ОБЛАСТЬ ТЕХНИКИ
[0001] Настоящее изобретение относится к промотору, проявляющему высокую экспрессионную активностью в клетках микроорганизмов, относящихся к роду Mortierella, к вектору, содержащему такой промотор, к нечеловеческому трансформанту, трансформированному таким промотором, а также к способу получения белков, липидов или жирных кислот с помощью такого промотора или трансформанта.
УРОВЕНЬ ТЕХНИКИ
[0002] Способы получения полезных соединений посредством микробного метаболизма (ферментационные способы в широком смысле) были разработаны и применяются на практике. Например, известно, что грибки рода Mortierella (например, Mortierella alpina) производят полиненасыщенные жирные кислоты, включая арахидоновую кислоту, и являются грибками, особенно пригодными для промышленных целей (Патентный документ 1).
Для применения этих грибков проводили селекцию, т.е. вносили модификации для улучшения генетических свойств полезных организмов (улучшение штамма). В частности, в ферментационных способах селекция становится очень важной с точки зрения улучшения эффективности производства полезных соединений микроорганизмами и снижения производственной себестоимости данных соединений и т.п.
Для выведения полезных организмов, обладающих более желательными свойствами, используют способы на основе трансформации. В этом случае фрагмент ДНК, кодирующий белок, которому необходимо придать желаемое свойство, делают экспрессирующимся под контролем подходящего промотора гена, а затем вводят в подходящий организм, подлежащий селекции (т.е. в хозяина) для получения популяции трансформантов. Из этой популяции отбирают желаемый штамм (линию). Эта процедура требует наличия промотора гена, который подходит для того типа организма, который служит хозяином, или для подлежащего модификации свойства.
Были опубликованы многие способы трансформации мицеллярных грибков, к которым относятся грибки рода Mortierella. Более того, что касается способности грибков рода Mortierella производить липиды, было получено много генов ферментов, входящих в системы синтеза липидов. Однако было сделано лишь несколько сообщений о промоторах генов, необходимых для введения этих полезных генов ферментов в грибки рода Mortierella и для стимуляции их экспрессии на высоком уровне.
[0003] Патентные документы
Патентный документ 1: JP 63-044891 A
РАСКРЫТИЕ ИЗОБРЕТЕНИЯ
[0004] В этих обстоятельствах существует необходимость выведения штаммов, которые эффективно производят полезные липиды. Для достижения этой цели требуется наличие промоторов генов, подходящих для грибков рода Mortierella.
[0005] В результате экстенсивных и интенсивных усилий авторам настоящего изобретения удалось клонировать промотор гена, экспрессирующегося на высоком уровне в Mortierella alpina (M. alpina), и таким образом завершить настоящее изобретение. То есть настоящее изобретение представляет полинуклеотид, экспрессионный вектор, трансформант и способ производства белков, липидов или жирных кислот с помощью такого полинуклеотида или трансформанта, как показано ниже.
[0006] Подробнее настоящее изобретение представляет собой следующее.
[1] Полинуклеотид, соответствующий любому, выбираемому из группы, состоящей из пунктов с (а) по (с), показанных ниже:
(а) полинуклеотид, который содержит любую нуклеотидную последовательность, выбираемую из группы, состоящей из SEQ ID NO: с 1 по 28;
(b) полинуклеотид, который содержит нуклеотидную последовательность с идентичностью равной 90% или более по отношению к любой нуклеотидной последовательности, выбираемой из группы, состоящей из SEQ ID NO: с 1 по 28, и который проявляет промоторную активность в клетках микроорганизмов, относящихся к роду Mortierella; и
(c) полинуклеотид, который может гибридизоваться в жестких условиях с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной любой нуклеотидной последовательности, выбираемой из группы, состоящей из SEQ ID NO: с 1 по 28, и который проявляет промоторную активность в клетках микроорганизмов, относящихся к роду Mortierella.
[2] Полинуклеотид по указанному выше [1], в котором промоторную активность подтверждают как активность белка GUS (глюкуронидаза, glucuronidase) равную по меньшей мере 500 нмоль/(мг⋅мин) при экспрессии репортерного гена GUS в клетках микроорганизмов, относящихся к роду Mortierella.
[3] Полинуклеотид по указанному выше [1], который содержит любую нуклеотидную последовательность, выбираемую из группы, состоящей из SEQ ID NO: с 1 по 28.
[4] Полинуклеотид по вышеперечисленным [1] или [2], который является ДНК.
[5] Вектор, содержащий полинуклеотид по любому из вышеперечисленных [1]-[4].
[6] Нечеловеческий трансформант, трансформированный полинуклеотидом по любому из вышеперечисленных [1]-[4].
[7] Нечеловеческий трансформант, трансформированный вектором по указанному выше [6].
[8] Трансформант по вышеперечисленным [7] или [8], в котором трансформант является продуцирующим липид грибком.
[9] Трансформант по указанному выше [8], в котором продуцирующий липид грибок является Mortierella alpina.
[0007] При применении в качестве промотора полинуклеотид согласно настоящему изобретению обеспечивает высоко эффективную экспрессию целевого гена в клетках микроорганизмов, относящихся к роду Mortierella.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
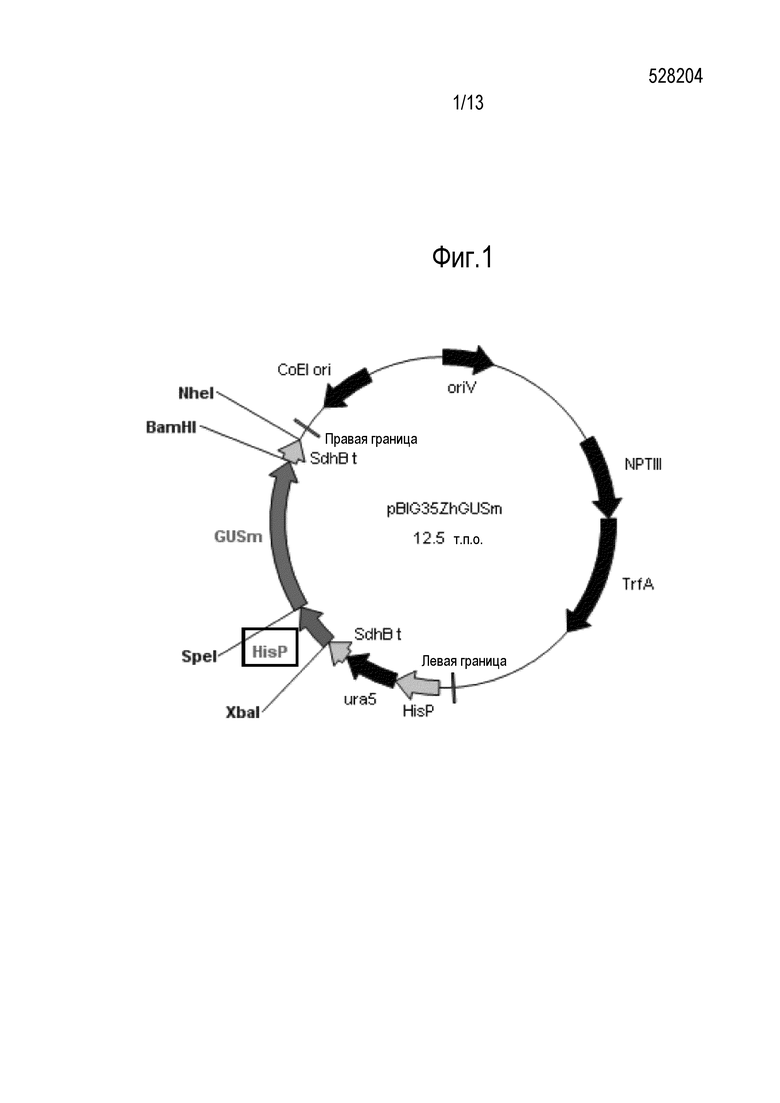
[0008] Фигура 1 показывает пример вектора для применения в оценке промотора согласно настоящему изобретению. Последовательность HisP заменена промотором согласно настоящему изобретению перед применением.
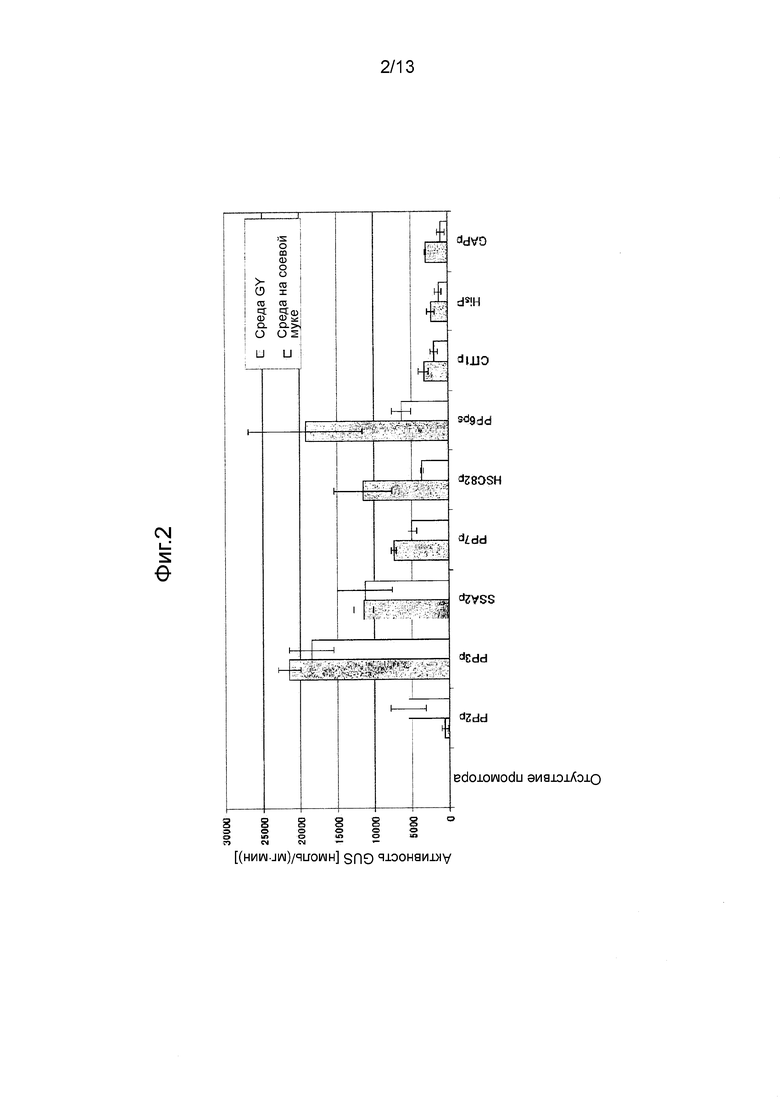
Фигура 2 показывает промоторную активность в трансформантах, трансформированных последовательностями промотора согласно настоящему изобретению, при культивировании в разных средах (серый столбец: среда GY, белый столбец: среда на соевой муке). Культивирование в каждой среде (10 мл) проводили при 28°C при 300 об/мин в течение 5 дней.
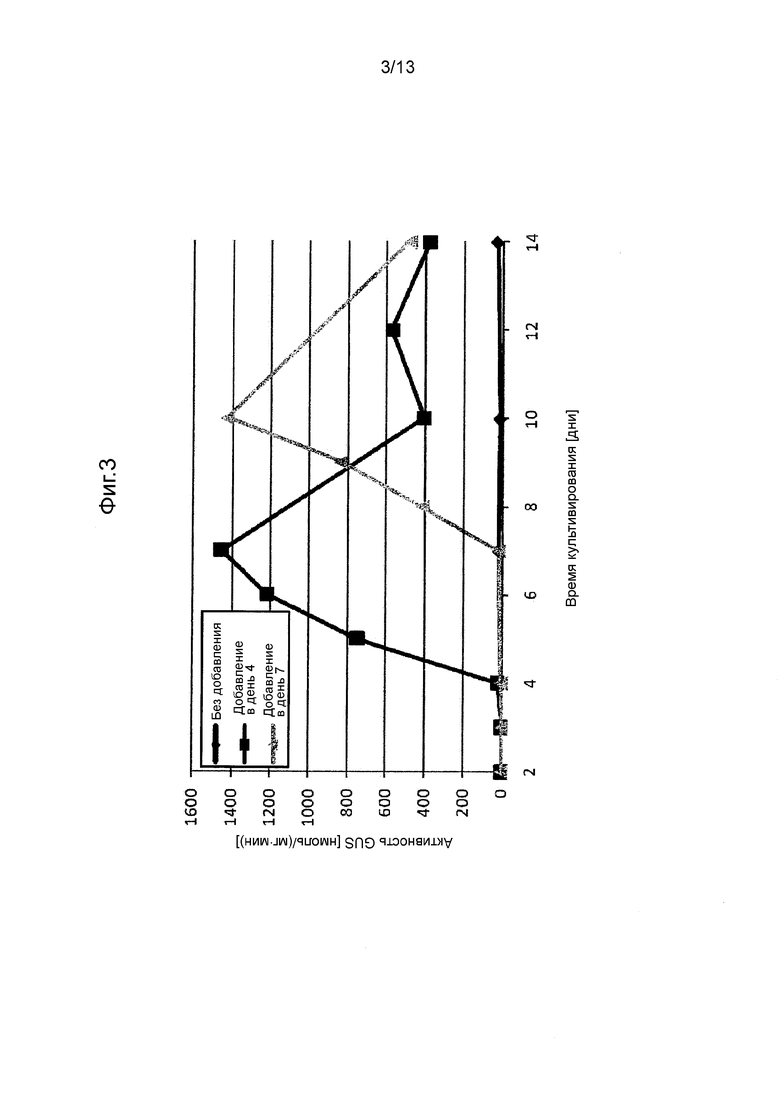
Фигура 3 показывает промоторную активность GAL10-2p. Эта фигура показывает промоторную активность, индуцированного при добавлении галактозы.
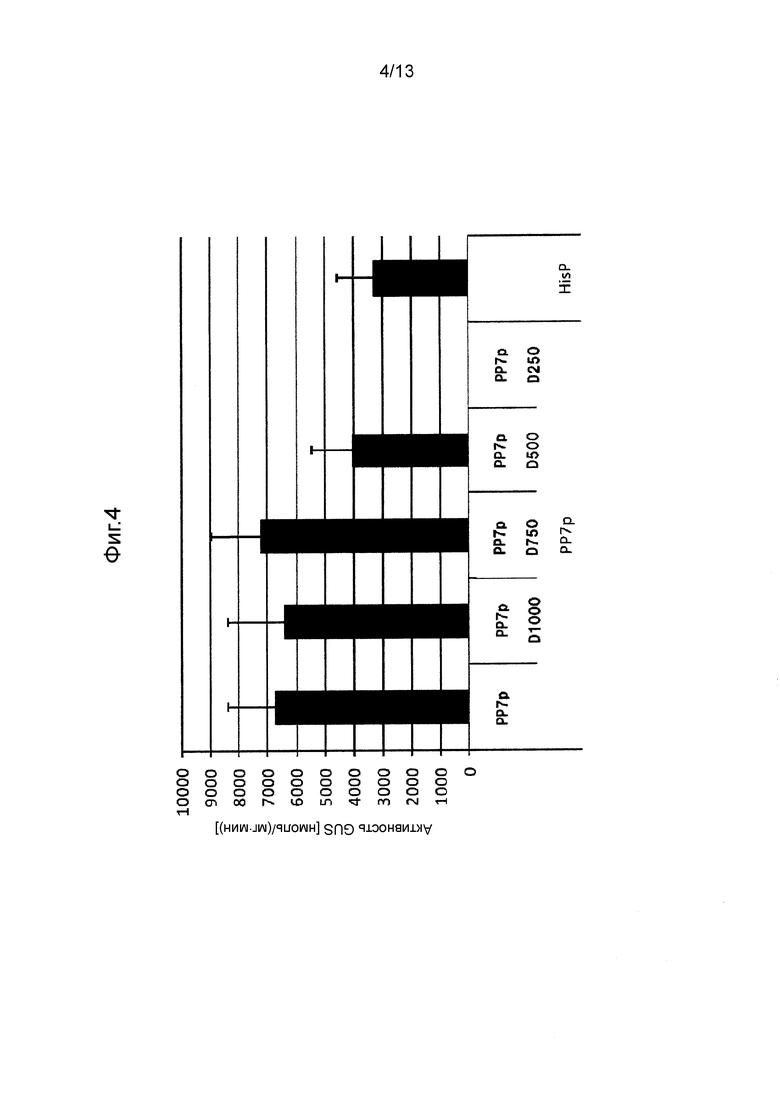
Фигура 4 показывает промоторную активность PP7p и его укороченных промоторов. Культивирование проводили в среде GY (10 мл) при 28°C при 300 об/мин в течение 5 дней.
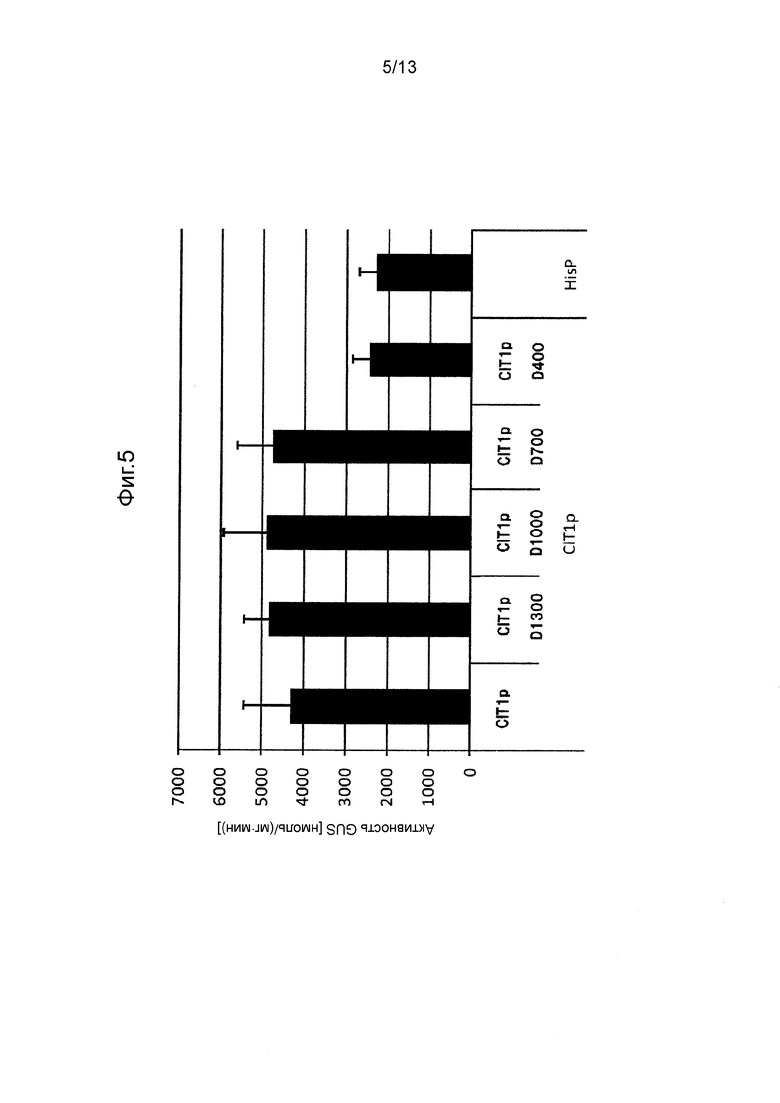
Фигура 5 показывает промоторную активность CIT1p и его укороченных промоторов. Культивирование проводили в среде GY (10 мл) при 28°C при 300 об/мин в течение 3 дней.
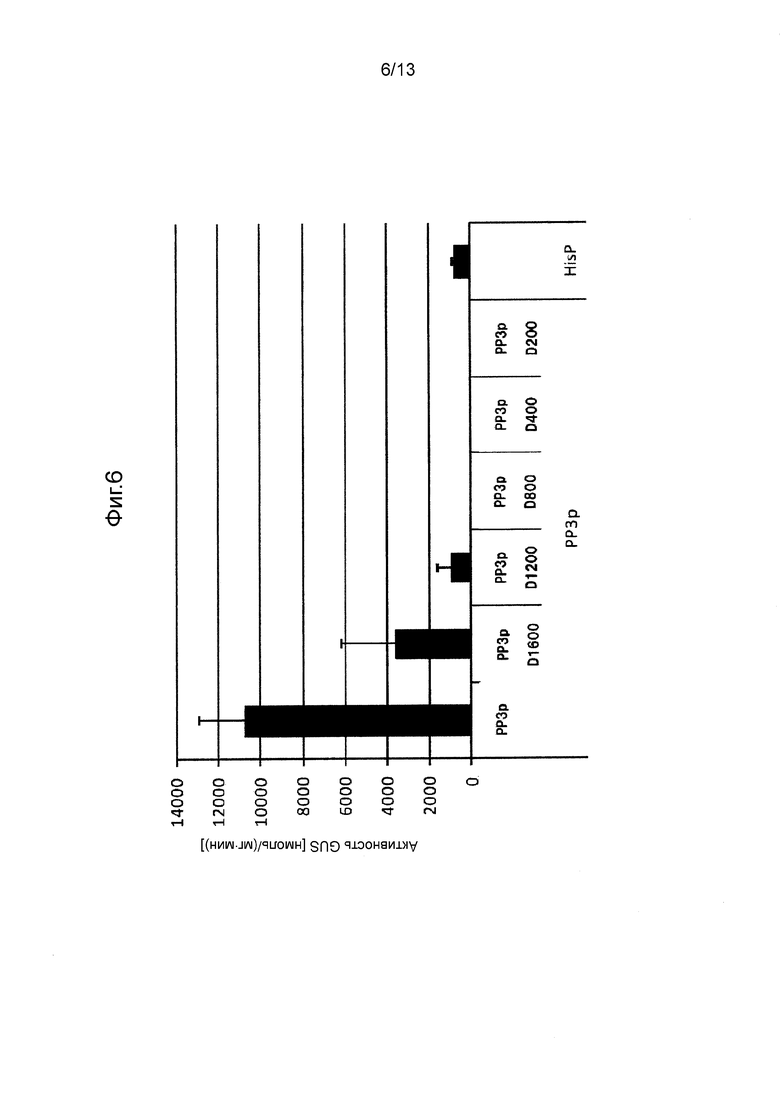
Фигура 6 показывает промоторную активность PP3p и его укороченных промоторов. Культивирование проводили в среде GY (10 мл) при 28°C при 300 об/мин в течение 10 дней.
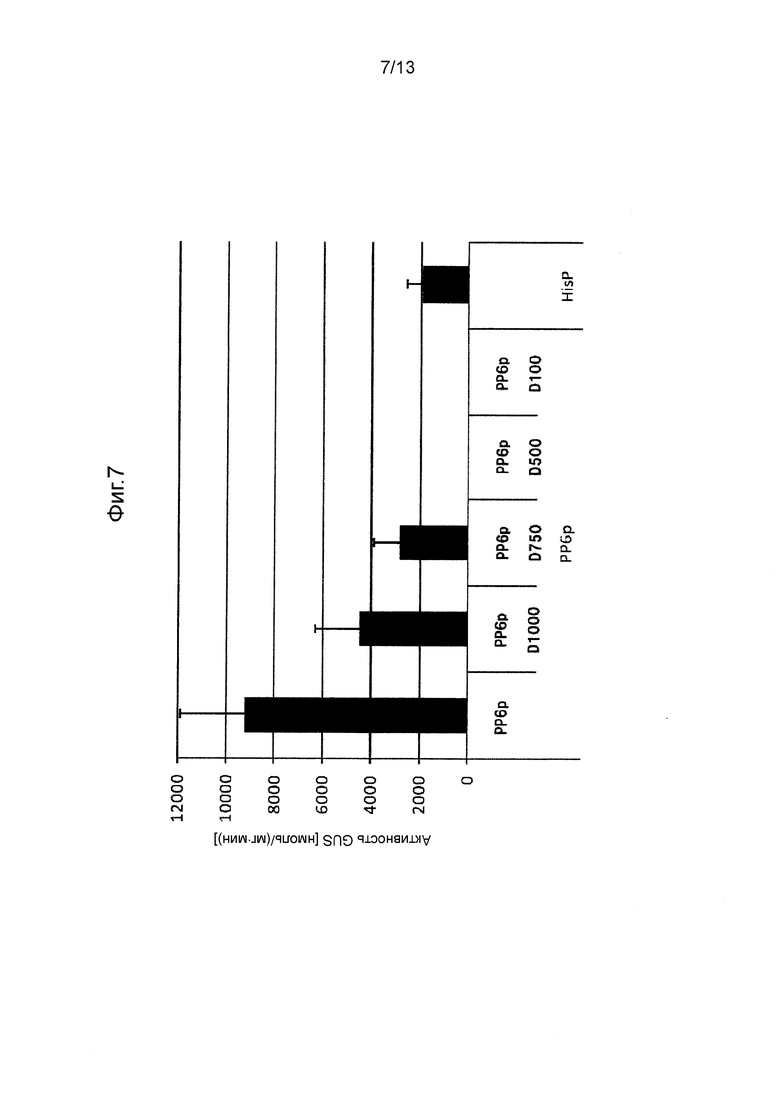
Фигура 7 показывает промоторную активность PP6p и его укороченных промоторов. Культивирование проводили в среде GY (10 мл) при 28°C при 300 об/мин в течение 5 дней.
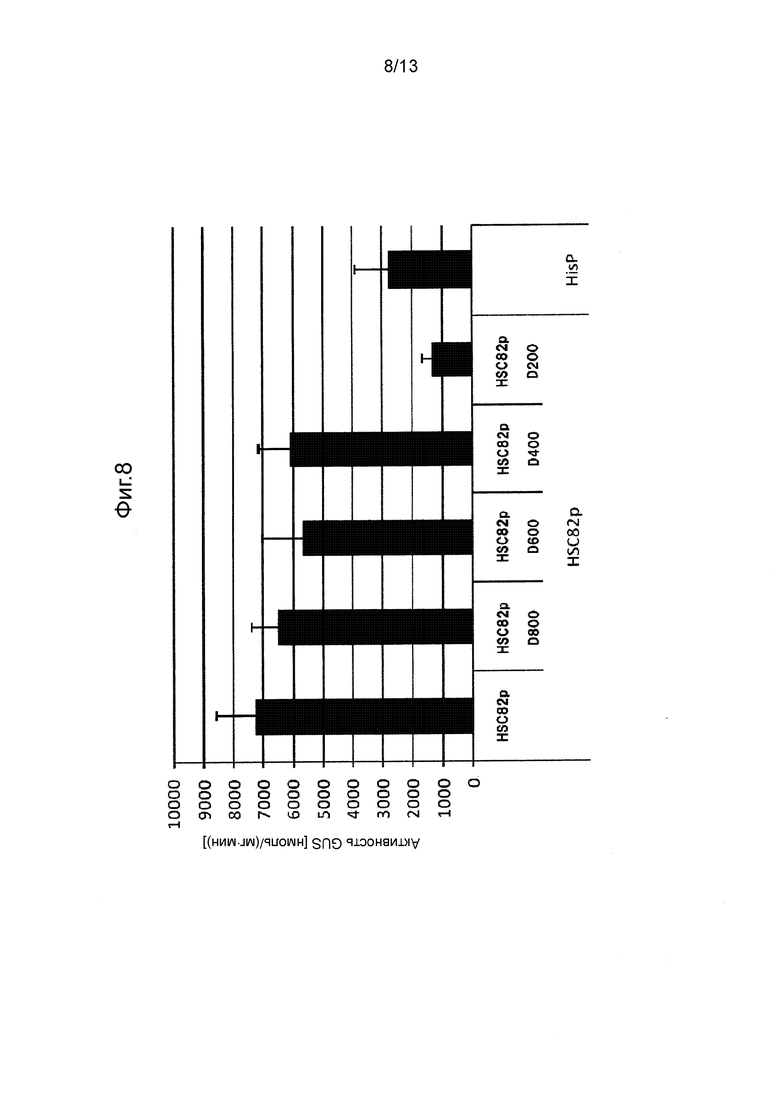
Фигура 8 показывает промоторную активность HSC82p и его укороченных промоторов. Культивирование проводили в среде GY (10 мл) при 28°C при 300 об/мин в течение 5 дней.
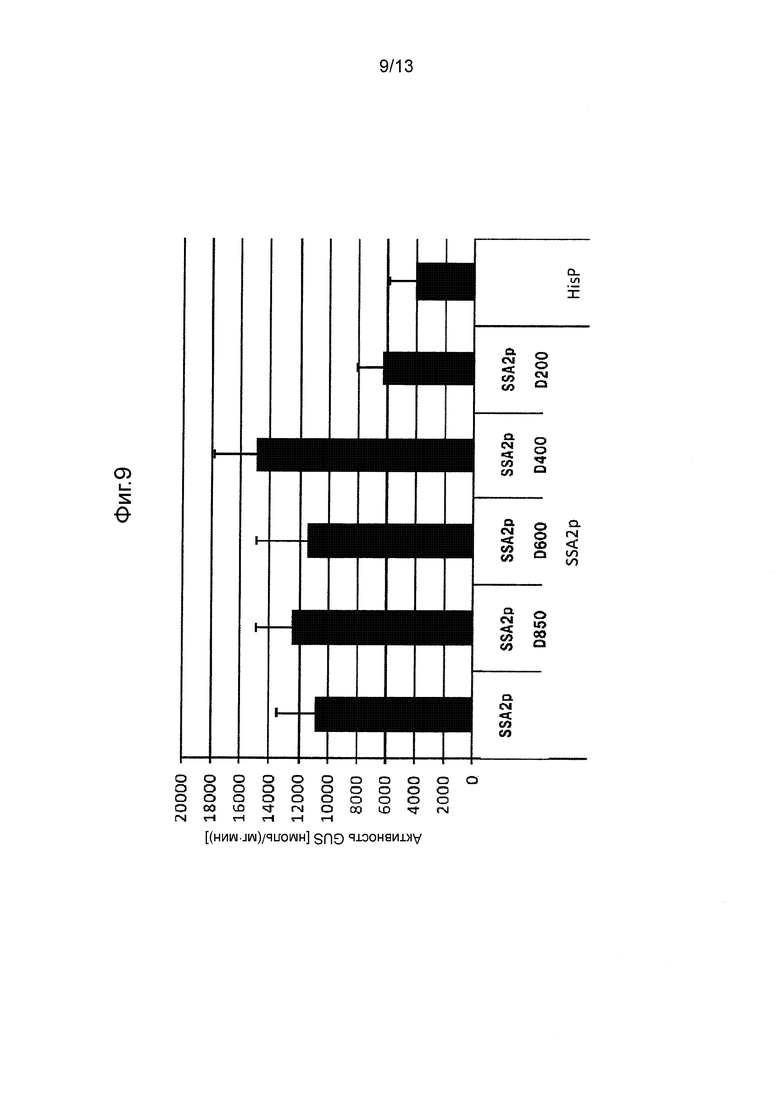
Фигура 9 показывает промоторную активность SSA2p и его укороченных промоторов. Культивирование проводили в среде GY (10 мл) при 28°C при 300 об/мин в течение 5 дней.
Фигура 10 показывает промоторную активность GAPp и его укороченных промоторов. Культивирование проводили в среде GY (10 мл) при 28°C при 300 об/мин в течение 5 дней.
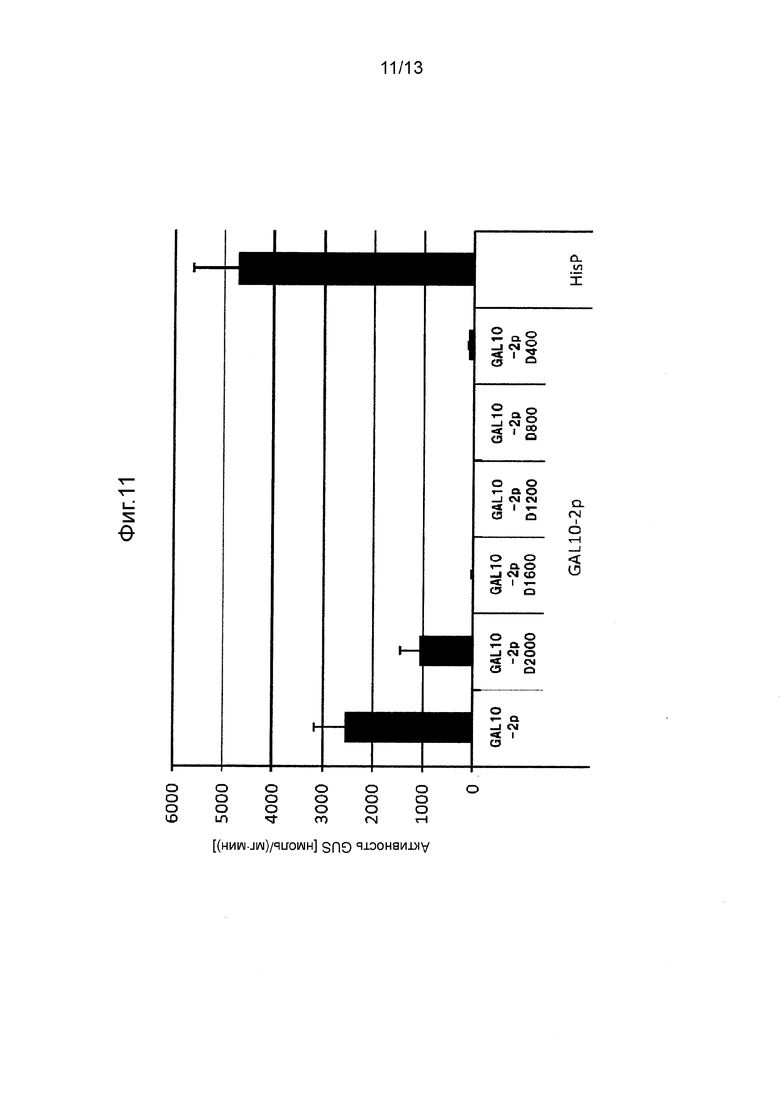
Фигура 11 показывает промоторную активность GAL10-2p и его укороченных промоторов.
Фигура 12A показывает выравнивание гена GUS из E. coli (последовательность кДНК: SEQ ID NO: 29, аминокислотная последовательность: SEQ ID NO: 30) и гена GUSm (последовательность кДНК: SEQ ID NO: 31, аминокислотная последовательность: SEQ ID NO: 32), который был модифицирован таким образом, чтобы адаптировать применение кодонов гена GUS из E. coli к микроорганизмам рода Mortierella.
Фигура 12B является продолжением Фигуры 12A.
НАИЛУЧШИЙ СПОСОБ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
[0009] Ниже настоящее изобретение будет описано более подробно. Следующие варианты осуществления настоящего изобретения приведены для описания настоящего изобретения, и не следует считать, что настоящее изобретение ограничивается только этими вариантами осуществления настоящего изобретения. Настоящее изобретение можно выполнить в разных режимах без отступления от сущности настоящего изобретения.
Следует отметить, что все публикации, процитированные в данной заявке, включая документы, касающиеся предшествующего уровня техники, патентные бюллетени и другие патентные документы, включены в настоящую заявку посредством ссылки. Более того, это описание изобретения включает содержимое, раскрытое в описании изобретения и чертежах заявки на патент Японии № 2013-066265 (поданной 27 марта 2013 г.), на основании которой по настоящей заявке испрашивается приоритет.
[0010] Если иное не определено в настоящей заявке, нуклеотидные последовательности показаны так, что их 5'-конец находится слева, а 3'-конец справа.
[0011] 1. Промоторы
Авторам настоящего изобретения удалось, опережая других, клонировать несколько типов промоторных последовательностей из продуцирующего липид грибка M. alpina, как описано более подробно далее в разделе Примеров. Более того, авторы настоящего изобретения также подтвердили, что белки, экспрессирующиеся под данными промоторами, проявляют биологическую активность.
Промоторами, в соответствии с настоящим изобретением, являются PP7p, CIT1p, PP3p, PP2p, PP6ps, HSC82p, SSA2p, GAL10-2p и/или их частичными последовательностями (укороченными последовательностями). Данные последовательности промоторных областей и их укороченные последовательности показаны в таблице ниже.
Любая последовательность, выбранная из нуклеотидных последовательностей, показанных в таблице, т.е. любая последовательность, выбранная из группы, состоящая из SEQ ID NO: с 1 по 28, здесь и далее называют «последовательностью промотора согласно настоящему изобретению».
[0012] [Таблица 1]
(SEQ ID NO)
(SEQ ID NO)
[0013] Таким образом, настоящее изобретение относится к следующему полинуклеотиду в качестве промотора, проявляющего высокую экспрессионную активность в клетках микроорганизмов, относящихся к роду Mortierella.
Полинуклеотид, соответствующий любому, выбираемому из группы, состоящей из пунктов с (а) по (с), показанных ниже:
(а) полинуклеотид, который содержит последовательность промотора согласно настоящему изобретению;
(b) полинуклеотид, который содержит нуклеотидную последовательность с идентичностью равной 90% или более по отношению к последовательности промотора согласно настоящему изобретению и который проявляет промоторную активность в клетках микроорганизмов, относящихся к роду Mortierella; и
(c) полинуклеотид, который может гибридизоваться в жестких условиях с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной последовательности промотора согласно настоящему изобретению, и который проявляет промоторную активность в клетках микроорганизмов, относящихся к роду Mortierella.
[0014] Каждый из указанных выше полинуклеотидов, показанных в п. с (а) по (с), здесь и далее называют «полинуклеотид согласно настоящему изобретению».
Более того, в контексте настоящего изобретения «обладающий» последовательностью промотора согласно настоящему изобретению означает «содержащий» последовательность промотора согласно настоящему изобретению. Таким образом, дополнительную последовательность (последовательности) (например, последовательность энхансера), отличную от последовательности промотора согласно настоящему изобретению, можно добавить до (с 5'-конца) или после (с 3'-конца) последовательности промотора согласно настоящему изобретению. Такую дополнительную последовательность можно добавить к последовательности промотора согласно настоящему изобретению через нуклеотидную последовательность длиной от 1 до 1000 п.о., от 1 до 900 п.о., от 1 до 800 п.о., от 1 до 700 п.о., от 1 до 600 п.о., от 1 до 500 п.о., от 1 до 400 п.о., от 1 до 300 п.о., от 1 до 200 п.о., от 1 до 100 п.о., от 1 до 75 п.о., от 1 до 50 п.о., от 1 до 25 п.о., от 1 до 10 п.о. или в качестве альтернативы можно напрямую добавить к последовательности промотора согласно настоящему изобретению (т.е. число нуклеотидных остатков, расположенных между последовательностью промотора согласно настоящему изобретению и дополнительной последовательностью, равно 0).
[0015] В рамках изобретения термин «полинуклеотид» предназначен для обозначения ДНК или РНК.
В рамках изобретения выражение «полинуклеотид, который может гибридизоваться в жестких условиях» предназначено для обозначения, например, полинуклеотида, который можно получить посредством, например, гибридизации колоний, гибридизации бляшек или Саузерн гибридизации, применяя в качестве зонда целый полинуклеотид или его часть, состоящий из нуклеотидной последовательности, комплементарной последовательности промотора согласно настоящему изобретению. Для гибридизации можно использовать способы, как описанные, например, в "Sambrook & Russell, Molecular Cloning: A Laboratory Manual Vol. 3, Cold Spring Harbor, Laboratory Press 2001" and "Ausubel, Current Protocols in Molecular Biology, John Wiley & Sons 1987-1997."
[0016] В рамках изобретения термин «жесткие условия» предназначен для обозначения, например, условий (1) 5 × цитратно-солевой буфер, 5 × раствор Денхардта, 0,5% додецилсульфата натрия, 50% формамида и 50°C, (2) 0,2 × цитратно-солевой буфер, 0,1% додецилсульфата натрия и 60°C, (3) 0,2 × цитратно-солевой буфер, 0,1% додецилсульфата натрия и 62°C, (4) 0,2 × цитратно-солевой буфер, 0,1% додецилсульфата натрия и 65°C или (5) 0,1 × цитратно-солевой буфер, 0,1% додецилсульфата натрия и 65°C, не ограничиваясь ими. В данных условиях ожидают, что ДНК, обладающую большей идентичностью последовательности, можно более эффективно получить при более высокой температуре. Однако на жесткость гибридизации будет влиять множество факторов, включая температуру, концентрацию зонда, длину зонда, ионную силу, время реакции, концентрацию соли и т.д. Специалисты в данной области техники смогут добиться такой же жесткости, должным образом подбирая данные факторы.
[0017] Следует отметить, что при использовании коммерческого набора для гибридизации в этих целях в качестве примера можно применять систему прямого мечения и детектирования Alkphos Direct Labelling and Detection System (GE Healthcare). В этом случае гибридизацию можно проводить в соответствии с протоколом, включенным в набор, т.е. мембрану можно инкубировать в течение ночи с меченным зондом, а затем промывать первичным промывочным буфером, содержащим 0,1% (вес/объем) додецилсульфата натрия, в условиях 55°C для детектирования гибридизованной ДНК. В качестве альтернативы, при применении коммерческого реактива (например, смеси для мечения ПЦР PCR labeling mix (Roche Diagnostics)) для мечения зонда дигоксигенином (DIG, digoxigenin) во время подготовки зонда на основе целой или части нуклеотидной последовательности, комплементарной последовательности промотора согласно настоящему изобретению можно применять набор для детектирования нуклеиновых кислот DIG nucleic acid detection kit (Roche Diagnostics) для детектирования гибридизации.
[0018] В дополнение к перечисленным выше другие способные к гибридизации полинуклеотиды включают полинуклеотиды и идентичностью равной 90% или более, 91% или более, 92% или более, 93% или более, 94% или более, 95% или более, 96% или более, 97% или более, 98% или более, 99% или более, 99,1% или более, 99,2% или более, 99,3% или более, 99,4% или более, 99,5% или более, 99,6% или более, 99,7% или более, 99,8% или более, или 99,9% или более по отношению к последовательности промотора согласно настоящему изобретению, по данным расчета с помощью программ поиска гомологии, таких как FASTA или BLAST с параметрами, заданными по умолчанию.
[0019] Следует отметить, что идентичность нуклеотидных последовательностей можно определить с помощью FASTA (Science 227 (4693): 1435-1441, (1985)) или алгоритма Карлин и Альтшуль (Karlin and Altschul), BLAST (Basic Local Alignment Search Tool) (Proc. Natl. Acad. Sci. USA 872264-2268, 1990; Proc Natl Acad Sci USA 90: 5873, 1993). На основании алгоритма BLAST, были разработаны программы, называемые blastn, blastx, tblastn и tblastx (Altschul SF, et al: J Mol Biol 215: 403, 1990). При применении blastn для анализа нуклеотидной последовательности можно задать параметры, например, такие вес = 100 и длина слова = 12. При применении программ BLAST и Gapped BLAST в каждой программе можно применять параметры, заданные по умолчанию.
[0020] В контексте настоящего изобретения термин «промоторная активность» предназначен для обозначения того, что при введении последовательности кодирующего белок гена (здесь и далее называемого «целевой ген») после промотора согласно настоящему изобретению получают продукта экспрессии данного гена.
В рамках изобретения термин «продукт экспрессии» предназначен для обозначения либо РНК (например, гяРНК (гетерогенной ядерной РНК), мРНК, миРНК (малой интерферирующей РНК) или микроРНК), которая является продуктом транскрипции данного гена, либо белка, который является продуктом трансляции данного гена, либо и РНК, и белка.
[0021] Введение целевого гена можно провести так, чтобы 5’-конец целевого гена был расположен в области в пределах 500 п.о., 400 п.о., 300 п.о., 200 п.о., 100 п.о., 50 п.о., 30 п.о. или 10 п.о. от 3’-конца последовательности промотора согласно настоящему изобретению.
[0022] При попытке подтвердить активность последовательности промотора согласно настоящему изобретению целевой ген никак не ограничивают, но предпочитают ген, кодирующий белок, чью активность можно измерить установленным способом.
[0023] Примеры такого гена включают, но не ограничиваются выбором селективных маркерных генов, таких как гена устойчивости к неомицину, гена гидромицин B фосфотрансферазы и т.д., а также экспрессионных репортерных генов, таких как генов LacZ, GFP (Green Fluorescence Protein, зеленого флуоресцентного белка) и люциферазы и пр.
[0024] Предпочтительно, подтверждать промоторную активность можно, применяя ген β-D-глюкуронидазы (GUS) для измерения активности GUS. В тех случаях, когда M. alpina применяют в качестве хозяина, предпочтительно, чтобы ген GUS был геном GUSm, частота применения кодонов которого адаптирована к M. alpina.
Активность GUS можно измерить, как описано ниже: применить последовательность промотора согласно настоящему изобретению, чтобы вызвать экспрессию гена GUS в клетках микроорганизмов, относящихся к роду Mortierella, затем запустить реакцию белка GUS, собранного с клеток, с п-нитрофенил-β-D-глюкуронидом и в реакционной смеси измерить поглощение в течение времени при длине волны 405 нм, а затем провести расчет на основании измеренных значений в соответствии со следующим уравнением:
Активность GUS (нмоль/(мг⋅мин)) =
1000 × [(величина наклона графика зависимости поглощения от времени для каждого образца)/(величина наклона калибровочного графика)]/[(концентрация белка в образце)/5]
[0025] Ген GUS, применяемый с этой целью, как правило, является геном GUS из E. coli (последовательность кДНК: SEQ ID NO: 29, аминокислотная последовательность: SEQ ID NO: 30). В тех случаях, когда последовательность промотора согласно настоящему изобретению применяют, чтобы вызвать экспрессию гена GUS в клетках микроорганизмов, относящихся к роду Mortierella, можно применять ген GUSm (последовательность кДНК: SEQ ID NO: 31, аминокислотная последовательность: SEQ ID NO: 32), который был модифицирован таким образом, чтобы адаптировать применение кодонов гена GUS из E. coli к микроорганизмам рода Mortierella.
Что касается примеров модификации кодонов, можно сослаться на выравнивание GUSm и GUS, показанное на Фигуре 12A и Фигуре 12B.
[0026] Промоторная активность в настоящем изобретении предпочтительно предназначена для обеспечения активности белка GUS равной по меньшей мере 500, 600, 700, 800, 900, 1000, 1200, 1400, 1600, 1800 или 2000 нмоль/(мг⋅мин) при экспрессии репортерного гена GUS в клетках микроорганизмов, относящихся к роду Mortierella, как описано выше.
Процедуры переноса генов в хозяйскую клетку описаны далее.
[0027] Упомянутый выше полинуклеотид согласно настоящему изобретению можно получить с помощью известных генно-инженерных процедур или известных процедур синтеза.
[0028] 2. Векторы и трансформанты
В другом варианте настоящее изобретение также представляет экспрессионный вектор, содержащий полинуклеотид согласно настоящему изобретению (здесь и далее называемый «вектор настоящего изобретения»).
Вектор настоящего изобретения, как правило, составлен так, что он содержит:
(i) промотор согласно настоящему изобретению; и
(ii) экспрессионную кассету, содержащую составные элементы, сигналы, которые функционируют в хозяйской клетке для терминации транскрипции и полиаденилирования молекулы РНК.
Составленный таким образом вектор вводят в хозяйские клетки. Примеры подходящих хозяйских клеток, применяемых в настоящем изобретении, включают продуцирующие липиды грибки, дрожжи и подобное.
[0029] В качестве продуцирующих липиды грибков можно применять штаммы, как описанные в MYCOTAXON, Vol. XLIV, No. 2, pp. 257-265 (1992). Конкретные примеры включают микроорганизмы, относящиеся к роду Mortierella, на примере микроорганизмов, относящихся к подроду Mortierella, таких как Mortierella elongata IFO8570, Mortierella exigua IFO8571, Mortierella hygrophila IFO5941, Mortierella alpina IFO8568, ATCC16266, ATCC32221, ATCC42430, CBS 219.35, CBS224.37, CBS250.53, CBS343.66, CBS527.72, CBS528.72, CBS529.72, CBS608.70, CBS754.68 и пр., а также микроорганизмов, относящихся к подроду Micromucor, таким как Mortierella isabellina CBS194.28, IFO6336, IFO7824, IFO7873, IFO7874, IFO8286, IFO8308, IFO7884, Mortierella nana IFO8190, Mortierella ramanniana IFO5426, IFO8186, CBS112.08, CBS212.72, IFO7825, IFO8184, IFO8185, IFO8287, Mortierella vinacea CBS236.82 и пр. Особенно предпочтительным является Mortierella alpina.
[0030] Такой вектор можно получить, начиная с существующего экспрессионного вектора, например, pDura5 (Appl. Microbiol. Biotechnol., 65, 419-425, (2004)), pBIG35 (Appl. Environ. Microbiol., (2009), vol. 75, p. 5529-5535), pD4 (Appl. Environ. Microbiol., November 2000, 66(11), p. 4655-4661), pDZeo (J. Biosci. Bioeng., December 2005, 100(6), p. 617-622), pDX vector (Curr. Genet., 2009, 55(3), p. 349-356) или pBIG3ura5 (Appl. Environ. Microbiol., 2009, 75, p. 5529-5535) посредством замены промоторной области в исходном экспрессионном векторе последовательностью промотора согласно настоящему изобретению, хотя исходный вектор не ограничен приведенными выше векторами.
[0031] Для трансформации хозяйских клеток можно применять селективный маркер, чтобы подтвердить, что вектор был введен. Примеры доступных для применения селективных маркеров включают ауксотрофные маркеры (ura5, niaD, trp1), маркеры лекарственной устойчивости (гигромицин, зеоцин), ген устойчивости к генетицину (G418r), ген устойчивости к меди (CUP1) (Marin et al., Proc. Natl. Acad. Sci. USA, 81, 337 1984), гены устойчивости к церуленину (fas2m, PDR4) (Junji Inokoshi et al., Biochemistry, vol. 64, p. 660, 1992; Hussain et al., gene, 101, 149, 1991) и подобные.
[0032] Примеры ауксотрофных маркеров включают, но не ограничиваются п. с (1) по (15), показанными ниже:
(1) метиониновый ауксотрофный маркер: met1, met2, met3, met4, met5, met6, met7, met8, met10, met13, met14 или met20;
(2) тирозиновый ауксотрофный маркер: tyr1 или изолейцин;
(3) валиновый ауксотрофный маркер: ilv1, ilv2, ilv3 или ilv5;
(4) фенилаланиновый ауксотрофный маркер: pha2;
(5) ауксотрофный маркер глутаминовой кислоты: glu3;
(6) треониновый ауксотрофный маркер: thr1 или thr4;
(7) ауксотрофный маркер аспарагиновой кислоты: asp1 или asp5;
(8) сериновый ауксотрофный маркер: ser1 или ser2;
(9) аргининовый ауксотрофный маркер: arg1, arg3, arg4, arg5, arg8, arg9, arg80, arg81, arg82 или arg84;
(10) урациловый ауксотрофный маркер: ura1, ura2, ura3, ura4, ura5 или ura6;
(11) адениновый ауксотрофный маркер: ade1, ade2, ade3, ade4, ade5, ade6, ade8, ade9, ade12 или ADE15;
(12) лизиновый ауксотрофный маркер: lys1, lys2, lys4, lys5, lys7, lys9, lys11, lys13 или lys14;
(13) триптофановый ауксотрофный маркер: trp1, trp2, trp3, trp4 или trp5;
(14) лейциновый ауксотрофный маркер: leu1, leu2, leu3, leu4 или leu5; и
(15) гистидиновый ауксотрофный маркер: his1, his2, his3, his4, his5, his6, his7 или his8.
[0033] Примеры маркеров лекарственной устойчивости включают, но не ограничиваются геном устойчивости к гигромицину (гигромицину B), геном устойчивости к блеомицину t (плеомицину) (Transformation of filamentous fungi based on hygromycin b and phleomycin resistance markers, Methods in Enzymology, Volume 216, 1992, Pages 447-457, Peter J. Punt, Cees A.M.J.J. van den Hondel), геном устойчивости к биалафосу (Avalos, J., Geever, R. F., and Case, M. E. 1989. Bialaphos resistance as a dominant selectable marker in Neurospora crassa. Curr. Genet. 16: 369-372), геном устойчивости к сульфонилмочевине (Zhang, S., Fan, Y., Xia, Y.X., and Keyhani, N.O. (2010) Sulfonylurea resistance as a new selectable marker for the entomopathogenic fungus Beauveria bassiana. Appl Microbiol Biotechnol 87: 1151-1156), геном устойчивости к беномилу (Koenraadt, H., S. C. Sommerville, and A. L. Jones. 1992. Characterization of mutations in the beta-tubulin gene of benomyl-resistant field strains of Venturia inaequalis and other pathogenic fungi. Mol. Plant Pathol. 82:1348-1354), геном ассимиляции ацетамида (ацетамидазой, AmdS) (Kelly, J. M. and Hynes, M. J. (1985). Transformation of Aspergillus niger by the Eamds gene of Aspergiilus nidulans. EMBO J. 4, 475-479) и подобным.
[0034] Для трансформации хозяйских клеток можно применять известные способы. Например, в случае с продуцирующими липиды грибками можно применять электропорацию (Mackenxie D. A. et al. Appl. Environ. Microbiol., 66, 4655-4661, 2000), способ доставки частиц (описанный в японском патенте JP 2005-287403 под названием “Breeding Method of Lipid Producing Fungi”) или способом с применением Agrobacterium, не ограничиваясь ими.
[0035] Кроме того, рассматривая стандартные способы клонирования, можно сослаться на "Sambrook & Russell, Molecular Cloning: A Laboratory Manual Vol. 3, Cold Spring Harbor Laboratory Press 2001" and "Methods in Yeast Genetics, A laboratory manual (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY)," и подобное.
[0036] 3. Способ получения белков, липидов или жирных кислот
В другом варианте применения настоящее изобретение также представляет способ получения белков, липидов или жирных кислот с помощью указанного выше трансформанта.
Целевой ген экспрессируется на высоком уровне в нечеловеческом трансформанте, трансформированном промотором согласно настоящему изобретению (здесь и далее называемом «трансформант настоящего изобретения»), особенно полученный с помощью микроорганизма, относящегося к роду Mortierella, в качестве хозяйской клетки. Таким образом, при применении трансформанта настоящего изобретения можно эффективно производить целевой белок.
[0037] Например, целевой ген можно эффективно ввести в вектор настоящего изобретения и культивировать трансформант, трансформированный данным вектором, в результате чего можно экспрессировать целевой белок с целевого гена в клетках трансформанта.
Экспрессированный целевой белок можно собрать, например, получив клеточный лизат из трансформанта и обработав этот лизат в соответствии с известными процедурами. При рассмотрении подробностей сбора целевого белка можно сослаться на "Sambrook & Russell, Molecular Cloning: A Laboratory Manual Vol. 3, Cold Spring Harbor Laboratory Press 2001" and "Methods in Yeast Genetics, A laboratory manual (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY)" и подобное.
[0038] Целевой ген никак не ограничен, но предпочтительно, чтобы он был геном, кодирующим липидсинтазу (здесь и далее называемым «ген липидсинтазы»). Примеры включают гены, кодирующие ацил-КоА-синтазу, глицерин-3-фосфатацилтрансферазу, диацилглицеринацилтрансферазу, элонгазу жирных кислот, ген Δ9-десатуразы жирных кислот, ген Δ12-десатуразы жирных кислот, ген Δ6-десатуразы жирных кислот, ген Δ5-десатуразы жирных кислот, ген Δ4-десатуразы жирных кислот, ген ω3-десатуразы жирных кислот, ген лизофосфолипилацилтрансферазы, ген фосфатазы фосфатидной кислоты, ген синтетазы жирных кислот, ген ацетил-КоА-карбоксилазы и ген АТФ:цитратлиазы.
[0039] При применении клеток, способных синтезировать липиды, например, продуцирующих липиды грибков или подобных, в качестве хозяина для экспрессии гена липидсинтазы, липидсинтаза, экспрессирующаяся с данного гена, вызывает синтез липидов и/или жирных кислот, которые впоследствии можно собрать. Таким образом, при культивировании трансформанта настоящего изобретения можно получить липиды и/или жирные кислоты с высокой эффективностью.
[0040] Липиды или жирные кислоты можно экстрагировать, как описано далее, из клеток, трансформированных в соответствии с настоящим изобретением. После культивирования трансформированный штамм организма (например, продуцирующего липиды грибка или дрожжей) обрабатывают стандартным способом, например, центрифугированием или фильтрацией, для получения культивируемых клеток. Клетки тщательно промывают водой и, предпочтительно, впоследствии высушивают. Высушивание можно провести посредством лиофилизации, высушивания на воздухе и т.п. Высушенные клетки при необходимости можно гомогенизировать, например, с помощью Dynomil или обработки ультразвуком, а затем экстрагировать органическим растворителем, предпочтительно в потоке азота. Органические растворители, применяемые для этой цели, включают простой эфир, гексан, метанол, этанол, хлороформ, дихлорметан, петролейный эфир и подобные. В качестве альтернативы хорошие результаты можно получить при чередующейся экстракции метанолом и петролейным эфиром или при экстракции однофазной системой растворителей хлороформ-метанол-вода. После отгонки органического растворителя из экстракта при пониженном давлении можно получить содержащие жирные кислоты липиды. Экстрагированные жирные кислоты можно перевести в соответствующие метиловые эфиры с помощью способа с применением соляной кислоты и метанола и т.п.
[0041] Более того, жирные кислоты можно отделить в виде смешанных жирных кислот или смешанных сложных эфиров жирных кислот от содержащих жирные кислоты липидов посредством концентрирования и разделения стандартным способом (например, добавлением мочевины, разделением при охлаждении, колоночной хроматографией).
ПРИМЕРЫ
[0042] Здесь настоящее изобретение будет описано подробнее с помощью следующих примеров, не предназначенных для ограничения объема настоящего изобретения.
[0043] Геномный анализ Mortierella alpina
Штамм M. alpina 1S-4 инокулировали в 100 мл среды GY2:1 (2% глюкоза, 1% дрожжевой экстракт, pH 6,0) и культивировали при 28°C в течение 2 дней при перемешивании. Клетки собирали фильтрацией, а их геномную ДНК получали с помощью DNeasy (QIAGEN).
Нуклеотидную последовательность указанной выше геномной ДНК определяли с помощью Roche 454 GS FLX Standard, проводя секвенирование нуклеотидов в два подхода для библиотеки фрагментов и в три подхода для библиотеки спаренных концов. Полученные в результате нуклеотидные последовательности объединяли для получения 300 суперконтигов.
[0044] Анализ экспрессии
Штамм M. alpina 1S-4 инокулировали в 100 мл среды (1,8% глюкоза, 1% дрожжевой экстракт, pH 6,0) и предварительно культивировали в течение 3 дней при 28ºC. 10-литровый культуральный сосуд (Able Co., Токио) заполняли 5 л среды (1,8% глюкозы, 1% соевой муки, 0,1% оливкового масла, 0,01% адеканола, 0,3% KH2PO4, 0,1% Na2SO4, 0,05% CaCl2⋅2H2O, 0,05% MgCl2⋅6H2O, pH 6,0) и инокулировали всем продуктом предварительного культивирования, а затем выращивали аэробную спинкультуру в условиях 300 об/мин, 1 об/об/мин (vvm) и 26ºC в течение 8 дней. В дни 1, 2 и 3 культивирования добавляли глюкозу в количестве, соответствующем 2%, 2% и 1,5%, соответственно. Клетки собирали на каждой стадии культивирования (в дни 1, 2, 3, 6 или 8) для получения общей РНК с помощью способа с применением гуанидингидрохлорида/CsCl. Применяя SOLiD™ Total RNA-Seq для полнотранскриптомных библиотек (Applied Biosystems), синтезировали кДНК на каждой стадии и секвенировали ее в SOLiD.
[0045] Клонирование промоторных областей
Клонирование проводили, как описано ниже, в промоторных областях генов, уровни экспрессии которых считали высокими в штамме M. alpina 1S-4 по результатам анализа экспрессии, или в промоторной области гомолога гена системы метаболизма галактозы.
Сначала праймеры, необходимые для ПЦР амплификации каждой промоторной области, составляли, как описано ниже. Следует отметить, что подчеркнутые части нуклеотидной последовательности праймеров, показанных ниже, представляют собой сайт узнавания рестрикционным ферментом. Праймеры составляли так, что последовательности узнавания XbaI и SpeI добавляли, соответственно, к обоим концам промоторной области. Однако только праймеры для GAL10-2p, имеющего последовательность узнавания SpeI в составе своей последовательности, составляли так, что последовательность узнавания XbaI добавляли на каждый конец. Символ «F» или «R», встречающийся в каждом названии праймера означает, что праймер является прямым (F, forward) или обратным (R, reverse), соответственно.
[0046] Промотор PP7p
PP7p F XbaI AATATCTAGATGACCGTGCGCTTTTTGAGAC (SEQ ID NO: 33)
PP7p R SpeI AGCAACTAGTCGTATATTTGTTGAAAGGTG (SEQ ID NO: 34)
Промотор CIT1p
CIT1p F XbaI ATTTTCTAGACACCTCAAAAACGTGCCTTG (SEQ ID NO: 35)
CIT1p R SpeI AATAACTAGTGGCGGATATGTGTATGGAG (SEQ ID NO: 36)
Промотор PP3p
PP3p F XbaI AACGTCTAGACGTGTTATCTTGCGCTGC (SEQ ID NO: 37)
PP3p R SpeI TCATACTAGTGATGATTTAGAGGTGTTGG (SEQ ID NO: 38)
Промотор PP2p
PP2p F XbaI AAGCTCTAGAGACTGTAAAGACGGAGGGG (SEQ ID NO: 39)
PP2p R SpeI AGTAACTAGTTGTGGATAGTGGGTAGTGG (SEQ ID NO: 40)
Промотор PP6ps
PP6ps F XbaI AAAGTCTAGACTGGCAATAGTTAGTGCACG (SEQ ID NO: 41)
PP6ps R SpeI ATCAACTAGTGATGGAGGTTTGTTTGAGAAG (SEQ ID NO: 42)
Промотор HSC82p
HSC82p F XbaI ATCATCTAGAGAGCTCAAGATGAAGGTGCTC (SEQ ID NO: 43)
HSC82p R SpeI AATAACTAGTGGTGTGTGTGGTTTGCGGG (SEQ ID NO: 44)
Промотор SSA2p
SSA2p F XbaI TTAGTCTAGAAAAGTGCTGCTTCGGAACC (SEQ ID NO: 45)
SSA2p R SpeI AGATACTAGTGATGTAGATGTGAGTGTGAG (SEQ ID NO: 46)
Промотор GAL10-2p
GAL10-2p F XbaI AATATCTAGAGGTTCCGAGAGGTGGATTTG (SEQ ID NO: 47)
GAL10-2p R XbaI ATAATCTAGATGGCTCCTGAAAGGACGAG (SEQ ID NO: 48)
Применяя геном штамма Mortierella alpina 1S-4 в качестве матрицы, клонировали каждую промоторную область с помощью ПЦР. Применяли полимеразу PrimeSTAR GXL (TaKaRa).
[0047] Создание вектора для оценки промотора
Ген GUSm (SEQ ID NO: 31), модифицированный таким образом, чтобы адаптировать применение кодонов гена GUS из E. coli (SEQ ID NO: 29) к микроорганизмам рода Mortierella (Фигура 12A и Фигура 12B), применяли в качестве репортерного гена.
GUSm лигировали в плазмиду pBIG35, содержащую гистоновый промотор (HisP), служащий конститутивным экспрессионным промотором (Appl. Environ. Microbiol., (2009), vol. 75, p. 5529-5535), для создания экспрессионной кассеты. Данную экспрессионную кассету впоследствии лигировали тандемно к гену урацилового ауксотрофного маркера (ura5) для создания бинарного вектора для трансформации pBIG35ZhGUSm (Фигура 1). Следует отметить, что ген GUSm, применяемый в этом векторе, является искусственно синтезированным геном β-D-глюкуронидазы, частота применения кодонов которого адаптирована к M. alpina. Ura5 является геном оротатфосфорибозилтрансферазы M. alpina. HisP является промотором гена гистона H4.1 M. alpina. SdhBt является терминатором гена сукцинатдегидрогеназы M. alpina. ColE1 ori является точкой начала репликации, NPTII является геном устойчивости к канамицину, TrfA является геном, отвечающим за амплификацию плазмиды, а левая и правая границы являются повторяющимися последовательностями для переноса гена.
Каждую их промоторных областей, клонированных, как описано выше, вырезали с помощью рестрикционных ферментов XbaI и SpeI или с помощью рестрикционного фермента XbaI, а затем вставляли на место HisP в разрезанный XbaI и SpeI вектор pBIG35ZhGUSm.
[0048] Трансформация Mortierella alpina
Ауксотрофный по урацилу штамм (Δura-3) индуцировали из штамма M. alpina 1S-4 в соответствии с процедурами, описанными в патентном документе (WO2005/019437), и культивировали на содержащей 0,05 мг/мл урацила агаризованной среде Чапека-Докса (Czapek-Dox) (3% сахароза, 0,2% NaNO3, 0,1% KH2PO4, 0,05% KC1, 0,05% MgSO4⋅7H2O, 0,001% FeSO4⋅7H2O, 2% агар, pH 6,0). Полученный таким образом продукт культивирования собирали и фильтровали через Miracloth (Calbiochem) для получения суспензии спор M. alpina Δura-3. Agrobacterium (Agrobacterium tumefaciens C58C1) трансформировали каждым из полученных векторов для оценки промотора посредством электропорации и культивировали при 28°C в течение 48 часов на агаризованной среде LB-Mg (1% триптон, 0,5% дрожжевой экстракт, 85 мМ NaCl, 0,5 мМ MgSO4⋅7H2O, 0,5 мМ NaOH, 1,5% агар, pH 7,0). Трансформанты Agrobacterium, несущие векторы, проверяли с помощью ПЦР. Трансформанты Agrobacterium, несущие векторы, культивировали при 28ºC при 120 об/мин в течение 2 дней при перемешивании в 100 мл минимальной среды MM (10 мМ K2HPO4, 10 мМ KH2PO4, 2,5 мМ NaCl, 2 мМ MgSO4⋅7H2O, 0,7 мМ CaCl2, 9 мкM FeSO4⋅7H2O, 4 мМ (NH4)2SO4, 10 мМ глюкоза, pH 7,0), центрифугировали при 5800 × g, а затем растворяли свежей средой IM (минимальной средой MM, содержащей 0,5% глицерин, 200 мкМ ацетосирингон и 40 мМ 2-(N-морфолино)этансульфоновая кислота (MES), и доведенной до pH 5,3) для получения суспензий. Данные суспензии культивировали на протяжении от 8 до 12 часов при 28°C при 300 об/мин при перемешивании до достижения оптической плотности OD 660 = от 0,4 до 3,7. Каждую из клеточных суспензий (100 мкл) смешивали с равным объемом указанной выше суспензии M. alpina Δura-3 (108 мл-1), распределяли по нитроцеллюлозной мембране (70 мм в диаметре; упрочненная малозольная уровня 50, Whatman), помещенной в среде сокультивирования (имеющей тот же состав, что и среда IM, за исключением того, что она содержит 5 мМ глюкозы вместо 10 мМ глюкозы и 1,5% агара), а затем культивировали при 23°C на протяжении от 2 до 5 дней. После сокультивирования мембрану переносили на не содержащую урацил и содержащую 0,03% нильский синий A (Sigma) агаризованную среду SC (5,0 г основы азотного агара без аминокислот и сульфата аммония (Difco), 1,7 г (NH4)2SO4, 20 г глюкозы, 20 мг аденина, 30 мг тирозина, 1,0 мг метионина, 2,0 мг аргинина, 2,0 мг гистидина, 4,0 мг лизина, 4,0 мг триптофана, 5,0 мг треонина, 6,0 мг изолейцина, 6,0 мг лейцина, 6,0 мг фенилаланина, 20 г/л агара) и культивировали при 28ºC в течение 5 дней. Гифы из видимых грибковых колоний переносили в не содержащую урацил среду SC. Перенос в свежую не содержащую урацил среду SC повторяли дважды, чтобы таким образом отобрать трансформанты, стабильно сохраняющие свои свойства.
[0049] Селекция экспрессирующихся на высоком уровне промоторов
Культивирование и сбор штаммов
Каждый трансформант культивировали при 28°C в течение 2 дней на агаризованной среде GY (2% глюкозы, 1% дрожжевого экстракта, 1,5% агара). После завершения культивирования клетки собирали, соскребая их вместе с агаром.
[0050] Экстракция белков из клеток
Собранные клетки смешивали с 500 мкл буфера для гомогенизации (100 мМ Трис-HCl (pH 8,0), 5 мМ 2-меркаптоэтанола) и гомогенизировали дважды при 5000 об/мин в течение 30 сек с помощью шарикового гомогенизатора TOMY beads shocker, применяя стеклянные шарики диаметром 0,1 мм. Гомогенат центрифугировали при 8000 × g в течение 10 минут и собранный супернатант впоследствии центрифугировали при 20400 × g в течение 10 минут для сбора супернатанта в виде раствора белков. В собранном растворе измеряли концентрацию белка и при желании разводили до любой концентрации буфером для гомогенизации. Все описанные выше операции проводили на льду.
[0051] Измерение активности GUS
Субстрат (п-нитрофенил-β-D-глюкуронид) растворяли в аналитическом буфере (21,7 мМ NaH2PO4, 33,9 мМ Na2HPO4, 1,11 мМ EDTA (pH 8,0)) для получения конечной концентрации равной 1,25 мМ. Этот раствор субстрата (160 мкл) и каждый образец белка (40 мкл) смешивали в 96-луночном микротитровальном планшете и измеряли поглощение при 405 нм в течение времени при 37ºC. Поглощение п-нитрофенола измеряли при 0,05 мМ, 0,1 мМ, 0,2 мМ и 0,5 мМ для получения калибровочной кривой, а значение активности GUS в каждом образце рассчитывали в соответствии со следующим уравнением:
Активность GUS (нмоль/(мг⋅мин)) =
1000 × [(величина наклона графика зависимости поглощения от времени для каждого образца)/(величина наклона калибровочного графика)]/[(концентрация белка в образце)/5]
[0052] Количество (нмоль) п-нитрофенил-β-D-глюкуронида, превращенного в п-нитрофенол под действием 1 мг/мл белка за 1 минуту определяют как 1 единицу активности GUS.
[0053] Селекция штаммов для оценки активности GUS
Стабильные трансформированные штаммы (30 штаммов), отобранные для оценки каждого промотора, культивировали на агаризованной среде GY, как описано выше, и измеряли в них активность GUS. Из этих 30 штаммов отобрали 10 штаммов, проявляющих умеренную активность GUS.
[0054] Оценка промоторной активности
Отобранные штаммы культивировали при 28ºC при 300 об/мин в течение 5 дней при перемешивании в жидкой среде GY (10 мл) или в среде на соевой муке (10 мл). После завершения культивирования клетки собирали фильтрованием и измеряли в них активность GUS. Оценивали среднее измеренных значений как промоторную активность. Полученные результаты показаны на Фигуре 2.
Было обнаружено, что оцененные промоторы обладают большей промоторной активностью, чем известные промоторы из Mortierella, HisP и GAPp, в среде GY medium и/или в среде на соевой муке.
[0055] Исследование времени культивирования и активности каждого промотора
Чтобы определить индуцированные временем культивирования изменения промоторной активности, штаммы, отобранные для каждого промотора, культивировали при 28°C при перемешивании в жидкой среде GY (10 мл) в течение 2 дней, 5 дней, 7 дней или 14 дней. После завершения культивирования клетки собирали фильтрованием и измеряли в них активность GUS. Полученные результаты показаны в таблице ниже.
[0056] [Таблица 2]
Количество дней культивирования и активность каждого промотора
[0057] Оценка индуцибельного промотора
Промотор GAL10-2p оценивали, как описано ниже.
Сначала стабильные трансформированные штаммы (30 штаммов) культивировали при 28°C в течение 3 дней на агаризованной среде SC + gal (агаризованная среда SC, содержащая 2% галактозы вместо 2% глюкозы) и измеряли в них активность GUS, как описано выше, чтобы таким образом отобрать 10 штаммов, проявляющих умеренную активность GUS. Эти штаммы инокулировали в жидкую среду GY, в которую добавляли галактозу в концентрации 2% в день 4 или 7. Условия культивирования задавали как 28ºC и 300 об/мин. Фигура 3 показывает активность GUS, измеренную между 2 и 14 днями после начала культивирования. Промотор GAL10-2p индуцировали к экспрессии при добавлении галактозы.
[0058] Исследование областей, необходимых для промоторной активности
Чтобы определить область, необходимую для промоторной активности каждого промотора получали фрагменты ДНК для каждого промотора, укорачивая расположенную выше область промотора, и оценивали промоторную активность.
Для получения таких фрагментов ДНК для каждого промотора получали следующие праймеры. Следует отметить, что каждая подчеркнутая часть представляют собой сайт узнавания рестрикционным ферментом.
[0059] PP7p
Праймер для амплификации промотора PP7p-D1000
PP7p D1000 F XbaI AGCATCTAGAAAAACTATTCAATAATGGGCG (SEQ ID NO: 49)
Праймер для амплификации промотора PP7p-D750
PP7p D750 F XbaI ATTTCTAGAATGGCGAGACGCAGGGGGTAG (SEQ ID NO: 50)
Праймер для амплификации промотора PP7p-D500
PP7p D500 F XbaI AATATCTAGAGAGTGGGCACTGAACTAAAAAG (SEQ ID NO: 51)
Праймер для амплификации промотора PP7p-D250
PP7p D250 F XbaI AATATCTAGAGACACTGCATGACGCGAAATC (SEQ ID NO: 52)
CIT1p
Праймер для амплификации промотора CIT1p-D1300
CIT1p D1300 F XbaI AAGTCTAGATGTCAATCATCTTTGCTGCTG (SEQ ID NO: 53)
Праймер для амплификации промотора CIT1p-D1000
CIT1p D1000 F XbaI TGCGTCTAGAATTATAATTATAATGAGGAAGTG (SEQ ID NO: 54)
Праймер для амплификации промотора CIT1p-D700
CIT1p D700 F XbaI TTATCTAGAGGCGAGTGGCGGACTGC (SEQ ID NO: 55)
Праймер для амплификации промотора CIT1p-D400
CIT1p D400 F XbaI TTGTCTAGACAATTGGCAAGGCTGGGTTG (SEQ ID NO: 56)
PP3p
Праймер для амплификации промотора PP3p-D1600
PP3p D1600 R XbaI AATATCTAGAGATCCTGGTCGAAAAAGACAG (SEQ ID NO: 57)
Праймер для амплификации промотора PP3p-D1200
PP3p D1200 R XbaI AATGTCTAGATGAGTTTCTGTTTTTTCCTTTTTGC (SEQ ID NO: 58)
Праймер для амплификации промотора PP3p-D800
PP3p D800 R XbaI AATATCTAGATGAACAATTCATGCAGCTTCACG (SEQ ID NO: 59)
Праймер для амплификации промотора PP3p-D400
PP3p D400 R XbaI AATATCTAGACGTCTAAGCGTTTACGTGCC (SEQ ID NO: 60)
Праймер для амплификации промотора PP3p-D200
PP3p D200 R XbaI AATATCTAGACTCGTTTTGATGGAGTTCTC (SEQ ID NO: 61)
PP2p
Праймер для амплификации промотора PP2p-D1200
PP2p D1200 F XbaI ATTTCTAGATGCATTTACAGGTGAATATTAC (SEQ ID NO: 62)
Праймер для амплификации промотора PP2p-D800
PP2p D800 F XbaI TTATCTAGACATAAAAGTGTCTGGAGCG (SEQ ID NO: 63)
Праймер для амплификации промотора PP2p-D400
PP2p D400 F XbaI TTATCTAGAACTAAGTGGTGTCTACTTTGG (SEQ ID NO: 64)
Праймер для амплификации промотора PP2p-D200
PP2p D200 F XbaI AATTCTAGAGGATACTCCATCCCCACCC (SEQ ID NO: 65)
Праймер для амплификации промотора PP6ps
PP6ps-D1000
PP6ps D1000 F XbaI AATTCTAGACAGTTACCGTGCGCCCACTG (SEQ ID NO: 66)
Праймер для амплификации промотора PP6ps-D750
PP6ps D750 F XbaI AATTCTAGACTTTCACAAATAGGCATCCTATC (SEQ ID NO: 67)
Праймер для амплификации промотора PP6ps-D500
PP6ps D500 F XbaI AATTCTAGAGGCTTTTTCGTTTATTGGATTG (SEQ ID NO: 68)
Праймер для амплификации промотора PP6ps-D100
PP6ps D100 F XbaI ACGTCTAGATATCCAATTCTCACCACTTC (SEQ ID NO: 69)
HSC82p
Праймер для амплификации промотора HSC82p-D800
HSC82p D800 F XbaI AATTCTAGATTTTACTACCGCATTCCCTTTTC (SEQ ID NO: 70)
Праймер для амплификации промотора HSC82p-D600
HSC82p D600 F XbaI ACGTCTAGACCTTTTCAGTAAACAATTTC (SEQ ID NO: 71)
Праймер для амплификации промотора HSC82p-D400
HSC82p D400 F XbaI ATTTCTAGACACAAAGAAGAAGGGTGTGTC (SEQ ID NO: 72)
Праймер для амплификации промотора HSC82p-D200
HSC82p D200 F XbaI ACGTCTAGAACTGTTTTCTTGAAACTTC (SEQ ID NO: 73)
SSA2p
Праймер для амплификации промотора SSA2p-D850
SSA2p D850 F SpeI AGTAACTAGTTGACGGCGTGTATATGTCAG (SEQ ID NO: 74)
Праймер для амплификации промотора SSA2p-D600
SSA2p D600 F SpeI AGGTACTAGTCCATTGTATCGATTTCTGAT (SEQ ID NO: 75)
Праймер для амплификации промотора SSA2p-D400
SSA2p D400 F SpeI AGTAACTAGTGCTATGCGAACGGTTCATTTTG (SEQ ID NO: 76)
Праймер для амплификации промотора SSA2p-D200
SSA2p D200 F SpeI AGGTACTAGTTTTTTTCTCTCTGGTGTGAACG (SEQ ID NO: 77)
GAL10-2p
Праймер для амплификации промотора GAL10-2p-D2000
GAL10-2p D2000 F XbaI AATTCTAGACGCAGAGTGATGGTCATTACC (SEQ ID NO: 78)
Праймер для амплификации промотора GAL10-2p-D1600
GAL10-2p D1600 F XbaI AATTCTAGACTCTATGGCAAGATTACGAG (SEQ ID NO: 79)
Праймер для амплификации промотора GAL10-2p-D1200
GAL10-2p D1200 F XbaI AATTCTAGATGCTCGTGAAGAGGGGCAC (SEQ ID NO: 80)
Праймер для амплификации промотора GAL10-2p-D800
GAL10-2p D800 F XbaI ACGTCTAGACATTTTTTGCCGCCAATTCTG (SEQ ID NO: 81)
Праймер для амплификации промотора GAL10-2p-D400
GAL10-2p D400 F XbaI ATTTCTAGACCCCCGCCTATTTTTTTTTTC (SEQ ID NO: 82)
[0060] Для получения укороченных промоторов каждого промотора полученный ранее вектор для оценки каждого промотора применяли в качестве матрицы в ПЦР с указанными выше праймерами и с обратными праймерами, применяемыми в примерах (PP7p R SpeI, CIT1p R SpeI, PP3p R SpeI, PP2p R SpeI, PP6ps R SpeI, HSC82p R SpeI, SSA2p R SpeI, GAL10-2p R XbaI), каждый из которых соответствует 3’-концу каждого промотора. Каждый из образованных в результате фрагментов ДНК разрезали рестрикционными ферментами XbaI и SpeI или рестрикционным ферментом XbaI, а затем вставляли в вектор для оценки промотора.
Так же, как описано в разделе «Трансформация Mortierella alpine» трансформировали M. alpina для отбора стабильных трансформированных штаммов. В этих штаммах измеряли активность GUS таким же способом, как и способ, применяемый в примерах. Следует отметить, что количество дней культивирования устанавливали равным 3 дням (CIT1p), 5 дням (PP7p, PP6p, HSC82p, SSA2p, GAPp) или 10 дням (PP3p), в зависимости от свойств каждого промотора. Полученные результаты показаны на Фигурах с 4 по 10.
[0061] В случае индуцируемого галактозой промотора стабильные трансформированные линии предварительно культивировали при 28°C в течение 3 дней на агаризованной среде SC + gal (агаризованная среда SC, содержащая 2% галактозы вместо 2% глюкозы) или предварительно культивировали при 28ºC при 300 об/мин в течение 4 дней в среде SC + raf (жидкая среда SC, содержащая 2% раффинозы вместо 2% глюкозы), после чего добавляли галактозу до конечной концентрации 2%. Культивирование продолжали дополнительно 1 день и измеряли активность GUS в клетках. Полученные результаты показаны на Фигуре 11.
[0062] Как видно на Фигурах с 4 по 10, было получено подтверждение того, что каждый из полноразмерных промоторов и укороченных промоторов, показанных в Таблице 1 выше, проявляет активность белка GUS равную 500 нмоль/(мг⋅мин) или выше.
[0063] ПРОМЫШЛЕННАЯ ПРИМЕНИМОСТЬ
Настоящее изобретение обеспечивает высокий уровень экспрессии целевых генов в продуцирующих липиды грибках и таким образом обеспечивает эффективный синтез и получение целевых белков, липидов и жирных кислот.
[0064] Список последовательностей в произвольной форме
SEQ ID NOs: с 31 по 82: синтетические ДНК
| название | год | авторы | номер документа |
|---|---|---|---|
| ГЛИЦЕРОЛ-3-ФОСФАТАЦИЛТРАНСФЕРАЗА | 2010 |
|
RU2499835C1 |
| ГЕНЫ ДИАЦИЛГЛИЦЕРОЛ-АЦИЛТРАНСФЕРАЗЫ И ИХ ИСПОЛЬЗОВАНИЕ | 2010 |
|
RU2514655C2 |
| СИНТЕТАЗА ЖИРНЫХ КИСЛОТ, КОДИРУЮЩИЙ ЕЕ ПОЛИНУКЛЕОТИД И ИХ ПРИМЕНЕНИЕ | 2007 |
|
RU2444572C2 |
| НОВЫЕ ГЕНЫ АТФ:ЦИТРАТЛИАЗЫ | 2008 |
|
RU2534560C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ | 2018 |
|
RU2697005C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ | 2014 |
|
RU2614258C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ | 2014 |
|
RU2651461C2 |
| ГОМОЛОГИ ФОСФАТАЗЫ ФОСФАТИДНОЙ КИСЛОТЫ И ИХ ПРИМЕНЕНИЕ | 2008 |
|
RU2507264C2 |
| ПОЛИНУКЛЕОТИД, КОДИРУЮЩИЙ ГОМОЛОГ АЦИЛ-СОА-СИНТЕТАЗЫ, И ЕГО ПРИМЕНЕНИЕ | 2011 |
|
RU2528248C2 |
| РЕГУЛИРУЮЩИЙ ВЫСОТУ РАСТЕНИЙ ГЕН И ЕГО ПРИМЕНЕНИЯ | 2008 |
|
RU2458132C2 |


































































Группа изобретений относится к биотехнологии. Предложен полинуклеотид для обеспечения высокой экспрессии целевого гена в микроорганизме, относящемся к роду Mortierella. Полинуклеотид соответствует любому полинуклеотиду, выбираемому из группы, состоящей из (а)-(с), где (а) полинуклеотид, который состоит из любой нуклеотидной последовательности, выбираемой из группы, состоящей из SEQ ID NO: с 22 по 26; (b) полинуклеотид, который имеет нуклеотидную последовательность с идентичностью 90% или более по отношению к любой нуклеотидной последовательности, выбираемой из группы, состоящей из SEQ ID NO: с 22 по 26, и который проявляет промоторную активность в клетках микроорганизма, относящегося к роду Mortierella; и (с) полинуклеотид, который может гибридизоваться в жестких условиях с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной любой из нуклеотидных последовательностей, выбираемых из группы, состоящей из SEQ ID NO: с 22 по 26, и который проявляет промоторную активность в клетках микроорганизма, относящегося к роду Mortierella. Также предложены вектор для трансформации Mortierella, содержащий указанный полинуклеотид; нечеловеческий трансформант, трансформированный указанным полинуклеотидом или трансформированный указанным вектором. Группа изобретений обеспечивает высокую экспрессию целевого гена. 4 н. и 4 з.п. ф-лы, 13 ил., 2 табл.
1. Полинуклеотид, соответствующий любому, выбираемому из группы, состоящей из (а)-(с), показанных ниже, для обеспечения высокой экспрессии целевого гена в микроорганизме, относящемся к роду Mortierella:
(а) полинуклеотид, который состоит из любой нуклеотидной последовательности, выбираемой из группы, состоящей из SEQ ID NO: с 22 по 26;
(b) полинуклеотид, который имеет нуклеотидную последовательность с идентичностью 90% или более по отношению к любой нуклеотидной последовательности, выбираемой из группы, состоящей из SEQ ID NO: с 22 по 26, и который проявляет промоторную активность в клетках микроорганизмов, относящихся к роду Mortierella; и
(с) полинуклеотид, который может гибридизоваться в жестких условиях с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной любой из нуклеотидных последовательностей, выбираемых из группы, состоящей из SEQ ID NO: с 22 по 26, и который проявляет промоторную активность в клетках микроорганизмов, относящихся к роду Mortierella.
2. Полинуклеотид по п.1, в котором промоторную активность подтверждают как активность белка GUS равную по меньшей мере 500 нмоль/(мг⋅мин) при экспрессии репортерного гена GUS в клетках микроорганизмов, относящихся к роду Mortierella.
3. Полинуклеотид по п.1, который состоит из любой нуклеотидной последовательности, выбираемой из группы, состоящей из SEQ ID NO: с 22 по 26.
4. Полинуклеотид по п.1, который представляет собой ДНК.
5. Вектор, содержащий полинуклеотид по любому из пп.1-4, для трансформации микроорганизма, относящегося к роду Mortierella.
6. Нечеловеческий трансформант, трансформированный полинуклеотидом по любому из пп.1-4, для обеспечения высокой экспрессии целевого гена, причем трансформант представляет собой микроорганизм, относящийся к роду Mortierella.
7. Нечеловеческий трансформант, трансформированный вектором по п.5, для обеспечения высокой экспрессии целевого гена, причем трансформант представляет собой микроорганизм, относящийся к роду Mortierella.
8. Трансформант по п.6 или 7, в котором микроорганизмом, относящимся к роду Mortierella, является Mortierella alpina.
| MACKENZIE D.A | |||
| ET AL | |||
| Isolation and use of a homologous histone promoter and a ribosomal DNA region in a transformation vector for the oilproducing fungus Mortierella alpine // APPL | |||
| ENVIRON | |||
| MICROBIOL., 2000, vol | |||
| Приспособление для соединения пучка кисти с трубкою или втулкою, служащей для прикрепления ручки | 1915 |
|
SU66A1 |
| Походная разборная печь для варки пищи и печения хлеба | 1920 |
|
SU11A1 |
| СПОСОБ И КЛАПАН ДЛЯ НЕПОСРЕДСТВЕННОГО ПОДОГРЕВА ВОДЫ ПАРОМ ДО НАПЕРЕД ЗАДАННОЙ ТЕМПЕРАТУРЫ | 1926 |
|
SU4655A1 |
| MULLER S | |||
| ET AL | |||
| Comparison of expression systems in the yeasts Saccharomyces cerevisiae, Hansenula polymorpha, Klyveromyces lactis, Schizosaccharomyces pombe and Yarrowia lipolytica | |||
| Cloning of two novel promoters from Yarrowia lipolytica // YEAST, 1998, vol | |||
| Паровоз для отопления неспекающейся каменноугольной мелочью | 1916 |
|
SU14A1 |
| Паровоз для отопления неспекающейся каменноугольной мелочью | 1916 |
|
SU14A1 |
| Поршень для двигателей внутреннего горения | 1924 |
|
SU1267A1 |
| US 5204250 A1, 20.04.1993 | |||
| СПОСОБ СЕЛЕКЦИИ ЛИПИД-ПРОДУЦИРУЮЩИХ ГРИБОВ | 2004 |
|
RU2340665C2 |

