ОБЛАСТЬ ТЕХНИКИ
[0001] Настоящее изобретение относится к вакцинам против кампилобактериоза. Более конкретно, настоящее изобретение относится к вакцинам против кампилобактериоза, содержащим клетки Escherichia coli, экспрессирующим гептасахарид гликан Campylobacter jejuni, образующийся в пути N-гликозилирования.
УРОВЕНЬ ТЕХНИКИ
[0002] Грамотрицательная бактерия Campylobacter является наиболее частым бактериальным возбудителем гастроэнтерита в Северной Америке и многих промышленно развитых странах. Также Campylobacter является значимым патогеном, вызывающим пищевые отравления у домашнего скота, включая птицу, что является основным источником кампилобактериоза у человека. Таким образом, контроль Campylobacter у домашней птицы на фермах мог бы снизить риск воздействия данного патогена на человека и оказать значительное влияние на безопасность продуктов питания и состояние здоровья населения.
[0003] Campylobacter является эндемичным во многих развивающихся странах, в основном вследствие плохих санитарных условий и близкого контакта человека с животными, которые являются резервуаром для данного патогена. Работа Katarzyna et al. (Expert Rev. Vaccines 8: 625-645, 2009) свидетельствует о том, что в Соединенных Штатах Америки инфекция, вызываемая Campylobacter, вызывает от 1,5 миллионов (данные Всемирной организации здравоохранения) до 2,4 миллионов (данные Государственного центра санитарно-эпидемического контроля США) случаев заболевания в год. Кроме того, согласно данным Всемирной организации здравоохранения приблизительно 1% населения Восточной Европы ежегодно инфицируется видами Campylobacter. Инфекцию у людей вызывают преимущественно два вида: С. coli и С. jejuni, которые обусловливают более чем 95% случаев камбилобактериоза. Клинические проявления инфекции, вызванной Campylobacter, могут варьировать от бессимптомных случаев до тяжелого гастроэнтерита, иногда сопровождающегося длительной диареей со слизистым, кровянистым или водянистым стулом.
[0004] В публикации Jun Lin "Novel Approaches for Campylobacter Control in Poultry" (FOODBORNE PATHOGENS AND DISEASE, Volume 6, Number 7, pp. 755-765, 2009), включенной в настоящую заявку посредством ссылки, обсуждаются разные стратегии со кращения инфицирования Campylobacter у домашней птицы. Lin предлагает три общие стратегии контролирования Campylobacter у домашней птицы на уровне ферм: (1) уменьшение проникновения в окружающую среду (меры биологической безопасности), (2) увеличение резистентности хозяев-птиц с целью снижения носительства Campylobacter в кишечнике (например, конкурентное исключение, вакцинация и генетическая селекция хозяина), и (3) применение противомикробных альтернативных средств для уменьшения и даже эрадикации Campylobacter из организма колонизированных куриц (например, терапия бактериофагами и лечение бактериоцином). Lin также утверждает, что за исключением мер биологической безопасности другие способы вмешательства не доступны для потребителя и все еще находятся в стадии разработки.
[0005] Эрадикация указанных патогенов у домашнего скота может способствовать снижению частоты возникновения инфекции у человека и предотвращению распространения среди сельскохозяйственных животных. Вакцинация на фермах также может уменьшать риск заражения людей при еде или обращении с продуктами животного происхождения, а также заражению вследствие распространения бактерий через навоз. Лечение кампилобактериоза антибиотиками также становится нарастающей проблемой, поскольку устойчивость Campylobacter к антибиотикам, в прошлом эффективным при борьбе с ним, становится более распространенной.
[0006] В свое время гликозилирование считалось явлением, специфически присущим эукариотам, но позже было показано, что оно широко распространено среди простейших и бактерий. Бактериальные О- и N-связи образуются в более широком диапазоне сахаров, а не только в сахарах в составе гликопротеинов эукариот. Обычный путь гликозилирования для белков у бактерий впервые был продемонстрирован у С. jejuni. (Szymanski et al. Molecular Microbiology 32: 1022-1030, 1999). Механизмы гликозилирования С. jejuni были охарактеризованы и даже были успешно перенесены в клетки Е. coli (Wacker et al. Science, 298: 1790-1793, 2002), и было продемонстрировано активное N-гликозилирование белков (Young et al. J Biol Chem, 277: 42530-42539, 2002; Wacker et al. Science, 298: 1790-1793, 2002). Локус гена С. jejuni, называемый pgl (для гликозилирования белка), участвует в гликозилировании многих белков. Молчащие мутации в нем приводят к утрате иммуногенности многих белков, среди многих биологических фенотипов.
[0007] В Публикации заявки на патент США 2006/0165728 А1, в настоящее время патент США №7598354, включенной в настоящую заявку посредством ссылки, описаны специфические и крайне иммуногенные гептасахариды, которые присутствуют во многих периплазматических и поверхностных гликопротеинах С. jejuni. Указанный гептасахарид распространен по меньшей мере у нескольких видов Campylobacter и целого ряда штаммов, которые являются значимыми медицинскими и ветеринарными патогенами (Nothaft et al. Mol. Cell. Proteomics 11: 1203-1219, 2012). Указанный гептасахарид соответствует следующей формуле (I): GalNAc-α1,4-GalNAc-α1,4-[Glc-β-1,3]GalNAc-α1,4-GalNAc-α1,4-GalNAc-α1,3-diNAcBac, где diNAcBac (также называемый ди-N-ацетилбациллосамин) - 2,4-диацетамидо-2,4-,6-тридезокси-D-глюкопираноза, GalNAc - N-ацелилгалактозамин, a Glc - глюкоза. Данная группа гликана является компонентом многих гликопротеинов. У С. jejuni N-гликан важен для взаимодействия С. jejuni с клетками-хозяевами. Мутации в механизме гликозилирования приводят к уменьшению колонизации кишечника у мышей и куриц. У С. jejuni N-гликан важен для присоединения и инвазии в клетки эпителия человека (Szymanski et al. Infect Immun 70: 2242-2244, 2002), колонизации кишечника мышей и куриц (Kelly et al. J Bacteriol 188: 2427-2434, 2006; Szymanski et al. Infect Immun 70: 2242-2244, 2002; Hendrixson & DiRita, Mol Microbiol 52: 471-484, 2004; Karlyshev et al. Microbiology 150: 1957-1964, 2004), природной компетентности у штаммов с системой секреции IV типа (Larsen et al. J Bacteriol 186: 6508-6514, 2004) и для связывания с лектинами С-типа макрофагов человека, MGL (van Sorge et al, Cell Microbiol 11: 1768-1781, 2009). Кроме того, было показано, что поверхностные N-гликаны Campylobacter играют защитную роль в отношении протеаз в кишечнике куриц, что приводит к повышению приспособляемости бактерий (Alemka et al. Infect Immun 81: 1674-82, 2013).
[0008] В Публикации заявки на патент США 2012/0100177 описан штамм Salmonella enterica, содержащий по меньшей мере один оперон pgl С. jejuni или его функциональное производное, и предъявляющий по меньшей мере один N-гликан С. jejuni, или производное такого гликана, на поверхности своей клетки. Была выдвинута гипотеза, что такая модифицированная S. enterica может быть применима в составе вакцины против инфекций, вызываемых Campylobacter, в частности у домашнего скота, такого как птица. Однако к сожалению, в последующих публикациях было показано, что хотя S. enterica, экспрессирующая N-гликан из Campylobacter на своей поверхности, способна создавать колонии в организме куриц, не вызывая заболевания, у куриц, вакцинированных против N-гликана Campylobacter, не было определимого гуморального иммунного ответа (Thommen "Campylobacter N-glycan presenting Salmonella Typhimurium: a new vaccine for broiler chickens?" Zurich Open Repository and Archive, University of Zurich, Dissertation, Vetsuisse Faculty, 2011). Кроме того, у вакцинированных куриц при инфекции, вызванной С. jejuni, колонизация С. jejuni не уменьшалась.
[0009] Потребность в эффективной вакцине для профилактики и/или лечения инфекций, вызываемых Campylobacter у людей и животных, в частности у домашнего скота, и в частности у птицы, сохраняется.
[0010] Данная информация об исходном уровне техники приведена в целях ознакомления со сведениями, которые по мнению заявителя могут объяснить потенциальную значимость настоящего изобретения. Нет необходимости признавать и не следует истолковывать, что любая предшествующая информация являет собой предшествующий уровень техники в сравнении с настоящим изобретением.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0011] Задача настоящего изобретения состоит в том, чтобы обеспечить вакцину против Campylobacter. Согласно одному аспекту предложена композиция вакцины, содержащая бактерии, модифицированные таким образом, что они экспрессируют по меньшей мере один N-гликан бактерии Campylobacter или производное такого гликана на своей поверхности; и один или более физиологический растворитель, вспомогательное вещество, адъювант или носитель. Согласно определенным вариантам реализации указанным видом Campylobacter является С. jejuni. Указанной бактерией может быть Escherichia coli или Salmonella, и указанная модифицированная бактерия экспрессирует на своей поверхности гептасахарид С. jejuni.
[0012] Согласно определенным вариантам реализации указанная композиция содержит живые, модифицированные клетки E. coli, или живые ослабленные, инактивированные или убитые модифицированные клетки Е. coli. Указанная композиция может содержать суспензию модифицированных бактерий в подходящем буферном растворителе, таком как фосфатно-солевой буфер, и может быть выполнена в форме для перорального введения, введения in ovo, парентерального введения (например, путем инъекции или инфузии), или, например, распыления. Также указанная композиция вакцины может быть выполнена в форме для добавления в корм домашнему скоту, пищевые добавки ил и вводу домашнему скоту, и для введения птице, например, курицам.
[0013] Согласно другому аспекту предложен способ вакцинации животных против Campylobacter, способ, включающий введение животному композиции вакцины, описанной в настоящей заявке, содержащей бактерии, модифицированные таким образом, что они экспрессируют по меньшей мере один N-гликан бактерии Campylobacter или производное такого гликана на своей поверхности; и один или более физиологический растворитель, вспомогательное вещество, адъювант или носитель.
[0014] Известно, что экспрессия N-гликана у сальмонеллы не приводит к индукции защитного иммунного ответа. Неожиданно авторы настоящего изобретения показали, что Е. coli - которая является бактерией, очень похожей на сальмонеллу, и, как ожидалось, должна была дать похожие результаты - действительно вызывала индукцию защитного иммунного ответа у куриц, когда она экспрессировала N-гликан.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0015] Для лучшего понимания настоящего изобретения, а также других аспектов и дополнительных его особенностей, дается ссылка на следующее описание, которое следует применять в сочетании с сопровождающими его фигурами, где:
[0016] На Фигуре 1 показаны лизаты клеток Е. coli, из E. coli, мутантных по полимеразе, обработанные протеиназой K.
[0017] На Фигуре 2 показана структура липид-А-N-гликана и эксперимент на основе ЯМР (ядерно-магнитного резонанса) для компонента N-гликана липида-А Campylobacter jejuni.
[0018] На Фигуре 3 показан эксперимент FACS с мутантом Е. coli по полимеразе.
[0019] На Фигурах 4А, В, С и D показаны эксперименты по вакцинации и провокации, которые описаны в Примере 2, и
[0020] На Фигурах 5А и В показаны специфические антительные ответы куриц на IgY (IgG) N-гликан (ELISA).
ПОДРОБНОЕ ОПИСАНИЕ
[0021] Определение
[0022] Если не указано другое, все технические и научные термины, применяемые в настоящей заявке, имеют те же значения, которые обычно подразумевают специалисты в той области техники, к которой относится настоящее изобретение.
[0023] В настоящем описании и формуле изобретения формы единственного числа включают множественное число, если из контекста явно не следует другое.
[0024] В настоящей заявке под термином «содержащий» следует понимать, что нижеследующий перечень не исчерпывающий, и может включать, а может и не включать любые другие дополнительные соответствующие термины, например, при необходимости одно или более дополнительных свойств, компонентов и/или ингредиентов.
[0025] Термины «гликан C. jejuni», «гептасахарид C. jejuni», «N-гликан гептасахарид», «N-гликан Campylobacter» и «гептасахарид» применяются в настоящей заявке взаимозаменяемо и относятся к компоненту-гликану, который присутствует в большинстве поверхностных гликопротеидов и свободных олигосахаридов во многих штаммах и видах Campylobacter. Данный гликан в случае C. jejuni и который в качестве примера приведен в настоящей заявке, имеет формулу: GalNAc-α1,4-GalNAc-α1,4-[Glc-β-1,3]GalNAc-α1,4-GalNAc-α1,4-GalNAc-α1,3-diNAcBac, где diNAcBac - 2,4-диацетамидо-2,4,6-тридезокси-D-глюкопираноза. Указанные термины могут относиться к гликозилированию либо путем N-гликозилирования, либо с применением сахаров, полученных из пути N-гликозилирования или других путей. В недавней работе авторов настоящего изобретения было продемонстрировано, что N-гликан C. jejuni и свободные олигосахариды являются достаточно консервативными в ряду термофильных видов Campylobacter (Nothaft et al. Mol. Cell. Proteomics 11: 1203-1219, 2012), причем некоторые виды вырабатывают гексасахаридное производное указанного гептасахарида, не содержащее боковой глюкозы. Применение альтернативных структур N-гликанов и свободных олигосахаридов, описанных в (Nothaft et al. Mol. Cell. Proteomics 11: 1203-1219, 2012), которые присутствуют у нетермофильных видов Campylobacter, таких как описаны в Публикации РСТ WO/2011/097733, также подразумевается и включено в настоящее изобретение.
[0026] В настоящей заявке термин «антиген» относится к химическим или биологическим категориям, которые вызывают иммунную реакцию у животных или человека. В системе, описываемой в настоящей заявке, антиген содержит гептасахарид бактерии Campylobacter jejuni или их производное в форме N-гликана. В настоящей заявке термин «производное N-гликана» относится к производным гептасахарида, которые вызывают иммунный ответ у животного, аналогичный или более выраженный, чем ответ, вызываемый самим указанным гептасахаридом. N-гликан может быть конъюгирован с носителями, например, такими как белки (как описано в Заявке РСТ №WO 2012/027850) и липиды (Nothaft et al. Mol. Cell. Proteomics 11: 1203-1219, 2012, van Sorge et al. Cell Microbiol 11: 1768-1781, 2009), или более короткими или более длинными сахаридными повторами гептасахарида в полисахариде.
[0027] В настоящей заявке термин «вакцина» относится к композиции для повышения иммунитета у животных или человека к определенным микроорганизмам. Описываемые в настоящей заявке вакцины можно применять у целого ряда животных, например, таких как птицы, например, домашние птицы, а также млекопитающие. Микроорганизмы, на которые направлена вакцина, описываемая в настоящей заявке, являются микроорганизмами из рода Campylobacter.
[0028] В настоящей заявке термин «Campylobacter» относится к роду бактерий, содержащих любой и все виды из рода Campylobacter. Разные виды Campylobacter из указанного рода включают без ограничений С. jejuni, С. hominis, С. rectus, С. lari, С. fetus, С. coli, С. upsaliensis, С. fetus subsp. venerealis, С. fetus subsp. fetus, С. peloridis, С. lari subsp. concheus, C. sputorum, C. gracilis, C. showae, C. lanienae, C. curvus, C. helveticus, C. hyointestinalis subsp. hyointestinalis, С. hyointestinalis subsp. lawsonii, С. mucosalis, C. sputorum bv. paraureolyticus, C. sputorum bv. fecalis, C. ureolyticus, C. insulaenigrae, С. concisus, C. subantarcticus, C. avium, С. cuniculorum и С. volucris.
[0029] Согласно настоящей заявке предложен гликан, и его иммунологически активные фрагменты, которые можно применять в качестве вакцин против инфекции, вызываемой Campylobacter у людей и животных. Такие вакцины могут быть применимы при профилактике или нейтрализации инфекций, вызываемых Campylobacter, у домашнего скота, что позволяет предотвратить поступление данного патогена в пищевую цепь человека. Согласно определенным вариантам реализации гептасахарид С. jejuni и его фрагменты, необязательно связанные с аминокислотой, олигопептидом, липидом, или другой подходящий конъюгат можно применять в качестве вакцины. Например, данную вакцину можно применять для любого животного, которое инфицировано кампилобактерами, для которых N-гликан может быть экспрессирован на поверхности клетки Е. coli в форме гибрида центральной частью липида А.
[0030] Композиция вакцины
[0031] В настоящей заявке предложена композиция вакцины, содержащая модифицированную E. coli, которая была создана так, чтобы экспрессировать по меньшей мере один N-гликан Campylobacter, или его гептасахаридное производное, на своей поверхности. Указанная модифицированная Е. coli является живой, мертвой и/или ослабленной.
[0032] Как было описано выше, гептасахарид Campylobacter является характерным по меньшей мере для нескольких видов Campylobacter и целого ряда штаммов, включая виды, которые являются значимыми медицинскими и ветеринарными патогенами. Он является компонентом многих гликопротеинов, например, включая гликопротеины С. jejuni, именуемые, Cj0200c, Cj0289c, Cj0367c и другие. Было показано, что данный компонент гликана также обладает высокой иммуногенностью, и как и сам данный гликан (и близкие его производные и гликопептиды, содержащие N-гликан или его производные) является хорошим кандидатом для применения в качестве антигена в составе вакцины для иммунизации против многих штаммов и видов Campylobacter у млекопитающих, включая людей, у домашнего скота, включая куриц (Патент США №7598354).
[0033] Е. coli является грамотрицательной бактерией, у которой наружная мембрана покрыта липополисахаридом (ЛПС), который способствует структурной целостности бактерии и создает физический барьер, защищающий мембраны. ЛПС состоит из трех основных компонентов: центральная часть липида А и О-антиген. Липид А заякоривает ЛПС в наружной мембране, а О-антиген является самой удаленной от центра частью ЛПС. Сердцевина представляет собой разветвленный олигосахарид, который соединяет компоненты липида А и О-антигена в ЛПС.
[0034] Штаммом Е. coli, пригодным для получения композиции вакцины, является любой штамм, который можно в достаточной степени ослабить для непатологического введения человеку и/или животным в живой или мертвой форме. Можно применять другие бактерии, такие как Salmonella или другие штаммы Е. coli, которые могут обеспечить достаточную экспрессию и повышение иммунного ответа.
[0035] В настоящей заявке термин «оперон pgl» относится к любому физиологически активному кластеру гликозилирования генов Campylobacter, способному к гликозилированию гомологичных или гетерологичных структур, вырабатываемых штаммом Е. coli, применяемым в составе композиции вакцины. Оперон pgl у С. jejuni кодирует все ферменты, необходимые для синтеза N-гликана гептасахарида С. jejuni, его транспорта через внутреннюю мембрану и переноса к белкам. PglD, Е, F кодируют ферменты, участвующие в биосинтезе ди-N-ацетилбациллосамина, PglC переносит UDP-ди-N-ацетилбациллосамин на андекарпенилфосфат, a PglA, Н и J добавляет остатки GalNAc. Боковая группа Glc присоединяется ферментом PglI. Перенос через внутреннюю мембрану готового гептасахарида происходит благодаря действию PglK, и олигосахарилтрансфераза PglB переносит N-гликан на белок, а также высвобождает гептасахарид в периплазму в его свободной форме.
[0036] Функциональное производное оперона pgl представляет собой кластер генов, полученный из любого оперона pgl Campylobacter, содержащий делеции, мутации и/или замены нуклеотида(ов) или целых генов, но все еще способный производить олиго- или полисахарид, который может соединяться с гомологичными или гетерологичными структурами, вырабатываемыми штаммом E. coli, применяемым в составе композиции вакцины. Один или более оперонов pgl или их производных можно интегрировать в хромосому штамма E. coli, или его/их можно ввести в составе по меньшей мере одной плазмиды. Обычно интеграция в хромосому предпочтительна, поскольку данный вариант более стабилен по сравнению с векторами на основе плазмид, утрата которых может происходить во время культивирования. Следует отметить, что указанный штамм Е. coli может содержать более одного оперона pgl или его производного, производящих один или более N-гликанов или их производных. Согласно определенным вариантам реализации указанная композиция вакцины содержит штамм Е. coli, содержащим опероны pgl более чем одного типа, что приводит к экспрессии на поверхности модифицированной E. coli более чем одной структуры гликанов. Это может благоприятствовать стимуляции более разнонаправленного иммунного ответа у человека или животного в отношении разных видов Campylobacter. Согласно альтернативному варианту реализации указанная композиция вакцины содержит штамм E. coli, содержащий оперон pgl единственного типа, что приводит к экспрессии на поверхности модифицированной E. coli гликанов одной структуры. Это может благоприятствовать стимуляции специфического иммунного ответа у человека или животного в отношении одного вида Campylobacter.
[0037] Необязательно, уровень экспрессии гликана С. jejuni можно регулировать посредством применения разных промоторов или других регуляторных элементов, располагая их до оперона pgl, включая без ограничений промоторы генов рибосомальных белков, а также промоторы из генов, кодирующих устойчивость к антибиотикам, таких как bla, или похожие и предпочтительно сильные промоторы. Данный тип регуляции доступен для кодируемых плазмидой или интегрированных в хромосому оперонов pgl. Кроме того, стабильность плазмиды можно необязательно увеличивать путем включения жизненно-важных генов в плазмиду, при условии делеции указанных генов в геноме штамма Е. coli, применяемого в составе композиции вакцины.
[0038] Согласно альтернативному варианту реализации ген pglB оперона pgl инактивироан, что означает, что соответствующая олигосахаридтрансфераза В либо не экспрессируется, либо по меньшей мере инактивирована как фермент. Продукт гена pglB переносит N-гликан на специфический акцепторный сайт специфического полипептида, который также описан ниже, и высвобождает гептасахарид в его свободной форме. Инактивация трансферазы ведет к тому, что N-гликан или производное N-гликана связывается исключительно липидом А сердцевины акцептора О-антигена в Е. coli, и приводит к обмену GlcNAc на diN-ацетилбациллосамин, поскольку О-антиген-лигаза Е. coli распознает только GlcNAc-содержащие гликаны в сайте присоединения (т.е., на восстанавливающем конце).
[0039] Согласно близкому варианту реализации производное pgl представлет собой такое производное, в котором один или более генов биосинтеза diN-ацетилбациллосамина, pglD, Е, F и переноса инактивированы, а также инактивирован ген pglB. Данный вариант реализации приводит к обмену GlcNAc на diN-ацетилбациллосамин. Включение такого производного pgl в клетку Salmonella приводит к увеличенному клеточному предъявлению и к переносу модифицированного гептасахарида на сердцевину липида А, а не на акцепторы полипептидов (см., Публикацию заявки на патент США №2012/0100177).
[0040] Указанный по меньшей мере один N-гликан С. jejuni, или его производное в форме гептасахарида, может быть любым N-гликаном, вырабатываемым любым опероном pgl Campylobacter, или его функциональным производным, при условии, что указанный гликан иммуногенен в том смысле, что он вызывает иммунный ответ, специфичный для вида Campylobacter.
[0041] Согласно специфическому варианту реализации указанный гликан представляет собой гептасахарид, соответствующий формуле (I), которая описана выше, т.е. GalNAc-α1,4-GalNAc-α1,4-[Glc-β-1,3]GalNAc-α1,4-GalNAc-α1,4-GalNAc-α1,3-diNAcBac, где diNAcBac (также называемый ди-N-ацетилбациллосамин) - 2,4-диацетамидо-2,4,6-тридезокси-D-глюкопираноза.
[0042] Альтернативным вариантом реализации, при котором оперон pgl, где гены для биосинтеза ди-N-ацетилбациллосамина инактивированы или в значительной степени или полностью делетированы, приводит к синтезу производного гептасахарида, соответствующего формуле (II), является GalNAc-α1,4-GalNAc-α1,4-[Glc-β-1,3]GalNAc-α1,4-GalNAc-α1,4-GalNAc-α1,3-GlcNAc.
[0043] Согласно определенному варианту реализации указанный N-гликан(ы) или производное(ые), полученные по меньшей мере с одного оперона pgl, или его производного, может быть связан по меньшей мере с одним гомологичным или гетерологичным полипептидом E. coli, который в итоге будет перенесен и предъявлен на поверхности клетки. Полипептид, связанный с N-гликаном (производным) может быть полипептидом любого типа, например, чистым полипептидом (только аминокислоты) или полипептидом с пострансляционными модификациями, например полипептид, связанный с липидом.
[0044] Согласно другому варианту реализации указанный гликан(ы) или его производное(ые) выделены из клеток природного хозяина, находятся в своей свободной олигосахаридной форме, а затем их химически конъюгируют с полипептидом или липидом-носителем.
[0045] Согласно специфическому варианту реализации по меньшей мере один гликан или его производное, полученные по меньшей мере с одного оперона pgl, или его производного, связаны с центральной частью липида A E. coli или его функционально эквивалентным производным. Центральная часть липида А Е. coli представляет собой олигосахаридную структуру, состоящую без ограничений из гексоз, гептоз и KDO (3-дезокси-D-маннооктулозоновой кислоты), соединенных двумя глюкозаминами с ацильными цепями, заякоривающими данную структуру в наружной мембране бактерии. Функционально эквивалентное производное центральной части липида А представляет собой производное, которое способно принимать один или более гликанов или их производных и предъявлять их на поверхности клетки. Следует отметить, что в данном случае указанный гептасахарид или его производное не является N-связанным, поскольку структура центральной части липида А E. coli не является полипептидом.
[0046] Необязательно по меньшей мере один гептасахарид или его производное заменяют собой боковые цепи О-антигена в ЛПС (липополисахариде). Внутренняя и наружная сторона центральной части липида A E. coli остаются неизменными, хотя биосинтез О-антигена устранятся посредством мутации, например, мутации wzy и/или других мутаций. Согласно определенным вариантам реализации по меньшей мере один гептасахарид, его производное или смесь их обоих экспрессируются одновременно с боковыми цепями О-антигена в ЛПС, что приводит к образованию гетерогенного ЛПС, содержащего и О-антиген, и указанный гептасахарид.
[0047] Предпочтительно, а для медицинских приложений весьма важно, чтобы штамм E. coli согласно настоящему изобретению не оказывал патогенного действия при его введении животному или человеку в живой и/или инактивированной форме. Специалисту в данной области техники известно много способов ослабления вирулентных видов E. coli посредством мутации. Например, мутации, которые ослабляют патогенную E. coli (1) мутант CarAB патогенной Escherichia coli O2 птиц ослаблен и пригоден в качестве живой пероральной вакцины против колибациллеза у индеек (Kwaga et al. Infect Immun. 62: 3766-3772, 1994); (2) мутация РНК-шаперона Hfq значительно снижает патогенность VTEC, ЕАЕС и UPEC в модели на нематоде (Bojer et al. Microbes Infect 14: 1034-1039, 2012); (3) мутации генов в фосфат-специфичной транспортной системе (Pst) ослабляют штаммы Е. coli (Buckles et al. Microbiology 152: 153-160, 2006; Daigle et al. Infect and Immun. 63: 4924-4927, 1995).
[0048] Согласно одному конкретному варианту реализации штамм Е. coli применяемый в композиции вакцины, ослабляют посредством частичной или полной инактивации экспрессии О-антигена, например, посредством мутации в гене wzy (приводящей к получению мутанта по полимеразе О-антигена) (Baba et al. Mol. Syst. Biol. 2: 2006).
[0049] Описанные выше штаммы Е. coli обладают высокой иммуногенностью и вызывают иммунный ответ, направленный против инфекции, вызванной Campylobacter, такого как С. jejuni. Кроме того, будучи полученными, они легко размножаются и нарабатываются в большом количестве. Их можно вводить в виде мертвых или живых вакцин, причем живые вакцины позволяют продолжить размножение и поддерживать длительный иммунный стимул в организме хозяина, а также получать полные иммунные ответы с адъювантом или без него.
[0050] Следовательно, настоящее изобретение также связано с медицинским применением живых или мертвых штаммов Е. coli, модифицированных так, чтобы они предъявляли один или более N-гликанов Camplyobacter или их производных, на своей поверхности, в частности для получения лекарственного препарата, предпочтительно вакцины.
[0051] Предпочтительно указанный лекарственный препарат применим при профилактике и/или лечении инфекции и/или колонизации, вызванной С. jejuni, предпочтительно у домашнего скота, более предпочтительно у крупного рогатого кота и птицы, более предпочтительно у птицы, такой как курицы, индюки, гуси и утки.
[0052] Согласно одному аспекту настоящего изобретения предложена композиция вакцины, которая является фармацевтической композицией, пищевой или кормовой добавкой, содержащими мертвую или живую Е. coli, модифицированную так, чтобы она предъявляла один или более N-гликанов Camplyobacter или их производных, на своей поверхности, и физиологически приемлемое вспомогательное вещество, растворитель или носитель. Необязательно указанная композиция вакцины включает дополнительные компоненты или вводится с дополнительными компонентами, такими как, например, адъювант. Согласно другому альтернативному аспекту композиция вакцины, описываемая в настоящей заявке, выполнена в форме для введения с другой композицией вакцины.
[0053] Адъюванты, как правило, содержат вещества, которые стимулируют иммунный ответ хозяина неспецифическим образом. В технике известно множество разных адъювантов. Примеры адъювантов включают полный и неполный адъювант Фрейнда, витамин Е, неионные блок-полимеры и полиамины, такие как сульфат декстрана, карбопол и пиран. Также подходят поверхностно-активные вещества, такие как Span, Tween, гексадециламин, лизолецитин, метоксигексадецилглицрин и сапонины (например, Quil А®). Кроме того, часто применяют пептиды, такие как мурамилпептиды, диметилглицин и тафцин. Наряду с перечисленными адъювантами можно с успехом применять иммуностимулирующие комплексы (ISCOMS), минеральное масло, например, Bayol® или Markol®, растительные масла или эмульсии и Diluvac® Forte.
[0054] Необязательно вакцины смешивают с одним или более стабилизаторами, например, для защиты склонных к распаду компонентов от разложения, чтобы увеличить срок хранения вакцины, или чтобы повысить эффективность лиофилизации. Применимые стабилизаторы включают, например, SPGA (Bovarnik et al. J. Bacteriology 59: 509, 1950), обезжиренное молоко, желатин, бычий сывороточный альбумин, углеводы, например, сорбитол, маннитол, трегалозу, крахмал, сахарозу, декстран или глюкозу, белки, такие как альбумин или казеин, или продукты их распада, и буферы, такие как фосфаты щелочных металлов.
[0055] Композиция вакцины может быть, например, в форме раствора, суспензии или лиофилизированной композиции, подходящей для восстановления перед введением.
[0056] Лиофилизация представляет собой эффективный способ сохранения композиции вакцины. Лиофилизированный материал можно сохранить стабильным в течение многих лет. Температуры хранения для лиофилизированного материала может быть значительно выше нуля градусов, что не наносит вреда материалу. Лиофилизацию можно проводить в соответствии с хорошо известными стандартными процедурами лиофилизации.
[0057] Способ иммунизации
[0058] В настоящей заявке предложен способ иммунизации животного от инфекции, вызываемой Campylobacter. Указанный способ включает этап введения животному композиции вакцины, содержащей E. coli, которая была сконструирована так, чтобы она предъявляла по меньшей мере один N-гликан Camplyobacter или его производное, на своей поверхности, как описано выше.
[0059] Кампилобактериоз представляет собой заболевание, вызываемое Campylobacter. Наиболее частыми симптомами являются диарея, боль в животе, лихорадка, головная боль, тошнота и/или рвота. Обычно данные симптомы сохраняются приблизительно только три-шесть дней. Однако в редких случаях инфекция, вызываемая Campylobacter, может вызывать длительные осложнения, такие как, например, синдром Джуллиана-Барр (GBS), артрит и бактериемию. Следовательно, вакцины, описываемые в настоящей заявке, применимы при иммунизации животных, включая человека и домашний скот, такой как курицы, которые являются основной причиной заболеваний, распространяющихся через пищеварительный тракт, у человека. Следовательно, в настоящей заявке также предложен способ профилактики или минимизации эффекта заболевания или расстройства, вызванного Campylobacter. Согласно особым вариантам реализации заболеванием или расстройством, вызванным Campylobacter, является кампилобактериоз, синдром Джуллиана-Барр (GBS) и/или артрит и/или бактериемия, хотя другие виды Campylobacter связаны с другими патологическими состояниями, такими как периодонтит и выкидыши.
[0060] Согласно варианту реализации, при котором композиция вакцины применяется для вакцинации домашнего скота, есть разные пути введения указанной композиции, которые могут быть удобны для массовой вакцинации. Введение можно проводить с питьевой водой, пищей или кормом, при помощи спрея/распылителя (например, курицам в возрасте одного дня в транспортных контейнерах, или животным в закрытой среде, например, в птичниках), в виде глазных капель, путем прокалывания или скарификации (кожный путь в перепонку крыла или стопу), путем инъекции (например, внутримышечной или подкожной) или введения in-ovo.
[0061] Необязательно животным вводят начальную дозу композиции вакцины, а затем одну или более бустер-доз с соответствующими интервалами времени. Специалист в данной области техники может легко определить объем дозы и режим дозирования, подходящие для конкретного приложения. В данном примере мы проводили начальную вакцинацию птицам в возрасте 1 неделя, вводя им 1×108 живых или фиксированных в формалине клеток Е. coli (или с любой другой гликоконъюгированной вакциной, т.е., N-гликан С. jejuni, связанный с ТохС). Мы вводили одну бустер-дозу с тем же количеством бактериальных клеток (или белка) через две недели. Провокация кампилобактером обычно проводилась еще через неделю, и птиц подвергали эвтаназии через 1 неделю после провокации (как описано ниже).
[0062] Чтобы улучшить понимание изобретения, описываемого в настоящей заявке, приведены следующие примеры. Следует понимать, что данные примеры приведены только в целях иллюстрации. Следовательно, они не должны ни коим образом ограничивать область настоящего изобретения.
ПРИМЕРЫ
[0063] Пример 1: Получение вакцины
[0064] N-гликан Campylobacter jejuni, присоединенный к белку
[0065] Экспрессия и очистка белка ToxC-GT, гликозилированного N-гликаном С.
jejuni: Белок ToxC-GT, гликозилированный N-гликаном С. jejuni экспрессировали в клетках E. coli BL21, экспрессирующих оперон pgl С. jejuni, и очищали посредством хроматографии с Ni-NTA (нитрилотрехуксусной кислотой), как описано в опубликованной заявке РСТ на международный патент №WO 2012/027850. Белок подвергали дальнейшей очистке при помощи ионообменной хроматографии с применением системы AEKTA FPLC, снабженной анионообменной колонки 2,5 мл MonoQ. Подвижной фазой был буфер 50 мМ Tris-HCl, рН 8,0 с градиентом NaCl в пределах 0-500 мМ NaCl в 30 объемах колонки. Фракции, содержащие глюкоконъюгат, анализировали в 12,5% полиакриламидном геле с додецил-сульфатом натрия (SDS PAGE), два раза пропускали через 1 г абсорбента для удаления липида (LRA, «Supelco»), проводили диализ с обменом на ФСБ и перед применением устанавливали концентрацию белка 0,5 мг/мл. Концентрацию белка определяли с применением стандартных способов (тест Бредфорда) с применением повышенных концентраций БСА в ФСБ для получения стандартной кривой.
[0066] N-гликан Campylobacter jejuni, присоединенный к структуре центральной части липида A E. coli: получение вакцины на основе E. coli
[0067] Клетки Е. coli, экспрессирующие гептасахарид С. jejuni, были описаны ранее (Nothaft et al. Molecular and Cellular Proteomics 11: 1203-1219). Клетки выращивали в жидком бульоне (2 × бульон YT (бульон из дрожжевого экстракта и триптона) при 37°С и интенсивном взбалтывании (220 об/мин) до достижения стационарной фазы. Клетки отбирали посредством центрифугирования и дважды промывали стерильным ФСБ. Количество клеток определяли посредством разлива в планшеты последовательных разведений суспензии клеток до оптической плотности OD600, равной 2,0, и применяли либо в таком виде, либо фиксированными в формалине.
[0068] Клетки Е. coli из ночной культуры, экспрессирующие локус pgl С. jejuni, отбирали посредством центрифугирования и дважды промывали стерильным ФСБ, как описано в Nothaft et al. Mol. Cell. Proteomics 11: 1203-1219, 2012. Затем 1 мл клеток, OD600 для которых при помощи стерильного ФСБ была доведена до 1,0, центрифугировали и повторно суспендировали в 100 мкл 1-кратного буфера Лэммли и нагревали в течение 10 мин до 95°С. Добавляли протеиназу до конечной концентрации 200 мкл/мл, и пробу инкубировали при 60°С в течение 1 ч, после чего инкубировали в течение 5 мин на льду и центрифугировали в течение 15 мин. Аликвоты надосадочной жидкости разделяли при помощи стандартного 12,5% SDS-PAGE. Гепатсахарид, присоединенный к структуре центральной части липида А, визуализировали при помощи Вестерн-блоттинга, как было описано (Nothaft et al. Molecular and Cellular Proteomics 11: 1203-1219), с применением N-гликан Campylobacter jejuni-специфической антисыворотки hR6, в качестве основного антитела, и антитела на белок кролика, конъюгированный со щелочной фосфатазы, в качестве дополнительного антитела (Фигура 1). На первой дорожке показаны обработанные протеиназой K лизаты клеток из мутанта Е. coli по О-антиген-полимеразе, экспрессирующего оперон гликозилирования белка C. jejuni с активным геном pglB (pACYC184pglBmut). Образование гибрида липид A-N-гликан отмечено стрелкой. На дорожке 2 показаны обработанные протеиназой K лизаты контрольных клеток из мутанта Е. coli по О-антиген-полимеразе с пустым вектором (pACYC184). Маркеры молекулярной массы (MW в килодальтонах, кДа) показаны слева. Полосы с более высокой молекулярной массой представляют собой компоненты Е. coli, которые в обоих препаратах проявляли перекрестную реактивность.
[0069] Ядерная магнитно-резонансная спектроскопия (ЯМР) очищенного компонента липид A-N-гликан. Гликолипиды получали из восьми литров культуры с OD600=1.0 мутанта Е. coli по О-антиген-полимеразе, экспрессирующего оперон гликозилирования белка C. jejuni с активным геном pglB (pACYC184pglBmut). ЛПС экстрагировали при смеси фенол-вода, проводили диализ, обрабатывали АсОН, чтобы осадить нуклеиновые кислоты, проводили диализ, высушивали, гидролизовали в 2% АсОН и разделяли на биогеле Р6. Фракции анализировали посредством ЯМР. Фракции, которые содержали сигналы от N-гликана С. jejuni, объединяли и разделяли на анионообменной колонке Hitrap с градиентом NaCl. Фракции анализировали посредством ЯМР. Фракции, которые содержали сигналы от N-гликана C. jejuni, подвергали обессоливанию посредством хроматографии на сефадексе G-15. Присоединение подтверждали при помощи спектроскопия ядерного эффекта Оверхаузера (NOESY) и гетероциклической корреляционной спектроскопии нескольких связей (НМВС). Можно было наблюдать специфические химические сдвиги для N-гликана С. jejuni, все 1-4-связи компонентов N-гликана С. jejuni давали ядерное усиление Оверхаузера 1:4 и 1:6, и заданные значения хорошо согласовывались с ранее опубликованными данными (Фигура 2 и Таблица 1, и Nothaft et al. Molecular and Cellular Proteomics 11: 1203-1219, 2012). Производное N-гликана С. jejuni с GlcNAc вместо diNAcBac в качестве восстановления конечного сахара (Фигура 2) присоединяли через O-7 L-глицеро-D-манногептозы центральной части липида А. Все сигналы от Hep (L) были обнаружены посредством анализа основного множества корреляций, и заданные значения для центральной части липида A E. coli согласовывались с опубликованными данными (Muller-Loennies et al. Journal of Biological Chemistry 278:, 34090-34101, 2003, и Таблица 1).

[0070] Анализ посредством флуоресцентной сортировки клеток (FACS). Сначала 1 мл клеток E. coli с OD600=1.0 осаждали посредством центрифугирования и суспендировали в 1 мл блокирующего раствора (ФСБ, 5% обезжиренного молока). Клетки исследовали с применением N-гликан С. jejuni-специфической антисыворотки hR6 и конъюгированной с Алекса-флор-546 антисывороткой на белок кролика, и анализировали посредством FACS. Данные FACS обрабатывали при помощи программы FACS Diva. Для идентификации и разблокировки интактных клеток применяли контрастную окраску DAPI (4',6-диамидин-2-фенилиндол). Анализ популяции из 2×104 клеток показал значительное увеличение флуоресценции для клеток E. coli, экспрессирующих N-гликан С. jejuni, по сравнению с контрольными клетками E. coli с пустым вектором (pACYC184), что подтверждает, что N-гликан C. jejuni присутствует на поверхности клеток (фигура 3). Появление пика и геометрия пика показывает, что каждая клетка E. coli предъявляла сопоставимое количество N-гликана С. jejuni на своей поверхности.
[0071] Пример 2: Вакцинация и провокация
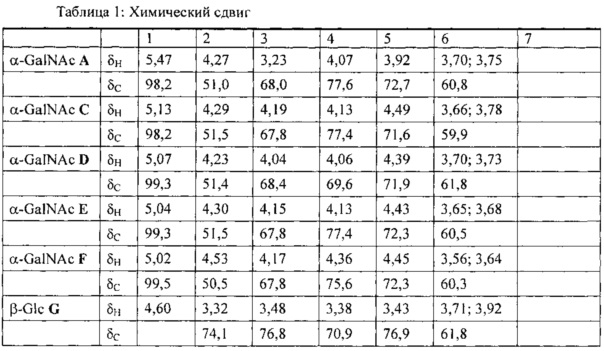
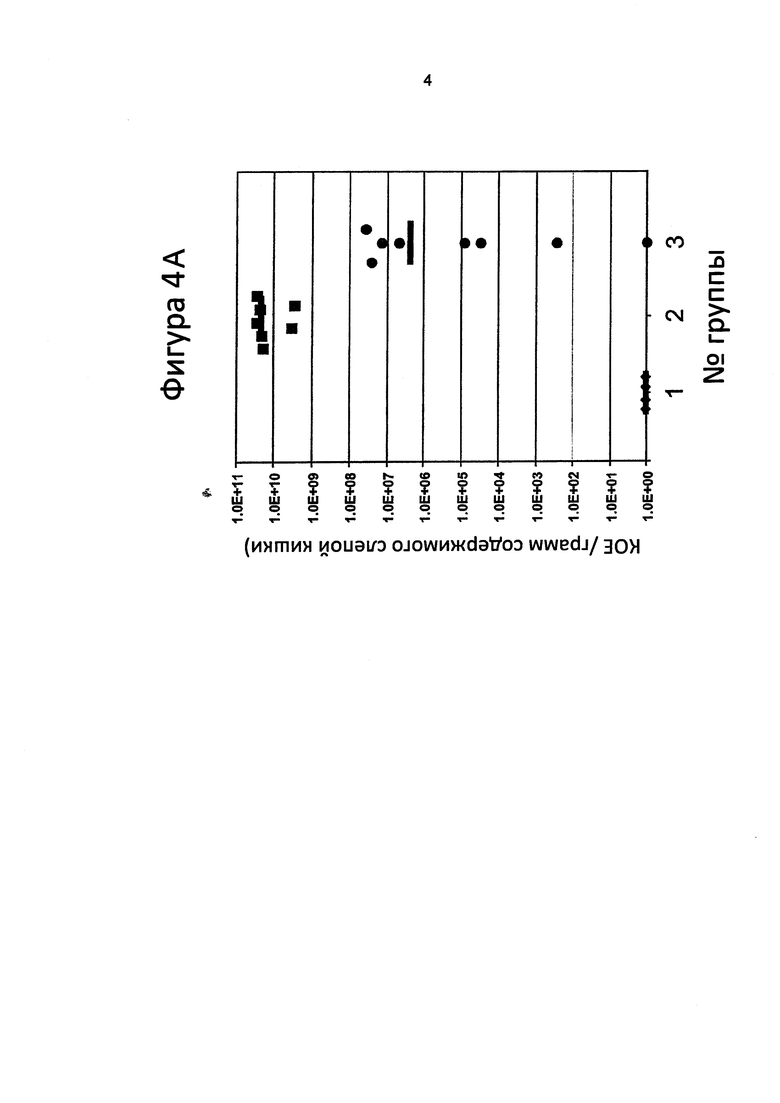
[0072] Введение курицам гликоконъюгата ToxC-GT посредством инъекций (Фигура 4D) или посредством перорального введения (Фигура 4А), а также живых штаммов (Фигура 4В и С) модифицированной E. coli, описанной в примере 1, приводило к значительному снижению содержания Campylobacter в слепой кишке куриц, подвергаемых провокации. Было проведено три эксперимента с вакцинацией куриц с применением Е. coli, экспрессирующей гептасахарид С. jejuni, чтобы продемонстрировать повышение иммунитета куриц против Campylobacter. Результаты указанных экспериментов показаны на Фигурах 4А, В и С.
[0073] В первом эксперименте с вакцинацией куриц провокацию проводили в о трех группах куриц. Контрольная группа с ФСБ включала четырех куриц, а в группе 2 и 3 было по восемь куриц. Условия для групп 1 и 2 приведены в таблице 2. Группе 3 через желудочный зонд вводили мертвые клетки E. coli, экспрессирующие на поверхности гептасахарид N-гликан C. jejuni, в дни 7 и 21. Впоследствии птиц повергали провокации следующим образом: группа 1 (контроль) получали через желудочный зонд 300 мкл ФСБ; группы 2 и 3 получали через желудочный зонд 300 мкл ФСБ, содержащие 102 клеток С. jejuni 81-176. В день 35 куриц повергали эвтаназии и определяли уровень колонизации посредством посева последовательных разведений содержимого слепой кишки каждой птицы на селективный агар Кармали. После инкубации чашек Петри в течение 48 часов при микроаэробных условиях определяли число колониеобразующих единиц (КОЕ). Результаты графически показаны на Фигуре 4А, и уровни колонизации показаны как КОЕ на грамм содержимого слепой кишки. Горизонтальные полоски соответствуют медиане для каждой группы. Более конкретно, результаты показывают, что вакцина на основе N-гликана уменьшает колонизацию Campylobacter у куриц. На чашках группы 1 не было обнаружено колониеобразующих единиц (контроль с ФСБ), тогда как у птиц из группы 2 была выявлена колонизация со средним значением 1010 клеток кампилобактера на грамм содержимого слепой кишки. В группе 3 колонизация уменьшалась приблизительно на 4 логарифмических единицы.
[0074] Во втором эксперименте с вакцинацией куриц провокацию проводили в отношении трех групп куриц. Группы 1 и 2 включали по 6 куриц, группа 3 включала 8 куриц. Условия для групп 1 и 2 приведены в таблице 2. Группе 3 через желудочный зонд вводили живые клетки E. coli, экспрессирующие гептасахарид N-гликан С. jejuni, в дни 7 и 21. Концентрации материала для провокации и уровень колонизации определяли, как описано в первом эксперименте. Результаты графически показаны на фигуре 4В, и уровни колонизации показаны как КОЕ на грамм содержимого слепой кишки. Горизонтальные полоски соответствуют медиане для каждой группы. Результаты показывают, что вакцина на основе N-гликана уменьшает колонизацию Campylobacter у куриц. На чашках группы 1 не было обнаружено колониеобразующих единиц (контроль с ФСБ), тогда как у птиц из группы 2 была выявлена колонизация со средним значением 1010 клеток Campylobacter на грамм содержимого слепой кишки. В группе 3 ни у одной из птиц колонизация была на неопределимом уровне.
[0075] В третьем эксперименте, показанном на Фигуре 4С, провокацию проводили в отношении 4 групп куриц. Группа 1 включала 6 куриц, группы 2, 3 и 4 включали по 8 куриц. Условия для групп 1 и 2 приведены в таблице 2. Группам 3 и 4 через желудочный зонд вводили вакцины в дни 7 и 21. Птицы из 3-ей группы получали живые клетки E. coli, не экспрессирующие на поверхности N-гликан, а птицы группы 4 получали живые клетки E. coli, экспрессирующие на своей поверхности гептасахарид N-гликан С. jejuni. Результаты графически показаны на фигуре 4С, и уровни колонизации показаны как КОЕ на грамм содержимого слепой кишки. Горизонтальные полоски соответствуют медиане для каждой группы. Результаты показывают, что вакцина на основе N-гликана воспроизводимо уменьшает колонизацию Campylobacter у куриц, и что клетки E. coli, не экспрессирующие N-гликан, не обладают пробиотическим эффектом, поскольку уровень колонизации Campylobacter после провокации был сходным с уровнем у птиц группы 2.
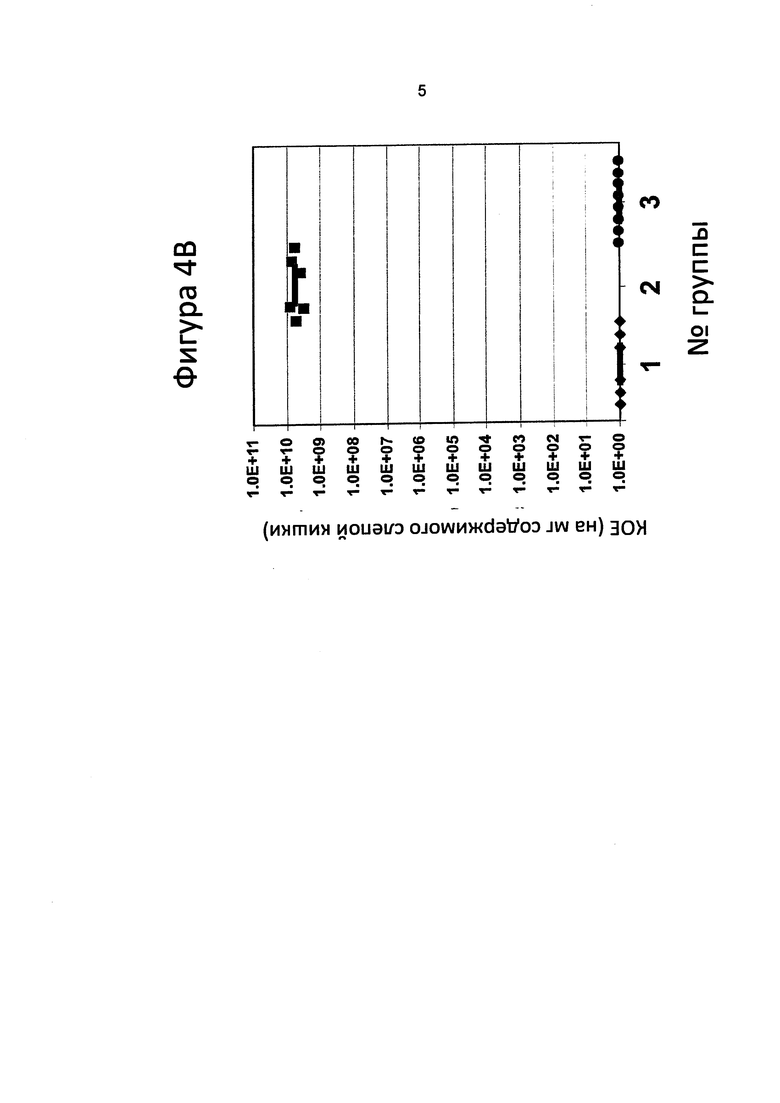
[0076] В четвертом эксперименте, который показан на Фигуре 4D, провокацию проводили в отношении шести групп куриц, в каждой из которых было по восемь куриц. Условия для каждой группы приведены в таблице 2.

[0077] Аналогично предыдущим экспериментам в день 1 у 10% птиц отбирали клоакальные мазки (5 случайно выбранных птиц) и завевали на селективный агар Кармали, чтобы подтвердить, что у птиц нет колонизации С. jejuni. После 48-часовой инкубации при микроаэробных условиях при 37°С колоний Campylobacter не наблюдалось.
[0078] Аналогично предыдущим экспериментам в день 7 у каждой птицы отбирали до 50 мкл крови (до иммунизации). Сыворотку готовили следующим образом: после хранения проб крови при 37°С в течение 1 ч после центрифугирования (5 мин, 18000xg, 4°С) надосадочную жидкость (сыворотку) переносили в новую пробирку и добавляли глицерин до конечной концентрации 10%. Сыворотку хранили при -20°С до дальнейшего применения. Последующее противомикробное лечение проводили следующим образом: Группа 1 (контроль ФСБ) и 2 (контроль колонизации) получали 300 мкл ФСБ с полным адъювантом Фрейнда в день 7 и такое же количество неполного адъюванта Фрейнда в день 21 (150 мкл ФСБ+150 мкл адъюванта), путем инъекции в два участка в грудную клетку со 150 мкл состава для вакцинации (без гликокнъюгата) на один участок. Группа 3 не получала антиген в день 7, но получала 1 дозу ToxC-GT, гликозилированного N-гликаном C. jejuni (100 мкг белка в 150 мкл ФСБ+150 мкл полного адъюванта Фрейнда) в день 21; группа 4 получала 1 дозу ToxC-GT, гликозилированного N-гликаном С. jejuni, с полным адъювантом Фрейнда в день 7 и не получала антиген в день 21. Группа 5 получала 2 дозы (в день 7 с полным адъювантом Фрейнда и в день 21 с неполным адъювантом Фрейнда) 100 мкг ToxC-GT, гликозилированного N-гликаном C. jejuni, в форме инъекции в заднюю конечность (150 мкл состава для вакцинации в каждую нижнюю конечность), а группа 6 получала 2 дозы через желудочный зонд (в день 7 и день 21) 300 мкл ФСБ, содержащих 108 живых клеток Е. coli, экспрессирующих на своей поверхности N-гликан C. jejuni.
[0079] Аналогично предыдущим экспериментам в день 28 у каждой птицы из групп 1-6 отбирали кровь (перед иммунизацией), и сыворотку готовили и хранили, как было описано выше. Впоследствии птиц подвергали провокации следующим образом: группа 1 (отрицательный контроль) перорально через зонд получала 300 мкл ФСБ; группы 2-6 перорально через зонд получали 300 мкл ФСБ, содержащие 102 клеток С. jejuni 81-176. В день 34 куриц подвергали эвтаназии, отбирали кровь (окончательный отбор крови) путем прокола сердца, сыворотку готовили и хранили, как было описано выше. Уровень колонизации определяли путем посева последовательных разведений содержимого слепой кишки каждой птицы на селективном агаре Кармали. Колониеобразующие единицы определяли после инкубации на чашках Петри в течение 48 часов при микроаэробных условиях.
[0080] Результаты графически показаны на фигуре 4D, и уровни колонизации показаны как КОЕ на грамм содержимого слепой кишки. Горизонтальные полоски соответствуют медиане для каждой группы. Более конкретно, результаты показывают, что вакцина на основе N-гликана уменьшает колонизацию Campylobacter у куриц, и что вакцины, содержащие живые клетки E. coli, экспрессирующие на своей поверхности N-гликан C. jejuni, у куриц проявляют себя лучше, чем вакцины на основе гликопротеина (см. ниже).
[0081] На чашках группы 1 не было обнаружено колониеобразующих единиц (контроль с ФСБ), тогда как у птиц из группы 2 была выявлена колонизация со средним значением 1010 клеток кампилобактера на грамм содержимого слепой кишки. Колонизация уменьшалась у группы 3, группы 4 и группы 5, где среднее количество КОЕ составило 2.2×104, 6.8×105 и 5.5×104 на грамм содержимого слепой кишки, соответственно. Колонизация в группе 6 была почти устранена, и среднее количество КОЕ составило 100 на грамм содержимого слепой кишки. Кроме того, 5 из 8 птиц не продемонстрировали никаких признаков колонизации С. jejuni. Это явно указывает на то, что лечение вакциной на основе белка и N-гликана С. jejuni приводит к уменьшению колонизации после провокации кампилобактером, независимо от времени проведения инъекции участка введения вакцины, и что пероральная вакцинация клетками E. coli, которые экспрессируют на своей поверхности гептасахарид, почти полностью упраздняла колонизацию кампилобактера. Кроме того, было продемонстрировано самоограничение штамма Е. coli, применяемого в составе вакцины, поскольку Е. coli обнаруживалась в содержимом слепой кишки данной группы, когда его засевали на селективную среду LB Kan-Cm. Во всех экспериментах наблюдалось устранение вакцинного штамма живой Е. coli из организма куриц.
[0082] Для анализа N-гликан-специфического иммунного ответа, более конкретно, IgY (IgG) N-гликан-специфического антительного ответа у куриц, проводили исследование на основе ELISA. Свободный олигосахарид (fOS) из С. jejuni готовили, как было описано ранее (Dwivedi et al. Biopolymers, 99: 772-7830, 2013) и соединяли с БСА посредством восстановительного аминирования, как было описано ранее (Nothaft et al. Mol. Cell. Proteomics 11: 1203-1219, 2012). Образование конъюгата БСА- Cj-N-гликана подтверждали посредством Вестерн-блоттинга с применением антисыворотки R1-4. После корректировки концентрации до 1 мг/мл при помощи ФСБ гликоконъюгат хранили при 4°С до дальнейшего применения. В десять планшетов на 96 лунок Maxisorb наносили 500 нг конъюгата БСА-Cj-N-гликана и оставляли на ночь при 4°С. После удаления несвязанного антигена планшет блокировали в течение 1 ч при комнатной температуре путем добавления 100 мкл ФСБ-Т, 5% обезжиренного молока при взбалтывании. После слива блокирующего раствора добавляли 100 мкл растворов антитела и инкубировали в течение 1, как описано выше. Растворы антитела содержали N-гликан-специфическую антисыворотку, разведенную в отношении 1:3000 в ФСБ-Т, 1% обезжиренного молока или сыворотки куриц (приготовленной из пробы крови из 2-го отбора (т.е. день 28) из экспериментов по вакцинации) и разводили в соотношении 1:50 в ФСБ-Т, 1% обезжиренного молока. Планшеты инкубировали в течение 1 ч при комнатной температуре, как было описано, и каждую лунку промывали 3 раза в течение 5 мин 100 мкл ФСБ-Т. После добавления 100 мкл раствора дополнительного антитела (либо на бело кролика АР (1:50) для контроля R1-4, либо на IgY цыпленка (1:500) для экспериментальных проб) инкубировали в течение 1 ч при комнатной температуре, затем дополнительное антитело удаляли и лунки промывали 4 раза в течение 5 мин 100 мкл ФСБ-Т. После последнего этапа отмывки оставшийся раствор для отмывки полностью удаляли из каждой лунки, и в планшет в качестве субстрата добавляли PNPP (р-нитрофенилфосфат). Иммунореактивность каждой сыворотки определяли после сканирования планшета при OD405 на планшетном ридере.
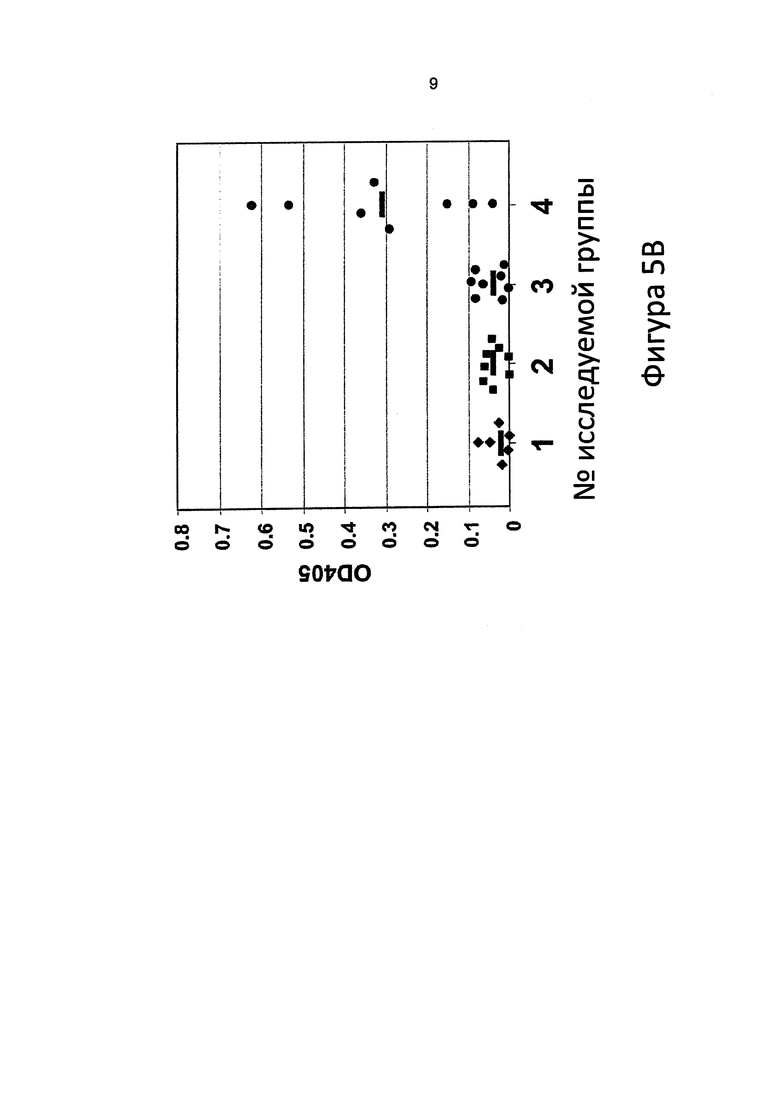
[0083] С. jejuni N-гликан-специфические антитела присутствовали (Фигура 5А) в сыворотке, приготовленной из крови, отобранной за 28 дней до провокации Campylobacter, у птиц, вакцинированных 1 дозой ToxC-GT с гликаном в день 21 (грудная клетка, в/м) (исследуемая группа 3), 2 дозами ToxC-GT с гликаном в день 7 и 21 (грудная клетка, в/м) (исследуемая группа 4), 2 дозами ToxC-GT с гликаном в день 7 и 21 (задняя конечность, в/м) (исследуемая группа 5) и 2 мертвых клеток E. coli (исследуемая группа 6). Антительный ответ (выражаемый как OD405) был максимальным в исследуемой группе 6, и большинство куриц в данной исследуемой группе не демонстрировали колонизации. Антительный ответ в группах положительного и отрицательного контроля колонизации (исследуемая группа 1 и 2) был ниже предела определения. Горизонтальные полоски показывают медиану для каждой группы.
[0084] С. jejuni N-гликан-специфические антитела присутствовали (Фигура 5В) в сыворотке, приготовленной из крови, отобранной за 28 дней до провокации Campylobacter, у птиц, вакцинированных живой E. coli, предъявляющей N-гликан на своей поверхности (исследуемая группа 4). Это соответствует эксперименту по вакцинации №3, показанному на Фигуре 4С. Антительный ответ (выражаемый как OD405) у птиц, вакцинированных живой E. coli, без гликана, (исследуемая группа 3) и из групп положительного и отрицательного контроля колонизации (исследуемая группа 1 и 2) был ниже предела определения. Горизонтальные полоски показывают медиану для каждой группы.
[0085] Наблюдалось следующее:
1) добавление в корм курицам мертвых E. coli, экспрессирующих гептасахарид С. jejuni, с последующей провокацией С. jejuni приводило к уменьшению колонизации С. jejuni в кишечнике куриц приблизительно на 4 логарифмических единицы (Фигура 4А); и
2) добавление в корм курицам живых E. coli, экспрессирующих гептасахарид C. jejuni, с последующей провокацией С. jejuni последовательно вызывало уменьшение колонизации С. jejuni в кишечнике куриц более чем на 7 логарифмических единиц (Фигура 4В, С и D).
[0086] Очевидно, что вакцины, содержащие мертвые или живые клетки Е. coli, экспрессирующие гептасахарид С. jejuni, способны значительно усиливать иммунитет куриц в отношении провокации C. jejuni.
[0087] Контрольный эксперимент
[0088] Чтобы продемонстрировать, что снижение колонизации С. jejuni является результатом вакцинации живыми Е. coli, экспрессирующими гептасахарид С. jejuni, а не пробиотического эффекта в связи с воздействием живых клеток E. coli, содержимое слепой кишки птиц, которых вакцинировали живым штаммом E. coli, также засевали на среду, селективную для E. coli, как упоминалось выше. Отсутствие колоний Е. coli у куриц, вакцинированных живыми Е. coli, указывает на то, что Е. coli выводились до окончания эксперимента.
[0089] Все публикации, патенты и заявки на патенты, упоминаемые в данном описании, указывают на уровень техники в той области, к которой относится данное изобретение, и включены в настоящую заявку посредством ссылки, так же как если бы каждая отдельная публикация, патент или заявка на патент были особо и индивидуально включены посредством ссылки.
[0090] Изобретение, описанное таким образом, очевидно, может варьировать и по-разному видоизменяться. Такие варианты не должны рассматриваться как отклонение от духа и области изобретения, и все такие модификации, которые должны быть очевидны специалистам в данной области техники, должны быть включены в область настоящего изобретения, охватываемую следующей формулой изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПРЕЗЕНТИРУЕМЫЙ В SALMONELLA ENTERICA N-ГЛИКАН ИЗ C. JEJUNI И ЕГО ПРОИЗВОДНЫЕ | 2010 |
|
RU2507253C2 |
| ПОВЕРХНОСТНЫЕ ГЛИКАНЫ CLOSTRIDIUM PERFRINGENS И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2798419C2 |
| СПОСОБ ПОЛУЧЕНИЯ СИАЛИЛИРОВАННЫХ ОЛИГОСАХАРИДОВ | 2007 |
|
RU2473695C2 |
| ПОЛИПЕПТИДЫ ГЕМАГГЛЮТИНИНОВ, А ТАКЖЕ СВЯЗАННЫЕ С НИМИ РЕАГЕНТЫ И СПОСОБЫ | 2011 |
|
RU2663718C2 |
| РЕКОМБИНАНТНАЯ ВАКЦИНА ПРОТИВ ВИРУСА АФРИКАНСКОЙ КАТАРАЛЬНОЙ ЛИХОРАДКИ | 2007 |
|
RU2446823C2 |
| КЛОСТРИДИАЛЬНЫЙ ТОКСИН NetB | 2008 |
|
RU2474587C2 |
| АДАПТАЦИЯ ВИРУСА ГРИППА Н5 К ЧЕЛОВЕКУ | 2014 |
|
RU2708447C2 |
| БАКТЕРИАЛЬНЫЕ ФАКТОРЫ ВИРУЛЕНТНОСТИ И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ | 2004 |
|
RU2465331C2 |
| УСОВЕРШЕНСТВОВАННАЯ КОРОНАВИРУСНАЯ ВАКЦИНА | 2022 |
|
RU2816182C2 |
| ЖИВЫЕ АТТЕНУИРОВАННЫЕ ВАКЦИНЫ | 2010 |
|
RU2556813C2 |





Группа изобретений относится к биотехнологии. Предложены композиция вакцины и способ вакцинации домашней птицы. Композиция вакцины против кампилобактериоза содержит модифицированные клетки Е. coli, экспрессирующие по меньшей мере один N-гликан или его производное на своей поверхности, полученные по меньшей мере с одного оперона pgl Campylobacter jejeni, и один или более физиологически приемлемый разбавитель, вспомогательное вещество, адъювант или носитель. Способ вакцинации предусматривает введение животному эффективной дозы композиции вакцины. Изобретения обеспечивают профилактику и/или лечение инфекций домашней птицы. 2 н. и 12 з.п. ф-лы, 9 ил., 2 табл., 2 пр.
1. Композиция вакцины против кампилобактериоза, содержащая: модифицированные клетки Е. coli, экспрессирующие по меньшей мере один N-гликан или его производное на своей поверхности, полученные по меньшей мере с одного оперона pgl Campylobacter jejeni; и один или более физиологически приемлемый разбавитель, вспомогательное вещество, адъювант или носитель.
2. Композиция вакцины по п. 1, отличающаяся тем, что указанная композиция содержит живую модифицированную Е. coli или живую ослабленную модифицированную Е. coli.
3. Композиция вакцины по п. 1, отличающаяся тем, что указанная композиция содержит инактивированные или убитые клетки модифицированной Е. coli.
4. Композиция вакцины по п. 1, отличающаяся тем, что указанная композиция содержит суспензию модифицированных бактерий в соответствующем буферном растворителе, таком как фосфатно-солевой буфер.
5. Композиция вакцины по п. 1, дополнительно содержащая адъювант, стабилизатор или консервант.
6. Композиция вакцины по п. 1, отличающаяся тем, что указанная композиция выполнена в форме для перорального введения, введения in ovo или парентерального введения (например, посредством инъекции или инфузии).
7. Композиция вакцины по п. 6, отличающаяся тем, что указанная композиция вакцины выполнена в форме для распыления или для добавления в корм, пищевые добавки или питьевую воду домашнему скоту.
8. Композиция вакцины по любому из пп. 1-7, выполненная в форме для введения домашней птице.
9. Композиция вакцины по п. 8, отличающаяся тем, что указанной домашней птицей являются курицы.
10. Способ вакцинации домашней птицы против Campylobacter, включающий введение животному композиции вакцины, содержащей эффективную дозу композиции вакцины по п. 1.
11. Способ по п. 10, отличающийся тем, что указанная домашняя птица представляет собой цыпленка.
12. Способ по п. 10, отличающийся тем, что указанную композицию вакцины вводят путем распыления или в составе фармацевтической композиции, корма, пищевой добавки или с питьевой водой.
13. Способ по п. 12, отличающийся тем, что указанная фармацевтическая композиция выполнена в форме для перорального введения, введения in ovo, введения путем инъекции или инфузии.
14. Способ по любому из пп. 10-13, включающий второй этап введения указанной композиции вакцины.
| Изложница с суживающимся книзу сечением и с вертикально перемещающимся днищем | 1924 |
|
SU2012A1 |
| MICHAEL WACKER et al | |||
| "N-linked glycosylation in Campylobacter jejuni and its functional transfer into E.coli".//SCIENCE, 29 NOVEMBER 2002, v | |||
| РАССЕИВАЮЩИЙ ТОПЛИВО МЕХАНИЗМ | 1920 |
|
SU298A1 |
| Лекало летной зоны | 1924 |
|
SU1790A1 |
| АССОЦИИРОВАННАЯ ИНАКТИВИРОВАННАЯ ВАКЦИНА ПРОТИВ ХЛАМИДИОЗА, КАМПИЛОБАКТЕРИОЗА, САЛЬМОНЕЛЛЕЗА И ЛЕПТОСПИРОЗА МЕЛКОГО РОГАТОГО СКОТА | 1994 |
|
RU2086259C1 |

