Область техники, к которой относится изобретение
Настоящее изобретение относится к полинуклеотиду, кодирующему новую глицерол-3-фосфатацилтрансферазу, и их применение.
Уровень техники
Жирные кислоты являются важными компонентами липидов, таких как фосфолипиды, триацилглицерины и т.д. Жирные кислоты, содержащие две или более ненасыщенных связей, в совокупности именуются полиненасыщенными жирными кислотами (ПНЖК), как известно, включают, в частности, арахидоновую кислоту, дигомо-γ-линоленовую кислоту, эйкозапентаеновую кислоту и докозагексаеновую кислоту и др. Некоторые из этих полиненасыщенных жирных кислот не могут синтезироваться в организме животного. Поэтому необходимо использовать такие полиненасыщенные жирные кислоты в качестве незаменимых аминокислот вместе с продуктами питания.
В организме животного полиненасыщенные жирные кислоты распределены по многим органам и тканям. Например, арахидоновая кислота была выделена из липидов, экстрагированных из надпочечников и печени животных. При этом полиненасыщенные жирные кислоты содержатся в малых количествах в органах животных, и экстракция и выделение полиненасыщенных жирных кислот из органов животных не достаточны для обеспечения большого количества полиненасыщенных жирных кислот. Поэтому были разработаны способы получения полиненасыщенных жирных кислот путем культивирования различных микроорганизмов. Среди этих микроорганизмов, микроорганизм Mortierella известен как микроорганизм, способный продуцировать липиды, содержащие полиненасыщенные жирные кислоты, такие как арахидоновая кислота и др. Кроме того, также предпринимались попытки продуцировать полиненасыщенные жирные кислоты в растениях. Полиненасыщенные жирные кислоты составляют запасные липиды, такие как триацилглицерины и т.д., и, как известно, накапливаются в клетках микроорганизмов или в семенах растений.
Триацилглицерины, которые представляют собой запасные липиды, продуцируются in vivo следующим образом. Передача ацила на глицерол-3-фосфат с образованием лизофосфатидной кислоты осуществляется при помощи глицерол-3-фосфатацилтрансферазы. Затем, передача ацила далее происходит при помощи лизофосфатидацилтрансферазы с образованием фосфатидной кислоты. Эта фосфатидная кислота, в свою очередь, дефосфорилируется при помощи фосфатидфосфатазы с образованием диацилглицерина. Наконец, передача ацила происходит при помощи диацилглицеринацилтрансферазы с образованием триацилглицерина.
В описанном выше триацилглицеридном пути биосинтеза или фосфолипидном пути биосинтеза известно, что реакция ацилирования глицерол-3-фосфата с образованием лизофосфолипидной кислоты опосредуется глицерол-3-фосфатацилтрансферазой (в дальнейшем именуемый иногда «GPAT»; EC 2.3.1.15).
До настоящего времени наличие генов GPAT отмечалось у нескольких организмов. В качестве генов GPAT млекопитающих были клонированы два типа генов: микросомальный (мембраносвязанный) и митохондриальный (мембраносвязанный) (непатентная литература 2). Аналогичным образом, в качестве генов GPAT растений также были клонированы три типа генов: микросомальный (мембраносвязанный), митохондриальный (мембраносвязанный) и хлоропластный (свободный) (непатентная литература 3).
В качестве генов GPAT, полученных из грибка Saccharomyces cerevisiae, были клонированы два типа генов: микросомальный (мембраносвязанный) GPT2/GAT1 (YKR067w) и SCT1/GAT2 (YBL011w); известно, что одновременное делетирование обоих генов приводит к летальности (непатентная литература 4). Для этих грибковых генов показано, что GPT2 имеет активность, чтобы использовать широкий спектр жирных кислот от пальмитиновой кислоты (16:0) до олеиновой кислоты (18:1) в качестве субстрата, принимая во внимание, тогда как SCT1 имеет сильную избирательность, используя 16-углеродные жирные кислоты, такие как пальмитиновая кислота (16:0) и пальмитолеиновая кислота (16:1)) в качестве субстрата (непатентная литература 4).
Кроме того, гены GPAT также были клонированы из многих других видов организмов. Прежде всего, о GPAT, полученном из микроорганизмов рода Mortierella, способных продуцировать липиды, сообщается следующее.
Для GPAT, полученного из Mortierella ramanniana, был изолирован микросомальный GPAT, и показано, что он используется в качестве донора ацил с избирательностью, в 5,4 раза большей для олеиновой кислоты (18:1), чем для пальмитиновой кислоты (16:0) (непатентная литература 5). Сообщается, что GPAT, полученный из Mortierella alpina (в дальнейшем именуемый иногда как «М. alpina»), обладает глицерол-3-фосфатацилтрансферазной активностью в своей микросомальной фракции (непатентная литература 6).
Показано, что в случае, когда GPAT, присутствующий в микросомах М. alpina (в мембранно-связанном состоянии), реагирует in vitro с различными ацил КоА, GPAT использует в качестве субстрата широкого спектра полиненасыщенные жирные кислоты, включая олеиновую кислоту (18:1), линолевую кислоту (18:2), дигомо-γ-линоленовую кислоту (DGLA) (20:3) и арахидоновую кислоту (20:4) (патентная литература 1).
Показано, что в случае, когда GPAT, клонированный из М. alpina (ATCC #16266) (в дальнейшем именуемый MaGPAT1 (ATCC#16266)), экспрессировали в трансформанте Yarrowia lipolytica, разработанном таким образом, чтобы обеспечить биосинтез для получения эйкозапентаеновой кислота (EPA) в общем количестве жирных кислотах, увеличенную композицию дигомо-γ-линоленовой кислоты (DGLA) (20:3) и уменьшенную композицию олеиновой кислоты (18:1). Результаты показывают, что полиненасыщенные жирные кислоты с более длинной цепью и высокой степенью ненасыщенности включаются избирательно (патентная литература 2).
В последние годы сообщалось, что гомолог GPAT, или MaGPAT2, был выделен из М. alpina (1S-4), и показал субстратную специфичность, отличную от MaGPAT1 (патентная литература 3). То есть, предполагается, что MaGPAT1 будет демонстрировать высокую специфичность к пальмитиновой кислоте и MaGPAT2 будет демонстрировать высокую специфичность к олеиновой кислоте.
РОДСТВЕННАЯ ОБЛАСТЬ
Патентая литература:
[Патентая литература 1] Инструкция к WO 2004/087902
[Патентая литература 2] США 2006/0094091
[Патентая литература 3] Инструкция к WO 2008/156026
Непатентная литература:
[Непатентная литература 1] Lipids, 39, 1147 (2004)
[Непатентная литература 2] Biochimica et Biophysica Acta, 1348, 17-26, 1997
[Непатентная литература 3] Biochimica et Biophysica Acta, 1348, 10-16, 1997
[Непатентная литература 4] The Journal of Biological Chemistry, 276 (45), 41710-41716, 2001
[Непатентная литература 5] The Biochemical Journal, 355, 315-322, 2001
[Непатентная литература 6] Biochemical Society Transactions, 28, 707-709, 2000
РАСКРЫТИЕ ИЗОБРЕТЕНИЯ
На основании приведенного выше желательно разработать новый гомолог GPAT, способствующий дальнейшей активизации и эффективному пути синтеза жирных кислот путем эффективной продукции лизофосфатидной кислоты и триацилглицеринов, образованных на их основе.
В результате обширных исследований, авторам настоящего изобретения удалось клонировать ген, кодирующий третий гомолог GPAT (MaGPAT3) липид-продуцирующего грибка М. alpina и усовершенствовать настоящее изобретение. То есть, настоящее изобретение относится к следующему: полинуклеотидам, белкам, векторам экспрессии, трансформантам, способу получения пищевых продуктов и т.д. с использованием трансформантов, пищевых продуктов и т.д., полученных способом, и так далее.
Таким образом, настоящее изобретение относится к следующим объектам.
[1] Полинуклеотиду, выбранному из группы, состоящей из нижеприведенных (a)-(e):
(a) полинуклеотиду, содержащему нуклеотидную последовательность с SEQ ID NO: 1 или 4;
(b) полинуклеотиду, кодирующему белок, состоящий из аминокислотной последовательности SEQ ID NO:2;
(c) полинуклеотиду, кодирующему белок, состоящий из последовательности аминокислот, где 1-100 аминокислот удалены, и заменены, встроены и/или добавлены в аминокислотную последовательность SEQ ID NO:2, и обладающему глицерол-3-фосфатацилтрансферазной активностью;
(d) полинуклеотиду, кодирующему белок с аминокислотной последовательностью, который по крайней мере на 85% гомологичен аминокислотной последовательности SEQ ID NO:2, и обладающему глицерол-3-фосфатацилтрансферазной активностью; и,
(e) полинуклеотиду, который гибридизуется с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной нуклеотидной последовательности SEQ ID NO:1 или 4 в жестких условиях, и который кодирует белок, обладающему глицерол-3-фосфатацилтрансферазной активностью.
[2] Полинуклеотиду по п. [1] выше, как определено в любом из пунктов (f) или (g) ниже:
(f) полинуклеотиду, кодирующему белок, состоящий из последовательности аминокислот, где 1-10 аминокислот удалены, заменены, встроены и/или добавлены в аминокислотную последовательность SEQ ID NO:2, и обладающему глицерол-3-фосфатацилтрансферазной активностью; и,
(g) полинуклеотиду, кодирующему белок с аминокислотной последовательностью, который по крайней мере на 90% гомологичен аминокислотной последовательности SEQ ID NO:2, и обладающему глицерол-3-фосфатацилтрансферазной активностью.
[3] Полинуклеотиду по п. [1] выше, содержащему нуклеотидную последовательность SEQ ID NO:1 или 4.
[4] Полинуклеотиду по п. [1] выше, кодирующему белок, состоящий из аминокислотной последовательности SEQ ID NO:2.
[5] Полинуклеотиду по любому из п.п. [1]-[4] выше, который представляет собой ДНК.
[6] Белку, кодируемому полинуклеотидом по любому из п.п. [1]-[5] выше.
[7] Вектору, содержащему полинуклеотид по любому из п.п. [1]-[5] выше.
[8] Трансформанту, не являющемуся трансформантом человека, в который введен полинуклеотид по любому из п.п. [1]-[5] выше.
[9] Трансформанту, не являющемуся трансформантом человека, в который введен вектор по п. [7] выше.
[10] Трансформанту по п. [8] или [9] выше, где трансформант представляет собой липид-продуцирующий грибок.
[11] Трансформанту по п. [10] выше, где липид-продуцирующий грибок представляет собой грибок Mortierella alpina.
[12] Способу получения липидной композиции или композиции на основе жирных кислот, который предусматривает сбор липидной композиции и композиции на основе жирных кислот из культуры трансформанта по любому из п.п. [8]-[11] выше.
[13] Пищевой продукт, фармацевтический препарат, косметический препарат или мыло, содержащие липидную композицию или композицию на основе жирных кислот, собранную при помощи способа производства по п. [12] выше.
Полинуклеотид по настоящему изобретению можно использовать для трансформации липид-продуцирующего грибка (например, М. alpina), дрожжей, растений и т.д., и трансформанты липид-продуцирующего грибка, трансформанты дрожжей, трансформанты растений и т.д., полученные таким образом, можно использовать для получения композиции на основе жирных кислот, пищевых продуктов, косметики, лекарственных препаратов, мыла и т.д.
Более конкретно, трансформанты по настоящему изобретению обеспечивают чрезвычайно высокую эффективность продукции триглицеридов, а большая часть жирных кислот, увеличенных в трансформантах, имеет место за счет увеличения жирных кислот в триглицеридах. Соответственно, эти трансформанты можно эффективно использовать для получения лекарственных препаратов и диетических пищевых продуктов.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
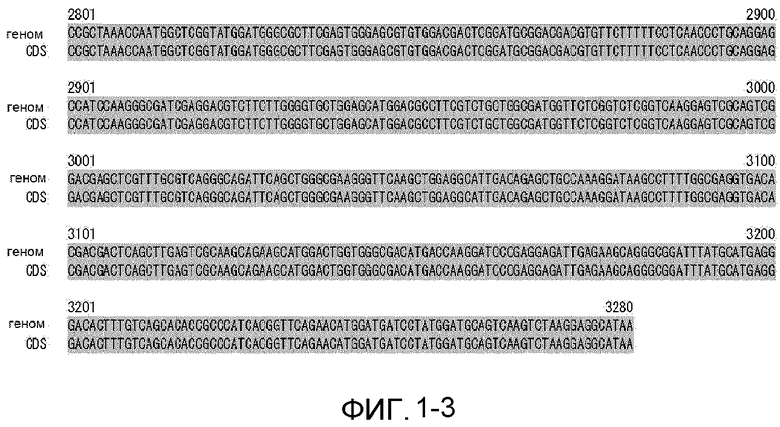
На фиг. 1-1 показано выравнивание геномной последовательности и CDS-последовательности MaGPAT3e.
Фиг. 1-2 является продолжением фиг. 1-1.
Фиг. 1-3 является продолжением фиг. 1-2.
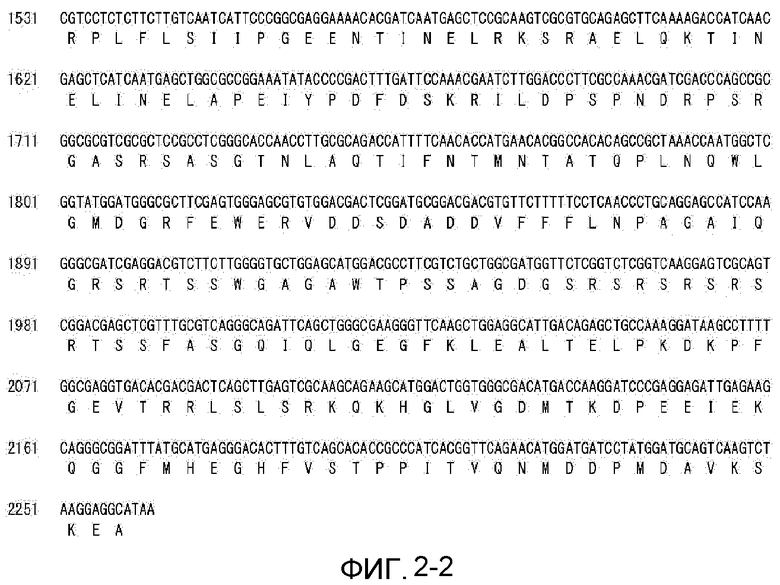
На фиг. 2-1 показана CDS-последовательность и предполагаемая аминокислотная последовательность MaGPAT3 (1S-4).
Фиг. 2-2 является продолжением фиг. 2-1.
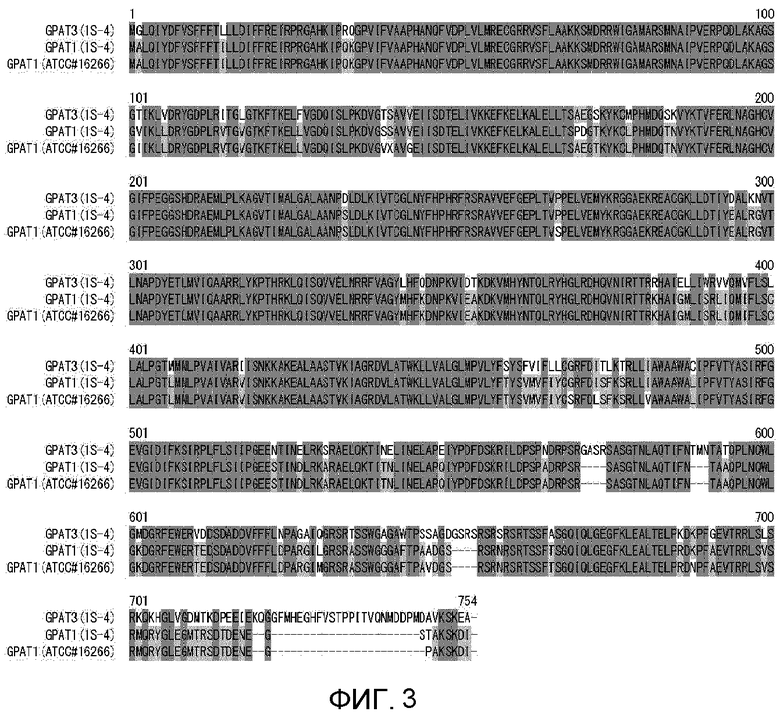
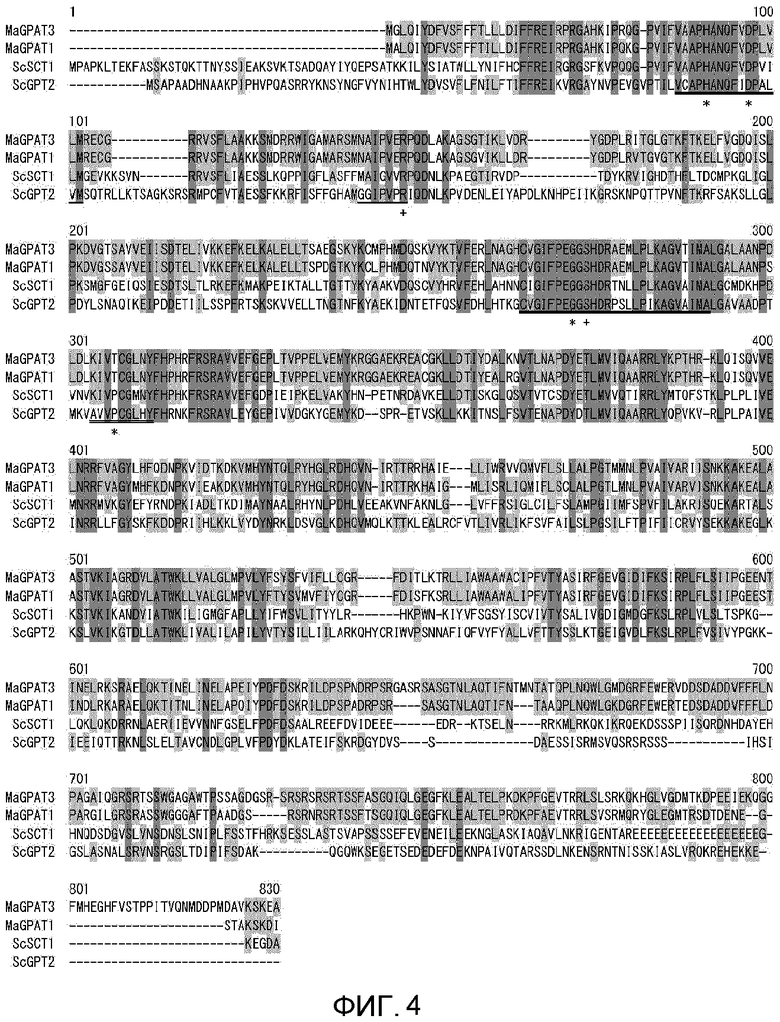
На фиг. 3 показано выравнивание аминокислотных последовательностей MaGPAT1 ((1S-4) и (ACTT#16266)) и MaGPAT3 (1S-4).
На фиг. 4 показано выравнивание аминокислотных последовательностей различных гомологичных белков GPAT (MaGPAT1, MaGPAT3, ScSCT1 и ScGPT2). Четыре домена, консервативные для GPAT-гомологов, и аминокислотные остатки (звездочки), которые считаются важными для GPAT-активности, и аминокислотные остатки (крестики), которые считаются важными для связывания с глицерол-3-фосфатом, в этих доменах были также сохранены в GPAT3.
НАЛУЧШИЙ СПОСОБ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Ниже настоящее изобретение описано более подробно. Приведенные ниже варианты осуществления предназначены, только для описания изобретение, а не для ограничения его только этими вариантами осуществления. Настоящее изобретение может быть осуществлено различными способами, не выходя за рамки его сущности.
Все публикации, опубликованные заявки на патенты, патенты и другая патентная литература, приведенные в данном приложении, включены в настоящем документе посредством ссылки в полном объеме. Это приложение настоящим включает посредством ссылки содержание спецификации и чертежей в японской патентной заявке No. 2009-217646, поданной 18 сентября 2009 года, из которой приоритет был востребован.
Авторам настоящего изобретения впервые удалось клонировать полноразмерную кДНК гена (MaGPAT3), кодирующего третий гомолог глицерол-3-фосфатацилтрансферазы липид-продуцирующего гриба M. alpine, как позднее будет подробно описано в ПРИМЕРАХ. Авторы настоящего изобретения также определили нуклеотидную последовательность геномной ДНК MaGPAT3 M. alpine и его предполагаемую аминокислотную последовательность. Последовательность ORF MaGPAT3, предполагаемая аминокислотная последовательность MaGPAT3, последовательность CDS MaGPAT3 и геномная последовательность MaGPAT3 представляют собой SEQ ID NO:1, SEQ ID NO:2, SEQ ID NO:3 и SEQ ID NO: 4, соответственно. Геномная последовательность MaGPAT1 M. alpine также показана в SEQ ID NO:5. Эти полинуклеотиды и ферменты можно получить с помощью способов, описанных в ПРИМЕРАХ, приведенных ниже, известных способов генной инженерии, известных способов синтеза и так далее.
1. Полинуклеотид по изобретению
Во-первых, настоящее изобретение относится к полинуклеотиду, выбранному из группы, состоящей из нижеприведенных (a)-(e):
(a) полинуклеотиду, содержащему нуклеотидную последовательность SEQ ID NO: 1 или 4;
(b) полинуклеотиду, кодирующему белок, состоящий из аминокислотной последовательности SEQ ID NO:2;
(c) полинуклеотиду, кодирующему белок, состоящий из последовательности аминокислот, где 1-100 аминокислот удалены, заменены, встроены и/или добавлены в аминокислотную последовательность SEQ ID NO:2, и обладающему глицерол-3-фосфатацилтрансферазной активностью;
(d) полинуклеотиду, кодирующему белок с аминокислотной последовательностью, который по крайней мере на 85% гомологичен аминокислотной последовательности SEQ ID NO:2, и обладающему глицерол-3-фосфатацилтрансферазной активностью; и,
(e) полинуклеотиду, который гибридизуется с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной нуклеотидной последовательности SEQ ID NO:1 или 4 в жестких условиях, и который кодирует белок, обладающий глицерол-3-фосфатацилтрансферазной активностью.
Как используется в настоящем документе, термин «полинуклеотид» означает ДНК или РНК.
Как используется в настоящем документе, термин «полинуклеотид, который гибридизуется в жестких условиях» относится к полинуклеотиду, полученному при помощи способа гибридизации на колониях, способа гибридизации на бляшках, способа гибридизации по Саузерну или подобных, с использованием в качестве зонда, например, полинуклеотида, состоящего из нуклеотидной последовательности, комплементарной нуклеотидной последовательности SEQ ID NO:1 или 4, или целой или части полинуклеотида, состоящего из нуклеотидной последовательности, кодирующей аминокислотную последовательность SEQ ID NO:2. В качестве способов гибридизации используются способы, описанные, например, в «Sambrook & Russell, Molecular Cloning; A Laboratory Manual Vol. 3, Cold Spring Harbor, Laboratory Press 2001» и «Ausubel, Current Protocols in Molecular Biology, John Wiley & Sons 1987-1997» и т.д.
Как используется в настоящем документе, термин «жесткие условия» может означать любые из слабо-жестких условий, умеренно-жестких условий или сильно-жестких условий. Термин «слабо-жесткие условия» представляет собой, например, 5x SSC, 5x раствор Денхардта, 0,5% SDS, 50% формамид при 32ºC. Термин «умеренно-жесткие условия» представляет собой, например, 5x SSC, 5x раствор Денхардта, 0,5% SDS, 50% формамид при 42ºC. Термин «сильно-жесткие условия» представляет собой, например, 5x SSC, 5x раствор Денхардта, 0,5% SDS, 50% формамид при 50ºC. При таких условиях, ДНК с более высокой гомологией, как ожидается, эффективно получаются при более высоких температурах, несмотря на то, что жесткость гибридизации определяется несколькими факторами, включая температуру, концентрацию зонда, длину зонда, ионную силу, время, концентрацию соли и другие, и специалист в рассматриваемой области может надлежащим образом подобрать эти факторы для достижения аналогичной жесткости.
При использовании коммерчески доступных наборов для гибридизации можно воспользоваться, например, Alkphos Direct Labeling and Detection System (GE Healthcare). В этом случае, в соответствии с прилагаемым протоколом, после инкубации с меченым зондом в течение ночи мембрану промывают первичным промывочным буфером, содержащем 0,1% (масса/объем) SDS при 55°C, детектируя тем самым гибридизовавшуюся ДНК.
В дополнение к описанным выше полинуклеотидам, другие полинуклеотиды, которые могут быть гибридизованы, включают ДНК, имеющие 74% или выше, 75% или выше, 76% или выше, 77% или выше, 78% или выше, 79% или выше, 80% или выше, 81% или выше, 82% или выше, 83% или выше, 84% или выше, 85% или выше, 86% или выше, 87% или выше, 88% или выше, 89% или выше, 90% или выше, 91% или выше, 92% и выше, 93% или выше, 94% и выше 95% или выше, 96% и выше, 97% или выше, 98% или выше, 99% или выше, 99,1% или выше, 99,2% или выше, 99,3% или выше, 99,4% или выше, 99,5% или выше, 99,6% или выше, 99,7% или выше, на 99,8% или выше, или 99,9% или выше гомологии с ДНК с SEQ ID NO:1 или 4, или с ДНК, кодирующей аминокислотную последовательность SEQ ID NO:2, как рассчитано при помощи программного обеспечения по поиску гомологий, такого как FASTA и BLAST, используя параметры по умолчанию.
Гомология между аминокислотными последовательностями или нуклеотидными последовательностями может быть определена с использованием алгоритма BLAST от Karlin and Altschul (Proc. Natl. Acad. Sci. USA, 87: 2264-2268, 1990; Proc. Nail Acad. Sci. USA, 90: 5873, 1993). Программы, называемые BLASTN и BLASTX, были разработаны на основе алгоритма BLAST (Altschul S. F. et al., J. Mol. Biol. 215: 403, 1990). В случае, когда нуклеотидную последовательность анализируют с использованием BLASTN, параметры составляют, например, оценка=100 и длина слова=12. В случае, когда нуклеотидную последовательность анализируют с использованием BLASTX, параметры составляют, например, оценка=50 и длина слова=3. При использовании программ BLAST и Gapped BLAST для каждой из программ применяют параметры по умолчанию.
Полинуклеотиды по настоящему изобретению, описанные выше, можно получить известными способами генной инженерии, известными способами синтеза и так далее.
2. Белки по изобретению
Настоящее изобретение относится к белкам, показанным ниже.
(i) Белку, кодируемому полинуклеотидом по любому из п.п. (a)-(e) выше.
(ii) Белку, включающему аминокислотную последовательность SEQ ID NO:2.
(iii) Белку, содержащему аминокислотную последовательность, где одна или более аминокислот удалены, заменены, встроены и/или добавлены в аминокислотной последовательности SEQ ID NO:2, и обладающему глицерол-3-фосфатацилтрансферазной активностью.
(iv) Белку с аминокислотной последовательностью, который по крайней мере на 85% гомологичен аминокислотной последовательности SEQ ID NO:2, и обладающему глицерол-3-фосфатацилтрансферазной активностью.
Белки, описанные выше в (iii) или (iv), как правило, являются мутантами природного белка SEQ ID NO:2 и включают те белки, которые могут быть искусственно получены с использованием сайт-направленного мутагенеза, описанного, например, в «Sambrook & Russell, Molecular Cloning: A Laboratory Manual, Vol. 3, Cold Spring Harbor Laboratory Press 2001», «Ausubel, Current Protocols in Molecular Biology, John Wiley & Sons 1987-1997», «Nuc. Acids. Res., 10, 6487 (1982)», «Proc. Natl. Acad. Sci. USA, 79, 6409 (1982)», «Gene, 34, 315 (1985)», «Nuc. Acids. Res., 13, 4431 (1985)», «Proc. Natl. Acad. Sci. USA, 82, 488 (1985)» и т.д.
Как используется в настоящем документе, «белок, содержащий аминокислотную последовательность, где одна или более аминокислот удалены, заменены, встроены и/или добавлены в аминокислотной последовательности SEQ ID NO:2, и обладающий глицерол-3-фосфатацилтрансферазной активностью», включает белки, содержащие аминокислотные последовательности, где, например, 1-100, 1-90, 1-80, 1-70, 1-60, 1-50, 1-40, 1-39, 1-38, 1-37, 1-36, 1-35, 1-34, 1-33, 1-32, 1-31, 1-30, 1-29, 1-28, 1-27, 1-26, 1-25, 1-24, 1-23, 1-22, 1-21, 1-20, 1-19, 1-18, 1-17, 1-16, 1-15, 1-14, 1-13, 1-12, 1-11, 1-10, 1-9 (от 1 до нескольких), 1-8, 1-7, 1-6, 1-5, 1-4, 1-3, 1-2, или одна аминокислота удалены, заменены, встроены и/или добавлены в аминокислотной последовательности SEQ ID NO:2, и обладающие глицерол-3-фосфатацилтрансферазной активностью. Как правило, число удалений, замен, вставок, и/или добавлений является, предпочтительно, меньшим.
Такие белки включают белки с аминокислотной последовательностью, которая гомологична приблизительно на 85% или выше, 86% или выше, 87% или выше, 88% или выше, 89% или выше, 90% или выше, 91% или выше, 92% и выше, 93% или выше, 94% и выше 95% или более, 96% и выше, 97% или выше, 98% или выше, 99% или выше, 99,1% или выше, 99,2% или более, 99,3% или выше, 99,4% или выше, 99,5% или выше, 99,6% или выше, 99,7% или выше, на 99,8% или выше, или 99,9% и выше, аминокислотной последовательности SEQ ID NO:2, и обладающие глицерол-3-фосфатацилтрансферазной активностью. Чем больше процент гомологий, описанный выше, тем обычно более предпочтительным является белок.
Глицерол-3-фосфатацилтрансферазную активность можно определять, например, при помощи способа, описанного в J.B.C. 276 (45), 41710-41716(2001). Для проверки GPAT-активности, существует эксперимент по комплементарности с использованием штаммов дрожжей Δgpt2 и Δsct1. Когда полинуклеотид, кодирующий фермент, экспрессируется в штаммах Δgpt2 и Δsct1 (одновременное делетирование GPT2 и SCT1 приводит к летальности) и штаммах Δgpt2 и Δsct1 способны расти, белок и пептид, кодируемый полинуклеотидом, как оказалось, обладает GPAT-активностью.
Удаление, замена, вставка и/или добавление одного или нескольких остатков аминокислот в аминокислотную последовательность белка по изобретению означает, что один или несколько остатков аминокислот удалены, заменены, встроены и/или добавлены в одной или нескольких положениях в той же последовательности аминокислот. Два или более типов делеций, замен, вставок и добавлений могут одновременно иметь место.
Примеры аминокислотных остатков, которые являются взаимозаменяемыми, приведены ниже. Аминокислотные остатки в одной группе являются взаимозаменяемыми. Группа А: лейцин, изолейцин, норлейцин, валин, норвалин, аланин, 2-аминобутаноевая кислота, метионин, o-метилсерин, t-бутилглицин, t-бутилаланин и циклогексилаланин; Группа B: аспарагиновая кислота, глутаминовая кислота, изоаспарагиновая кислота, изоглутаминовая кислота, 2- аминоадипиновая кислота и 2-аминопробковая кислота; Группа C: аспарагин и глютамин, Группа D: лизин, аргинин, орнитин, 2,4- диаминобутановая кислота и 2,3-диаминопропионовая кислота; Группа E: пролин, 3-гидроксипролин и 4-гидроксипролин; Группа F: Серин, треонин и гомосерин; и Группа G: фенилаланин и тирозин.
Белок по настоящему изобретению также может быть получен при помощи способов химического синтеза, таких, как способ Fmoc (флюоренилметилоксикарбониловым способ), способом tBoc (t-бутилоксикарбониловым способ) и т.д. Кроме того, пептидные синтезаторы, доступные от Advanced Automation Peptide Protein Technologies, Perkin Elmer, Protein Technology Instrument, PerSeptive, Applied Biosystems, SHIMADZU Corp. и т.д., также могут использоваться для химического синтеза.
3. Вектор по изобретению и трансформант с введенным вектором
В другом варианте осуществления, настоящее изобретение также относится к вектору экспрессии, содержащему полинуклеотид по изобретению.
Вектор по изобретению обычно конструируют так, чтобы он содержал экспрессионную кассету, содержащую:
(i) промотор, который может транскрибироваться в клетке-хозяине;
(ii) любой из полинуклеотидов, описанных в пунктах (a) -(g) выше, который связан с промотором; и,
(iii) сигнал, который действует в клетке-хозяине в отношении терминации транскрипции и полиаденилирования молекулы РНК. Таким образом сконструированный вектор вводят в клетку-хозяин. Примеры клеток-хозяев, которые можно соответствующим образом использовать в настоящем изобретении, включают липид-продуцирующие грибки, дрожжи и тому подобное.
Липид-продуцирующие грибки, которые можно использовать, представляют собой штаммы, описанные, например, MYCOTAXON, Vol. XLIV, No. 2, стр.257-265(1992). Конкретные примеры включают микроорганизмы, принадлежащих роду Mortierella в том числе микроорганизмы, принадлежащие подроду Mortierella, например, Mortierella elongata IFO8570, Mortierella exigua IFO8571, Mortierella hygrophila IFO5941, Mortierella alpina IFO8568, ATCC16266, ATCC32221, ATCC42430, CBS219.35, CBS224.37, CBS250.53, CBS343.66, CBS527.72, CBS528.72, CBS529.72, CBS608.70 и CBS754.68, и т.д., или микроорганизмы, принадлежащие подроду Micromucor, например, Mortierella isabellina CBS194.28, IFO6336, IFO7824, IFO7873, IFO7874, IFO8286, IFO8308 и IFO7884, Mortierella nana IFO8190, Mortierella ramanniana IFO5426, IFO8186, CBS112.08, CBS212.72, IFO7825, IFO8184, IFO8185 и IFO8287, Mortierella vinacea CBS236.82, и т.д. Среди прочего, Mortierella alpina является предпочтительным.
Примерами дрожжей являются Saccharomyces cerevisiae NBRC1951, NBRC1952, NBRC1953, NBRC1954 и т.д.
Введение вектора по изобретению в дрожжи и тестирование глицерол-3-фосфатацилтрансферазной активности белка GPAT3, кодируемого вектором, отсутствие генов GPAT (Gpt2p и Sct1p) дрожжей, используемых в качестве клетки-хозяина, позволяет оценить ферментативную активность только белка GPAT3. Соответственно, в варианте осуществления изобретения дрожжи в качестве клетки-хозяина, предпочтительно, являются дефицитными по гену Gpt2p и гену Sct1p.
Эти клетки-хозяева, трансформированные вектором по изобретению, продуцируют большее количество триглицеридов, чем клетки-хозяева, которые не трансформированы вектором по изобретению. Большая часть жирных кислот представляют собой жирные кислоты, которые составляют триглицериды, увеличенные путем введения вектора по изобретению.
Векторы, используемые для введения в липид-продуцирующие грибки, включают, в качестве неограничивающего примера, pDura5 (Appl. Microbiol. Biotechnol., 65, 419-425, (2004)).
В качестве векторов, которые можно использовать для введения в дрожжи, подходит любой вектор без особенных ограничений, при условии, что вектор обладает активностью экспресировать вставку в клетках дрожжей, и включает, например, pYE22m (Biosci. Biotech. Biochem., 59, 1221-1228, 1995).
Промоторы/терминаторы для регулирования экспрессии генов в клетках-хозяевах, могут быть в любой комбинации, при условии, что они функционируют в клетках-хозяевах. Например, можно использовать промотор гена гистона H4.1, промотор глицеральдегид-3-фосфатдегидрогеназы и т.д.
В качестве селективного маркера, используемого для трансформации, можно использовать ауксотрофные маркеры (ura5, niaD), маркеры химической устойчивости (гигромицин, зеоцин), ген устойчивости к генетицину (G418r), ген устойчивости к меди (CUP1) (Marin et al., Proc. Natl. Acad. Sci. USA, 81, 337 1984), ген устойчивости к церуленину (fas2m, PDR4) (Junji Inokoshi, et al., Biochemistry, 64, 660, 1992; и Hussain et al., Gene, 101: 149, 1991, соответственно).
Для трансформации клеток-хозяев, как правило, можно использовать известные способы. Например, способы, которые можно использовать, включают, в качестве неограничивающих примеров, способ электропорации (Mackenxie D. A. et al., Appl. Environ. Microbiol., 66, 4655-4661, 2000), способ доставки частиц (способ, описанный в JPA 2005-287403 «Method of Breeding Lipid-Producing Fungus»), способ сферопластов (Proc. Natl. Acad. Sci. USA, 75 p1929 (1978)), литийацетатный способ (J. Bacteriology, 153 p163 (1983)) и способы, описанные в Proc. Natl. Acad. Sci. USA, 75 p1929 (1978), Methods in yeast genetics, 2000 Edition: A Cold Spring Harbor Laboratory Course Manual и т.д.
Кроме того, может быть сделана ссылка на «Sambrook & Russell, Molecular Cloning: A Laboratory Manual Vol. 3, Cold Spring Harbor Laboratory Press 2001», «Methods in Yeast Genitics, A laboratory manual (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY)» и т.д., для общих способов клонирования.
4. Способ получения липидной композиции или композиции на основе жирных кислот по изобретению
В другом варианте осуществления, настоящее изобретение также относится к способу получения липидной или композиции на основе жирных кислот, который предусматривает использование трансформанта липид-продуцирующего грибка или дрожжей, описанного выше.
Как используется в настоящем документе, термин «липидный» предназначается для обозначения простого липида, включая соединение (например, глицерид), которое состоит из жирных кислот и спирта, соединенного с помощью сложноэфирной связи, или его аналога (например, холестероловый эфир), и т.д.; сложного липида, в котором фосфорная кислота, аминокислоты(ы), сахарид(ы) или подобное присоединены к части простого липида; или производному липида, которое является гидролизатом вышеприведенного липида и нерастворимым в воде.
Как используется в настоящем документе, термин «масла и жир» предназначается для обозначения сложного эфира глицерина и жирной кислоты (глицерида).
Как используется в настоящем документе, термин «жирная кислота» означает алифатическую монокарбоновую кислоту (карбоновую кислоту, имеющую одну карбоксильную группу и атомы углерода, соединенные друг с другом в цепь), представленную общей формулой RCOOH (где R представляет собой алкил). Жирная кислота включает насыщенную жирную кислоту, не имеющую двойных связей, и ненасыщенную жирную кислоту, содержащую двойные связи в углеводородной цепи.
Липидная композиция или композиция на основе жирных кислот по настоящему изобретению может быть получена, как следует, из клеток, трансформированных в соответствии с настоящим изобретением. После окончания инкубации, трансформанты организма (например, липид-продуцирующий грибок и дрожжи) обрабатываются обычным способом, включая центрифугирование, фильтрацию и т.д. для сбора клеток культуры. Клетки тщательно промывают водой и, желательно, высушивают. Высушивание может осуществляться при помощи лиофилизации, воздушной сушки и т.д. Высушенные клетки разрушают в мельнице Dyno или при помощи обработки ультразвуком и т.д., если необходимо, и затем экстрагируют органическим растворителем, предпочтительно, в струе азота. Органический растворитель, который можно использовать, включает эфир, формальдегид, метанол, этанол, хлороформ, дихлорметан, петролейный эфир и др. Кроме того, хорошие результаты можно также получить путем попеременного экстрагирования метанолом и петролейным эфиром, однофазной экстракцией хлороформ-метанол-вода. При удалении органического растворителя путем перегонки при пониженном давлении можно получить липид, содержащий жирные кислоты. Экстрагированные жирные кислоты также можно этерифицировать способом соляной кислоты-метанола и т.д.
Кроме того, выделение жирных кислот из липидов, содержащих жирные кислоты, описанное выше, можно проводить путем концентрирования и разделения обычным образом (например, способом с добавлением мочевины, способом разделения с охлаждением, колоночной хроматографией и др.) при насыщении смешанных жирных кислот или смешанных сложных эфиров жирных кислот.
Липидная композиция или композиция на основе жирных кислот, полученная способом получения по настоящему изобретению, может быть использована для получения, например, пищи, фармацевтической продукции, промышленных материалов (сырья для косметической продукции, мыла и т.д.), содержащих масла и жиры, и тому подобное.
В еще одном варианте осуществления, настоящее изобретение относится к способу получения пищевых продуктов, косметики, фармацевтической продукции, мыла и т.д., с использованием трансформанта липид-продуцирующего грибка или дрожжей по настоящему изобретению. Способ предусматривает стадию формирования липидов и жирных кислот с помощью трансформанта липид-продуцирующего грибка или трансформанта дрожжей по настоящему изобретению. Продукты питания, косметические препараты, фармацевтические препараты, мыло и прочие, содержащие образованные липиды или жирные кислоты, подготавливают в традиционной манере. Таким образом, продукты питания, косметические препараты, фармацевтические препараты, мыло и прочие, подготовленные способом по настоящему изобретению, содержат липиды и жирные кислоты, образованные с применением трансформанта липид-продуцирующего грибка или трансформанта дрожжей по настоящему изобретению. Настоящее изобретение также относится к приготовленным способом продуктам питания, косметическим препаратам, фармацевтическим препаратам, мылу и прочему.
Форма косметической (композиции) или фармацевтической (композиции) по настоящему изобретению особенно не ограничена и может быть в любом виде, включая состояние раствора, пасты, геля, сухого вещества или порошка. Также, косметическая композиция или фармацевтическая композиция по настоящему изобретению могут применяться в качестве косметики или местных агентов для кожи, включая масло, лосьон, крем, эмульсию, гель, шампунь, ополаскиватель для волос, кондиционер для волос, эмаль, крем-основу, помаду, пудру для лица, маску для лица, мази, духи, пудру, одеколон, зубную пасту, мыло, аэрозоль, очищающую пену и т.д., средство по уходу за кожей, замедляющее старение, противовоспалительное средство для кожи, средство для ванн, лечебный тоник, эссенция для придания красоты коже, солнцезащитное средство, или защитное и улучшающее средство при кожных заболеваниях, вызванных травмой, растрескиванием или трещиной кожи и т.д.
Косметическая композиция по настоящему изобретению также может быть составлена соответствующим образом с другими маслами и жирами и/или красящими веществами, ароматизаторами, консервантами, поверхностно-активными веществами, красителями, антиоксидантами и т.д., если это необходимо. Кроме того, фармацевтическая композиция по настоящему изобретение также может содержать другие фармацевтически активные компоненты (например, противовоспалительные компоненты) или вспомогательные компоненты (например, компоненты смазки или носителя). Примеры других компонентов, которые широко используются в косметических препаратах или препаратах для кожи для наружного применения, включают средство от угрей, средство для профилактики перхоти и зуда, дезодорантное и антиперспирантное средство, средство от ожогов, средство против клещей и вшей, средство для смягчения кератина, агент против сухости кожи, противовирусное вещество, вещество, способствующее чрескожной абсорбции, и тому подобное.
Продукт питания по настоящему изобретению включает диетическую добавку, лечебное питание, функциональное питание, питание для детей, питание для младенцев, модифицированное молоко для младенцев, модифицированное молоко для недоношенных младенцев, гериатрическое питание и т.д. Как используется в настоящем документе, под пищей подразумевается твердая, текучая и жидкая пища, а также их смесь, и в совокупности означает съедобный продукт.
Термин «пищевая добавка» относится к пищевым продуктам, обогащенным конкретными пищевыми ингредиентами. Термин «диетический продукт» относится к продуктам питания, которые полезны и благоприятны для здоровья, и включает в себя диетические добавки, натуральные продукты питания и диетические пищевые продукты. Термин «функциональная пища» относится к пищевому продукту для компенсирования питательных компонентов, которые способствуют регулирующим функциям организма. «Функциональные пищевые продукты» являются синонимами продуктов для указанного медицинского применения. Термин «пищевой продукт для маленьких детей» относится к пищевому продукту, предназначенному детям до 6 лет. Термин «гериатрический пищевой продукт» относится к пищевому продукту, обработанному для того, чтобы облегчить переваривание и усвоение по сравнению с необработанным пищевым продуктом. Термин «модифицированное молоко для младенцев» относится к модифицированному молоку, предназначенному для детей до одного года. Термин «модифицированное молоко для недоношенных младенцев» относится к модифицированному молоку, предназначенному для недоношенных младенцев в возрасте до 6 месяцев после родов.
Эти продукты включают природные продукты (обработанные жирами и маслами), такие как мясо, рыба и орехи; продукты с добавлением жиров и масел во время приготовления, например, китайские продукты, китайская лапша, супы и т.д.; пищевые продукты, приготовленные с применением жиров и масел в качестве нагретой среды, например, темпура или жареные во фритюре рыба и овощи, жареные во фритюре продукты, жареный соевый творог, китайский жареный рис, пончики, японское жареное печение из теста или каринто; продукты на основе жира и масла или переработанные продукты с добавлением жиров и масел в процессе переработки, например, сливочное масло, маргарин, майонез, соус, шоколад, макароны быстрого приготовления, карамель, бисквит, печенье, пирожное, мороженое; и пищевые продукты с напылением или покрытием жирами и маслами после окончания приготовления, например, рисовые крекеры, жесткое печенье, хлеб со сладкой бобовой пастой и т.д. Однако, продукт питания не ограничивается продуктами, содержащими жиры и масла, и другие примеры включают продукты сельского хозяйства, такие как хлебобулочные изделия, лапша, вареный рис, сладости (например, конфеты, жевательная резинка, клейкие массы, таблетки, японские сладости), соевый творог и продукты переработки этого; ферментированные пищевые продукты, такие, как японское рисовое вино или сакэ, лекарственные ликеры, Шерри десертного приготовления (мирин), уксус, соевый соус и мисо или бобовые пасты и т.д.; пищевые продукты животноводства, такие как йогурт, ветчина, бекон, колбаса и т.д.; морепродукты, такие как рубленые и приготовленные на пару рыбные котлеты или камабоко, жареные во фритюре рыбные лепешки или агетен и взошедшие рыбные котлеты или хенпен и т.д., а также фруктовые напитки, безалкогольные напитки, напитки для спортсменов, алкогольные напитки, чай и т.д.
Пищевой продукт по настоящему изобретению также может быть в виде фармацевтических препаратов, таких как капсулы и т.п., или в виде переработанного продукта питания, такого как природное жидкое питание, питание с определенным составом и элементарное питание, составленное с маслом и жиром по настоящему изобретению вместе с белками, сахарами, микроэлементами, витаминами, эмульгаторами, синтетическими душистыми веществами и т.п., целебных напитков, энтеральных питательных веществ и тому подобное.
Как описано выше, жирные кислоты можно эффективно изготавливать путем экспрессии гена GPAT3 по настоящему изобретению в клетке-хозяине.
Кроме того, уровень экспрессии гена можно использовать в качестве показателя для изучения условий культивирования, контроля инкубации и т.д. для эффективной продукции жирных кислот.
ПРИМЕРЫ
Далее настоящее изобретение описано более подробно со ссылкой на ПРИМЕРЫ, но без намерения ограничить объем изобретения этими ПРИМЕРАМИ.
[ПРИМЕР 1]
Анализ генома М. alpina
Штамм М. alpina 1S-4 засевали в 100 мл среды GY2:1 (2% глюкозы, 1% дрожжевого экстракта, pH 6,0), затем культивировали с перемешиванием при 28°C в течение 2 дней. Клетки грибков собирали путем фильтрации, и геномная ДНК была получена с использованием DNeasy (QIAGEN).
Нуклеотидную последовательность описанной выше геномной ДНК определяли с использованием Roche 454 Genome Sequencer FLX Standard. Эта процедура включала два раунда определения нуклеотидной последовательности библиотеки фрагментов и три раунда определения нуклеотидной последовательности совмещенной парной библиотеки. Полученные нуклеотидные последовательности были собраны в 300 суперконтигов.
Синтез кДНК штамма М. alpina Штамм 1S-4
Штамм М. alpina 1S-4 засевали в 4 мл среды (2% глюкозы, 1% дрожжевой экстракт, pH 6,0) и культивировали течение 4 дней при 28°C. Клетки собирали при помощи фильтрации, и РНК экстрагировали при помощи RNeasy Plant Kit (QIAGEN). кДНК синтезировали с помощью SuperScript First Strand System при помощи RT-PCR (Invitrogen).
Поиск гомологов GPAT
Был проведен поиск аминокислотной последовательности MaGPAT1 (ATCC#16266) для сравнения с геномной нуклеотидной последовательностью штамма М. alpina 1S-4 с помощью tblastn. В результате, были определены суперконтиги, содержащие последовательности, приведенные в SEQ ID NO:4 и SEQ ID NO:5. Считалось, что SEQ ID NO:5 представляет собой геномную последовательность GPAT1, полученного из штамма M. alpina 1S-4 (в дальнейшем именуемый MaGPAT1). С другой стороны, считалось, что SEQ ID NO:4 представляет собой последовательность, кодирующую новый гомолог GPAT, на основании наличия инициирующего кодона или терминирующего кодона и сравнения с MaGPAT1. Кроме того, было принято, что остатки с первого по третий и 3278-3280 в SEQ ID NO:4 являются инициирующим кодоном и терминирующим кодоном этого гомолога, соответственно. Этот ген был назван MaGPAT3.
Клонирование гена MaGPAT3
Для клонирования CDS гена MaGPAT3 были получены следующие праймеры.
Eco-MaGPAT3-F: 5'-GAATTCATGGGTCTCCAGATCTATGACTTCGTCTC-3' (SEQ ID NO:6)
Sal-MaGPAT3-R: 5'-GTCGACTTATGCCTCCTTAGACTTGACTGCATCC-3' (SEQ ID NO:7)
С помощью вышеупомянутой кДНК в качестве матрицы, была выполнена ПЦР-амплификация с KOD-Plus (TOYOBO) с использованием праймеров, Есо-MaGPAT3-F и Sal-MaGPAT3-R, и был амплифицирован фрагмент ДНК примерно 2,2 т.п.н. Фрагмент был клонирован с использованием Zero Blunt TOPO PCR Cloning Kit (Invitrogen), и полученная в результате плазмида был назван pCR-MAGPAT3. Последовательность вставки этой плазмиды, т.е., CDS-последовательность гена MaGPAT3, показана в SEQ ID NO:3. ORF-последовательность гена MaGPAT3 представлена также в SEQ ID NO:1.
Анализа нуклеотидной последовательность
Из сравнения геномной последовательности (SEQ ID NO:4) и CDS-последовательности (SEQ ID NO:3) гена MaGPAT3 предполагается, что геномная последовательность гена должна содержать 7 экзонов и 6 интронов и кодировать белок, состоящий из 753 аминокислотных остатков (фиг. 1 и 2). Последовательность MaGPAT3 сравнивали с последовательностью известного GPAT-гомолога М. alpina. CDS-последовательность MaGPAT3 показала гомологию 73,7% с GPAT1 (ATCC#16266) и 73,2% с GPAT1 (1S-4). Аминокислотная последовательность MaGPAT3 показала гомологию 14,4% с GPAT2 (1S-4).
Предполагаемая аминокислотная последовательность (SEQ ID NO:2) MaGPAT3 была представлена для анализа гомологий с аминокислотной последовательностью, зарегистрированной в GENEBANK nr, при помощи BLASTp. В результате, аминокислотная последовательность с самым низким E-значением для последовательности, а именно, аминокислотная последовательность с высокой гомологией, представляла собой полученный из Cryptococcus neoformans var. neoformans JEC21 GPAT-гомолог (инвентарный номер GENEBANK No. XP_569487), и гомология аминокислотной последовательности, как оказалось, составляла 40,3%. Кроме того, предполагаемая аминокислотная последовательность MaGPAT3 показала 33,3% гомологии с аминокислотной последовательностью Sct1p, которая представляет собой полученный из дрожжей S. cerevisiae GPAT, и 31,5% с аминокислотной последовательностью Gpt2p. Сравнивались аминокислотные последовательности GPAT3 и GPAT1, полученных из М. alpina 1S-4, и полученных из S. cerevisiae GPAT Sct1 и Gpt2 (фиг. 4). Четырех домена, консервативные в GPAT-гомологах, и аминокислотные остатки (звездочки на фиг. 4), считающиеся важными для GPAT-активности, и аминокислотные остатки (крестики на фиг. 4), считающиеся важными для связывания глицерол-3-фосфата, в этих областях, также были консервативными в GPAT3.
[ПРИМЕР 2]
Комплементарный эксперимент для дрожжей S. cerevisiae (Δsct1, Δgpt2)
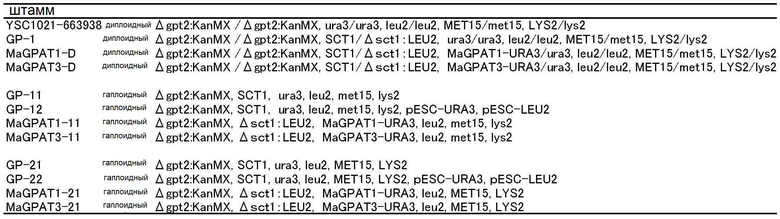
В дрожжах S. cerevisiae, SCT1 и GPT2 известны как гены, ответственные за GPAT-активность; известно, что одновременное удаление этих генов приводит к летальности. Для того, чтобы подтвердить, обладают ли белки, кодируемые MaGPAT1 и MaGPAT3 М. alpina, GPAT-активностью, был проведен дополнительный эксперимент с Δsct1 и Δgpt2. Генотипы штаммов, полученных в этом эксперименте, сведены в таблицу 1.
Получение штамма GP-1
Ген SCT1 Δgpt2-гомозиготных диплоидных дрожжей (Номер по каталогу No. YSC1021-663938) из Коллекции штаммов дрожжей с нокаутами генов (Open Biosystems) был разрушен следующим способом. Сначала, ДНК экстрагировали из клеток штамма S. cerevisiae S288C с помощью Gen Toru-Kun (для дрожжей) (TAKARA BIO). Используя ДНК в качестве матрицы, частичная последовательность гена SCT1 была амплифицирована при помощи ПЦР с KOD-Plus (TOYOBO) с использованием праймера Xba1-Des-SCT1-F: 5'-TCTAGAATGCCTGCACCAAAACTCAC-3 (SEQ ID NO:8) и праймера Xba1-Des-SCT1-R: 5'-TCTAGACCACAAGGTGATCAGGAAGA-3' (SEQ ID NO:9). Амплифицированный фрагмент ДНК примерно в 1,3 т.п.н. был клонирован с использованием Zero Blunt TOPO PCR Cloning Kit (Invitrogen), и получившаяся в результате плазмида была названа pCR-SCT1P. Затем, плазмиду YEp13 расщепляли рестрикционными ферментами SalI и XhoI. Получившийся в результате фрагмент ДНК размером около 2,2 т.п.н., содержащий CDS гена LEU2, был лигирован с ДНК-фрагментом размером около 4,4 т.п.н., полученным путем расщепления плазмиды pCR-SCT1P SalI, с использованием Ligation High (TOYOBO), чтобы сконструировать плазмиду с геном LEU2, встроенном в противоположном направлении по отношению к гену SCT1, и эта плазмида была названа pCR-Δsct1:LEU2. pCR-Δsct1: LEU2 была обработана с ограничением ферментом рестрикции XbaI, и вышеописанные Δgpt2 гомозиготные диплоидные дрожжи трансформировали при помощи литийацетатного способа. Трансформанты селектировали по способности расти на агаризованной среде SD-Leu (2% агар) (6,7 г дрожжей на основе азота без аминокислот (DIFCO), 20 г глюкозы и 1,3 г аминокислот в виде порошков (смесь 1,25 г аденинсульфата, 0,6 г аргинина, 3 г аспарагиновой кислоты, 3 г глутаминовой кислоты, 0,6 г гистидина, 0,9 г лизина, 0,6 г метионина, 1,5 г фенилаланина, 11,25 г серина, 0,9 г тирозина, 4,5 г валина, 6 г треонина, 1,2 г триптофана и 0,6 г урацила) на 1л). Из клеток полученных трансформантов экстрагировали ДНК при помощи способа, описанного выше. Используя (1) пару из праймера SCT1outORF-F: 5'-AGTGTAGGAAGCCCGGAATT-3 (SEQ ID NO:10) и праймера SCT1inORF-R: 5'-GCGTAGATCCAACAGACTAC-3' (SEQ ID NO:11) (0,5 т.п.н.), и (2) пару из праймера SCT1outORF-F1 и праймера LEU2inORF-F: 5'-TTGCCTCTTCCAAGAGCACA-3' (SEQ ID NO:12) (1,2 т.п.н.), выполняли ПЦР-амплификацию, чтобы проверить генотип, т.е., SCT1/Δsct1: LEU2, и трансформированная клетка дрожжей, имеющая этот генотип, была названа штаммом GP-1.
Конструирование штамма MaGPAT1-D и штамма MaGPAT3-D
Для того, чтобы встроить GPAT-гены MaGPAT1 и MaGPAT3 М. alpina в хромосомы дрожжей, плазмида pUC-URA3-MAGPAT1 и плазмида pUC-URA3-MAGPAT3 были сконструированы следующим образом.
Плазмида pUC-URA3-MAGPAT1
Множественный сайт для клонирования плазмиды pUC18 был модифицирован таким образом, чтобы содержать лишь сайт HindIII. ДНК-фрагмент размером около 1,2 т.п.н., который был получен расщеплением pURA34 (WO0131000) HindIII, был встроен в сайт HindIII, чтобы сконструировать плазмиды pUC-URA3. После расщепления pYE-MAGPAT1 (WO2008156026) ферментом рестрикции HindIII концы затупляли при помощи Blunting Kit (TAKARA BIO). В результате фрагмент ДНК около 3,5 т.п.н.был встроен в сайт SmaI плазмиды pUC-URA3, чтобы сконструировать плазмиду с геном GPAT1 и геном URA3, встроенными в одном и том же направлении. Эта плазмида была названа pUC-URA3-MAGPAT1.
Плазмида pUC-URA3-MAGPAT3
Плазмида pCR-MAGPAT3 была обработана рестрикционными ферментами EcoRI и SalI, и полученный в результате ДНК-фрагмент размером около 2,3 т.п.н. был встроен в сайты EcoRI/SalI вектора pYE22m (Biosci. Biotech. Biochem., 59, 1221-1228, 1995) для экспрессии в дрожжах, чтобы сконструировать плазмиду pYE-MAGPAT3. Далее, плазмида pYE-MAGPAT3 была обработана с рестрикционным ферментом HindIII, и затем концы были затуплены при помощи Blunting Kit (TAKARA BIO). Полученный в результате фрагмент ДНК примерно 3,6 т.п.н. был встроен в сайт SmaI плазмиды pUC-URA3, чтобы сконструировать плазмиду с геном GPAT3 и геном URA3, встроенных в одном и том же направлении. Эта плазмида была названа pUC-URA3-MAGPAT3.
Штамм GP-1 был трансформирован плазмидой pUC-URA3-MAGPAT1 и плазмидой pUC-URA3-MAGPAT3 с рестрикцией ферментом HindIII, соответственно, с помощью литийацетатного способа. Трансформанты селектировали по способности расти на агаризованной среде SD-Ura (2% агар) (6,7 г дрожжей на основе азота без аминокислот (DIFCO), 20 г глюкозы и 1,3 г аминокислот в виде порошков (смесь 1,25 г аденинсульфата, 0,6 г аргинина, 3 г аспарагиновой кислоты, 3 г глутаминовой кислоты, 0,6 г гистидина, 0,9 г лизина, 0,6 г метионина, 1,5 г фенилаланина, 11,25 г серина, 0,9 г тирозина, 4,5 г валина, 6 г треонина, 1,2 г триптофана и 1,8 г лейцина) на 1 л). ДНК экстрагировали из штаммов, необязательно выбранных из полученных трансформантов, используя Gen Toru-Kun (для дрожжей) (TAKARA BIO). В трансформант вставлены в URA3-MAGPAT1, геномную ДНК амплифицировали ПЦР, используя (3) пару из праймера GPAT1-f1: 5'-GTCAAGAAGGAATTCAAGGAGCTCAAG-3' (SEQ ID NO:13) и праймера GPAT1-r2: 5'-CCTGGGATGATGGACAAGAACAATG-3' (SEQ ID NO:14) (1,1 т.п.н.). В трансформанте с встроенной URA3-MAGPAT3 геномную ДНК амплифицировали при помощи ПЦР, используя (4) пару из праймера MaGPAT3-2F: 5'-TTTTGAACGCTTAAATGCTGGC-3' (SEQ ID NO:15) и праймера 5'-GGTCTTTTGAAGCTCTGCACGCGAC-3' (SEQ ID NO:16) (1,1 т.п.н.). Трансформанты, в которых вставка экспрессионной кассеты MaGPAT1 или экспрессионной кассеты MaGPAT3 в геном могла быть подтверждена, были названы штаммом MaGPAT1-D или штаммом MaGPAT3-D.
Споруляция и тетрадный анализ
Штаммы MaGPAT1-D и MaGPAT3-D высевали на агаризованную среду YPD, соответственно, и инкубируют при 30°C в течение 2 дней. Выращенные клетки засевали на агаризованную среду для споруляции (0,5% ацетат калия, 2% агар) и инкубировали при 25°C в течение 4 дней. Соответствующее количество спорулирующих клеток соскребали и суспендировали в 100 мкл раствора Zymolyase (0,125 мг/мл Zymolyase 100Т, 1М сорбитол, 40 мМ калийфосфатный буфер (pH 6,8)). После инкубации при комнатной температуре в течение 30 минут, к пробиркам добавляли раствор Zymolyase, и клетки переносили на лед. После того, как образование аскоспор было микроскопически подтверждено, четыре аскоспоры изолировали на агаризованной среде YPD при помощи микроманипулятора и инкубировали при 30°C в течение 2 дней, чтобы отобрать колонии, полученные из соответствующих спор. Полученные из спор клоны отпечатывали на агаризованную среду SD-Ura и агаризованную среду SD-Leu. Инкубацию проводили при 30°C в течение 3 дней, чтобы выявить ауксотроф по урацилу и ауксотроф по лейцину. Наличие или отсутствие роста на каждой чашке и число клонов приведены в таблице 2.
В споровых клонах от обоих штаммов, соотношение прототрофных по урацилу и прототрофных по лейцину штаммов: прототрофных по урацилу и ауксотрофных по лейцину штаммов: ауксотрофных по урацилу и ауксотрофных по лейцину штаммов составило примерно 1:1:1. Штамм, ауксотрофный по урацилу и прототрофный по лейцину, получен не был. Далее, в целях определения генотипа штамма, прототрофного по урацилу и прототрофного по лейцину, и штамма, ауксотрофного по урацилу и ауксотрофного по лейцин, полученных от штамма MaGPAT1-D и штамма MaGPAT3-D, соответственно, ДНК экстрагировали из клеток таким же образом, как описано выше, и ПЦР проводили, используя (1) пару из праймера SCT1outORF-F и праймера SCT1inORF-R и (2) пару из праймера SCT1outORF-F1 и праймера LEU2in ORF-F, и в штамме, полученном из MaGPAT1-D, используя (3) пару из праймера GPAT1-f1 и праймера GPAT1-r2, и в штамме, полученном из MaGPAT3-D, (4) - пару из праймера MaGPAT3-2F и праймера MaGPAT3-3R.
Штаммы, прототрофные по урацилу и прототрофные по лейцину, не продемонстрировали какой-либо амплификации с помощью ПЦР с парой (1), однако продемонстрировали амплификацию с парой (2). Поэтому было продемонстрировано, что эти штаммы, как выяснилось, являются Δsct1:LEU2. Кроме того, поскольку пары (3) или (4) показали амплификацию, было продемонстрировано, что MaGPAT1 или MaGPAT3 вставлены в эти штаммы.
Вышеизложенные результаты показали, что мутации Δgpt2 и Δsct1 в штаммах вызывают летальность у S. cerevisiae, однако штаммы становятся жизнеспособными при экспрессии MaGPAT1 или MaGPAT3, полученных от M. alpina. То есть, MaGPAT1 или MaGPAT3, полученный от M. alpina, был способен комплементировать мутации Δgpt2 и Δsct1 в штаммах дрожжей. Это говорит о том, что белки, кодируемые MaGPAT1 или MaGPAT3, полученный от M. alpina, могли бы обладать GPAT-активностью.
С другой стороны, в штаммах, ауксотрофных по урацилу и ауксотрофных по лейцину, амплификация при ПЦР наблюдалась с парой (1), но не наблюдалась с парой (2), указавая, что эти штаммы представляли собой SCT1. Отсутствие амплификации с парой (3) или парой (4) показало, что MaGPAT1 или MaGPAT3, полученный от M. alpina, не был встроен.
Кроме того, эти штаммы были отпечатаны на агаризованную среду SD-Met (2% агар) (6,7 г дрожжей на основе азота без аминокислот (DIFCO), 20 г глюкозы и 1,3 г аминокислот в порошках (смесь 1,25 г аденинсульфата, 0,6 г аргинина, 3 г аспарагиновой кислоты, 3 г глутаминовой кислоты, 0,6 г гистидина, 0,9 г лизина, 1,5 г фенилаланин, 11,25 г серина, 0,9 г тирозина, 4,5 г валина, 6 г треонина, 1,2 г триптофана, 1,8 г лейцина, 0,6 г урацила) и агаризованную среду SD-Lys (2% агар) (6,7 г дрожжей на основе азота без аминокислот (DIFCO), 20 г глюкозы и 1,3 г аминокислот в порошках (смесь 1,25 г аденинсульфата, 0,6 г аргинина, 3 г аспарагиновой кислоты, 3 г глутаминовой кислоты, 0,6 г гистидина, 0,6 г метионина, 1,5 г фенилаланина, 11,25 г серина, 0,9 г тирозина, 4,5 г валина, 6 г треонина, 1,2 г триптофана, 1,8 г лейцина и 0,6 г урацила) на 1 литр). Инкубацию проводили при 30°C в течение 3 дней, чтобы определить ауксотрофов по метионину и ауксотрофов по лизину. На основании результатов, GP-11 (#SC-1), MaGPAT1-11 (#3b, #4a, #8a), MaGPAT3-11 (#2d, #19a, #32a), GP-21 (#SC-2), MaGPAT1-21 (#1a, #2d, #13a) и MaGPAT3-21 (#10a, #20c, #26b), выбранные из штаммов для соответствующих генотипов, показанных в ТАБЛИЦЕ 1, были представлены для следующих проверок.
Сравнивали способности продуцировать жирные кислоты штамма только с SCT1, полученным из дрожжей, штамма только с GPAT1, полученным из М. alpina, штамма только с GPAT3, полученными из М. alpina, в качестве гена GPAT. Описанные выше штаммы GP-11 и GP-21, содержащие только SCT1, полученный из дрожжей, являются как ura3, так и leu2 и, таким образом, нуждаются в урациле и лейцине. Для комплементации ауксотрофности к урацилу и лейцину, GP-11 и GP-21 были совместно трансформированы плазмидой pESC-URA3 и плазмидой pESC-LEU2, соответственно. Штаммы были выбраны как трансформанты со способностью расти на агаризованной среде SD-Ura,Leu (2% агар) (6,7 г дрожжей на основе азота без аминокислот (DIFCO), 20 г глюкозы и 1,3 г аминокислот в порошке (смесь 1,25 г аденинсульфата, 0,6 г аргинина, 3 г аспарагиновой кислоты, 3 г глутаминовой кислоты, 0,6 г гистидина, 0,9 г лизина, 0,6 г метионина, 1,5 г фенилаланин, 11,25 г серина, 0,9 г тирозина, 4,5 г валина, 6 г треонина и 1,2 г триптофана) на 1 л). Дополнительно отобранные штаммы GP-12 (#1, 2, 3) и GP-22 (#1, #2, #3) были предоставлены для последующих проверок.
[ПРИМЕР 3]
Анализ жирных кислот дрожжей
Штаммы GP-12 (#1, 2, 3), MaGPAT1-11 (#3b, #4a, #8a), MaGPAT3-11 (#2d, #19a, #32a), GP-22 (#1, #2, #3), MaGPAT1-21 (#1a, #2d, #13a) и MaGPAT3-21 (#10a, #20c, #26b), полученные как описано выше, инкубировали следующим образом.
Одну платиновую петлю каждого штамма засевали в 10 мл жидкой среды SD-Ura, Leu и культивировали с перемешиванием при 30°C в течение 1 дня. Затем 100 мкл полученной культуры переносили в 10 мл жидкой среды SD-Ura, Leu, после чего культивировали с перемешиванием при 30°C в течение 2 дней. Дрожжевые культуры центрифугировали для сбора клеток. Клетки промывали 10 мл стерильной воды и центрифугировали снова для сбора клеток. Клетки лиофилизировали. Жирные кислоты в клетках преобразовывали в метиловый эфир способом соляной кислоты-метанола. Эфиры экстрагировали гексаном. После отгонки гексана был проведен анализ с помощью газовой хроматографии. Результаты анализа композиции на основе жирных кислот представлены в таблицах 3-8.
Таблица 3 показывает клеточную композицию на основе жирных кислот (%) дрожжей met15, lys2. Числовые значения выражены при помощи среднего ± стандартное отклонение (SD).
Таблица 4 показывает плотность клеток дрожжей met15, lys2 после инкубации. Числовые значения выражены при помощи среднего ±SD.
Таблица 5 показывает уровень продукции жирных кислот в дрожжах met15, lys2. Числовые значения выражены при помощи среднего ±SD.
Таблица 6 показывает композицию клеточных жирных кислот (%) дрожжей MET15,LYS2. Числовые значения выражены при помощи среднего ±SD.
Таблица 7 показывает плотность клеток дрожжей MET15,LYS2 после инкубации. Числовые значения выражены посредством среднего±SD.
Таблица 8 показывает уровень продукции жирных кислот дрожжей MET15,LYS2. Числовые значения выражены при помощи среднего ±SD.
В соответствующих клетках, плотность клеток зависела от ауксотрофности. Клеточная плотность дрожжей MET15,LYS2 была в 5 раз выше, чем клеточная плотность дрожжей met15,lys2, но какого-либо воздействия гена GPAT на его композицию на основе жирных кислот не наблюдалось (Таблицы 3 и 6). Далее, в дрожжах MET15,LYS2, также не было отмечено сколько-нибудь заметной разницы в способности продуцировать жирные кислоты, на основании различий в гене GPAT.
С другой стороны, в дрожжах met15,lys2 уровень продукции жирных кислот был выше примерно в 1,5 раза в штамме MaGPAT1-11 и выше примерно в 2,2 раза в штамме MaGPAT3-11 с полученным из M. alpine геном GPAT, по сравнению со штаммом GP-12, имеющим только SCT1 в качестве гена GPAT. Кроме того, уровень продукции жирных кислот штамма MaGPAT3-11 было выше примерно в 1,4 раза, чем у штамма MaGPAT1-11. Дрожжи met15,lys2 подавляли пролиферацию по сравнению с дрожжами MET15,LYS2, и оказалось, что экспрессия полученного из M. alpine гена GPAT1 или гена GPAT3 значительно повышала уровень продукции жирных кислот (таблицы 4 и 5). Сравнение композиции на основе жирных кислот показали увеличение соотношения насыщенной жирной кислоты пальмитиновой кислоты (16:0) или стеариновой кислоты (18:0) в штамме MaGPAT3-11, по сравнению со штаммами G-12 и MaGPAT1-11 (таблица 3).
[ПРИМЕР 4]
Анализ липидов дрожжей
Уровень каждого липида и композиции на основе жирных кислот проверяли для дрожжей met15,lys2 GP-12 (#1, 2, 3), MaGPAT1-11 (#3b, #4a, #8a) и MaGPAT3-11 (#2d, #19a и #32a), что показало различия в уровне продукции жирных кислот.
Одну платиновую петлю каждого штамма засевали в 10 мл жидкой среды SD-Ura,Leu, и культивировали с перемешиванием при 30ºC в течение 1 дня. Затем 1 мл полученной культуры каждого из 2 штаммов засевали в 10 мл жидкой среды SD-Ura,Leu, после чего культивировали с перемешиванием при 30ºC в течение 1 дня. Дрожжевую культуру центрифугировали для сбора клеток. Клетки промывали 10 мл стерильной воды и центрифугировали снова, чтобы собрать клетки. Клетки лиофилизировали. Жирные кислоты в клетках на один штамм преобразовывали в метиловые эфиры способом соляной кислоты-метанола. Эфиры экстрагировали гексаном. После удаления гексана перегонкой анализ проводили с помощью газовой хроматографии.
Кроме того, липиды экстрагировали из каждого из штаммов следующим образом. То есть, добавляли 1 мл хлороформа:метанола (2:1) и стеклянные бусы, и клетки разрушали при помощи устройства для разрушения бусами и центрифугировали, чтобы отобрать супернатант. К оставшимся клеткам дополнительно добавляли 1 мл хлороформа:метанола (2:1), и супернатант отобрали, как описано выше, что повторяли для того, чтобы получить липиды в итоге с 4 мл хлороформа:метанола (2:1). Растворитель удаляли путем перегонки, используя прибор speed-vac, а остаток растворяли в небольшом количестве хлороформа. Тонкослойную хроматографию проводили в условиях Silica Gel 60 Plate (Merck) и проявляющего растворителя гексан:диэтиловый эфир:уксусная кислота = 70:30:1 для того, чтобы фракционировать липиды. Липиды детектировали путем распыления раствора примулина и облучения УФ-лучами. Фракцию триглицеридов и фракцию фосфолипидов соскребали, соответственно, и собирали в пробирки. Жирные кислоты преобразовывали в метиловые эфиры способом соляной кислоты-метанола. Анализ жирных кислот выполняли с помощью газовой хроматографии.
Результаты представлены в таблицах ниже.
Плотность клеток после инкубации дрожжей met15,lys2 (среднее +SD)
Общий уровень жирных кислот у дрожжей met15,lys2 (среднее ±SD)
Уровень жирных кислот в фосфолипидной фракции дрожжей met15,lys2 (среднее ±SD)
Композиция на основе жирных кислот в фосфолипидной фракции дрожжей met15,lys2 (среднее ±SD)
Уровень жирных кислот в триглицеридной фракции дрожжей met15,lys2 (среднее ±SD)
Композиция на основе жирных кислот в триглицеридной фракции дрожжей met15,lys2 (среднее ± SD)
На клеточную плотность гены GPAT не оказывали воздействия. По уровням продукции жирных кислот в пересчете на среду, штамм MaGPAT1-11 с геном GPAT, полученным из Mortierella alpina, показал увеличение примерно в 1,5 раза, а штамм MaGPAT3-11 - увеличение приблизительно в 2 раза, чем штамм GP-12, имеющий только SCT1 в качестве гена GPAT.
На уровень жирных кислот и композицию на основе жирных кислот фосфолипидов гены GPAT не оказывали воздействия. С другой стороны, уровень триглицеридов был выше в 1,6 раза в штамме MaGPAT1-11 и в 2 раза выше в штамме MaGPAT3-11 с геном GPAT, полученным из Mortierella alpina, чем в штамме GP-12, имеющем только SCT1 в качестве гена GPAT.
Выяснилось, что увеличенный общей уровень продукции жирных кислот в штамме MaGPAT1-11 и штамме MaGPAT3-11, в основном, связан с увеличением уровня триглицеридов. Сравнение композиции на основе жирных кислот в триглицеридах выявило увеличение соотношения насыщенных жирных кислот в штамме MaGPAT3-11, по сравнению со штаммом GP-12.
Было установлено, что способность продуцировать триглицериды, в частности, может быть улучшена путем экспрессии гена MaGPAT3.
Промышленная применимость
Путем экспрессии полинуклеотида по настоящему изобретению в соответствующей клетке-хозяине можно эффективно продуцировать триглицериды независимо от скорости роста клетки-хозяина. Кроме того, увеличение триглицеридов также приводит к увеличению жирных кислот, которые образуют триглицериды. В соответствии с настоящим изобретением, триглицериды и жирные кислоты могут быть использованы для производства пищевых продуктов, косметических препаратов, лекарств, мыла и так далее.
















Изобретение относится к биотехнологии, в частности к новым глицерол-3-фосфатацилтрансферазам и полинуклеотидам, их кодирующим. Настоящее изобретение относится также к вектору экспрессии и трансформанту, содержащему полинуклеотид, а также способу получения липидной композиции или композиции на основе жирных кислот с использованием трансформанта. Изобретение позволяет получить новый фермент с улучшенными свойствами,. обладающий глицерол-3-фосфатацилтрансферазной активностью. 6 н. и 5 з.п. ф-лы, 4 ил., 14 табл., 4 пр.
1. Полинуклеотид, выбранный из группы, состоящей из нижеприведенных (a)-(e), который кодирует белок с глицерин-3-фосфатацилтрансферазной активностью:
(a) полинуклеотид, содержащий нуклеотидную последовательность SEQ ID NO: 1 или 4;
(b) полинуклеотид, кодирующий белок, состоящий из аминокислотной последовательности SEQ ID NO: 2;
(c) полинуклеотид, кодирующий белок, состоящий из последовательности аминокислот, где 1-10 аминокислот удалены, заменены, встроены и/или добавлены в аминокислотную последовательность SEQ ID NO: 2, и обладающий глицерол-3-фосфатацилтрансферазной активностью;
(d) полинуклеотид, кодирующий белок с аминокислотной последовательностью, гомологичной по меньшей мере на 90% аминокислотной последовательности SEQ ID NO: 2, и обладающий глицерол-3-фосфатацилтрансферазной активностью; и,
(e) полинуклеотид, который гибридизуется с полинуклеотидом, состоящим из нуклеотидной последовательности, комплементарной нуклеотидной последовательности SEQ ID NO: 1 или 4 в жестких условиях, и который кодирует белок, обладающий глицерол-3-фосфатацилтрансферазной активностью.
2. Полинуклеотид по п.1, содержащий нуклеотидную последовательность SEQ ID NO: 1 или 4.
3. Полинуклеотид по п.1, кодирующий белок, состоящий из аминокислотной последовательности SEQ ID NO: 2.
4. Полинуклеотид по любому из пп.1-3, который представляет собой ДНК.
5. Белок, кодируемый полинуклеотидом по любому из пп.1-4, который используется для получения липидной композиции или композиции жирных кислот.
6. Вектор, содержащий полинуклеотид по любому из пп.1-4, который используется для экспрессии глицерин-3-фосфатацилтрасферазы.
7. Трансформант, не являющийся трансформантом человека, в который введен полинуклеотид по любому из пп.1-4, который используется для получения липидной композиции или композиции жирных кислот, где трансформант является дрожжами или липид-продуцирующим грибом.
8. Трансформант, не являющийся трансформантом человека, в который введен вектор по п.6, который используется для получения липидной композиции или композиции жирных кислот, где трансформант является дрожжами или липид-продуцирующим грибом.
9. Трансформант по п.7 или 8, где трансформант представляет собой липид-продуцирующий грибок.
10. Трансформант по п.9, где липид-продуцирующий грибок представляет собой Mortierella alpina.
11. Способ получения липидной композиции или композиции на основе жирных кислот, который предусматривает сбор липидной композиции или композиции на основе жирных кислот из культуры трансформанта по любому из пп.7-10.
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| 0 |
|
SU78974A1 | |
| CHATRATTANAKUNCHAI S | |||
| et al | |||
| Oil biosynthesis in microsomal membrane preparations from Mortierella alpina | |||
| Biochem Soc Trans | |||
| ЩИТОВОЙ ДЛЯ ВОДОЕМОВ ЗАТВОР | 1922 |
|
SU2000A1 |
| RU 2001129499 A, 27.08.2003. | |||

