Область техники, к которой относится изобретение
Данное изобретение относится к экспериментальной биологии и медицины и касается способов доставки защитной изоформы АроЕ в центральную нервную систему млекопитающего. Способы включают введение в спинномозговую жидкость млекопитающего частицы rAAV, содержащей капсидный белок AAV и вектор, содержащий нуклеиновую кислоту, кодирующую изоформу АроЕ.
Уровень техники
Перенос генов в настоящее время широко признан в качестве мощного инструмента анализа биологических явлений и процессов заболеваний как на клеточном, так и на молекулярной уровне. Совсем недавно значительное внимание получило применение генной терапии для лечения человеческих заболеваний, либо наследуемых (например, дефицита ADA), либо приобретенных (например, злокачественной опухоли или инфекционного заболевания). С появлением усовершенствованных способов переноса генов и постоянно расширяющейся библиотеки идентифицированных заболеваний, связанных с дефектными генами, генная терапия быстро перешла из теории лечения в практическую реальность.
Традиционно, генную терапию определяли как процедуру, в которой экзогенный ген вводится в клетки пациента для того, чтобы исправить врожденную генетическую ошибку. Совсем недавно генная терапия была определена шире как коррекция фенотипа заболевания через введение новой генетической информации в пострадавший организм. В in vivo генной терапии перенесенный ген вводится в клетки реципиентного организма in situ то есть в реципиенте. In vivo генная терапия была проверена на животных моделях. Сообщалось о возможности прямого генного переноса in situ в органы и ткани, такие как мышца, гематопоэтические стволовые клетки, артериальная стенка, нервная система и легкое. Также сообщалось о получении детектируемого уровня экспрессии введенного геннного продукта (ов) in vivo с помощью прямой инъекции ДНК в скелетную мышцу, сердечную мышцу и инъекции ДНК-липидных комплексов в сосудистую систему.
Лечение заболеваний центральной нервной системы (ЦНС), например, генетических заболеваний головного мозга, таких как болезнь Альцгеймера, остается трудноразрешимой проблемой. Основная проблема с лечением заболеваний головного мозга заключается в том, что терапевтические белки, доставляемые внутривенно, не пересекают гематоэнцефалический барьер, а доставляемые непосредственно в мозг, не распространяются широко. Таким образом, необходимо разработать способы лечения болезни Альцгеймера.
Раскрытие изобретения
В некоторых воплощениях настоящее изобретение относится к способу лечения болезни Альцгеймера у млекопитающих, включающий введение в спинномозговую жидкость (CSF) млекопитающего частицы rAAV, содержащей капсидный белок AAV и вектор, содержащий нуклеиновую кислоту, кодирующую защитную изоформу АроЕ, вставленную между парой инвертированных концевых повторов AAV таким образом, который является эффективным для инфекции эпендимных клеток в млекопитающем, не являющемся грызуном, где эпендимные клетки секретируют АроЕ для лечения заболевания. В данном документе, термин «защитная изоформа АроЕ» используется для того, чтобы отличить изоформы АроЕ, которые снижают риск болезни Альцгеймера, по меньшей мере, на 5%, например, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100% или более.
В некоторых воплощениях настоящее изобретение относится к способу доставки защитной изоформы АроЕ в центральную нервную систему млекопитающего, не являющегося грызуном, включающему введение в спинномозговую жидкость (CSF) млекопитающего, не являющегося грызуном, частицы rAAV, содержащей капсидный белок AAV и вектор, содержащий нуклеиновую кислоту, кодирующую защитную изоформу АроЕ, вставленную между парой инвертированных концевых повторов AAV таким образом, который является эффективным для инфекции эпендимных клеток в млекопитающем, не являющемся грызуном, например, эпендимных клеток, секретирующих АроЕ в CSF млекопитающего. В некоторых воплощениях частица rAAV является частицей rAAV2, которая заражает эпендимную клетку негрызунов со скоростью, которая более чем на 20% больше, чем скорость инфицирования AAV4, например, со скоростью, которая на 50% или 100%, 1000% или 2000% больше, чем скорость инфицирования AAV4.
В некоторых воплощениях настоящее изобретение обеспечивает способ лечения заболевания у млекопитающего, не являющегося грызуном, включающий введение в эпендимные клетки млекопитающего частицы rAAV, содержащей капсидный белок AAV и вектор, содержащий нуклеиновую кислоту, кодирующую защитную изоформу АроЕ, вставленную между парой инвертированных концевых повторов AAV, тем самым доставляя нуклеиновую кислоту в эпендимную клетку, где эпендимная клетка секретирует белок АроЕ для лечения заболевания. Настоящее изобретение относится к способу доставки нуклеиновой кислоты в эпендимную клетку у млекопитающего, включающему введение млекопитающему частицы AAV, содержащей нуклеиновую кислоту, вставленную между парой инвертированных концевых повторов AAV, тем самым доставляя нуклеиновую кислоту в эпендимную клетку в млекопитающем.
В некоторых воплощениях, данное изобретение обеспечивает способ доставки нуклеиновой кислоты, кодирующей защитную изоформу АроЕ в эпендимную клетку млекопитающего, включающее введение в эпендимную клетку частицы rAAV, содержащей капсидный белок AAV и вектор, включающий нуклеиновую кислоту, вставленную между парой инвертированных концевых повторов AAV, тем самым доставляя нуклеиновую кислоту в эпендимную клетку.
В некоторых воплощениях настоящее изобретение относится к способу доставки нуклеиновой кислоты, кодирующей защитную изоформу АроЕ, в млекопитающего, включающему введение в эпендимную клетку млекопитающего частицы rAAV, содержащей капсидный белок AAV и вектор, содержащий нуклеиновую кислоту, вставленную между парой инвертированных концевых повторов AAV, и возвращение эпендимной клетки в млекопитающего, посредством чего осуществляется доставка нуклеиновой кислоты млекопитающему.
В некоторых воплощениях настоящее изобретение относится к способу доставки нуклеиновой кислоты, кодирующей защитную изоформу АроЕ, в эпендимную клетку в млекопитающем, включающий введение млекопитающему частицы rAAV, содержащей капсидный белок AAV и вектор, содержащий нуклеиновую кислоту, вставленную между парой инвертированных концевых повторов AAV, тем самым доставляя нуклеиновую кислоту в эпендимную клетку в млекопитающем.
В некоторых воплощениях настоящее изобретение относится к способу трансфекции эпендимной клетки в мозге млекопитающего, включающему введение в спинномозговую жидкость (CSF) млекопитающего частицы rAAV, содержащей капсидный белок AAV и вектор, содержащий нуклеиновую кислоту, кодирующую защитную изоформу АроЕ, вставленную между парой инвертированных концевых повторов AAV таким образом, который является эффективным для инфекции эпендимных клеток в млекопитающем, так что эпендимные клетки секретируют агент в CSF млекопитающего.
В некоторых воплощениях млекопитающее представляет собой млекопитающее, не являющееся грызуном. В некоторых воплощениях млекопитающее, не являющееся грызуном, представляет собой примата, лошадь, овцу, козу, свинью или собаку. В некоторых воплощениях, примат является человеком.
В некоторых воплощениях, защитная изоформа АроЕ имеет, по меньшей мере, приблизительно 80% гомологию с АроЕ ε2. В некоторых воплощениях, защитная изоформа АроЕ имеет 100% гомологию с АроЕ ε2.
В некоторых воплощениях, частица AAV является частицей rAAV4. В некоторых воплощениях частица AAV является частицей rAAV2. В некоторых воплощениях капсид rAAV2 имеет, по меньшей мере, 80% гомологию с капсидным белком VP1, VP2 и/или VP3 из AAV2. В некоторых воплощениях капсида rAAV2 имеет 100% гомологию с VP1, VP2 и/или VP3 капсида AAV2.
В некоторых воплощениях настоящее изобретение относится к частице rAAV, содержащей вектор, содержащий нуклеиновую кислоту, кодирующую защитную изоформу АроЕ, вставленную между парой инвертированных концевых повторов AAV, для использования при трансфекции эпендимных клеток у млекопитающего для получения терапевтического результата.
В некоторых воплощениях, данное изобретение обеспечивает применение частицы rAAV, содержащей вектор, включающий нуклеиновую кислоту, кодирующую защитную изоформу АроЕ, вставленную между парой инвертированных концевых повторов AAV для изготовления лекарственного средства, полезного при лечении или профилактике болезни Альцгеймера у животного, такого как человек.
Настоящее изобретение относится к клетке, как описано выше, для применения при лечении или диагностике.
Настоящее изобретение относится к применению клетки, как описано выше, для получения лекарственного средства полезного при лечении болезни Альцгеймера у млекопитающего.
В некоторых воплощениях в настоящем изобретении предлагается набор, содержащий комбинацию частицы rAAV, содержащей вектор, включающий нуклеиновую кислоту, кодирующую защитную изоформу АроЕ, вставленную между парой инвертированных концевых повторов AAV, контейнера и вкладыша или этикетки, предписывающих введение частицы AAV в CSF для лечения болезни Альцгеймера у животного.
Краткое описание чертежей
Фигуры 1А-1В. Внутрижелудочковая инъекция AAV4-ApoE приводит к стабильной экспрессии huAPOE и устойчивому обнаружению рекомбинантного белка huApoE в головном мозге.
Фигура 2. Избыточная экспрессия каждой изоформы АроЕ дифференциально влияет на прогрессию амилоидоза.
Фигура 3. Размеры амилоидных бляшек варьируют в соответствии с каждой изоформой АроЕ.
Фигуры 4А-4В. Посмертная оценка амилоидной нагрузки подтверждает влияние ароЕ2 и ароЕ4 на отложение амилоида.
Фигуры 5A-5D. Каждая изоформа АроЕ дифференциально влияет на синаптическую плотность вокруг амилоидных отложений.
Фигура 6А является выравниванием белков AAV2 (SEQ ID NO: 1) и AAV4 (SEQ ID NO: 2), а Фигура 6В является и выравниванием нуклеотидов AAV2 (SEQ ID NO: 3) и AAV4 (SEQ ID NO: 4) на основании последовательности AAV2 (NC_001401) и AAV4 (NC_001829).
Фигура 7 демонстрирует повышенную активность ТРР1 в различных областях головного мозга.
Фигура 8 демонстрирует результаты прохождения Т-образного лабиринта контрольными и обработанными собаками. Светлые круги для пораженных собак; темные квадраты для нормальных собак, а темные круги для ТРР-/- собак, обработанных AAV2-CLN2.
Фигуры 9А-9В. Фигуры 9А-9В. Подтверждение подхода с переносом гена АРОЕ путем внутрижелудочкового введения AAV серотипа 4. Иммуногистологическое мечение GFP или АроЕ показало наличие GFP или человеческого белка АроЕ в эпендиме и в хориоидном сплетении. (А) Использование видоспецифического тИФА для количественного определения концентрации рекомбинантного человеческого белка АроЕ в мозговых гомогенатах инъецированных мышей. (В) Оценка процента человеческого белка АроЕ по сравнению с эндогенным ароЕ на мышь. Отношение человеческого АроЕ и мышиного эндогенного ароЕ рассчитывали для каждого животного. С помощью специфического анти-человеческого антитела ЗН1 АроЕ, наличие рекомбинантного белка может быть выявлено вокруг некоторых амилоидных отложений, где он имеет тенденцию к накоплению, в кортикальной паренхиме инъецированных APP/PS1 мышей. Обнаружение АроЕ вестерн-блоттингом в ISF образце нокаутных по ароЕ мышей инъецированных вектором AAV4-apoE4. Высоко чувствительное (но не видоспецифичное) козье антитело против ароЕ от «Millipore» (АВ947) было использовано в качестве детектирующего антитела. Альбумин использовали в качестве контроля, n=4-6 животных на группу. *<0,05.
Фигуры 10A-10D. Уровни Αβ пептидов и плотность амилоидных отложений модулируются гиперэкспрессией разных аллелей АРОЕ. (А) Анализ плотности амилоидных отложений в коре (слева) и гиппокампе (справа) инъецированных трансгенных мышей. Аналогичную тенденцию можно наблюдать между обеими церебральными областями, но данные достигли статистической значимости только в коре головного мозга. (В) определение с помощью тИФА концентраций пептидов Аβ40 и β42 в фракции муравьиной кислоты (FA). (С) Количественная оценка с помощью тИФА уровней пептидов Аβ40 и Αβ42 в растворимой фракции TBS через 5 месяцев после внутрижелудочкового введения каждого AAV. (D) Количественная оценка уровней в плазмы петидов Аβ40, через 5 месяцев после внутрижелудочкового введения APP/PS1 мышам векторов AAV-GFP и AAV-apoE2/3/4. n=4-7 животных на группу. *p<0,05
Фигуры 11 А-11 В. Сверхэкспрессия каждого варианта АРОЕ дифференциально модулирует прогресс амилоидоза in vivo. Были получены in vivo двухфотонные изображения осаждения амилоида в APP/PS1 мышах за одну неделю (ТО), один месяц (Т1) и два месяца (Т2) после интрацеребровентрикулярной инъекции векторов AAV-GFP, АРОЕ-2, -3 или -4. Внутривенное введение декстрана, техасского красного (70000 Да) проводили перед имиджингом, для того, чтобы одни и те же поля зрения можно было отслеживать в течение долгого периода времени. В течение двухмесячного периода могут быть обнаружены несколько новых амилоидных бляшек, в то время как случайные отложения, первоначально видные, не обнаруживались уже через один-два месяца. (А) Оценка объемной кортикальной плотности амилоидных отложений в течение двухмесячного периода после внутрижелудочковой инъекции AAV-GFP, -АРОЕ2, -АРОЕ3 или АРОЕ4 в 7-месячных APP/PS1 мышей. От шести до восьми полей зрения зафиксировали в продольном направлении для каждого животного, рассчитывали плотность бляшек на объем коры и представляли относительно начального значения для каждого животного на исходном уровне (ТО). Общий прогресс 0,23 плотности амилоидных отложений наблюдался в зависимости от времени (Т2/Т1, p<0,011). Кроме того, АроЕ2 значительно снижает плотность относительно GFP на 0,66 (SE=0,21, p=0,002), относительно АроЕ3 на 0,67 (SE=0,17, p<0,0001) и относительно АроЕ4 на 0,74 (SE=0,17, p<0,0001). (В) Линейнорегрессионный анализ прогрессирования амилоидоза в течение 2 месяцев после переноса генов в APP/PS1 мыши показал, что только AAV-АРОЕ4 индуцирует значительный положительный угловой коэффициент в течение этого периода времени, n=4-6 животных на группу. *p<0,05.
Фигура 12. Эволюция размера амилоидных отложений через один и два месяца после инфузии АроЕ2, -3 и -4. Точечные диаграммы рассеяния, представляющие соотношение размеров бляшек между Т1 и Т0, показали, что с АроЕ4 был связан повышенный рост бляшек по сравнению как с ароЕ2, так и с АроЕ3 после одного месяца. Этот эффект не поддерживается по истечении двух месяцев. n>50 бляшек измеряли на группу из 3-4 животных. *p<0,05.
Фигуры 13А-13С. Каждый вариант АРОЕ дифференциально влияет на нейропатологические изменения, связанные с амилоидными отложениями. Были получены изображения массива томографических срезов, иммуноокрашенных по PSD95 (пост-синаптической элемент) и амилоидным отложениям в APP/PS1 мышах через 2 месяца после внутрижелудочковой инъекции AAV-GFP, -АРОЕ2, -АРОЕ3 и -АРОЕ4. Амилоидные отложения метили с помощью антитела NAB61, которое, как было показано ранее, преимущественно метит токсичные олигомерные виды Αβ (А) значительно более высокая потеря маркера синапсина-1 наблюдается в непосредственной близости от амилоидных бляшек, при экспрессии как АРОЕ3, так и АРОЕ4, по сравнению с GFP или АРОЕ2. (В) Аналогичный эффект наблюдался, при количественной оценке постсинаптических элементов, так что плотность PSD95, окружающих отложения, снижалась через 2 месяца после внутрижелудочковой инъекции AAV4-APOE4. В качестве дополнительного параметра нейропатологического изменения, оценивали количество нейритных дистрофий на амилоидную бляшку в мозге инъецированных APP/PS1 мышей, после иммуноокрашивания ThioS и аксональным маркером SMI312. (С) Значимый сдвиг в сторону более высокого количества дистрофий наблюдали при инфузии мышей АроЕ4 по сравнению с группами, инфузированными АроЕЗ и АроЕ2, что позволяет предположить, что АроЕ4 может иметь вредные эффекты помимо формирования амилоидных бляшек и может модулировать нейротоксический потенциал меньших олигомерных амилоидных агрегатов. n=4-6 животных на группу. *p<0,05.
Фигура 14. Ранние изменения в содержании видов олигомерных Αβ наблюдаются в ISF после интрацеребровентрикулярной инъекции AAV4-APOE2, -3, -4 в мышей Tg2576. Количественная оценка содержания ISF в οΑβ с помощью 82Е1/82Е1 тИФА показывает, что существует высокая концентрация видов олигомерного амилоида β после инъекции AAV4-APOE4 по сравнению с AAV4-APOE2 и -GFP, в то время как у мышей, инъецированных AAV4-APOE3, достигает среднего уровня, n=3-6 животных на группу. *p<0,05.
Фигура 15А-15В. Обнаружение человеческих и эндогенных мышиных АРОЕ мРНК и белка после внутрижелудочкового введения AAV4 в мышей APP/PS1. (А) Коробчатые диаграммы, представляющие количества эндогенного мышиного белка ароЕ в мозге инъецированных мышей. (В) Сравнение уровней белка АроЕ через 2 и 5 месяцев после интрацеребровентрикулярной инъекции AAV4 в мышей APP/PS1 (образцы из всех АроЕ-инъецированных мышей объединяли вместе через 2 и 5 месяцев, без дискриминации по варианту АРОЕ). n=4-6 животных на группу. *p<0,05.
Фиг. 16А-16В. Влияние на Αβ ассоциировано с каждой изоформой АроЕ после короткого (2 месячного) воздействия. Были получены изображения амилоидного отложения в APP/PS1 мышах через 2 месяца после инъекции. Иммуноокрашивание как с помощью антитела Bam10, так и с помощью ThioS использовали для окрашивания всех амилоидных отложений или бляшек с плотным ядром, соответственно. (А) Стереологический анализ плотности амилоидных отложений в коре головного мозга показал, что избыточная экспрессия АРОЕ4 привела к увеличению количества бляшек уже через 2 месяца после инъекции, в то время как никаких отличий не наблюдалось между другими экспериментальными группами. (В) Соотношение между окрашиванием Bam10 и ThioS, с другой стороны, осталось неизменным среди всех групп.(С) Определение концентраций пептидов Аβ40 (левые панели) и Αβ42 (правые панели) в нерастворимых экстрактах муравьиной кислотой после короткого воздействия различными вариантами АроЕ. n=3-5 животных на группу. *p<0,05.
Фигуры 17А-17В. Изменения в растворимых и нерастворимых видах Αβ, обнаруженные через 3 месяца после инъекции в мышей Tg2576. (А) Количественная оценка с помощью тИФА содержания ISF в Аβ40 и Αβ42 (В) показала, что существует тенденция к более высокой концентрации растворимых пептидов амилоида β после инъекции AAV4-APOE4 по сравнению с AAV4-apoE2, -АРОЕ3 и -GFP. (Б) Как ранее наблюдалось у мышей APP/PS1, более сильный эффект наблюдался с АроЕ4, который дает значительно более высокие количества Аβ42 во фракции муравьиной кислоты из мышей Tg2576. n=3-5 животных на группу. *p<0,05.
Подробное описание
Существует несколько различных изоформ человеческого аполипопротеина Ε (АроЕ), присутствие некоторых из этих изоформ в мозге повышает риск развития болезни Альцгеймера (AD), в то время как присутствие других изоформ снижает риск AD. Наличие ε4-изоформы АроЕ представляет собой сильный генетический фактор риска для спорадической AD с поздним началом (Casellano et al., Sci Transi Med, 3(89):89ra57 (29 June 2011).). ε4-Аллель АроЕ сильно повышает риск AD и снижает возраст возникновения. С другой стороны, наличие аллеля ε2 АроЕ по-видимому, снижает риск AD. Это позволяет предположить, что человеческие изоформы АроЕ по-разному влияют на выведение или синтез амилоида-β (Αβ) in vivo.
Аденоассоциированный вирус (AAV) представляет собой небольшой непатогенный вирус семейства Parvoviridae. AAV отличается от других представителей этого семейства своей зависимостью при репликации от вируса-помощника. В отсутствие вируса-помощника, AAV может интегрироваться локус-специфичным образом в q-плечо хромосомы 19. Геном AAV длиной приблизительно 5 тыс. п.о. состоит из одного сегмента одноцепочечной ДНК либо плюс, либо минус полярности. Концами генома являются короткие инвертированные концевые повторы, которые могут складываться в шпилечные структуры и служить в качестве начала репликации вирусной ДНК. Физически, вирион парвовируса является безоболочечным и его икосаэдрический капсид имеет диаметр около 20 нм.
На сегодняшний день были определены восемь серологически отличных AAV, а пять были выделены из человека или приматов и их обозначают как типы AAV 1-5. Govindasamy et al., «Structurally Mapping the Diverse Phenotype of Adeno-Associated Virus Serotype 4,» J. Vir., 80 (23): 11556-11570 (2006). Геном AAV2 имеет 4680 нуклеотидов в длину и содержит две открытые рамки считывания (ORF). Левая ORF кодирует неструктурные белки Rep, Rep 40, Rep 52, Rep 68 и Rep 78, которые участвуют в регуляции репликации и транскрипции в дополнение к производству одноцепочечных геномов потомков. Кроме того, два из белков Rep были ассоциированы с преференциальной интеграцией геномов AAV в области q-плеча хромосомы человека 19. Также было показано, что Rep68/78, обладают NTP-связывающей активностью, а также ДНК- и РНК- геликазной активностью. Белки Rep обладают сигналом ядерной локализации, а также несколькими потенциальными местами фосфорилирования. Мутация одного из этих киназных сайтов приводит к потере активности репликации.
Концы генома являются короткими инвертированными концевыми повторами (ITR), которые имеют возможность складываться в Т-образные шпилечные структуры и служить в качестве начала репликации вирусной ДНК. В регионе ITR были описаны два элемента, которые являются центральными в функции ITR, мотив повтора GAGC и сайт концевого разрешения (TRS). Было показано, что мотив повтора связывает Rep, когда ITR находится либо в линейной, либо в шпилечной конформации. Это связывание служит для позиционирования Rep68/78 при расщеплении TRS, которое происходит сайт- и нить-специфическим образом. В дополнение к их роли в репликации, эти два элемента, как представляется, играют центральную роль в вирусной интеграции. Содержащийся в хромосоме 19 локус интеграции является сайтом связывания Rep с соседними TRS. Было показано, что эти элементы являются функциональным и необходимы для локус-специфической интеграции.
Вирион AAV2 является безоболочечной, икосаэдрической частицей примерно 25 нм в диаметре, состоящий из трех родственных белков, называемых VP1, VP2 и VP3. Правый ORF кодирует белки капсида VP1, VP2 и VP3. Обнаружено, что эти белки находятся в соотношении 1:1:10, соответственно, и все они получены из правой ORF. Белки капсида отличаются друг от друга из-за применения альтернативного сплайсинга и необычного стартового ко дона. Делеционный анализ показал, что удаление или изменение VP1, который сплайсйруется с альтернативно-сплайсированного носителя, приводит в снижению выхода инфекционных частиц. Мутации в пределах кодирующей области VP3 приводят к неспособности продуцировать какую-либо одноцепочечную ДНК потомства или инфекционные частицы. Частица AAV2 является вирусной частицей, содержащей капсидный белок AAV2. Полипептид капсида AAV2 может кодировать все полипептиды VP1, VP2 и VP3. Частица может быть частицей, содержащей AAV2 и другие капсидные белки AAV (т.е. химерный белок, такой как AAV4 и AAV2). Вариации в аминокислотной последовательности капсидного белка AAV2 рассматриваются в данном описании, при условии, что в результате вирусная частица, содержащая капсид AAV2, остается антигенно или иммунологически отличной от AAV4, что можно определить обычным образом с помощью стандартных методов. В частности, например, тИФА и Вестерн-блоттинг можно использовать для определения, является ли вирусная частица антигенно или иммунологически отличной от AAV4. Кроме того, вирусная частица AAV2 предпочтительно сохраняет тропизм к ткани, отличный от AAV4.
Частица AAV2 является вирусной частицей, содержащей капсидный белок AAV2. Капсидный полипептид AAV2, кодирующий все полипептиды VP1, VP2, VP3, может в целом иметь, по меньшей мере, около 63% гомологии (или идентичности) по отношению к полипептиду, имеющему аминокислотную последовательность, кодируемую нуклеотидами, представленными в SEQ ID NO: 1 (капсидный белок AAV2). Капсидный белок может иметь около 70% гомологии, около 75% гомологии, 80% гомологии, 85% гомологии, 90% гомологии, 95% гомологии, 98% гомологии, 99% гомологию, или даже 100% гомологию по отношению к белку, представленному в SEQ ID NO: 1. Капсидный белок может иметь около 70% идентичности, 75% идентичности, 80% идентичности, 85% идентичности, 90% идентичности, 95% идентичности, 98% идентичности, 99% идентичности, или даже 100% идентичности по отношению к белку, представленному в SEQ ID NO: 1. Частица может быть частицей, содержащей как AAV4- так и AAV2-капсидный белок, т.е. химерный белок. Изменения в аминокислотной последовательности капсидного белка AAV2 рассматриваются в данном описании, при условии, что конечная вирусная частица, содержащая капсид AAV2 остается антигенно или иммунологически отличной от AAV4, что можно определить обычным образом с помощью стандартных способов. В частности, например, можно использовать тИФА и Вестерн-блоттинг для того, чтобы определить, являются ли вирусные частицы антигенно или иммунологически отличными от AAV4. Кроме того, вирусные частицы AAV2 предпочтительно сохраняют различие от AAV4 по тропизму к ткани, как проиллюстрировано, например, в приведенных ниже примерах, хотя химерная частица AAV2 содержащая, по меньшей мере, один белок оболочки AAV2 может иметь тропизм к ткани, отличный от такового для частицы AAV2, состоящей только из белков оболочки AAV2.
Как показано в Фигурах 6А и 6В, последовательности капсида AAV2 и последовательности капсида AAV4 имеют около 60% гомологии. В некоторых воплощениях капсид AAV2 содержит (или состоит из) последовательности, по меньшей мере, на 65% гомологичной аминокислотной последовательности, представленной в SEQ IDNO: 1.
В некоторых воплощениях изобретение также относится к частице AAV2 содержащей, например, инкапсулирующей, вектор, содержащий пару инвертированных концевых повторов AAV2. Нуклеотидная последовательность ITR из AAV2 известна в данной области. Кроме того, частица может быть частицей, содержащей как капсидный белок AAV4, так и капсидный белок AAV2, т.е. химерный белок. Кроме того, частицы могут быть частицей, инкапсулирующей вектор, содержащий пару инвертированных концевых повторов AAV из других AAV (например, AAV1-AAV8). Вектор, инкапсулированный в частице может дополнительно содержать экзогенную нуклеиновую кислоту, вставленную между инвертированными концевыми повторами.
Нижеупомянутые особенности AAV сделали его привлекательным вектором для переноса генов. Было показано in vitro, что векторы AAV стабильно интегрируются в клеточный геном; обладают широким спектром хозяев; трансдуцируют как делящиеся, так и неделящиеся клетки в пробирке in vitro и in vivo и поддерживают высокие уровни экспрессии трансдуцированных генов. Вирусные частицы термостабильны, устойчивы к действию растворителей, детергентов, изменений pH, температуры, и могут быть сконцентрированы в градиентах CsCl. Интеграция провируса AAV не связана с какими-либо долгосрочными негативными воздействиями на рост или дифференцировку клеток. Было показано, что в ITR только цис-элементы необходимы для репликации, упаковки и интеграции и могут включать некоторую промоторную активность.
Настоящее изобретение относится к способам введения частиц AAV, рекомбинантных векторов AAV и рекомбинантных вирионов AAV. Например, частица AAV2 является вирусной частицей, содержащей капсидный белок AAV2 или частица AAV4 является вирусной частицей, содержащей капсидный белок AAV4. Рекомбинантный вектор AAV2 представляет собой конструкцию нуклеиновой кислоты, которая содержит, по меньшей мере, одну уникальную нуклеиновую кислоту AAV2. Рекомбинантный вирион AAV2 является частицей, содержащей рекомбинантный вектор AAV2. Чтобы рассматриваться в пределах термина «AAV2 ITR» последовательность нуклеотидов должна сохранить один или оба признака, описанных здесь, которые отличают AAV2 ITR от AAV4 ITR: (1) три (а не четыре, как в AAV4) «GAGC» повтора и (2) в сайте связывания Rep в AAV2 ITR четвертый нуклеотид в первых двух «GAGC» повторов C, а не Т.
Промотор может быть любым желаемым промотором, выбранным исходя из известных соображений, таких, как уровень экспрессии нуклеиновой кислоты, функционально связанной с промотором и тип клеток, в которых вектор должен быть использован. Промоторы могут быть экзогенным или эндогенным промотором. Промоторы могут включать в себя, например, известные сильные промоторы, такие как SV40 или индуцируемый металлотионеиновый промотор, или AAV промотор, такой как промотор AAV р5. Дополнительные примеры промоторов включают промоторы, полученные из генов актина, генов иммуноглобулина, цитомегаловируса (CMV), аденовируса, вируса папилломы крупного рогатого скота, аденовирусных промоторов, таких как аденовирусный главный поздний промотор, индуцируемого промотора теплового шока, респираторно-синцитиального вируса, вирус саркомы Рауса (RSV), и т.п. В частности, промотор может быть AAV2 Р5 промотором или AAV4 Р5 промотором. Кроме того, более мелкие фрагменты р5 промотора, которые сохраняют активность промотора, могут быть легко определены с помощью стандартных процедур, включая, например, создание серии делеций в промоторе р5, связывание делеции с репортерным геном, и определение экспрессии гена-репортера, т.е., транскрипции и/или трансляции.
Вектор AAV может дополнительно содержать экзогенную (гетерологичную) нуклеиновую кислоту, функционально связанную с промотором. Под «гетерологичной нуклеиновой кислоты» подразумевается, что любая гетерологичная или экзогенная нуклеиновая кислота может быть встроена в вектор для переноса в клетки, ткани или организм. Например, в некоторых воплощениях гетерологичная нуклеиновая кислота кодирует защитную изоформу АроЕ. Под «функционально связанный» подразумевается, например, что промотор может способствовать экспрессии гетерологичной нуклеиновой кислоты, как известно в данной области, например, в соответствующей ориентации промотора по отношению к гетерологичной нуклеиновой кислоте. Кроме того, гетерологичная нуклеиновая кислота предпочтительно имеет все необходимые последовательности для экспрессии нуклеиновой кислоты, как известно в данной области, для того чтобы кодировать функционально, т.е. позволять нуклеиновой кислоте экспрессироваться. Нуклеиновая кислота может включать, например, контролирующие экспрессию последовательности, такие как энхансер, и необходимые сайты обработки информации, такие как сайты связывания рибосом, сплайсинга РНК, сайты полиаденилирования, и последовательности терминации транскрипции. Нуклеиновая кислота может кодировать более чем один генный продукт, ограничиваясь только размером нуклеиновой кислоты, которая может быть упакована.
Частица AAV2 является вирусной частицей, содержащей капсидный белок AAV2. Изменения в аминокислотной последовательности капсидного белка AAV2 рассматриваются в данном описании, при условии, что в результате вирусная частица, содержащая капсид AAV2, остается антигенно или иммунологически отличной от AAV4, что можно определить обычным способом с помощью стандартных методов. В частности, например, тИФА и Вестерн-блоттинг можно использовать для того, чтобы определить, является ли вирусная частица антигенно или иммунологически отличной от других серотипов AAV.
AAV4 является уникальным представителем семейства AAV. Обсуждение AAV4 приводится в патенте США №6468524, который включен в данное описание ссылкой. Данные гибридизации ДНК указывают на аналогичный уровень гомологии для AAV1-4. Тем не менее, в отличие от других AAV, в AAV4 был идентифицирован только один ORF, соответствующий капсидным белкам, и ни одного ORF не было обнаружено для белков Rep. Настоящее изобретение относится к вектору, содержащему вирус AAV4, а также вирусные частицы AAV4. Хотя AAV4 аналогичен AAV2, два вируса как установлено в данном документе, физически и генетически различны. Эти различия придают AAV4 некоторые уникальные преимущества, которые делают его более подходящим в качестве вектора для генной терапии. Например, геном AAV4 дикого типа больше, чем AAV2, что позволяет эффективно инкапсулировать больший рекомбинантный геном. Кроме того, частицы AAV4 дикого типа имеют большую плавучую плотность, чем частицы AAV2 и, следовательно, более легко отделяется от загрязняющего вспомогательного вируса и пустых частиц AAV, чем частицы на основе AAV2. Кроме того, в отличие от AAV1, 2, и 3, AAV4 способен гемагглютинировать эритроциты человека, морских свинок и овец.
В некоторых воплощениях настоящее изобретение относится к вектору, содержащему вирус AAV5 или вектор, содержащий составные части вируса, а также вирусные частицы AAV5. Обсуждение AAV5 приводится в патенте США №6855314, который включен в данное описание ссылкой. Хотя AAV5 аналогичен AAV2, два вируса, как установлено в данном документе, физически и генетически различны. Эти различия придают AAV5 некоторые уникальные преимущества, которые делают его более подходящим в качестве вектора для генной терапии. Например, одна из характеристик ограничивающих использование AAV2 в качестве вектора для генной терапии заключается в выработке больших количеств вируса. Используя стандартные методы производства, AAV5 продуцируется в 10-50 раз на более высоком уровне по сравнению с AAV2. Благодаря своему уникальному сайту TRS и rep белкам, AAV5 также должен иметь индивидуальный локус интеграции по сравнению с AAV2.
Более того, капсидный белок AAV5, опять же, отличается от капсидного белка AAV2 и имеет различный тканевой тропизм, что делает частицы, содержащие капсид AAV5, подходящими для трансдукции типов клеток, для которых AAV2 не подходит или подходит менее хорошо. AAV2 и AAV5, как было показано, серологически отличаются и таким образом, в генно-терапевтическом применении, AAV5 и векторы на основе AAV5 позволят трансдуцировать пациента, который уже обладает нейтрализующими антителами к AAV2 либо в результате естественной иммунологической защиты либо из-за предварительного воздействия AAV2-векторами. Еще одно преимущество AAV5 заключается в том, что AAV5 не может быть восстановлен другими серотипами. Только AAV5 может восстановить интегрированный геном AAV5 и осуществить репликацию, что позволяет избежать непреднамеренной репликации AAV5, вызванной другими серотипами AAV.
Термин «полипептид», при использовании в данном документе, относится к полимеру аминокислот и включает в себя полноразмерные белки и их фрагменты. Таким образом, «белок», «полипептид» и «пептид» часто используются взаимозаменяемо в данном документе. Замены могут быть выбраны нейтральными согласно известным параметрам. Как будет понятно специалистам в данной области, изобретение также включает те полипептиды, которые имеют небольшие изменения аминокислотных последовательностей или других свойств. Такие изменения могут возникнуть естественно, как аллельные вариации (например, из-за генетического полиморфизма) или могут быть получены путем вмешательства человека (например, мутагенеза клонированных последовательностей ДНК), таких как индуцированные точечные, делеционные, вставочные и замещающие мутанты. Незначительные изменения в аминокислотной последовательности, как правило, являются предпочтительными, например, консервативные аминокислотные замены, небольшие внутренние делеции или вставки и добавления или делеции на концах молекул. Эти модификации могут привести к изменениям в аминокислотной последовательности, дать молчащие мутации, изменить сайт рестрикции, или дать другие специфические мутации.
Данный способ обеспечивает способ доставки нуклеиновой кислоты в клетку, включающий введение в клетку частицы AAV, содержащую вектор, содержащий нуклеиновую кислоту, вставленную между парой инвертированных концевых повторов AAV, на основании чего осуществляется доставка нуклеиновой кислоты в клетку. Введение в клетку может быть осуществлено с помощью любых средств, в том числе простого контакта частиц, необязательно содержащихся в желаемой жидкости, такой как тканевая культуральная среда, или буферный солевой раствор, с клетками. Частице может быть позволено оставаться в контакте с клетками в течение любого желаемого периода времени, и обычно частицы вводят и оставляют на неопределенный срок. Для таких методов in vitro, вирус может быть введен в клетки с помощью стандартных способов вирусных трансдукции, как известно в данной области техники и как приведено в качестве примера в данном описании. Титры вируса для введения могут варьировать, в частности, в зависимости от типа клеток, но типично будут такими, которые используются для трансдукции AAV в большинстве случаев. Кроме того могут использоваться титры, которые применяются для трансдукции конкретных клеток в настоящих примерах. Клетки могут включать любую желаемую клетку человека, а также других крупных (не грызунов) млекопитающих, таких как приматы, лошади, овцы, козы, свиньи, и собаки.
Более конкретно, настоящее изобретение относится к способу доставки нуклеиновой кислоты в эпендимную клетку, включающему введение в эпендимную клетку частицы AAV, включающей вектор, содержащий нуклеиновую кислоту, вставленную между парой инвертированных концевых повторов AAV, на основании чего осуществляется доставка нуклеиновой кислоты в эпендимные клетки.
Настоящее изобретение также относится к способу доставки нуклеиновой кислоты объекту, включающий введение в клетку из объекта частицы AAV, содержащей нуклеиновую кислоту, вставленную между парой инвертированных концевых повторов AAV, и возвращение клетки объекту, на основании чего осуществляется доставка нуклеиновой кислоты в объект. В некоторых воплощениях AAV ITR может быть AAV2 ITR. Для такого ex vivo введения, клетки выделяют из объекта с помощью стандартных средств в зависимости от типа клеток и помещают в соответствующую культуральную среду, опять же в зависимости от типа клеток. Вирусные частицы затем контактируют с клетками, как описано выше, и вирусу позволяют трансфецировать клетки. Клетки затем могут быть пересажены обратно в тело объекта, опять же с помощью стандарта для этого типа клеток и тканей. При необходимости перед трансплантацией с помощью известных способов обнаружения и, как описано в данном документе, может быть изучена степень трансфецируемости клеток вирусом.
Настоящее изобретение также относится к способу доставки нуклеиновой кислоты в клетку объекта, включающему введение объекту частицы AAV, содержащей нуклеиновую кислоту, вставленную между парой инвертированных концевых повторов AAV, на основании чего осуществляется доставка нуклеиновой кислоты в клетку объекта. Введение может быть ex vivo введением непосредственно в клетку, удаленную из объекта, например, в любую из клеток, перечисленных выше, с последующей реплантацией клетки обратно в объект, или введение может быть введением in vivo в клетку объекта. Для ех vivo введения, клетки выделяют из объекта с помощью стандартных средств в зависимости от типа клеток, и помещают в соответствующую культуральную среду, опять же в зависимости от типа клеток. Вирусные частицы затем контактируют с клетками, как описано выше, и вирусу позволяют трансфецировать клетки. Клетки затем могут быть трансплантированы обратно в организм объекта, опять же с помощью стандарта для этого типа клеток и тканей. При необходимости перед трансплантацией с помощью известных способов обнаружения и как описано в данном документе может быть изучена степень трансфецируемости клеток вирусом.
Также предлагается способ доставки нуклеиновой кислоты в эпендимную клетку объекта, включающий введение объекту частицы AAV, содержащую нуклеиновую кислоту, вставленную между парой инвертированных концевых повторов AAV, на основании чего осуществляется доставка нуклеиновой кислоты в эпендимную клетку объекта.
В некоторых воплощениях аминокислотная последовательность, которая нацелена на сосудистый эндотелий головного мозга, нацелена на сосудистый эндотелий головного мозга у объекта, который страдает от заболевания, например, болезни Альцгеймера.
В некоторых воплощениях аминокислотная последовательность, которая нацелена на сосудистый эндотелий головного мозга, нацелена на сосудистый эндотелий головного мозга у объекта, который не страдает от болезни Альцгеймера.
В некоторых воплощениях, вирусный вектор содержит последовательность нуклеиновой кислоты, кодирующую терапевтический агент. В некоторых воплощениях терапевтический агент представляет собой защитную изоформу АроЕ.
Некоторые воплощения настоящего описания обеспечивают клетку, содержащую вирусный вектор, описанный выше.
В некоторых воплощениях клетка представляет собой клетку млекопитающих, не являющихся грызунами. В некоторых воплощениях клетка представляет собой клетку приматов. В некоторых воплощениях клетка представляет собой клетку человека. В некоторых воплощениях клетка представляет собой нечеловеческую клетку. В некоторых воплощениях клетка находится in vitro. В некоторых воплощениях клетка находится in vivo. В некоторых воплощениях клетка представляет собой эпендимную клетку.
В некоторых воплощениях настоящего описания предлагается способ лечения заболевания у млекопитающего, включающий введение млекопитающему вирусного вектора или клетки, как описано в данном документе.
В некоторых воплощениях млекопитающее является человеком.
В некоторых воплощениях настоящего описания предлагается способ доставки агента в центральную нервную систему объекта, включающий введение в спинномозговую жидкость вирусного вектора, описанного в данном документе так, чтобы трансдуцированные эпендимные клетки экспрессировали терапевтический агент и доставляли агент в центральную нервную систему объекта. В некоторых воплощениях вирусный вектор трансдуцирует эпендимные клетки.
В некоторых воплощениях настоящего описания предлагается вирусный вектор или клетка, описанные в данном документе, для применения при оказании медицинской помощи.
В некоторых воплощениях настоящего описания предлагается применение вирусного вектора или клетки, как описано выше, для получения лекарственного средства, пригодного для лечения заболевания у млекопитающего, например, болезни Альцгеймера.
Вектор может дополнительно содержать защитную изоформу белка АроЕ. При использовании в данном документе, термин «секретируемый белок» включает в себя любой секретируемый белок, секретируемый естественным образом или модифицированный путем включения сигнальной последовательности, для обеспечения секретируемости.
Нуклеиновая кислота является «функционально связанной», когда он находится в функциональной связи с другой последовательностью нуклеиновой кислоты. Как правило, «функционально связанный» означает, что последовательности ДНК, будучи связанными, являются смежными. Однако, энхансеры не должны быть смежными. Связывание осуществляют лигированием по удобным сайтам рестрикции. Если такие сайты не существуют, используют синтетические олигонуклеотидные адаптеры или линкеры в соответствии с обычной практикой. Кроме того, множественные копии нуклеиновой кислоты, кодирующей ферменты, могут быть связаны друг с другом в экспрессирующем векторе. Такие множественные нуклеиновые кислоты могут быть отделены друг друга линкерами.
В настоящем описании также предлагается клетка млекопитающего, содержащая вектор, описанный в данном документе. Клетка может быть человеческой, и может быть из мозга. Тип клеток может представлять собой популяцию стволовых клеток или клеток-предшественников.
Настоящее изобретение относится к способу лечения заболевания, такого как генетическое заболевание или злокачественная опухоль у млекопитающего путем введения полинуклеотида, полипептида, экспрессирующего вектора, или клетки, описанных в данном документе. Генетическое заболевание может быть нейродегенеративным заболеванием, таким как болезнь Альцгеймера.
Некоторые аспекты изобретения относятся к полинуклеотидам, полипептидам, векторам, и генно-инженерным клеткам (модифицированным in vivo), а также к их применению. В частности, изобретение относится к способу генной или белковой терапии, которая приспособлена к системной доставке терапевтически эффективной дозы терапевтического агента.
Согласно одному аспекту, предлагается экспрессирующая клеточная система для экспрессии терапевтического агента в реципиенте-млекопитающем. Экспрессирующая система (также называемая в данном документе как «генетически модифицированная клетка») включает в себя клетку и экспрессирующий вектор для экспрессии терапевтического агента. Экспрессирующие векторы включают, без ограничения перечисленным, вирусы, плазмиды и другие носители для доставки гетерологичного генетического материала в клетки. Соответственно, термин «экспрессирующий вектор», при использовании в данном документе, относится к носителю для доставки гетерологичного генетического материала в клетки. В частности, экспрессирующий вектор представляет собой вектор на основе рекомбинантных аденовируса, аденоассоциированного вируса или лентивируса или ретровируса.
Экспрессирующий вектор дополнительно содержит промотор для контроля транскрипции гетерологичного гена. Промотор может быть индуцируемым промотором (описано ниже). Экспрессирующая система подходит для введения реципиенту-млекопитающему. Экспрессирующая система может включать в себя множество неиммортализованных генетически модифицированных клеток, где каждая клетка, содержит, по меньшей мере, один рекомбинантный ген, кодирующий, по меньшей мере, один терапевтический агент.
Экспрессирующая клеточная система может быть сформирована in vivo. В соответствии с еще одним аспектом предлагается способ лечения реципиента-млекопитающего in vivo. Способ включает в себя введение экспрессирующего вектора для экспрессии гетерологичного генного продукта в клетке пациента in situ, например, с помощью внутривенного введения. Для формирования экспрессирующей системы in vivo, экспрессирующий вектор для экспрессии терапевтического агента вводят in vivo в реципиента-млекопитающего, где вектор мигрирует через сосудистую систему в мозг.
В соответствии с еще одним аспектом, предлагается способ лечения реципиента-млекопитающего in vivo. Способ включает введение целевого белка в пациента in vivo.
Экспрессирующий вектор для экспрессии гетерологичного гена может включать индуцируемый промотор для контроля транскрипции гетерологичного генного продукта. Соответственно, доставки терапевтического агента in situ контролируется воздействием на клетку in situ условиями, которые индуцируют транскрипцию гетерологичного гена.
Реципиент-млекопитающее может иметь состояние, которое поддается генозамещающей терапии. При использовании в данном документе, «генозамещающая терапия» относится к введению реципиенту экзогенного генетического материала, кодирующего терапевтический агент и последующей экспрессии введенного генетического материала in situ. Таким образом, фраза «состояние поддается генозамещающей терапии» включает в себя такие состояния, как генетические заболевания (например, болезненное состояние, которому свойственны один или несколько генных дефектов), приобретенные патологии (т.е. патологическое состояние, которому несвойственны врожденные дефекты), злокачественные опухоли и профилактические процессы (т.е., предотвращение болезни или нежелательного медицинского состояния). Соответственно, при использовании в данном документе, термин «терапевтический агент» относится к любому агенту или материалу, который оказывает благоприятное воздействие на реципиента-млекопитающего. Таким образом, «терапевтический агент» охватывает как терапевтические, так и профилактические молекулы, имеющие нуклеотидные или белковые компоненты.
Согласно одному варианту осуществления, реципиент-млекопитающее имеет генетическое заболевание, а экзогенный генетический материал содержит гетерологичный ген, кодирующий терапевтический агент для лечения болезни. В еще одном воплощении реципиент-млекопитающее имеет приобретенную патологию, а экзогенный генетический материал содержит гетерологичный ген, кодирующий терапевтический агент для лечения патологии. Согласно другому воплощению, пациент имеет злокачественную опухоль, а экзогенный генетический материал содержит гетерологичный ген, кодирующий противоопухолевый агент. В еще одном воплощении пациент имеет нежелательное медицинское состояние, а экзогенный генетический материал содержит гетерологичный ген, кодирующий терапевтический агент для лечения состояния.
При использовании в данном документе, термин «защитная изоформа АроЕ» включает варианты или биологически активные или неактивные фрагменты этого полипептида. «Вариант» одного из полипептидов представляет собой полипептид, который не полностью идентичен нативному белку. Такой вариантный белок может быть получено путем изменения аминокислотной последовательности вставкой, делецией или заменой одной или нескольких аминокислот. Аминокислотная последовательность белка модифицирована, например, путем замещения, для создания полипептида, имеющего, по существу, одни и те же или улучшенные свойства по сравнению с нативным полипептидом. Замена может быть консервативной заменой. «Консервативная замена» является заменой аминокислоты другой аминокислотой, имеющей сходную боковую цепь. Консервативная замена будет заменой аминокислотой, которая дает наименьшее возможное изменение заряда аминокислоты или размера боковой цепи аминокислоты (в ином случае, размера, заряда или типа химической группы в боковой цепи), так что общий пептид сохраняет свою пространственную конформацию, но имеет измененную биологическую активность. Например, распространенными консервативными изменениями могут быть Asp на Glu, Asn или Gin; His на Lys, Arg или Phe; Asn на Gin, Asp или Glu и Ser на Cys, Thr или Gly. Алании обычно используется для замены других аминокислот. 20 незаменимых аминокислот могут быть сгруппированы следующим образом: аланин, валин, лейцин, изолейцин, пролин, фенилаланин, триптофан и метионин, имеющие неполярные боковые цепи; глицин, серии, треонин, цистеин, тирозин, аспарагин и глутамин, имеющие незаряженные полярные боковые цепи; аспартат и глутамат, имеющие кислотные боковые цепи; и лизин, аргинин, гистидин и имеющие основные боковые цепи.
Аминокислотные замены достигаются путем изменения кодонов соответствующей последовательности нуклеиновой кислоты. Известно, что такие полипептиды могут быть получены на основе замен определенных аминокислот другими аминокислотами в полипептидной структуре для изменения или улучшения биологической активности. Например, посредством замены на альтернативные аминокислоты, полипептиду могут быть приданы небольшие конформационные изменения, которые приводят к повышению активности. В ином случае, аминокислотные замены в определенных полипептидах могут быть использованы для обеспечения остатков, которые затем могут быть связаны с другими молекулами, для получения пептид-молекулярных конъюгатов, которые, сохраняют свойства исходного полипептида достаточные для того, чтобы быть полезными для других целей.
Можно использовать индекс гидрофобности аминокислот для придании им интерактивной биологической функции на полипептиде, поскольку было обнаружено, что некоторые аминокислоты могут быть заменены другими аминокислотами, имеющими аналогичные индексы гидрофобности и при этом сохраняют схожую биологическую активность. В ином случае, замена подобных аминокислот может быть осуществлена на основе гидрофильности, особенно в случае, когда в полипептиде желательны биологические функции, предназначенные для использования в иммунологических воплощениях. Наибольшая локальная средняя гидрофильность «белка», регулируемая гидрофильностью смежных аминокислот, коррелирует с его иммуногенностью. Соответственно, следует отметить, что замены могут быть осуществлены на основании гидрофильности, назначенной для каждой аминокислоты.
При использовании либо индекса гидрофильности, либо индекса гидрофобности, которые присваивают значения каждой аминокислоте, предпочтительно проводить замену аминокислот, когда эти значения составляют ± 2, а значения ± 1 являются особенно предпочтительными, а значения в пределах ± 0,5 является наиболее предпочтительными заменами.
Вариантный белок имеет, по меньшей мере, 50%, по меньшей мере, приблизительно 80% или даже, по меньшей мере, примерно 90%, но менее 100%, гомологии непрерывной аминокислотной последовательности или идентичности по отношению к аминокислотной последовательности соответствующего нативного белка.
Аминокислотная последовательность вариантного полипептида по существу соответствует аминокислотной последовательности нативного полипептида. При использовании в данном документе термин «по существу соответствует» относится к полипептидной последовательности, которая будут вызывать биологический ответ, по существу такой же, как ответ, вызываемый нативным белком. Такой ответ может составлять, по меньшей мере, 60% от уровня, вызываемого нативным белком, и может составлять даже, по меньшей мере, 80% от уровня, вызываемого нативным белком.
Вариант может включать аминокислотные остатки, не присутствующие в соответствующем нативном белке или включать делеции относительно соответствующего нативного белка. Вариант также может быть усеченным «фрагментом», по сравнению с соответствующим нативным белком, то есть может быть только частью полноразмерного белка. Варианты белков также включают пептиды, имеющие, по меньшей мере, одну D-аминокислоту.
Вариантный белок может быть экспрессирован с выделенной последовательности ДНК, кодирующей вариантный белок. «Рекомбинантный» определяется как пептид или нуклеиновая кислота, полученная с помощью способов генной инженерии. Следует отметить, что в данной области хорошо известно, что из-за избыточности генетического кода, отдельные нуклеотиды могут быть легко заменены в кодоне и при этом привести к идентичной аминокислотной последовательности. Термины «белок», «пептид» и «полипептид» используются здесь взаимозаменяемо.
Настоящее изобретение относится к способам лечения заболевания у млекопитающего путем введения экспрессирующего вектора в клетку или пациента. Для способов генной терапии, специалист в данной области молекулярной биологии и генной терапии в состоянии определить, без проведения чрезмерных экспериментов, соответствующие дозировки и способы введения экспрессирующего вектора, используемого в новых способах по настоящему изобретению.
Согласно одному варианту осуществления, клетки трансформированы или иным генетически модифицированы in vivo. Клетки из реципиента-млекопитающего трансформируются (например, трансдуцируются или трансфецируются) in vivo вектором, содержащим экзогенный генетический материал для экспрессии гетерологичного (например, рекомбинантного) гена, кодирующего терапевтический агент и терапевтический агент доставляется in situ.
При использовании в данном документе, «экзогенный генетический материал» относится к нуклеиновой кислоте или олигонуклеотиду, либо природному, либо синтетическому, который, естественным образом не обнаруживается в клетках; или если он естественным образом содержится в клетках, то не транскрибируется или не экспрессируется клетками в биологически значимых уровнях. Таким образом, «экзогенный генетический материал» включает, например, неприродные нуклеиновой кислоты, которые могут быть транскрибированы в антисмысловую РНК, а также «гетерологичный ген» (то есть, ген, кодирующий белок, который не экспрессирован или экспрессирован на биологически незначительных уровнях в природной клетке того же типа).
В некоторых воплощениях реципиент-млекопитающее имеет состояние, которое поддается генно-заместительной терапии. При использовании в данном документе, «геннозамещающая терапия» относится к введению реципиенту экзогенного генетического материала, кодирующего терапевтический агент и к последующей экспрессии введенного генетического материала in situ. Таким образом, фраза «состояние поддается генно-заместительной терапии» включает в себя такие состояния, как генетические заболевания (например, болезненное состояние, которому свойственны один или несколько генных дефектов), приобретенные патологии (т.е. патологическое состояние, которому несвойственны врожденные дефекты), злокачественные опухоли и профилактические процессы (т.е., предотвращение болезни или нежелательного медицинского состояния). Соответственно, при использовании в данном документе, термин «терапевтический агент» относится к любому агенту или материалу, который оказывает благоприятное воздействие на реципиента-млекопитающего. Таким образом, «терапевтический агент» охватывает как терапевтические и профилактические молекулы, состоящие из нуклеиновой кислоты (например, антисмысловой РНК), и/или белковых компонентов.
С другой стороны, состояние, поддающееся генно-заместительной терапии является профилактическим процессом, т.е., способом профилактики заболевания или нежелательного состояния здоровья. Таким образом, настоящее описание охватывает системы экспрессии клеток для доставки терапевтического агента, который имеет профилактическую функцию (т.е. профилактический агент), реципиенту-млекопитающему.
Таким образом, термин «терапевтический агент» включает, без ограничения перечисленным, агенты, связанные с перечисленными выше состояниями, а также их функциональные эквиваленты. При использовании в данном документе, термин «функциональный эквивалент» относится к молекуле (например, пептиду или белку), который оказывает такое же или улучшенное благоприятное воздействие на реципиента-млекопитающего,, как и терапевтический агент, относительного которого подразумевается функциональный эквивалент.
Вышеописанные терапевтические агенты и состояния, поддающиеся генно-заместительной терапии, являются всего лишь иллюстративными и не предназначены для ограничения объема настоящего изобретения. Выбор подходящего терапевтического средства для лечения известного состояния без проведения излишних экспериментов подразумевается в рамках квалификации специалиста в данной области.
Векторы AAV
В одном воплощении вирусный вектор изобретения является вектором AAV. Вектор «AAV» относится к аденоассоциированному вирусу, и может быть использован для обозначения природного вируса дикого типа или его производных. Термин охватывает все подтипы, серотипы и псевдотипы, как естественные, так и рекомбинантные формы, за исключением случаев, когда требуется иное. При использовании в данном документе, термин «серотип» относится к AAV, который идентифицируется с помощью, и отличается от других AAV на основании реактивности капсидного белка с определенной антисывороткой, например, существует восемь известных серотипов AAV приматов, AAV-1-AAV-8. Например, серотип AAV2 используется для обозначения AAV, который содержит капсидные белки, кодируемые геном cap из AAV2 и геном, содержащий 5' и 3' ITR последовательности из того же серотипа AAV2. При использовании в данном документе, например, rAAV может быть использован для обозначения AAV, имеющего как капсидные белки и 5'-3' ITR из того же серотипа или может быть использован для обозначения AAV, имеющего белки капсида из одного серотипа и 5'-3' ITR из другого серотипа AAV, например, капсида от AAV серотипа 2 и ITR от AAV серотипа 5. Для каждого примера, показанного здесь, описание разработки и производства вектора описывает серотип капсида и 5-3 'ITR последовательности. Аббревиатура «rAAV» относится к рекомбинантному адено-ассоциированному вирусу, также называемому рекомбинантным AAV-вектором (или «rAAV вектором").
"Вирус AAV» или «вирусная частица AAV» относится к вирусной частице, состоящей, по меньшей мере, из одного капсидного белка AAV (предпочтительно из всех белков капсида AAV дикого типа) и инкапсулированного полинуклеотида. Если частица содержит гетерологичный полинуклеотид (т.е. полинуклеотид, отличный от генома AAV дикого типа, такой как трансген, доставляемый в клетку млекопитающего), то она, как правило, называется «rAAV".
В одном воплощении экспрессирующие векторы AAV созданы с использованием известных методов обеспечивающих, по меньшей мере, в виде функционально связанных компонентов в направлении транскрипции, контрольные элементы, в том числе в области инициации транскрипции, представляющую интерес ДНК и область терминации транскрипции. Контрольные элементы выбираются так, чтобы быть функциональными в клетке млекопитающего. Полученный конструкт, который содержит функционально связанные компоненты, примыкает (5' и 3') к функциональным последовательностям AAV ITR.
Под «инвертированными концевыми повторами аденоассоциированного вируса» или «AAV ITR» понимаются известные в данной области участки, обнаруженные на каждом конце AAV генома, функционирующие в cis-положении в качестве начала репликации ДНК и в качестве упаковочных сигналов для вируса. AAV ITR, вместе с кодирующей областью AAV Rep, обеспечивают эффективное удаление и освобождение и интеграцию нуклеотидной последовательности, расположенной между двумя фланговыми ITR в геноме клетки млекопитающего.
Нуклеотидные последовательности участков AAV ITR известны. При использовании в данном документе, «AAV ITR» не должен иметь нуклеотидную последовательность изображенного дикого типа, но может быть изменен, например, путем вставки, делеции или замены нуклеотидов. Кроме того, AAV ITR могут быть получены из любого из нескольких серотипов AAV, в том числе, без ограничения перечисленным, AAV1, AAV2, AAV3, AAV4, AAV5, AAV7 и т.д. Кроме того, 5' и 3' ITR, которые фланкируют нуклеотидную последовательность, выбранную в векторе AAV не обязательно должны быть идентичными или получены из одного и той же серотипа или изолята AAV, при условии, что они функционируют надлежащим образом, то есть, для обеспечения удаления и освобождения интересующей последовательности из генома клетки-хозяина или вектора, и, чтобы позволить интеграцию гетерологичной последовательности в геном клетки-реципиента, когда генные продукты AAV Rep присутствуют в клетке.
В одном воплощении, AAV ITR могут быть получены из любого из нескольких серотипов AAV, в том числе, без ограничения перечисленным, AAV1, AAV2, AAV3, AAV4, AAV5, AAV7 и т.д. Кроме того, 5' и 3' ITR, которые фланкируют нуклеотидную последовательность, выбранную в экспрессирующем векторе AAV не обязательно должны быть идентичными или получены из одного и того же серотипа или изолята AAV, при условии, что они функционируют надлежащим образом, то есть, для обеспечения удаления и освобождения интересующей последовательности из генома клетки-хозяина или вектора, и для того, чтобы позволить интеграцию молекулы ДНК в геном клетки-реципиента, когда генные продукты AAV Rep присутствуют в клетке.
В одном воплощении, капсид AAV может быть получен из AAV2. Подходящие молекулы ДНК для использования в AAV векторах будут меньше, чем около 5 тысяч пар нуклеотидов (тыс. п.н.), меньше чем около 4,5 тыс. п.н., меньше чем около 4 тыс.п.н., меньше чем около 3,5 тыс. п.н., меньше чем около 3 тыс.п.н., меньше чем около 2,5 тыс. п.н. и известны в данной области.
В одном воплощении выбранная нуклеотидная последовательность функционально связана с регуляторными элементами, которые направляют транскрипцию или экспрессию их в объекте in' vivo. Такие регуляторные элементы могут содержать регуляторные последовательности, как правило, связанные с выбранным геном. В ином случае, можно использовать гетерологичные регуляторные последовательности. Полезные последовательности гетерологичных регуляторных последовательностей, как правило, включают те, которые получены из последовательностей, кодирующих гены млекопитающих · или вирусные гены. Примеры включают, без ограничения перечисленным, ранний промотор SV40, промотор LTR вируса опухоли молочной железы мыши; основной поздний промотор аденовируса (Ad MLP); промотор вируса простого герпеса (HSV), промотор цитомегаловируса (CMV) промотор, такой как предранний участок промотора CMV (CMVIE), промотор вируса саркомы Рауса (RSV), промоторы Pol II, промоторы Pol. III, синтетические промоторы, гибридные промоторы, и тому подобное. Кроме того, последовательности, полученные из невирусных генов, таких как ген мышиного металлотионеина, также могут найти применение в настоящем документе. Такие промоторные последовательности являются коммерчески доступными, например, в «Stratagene» (Сан-Диего, Калифорния).
В одном воплощении, как гетерологичные промоторы, так и другие регуляторные элементы, такие как ЦНС-специфические и индуцируемые промоторы, энхансеры и т.п., будут особенно полезны. Примеры гетерологичных промоторов включают промотор CMV. Примеры ЦНС-специфических промоторов включают промоторы, выделенные из генов основного белка миелина (МВР), глиального фибриллярного кислого белка (GFAP), и нейрон-специфичной энолазы (NSE). Примеры индуцируемых промоторов включают респонсивные элементы ДНК для экдизона, тетрациклина, гипоксии и ауфина.
В одном " воплощении экспрессирующий вектор AAV, который несет представляющую интерес молекулу ДНК, ограниченную AAV ITR, может быть построен путем непосредственного добавления выбранной последовательности (ей) в геном AAV, в котором вырезаны основные открытые рамки считывания AAV («ORF»). Другие части генома AAV также могут быть удалены, при условии, что достаточная часть ITR остается, для обеспечения функций репликации и упаковки. Такие конструкты могут быть разработаны с использованием методов, хорошо известных в данной области.
Кроме того, AAV ITR могут быть вырезаны из вирусного генома или из AAV вектора, содержащего то же самое и объединены с 5' и 3' выбранного нуклеотидного конструкта, который находится в другом векторе с использованием стандартных методов лигирования. Например, лигирование может быть осуществлено в 20 мМ Трис-Cl, pH 7,5, 10 мМ MgCl2, 10 мМ DTT, 33 мкг/мл БСА, 10 мМ-50 мМ NaCl и либо 40 мкМ АТФ, 0,01-0,02 (Weiss) единицы Т4 ДНК-лигазы при 0°C (для лигирования по «липким концам») или 1 мМ АТР, 0,3-0,6 (Weiss) единицы Т4 ДНК-лигазы при 14°C (для лигирования по «тупым концам»). Межмолекулярные лигирования по «липким концам», как правило, осуществляются при общей концентрации ДНК 30-100 мкг/мл (5-100 нМ общей концентрации концов). AAV - векторы, которые содержат ITR.
Кроме того, химерные гены могут быть получены синтетически, путем включения последовательностей AAV ITR расположенных 5' и 3' относительно одной или нескольких выбранных последовательностей нуклеиновых кислот. Для экспрессии химерной последовательности гена в клетках ЦНС млекопитающих могут быть использованы предпочтительные кодоны. Полная химерная последовательность собирается из перекрывающихся олигонуклеотидов полученных с помощью стандартных способов.
Для того чтобы получить вирионы rAAV, экспрессирующий вектор AAV вводили в подходящую клетку-хозяина с использованием известных методов, например, путем трансфекции. Несколько методов трансфекции широко известны в данной области. См., например, Sambrook et al. (1989) Molecular Cloning, a laboratory manual, Cold Spring Harbor Laboratories, New York. Особенно подходящие способы трансфекции включают копреципитацию фосфатом кальция, прямую микроинъекцию в культивируемые клетки, электропорацию, липосома-опосредованный перенос генов, липид-опосредованную трансдукцию и доставку нуклеиновой кислоты с использованием микрочастицами высокой скорости.
В одном воплощении, подходящие клетки-хозяева для получения вирионов rAAV, включают микроорганизмы, дрожжевые клетки, клетки насекомых и клетки млекопитающих, которые могут быть или использовались в качестве реципиентов гетерологичной молекулы ДНК. Этот термин включает потомство исходной клетки, которая была трансфецирована. Таким образом, «клетка-хозяин», при использовании в данном документе, как правило, относится к клетке, которая была трансфецирована экзогенной последовательностью ДНК. Клетки из стабильной линии клеток человека, 293 (легко доступны, например, в Американской коллекции типовых культур под каталожным номером АТСС CRL1573) могут быть использованы при практическом осуществлении настоящего изобретения. В частности, линии клеток человека 293 является эмбриональной линией клеток почки человека, которая была трансформирована ДНК-фрагментами аденовируса типа 5, и экспрессирует аденовирусные гены E1A и E1B. Клеточная линия 293 легко трансфицируется, и дает особенно удобную платформу для получения вирионов rAAV.
Под «кодирующей областью AAV Rep» в данной области понимается область генома AAV, которая кодирует белки репликации Rep 78, Rep 68, Rep 52 и Rep 40. Эти продукты экспрессии Rep, как было показано, обладают множеством функций, включая распознавание, связывание и надрезание начала репликации ДНК AAV, ДНК-геликазную активность и модуляцию транскрипции с AAV (или других гетерологических) промоторов. Продукты экспрессии Rep в совокупности необходимы для репликации генома AAV. Подходящие гомологи с кодирующей областью AAV rep включают rep ген человеческого вируса герпеса 6 (HHV-6), который также известен в качестве посредника репликации ДНК AAV2.
Под «кодирующей областью AAV cap» в данной области понимается область генома AAV, которая кодирует белки капсида VP1, VP2 и VP3, или их функциональные гомологи. Эти продукты экспрессии Сар осуществляют упаковочные функции, которые в совокупности, необходимы для упаковки вирусного генома.
В одном воплощении, хелперные функции для AAV вводят в клетку-хозяина путем трансфекции клетки-хозяина хелперным конструктом для AAV либо до, либо одновременно с трансфекцией экспрессирующим вектором AAV. Хелперные конструкты для AAV, таким образом, используется для обеспечения, по меньшей мере, временной экспрессии генов AAV rep и/или cap для комплементации отсутствующих функций AAV, которые необходимы для продуктивной инфекции AAV. Хелперные конструкты AAV утратили AAV ITR и не могут ни копировать, ни упаковывать себя. Эти конструкты могут быть в форме плазмиды, фага, космиды, транспозона, вируса или вириона. Несколько хелперных конструктов AAV были описаны, например, широко используемая плазмида pAAV/AD и pIM29+45, которые кодируют продукты экспрессии как Rep, так и Сар. Был описан ряд других векторов, которые кодируют продукты экспрессии Rep и/или Сар.
Способы доставки вирусных векторов включают инъекции AAV в CSF. Как правило, вирионы rAAV могут быть введены в клетки ЦНС с использованием либо in vivo либо in vitro методов трансдукции. При трансдукции in vitro, искомая клетка-реципиент удаляется из объекта, трансдуцируется вирионами rAAV и повторно вводится объекту. В ином случае, могут быть использованы сингенные или ксеногенные клетки, когда эти клетки не будут генерировать несоответствующий иммунный ответ в объекте.
Были описаны подходящие способы доставки и внедрения трансдуцированных клеток в объект. Например, клетки могут быть трансдуцированы in vitro путем объединения рекомбинантных AAV вирионов с клетками ЦНС, например, в соответствующей среде, и скрининг клеток, несущих представляющую интерес ДНК может быть проведен с использованием обычных методов, таких как Саузерн-блоттинг и/или ПЦР, или с помощью селективных маркеров. Трансдуцированные клетки могут быть приготовлены в виде фармацевтических композиций, как более подробно описано ниже, и композиция вводится объекту различными методами, например, путем прививки, внутримышечной, внутривенной, подкожной и внутрибрюшинной инъекции.
В одном воплощении фармацевтические композиции будут содержать достаточное количество генетического материала для того, чтобы произвести терапевтически эффективное количество представляющей интерес нуклеиновой кислоты, т.е. количество, достаточное для снижения или облегчения симптомов рассматриваемого болезненного состояния или количества, достаточного для придания желаемого эффекта. Фармацевтические композиции также будут содержать фармацевтически приемлемый наполнитель. Такие наполнители включают любой фармацевтический агент, который сам по себе не индуцирует выработку антител, вредных для объекта, получающего композицию, и который может быть введен без чрезмерной токсичности. Фармацевтически приемлемые наполнители включают, без ограничения перечисленным, сорбит, Tween80, и жидкости, такие как вода, физиологический раствор, глицерин и этанол. Фармацевтически приемлемые соли могут быть включены в них, например, соли минеральных кислот, такие как гидрохлориды, гидробромиды, фосфаты, сульфаты и тому подобное; и соли органических кислот, такие как ацетаты, пропионаты, малонаты, бензоаты и тому подобное. Кроме того, вспомогательные вещества, такие как смачивающие или эмульгирующие агенты, pH-буферные вещества и т.п., могут присутствовать в таких носителях. Подробное обсуждение фармацевтически приемлемых наполнителей доступно в Remington's Pharmaceutical Sciences (Mack Pub. Co., N. J. 1991).
Как очевидно специалистам в данной области при рассмотрении излагаемого в данном описании, эффективное количество вирусного вектора, который должен быть добавлен, может быть определено эмпирически. Введение может быть осуществлено в одной дозе, непрерывно или периодически в течение всего курса лечения. Способы определения наиболее эффективных средств и доз введения хорошо известны специалистам в данной области и будут зависеть от вирусного вектора, состава терапии, клеток-мишеней, и объекта, подвергаемого лечению. Одно- и многократные введения могут быть осуществлены с уровнем дозы и паттерном, выбранными лечащим врачом.
Следует понимать, что с помощью доставленного вирусного вектора может быть экспрессирован более чем один трансген. В ином случае, отдельные векторы, каждый экспрессирующий один или несколько различных трансгенов, также могут быть доставлены в ЦНС, как описано в данном документе. Кроме того также предполагается, что вирусные векторы, доставляемые способами по настоящему изобретению, могут быть объединены с другими подходящими композициями и терапиями.
Способы введения генетического материала в клетки
Экзогенный генетический материал (например, кДНК, кодирующая один или несколько терапевтических белков) вводят в клетку ex vivo или in vivo с помощью способов переноса генов, таких как трансфекция или трансдукция, для обеспечения генетически модифицированных клеток. Различные экспрессирующие векторы (т.е., носители для облегчения доставки экзогенного генетического материала в клетку-мишень), как известны любому специалисту в данной области.
При использовании в данном документе «трансфекция клеток» относится к приобретению клеткой нового генетического материала путем включения добавленной ДНК. Таким образом, к трансфекции относится вставка нуклеиновой кислоты в клетку с помощью физических или химических способов. Несколько методов трансфекции известны специалистам в данной области, включая: копреципитацию ДНК фосфатом кальция; DEAE-декстран; электропорацию; опосредованную катионными липосомами трансфекцию; и облегченную вольфрамовыми частицами бомбардировку микрочастицами. Копреципитация ДНК фосфатом стронция является еще одним возможным способом трансфекции.
Напротив, «трансдукция клеток» относится к процессу передачи нуклеиновой кислоты в клетку с помощью ДНК- или РНК-вируса. РНК-вирус (т.е. ретровирус) для передачи нуклеиновой кислоты в клетку, называется в данном документе трансдуцирующим химерным ретровирусом. Экзогенный генетический материал, содержащийся в ретровирусе, включается в геном трансдуцированной клетки. Клетка, которая была трансдуцирована химерным ДНК-вирусом (например, аденовирусом, несущий кДНК, кодирующую терапевтический агент), не имеет экзогенного генетического материала, включенного в ее геном, но будет способна экспрессировать экзогенный генетический материал, который удерживается внутри клетки экстрахромосомально.
Как правило, экзогенный генетический материал включает гетерологичный ген (обычно в виде кДНК, содержащей экзоны, кодирующей терапевтический белок) вместе с промотором, для контроля транскрипцию нового гена. Промотор обычно имеет специфическую нуклеотидную последовательность, необходимую для инициации транскрипции. Необязательно, экзогенный генетический материал дополнительно включает в себя дополнительные последовательности (например, энхансеры), необходимые для получения транскрипционной активности искомого гена. Для целей настоящего обсуждения «энхансер» просто является какой-либо нетрансляционной последовательностью ДНК, которая работает смежно с кодирующей последовательностью (в cis-положении), для того, чтобы изменить исходный уровень транскрипции обусловленный промотором. Экзогенный генетический материал может вводиться в геном клетки сразу же после промотора, так что промотор и кодирующая последовательность функционально связаны так, чтобы обеспечить транскрипцию кодирующей последовательности. Ретровирусный экспрессирующий вектор может включать в себя экзогенный промоторный элемент, для регуляции транскрипции вставленного экзогенного гена. Такие экзогенные промоторы включают как конститутивные, так и индуцируемые промоторы.
Природные конститутивные промоторы контролируют экспрессию важных клеточных функций. В результате, ген под контролем конститутивного промотора экспрессируется во всех условиях роста клеток. Примеры конститутивных промоторов включают промоторы для следующих генов, которые кодируют определенные конститутивные функции или функции «домашнего хозяйства»: гипоксантин фосфорибозилтрансферазы (HPRT), дигидрофолатредуктазы (DHFR), аденозиндезаминазы, фосфоглицеринкиназы (PGK), пируваткиназы, фосфоглицеринмутазы, промотор актина и другие конститутивные промоторы, известные специалистам в данной области. Кроме того, многие вирусные промоторы функционируют конститутивно в эукариотических клетках. К ним относятся: ранние и поздние промоторы SV40; длинные концевые повторы (LTR) из вируса лейкоза Молони и других ретровирусов; и промотор тимидинкиназы вируса простого герпеса, среди прочего. Соответственно, любой из упомянутых выше конститутивных промоторов может быть использован для контроля транскрипции вставки гетерологичного гена.
Гены, которые находятся под контролем индуцируемых промоторов, экспрессируются только или в большей степени, в присутствии индуцирующего агента (например, транскрипция под контролем промотора металлотионеина значительно возрастает в присутствии определенных ионов металлов). Индуцируемые промоторы включают респонсивные элементы (RE), которые стимулируют транскрипцию, когда с ними связываются индуцирующие факторы. Например, существуют RE для сывороточных факторов, стероидных гормонов, ретиноевой кислоты и циклического АМФ. Промоторы, содержащие конкретный RE, могут быть выбраны для того, чтобы получить индуцируемый ответ в определенных случаях, сам RE может быть прикреплен к другому промотору, тем самым придавая индуцибельность рекомбинантному гену. Таким образом, выбрав соответствующий промотор (конститутивный по сравнению с индуцируемым, сильный по сравнению со слабым), можно контролировать как наличие, так и уровень экспрессии терапевтического агента в генетически модифицированной клетке. Если ген, кодирующий терапевтический агент, находится под контролем индуцируемого промотора, доставка терапевтического агента in situ срабатывает при воздействии на генетически модифицированную клетку in situ условиями, разрешающими транскрипцию терапевтического агента, например, путем внутрибрюшинной инъекции специфических индукторов индуцируемых промоторов, которые контролируют транскрипцию агента. Например, in situ экспрессия генетически модифицированными клетками терапевтического агента, кодируемого геном под контролем промотора металлотионеина, усиливается при контакте генетически модифицированных клеток с раствором, содержащим соответствующие (т.е. индукцирующие) ионы металлов in situ.
Соответственно, количество терапевтического агента, который поставляется in situ регулируется путем контроля таких факторов, как: (1) природа промотора, используемого для прямой транскрипции введенного гена, (то есть, является ли промотор конститутивным или индуцибельным, сильным или слабым); (2) количество копий экзогенного гена, которое вставлено в клетку; (3) количество трансдуцированных/трансфецированных клеток, которые вводят (например, имплантируют) пациенту; (4) размер имплантата (например, имплантата или инкапсулированной системы экспрессии); (5) количество имплантатов; (6) длительность с которой трансдуцированные/трансфецированные клетки или имплантаты остаются in situ; и (7) скорость производства терапевтического агента генетически модифицированными клетками. Выбор и оптимизация этих факторов для доставки терапевтически эффективной дозы конкретного терапевтического агента, считаются находящимися в компетенции любого специалиста в данной области без проведения излишних экспериментов с учетом раскрытых выше факторов и клинического профиля пациента.
В дополнение, по меньшей мере, к одному промотору и, по меньшей мере, одной гетерологичной нуклеиновой кислоте, кодирующей терапевтический агент, экспрессирующий вектор может включать селективный ген, например, ген устойчивости к неомицину, для облегчения отбора клеток, которые были трансфецированы или трансдуцированы с помощью экспрессирующего вектора. В ином случае, клетки трансфицируются двумя или несколькими экспрессирующими векторами, по меньшей мере, одним вектором, содержащим ген(ы), кодирующий терапевтический агент(ы), и другим вектором, содержащим селективный ген. Выбор подходящего промотора, энхансера, селективного гена и/или сигнальной последовательностью (описано ниже), считается находящимся в рамках компетенции обычного специалиста в данной области, без проведения излишних экспериментов.
Терапевтический агент может быть направлен на доставку во внеклеточное, внутриклеточное или мембранное расположение. Если необходимо, чтобы продукт гена, секретировался из клеток, экспрессирующий вектор разрабатывается так, чтобы включать в себя соответствующую секреционную «сигнальную» последовательность для секреции терапевтического генного продукта из клетки во внеклеточную среду. Если необходимо, чтобы продукт гена сохранялся в клетке, эту последовательность сигнала секреции отбрасывают. Аналогичным образом, экспрессирующий вектор может быть сконструирован так, чтобы включать в себя «удерживающие» сигнальные последовательности для закрепления терапевтического агента в плазматической мембраны клетки. Например, все мембранные белки имеют гидрофобные трансмембранные участки, останавливающие перемещение белка в мембране и не позволяют белку секретироваться. Конструирование экспрессирующего вектора, в том числе сигнальных последовательностей для нацеливания генного продукта в конкретное место, считается в пределах компетенции любого специалиста в данной области без необходимости проведения чрезмерных экспериментов.
Пример 1
Изменения в прогрессии амилоидных отложений
В этом примере изучаются изменения прогрессии отложения амилоидов в мышах app/ps после избыточной экспрессии различных изоформ АроЕ посредством внутрижелудочкового введения аденоассоциированного вируса серотипа 4 (AAV4).
Аллель эпсилон 4 АроЕ (АроЕ ε4) является первым генетическим фактором риска развития болезни Альцгеймера (AD), тогда как наследование редкого аллеля эпсилон 2 АроЕ (АроЕ ε2) снижает этот риск примерно в два раза. Однако, несмотря на открытие этих сильных генетических ключей почти 17 лет назад, механизмы, посредством которого АроЕ определяет риск развития остаются неопределенными.
Для того, чтобы расшифровать, как различные изоформы (АроЕ ε2, ε3 и ε4) воздействуют на формирование и стабильность фибриллярных амилоидных бляшек, векторы AAV4, кодирующие изоформы каждого АроЕ, инъецировали в желудочек 7 месячных мышей APP/PS. С помощью in vivo мультифотонного имиджинга, популяции амилоидных отложений были записаны на исходном уровне и после экспонирования АроЕ в течение двухмесячного интервала, получили динамическое представление о прогрессии амилоидоза в живом животном.
Кинетика отложения амилоидных бляшек наблюдали как вариабельную в зависимости от каждой изоформы, при том, что мыши, инъецированные АроЕ ε4, имели 38% увеличение сенильных бляшек, тогда как мыши, получавшие АроЕ ε2 демонстрировали 15% снижение количества амилоидных отложений в сравнению с АроЕ ε3 после 2 месяцев. Посмертный анализ подтвердил эти результаты и выявил наличие белков человека АроЕ, декорирующих бляшки в коре головного мозга, что свидетельствует о большой диффузии белка через паренхиму и его фокальное накопление, где депонируются пептиды АВ. Важно отметить, что это повышенное содержание белка АроЕ ε4 также было связано с более тяжелой потерей синапсов вокруг амилоидных отложений.
В целом, имеющиеся данные показали, что перепроизводство различных изоформ АроЕ было способно влиять на развитие болезни и может модулировать степень потери синапсов, один из параметров, который коррелирует с лучшим когнитивным нарушением у пациентов с AD.
1. Внутрижелудочковая инъекция AAV4-ApoE приводит к стабильной экспрессии huApoE и устойчивому обнаружению рекомбинантного человеческого белка АроЕ (huApoE) в головном мозге.
Вкратце, GFP и huApoE иммунодетектировали у мышей APP/PS, инъецированных векторами AAV4. Сигнал GFP можно было наблюдать во всей области желудочка (верхняя панель) и в клетках, выстилающих желудочек, также как и человеческий АРОЕ.
Для того чтобы оценить данный подход, AAV4-Venus (контроль), -АроЕ2, -АроЕ3 и -АроЕ4 инъецировали в желудочек мышей дикого типа. Через два месяца после инъекции человеческие белки АроЕ могут быть обнаружены в корковой паренхиме вокруг амилоидных отложений (обратите внимание на антитело 3Н1, в мышах, инъецированных AAV4-GFP, наблюдали только неспецифический фон). Таким образом, значительный уровень человеческого АроЕ был обнаружен с помощью тИФА в головном мозге, а иммуногистологическое окрашивание Venus и АроЕ подтвердило экспрессию различных трансгенов в клетках, выстилающих желудочек.
Эксперименты количественного ОТ-ПЦР проводили с целью оценки уровней мРНК трансгена. Стандартная кривая позволила нам определить концентрации huApoE мРНК относительно уровня эндогенного GAPDH. Были включены образцы из мышей, которые подвергались воздействию в течение 2 или 5 месяцев. тИФА, разработанный для специфической детекции человеческого АРОЕ проводили на гомогенатах мозга (Фиг. 1 А). Низкие уровни рекомбинантных белков могут быть обнаружены в мышах, которым инъецировали AAV4-APOE по сравнению с животными, обработанными AAV4-GFP, что количественно определено тИФА, специфическим для человеческого АРОЕ (Фиг. 1В) и подтверждено Вестерн-блоттингом.
2. Сверхэкспрессия каждой изоформы АРОЕ дифференциально влияет на прогресс амилоидоза.
In vivo двухфотонный имиджинг использовали для того, чтобы исследовать отложение амилоида с течением времени в живом животном. Вкратце, мышам APP/PS (7 месячным) стереотаксически инъецировали AAV4 векторы, кодирующие АроЕ2, АроЕ3, АроЕ4 и Venus. - После 1 недели, имплантировали черепное окошко и получали изображения отложений амилоида с течением времени после краниотомии. Через 2 месяца животных умерщвляли и проводили посмертные анализы.
Были получены 2-фотонные изображения мышей APP/PS, которым вводили AAV4-АроЕ2, AAV4-ApoE3 или AAV4-ApoE4. Амилоидные бляшки могут быть обнаружены после внутрибрюшинного введения метокси-Х04 (5 мг/кг) и Texas Red-декстрана (молекулярная масса 70000 Да; 12,5 мг/мл в стерильном PBS) вводили в боковую хвостовую вену для получения флуоресцентной ангиограммы. Изображения были получены через одну неделю (=Т0), один месяц и через два месяца после инъекции. С тех же полей получали изображения в течение периода времени для отслеживания прогресса поражений. Несколько новых амилоидных отложений появились, в то время как некоторые из них не обнаруживались более в течение двухмесячного периода.
Полный анализ изображений in vivo показывает, что количество амилоидных отложений значительно увеличивается более быстрыми темпами в мышах APP/PS, инъецированных AAV4-apoE4 по сравнению с животными, обработанными как AAV4-АроЕ3, так и AAV4-Venus. С другой стороны, небольшое, но значимое снижение плотности бляшек выявили при использовании AAV4-ApoE2 (Фиг. 2). Тенденция к более крупным бляшкам наблюдается у мышей APP/PS, которых инъецировали AAV4-ApoE4 (p<0,06), но в целом размер бляшек остается постоянным. Обобщенные данные in vivo показывают, что избыточная экспрессия каждой изоформы АРОЕ дифференциально влияет на прогресс амилоидных отложений in vivo. Инъекция AAV4-ApoE2 приводит к незначительному снижению амилоидной плотности с течением времени, в то время как инъекция AAV4-apoE4 усугубляет амилоидоз.
3. Размер амилоидных бляшек варьирует в зависимости от каждой изоформы АроЕ.
In vivo двухфотонный имиджинг позволил отследить изменения размера каждого амилоидного отложения в течение 2-х месячного периода. Размер бляшек может оставаться стабильным, увеличиваться или уменьшаться с течением времени. Распределения соотношений размеров между T1/Т0 и Т2/Т1 показывают, что существует сдвиг в сторону более крупных амилоидных бляшек у мышей, инъецированных AAV4-ароЕ4 по сравнению с другими группами (Фиг. 3).
4. Посмертная оценка амилоидной нагрузки подтверждает влияние ароЕ2 и ароЕ4 на отложение амилоида.
Через два месяца после инъекции AAV4, посмертная стереологическая оценка показала, что животные, которым инъецировали AAV4-ApoE4, имеют более высокую плотность амилоидных бляшек в коре головного мозга, в то время как никакой разницы не было обнаружено между другими группами (Фиг. 4А). Это увеличенное количество амилоидных отложений наблюдалось, когда бляшки были помечены ThioS или Bam10. Тем не менее, никаких изменений в соотношении между Bam10 и ThioS не было обнаружено. Через пять месяцев после инъекции эффекты каждой изоформы АроЕ были более выраженными по сравнению с двухмесячными (Фиг. 4В). Значительное увеличение плотности отложений наблюдалось, когда мышам инъецировали AAV4-ApoE4, а обратный эффект детектировали при использовании ароЕ2. Опять же, никаких изменений соотношения между Bam10 и ThioS не было обнаружено.
5. Каждая изоформа АроЕ дифференциально влияет на синаптическую плотность вокруг амилоидных отложений.
Array-томография используется для точного определения плотности пре- и постсинаптических элементов вокруг амилоидных отложений. Этот новый способ визуализации предлагает возможности изображения с высоким разрешением молекулярной архитектуры тканей. Array-томография основана на ультратонких срезах образца (70 нм), иммуноокрашивании и 3D-реконструкции. Характерные изображения образцов array-томографии окрашивали по амилоидным бляшкам и постсинаптическому маркеру PSD95. Array-томография показывает, что уменьшение количества постсинаптического маркера PSD95 наблюдается вокруг амилоидных отложений, но этот эффект исчезал вдалеке от бляшек. Количественное определение пре- (синапсин-1) и постсинаптических маркеров в непосредственной близости или далеко от бляшек было осуществлено в каждой группе мышей, которым вводили AAV4 (Фиг. 5A-D). Обширное количественное определение пре- и постсинаптических элементов подтвердили, что уменьшенная плотность Синапсина 1 и PSD95 была связан с амилоидными бляшками, причем этот эффект значительно усиливается, когда АроЕ4 избыточно экспрессируется в головном мозге мышей APP/PS1 (Фиг. 5С, Фиг. 5D). Сверхэкспрессия АроЕ4 связана с повышенной потерей в позвоночнике по сравнению с другими группами в непосредственной близости от амилоидных отложений. Напротив, вокруг бляшек плотность пятна синапсина выше в животных, обработанных АроЕ2.
Заключение
Интравентрикулярные инъекции вируса AAV серотипа 4 привели к устойчивому и длительному сверхпродуцированию растворимых рекомбинантных белков по всей головной паренхиме. Сверхэкспрессия АроЕ2, АроЕ3 и АроЕ4 дифференциально повлияла на ход патологии в мышах APP/PS, так что прогрессия амилоидной нагрузки значительно возрастает, когда АроЕ4 инъецируется по сравнению с АроЕ3. С другой стороны АроЕ2 был ассоциирован с защитными эффектами и несколько амилоидных отложений не детектировалось, больше через два месяца после инъекции. Посмертное иммуногистологический анализ подтвердил отрицательное влияние АроЕ4. Устойчивое перепроизводство АроЕ4 усугубляет потерю синапсов, наблюдаемую вокруг амилоидных отложений по сравнению с АроЕ3, в то время как АроЕ2 оказывает умеренное воздействие. Настоящее исследование показало прямую связь между АроЕ изоформой, прогрессом амилоидоза и потерей синапсов in vivo.
Пример 2
Лечение расстройств центральной нервной системы через спинно-мозговую жидкость (ликвор) у крупных млекопитающих
Для осуществления генной терапии заболеваний мозга, таких как болезнь Альцгеймера, необходимо определить, можно ли достичь долгосрочных, стационарных уровней терапевтических ферментов у млекопитающего. Было обнаружено, что эпендимные клетки (клетки, которые выстилают желудочки головного мозга) могут быть трансдуцированы и секретировать целевой фермент в спинно-мозговую жидкость (CSF). Было установлено, что аденоассоциированный вирус (AAV4) может трансдуцировать эпендиму в мышиной модели с высокой эффективностью. (Davidson et al., PNAS, 28: 3428-3432, 2000.) У мышей наблюдалась нормализация уровней запасенного субстрата в заболевшем мозге после обработки AAV4.
Исследовали возможность эффективного осуществления глобальной доставки вектора, для достижения устойчивых уровней фермента в CSF. Во-первых, необходимо найти вектор, который может трансдуцировать эпендимные клетки (клетки, которые выстилают желудочки) в головном мозге больших млекопитающих. Исследования проводились в собачьей модели LINCL и модели LINCL на приматах, не являющихся человеком. Собаки LINCL нормальны при рождении, но у них развиваются неврологические симптомы в возрасте около 7 месяцев, тестируемые когнитивные расстройства в возрасте ~5-6 месяцев, судороги в возрасте 10-11 месяцев, и прогрессирующая потеря зрения.
Аденоассоциированный вирус (AAV) был выбран в качестве вектора из-за его малого размера (20 нм), из-за того, что большая часть его генетического материала может быть удалена («вычищена»), так что вирусные гены отсутствуют, и из-за чего он является репликативно некомпетентным. Ранее было протестировано, может ли аденоассоциированный вирус типа 4 (AAV4) быть посредником глобальных функциональных и патологических улучшений в мышиной модели мукополисахаридоза типа VII (MPS VII), вызванных дефицитом бета-глюкуронидазы (Liu et al., J. Neuroscience, 25 (41): 9321-9327, 2005). Рекомбинантные векторы AAV4, кодирующие бета-глюкуронидазу, инъецировали в одностороннем порядке в боковой желудочек мышей MPS VII с развившимся заболеванием. Трансдуцированная эпендима экспрессирует высокие уровни рекомбинантного фермента, при этом секретируемый фермент проникает внутрь структур головного мозга и мозжечка, а также ствол мозга. Иммуногистохимические исследования показали тесную связь рекомбинантного фермента и микроциркуляторной части сосудистого русла головного мозга, указывая на то, что бета-глюкуронидаза достигает паренхиму мозга через периваскулярные пространства, выстилающие кровеносные сосуды. Аверсивное ассоциативное обучение было проверено в контексте выработки условного рефлекса страха. По сравнению с подобранными по возрасту гетерозиготными контролями, пораженные мыши показали ослабленный условный рефлекс страха и контекстную дискриминацию. Такой поведенческий дефицит был обращен через 6 недель после генного переноса в мышей MPS VII, обработанных AAV4 с бета-глюкуронидазой. Данные показывают, что эпендимные клетки могут служить в качестве источника секреции ферментов в окружающей паренхиме головного мозга и CSF.
Удивительно, но, однако, когда эти исследования были продолжены на крупных млекопитающих (т.е. собак и приматах), AAV4-векторы не были эффективными при направленном воздействии на эпендиму в этих животных. Вместо этого необходимо использовать вектор AAV2. Вкратце, получали rAAV2, кодирующий ТРР1 (AAV2-CLN2), и инъецировали интравентрикулярно для трансдукции эпендимы (Liu et al., J. Neuroscience, 25 (41): 9321-9327, 2005). TPP1 является ферментом, дефицитным при LINCL. Данные показали, что эпендимная трансдукция в NHP-мозг приводила к значительному увеличению фермента в CSF. Результаты показали повышенные уровни активности ТРР1 в различных участках мозга, где вертикальные оси показывают % контроля активности (Фигура 7).
У первой обработанной собаки, доставка вектора был субоптимальной, но все еще демонстрировала активность CLN2 в головном мозге. Последующие собаки подверглись ICV-доставке со стереотаксией. Было обнаружено, что когнитивные способности обработанных собак были значительно улучшены по сравнению с необработанной собакой, что было измерено по эффективности в Т-образном лабиринте (Фигура 8). Кроме того, эффекты ICV-доставки AAV2-CLN2 в собачьей модели LINCL были весьма выраженными. У необработанного (-/-) животного, присутствуют большие желудочки, тогда как в мозге необработанного контроля и обработанных животных не было обнаружено желудочков. После доставки AAV. TPP1 в желудочки LINCL-собак, обнаруживаемая активность фермента была отмечена в различных областях головного мозга, в том числе в мозжечке и верхней части спинного мозга. В двух дополнительных живых пораженных собаках, атрофия мозга была значительно ослаблена, продолжительность жизни была увеличена, а когнитивная функция была улучшена. Наконец, в NHP, мы показали, что этот способ может помочь достичь уровней активности ТРР1 в 2-5 раз выше, чем у дикого типа.
Несколько векторов AAV были созданы и испытаны для определения оптимального сочетания ITR и капсида. Были получены пять различных комбинаций, и было определено, что AAV2 ITR является наиболее эффективной: AAV2/1 (то есть, AAV2ITR и капсид AAV1), AAV2/2, AAV2/4, AAV2/5, и AAV2/8. Было обнаружено, что AAV2/2 работала намного лучше в крупных млекопитающих (собаки и NHP), а за ней следовали AAV2/8, AAV2/5, AAV2/1 и AAV2/4. Это было довольно неожиданно, поскольку порядок эффективности вирусных векторов противоположен тому, что наблюдался у мышей.
Таким образом, данная работа показала, что выстилающие желудочек клетки могут быть источником рекомбинантного фермента в CSF для распределения по всему мозгу, и что AAV2/2 является эффективным средством введения терапевтических агентов, таких как ген, кодирующий CLN2 (ТРР1), в собаку и приматов.
Пример 3
Человеческие изоформы АроЕ, доставляемые переносом генов, дифференциально модулируют болезнь Альцгеймера, влияя на отложение амилоидов, очистку и нейротоксичность
Болезнь Альцгеймера (AD) является наиболее частым связанным с возрастом нейродегенеративным расстройством и превратилась в серьезную проблему общественного здравоохранения. Среди генов восприимчивости, связанных с поздним возникновением спорадической формы AD, аллель аполипопротеина Ε ε4 (АРОЕ - ген; АроЕ - белок) на сегодняшний день является наиболее значимым фактором генетического риска. Наличие одной копии АРОЕ ε4 существенно повышает риск развития заболевания на 3 по сравнению с наиболее распространенными аллелем АРОЕ ε3, в то время как две копии приводят к 12-кратному повышению риска. Интересно, что АРОЕ ε2 имеет противоположный эффект и является защитным фактором, так что наследование этого специфического аллеля снижает стандартизированный по возрасту риск AD примерно вдвое по сравнению с АРОЕ3/3. Средний возраст начала деменции также соответствует этому профилю риска, у носителей ароЕ4/4 это значение приходится на середину десятилетия после 60 лет, у носителей ароЕ2/3 - это значение приходится на начало десятилетия после 90 лет, со сдвигом почти в 3 десятилетия, в то время как у индивидуумов с АРОЕ3/3 возраст начала заболевания приходится на промежуток - середину десятилетия после 70 лет.
Механизм, посредством которого АроЕ воздействует на AD, является спорным. Накопление Αβ, содержащего сенильные бляшки в гиппокампе и коре пациентов, как полагают, играет центральную роль в AD, потому что все известные гены, ответственные за редкие аутосомно-доминантные формы заболевания, участвуют в выработке пептидов Aβ. Интересно, что было показано, что генотип АРОЕ сильно влияет на степень отложения амилоида у больных с AD, а также количество нейротоксического растворимого олигомерного Αβ, обнаруженного в аутопсии образцов. Было высказано предположение, что изоформы АроЕ дифференциально влияют на цереброваскулярную целостность и влияют на выведение пептидов Αβ через гематоэнцефалический барьер, таким образом, модулируя накопление амилоидных агрегатов вокруг кровеносных сосудов (церебральной амилоидной ангиопатии или CAA). Кроме того, АроЕ также вовлечен непосредственно в нейродегенерацию и нейрональную пластичность. Последствия АроЕ2 были относительно мало исследованы в этих контекстах.
Генетически сконструированные животные, экспрессирующие человеческий АРОЕ2, -Е3 и - Е4 имеют ранговый порядок амилоидной нагрузки как у людей, что соответствует гипотезе о том, что различные изоформы АроЕ влияют на инициацию и/или рост бляшек. Тем не менее, необходимы дальнейшие исследования для того, чтобы проанализировать механизмы АроЕ-опосредованных эффектов на существующие амилоидные отложения и на сохранившуюся нейродегенерацию. Для того, чтобы преодолеть этот пробел в знаниях, мы использовали подход переноса генов, в котором аденоассоциированный вирусный вектор, экспрессирующие различные аллели АРОЕ (или контроль с GFP), инъецировали в боковой желудочек для первичной трансдукции эпендимы, которая затем будет действовать в качестве биологического завода по доставке АроЕ в спинномозговую и интерстициальную жидкости. Затем мы использовали прижизненную многофотонную микроскопию для отслеживания влияния различных изоформ АроЕ на образование, рост, и в случае ароЕ2, растворения бляшек, а также in vivo микродиализных подходов для отслеживания биохимические переменных АроЕ и Αβ в ISF, и array-томографии для оценки изменений в Αβ-ассоциированной нейротоксичности. Мы обнаружили, что изоформы АроЕ воздействуют на уровни растворимого олигомерного Aβ в ISF, темпы Αβ фибрилизации и осаждения, стабильность амилоидных отложений после формирования, их удаление, и степень околобляшечных нейротоксических эффектов. Действительно, мыши с AD, обработанные ароЕ4, показали увеличенное количество растворимого Aβ, более высокую плотность фибриллярных бляшек, рецидив потери синаптических элементов и увеличение количества нейритных дистрофий вокруг каждого отложения, в то время как относительный защитный эффект наблюдался при АроЕ2. Эти данные подтверждают гипотезу о том, что аллели АРОЕ опосредуют их влияние на AD, прежде всего, через Aβ, и привлекают внимание к АРОЕ как к терапевтической мишени.
Результаты
Внутрижелудочковая инъекция AAV4-APOE приводит к стабильной экспрессии АРОЕ и устойчивому производству человеческого АроЕ в головном мозге.
Аполипопротеин Е, является естественно секретируемым белком, выделяемым главным образом, астроцитами и клетками микроглии и может диффундировать по всей церебральной паренхиме. Мы воспользовались преимуществом этого свойства путем инъекции AAV серотипа 4, кодирующего GFP (контроль) или каждый из аллелей АроЕ в боковые желудочки головного мозга в 7 месячных мышей APP/PS1. Учитывая значительные области, пострадавшие от характерных повреждений AD, эта стратегия дает большое преимущество по сравнению с несколькими интрапаренхимальными инъекциями.
Через два месяца после инъекции, трансдуцированные клетки были обнаружены в хориоидном сплетении и эпендиме, выстилающей желудочек, что подтверждает функциональность векторов AAV4. При использовании антител, специфичных для обоих видов с помощью тИФА и вестерн-блоттинга были обнаружены человеческий и мышиный белки АроЕ (Фиг. 9А, 9В и 15А). Мы наблюдали, что концентрация человеческого аполипопротеина Ε достигла 20 цг/мг общего белка в среднем (Фиг. 9А), что составляет около 10% от эндогенного мышиного ароЕ (Фиг. 9В). Наличие этой скромного дополнительного. количества человеческого АроЕ не изменяло детектируемо уровни эндогенного мышиного белка ароЕ (Фиг. 15А). Небольшое, но статистически значимое снижение наблюдалось между 2 и 5 месяцами после инъекции AAV4 (Фиг. 15В). Тем не менее, уровни человеческого белка оставались детектируемыми по сравнению с контрольной группой, что указывает на ААУ4-опосредованную трансдукцию, обеспечиваемую платформой для устойчивой выработки рекомбинантного белка, секретируемого по всей паренхиме. Действительно, человеческие белки АроЕ могут быть обнаружены вокруг амилоидных отложений у мышей APP/PS1 по всей корковой мантии, где, как известно, накапливается эндогенный мышиный белок ароЕ.
Далее, мы оценивали наличие человеческого АроЕ в межклеточной жидкости (ISF), внеклеточном компартменте, который также содержит биологически высокоактивные растворимые виды Aβ. Из-за относительно небольшого количества АроЕ, обнаруженного в лизате целого мозга, мы инъецировали несколько мышей, нокаутных по ароЕ, каждым вектором AAV4-APOE и отслеживали присутствие человеческого белка с использованием высокочувствительных, но не видоспецифических антител. Используя технику микродиализа, мы подтвердили наличие АроЕ в ISF инъецированных животных, нокаутных по ароЕ.
В целом, эти данные подтверждают, что одиночной интрацеребровентрикулярной инъекции AAV4 было достаточно, чтобы привести к устойчивой выработке представляющего интерес белка по всей паренхиме головного мозга и в ISF, и что эпендима/хориоидное сплетение могут быть использованы в качестве «биологического насоса» для того, чтобы доставить потенциально терапевтические белки в мозг.
Инфузия изоформ АроЕ дифференциально влияет на амилоидные пептиды и отложение бляшек
Мышей APP/PS 1 трансдуцировали векторами, которые экспрессировали GFP или различные изоформы АроЕ в течение 5 месяцев перед эвтаназией. Анализ нагрузки амилоидными бляшками показал, что через 5 месяцев наблюдалось значительное увеличение плотности амилоидных отложений в коре головного мозга животных, инъецированных AAV4-APOE4, по сравнению с теми, что экспрессируют АРОЕ2. Плотность бляшек в мышах, обработанных AAV4-GFP и AAV4-APOE3, не отличалась друг от друга на промежуточном уровне (Фиг. 16А).
Концентрации пептидов Aβ40 и Αβ42, измеренных в экстрактах муравьиной кислотой, имитировали изменения, наблюдаемые в содержимом амилоидных бляшек, при этом повышение концентрации амилоидных пептидов было обнаружено у мышей, экспрессирующих аллель АроЕ4 (Фиг. 16В), а противоположный эффект был обнаружен при использовании АРОЕ2 через 5 месяцев. На содержание пептидов Aβ40 и Αβ42 в TBS-растворимой фракции также оказывали влияние инъекции каждого из AAV-APOE (Фиг. 16С). Кроме того, соотношение между агрегированным и растворимым пептидами Aβ осталось неизменным под воздействием АроЕ, что, таким образом, позволяет предположить, что сверхэкспрессия каждой отдельной изоформы человеческого АроЕ одновременно модулирует как фибриллярные, так и растворимые виды амилоида.
Сверхэкспрессия каждой изоформы АроЕ в течение всего 2 месяцев приводит к меньшим эффектам, чем наблюдаемые в 5-месячном исследовании. Тем не менее, наблюдалось значительное увеличение плотности амилоидных бляшек в кортикальной области мышей, инъецированных AAV4-APOE4, по сравнению с другими экспериментальными группами (Фиг. 16А). Это соответствовало количеству Aβ, содержащемуся в фракции муравьиной кислоты (Фиг. 16С), что свидетельствует о преобладающем воздействие этого конкретного варианта. TBS-растворимые виды Аβ40/42 показали тенденцию к тому, чтобы быть ниже или выше, чем AAV4-APOE2 или AAV4-АРОЕ4, соответственно, экспрессируемых в течение 2 месяцев (данные не показаны).
Для того, чтобы определить, может ли наличие человеческих изоформ АроЕ отражать раннее изменение в степени фибриллизации Aβ, мы также измерили соотношение между надежным иммуноокрашиванием Aβ с помощью Bam10 (который метит все амилоидные отложения) и тио-S (который окрашивает только плотное ядро) через 2 месяца после инъекции. Никаких изменений не было обнаружено среди 3 изоформ, что указывает на то, что нет дифференциального воздействия на распределение популяций плотных и диффузных амилоидных отложений среди экспериментальных групп в этих временных рамках (Фиг. 16В). Эти данные указывают на то, что длительная экспозиция вариантами АроЕ оказывает более сильное воздействие на отложение амилоида, чем более короткая экспозиция.
Было высказано предположение, что АроЕ играет важную роль в транспорте Aβ через гематоэнцефалический барьер. Для того, чтобы проверить, может ли экспонирование изоформами АроЕ модулировать отток пептидов Aβ через гематоэнцефалический барьер, концентрации Аβ40 измеряли в плазме каждого инъецированного животного. Мы заметили, что содержание в плазме человеческого Aβ интрацеребровентрикулярно введенного как в мышей AAV4-АРОЕ3, так и мышей AAV4-АРОЕ4 было ниже по сравнению с AAV4-APOE2 и AAV4-GFP (Фиг. 10D). Это говорит о том, что варианты ЕЗ и Е4 помогают сохранить Aβ в компартменте центральной нервной системы, в соответствии с наблюдаемыми относительными повышенными концентрациями Aβ в церебральной паренхиме и в соответствии с предыдущими данными, предполагающими повышенный период полужизни Aβ из-за АроЕ.
Носители АРОЕ4 являются более восприимчивыми к нервно-сосудистого дисфункции, а недавно было показано, что нарушение гематоэнцефалического барьера преимущественно проходит в трансгенных мышах с АроЕ4, даже в отсутствие отложения амилоидов. Для оценки того, может ли внутрижелудочковая инъекция AAV4-APOE в APP/PS поставить под угрозу целостность ВВВ, было осуществлено посмертное окрашивание с помощью прусского синего. Несмотря на наличие нескольких положительных по гемосидерину фокальных областей, редко разбросанных по всему мозгу во всех группах, никаких очевидных различий не наблюдалось между любыми экспериментальными группами животных.
Экспрессия изоформ АроЕ модулирует кинетику прогрессии амилоидоза
АроЕ4 было связан с увеличением плотности амилоидных отложений, в то время как с АроЕ2 после 5 месяцев наблюдали противоположный эффект. Это может отражать изменения в скоростях отложения амилоидов β, очистки или и того, и другого. Для оценки того, как варианты АроЕ оказывают влияние на динамическое развитие амилоидоза, мы использовали in vivo двухфотонный имиджинг с последующим изучением кинетики образования и удаления амилоидных бляшек. Мышам в возрасте 7 месяцев интравентрикулярно инъецировали векторы AAV4, после чего через одну неделю после инъекции имплантировали черепное окошко для того, чтобы получить первую сессию изображений (Т0). Через 1 (Т1) и 2 месяца (Т2), получали изображения амилоидных отложений в тех же полях зрения. Мышей подвергали эвтаназии для посмертного анализа после второй сессии изображений.
Подавляющее большинство амилоидных отложений оставалось стабильным, хотя в течение двухмесячного периода время от времени обнаруживались новые бляшки в небольшом объеме изображений. Кроме того, в редких случаях, метоксиположительные бляшки, отображенные в начале эксперимента, не обнаруживались через один или два месяца, что позволяет предположить, что некоторые бляшки могут быть устранены. С течением времени мы наблюдали общее увеличение объемной плотности амилоидных отложений, с плотностью в Т2 в среднем на 23% больше, чем в Т1. Скорость прогрессии амилоида была быстрее в мышах APP/PS1, обработанных АроЕ4, в то время как животные, экспонированные АроЕ2, через 2 месяца имели значительно меньшую плотность отложения амилоида относительно GFP (0,66), АроЕ3 (0,67) и ароЕ4 (0,74) (Фиг. 11А, 11В). Важно отметить, что эти изменения АроЕ2 отражают снижение по сравнению с исходной линией, демонстрируя напрямую и впервые неимунное активное устранение бляшек. В отличие от данных, полученных на АРОЕ-трансгенных животных, эти результаты демонстрируют, что индукция умеренного увеличения количества АроЕ, может повлиять на текущий амилоидогенный процесс даже после того, как отложение амилоида уже началось.
Затем мы оценивали одиночный рост амилоидных бляшек, путем измерения соотношения площади поперечного сечения индивидуальных отложений между Т1/Т0 и Т2/Т1. Различия были обнаружены в группах в Т1 (соотношение Т1/Т0), но не в Т2 (соотношение Т2/Т1, Фиг. 12), что свидетельствует о том, что наличие вариантов человеческого АроЕ, главным образом, влияет на рост бляшек в течение первого месяца после экспозиции, но этот параметр не отличается впоследствии. В частности, размер амилоидных отложений вырос значительней у мышей, обработанных АроЕ4, по сравнению как с АроЕ2, так и с АроЕ3, что позволяет предположить, что этим аллелем усугубляется не только количество бляшек, но и их размер. Поэтому АроЕ4 влияет как на рассев пептидов Aβ, так и на размер уже существующих бляшек.
Изоформы АроЕ3 и АроЕ4 ухудшают синоптическую плотность вокруг амилоидных отложений по сравнению с АроЕ2
Потеря синапсов является параметром, который наилучшим образом коррелирует с когнитивными нарушениями. Недавно мы показали, что присутствие АроЕ4 связано с более высоким уровнем синаптического олигомерного Aβ в мозге пациентов-людей с болезнью Альцгеймера и приводит к значительно снижению плотности синапсов вокруг амилоидных бляшек по сравнению с АроЕ3 (R.M. Koffie et al., Apolipoprotein E4 effects in Alzheimer's disease are mediated by synaptotoxic oligomeric amyloid-beta. Brain 135, 2155 (Jul, 2012); T. Hashimoto et al., Apolipoprotein Ε, Especially Apolipoprotein E4, Increases the Oligomerization of Amyloid beta Peptide. J Neurosci 32, 15181 (Oct 24, 2012)). Кроме того, недавно полученные in vitro данные показали, что АроЕ4 не может защитить от Aβ индуцированной потери синапсов (M. Buttini et al., Modulation of Alzheimer-like synaptic and cholinergic déficits in transgenic mice by human apolipoprotein Ε depends on isoform, aging, and overexpression of amyloid beta peptides but not on plaque formation. J Neurosci 22, 10539 (Dec 15, 2002); A. Sen, D. L. Alkon, T. J. Nelson, Apolipoprotein E3 (АроЕ3) but not ApoE4 protects against synaptic loss through increased expression of protein kinase С epsilon. J Biol Chem 287, 15947 (May 4, 2012)). Поэтому мы предположили, что непрерывное и диффузное распределение каждой изоформы АроЕ может не только по-разному влиять на кинетику отложения и удаления Aβ в мозге мышей APP/PS, но и на целостность синапсов, окружающих амилоидные отложения.
Плотности пре- и постсинаптических элементов (соответственно синапсина-1 и PSD95) были определены с использованием array-томографии, метода высокого разрешения, основанного на иммунофлуоресцентном окрашивании ультратонких срезов тканей (К.D. Micheva, S.J. Smith, Array tomography: a new tool for imaging the molecular architecture and ultrastructure of neural circuits. Neuron 55, 25 (Jul 5, 2007); R.M. Koffie et al., Oligomeric amyloid beta associates with postsynaptic densities and correlates with excitatory synapse loss near senile plaques. Proc Natl Acad Sci USA 106, 4012 (Mar 10, 2009)). Поскольку было, показано, что олигомерные виды сильно сконцентрированы в непосредственной близости от амилоидных отложений, пятна синапсина-1 и PSD95 определяли количественно либо далеко (>50 μм), либо близко (<50 μм) от бляшек, с использованием ранее разработанных протоколов (R.М. Koffie et al., Oligomeric amyloid beta associates with postsynaptic densities and correlates with excitatory synapse loss near senile plaques. Proc Natl Acad Sci USA 106, 4012 (Mar 10, 2009)). Мы заметили, что потеря пресинаптических элементов вблизи бляшек усугубляется, когда экспрессируется либо АРОЕ3, либо АРОЕ4, чего не было после инъекции AAV4-APOE2 или AAV4-GFP (Фиг. 13А). В отличие от этого, плотность постсинаптических пятен осталась неизменной между мышами, которым вводили GFP, АроЕ2 и АроЕ3, тогда как животные, обработанные АроЕ4 показали значительную потерю PSD95 вокруг амилоидных отложений, что усиливает вредное воздействие АроЕ4 на нейротоксичные эффекты Aβ (Фиг. 13С). Когда плотность синаптических элементов оценивали в областях, расположенных далеко от амилоидных отложений (>50 μм), между группами не было обнаружено никакой разницы, что позволяет предположить, что нет никакого влияния вариантов человеческого АроЕ per se на синаптическую плотность, но есть важное влияние изоформ АроЕ на Aβ-индуцированную нейротоксичность. Относительная синаптическая потеря, наблюдаемая с АроЕ3 и АроЕ4, следовательно, напрямую связана с наличием пептидов Aβ, окружающих каждую бляшку (на расстоянии <50 μм от ее края).
В качестве дополнительного нейропатологического параметра мы также оценивали количество аксональных дистрофий, связанных с амилоидными отложениями в мышах APP/PS1, инъецированных AAV4. В дополнение к снижению плотности шипов вокруг них, сенильные бляшки также вызывают более общее изменение нейропиля с увеличением кривизны аксонов и появлением набухших дистрофий. Эти патологические изменения, вероятно, связано с растворимыми олигомерными видами Aβ, которыми обогащены области в пределах 50 мкм от поверхности бляшек. Мы заметили, что сверхэкспрессия ароЕ4 усугубляет образование SMI312-положительных аксональных дистрофий, связанных с амилоидными отложениями по сравнению с GFP, АроЕ2 и АроЕ3 (Фиг. 13С). Этот результат подтверждает наблюдение, что изоформа АроЕ4 оказывает сильнейшее влияние не только модуляцию образования бляшек, но и влияет на амилоид-ассоциированную нейротоксичность.
Белки человеческого АроЕ модифицируют количество олигомерных частиц Aβ, содержащихся в межклеточной жидкости, в другой мышиной модели AD
Далее мы рассмотрели вопрос о том, может ли наличие различных изоформ АроЕ в ISF изменить количество видов растворимого амилоида в том же самом внеклеточном компартменте. Мы решили инъецировать другую модель AD, мышей Tg2576, с целью подтверждения наших предыдущих выводов в другой трансгенной линии мышей. Мыши Tg2576 сверхэкспрессируют мутантную форму АРР, которая содержит шведскую мутацию, и в данном возрасте представляют собой гораздо более умеренный фенотип, чем мыши APP/PS1. Мы инъецировали когорты 16-18 месячных животных, так что амилоидные отложения уже присутствовали в момент трансдукции AAV4-APOE. Через три месяца после генного переноса, в гиппокамп вставляли микродиализный зонд и собирали образцы для описания ранних изменений, связанных с каждым вариантом АРОЕ в пределах ISF.
Мы наблюдали, что концентрация олигомерных частиц Aβ, измеренных с использованием конкретного 82Е1/82Е1 тИФА, была значительно выше (на 42±7%) после инъекции AAV4-APOE4 по сравнению с AAV4-APOE2 (Фиг. 14), что свидетельствует о том, что присутствие АроЕ может модулировать характер амилоидных агрегатов в этом внеклеточном компартменте. Кроме того, когда в ISF оценивали общие Aβ40 и Αβ42, наблюдались одни и те же тенденции, но без достижения значимости (Фиг. 17А), что свидетельствует о том, что наличие различных изоформ АроЕ в ISF несколько больше влияет на состояние агрегации амилоидных пептидов, чем на общее количество.
Как и ожидалось, посмертные биохимические анализы мозга мышей Tg2576, подвергнутых воздействию различных изоформ АроЕ, показали, что концентрация Αβ42 во фракции муравьиной кислоты была значительно увеличена в животных, обработанных АроЕ4 (Фиг. 17В), что подтверждает во второй трансгенной модели наши наблюдения в мышах APP/PS1.
В совокупности эти биохимические критерии подтверждают, что экспрессия АроЕ в мышах Tg2576 вызывает изменения в амилоидной биологии, схожие с наблюдаемыми в мышах APP/PS1. Важно отметить, что наблюдались ранние изменения содержания οAβ в ISF, где эти нейротоксичные виды могут непосредственно взаимодействовать с синаптическими окончаниями.
Обсуждение
Убедительная связь между наследованием аллелей АРОЕ4, которые повышают риск развития AD, и аллелями АРОЕ2, которые оказывают значимый противоположный эффект, привела к появлению множества предложений о том, как опосредуется этот риск. АроЕ вовлечен в качестве Aβ-связывающего белка в участие в удаление Aβ. Тем не менее, исследования на мышах, нокаутных по ароЕ, к удивлению показали, что отложения Aβ были значительно ниже в отсутствие ароЕ. Замена на человеческий АРОЕ2, АРОЕ3 или АРОЕ4 привела к увеличению отложений амилоида того же порядка, что и у пациентов с AD, что постулируется как происходящее через влияние на инициацию бляшек или образование фибрилл. Альтернативные гипотезы сосредотачиваются на дифференциальных эффектах на разрастание аксонов, или даже предполагают, что эффект АРОЕ генотипа на фенотип болезни Альцгеймера является следствием другого гена в генетическом нарушенном равновесии с АРОЕ на хромосоме 19.
Наши данные, полученные при исследовании 2-х различных мышиных моделей с использованием подхода, ранее протестировано в условиях лизосомной болезни накопления и болезнь Хантингтона, непосредственно отвечают на эти вопросы, используя комбинацию из исследований с помощью in vivo многофотонного имиджинга, стандартной количественной иммуногистопатологии, array-томографии синаптической структуры и новых высокомолекулярных микродиализных подходов, которые позволяют исследовать олигомерные Aβ. Мы показали, что изменение микросреды ISF АроЕ у животных с развившимся заболеванием оказывает поразительное и быстрое аллель-специфическое влияние на сбережение Aβ. Наше исследование показало, что даже умеренное (~10%) увеличение уровней АроЕ4, доставленных в ISF, заметно влияет на Aβ-фенотип и кинетику выведения, с ассоциированным с АроЕ4 удержанием повышенного растворимого Aβ, а также фибриллярных и экстрагируемых муравьиной кислотой форм и повышает нейротоксичность вокруг бляшек, отмеченных синаптической потерей и повышенной аксональной дистрофией. И наоборот, ароЕ2 уменьшает Aβ, и оказывает заметное нейропротекторное действие.
Поскольку небольшие изменения в уровнях ISF АроЕ имеют такие драматические последствия, эти результаты могут привести к пониманию относительно влияния широкого спектра факторов окружающей среды и генетических факторов, которые могут изменить риск или прогрессию AD, путем воздействия на экспрессию АРОЕ. Увеличение АроЕ, по существу больше, чем величина, которая, как мы продемонстрировали, может быть произойти после травмы, эпилепсии, ишемии и диет с высоким уровнем холестерина, с которыми связаны повышенные церебральные уровни Aβ. Кроме того, на экспрессию АРОЕ влияют промоторные полиморфизмы, которые ранее были обнаружены в генетическом дисбалансе аллеля АРОЕ4.
Другие манипуляции, которые влияют на гомеостаз АроЕ или АроЕ-липопротеинов в ЦНС, четко изменяют отложение Aβ. Например, в экспериментах с фокальным переносом генов с помощью АРОЕ-лентивирусов (прежде всего в нейронах гиппокампа), сверхэкспрессия АРОЕ4 оказывает более сильное воздействие на амилоид по сравнению с АРОЕ3. Предыдущие исследования также показали, что агонисты RXR, которые имеют множественные эффекты, включая повышение эндогенного синтеза ароЕ, приводят к выведению Aβ из мозга, возможно, путем влияния на выведение через гематоэнцефалический барьер. Кроме того, трансдукция мозга с помощью CYP46A, который метаболизирует холестерин в ЦНС и снижает его уровень, уменьшает отложение Aβ, поскольку повышается уровень LDL-R в головном мозге, который, как известно, снижает уровни ароЕ. Наконец, генетические манипуляции позволяют предположить, что меняя экспрессию АроЕ наполовину, можно повлиять на Aβ-фенотип. Интересно, что наши результаты позволяют предположить, что более умеренные изменения также могут иметь драматические последствия.
Наши данные непосредственно касаются четырех других важных областей противоречий в литературе по АРОЕ-Альцгеймеру. 1) Мы продемонстрировали четкое влияние изоформы АроЕ на нейротоксичность, оцениваемую по потере синапсов и аксональной дистрофии, которые, вероятно, связаны с нарушениями функции нервной системы. Так как эти эффекты были очевидны в непосредственной близости от бляшек, но не в зонах, удаленных от бляшек, синапс-защитная природа АроЕ2 по сравнению с АроЕ4, вероятно, опосредуется влиянием на пери-бляшечный Aβ, а не непосредственным влиянием АроЕ на синаптическую устойчивость. 2) Прямое наблюдение кинетики отложения и росте бляшек с использованием продольного многофотонного in vivo имиджинга, показывает, что АроЕ4 усиливает отложение и рост бляшек, в то время как АроЕ2 на самом деле связан с растворением бляшек - что указывает на то, что изоформы АроЕ оказывают мощное воздействие на темп и прогресс заболевания за рамками первоначального воздействия на фибриллярное формирование бляшек. Результаты подкрепляют идею о том, что АроЕ4 может ускорить прогресс болезни как по показателю отложения амилоидов, так и по показателю нейротоксичности (и, следовательно, является причиной более раннего возраста возникновения заболевания), а АроЕ2 оказывает противоположное воздействие, что повышает вероятность того, что введение АроЕ2 (или миметика АроЕ2) в ЦНС может иметь терапевтическое значение, даже после развития болезни. 3) были предложены различные механизмы, которыми АроЕ выводит Aβ из мозга или служит в качестве молекулы удержания, которая повышает период полужизни выведения; наши текущие результаты показывают, что введения умеренных количеств АроЕ в CNS достаточно для повышения удержания Aβ в ЦНС, если это не АроЕ2. 4) механизмы значимого защитного действия ароЕ2 при AD были не ясны в течение длительного времени, в частности, поскольку АроЕ2 связывает рецепторы АроЕ относительно слабо. Наши текущие данные свидетельствуют о том, что АроЕ2 имеет усиление функции - способен на самом деле обратить развитие сформировавшихся отложений Aβ, а также осуществить поддержку синаптической и аксональной пластичности - в дополнение к вероятно нейтральному или нулевому воздействию на выведение Aβ в плазму. Это позволяет предположить, что десятилетняя разница в возрасте возникновения заболевания между пациентами, которые наследуют аллели АРОЕ4 и АРОЕ2, может отражать разные точки инициации и обширные различия в кинетике отложения и выведения Aβ, а также аллель-специфические различия по степени нейротоксичности, связанной с отложениями. Эта двойственная функция АроЕ2 может привести к терапевтическим подходам, направленным на имитацию выведения бляшек и способности синаптического восстановления.
Эти результаты согласуются с моделью, в которой ароЕ действует в качестве каркаса для олигомеризации Aβ, с эффективностью формирования и стабилизации олигомерных Aβ АроЕ4>АроЕ3>АроЕ2, и с нашими недавними наблюдениями которые заключаются в том, что содержание олигомерных Aβ повышено в ЦНС ароЕ4>АроЕ3 пациентов-людей с AD (даже тогда, когда нагрузка бляшками нормализуется по всем случаям). Если АроЕ, особенно АроЕ4, опосредует образование нейротоксического олигомерного Aβ, мы прогнозировали, что повышение АроЕ4 приведет к усилению синаптических и аксональных изменений, что очевидно из текущих данных. На основании этих результатов следует проявлять осторожность в отношении агентов, которые могут повысить уровень АроЕ в мозге AD-пациентов, унаследовавших аллель АРОЕ4.
Наконец, наши данные подтверждают силу AAV-опосредованной трансдукции эпендимы, для доставки секретируемых белков в мозг и по всей кортикальной мантии в нем. Генный перенос или другие подходы, которые снижают ароЕ4 или повышают ароЕ2, являются мощными средствами, влияющими на прогресс AD.
Материалы и методы
Животные.
Эксперименты проводились с использованием мышей, трансгенных по двух генам APPswe/PS1dE9 (APP/PS1) (D.R. Borchelt et al., Accelerated amyloid deposition in the brains of transgenic mice coexpressing mutant presenilin 1 and amyloid precursor proteins. Neuron 19, 939 (Oct, 1997)) (получены из Jackson laboratory, Бар-Харбор, Мэн) и мышей Tg2576 (К. Hsiao et al., Correlative memory deficits, Abeta elevation, and amyloid plaques in transgenic mice. Science 274, 99 (Oct 4, 1996)). Ген человеческого мутантного белка-предшественника амилоида, содержащий шведскую двойную мутацию K594N/M595L вставляли в геном этих двух линий мышей под контролем промотора прионного белка. Кроме того, мышиная модель APP/PS1 сверхэкспрессирует вариант гена пресенилина 1 с удаленным экзоном 9 (под управлением того же промотора). Сопутствующая избыточная экспрессия APPswe и PSEN1 в мышах APP/PS1 приводит к более тяжелым фенотипам, со значительным отложением амилоида, видимым уже в 6-ти месячном возрасте. С другой стороны, мышиная линии Tg2576 является гораздо более умеренной моделью, у которой амилоидные бляшки развиваются в возрасте около одного года. Чтобы определить, как введение различных изоформ АроЕ влияет на прогресс заболевания, мы соответственно инъецировали 7-месячных и 16-месячных мышей APP/PS1 (от 4 до 7 животных на состояние) и Tg2576 (от 3 до 5 животных на состояние). Также использовали АРОЕ-дефицитных мышей (АроЕ-КО, the Jackson Laboratory, Бар-Харбор, Мэн). Эксперименты проводили согласно NIH и установленным принципам.
Конструирование и создание вирусных векторов
кДНК АРОЕ-2, -3 и -4 любезно предоставлена доктором LaDu из университета штата Иллинойс (Чикаго). После амплификации с помощью ПЦР, каждую из них гидролизовали с помощью BamHI и вставляли в остов AAV2-PCMV-hrGFP. Высокие титры векторов AAV серотипа 4 (AAV4-apoE2, AAV4-АРОЕ3, AAV4-apoE4 и AAV4-GFP) получали с использованием бакуловирусной системы при содействии подразделения Gene Transfer Vector Core Университета штата Айова, Айова-Сити. Вирусы титровали с помощью количественной ПЦР.
Стереотаксические интравентрикулярные инъекции. Стереотаксические внутрижелудочковые инъекции векторов AAV серотипа 4 проводили, как описано ранее (T.L. Spires et al., Dendritic spine abnormalities in amyloid precursor protein transgenic mice demonstrated by gene transfer and intravital multiphoton microscopy. JNeurosci 25, 7278 (Aug 3, 2005); G. Liu, I.H. Martins, J.A. Chiorini, B.L. Davidson, Adeno-associated virus type 4 (AAV4) targets ependyma and astrocytes in the subventricular zone and RMS. Gene Ther 12, 1503 (Oct, 2005)). Животных анестезировали с помощью интраперитонеальной инъекции кетамина/ксилазина (100 мг/кг и 50 мг/кг массы тела, соответственно), и помещали на стереотаксическую раму (David Kopf Instruments, Туюнга, Калифорния). Инъекции векторов осуществляли в каждый боковой желудочек с использованием 5 ил вирусного препарата (титр 2×1012 vг/мл) с использованием острой микропипетки 33 калибра, прикрепленной к 10-мкл шприцу Гамильтона (Hamilton Medical, Рено, Невада) со скоростью 0,25 мкл/мин. Стереотаксические координаты участков для инъекции рассчитывали от темени (фронтально + 0,3 мм, саггитально ± 1 мм и вертикально - 2 мм).
Черепно-мозговая имплантация и мультифотонный имиджинг. Через неделю после внутрижелудочковой инъекции мышей анестезировали изофлураном (1,5%) и имплантировали черепные окошки путем удаления куска черепа и замены его на покровное стекло диаметром 8 мм (как описано ранее, Т. L. Spires et al., Dendritic spine abnormalities in amyloid precursor protein transgenic mice demonstrated by gene transfer and intravital multiphoton microscopy. J Neurosci 25, 7278 (Aug 3, 2005)). Для обработки изображений, вдоль границы окна было построено восковое кольцо для того, чтобы создать колодец с водой для объектива (20 * объектив, числовая апертура 0,95, Olympus). Для визуализации амилоидных отложений, трансгенные животные за 24 часа до операции получали внутрибрюшинную инъекцию метокси-Х04 (5 мг/кг), флуоресцентного соединения, которое пересекает гематоэнцефалический барьер и связывается с амилоидными отложениями (В.J. Bacskai, W.Ε. Klunk, С.A. Mathis, В.T. Hyman, Imaging amyloid-beta deposits in vivo. J Cereb Blood Flow Metab 22, 1035 (Sep, 2002)). До обработки изображений, в боковую хвостовую вену вводили Texas Red декстран (70000 Да молекулярная масса; 12,5 мг/мл в стерильном PBS; Molecular Probes, Юджин, Орегон), для получения флуоресцентной ангиограммы, так чтобы форма сосудистой сети могла бы использоваться в качестве ориентира для того, чтобы исследовать одни и те же поля зрения с течением времени. Мышей обследовали через неделю после инъекции AAV для того, чтобы оценить базовый уровень амилоидных отложений, а затем через один и два месяца после инъекции.
Тксапфировый лазер со сфазированными модами (Maitai, Spectra-Physics, Маунтин-Вью, Калиформния), установленный в многофотонной системе визуализации (Bio-Rad 1024ES, Bio-Rad, Геркулес, Калифория), генерировал 860 нм двухфотонный свет возбуждения флуоресценции. Излучение света была собрано с помощью специально построенного внешнего детектора, содержащего три фотоэлектронных умножителей (Hamamatsu Photonics, Бриджуотер, Нью-Джерси), в диапазоне 380-480, 500-540 и 560-650 нм. 2-цветные изображения получали для бляшек и ангиографии одновременно. Получали in vivo изображения со слабым увеличением (615×615 мкм; z-step, 2 мкм, глубина ~200 мкм) и собирали изображения 6-8 полей зрения для покрытия большей области коры.
Обработка изображений и анализ. Плотность бляшек в каждом поле зрения количественно оценивали с помощью «Image J», путем получения общего количества амилоидных отложений на объем отображаемой коры. Мы рассмотрели корковой объем, начиная с первого среза Z-стека на поверхности до последнего среза, в котором может быть обнаружено амилоидное отложение. Размер амилоидных отложений оценивали в зависимости от времени путем измерения области их поперечного сечения от максимальной интенсивности, после двумерной проекции. Для каждой бляшки рассчитывали отношение площади между начальной временной точкой и первым месяцем (Т1/Т0), или между вторым и первым месяцами (Т2/Т1).
Настройки мультифотонного микроскопа (мощности лазера и РМТ) сохраняли неизменными на протяжении различных сессий получения изображений в течение всего эксперимента.
In vivo сбор образцов для микродиализа. In vivo сбор образцов для микродиализа интерстициального Aβ и АроЕ мозга проводили на мышах Tg2576, через 3 месяца после интрацеребровентрикулярной инъекции каждого AAV4 (S. Takeda et al., Novel microdialysis method to assess neuropeptides and large molecules in free-moving mouse. Neuroscience 186, 110 (Jul 14, 2011)). Микродиализный зонд имел 4 мм стержень с 3,0 мм полиэтиленовой (ПЭ) мембраной с отсечением по молекулярной массе 1000 кДа (MWCO) (РЕР-4-03, Eicom, Киото, Япония). Перед использованием зонд предварительно обрабатывали коротким погружением его в этанол, с последующей промывкой перфузионным буфером - искусственной спинномозговой жидкостью (aCSF) (в мМ: 122 NaCl, 1,3 CaCl2, MgCl2 1,2, 3,0 KH2PO4, 25,0 NaHCO3), который фильтровали через мембрану с размером пор 0,2 мкм. Входное и выходное отверстие предварительно обработанного зонда соединяли с перистальтическим насосом (ERP-10, Eicom, Киото, Япония) и насосом-микрошприцем (ESP-32, Eicom, Kyoto, Japan), соответственно, с использованием фторированной этилен-пропиленовой (FEP) трубки (внутренний диаметр φ 250 мкм).
Имплантацию зонда проводили, как описано ранее (S. Takeda et al., Novel microdialysis method to assess neuropeptides and large molecules in free-moving mouse. Neuroscience 186, 110 (Jul 14, 2011; J.R. Cirrito et al., In vivo assessment of brain interstitial fluid with microdialysis reveals plaque-associated changes in amyloid-beta metabolism and half-life. J Neurosci 23, 8844 (Oct 1, 2003)), с небольшими модификациями. Вкратце, наркотизированных животных (1,5% изофлурана) стереотаксически имплантировали с помощью направляющей канюли (ПЭГ-4, Eicom, Киото, Япония) в гиппокамп (брегма -3,1 мм, -2,5 мм латерально к средней линии, -1,2 мм вентрально к твердой мозговой оболочке). Направляющую фиксировали к черепу, используя бинарный стоматологический цемент.
Через четыре дня после имплантации направляющей канюли мышей помещали в стандартную микродиализную клетку и вставляли зонд в направляющую. После введения зонда, для того, чтобы получить стабильные записи, зонд и соединительные трубки перфузировали ACSF в течение 240 мин при скорости потока 10 мкл/мин перед отбором проб. Образцы собирали при скорости потока 0,25 (для количественного определения Aβ) и 0,1 мкл/мин (для обнаружения АроЕ). Образцы хранили при 4°C в полипропиленовых пробирках. Во время сбора микродиализных образцов, мыши бодрствовали и свободно перемещались в микродиализной клетке, разработанной для того, чтобы позволить беспрепятственное перемещение животных без применения давления на зонд в сборе (AtmosLM microdialysis system, Eicom, Киото, Япония).
Иммуногистологический анализ. Мышей APP/PS 1 подвергали эвтаназии ингаляцией CO2 через 2 или 5 месяцев после внутрижелудочковой инъекции (краткосрочного и долгосрочного воздействия), в то время как животных Tg2576 умерщвляли через 3 месяца. Одно целое полушарие фиксировали в 4% параформальдегиде в фосфатном буферном солевом растворе для иммуногистохимического анализа и заливали в парафин. Осуществляли 1 мм коронарный срез через лобную кору для анализа array-томографией, а вторую половину мозга быстро замораживали для выполнения биохимических и биомолекулярных анализов.
Для детекции амилоидных отложений, АРОЕ и GFP, парафиновые срезы (10 мкм) последовательно депарафинизировали в ксилоле, регидратировали в этаноле, обрабатывали в цитратном буфере (10 мМ цитрат натрия, 0,05% Твин 20, pH 6,0), пермеабилизовали в PBS с 0,5% Тритон и блокировали в PBS с 3% БСА в течение 2 часов при комнатной температуре. Инкубацию с первичными антителами проводили в течение ночи при 4°C: Bam10 (Sigma 1: 1000) и R1282 (1: 500, предоставлены д-р Dennis Selkoe) для амилоидных бляшек, мышиные моноклональные антитела 3Н1 (Ottawa Heart Institute) для человеческого АроЕ, куриные анти-GFP (1: 500, Aves) и SMI-312 (Covanee) для аксональных дистрофий. Инкубацию со вторичным антителом проводили в течение 2 ч при комнатной температуре на следующий день. Амилоидные бляшки с плотным ядром метили путем инкубации срезов в течение 8 минут в растворе Thio-S (Sigma, Сент-Луис, штат Миссури) 0,05% в 50%-ном этаноле перед заливкой.
Подготовка образцов, иммунноокрашивание и анализ изображений для array-томографии
Анализ array-томографии для пре- и постсинаптических элементов проводили, как описано ранее (R.М. Koffie et al., Oligomeric amyloid beta associates with postsynaptic densities and correlates with excitatory synapse loss near senile plaques. Proc Natl Acad Sci USA 106, 4012 (Mar 10, 2009)). Вкратце, кусок коры ткани головного мозга (1 мм3), примыкающей к области желудочка иссекали и фиксировали в течение 3 ч в 4% параформальдегиде, 2,5% сахарозе в 0,01 M PBS. После дегидратации в этаноле, образцы инкубировали в «LR White resin» (Electron Microscopy Sciences) в течение ночи при 4°C до полимеризации при 53°C. Ленты срезов (70 нм) затем разрезали на микротоме «ULTRACUT» (Leica) с помощью ножа «Jumbo Histo Diamond» (Diatome).
После регидратации в 50 мМ глицине в TBS в течение 5 минут, срезы блокировали в 0,05% Tween и 0,1% БСА в Tris в течение 5 минут, и наносили первичные антитела 1:50 в блокирующем буфере в течение 2 часов (PSD95 Abeam Ab 12093, Синапсин I Millipore АВ1543 и NAB61 от д-ра Вирджинии Ли, которые преимущественно окрашивают виды олигомерного Aβ, Ε. В. Lee et al., Targeting amyloid-beta peptide (Abeta) oligomers by passive immunization with a conformation-selective monoclonal antibody improves learning and memory in Abeta precursor protein (APP) transgenic mice. J Biol Chem 281, 4292 (Feb 17, 2006)). Срезы промывали TBS и наносили вторичные антитела (анти козьи-Alexa Fluor 488, анти-мышиные Су3, или анти-мышиные Alexa Fluor 488 Invitrogen). Изображения получали на 7-30 серийных срезов сквозь лобную кору с помощью конфокального/многофотонного микроскопа Zeiss Axioplan LSM510 (план апохроматический масляный объектив с 63х числовой апертурой).
Изображения анализировали, как описано выше, используя «Image J» (открытое программное обеспечение Национальных институтов здравоохранения) и «MATLAB» (Mathworks) (R.M. Koffie et al., Oligomeric amyloid beta associates with postsynaptic densities and correlates with excitatory synapse loss near senile plaques. Proc Natl Acad Sci USA 106, 4012 (Mar 10, 2009)). Каждый набор изображений был преобразован в стеки, и выровнен с помощью плагинов «MultiStackReg» и «StackReg» из «Image J» (любезно предоставлены Brad Busse и P. Thevenaz, Стэнфордский университет). Известные объемы были отобраны и автоматизированы, пороговая программа детекции была использована для подсчета пятен как PSD95, так и Синапсина, которые появились в более чем одном последовательном срезе (программа Watershed, представленная Brad Busse, Stephen Smith, и Kristina Micheva, Стэнфордский университет). Экспортированый из «Watershed» стэк изображений с заданными пределами (отдельно для каждого канала) демонстрировал пятна, которые присутствовали в более чем одном срезе массива. Отбирали по несколько участков в коре головного мозга на мышь, и измеряли расстояния от края бляшек.
Количественная оценка Aβ
Концентрации Аβ40 и Αβ42 в TBS-растворимой фракции, фракции муравьиной кислоты, а также- в микродиализате определяли с помощью ΒΝΤ-77/ΒΑ-27 (для Аβ40) и BNT-77/BC-05 (для Αβ42) сэндвич тИФА (Wako Pure Chemical Industries, Осака, Япония), в соответствии с инструкциями изготовителя. Количество олигомеров Aβ в образце определяли с помощью 82Е1/82Е1 сэндвич тИФА (Immuno-Biological Laboratories, Inc, Гамбург, Германия), в которых одни и те же N-концевые (остатки 1-16) антитела были использованы как для захвата, так и обнаружения (W. Xia et al., A specific enzyme-linked immunosorbent assay for measuring beta-amyloid protein oligomers in human plasma and brain tissue of patients with Alzheimer disease. Arch Neurol 66,190 (Feb, 2009)).
Анализ иммуноблоттингом
Мозговые TBS-растворимые фракции и микродиализаты (20 мкг белка), подвергали электрофорезу на гелях 4-12% «Novex Bis-Tris» (Invitrogen) в электродном буфере с MOPS для ДСН-ПААГ (Invitrogen). Гели переносили на PVDF-мембрану и блокировали в течение 60 мин. при комнатной температуре в 5% молоке/TBS-T. Мембраны исследовали с помощью зонда, козьего анти-АроЕ антитела (1: 1000, Millipore, АВ947) для обнаружения небольшого количества АРОЕ в ISF из АРОЕ-нулевых животных, в то время как альбумин детектировали в качестве контроля. Блоты для человеческого и мышиного АроЕ, соответственно, исследовали с помощью антитела ЕР 1373Y (1: 1000, No vus Biologicals, NB110-55467) и кроличьего поликлонального антитела к ароЕ (1: 1000, Abeam, ab20874). Инкубация с HRP-сопряженными козьими IgG антителами (Vector) осуществляли в течение 2 часов. Иммунореактивные белки исследовали с помощью набора «ECL» (Western Lightning, PerkinElmer) и детектировали на «Hyperfilm ECL» (GE Healthcare).
кОТ-ПЦР
Общую РНК из образцов мозга извлекали с помощью реактива «TRIzol®» (Life Technologies, 15596-026), а затем синтезировали кДНК согласно инструкциям производителя системы «SuperScript® III One-Step RT-PCR» (Life Technologies, 12574-018). ПЦР-праймеры специально разрабатывали для амплификации мРНК рекомбинантного человеческого АРОЕ и мРНК эндогенного АроЕ и GAPDH (АроЕ прямой: 5'-AGCTCCCAAGTCACACAAGA; АроЕ обратный: 5'GTTGCGTAGATCCTCCATGT; АРОЕ прямой: 5'CCAGCGACAATCACTGAAC; АРОЕ обратный: 5'- GCGCGTAATACGACTCACTA; GAPDH прямой: 5 ' ATGAC АТС AAGA AGGTGGTG и GAPDH обратный: 5'CATACCAGGAAATGAGCTTG). тИФААРОЕ
Специфический тИФА использовали для обнаружения как человеческого, так и эндогенного мышиного белка АРОЕ. Вкратце, тИФА-планшеты покрывали в течение ночи с помощью 1,5 мкг/мл козьего анти-АРОЕ антитела (для обнаружения мышиного АРОЕ) или 1,5 мкг/мл антитела WUE4 (для детекции человеческого АРОЕ) и блокировали 1% обезжиренным молоком, разведенным в PBS в течение 1,5 часа при 37°C. Используемые в качестве стандартов человеческие рекомбинантные белки ароЕ (для человек-специфичного анализа, «BioVision») или стандарт белков домашнего хозяйства мыши из экстракта головного мозга (для мышь-специфичного анализа) и образцы разводили в буфере для тИФА (0,5% BSA и 0,025% Твин-20 в PBS) и инкубировали в течение ночи. После промывки использовали детектирующие антитела, специфичные для человека (козьи - АроЕ Millipore; 1: 10000) или мыши (Abeam ab20874; 1: 2000), соответственно, с последующей 1,5 ч инкубацией с соответствующим HRP-конъюгированным вторичным антителом. Обнаружение сигнала осуществляли с использованием субстрата ТМВ до остановки раствора с использованием Н3РО4. Колориметрические результаты измеряли при длине волны 450 нм.
Статистические анализы
Статистический анализ проводился с использованием программного обеспечения «Prism». Из-за небольшого размера выборок, нормальность не может быть принята для большинства анализов. Для всех посмертных анализов был осуществлен непараметрический критерий Крускала-Уоллиса с последующим критерием множественных сравнений Данна для оценки влияния каждого инъецированного вектора. Данные in vivo имиджинга анализировали с помощью модели смешанных эффектов, визуализации данных прогрессии амилоида с использованием случайного эффекта для мыши, и фиксированных эффектов для вектора, времени и исходной объемной плотности. В этом анализе учитывали взаимодействие между временем и вектором, но оно не было значимым. Для анализа размера бляшек в зависимости от времени, две модели смешанных эффектов были подогнаны для логарифма отношения двух последовательных временных точек, со случайными эффектами для мыши и фиксированными эффектами для логарифма исходного размера (t0 в первом анализе, t1 во втором анализе).
Все публикации, патенты и патентные заявки включены в настоящее описание ссылкой. Хотя в вышеуказанном описании данное изобретение было представлено в отношении некоторых предпочтительных вариантов осуществления, и многие подробности были изложены в иллюстративных целях, специалистам в данной области техники должно быть понятно, что изобретение допускает дополнительные варианты осуществления, и что некоторые подробности, описанные здесь, могут значительно варьировать без отхода от основных принципов изобретения.
Термины в контексте описания изобретения должны толковаться как охватывающее как единственное, так и множественное число, если не указано иное или если есть явное противоречие контексту. Термины «содержащий», «имеющий», «включающий» и «охватывающий» должны быть истолкованы как неограничивающие термины (т.е. означающие «включающий, без ограничения перечисленным»), если не указано иное. Указание диапазонов значений в данном документе предназначены только, чтобы служить кратким способом отсылки индивидуально на каждое отдельное значение, попадающее в диапазон, если здесь не указано иное, и каждое отдельное значение включено в описание, как если бы оно было индивидуально указано в данном документе. Все описанные в данном документе способы могут быть выполнены в любом подходящем порядке, если здесь не указано иное или если есть явное противоречие контексту. Использование любых и всех примеров или примерных формулировок (например, «такой как») в данном документе, предназначено только для лучшего освещения изобретения и не накладывает ограничения на объем изобретения, если не заявлено иное. Ни одна формулировка в описании не должна быть истолкована как указание на любой незаявленный элемент как существенный при практическом осуществлении изобретения.
Воплощения настоящего изобретения описаны в данном документе, в том числе лучший способ, известный изобретателям для осуществления изобретения. Варианты этих воплощений могут стать очевидными для специалистов в данной области техники после прочтения приведенного выше описания. Изобретатели ожидают, что специалист в данной области использует такие вариации соответствующим образом, и авторы изобретения подразумевают, что изобретение будет осуществлено на практике иначе, чем конкретно описано в данном документе. Соответственно, данное изобретение включает все модификации и эквиваленты объекта изобретения, изложенного в прилагаемой формуле изобретения в соответствии с действующим законодательством. Кроме того, изобретением охватывается любая комбинация вышеописанных элементов во всех возможных вариантах, если иное не указано в данном описании, или если иное явно не противоречит контексту.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНЕЙ МОЗГА | 2014 |
|
RU2664471C2 |
| ВЕКТОРНЫЕ КОМПОЗИЦИИ С МОДИФИЦИРОВАННЫМ АДЕНОАССОЦИИРОВАННЫМ ВИРУСОМ | 2013 |
|
RU2679843C2 |
| КОМПОЗИЦИИ, СПОСОБЫ И ПРИМЕНЕНИЯ ПЕРЕНОСА ГЕНОВ ДЛЯ ЛЕЧЕНИЯ НЕЙРОДЕГЕНЕРАТИВНЫХ ЗАБОЛЕВАНИЙ | 2017 |
|
RU2805606C2 |
| ИНТРАТЕКАЛЬНОЕ ВВЕДЕНИЕ ВЕКТОРОВ НА ОСНОВЕ АДЕНОАССОЦИИРОВАННЫХ ВИРУСОВ ДЛЯ ГЕННОЙ ТЕРАПИИ | 2016 |
|
RU2775138C2 |
| ВАРИАНТ СРЕДСТВА ДЛЯ RNAi | 2018 |
|
RU2789647C2 |
| АДЕНОАССОЦИИРОВАННЫЕ ВИРУСНЫЕ ВЕКТОРЫ ДЛЯ ЛЕЧЕНИЯ МИОЦИЛИНОВОЙ (MYOC) ГЛАУКОМЫ | 2015 |
|
RU2746991C2 |
| ТЕРАПЕВТИЧЕСКИЕ СОЕДИНЕНИЯ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ХАНТИНГТОНА | 2015 |
|
RU2711147C2 |
| AAV ВЕКТОРЫ С УСОВЕРШЕНСТВОВАННЫМИ Rep-КОДИРУЮЩИМИ ПОСЛЕДОВАТЕЛЬНОСТЯМИ, ИСПОЛЬЗУЕМЫМИ В СИСТЕМАХ ПРОДУКЦИИ НА ОСНОВЕ КЛЕТОК НАСЕКОМЫХ | 2007 |
|
RU2457252C2 |
| АДЕНОАССОЦИИРОВАННЫЕ ВИРУСНЫЕ ВЕКТОРЫ ДЛЯ ЛЕЧЕНИЯ МИОЦИЛИНОВОЙ (MYOC) ГЛАУКОМЫ | 2015 |
|
RU2718047C2 |
| ВЕКТОРЫ НА ОСНОВЕ АДЕНОАССОЦИИРОВАННОГО ВИРУСА ДЛЯ ЛЕЧЕНИЯ МУКОЛИПИДОЗА ТИПА II | 2016 |
|
RU2742612C2 |










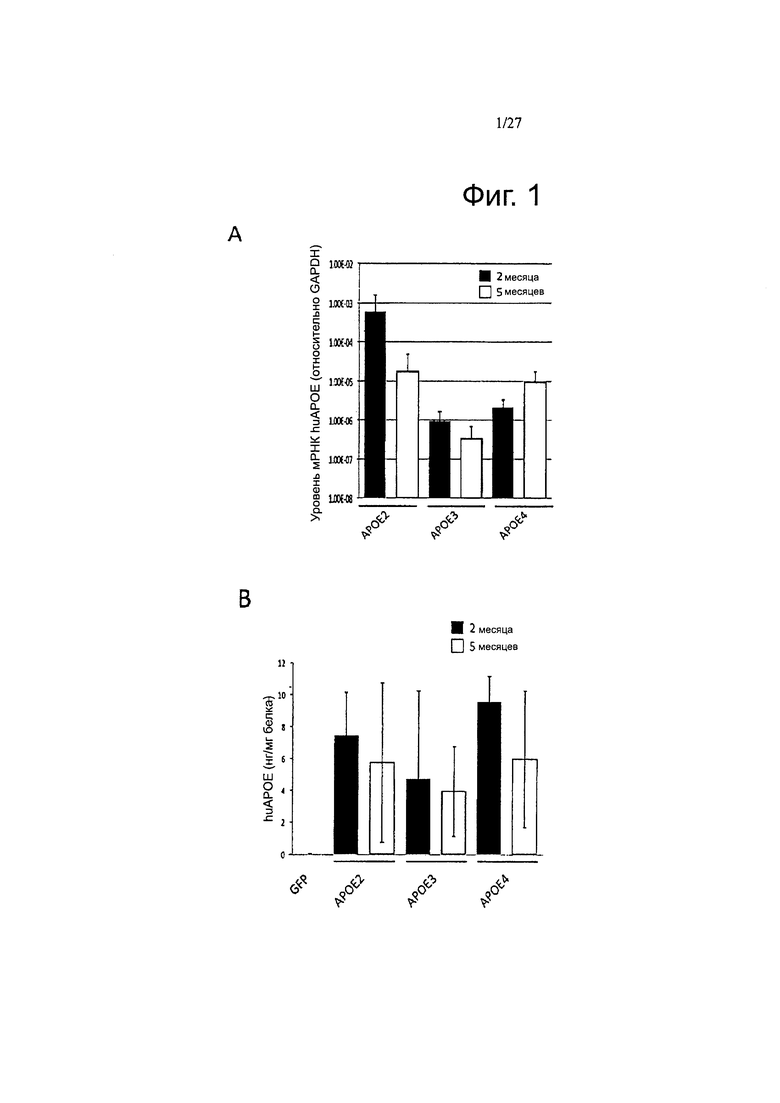
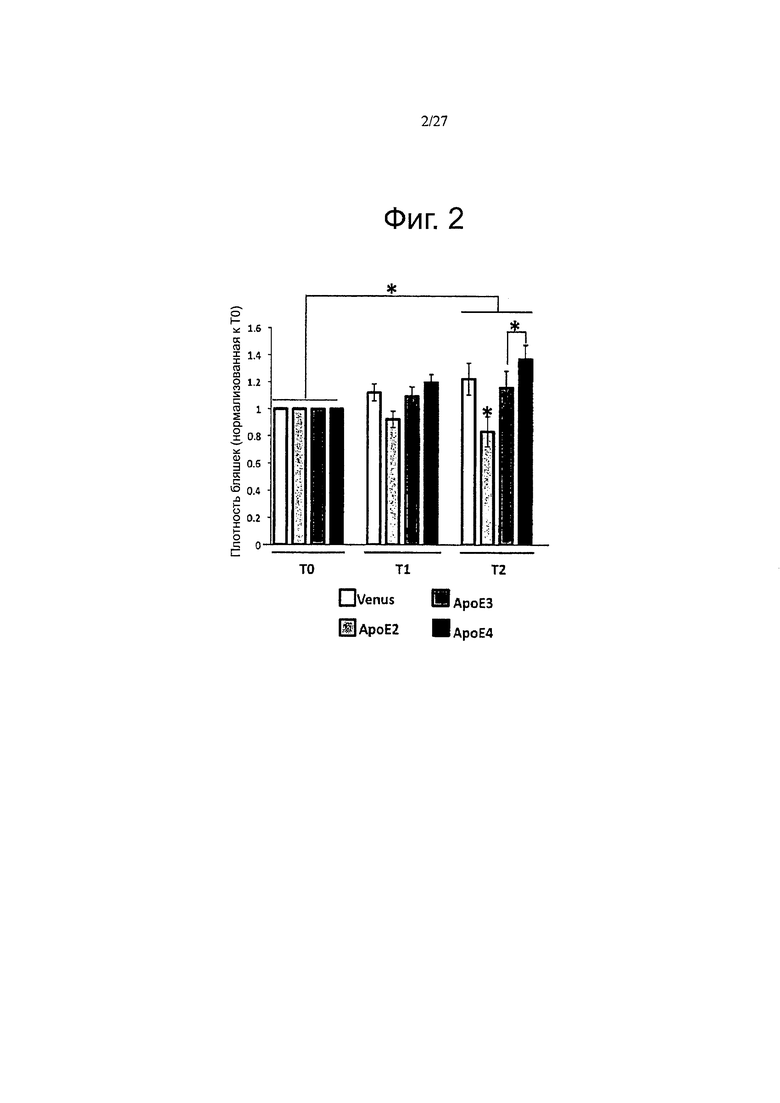
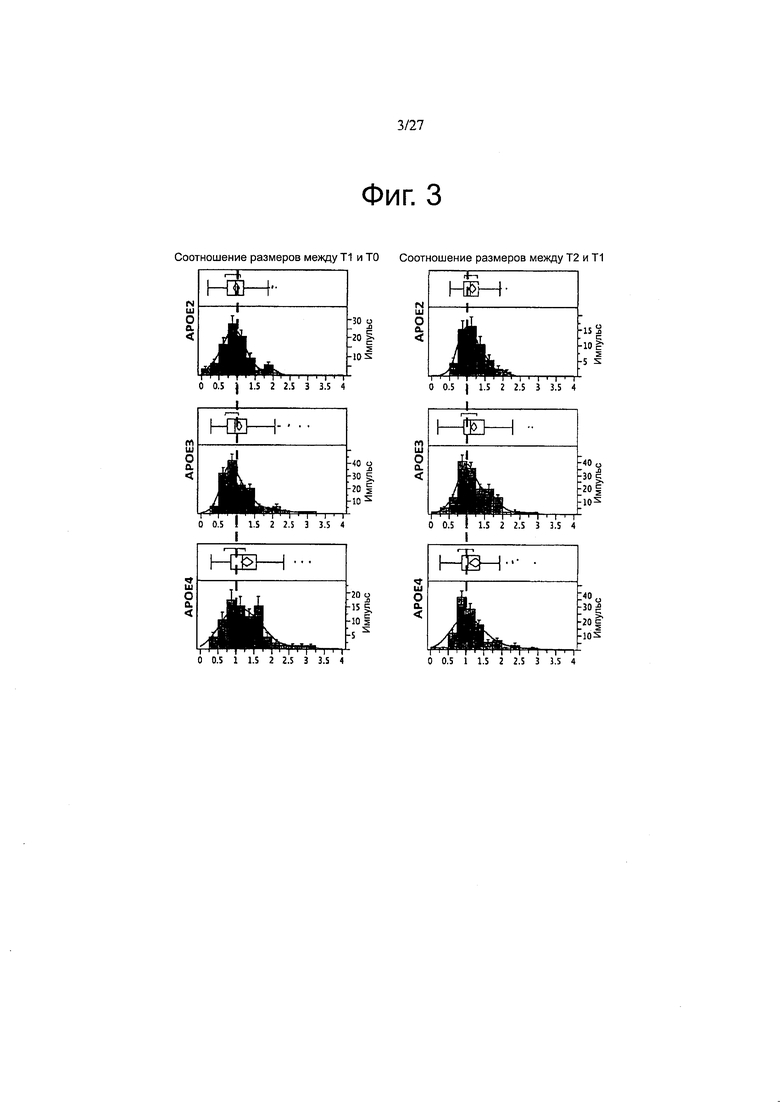
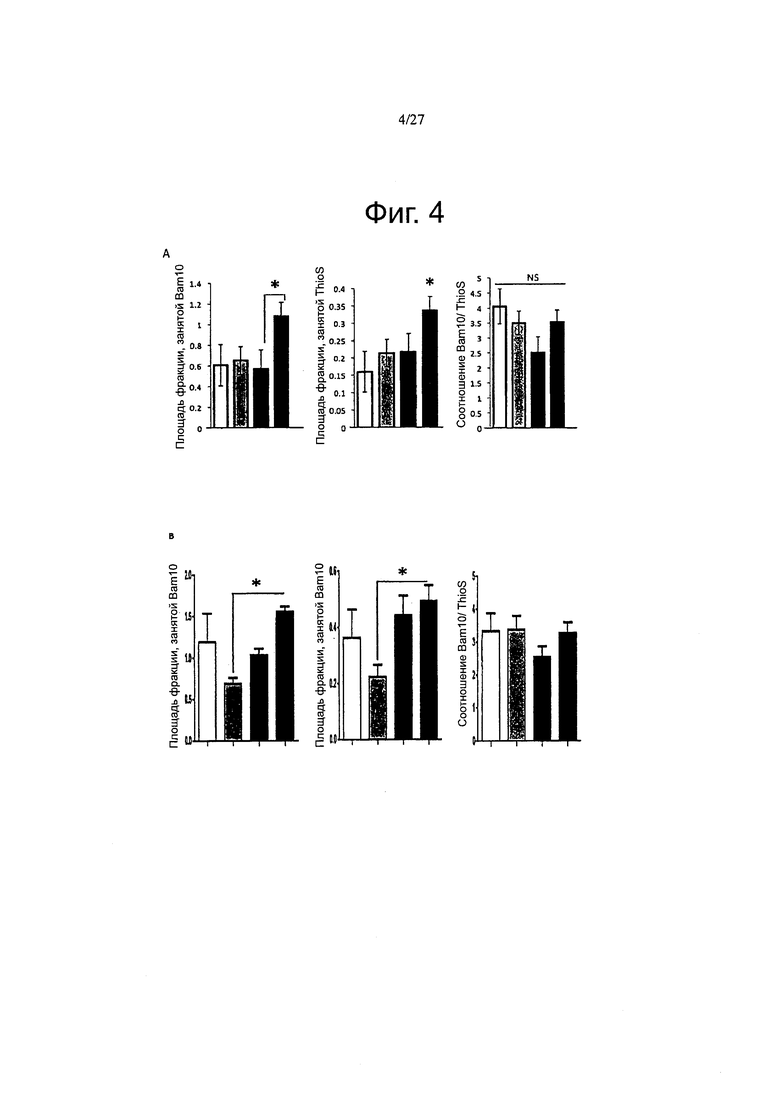
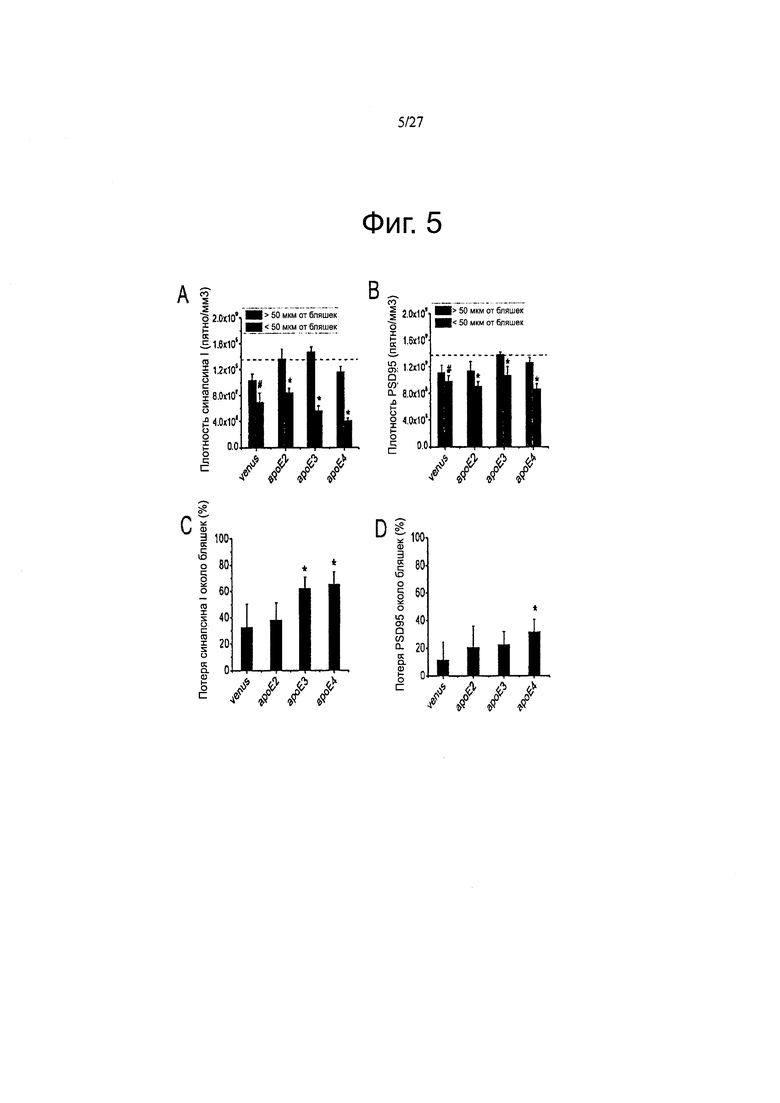










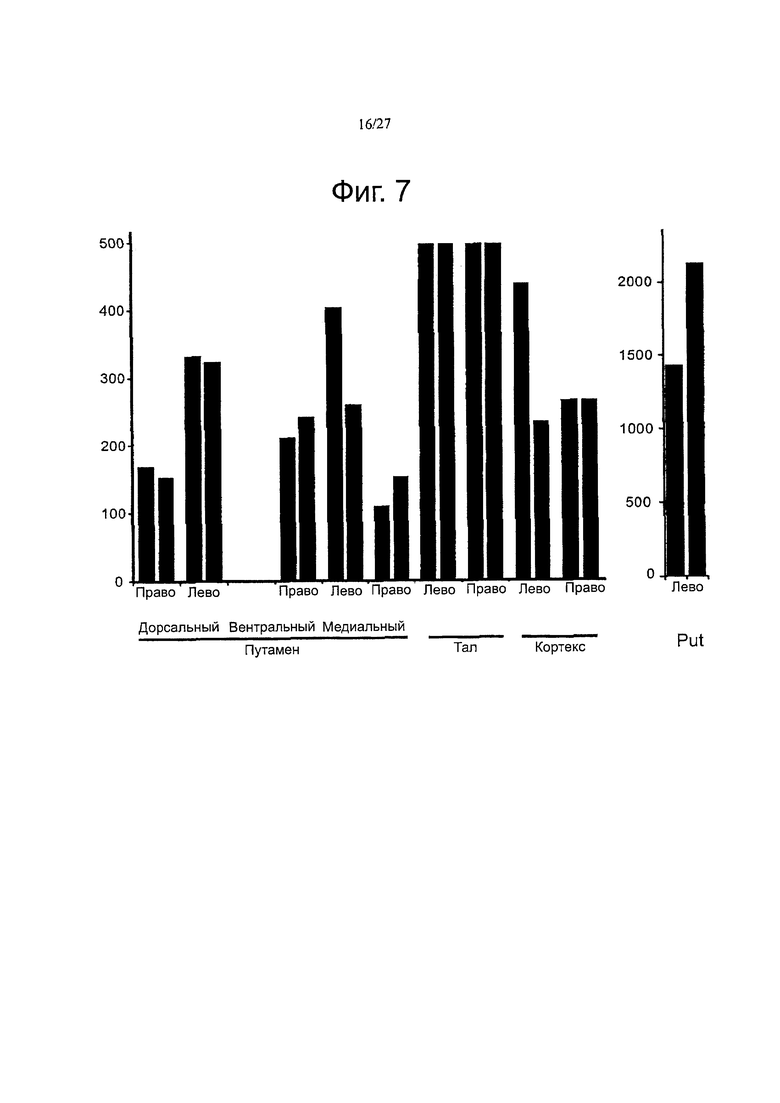
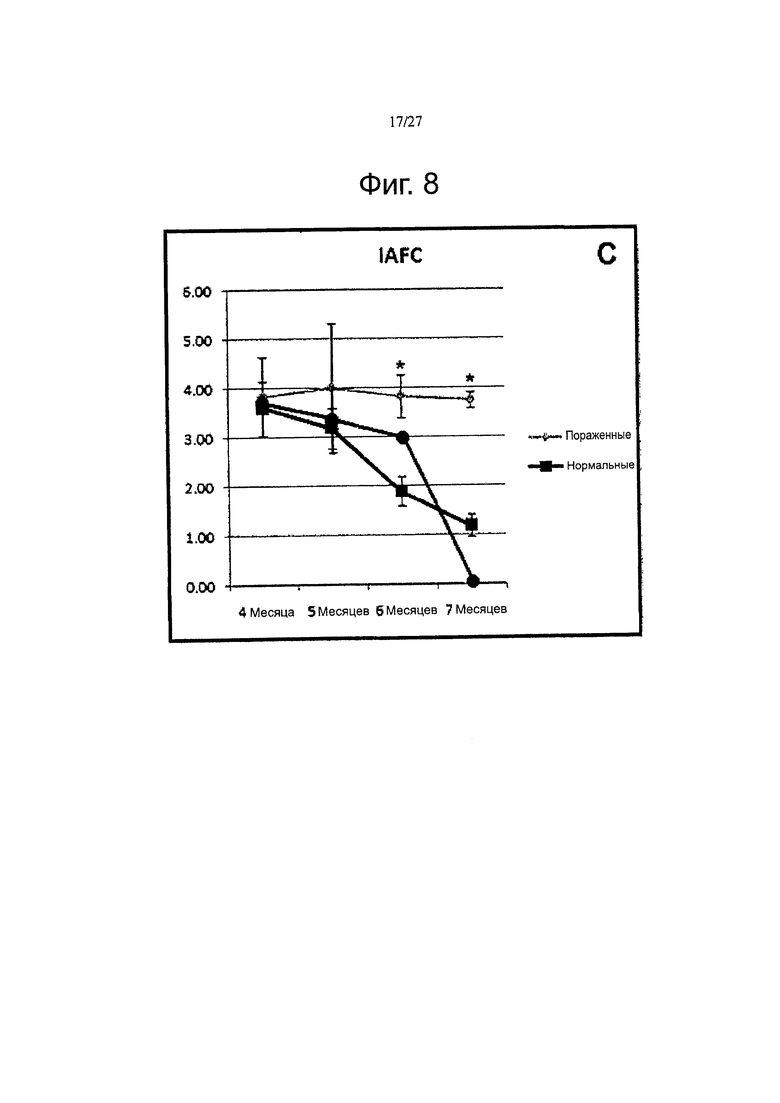
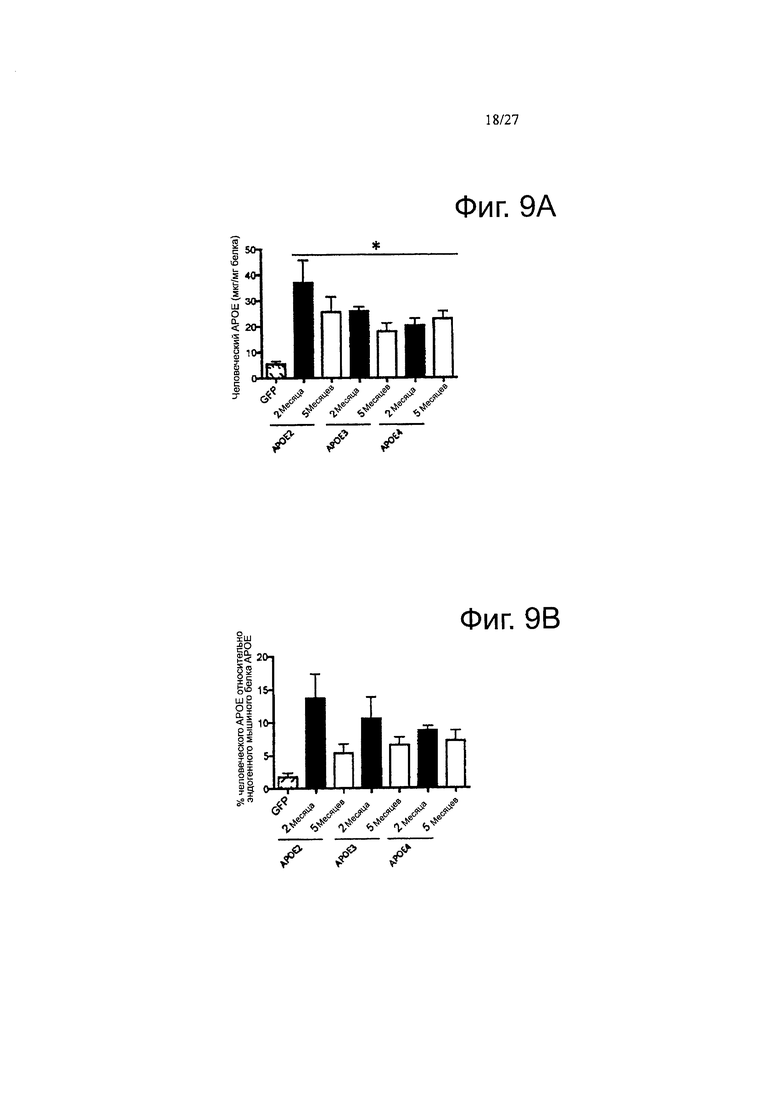
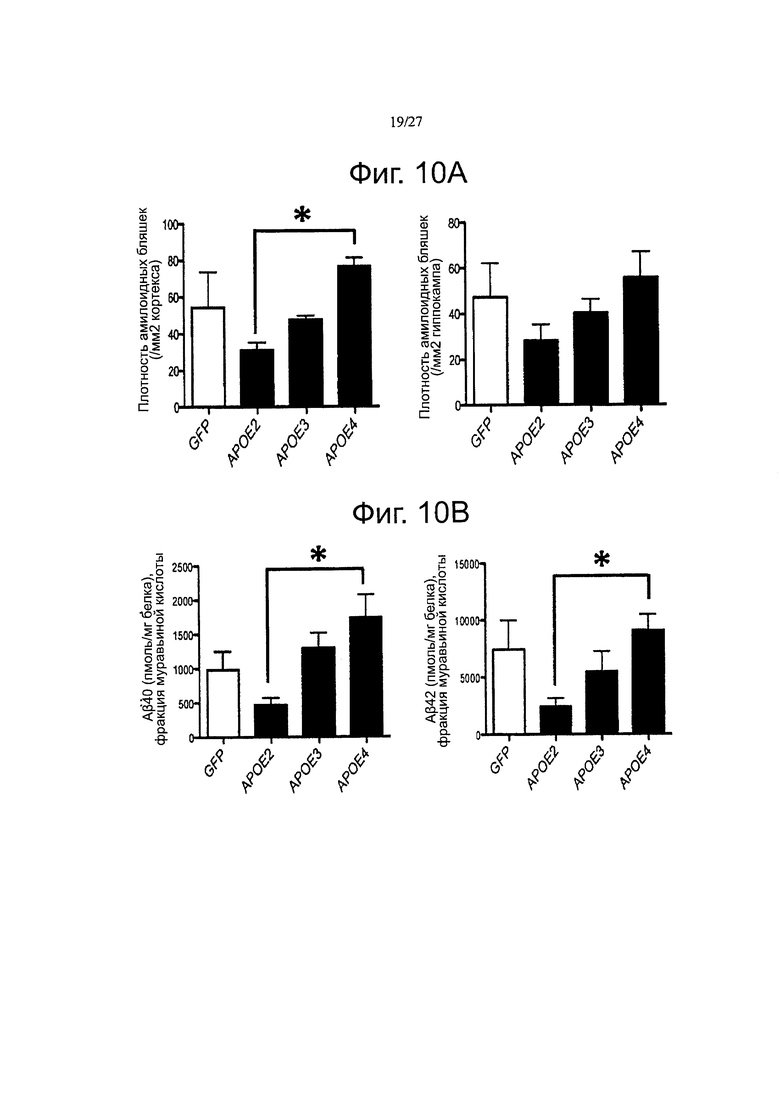
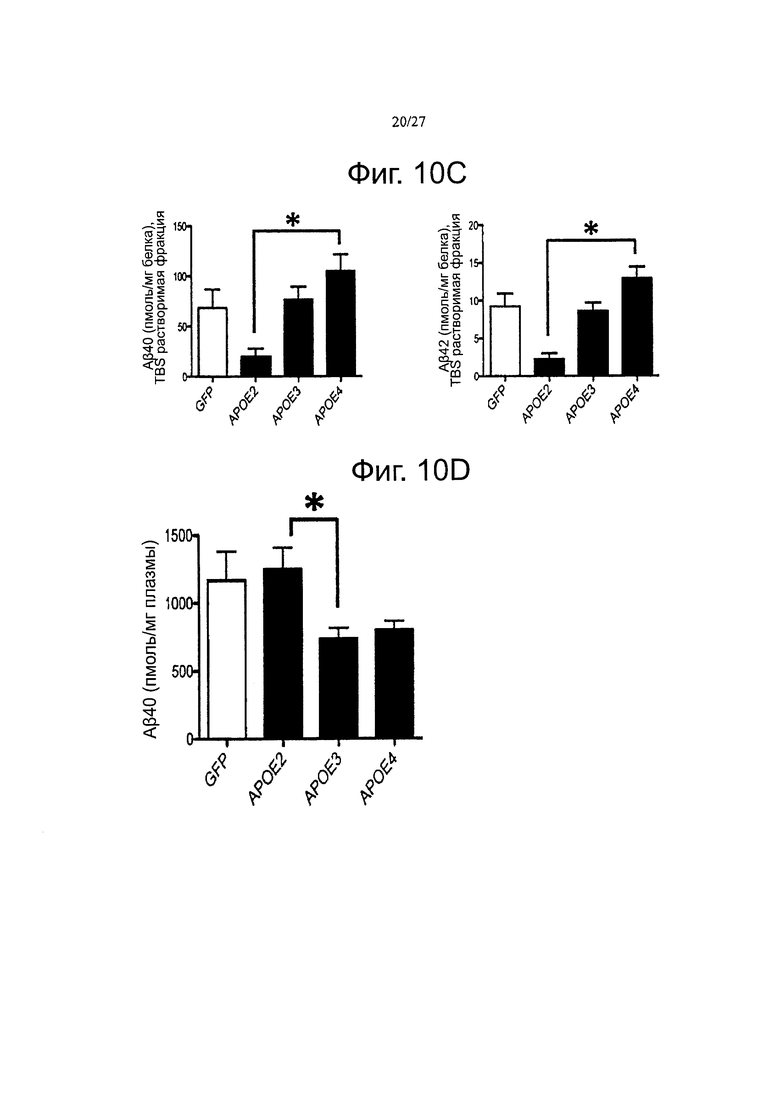
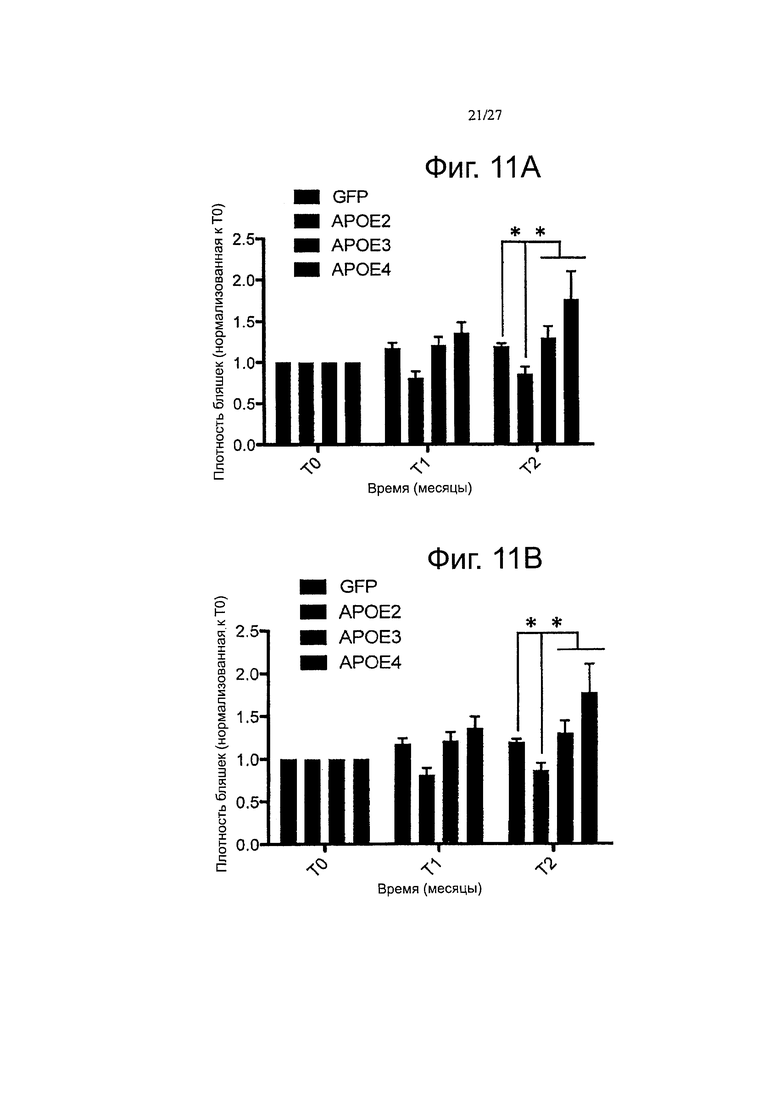
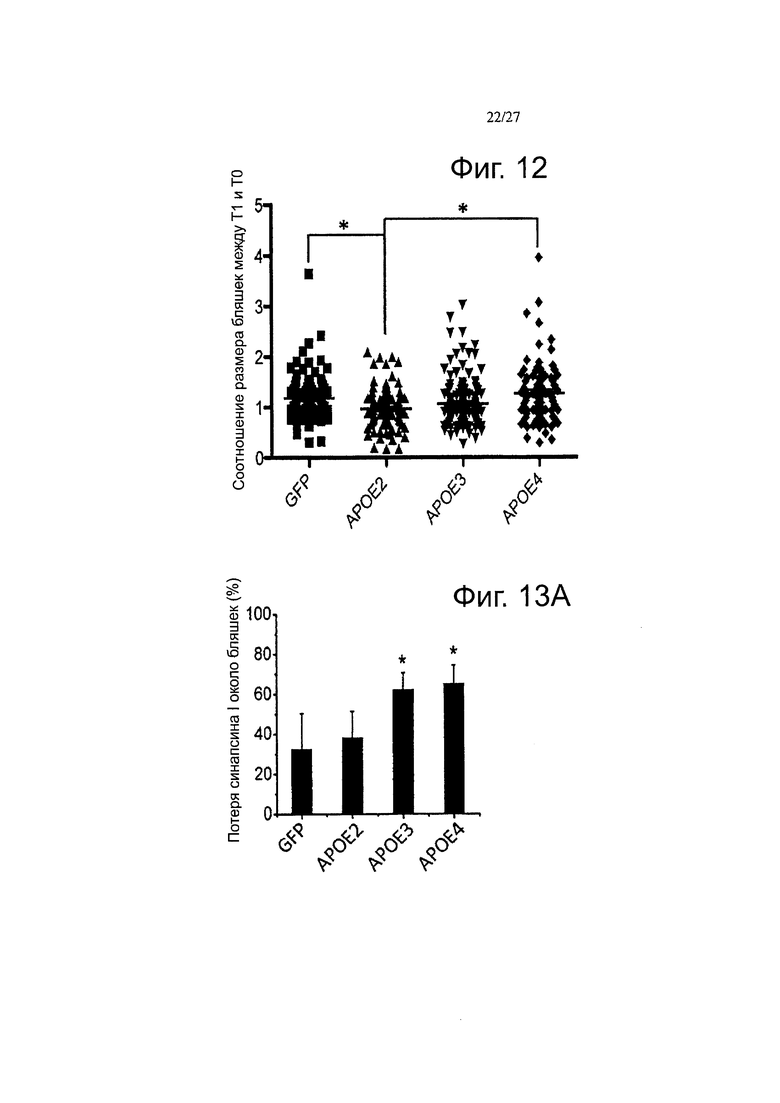
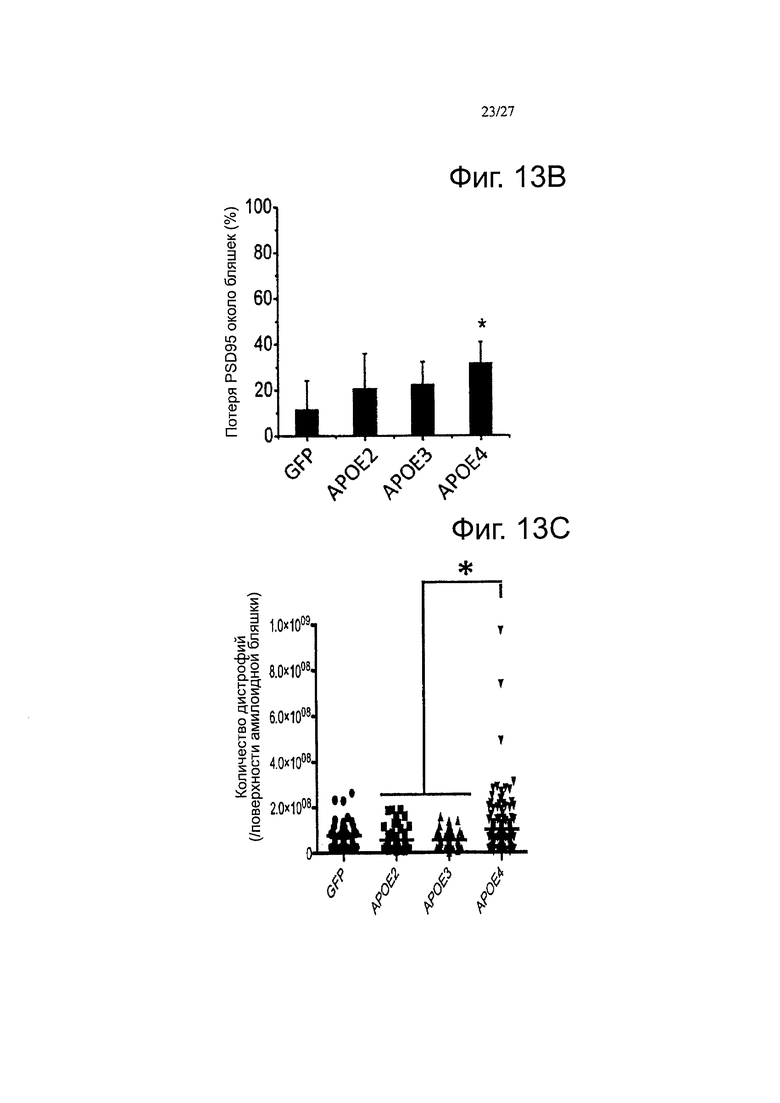
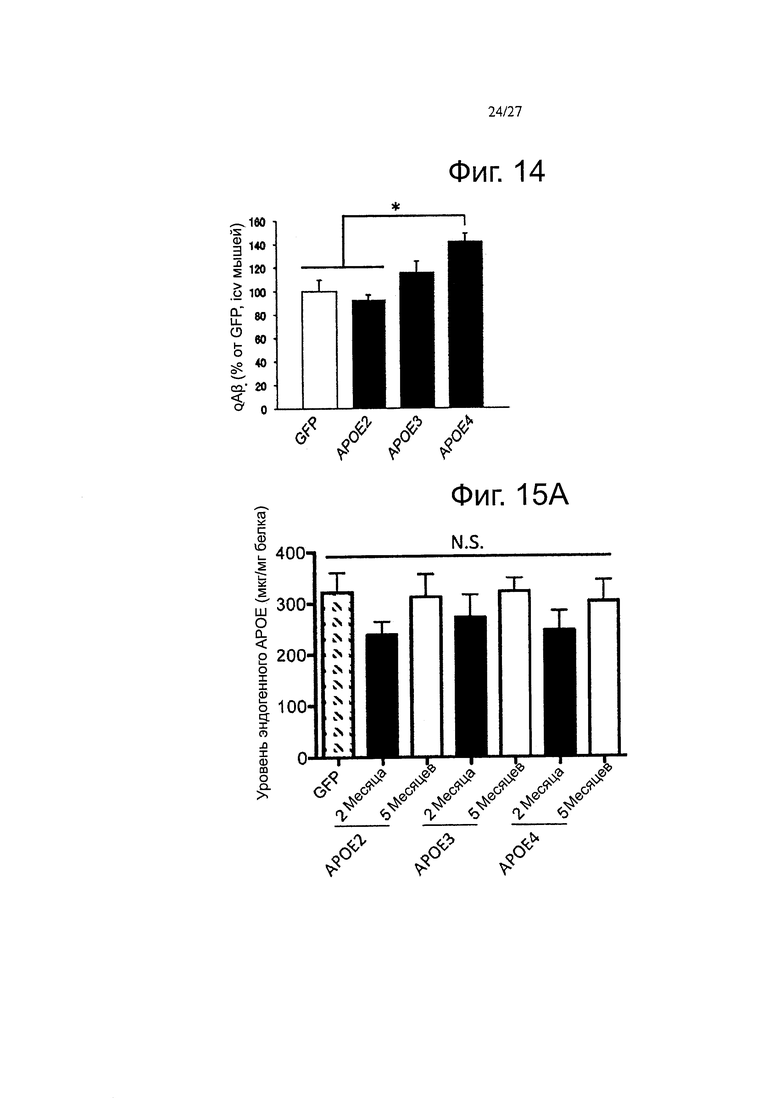
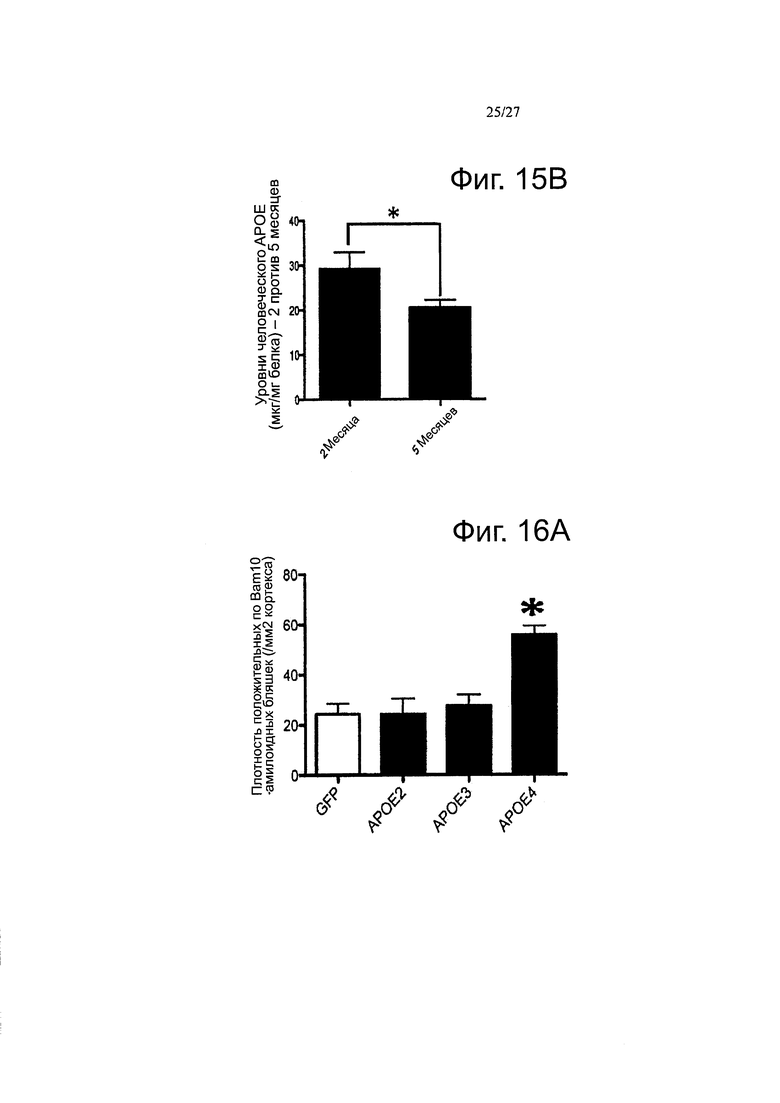
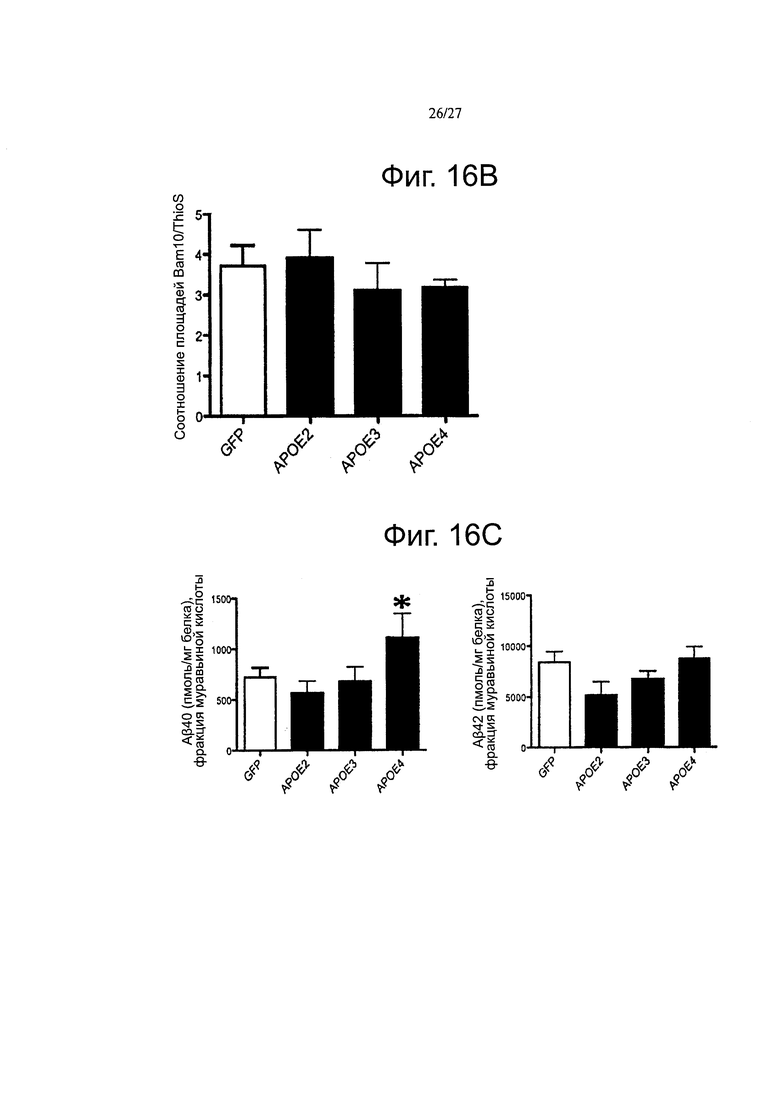
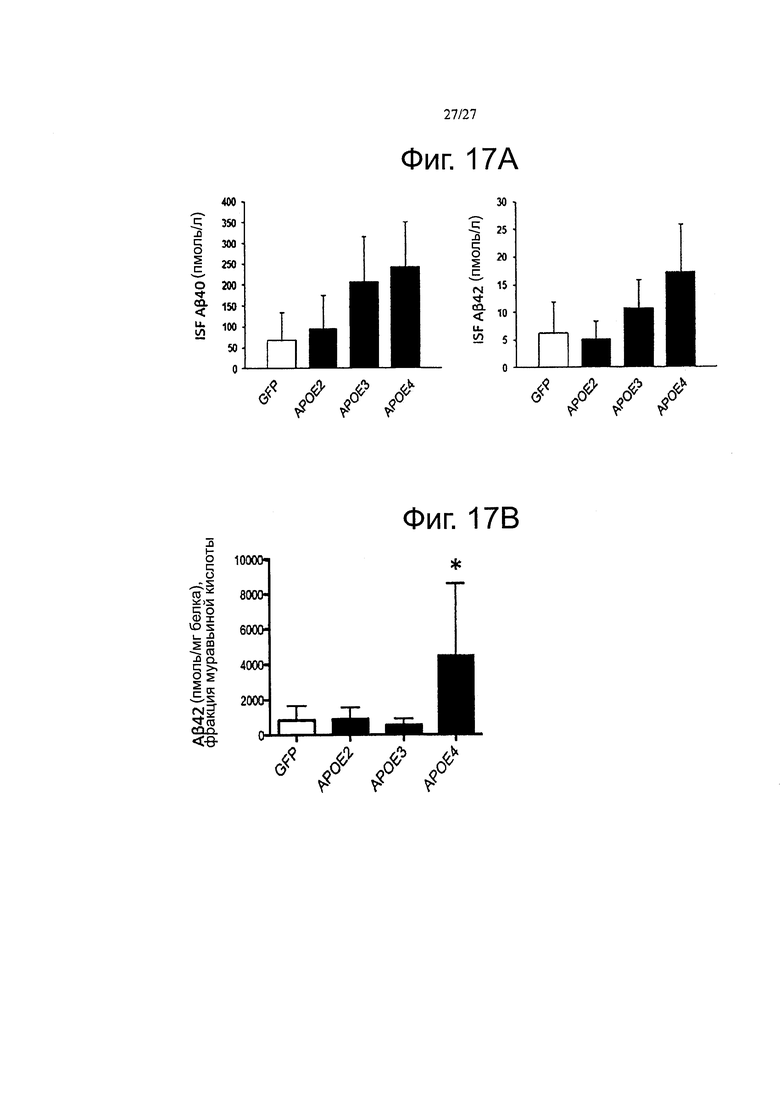
Предложенная группа изобретений относится к области медицины. Предложены способы доставки защитной изоформы ApoE и кодирующей ее нуклеиновой кислоты в центральную нервную систему млекопитающего. Предложены частица rAAV2, содержащая вектор, включающий нуклеиновую кислоту, кодирующую защитную изоформу ApoE ε2, и фармацевтическая композиция, содержащая указанную частицу, для лечения болезни Альцгеймера. Предложенная группа изобретений обеспечивает снижение риска развития болезни Альцгеймера и ее эффективное лечение. 4 н. и 10 з.п. ф-лы, 17 ил., 3 пр.
1. Способ доставки защитной изоформы ApoE в центральную нервную систему млекопитающего, не являющегося грызуном, для лечения болезни Альцгеймера, включающий введение в спинномозговую жидкость (CSF) млекопитающего, не являющегося грызуном, частицы rAAV, содержащей капсидный белок AAV2 и вектор, содержащий нуклеиновую кислоту, кодирующую защитную изоформу ApoE ε2, вставленную между парой инвертированных концевых повторов AAV таким образом, который является эффективным для инфекции эпендимных клеток в млекопитающем, не являющемся грызуном, секретирующих защитную форму ApoE ε2 в CSF млекопитающего, не являющегося грызуном.
2. Способ по п. 1, в котором животное, не являющееся грызуном, представляет собой примата, лошадь, овцу, козу, свинью или собаку.
3. Способ по п. 2, в котором приматом является человек.
4. Способ доставки нуклеиновой кислоты, кодирующей защитную изоформу белка ApoE ε2 в эпендимную клетку млекопитающего, не являющегося грызуном, для лечения болезни Альцгеймера, включающий введение в эпендимную клетку частицы rAAV, содержащей капсидный белок AAV2 и вектор, содержащий нуклеиновую кислоту, вставленную между парой инвертированных концевых повторов AAV с образованием трансфецированной эпендимальной клетки.
5. Способ по п. 4, который дополнительно включает доставку этой трансфецированной клетки назад в млекопитающее, не являющееся грызуном.
6. Способ по п. 4, в котором млекопитающее, не являющееся грызуном, является приматом, лошадью, овцой, козой, свиньей или собакой.
7. Способ по п. 6, в котором примат является человеком.
8. Способ по п. 4, в котором эпендимальная клетка находится в млекопитающем.
9. Способ по п. 4, в котором эпендимальная клетка в млекопитающем, не являющемся грызуном, секретирует защитную изоформу АроЕ ε2 в CSF животного, не являющегося грызуном.
10. Частица rAAV2, содержащая вектор, включающий нуклеиновую кислоту, кодирующую защитную изоформу ApoE ε2, вставленную между парой инвертированных концевых повторов AAV для трансфекции эпендимных клеток в млекопитающем, не являющемся грызуном, для лечения болезни Альцгеймера.
11. Частица rAAV2 по п. 10, где млекопитающее, не являющееся грызуном, является приматом, лошадью, овцой, козой, свиньей или собакой.
12. Частица rAAV2 по п. 11, где примат является человеком.
13. Частица по п. 10, в которой вектор дополнительно содержит ITR и/или регуляторный элемент.
14. Фармацевтическая композиция для лечения болезни Альцгеймера, содержащая rAAV2 частицу по п. 10.
| US 6468524 B1, 22.10.2002 | |||
| US 2011038851 A1, 17.02.2011 | |||
| НОВОЕ ПРИМЕНЕНИЕ | 2004 |
|
RU2416426C2 |
| WARRINGTON K.H.Jr | |||
| et al | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| J Virol | |||
| Способ приготовления мыла | 1923 |
|
SU2004A1 |

