Уровень техники
Рак поджелудочной железы является одним из самых летальных видов рака. Смертность достигает 100% из-за склонности к раннему метастазированию и вследствие того, что болезнь очень устойчива к лучевой и химиотерапии. Учитывая, что каждый год диагностируется 27000 новых случаев в Северной Америке и 68000 в Европе, существует настоятельная потребность в разработке новых стратегий лечения для снижения смертности у больных раком поджелудочной железы.
Сплайс-вариант 2 молекулы плотных контактов клаудина 18 (claudin 18.2, или CLDN18.2) входит в семейство белков плотных контактов - клаудинов. CLDN18.2 - трансмембранный белок в 27,8 кДа, содержащий четыре трансмембранных домена с двумя небольшими внеклеточными петлями. В нормальных тканях экспрессия CLDN18.2 методом ОТ-ПЦР не обнаружена, за исключением желудка. Иммуногистохимия с помощью специфичных к CLDN18.2 антител показала, что положительную реакцию дает единственная ткань - желудок. CLDN18.2 является весьма избирательным антигеном желудочной линии, которая экспрессируется исключительно в короткоживущих дифференцированных эпителиальных клетках желудка. CLDN18.2 сохраняется при злокачественной трансформации и поэтому часто выступает на поверхности раковых клеток желудка человека. Кроме того, этот пан-опухолевый антиген активируется эктопически на существенном уровне в клетках аденокарциномы пищевода, поджелудочной железы и легких.
Фирмой Ganymed Pharmaceuticals AG было разработано химерное антитело IMAB362 типа IgG1, направленное против CLDN18.2. IMAB362 распознает первый внеклеточный домен (ECD1) CLDN18.2 с высоким сродством и специфичностью. IMAB362 не связывается с другими представителями семейства, включая близкородственный сплайс-вариант 1 клаудина 18 (CLDN18.1). IMAB362 проявляет сильную специфичность к опухолевым клеткам и в нем сочетаются четыре независимых высокоактивных механизма действия. После связывания с мишенью IMAB362 опосредует уничтожение клеток посредством ADCC, CDC и индукции апоптоза, вызванного перекрестными сшивками мишени на поверхности опухолевых клеток и непосредственным ингибированием пролиферации. При этом IMAB362 эффективно вызывает лизис CLDN18.2-положительных клеток, в том числе клеток раковых линий желудка человека in vitro и in vivo.
Токсичность и PK/TK-профиль IMAB362 тщательно исследовались на мышах и яванских макаках, включая исследования по выявлению дозового диапазона, 28-дневные исследования по токсичности повторных доз на макаках и 3-месячные исследования по токсичности повторных доз на мышах. Как мыши (наибольшая продолжительность обработки при еженедельном введении - 3 месяца, максимальный уровень доз - 400 мг/кг), так и яванские макаки (до 5 еженедельных введений вплоть до 100 мг/кг) хорошо переносили повторные дозы IMAB362 в/в. Не отмечалось никаких признаков системной или местной токсичности. В частности, ни в одном исследовании по токсичности не отмечалось желудочной токсичности. IMAB362 не вызывает иммунной активации и высвобождения цитокинов. Не отмечалось никаких отрицательных эффектов на репродуктивные органы у самцов или самок. IMAB362 не связывается с такими тканями, в которых нет мишени. Исследования по биораспределению на мышах показали, что отсутствие желудочной токсичности, скорее всего, обусловлено компартментализацией плотных контактов на люминальной стороне нормального эпителия желудка, что сильно затрудняет доступ к эпитопам IMAB362.
IMAB362 находится в начале клинических испытаний. Проводилась I фаза клинических испытаний на людях. По 3 пациента в 5 дозовых группах (33 мг/м2, 100 мг/м2, 300 мг/м2, 600 мг/м2, 1000 мг/м2) получали внутривенно однократные дозы IMAB362 и наблюдались в течение 28 дней. IMAB362 очень хорошо переносился, при этом не было существенных замечаний по безопасности у пациентов. У одного пациента все измеряемые опухолевые маркеры значительно уменьшались на протяжении 4 недель после лечения. В текущей фазе IIа клинических испытаний IMAB362 вводится неоднократно.
Далее здесь приводятся данные, свидетельствующие о том, что химиотерапевтические средства могут стабилизировать или усиливать экспрессию CLDN18.2 на поверхности раковых клеток поджелудочной железы, приводя к повышению доступности CLDN18.2 для антител против CLDN18.2 типа IMAB362. Наблюдалось синергическое действие антител против CLDN18.2 типа IMAB362 с определенными химиотерапевтическими препаратами, в частности, с химиотерапевтическими препаратами, используемыми для лечения рака поджелудочной железы. При химиотерапевтической обработке раковые клетки человека становятся более восприимчивыми к индуцируемой антителами мишень-специфичного уничтожения. На моделях опухолей у мышей контролирование опухолей с помощью антител против CLDN18.2 плюс химиотерапии превосходит таковое с помощью одних лишь антител против CLDN18.2 как единственного средства.
Сущность изобретения
Настоящим изобретением в общем предусмотрена комбинированная терапия для эффективного лечения и/или профилактики заболеваний, связанных с клетками, экспрессирующими CLDN18.2, включая такие раковые заболевания, как рак желудка, рак пищевода, рак поджелудочной железы, рак легких типа немелкоклеточного рака легких (NSCLC), рак яичников, рак толстой кишки, рак печени, рак головы и шеи и рак желчного пузыря, а также их метастазов, в частности метастазов рака желудка типа опухолей Крукенберга, метастазов в брюшине и метастазов в лимфатических узлах. Особенно предпочтительными раковыми заболеваниями являются рак поджелудочной железы и его метастазы.
В одном аспекте настоящего изобретения предусмотрен способ лечения или профилактики рака поджелудочной железы у пациентов, включающий введение пациентам (i) антитела, обладающего способностью к связыванию с CLDN18.2, и (ii) средства, стабилизирующего или усиливающего экспрессию, т.е. уровень CLDN18.2. Экспрессия CLDN18.2 предпочтительно происходит на клеточной поверхности раковых клеток. Средство, стабилизирующее или усиливающее экспрессию CLDN18.2, может вводиться до, одновременно с или после введения антитела, обладающего способностью к связыванию с CLDN18.2, или в комбинации с ним.
Средство, стабилизирующее или усиливающее экспрессию CLDN18.2, может быть цитотоксическим и/или цитостатическим средством. В одном воплощении средство, стабилизирующее или усиливающее экспрессию CLDN18.2, включает средство, которое индуцирует остановку клеточного цикла или накопление клеток в одной или нескольких фазах клеточного цикла, предпочтительно в одной или нескольких фазах клеточного цикла, отличных от фазы G1, типа фазы S, фазы G2 или их комбинации либо комбинации фазы S или фазы G2 с фазой G1. Средство, стабилизирующее или усиливающее экспрессию CLDN18.2, может включать средство, выбранное из группы, состоящей из аналогов нуклеозидов, соединений платины, аналогов камптотецина и таксанов, их пролекарственных форм, солей и комбинаций. Аналоги нуклеозидов можно выбирать из группы, состоящей из гемцитабина, 5-фторурацила, их пролекарственных форм и солей. Соединения платины можно выбирать из группы, состоящей из оксалиплатина, цисплатина, их пролекарственных форм и солей. Аналоги камптотецина можно выбирать из группы, состоящей из иринотекана, топотекана, их пролекарственных форм и солей. Таксаны можно выбирать из группы, состоящей из паклитакселя, доцетакселя, их пролекарственных форм и солей. Средство, стабилизирующее или усиливающее экспрессию CLDN18.2, может включать средство, выбранное из группы, состоящей из гемцитабина, 5-фторурацила, оксалиплатина, иринотекана, паклитакселя, их пролекарственных форм, солей и комбинаций. Средство, стабилизирующее или усиливающее экспрессию CLDN18.2, может включать комбинацию из оксалиплатина и 5-фторурацила или их пролекарственных форм, комбинацию из цисплатина и 5-фторураггила или их пролекарственных форм, комбинацию из по меньшей мере одного таксана и оксалиплатина, комбинацию из по меньшей мере одного таксана и цисплатина, комбинацию из по меньшей мере одного таксана и 5-фторурацила или его пролекарственных форм, либо комбинацию из по меньшей мере одного аналога камптотецина и 5-фторурацила или его пролекарственных форм. Средство, стабилизирующее или усиливающее экспрессию CLDN18.2, может включать комбинацию из гемцитабина и оксалиплатина, комбинацию из гемцитабина и цисплатина, комбинацию из гемцитабина и карбоплатина или комбинацию из оксалиплатина, 5-фторурацила или его пролекарственных форм и иринотекана. Соответственно, способ по изобретению может включать введение комбинации из гемцитабина и оксалиплатина, комбинации из гемцитабина и цисплатина, комбинации из гемцитабина и карбоплатина или комбинации из оксалиплатина, 5-фторурацила или его пролекарственных форм и иринотекана. В одном воплощении способ по изобретению включает введение фолиновой кислоты, 5-фторурацила или его пролекарственных форм, иринотекана и оксалиплатина. Средство, стабилизирующее или усиливающее экспрессию CLDN18.2, может включать средство, индуцирующее иммуногенную гибель клеток. Средство, индуцирующее иммуногенную гибель клеток, может включать оксалиплатин.
В следующем аспекте настоящего изобретения предусмотрен способ лечения или профилактики рака у пациентов, включающий введение пациентам (i) антитела, обладающего способностью к связыванию с CLDN18.2, и (ii) гемцитабина. В одном воплощении рак выбирают из группы, состоящей из рака желудка, рака пищевода, рака поджелудочной железы, рака легких, рака яичников, рака толстой кишки, рака печени, рака головы и шеи, рака желчного пузыря и их метастазов. Раковое заболевание может быть представлено опухолью Крукенберга, метастазами в брюшине и/или метастазами в лимфатических узлах. В одном воплощении рак представлен аденокарциномой, в частности, поздней стадией аденокарциномы. В одном воплощении рак представлен раком поджелудочной железы.
В одном воплощении способ по изобретению дополнительно включает введение средства, стимулирующего Т-клетки γδ. В одном воплощении Т-клетки γδ представлены Т-клетками Vγ9Vδ2. В одном воплощении средство, стимулирующее Т-клетки γδ, представлено бисфосфонатом типа азотсодержащего бисфосфоната (аминобисфосфоната). В одном воплощении средство, стимулирующее Т-клетки γδ, выбирается из группы, состоящей из золедроновой кислоты, клодроновой кислоты, ибандроновой кислоты, памидроновой кислоты, ризедроновой кислоты, минодроновой кислоты, олпадроновой кислоты, алендроновой кислоты, инкадроновой кислоты и их солей. В одном воплощении средство, стимулирующее Т-клетки γδ, вводится в сочетании с интерлейкином-2.
Способ по изобретению может дополнительно включать введение по меньшей мере еще одного химиотерапевтического средства, которое может быть цитотоксическим средством.
Антитело, обладающее способностью к связыванию с CLDN18.2, может связываться с нативными эпитопами CLDN18.2, находящимися на поверхности живых клеток. В одном воплощении антитело, обладающее способностью к связыванию с CLDN18.2, связывается с первой внеклеточной петлей CLDN18.2. В одном воплощении антитело, обладающее способностью к связыванию с CLDN18.2, опосредует уничтожение клеток по одному или нескольким механизмам из числа опосредованного комплементзависимой цитотоксичностью (CDC) лизиса, опосредованного антителозависимой клеточной цитотоксичностью (ADCC) лизиса, индукции апоптоза и ингибирования пролиферации. В одном воплощении антитело, обладающее способностью к связыванию с CLDN18.2, представляет собой моноклональное, химерное или гуманизированное антитело либо фрагмент антитела. В одном воплощении антитело опосредует уничтожение клеток при связывании с клеточным CLDN18.2, в частности, с CLDN18.2, который экспрессируется клетками на их клеточной поверхности, причем клетки предпочтительно представлены раковыми клетками типа описанных здесь раковых клеток. В одном воплощении антитело, обладающее способностью к связыванию с CLDN18.2, представляет собой антитело, выбранное из группы, состоящей из (i) антител, вырабатываемых и/или получаемых из клона, депонированного под номером доступа DSM АСС2737, DSM АСС2738, DSM АСС2739, DSM АСС2740, DSM АСС2741, DSM АСС2742, DSM АСС2743, DSM АСС2745, DSM АСС2746, DSM АСС2747, DSM АСС2748, DSM АСС2808, DSM АСС2809 или DSM АСС2810, (ii) антител, представляющих собой химеризованные или гуманизованные формы антител по п. (i), (iii) антител, обладающих специфичностью антител по п. (i), и (iv) антител, содержащих антигенсвязывающий участок или антигенсвязывающий сайт, в частности, вариабельную область антител по п. (i), и предпочтительно обладающих специфичностью антител по п. (i). В одном воплощении антитело конъюгировано с терапевтическим средством, как-то токсином, радиоизотопом, лекарственным препаратом или цитотоксическим средством.
В одном воплощении способ по изобретению включает введение антитела, обладающего способностью к связыванию с CLDN18.2, в дозе вплоть до 1000 мг/м2. В одном воплощении способ по изобретению включает неоднократное введение антитела, обладающего способностью к связыванию с CLDN18.2, в дозе от 300 до 600 мг/м2.
В соответствии с изобретением, CLDN18.2 предпочтительно имеет аминокислотную последовательность согласно SEQ ID NO: 1.
В одном воплощении описанный здесь рак является CLDN18.2-положительным. В одном воплощении раковые клетки описанного здесь рака являются CLDN18.2-положительными. В одном воплощении раковые клетки описанного здесь рака экспрессируют CLDN18.2 на своей клеточной поверхности.
В одном воплощении описанный здесь рак поджелудочной железы включает первичный рак, рак на поздних стадиях или метастатический рак либо их комбинации типа сочетания первичного рака поджелудочной железы и метастатического рака. В одном воплощении способы по изобретению предназначены для одновременного лечения первичного рака и метастазирующего рака, как-то первичного рака поджелудочной железы и метастатического рака поджелудочной железы. В одном воплощении метастазирующий рак включает метастазы в лимфатических узлах, яичниках, печени или легких либо их комбинации. В одном воплощении рак поджелудочной железы включает рак протоков поджелудочной железы. В одном воплощении рак поджелудочной железы включает аденокарциному или карциному либо их комбинации. В одном воплощении рак поджелудочной железы включает аденокарциному протоков, слизистую аденокарциному, нейроэндокринную карциному или ацинозно-клеточную карциному либо их комбинации. В одном воплощении рак поджелудочной железы частично или полностью не поддается лечению гемцитабином типа монотерапии гемцитабином. В одном воплощении профилактика рака поджелудочной железы включает предотвращение рецидива рака поджелудочной железы.
В одном воплощении у пациента, подлежащего лечению по изобретению, была операция по поводу рака поджелудочной железы. В одном воплощении у пациента есть предраковые поражения поджелудочной железы, в частности, предраковые поражения поджелудочной железы, включающие возникновение злокачественных гистологических изменений в протоках поджелудочной железы. В этих воплощениях способы по изобретению предпочтительно направлены на предотвращение развития злокачественного рака поджелудочной железы.
В следующем аспекте настоящего изобретения предусмотрены лекарственные препараты для лечения или профилактики рака поджелудочной железы, включающие: (i) антитело, обладающее способностью к связыванию с CLDN18.2, и (ii) средство, стабилизирующее или усиливающее экспрессию CLDN18.2. Лекарственные препараты настоящего изобретения могут дополнительно содержать средство, стимулирующее Т-клетки γδ. Антитело, обладающее способностью к связыванию с CLDN18.2, и средство, стабилизирующее или усиливающее экспрессию CLDN18.2, а также, необязательно, средство, стимулирующее Т-клетки γδ, могут находиться в лекарственном препарате в виде смеси или отдельно друг от друга. Лекарственный препарат может быть представлен в виде набора, включающего первый контейнер, содержащий антитело, обладающее способностью к связыванию с CLDN18.2, и второй контейнер, содержащий средство, стабилизирующее или усиливающее экспрессию CLDN18.2, а также, необязательно, контейнер, содержащий средство, стимулирующее Т-клетки γδ. Лекарственный препарат может дополнительно включать в себя печатные инструкции по применению препарата для лечения или профилактики рака поджелудочной железы, в частности, по применению препарата в способе по изобретению. Различные воплощения лекарственных препаратов, в частности, антител, обладающих способностью к связыванию с CLDN18.2, средств, стабилизирующих или усиливающих экспрессию CLDN18.2, и средств, стимулирующих Т-клетки γδ, уже описаны выше для способов по изобретению.
В отдельном аспекте настоящего изобретения предусмотрены лекарственные препараты, включающие: (i) антитело, обладающее способностью к связыванию с CLDN18.2, и (ii) гемцитабин. Лекарственные препараты настоящего изобретения могут дополнительно содержать средство, стимулирующее Т-клетки γδ. Антитело, обладающее способностью к связыванию с CLDN18.2, и гемцитабин, а также, необязательно, средство, стимулирующее Т-клетки γδ, могут находиться в лекарственном препарате в виде смеси или отдельно друг от друга. Лекарственный препарат может служить для лечения или профилактики рака, как-то рака поджелудочной железы. Лекарственный препарат может быть представлен в виде набора, включающего первый контейнер, содержащий антитело, обладающее способностью к связыванию с CLDN18.2, и второй контейнер, содержащий гемцитабин, а также, необязательно, контейнер, содержащий средство, стимулирующее Т-клетки γδ. Лекарственный препарат может дополнительно включать в себя печатные инструкции по применению препарата для лечения или профилактики рака типа рака поджелудочной железы, в частности, по применению препарата в способе по изобретению. Различные воплощения лекарственных препаратов, в частности, антител, обладающих способностью к связыванию с CLDN18.2, средств, стабилизирующих или усиливающих экспрессию CLDN18.2, и средств, стимулирующих Т-клетки γδ, уже описаны выше для способов по изобретению.
Настоящим изобретением также предусмотрены описанные здесь средства, такие как антитела, обладающие способностью к связыванию с CLDN18.2, и/или средства, стабилизирующие или усиливающие экспрессию CLDN18.2, для применения в описанных здесь способах. Например, настоящим изобретением также предусмотрены антитела, обладающие способностью к связыванию с CLDN18.2, для введения в сочетании со средством типа гемцитабина, стабилизирующим или усиливающим экспрессию CLDN18.2, а также, необязательно, средством, стимулирующим Т-клетки γδ.
Другие особенности и преимущества настоящего изобретения станут понятными из следующего подробного описания и формулы изобретения.
Краткое описание фигур
Фиг. 1. Лентивирусный вектор, используемый для трансдукции клеток раковых линий поджелудочной железы. CLDN18.2 человека клонировали ниже промотора EF1α. Экспрессионную кассету вставляли между длинными концевыми повторами (5' и 3-LTR), которые способствуют упаковке и обратной транскрипции вирусной мРНК. RSV: вирус саркомы Rous обеспечивает Tat-независимое получение вирусной мРНК. Amp: ген устойчивости к ампициллину. PGKp: промотор бластицидина. WPRE: посттранскрипционный регуляторный элемент woodchuck; повышает экспрессию трансгена. LTR: длинный концевой повтор, способствует упаковке вируса. SV40A способствует терминации транскрипции и полиаденилированию мРНК. pUC: остов бактериального вектора. Bla: промотор ампициллина.
Фиг. 2. Анализ метастазирования клеток поджелудочной железы в легких мыши. Схема вскрытия легких у мышей после в/в введения им раковых клеток поджелудочной железы.
Фиг. 3. Экспрессия CLDN18.2 в нормальных и раковых тканях поджелудочной железы. Окрашивание фиксированной формалином и залитой в парафин (FFPE) нормальной ткани поджелудочной железы (А) и ткани аденокарциномы поджелудочной железы (В) с помощью моноклонального антитела 35-22А мыши (0,2 мкг/мл). Контрастное окрашивание гематоксилином (2:00 мин). Увеличение ×200.
Фиг. 4. Экспрессия CLDN18.2 в нормальных и предраковых тканях поджелудочной железы. Окрашивание с помощью 43-14А различных предраковых структур в норме (А) и PanIN1; PanIN2 (В); PanIN3 (С). Увеличение ×200.
Фиг. 5. Пилотное исследование. Корреляция между интенсивностью сигнала CLDN18.2 и количеством положительных опухолевых клеток при анализе первичных опухолей поджелудочной железы. Каждая точка представляет случай первичного рака поджелудочной железы при анализе путем окрашивания FFPE-срезов с помощью моноклонального антитела 35-22А мыши (0,2 мкг/мл). Пунктирной линией отмечено значение в 10%.
Фиг. 6. Пилотное исследование. Экспрессия CLDN18.2 в тканях первичных и метастатических опухолей поджелудочной железы. Окрашивание FFPE-срезов ткани (3 мкм) с помощью моноклонального антитела 35-22А мыши в первичной опухоли аденокарциномы (А) и метастазах в лимфатических узлах (В). Контрастное окрашивание гематоксилином (Mayers).
Фиг. 7. Основное исследование. Корреляция между интенсивностью сигнала CLDN18.2 и количеством положительных опухолевых клеток при анализе первичных опухолей поджелудочной железы. Каждая точка представляет случай первичной опухоли аденокарциномы протоков поджелудочной железы (черные кружочки) или первичной нейроэндокринной опухоли (светлые кружочки) при анализе путем окрашивания FFPE-срезов с помощью моноклонального антитела 43-14А мыши (0,2 мкг/мл).
Фиг. 8. Корреляция между интенсивностью сигнала CLDN18.2 и количеством положительных опухолевых клеток при анализе метастазов поджелудочной железы. Каждая точка представляет случай метастазов рака поджелудочной железы в лимфатических узлах (черные кружочки) или печени (светлые кружочки) при анализе путем окрашивания FFPE-срезов с помощью моноклонального антитела 43-14А мыши (0,2 мкг/мл). Пунктирной линией отмечено значение в 10%.
Фиг. 9. Экспрессия CLDN18.2 в тканях первичных и метастатических опухолей поджелудочной железы. Окрашивание FFPE-срезов ткани (3 мкм) с помощью моноклонального антитела 43-14А мыши в первичных опухолях аденокарциномы (А, С, Е) и их метастазах в лимфатических узлах (В, D, F). Контрастное окрашивание срезов гематоксилином (Mayers).
Фиг. 10. Графический анализ. Экспрессия CLDN18.2 в тканях первичных опухолей поджелудочной железы и соответствующих метастазов в лимфатических узлах.
Фиг. 11. Экспрессия CLDN18.2 в тканях первичных опухолей поджелудочной железы и соответствующих метастазов. Окрашивание FFPE-срезов ткани (3 мкм) из первичной аденокарциномы (А), метастазов в печени (В) и метастазов в лимфатических узлах (С) с помощью моноклонального антитела 43-14А мыши. Контрастное окрашивание срезов гематоксилином (Mayers). Увеличение ×200.
Фиг. 12. Уровень мРНК CLDN18.2 в клеточных линиях карциномы поджелудочной железы. (А) Анализ экспрессии методом количественной ПЦР в различных линиях клеток СА поджелудочной железы, трансдуцированных лентивирусом (LVT) линиях клеток (серые столбики), линии клеток рака желудка КАТО-III (положительный контроль) и линии клеток рака молочной железы SKBR-3 (отрицательный контроль). Транскрипты CLDN18.2 амплифицировали с помощью геноспецифичных праймеров. Эндогенные линии клеток, дающие относительный уровень экспрессии более 1×105, оценивали как CLDN18.2-положительные (заштрихованные столбики). NTC: контрольный образец с Н2О. Планки погрешностей: среднее + SD. (B-D) Анализ зависящей от пассажа экспрессии CLDN18.2 в клетках Patu8988S (В), Panc05.04 (С) и указанных линиях LVT-клеток (D). Номер пассажа указан под каждым столбиком.
Фиг. 13. Уровень белка CLDN18.2 в клеточных лизатах линий клеток карциномы поджелудочной железы. Белки разделяли на 12,5% SDS-PAGE. Вестерн-блоттинг проводили с помощью антитела CLDN18, распознающего С-концевой участок CLDN18.1 и CLDN18.2 (Zymed-MID), и с помощью контрольного антитела, распознающего β-актин. Время экспозиции - 140 сек (Pierce SuperSignal West Dura) и 20 сек (Pierce SuperSignal West Pico), соответственно. (А) Выявление CLDN18 в лизатах клеточных линий поджелудочной железы, лизатах клеток положительного контроля (HEK293-р740) и отрицательного контроля (SKBR-3). (В) Сравнение экспрессии CLDN18.2 между лизатами нетрансдуцированных исходных клеток и лизатами трансдуцированных лентивирусом (LVT) клеточных линий. В качестве положительного и отрицательного контроля, соответственно, добавляли клетки Patu8988S и SKBR-3.
Фиг. 14. Выявление и клеточная локализация экспрессии CLDN18 в клетках раковых линий поджелудочной железы. Окрашивание клеток раковых линий поджелудочной железы, выращенных на покровных стеклах. Антитело: 35-22А (увеличение ×20, время экспозиции указано под каждым снимком). Для окрашивания ядер использовали DAPI (синие). A: AsPC1; В: ВхРС3; С: CFPAC; D: DANG; Е: HPAF-II; F: HUP-T3; G: HUP-T4; Н: KCI-M3; I: Panc1; J: Panc05.04; K: Panc02.04; L: Panc04.03; M: Patu8902; N: Patu8988S; O: Su86.86; P: Suit-2, Q: SW-1990; R: YAPC; S: контрольная линия раковых клеток желудка KATO-III.
Фиг. 15. Выявление и клеточная локализация экспрессии CLDN18 в трансдуцированных CLDN18.2 клетках раковых линий поджелудочной железы. Детектирование CLDN18 в трансдуцированных лентивирусом (LVT) клетках раковых линий поджелудочной железы с помощью антитела 35-22А после фиксации и пермеабилизации. Для детектирования использовали меченые Alexa488 или Alexa555 вторичные антитела. А: ВхРС3-LVT; В: CAPAN1-LVT; С: DANG-LVT; D: HPAC-LVT; Е: MiaPaCa2-LVT; F: Patu8902-LVT; G: Suit-2-CKO; Н: YAPC-LVT.
Фиг. 16. Связывание IMAB362 с клеточной поверхностью CLDN18.2-положительных линий клеток СА поджелудочной железы (фармакодинамика). IF-анализ клеток раковых линий поджелудочной железы (А, В, D, Е), трансдуцированных лентивирусом клеточных линий поджелудочной железы (GL) и контрольных раковых клеток желудка KATO-III (С, F), экспрессирующих CLDN18.2. Клетки окрашивали IMAB362 в нативных условиях (D-E), а для сравнения - 35-22А после фиксации и пермеабилизации клеток (А-С). Для окрашивания ядер использовали DAPI. Время экспозиции указано на каждой панели. G: ВхРС3-LVT; Н: CAPAN1-LVT; I: DANG-LVT; J: MiaPaCa2-LVT; K: Patu8902-LVT; L: Suit2-LVT.
Фиг. 17. Экспрессия CLDN18.2 в привитых опухолях из различных клеточных линий. Экспрессия CLDN18.2 в привитых опухолях CAPAN1-LVT (А, В), ВхРС3-LVT (С, D), PATU8988S-LVT (Е, F), MiaPaCa2-LVT (G, Н), YAPC-LVT (J, K) и DANG-LVT (L, М). Проводили окрашивание тканей с помощью антитела Zymed-MID. Увеличение объектива ×10 (А, С, Е, G, J, L) и ×20 (В, D, F, Н, K, М).
Фиг. 18. Проверка приживления клеток раковых линий Suit-2 и MiaPaCa2 поджелудочной железы. Клетки вводили в хвостовую вену мышей nude. Животных забивали через 45 (А), 52 (В), 59 (С) дней после введения Suit-2 (А-С) или через 59 (D), 66 (Е), 73 (F) дней после введения MiaPaCa2 (D-F). Легкие препарировали и окрашивали с помощью антител к МНС класса I (против МНС I человека, клон EPR1394Y) для выявления клеток человека в тканях мыши.
Фиг. 19. Анализ приживления метастазов Patu8988S. Клетки Patu8988S вводили мышам Nu/Nu внутривенно по 1×106 или 2×106 клеток и в различные моменты времени у мышей извлекали легкие (А) и печень (В), как указано под осью х. Для вычисления % присутствия ДНК человека в каждом препарате ткани строили стандартную кривую, смешивая ДНК человека и мыши и делая 7×5-кратных разведений, получая от 100% (1) до 0,0064% (7) ДНК человека.
Фиг. 20. Анализ методом IHC метастазов Patu8988S в тканях легких мыши. Мышам вводили в хвостовую вену клетки Patu8988S, а через различные промежутки времени (A-D=70 дней, Е-Н=86 дней) их забивали, выделяли ткани легких и окрашивали с помощью антитела к MHC-I (EPR1394Y) (А, В, Е, F) в разведении 1:1000 или против клаудина 18 (Zymed-Mid) (С, D, G, Н) при 0,2 мкг/мл. Увеличение: А, С, Е, G = ×10 и В, D, F, Н = ×20.
Фиг. 21. Опосредованный IMAB362 апоптоз обработанных гемцитабином раковых клеток поджелудочной железы. Апоптоз индуцировали сшиванием CLDN18.2 на клетках BxPC3~CLDN18 через 48 часов. Клетки BxPC3~CLDN18 культивировали в среде без или +100 нг/мл гемцитабина. Доля апоптозных клеток во фракции мононуклеарных клеток смещалась. Аналогичное смещение наблюдалось при инкубации опухолевых клеток с камптотецином.
Фиг. 22. Выраженность IMAB362-индуцированной ADCC-активности на раковых клетках поджелудочной железы. (A) ADCC проводили с CLDN18.2-положительными клетками раковых линий поджелудочной железы, используя РВМС от разных доноров. (B-F) ADCC проводили с LVT-линиями раковых клеток поджелудочной железы, эктопически экспрессирующими CLDN18.2, и соответствующими исходными клетками. (G): точечный график.
Фиг. 23. Выраженность IMAB362-индуцированной CDC-активности на раковых клетках поджелудочной железы. (A) CDC проводили со сборной сывороткой здорового человека в качестве источника комплемента, IMAB362 и CLDN18.2-положительными контрольными клетками CDOK1-p740 поджелудочной железы в 4-х независимых экспериментах. (В) CDC проводили с CLDN18.2-положительными (Patu8988S, DANG, Panc05.04) и CLDN18.2-отрицательными (CAPAN1, Suit2, ВхРС3, YAPC) линиями клеток поджелудочной железы. (С) CDC проводили с эктопически экспрессирующими LVT-линиями клеток. (D) Точечный график, показывающий концентрации IMAB362, вызывающие полумаксимальную степень лизиса (ЕС50) клеток раковых линий поджелудочной железы. (Е) Максимальная степень уничтожения клеток, полученная с IMAB362 на клетках раковых линий поджелудочной железы.
Фиг. 24. Влияние обработки IMAB362 на подкожные ксенотрансплантаты MiaPaCa2-LVT. Опухоли-ксенотрансплантаты MiaPaCa2-LVT прививали путем инъекции 1×107 клеток MiaPaCa2-LVT подкожно в бок 15 самкам бестимусных мышей nude Hsd:Foxnlnu на каждую группу обработки. На третий день после введения раковых клеток начинали обработку по 200 мкг IMAB362 или контролей, соответственно. Обработку продолжали по 2 раза в неделю, чередуя в/б и в/в введение вплоть до забоя животных. (А) Влияние обработки IMAB362 на рост опухолей. Размер п/к опухолей измеряли 2 раза в неделю (среднее + SEM). (В) Графики выживаемости Kaplan-Meier. Мышей забивали, когда объем опухолей достигал 1400 мм3 или же опухоли становились изъязвленными.
Фиг. 25. Обработка IMAB362 подкожных ксенотрансплантатов ВхРС3-LVT. Опухоли-ксенотрансплантаты ВхРС3-LVT прививали путем инъекции 1×107 клеток ВхРС3-LVT подкожно в бок 15 самкам бестимусных мышей nude Hsd:Foxnlnu на каждую группу обработки. На третий день после введения раковых клеток начинали обработку по 200 мкг IMAB362 или контролей, соответственно. Обработку продолжали по 2 раза в неделю, чередуя в/б и в/в введение вплоть до забоя животных. (А) Влияние обработки IMAB362 на рост опухолей. Размер п/к опухолей измеряли 2 раза в неделю (среднее + SEM). (В) Графики выживаемости Kaplan-Meier. Мышей забивали, когда объем опухолей достигал 1400 мм3 или же опухоли становились изъязвленными.
Фиг. 26. Влияние обработки IMAB362 на рост панкреатических метастазов Suit2-LVT. По 2×106 раковых клеток Suit2-LVT вводили внутривенно в хвостовую вену 12 самкам бестимусных мышей nude Hsd:Foxnlnu на каждую группу обработки. На третий день после введения раковых клеток начинали обработку по 200 мкг IMAB362, 200 мкг изотипного контроля или равным объемом PBS. Животных забивали на 42-й день после трансплантации. (А) Анализ методом количественной ПЦР (среднее из 2-4 реакций на 1 образец) для определения процентного содержания ДНК человека в образцах легких у мышей. (В) Процент покрытой клетками человека поверхности легких у мышей по данным планиметрии. Иммуногистохимическое окрашивание клеток человека на срезах тканей с помощью антитела против МНС класса I человека. * р<0,05 (тест Kruskal-Wallis). Планки погрешностей: среднее ± SD.
Фиг. 27. Анализ методами количественной ПЦР и IHC метастазов Patu8988S в легких. Мышам вводили по 2×106 клеток Patu8988S на мышь. Через 65 дней животных забивали. Светлые кружочки: мышей забивали через 63 дня. (А) Мышей обрабатывали по 2 раза в неделю 200 мкг IMAB362 или контрольным физраствором. Количество ДНК человека (нг) при определении методом количественной ПЦР рассчитывали по значениям Ct. (В) Повтор эксперимента по к-ПЦР, как описано в А. При этом процентное содержание ДНК человека в ДНК мыши рассчитывали по значениям Ct. (С) Мышей обрабатывали IMAB362 и контрольным антителом того же изотипа (ритуксимаб). Процентное содержание ДНК человека в легких мышей рассчитывали по значениям Ct. У группы DVLAB362 обнаружился один выброс (светлый треугольник). Значимость приводится как с включением, так и с исключением значений выброса. (D/E) Такой же эксперимент, как и в С. При этом площадь метастазов определяли с помощью программы Image J. На точечных графиках значимость ингибирования IMAB362 представлена как с включением (D), так и с исключением (Е) значения выброса. Значения р: непарный t-критерий. Планки погрешностей: среднее ± SD.
Фиг. 28. Кривые доза-ответ для гемцитабина. Клетки раковых линий поджелудочной железы проявляют очень большие различия в чувствительности к гемцитабину. Клеточные линии подвергали воздействию различных концентраций гемцитабина на протяжении 4 дней и определяли ингибирование пролиферации посредством анализа на жизнеспособность.
Фиг. 29. Кривые доза-ответ для оксалиплатина. Клетки раковых линий поджелудочной железы проявляют очень большие различия в чувствительности к оксалиплатину. Клеточные линии подвергали воздействию различных концентраций оксалиплатина на протяжении 4 дней и определяли ингибирование пролиферации посредством анализа на жизнеспособность.
Фиг. 30. Влияние обработки химиотерапевтическими средствами на экспрессию CLDN18.2 (РНК). РНК у необработанных и предварительно обработанных Gem (1 нг/мл) или GemOx (Gem 1 нг/мл + Ох 10 нг/мл) клеток DANG (2 дня) (А) или Patu8988S (В) при обработке в течение 3 дней Gem (10 нг/мл или GemOx (Gem 10 нг/мл + Ох 100 нг/мл). РНК превращали в кДНК и определяли уровень транскриптов CLDN18.2 количественным методом ПНР в реальном времени. Результаты представлены в относительных единицах по сравнению с уровнем транскрипта гена домашнего хозяйства HPRT.
Фиг. 31. Влияние химиотерапии на уровень белка CLDN18.2 в клетках карциномы поджелудочной железы. Анализу на экспрессию CLDN18.2 подвергали общий белок из клеточных лизатов необработанных (med) и предварительно обработанных Gem (1 нг/мл) или GemOx (GEM 1 нг/мл + Ох 10 нг/мл) клеток DANG (А) или Patu8988S (В) при детектировании с помощью поликлональной антисыворотки Zymed C-term. Для проверки одинакового нанесения белков использовали актин.
Фиг. 32. Анализ методом FACS экспрессии CLDN18.2 на клеточной поверхности. Экспрессия CLDN18 (темная гистограмма) при культивировании Patu8988S в среде (слева) и обработке Gem (справа) представлена в виде наложения на изотипный контроль. Клетки Patu8988S обрабатывали гемцитабином (10 нг/мл) в течение 3 дней.
Фиг. 33. Анализ клеточного цикла у клеток DANG, обработанных или не обработанных гемцитабином (GEM; 2 нг/мл) или гемцитабином + оксалиплатином (GemOx; 1 нг/мл + 10 нг/мл) в течение двух дней. (А) Обработка гемцитабином приводит к остановке клеточного цикла клеток в фазе S. Площадь каждого столбика разделена с тем, чтобы показать процент клеток в фазах G0/G1, S и G2. (В) Вестерн-блоттинг показал положительную регуляцию CLDN18 после обработки Gem.
Фиг. 34. Влияние гемцитабина на клеточный цикл (А) и экспрессию CLDN18.2 (В, С) в клетках Patu8988S. Клетки Patu8988S обрабатывали либо не обрабатывали гемцитабином (10 нг/мл) в течение 2 дней. (А) Площадь каждого столбика разделена с тем, чтобы показать процент клеток в фазах G0/G1, S и G2. На графике представлена плотность CLDN18.2 (по оси x) относительно количества клеток (по оси y). (В) Экспрессия CLDN18.2 в отсутствие обработки (пунктирная линия) в сравнении с обработкой Gem (сплошная линия). (С) Экспрессия CLDN18.2 в обработанных Gem клетках Patu8988S в фазе G0/G1 (пунктирная линия) в сравнении с клетками в фазе S (сплошная линия).
Фиг. 35. Влияние химиотерапии на раковые клетки желудка. Культивирование клеток KATO-III в течение 96 ч приводит к остановке клеточного цикла в фазе G0/G1 (а) и отрицательной регуляции CLDN18.2 (с). Цитостатические соединения, вызывающие остановку клеточного цикла в различных фазах клеточного цикла, стабилизируют экспрессию CLDN18.2 (с).
Фиг. 36. Влияние химиотерапии на раковые клетки желудка. Цитостатические соединения, вызывающие остановку клеточного цикла в различных фазах клеточного цикла: фазе S/G2 (иринотекан) или фазе G2 (доцетаксель). Площадь каждого столбика разделена с тем, чтобы показать процент клеток в фазах G0/G1, S и G2.
Фиг. 37. Кривые доза-ответ для опосредованной IMAB362 ADCC после химиотерапевтической обработки клеток DANG. (А) Кривые доза-ответ у одного репрезентативного донора после предварительной обработки раковых клеток DANG поджелудочной железы с помощью Gem или GemOx в течение 40 ч. (В) Значения ЕС50 (средние) для опосредованной IMAB362 ADCC. Значения р: непарный t-критерий.
Фиг. 38. Влияние химиотерапии на раковые клетки желудка, а: Клетки, обработанные иринотеканом, доцетакселем или цисплатином, проявляют более низкий уровень жизнеспособных клеток по сравнению с культивированными в среде клетками мишени, c/d: Экспрессия CLDN18.2 в клетках, обработанных иринотеканом, доцетакселем или цисплатином, повышалась по сравнению с клетками, культивированными в среде, c/d: Обработка клеток иринотеканом, доцетакселем или цисплатином усиливает способность IMAB362 индуцировать ADCC.
Фиг. 39. Влияние химиотерапевтических средств на опосредованную IMAB362 CDC у клеток MiaPaCa2-LVT. Кривые доза-ответ по 2 независимым опытам. MiaPaCa2-LVT культивировали в среде либо в среде с Gem (10 нг/мл) или GemOx (10 нг/мл Gem + 100 нг/мл Ох) в течение 70 ч.
Фиг. 40. Влияние химиотерапии на индуцированную IMAB362 CDC.
Фиг. 41. Влияние обработки IMAB362 в сочетании с Gem или GemOx на ксенотрансплантаты ВхРС3-LVT. Опухоли-ксенотрансплантаты ВхРС3-LVT прививали путем инъекции 8,5×106 клеток ВхРС3-LVT подкожно в бок 10 самкам бестимусных мышей nude Hsd:Foxnlnu на каждую группу обработки. На третий день после введения раковых клеток начинали применение химиотерапии (50 мг/кг гемцитабина в/б либо 50 мг/кг гемцитабина плюс 5 мг/кг оксалиплатина в/б, соответственно) и продолжали ее раз в неделю на протяжении 6 недель. Через 24 ч после введения химиотерапевтических средств вводили внутривенно в хвостовую вену 800 мкг IMAB362 или контроля. Обработку IMAB362 продолжали раз в неделю вплоть до забоя мышей. (А) Кривые роста подкожных ксенотрансплантатов ВхРС3-LVT. Размер п/к опухолей измеряли два раза в неделю (среднее + SEM). (В) Кривые выживаемости Kaplan-Meier. Мышей забивали, когда объем опухолей достигал 1400 мм3 или же опухоли становились изъязвленными.
Фиг. 42. Повышение противоопухолевой эффективности при комбинировании гемцитабина с IMAB362. Опухоли-ксенотрансплантаты ВхРС3-LVT прививали путем инъекции 8,5×106 клеток ВхРС3-LVT подкожно в бок 10 самкам бестимусных мышей nude Hsd:Foxnlnu на каждую группу обработки. На третий день после введения раковых клеток начинали применение химиотерапии (100 мг/кг гемцитабина в/б либо 100 мг/кг гемцитабина плюс 5 мг/кг оксалиплатина в/б, соответственно) и продолжали ее раз в неделю на протяжении 6 недель. Через 24 ч после введения химиотерапевтических средств вводили внутривенно в хвостовую вену 200 мкг (1/2 дозы) или 400 мкг (полная доза) IMAB362. Обработку IMAB362 продолжали по 2 раза в неделю, чередуя в/б и в/в введение вплоть до забоя мышей. (А) Кривые роста подкожных ксенотрансплантатов ВхРС3-LVT. Размер п/к опухолей измеряли два раза в неделю (среднее + SEM). (В) Кривые выживаемости Kaplan-Meier. Мышей забивали, когда объем опухолей достигал 1400 мм3 или же опухоли становились изъязвленными.
Фиг. 43. Влияние обработки IMAB362 в сочетании с гемцитабином на ксенотрансплантаты MiaPaCa2-LVT. Опухоли-ксенотрансплантаты MiaPaCa2-LVT прививали путем инъекции 5×106 клеток MiaPaCa2-LVT подкожно в бок 10 самкам бестимусных мышей nude Hsd:Foxnlnu на каждую группу обработки. Через 4 дня после введения раковых клеток начинали применение химиотерапии (50 мг/кг гемцитабина в/б) и продолжали ее раз в неделю на протяжении 6 недель. Через 24 ч после введения химиотерапевтических средств вводили внутривенно в хвостовую вену 200 мкг IMAB362 или контроля. Обработку IMAB362 продолжали по 2 раза в неделю, чередуя в/б и в/в введение вплоть до забоя мышей. (А) Рост подкожных ксенотрансплантатов. Размер опухолей измеряли два раза в неделю (среднее + SEM). (В) Кривые выживаемости Kaplan-Meier. Мышей забивали, когда объем опухолей достигал 1400 мм3 или же опухоли становились изъязвленными.
Фиг. 44. Влияние обработки IMAB362 в сочетании с гемцитабином на привитые опухоли-ксенотрансплантаты MiaPaCa2-LVT. Опухоли-ксенотрансплантаты MiaPaCa2-LVT прививали путем инъекции 1×107 клеток MiaPaCa2-LVT подкожно в бок самкам бестимусных мышей nude Hsd:Foxnlnu. Через 9 дней после подкожной инокуляции несущих опухоли мышей разбивали на однородные группы обработки по 8 животных в группе и начинали обработку. Мышам вводили 150 мг/кг гемцитабина в/б по 2 раза в неделю на протяжении 4 недель. Через 24 ч после введения гемцитабина вводили внутривенно в хвостовую вену 200 мкг IMAB362 или контроля. Обработку IMAB362 продолжали по 2 раза в неделю, чередуя в/б и в/в введение вплоть до забоя мышей. (А) Размер подкожных опухолей измеряли два раза в неделю (среднее + SEM; ** = р<0,01). (В) Кривые выживаемости Kaplan-Meier. Мышей забивали, когда объем опухолей достигал 1400 мм3 или же опухоли становились изъязвленными (лог-ранговый критерий Mantel-Сох; **=р 0,01).
Фиг. 45. Влияние IMAB362 в сочетании с гемцитабином на метастазы в легких на модели ксенотрансплантации Patu8988S. Вводили внутривенно 2×106 раковых клеток Patu8988S в хвостовую вену 12 самкам бестимусных мышей nude Hsd:Foxnlnu на каждую группу обработки. Через 2 недели после внутривенного введения раковых клеток начинали обработку 200 мкг IMAB362 по 2 раза в неделю (чередуя в/в-в/б) в сочетании с введением 100 мг/кг гемцитабина в/б по 2 раза в неделю на протяжении 4 недель. Контрольная группа получала по 200 мкг контрольного антитела того же изотипа в сочетании с 100 мг/кг гемцитабина по 2 раза в неделю. Животных забивали на 70-й день после трансплантации. (А) Количественное определение методом ПЦР (среднее из 3 реакций на 1 образец) ДНК человека в образцах легких у получавших IMAB362 и изотипное антитело мышей. (В) Процент покрытой клетками человека поверхности легких у мышей определяли на основе компьютерного анализа. Проводили иммуногистохимическое окрашивание с помощью антитела против MHC-I человека (клон EPR1394Y) на заключенных в парафин тканях легких (среднее ± SEM; р=0,0003, критерий Mann-Whitney). С и D: Примеры иммуногистохимического окрашивания с помощью антитела против MHC-I человека метастазов Patu8988S в легких у мышей, получавших IMAB362 + гемцитабин (С) или изотипное антитело + гемцитабин (D).
Раскрытие сущности изобретения
Несмотря на то, что настоящее изобретение подробно описано ниже, следует иметь в виду, что оно не ограничивается конкретными методами, методиками и реагентами, описанными здесь, поскольку они могут варьироваться. Также следует иметь в виду, что используемая здесь терминология служит только для описания конкретных воплощений и не должна ограничивать объем настоящего изобретения, который ограничивается лишь прилагаемой формулой изобретения. Если не указано иначе, все технические и научные термины, используемые здесь, имеют такие значения, которые широко известны рядовым специалистам в данной области.
Далее будут описаны все элементы настоящего изобретения. Эти элементы приводятся вместе с конкретными воплощениями, однако следует иметь в виду, что их можно комбинировать любым способом и в любом количестве для создания дополнительных воплощений. Приведенные примеры и предпочтительные воплощения не должны рассматриваться как ограничивающие настоящее изобретение только явно описанными воплощениями. Такое описание следует понимать как поддерживающее и охватывающее такие воплощения, в которых явно описанные воплощения комбинируются с любым количеством приведенных и/или предпочтительных элементов. Более того, любые перестановки и комбинации всех описанных элементов в настоящей заявке следует рассматривать как раскрытые описанием в настоящей заявке, если из контекста не следует иное.
Предпочтительно используемые здесь термины определяются так, как описано в "A multilingual glossary of biotechnological terms: (IUPAC Recommendations)", H.G.W. Leuenberger, B. Nagel, and H. 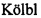 , Eds., Helvetica Chimica Acta, CH-4010 Basel, Switzerland (1995).
, Eds., Helvetica Chimica Acta, CH-4010 Basel, Switzerland (1995).
При практическом применении настоящего изобретения должны применяться, если не указано иначе, стандартные методы химии, биохимии, клеточной биологии, иммунологии и технологии рекомбинантной ДНК, которые описаны в литературе по данной области (например, см. Molecular Cloning: A Laboratory Manual, 2nd Edition, J. Sambrook et al., eds., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, 1989).
По всему описанию и следующей за ним формуле изобретения, если из контекста не следует иное, слово "включать" и такие его варианты, как "включает" и "включающий", следует понимать как означающие включение указанного элемента, числа или стадии либо группы элементов, чисел или стадий, но не исключение любого другого элемента, числа или стадии либо группы элементов, чисел или стадий, хотя в каких-то воплощениях такие элементы, числа или стадии либо группы элементов, чисел или стадий могут исключаться, т.е. этот вопрос заключается во включении указанного элемента, числа или стадии либо группы элементов, чисел или стадий. Формы единственного числа при описании изобретения (особенно в контексте формулы) следует понимать как охватывающие и единственное, и множественное число, если иное не указано прямо или явно противоречит контексту. Указание диапазонов значений здесь всего лишь служит кратким способом индивидуального указания всех отдельных значений, попадающих в этот диапазон. Если не указано иначе, каждое индивидуальное значение включается в описание, как если бы оно было указано отдельно. Все описанные здесь методы могут выполняться в любом подходящем порядке, если это не указано иначе или же явно противоречит контексту. Использование каких бы то ни было примеров или типичных выражений (например, "таких как"), приведенных здесь, служит только лишь для лучшего раскрытия изобретения и не накладывает ограничений на объем изобретения, заявленный в формуле. Никакие выражения в описании не следует понимать как указывающие на какой-либо незаявленный элемент, необходимой для практического применения изобретения.
В тексте настоящего описания приводятся некоторые документы. Каждый из приведенных здесь документов (включая все патенты, патентные заявки, научные публикации, спецификации производителя, инструкции и т.п.), будь-то выше или ниже, тем самым включаются сюда путем ссылки во всей полноте. При этом ничто не должно толковаться как допущение того, будто изобретение не может предвосхищать такие описания ввиду предшествующего ему изобретения.
Термин "CLDN18" относится к клаудину 18 и включает в себя любые варианты, включая сплайсинг-вариант 1 клаудина 18 (клаудин 18.1 (CLDN18.1)) и сплайсинг-вариант 2 клаудина 18 (клаудин 18.2 (CLDN18.2)).
Термин "CLDN18.2" предпочтительно относится к CLDN18.2 человека, в частности, к белку, содержащему, предпочтительно состоящему из аминокислотной последовательности по SEQ ID NO: 1 из перечня последовательностей или варианта данной аминокислотной последовательности.
Термин "CLDN18.1" предпочтительно относится к CLDN18.1 человека, в частности, к белку, содержащему, предпочтительно состоящему из аминокислотной последовательности по SEQ ID NO: 2 из перечня последовательностей или варианта данной аминокислотной последовательности.
Термин "вариант" в соответствии с изобретением относится, в частности, к мутантам, сплайс-вариантам, конформациям, изоформам, аллельным вариантам, видовым вариантам и видовым гомологам, в частности тем, что встречаются в природе. Аллельный вариант означает такое изменение в нормальной последовательности гена, значение которого зачастую неясно. При полном секвенировании гена часто идентифицируются многочисленные аллельные варианты данного гена. Видовой гомолог означает то, что нуклеотидная или аминокислотная последовательность происходит из другого вида, чем данная нуклеотидная или аминокислотная последовательность. Термин "вариант" также охватывает любые подвергшиеся посттрансляционной модификации варианты и конформационные варианты.
В соответствии с изобретением, термин "CLDN18.2-положительный рак" означает рак с участием раковых клеток, экспрессирующих CLDN18.2, предпочтительно на поверхности данных раковых клеток.
"Клеточная поверхность" используется в соответствии со своим обычным значением в данной области, так что она включает в себя наружную поверхность клетки, которая доступна для связывания с белками и другими молекулами. Например, трансмембранный белок, имеющий один или несколько внеклеточных участков, считается экспрессирующимся на клеточной поверхности.
CLDN18.2 экспрессируется на клеточной поверхности, если он располагается на поверхности данных клеток и доступен для связывания с CLDN18.2-специфичными антителами при добавлении их к клеткам.
В соответствии с изобретением, CLDN18.2 практически не экспрессируется в клетках, если его уровень экспрессии ниже по сравнению с экспрессией в клетках желудка или ткани желудка. Предпочтительно уровень экспрессии составляет менее 10%, предпочтительно менее 5%, 3%, 2%, 1%, 0,5%, 0,1% или 0,05% от уровня экспрессии в клетках желудка или ткани желудка либо еще ниже. Предпочтительно CLDN18.2 практически не экспрессируется в клетках, если его уровень экспрессии превышает уровень экспрессии в другой нераковой ткани, кроме желудка, не более чем в 2 раза, предпочтительно в 1,5 раза, а предпочтительно не превышает уровень экспрессии в данной нераковой ткани. Предпочтительно CLDN18.2 практически не экспрессируется в клетках, если его уровень экспрессии ниже предела обнаружения и/или если уровень экспрессии будет слишком низким для связывания CLDN18.2-специфичных антител при добавлении их к клеткам.
В соответствии с изобретением, CLDN18.2 экспрессируется в клетках, если его уровень экспрессии превышает уровень экспрессии в другой нераковой ткани, кроме желудка, предпочтительно более чем в 2 раза, предпочтительно в 10 раз, 100 раз, 1000 раз или 10000 раз. Предпочтительно CLDN18.2 экспрессируется в клетках, если его уровень экспрессии выше предела обнаружения и/или если уровень экспрессии будет достаточно высоким для связывания CLDN18.2-специфичных антител при добавлении их к клеткам. Предпочтительно при экспрессировании CLDN18.2 в клетках он экспрессируется или выступает на поверхности данных клеток.
В соответствии с изобретением термин "заболевание" означает любое патологическое состояние, в том числе рак, в особенности формы рака, описанные здесь. Любые упоминания рака или определенных форм рака также включают и их метастазы. В предпочтительном воплощении в заболевании, подлежащем лечению в соответствии с настоящей заявкой, участвуют клетки, экспрессирующие CLDN18.2.
"Заболевания, связанные с клетками, экспрессирующими CLDN18.2" или аналогичные выражения означают в соответствии с изобретением, что в клетках пораженных тканей или органов экспрессируется CLDN18.2. В одном воплощении экспрессия CLDN18.2 в клетках пораженных тканей или органов возрастает по сравнению со здоровой тканью или органом. Возрастание означает повышение по меньшей мере на 10%, в частности, по меньшей мере на 20%, на 50%, на 100%, на 200%, на 500%, на 1000%, на 10000% или еще больше. В одном воплощении экспрессия происходит только в пораженной ткани, тогда как в соответствующей здоровой ткани экспрессия подавляется. Например, CLDN18.2 экспрессируется в раковой ткани поджелудочной железы, но он не выявляется в нераковой ткани поджелудочной железы. В соответствии с изобретением, заболевания, связанные с клетками, экспрессирующими CLDN18.2, включают раковые заболевания. Кроме того, в соответствии с изобретением, раковые заболевания предпочтительно представлены теми, при которых в раковых клетках экспрессируется CLDN18.2.
В настоящем изобретении "раковое заболевание" или "рак" включает такие заболевания, которые характеризуются аномальной регуляцией роста, пролиферации, дифференцировки, адгезии и/или миграции клеток. Под "раковыми клетками" имеются в виду аномальные клетки, которые растут в процессе быстрой, неконтролируемой пролиферации клеток и продолжают расти после прекращения тех стимулов, которые вызвали рост новообразования. Предпочтительно "раковое заболевание" характеризуется наличием клеток, экспрессирующих CLDN18.2, причем раковые клетки экспрессируют CLDN18.2. Клетки, экспрессирующие CLDN18.2, предпочтительно представляют собой раковые клетки, предпочтительно описанных здесь видов рака.
В соответствии с изобретением, "карцинома" означает злокачественную опухоль, происходящую из эпителиальных клеток.
"Аденокарцинома" означает такой рак, который возникает в железистой ткани. Такие ткани также входят в более крупную категорию тканей, известных как эпителиальные ткани. Эпителиальные ткани включает в себя кожу, железы и целый ряд других тканей, выстилающих полости и органы тела. Эмбриологически эпителий происходит из эктодермы, эндодермы и мезодермы. Для классификации в качестве аденокарциномы клетки не обязательно должны быть частью железы, если только они обладают секреторными свойствами. Эта форма карциномы может возникать у некоторых высших млекопитающих, включая человека. Хорошо дифференцированная аденокарцинома обычно похожа на ту железистую ткань, из которой она происходит, тогда как плохо дифференцированная может и не быть. По окрашиванию клеток из биопсии патологоанатом должен определить, является ли опухоль аденокарциномой или принадлежит к какому-то другому типу рака. Аденокарцинома может возникать во многих тканях организма вследствие повсеместной природы желез в организме. Хотя не каждая железа может секретировать одно и то же вещество, однако, если клетки выполняют экзокринные функции, то они считаются железистыми, а их злокачественные формы при этом называются аденокарциномами. Злокачественные аденокарциномы проникают в другие ткани и зачастую дают метастазы, если у них будет достаточно времени для этого.
Поджелудочная железа - орган эндодермального происхождения, она является ключевым регулятором расщепления белков и углеводов и гомеостаза глюкозы. Экзокринная часть поджелудочной железы (80% от массы всего органа) состоит из разветвленной сети ацинарных клеток и клеток протоков, которые вырабатывают и доставляют пищеварительные ферменты в желудочно-кишечный тракт. Ацинарные клетки, которые образуют функциональные единицы вдоль сети протоков, синтезируют и секретируют ферменты в просвет протоков в ответ на сигналы из желудка и двенадцатиперстной кишки. В ацинарных звеньях вблизи протоков находятся центроацинарные клетки. Эндокринная часть поджелудочной железы, которая регулирует метаболизм и гомеостаз глюкозы посредством выделения гормонов в кровь, состоит из четырех специализированных типов эндокринных клеток, собранных в кластеры, называемые островками Лангерганса.
Рак поджелудочной железы является злокачественным новообразованием, происходящим из трансформированных клеток, возникающих в тканях, составляющих поджелудочную железу. Рак поджелудочной железы занимает четвертое место по смертности от рака в США, а во всем мире - восьмое. Вначале рак поджелудочной железы зачастую не вызывает симптомов, а более поздние симптомы, как правило, неспецифичны и разнообразны. Поэтому рак поджелудочной железы часто не выявляется вплоть до поздних стадий. Рак поджелудочной железы имеет плохой прогноз: на всех стадиях вместе коэффициент выживаемости на 1 год и на 5 лет составляет 25% и 6%, соответственно. При локализованном заболевании 5-летняя выживаемость составляет примерно 20%, тогда как средняя выживаемость при локализованном запущенном и при метастатическом заболевании, которые в совокупности составляют более 80% больных, - около 10 и 6 месяцев, соответственно.
Рак поджелудочной железы включает аденокарциномы (опухоли, проявляющие железистую архитектуру), возникающие в экзокринной части поджелудочной железы, и нейроэндокринные карциномы, возникающие из островковых клеток.
Наиболее распространенная форма рака поджелудочной железы - аденокарцинома протоков, как правило, характеризуется умеренно или слабо дифференцированными железистыми структурами при микроскопическом исследовании. Аденокарцинома протоков поджелудочной железы (PDAC) обычно возникает в головке поджелудочной железы с инфильтрацией в окружающие ткани, в том числе лимфатические узлы, селезенку и брюшную полость, а также с метастазами в печени и легких. PDAC главным образом проявляет железистый профиль с протокоподобными структурами и разной степенью атипии и дифференцировки клеток. Менее распространенные подтипы PDAC включают коллоидную, железисто-плоскоклеточную или саркоматоидную гистологию. Зачастую в индивидуальной опухоли существуют региональные различия по гистологии, степени злокачественности и степени дифференцировки. Даже мельчайшие первичные поражения обычно проявляют периневральное и лимфо-сосудистое инвазирование, что свидетельствует о склонности к раннему и дальнему распространению.
Вторым по распространенности типом рака экзокринной поджелудочной железы является слизистый. Слизистая аденокарцинома вырабатывает большой объем муцина, что приводит к кистозному виду при томографии.
Нейроэндокринные опухоли поджелудочной железы образуются из вырабатывающих гормоны клеток (островковых клеток) поджелудочной железы. Ацинозно-клеточные новообразования возникают из ацинарных клеток поджелудочной железы.
В соответствии с изобретением, термин "рак" также включает метастазы рака из первичных опухолей типа первичного рака поджелудочной железы. Так, если, к примеру, упоминается рак поджелудочной железы, то это также включает метастазы рака поджелудочной железы, к примеру, метастазы в легких, печени и/или лимфатических узлах.
Под "метастазированием" понимается распространение раковых клеток из своего исходного места в другие части тела. Образование метастазов является очень сложным процессом и зависит от отделения злокачественных клеток из первичной опухоли, инвазии во внеклеточный матрикс, проникновения через базальные мембраны эндотелия для попадания в полости тела и сосуды, а затем, после переноса с кровью, инфильтрации органов-мишеней. Наконец, рост новых опухолей на месте мишени зависит от ангиогенеза. Метастазирование опухолей часто происходит даже после удаления первичной опухоли, так как опухолевые клетки или их компоненты могут оставаться и создавать метастатический потенциал. В одном воплощении термин "метастазирование" по изобретению относится к "удаленным метастазам", что означает метастазы, удаленные от первичной опухоли и региональной системы лимфатических узлов. В одном воплощении термин "метастазы" по изобретению относится к метастазам в лимфатических узлах. Один конкретный вид метастазов, который можно лечить с помощью терапии по изобретению, представлен метастазами, происходящими из рака поджелудочной железы в качестве первичного сайта. В предпочтительных воплощениях такие метастазы рака поджелудочной железы представлены метастазами в лимфатических узлах, метастазами в легких и/или метастазами в печени.
Опухоль Крукенберга - редко встречающаяся метастатическая опухоль яичников, составляющая от 1% до 2% от всех опухолей яичников. Опухоль Крукенберга является метастатической аденокарциномой перстневидных клеток яичников. В большинстве случаев опухолей Крукенберга (70%) первичным участком является желудок. Следующими по распространенности первичными участками являются карциномы толстой кишки, аппендикса и молочной железы (в основном инвазивная дольковая карцинома). Отмечены редкие случаи опухолей Крукенберга, происходящих из карциномы желчного пузыря, желчевыводящих путей, поджелудочной железы, тонкой кишки, фатеровой ампулы, шейки матки и мочевого пузыря/мочевого протока.
Рефрактерным является такое раковое заболевание, для которого определенное лечение неэффективно, причем оно либо изначально не поддается лечению, либо перестает поддаваться лечению со временем.
Под "лечением" понимается введение субъекту соединения или композиции либо комбинации соединений или композиций для предотвращения или устранения заболевания, включая уменьшение размера опухоли или количества опухолей у субъекта; прекращение или замедление болезни у субъекта; подавление или замедление развития нового заболевания у субъекта; уменьшение частоты или тяжести симптомов и/или рецидивов у субъекта, у которого сейчас или ранее было заболевание; и/или продление, т.е. увеличение продолжительности жизни у субъекта.
В частности, термин "лечение заболевания" включает в себя лечение, сокращение продолжительности, ослабление, предотвращение, замедление или подавление развития или ухудшения либо предотвращение или задержку возникновения заболевания или его симптомов.
Термин "пациент" в соответствии с изобретением означает подлежащего лечению субъекта, в частности, больных субъектов, включая человека, приматов или других животных, в особенности таких млекопитающих, как коровы, лошади, свиньи, овцы, козы, собаки, кошки или такие грызуны, как мыши и крысы. В особенно предпочтительном воплощении пациентом является человек.
Термин "средство, стабилизирующее или повышающее экспрессию CLDN18.2" относится к таким средствам или комбинациям средств, предоставление которых клеткам приводит к повышению уровня РНК и/или белка CLDN18.2 в данных клетках, предпочтительно к повышению уровня белка CLDN18.2 на поверхности клеток по сравнению с ситуацией, когда клетки не получают этого средства или комбинации средств. Предпочтительно клетки представлены раковыми клетками, в частности раковыми клетками, экспрессирующими CLDN18.2, поэтому они являются мишенью для антител, связывающих CLDN18.2, как-то клетки описанных здесь типов рака, в частности, рака поджелудочной железы. Термин "средство, стабилизирующее или повышающее экспрессию CLDN18.2", в частности, относится к таким средствам или комбинациям средств, предоставление которых клеткам приводит к повышению плотности CLDN18.2 на поверхности данных клеток по сравнению с ситуацией, когда клетки не получают этого средства или комбинации средств. "Стабилизация экспрессии CLDN18.2", в частности, включает в себя ситуации, когда средство или комбинация средств предотвращает снижение или уменьшает снижение экспрессии CLDN18.2, например, экспрессия CLDN18.2 будет снижаться без предоставления средства или комбинации средств, а предоставление средства или комбинации средств предотвращает такое снижение или уменьшает такое снижение экспрессии CLDN18.2. "Повышение экспрессии CLDN18.2", в частности, включает ситуации, когда средство или комбинация средств повышает экспрессию CLDN18.2, например, экспрессия CLDN18.2 будет снижаться, оставаться практически постоянной или повышаться без предоставления средства или комбинации средств, а предоставление средства или комбинации средств повышает экспрессию CLDN18.2 по сравнению с ситуацией без предоставления средства или комбинации средств, в результате чего экспрессия будет выше по сравнению с ситуацией, когда экспрессия CLDN18.2 могла бы снижаться, оставаться практически постоянной или повышаться без предоставления средства или комбинации средств.
В соответствии с изобретением, термин "средство, стабилизирующее или повышающее экспрессию CLDN18.2" включает химиотерапевтические средства или комбинации таких химиотерапевтических средств, как цитостатики. Химиотерапевтические средства могут влиять на клетки одним из следующих способов: (1) повреждать ДНК в клетках, так что они больше не смогут воспроизводиться, (2) ингибировать синтез новых цепей ДНК, так что репликация клеток станет невозможной, (3) останавливать митотические процессы в клетках, так что клетки не смогут делиться на две клетки.
В соответствии с изобретением, термин "средство, стабилизирующее или повышающее экспрессию CLDN18.2" предпочтительно относится к таким средствам или комбинациям средств, как цитостатические соединения или комбинации цитостатических соединений, предоставление которых клеткам, в частности раковым клеткам, приводит к тому, что клетки останавливаются или накапливаются в одной или нескольких фазах клеточного цикла, предпочтительно в одной или нескольких других фазах клеточного цикла, чем фазы G1 и G0, предпочтительно других, чем фаза G1, предпочтительно в одной или нескольких из фаз G2 или S клеточного цикла, как-то фаз G1/G2, S/G2, G2 или S клеточного цикла. Термин "клетки останавливаются или накапливаются в одной или нескольких фазах клеточного цикла" означает то, что возрастает процент клеток, находящихся в одной или нескольких фазах клеточного цикла. Каждая клетка для своего воспроизведения проходит через цикл, состоящий из четырех фаз. Первая фаза называется G1, когда клетка готовится к воспроизведению своих хромосом. Второй этап называется S, и в этой фазе происходит синтез ДНК и дупликация ДНК. Следующая фаза - фаза G2, когда происходит дупликация РНК и белков. Заключительная стадия - стадия М, которая заключается в реальном делении клеток. На этой заключительной стадии прошедшие дупликацию ДНК и РНК разделяются и движутся к разным полюсам клетки, а клетка в самом деле делится на две одинаковые функциональные клетки. Химиотерапевтические средства, которые вызывают повреждения ДНК, обычно приводят к накоплению клеток в фазе G1 и/или G2. Химиотерапевтические средства, которые блокируют рост клеток, препятствуя синтезу ДНК, типа антиметаболитов, обычно вызывают накопление клеток в фазе S. Примеры таких препаратов - гемцитабин, 6-меркаптопурин и 5-фторурацил.
В соответствии с изобретением, термин "средство, стабилизирующее или повышающее экспрессию CLDN18.2" включает аналоги нуклеозидов, такие как гемцитабин, 5-фторурацил или их пролекарственные формы, соединения платины, такие как цисплатин и оксалиплатин, таксаны, такие как паклитаксель и доцетаксель, и аналоги камптотецина, такие как иринотекан и топотекан, и комбинированные препараты, как-то комбинированные препараты, включающие один или несколько из числа гемцитабина, оксалиплатина и 5-фторурацила, как-то комбинированные препараты, содержащие гемцитабин и оксалиплатин, гемцитабин и 5-фторурацил, оксалиплатин и 5-фторурацил или другие описанные здесь комбинации препаратов. В соответствии с изобретением, ссылки на средство, стабилизирующее или повышающее экспрессию CLDN18.2, как-то ссылки на аналог нуклеозида, соединение платины либо аналог камптотецина или таксан, к примеру, ссылки на гемцитабин, 5-фторурацил, оксалиплатин, иринотекан или паклитаксель, должны включать и их пролекарственные формы, такие как сложные эфиры, соли или производные типа конъюгатов данных средств. Примерами их являются конъюгаты данных средств с веществами-носителями, например, связанный с белком паклитаксель типа связанного с альбумином паклитакселя. Предпочтительно соли данных средств являются фармацевтически приемлемыми.
В одном предпочтительном воплощении "средство, стабилизирующее или повышающее экспрессию CLDN18.2" представляет собой или включает в себя "средство, индуцирующее иммуногенную гибель клеток".
В определенных обстоятельствах раковые клетки могут вступать на летальный стрессовый путь, связанный с подачей заданной в пространстве и времени комбинации сигналов, которая декодируется иммунной системой для активации опухолеспецифичных иммунных ответов (Zitvogel L. et al. (2010) Cell 140: 798-804). По такому сценарию раковые клетки активируются и подают сигналы, которые воспринимаются врожденными иммунными эффекторами типа дендритных клеток и запускают когнатный иммунный ответ, который включает Т-клетки CD8+ и сигнализацию IFN-γ с тем, чтобы гибель раковых клеток могла вызвать продуктивный противораковый иммунный ответ. Эти сигналы включают преапоптотическое воздействие на шаперон эндоплазматического ретикулума (ER) - кальретикулин (CRT) на клеточной поверхности, преапоптотическую секрецию АТФ и постапоптотическое высвобождение ядерного белка HMGB1. В целом эти процессы составляют молекулярные детерминанты иммуногенной гибели клеток (ICD). Антрациклины, оксалиплатин, и γ-облучение способны индуцировать все сигналы, которые определяют ICD, тогда как для цисплатина, к примеру, который не способен индуцировать транслокацию CRT из ER на поверхность гибнущих клеток, т.е. процесс, требующий ER-стресса, необходимо содействие тапсигаргина, индуктора ER-стресса.
В соответствии с изобретением, термин "средство, индуцирующее иммуногенную гибель клеток" относится к таким средствам или комбинациям средств, которые при предоставлении их клеткам, в частности раковым клеткам, способны индуцировать вступление клеток на летальный стрессовый путь, который в конечном счете вызывает опухолеспецифичные иммунные ответы. В частности, средства, индуцирующие иммуногенную гибель клеток, при предоставлении их клеткам вызывают подачу клетками заданной в пространстве и времени комбинации сигналов, включающих, в частности, преапоптотическое воздействие на шаперон эндоплазматического ретикулума (ER) - кальретикулин (CRT) на клеточной поверхности, преапоптотическую секрецию АТФ и постапоптотическое высвобождение ядерного белка HMGB1.
В соответствии с изобретением, термин "средство, индуцирующее иммуногенную гибель клеток", включает в себя антрациклины и оксалиплатин.
Термин "аналог нуклеозида" относится к структурным аналогам нуклеозидов - категории, которая включает и аналоги пуринов, и аналоги пиримидинов.
Термин "гемцитабин" означает соединение, которое является аналогом нуклеозида по следующей формуле:
 .
.
В частности, этот термин обозначает соединение 4-амино-1-(2-дезокси-2,2-дифтор-β-D-эритро-пентофуранозил)пиримидин-2(1Н)-он или 4-амино-1-[(2R,4R,5R)-3,3-дифтор-4-гидрокси-5-(гадроксиметил)оксолан-2-ил]-1,2-дигидрогшримидин-2-он.
В соответствии с изобретением, гемцитабин предпочтительно вводится внутривенно. Предпочтительно гемцитабин вводится в дозах от 0,5 до 2 г/м2, предпочтительно от 0,8 до 1,5 г/м2, более предпочтительно от 1 до 1,2 г/м2 площади поверхности тела. Например, гемцитабин можно вводить в дозе 1000 мг на квадратным метр раз в неделю на протяжении 7 из 8 недель, а затем раз в неделю на протяжении 3 из 4 недель.
Термин "аналог нуклеозида" включает такие производные фторпиримидинов, как фторурацил и его пролекарственные формы. Термин "фторурацил" или "5-фторурацил" (5-FU или f5U) (в продаже под торговыми марками Adrucil, Carac, Efudix, Efudex и Fluoroplex) означает соединение, которое является аналогом пиримидина по следующей формуле:
 .
.
В частности, этот термин относится к соединению 5-фтор-1Н-пиримидин-2,4-дион.
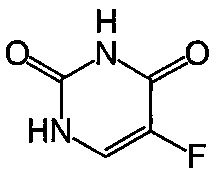
Термин "капецитабин" (Xeloda, Roche) относится к химиотерапевтическому средству, которое является пролекарством, которое в тканях превращается в 5-FU. Капецитабин, который можно вводить перорально, имеет следующую формулу:
 .
.
В частности, этот термин относится к соединению пентил-[1-(3,4-дигидрокси-5-метилтетрагидрофуран-2-ил)-5-фтор-2-оксо-1Н-пиримидин-4-ил]карбамат.
В соответствии с изобретением, термин "соединение платины" относится к соединениям, содержащим платину в своей структуре типа комплексов платины, и включает такие соединения, как цисплатин, карбоплатин и оксалиплатин.
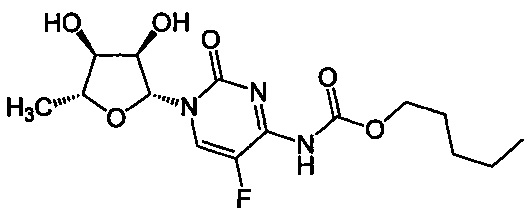
Термин "цисплатин" или "цисплатина" относится к соединению цис-диаминхлорплатина(II) (CDDP) следующей формулы:
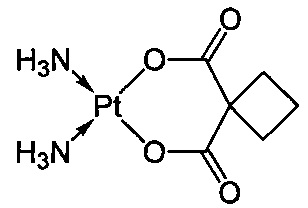
Термин "карбоплатин" относится к соединению цис-диамин-(1,1-циклобутандикарбоксилато)платина(II) следующей формулы:
Термин "оксалиплатин" относится к соединению, которое является соединением платины, образующим комплекс с несущим диаминоциклогексан лигандом по следующей формуле:
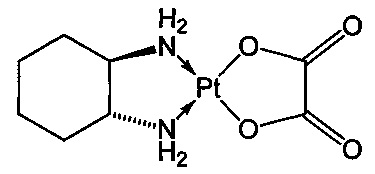
В частности, термин "оксалиплатин" относится к соединению [(1R,2R)-циклогексан-1,2-диамин](этандиоато-O,O')платина(II). Оксалиплатин для инъекций также поступает в продажу под торговым названием Eloxatine.
Таксаны представляют собой класс дитерпеновых соединений, которые были впервые получены из таких природных источников, как растения из рода Taxus, но некоторые были синтезированы искусственно. Основным механизмом действия препаратов класса таксанов является нарушение функции микротрубочек, что ингибирует процесс деления клеток. Таксаны включают доцетаксель (Taxotere) и паклитаксель (Taxol).
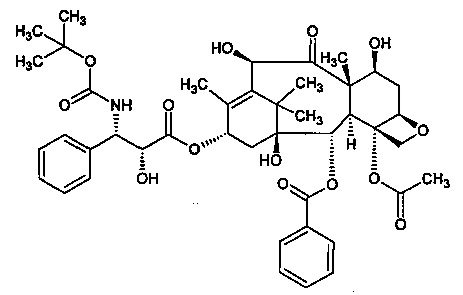
В соответствии с изобретением, термин "доцетаксель" относится к соединению, имеющему следующую формулу:
 .
.
В частности, термин "доцетаксель" относится к соединению 1,7β,10β-тригидрокси-9-оксо-5β,20-эпокситакс-11-ен-2α,4,13α-триил-4-ацетат-2-бензоат-13-{(2R,3S)-3-[(трет-бутоксикарбонил)амино]-2-гидрокси-3-фенилпропаноат}.
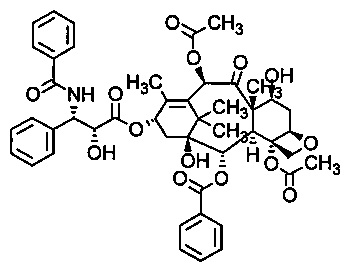
В соответствии с изобретением, термин "паклитаксель" относится к соединению, имеющему следующую формулу:
 .
.
В частности, термин "паклитаксель" относится к соединению (2α,4α,5β,7β,10β,13α)-4,10-бис-(ацетилокси)-13-{[(2R,3S)-3-(бензоиламино)-2-гидрокси-3-фенилпропаноил]окси}-1,7-дигидрокси-9-оксо-5,20-эпокситакс-11-ен-2-ил-бензоат.
В соответствии с изобретением, термин "аналог камптотецина" относится к производным соединения камптотецина (СРТ; (S)-4-этил-4-гидрокси-1Н-пирано[3',4':6,7]индолизино-[1,2-b]хинолин-3,14-(4Н,12Н)-дион). Предпочтительно термин "аналог камптотецина" относится к соединениям, содержащим следующую структуру:
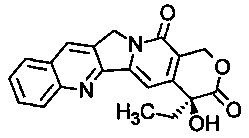
В соответствии с изобретением, предпочтительными аналогами камптотецина являются ингибиторы фермента ДНК-топоизомеразы I (topo I). Предпочтительными аналогами камптотецина по изобретению являются иринотекан и топотекан.
Иринотекан - препарат, препятствующий раскручиванию ДНК путем ингибирования топоизомеразы I. С химической точки зрения, он является полусинтетическим аналогом природного алкалоида камптотецина, имеющего следующую формулу:
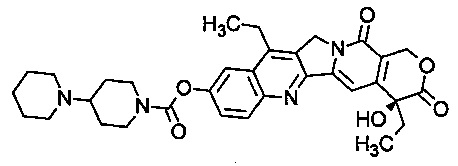
В частности, термин "иринотекан" относится к соединению (S)-4,11-диэтил-3,4,12,14-тетрагидро-4-гидрокси-3,14-диоксо-1Н-пирано[3',4':6,7]индолизино[1,2-b]хинолин-9-ил-[1,4'-бипиперидин]-1'-карбоксилат.
Топотекан является ингибитором топоизомеразы по формуле:

В частности, термин "топотекан" относится к соединению (S)-10-[(диметиламино)метил]-4-этил-4,9-дигидрокси-1Н-пирано[3',4':6,7]индолизино[1,2-b]хинолин-3,14(4Н,12Н)-дионмоногидрохлорид.
Антрациклины составляют класс лекарственных препаратов, которые широко применяются при химиотерапии рака, а также являются антибиотиками. По структуре все антрациклины содержат общую 4-кольцевую структуру 7,8,9,10-тетрагидротетрацен-5,12-хинона и обычно требуют гликозилирования по определенным сайтам.
Антрациклины предпочтительно действуют по одному или нескольким из следующих механизмов действия: 1) ингибирование синтеза ДНК и РНК путем интеркаляции между парами оснований в нити ДНК/РНК, тем самым предотвращая репликацию быстрорастущих раковых клеток; 2) ингибирование фермента топоизомеразы II, что предотвращает раскручивание сверхпиральной ДНК и тем самым блокирует транскрипцию и репликацию ДНК; 3) опосредованная железом генерация свободных радикалов кислорода, которые повреждают ДНК и клеточные мембраны.
В соответствии с изобретением, термин "антрациклин" предпочтительно относится к таким средствам, предпочтительно противораковым средствам, которые индуцируют апоптоз, предпочтительно путем ингибирования раскручивания ДНК топоизомеразой II.
Предпочтительно, согласно изобретению, термин "антрациклин" вообще относится к классу соединений, имеющих следующую кольцевую структуру
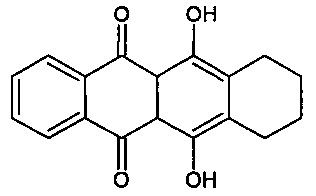
включая их аналоги и производные, фармацевтические соли, гидраты, эфиры, конъюгаты и пролекарственные формы.
Примеры антрациклинов и аналогов антрациклинов включают, без ограничения, даунорубицин (дауномицин), доксорубицин (адриамицин), эпирубицин, идарубицин, родомицин, пирарубицин, вальрубицин, N-трифтор-ацетил-доксорубицин-14-валерат, аклациномицин, морфолинодоксорубицин (морфолино-DOX), цианоморфолинодоксорубицин (цианоморфолино-DOX), 2-пирролинодоксорубицин (2-PDOX), 5-иминодауномицин, митоксантрон и аклациномицин (акларубицин). Митоксантрон является представителем класса соединений антрацендионов, которые являются аналогами антрациклина, не содержащими сахаридной части антрациклинов, но сохраняют планарную структуру полициклического ароматического кольца, которая позволяет интеркаляцию в ДНК.
Особенно предпочтительными антрациклинами по изобретению являются соединения следующей формулы:
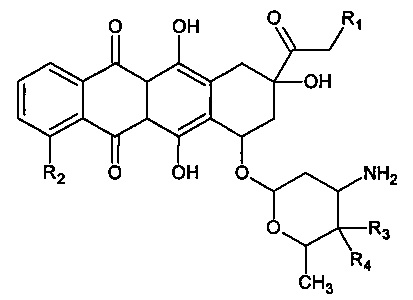
где: R1 выбран из группы, состоящей из Н и ОН, R2 - из группы, состоящей из Н и ОМе, R3 - из группы, состоящей из Н и ОН, a R4 - из группы, состоящей из Н и ОН.
В одном воплощении R1 означает Н, R2 - ОМе, R3 - Н, a R4 - ОН. В другом воплощении R1 означает ОН, R2 - ОМе, R3 - Н, a R4 - ОН. В другом воплощении R1 означает ОН, R2 - ОМе, R3 - ОН, a R4 - Н. В другом воплощении R1 означает Н, R2 - Н, R3 - H, a R4 - OH.
Особенно предпочтительным антрациклином в настоящем изобретении является эпирубицин - антрациклиновый препарат, который имеет следующую формулу:
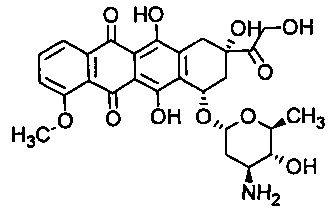
и поступает в продажу под торговой маркой Ellence в США и Pharmorubicin или Epirubicin Ebewe в других странах. В частности, термин "эпирубицин" относится к соединению (8R,10S)-10-[(2S,4S,5R,6S)-4-амино-5-гадрокси-6-метил-оксан-2-ил]окси-6,11-дигидрокси-8-(2-гидроксиацетил)-1-метокси-8-метил-9,10-дигидро-7Н-тетрацен-5,12-дион. Эпирубицин в некоторых схемах химиотерапии предпочтителен перед доксорубицином, наиболее популярным антрациклином, так как он вызывает меньше побочных эффектов.
В соответствии с изобретением, средство, стабилизирующее или повышающее экспрессию CLDN18.2, может представлять собой химиотерапевтическое средство, в частности, химиотерапевтическое средство, признанное для лечения рака, которое может входить в комбинации препаратов типа комбинаций препаратов, признанных для применения при лечении рака. Такие комбинации препаратов могут представлять собой комбинации препаратов, используемые в химиотерапии, как-то комбинации препаратов, которые применяются в режиме химиотерапии FOLFIRINOX.
Комбинация препаратов, используемая при химиотерапии FOLFIRINOX, включает лейковорин, фторурацил, иринотекан (как-то иринотекан гидрохлорид) и оксалиплатин. Оксалиплатин может вводиться в дозе 85 мг/м2 площади поверхности тела; иринотекан в дозе 180 мг/м2; лейковорин в дозе 400 мг/м2; и фторурацил в дозе 400 мг/м2 в виде болюса с последующим введением 5-фторурацила в дозе 2400 мг/м2 в виде непрерывной инфузии предпочтительно на протяжении 46 ч, предпочтительно каждые 2 недели.
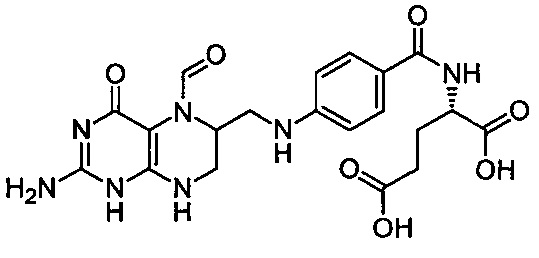
Термин "фолиновая кислота" или "лейковорин" относится к соединению, которое применяется в синергической комбинации с химиотерапевтическим средством 5-фторурацилом. Так, если здесь упоминается введение 5-фторурацила или его пролекарственной формы, то такое введение в одном воплощении может включать введение в сочетании с фолиновой кислотой. Фолиновая кислота имеет следующую формулу:
 .
.
В частности, этот термин относится к соединению (2S)-2-{[4-[(2-амино-5-формил-4-оксо-5,6,7,8-тетрагидро-1Н-птеридин-6-ил)метиламино]бензоил]амино}пентадионовая кислота.
Т-клетки γδ (Т-клетки гамма-дельта) составляют небольшое подмножество Т-клеток, обладающих особыми Т-клеточными рецепторами (TCR) на своей поверхности. У большинства Т-клеток TCR состоят из двух гликопротеидных цепей, называемых α- и β-цепи TCR. В противоположность этому, у Т-клеток γδ TCR состоят из одной γ-цепи и одной δ-цепи. Эта группа Т-клеток обычно гораздо менее распространена, чем Т-клетки αβ. Т-клетки γδ человека играют важную роль в реакциях стрессового надзора типа инфекционных заболеваний и аутоиммунитета. Также предполагается, что вызванные трансформацией изменения в опухолях вызывают реакции стрессового надзора, опосредованные Т-клетками γδ, и усиливают противоопухолевый иммунитет. Важно отметить, что активированные после воздействия антигена Т-клетки γδ в очаге повреждения обеспечивают цитокины (например, INFγ, TNFα) и/или хемокины, опосредующие привлечение (рекрутмент) других эффекторных клеток, и проявляют непосредственные эффекторные функции, такие как цитотоксичность (через рецепторы смерти и пути цитолитических гранул) и ADCC.
Большинство Т-клеток γδ в периферической крови экспрессируют Т-клеточный рецептор Vγ9Vδ2 (TCRγδ). Т-клетки Vγ9Vδ2 уникальны для человека и приматов и предполагается, что они играют раннюю и важнейшую роль в восприятии "опасности" при вторжении патогенов, так как их количество сильно возрастает при многих острых инфекциях и за несколько дней может превзойти все другие лимфоциты, например, при туберкулезе, сальмонеллезе, эрлихиозе, бруцеллезе, туляремии, листериозе, токсоплазмозе и малярии.
Т-клетки γδ реагируют на небольшие непептидные фосфорилированные антигены (фосфоантигены) типа пирофосфатов, синтезирующихся в бактериях, и изопентенил-пирофосфатов (IPP), вырабатывающихся в клетках млекопитающих через мевалонатный путь. В то время как продукция IPP в нормальных клетках недостаточна для активации Т-клеток γδ, нарушение регуляции мевалонатного пути в опухолевых клетках приводит к накоплению IPP и активации Т-клеток γδ. Уровень IPPs также может и терапевтически повышаться аминобисфосфонатами, которые ингибируют фермент мевалонатного пути - фарнезил-пирофосфатсинтазу (FPPS). Среди прочего, такие аминобисфосфонаты представлены золедроновой кислотой (ZA, золедронат, Zometa™, Novartis), которая уже применяется клинически на пациентах для лечения остеопороза и метастатических заболеваний костей. После обработки PBMCs in vitro ZA в особенности поглощается моноцитами. IPP накапливаются в моноцитах, которые дифференцируются в антиген-презентирующие клетки, стимулирующие развитие Т-клеток γδ. При этом предпочтительно добавление интерлейкина-2 (IL-2) в качестве фактора роста и выживания для активированных Т-клеток γδ. Наконец, были описаны некоторые алкилированные амины, активирующие Т-клетки Vγ9Vδ2 in vitro,no только в миллимолярных концентрациях.
В соответствии с изобретением, термин "средство, стимулирующее Т-клетки γδ" относится к соединениям, стимулирующим развитие Т-клеток γδ, в частности Т-клеток Vγ9Vδ2, in vitro и/или in vivo, в частности, вызывая активацию и экспансию Т-клеток γδ. Предпочтительно этот термин относится к соединениям, которые in vitro и/или in vivo повышают продукцию изопентенил-пирофосфатов (IPP), вырабатывающихся в клетках млекопитающих, предпочтительно путем ингибирования фермента мевалонатного пути - фарнезил-пирофосфатсинтазы (FPPS).
Одной из предпочтительных групп соединений, стимулирующих Т-клетки γδ, являются бисфосфонаты, в особенности азотсодержащие бисфосфонаты (N-бисфосфонаты; аминобисфосфонаты).
Например, подходящие бисфосфонаты для применения в изобретении могут включать одно или несколько из следующих соединений, включая их аналоги и производные, фармацевтические соли, гидраты, эфиры, конъюгаты и пролекарственные формы:
[1-гадрокси-2-(1Н-имидазол-1-ил)этан-1,1-диил]бис(фосфоновая кислота), т.е. золедроновая кислота, например, золедронат;
(дихлорфосфоно-метил)фосфоновая кислота, например, клодронат;
{1-гидрокси-3-[метил(пентил)амино]пропан-1,1-диил}бис(фосфоновая кислота), т.е. ибандроновая кислота, например, ибандронат;
(3-амино-1-гидроксипропан-1,1-диил)бис(фосфоновая кислота), т.е. памидроновая кислота, например, памидронат;
(1-гидрокси-1-фосфоно-2-пиридин-3-ил-этил)фосфоновая кислота, т.е. ризедроновая кислота, например, ризедронат;
(1-гидрокси-2-имидазо[1,2-а]пиридин-3-ил-1-фосфоноэтил)фосфоновая кислота, т.е. минодроновая кислота;
[3-(диметиламино)-1-гидроксипропан-1,1-диил]бис(фосфоновая кислота), т.е. олпадроновая кислота;
[4-амино-1-гидрокси-1-(гидрокси-оксидофосфорил)бутил]фосфоновая кислота, т.е. алендроновая кислота, например, алендронат;
[(циклогептиламино)метилен]бис(фосфоновая кислота), т.е. инкадроновая кислота;
(1-гидроксиэтан-1,1-диил)бис(фосфоновая кислота), т.е. этидроновая кислота, например, этидронат; и
{[(4-хлорфенил)тио]метилен}бис(фосфоновая кислота), т.е. тилудроновая кислота.
В соответствии с изобретением, особенно предпочтительным бисфосфонатом является золедроновая кислота (INN) или золедронат (продается фирмой Novartis под торговыми марками Zometa, Zomera, Aclasta и Reclast). Zometa применяется для предотвращения скелетных переломов у больных раком типа множественной миеломы и рака простаты, а также для лечения остеопороза. Она также может применяться для лечения гиперкальциемии при раке и может быть полезной для лечения боли при метастазах в костях.
В одном особенно предпочтительном воплощении средство, стимулирующее Т-клетки γδ по изобретению, вводится в комбинации с IL-2. Такая комбинация, как было показано, особенно эффективно опосредует экспансию и активацию Т-клеток γ9δ2.
Интерлейкин-2 (IL-2) представляет собой интерлейкин, сигнальную молекулу цитокинового типа в иммунной системе. Это белок, который привлекает лимфоциты и является частью природного ответа организма на микробные инфекции, а также участвует в различении между чужими (не своими) и собственными антигенами. IL-2 опосредует свои эффекты путем связывания с рецепторами IL-2, которые экспрессируются лимфоцитами.
При применении по изобретению IL-2 может представлять собой любой IL-2, поддерживающий или способствующий стимуляции Т-клеток γδ, и может происходить из любого вида, предпочтительно человека. IL-2 может представлять собой выделенный, полученный рекомбинантно или синтетический IL-2, а также может быть природного происхождения или модифицированный IL-2.
Термин "антиген" относится к таким веществам, как белки или пептиды, содержащие эпитоп, против которого направлен и/или должен быть направлен иммунный ответ. В предпочтительном воплощении антиген представляет собой опухолевый антиген типа CLDN18.2, т.е. компонент раковых клеток, который может происходить из цитоплазмы, клеточной поверхности или клеточного ядра, в частности, такой антиген, который вырабатывается, предпочтительно в большом количестве, внутри клетки или как поверхностный антиген на раковых клетках.
В настоящем изобретении термин "ассоциированный с опухолью (опухолевый) антиген" предпочтительно относится к таким белкам, которые в нормальных условиях специфически экспрессируются в ограниченном числе тканей и/или органов или на определенных этапах онтогенеза и экспрессируются или аномально экспрессируются в одной или нескольких опухолевых или раковых тканях. В настоящем изобретении опухолевые антигены предпочтительно связаны с клеточной поверхностью раковых клеток и предпочтительно не экспрессируются или редко экспрессируются в нормальных тканях.
Термин "эпитоп" относится к антигенным детерминантам в молекулах, т.е. той части молекул, которая распознается иммунной системой, к примеру, распознается антителом. Например, эпитопы представляют собой дискретные, трехмерные сайты на антигенах, которые распознаются иммунной системой. Как правило, эпитопы состоят из химически активных поверхностных группировок молекул типа аминокислот или боковых цепей Сахаров и обычно имеют специфические трехмерные структурные характеристики, а также специфические характеристики зарядов. Конформационные и неконформационные эпитопы отличаются тем, что связывание с первыми, но не последними утрачивается в присутствии денатурирующих растворителей. Эпитопы таких белков, как CLDN18.2, предпочтительно включают в себя непрерывный или прерывистый участок данного белка длиной предпочтительно от 5 до 100, предпочтительно от 5 до 50, более предпочтительно от 8 до 30, наиболее предпочтительно от 10 до 25 аминокислот, например, эпитоп может быть длиной предпочтительно в 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 аминокислот.
Термин "антитело" относится к гликопротеинам, содержащим по меньшей мере две тяжелые (Н) цепи и две легкие (L) цепи, связанные между собой дисульфидными связями, и включает в себя любые молекулы, содержащие антигенсвязывающие участки. Термин "антитело" включает в себя моноклональные антитела и фрагменты или производные антител, в том числе, без ограничения, человеческие антитела, гуманизованные антитела, химерные антитела, одноцепочечные антитела, например, scFv, и антиген-связывающие фрагменты антител, такие как фрагменты Fab и Fab', а также включает в себя все рекомбинантные формы антител, например, антитела, экспрессированные в прокариотах, негликозилированные антитела, а также любые антигенсвязывающие фрагменты и производные антител, как описано здесь. Каждая тяжелая цепь состоит из вариабельной области тяжелой цепи (сокращенно VH) и константной области тяжелой цепи. Каждая легкая цепь состоит из вариабельной области легкой цепи (сокращенно VL) и константной области легкой цепи. Области VH и VL могут еще дальше подразделяться на участки гипервариабельности, именуемые определяющими комплементарность участками (CDR), которые перемежаются с более консервативными участками, именуемыми каркасными участками (FR). Каждая область VH и VL состоит из трех CDR и четырех FR, располагающихся от N-конца к С-концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Вариабельные области тяжелой и легкой цепей содержат связывающий домен, который взаимодействует с антигеном. Константные области антител могут опосредовать связывание иммуноглобулина с тканями или факторами хозяина, включая различные клетки иммунной системы (например, эффекторные клетки) и первый компонент (Clq) классической системы комплемента.
Описанные здесь антитела могут быть человеческими антителами. Термин "человеческое антитело" в настоящем изобретении служит для обозначения антител, имеющих вариабельные и константные области, происходящие из последовательностей иммунолобулинов человека зародышевой линии. Описанные здесь человеческие антитела могут содержать остатки аминокислот, не кодируемых последовательностями иммунолобулинов человека зародышевой линии (например, мутации, введенные случайным образом или методом сайт-специфичного мутагенеза in vitro или посредством соматической мутации in vivo).
Термин "гуманизованное антитело" относится к молекулам, у которых антигенсвязывающие сайты в основном происходят из иммуноглобулинов других видов, чем человек, тогда как остальная иммуноглобулиновая структура молекулы основывается на структуре и/или последовательности иммуноглобулина человека. Антигенсвязывающие сайты могут либо содержать полные вариабельные домены, встроенные в константные домены, либо только гипервариабельные участки (CDR), встроенные в соответствующие каркасные участки вариабельных доменов. Антигенсвязывающие участки могут быть дикого типа или модифицированные заменой одной или нескольких аминокислот, например, модифицированы так, чтобы иметь большее сходство с иммуноглобулинами человека. У некоторых форм гуманизованных антител сохраняются все последовательности CDR (к примеру, гуманизованное антитело мыши, которое содержит все шесть CDR из антитела мыши). У других форм один или несколько участков CDR подвергались изменениям по сравнению с исходным антителом.
Термин "химерное антитело" относится к таким антителам, у которых одна часть каждой из аминокислотных последовательностей тяжелых и легких цепей гомологична соответствующим последовательностям у антител, происходящих из определенного вида или принадлежащих к определенному классу, а остальные сегменты цепи гомологичны соответствующим последовательностям у других. Как правило, вариабельные области легких и тяжелых цепей воспроизводят вариабельные области антител, происходящих из одного вида млекопитающих, а константные области гомологичны последовательности антител, происходящих из другого. Одним из очевидных преимуществ таких химерных форм является то, что вариабельная область может легко быть получена из известных в настоящее время источников с помощью легкодоступных В-клеток или гибридом из других организмов, чем человек, в сочетании с константными областями, полученными, к примеру, из препаратов клеток человека. В то время, как вариабельная область имеет преимущество в легкости получения, а специфичность ее не зависит от источника, то константная область, полученная из человека, с меньшей вероятностью будет вызывать иммунный ответ у субъекта-человека при введении антител, чем если бы константная область происходила другого из источника, чем человек. Однако это определение не ограничивается данным конкретным примером.
Термины "антигенсвязывающая часть" антитела (или просто "связывающая часть") или "антигенсвязывающий фрагмент" антитела (или просто "связывающий фрагмент") или подобные термины относятся к одному или нескольким фрагментам, сохраняющим способность антитела к специфичному связыванию с антигеном. Было показано, что антигенсвязывающая функция антител может выполняться фрагментами полноразмерных антител. Примеры связывающих фрагментов, охватываемые термином "антигенсвязывающая часть" антитела, включают в себя: (i) Fab-фрагменты, моновалентные фрагменты, состоящие из доменов VL, VH, CL и CH; (ii) F(ab')2-фрагменты, бивалентные фрагменты, содержащие два двухвалентных Fab-фрагмента, соединенных дисульфидным мостиком в шарнирной области; (iii) Fd-фрагменты, состоящие из доменов VH и CH; (iv) Fv-фрагменты, состоящие из доменов VL и VH на одном плече антитела; (v) dAb-фрагменты (Ward et al. (1989) Nature 341: 544-546), которые состоят из домена VH; (vi) выделенные участки, определяющие комплементарность (CDR); и (vii) комбинации из двух или нескольких выделенных CDR, которые необязательно могут соединяться через синтетический линкер. Кроме того, хотя два домена Fv-фрагмента, VL и VH, кодируются отдельными генами, их можно соединить, рекомбинантными методами, при помощи синтетического линкера, который позволяет им составлять одну белковую цепь, в которой сочетаются области VL и VH с образованием моновалентных молекул (известных как одноцепочечный Fv (scFv), например, см, Bird et al. (1988) Science 242: 423-426; и Huston et al. (1988) Proc. Natl. Acad. Sci. USA 85: 5879-5883). Такие одноцепочечные антитела тоже охватываются термином "антигенсвязывающий фрагмент" антитела. Следующий пример - слитые белки связывающего домена иммуноглобулина, включающие: (i) полипептид связывающего домена, слитый с полипептидом шарнирного участка иммуноглобулина, (ii) константную область CH2 тяжелой цепи иммуноглобулина, слитую с шарнирным участком, и (iii) константную область CH3 тяжелой цепи иммуноглобулина, слитую с константной областью CH2. Полипептид связывающего домена может представлять собой вариабельную область тяжелой цепи или вариабельную область легкой цепи. Слитые белки связывающего домена иммуноглобулина более подробно раскрыты в US 2003/0118592 и US 2003/0133939. Эти фрагменты антител получают стандартными методами, известными специалистам в данной области, и фрагменты подвергают скринингу на применимость таким же образом, как и интактные антитела.
Термин "биспецифичные молекулы" служит для обозначения таких агентов, например, белков, пептидов либо белковых или пептидных комплексов, которые обладают двумя различными специфичностями связывания. Например, молекула может связываться или взаимодействовать с (а) антигеном на клеточной поверхности и (b) Fc-рецептором на поверхности эффекторной клетки. Термин "полиспецифичная молекула" или "гетероспецифичная молекула" служит для обозначения таких агентов, например, белков, пептидов либо белковых или пептидных комплексов, которые обладают более чем двумя различными специфичностями связывания. Например, молекула может связываться или взаимодействовать с (а) антигеном на клеточной поверхности, (b) Fc-рецептором на поверхности эффекторной клетки и (с) по меньшей мере еще с одним компонентом. Соответственно, изобретение включает, без ограничения, биспецифичные, триспецифичные, тетраспецифичные и другие мультиспецифичные молекулы, которые направлены на CLDN18.2 и другие мишени, как-то Fc-рецепторы на эффекторных клетках. Термин "биспецифичные антитела" также включает в себя диатела. Диатела представляют собой бивалентные биспецифичные антитела, у которых домены VH и VL экспрессируются на одной и той же полипептидной цепи, но с помощью такого линкера, который слишком короткий для образования связи между двумя доменами на одной и той же цепи, что вынуждает домены связываться с комплементарными доменами на другой цепи, образуя два антигенсвязывающих сайта (например, см. Holliger P. et al. (1993) Proc. Natl. Acad. Sci. USA 90: 6444-6448; Poljak R.J. et al. (1994) Structure 2: 1121-1123).
Антитело можно конъюгировать с терапевтической молекулой или средством, как-то цитотоксином, лекарственным препаратом (например, иммунодепрессантом) или радиоизотопом. Цитотоксины или цитотоксические средства включает в себя любые средства, которые приносят вред и, в частности, убивают клетки. Примеры включают таксол, цитохалазин В, грамицидин D, этидия бромид, эметин, митомицин, этопозид, винкристин, тенопозид, винбластин, колхицин, доксорубицин, даунорубицин, дигидроксиантрациндион, митоксантрон, митрамицин, актиномицин D, 1-дегидротестостерон, глюкокортикоиды, прокаин, тетракаин, лидокаин, пропранолол и пуромицин и их аналоги или гомологи. Подходящие терапевтические средства для получения конъюгатов антител включают, без ограничения, антиметаболиты (например, метотрексат, 6-меркаптопурин, 6-тиогуанин, цитарабин, флударабин, 5-фторурацил, дакарбазин), алкилирующие средства (например, мехлорэтамин, тиоэпа, хлорамбуцил, мелфалан, кармустин (BSNU) и ломустин (CCNU), циклофосфамид, бусульфан, дибромманнитол, стрептозотоцин, митомицин С и цис-дихлордиаминплатина(II) (DDP, цисплатин), антрациклины (например, даунорубицин (прежде дауномицин) и доксорубицин), антибиотики (например, дактиномицин (прежде актиномицин), блеомицин, митрамицин и антрамицин (АМС) и анти-митотические средства (например, винкристин и винбластин). В предпочтительном воплощении терапевтическое средство представлено цитотоксическим средством или радиотоксическим средством. В другом воплощении терапевтическое средство представлено иммунодепрессантом. В следующем воплощении терапевтическое средство представлено GM-CSF. В предпочтительном воплощении терапевтическим средством служит доксорубицин, цисплатин, блеомицин сульфат, кармустин, хлорамбуцил, циклофосфамид или рицин А.
Антитела также можно конъюгировать с радиоизотопами, например, с йодом-131, иттрием-90 или индием-111, получая цитотоксические радиофармпрепараты.
Конъюгаты антител по изобретению могут применяться для модификации заданного биологического ответа, а молекулы лекарственных средств не должны ограничиваться классическими химическими терапевтическими средствами. Например, молекула лекарственного средства может быть белком или полипептидом, обладающим требуемой биологической активностью. Такие белки могут включать в себя, к примеру, ферментативно активные токсины или их активные фрагменты, как-то абрин, рицин А, экзотоксин Pseudomonas или дифтерийный токсин; такие белки, как фактор некроза опухолей или γ-интерферон; либо модификаторы биологических ответов, такие, к примеру, как лимфокины, интерлейкин-1 ("IL-1"), интерлейкин-2 ("IL-2"), интерлейкин-6 ("IL-6"), гранулоцитарно-макрофагальный колониестимулирующий фактор ("GM-CSF"), гранулоцитарный колониестимулирующий фактор ("G-CSF") или другие факторы роста.
Методы конъюгирования таких терапевтических молекул с антителами хорошо известны, например, см. Arnon et al., "Monoclonal antibodies for immunotargeting of drugs in cancer therapy", in Monoclonal Antibodies and Cancer Therapy, Reisfeld et al. (eds.), pp. 243-56 (Alan R. Liss, Inc. 1985); Hellstrom et al., "Antibodies for drug delivery", in Controlled Drug Delivery (2nd Ed.), Robinson et al. (eds.), pp. 623-53 (Marcel Dekker, Inc. 1987); Thorpe, "Antibody carriers of cytotoxic agents in cancer therapy: a review", in Monoclonal Antibodies '84: Biological and Clinical Applications, Pincheraet al. (eds.), pp. 475-506 (1985); "Analysis, results, and future prospective of the therapeutic use of radiolabeled antibodies in cancer therapy", in Monoclonal Antibodies for Cancer Detection and Therapy, Baldwin et al. (eds.), pp. 303-16 (Academic Press 1985), и Thorpe et al., "The Preparation and cytotoxic properties of antibody-toxin conjugates", Immunol. Rev., 62: 119-58 (1982).
В настоящем изобретении антитело "происходит из" определенной последовательности зародышевой линии, если оно получено из системы путем иммунизации животного или скрининга библиотеки иммуноглобулиновых генов, причем выбранное антитело по аминокислотной последовательности по меньшей мере на 90%, более предпочтительно 95%, еще более предпочтительно по меньшей мере на 96%, 97%, 98% или на 99% идентично аминокислотной последовательности, кодируемой иммуноглобулиновым геном зародышевой линии. Как правило, антитело, происходящее из определенной последовательности зародышевой линии, должно проявлять отличия не более чем по 10 аминокислотам, более предпочтительно не более чем по 5 или еще более предпочтительно не более чем по 4, 3, 2 или по 1 аминокислоте от аминокислотной последовательности, кодируемой иммуноглобулиновым геном зародышевой линии.
В настоящем изобретении термин "гетероантитела" относится к соединенным друг с другом двум или нескольким антителам, их производным или антигенсвязывающим участкам, причем по крайней мере два из них имеют различные специфичности. Эти различные специфичности включают специфичность связывания для Fc-рецепторов на эффекторных клетках и специфичность связывания с антигеном или эпитопом на клетках мишени, например, опухолевых клетках.
Описанные здесь антитела могут быть моноклональными антителами. Термин "моноклональное антитело" в настоящем изобретении относится к препаратам молекул антител одного молекулярного состава. Моноклональное антитело проявляет лишь одну специфичность связывания и сродство. В одном воплощении моноклональные антитела вырабатываются гибридомой, которая включает В-клетки, полученные из животного, а не человека, например, мыши, слитые с иммортализованными клетками.
Описанные здесь антитела могут быть рекомбинантными антителами. Термин "рекомбинантное антитело" в настоящем изобретении включает все антитела, которые получены, экспрессированы, созданы или выделены рекомбинантными методами, как-то (а) антитела, выделенные из таких животных (например, мышей), которые трансгенны или трансхромосомны по отношению к генам иммуноглобулинов или гибридоме, из которой они получены, (b) антитела, выделенные из клеток хозяина, трансформированных для экспрессии антител, например, из трансфектомы, (с) антитела, выделенные из рекомбинантной или комбинаторной библиотеки антител, и (d) антитела, полученные, экспрессированные, созданные или выделенные любыми другими методами, которые включают сплайсинг последовательностей генов иммуноглобулинов с другими последовательностями ДНК.
Описанные здесь антитела могут происходить из различных видов, в том числе, без ограничения, мышей, крыс, кроликов, морских свинок и человека.
Описанные здесь антитела включают поликлональные и моноклональные антитела и охватывают антитела типа IgA, как-то IgA1 или IgA2, IgG1, IgG2, IgG3, IgG4, IgE, IgM и IgD. В различных воплощениях антитела представляют собой антитела типа IgG1, более предпочтительно антитела типа IgG1 изотипа каппа или лямбда (то есть IgG1, κ, λ), антитела типа IgG2a (например, IgG2a, κ, λ), антитела типа IgG2b (например, IgG2b, κ, λ), антитела типа IgG3 (например, IgG3, κ, λ) или антитела типа IgG4 (например, IgG4, κ, λ).
Термин "трансфектома" в настоящем изобретении охватывает рекомбинантные эукариотические клетки хозяина, экспрессирующие антитела, как-то клетки СНО, клетки NS/0, клетки HEK293, клетки HEK293T, растительные или грибковые клетки, в том числе дрожжевые клетки.
В настоящем изобретении термин "гетерологичное антитело" определяется по отношению к трансгенным организмам, вырабатывающим такие антитела. Этот термин относится к антителам, у которых аминокислотная последовательность или кодирующая последовательность нуклеиновой кислоты соответствует той, что находится у организма, не являющегося трансгенным, и обычно происходит из другого вида, чем трансгенный организм.
В настоящем изобретении "гетерогибридное антитело" означает такое антитело, у которого легкие и тяжелые цепи происходят из различных организмов. Например, антитело, у которого тяжелая цепь человека связана с легкой цепью мыши, является гетерогибридным антителом.
Изобретение охватывает все описанные здесь антитела и производные антител, которые для целей настоящего изобретения охватываются термином "антитело". Термин "производные" антител относится к любым модифицированным формам антител, например, конъюгатам антител с другими молекулами или антителами или фрагментами антител.
Описанные здесь антитела предпочтительно являются выделенными. Термин "выделенное антитело" в настоящем изобретении служит для обозначения такие антител, которые практически свободны от других антител, обладающих другой антигенной специфичностью (например, выделенное антитело, которое специфически связывается с CLDN18.2, практически свободно от антител, специфически связывающихся с другими антигенами, чем CLDN18.2). Однако выделенное антитело, которое специфически связывается с эпитопом, изоформой или вариантом CLDN18.2 человека, может обладать перекрестной реактивностью с другими родственными антигенами, например, из другого вида (например, с видовыми гомологами CLDN18.2). Более того, выделенное антитело может быть практически свободным от другого клеточного материала и/или химических веществ. В одном воплощении изобретения комбинация "выделенных" моноклональных антител относится к антителам, обладающим различной специфичностью и объединенным в четко определенные композиции или смеси.
Термин "связывание" в соответствии с изобретением предпочтительно относится к специфическому связыванию.
В соответствии с настоящим изобретением, антитело способно связываться с заданной мишенью, если оно обладает значительным сродством к этой заданной мишени и связывается с ней при стандартных анализах. "Сродство" или "сродство связывания" часто измеряется по равновесной константе диссоциации (KD). Предпочтительно термин "существенное сродство" означает связывание с заданной мишенью с константой диссоциации (KD) 10-5 М или меньше, 10-6 М или меньше, 10-7 М или меньше, 10-8 М или меньше, 10-9 М или меньше, 10-10 М или меньше, 10-11 М или меньше или 10-12 М или меньше.
Антитело (практически) не способно связываться с мишенью, если оно не обладает существенным сродством к данной мишени и практически не связывается, в частности, связывание с данной мишенью не выявляется при стандартных анализах. Предпочтительно, связывание антитела с данной мишенью не выявляется, если оно присутствует в концентрации вплоть до 2, предпочтительно 10, более предпочтительно 20, в частности, 50 или 100 мкг/мл или больше. Предпочтительно антитело не обладает существенным сродством к мишени, если оно связывается с данной мишенью с KD, которая по меньшей мере в 10 раз, 100 раз, 103 раз, 104 раз, 105 раз или 106 раз выше, чем KD для связывания с такой мишенью, с которой антитело способно связываться. Например, если KD для связывания антитела с такой мишенью, с которой антитело способно связываться, составляет 10-7 М, то KD для связывания с мишенью, к которой антитело не обладает существенным сродством, будет составлять не менее 10-6 М, 10-5 М, 10-4 M, 10-3 M, 10-2 M или 10-1 М.
Антитело является специфичным для заданной мишени, если оно способно связываться с этой заданной мишенью, но не способно связываться с другими мишенями, т.е. не обладает существенным сродством к другим мишеням и не связывается с другими мишенями существенно при стандартных анализах. В соответствии с изобретением, антитело является специфичным для CLDN18.2, если оно способно связываться с CLDN18.2, но (практически) не способно связываться с другими мишенями. Предпочтительно антитело является специфичным для CLDN18.2, если сродство к или связывание с такими другими мишенями существенно не превышает сродства к или связывания с неродственными CLDN18.2 белками, такими как бычий сывороточный альбумин (BSA), казеин, сывороточный альбумин человека (HSA), или же с другими трансмембранными белками, чем клаудин, типа молекул МНС или рецепторов трансферрина, или с любыми другими такими полипептидами. Предпочтительно антитело является специфичным для заданной мишени, если оно связывается с данной мишенью с KD, которая по меньшей мере в 10 раз, 100 раз, 103 раз, 104 раз, 105 раз или 106 раз ниже, чем KD для связывания с такой мишенью, для которой оно не является специфичным. Например, если KD для связывания антитела с такой мишенью, для которой оно является специфичным, составляет 10 М, то KD для связывания с мишенью, для которой оно не является специфичным, будет составлять не менее 10-6 М, 10-5 М, 10-4 М, 10-3 М, 10-2 М или 10-1 М.
Связывание антитела с мишенью можно определить экспериментально любым подходящим методом, к примеру, см. Berzofsky et al., "Antibody-Antigen Interactions" in Fundamental Immunology, Paul W.E., ed., Raven Press, New York, NY (1984); Kuby & Janis, Immunology, W.H. Freeman and Company, New York, NY (1992); а также описанными здесь методами. Сродство легко определяется стандартными методами, такими как метод равновесного диализа; с помощью прибора BIAcore 2000, используя общие процедуры, изложенные изготовителем; методом радиоиммуноанализа с использованием радиоактивного антигена-мишени; или другим методом, известным специалистам в данной области. Данные по сродству можно анализировать, к примеру, по методу Scatchard et al., Ann N.Y. Acad. Sci, 51: 660 (1949). Измеренное сродство конкретного взаимодействия антитело-антиген может варьироваться при измерении в различных условиях, например, концентрации соли, рН. Поэтому измерения сродства и других параметров связывания с антигеном, например, KD, IC50, предпочтительно проводятся со стандартными растворами антитела и антигена и стандартизованным буфером.
В настоящем изобретении термин "изотип" означает класс антител (например, IgM или IgG1), который кодируется генами константной области тяжелой цепи.
В настоящем изобретении термин "переключение изотипа" относится к явлению, при котором класс или изотип антитела меняется из одного класса Ig на один из других классов Ig.
Термин "встречающийся в природе" или "природный" в настоящем изобретении применительно к объекту означает то, что объект встречается в природе. Например, последовательность полипептида или полинуклеотида, присутствующая в организме (включая вирусы), которая может быть выделены из природного источника и не была преднамеренно модифицирована человеком в лаборатории, является природной.
Термин "перестановка" в настоящем изобретении относится к такой конфигурации локуса тяжелой цепи или легкой цепи иммуноглобулина, при которой V-сегмент располагается непосредственно рядом с сегментом D-J или J в конформации, кодирующей практически полный домен VH или VL, соответственно. Перестройка локуса гена иммуноглобулина (антитела) выявляется при сравнении с зародышевой ДНК; при перестройке локус будет содержать по крайней мере один рекомбинированный элемент гомологии типа гептамера/нонамера.
Термин "неперестроенная" или "зародышевая конфигурация" в настоящем изобретении по отношению к V-сегменту относится к такой конфигурации, при которой V-сегмент не подвергался рекомбинации и поэтому не находится непосредственно рядом с сегментом D или J.
В соответствии с изобретением антитело, обладающее способностью к связыванию с CLDN18.2, представлено антителом, способным связываться с эпигоном, присутствующим в CLDN18.2, предпочтительно с эпитопом, расположенным во внеклеточных доменах CLDN18.2, в частности, в первом внеклеточном домене, предпочтительно в положении аминокислот от 29 до 78 в CLDN18.2. В отдельных воплощениях антитело, обладающее способностью к связыванию с CLDN18.2, представлено антителом, способным связываться с (i) таким эпитопом на CLDN18.2, которого нет на CLDN18.1, предпочтительно SEQ ID NO: 3, 4 и 5, (ii) эпитопом, расположенным на петле 1 CLDN18.2, предпочтительно SEQ ID NO: 8, (iii) эпитопом, расположенным на петле 2 CLDN18.2, предпочтительно SEQ ID NO: 10, (iv) эпитопом, расположенным на петле D3 CLDN18.2, предпочтительно SEQ ID NO: 11, (v) эпитопом, охватывающим петлю 1 CLDN18.2 и петлю D3 CLDN18.2, или (vi) негликозилированным эпитопом, расположенным на петле D3 CLDN18.2, предпочтительно SEQ ID NO: 9.
В соответствии с изобретением антитело, обладающее способностью к связыванию с CLDN18.2, предпочтительно представлено антителом, обладающим способностью к связыванию с CLDN18.2, но не CLDN18.1. Предпочтительно антитело, обладающее способностью к связыванию с CLDN18.2, специфично к CLDN18.2. Предпочтительно антитело, обладающее способностью к связыванию с CLDN18.2, представлено антителом, обладающим способностью к связыванию с CLDN18.2, экспрессированым на клеточной поверхности. В особенно предпочтительных воплощениях антитело, обладающее способностью к связыванию с CLDN18.2, связывается с нативными эпитопами CLDN18.2, присутствующими на поверхности живых клеток. Предпочтительно антитело, обладающее способностью к связыванию с CLDN18.2, связывается с одним или несколькими пептидами, выбранными из группы, состоящей из SEQ ID NOs: 1, 3-11, 44, 46, 48-50. Предпочтительно антитело, обладающее способностью к связыванию с CLDN18.2, специфично к вышеупомянутым белкам, пептидам либо их иммуногенным фрагментам или производным. Антитело, обладающее способностью к связыванию с CLDN18.2, может быть получено способом, включающим стадию иммунизации животного белком или пептидом, содержащим аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 1, 3-11, 44, 46 и 48-50, либо нуклеиновой кислотой или клетками хозяина, экспрессирующими данный белок или пептид. Предпочтительно антитело связывается с раковыми клетками, в частности клетками приведенных выше типов рака, и предпочтительно практически не связывается с нераковыми клетками.
Предпочтительно связывание антитела, обладающего способностью к связыванию с CLDN18.2, с клетками, экспрессирующими CLDN18.2, индуцирует или опосредует уничтожение клеток, экспрессирующих CLDN18.2. Клетки, экспрессирующие CLDN18.2, предпочтительно представлены раковыми клетками, в частности, выбранными из группы, состоящей из опухолевых клеток рака желудка, пищевода, поджелудочной железы, легких, яичников, толстой кишки, печени, головы и шеи и желчного пузыря. Предпочтительно антитело индуцирует или опосредует уничтожение клеток путем индукции одного или нескольких из числа опосредованного комплементзависимой цитотоксичностью (CDC) лизиса, опосредованного антителозависимой клеточной цитотоксичностью (ADCC) лизиса, апоптоза и ингибирования пролиферации клеток, экспрессирующих CLDN18.2. Предпочтительно опосредованный ADCC лизис клеток происходит в присутствии эффекторных клеток, которые в отдельных воплощениях выбирают из группы, состоящей из моноцитов, мононуклеаров, NK-клеток и PMNs. Ингибирование пролиферации клеток можно измерять in vitro путем определения пролиферации клеток методом с использованием бромдезоксиуридина (5-бром-2-дезоксиуридина, BrdU). BrdU - синтетический нуклеозид, который является аналогом тимидина и может включаться в новосинтезированную ДНК при репликации клеток (во время фазы S клеточного цикла) вместо тимидина во время репликации ДНК. Обнаружение его включения, к примеру, с помощью антител, специфичных к BrdU, указывает на то, что в клетках происходила активная репликация ДНК.
В предпочтительных воплощениях описанные здесь антитела характеризуются одним или несколькими из следующих свойств:
a) специфичность к CLDN18.2;
b) сродство связывания с CLDN18.2 составляет около 100 нм или меньше, предпочтительно 5-10 нМ или меньше, более предпочтительно 1-3 нм или меньше;
c) способность индуцировать или опосредовать CDC на CLDN18.2-положительных клетках;
d) способность индуцировать или опосредовать ADCC на CLDN18.2-положительных клетках;
e) способность ингибировать рост CLDN18.2-положительных клеток;
f) способность индуцировать апоптоз CLDN18.2-положительных клеток.
В особенно предпочтительном воплощении антитело, обладающее способностью к связыванию с CLDN18.2, вырабатывается гибридомой, депонированной в DSMZ (Mascheroder Weg 1b, 31824 Braunschweig, Германия; новый адрес: Inhoffenstr. 7В, 31824 Braunschweig, Германия) и имеющей следующее обозначение и номер доступа:
a). 182-D1106-055, номер доступа DSM АСС2737, депонирована 19 октября 2005 г.,
b). 182-D1106-056, номер доступа DSM АСС2738, депонирована 19 октября 2005 г.,
c). 182-D1106-057, номер доступа DSM АСС2739, депонирована 19 октября 2005 г.,
d). 182-D1106-058, номер доступа DSM АСС2740, депонирована 19 октября 2005 г.,
e). 182-D1106-059, номер доступа DSM АСС2741, депонирована 19 октября 2005 г.,
f). 182-D1106-062, номер доступа DSM АСС2742, депонирована 19 октября 2005 г.,
g). 182-D1106-067, номер доступа DSM АСС2743, депонирована 19 октября 2005 г.,
h). 182-D758-035, номер доступа DSM АСС2745, депонирована 17 ноября 2005 г.,
i). 182-D758-036, номер доступа DSM АСС2746, депонирована 17 ноября 2005 г.,
j). 182-D758-040, номер доступа DSM АСС2747, депонирована 17 ноября 2005 г.,
k). 182-D1106-061, номер доступа DSM АСС2748, депонирована 17 ноября 2005 г.,
l). 182-D1106-279, номер доступа DSM АСС2808, депонирована 26 октября 2006 г.,
m). 182-D1106-294, номер доступа DSM АСС2809, депонирована 26 октября 2006 г.,
n). 182-D1106-362, номер доступа DSM АСС2810, депонирована 26 октября 2006 г.
Предпочтительными антителами по изобретению являются антитела, которые вырабатываются и их получают из вышеописанных гибридом, т.е. 37G11 в случае 182-D1 106-055, 37H8 в случае 182-D1106-056, 38G5 в случае 182-D1106-057, 38Н3 в случае 182-D1106-058, 39F11 в случае 182-D1106-059, 43А11 в случае 182-D1106-062, 61С2 в случае 182-D1106-067, 26В5 в случае 182-D758-035, 26D12 в случае 182-D758-036, 28D10 в случае 182-D758-040, 42Е12 в случае 182-D1106-061, 125Е1 в случае 182-D1106-279, 163Е12 в случае 182-D1106-294 и 175D10 в случае 182-D1106-362; и их химерные и гуманизованные формы.
Предпочтительные химерные антитела и их последовательности представлены в следующей таблице.
В предпочтительных воплощениях антитела, в частности химеризованные формы антител по изобретению, включают антитела, содержащие константную область тяжелой цепи (CH), содержащую аминокислотную последовательность, происходящую из константной области тяжелой цепи человека, как-то аминокислотную последовательность, представленную SEQ ID NO: 13, или ее фрагмент. В других предпочтительных воплощениях антитела, в частности химеризованные формы антител по изобретению, включают в себя антитела, содержащие константную область легкой цепи (CL), содержащую аминокислотную последовательность, происходящую из константной области легкой цепи человека, как-то аминокислотную последовательность, представленную SEQ ID NO: 12, или ее фрагмент. В особенно предпочтительном воплощении антитела, в частности химеризованные формы антител по изобретению, включают антитела, которые содержат CH, содержащую аминокислотную последовательность, происходящую из CH человека, как-то аминокислотную последовательность, представленную SEQ ID NO: 13, или ее фрагмент, а также содержат CL, содержащую аминокислотную последовательность, происходящую из CL человека, как-то аминокислотную последовательность, представленную SEQ ID NO: 12, или ее фрагмент.
В одном воплощении антитело, обладающее способностью к связыванию с CLDN18.2, представлено химерным моноклональным антителом мыши/человека типа IgG1, включающим вариабельную область легкой каппа-цепи мыши, константную область легкой каппа-цепи аллотипа Km(3) человека, вариабельную область тяжелой цепи мыши, и константную область IgG1 аллотипа G1m(3) человека.
В некоторых предпочтительных воплощениях химеризованные формы антител включают антитела, содержащие тяжелую цепь, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 14, 15, 16, 17, 18, 19 и их фрагментов, и/или содержащие легкую цепь, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NOs: 20, 21, 22, 23, 24, 25, 26, 27, 28 и их фрагментов.
В некоторых предпочтительных воплощениях химеризованные формы антител включают антитела, содержащие комбинацию тяжелых цепей и легких цепей, выбранную из следующих возможных вариантов от (i) до (ix):
(i) тяжелая цепь содержит аминокислотную последовательность, представленную SEQ ID NO: 14, или ее фрагмент, а легкая цепь содержит аминокислотную последовательность, представленную SEQ ID NO: 21, или ее фрагмент,
(ii) тяжелая цепь содержит аминокислотную последовательность, представленную SEQ ID NO: 15, или ее фрагмент, а легкая цепь содержит аминокислотную последовательность, представленную SEQ ID NO: 20, или ее фрагмент,
(iii) тяжелая цепь содержит аминокислотную последовательность, представленную SEQ ID NO: 16, или ее фрагмент, а легкая цепь содержит аминокислотную последовательность, представленную SEQ ID NO: 22, или ее фрагмент,
(iv) тяжелая цепь содержит аминокислотную последовательность, представленную SEQ ID NO: 18, или ее фрагмент, а легкая цепь содержит аминокислотную последовательность, представленную SEQ ID NO: 25, или ее фрагмент,
(v) тяжелая цепь содержит аминокислотную последовательность, представленную SEQ ID NO: 17, или ее фрагмент, а легкая цепь содержит аминокислотную последовательность, представленную SEQ ID NO: 24, или ее фрагмент,
(vi) тяжелая цепь содержит аминокислотную последовательность, представленную SEQ ID NO: 19, или ее фрагмент, а легкая цепь содержит аминокислотную последовательность, представленную SEQ ID NO: 23, или ее фрагмент,
(vii) тяжелая цепь содержит аминокислотную последовательность, представленную SEQ ID NO: 19, или ее фрагмент, а легкая цепь содержит аминокислотную последовательность, представленную SEQ ID NO: 26, или ее фрагмент,
(viii) тяжелая цепь содержит аминокислотную последовательность, представленную SEQ ID NO 19, или ее фрагмент, а легкая цепь содержит аминокислотную последовательность, представленную SEQ ID NO: 27, или ее фрагмент, и
(ix) тяжелая цепь содержит аминокислотную последовательность, представленную SEQ ID NO: 19, или ее фрагмент, а легкая цепь содержит аминокислотную последовательность, представленную SEQ ID NO: 28, или ее фрагмент.
"Фрагмент" или "фрагмент аминокислотной последовательности", приводимые выше, относятся к части последовательности антитела, т.е. последовательности, которая представляет собой последовательность антитела, укороченную на N- и/или С-конце, у которой, когда она заменяет данную последовательность у антитела, сохраняется связывание данного антитела с CLDN18.2 и предпочтительно сохраняются функции данного антитела, описанные здесь, например, опосредованного CDC лизиса или опосредованного ADCC лизиса. Предпочтительно фрагмент аминокислотной последовательности содержит по меньшей мере 80%, предпочтительно по меньшей мере 90%, 95%, 96%, 97%, 98% или 99% аминокислотных остатков из данной аминокислотной последовательности. Фрагмент аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NOs: 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27 и 28, предпочтительно означает то, что из данной последовательности удалено 17, 18, 19, 20, 21, 22 или 23 аминокислоты на N-конце.
В предпочтительном воплощении антитело, обладающее способностью к связыванию с CLDN18.2, содержит вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 29, 30, 31, 32, 33, 34 и их фрагментов.
В предпочтительном воплощении антитело, обладающее способностью к связыванию с CLDN18.2, содержит вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 35, 36, 37, 38, 39, 40, 41, 42, 43 и их фрагментов.
В некоторых предпочтительных воплощениях антитело, обладающее способностью к связыванию с CLDN18.2, содержит комбинацию вариабельной области тяжелой цепи (VH) и вариабельной области легкой цепи (VL), выбранную из следующих возможных вариантов от (i) до (ix):
(i) VH содержит аминокислотную последовательность, представленную SEQ ID NO: 29, или ее фрагмент, a VL содержит аминокислотную последовательность, представленную SEQ ID NO: 36, или ее фрагмент,
(ii) VH содержит аминокислотную последовательность, представленную SEQ ID NO: 30, или ее фрагмент, a VL содержит аминокислотную последовательность, представленную SEQ ID NO: 35, или ее фрагмент,
(iii) VH содержит аминокислотную последовательность, представленную SEQ ID NO: 31, или ее фрагмент, a VL содержит аминокислотную последовательность, представленную SEQ ID NO: 37, или ее фрагмент,
(iv) VH содержит аминокислотную последовательность, представленную SEQ ID NO: 33, или ее фрагмент, a VL содержит аминокислотную последовательность, представленную SEQ ID NO: 40, или ее фрагмент,
(v) VH содержит аминокислотную последовательность, представленную SEQ ID NO: 32, или ее фрагмент, a VL содержит аминокислотную последовательность, представленную SEQ ID NO: 39, или ее фрагмент,
(vi) VH содержит аминокислотную последовательность, представленную SEQ ID NO: 34, или ее фрагмент, a VL содержит аминокислотную последовательность, представленную SEQ ID NO: 38, или ее фрагмент,
(vii) VH содержит аминокислотную последовательность, представленную SEQ ID NO: 34, или ее фрагмент, a VL содержит аминокислотную последовательность, представленную SEQ ID NO: 41, или ее фрагмент,
(viii) VH содержит аминокислотную последовательность, представленную SEQ ID NO: 34, или ее фрагмент, a VL содержит аминокислотную последовательность, представленную SEQ ID NO: 42, или ее фрагмент, и
(ix) VH содержит аминокислотную последовательность, представленную SEQ ID NO: 34, или ее фрагмент, a VL содержит аминокислотную последовательность, представленную SEQ ID NO: 43, или ее фрагмент.
В предпочтительном воплощении антитело, обладающее способностью к связыванию с CLDN18.2, содержит область VH, включающую комплект определяющих комплементарность участков CDR1, CDR2 и CDR3, выбранный из следующих вариантов от (i) до (vi):
(i) CDR1: положения 45-52 в SEQ ID NO: 14, CDR2: положения 70-77 в SEQ ID NO: 14, CDR3: положения 116-125 в SEQ ID NO: 14,
(ii) CDR1: положения 45-52 в SEQ ID NO: 15, CDR2: положения 70-77 в SEQ ID NO: 15, CDR3: положения 116-126 в SEQ ID NO: 15,
(iii) CDR1: положения 45-52 в SEQ ID NO: 16, CDR2: положения 70-77 в SEQ ID NO: 16, CDR3: положения 116-124 в SEQ ID NO 16,
(iv) CDR1: положения 45-52 в SEQ ID NO: 17, CDR2: положения 70-77 в SEQ ID NO: 17, CDR3: положения 116-126 в SEQ ID NO: 17,
(v) CDR1: положения 44-51 в SEQ ID NO: 18, CDR2: положения 69-76 в SEQ ID NO: 18, CDR3: положения 115-125 в SEQ ID NO 18, и
(vi) CDR1: положения 45-53 в SEQ ID NO: 19, CDR2: положения 71-78 в SEQ ID NO 19, CDR3: положения 117-128 в SEQ ID NO 19.
В предпочтительном воплощении антитело, обладающее способностью к связыванию с CLDN18.2, содержит область VL, включающую комплект определяющих комплементарность участков CDR1, CDR2 и CDR3, выбранных из следующих вариантов от (i) до (ix):
(i) CDR1: положения 47-58 в SEQ ID NO: 20, CDR2: положения 76-78 в SEQ ID NO: 20, CDR3: положения 115-123 в SEQ ID NO 20,
(ii) CDR1: положения 49-53 в SEQ ID NO: 21, CDR2: положения 71-73 в SEQ ID NO: 21, CDR3: положения 110-118 в SEQ ID NO: 21,
(iii) CDR1: положения 47-52 в SEQ ID NO: 22, CDR2: положения 70-72 в SEQ LD NO: 22, CDR3: положения 109-117 в SEQ LD NO: 22,
(iv) CDR1: положения 47-58 в SEQ ID NO: 23, CDR2: положения 76-78 в SEQ LD NO: 23, CDR3: положения 115-123 в SEQ ID NO: 23,
(v) CDR1: положения 47-58 в SEQ ID NO: 24, CDR2: положения 76-78 в SEQ ID NO: 24, CDR3: положения 115-123 в SEQ ID NO: 24,
(vi) CDR1: положения 47-58 в SEQ ID NO: 25, CDR2: положения 76-78 в SEQ ID NO: 25, CDR3: положения 115-122 в SEQ ID NO: 25,
(vii) CDR1: положения 47-58 в SEQ LD NO: 26, CDR2: положения 76-78 в SEQ ID NO: 26, CDR3: положения 115-123 в SEQ ID NO: 26,
(viii) CDR1: положения 47-58 в SEQ ID NO: 27, CDR2: положения 76-78 в SEQ ID NO: 27, CDR3: положения 115-123 в SEQ ID NO 27, и
(ix) CDR1: положения 47-52 в SEQ ID NO: 28, CDR2: положения 70-72 в SEQ ID NO: 28, CDR3: положения 109-117 в SEQ ID NO: 28.
В предпочтительном воплощении антитело, обладающее способностью к связыванию с CLDN18.2, содержит комбинацию VH и VL, которая включает комплект определяющих комплементарность участков CDR1, CDR2 и CDR3, выбранный из следующих воплощений от (i) до (ix):
(i) VH: CDR1: положения 45-52 в SEQ ID NO: 14, CDR2: положения 70-77 в SEQ ID NO: 14, CDR3: положения 116-125 в SEQ ID NO: 14; VL: CDR1: положения 49-53 в SEQ ID NO: 21, CDR2: положения 71-73 в SEQ ID NO: 21, CDR3: положения 110-118 в SEQ ID NO: 21;
(ii) VH: CDR1: положения 45-52 в SEQ ID NO: 15, CDR2: положения 70-77 в SEQ ID NO: 15, CDR3: положения 116-126 в SEQ ID NO: 15; VL: CDR1: положения 47-58 в SEQ ID NO: 20, CDR2: положения 76-78 в SEQ ID NO: 20, CDR3: положения 115-123 в SEQ ID NO: 20;
(iii) VH: CDR1: положения 45-52 в SEQ ID NO: 16, CDR2: положения 70-77 в SEQ ID NO: 16, CDR3: положения 116-124 в SEQ ID NO: 16; VL: CDR1: положения 47-52 в SEQ ID NO: 22, CDR2: положения 70-72 в SEQ ID NO: 22, CDR3: положения 109-117 в SEQ ID NO: 22;
(iv) VH: CDR1: положения 44-51 в SEQ ID NO: 18, CDR2: положения 69-76 в SEQ ID NO: 18, CDR3: положения 115-125 в SEQ ID NO: 18; VL: CDR1: положения 47-58 в SEQ ID NO: 25, CDR2: положения 76-78 в SEQ ID NO: 25, CDR3: положения 115-122 в SEQ ID NO: 25;
(v) VH: CDR1: положения 45-52 в SEQ ID NO: 17, CDR2: положения 70-77 в SEQ ID NO: 17, CDR3: положения 116-126 в SEQ ID NO: 17; VL: CDR1: положения 47-58 в SEQ ID NO: 24, CDR2: положения 76-78 в SEQ ID NO: 24, CDR3: положения 115-123 в SEQ ID NO: 24;
(vi) VH: CDR1: положения 45-53 в SEQ ID NO: 19, CDR2: положения 71-78 в SEQ ID NO: 19, CDR3: положения 117-128 в SEQ ID NO: 19; VL: CDR1: положения 47-58 в SEQ ID NO: 23, CDR2: положения 76-78 в SEQ ID NO: 23, CDR3: положения 115-123 в SEQ ID NO: 23;
(vii) VH: CDR1: положения 45-53 в SEQ ID NO: 19, CDR2: положения 71-78 в SEQ ID NO: 19, CDR3: положения 117-128 в SEQ ID NO: 19; VL: CDR1: положения 47-58 в SEQ ID NO: 26, CDR2: положения 76-78 в SEQ ID NO: 26, CDR3: положения 115-123 в SEQ ID NO: 26;
(viii) VH: CDR1: положения 45-53 в SEQ ID NO: 19, CDR2: положения 71-78 в SEQ ID NO: 19, CDR3: положения 117-128 в SEQ ID NO: 19; VL: CDR1: положения 47-58 в SEQ ID NO: 27, CDR2: положения 76-78 в SEQ ID NO: 27, CDR3: положения 115-123 в SEQ ID NO: 27; и
(ix) VH: CDR1: положения 45-53 в SEQ ID NO: 19, CDR2: положения 71-78 в SEQ ID NO: 19, CDR3: положения 117-128 в SEQ ID NO: 19; VL: CDR1: положения 47-52 в SEQ ID NO: 28, CDR2: положения 70-72 в SEQ ID NO: 28, CDR3: положения 109-117 в SEQ ID NO: 28.
В других предпочтительных воплощениях антитело, обладающее способностью к связыванию с CLDN18.2, предпочтительно содержит один или несколько участков, определяющих комплементарность (CDR), предпочтительно по крайней мере участок CDR3 из вариабельной области тяжелой цепи (VH) и/или вариабельной области легкой цепи (VL) моноклонального антитела против CLDN18.2, предпочтительно описанного здесь моноклонального антитела против CLDN18.2, а предпочтительно оно содержит один или несколько определяющих комплементарность участков (CDR), предпочтительно по крайней мере участок СDR3 из описанных здесь вариабельных областей тяжелой цепи (VH) и/или вариабельных областей легкой цепи (VL). В одном воплощении данные один или несколько участков, определяющих комплементарность (CDR), выбирают из описанного здесь комплекта определяющих комплементарность участков CDR1, CDR2 и CDR3. В особенно предпочтительном воплощении антитело, обладающее способностью к связыванию с CLDN18.2, предпочтительно содержит определяющие комплементарность участки CDR1, CDR2 и CDR3 из вариабельной области тяжелой цепи (VH) и/или вариабельной области легкой цепи (VL) моноклонального антитела против CLDN18.2, предпочтительно описанного здесь моноклонального антитела против CLDN18.2, а предпочтительно оно содержит определяющие комплементарность участки CDR1, CDR2 и CDR3 из описанных здесь вариабельных областей тяжелой цепи (VH) и/или вариабельных областей легкой цепи (VL).
В одном воплощении антитело, содержащее один или несколько участков CDR, комплект CDR или комбинацию комплектов CDR, как описано здесь, содержит данные CDRs вместе с перемежающимися с ними каркасными участками. Предпочтительно эта часть также должна включать по меньшей мере 50% одного из каркасных участков - первого и четвертого или обоих, причем эти 50% составляют С-концевые 50% первого каркасного участка и N-концевые 50% четвертого каркасного участка. Конструирование антител методами рекомбинантной ДНК может привести к включению остатков со стороны N- или С-конца в вариабельные области, кодируемые линкерами, введенными для облегчения клонирования или других стадий обработки, включая введение линкеров для соединения вариабельных областей по изобретению с последовательностями других белков, в том числе тяжелых цепей иммуноглобулинов, других вариабельных доменов (к примеру, при получении диател) или белковых меток.
В одном воплощении антитело, содержащее один или несколько участков CDR, комплект CDR или комбинацию комплектов CDR, как описано здесь, содержит данные CDRs в каркасе человеческого антитела.
Упоминание здесь антител, содержащих по отношению к своей тяжелой цепи определенную цепь либо определенную область или последовательность, предпочтительно относится к такой ситуации, когда все тяжелые цепи данного антитела содержат данную конкретную цепь, область или последовательность. Это также относится и к легкой цепи антител, соответственно.
Термин "нуклеиновая кислота" в настоящем изобретении служит для обозначения ДНК и РНК. Нуклеиновая кислота может быть одноцепочечной или двухцепочечной, но предпочтительно это двухцепочечная ДНК.
В соответствии с изобретением, термин "экспрессия" применяется в его самом общем значении и включает продукцию РНК либо РНК и белка/пептида. Он также включает частичную экспрессию нуклеиновых кислот. Кроме того, экспрессия может осуществляться временно или стабильно.
Приведенные здесь сведения в отношении определенных аминокислотных последовательностей, например, представленных в перечне последовательностей, следует понимать так, что они также относятся и к таким вариантам данных конкретных последовательностей, у которых последовательности функционально эквивалентны данной конкретной последовательности, например, их аминокислотные последовательности проявляют свойства, идентичные или близкие таковым у определенных аминокислотных последовательностей. Одним из важных свойств является сохранение связывания антитела со своей мишенью или поддержание эффекторных функций антитела. Предпочтительно последовательность, которая является вариантом по отношению к определенной последовательности, когда она заменяет эту конкретную последовательность у антитела, сохраняет связывание данного антитела с CLDN18.2 и предпочтительно функции данного антитела, как описано здесь, например, опосредованный CDC лизис или опосредованный ADCC лизис.
Специалистам должно быть известно то, в частности, что последовательности участков CDR, гипервариабельных и вариабельных областей можно модифицировать без потери способности к связыванию с CLDN18.2. Например, участки CDR должны быть либо идентичными, либо сильно гомологичными приведенным здесь участкам антител. Под "высокой гомологичностью" подразумевается то, что в CDR может быть от 1 до 5, предпочтительно от 1 до 4, как-то от 1 до 3 либо 1 или 2 замены. Кроме того, гипервариабельные и вариабельные области можно модифицировать так, чтобы они проявляли существенную гомологию с конкретными приведенными здесь участками антител.
В настоящем изобретении "варианты" аминокислотной последовательности включают варианты со вставкой аминокислот, варианты с добавлением аминокислот, варианты с делецией аминокислот и/или варианты с заменой аминокислот. Варианты с делецией аминокислот, включающие делении на N-конце и/или С-конце белка, также называются N-концевыми и/или С-концевыми усеченными вариантами.
Варианты со вставкой аминокислот включают вставки одной или двух или нескольких аминокислот в определенной аминокислотной последовательности. В случае вариантов аминокислотной последовательности со вставками в определенное место в аминокислотной последовательности вставляется один или несколько аминокислотных остатков, хотя также возможны и случайные вставки с соответствующим скринингом полученного продукта.
Варианты с добавлением аминокислот включают присоединение по N- и/или С-концам одной или нескольких аминокислот, как-то 1, 2, 3, 5, 10, 20, 30, 50 или больше аминокислот.
Варианты с делецией аминокислот характеризуются удалением из последовательности одной или нескольких аминокислот, как-то удалением 1, 2, 3, 5, 10, 20, 30, 50 или больше аминокислот. Делеции могут быть в любом положении белка.
Варианты с заменой аминокислот характеризуются тем, что по меньшей мере один остаток в последовательности удаляется, а на его место вставляется другой остаток. Предпочтение отдается модификациям, приходящимся на такие положения в аминокислотной последовательности, которые не сохраняются между гомологичными белками или пептидами, и/или заменам аминокислот на другие с аналогичными свойствами. Предпочтительно замены аминокислот в белковых вариантах являются консервативными заменами, т.е. это замены аналогичных заряженных или незаряженных аминокислот. Консервативные аминокислотные замены включают замены в рамках одного семейства аминокислот, сходных по своим боковым цепям. Встречающиеся в природе аминокислоты обычно подразделяются на четыре семейства: кислые (аспартат, глутамат), основные (лизин, аргинин, гистидин), неполярные (аланин, валин, лейцин, изолейцин, пролин, фенил аланин, метионин, триптофан) и незаряженные полярные (глицин, аспарагин, глутамин, цистеин, серии, треонин, тирозин) аминокислоты. Фенил аланин, триптофан и тирозин иногда классифицируются в совокупности как ароматические аминокислоты.
Предпочтительно степень сходства, предпочтительно идентичности между данной аминокислотной последовательностью и аминокислотной последовательностью, которая является вариантом данной аминокислотной последовательности, должна составлять по меньшей мере 60%, 65%, 70%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%. Степень сходства или идентичности предпочтительно приводится для аминокислотного участка, составляющего по меньшей мере 10%, по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% или около 100% от всей длины контрольной аминокислотной последовательности. Например, если контрольная аминокислотная последовательность состоит из 200 аминокислот, то степень сходства или идентичности предпочтительно приводится по меньшей мере для 20, по меньшей мере 40, по меньшей мере 60, по меньшей мере 80, по меньшей мере 100, по меньшей мере 120, по меньшей мере 140, по меньшей мере 160, по меньшей мере 180 или около 200 аминокислот, предпочтительно непрерывных аминокислот. В предпочтительном воплощении степень сходства или идентичности приводится для всей длины контрольной аминокислотной последовательности. Совмещение последовательностей для определения сходства, предпочтительно идентичности последовательностей проводится с помощью известных инструментов, предпочтительно методом наилучшего совмещения последовательностей, например, с помощью Align, используя стандартные настройки, предпочтительно EMBOSS::needle, Matrix: Blosum62, Gap Open 10.0, Gap Extend 0.5.
"Сходство последовательностей" означает процент аминокислот, которые либо идентичны, либо представляют консервативные замены аминокислот. "Идентичность последовательностей" между двумя аминокислотными последовательностями означает процент аминокислот, идентичных между этими последовательностями.
Термин "степень идентичности" служит для обозначения процента идентичных аминокислотных остатков между двумя сравниваемыми последовательностями, полученного после наилучшего выравнивания, причем этот процент является чисто статистическим, а отличия между двумя последовательностями распределяются случайным образом и по всей длине. Сравнение последовательностей между двумя аминокислотными последовательностями обычно проводится путем сравнения этих последовательностей после их оптимального выравнивания, причем такое сравнение проводится по сегментам либо по "окнам сравнения" с целью выявления и сравнения локальных участков сходства последовательностей. Оптимальное выравнивание последовательностей для их сравнения может проводиться вручную или же с помощью алгоритма локальной гомологии Smith and Waterman, 1981, Ads. App. Math. 2, 482, с помощью алгоритма локальной гомологии Neddleman and Wunsch, 1970, J. Mol. Biol. 48, 443, методом поиска сходства Pearson and Lipman, 1988, Proc. Natl. Acad. Sci. USA 85, 2444, или при помощи компьютерных программ, в которых используются эти алгоритмы (GAP, BESTFIT, FASTA, BLASTP, BLASTN и TFASTA в пакете программ Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Drive, Madison, Wis.).
Степень идентичности вычисляется путем определения числа идентичных положений между двумя сравниваемыми последовательностями, деления этого количества на число сравниваемых положений и умножения полученного результата на 100, получая процент идентичности между этими двумя последовательностями.
Термин "трансгенное животное" относится к таким животным, у которых геном содержит один или несколько трансгенов, предпочтительно трансгенов тяжелых и/или легких цепей либо трансхромосом (встроенных или не встроенных в природную геномную ДНК животного), которые предпочтительно способны экспрессировать эти трансгены. Например, трансгенная мышь может содержать трансген легкой цепи человека и либо трансген тяжелой цепи человека, либо трансхромосому тяжелой цепи человека, так что мышь будет вырабатывать человеческие антитела против CLDN18.2 при иммунизации антигеном CLDN18.2 и/или клетками, экспрессирующими CLDN18.2. Трансген тяжелой цепи человека может быть встроен в хромосомную ДНК мыши, как в случае трансгенных мышей, например, мышей HuMAb, как-то мышей НСо7 или НСо12, или же трансген тяжелой цепи человека может содержаться вне хромосом, как в случае трансхромосомных мышей (например, мышей KM), как описано в WO 02/43478. Такие трансгенные и трансхромосомные мыши могут вырабатывать несколько изотопов человеческих моноклональных антител к CLDN18.2 (например, IgG, IgA и/или IgE) путем прохождения через рекомбинацию V-D-J и переключение изотипа.
"Уменьшение", "снижение" или "ингибирование" в настоящем изобретении означает общее снижение или способность вызывать общее снижение уровня, предпочтительно на 5% или больше, 10% или больше, 20% или больше, более предпочтительно на 50% или больше и наиболее предпочтительно на 75% или больше, например, уровня экспрессии или уровня пролиферации клеток.
Термины типа "повышение" или "увеличение" предпочтительно означают повышение или увеличение по меньшей мере на 10%, предпочтительно по меньшей мере на 20%, предпочтительно по меньшей мере на 30%, более предпочтительно по меньшей мере на 40%, более предпочтительно по меньшей мере на 50%, еще более предпочтительно по меньшей мере на 80% и наиболее предпочтительно по меньшей мере на 100%, на 200%, на 500%, на 1000%, на 10000% или еще больше.
Механизмы действия mAb
Хотя далее представлены соображения относительно механизма, лежащего в основе терапевтической эффективности антител по изобретению, однако это никоим образом не следует рассматривать как ограничение изобретения.
Описанные здесь антитела предпочтительно взаимодействуют с компонентами иммунной системы, предпочтительно через ADCC или CDC. Описанные здесь антитела также могут применяться для направленной доставки полезных грузов (например, радиоизотопов, лекарственных препаратов или токсинов) для непосредственного уничтожения опухолевых клеток или же могут применяться синергично с традиционными химиотерапевтическими средствами, атакующими опухоли через дополнительные механизмы действия, которые могут включать противоопухолевые иммунные ответы, оказавшиеся нарушенными из-за цитотоксических побочных эффектов химиотерапевтических препаратов на Т-лимфоциты. Однако описанные здесь антитела также могут оказывать действие просто путем связывания с CLDN18.2 на клеточной поверхности, тем самым, например, блокируя пролиферацию клеток.
Антителозависимая клеточная питотоксичность (ADCC)
ADCC означает способность к уничтожению клеток у эффекторных клеток, как описано здесь, в частности, у лимфоцитов, для чего предпочтительно требуется, чтобы клетки мишени были помечены антителом.
ADCC предпочтительно происходит тогда, когда антитела связываются с антигенами на опухолевых клетках, a Fc-домены антител задействуют Fc-рецепторы (FcR) на поверхности иммунных эффекторных клеток. Идентифицировано несколько семейств Fc-рецепторов, причем определенные популяции клеток экспрессируют характерные для них Fc-рецепторы. ADCC может рассматриваться в качестве механизма непосредственной индукции в различной степени немедленного разрушения опухоли, что приводит к презентации антигена и индукции направленных на опухоль Т-клеточных ответов. Предпочтительно индукция ADCC in vivo должна вызывать направленные на опухоль Т-клеточные ответы и антительные ответы организма.
Комплементзависимая питотоксичность (CDC)
CDC является еще одним методом уничтожения клеток, который запускается антителами. Наиболее эффективным изотипом для активации комплемента является IgM. Также IgG1 и IgG3 оба очень эффективно запускают CDC по классическому пути активации комплемента. Предпочтительно в этом каскаде образование комплексов антиген-антитело приводит к демаскированию нескольких ближайших сайтов связывания C1q на доменах CH2 задействованных молекул антител типа молекул IgG (C1q является одним из трех субкомпонентов комплемента С1). Предпочтительно эти демаскированные сайты связывания C1q превращают прежде низкоаффинное взаимодействие C1q-IgG во взаимодействие с высоким сродством, которое запускает каскад событий с участием ряда других белков комплемента и приводит к протеолитическому высвобождению хемотаксических/активирующих эффекторные клетки молекул С3а и С5а. Предпочтительно каскад комплемента заканчивается образованием мембраноатакующего комплекса, который создает поры в клеточной мембране, способствующие свободному прохождению воды и растворенных веществ в клетку и из клетки.
Получение и тестирование антител
Описанные здесь антитела могут быть получены различными способами, включая стандартные методы моноклональных антител, например, стандартный метод гибридизации соматических клеток Kohler and Milstein, Nature 256: 495 (1975). Хотя методы гибридизации соматических клеток и предпочтительны, однако в принципе можно использовать и другие методы получения моноклональных антител, например, вирусной или онкогенной трансформации В-лимфоцитов или методы фагового дисплея с использованием библиотек генов антител.
Предпочтительной животной системой для получения гибридом, секретирующих моноклональные антитела, является мышиная система. Получение гибридом на мышах является очень хорошо разработанной процедурой. Известны методики иммунизации и методы выделения иммунизованных спленоцитов для слияния. Также известны партнеры для слияния (например, миеломные клетки мыши) и процедуры слияния.
Другими предпочтительными системами на животных для получения гибридом, секретирующих моноклональные антитела, являются системы на крысах и кроликах (например, описанные в Spieker-Polet et al., Proc. Natl. Acad. Sci. USA 92: 9348 (1995), также см. Rossi et al., Am. J. Clin. Pathol. 124: 295 (2005)).
В еще одном предпочтительном воплощении моноклональные антитела человека могут быть получены с использованием трансгенных или трансхромосомных мышей, несущих части иммунной системы человека, а не системы мыши. Эти трансгенные и трансхромосомные мыши включают мышей, известных как мыши HuMAb и KM мыши, соответственно, и собирательно именуются здесь как "трансгенные мыши". Получение человеческих антител в таких трансгенных мышах может проводиться, как описано подробно для CD20 в WO 2004/035607.
Еще одна стратегия для получения моноклональных антител состоит в том, чтобы непосредственно выделить гены, кодирующие антитела, из лимфоцитов, вырабатывающих антитела определенной специфичности, например, см. Babcock et al., 1996: A novel strategy for generating monoclonal antibodies from single, isolated lymphocytes producing antibodies of defined specificities. Подробно о конструировании рекомбинантных антител см. также Welschof and Kraus, Recombinant antibodies for cancer therapy: ISBN-0-89603-918-8; и Benny K.C. Lo, Antibody Engineering: ISBN 1-58829-092-1.
Для получения антител можно иммунизировать мышей конъюгированными с носителями пептидами, полученными из последовательности антигена, т.е. последовательности, против которой должны быть направлены антитела, обогащенным препаратом рекомбинантно экспрессированного антигена или его фрагментов и/или клетками, экспрессирующими антиген, как описано. С другой стороны, можно иммунизировать мышей ДНК, кодирующей антиген или его фрагменты. В том случае, когда иммунизация очищенным или обогащенным препаратом антигена не приводит к образованию антител, мышей также можно иммунизировать клетками, экспрессирующими антиген, например, клеточной линии, чтобы усилить иммунные ответы.
Иммунный ответ можно отслеживать по ходу иммунизации с помощью образцов плазмы и сыворотки, полученных из хвостовой вены или заглазничных проб крови. Мышей с достаточным титром иммуноглобулина можно использовать для слияния. Для повышения процента гибридом, секретирующих определенное антитело, мышей можно ревакцинировать внутрибрюшинно или внутривенно клетками, экспрессирующими антиген, за 3 суток до забоя и извлечения селезенки.
Для получения гибридом, продуцирующих моноклональные антитела, можно выделить спленоциты и клетки лимфатических узлов из иммунизированных мышей и слить их с соответствующей иммортализованной линией клеток типа линии клеток миеломы мыши. Полученные гибридомы можно затем подвергнуть скринингу на выработку антиген-специфичных антител. Затем можно провести скрининг индивидуальных лунок методом ELISA на предмет гибридом, секретирующих антитела. Используя клетки, экспрессирующие антиген, можно идентифицировать антитела со специфичностью к антигену по иммунофлуоресценции и методом FACS. Секретирующие антитела гибридомы можно пересеять, снова провести скрининг и, если они по-прежнему будут положительными на наличие моноклональных антител, субклонировать методом серийных разведений. Затем можно культивировать стабильные субклоны in vitro, чтобы получить антитела в среде для культуры ткани для изучения.
Антитела также можно получить в клетках трансфектомы, например, сочетанием методов рекомбинантной ДНК и методов генной трансфекции, которые хорошо известны в данной области (Morrison S. (1985) Science 229: 1202).
Например, в одном воплощении, нужные гены, к примеру, гены антитела, можно лигировать в экспрессионный вектор типа экспрессирующей плазмиды для эукариот типа той, что используется в системе экспрессии генов GS, раскрытой в WO 87/04462, WO 89/01036 и ЕР 338 841, или другой системы экспрессии, хорошо известной в данной области. Очищенную плазмиду с клонированными генами антитела можно ввести в эукариотические клетки хозяина, как-то клетки СНО, клетки NS/0, клетки HEK293T или клетки HEK293 или же в другие эукариотические клетки типа растительных клеток, грибковых или дрожжевых клеток. Для введения этих генов можно использовать уже описанные методы, такие как электропорация, липофекция, липофектамин или другие. После введения этих генов антитела в клетки хозяина можно идентифицировать и отобрать клетки, экспрессирующие антитело. Эти клетки представляют собой трансфектомы, которые можно затем размножить до нужного уровня экспрессии и получения антител. Из супернатантов этих культур и/или клеток можно выделить и очистить рекомбинантные антитела.
С другой стороны, клонированные гены антитела можно экспрессировать в других системах экспрессии, включая прокариотические клетки типа микроорганизмов, например, Е. coli. Более того, антитела можно получать и в трансгенных животных, например, в молоке из овец и кроликов или в яйцах из кур, либо в трансгенных растениях; например, см. Verma R. et al. (1998) J. Immunol. Meth. 216: 165-181; Pollock et al. (1999) J. Immunol. Meth. 231:147-157; и Fischer R. et al. (1999) J. Biol. Chem. 380: 825-839.
Химеризация
Мышиные моноклональные антитела можно использовать в качестве терапевтических антител для человека, если пометить их токсинами или радиоактивными изотопами. Немеченые мышиные антитела сильно иммуногенны для человека, что при повторном применении ведет к снижению терапевтического эффекта. В основном иммуногенность опосредована константными областями тяжелой цепи. Иммуногенность мышиных антител для человека можно уменьшить или полностью устранить, если соответствующие антитела подвергнуть химеризации или гуманизации. Химерные антитела - это такие антитела, различные части которых происходят от разных видов животных, как-то вариабельная область происходит из антитела мыши, а константная область - из иммуноглобулина человека. Химеризация антител осуществляется путем присоединения вариабельных областей тяжелой и легкой цепи мышиного антитела к константной области тяжелой и легкой цепи человека (например, как описано Kraus et al., in Methods in Molecular Biology series, Recombinant antibodies for cancer therapy ISBN-0-89603-918-8). В предпочтительном воплощении химерные антитела получают путем соединения константной области легкой каппа-цепи человека с вариабельной областью легкой цепи мыши. В другом предпочтительном воплощении химерные антитела можно получить путем соединения константной области легкой лямбда-цепи человека с вариабельной областью легкой цепи мыши. Для получения химерных антител предпочтительны константные области тяжелой цепи типа IgG1, IgG3 и IgG4. Другие предпочтительные константные области тяжелой цепи для получения химерных антител IgG2, IgA, IgD и IgM.
Гуманизация
Антитела взаимодействуют с антигенами мишени преимущественно через те аминокислотные остатки, которые находятся в шести определяющих комплементарность участках (CDR) тяжелой и легкой цепи. По этой причине аминокислотные последовательности в пределах CDR более разнообразны между отдельными антителами, чем последовательности вне CDR. Поскольку последовательности CDR ответственны за большую часть взаимодействий антитело-антиген, то можно экспрессировать рекомбинантные антитела, имитирующие свойства определенных природных антител, путем конструирования экспрессирующих векторов, включающих последовательности CDR из данного природного антитела, привитые на каркасные последовательности из другого антитела с другими свойствами (например, см. Riechmann L. et al. (1998) Nature 332: 323-327; Jones P. et al. (1986) Nature 321: 522-525; и Queen C. et al. (1989) Proc. Natl. Acad. Sci. USA 86: 10029-10033). Такие каркасные последовательности можно получить из общедоступных баз данных по ДНК, содержащих последовательности генов антител зародышевой линии. Эти последовательности зародышевой линии будут отличаться от зрелых последовательностей генов антител, потому что они не включают полностью собранные гены вариабельной области, которые образуются при соединении V-(D)-J в процессе созревания В-клеток. Последовательности зародышевой линии генов также отличаются от последовательностей антител с высоким сродством из вторичного репертуара по отдельным точкам равномерно по всей вариабельной области.
Способность антител к связыванию с антигеном можно определить стандартными методами анализа связывания (например, методами ELISA, Вестерн-блот, иммунофлуоресценции и проточной цитометрии).
Для очистки антител можно выращивать отобранные гибридомы в двухлитровых вращающихся колбах для очистки моноклональных антител. С другой стороны, антитела можно получать в диализных биореакторах. Супернатанты фильтруют, а при необходимости и концентрируют перед проведением аффинной хроматографии на сефарозе с белком G или сефарозе с белком А. Элюированный IgG подвергают проверке на чистоту методами гель-электрофореза и высокоэффективной жидкостной хроматографии. Буферный раствор меняют на PBS, а концентрацию определяют по OD280, используя коэффициент экстинкции 1,43. Моноклональные антитела разделяют на порции и хранят при -80°С.
Чтобы определить, связываются ли выбранные моноклональные антитела с уникальными эпитопами, можно использовать сайт-направленный или множественный сайт-направленный мутагенез.
Для определения изотипа антител можно провести ELISA на изотип с помощью различных коммерческих наборов (например, Zymed, Roche Diagnostics). Лунки микропланшета покрывают антителом против Ig мыши. После блокирования планшет подвергают реакции с моноклональными антителами или очищенными контролями на изотип, при комнатной температуре в течение двух часов. Затем лунки подвергают реакции с конъюгированными с пероксидазой зондами, специфичными к IgG1, IgG2a, IgG2b или IgG3 мыши, IgA или IgM мыши. После отмывки планшет проявляют с помощью субстрата ABTS (1 мг/мл) и анализируют по OD405-650. С другой стороны, можно использовать набор IsoStrip Mouse Monoclonal Antibody Isotyping Kit (Roche, кат. №1493027), как описано производителем.
Для того, чтобы показать наличие антител в сыворотке иммунизованных мышей или связывание моноклональных антител с живыми клетками, экспрессирующими антиген, можно использовать проточную цитометрию. Клеточные линии, экспрессирующие антиген естественным образом или после трансфекции, и отрицательные контроли, не экспрессирующие антиген (выращенные в стандартных условиях роста), смешивают с различными концентрациями моноклональных антител в супернатантах гибридом или в PBS, содержащем 1% FBS, и инкубируют при 4°С в течение 30 мин. После отмывки проводят связывание меченных Аре или Alexa647 антител против IgG со связавшимся с антигеном моноклональным антителом в таких же условиях, как и при связывании первичного антитела. Образцы анализируют методом проточной цитометрии на приборе FACS с настройкой канала на одиночные живые клетки по свойствам прямого и бокового светорассеяния. Для того, чтобы отличить антиген-специфичные моноклональные антитела от неспецифического связывания при единичном измерении, можно использовать метод котрансфекции. Клетки, прошедшие временную трансфекцию плазмидами, кодирующими антиген и флуоресцентный маркер, подвергают окрашиванию, как описано выше. Трансфецированные клетки детектируют на другом канале флуоресценции, чем окрашенные антителом клетки. Поскольку большинство трансфецированных клеток экспрессируют оба трансгена, то антиген-специфичные моноклональные антитела преимущественно связываются с клетками, экспрессирующими флуоресцентный маркер, тогда как неспецифические антитела связываются в сопоставимом соотношении с не-трансфецированными клетками. Наряду с методом проточной цитометрии или вместо него можно использовать альтернативный метод с использованием флуоресцентной микроскопии. Клетки подвергают окрашиванию точно так же, как описано выше, и анализируют с помощью флуоресцентной микроскопии.
Для того, чтобы показать наличие антител в сыворотке иммунизованных мышей или связывание моноклональных антител с живыми клетками, экспрессирующими антиген, можно использовать метод иммунофлуоресцентной микроскопии. Например, линии клеток, экспрессирующих антиген спонтанно или после трансфекции, и отрицательные контроли, не экспрессирующие антиген, культивируют на предметных стеклах с лунками в стандартных условиях роста в среде DMEM/F12 с добавлением 10% эмбриональной телячьей сыворотки (FCS), 2 мМ L-глутамина, 100 МЕ/мл пенициллина и 100 мкг/мл стрептомицина. Затем клетки фиксируют в метаноле или параформальдегиде либо оставляют необработанными. Затем клетки подвергают реакции с моноклональными антителами против антигена в течение 30 мин при 25°С. После отмывки клетки подвергают реакции с меченным Alexa555 вторичным антителом против IgG мыши (Molecular Probes) в тех же условиях. Затем клетки исследуют методом флуоресцентной микроскопии.
Можно приготовить клеточные экстракты из клеток, экспрессирующих антиген, и соответствующих отрицательных контролей, и подвергнуть их электрофорезу в полиакриламидном геле с додецилсульфатом натрия (SDS). После электрофореза разделенные антигены переносят на нитроцеллюлозные мембраны, блокируют и зондируют моноклональными антителами, подлежащими тестированию. Связывание с IgG детектируют с помощью конъюгированного с пероксидазой антитела против IgG мыши и проявляют с помощью субстрата ECL.
Антитела также можно тестировать на реактивность с антигеном методом иммуногистохимии, хорошо известным специалистам, например, используя фиксированные параформальдегидом или ацетоном криосрезы или заключенные в парафин и фиксированные параформальдегидом срезы тканей из образцов нераковой ткани или раковой ткани, полученных от пациентов при рутинных хирургических процедурах или из мышей, несущих привитые опухоли из клеточных линий, экспрессирующих антиген спонтанно или после трансфекции. Для иммуноокрашивания их инкубируют с реагирующими с антигеном антителами, а затем с конъюгированными с пероксидазой хрена козьими антителами против мыши или козьими антителами против кролика (DAKO) в соответствии с инструкциями производителя.
Антитела можно протестировать на их способность опосредовать фагоцитоз и уничтожение клеток, экспрессирующих CLDN18.2. Тестирование активности моноклональных антител in vitro обеспечивает первоначальный скрининг перед тестированием на моделях in vivo.
Антителозависимая клеточная питотоксичность (ADCC)
Вкратце, полиморфноядерные клетки (PMNs), NK-клетки, моноциты, мононуклеары или другие эффекторные клетки от здоровых доноров подвергают очистке с помощью центрифугирования в градиенте плотности Ficoll Hypaque с последующим лизисом примесных эритроцитов. Промытые эффекторные клетки суспендируют в среде RPMI с добавлением 10% инактивированной нагреванием эмбриональной телячьей сыворотки либо 5% инактивированной нагреванием человеческой сыворотки и смешивают с меченными 51Cr клетками мишени, экспрессирующими CLDN18.2, при различных соотношениях эффекторных клеток и клеток мишени. В качестве альтернативы клетки мишени можно пометить усиливающим флуоресценцию лигандом (BATDA). Сильно флуоресцирующий хелат европия с усиливающим лигандом, который высвобождается из мертвых клеток, измеряют на флуориметре. В другом альтернативном методе применяется трансфекция клеток мишени люциферазой. При добавлении Lucifer Yellow он может быть окислен только жизнеспособными клетками. Затем добавляют очищенный IgG против CLDN18.2 в различных концентрациях. В качестве отрицательного контроля используют посторонний IgG человека. Анализы проводятся в течение 4-20 часов при 37°С в зависимости от типа используемых эффекторных клеток. Образцы исследуют на цитолиз по измерению высвобождения 51Cr или присутствия хелата EuTDA в супернатанте культуры. С другой стороны, мерой жизнеспособных клеток может служить люминесценция в результате окисления Lucifer Yellow.
Моноклональные антитела против CLDN18.2 также можно протестировать в различных комбинациях, чтобы определить, будет ли усиливаться цитолиз при нескольких моноклональных антителах.
Комплементзависимая цитотоксичность (CDC)
Моноклональные антитела против CLDN18.2 можно протестировать на их способность опосредовать CDC различными известными методами. Например, можно получить сыворотку для комплемента из крови известным специалистам способом. Для определения CDC-активности mAbs можно использовать различные методы. Например, можно измерять высвобождение 51Cr или повышение проницаемости мембраны методом анализа исключения пропидия йодида (PI). Вкратце, клетки мишени промывают и инкубируют при 5×105/мл с различными концентрациями mAb в течение 10-30 мин при комнатной температуре или при 37°С. Затем добавляют сыворотку или плазму до конечной концентрации 20% (об./об.) и инкубируют клетки при 37°С в течение 20-30 мин. Все клетки из каждого образца вносят в раствор PI в пробирке для FACS. Затем смесь сразу же подвергают анализу методом проточной цитометрии с использованием FACSArray.
В альтернативном методе определяется индукция CDC на адгезивных клетках. В одном воплощении этого метода клетки высеивают за 24 ч до анализа при 3×104/лунку в плоскодонные микропланшеты для тканевых культур. На следующий день среду роста удаляют, а клетки инкубируют в трех повторах с антителами. Контрольные клетки инкубируют в среде роста или ростовой среде, содержащей 0,2% сапонина, для определения фонового лизиса и максимального лизиса, соответственно. После инкубации 20 мин при комнатной температуре супернатант удаляют, а к клеткам добавляют 20% (об./об.) плазмы или сыворотки человека в DMEM (нагретой до 37°С) и инкубируют еще 20 мин при 37°С. Все клетки из каждого образца вносят в раствор пропидия йодида (10 мкг/мл). Затем супернатанты заменяют на PBS, содержащий 2,5 мкг/мл этидия бромида, и измеряют флуоресценцию при 600 нм с возбуждением при 520 нм на приборе Tecan Safire. Специфический лизис в процентах рассчитывают следующим образом: специфический лизиса (%) = (флуоресценция образца - фоновая флуоресценция) / (флуоресценция при максимальном лизисе - фоновая флуоресценция) × 100.
Индукция апоптоза и ингибирование клеточной пролиферации моноклональными антителами
Для тестирования на способность запускать апоптоз моноклональные антитела против CLDN18.2 можно, к примеру, инкубировать с CLDN18.2-положительными раковыми клетками, например, трансфецированными CLDN18.2 раковыми клетками SNU-16, DANG или KATO-III, при 37°С в течение примерно 20 часов. Клетки собирают, промывают в буфере для связывания с Annexin-V (BD Biosciences) и инкубируют с Annexin V, конъюгированным с FITC или АРС (BD Biosciences), в течение 15 мин в темноте. Все клетки из каждого образца вносят в раствор PI (10 мкг/мл в PBS) в пробирке для FACS и сразу же оценивают методом проточной цитометрии (как описано выше). С другой стороны, можно детектировать общее ингибирование пролиферации клеток моноклональными антителами с помощью коммерчески доступных наборов. Набор DELFIA Cell Proliferation Kit (Perkin-Elmer, кат. № AD0200) представляет собой неизотопный иммуноанализ на основе измерения включения 5-бром-2'-дезоксиуридина (BrdU) во время синтеза ДНК в пролиферирующих клетках на микропланшетах. Включение BrdU определяется с помощью меченных европием моноклональных антител. Для детектирования антител клетки фиксируют и денатурируют ДНК с помощью раствора Fix. Несвязанные антитела вымываются и добавляется DELFIA Inducer, который вытесняет ионы европия из меченного антитела в раствор, где они образуют сильнофлуоресцирующие хелаты с компонентами DELFIA Inducer. Измеряют флуоресценцию - методом флуорометрии с временным разрешением, - которая пропорциональна синтезу ДНК в клетках по каждой лунке.
Доклинические исследования
Моноклональные антитела, которые связываются с CLDN18.2, также можно протестировать на модели in vivo (например, на иммунодефицитных мышах, несущих привитые опухоли из клеточных линий, экспрессирующих CLDN18.2, например, DANG, SNU-16 или KATO-III, либо после трансфекции, например, HEK293), чтобы определить их эффективность в контролировании роста экспрессирующих CLDN18.2 опухолевых клеток.
Исследования in vivo после ксенотрансплантации экспрессирующих CLDN18.2 опухолевых клеток мышам или другим животным с ослабленным иммунитетом могут проводиться с использованием описанных здесь антител. Антитела можно вводить лишенным опухолей мышам с последующим введением опухолевых клеток, чтобы измерить эффекты антител в предотвращении образования опухолей или связанных с ними симптомов. Антитела можно вводить несущим опухоли мышам, чтобы определить терапевтическую эффективность соответствующих антител в уменьшении роста, метастазирования опухолей или связанных с ними симптомов. Применение антител можно сочетать с применением других веществ типа цитостатических препаратов, ингибиторов факторов роста, блокаторов клеточного цикла, ингибиторов ангиогенеза или других антител, чтобы определить синергическую эффективность и потенциальную токсичность комбинаций. Для анализа токсических побочных эффектов, опосредованных антителами, можно инокулировать животным антитела или контрольные реагенты и тщательно исследовать на наличие симптомов, возможно связанных с терапией антителами к CLDN18.2. Возможные побочные эффекты применения in vivo антител к CLDN18.2 включают, в частности, токсичность в экспрессирующих CLDN18.2 тканях, включая желудок. Антитела, распознающие CLDN18.2 у человека и других видов, например, мыши, особенно полезны для прогнозирования возможных побочных эффектов, опосредованных применением моноклональных антител к CLDN18.2 на людях.
Картирование эпитопов, распознаваемых антителами, может проводиться так, как подробно описано в "Epitope Mapping Protocols" (Methods in Molecular Biology) by Glenn E. Morris, ISBN-089603-375-9, и в "Epitope Mapping: A Practical Approach" in Practical Approach Series, 248 by Olwyn M.R. Westwood, Frank C. Hay.
Описанные здесь соединения и средства можно вводить в виде любой подходящей фармацевтической композиции.
Фармацевтические композиции обычно представлены в стандартной дозовой форме и могут быть получены известными per se способами. Фармацевтические композиции, например, могут быть в виде раствора или суспензии.
Фармацевтические композиции могут содержать соли, буферные вещества, консерванты, носители, разбавители и/или наполнители, которые все предпочтительно являются фармацевтически приемлемыми. Термин "фармацевтически приемлемый" означает отсутствие токсичности у материала, который не влияет на действие активного компонента фармацевтической композиции.
Соли, которые не являются фармацевтически приемлемыми, могут применяться для получения фармацевтически приемлемых солей и включены в изобретение. Фармацевтически приемлемые соли такого типа включают, без ограничения, соли, полученные из следующих кислот: соляной, бромистоводородной, серной, азотной, фосфорной, малеиновой, уксусной, салициловой, лимонной, муравьиной, малоновой, янтарной кислоты и др. Фармацевтически приемлемые соли также могут быть получены в виде солей щелочных металлов или солей щелочноземельных металлов, как-то солей натрия, солей калия или солей кальция.
Подходящие буферные вещества для применения в фармацевтических композициях включают уксусную кислоту в виде соли, лимонную кислоту в виде соли, борную кислоту в виде соли и фосфорную кислоту в виде соли.
Подходящими консервантами для применения в фармацевтических композициях являются бензалкония хлорид, парабен, хлорбутанол и тимеросал.
Формы для инъекций могут содержать фармацевтически приемлемые наполнители типа лактата для Рингера.
Термин "носитель" относится к таким органическим или неорганическим компонентам, природного или синтетического происхождения, с которыми сочетается активный компонент для того, чтобы облегчить, усилить или сделать возможным применение. В соответствии с изобретением, термин "носитель" также включает один или несколько совместимых твердых или жидких наполнителей, разбавителей или инкапсулирующих веществ, которые подходят для введения пациентам.
Возможными веществами-носителями для парентерального введения являются, к примеру, стерильная вода, раствор Рингера, раствор Рингера с лактатом, стерильный раствор хлорида натрия, полиалкиленгликоли, гидрогенизированные нафталины и в особенности биосовместимые полимеры лактидов, сополимеры лактидов/гликолидов или сополимеры полиоксиэтилена/полиоксипропилена.
Термин "наполнитель" в настоящем изобретении служит для обозначения всех веществ, которые могут присутствовать в фармацевтической композиции, но не являются активными ингредиентами, таких, например, как носители, связующие вещества, смазывающие вещества, загустители, поверхностно-активные вещества, консерванты, эмульгаторы, буферы, ароматизаторы или красители.
Описанные здесь средства и композиции могут вводиться любым стандартным способом, как-то посредством парентерального введения, в том числе посредством инъекций или инфузий. Предпочтительным является парентеральное введение, например, внутривенно, внутриартериально, подкожно, внутрикожно или внутримышечно.
Композиции, подходящие для парентерального введения, обычно содержат стерильный водный или неводный препарат активного соединения, который предпочтительно является изотоническим для крови реципиента. Примерами совместимых носителей и растворителей служат раствор Рингера и изотонический раствор хлорида натрия. Кроме того, в качестве среды для растворов или суспензий используют нелетучие масла, обычно стерильные.
Описанные здесь средства и композиции вводятся в эффективных количествах. Термин "эффективное количество" означает такое количество, которое вызывает нужную реакцию или требуемый эффект само по себе или вместе с последующими дозами. В случае лечения определенного заболевания или состояния желательная реакция предпочтительно означает торможение развития заболевания. Это включает замедление развития заболевания, в частности, прекращение или обращение прогрессирования заболевания. При лечении заболевания или состояния желательной реакцией может быть задержка возникновения или предотвращение возникновения данного заболевания или состояния.
Эффективное количество описанного здесь средства или композиции будет зависеть от подлежащего лечению заболевания, степени тяжести заболевания, индивидуальных параметров пациента, таких как возраст, физиологическое состояние, размер и вес, продолжительности лечения, типа попутной терапии (если она есть), конкретного способа введения и тому подобных факторов. Соответственно, вводимые дозы описанных здесь средств могут зависеть от этих различных параметров. В случае, когда реакция пациента недостаточна при начальной дозе, можно применять более высокие дозы (или эффективно более высокие дозы, достигаемые другим, более локализованным способом введения).
Описанные здесь средства и композиции можно вводить пациентам, например, in vivo, для лечения или профилактики различных заболеваний типа тех, что описаны здесь. Предпочтительными пациентами являются больные люди с такими заболеваниями, которые могут быть исправлены или облегчены при введении описанных здесь средств и композиций. Это включает в себя заболевания с участием клеток, характеризующихся изменением профиля экспрессии CLDN18.2.
Например, в одном воплощении описанные здесь антитела могут применяться для лечения больных раковыми заболеваниями, например, раковыми заболеваниями типа описанных здесь, характеризующихся наличием раковых клеток, экспрессирующих CLDN18.2.
Фармацевтические композиции и способы лечения, описанные в соответствии с изобретением, также могут применяться при иммунизации или вакцинации для предотвращения описанных здесь заболеваний.
Далее настоящее изобретение раскрывается на следующих примерах, которые не должны восприниматься как ограничивающие объем изобретения.
Примеры
Пример 1. Материалы и методы
1. Антитела

2. Иммуногистохимия (IHC)
Срезы тканей (толщиной 4 мкм) хранили при 2-8°С до использования.
Перед процессом депарафинизации срезы инкубировали при 58-60°С в сушильном шкафу в течение 1 часа, чтобы расплавить парафин и количественно удалить воду, тем самым улучшая адгезию тканей к стеклам ("пропекание").
Депарафинизапия
После расплавления и сушки стекла подвергали депарафинизации ксилолом в две стадии (по 5 мин) и регидратации с использованием убывающего ряда растворов спирта (при температуре окружающей среды 20-27°С):
• 5 (±1) мин в ксилоловой ванне;
• еще раз в свежей ксилоловой ванне;
• удалить лишнюю жидкость;
• 5 (±1) мин в абсолютном этаноле;
• еще раз в свежей ванне;
• удалить лишнюю жидкость;
• 5 (±1) мин в 96% этаноле;
• еще раз в свежей ванне;
• удалить лишнюю жидкость;
• 5 (±1) мин в 80% этаноле;
• удалить лишнюю жидкость;
• 5 (±1) мин в 70% этаноле;
• удалить лишнюю жидкость;
• 5 мин в дистиллированной или деионизованной воде.
Воспроизведение эпитопов и гашение
После удаления парафина проводили извлечение эпитопов мишени по методике термоиндуцированного извлечение эпитопов. Для этого стекла помещали в сосуд для окрашивания, заполненный 200 мл буфера для воспроизведения (10 мМ цитратный буфер, 0,05% Tween-20, рН 6), и инкубировали в скороварке (PASCAL, Dako) при 120°С в течение 10 мин. После этого сосуд вынимали из скороварки и оставляли охлаждаться в растворе для воспроизведения эпитопов на 10 (±1) мин при комнатной температуре. Стекла промывали промывочным буфером (1×PBS).
После охлаждения срезы помещали в сосуд для окрашивания, заполненный 200 мл гасящего раствора (0,3% пероксидазы в 1×PBS), и инкубировали 15 мин при комнатной температуре, а затем промывали 2×5 мин свежим промывочным буфером.
Блокирование и инкубация антител
Удаляли избыток промывочного буфера, покрывали стекла 200 мкл блокирующего буфера (10% козьей сыворотки в 1×PBS) и инкубировали при комнатной температуре в течение 30 мин. Блокирующий буфер удаляли и заменяли на 200 мкл разведенного раствора антитела (разведение в блокирующем буфере). Предметные стекла инкубировали с первичным антителом в течение ночи при 2-8°С:
На следующий день удаляли раствор первичного антитела и промывали срезы 3×5 мин промывочным буфером. После этого избыток промывочного буфера удаляли и добавляли 200 мкл готового к использованию раствора вторичного антитела (Power Vision козье-против-мыши с HRP; Immunologic, NL). Предметные стекла инкубировали 30 мин при комнатной температуре. Избыток жидкости удаляли, а стекла промывали 3×5 мин свежим промывочным буфером.
Реакция с субстратом и контрастное окрашивание
После удаления излишка промывочного буфера срезы покрывали ок. 50-150 мкл свежеприготовленного раствора субстрата-хромогена (VectorRed; Vector Labs) в течение 2 мин. Излишек субстрата удаляли, а предметные стекла инкубировали в сосудах с деионизированной водой в течение 1-5 мин.
После этого проводили контрастное окрашивание погружением срезов в сосуды, содержащих по 200 мл гематоксилина Mayer, на 2 мин. После этого срезы помещали в водопроводную воду на 5-10 мин для подсинивания ядер.
Обезвоживание и заключение препаратов
После контрастного окрашивания срезы обезвоживали с помощью возрастающего ряда растворов спирта:
• погружение в 70%-й этанол (на 5-10 сек);
• погружение в 80%-й этанол (на 5-10 сек);
• погружение в 96%-й этанол (на 5-10 сек);
• погружение в 96%-й этанол (на 5-10 сек);
• погружение в абсолютный этанол (на 5-10 сек);
• 5 мин в ксилоле;
• 5 мин в ксилоле.
Для заключения препаратов использовали неводную заливочную среду (X-TRA-Kit, Medite). Предметные стекла заливали прямо сразу после последнего заполненного ксилолом сосуда и сушили на воздухе при комнатной температуре.
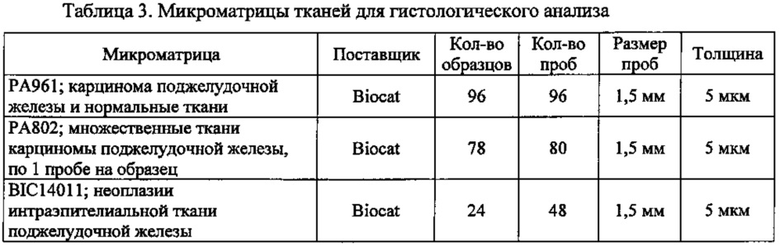
3. Культивирование клеток
Все раковые линии клеток поджелудочной железы и дополнительные контрольные линии клеток, использовавшиеся для представленных здесь экспериментов, культивировали в средах согласно исходным спецификациям и по стандартным методикам для тканевых культур. Условия приведены в таблице 4. Для всех вновь полученных клеточных линий проверяли клетки на загрязнение микоплазмой и готовили мастер-банк клеток.
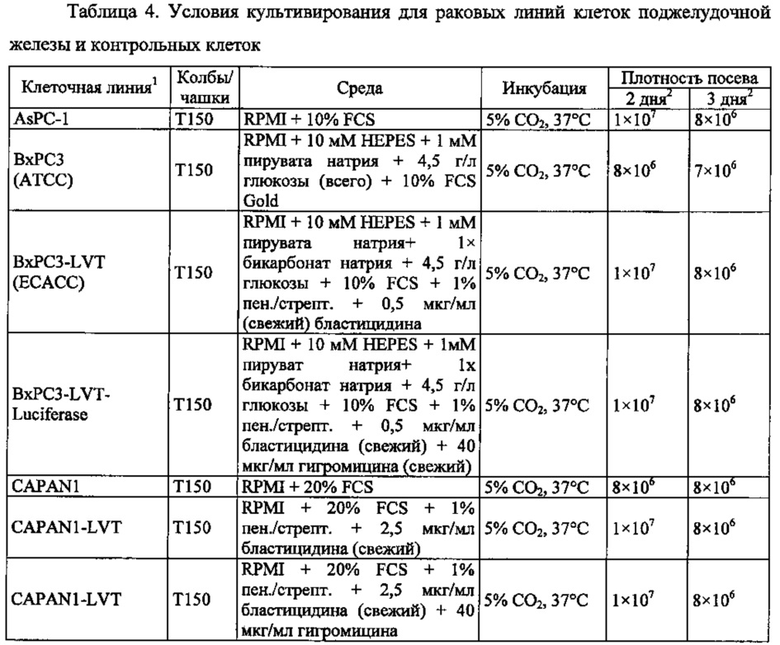
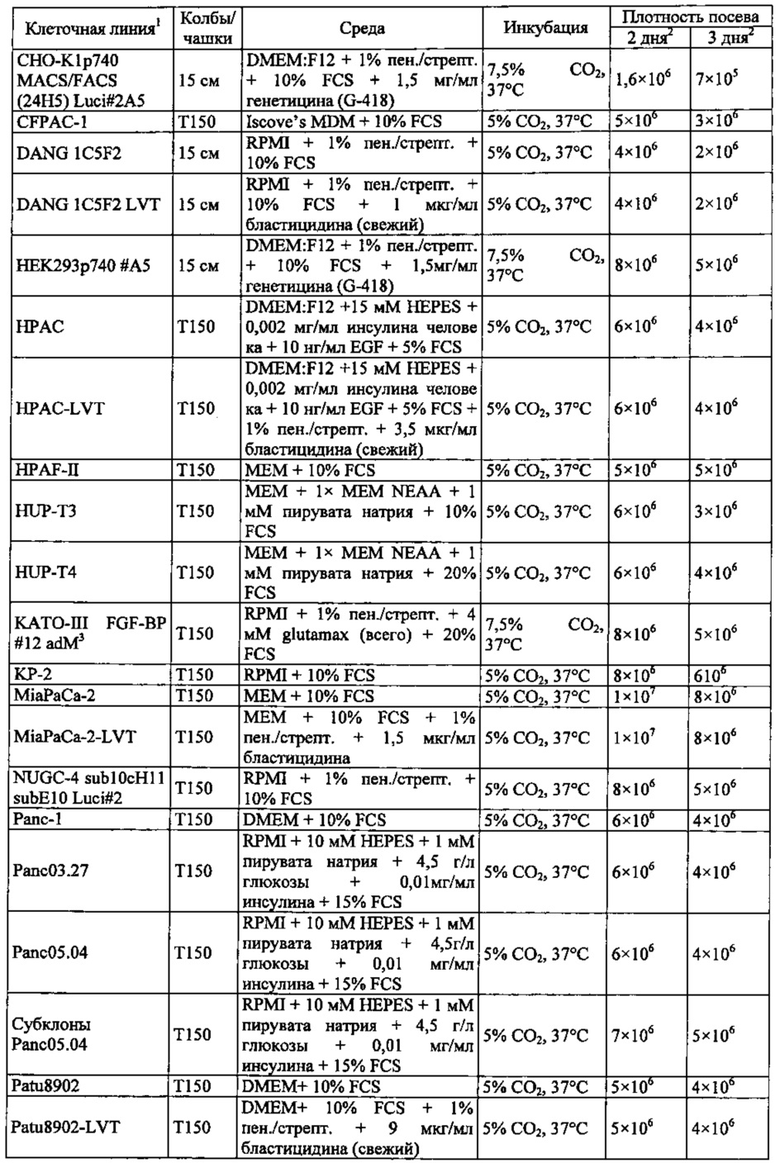
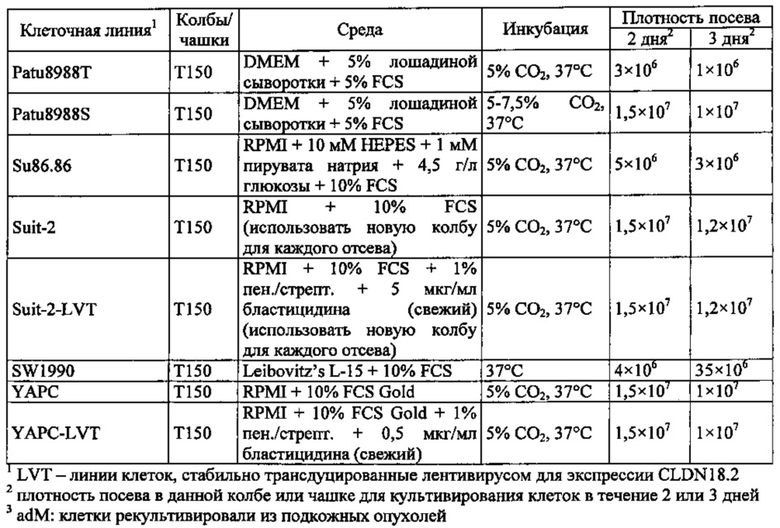
4. Трансфекции люциферазой клеточных линий поджелудочной железы
Для анализа ADCC клетки раковых линий поджелудочной железы трансфецировали РНК люциферазы. РНК люциферазы (вектор Pst1-luc2mut-2hBgUTR-A121-EciI (pST1-109)) получали с кэпом ARCA и растворяли в Н2О. Эту РНК хранили порциями по 22 мкл при -80°С. Для всех клеточных линий поджелудочной железы определяли оптимальные условия электропорации, дающие самые высокие показатели трансфекции и жизнеспособности клеток. При каждом анализе клетки отделяли с помощью PBS/5 мМ ЭДТА, растворяли 2,5×106 клеток в 250 мкл Х-Vivo и смешивали в охлаждаемых льдом кюветах с 10 мкг РНК. Клетки сразу же подвергали электропорации (GenePulser Xcell, BioRad) и ресуспендировали в подогретой среде для анализа, доводя их концентрацию до 5×105 клеток/мл. Проверенные условия электропорации для всех клеточных линий были таковы:
ЕР1: 250 В, 475 мкФ
ЕР2: 200 В, 300 мкФ
ЕР3: 150 В, 300 мкФ
ЕР4: 200 В, 400 мкФ
ЕР5: 250 В, 950 мкФ
контроль: 0 В, 0 мкФ
Жизнеспособность клеток определяли сразу после электропорации с помощью CASY или окрашиванием клеток трипановым синим и определением процента мертвых клеток в камере Neubauer. Клетки высеивали в четырех повторах в белые 96-луночные планшеты (2,5×104 клеток на лунку) и инкубировали в течение 24 ч. После этого измеряли люциферазную активность в лиминометре (Tecan Infinite200) после добавления смеси люциферина в течение 90 мин. Трансфекция была успешной, a ADCC измеримой, если значения RLU составляли >1000.
5. Количественная реакция ПЦР в реальном времени (количественная ПЦР)
Для выделения РНК из клеток раковых линий поджелудочной железы клетки высеивали в чашки на 10 см и культивировали 2-3 дня, до 80%-й конфлюентности. РНК выделяли в соответствии с инструкциями, прилагаемыми к набору RNeasy® Mini Kit (Qiagen). Получение кДНК проводили в соответствии с инструкциями производителя, входящими в набор Superscript® III First Strand Kit (Invitrogen). Образцы РНК и кДНК хранили при -80°С.
Количественный анализ транскриптов CLDN18.2 проводили путем амплификации олиго(dT)-примированной кДНК в реакции ПЦР на 40 циклов с использованием ПЦР-праймеров #5054s (5'-AGAGAGCTCTGGCTTCACCGAGTG-3') и #5060as (5'-CCAGAAGTTAGTCACCAGCATGTTGG-3'), отличающих изоформы CLDN18.1 и CLDN18.2. Реакционную смесь готовили с красителем SYBR Green (QuantiTect SYBR Green PCR Kit, Qiagen), который интеркалирует в двухцепочечную ДНК. Реакции и измерения проводили на приборе ABI-PRISM7900 Sequence Detection System с использованием программного обеспечения (Applied Biosystems).
Относительные уровни экспрессии транскриптов CLDN18 вычисляли по значениям ΔΔСТ относительно гена домашнего хозяйства HPRT.
6. Анализ методом Вестерн-блот
Для выделения белков из клеток раковых линий поджелудочной железы клетки высеивали в чашки на 10 см и культивировали 2-3 дня, до 80%-й конфлюентности. Клетки лизировали добавлением 800 мкл 4×буфера с SDS для образцов (34% глицина, 250 мМ трис рН 6,8, 5% β-меркаптоэтанола, 8,2% SDS). Для разрушения геномной ДНК образцы белка обрабатывали ультразвуком при следующих условиях: выходная мощность: level 1, рабочий цикл: 70% в течение 20-25 сек. Концентрацию белка измеряли на спектрофотометре (поглощение при 280 нм), а образцы хранили при -80°С до использования.
Для выявления экспрессии CLDN18.2 на вестерн-блотах готовили 12,5% полиакриламидный гель для разделения (для 2 небольших гелей: 4,1 мл смеси акриламид/бисакриламид 29:1, 100 мкл 10% SDS, 2,5 мл трис рН 8,8, 3,2 мл H2O, 100 мкл APS, 10 мкл TEMED) между двумя с фиксированными стеклянными пластинами. После полимеризации на этот гель наслаивали концентрирующий гель (1,5 мл смеси акриламид/бисакриламид 29:1, 100 мкл 10% SDS, 2,5 мл трис рН 6,8, 5,8 мл H2O, 100 мкл APS, 10 мкл TEMED) и между стеклянными пластинами вставляли в него гребенку. После полимеризации на гель наносили по 75 мкг каждого образца белка, приготовленного добавлением (1:20) 4×буфера с SDS для образцов (250 мМ трис-HCl, 34% глицерина, 8,2% SDS, рН 6,8) и 7,5 мкл смеси маркеров размера (1,5 мкл Magic Mark ХР Western Standard вместе с 6 мкл SeaBlue Plus2 Prestained Standard). Гели разгоняли в 1× рабочем буфере с SDS (25 мМ трис, 0,192 М глицина, 0,1% SDS) по 30 мин при 80 В и 60 мин при 180 В. Полусухой блоттинг геля на нитроцеллюлозную мембрану проводили в течение 90 мин при 160 мА в 1× буфере для переноса (25 мМ трис, 0,192 мМ глицина, 20% метанола). Блоты сначала блокировали в 5% растворе сухого молока в PBS, а затем добавляли первичные антитела (0,25 мкг/мл против клаудина 18 (С-конец) или 0,1 мкг/мл против β-актина) в 1% растворе сухого молока в PBS. Блоты инкубировали в течение ночи при 4°С, промывали 3 раза по 10 мин в 1×PBS/0,05% Tween 20, а затем инкубировали 1 час с мечеными вторичными антителами при комнатной температуре в 1% растворе сухого молока в PBS (козьи против IgG кролика (FC) в разведении 1:1000). Блоты опять промывали 3 раза по 10 мин в 1×PBS/0,05% Tween 20, а затем проводили детектирование добавлением 1-3 мл раствора для детектирования (Pico and Dura Detection System (Pierce) на 1 мин и сканированием блотов в детекторной камере LAS-3000 (инкремент: 10 сек, интервальное время: 10 сек, чувствительность: высокая) в соответствии с GA_056_Chemolumineszenzentwickler LAS3000.
7. Проточная цитометрии (FACS)
Клетки из экспоненциально растущей культуры при конфлюэнтности 70-85% собирали с помощью PBS с 5 мМ ЭДТА или трипсина с ЭДТА. Клетки подсчитывали, центрифугировали 5 мин (468×g), а осадок ресуспендировали в буфере FACS (2% FCS, 0,1% азида натрия в PBS), доводя концентрацию до 2×106/мл. По 100 мкл клеток высеивали в круглодонные 96-луночные планшеты и опять центрифугировали (5 мин, 468×g). Делали серийные разведения IMAB362 (или изотопического контроля - ритуксимаб) от 0,1 до 200 мкг/мл (11 ступеней разведения + контроль без антител) в 50 мкл буфера FACS и добавляли к клеткам на 30 мин при 4°С. Затем в каждую лунку добавляли 200 мкл буфера FACS и центрифугировали планшеты (5 мин, 468×g). Удаляли супернатанты и повторяли промывку. Вторичные козьи антитела против человека (FC-специфичные, F(ab')2, конъюгированные с АРС (Dianova)) разводили 1:100 в буфере FACS и добавляли по 30 мкл в каждую лунку. Планшеты инкубировали 30 мин при 4°С. После инкубации планшеты снова дважды промывали по 200 мкл буфера FACS, а осадок окончательно ресуспендировали в 100 мкл буфера FACS для измерения на FACS Array Bioanalyzer (BD) в соответствии с GA_018_BD FACS Array Bioanalyzer.
8. Лентивирусная трансдукция
Конструирование лентивирусного вектора. Лентивирусы относятся к РНК-вирусам, которые стабильно встраиваются в геномную ДНК человека как делящихся, так и не делящихся клеток. За основу был принят вектор pLenti6.4 (Invitrogen). Он содержит ген бластицидина для отбора положительно трансдуцированных клеток. В рекомбинантную область вектора клонировали CLDN18.2, слитый с промотором EF1α, получая pL64B42E (EF1α-hCLaudin18.2)-Blasticidin (фиг. 1).
Выбор клеточных линий. Клеточные линии выбирали по данным литературы либо из тех, что тестировались ранее in vivo. Критерии отбора включали однородный подкожный рост у мышей nude и терапевтическое окно в 20-100 дней. Были включены три клеточные линии (DANG, YAPC и ВхРС3), которые уже показали слабую экспрессию мРНК CLDN18.2, и еще три (MiaPaCa-2, Patu8902 и Suit-2), способные к метастазированию по данным литературы. Две другие клеточные линии (которые растут в виде однородных подкожных опухолей in vivo) были выбраны случайным образом (HPAC, CAPAN1).
Определение условий отбора по бластицидину. Для всех клеточных линий перед трансдукцией определяли, какая концентрация бластицидина необходима для отбора клеток после лентивирусной трансдукции. Раковые клетки поджелудочной железы высеивали в 6-луночные планшеты при высокой плотности, дающей конфлюентность в 80-90% через 24 часа. В лунки добавляли бластицидин (исходно: 10 мг/мл, Invitrogen) в возрастающих концентрациях от 0,5 до 12 мкг/мл (5 ступеней разведения + контроль без бластицидина). Среду меняли каждые 3-4 дня, а клетки анализировали под микроскопом перед удалением среды. Отмечали количество мертвых клеток и состояние живых клеток. Клетки культивировали в течение 14 дней. Для отбора трансдуцированных лентивирусом клеток использовали наименьшую концентрацию бластицидина, вызывающую 100%-й апоптоз клеток через 14 дней. Необходимые концентрации бластицидина для каждой из полученных LVT-линий клеток приведены в таблице 4.
Выбор оболочки. Для лентивирусной трансдукции контрольный GFP-лентивирусный вектор pL64B42E-(EF1a-GFP)-blast упаковывали в частицы с различными оболочками (VSV-G, GALV, RD114, Mokola-G и Rabies-G). В зависимости от белков, присутствующих в оболочке, и состава клеточной мембраны, прикрепление к раковым клеткам мишени было более или менее эффективным. Для всех раковых линий клеток поджелудочной железы оболочка VSV-G давала самую высокую эффективность трансдукции (68,5-91,2%) (таблица 5). Поэтому экспрессирующий CLDN18.2 вектор pL64B42E (EF1α-hCLauclin18.2)-Blasticidin упаковывали в оболочки VSV-G. Инфицировали клетки-продуценты и выделяли вирус из среды при высоком титре (3,86×107 частиц/мл). Вирусные супернатанты хранили при -80°С.
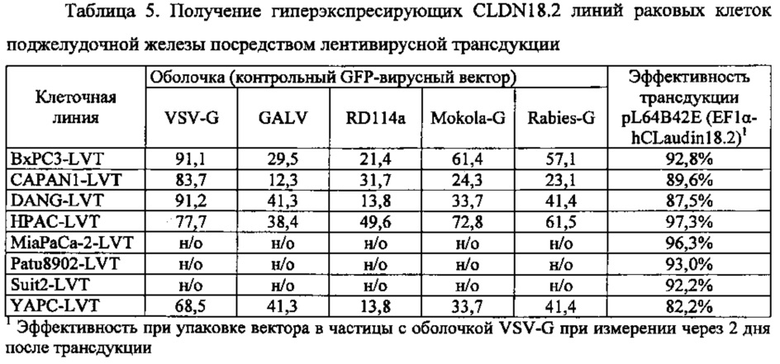
Лентивирусная трансдукции клеток раковых линий поджелудочной железы. Для инфицирования клеток намеченных раковых линий поджелудочной железы 24-луночный планшет покрывали 200 мкл 1×retronectin® (20 мкг/мл, Takara Inc.), плотно закрывали его парафильмом и инкубировали 3-16 ч при 4°С. Планшеты промывали 200 мл PBS и блокировали PBS/2% BSA в течение 30 мин при комнатной температуре. Планшеты опять промывали и наносили по 300 мкл вирусного супернатанта с центрифугированием 25 мин при 2500 об/мин при 15°С. Супернатант удаляли и повторяли нанесение 3 раза. Наконец планшеты промывали один раз PBS и в каждую лунку высеивали намеченные клетки с низким количеством пересевов. Для всех линий раковых клеток поджелудочной железы высеивали по 5×105-1×107 клеток в каждую из 24 лунок. Планшеты инкубировали 2 дня при 37°С. После этого клетки отделяли и определяли эффективность трансдукции методом FACS с использованием меченного FITC антитела IMAB362. Клетки подвергали экспансии и для каждой клеточной линии готовили мастер-банк клеток.
9. Анализ ADCC
Намеченные раковые клетки поджелудочной железы высеивали в колбы заранее за два дня, чтобы получить конфлюентные на 80-90% культуры в день начала ADCC. Раковые клетки поджелудочной железы трансфецировали РНК люциферазы и высеивали в белые 96-луночные планшеты при плотности 1×104 клеток на лунку в 50 мкл среды определения (культуральная среда, приведенная в таблице 4, с 20 мМ HEPES). Кроме того, высеивали клетки NUGC-4 sub10cH11 subE10 Luci#2 (8000 клеток на лунку) в качестве положительного контроля при всех анализах. Клетки культивировали 4-6 ч перед добавлением антител и очищенных PBMCs.
PBMCs получали из свежей лейкоцитной пленки человека, полученной от здоровых доноров. Три порции крови по 20-25 мл разводили PBS (1:2) и осторожно наслаивали на 4 порции по 15 мл Ficol-Paque Plus (GE Healthcare) в пробирках Falcon на 50 мл. Градиенты центрифугировали (25 мин, 700×g). После центрифугирования из интерфазы собирали мононуклеарные клетки периферической крови (РВМС), промывали в PBS с 2 мМ ЭДТА, центрифугировали (5 мин, 468×g), опять суспендировали в PBS с 2 мМ ЭДТА и центрифугировали (10 мин, 208×g), чтобы удалить тромбоциты. Осадки ресуспендировали в 50 мл PBS с 2 мМ ЭДТА и проводили подсчет клеток. РВМС центрифугировали (5 мин, 468×g) и ресуспендировали в культуральной среде X-Vivo-15 при концентрации 1,6×107 клеток/мл для добавления к клеткам поджелудочной железы или 1,28×107 клеток/мл для добавления к клеткам NUGC-4 sub 10сН11 sub Е10 Luci#2.
Антитела (IMAB362 и антитело ch78H11-1Н6 из изотопического контроля) подвергали серийному (4,5-кратному) разведению по 10 раз, получая концентрации от 200 мкг/мл до 0,26 нг/мл. Из каждого разведения по 25 мкл добавляли в четырех повторах к намеченным клеткам. В лунки с контролем на среду и на лизис добавляли PBS без антител. После этого в каждую лунку добавляли 25 мкл РВМС (соотношение Е : Т = 40:1) и инкубировали планшеты в течение 24 ч ± 1 ч при 37°С, 5% CO2.
На следующий день в лунки с контролем на лизис добавляли по 10 мкл 8% раствора Triton X100/PBS, а во все другие лунки - по 10 мкл PBS. Наконец, в каждую лунку добавляли 50 мкл свежеприготовленного раствора люциферина (160 мМ HEPES, 1×PBS, 3,84 мг/мл D-люциферина (BD Biosciences)) и инкубировали планшеты 80 мин при комнатной температуре в темноте. Измеряли люминесценцию в результате окисления Lucifer Yellow люциферазой жизнеспособных клеток на считывающем устройстве для микропланшетов (Infinite200, Tecan, Швейцария). Степень клеточной цитотоксичности рассчитывали по следующей формуле:
специфическое уничтожение (%) = 100 - [(RLUпроба - RLUTriton) / (RLUконтр.на среду - RLUTriton) × 100].
10. CDC
Анализ CDC проводили следующим образом.
Намеченные клетки (СНО-K1 р740 MACS/FACS (24Н5) р3151 Luci#2A5) высеивали в 50 мкл среды определения в белые 96-луночные планшеты для анализа (10000 клеток на лунку) и культивировали в течение 24 ч + 20 мин при 37°С, 7,5% CO2 при относительной влажности 95% перед добавлением образцов. Каждый 96-луночный планшет содержал в общей сложности 3 различных отрицательных контроля (инактивированная нагреванием сыворотка, сыворотка с IMAB362 и без него и сыворотка с антителом из изотопического контроля (Rituximab)) и положительный контроль из сборной сыворотки здорового человека (lot #31032011) с 500 нг/мл IMAB362. Дополнительный положительный контроль получали по окончании реакции добавлением 0,8% Triton X100 во вторую лунку контроля на среду, что вызывает полный лизис. Один из 96-луночных планшетов содержал функциональный положительный контроль, полученный путем 7 серийных 3,16-кратных разведениях IMAB362 (от 10000 до 31,8 нг/мл). Этот контроль давал сигмовидную зависимость лизиса клеток от дозы. Все образцы готовили (по 200 мкл) в одно и то же время в 96-луночном планшете с углубленными лунками для разведения. Из каждой лунки брали пробы по 3 раза пипеткой, чтобы получить по три повтора в планшетах для анализа. После внесения по 50 мкл исследуемых и контрольных проб в планшеты для анализа их инкубировали в течение 80+5 мин при 37°С, 7,5% СО2 при относительной влажности 95%.
В каждую лунку добавляли по 10 мкл PBS, за исключением лунок контроля на лизис с тритоном. В каждую лунку контроля на лизис с тритоном добавляли по 10 мкл 0,8% раствора Triton/PBS. Готовили раствор субстрата - люциферина (6114 мкл бидист. воды, 2496 мкл HEPES (1М), 1998 мкл 1×DPBS, 4992 мкл исходного раствора D-люциферина (12 мг/мл)). В каждую лунку добавляли 50 мкл раствора субстрата - люциферина. Планшеты инкубировали при 37°С, 7,5% CO2 при относительной влажности 95% в течение 45 мин. Планшеты измеряли на считывающем устройстве для микропланшетов.
Комплемент-зависимый лизис рассчитывали по следующей формуле:
специфический лизис (%) = 100 - [(RLUпроба - RLUTriton) / (RLUHSCM - RLUTriton) × 100]
Модификации для тестирования раковых линий клеток поджелудочной железы:
• раковые клетки поджелудочной железы трансфецировали РНК люциферазы при оптимизированных условиях; для каждой тестируемой клеточной линии высеивали по 1,5×104 клеток на лунку;
• поскольку большинство раковых линий клеток поджелудочной железы с трудом отделяются и образуют одиночные клетки, то в 1-й день использовали трипсин;
• раковые клетки поджелудочной железы культивировали в планшетах для анализа при 37°С, 5% CO2;
• анализ CDC с предобработкой клеток химиотерапевтическими средствами проводили при следующих концентрациях IMAB362 или в качестве изотопического контроля - антитела ch78H11 1Н6: 640000, 160000, 40000, 10000, 2500, 625, 156 и 39 нг/мл.
11. Ингибирование пролиферации
Для анализа кривых доза-ответ для каждого химиотерапевтического средства проводили анализ пролиферации. Анализировали ингибирование пролиферации после применения гемцитабина или оксалиплатина в различных концентрациях для каждой линии раковых клеток поджелудочной железы.
Клетки высеивали в 96-луночные планшеты и через 4-6 часов добавляли гемцитабин или оксалиплатин в следующих концентрациях: 1000, 500, 250, 100 и 20 нг/мл. Для анализа пролиферации клетки инкубировали 4 дня при 37°С и 5% CO2. Добавляли 50 мкл полного реагента ХТТ (50 частей ХТТ + 1 часть сшивающего реагента) и инкубировали при 37°С. Через 3 ч и 4 ч проводили измерение поглощения (клетки плюс супернатант) на Tecan Safire. Рассчитывали ингибирование пролиферации по сравнению с контролем на среду, принятым за 100%. Значения ЕС50 для гемцитабина и оксалиплатина рассчитывали в программе GraphPad Prism.
12. Культивирование клеток раковых линий поджелудочной железы с химиотерапевтическими препаратами для ADCC или CDC
Линия DANG: высеивали 4-6×106 клеток и культивировали 2 дня в среде или в среде + 1 нг/мл гемцитабина или 1 нг/мл гемцитабина + 10 нг/мл оксалиплатина. Линия Patu8988S: высеивали 1-1,4×107 клеток и культивировали в отсутствие или в присутствии 10 нг/мл гемцитабина или 10 нг/мл гемцитабина в сочетании со 100 нг/мл оксалиплатина.
В день начала ADCC следовали вышеописанной методике и определяли экспрессию CLDN18 на клеточной поверхности методом FACS, как описано выше.
13. Анализ клеточного цикла
Клетки засеивали в 6-луночные планшеты и через 5-6 часов добавляли химиотерапевтические средства на 24 ч, 48 ч или 3 дня. Свободноплавающие в среде клетки объединяли с клетками адгезионного слоя, который подвергали трипсинизации. Клетки промывали. Анализ клеточного цикла либо начинали сразу же, либо сначала проводили окрашивание клеточной поверхности, как описано выше. Клетки ресуспендировали в 1 мл PBS и вносили в 3 мл 4% PFA. После фиксации клеток 15 мин при комнатной температуре клетки осаждали и промывали. Для обработки РНКазой клетки ресуспендировали в 200 мкл РНКазы (10000 Ед/мл) плюс 0,05% Triton Х-100 и инкубировали 30 мин при 37°С. Добавляли 1 мл PBS, образцы центрифугировали и ресуспендировали в 200 мкл PBS с 50 мкг/мл PJ. Уже через 30 минут образцы были готовы для анализа методом проточной цитометрии. Распределение фаз клеточного цикла определяли с помощью программного обеспечения FlowJo в виде гистограмм содержания ДНК.
14. Анализ апоптоза
После указанных процедур измеряли апоптоз по связыванию аннексина V (набор для обнаружения I) или по фрагментации ДНК (Apo-Direct) согласно рекомендациям изготовителя (Pharmingen, San Diego, СА). Вкратце, свободноплавающие в супернатанте клетки объединяли с клетками адгезионного слоя, который подвергали трипсинизации, а затем промывали. Порции по 5×105 клеток инкубировали с аннексином V-APC и PI в течение 15 мин при комнатной температуре в темноте. Клетки сразу же анализировали методом проточной цитометрии. Жизнеспособные клетки не содержат ни аннексина V-АРС, ни PI. В ранней стадии апоптоза клетки положительны на аннексии V-APC и отрицательны на PI, тогда как клетки, потерявшие жизнеспособность вследствие апоптотической или некротической смерти клеток, дают положительное окрашивание и на аннексии V, и на PI. Процент окрашенных клеток в каждом квадранте количественно определяли с помощью программного обеспечения FlowJo (BD Biosciences, Franklin Lakes, NJ).
Анализ апоптоза по фрагментации ДНК проводили следующим образом. Обработанные клетки (прикрепленные и плавающие) фиксировали в 70% ледяном этаноле в течение ночи. После промывки инкубировали 106 фиксированных клеток с ферментом терминальная дезоксинуклеотидилтрансфераза (TdT) и FITC-dUTP в течение 90 мин при 37°С, чтобы пометить разрывы ДНК. Клетки промывали, инкубировали с РНКазой А и йодидом пропидия в темноте 30 мин при комнатной температуре для окрашивания тотальной ДНК, а затем анализировали методом проточной цитометрии. Двойные и слипшиеся клетки исключались из анализа при настройке каналов.
15. Исследования in vivo
Все эксперименты in vivo проводились в соответствии с национальными правилами и этическими принципами для экспериментальных исследований на животных.
15.1. Обработка ксенотрансплантатов
Ксенотрансплантаты опухолей прививали путем подкожной инъекции раковых клеток в 200 мкл PBS в бок самкам бестимусных мышей nude Hsd:Foxnlnu. Несущие опухоли мыши получали 0 мкг, 200 мкг, 400 мкг или 800 мкг антител путем в/в инъекции раз в неделю или же чередуя в/в и в/б введение 2 раза в неделю. Химиотерапевтические средства вводили в/б раз в неделю или 2 раза в неделю. Размер опухолей и состояние животных отслеживали по 2 раза в неделю. По окончании химиотерапии введение антител продолжалось до тех пор, пока размер опухолей не достигал >1400 мм3 или же опухоли становились изъязвленными. Образцы опухолей хранили замороженными или фиксированными в 4% формалине для последующего анализа.
15.2. Анализ метастазирования
Различные линии раковых клеток поджелудочной железы сначала подвергали анализу на их способность к образованию метастазов после внутривенного введения этих клеток мышам nude. Для анализа их приживления группам из 5-10 мышей вводили 1×106 и/или 2×106 клеток и в различные моменты времени забивали по одной мыши, чтобы найти моменты времени приживления и роста метастазов.
Обработку метастазов проводили на 10-12 бестимусных мышах nude Hsd:Foxnlnu в 1 группе обработки. Им вводили по 2×106 клеток (Patu8988S или Suit2-LVT) внутривенно. Всех мышей забивали в одно и то же время, как только появлялись первые симптомы метастазирования (потеря веса, слабость, одышка) или же умирала первая мышь.
Препарирование тканей. При исследовании приживления мышей забивали в различные моменты времени либо как только у них появлялись четкие физиологические признаки метастазирования (потеря веса, слабость, одышка). Все органы у них подвергали макроскопическому анализу на метастазы. Только при клетках Patu8988S и Suit-2 в легких и легких/печени проявлялись макроскопически заметные метастазы, соответственно. Эти органы иссекали на 4 равных кусочка, из которых два (легкие: верхняя правая и нижняя левая доли) хранили для выделения геномной ДНК. Два других кусочка фиксировали в формалине и хранили для IHC-анализа (фиг. 2).
Получение геномной ДНК и стратегия количественной ПЦР. Геномную ДНК экстрагировали из ткани легких или печени. В качестве контроля геномную ДНК также выделяли из раковых клеток поджелудочной железы Patu8988S человека, а также из не получавших инъекций мышей отрицательного контроля.
Стратегия количественной ПЦР основывается на амплификации ДНК человека, присутствующей в метастазах. Относительный уровень ДНК человека, обнаруженной в образцах легких мыши, прямо коррелирует с количеством и/или размером метастазов. Поскольку этот метод подвержен смещению потому, что метастазы в легких распределяются неравномерно и иногда одна доля поражается больше, чем другая, то для одного препарата ДНК смешивали два различных участка легких (фиг. 2).
Реакции количественной ПЦР проводили с парой праймеров #5861: 5'-GGGATAATTTCAGCTGACTAAACAG-3' и #5862: 5'-TTCCGTTTAGTTAGGTGCAGTTATC-3', которые специфически амплифицируют альфа-сателлитную ДНК, присутствующую в 17-й хромосоме человека, но не в ДНК мыши. Для составления стандартной кривой и в качестве положительного контроля ДНК Patu8988S смешивали с ДНК мыши и готовили 5-кратные разведения, получая 100%, 20%, 4%, 0,8%, 0,16%, 0,032% и 0,0064% ДНК человека в ДНК мыши. Эту кривую использовали для вычисления (линейная регрессия) количества ДНК метастазов человека в ткани легких мыши. Реакции количественной ПЦР проводили в конечном объеме 50 мкл, содержащем 20 мкл (200 нг) ДНК легких мыши, 25 мкл SYBR Green (Qiagen), 1,6 мкл смыслового праймера (10 мкМ) и 1,6 мкл антисмыслового праймера и 1,8 мкл Н2О.
Пример 2. Экспрессия CLDN18.2 в нормальных и опухолевых тканях поджелудочной железы человека
Для анализа уровня и профиля экспрессии CLDN18.2 в нормальных и опухолевых тканях поджелудочной железы проводили гистологическое окрашивание FFPE-срезов с помощью двух мышиных моноклональных антител (фиг. 3).
Поисковые пилотные эксперименты проводили с использованием первоначального антитела 35-22А на микроматрицах тканей (TMAs). Главным недостатком TMAs является вариабельное качество нанесенных тканей и малый размер пятен, то есть нерепрезентативность проб. Наряду с не полностью оптимизированной методикой окрашивания, это могло приводить к недооценке положительных случаев.
Основные эксперименты проводили с антителом 43-14А. При этом окрашивание проводили на срезах тканей, которые (по сравнению с TMAs) были крупнее и подвергались предварительной оценке на наличие опухолевых клеток.
Предраковые очаги поражения, происходящие из протоков поджелудочной железы, можно ранжировать в соответствии с международной системой интраэпителиалъной неоплазии поджелудочной железы (pancreas intraepithelial neoplasia, PanIN) (подтипы PanIN-1A, -1B, -2, -3).
Очаги типа PanIN-1 (фиг. 4А) плоские, состоят из высоких столбчатых клеток с базально расположенными ядрами и обильным супрануклеарным муцином. Ядра маленькие и круглой или овальные формы, ориентированы перпендикулярно базальной мембране. Существует гистологическое перекрывание между плоскими неопухолевыми гиперпластическими очагами и плоскими опухолевыми очагами без атипии.
Очаги подтипа PanIN-1В имеют папиллярную, микропапиллярную или базальую псевдомногослойную архитектуру и в остальном идентичны PanIN-1A (Hruban et al., Am J Surg Pathol. 2001, 25(5): 579-86).
Очаги типа PanIN-2 (фиг. 4B) плоские или папиллярные, имеют характерные ядерные аномалии, включая некоторую потерю полярности, уплотнение, увеличение размеров, псевдостратификацию и гиперхроматизм ядер. Митозы встречаются редко, но если они есть, то не люминальные (не апикальные) и не атипичные (Hruban et al., Am J Surg Pathol. 2001, 25(5): 579-86).
Очаги типа PanIN-3 (фиг. 4C) обычно папиллярные или микропапиллярные, однако изредка они могут быть плоскими. Истинная решетчатость, отпочковывание небольших кластеров эпителиальных клеток в просвет и люминальный некроз свидетельствуют о диагнозе PanIN-3. Очаги характеризуются потерей полярности ядер, дистрофическими бокаловидными клетками (бокаловидные клетки с ядрами, ориентированными в просвет, и мукоидной цитоплазмой, ориентированной к базальной мембране), митозами, которые иногда могут быть аномальными, неоднородностью ядер и вьщеляющимися (макро)-ядрышками (Hruban et al., Am J Surg Pathol. 2001, 25(5): 579-86).
Экспрессию CLDN18.2 в предраковых тканях анализировали с помощью антитела 43-14А на образцах тканей из различных источников. CLDN18.2 часто обнаруживался в структурах PanIN подтипов PanIN-1, -2 и -3, что свидетельствует о ранней экспрессии CLDN18.2 в предраковых очагах (фиг. 4), которая сохраняется и на более поздних стадиях. Напротив, экспрессия не наблюдалась в образцах нормальных тканей поджелудочной железы, включая протоки поджелудочной железы. Таким образом, CLDN18.2 является ранним маркером начала злокачественных гистологических изменений в протоках поджелудочной железы.
Для оценки экспрессии CLDN18.2 при первичном раке поджелудочной железы проводились два исследования. В пилотном исследовании несколько тканевых микроматриц (TMAs), включающих в общей сложности 141 случай первичного рака поджелудочной железы, подвергали окрашиванию специфичным к CLDNA18.2 моноклональным антителом 35-22А. В целом качество анализируемых TMAs было неудовлетворительным. Многие пятна были частично потеряны во время воспроизведения эпитопов, а неоднородное контрастное окрашивание гематоксилином свидетельствует о субоптимальной FFPE-обработке тканей.
В целом >48,9% случаев окрашивания были положительными на CLDN18.2, в том числе 49,2% (65/132) аденокарцином протоков, 50% (1/2) ацинозно-клеточных карцином и 3 из 7 нейроэндокринных карцином (таблица 7). Мембраны опухолевых клеток окрашивались без какого-либо фона на других типах клеток (фиг. 6). Более того, наблюдалась корреляция между интенсивностью экспрессии CLDN18.2 и содержанием окрашенных опухолевых клеток в опухоли (таблица 8, фиг. 5).
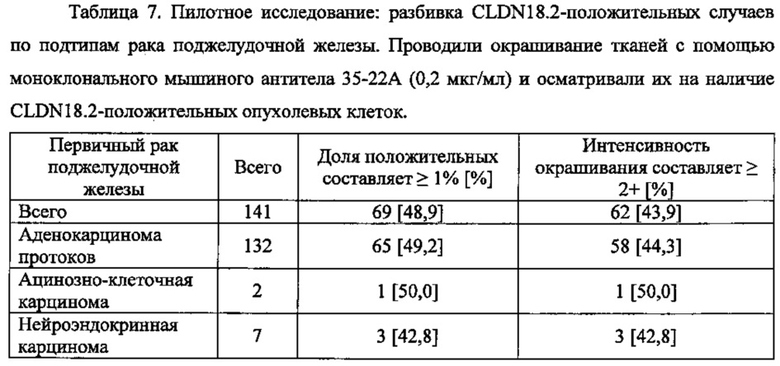
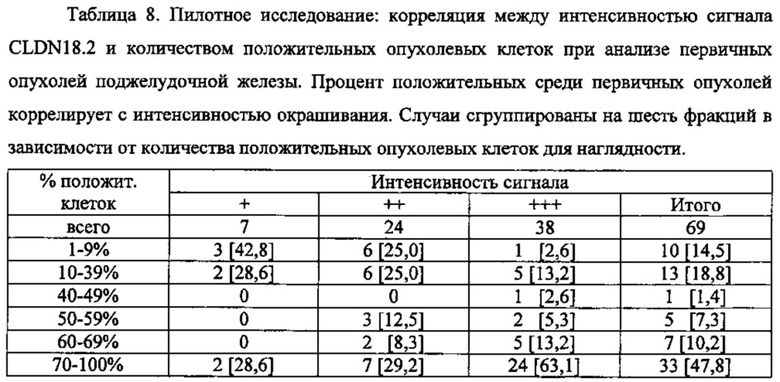

Второе исследование проводили по оптимизированной методике окрашивания с высокочувствительным антителом 43-14А, используя проверенные на качество срезы тканей.
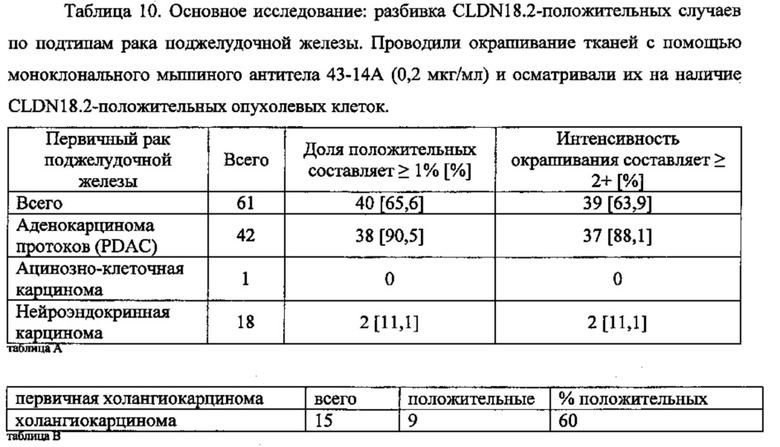
В общей сложности проанализировали 42 образца первичного рака протоков поджелудочной железы. Около 90% (38 из 42 случаев) из них были положительными на CLDN18.2 (таблица 10), причем большинство (>60%) проявляло большую интенсивность сигнала +++ (фиг. 7, таблица 11). При этом также наблюдалась корреляция между уровнем экспрессии CLDN18.2 и содержанием положительных опухолевых клеток. Большинство анализируемых случаев (62%) составляли опухоли 3-й степени (таблица 12).
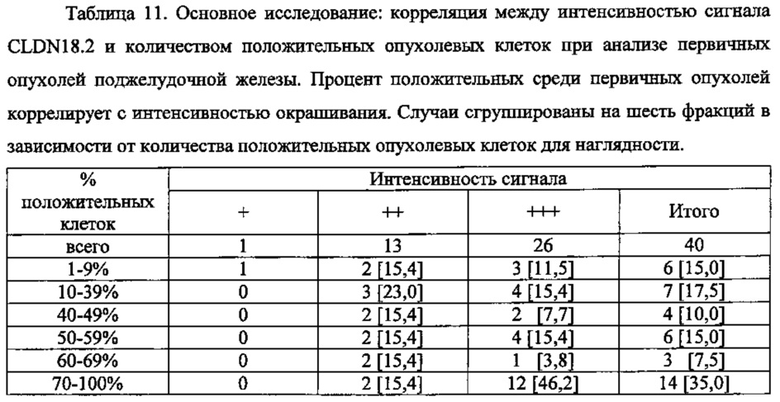
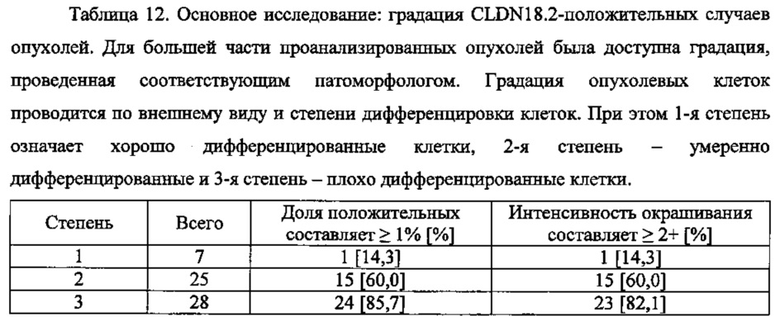
У большинства пациентов рак поджелудочной железы диагностируется на поздней стадии. Опухоли у пациентов уже дали метастазы в лимфатические узлы и другие органы, в частности, в печень. В основном исследовании подвергали анализу 79 FFPE-образцов тканей лимфатических узлов и печени с метастазами рака поджелудочной железы методом иммуногистохимии, используя специфичное к CLDN18.2 антитело 43-14А.
У 70,5% метастазов в лимфатических узлах (31/44 случаев) и 68,6% отдаленных метастазов в печени (24/35 случаев) проявлялось четкое окрашивание опухолевых клеток на CLDN18.2 (таблица 13). Профиль окрашивания у положительных опухолевых клеток был мембранным, в некоторых случаях с дополнительными слабыми цитоплазматическими сигналами (фиг. 9). В соответствии с результатами анализа первичных опухолей была обнаружена корреляция между уровнем экспрессии CLDN18.2 и содержанием CLDN18.2-положительных опухолевых клеток в образцах метастазов (фиг. 8).
Не обнаружено никакой корреляции между градацией анализируемых опухолей и уровнем экспрессии CLDN18.2 или содержанием положительных опухолевых клеток.
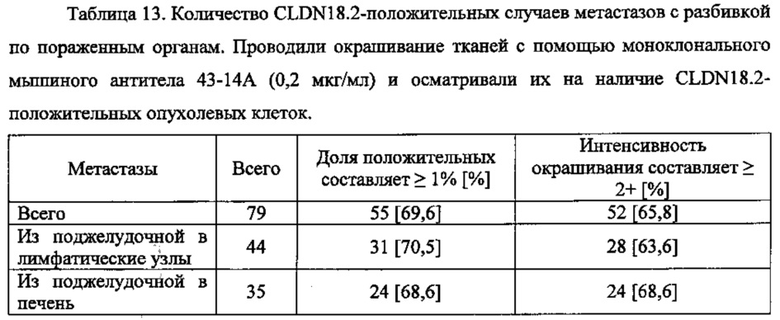
Чтобы проверить, сохраняется ли экспрессия CLDN18.2 в метастазах из положительных первичных опухолей у того же пациента, проводили скрининг парных образцов первичного рака и метастазов в лимфатических узлах с помощью антитела 43-14А.
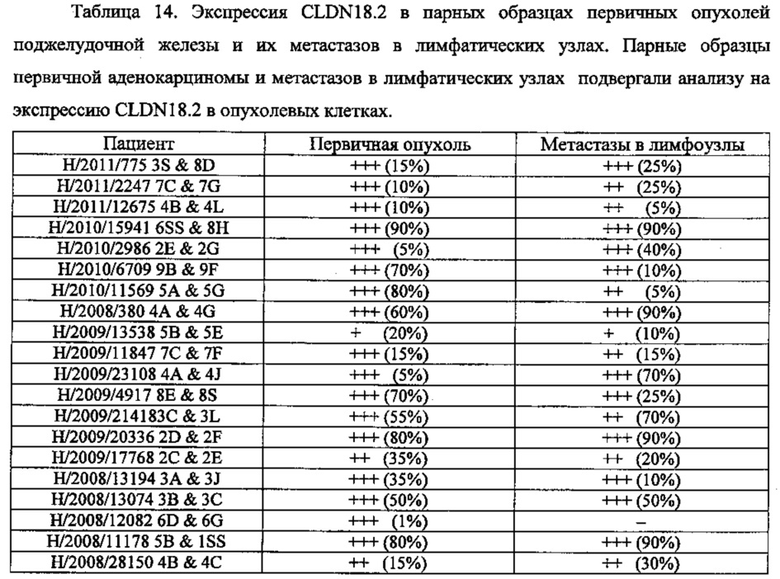

В 25 (92,5%) из 27 проанализированных случаев и первичная опухоль, и парные ей лимфатические узлы были положительными на CLDN18.2. В одном случае обе ткани были отрицательными, а в одном другом случае первичная опухоль была положительной на CLDN18.2, тогда как метастазы были отрицательными.
В 21 (80,7%) из 26 положительных парных образцов интенсивность сигнала у первичной опухоли и метастатических опухолевых клеток была одинаковой. В 5 случаях интенсивность сигнала снижалась с +++ до ++. В 11 (44%) из 25 парных образцов тканей число положительных опухолевых клеток в метастазах было меньше, чем у первичной опухоли (таблица 14).
Таким образом, экспрессия CLDN18.2 сохраняется при переходе клеток первичной опухоли в метастазирующую стадию. Общая интенсивность и доля положительных опухолевых клеток у метастазов в лимфатических узлах была лишь немного меньше, чем у первичной опухоли (фиг. 10).
У небольшого количества пациентов были доступны образцы тканей, полученные из первичной опухоли, метастазов в лимфатических узлах и метастазов в печени. Эти тройные пары подвергали окрашиванию для проверки сохранности экспрессии CLDN18.2 в отдаленных метастазах. Шесть таких тройных пар подвергали анализу с помощью антитела 43-14А.
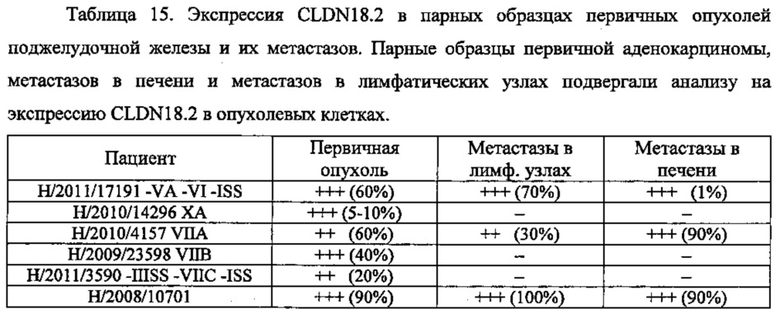
У 3 из 6 тройных пар все три образца ткани были сопоставимы в отношении их положительности на CLDN18.2 (фиг. 11). В трех случаях часть опухолевых клеток в первичной опухоли была положительной на CLDN18.2, тогда как метастатические очаги не проявляли окрашивания на CLDN18.2 (таблица 15).
Пример 3. Экспрессия мишени в клетках раковых линий поджелудочной железы человека, использовавшихся для моделей in vitro и in vivo и моделей рака поджелудочной железы
Источники клеточных линий
Главной целью данного доклинического исследования был анализ ингибирующих эффектов применения IMAB362 на подходящих модельных системах. Для идентификации CLDN18.2-положительных клеточных линий, которые могут использоваться для характеристики эффектов IMAB362 in vitro и in vivo, проводили скрининг комплекта из 26 коммерчески доступных раковых линий поджелудочной железы на экспрессию CLDN18.2 и их подробную характеристику. Для каждой линии клеток сразу по прибытии готовили банк клеток для экспериментального использования. Они происходили из первичных аденокарцином поджелудочной железы (10, из них 6 - слизистые аденокарциномы), первичных карцином (4), метастазов аденокарциномы поджелудочной железы в печени (5) или селезенке (1), или были выделены из асцитной жидкости (5) (см. таблицу 16). Некоторые из этих клеточных линий (8) подвергались лентивирусной трансдукции для экспрессии CLDN18.2.
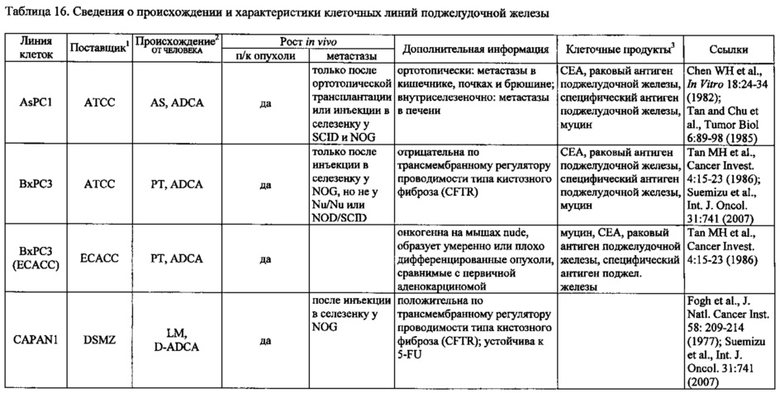
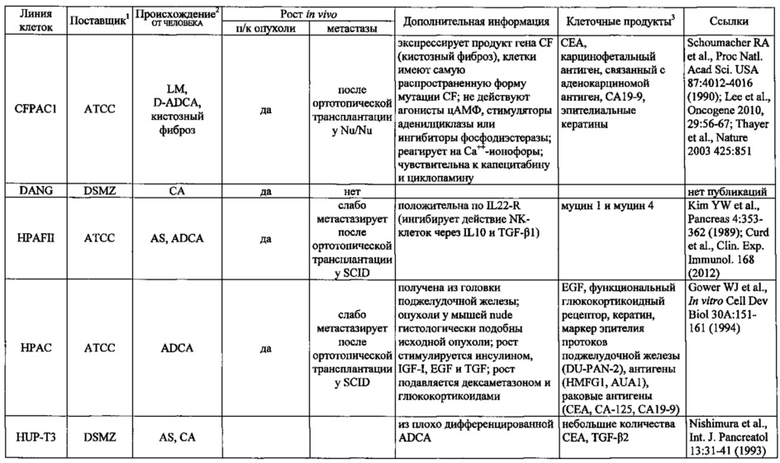

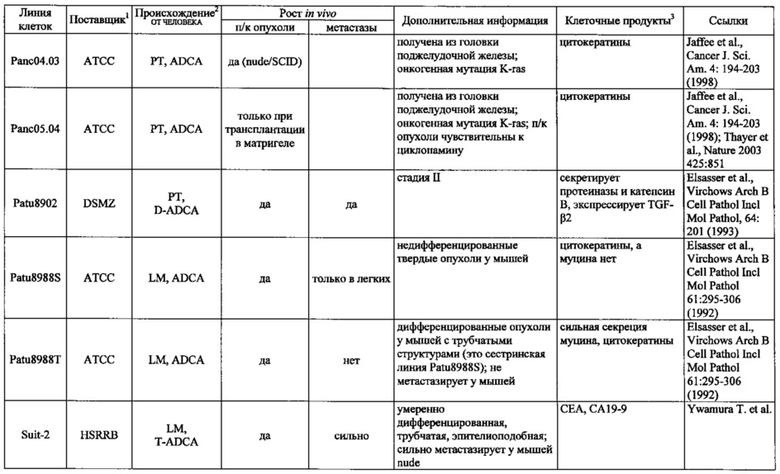
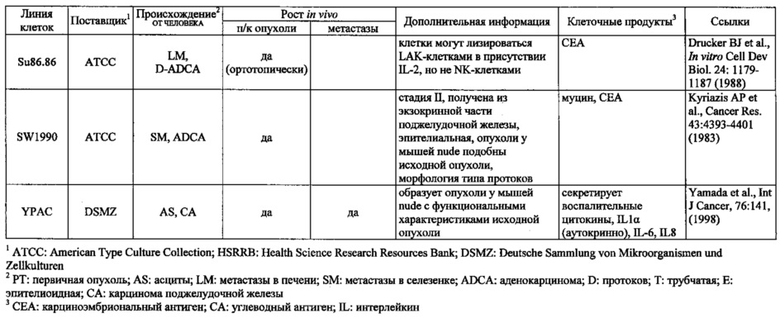
Экспрессия транскрипта CLDN18.2 в раковых линиях клеток поджелудочной железы человека
Для идентификации экспрессирующих CLDN18.2 линий клеток поджелудочной железы определяли уровни транскриптов методом количественной ПЦР в реальном времени (RT-PCR), используя прямой праймер, связывающийся с экзоном-1 CLDN18.2, и обратный праймер, связывающийся с экзоном-3 CLDN18. В качестве положительного и отрицательного контроля, соответственно, включали эндогенно экспрессирующую CLDN18.2 линию клеток карциномы желудка человека KATO-III и отрицательную по CLDN18.2 линию раковых клеток молочной железы SKBR-3. ПЦР в реальном времени показал четкую эндогенную экспрессию CLDN18.2 в раковых линиях клеток поджелудочной железы DANG, Panc03.27, Panc05.04, Patu8988S и YAPC, при этом относительный уровень превышал 1×105. Интересно, что клетки Patu8988S проявляли уровень экспрессии CLDN18.2 (~1×108), сравнимый с клетками СА желудка KATO-III (фиг. 12А). Таким образом, мы обнаружили сильную экспрессию CLDN18.2 у 5 из 22 линий раковых клеток поджелудочной железы.
Наряду с эндогенными линиями клеток, анализировали на уровне транскрипта LVT-линии клеток, эктопически экспрессирующие CLDN18.2 (фиг. 12А). У 6 из 8 LVT-линий клеток относительные уровни экспрессии CLDN18.2 составляли более 1×108. Только в клетках HAPC-LVT и Suit2-LVT уровень экспрессии был выше 1×105.
Далее исследовали, будет ли экспрессия CLDN18.2 стабильной при культивировании in vitro. Клетки Patu8988S, Panc05.04 и трансдуцированные лентивирусом линии клеток Suit2-LVT, MiaPaCa2-LVT и Patu8902-LVT пересеивали вплоть до 15 раз и проводили анализ на траскрипты CLDN18.2 (фиг. 12B-D). При большом числе пересевов как в эндогенных, так и в трансдуцированных клетках наблюдалась потеря экспрессии CLDN18.2. Потеря экспрессии была самой высокой в трансдуцированных клетках. Поэтому для экспериментов in vitro использовали ранние пересевы, где это возможно, а в приведенных ниже экспериментах по приживлению проверяли экспрессию CLDN18.2 в привитых опухолях.
Экспрессия белка CLDN18.2 в раковых линиях клеток поджелудочной железы человека
Выявление CLDN18.2 в лизатах клеток
В дополнение к анализу транскриптов, анализировали экспрессию CLDN18.2 на уровне белка методами вестерн-блоттинга и IF. При вестерн-блоттинге исследовали клеточные лизаты 26 линий клеток поджелудочной железы методом вестерн-блоттинга (WB) с использованием специфичного к CLDN18 антитела против клаудина 18 (С-конец). Опять же в качестве отрицательного контроля использовали лизаты клеток SKBR-3, а в качестве положительного контроля - лизаты клеток HEK293, стабильно трансфецированных CLDN18.2 (HEK293-р740). При этом отмечалась сильная экспрессия белка в клетках Patu8988S, DANG и Panc05.04, что подтверждает данные по РНК. В лизатах клеток Panc03.27 и ВхРС3 проявлялись слабые полосы. Клетки YAPC, которые были положительными на уровне РНК, проявляли слабую полосу меньшего размера на вестерн-блотах. Все остальные клеточные линии были отрицательными (фиг. 13).
Клеточная экспрессия CLDN18 в раковых клетках поджелудочной железы
Для получения данных, подтверждающих экспрессию белка, клеточные линии карциномы поджелудочной железы исследовали методом иммунофлуоресценции (IF) после фиксации и пермеабилизации клеток, используя для детектирования антитело 35-22А. Анализ методом LF подтвердил прежние данные по РНК и белку, показывающие, что большинство раковых линий клеток поджелудочной железы отрицательны по окрашиванию на CLDN18.2 (фиг. 14). В некоторых клеточных линиях (типа AsPC1, DANG, HUP-T3, T4-HUP, Panc01) наблюдались пятнышка в ядрах, которые, скорее всего, являются артефактами окрашивания. Клетки DANG, Panc03.27 и ВхРС3, которые проявляли низкие уровни CLDN18.2 на уровне РНК и/или белка, были отрицательными и при IF-анализе, который обладает меньшей чувствительностью. Напротив, у контрольных клеток карциномы желудка Panc05.04, Patu8988S и KATO-III мембраны и цитоплазма давали сильное положительное окрашивание на CLDN18.2. Интенсивность окрашивания была различной у разных клеток, а в популяции также отмечались отрицательные клетки (фиг. 14J и N). У LVT-линий клеток сильное окрашивание мембран отмечалось у более чем 80% клеток.
Подтверждение экспрессии CLDN18.2 в раковых клетках поджелудочной железы
Для подтверждения экспрессии CLDN18.2 и оценки количества этой мишени на клеточной поверхности, эндогенные линии клеток Panc05.04 и Patu8988S, а также LVT-линии клеток подвергали окрашиванию с помощью IMAB362 по нативной методике окрашивания. Хотя окрашивание IMAB362 контрольных раковых клеток желудка Patu8988S, Panc05.04 и KATO-III было менее интенсивным и процент положительных клеток снижался по сравнению с окрашиванием клеток 35-22А (фиг. 16A-F), анализ методом IF подтвердил, что CLDN18.2 экспрессируется на поверхности раковых клеток поджелудочной железы. У 8 LVT-линий раковых клеток поджелудочной железы, эктопически экспрессирующих CLDN18.2, наблюдалось четкое окрашивание мембран почти на всех клетках (как это видно у 6 LVT-линий клеток на фиг. 16G-L).
Таким образом, анализ экспрессии CLDN18.2 привел к идентификации эндогенно экспрессирующих раковых линий клеток поджелудочной железы Panc05.04 и Patu8988S и всех 8 трансдуцированных лентивирусом клеточных линий ВхРС3-LVT, CAPAN1-LVT, DANG-LVT, MiaPaCa-2-LVT, Suit2-LVT, Patu8902-LVT и YAPC-LVT как подходящих для моделирования систем CLDN18.2-положительных клеток.
Разработка моделей ксенотрансплантации и метастазирования рака поджелудочной железы
Исследования по приживлению для идентификации подходящих моделей подкожных опухолей при раке поджелудочной железы
В общей сложности проводили 37 исследований по приживлению с различными линиями раковых клеток поджелудочной железы для идентификации подходящих подкожных моделей ксенотрансплантации для тестирования эффективности IMAB362 in vivo. Из всех исследованных клеточных линий для подкожных моделей ксенотрансплантации были отобраны линии клеток ВхРС3-LVT, CAPAN1-LVT, MiaPaCa-2-LVT, HPAC-LVT, DANG-LVT и YAPC-LVT с эктопической экспрессией CLDN18.2, проявляющие высокую степень приживления и однородный рост опухолей. Кроме того, для тестирования эффективности IMAB362 in vivo были выбраны подкожные модели ксенотрансплантации с эндогенно экспрессирующими CLDN18.2 линиями клеток Patu8988S и DANG. Подкожное введение клеток Panc05.04 не приводило к образованию подкожных опухолей.
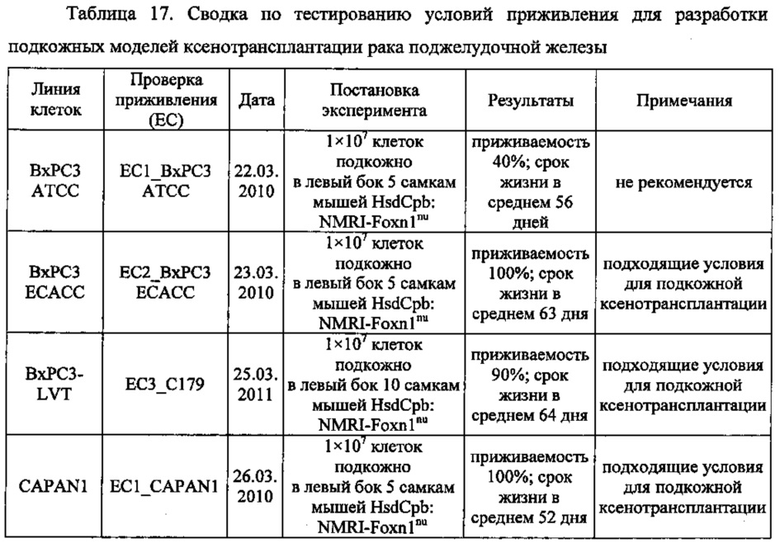
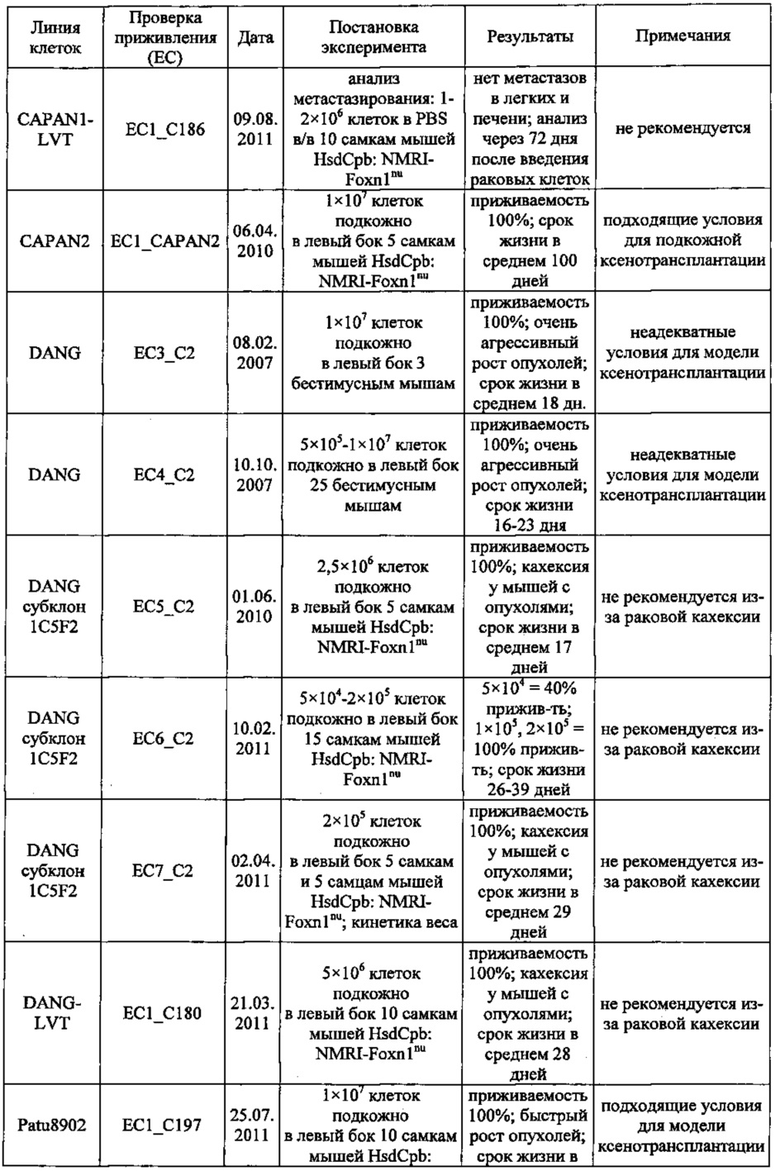
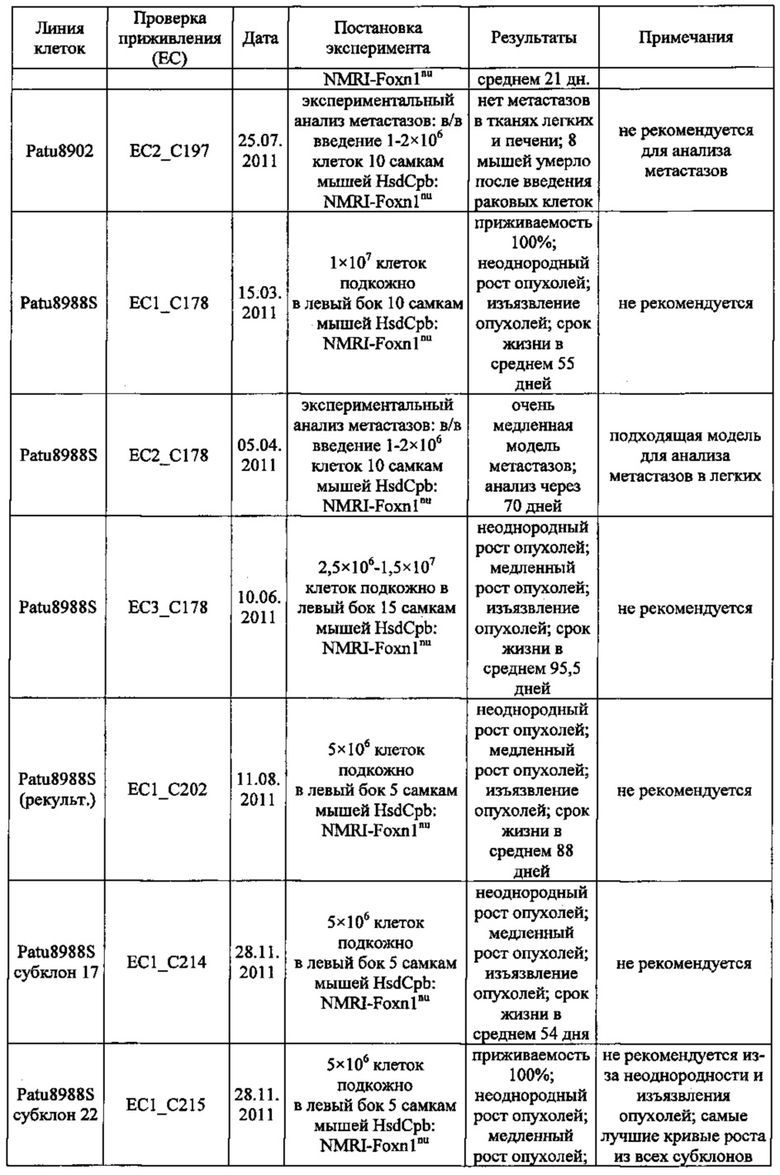

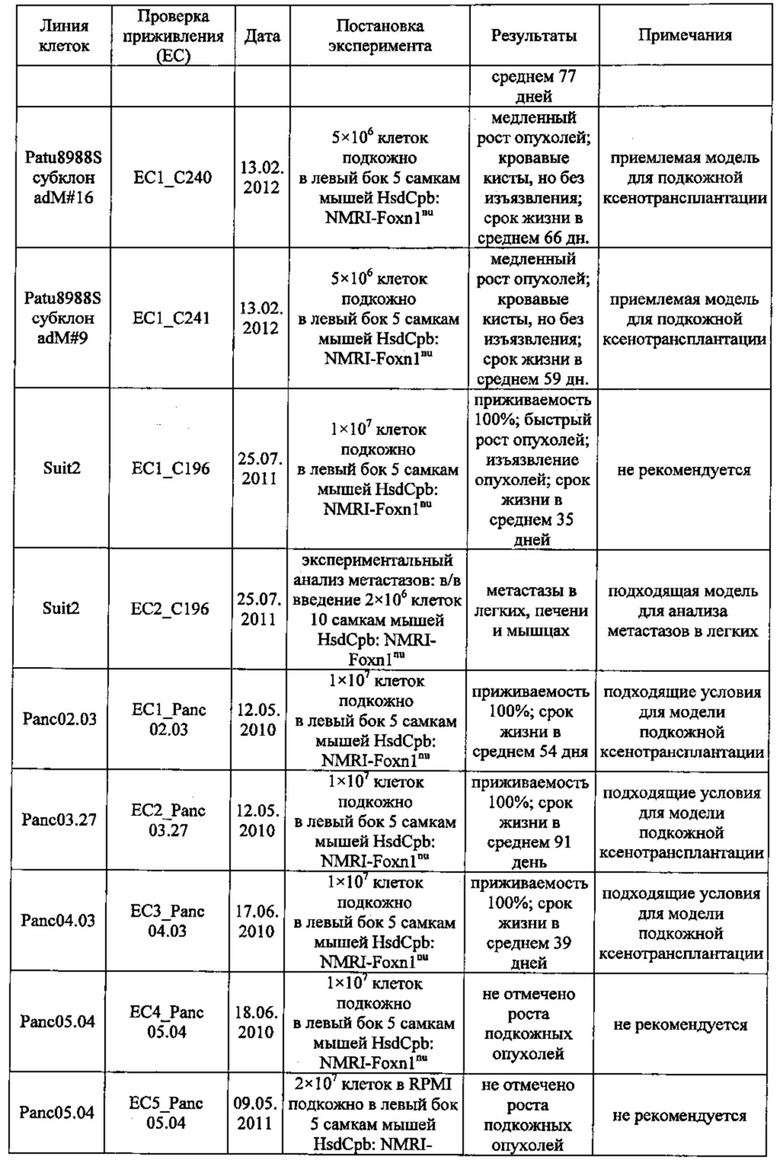
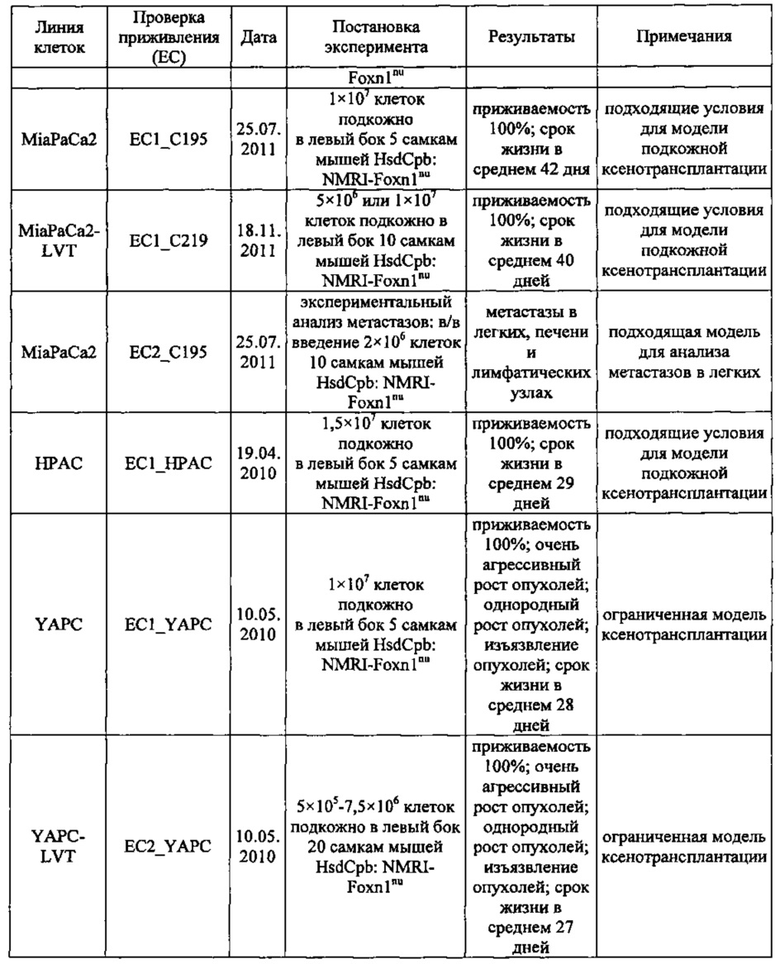
Исследования по приживлению для идентификации подходящих моделей метастазирования
Для изучения эффектов IMAB362 на образование метастазов были поставлены модели метастазирования рака на мышах nude. Клетки раковых линий поджелудочной железы подвергали анализу на их способность к метастазированию после в/в введения. Клетки CAPAN1-LVT, MiaPaCa-2, Patu8988S, Patu8902 и Suit-2 вводили в хвостовую вену мышей nude, как описано в Mohanty and Xu, 2010. Для определения момента времени приживления и скорости роста метастазов мышей забивали в различные моменты времени (таблица 18).
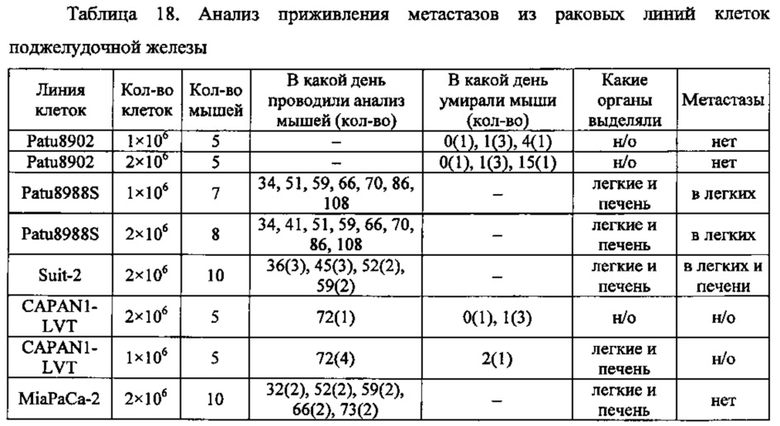
Анализ приживления клеток Patu8902 и CAPAN1-LVT был невозможен, так как большинство мышей погибало почти сразу. В легких и печени 5 выживших мышей, получавших клетки CAPAN-LVT, через 72 дня не было обнаружено макроскопически видимых метастазов. Напротив, введение клеток Suit-2 и MiaPaCa-2 хорошо переносилось. Ткани легких этих мышей подвергали анализу методом IHC в различные моменты времени после инъекции. У мышей, получавших клетки MiaPaCa-2, не отмечалось метастазов в легких и через 73 дня, поэтому эта линия клеток не была выбрана в качестве модели для обработки IMAB362. Раковые клетки Suit-2 давали метастазы в легкие мышей. Отмечались множественные очаги по всей ткани. Именно поэтому трансдуцированная лентивирусным CLDN18.2 линия клеток Suit-2-LVT была выбран в качестве модельной системы для анализа эффектов применения IMAB362 на образование метастазов.
Наряду с Suit-2, также анализировали способность к образованию метастазов у клеток Patu8988S, эндогенно экспрессирующих CLDN18.2. Проверку на приживление проводили при внутривенном введении мышам 2 различных количеств клеток (1×106, 2×106). В различные моменты времени выделяли легкие и печень, как указано в таблице 18. Сначала различные полученные ткани подвергали анализу методом к-ПЦР. Легкие и печень, полученные до 70-го дня, анализировали путем амплификации α-сателлитной ДНК из хромосомы 17 человека. Результаты на легких показали четкое возрастание содержания ДНК человека в легких мыши по времени, которое не зависело от количества введенных клеток. При внутривенном введении 1×106 или 2×106 клеток через 70 дней отмечалось 5,8% и 3,7% ДНК человека, соответственно (фиг. 19). В печени ДНК человека почти не амлифицировалась. Через 70 дней ее содержание слегка повышалось, но все еще было менее 0,005%.
Для проверки экспрессии CLDN18.2 в метастазах Patu8988S подвергали ткани легких иммуногистохимическому окрашиванию с помощью антител против МНС класса I человека для выявления клеток человека в тканях мыши, а также антитела против клаудина 18 (Mid). Окрашивание на MHC-I показало, что у мышей отмечаются четкие очаги метастазов на срезах ткани легких, но не на срезах печени (фиг. 20). Более того, мембраны клеток в этих очагах окрашивались антителом против клаудина 18 (Mid), что четко свидетельствует об экспрессии белка мишени IMAB362 в этих клетках. Поэтому эта модель эндогенных метастазов была выбрана для исследования применения IMAB362 наряду с моделью Suit2-LVT.
Пример 4. Опосредованные IMAB362 эффекты уничтожения клеток
Перекрестное сшивание IMAB362 вызывает эффективный апоптоз
Связывание антител с клеточной поверхностью мишени может запускать аномальную передачу сигналов, приводящую непосредственно к гибели клеток. Такие сигнальные события могут зависеть от эпитопа мишени, валентности связывания и того, ассоциировано ли связывание с перекрестным сшиванием мишени. Например, индукция апоптоза ритуксимабом у нескольких CD20-положительных клеточных линий лимфомы наблюдается только при условии перекрестного сшивания. Такое сшивание может происходить in vivo при высокоаффинном взаимодействии положительных по Fc-рецепторам иммунных клеток с покрытыми антителом опухолевыми клетками.
Перекрестное сшивание IMAB362 вызывает апоптоз прямо в пределах 18-42 часов в раковых клетках желудка человека NUGC-4 и KATO-III при определении методом TUNEL. Степень апоптоза коррелирует с дозой антитела и уровнем экспрессии мишени в раковых клетках. Обработка гемцитабином приводит к остановке клеточного цикла раковых клеток с последующим апоптозом клеток. Апоптоз обработанных гемцитабином опухолевых клеток поджелудочной железы представлен на фиг. 21.
Опосредованная IMAB362 активность ADCC против раковых клеток поджелудочной железы
IMAB362 очень действенно вызывает рекрутинг и активацию положительных по Fcγ-рецептору иммунных эффекторных клеток, таких как клетки нормальных киллеров. Связывание IMAB362 с клетками мишени вызывает антителозависимую клеточную цитотоксичность (ADCC) под действием гранзимов и перфоринов, секретируемых эффекторными клетками при связывании их Fcγ-рецепторов с антителом. Ранее было показано влияние этого механизма действия на положительные по люциферазе и CLDN18.2 клетки СА желудка (типа NUGC-4 и KATO-III) при инкубации с IMAB362 в течение 24 часов в присутствии мононуклеаров периферической крови человека (РВМС) (соотношение эффектор : мишень = 40:1). Применение до 200 мкг/мл IMAB362 давало максимальную степень лизиса в 80-100%.
Здесь мы определяли ADCC-активность IMAB362 против раковых линий клеток поджелудочной железы. IMAB362 в возрастающих концентрациях инкубировали с различными линиями клеток при соотношении Е : Т = 40:1. В каждом эксперименте добавляли PBMCs от различных доноров. Результаты для всех клеточных линий приведены в таблице 19. Из 5 первоначально выявленных CLDN18.2-положительных линий клеток поджелудочной железы только Patu8988S, Panc05.04 и DANG эффективно погибали при добавлении IMAB362 и PBMCs (фиг. 22А). Хотя поверхностная экспрессия CLDN18.2 у Panc05.04 и DANG не выявлялась методом FACS, уровень экспрессии был достаточно значительным, чтобы вызвать зависимое от эффекторных клеток уничтожение (ЕС50: Patu8988S: 0,01-1,4 мкг/мл, DANG/Pan05.04: 0,1-38 мкг/мл). Из этих данных можно сделать вывод, что эффективно лизируются только клетки, экспрессирующие РНК на относительном уровне >5,5×105.
Анализ ADCC также проводили с LVT-линиями раковых клеток поджелудочной железы и соответствующими исходными линиями клеток (фиг. 22B-F). ADCC строго зависит от специфического связывания IMAB362 с мишенью, так как только CLDN18.2-положительные клетки мишени уничтожаются IMAB362 и PBMCs. Полумаксимальное уничтожение и максимальная степень уничтожения раковых клеток поджелудочной железы человека под действием IMAB362 варьировалась между донорами РВМС, а также зависела от количества пересевов клеток, влияющего на уровень экспрессии CLDN18.2.
Концентрации IMAB362, вызывающие полумаксимальное уничтожение клеток мишени, а также максимальную степень уничтожения представлены на фиг. 22G-H. LVT-линии клеток СА поджелудочной железы погибали при добавлении небольших количеств антитела в большой степени, тогда как для DANG и Panc05 требуются самые высокие концентрации антитела для достижения степени уничтожения ~50% от максимальной. На фигуре также представлены результаты для Panc05.04, полученные с субклоном 15D3 (CLDN18.2-положительный клон, отобранный методом серийных разведений Panc05.04 и FACS), который проявлял сравнимую с LVT-линиями клеток степень вызванного ADCC лизиса. К сожалению, экспрессия CLDN18.2 у этого клона быстро затихала in vitro после пересева клеток, поэтому этот клон не использовался для дальнейших экспериментов.
Опосредованная IMAB362 активность CDC против раковых клеток поджелудочной железы
Раковые клетки поджелудочной железы, которые уничтожались IMAB362 при анализе ADCC, подвергали анализу на их чувствительность к комплементзависимой литической активности IMAB362. Кроме того, тестировали на CDC и LVT-линии клеток и их исходные штаммы.
Активность CDC активируется комплексами антигена с антителами типа IgM или IgG (классический путь) или поверхностями микроорганизмов (альтернативный путь). В классическом пути комплемент С5 превращается в C5b. При последовательном связывании C5b, С7, С8 и С9 высвобождаются анафилотоксины С3а, С4а и C5b и образуется мембраноатакующий комплекс (MAC). Этот путь подавляется как растворимыми, так и мембраносвязанными белками (например, CR1, DAF, МСР, CD59, CD55, CD46), защищающими свои ткани.
В качестве положительных контролей при каждом анализе использовали клетки СНО-K1, стабильно трансфецированные CLDN18.2 (р740) и люциферазой (фиг. 23А). Линии клеток DANG, ВхРС3, YAPC, Patu8988S, Panc05.04, CAPAN1 и Suit-2 не подвергались лизису при добавлении IMAB362 и сборной сыворотки здорового человека (фиг. 23В). Хотя клетки DANG, Patu8988S и Panc05.04 являются CLDN18.2-положительными, как было показано во всех предыдущих экспериментах, однако они не лизируются комплементзависимым образом. Скорее всего, это связано с тем, что раковые клетки гиперэкспрессируют один или несколько белков, ингибирующих мембраносвязанный комплемент (например, CD46, CD55 и CD59) (Geis et al., Curr Cancer Drag Targets 2010, 10: 922-931). Однако все еще остается неясным, влияет ли экспрессия этих ингибиторных белков в опухолевых клетках на клинический исход терапии антителами (Dzietczenia et al., Med. Oncol. 2010, 27: 743-6; Weng and Levy et al., Blood 2001, 98: 1352-7).
Наряду с эндогенными линиями клеток, тестировали на CDC и все LVT-линии клеток. Как видно из фиг. 23, добавление IMAB362 и сыворотки к клеткам MiaPaCa-2-LVT, Suit2-LVT и CAPAN1-LVT вызывало дозозависимый лизис со значениями ЕС50 от 0,3 до 2,6 мкг/мл.
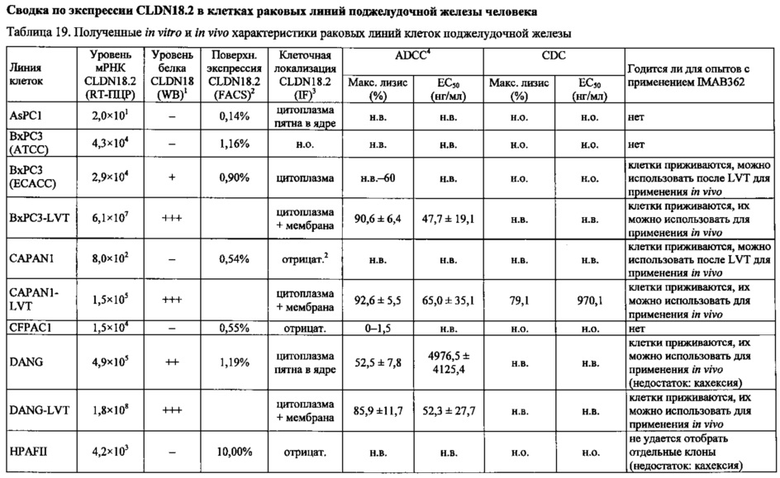
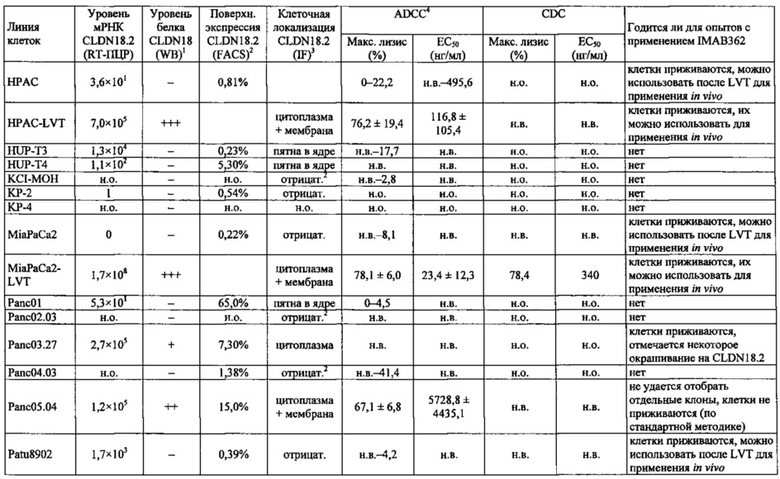
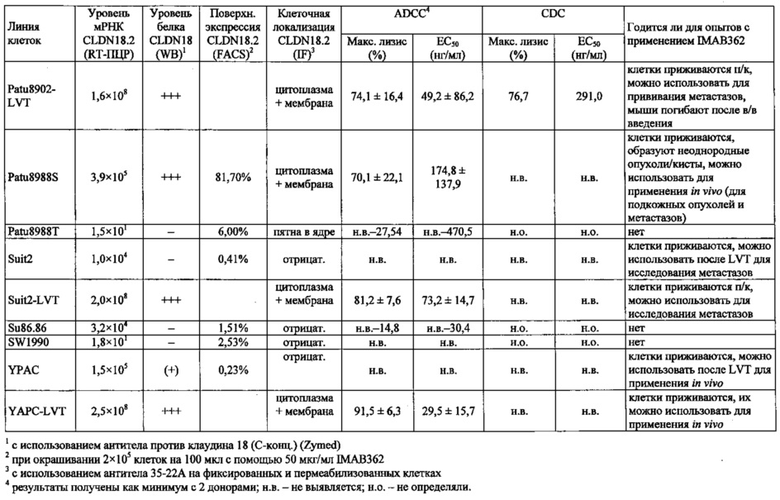
Пример 5. Эффективность IMAB362 на моделях ксенотрансплантации рака поджелудочной железы
Для исследования эффективности IMAB362 in vivo было выбрано 10 из 41 проверенной модели ксенотрансплантации рака поджелудочной железы. На моделях ксенотрансплантатов поджелудочной железы с высокой экспрессией CLDN18.2, обработка IMAB362 показала высокий противоопухолевый эффект. Его исследовали путем обработки мышей, несущих в левом боку подкожные ксенотрансплантаты ВхРС3-LVT или MiaPaCa-2-LVT. Обработку начинали через 3 дня после прививания опухолей введением 200 мкг IMAB362 по 2 раза в неделю. Получавшие IMAB362 мыши проявляли существенное ингибирование роста опухолей по сравнению с мышами, получавшими контрольный солевой раствор. Кроме того, подавление роста опухолей у получавших IMAB362 мышей приводило к удлинению среднего срока жизни (фиг. 24 и фиг. 25). Эффективность IMAB362 коррелирует с продолжительностью обработки. Если обработка IMAB362 начинается в ранних временных точках, то ингибирование роста опухолей повышается больше, чем при позднем начале обработки для исследования действия на прижившиеся опухоли. Более того, противоопухолевый эффект IMAB362 зависел от степени экспрессии CLDN18.2 в мишени. У опухолей с низкой экспрессией CLDN18.2 типа ксенотрансплантатов DANG и Patu8988S опосредованное IMAB362 ингибирование роста снижалось по сравнению с ингибированием роста опухолей при ксенотрансплантации опухолей с высокой экспрессией CLDN18.2.
Пример 6. Обработка на моделях метастазов рака поджелудочной железы у мышей


Модель метастазов Suit2-LVT
Мышам вводили внутривенно 2×106 клеток Suit2-LVT и обрабатывали 200 мкг IMAB362, антитела из изотопического контроля (IMAB027) или PBS, как указано в таблице 20. Через 35 дней умерла первая мышь (группа изотопического контроля). После этого всех мышей забивали на 42-й день и извлекали легкие и печень для анализа методами IHC и количественной ПЦР.
Анализ ДНК человека в легких мышей методом количественной ПЦР повторяли по меньшей мере дважды в трех повторах. Расчет содержания ДНК человека по полученным значениям Ct показал значительное снижение (р<0,05) метастазов Suit2-LVT, обнаруженных в легких, при обработке мышей IMAB362 (фиг. 26А) по сравнению с обработкой PBS и изотопическим контролем. Для проверки этих результатов готовили срезы из образцов ткани легких и проводили окрашивание с помощью антител к MHC-I. Рассчитывали площадь поверхности положительно окрашенных клеток на срезах легких с помощью программы ImageJ. При обработке IMAB362 наблюдалось значительное ингибированию (р<0,05) по сравнению с обработкой PBS, подтверждая результаты, полученные методом к-ПЦР. Однако в случае антитела из изотопического контроля отличия не были значимы (фиг. 26В). Это расхождение, скорее всего, вызвано различиями в обработке тканей: обработка срезов тканей для IHC обеспечивает доступ только в очень небольшой участок легких по сравнению с методом к-ПЦР, для которого экстрагируется геномная ДНК из половины ткани.
Наряду с обработкой тканей, возможно, что этот результат представляет неожиданные ингибирующие эффекты антитела из изотопического контроля против CLDN6. Для исследования этой возможности клетки Suit2-LVT подвергали анализу на экспрессию CLDN6 и связывание IMAB027 методом FACS. При добавлении 200 мкг/мл IMAB362 к клеткам Suit2-LVT проявлялось сильное связывание с клетками, тогда как добавление 200 мкг/мл IMAB027 давало слабое связывание антитела с этими клетками мишени, указывая на то, что CLDN6 действительно слабо экспрессируется в этих клетках. Эти результаты свидетельствуют, что по меньшей мере два фактора (обработка ткани и слабое ингибирование IMAB027) вызывали наблюдавшиеся расхождения с антителом из изотопического контроля.
Модель метастазов Patu8988S
Для анализа эффекта обработки IMAB362 на развитие и рост метастазов Patu8988S in vivo группам по 10 мышей вводили 2×106 клеток Patu8988S. Первый эксперимент проводили путем сравнения обработки IMAB362 с обработкой мышей PBS. В каждой группе 1 мышь умирала сразу же после инъекции клеток. У других 18 мышей метастазы возникали очень быстро по сравнению с экспериментами по приживлению. Через 63 дня забивали первых 2 мышей в группе PBS из-за плохого состояния здоровья. Всех других мышей забивали через 65 дней. Оптический анализ легких показал крупные метастазы по всей ткани легких. Степень метастазирования анализировали методом количественной ПЦР (фиг. 27). Результаты показывают, что IMAB362 ингибирует рост метастазов в ткани легких.
Второй эксперимент по 11 мышей в каждой группе проводили путем сравнения обработки IMAB362 с обработкой изотопическим контролем (ритуксимаб). В этом эксперименте метастазы развивались медленно, как это наблюдалось при приживлении. Тем не менее, для сопоставимости данных этот второй эксперимент тоже прекращали через 65 дней. Опять проводили анализ тканей легких методом количественной ПЦР и опять IMAB362 снижал рост метастазов. Одна мышь из группы IMAB362 была установлена как выброс, и исключение этого выброса давало почта значимое (р=0,0588) ингибирование. Эта данные проверяли по площади поверхности методом IHC, как описано для эксперимента с метастазами Suit2-LVT. При этом проявлялся тот же самый выброс, и при пропуске этого значения при проверке по t-критерию ингибирование IMAB362 также оказалось на грани значимости (р=0,0691), как и значения этой мыши.
Пример 7. Первичная фармакодинамика IMAB362 в сочетании с химиотерапией
Чувствительность клеток карциномы поджелудочной железы к гемцитабину и оксалиплатину
Для изучения механизмов действия IMAB362 в сочетании с химиотерапевтическими средствами оксалиплатином или гемцитабином использовали клетки раковых линий поджелудочной железы, конституитавно экспресирующие CLDN18.2 (DANG, Patu8988S), и клетки, стабильно трансдуцированные CLDN18.2 (MiaPaCa-2-LVT, ВхРС3-LVT).
В химическом отношении гемцитабин (Gemzar, выпускается фирмой Eli Lilly & Со) является аналогом нуклеозидов. Как и 5-фторурацил (5-FU) и другие аналоги пиримидинов, трифосфатный аналог гемцитабина заменяет собой один из строительных блоков нуклеиновых кислот при репликации ДНК. Этот процесс останавливает рост опухолей, так как к "неправильному" нуклеозиду может присоединяться только еще один нуклеозид, что приводит к апоптозу.
Оксалиплатин функционирует путем образования межцепочечных и внутрицепочечных перекрестных сшивок в ДНК. Перекрестные сшивки в ДНК предотвращают репликацию и транскрипцию ДНК, что приводит к гибели клеток (Graham J., Mushin М., Kirkpatrick Р. (2004) "Oxaliplatin" Nature Reviews Drug Discovery 3(1): 11-2).
Кривые доза-ответ у гемцитабина и оксалиплатина свидетельствуют о различной чувствительности исследуемых раковых линий клеток поджелудочной железы (фиг. 28 и фиг. 29).
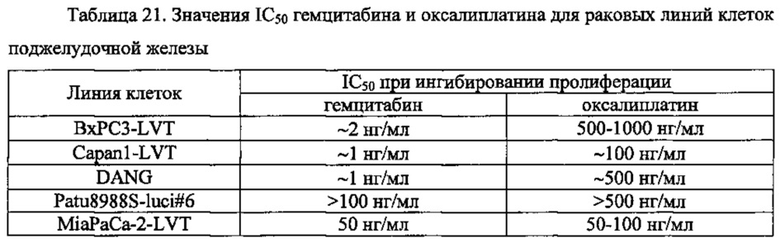
Для ингибирования пролиферации клеток Patu8988S необходимы высокие концентрации гемцитабина (IC50>100 нг/мл) или оксалиплатина (IC50>500 нг/мл). Клетки DANG и ВхРС3-LVT очень чувствительны к гемцитабину, но не к оксалиплатину. Клетки MiaPaCa-2-LVT наиболее чувствительны к оксалиплатину, но менее чувствительны к обработке гемцитабином (фиг. 28, фиг. 29 и таблица 21).
Влияние химиотерапевтических средств на экспрессию CLDN18.2 в клеточных линиях карциномы поджелудочной железы
Способ действия, запускаемого связыванием IMAB362, строго зависит от наличия и плотности на поверхности клеток его мишени - CLDN18.2. Предварительная обработка клеток DANG и Patu8988S гемцитабином (GEM), а также гемцитабином в комбинации с оксалиплатином (GemOx), приводила к повышению уровней мРНК и белка CLDN18.2 по данным анализа методом ОТ-ПЦР (фиг. 30) и вестерн-блоттинга (фиг. 31) клеток без обработки и клеток с предварительной химиотерапией. Следовательно, количество белка CLDN18.2, доступного для IMAB362 на поверхности предварительно обработанных Gem или GemOx клеток раковых линий поджелудочной железы, возрастало, как показала проточная цитометрия (фиг. 32).
Обработка DANG и Patu8988S гемцитабином приводит к повышению уровня CLDN18.2. Patu8988S проявляет сильную повышающую регуляцию CLDN18.2 при воздействии Gem и меньшую при воздействии GemOx.
Влияние химиотерапевтических препаратов на клеточный цикл и экспрессию CLDN18.2
Прохождение клеточного цикла относится к последовательности событий от одного митотического деления к другому в клетке. За фазой покоя (G0/G1) следует фаза синтеза ДНК (S), а затем фаза роста клеток (G2) и репликации ДНК (М) с последующим делением клетки на две дочерние клетки. Всякое вмешательство в клеточную машину может препятствовать прохождению всего цикла в любой фазе клеточного цикла. Например, определенные химиотерапевтические средства могут блокировать его в фазе G2 или М либо одновременно в G2 и М (G2/M).
Обработка гемцитабином клеток DANG или Patu8988S приводит к остановке клеточного цикла в фазе S (фиг. 33, фиг. 34). Проводили анализ Patu8988S при культивировании с Gem. Обработка гемцитабином не только приводит к остановке клеточного цикла, но также изменяет экспрессию CLDN18.2 (фиг. 34В). Изменение плотности CLDN18.2 после обработки гемцитабином было еще выше, если сравнивать пролиферирующие клетки в фазе S и покоящиеся клетки в фазе G0/G1 (фиг. 34С). В клетках Patu8988S CLDN18.2 экспрессируется на всех фазах клеточного цикла. После обработки гемцитабином его экспрессия еще повышается, а самые высокие уровни CLDN18.2 в клетках отмечаются в популяции клеток фазы S.
Такое искажение фенотипа опухолевых клеток оказывает значительное влияние на биологическую эффективность терапевтических антител. ADCC и CDC зависят от дозы, поэтому возрастание уровня мишени - CLDN18.2 обеспечивает синергическую пользу при стандартных режимах химиотерапии.
Клетки KATO-III, линии раковых клеток желудка человека, культивировали в среде RPMI 1640 (Invitrogen), содержащей 20% FCS (Perbio) и 2 мМ Glutamax (Invitrogen) при 37°С и 5% СО2, с добавлением или без цитостатических соединений. 5-FU (Neofluor фирмы NeoCorp AG) тестировали в концентрации 10 или 100 нг/мл, а оксалиплатин (Hospira) тестировали в концентрации 50 или 500 нг/мл. Культивировали 8×105 клеток KATO-III в течение 96 часов без замены среды либо 72 часов с последующим культивированием еще 24 часа в стандартной среде, чтобы снять остановку клеточного цикла, в 6-луночном планшете для культивирования тканей при 37°С, 5% CO2. Клетки собирали с помощью трипсина/EDTA, промывали и анализировали.
Для внеклеточного выявления CLDN18.2 проводили окрашивание клеток с помощью моноклонального антитела IMAB362 против CLDN18.2 (Ganymed) или контрольного антитела того же изотипа (Ganymed). В качестве вторичного реагента использовали козье антитело против huIgG-APC фирмы Dianova.
Стадии клеточного цикла определяли по измерению содержания ДНК в клетках. Это позволяет различать клетки в фазе G1, S или G2 клеточного цикла. В фазе S происходит удвоение ДНК, тогда как в фазе G2 клетки растут и готовятся к митозу. Анализ клеточного цикла проводили с помощью набора CycleTest Plus DNA Reagent Kit фирмы BD Biosciences по методике производителя. Получение и анализ данных проточной цитометрии проводили на установке BD FACS CantoII (BD Biosciences) с помощью программы FlowJo (Tree Star).
На фиг. 35а и b столбики показывают процент клеток в фазе G1, S или G2 клеточного цикла, соответственно. При культивировании клеток KATO-III в среде происходит остановка клеточного цикла, преимущественно в фазе G1. При обработке клеток 5-FU они блокируются преимущественно в фазе S. При обработке клеток KATO-III оксалиплатином происходит обогащение клетками преимущественно в фазах G1 и G2. Как видно из фиг. 35с, остановка клеточного цикла в фазе S или фазе G2 приводит к стабилизации или повышению уровня CLDN18.2. Как только клетки выходят из какой-либо фазы клеточного цикла (фиг. 35b), экспрессия CLDN18.2 на поверхности клеток KATO-III усиливается (фиг. 35d).
Клетки KATO-III подвергали предварительной обработке иринотеканом или доцетакселем в течение 4 дней и проводили анализ на экспрессию CLDN18.2 и остановку клеточного цикла. Обработка клеток иринотеканом приводила к дозозависимому ингибированию роста клеток и остановке клеточного цикла в фазе S/G2 (фиг. 36). Обработка клеток доцетакселем приводила к дозозависимому ингибированию роста клеток и остановке клеточного цикла в фазе G2 (фиг. 36).
Влияние химиотерапии на индуцированную IMAB362 антителозависимую клеточную питотоксичность (ADCC)
Проводили ряд экспериментов с конститутивно экспрессирующими CLDN18.2 клетками раковых линий поджелудочной железы Patu8988S и DANG для исследования эффектов гемцитабина (Gem) или гемцитабина + оксалиплатина (GemOx) на IMAB362-опосредованную ADCC. Сравнивали кривые доза-ответ для опосредованного IMAB362 лизиса клеток при предварительной обработке клеток и при культивировании в среде.
Кривые доза-ответ у предварительно обработанных Gem (1 нг/мл) или GemOx (Gem 1 нг/мл + Ох 10 нг/мл) клеток DANG (2 дня) смещались вверх и влево по сравнению с необработанными клетками мишени (фиг. 37А). Обработка раковых клеток Gem или GemOx приводила к повышению уровня CLDN18.2 и большей подверженности IMAB362-опосредованной ADCC. После обработки химиотерапевтическими средствами у клеток DANG наблюдалось снижение значений ЕС50 и повышение максимального лизиса клеток при IMAB362-опосредованной ADCC (фиг. 37В).
Очищали мононуклеарные клетки периферической крови (РВМС), включая NK-клетки, моноциты, мононуклеары или другие эффекторные клетки от здоровых доноров, центрифугированием в градиенте плотности Ficoll Hypaque. Промытые эффекторные клетки высеивали в среду Х-Vivo. При этом в качестве клеток мишени использовали клетки KATO-III, эндогенно экспрессирующие CLDN18.2 и происходящие из желудка. Клетки мишени стабильно экспрессировали люциферазу, a Lucifer желтый окисляется только жизнеспособными клетками. Добавляли очищенное антитело IMAB362 против CLDN18.2 в различных концентрациях, а в качестве антитела для изотопического контроля использовали постороннее химерное антитело против huIgG1. Образцы анализировали на цитолиз по измерению люминесценции в результате окисления Lucifer желтого, которая является мерой количества жизнеспособных клеток, оставшихся после индуцированной IMAB362 цитотоксичности. KATO-III, предварительно обработанные в течение 3 дней иринотеканом (1000 нг/мл), доцетакселем (5 нг/мл) или цисплатином (2000 нг/мл), сравнивали с необработанными клетками мишени, культивировавшимися в среде, и проводили количественное определение IMAB362-индуцированной ADCC.
При предварительной обработке в течение 3 дней иринотеканом, доцетакселем или цисплатином клетки KATO-III проявляли снижение уровня жизнеспособных клеток по сравнению с клетками мишени, культивировавшимися в среде (фиг. 38а), а экспрессия клаудина 18.2 в клетках при предварительной обработке иринотеканом, доцетакселем или цисплатином повышалась по сравнению с клетками, культивировавшимися в среде (фиг. 38b).
Кроме того, предварительная обработка клеток KATO-III иринотеканом, доцетакселем или цисплатином усиливала способность IMAB362 индуцировать ADCC (фиг. 38с, d).
Влияние химиотерапии на IMAB362-индуцированную CDC
Действие IMAB362 на CDC исследовали путем инкубации с клетками мишени в присутствии сыворотки человека в качестве источника комплемента.
Культивировавшиеся в среде клетки MiaPaCa-2-LVT проявляли значения ЕС50 для IMAB362-специфичного лизиса в 7665 нг/мл. Обработка Gem приводила к снижению ЕС50 до 4677 нг/мл наряду с повышением максимальной степени лизиса (фиг. 39).
Действие химиотерапевтических средств на IMAB362-индуцированную CDC анализировали при предварительной обработке раковых клеток желудка KATO-III с 10 нг/мл 5-FU и 500 нг/мл оксалиплатина (5-FU + ОХ) в течение 48 часов. Репрезентативные кривые доза-ответ по IMAB362-индуцированную CDC при предварительной химиотерапевтической обработке клеток KATO-III представлены на фиг. 40. Предварительная обработка раковых клеток в течение 48 часов усиливала способность IMAB362 индуцировать ADCC, что приводило к повышению максимальной степени лизиса у предварительно обработанных раковых клеток по сравнению с необработанными клетками.
Пример 8. Эффективность IMAB362 в сочетании с химиотерапией на моделях опухолей у мышей
Исследовали противоопухолевую активность IMAB362 в сочетании с Gem или GemOx на подкожных моделях ксенотрансплантации карциномы поджелудочной железы, которые ранее использовались для тестирования эффективности IMAB362 в качестве единственного средства.
У обработанных IMAB362 мышей nude с опухолями ВхРС3-LVT или MiaPaCa-2-LVT проявлялось значительное замедление роста опухолей по сравнению с контрольными мышами, получавшими контрольный солевой раствор. Химиотерапия гемцитабином вплоть до 100 мг/кг без дополнительной обработки IMAB362 не оказывала существенного терапевтического влияния на ксенотрансплантаты ВхРС3-LVT или MiaPaCa-2-LVT. Напротив, комбинированная обработка 50-100 мг/кг гемцитабина плюс IMAB362 приводила к значительному усилению ингибирования роста опухолей и продлению срока жизни у мышей с опухолями по сравнению с мышами, получавшими только химиотерапию (фиг. 41, фиг. 42, фиг. 43). Эти наблюдения указывают на существование синергических терапевтических эффектов при комбинировании гемцитабина и иммунотерапии IMAB362.
При использовании высоких доз гемцитабина 2×150 мг/кг в неделю у привитых ксенотрансплантатов опухолей MiaPaCa-2-LVT сильно ингибировался рост опухолей независимо от обработки IMAB362 (фиг. 44А). Однако мыши, получавшие комбинированную терапию IMAB362 и гемцитабином, проявляли значительно более длительный срок жизни по сравнению с мышами, получавшими гемцитабин в качестве единственного средства (фиг. 44В).
Пример 9. Обработка ZA/IL-2 приводит к экспансии большого количества Т-клеток Vγ9Vδ2
PBMCs культивировали в течение 14 дней в среде RPMI с добавлением 300 Ед/мл IL-2 и 1 мкМ золедроновой кислоты (ZA) или без нее. В день 0 и день 14 определяли процент Т-клеток Vγ9+ Vδ2+ в популяции лимфоцитов CD3+ и процент клеток CD16+ в популяции Т-клеток CD3+ Vγ9+ Vδ2+ многоцветным методом FACS.
Для выживания и роста лимфоцитов требуется добавление IL-2 в культуры PBMCs. Они эффективно экспандируют в культурах при добавлении 300 Ед/мл IL-2. Методом FACS с использованием специфичных к Vγ9 и Vδ2 антител показали, что добавление ZA/IL-2 специфически индуцирует накопление Т-клеток Vγ9Vδ2. Через 14 дней популяция лимфоцитов CD3+ может содержать до 80% Т-клеток Vγ9Vδ2. Часть Т-клеток Vγ9Vδ2 экспрессирует CD 16, причем обогащение этими клетками в популяции лимфоцитов CD3+ составляет 10-700 раз, в зависимости от донора. Обогащение Т-клетками CD16+ Vγ9+ Vδ2+ в культурах составляет 10-600 раз по сравнению с культурами, растущими без ZA. Делаем вывод, что обработка PBMCs in vitro с помощью ZA/IL-2 приводит к повышающей регуляции опосредующего ADCC FcγIII-рецептора CD16 у значительной части Т-клеток γδ.
Подобно NK-клеткам, при вызванной ZA/IL-2 экспансии Т-клетки Vγ9Vδ2 будут положительными по CD16 - рецептору FcγRIII, через которые связанное с клетками антитело запускает ADCC. Чтобы определить, будут ли Т-клетки Vγ9Vδ2 способны индуцировать сильную ADCC в сочетании с IMAB362, проводили ряд экспериментов.
PBMCs, полученные от 2 различных доноров (#1 и #2), культивировали в среде с 300 Ед/мл IL-2 и 1 мкМ ZA или без нее. Через 14 дней клетки собирали и добавляли их при возрастающих концентрациях (0,26 нг/мл - 200 мкг/мл) IMAB362 к клеткам NUGC-4, экспрессирующим CLDN18.2. Определяли специфическую гибель клеток люциферазным методом. Проводили анализ ADCC с 27 донорами при культивировании в присутствии 300 Ед/мл IL-2 с ZA или без нее, при этом клетки NUGC-4 служили в качестве мишени. Для каждого донора вычисляли значения ЕС50 из кривых доза-ответ и оценивали максимальную степень специфической гибели при дозе в 200 мкг/мл IMAB362 на точечных графиках.
При использовании PBMCs, культивировавшихся 14 дней с ZA/IL-2, наблюдалась сильная IMAB362-зависимая активность ADCC против CLDN18.2-положительных клеток NUGC-4. При использовании обработанных ZA/IL-2 культур PBMCs активность ADCC зависит от присутствия Т-клеток Vγ9Vδ2. При культивировании клеток без ZA активность ADCC снижается у большинства доноров. В этих культурах остаточная активность ADCC зависит от NK-клеток. При тестировании более 20 доноров анализ ADCC показывает, что обработка PBMCs с помощью ZA/IL-2 улучшает значения ЕС50 и максимальной степени специфической гибели по сравнению с культивированием PBMCs только с одним IL-2.
Пример 10. Эффективность IMAB362 в сочетании с гемцитабином на модели метастазов у мышей
Для анализа влияния обработки IMAB362 в комбинации с гемцитабином на метастазы Patu8988S в легких in vivo, по 12 бестимусных мышей nude Hsd:Foxnlnu на группу получали внутривенную инъекцию 2×106 клеток Patu8988S в хвостовую вену. Через 14 дней после введения опухолевых клеток мыши получали 200 мкг IMAB362 или PBS в качестве контроля (в/в-в/б) по 2 раза в неделю плюс раз в неделю дозу в 100 мг/кг гемцитабина в/б на протяжении 4 недель. Обработку IMAB362 или PBS продолжали до забоя мышей на 70-й день после введения опухолевых клеток. Проводили анализ нагруженности легких ксенотрансплантированными опухолями методом количественной ПЦР на ДНК человека в препаратах легких и оптическим методом иммуногистологического окрашивания с помощью антитела против MHC-I человека (клон EPR1394Y). Результаты показывают, что у мышей, получавших IMAB362 плюс гемцитабин, значительно снижалось количество ДНК человека в легких (фиг. 45А), а площадь поверхности, окрашенной комплексом против MHC-I человека на срезах легких, была значительно меньше, чем в легких у мышей, получавших постороннее антитело плюс гемцитабин (фиг. 45В). Оба метода показали снижение нагруженности ксенотрансплантатами опухолей Patu8988s в легких у получавших IMAB362 плюс гемцитабин мышей, свидетельствуя, что комбинирование с IMAB362 значительно превосходит монотерапию гемцитабином.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМБИНИРОВАННАЯ ТЕРАПИЯ С ИСПОЛЬЗОВАНИЕМ АНТИТЕЛ ПРОТИВ КЛАУДИНА 18.2 ДЛЯ ЛЕЧЕНИЯ РАКА | 2014 |
|
RU2792932C2 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ С ИСПОЛЬЗОВАНИЕМ АНТИТЕЛ К КЛАУДИНУ 18.2 ДЛЯ ЛЕЧЕНИЯ РАКА | 2013 |
|
RU2789478C2 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ С ИСПОЛЬЗОВАНИЕМ АНТИТЕЛ К КЛАУДИНУ 18.2 ДЛЯ ЛЕЧЕНИЯ РАКА | 2013 |
|
RU2665321C2 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ С ИСПОЛЬЗОВАНИЕМ АНТИТЕЛ К КЛАУДИНУ 18.2 ДЛЯ ЛЕЧЕНИЯ РАКА | 2013 |
|
RU2662066C2 |
| КОНЪЮГАТЫ ЛЕКАРСТВЕННЫХ СРЕДСТВ, СОДЕРЖАЩИЕ АНТИТЕЛА ПРОТИВ КЛАУДИНА 18.2 | 2016 |
|
RU2841168C2 |
| АНТИТЕЛА ПРОТИВ КЛАУДИНА 18.2 ДЛЯ ДИАГНОСТИКИ РАКА | 2013 |
|
RU2661772C2 |
| ТЕРАПИЯ ДЛЯ ЛЕЧЕНИЯ РАКА, ВКЛЮЧАЮЩАЯ АНТИТЕЛА ПРОТИВ КЛАУДИНА 18.2 | 2014 |
|
RU2678700C2 |
| ТЕРАПИЯ ДЛЯ ЛЕЧЕНИЯ РАКА, ВКЛЮЧАЮЩАЯ АНТИТЕЛА ПРОТИВ КЛАУДИНА 18.2 | 2014 |
|
RU2832015C2 |
| АГЕНТЫ ДЛЯ ЛЕЧЕНИЯ ЭКСПРЕССИРУЮЩИХ КЛАУДИН РАКОВЫХ ЗАБОЛЕВАНИЙ | 2013 |
|
RU2678127C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ПРОГНОЗИРОВАНИЯ ТЕРАПЕВТИЧЕСКОЙ ЭФФЕКТИВНОСТИ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ И ПРОГНОЗА ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2016 |
|
RU2785291C2 |







































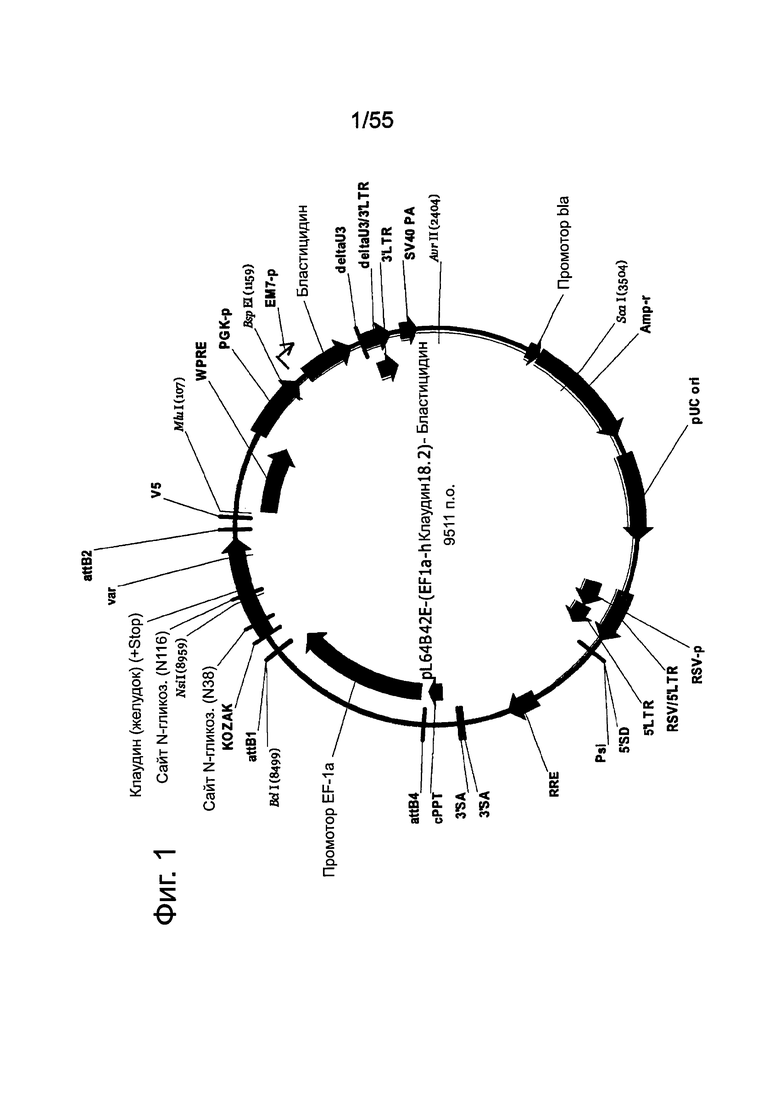

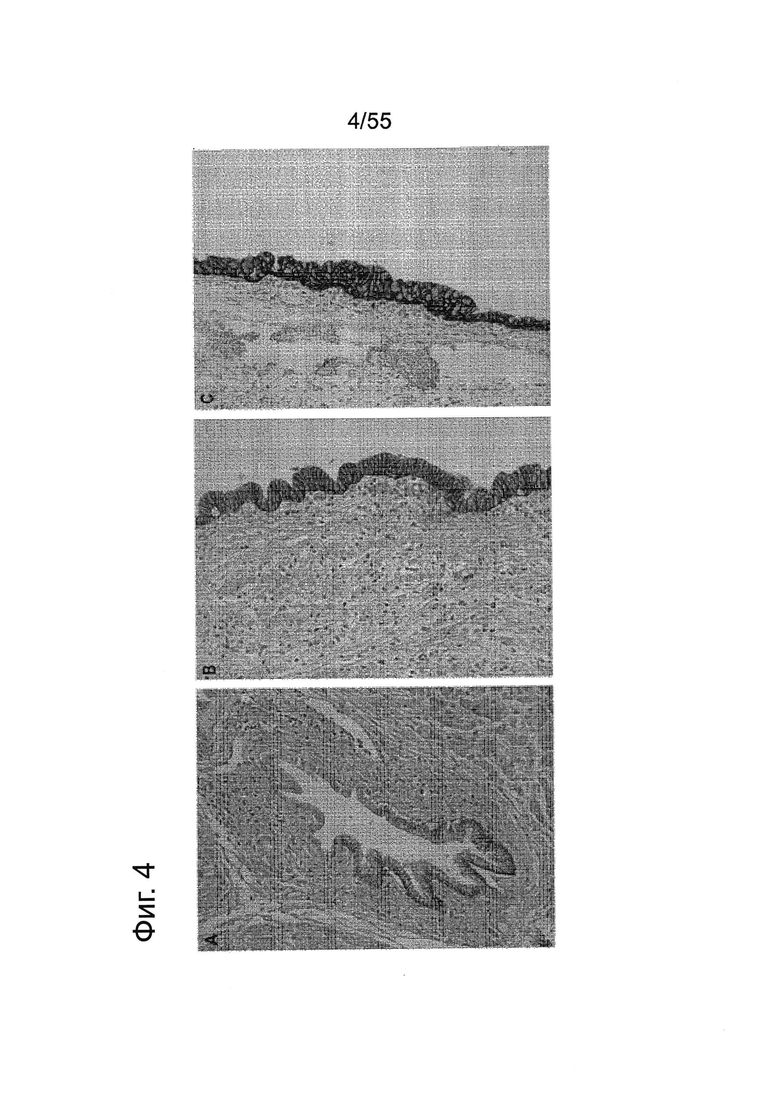
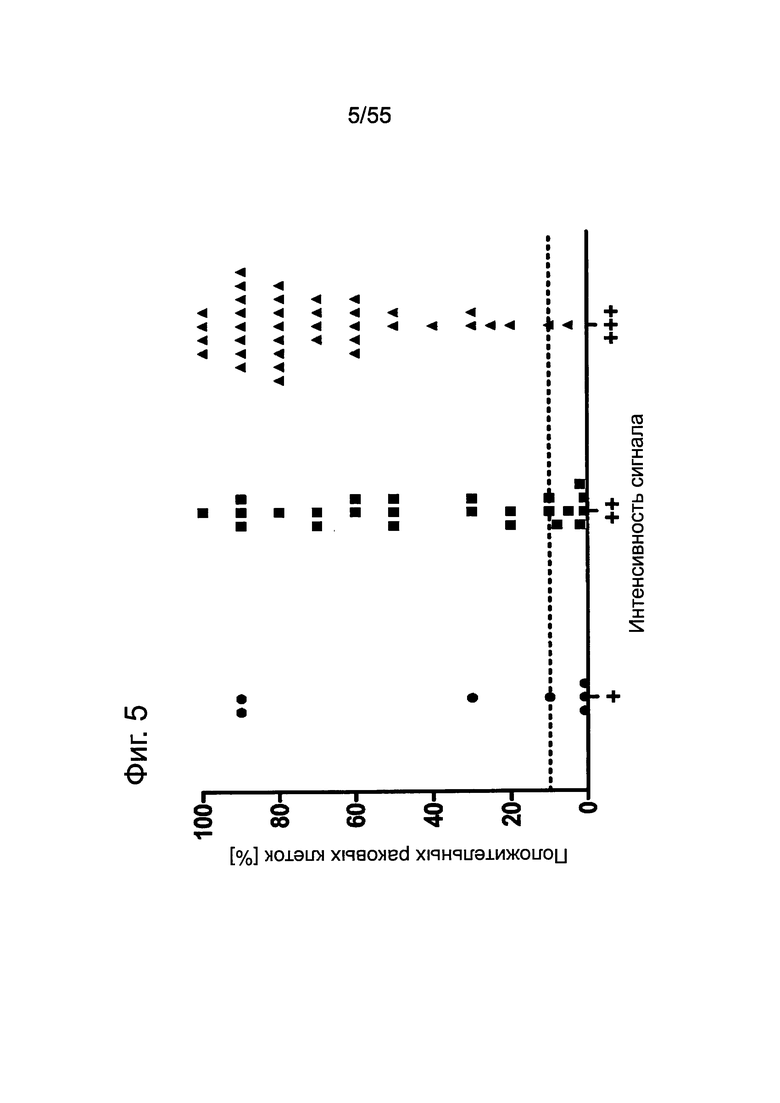
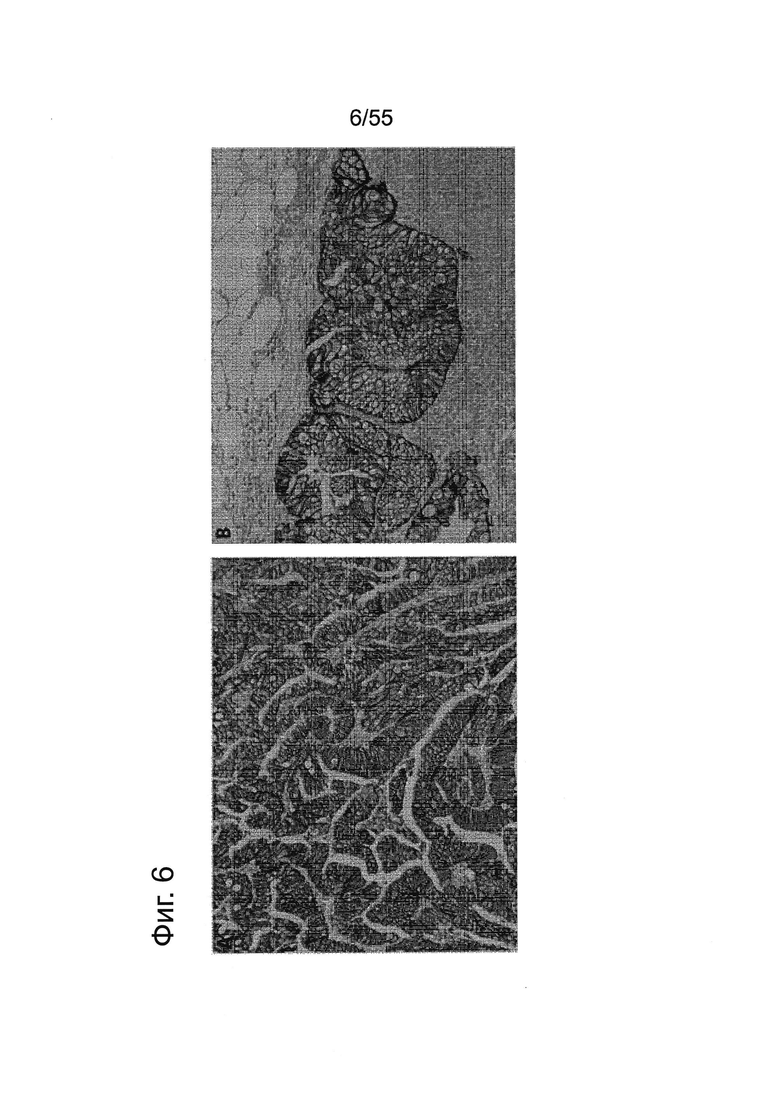
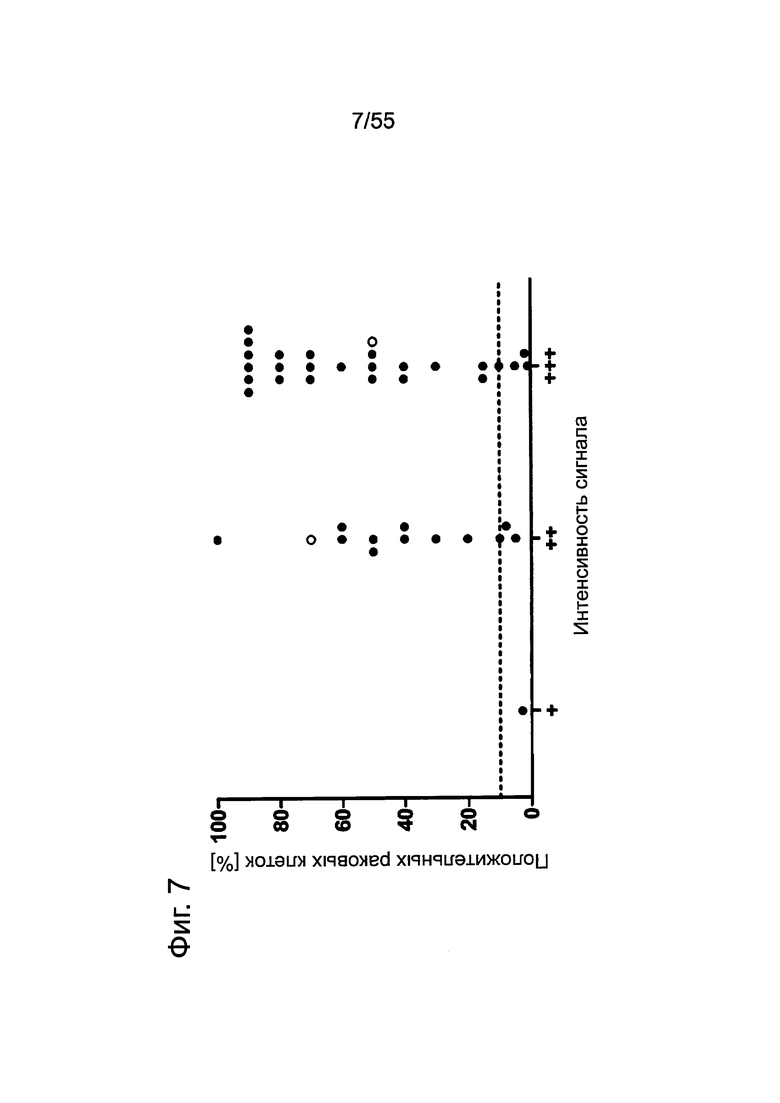
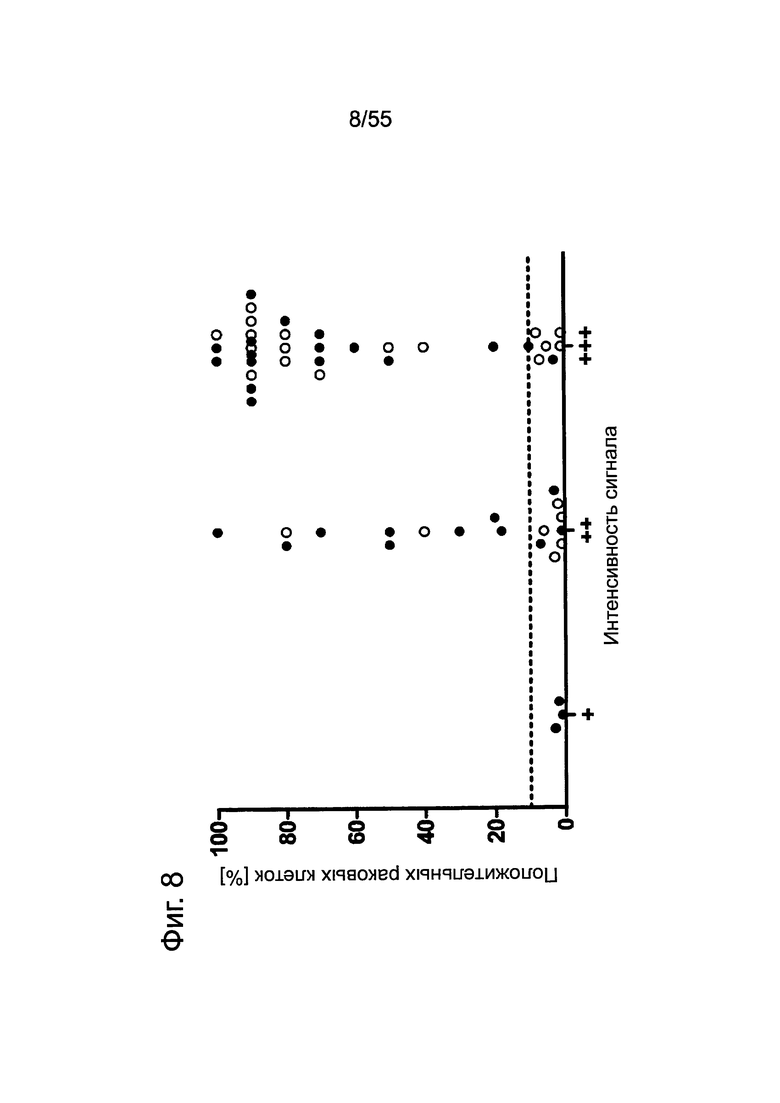
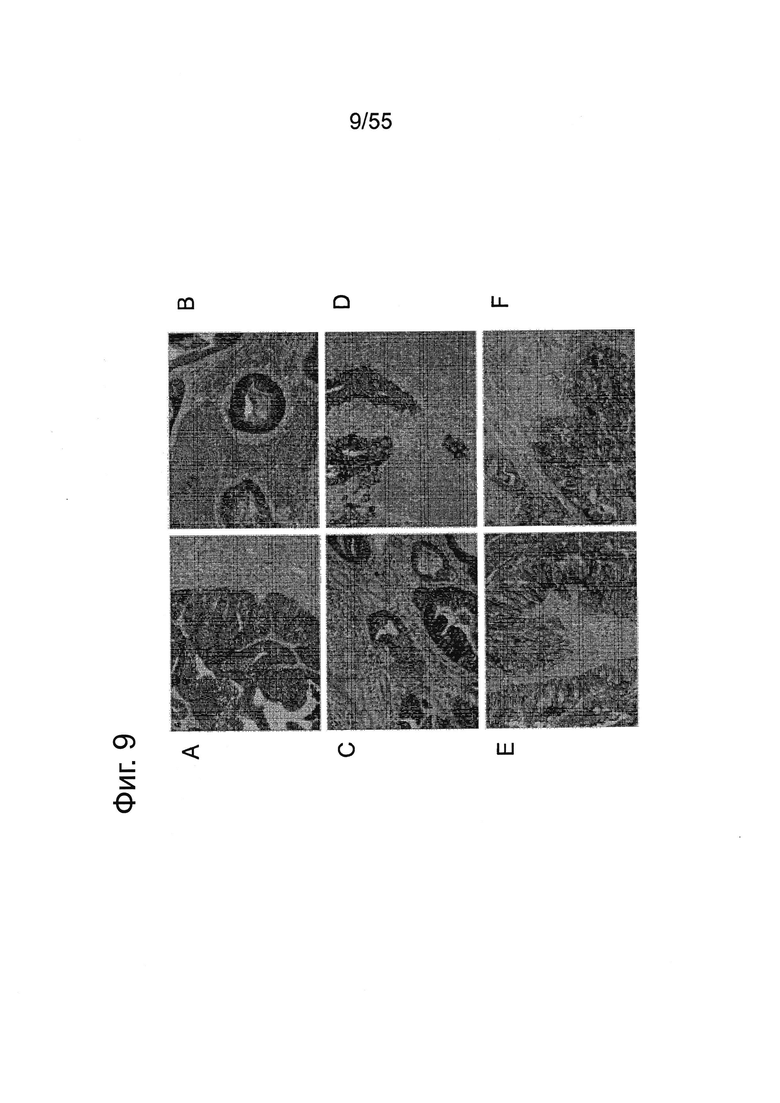
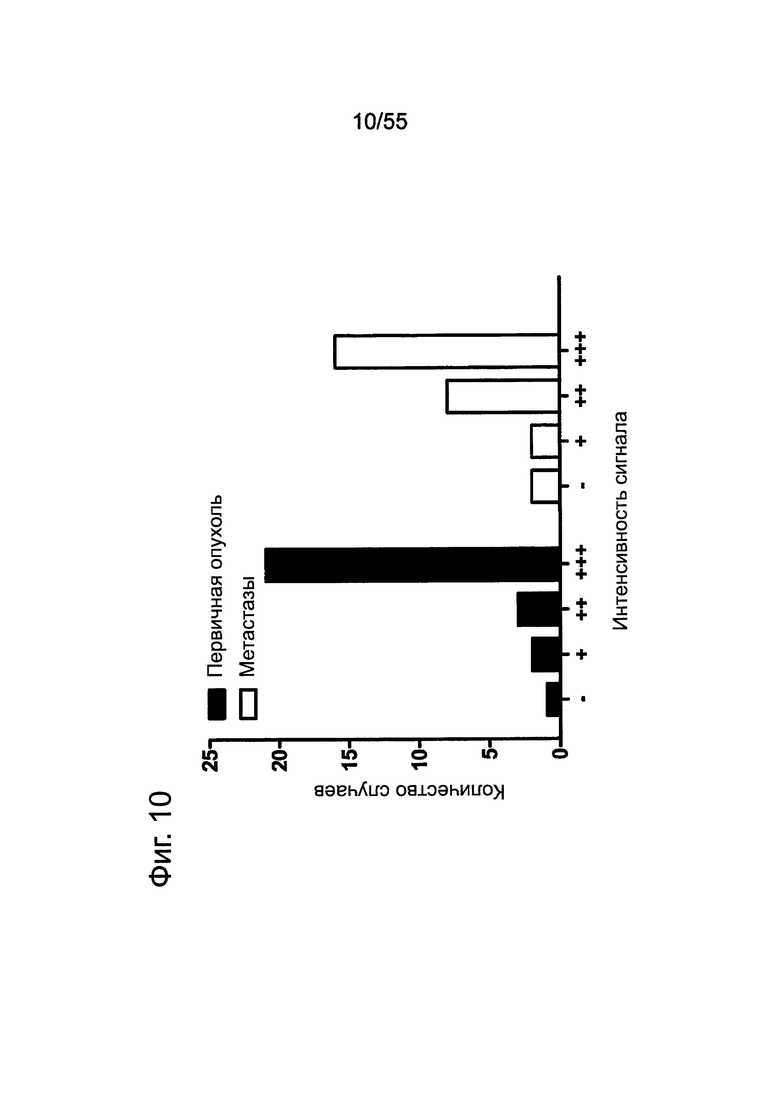
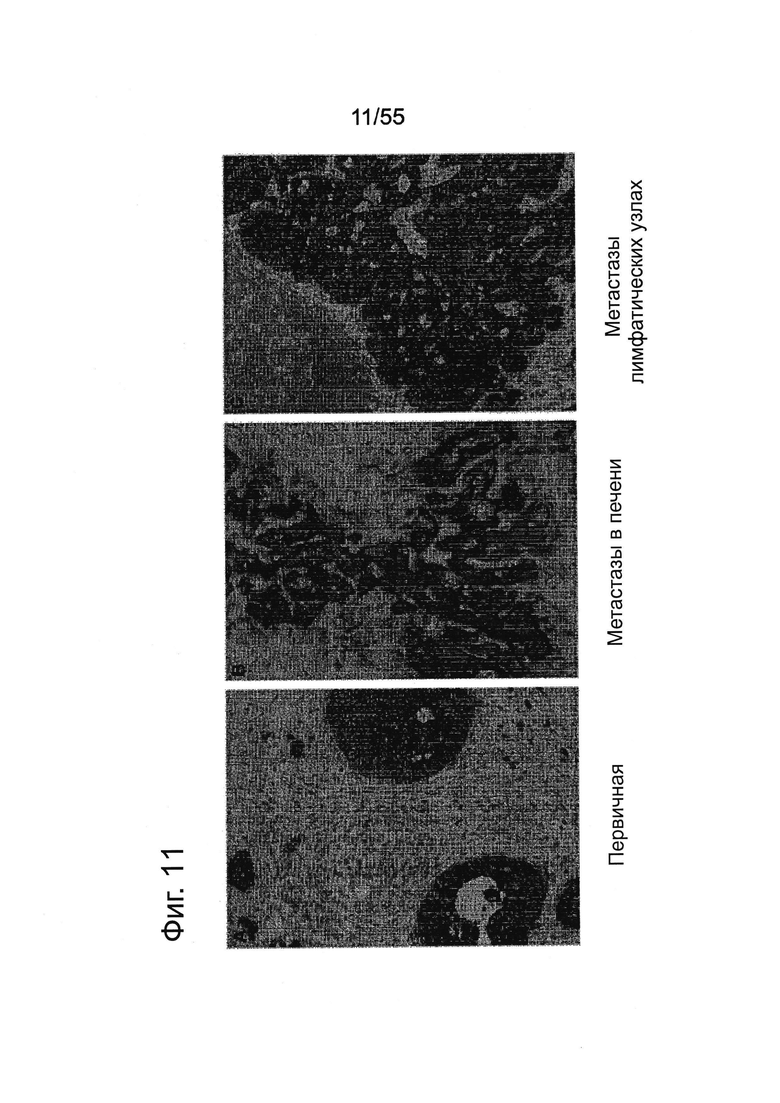
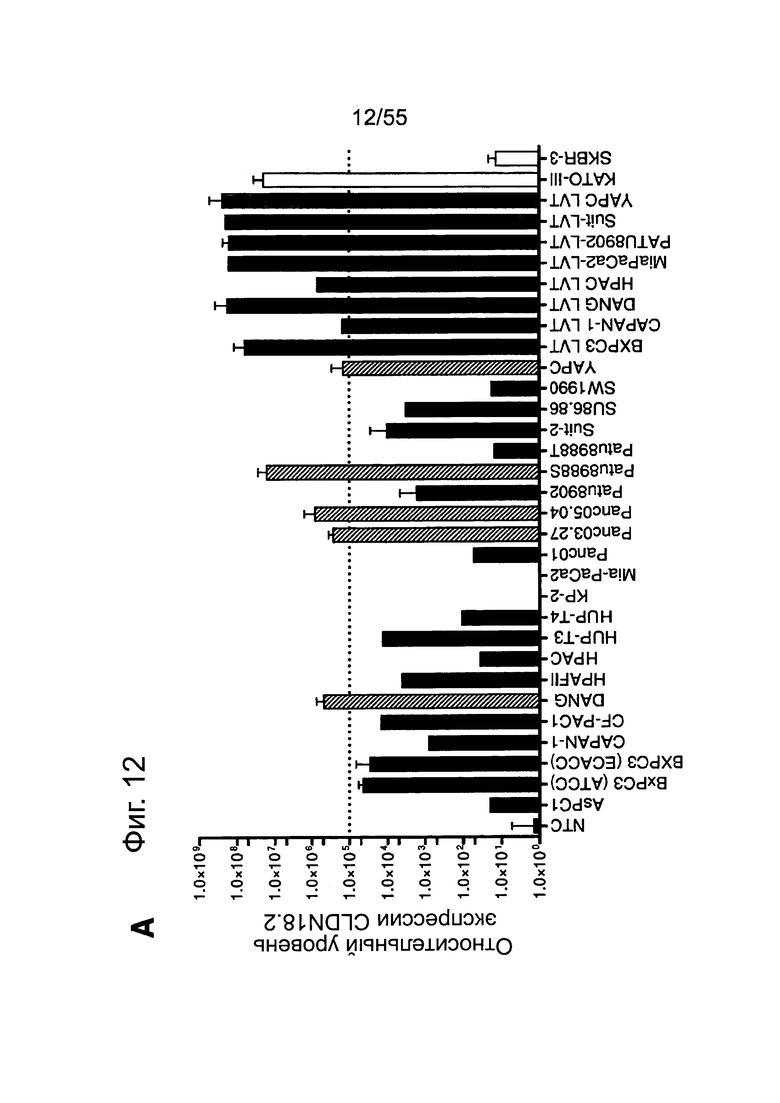
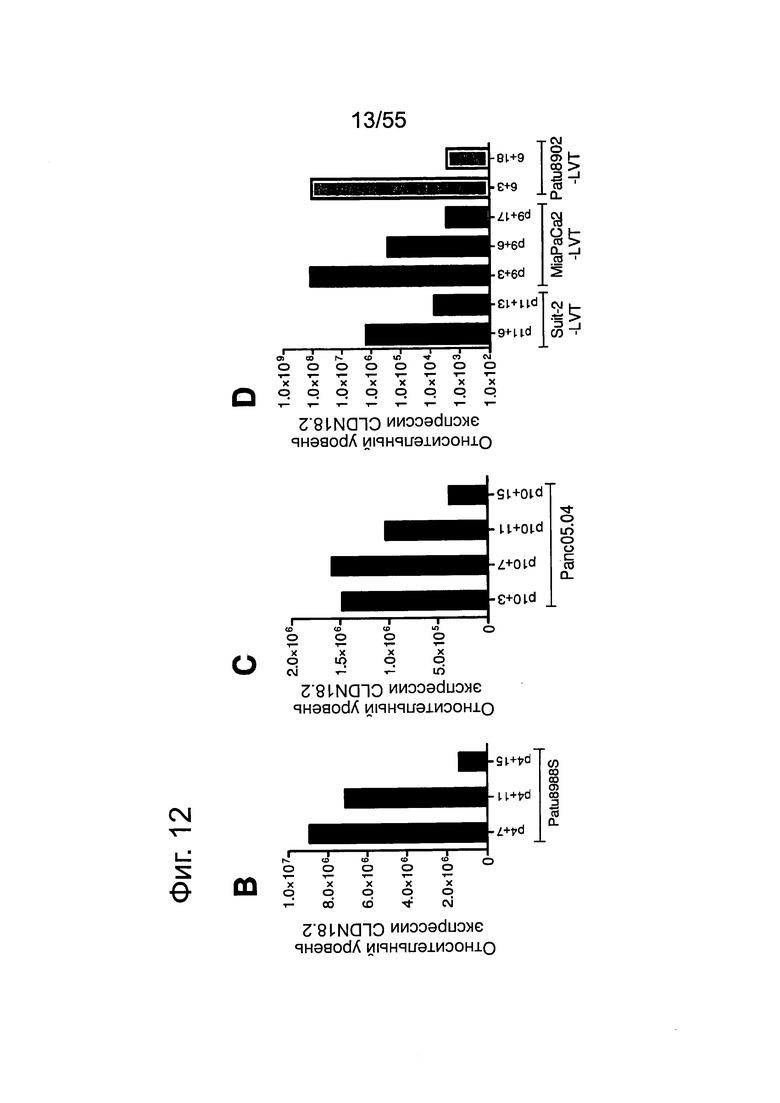
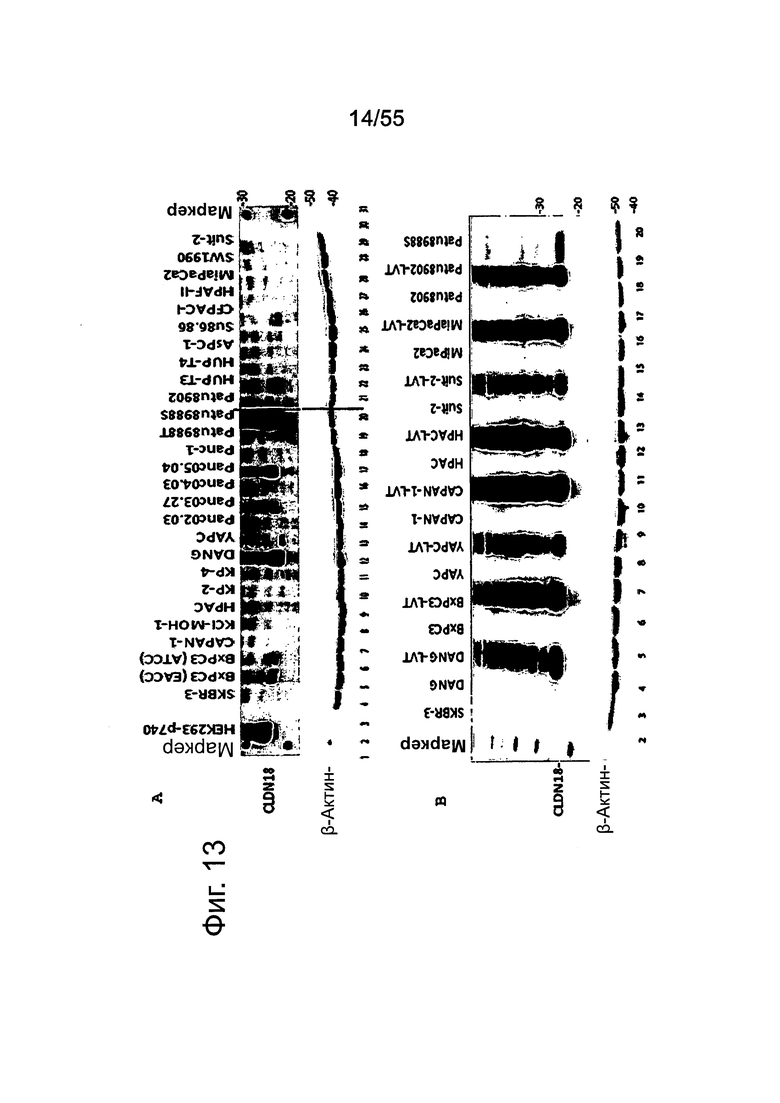

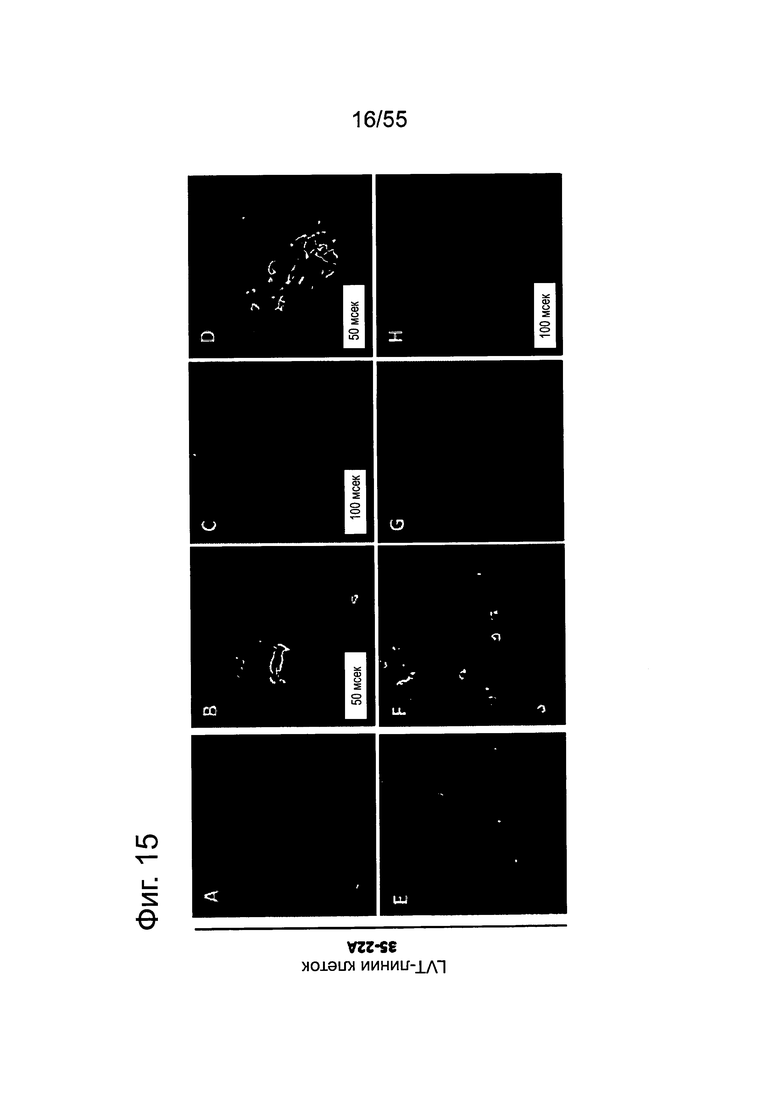
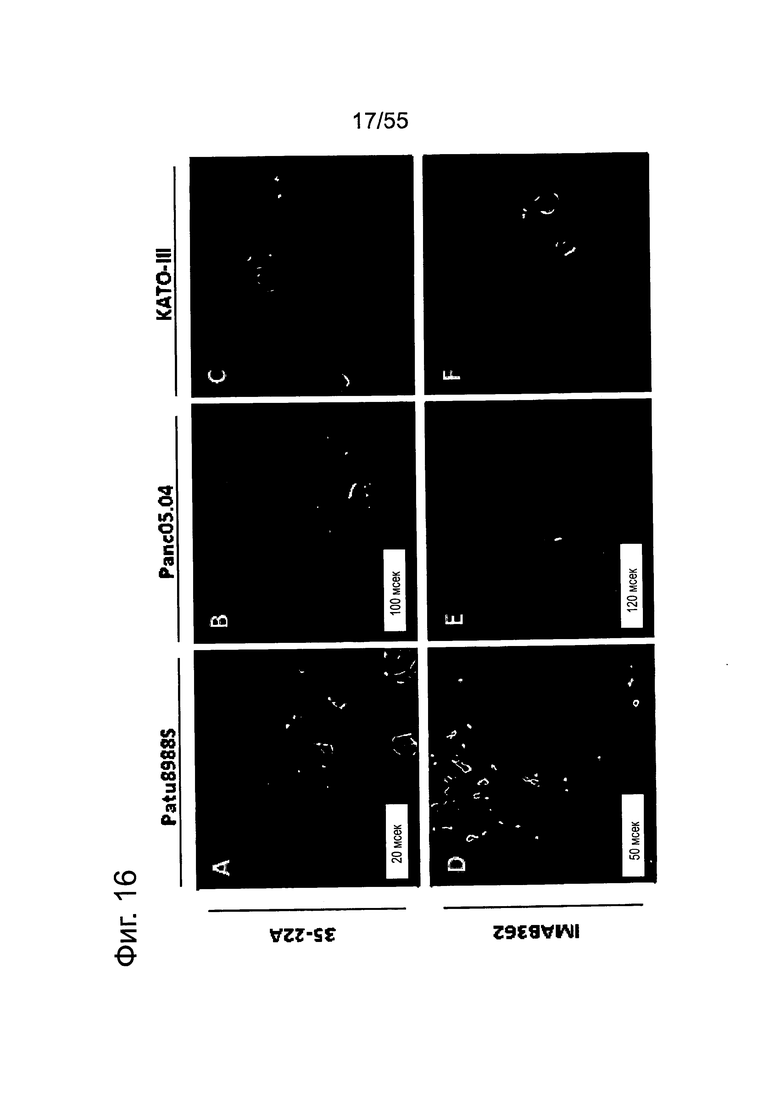

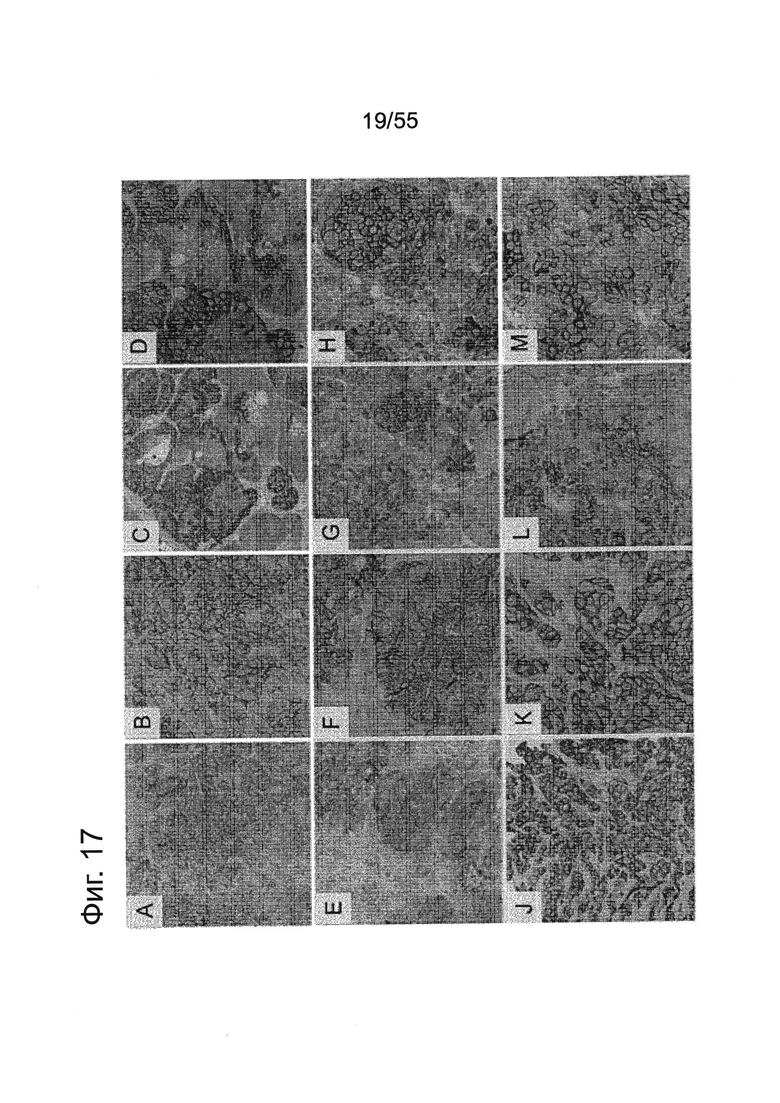

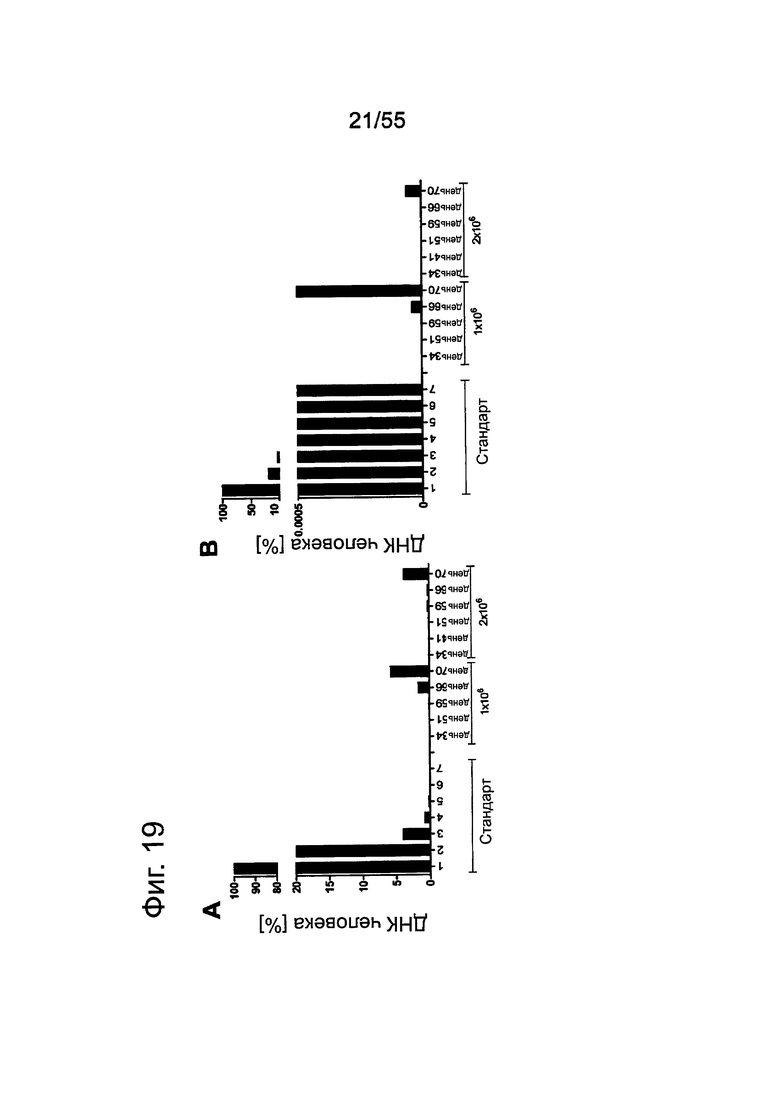
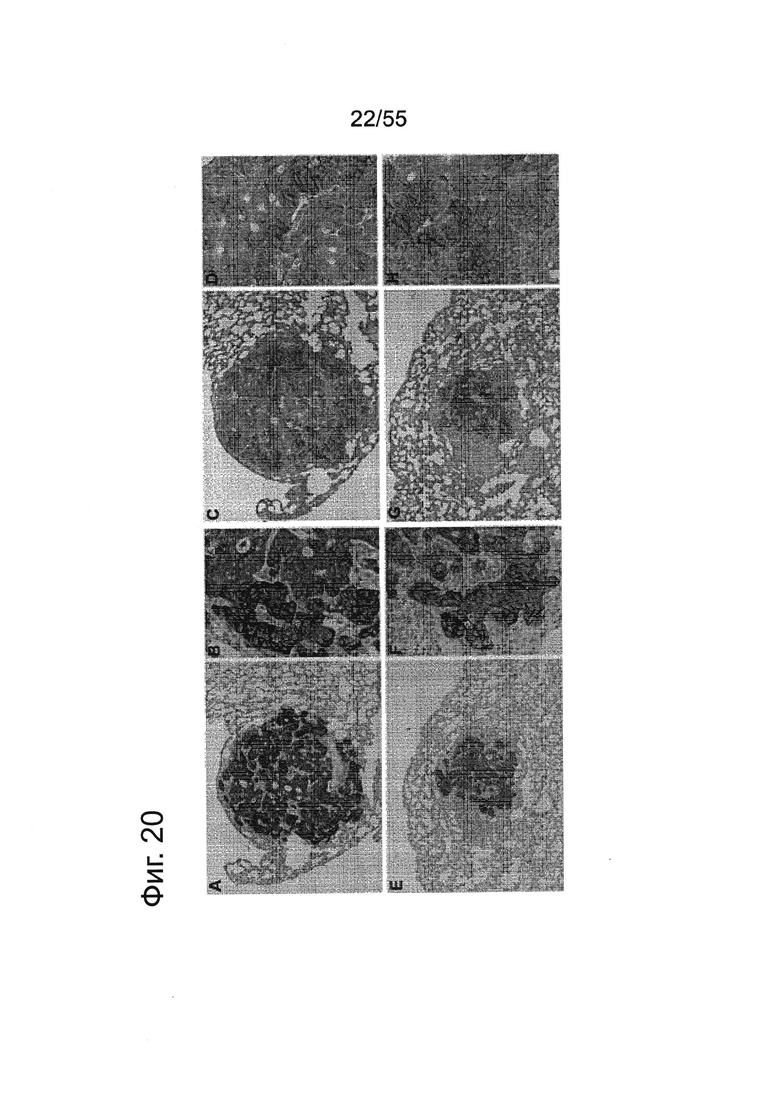
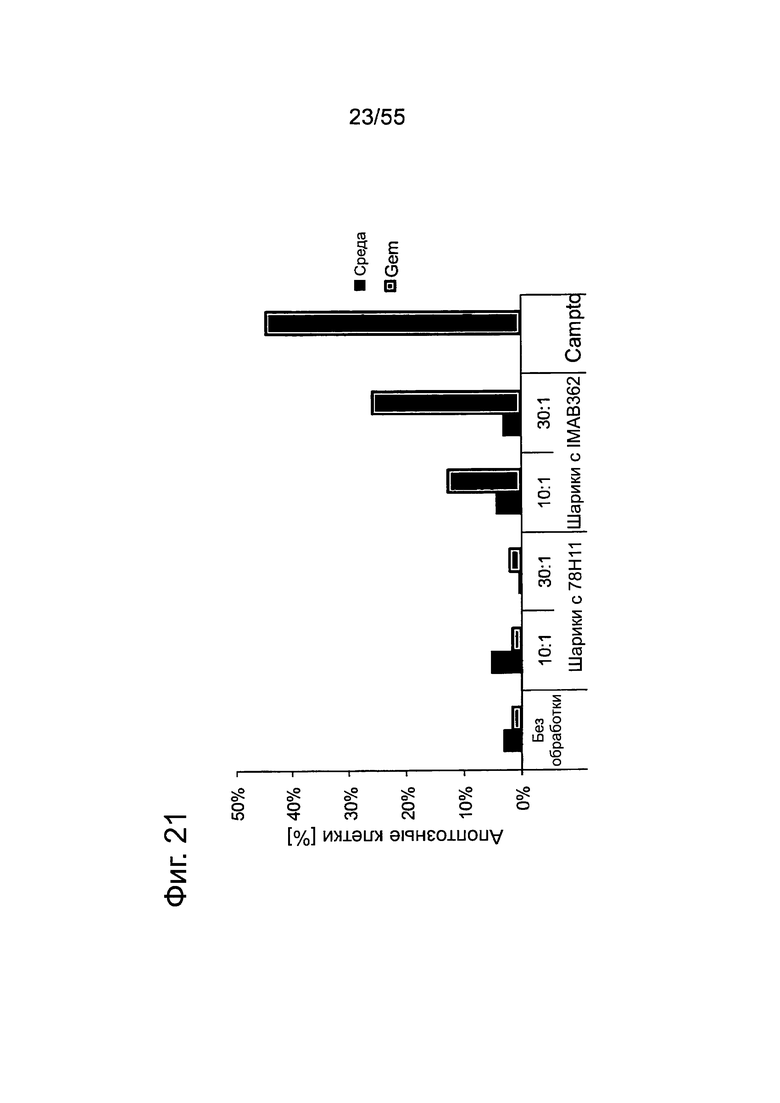
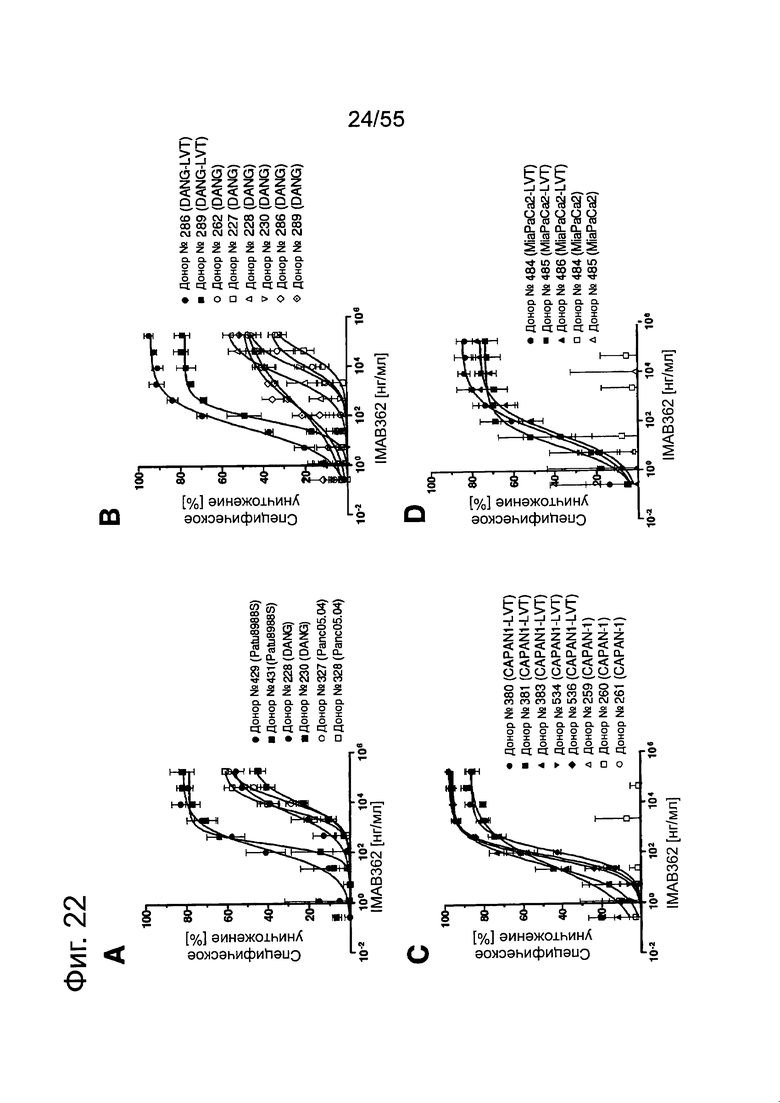
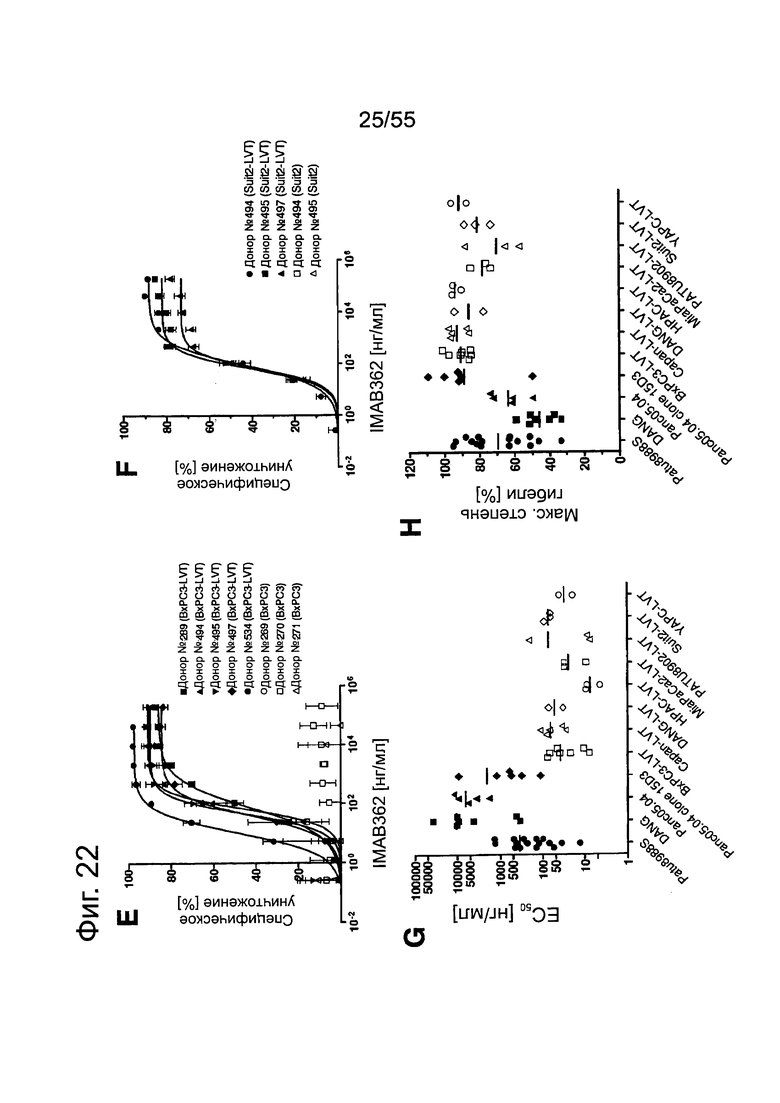
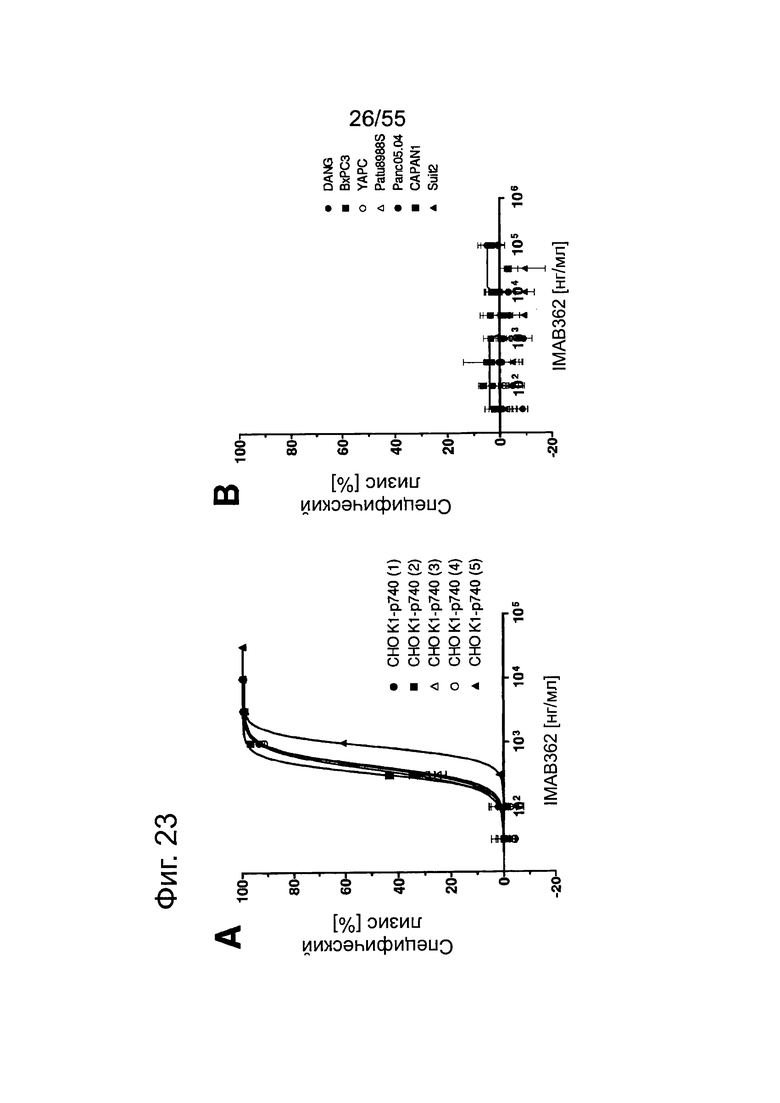
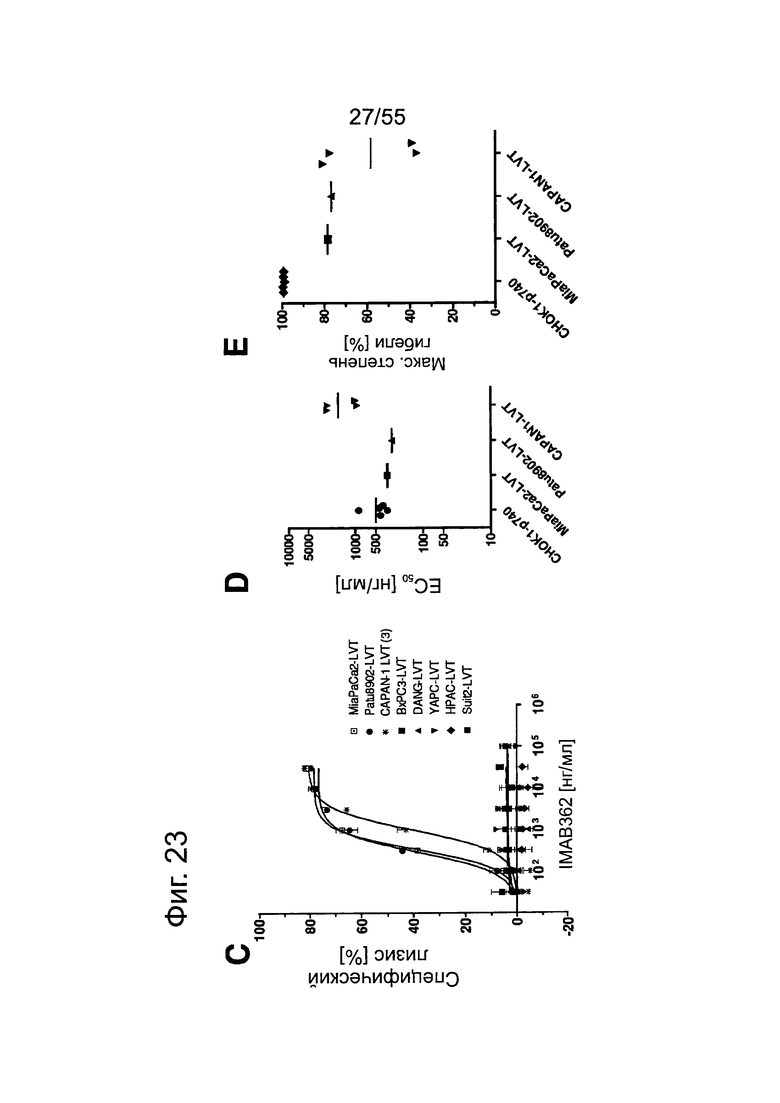
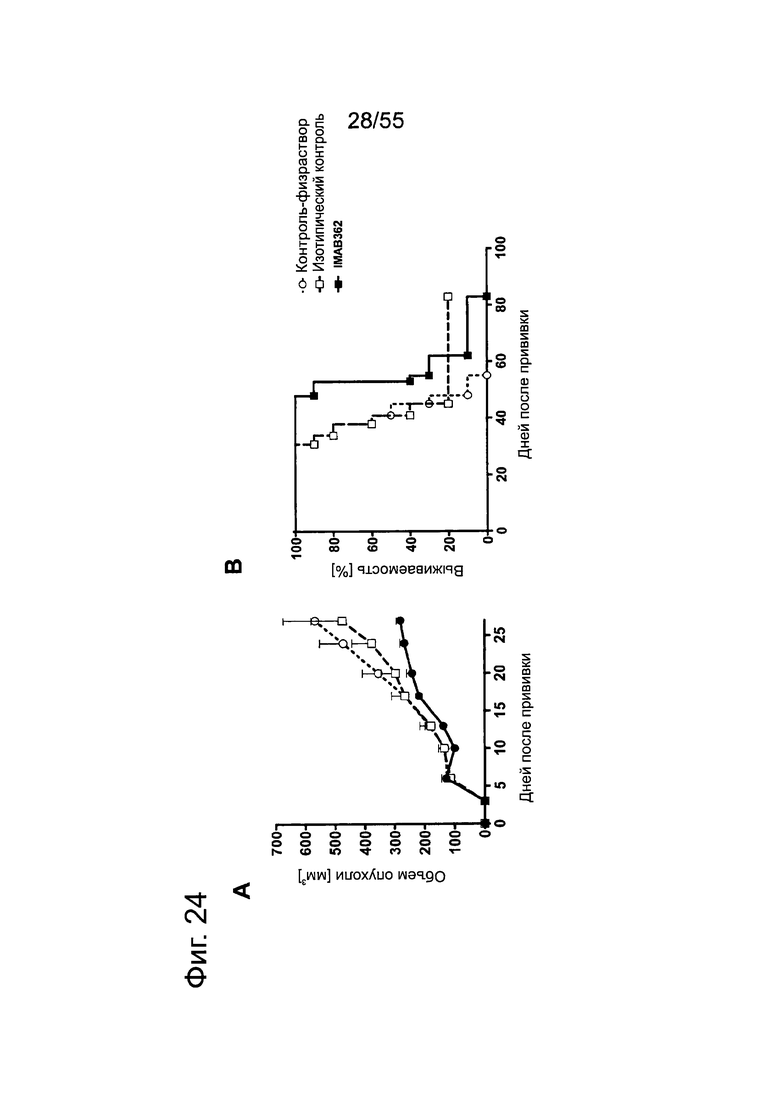
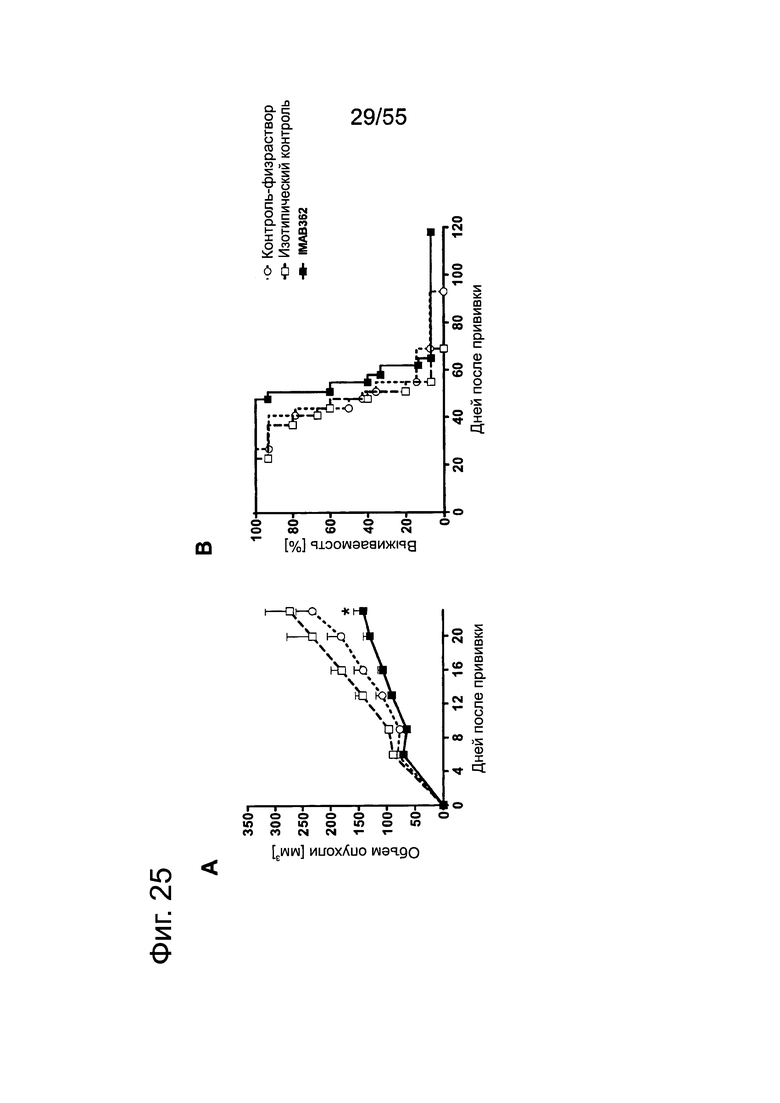
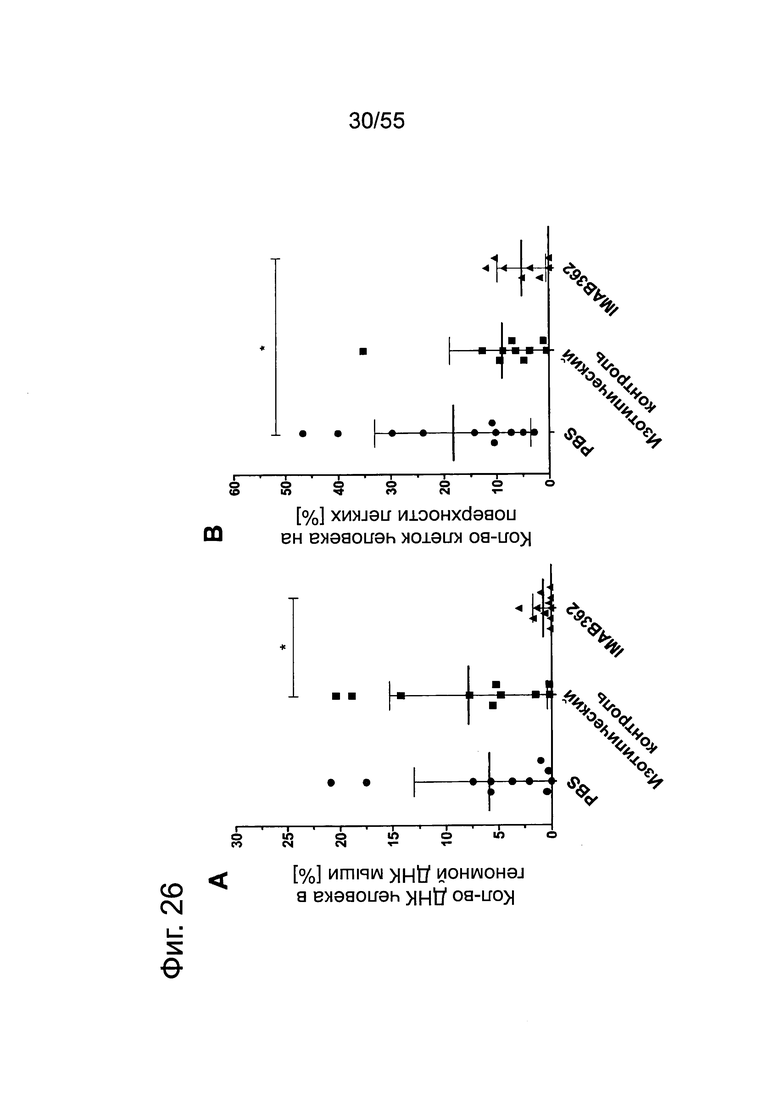
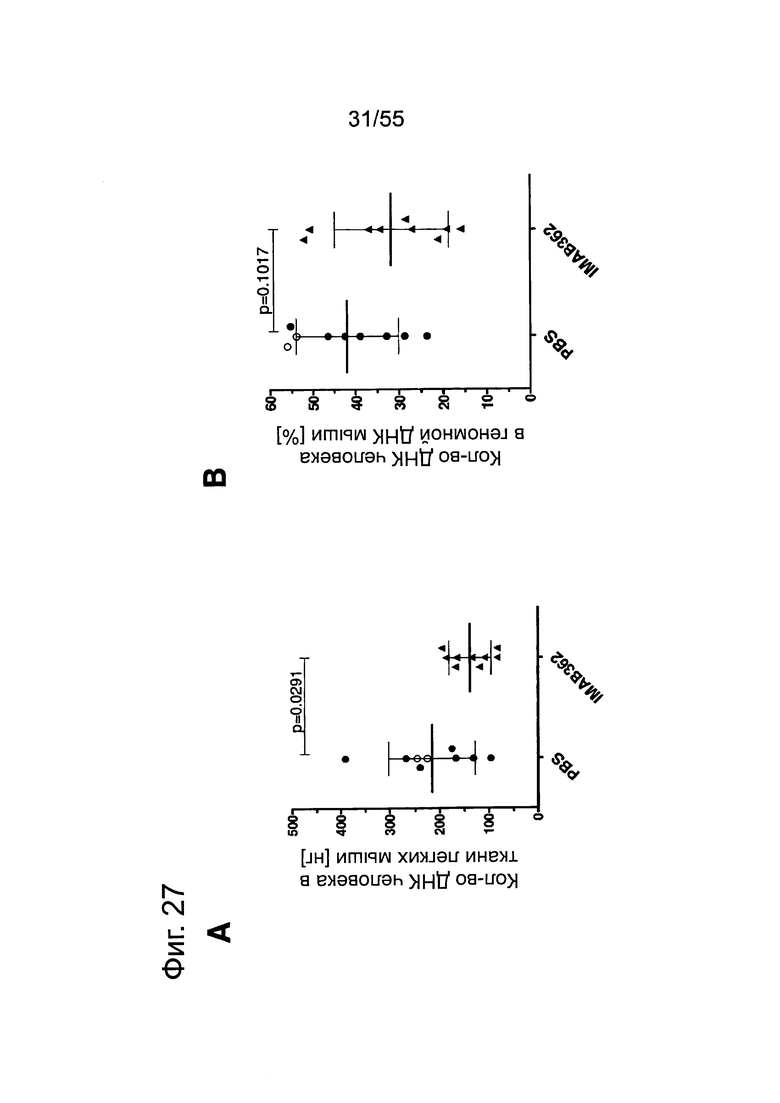
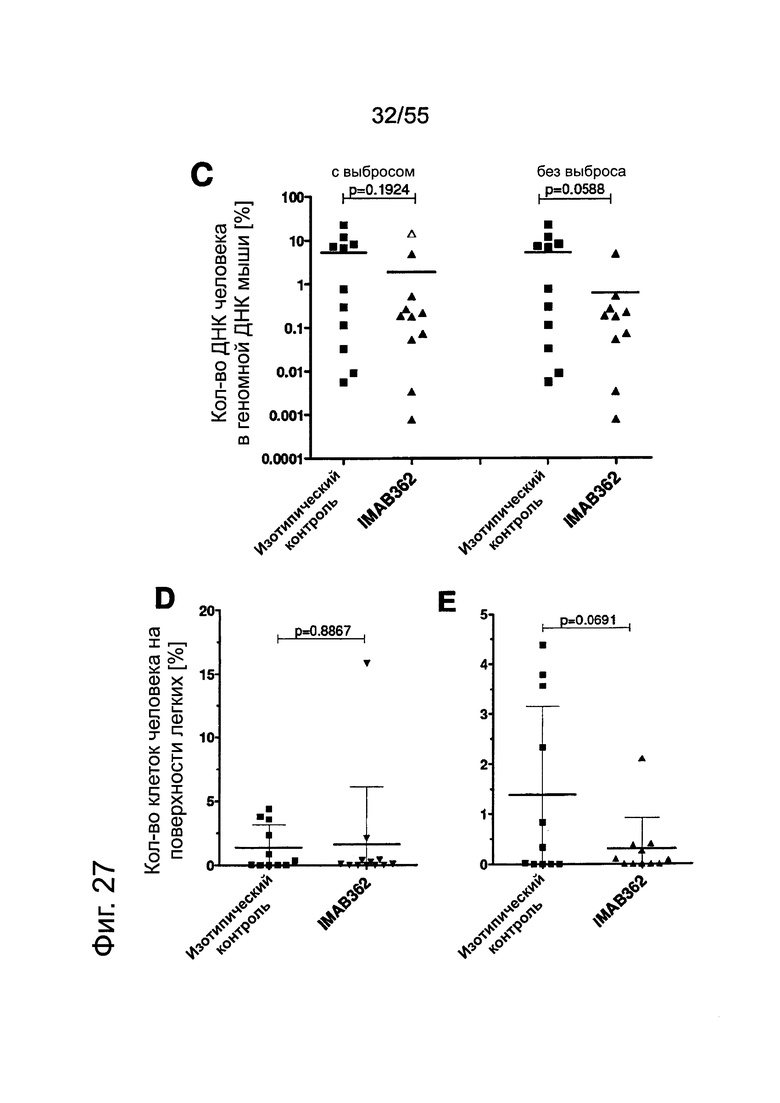
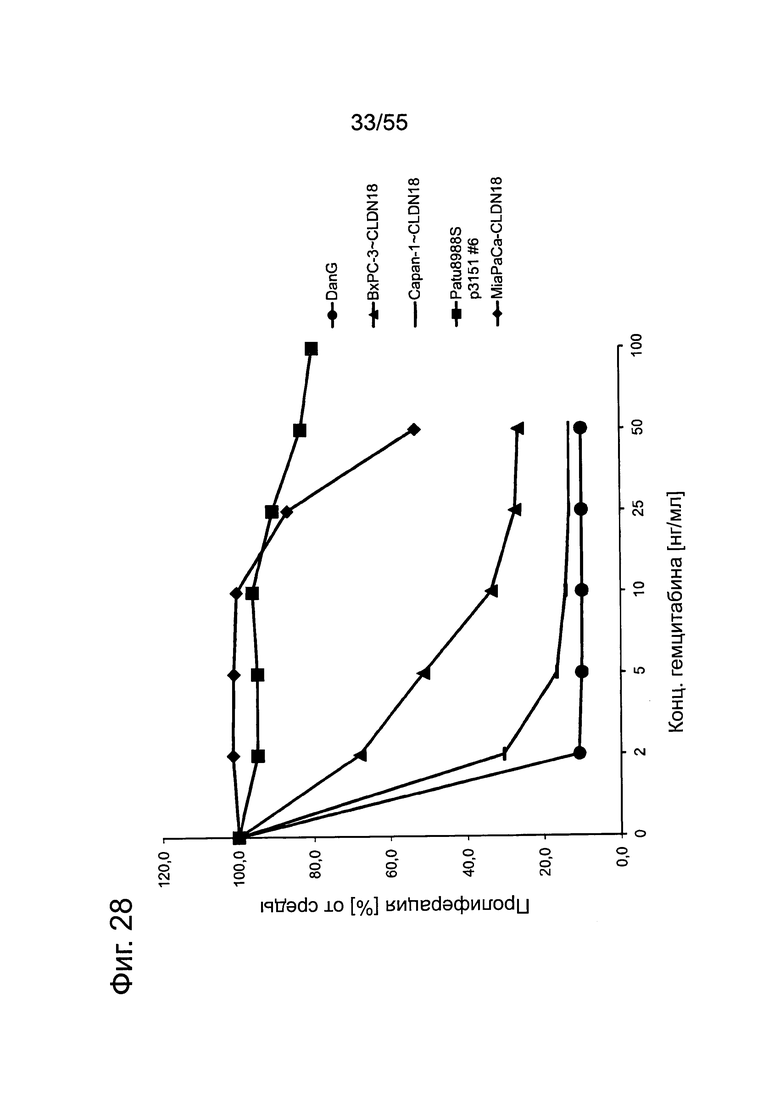
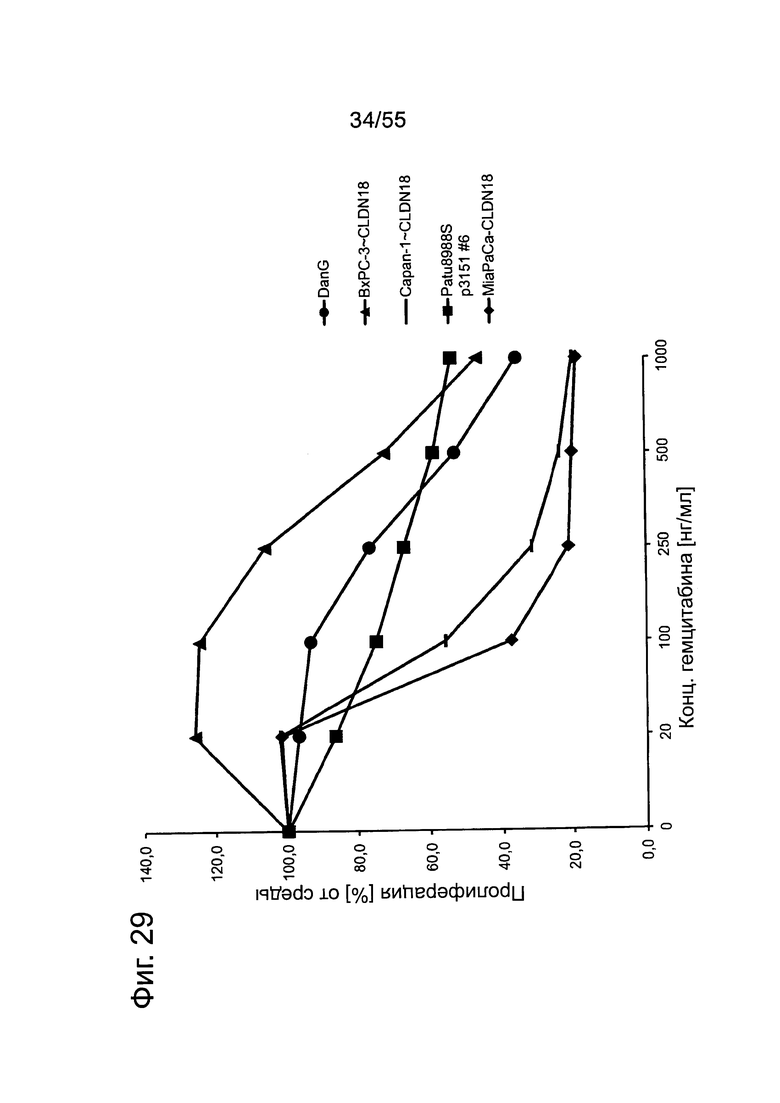
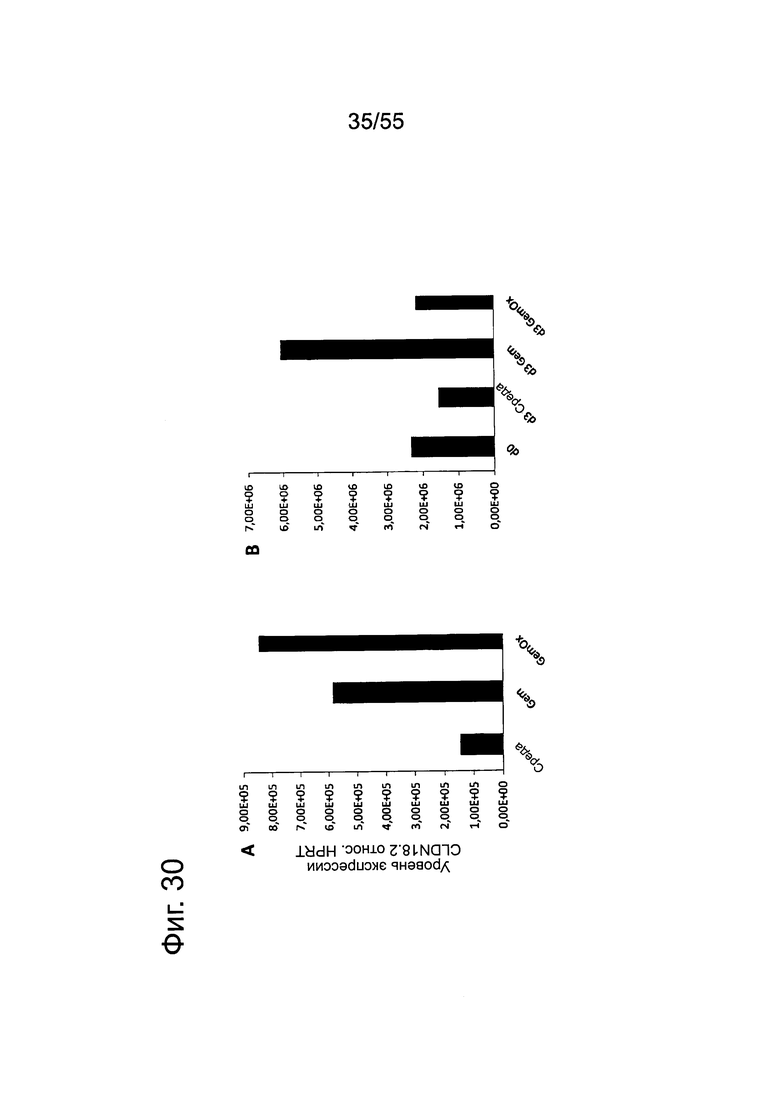
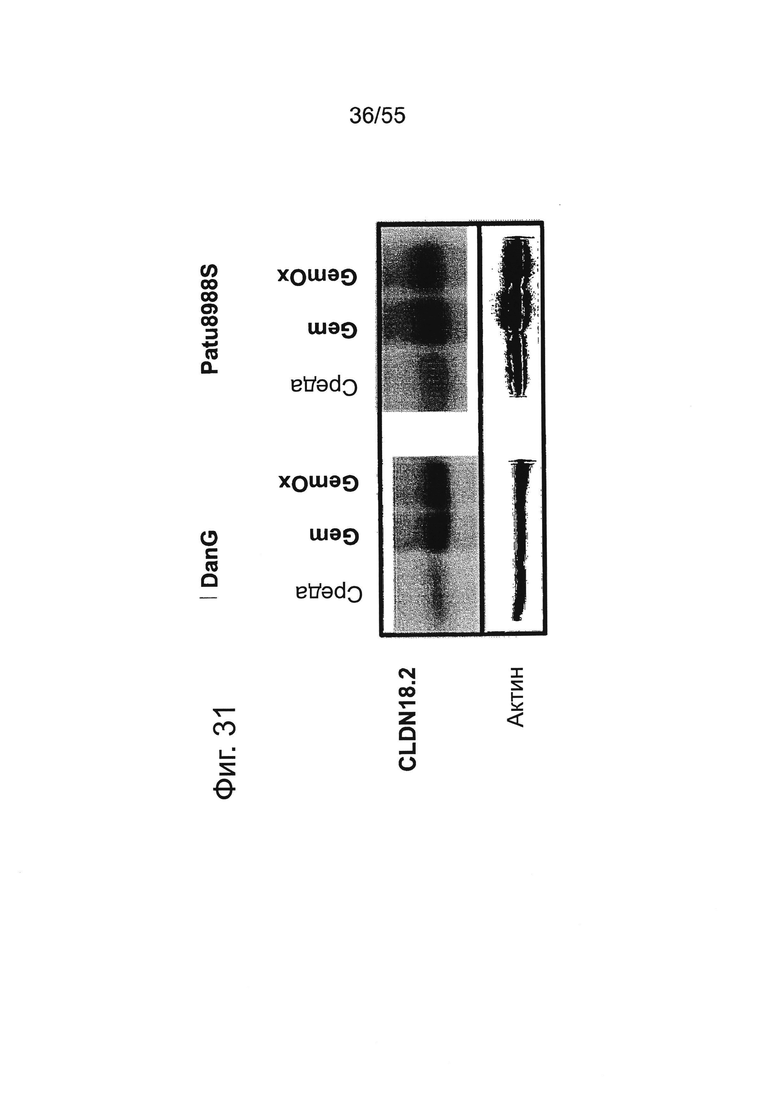
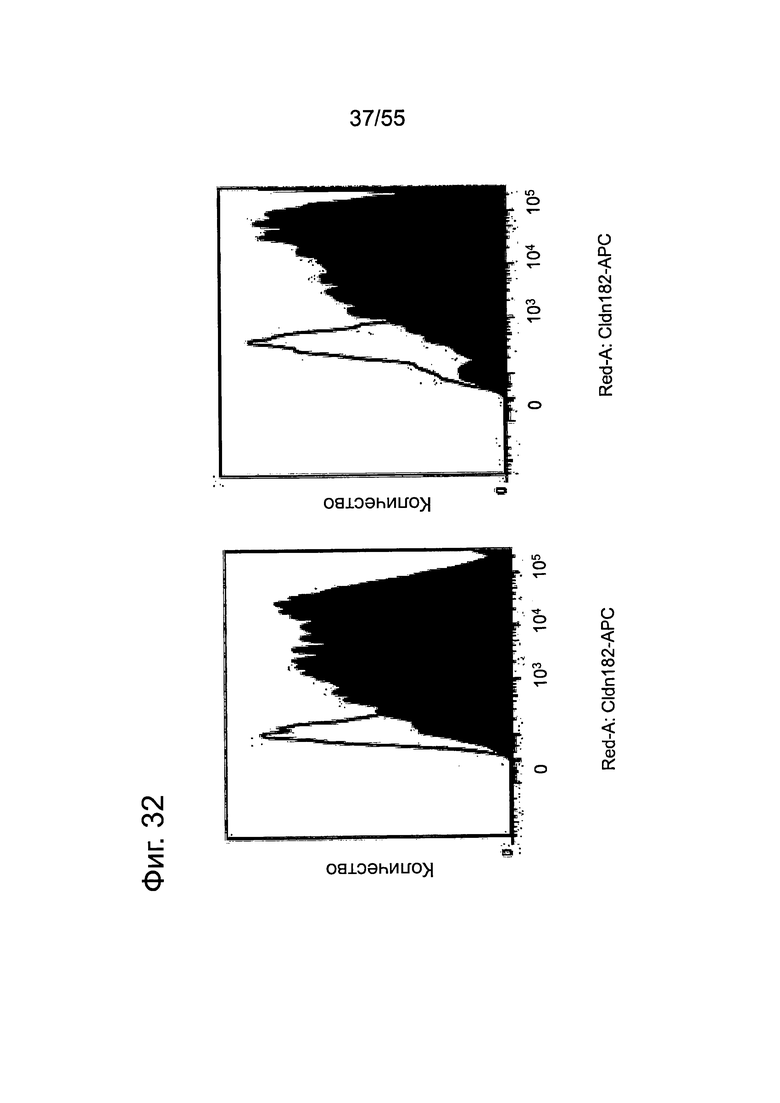
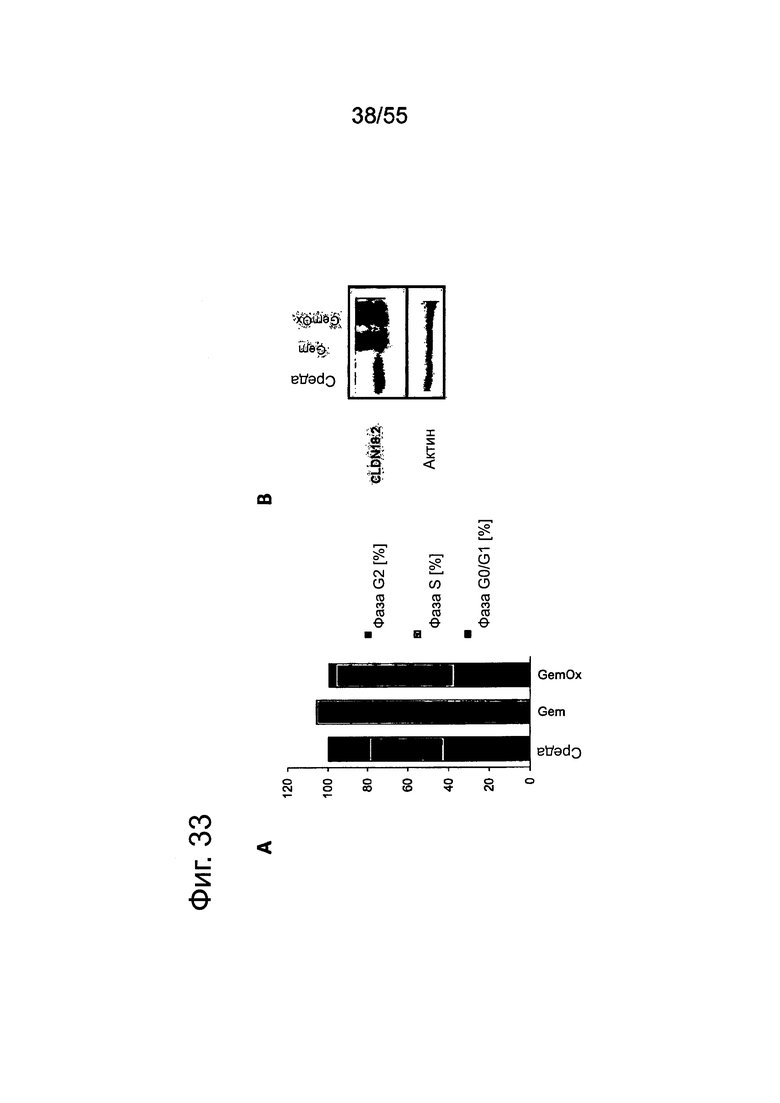
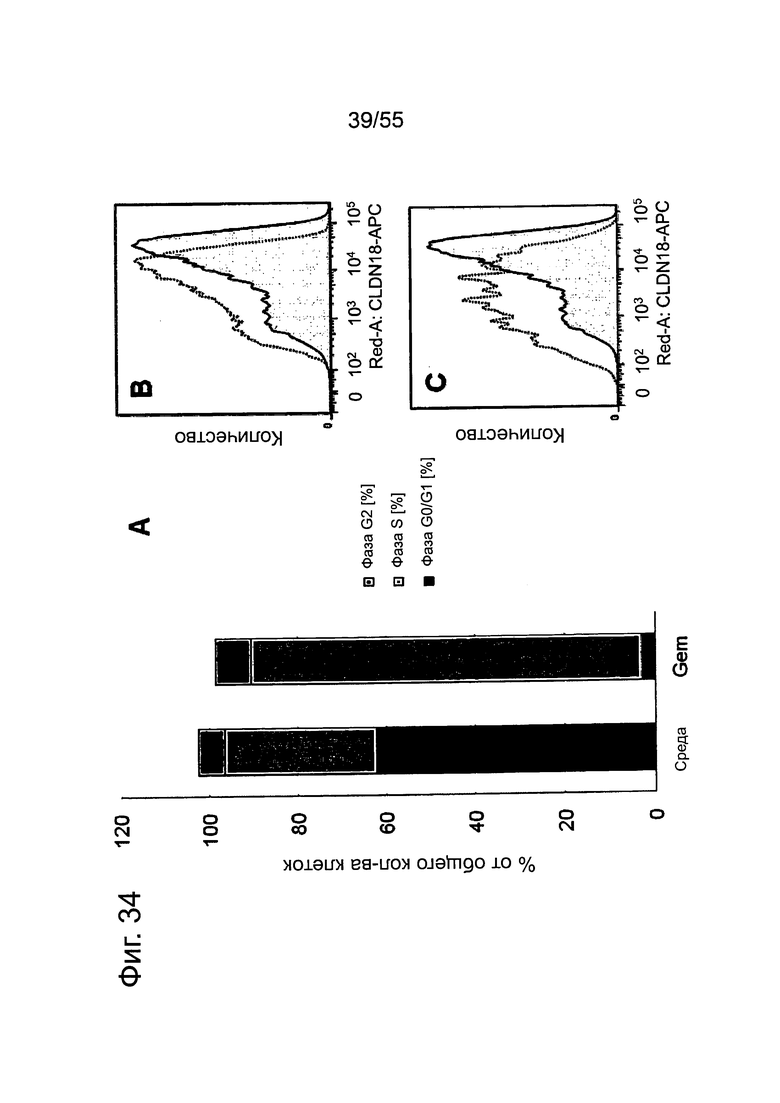
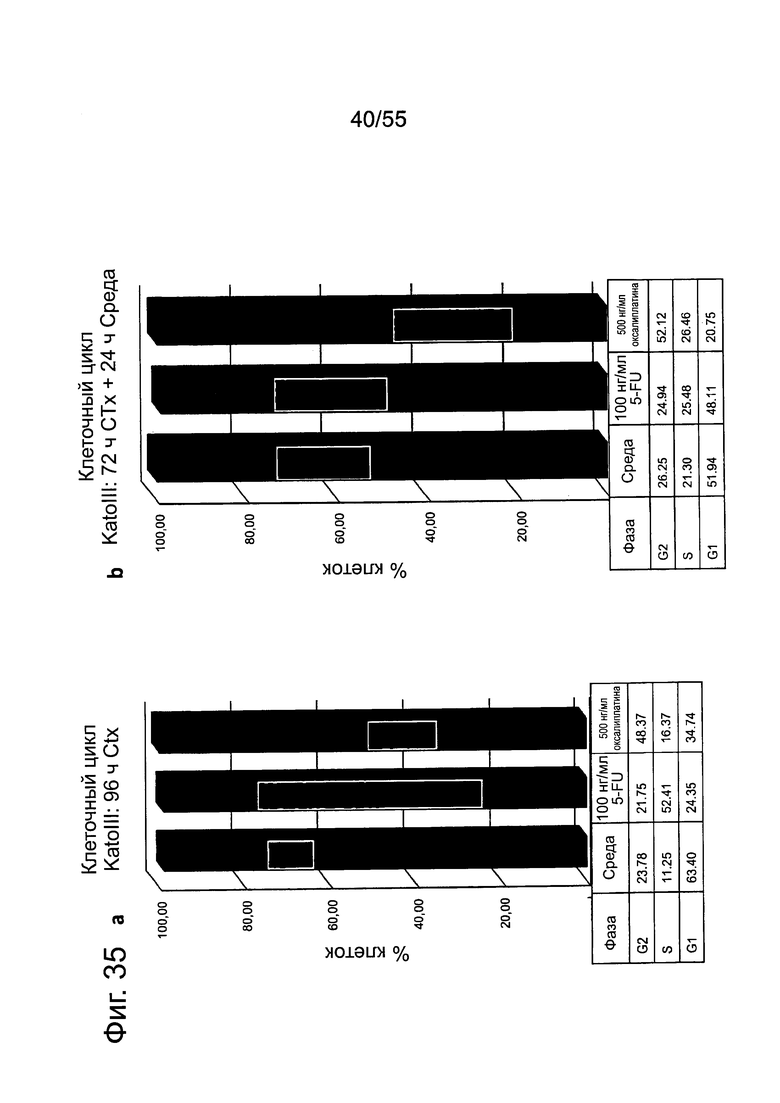
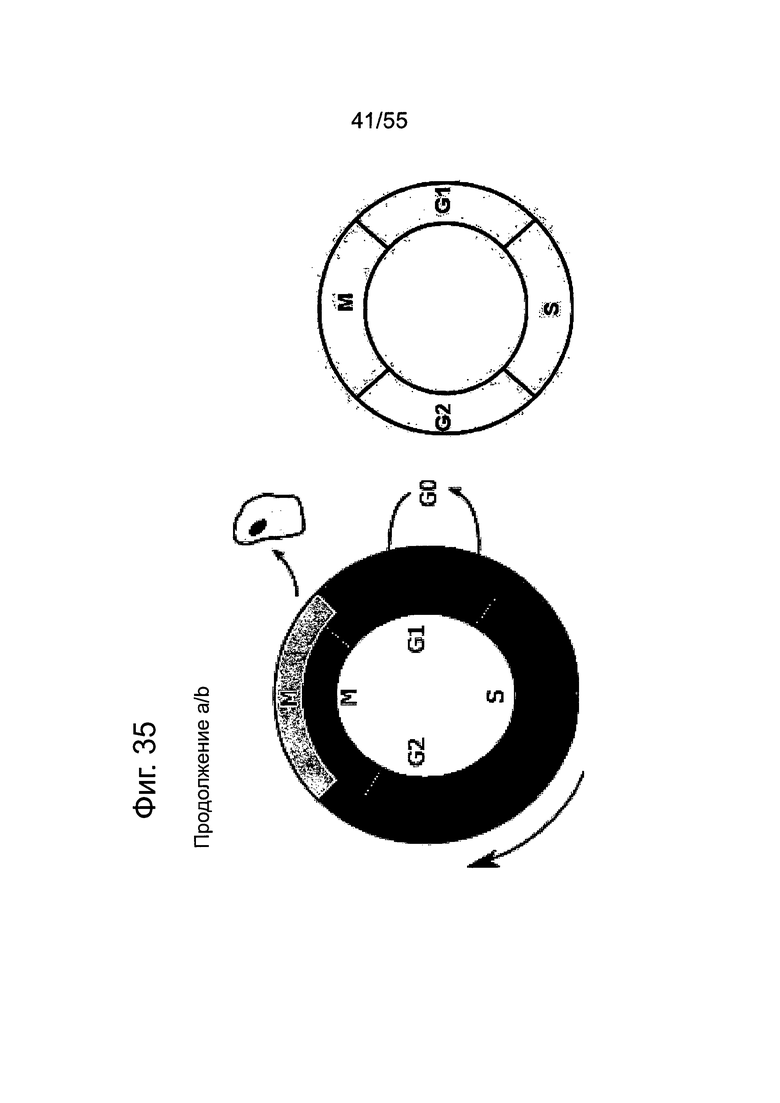
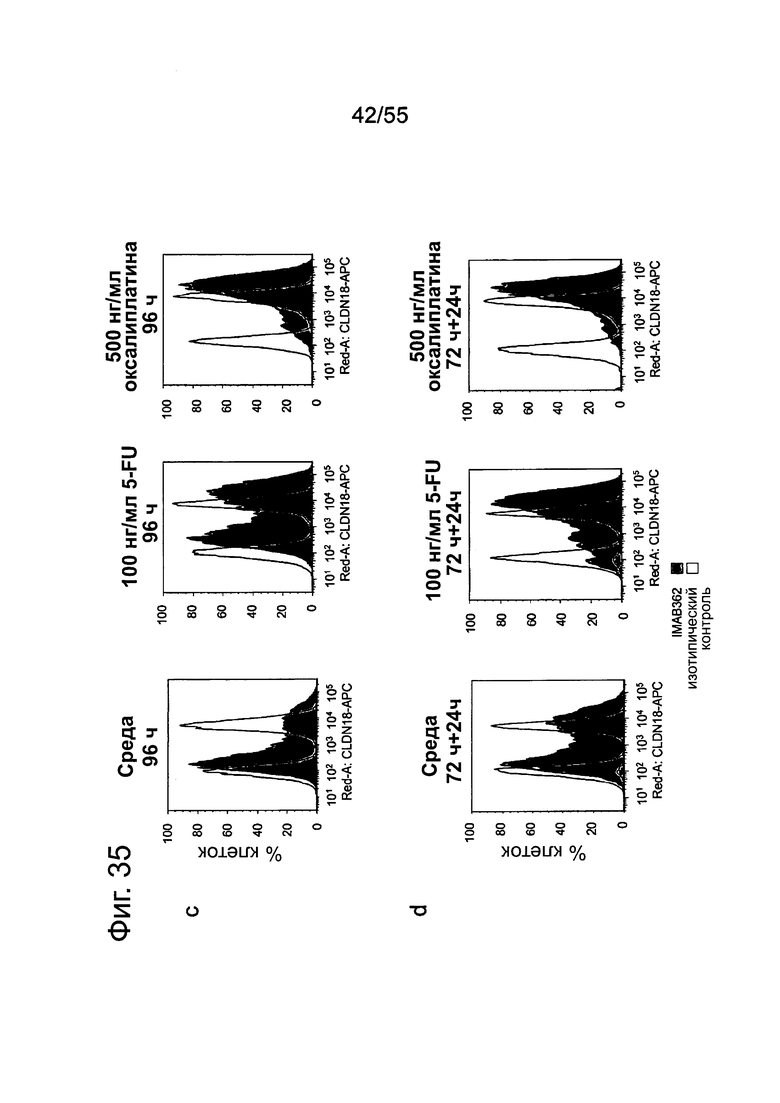

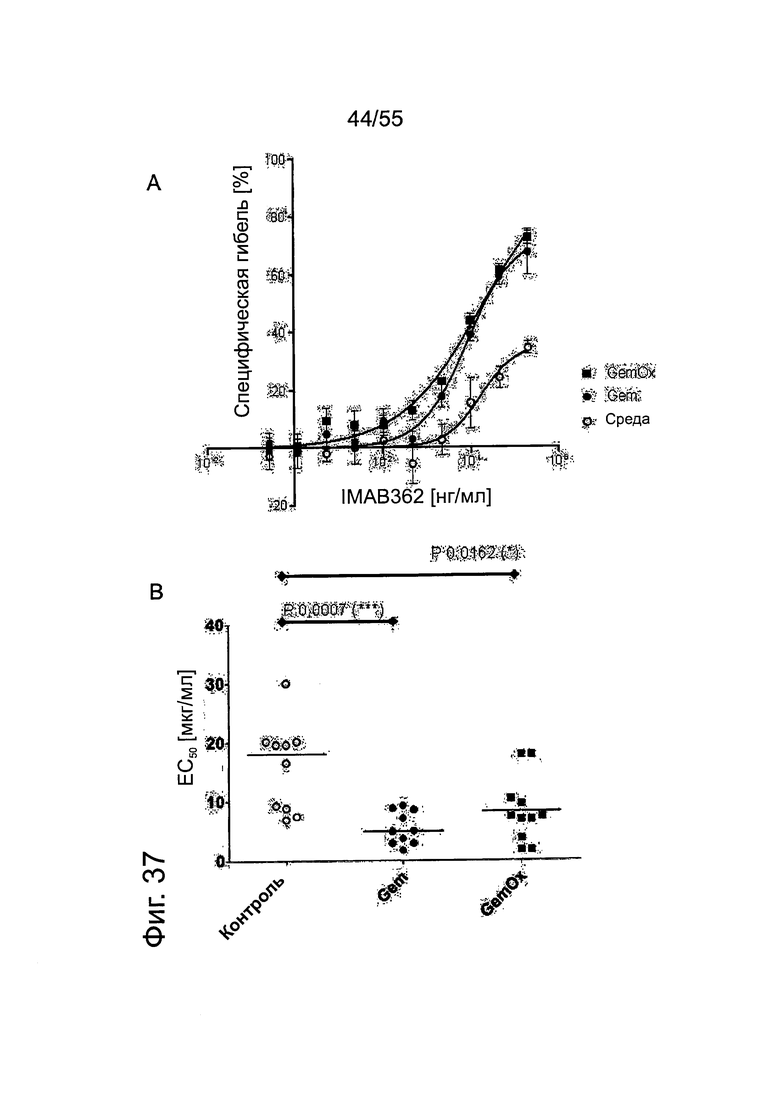
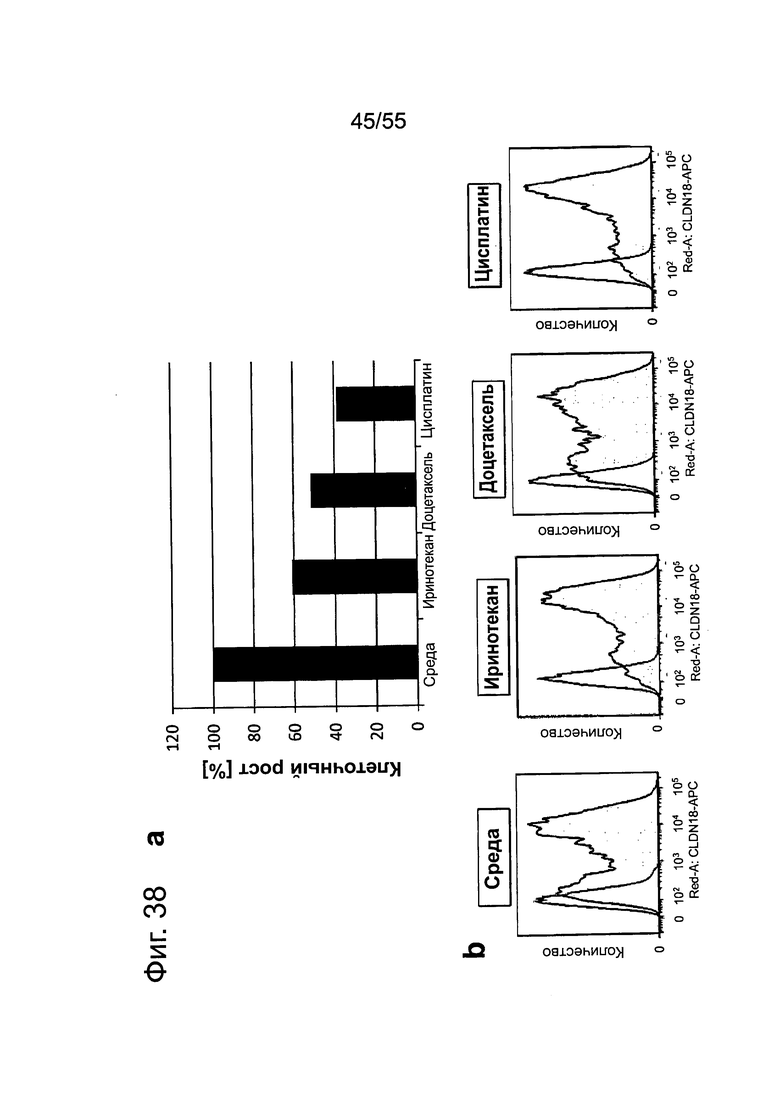
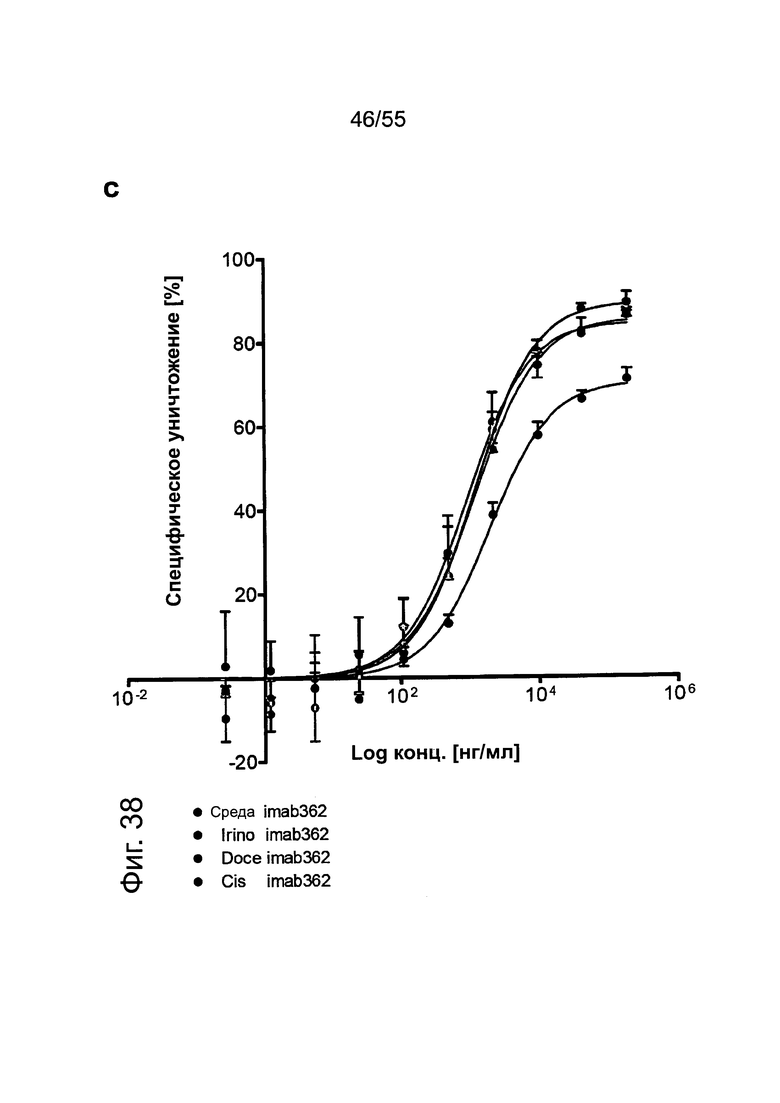
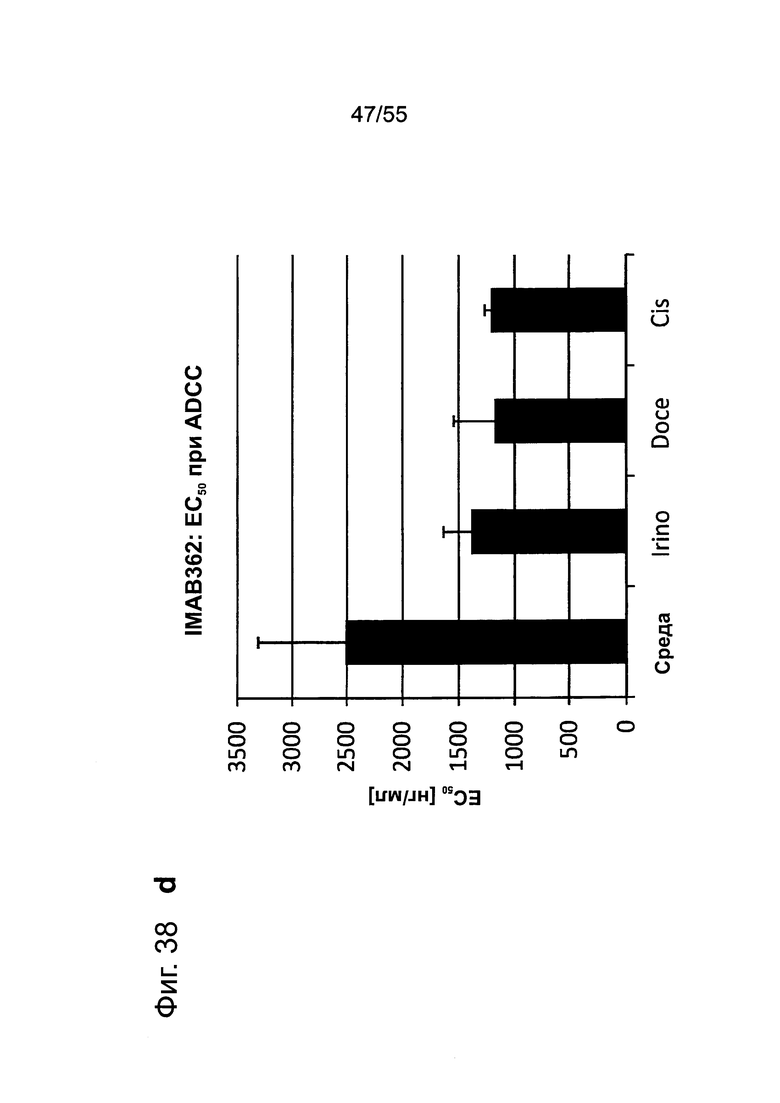
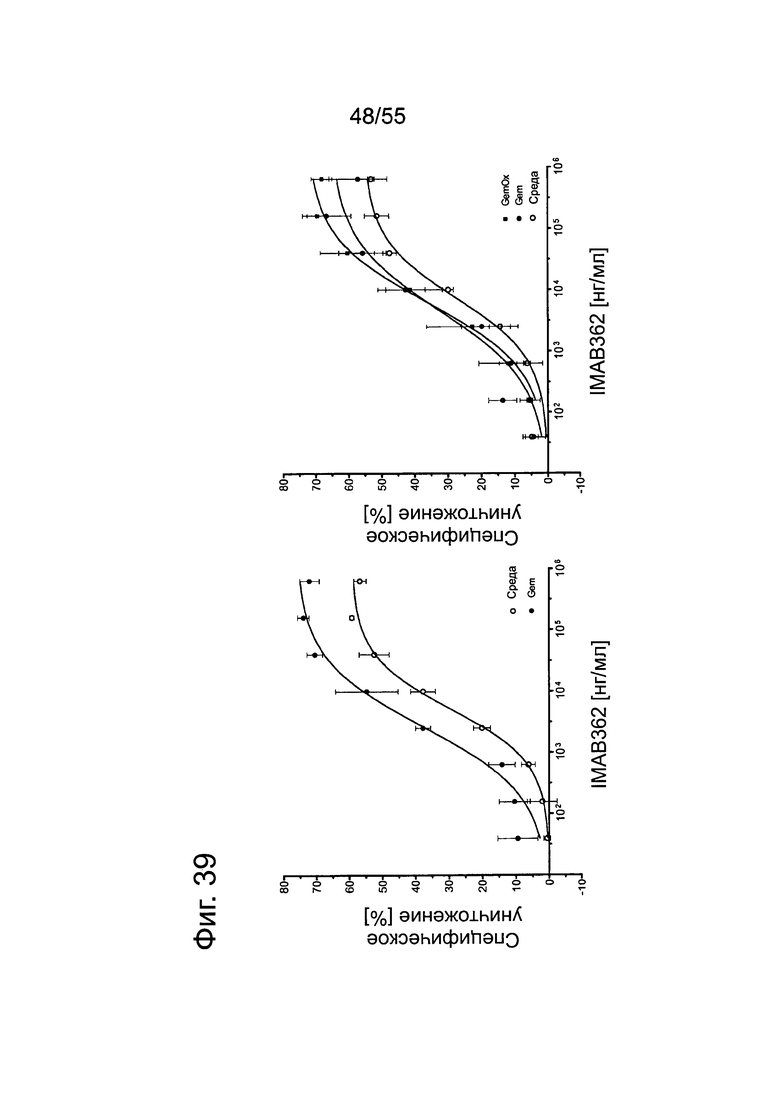
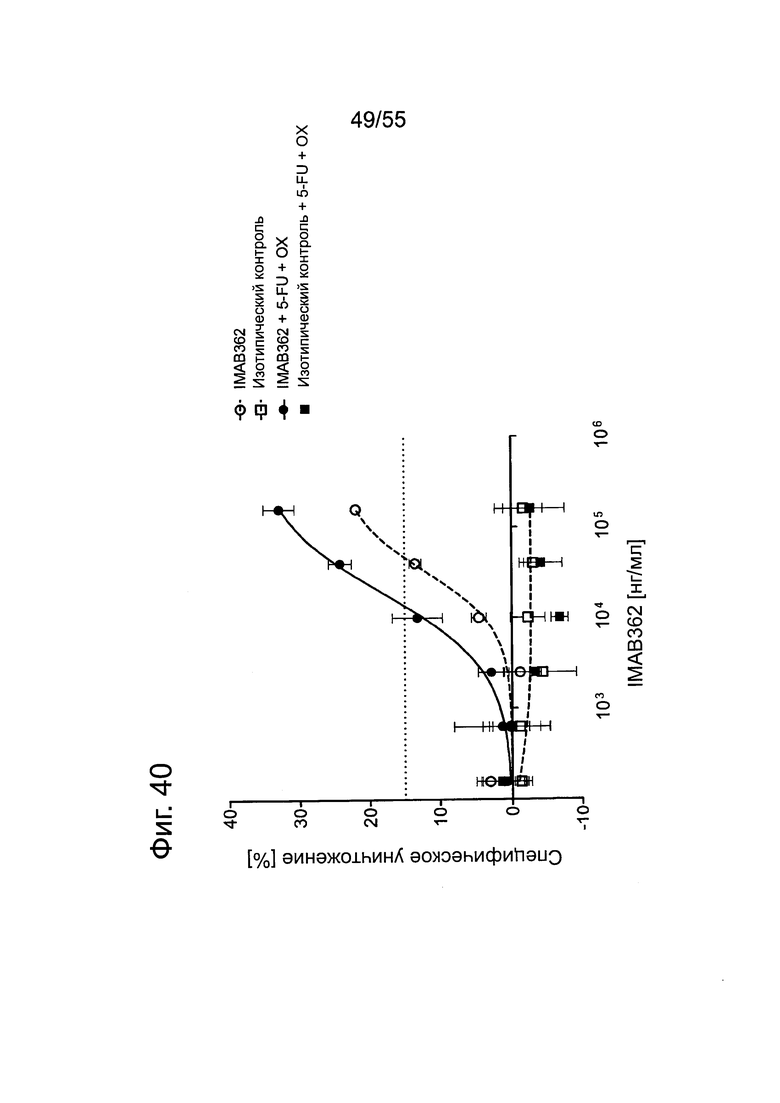
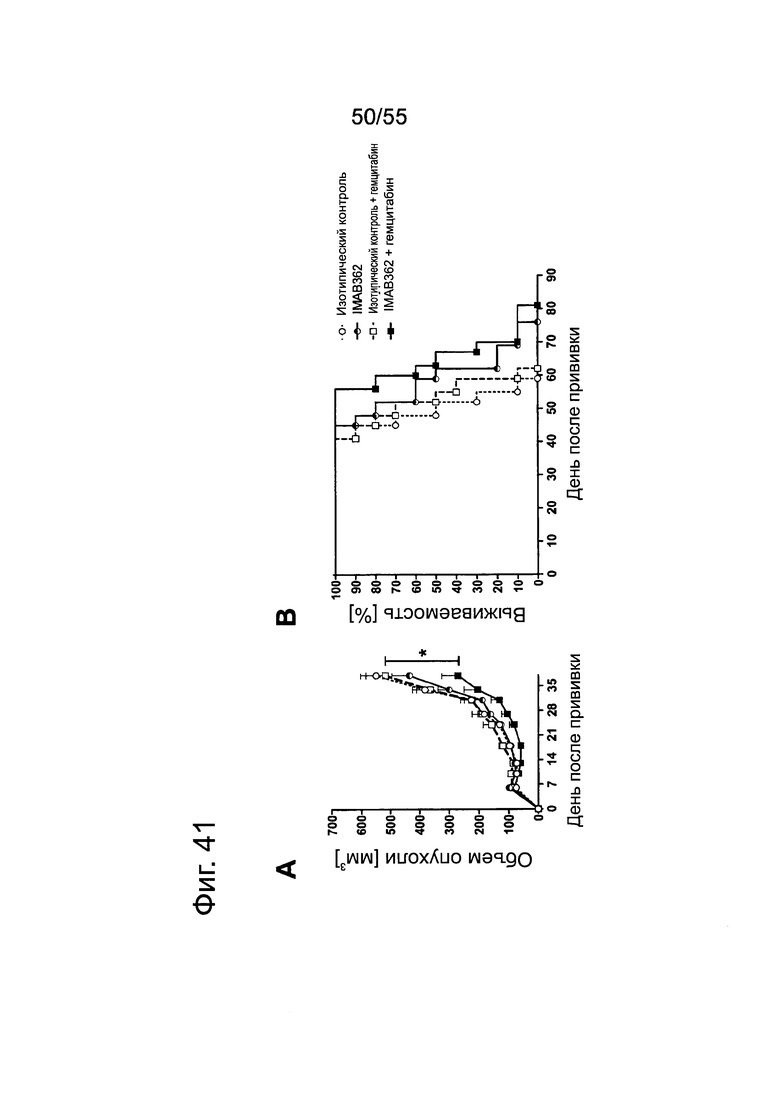
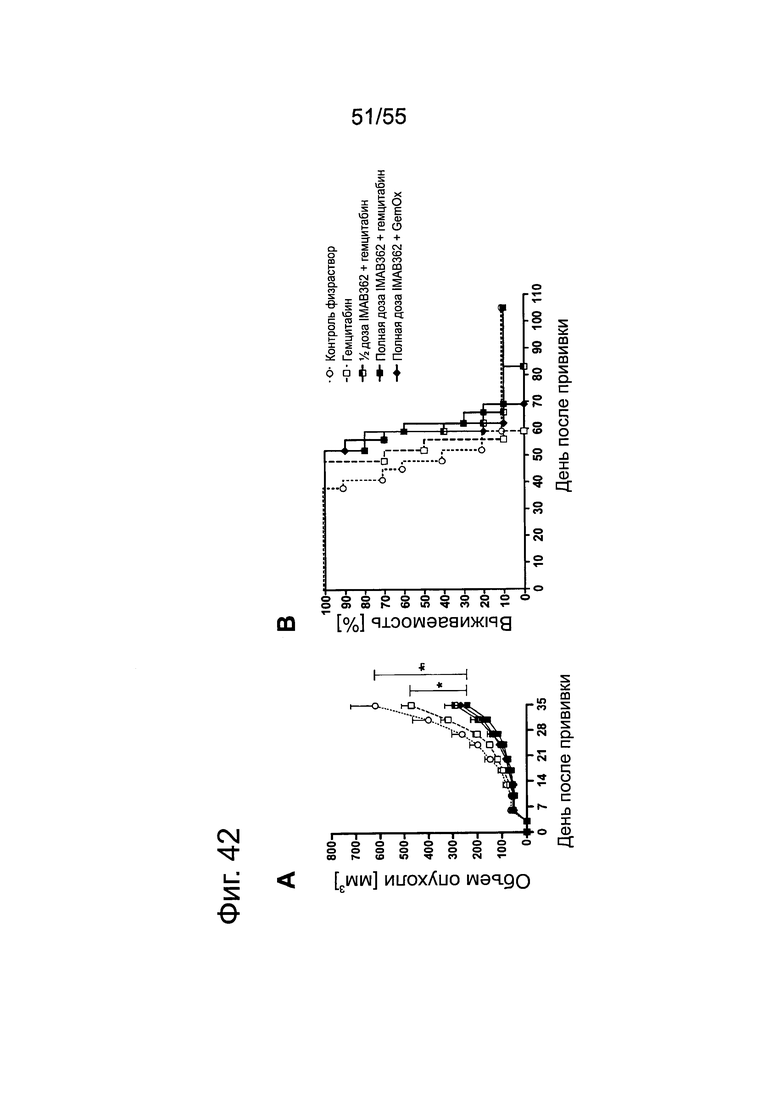
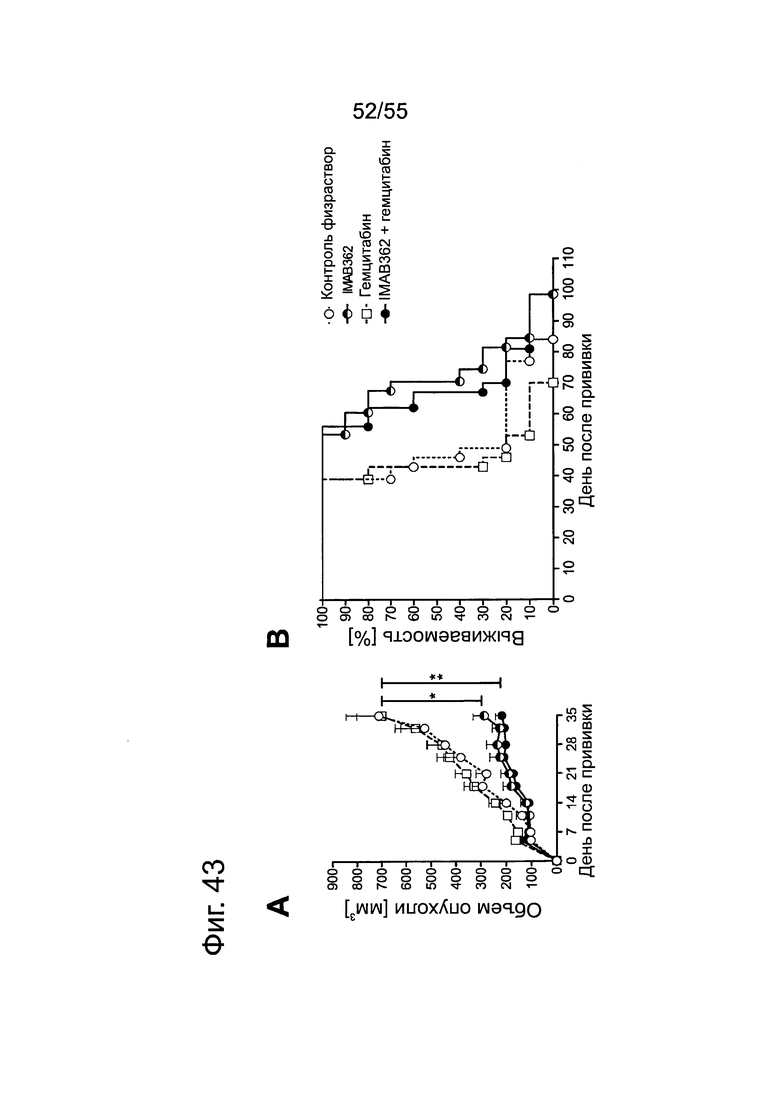
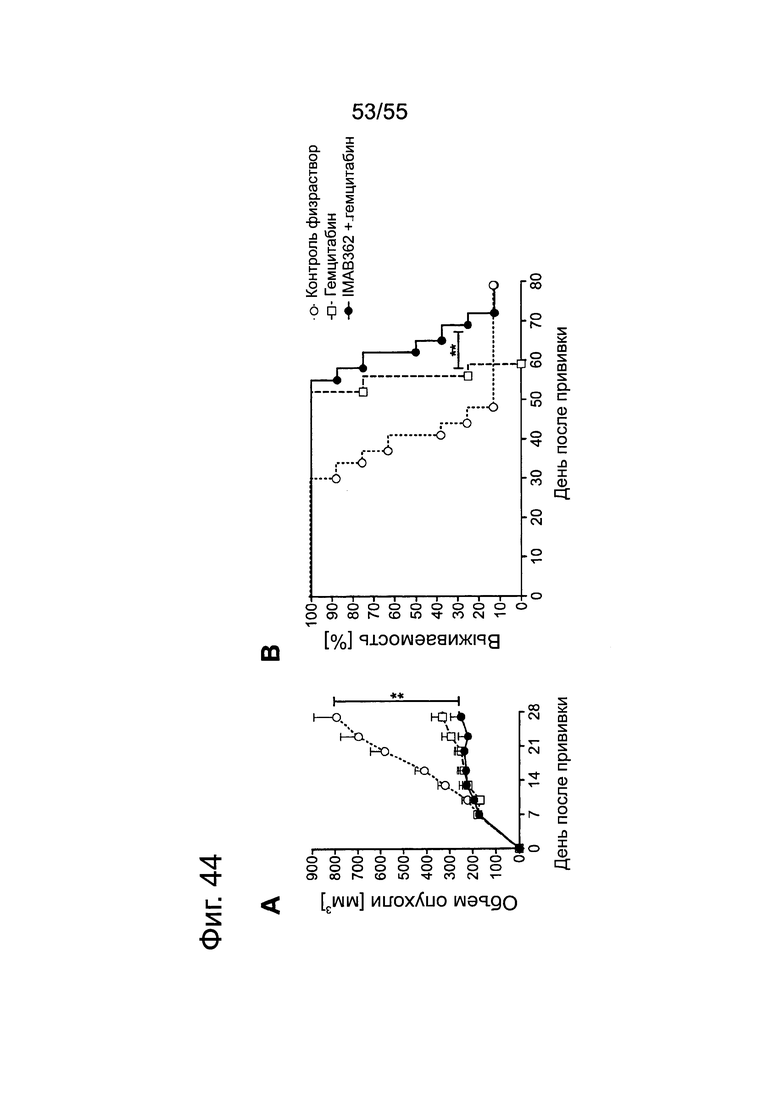

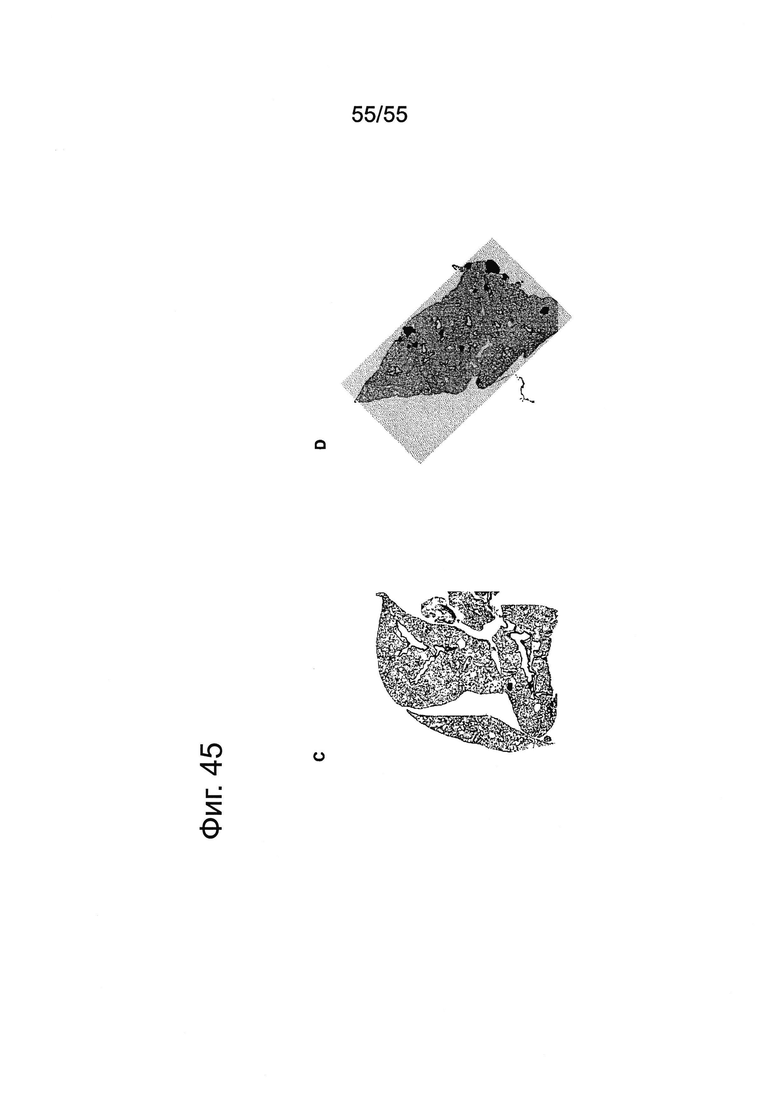
Группа изобретений относится к медицине и может быть использована для лечения рака поджелудочной железы у пациента. Для этого пациенту вводят (i) антитело, обладающее способностью к связыванию с CLDN18.2, и (ii) средство, стабилизирующее или усиливающее экспрессию CLDN18.2, где указанный рак характеризуется раковыми клетками, экспрессирующими CLDN18.2, и где (а) антитело связывается с CLDN18.2 и обеспечивает уничтожение клеток, экспрессирующих CLDN18.2, где антитело содержит тяжелую цепь антитела, содержащую аминокислотную последовательность, представленную SEQ ID NO: 32, и легкую цепь антитела, содержащую аминокислотную последовательность, представленную SEQ ID NO: 39, и (б) средство представляет собой нуклеозидный аналог, выбранный из группы, состоящей из гемцитабина и его эфира или соли. Группа изобретений обеспечивает эффективное лечение рака поджелудочной железы. 2 н. и 16 з.п. ф-лы, 55 ил., 21 табл., 10 пр.
1. Способ лечения рака поджелудочной железы у пациента, включающий введение пациенту (i) антитела, обладающего способностью к связыванию с CLDN18.2, и (ii) средства, стабилизирующего или усиливающего экспрессию CLDN18.2, где указанный рак характеризуется раковыми клетками, экспрессирующими CLDN18.2, и где
(а) антитело связывается с CLDN18.2 и обеспечивает уничтожение клеток, экспрессирующих CLDN18.2, где антитело содержит тяжелую цепь антитела, содержащую аминокислотную последовательность, представленную SEQ ID NO: 32, и легкую цепь антитела, содержащую аминокислотную последовательность, представленную SEQ ID NO: 39, и
(б) средство представляет собой нуклеозидный аналог, выбранный из группы, состоящей из гемцитабина и его эфира или соли.
2. Способ по п. 1, где экспрессия CLDN18.2 происходит на клеточной поверхности раковой клетки.
3. Способ по п. 1 или 2, где способ включает введение средства, выбранного из группы, состоящей из соединений платины, таксанов, их солей и комбинаций.
4. Способ по любому из пп. 1 или 2, где способ включает введение средства, выбранного из группы, состоящей из 5-фторурацила, оксалиплатина, иринотекана, паклитакселя, связанного с альбумином паклитакселя, их солей и комбинаций.
5. Способ по любому из пп. 1 или 2, где способ включает введение комбинации из гемцитабина и оксалиплатина, комбинации из гемцитабина и цисплатина, комбинации из гемцитабина и карбоплатина или комбинации паклитакселя или связанного с альбумином паклитакселя и гемцитабина.
6. Способ лечения рака поджелудочной железы у пациента, включающий введение пациенту (i) антитела, обладающего способностью к связыванию с CLDN18.2, и (ii) гемцитабина, где указанный рак характеризуется раковыми клетками, экспрессирующими CLDN18.2, и где антитело связывается с CLDN18.2 и обеспечивает уничтожение клеток, экспрессирующих CLDN18.2, где антитело содержит тяжелую цепь антитела, содержащую аминокислотную последовательность, представленную SEQ ID NO: 32, и легкую цепь антитела, содержащую аминокислотную последовательность, представленную SEQ ID NO: 39, и где способ включает введение бисфосфоната пациенту.
7. Способ п. 6, где бисфосфонат является азотсодержащим бисфосфонатом (аминобисфосфонатом).
8. Способ по п. 6, где бисфосфонат выбирают из группы, состоящей из золедроновой кислоты, клодроновой кислоты, ибандроновой кислоты, памидроновой кислоты, ризедроновой кислоты, минодроновой кислоты, олпадроновой кислоты, алендроновой кислоты, инкадроновой кислоты и их солей.
9. Способ по п.6, где бисфосфонат вводят в сочетании с интерлейкином-2.
10. Способ по любому из пп. 1-9, где способ включает введение антитела, обладающего способностью к связыванию с CLDN18.2, в дозе вплоть до 1000 мг/м2.
11. Способ по любому из пп. 1-9, где способ включает введение антитела, обладающего способностью к связыванию с CLDN18.2, неоднократно в дозе от 300 до 600 мг/м2.
12. Способ по любому из пп. 1-11, где рак поджелудочной железы включает первичный рак, рак на поздних стадиях или метастатический рак либо их комбинации типа сочетания первичного рака и метастатического рака поджелудочной железы.
13. Способ по пп. 1-11, где рак поджелудочной железы включает в себя рак протоков поджелудочной железы.
14. Способ по любому из пп. 1-11, где рак поджелудочной железы включает в себя аденокарциному или карциному, либо их комбинации.
15. Способ по любому из пп. 1-11, где рак поджелудочной железы включает в себя аденокарциному протоков, слизистую аденокарциному, нейроэндокринную карциному или ацинозно-клеточную карциному, либо их комбинации.
16. Способ по любому из пп. 1-11, где рак поджелудочной железы частично или полностью не поддается лечению гемцитабином типа монотерапии гемцитабином.
17. Способ по любому из пп. 1-16, где пациент подвергался операции по поводу рака поджелудочной железы.
18. Способ по любому из пп. 1-16, где у пациента есть предраковые поражения поджелудочной железы, в частности предраковые поражения поджелудочной железы, включающие возникновение злокачественных гистологических изменений в протоках поджелудочной железы.
| WO2007059997 А1, 31.05.2007 | |||
| ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ, НАПРАВЛЕННЫЕ НА РЕЦЕПТОРЫ ErbB1 | 2003 |
|
RU2354402C2 |
| CARTWRIGHT THOMAS ET AL, Cancer of the pancreas: are we making progress? A review of studies in the US Oncology Research Network.// CANCER CONTROL : JOURNAL OF THE MOFFITT CANCER CENTER OCT 2008, vol | |||
| Прибор для нагревания перетягиваемых бандажей подвижного состава | 1917 |
|
SU15A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| SAHIN UGUR ET AL, // Claudin-18 splice variant 2 is a pan-cancer target suitable for therapeutic antibody development", CLINICAL CANCER RESEARCH, THE AMERICAN ASSOCIATION FOR CANCER RESEARCH, US, (20081201), vol | |||
| Паровоз для отопления неспекающейся каменноугольной мелочью | 1916 |
|
SU14A1 |
| Прибор для равномерного смешения зерна и одновременного отбирания нескольких одинаковых по объему проб | 1921 |
|
SU23A1 |

