% агарозном геле. Наконец, получали штамм с делегированным геном metB. Полученный ауксотрофный по метионину штамм был назван CC03-0132.
1-2. Деления гена thxB
Для повышения продукции O-сукцинилгомосерина из гомосерина делегировали ген thrB, который является геном, кодирующим гомосеринкиназу. В частности, при использовании треонин-продуцирующего штамма, делеция гена thrB является необходимой, поскольку активность утилизации гомосерина очень высока. Для делеции гена thrB в штамме CC03-0132, полученном в ссылочном примере 1-1, осуществляли метод делеции с использованием FRT-одностадийной ПЦР (PNAS (2000), Vol. 97, p. 6640-6645). Для делеции гена thrB ПЦР проводили с использованием праймеров SEQ ID NO: 5 и 6 и вектора pKD3 (PNAS (2000) Vol. 97, p. 6640-6645) в качестве матрицы для получения делеционной кассеты.
SEQIDNO: 5:
5'-CATGGTTAAAGTTTATGCCCCGGCTTCCAGTGCCAATATGAGCGTCGGGTGTGTAGGCTGGAGCTGCTTC-3'
SEQ ID NO: 6:
5'-GGAGATACCGCTCGCTACCGCGCCGATTTCCGCGACCGCCTGCCGCGCCTCATATGAATATCCTCCTTAG-3'
ПЦР проводили в следующих условиях: 30 циклов, состоящих из денатурации при 95°C в течение 30 секунд, отжига при 55°C в течение 30 секунд и удлинения при 72°C в течение 1 минуты. Полученный таким образом продукт ПЦР подвергали электрофорезу на 1,0% агарозном геле, с последующим элюированием и очисткой полосы размером 1,1 т.п.н. Полученный таким образом фрагмент ДНК электропорировали в штамм СС03-0132, предварительно трансформированный вектором pKD46 (PNAS (2000) Vol. 97, p. 6640-6645). Для электропорации штамм СС03-0132, трансформированный pKD46, культивировали при 30°C в среде LB, содержащей 200 мкг/л ампициллина и 5 мМ L-арабинозы до тех пор, пока OD600 не достигала 0,5. Затем штамм промывали трижды 10% глицерином. Электропорацию проводили при 2500 B. Полученный штамм сеяли штрихом в среду LB для чашек Петри, содержащую 30 мкг/л хлорамфеникола, с последующим культивированием при 37°C в течение 1-2 суток. Затем отбирали штамм, проявляющий устойчивость.
ПЦР проводили с использованием отобранного штамма в качестве матрицы и праймеров SEQ ID NO: 7 и 8 в описанных выше условиях. Делецию гена thrB подтверждали идентификацией гена размером 1,5 т.п.н на 1,0% агарозном геле.
SEQ ID NO: 7: 5'-ACTCGACGATCTCTTTGCC-3'
SEQ ID NO: 8: 5'-ACGCCGAGAGGATCTTCGCAG-3'
Подтвержденный таким образом штамм затем трансформировали вектором pCP20 (PNAS (2000) vol. 97, p. 6640-6645) и культивировали в среде LB, содержащей 100 мкг/л ампицилина. Затем ПЦР проводили в тех же самых условиях, и элиминирование хлорамфеникольного маркера подтверждали наблюдением меньшего по размерам ПЦР-продукта на 1,0% агарозном геле. Наконец, получали штамм с делегированным геном thrB. Полученный таким образом штамм был назван СС03-0133.
1-3. Деления гена metA
Для введения устойчивого к регуляции по типу обратной связи гена metA, в хромосому, аутентичный хромосомный ген metA на основе штамма СС03-0133, который получали с помощью делеции генов metB и thrB в штамме W3110 Е. coli (К 12), делегировали. Для делеции гена metA осуществляли метод делеции с использованием FRT-одностадийной ПЦР (PNAS (2000), Vol. 97, p. 6640-6645). Для делеции гена metA ПЦР проводили с использованием праймеров SEQ ID NO: 9 и 10 и вектора pKD3 (PNAS (2000) Vol. 97, pp. 6640-6645) в качестве матрицы для получения делеционной кассеты.
SEQ ID NO: 9:
5'-TCAGCTGTTGCGCATCGATTCCCGTGAATCGCGCAACACGCCCGCAGAGCGTGTAGGCTGGAGCTGCTTC-3'
SEQ ID NO: 10:
5'-CCGTCACAAAGGCAATGCGCTTATCTTTACTGGCAAACAGATATGCATCCCATATGAATATCCTCCTTAG-3'
ПЦР проводили в следующих условиях: 30 циклов, состоящих из денатурации при 95°C в течение 30 секунд, отжига при 55°C в течение 30 секунд и удлинения при 72°C в течение 1 минуты. Полученный таким образом продукт ПЦР подвергали электрофорезу на 1,0% агарозном геле, с последующим элюированием и очисткой полосы размером 1,1 т.п.н. Полученный таким образом фрагмент ДНК электропорировали в штамм СС03-0133, предварительно трансформированный вектором pKD46 (PNAS (2000) Vol. 97, p. 6640-6645). Для электропорации штамм СС03-0133, трансформированный pKD46, культивировали при 30°C в среде LB, содержащей 200 мкг/л ампициллина и 5 мМ L-арабинозы до тех пор, пока OD600 не достигала 0,5. Затем штамм промывали трижды 10% глицерином. Электропорацию проводили при 2500 В. Полученный штамм сеяли штрихом в среду LB для чашек Петри, содержащую 30 мкг/л хлорамфеникола, с последующим культивированием при 37°C в течение 1-2 суток. Затем отбирали штамм, проявляющий устойчивость.
ПЦР проводили с использованием отобранного штамма в качестве матрицы и праймеров SEQ ID NO: 11 и 12 в описанных выше условиях. Делецию гена metA подтверждали идентификацией гена размером 1,5 т.п.н. на 1,0% агарозном геле.
SEQ ID NO: 11: 5'-CTCATTAACGTTGGTTGTCA-3'
SEQ ID NO: 12: 5'-TATCTTGCTGCTGCTGAATG-3'
Подтвержденный таким образом штамм затем трансформировали вектором pCP20 (PNAS (2000) vol. 97, p. 6640-6645) и культивировали в среде LB, содержащей 100 мкг/л ампицилина. Затем ПЦР проводили в тех же самых условиях, и элиминирование хлорамфеникольного маркера подтверждали наблюдением меньшего по размерам ПЦР-продукта на 1,0% агарозном геле. Наконец, получали штамм с делегированным геном metA. Полученный таким образом штамм был назван СС03-0134.
1.4. Инсерция гена metA11
- Получение вектора pSG для инсерции metA11
Большая часть активности гомосеринсукцинилтрансферазы регулируется ингибированием по типу обратной связи небольшим количеством метионина, добавляемого в среду, и, поэтому, мутант, в котором была устранена регуляция метионином по типу обратной связи, был замещен для повышенного образования предшественника L-метионина, O-сукцинилгомосерина. Для замещения хромосомного гена metA дикого типа, кодирующего гомосеринсукцинилтрансферазу E. coli, на metA11 (SEQ ID NO: 27), кодирующий мутант, в котором устранена регуляция метионином по типу обратной связи, получали pSG-metA11 вектор для инсерции. Согласно корейской патентной публикации №2009-0106365 была получена информация о последовательности оснований гена metA11, и на основе данной последовательности оснований праймеры (SEQ ID NO: 13 и 14), включающие от ATG до ORF гена metA11 и сайты узнавания для рестрикционных ферментов EcoRI и SacI, были синтезированы. ПЦР проводили с использованием плазмиды pMetA11-CL в качестве матрицы (корейская патентная публикация №2009-0106365), которая была получена путем лигирования гена metA11 вектором pCL1920, и праймеров следующих SEQ ID NO.
SEQ ID NO: 13: 5'-ggccgaattcatgccgattcgtgtgccgga-3'
SEQ ID NO: 14: 5'-ggccgagctcgttaatccagcgttggattca-3'
ПЦР проводили с использованием pfu-X ДНК-полимеразы (SolGent; SPX16-R250), и условиями ПЦР были: 30 циклов, состоящих из денатурации при 95°C в течение 30 секунд, отжига при 55°C в течение 30 секунд и удлинения при 72°C в течение 2 минут. В результате, был получен продукт ПЦР амплифицированного metA11 ORF, включающий сайты узнавания для рестрикционных ферментов EcoRI и SacI на обоих концах. Полученный в результате ПЦР ген metA11 обрабатывали рестрикционными ферментами EcoRI и SacI и лигировали вектором pSG76-C (Nucleic Acids Res. 1999 Nov 15; 27(22):4409-15), который обрабатывали рестрикционными ферментами EcoRI и SacI для клонирования гена. Наконец, получали рекомбинантный вектор pSG-metA11 клонированного гена metA11.
- Получение штамма с инсертированным геном metA11
Штамм СС03-0134, полученный в ссылочном примере 1-3, трансформировали с получением pSG-metA11, который был вектором для инсерции гена metA11, и культивировали в среде LB Cm (10 г/л дрожжевого экстракта, 5 г/л NaCl, 10 г/л триптона и 30 мкг/л хлорамфеникола). Затем выбирали колонию, проявляющую устойчивость к хлорамфениколу. Выбранный трансформант представлял собой штамм, в котором вектор pSG-metA11 был преимущественно инсертирован в хромосомный metA. Штамм с инсертированным геном metA11 трансформировали вектором pST76-ASceP (Nucleic Acids Res. 1999 Nov 15; 27(22): 4409-15), экспрессирующим I-SceI, который является рестрикционным ферментом, расщепляющим I-SceI в векторе pSG, и штамм, выращиваемый в LB-Amp (10 г/л дрожжевого экстракта, 5 г/л NaCl, 10 г/л триптона и 100 мкг/л хлорамфеникола), отбирали. Этот штамм, отобранный таким путем, являлся штаммом, в котором metA дикого типа был замещен на metA11 и инсертированный вектор pSG76-C был удален. Полученный штамм был назван Е. coli СС03-0038.
Пример 1. Ослабление и деления гена zwf
1-1. Ослабление гена zwf
- Получение вектора pSG для замещения инициирующего кодона гена zwf
Для ослабления гена zwf в штамме СС03-0038, полученном в ссылочном примере 1-4, применяли способ замещения инициирующего кодона ATG области ORF гена zwf на GTG. ПЦР проводили с использованием праймеров SEQ ID NO: 15 и 16 и SEQ ID NO: 17 и 18, и генома Е. coli W3110 в качестве матрицы.
SEQ ID NO: 15: 5'-ggccgaattcctgaaagaaatcgaaatgcag-3'
SEQ ID NO: 16: 5'-cacgtcattctccttaagaattc-3'
SEQ ID NO: 17: 5'-gaattcttaaggagaatgacgtg-3'
SEQ ID NO: 18: 5'-ggccgagctcgggcatggcaaagtagttaatg-3'
ПЦР проводили с использованием pfu-X ДНК-полимеразы (SolGent; SPX16-R250), и ПЦР проводили в следующих условиях: 30 циклов, состоящих из денатурации при 95°C в течение 30 секунд, отжига при 55°C в течение 30 секунд и удлинения при 72°C в течение 1 минуты. ПЦР со сплайсингом перекрывающимися расширениями (SOE, Splicing by Overlap Extension) проводили с использованием двух фрагментов, полученных таким образом в качестве матриц. В результате, были получены область zwf, включающая сайты узнавания для рестрикционных ферментов EcoRI и SacI на обоих концах, и инициирующий кодон GTG. Рестрикционные ферменты EcoRI и SacI были обработаны на концах полученного фрагмента, включая сайты узнавания для ферментов, и клонированы в вектор pSG76-C (Nucleic Acids Res. 1999 Nov 15; 27(22): 4409-15), обработанный рестрикционными ферментами EcoRI и SacI путем лигирования. Наконец, получали рекомбинантный вектор pSG-zwf(GTG).
- Получение штамма с замещением инициирующего кодона гена zwf
pSG-zwf(GTG), который был получен для замещения инициирующего кодона гена zwf, как описано выше, трансформировали в штамм СС03-0038, полученный в ссылочном примере 1-4, и культивировали в среде LB_Cm (10 г/л дрожжевого экстракта, 5 г/л NaCl, 10 г/л триптона и 30 мкг/л хлорамфеникола). Затем выбирали колонию, проявляющую устойчивость к хлорамфениколу. Выбранный трансформант представлял собой штамм, в котором вектор pSG-zwf(GTG) был преимущественно инсертирован в хромосомный zwf. Выбранный штамм трансформировали pST76-ASceP (Nucleic Acids Res. 1999 Nov 15; 27(22): 4409-15), экспрессирующим I-SceI, который является рестрикционным ферментом, расщепляющим I-SceI на вектор pSG, и штамм, выращиваемый в LB-Amp (10 г/л дрожжевого экстракта, 5 г/л NaCl, 10 г/л триптона и 100 мкг/л хлорамфеникола), отбирали. Этот штамм, отобранный таким образом, являлся штаммом, в котором ген zwf был ослаблен замещением инициирующего кодона ATG гена zwf на GTG, и затем инсертированный вектор pSG76-C был удален. Полученный штамм был назван СС03-0038zwfGTG.
1-2. Деления гена zwf
Для делеции гена zwf в штамме СС03-0038, полученном в ссылочном примере 1-4, осуществляли метод делеции с использованием FRT-одностадийной ПЦР (PNAS (2000), Vol. 97, p. 6640-6645). Для делеции гена zwf ПЦР проводили с использованием праймеров SEQ ID NO: 19 и 20 и вектора pKD3 (PNAS (2000) Vol. 97, p. 6640-6645) в качестве матрицы для получения делеционной кассеты.
SEQ ID NO: 19:
5'-CAAGTATACCCTGGCTTAAGTACCGGGTTAGTTAACTTAAGGAGAATGACGTGTAGGCTGGAGCTGCTTC-3'
SEQ ID NO: 20:
5'-CTGCGCAAGATCATGTTACCGGTAAAATAACCATAAAGGATAAGCGCAGATACATATGAATATCCTCCTTAG-3'
ПЦР проводили в следующих условиях: 30 циклов, состоящих из денатурации при 95°C в течение 30 секунд, отжига при 55°C в течение 30 секунд и удлинения при 72°C в течение 1 минуты. Полученный таким образом продукт ПЦР подвергали электрофорезу на 1,0% агарозном геле, с последующим элюированием и очисткой полосы размером 1,1 т.п.н. Полученный таким образом фрагмент ДНК электропорировали в штамм СС03-0038, предварительно трансформированный вектором pKD46 (PNAS (2000) Vol. 97, p. 6640-6645). Для электропорации штамм СС03-0038, трансформированный pKD46, культивировали при 30°C в среде LB, содержащей 200 мкг/л ампициллина и 5 мМ арабинозы, до тех пор, пока OD600 не достигала 0,5. Затем штамм промывали трижды 10% глицерином. Электропорацию проводили при 2500 В. Полученный штамм сеяли штрихом в среду LB для чашек Петри, содержащую 30 мкг/л хлорамфеникола, с последующим культивированием при 37°C в течение 1-2 суток. Затем отбирали штамм, проявляющий устойчивость.
ПЦР проводили с использованием отобранного штамма в качестве матрицы и праймеров SEQ ID NO: 21 и 22 в описанных выше условиях. Делецию гена zwf подтверждали идентификацией гена размером 2 т.п.н. на 1,0% агарозном геле.
SEQ ID NO: 21: 5'-CATAACATGATCAGTGTCAGAT-3'
SEQ ID NO: 22: 5'-CGCGTAACAATTGTGGATTCAT-3'
Идентифицированный таким образом штамм был назван СС03-0156.
1-3. Получение O-сукцинилгомосерин-продуцирующего штамма на основе треонин-продуцирующего штамма
Треонин-продуцирующий штамм, E. coli KCCM 10541P, описанный в международной патентной публикации WO 2005/075625, использовали для делеции генов metB, thrB и метА таким же образом, как описано в ссылочном примере 1, и затем устойчивый к регуляции по типу обратной связи ген metA11 вводили для получения штамма для продуцирования O-сукцинилгомосерина, который был назван CJM2-A11. Кроме того, штамм с делегированным геном zwf получали таким же образом, как описано в примере 1-2, и назвали CJM2-A11Z.
Пример 2. Ферментация для получения O-сукцинилгомосерина
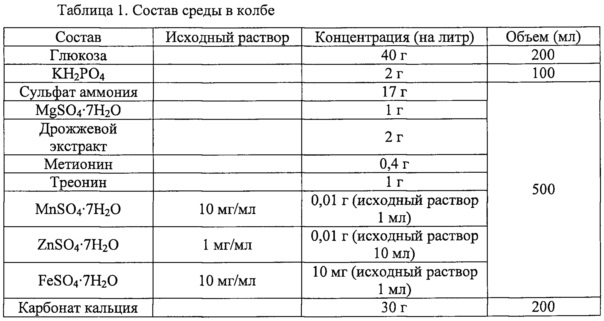
Для изучения влияния делеции гена zwf в штамме, полученном в примере 1, проводили культивирование в колбе Эрленмейера. Состав среды в колбе был таким, как в приведенной ниже таблице 1.
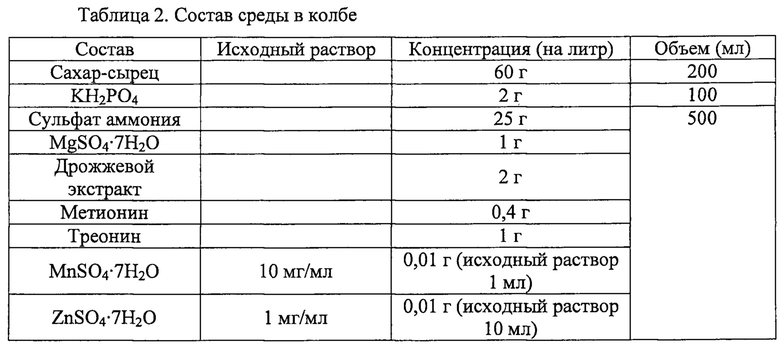
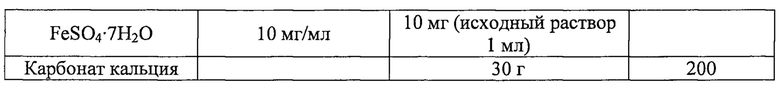
Далее, плазмиду pAscrSM (SEQ ID NO: 28), имеющую последовательность scrKYABR (ниже называется «scrO»), описанную в корейской патентной публикации №2009-0018128, вводили для получения штамма, способного утилизировать сахар-сырец, с последующим культивированием в колбе. Состав среды в колбе был таким, как в приведенной ниже таблице 2. Штаммы, полученные указанным выше способом, были названы CC03-0038/pAscrSM, CC03-0038zwfGTG/pAscrSM, CC03-0156/pAscrSM, CJM2-A11/pAscrSM и CJM2-A11Z/pAscrSM, соответственно.
Каждый из штаммов СС03-0038, CC03-0038zwfGTG, СС03-0156, CJM2-A11, CJM2-A11Z, CC03-0038/pAscrSM, CC03-0038zwfGTG/pAscrSM, CC03-0156/pAscrSM, CJM2-A11/pAscrSM и CJM2-A11Z/pAscrSM инокулировали в среды LB для чашек Петри и культивировали при 33°C в течение ночи. Одиночную колонию инокулировали в 2 мл среды LB, с последующим культивированием при 33°C в течение 2 ч. Культуру инокулировали в 250 мл колбе Эрленмейера, содержащей 25 мл среды в колбе при OD600=0,5, с последующим культивированием при 33°C, 200 об/мин в течение 48 ч. Для сравнения продуцирования O-сукцинилгомосерина проводили ВЭЖХ. Результаты приведены в таблице 3 ниже.
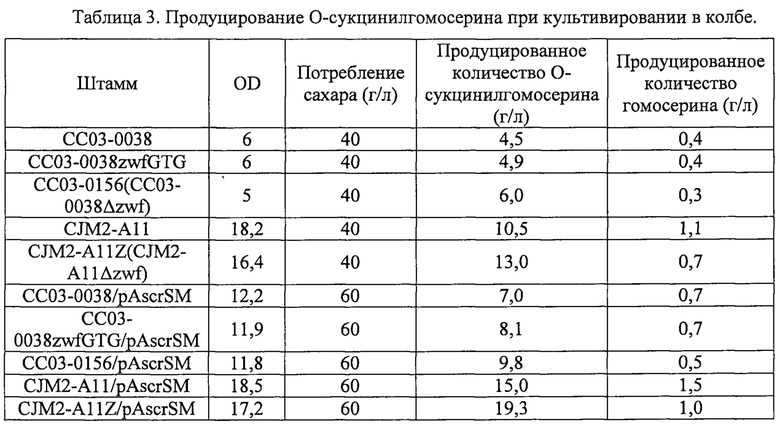
Результаты культивирования в колбе показали, что штаммы с ослабленным или делегированным геном zwf CC03-0038zwfGTG, СС03-0156, CJM2-A11Z, СС03-0038zwfGTG/pAscrSM, СС03-0156/pAscrSM, и CJM2-A11Z/pAscrSM демонстрируют продукцию O-сукцинилгомосерина, которая на 8,9-40,0% выше, чем для соответствующих контрольных штаммов. Кроме того, было отмечено уменьшение образования гомосерина, что позволяет предположить, что образование сукцинил-КоА повысилось за счет усиления цикла TCA.
Авторы настоящего изобретения подтвердили, что O-сукцинилгомосерин может быть получен с высоким выходом путем ослабления или делеции гена zwf в штамме СС03-0038 и штамме CJM2-A11. Штамм СС03-0156 депонировали в соответствии с условиями Будапештского договора в Корейском центре культур микроорганизмов (КССМ) 22 ноября 2013 года под номером доступа KCCM11487P.
Что касается применения гена zwf для усиления цикла TCA, следует понимать, что ослабление или делеция гена zwf не ограничивается Е. coli, но точно так же применимо для всех микроорганизмов, имеющих цикл TCA, включая род Escherichia, род Corynebacterium, дрожжи и т.д., для получения высокого выхода O-сукцинилгомосерина.
















Группа изобретений относится к микроорганизмам рода Escherichia для продуцирования О-сукцинилгомосерина и способу получения O-сукцинилгомосерина с использованием указанного микроорганизма. В предложенном микроорганизме рода Escherichia активность глюкозо-6-фосфат-1-дегидрогеназы ослаблена или элиминирована по сравнению с ее эндогенной активностью, активности одной или более из цистатионин-гамма-синтазы и гомосеринкиназы элиминированы по сравнению с их эндогенными активностями у микроорганизма дикого типа, активность гомосерин-O-сукцинилтрансферазы усилена по сравнению с ее эндогенной активностью у микроорганизма дикого типа, и продукция О-сукцинилгомосерина увеличена по сравнению с продукцией у микроорганизма дикого типа. Предложен также способ получения О-сукцинилгомосерина, включающий культивирование вышеуказанного микроорганизма в среде и извлечение О-сукцинилгомосерина из культуральной среды или культивируемого микроорганизма. Изобретение обеспечивает получение О-сукцинилгомосерина с высоким выходом. 2 н. и 8 з.п. ф-лы, 3 табл., 2 пр.
1. Микроорганизм рода Escherichia для продуцирования О-сукцинилгомосерина, в котором:
- активность глюкозо-6-фосфат-1-дегидрогеназы ослаблена или элиминирована по сравнению с ее эндогенной активностью,
- активности одной или более из цистатионин-гамма-синтазы и гомосеринкиназы элиминированы по сравнению с их эндогенными активностями у микроорганизма дикого типа,
- активность гомосерин-O-сукцинилтрансферазы усилена по сравнению с ее эндогенной активностью у микроорганизма дикого типа, и
- продукция О-сукцинилгомосерина увеличена по сравнению с продукцией у микроорганизма дикого типа.
2. Микроорганизм рода Escherichia по п. 1, в котором глюкозо-6-фосфат-1-дегидрогеназа имеет аминокислотную последовательность SEQ ID NO: 23.
3. Микроорганизм рода Escherichia по п. 1, в котором цистатионин-гамма-синтаза имеет аминокислотную последовательность SEQ ID NO: 24 и гомосеринкиназа имеет аминокислотную последовательность SEQ ID NO: 25.
4. Микроорганизм рода Escherichia по п. 1, в котором гомосерин-O-сукцинилтрансфераза имеет аминокислотную последовательность SEQ ID NO: 26.
5. Микроорганизм рода Escherichia по любому из пп. 1-4, который является Escherichia coli (E.coli).
6. Способ получения О-сукцинилгомосерина, включающий:
культивирование микроорганизма по п. 1 в среде, в результате чего продукция О-сукцинилгомосерина увеличивается по сравнению с продукцией у микроорганизма дикого типа, и
извлечение О-сукцинилгомосерина из культуральной среды или культивируемого микроорганизма.
7. Способ получения О-сукцинилгомосерина по п. 6, в котором микроорганизмом является E.coli.
8. Микроорганизм рода Escherichia по п. 1, в котором ген, кодирующий глюкозо-6-фосфат-1-дегидрогеназу, представляет собой ген zwf.
9. Микроорганизм рода Escherichia по п. 1, в котором ген, кодирующий цистатионин-гамма-синтазу, представляет собой ген metB и ген, кодирующий гомосеринкиназу, представляет собой ген thrB.
10. Микроорганизм рода Escherichia по п. 1, в котором ген, кодирующий гомосерин-О-сукцинилтрансферазу, представляет собой ген metA, и активность гомосерин-O-сукцинилтрансферазы увеличена путем замены гена metA на ген metA11.
| WO 2008013432 A1, 31.01.2008 | |||
| ZHAO J | |||
| ET AL | |||
| Effect of zwf gene knockout on the metabolism of Escherichia coli grown on glucose or acetate // Metabolic Engineering, 2004, 6, pp | |||
| Способ получения суррогата олифы | 1922 |
|
SU164A1 |
| NICOLAS C ET AL | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| СПОСОБ ИССЛЕДОВАНИЯ И ПРОГНОЗИРОВАНИЯ ПУТИ БИОСИНТЕЗА МЕТИОНИНА В РОДСТВЕННЫХ ГЕНОМАХ КОРИНЕБАКТЕРИЙ | 2006 |
|
RU2307169C1 |
| СПОСОБ ПОЛУЧЕНИЯ АРОМАТИЧЕСКОЙ L-АМИНОКИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН ydiB, СПОСОБ ПОЛУЧЕНИЯ СЛОЖНОГО ЭФИРА НИЗШИХ АЛКИЛОВ АЛЬФА-L-АСПАРТИЛ-L-ФЕНИЛАЛАНИНА | 2007 |
|
RU2355759C1 |
| PAKULA A.A | |||
| Genetic analysis of protein stability and function | |||
| Anna | |||
| Rev | |||
| Genet., 1989, 23:289-310, с | |||
| Держатель для поленьев при винтовом колуне | 1920 |
|
SU305A1 |

