В последнее десятилетие антитела успешно внедряются в клиническую практику для применения при лечении рака; они представляются наиболее многообещающим терапевтическим средством в онкологии. По сравнению с обычными лекарственными средствами для лечения рака терапевтические методы, основанные на антителах, обладают потенциально более высокой специфичностью и меньшим спектром побочных эффектов, потому что антитела точно различают нормальные и неопластические клетки и их действие базируется на менее токсичных иммунологических противоопухолевых механизмах, например активации комплемента и мобилизации цитотоксических клеток иммунной системы.
Клаудины - это интегральные мембранные белки, располагающиеся в местах плотных контактов между эпителием и эндотелием. Считается, что в клаудиновой молекуле четыре трансмембранных сегмента с двумя внеклеточными «петлями», a N- и С-концы находятся в цитоплазме. Трансмембранные белки семейства клаудинов (CLDN) играют ключевую роль в поддержании плотных контактов между эпителием и эндотелием, а также, возможно, участвуют в поддержании цитоскелета и в клеточных сигнальных механизмах. Экспрессия этих белков существенно различается в нормальных клетках и в опухолевых, что, наряду с их мембранной локализацией, делает их заманчивой мишенью иммунотерапии рака; применение терапевтических методов, основанных на антителах, для прицельного воздействия на клаудины при лечении рака сулит высокую специфичность терапевтического воздействия.
Однако клиническое применение терапевтических воздействий, нацеленных на клаудины, сталкивается с рядом препятствий. В силу повсеместного присутствия клаудинов в организме и их важной роли в поддержании плотных контактов необходима высокая специфичность нацеленных на клаудины терапевтических воздействий, чтобы обеспечить максимальную специфичность лечения и свести к минимуму системную токсичность.
Публикация WO 2009/087978 относится к антителам, направленным против калудина-6 и их применению в качестве противораковых агентов. В частности описываются моноклональные антитела, обозначенные АВ3-1, АЕ1-16, АЕ49-11 и АЕ3-20. Однако, как показал анализ методом флуоресцентно-активируемой клеточной сортировки (FACS), описанный в примере 5, ни одно из этих антител не было специфичным в отношении клаудина-6. Антитела АЕ3-20 взаимодействовали с CLDN9, тогда как антитела АЕ1-16 и АЕ49-11 проявляли значительную реактивность в отношении CLDN9 и также взаимодействовали с CLDN4. Прочность связывания антител АВ3-1 с CLDN6 была такой же, как с CLDN9. В примере 7 описывается, что введение антител АЕ49-11 мышам с модельными опухолями подавляло опухолевый рост и продлевало жизнь. Но с учетом неспецифичности использованных в этом примере антител остается неясным, действительно ли наблюдавшийся эффект был обусловлен связыванием этих антител с CLDN6.
Таким образом, до сих пор не были описаны антитела, специфичные в отношении CLDN6, которые бы избирательно связывались с поверхностью клеток, в которых экспрессируется CLDN6. А для основанных на антителах терапевтических воздействий, нацеленных на CLDN6, требуются как раз специфичные антитела.
Показанное на фигуре 1 сравнение аминокислотных последовательностей CLDN3, CLDN4, CLDN6 и CLDN9 демонстрирует высокую консервативность CLDN6 относительно других клаудинов. Высокая степень гомологии CLDN6 с другими клаудинами, в особенности с CLDN9 и CLDN4, и тот факт, что в работе WO 2009/087978 не удалось получить антитела, специфичные к CLDN6, заставляют предполагать, что невозможно получить антитела, специфически связывающиеся с CLDN6.
Раскрытие изобретения
Описываемые в настоящем документе результаты экспериментов подтверждают, что у человека CLDN6 экспрессируется в различных раковых клетках, тогда как в нормальных тканях его экспрессия ограничена плацентой.
В данном изобретении впервые описывается успешное получение антител, специфичных в отношении CLDN6 и способных связываться с поверхностью интактных клеток, в которых экспрессируется CLDN6. Исследование интактных клеток, в которых экспрессируется CLDN6, методом флуоресцентно-активируемой клеточной сортировки (FACS) продемонстрировало специфическое связывание антител против CLDN6 в то время как в случае клеток, в которых экспрессируются другие клаудиновые белки, в частности CLDN3, CLDN4 and CLDN9, или клеток, в которых не образуются ни один из этих белков, связывание вообще не наблюдалось. Таким образом, настоящее изобретение неожиданно показало, что можно получить антитела, специфически взаимодействующие с CLDN6 на поверхности клеток, в которых экспрессируется CLDN6, но не проявляющие существенной реактивности в отношении других высокогомологичных клаудинов как антигенов.
В общем, настоящим изобретением предлагаются антитела, полезные в качестве терапевтических средств для лечения и/или предотвращения заболеваний, при которых играют ту или иную роль клетки экспрессирующие CLDN6, у которых CLDN6 связан с их клеточной поверхностью, включая заболевания, сопровождающиеся образованием опухолей, в частности рак, например рак яичника, в том числе аденокарциному яичника и тератокарциному яичника, рак легких, в том числе мелкоклеточный рак легких (SCLC) и немелкоклеточный рак легких (NSCLC), в особенности плоскоклеточную карциному легких и аденокарциному, рак желудка, рак молочной железы, рак печени, рак поджелудочной железы, рак кожи, в частности базальноклеточную карциному и плоскоклеточную карциному, злокачественную меланому, рак головы и шеи, в том числе злокачественную полиморфную аденому, саркому, в частности синовиальную саркому и карциносаркому, рак желчного протока, рак мочевого пузыря, в частности переходноклеточная карцинома и папиллярная карцинома, рак почек, в частности почечно-клеточная карцинома, включая светлоклеточную почечно-клеточную карциному и папиллярную почечно-клеточную карциному, рак толстого кишечника, рак тонкого кишечника, включая рак подвздошной кишки, в частности аденокарциному тонкого кишечника и аденокарциному подвздошной кишки, эмбриональную карциному яичка, хориокарциному плацентарной площадки, рак шейки матки, рак яичка, в частности семиному яичка, тератому яичка и эмбриональный рак яичка, рак матки, герминогенные опухоли, например тератокарциному или эмбриональную карциному, в частности герминогенную опухоль яичка, и их метастазирующие формы.
Один из вариантов данного изобретения относится к антителам, способным связываться с CLDN6, ассоциированным с поверхностью клеток, в которых экспрессируется CLDN6. Предпочтительно, эти антитела не способны в основном связываться с CLDN9, ассоциированным с поверхностью клеток, в которых экспрессируется CLDN9. Предпочтительно, эти антитела не способны в основном связываться с CLDN4, ассоциированным с поверхностью клеток, в которых экспрессируется CLDN4 и/или не способны в основном связываться с CLDN3, ассоциированным с поверхностью клеток, в которых экспрессируется CLDN3. Наиболее предпочтительно, эти антитела не способны связываться ни с каким другим клаудином, кроме CLDN6, ассоциированным с поверхностью клеток, в которых экспрессируется указанный клаудин, и специфичны в отношении CLDN6. Предпочтительно, указанные клетки, в которых экспрессируется указанный клаудин, являются интактными, в частности с ненарушенной проницаемостью клеточной мембраны, и указанный клаудин, ассоциированный с поверхностью клетки, обладает нативной конформацией т.е. не денатурирован. Предпочтительно, эти антитела способны связываться с одним или более эпитопами CLDN6 в их нативной конформации.
В одном из воплощений данного изобретения предлагаемые антитела способны связываться с эпитопом, находящимся в пределах внеклеточной части CLDN6, причем указанная часть CLDN6 предпочтительно содержит какую-либо аминокислотную последовательность из числа следующих: SEQ ID NO:6, SEQ ID NO:7, SEQ ID NO:14 и SEQ ID NO:15, предпочтительно аминокислотную последовательность SEQ ID NO:6 или SEQ ID NO:7, более предпочтительно аминокислотную последовательность SEQ ID NO:6. Предпочтительно, предлагаемые антитела способны связываться с эпитопом, находящимся в пределах одной из следующих аминокислотных последовательностей: SEQ ID NO:6, SEQ ID NO:7, SEQ ID NO:14 и SEQ ID NO:15, предпочтительно аминокислотной последовательности SEQ ID NO:6 или SEQ ID NO:7.
В одном из воплощений данного изобретения предлагаемые антитела способны связываться с CLDN6 путем взаимодействия с по меньшей мере одним, предпочтительно более чем с одним, например с двумя, тремя, четырьмя или пятью, предпочтительно со всеми аминокислотными остатками, выбираемыми из группы, состоящей из Thr33, Phe35, Gly37, Ser39, Ile40 и Leu151, предпочтительно путем взаимодействия с по меньшей мере одним, предпочтительно с более чем одним, предпочтительно со всеми аминокислотными остатками, выбираемыми из группы, состоящей из Thr33, Phe35, Gly37, Ser39 и Ile40, более предпочтительно путем взаимодействия с по меньшей мере одним, предпочтительно с более чем одним, предпочтительно со всеми аминокислотными остатками, выбираемыми из группы, состоящей из Phe35, Gly37, Ser39 и Ile40, или состоящей из Thr33, Phe35, Gly37 и Ser39, и в особенности путем взаимодействия с по меньшей мере одним, предпочтительно с более чем одним, предпочтительно со всеми аминокислотными остатками, выбираемыми из группы, состоящей из Phe35, Gly37, и Ser39. Предпочтительно, предлагаемые антитела не взаимодействуют с одним или более, предпочтительно всеми аминокислотными остатками, выбираемыми из группы, состоящей из Glu154, Ala155, Arg158 и Gly161, и предпочтительно не взаимодействуют с одним или более, предпочтительно всеми аминокислотными остатками, выбираемыми из группы, состоящей из Arg158 и Gly161.
Взаимодействие между антителами и CLDN6, в частности в его нативной конформации, можно изучить путем мутагенеза на основе сканирования аланином. Мутантные CLDN6 можно характеризовать по их способности связываться со специфичными моноклональными антителами. Нарушение связывания специфичного моноклонального антитела с мутантным CLDN6 позволяет полагать, что аминокислотный остаток, по которому имела место мутация, важен для контакта с антителом. Определять связывание можно, например, путем проточной цитометрии.
В одном воплощении данного изобретения предлагаемые антитела получают способом, включающим стадию иммунизации животного пептидом с аминокислотной последовательностью, являющейся какой-либо из следующих: SEQ ID NO:6, SEQ ID NO:7, SEQ ID NO:14 и SEQ ID NO:15, предпочтительна аминокислотная последовательность SEQ ID NO:6 или SEQ ID NO:7 или иммунологически эквивалентный пептид, или нуклеотидная последовательность, или клетка-хозяин, в которой экспрессируется указанный пептид.
В других воплощениях данного изобретения CLDN6, с которым способно связываться предлагаемое антитело, имеет аминокислотную последовательность SEQ ID NO:2 или аминокислотную последовательность SEQ ID NO:8. Особенно предпочтительно, чтобы предлагаемое антитело было способно связываться с CLDN6, имеющем аминокислотную последовательность SEQ ID NO:2, и с CLDN6, имеющем аминокислотную последовательность SEQ ID NO:8.
В одном из воплощений настоящего изобретения предлагаемое антитело содержит тяжелую цепь, включающую по меньшей мере одну, предпочтительно две, более предпочтительно все три последовательности CDR (области, определяющие комплементарность) из последовательности тяжелой цепи антитела, выбираемые из SEQ ID NO:34, 36, 38 и 40 или их вариантов. На фигуре 25 последовательности CDR в упомянутых выше последовательностях тяжелой цепи антитела выделены квадратной рамочкой.
В одном из воплощений настоящего изобретения предлагаемое антитело содержит тяжелую цепь, включающую последовательность CDR3 Xaa1 Gly Xaa2 Val Хаа3, где Xaa1 представляет любую аминокислоту, предпочтительно ароматическую аминокислоту, более предпочтительно Phe или Tyr, наиболее предпочтительно Tyr, Xaa2 представляет любую аминокислоту, предпочтительно ароматическую аминокислоту, более предпочтительно Phe или Tyr, наиболее предпочтительно Tyr, и Хаа3 представляет любую аминокислоту, предпочтительно Leu или Phe, более предпочтительно Leu. В одном из воплощений настоящего изобретения предлагаемое антитело содержит тяжелую цепь, включающую последовательность CDR3 Asp Xaa1 Gly Xaa2 Val Хаа3 или Xaa1 Gly Xaa2 Val Хаа3 Asp, где Xaa1, Xaa2 и Хаа3 представляют аминокислоты, указанные выше. В одном из воплощений настоящего изобретения предлагаемое антитело содержит тяжелую цепь, включающую последовательность CDR3 Asp Xaa1 Gly Xaa2 Val Хаа3 Asp, где Xaa1, Хаа2 и Хаа3 представляют аминокислоты, указанные выше. В одном из воплощений настоящего изобретения предлагаемое антитело содержит тяжелую цепь, включающую последовательность CDR3 Ala Arg Asp Xaa1 Gly Хаа2 Val Хаа3 Asp Tyr, где Xaa1, Хаа2 and Хаа3 представляют аминокислоты, указанные выше. В одном из воплощений настоящего изобретения предлагаемое антитело по указанным выше воплощениям содержит последовательность CDR1, соответствующую SEQ ID NO:47 или ее варианту, и/или последовательность CDR2, соответствующую SEQ ID NO:48 или ее варианту.
В одном из воплощений настоящего изобретения предлагаемое антитело содержит тяжелую цепь, включающую последовательность из антительной тяжелой цепи, выбираемую из SEQ ID NO:34, 36, 38 и 40 или их вариантов.
В одном из воплощений настоящего изобретения предлагаемое антитело содержит легкую цепь, включающую по меньшей мере одну, предпочтительно две, более предпочтительно все три последовательности CDR из последовательности легкой цепи антитела, выбираемые из SEQ ID NO:35, 37, 39 и 41 или их вариантов. На фигуре 26 последовательности CDR в упомянутых выше последовательностях легкой цепи антитела выделены квадратной рамочкой.
В одном из воплощений настоящего изобретения предлагаемое антитело содержит легкую цепь, включающую последовательность CDR3 Arg Xaa1 Хаа2 Хаа3 Pro, где Xaa1 представляет любую аминокислоту, предпочтительно Ser или Asn, наиболее предпочтительно Ser, Хаа2 представляет любую аминокислоту, предпочтительно Tyr, Ser, Ile, Asn или Thr, более предпочтительно Tyr, Ser или Asn, наиболее предпочтительно Asn, и Хаа3 представляет любую аминокислоту, предпочтительно Ser или Tyr, более предпочтительно Tyr. В одном из воплощений настоящего изобретения предлагаемое антитело содержит легкую цепь, включающую последовательность CDR3 Gln Arg Xaa1 Хаа2 Хаа3 Pro Pro, где Xaa1, Хаа2 и Хаа3 представляют аминокислоты, указанные выше. В одном из воплощений настоящего изобретения предлагаемое антитело содержит легкую цепь, включающую последовательность CDR3 Gln Gln Arg Xaa1 Хаа2 Хаа3 Pro Pro Trp Thr, где Xaa1, Хаа2 и Хаа3 представляют аминокислоты, указанные выше. В одном из воплощений настоящего изобретения предлагаемое антитело по указанным выше воплощениям содержит содержит легкую цепь, включающую последовательность CDR1, соответствующую SEQ ID NO:52 или ее варианту, и/или последовательность CDR2, соответствующую SEQ ID NO:53 или ее варианту.
В одном из воплощений настоящего изобретения предлагаемое антитело содержит легкую цепь, включающую последовательность легкой цепи антитела, выбираемую из SEQ ID NO:35, 37, 39, 41, 54 и 55 или их вариантов.
В различных воплощениях настоящего изобретения предлагаемое антитело содержит тяжелую цепь, как указано выше, и легкую цепь, как указано выше.
В одном из воплощений настоящего изобретения предлагаемое антитело содержит:
(i) тяжелую цепь, включающую по меньшей мере одну, предпочтительно две, более предпочтительно все три последовательности CDR из последовательности тяжелой цепи антитела SEQ ID NO: x или ее варианта, и
(ii) легкую цепь, включающую по меньшей мере одну, предпочтительно две, более предпочтительно все три последовательности CDR легкой цепи антитела SEQ ID NO: x+1 или ее варианта;
где x выбирают из 34, 36, 38 и 40.
На фигурах 25 и 26 соответственно указанные последовательности CDR в упомянутых выше последовательностях тяжелой и легкой цепей антитела выделены квадратными рамочками.
В одном из воплощений настоящего изобретения предлагаемое антитело содержит:
(i) тяжелую цепь, включающую последовательность CDR3, выбираемую из группы, состоящей из Xaa1 Gly Xaa2 Val Хаа3, Asp Xaa1 Gly Xaa2 Val Xaa3, Xaa1 Gly Xaa2 Val Xaa3 Asp, Asp Xaa1 Gly Xaa2 Val Xaa3 Asp и Ala Arg Asp Xaa1 Gly Xaa2 Val Xaa3 Asp Tyr, где Xaa1 представляет любую аминокислоту, предпочтительно ароматическую аминокислоту, более предпочтительно Phe или Tyr, наиболее предпочтительно Tyr, Xaa2 представляет любую аминокислоту, предпочтительно ароматическую аминокислоту, более предпочтительно Phe или Tyr, наиболее предпочтительно Tyr, и Xaa3 представляет любую аминокислоту, предпочтительно Leu или Phe, более предпочтительно Leu.
(ii) легкую цепь антитела, включающую последовательность CDR3, выбираемую из группы, состоящей из Arg Xaa1 Xaa2 Xaa3 Pro, Gln Arg Xaa1 Xaa2 Xaa3 Pro Pro, Gln Gln Arg Xaa1 Xaa2 Xaa3 Pro Pro Trp Thr, где Xaa1 представляет любую аминокислоту, предпочтительно Ser или Asn, наиболее предпочтительно Ser, Xaa2 представляет любую аминокислоту, предпочтительно Tyr, Ser, Ile, Asn или Thr, более предпочтительно Tyr, Ser или Asn, наиболее предпочтительно Asn, и Xaa3 представляет любую аминокислоту, предпочтительно Ser или Tyr, более предпочтительно Tyr.
В одном из воплощений настоящего изобретения предлагаемое антитело по одному из упомянутых выше воплощений содержит: (i) тяжелую цепь антитела, включающую последовательность CDR1, соответствующую SEQ ID NO:47 или ее вариант и/или последовательность CDR2, соответствующую SEQ ID NO:48 или ее вариант и/или (ii) легкую цепь антитела, включающую последовательность CDR1, соответствующую SEQ ID NO:52, или ее вариант и/или последовательность CDR2, соответствующую SEQ ID NO:53, или ее вариант.
В одном из воплощений настоящего изобретения предлагаемое антитело содержит:
(i) тяжелую цепь, включающую последовательность тяжелой цепи, выбираемую из SEQ ID NO 34, 36, 38 и 40 или их вариантов, предпочтительно SEQ ID NO:36 или ее вариант, и
(ii) легкую цепь, включающую последовательность легкой цепи антитела, выбираемую из SEQ ID NO 35, 37, 39, 41, 54 и 55 или их вариантов, предпочтительно SEQ ID NO:35 или ее вариант.
В одном из воплощений настоящего изобретения предлагаемое антитело содержит:
(i) тяжелую цепь, включающую последовательность тяжелой цепи SEQ ID NO:36 или ее вариант, и
(ii) легкую цепь, включающую последовательность легкой цепи антитела, выбираемую из SEQ ID NO 35, 54 и 55 или их вариантов.
В одном из воплощений настоящего изобретения предлагаемое антитело содержит:
(i) тяжелую цепь, включающую последовательность тяжелой цепи SEQ ID NO:36 или ее вариант, и
(ii) легкую цепь, включающую последовательность легкой цепи антитела, выбираемую из SEQ ID NO 54 и 55 или их вариантов.
В одном из воплощений настоящего изобретения предлагаемое антитело содержит:
(i) тяжелую цепь, включающую последовательность тяжелой цепи SEQ ID NO:36 или ее вариант, и
(ii) легкую цепь, включающую последовательность легкой цепи антитела SEQ ID NO 35 или ее вариант.
В одном из воплощений настоящего изобретения предлагаемое антитело содержит:
(i) тяжелую цепь, включающую последовательность тяжелой цепи SEQ ID NO: x или ее вариант, и
(ii) легкую цепь, включающую последовательность легкой цепи антитела SEQ ID NO x+1 или ее вариант,
где x выбирают из 34, 36, 38 и 40.
В предпочтительных воплощениях настоящего изобретения предлагаемое антитело содержит тяжелую цепь, включающую константную область гамма-1 тяжелой цепи, предпочтительно константную область гамма-1 тяжелой цепи антитела человека, например такую последовательность, как SEQ ID NO:25, и/или содержит легкую цепь, включающую константную область каппа легкой цепи антитела, предпочтительно константную область каппа легкой цепи антитела человека, например такую последовательность, как SEQ ID NO:27.
В предпочтительных воплощениях настоящего изобретения предлагаемые антитела обладают одной или более из следующих активностей: (i) уничтожают клетки, в которых экспрессируется CLDN6, (ii) ингибируют пролиферацию клеток, в которых экспрессируется CLDN6, (iii) ингибируют образование колоний клетками, в которых экспрессируется CLDN6, (iv) опосредуют ремиссию, т.е. уменьшение размеров, предпочтительно полную ремиссию, т.е. полное исчезновение сформировавшихся опухолей, (v) предотвращают образование или восстановление ранее существовавших опухолей и (vi) подавляют метастазирование клеток, в которых экспрессируется CLDN6. Соответственно, антитела по данному изобретению можно применять для достижения одного или более из вышеуказанных явлений, в частности при введении пациенту. Уничтожение клеток и/или ингибирование одной или более из их активностей можно использовать в терапевтических целях, как описано в настоящем документе. В частности, уничтожение клеток, ингибирование пролиферации клеток и/или ингибирование образования колоний клеток можно использовать для лечения или предотвращения рака, включая образование метастазов. Ингибирование пролиферации, образования колоний и/или метастазирования клеток можно использовать, в частности, для лечения или предотвращения образования метастазов при раке и метастатического распространения раковых клеток. Предпочтительно антитела по данному изобретению опосредуют уничтожение клеток, вызывая лизис, опосредуемый комплемент-зависимой цитотоксичностью (CDC), лизис, опосредуемый антителозависимой клеточной цитотоксичностью (ADCC), апоптоз, гомотипическую адгезию и/или фагоцитоз, предпочтительно вызывая лизис, опосредуемый комплемент-зависимой цитотоксичностью (CDC), и/или лизис, опосредуемый антителозависимой клеточной цитотоксичностью (ADCC). Настоящее изобретение также включает воплощения, в которых предлагаемые антитела проявляют свои описанные здесь активности, например уничтожение клеток и/или ингибирование одной или более клеточных активностей, например пролиферации клеток и/или образования колоний, не вызывая лизис, опосредуемый комплемент-зависимой цитотоксичностью (CDC), лизис, опосредуемый антителозависимой клеточной цитотоксичностью (ADCC), апоптоз, гомотипическую адгезию и/или фагоцитоз. Так, антитела по данному изобретению могут оказывать свое действие, просто связываясь с CLDN6 на клеточной поверхности и таким образом блокируя пролиферацию клеток, например. В одном из воплощений настоящего изобретения предлагаемые антитела не вызывают лизис клеток, опосредуемый комплемент-зависимой токсичностью.
Предпочтительно лизис, опосредуемый антителозависимой клнточной цитотоксичностью, происходит в присутствии эффекторных клеток, которые в конкретных воплощениях данного изобретения выбирают из группы, состоящее из моноцитов, мононуклеарных клеток, NK-клеток (естественных киллеров) и полиморфноядерных лейкоцитов (PMN), а фагоцитоз осуществляется макрофагами.
Активность, состоящую в ингибировании или уменьшении пролиферации клеток, в которых экспрессируется CLDN6, предпочтительно раковых клеток, можно измерить in vitro, определяя пролиферацию раковых клеток, в которых экспрессируется CLDN6, с помощью аналитического метода с использованием бромдезоксиуридина (5-бром-2-дезоксиуридина, BrdU). Бромдезоксиуридин представляет собой синтетический нуклеотид, являющийся аналогом тимидина, который может включаться в новосинтезированную ДНК реплицирующейся клетки (во время S-фазы клеточного цикла), замещая собой тимидин в процессе репликации ДНК. Определяя количество включившегося бромдезкосиуридина, например при помощи специфичных к нему антител, можно выявить клетки, в которых происходит активная репликация ДНК.
Активность, состоящую в ингибировании или сокращении образования колоний клетками, в которых экспрессируется CLDN6, предпочтительно раковых клеток, можно измерить in vitro с помощью клоногенного теста. Клоногенный тест - это микробиологический метод для изучения эффективности влияния различных агентов на жизнеспособность и пролиферацию клеток. Его часто используют в онкологических исследованиях для определения воздействия лекарственных препаратов и облучения на размножающиеся опухолевые клетки. Такой эксперимент включает три основных этапа: (i) осуществление изучаемого воздействия на образец клеток, в частности раковых клеток, (ii) высевание клеток в сосуд для культуры ткани и (in) рост клеток. Образующиеся колонии фиксируют, окрашивают и подсчитывают. Образование колоний важно потому, что оно связано со способностью отдельных опухолевых клеток, заселять органы, формируя метастазы. Ингибирующая активность антител в отношении образования колоний отражает их потенциальную эффективность в ингибировании метастазов. Антитела, обладающие такой активностью, т.е способные ингибировать или сокращать образование колоний в клоногенном тесте особенно полезны для лечения или предотвращения метастазов и метастатического распространения раковых клеток, в частности тех типов, которые обсуждаются в настоящем документе.
В предпочтительных воплощениях настоящего изобретения предлагаемые антитела проявляют одну или более иммуноэффекторных функций против клеток, несущих CLDN6 в его нативной конформации, причем эти одна или более эффекторные функции предпочтительно выбирают из группы, состоящей из комплемент-зависимой цитотоксичности (CDC), антителозависимой клеточноопосредованной цитотоксичности (ADCC), индукции апоптоза и ингибирования пролиферации; предпочтительными эффекторными функциями являются антителозависимая клеточно-опосредованная цитотоксичность и/или комплемент-зависимая цитотоксичность.
Предпочтительно указанные одна или более активности или одна или более иммуноэффекторные функции, проявляемые указанными антителами индуцируются связыванием указанных антител с CLDN6, предпочтительно с эпитопом, расположенным в пределах внеклеточной части CLDN6, причем указанная внеклеточная часть CLDN6 предпочтительно содержит любую аминокислотную последовательность из следующих: SEQ ID NO:6, SEQ ID NO:7, SEQ ID NO:14 и SEQ ID NO:15, предпочтительна аминокислотная последовательность SEQ ID NO:6 или SEQ ID NO:7, более предпочтительна аминокислотная последовательность SEQ ID NO:6.
По данному изобретению клетки, в которых экспрессируется CLDN6, предпочтительно отличаются тем, что CLDN6 связан с их поверхностью. Клетки, в которых экспрессируется CLDN6 или клетки, несущие на своей поверхности CLDN6 в его нативной конформации, предпочтительно являются опухолевыми клетками, например раковыми клетками, предпочтительно раковыми клетками при раковых заболеваниях, выбираемых из группы, состоящей из рака яичника, в том числе аденокарциномы яичника и тератокарциномы яичника, рака легких, в том числе мелкоклеточного рака легких (SCLC) и немелкоклеточного рака легких (NSCLC), в особенности плоскоклеточной карциномы легких и аденокарциномы, рака желудка, рака молочной железы, рака печени, рака поджелудочной железы, рака кожи, в частности базальноклеточной карциномы и плоскоклеточной карциномы, злокачественной меланомы, рака головы и шеи, в том числе злокачественной полиморфной аденомы, саркомы, в частности синовиальной саркомы и карциносаркомы, рака желчного протока, рака мочевого пузыря, в частности переходноклеточной карциномы и папиллярной карциномы, рака почек, в частности почечно-клеточной карциномы, включая светлоклеточную почечно-клеточную карциному и папиллярную почечно-клеточную карциному, рака толстого кишечника, рака тонкого кишечника, включая рак подвздошной кишки, в частности аденокарциному тонкого кишечника и аденокарциному подвздошной кишки, эмбриональной карциномы яичка, хориокарциномы плацентарной площадки, рака шейки матки, рака яичка, в частности семиномы яичка, тератомы яичка и эмбрионального рака яичка, рака матки, герминогенных опухолей, например тератокарциномы или эмбриональной карциномы, в частности герминогенной опухоли яичка, и их метастазирующих форм.
К антителам по данному изобретению можно присоединять один или более терапевтических эффекторов, например радиоактивные метки, цитотоксины, ферменты терапевтического назначения, агенты, вызывающие апоптоз и т.п., чтобы обеспечить прицельную цитотоксичность, т.е. уничтожение опухолевых клеток.
В одном из воплощений данного изобретения предлагаемые антитела (i) связываются с клетками, в которых экспрессируется CLDN6 и которые отличаются тем, что CLDN6 ассоциирован с клеточной поверхностью и (ii) не связываются с клетками, в которых не экспрессируется CLDN6 и у которых нет CLDN6, ассоциированного с клеточной поверхностью. Антитела по данному изобретению предпочтительно (i) опосредуют уничтожение и/или ингибируют пролиферацию клеток, в которых экспрессируется CLDN6 и которые отличаются тем, что CLDN6 ассоциирован с клеточной поверхностью, и (ii) не опосредует уничтожение и/или не ингибирует пролиферацию клеток, в которых экспрессируется CLDN6 и у которых нет CLDN6, ассоциированного с клеточной поверхностью.
В особенно предпочтительном воплощении настоящего изобретения предлагаемые антител связываются с нативными эпитопами CLDN6, находящимися на поверхности живых клеток, например с последовательностью SEQ ID NO:6 или 7. В других предпочтительных воплощениях данного изобретения предлагаемые антитела специфичны в отношении раковых клеток, в которых экспрессируется CLDN6 и не связываются с раковыми клетками, в которых не экспрессируется CLDN6.
Антитела по данному изобретению можно получить из различных видов живых организмов, включая (здесь перечислены не ограничивающие примеры) мышь, крысу, кролика, морскую свинку и человека. Антитела по данному изобретению включают также гибридные молекулы, в которых константная область, происходящая от одного вида живых организмов, предпочтительно человека, объединена с участком связывания антигена, происходящим от другого вида. Также антитела по данному изобретению включают гуманизированные молекулы, в которых участки связывания антигена, происходящие не от человека, объединены с константными и каркасными областями человеческого происхождения.
Антитела по данному изобретению включают поликлональные и моноклональные антитела и антитела IgG2a (например, IgG2a, κ, λ), IgG2b (например, IgG2b, κ, λ), IgG3 (например, IgG3, κ, λ) и IgM. Но другие изотипы антител тоже входят в данное изобретение, включая IgG1, IgA1, IgA2, секреторный IgA, IgD, IgE. Предлагаемые антитела могут представлять собой полноразмерные антитела или же их антиген связывающие фрагменты, включая, например, Fab, F(ab')2, Fv, одноцепочечные фрагменты Fv или биспецифичные антитела. Кроме того, антиген связывающие фрагменты,, включают связывающий домен гибридных иммуноглобулинов, содержащий (i) полипептид, который формирует связывающий домен (например, вариабельная область тяжелой цепи или вариабельная область легкой цепи) который соединен с полипептидом, образующим шарнирную область имуноглобулиновой молекулы, (ii) константная область СН2 тяжелой цепи иммуноглобулина, соединенная с шарнирной областью, и (iii) константная область СН3 тяжелой цепи иммуноглобулина, соединенная с константной областью СН2. Такие гибридные белки также описываются в заявках на патент США №№2003/0118592 и 2003/0133939.
Антитела по данному изобретению являются предпочтительно моноклональными, гибридными, человеческими или гуманизированными антителами или фрагментами антител. Антитела по данному изобретению включают полностью человеческие антитела. Такие антитела могут быть выработаны трансгенными животными (не человеком), например трансгенными мышами, способными производить множественные изотипы человеческих моноклональных антител против CLDN6, путем V-D-J рекомбинации и переключения изотипа. Таким трансгенным животным для производства поликлональных антител, например таких, какие описаны в заявке на патент США №2003/0017534, может также быть кролик.
Антитела по данному изобретению предпочтительно высвобождаются из комплекса с CLDN6 с константой диссоциации (KD) приблизительно 1-100 нМ или меньше. Предпочтительно, антитела по данному изобретению не проявляют перекрестного взаимодействия (кросс-реактивности) с близкими антигенами клеточной поверхности и тем самым не ингибируют их функцию.
В предпочтительных воплощениях данного изобретения предлагаемые антитела отличаются одним или более из следующих свойств:
a) специфичность в отношении CLDN6;
b) сродство связывания с CLDN6 около 100 нМ или меньше, предпочтительно около 5-10 нМ или меньше и более предпочтительно около 1-3 нМ или меньше;
c) способность вызывать комплемент-зависимую цитотоксичность в отношении клеток, в которых экспрессируется CLDN6 и которые отличаются тем, что CLDN6 связан с их поверхностью;
d) способность ингибировать пролиферацию клеток, в которых экспрессируется CLDN6 и которые отличаются тем, что CLDN6 связан с их поверхностью;
e) способность вызывать апоптоз клеток, в которых экспрессируется CLDN6 и которые отличаются тем, что CLDN6 связан с их поверхностью;
f) способность вызывать гомотипическую адгезию клеток, в которых экспрессируется CLDN6 и которые отличаются тем, что CLDN6 связан с их поверхностью;
g) способность вызывать антителозависимую клеточно-опосредованную цитотоксичность в отношении клеток, в которых экспрессируется CLDN6 и которые отличаются тем, что CLDN6 связан с их поверхностью, в присутствии эффекторных клеток;
h) способность продлевать жизнь пациента, имеющего опухолевые клетки, в которых экспрессируется CLDN6 и которые отличаются тем, что CLDN6 связан с их поверхностью;
i) способность уменьшать количество клеток, в которых экспрессируется CLDN6 и которые отличаются тем, что CLDN6 связан с их поверхностью;
j) способность агрегировать CLDN6 на поверхности живых клеток.
Предпочтительные антитела, описанные в настоящем документе, - это антитела, произведенные клетками гибридомы (или полученные из них), хранящимися в Коллекции микроорганизмов и культур клеток Германии Института им. Лейбница (DSMZ) (Инхоффенштрассе, 7B, 38124 Брауншвейг, Германия, Germany) и имеющими следующие обозначения и регистрационные номера:
1. GT512muMAB 59A, регистрационный номер DSM ACC3067, депонирована 21.06.2010;
2. GT512muMAB 60А, регистрационный номер DSM ACC3068, депонирована 21.06.2010;
3. GT512muMAB 61D, регистрационный номер DSM ACC3069, депонирована 21.06.2010;
4. GT512muMAB 64А, регистрационный номер DSM ACC3070, депонирована 21.06.2010;
5. GT512muMAB 65А, регистрационный номер DSM ACC3071, депонирована 21.06.2010;
6. GT512muMAB 66B, регистрационный номер DSM ACC3072, депонирована 21.06.2010;
7. GT512muMAB 67А, регистрационный номер DSM ACC3073. депонирована 21.06.2010;
8. GT512muMAB 55А, регистрационный номер DSM ACC3089, депонирована 31.07.2010; или
9. GT512muMAB 89A, регистрационный номер DSM ACC3090, депонирована 31.07.2010.
В обозначениях антител по данному изобретению в настоящем документе отражены обозначение самого антитела и/или клона, его производящего, например muMAB 59A.
Также предпочтительны антитела обладающие специфичностью антител, вырабатываемых описанными выше гибридомами или получаемых из них, в частности тех, которые содержат антигенсвязывающий участок или антигенсвязывающий сайт, в частности вариабельную область, идентичную таковой антител, вырабатываемых описанными выше гибридомами или получаемых из них, или имеющую высокую степень гомологии с ними. Имеется в виду, что предпочтительные по данному изобретению антитела имеют область CDR, идентичную соответствующей области антител вырабатываемых описанными выше гибридомами или получаемых из них. Под высокой степенью гомологии понимается, что в каждой области CDR может быть от 1 до 5, предпочтительно от 1 до 4, например от 1 до 3 или от 1 до 2 замен, особенно предпочтительны антитела, которые являются гибридными и гуманизированными формами антител, вырабатываемых описанными выше гибридомами или получаемых из них.
Таким образом, антитела по данному изобретению могут быть выбраны из группы, состоящей из i) антител, вырабатываемых клонами, депонированными под регистрационным номером DSM ACC3067 (GT512muMAB 59A), DSM ACC3068 (GT512muMAB 60А), DSM ACC3069 (GT512muMAB 61D), DSM ACC3070 (GT512muMAB 64А), DSM ACC3071 (GT512muMAB 65А), DSM ACC3072 (GT512muMAB 66В), DSM ACC3073 (GT512muMAB 67А), DSM ACC3089 (GT512muMAB 55А) или DSM ACC3090 (GT512muMAB 89A), либо получаемых из них; (ii) антител, являющихся гибридными или гуманизированными формами антител по пункту (i); (iii) антитела, обладающие специфичностью антител по пункту (i); (iv) антитела, содержащие участок связывания антигена или сайт связывания антигена, как у антител по пункту (i). Участок связывания антигена или сайт связывания антигена, как у антител по пункту (i) может включать вариабельную область антител по пункту (i).
Данное изобретение относится также к таким клеткам, как клетки гибридомы, вырабатывающими описанные в настоящем документе антитела.
Предпочтительны клетки гибридомы, имеющиеся в коллекции DSMZ (Inhoffeenstr. 7B, 38124 Braunschweig, Germany) с каким-либо из следующих обозначений и регистрационных номеров:
1. GT512muMAB 59A, регистрационный номер DSM ACC3067, депонирована 21.06.2010;
2. GT512muMAB 60А, регистрационный номер DSM ACC3068, депонирована депонирована 21.06.2010;
3. GT512muMAB 61D, регистрационный номер DSM ACC3069, депонирована 21.06.2010;
4. GT512muMAB 64А, регистрационный номер DSM ACC3070, депонирована 21.06.2010;
5. GT512muMAB 65А, регистрационный номер DSM ACC3071, депонирована 21.06.2010;
6. GT512muMAB 66B, регистрационный номер DSM ACC3072, депонирована 21.06.2010;
7. GT512muMAB 67А, регистрационный номер DSM ACC3073. депонирована 21.06.2010;
8. GT512muMAB 55А, регистрационный номер DSM ACC3089, депонирована 31.08.2010; или
9. GT512muMAB 89А, регистрационный номер DSM ACC3090, депонирована 31.08.2010.
Антитела против CLDN6 можно модифицировать, соединить или обеспечить совместную экспрессию с носителями иных специфичностей. В одном конкретном воплощении данного изобретения предлагаются биспецифичные или полиспецифичные антитела, обладающие по меньшей мере одной первой специфичностью в отношении CLDN6 (например, антитело против CLDN6 или его миметик) и второй специфичностью в отношении эффекторной клетки, например специфичностью связывания с рецептором Fc (например, рецептором Fc-γ, в том числе Fc-γRI, или любым другим рецептором Fc) или с рецептором на Т-клетках, например CD3.
Соответственно сказанному выше данное изобретение включает биспецифичные и полиспецифичные молекулы, связывающиеся как с CLDN6, так и с рецептором Fc или с рецептором на Т-клетках, например CD3. Примерами рецепторов Fc являются рецептор IgG, рецептор Fcγ, например, FcγRI (CD64), FcγRII (CD32) и FcγRIII (CD16). Другие рецепторы Fc, например рецепторы IgA (например, FcαRI) тоже могут служить мишенями. Рецептор Fc предпочтительно располагается на поверхности эффекторных клеток, например моноцитов, макрофагов или активированных мононуклеаров. В одном из предпочтительных воплощений данного изобретения биспецифичные и полиспецифичные молекулы связываются с рецептором Fc в сайте, отличном от сайта связывания этим рецептором иммуноглобулинов (например, IgG или IgA), так что с ввязывание биспецифичных и полиспецифичных молекул не блокируется при физиологических уровнях иммуноглобулинов.
В еще одном варианте настоящего изобретения предлагаемые антитела против CLDN6 модифицируют, присоединяют или обеспечивают совместную экспрессию с другими функциональными молекулами, например иными пептидами или белками (например, Fab'-фрагментами). Так, антитела по данному изобретению можно функционально соединить (например, путем химического присоединения, слияния генетического материала, нековалентной ассоциации и проч.) с одной или более других молекулярных структур, например с другими антителами (например, для получения биспецифичных или полиспецифичных антител), цитотоксическими агентами, клеточными лигандами или антигенами (например, для получения иммуноконъюгата, например иммунотоксина). Антитела по данному изобретению можно соединить с другими терапевтическими агентами, например радиоактивными изотопами, лекарственными низкомолекулярными соединениями противоракового действия, рекомбинантными цитокинами или хемокинами. Таким образом, настоящее изобретение охватывает множество разнообразных конъюгатов антител, биспецифичных и полиспецифичных молекул и гибридных белков, которые все способны связываться с клетками, в которых экспрессируется CLDN6 и/или с клетками, отличающимися тем, что CLDN6 ассоциирован с их поверхностью; все эти агенты можно использовать для прицельного воздействия на указанные клетки.
В целом в контексте данного изобретения все производные антител, например их конъюгаты, биспецифичные и полиспецифичные молекулы и гибридные белки, в настоящем документе включаются в термин «антитела».
В другом своем варианте настоящее изобретение предусматривает белки, связывающие CLDN6, которые происходят не из иммуноглобулиновых доменов, в том числе одноцепочечные белки. Такие связывающие белки и способы их получения описаны, например, в работе Binz et al. (2005) Nature Biotechnology 23(10):1257-1268, включенной в настоящий документ путем отсылки. Следует учесть, что все сказанное в настоящем документе относительно иммуноглобулинов или происходящих из них связывающих молекул соответственно применимо к связывающим молекулам, происходящим из неиммуноглобулиновых доменов. В частности, используя такие связывающие молекулы, происходящие из неиммуноглобулиновых доменов, можно блокировать CLDN6 в клетках, в которых экспрессируется указанная мишень и которые отличаются тем, что указанная мишень ассоциирована с их поверхностью; таким образом можно достичь терапевтического эффекта, описанного в настоящем документе применительно к антителам по данному изобретению, в частности добиться ингибирования одной или более из множества активностей опухолевых клеток, описанных в настоящем документе, например пролиферации. Возможно также (хотя и не обязательно) придать таким неиммуноглобулиновым связывающим молекулам эффекторные функции антител путем, например, соединения их с Fc-областью антител.
В целом данное изобретение охватывает лечение и/или диагностирование заболеваний, в частности онкологических заболеваний, путем прицельного воздействия на CLDN6, который экспрессируется в клетках и ассоциирован с их поверхностью. Этот подход обеспечивает избирательное определение и/или ликвидацию таких клеток и тем самым сведение к минимуму негативного воздействия на нормальные клетки, в которых не экспрессируется CLDN6 и которым не свойственна связь CLDN6 с их поверхностью. Предпочтительными заболеваниями для лечения или диагностирования являются те, при которых задействованы клетки, в которых экспрессируется CLDN6 и которые отличаются связью CLDN, например онкологические заболевания, в частности раковые заболевания типа тех, что описаны в настоящем документе.
В одном из вариантов настоящего изобретения предлагаются композиции, например фармацевтические и диагностические композиции/наборы, включающие антитела или комбинации антител по данному изобретению. Фармацевтическая композиция по данному изобретению может содержать фармацевтически приемлемый носитель и, при необходимости, один или более вспомогательных компонентов, стабилизирующих агентов и др. В одном конкретном воплощении данного изобретения такая композиция включает комбинацию антител, которые связываются с отдельными эпитопами или которые имеют различные функциональные свойства, например способны вызывать цитотоксичность, зависимую от комплемента и/или антителозависимую клеточно-опосредованную цитотоксичность и способность вызывать апоптоз. В этом воплощении данного изобретения антитела могут использоваться в сочетании, например в виде фармацевтической композиции, содержащей два или более моноклональных антитела против CLDN6. Например, моноклональные антитела против CLDN6, обладающие различными, но дополняющими друг друга активностями, можно сочетать в одном методе лечения для достижения желаемого терапевтического эффекта. В одном предпочтительном воплощении данного изобретения предлагаемая композиция включает антитела против CLDN6, которые опосредуют цитотоксичность, зависимую от комплемента, вместе с другими антителами против CLDN6. которые опосредуют апоптоз. В другом воплощении данного изобретения предлагаемая композиция включает антитела против CLDN6, которые опосредуют высоко эффективное уничтожение клеток-мишеней в присутствии эффекторных клеток, вместе с другими антителами против CLDN6. которые подавляют пролиферацию клеток с экспрессией CLDN6, отличающихся тем, что CLDN6 связан с их поверхностью.
Настоящее изобретение также включает одновременное или последовательное введение двух или более антител против CLDN6, из которых по меньшей мере одно является гибридным антителом против CLDN6 и по меньшей мере одно другое антитело является человеческим антителом против CLDN6, причем эти антитела связываются с одними и теми же либо разными эпитопами CLDN6. Предпочтительно сначала пациенту вводят гибридные антитела против CLDN6, а затем человеческие антитела против CLDN6, причем человеческие антитела против CLDN6 предпочтительно вводят на протяжении некоторого (продолжительного) периода времени, например в качестве поддерживающей терапии.
Антитела, конъюгаты, биспецифические/полиспецифические молекулы и композиции по данному изобретению можно использовать в различных способах для ингибирования пролиферации клеток, в которых экспрессируется CLDN6 и которые отличаются теми, что CLDN6 связан с их поверхностью, и/или для избирательного уничтожения клеток, в которых экспрессируется CLDN6 и которые отличаются теми, что CLDN6 связан с их поверхностью, путем воздействия на эти клетки эффективного количества указанных антител, конъюгатов, биспецифических/полиспецифических молекул и композиций таким образом, что пролиферация указанных клеток подавляется и/или они гибнут. В одном и воплощений данного изобретения такой способ включает уничтожение клеток, в которых экспрессируется CLDN6 и которые отличаются теми, что CLDN6 связан с их поверхностью, при необходимости в присутствии эффекторных клеток, например путем цитотоксичности, зависимой от комплемента, апоптоза, цитотоксичности, зависимой от антител, фагоцитоза или путем комбинированного действия этих механизмов. К клеткам с экспрессией CLDN6, которые отличаются теми, что CLDN6 связан с их поверхностью и которые могут быть ингибированы или убиты с помощью антител по данному изобретению, относятся раковые клетки.
Антитела, конъюгаты, биспецифические/полиспецифические молекулы и композиции по данному изобретению можно использовать для лечения и/или предотвращения различных заболеваний, при которых задействованы клетки, в которых экспрессируется CLDN6 и которые отличаются теми, что CLDN6 связан с их поверхностью, путем введения указанных антител пациентам, страдающим от этих заболеваний. Примеры заболеваний, которые можно лечить (например, облегчать течение болезни) или предотвращать таким образом, включают (но не ограничиваются перечисленным здесь) заболевания, при которых образуются опухоли. Примеры заболеваний, при которых образуются опухоли и которые можно лечить и/или предотвращать указанным образом, включают раковые заболевания, например рак яичника, в частности аденокарциному яичника и тератокарциному яичника, рак легких, в том числе мелкоклеточный рак легких (SCLC) и немелкоклеточный рак легких (NSCLC), в частности плоскоклеточную карциному легких и аденокарциному, рак желудка, рак молочной железы, рак печени, рак поджелудочной железы, рак кожи, в частности базальноклеточную карциному и плоскоклеточную карциному, злокачественную меланому, рак головы и шеи, в том числе злокачественную полиморфную аденому, саркому, в частности синовиальную саркому и карциносаркому, рак желчного протока, рак мочевого пузыря, в частности переходноклеточную карциному и папиллярную карциному, рак почек, в частности почечно-клеточную карциному, включая светлоклеточную почечно-клеточную карциному и папиллярную почечно-клеточную карциному, рак толстого кишечника, рак тонкого кишечника, включая рак подвздошной кишки, в частности аденокарциному тонкого кишечника и аденокарциному подвздошной кишки, эмбриональную карциному яичка, хориокарциному плацентарной площадки, рак шейки матки, рак яичка, в частности семиному яичка, тератому яичка и эмбриональный рак яичка, рак матки, герминогенные опухоли, например тератокарциному или эмбриональную карциному, в частности герминогенную опухоль яичка, и их метастазирующие формы.
В другом своем аспекте настоящее изобретение относится к способу лечения или предотвращения заболеваний или расстройств, при которых задействованы клетки, в которых экспрессируется CLDN6 и которые отличаются теми, что CLDN6 связан с их поверхностью, включающий введение пациенту антител, конъюгатов, биспецифичных/полиспецифичных молекул и композиций по данному изобретению. Предпочтительно эти болезни и расстройства являются такими, при которых образуются опухоли, и в конкретных воплощениях данного изобретения их выбирают из группы, состоящей из рака яичника, в том числе аденокарциномы яичника и тератокарциномы яичника, рака легких, в том числе мелкоклеточного рака легких (SCLC) и немелкоклеточного рака легких (NSCLC), в особенности плоскоклеточной карциномы легких и аденокарциномы, рака желудка, рака молочной железы, рака печени, рака поджелудочной железы, рака кожи, в частности базальноклеточной карциномы и плоскоклеточной карциномы, злокачественной меланомы, рака головы и шеи, в том числе злокачественной полиморфной аденомы, саркомы, в частности синовиальной саркомы и карциносаркомы, рака желчного протока, рака мочевого пузыря, в частности переходноклеточной карциномы и папиллярной карциномы, рака почек, в частности почечно-клеточной карциномы, включая светлоклеточную почечно-клеточную карциному и папиллярную почечно-клеточную карциному, рака толстого кишечника, рака тонкого кишечника, включая рак подвздошной кишки, в частности аденокарциному тонкого кишечника и аденокарциному подвздошной кишки, эмбриональной карциномы яичка, хориокарциномы плацентарной площадки, рака шейки матки, рака яичка, в частности семиномы яичка, тератомы яичка и эмбрионального рака яичка, рака матки, герминогенных опухолей, например тератокарциномы или эмбриональной карциномы, в частности герминогенной опухоли яичка, и их метастазирующих форм. Предпочтительно CLDN6 экспрессируется на поверхности указанных клеток.
Данное изобретение включает применение описанных в настоящем документе агентов и композиций для профилактики и лечения онкологических заболеваний, т.е. для лечения пациентов, которые страдают каким-либо онкологическим заболеванием или для которых повышен риск развития такого заболевания. В одном из аспектов данного изобретения предлагаются способы ингибирования опухолевого роста, включающие введение одного (одной) или более описанных в настоящем документе агентов и композиций.
Предпочтительно, описанные в настоящем документе агенты и композиции вводят пациентам таким образом, что терапевтически активное вещество не попадает или в основном не попадает в ткани или органы, где нет опухолей, в клетках которых экспрессируется CLDN6 и которые отличаются тем, что CLDN6 связан с их поверхностью, например плацентарная ткань или плацента. Поэтому описанные в настоящем документе агенты и композиции можно вводить локально.
В одном аспекте данного изобретения предлагаются антитела, описанные в настоящем документе, для применения в способах лечения, описанных в настоящем документе. В одном из воплощений данного изобретения предлагается описанная в настоящем документе фармацевтическая композиция для применения в способах лечения, описанных в настоящем документе.
В одном конкретном воплощении и данного изобретения пациент, которому вводят предлагаемые антитела, дополнительно получает химио- или лучевую терапию или агенты, которые модулируют, т.е. усиливают или ингибируют, экспрессию либо активность рецепторов Fc, например рецепторов Fey, например цитокинов. Цитокины для введения в ходе указанного лечения включают гранулоцитарный колониестимулирующий фактор (G-CSF), гранулоцитарно-макрофагальный колониестимулирующий фактор (GM-CSF), гамма-интерферон (IFN-γ) и фактор некроза опухолей (TNF). Химиотерапевтические агенты для введения в ходе указанного лечения включают, помимо прочих, антинеопластические агенты, например доксорубицин, цисплатин, таксотер, 5-фторурацил. метотрексат, гемцитабин и циклофосфамид.
В другом своем аспекте данное изобретение относится к иммунизации животных, например мышей человеческим CLDN6 или пептидом, являющимся его фрагментом, для получения антител. Предпочтительными для иммунизации являются пептиды, выбираемые из группы, состоящей из SEQ ID NO:6, SEQ ID NO:7, SEQ ID NO:14 и SEQ ID NO:15 и иммунологически эквивалентных им пептидов.
Животных дикого типа, а также трансгенных животных (которые не являются человеком), можно иммунизировать очищенными или обогащенными препаратами антигенов CLDN6 или пептидов, являющихся их фрагментами, и/или нуклеиновыми кислотами и/или клетками, в которых экспрессируется CLDN6 или его фрагмент. Предпочтительно, для этой цели трансгенные животные (которые не являются человеком) должны быть способны производить множественные изотипы моноклональных человеческих антител против CLDN6 (например, IgG, IgA или IgM) путем V-D-J рекомбинации и переключения изотипа. Переключение изотипа может происходить классическим либо неклассическим путем.
В другом аспекте данного изобретения предлагаются В-клетки животных (не человека), как описано выше. Выделенные В-клетки можно затем иммортализировать путем слияния с уже иммортализованными клетками, чтобы создать источник (например, гибридому) антител по данному изобретению. Такие гибридомы, производящие антитела по данному изобретению, также включаются в его объем.
В другом своем аспекте данное изобретение относится к способам диагностирования, выявления и наблюдения за течением онкологического заболевания, включающим выявление и/или определение количества CLDN6 или клеток, в которых экспрессируется CLDN6 и которые отличаются тем, что CLDN6 связан с их поверхностью, в образцах биологического материала, взятых у пациента с использованием антител по данному изобретению. Образцы биологического материала могут быть взяты у пациента с начавшимся или развившимся онкологическим заболеванием либо с подозрением на такое заболевание, либо с повышенным риском такого заболевания.
В одном из воплощений способа диагностирования, выявления и наблюдения за течением онкологического заболевания по данному изобретению образец биологического материала и/или контрольный образец/образец для сравнения берут из ткани или органа, соответствующих ткани или органу, подлежащим диагностированию, выявлению или наблюдению в отношении поражения онкологическим заболеванием; например, онкологическое заболевание, которое должно быть диагностировано, выявлено или отслежено, является раком яичника и образец биологического материала и/или контрольный образец/образец для сравнения берут из ткани яичника. Такие органы и ткани описаны в настоящем документе, например, в связи с различными онкологическими и раковыми заболеваниями.
В одном из воплощений способа диагностирования, выявления и наблюдения за течением онкологического заболевания по данному изобретению образец биологического материала и/или контрольный образец/образец для сравнения берут из ткани или органа, в клетках которых в отсутствие опухоли не экспрессируется CLDN6 и они не отличаются выраженным присутствием CLDN6 на своей поверхности. Предпочтительно указанная ткань не является плацентарной.
Как правило, уровень молекул-мишеней в образце биологического материала сравнивают с каким-либо уровнем, принятым за контрольный или базовый, и тогда отклонение от этого последнего считается указанием на наличие и/или стадию онкологического заболевания у данного пациента. Уровень, взятый для сравнения, может быть тем, который имеет место в контрольном образце (например, в образце здоровой ткани того же пациента) или средней величиной, установленной по результатам определения у ряда здоровых индивидов. Под отклонением от указанного уровня, взятого для сравнения, понимается любое существенное изменение, например уменьшение или увеличение не менее чем на 10%, 20% или 30%, предпочтительно не менее чем на 40% или 50% или даже больше. Предпочтительно, если в образце биологического материала присутствуют CLDN6 или клетки, в которых экспрессируется CLDN6 и которые отличаются тем, что CLDN6 связан с их поверхностью, или количество CLDN6 или клеток, в которых экспрессируется CLDN6 и которые отличаются тем, что CLDN6 связан с их поверхностью увеличено по сравнению с уровнем, взятым для сравнения, то это свидетельствует о наличии онкологического заболевания.
Как правило, выявление и/или определение количества в способах по данному изобретению включает использование меченых антител, специфически связывающихся с молекулами-мишенями.
В одном особом аспекте данное изобретение относится к способу выявления, т.е. определения локализации или распространения в организме онкологического заболевания, например какие ткани или органы поражены, который включает введение пациенту антител по данному изобретению, соединенных с детектируемой меткой. Мечение органа или ткани у указанного пациента может свидетельствовать о наличии или повышенном риске развития опухоли в этой ткани или органе.
Как отмечалось выше, антитела по данному изобретению могут быть получены непосредственно из клеток гибридомы, в которых эти антитела экспрессируются, или путем клонирования и рекомбинантной экспрессии в клетке-хозяине (например, в клетках СНО или лимфоцитах). К числу примеров клетки-хозяина относятся также такие микроорганизмы, как Escherichia coli и грибы, например дрожжи. Или же указанные антитела могут вырабатываться рекомбинантным путем у трансгенных животных (которые не являются человеком) или растений. Настоящее изобретение включает также воплощения, в которых указанные антитела производятся путем иммунизации или вакцинации in situ (в организме пациента) с помощью описанных в настоящем документе методов иммунизации.
Данное изобретение относится также к нуклеиновым кислотам, включающим гены или нуклеотидные последовательности, кодирующие антитела или их фрагменты, например какую-либо цепь молекулы антитела, как описано в настоящем документе. Эти нуклеиновые кислоты могут быть включены в вектор, например в плазмиду, космиду, вирус, бактериофаг или иной вектор с помощью, например, обычно применяемых для таких целей методов генетической инженерии. Вектор может включать также другие гены, например маркерные гены, которые позволят вести отбор в подходящих клетках-хозяевах и в подходящих условиях. Кроме того, вектор может включать регуляторные элементы, обеспечивающие должную экспрессию кодирующих последовательностей в подходящей клетке-хозяине. Специалистам в данной области техники известны такие регуляторные элементы, включая промотор, кассеты экзонов, подлежащие альтернативному сплайсингу, и кодоны инициации трансляции.
Предпочтительно нуклеиновые кислоты по данному изобретению функционально соединены с упомянутыми выше последовательностями, обеспечивающими регуляцию экспрессии в эукариотических или прокариотических клетках. Регуляторные элементы, обеспечивающие экспрессию в эукариотических или прокариотических клетках, хорошо известны специалистам в данной области техники.
Методы создания молекул нуклеиновых кислот по данному изобретению и векторов, включающих эти молекулы, введения указанных векторов в соответственно выбранные клетки-хозяева и обеспечения экспрессии нужных последовательностей, хорошо известны в данной области техники.
В еще одном своем аспекте данное изобретение относится к клеткам-хозяевам, несущим описанные в настоящем документе нуклеиновые кислоты или векторы.
Другие признаки и преимущества настоящего изобретения явствуют из нижеследующего подробного описания и формулы изобретения.
Краткое описание чертежей
Фигура 1. Выравнивание последовательностей CLDN3, CLDN4, CLDN6 и CLDN9
Фигура 2. Иммунофлуоресцентный анализ сыворотки крови, взятой у мышей, иммунизированных для получения антител, специфичных против CLDN6
(А) Нефиксированные клетки СНО-K1, ко-трансфицированные нуклеиновой кислотой, кодирующей CLDN6, вместе с флуоресцентным белком GFP, тестировали с помощью мышиных моноклональных антител против CLDN6 (R&D Systems, MAB3656). CLDN6 локализован на плазматической мембране трансфицированных клеток и на живых клетках может служить мишенью для специфичных антител. (В) Сыворотка мышей, использованных для получения гибридом F3-6C3-H8, содержала антитела, связывающиеся с CLDN6 на поверхности нефиксированных клеток СНО-K1, ко-трансфицированных нуклеиновой кислотой, кодирующей CLDN6, вместе с флуоресцентным белком GFP.
Фигура 3. Определение эндогенной экспрессии клаудинов в клетках HEK293T методом Вестерн-блоттинга
Белковый лизат клеток HEK293T, трансфицированных нуклеиновыми кислотами, кодирующими CLDN3, CLDN4, CLDN6 и CLDN9, соответственно, или контрольных клеток (с которыми проделали все те же манипуляции, но без нуклеиновой кислоты) анализировали методом Вестерн-блоттинга, используя готовые имеющиеся в продаже антитела против CLDN3(A) (Invitrogen, Cat No. 34-1700), против CLDN4(A) (Zymed, 32-9400), против CLDN6(A) (ARP, 01-8865) и против CLDN9(A) (Santa Cruz, sc-17672). Этими антителами выявлялась экспрессия соответствующих мишеней только в соответствующим образом трансфицированных клетках HEK293T. В нетрансфицированных клетках HEK293T не наблюдалось эндогенной экспрессии ни одного из перечисленных клаудинов.
Фигура 4. Определение специфичности готовых имеющихся в продаже антител против CLDN методом проточной цитометрии
Методом проточной цитометрии определяли связывание готовых имеющихся в продаже антител против CLDN с клетками HEK293T, трансфицированными нуклеиновыми кислотами, кодирующими CLDN3, CLDN4, CLDN6 и CLDN9, соответственно, и с нетрансфицированными клетками HEK293T. Специфичными к своей мишени оказались только готовые антитела против CLDN3.
Фигура 5. Определение специфичности антител против CLDN, полученных по данному изобретению, методом проточной цитометрии
Методом проточной цитометрии определяли связывание антител в супернатантах моноклональных субклонов гибридом с клетками HEK293T, трансфицированных одновременно двумя векторами, один из которых кодировал CLDN6, CLDN3, CLDN4 или CLDN9, а другой - флуоресцентный маркер.
(A) Антитела в супернатанте моноклонального субклона гибридом F3-6C3-H8 специфично связывались с клетками, трансфицированными CLDN6, но не с клетками, трансфицированными CLDN3, CLDN4 и CLDN9, соответственно. В противоположность этому антитела в супернатанте моноклонального субклона гибридом F4-4F7-F2 связывались с клетками, трансфицированными CLDN6 или CLDN9. Антитела в супернатанте моноклонального субклона гибридом F3-6C3-H8 также связывались с клетками, трансфицированными вариантом CLDN6 (I143V)-SNP.
(B) Антитела в из супернатанта моноклонального субклона гибридом F3-7B3-B4 связывались с клетками, трансфицированными CLDN6, CLDN3 или CLDN9. Моноклональные антитела в супернатанте субклона гибридом F3-3F7-A5 связывались с клетками, трансфицированными CLDN6, CLDN4 или CLDN9
Фигура 6. Специфичность связывания мышиных мноклональных антител против CLDN6 muMAB 59А, 60А, 61D, 64А, 65А, 66В и 67А
Связывание антител против CLDN6 с человеческими белками CLDN6, 3, 4, 9 и CLDN6 SNP (однонуклеотидный полиморфизм) вариант I143V определяли путем проточной цитометрии, используя клетки HEK293T с транзиентной (временной) экспрессией соответствующего человеческого клаудина. Эти клетки одновременно трансфицировали флуоресцентным маркером, чтобы отличать нетрансфицированные клетки (популяции Q1 и Q3) от трансфицированных (популяции Q2 и Q4). Концентрацию антител брали насыщающей места связывания с CLDN6 (25 мкг/мл). Экспрессию человеческих белков CLDN6, 3, 4, 9 и CLDN6-SNP(I143V) подтверждали с помощью готовых имеющихся в продаже моноклональных антител против клаудина-6 человека (R&D Systems, MAB3656), клаудина-3 человека (R&D Systems, MAB4620) и клаудина-4 человека (R&D Systems, MAB 4219).
Фигура 7. Относительная аффинность мышиных монолкональных антител против CLDN6 muMAB 59А, 60А, 61D, 64А, 65А, 66В и 67А
Для определения относительной аффинности анализировали связывание антител против CLDN6 с человеческим белком CLDN6, стабильно экспрессирующимся на поверхности клеток HEK293, методом проточной цитометрии. По данным о насыщении мест связывания строили график зависимости сигнала FACS (средней интенсивности флуоресценции) от концентрации антител. Рассчитывали ЕС50 (концентрацию антител, при которой в равновесии занята половина мест их связывания), используя метод нелинейной регрессии. Для специфичных в отношении CLDN6 антител muMAB 59A, 60А, 61D, 64A, 65А, 66В и 67А получались очень низкие значения ЕС50 (200-500 нг/мл) и насыщение мест связывания достигалось при низких концентрациях.
Фигура 8. Активность, проявляющаяся комплемент-зависимой цитотоксичностью (CDC), мышиных моноклональных антител против CLDN6 muMAB 59A, 60А, 61D, 64A, 65А, 66B и 67А
Активность, проявляющуюся комплемент-зависимой цитотоксичностью, антител против CLDN6 определяли в люциферазной системе, выявляющей эндогенный аденозинтрифосфат в нелизированных клетках. Клетки СНО-K1 со стабильной экспрессией человеческого CLDN6 инкубировали с различными концентрациями антител muMAB 59A, 60А, 61D, 64A, 65А, 66B и 67А. В случае антител MuMAB 59A, 60А, 61D, 64A, 65А, 66B и 67А наблюдалась зависимая от дозы цитотоксичность, зависимая от комплемента, причем она возникала при низких концентрациях антител.
Фигура 9. Активность, проявляющаяся комплемент-зависимой цитотоксичностью (CDC), мышиных моноклональных антител против CLDN6 muMAB 65А и 66B в отношении клеток NEC8 с эндогенной экспрессией CLDN6 и клеток NEC8 LVTS2 54 (нокдаунных по CLDN6)
Мышиные моноклональные антитела против CLDN6 muMAB 65А и 66B вызывали цитотоксичность в отношении клеток NEC8 в зависимости от дозы. Специфичность muMAB 65А и 66B в отношении мишени подтверждалась в опыте с клетками NEC8 LVTS2 54 (нокдаунными по CLDN6).
Фигура 10. Терапевтический эффект мышиных моноклональных антител против CLDN6 muMAB 59A, 60А, 61D, 64A, 65А, 66B и 67А на ранних стадиях развития ксенотрансплантата опухоли у мышей, где мышам прививали линию опухолевых клеток NEC8
Бестимусным мышам линии Nude-Foxn1nu вводили клетки NEC8 с эндогенной экспрессией CLDN6. Мышиные моноклональные антитела против CLDN6 muMAB 59A, 60А, 61D, 64A, 65А, 66B и 67А подавляли опухолевый рост по сравнению с особями контрольной группы, которые получали вместо антител физиологический раствор.
Фигура 11. Специфичность связывания гибридных моноклональных антител против CLDN6 chimAB 61D, 64А, 67А и 89А
Методом проточной цитометрии определяли связывание антител против CLDN6 с человеческими белками CLDN6, 3, 4 и 9, используя клетки HEK293 со стабильной экспрессией соответствующего человеческого клаудина. Концентрация антител была насыщающей места связывания (25 мкг/мл). Экспрессию человеческих белков CLDN3, 4, 6 и 9 подтверждали с помощью готовых имеющихся в продаже моноклональных антител против человеческого клаудина-3 (R&D Systems, MAB4620) и человеческого клаудина-4 (R&D Systems, MAB 4219), и CLDN6/9-реактивных мышиных моноклональных антител muMAB 5F2D2, соответственно. Для отрицательного контроля проделывали те же манипуляции в тех же условиях, но без первых антител.
Фигура 12. Относительное сродство моноклональных гибридных антител против CLDN6 chimAB 61D, 64А, 67А и 89А к клеткам HEK293-CLDN6
Методом проточной цитометрии определяли связывание антител против CLDN6 с человеческим CLDN6, стабильно экспрессирующимся на поверхности клеток HEK293. Результаты опыта с насыщением мест связывания представляли в виде графика, откладывая по оси ординат сигнал FACS (среднюю интенсивность флуоресценции), а по оси абсцисс - концентрацию антител. Рассчитывали ЕС50 (концентрацию антител, при которой в равновесии занята половина мест их связывания), используя метод нелинейной регрессии. Для специфичных в отношении CLDN6 антител chimAB 64A и 89А получались очень низкие значения ЕС50 (450-600 нг/мл) и насыщение мест связывания достигалось при низких концентрациях. Антитела chimAB 67А и 61D характеризовались низким (1000 нг/мл) и средним (2300 ng/ml) значениями ЕС50 соответственно.
Фигура 13. Относительное сродство моноклональных гибридных антител против CLDN6 chimAB 61D, 64А, 67А и 89А к клеткам NEC8
Чтобы установить сродство антител против CLDN6 к опухолевым клеткам, в которых эндогенно экспрессируется человеческий CLDN6, методом проточной цитометрии определяли связывание этих антител с клетками линии NEC8 (рак яичка). Для специфичных в отношении CLDN6 антител chimAB 64А и 89А получались очень низкие значения ЕС50 (600-650 нг/мл) и насыщение мест связывания достигалось при низких концентрациях. Антитела chimAB 61D и 67А характеризовались средним (1700 нг/мл) и высоким (6100 нг/мл) значениями ЕС50 соответственно.
Фигура 14. Относительное сродство моноклональных гибридных антител против CLDN6 chimAB 61D, 64А, 67А и 89А к клеткам OV90
Чтобы установить сродство антител против CLDN6 к опухолевым клеткам, в которых эндогенно экспрессируется человеческий CLDN6, методом проточной цитометрии определяли связывание этих антител с клетками линии OV90 (рак яичника). Для специфичных в отношении CLDN6 антител chimAB 64A и 89А получались очень низкие значения ЕС50 (550-600 нг/мл) и насыщение мест связывания достигалось при низких концентрациях. Антитела chimAB 61D и 67А характеризовались средними значениями ЕС50 (1500 нг/мл и 2300 нг/мл, соответственно).
Фигура 15. Активность, проявляющаяся комплемент-зависимой цитотоксичностью (CDC), моноклональных гибридных антител против CLDN6 chimAB 61D, 64A, 67А и 89А в отношении клеток NEC8 дикого типа и нокдаунных
Активность, проявляющуюся комплемент-зависимой цитотоксичностью, антител против CLDN6 определяли в люциферазной системе, выявляющей эндогенный аденозинтрифосфат в нелизированных клетках. Клетки NEC8 дикого типа (NEC8 LVTS2 77) с эктопической экспрессией люциферазы инкубировали с различными концентрациями антител chimAB 61D, 64A, 67А и 89А. Антитела chimAB 61D, 64A, 67А и 89А влияли на клетки NEC8 зависимым от дозы образом, тогда как в случае клеток NEC8, нокдаунных по CLDN6 (NEC8 LVTS2 54), ни одно из указанных антител не вызывало неспецифического лизиса. Эти результаты демонстрируют специфичные в отношении мишени эффекторные функции chimAB 61D, 64A, 67А и 89А.
Фигура 16. Активность, проявляющаяся антителозависимой клеточно-опосредованной цитотоксичностью (ADDC), моноклональных гибридных антител против CLDN6 chimAB 61D, 64A, 67А и 89А в отношении клеток NEC8 дикого типа и нокдаунных
Активность, проявляющуюся зависимой от антител клеточно-опосредованной цитотоксичностью (ADDC), антител против CLDN6 определяли в люциферазной системе, выявляющей эндогенный аденозинтрифосфат в нелизированных клетках. Клетки NEC8 дикого типа (NEC8 LVTS2 77) инкубировали с различными концентрациями антител chimAB 61D, 64A, 67А и 89А. Активность антител chimAB 61D, 64A, 67А и 89А, проявляющаяся ADDC, зависила от дозы и возникала даже при низких концентрациях антител. Чтобы продемонстрировать специфичность антител в отношении мишени, использовали клетки NEC8 стабильно нокдаунные по CLDN6 (NEC8 LVTS2 54).
Фигура 17. Долгосрочный терапевтический эффект мышиных моноклональных антител против CLDN6 muMAB 61D, 64A и 67А у мышей с ксенографтами опухолевых клеток линии NEC8 при воздействии на ранних стадиях развития опухолей
Бестимусные мыши Nude-Foxn1nu с ксенографтами клеток NEC8, в которых эндогенно экспрессируется CLDN6, в течение 46 суток получали антитела, специфичные к CLDN6. После этого воздействия на протяжении 60 суток отслеживали опухолевый рост. Даже после прекращения этой иммунотерапии у мышей, получавших мышиные моноклональные антитела muMAB 61D, 64A и 67А роста опухолей не наблюдалось.
Фигура 18. Терапевтический эффект мышиных моноклональных антител против CLDN6 muMAB 89A у мышей с ксенографтами опухолевых клеток линии NEC8 при воздействии на ранних стадиях развития опухолей
Использовали бестимусные мышей Nude-Foxn1nu с ксенографтами клеток NEC8, в которых эндогенно экспрессируется CLDN6. Разбросанные кружочки представляют значения объема опухолей в различные моменты времени в ходе лечения антителами в ранний период развития ксенографтов NEC8 у бестимусных мышей Nude-Foxn1nu. По сравнению с контрольными особями у мышей, получавших muMAB 89A, наблюдалось ингибирование опухолевого роста (А). Введение антител, специфичных к CLDN6, либо PBS (контроль) продолжалось 47 суток, в течение которых и на протяжении последующих 51 суток регистрировали опухолевый рост. К концу ксперимента у мышей, получавших muMAB 89A, опухолей не обнаруживалось - в отличие от контрольных особей, получавших PBS (В).
Фигура 19. Терапевтический эффект мышиных моноклональных антител против CLDN6 muMAB 64A у мышей с ксенографтами опухолевых клеток линии NEC8 при воздействии на поздних стадиях развития опухолей
Разбросанные кружочки представляют значения объема опухолей в различные моменты времени в ходе лечения антителами в поздний период развития ксенографтов NEC8 у бестимусных мышей Nude-Foxn1nu. При иммунотерапии мышиными моноклональными антителами против CLDN6 muMAB 64A наблюдалось ингибирование роста солидных опухолей, развившихся из ксенографтов NEC8, по сравнению с обеими контрольными группами, в одной из которых мыши получали PBS без антител, а во второй - неспецифичные антитела.
Фигура 20. Долгосрочный терапевтический эффект мышиных моноклональных антител против CLDN6 muMAB 64A у мышей с ксенографтами опухолевых клеток линии NEC8 при воздействии на поздних стадиях развития опухолей
Через 15 суток после введения опухолевых клеток 15 мыши в течение 45 суток получали специфичные к CLDN6 антитела muMAB 64A. На протяжении еще 49 суток регистрировали опухолевый рост (А). График, представляющий выживание мышей, показывает, что животные, получавшие специфичные к CLDN6 антитела muMAB 64A жили дольше (В).
Фигура 21. Терапевтический эффект мышиных моноклональных антител против CLDN6 muMAB 61D, 67А и 89А у мышей с ксенографтами опухолевых клеток линии NEC8 при воздействии на поздних стадиях развития опухолей
Разбросанные кружочки, треугольники и квадратики представляют значения объема опухолей в различные моменты времени в ходе лечения антителами в поздний период развития ксенографтов NEC8. Мышиные моноклональные антитела против CLDN6 muMAB 61D, 67А и 89А подавляли опухолевый рост - в отличие от контрольных воздействий (PBS без антител и неспецифичные антитела.
Фигура 22. Долгосрочный терапевтический эффект мышиных моноклональных антител против CLDN6 muMAB 61D, 67А и 89А у мышей с ксенографтами опухолевых клеток линии NEC8 при воздействии на поздних стадиях развития опухолей
Через 17 суток после введения опухолевых клеток мыши на протяжении 42 суток получали специфичные к CLDN6 антитела muMAB 61D, 67А и 89А. В период иммунотерапии и на протяжении еще 49 суток регистрировали опухолевый рост (А). График, представляющий выживание мышей, показывает, что животные, получавшие специфичные к CLDN6 антитела muMAB 61D и 67А жили дольше (В).
Фигура 23. Терапевтический эффект мышиных моноклональных антител против CLDN6 muMAB 64A и 89А у мышей с ксенографтами опухолевых клеток NEC8 дикого типа и клеток NEC8, стабильно нокдаунных по CLDN6
Антитела muMAB 64A и 89А оказывали терапевтическое действие только у мышей с ксенографтами NEC8 дикого типа, а у животных, которым были введены клетки NEC8, нокдаунные по CLDN6, такого эффекта не наблюдалось, что демонстрирует специфичность этих антител в отношении мишени in vivo.
Фигура 24. Картирование с высоким разрешением эпитопов, с которыми взаимодействуют антитела chimAB 61D, 64A, 67А и 89А
Аланиновые мутантные варианты обозначенные « остаток дикого типа номер аланин» или «остаток дикого типа номер)глицин» в случаях наличия в данном положении аланина в белке дикого типа; аминокислоты названы однобуквенным обозначениями. Для взаимодействия со специфичными к CLDN6 гибридными антителами chimAB 61D, 64A, 67А и 89А важны аминокислотные остатки F35, G37, S39 и, вероятно, Т33 первого внеклеточного домена CLDN6. Аминокислотный остаток 140 имеет существенное значение для связывания chimAB 89А и участвует в связывании chimAB 61D and 67A. Кроме того, аминокислотный остаток L151 второго внеклеточного домена участвует во взаимодействии с антителами chimAB 67A. Хотя иммунофлуоресцентный анализ подтвердил экспрессию мутантных вариантов CLDN6 Р28А, W30A, G49A, L50A, W51A, С54А и С64А, они не давали окрашивания мембран. Поэтому нельзя исключить взаимодействия антител по данному изобретению с этими аминокислотами. В целом идентифицированные эпитопы согласуются с теми ДНК и пептидами домена ЕС1 CLDN6, которые использовались для иммунизации.
Фигура 25. Выравнивание аминокислотных последовательностей вариабельной области тяжелой цепи специфичных к CLDN6 антител по данному изобретению
Участки, определяющие комплементарность (HCDR1, HCDR2 и HCDR3) показаны рамкой.
Фигура 26. Выравнивание аминокислотных последовательностей вариабельной области легкой цепи специфичных к CLDN6 антител по данному изобретению
Участки, определяющие комплементарность (LCDR1, LCDR2 и LCDR3) показаны рамкой.
Фигура 27. Специфичность связывания моноклональных гибридных антител против CLDN6 chimAB 64A, mAb206-LCC, mAb206-SUBG и mAb206-SUBS
Специфичность связывания антител против CLDN6 определяли методом проточной цитометрии, используя клетки HEK293T, временно трансфицированные человеческими CLDN6, 3, 4 и 9,соответственно. Чтобы отличать нетрансфицированные клетки от трансфицированных, клетки одновременно трансфицировали флуоресцентным маркером. Концентрация антител была насыщающей места связывания (100 мкг/мл). Экспрессию человеческих белков CLDN3, 4, 6 и 9 подтверждали с помощью готовых имеющихся в продаже моноклональных антител против человеческого клаудина-3 (R&D Systems, MAB4620) и человеческого клаудина-4 (R&D Systems, MAB 4219), и CLDN6/9-реактивных мышиных моноклональных антител muMAB 5F2D2, соответственно.. Моноклональные гибридные антитела chimAB 64A, mAb206-LCC, mAb206-SUBG и mAb206-SUBS связывались с CLDN6, но не взаимодействовали с CLDN3, 4 и 9, соответственно.
Фигура 28. Относительное сродство связывания (аффинность) моноклональных гибридных антител против CLDN6 chimAB 64A, mAb206-LCC, mAb206-SUBG и mAb206-SUBS с клетками HEK293-CLDN6
Для оценки относительного сродства (аффинности) методом проточной цитометрии определяли связывание антител против CLDN6 с человеческим CLDN6, стабильно экспрессирующимся на поверхности клеток HEK293. По результатам опыта с насыщением мест связывания строили график зависимости сигнала FACS (средней интенсивности флуоресценции) от концентрации антител. Рассчитывали ЕС50 (концентрацию антител, при которой в равновесии занята половина мест их связывания), используя метод нелинейной регрессии. Специфичные к CLDN6 антитела характеризовались сходными низкими значениями ЕС50, и насыщение мест связывания достигалось при низких концентрациях.
Фигура 29. Относительное сродство связывания (аффинность) моноклональных гибридных антител против CLDN6 chimAB 64A, mAb206-LCC, mAb206-SUBG и mAb206-SUBS с клетками NEC8
Для оценки относительного сродства (аффинности) методом проточной цитометрии определяли связывание антител против CLDN6 с опухолевыми клетками, в которых эндогенно экспрессируется человеческий CLDN6, а именно с клетками линии NEC8 (рак яичка). По сравнению со специфичными к CLDN6 антителами chimAB 64A, mAb206-SUBG и mAb206-SUBS вариант с комбинацией легких цепей mAb206-LCC проявлял втрое большую аффинность по отношению к клеткам NEC8. Во всех случаях насыщение мест связывания достигалось при низких концентрациях.
Фигура 30. Относительное сродство связывания (аффинность) моноклональных гибридных антител против CLDN6 chimAB 64A, mAb206-LCC, mAb206-SUBG и mAb206-SUBS с клетками OV90
Методом проточной цитометрии определяли сродство связывания (аффинность) антител против CLDN6 с человеческими клетками линии OV90 (рак яичника). Антитела, специфичные к CLDN6, характеризовались сходными низкими значениями ЕС50. Вариант с комбинацией легких цепей mAb206-LCC проявлял наибольшую прочность связывания с клетками OV90.
Фигура 31а. Активность моноклональных гибридных антител против CLDN6 chimAB 64A, mAb206-LCC и mAb206-SUBG, проявляющаяся комплемент-зависимой цитотоксичностью (CDC) в отношении клеток NEC8 дикого типа и нокдаунных клеток NEC8
Активность, проявляющуюся комплемент-зависимой цитотоксичностью, антител против CLDN6 определяли в люциферазной системе, выявляющей эндогенный аденозинтрифосфат в нелизированных клетках. Клетки NEC8 дикого типа с эктопической экспрессией люциферазы инкубировали с различными концентрациями антител chimAB 64A, mAb206-LCC и mAb206-SUBG. Эти антитела влияли на клетки NEC8 зависимым от дозы образом, тогда как в случае клеток NEC8, нокдаунных по CLDN6 (NEC8 LVTS2 54), ни одно из них не вызывало неспецифического лизиса. Эти результаты демонстрируют специфичные в отношении мишени эффекторные функции chimAB 64A, mAb206-LCC и mAb206-SUBG.
Фигура 31b. Активность моноклональных гибридных антител против CLDN6 chimAB 64A, mAb206-LCC, mAb206-SUBG и mAb206-SUBS, проявляющаяся комплемент-зависимой цитотоксичностью (CDC) в отношении клеток NEC8
Эти антитела вызывали CDC в отношении клеток NEC8 зависимым от дозы образом. По сравнению с антителами chimAB 64A варианты с аминокислотными заменами mAb206-SUBG и mAb206-SUBS обладали сходной активностью.
Фигура 32а. Активность моноклональных гибридных антител против CLDN6 chimAB 64A, mAb206-LCC, mAb206-SUBG и mAb206-SUBS, проявляющаяся антителозависимой клеточно-опосредованной цитотоксичностью (ADDC), в отношении клеток NEC8 и OV90
Активность антител против CLDN6, проявляющуюся антителозависимой клеточно-опосредованной цитотоксичностью, определяли в люциферазной системе, выявляющей эндогенный аденозинтрифосфат в нелизированных клетках. Клетки NEC8 и OV90 инкубировали с различными концентрациями гибридных антител против CLDN6. В обеих линий опухолевых клеток все указанные антитела вызывали ADCC даже в низких концентрациях, причем эффект ADDC зависел от дозы.
Фигура 32b. Активность моноклональных гибридных антител против CLDN6 chimAB 64A, mAb206-LCC и mAb206-SUBG, проявляющаяся антителозависимой клеточно-опосредованной цитотоксичностью (ADDC), в отношении клеток NEC8 дикого типа и нокдаунных клеток NEC8
Чтобы продемонстрировать специфичность антител в отношении мишени, использовали клетки NEC8, стабильно нокдаунные по CLDN6,
Фигура 33. Терапевтический эффект моноклональных гибридных антител против CLDN6 mAb206-LCC, mAb206-SUBG и mAb206-SUBS у мышей с ксенографтами опухолевых клеток линии NEC8 при воздействии на поздних стадиях развития опухолей
Использовали бестимусных мышей линии Nude-Foxn1nu с ксенографтами клеток NEC8. Через 17 суток после введения опухолевых клеток мышам вводили специфичные к CLDN6 антитела mAb206-LCC, mAb206-SUBG и mAb206-SUBS и затем регистрировали опухолевый рост. Разбросанные кружочки, треугольники и квадратики представляют значения объема опухолей в различные моменты времени в ходе лечения антителами в поздний период развития ксенографтов NEC8. По сравнению с контролем (мыши получали неспецифичные антитела) моноклональные гибридные антитела против CLDN6 mAb206-LCC, mAb206-SUBG и mAb206-SUBS подавляли опухолевый рост.
Фигура 34. Терапевтический эффект моноклональных гибридных антител против CLDN6 mAb206-LCC, mAb206-SUBG и mAb206-SUBS у мышей с ксенографтами опухолевых клеток линии NEC8 при воздействии на поздних стадиях развития опухолей
Для сравнения с фиг.33 данные по опухолевому росту во времени представлены более подробно. Видно, что моноклональные гибридные антитела против CLDN6 способны подавлять опухолевый рост (А). Кривая выживания показывает, что мыши, получавшие специфичные к CLDN6 антитела, жили дольше (В).
Фигура 35. Картирование с высоким разрешением эпитопов, с которыми взаимодействуют антитела chimAB 64A, mAb206-LCC, mAb206-SUBG and mAb206-SUBS
Аланоновые мутантные варианты названы «остаток дикого типа номер аланин»; аминокислоты названы однобуквенным обозначениями. Для взаимодействия со специфичными к CLDN6 гибридными антителами важны аминокислотные остатки F35, G37, S39 и, вероятно, Т33 первого внеклеточного домена CLDN6. Антитела chimAB 64A, mAb206-LCC, mAb206-SUBG и mAb206-SUBS характеризовались одинаковыми особенностями связывания.
Фигура 36. Терапевтический эффект мышиных моноклональных антител против CLDN6 muMAB 64A в отношении метастазирования у мышей с ксенографтами опухолевых клеток линии NEC8
Бестимусным мышам линии Nude-Foxn1nu вводили в хвостовую вену клетки NEC8, а через трое суток эти животные получали специфичные к CLDN6 антитела muMAB 64A. Спустя 8 недель определяли опухолевую нагрузку в ткани легких с помощью полимеразной цепной реакции. Мышиные моноклональные антитела против CLDN6 muMAB 64A по сравнению с контролем (PBS без антител) вызывали отчетливое ингибирование опухолевого роста.
Фигура 37. Иммуногистохимическое окрашивание раковых и нормальных тканей человека с использованием моноклональных антител muMAB 64A, mAb206-LCC и mAb206-SUBG
В противоположность нормальным тканям в срезах опухолей яичников и яичек наблюдалось выраженное равномерное окрашивание. Очень интенсивное окрашивание мембран имело место в злокачественных эпителиальных клетках, тогда как соседние стромальные клетки и незлокачественные эпителиальные клетки не окрашивались. Эти результаты с очевидностью свидетельствуют, что специфичные к CLDN6 антитела по данному изобретению специфично связываются со злокачественными клетками, полученными от пациентов с опухолями. (Пояснение: число тканей, окрашивающихся антителами/ число изученных тканей).
Определения терминов
Чтобы облегчить понимание настоящего изобретения мы вначале дадим определение некоторым терминам. Дополнительные определения даются по ходу раскрытия изобретения.
В нижеследующем описании и в прилагаемой формуле изобретения слова «включает» и «содержит» и их производные (и синонимы, например «в их число входит») следует понимать так (если не оговорено иного), что указанные члены, целые числа или стадии или группы членов, целых чисел или стадий входят в рассматриваемую группу, но не исключается, что в нее входят и другие члены, целые числа или стадии или группы членов, целых чисел или стадий. Определенный и неопределенный артикль и их аналоги в описании данного изобретения и особенно в формуле изобретения подразумевают как единственное, так и множественное число, если не оговорено иного или если это явно не противоречит конкретному контексту. Диапазоны значений приводятся здесь для сокращенного указания всех значений, входящих в данный диапазон, вместо того, чтобы указывать их каждое в отдельности. И, если не оговорено иного, каждое отдельное значение включено в настоящее описание, как если бы оно было указано в отдельности. Все описанные здесь способы могут быть осуществлены в любом нужном порядке, если не оговорено иного или если это явно не противоречит конкретному контексту. Любой приведенный здесь пример и все они вместе взятые, а также употребление здесь выражений типа «такой/такие, как» или «например» или «в том числе» служат только для иллюстрации данного изобретения и не накладывают ограничений на объем изобретения, если не указано иного. Никакие словесные конструкции в настоящем описании не следует истолковывать как указывающие на какой-либо незаявленный элемент, существенный для практического осуществления данного изобретения.
Клаудины - это семейство белков, которые являются очень важными компонентами плотных контактов, где они образуют околоклеточный барьер, регулирующий поток веществ в межклеточном пространстве эпителия. Клаудины представляют собой трансмембранные белки; молекулы клаудина пересекает мембрану четыре раза, причем и N-конец, и С-конец находятся в цитоплазме. Первая внеклеточная петля состоит из в среднем 53 аминокислотных остатков, вторая - из около 24 аминокислотных остатков. В семействе клаудинов белки CLDN6 и CLDN9 наиболее сходны между собой. В настоящем документе сокращение "CLDN" означает «клаудин» (claudin) и включает белки CLDN6, CLDN9, CLDN4 и CLDN3. Предпочтительно, CLDN является человеческим клаудином.
Обозначение "CLDN6" относится предпочтительно к человеческому CLDN6, в частности к (i) нуклеиновой кислоте, содержащей нуклеотидную последовательность, кодирующую аминокислотную последовательность SEQ ID NO:2 или кодирующую аминокислотную последовательность SEQ ID NO:8, например, к нуклеиновой кислоте, содержащей нуклеотидную последовательность8ЕО ID NO:1, или (ii) к белку, содержащему аминокислотную последовательность SEQ ID NO:2 или содержащему аминокислотную последовательность SEQ ID NO:8. Первая внеклеточная петля CLDN6 предпочтительно содержит аминокислотные остатки с 28 по 80, более предпочтительно аминокислотные остатки с 28 по 76 аминокислотной последовательности, показанной в SEQ ID NO:2, или аминокислотной последовательности, показанной в SEQ ID NO:8, например аминокислотной последовательности, показанной в SEQ ID NO:7. Вторая внеклеточная петля CLDN6 предпочтительно содержит аминокислотные остатки с 138 по 160, предпочтительно аминокислотные остатки с 141 по 159, более предпочтительно аминокислотные остатки с 145 по 157 аминокислотной последовательности, показанной в SEQ ID NO:2, или аминокислотной последовательности, показанной в SEQ ID NO:8, например аминокислотной последовательности, показанной в SEQ ID NO:6. Указанные первая и вторая внеклеточные петли предпочтительно формируют внеклеточную часть CLDN6.
Обозначение "CLDN9" относится предпочтительно к человеческому CLDN9, в частности к (i) нуклеиновой кислоте, содержащей нуклеотидную последовательность, кодирующую аминокислотную последовательность SEQ ID NO:9 или (ii) к белку, содержащему аминокислотную последовательность SEQ ID NO:9. Первая внеклеточная петля CLDN9 предпочтительно содержит аминокислотные остатки с 28 по 76, аминокислотной последовательности показанной в SEQ ID NO:9. Вторая внеклеточная петля CLDN9 предпочтительно содержит аминокислотные остатки с 141 по 159 аминокислотной последовательности, показанной в SEQ ID NO:9. Указанные первая и вторая внеклеточные петли предпочтительно формируют внеклеточную часть CLDN9.
Обозначение "CLDN4" относится предпочтительно к человеческому CLDN4, в частности к (i) нуклеиновой кислоте, содержащей нуклеотидную последовательность, кодирующую аминокислотную последовательность SEQ ID NO:10 или (ii) к белку, содержащему аминокислотную последовательность SEQ ID NO:10. Первая внеклеточная петля CLDN4 предпочтительно содержит аминокислотные остатки с 28 по 76 аминокислотной последовательности, показанной в SEQ ID NO:10. Вторая внеклеточная петля CLDN4 предпочтительно содержит аминокислотные остатки с 141 по 159 аминокислотной последовательности, показанной в SEQ ID NO:10. Указанные первая и вторая внеклеточные петли предпочтительно формируют внеклеточную часть CLDN4.
Обозначение "CLDN3" относится предпочтительно к человеческому CLDN3, в частности к (i) нуклеиновой кислоте, содержащей нуклеотидную последовательность, кодирующую аминокислотную последовательность SEQ ID NO:11 или (ii) к белку, содержащему аминокислотную последовательность SEQ ID NO:11. Первая внеклеточная петля CLDN3 предпочтительно содержит аминокислотные остатки с 27 по 75 аминокислотной последовательности, показанной в SEQ ID NO:11. Вторая внеклеточная петля CLDN9 предпочтительно содержит аминокислотные остатки с 140 по 158 аминокислотной последовательности, показанной в SEQ ID NO:11. Указанные первая и вторая внеклеточные петли предпочтительно формируют внеклеточную часть CLDN3.
Описанные выше последовательности CLDN включают любые варианты указанных последовательностей, в частности мутантные формы, варианты сплайсинга, конформации, изоформы, аллельные варианты, варианты и гомологичные формы различного видового происхождения, в том числе те, которые встречаются в природе. Аллельные варианты относятся к изменениям нормальной последовательности гена, значение которых часто остается неясным. При определении полной нуклеотидной последовательности гена часто выявляются многочисленные варианты этого гена. Гомологичные формы различного видового происхождения - это соответственные нуклеотидные или аминокислотные последовательности, имеющиеся у разных видов. Термин «клаудин» (обозначаемый "CLDN") охватывает (i) варианты сплайсинга CLDN, (ii) варианты посттрансляционной модификации CLDN, в частности различные варианты гликозилирования, например гликозилирования N-концевого участка полипептидной цепи, (iii) конформационные варианты CLDN, (iv) варианты CLDN, связанные с раком, и несвязанные с раком. Предпочтительно, чтобы CLDN присутствовал в своей нативной конформации.
Экспрессия CLDN6 обнаружена, например, в опухолевой ткани при раке яичника, раке легких, желудка, молочной железы, печени, поджелудочной железы, кожи, при меланомах, раке головы и шеи, саркомах, раке желчного протока, почечноклеточном раке и раке мочевого пузыря. CLDN6 представляет особенно предпочтительную мишень для лечения и/или предотвращения рака яичника, в частности аденокарциномы яичника и тератокарциномы яичника, а также рака легких, включая мелкоклеточный рак легких (SCLC) и немелкоклеточный рак легких (NSCLC), в том числе плоскоклеточную карциному и аденокарциному легких, рак желудка, молочной железы, печени, поджелудочной железы, кожи, в частности базальноклеточную карциному и плоскоклеточную карциному, злокачественную меланому, рак головы и шеи, в том числе злокачественную полиморфную аденому, саркому, в частности синовиальную саркому и карциносаркому, рак желчного протока, рак мочевого пузыря, в частности переходноклеточную карциному и папиллярную карциному, рак почек, в частности почечно-клеточную карциному, включая светлоклеточную почечно-клеточную карциному и папиллярную почечно-клеточную карциному, рак толстого кишечника, рак тонкого кишечника, включая рак подвздошной кишки, в частности аденокарциному тонкого кишечника и аденокарциному подвздошной кишки, эмбриональную карциному яичка, хориокарциному плацентарной площадки, рак шейки матки, рак яичка, в частности семиному яичка, тератому яичка и эмбриональный рак яичка, рак матки, герминогенные опухоли, например тератокарциному или эмбриональную карциному, в частности герминогенную опухоль яичка, и их метастазирующие формы. В одном из воплощений данного изобретения раковое заболевание, связанное с экспрессией CLDN6, выбирают из группы, состоящей из рака яичника, рака легких, метастазирующего рака яичника и метастазирующего рака легких, предпочтительно рак яичника является карциномой или аденокарциномой. Предпочтительно, рак легких является карциномой или аденокарциномой, предпочтительно бронихиолярным раком, например бронихиолярной карциномой или бронихиолярной аденокациномой. В одном из воплощений данного изобретения опухолевые клетки, связанные с экспрессией CLDN6, являются раковыми клетками указанных заболеваний.
Термин «участок» относится к части. Применительно к таким конкретным структурам, как аминокислотная последовательность или белок термин «участок» может означать непрерывную или составную часть указанной структуры. Предпочтительно участок аминокислотной последовательности содержит по меньшей мере 1%, по меньшей мере 5%, по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, предпочтительно по меньшей мере 40%, предпочтительно по меньшей мере 50%, более предпочтительно по меньшей мере 60%, более предпочтительно по меньшей мере 70%, еще более предпочтительно по меньшей мере 80%, и наиболее предпочтительно по меньшей мере 90% аминокислотных остатков указанной аминокислотной последовательности. Предпочтительно, если участок является составной частью указанная составная часть состоит из 2, 3, 4, 5, 6, 7, 8 или более частей структуры, причем каждая часть является непрерывным элементом этой структуры. Например, составная часть аминокислотной последовательности может быть образована из 2, 3, 4, 5, 6, 7, 8 или более, предпочтительно не более 4 частей указанной аминокислотной последовательности, причем каждая часть предпочтительно содержит по меньшей мере 5 непрерывно расположенных аминокислотных остатков, по меньшей мере 10 непрерывно расположенных аминокислотных остатков, предпочтительно по меньшей мере 20 непрерывно расположенных аминокислотных остатков, предпочтительно по меньшей мере 30 непрерывно расположенных аминокислотных остатков данной аминокислотной последовательности.
Термины «часть» и «фрагмент» в настоящем документе употребляются как синонимы и относятся к непрерывным элементам. Например, часть такой структуры, как аминокислотная последовательность или белок, относится к непрерывному элементу указанной структуры. В участке, части или фрагменте структуры предпочтительно заключено одно или более функциональных свойств указанной структуры. Например, участок, часть или фрагмент эпитопа или пептида предпочтительно иммунологически эквивалентны эпитопу или пептиду, из которого они происходят.
Термин «внеклеточный участок CLDN" в контексте данного изобретения относится к части клаудина, обращенной во внеклеточное пространство и предпочтительно доступной извне указанной клетки, например для находящихся снаружи клетки антител. Предпочтительно, данный термин относится к одной или более внеклеточных петель или их частей или к любой другой внеклеточной части CLDN, которая предпочтительно специфична для указанного CLDN. предпочтительно, указанная часть содержит по меньшей мере 5, по меньшей мере 8, по меньшей мере 10, по меньшей мере 15, по меньшей мере 20, по меньшей мере 30 или по меньшей мере 50 аминокислотных остатков или больше.
Термин «CLDN, связанный/ассоциированный с поверхностью клетки» следует понимать как относящийся к нативному CLDN, т.е. CLDN в его не денатурированном виде, предпочтительно в естественной конформации. Предпочтительно термин «CLDN, связанный/ассоциированный с поверхностью клетки» означает, что между этим белком и плазматической мембраной существует связь и что он располагается на клеточной мембране, где по меньшей мере часть CLDN, предпочтительно внеклеточный участок, обращен во внеклеточное пространство указанной клетки и доступен снаружи от нее, например для антител, находящихся вне клетки. Связь может быть прямой либо непрямой. Например, связь может осуществляться за счет одного или более трансмембранных доменов, одной или более липидных заякоривающих структур и/или благодаря взаимодействию с каким-либо другим белком, липидом, сахаридом или другой структурой, находящейся на наружном слое плазматической мембраны клетки. Например, CLDN, связанный с поверхностью клетки, может быть трансмембранным белком, т.е. белком, встроенным в мембрану (интегральным мембранным), имеющим внеклеточный участок, или CLDN может быть белком, связанным с клеточной поверхностью за счет взаимодействия с другим белком, являющимся трансмембранным.
CLDN6 связан с поверхностью клетки, если он располагается на ее поверхности и доступен для связывания с добавленными к таким клеткам антителами, специфичными к CLDN6.. В предпочтительных воплощениях данного изобретения клетки, отличающиеся тем, что CLDN6 ассоциирован с их поверхностью, - это клетки, в которых CLDN6 экспрессируется. Следует подчеркнуть, что если в клетке имеет место экспрессия CLDN6, то CLDN6, связанный с клеточной поверхностью, может быть только из числа этих белковых молекул.
Выражение «клетка, несущая CLDN» предпочтительно означает, что указанная клетка имеет на своей поверхности молекулы клаудина, т.е. он связан с поверхностью указанной клетки.
Словосочетания «клеточная поверхность» или «поверхность клетки» используются в их обычном для данной области техники смысле, т.е. включают наружную поверхность клетки, доступную для связывания с белками и другими веществами извне.
Выражение «CLDN, экспрессируемый на поверхности клетки» означает, что клаудин, образующийся в данной клетке, оказывается связан с ее поверхностью.
По данному изобретению считается, что в клетках нет существенной экспрессии CLDN6 и на них нет существенного количества этого белка, связанного с клеточной поверхностью, если уровень экспрессии и количество CLDN6 на клеточной поверхности меньше, чем в плацентарных клетках или в ткани плаценты. Предпочтительно уровень экспрессии и количество CLDN6 на клеточной поверхности должны быть менее 10%, предпочтительно менее 5%, 3%, 2%, 1%, 0,5%, 0,1% или 0,05% от уровня экспрессии и количества этого белка на клеточной поверхности в плацентарных клетках или в ткани плаценты (или даже ниже). Предпочтительно, в клетках нет существенной экспрессии CLDN6 и на них нет существенного количества этого белка, связанного с клеточной поверхностью, если уровень экспрессии и количество CLDN6 на клеточной поверхности превышают таковые в неопухолевых нераковых тканях, отличных от ткани плаценты, не более чем в 2 раза, предпочтительно не более чем в 1,5 раза и предпочтительно не превышают уровень экспрессии и количество CLDN6 на клеточной поверхности в указанных неопухолевых нераковых тканях. Предпочтительно, в клетках нет существенной экспрессии CLDN6 и на них нет существенного количества этого белка, связанного с клеточной поверхностью, если уровень экспрессии и количество CLDN6 на клеточной поверхности ниже предела чувствительности метода определения и/или если уровень экспрессии и количество CLDN6 на клеточной поверхности слишком низкие для того, чтобы происходило связывание специфичных к CLDN6 антител, добавленных к этим клеткам.
По данному изобретению считается, что в клетках экспрессируется CLDN6 и этот белок имеется на клеточной поверхностью, если уровень экспрессии и количество CLDN6 на клеточной поверхности превышают таковые в неопухолевых нераковых тканях, отличных от ткани плаценты, предпочтительно более чем в 2 раза, предпочтительно в 10, 100, 1000 или 10000 раз. Предпочтительно в клетках экспрессируется CLDN6 и этот белок имеется на клеточной поверхностью, если уровень экспрессии и количество CLDN6 на клеточной поверхности превышают предел чувствительности метода определения и/или если уровень экспрессии и количество CLDN6 на клеточной поверхности достаточны для того, чтобы происходило связывание специфичных к CLDN6 антител, добавленных к этим клеткам. Предпочтительно, экспрессируемый в клетках CLDN6 экспонируется на их клеточной поверхности.
Термин «рафт (плотик)» означает микродомен липидного бислоя клеточной мембраны, обогащенный холестерином и сфинголипидами, который расположен во внешнем слое плазматической мембраны клетки. Некоторые белки способны связываться в такие домены, образовывать агрегаты или участки фокальной адгезии, что влияет на функционирование белков. Например, перенос молекул CLDN6 в такие структуры после того как этот белок связался с антителами по данному изобретению, приводит к образованию комплексов высокой плотности антиген (CLDN6)-антитело в плазматической мембране. Высокая плотность комплексов антиген (CLDN6)-антитело имеет значение для эффективной активации системы комплемента при комплемент-зависимой цитотоксичности.
По данному изобретению термин «заболевание/болезнь» относится к любому патологическому состоянию, включая рак, в частности к тем раковым заболеваниям, которые упоминаются в настоящем документе.
Выражение «заболевания с участием клеток, в которых экспрессируется CLDN6 и которые отличаются тем, что CLDN6 связан с их поверхностью», по данному изобретению означает, что экспрессия и количество этого белка на клеточной поверхности в пораженной ткани или органе предпочтительно увеличены по сравнению с состоянием в здоровом органе или ткани. Под увеличением понимается увеличение по меньшей мере на 10%, в частности по меньшей мере на 20%, по меньшей мере на 50%, по меньшей мере на 100%, по меньшей мере на 200%, по меньшей мере на 500%, по меньшей мере на 1000%, по меньшей мере на 10 000% или даже больше. В одном из воплощений данного изобретения экспрессия CLDN6 и его наличие на клеточной поверхности имеют место только в пораженной ткани, а в здоровых тканях его экспрессия подавлена. По данному изобретению заболевания с участием клеток, в которых экспрессируется CLDN6 и которые отличаются тем, что CLDN6 связан с их поверхностью, включают онкологические заболевания, например раковые заболевания. Также по данному изобретению онкологические заболевания, например раковые заболевания, - это предпочтительно те, при которых в опухолевых клетках или раковых клетках экспрессируется CLDN6 и этот белок связан с их клеточной поверхностью.
В настоящем документе термины «онкологическое заболевание», «заболевание, ассоциированное с опухолью» или «онкогенное заболевание» включают заболевания, отличающиеся нарушением регуляции клеточного роста, пролиферации, дифференцировки, адгезии и/или миграции, что может приводить к образованию опухолей и/или метастазов или к возникновению тенденции к их образованию. Под «опухолевыми клетками» понимаются аномальные клетки, которым свойственна быстрая бесконтрольная пролиферация и продолжение клеточного роста и которые продолжают размножаться и после того, как фактор, инициировавший клеточный рост, прекратил свое действие.
Под «опухолью» понимается аномальная группа клеток или ткань, быстро растущая за счет бесконтрольной пролиферации и продолжающая расти после прекращения действия фактора, вызвавшего клеточный рост. Опухолям свойственно частичное или полное отсутствие структурной организации и функциональной координации с нормальной тканью; обычно опухоль представляет собой отдельную массу ткани, которая может быть доброкачественной, предраковой или злокачественной.
Предпочтительно «онкологическое заболевание», «заболевание, ассоциированное с опухолью» или «онкогенное заболевание» по данному изобретению являются раковыми заболеваниями, т.е. злокачественными заболеваниями, а опухолевая клетка является раковой клеткой. Предпочтительно «онкологическое заболевание», «заболевание, ассоциированное с опухолями» или «онкогенное заболевание» отличаются тем, что имеются клетки, в которых экспрессируется CLDN6 и он связан с их поверхностью, и в опухолевых клетках экспрессируется CLDN6 и он связан с их поверхностью.
Клетки, в которых экспрессируется CLDN6 и которые отличаются тем, что этот белок связан с их поверхностью, предпочтительно являются опухолевыми клетками или раковыми клетками, предпочтительно тех опухолей и раковых заболеваний, которые описаны в настоящем документе. Предпочтительно такие клетки не являются плацентарными клетками.
Предпочтительные раковые заболевания и злокачественные опухоли по данному изобретению выбирают из группы, состоящей из рака яичника, в том числе аденокарциномы яичника и тератокарциномы яичника, рака легких, в том числе мелкоклеточного рака легких (SCLC) и немелкоклеточного рака легких (NSCLC), в особенности плоскоклеточной карциномы легких и аденокарциномы, рака желудка, рака молочной железы, рака печени, рака поджелудочной железы, рака кожи, в частности базальноклеточной карциномы и плоскоклеточной карциномы, злокачественной меланомы, рака головы и шеи, в том числе злокачественной полиморфной аденомы, саркомы, в частности синовиальной саркомы и карциносаркомы, рака желчного протока, рака мочевого пузыря, в частности переходноклеточной карциномы и папиллярной карциномы, рака почек, в частности почечно-клеточной карциномы, включая светлоклеточную почечно-клеточную карциному и папиллярную почечно-клеточную карциному, рака толстого кишечника, рака тонкого кишечника, включая рак подвздошной кишки, в частности аденокарциному тонкого кишечника и аденокарциному подвздошной кишки, эмбриональной карциномы яичка, хориокарциномы плацентарной площадки, рака шейки матки, рака яичка, в частности семиномы яичка, тератомы яичка и эмбрионального рака яичка, рака матки, герминогенных опухолей, например тератокарциномы или эмбриональной карциномы, в частности герминогенной опухоли яичка, и их метастазирующих форм.
Основными типами рака легкого являются мелкоклеточная карцинома легкого (SCLC) и немелкоклеточная карцинома легкого (NSCLC). Немелкоклеточную карциному легкого подразделяют на три основных подтипа: плоскоклеточная карцинома легкого, аденокарцинома и крупноклеточная карцинома легкого. На долю аденокарциномы приходится приблизительно 10% всех случаев рака легких. Раковые поражения этого вида обычно заметны на периферии легких - в противоположность мелкоклеточному и плоскоклеточному раку, которым свойственна более центральная локализация.
Рак кожи - это злокачественные новообразования в коже. Наиболее распространенными раковыми заболеваниями кожи являются базальноклеточный рак, плоскоклеточный рак и меланома. Злокачественная меланома - весьма агрессивное заболевание. Оно обусловлено неконтролируемым ростом пигментных клеток, называемых меланоцитами.
По данному изобретению карцинома (рак) - это злокачественная опухоль, происходящая из эпителиальных клеток того или иного органа.
Бронхиолярная карцинома - это рак легкого, происходящий, как полагают, из эпителия мелких дыхательных путей (бронхиол); при этом заболевании неопластическая ткань простирается вдоль стенок альвеол и прорастает небольшими массами внутри альвеол. В некоторых клетках обнаруживается муцин, который присутствует и в альвеолах, где также имеются денудированные клетки.
Аденокарцинома - это опухоль, происходящая из железистой ткани. Железистая ткань относится к эпителиальной ткань, которая включат кожу, железы и разнообразные выстилки полостей и органов. В процессе эмбрионального развития эпителий происходит из эктодермы, эндодермы или мезодермы. Для того, чтобы клетки можно было назвать аденокарциномными, они не обязательно должны быть частью железы - достаточно того, что они способны к секреции. Такая карцинома встречается у высших животных и человека. Высоко дифференцированная аденокарцинома похожа на железистую ткань, из которой она происходит, тогда как мало дифференцированная аденокарцинома может быть на нее вовсе не похожа. Путем окрашивания клеток в материале биопсии патологоанатом может определить, является рассматриваемая опухоль аденокарциномой или же иным злокачественным образованием. Аденокарциномы могут возникать в различных тканях организма, поскольку те или иные железы имеются в нем повсюду. У каждой железы свой специфический состав секрета, но всякая клетка, обладающая экзокринной функцией, считается железистой, и ее злокачественная форма называется аденокарциномной. Злокачественные аденокарциномы прорастают в другие ткани и часто дают метастазы, если развивались достаточно долго. Наиболее частый тип карциномы яичника - аденокарцинома. Аденокарцинома яичника включает серозную, муцинозную, светлоклеточную и эндометриоидную аденокарциномы.
Цистаденокарцинома - это злокачественная форма поверхностной эпителиально-стромальной опухоли, разновидность рака яичника.
Поверхностные эпителиально-стромальные опухоли - это группа неопластических образований в яичнике, которые, как считается, происходят из поверхностного эпителия яичника (видоизмененного перитонеума) или из эктопической эндометриоидной ткани либо ткани фаллопиевой трубы. Такие опухоли составляют большинство всех онкологических поражений яичника.
Тератокарциномой называют герминогенную опухоль, представляющую собой смесь тератомы и эмбриональной карциномы, или хориокарциномы, или той и другой. Хориокарцинома - это злокачественная трофобластическая опухоль, весьма агрессивная, обычно развивающаяся из плаценты. Для нее характерно раннее распространение через кровь в легкие.
Саркома - это рак соединительной ткани (костной, хрящевой, жировой), при котором происходит пролиферация клеток, происходящих из мезодермы, в отличие от карцином, происхождение которых эпителиальное. Синовиальная саркома - это довольно редко встречающееся раковое поражение мягких тканей в области суставов конечностей.
Почечноклеточная карцинома, известная также под названиями «почечноклеточный рак» или «почечноклеточная аденокарцинома», - это рак почек, развивающийся и выстилки проксимальных извитых канальцев, где происходит фильтрация крови и удаляются отходы. На сегодняшний день почечноклеточная карцинома является наиболее распространенным раковым поражением почек у взрослых; из всех опухолей мочеполовой системы это раковое заболевание отличается наивысшей летальностью. Почечноклеточная карцинома подразделяется на светлоклеточную и папиллярную. Светлоклеточная почечноклеточная карцинома встречается чаще других форм почечноклеточной карциномы. Клетки светлоклеточной почечноклеточной карциномы под микроскопом выглядят очень бледными или прозрачными. Также часто встречается папиллярная почечноклеточная карцинома. При этом заболевании в большинстве случаев опухоль образует характерные небольшие пальцевидные выросты (папиллы).
Герминогенная опухоль - это новообразование, происходящее из клеток зародышевой линии. Герминогенные опухоли могут быть злокачественными (раковыми) либо доброкачественными. Клетки зародышевой линии в норме находятся в гонадах (яичниках или семенниках). Герминогенные опухоли, возникающие вне гонад (например, в области головы и шеи, во рту, в тазовой области; в эмбрионах, у новорожденных и маленьких детей - чаще всего по средней линии тела, в частности на конце копчика) могут быть врожденными дефектами, возникшими вследствие ошибок в ходе индивидуального развития.
Различают два основных класса герминогенных опухолей - семиномы и несеминомные опухоли. Последние включают тератокарциномы, эмбриональные карциномы, опухоли желточного мешка, хориокарциномы и дифференцированные тератомы. Большинство клеточных линий, полученные из несеминомных опухолей, эквивалентны эмбриональным карциномам, т.е. они состоят полностью из стволовых клеток, не дифференцирующихся при обычных условиях, хотя некоторые такие опухоли могут реагировать на индукторы дифференцировки, например на ретиноевую кислоту.
Метастаз - это результат распространения раковых клеток из места первоначального злокачественного поражения в другие части тела. Метастазирование, т.е. образование метастазов, представляет собой очень сложный процесс, зависящий от отделения злокачественных клеток от первичной опухоли, их внедрения в межклеточный матрикс, проникновения через базальную мембрану и эндотелий в полости тела и в просвет сосудов, переноса кровью и включения в органы-мишени. Когда образовалась новая опухоль, ее рост зависит от прорастания ее сосудами. Метастазы нередко образуются даже после удаления первичной опухоли, потому что в организме могут остаться опухолевые клетки или компоненты, способные к метастазированию. В одном из воплощений данного изобретения термин «метастаз» относится к отдаленным метастазам, т.е. находящимся вне области распространения от первичной опухоли по лимфатическим сосудам и узлам в регионе пораженного органа.
Клетки вторичной опухоли (метастаза) такие же, как в первичной опухоли. Это означает, например, что если рак яичника метастазирует в печень, то вторичная опухоль состоит из аномальных клеток яичника, а не из аномальных клеток печени. Соответственно, эта опухоль в печени называется метастатическим раком яичника, а не раком печени.
Термин «лечение» означает введение пациенту описанного в настоящем документе вещества или композиции с целью предотвратить или ликвидировать болезнь, включая уменьшение опухоли в размерах или числа опухолей у данного больного; приостановление или замедление прогрессирования заболевания; ингибирование или замедление развития нового заболевания у данного пациента; снижение частоты проявления или тяжести симптомов и/или рецидивирования у пациента, у которого на данный момент имеет место рак или же он был ранее; и/или продление, т.е. увеличение продолжительности, жизни больного.
Термин «лечение заболевания» включает излечение, сокращение продолжительности болезненного состояния, ослабление его тяжести, предотвращение развития или ингибирование прогрессирования патологического процесса или ухудшения состояния больного или задержка начала развития заболевания или его симптомов.
Выражения «находиться в группе риска», «существует риск развития заболевания», «повышен риск развития заболевания» означают, что для данного индивида (больного, пациента) вероятность развития того или иного заболевания, в частности рака, выше «нормальной», т.е. средней, в данной популяции. Кроме того, риск развития заболевания, в частности рака, повышен для индивидов, у которых было данное заболевание в прошлом или оно имеется в настоящее время, так как у такого индивида возможно продолжение развития этого заболевания. Для индивидов, у которых был или имеется в настоящее время рак, также повышена вероятность метастазирования рака.
Термин «иммунотерапия» относится к лечению, при котором используются те или иные иммунологические процессы. В контексте настоящего изобретения термины «защищать», «предотвращать», «профилактический», «превентивный» или «защитный» относятся к предотвращению или лечению, или к тому и другому вместе возникновения и/или распространения опухоли у индивида. В контексте настоящего изобретения термин «иммунотерапия» предпочтительно относится к активной иммунизации или вакцинации опухоли. Профилактическое применение иммунотерапии, например введение композиции по данному изобретению, предпочтительно защищает пациента от развития опухолевого роста. Терапевтическое применение иммунотерапии, например терапевтическое введение композиции по данному изобретению, может вести к ингибированию развития/роста опухоли, включающему замедление развития/роста опухоли, в частности к прерыванию прогрессирования опухоли, что предпочтительно ведет к ее ликвидации. Применение иммунотерапии в лечебных целях может защитить индивида, например, от распространения или метастазирования существующей опухоли.
Термин «иммунизация» или «вакцинация» означает процесс введения индивиду антигена для того, чтобы вызывать у него иммунный ответ с целью лечения или профилактики.
Термины «индивид», «особь», «больной», «пациент», «объект лечения/профилактики» или «организм» употребляются в настоящем документе как синонимы и относятся к позвоночным, предпочтительно к млекопитающим. Например, млекопитающие в контексте данного изобретения - это люди и другие приматы, домашние животные (собаки, кошки, овцы, крупный рогатый скот, козы, свиньи, лошади и проч.), лабораторные животные (мыши, крысы, кролики, морские свинки и др.), а также животные, содержащиеся в неволе (например, в зоопарке). Термин «животное» в настоящем документе включает также человека. Термины «индивид», «особь», «объект лечения/профилактики» могут также включать значение «пациент/больной», т.е. животное, предпочтительно человек, страдающий от той или иной болезни, предпочтительно от болезни, связанной с экспрессией CLDN6, предпочтительно от онкогенного заболевания, например рака.
Термин «адъювант» относится к веществам, которые продлевают или усиливают или ускоряют иммунный ответ. Композиции по данному изобретению предпочтительно оказывают свое действие без добавления адъювантов. Тем не менее, композиции по данной заявке на патент могут содержать любой известный адъювант. К адъювантам относятся различные вещества, например масляные эмульсии (например, адъювант Фрейнда и его модификации), минеральные вещества (например, квасцы), продукты бактериального происхождения (например, токсин Bordetella pertussis), липосомы, иммуностимулирующие комплексы. Примеры адъювантов: монофосфорил-липид-А (MPL SmithKline Beecham), сапонины, например QS21 (SmithKline Beecham), DQS21 (SmithKline Beecham; WO 96/33739), QS7, QS17, QS18, и QS-L1 (So et al., 1997, Mol. Cells 7:178-186), неполные адъюванты Фрейнда, полные адъюванты Фрейнда, витамин Е, монтанид, квасцы, CpG-олигонуклеотиды (Krieg et al., 1995, Nature 374:546-549) и различные эмульсии типа «вода в масле», получаемые из масел, подлежащих биологическому расщеплению, например скваленов и/или токоферола.
По данному изобретению образец может быть любым, какой можно использовать соответственно настоящему описанию, в частности биологическим образцом, например образцом ткани, включая жидкости тела и/или клеточные образцы, и который может быть получен обычными методами, например путем биопсии, включая пункционную биопсию, путем забора крови, бронхиального аспирата, мокроты, мочи, кала или иных жидкостей тела. По данному изобретению термин «биологический образец» также включает фракции биологических образцов.
Термин «антитело» относится к гликопротеинам, содержащим по меньшей мере две тяжелых (Н) цепи и две легких (L) цепи, соединенных между собой дисульфидными связями, и включает любое вещество/молекулу, содержащую участок, связывающий антиген. Термин «антитела» включает моноклональные антитела и их фрагменты или производные, в том числе (перечисленные здесь примеры не являются ограничивающими): человеческие моноклональные антитела, гуманизированные моноклональные антитела, гибридные моноклональные антитела, одноцепочечные антитела, например scFv, и фрагменты антител, связывающие антиген, например фрагменты Fab и Fab'; также этот термин включает все рекомбинантные формы антител, (например, антитела, экспрессирующиеся у прокариот), дегликозилированные антитела и любые антиген-связывающие фрагменты и производные, описанные в настоящем документе. Каждая тяжелая цепь антитела состоит из вариабельной области (обозначается сокращением VH) и константной области. Каждая легкая цепь состоит из вариабельной области (сокращенно VL) и константной области. Области VH и VL можно далее подразделить на гипервариабельные участки, называемые участками, определяющими комплементарность (CDR), между которыми имеются более консервативные по структуре участки, называемые каркасными (FR). Каждая вариабельная область как легкой, так и тяжелой цепи образована тремя CDR и четырьмя FR, располагающимися в следующем порядке (считая от N-конца к C-концу полипептидной цепи): FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Вариабельные области легких и тяжелых цепей содержат связывающий домен, который взаимодействует с антигеном. Константные области антител могут опосредовать связывание молекулы иммуноглобулина с тканями или факторами собственного организма, включая различные клетки иммунной системы (например, эффекторные клетки) и первый компонент (C1q) системы комплемента.
По данному изобретению выражение «по меньшей мере одна из последовательностей CDR» предпочтительно означает по меньшей мере одну последовательность CDR3. Выражение «последовательности CDR цепи антитела» предпочтительно относится к участкам CDR1, CDR2 и CDR3 тяжелой или легкой цепи антитела.
По данному изобретению если указывается, что какая-либо цепь антитела содержит конкретную последовательность CDR, например последовательность CDR3, это означает, что указанная конкретная последовательность CDR либо образует участок CDR, например участок CDR3 указанной цепи антитела (то есть участок CDR состоит из указанной конкретной последовательности CDR), либо образует часть участка CDR, например участка CDR3 указанной цепи антитела (то есть участок CDR содержит указанную конкретную последовательность CDR).
Если по данному изобретению указывается, что антитело содержит конкретную тяжелую цепь и/или конкретную легкую цепь, например цепь, содержащую определенные последовательности CDR, то предпочтительно, чтобы обе тяжелые цепи и/или обе легкие цепи данного антитела каждая состояли из определенной тяжелой цепи и/или определенной легкой цепи.
Термин «гуманизированные антитела» относится к молекулам, в которых имеется антиген-связывающий участок, происходящий в основном из нечеловеческого иммуноглобулина, а остальная часть молекулы имеет основой последовательность и/или структуру человеческого иммуноглобулина. У такого антитела антиген-связывающий участок может содержать либо полностью вариабельные домены, присоединенные с константным доменам, либо только области, определяющие комплементарность (CDR), присоединенные к каркасным участкам вариабельных доменов. Участки, связывающие антиген, при этом могут быть нативными или же модифицированными путем одной или более аминокислотных замен, например модифицированными таким образом, чтобы было больше сходство с иммуноглобулинами человека. В некоторых формах гуманизированных антител сохраняются все последовательности CDR (например, гуманизированное мышиное антитело, которое содержит все шесть последовательностей CDR из мышиного антитела). В других формах гуманизированных антител имеется одна или более последовательностей CDR, измененных по сравнению с исходным антителом.
Термин «гибридные антитела» относится к таким антителам, в которых одна часть каждой аминокислотной последовательности как легких, так и тяжелых цепей гомологична соответствующим последовательностям антител, происходящих от определенного вида животных или принадлежащим к определенному классу, в то время как остальная часть той же гомологична соответствующим последовательностям других антител. Обычно вариабельные области как легких, так и тяжелых цепей походят на вариабельные области антител, происходящих от одного вида млекопитающих, а константные части гомологичны последовательностям антител, происходящих от другого вида. Одно из явных преимуществ таких гибридных форм состоит в том, что вариабельная область может происходить из известных источников, что удобно, так как можно использовать легко доступные В-клетки или гибридомы от других (не от человека) организмов в сочетании с константными областями, происходящими из, например, препаратов человеческих клеток. Если вариабельные области сравнительно легко получить и их специфичность не зависит от источника, то константные области, будучи человеческими, с меньшей вероятностью вызовут иммунный ответ у человека при введении таких антител, чем вызывали бы константные области, происходящие от другого вида живых организмов. Впрочем, данное определение не ограничивается этим конкретным примером.
Термин «участок, связывающий антиген» (антиген-связывающий участок) антитела (или просто - «участок связывания») в настоящем документе относится к одному или более фрагментам антитела, которые сохраняют способность специфично связываться с антигеном. Показано, что функция связывания антигена может осуществляться фрагментами антитела, т.е. вся его последовательность для этого не требуется. Примеры связывающих фрагментов, охватываемых термином «антиген-связывающий участок», включают: (i) фрагменты Fab - одновалентные фрагменты, состоящие из доменов VL, VH, CL и СН; (ii) фрагменты F(ab')2 - двухвалентные фрагменты, содержащие два фрагмента Fab, соединенных дисульфидным «мостиком» в шарнирной области; (iii) фрагменты Fd, состоящие из доменов VH и СН; (iv) фрагменты Fv, состоящие из доменов VL и VH одного «плеча» молекулы антитела, (v) фрагменты dAb (Ward et al., (1989) Nature 341:544-546), состоящие из домена VH; (vi) изолированные участки, определяющие комплементарность (CDR) и (vii) комбинации двух или более изолированных CDR, которые при необходимости могут быть соединены синтетической линкерной структурой. Также, хотя два домена фрагмента Fv - VL и VH - кодируются отдельными генами, их можно соединить, применяя технологию рекомбинантной ДНК, с помощью линкерной структуры, что позволяет получать их как единую полипептидную цепь, в которой области VL и VH составляют пару, образующую одновалентную молекулу (называемую одноцепочечным Fv, сокращенно scFv); см., например, Bird et al. (1988) Science 242:423-426; Huston et al. (1988) Proc. Natl. Acad. Sci. USA 85:5879-5883). Такие одноцепочечные антитела тоже охватываются термином «антиген-связывающий участок» антитела. Другой пример - гибридные белки типа «связывающий домен-иммуноглобулин», содержащие (i) полипептид, являющийся связывающим доменом и присоединенный к полипептиду, представляющему собой шарнирную область молекулы иммуноглобулина, (ii) константную область тяжелой цепи иммуноглобулиновой молекулы СН2, присоединенную к шарнирной области, и (iii) константную область тяжелой цепи иммуноглобулиновой молекулы СН3, присоединенную к константной области СН2. Полипептид, формирующий связывающий домен, может быть вариабельной областью тяжелой цепи или вариабельной областью легкой цепи. Гибридные белки типа «связывающий домен-иммуноглобулин» также описаны в заявках на патент США 2003/0118592 и 2003/0133939. Эти фрагменты антител получены с применением обычных методов, известных специалистам в данной области техники; возможность практического использования таких фрагментов выявляется таким же образом, как интактных антител.
Термин «эпитоп» относится к антигенной детерминанте молекулы, то есть к той части молекулы, которая распознается иммунной системой, например, которая распознается антителами. Например, эпитопы являются дискретными пространственными структурами на антигенной молекуле, распознаваемыми иммунной системой. В контексте настоящего изобретения эпитоп предпочтительно происходит из белка CLDN. Эпитопы обычно состоят из химически активных поверхностных группировок, например аминокислотных остатков или боковых цепей Сахаров, и обладают специфическими свойствами в отношении пространственной структуры и электрического заряда. Различают конформационные и неконформационные эпитопы: первые утрачивают способность к связыванию в присутствии денатурирующих растворителей, а вторые нет. Эпитопы таких белков, как клаудины, предпочтительно содержат непрерывные (секвенциальные) или составные (прерывистые, комбинированные) участки указанных белков и содержат предпочтительно от 5 до 100, предпочтительно от 5 до 50, более предпочтительно от 8 до 30, наиболее предпочтительно от 10 до 25 аминокислотных остатков; например, эпитоп может быть длиной предпочтительно 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 аминокислотных остатков.
Термин «прерывистый (составной, комбинированный) эпитоп» в настоящем документе означает конформационный эпитоп на белковой антигенной молекуле, сформированный по меньшей мере двумя раздельными участками первичной структуры (аминокислотной последовательности) этого антигенного белка.
Термин «молекула с двойной специфичностью» («биспецифичная молекула») подразумевает любой агент, например белок, пептид или белковый или пептидный комплекс, который обладает двумя различными специфичностями связывания. Например, такая молекула может связываться (взаимодействовать) с (а) антигеном клеточной поверхности и (b) рецептором Fc на поверхности эффекторной клетки. Термин «полиспецифичная молекула» или «гетероспецифичная молекула» подразумевает любой агент, например белок, пептид или белковый или пептидный комплекс, который обладает более чем двумя различными специфичностями связывания. Например, такая молекула может связываться (взаимодействовать) с (а) антигеном клеточной поверхности и (b) рецептором Fc на поверхности эффекторной клетки (b) и (с) еще по меньшей мере с одним иным компонентом. Соответственно, данное изобретение включает (но не ограничивается перечисленным здесь) молекулы с двумя, тремя, четырьмя и многими специфичностями связывания, направленные CLDN6 и против других мишеней, например рецепторов Fc на эффекторных клетках. Термин «биспецифичные антитела» также включает диантитела. Диантитела представляют собой бивалентные биспецифичные антитела, в которых домены VH и VL синтезируются как единая полипептидная цепь, но с использованием линкерной структуры, слишком короткой, чтобы допустить объединение двух доменов одной и той же цепи, тем самым вынуждая соединяться комплементарные домены различных цепей, так что создаются два участка связывания антигена (см., например, Holliger, P., et al. (1993) Proc. Natl. Acad. Sci. USA 90:6444-6448; Poljak, R. J., et al. (1994) Structure 2:1121-1123).
В настоящем документе термин «гетероантитела» относится к двум или более антителам, производные или антиген-связывающие участки которых соединены вместе, причем по меньшей мере два из них обладают различной специфичностью. Эти различные специфичности включают специфичность связывания с рецептором Fc на эффекторных клетках и специфичность связывания с антигеном или эпитопом на клетке-мишени, например на опухолевой клетке.
Антитела, описываемые в настоящем документе, могут быть человеческими. Термин «человеческое антитело» в данном описании подразумевает антитела, имеющие вариабельные и константные области, происходящие из иммуноглобулиновых последовательностей зародышевой линии человека. Человеческие антитела по данному изобретению могут включать аминокислотные остатки, не кодируемые иммуноглобулиновыми последовательностями зародышевой линии человека (например, мутации, возникшие путем случайного или же направленного (сайт-специфического) мутагенеза in vitro или путем соматических мутаций in vivo.
Термин «моноклональные антитела» в настоящем документе относится к препаратам антител, включающим молекулы одного состава и строения. Моноклональное антитело обладает единственной специфичностью связывания и сродством к определенному эпитопу. В одном из воплощений данного изобретения моноклональные антитела производятся гибридомами, включающими В-клетки, полученные от животных (не человека), например мыши, слитые с иммортализованными клетками.
Термин «рекомбинантные антитела» в настоящем документе включает все антитела, полученные, экспрессируемые, синтезируемые или выделяемые с помощью методов, использующих рекомбинацию, например (а) антитела, выделенные у животного (например, у мыши), трансгенного или трансхромосомного в отношении генов иммуноглобулинов, или из полученной из него гибридомы, (b) антитела,выделенные из клетки-хозяина, трансформированной таким образом, чтобы в ней экспрессировалось данное антитело, например из трансфектом, (с) антитела, выделенные из рекомбинантной комбинаторной библиотеки антител, и (d) антитела, полученные, экспрессируемые, синтезированные или выделенные любыми другими способами с использованием сплайсинга генов иммуноглобулинов с другими последовательностями ДНК.
Термин «трансфектома» в настоящем документе включает рекомбинантные эукариотические клетки-хозяева, в которых экспрессируются антитела, например клетки СНО, NS/0, HEK293, HEK293T, растительные клетки или грибы, включая дрожжи.
Термин «гетерологичные антитела» в настоящем документе определяется относительно трансгенного организма, производящего такие антитела. Этот термин относится к антителам, обладающим аминокислотной последовательностью или кодирующей нуклеотидной последовательностью, соответствующей таковым у организма, не являющегося трансгенным, и происходящим, как правило, из организма не того вида, к которому относится трансгенный организм.
Термин «гетерогибридные антитела» в настоящем документе относится к антителам, в которых легкие цепи и тяжелые цепи происходят от разных организмов. Например, антитела, в которых тяжелая цепь человеческая, а легкая цепь мышиная, являются гетерогибридными.
Данное изобретение включает все антитела и производные антител, описанные в настоящем документе, которые в целях данного изобретения охватываются термином «антитела». Термин «производные антител» относится к любым модифицированным формам антител, например к конъюгатам антител и иного агента или антитела, или фрагмента антитела.
Антитела, описываемые в настоящем документе, являются предпочтительно изолированными. Под изолированными антителами в настоящем документе подразумеваются антитела в основном без примеси других антител с иными антигенными специфичностями (например, изолированные антитела, специфично связывающиеся с CLDN6, в основном не содержат антител специфично связывающихся с антигенами, отличными от CLDN6). Изолированные антитела, специфично связывающиеся с эпитопом, изоформой или вариантом человеческого CLDN6, могут, однако, проявлять перекрестную реактивность в отношении других, родственных антигенов, например от других видов живых организмов (например, гомологичных CLDN6). Кроме того, изолированное антитело в основном не содержит другой клеточный материал и/или химические вещества. В одном из воплощений данного изобретения «комбинация изолированных моноклональных антител» означает антитела с разными специфичностями, объединенные в определенной композиции.
По данному изобретению антитела способны связываться с предопределенной мишенью, если они имеют значительное сродство к указанной предопределенной мишени и связываются с указанной предопределенной мишенью в стандартных аналитических системах наподобие описанных в настоящем документе. Предпочтительно антитела способны связываться с некоторой мишенью, если связывание указанных антител с указанной мишенью на поверхности интактных клеток можно выявить и измерить методом проточной цитометрии (FACS). Предпочтительно связывание антител с указанной мишенью можно выявить и измерить при концентрации антител 10 мкг/мл или меньше, 5 мкг/мл или меньше, либо 2 мкг/мл или меньше. Предпочтительно связывание антител с указанной мишенью можно выявить и измерить при концентрации антител 50 нМ или меньше, 30 нМ или меньше, либо 15 нМ или меньше. Сродство (сродство связывания) часто количественно определяют константой равновесия реакции диссоциации (KD). Предпочтительно термин «значительное сродство» относится к связыванию с предопределенной мишенью, для которого (KD)=10-5 М или меньше, 10-6 М или меньше, 10-7 М или меньше, 10-8 М или меньше, 10-9 М или меньше, 10-10 М или меньше, 10-11 М или меньше, or 10-12 M или меньше. Для антител по данному изобретению предпочтительно величина ЕС50 при связывании с CLDN6 составляет 6500 нг/мл или меньше, 3000 нг/мл или меньше, 2500 нг/мл или меньше, 2000 нг/мл или меньше, 1500 нг/мл или меньше, 1000 нг/мл или меньше, 500 нг/мл или меньше, 400 нг/мл или меньше, 300 нг/мл или меньше, 200 нг/мл или меньше либо 100 нг/мл или меньше.
Антитела не способны (практически) связываться с данной мишенью, если отсутствует значительное сродство к указанной мишени и при использовании стандартных аналитических систем не выявляется значительного связывания этих антител с указанной мишенью. Предпочтительно, антитела не способны (практически0 связываться с данной мишенью, если при анализе методом проточной цитометрии (FACS) не выявляется связывания с указанной мишенью на поверхности интактных клеток. Предпочтительно связывание антител с указанной мишенью не выявляется при концентрации антител до 2 мкг/мл, предпочтительно до 5 мкг/мл, предпочтительно до 10 мкг/мл, предпочтительно до 20 мкг/мл, более предпочтительно до 50 мкг/мл, в особенности до 100 мкг/мл или до 150 мкг/мл, до 200 мкг/мл или более. Предпочтительно связывание антител с указанной мишенью не выявляется при концентрации антител до 15 нМ, предпочтительно до 30 нМ, предпочтительно до 50 н, предпочтительно до 100 нм, предпочтительно до 150 нм или до 170 нм, до 300 нм, до 600 нм, до 1000 нм, до 1300 нм или более. Предпочтительно связывание антител с указанной мишенью не выявляется при насыщающей концентрации антител для связывания с данной мишенью, т.е. с CLDN6. Предпочтительно значительное сродство антител к данной мишени отсутствует, если KD по меньшей мере в 10, 100, 103, 104, 105 или 106 раз больше, чем KD для связывания с предопределенной мишенью, с которой данные антитела способны связываться. Например, если KD для связывания данных антител с предопределенной мишенью, с которой они способны связываться, составляет 10-7 М, то KD для связывания этих антител с мишенью, к которой нет значительного сродства, будет по меньшей мере 10-6 М, 10-5 М, 10-4 М, 10-3 М, 10-2 М или 10-1 М.
Антитела являются специфичными к предопределенной мишени, если они способны связываться с указанной предопределенной мишенью, но не способны связываться с другими мишенями, т.е. не имеют значительного сродства к другим мишеням и в стандартных аналитических системах не выявляется значительного связывания с другими мишенями. По данному изобретению антитела являются специфичными в отношении CLDN6, если они способны связываться с CLDN6, но не способны связываться с другими мишенями, в частности с другими клаудиновыми белками, отличными от CLDN6, например с CLDN9, CLDN4, CLDN3 и CLDN1. Предпочтительно, антитела являются специфичными в отношении CLDN6, если сродство к другим клаудиновым белкам, отличным от CLDN6, например к CLDN9, CLDN4, CLDN3 и CLDN1, и связывание с ними не превышают существенно сродство и связывание с белками, не родственными клаудинам, например с бычьим сывороточным альбумином (BSA), казеином, человеческим сывороточным альбумином (HSA) или с неклаудиновыми трансмембранными белками, например с белками МНС или с рецептором трансферрина или с любым другим известным полипептидом. Предпочтительно антитела являются специфичными к предопределенной мишени, если они связываются с указанной мишенью с KD, величина которой по меньшей мере в 10, 100, 103, 104, 105 или 106 раз меньше, чем KD для связывания с мишенью, в отношении которой эти антитела не специфичны. Например, если KD для связывания данных антител с предопределенной мишенью, в отношении которой они специфичны, составляет 10-7 М, то KD для связывания этих антител с мишенью, к которой они не специфичны, будет по меньшей мере 10-6 М, 10-5 М, 10-4 М, 10-3 М, 10-2 М или 10-1 М.
Связывание антител с мишенью можно определить экспериментальным путем с помощью любого подходящего метода; см., например, Berzofsky et al., "Antibody-Antigen Interactions" In Fundamental Immunology, Paul, W. E., Ed., Raven Press New York, N Y (1984), Kuby, Janis Immunology, W. H. Freeman and Company New York, N Y (1992), и способов, описанных в настоящем документе. Сродство можно легко определить с помощью обычно применяемых для этой цели методов, например путем равновесного диализа; с помощью биосенсоров BIAcore 2000 путем стандартной процедуры согласно инструкциям производителя; путем радиоиммунологического анализа с использованием радиоактивно меченного антигена-мишени или любым другим путем, известным специалистам в данной области техники. Анализ данных по сродству можно проводить, например, по методу, описанному в работе Scatchard et al., Ann N.Y. Acad. ScL, 51:660 (1949). Численные значения параметров сродства для данного взаимодействия антитело-антиген могут варьировать в зависимости от условий их определения, например от концентрации солей, величины рН. Таким образом, определенные экспериментальным путем значения сродства иди других параметров, характеризующих взаимодействие антитела с антигеном, могут варьировать в зависимости от условий определения, например от концентрации солей и величины рН. Таким образом, при измерениях сродства и других параметров, характеризующих взаимодействие антитела с антигеном, например KD и IC50, предпочтительно использовать стандартизованные растворы антител и антигенов и стандартные буферные растворы.
Уникальным свойством антител по данному изобретению является их способность связывать клаудин 6 клеточной поверхности. Это демонстрируется результатами анализа клеток, в которых экспрессируется клаудин 6, методом проточной цитометрии.
Для того, чтобы определить связывание моноклональных антител с живыми клетками, в которых экспрессируются клаудины, можно применить метод проточной цитометрии. Вкратце, поступают следующим образом. Клетки нужных линий, в которых экспрессируются связанные с мембраной клаудины, (выращенные в стандартных условиях культивирования) смешивают с раствором антител (в различной концентрации) в PBS, содержащем 2% инактивированной нагреванием FCS и 0,1% NaN3 при 4°С в течение 30 мин. Затем клетки промывают, после чего они взаимодействуют с флуоресцентно меченными вторыми антителами в тех же условиях, в каких проводилась реакция с первыми антителами. Образцы анализируют путем FACS, регистрируя прямое и боковое светорассеяние каждой отдельной клетки и определяют связывание меченых антител.
Термин «связывание» по данному изобретению относится предпочтительно к специфичному связыванию, определение которому дано выше.
В настоящем документе термин «изотип» относится к классу антител (например, IgM или IgG1), которые кодируются генами константной области тяжелой цепи.
В настоящем документе термин «переключение изотипа» относится к явлению, состоящему в том, что синтез иммуноглобулинов переключается с одного изотипа (класса) на другой.
Термин «природный»/«встречающийся в природе» в настоящем документе употребляется применительно к объектам, которые можно найти в естественных условиях (в природе). Например, это полипептид или нуклеотидная последовательность, имеющиеся у какого-либо организма (включая вирусов), которые могут быть выделены из природного источника, которые не подвергались намеренной модификации человеком в лабораторных условиях и которые встречаются в природе.
Термин «перестроенный» в настоящем документе относится к иммуноглобулинам с такой конфигурацией тяжелой или легкой цепи, когда сегмент V располагается непосредственно рядом с сегментом D-J или J, что соответствует практически полному домену VH или VL соответственно. Перестроенный локус генов иммуноглобулинов (антител) можно идентифицировать путем сравнения с ДНК клеток зародышевой линии: перестроенный локус должен иметь по меньшей мере один рекомбинировавший гомологичный элемент гептамер/нонамер.
Термин «неперестроенный» или «конфигурация зародышевой линии» в настоящем документе применительно к сегменту V относится к конфигурации, в которой сегмент V не рекомбинировал таким образом, чтобы оказаться в непосредственном соседстве к сегменту D или J.
Термин «молекула нуклеиновой кислоты» в настоящем документе подразумевает молекулы ДНК и РНК. Молекула нуклеиновой кислоты может быть одноцепочечной или двухцепочечной, предпочтительно это двухцепочечная (двуспиральная) ДНК. Молекулы нуклеиновых кислот могут использоваться для введения в клетки (трансфекции), например в форме РНК, которую можно получить in vitro путем транскрипции с ДНК-матрицы. Перед применением РНК может быть модифицирована путем стабилизации последовательности, кэпирования и полиаденилирования.
Нуклеиновые кислоты по данному изобретению, предпочтительно являются изолированными. Термин «изолированная нуклеиновая кислота» по данному изобретению означает, что данная нуклеиновая кислота была (i) амплифицирована in vitro, например с помощью полимеразной цепной реакции (PCR), (ii) произведена рекомбинантным образом путем клонирования, (iii) очищена, например, путем расщепления и фракционирования гель-электрофорезом или (iv) синтезирована, например химическим путем. Изолированная нуклеиновая кислота служит для использования в методах рекомбинантной ДНК.
Нуклеиновые кислоты по данному изобретению могут использоваться по отдельности или в сочетании с другими нуклеиновыми кислотами, которые могут быть гомологичными или гетерологичными. В предпочтительных воплощениях данного изобретения нуклеиновые кислоты функционально связаны с последовательностями. регулирующими экспрессию, которые могут быть гомологичными либо гетерологичными по отношению к данной нуклеиновой кислоте, причем термин «гомологичная» означает, что данная нуклеиновая кислота также естественным образом (от природы) функционально связана с последовательностями, регулирующими экспрессию, а термин «гетерологичная» означает, что данная нуклеиновая кислота функционально не связана естественным образом (от природы) с последовательностями, регулирующими экспрессию.
Нуклеиновая кислота, например нуклеиновая кислота для экспрессии РНК и/или белка или пептида, и последовательности, регулирующие экспрессию, являются функционально связанными друг с другом, если они ковалентно соединены таким образом, что экспрессия или транскрипция данной нуклеиновой кислоты оказывается под контролем или под влиянием данных регуляторных последовательностей. Если данная нуклеиновая кислота должна транслироваться с образованием функционального белка, то при наличии последовательности, регулирующей экспрессию, которая функционально связана с кодирующей последовательностью, индукция данной регуляторной последовательности приводит к транскрипции данной нуклеиновой кислоты без сдвига рамки считывания в этой кодирующей последовательности или данная кодирующая последовательность не может транслироваться с образованием желаемого белка или пептида.
Термин «последовательность, регулирующая экспрессию» (регуляторная последовательность) по данному изобретению включает промоторы, участки связывания с рибосомой, энхансеры и другие регуляторные элементы, влияющие на транскрипцию гена или трансляцию матричной РНК. В некоторых конкретных воплощениях данного изобретения последовательности, регулирующие экспрессию, могут в свою очередь регулироваться. Структура последовательностей, регулирующих экспрессию, может быть различной: она зависит от вида живого организма или типа клетки, но как правило в ней имеются 5'-не транскрибируемая и 5'- и 3'- не транслируемые последовательности, которые участвуют в инициации транскрипции и трансляции, соответственно, например ТАТА-бокс, кэп-последовательность, последовательность СААТ и проч. В частности, 5'-не транскрибируемая регуляторная последовательность содержит промоторный участок, который включает промоторную последовательность для регуляции транскрипции функционально сопряженной нуклеиновой кислоты. Последовательности, регулирующие экспрессию, могут также содержать энхансерные последовательности или вышерасположенные активирующие последовательности (UAS).
По данному изобретению термин «промотор» или «промоторная область»» относится к нуклеотидной последовательности, расположенной «выше», т.е. в направлении 5' от нуклеотидной последовательности, которая экспрессируется. и регулирующей экспрессию этой последовательности, содержа сайт узнавания и связывания с РНК-полимеразой. Промоторная область может включать другие сайты распознавания и связывания для других факторов, участвующих в регуляции транскрипции данного гена. Промотор может регулировать транскрипцию прокариотического или эукариотического гена. Также промотор может быть индуцируемым и инициировать транскрипцию в ответ на индуцирующий агент или же промотор может быть конститутивным (не индуцируемым) и тогда транскрипция не регулируется индуцирующим агентом. Ген, регулируемый индуцируемым промотором, в отсутствие индуцирующего агента не экспрессируется или экспрессируется лишь в небольшой степени. В присутствии индуцирующего агента ген «включается» или увеличивается уровень транскрипции. Это опосредуется, как правило, связыванием специфического фактора транскрипции.
Промоторы, предпочтительные по данному изобретению, включают промоторы для полимераз SP6, Т3 и Т7, человеческий промотор U6, промотор CMV и искусственные гибридные промоторы (например, CMV), в которых какая-то часть (или части) соединена с частью (или частями) промоторов генов, кодирующих другие клеточные белки, например человеческую глицеральдегид-3-фосфатдегидрогеназу (GAPDH), включая или не включая дополнительный интрон (интроны).
По данному изобретению термин «экспрессия» употребляется в его наиболее общем значении и включает образование РНК или РНК и белка/пептида. Он также включает частичную экспрессию нуклеиновых кислот. Экспрессия может осуществляться временно (транзиентная экспрессия) или постоянно (стабильная экспрессия). По данному изобретению термин «экспрессия» также включает понятия «аберрантная экспрессия» и «аномальная экспрессия».
«Аберрантная экспрессия» или «аномальная экспрессия» по данному изобретению означает измененную экспрессию, предпочтительно усиленную, по сравнению с уровнем, принятым за нормальный, предпочтительно по сравнению с уровнем экспрессии в нормальных клетках (не онкогенных) или у здорового индивида. Усилением экспрессии считается повышение ее уровня на по меньшей мере 10%, в частности по меньшей мере на 20%, по меньшей мере на 50% или по меньшей мере на 100%. В одном из воплощений настоящего изобретения экспрессия имеет место только в пораженной ткани, тогда как в нормальной ткани она подавлена.
В предпочтительном воплощении настоящего изобретения молекула нуклеиновой кислоты присутствует в векторе, где снабжена промотором, регулирующим экспрессию данной нуклеиновой кислоты. Термин «вектор» в настоящем документе употребляется в его наиболее общем значении и включает любое вспомогательное средство, позволяющее указанной нуклеиновой кислоте, например, проникнуть в прокариотические и/или эукариотические клетки и, если нужно, встроиться в геном. Такие векторы предпочтительно реплицируются и/или экспрессируются в клетках. Векторы включают плазмиды, фагмиды, бактериофаги или вирусные геномы. Термин «плазмида» в настоящем документе употребляется, как правило, применительно к конструкциям из внехромосомного генетического материала, обычно кольцевым двуспиральным ДНК, которые могут реплицироваться независимо от хромосомной ДНК.
В качестве вектора для экспрессии антител можно использовать такие векторы, с которыми тяжелая цепь и легкая цепь антитела представлены разными векторами или же такие векторы, с которыми тяжелая цепь и легкая цепь антитела представлены в одном векторе.
Содержащиеся в настоящем документе утверждения относительно конкретных нуклеиновых кислот и аминокислотных последовательностей, например приведенных в Списке последовательностей, подразумевают также модификации, то есть варианты, указанных конкретных последовательностей, в результате которых получаются последовательности, функционально эквивалентные указанным конкретным последовательностям, например аминокислотные последовательности, обладающие свойствами, идентичными таковым (или сходными с ними) данных конкретных последовательностей и нуклеотидных последовательностей, кодирующих эти аминокислотные последовательности, обладающие свойствами, идентичными таковым (или сходными с ними) аминокислотных последовательностей, кодируемых этими конкретными нуклеотидными последовательностями. Одно из важных свойств состоит в сохранении способности связываться со своей мишенью или поддерживать эффекторные функции антител, например комплемент-зависимую цитотоксичность (CDC), и/или антителозависимую клеточно-опосредованную цитотоксичность (ADCC). Предпочтительно, когда последовательность, модифицированная по сравнению с какой-либо конкретной последовательностью, заменяет эту конкретную последовательность в молекуле антитела, сохраняются способность указанного антитела связываться с соответствующей мишенью и предпочтительно функции указанного антитела, как это описано в настоящем документе.
Аналогично, содержащиеся в настоящем документе утверждения относительно конкретных антител или гибридом, производящих определенные антитела, следует относить также к антителам, отличающимся аминокислотной последовательностью и/или кодирующей ее нуклеотидной последовательностью, модифицированной по сравнению с аминокислотной последовательностью и/или кодирующей ее нуклеотидной последовательностью этих конкретных антител, но функционально им эквивалентным. Важным свойством является сохранение связывания антител с их мишенью или сохранение их эффекторной функции. Предпочтительно последовательность, модифицированная по сравнению с какой-либо конкретной последовательностью, заменяя последнюю в молекуле антитела, сохраняет способность связывания с данного антитела с его мишенью и предпочтительно функции данного антитела, например лизис, опосредованный CDC или ADCC.
Специалистам в данной области техники понятно, что, в частности, последовательности CDR, гипервариабельные и вариабельные области можно модифицировать без потери способности связываться с мишенью. Например, области CDR окажутся идентичны или высоко гомологичны областям антител, описанным в настоящем документе. Термин «высоко гомологичный» подразумевает, что в CDR может быть произведено от 1 до 5, предпочтительно от 1 до 4, например от 1 до 3 или 1, или 2, замен. Кроме того, гипервариабельные и вариабельные области могут быть модифицированы таким образом, что они будут в значительной степени гомологичны определенным областям конкретных антител, описанных в настоящем документе.
Следует учитывать, что описанные в настоящем документе конкретные нуклеиновые кислоты также включают нуклеиновые кислоты, модифицированные ради оптимизации частоты использования кодонов в определенных хозяйских клетках или организме. Различия в частоте использования кодонов между организмами могут приводить к разнообразным проблемам, касающимся гетерологичной генной экспрессии. В результате оптимизации кодонов путем изменения одного или более нуклеотидов в исходной последовательности возможна оптимизация экспрессии данной нуклеиновой кислоты, в частности оптимизация эффективности трансляции, в гомологичном или гетерологичном организме-хозяине, у которого должна экспрессироваться данная нуклеиновая кислота.
По данному изобретению вариант, производное, модифицированная форма или фрагмент нуклеотидной последовательности, аминокислотной последовательности или пептида предпочтительно обладает функциональным свойством той нуклеотидной последовательности, аминокислотной последовательности или пептида, от которых они происходят. Такие функциональные свойства включают взаимодействие или связывание с другими молекулами. В одном из воплощений данного изобретения вариант, производное, модифицированная форма или фрагмент нуклеотидной последовательности, аминокислотной последовательности или пептида иммунологически эквивалентны той нуклеотидной последовательности, аминокислотной последовательности или пептида, от которых они происходят.
Предпочтительно степень идентичности какой-либо нуклеотидной последовательности и нуклеотидной последовательности, модифицированной по сравнению с данной конкретной последовательностью, или являющейся ее вариантом, должна составлять по меньшей мере 70%, предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, еще более предпочтительно по меньшей мере 90% или наиболее предпочтительно по меньшей мере 95%, 96%, 97%, 98% или 99%.
Что касается вариантов нуклеотидной последовательности, соответствующей CLDN6, то степень идентичности предпочтительно берется для области протяженностью по меньшей мере около 300, по меньшей мере около 400, по меньшей мере около 450, по меньшей мере около 500, по меньшей мере около 550, по меньшей мере около 600 или по меньшей мере около 630 нуклеотидов. В предпочтительных воплощениях данного изобретения степень идентичности берется для всей длины нуклеотидной последовательности, с которой проводится сравнение, например тех нуклеотидных последовательностей, которые приведены в Списке последовательностей. Предпочтительно, две нуклеотидные последовательности способны гибридизоваться друг с другом и образовывать стабильную двойную спираль предпочтительно, если это происходит в условиях, в которых возможна специфическая гибридизация полинуклеотидов (жесткие условия). Жесткие условия гибридизации описаны, например, в работах Molecular Cloning: A Laboratory Manual, J. Sambrook et al., Editors, 2nd Edition, Cold Spring Harbor Laboratory press, Cold Spring Harbor, New York, 1989 или Current Protocols in Molecular Biology, F.M. Ausubel et al., Editors, John Wiley & Sons, Inc., New York и относятся, например, к гибридизации при температуре 65°C в буферном растворе (3,5×SSC, 0,02% фикол, 0,02% поливинилпирролидон, 0,02% бычий сывороточный альбумин, 2,5 мМ NaH2PO4 (рН 7), 0,5% SDS, 2 мМ EDTA). Аббревиатурой SSC обозначен раствор 0,15 М хлорид натрия//0,15 М цитрат натрия, рН 7. После гибридизации мембрану, на которую шел перенос ДНК, промывают, например, дважды SSC при комнатной температуре и затем в 0,1-0,5×SSC/0,1×SDS при температуре до 68°С.
Термин «вариант» по данному изобретению также включает мутантов, варианты сплайсинга, конформации, изоформы, аллельные варианты, видовые варианты и видовые гомологи, в частности существующие в природе. Под аллельным вариантом понимается изменение в нормальной нуклеотидной последовательности гена, причем значение этого изменения зачастую остается неясным. При определении полной последовательности ген нередко выявляются многочисленные аллельные варианты данного гена. Видовый гомолог - это нуклеотидная либо аминокислотная последовательность иного видового происхождения, нежели таковое данной нуклеотидной или аминокислотной последовательности.
Для целей данного изобретения варианты аминокислотной последовательности включают варианты вследствие инсерций либо делеций, добавления и/или замен аминокислотных остатков. Возникшие в результате делеций варианты, включающие делецию на N-конце и/или на С-конце белка, называются также укороченными по N-либо С-концу вариантами.
Варианты, возникшие в результате инсерций аминокислотных остатков включают инсерций одного или двух или больше аминокислотных остатков в определенной аминокислотной последовательности. В случае вариантов аминокислотной последовательности, содержащих инсерцию, один или более аминокислотных остатков встраиваются в определенное место аминокислотной последовательности, хотя возможны также инсерций в случайные места с должным скринингом получающегося продукта.
Варианты, возникшие в результате добавления аминокислотных остатков включают присоединение одного или больше аминокислотных остатков (например, 1, 2, 3, 5, 10, 20, 30, 50 или более) к амино- и/или карбоксильному концу полипептидной цепи.
Варианты, возникшие в результате делеций аминокислотных остатков отличаются тем, что один или больше аминокислотных остатков (например, 1, 2, 3, 5, 10, 20, 30, 50 или более) удалены из полипептидной цепи. Делеций возможны в любом положении полипептидной цепи.
Варианты, возникшие в результате замены аминокислотных остатков отличаются тем, что по меньшей мере один аминокислотный остаток удален из полипептидной цепи, а на его место введен другой аминокислотный остаток. Предпочтение отдается изменениям в таких положениях аминокислотной последовательности, которые не являются консервативными среди гомологичных белков или пептидов и/или предпочтительно, чтобы заменяемые и заменяющие аминокислотные остатки обладали сходными свойствами. Предпочтительно, чтобы аминокислотные замены в вариантах белка были консервативными, то есть чтобы заменяемый и заменяющий аминокислотные остатки имели сходный электрический заряд. Консервативная аминокислотная замена подразумевает, что заменяющая аминокислота относится к той же группе по свойствам боковых цепей, что и заменяемая. Существующие в природе аминокислоты в целом подразделяются на четыре группы: кислотные (аспарагиновая кислота, глутаминовая кислота), основные (лизин, аргинин, гистидин), неполярные (аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан) и полярные, несущие заряд (глицин, аспарагин, глутамин, цистеин, серии, треонин, тирозин). Фенилаланин, триптофан и тирозин иногда выделяют в группу ароматических аминокислот.
В отношении аминокислотной последовательности, соответствующей SEQ ID NO:37, термин «вариант» относится в частности к последовательности, в которой остаток цистеина в положении 436 заменен на остаток другой аминокислоты из числа перечисленных выше, предпочтительно на глицин, аланин, серин, треонин, валин или лейцин.
Степень сходства, предпочтительно идентичности между данной аминокислотной последовательностью и последовательностью, модифицированной по сравнению с данной, или являющейся ее вариантом, например между значительно гомологичными аминокислотными последовательностями, предпочтительно составляет по меньшей мере 70%, более предпочтительно по меньшей мере 80%, еще более предпочтительно по меньшей мере 90% или наиболее предпочтительно по меньшей мере 95%, 96%, 97%, 98% или 99%. Степень сходства или идентичности берется предпочтительно для участка полипептидной цепи, составляющего по меньшей мере около 10%, по меньшей мере около 20%, по меньшей мере около 30%, по меньшей мере около 40%, по меньшей мере около 50%, по меньшей мере около 60%, по меньшей мере около 70%, по меньшей мере около 80%, по меньшей мере около 90% или около 100% все длины аминокислтной последовательности, с которой проводится сравнение. Например, если аминокислтная последовательность, с которой проводится сравнение, состоит из 200 аминокислотных остатков, то степень сходства или идентичности берется предпочтительно для по меньшей мере около 20, по меньшей мере около 40, по меньшей мере около 60, по меньшей мере около 80, по меньшей мере около 100, по меньшей мере около 120, по меньшей мере около 140, по меньшей мере около 160, по меньшей мере около 180 или около 200 аминокислотных остатков, предпочтительно расположенных непрерывно. Что касается вариантов полипептидной цепи CLDN6, то степень сходства или идентичности берется предпочтительно для участка, состоящего из по меньшей мере около 100, по меньшей мере около 120, по меньшей мере около 140, по меньшей мере около 160, по меньшей мере около 180, по меньшей мере около 200 или по меньшей мере около 210 аминокислотных остатков. В предпочтительных воплощениях данного изобретения степень сходства или идентичности берется для всей длины аминокислотной последовательности, с которой проводится сравнение, например аминокислотных последовательностей, приведенных в Списке последовательностей. Для выравнивания последовательностей с целью определения их сходства, предпочтительно идентичности, можно применять средства, известные в данной области техники, предпочтительно с использованием наилучшего выравнивания, например при помощи программ Align, предпочтительно EMBOSS::needle, Matrix: Blosum62, Gap Open 10.0, Gap Extend 0.5.
«Сходство последовательностей» указывает доля аминокислотных остатков (в процентах), которые идентичны либо представляют консервативные аминокислотные замены. «Идентичность последовательностей» при сравнении двух полипептидов или нуклеотидных последовательностей указывает долю аминокислотных остатков или нуклеотидов, которые в сравниваемых последовательностях одинаковы.
«Идентичность (в процентах)» получают после наилучшего выравнивания, причем эта величина чисто статистическая и различия между двумя сравниваемыми последовательностями распределяются равномерно по всей длине последовательностей. Сравнение двух нуклеотидных или аминокислотных последовательностей обычно осуществляется после оптимального выравнивания, причем сравнение проводят с помощью сегмента или окна сравнения, чтобы идентифицировать и сравнить локальные области сходства последовательностей. Оптимальное выравнивание последовательностей для сравнения можно проделать, помимо «вручную», с помощью алгоритма локальной гомологии Смита и Уотермана (см. Smith, Waterman, 1981, Ads App. Math. 2, 482), CC помощью алгоритма локальной гомологии Нидлмана и Уанча (Neddleman, Wunsch, 1970, J. Mol. Biol. 48, 443), с помощью метода поиска подобия Пирсона и Липмана (Pearson, Lipman, 1988, Proc. Natl Acad. Sci. USA 85, 2444) или с помощью компьютерных программ, в которых используются эти алгоритмы (GAP, BESTFIT, FASTA, BLAST P, BLAST N, TFASTA, Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Drive, Мадисон, шт. Висконсин, США).
Идентичность (в процентах) рассчитывается путем определения числа одинаковых позиций в двух сравниваемых последовательностях и деления его на число сравненных позиций и умножения полученного результата на 100, так что в итоге получается процентная идентичность двух последовательностей.
Консервативные замены можно сделать, например, исходя из сходства полярности, заряда, растворимости, гидрофобности, гидрофильности и/или амфипатической природы рассматриваемых аминокислотных остатков. Например: (а) неполярные (гидрофобные) аминокислоты включают аланин, лейцин, изолейцин, валин, пролин, фенилаланин, триптофан и метионин); (b) полярные нейтральные аминокислоты включают глицин, серин, треонин, цистеин, тирозин, аспарагин и глутамин; (с) положительно заряженные (основные) аминокислоты включают аргинин, лизин и гистидин; и (d) отрицательно заряженные (кислотные) аминокислоты включают аспарагиновую кислоту и глутаминовую кислоту. Как правило, замены осуществляются в пределах групп (a)-(d). Кроме того, глицин и пролин могут заменять друг друга, так как и та и другая аминокислоты способны нарушать правильную альфа-спиральную структуру полипептидной цепи. Некоторые предпочтительные замены делаются в пределах следующих групп: (i) S и Т; (ii) Р и G; и (iii) А, V, L и I. Зная генетический код и располагая технологией рекомбинантной ДНК и возможностями ее синтеза, специалист в данной области техники вполне может создать ДНК, кодирующую консервативные варианты аминокислотных последовательностей.
В настоящее изобретение входят антитела, в которых сделаны изменении, затрагивающие область Fc, с целью изменить функциональные или фармакокинетические свойства этих антител. Такие изменения могут приводить к ослаблению или усилению связывания C1q и к комплемент-зависимой цитотоксичности или к усилению либо ослабления связывания FcγR и к антителозависимой клеточной (клеточно-опосредованной) цитотоксичности. Возможны, например, замены одного или более аминокислотных остатков в константной области тяжелой цепи, что должно вызвать изменение эффекторной функции при сохранении способности к связыванию антигена по сравнению с модифицированными антителами (см. для сравнения патенты США 5,624,821 и 5,648,260).
Время жизни (полужизни) антител in vivo можно увеличить, модифицируя эпитоп связывания рецептора реутилизации в константном домене иммуноглобулина или в подобном домене таким образом, чтобы молекула не содержала интактного домена СН2 или интактной области Fc (см. для сравнения патенты США 6,121,022 и 6,194,551. Время жизни (полужизни) антител может быть также увеличено путем мутаций в области Fc, например путем замены треонина в положении 252 на лейцин, треонина в положении 254 на серии или треонина в положении 256 на фенилаланин (см. для сравнения патент США 6,277,375).
Для изменения эффекторной функции антител можно также модифицировать паттерн гликозилирования молекул антител. Например, экспрессию антител можно осуществлять в трансфектомах, где не происходит присоединения фукозы, которая в норме присоединяется к остатку аспарагина в положении 297 области; в результате увеличится сродство области Fc к рецептору Fc, что в свою очередь приведет к усилению антителозависимой клеточной цитотоксичности в присутствии естественных киллеров (NK-клеток); см. для сравнения Shield et al. (2002) JBC, 277:26733. Для модифицирования комплемент-зависимой цитотоксичности можно проводить модификацию галактозилирования.
В другом воплощении данного изобретения мутации вызывают случайным образом по всей последовательности, кодирующей антитело против CLDN6, или в какой-то ее части, например, путем насыщающего мутагенеза, и получающиеся в результате модифицированные антитела против CLDN6 можно проверить на активность связывания с мишенью.
По данному изобретению термины «клетка» и «клетка-хозяин» предпочтительно относятся к интактной клетке, то есть к клетке с неповрежденной и неизмененной мембраной, которая не теряла свои в норме внутриклеточные компоненты, например ферменты, органеллы или генетический материал. Интактная клетка является предпочтительно жизнеспособной, то есть живой клеткой, способной нормально осуществлять свойственные ей метаболические функции. Предпочтительно указанные термины по данному изобретению относятся к любой клетке, которая может быть трансформирована или трансфецирована с использованием экзогенной нуклеиновой кислоты. Термин «клетка» по данному изобретению включает прокариотические клетки (например, Ecsherichia coli) или эукариотические клетки (например, дендритные клетки, В-клетки, клетки СНО, COS, K562, HEK293, HELA, дрожжевые клетки и клетки насекомых). Экзогенная нуклеиновая кислота внутри такой клетки может находиться (i) сама по себе, в свободно распределенном состоянии, (ii) включенная в рекомбинантный вектор или (iii) интегрированная в геном клетки-хозяина или в митохондриальную ДНК. Особенно предпочтительны клетки млекопитающих, например мыши, хомячка, свиньи, козы и приматов, включая человека. Клетки по данному изобретению могут происходить из различных типов ткани и включают эмбриональные клетки и клеточные линии. Конкретные примеры включают кератиноциты, лейкоциты периферической крови, стволовые клетки костного мозга и эмбриональные стволовые клетки. В других воплощениях данного изобретения клетки являются представляющими антиген, например это дендритные клетки, моноциты или макрофаги. Термин «клетка-хозяин» в настоящем документе предпочтительно относится к клеткам, в которые введен рекомбинантный вектор экспрессии.
В клетке, содержащей молекулу нуклеиновой кислоты, предпочтительно экспрессируется пептид или белок, кодируемый этой нуклеиновой кислотой.
Термин «трансгенное животное» относится к животным, геном которых содержит один или более чужеродных генов (трансгенов), предпочтительно генов тяжелых и/или легких цепей антител, или чужеродных хромосом (трансхромосом), интегрированных или не интегрированных в природную геномную ДНК данного животного, и которые предпочтительно способны к экспрессии этих трансгенов. Например, у трансгенной мыши может иметься трансген или трансхромосома - человеческий ген легкой цепи антитела и человеческий ген тяжелой цепи или человеческая хромосома с нуклеотидной последовательностью, кодирующей тяжелую цепь, так что у этой мыши после иммунизации антигеном CLDN6 и/или клетками, в которых экспрессируется CLDN6, образуются человеческие антитела против CLDN6. Трансген человеческой тяжелой цепи может быть интегрирован в мышиную хромосомную ДНК, как в случае трансгенных мышей, например мышей HuMAb, HCo7 или НСо12, или трансген человеческой тяжелой цепи может существовать вне хромом, как в случае трансхромосомных мышей (например, КМ), описанных в публикации WO 02/43478. Такие трансгенные и трансхромосомные мыши могут быть способными к образованию множества изотипов человеческих моноклональных антител против CLDN6 (например, IgG, IgA и/или IgE) за счет рекомбинации V-D-J и переключения изотипов.
Слова «снизить» или «ингибировать» в настоящем документе означают способность вызвать в целом уменьшение уровня, например пролиферации клеток, предпочтительно на 5% или больше, 10% или больше, 20% или больше, более предпочтительно на 50% или больше, наиболее предпочтительно на 75% или больше. Термин «ингибировать» (или подобные выражения) включает полное или практически полное ингибирование, то есть снижение до нуля или практически до нуля.
Слова «увеличение» или «усиление» предпочтительно относятся к увеличению или усилению на по меньшей мере 10%, предпочтительно на по меньшей мере 20%, предпочтительно на по меньшей мере 30%, более предпочтительно на по меньшей мере 40%, более предпочтительно на по меньшей мере 50%, еще более предпочтительно на по меньшей мере 80% и наиболее предпочтительно на по меньшей мере 100%. Эти слова могут также относиться к обстоятельствам, когда в момент времени «ноль» отсутствует определяемый/детектируемый сигнал для определенного вещества или состояния и в некоторый момент времени позже нулевого имеется детектируемый сигнал для данного вещества или состояния.
Термин «иммунологически эквивалентный» означает, что иммунологически эквивалентный агент (молекула), например иммунологически эквивалентная аминокислотная последовательность, проявляет такие же или практически такие же иммунологические свойства и/или имеет такие же или практически такие же иммунологические эффекты, например в отношении типа иммунологического эффекта (индукция гуморального и/или клеточного иммунного ответа, интенсивность и/или продолжительность вызванной иммунологической реакции) или в отношении специфичности вызванной иммунологической реакции. В контексте настоящего изобретения термин «иммунологически эквивалентный» предпочтительно употребляется применительно к иммунологическим свойствам или эффектам пептида или варианта пептида, взятого для иммунизации. В частности иммунологическим свойством является способность связываться с антителами и вызывать иммунный ответ, предпочтительно путем стимуляции образования антител. Например, аминокислотная последовательность является иммунологически эквивалентной другой последовательности, взятой для сравнения, если указанная аминокислотная последовательность, предъявленная иммунной системе индивида, вызывает иммунологическую реакцию, предпочтительно образование антител, специфично реагирующих с последовательностью, взятой для сравнения, например с аминокислотной последовательностью, формирующей часть CLDN6.
Термин «иммунные эффекторные функции» в контексте настоящего изобретения включает любые функции, опосредуемые компонентами иммунной системы и приводящие к ингибированию опухолевого роста и/или ингибированию развития опухоли, включая ингибирование диссеминации опухоли и метастазирования. Предпочтительно иммунные эффекторные функции приводят к уничтожению опухолевых клеток. Предпочтительно иммунные эффекторные функции в контексте данного изобретения опосредуются антителами. Такие функции включают комплемент-зависимую цитотоксичность (CDC), антителозависимую клеточную цитотоксичность (ADCC), индукцию апоптоза в клетках, несущих антигены, ассоциированные с опухолями, например путем связывания антител с поверхностным антигеном, и/или ингибирование пролиферации клеток, несущих антигены, ассоциированные с опухолями, предпочтительно ADCC и/или CDC. Таким образом, антитела, способные опосредовать одну или более иммунных эффекторных функций, предпочтительно способны опосредовать уничтожение клеток, вызывая лизис, опосредованный CDC, лизис, опосредованный ADCC, апоптоз, гомотипическую адгезию и/или фагоцитоз, предпочтительно вызывая лизис, опосредованный CDC, и/или лизис, опосредованный ADCC. Эффект антител может также осуществляться просто за счет связывания с антигенами, ассоциированными с опухолями, на поверхности опухолевых клеток. Например, антитела могут блокировать функцию антигенов, ассоциированных с опухолями, в результате связывания с антигенами, ассоциированными с опухолями, на поверхности опухолевых клеток.
Подробное описание изобретения
Механизм действия mAb
Хотя в изложенном ниже предлагаются рассуждения, касающиеся механизма, лежащего в основе лечебного действия антител, их не следует считать как-либо ограничивающими данное изобретение.
Антитела, описанные в настоящем документе, могут взаимодействовать с компонентами иммунной системы, предпочтительно через цитотоксичность - ADCC или CDC. Антитела по данному изобретению можно также использовать для нацеливания нужного агента (например, радиоактивного изотопа, лекарственного вещества или токсина) с целью непосредственного уничтожения опухолевых клеток или применять в сочетании, предполагающем синергизм действия, с обычными химиотерапевтическими агентами, воздействуя на опухоль взаимодополняющими путями, которые могут включать противоопухолевый иммунный ответ, возможно, ослабленный вследствие цитотоксических побочных эффектов химиотерапевтических препаратов, влияющих на Т-лимфоциты. Однако эффект антител по данному изобретению может также осуществляться просто за счет связывания с CLDN6 на поверхности клеток, что блокирует пролиферацию клеток.
Антителозависимая клеточно-опосредованная цитотоксичность
ADCC характеризует способность описанных в настоящем документе эффекторных клеток, в частности лимфоцитов, уничтожать другие клетки, для чего предпочтительно требуется, чтобы клетки-мишени были помечены антителами.
Предпочтительно ADCC осуществляется, когда антитела связываются с антигеном на опухолевых клетках и домен Fc молекулы антитела связывается с рецептором Fc (FcR) на поверхности эффекторной клетки. Идентифицированы несколько семейств рецепторов Fc; определенным клеточным популяциям свойственна экспрессия определенных рецепторов Fc. ADCC можно рассматривать как механизм, предназначенный для того, чтобы непосредственно вызывать - в той или иной степени - немедленное разрушение опухоли, и ведущий к представлению антигена и индукции противоопухолевого Т-клеточного ответа. Предпочтительно индукция ADCC in vivo должна приводить к противоопухолевому Т-клеточному ответу и антительному ответу с участием собственных антител организма.
Комплемент-зависимая цитотоксичность
CDC - это еще один опосредованный антителами путь уничтожения клеток. Наиболее эффективным для активации комплемента изотипом антител являются IgM. IgG1 и IgG3 тоже очень эффективно направляют CDC по классическому пути активации комплемента. В этом каскаде реакций предпочтительно образование комплексов антиген-антитело приводит к доступности множества участков связывания C1q поблизости от домена CH2 задействованных молекул антител, например IgG (C1q - это один из трех белков, комплекс которых представляет собой компонент С1 системы комплемента). Предпочтительно, когда эти участки связывания с C1q становятся доступными, сродство IgGk C1q, до того низкое, существенно возрастает, что инициирует каскад реакций, включающий ряд других белков системы комплемента и ведущий к протеолитическому высвобождению из эффекторной клетки хемотаксических/активирующих агентов С3а и С5а. Предпочтительно каскад системы комплемента завершается формированием комплекса, атакующего мембрану клетки-мишени и создающего в ней поры, способствующие свободному входу воды и растворенных веществ внутрь клетки и наружу.
Образование антител
Антитела по данному изобретению можно производить различными методами, включая обычную технологию моноклональных антител, например стандартный метод гибридизации соматических клеток по Келеру-Мильштейну (см. G. Kohler, C. Milstein, Nature 256:495, 1975). Хотя метод гибридизации соматических клеток предпочтителен для получения антител по данному изобретению, в принципе можно применять и другие методы производства моноклональных антител, например вирусную или онкогенную трансформацию В-лимфоцитов или метод фагового дисплея с использованием библиотек генов антител.
Предпочтительной животной системой для получения гибридом, производящих моноклональные антитела, является мышиная система. Получение гибридом у мышей - налаженная лабораторная процедура. Протоколы иммунизации мышей и способы выделения активированных спленоцитов для гибридизации известны в данной области техники. Известны также пары для гибридизации (например, мышиные миеломные клетки) и методики слияния клеток.
К числу предпочтительных по данному изобретению животных систем для получения гибридом, секретирующих моноклональные антитела, также относятся крысиная и кроличья системы (например, описанные в работе Spieker-Polet et al., Proc. Natl. Acad. Sci. U.S.A. 92:9348 (1995); см. также Rossi et al., Am. J. Clin. Pathol. 124:295 (2005)).
В еще одном предпочтительном воплощении данного изобретения человеческие моноклональные антитела, направленные против CLDN6, можно получать с использованием трансгенных или трансхромосомных мышей, несущих элементы человеческой иммунной системы вместо соответствующих мышиных. К таким трансгенным и трансхромосомным мышам относятся животные, обозначаемые соответственно HuMAb и KM; в настоящем документе тех и других вместе называют трансгенными. Получение человеческих антител с помощью таких трансгенных мышей можно проделывать так, как подробно описано для CD20 в публикации WO2004 035607.
Другой подход к получению моноклональных антител - непосредственное выделение генов, кодирующих антитела, из лимфоцитов, производящих определенные антитела (см., например, Babcock et al., 1996; A novel strategy for generating monoclonal antibodies from single, isolated lymphocytes producing antibodies of defined strategy). Подробности получения рекомбинантных антител можно найти также в работах M. Welschof, J. Kraus, Recombinant antibodes for cancer therapy ISBN-0-89603-918-8 and Benny K.C. Lo Antibody Engineering ISBN 1-58829-092-1.
Иммунизация
Для получения антител против CLDN6, мышей можно иммунизировать конъюгированными с носителем пептидами, происходящими из аминокислотной последовательности CLDN6, обогащенным препаратом антигена CLDN6, экпрессирующегося как рекомбинантный белок, или их фрагментами и/или клетками, в которых экспрессируется CLDN или его фрагменты, как описано в настоящем документе. Или же мышей можно иммунизировать ДНК, кодирующей полную полипептидную цепь человеческого или его фрагменты. В случае, если иммунизация с использованием очищенного или обогащенного препарата антигена CLDN6 не приводит к образованию нужных антител, мышей для стимуляции иммунного ответа можно иммунизировать клетками, в которых экспрессируется CLDN6, например взятых из культуры клеток подходящей линии.
Иммунный ответ можно отслеживать в ходе иммунизации по образцам плазмы и сыворотки крови из хвостовой вены или ретроорбитального венозного синуса. Для слияния клеток отбирают животных с достаточно высоким титром иммуноглобулинов против CLDN6. Чтобы увеличить интенсивность образования специфических антител гибридомами, мышам вводят клетки, в которых экспрессируется CLDN6, путем внутрибрюшинной или внутривенной инъекции за 3-5 дней до умерщвления и взятия селезенки.
Создание гибридом, продуцирующих моноклональные антитела
Чтобы получить гибридомы, продуцирующие моноклональные антитела против CLDN6, выделяют клетки из лимфатических узлов или селезенки, взятых у иммунизированных мышей, и сливают их с иммортализованными клетками подходящей линии, например с мышиными миеломными клетками. Получаемые в результате гибридомы проверяют на образование антиген-специфичных антител. Содержимое отдельных ячеек исследуют методом твердофазного иммуноферментного анализа (ELISA) на наличие гибридом, секретирующих антитела. Антитела, специфичные в отношении CLDN6, можно идентифицировать, применяя иммунофлуоресцентный анализ и метод FACS с использованием клеток, в которых экспрессируется CLDN6. Гибридомы, продуцирующие нужные антитела, пересевают, вновь проверяют и те, в которых образуются моноклональные антитела против CLDN6, можно субклонировать методом серийных разведении. Затем стабильные субклоны культивируют in vitro, чтобы получить антитела в культуральной среде и охарактеризовать их.
Создание трансфертом, продуцирующих моноклональные антитела
Антитела по данному изобретению можно также получать в трансфектомах клеток-хозяев, применяя, например, сочетание технологии рекомбинантной ДНК и методов трансфекции генов, хороши известных в данной области техники (Morrison, S. (1985) Science 229:1202).
Например, в одном из воплощений данного изобретения нужный ген (гены), например гены антител, встраивают в вектор экспрессии, например в плазмиду эукариотической клетки, как описано для системы экспрессии гена GS в публикациях WO 87/04462 и WO 89/01036, а также в европейском патенте 338841, или используют другую систему экспрессии, которые хорошо известны в данной области техники. Очищенные плазмиды, содержащие клонированные гены антител, вводят в эукариотические клетки-хозяева, например в клетки СНО, NS/0, HEK293T или HEK293, либо в иные эукариотические клетки, например в клетки растительного происхождения или в клетки грибов, включая дрожжи. Для введения генов по данному изобретению можно применять методы, описанные в литературе по данной области техники, например электропорез, использование липофектина, липофектамина и др. Клетки-хозяева, в которые введенные гены антител экспрессируются, можно выявить и отобрать. Уровень экспрессии генов антител в трансфицированных клетках (трансфектомах) можно повысить, затем размножить эти клетки и получать в нужном количестве продуцируемые ими антитела. Из супернатанта культуральной среды и/или клеток выделяют и очищают рекомбинантные антитела.
Или же экспрессию клонированных генов антител по данному изобретению можно получить в других экспрессионных системах, включая прокариотические клетки, например микроорганизмы, включая Escherichia coli. Также антитела по данному изобретению могут производиться трансгенными животными, например в составе молока у овец и кроликов или в составе яиц у кур, или в трансгенных растениях; см., например, Verma, R., et al. (1998) J. Immunol. Meth. 216:165-181; Pollock, et al. (1999) J. Immunol. Meth. 231:147-157; and Fischer, R., et al. (1999) Biol. Chem. 380:825-839.
Экспрессия интактных антител с использованием частичных последовательностей (гуманизация, химеризация)
а) Химеризация
Для лечения людей можно использовать мышиные моноклональные антитела, меченные токсическими агентами или радиоактивными изотопами. Немеченые мышиные антитела при многократном применении сильно иммуногенны для человека, что снижает терапевтический эффект. Иммуногенность обусловлена в основном константными областями тяжелых цепей молекул антител. Иммуногенность мышиных антител для человека можно снизить или вообще снять, если эти антитела подвергнуть химеризации или гуманизации. Гибридные (или химерные) антитела - это антитела, в молекулах которых различные участки происходят от животных разных видов, например вариабельная область - от мышиного антитела, а константная - от человеческого иммуноглобулина. Химеризация антител достигается путем соединения тяжелой и легкой цепей вариабельной области мышиного антитела с тяжелой и легкой цепями константной области человеческого антитела (например, как описано в работе Kraus et al., in Methods in Molecular Biology series, Recombinant antibodies for cancer therapy ISBN-0-89603-918-8). В одном из предпочтительных воплощений данного изобретения гибридные антитела сочетают в себе константную область человеческой легкой каппа-цепи и вариабельную область мышиной легкой цепи. В другом предпочтительном воплощении данного изобретения гибридные антитела образованы в результате присоединения константной области человеческой легкой лямбда-цепи к вариабельной области мышиной легкой цепи. Для получения гибридных антител предпочтительны константные области тяжелых цепей IgG1, IgG3 и IgG4. Для получения гибридных антител предпочтительны также константные области тяжелых цепей IgG2, IgA, IgD и IgM.
b) Гуманизация
Антитела взаимодействуют с антигенами-мишенями преимущественно аминокислотными остатками, расположенными в шести участках, определяющих комплементарность (CDR). Поэтому аминокислотные последовательности в пределах CDR сильнее различаются у разных антител, чем последовательности вне CDR. Поскольку последовательности CDR обеспечивают большинство взаимодействий между антителом и антигеном, возможна экспрессия рекомбинантных антител, имитирующих свойства определенных природных антител, путем создания векторов экспрессии, которые включают последовательности CDR от определенных природных антител, объединенные с каркасными последовательностями от других антител с другими свойствами (см., например, Riechmann, L. et al. (1998) Nature 332:323-327; Jones, P. et al. (1986) Nature 321:522-525; and Queen, С. et al. (1989) Proc. Natl. Acad. Sci. U.S.A. 86:10029-10033). Такие каркасные последовательности можно получить из общедоступных баз данных по ДНК, включающих нуклеотидные последовательности генов антител «зародышевой линии». Эти гаметные последовательности отличаются от последовательностей зрелых генов антител, потому что в них нет полностью собранных вариабельных последовательностей, которые образуются путем соединения сегментов V (D) J в ходе созревания В-клеток. Гаметные нуклеотидные последовательности также отличаются от последовательностей, соответствующих высокоаффинным антителам вторичного репертуара по всей вариабельной области. Например, соматические мутации относительно редки вблизи N-конца каркасного участка 1 и вблизи С-конца каркасного участка 4. Кроме того, многие соматические мутации существенно не влияют на связывание антител. По этой причине для того, чтобы воссоздать интактное рекомбинантное антитело, обладающее свойствами связывания, как у исходного антитела, нет необходимости получать полную последовательность ДНК данного антитела (см. WO 99/45962). Для этой цели обычно достаточен охват участков CDR частичных последовательностей тяжелых и легких цепей. Частичная последовательность используется, чтобы определить, какие вариабельные и соединяющие сегменты гена зародышевой линии (гаметной последовательности) участвуют в образовании вариабельных генов рекомбинированного антитела. Гаметная последовательность затем используется для заполнения недостающих частей вариабельных областей. Лидерные последовательности тяжелой и легкой цепи отщепляются в процессе созревания белка и не вносят вклад в формирование свойств конечного антитела. Для восполнения недостающих последовательностей клонированные последовательности кДНК можно комбинировать с синтетическими олигонуклеотидами путем лигирования или амплификации с помощью полимеразной цепной реакции (ПЦР). Или же можно синтезировать целую вариабельную область как набор коротких перекрывающихся олигонуклеотидов и объединить их посредством ПЦР-амплификации, чтобы создать полностью синтетический клон вариабельной области. Такой процесс обладает рядом преимуществ: элиминируются или включаются определенные сайты рестрикции либо оптимизируются определенные кодоны.
Транскрипты нуклеотидных последовательностей тяжелых и легких цепей из гибридом используются для составления набора перекрывающихся синтетических олигонуклеотидов для создания синтетических V-последовательностей, способных кодировать такие же аминокислотные последовательности, как природные нуклеотидные последовательности. Синтетические последовательности тяжелых и каппа-цепей могут отличаться от природных последовательностей в трех аспектах: вереницы повторяющихся нуклеотидов прерывают для облегчения синтеза олигонуклеотидов и амплификации методом ПЦР; включают оптимальные сайты инициации трансляции по Козаку (Kozak, 1991, J. Biol. Chem. 266:19867-19870); сайты рестрикции эндонуклеазой HindIII располагают в сторону 5'-конца нуклеотидной последовательности от сайтов инициации трансляции.
Для вариабельных областей как легких, так и тяжелых цепей оптимизированные кодирующие и соответствующие некодирующие последовательности разбивают на сегменты из 30-50 нуклеотидов примерно посередине соответствующего некодирующего олигонуклеотида. Таким образом для каждой цепи олигонуклеотиды можно собрать в перекрывающиеся двухцепочечные наборы, охватывающие сегменты из 150-400 нуклеотидов. Эти пулы затем используют как матрицы для получения продуктов амплификации методом ПЦР из 150-400 нуклеотидов. Как правило, из одного набора олигонуклеотидов вариабельной области получается два пула, которые амплифицируют по отдельности, чтобы получить два перекрывающихся продукта ПЦР. Эти перекрывающиеся продукты затем объединяют путем амплификации методом ПЦР, чтобы сформировать полную вариабельную область. Может оказаться желательным включить в амплификацию методом ПЦР также перекрывающийся фрагмент константной области тяжелой или легкой цепи, чтобы получить фрагменты, легко клонируемые в экспрессионных векторных конструкциях.
Затем сконструированные гибридные или гуманизированные вариабельные области тяжелых и легких цепей объединяют с клонированными промоторной и лидерной последовательностями, сайтом инициации трансляции, константной областью, 3'-нетранслируемой последовательностью, сайтом полиаденилирования и сайтом терминации транскрипции, чтобы сформировать экспрессионную векторную конструкцию. Экспрессионные конструкции для тяжелой и легкой цепей можно объединить в одном векторе, и провести совместную, последовательную или раздельную трансфекцию в клетки-хозяева, которые затем сливают, чтобы получить клетку-хозяин, в которой экспрессируются обе цепи. Известны плазмиды для использования при конструировании векторов экспрессии для человеческого IgGK. Плазмиды можно сконструировать таким образом, что амплифицированные методом ПЦР последовательности кДНК вариабельных областей тяжелой цепи и легкой каппа-цепи можно использовать для воссоздания минигенов полных тяжелой и легкой цепей. Эти плазмиды можно использовать для экспрессии полностью человеческих или же гибридных IgG1, каппа или IgG4, каппа-антител. Подобные плазмиды можно сконструировать для экспрессии других изотипов тяжелых цепей или для экспрессии антител, содержащих легкие лямбда-цепи.
Таким образом, в другом аспекте данного изобретения структурные свойства антител против CLDN6 используются для создания структурно родственных гуманизированных антител против CLDN6, сохраняющих по меньшей мере одно функциональное свойство антител по данному изобретению, например связывание с CLDN6. Более конкретно, один или более участков CDR мышиных моноклональных антител можно объединить рекомбинантным путем с известными человеческими каркасными участками и участками CDR, чтобы получить дополнительные рекомбинантные гуманизированные антитела против CLDN6 по данному изобретению.
Связывание с клетками, в которых экспрессируется антиген
Способность антител по данному изобретению связываться с CLDN6 можно определить с помощью стандартных аналитических методов наподобие тех, которые представлены в примерах - например, твердофазного иммуноферментного анализа (ELISA), Вестерн-блоттинга, иммунофлуореценции, проточной цитометрии.
Выделение и определение свойств антител
Для очистки антител против CLDN6 отобранные гибридомы выращивают в двухлитровых роллерных колбах для очистки моноклональных антител. Или же антитела против CLDN6 получают в биореакторах с диализом. Супернатант фильтруют и, при необходимости, концентрируют, после чего проводят аффинную хроматографию с белком G или белком А, иммобилизованным на сефарозе. Чистоту элюированного IgG проверяют методом гель-электрофореза и высокоэффективной жидкостной хроматографии. Буферный раствор обменивается на PBS, концентрацию определяют по оптической плотности на длине волны 280 нм (коэффициент экстинкции 1,43). Аликвоты моноклональных антител хранят при температуре -80°С.
Чтобы определить, связываются ли отобранные моноклональные антитела против CLDN6 с данным эпитопом, можно использовать направленный (сайт-специфический) мутагенез и множественный сайт-специфический мутагенез.
Определение изотипа
Чтобы определить изотип очищенных антител, можно применить твердофазный иммуноферментный анализ (ELISA), используя различные готовые имеющиеся в продаже наборы (например, Zymed, Roche Diagnostics). Лунки титрационного микропланшета покрывают антимышиным Ig. После блокирования, проводят реакцию с моноклональными антителами или очищенным контрольным (по изотипу) образцом при комнатной температуре в течении 2 часов. Затем в ячейки вносят мышиные IgG1, IgG2a, IgG2b или IgG3, IgA, либо мышиные специфичные к IgM конъюгаты с пероксидазой. После промывания планшеты обрабатывают 2,2'-азино-бис-(3-этилбензотиазолин)-6-судьфоновой кислотой (ABTS) в качестве субстрата (1 мг/мл) и определяют оптическую плотность при 405-650 нм. Или же используют готовый набор IsoStrip Mouse Monoclonal Antibody Isotyping Kit (Roche, Cat. No. 1493027) согласно инструкции производителя.
Проточная цитометрия
Чтобы продемонстрировать присутствие антител против CLDN6 в сыворотке крови иммунизированных мышей или связывание моноклональных антител с живыми клетками, в которых экспрессируется CLDN6, можно применять метод проточной цитометрии. Образцы линий клеток, в которых CLDN6 экспрессируется от природы или после трансфекции, и отрицательный контроль (клетки без экспрессии CLDH6, выращенные в стандартных условиях культивирования) смешивают с моноклональными антителами в различной концентрации в супернатанте гибридом или в PBS, содержащем 1% FBS, и инкубируют при температуре 4°С в течение 30 мин. После промывания проводят связывание моноклональных антител, связавших CLDH6, с антителами против IgG, меченными аллофикоцианином (АРС) или Alexa647, в тех же условиях, в которых проводили окрашивание первичных антител. Образцы анализируют с помощью проточного цитофлуориметра FACS, определяя параметры прямого(переднего) и бокового светорассеяния единичных живых клеток. Чтобы отличить связывание моноклональных антител, специфичных в отношении CLDN6, от неспецифического связывания в каждом отдельном измерении, можно применять метод котрансфекции. Клетки временно трансфецируют плазмидами, кодирующими CLDN6 и флуоресцентный маркер, и окрашивают, как описано выше. Трансфицированные клетки выявляют в канале регистрации флуоресценции, отличном от такового для клеток, окрашенных антителами. Поскольку в большинстве траснфецированных клеток экспрессируются оба трансгена, моноклональные антитела, специфичные к CLDN6, связываются предпочтительно с клетками, в которых экспрессируется флуоресцентный маркер, тогда как неспецифичные антитела связываются в сравнимом количестве с нетрансфицированными клетками. В дополнение к проточной цитометрии или вместо нее можно применять флуоресцентную микроскопию. Для использования флуоресцентного микроскопа клетки окрашивают так же, как описано выше.
Иммунофлуоресцентная микроскопия
Чтобы продемонстрировать присутствие антител против CLDN6 в сыворотке крови иммунизированных мышей или связывание моноклональных антител с живыми клетками, в которых экспрессируется CLDN6, можно применять метод иммунофлуоресцентной микроскопии. Например, образцы линий клеток, в которых CLDN6 экспрессируется спонтанно или после трансфекции, и отрицательный контроль (клетки без экспрессии CLDH6) выращивают в слайд-камерах при стандартных условиях культивирования в среде DMEM/F12, содержащей 10% эмбриональной телячьей сыворотки (FCS), 2 мМ L-глутамина, 100 МЕ/мл пенициллина и 100 мкг/мл стрептомицицна. Затем клетки фиксируют метиловым спиртом или параформальдегидом либо ничем не обрабатывают. Затем в течение 30 мин клетки взаимодействуют с моноклональными антителами против CLDN6 при 25°С. После промывания проводят связывание клеток с вторичными антителами против мышиного IgG, меченными Alexa555 (Molecular Probes) в тех же условиях. Затем клетки исследуют методом флуоресцентной микроскопии.
Суммарный уровень CLDN6 в клетках можно определить, когда клетки фиксированы метиловым спиртом или параформальдегидом и обработаны детергентом Triton Х-100 для увеличения проницаемости. В живых клетках и в обработанных параформальдегидом, но с нормальной проницаемостью можно изучить локализацию CLDN6. Кроме того, можно изучить связывание CLDN6 в участках плотного контакта путем окрашивания маркерами плотных контактов, например ZO-1. Также можно изучить эффекты связывания антител и локализацию CLDN6 в клеточной мембране.
Вестерн-блоттинг
Взаимодействие иммуноглобулинов G против CLDN6 с антигеном CLDN6 можно также проверить с помощью метода вестерн-блоттинга. Вкратце делают следующее. Получают экстракты клеток, в которых экспрессируется CLDN6, и соответствующие отрицательные контроли и подвергают их электрофорезу в полиакриламидном геле с додецилсульфатом натрия (SDS) Разделенные в результате электрофореза антигены переносят на нитроцеллюлозные мембраны, блокируют и детектируют моноклональными антителами, подлежащими проверке. Связывание IgG можно выявить при помощи антимышиных IgG и пероксидазы, окрашивая субстратом для усиления хемилюминисценции (ECL).
Иммуноцитохимический анализ
Взаимодействие иммуноглобулинов G против CLDN6 с антигеном CLDN6 можно также проверить иммуноцитохимическим методом, способы которого хорошо известны специалистам в данной области техники. Например, используют замороженные срезы ткани, фиксированной параформальдегидом или ацетоном, или заключенные в парафин срезы ткани, фиксированной параформальдегидом, образцы которой берут из опухоли или здоровой ткани пациента в ходе хирургического вмешательства или у мышей с ксенографтами опухолей, инокулированных клетками линий со спонтанной или посттрансфекционной экспрессией CLDN6. Для иммунологического окрашивания, выявляющего антитела, способные взаимодействовать с CLDN6, можно использовать конъюгаты пероксидазы хрена с козьими антимышиными или антикроличьими антителами (производства DAKO) по инструкциям производителя
Активность антител in vitro: фагоцитоз и гибель клеток-мишеней
Помимо специфичного связывания с CLDN6, у антител против этого белка проверяют способность опосредовать фагоцитоз и гибель клеток, в которых экспрессируется CLDN6 и которые отличаются тем, что CLDN6 связан с их поверхностью. Результаты проверки активности моноклональных антител in vitro служат для первоначального скрининга, предшествующего проверке на моделях in vivo.
Антителозависимая клеточно-опосредованная цитотоксичность (ADCC):
Вкратце, проделывают следующее, полиморфноядерные клетки (PMN), естественные киллеры (NK-клетки), моноциты, мононуклеарные клетки или иные эффекторные клетки очищают путем центрифугирования в градиенте плотности в среде Ficoll Hypaque, после чего осуществляют лизис эритроцитов. Промытые эффекторные клетки суспендируют в среде RPMI, содержащей 10% инактивированной нагреванием эмбриональной телячьей сыворотки или же 5% инактивированной нагреванием человеческой сыворотки и смешивают с меченными 51Cr клетками-мишенями, в которых экспрессируется CLDN6 и которые отличаются тем, что CLDN6 связан с их поверхностью, при различных количественных соотношениях эффекторных клеток и клеток-мишеней. Или же клетки-мишени метят лигандом, обеспечивающим усиление флуоресценции (BATDA). С помощью флуориметра измеряют флуоресценцию комплекса хелата европия с усиливающим лигандом. Или же можно применить трансфекцию клеток-мишеней люциферазой. Добавленный краситель люциферовый желтый будет окисляться только жизнеспособными клетками. Затем добавляют очищенные иммуноглобулины G против CLDN6 в различных концентрациях. В качестве отрицательного контроля можно использовать другие человеческие IgG. Инкубация (при температуре 37°С) занимает от 4 до 20 часов в зависимости от типа взятых эффекторных клеток. Цитолиз в образцах определяют, измеряя высвобождение 51Cr в присутствии хелата EuTDA в культуральном супернатанте. Или же мерой количества жизнеспособных клеток может служить люминесценция в результате окисления люциферового желтого.
Проверяют также, усиливают ли моноклональные антитела против CLDN6 цитолиз в различных комбинациях с другими антителами.
Комплемент-зависимая цитотоксичностъ Моноклональные антитела против CLDN6 проверяют на способность опосредовать комплемент-зависимую цитотоксичность, для чего применяют разнообразные известные в данной области техники методы. Например, известными специалистам в данной области техники способами получают сыворотку крови, содержащую комплемент. Для определения активности моноклональных антител в отношении комплемент-зависимой цитотоксичности можно использовать разные методы. Например, можно измерять высвобождение 5lCr или оценивать изменение проницаемости мембраны с использованием пропидий иодида (PI). Вкратце проделывают следующее. Клетки-мишени промывают и 5×105/мл инкубируют с различными концентрациями моноклональных антител в течение 10-30 мин при комнатной температуре или при 37°С. Добавляют сыворотку или плазму крови до конечной концентрации 20% (объем/объем) и клетки инкубируют при температуре 37°С в течение 20-30 мин. Из каждого образца все клетки помещают в раствор пропидий иодида в пробирки для цитофлуориметра FACS и сразу же проводят анализ методом проточной цитометрии, используя FACS-планшет.
Или же индукцию комплемент-зависимой цитотоксичности можно определять на адгезивных клетках. В одном из вариантов осуществления этого подхода за 24 часа до опыта клетки высевают на плоскодонные планшеты для микротитрования с плотностью 3×104/на лунку. На следующий день удаляют культуральную среду и клетки инкубируют с антителами (в трех параллельных опытах). Контрольные клетки инкубируют с культуральной средой или с культуральной средой, содержащей 0,2% сапонина для определения фонового лизиса и максимального лизиса соответственно. Через 20 мин инкубации при комнатной температуре удаляют супернатант, а к клеткам прибавляют 20% (объем/объем) человеческой сыворотки или плазмы крови в среде DMEM (подогретой до 37°С) и инкубируют еще 20 мин при температуре 37°С. Все клетки из каждого образца помещают в раствор пропидия йодида (10 мкг/мл). Затем супернатанты заменяют на PBS, содержащий 2,5 мкг/мл этидия бромида и измеряют испускание флуоресценции при 600 нм (длина волны возбуждении 520 нм) с помощью микропланшетного ридера Tecan Safire. Долю (в процентах) специфичного лизиса рассчитывают следующим образом % специфичного лизиса = (флуоресценция образца - фоновая флуоресценция)/(флуоресценция при максимальном лизисе - фоновая флуоресценция) × 100.
Ингибирование пролиферации клеток моноклоналъными антителами
Чтобы проверить способность моноклональных антител против CLDN6 вызывать апоптоз, их можно, например, инкубировать с опухолевыми клетками, несущими CLDN6, или с опухолевыми клетками, трансфицированными CLDN6. Инкубацию проводят при температуре 37°С в течение 20 часов. Затем клетки собирают, промывают буферным раствором для связывания аннексина V (производство BD biosciences) и инкубируют с аннексином V, конъюгированным с флуоресцеинизотиоцианатом (FITC) или аллофикоцианином (АРС) (BD biosciences), в течение 15 мин в темноте. Из каждого образца все клетки помещают в раствор йодистого пропидия (PI) (10 мкг/мл в PBS) в пробирки для цитофлуориметра FACS и сразу же проводят проточную цитометрию (как описано выше). Можно также определять общее ингибирование пролиферации клеток моноклональными антителами при помощи готовых имеющихся в продаже наборов. Так, набор DELFIA Cell Proliferation Kit (Perkin-Elmer, Cat. No. AD0200) позволяет осуществить иммунологический анализ без использования радиоактивных изотопов на основе количественного определения включения 5-бром-2'-дезоксиуридина (BrdU) в синтезируемую ДНК пролиферирующих клеток на микропланшетах. Включение BrdU выявляют с помощью меченных европием моноклональных антител. Для выявления антител клетки фиксируют и денатурируют ДНК, используя фиксирующий раствор. Не связавшиеся антитела отмывают и прибавляют индуктор DELFIA для диссоциации ионов европия, которые переходят из меченых антител в раствор, где образуют интенсивно флуоресцирующие хелатные комплексы с компонентами индуктора DELFIA. Получаемая в результате измерения (с временным разрешением) величина, характеризующая флуоресценцию, в каждой ячейке пропорциональна интенсивности синтеза ДНК в клетках.
Доклинические исследования
Моноклональные антитела, связывающиеся с CLDN6, также можно проверять на моделях in vivo (например, у мышей с иммунодефицитом, имеющих опухоли-ксенографты, в которые инокулированы клетки с экспрессией CLDN6, возможно, после трансфекции), с целью определить их эффективность для регуляции пролиферации опухолевых клеток, в которых экспрессируется CLDN6.
В исследованиях in vivo с использованием мышей с ослабленной иммунной системой и ксенографтами опухолевых клеток, в которых экспрессируется CLDN6, можно применять антитела по данному изобретению. Эти антитела вводят мышам без опухолей, а затем этим животным вводят опухолевые клетки и оценивают производимый антителами эффект предотвращения образования опухолей или связанных с опухолями симптомов. Также изучаемые антитела вводят мышам с опухолями, чтобы оценить их лечебный эффект, т.е. ослабление опухолевого роста, метастазирования или связанных с опухолями симптомов. Применение антител по данному изобретению можно сочетать с применением других веществ, используемых в качестве цитостатических препаратов, ингибиторов факторов роста, агентов, блокирующих клеточный цикл или ангиогенез, или с другими антителами, чтобы оценить синергический эффект и потенциальную токсичность таких комбинаций. Для изучения токсических побочных эффектов, опосредуемых антителами по данному изобретению, животным вводят соответствующие антитела или регуляторные агенты и детально оценивают симптомы, которые могут быть связаны с лечебным влиянием антител против CLDN6. Возможные побочные эффекты применения антител против in vivo включают, в частности, токсичность для тканей, в которых экспрессируется CLDN6, включая плаценту. Антитела, распознающие CLDN6 у человека и других видов, например мыши, особенно полезны для предсказания возможных побочных эффектов, опосредуемых применением моноклональных антител против CLDN6 у человека.
Картирование эпитопов
Картирование эпитопов, узнаваемых антителами по данному изобретению, может быть проделано, как подробно описано в работах Glenn E. Morris "Epitope Mapping Protocols (Methods in Molecular Biology), ISBN-089603-375-9 и Olwyn M. R. Westwood, Frank C. Hay "Epitope Mapping: A Practical Approach" Practical Approach Series, 248.
I. Биспецифичные/полиспецифичные вещества, связывающиеся с CLDN6
В другом воплощении настоящего изобретения антитела против CLDN6 модифицируют или присоединяют к иным функциональным молекулам, например пептидам или белкам (например, фрагментам Fab'), чтобы создать биспецифичные или полиспецифичные молекулы, связывающиеся со многими участками связывания или эпитопами мишени. Например, антитело по данному изобретению может быть функционально соединено (например, путем химического сопряжения, объединения генетического материала, нековалентной связи или как-то иначе) с одним или более других связывающих молекул, например другим антителом, пептидом или миметиком по связыванию.
Соответственно, настоящее изобретение включает биспецифичные и полиспецифичные вещества, содержащие по меньшей мере одну первую специфичность связывания для CLDN6 и вторую специфичность связывания для второго эпитопа мишени. В одном конкретном воплощении данного изобретения второй эпитоп мишени является рецептором Fc, например человеческими рецепторами Fc-γRI (CD64) или Fc-a (CD89), или рецептором Т-клеток, например CD3. Таким образом, данное изобретение включает биспецифичные и полиспецифичные вещества, способные связываться как с эффекторными клетками (например, моноцитами, макрофагами или полиморфноядерными клетками), в которых экспрессируется Fc-γR, Fc-αR или Fc-εR, так и с клетками-мишенями, в которых экспрессируется CLDN6 и которые отличаются том, что CLDN6 связан с их поверхностью. Эти биспецифичные и полиспецифичные вещества делают клетки, в которых экспрессируется CLDN6 и которые отличаются том, что CLDN6 связан с их поверхностью, мишенью для эффекторных клеток и инициируют активность эффекторных клеток, опосредованную рецепторами Fc, например фагоцитоз клеток, в которых экспрессируется CLDN6 и которые отличаются том, что CLDN6 связан с их поверхностью, антителозависимую клеточно-опосредованную цитотоксичность, высвобождение цитокинов или образование супероксид-аниона.
Биспецифичные и полиспецифичные агенты по данному изобретению могут также обладать третьей специфичностью связывания, в дополнение к специфичности связывания Fc и специфичности связывания CLDN6. В одном из воплощений данного изобретения третья специфичность связывания относится части направленной на фактор усиления (EF), например веществу, которое связывается с белком клеточной поверхности, участвующим в цитотоксической активности и таким образом усиливает иммунный ответ, направленный на клетки-мишени. Часть молекулы полиспецифичного агента, связывающаяся с фактором усиления, может быть антителом, функциональным фрагментом антитела или лигандом, связывающимся с данным веществом, например с антигеном или с рецептором, и таким образом вызывающая усиление эффекта детерминант связывания для рецептора Fc или антигена клетки-мишени. Часть молекулы полиспецифичного агента, связывающаяся с фактором усиления, может связываться с рецептором Fc или антигеном клетки-мишени. Или же часть молекулы полиспецифичного агента, связывающаяся с фактором усиления, может связываться со структурой, отличной от тех двух, к которым относятся первая и вторая специфичности данного агента. Например, часть молекулы полиспецифичного агента, связывающаяся с фактором усиления, может связываться с цитотоксической Т-клеткой (например, через CD2, CD3, CD8, CD28, CD4, CD40, ICAM-1 или с другой клеткой иммунной системы, что ведет к усилению иммунного ответа, направленного против клетки-мишени).
В одном из воплощений данного изобретения биспецифичные и полиспецифичные агенты содержат по меньшей мере одно антитело, обеспечивающее специфичность связывания, включая, например. Fab, Fab', F(ab')2, Fv или одноцепочечный Fv. Это антитело также может быть димером легкой или тяжелой цепи или любым их минимальным фрагментом, например Fv или одноцепочечной конструкцией, как описано в работе Ladner et al., патент США 4,946,778. Это антитело может также быть связывающим доменом гибридного иммуноглобулина, как описано в заявках на патент США 2003/0118592 и 2003/0133939.
В одном из воплощений данного изобретения биспецифичные и полиспецифичные агенты обладают специфичностью связывания с Fc-γR или Fc-αR, присутствующих на поверхности эффекторных клеток, и второй специфичностью связывания с антигеном клетки-мишени, например CLDN6.
В одном из воплощений данного изобретения специфичность связывания с рецептором Fc обеспечивается моноклональным антителом связывание которого не блокируется человеческим иммуноглобулином G (IgG). В настоящем документе термин «рецептор IgG» относится к любому из восьми генов гамма-цепей, находящихся в хромосоме 1. Эти гены кодируют в совокупности двенадцать трансмембранных или растворимых изоформ рецептора, которые группируются в три класса рецепторов Fc-γ: Fc-γRI (CD64), Fc-γRII (CD32) и Fc-γRIII (CD16). В одном из предпочтительных воплощений данного изобретения рецептор Fc-γ является человеческим высокоаффинным Fc-γRI.
В другом предпочтительном воплощении данного изобретения специфичность связывания с рецептором Fc обеспечивается антителом, которое связывается с человеческим рецептором IgA, например с рецептором Fc-α (Fc-αRI (CD89)), причем это связывание предпочтительно не блокируется человеческим иммуноглобулином A (IgA). Термин «рецептор IgA» подразумевает продукт гена одного α-гена (Fc-αRI), расположенного в хромосоме 19. Известно, что этот ген кодирует несколько образующихся в результате альтернативного сплайсинга изоформ с молекулярной массой от 55 до 110 кДа. Fc-αRI (CD89) постоянно экспрессируется на моноцитах/макрофагах, гранулоцитах (эозинофилах и нейтрофилах), но не на эффекторных клетках. Fc-αRI обладает умеренным сродством к IgA1 и IgA2, которое увеличивается под действием цитокинов, например гранулоцитарного колониестимулирующего фактора (G-CSF) или гранулоцитарно-макрофагального колониестимулирующего фактора (GM-CSF) (Morton, Н.С. et al. (1996) Critical Reviews in Immunology 16:423-440). Идентифицировано четыре моноклональных антитела, специфичных к Fc-αRI (обозначаются A3, А59, А62 и А77), которые связываются с Fc-αRI вне домена, обеспечивающего связывание с IgA (Monteiro, R.С. et al. (1992) J. Immunol. 148:1764).
В другом воплощении настоящего изобретения биспецифичные агенты содержат два моноклональных антитела по данному изобретению, обладающих комплементарными активностями, например одно антитело действует преимущественно путем индуцирования зависимой от комплемента цитотоксичности, а другое преимущественно вызывает апоптоз.
Термин «антитела, специфичные к эффекторным клеткам» в настоящем документе относится к антителам или функциональным фрагментам антител, которые связываются с рецептором Fc эффекторных клеток. Предпочтительно по данному изобретению используются антитела связываются с рецептором Fc эффекторных клеток в участке, который не связывается с эндогенными иммуноглобулинами.
В настоящем документе термин «эффекторная клетка» употребляется применительно к клеткам иммунной системы, участвующим в эффекторной фазе иммунного ответа, - в отличие от стадии распознавания антигена и стадии активации эффекторных элементов. Примеры клеток иммунной системы, участвующих в эффекторной фазе иммунного ответа, включают клетки миелоидного и лимфоидного происхождения, например лимфоциты (например, В-клетки и Т-клетки, включая цитолитические Т-клетки (CTL)), киллерные клетки, естественные киллерные клетки (NK-клетки), макрофаги, моноциты, эозинофилы, нейтрофилы, полиморфноядерные клетки, гранулоциты, тучные клетки и базофилы. В некоторых эффекторных клетках экспрессируются специфические рецепторы Fc и эти клетки выполняют специфические функции. В предпочтительных воплощениях данного изобретения эффекторные клетки способны вызывать антителозависимую клеточно-опосредованную цитотоксичность (ADCC), например нейтрофилы могут вызывать ADCC. К примеру, моноциты и макрофаги, в которых экспрессируется FcR, участвуют в специфичном уничтожении клеток-мишеней и в представлении антигена другим компонентам иммунной системы или в связывании клеток, представляющих антигены. В других воплощениях данного изобретения эффекторные клетки могут поглощать путем фагоцитоза антиген-мишень, клетку-мишень или микроорганизм. Экспрессия конкретных FcR на эффекторных клетках может регулироваться гуморальными факторами, например цитокинами. Так, обнаружено, что экспрессия Fc-γRI регулируется, а именно усиливается, γ-интерфероном. В результате этого усиления экспрессии увеличивается направленная против клеток-мишеней цитотоксическая активность клеток, несущих Fc-γRI. Эффекторные клетки могут осуществлять фагоцитоз или лизис антигена-мишени или клеток-мишеней.
Термин «клетка-мишень» в настоящем документе означает любую нежелательную клетку у индивида (например, человека или животного), которая может быть мишенью для антител по данному изобретению. В предпочтительных воплощениях данного изобретения клетками-мишенями являются клетки, в которых экспрессируется или чрезмерно экспрессируется CLDN6 и которые отличаются тем, что CLDN6 связан с их поверхностью. Клетки, в которых экспрессируется CLDN6 и которые отличаются тем, что CLDN6 связан с их поверхностью, как правило, включают опухолевые клетки.
II. Иммуноконъюгаты
В другом своем аспекте данное изобретение предлагает антитела против CLDN6, конъюгированные с играющей терапевтическую роль структурой или агентом, например, цитотоксином, лекарственным веществом (например, иммуносупрессором) или радиоактивным изотопом. Такие конъюгаты в настоящем документе называются иммуноконъюгатами. Иммуноконъюгаты, включающие один или более цитотоксинов, называются здесь иммунотоксинами. Цитотоксины или цитотоксические агенты включают любые агенты, губительные для живых клеток, в частности убивающие их. Примеры таких веществ включают таксол, цитохалазин В, грамицидин D, этидия бромид, эметин, митомицин, этопозид, тенопозид, винкристин, винбластин, колхицин, доксорубицин, даунорубицин, дигидроксиантрациндион, митоксантрон, митрамицин, актиномицин D, 1-дегидротестостерон, глюкокортикоиды, прокаин, тетракаин, лидокаин, пропранолол, пуромицин и их аналоги или гомологичные им вещества.
Терапевтические агенты, подходящие для образования иммуноконъюгатов по данному изобретению, включают (не ограничиваясь приведенными здесь примерами) антиметаболиты (например, метотрексат, 6-меркаптопурин, 6-тиогуанин, цитарабин, флударабин, 5-фторурацил, декарбазин), алкилирующие агенты (например, меклоретамин, тиотепа, хлорамбуцил, мелфалан, кармустин (BSNU) и ломустин (CCNU), циклофосфамид, бусульфан, дибромманнит, стрептозотоцин, митомицин С и цис-дихлордиаминплатина (II) (DDP) цисплатин), антрациклины (например, даунорубицин, ранее известный под названием дауномицина, и доксорубицин), антибиотики (например, дактиномицин, ранее известный под название актиномицина, блеомицин, митрамицин и антрамицин) и агенты, подавляющие митоз (например, винкристин и винбластин). В одном из предпочтительных воплощений данного изобретения терапевтическим агентом является цитотоксический или радиотоксический агент. В другом воплощении данного изобретения терапевтическим агентом является иммуносупрессор. В еще одном воплощении данного изобретения терапевтическим агентом является гранулоцитарно-макрофагальный колониестимулирующий фактор (GM-CSF). В одном из предпочтительных воплощений данного изобретения терапевтическим агентом является доксорубицин, цисплатин, блеомицин, сульфат, кармустин, хлорамбуцил, циклофосфамид или рицин А.
Антитела по данному изобретению можно также конъюгировать с радиоактивными изотопами, например, иодом-131, иттрием-90 или индием-111, для создания цитотоксических радиоактивных лекарств для лечения расстройств, связанных с CLDN6, например рака. Конъюгаты антител по данному изобретению можно использовать для модификации той или иной биологической реакции, и обладающую лечебным эффектом структуру не следует считать ограниченной классическими химиотерапевтическими агентами. Например, этой структурой может быть белок или полипептид, обладающий желаемой биологической активностью. Такие белки могут включать, например, ферментативно активные токсины или их активные фрагменты, например абрин, рицин А, экзотоксин Pseudomonas или дифтерийный токсин; белок, например фактор некроза опухолей или γ-интерферон; или модификаторы биологических реакций, например лимфокины, интерлейкин-1, интерлейкин-2, интерлейкин-6, гранулоцитарно-макрофагальный колониестимулирующий фактор, гранулоцитарный колониестимулирующий фактор или другие факторы роста.
Методики конъюгирования подобных структур, обладающих терапевтическим эффектом, с антителами хорошо известны в данной области техники; см., например, Arnon et al., "Monoclonal Antibodies For Immunotargeting Of Drugs In Cancer Therapy", in Monoclonal Antibodies And Cancer Therapy, Reisfeld et al. (eds.), pp.243-56 (Alan R. Liss, Inc. 1985); Hellstrom et al., "Antibodies For Drug Delivery", in Controlled Drug Delivery (2nd Ed.), Robinson et al. (eds.), pp.623-53 (Marcel Dekker, Inc. 1987); Thorpe, "Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review", in Monoclonal Antibodies '84: Biological And Clinical Applications, Pincheraet al. (eds.), pp.475-506 (1985); "Analysis, Results, And Future Prospective Of The Therapeutic Use Of Radiolabeled Antibody In Cancer Therapy", in Monoclonal Antibodies For Cancer Detection And Therapy, Baldwin et al. (eds.), pp.303-16 (Academic Press 1985), Thorpe et al., "The Preparation And Cytotoxic Properties Of Antibody-Toxin Conjugates", Immunol. Rev., 62:119-58 (1982).
В еще одном воплощении данного изобретения антитела присоединяются к хелатообразующей линкерной структуре (линкеру-хелатору), например тиуксетану, которая позволяет присоединить к антителу радиоактивный изотоп.
III. Фармацевтические композиции
В другом аспекте настоящего изобретения предлагаются композиции, например фармацевтические композиции, содержащие один вариант антител или комбинацию антител по данному изобретению. В состав этих фармацевтических композиций могут входить фармацевтически приемлемые носители и разбавители, а также любые другие известные дополнительные компоненты и эксипиенты в соответствии с общепринятыми методиками, описанными в работе Remington: The Science and Practice of Pharmacy, 19th Edition, Gennaro, Ed., Mack Publishing Co., Easton, PA, 1995. В одном из воплощений данного изобретения предлагаемые композиции включают комбинацию множества (например, двух или более) изолированных антител по данному изобретению, действующих различными путями, например одно антитело, которое преимущественно вызывает зависимую от комплемента цитотоксичность, сочетают с другим антителом, которое преимущественно вызывает апоптоз.
Фармацевтические композиции по данному изобретению можно использовать как часть комбинированной терапии, то есть сочетать с другими лечебными агентами. Например, такая комбинированная терапия может включать композицию по данному изобретению вместе с по меньшей мере одним противовоспалительным агентом или с по меньшей мере одним иммуносупрессивным агентом. В одном из воплощений данного изобретения такие терапевтические агенты включают один или более противовоспалительных агентов, например стероидные препараты или нестероидные противовоспалительные средства. Предпочтительные агенты включают, например аспирин и другие салицилаты, ингибиторы циклоокисгеназы-2 (Сох-2), например рофекоксиб (Vioxx), целекоксиб (Celebrex), нестероидные противовоспалительные средства, например ибупрофен (Motrin, Advil), фенопрофен (Nalfon), напроксен (Naprosyn), сулиндак (Clinoril), диклофенак (Voltaren), пироксикам (Feldene), кетопрофен (Orudis), дифлунизал (Dolobid), набуметон (Relafen), этодолак (Lodine), оксапрозин (Daypro) и индометацин (Indocin).
В другом воплощении данного изобретения такие терапевтические агенты включают агенты, вызывающие истощение или функциональную инактивацию регуляторных Т-клеток; это циклофосфамид в малых дозах, антитела против CTLA4, против интерлейкина-2 или против рецептора интерлейкина-2.
В еще одном воплощении настоящего изобретения такие терапевтические агенты включают химиотерапевтические препараты, например производные таксола, таксотер, гемцитабин, 5-фторурацил, доксорубицин (Adriamycin), цисплатин (Platinol), циклофосфамид (Cytoxan, Procytox, Neosar). В другом воплощении данного изобретения предлагаемые антитела можно вводить индивиду в сочетании с химиотерапевтическими агентами, которые предпочтительно проявляют терапевтическую эффективность у больных раком, например раковыми заболеваниями, описываемыми в настоящем документе.
В еще одном воплощении настоящего изобретения предлагаемые антитела можно вводить индивиду параллельно лучевой терапии и/или аутологичной трансплантации стволовых клеток периферической крови или костного мозга.
В настоящем документе термин «фармацевтически приемлемый носитель» включает любой растворитель, дисперсионную среду, оболочку, антибактериальные или противогрибковые агенты, агенты, обеспечивающие нужное осмотическое давление и агенты, задерживающие абсорбцию, а также иные агенты, являющиеся физиологически совместимыми. Предпочтительно носитель по данному изобретению подходит для внутривенного, внутримышечного, подкожного, парентерального, спинального или эпидермального введения (например, путем инъекции или инфузии). В зависимости от пути введения активное вещество, например антитела, биспецифичные и полиспецифичные соединения, может быть покрыто материалом для защиты активного вещества от воздействия кислот и других естественных факторов, которые могут инактивировать активное вещество.
Термин «фармацевтически приемлемая соль» в настоящем документе относится к солям, которые сохраняют биологическую активность исходного соединения и не привносят каких-либо нежелательных токсических эффектов (см., например, Berge, S.М., et al. (1977) J. Pharm. Sci. 66:1-19).
Примеры таких солей включают кислотные аддукты солей и щелочные аддукты солей. Кислотные аддукты солей включают производные нетоксичных неорганических кислот, например соляной, азотной, фосфорной, серной, бромистоводородной, йодистоводородной, фосфористой кислот и проч., а также производные нетоксичных органических кислот, например алифатических моно- и дикарбоновых кислот, фенил-замещенных алкановых кислот, гидроксиалкановых кислот, ароматических кислот, алифатических и ароматических сульфоновых кислот и проч. Щелочные аддукты солей включают производные щелочных и щелочноземельных металлов, например натрия, калия, магния, кальция и проч., а также производные нетоксичных органических аминов, например N,N'-дибензилэтилендиамина, N-метилглюкамина, хлорпрокаина, холина, диэтаноламина, этилендиамина, прокаина и проч.
Композиции по настоящему изобретению можно вводить индивиду различными методами, известными в данной области техники, специалистам в которой понятно, что путь и/или способ введения будут варьировать в зависимости от желаемых результатов. Активное вещество может быть объединено с носителем, который предотвратит его быстрое высвобождение, например активное вещество может входить в состав лекарственной формы с контролируемым высвобождением, включая имплантаты, пластыри для трансдермального введения и системы на основе микрокапсул, в которые заключается активное вещество. Здесь могут использоваться подлежащие биологической деградации и биологически совместимые полимеры, например этиленвинилацета, полиангидриды, полигликолевая кислота, коллаген, полиортоэфиры и полимолочная кислота. Методы изготовления таких препаратов общеизвестны в данной области техники; см., например, Sustained and Controlled Release Drug Delivery Systems, J. R. Robinson, ed., Marcel Dekker, Inc., New York, 1978.
Для применения некоторых способов введения активного вещества в организм может потребоваться покрыть это вещество материалом, предотвращающим его инактивацию, или вводить вместе с таким материалом. Например, активное вещество может вводиться индивиду в подходящем носителе, например в липосомах, или в разбавителе. Фармацевтически приемлемые разбавители включают физиологический раствор и водные буферные растворы. Липосомы включают эмульсии типа «вода в масле в воде» (CGF Inc.), а также обычные липосомы (Strejan et al. (1984) J. Neuroimmunol. 7:27).
Фармацевтически приемлемые носители включают стерильные водные растворы или дисперсии и стерильные порошки для приготовления инъецируемого раствора или дисперсии непосредственно перед введением. Применение таких сред и агентов для фармацевтически активных веществ известно в данной области техники. За исключением случаев несовместимости обычных сред или агентов с активным веществом по данному изобретению, любые среды и агенты могут использоваться в фармацевтических композициях по данному изобретению. В эти композиции могут входить также дополнительные активные вещества.
Как правило, терапевтические композиции должны быть стерильными и стабильными в условиях изготовления и хранения. Композиции по данному изобретению могут быть представлены раствором, микроэмульсией, липосомами или другими упорядоченными структурами, подходящими для высокой концентрации лекарственного препарата. Носителем может быть растворитель или дисперсионная среда, содержащие, например, воду, этиловый спирт, многоатомный спирт (Например, глицерин, пропиленгликоль, жидкий полиэтиленгликоль и т.п.) и их подходящие смеси. Нужная текучесть поддерживается, например, при помощи кроющего материала, например лецитина, путем достижения требующегося размера частиц (в случае дисперсий) и при помощи поверхностно-активных агентов. Во многих случаях предпочтительно включать в состав композиции агенты, обеспечивающие нужное осмотическое давление, например сахара, многоатомные спирты (например, манит, сорбит) или хлорид натрия. Продление абсорбции композиции, вводимой путем инъекции, достигается включением в ее состав агента, задерживающего абсорбцию, например однозамещенных солей стеариновой кислоты и желатина.
Стерильные растворы для инъекций можно готовить путем объединения требующегося количества активного вещества в подходящем растворителе с одним из упомянутых выше ингредиентов или с их комбинацией и последующей стерилизацией микрофильтрацией.
Как правило, дисперсии получают путем включения активного вещества в стерильный носитель, содержащий основную дисперсионную среду и другие необходимые ингредиенты из числа упомянутых выше. В случае стерильных порошков для приготовления стерильных растворов для инъекций, то предпочтительными методами их получения являются высушивание в вакууме и высушивание замораживанием (лиофилизация), которые позволяют получить в виде порошка активное вещество вместе с любым дополнительным желаемым ингредиентом из их раствора, предварительно стерилизованного фильтрованием.
Дозировка и режим введения подбираются так, чтобы обеспечить оптимальную желаемую реакцию (например, лечебный эффект). Например, можно вводить некоторую дозу однократно или же в несколько приемов на протяжении некоторого промежутка времени или доза может быть пропорционально уменьшена или увеличена в зависимости от терапевтических показаний в конкретной ситуации. Особенно предпочтительно составлять композицию для парентерального введения как единичную лекарственную форму - это облегчает введение и обеспечивает единообразие дозировки. В настоящем документе термин единичная лекарственная форма означает физически отдельные единицы, удобные для разового введения нуждающемуся в том индивиду; каждая единица содержит заранее определенное количество активного вещества, рассчитанное таким образом, чтобы достичь желаемого терапевтического эффекта, вместе с нужным фармацевтическим носителем.
Примеры фармацевтически приемлемых антиоксидантов включают: (1) водорастворимые вещества, например аскорбиновая кислота, цистеина гидрохлорид, бисульфат натрия, метабисульфит натрия, сульфит натрия и проч.; (2) жирорастворимые вещества, например аскорбилпальмитат, гидроксианизола бутилат (ВНА), гидрокситолуола бутилат (ВНТ), лецитин, пропилгаллат, α-токоферол и проч.; (3) агенты, образующие комплексы с металлами (хелатирующие), например, лимонная кислота, этилендиаминтетраацетат (EDTA), сорбит, винная кислота, фосфорная кислота и проч.
Составы терапевтических композиций по данному изобретению включают предназначенные для перорального, назального, местного (включая буккальное и подъязычное), ректального, вагинального и/или парентерального введения. Эти препараты могут быть для удобства представлены как единичная лекарственная форма и могут быть изготовлены любым методом, известным в области фармацевтики. Количество активного ингредиента, объединяемое с материалом носителя для получения единичной лекарственной формы варьирует в зависимости от индивидуальных потребностей пациента и конкретного способа введения препарата. Как правило, количество активного ингредиента, объединяемое с материалом носителя для получения единичной лекарственной формы, должно быть таким, чтобы вызвать терапевтический эффект.
Препараты по данному изобретению, пригодные для вагинального введения, включают также пессарии, тампоны, кремы, гели, пасты, пенящиеся или аэрозольные составы, содержащие подходящие носители, известные в данной области техники. Лекарственные формы для местного и трансдермального введения композиций по данному изобретению включают порошки, спреи, мази, пасты, кремы, лосьоны, гели, растворы, пластыри и вдыхаемые препараты. Активное вещество можно смешивать в условиях стерильности с фармацевтически приемлемым носителем и при необходимости с любыми консервантами, забуферивающими агентами или пропеллентами.
Термин «парентеральное введение» и выражение «вводится парентерально» в настоящем документе означают способы введения препарата в организм помимо энтерального и местного путей; обычно они подразумевают инъекции и включают (не ограничиваясь перечисленным здесь) внутривенные, внутримышечные, интраартериальные, интратекальные, интракапсулярные, интраорбитальные, интракардиальные, интрадермальные, интраперитонеальные, транстрахеальные, подкожные, субкутикулярные, внутрисуставные, субкапсулярные, субаракноидальные, интраспинальные, эпидуральные и интрастернальные инъекции и инфузии.
Примеры водных и неводных носителей, пригодных для использования в фармацевтических композициях по данному изобретению, включают воду, этиловый спирт, многоатомные спирты (например, глицерин, пропиленгликоль, полиэтиленгликоль и т.п.) и подходящие их смеси, растительные масла (например, оливковое масло), пригодные для инъекций органические эфиры (например, этилолеат). Должная текучесть поддерживается, например, при помощи кроющих материалов, например лецитина, путем достижения требующегося размера частиц (в случае дисперсий) и при помощи поверхностно-активных агентов.
Композиции по данному изобретению могут также содержать дополнительные компоненты, например консерванты, увлажняющие агенты, эмульгирующие и диспергирующие агенты. Предотвратить присутствие микроорганизмов можно как путем стерилизации тем или иным способом, так и с помощью включения в состав композиции антибактериальных и противогрибковых агентов, например парабенов, хлорбутанола, фенолсорбиновой кислоты и т.п. Может быть также желательно включить в состав композиции агенты, обеспечивающие нужное осмотическое давление, например сахара, хлорид натрия и проч. Кроме того, продление абсорбции фармацевтической формы, вводимой путем инъекции, достигается включением в ее состав агентов, задерживающих абсорбцию, например однозамещенного стеарата алюминия и желатина.
Независимо от выбранного пути введения соединения по данному изобретению, которые можно использовать в подходящей гидратированной форме, и/или фармацевтические композиции по данному изобретению, включаются в состав фармацевтически приемлемых лекарственных форм при помощи методов, известных специалистам в данной области техники.
Конкретное содержание активных ингредиентов в фармацевтических композициях по данному изобретению и дозировка препарата могут варьировать таким образом, чтобы активный ингредиент присутствовал в эффективном количестве, вызывающем желаемый терапевтический эффект у конкретного пациента при конкретном составе препарата и при выбранном способе его введения, не оказывая в то же время токсического действия на данного индивида. Выбор концентрации активного ингредиента в препарате и дозировки препарата определяются рядом фармакокинетических факторов, включая активность конкретной применяемой композиции по данному изобретению, способ и режим введения препарата, скорость выведения данного соединения из организма, продолжительность курса лечения, другие лекарственные препараты, вещества и/или материалы, используемые в сочетании с данными композициями, возраст, пол, массу тела, общее состояние здоровья и конкретное состояние пациента, его историю болезни, а также другие факторы, хорошо известные в области медицины.
Врачам и ветеринарам, ориентирующимся в данной области техники, не составит труда определить и назначить эффективное в конкретном случае количество нужной фармацевтической композиции. Например, применение соединений по данному изобретению в составе фармацевтических композиций у человека или животного можно начать с доз ниже требующихся для достижения желаемого лечебного эффекта и постепенно увеличивать дозировку до тех пор, пока желаемый эффект не будет достигнут. Вообще говоря, подходящей суточной дозой композиции по данному изобретению будет такая, при которой количество активного вещества минимально для эффективного оказания лечебного действия. Такая эффективная доза, как правило, зависит от рассмотренных выше факторов. Предпочтительные пути введения - внутривенный, внутримышечный, внутрибрюшинный или подкожный, причем место введения предпочтительно проксимально по отношению к локализации мишени. При желании эффективную суточную дозу терапевтической композиции можно вводить, разделив ее на два, три, четыре, пять, шесть или более приемов, распределенных с соответствующими интервалами на протяжении суток, при необходимости используя единичные лекарственные формы. Хотя соединение по данному изобретению возможно вводить само по себе, предпочтительно вводить его в составе фармацевтического препарата (композиции).
В одном из воплощений данного изобретения предлагаемые антитела вводят пациенту путем инфузии, предпочтительно медленно непрерывно на протяжении продолжительного времени, например долее 24 часов, чтобы уменьшить токсические побочные эффекты. Введение путем непрерывной инфузии можно также осуществлять за период от 2 до 24 часов, например от 2 до 12 часов, повторяя, если нужно, еще один или несколько раз через, например, 6 или 12 месяцев. Дозировку можно определить или нужным образом изменить, измеряя количество моноклональных антител против CLDN6 в кровотоке после введения биологического образца с помощью антиидиотипических антител, мишенью которых являются антитела против CLDN6.
В другом воплощении данного изобретения предлагаемые антитела вводят в режиме поддерживающей терапии, например раз в неделю на протяжении 6 месяцев или дольше.
В еще одном воплощении данного изобретения предлагаемые антитела вводят по схеме, включающей одну инфузию антител против CLDN6, после которой проводят инфузию антител против CLDN6, конъюгированных с радиоактивным изотопом. Эту схему можно повторить, например 7-9 суток спустя.
В одном из воплощений данного изобретения вещества терапевтического назначения включают в липосомы. В более предпочтительном воплощении данного изобретения липосомы несут нацеливающую структуру. В наиболее предпочтительном воплощении вещества терапевтического назначения в липосомах вводят в ударной дозе, причем место введения проксимально по отношению к нужному участку тела, например к месту расположения опухоли. Эта композиция должна быть достаточно жидкой, чтобы ее легко было вводить шприцем. И она должна быть стабильной в условиях изготовления и хранения, защищенной от загрязнения микроорганизмами, например бактериями и грибами.
В еще одном воплощении данного изобретения предлагаемые антитела обработаны таким образом, чтобы предотвратить или сократить их перенос через плаценту. Это достигается известными в данной области техники способами, например путем ПЭГилирования антител или при помощи фрагментов F(ab)2'. Можно также сослаться на работы Cunningham-Rundles С, Zhuo Z, Griffith В, Keenan J. (1992) Biological activities of polyethylene-glycol immunoglobulin conjugates. Resistance to enzymatic degradation. J. Immunol. Methods, 152:177-190; "Landor M. (1995) Maternal-fetal transfer of immunoglobulins, Ann. Allergy Asthma Immunol. 74:279-283.
Терапевтически эффективную дозу при онкотерапии можно определить по объективной реакции опухоли, которая может быть полной или же частичной. Полная реакция (CR) - это отсутствие клинических, радиологических или каких-либо иных признаков болезни. Частичная реакция (PR) - это сокращение размеров опухолевого образования более чем на 50%. Продолжительность объективной реакции опухоли характеризуется средним временем до прогрессирования.
Терапевтически эффективную дозу препарата при онкотерапии можно также определить по его способности стабилизировать прогрессирование болезни. Способность вещества подавлять рак можно оценить с помощью животных моделей, по которым можно предсказать эффективность препарата в отношении опухолей человека. Или же это свойство композиции можно оценить, изучая способность данного вещества подавлять пролиферацию клеток или апоптоз методами in vitro, известными специалистам в данной области техники. Терапевтически эффективное количество вещества лечебного назначения может обеспечить сокращение размеров опухоли или как-либо иначе ослабить симптомы болезни у пациента. Рядовой специалист в данной области техники вполне сможет определить это количество на основании таких факторов, как масса тела пациента, выраженность симптомов, конкретный состав композиции и выбранный способ введения препарата.
Композиция по данному изобретению должна быть стерильной и достаточно жидкой, чтобы ее можно было вводить с помощью шприца. Помимо воды носитель может быть изотоническим буферным солевым раствором, этиловым спиртом, многоатомным спиртом (например, глицерином, пропиленгликолем, жидким полиэтиленгликолем и т.п.) и их подходящими смесями. Должная текучесть поддерживается, например, при помощи кроющих материалов, например лецитина, путем достижения требующегося размера частиц (в случае дисперсий) и при помощи поверхностно-активных агентов. Во многих случаях предпочтительно включать в состав композиции агенты, обеспечивающие нужное осмотическое давление, например сахара, многоатомные спирты (например, маннит или сорбит) и хлорид натрия. Длительная абсорбция композиции, вводимой путем инъекции, достигается включением в ее состав агентов, задерживающих абсорбцию, например однозамещенного стеарата алюминия или желатина.
Когда активное вещество должным образом защищено, как описано выше, его можно вводить перорально, например с инертным разбавителем или усваиваемым съедобным носителем.
IV. Применения и способы по данному изобретению
Антитела по данному изобретению (включая иммуноконъюгаты, биспецифичные/полиспецифичные агенты, композиции и другие производные, описанные в настоящем документе) находят множество применений в терапевтических целях, имеющих отношение к лечению расстройств, затрагивающих клетки, в которых экспрессируется CLDN6 и которые отличаются тем, что CLDN6 связан с их поверхностью. Например, антитела по данному изобретению можно вводить в культивируемые клетки, например in vitro или ex vivo, или пациентам (людям), например in vivo, для лечения или предотвращения различных расстройств, описанных в настоящем документе. Предпочтительные объекты такого воздействия включают пациентов (людей) с расстройствами, которые подлежат коррекции или при которых возможно облегчение состояния пациента путем уничтожения пораженных клеток, в частности клеток, отличающихся измененной по сравнению с нормальными клетками экспрессией CLDN6 и/или измененной связью CLDN6 с их поверхностью.
Например, в одном из воплощений настоящего изобретения предлагаемые антитела используются для лечения пациента с онкогенным расстройством, например с расстройством, при котором характерно присутствие клеток, в которых экспрессируется CLDN6 и которые отличаются тем, что CLDN6 связан с их поверхностью. Примерами онкогенных заболеваний, которые можно лечить и/или предотвращать, являются все раковые заболевания и опухоли с экспрессией CLDN6, включая описанные в настоящем документе.
Фармацевтические композиции и способы лечения по данному изобретению можно также использовать для иммунизации или вакцинации для предотвращения заболеваний, описанных в настоящем документе.
В другом воплощении данного изобретения предлагаемые антитела используются для определения уровня CLDN6 или конкретных форм CLDN6, или количества клеток, содержащих CLDN6 на поверхности своей мембраны, и полученные таким образом величины затем связывают с определенными заболеваниями или симптомами наподобие описанных выше. Или же антитела по данному изобретению используют для истощения или нарушения функции клеток, в которых экспрессируется CLDN6 и которые отличаются тем, что CLDN6 связан с их поверхностью; тем самым выявляется важное участие этих клеток в развитии заболевания. Это реализуется путем инкубации изучаемого и контрольного образцов с антителами против CLDN6 в условиях, при которых возможно образование комплекса между взятыми антителами и CLDN6. Выявляются любые комплексы, образовавшиеся в результате взаимодействия взятых антител с CLDN6, и по ним сравниваются опытный и контрольный (референсный) образцы.
Вначале у антител по данному изобретению проверяют их связывающую активность, имеющую отношение к терапевтическим и диагностическим применениям, in vitro. Например, для этого можно применить метод проточной цитометрии, как описано в настоящем документе.
С помощью антител по данному изобретению можно вызывать in vivo или in vitro одну или более из следующих биологических активностей: ингибирование пролиферации и/или дифференцировки клеток, в которых экспрессируется CLDN6 и которые отличаются тем, что CLDN6 связан с их поверхностью; уничтожение клеток, в которых экспрессируется CLDN6 и которые отличаются тем, что CLDN6 связан с их поверхностью; опосредование фагоцитоза клеток, в которых экспрессируется CLDN6 и которые отличаются тем, что CLDN6 связан с их поверхностью, или направленной на них антителозависимой клеточной цитотоксичности в присутствии эффекторных клеток; опосредование комплемент-зависимой цитотоксичности, направленной на клетки, в которых экспрессируется CLDN6 и которые отличаются тем, что CLDN6 связан с их поверхностью, в присутствии комплемента; опосредование апоптоза клеток, в которых экспрессируется CLDN6 и которые отличаются тем, что CLDN6 связан с их поверхностью; индуцирование гомотипической адгезии; и/или индуцирование переноса в липидные рафты после связывания CLDN6.
В одном конкретном воплощении данного изобретения антитела используют in vivo или in vitro для лечения, предотвращения или диагностики различных заболеваний, ассоциированных с CLDN6. Примеры заболеваний, ассоциированных с CLDN6, включают помимо прочего раковые заболевания, например описанные в настоящем документе.
Как говорилось выше, антитела против CLDN6 по данному изобретению можно вводить пациенту вместе с одним или более других терапевтических агентов, например цитотоксических агентов, радиоактивных агентов, агентов, препятствующих ангиогенезу, и/или иммуносупрессоров, для того, чтобы ослабить индукцию иммунного ответа против антител по данному изобретению. Эти антитела могут быть соединены с тем или иным агентом, образуя иммунокомплекс, или их вводят отдельно от взятого агента. В последнем случае (раздельное введение) антитела могут быть введены до, после или одновременно с дополнительным агентом или могут комбинироваться с другими известными терапевтическими подходами, например, с противораковой терапией, например с лучевой терапией. Такие терапевтические агенты включают, помимо прочего, агенты, препятствующие неоплазии, например упомянутые выше. Введение антител против CLDN6 по данному изобретению вместе с химиотерапевтическими агентами дает двоякое противораковое действие посредством различных механизмов, приводя к цитотоксическому эффекту в отношении опухолевых клеток. Такое совместное введение может решить проблемы развития невосприимчивости к препаратам или изменения антигенных свойств опухоли, что делает ее не способной взаимодействовать с данными антителами.
Композиции (например, антитела, полиспецифичные и биспецифичные агенты, иммуноконъюгаты) по данному изобретению, содержащие места связывания комплемента, например участки в молекулах IgG1, -2 или -3 или IgM, связывающие комплемент, также можно использовать в присутствии комплемента. В одном из воплощений данного изобретения обработка популяции клеток, включающей клетки-мишени, ех vivo связывающим агентом по данному изобретению и соответствующими эффекторными клетками дополняется добавлением комплемента или сыворотки, содержащей комплемент. Связывание белков комплемента способствует фагоцитозу клеток-мишеней, несущих на своей поверхности связывающий агент по данному изобретению. В другом воплощении данного изобретения клетки, несущие на своей поверхности композиции по данному изобретению, подвергаются лизису при участии комплемента. В еще одном воплощении данного изобретения предлагаемые композиции не активируют комплемент.
Композиции по данному изобретению можно вводить в организм вместе с комплементом. Соответственно, в объем данного изобретения входят композиции, содержащие антитела, полиспецифичные и биспецифичные агенты и сыворотку или комплемент. Эти композиции обладают тем преимуществом, что комплемент оказывается в тесной близости от антител, биспецифичных и полиспецифичных молекул.
Или же антитела, полиспецифичные и биспецифичные агенты по данному изобретению и сыворотку или комплемент вводят по отдельности Связывание композиций по данному изобретению с клетками-мишенями может вызывать перенос комплекса антиген CLDN6-антитело в липидный рафт («плотик») клеточной мембраны. Благодаря такому переносу создается высокая плотность комплексов антиген-антитело, что может эффективно активировать и/или усиливать цитотоксичность, зависимую от комплемента.
В объем данного изобретения входят также наборы, включающие композиции на основе антител (например, антитела и иммуноконъюгаты) и инструкции по их применению. Такой набор может также содержать один или более дополнительных реагентов, например иммуносупрессор, цитотоксический агент или радиоактивно токсический агент, или одно или более дополнительных антител по данному изобретению (например, антитела с комплементарной активностью).
Соответственно, пациентам, получающим композиции на основе антител по данному изобретению, можно дополнительно вводить (до, одновременно с или после введения антител) другой терапевтический агент, например, цитотоксический или радиоактивно токсический агент, который усиливает или продлевает терапевтический эффект антител по данному изобретению.
В других воплощениях данного изобретения пациент может дополнительно получать агент, модулирующий, например усиливающий или ингибирующий, экспрессию или активность рецепторов Fc-γ или Fc-α путем; таким агентом может быть, например, цитокин. Предпочтительные в этом отношении цитокины включают гранулоцитарный колониестимулирующий фактор (G-CSF), гранулоцитарнов-макрофагальный колониестимулирующий фактор (GM-CSF), γ-интерферон (IFN-γ) и фактор некроза опухолей (TNF). К числу важных агентов для увеличения терапевтической эффективности антител и фармацевтических композиций, описанных в настоящем документе, относятся β-глюканы, которые представляют собой разветвленные гомополисахариды, состоящие из остатков глюкозы; они образуются у различных растений (например, в семенах злаков) и микроорганизмов, например бактерий, водорослей и грибов (включая дрожжи). Можно использовать также фрагменты молекул β-глюканов, производимых живыми организмами. Предпочтительны β-глюканы, являющиеся полимерами β(1,3) глюкозы, в которых по крайней мере некоторые из остатков глюкозы главной цепи, например 3-6% остатков глюкозы главной цепи, имеют β(1,6) ответвления.
В одном из конкретных воплощений данного изобретения предлагаются способы для выявления присутствия антигена CLDN6 в образце или измерения количества антигена CLDN6, включающие обработку анализируемого образца и контрольного образца антителами, специфично связывающимися с CLDN6, в условиях, обеспечивающих возможность образования комплекса данного антитела или его участка и CLDN6. Образование такого комплекса выявляется, и разница в соответствующем показателе между анализируемым и контрольным образцами служит мерой присутствия антигена CLDN6 в образце.
В еще одном воплощении данного изобретения предлагается способ выявления присутствия или измерения количества клеток, в которых экспрессируется CLDN6 и которые отличаются тем, что CLDN6 связан с их поверхностью, in vivo или in vitro. Этот способ включает (i) введение в объект анализа (пациенту) композиции по данному изобретению, конъюгированной с выявляемым маркером; (ii) воздействие на объект анализа (пациента) средством для выявления указанного маркера с целью установить области, содержащие клетки, в которых экспрессируется CLDN6 и которые отличаются тем, что CLDN6 связан с их поверхностью.
Упомянутые выше способы можно использовать, в частности, для диагностирования заболеваний, при которых имеет значение CLDN6, и/или определения локализации поражений при связанных с CLDN6 заболеваниях, например при раковых заболеваниях. Предпочтительно указанием на наличие у индивида, в частности у человека, заболевания, связанного с CLDN6, является присутствие CLDN6 в анализируемом образце, взятом у указанного индивида, в количестве выше, чем в контрольном образце.
При использовании в способах, упомянутых выше, антитела, описанные в настоящем документе, могут нести метку, которая может выполнять следующие функции: (i) обеспечивать детектируемый сигнал; (ii) взаимодействовать со второй меткой для модификации детектируемого сигнала, обеспечиваемого первой или второй меткой, например при резонансном переносе энергии флуоресценции (FRET); (iii) влиять на подвижность, например на электрофоретическую подвижность, изменяя заряд, гидрофобность, форму или другие физические свойства, или (iv) предоставлять/создавать связывающую структуру, например обеспечивать аффинность, взаимодействие антиген/антитело или образование ионного комплекса. В качестве таких меток пригодны, например, следующие структуры: флуоресцентные метки, люминисцентные метки, хромофорные метки, радиоактивные метки, изотопные метки (предпочтительно стабильные), изобарные метки, ферментные метки, корпускулярные метки (в частности частицы металлов), магнитные корпускулярные метки, полимерные корпускулярные метки, низкомолекулярные органические соединения (например, биотин), лиганды рецепторов или связывающие соединения (например, адгезивные белки или лектины), последовательности, включающие нуклеиновые кислоты и/или остатки аминокислот, которые могут быть выявлены с помощью связывающих агентов, и проч. Метки включают (пне ограничиваясь перечисленным здесь) сульфат бария, йоцетамовая кислота, йопаноевая кислота, йоподат кальция, диатризоат натрия, диатризоат меглумина, метризамид, тиропаноат натрия и средства лучевой диагностики, включая источники позитронов (например, фтор-18 и углерод-11), источники γ-излучения (например, иод-123, технеций-99m, йод-131 и индий-111), нуклиды для ядерного магнитного резонанса (например, фтор и гадолиний).
В еще одном воплощении данного изобретения для нацеливания соединений (например, терапевтических агентов, меток, цитокинов, радиоактивных веществ, иммуносупрессоров и проч.) на клетки, несущие на своей поверхности CLDN6, используются иммуноконъюгаты: такие соединения присоединяют к антителам. Таким образом, данным изобретением предлагаются также способы для выявления ex vivo или in vitro расположения клеток, в которых экспрессируется CLDN6 и которые отличаются тем, что CLDN6 связан с их поверхностью, например циркулирующих в кровотоке опухолевых клеток.
Далее настоящее изобретение иллюстрируется примерами, которые не следует считать ограничивающими объем данного изобретения.
Примеры
Методики и способы, использованные в данном изобретении, осуществлялись или описаны в настоящем документе известным по существу образом и так, как описывается, например, в работе Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd Edition (1989) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. Все методы, включая использование наборов и реагентов, осуществлялись согласно информации, предоставленной производителем, если не оговорено иного.
Пример 1: Количественная оценка экспрессии CLDN6 в нормальных тканях, раковых тканях и линиях клеток с помощью полимеразной цепной реакции в режиме реального времени (RT-PCR)
Из замороженных образцов ткани и линий раковых клеток выделяли тотальную клеточную РНК, используя набор RNeasy Mini Kit (Qiagen); для примирования использовали олиго(dT18), для обратной транскрипции Superscript II (GIBCO/Lifetech) согласно инструкциям производителя. Целостность полученной кДНК проверяли путем амплификации транскрипта р53 в 30 циклах PCR. Результаты нормализовали нормализовали относительно гена HPRT и рассчитывали величину экспрессии CLDN6, используя ΔΔСТ.
Для каждой нормальной ткани исследовали образцы, взятые у трех индивидов. В нормальных тканях после 40 циклов RT-PCR обнаруживались лишь следовые количества транскриптов CLDN6. Единственной нормальной тканью с экспрессией CLDN6, несколько превышающей порог детекции, была плацента.
Мы обнаружили, что в отличие от нормальных тканей в образцах из опухолевой ткани уровень экспрессии CLDN6 высокий при раке яичника (аденокарциноме), раке легкого (немелкоклеточном, с самой высокой частотой и уровнем экспрессии в аденокарциномах), раке желудка, раке молочной железы, раке печени, раке поджелудочной железы, раке кожи (базальноклеточной карциноме и плоскоклеточной карциноме), злокачественной меланоме, раке головы и шеи (злокачественной плеоморфной аденоме), саркоме (синовиальной саркоме и карциносаркоме), раке желчного протока, почечно-клеточном раке (светлоклеточной карциноме и папиллярной карциноме) и раке матки, а также в линиях клеток А2780 (рак яичника), NIH-OVCAR3 (рак яичника), НСТ-116 (рак толстой кишки), EFO-27 (рак яичника), CPC-N (SCLC), NCI-Н552 (NSCLC), SNU-1 (рак желудка), KATOIII (рак желудка), YAPC (рак поджелудочной железы), AGS (рак желудка), FU97 (рак желудка), MKN7 (рак желудка).
Пример 2. Количественная оценка экспрессии CLDN6 в нормальных тканях, раковых тканях и линиях клеток с помощью Вестерн-блоттинга
Для проведения анализа методом Вестерн-блоттинга брали 20 мкг тотального белка, выделенного из клеток, лизированных буфером Лемли для лизиса (Lemley P.V). Экстракты разбавляли восстанавливающим буферным раствором для образцов (фирма Roth), проводили электрофорез в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE), после чего осуществляли электроблоттинг на мембрану из поливинилиденфторида (PVDF, корпорация Pall). Иммунологическое окрашивание проводили с поликлональными антителами против CLDN6 (фирма ARP) и β-актином (фирма Abcam), затем определяли первые антитела с помощью пероксидазы хрена, конъюгированной с козьими антимышиными и козьими антикроличьими вторыми антителами (фирма Dako).
Для каждой нормальной ткани исследовали тканевые лизаты от пяти индивидов. Ни в одной из исследованных нормальных тканей не выявлялась экспрессия белка CLDN6. В противоположность нормальным тканям наблюдался высокий уровень экспрессии белка CLDN6 в образцах опухолевой ткани при раке яичника и раке легкого. Экспрессия CLDN6 была обнаружена в клетках линий NIH-OVCAR3 (рак яичника), MKN7 (рак желудка), AGS (рак желудка), CPC-N (мелкоклеточный рак легкого), НСТ-116 (рак толстой кишки), FU97 (рак желудка), NEC8 (эмбриональная карцинома яичка), JAR (хориокарцинома плаценты), JEG3 (хориокарцинома плаценты), BEWO (хориокарцинома плаценты), and PA-1 (тератокарцинома яичника).
Пример 3. Иммуногистохимический анализ (IHC) экспрессии CLDN6 в нормальных тканях и раковых тканях
Залитые парафином срезы ткани (4 мкм) инкубировали в течение 1 часа при температуре 58°С на нагревательном столике (HI 1220, Leica). Парафин из срезов удаляли путем инкубации стекол со срезами в растворителе Roticlear (Roth) два раза по 10 мин при комнатной температуре. Затем срезы регидратировали в спирте убывающей концентрации (99%, 2×96%, 80% и 70%, по 5 мин в каждом). Для демаскировки антигенных детерминант препараты кипятили при температуре 120°С, давление 15 psi (фунт/дюйм2) в течение 15 мин в цитратном буферном растворе (10 мМ, рН 6,0) с 0,05% Tween-20. Тотчас после кипячения препараты инкубировали в фосфатном буферном растворе (PBS) в течение 5 мин. Эндогенную пероксидазную активность блокировали с помощью перекиси водорода (0,3%) в метиловом спирте в течение 15 мин при комнатной температуре. Чтобы избежать неспецифического связывания препараты блокировали козьей сывороткой (10%) в PBS в течение 30 мин при комнатной температуре. Затем препараты инкубировали с поликлональными антителами, специфичными в отношении CLDN6 (1 мкг/мл) (ARP) в течение ночи при температуре 4°С. На следующий день препараты промывали PBS при комнатной температуре 3 раза по 5 мин, после чего инкубировали с 100 мкл вторых антител (Power-Vision готовые к использованию антикроличьи поли-HRP IgG (ImmunoLogic)) в течение 1 часа при комнатной температуре. Затем препараты промывали PBS при комнатной температуре 3 раза по 5 мин. Окончательное окрашивание осуществляли с использованием набора VECTOR NovaRED Substrate Kit SK-4800 производства Vector Laboratories (Берлингейм, шт. Калифорния, США). Контрастное окрашивание срезов проводили гематоксилином в течение 90 секунд при комнатной температуре. Препараты обезвоживали в этиловом спирте возрастающей концентрации (70%, 80%, 2×96% и 99%, по 5 мин в каждом) и 10 мин инкубировали в ксилоле, после чего заливали X-tra Kit (Medite Histotechnic).
В нормальных тканях легких, яичников, желудка, толстой кишки, поджелудочной железы, печени, двенадцатиперстной кишки и почек экспрессия белка CLDN6 не обнаруживалась. В отличие от нормальных тканей в срезах опухолевой ткани наблюдалось интенсивное или по меньшей мере заметное окрашивание при раке яичника, легкого, кожи, поджелудочной железы, желудка, молочной железы, мочевого пузыря (переходноклеточной карциноме), шейки матки, яичка (семиноме) и матки. Окрашивание было явно сосредоточено на плазматической мембране злокачественных эпителиальных клеток, тогда как соседние стромальные клетки и незлокачественные эпителиальные клетки не окрашивались. Эти результаты свидетельствуют, что белок CLDN6 локализован на плазматической мембране злокачественных клеток.
Пример 4. Получение мышиных антител против CLDN6
а. Получение экспрессионных векторов, кодирующих полный белок CLDN6 и фрагменты CLDN6
Неприродные последовательности ДНК с оптимизированными кодонами (SEQ ID NO:3) кодирующие полную полипептидную цепь CLDN6 (регистрационный номер Национального центра биотехнологической информации США (NCBI) NP_067018.2, SEQ ID NO:2) получали путем химического синтеза (GENEART AG, Германия) и клонировали в векторе pcDNA3.1/myc-His (Invitrogen, США), в результате получали вектор р3953. Инсерция стоп-кодона позволяла белку CLDN6 экспрессироваться без слияния с кодируемой этим вектором меткой myc-His. Экспрессию CLDN6 проверяли путем Вестерн-блоттинга, проточной цитометрии и иммунофлуоресцентного анализа, используя готовые имеющиеся в продаже антитела против CLDN6 (ARP, 01-8865; R&D Systems, MAB3656).
Кроме того получали последовательность ДНК с оптимизированными кодонами (SEQ ID NO:4), кодирующую предполагаемый фрагмент внеклеточного домена 2 (ЕС2) CLDN6 (SEQ ID NO:6) как объединение с сигнальным пептидом, происходящим из N-концевой лидерной последовательности иммуноглобулиновой κ-цепи, за которой следуют 4 дополнительных аминокислотных остатка для обеспечения правильного сайта расщепления сигнальной пептидазой (SEQ ID NO:5) и клонировали ее в векторе pcDNA3.1/myc-His, что давало вектор р3974. До иммунизации экспрессию фрагмента ЕС2 подтверждали путем иммунофлуоресцентной микроскопии на временно трансфицированных и фиксированных параформальдегидом (PFA) клетках СНО-K1, используя готовые имеющиеся в продаже антитела против туе (Cell Signaling, MAB 2276).
b. Получение линий клеток со стабильной экспрессией CLDN6
Клетки линий HEK293 и P3X63Ag8U.1, в которых стабильно экспрессировался CLDN6, получали по стандартной методике, используя вектор р3953.
с. Иммунизация
Мышей линии Balb/c иммунизировали ДНК плазмиды р3974 в количестве 25 мкг вместе с 4 мкл полиэтилениминманнозы (PEI-Man; in vivo-jetPEI™-Man, производство компании PolyPlus Transfection) (водный раствор PEI-Man150 мМ с 5% глюкозы) путем внутрибрюшинной инъекции в дни 0, 16 и 36. В дни 48 и 62 животных иммунизировали (путем внутрибрюшинной инъекции) миеломными клетками P3X63Ag8U.1, трансфицированными вектором р3953 для стабильной экспрессии CLDN6. Клетки, которые вводили мышам в день 62, перед инъекцией подвергали воздействию радиоактивного излучения (3000 рад). Присутствие антител против CLDN6 в сыворотке крови подопытных животных определяли путем иммунофлуоресцентной микроскопии в период с 20-го по 70-й день, используя клетки СНО-K1, трансфицированные нуклеиновой кислотой, кодирующей CLDN6 вместе с флуоресцентным белком GFP. Для этого через 24 часа после трансфекции клетки, фиксированные параформальдегидом и нефиксированные клетки инкубировали с сывороткой (разведение 1:100) иммунизированных мышей dilution в течение 45 мин при комнатной температуре. Клетки промывали, инкубировали с антимышиными иммуноглобулинами, меченными флуорофором Alexa555 (Molecular Probes) и анализировали с помощью флуоресцентного микроскопа.
Антитела, специфичные в отношении CLDN6 определяли в образцах сыворотки крови, взятой у мышей, которых использовали для получения гибридом F3-6C3-H8; см. фиг.2.
Для образования моноклональных антител мышей с заметным иммунным ответом против CLDN6 за 4 дня до взятия селезенки иммунизировали (путем внутрибрюшинной инъекции) клетками HEK293 (2×107), стабильно трансфицированными вектором р3953.
d. Получение гибридом, продуцирующих мышиные моноклональные антитела против CLDN6
Спленоциты, выделенные у иммунизированных мышей в количестве 6×107 клеток сливали с мышиными миеломными клетками линии P3X63Ag8.653 (ATCC, CRL 1580) в количестве 3×107, используя PEG 1500 (Roche, CRL 10783641001). Клетки высевали на плоскодонные планшеты для микротитрования (приблизительно 5×104 на лунку) и культивировали в течение около двух недель в элективной среде RPMI, содержавшей 10% инактивированной нагреванием эмбриональной телячьей сыворотки, 1% добавки для слияния и клонирования (HFCS, Roche, CRL 11363735), 10 мМ HEPES, 1 мМ пируват натрия, 4,5%глюкоза, 0,1 мМ 2-меркаптоэтанол, 1 × пенициллин/стрептомицин и 1 × HAT (гипоксантин, аминоптерин, тимидин) (Invitrogen, CRL 21060). спустя 10-14 суток каждую лунку проверяли методом проточной цитометрии на присутствие моноклональных антител против CLDN6. Гибридомы, секретирующие нужные антитела, субклонировали методом серийных разведении и вновь проверяли на наличие моноклональных антител против CLDN6. Стабильные субклоны культивировали, чтобы получить небольшие количества антител в культуральной среде и охарактеризовать их. Отбирали по меньшей мере один клон от каждой гибридомы, сохранявший активность исходных клеток (проверяли методом проточной цитометрии). Для каждого клона делали запас из девяти ампул, которые хранили в жидком азоте.
Пример 5. Связывающие свойства супернатантов гибридом и моноклональных антител
а. Качественный контроль временно трансфицированныхклеток НЕК293Т методами (i) Вестерн-блоттинга и (ii) проточной цитометрии
(i) Клетки HEK293T трансфицировали нуклеиновой кислотой, кодирующей CLDN3, CLDN4, CLDN6 и CLDN9 соответственно или для контроля проделывали все манипуляции без нуклеиновой кислоты. Экспрессию CLDN3, CLDN4, CLDN6 или CLDN9 в клетках HEK293T определяли путем Вестерн-блоттинга. Для этого через 24 часа после трансфекции клетки собирали и подвергали лизису. С лизатом проделывали электрофорез в полиакриламидном геле с додецилсульфатом натрия, переносили (с помощью блота) на нитроцеллюлозную мембрану и окрашивали антителами против CLDN3 (A) (Invitrogen, 34-1700), против CLDN4(A) (Zymed, 32-9400), против CDLN6(A) (ARP, 01-8865) или против CLDN9(A) (Santa Cruz, sc-17672), специфично связывающимися с C-концевым участком соответствующего клаудина в денатурирующих условиях. Инкубировали с вторыми антителами, меченными пероксидазой, проявляли электрохемилюминофором (ECL-реагентом), для визуализации использовали спектрометр LAS-3000 (Fuji). Полосы, соответствующие ожидаемым молекулярным массам CLDN3, CLDN4, CLDN6 и CLDN9, наблюдались только в случае трансфицированных клеток, но не в контроле (фиг.3); это демонстрирует, что клеткам HEK293T не свойственна эндогенная экспрессия ни одного из взятых клаудинов и, таким образом, они являются удобным инструментом для определения перекрестной реактивности антител против CLDN6.
(ii) Клетки HEK293T по (i) также анализировали методом проточной цитометрии, используя антитела против CLDN, распознающие нативные эпитопы (мышиные IgG2a против CLDN3 (R&D, MAB4620), мышиные IgG2a против CLDN4 (R&D, МАВ4219), мышиные IgG2b против CLDN6 (R&D, MAB3656)). Эти антитела, производства фирмы Sigma (продукты под номерами М9144 и М8894) служили изотипическими контролями. Специфичность этих антиклаудиновых антител определяли с помощью клеток HEK293T, временно трансфицированных нуклеиновыми кислотами, кодирующими соответственно CLDN3, CLDN4, CLDN6 и CLDN9. У антител против CLDN6 наблюдалась перекрестная реактивность с CLDN3, CLDN4 и CLDN9. У антител против CLDN4 наблюдалась перекрестная реактивность с CLDN3, CLDN6 и CLDN9. Антитела против CLDN3 связывались специфично с CLDN3 (Фиг.4).
b. Определение специфичности моноклональных антител, полученных по данному изобретению, методом проточной цитометрии
Клетки HEK293T трансфицировали одновременно двумя векторами, из которых один кодировал различные клаудины, а другой - флуоресцентный маркер. Через 24 часа после трансфекции клетки собирали, используя раствор трипсин (0,05%)/EDTA и промывали буферным раствором для FACS (PBS, содержащий 2% FCS и 0,1% азида натрия). Клетки переносили на планшеты для микротитрования с U-образным дном в количестве 2×105 клеток на лунку и инкубировали с гибридомными супернатантами в течение 60 мин при температуре 4°С. Затем клетки промывали буферным раствором для FACS и инкубировали со специфичными вторыми антителами (антимышиные IgG 1+2a+2b+3, Dianova, 115-135-164), конъюгированными с аллофикоцианином (АРС). После этого клетки дважды промывали и определяли связывание при помощи проточного цитофлуориметра-биоанализатора BD FACSArray (фиг.5). Экспрессия флуоресцентного маркера отложена по горизонтальной оси, а связывание антител - по вертикальной. Положительным контролем служили готовые имеющиеся в продаже мышиные антитела IgG2b против CLDN6 (R&D, MAB3656); изотипическим контролем служили антитела производства Sigma (каталожный номер продукта М8894).
Моноклональные антитела в супернатантах от субклонов гибридом F3-6C3-H2, F3-6С3-Н8, F3-6C3-H9, F3-6C3-D8 и F3-6C3-G4 (все происходят от гибридомы F3-6C3) были специфичны в отношении CLDN6 и не связывались с CLDN9, CLDN3 и CLDN4. На фиг.5А приведены примеры результатов для субклона гибридом F3-6C3-H8. Моноклональные антитела в супернатанте от субклона гибридом F3-6C3-H8 также связывались с клетками, трансфицированными вариантом CLDN6 (I143V)-SNP. Моноклональные антитела в супернатанте от субклона гибридом F4-4F7-F2 связывались как с CLDN6, так и с CLDN9 (фиг.5А). Моноклональные антитела в супернатанте от субклона гибридом F3-7B3-B4 связывались с CLDN6, CLDN3 и CLDN9 (фиг.5B). Моноклональные антитела в супернатанте от субклона гибридом F3-3F7-A5 связывались с CLDN6, CLDN4 и CLDN9 (фиг.5B).
Пример 6. Получение и проверка моноклональных антител против CLDN6
а. Получение экспрессионных векторов, кодирующих внеклеточный домен 1 CLDN6
Получали последовательность ДНК с оптимизированными кодонами (SEQ ID NO:12), кодирующая фрагмент CLDN6, предположительно соответствующий внеклеточному домену 1 (ЕС1) этого белка (SEQ ID NO:7), как объединение с сигнальным пептидом, происходящим из N-концевой лидерной последовательности иммуноглобулиновой κ-цепи, за которой следуют 4 дополнительных аминокислотных остатка для обеспечения правильного сайта расщепления сигнальной пептидазой (SEQ ID NO:13) и клонировали ее в векторе pcDNA3.1/myc-His, что давало вектор р3974. До иммунизации экспрессию фрагмента ЕС1 подтверждали путем иммунофлуоресцентной микроскопии на временно трансфицированных и фиксированных параформальдегидом (PFA) клетках СНО-K1, используя готовые имеющиеся в продаже антитела против туе (Cell Signaling, MAB 2276).
b. Иммунизация
Мышей линии Balb/c иммунизировали ДНК плазмиды р3973 в количестве 25 мкг вместе с 4 мкл полиэтилениминманнозы (PEI-Man; in vivo-jetPEI™-Man, производство компании PolyPlus Transfection) (водный раствор PEI-Man 150 мМ с 5% глюкозы) путем внутрибрюшинной инъекции в дни 0 и 14. В дни 28 и 44 животных иммунизировали (путем подкожной инъекции) конъюгированными с белком KLH пептидами SEQ ID NO:14 и SEQ ID NO:15 (100 мкг каждый в PBS, производство JPT Peptide Technologies GmbH, Германия) вместе с очищенным путем ВЭЖХ олигонуклеотидом (PTO-CpG-ODN, 25 мкг в PBS; 5'-TCCATGACGTTCCTGACGTT; производство Eurofins MWG Operon, Германия). В дни 64, 77 и 97 животных иммунизировали (путем внутрибрюшинной инъекции) миеломными клетками P3X63Ag8U.1 (в количестве 2×107), трансфицированными вектором р3953 для стабильной экспрессии CLDN6. Перед иммунизацией клетки обрабатывали митомицином-C (2,5 мкг/мл, Sigma-Aldrich, M4287). В дни 64 и 97 клетки вводили вместе с очищенным путем ВЭЖХ олигонуклеотидом PTO-CpG-ODN (50 мкг в PBS), в день 77 - вместе с неполным адъювантом Фрейнда.
Для образования моноклональных антител мышей с заметным иммунным ответом против CLDN6 за четыре дня до взятия селезенки иммунизировали (путем внутрибрюшинной инъекции) клетками HEK293 (в количестве 2×107), стабильно трансфицированными вектором р3953.
c. Проверка моноклональных антител против CLDN6
Проточная цитометрия
Для проверки связывания моноклональных антител с CLDN6 и его гомологами, клетки HEK293T временно трансфицировали соответствующей плазмидой, кодирующей тот или иной клаудин, и определяли его экспрессию путем проточной цитометрии. Чтобы отличать трансфицированные клетки от нетрансфецированных, клетки НЕК293Т были одновременно трансфицированы флуоресцентным маркером. Через 24 часа после трансфекции клетки собирали с помощью раствора трипсина (0,05%)/EDTA, промывали буферным раствором для FACS (PBS, содержащий 2% FCS и 0,1% азида натрия) и ресуспендировали в буферном растворе для FACS в концентрации 2×106 клеток/мл. Эту суспензию клеток (100 мкл) инкубировали с соответствующими антителами в указанной концентрации в течение 30 мин при температуре 4°С. Для определения экспрессии CLDN6 и CLDN9 использовали перекрестно-реактивные антитела. В качестве положительного контроля служили готовые имеющиеся в продаже мышиные антитела против CLDN3 (R&D, MAB4620) и против CLDN4 (R&D, MAB4219); изотопическим контролем служили мышиные IgG2a (Sigma, M9144) и IgG2b (Sigma, M8894), соответственно. Клетки промывали три раза буферным раствором для FACS и инкубировали со специфичными вторыми антителами (антимышиные IgG 1+2a+2b+3, Dianova, 115-135-164), конъюгированными с аллофикоцианином (АРС) в течение 30 мин при температуре 4°С. После этого клетки дважды промывали и ресуспендировали в буферном растворе для FACS. Определяли связывание при помощи проточного цитофлуориметра-биоанализатора BD FACSArray. Экспрессия флуоресцентного маркера отложена по горизонтальной оси, а связывание антител - по вертикальной.
Комплемент-зависимая цитотоксичность (CDC)
Комплемент-зависимую цитотоксичность (CDC), определяли путем измерения содержания внутриклеточного аденозинтрифосфата (АТР) в нелизированных клетках после добавления человеческого комплемента к клеткам-мишеням, инкубированным с антителами против CLDN6. Для количественного определения АТР применяли высокочувствительный метод с использованием люминесцентной реакции, катализируемой люциферазой.
Клетки СНО-K1, стабильно трансфицированные CLDN6 (СНО-K1-CLDN6), собирали с помощью раствора трипсина (0,05%)/EDTA, два раза промывали средой X-Vivo 15 (Lonza, BE04-418Q) и суспендировали в концентрации 1×107 клеток/мл в среде X-Vivo 15. Эту суспензию клеток (250 мкл) переносили в электропорационную кювету (0,4 см) и смешивали с 7 мкг РНК, траскрибированной in vitro и кодирующей люциферазу. Клетки подвергали электропорации при напряжении 200 В и емкости 300 мкФ, используя электропорационную систему Gene Pulser Xcell (Bio Rad). После электропорации клетки ресуспендировали в 2,4 мл предварительно подогретой среды D-MEM/F12 (1:1) со средой GlutaMax-I (Invitrogen, 31331-093), содержащей 10% (объем/объем) FCS, 1% (объем/объем) пенициллин/стрептомицин и 1,5 мг/мл селективного антибиотика G418. Суспензию клеток помещали в лунки (50 мкл на лунку) 96-луночного белого полипропиленового планшета и инкубировали при температуре 37°С в атмосфере с 7,5% CO2. Через 24 часа после электропорации к клеткам прибавляли 50 мкл моноклональных мышиных антител против CLDN6 (в указанных концентрациях) в смеси 60% среды RPMI (содержащей 20 мМ HEPES) и 40% человеческой сыворотки крови (от шести здоровых доноров). Для контроля «тотальный лизис» добавляли 10 мкл на лунку смеси 8% (объем/объем) Triton Х-100 в PBS; для контроля «максимальное количество жизнеспособных клеток» добавляли 10 мкл на лунку PBS; к опытным образцам добавляли также 10 мкл на лунку PBS. Инкубировали в течение 80 мин при температуре 37°С в атмосфере с 7,5% CO2, затем прибавляли 50 мкл на лунку «люцифериновой смеси» (3,84 мг/мл D-люциферин, 0,64 Ед/мл аденозинтрифосфатаза, 160 мМ HEPES в дважды дистиллированной воде). Планшет инкубировали в темноте в течение 45 мин при комнатной температуре. Измеряли люминесценцию при помощи люминометра Infinite М200 (TECAN). Данные представлены как результаты цифрового интегрирования сигнала в относительных световых единицах (RLU).
Клетки NEC8 подвергали электропорации при напряжении 200 В и емкости 400 мкФ и культивировали в смеси сред RPMI 1640 и GlutaMAX-I (Invitrogen, 61870), содержащей 10% (объем/объем) FCS.
Специфичный лизис рассчитывали следующим образом:

максимальное количество жизнеспособных клеток: 10 мкл PBS, без антител тотальный лизис: 10 мкл 8% (объем/объем) Triton X-100 в PBS, без антител.
Воздействие антителами на ранних стадиях
Для воздействия антителами на ранних стадиях бестимусным мышам Nude-Foxn1nu вводили подкожно в бок клетки NEC8 в количестве 2×107 в 200 мкл PBS. В каждой группе было по десять самок возрастом 6-8 недель. Через трое суток после этой инокуляции на протяжении 46 дней два раза в неделю животным вводили (чередуя внутривенные и внутрибрюшинные инъекции) по 200 мкг очищенных моноклональных мышиных антител muMAB 59A, 60А, 61D, 64A, 65А, 66В и 67А. Отрицательным контролем служили особи, которым вводили PBS. Дважды в неделю определяли размеры опухолей: Объем опухоли (TV) = (длина × ширина2)/2. TV выражали в кубических миллиметрах (мм3) и строили кривую роста опухоли во времени. Когда опухоль превышала 1500 мм3, животное умерщвляли.
Результаты
Мышиные моноклональные антитела muMAB 59A, 60А, 61D, 64A, 65А, 66В и 67А прочно связывались с человеческими белками CLDN6 и CLDN6 SNP (однонуклеотидный полиморфизм) вариант I143V, но связывания с CLDN3, 4 и 9 не наблюдалось (фиг.6).
Для антител MuMAB 59A, 60А, 61D, 64A, 65А, 66В и 67А получились очень низкие значения ЕС50 (200-500 нг/мл) и насыщение мест связывания достигалось при низких концентрациях (фиг.7).
В случае антител МиВ 59A, 60А, 61D, 64A, 65А, 66В и 67А наблюдалась зависимая от дозы CDC, причем она возникала при низких концентрациях антител (фиг.8). Антитела против CLDN6 muMAB 65А и 66В вызывали CDC в отношении клеток NEC8 зависимым от дозы образом (фиг.9). Специфичность muMAB 65А и 66B в отношении мишени подтверждалась опытом с клетками NEC8 LVTS2 54 (нокдаунными по CLDN6).
Также антитела muMAB 59A, 60А, 61D, 64A, 65А, 66B и 67А подавляли рост опухолей у мышей, которым ввели клетки NEC8 (фиг.10).
Пример 7. Получение и проверка гибридных моноклональных антител против CLDN6
а. Получение гибридных мышино-человеческих моноклональных антител
Чтобы создать гибридные антитела, нуклеотидные последовательности, кодирующие вариабельные области тяжелой и легкой цепей мышиных антител, включая лидерные последовательности, амплифицировали путем полимеразной ПЦР, используя праймеры, указанные в приведенной ниже таблице.
Нуклеотидные последовательности, соответствующие мышиным тяжелым цепям, соединялись в сайте рестрикции ApaI (5'-GGGCCC-3') с нуклеотидной последовательностью, соответствующей N-концевой части человеческой цепи Fcγ1, в экспрессионном векторе. Нуклеотидные последовательности, соответствующие вариабельным областям мышиной κ-цепи, включая лидерную последовательность, клонировали перед константной областью, используя сайт рестрикции BsiWI. Правильность полученной конструкции и ориентации для предшествующего промотора в векторе последовательности, соответствующей константной области, проверяли путем секвенирования. Вследствие положения сайта рестрикции ApaI амплификация вариабельной области, включая лидерную последовательность в любом случае включает первые 11 нуклеотидов последовательности, кодирующей человеческую константную область γ1, вместе с последовательностью сайта ApaI. Нуклеотидная последовательность, соответствующая константной области человеческой тяжелой цепи γ1, обозначена SEQ ID NO:24; аминокислотная последовательность экспрессирующейся таким образом человеческой константной области γ1, обозначена SEQ ID NO:25. Нуклеотидная последовательность, кодирующая константную часть легкой κ-цепи, обозначена SEQ ID NO:26; соответствующая аминокислотная последовательность обозначена SEQ ID NO:27.
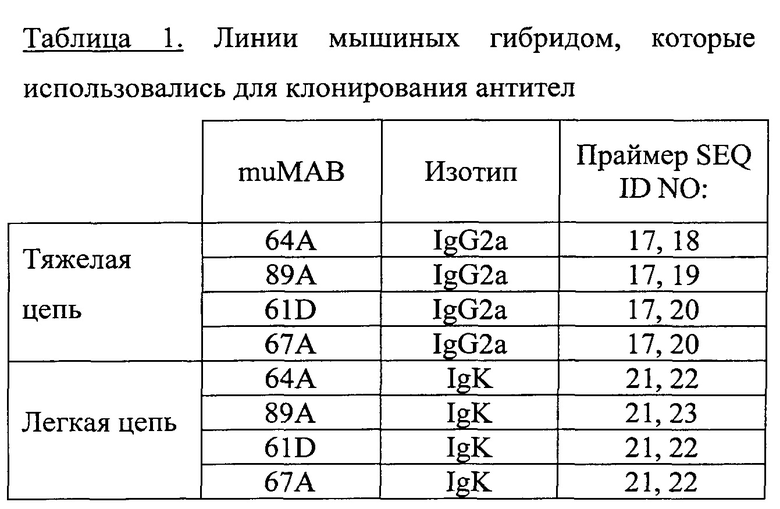
Соответствующие мышиные участки гибридных моноклональных антител обозначены с приставкой "chim" (от англ. chimeric - гибридный), например chimAB 64A.
Амплификацию нуклеотидных последовательностей, соответствующих мышиным вариабельным областям легкой и тяжелой цепей, включая лидерные последовательности, осуществляли методом ПЦР с быстрой амплификацией концов кДНК («step-out»), описанный в работе Matz et al. (Nucleic Acids Research, 1999, Vol.27, No. 6). Для этого получали тотальную РНК из гибридомных клеточных линий (см. таблицу 1) стандартными методами, известными специалистам в данной области техники, например с использованием набора RNeasy Mini Kit (Qiagen). Одноцепочечную кДНК получали методом со сменой матриц, также описанным в работе Matz et al. (Nucleic Acids Research, 1999, Vol.27, No. 6, 1558). В дополнение к олигомеру (dT)30 (SEQ ID NO:28) использовался гибридный олигомер ДНК/РНК (SEQ ID NO:29), служивший 5'-адаптером для смены матриц в процессе полимеризации цепи кДНК. В этом адапторном олигомере последние три нуклеотида были не дезоксирибо-, а рибонуклеотидами. В последующей "step-out" ПЦР использовался антисмысловой олигомер, мишенью которого были константная область мышиной κ-цепи или константной области подкласса 2а γ-цепи (SEQ ID NO:30 и 31, соответственно). Подкласс IgG мышиных моноклональных антител, продуцируемых гибридомными клеточными линиями заранее определяли с помощью набора IsoStrip (Roche), и соответственно выбирали подходящий антисмысловой олигомер (см. таблицу 1). Смысловым олигомером в «step-out» ПЦР, включавшим два олигомера (SEQ ID NO:32 и 33), была смесь праймеров.
Затем идентифицированные нуклеотидные последовательности, соответствующие мышиным вариабельным областям, включая лидерные последовательности, амплифицировали путем ПЦР исключая 5'-концевую нетранслируемую область (5'-UTR) и мышиную константную 3'-область, добавляя сайты рестрикции на концах, что давало возможность субклонирования в подготовленных экспрессионных векторах, несущих нуклеотидные последовательности, соответствующие человеческим константным областям. Кроме того, смысловые олигомеры обеспечивали консенсусную последовательность Козак (5'-GCCGCCACC-3'), а антисмысловые олигомеры для вариабельных областей тяжелых цепей включали первые 11 нуклеотидов константной области человеческой тяжелой цепи γ1 в дополнение к сайту рестрикции ApaI (см. таблицу 1, SEQ ID NO:17-23). Нуклеотидные последовательности, соответствующие вариабельным областям легких κ-цепей, включая лидерные последовательности, клонировали, используя рестрикционные эндонуклеазы HindIII и BsiWI; для вариабельных областей тяжелых γ-цепей требовались ферменты HindIII и ApaI.
Также были амплифицированы нуклеотидные последовательности. соответствующие другим вариабельным областям легких и тяжелых цепей мышиных антител и получены другие гибридные моноклональные антитела против CLDN6 согласно описанному выше.
b. Получение гибридных моноклональных антител против CLDN6
Гибридные моноклональные антитела временно экспрессировались в клетках HEK293T (АТСС CRL-11268), трансфицированных плазмидной ДНК, кодирующей легкие и тяжелые цепи соответствующих антител. За 24 часа до трансфекции клетки в количестве 8×107 высевали на культуральные чашки (145 мм) и растили в 25 мл среды HEK293T (DMEM/F12 + GlutaMAX - I, 10% FCS, 1% пенициллин/стрептомицин). Плазмидную ДНК растворяли в среды HEK293T без добавок (20 мкг в 5 мл на одну чашку). Добавляли 75 мкл полиэтиленимина (PEI) (1 mg/ml) (Polyscience, 23966) и смесь DNA:PEI инкубировали в течение 15 мин при комнатной температуре. Затем эту трансфекционную смесь прибавляли по каплям к клеткам. Через 24 часа после трансфекции среду HEK293T-заменяли на среду Pro293a (Lonza, BE12-764Q), содержащую пенициллин/стрептомицин (1%). Для оптимальной экспрессии трансфицированные клетки культивировали в течение 96-120 часов при температуре 37°С в атмосфере, сродержащей 7,5% СО2). Собирали супернатант и очищали гибридные антитела методом быстрой жидкостной хроматографии белков (FPLC, используя колонки с белком А. Определяли концентрацию антител и проверяли их чистоту методом электрофореза в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE).
c. Исследование свойств гибридных моноклональных антител против CLDN6
Проточная цитометрия
Для определения специфичности и аффинности гибридных моноклональных антител, специфичных в отношении CLDN6, методом проточной цитометрии изучали их связывание с клетками HEK293, стабильно трансфицированными соответственно CLDN3, 4, 6 или 9, и с опухолевыми клетками линий с эндогенной экспрессией CLDN6. Клетки собирали с помощью раствора 0,05% трипсин/EDTA, промывали буферным раствором для FACS (PBS, содержащий 2% FCS и 0,1% азида натрия) и ресуспендировали в буферном растворе для FACS в концентрации 2×106 клеток/мл. Эту клеточную суспензию в количестве 100 мкл инкубировали с соответствующими антителами в указанных концентрациях в течение 60 мин при температуре 4°С. Для определения экспрессии CLDN6 и CLDN9 А использовали гибридные перекрестно-реактивные антитела (chimAB 5F2D2). Положительным контролем служили готовые имеющиеся в продаже мышиные антитела против CLDN3 (R&D, MAB4620) и против CLDN4 (R&D, MAB4219); отрицательным контролем служили человеческие IgG1-κ (Sigma, I5154). Клетки промывали три раза буферным раствором для FACS и инкубировали в течение 30 мин при температуре 4°С со специфичными вторыми антителами - соответственно козьими античеловеческими IgG Fc-γ, конъюгированными с аллофикоцианином (АРС) (Dianova, 109-136-170), или с антимышиными IgG 1+2a+2b+3a, конъюгированными с АРС (Dianova, 115-135-164). Клетки дважды промывали и ресуспендировали в буферном растворе для FACS. Связывание определяли при помощи проточного цитофлуориметра-биоанализатора BD FACSArray.
Комплемент-зависимая цитотоксичность (СРС)
Комплемент-зависимую цитотоксичность (CDC), определяли путем измерения содержания внутриклеточного аденозинтрифосфата (АТР) в нелизированных клетках после добавления человеческого комплемента к клеткам-мишеням, инкубированным с антителами против CLDN6. Для количественного определения АТР применяли высокочувствительный метод с использованием люминесцентной реакции, катализируемой люциферазой.
Для этого исследования брали клетки NEC 8 дикого типа (несущие CLDN6) и клетки NEC8, нокдаунные по этому белку (не имеющие CLDN6); и те, и другие были стабильно трансдуцированы люциферазной экспрессионной конструкцией. Клетки собирали с помощью раствора 0,05% трипсин/EDTA, разбавляли до концентрации 2×105 клеток/мл смесью сред RPMI и GlutaMax-I (Invitrogen, 61870-010), содержащей 10% (объем/объем) FCS. Клетки в количестве 1×104 высевали на белый полипропиленовый 96-луночный планшет и инкубировали 24 часа при температуре 37°С в атмосфере с 5% CO2. После этого к клеткам прибавляли 50 мкл моноклональных гибридных антител против CLDN6 (в указанных концентрациях) в смеси среды RPMI, содержащей 20 мМ HEPES (60%) и человеческой сыворотки крови, полученной от шести здоровых доноров (40%). Для контроля на тотальный лизис добавляли 10 мкл на лунку 8%-ного (объем/объем) раствора Triton Х-100 в PBS; для контроля на максимальное количество жизнеспособных клеток добавляли 10 мкл на лунку PBS; в опытные образцы также добавляли 10 мкл на лунку PBS. Инкубировали в течение 80 мин при температуре 37°С в атмосфере с 7,5% CO2, затем прибавляли 50 мкл на лунку «люцифериновой смеси» (3,84 мг/мл D-люциферин, 0,64 Ед/мл аденозинтрифосфатаза, 160 мМ HEPES в дважды дистиллированной воде). Планшет инкубировали в темноте в течение 45 мин при комнатной температуре. Измеряли люминесценцию при помощи люминометра Infinite М200 (TECAN). Данные представлены как результаты цифрового интегрирования сигнала в относительных световых единицах (RLU).
Специфичный лизис рассчитывали следующим образом:

максимальное количество жизнеспособных клеток: 10 мкл PBS, без антител
тотальный лизис: 10 мкл 8% (объем/объем) Triton Х-100 в PBS, без антител.
Антитело-завивимая клеточно-опосредованная цитотоксичность (ADCC)
Антитело-зависимую клеточно-опосредованную цитотоксичность (ADDC), определяли путем измерения содержания внутриклеточного аденозинтрифосфата (АТР) в нелизированных клетках после добавления человеческих мононуклеаров периферической крови (РВМС) к клеткам-мишеням, инкубированным с антителами против CLDN6. Для количественного определения АТР применяли высокочувствительный метод с использованием люминесцентной реакции, катализируемой люциферазой.
Для этого исследования брали клетки NEC8 дикого типа (несущие CLDN6) и клетки NEC8, нокдаунные по этому белку (не имеющие CLDN6); и те, и другие были стабильно трансдуцированы люциферазной экспрессионной конструкцией. Клетки собирали с помощью раствора 0,05% трипсин/EDTA, разбавляли до концентрации 2×105 клеток/мл смесью сред RPMI и GlutaMax-I (Invitrogen, 61870-010), содержащей 10% (объем/объем) FCS и 20 мМ Hepes. Клетки в количестве 1×104 высевали на белый полипропиленовый 96-луночный планшет и инкубировали 4 часа при температуре 37°С в атмосфере с 5% СО2.
Мононуклеары периферической крови выделяли из человеческой донорской крови путем центрифугирования в градиенте плотности, используя фиколл/гипак (GE Healthcare, 17144003). Сбирали слой, содержащий РВМС, и клетки промывали два раза раствором PBS/EDTA (2 мМ). РВМС в количестве 1×108 клеток вносили в 50 мл среды X-Vivo 15 (Lonza, BE04-418Q), содержащей 5% инактивированной нагреванием человеческой сыворотки крови (Lonza, US14-402E) и инкубировали в течение 2 часов при температуре 37°С в атмосфере с 5% CO2.
Через 4 часа после этой манипуляции с клетками-мишенями (NEC-8) к клеткам прибавляли моноклональные гибридные антитела против CLDN6 в растворе PBS в указанных концентрациях. Собирали неадгезивные РВМС, которые за 2 часа инкубации отделялись от адгезивных моноцитов, и разводили их средой X-vivo 15 до концентрации 8×106 клеток/мл. Эту клеточную суспензию (25 мкл) прибавляли к клеткам-мишеням и моноклональным гибридным антителам против CLDN6. Планшеты инкубировали в течение 24 часов при температуре 37°С в атмосфере с 5% CO2.
Через 24 часа инкубации для контроля «тотальный лизис» прибавляли раствор Triton Х-100 (8% объем/объем) в PBS в количестве 10 мкл на лунку; для контроля «максимальное количество жизнеспособных клеток» прибавляли PBS 10 мкл на лунку; к опытным образцам также прибавляли PBS 10 мкл на лунку. Затем прибавляли 50 мкл на лунку «люцифериновой смеси» (3,84 мг/мл D-люциферин, 0,64 Ед/мл аденозинтрифосфатаза, 160 мМ HEPES в дважды дистиллированной воде). Планшет инкубировали в темноте в течение 30 мин при комнатной температуре. Измеряли люминесценцию при помощи люминометра Infinite M200 (TECAN). Данные представлены как результаты цифрового интегрирования сигнала в относительных световых единицах (RLU).
Специфичный лизис рассчитывали следующим образом:

максимальное количество жизнеспособных клеток: 10 мкл PBS, без антител
тотальный лизис: 10 мкл 8% (объем/объем) Triton Х-100 в PBS, без антител.
d. Результаты
Гибридные моноклональные антитела против CLDN6 chimAB 61D, 64A, 67А и 89А прочно связывались с человеческим CLDN6, а связывания с CLDN3, 4 и 9 не наблюдалось (фиг.11).
При связывании с человеческим CLDN6, стабильно экспрессирующимся на поверхности клеток HEK293, гибридные моноклональные антитела против CLDN6 chimAB 64A и 89А характеризовались очень низкими значениями ЕС50 (450-600 нг/мл) и насыщение мест связывания достигалось при низких концентрациях. Антитела chimAB 67А и 61D характеризовались низким (1000 нг/мл) и средним (2300 нг/мл) значениями ЕС50 соответственно (фиг.12).
При связывании с CLDN6, эндогенно экспрессирующимся в клетках NEC8, гибридные моноклональные антитела против CLDN6 chimAB 64A и 89А характеризовались очень низкими значениями ЕС50 (600-650 нг/мл) и насыщение мест связывания достигалось при низких концентрациях. Антитела chimAB 61D и 67А характеризовались средним (1700 нг/мл) и высоким (6100 нг/мл) значениями ЕС50 соответственно (фиг.13).
При связывании с CLDN6, эндогенно экспрессирующимся в клетках OV90, гибридные моноклональные антитела против CLDN6 chimAB 64A и 89А характеризовались очень низкими значениями ЕС50 (550-600 нг/мл) и насыщение мест связывания достигалось при низких концентрациях. Антитела chimAB 61D и 67А характеризовались средними значениями ЕС50 (1500 нг/мл 2300 нг/мл соответственно) (фиг.14).
Гибридные моноклональные антитела против CLDN6 chimAB 61D, 64A, 67А и 89А проявляли активность, выражающуюся в зависимой от комплемента цитотоксичности в отношении клеток NEC-8, причем эта активность зависела от дозы (фиг.15).
Гибридные моноклональные антитела против CLDN6 chimAB 61D, 64A, 67А и 89А проявляли активность, выражающуюся в зависимой от антитело-зависимой клеточно-опосредованной цитотоксичности в отношении клеток NEC-8, причем эта активность зависела от дозы и цитотоксичность возникала даже при низких концентрациях антител (фиг.16).
Эти результаты ясно демонстрируют специфичность указанных гибридных моноклональных антител в отношении CLDN6.
Пример 8. Лечение с использованием моноклональных антител против CLDN6
Лечение на ранних стадиях
Для воздействия антителами на ранних стадиях бестимусным мышам линии Nude-Foxn1nu вводили путем подкожной инъекции в бок клетки NEC8 в количестве 2×107 в 200 мкл среды RPMI (Gibco). В каждой группе было по десять самок возрастом 6-8 недель. Через трое суток после инокуляции опухолевых клеток животным на протяжении семи недель дважды в неделю вводили путем внутривенной или внутрибрюшинной инъекции (чередовали) 200 мкг очищенных мышиных моноклональных антител muMAB 89A. Отрицательным контролем служила группа особей, которым вводили такой же объем PBS. Два раза в неделю определяли размеры опухолей: объем опухоли (TV) = (длина × ширина2)/2). TV выражали в мм3 и строили графики роста опухолей во времени. Когда размеры опухоли превышали 1500 мм3, животное умерщвляли.
Лечение на поздних стадиях
Для воздействия антителами на поздних стадиях развития опухолей-ксенографтов бестимусным мышам-самкам возрастом 6-8 недель линии Nude-Foxn1nu вводили путем подкожной инъекции в бок клетки NEC8 в количестве 2×107 в 200 мкл среды RPMI (Gibco). Два раза в неделю определяли размеры опухолей: объем опухоли (TV) = (длина × ширина2)/2). TV выражали в мм3 и строили графики роста опухолей во времени. Через 15-17 суток после инокуляции опухолевых клеток животных разделяли на группы для воздействия; в каждой группе было 8 особей с опухолями одинаковых размеров - более 80 мм. На протяжении пяти недель дважды в неделю мышам вводили путем внутривенной или внутрибрюшинной инъекции (чередовали) 200 мкг очищенных мышиных моноклональных гибридных антител. Отрицательным контролем служила группа особей, которым вводили неспецифичные антитела. Когда размеры опухоли превышали 1500 мм3, животное умерщвляли.
При воздействии на мышей с ксенографтами опухолевых клеток линии NEC8 мышиными моноклональными антителами muMAB 61D, 64A и 67А на ранних стадиях развития опухолей роста опухолей не наблюдалось даже после прекращения иммунотерапии (фиг.17).
При воздействии на мышей с ксенографтами опухолевых клеток линии NEC8 мышиными моноклональными антителами muMAB 89A на ранних стадиях развития опухолей рост опухолей подавлялся и в конце исследования у мышей, получавших muMAB89A, заметных опухолей не было (фиг.18).
При воздействии на мышей с ксенографтами опухолевых клеток линии NEC8 мышиными моноклональными антителами muMAB 64A на поздних стадиях развития опухолей рост опухолей подавлялся (фиг.19).
При воздействии на мышей с ксенографтами опухолевых клеток линии NEC8 мышиными моноклональными антителами muMAB 64A на поздних стадиях развития продолжительность жизни подопытных особей увеличивалась (фиг.20).
При воздействии на мышей с ксенографтами опухолевых клеток линии NEC8 мышиными моноклональными антителами muMAB 61D, 67A и 89А на поздних стадиях развития опухолей достигалось ингибирование опухолевого роста опухолей (фиг.21).
При воздействии на мышей с ксенографтами опухолевых клеток линии NEC8 особи, получавшие специфичные антитела против CLDN6 muMAB 61D или 67A на поздних стадиях развития опухолей, жили дольше (фиг.22).
При позднем воздействии на мышей с ксенографтами клеток линии NEC8 дикого типа и со стабильным нокдауном по CLDN6 антитела muMAB 64A и 89А оказывали терапевтическое действие только у мышей с ксенографтами клеток NEC8 дикого типа, а у мышей с ксенографтами клеток NEC8, нокдаунных по CLDN6, терапевтического эффекта этих антител не было, что демонстрирует специфичность антител против CLDN6 in vivo (фиг.23).
Пример 9. Картирование с высоким разрешением эпитопов моноклональных антител против CLDN6
При картировании эпитопов с помощью метода твердофазного иммуноферментного анализа (ELISA) специфичные моноклональные антитела против CLDN6 лишь очень слабо связывались с линейными эпитопами, что указывало на то, что они имеют конформационные эпитопы. Чтобы изучить взаимодействие описанных в настоящем документе антител с CLDN6 в его нативной конформации, для эпитопного картирования был применен метод сайт-специфического мутагенеза в культурах клеток млекопитающих. Был проведен сканирующий аланином мутагенез аминокислотных остатков 27-81 и 137-161 в первом и втором внеклеточных доменах, соответственно. После осуществления временной экспрессии в клетках HEK293T у мутантных вариантов CLDN6 определяли способность связываться со специфичными моноклональными антителами. Нарушение связывания специфичного моноклонального антитела с мутантным вариантом CLDN6 указывало на то, что затронутый мутацией аминокислотный остаток важен для контакта и/или пространственной структуры. Связывание определяли методом проточной цитометрии. Чтобы отличать трансфицированные клетки от нетрансфицированных трансфекцию проводили одновременно с флуоресцентным маркером.
В белке CLDN6 путем сканирования аланином были идентифицированы аминокислотные остатки, важные для взаимодействия со специфичными гибридными антителами против CLDN6. Путем сайт-специфического мутагенеза были получены замены на аланин и глицин (GENEART AG, Германия). Для проверки связывания моноклональных гибридных антител с CLDN6 дикого типа и его мутантными вариантами, клетки HEK293T временно трансфицировали соответствующими плазмидами, кодирующими тот или иной клаудин, и определяли экспрессию методом проточной цитометрии. Чтобы отличать трансфицированные клетки от нетрансфицированных, проводили одновременную трансфекцию клеток HEK293T флуоресцентным маркером. Через 24 часа после трансфекции клетки собирали с помощью раствора трипсина (0,05%)/EDTA, промывали буферным раствором для FACS (PBS, содержащий 2% FCS и 0,1% азида натрия) и ресуспендировали в буферном растворе для FACS в концентрации 2×106 клеток/мл. Эту суспензию клеток (100 мкл) инкубировали с антителами в концентрации 10 мкг/мл в течение 45 мин при температуре 4°С. В качестве контроля для определения экспрессии мутантных вариантов CLDN6 на поверхности клеток использовали готовые имеющиеся в продаже мышиные антитела против CLDN6 (R&D, МАВ3656). Клетки промывали три раза буферным раствором для FACS и инкубировали со специфичными вторыми антителами - козьими античеловеческими IgG Fc-γ (Dianova, 109-136-170) или антимышиными IgG 1+2a+2b+3a (Dianova, 115-135-164), - конъюгированными с аллофикоцианином (АРС) в течение 30 мин при температуре 4°С. После этого клетки дважды промывали и ресуспендировали в буферном растворе для FACS. Определяли связывание с трансфицированными клетками при помощи проточного цитофлуориметра-биоанализатора BD FACSArray. Экспрессия флуоресцентного маркера откладывали по горизонтальной оси, связывание антител - по вертикальной. Среднюю интенсивность сигнала для связывания моноклональных гибридных антител, специфичных к CLDN6, с мутантными вариантами CLDN6 выражали как процент относительно связывания с CLDN6 дикого типа. Если аминокислотный остаток имел ключевое значение для связывания, то после его мутации связывания не происходило; если аминокислотный остаток имел поддерживающее значение, то после мутации связывание лишь снижалось по сравнению со связыванием CLDN6 дикого типа.
Картирование эпитопов с высоким разрешением показало, что для взаимодействия со специфичными гибридными антителами против CLDN6 chimAB 61D, 64A, 67А и 89А важны аминокислотные остатки F35, G37, S39 и, вероятно, Т33 первого внеклеточного домена CLDN6. Аминокислотный остаток 140 существенно важен для связывания с антителами chimAB 89A и участвует в связывании с chimAB 61D и 67А. Кроме того, аминокислотный остаток L151 второго внеклеточного домена CLDN6 важен для взаимодействия с chimAB 67А (фиг.24).
Пример 10. Получение и проверка усовершенствованных моноклональных антител против CLDN6
Оказалось, что антитела 64A, обладая превосходными качествами в отношении связывания с CLDN6 и эффективного лечения опухолей, содержат свободный остаток цистеина в положении 46 легкой цепи, который существенно ухудшает такие свойства белка, как растворимость, стабильность и способность к образованию агрегатов. Свободный остаток цистеина нежелателен также по другим причинам, например для регуляции. Поэтому мы попытались создать антитела на основе 64A, не содержащие свободного остатка цистеина в положении 46, не теряя при этом их желательных свойств.
Были получены гибридные антитела mAb206-LCC, в которых объединены тяжелая цепь chimAB 64A и легкая цепь chimAB 61D. В результате аминокислотных замен были получены MAb206-SUBG и mAb206-SUBS, в которых цистеин в положении 46 легкой цепи chimAB 64A был заменен на глицин и серии, соответственно.
Получение моноклональных гибридных антител против CLDN6 в клетках HEK293T
Моноклональные гибридные антитела временно экспрессировались в клетках HEK293T (АТСС CRL-11268), трансфицированных плазмидной ДНК, кодирующей легкие и тяжелые цепи соответствующих антител, как описано в разделе b примера 7.
Получение моноклональных гибридных антител против CLDN6 клеток СНО, адаптированных к росту в суспензии
Адаптированные к росту в суспензии клетки СНО субкультивировали в бессывороточной среде в CO2-инкубаторе с шейкером и увлажнением рабочей камеры. За сутки до трансфекции клетки высевали в бессывороточную среду в шейкерные колбы.
В день трансфекции клетки центрифугировали (5 мин при 200 g) и ресуспендировали в свежей среде DMEM (Invitrogen, 41965-039) в шейкерных колбах. К клеткам прибавляли ДНК и трансфекционный реагент и осторожно перемешивали встряхиванием. После накопительного культивирования в CO2-инкубаторе клетки разводили бессывороточной питательной средой и культивировали далее в шейкере-инкубаторе. В дни 0, 2, 4 и 6 клетки подпитывали готовым препаратом СНО CD EfficientFeed™ А и В, соответственно (Invitrogen, A 1023401 и А 1024001). Когда жизнеспособность клеток становилась ниже 90% собирали гибридные антитела. Полученные антитела очищали путем жидкостной экспресс-хроматографии белков (FPLC), используя колонки с белком А. Концентрацию антител определяли по поглощению при 280 нм, чистоту проверяли методом электрофореза в полиакриламидном геле с додецилсульфатом натрия.
Проточная цитометрия
Связывание специфичных гибридных моноклональных антител против CLDN6 с клетками, в которых экспрессируется их мишень, определяли методом проточной цитометрии. Клетки собирали с помощью раствора 0,05% трипсин/EDTA, промывали буферным раствором для FACS (PBS, содержащий 2% FCS и 0,1% азида натрия) и ресуспендировали в буферном растворе для FACS в концентрации 2×106 клеток/мл. Эту клеточную суспензию в количестве 100 мкл инкубировали с соответствующими антителами в указанных концентрациях в течение 30 мин при температуре 4°С. Для определения экспрессии CLDN6 и CLDN9 использовались антитела ChimAB 5F2D2. Положительным контролем служили готовые имеющиеся в продаже мышиные антитела против CLDN3 (R&D, MAB4620) и против CLDN4 (R&D, MAB4219); отрицательным контролем служили человеческие IgG1-κ (Sigma, I5154). Клетки промывали три раза буферным раствором для FACS и инкубировали в течение 30 мин при температуре 4°С со специфичными вторыми антителами - соответственно козьими античеловеческими IgG Fc-γ, конъюгированными с аллофикоцианином (АРС) (Dianova, 109-136-170), или с антимышиными IgG 1+2a+2b+3a, конъюгированными с АРС (Dianova, 115-135-164). Клетки дважды промывали и ресуспендировали в буферном растворе для FACS. Связывание определяли при помощи проточного цитофлуориметра-биоанализатора BD FACSArray.
Комплемент-зависимая цитотоксичность (CDC)
Комплемент-зависимую цитотоксичность (CDC), определяли путем измерения содержания внутриклеточного аденозинтрифосфата (АТР) в нелизированных клетках после добавления человеческого комплемента к клеткам-мишеням, инкубированным с антителами против CLDN6, как описано в примере 7.
Антитиело-зависимая клеточно-опосредованная цитотоксичность
Антитело-зависимую клеточно-опосредованную цитотоксичность (ADDC), определяли путем измерения содержания внутриклеточного аденозинтрифосфата (АТР) в нелизированных клетках после добавления человеческих мононуклеаров периферической крови (РВМС) к клеткам-мишеням, инкубированным с антителами против CLDN6. Для количественного определения АТР применяли высокочувствительный метод с использованием люминесцентной реакции, катализируемой люциферазой.
Для этого исследования брали клетки NEC8 дикого типа (несущие CLDN6) и клетки NEC8, нокдаунные по этому белку (не имеющие CLDN6); и те, и другие были стабильно трансдуцированы люциферазной РНК, а клетки OV90 временно трансфицировали транскрибированной in vitro РНК, кодирующей люциферазу. Клетки собирали с помощью раствора 0,05% трипсин/EDTA, разбавляли до концентрации 2×105 клеток/мл (NEC-8 дикого типа и нокдаунные по CLD6) или 1×106 клеток/мл (OV90) соответствующей питательной средой, содержащей дополнительно 20 ммМ Hepes. Клетки в количестве, соответственно, 1×104 или 5×104 высевали на белый полипропиленовый 96-луночный планшет и инкубировали при температуре 37°С в атмосфере с 5% CO2.
Мононуклеары периферической крови выделяли из человеческой донорской крови путем центрифугирования в градиенте плотности, используя фиколл/гипак (GE Healthcare, 17144003). Собирали слой, содержащий РВМС, и клетки промывали два раза раствором PBS/EDTA (2 мМ). РВМС в количестве 1×108 клеток вносили в 50 мл среды X-Vivo 15 (Lonza, BE04-418Q), содержащей 5% человеческой сыворотки крови (от шести здоровых доноров) и инкубировали в течение 2 часов при температуре 37°С в атмосфере с 5% CO2. Через 4 часа после этой манипуляции к клеткам прибавляли моноклональные гибридные антитела против CLDN6 (25 мкл) в растворе PBS в указанных концентрациях. Собирали неадгезивные РВМС, которые за 2 часа инкубации отделялись от адгезивных моноцитов, и разводили их средой X-vivo 15 до концентрации 1,6×107 клеток/мл (клетки NEC-8 дикого типа или нокдаунные по CLDN6) или 4×107 клеток/мл (клетки OV90). Эту клеточную суспензию (25 мкл) прибавляли к клеткам-мишеням и моноклональным гибридным антителам против CLDN6. Планшеты инкубировали в течение 24 часов при температуре 37°С в атмосфере с 5% CO2.
Через 24 часа инкубации для контроля «тотальный лизис» прибавляли раствор Triton Х-100 (8% объем/объем) в PBS в количестве 10 мкл на лунку; для контроля «максимальное количество жизнеспособных клеток» прибавляли PBS 10 мкл на лунку; к опытным образцам также прибавляли PBS 10 мкл на лунку. Затем прибавляли 50 мкл на лунку «люцифериновой смеси» (3,84 мг/мл D-люциферин, 0,64 Ед/мл аденозинтрифосфатаза, 160 мМ HEPES в дважды дистиллированной воде). Планшет инкубировали в темноте в течение 30 мин при комнатной температуре. Измеряли люминесценцию при помощи люминометра Infinite M200 (TECAN). Данные представлены как результаты цифрового интегрирования сигнала в относительных световых единицах (RLU).
Специфичный лизис рассчитывали следующим образом:

максимальное количество жизнеспособных клеток: 10 мкл PBS, без антител
тотальный лизис: 10 мкл 8% (объем/объем) Triton Х-100 в PBS, без антител.
Лечение на поздних стадиях
Для воздействия антителами на поздние стадии развития опухолей, возникших из - ксенографтов опухолевых клеток, бестимусным мышам-самкам линии Nude-Foxn1nu возрастом 6-8 недель вводили путем подкожной инъекции в бок 2×107 клеток NEC8 в 200 мкл среды RPMI (Gibco). Два раза в неделю определяли размеры опухолей: объем опухоли (TV) = (длина × ширина2)/2). TV выражали в мм3 и строили графики роста опухолей во времени. Через 17 суток после инокуляции опухолевых клеток животных разделяли на группы для воздействия; в каждой группе было 8 особей с опухолями одинаковых размеров - более 80 мм3. На протяжении пяти недель дважды в неделю мышам вводили путем внутривенной или внутрибрюшинной инъекции (чередовали) 200 мкг очищенных мышиных моноклональных гибридных антител. Отрицательным контролем служила группа особей, которым вводили неспецифичные антитела. Когда размеры опухоли превышали 1500 мм3, животное умерщвляли.
Исследование метастазирования
Для изучения метастазирования клетки NEC8 собирали, фильтровали через сетку (70 мкм), чтобы удалить крупные клеточные агрегаты. Бестимусным мышам-самкам линии Nude-Foxn1nu возрастом 6 недель вводили путем инъекции в хвостовую вену 4×106 клеток NEC8. Через трое суток после введения опухолевых клеток два раза в неделю животным вводили путем внутривенной или внутрибрюшинной инъекции (чередовали) очищенные мышиные моноклональные антитела muMAB 64A в количестве 200 мкг в PBS либо PBS без антител. Через 8 недель мышей умерщвляли, извлекали легкие и замораживали образцы в жидком азоте.
Из замороженной ткани выделяли геномную ДНК согласно инструкциям "Blood & Cell Culture DNA Midi Kit" (Qiagen, 13343). Легочную ткань гомогенизировали при помощи диспергатора Ultra Torax Т8.10 (IKA-Werke). Чтобы избежать загрязнения человеческой геномной ДНК, легочную геномную ДНК получали в стерильных условиях, используя количественную полимеразную цепную реакцию (qPCR). Чтобы определить опухолевую нагрузку человеческих клеток NEC8 в образцах мышиной легочной ткани, строили стандартную кривую с определенными количествами человеческой геномной ДНК клеток NEC8 и мышиной легочной геномной ДНК. Использовали следующие серии разведении (ДНК клеток NEC8/мышиная легочная ДНК): 200/0, 40/160, 8/192, 1,6/198,4, 0,32/199,68, 0,064/199,94, 0,013/200 и 0/200 нг. Для qPCR брали 200 нг геномной ДНК, краситель 2xSYBR Green (Qiagen, 204145), 16 нмоль смысловой праймер (GGGATAATTTCAGCTGACTAAACAG, Eurofins) и 16 нмоль антисмысловой праймер (TTCCGTTTAGTTAGGTGCAGTTATC, Eurofins) в суммарном объеме 50 мкл; для анализа использовали систему ПЦР в режиме реального времени 7300 Real Time PCR-System (Applied Biosystems, США). Для отрицательного контроля вместо ДНК брали воду. Количественную ПЦР проводили при следующих условиях: 15 мин при 95°С (1 цикл), 30 с при 95°С/30 с при 62°С/30 с при 72°С (40 циклов), 15 с при 95°С/30 с при 60°С/15 с при 95°С (1 цикл). Все реакции qPCR проводили в трех повторах. Опухолевую нагрузку определяли методом стандартных кривых и рассчитывали с помощью прилагаемой к прибору 7300 System (Applied Biosystems) программы.
Картирование эпитопов с высоким разрешением
Чтобы изучить взаимодействие антител по данному изобретению с CLDN6 в его нативной конформации применяли метод сайт-специфического мутагенеза в культурах клеток млекопитающих для картирования эпитопов. Проводили сканирование аланином аминокислотных остатков 27-81 и 137-161 первого и второго внеклеточных доменов CLDN6, соответственно. Замены на аланин осуществляли методом сайт-специфического мутагенеза (GENEART AG, Германия). После осуществления временной экспрессии в клетках HEK293T у мутантных вариантов CLDN6 определяли способность связываться со специфичными моноклональными антителами. Нарушение связывания специфичного моноклонального антитела с мутантным вариантом CLDN6 указывало на то, что затронутый мутацией аминокислотный остаток важен для контакта и/или пространственной структуры. Связывание определяли методом проточной цитометрии. Чтобы отличать трансфицированные клетки от нетрансфицированных, трансфекцию проводили одновременно с флуоресцентным маркером. Через 24 часа после трансфекции клетки собирали с помощью раствора трипсина (0,05%)/EDTA, промывали буферным раствором для FACS (PBS, содержащий 2% FCS и 0,1% азида натрия) и ресуспендировали в буферном растворе для FACS в концентрации 2×106 клеток/мл. Эту суспензию клеток (100 мкл) инкубировали с антителами в концентрации 10 мкг/мл в течение 30 мин при температуре 4°С. В качестве контроля для определения экспрессии мутантных вариантов CLDN6 на поверхности клеток использовали готовые имеющиеся в продаже мышиные антитела против CLDN6 (R&D, MAB3656). Клетки промывали три раза буферным раствором для FACS и инкубировали со специфичными вторыми антителами - козьими античеловеческими IgG Fc-γ (Dianova, 109-136-170) или антимышиными IgG 1+2a+2b+3a (Dianova, 115-135-164), - конъюгированными с аллофикоцианином (АРС) в течение 30 мин при температуре 4°С. После этого клетки дважды промывали и ресуспендировали в буферном растворе для FACS. Определяли связывание с трансфицированными клетками при помощи проточного цитофлуориметра-биоанализатора BD FACSArray. Экспрессия флуоресцентного маркера откладывали по горизонтальной оси, связывание антител - по вертикальной. Среднюю интенсивность сигнала для связывания моноклональных гибридных антител, специфичных к CLDN6, с мутантными вариантами CLDN6 выражали как процент относительно связывания с CLDN6 дикого типа. Если аминокислотный остаток имел ключевое значение для связывания, то после его мутации связывания не происходило; если аминокислотный остаток имел поддерживающее значение, то после мутации связывание лишь снижалось по сравнению со связыванием CLDN6 дикого типа.
Иммуногистохимический анализ
Криосрезы ткани (4 мкм) сразу после изготовления фиксировали ацетоном в течение 10 мин при температуре -20°С. Затем срезы высушивали в течение 10 мин при комнатной температуре и хранили при -80°С. Перед использованием срезы согревали (10 мин при комнатной температуре) и регидратировали в PBS в течение 5 мин. Блокировали эндогенную пероксидазную активность 0,3%-ной перекисью водорода в PBS в течение 15 мин при комнатной температуре. Чтобы избежать неспецифичного связывания препараты инкубировали с козьей сывороткой (10%) в PBS в течение 30 мин при комнатной температуре. После этого препараты инкубировали с мышиными моноклональными антителами, специфичными к CLDN6, muMAB 64A (0,2 мкг/мл в PBS, содержащем 10% козьей сыворотки), в течение ночи при температуре 4°С. На следующий день препараты промывали PBS (3 раза по 5 мин) при комнатной температуре и инкубировали с вторыми антителами (100 мкл готовых имеющихся в продаже антимышиных IgG (ImmunoLogic), конъюгированных с комплексом полимер-пероксидаза хрена (PowerVision)) в течение часа при комнатной температуре. Затем препараты промывали PBS (3 раза по 5 мин) при комнатной температуре. Окончательное окрашивание осуществляли в течение полутора минут, используя набор VECTOR NovaRED Substrate Kit SK-4800 производства Vector Laboratories (Берлингейм, шт. Калифорния, США). Контрастное окрашивание срезов проводили гематоксилином в течение 90 секунд при комнатной температуре. Препараты обезвоживали в этиловом спирте возрастающей концентрации (70%, 80%, 2×96% и 99%, по 5 мин в каждом) и 10 мин инкубировали в ксилоле, после чего заливали X-tra Kit (Medite Histotechnic).
Чтобы использовать моноклональные гибридные антитела mAb206-LCC и mAb206-SUBG в человеческих тканях, их метили флуоресцеинизотиоцианатом (FITC, Squarix GmbH, Германия). Изготавливали криосрезы и обрабатывали их, как описано выше (фиксация, блокирование эндогенной пероксидазы, блокирование участков неспецифичного связывания). Затем препараты инкубировали с антителами mAb206-LCC-FITC и mAb206-SUBG-FITC (1 мкг/мл в PBS, содержащем 10% козьей сыворотки) в течение часа при комнатной температуре в темноте. После этого препараты промывали PBS (3 раза по 5 мин) при комнатной температуре и инкубировали с 200 мкл кроличьих анти-FITC-HRP антител (AbD Serotec, разведенные 1:300 в PBS, содержащем 10% козьей сыворотки) в течение 30 мин при комнатной температуре. Промывали PBS (3 раза по 5 мин) при комнатной температуре, после чего осуществляли реакцию с субстратом в течение 2 мин 30 используя набор VECTOR NovaRED Substrate Kit SK-4800 производства Vector Laboratories (Берлингейм, шт. Калифорния, США). Контрастное окрашивание, обезвоживание и заключение в среду осуществляли, как описано выше.
Изучение специфичности связывания моноклональных гибридных антител против CLDN6 методом проточной цитометрии с использованием клеток HEK293T, временно трансфицированных человеческими CLDN6, 3, 4 и 9, показало, что моноклональные гибридные антитела chimAB 64A, mAb206-LCC, mAb206-SUBG и mAb206-SUBS связываются с CLDN6, но не взаимодействуют с CLDN3, 4 и 9, соответственно (фиг.27).
Что касается связывания с человеческим CLDN6, стабильно экспрессирующимся на поверхности клеток HEK293, то моноклональные гибридные антитела против CLDN6 chimAB 64A, mAb206-LCC, mAb206-SUBG и mAb206-SUBS характеризовались сходно низкими значениями ЕС50 и насыщение мест связывания достигалось при низких концентрациях (фиг.28).
Сродство связывания (аффинность) моноклональных гибридных антител против CLDN6 chimAB 64A, mAb206-LCC, mAb206-SUBG и mAb206-SUBS с опухолевыми клетками, в которых эндогенно экспрессируется человеческий CLDN6, оценивали по связыванию с опухолевыми клетками линии NEC8 (рак яичка), которое определяли методом проточной цитометрии. По сравнению со специфичными к CLDN6 антителами chimAB 64A, mAb206-SUBG и mAb206-SUBS вариант с комбинацией легких цепей mAb206-LCC проявлял втрое большую аффинность по отношению к клеткам NEC8. Во всех случаях насыщение мест связывания достигалось при низких концентрациях (фиг.29).
Изучение специфичности связывания моноклональных гибридных антител против CLDN6 chimAB 64A, mAb206-LCC, mAb206-SUBG и mAb206-SUBS с человеческими клетками линии OV90 (рак яичника) методом проточной цитометрии показало, что антитела, специфичные к CLDN6, характеризовались сходными низкими значениями ЕС50. Вариант с комбинацией легких цепей mAb206-LCC проявлял наибольшую прочность связывания с клетками OV90 (фиг.30).
В отношении клеток NEC-8 моноклональные гибридные антитела против CLDN6 chimAB 64A, mAb206-LCC и mAb206-SUBG обладали активностью, проявляющейся комплемент-зависимой цитотоксичностью (CDC), причем эффект зависел от дозы. А с клетками NEC-8, нокдаунными по CLDN6, ни одно из указанных антител не вызывало неспецифический лизис. Этот результат демонстрирует специфичную к мишени эффекторную функцию антител chimAB 64A, mAb206-LCC и mAb206-SUBG (фиг.31а).
Активность антител chimAB 64A, mAb206-LCC, mAb206-SUBG и mAb206-SUBS, проявляющаяся CDC в отношении клеток NEC8, зависела от дозы. По сравнению с chimAB 64A варианты с аминокислотными заменами mAb206-SUBG и mAb206-SUBS обладали сходной активностью, проявляющейся CDC в отношении клеток NEC8 (фиг.31b).
Моноклональные гибридные антитела против CLDN6 chimAB 64A, mAb206-LCC, mAb206-SUBG и mAb206-SUBS обладали зависимой от дозы активностью, проявляющейся антителозависимой клеточно-опосредованной цитотоксичностью (ADCC), и вызывали ADCC в отношении опухолевых клеток линий NEC8 и OV90 даже в низких концентрациях (фиг.32а).
Фиг.32b демонстрирует активность, проявляющуюся антителозависимой клеточно-опосредованной цитотоксичностью (ADCC), моноклональных гибридных антител против CLDN6 chimAB 64A, mAb206-LCC и mAb206-SUBG в отношении клеток NEC8 дикого типа и нокдаунных клеток NEC8. Чтобы показать специфичность антител к мишени, использовали клетки NEC8, стабильно нокдаунные по CLDN6.
В опытах с ксенографтами опухолевых клеток линии NEC8 у мышей воздействие специфичными к CLDN6 антителами mAb206-LCC, mAb206-SUBG и mAb206-SUBS на поздних стадиях развития опухолей подавляло опухолевый рост по сравнению с контрольными воздействиями (фиг.33).
Кривые опухолевого роста, представленные на фиг.34а, показывают, что моноклональные гибридные антитела против CLDN6 mAb206-LCC, mAb206-SUBG и mAb206-SUBS способны подавлять опухолевый рост. График выживания (фиг.34b) демонстрирует увеличение продолжительности жизни мышей, получавших антитела, специфичные к CLDN6.
Картирование эпитопов с высоким разрешением показало, что для взаимодействия специфичных к CLDN6 гибридных антител chimAB 64A, mAb206-LCC, mAb206-SUBG и mAb206-SUBS с CLDN6 важны аминокислотные остатки F35, G37 и S39, а также, вероятно, Т33 первого внеклеточного домена CLDN6. Особенности связывания chimAB 64A, mAb206-LCC, mAb206-SUBG и mAb206-SUBS были одинаковыми (фиг.35).
Для изучения терапевтического эффекта мышиных моноклональных антител против CLDN6 muMAB 64A в отношении метастазирования использовали модель с ксенографтами опухолевых клеток: бестимусным мышам линии Nude-Foxn1nu вводили в хвостовую вену клетки NEC8, а через трое суток эти животные получали специфичные к CLDN6 антитела muMAB 64A. Спустя 8 недель определяли опухолевую нагрузку в ткани легких с помощью полимеразной цепной реакции. Мышиные моноклональные антитела против CLDN6 muMAB 64A по сравнению с контролем (PBS без антител) вызывали отчетливое ингибирование опухолевого роста (фиг.36).
Иммуногистохимическое окрашивание раковых и нормальных тканей человека с использованием моноклональных антител muMAB 64A, mAb206-LCC и mAb206-SUBG показало, что в противоположность нормальным тканям в срезах опухолей яичников и яичек наблюдалось выраженное равномерное окрашивание. Очень интенсивное окрашивание мембран имело место в злокачественных эпителиальных клетках, тогда как соседние стромальные клетки и незлокачественные эпителиальные клетки не окрашивались (фиг.37). Эти результаты с очевидностью свидетельствуют, что специфичные к CLDN6 антитела по данному изобретению специфично связываются со злокачественными клетками, полученными от пациентов с опухолями. (Пояснение: число тканей, окрашивающихся антителами/число изученных тканей).
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА ДЛЯ ЛЕЧЕНИЯ РАКОВЫХ ЗАБОЛЕВАНИЙ, ПРИ КОТОРЫХ ЭКСПРЕССИРУЕТСЯ КЛАУДИН 6 | 2012 |
|
RU2816850C2 |
| АНТИТЕЛА, СПЕЦИФИЧНЫЕ К КЛАУДИНУ 6 (CLDN6) | 2010 |
|
RU2675997C2 |
| ЛЕЧЕНИЕ РАКА ПРИ ПОМОЩИ НАПРАВЛЕННЫХ НА МИШЕНЬ АНТИТЕЛ IN VIVO | 2011 |
|
RU2642305C2 |
| АНТИТЕЛА, ПРИГОДНЫЕ В ДИАГНОСТИКЕ РАКА | 2018 |
|
RU2815883C1 |
| АНТИТЕЛА ПРОТИВ КЛАУДИНА 18.2 ДЛЯ ДИАГНОСТИКИ РАКА | 2013 |
|
RU2661772C2 |
| Модифицированные клетки с химерным антигеновым рецептором для лечения рака, экспрессирующего CLDN6 | 2020 |
|
RU2826058C2 |
| БИСПЕЦИФИЧЕСКИЕ ТРЕХВАЛЕНТНЫЕ АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С КЛАУДИНОМ 6 ИЛИ КЛАУДИНОМ 18.2, И CD3, ДЛЯ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ С ЭКСПРЕССИЕЙ КЛАУДИНА | 2017 |
|
RU2798988C2 |
| АГЕНТЫ ДЛЯ ЛЕЧЕНИЯ ЭКСПРЕССИРУЮЩИХ КЛАУДИН РАКОВЫХ ЗАБОЛЕВАНИЙ | 2013 |
|
RU2678127C2 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ С ИСПОЛЬЗОВАНИЕМ АНТИТЕЛ ПРОТИВ КЛАУДИНА 18.2 ДЛЯ ЛЕЧЕНИЯ РАКА | 2014 |
|
RU2699549C2 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ПРОТИВ КЛАУДИНА-18 ДЛЯ ЛЕЧЕНИЯ РАКА | 2008 |
|
RU2571923C2 |



























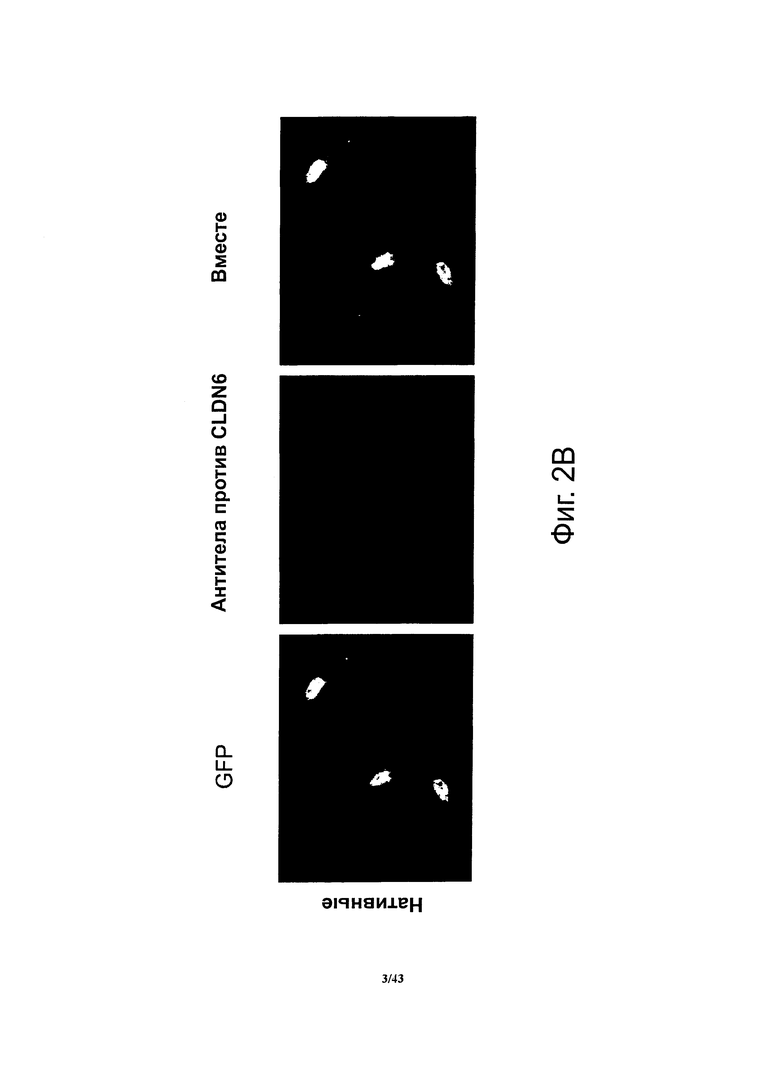
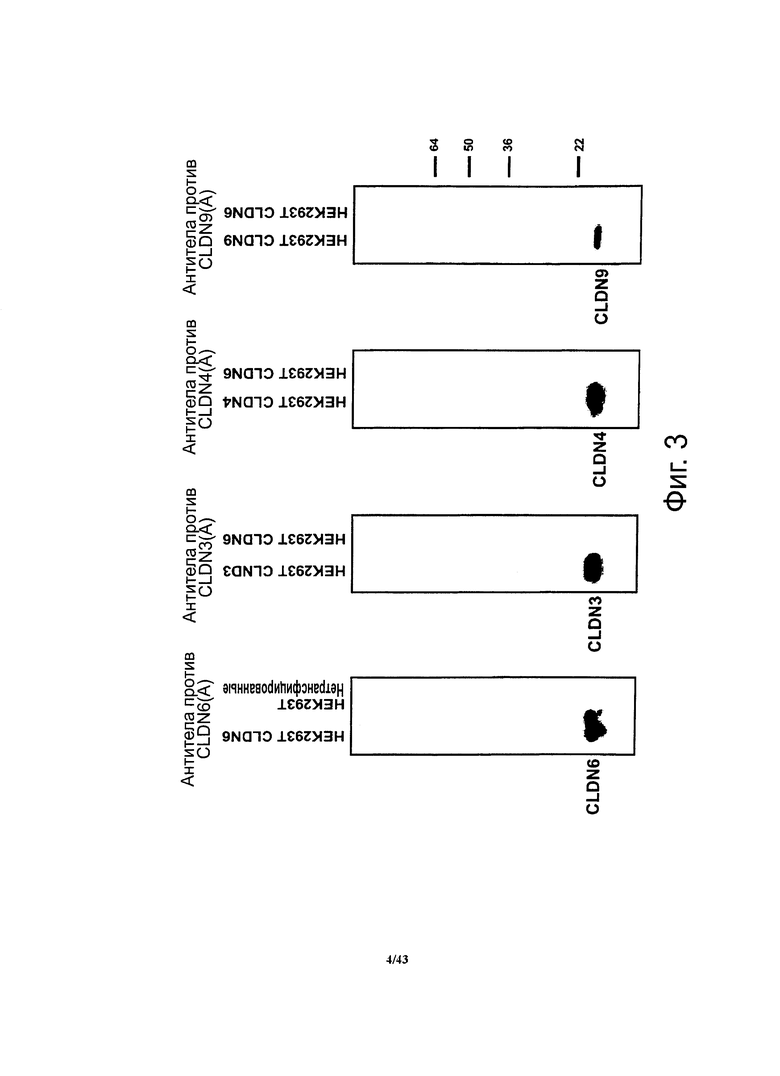
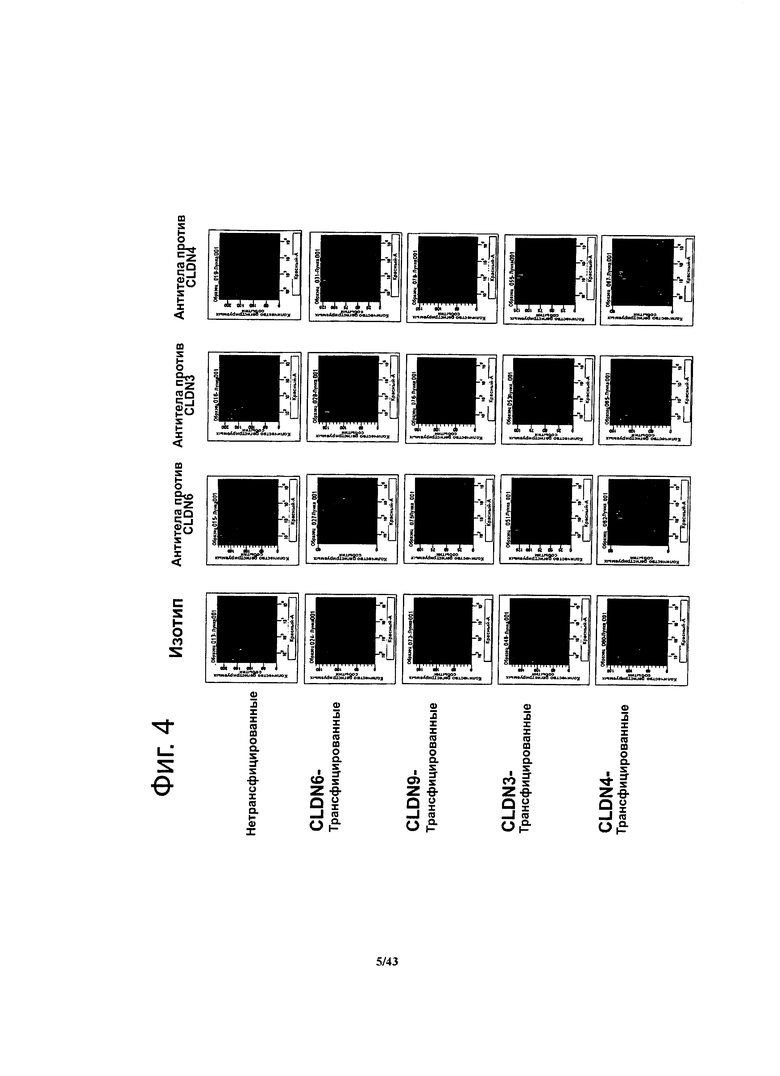
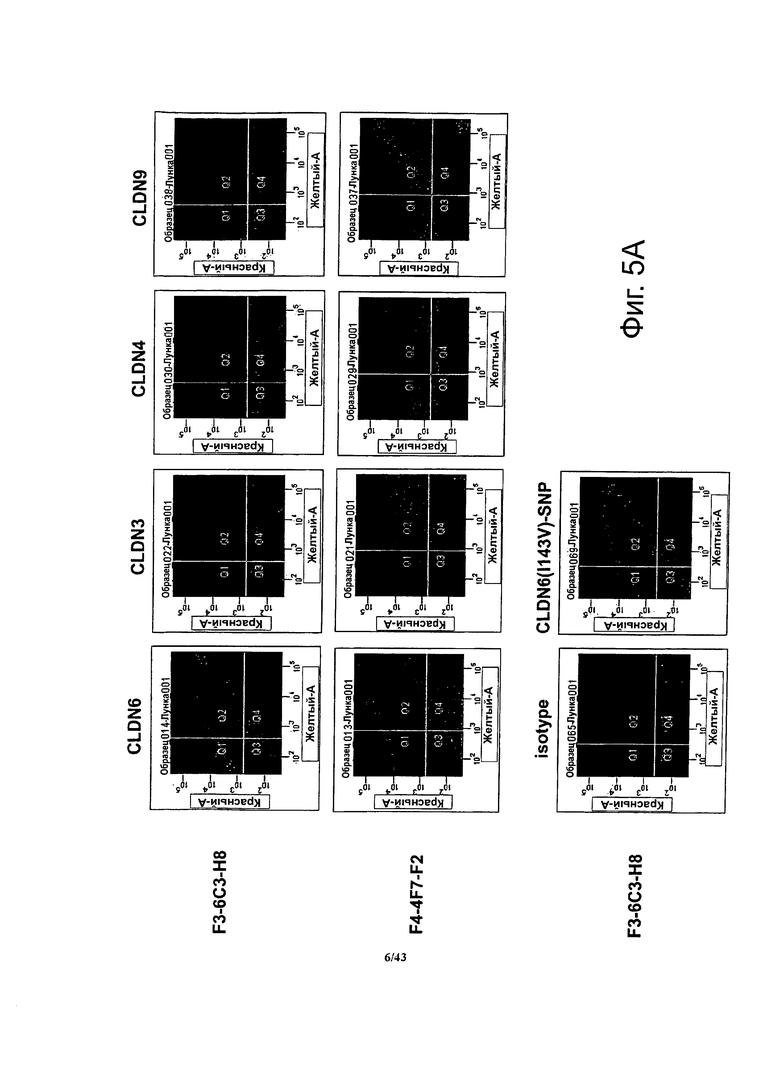
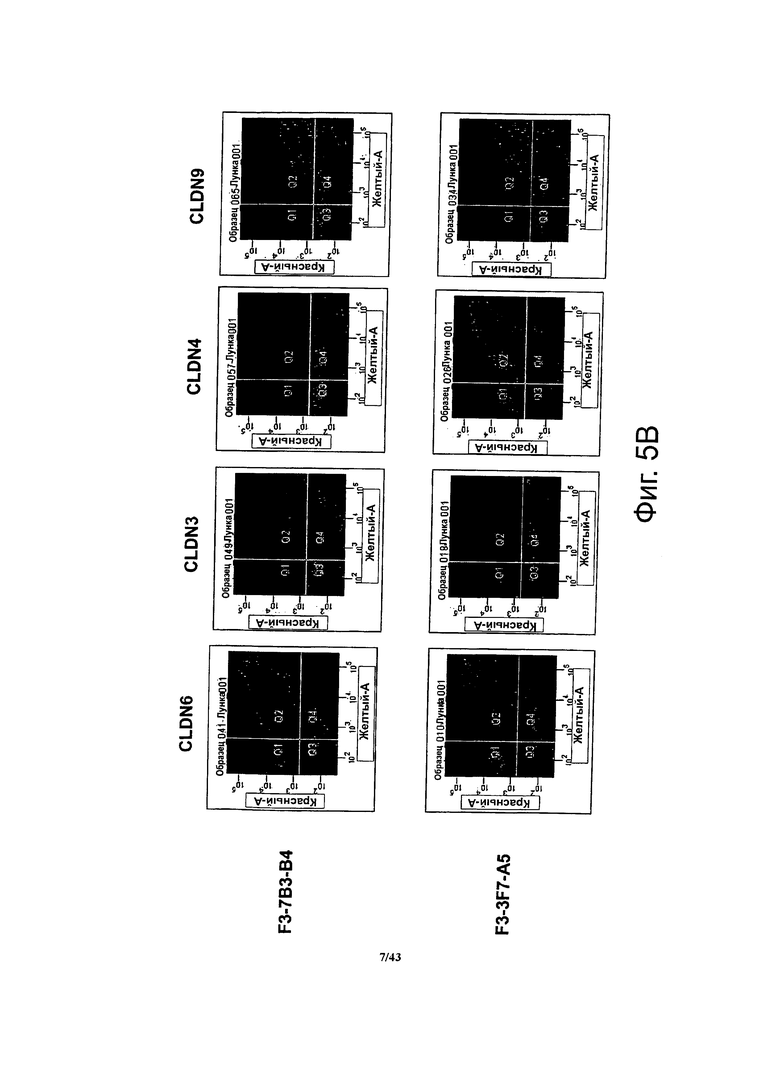

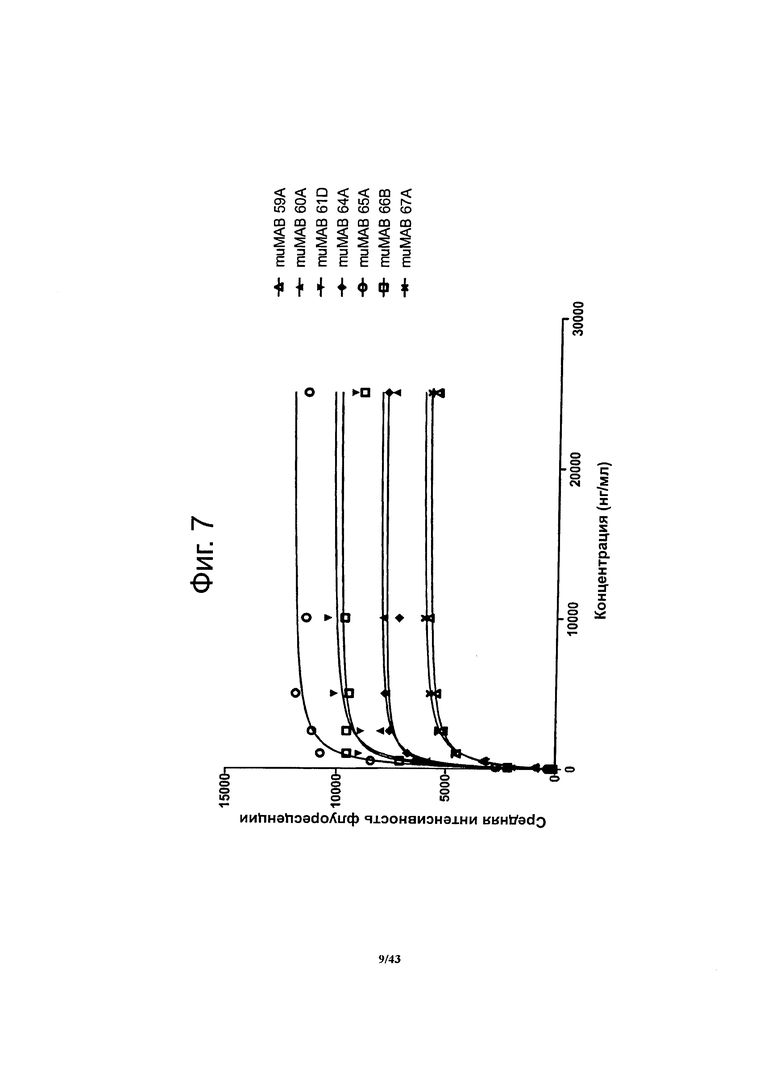
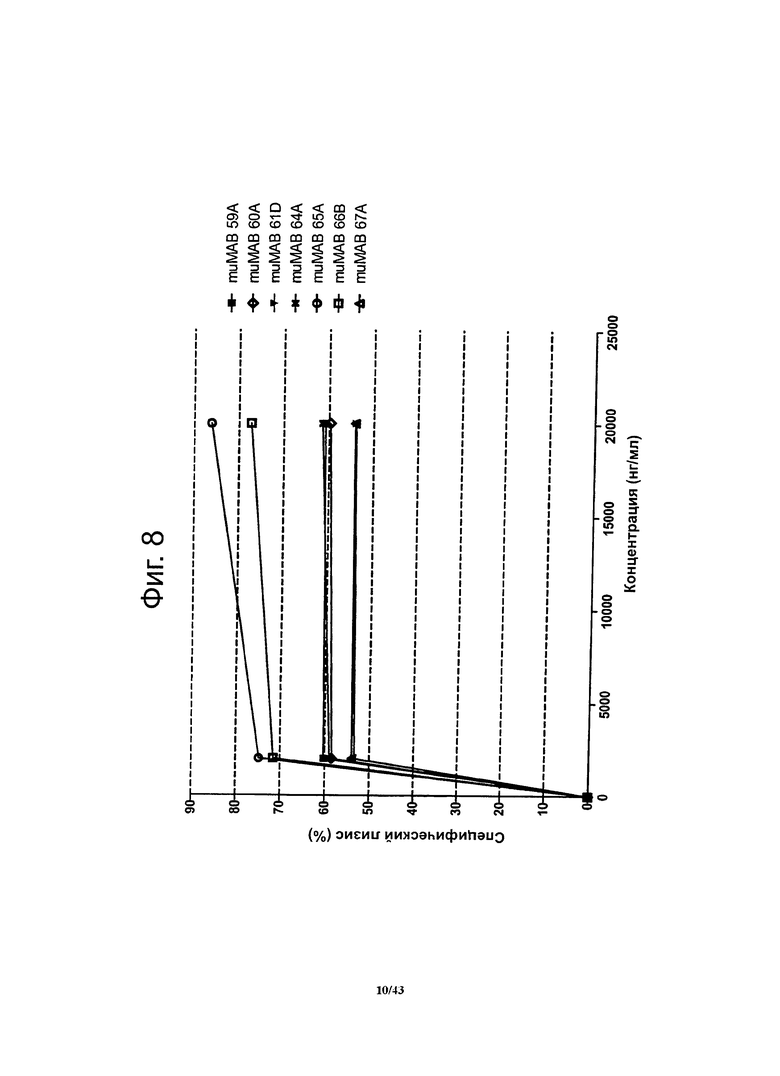
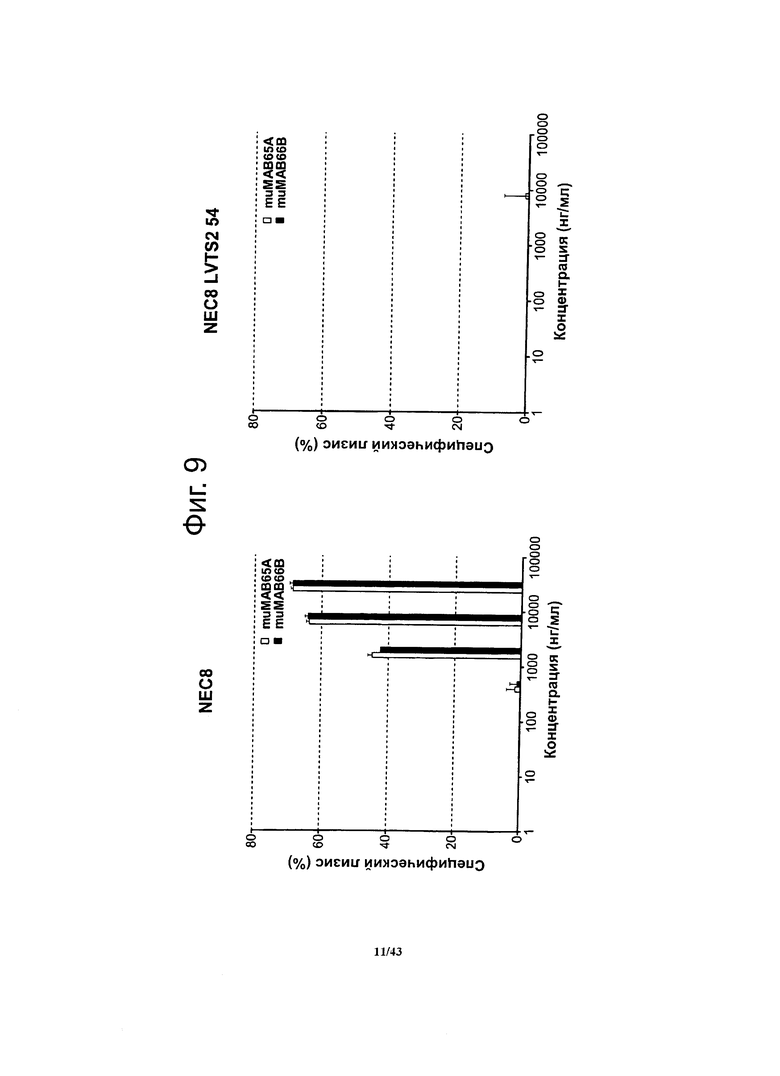
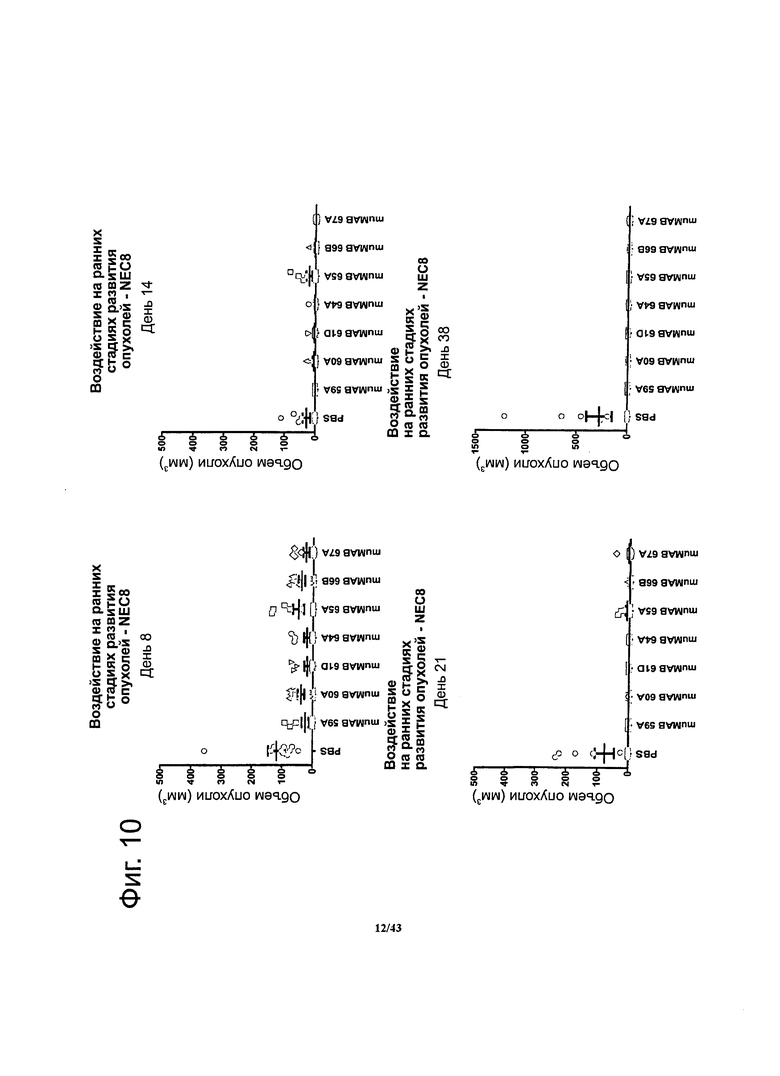
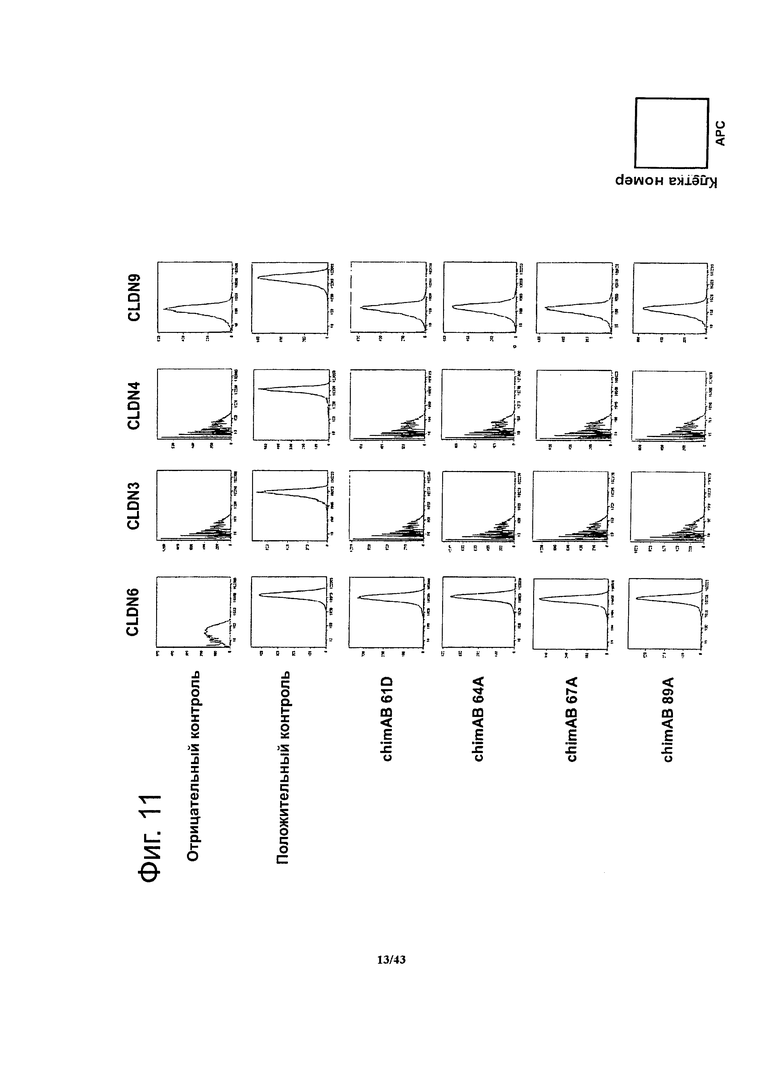
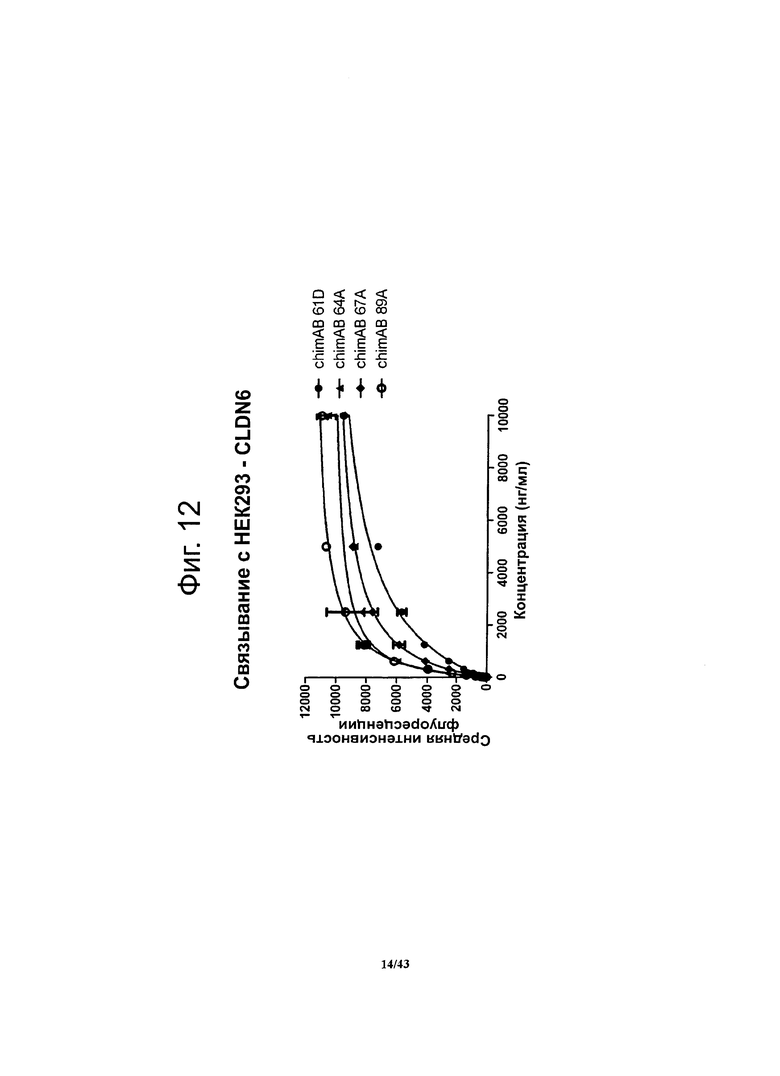
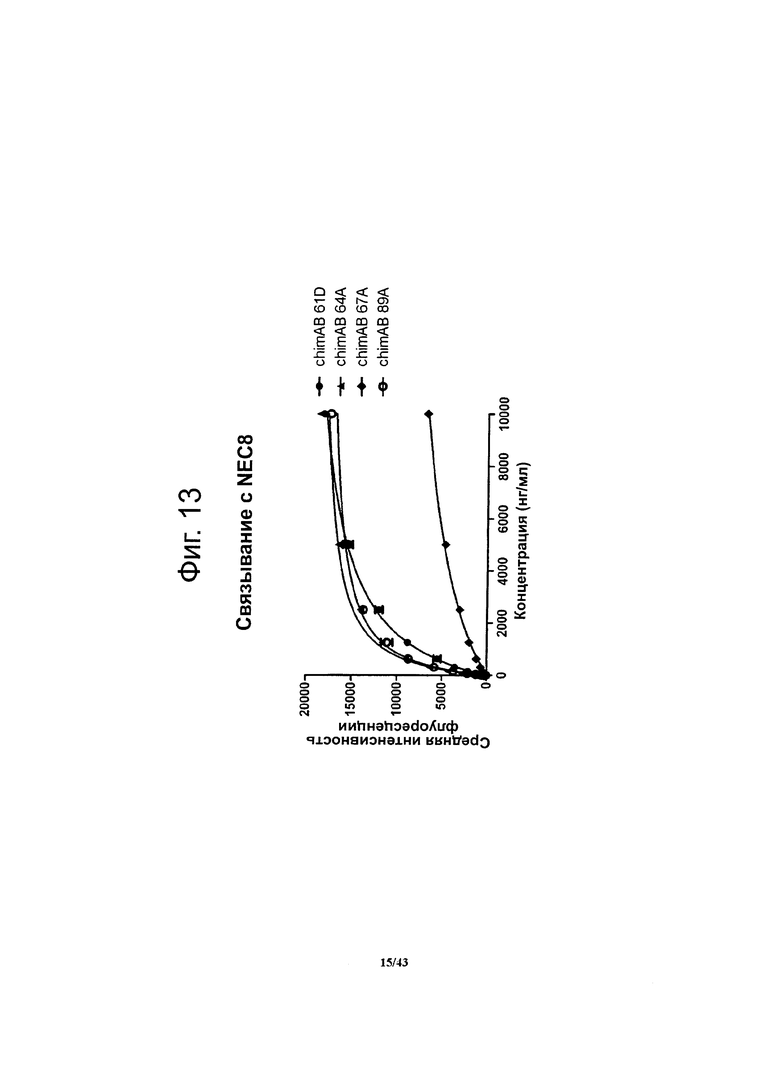
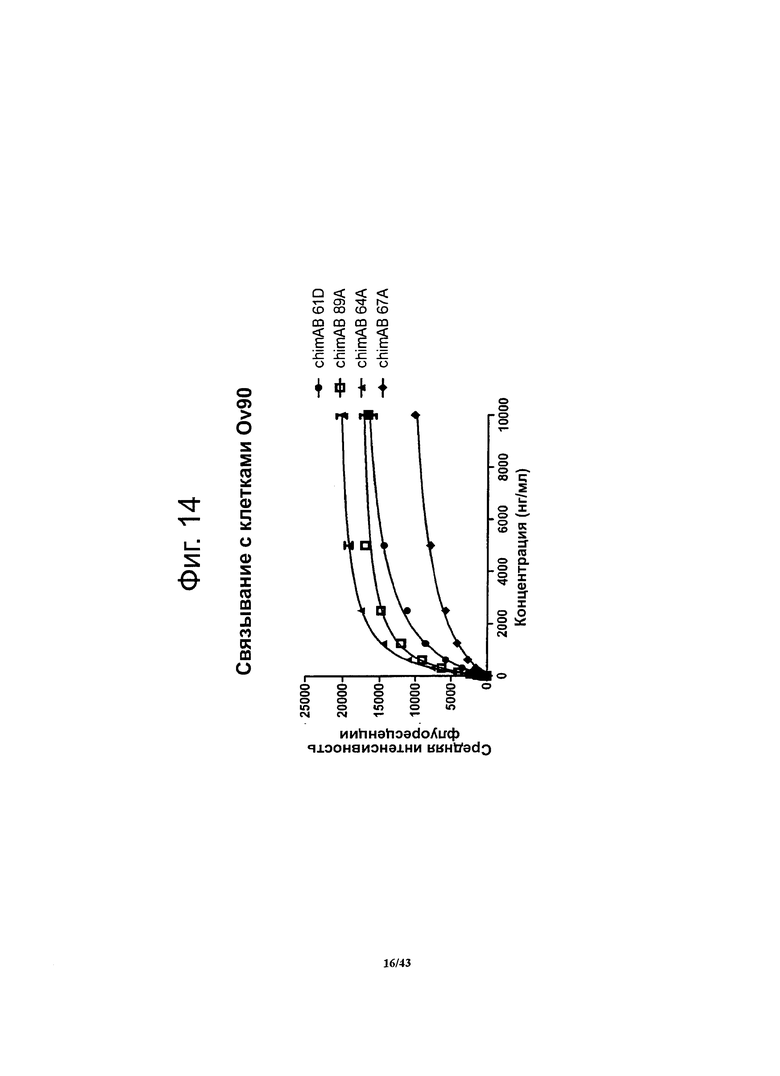
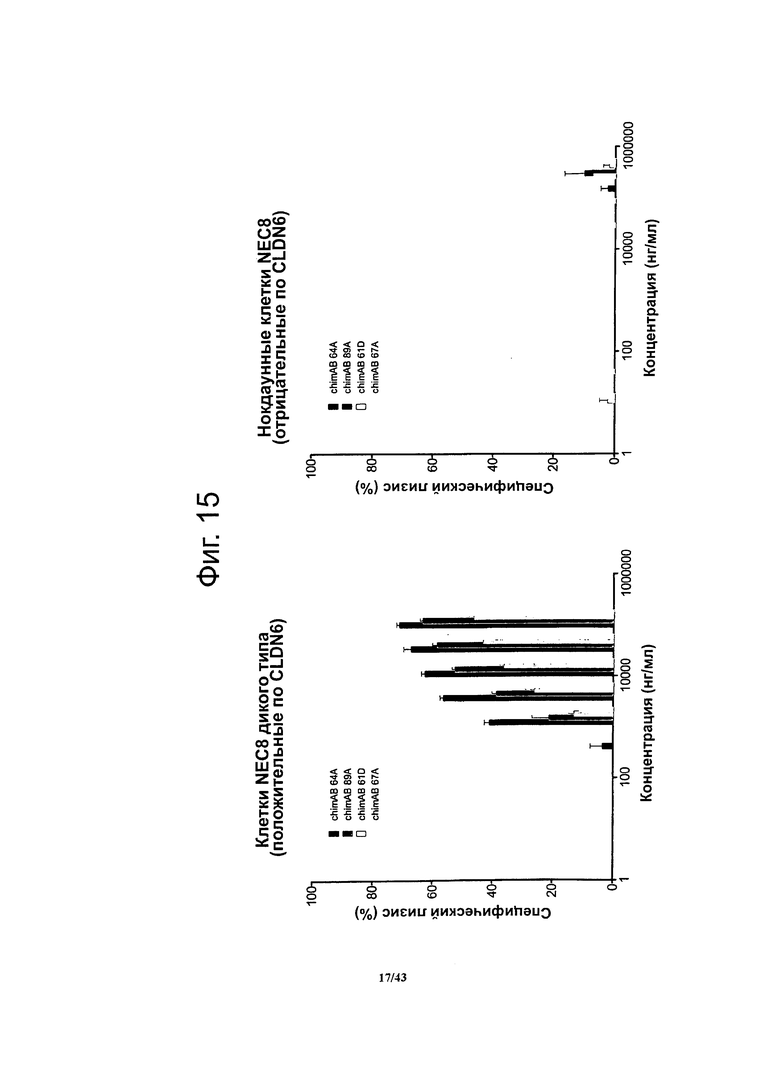
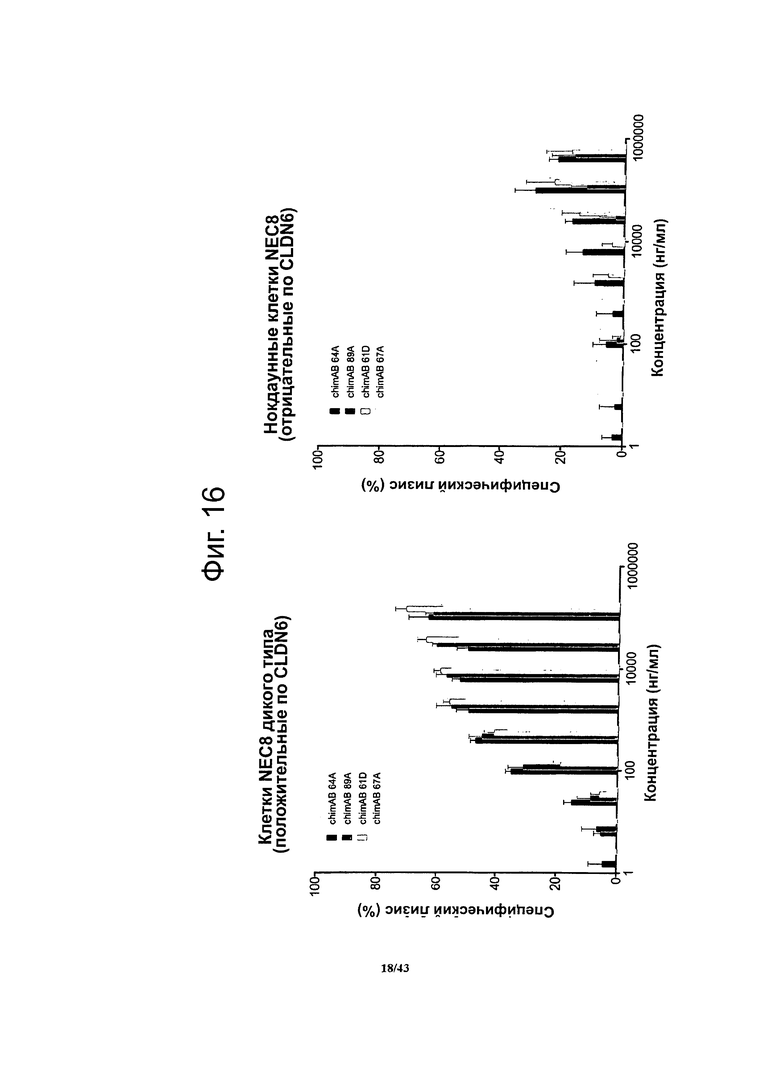
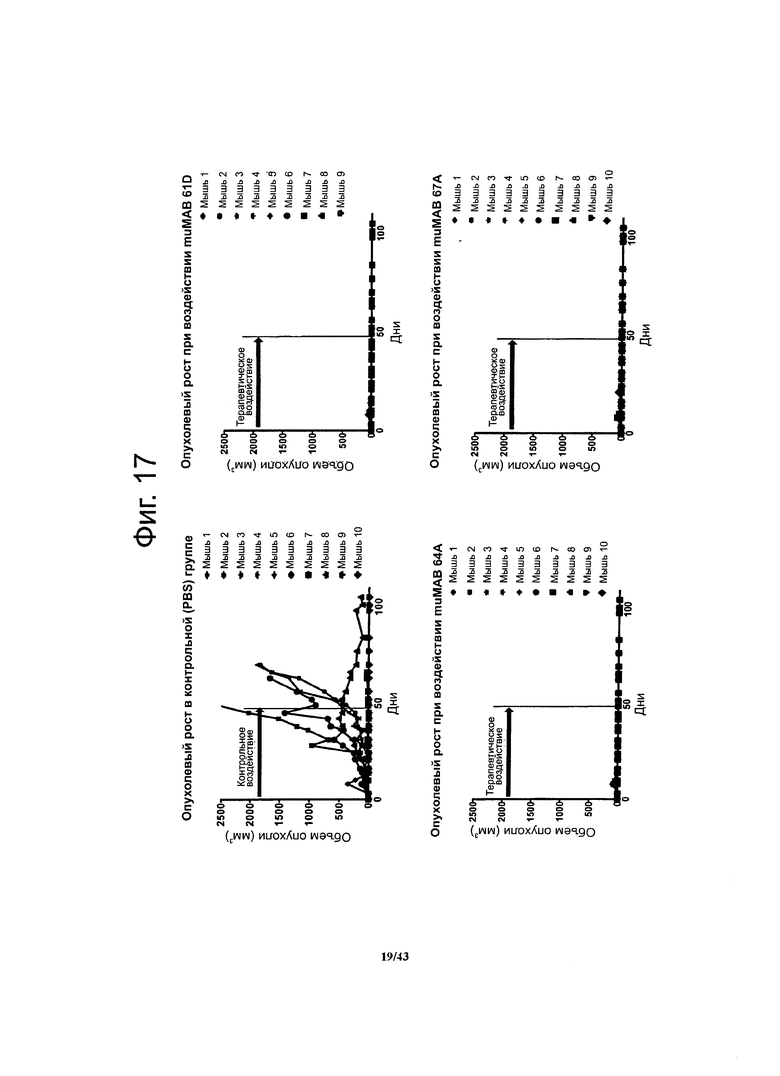
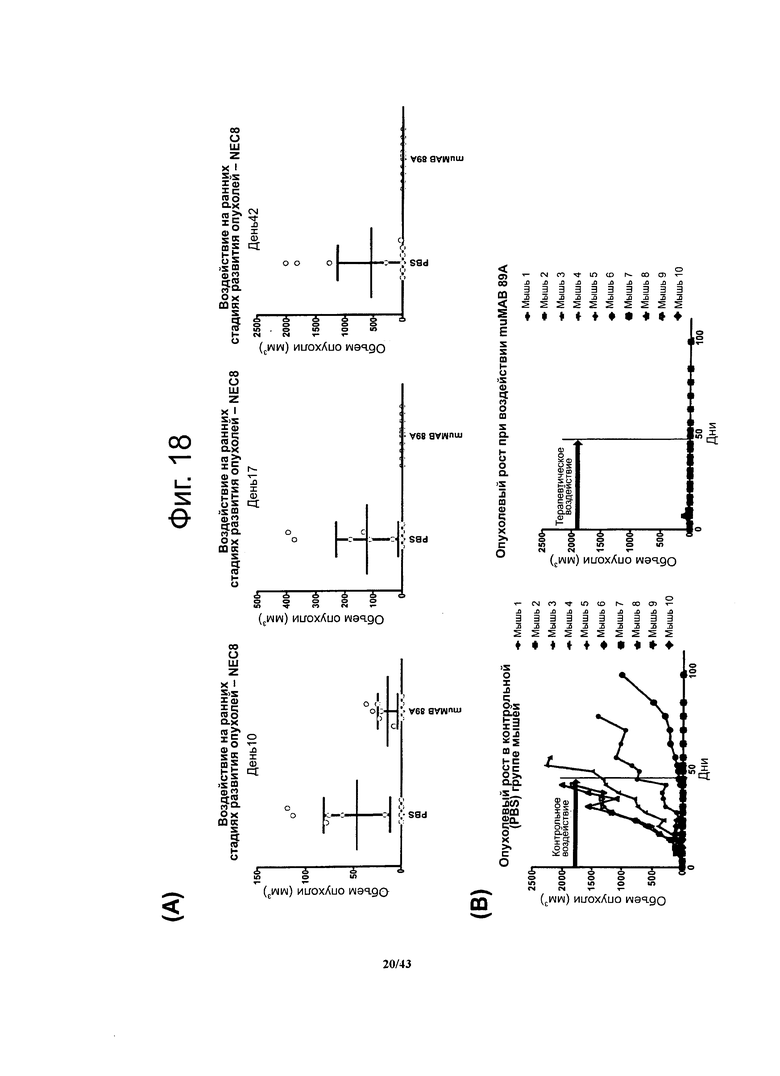

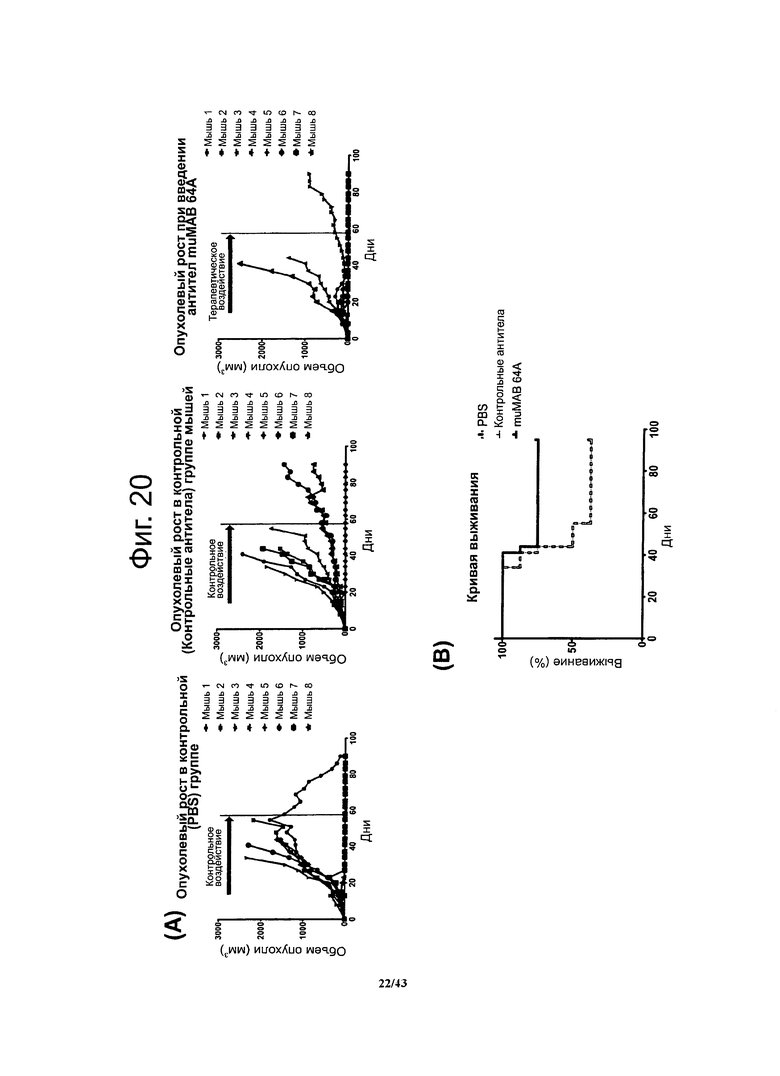

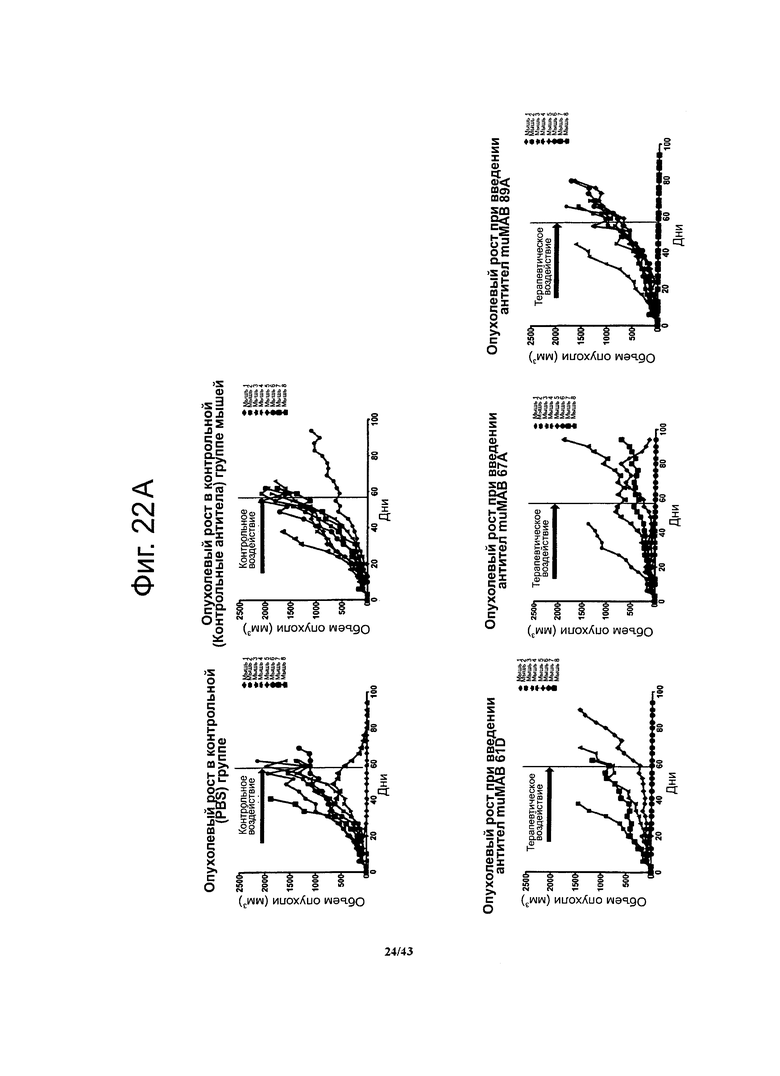
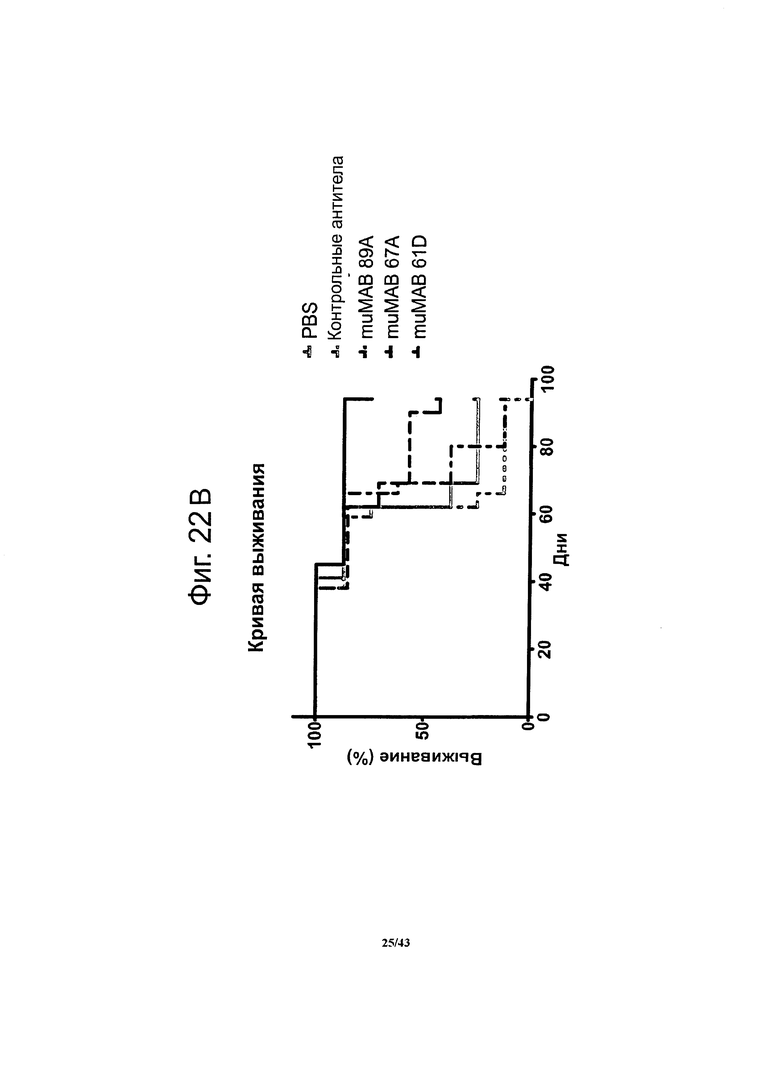

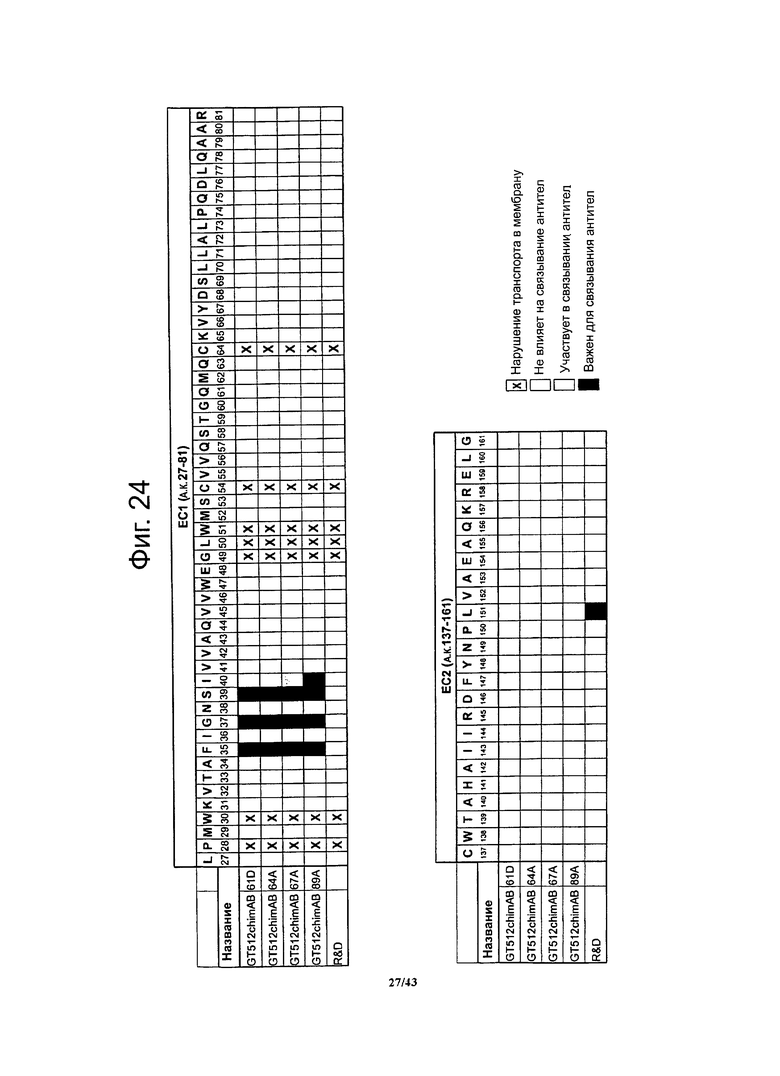
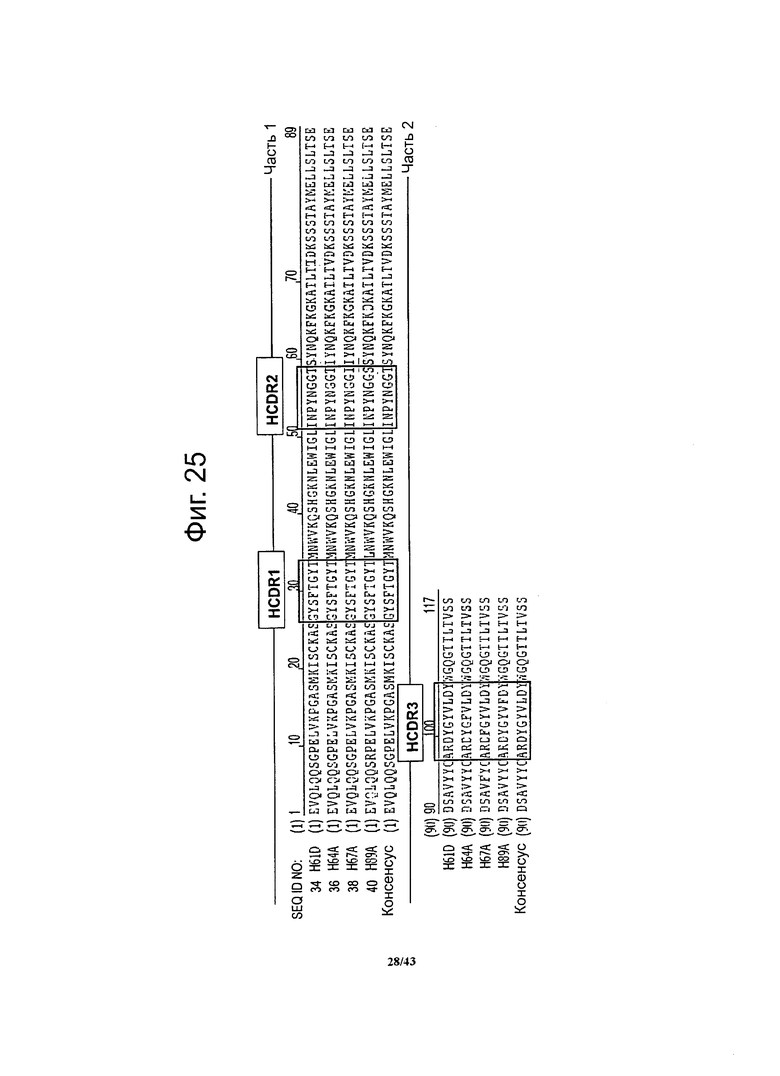
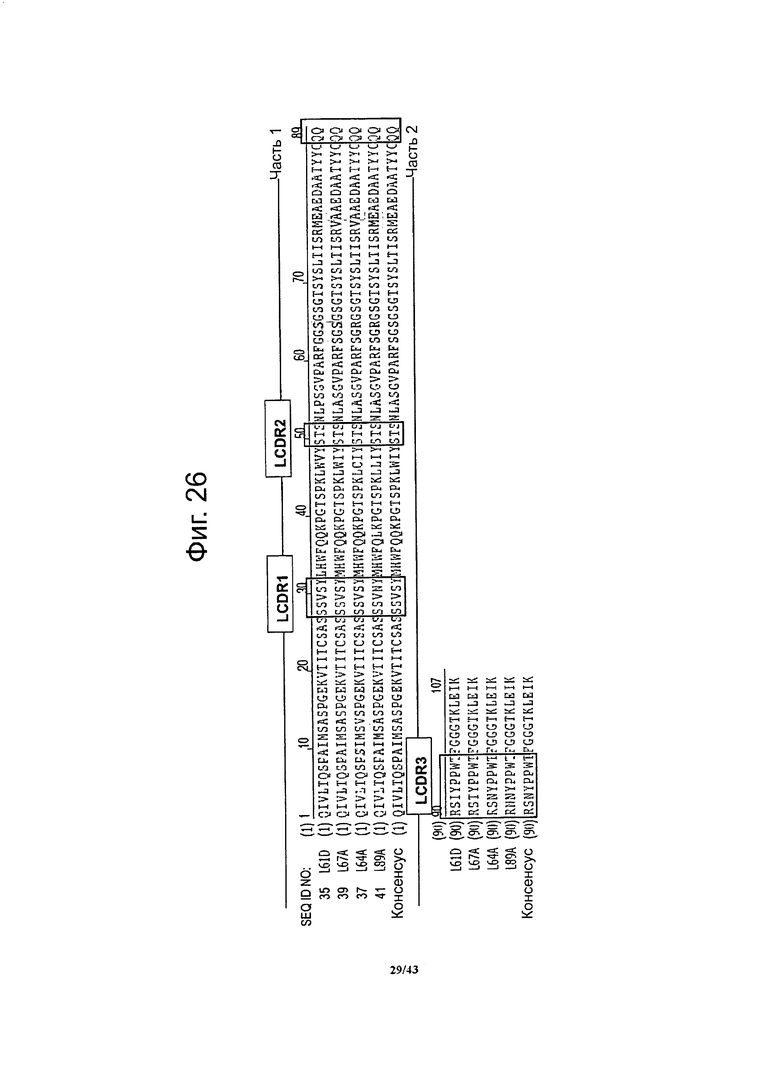
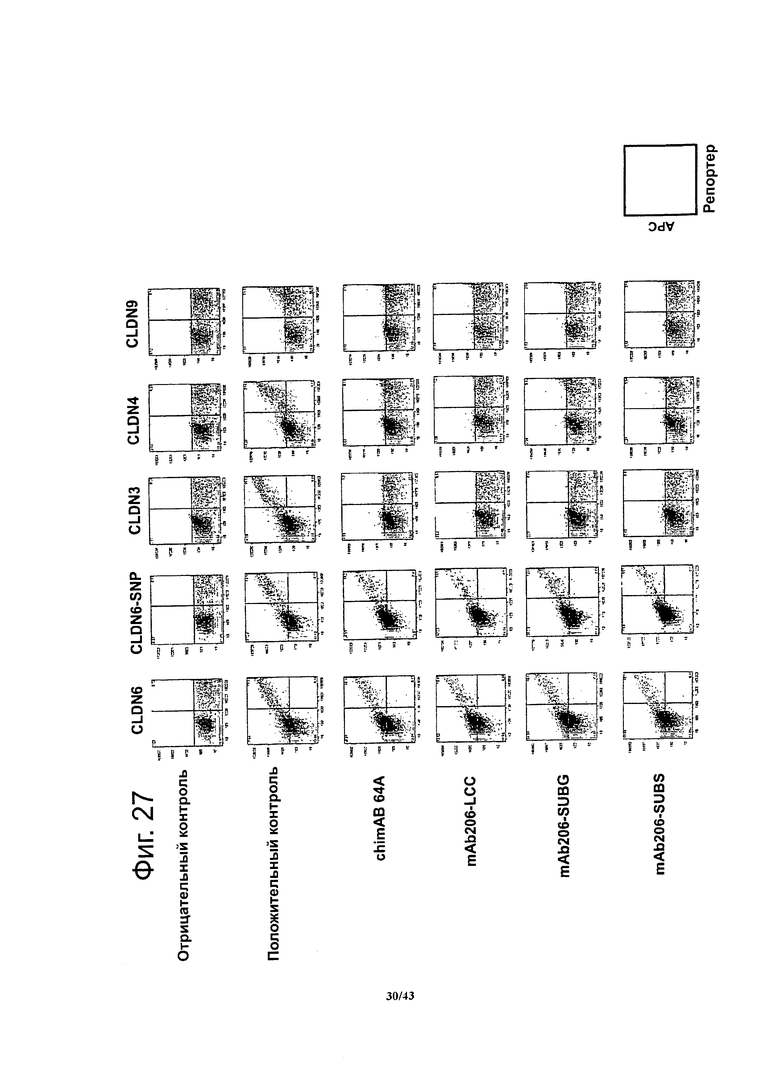

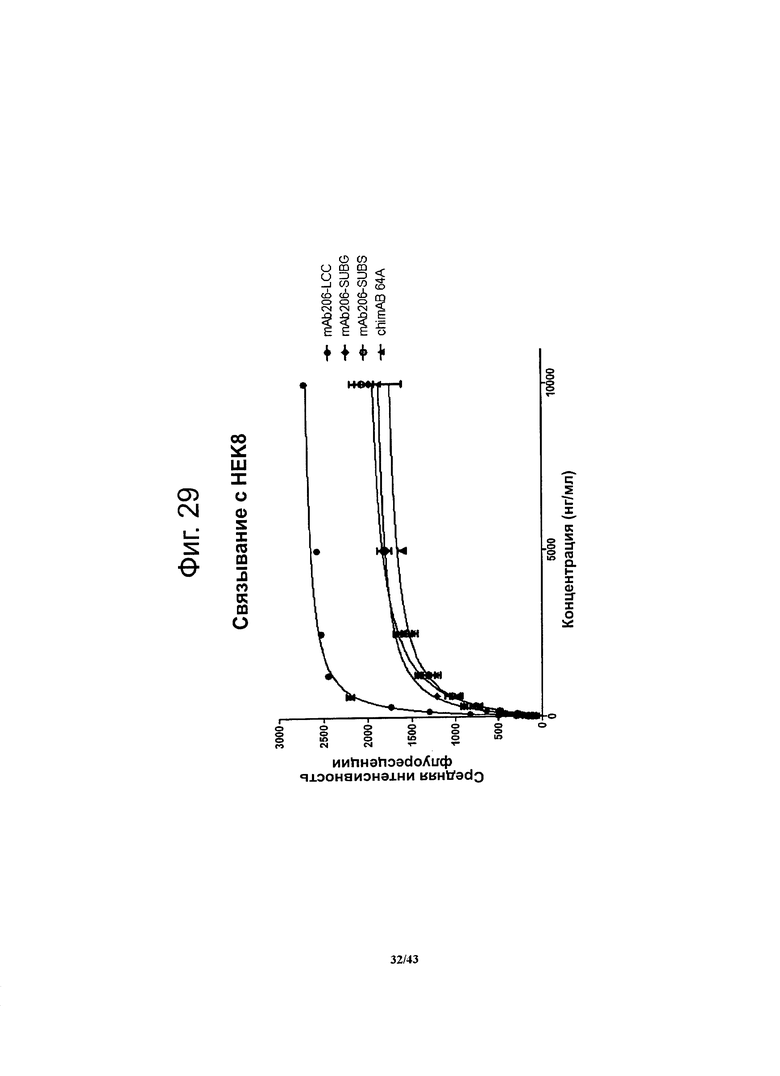
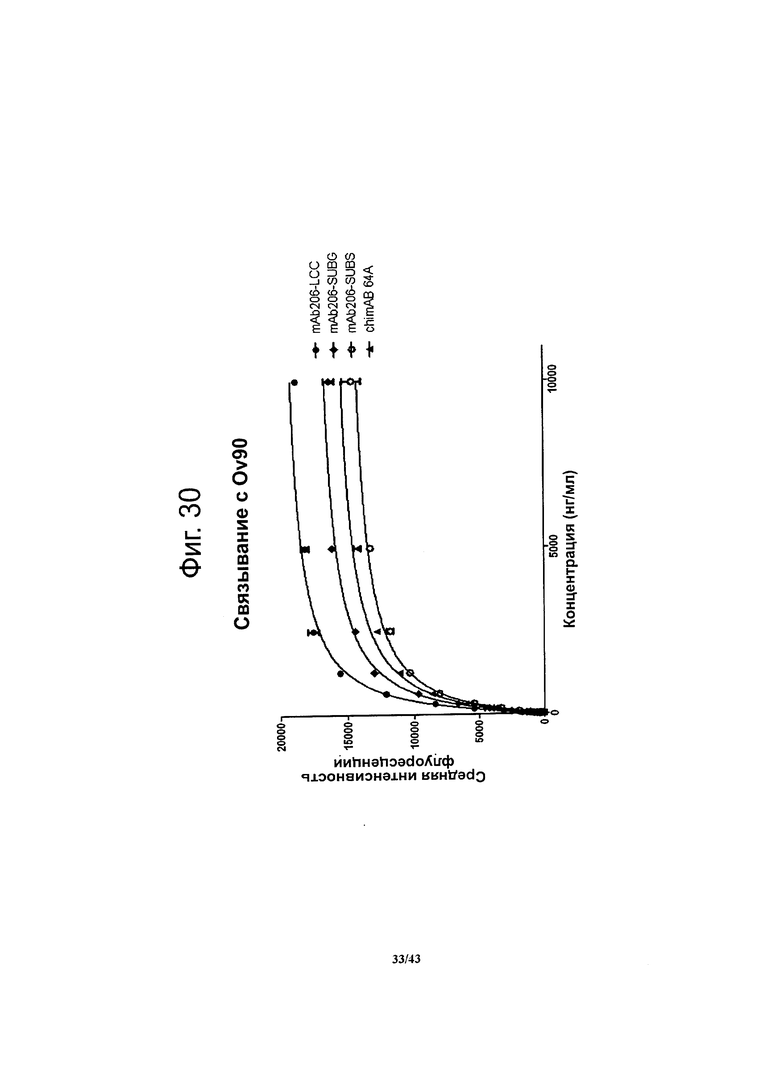
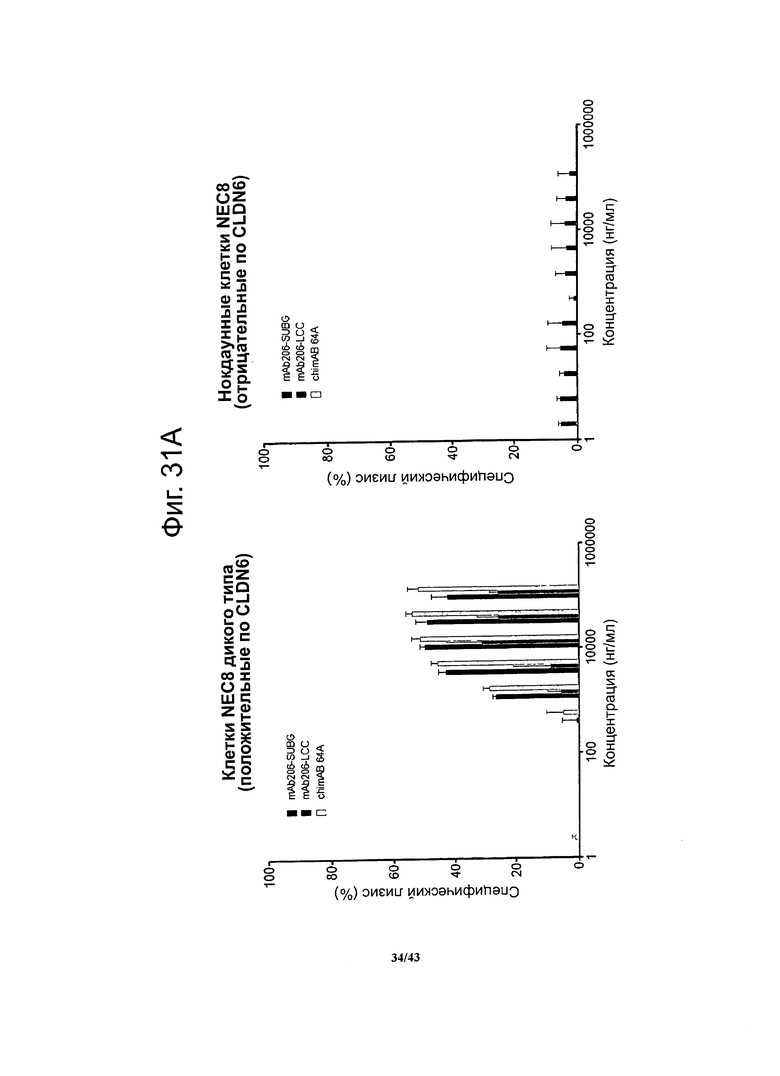
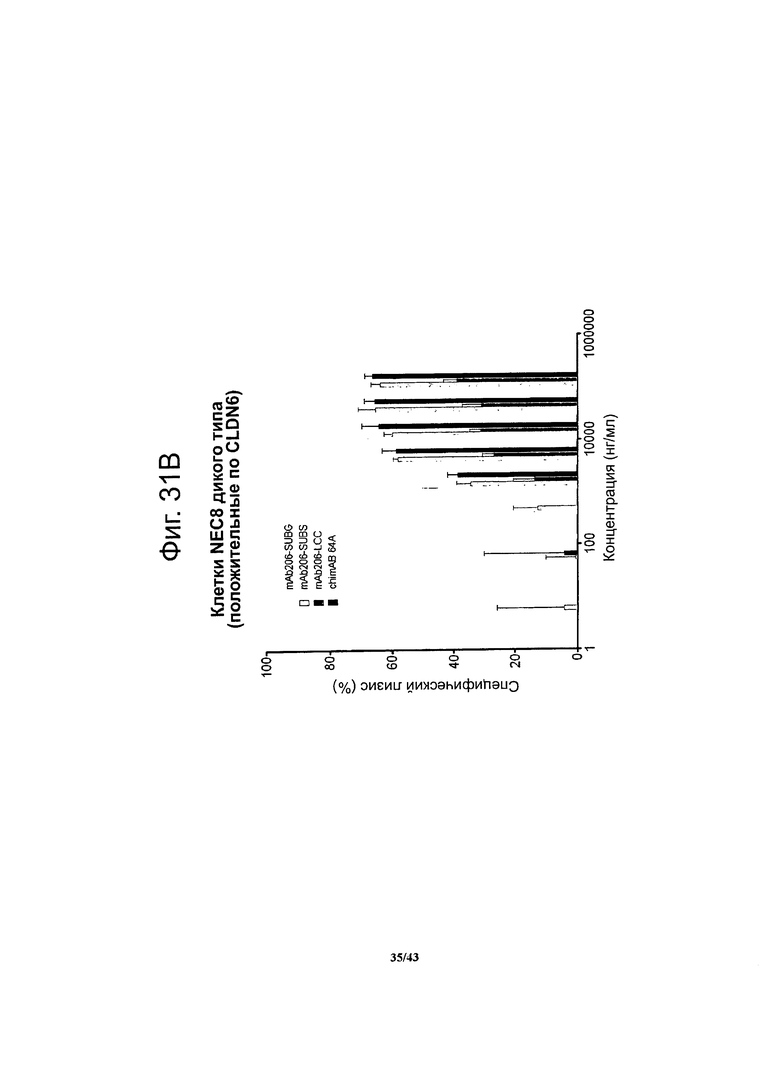
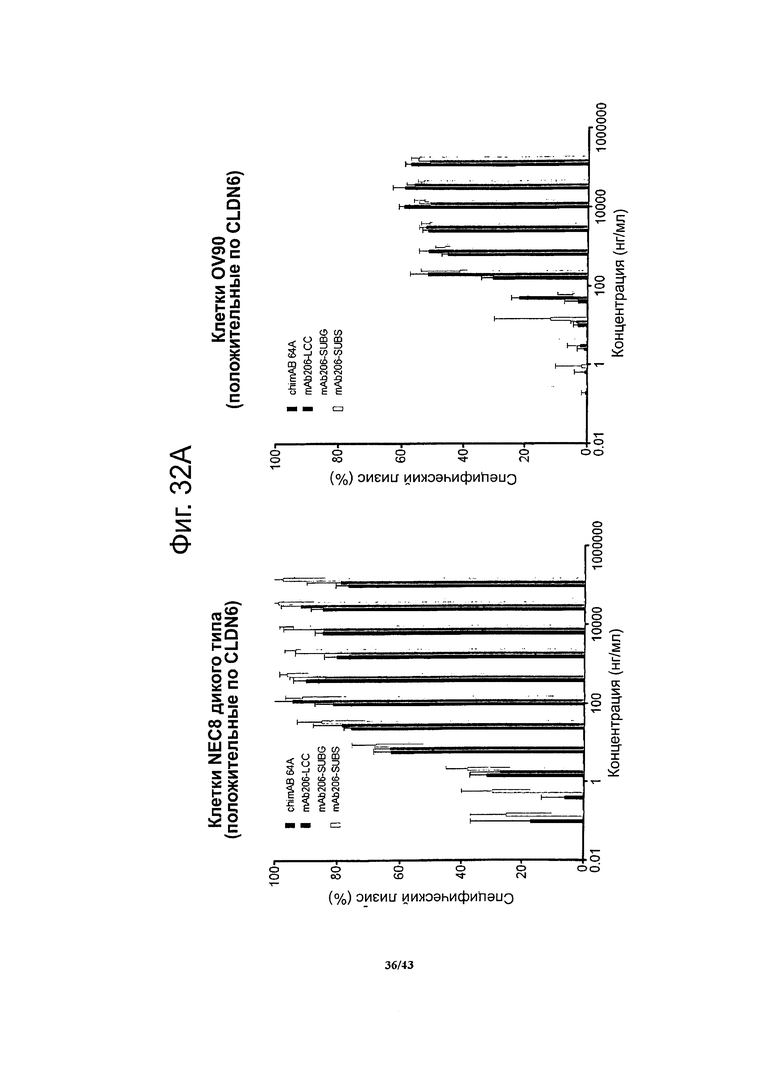
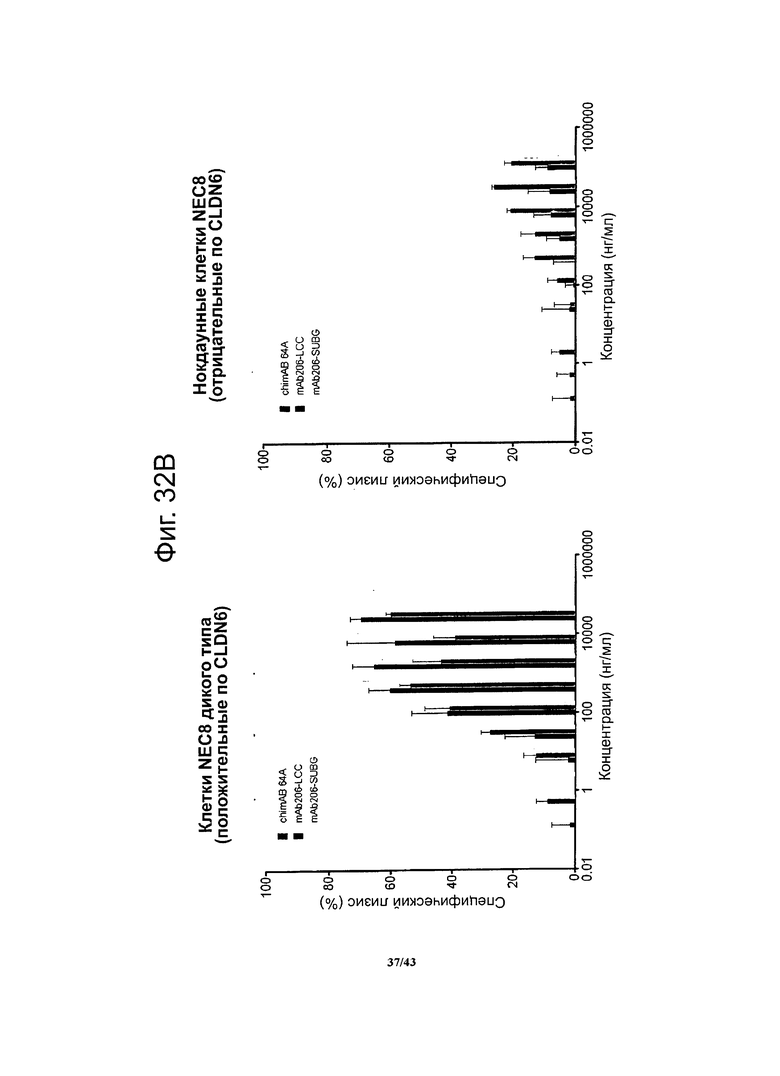

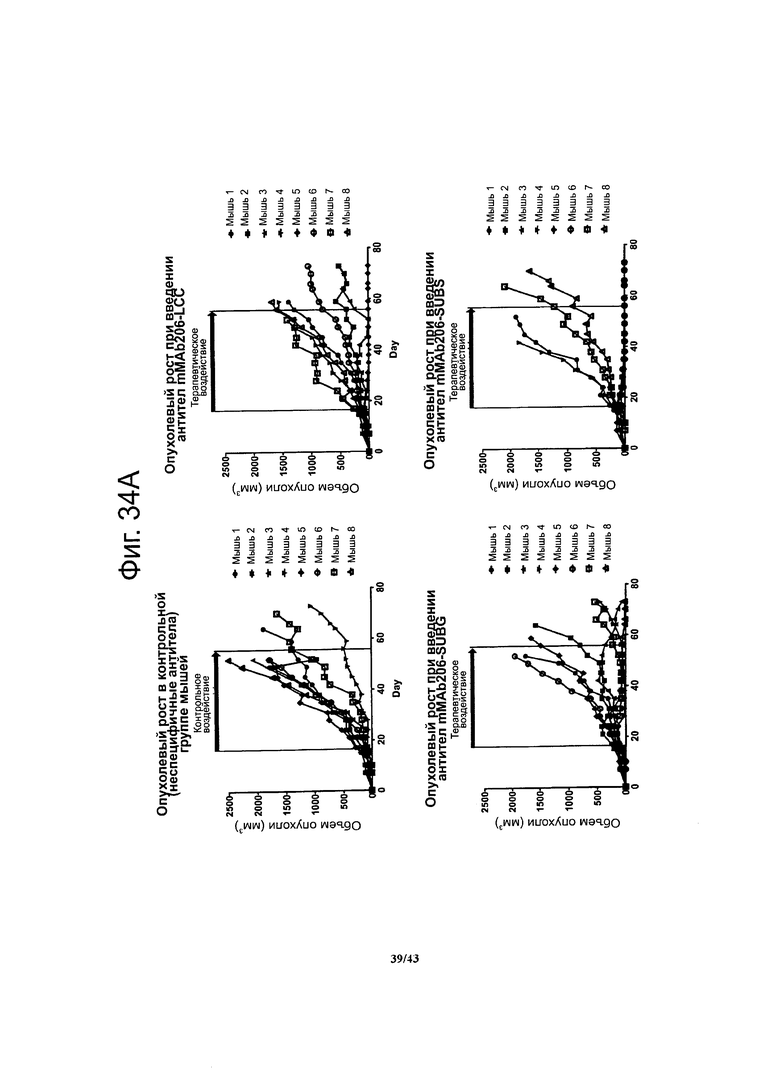
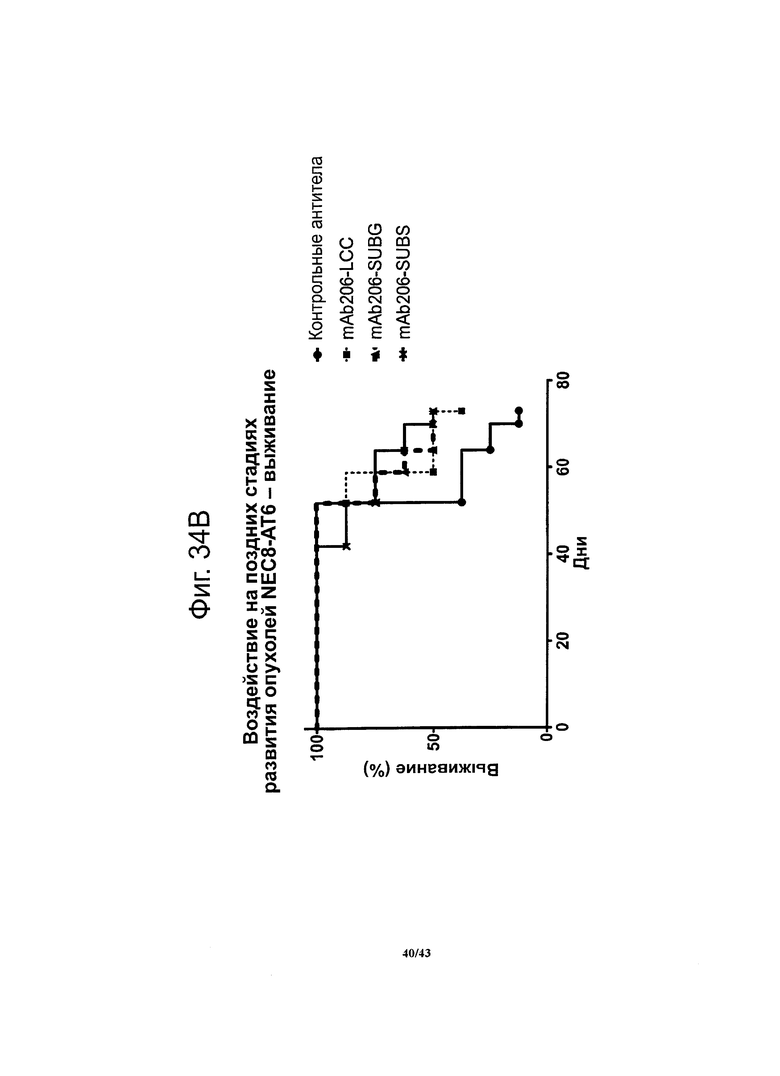
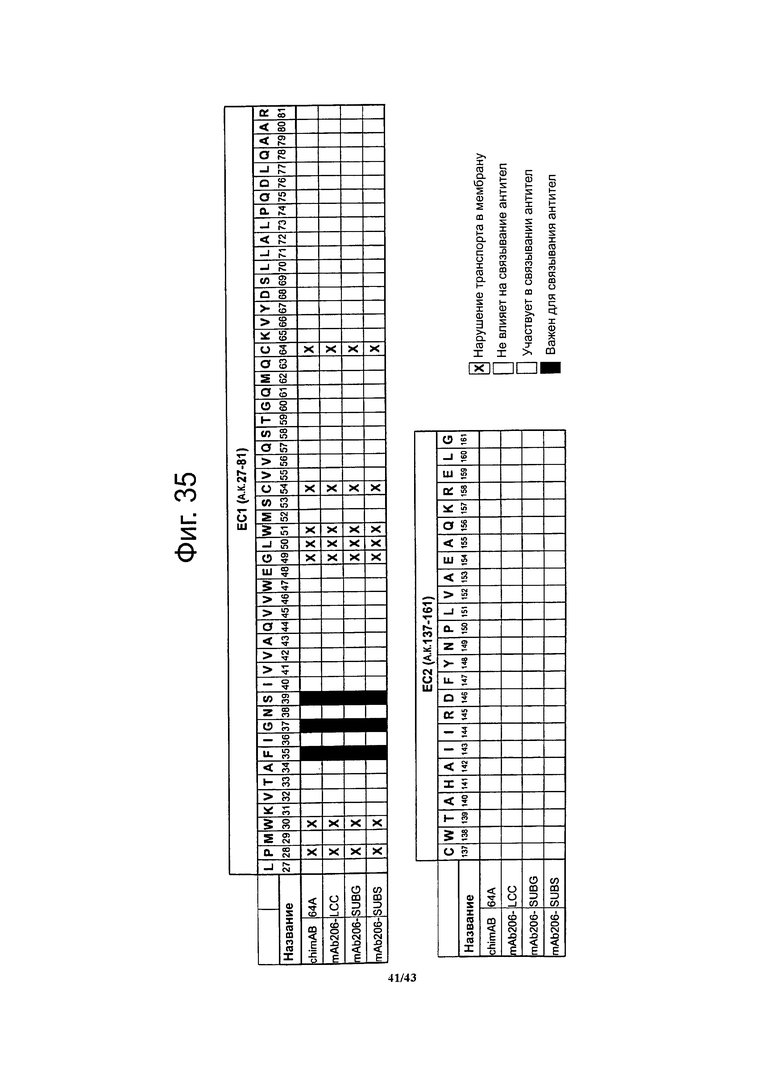
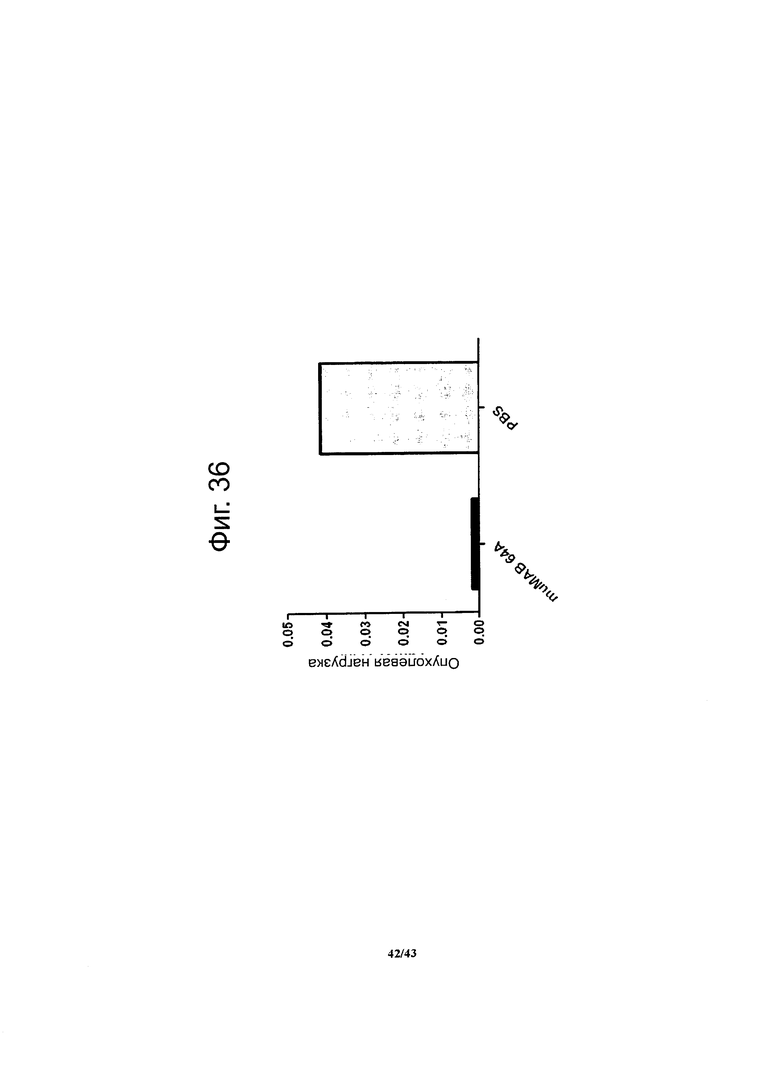
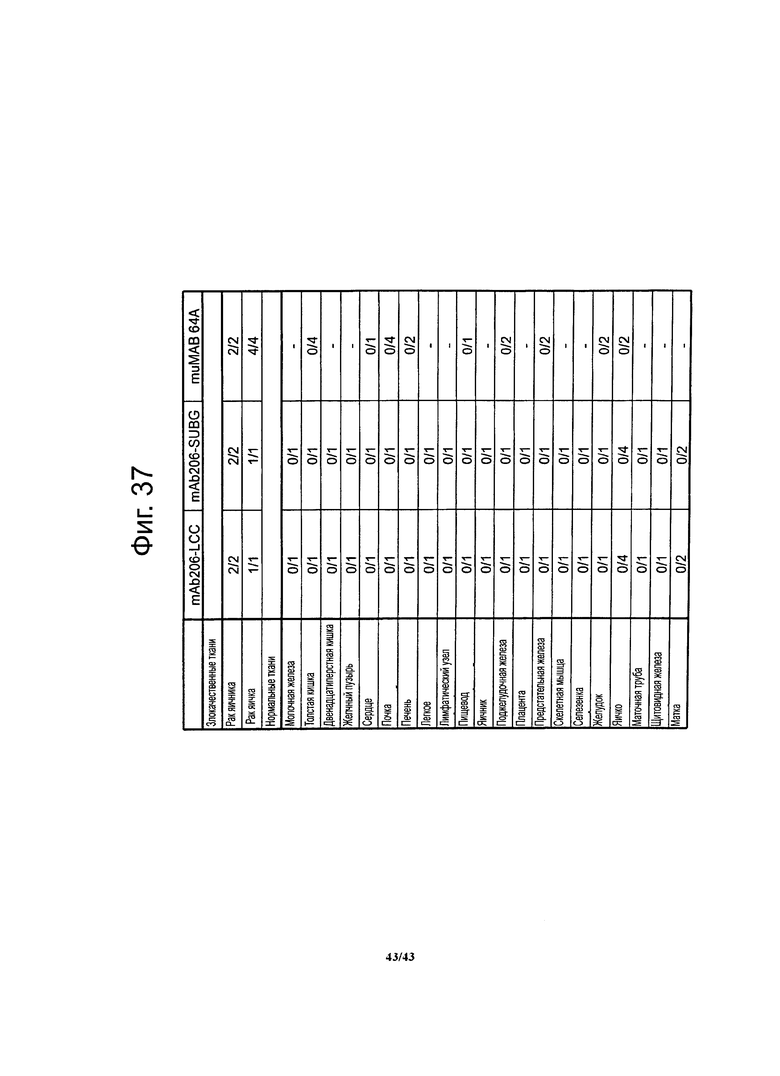
Изобретение относится к области биохимии, в частности к антителу, связывающемуся с CLDN6. Также раскрыты: способ ингибирования роста клеток, способ уничтожения клетки, способ лечения или предотвращения заболевания или расстройства, способ ингибирования метастатического распространения клеток – с помощью указанного антитела. Раскрыты конъюгат, включающий указанное антитело, фармацевтическая композиция, содержащая указанное антитело. Изобретение позволяет эффективно лечить заболевания, ассоциированные с CLDN6. 9 н. и 21 з.п. ф-лы, 43 ил., 1 табл., 10 пр.
1. Антитело, специфически связывающееся с CLDN6, ассоциированным с поверхностью клетки, в которой экспрессируется CLDN6, содержащее:
(i) тяжелую цепь антитела, включающую последовательность тяжелой цепи антитела, выбираемую из SEQ ID NO: 34, 36, 38 и 40, предпочтительно SEQ ID NO: 36, и
(ii) легкую цепь антитела, включающую последовательность легкой цепи антитела, выбираемую из SEQ ID NO: 35, 37, 39, 41, 54 и 55, предпочтительно SEQ ID NO: 35,
где указанное антитело связывается с указанным CLDN6 с KD, которая по меньшей мере в 10 раз ниже, чем KD при связывании с мишенью, для которой антитело не является специфическим.
2. Антитело по п. 1, содержащее:
(i) тяжелую цепь антитела, включающую последовательность тяжелой цепи антитела SEQ ID NO: 36 и
(ii) легкую цепь антитела, включающую последовательность легкой цепи антитела, выбираемую из SEQ ID NO: 35, 54 и 55.
3. Антитело по п. 1, содержащее:
(i) тяжелую цепь антитела, включающую последовательность тяжелой цепи антитела SEQ ID NO: 36 и
(ii) легкую цепь антитела, включающую последовательность легкой цепи антитела, выбираемую из SEQ ID NO: 54 и 55.
4. Антитело по п. 1, содержащее:
(i) тяжелую цепь антитела, включающую последовательность тяжелой цепи антитела SEQ ID NO: 36 и
(ii) легкую цепь антитела, включающую последовательность легкой цепи антитела, выбираемую из SEQ ID NO: 35.
5. Антитело по любому из пп. 1-4, способное связываться с CLDN6, ассоциированным с поверхностью клетки, в которой экспрессируется CLDN6, и в основном не способное связываться с CLDN9, ассоциированным с поверхностью клетки, в которой экспрессируется CLDN9.
6. Антитело по п. 5, в основном не способное связываться с CLDN4, ассоциированным с поверхностью клетки, в которой экспрессируется CLDN4, и/или в основном не способное связываться с CLDN3, ассоциированным с поверхностью клетки, в которой экспрессируется CLDN3.
7. Антитело по п. 5, отличающееся тем, что указанные клетки являются интактными, в частности непермеабилизированными.
8. Антитело по любому из пп. 1-4, способное связываться с эпитопами, расположенными во внеклеточной части CLDN6.
9. Антитело по п. 5, отличающееся тем, что CLDN6 имеет аминокислотную последовательность SEQ ID NO: 2 или аминокислотную последовательность SEQ ID NO: 8.
10. Антитело по п. 1, отличающееся тем, что CLDN6 имеет аминокислотную последовательность SEQ ID NO: 2 или аминокислотную последовательность SEQ ID NO: 8.
11. Антитело по п. 8, отличающееся тем, что CLDN6 имеет аминокислотную последовательность SEQ ID NO: 2 или аминокислотную последовательность SEQ ID NO: 8.
12. Антитело по любому из пп. 1-4, обладающее одной или более из следующих активностей:
(i) вызывает уничтожение клетки, в которой экспрессируется CLDN6,
(ii) ингибирует пролиферацию клетки, в которой экспрессируется CLDN6,
(iii) ингибирует образование колонии клеток, в которых экспрессируется CLDN6, и
(vi) ингибирует метастазирование клетки, в которой экспрессируется CLDN6.
13. Антитело по любому из пп. 1-4, которое способно ингибировать пролиферацию или опосредовать уничтожение клетки, несущей CLDN6 на ее поверхности в его нативной конформации, причем указанное уничтожение индуцируется комплементзависимой цитотоксичностью (CDC) или антителозависимой клеточно-опосредованной цитотоксичностью (ADCC).
14. Антитело по п. 12, отличающееся тем, что указанное ингибирование пролиферации или уничтожение индуцируется связыванием указанного антитела с эпитопом, расположенным во внеклеточной части CLDN6.
15. Антитело по п. 5, отличающееся тем, что указанная клетка с экспрессией CLDN6 является опухолевой или раковой клеткой.
16. Антитело по п. 13, отличающееся тем, что указанная клетка, несущая CLDN6 в его нативной конформации, является опухолевой или раковой клеткой.
17. Антитело по п. 15 или 16, отличающееся тем, что указанная опухолевая или раковая клетка происходит из опухоли или рака, выбираемого из группы, состоящей из рака яичника, в том числе аденокарциномы яичника и тератокарциномы яичника, рака легких, в том числе мелкоклеточного рака легких (SCLC) и немелкоклеточного рака легких (NSCLC), в частности плоскоклеточной карциномы легких и аденокарциномы, рака желудка, рака молочной железы, рака печени, рака поджелудочной железы, рака кожи, в частности базальноклеточной карциномы и плоскоклеточной карциномы, злокачественной меланомы, рака головы и шеи, в том числе злокачественной полиморфной аденомы, саркомы, в частности синовиальной саркомы и карциносаркомы, рака желчного протока, рака мочевого пузыря, в частности переходноклеточной карциномы и папиллярной карциномы, рака почек, в частности почечно-клеточной карциномы, включая светлоклеточную почечно-клеточную карциному и папиллярную почечно-клеточную карциному, рака толстого кишечника, рака тонкого кишечника, включая рак подвздошной кишки, в частности аденокарциному тонкого кишечника и аденокарциному подвздошной кишки, эмбриональной карциномы яичка, хориокарциномы плацентарной площадки, рака шейки матки, рака яичка, в частности семиномы яичка, тератомы яичка и эмбрионального рака яичка, рака матки, герминогенных опухолей, например тератокарциномы или эмбриональной карциномы, в частности герминогенной опухоли яичка, и их метастазирующих форм.
18. Антитело по любому из пп. 1-4, являющееся моноклональным антителом.
19. Конъюгат, включающий антитело по любому из пп. 1-18, объединенное с терапевтическим агентом, для лечения или предотвращения заболевания или расстройства, характеризующегося наличием клетки, экспрессирующей CLDN6, у которой CLDN6 связан с ее клеточной поверхностью, у субъекта.
20. Конъюгат, включающий антитело по любому из пп. 1-18, объединенное с терапевтическим агентом, для ингибирования пролиферации клетки, экспрессирующей CLDN6 и отличающейся тем, что CLDN6 связан с ее поверхностью.
21. Конъюгат по п. 19 или 20, в котором терапевтический агент является токсином, радиоактивным изотопом, лекарственным веществом или цитотоксическим агентом.
22. Фармацевтическая композиция, содержащая терапевтически эффективное количество антитела по любому из пп. 1-18 и/или конъюгат по любому из пп. 19-21 и фармацевтически приемлемый носитель, для лечения или предотвращения заболевания или расстройства, характеризующегося наличием клетки, экспрессирующей CLDN6, у которой CLDN6 связан с ее клеточной поверхностью, у субъекта.
23. Фармацевтическая композиция, содержащая терапевтически эффективное количество антитела по любому из пп. 1-18 и/или конъюгат по любому из пп. 19-21 и фармацевтически приемлемый носитель, для ингибирования пролиферации клетки, экспрессирующей CLDN6 и отличающейся тем, что CLDN6 связан с ее поверхностью
24. Способ ингибирования роста клетки, в которой экспрессируется CLDN6 и которая отличается тем, что CLDN6 связан с ее поверхностью, включающий контакт указанной клетки с антителом по любому из пп. 1-18 и/или конъюгатом по любому из пп. 19-21.
25. Способ уничтожения клетки, в которой экспрессируется CLDN6 и которая отличается тем, что CLDN6 связан с ее поверхностью, включающий контакт указанной клетки с антителом по любому из пп. 1-18 и/или конъюгатом по любому из пп. 19-21.
26. Способ лечения или предотвращения заболевания или расстройства, затрагивающего клетку, в которой экспрессируется CLDN6 и которая отличается тем, что CLDN6 связан с ее поверхностью, включающий введение указанному индивиду антитела по любому из пп. 1-18, конъюгата по любому из пп. 19-21 или фармацевтической композиции по п. 22 или 23.
27. Способ по п. 26, в котором указанное заболевание или расстройство является заболеванием, связанным с образованием опухолей.
28. Способ по п. 27, в котором заболевание, связанное с образованием опухолей, является раком.
29. Способ по п. 28, в котором раковое заболевание выбирают из группы, состоящей из рака яичника, в том числе аденокарциномы яичника и тератокарциномы яичника, рака легких, в том числе мелкоклеточного рака легких (SCLC) и немелкоклеточного рака легких (NSCLC), в частности плоскоклеточной карциномы легких и аденокарциномы, рака желудка, рака молочной железы, рака печени, рака поджелудочной железы, рака кожи, в частности базальноклеточной карциномы и плоскоклеточной карциномы, злокачественной меланомы, рака головы и шеи, в том числе злокачественной полиморфной аденомы, саркомы, в частности синовиальной саркомы и карциносаркомы, рака желчного протока, рака мочевого пузыря, в частности переходноклеточной карциномы и папиллярной карциномы, рака почек, в частности почечно-клеточной карциномы, включая светлоклеточную почечно-клеточную карциному и папиллярную почечно-клеточную карциному, рака толстого кишечника, рака тонкого кишечника, включая рак подвздошной кишки, в частности аденокарциному тонкого кишечника и аденокарциному подвздошной кишки, эмбриональной карциномы яичка, хориокарциномы плацентарной площадки, рака шейки матки, рака яичка, в частности семиномы яичка, тератомы яичка и эмбрионального рака яичка, рака матки, герминогенных опухолей, например тератокарциномы или эмбриональной карциномы, в частности герминогенной опухоли яичка, и их метастазирующих форм.
30. Способ ингибирования метастатического распространения клеток, в которых экспрессируется CLDN6 и которые отличаются тем, что CLDN6 связан с их поверхностью, включающий контакт указанных клеток с антителами по любому из пп. 1-18 и/или конъюгатом по любому из пп. 19-21.
| WO 2010094499 A1, 28.08.2010 | |||
| ARABZADEH A | |||
| et al., Changes in the distribution pattern of Claudin tight junction proteins during the progression of mouse skin tumorigenesis, BMC CANCER, 2007, vol.7:196, doi:10.1186/1471-2407-7-196 | |||
| КОМБИНИРОВАННЫЙ ИГЛОУПРОЧНЯЮЩИЙ ИНСТРУМЕНТ | 2003 |
|
RU2241578C1 |
| RU 2008125324 A, 27.12.2009. | |||

