Родственные заявки
Настоящая заявка на изобретение представляет собой частичное продолжение заявки на патент США №10/985847, поданной 10 ноября 2004 г., представляющей собой:
1) частичное продолжение международной заявки на патент № PCT/GB2004/004253, которая указывает Соединенные Штаты Америки и подана 8 октября 2004 г.; и
2) частичное продолжение международной заявки на патент № PCT/GB2003/005646, которая указывает Соединенные Штаты Америки, подана 24 декабря 2003 г. и заявляет приоритет заявки на патент Великобритании № GB 0230202.4, поданной 27 декабря 2002 г., и заявки на патент Великобритании № GB 0327706.8, поданной 28 ноября 2003 г., представляющей собой:
частичное продолжение международной заявки на патент № PCT/GB2003/002804, которая указывает Соединенные Штаты Америки, подана 30 июня 2003 г. и заявляет приоритет заявки на патент Великобритании № GB 0230202.4, поданной 27 декабря 2002 г., представляющей собой:
частичное продолжение международной заявки на патент № PCT/GB02/03014, которая указывает Соединенные Штаты Америки и подана 28 июня 2002 г.
Полное содержание вышеупомянутых заявок на изобретения включены в настоящее описание путем ссылки.
Предшествующий уровень техники
Антиген-связывающий домен антитела включает две отдельные области: вариабельный домен тяжелой цепи (VH) и вариабельный домен легкой цепи (VL, который может представлять собой либо VK, либо Vλ). Антиген-сайт связывания сам по себе образован шестью полипептидными петлями: три из VH-домена (Н1, Н2 и Н3) и три из VL-домена (L1, L2 и L3). Разнообразный первоначальный репертуар V-генов, кодирующих VH- и VL-домены, формируется в результате комбинаторной перегруппировки генных сегментов. Ген VH формируется в результате рекомбинации трех генных сегментов VH, D и JH. У людей существует примерно 51 функциональный сегмент VH (Cook and Tomlinson (1995) Immunol Today, 16: 237), 25 функциональных сегментов D (Corbett et al (1997) J. Mol. Biol., 268: 69) и 6 функциональных сегментов JH (Ravetch et al. (1981) Cell, 27: 583) в зависимости от гаплотипа. Сегмент VH кодирует область полипептидной цепи, которая образует первую и вторую антиген-связывающие петли VH-домена (Н1 и Н2), тогда как сегменты VH, D и JH объединяются с образованием третьей антиген-связывающей петли VH-домена (Н3). Ген VL формируется в результате рекомбинации лишь двух генных сегментов VL и JL. У людей существует примерно 40 функциональных сегментов VK (Schable and Zachau (1993) Biol. Chem. Hoppe-Seyler, 374: 1001), 31 функциональный сегмент VK (Williams et al. (1996) J. Mol. Biol., 264: 220; Kawasaki et al. (1997) Genome Res., 7: 250), 5 функциональных сегментов JK (Hieter et al. (1982) J. Biol. Chem., 257: 1516) и 4 функциональных сегмента Jλ (Vasicek and Leder (1990) J. Exp. Med., 172: 609), в зависимости от гаплотипа. Сегмент VL кодирует область полипептидной цепи, которая образует первую и вторую антиген-связывающие петли VL-домена (L1 и L2), тогда как сегменты VL и JL объединяются с образованием третьей антиген-связывающей петли VL-домена (L3). Полагают, что антитела, выбранные из этого первоначального репертуара, достаточно разнообразны, чтобы связывать почти все антигены по меньшей мере с умеренной аффинностью. Высокоаффинные антитела продуцируются путем "аффинного созревания" перегруппированных генов, в которых образуются точечные мутации, и отбираются иммунной системой на основе улучшенного связывания.
Анализ структур и последовательностей антител демонстрирует, что пять из шести антиген-связывающих петель (Н1, Н2, L1, L2, L3) обладают ограниченным количеством конформаций основной цепи или канонических структур (Chothia and Lesk (1987) J. Mol. Biol, 196: 901; Chothia et al. (1989) Nature, 342: 877). Конформации основной цепи определяются (1) длиной антиген-связывающей петли и (2) конкретными остатками или типами остатка, находящимися в определенном ключевом положении в антиген-связывающей петле и каркасе антитела. Анализ длин петель и ключевых остатков позволил авторам изобретрения предсказать конформации основных цепей Н1, Н2, L1, L2 и L3, кодируемых большинством последовательностей человеческих антител (Chothia et al (1992) J. Mol. Biol, 227: 799; Tomlinson et al (1995) EMBO J., 14: 4628; Williams et al (1996) J. Mol. Biol, 264: 220). Хотя область Н3 гораздо более разнообразна с точки зрения последовательности, длины и структуры (вследствие применения сегментов D), она также образует ограниченное количество конформаций основной цепи для коротких петель, зависящих от длины и присутствия конкретных остатков или типов остатка, находящихся в ключевых положениях в петле и каркасе антитела (Martin et al (1996) J. Mol. Biol, 263: 800; Shirai et al (1996) FEBS Letters, 399:1.
В данной области известны биспецифические антитела, включающие комплементарные пары VH - и VL-областей. Эти биспецифические антитела должны включать две пары VH и VL, причем каждая пара VH/VL связывается с одним антигеном или эпитопом. Описанные способы включают гибридные гибридомы (Milstein & Cuello AC, Nature 305:537-40), мини-антитела (Hu et al, (1996) Cancer Res 56:3055-3061;), диа-антитела (Holliger et al, (1993) Proc. Natl. Acad. Sci. USA 90, 6444-6448; WO 94/13804), хелатообразующие рекомбинантные антитела (CRAbs; (Neri et al, (1995) J. Mol. Biol. 246, 367-373), biscFv (например, Atwell et al, (1996) Mol. Immunol. 33, 1301-1312), антитела, стабилизированные по механизму "knobs in holes" ("выступы в углубления") (Carter et al, (1997) Protein Sci. 6, 781-788). В каждом случае каждая разновидность антитела включает два антиген-связывающих сайта, каждый из которых моделируется комплементарной парой доменов VH и VL. Таким образом, каждое антитело способно связываться с двумя различными антигенами или эпитопами одновременно, причем связывание с КАЖДЫМ антигеном или эпитопом опосредуется VH и его комплементарным VL-доменом. Каждый из этих способов имеет свои конкретные недостатки; например, в случае гибридных гибридом неактивные пары VH/VL могут значительно уменьшать фракцию биспецифического IgG. Кроме того, большая часть биспецифических подходов основана на ассоциации различных пар VH/VL или ассоциации VH- и VL-цепей для воссоздания двух различных VH/VL-связывающих сайтов. Поэтому невозможно контролировать соотношение связывающих сайтов для каждого антигена или эпитопа в собранной молекуле, и, таким образом, множество собранных молекул будет связываться с одним антигеном или эпитопом, а не с другим. В некоторых случаях можно конструировать тяжелые или легкие цепи на поверхности раздела субъединиц (Carter et al, 1997) с целью повышения количества молекул, которые имеют сайты связывания с обоими антигенами или эпитопами, но это никогда не приведет к тому, что все молекулы будут связываться с обоими антигенами или эпитопами.
Существует доказательство того, что специфичности связывания двух различных антител могут быть включены в один и тот же сайт связывания, но они, как правило, представляют две или более чем две специфичности, которые соответствуют структурно родственным антигенам или эпитопам, или антителам, которые обладают широкой перекрестной реактивностью. Например, описаны перекрестно реактивные антитела, обычно когда два антигена имеют родственную последовательность и структуру, такие как лизоцим белка куриного яйца и лизоцим индюшки (McCafferty et al., WO 92/01047), или свободный гаптен и гаптен, конъюгированный с носителем (Griffiths AD et al. EMBO J 1994 13:14 3245-60). В еще одном примере WO 02/02773 (Abbott Laboratories) описаны молекулы антитела с "двойной специфичностью". Молекулы антитела, названные антителами, образуются или отбираются против множества антигенов, так что их специфичность распространяется более чем на один антиген. Каждая комплементарная пара VH/VL в антителах WO 02/02773 обуславливает единичную связывающую специфичность в отношении двух или более чем двух структурно родственных антигенов; каждый из VH- и VL-доменов в таких комплементарных парах не обладает отдельной специфичностью. Таким образом, антитела обладают широкой единичной специфичностью, охватывающей два структурно родственных антигена. Кроме того, описаны природные аутоантитела, которые являются полиреактивными (Casali & Notions, Ann. Rev. Immunol. 7, 515-531), вступающими в реакцию по меньшей мере с двумя (обычно более чем с двумя) различными, структурно не родственными антигенами или эпитопами. Кроме того, продемонстрировано, что выбор произвольных пептидных репертуаров с использованием технологии фагового дисплея на моноклональном антителе будет определять диапазон пептидных последовательностей, которые соответствуют антиген-связывающему сайту. Некоторые из последовательностей являются высоко родственными, соответствуя консенсусной последовательности, тогда как другие весьма различны и названы мимитопами (Lane & Stephen, Current Opinion in Immunology, 1993, 5, 268-271). Поэтому очевидно, что природное четырехцепочечное антитело, включающее ассоциированные и комплементарные VH- и VL-домены, обладают возможностью связываться с множеством различных антигенов из большой совокупности известных антигенов. В меньшей степени понятно, как конструировать сайт связывания для двух данных антигенов в одном и том же антителе, в частности антигенов, которые не обязательно структурно родственны.
Предложены способы белковой инженерии, которые могут быть основаны на этом. Например, также было предложено, что может быть сконструировано каталитическое антитело со связывающей активностью к иону металла, опосредованной одним вариабельным доменом, и к гаптену (субстрату), опосредованной контактами с ионом металла и комплементарным вариабельным доменом (Barbas et al, 1993 Proc. Natl. Acad. Sci USA 90, 6385-6389). Однако в этом случае предложено связывание и катализ субстрата (первый антиген), необходимые для связывания с ионом металла (второй антиген). Таким образом, связывание с парами VH/VL относится к единичному, но многокомпонентному антигену.
Описаны способы создания биспецифических антител из единичных доменов тяжелой цепи антитела Camelid, в которых связывающие контакты для одного антигена созданы в одном вариабельном домене, а для второго антигена - во втором вариабельном домене. Однако вариабельные домены не были комплементарными. Таким образом, первый вариабельный домен тяжелой цепи выбран против первого антигена, а второй вариабельный домен тяжелой цепи - против второго антигена, и затем оба домена связывают вместе на одной и той же цепи с получением фрагмента биспецифического антитела (Conrath et al., J. Biol. Chem. 270, 27589-27594). Однако единичные домены тяжелой цепи антитела Camelid не обычны тем, что они получены из природных антител Camelid, которые не имеют легких цепей, и действительно единичные домены тяжелой цепи не способны ассоциироваться с легкими цепями антитела Camelid с образованием комплементарных пар VH и VL.
Также описаны единичные вариабельные домены тяжелой цепи, полученные из природных антител, которые обычно ассоциированы с легкими цепями (из моноклональных антител или из репертуаров доменов; смотри ЕР-А-0368684). Показано, что эти вариабельные домены тяжелой цепи специфически взаимодействуют с одним или более чем одним родственным антигеном, но не комбинируются с другими вариабельными доменами тяжелой или легкой цепей с образованием лиганда, обладающего специфичностью в отношении двух или более чем двух различных антигенов. Кроме того, было показано, что эти единичные домены обладают очень коротким периодом полувыведения in vivo. Таким образом, такие домены обладают ограниченной терапевтической ценностью.
Предложено получать фрагменты биспецифического антитела путем связывания вместе вариабельных доменов тяжелой цепи, обладающих различными специфичностями (как описано выше). Недостаток этого подхода заключается в том, что вариабельные домены выделенного антитела могут обладать гидрофобной поверхностью контакта, которая в норме обеспечивает взаимодействия с легкой цепью и доступна для растворителя, и может быть "липкой", что позволяет единичному домену связываться с гидрофобными поверхностями. Кроме того, в отсутствие легкой цепи-партнера комбинация двух или более чем двух различных вариабельных доменов тяжелой цепи и их ассоциация, возможно через их гидрофобные поверхности контакта, может предотвращать их связывание с одним, а не с обоими лигандами, с которыми они способны связываться при выделении. Кроме того, в этом случае вариабельные домены тяжелой цепи могут быть не ассоциированы с комплементарными вариабельными доменами легкой цепи и, таким образом, могут быть менее стабильными и легко разворачивающимися (Worn & Pluckthun, 1998 Biochemistry 37, 13120-7).
Краткое изложение сущности изобретения
Изобретение относится к антагонистам фактора некроза опухоли 1 (TNFR1, р55, CD120a, P60, член 1А суперсемейства рецепторов TNF, TNFRSF1A) и способам применения этих антагонистов. Предпочтительные антагонисты обладают эффективностью при лечении, подавлении или предупреждении хронического воспалительного заболевания и по существу не оказывают антагонистического действия на фактор некроза опухоли 2 (TNFR2, Р75, Р80, CD120b, член 1 В суперсемейства рецепторов TNF, TNFRSF1B). В некоторых воплощениях антагонист является моновалентным.
В других воплощениях антагонист представляет собой антитело или его антиген-связывающий фрагмент, такой как моновалентный антиген-связывающий фрагмент (например, scFv, Fab, Fab', dAb), который обладает связывающей специфичностью в отношении TNFR1.
Другие предпочтительные антагонисты представляют собой описанные в данном изобретении лиганды, которые связывают TNFR1. Лиганды включают в себя единичный вариабельный домен иммуноглобулина или доменное антитело (dAb), которые обладают связывающей специфичностью в отношении TNFR1, или области, определяющие комплементарность, такого dAb в подходящем формате. В некоторых воплощениях лиганд представляет собой мономер dAb, который состоит по существу или состоит из единичного вариабельного домена иммуноглобулина или dAb, обладающего связывающей специфичностью в отношении TNFR1. В других воплощениях лиганд представляет собой полипептид, который включает dAb (или CDR (области, определяющие комплементарность) dAb) в подходящем формате, таком как формат антитела.
В некоторых воплощениях лиганд представляет собой лиганд с двойной специфичностью, который включает первое dAb, которое связывает TNFR1, и второе dAb, которое имеет связывающую специфичность, отличающуюся от связывающей специфичности первого dAb. В одном из примеров лиганд с двойной специфичностью включает первое dAb, которое связывает первый эпитоп на TNFR1, и второе dAb, которое связывает эпитоп на отличающейся мишени. В одном из примеров второе dAb связывает эпитоп на сывороточном альбумине.
В других воплощениях лиганд представляет собой полиспецифический лиганд, который включает первый эпитоп-связывающий домен, обладающий связывающей специфичностью в отношении TNFR1, и по меньшей мере один другой эпитоп-связывающий домен, обладающий связывающей специфичностью, отличающейся от связывающей специфичности первого эпитоп-связывающего домена. Например, первый эпитоп-связывающий домен может представлять собой dAb, которое связывает TNFR1, или может представлять собой домен, который включает CDR dAb, связывающего TNFR1 (например, CDR, перенесенные на подходящий белковый каркас или скелет, например аффитело, каркас SpA, домен класса А рецептора LDL (липопротеинов низкой плотности) или домен EGF (эпидермального фактора роста)) или может представлять собой домен, который связывает TNFR1, где домен выбран из антитела, домена SpA, домена класса А рецептора LDL или домена EGF).
В некоторых воплощениях лиганд или мономер dAb характеризуется одним или более чем одним из следующих: 1) диссоциирует из человеческого TNFR1 с константой диссоциации (Kd) от 50 нМ до 20 пМ, и константой скорости диссоциации Koff от 5×10-1 до 1×10-7 с-1; 2) ингибирует связывание фактора некроза опухоли альфа (TNFα) с TNFR1 с IC50 (50% ингибирующая концентрация) от 500 нМ до 50 пМ; 3) нейтрализует человеческий TNFR1 в стандартном анализе клеток L929 с ND50 (50% нейтрализующая доза) от 500 нМ до 50 пМ; 4) оказывает антагонистическое действие на активность TNFR1 в стандартном клеточном анализе с ND50 не более 100 нМ, и в концентрации не более 10 мкМ dAb оказывает агонистическое действие на активность TNFR1 не более чем 5% в анализе; 5) ингибирует смертность мышей в модели септического шока, индуцированного ЛПС (липополисахаридом)/D-галактозамином; 6) противодействует агрегации; 7) секретируется в количестве, составляющем по меньшей мере примерно 0,5 мг/л при экспрессии в Е. coli или видах Pichia (например, Р. pastoris); 8) обратимо разворачивается; 9) обладает эффективностью в модели хронического воспалительного заболевания, выбранной из группы, состоящей из модели коллаген-индуцированного артрита у мышей, модели артрита у мышей ДАРЕ, модели воспалительного заболевания кишечника у мышей AARE, модели воспалительного заболевания кишечника у мышей, индуцированного декстрансульфатом натрия, модели хронического обструктивного заболевания легких у мышей, индуцированного табачным дымом, и подходящих моделей у приматов (например, модели коллаген-индуцированного артрита у приматов); и/или 10) обладает эффективностью при лечении, подавлении или предупреждении хронического воспалительного заболевания.
В конкретных воплощениях лиганд или мономер dAb диссоциирует из человеческого TNFR1 с константой диссоциации (Kd) от 50 нМ до 20 пМ и константой скорости диссоциации Koff от 5×10-1 до 1×10-7 с-1; ингибирует связывание фактора некроза опухоли альфа (TNFα) с TNFR1 с IC50 от 500 нМ до 50 пМ; и нейтрализует человеческий TNFR1 в стандартном анализе клеток L929 с ND50 от 500 нМ до 50 пМ. В других конкретных воплощениях лиганд или мономер dAb диссоциирует из человеческого TNFR1 с константой диссоциации (Kd) от 50 нМ до 20 пМ и константой скорости диссоциации Koff от 5×10-1 до 1×10-7 с-1; ингибирует связывание фактора некроза опухоли альфа (TNFα) с TNFR1 с IC50 от 500 нМ до 50 пМ; и обладает эффективностью в модели хронического воспалительного заболевания, выбранной из группы, состоящей из модели коллаген-индуцированного артрита у мышей, модели артрита у мышей AARE, модели воспалительного заболевания кишечника у мышей AARE, модели воспалительного заболевания кишечника у мышей, индуцированного декстрансульфатом натрия, модели хронического обструктивного заболевания легких у мышей, индуцированного табачным дымом, и подходящих моделей у приматов (например, модели коллаген-индуцированного артрита у приматов). В других конкретных воплощениях лиганд или мономер dAb диссоциирует из человеческого TNFR1 с константой диссоциации (Kd) от 50 нМ до 20 пМ и константой скорости диссоциации Koff от 5×10-1 до 1×10-7 с-1; нейтрализует человеческий TNFR1 в стандартном анализе клеток L929 с ND50 от 500 нМ до 50 пМ; и оказывает антагонистическое действие на активность TNFR1 в стандартном клеточном анализе с ND50 не более 100 нМ, и в концентрации не более 10 мкМ dAb оказывает агонистическое действие на активность TNFR1 не более чем 5% в анализе.
В более конкретном воплощении лиганд или мономер dAb содержит аминокислотную последовательность, которая по меньшей мере примерно на 90% гомологична аминокислотной последовательности dAb, выбранной из группы, состоящей из TAR2h-12 (SEQ ID NO:32), TAR2h-13 (SEQ ID NO:33), TAR2h-14 (SEQ ID NO:34), TAR2h-16 (SEQ ID NO:35). TAR2h-17 (SEQ ID NO:36), TAR2h-18 (SEQ ID NO:37), TAR2h-19 (SEQ ID NO:38), TAR2h-20 (SEQ ID NO:39), TAR2h-21 (SEQ ID NO:40), TAR2h-22 (SEQ ID NO:41), TAR2h-23 (SEQ ID NO:42), TAR2D-24 (SEQ ID NO:43), TAR2h-25 (SEQ ID NO:44), TAR2h-26 (SEQ ID NO:45), TAR2h-27 (SEQ ID NO:46), TAR2h-29 (SEQ ED NO:47), TAR2h-30 (SEQ ID NO:48), TAR2h-32 (SEQ ID NO:49), TAR2h-33 (SEQ ID NO:50), TAR2h-10-1 (SEQ ID NO:51), TAR2h-10-2 (SEQ ID NO:52), TAR2h-10-3 (SEQ ID NO:53), TAR2h-10-4 (SEQ ID NO:54), TAR2h-10-5 (SEQ ID NO:55), TAR2h-10-6 (SEQ ID NO:56), TAR2h-10-7 (SEQ ID NO:57), TAR2h-10-8 (SEQ ID NO:58), TAR2h-10-9 (SEQ ID NO:59), TAR2h-10-10 (SEQ ID NO:60), TAR2h-10-11 (SEQ ID NO:61), TAR2h-10-12 (SEQ ID NO:62), TAR2h-10-13 (SEQ ID NO:63), TAR2h-10-14 (SEQ ID NO:64), TAR2h-10-15 (SEQ ID NO:65), TAR2h-10-16 (SEQ ID NO:66), TAR2h-10-17 (SEQ ID NO:67), TAR2h-10-18 (SEQ ID NO:68), TAR2h-10-19 (SEQ ID NO:69), TAR2h-10-20 (SEQ ID NO:70), TAR2h-10-21 (SEQ ID NO:71), TAR2h-10-22 (SEQ ID NO:72), TAR2h-10-27 (SEQ ID NO:73), TAR2h-10-29 (SEQ ID NO:74), TAR2h-10-31 (SEQ ID NO:75), TAR2h-10-35 (SEQ ID NO:76), TAR2h-10-36 (SEQ ID NO:77), TAR2h-10-37 (SEQ ID NO:78), TAR2h-10-38 (SEQ ID NO:79), TAR2h-10-45 (SEQ ID NO:80), TAR2h-10-47 (SEQ ID NO:81), TAR2h-10-48 (SEQ ID NO:82), TAR2h-10-57 (SEQ ID NO:83), TAR2h-10-56 (SEQ ID NO:84), TAR2h-10-58 (SEQ ID NO:85), TAR2h-10-66 (SEQ ID NO:86), TAR2h-10-64 (SEQ ID NO:87), TAR2h-10-65 (SEQ ID NO:88), TAR2h-10-68 (SEQ ID NO:89), TAR2h-10-69 (SEQ ID NO:90), TAR2h-10-67 (SEQ ID NO:91), TAR2h-10-61 (SEQ ID NO:92), TAR2h-10-62 (SEQ ID NO:93), TAR2h-10-63 (SEQ ID NO:94), TAR2h-10-60 (SEQ ID NO:95), TAR2h-10-55 (SEQ ID NO:96), TAR2h-10-59 (SEQ ID NO:97), TAR2h-10-70 (SEQ ID NO:98), TAR2h-34 (SEQ ID NO:373), TAR2h-35 (SEQ ID NO:374), TAR2h-36 (SEQ ID NO:375), TAR2h-37 (SEQ ID NO:376), TAR2h-38 (SEQ ID NO:377), TAR2h-39 (SEQ ID NO:378), TAR2h-40 (SEQ ID NO:379), TAR2h-41 (SEQ ID NO:380). TAR2h-42 (SEQ ID NO:381), TAR2h-43 (SEQ ID NO:382), TAR2h-44 (SEQ ID NO:383), TAR2h-45 (SEQ ID NO:384), TAR2h-47 (SEQ ID NO:385), TAR2h-48 (SEQ ID NO:386), TAR2h-50 (SEQ ID NO:387), TAR2h-51 (SEQ ID NO:388), TAR2h-66 (SEQ ID NO:389), TAR2h-67 (SEQ ID NO:390), TAR2h-68 (SEQ ID NO:391), TAR2h-70 (SEQ ID NO:392), TAR2h-71 (SEQ ID NO:393), TAR2h-72 (SEQ ID NO:394), TAR2h-73 (SEQ ID NO:395), TAR2h-74 (SEQ ID NO:396), TAR2h-75 (SEQ ID NO:397), TAR2h-76 (SEQ ID NO:398), TAR2h-77 (SEQ ID NO:399), TAR2h-78 (SEQ ID NO:400), TAR2h-79 (SEQ ID NO:401) и TAR2h-15 (SEQ ID NO:431).
В дополнительных воплощениях лиганд или мономер dAb содержит аминокислотную последовательность, которая по меньшей мере примерно на 90% гомологична аминокислотной последовательности dAb, выбранной из группы, состоящей из TAR2h-131-8 (SEQ ID NO:433), TAR2h-131-24 (SEQ ID NO:434), TAR2h-15-8 (SEQ ID NO:435), TAR2h-15-8-1 (SEQ ID NO:436), TAR2h-15-8-2 (SEQ ID NO:437), TAR2h-185-23 (SEQ ID NO:438), TAR2h-154-10-5 (SEQ ID NO:439), TAR2h-14-2 (SEQ ID NO:440), TAR2h-151-8 (SEQ ID NO:441), TAR2h-152-7 (SEQ ID NO:442), TAR2h-35-4 (SEQ ID NO:443), TAR2h-154-7 (SEQ ID NO:444), TAR2h-80 (SEQ ID NO:445), TAR2h-81 (SEQ ID NO:446), TAR2h-82 (SEQ ID NO:447), TAR2h-83 (SEQ ID NO:448), TAR2h-84 (SEQ ID NO:449), TAR2h-85 (SEQ ID NO:450), TAR2h-86 (SEQ ID NO:451), TAR2h-87 (SEQ ID NO:452), TAR2h-88 (SEQ ID NO:453), TAR2h-89 (SEQ ID NO:454), TAR2h-90 (SEQ ID NO:455), TAR2h-91 (SEQ ID NO:456), TAR2h-92 (SEQ ID NO:457), TAR2h-93 (SEQ ID NO:458), TAR2h-94 (SEQ ID NO:459), TAR2h-95 (SEQ ID NO:460), TAR2h-96 (SEQ ID NO:461), TAR2h-97 (SEQ ID NO:462), TAR2h-99 (SEQ ID NO:463), TAR2h-100 (SEQ ID NO:464), TAR2h-101 (SEQ ID NO:465), TAR2h-102 (SEQ ID NO:466), TAR21h-103 (SEQ ID NO:467), TAR2h-104 (SEQ ID NO:468), TAR2h-105 (SEQ ID NO:469), TAR2h-106 (SEQ ID NO:470), TAR2h-107 (SEQ ID NO:471), TAR2h-108 (SEQ ID NO:472), TAR2h-109 (SEQ ID NO:473), TAR2h-110 (SEQ ID NO:474), TAR2h-111 (SEQ ID NO:475), TAR2h-112 (SEQ ID NO:476), TAR2h-113 (SEQ ID NO:477), TAR2h-114 (SEQ ID NO:478), TAR2h-115 (SEQ ID NO:479), TAR2h-116 (SEQ ID NO:480), TAR2h-117 (SEQ ID NO:481), TAR2h-118 (SEQ ID NO:482), TAR2h-119 (SEQ ID NO:483), TAR2h-120 (SEQ ID NO:484), TAR2h-121 (SEQ ID NO:485), TAR2h-122 (SEQ ID NO:486), TAR2h-123 (SEQ ID NO:487), TAR2h-124 (SEQ ID NO:488), TAR2h-125 (SEQ ID NO:489), TAR2h-126 (SEQ ID NO:490), TAR2h-127 (SEQ ID NO:490), TAR2h-128 (SEQ ID NO:492), TAR2h-129 (SEQ ID NO:493), TAR2h-130 (SEQ ID NO:494), TAR2h-131 (SEQ ID NO:495), TAR2h-132 (SEQ ID NO:496), TAR2h-133 (SEQ ID NO:497), TAR2h-151 (SEQ ID NO:498), TAR2h-152 (SEQ ID NO:499), TAR2h-153 (SEQ ID NO:500), TAR2h-154 (SEQ ID NO:501), TAR2h-159 (SEQ ID NO:502), TAR2h-165 (SEQ ID NO:503), TAR2h-166 (SEQ ID NO:504), TAR2h-168 (SEQ ID NO:505), TAR2h-171 (SEQ ID NO:506), TAR2h-172 (SEQ ID NO:507), TAR2h-173 (SEQ ID NO:508), TAR2h-174 (SEQ ID NO:509), TAR2h-176 (SEQ ID NO:510), TAR2h-178 (SEQ ID NO:511), TAR2h-201 (SEQ ID NO:512), TAR2h-202 (SEQ ID NO:513), TAR2h-203 (SEQ ID NO:514), TAR2h-204 (SEQ ID NO:515), TAR2h-185-25 (SEQ ID NO:516), TAR2h-154-10 (SEQ ID NO:517) и TAR2h-205 (SEQ ID NO:627).
Изобретение относится к антагонисту фактора некроза опухоли I (TNFR1), связывающему фактор некроза опухоли I (TNFR1) и ингибирующему передачу сигнала через TNFR1, где указанный антагонист не ингибирует связывание TNFα с TNFR1. В некоторых воплощениях антагонист включает мономер первого доменного антитела (dAb) и мономер второго dAb, где указанный мономер первого dAb связывает домен TNFR1, выбранным из группы, состоящей из домена 1, домена 2, домена 3 и домена 4, и указанный мономер второго dAb связывает домен TNFR1, выбранный из группы, состоящей из домена 1, домена 2, домена 3 и домена 4, где указанный антагонист не оказывает агонистического действия на TNFR1, когда присутствует в концентрации примерно 1 мкМ в стандартном анализе цитотоксичности в отношении L929 или стандартном анализе IL-8 в HeLa.
В некоторых воплощениях изобретение представляет собой мономер доменного антитела (dAb) или лиганд, содержащий dAb, которое связывает фактор некроза опухоли I (TNFR1) и ингибирует передачу сигнала через TNFR1, где указанный мономер dAb не ингибирует связывание TNFα с TNFR1.
В других воплощениях изобретение представляет собой мономер доменного антитела (dAb) или лиганд, содержащий dAb, которое связывает фактор некроза опухоли I (TNFR1), где указанное dAb связывает домен 1 TNFR1 и конкурирует с TAR2m-21-23 за связывание с мышиным TNFR1 или конкурирует с TAR2h-205 за связывание с человеческим TNFR1.
В других воплощениях изобретение представляет собой мономер доменного антитела (dAb) или лиганд, содержащий dAb, которое связывает фактор некроза опухоли I (TNFR1), где указанное dAb связывает домен 3 TNFR1 и конкурирует с TAR2h-131-8, TAR2h-15-8, TAR2h-35-4, TAR2h-154-7, TAR2h-154-10 или TAR2h-185-25 за связывание с человеческим TNFR1.
Изобретение также относится к антителу или его антиген-связывающему фрагменту, обладающему связывающей специфичностью в отношении TNFR1 и эффективному при лечении, подавлении или предупреждении хронического воспалительного заболевания. В некоторых воплощениях антитело или антиген-связывающий фрагмент представляет собой моновалентный антиген-связывающий фрагмент.
В изобретении также предложен мономер dAb и лиганды, включающие мономер dAb, который обладает связывающей специфичностью в отношении TNFR1 и ингибирует TNFR-1-опосредованную передачу сигнала, но по существу не ингибирует связывание TNFα с TNFR1. В некоторых воплощениях мономер dAb ингибирует TNFα-индуцированное перекрестное связывание или кластеризацию TNFR1 на поверхности клетки.
В изобретении также предложены выделенные и/или рекомбинантные молекулы нуклеиновой кислоты, которые кодируют лиганды по изобретению, и векторы, которые включают рекомбинантные молекулы нуклеиновой кислоты. Также предложены клетки-хозяева, включающие рекомбинантные молекулы нуклеиновой кислоты или векторы по изобретению, и способы получения лигандов.
Изобретение также относится к фармацевтическим композициям, содержащим антагонист или лиганд по изобретению и фармакологически, физиологически или фармацевтически приемлемый носитель.
Изобретение также относится к способам лечения, подавления или предупреждения заболевания или расстройства (например, хронического воспалительного заболевания, аутоиммунного расстройства, воспалительного заболевания, артрита, рассеянного склероза, воспалительного заболевания кишечника, хронического обструктивного заболевания легких, пневмонии, септического шока), включающим введение млекопитающему, нуждающемуся в этом, терапевтически эффективного количества или дозы антагониста или лиганда по изобретению.
Изобретение также относится к антагонисту или лиганду по изобретению для применения в терапии или диагностике, и к применению антагониста или лиганда по изобретению для изготовления лекарственного средства для лечения, подавления или предупреждения заболевания или расстройства, как описано в данном изобретении (например, хронического воспалительного заболевания, аутоиммунного расстройства, воспалительного заболевания, артрита, рассеянного склероза, воспалительного заболевания кишечника, хронического обструктивного заболевания легких, пневмонии или септического шока. В других воплощениях заболевание может представлять собой муковисцидоз или тяжелую стероидорезистентную астму).
Изобретение также относится к фармацевтической композиции для лечения, подавления или предупреждения заболевания или расстройства, как описано в данном изобретении (например, хронического воспалительного заболевания, аутоиммунного расстройства, воспалительного заболевания, артрита, рассеянного склероза, воспалительного заболевания кишечника, хронического обструктивного заболевания легких, пневмонии или септического шока. В других воплощениях заболевание может представлять собой муковисцидоз или тяжелую стероидорезистентную астму), содержащей антагонист или лиганд по изобретению в качестве активного ингредиента.
Единичные вариабельные домены или доменные антитела (dAb), которые обладают связывающей специфичностью в отношении TNFR1, и лиганды, включающие эти единичные вариабельные домены или dAb, имеют несколько преимуществ. Например, описанные в данном изобретении единичные вариабельные домены или dAb, которые обладают связывающей специфичностью в отношении TNFR1, обладают антагонистическим эффектом на TNFR1. Соответственно, могут быть введены терапевтические агенты, которые включают единичный вариабельный домен иммуноглобулина против TNFR1 или dAb по изобретению (например, для терапевтических, диагностических или профилактических целей) с существенно сниженным риском побочных эффектов, вызванных связыванием и/или антагонистическим действием в отношении TNFR2 (например, иммуносупрессия). Терапевтические агенты, нацеленные на TNF-альфа, такие как ENBREL® (entarecept; Immunex Corporation) оказывают антагонистическое действие на TNFR1 и TNFR2, и введение таких агентов может вызывать иммуносупрессию и родственные побочные эффекты (например, тяжелые инфекции). Эти побочные эффекты могут ограничить применение таких агентов, в частности при хронических заболеваниях, при которых агент вводят в течение длительного периода времени (Kollias G. and Kontoyiannis D., Cytokine Growth Factor Rev., 73(4-5):315-321 (2002).) Напротив, поскольку лиганды по изобретению оказывают специфический антагонистическое действие на TNFR1, то их можно вводить в течение длительных периодов при хронических заболеваниях со сниженным риском побочных эффектов и обеспечивают преимущества при лечении воспалительных состояний и хронических воспалительных состояний (включая продолжительные заболевания, характеризующиеся периодами покоя и периодами активного воспаления, такие как воспалительное заболевание кишечника и артрит).
Краткое описание графических материалов
На Фиг.1 показано разнообразие VH/HAS (человеческий сывороточный альбумин) в положениях Н50, Н52, Н52а, Н53, Н55, Н56, Н58, Н95, Н96, Н97, Н98 (кодируемые соответственно DVT или NNK), которые располагаются в антиген-связывающем сайте VH HSA. (SEQ ID NO:1, нуклеотидная последовательность; SEQ ID NO:2, аминокислотная последовательность). Последовательность VK варьируется в положениях L50, L53.
На Фиг.2 представлено схематическое изображение структуры плазмиды pIT1/pIT2, используемой для получения библиотек одноцепочечных Fv (scFv), и показана нуклеотидная последовательность плазмиды от областей, регулирующих экспрессию и клонирование (SEQ ID NO:3), до кодируемой аминокислотной последовательности (SEQ ID NO:4). Плазмиду использовали для получения:
Библиотеки 1: Герминальный VK/DVT VH,
Библиотеки 2: Герминальный VK/NNK VH,
Библиотеки 3: Герминальный VH/DVT VK и
Библиотеки 4: Герминальный VH/NNK VK в формате фагового дисплея/ScFv.
Эти библиотеки предварительно подвергали селекции на связывание с типичными лигандами белком А и белком L, так чтобы большая часть клонов и выбранных библиотек была функциональной. Библиотеки подвергали селекции на HSA (первый раунд) и β-gal (второй раунд) или HSA β-gal, или на β-gal (первый раунд) и HSA (второй раунд) или β-gal HSA. Растворимый scFv из этих клонов ПЦР последовательно амплифицируют.Один из клонов, кодирующих антитело К8 с двойной специфичностью, выбрали для дальнейшей работы.
На Фиг.3 показано выравнивание VH-цепей (пустой VH (SEQ ID NO: 5), К8 (SEQ ID NO:6), VH2 (SEQ ID NO:7), VH4 (SEQ ID NO:8), VHC11 (SEQ ID NO:9), VHA-10sd (SEQ ID NO: 10), VHA-1sd (SEQ ID NO:11), VHA5sd (SEQ ID NO: 12), VHC5sd (SEQ ID NO:13), VHC11sd (SEQ ID NO: 14), VHC11sd (SEQ ID NO: 15)) и VK-цепей (пустой VK (SEQ ID NO: 16), K8 (SEQ ID NO: 17), E5sc (SEQ ID NO: 18), C3 (SEQ ID NO: 19)).
На Фиг.4 представлена характеристика связывающих свойств антитела K8, связывающие свойства антитела K8, охарактеризованные моноклональным фаговым иммуноферментным анализом (ИФА). Обнаружено, что антитело K8 с двойной специфичностью связывает HSA и β-gal и представлено на поверхности фага с сигналами абсорбции более 1,0. Не обнаружено перекрестной реактивности с другими белками.
На Фиг.5 показан ИФА растворимого scFv, осуществляемый с использованием известных концентраций фрагмента антитела K8. 96-луночный планшет покрывали 100 мкг HSA, BSA (бычий сывороточный альбумин) и β-gal в концентрации 10 мкг/мл и 100 мкг/мл белка А в концентрации 1 мкг/мл. Наносили по 50 мкг серийных разведении scFv K8 и фрагменты связанного антитела обнаруживали с использованием белка L, конъюгированного с HRP (пероксидаза хрена). Результаты ИФА подтверждают двойную специфическую природу антитела K8.
На Фиг.6 показаны связывающие характеристики клона К8VK/пустой VH, проанализированные с использованием ИФА растворимого scFv. Получение растворимых фрагментов scFv индуцировали с помощью IPTG (изопропилтиогалакгозид), как описано Harrison et al. Methods Enzymol. 1996;267:83-109, и непосредственно анализировали супернатант, содержащий scFv. ИФА растворимого scFv осуществляют, как описано в примере 1, и связанные scFv обнаруживают с помощью белка L, конъюгированного с HRP. Результаты ИФА показали, что этот клон способен связывать β-gal, тогда как связывание BSA отменялось.
На Фиг.7 показана последовательность (SEQ ID NO:2 и SEQ ID NO:3) векторов 1 и 2 для вариабельных доменов.
На Фиг.8 представлена карта вектора CH, используемого для конструирования полиспецифического лиганда VH1/VH2.
На Фиг.9 представлена карта вектора CK, используемого для конструирования полиспецифического лиганда VK1/VK2.
На Фиг.10 представлен анализ рецептора TNF, сравнивающий димер 4 TAR1-5, димер 4 TAR1-5-19 и мономер TAR1-5-19.
На Фиг.11 представлен анализ рецептора TNF, сравнивающий димеры 1-6 TAR1-5. Все димеры были очищены с помощью FPLC (быстрая жидкостная хроматография пептидов), и результаты представлены для оптимальных видов димеров.
На Фиг.12 представлен анализ рецептора TNF для гомодимеров TAR1-5 19 в различных форматах: формат dAb-линкер-dAb с линкером 3U, 5U или 7U, формат Fab и формат, представляющий собой цистеиновый шарнир-линкер.
На Фиг.13 показана пустая последовательность VH для библиотеки 1 (аминокислотная последовательность ((SEQ ID NO:5; нуклеотидные последовательности: кодирующая цепь (SEQ ID NO:20), некодирующая цепь (SEQ ID NO:21)). Последовательность каркаса VH, основанная на герминальной последовательности DP47-JH4b. Положения, где рандомизация NNK (N=нуклеотиды А или Т, или С, или G; К=нуклеотиды G или Т) включена в библиотеку 1, указаны жирным подчеркнутым текстом.
На Фиг.14 показана пустая последовательность VH для библиотеки 2 (аминокислотная последовательность ((SEQ ID NO:22; нуклеотидные последовательность: кодирующая цепь (SEQ ID NO:23), некодирующая цепь (SEQ ID NO:24). Последовательность каркаса VH, основанная на герминальной последовательности DP47-JH4b. Положения, где рандомизация NNK (N=нуклеотиды А или Т, или С, или G; К=нуклеотиды G или Т) включена в библиотеку 2, указаны жирным подчеркнутым текстом.
На Фиг.15 показана пустая последовательность VK для библиотеки 3. (аминокислотная последовательность ((SEQ ID NO:16; нуклеотидные последовательность: кодирующая цепь (SEQ ID NO:25), некодирующая цепь (SEQ ID NO:26). Последовательность каркаса VK, основанная на герминальной последовательности DPK9-JK1. Положения, где рандомизация NNK (N=нуклеотиды А или Т, или С, или G; К=нуклеотиды G или Т) включена в библиотеку 3, указаны жирным подчеркнутым текстом.
На Фиг.16 показана нуклеотидная и аминокислотная последовательность dAb против MSA (мышиный сывороточный альбумин) MSA 16 (нуклеотидная последовательность (SEQ ID NO:27), аминокислотная последовательность (SEQ ID NO:28) и MSA 26 (нуклеотидная последовательность (SEQ ID NO:29), аминокислотная последовательность (SEQ ID NO:30).
На Фиг.17 показано biacore ингибирование MSA 16 и 26. Очищенные dAb MSA16 и MSA26 анализировали с помощью biacore ингибирования для определения Кd. Кратко, dAb тестировали для определения концентрации dAb, необходимой для достижения ответа 200 единиц на biacore чипе СМ5, покрытом MSA с высокой плотностью. После того как необходимые концентрации dAb были определены, антиген MSA в диапазоне концентраций около ожидаемой Кd предварительно смешивали с dAb и инкубировали в течение ночи. Связывание dAb с biacore чипом, покрытым MSA, в каждой из предварительных смесей затем измеряли при высокой скорости потока 30 мкл/мин.
На Фиг.18 показаны уровни MSA16 в сыворотке крови после инъекции. У мышей определяли период полувыведения dAb MSA16 из сыворотки крови. MSA16 дозировали в виде разовых внутривенных (в/в) инъекций в концентрации приблизительно 1,5 мг/кг мышам CD1. Моделирование с использованием 2-компартментной модели продемонстрировало, что MSA16 имел t1/2α 0,98 ч, t1/2β 36,5 ч и AUC (площадь под кривой) 913 ч.мг/мл. MSA16 имел сравнительно удлиненный период полувыведения по сравнению с HEL4 (dAb против лизоцима белка куриного яйца), имеющего t1/2α 0,06 ч и t1/2β 0,34 ч.
На Фиг.19а-19в показан ИФА (Фиг.19а) и анализ рецептора TNF (Фиг.196, 19в), демонстрирующий ингибирование связывания TNF с Fab-подобным фрагментом, включающим MSA26Ck и TAR1-5-19CH. Добавление MSA с Fab-подобным фрагментом уменьшает уровень ингибирования. Планшет для ИФА, покрытый 1 мкг/мл TNFα, зондировали обладающим двойной специфичностью VKCH и VKCK Fab-подобным фрагментом, а также контрольным TNFα-связывающим dAb в концентрации, рассчитанной для получения похожего сигнала на ИФА. Как обладающее двойной специфичностью, так и контрольное dAb использовали для зондирования планшета для ИФА в присутствии и в отсутствие 2 мг/мл MSA. Сигнал в лунке с антителами с двойной специфичностью уменьшался более чем на 50%, а сигнал в лунке с dAb не уменьшался вовсе (смотри Фиг.19а). Тот же самый белок с двойной специфичностью также вводили в анализ рецептора с MSA и без него, и также показана конкуренция с MSA (смотри Фиг.19в). Это демонстрирует, что связывание MSA с антителами с двойной специфичностью конкурирует со связыванием с TNFα.
На Фиг.20 представлен анализ рецептора TNF, демонстрирующий ингибирование связывания TNF со связанным дисульфидными связями гетеродимером dAb TAR1-5-19 и dAb MSA16. Добавление MSA с димером уменьшает уровень ингибирования дозозависимым образом. Анализ рецептора TNF (Фиг.19(б)) проводили в присутствии постоянной концентрации гетеродимера (18 нМ) и серий разведении MSA и HSA. Присутствие HSA в диапазоне концентрций (до 2 мг/мл) не вызывало уменьшения способности димера ингибировать TNFα. Однако добавление MSA вызывало дозозависимое уменьшение способности димера ингибировать TNFα (Фиг.19а). Это демонстрирует, что MSA и TNFα конкурируют за связывание с cys связанным димером TAR1-5-19, MSA16. MSA и HSA сами по себе не оказывали эффекта на уровень связывания TNF в анализе.
На Фиг.21А-21М показаны аминокислотные последовательности (SEQ ID NO:31-98 и SEQ ID NO:373-401 и 431) нескольких вариабельных доменов человеческого иммуноглобулина, обладающих связывающей специфичностью в отношении человеческого TNFR1. Представленные аминокислотные последовательности являются непрерывными без пропусков; символ ~ вставлен в последовательности для указания мест расположения областей, определяющих комплементарность (CDR). CDR1 фланкирована ~, CDR2 фланкирована ~~, и CDR3 фланкирована ~~~.
На Фиг.22А-22Т показаны нуклеотидные последовательности (SEQ ID NO: 99-166 и SEQ ID NO:402-430 и 432) нескольких нуклеиновых кислот, которые кодируют вариабельные домены человеческого иммуноглобулина, представленные на Фиг.21А-21М. Представленные нуклеотидные последовательности являются непрерывными без пропусков; символ ~ вставлен в последовательности для указания мест расположения областей, определяющих комплементарность (CDR). CDR1 фланкирована ~, CDR2 фланкирована ~~, и CDR3 фланкирована ~~~.
На Фиг.23А-23 В показаны аминокислотные последовательности (SEQ ID NO: 167-179) нескольких вариабельных доменов человеческого иммуноглобулина, которые обладают связывающей специфичностью в отношении мышиного TNFR1. Представленные аминокислотные последовательности являются непрерывными без пропусков. В некоторых последовательностях символ ~ вставлен для указания на место расположения областей, определяющих комплементарность (CDR). CDR1 фланкирована ~, CDR2 фланкирована ~~, и CDR3 фланкирована ~~~.
На Фиг.24А-24С показаны нуклеотидные последовательности (SEQ ID NO:180-192 и 626) нескольких нуклеиновых кислот, кодирующих вариабельные домены человеческого иммуноглобулина, представленные на Фиг.23А-23 В. И SEQ ID NO: 186, и SEQ ID NO:626 кодирует аминокислотную последовательность SEQ ID NO: 173. Последовательности SEQ ID NO:626, кодирующие CDR1, фланкированы "-, последовательности, кодирующие CDR2, фланкированы ~~, и последовательности, кодирующие CDR3, фланкированы ~~~.
На Фиг.25A-25L показаны нуклеотидные последовательности, кодирующие несколько вариабельных доменов человеческого иммуноглобулина, и аминокислотные последовательности кодируемых вариабельных доменов человеческого иммуноглобулина (SEQ ID NO:193-198 и 200-295).
На Фиг.26 представлен график, демонстрирующий, что форматы dAb против TNFR1 по существу не оказывают агонистического действия на TNFR1 в анализе L929. Клетки L929 культивировали в средах, которые содержали диапазон концентраций мономера dAb против TNFR1 (TAR2m-21-23), мономера TAR2m-21-23, перекрестно связанного имеющимся в продаже антителом против myc (9E10), dAb с двойной специфичностью против TNFR1/dAb против SA (TAR2m-21-23 3D TAR7m-16), или пэгилированного мономера dAb против TNFR1 (TAR2m-21-23 40К PEG). В случае мономера TAR2m-21-23, перекрестно связанного антителом против myc, dAb и антитело смешивали в отношении 2:1 и предварительно инкубировали в течение одного часа при комнатной температуре для моделирования эффектов иммунного перекрестного связывания in vivo перед культивированием (Мономер TAR2m-21-23 включает эпитоп myc.) Мономер TAR2m-21-23 инкубировали с клетками L929 в концентрации 3000 нМ. Мономер TAR2m-21-23 и антитело против Мус инкубировали при концентрации dAb 3000 нМ. TAR2m-21-23 3U TAR7m-16 инкубировали с клетками в концентрациях 25 нМ, 83,3 нМ, 250 нМ, 833 нМ и 2500 нМ. TAR2m-21-23 40К PEG инкубировали с клетками в концентрациях 158,25 нМ, 527,5 нМ, 1582,5 нМ, 5,275 нМ и 15,825 нМ. После инкубации в течение ночи оценивали жизнеспособность клеток. Результаты показали, что инкубация клеток L929 с 10 нМ, 1 нМ или 0,1 нМ имеющегося в продаже IgG антитела против TNFR1, которое перекрестно связывает и оказывает агонистическое действие на TNFR1 (кат. №AF-425-PB; R&D Systems, Minneapolis, MN), привели в результате к дозозависимому увеличению нежизнеспособных клеток, таким образом демонстрируя чувствительность этих клеток к агонистам TNFR1. В отличие от этого, инкубация с различными количествами форматов антител против TNFR1 не оказывала антагонистического действия на TNFR1 и не приводила к увеличению количества нежизнеспособных клеток в культурах даже при использовании более чем 1000-кратной концентрации имеющегося в продаже IgG антитела против TNFR1.
На Фиг.27A-27I показаны аминокислотные последовательности (SEQ ID NO:433-517 и 627) нескольких вариабельных доменов человеческого иимуноглобулина, обладающего связывающей специфичностью в отношении человеческого TNFR1. Представленные аминокислотные последовательности являются непрерывными без пропусков; символ ~ вставлен в последовательности для указания мест расположения областей, определяющих комплементарность (CDR). CDR1 фланкирована ~, CDR2 фланкирована ~~, и CDR3 фланкирована ~~~.
На Фиг.28А-280 показаны нуклеотидные последовательности (SEQ ID NO:518-602 и 628) нескольких нуклеиновых кислот, кодирующих вариабельные домены человеческого иммуноглобулина, представленные на Фиг.27А-27Н. Представленные нуклеотидные последовательности являются непрерывными без пропусков; символ ~ вставлен в последовательности для указания мест расположения областей, определяющих комплементарность (CDR). Последовательности, кодирующие CDR1, фланкированы последовательности, кодирующие CDR2, фланкированы ~~, и последовательности, кодирующие CDR3, фланкированы ~~~.
Подробное описание изобретения
В этом описании воплощения раскрыты таким образом, что обеспечивают написание ясного и точного описания, но предполагается и будет понятно, что воплощения могут быть различным образом скомбинированы или разделены без отклонения от объема изобретения.
Определения
"Комплементарный". Два иммуноглобулиновых домена "комплементарны", когда они относятся к семействам структур, которые образуют когнатные пары или группы, или происходят из таких семейств и сохраняют это свойство. Например, VH-домен и VL-домен антитела комплементарны; два VH -домена не комплементарны, и два домена VL не комплементарны. Комплементарные домены могут быть обнаружены у других членов суперсемейства иммуноглобулинов, такие как домены Vα и Vβ (или γ и δ) Т-клеточного рецептора. В контексте второй конфигурации по настоящему изобретению некомплементарные домены не связываются кооперативно с молекулой-мишенью, а действуют независимо на различные эпитопы-мишени, которые могут находиться на одной и той же или различных молекулах. Домены, которые являются искусственными, такие как домены на основе белковых каркасов, которые не связываются с эпитопами, если только не сконструированы для этого, являются некомплементарными. Аналогично, два домена, основанные (например) на иммуноглобулиновом домене и фибронектиновом домене, не комплементарны.
"Иммуноглобулин". Относится к семейству полипептидов, которые сохраняют характеристики складывания иммуноглобулина для молекул антител, содержащих два β-слоя и обычно консервативную дисульфидную связь. Члены суперсемейства иммуноглобулинов вовлечены во многие аспекты клеточных и неклеточных взаимодействий in vivo, включая широко распространенные роли в иммунной системе (например, антитела, Т-клеточные рецепторные молекулы и т.п.), вовлечены в клеточную адгезию (например, молекулы ICAM (молекулы межклеточной адгезии)) и внутриклеточную передачу сигнала (например, рецепторные молекулы, такие как рецептор PDGF (тромбоцитарного фактора роста)). Настоящее изобретение применимо ко всем молекулам суперсемейства иммуноглобулинов, которые имеют связывающие домены. Предпочтительно, настоящее изобретение относится к антителам.
"Комбинирование". Вариабельные домены по изобретению комбинируют с образованием группы доменов; например, можно комбинировать комплементарные домены, такие как VL-домены, комбинированные с VH-доменами. Также можно комбинировать некомплементарные домены. Домены можно комбинировать многими путями, включая ковалентное и нековалентное связывание доменов.
"Домен". Домен представляет собой свернутую белковую структуру, которая сохраняет свою третичную структуру независимо от остальной части белка. Как правило, домены ответственны за дискретные функциональные свойства белков, и во многих случаях могут быть добавлены, удалены или перенесены на другие белки без утраты функции оставшейся части белка и/или домена. Под единичным вариабельным доменом антитела понимают свернутый полипептидный домен, включающий последовательности, характерные для вариабельных доменов антитела. Поэтому он включает вариабельные домены и модифицированные вариабельные домены полного антитела, например, в которых одна или более чем одна петля заменена на последовательности, которые не характерны для вариабельных доменов антитела, или вариабельные домены антитела, которые укорочены или включают N- или С-концевые удлинения, а также свернутые фрагменты вариабельных доменов, которые сохраняют по меньшей мере частично связывающую активность и специфичность полноразмерного домена.
"Репертуар". Коллекция различных вариантов, например полипептидных вариантов, которые отличаются своей первичной последовательностью. Библиотека, используемая в настоящем изобретении, охватывает репертуар полипептидов, включающих по меньшей мере 1000 членов.
"Библиотека". Термин библиотека относится к смеси гетерогенных полипептидов или нуклеиновых кислот. Библиотека состоит из членов, каждый из которых имеет последовательность одного полипептида или нуклеиновой кислоты. В этом смысле библиотека является синонимом репертуара. Различия последовательностей между членами библиотеки ответственны за разнообразие, присутствующее в библиотеке. Библиотека может принимать форму простой смеси полипептидов или нуклеиновых кислот, или может находиться в форме организмов или клеток, например бактериальных клеток, вирусов, животных или растительных клеток и т.п, трансформированных библиотекой нуклеиновых кислот. Предпочтительно, каждый индивидуальный организм или клетка содержит только один или ограниченное количество членов библиотеки. Предпочтительно, нуклеиновые кислоты включены в экспрессирующие векторы для того, чтобы обеспечить возможность экспрессии полипептидов, кодируемых нуклеиновыми кислотами. Поэтому в предпочтительном аспекте библиотека может принимать форму популяции организмов-хозяев, где каждый организм содержит одну или более чем одну копию экспрессирующего вектора, содержащего один член библиотеки в форме нуклеиновой кислоты, который может экспрессироваться с получением соответствующего ему полипептидного члена. Таким образом, популяция организмов-хозяев имеет потенциал для кодирования большого репертуара генетически различных полипептидных вариантов.
"Полиспецифический лиганд с закрытой конформацией" описывает полиспецифический лиганд, как определено в данном описании, включающий по меньшей мере два эпитоп-связывающих домена, как определено в данном описании. Термин 'закрытая конформация' (полиспецифический лиганд) означает, что эпитоп-связывающие домены лиганда расположены таким образом, что связывание эпитопа с одним эпитоп-связывающим доменом, конкурирует со связыванием эпитопа другим эпитоп-связывающим доменом. То есть, когнатные эпитопы могут быть связаны каждым эпитоп-связывающим доменом индивидуально, а не одновременно. Закрытой конформации лиганда можно достичь с использованием раскрытых в данном описании способов.
"Антитело". Антитело (например, IgG, IgM, IgA, IgD или IgE) или фрагмент (такой как Fab, F(ab')2, Fv, дисульфид-связанные Fv, scFv, полиспецифическое антитело с закрытой конформацией, дисульфид-связанный scFv, диа-антитело), полученное из видов, естественно продуцирующих антитело, или созданное с использованием технологии рекомбинантных ДНК; выделенное из сыворотки крови, В-клеток, гибридом, трансфектом, дрожжей или бактерий).
"Лиганд с двойной специфичностью". Лиганд, содержащий единичный вариабельный домен первого иммуноглобулина и единичный вариабельный домен второго иммуноглобулина, как описано в данном изобретении, где вариабельные области способны связываться с двумя различными антигенами или двумя эпитопами на одном и том же антигене, которые обычно не связываются моноспецифическим иммуноглобулином. Например, два эпитопа могут располагаться на одном и том же гаптене, но не являются одним и тем же эпитопом или достаточно близки для того, чтобы быть связанными моноспецифическим лигандом. Лиганды с двойной специфичностью по изобретению состоят из вариабельных доменов, обладающих различными специфичностями, и не содержат пары взаимно комплементарных вариабельных доменов, которые обладают той же самой специфичностью.
"Антиген". Молекула, которая связана с лигандом по настоящему изобретению. Как правило, антигены связаны с лигандами антитела и способны приводить к увеличению антительного ответа in vivo. Он может представлять собой полипептид, белок, нуклеиновую кислоту или другую молекулу. Как правило, лиганды с двойной специфичностью по изобретению отбирают на целевую специфичность против конкретного антигена. В случае стандартных антител и их фрагментов сайт связывания антитела, определенный вариабельными петлями (L1, L2, L3 и Н1, Н2, Н3), способен связываться с антигеном.
"Эпитоп". Единица структуры, обычно связанная иммуноглобулиновой парой VH/VL. Эпитопы определяют минимальный сайт связывания антитела и таким образом демонстрируют мишень для специфичности антитела. В случае однодоменного антитела эпитоп представляет единицу структуры, связанной с вариабельным доменом при выделении.
"Общий лиганд". Лиганд, который связывается со всеми членами репертуара. Как правило, не связан через антиген-связывающий сайт, как определено выше. Не ограничивающие примеры включают белок А, белок L и белок G.
"Селекция". Получают путем скрининга или в результате дарвиновского процесса селекции, при котором связывающие взаимодействия образуются между доменом и антигеном или эпитопом, или между антителом и антигеном или эпитопом. Таким образом, первый вариабельный домен может быть выбран для связывания с антигеном или эпитопом в присутствии или в отсутствие комплементарного вариабельного домена.
"Универсальный каркас". Последовательность каркаса единичного антитела, соответствующая областям антитела, консервативным по последовательности, как определено Kabat ("Sequences of Proteins of Immunological Interest", US Department of Health and Human Services) или соответствующая человеческому герминальному репертуару иммуноглобулинов или структуре, как определено Chothia и Lesk, (1987) J. Mol. Biol. 196:910-917. В изобретении предложен для применения единичной каркас или набор таких каркасов, которые, как было обнаружено, дают возможность получать фактически любую связывающую специфичность путем вариации только в гипервариабельных областях.
"Период полувыведения". Время, которое необходимо для уменьшения концентрации лиганда в сыворотке крови in vivo на 50%, например вследствие разрушения лиганда и/или клиренса, или секвестрирования лиганда природными механизмами. Лиганды по изобретению стабилизированы in vivo и их период полувыведения увеличен путем связывания с молекулами, которые устойчивы к разрушению и/или клиренсу или секвестрированию. Как правило, такие молекулы представляют собой встречающиеся в природе белки, которые сами по себе имеют длительный период полувыведения in vivo. Период полувыведения лиганда увеличивается, если сохраняется его функциональная активность in vivo в течение более длительного периода по сравнению с похожим лигандом, который не специфичен в отношении молекулы, увеличивающей период полувыведения. Таким образом, лиганд, специфичный в отношении HSA и молекулы-мишени, сравнивают с тем же самым лигандом, у которого отсутствует специфичность в отношении HSA, то есть он не связывает HSA, а связывает другую молекулу. Например, он может связывать второй эпитоп на молекуле-мишени. Как правило, период полувыведения увеличивается на 10%, 20%, 30%, 40%, 50% или более процентов. Возможны увеличения периода полувыведения в 2х, 3х, 4х, 5х, 10х, 20х, 30х, 40х, 50х или более раз. Альтернативно или дополнительно возможны увеличения периода полувыведения, находящиеся в диапазоне до 30х, 40х, 50х, 60х, 70х, 80х, 90х, 100, 150х.
"Гомогенный иммуноанализ". Иммуноанализ, в котором аналит обнаруживают без необходимости стадии разделения связанных и несвязанных реагентов.
"По существу идентичный (или "по существу гомогенный")". Первая аминокислотная или нуклеотидная последовательность, которая содержит достаточное количество аминокислотных остатков или нуклеотидов, идентичных или эквивалентных (например, с похожей боковой цепью, например консервативные замены аминокислот) второй аминокислотной или нуклеотидной последовательности, так что первая и вторая аминокислотная или нуклеотидная последовательности имеют похожие активности. В случае антител, второе антитело имеет ту же самую связывающую специфичность и имеет по меньшей мере 50% аффинности.
Используемые в описании термины "низкая жесткость", "средняя жесткость", "высокая жесткость" или "условия с очень высокой жесткостью" описывают условия гибридизации и промывки нуклеиновых кислот. Руководство по проведению реакций гибридизации можно найти в Current Protocols in Molecular Biology, John Wiley & Sons, N. Y. (1989), 6.3.1-6.3.6, которая включена в данное описание путем ссылки. Водные и неводные способы описаны в этой ссылке и любой из них может быть использован. Упомянутые в данном описании специфические условия гибридизации представляют собой следующие: (1) условия гибридизации с низкой жесткостью в смеси 6Х хлорид натрия/цитрат натрия (SSC) при примерно 45°С с последующими двумя промывками в 0,2Х SSC, 0,1% SDS (додецилсульфат натрия) по меньшей мере при 50°С (температура промывок может быть увеличена до 55°С для условий с низкой жесткостью); (2) условия гибридизации со средней жесткостью в 6Х SSC при примерно 45°С с последующей одной или более промывками в 0,2Х SSC, 0,1% SDS при 60°С; (3) условия гибридизации с высокой жесткостью в 6Х SSC при примерно 45°С с последующей одной или более промывками в 0,2Х SSC, 0,1% SDS при 65°С; и предпочтительно (4) условия гибридизации с очень высокой жесткостью представляют собой 0,5М фосфат натрия, 7% SDS при 65°С с последующей одной или более промывками в 0,2Х SSC, 1% SDS при 65°С. Условия с очень высокой жесткостью (4) представляют собой предпочтительные условия и должны быть использованы, если не указано иначе.
Определенный в данном описании термин "закрытая конформация" (полиспецифический лиганд) означает, что эпитоп-связывающие домены лиганда присоединены или ассоциированы друг с другом, возможно с помощью белкового скелета, так что связывание эпитопа с одним из эпитоп-связывающих доменов конкурирует со связыванием эпитопа с другим эпитоп-связывающим доменом. То есть когнатные эпитопы могут быть связаны каждым из эпитоп-связывающих доменов индивидуально, а не одновременно. Закрытой конформации лиганда можно достичь, используя описанные в данном изобретении способы.
"Открытая конформация" означает, что эпитоп-связывающие домены лиганда присоединены друг к другу или ассоциированы друг с другом, возможно с помощью белкового скелета, так что связывание эпитопа с одним из эпитоп-связывающих доменов не конкурирует со связыванием эпитопа с другим эпитоп-связывающим доменом.
Используемый в данном описании термин "конкурирует" означает, что связывание первого эпитопа с когнатным ему эпитоп-связывающим доменом ингибируются, когда второй эпитоп связан с когнатным ему эпитоп-связывающим доменом. Например, связывание может ингибироваться стерически, например путем физического блокирования связывающего домена или путем изменения структуры или окружения связывающего домена таким образом, что его аффинность или авидность в отношении эпитопа уменьшается.
Фраза "единичный вариабельный домен иммуноглобулина" относится к вариабельной области антитела (VH, VHH, VL), которая специфически связывает антиген или эпитоп независимо от других V областей или доменов; однако используемый в данном описании термин "единичный вариабельный домен иммуноглобулина" может быть представлен в формате (например, гомо- или гетеромультимера) с другими вариабельными областями или вариабельными доменами, где другие области или домены не требуются для связывания антигена с единичным вариабельным доменом иммуноглобулина (т.е. когда единичный вариабельный домен иммуноглобулина связывает антиген независимо от дополнительных вариабельных доменов). "Единичный вариабельный домен иммуноглобулина" охватывает не только полипептид единичного вариабельного домена выделенного антитела, но также более крупные полипептиды, которые включают один или более чем один мономер полипептидной последовательности единичного вариабельного домена. "Доменное антитело" или "dAb" представляет собой то же самое, что и используемый в данном описании термин полипептид "единичного вариабельного домена иммуноглобулина". Используемый в данном описании полипептид единичного вариабельного домена иммуноглобулина относится к полипептиду единичного вариабельного домена иммуноглобулина млекопитающего, предпочтительно человека, но также включает грызунов (например, как описано в WO 00/29004, содержание которой включено в данное описание путем ссылки) или VHH dAb Camelid. dAb Camelid представляют собой полипептиды единичного вариабельного домена иммуноглобулина, которые происходят из видов, включающих Camelid, ламу, альпака, дромадера и гуанако, и включают антитела, состоящие из тяжелой цепи, естественно лишенные легкой цепи: молекулы VHH. Молекулы VHH примерно в десять раз меньше, чем молекулы IgG, и в виде единичных полипептидов они очень стабильны, устойчивы к экстремальным условиям рН и температуры.
Используемый в данном описании термин "антагонист рецептора фактора некроза опухоли 1 (TNFR1)" относится к агенту (например, молекуле, соединению), который связывает TNFR1 и может ингибировать (т.е. одну или более чем одну) функцию TNFR1. Например, антагонист TNFR1 может ингибировать связывание TNFα с TNFR1 и/или ингибировать передачу сигнала, опосредованную через TNFR1. Соответственно, TNFR1-опосредованные способы и клеточные ответы (например, TNFα-индуцированная гибель клеток в стандартном анализе цитотоксичности в отношении L929) можно ингибировать антагонистом TNFR1. Антагонист TNFR1 может представлять собой, например, небольшую органическую молекулу, природный продукт, белок, пептид или пептидомиметик. Антагонисты TNFR1 могут быть идентифицированы, например, путем скрининга библиотек или коллекций молекул, таких как Химический депозитарий Национального института рака (Chemical Repository of the National Cancer Institute), как описано в данном изобретении, или с использованием других подходящих способов. Предпочтительные антагонисты TNFR1 представляют собой описанные в данном изобретении антитела, антиген-связывающие фрагменты антител, лиганды и мономеры dAb.
Последовательности, похожие или гомологичные (например, по меньшей мере примерно 70% идентичности последовательности) последовательностям, раскрытым в данном изобретении, также представляют собой часть изобретения. В некоторых воплощениях идентичность последовательности на аминокислотном уровне может составлять примерно 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или выше. На уровне нуклеиновых кислот идентичность последовательности может составлять примерно 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или выше. Альтернативно, существует значительная идентичность, когда сегменты нуклеиновых кислот гибридизуются в селективных условиях гибридизации (например, условия гибридизации с очень высокой жесткостью) с комплементарной цепью. Нуклеиновые кислоты могут присутствовать в целых клетках, в клеточном лизате или в частично очищенной или по существу чистой форме.
Расчеты "гомологии" или "идентичности последовательности" или "сходства" между двумя последовательностями (в данном изобретении термины используются взаимозаменяемо) осуществляют в соответствии со следующим. Последовательности выравнивают для целей оптимального сравнения (например, могут быть введены пробелы в одной из или обеих первой и второй аминокислотных последовательностей или последовательностей нуклеиновой кислоты для оптимального выравнивания, а негомологичные последовательности могут быть проигнорированы для целей сравнения). В предпочтительном воплощении длина ссылочной последовательности, выровненной для целей сравнения, составляет по меньшей мере 30%, предпочтительно по меньшей мере 40%, более предпочтительно по меньшей мере 50%, еще более предпочтительно по меньшей мере 60%, и еще более предпочтительно по меньшей мере 70%, 80%, 90%, 100% от длины ссылочной последовательности. Затем сравнивают аминокислотные остатки или нуклеотиды в соответствующих аминокислотных положениях или нуклеотидных положениях. Когда положение в первой последовательности занято одним и тем же аминокислотным остатком или нуклеотидом, как в соответствующем положении во второй последовательности, то молекулы идентичны по этому положению (используемая в данном изобретении "гомология" аминокислот или нуклеиновых кислот эквивалентна "идентичности" аминокислот или нуклеиновых кислот). Процентная идентичность между двумя последовательностями представляет собой функцию количества идентичных положений, разделяемых последовательностями, с учетом количества пробелов, и длину каждого пробела, которые необходимо вводить для оптимального выравнивания двух последовательностей.
Предпочтительно, алгоритм BLAST (версия 2.0) используют для выравнивания последовательностей со значениями параметров, установленными по умолчанию. Алгоритм BLAST подробно описан на сайте в Интернете ("www") Национального центра биотехнологической информации (National Center for Biotechnology Information) (".ncbi") Национальных институтов здоровья (National Institutes of Health) ("nih") правительства США (".gov") в директории "/Blast/", в файле "blastjielp.html". Параметры поиска определены следующим образом и являются предпочтительными для установления определенных параметров по умолчанию.
BLAST (Basic Local Alignment Search Tool) представляет собой эвристический алгоритм поиска, используемый в программах blastp, blastn, blastx, tbiastn, и tbiastx; эти программы приписывают важность своего поиска использованию статистических способов Karlin and Altschul, 1990, Proc. Natl. Acad. Sci. USA 87(6):2264-8 (смотри файл "blast_help.html", как описано выше) с несколькими улучшениями. Программы BLAST разработаны для поиска сходства последовательностей, например для идентификации гомологии с интересующей последовательностью. Данные программы обычно не подходят для поиска мотивов. Обсуждение основных проблем поиска сходства в базах данных последовательностей смотри в Altschul era/. (1994).
Пять программ BLAST, доступных на сайте в Интернете National Center for Biotechnology Information, решают следующие задачи:
"blastp" сравнивает интересующую аминокислотную последовательность с базой данных белковых последовательностей;
"blastn" сравнивает интересующую нуклеотидную последовательность с базой данных нуклеотидных последовательностей;
"blastx" сравнивает умозрительные продукты трансляции интересующей нуклеотидной последовательности (обе цепи) с шести рамок с базой данных белковых последовательностей;
"tbiastn" сравнивает интересующую белковую последовательность с базой данных нуклеотидных последовательностей, динамически транслируемых во всех шести рамках считывания (обе цепи);
"tbiastx" сравнивает продукты трансляции интересующей нуклеотидной последовательности с шести рамок с продуктами трансляции базы данных нуклеотидных последовательностей с шести рамок.
BLAST использует следующие параметры поиска:
ГИСТОГРАММА. Демонстрирует гистограмму оценок для каждого поиска;
по умолчанию "да". (Смотри параметр Н в руководстве пользователя BLAST).
ОПИСАНИЯ. Ограничивает количество коротких описаний совпадающих последовательностей, сообщенных для указанного количества; предел по умолчанию 100 описаний (смотри параметр V в руководстве пользователя). Смотри также ОЖИДАЕМОЕ и ОТСЕЧЕНИЕ.
ВЫРАВНИВАНИЯ. Ограничивает последовательности из базы данных до указанного количества, для которого сообщены пары сегментов с высоким сходством (HSP от англ. high-scoring segment pair); предел по умолчанию составляет 50. Если в базе данных больше последовательностей по сравнению с указанным удовлетворяют порогу статистической значимости для приведенного (смотри ОЖИДАЕМОЕ и ОТСЕЧЕНИЕ ниже), то приводятся лишь сочетания, которым приписывают наибольшую статистическую значимость (смотри параметр В в руководстве пользователя BLAST).
ОЖИДАЕМОЕ. Порог статистической значимости для приведенных сочетаний в отношении последовательностей из базы данных; значение по умолчанию составляет 10, так что 10 сочетаний, как ожидают, должны обнаруживаться лишь случайно, в соответствии со стехиометрической моделью Karlin and Altschul (1990). Если статистическая значимость, приписываемая сочетанию, больше ОЖИДАЕМОГО порога, сочетание не приводят. Меньшие ОЖИДАЕМЫЕ пороги являются более строгими, что приводит к тому, что сообщают о менее случайных сочетаниях. Дробные значения являются приемлемыми (смотри параметр Е в руководстве пользователя BLAST).
ОТСЕЧЕНИЕ. Оценка отсечения для приведенных пар сегментов с максимальным сходством. Значение по умолчанию рассчитывают из ОЖИДАЕМОГО значения (смотри выше). HSP приведены для последовательности из базы данных, только если приписываемая ей статистическая значимость по меньшей мере столь же высока как приписываемая единственной HSP, имеющей оценку, равную значению ОТСЕЧЕНИЯ. Более высокие значения ОТСЕЧЕНИЯ являются более строгими, что приводит к меньшей вероятности сочетаний (смотри параметр S в руководстве пользователя BLAST). Как правило, значительными порогами можно более интуитивно управлять, используя ОЖИДАЕМОЕ.
МАТРИЦА. Указывает на альтернативную матрицу оценок для BLASTP, BLASTX, TBLASTN и TBLASTX. Матрица по умолчанию представляет собой BLOSUM62 (Henikoff & Henikoff, 1992, Proc. Natl. Acad. Sci. USA 89(22): 10915-9). Действительные альтернативные выборы включают: РАМ40, РАМ120, РАМ250 и IDENTITY. Для BLASTN отсутствуют альтернативные матрицы оценки; указывает на то, что команда MATRIX в запросе BLASTN предлагает возврат к ошибочному ответу.
ЦЕПЬ. Ограничивает поиск TBLASTN лишь верхней или нижней цепью в последовательностях из базы данных; или ограничивает поиск BLASTN, BLASTX или TBLASTX лишь рамками считывания на верхней или нижней цепи запрашиваемой последовательности.
ФИЛЬТР. Маскирует сегменты запрашиваемой последовательности, которые обладают низкой композиционной сложностью, как определено с помощью программы SEG Wootton & Federhen (1993) Computers and Chemistry 17:149-163, или сегменты, состоящие из внутренних повторов с короткой периодичностью, как определено с использованием программы XNU Claverie & States, 1993, Computers and Chemistry 17:191-201, или для BLASTN с использованием программы DUST Tatusov and Lipman (смотри сайт в Интернете NCBI). Фильтр может устранять статистически значимые, но биологические не интересные сообщения, выдаваемые blast (например, против обычных кислых, основных или пролин-богатых областей), оставляя более биологически интересные области запрашиваемой последовательности, доступные для специфического совпадения с последовательностями из базы данных.
Последовательность с низкой сложностью, обнаруженная фильтрующей программой, замещают с использованием символа "N" в нуклеотидной последовательности (например, "N", повторенная 13 раз) и буквой "X" в белковых последовательностях (например, "X", повторенная 9 раз).
Фильтрование применимо только к запрашиваемой последовательности (или продуктам ее трансляции), а не к последовательностям из базы данных. Фильтрование по умолчанию представляет собой DUST для BLASTN, SEG для других программ.
Не является необычным, когда ничего вовсе не маскируется с помощью SEG, XNU или и той, и другой программы при применении к последовательностям в SWISS-PROT, таким образом, не следует ожидать, что фильтрование всегда будет эффективно. Кроме того, в некоторых случаях последовательности маскируются полностью, что указывает на то, что статистическую значимость любых сообщаемых совпадений относительно не подвергнутой фильтрованию интересующей последовательности следует подвергать сомнению.
NCBI-gi приводит к тому, что идентификаторы NCBI gi должны быть представлены на выходе, дополнительно к номеру доступа и/или имени локуса.
Наиболее предпочтительно, сравнения последовательностей проводят с использованием простого алгоритма поиска BLAST, предложенного на описанном выше сайте NCBI в Интернете, в директории "/BLAST".
Если не определено иначе, все используемые в данном изобретении технические и научные термины имеют такое же значение, которое обычно понятно специалисту в данной области (например, в культивировании клеток, молекулярной генетике, химии нуклеиновых кислот, способах гибридизации и биохимии). Стандартные способы используют для молекулярных, генетических и биохимических способов (смотри в основном Sambrook et al, Molecular Cloning: A Laboratory Manual, 2d ed. (1989) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N. Y. и Ausubel et al. Short Protocols in Molecular Biology (1999) 4th Ed, John Wiley & Sons, Inc., которые включены в данное описание путем ссылки), и химических способов.
TNFR1 представляет собой трансмембранный рецептор, содержащий внеклеточную область, которая связывает лиганд, и внутриклеточный домен, который лишен присущей ему активности в отношении передачи сигнала, но который может связываться с молекулами, передающими сигнал. Комплекс TNFR1 со связанным TNF содержит три цепи TNFR1 и три цепи TNF. (Banner et al, Cell, 73(3) 431-445 (1993).) Лиганд TNF представлен в виде тримера, который связан тремя цепями TNFR1 (там же). Три цепи TNFR1 плотно кластеризованы вместе в рецептор-лигандном комплексе, и эта кластеризация является предпосылкой для TNFR1-опосредованной передачи сигнала. Фактически, поливалентные агенты, которые связывают TNFR1, такие как антитела против TNFR1, могут индуцировать кластеризацию TNFR1 и передачу сигнала в отсутствие TNF и обычно используются в качестве агонистов TNFR1. (Смотри, например, Belka et al, EMBO, 14(6): 1156-1165 (1995); Mandik-Nayak et al, J. Immunol, 767:1920-1928 (2001).) Соответственно, поливалентные агенты, которые связывают TNFR1, обычно не являются эффективными антагонистами TNFR1, даже если они блокируют связывание TNFα с TNFR1.
Внеклеточная область TNFR1 включает аминоконцевой сегмент из тринадцати аминокислот (аминокислоты 1-13 SEQ ID NO: 603 (человек); аминокислоты 1-13 SEQ ID NO:604 (мышь)), домен 1 (аминокислоты 14-53 SEQ ID NO:603 (человек); аминокислоты 14-53 SEQ ID NO:604 (мышь)), домен 2 (аминокислоты 54-97 SEQ ID NO: 603 (человек); аминокислоты 54-97 SEQ ID NO:604 (мышь)), домен 3 (аминокислоты 98-138 SEQ ID NO: 603 (человек); аминокислоты 98-138 SEQ ED NO:604 (мышь)) и домен 4 (аминокислоты 139-167 SEQ ID NO:603 (человек); аминокислоты 139-167 SEQ ID NO:604 (мышь)), за которыми следует проксимальная к мембране область (аминокислоты 168-182 SEQ ID NO:603 (человек); аминокислоты 168-183 SEQ ID NO: 604 (мышь)). (Смотри Banner et al, Cell 73(3) 431-445 (1993) и Loetscher et al, Cell 61(2) 351-359 (1990)). домены 2 и 3 образуют контакт со связанным лигандом (TNFp, TNFα). (Banner et al. Cell, 73(3) 431-445 (1993)). Внеклеточная область TNFR1 также содержит область, названную прелиганд-связывающий домен сборки или PLAD (от англ. pre-ligand binding assembly domain) домен (аминокислоты 1-53 SEQ ID NO:603 (человек); аминокислоты 1-53 SEQ ID NO:604 (мышь)) (The Government of the USA, WO 01/58953; Deng et al, Nature Medicine, doi: 10.1038/nm1304 (2005)).
TNPR1 сбрасывается с поверхности клетки in vivo посредством способа, который включает протеолиз TNFR1 в домене 4 или в проксимальной к мембране области (аминокислоты 168-182 SEQ ID NO:603; аминокислоты 168-183 SEQ ID NO:604) с получением растворимой формы TNFR1. Растворимый TNFR1 сохраняет способность связывать TNFα, и таким образом действует в качестве эндогенного ингибитора активности TNFα.
Изобретение относится к антителу или его антиген-связывающему фрагменту (например, dAb) или лиганду, который связывает TNFR1, но не конкурирует с TNF за связывание с TNFR1. Например, антитело или его антиген-связывающий фрагмент (например, dAb) или лиганд может связывать домен 1 TNFR1 или домен 4 TNFR1. Такое антитело или его антиген-связывающий фрагмент (например, dAb) или лиганд обеспечивают преимущества в качестве диагностических агентов и могут быть использованы для связывания и обнаружения, количественной оценки или измерения TNFR1 в образце, но не конкурирует с TNF в образце за связывание с TNFR1. Соответственно, может быть получено точное определение того, присутствует ли TNFR1 в образце или сколько TNFR1 в образце. В некоторых воплощениях антитело или его антиген-связывающий фрагмент (например, dAb) или лиганд, который связывает TNFR1, но не конкурирует с TNF за связывание с TNFR1, представляет собой антагонист TNFR1, как описано в данном изобретении.
Изобретение также относится к диагностическому набору для определения, присутствует ли в образце TNFR1 или сколько TNFR1 присутствует в образце, включающему антитело или его антиген-связывающий фрагмент (например, dAb), или лиганд, который связывает TNFR1, но не конкурирует с TNF за связывание с TNFR1, и инструкции по применению (например, для определения присутствия и/или количества TNFR1 в образце). В некоторых воплощениях набор дополнительно включает один или более чем один дополнительный реагент, такой как подходящий буфер или подходящий агент для обнаружения (например, поддающееся обнаружению меченое антитело или его антиген-связывающий фрагмент, который связывает антитело или его антиген-связывающий фрагмент (например, dAb), или лиганд, который связывает TNFR1, но не конкурирует с TNF за связывание с TNFR1).
Изобретение также относится к устройству, включающему твердую поверхность, на которой иммобилизовано антитело или его антиген-связывающий фрагмент (например, dAb) или лиганд, который связывает TNFR1, но не конкурирует с TNF за связывание с TNFR1, так что иммобилизованное антитело или его антиген-связывающий фрагмент (например, dAb), или лиганд связывает TNFR1. Могут быть использованы любые подходящие твердые поверхности, на которых может быть иммобилизовано антитело, или его антиген-связывающий фрагмент (например, dAb), или лиганд, например стекло, пластик, углеводы (например, агарозные смолы). При желании, носитель может содержать или быть модифицирован таким образом, чтобы содержать желаемые функциональные группы для облегчения иммобилизации антитела, или его антиген-связывающего фрагмента (например, dAb), или лиганда. Устройство и/или носитель может иметь любую подходящую форму, например лист, стержень, лента, пластина, предметное стекло, бусина, гранула, диск, гель, трубка, сфера, чип, планшет или чашка и т.п. В некоторых воплощениях устройство представляет собой полоску (dipstick).
Изобретение относится к антагонистам TNFR1 (например, лигандам, описанным в данном изобретении), которые обладают связывающей специфичностью в отношении рецептора фактора некроза опухоли 1 (TNFR1; р55; CD120a). Предпочтительно, антагонисты по изобретению не обладают связывающей специфичностью в отношении фактора некроза опухоли 2 (TNFR2), или по существу не оказывают антагонистического действия на TNFR2. Антагонист TNFR1 по существу не оказывает антагонистического действия на TNFR2, когда данный антагонист (1 нМ, 10 нМ, 100 нМ, 1 мкМ, 10 мкМ или 100 мкМ) дает в результате не более чем примерно 5% ингибирования TNFR2-опосредованной активности, индуцированной TNFα (100 пг/мл), в стандартном клеточном анализе. Особенно предпочтительные антагонисты TNFR1 представляют собой эффективные терапевтические вещества для лечения хронического воспалительного заболевания (эффективны, обладают терапевтической эффективностью). Например, в некоторых воплощениях антагонист TNFR1 эффективен в модели хронического воспалительного заболевания, такой как модель коллаген-индуцированного артрита у мышей, модели артрита у мышей ΔАРЕ, модели воспалительного заболевания кишечника у мышей, индуцированного декстрансульфатом натрия, модели воспалительного заболевания кишечника у мышей ΔАРЕ, модели хронического обструктивного заболевания легких у мышей, индуцированного табачным дымом, или подходящих моделей у приматов (например, модели коллаген-индуцированного артрита у приматов).
Антагонисты TNFR1 могут быть моновалентными или поливалентными. В некоторых воплощениях антагонист является моновалентным и содержит один сайт связывания, который взаимодействует с TNFR1. Моновалентные антагонисты связывают один TNFR1 и не вызывают перекрестного связывания или кластеризации TNFR1 на поверхности клеток, которые могут приводить к активации рецептора и передаче сигнала. В конкретных воплощениях моновалентный антагонист TNFR1 связывает домен 1 TNFR1. В более конкретных воплощениях моновалентный антагонист TNFR1 связывает домен 1 TNFR1 и конкурирует с TAR2m-21-23 за связывание с мышиным TNFR1 или конкурирует с TAR2h-205 за связывание с человеческим TNFR1.
В других воплощениях антагонист TNFR1 является поливалентным. Поливалентные антагонисты TNFR1 могут содержать две или более копий конкретного сайта связывания TNFR1 или содержат два или более различных сайтов связывания, которые связывают TNFR1. Например, как описано в данном изобретении, антагонист TNFR1 может представлять собой димер, тример или мультимер, включающий две или более копий конкретного dAb, который связывает TNFR1, или два или более различных dAb, которые связывают TNFR1. Предпочтительно, поливалентный антагонист TNFR1 по существу не оказывает агонистического действия на TNFR1 (действует как агонист TNFR1) в стандартном клеточном анализе (т.е. когда присутствует в концентрации 1 нМ, 10 нМ, 100 нМ, 1 мкМ, 10 мкМ, 100 мкМ, 1000 мкМ или 5000 мкМ, дает не более чем примерно 5% от TNFR1-опосредованной активности, индуцированной TNFα (100 пг/мл) в анализе).
В некоторых воплощениях поливалентный антагонист TNFR1 содержит два или более сайтов связывания для желаемого эпитопа или домена TNFR1. Например, поливалентный антагонист TNFR1 может включать два или более сайтов связывания, которые связывают такой же эпитоп в домене 1 TNFR1.
В других воплощениях поливалентный антагонист TNFR1 содержит два или более сайтов связывания, которые связываются с различными эпитопами или доменами TNFR1. В одном из примеров поливалентный антагонист TNFR1 включает первый сайт связывания, который связывает первый эпитоп в домене 1 TNFR1, и второй сайт связывания, который связывает второй отличающийся эпитоп в домене 1. В других примерах поливалентный антагонист TNFR1 может включать сайты связывания, которые связывают два или более желаемых эпитопов или доменов TNFR1. Например, поливалентные антагонисты TNFR1 могут включать сайты связывания для доменов 1 и 2, доменов 1 и 3, доменов 1 и 4, доменов 2 и 3, доменов 2 и 4 или доменов 3 и 4 TNFR1. Например, поливалентные антагонисты TNFR1 могут включать сайты связывания для доменов 1, 2 и 3, сайты связывания для доменов 1, 2 и 4, или сайты связывания для доменов 1, 3 и 4 TNFR1. В некоторых воплощениях антагонист TNFR1 представляет собой лиганд с двойной специфичностью, включающий dAb, которое связывает домен 1 TNFR1, и dAb, которое связывает домен 3 TNFR1. Предпочтительно, такие поливалентные антагонисты не оказывают антагонистического действия на TNFR1, когда присутствуют в концентрации примерно 1 нМ, или примерно 10 нМ, или примерно 100 нМ, или примерно 1 мкМ, или примерно 10 мкМ, в стандартном анализе цитотоксичности в отношении L929 или в стандартном анализе IL-8 в HeLa, как описано в данном изобретении.
Некоторые антагонисты TNFR1 связывают TNFR1 и ингибируют связывание TNFα с TNFR1. В некоторых воплощениях такой антагонист TNFR1 связывает домен 2 и/или домен 3 TNFR1. В конкретных воплощениях антагонист конкурирует с TAR2h-10-27, TAR2h-131-8, TAR2h-15-8, TAR2D-35-4, TAR2h-154-7, TAR2h-154-10 или TAR2h-185-25 за связывание с TNFR1.
Другие лиганды (которые в предпочтительных воплощениях представляют собой антагонисты TNFR1) не ингибируют связывание TNFα с TNFR1. Такие лиганды (и антагонисты) обеспечивают преимущества в качестве диагностических агентов, поскольку они могут быть использованы для связывания и обнаружения, количественного определения или измерения TNFR1 в образце и не будут конкурировать с TNF в образце за связывание с TNFR1. Соответственно, может быть проведено точное определение того, присутствует ли TNFR1 или сколько TNFR1 присутствует в образце.
Некоторые антагонисты TNFR1 не ингибируют связывание TNFα с TNFR1, но ингибируют передачу сигнала, опосредованную через TNFR1. Например, антагонист TNFR1 может ингибировать TNFα-индуцированную кластеризацию TNFR1, которая предшествует передаче сигнала через TNFR1. Такие антагонисты обеспечивают некоторые преимущества. Например, в присутствии такого антагониста, TNFα может связывать TNFR1, экспрессирующийся на поверхности клеток и удаляться из клеточного окружения, но TNFR1-опосредованная передача сигнала не будет активироваться. Таким образом, TNFR1 сигнал-индуцированная продукция дополнительного количества TNFα и других медиаторов воспаления будет ингибироваться. Аналогично, антагонисты TNFR1, которые связывают TNFR1 и ингибируют передачу сигнала, опосредованную через TNFR1, но не ингибируют связывание TNFα с TNFR1, не будут ингибировать связывание TNFα и ингибирующую активность эндогенно продуцируемого растворимого TNFR1. Соответственно, введение такого антагониста млекопитающему, нуждающемуся в этом, может дополнить эндогенные регуляторные пути, которые ингибируют активность TNFα и активность TNFR1 in vivo. Изобретение также относится к лигандам, которые (1) связывают TNFR1 (например, в домене 1), (2) не оказывают антагонистического действия на активацию TNFR1-опосредованной передачи сигнала, (3) не ингибируют связывание TNFα с TNFR1. Такой лиганд связывает растворимый TNFR1 и не предотвращает связывание растворимого рецептора с TNFα, и поэтому введение такого антагониста млекопитающему, нуждающемуся в этом, может дополнить эндогенные регуляторные пути, которые ингибируют активность TNFα in vivo путем увеличения периода полувыведения растворенного рецептора из сыворотки крови. Эти преимущества особенно касаются лигандов, которые были представлены в формате, имеющем более крупный гидродинамический размер, например путем присоединения ПЭГ (полиэтиленгликоль) группы, сывороточного альбумина, трансферрина, рецептора трансферрина или по меньшей мере его трансферрин-связывающего участка, Fc-области антитела, или путем конъюгирования с доменом антитела. Например, агент (например, полипептид), который 1) связывает TNFR1 (например, в домене 1), (2) не оказывает антагонистического действия на активацию TNFR1-опосредованной передачи сигнала, и (3) не ингибируют связывание TNFα с TNFR1, такой как мономер dAb, может быть представлен в виде более крупного антиген-связывающего фрагмента антитела или в виде антитела (например, в формате Fab, Fab', F(ab)2, F(ab')2, IgG, scFv). Гидродинамический размер лиганда и период его полувыведения из сыворотки крови также может быть увеличен путем конъюгирования или связывания TNFR1-связывающего агента со связывающим доменом (например, антителом или фрагментом антитела), который связывает антиген или эпитоп, увеличивающий период полувыведения in vivo, как описано в данном изобретении (смотри Приложение 1). Например, TNFR1-связывающий агент (например, полипептид) может быть конъюгирован или связан с антителом или фрагментом антитела против сывороточного альбумина или неонатального рецептора Fc, например dAb, Fab, Fab' или scFv против SA, или против неонатального рецептора Fc, или с аффителом против SA, или аффителом против неонатального рецептора Fc.
Примеры подходящего альбумина, фрагментов альбумина или вариантов альбумина для применения в TNFR1-связывающем лиганде по изобретению описаны в WO2005/077042A2, которая включена в данное описание путем ссылки во всей своей полноте. В частности, в настоящем изобретении могут быть использованы следующие альбумин, фрагменты альбумина или варианты альбумина:
- SEQ ID NO: 1 (как раскрыто в WO 2005/077042A2, причем эта последовательность непосредственно включена в настоящее описание путем ссылки);
- Фрагмент или вариант альбумина, включающий или состоящий из аминокислот 1-387 SEQ ID NO:1 в WO 2005/077042A2;
- Альбумин или его фрагмент или вариант, включающий аминокислотную последовательность, выбранную из группы, состоящей из: (а) аминокислот 54-61 SEQ ID NO: 1 в WO 2005/077042A2; (б) аминокислот 76-89 SEQ ID NO: 1 в WO 2005/077042A2; (в) аминокислот 92-100 SEQ ID NO:1 в WO 2005/077042A2; (г) аминокислот 170-176 SEQ ID NO:1 в WO 2005/077042A2; (д) аминокислот 247-252 SEQ ID NO:1 в WO 2005/077042A2; (e) аминокислот 266-277 SEQ ID NO:1 в WO 2005/077042A2; (ж) аминокислот 280-288 SEQ ID NO: 1 в WO 2005/077042A2; (з) аминокислот 362-368 SEQ ID NO:1 в WO 2005/077042A2; (и) аминокислот 439-447 SEQ ID NO:1 в WO 2005/077042A2 (к) аминокислот 462-475 SEQ ID NO:1 в WO 2005/077042A2; (л) аминокислот 478-486 SEQ ID NO:1 в WO 2005/077042A2; и (м) аминокислот 560-566 SEQ ID NO:1 в WO 2005/077042A2.
Дополнительные примеры подходящего альбумина, фрагментов и аналогов для применения в TNFR1-связывающем лиганде по изобретению описаны в WO 03/076567 А2, которая включена в данное описание путем ссылки. В частности, в настоящем изобретении могут быть использованы следующие альбумин, фрагменты или варианты:
- человеческий сывороточный альбумин, описанный в WO 03/076567А2, например на Фиг.3 (причем информация об этой последовательности непосредственно включена в настоящее описание путем ссылки);
- человеческий сывороточный альбумин (HSA), состоящий из единичной негликозилированной полипептидной цепи из 585 аминокислот с молекулярной массой 66500 (смотри Meloun, et at, FEBS Letters 58:136 (1975); Behrens, et al. Fed. Proc. 34:591 (1975); Lawn, et al. Nucleic Acids Research 9:6102-6114 (1981); Minghetti, etal, J. Biol Chem. 261:6747 (1986));
- полиморфный вариант или аналог, или фрагмент альбумина, как описано BWeitkamp, et al, Ann. Hum. Genet. 37:219 (1973);
- фрагмент или вариант альбумина, как описано в ЕР 322094, например НА(1-373), НА(1-388), НА(1-389), НА(1-369) и НА(1-419) и фрагменты между 1-369 и 1-419;
- фрагмент или вариант альбумина, как описано в ЕР 399666, например НА(1-177) и НА(1-200) и фрагменты между НА (1-Х), где Х означает число от 178 до 199.
Когда (одну или более чем одну) группировку, увеличивающую период полувыведения (например, альбумин, трансферрин и их фрагменты и аналоги), используют в TNFR1-связывающих лигандах по изобретению, тогда она может быть конъюгирована с помощью любого подходящего способа, такого как прямое слияние с TNFR1-связывающей группировкой (например, dAb или фрагмент антитела против TNFR1), например с использованием единичной нуклеотидной конструкции, которая кодирует слитый белок, где слитый белок кодируется в виде единичной полипептидной цепи с группировкой, увеличивающей период полувыведения, расположенной на N- или С-конце относительно TNFR1-связывающей группировки. Альтернативно, конъюгирования можно достичь путем использования пептидного линкера между группировками, например пептидного линкера, как описано в WO 03/076567 А2 или WO 2004/003019 (описания этого линкера включены путем ссылки в настоящее описание для обеспечения примеров для применения в настоящем изобретении).
В более конкретных воплощениях антагонист TNFR1, который связывает TNFR1 и ингибирует передачу сигнала, опосредованную через TNFR1, но не ингибирует связывание TNFα с TNFR1, связывает домен 1 TNFR1 или домен 4 TNFR1. В некоторых воплощениях такой антагонист TNFR1 представляет собой мономер dAb или лиганд, который связывает домен 1 TNFR1 или домен 4 TNFR1.
В конкретном воплощении антагонист TNFR1 (например, мономер или лиганд dAb) связывает домен 1 TNFR1 и ингибирует передачу сигнала, опосредованную через TNFR1 при связывании TNFα. Такой антагонист может ингибировать передачу сигнала через TNFR1, но не ингибирует связывание TNFα с TNFR1 и/или шеддинг (сбрасывание от англ. shedding) TNFR1 с получением растворимого TNFR1. Соответственно, введение такого антагониста млекопитающему, нуждающемуся в этом, может дополнять эндогенные регуляторные пути, которые ингибируют активность TNFα и активность TNFR1 in vivo.
Другие антагонисты TNFR1 связывают TNFR1, но не связывают домен 4. Такие антагонисты ингибируют функцию TNFR1, но не ингибируют шеддинг растворимого TNFR1. Соответственно, введение такого антагониста млекопитающему, нуждающемуся в этом, может дополнять эндогенные регуляторные пути, которые ингибируют активность TNFα и активность TNFR1 in vivo.
В некоторых воплощениях антагонист (например, химическое соединение, новый химический структурный элемент, мономер dAb, лиганд) связывает домен 1 TNFR1 и конкурирует с TAR2m-21-23 за связывание с мышиным TNFR1 или конкурирует с TAR2h-205 за связывание с человеческим TNFR1. В других воплощениях антагонист (например, химическое соединение, новый химический структурный элемент, лиганд) связывает домен 2 или домен 4 TNFR1. В других воплощениях антагонист (например, химическое соединение, новый химический структурный элемент, мономер dAb, лиганд) связывает домен 3 TNFR1 и конкурирует с TAR2h-131-8, TAR2h-15-8, TAR2h-35-4, TAR2h-154-7, TAR2h-154-10, TAR2h-185-25 или TAR2h-27-10 за связывание с TNFR1 (например, человеческим и/или мышиным TNFR1).
Некоторые лиганды (которые в предпочтительных воплощениях представляют собой антагонисты TNFR1) связывают человеческий TNFR1 и мышиный TNFR1. Такие лиганды (например, антагонисты, мономеры dAb) обеспечивают преимущество, дающее возможность проводить доклинические и клинические испытания с использованием одного и того же лиганда, и устраняют необходимость в проведении доклинических испытаний с подходящим суррогатным лигандом.
В других воплощениях антагонист или лиганд представляет собой антитело, которое обладает связывающей специфичностью в отношении TNFR1, или его антиген-связывающий фрагмент, такой Fab-фрагмент, Fab'-фрагмент, F(ab')2-фрагмент или Fv-фрагмент (например, scFV). В других воплощениях антагонист или лиганд является моновалентным, таким как dAb или моновалентный антиген-связывающий фрагмент антитела, такой как Fab-фрагмент, Fab'-фрагмент или Fv-фрагмент.
В других воплощениях изобретения, раскрытых в этом описании, вместо применения "dAb" в антагонисте или лиганде по изобретению, ожидают, что специалист в данной области может использовать домен, который включает CDR (области, определяющие комплементарность) dAb, которое связывает TNFR1 (например, CDR, перенесенные на подходящий белковый каркас или скелет, например аффитело, каркас SpA, домен класса А рецептора LDL или домен EGF) или может представлять собой белковый домен, включающий сайт связывания для TNFR1, например, где домен выбран из аффитела, домена SpA, домена класса А рецептора LDL или домена EGF. Описание в целом следует интерпретировать соответственно для обеспечения описания антагонистов, лигандов и способов с использованием таких доменов вместо dAb.
Предпочтительно, антагонист TNFR1 представляет собой лиганд, как описано в данном изобретении. Лиганды включают единичный вариабельный домен иммуноглобулина или доменное антитело (dAb), которые обладают связывающей специфичностью в отношении TNFR1 или определяющих комплементарность областей такого dAb в подходящем формате. Лиганд может представлять собой полипептид, который состоит из такого dAb, или состоит по существу из такого dAb. Лиганд также может представлять собой полипептид, который включает dAb (или CDR dAb) в подходящем формате, таком как формат антитела (например, IgG-подобный формат, scFv, Fab, Fab', F(ab')2), лиганд с двойной специфичностью, который включает dAb, связывающее TNFR1, и второе dAb, связывающее другой белок-мишень, антиген или эпитоп (например, сывороточный альбумин), или полиспецифический лиганд, как описано в данном изобретении.
Антагонисты TNFR1, включая лиганды по любому аспекту настоящего изобретения, а также мономера dAb, полезные при конструировании таких лигандов, могут диссоциировать предпочтительно от их когнатной мишени(ей) с Kd от 300 нМ до 5 пМ (т.е. от 3×10-7 до 5×10-12 М), предпочтительно от 50 нМ до 20 пМ, или от 5 нМ до 200 пМ, или от 1 нМ до 100 пМ, 1×10-7 М или меньше, 1×10-8 М или меньше, 1×10-9 М или меньше, 1×10-10 М или меньше, 1×10-11 М или меньше; и/или с константой скорости диссоциации Koff от 5×10-1 с-1 до 1×10-7 с-1, предпочтительно от 1×10-2 с-1 до 1×10-6 с-1, или от 5×10-3 с-1 до 1×10-5 с-1, или 5×10-1 с-1 или меньше, или 1×10-2 с-1 или меньше, или 1×10-3 с-1 или меньше, или 1×10-4 с-1 или меньше, или 1×10-5 с-1 или меньше, или 1×10-6 с-1 или меньше, как определено методом поверхностного плазмонного резонанса. Константу скорости Kd определяют как Koff/Kon.
В других воплощениях антагонист связывает TNFR1 и ингибирует (т.е. одну или более чем одну) функцию TNFR1 (например, кластеризацию рецепторов, передачу сигнала с рецептора или связывание TNFα с TNFR1), а также связывается с другим членом суперсемейства рецепторов TNF. Предпочтительно, этот тип антагониста также ингибирует функцию (например, кластеризацию членов, передачу сигнала или связывание члена с его когнатным лигандом) другого члена суперсемейства рецепторов TNF. Суперсемейство рецепторов TNF представляет собой известную в данной области техники группу белков, которая включает TNFR1 (р55, CD120a, p60, член 1А суперсемейства рецепторов TNF, TNFRSF1A), TNFR2 (р75, р80, CD120D, член 1 В суперсемейства рецепторов TNF, TNFRSF1B), CD18 (TNFRSF3, LTBR, TNFR2-RP, TNFR-RP, TNFCR, TNF-R-III), 0Х40 (TNFRSF4, АСТ35, TXGPIL), CD40 (TNFRSF5, р50, Вр50), Fas (CD95, TNFRSF6, АРО-1, APTI), DcR3 (TNFRSF6B), CD27 (TNFRSF7, Тр55, S152), CD30 (TNFRSF8, Ki-1, D1S166E), CD137 (TNFRSF9, 4-1 ВВ, ILA), TRAILR-1 (TNFRSFIOA, DR4, Аро2), TRAIL-R2 (TNFRSF10B, DR5, KILLER, TRICK2A, TRICKB), TRAILR3 (TNFRSF10C, DcR1, LIT, TRID), TRAILR4 (TNFRSFIOD, DcR2, TRUNDD), RANK (TNFRSF11A), OPG (TNFRSF11B, OCIF, TR1), DR3 (TNFRSF12, TRAMP, WSL-1, LARD, WSL-LR, DDR3, TR3, APO-3), DR3L (TNFRSF12L), TACI (TNFRSF13B), BAFFR (TNFRSF13C), HVEM (TNFRSF14, ATAR, TR2, LIGHTR, HVEA), NGFR (TNFRSF16), BCMA (TNFRSF17, BCM), AITR (TNFRSF18, GITR), TNFRSF19, FLJ14993 (TNFRSF19L, RELT), DR6 (TNFRSF21), SOBa (TNFRSF22, Tnfrh2, 2810028K06Rik), mSOB (THFRSF23, Tnfrhl). В некоторых воплощениях антагонист включает первое dAb, которое связывает TNFR1 и ингибирует функцию TNFR1, и второе dAb, которое связывает другой член суперсемейства рецепторов TNF, такой как TNFR2 (CD 120b), OX40, CD40, Fas (CD95), TRAILR-1, TRAILR-2, TAC1, BCMA и т.п., как перечислено выше. В еще одном воплощении антагонист включает мономер dAb, который связывает TNFR1 и ингибирует функцию (например, кластеризацию рецепторов, передачу сигнала с рецептора или связывание TNFα с TNFR1) TNFR1, и также связывает другой член суперсемейства рецепторов TNF, такой как TNFR2 (CD 120b), OX40, CD40, Fas (CD95), TRAILR-1, TRAILR-2, TAC1, BCMA и т.п., как перечислено выше.
Лиганды и мономеры dAb, которые связывают TNFR1
В изобретении предложены лиганды, которые включают мономер dAb против TNFR1 (например, лиганд с двойной специфичностью, содержащий такое dAb), которые связывают рецептор I TNF с Kd от 300 нМ до 5 пМ (т.е. от 3×10-7 до 5×10-12 М), предпочтительно от 50 нМ до 20 пМ, более предпочтительно от 5 нМ до 200 пМ и наиболее предпочтительно от 1 нМ до 100 пМ, например 1×10-7 М или меньше, предпочтительно 1×10-8 М или меньше, более предпочтительно 1×10-9 М или меньше, предпочтительно 1×10-10 М или меньше и наиболее предпочтительно 1×10-11 М или меньше; и/или константу скорости Koff от 5×10-1 с-1 до 1×10-7 с-1, предпочтительно от 1×10-2 с-1 до 1×10-6 с-1, более предпочтительно от 5×10-3 с-1 до 1×10-5 с-1, например 5×10-1 с-1 или меньше, предпочтительно 1×10-2 с-1 или меньше, предпочтительно 1×10-3 с-1 или меньше, более предпочтительно 1×10-4 с-1 или меньше, еще более предпочтительно 1×10-5 с-1 или меньше, и наиболее предпочтительно 1×10-6 с-1 или меньше, как определено методом поверхностного плазменного резонанса.
Предпочтительно, лиганд или мономер dAb ингибирует связывание TNF-альфа с рецептором I TNF-альфа (рецептор р55) с 50% ингибирующей концентрацией (IC50) от 500 нМ до 50 пМ, предпочтительно от 100 нМ до 50 пМ, более предпочтительно от 10 нМ до 100 пМ, предпочтительно от 1 нМ до 100 пМ; например 50 нМ или меньше, предпочтительно 5 нМ или меньше, более предпочтительно 500 пМ или меньше, предпочтительно 200 пМ или меньше, и наиболее предпочтительно 100 пМ или меньше. Предпочтительно, мишенью для рецептора TNF I является человеческий TNFα.
Предпочтительно, лиганд или dAb связывает человеческий TNFR1 и ингибирует связывание человеческого TNF-альфа с человеческим TNFR1, или ингибирует передачу сигнала через TNFR1 в ответ на связывание TNF-альфа. Например, в некоторых воплощениях лиганд или мономер dAb может связывать TNFR1 и ингибировать TNFR1-опосредованную передачу сигнала, но по существу не ингибирует связывание TNFα с TNFR1. В некоторых воплощениях лиганд или мономер dAb ингибирует TNFα-индуцированное перекрестное связывание или кластеризацию TNFR1 на поверхности клетки. Такие лиганды или dAb (например, описанный в данном изобретении TAR2m-21-23) предпочтительны, поскольку они могут оказывать антагонистическое действие на TNFR1 клеточной поверхности, но по существу не уменьшают ингибирующую активность эндогенного растворимого TNFR1. Например, лиганд или dAb может связывать TNFR1, но ингибировать связывание TNFα с TNFR1 в анализе связывания рецепторов не более чем примерно на 10%, не более чем примерно на 5%, не более чем примерно на 4%, не более чем примерно на 3%, не более чем примерно на 2% или не более чем примерно на 1%. Кроме того, в этих воплощениях лиганд или dAb ингибирует TNFα-индуцированное перекрестное связывание TNFR1 и/или TNFR1-опосредованную передачу сигнала в стандартном клеточном анализе по меньшей мере примерно на 10%, по меньшей мере примерно на 20%, по меньшей мере примерно на 30%, по меньшей мере примерно на 40%, по меньшей мере примерно на 50%, по меньшей мере примерно на 60%, по меньшей мере примерно на 70%, по меньшей мере примерно на 80%, по меньшей мере примерно на 90%, по меньшей мере примерно на 95% или по меньшей мере примерно на 99%.
Лиганд может быть моновалентным (например, мономер dAb) или поливалентным (например, с двойной специфичностью, полиспецифическим), как описано в данном изобретении. В конкретных воплощениях лиганд представляет собой мономер dAb, который связывает домен 1 TNFR1. Мономеры доменного антитела, которые связывают домен 1 TNFR1, имеют небольшой след по сравнению с другими связывающими форматами, такими как, например, моноклинальное антитело. Таким образом, такой мономер dAb может избирательно блокировать домен 1, но не влиять на функцию других доменов TNFR1. Например, мономер dAb, который связывает домен 1 TNFR1, может оказывать антагонистическое действие на TNFR1, но не ингибировать связывание TNFα с TNFR1 или шеддинг TNFR1.
В более конкретных воплощениях лиганд представляет собой мономер dAb, который связывает домен 1 TNFR1 и конкурирует с TAR2m-21-23 за связывание с мышиным TNFR1 или конкурирует с TAR2h-205 за связывание с человеческим TNFR1.
В других воплощениях лиганд является поливалентным и включает два или более мономеров dAb, которые связывают TNFR1. Поливалентные лиганды могут содержать две или более копий конкретного dAb, которое связывает TNFR1, или содержать два или более dAb, которые связывают TNFR1. Например, как описано в данном изобретении, лиганд может представлять собой димер, тример или мультимер, включающий две или более копий конкретного dAb, которое связывает TNFR1, или два или более различных dAb, которые связывают TNFR1. В некоторых примерах лиганд представляет собой гомодимер или гомотример, который включает две или три копии конкретного dAb, которое связывает соответственно TNFR1. Предпочтительно, поливалентный лиганд по существу не оказывает агонистического действия на TNFR1 (действует как агонист TNFR1) в стандартном клеточном анализе (т.е. когда присутствует в концентрации 1 нМ, 10 нМ, 100 нМ, 1 мкМ, 10 мкМ, 100 мкМ, 1000 мкМ или 5000 мкМ, дает не более чем примерно 5% от TNFR1-опосредованной активности, индуцированной TNFα (100 пг/мл) в анализе).
В некоторых воплощениях поливалентный лиганд содержит два или более dAb, которые связывают желаемый эпитоп или домен TNFR1. Например, поливалентный лиганд может включать две или более копий dAb, которое связывает желаемый эпитоп в домене 1 TNFR1.
В других воплощениях поливалентный лиганд содержит два или более dAb, которые связывают различные эпитопы или домены TNFR1. В одном из примеров поливалентный лиганд включает первое dAb, которое связывает первый эпитоп в домене 1 TNFR1, и второе dAb, которое связывает второй отличающийся эпитоп в домене 1 TNFR1. В других примерах поливалентный лиганд включает dAb, которые связывают два или более желаемых эпитопов или доменов TNFR1. Например, поливалентный лиганд может включать dAb, которые связывают домены 1 и 2, домены 1 и 3, домены 1 и 4, домены 2 и 3, домены 2 и 4 или домены 3 и 4 TNFR1.
В некоторых воплощениях поливалентный лиганд представляет собой лиганд с двойной специфичностью, включающий dAb, которое связывает домен 1 TNFR1, и dAb, которое связывает домен 3 TNFR1. Лиганды этого типа могут связывать TNFR1 с высокой авидностью и быть более избирательными в отношении связывания с клетками, которые сверхэкспрессируют TNFR1 или экспрессируют TNFR1 на своей поверхности с высокой плотностью по сравнению с другими форматами лиганда, такими как мономеры dAb.
В других конкретных воплощениях поливалентный лиганд включает два или более dAb, или две или более копий конкретного dAb, которое связывает домен 1 TNFR1. Поливалентные лиганды этого типа могут связывать мономеры TNFR1 с низкой аффинностью, но связывать мультимеры рецептора (например, тримеры смотри в рецептор-лигандном комплексе) с высокой авидностью. Таким образом, лиганды, имеющие этот формат, можно вводить для эффективного нацеливания на рецепторы, которые были кластеризованы или ассоциированы друг с другом, и/или лиганд (например, TNFα), который требуется для TNFR1-опосредованной передачи сигнала.
Некоторые лиганды или мономеры dAb связывают TNFR1 и ингибируют связывание TNFα с TNFR1. В некоторых воплощениях такой лиганд или мономер dAb связывает домен 2 и/или домен 3 TNFR1. В конкретных воплощениях лиганд или мономер dAb связывает домен 3 TNFR1. В более конкретных воплощениях лиганд или мономер dAb связывает домен 3 TNFR1 и конкурирует с TAR2h-10-27, TAR2h-131-8, TAR2h-15-8, TAR2h-35-4, TAR2h-154-7, TAR2h-154-10 или TAR2h-185-25 за связывание с TNFR1.
Другие лиганды или мономеры dAb не ингибируют связывание TNFα с TNFR1. Такие антагонисты обеспечивают преимущества в качестве диагностических агентов, поскольку они могут быть использованы для связывания и обнаружения, количественного определения или измерения TNFR1 в образце и не будут конкурировать с TNF в образце за связывание с TNFR1. Соответственно, можно точно определить, присутствует ли TNFR1 в образце или в каком количестве присутствует TNFR1 в образце.
Некоторые лиганды и мономеры dAb не ингибируют связывание TNFα с TNFR1, но ингибируют передачу сигнала, опосредованную через TNFR1. Например, лиганд или мономер dAb может ингибировать TNFα-индуцированную кластеризацию TNFR1, которая предшествует передаче сигнала через TNFR1. Такие лиганды или мономеры dAb обеспечивают некоторые преимущества, как обсуждается в данном изобретении в отношении антагонистов, обладающих этими свойствами. В конкретных воплощениях лиганд или мономер dAb этого типа связывает домен 1 TNFR1 или домен 4 TNFR1. В некоторых воплощениях лиганд представляет собой мономер dAb, который связывает домен 1 TNFR1 или домен 4 TNFR1.
В конкретном воплощении лиганд или мономер dAb связывает домен 1 TNFR1 и ингибирует передачу сигнала, опосредованную через TNFR1 после связывания с TNFα. Такой лиганд или мономер dAb может ингибировать передачу сигнала через TNFR1, но не ингибировать связывание TNFα с TNFR1 и/или шеддинг TNFR1 с получением растворимого TNFR1. Соответственно, введение такого лиганда или мономера dAb млекопитающему, нуждающемуся в этом, может дополнять эндогенные регуляторные пути, ингибирующие активность TNFα активность TNFR1 in vivo.
Другие лиганды или мономеры dAb связывают TNFR1, но не связывают домен 4. Такой лиганд или мономер dAb ингибирует функцию TNFR1, но не ингибирует шеддинг растворимого TNFR1. Соответственно, введение такого антагониста млекопитающему, нуждающемуся в этом, может дополнять эндогенные регуляторные пути, которые ингибируют активность TNF(и активность TNFR1 in vivo.
Предпочтительно, лиганд или мономер dAb нейтрализует (ингибирует активность) TNFα или TNFR1 в стандартном анализе (например, стандартный анализ L929 или стандартный анализ IL-8 в HeLa, как описано в данном изобретении) с 50% нейтрализующей дозой (ND50) от 500 нМ до 50 пМ, предпочтительно от 100 нМ до 50 пМ, более предпочтительно от 10 нМ до 100 пМ, предпочтительно от 1 нМ до 100 пМ; например 50 нМ или меньше, предпочтительно 5 нМ или меньше, более предпочтительно 500 пМ или меньше, предпочтительно 200 пМ или меньше и наиболее предпочтительно 100 пМ или меньше.
В некоторых воплощениях лиганд или мономер dAb специфически связывает человеческий рецептор фактора некроза опухоли 1 (TNFR1; р55) и диссоциирует из TNFR1 с константой диссоциации (Kd) от 50 нМ до 20 пМ и константой скорости диссоциации Koff от 5×10-1 с-1 до 1×10-7 с-1, как определено методом поверхностного плазменного резонанса.
В других воплощениях лиганд или мономер dAb специфически связывает TNFR1 с описанной в данном изобретении Kd и ингибирует летальность в стандартной модели септического шока у мышей, индуцированного ЛПС (липополисахаридом)/D-галактозамином (т.е. предотвращает летальность или уменьшает летальность по меньшей мере примерно на 10% по сравнению с соответствующим контролем). Предпочтительно, мономер dAb ингибирует летальность по меньшей мере примерно на 25% или по меньшей мере примерно на 50% по сравнению с соответствующим контролем в стандартной модели септического шока у мышей, индуцированного ЛПС/D-галактозамином при введении в концентрации примерно 5 мг/кг или более предпочтительно примерно 1 мг/кг.
В других воплощениях лиганд или мономер dAb связывает TNFR1 и оказывает антагонистическое действие на активность TNFR1 в стандартном клеточном анализе с ND50 не более 100 нМ, и в концентрации не более 10 мкМ dAb оказывает агонистическое действие на активность TNFR1 не более чем 5% в анализе.
В конкретных воплощениях лиганд или мономер dAb по существу не оказывает агонистического действия на TNFR1 (действует как агонист TNFR1) в стандартном клеточном анализе (т.е. когда присутствует в концентрации 1 нМ, 10 нМ, 100 нМ, 1 мкМ, 10 мкМ, 100 мкМ, 1000 мкМ или 5000 мкМ, дает не более чем примерно 5% от TNFR1-опосредованной активности, индуцированной TNF((100 пг/мл) в анализе).
В некоторых воплощениях лиганд или мономер dAb по существу устойчив к агрегации. Например, в некоторых воплощениях меньше чем примерно 10%, меньше чем примерно 9%, меньше чем примерно 8%, меньше чем примерно 7%, меньше чем примерно 6%, меньше чем примерно 5%. меньше чем примерно 4%, меньше чем примерно 3%, меньше чем примерно 2% или меньше чем примерно 1% лиганда или мономера dAb агрегирует, когда 1-5 мг/мл, 5-10 мг/мл, 10-20 мг/мл, 20-50 мг/мл, 50-100 мг/мл, 100-200 мг/мл или 200-500 мг/мл раствора лиганда или dAb в растворителе, который стандартно используют для изготовления лекарства, такого как физиологический раствор, забуференный физиологический раствор, забуференный цитратом физиологический раствор, вода, эмульсия и любой из этих растворителей с приемлемым эксципиентом, таким как одобренный Управлением по санитарному надзору за качеством пищевых продуктов и медикаментов США (FDA), поддерживают при примерно 22°С, 22-25°С, 25-30°С, 30-37°С, 37-40°С, 40-50°С, 50-60°С, 60-70°С, 70-80°С, 15-20°С, 10-15°С, 5-10°С, 2-5°С, 0-2-С, от -10°С до 0°С, от -20°С до -10°С, от -40°С до -20°С, от -60°С до -40°С или от -80°С до -60°С, в течение периода времени, например 10 минут, 1 часа, 8 часов, 24 часов, 2 суток, 3 суток, 4 суток, 1 недели, 2 недель, 3 недель, 1 месяца, 2 месяцев, 3 месяцев, 4 месяцев, 6 месяцев, 1 года или 2 лет.
Агрегация может быть оценена с использованием любого подходящего способа, такого как микроскопия, оценка мутности раствора с помощью визуальной оценки или спектроскопии, или любого другого подходящего способа. Предпочтительно, агрегацию оценивают с помощью динамического светорассеяния. Лиганды или мономеры dAb, которые устойчивы к агрегации, обеспечивают некоторые преимущества. Например, такие лиганды или мономеры dAb можно легко получить с высоким выходом в виде растворенных белков путем экспрессии с использованием подходящей биологической продуцирующей системы, такой как E. coli, и можно приготавливать и/или хранить в более высоких концентрациях, чем стандартные полипептиды, и с меньшей агрегацией и потерей активности.
Кроме того, лиганды или мономеры dAb, которые устойчивы к агрегации, могут быть получены с большей экономией, чем другие антиген- или эпитоп-связывающие полипептиды (например, стандартные антитела). Например, как правило, получение антиген- или эпитоп-связывающих полипептидов, предназначенных для применений in vivo, включает способы (например, гель-фильтрацию), которые удаляют агрегированные полипептиды. Невозможность удалить такие агрегаты может дать в результате препарат, не подходящий для применений in vivo, поскольку, например, агрегаты антиген-связывающего полипептида, который предназначен действовать в качестве антагониста, могут действовать в качестве агониста путем индукции перекрестного связывания или кластеризации антигена-мишени. Белковые агрегаты также могут уменьшать эффективность терапевтического полипептида путем индукции иммунного ответа у субъекта, которому их вводят.
В отличие от этого, лиганды или мономеры dAb по изобретению, устойчивые к агрегации, могут быть получены для применений in vivo без необходимости включать стадии способа, удаляющие агрегаты, и могут быть использованы в применениях in vivo без вышеупомянутых недостатков, вызванных полипептидными агрегатами.
В некоторых воплощениях лиганд или мономер dAb обратимо разворачивается при нагревании до температуры (Ts) и охлаждении до температуры (Ts), где Ts больше температуры плавления (Tm) dAb, а Те меньше температуры плавления dAb. Например, мономер dAb может обратимо разворачиваться при нагревании до 80°С и охлаждении до примерно комнатной температуры. Полипептид, который обратимо разворачивается, теряет функцию после разворачивания, но восстанавливает функцию после рефолдинга (повторного сворачивания). Такие полипептиды отличаются от полипептидов, которые агрегируют после разворачивания или неправильного рефолдинга (неправильно свернутые полипептиды), т.е. не восстанавливают функцию.
Разворачивание и рефолдинг полипептидов можно оценить, например, путем непосредственного или опосредованного обнаружения полипептидной структуры с использованием подходящего способа. Например, полипептидная структура может быть обнаружена с помощью кругового дихроизма (CD) (например, CD в далеком ультрафиолетовом излучении (far-UV CD), CD в близком ультрафиолетовом излучении (near-UV CD)), флуоресценции (например, флуоресценции боковых цепей триптофана), чувствительности к протеолизу, ядерного магнитного резонанса (ЯМР) или путем обнаружения или измерения функции полипептида, которая зависит от правильного сворачивания (например, связывание с лигандом-мишенью, связывание с когнатным лигандом). В одном из примеров разворачивание полипептидов оценивают с использованием функционального анализа, в котором потеря связывающей функции (например, связывания с когнатным лигандом и/или лигандом-мишенью, связывания с субстратом) указывает на то, что полипептид развернут.
Степень разворачивания и рефолдинга лиганда или мономера dAb может быть определена с использованием кривой разворачивания или денатурации. Кривая разворачивания может быть построена путем нанесения на график температуры в виде ординаты и относительной концентрации свернутого полипептида в виде абсциссы. Относительная концентрация свернутого лиганда или мономера dAb может быть определена непосредственно или опосредованно с использованием любого подходящего способа (например, CD, флуоресценции, анализа связывания). Например, может быть приготовлен раствор лиганда или мономера dAb, и эллиптичность раствора определяют с помощью CD. Полученное значение эллиптичности представляет относительную концентрацию свернутого лиганда или мономера dAb из 100%. Лиганд или мономер dAb в растворе затем разворачивают путем постепенного увеличения температуры раствора, и эллиптичность определяют при подходящих приращениях (например, после каждого увеличения температуры на один градус). Лиганд или мономер dAb в растворе затем разворачивают путем постепенного уменьшения температуры раствора, и эллиптичность определяют при подходящих приращениях. Данные могут быть нанесены на график с получением кривой разворачивания и кривой рефолдинга. Кривые разворачивания и рефолдинга имеют характерную сигмоидную форму, которая включает участок, на котором молекулы лиганда или мономера dAb свернуты, участок перехода разворачивание/рефолдинг, на котором молекулы лиганда или мономера dAb развернуты в различной степени, и участок, на котором молекулы лиганда или мономера dAb развернуты. Отрезок кривой рефолдинга по оси у представляет собой относительное количество свернутого лиганда или восстановленного мономера dAb. Восстановление по меньшей мере примерно на 50% или по меньшей мере примерно на 60%, или по меньшей мере примерно на 70%, или по меньшей мере примерно на 75%, или по меньшей мере примерно на 80%, или по меньшей мере примерно на 85%, или по меньшей мере примерно на 90%, или по меньшей мере примерно на 95% указывает на то, что лиганд или мономер dAb разворачивается обратимо.
В предпочтительном воплощении обратимость разворачивания лиганда или мономера dAb определяют путем приготовления раствора лиганда или мономера dAb и нанесения на график кривых теплового разворачивания и рефолдинга. Раствор лиганда или мономера dAb может быть приготовлен в любом подходящем растворителе, таком как водный буфер, который имеет рН, подходящий для того, чтобы обеспечить растворение лиганда или мономера dAb (например, рН, который составляет примерно 3 единицы выше или ниже изоэлектрической точки (р1)). Раствор лиганда или мономера dAb концентрируют в достаточной степени для того, чтобы обеспечить возможность обнаружения разворачивания/фолдинга. Например, раствор лиганда или мономера dAb может составлять от примерно 0,1 мкМ до примерно 100 мкМ, или предпочтительно от примерно 1 мкМ до примерно 10 мкМ.
Если известна температура плавления (Tm) лиганда или мономера dAb, то раствор может быть нагрет до примерно десяти градусов ниже Тт (Тт-10), и фолдинг можно оценить с помощью эллиптичности или флуоресценции (например, сканирование far-UV CD от 200 нм до 250 нм, фиксированная длина волны CD при 235 нМ или 225 нМ; спектры флуоресцентной эмиссии триптофана при 300-450 нМ с возбуждением при 298 нМ) с получением 100% относительно свернутого лиганда или мономера dAb. Раствор затем нагревают до температуры, по меньшей мере десяти на градусов выше Тт (Тт+10) с заданными приращениями (например, увеличения от примерно 0,1 до примерно 1 градуса), и эллиптичность или флуоресценцию определяют при каждом приращении. Затем лиганд или мономер dAb подвергают рефолдингу путем охлаждения по меньшей мере до Тт-10 с заданными приращениями, и эллиптичность или флуоресценцию определяют при каждом приращении. Если температура плавления лиганда или мономера dAb не известна, то раствор может быть подвергнут разворачиванию путем постепенного нагревания от примерно 25°С до примерно 100°С и затем подвергнут рефолдингу путем постепенного охлаждения до по меньшей мере примерно 25°С, и при каждом приращении нагревания и охлаждения определяют эллиптичность или флуоресценцию. Полученные данные могут быть нанесены на график с получением кривой разворачивания и кривой рефолдинга, в которых отрезок кривой рефолдинга на оси у представляет собой относительное количество восстановленного белка, подвергшегося рефолдингу.
В некоторых воплощениях лиганды или мономеры dAb по изобретению эффективны в моделях хронических воспалительных заболеваний при введении эффективного количества. Как правило, эффективное количество составляет от примерно 1 мг/кг до примерно 10 мг/кг (например, примерно 1 мг/кг, примерно 2 мг/кг, примерно 3 мг/кг, примерно 4 мг/кг, примерно 5 мг/кг, примерно 6 мг/кг, примерно 7 мг/кг, примерно 8 мг/кг, примерно 9 мг/кг или примерно 10 мг/кг). Описанные в данном изобретении модели хронического воспалительного заболевания признаны специалистами в данной области как предпосылки терапевтической эффективности у людей. В предшествующем уровне техники не предложено применение антагонистов TNFR1 (например, моновалентных антагонистов, лигандов, как описано в данном изобретении) в этих моделях или то, что они могут быть эффективны.
В конкретных воплощениях лиганд или мономер dAb эффективен в стандартной модели коллаген-индуцированного артрита у мышей (Пример 15А). Например, введение эффективного количества лиганда может уменьшить среднюю оценку артрита в результате суммирования результатов для четырех конечностей в стандартной модели коллаген-индуцированного артрита у мышей, например, от примерно 1 до примерно 16, от примерно 3 до примерно 16, от примерно 6 до примерно 16, от примерно 9 до примерно 16 или от примерно 12 до примерно 16 по сравнению с соответствующим контролем. В еще одном примере введение эффективного количества лиганда может замедлить начало развития симптомов артрита в стандартной модели коллаген-индуцированного артрита у мышей, например, примерно на 1 сутки, примерно на 2 суток, примерно на 3 суток, примерно на 4 суток, примерно на 5 суток, примерно на 6 суток, примерно на 7 суток, примерно на 10 суток, примерно на 14 суток, примерно на 21 сутки или примерно на 28 суток по сравнению с соответствующим контролем. В еще одном примере введение эффективного количества лиганда может привести в результате к средней оценке артрита в результате суммирования результатов для четырех конечностей в стандартной модели коллаген-индуцированного артрита у мышей от 0 до примерно 3, от примерно 3 до примерно 5, от примерно 5 до примерно 7, от примерно 7 до примерно 15, от примерно 9 до примерно 15, от примерно 10 до примерно 15, от примерно 12 до примерно 15 или от примерно 14 до примерно 15.
В других воплощениях лиганд или мономер dAb эффективен в модели артрита у мышей ΔARE (Пример 15 В). Например, введение эффективного количества лиганда может уменьшить среднюю оценку артрита в модели артрита у мышей ΔARE, например, от примерно 0,1 до примерно 2,5, от примерно 0,5 до примерно 2,5, от примерно 1 до примерно 2,5, от примерно 1,5 до примерно 2,5 или от примерно 2 до примерно 2,5 по сравнению с соответствующим контролем. В еще одном примере введение эффективного количества лиганда может замедлить начало развития симптомов артрита в модели артрита у мышей ДАРЕ, например, примерно на 1 сутки, примерно на 2 суток, примерно на 3 суток, примерно на 4 суток, примерно на 5 суток, примерно на 6 суток, примерно на 7 суток, примерно на 10 суток, примерно на 14 суток, примерно на 21 сутки или примерно на 28 суток по сравнению с соответствующим контролем. В еще одном примере введение эффективного количества лиганда может привести в результате к средней оценке артрита в модели артрита у мышей ΔARE от 0 до примерно 0,5, от примерно 0,5 до примерно 1, от примерно 1 до примерно 1,5, от примерно 1,5 до примерно 2, или от примерно 2 до примерно 2,5.
В других воплощениях лиганд или мономер dAb эффективен в модели воспалительного заболевания кишечника у мышей ΔARE (IBD) (Пример 15 В). Например, введение эффективного количества лиганда может уменьшить среднюю оценку острого и/или хронического воспаления в модели IBD у мышей ΔARE, например, от примерно 0,1 до примерно 2,5, от примерно 0,5 до примерно 2,5, от примерно 1 до примерно 2,5, от примерно 1,5 до примерно 2,5 или от примерно 2 до примерно 2,5 по сравнению с соответствующим контролем. В еще одном примере введение эффективного количества лиганда может замедлить начало развития симптомов IBD в модели IBD у мышей ΔARE, например, примерно на 1 сутки, примерно на 2 суток, примерно на 3 суток, примерно на 4 суток, примерно на 5 суток, примерно на 6 суток, примерно на 7 суток, примерно на 10 суток, примерно на 14 суток, примерно на 21 сутки или примерно на 28 суток по сравнению с соответствующим контролем. В еще одном примере введение эффективного количества лиганда может привести в результате к средней оценке острого и/или хронического воспаления в модели IBD у мышей ΔARE от 0 до примерно 0,5, от примерно 0,5 до примерно 1, от примерно 1 до примерно 1,5, от примерно 1,5 до примерно 2 или от примерно 2 до примерно 2,5.
В других воплощениях лиганд или мономер dAb эффективен в модели IBD у мышей, индуцированного декстрансульфатом натрия (ДСН) (Пример 15 В). Например, введение эффективного количества лиганда может уменьшить среднюю оценку тяжести в модели IBD у мышей, индуцированного ДСН, например, от примерно 0,1 до примерно 2,5, от примерно 0,5 до примерно 2,5, от примерно 1 до примерно 2,5, от примерно 1,5 до примерно 2,5, или от примерно 2 до примерно 2,5 по сравнению с соответствующим контролем. В еще одном примере введение эффективного количества лиганда может замедлить начало развития симптомов IBD в модели IBD у мышей, индуцированного ДСН, например, примерно на 1 сутки, примерно на 2 суток, примерно на 3 суток, примерно на 4 суток, примерно на 5 суток, примерно на 6 суток, примерно на 7 суток, примерно на 10 суток, примерно на 14 суток, примерно на 21 сутки или примерно на 28 суток по сравнению с соответствующим контролем. В еще одном примере введение эффективного количества лиганда может привести в результате к средней оценке тяжести в модели IBD у мышей, индуцированного ДСН, от 0 до примерно 0,5, от примерно 0,5 до примерно 1, от примерно 1 до примерно 1,5, от примерно 1,5 до примерно 2 или от примерно 2 до примерно 2,5.
В конкретных воплощениях лиганд или мономер dAb эффективен в модели хронического обструктивного заболевания легких (ХОЗЛ) у мышей, индуцированного табачным дымом (Пример 15Г). Например, введение эффективного количества лиганда может уменьшить или замедлить начало развития симптомов ХОЗЛ по сравнению с соответствующим контролем.
В конкретных воплощениях лиганд или мономер dAb специфически связывает TNFR1 и включает аминокислотную последовательность TAR2-10 (SEQ ID NO:31) или последовательность, которая по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%. по меньшей мере на 98% или по меньшей мере на 99% гомологична ей.
В конкретных воплощениях лиганд или мономер dAb специфически связывает TNFR1 и включает аминокислотную последовательность TAR2-5 (SEQ ID NO: 195) или последовательность, которая по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% гомологична ей.
В других воплощениях лиганд включает мономер доменного антитела (dAb), который специфически связывает рецептор фактора некроза опухоли I (TNFR1, р55, CD120a) с Kd от 300 нМ до 5 пМ и содержит аминокислотную последовательность, которая по меньшей мере примерно на 80%, по меньшей мере примерно на 85%, по меньшей мере примерно на 90%, по меньшей мере примерно на 91%, по меньшей мере примерно на 92%, по меньшей мере примерно на 93%, по меньшей мере примерно на 94%, по меньшей мере примерно на 95%, по меньшей мере примерно на 96%, по меньшей мере примерно на 97%, по меньшей мере примерно на 98% или по меньшей мере примерно на 99% гомологична аминокислотной последовательности или dAb, выбранному из группы, состоящей из TAR2h-12 (SEQ ID NO:32), TAR2h-13 (SEQ ID NO:33), TAR2h-14 (SEQ ID NO:34), TAR2h-16 (SEQ ID NO:35), TAR2h-1 7 (SEQ ID NO:36), TAR2h-1 8 (SEQ ID NO:37), TAR2h-1 9 (SEQ ID NO:38), TAR2h-20 (SEQ ID NO:39), TAR2h-21 (SEQ ID NO:40), TAR2h-22 (SEQ ID NO:41), TAR2h-23 (SEQ ID NO:42), TAR2h-24 (SEQ ID NO:43), TAR2h-25 (SEQ ID NO:44), TAR2h-26 (SEQ ID NO:45), TAR2h-27 (SEQ ID NO:46), TAR2h-29 (SEQ ID NO:47), TAR2h-30 (SEQ ID NO:48), TAR2h-32 (SEQ ID NO:49), TAR2h-33 (SEQ ID NO:50), TAR2h-10-1 (SEQ ID NO:51), TAR2h-10-2 (SEQ ID NO:52), TAR2h-10-3 (SEQ ID NO:53), TAR2h-10-4 (SEQ ID NO:54), TAR2h-10-5 (SEQ ID NO:55), TAR2h-10-6 (SEQ ID NO:56), TAR2h-10-7 (SEQ ID NO:57), TAR2h-10-8 (SEQ ID NO:58), TAR2h-10-9 (SEQ ID NO:59), TAR2h-10-10 (SEQ ID NO:60), TAR2h-10-11 (SEQ ID NO:61), TAR2h-10-12 (SEQ ID NO:62), TAR2h-10-13 (SEQ ID NO:63), TAR2h-10-14 (SEQ ID NO:64), TAR2h-10-15 (SEQ ID NO:65), TAR2h-10-16 (SEQ ID NO:66), TAR2h-10-17 (SEQ ID NO:67), TAR2h-10-18 (SEQ ID NO:68), TAR2h-10-19 (SEQ ID NO:69), TAR2h-10-20 (SEQ ID NO:70), TAR2h-10-21 (SEQ ID NO:71), TAR2h-10-22 (SEQ ID NO:72), TAR2h-10-27 (SEQ ID NO:73), TAR2h-10-29 (SEQ ID NO:74), TAR2h-10-31 (SEQ ID NO:75), TAR2h-10-35 (SEQ ID NO:76), TAR2h-10-36 (SEQ ID NO:77), TAR2h-10-37 (SEQ ID NO:78), TAR2h-10-38 (SEQ ID NO:79), TAR2h-10-45 (SEQ ID NO:80), TAR2h-10-47 (SEQ ID NO:81), TAR2h-10-48 (SEQ ID NO:82), TAR2h-10-57 (SEQ ID NO:83), TAR2h-10-56 (SEQ ID NO:84), TAR2h-10-58 (SEQ ID NO:85), TAR2h-10-66 (SEQ ID NO:86), TAR2h-10-64 (SEQ ID NO:87), TAR2h-10-65 (SEQ ID NO:88), TAR2h-10-68 (SEQ ID NO:89), TAR2h-10-69 (SEQ ID NO:90), TAR2h-10-67 (SEQ ID NO:91), TAR2h-10-61 (SEQ ID NO:92), TAR2h-10-62 (SEQ ID NO:93), TAR2h-10-63 (SEQ ID NO:94), TAR2h-10-60 (SEQ ID NO:95), TAR2h-10-55 (SEQ ID NO:96), TAR2h-10-59 (SEQ ID NO:97), TAR2h-10-70 (SEQ ID NO:98), TAR2h-34 (SEQ ID NO:373), TAR2h-35 (SEQ ID NO:374), TAR2h-36 (SEQ ID NO:375), TAR2h-37 (SEQ ID NO:376), TAR2h-38 (SEQ ID NO:377), TAR2h-39 (SEQ ID NO:378), TAR2h-40 (SEQ ID NO:379), TAR2h-41 (SEQ ID NO:380), TAR2h-42 (SEQ ID NO:381), TAR2h-43 (SEQ ID NO:382), TAR2h-44 (SEQ ID NO:383), TAR2h-45 (SEQ ID NO:384), TAR2h-47 (SEQ ID NO:385), TAR2h-48 (SEQ ID NO:386), TAR2h-50 (SEQ ID NO:387), TAR2h-51 (SEQ ED NO:388), TAR2h-66 (SEQ ID NO:389), TAR2h-67 (SEQ ID NO:390), TAR2h-68 (SEQ ID NO:391), TAR2h-70 (SEQ ID NO:392), TAR2h-71 (SEQ ID NO:393), TAR2h-72 (SEQ ID NO:394), TAR2h-73 (SEQ ID NO:395), TAR2h-74 (SEQ ID NO:396), TAR2h-75 (SEQ ID NO:397), TAR2h-76 (SEQ ID NO:398), TAR2h-77 (SEQ ID NO:399), TAR2h-78 (SEQ ID NO:400), TAR2h-79 (SEQ ID NO:401) и TAR2h-15 (SEQ ED NO:431).
В дополнительных воплощениях лиганд включает мономер доменного антитела (dAb), который специфически связывает рецептор фактора некроза опухоли I (TNFR1, р55, CD120a) с Кd от 300 нМ до 5 пМ и содержит аминокислотную последовательность, которая по меньшей мере примерно на 80%, по меньшей мере примерно на 85%, по меньшей мере примерно на 90%, по меньшей мере примерно на 91%, по меньшей мере примерно на 92%, по меньшей мере примерно на 93%, по меньшей мере примерно на 94%, по меньшей мере примерно на 95%, по меньшей мере примерно на 96%, по меньшей мере примерно на 97%, по меньшей мере примерно на 98%, или по меньшей мере примерно на 99% гомологична аминокислотной последовательности или dAb, выбранному из группы, состоящей из TAR2h-131-8 (SEQ ID NO:433), TAR2h-131-24 (SEQ ID NO:434), TAR2h-15-8 (SEQ ID NO:465), TAR2h-15-8-1 (SEQ ID NO:436), TAR2h-15-8-2 (SEQ ID NO:437), TAR2h-185-23 (SEQ ID NO:438), TAR2h-154-10-5 (SEQ ID NO:439), TAR2h-14-2 (SEQ ED NO:440), TAR2h-151-8 (SEQ ID NO:441), TAR2h-152-7 (SEQ ED NO:442), TAR2h-35-4 (SEQ ID NO:443), TAR2h-154-7 (SEQ ID NO:444), TAR2h-80 (SEQ ID NO:445), TAR2D-81 (SEQ ID NO:446), TAR2h-82 (SEQ ED NO:447), TAR2h-83 (SEQ ID NO:448), TAR2h-84 (SEQ ID NO:449), TAR2h-85 (SEQ ID NO:450), TAR2h-86 (SEQ ID NO:45 1), TAR2h-87 (SEQ ID NO:452), TAR2h-88 (SEQ ID NO:453), TAR2h-89 (SEQ ID NO:454), TAR2h-90 (SEQ ID NO:455), TAR2h-91 (SEQ ID NO:456), TAR2h-92 (SEQ ID NO:457), TAR2h-93 (SEQ ID NO:458), TAR2h-94 (SEQ ID NO:459), TAR2h-95 (SEQ ID NO:460), TAR2h-96 (SEQ ID NO:461), TAR2h-97 (SEQ ID NO:462), TAR2h-99 (SEQ ID NO:463), TAR2h-100 (SEQ ID NO:464), TAR2h-101 (SEQ ID NO:465), TAR2h-102 (SEQ ID NO:466), TAR2h-103 (SEQ ID NO:467), TAR2h-104 (SEQ ID NO:468), TAR2h-105 (SEQ ID NO:469), TAR2h-106 (SEQ ID NO:470), TAR2h-107 (SEQ ID NO:471), TAR2h-108 (SEQ ID NO:472), TAR2h-109 (SEQ ID NO:473), TAR2h-110 (SEQ ID NO:474), TAR2h-111 (SEQ ID NO:475), TAR2h-112 (SEQ ID NO:476), TAR2h-113 (SEQ ID NO:477), TAR2h-114 (SEQ ID NO:478), TAR2h-115 (SEQ ID NO:479), TAR2h-116 (SEQ ID NO:480), TAR2h-117 (SEQ ID NO:481), TAR2h-118 (SEQ ID NO:482), TAR2h-119 (SEQ ID NO:483), TAR2h-120 (SEQ ID NO:484), TAR2h-121 (SEQ ID NO:485), TAR2h-122 (SEQ ID NO:486), TAR2h-123 (SEQ ID NO:487), TAR2h-124 (SEQ ID NO:488), TAR2h-125 (SEQ ID NO:489), TAR2h-126 (SEQ ID NO:490), TAR2h-127 (SEQ ID NO:490), TAR2h-128 (SEQ ID NO:492), TAR2h-129 (SEQ ID NO:493), TAR2h-130 (SEQ ID NO:494), TAR2h-131 (SEQ ID NO:495), TAR2h-132 (SEQ ID NO:496), TAR2h-133 (SEQ ID NO:497), TAR2h-151 (SEQ ID NO:498), TAR2h-152 (SEQ ID NO:499), TAR2h-153 (SEQ ID NO:500), TAR2h-154 (SEQ ID NO:501), TAR2h-159 (SEQ ID NO:502), TAR2h-165 (SEQ ID NO:503), TAR2h-166 (SEQ ID NO:504), TAR2h-168 (SEQ ID NO:505), TAR2h-171 (SEQ ID NO:506), TAR2h-172 (SEQ ID NO:507), TAR2h-173 (SEQ ID NO:508), TAR2h-174 (SEQ ID NO:509), TAR2h-176 (SEQ ID NO:510), TAR2h-178 (SEQ ID NO:511), TAR2h-201 (SEQ ID NO:512), TAR2h-202 (SEQ ID NO:513), TAR2h-203 (SEQ ID NO:514), TAR2h-204 (SEQ ID NO:515), TAR2h-185-25 (SEQ ID NO:516), TAR2h-154-10 (SEQ ID NO:517) и TAR2h-205 (SEQ ID NO:627).
В предпочтительных воплощениях лиганды или dAb включают аминокислотную последовательность, которая по меньшей мере примерно на 90%, по меньшей мере примерно на 91%, по меньшей мере примерно на 92%, по меньшей мере примерно на 93%, по меньшей мере примерно на 94%, по меньшей мере примерно на 95%, по меньшей мере примерно на 96%, по меньшей мере примерно на 97%, по меньшей мере примерно на 98% или по меньшей мере примерно на 99% гомологична аминокислотной последовательности dAb, выбранной из группы, состоящей из TAR2h-10-27 (SEQ ID NO:73), TAJR2h-10-57 (SEQ ID NO:83), TAR2h-10-56 (SEQ ID NO:84), TAR2h-10-58 (SEQ ID NO:85), TAR2h-10-66 (SEQ ID NO:86), TAR2h-10-64 (SEQ ID NO:87), TAR2h-10-65 (SEQ ID NO:88), TAR2h-10-68 (SEQ ID NO:89), TAR2h-10-69 (SEQ ID NO:90), TAR2h-10-67 (SEQ ID NO:91), TAR2h-10-61 (SEQ ID NO:92), TAR2h-10-62 (SEQ ID NO:93), TAR2h-10-63 (SEQ ID NO:94), TAR2h-10-60 (SEQ ID NO:95), TAR2h-10-55 (SEQ ID NO:96), TAR2h-10-59 (SEQ ID NO:97) и TAR2h-10-70 (SEQ ID NO:98).
Особенно предпочтительные лиганды или dAb включают аминокислотную последовательность, которая по меньшей мере примерно на 90%, по меньшей мере примерно на 91%, по меньшей мере примерно на 92%, по меньшей мере примерно на 93%, по меньшей мере примерно на 94%, по меньшей мере примерно на 95%, по меньшей мере примерно на 96%, по меньшей мере примерно на 97%, по меньшей мере примерно на 98% или по меньшей мере примерно на 99% гомологична аминокислотной последовательности SEQ ID NO: 73.
В других воплощениях лиганд включает мономер доменного антитела (dAb), который специфически связывает рецептор фактора некроза опухоли I (TNFR1, р55, CD 120a) с Ка от 300 нМ до 5 пМ и содержит аминокислотную последовательность, которая по меньшей мере примерно на 80%, по меньшей мере примерно на 85%, по меньшей мере примерно на 90%, по меньшей мере примерно на 91%, по меньшей мере примерно на 92%, по меньшей мере примерно на 93%, по меньшей мере примерно на 94%, по меньшей мере примерно на 95%, по меньшей мере примерно на 96%, по меньшей мере примерно на 97%, по меньшей мере примерно на 98% или по меньшей мере примерно на 99% гомологична аминокислотной последовательности или dAb, выбранному из группы, состоящей из TAR2m-14 (SEQ ID NO:167), TAR2m-15 (SEQ ID NO:168), TAR2m-19 (SEQ ID NO:169), TAR2m-20 (SEQ ID NO:170), TAR2m-21 (SEQ ID NO:171), TAR2m-24 (SEQ ID NO:172), TAR2m-21-23 (SEQ ID NQ:173), TAR2m-21-07 (SEQ ID NO:174), TAR2m-21-43 (SEQ ID NO:175), TAR2m-21-48 (SEQ ID NO:176), TAR2m-21-10 (SEQ ID NO:177), TAR2m-21-06 (SEQ ID NO:178), TAR2m-21-17 (SEQ ID NO: 179).
В некоторых воплощениях лиганд включает мономер dAb, который связывает TNFR1 и конкурирует с любым из описанных в данном изобретении dAb за связывание с TNFR1 (например, мышиный и/или человеческий TNFR1).
Предпочтительно, лиганд или мономер dAb секретируется в количестве, составляющем по меньшей мере примерно 0,5 мг/л при экспрессии в Е. coli или видах Pichia (например, Р. pastoris). В других предпочтительных воплощениях мономер dAb выбран в количестве, составляющем по меньшей мере примерно 0,75 мг/л, по меньшей мере примерно 1 мг/л, по меньшей мере примерно 4 мг/л, по меньшей мере примерно 5 мг/л, по меньшей мере примерно 10 мг/л, по меньшей мере примерно 15 мг/л, по меньшей мере примерно 20 мг/л, по меньшей мере примерно 25 мг/л, по меньшей мере примерно 30 мг/л, по меньшей мере примерно 35 мг/л, по меньшей мере примерно 40 мг/л, по меньшей мере примерно 45 мг/л или по меньшей мере примерно 50 мг/л, или по меньшей мере примерно 100 мг/л, или по меньшей мере примерно 200 мг/л, или по меньшей мере примерно 300 мг/л, или по меньшей мере примерно 400 мг/л, или по меньшей мере примерно 500 мг/л, или по меньшей мере примерно 600 мг/л, или по меньшей мере примерно 700 мг/л, или по меньшей мере примерно 800 мг/л, по меньшей мере примерно 900 мг/л, или по меньшей мере примерно 1 г/л при экспрессии в Е. со// или в видах Pichia (например, Р. pastoris). В других предпочтительных воплощениях мономер dAb секретируется в количестве по меньшей мере от примерно 1 мг/л по меньшей мере до примерно 1 г/л, по меньшей мере от примерно 1 мг/л по меньшей мере до примерно 750 мг/л, по меньшей мере от примерно 100 мг/л по меньшей мере до примерно 1 г/л, по меньшей мере от примерно 200 мг/л по меньшей мере до примерно 1 г/л, по меньшей мере от примерно 300 мг/л по меньшей мере до примерно 1 г/л, по меньшей мере от примерно 400 мг/л по меньшей мере до примерно 1 г/л, по меньшей мере от примерно 500 мг/л по меньшей мере до примерно 1 г/л, по меньшей мере от примерно 600 мг/л по меньшей мере до примерно 1 г/л, по меньшей мере от примерно 700 мг/л по меньшей мере до примерно 1 г/л, по меньшей мере от примерно 800 мг/л по меньшей мере до примерно 1 г/л, или по меньшей мере от примерно 900 мг/л по меньшей мере до примерно 1 г/л при экспрессии в E. coli' или видах Pichia (например, Р. pastoris). Хотя описанные в данном изобретении лиганды и мономер dAb могут секретироваться при экспрессии в Е. coli или видах Pichia (например, Р. pastoris), они могут быть получены с использованием любого подходящего способа, такого как способы химического синтеза или способы биологической продукции, в которых не используют Е. со//' или виды Pichia.
Мономер dAb может включать любой подходящий вариабельный домен иммуноглобулина и предпочтительно включает человеческий вариабельный домен или вариабельный домен, который включает человеческие каркасные области. В некоторых воплощениях мономер dAb содержит универсальный каркас, как описано в данном изобретении.
Универсальный каркас может представлять собой каркас VL (Vλ или VK), такой как каркас, включающий каркасные аминокислотные последовательности, кодируемые герминальным генным сегментом DPK1, DPK2, DPK3, DPK4, DPK5, DPK6, DPK7, DPK8, DPK9, DPK10, DPK12, DPK13, DPK15, DPK16, DPK18, DPK19, DPK20, DPK21, DPK22, DPK23, DPK24, DPK25, DPK26 или DPK 28 человеческого иммуноглобулина. При желании, каркас VL может дополнительно включать каркасную аминокислотную последовательность, кодируемую герминальным генным сегментом JK1, JK2, JK3, JK4 или JK5 человеческого иммуноглобулина.
В других воплощениях универсальный каркас может представлять собой каркас VH, такой как каркас, который включает каркасные аминокислотные последовательности, кодируемые герминальным генным сегментом DP4, DP7, DP8, DP9, DP10, DP31, DP33, DP38, DP45, DP46, DP47, DP49, DP50, DP51, DP53, DP54, DP65, DP66, DP67, DP68 или DP69 человеческого иммуноглобулина. При желании, каркас VH может дополнительно включать каркасную аминокислотную последовательность, кодируемую герминальным генным сегментом JH1, JH2, JH3, JH4, JH4b, JH5 и JH6 человеческого иммуноглобулина.
В конкретных воплощениях лиганд, представляющий собой мономер dAb, содержит каркас DPK9 VL или каркас VH, выбранный из группы, состоящей из DP47, DP45 и DP38.
Мономер dAb может включать сайт связывания общего лиганда, такого как белок А, белок L и белок G.
В некоторых воплощениях мономер dAb содержит одну или более каркасных областей, включающих аминокислотную последовательность, то есть такую же, как аминокислотная последовательность соответствующей каркасной области, кодируемой герминальным генным сегментом человеческого антитела, или аминокислотные последовательности одной или более чем одной из указанных каркасных областей вместе содержат до 5 аминокислотных различий относительно аминокислотной последовательности указанной соответствующей каркасной области, кодируемой герминальным генным сегментом человеческого антитела.
В других воплощениях аминокислотные последовательности FW1, FW2, FW3 и FW4 мономера dAb являются такими же, как аминокислотные последовательности соответствующих каркасных областей, кодируемых герминальным генным сегментом человеческого антитела, или аминокислотные последовательности FW1, FW2, FW3 и FW4 вместе содержат до 10 аминокислотных различий относительно аминокислотных последовательностей соответствующих каркасных областей, кодируемых указанным герминальным генным сегментом человеческого антитела.
В других воплощениях мономер dAb содержит области FW1, FW2 и FW3, и аминокислотная последовательность указанных областей FW1, FW2 и FW3 являются такими же, как аминокислотные последовательности соответствующих каркасных областей, кодируемых герминальными генными сегментами человеческого антитела.
В некоторых воплощениях мономер dAb не включает вариабельный домен иммуноглобулина Camelid, или одну или более каркасных аминокислот, которые являются уникальными для вариабельных доменов иммуноглобулина, кодируемых герминальными генными сегментами антитела Camelid.
Лиганды и мономеры dAb, которые связывают сывороточный альбумин
В изобретении предложен лиганд или мономер dAb (например, лиганд с двойной специфичностью, включающий такое dAb), который связывается с сывороточным альбумином (SA) с Ка от 1 нМ до 500 мкМ (т.е. от ×10-9 до 5×10-4), предпочтительно от 100 нМ до 10 мкМ. Предпочтительно, для лиганда с двойной специфичностью, включающего первое dAb против SA и второе dAb против другой мишени, аффинность (например, Ка и/или Код, измерененные методом поверхностного плазменного резонанса, например с использованием BiaCore) второго dAb в отношении его мишени составляет от 1 до 100000 раз (предпочтительно от 100 до 100000, более предпочтительно от 1000 до 100000, или от 10000 до 100000 раз) относительно аффинности первого dAb в отношении SA. Например, первое dAb связывает SA с аффинностью, составляющей примерно 10 мкМ, тогда как второе dAb связывает свою мишень с аффинностью, составляющей 100 пМ. Предпочтительно, сывороточный альбумин представляет собой человеческий сывороточный альбумин (HSA). В одном из воплощений первое dAb (или мономер dAb) связывает SA (например, HSA) с Kd примерно 50, предпочтительно 70, и более предпочтительно 100, 150 или 200 нМ.
В некоторых воплощениях мономер dAb, специфичный в отношении SA, включает аминокислотную последовательность MSA-16 (SEQ ID NO:28) или последовательность, которая по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% гомологична ей.
В других воплощениях мономер dAb, специфичный в отношении SA, включает аминокислотную последовательность MSA-26 (SEQ ID NO:30) или последовательность, которая по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% гомологична ей.
В некоторых воплощениях мономер dAb, который связывает SA, противодействует агрегации, обратимо разворачивается и/или включает каркасную область, как описано выше для мономеров dAb, которые связывают TNFR1.
Молекулы нуклеиновых кислот, векторы и клетки-хозяева
В изобретении также предложены выделенные и/или рекомбинантные молекулы нуклеиновых кислот, которые кодируют лиганды и мономеры dAb, описанные в данном изобретении. В некоторых воплощениях выделенная и/или рекомбинантная нуклеиновавя кислота включает нуклеотидную последовательность, которая кодирует мономер доменного антитела (dAb), который специфически связывает рецептор фактора некроза опухоли I (TNFR1), где указанная нуклеотидная последовательность по меньшей мере примерно на 80%, по меньшей мере примерно на 85%, по меньшей мере примерно на 90%, по меньшей мере примерно на 91%, по меньшей мере примерно на 92%, по меньшей мере примерно на 93%, по меньшей мере примерно на 94%, по меньшей мере примерно на 95%, по меньшей мере примерно на 96%, по меньшей мере примерно на 97%, по меньшей мере примерно на 98% или по меньшей мере примерно на 99% гомологична нуклеотидной последовательности, выбранной из группы, состоящей из TAR2h-12 (SEQ ID NO:32), TAR2h-13 (SEQ ID NO:33), TAR2h-14 (SEQ ID NO:34), TAR2h-16 (SEQ ID NO:35), TAR2h-17 (SEQ ID NO:36), TAR2h-18 (SEQ ID NO:37), TAR2h-19 (SEQ ID NO:38), TAR2h-20 (SEQ ID NO:39), TAR2h-21 (SEQ ID NO:40), TAR2h-22 (SEQ ID NO:41), TAR2h-23 (SEQ ID NO:42), TAR2h-24 (SEQ ID NO:43), TAR2h-25 (SEQ ID NO:44), TAR2h-26 (SEQ ID NO:45), TAR2h-27 (SEQ ID NO:46), TAR2h-29 (SEQ ID NO:47), TAR2h-30 (SEQ ID NO:48), TAR2h-32 (SEQ ID NO:49), TAR2h-33 (SEQ ID NO:50), TAR2h-10-1 (SEQ ID NO:51), TAR2h-10-2 (SEQ ID NO:52), TAR2h-10-3 (SEQ ID NO:53), TAR2h-10-4 (SEQ ID NO:54), TAR2h-10-5 (SEQ ID NO:55), TAR2h-10-6 (SEQ ID NO:56), TAR2h-10-7 (SEQ ID NO:57), TAR2h-10-8 (SEQ ID NO:58), TAR2h-10-9 (SEQ ID NO:59), TAR2h-10-10 (SEQ ID NO:60), TAR2h-10-11 (SEQ ID NO:61), TAR2h-10-12 (SEQ ID NO:62), TAR2h-10-13 (SEQ ID NO:63), TAR2h-10-14 (SEQ ID NO:64), TAR2h-10-15 (SEQ ID NO:65), TAR2h-10-16 (SEQ ID NO:66), TAR2h-10-17 (SEQ ID NO:67), TAR2h-10-18 (SEQ ID NO:68), TAR2h-10-19 (SEQ ID NO:69), TAR2h-10-20 (SEQ ID NO:70), TAR2h-10-21 (SEQ ID NO:71), TAR2h-10-22 (SEQ ID NO:72), TAR2h-10-27 (SEQ ID NO:73), TAR2h-10-29 (SEQ ID NO:74), TAR2h-10-31 (SEQ ID NO:75), TAR2h-10-35 (SEQ ID NO:76), TAR2h-10-36 (SEQ ID NO:77), TAR2h-10-37 (SEQ ID NO:78), TAR2h-10-38 (SEQ ID NO:79), TAR2h-10-45 (SEQ ID NO:80), TAR2h-10-47 (SEQ ID NO:81), TAR2h-10-48 (SEQ ID NO:82), TAR2h-10-57 (SEQ ID NO:83), TAR2h-10-56 (SEQ ID NO:84), TAR2h-10-58 (SEQ ID NO:85), TAR2h-10-66 (SEQ ID NO:86), TAR2h-10-64 (SEQ ID NO:87), TAR2h-10-65 (SEQ ID NO:88), TAR2h-10-68 (SEQ ID NO:89), TAR2h-10-69 (SEQ ID NO:90), TAR2h-10-67 (SEQ ID NO:91), TAR2h-10-61 (SEQ ID NO:92), TAR2h-10-62 (SEQ ID NO:93), TAR2h-10-63 (SEQ ID NO:94), TAR2h-10-60 (SEQ ID NO:95), TAR2h-10-55 (SEQ ID NO:96). TAR2h-10-59 (SEQ ID NO:97), TAR2h-10-70 (SEQ ID NO:98), TAR2h-34 (SEQ ID NO:402), TAR2h-35 (SEQ ID NO:403), TAR2h-36 (SEQ ID NO:404), TAR2h-37 (SEQ ID NO:405), TAR2h-38 (SEQ ID NO:406), TAR2h-39 (SEQ ID NO:407), TAR2h-40 (SEQ ID NO:408), TAR2h-41 (SEQ ID NO:409), TAR2h-42 (SEQ ID NO:410), TAR2h-43 (SEQ ID NO:411), TAR2h-44 (SEQ ID NO:412), TAR2h-45 (SEQ ID NO:413), TAR2h-47 (SEQ ID NO:414), TAR2h-48 (SEQ ID NO:415), TAR2h-50 (SEQ ID NO:416), TAR2h-51 (SEQ ID NO:417), TAR2h-66 (SEQ ID NO:418), TAR2h-67 (SEQ ID NO:419), TAR2h-68 (SEQ ID NO:420), TAR2h-70 (SEQ ID NO:421), TAR2h-71 (SEQ ID NO:422), TAR2h-72 (SEQ ID NO:423), TAR2h-73 (SEQ ID NO:424), TAR2h-74 (SEQ ID NO:425), TAR2h-75 (SEQ ID NO:426), TAR2h-76 (SEQ ID NO:427), TAR2h-77 (SEQ ID NO:428), TAR2h-78 (SEQ ID NO:429), TAR2h-79 (SEQ ID NO:430) и TAR2h-15 (SEQ ID NO:432).
В других воплощениях выделенная и/или рекомбинанатная нуклеиновая кислота включает нуклеотидную последовательность, кодирующую мономер доменного антитела (dAb), который специфически связывает рецептор фактора некроза опухоли I (TNFR1), где указанная нуклеотидная последовательность по меньшей мере примерно на 80%, по меньшей мере примерно на 85%, по меньшей мере примерно на 90%, по меньшей мере примерно на 91%, по меньшей мере примерно на 92%, по меньшей мере примерно на 93%, по меньшей мере примерно на 94%, по меньшей мере примерно на 95%, по меньшей мере примерно на 96%, по меньшей мере примерно на 97%, по меньшей мере примерно на 98% или по меньшей мере примерно на 99% гомологична нуклеотидной последовательности, выбранной из группы, состоящей из TAR2h-131-8 (SEQ ID NO:518), TAR2h-131-24 (SEQ ID NO:519), TAR2h-15-8 (SEQ ID NO:520), TAR2h-15-8-1 (SEQ ID NO:521), TAR2h-15-8-2 (SEQ ID NO:522), TAR2h-185-23 (SEQ ID NO:523), TAR2h-154-10-5 (SEQ ID NO:524), TAR2h-14-2 (SEQ ID NO:525), TAR2h-151-8 (SEQ ID NO:526), TAR2h-152-7 (SEQ ID NO:527), TAR2h-35-4 (SEQ ID NO:528), TAR2h-154-7 (SEQ ID NO:529), TAR2h-80 (SEQ ID NO:530), TAR2h-81 (SEQ ID NO:531), TAR2h-82 (SEQ ID NO:532), TAR2h-83 (SEQ ID NO:533), TAR2h-84 (SEQ ID NO:534), TAR2h-85 (SEQ ID NO:535), TAR2h-86 (SEQ ID NO:536), TAR2h-87 (SEQ ID NO:537), TAR2h-88 (SEQ ID NO:538), TAR2h-89 (SEQ ID NO:539), TAR2h-90 (SEQ ID NO:540), TAR2h-91 (SEQ ID NO:541), TAR2h-92 (SEQ ID NO:542), TAR2h-93 (SEQ ID NO:543), TAR2h-94 (SEQ ID NO:544), TAR2h-95 (SEQ ID NO:545), TAR2h-96 (SEQ ID NO:546), TAR2h-97 (SEQ ID NO:547), TAR2h-99 (SEQ ID NO:548), TAR2h-100 (SEQ ID NO:549), TAR2h-101 (SEQ ID NO:550), TAR2h-102 (SEQ ID NO:551), TAR2h-103 (SEQ ID NO:552), TAR2h-104 (SEQ ID NO:553), TAR2h-105 (SEQ ID NO:554), TAR2h-106 (SEQ ID NO:555), TAR2h-107 (SEQ ID NO:556), TAR2h-108 (SEQ ID NO:557), TAR2h-109 (SEQ ID NO:558), TAR2h-110 (SEQ ID NO:559), TAR2h-111 (SEQ ID NO:560), TAR2h-112 (SEQ ID NO:561), TAR2h-113 (SEQ ID NO:562), TAR2h-114 (SEQ ID NO:563), TAR2h-115 (SEQ ID NO:564), TAR2h-116 (SEQ ID NO:565), TAR2h-117 (SEQ ID NO:566), TAR2h-118 (SEQ ID NO:567), TAR2h-119 (SEQ ID NO:568), TAR2h-120 (SEQ ID NO:569), TAR2h-121 (SEQ ID NO:570), TAR2h-122 (SEQ ID NO:571), TAR2h-123 (SEQ ID NO:572), TAR2h-12 (SEQ ID NO:573), TAR2h-125 (SEQ ID NO:574), TAR2h-126 (SEQ ID NO:575), TAR2h-127 (SEQ ID NO:576), TAR2h-128 (SEQ ID NO:577), TAR2h-129 (SEQ ID NO:578), TAR2h-130 (SEQ ID NO:579), TAR2h-131 (SEQ ID NO:580), TAR2h-132 (SEQ ID NO:581), TAR2h-133 (SEQ ID NO:582), TAR2h-151 (SEQ ID NO:583), TAR2h-152 (SEQ ID NO:584), TAR2h-153 (SEQ ID NO:585), TAR2h-154 (SEQ ID NO:586), TAR2h-159 (SEQ ID NO:587), TAR2h-165 (SEQ ID NO:588), TAR2h-166 (SEQ ID NO:589), TAR2h-168 (SEQ ID NO:590), TAR2h-171 (SEQ ID NO:591), TAR2h-172 (SEQ ID NO:592), TAR2h-173 (SEQ ID NO:593), TAR2h-174 (SEQ ID NO:594), TAR2h-176 (SEQ ID NO:595), TAR2h-178 (SEQ ID NO:596), TAR2h-201 (SEQ ID NO:597), TAR2h-202 (SEQ ID NO:598), TAR2h-203 (SEQ ID NO:599), TAR2h-204 (SEQ ID NO:600), TAR2h-185-25 (SEQ ID NO:601), TAR2h-154-10 (SEQ ID NO:602) и TAR2h-205 (SEQ ID NO:628).
В предпочтительном воплощении выделенная и/или рекомбинантная нуклеиновая кислота включает нуклеотидную последовательность, которая по меньшей мере примерно на 90%, по меньшей мере примерно на 91%, по меньшей мере примерно на 92%, по меньшей мере примерно на 93%, по меньшей мере примерно на 94%, по меньшей мере примерно на 95%, по меньшей мере примерно на 96%, по меньшей мере примерно на 97%, по меньшей мере примерно на 98% или по меньшей мере примерно на 99% гомологична нуклеотидной последовательности, выбранной из группы, состоящей из TAR2h-10-27 (SEQ ID NO:141), TAR2h-10-57 (SEQ ID NO:151), TAR2h-10-56 (SEQ ID NO:152), TAR2h-10-58 (SEQ ID NO: 153), TAR2h-10-66 (SEQ ID NO: 154), TAR2h-10-64 (SEQ ID NO:155), TAR2h-10-65 (SEQ ID NO:156), TAR2h-10-68 (SEQ ID NO:157), TAR2h-10-69 (SEQ ID NO:158), TAR2h-10-67 (SEQ ID NO:159), TAR2h-10-61 (SEQ ID NO: 160), TAR2h-10-62 (SEQ ID NO:161), TAR2h-10-63 (SEQ ID NO:162), TAR2h-10-60 (SEQ ID NO:163), TAR2h-10-55 (SEQ ID NO:164), TAR2h-10-59 (SEQ ID NO:165) и TAR2h-10-70 (SEQ ID NO:166).
В других воплощениях выделенная и/или рекомбинантная нуклеиновая кислота включает нуклеотидную последовательность, кодирующую мономер доменного антитела (dAb), который специфически связывает рецептор фактора некроза опухоли I (TNFR1), где указанная нуклеотидная последовательность по меньшей мере примерно на 80%, по меньшей мере примерно на 85%, по меньшей мере примерно на 90%, по меньшей мере примерно на 91%, по меньшей мере примерно на 92%, по меньшей мере примерно на 93%, по меньшей мере примерно на 94%, по меньшей мере примерно на 95%, по меньшей мере примерно на 96%, по меньшей мере примерно на 97%, по меньшей мере примерно на 98% или по меньшей мере примерно на 99% гомологична нуклеотидной последовательности, выбранной из группы, состоящей из TAR2m-14 (SEQ ID NO: 180), TAR2m-15 (SEQ ID NO:181), TAR2m-19 (SEQ ID NO:182), TAR2m-20 (SEQ ID NO:183), TAR2m-21 (SEQ ID NO:184), TAR2m-24 (SEQ ID NO:185), TAR2m-21-23 (SEQ ID NO:186), TAR2m-21-07 (SEQ ID NO:187), TAR2m-21-43 (SEQ ID NO:188), TAR2m-21-48 (SEQ ID NO:189), TAR2m-21-10 (SEQ ID NO:190), TAR2m-21-06 (SEQ ID NO:191), TAR2m-21-17 (SEQ ID NO: 192) и TAR2m-21-23a (SEQ ID NO:626).
В изобретении также предложен вектор, содержащий рекомбинантную молекулу нуклеиновой кислоты по изобретению. В некоторых воплощениях вектор представляет собой экспрессирующий вектор, содержащий один или более элементов, регулирующих экспрессию, или последовательности, которые функционально связаны с рекомбинантной нуклеиновой кислотой по изобретению. Подходящие векторы (например, плазмиды, фагмиды) и элементы, регулирующие экспрессию, дополнительно описаны ниже.
В изобретении также предложена рекомбинантная клетка-хозяин, включающая молекулу рекомбинантной нуклеиновой кислоты или вектор по изобретению. Подходящие клетки-хозяева и способы получения рекомбинантной клетки-хозяина по изобретению дополнительно описаны ниже.
В изобретении также предложен способ получения лиганда (например, мономера dAb, лиганда с двойной специфичностью, полиспецифического лиганда) по изобретению, включающий поддержание рекомбинантной клетки-хозяина, включающей рекомбинантную нуклеиновую кислоту по изобретению в условиях, подходящих для экспрессии рекомбинантной нуклеиновой кислоты, в результате чего рекомбинантная экспрессируется нуклеиновая кислота и продуцируется лиганд. В некоторых воплощениях способ дополнительно включает выделение лиганда.
Форматы лиганда
Лиганды по изобретению могут иметь форматы моно- или полиспецифических антител или фрагментов антител или моно- или полиспецифических неиммуноглобулиновых структур. Подходящие форматы включают любую подходящую полипептидную структуру, в которой вариабельный домен антитела или его CDR могут быть включены таким образом, чтобы придавать связывающую специфичность в отношении антигена на структуре. В данной области известно множество подходящих форматов антител, таких как lgG-подобные форматы, химерные антитела, гуманизированные антитела, человеческие антитела, одноцепочечные антитела, биспецифические антитела, тяжелые цепи антитела, легкие цепи антитела, гомодимеры и гетеродимеры тяжелых цепей и/или легких цепей антитела, антиген-связывающие фрагменты любого из вышеприведенного (например, Fv-фрагмент (например, одноцепочечный Fv (scFv), связанный дисульфидными связями Fv), Fab-фрагмент, Fab'-фрагмент, Р(ab')2-фрагмент), единичный вариабельный домен (например, VH, VL), dAb, и модифицированные варианты любого из вышеперечисленного (например, модифицированные путем ковалентного присоединения полиалкиленгликоля (например, полиэтиленгликоля, полипропиленгликоля, полибутиленгликоля) или другого подходящего полимера). Смотри PCT/GB03/002804, поданую 30 июня 2003 г., которая указывает США (WO 2004/081026), в отношении пэгилирования единичных вариабельных доменов и dAb, подходящие способы их получения, увеличенный период полувыведения in vivo пэгилированных единичных вариабельных доменов и мономеров и мультимеров dAb, подходящие ПЭГ, предпочтительные гидродинамические размеры ПЭГ и предпочтительные гидродинамические размеры пэгилированных единичных вариабельных доменов и мономеров и мультимеров dAb. Полное содержание PCT/GB03/002804 (WO 2004/081026), включая упомянутые выше фрагменты включены в данное описание путем ссылки.
В конкретных воплощениях лиганд (например, мономер, димер или мультимер dAb, двойной специфический формат, полиспецифический формат) пэгилирован и связывает домен 1 TNFR1 и возможно ингибирует функцию TNFR1. Предпочтительно, пэгилированный лиганд ингибирует передачу сигнала через TNFR1. Предпочтительно, пэгилированный лиганд связывает домен 1 TNFR1, по существу с такой же аффинностью как такой же лиганд, который не пэгилирован. Например, в одном из воплощений лиганд представляет собой пэгилированный мономер dAb, который связывает домен 1 TNFR1 и ингибирует передачу сигнала через TNFR1, где пэгилированный мономер dAb связывает домен 1 TNFR1 с аффинностью, которая отличается от аффинности dAb в непэгилированной форме не более чем на множитель, составляющий примерно 1000, предпочтительно не более чем на множитель, составляющий примерно 100, более предпочтительно не более чем на множитель, составляющий примерно 10, или по существу с неизменной аффинностью относительно непэгилированной формы.
Лиганд по изобретению, который связывает домен 1 мембранно-связанной (трансмембранной) и растворимой форм TNFR1, но не ингибирует связывание TNF(с рецепторными формами, может быть полезен для блокирования домена 1 на мембранно-связанном рецепторе (например, для ингибирования кластеризации рецептора и/или ингибирования проведения сигнала), а также для связывания с растворимой формой для увеличения периода полувыведения, особенно при присоединении к ПЭГ, или как иначе описано выше. В предпочтительном воплощении лиганд не связывает домен 2 мембранно-связанной (трансмембраной) и растворимой форм TNFR1. В еще одном предпочтительном воплощении лиганд не связывает домен 3 мембранно-связанной (трансмембранной) и растворимой форм TNFR1. В еще одном предпочтительном воплощении лиганд не связывает домен 2 или домен 3 мембранно-связанной (трансмембранной) и растворимой форм TNFR1.
В некоторых воплощениях лиганд находится в IgG-подобном формате. Такие форматы имеют стандартную четырехцепочечную структуру молекулы IgG (2 тяжелые цепи и две легкие цепи), в которой одна или более вариабельных областей (VH и/или VL) заменены на dAb или единичный вариабельный домен, имеющий желаемую специфичность. Предпочтительно, каждая из вариабельных областей (2 области VH и 2 области VL) заменена на dAb или единичный вариабельный домен. dAb или единичный вариабельный домен(ы), которые включены в IgG-подобный формат, могут иметь одинаковую специфичность или разные специфичности. В некоторых воплощениях IgG-подобный формат является тетравалентным и может иметь одну, две, три или четыре специфичности. Например, IgG-подобный формат может быть моноспецифическим и включать 4 dAb, которые имеют одинаковую специфичность; биспецифическим и включать 3 dAb, которые имеют одинаковую специфичность, и другое dAb, которое имеет другую специфичность; биспецифическим и включать два dAb, которые имеют одинаковую специфичность, и два dAb, которые имеют общую, но отличающуюся специфичность; триспецифическим и включать первое и второе dAb, которые имеют одинаковую специфичность, третье dAb с другой специфичностью и четвертое dAb с другой специфичностью, отличающейся от первого, второго и третьего dAb; или тетраспецифическим и включать четыре dAb, каждое из которых имеет отличающуюся специфичность. Могут быть получены антиген-связывающие фрагменты IgG-подобных форматов (например, Fab, F(ab')2, Fab', Fv, scFv). Предпочтительно, IgG-подобные форматы или их антиген-связывающие фрагменты не связывают перекрестно TNFR1.
Лиганды по изобретению, включающие мономеры, димеры и тримеры dAb, могут быть связаны с областью антитела Fc, включающей один или оба домена CH2 и CH3, и возможно шарнирную область. Для получения таких полипептидов могут быть использованы, например, векторы, кодирующие лиганды, связанные в виде единичной нуклеотидной последовательности с областью Fc.
В изобретении также предложены димеры, тримеры и полимеры вышеупомянутых мономеров dAb в соответствии со следующими аспектами настоящего изобретения.
Лиганды по изобретению могут быть скомбинированы и/или принимать формат неиммуноглобулиновых мультилигандных структур с образованием поливалентных комплексов, которые связывают молекулы-мишени с одним и тем же антигеном, таким образом обеспечивая превосходную авидность. В некоторых воплощениях по меньшей мере один вариабельный домен связывает антиген для увеличения периода полувыведения мультимера. Например, природные бактериальные рецепторы, такие как SpA, были использованы в качестве каркасов для присоединения CDR с образованием лигандов, которые специфически связываются с одним или более чем одним эпитопом. Детали этого способа описаны в US 5831012. Другие подходящие каркасы включают каркасы, основанные на фибронектине, и аффитела. Детали подходящих способов описаны в WO 98/58965. Другие подходящие каркасы включают липокаллин и CTLA4, как описано в van den Beuken et al, J. Mol. Biol. (2001) 310, 591-601, и такие каркасы, как описано в WO 00/69907 (Medical Research Council), которые основаны, например, на кольцевой структуре бактериального GroEL или других шаперонных полипептидах.
Белковые каркасы могут быть скомбинированы; например, CDR могут быть пришиты к каркасу CTLA4 и использованы вместе с иммуноглобулиновыми доменами VH или VL с образованием лиганда. Аналогично, могут быть скомбинированы фибронектиновые, липокаллиновые и другие каркасы.
Лиганды с двойной специфичностью и полиспецифические лиганды
Авторы изобретения описали в находящейся на одновременном рассмотрении международной заявке на патент WO 03/002609, а также в находящейся на одновременном рассмотрении неопубликованной заявке на патент Великобритании 0230203.2, иммуноглобулиновые лиганды с двойной специфичностью, которые включают единичные вариабельные домены иммуноглобулина, каждый из которых имеет различную специфичность. Эти домены могут конкурировать друг с другом или независимо связываться с антигенами или эпитопами на молекулах-мишенях.
Лиганды с двойной специфичностью по настоящему изобретению предпочтительно включают комбинации доменов тяжелой и легкой цепи. Например, лиганд с двойной специфичностью может включать VH-домен и VL-домен, которые могут быть связаны вместе в форме scFv. Кроме того, лиганды могут включать один или более чем один домен CH или CL. Например, лиганды могут включать домен CH1, CH2 или CH3 и/или домен CL, домены Cµ1, Cµ2, Сµ3 или Сµ4 или любую их комбинацию. Домен шарнирной области также может быть включен. Такие комбинации доменов могут, например, имитировать природные антитела, такие как IgG или IgM, или их фрагменты, такие как молекулы Fv, scFv, Fab или F(ab')2. Предусматриваются и другие структуры, такие как единичное плечо молекулы IgG, включающее домены VH, VL, CH1 и CL.
В предпочтительном воплощении изобретения вариабельные области выбраны из генных репертуаров единичного домена V. Как правило, репертуар единичных доменов антитела представлен на поверхности филаментного бактериофага. В предпочтительном воплощении каждый домен единичного антитела выбирают путем связывания фагового репертуара с антигеном.
В предпочтительном воплощении изобретения каждый единичный вариабельный домен может быть выбран для связывания с его целевым антигеном или эпитопом в отсутствие комплементарной вариабельной области. В альтернативном воплощении единичные вариабельные домены могут быть выбраны для связывания с его целевым антигеном или эпитопом в присутствии комплементарной вариабельной области. Таким образом, первый единичный вариабельный домен может быть выбран в присутствии третьего комплементарного вариабельного домена, а второй вариабельный домен может быть выбран в присутствии четвертого комплементарного вариабельного домена. Третий или четвертый комплементарный вариабельный домен может представлять собой природный когнатный вариабельный домен, имеющий такую же специфичность как единичный тестируемый домен, или некогнатный комплементарный домен - такой как "пустой" вариабельный домен.
Предпочтительно, лиганд с двойной специфичностью по изобретению включает лишь два вариабельных домена, хотя несколько таких лигандов может быть включено вместе в один и тот же белок, например два таких лиганда могут быть включены в IgG или мультимерный иммуноглобулин, такой как IgM. Альтернативно, в еще одном воплощении множество лигандов с двойной специфичностью комбинируют с образованием мультимера. Например, два разных лиганда с двойной специфичностью комбинируют с получением тетраспецифической молекулы.
Специалисту в данной области будет понятно, что вариабельные области легких и тяжелых цепей лиганда с двойной специфичностью, получаемого с использованием способа по настоящему изобретению, могут находиться на одной и той же полипептидной цепи, или альтернативно, на разных полипептидных цепях. В случае, когда вариабельные области находятся на разных полипептидных цепях, они могут быть связаны через линкер, обычно гибкий линкер (такой как полипептидная цепь), химическую связывающую группу или любой другой способ, известный в данной области.
В первой конфигурации настоящего изобретения предложено дополнительное улучшение лигандов с двойной специфичностью, разработанное авторами настоящего изобретения, в которых одна специфичность лиганда направлена на белок или полипептид, присутствующий in vivo в организме, которое может действовать с целью увеличения периода полувыведения лиганда путем связывания с ним.
Соответственно, в первом аспекте предложен лиганд с двойной специфичностью, включающий первый единичный вариабельный домен иммуноглобулина, обладающий связывающей специфичностью в отношении первого антигена или эпитопа, и второй комплементарный единичный вариабельный домен иммуноглобулина, обладающий связывающей активностью в отношении второго антигена или эпитопа, где один или оба указанных антигена или эпитопа действуют для увеличения периода полувыведения лиганда in vivo, и где указанный первый и второй домены лишены взаимно комплементарных доменов, которые имеют одинаковую специфичность, при условии, что указанный лиганд с двойной специфичностью не состоит из VH-домена антитела против HSA и VK-домена против β-галактозидазы. Предпочтительно, чтобы ни первый, ни второй вариабельный домен не связывались с человеческим сывороточным альбумином (HSA).
Антигены или эпитопы, которые увеличивают период полувыведения лиганда, как описано в данном изобретении, предпочтительно присутствуют на белках или полипептидах, обнаруженных в организме in vivo. Примеры включают в себя белки внеклеточного матрикса, белки крови и белки, присутствующие в различных тканях в организме. Белки действуют для уменьшения скорости клиренса лиганда из крови, например, путем действия в качестве наполнителей, или путем заякоривания лиганда в желаемом сайте действия. Примеры антигенов/эпитопов, которые увеличивают период полувыведения in vivo, приведены в Приложении 1 ниже.
Увеличенный период полувыведения полезен в применениях иммуноглобулинов in vivo, в особенности антител и в особенности фрагментов антител, имеющих небольшой размер. Такие фрагменты (Fv, связанные дисульфидными связями Fv, Fab, scFv, dAb) страдают от быстрого клиренса из организма; таким образом, хотя они способны быстро достигать большинства частей организма и могут быть быстро получены и с ними легче обращаться, их применения in vivo ограничены лишь кратким их присутствием in vivo. Изобретение решает эту проблему путем обеспечения увеличенного периода полувыведения лигандов in vivo и, следовательно, более длительного поддержания в организме функциональной активности лиганда.
Способы фармакокинетического анализа и определения периода полувыведения лиганда хорошо известны специалистам в данной области. Детали можно найти в Kenneth, A et al: Chemical Stability of Phamnaceuticals: A Handbook for Pharmacists и в Peters et al, Pharmacokinetc analysis: A Practical Approach (1996). Также сделана ссылка на "Pharmacokinetics", M Gibaldi & D Perron, опубликованную Marcel Dekker, 2nd Rev. ex edition (1982), в котором описаны фармакокинетические параметры, такие как периоды полувыведения t-альфа и t-бета и площадь под кривой (AUC).
Периоды полувыведения (t1/2-альфа и t1/2-бета) и AUC могут быть определены из кривой зависимости концентрации лиганда в сыворотке крови от времени. Пакет для анализа WinNonlin (поставляется Pharsight Corp., Mountain View, CA94040, USA) может быть использован, например, для моделирования кривой. В первой фазе (альфа-фаза) лиганд подвергается в основном распределению в организме пациента с некоторым выведением. Вторая фаза (бета-фаза) представляет собой конечную фазу, когда лиганд распределен, и канцентрация в сыворотке крови уменьшается, поскольку лиганд подвергается клиренсу из организма пациента. Период полувыведения t-альфа представляет собой период полувыведения в первой фазе, а период полувыведения t-бета представляет собой период полувыведения во второй фазе. Таким образом, предпочтительно, в настоящем изобретении предложен лиганд или композиция, содержащая лиганд по изобретению, имеющая период полувыведения tα в диапазоне от 15 минут или более. В одном из воплощений нижняя граница диапазона составляет 30 минут, 45 минут, 1 час, 2 часа, 3 часа, 4 часа, 5 часов, 6 часов, 7 часов, 10 часов, 11 часов или 12 часов. Дополнительно или альтернативно, лиганд или композиция по изобретению будет иметь период полувыведения tα в диапазоне вплоть до и включая 12 часов. В одном из воплощений верхняя граница диапазона составляет 11, 10, 9, 8, 7, 6 или 5 часов. Пример подходящего диапазона составляет от 1 до 6 часов, от 2 до 5 часов или от 3 до 4 часов.
Предпочтительно, в настоящем изобретении предложен лиганд или композиция, содержащая лиганд по изобретению, имеющая период полувыведения tβ в диапазоне от 2,5 часов и более. В одном из воплощений нижняя граница диапазона составляет 3 часа, 4 часа, 5 часов, 6 часов, 7 часов, 10 часов, 11 часов или 12 часов. Дополнительно или альтернативно, лиганд или композиция по изобретению имеет период полувыведения tβ в диапазоне вплоть до и включая 21 сутки. В одном из воплощений верхняя граница диапазона составляет 12 часов, 24 часа, 2 суток, 3 суток, 5 суток, 10 суток, 15 суток или 20 суток. Предпочтительно, лиганд или композиция по изобретению имеет период полувыведения tβ в диапазоне от 12 до 60 часов. В другом воплощении он находится в диапазоне от 12 до 48 часов. В еще одном воплощении он находится в диапазоне от 12 до 26 часов.
Дополнительно или альтернативно к вышеприведенным критериям в настоящем изобретении предложен лиганд или композиция, содержащая лиганд по изобретению, имеющие значение AUC (площадь под кривой) в диапазоне от 1 мг·мин/мл или более. В одном из воплощений нижняя граница диапазона составляет 5, 10, 15, 20, 30, 100, 200 или 300 мг·мин/мл. Дополнительно или альтернативно лиганд или композиция по изобретению имеет AUC в диапазоне вплоть до 600 мг·мин/мл. В одном из воплощений верхняя граница диапазона составляет 500, 400, 300, 200, 150, 100, 75 или 50 мг·мин/мл. Предпочтительно, лиганд по изобретению имеет AUC в диапазоне, выбранном из группы, состоящей из следующих: от 15 до 150 мг·мин/мл, от 15 до 100 мг·мин/мл, от 15 до 75 мг·мин/мл и от 15 до 50 мг·мин/мл.
В первом воплощении лиганд с двойной специфичностью включает два комплементарных вариабельных домена, т.е. два вариабельных домена, которые в их природном окружении способны функционировать вместе в виде когнатной пары или группы, даже если в контексте настоящего изобретения они связываются раздельно с их когнатными эпитопами. Например, комплементарные вариабельные домены могут представлять собой тяжелую цепь иммуноглобулина и вариабельные домены легких цепей (VH и VL). Домены VH и VL предпочтительно обеспечиваются фрагментами scFv или Fab антитела. Вариабельные домены могут быть связаны вместе с образованием поливалентных лигандов, например, посредством: обеспечения шарнирной области на С-конце каждого V-домена и дисульфидного связывания между цистеинами в шарнирных областях; или обеспечения dAb, каждый из которых имеет цистеин на С-конце домена, причем эти цистеины связаны вместе дисульфидной связью; или продукцией V-CH и V-CL с получением формата Fab; или применения пептидных линкеров (например, Gly4Ser линкеров, обсуждаемых ниже) с получением димеров, тримеров и последующих мультимеров.
Авторы изобретения обнаружили, что применение комплементарных вариабельных доменов делает возможным совместную укладку поверхностей двух доменов и отделение от растворителя. Кроме того, комплементарные домены способны стабилизировать друг друга. Кроме того, оно позволяет создавать IgG антитела с двойной специфичностью без недостатков гибридных гибридом, используемых в предшествующем уровне техники, или необходимости конструировать тяжелые или легкие цепи на поверхностях контакта субъединиц. Лиганды с двойной специфичностью, в соответствии с первым аспектом настоящего изобретения, имеют по меньшей мере одну пару VH/VL. Биспецифические IgG по изобретению, таким образом, включают две такие пары, одна пара на каждом плече Y-образной молекулы. Поэтому в отличие от стандартных биспецифических антител или диа-антител, когда отношение используемых цепей является определяющим для успеха их получения и приводит к практическим сложностям, лиганды с двойной специфичностью по изобретению свободны от проблем баланса цепей. Дисбаланс цепей в стандартных биспецифических антителах возникает в результате ассоциации двух различных цепей VL с двумя различными цепями VH, где VL цепь 1 вместе с VH цепью 1 способна связываться с антигеном или эпитопом 1, и VL цепь 2 вместе с VH цепью 2 способна связываться с антигеном или эпитопом 2, и две корректные пары каким-то образом связаны друг с другом. Таким образом, только когда VL цепь 1 спарена с VH цепью 1, и VL цепь 2 спарена с VH цепью 2, в единой молекуле возникает биспецифичность. Такие биспецифические молекулы могут быть получены двумя различными путями. Во-первых, они могут быть получены путем ассоциации двух существующих пар VH/VL, каждая из которых связывается с различным антигеном или эпитопом (например, в биспецифическом IgG). В этом случае пары VH/VL должны вместе приводить к соотношению 1:1 для создания популяции молекул, все из которых являются биспецифическими. Этого никогда не наблюдается (даже когда комплементарный СН домен усиливается за счет конструирования по принципу "knobs into holes"), приводя к смеси биспецифических молекул и молекул, которые способны связываться только с одним антигеном или эпитопом, но не с другим. Второй путь создания биспецифического антитела осуществляют путем одновременной ассоциации двух различных цепей VH с двумя различными цепями VL (например, в биспецифическом диа-антителе). В этом случае, хотя существует тенденция в предпочтительном спаривании VL цепи 1 с VH цепью 1 и спаривании VL цепи 2 с VH цепью 2 (которое может усиливаться за счет конструирования VL и VH доменов по принципу "knobs into holes"), это спаривание никогда не достигается во всех молекулах, что приводит к смешанному препарату, в котором имеются некорректные пары, которые не способны связываться с антигеном или эпитопом.
Биспецифические антитела, сконструированные в соответствии с подходом с применением лиганда с двойной специфичностью, согласно первому аспекту настоящего изобретения позволяют преодолеть все эти проблемы, поскольку связывание с антигеном или эпитопом 1 происходит в домене VH или VL, а связывание с антигеном или эпитопом 2 происходит соответственно с комплементарным доменом VL или VH. Поскольку домены VH и VL спариваются в соотношении 1:1, все спаривания VH/VL являются биспецифическими, и, таким образом, все форматы, сконструированные с использованием этих спариваний VH/VL (Fv, scFv, Fab, мини-антитела, IgG и т.д.) имеют 100%-ную биспецифическую активность.
В контексте настоящего изобретения понятно, что первый и второй "эпитопы" представляют собой эпитопы, которые не являются одинаковыми и не связаны единым моноспецифическим лигандом. В первой конфигурации по изобретению они предпочтительно находятся на разных антигенах, один из них действует для увеличения периода полувыведения лиганда in vivo. Аналогично, первый и второй антигены предпочтительно не являются одинаковыми.
Лиганды с двойной специфичностью по изобретению не включают лиганды, описанные в WO 02/02773. Таким образом, лиганды по настоящему изобретению не включают комплементарные пары VH/VL, которые совместно связывают любой один или более антигенов или эпитопов. Наоборот, лиганды согласно первому аспекту изобретения включают комплементарную пару VH/VL, где домены V имеют различные специфичности. Кроме того, лиганды согласно первому аспекту изобретения включают комплементарные пары VH/VL, имеющие различные специфичности в отношении структурно неродственных эпитопов или антигенов. Структурно родственные эпитопы или антигены представляют собой эпитопы или антигены, которые имеют достаточное структурное сходство для того, чтобы связываться стандартной комплементарной парой VH/VL, которая действует совместным образом для связывания антигена или эпитопа; в случае структурно родственных эпитопов, эти эпитопы достаточно похожи по структуре в том, что они "подходят" к тому же связывающему карману, образованному в антиген-связывающем сайте димера VH/VL.
Во втором аспекте настоящего изобретения предложен лиганд, содержащий первый вариабельный домен иммуноглобулина, обладающий связывающей специфичностью в отношении первого антигена или эпитопа, и второй вариабельный домен иммуноглобулина, обладающий связывающей специфичностью в отношении второго антигена или эпитопа, где один или оба из указанных первых и вторых вариабельных доменов связываются с антигеном, что увеличивает период полувыведения лиганда in vivo, и вариабельные домены не комплементарны друг другу.
В одном из воплощений связывание через один вариабельный домен модулирует связывание лиганда через второй вариабельный домен.
В этом воплощении вариабельные домены могут паредставлять собой, например, пары доменов VH или пары доменов VL. Связывание антигена в первом сайте может модулировать, например усиливать или ингибировать, связывание антигена во втором сайте. Например, связывание в первом сайте по меньшей мере частично ингибирует связывание антигена во втором сайте. В таком воплощении лиганд может, например, поддерживаться в организме субъекта in vivo путем связывания с белком, который увеличивает период полувыведения лиганда до такого времени, когда он становится связанным со вторым антигеном-мишенью и диссоциирует из белка, увеличивающего период полувыведения.
Модуляцию связывания в указанном выше контексте достигают в виде структурной близости антиген-связывающих сайтов относительно друг друга. Такая структурная близость может быть достигнута за счет природы структурных компонентов, связывающих два или более антиген-связывающих сайтов, например путем обеспечения лиганда с относительно жесткой структурой, которая удерживает антиген-связывающие сайты в непосредственной близости. Предпочтительно, два или более антиген-связывающих сайтов находятся физически очень близко друг к другу, так что один сайт модулирует связывание антигена в другом сайте способом, который включает стерическое ограничение и/или конформационные изменения в молекуле иммуноглобулина.
Первый и второй антиген-связывающие домены могут быть связаны ковалентно или нековалентно. В случае, когда домены связаны ковалентно, то связывание может опосредоваться, например дисульфидными связями или полипептидным линкером, таким как (Gly4Ser)n, где n=от 1 до 8, например 2, 3, 4, 5 или 7.
В случае когда вариабельные домены выбраны из репертуаров V-гена, выбранных, например, с использованием технологии фагового дисплея, как описано в данном изобретении, тогда эти вариабельные домены могут включать универсальную каркасную область, так что они могут распознаваться специфическим общим лигандом, как определено в данном изобретении. Применение универсальных каркасов, общих лигандов и т.п. описано в WO 99/20749. В настоящем изобретении ссылка на фаговый дисплей включает применение фага и/или фагмид.
Когда используют репертуары V-генов, тогда вариация полипептидной последовательности предпочтительно располагается в структурных петлях вариабельных доменов. Полипептидные последовательности любого вариабельного домена могут быть изменены путем перестройки ДНК или с помощью мутации для усиления взаимодействия каждого вариабельного домена с его комплементарной парой. Перестройка ДНК известна в данной области и изложена, например в Stemmer, Nature 370:389-391 (1994) и патенте США №6297053, которые оба включены путем ссылки. Другие способы мутагенеза хорошо известны специалистам в данной области.
В предпочтительном воплощении изобретения 'лиганд с двойной специфичностью' представляет собой одноцепочечный Fv-фрагмент.В альтернативном воплощении изобретения 'лиганд с двойной специфичностью' состоит из Fab-области антитела. Термин "Fab-область" включает Fab-подобную область, в которой использованы два VH-домена или два VL-домена.
Вариабельные области могут быть получены из антител, направленных против целевых антигенов или эпитопов. Альтернативно, они могут быть получены из репертуара единичных доменов антител, таких как те, которые экспрессируются на поверхности филаментного бактериофага. Селекцию можно осуществлять, как описано ниже.
В еще одном аспекте настоящего изобретения предложена одна или более чем одна молекула нуклеиновой кислоты, кодирующая по меньшей мере лиганд с двойной специфичностью, как определено в данном изобретении. Лиганд с двойной специфичностью может кодироваться на одной молекуле нуклеиновой кислоты; альтернативно, каждый домен может кодироваться разными молекулами нуклеиновых кислот. Когда лиганд кодируется одной молекулой нуклеиновой кислоты, тогда домены могут экспрессироваться в виде слитого полипептида, в виде scFv молекулы, или могут экспрессироваться раздельно и затем связываться вместе, например с использованием химических сшивающих агентов. Лиганды, экспрессирующиеся из отдельных нуклеиновых кислот, сшиваются вместе подходящими способами.
Нуклеиновая кислота, кроме того, может кодировать сигнальную последовательность для экспорта полипептидов из клетки-хозяина при экспрессии и может быть слита с поверхностным компонентом филаментной частицы бактериофага (или другим компонентом системы селективного дисплея) после экспрессии.
В еще одном аспекте настоящего изобретения предложен вектор, содержащий нуклеиновую кислоту, кодирующую лиганд с двойной специфичностью по настоящему изобретению.
В еще одном аспекте настоящего изобретения предложена клетка-хозяин, трансфицированная вектором, кодирующим лиганд с двойной специфичностью по настоящему изобретению.
Экспрессия с такого вектора может иметь конфигурацию для получения, например, на поверхности частицы бактериофага, вариабельных доменов для селекции. Это дает возможность для селекции представленных вариабельных областей и таким образом для селекции 'лигандов с двойной специфичностью' с использованием способа по настоящему изобретению.
В настоящем изобретении дополнительно предложен набор, включающий по меньшей мере лиганд с двойной специфичностью по настоящему изобретению.
В третьем аспекте изобретения предложен способ получения лиганда, содержащего первый единичный вариабельный домен иммуноглобулина, имеющий первую связывающую специфичность, и второй единичный вариабельный домен иммуноглобулина, имеющий вторую (отличающуюся) связывающую специфичность, причем одна или обе связывающие специфичности являются специфичными в отношении антигена, который увеличивает период полувыведения лиганда in vivo, включающий стадии:
а) селекции первого вариабельного домена по его способности связываться с первым эпитопом,
б) селекции второй вариабельной области по ее способности связываться со вторым эпитопом,
в) комбинирования вариабельных доменов; и
г) селекции лиганда по его способности связываться с указанным первым эпитопом и с указанным вторым эпитопом.
Лиганд может связываться с первым и вторым эпитопами либо одновременно, либо когда существует конкуренция между связывающими доменами за связывание с эпитопом, тогда связывание одного домена может препятствовать связыванию другого домена с когнатным эпитопом. Таким образом, в одном из воплощений стадия (г) выше требует одновременного связывания с обоими первым и вторым (и возможно дополнительным) эпитопами; в еще одном воплощении связывание с первым и вторым эпитопами происходит не одновременно.
Эпитопы предпочтительно находятся на отдельных антигенах.
Лиганды предпочтительно включают комбинации VH/VL, или комбинации VH/VH, или VL/VL вариабельных доменов иимуноглобулина, как описано выше. Лиганды могут также включать Camelid домены VHH, при условии, что VHH-домен, специфичный в отношении антигена, который увеличивает период полувыведения лиганда in vivo, не связывает лизоцим белка куриного яйца (HEL), панкреатическую альфа-амилазу свиньи или NmC-A; hcg, BSA-связанный азокраситель RR6 или клетки HG982 S. mutans, как описано в Conrath et al, (2001) JBC 276:7346-7350 и W099/23221, ни один из которых не описывает применение специфичности в отношении антигена, который увеличивает период полувыведения, для увеличения периода полувыведения лиганда in vivo.
В одном из воплощений указанный первый вариабельный домен выбран для связывания с указанным первым эпитопом в отсутствие комплементарного вариабельного домена. В еще одном воплощении указанный первый вариабельный домен выбран для связывания с указанным первым эпитопом/антигеном в присутствии третьего вариабельного домена, в котором указанный третий вариабельный домен отличается от указанного второго вариабельного домена и комплементарен первому домену. Аналогично, второй домен может быть выбран в отсутствие или в присутствии комплементарного вариабельного домена.
Антигены или эпитопы, являющиеся мишенями для лигандов по изобретению, дополнительно к белку, увеличивающему период полувыведения, могут представлять собой любой антиген или эпитоп, но предпочтительно представляет собой антиген или эпитоп, который является мишенью с терапевтической пользой. В изобретении предложены лиганды, включающие лиганды с открытой конформацией, закрытой конформацией и лиганды на основе выделенного мономера dAb, специфичные в отношении любой такой мишени, в частности мишеней, дополнительно идентифицированных в данном изобретении. Такие мишени могут представлять собой полипептиды, белки или нуклеиновые кислоты, которые могут быть природными или синтетическими, или являются их частью. В этом отношении лиганд по изобретению может связывать эпитоп или антиген и действовать в качестве антагониста или агониста (например, агонист рецептора ЕРО). Специалисту в данной области понятно, что выбор огромен и варьирует.Они могут представлять собой, например человеческие или животные белки, цитокины, рецепторы цитокинов, ферменты, кофакторы ферментов или ДНК-связывающие белки. Подходящие цитокины и факторы роста включают: АроЕ (аполипопротеин Е), Apo-SAA, BDNF (нейротрофический фактор головного мозга), кардиотрофин-1, EGF (эпидермальный фактор роста), рецептор EGF, ENA-78, эотаксин, эотаксин-2, Exodus-2, EpoR, FGF-кислый (фактор роста фибробластов), FGF-основный, фактор роста фибробластов-10, лиганд FLT3, фракталкин (СХЗС), GDNF (нейротрофический фактор глиальных клеток), G-CSF (гранулоцитарный колониестимулирующий фактор), GM-CSF (колониестимулирующий фактор гранулоцитов и макрофагов), GF-β1, инсулин, IFN-γ, IGF-I (инсулиноподобный фактор роста), IGF-II, IL-1α, IL-1β, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8 (72 аминокислоты), IL-8 (77 аминокислот), IL-9, IL-10, IL-11, IL-12, IL-13, IL-15, IL-16, IL-17, IL-18 (IGIF), ингибин α, ингибин β, IP-10, KGF-2 (фактор роста кератиноцитов-2), KGF, лептин, LIF, лимфотактин, ингибитор Мюллера, моноцитарный колониеингибирующий фактор, белок-аттрактант моноцитов, М-CSF, MDC (67 аминокислот), MDC (69 аминокислот), МСР-1 (моноцитарный белок-хемоаттрактант) (MCAF (хемотактический и активирующий фактор моноцитов)), МСР-2, МСР-3, МСР-4, MDC (67 аминокислот), MDC (69 аминокислот), MIG, MIP-1α, MIP-1β, MIP-3α, MIP-3β, MDP-4, фактор-ингибитор миелоидных предшественников-1 (MPIF-1), NAP-2 (нейтрофил-активирующий белок), нейртурин, фактор роста нервов, β-NGF, NT-3, NT-4, онкостатин М, тромбоцитарный фактор роста PDGF-AA, PDGF-AB, PDGF-BB, тромбоцитарный фактор PF-4, RANTES, SDF1α (фактор стромальных клеток), SDF1β, SCF, фактор роста стволовых клеток SCGF, фактор стволовых клеток (SCF), TARC (хемокины, регулируемые тимусом и активацией), TGF-α (трансформирующий фактор роста), TGF-β, TGF-β2, TGF-β3, TNF (фактор некроза опухоли), TNF-α, TNF-β, рецептор TNF I, рецептор TNF II. TNIL-1, ТРО, VEGF (фактор роста эндотелия сосудов), рецептор VEGF 1, рецептор VEGF 2, рецептор VEGF 3, GCP-2 (хемотактический белок гранулоцитов), GRO/MGSA, GRO-β, GRO-γ, НСС1, 1-309, HER 1, HER 2, HER 3 и HER 4, но не ограничиваются ими.
Рецепторы цитокинов включают рецепторы вышеуказанных цитокинов. Понятно, что этот перечень никоим образом не является исчерпывающим.
В одном из воплощений изобретения вариабельные домены получены из соответствующего антитела, направленного против антигена или эпитопа. В предпочтительном воплощении вариабельные домены получены из репертуара единичных вариабельных доменов антитела.
Во второй конфигурации в настоящем изобретении предложены полиспецифические лиганды. По настоящему изобретению термин "полиспецифический лиганд" относится к лиганду, который обладает более чем одной эпитоп-связывающей специфичностью, как определено в данном изобретении. Как правило, полиспецифический лиганд включает два или более эпитоп-связывающих доменов.
Эпитоп-связывающие домены по настоящему изобретению включают белковый каркас и сайты взаимодействия с эпитопом (которые предпочтительно находятся на поверхности белкового каркаса). Согласно настоящему изобретению предпочтительно, чтобы каждый эпитоп-связывающий домен имел отличающуюся эпитоп-связывающую специфичность.
В контексте настоящего изобретения понятно, что первый и второй "эпитопы" представляют собой эпитопы, которые не являются одинаковыми и не связаны единым моноспецифическим лигандом. Они могут располагаться на различных антигенах или на одном и том же антигене, но разделены достаточным расстоянием, так что они не образуют единую структуру, которая могла бы быть связана единой моноспецифической связывающей парой VH/VL стандартного антитела. Экспериментально, если оба индивидуальных вариабельных домена в форме одноцепочечного антитела (доменные антитела или dAb) отдельно конкурируют с моноспецифическим лигандом VH/VL против двух эпитопов, то эти два эпитопа не достаточно удалены для того, чтобы рассматриваться как отдельные эпитопы согласно настоящему изобретению.
В соответствии с настоящим изобретением предпочтительно, чтобы каждый эпитоп-связывающий домен включал вариабельный домен иммуноглобулина. Более предпочтительно, чтобы каждый вариабельный домен иммуноглобулина представлял собой вариабельный домен легкой цепи (VL) или вариабельный домен тяжелой цепи VH. Во второй конфигурации настоящего изобретения домены иммуноглобулина, представленные на лиганде по настоящему изобретению, не комплементарны, то есть они не ассоциируются с образованием VH/VL антиген-связывающего сайта. Таким образом, полиспецифические лиганды, как определено во второй конфигурации изобретения, включают иммуноглобулиновые домены того же самого подтипа, то есть либо вариабельные домены легкой цепи (VL), либо вариабельные домены тяжелой цепи (VH). Кроме того, когда лиганд по изобретению находится в закрытой конформации, иммуноглобулиновые домены могут иметь VHH Camelid типа.
В альтернативном воплощении лиганд(ы) по изобретению не включают Camelid Vнн-домен. Более конкретно, лиганд(ы) по изобретению не включают один или более чем один аминокислотный остаток, специфичный в отношении Camelid VHH-доменов по сравнению с человеческими VH-доменами.
Предпочтительно, единичные вариабельные домены получены из антитела, выбранного для связывающей активности против различных антигенов или эпитопов. Например, вариабельные домены могут быть выделены по меньшей мере частично путем иммунизации людей. В данной области известны альтернативные способы, включающие выделение из библиотек человеческих антител и синтез генов искусственных антител.
Вариабельные домены предпочтительно связывают суперантигены, такие как белок А или белок L. Связывание с суперантигенами представляет собой свойство правильно свернутых вариабельных доменов антител и позволяет выделить такие домены, например, из библиотек рекомбинантных или мутантных доменов.
Эпитоп-связывающие домены также могут быть основаны на белковых каркасах или скелетах, отличающихся от иммуноглобулиновых доменов. Например, природные бактериальные рецепторы, такие как SpA, используют в качестве каркасов для присоединения CDR с получением лигандов, которые специфически связываются с одним или более эпитопами. Детали этого способа описаны в US 5831012. Другие подходящие каркасы включают каркасы на основе фибронектина и аффител. Детали подходящих способов описаны в WO 98/58965. Другие подходящие каркасы включают липокаллин и CTLA4, как описано в van den Beuken etal, J. Mol. Biol. (2001) 310, 591-601, и каркасы, такие как описано в WO 0069907 (Medical Research Council), которые основаны, например, на кольцевой структуре бактериального GroEL или других шаперонных полипептидов.
Белковые каркасы можно комбинировать; например CDR можно присоединять к каркасу CTLA4 и использовать вместе с иммуноглобулиновыми доменами VH или VL с образованием поливалентного лиганда. Аналогично, можно комбинировать фибронектин, липокаллин и другие каркасы.
В одном из воплощений полиспецифический лиганд включает эпитоп-связывающий домен, который включает CDR единичного иммуноглобулинового домена (dAb), который связывает описанный в данном изобретении TNFR1, присоединенный к подходящему белковому каркасу или скелету.
Специалисту в данной области понятно, что эпитоп-связывающие домены полиспецифического лиганда с закрытой конформацией, полученного с использованием способа по настоящему изобретению, могут находиться на одной и той же полипептидной цепи или, альтернативно, на разных полипептидных цепях. В случае, когда вариабельные области располагаются на разных полипептидных цепях, они могут быть связаны через линкер, предпочтительно гибкий линкер (такой как полипептидная цепь), химическая связывающая группа, или любым другим способом, известным в данной области.
Первый и второй эпитоп-связывающие домены могут быть ассоциированы ковалентно или нековалентно. В случае, когда домены ассоциированы ковалентно, ассоциация может быть опосредована, например, дисульфидными связями.
В некоторых воплощениях второй конфигурации по изобретению эпитопы могут замещать друг друга при связывании. Например, первый эпитоп может быть представлен на антигене, который после связывания с его когнатным первым связывающим доменом, вызывает стерическое ограничение для второго связывающего домена или конформационное изменение в нем, которое замещает эпитоп, связанный со вторым связывающим доменом.
Предпочтительно, связывание уменьшается на 25% или более, предпочтительно на 40%, 50%, 60%, 70%, 80%, 90% или, и предпочтительно вплоть до 100% или около 100%, так что связывание полностью ингибируются. Связывание эпитопов можно измерить с помощью стандартных анализов связывания антигена, таких как ИФА, флуоресцентные методы, включая FRET (резонансный перенос энергии флуоресценции), или способов, таких как поверхностный плазменный резонанс, который измеряет массу молекул.
Кроме того, в изобретении предложен полиспецифический лиганд с закрытой конформацией, включающий первый эпитоп-связывающий домен, обладающий первой эпитоп-связывающей специфичностью, и некомплементарный второй эпитоп-связывающий домен, обладающий второй эпитоп-связывающей специфичностью, где первая и вторая связывающие специфичности конкурируют за связывание с эпитопом, так что полиспецифический лиганд с закрытой конформацией может не связывать оба эпитопа одновременно.
Полиспецифические лиганды с закрытой конформацией по изобретению не включают лиганды, описанные в WO 02/02773. Таким образом, лиганды по настоящему изобретению не включают комплементарные пары VH/VL, которые кооперативно связывают любой один или более антигенов или эпитопов. Вместо этого, лиганды по изобретению предпочтительно включают некомплементарные пары VH-VH или VL-VL. Предпочтительно, каждый домен VH или VL в каждой паре VH-VH или VL-VL имеет разную эпитоп-связывающую специфичность, и сайты связывания эпитопов расположены таким образом, что связывание эпитопа в одном сайте конкурирует со связыванием эпитопа в другом сайте.
Предпочтительно, полиспецифический лиганд с закрытой конформацией может включать первый домен, способный связываться с молекулой-мишенью, и второй домен, способный связываться с молекулой или группой, которая увеличивает период полувыведения лиганда. Например, молекула или группа может представлять собой объемный агент, такой как HSA или белок клеточного матрикса. Используемая в данном изобретении фраза "молекула или группа, которая увеличивает период полувыведения лиганда" относится к молекуле или химической группе, которая после связывания с лигандом с двойной специфичностью, как описано в данном изобретении, увеличивает период полувыведения in vivo такого лиганда с двойной специфичностью при введении животному относительно лиганда, который не связывает эту молекулу или группу. Примеры молекул или групп, которые увеличивают период полувыведения лиганда, описаны ниже. В предпочтительном воплощении полиспецифический лиганд с закрытой конформацией может связывать молекулу-мишень только после замещения молекулы или группы, увеличивающей период полувыведения. Таким образом, например, полиспецифический лиганд с закрытой конформацией поддерживается в системе кровообращения субъекта с помощью объемной молекулы, такой как HSA. При столкновении с молекулой-мишенью конкуренция между связывающими доменами полиспецифического лиганда с закрытой конформацией приводит в результате к замещению HSA и связыванию с мишенью.
В предпочтительном воплощении второй конфигурации по изобретению эпитоп-связывающие домены представляют собой вариабельные области иммуноглобулина и выбраны из генных репертуаров единичного домена V. Как правило, репертуар единичных доменов антитела представлен на поверхности филаментного бактериофага. В предпочтительном воплощении каждый единичный домен антитела выбран путем связывания фагового репертуара с антигеном. В предпочтительном воплощении второй конфигурации по изобретению эпитоп-связывающие домены представляют собой вариабельные области иммуноглобулина и выбраны из генных репертуаров единичного домена V. Как правило, репертуар единичных доменов антитела представлен на поверхности филаментного бактериофага. В предпочтительном воплощении каждый единичный домен антитела выбран путем связывания фагового репертуара с антигеном.
В одном из аспектов полиспецифический лиганд включает по меньшей мере два некомплементарных вариабельных домена. Например, лиганды могут включать пару VH-доменов или пару VL-доменов. Предпочтительно, домены происходят не из Camelid; предпочтительно они представляют собой человеческие домены или включают человеческие каркасные области (FW) и одну или более гетерологичных CDR. CDR и каркасные области представляют собой области вариабельного домена иммуноглобулина, как определено в базе данных последовательностей белков, представляющих иммунологический интерес (Kabat база данных).
Предпочтительные человеческие каркасные области представляют собой области, которые кодируются герминальными генными сегментами DP47 и DPK9. Предпочтительно, FW1, FW2 и FW3 в домене VH или VL имеют последовательность FW1, FW2 или FW3 из DP47 или DPK9. Человеческие каркасные области могут содержать мутации, например до примерно 5 аминокислотных замен или до примерно 10 аминокислотных замен вместе в человеческих каркасных областях, используемых в лигандах по изобретению.
Вариабельные домены в полиспецифических лигандах в соответствии со второй конфигурацией по изобретению могут быть расположены в открытой или закрытой конформации; то есть, они могут быть расположены таким образом, что вариабельные домены могут связывать их когнатные лиганды независимо и одновременно, или таким образом, что только один из вариабельных доменов может связывать свой когнатный лиганд в любой момент времени.
Авторы изобретения понимают, что при некоторых структурных условиях некомплементарные вариабельные домены (например, два вариабельных домена легких цепей или два вариабельных домена тяжелых цепей) могут быть представлены в лиганде, так что связывание первого эпитопа с первым вариабельным доменом ингибирует связывание второго эпитопа со вторым вариабельным доменом, даже если такие некомплементарные домены не функционируют вместе как когнатная пара.
Предпочтительно, лиганд включает две или более пар вариабельных доменов; то есть он включает по меньшей мере четыре вариабельных домена. Предпочтительно, четыре вариабельных домена включают каркасные области человеческого происхождения.
В предпочтительном воплощении каркасные области человеческих антител идентичны каркасным областям человеческих герминальных последовательностей.
Авторы настоящего изобретения считают, что такие антитела имеют конкретное применение в анализах связывания лигандов для терапевтического и других применений.
В одном из воплощений второй конфигурации по изобретению вариабельные домены получены из антитела, направленного против первого и/или второго антигена или эпитопа. В предпочтительном воплощении вариабельные домены получены из репертуара единичных вариабельных доменов антител. В одном из примеров репертуар представляет собой репертуар, который не образуется в организме животного, или синтетический репертуар. В другом примере единичные вариабельные домены не выделены (по меньшей мере частично) путем иммунизации животного. Таким образом, единичные домены могут быть выделены из наивной библиотеки.
Вторая конфигурация по изобретению в еще одном аспекте предлагает полиспецифический лиганд, содержащий первый эпитоп-связывающий домен, обладающий первой эпитоп-связывающей специфичностью, и некомплементарный второй эпитоп-связывающий домен, обладающий второй эпитоп-связывающей специфичностью. Первая и вторая связывающие специфичности могут быть одинаковыми или разными.
В еще одном аспекте настоящего изобретения предложен полиспецифический лиганд с закрытой конформацией, включающий первый эпитоп-связывающий домен, обладающий первой эпитоп-связывающей специфичностью, и некомплементарный второй эпитоп-связывающий домен, обладающий второй эпитоп-связывающей специфичностью, где первая и вторая связывающие специфичности способны конкурировать за связывание эпитопа, так что полиспецифический лиганд с закрытой конформацией не может связывать оба эпитопа одновременно.
В еще одном аспекте изобретения предложены лиганды с открытой конформацией, включающие некомплементарные связывающие домены, где домены являются специфичными в отношении различных эпитопов на той же самой мишени. Такие лиганды связываются с мишенями с повышенной авидностью. Аналогично, в изобретении предложены поливалентные лиганды, включающие некомплементарные связывающие домены, специфичные в отношении одного и того же эпитопа и направленные в отношении мишеней, которые включают множественные копии указанного эпитопа, таких как IL-5, PDGF-AA, PDGF-BB, TGF-бета, TGF-бета 2, TGF-бета 3 и TNFα, например человеческий рецептор TNF 1 и человеческий TNFα.
В аналогичном аспекте лиганды по изобретению могут быть подвергнуты конфигурации для связывания с индивидуальными эпитопами с низкой аффинностью, так что связывание с индивидуальными эпитопами не является терапевтически значимым; но повышенная авидность, возникающая в результате связывания с двумя эпитопами, обеспечивает терапевтическую пользу. В конкретном примере в качестве мишеней могут выступать эпитопы, представленные индивидуально на нормальных типах клеток, но представленные вместе только на аномальных типах клеток или клетках, пораженных заболеванием, таких как опухолевые клетки. В такой ситуации только аномальные или пораженные заболеванием клетки являются эффективными мишенями для биспецифических лигандов по изобретению.
Лиганд, специфичный в отношении множества копий одного и того же эпитопа или соседних эпитопов на той же самой мишени (известный как хелатообразующие dAb) также может быть тримерным или полимерным (тетрамерным или более чем тетрамерным) лигандом, включающим три, четыре или более четырех некомплементарных связывающих доменов. Например, могут быть сконструированы лиганды, включающие три или четыре VH-домена или VL-домена.
Кроме того, предложены лиганды, которые связываются с мультисубъединичными мишенями, где каждый связывающий домен является специфичным в отношении субъединицы указанной мишени. Лиганд может быть димерным, тримерным или полимерным.
Предпочтительно, полиспецифические лиганды в соответствии с вышеизложенными аспектами изобретения могут быть получены с использованием способа в соответствии с первым аспектом изобретения.
В соответствии с вышеизложенным первым аспектом второй конфигурации по изобретению, предпочтительно первый эпитоп-связывающий домен и второй эпитоп-связывающий домен представляют собой некомплементарные вариабельные домены иимуноглобулина, как определено в данном изобретении. То есть вариабельные домены VH-VH или VL-VL.
Хелатообразующие dAb, в частности, могут быть получены в соответствии с предпочтительным аспектом изобретения, а именно путем применения заякоривающих dAb, при котором конструируют библиотеку димерных, тримерных или мультимерных dAb с использованием вектора, который содержит константную область dAb выше или ниже линкерной последовательности, где репертуар второго, третьего и других dAb встроен в другой конец линкера. Например, заякоривающие или направляющие dAb могут представлять собой TAR1-5 (VK), TAR1-27 (VK), TAR2h-5 (VH) или TAR2h-6 (VK).
В альтернативных методологиях применения линкеров можно избежать, например путем использования нековалентного связывания или природной аффинности между связывающими доменами, такими как VH и VK. В изобретении соответственно предложен способ получения хелатообразующего мультимерного лиганда, включающий стадии:
(а) получения вектора, содержащего последовательность нуклеиновой кислоты, кодирующей единичный связывающий домен, специфичный в отношении первого эпитопа на мишени;
(б) получения вектора, кодирующего репертуар, включающий вторые связывающие домены, специфичные в отношении второго эпитопа на указанной мишени, где эпитоп может быть тем же самым или отличающимся от первого эпитопа, причем указанный второй эпитоп является соседним по отношению к указанному первому эпитопу; и
(в) экспрессии указанных первого и второго связывающих доменов; и
(г) выделения этих комбинаций первого и второго связывающих доменов, которые комбинируют вместе с получением димера, связывающего мишень.
Первый и второй эпитопы являются соседними, так что мультимерный лиганд способен связываться с обоими эпитопами одновременно. Это позволяет получить лиганд, обладающий преимуществами повышенной авидности при связывании. Когда эпитопы являются одинаковыми, добиваются повышенной авидности с помощью присутствия множества копий эпитоп на мишени, давая возможность для одновременного связывания по меньшей мере двум копиям для достижения эффекта повышенной авидности.
Связывающие домены могут быть ассоциированы несколькими способами, а также с использованием линкеров. Например, связывающие домены могут включать остатки цистеина, авидиновые и стрептавидиновые группы или другие средства для нековалентного присоединения после синтеза; выделяют такие комбинации, которые эффективно связываются с мишенью. Альтернативно, линкер может присутствовать между первым и вторым связывающими доменами, которые экспрессируются в виде единичного полипептида из единичного вектора, который содержит первый связывающий домен, линкер и репертуар вторых связывающих доменов, например, как описано выше.
В предпочтительном аспекте первый и второй связывающие домены в природе ассоциируются при связывании с антигеном; например домены VH и VK, когда они связаны с соседними эпитопами, в природе ассоциируются путем трехстороннего взаимодействия с образованием стабильного димера. Такие ассоциированные белки могут быть выделены в анализе связывания мишени. Преимущество этого способа заключается в том, что только связывающие домены, которые связываются с близкими соседними эпитопами, находящимися в правильной конформации, будут ассоциировать и, таким образом, будут выделены в результате их повышенной авидности в отношении мишени.
В альтернативном воплощении вышеприведенного аспекта второй конфигурации по изобретению, по меньшей мере один эпитоп-связывающий домен включает неиммуноглобулиновый "белковый каркас" или "белковый скелет", как определено в данном изобретении. Подходящие неиммуноглобулиновые белковые каркасы включают каркасы, выбранные из группы, состоящей из: SpA, фибронектина, GroEL и других шаперонов, липокаллина, CCTLA4 и аффител, как указано выше, но не ограничиваются ими.
В соответствии с вышеприведенным аспектом второй конфигурации по изобретению предпочтительно, чтобы эпитоп-связывающие домены были присоединены к белковому скелету. Предпочтительно, белковый скелет по изобретению представляет собой иммуноглобулиновый скелет.
Согласно настоящему изобретению термин "иммуноглобулиновый скелет" относится к белку, который включает по меньшей мере одну иммуноглобулиновую складку и который действует в качестве ядра для одного или более чем одного эпитоп-связывающего домена, как определено в данном изобретении.
Предпочтительные иммуноглобулиновые скелеты, как определено в данном изобретении, включают любой один или более чем один из следующих: молекула иммуноглобулина, включающая по меньшей мере (1) CL-домен антитела (подкласс каппа или лямбда); или (2) СН1-домен тяжелой цепи антитела; молекула иммуноглобулина, включающая СН1- и СН2-домены тяжелой цепи антитела; молекула иммуноглобулина, включающая СН1-, СН2- и СН3-домены тяжелой цепи антитела; или любой из подгруппы (2) в комбинации с CL-доменом (подкласс каппа или лямбда) антитела. Также может быть включен домен шарнирной области. Такие комбинации доменов могут, например, имитировать природные антитела, такие как IgG или IgM, или их фрагменты, такие как молекулы Fv, scFv, Fab или F(ab')2. Специалист в данной области понимает, что этот перечень не следует рассматривать как исчерпывающий.
Связывание скелета с определенными в данном изобретении эпитоп-связывающими доменами может быть достигнуто на полипептидном уровне, то есть после экспрессии нуклеиновой кислоты, кодирующей скелет и/или эпитоп-связывающие домены. Альтернативно, стадия связывания может быть осуществлена на уровне нуклеиновой кислоты. Способы связывания белкового скелета по настоящему изобретению с одним или более чем одним эпитоп-связывающим доменом включают применение способов белковой химии и/или молекулярной биологии, которые знакомы специалистам в данной области и описаны в данном изобретении.
Предпочтительно, полиспецифический лиганд с закрытой конформацией может включать первый домен, способный связывать молекулу-мишень, и второй домен, способный связывать молекулу или группу, увеличивающую период полувыведения лиганда. Например, молекула или группа может представлять собой объемный агент, такой как HSA или белок клеточного матрикса. Используемая в данном изобретении фраза "молекула или группа, увеличивающая период полувыведения лиганда" относится к молекуле или химической группе, которая при связывании с лигандом с двойной специфичностью, как описано в данном изобретении, увеличивает период полувыведения in vivo такого лиганда с двойной специфичностью при введении животному относительно лиганда, который не связывает данную молекулу или группу. Примеры молекул или групп, которые увеличивают период полувыведения лиганда, описаны ниже. В предпочтительном воплощении полиспецифический лиганд с закрытой конформацией может связывать молекулу-мишень только после замещения молекулы или группы, увеличивающей период полувыведения. Таким образом, например, полиспецифический лиганд с закрытой конформацией поддерживается в системе кровообращения субъекта с помощью объемной молекулы, такой как HSA. При столкновении с молекулой-мишенью конкуренция между связывающими доменами полиспецифического лиганда с закрытой конформацией приводит в результате к замещению HSA и связыванию мишени.
В еще одном аспекте второй конфигурации по изобретению в настоящем изобретении предложена одна или более чем одна молекула нуклеиновой кислоты, кодирующая по меньшей мере полиспецифический лиганд, как определено в данном изобретении. В одном из воплощений лиганд представляет собой лиганд с закрытой конформацией. В еще одном воплощении он представляет собой лиганд с открытой конформацией. Полиспецифический лиганд может кодироваться на одной молекуле нуклеиновой кислоты; альтернативно, каждый эпитоп-связывающий домен может кодироваться отдельной молекулой нуклеиновой кислоты. Когда лиганд кодируется одной молекулой нуклеиновой кислоты, тогда домены могут экспрессироваться в виде слитого полипептида или могут экспрессироваться раздельно и затем связываться вместе, например, с использованием химических связывающих агентов. Лиганды, экспрессирующиеся с различных нуклеиновых кислот, связаны вместе с использованием подходящих средств.
Нуклеиновая кислота может дополнительно кодировать сигнальную последовательность для экспорта полипептидов из клетки-хозяина после экспрессии и может быть слита с поверхностным компонентом частицы филаментного бактериофага (или другого компонента дисплейной системы для селекции) после экспрессии. Лидерные последовательности, которые могут быть использованы в бактериальной экспрессии и/или фаговом или фагмидном дисплее, включают pelB, stll, ompA, phoA, bla и pelA.
В еще одном аспекте второй конфигурации по изобретению в настоящем изобретении предложен вектор, содержащий нуклеиновую кислоту по настоящему изобретению.
В еще одном аспекте настоящего изобретения предложена клетка-хозяин, трансфицированная вектором по настоящему изобретению.
Экспрессия с такого вектора может быть сконфигурирована для получения, например, на поверхности частицы бактериофага эпитоп-связывающих доменов для селекции. Это дает возможность для селекции представленных доменов и таким образом для селекции 'полиспецифических лигандов' с использованием способа по настоящему изобретению.
В настоящем изобретении дополнительно предложен набор, включающий по меньшей мере полиспецифический лиганд по настоящему изобретению, который может представлять собой лиганд с открытой конформацией или закрытой конформацией. Наборы по изобретению могут представлять собой, например, диагностические наборы, терапевтические наборы, наборы для обнаружения химических или биологических видов и т.п.
В настоящем изобретении предложен способ получения полиспецифического лиганда, включающий стадии:
а) селекции первого эпитоп-связывающего домена по его способности связываться с первым эпитопом,
б) селекции второго эпитоп-связывающего домена по его способности связываться со вторым эпитопом,
в) комбинирования эпитоп-связывающих доменов; и
г) селекции полиспецифического лиганда с закрытой конформацией по его способности связываться с указанным первым вторым эпитопом и указанным вторым эпитопом.
В еще одном аспекте второй конфигурации в изобретении предложен способ получения полиспецифического лиганда с закрытой конформацией, включающего первый эпитоп-связывающий домен, обладающий первой эпитоп-связывающей специфичностью, и некомплементарный второй эпитоп-связывающий домен, обладающий второй эпитоп-связывающей специфичностью, где первая и вторая связывающие специфичности конкурируют за связывание с эпитопом, так что полиспецифический лиганд с В предпочтительном воплощении изобретения вариабельные области выбраны из генных репертуаров единичного домена V. Как правило, репертуар единичных доменов антитела представлен на поверхности филаментного бактериофага. В предпочтительном воплощении каждый домен единичного антитела выбирают путем связывания фагового репертуара с антигеном.
В предпочтительном воплощении изобретения каждый единичный вариабельный домен может быть выбран для связывания с его целевым антигеном или эпитопом в отсутствие комплементарной вариабельной области. В альтернативном воплощении единичные вариабельные домены могут быть выбраны для связывания с его целевым антигеном или эпитопом в присутствии комплементарной вариабельной области. Таким образом, первый единичный вариабельный домен может быть выбран в присутствии третьего комплементарного вариабельного домена, а второй вариабельный домен может быть выбран в присутствии четвертого комплементарного вариабельного домена. Третий или четвертый комплементарный вариабельный домен может представлять собой природный когнатный вариабельный домен, имеющий такую же специфичность как единичный тестируемый домен, или некогнатный комплементарный домен - такой как "пустой" вариабельный домен.
Предпочтительно, лиганд с двойной специфичностью по изобретению включает лишь два вариабельных домена, хотя несколько таких лигандов может быть включено вместе в один и тот же белок, например два таких лиганда могут быть включены в IgG или мультимерный иммуноглобулин, такой как IgM. Альтернативно, в еще одном воплощении множество лигандов с двойной специфичностью комбинируют с образованием мультимера. Например, два разных лиганда с двойной специфичностью комбинируют с получением тетраспецифической молекулы.
Специалисту в данной области будет понятно, что вариабельные области легких и тяжелых цепей лиганда с двойной специфичностью, получаемого с использованием способа по настоящему изобретению, могут находиться на одной и той же полипептидной цепи, или альтернативно, на разных полипептидных цепях. В случае, когда вариабельные области находятся на разных полипептидных цепях, они могут быть связаны через линкер, обычно гибкий линкер (такой как полипептидная цепь), химическую связывающую группу или любой другой способ, известный в данной области.
В первой конфигурации настоящего изобретения предложено дополнительное улучшение лигандов с двойной специфичностью, разработанное авторами настоящего изобретения, в которых одна специфичность лиганда направлена на белок или полипептид, присутствующий in vivo в организме, которое может действовать с целью увеличения периода полувыведения лиганда путем связывания с ним.
Соответственно, в первом аспекте предложен лиганд с двойной специфичностью, включающий первый единичный вариабельный домен иммуноглобулина, обладающий связывающей специфичностью в отношении первого антигена или эпитопа, и второй комплементарный единичный вариабельный домен иммуноглобулина, обладающий связывающей активностью в отношении второго антигена или эпитопа, где один или оба указанных антигена или эпитопа действуют для увеличения периода полувыведения лиганда in vivo, и где указанный первый и второй домены лишены взаимно комплементарных доменов, которые имеют одинаковую специфичность, при условии, что указанный лиганд с двойной специфичностью не состоит из VH-домена антитела против HSA и VK-домена против β-галактозидазы. Предпочтительно, чтобы ни первый, ни второй вариабельный домен не связывались с человеческим сывороточным альбумином (HSA).
Антигены или эпитопы, которые увеличивают период полувыведения лиганда, как описанл в данном изобретении, предпочтительно присутствуют на белках или полипептидах, обнаруженных в организме in vivo. Примеры включают в себя белки внеклеточного матрикса, белки крови и белки, присутствующие в различных тканях в организме. Белки действуют для уменьшения скорости клиренса лиганда из крови, например, путем действия в качестве наполнителей, или путем заякоривания лиганда в желаемом сайте действия. Примеры антигенов/эпитопов, которые увеличивают период полувыведения in vivo, приведены в Приложении 1 ниже.
Увеличенный период полувыведения полезен в применениях иммуноглобулинов in vivo, в особенности антител и в особенности фрагментов антител, имеющих небольшой размер. Такие фрагменты (Fv, связанные дисульфидными связями Fv, Fab, scFv, dAb) страдают от быстрого клиренса из организма; таким образом, хотя они способны быстро достигать большинства частей организма и могут быть быстро получены и с ними легче обращаться, их применения in vivo ограничены лишь кратким их присутствием in vivo. Изобретение решает эту проблему путем обеспечения увеличенного периода полувыведения лигандов in vivo и, следовательно, более длительного поддержания в организме функциональной активности лиганда.
Способы фармакокинетического анализа и определения периода полувыведения лиганда хорошо известны специалистам в данной области. Детали можно найти в Kenneth, A et al: Chemical Stability of Phamnaceuticals: A Handbook for Pharmacists и в Peters et al, Pharmacokinetc analysis: A Practical Approach (1996). Также сделана ссылка на "Pharmacokinetics", M Gibaldi & D Perron, опубликованную Marcel Dekker, 2nd Rev. ex edition (1982), в котором описаны фармакокинетические параметры, такие как периоды полувыведения t-альфа и t-бета и площадь под кривой (AUC).
Периоды полувыведения (t1/2-альфа и t1/2-бета) и AUC могут быть определены из кривой зависимости концентрации лиганда в сыворотке крови от времени. Пакет для анализа WinNonlin (поставляется Pharsight Corp., Mountain View, CA94040, USA) может быть использован, например, для моделирования кривой. В первой фазе (альфа-фаза) лиганд подвергается в основном распределению в организме пациента с некоторым выведением. Вторая фаза (бета-фаза) представляет собой конечную фазу, когда лиганд распределен, и канцентрация в сыворотке крови уменьшается, поскольку лиганд подвергается клиренсу из организма пациента. Период полувыведения t-альфа представляет собой период полувыведения в первой фазе, а период полувыведения t-бета представляет собой период полувыведения во второй фазе. Таким образом, предпочтительно, в настоящем изобретении предложен лиганд или композиция, содержащая лиганд по изобретению, имеющая период полувыведения tα в диапазоне от 15 минут или более. В одном из воплощений нижняя граница диапазона составляет 30 минут, 45 минут, 1 час, 2 часа, 3 часа, 4 часа, 5 часов, 6 часов, 7 часов, 10 часов, 11 часов или 12 часов. Дополнительно или альтернативно, лиганд или композиция по изобретению будет иметь период полувыведения tα в диапазоне вплоть до и включая 12 часов. В одном из воплощений верхняя граница диапазона составляет 11, 10, 9, 8, 7, 6 или 5 часов. Пример подходящего диапазона составляет от 1 до 6 часов, от 2 до 5 часов или от 3 до 4 часов.
Предпочтительно, в настоящем изобретении предложен лиганд или композиция, содержащая лиганд по изобретению, имеющая период полувыведения tβ в диапазоне от 2,5 часов и более. В одном из воплощений нижняя граница диапазона составляет 3 часа, 4 часа, 5 часов, 6 часов, 7 часов, 10 часов, 11 часов или 12 часов. Дополнительно или альтернативно, лиганд или композиция по изобретению имеет период полувыведения tβ в диапазоне вплоть до и включая 21 сутки. В одном из воплощений верхняя граница диапазона составляет 12 часов, 24 часа, 2 суток, 3 суток, 5 суток, 10 суток, 15 суток или 20 суток. Предпочтительно, лиганд или композиция по изобретению имеет период полувыведения tβ в диапазоне от 12 до 60 часов. В другом воплощении он находится в диапазоне от 12 до 48 часов. В еще одном воплощении он находится в диапазоне от 12 до 26 часов.
Дополнительно или альтернативно к вышеприведенным критериям в настоящем изобретении предложен лиганд или композиция, содержащая лиганд по изобретению, имеющие значение AUC (площадь под кривой) в диапазоне от 1 мг.мин/мл или более. В одном из воплощений нижняя граница диапазона составляет 5, 10, 15, 20, 30, 100, 200 или 300 мг·мин/мл. Дополнительно или альтернативно лиганд или композиция по изобретению имеет AUC в диапазоне вплоть до 600 мг·мин/мл. В одном из воплощений верхняя граница диапазона составляет 500, 400, 300, 200, 150, 100, 75 или 50 мг.мин/мл. Предпочтительно, лиганд по изобретению имеет AUC в диапазоне, выбранном из группы, состоящей из следующих: от 15 до 150 мг·мин/мл, от 15 до 100 мг·мин/мл, от 15 до 75 мг·мин/мл и от 15 до 50 мг·мин/мл.
В первом воплощении лиганд с двойной специфичностью включает два комплементарных вариабельных домена, т.е. два вариабельных домена, которые в их природном окружении способны функционировать вместе в виде когнатной пары или группы, даже если в контексте настоящего изобретения они связываются раздельно с их когнатными эпитопами. Например, комплементарные вариабельные домены могут представлять собой тяжелую цепь иммуноглобулина и вариабельные домены легких цепей (VH и VL). Домены VH и VL предпочтительно обеспечиваются фрагментами scFv или Fab антитела. Вариабельные домены могут быть связаны вместе с образованием поливалентных лигандов, например, посредством: обеспечения шарнирной области на С-конце каждого V-домена и дисульфидного связывания между цистеинами в шарнирных областях; или обеспечения dAb, каждый из которых имеет цистеин на С-конце домена, причем эти цистеины связаны вместе дисульфидной связью; или продукцией V-CH и V-CL с получением формата Fab; или применения пептидных линкеров (например, Gly4Ser линкеров, обсуждаемых ниже) с получением димеров, тримеров и последующих мультимеров.
Авторы изобретения обнаружили, что применение комплементарных вариабельных доменов делает возможным совместную укладку поверхностей двух доменов и отделение от растворителя. Кроме того, комплементарные домены способны стабилизировать друг друга. Кроме того, оно позволяет создавать IgG антитела с двойной специфичностью без недостатков гибридных гибридом, используемых в предшествующем уровне техники, или необходимости конструировать тяжелые или легкие цепи на поверхностях контакта субъединиц. Лиганды с двойной специфичностью, в соответствии с первым аспектом настоящего изобретения, имеют по меньшей мере одну пару VH/VL. Биспецифические IgG по изобретению, таким образом, включают две такие пары, одна пара на каждом плече Y-образной молекулы. Поэтому в отличие от стандартных биспецифических антител или диа-антител, когда отношение используемых цепей является определяющим для успеха их получения и приводит к практическим сложностям, лиганды с двойной специфичностью по изобретению свободны от проблем баланса цепей. Дисбаланс цепей в стандартных биспецифических антителах возникает в результате ассоциации двух различных цепей VL с двумя различными цепями VH, где VL цепь 1 вместе с VH цепью 1 способна связываться с антигеном или эпитопом 1, и VL цепь 2 вместе с VH цепью 2 способна связываться с антигеном или эпитопом 2, и две корректные пары каким-то образом связаны друг с другом. Таким образом, только когда VL цепь 1 спарена с VH цепью 1, и VL цепь 2 спарена с VH цепью 2, в единой молекуле возникает биспецифичность. Такие биспецифические молекулы могут быть получены двумя различными путями. Во-первых, они могут быть получены путем ассоциации двух существующих пар VH/VL, каждая из которых связывается с различным антигеном или эпитопом (например, в биспецифическом IgG). В этом случае пары VH/VL должны вместе приводить к соотношению 1:1 для создания популяции молекул, все из которых являются биспецифическими. Этого никогда не наблюдается (даже когда комплементарный СН домен усиливается за счет конструирования по принципу "knobs into holes"), приводя к смеси биспецифических молекул и молекул, которые способны связываться только с одним антигеном или эпитопом, но не с другим. Второй путь создания биспецифического антитела осуществляют путем одновременной ассоциации двух различных цепей VH с двумя различными цепями VL (например, в биспецифическом диа-антителе). В этом случае, хотя существует тенденция в предпочтительном спаривании VL цепи 1 с VH цепью 1 и спаривании VL цепи 2 с VH цепью 2 (которое может усиливаться за счет конструирования VL и VH доменов по принципу "knobs into holes"), это спаривание никогда не достигается во всех молекулах, что приводит к смешанному препарату, в котором имеются некорректные пары, которые не способны связываться с антигеном или эпитопом.
Биспецифические антитела, сконструированные в соответствии с подходом с применением лиганда с двойной специфичностью, согласно первому аспекту настоящего изобретения позволяют преодолеть все эти проблемы, поскольку связывание с антигеном или эпитопом 1 происходит в домене VH или VL, а связывание с антигеном или эпитопом 2 происходит соответственно с комплементарным доменом VL или VH. Поскольку домены VH и VL спариваются в соотношении 1:1, все спаривания VH/VL являются биспецифическими, и, таким образом, все форматы, сконструированные с использованием этих спариваний VH/VL (Fv, scFv, Fab, мини-антитела, IgG и т.д.) имеют 100%-ную биспецифическую активность.
В контексте настоящего изобретения понятно, что первый и второй "эпитопы" представляют собой эпитопы, которые не являются одинаковыми и не связаны единым моноспецифическим лигандом. В первой конфигурации по изобретению они предпочтительно находятся на разных антигенах, один из них действует для увеличения периода полувыведения лиганда in vivo. Аналогично, первый и второй антигены предпочтительно не являются одинаковыми.
Лиганды с двойной специфичностью по изобретению не включают лиганды, описанные в WO 02/02773. Таким образом, лиганды по настоящему изобретению не включают комплементарные пары VH/VL, которые совместно связывают любой один или более антигенов или эпитопов. Наоборот, лиганды согласно первому аспекту изобретения включают комплементарную пару VH/VL, где домены V имеют различные специфичности. Кроме того, лиганды согласно первому аспекту изобретения включают комплементарные пары VH/VL, имеющие различные специфичности в отношении структурно неродственных эпитопов или антигенов. Структурно родственные эпитопы или антигены представляют собой эпитопы или антигены, которые имеют достаточное структурное сходство для того, чтобы связываться стандартной комплементарной парой VH/VL, которая действует совместным образом для связывания антигена или эпитопа; в случае структурно родственных эпитопов, эти эпитопы достаточно похожи по структуре в том, что они "подходят" к тому же связывающему карману, образованному в антиген-связывающем сайте димера VH/VL.
Во втором аспекте настоящего изобретения предложен лиганд, содержащий первый вариабельный домен иммуноглобулина, обладающий связывающей специфичностью в отношении первого антигена или эпитопа, и второй вариабельный домен иммуноглобулина, обладающий связывающей специфичностью в отношении второго антигена или эпитопа, где один или оба из указанных первых и вторых вариабельных доменов связываются с антигеном, что увеличивает период полувыведения лиганда in vivo, и вариабельные домены не комплементарны друг другу.
В одном из воплощений связывание через один вариабельный домен модулирует связывание лиганда через второй вариабельный домен.
В этом воплощении вариабельные домены могут паредставлять собой, например, пары доменов VH или пары доменов VL. Связывание антигена в первом сайте может модулировать, например усиливать или ингибировать, связывание антигена во втором сайте. Например, связывание в первом сайте по меньшей мере частично ингибирует связывание антигена во втором сайте. В таком воплощении лиганд может, например, поддерживаться в организме субъекта in vivo путем связывания с белком, который увеличивает период полувыведения лиганда до такого времени, когда он становится связанным со вторым антигеном-мишенью и диссоциирует из белка, увеличивающего период полувыведения.
Модуляцию связывания в указанном выше контексте достигают в виде структурной близости антиген-связывающих сайтов относительно друг друга. Такая структурная близость может быть достигнута за счет природы структурных компонентов, связывающих два или более антиген-связывающих сайтов, например путем обеспечения лиганда с относительно жесткой структурой, которая удерживает антиген-связывающие сайты в непосредственной близости. Предпочтительно, два или более антиген-связывающих сайтов находятся физически очень близко друг к другу, так что один сайт модулирует связывание антигена в другом сайте способом, который включает стерическое ограничение и/или конформационные изменения в молекуле иммуноглобулина.
Первый и второй антиген-связывающие домены могут быть связаны ковалентно или нековалентно. В случае, когда домены связаны ковалентно, то связывание может опосредоваться, например дисульфидными связями или полипептидным линкером, таким как (Gly4Ser)n, где n=от 1 до 8, например 2, 3, 4, 5 или 7.
В случае когда вариабельные домены выбраны из репертуаров V-гена, выбранных, например, с использованием технологии фагового дисплея, как описано в данном изобретении, тогда эти вариабельные домены могут включать универсальную каркасную область, так что они могут распознаваться специфическим общим лигандом, как определено в данном изобретении. Применение универсальных каркасов, общих лигандов и т.п. описано в WO 99/20749. В настоящем изобретении ссылка на фаговый дисплей включает применение фага и/или фагмид.
Когда используют репертуары V-генов, тогда вариация полипептидной последовательности предпочтительно располагается в структурных петлях вариабельных доменов. Полипептидные последовательности любого вариабельного домена могут быть изменены путем перестройки ДНК или с помощью мутации для усиления взаимодействия каждого вариабельного домена с его комплементарной парой. Перестройка ДНК известна в данной области и изложена, например в Stemmer, Nature 370:389-391 (1994) и патенте США №6297053, которые оба включены путем ссылки. Другие способы мутагенеза хорошо известны специалистам в данной области.
В предпочтительном воплощении изобретения 'лиганд с двойной специфичностью' представляет собой одноцепочечный Fv-фрагмент. В альтернативном воплощении изобретения 'лиганд с двойной специфичностью' состоит из Fab-области антитела. Термин "Fab-область" включает Fab-подобную область, в которой использованы два VH-домена или два VL-домена.
Вариабельные области могут быть получены из антител, направленных против целевых антигенов или эпитопов. Альтернативно, они могут быть получены из репертуара единичных доменов антител, таких как те, которые экспрессируются на поверхности филаментного бактериофага. Селекцию можно осуществлять, как описано ниже.
В еще одном аспекте настоящего изобретения предложена одна или более чем одна молекула нуклеиновой кислоты, кодирующая по меньшей мере лиганд с двойной специфичностью, как определено в данном изобретении. Лиганд с двойной специфичностью может кодироваться на одной молекуле нуклеиновой кислоты; альтернативно, каждый домен может кодироваться разными молекулами нуклеиновых кислот. Когда лиганд кодируется одной молекулой нуклеиновой кислоты, тогда домены могут экспрессироваться в виде слитого полипептида, в виде scFv молекулы, или могут экспрессироваться раздельно и затем связываться вместе, например с использованием химических сшивающих агентов. Лиганды, экспрессирующиеся из отдельных нуклеиновых кислот, сшиваются вместе подходящими способами.
Нуклеиновая кислота, кроме того, может кодировать сигнальную последовательность для экспорта полипептидов из клетки-хозяина при экспрессии и может быть слита с поверхностным компонентом филаментной частицы бактериофага (или другим компонентом системы селективного дисплея) после экспрессии.
В еще одном аспекте настоящего изобретения предложен вектор, содержащий нуклеиновую кислоту, кодирующую лиганд с двойной специфичностью по настоящему изобретению.
В еще одном аспекте настоящего изобретения предложена клетка-хозяин, трансфицированная вектором, кодирующим лиганд с двойной специфичностью по настоящему изобретению.
Экспрессия с такого вектора может иметь конфигурацию для получения, например, на поверхности частицы бактериофага, вариабельных доменов для селекции. Это дает возможность для селекции представленных вариабельных областей и таким образом для селекции 'лигандов с двойной специфичностью' с использованием способа по настоящему изобретению.
В настоящем изобретении дополнительно предложен набор, включающий по меньшей мере лиганд с двойной специфичностью по настоящему изобретению.
В третьем аспекте изобретения предложен способ получения лиганда, содержащего первый единичный вариабельный домен иммуноглобулина, имеющий первую связывающую специфичность, и второй единичный вариабельный домен иммуноглобулина, имеющий вторую (отличающуюся) связывающую специфичность, причем одна или обе связывающие специфичности являются специфичными в отношении антигена, который увеличивает период полувыведения лиганда in vivo, включающий стадии:
а) селекции первого вариабельного домена по его способности связываться с первым эпитопом,
б) селекции второй вариабельной области по ее способности связываться со вторым эпитопом,
в) комбинирования вариабельных доменов; и
г) селекции лиганда по его способности связываться с указанным первым эпитопом и с указанным вторым эпитопом.
Лиганд может связываться с первым и вторым эпитопами либо одновременно, либо когда существует конкуренция между связывающими доменами за связывание с эпитопом, тогда связывание одного домена может препятствовать связыванию другого домена с когнатным эпитопом. Таким образом, в одном из воплощений стадия (г) выше требует одновременного связывания с обоими первым и вторым (и возможно дополнительным) эпитопами; в еще одном воплощении связывание с первым и вторым эпитопами происходит не одновременно.
Эпитопы предпочтительно находятся на отдельных антигенах.
Лиганды предпочтительно включают комбинации VH/VL, или комбинации VH/VH или VL/VL вариабельных доменов иимуноглобулина, как описано выше. Лиганды могут также включать Camelid домены VHH, при условии, что VHH-домен, специфичный в отношении антигена, который увеличивает период полувыведения лиганда in vivo, не связывает лизоцим белка куриного яйца (HEL), панкреатическую альфа-амилазу свиньи или NmC-A; hcg, BSA-связанный азокраситель RR6 или клетки HG982 S. mutans, как описано в Conrath et al, (2001) JBC 276:7346-7350 и W099/23221, ни один из которых не описывает применение специфичности в отношении антигена, который увеличивает период полувыведения, для увеличения периода полувыведения лиганда in vivo.
В одном из воплощений указанный первый вариабельный домен выбран для связывания с указанным первым эпитопом в отсутствие комплементарного вариабельного домена. В еще одном воплощении указанный первый вариабельный домен выбран для связывания с указанным первым эпитопом/антигеном в присутствии третьего вариабельного домена, в котором указанный третий вариабельный домен отличается от указанного второго вариабельного домена и комплементарен первому домену. Аналогично, второй домен может быть выбран в отсутствие или в присутствии комплементарного вариабельного домена.
Антигены или эпитопы, являющиеся мишенями для лигандов по изобретению, дополнительно к белку, увеличивающему период полувыведения, могут представлять собой любой антиген или эпитоп, но предпочтительно представляет собой антиген или эпитоп, который является мишенью с терапевтической пользой. В изобретении предложены лиганды, включающие лиганды с открытой конформацией, закрытой конформацией и лиганды на основе выделенного мономера dAb, специфичные в отношении любой такой мишени, в частности мишеней, дополнительно идентифицированных в данном изобретении. Такие мишени могут представлять собой полипептиды, белки или нуклеиновые кислоты, которые могут быть природными или синтетическими, или являются их частью. В этом отношении лиганд по изобретению может связывать эпитоп или антиген и действовать в качестве антагониста или агониста (например, агонист рецептора ЕРО). Специалисту в данной области понятно, что выбор огромен и варьирует.Они могут представлять собой, например человеческие или животные белки, цитокины, рецепторы цитокинов, ферменты, кофакторы ферментов или ДНК-связывающие белки. Подходящие цитокины и факторы роста включают: АроЕ (аполипопротеин Е), Apo-SAA, BDNF (нейротрофический фактор головного мозга), кардиотрофин-1, EGF (эпидермальный фактор роста), рецептор EGF, ENA-78, эотаксин, эотаксин-2, Exodus-2, EpoR, FGF-кислый (фактор роста фибробластов), FGF-основный, фактор роста фибробластов-10, лиганд FLT3, фракталкин (СХЗС), GDNF (нейротрофический фактор глиальных клеток), G-CSF (гранулоцитарный колониестимулирующий фактор), GM-CSF (колониестимулирующий фактор гранулоцитов и макрофагов), GF-β1, инсулин, IFN-γ, IGF-I (инсулиноподобный фактор роста), IGF-II, IL-1α, IL-1β, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8 (72 аминокислоты), IL-8 (77 аминокислот), IL-9, IL-10, IL-11, IL-12, IL-13, IL-15, IL-16, IL-17, IL-18 (IGIF), ингибин α, ингибин β, IP-10, KGF-2 (фактор роста кератиноцитов-2), KGF, лептин, LIF, лимфотактин, ингибитор Мюллера, моноцитарный колониеингибирующий фактор, белок-аттрактант моноцитов, М-CSF, MDC (67 аминокислот), MDC (69 аминокислот), МСР-1 (моноцитарный белок-хемоаттрактант) (MCAF (хемотактический и активирующий фактор моноцитов)), МСР-2, МСР-3, МСР-4, MDC (67 аминокислот), MDC (69 аминокислот), MIG, MIP-1α, MIP-1β, MIP-3α, MIP-3β, MDP-4, фактор-ингибитор миелоидных предшественников-1 (MPIF-1), NAP-2 (нейтрофил-активирующий белок), нейртурин, фактор роста нервов, β-NGF, NT-3, NT-4, онкостатин М, тромбоцитарный фактор роста PDGF-AA, PDGF-AB, PDGF-BB, тромбоцитарный фактор PF-4, RANTES, SDF1α (фактор стромальных клеток), SDF1β, SCF, фактор роста стволовых клеток SCGF, фактор стволовых клеток (SCF), TARC (хемокины, регулируемые тимусом и активацией), TGF-α (трансформирующий фактор роста), TGF-β, TGF-β2, TGF-β3, TNF (фактор некроза опухоли), TNF-α, TNF-β, рецептор TNF I, рецептор TNF II. TNIL-1, ТРО, VEGF (фактор роста эндотелия сосудов), рецептор VEGF 1, рецептор VEGF 2, рецептор VEGF 3, GCP-2 (хемотактический белок гранулоцитов), GRO/MGSA, GRO-β, GRO-γ, НСС1, 1-309, HER 1, HER 2, HER 3 и HER 4, но не ограничиваются ими.
Рецепторы цитокинов включают рецепторы вышеуказанных цитокинов. Понятно, что этот перечень никоим образом не является исчерпывающим.
В одном из воплощений изобретения вариабельные домены получены из соответствующего антитела, направленного против антигена или эпитопа. В предпочтительном воплощении вариабельные домены получены из репертуара единичных вариабельных доменов антитела.
Во второй конфигурации в настоящем изобретении предложены полиспецифические лиганды. По настоящему изобретению термин "полиспецифический лиганд" относится к лиганду, который обладает более чем одной эпитоп-связывающей специфичностью, как определено в данном изобретении. Как правило, полиспецифический лиганд включает два или более эпитоп-связывающих доменов.
Эпитоп-связывающие домены по настоящему изобретению включают белковый каркас и сайты взаимодействия с эпитопом (которые предпочтительно находятся на поверхности белкового каркаса). Согласно настоящему изобретению предпочтительно, чтобы каждый эпитоп-связывающий домен имел отличающуюся эпитоп-связывающую специфичность.
В контексте настоящего изобретения понятно, что первый и второй "эпитопы" представляют собой эпитопы, которые не являются одинаковыми и не связаны единым моноспецифическим лигандом. Они могут располагаться на различных антигенах или на одном и том же антигене, но разделены достаточным расстоянием, так что они не образуют единую структуру, которая могла бы быть связана единой моноспецифической связывающей парой VH/VL стандартного антитела. Экспериментально, если оба индивидуальных вариабельных домена в форме одноцепочечного антитела (доменные антитела или dAb) отдельно конкурируют с моноспецифическим лигандом VH/VL против двух эпитопов, то эти два эпитопа не достаточно удалены для того, чтобы рассматриваться как отдельные эпитопы согласно настоящему изобретению.
В соответствии с настоящим изобретением предпочтительно, чтобы каждый эпитоп-связывающий домен включал вариабельный домен иммуноглобулина. Более предпочтительно, чтобы каждый вариабельный домен иммуноглобулина представлял собой вариабельный домен легкой цепи (VL) или вариабельный домен тяжелой цепи VH. Во второй конфигурации настоящего изобретения домены иммуноглобулина, представленные на лиганде по настоящему изобретению, не комплементарны, то есть они не ассоциируются с образованием VH/VL антиген-связывающего сайта. Таким образом, полиспецифические лиганды, как определено во второй конфигурации изобретения, включают иммуноглобулиновые домены того же самого подтипа, то есть либо вариабельные домены легкой цепи (VL), либо вариабельные домены тяжелой цепи (VH). Кроме того, когда лиганд по изобретению находится в закрытой конформации, иммуноглобулиновые домены могут иметь VHH Camelid типа.
В альтернативном воплощении лиганд(ы) по изобретению не включают Camelid Vнн-домен. Более конкретно, лиганд(ы) по изобретению не включают один или более чем один аминокислотный остаток, специфичный в отношении Camelid VHH-доменов по сравнению с человеческими VH-доменами.
Предпочтительно, единичные вариабельные домены получены из антитела, выбранного для связывающей активности против различных антигенов или эпитопов. Например, вариабельные домены могут быть выделены по меньшей мере частично путем иммунизации людей. В данной области известны альтернативные способы, включающие выделение из библиотек человеческих антител и синтез генов искусственных антител.
Вариабельные домены предпочтительно связывают суперантигены, такие как белок А или белок L. Связывание с суперантигенами представляет собой свойство правильно свернутых вариабельных доменов антител и позволяет выделить такие домены, например, из библиотек рекомбинантных или мутантных доменов.
Эпитоп-связывающие домены также могут быть основаны на белковых каркасах или скелетах, отличающихся от иммуноглобулиновых доменов. Например, природные бактериальные рецепторы, такие как SpA, используют в качестве каркасов для присоединения CDR с получением лигандов, которые специфически связываются с одним или более эпитопами. Детали этого способа описаны в US 5831012. Другие подходящие каркасы включают каркасы на основе фибронектина и аффител. Детали подходящих способов описаны в WO 98/58965. Другие подходящие каркасы включают липокаллин и CTLA4, как описано в van den Beuken etal, J. Mol. Biol. (2001) 310, 591-601, и каркасы, такие как описано в WO 0069907 (Medical Research Council), которые основаны, например, на кольцевой структуре бактериального GroEL или других шаперонных полипептидов.
Белковые каркасы можно комбинировать; например CDR можно присоединять к каркасу CTLA4 и использовать вместе с иммуноглобулиновыми доменами VH или VL с образованием поливалентного лиганда. Аналогично, можно комбинировать фибронектин, липокаллин и другие каркасы.
В одном из воплощений полиспецифический лиганд включает эпитоп-связывающий домен, который включает CDR единичного иммуноглобулинового домена (dAb), который связывает описанный в данном изобретении TNFR1, присоединенный к подходящему белковому каркасу или скелету.
Специалисту в данной области понятно, что эпитоп-связывающие домены полиспецифического лиганда с закрытой конформацией, полученного с использованием способа по настоящему изобретению, могут находиться на одной и той же полипептидной цепи или, альтернативно, на разных полипептидных цепях. В случае, когда вариабельные области располагаются на разных полипептидных цепях, они могут быть связаны через линкер, предпочтительно гибкий линкер (такой как полипептидная цепь), химическая связывающая группа, или любым другим способом, известным в данной области.
Первый и второй эпитоп-связывающие домены могут быть ассоциированы ковалентно или нековалентно. В случае, когда домены ассоциированы ковалентно, ассоциация может быть опосредована, например, дисульфидными связями.
В некоторых воплощениях второй конфигурации по изобретению эпитопы могут замещать друг друга при связывании. Например, первый эпитоп может быть представлен на антигене, который после связывания с его когнатным первым связывающим доменом вызывает стерическое ограничение для второго связывающего домена или конформационное изменение в нем, которое замещает эпитоп, связанный со вторым связывающим доменом.
Предпочтительно, связывание уменьшается на 25% или более, предпочтительно на 40%, 50%, 60%, 70%, 80%, 90% или, и предпочтительно вплоть до 100% или около 100%, так что связывание полностью ингибируются. Связывание эпитопов можно измерить с помощью стандартных анализов связывания антигена, таких как ИФА, флуоресцентные методы, включая FRET (резонансный перенос энергии флуоресценции), или способов, таких как поверхностный плазменный резонанс, который измеряет массу молекул.
Кроме того, в изобретении предложен полиспецифический лиганд с закрытой конформацией, включающий первый эпитоп-связывающий домен, обладающий первой эпитоп-связывающей специфичностью, и некомплементарный второй эпитоп-связывающий домен, обладающий второй эпитоп-связывающей специфичностью, где первая и вторая связывающие специфичности конкурируют за связывание с эпитопом, так что полиспецифический лиганд с закрытой конформацией может не связывать оба эпитопа одновременно.
Полиспецифические лиганды с закрытой конформацией по изобретению не включают лиганды, описанные в WO 02/02773. Таким образом, лиганды по настоящему изобретению не включают комплементарные пары VH/VL, которые кооперативно связывают любой один или более антигенов или эпитопов. Вместо этого, лиганды по изобретению предпочтительно включают некомплементарные пары VH-VH или VL-VL. Предпочтительно, каждый домен VH или VL в каждой паре VH-VH или VL-VL имеет разную эпитоп-связывающую специфичность, и сайты связывания эпитопов расположены таким образом, что связывание эпитопа в одном сайте конкурирует со связыванием эпитопа в другом сайте.
Предпочтительно, полиспецифический лиганд с закрытой конформацией может включать первый домен, способный связываться с молекулой-мишенью, и второй домен, способный связываться с молекулой или группой, которая увеличивает период полувыведения лиганда. Например, молекула или группа может представлять собой объемный агент, такой как HSA или белок клеточного матрикса. Используемая в данном изобретении фраза "молекула или группа, которая увеличивает период полувыведения лиганда" относится к молекуле или химической группе, которая после связывания с лигандом с двойной специфичностью, как описано в данном изобретении, увеличивает период полувыведения in vivo такого лиганда с двойной специфичностью при введении животному относительно лиганда, который не связывает эту молекулу или группу. Примеры молекул или групп, которые увеличивают период полувыведения лиганда, описаны ниже. В предпочтительном воплощении полиспецифический лиганд с закрытой конформацией может связывать молекулу-мишень только после замещения молекулы или группы, увеличивающей период полувыведения. Таким образом, например, полиспецифический лиганд с закрытой конформацией поддерживается в системе кровообращения субъекта с помощью объемной молекулы, такой как HSA. При столкновении с молекулой-мишенью конкуренция между связывающими доменами полиспецифического лиганда с закрытой конформацией приводит в результате к замещению HSA и связыванию с мишенью.
В предпочтительном воплощении второй конфигурации по изобретению эпитоп-связывающие домены представляют собой вариабельные области иммуноглобулина и выбраны из генных репертуаров единичного домена V. Как правило, репертуар единичных доменов антитела представлен на поверхности филаментного бактериофага. В предпочтительном воплощении каждый единичный домен антитела выбран путем связывания фагового репертуара с антигеном. В предпочтительном воплощении второй конфигурации по изобретению эпитоп-связывающие домены представляют собой вариабельные области иммуноглобулина и выбраны из генных репертуаров единичного домена V. Как правило, репертуар единичных доменов антитела представлен на поверхности филаментного бактериофага. В предпочтительном воплощении каждый единичный домен антитела выбран путем связывания фагового репертуара с антигеном.
В одном из аспектов полиспецифический лиганд включает по меньшей мере два некомплементарных вариабельных домена. Например, лиганды могут включать пару VH-доменов или пару VL-доменов. Предпочтительно, домены происходят не из Camelid; предпочтительно они представляют собой человеческие домены или включают человеческие каркасные области (FW) и одну или более гетерологичных CDR. CDR и каркасные области представляют собой области вариабельного домена иммуноглобулина, как определено в базе данных последовательностей белков, представляющих иммунологический интерес (Kabat база данных).
Предпочтительные человеческие каркасные области представляют собой области, которые кодируются герминальными генными сегментами DP47 и DPK9. Предпочтительно, FW1, FW2 и FW3 в домене VH или VL имеют последовательность FW1, FW2 или FW3 из DP47 или DPK9. Человеческие каркасные области могут содержать мутации, например до примерно 5 аминокислотных замен или до примерно 10 аминокислотных замен вместе в человеческих каркасных областях, используемых в лигандах по изобретению.
Вариабельные домены в полиспецифических лигандах в соответствии со второй конфигурацией по изобретению могут быть расположены в открытой или закрытой конформации; то есть они могут быть расположены таким образом, что вариабельные домены могут связывать их когнатные лиганды независимо и одновременно, или таким образом, что только один из вариабельных доменов может связывать свой когнатный лиганд в любой момент времени.
Авторы изобретения понимают, что при некоторых структурных условиях некомплементарные вариабельные домены (например, два вариабельных домена легких цепей или два вариабельных домена тяжелых цепей) могут быть представлены в лиганде, так что связывание первого эпитопа с первым вариабельным доменом ингибирует связывание второго эпитопа со вторым вариабельным доменом, даже если такие некомплементарные домены не функционируют вместе как когнатная пара.
Предпочтительно, лиганд включает две или более пар вариабельных доменов; то есть он включает по меньшей мере четыре вариабельных домена. Предпочтительно, четыре вариабельных домена включают каркасные области человеческого происхождения.
В предпочтительном воплощении каркасные области человеческих антител идентичны каркасным областям человеческих герминальных последовательностей.
Авторы настоящего изобретения считают, что такие антитела имеют конкретное применение в анализах связывания лигандов для терапевтического и других применений.
В одном из воплощений второй конфигурации по изобретению вариабельные домены получены из антитела, направленного против первого и/или второго антигена или эпитопа. В предпочтительном воплощении вариабельные домены получены из репертуара единичных вариабельных доменов антител. В одном из примеров репертуар представляет собой репертуар, который не образуется в организме животного, или синтетический репертуар. В другом примере единичные вариабельные домены не выделены (по меньшей мере частично) путем иммунизации животного. Таким образом, единичные домены могут быть выделены из наивной библиотеки.
Вторая конфигурация по изобретению в еще одном аспекте предлагает полиспецифический лиганд, содержащий первый эпитоп-связывающий домен, обладающий первой эпитоп-связывающей специфичностью, и некомплементарный второй эпитоп-связывающий домен, обладающий второй эпитоп-связывающей специфичностью. Первая и вторая связывающие специфичности могут быть одинаковыми или разными.
В еще одном аспекте настоящего изобретения предложен полиспецифический лиганд с закрытой конформацией, включающий первый эпитоп-связывающий домен, обладающий первой эпитоп-связывающей специфичностью, и некомплементарный второй эпитоп-связывающий домен, обладающий второй эпитоп-связывающей специфичностью, где первая и вторая связывающие специфичности способны конкурировать за связывание эпитопа, так что полиспецифический лиганд с закрытой конформацией не может связывать оба эпитопа одновременно.
В еще одном аспекте изобретения предложены лиганды с открытой конформацией, включающие некомплементарные связывающие домены, где домены являются специфичными в отношении различных эпитопов на той же самой мишени. Такие лиганды связываются с мишенями с повышенной авидностью. Аналогично, в изобретении предложены поливалентные лиганды, включающие некомплементарные связывающие домены, специфичные в отношении одного и того же эпитопа и направленные в отношении мишеней, которые включают множественные копии указанного эпитопа, таких как IL-5, PDGF-AA, PDGF-BB, TGF-бета, TGF-бета 2, TGF-бета 3 и TNFα, например человеческий рецептор TNF 1 и человеческий TNFα.
В аналогичном аспекте лиганды по изобретению могут быть подвергнуты конфигурации для связывания с индивидуальными эпитопами с низкой аффинностью, так что связывание с индивидуальными эпитопами не является терапевтически значимым; но повышенная авидность, возникающая в результате связывания с двумя эпитопами, обеспечивает терапевтическую пользу. В конкретном примере в качестве мишеней могут выступать эпитопы, представленные индивидуально на нормальных типах клеток, но представленные вместе только на аномальных типах клеток или клетках, пораженных заболеванием, таких как опухолевые клетки. В такой ситуации только аномальные или пораженные заболеванием клетки являются эффективными мишенями для биспецифических лигандов по изобретению.
Лиганд, специфичный в отношении множества копий одного и того же эпитопа или соседних эпитопов на той же самой мишени (известный как хелатообразующие dAb), также может быть тримерным или полимерным (тетрамерным или более чем тетрамерным) лигандом, включающим три, четыре или более четырех некомплементарных связывающих доменов. Например, могут быть сконструированы лиганды, включающие три или четыре VH-домена или VL-домена.
Кроме того, предложены лиганды, которые связываются с мультисубъединичными мишенями, где каждый связывающий домен является специфичным в отношении субъединицы указанной мишени. Лиганд может быть димерным, тримерным или полимерным.
Предпочтительно, полиспецифические лиганды в соответствии с вышеизложенными аспектами изобретения могут быть получены с использованием способа в соответствии с первым аспектом изобретения.
В соответствии с вышеизложенным первым аспектом второй конфигурации по изобретению, предпочтительно первый эпитоп-связывающий домен и второй эпитоп-связывающий домен представляют собой некомплементарные вариабельные домены иимуноглобулина, как определено в данном изобретении. То есть вариабельные домены VH-VH или VL-VL.
Хелатообразующие dAb, в частности, могут быть получены в соответствии с предпочтительным аспектом изобретения, а именно путем применения заякоривающих dAb, при котором конструируют библиотеку димерных, тримерных или мультимерных dAb с использованием вектора, который содержит константную область dAb выше или ниже линкерной последовательности, где репертуар второго, третьего и других dAb встроен в другой конец линкера. Например, заякоривающие или направляющие dAb могут представлять собой TAR1-5 (VK), TAR1-27 (VK), TAR2h-5 (VH) или TAR2h-6 (VK).
В альтернативных методологиях применения линкеров можно избежать, например путем использования нековалентного связывания или природной аффинности между связывающими доменами, такими как VH и VK. В изобретении соответственно предложен способ получения хелатообразующего мультимерного лиганда, включающий стадии:
(а) получения вектора, содержащего последовательность нуклеиновой кислоты, кодирующей единичный связывающий домен, специфичный в отношении первого эпитопа на мишени;
(б) получения вектора, кодирующего репертуар, включающий вторые связывающие домены, специфичные в отношении второго эпитопа на указанной мишени, где эпитоп может быть тем же самым или отличающимся от первого эпитопа, причем указанный второй эпитоп является соседним по отношению к указанному первому эпитопу; и
(в) экспрессии указанных первого и второго связывающих доменов; и
(г) выделения этих комбинаций первого и второго связывающих доменов, которые комбинируют вместе с получением димера, связывающего мишень.
Первый и второй эпитопы являются соседними, так что мультимерный лиганд способен связываться с обоими эпитопами одновременно. Это позволяет получить лиганд, обладающий преимуществами повышенной авидности при связывании. Когда эпитопы являются одинаковыми, добиваются повышенной авидности с помощью присутствия множества копий эпитоп на мишени, давая возможность для одновременного связывания по меньшей мере двум копиям для достижения эффекта повышенной авидности.
Связывающие домены могут быть ассоциированы несколькими способами, а также с использованием линкеров. Например, связывающие домены могут включать остатки цистеина, авидиновые и стрептавидиновые группы или другие средства для нековалентного присоединения после синтеза; выделяют такие комбинации, которые эффективно связываются с мишенью. Альтернативно, линкер может присутствовать между первым и вторым связывающими доменами, которые экспрессируются в виде единичного полипептида из единичного вектора, который содержит первый связывающий домен, линкер и репертуар вторых связывающих доменов, например, как описано выше.
В предпочтительном аспекте первый и второй связывающие домены в природе ассоциируются при связывании с антигеном; например домены VH и VK, когда они связаны с соседними эпитопами, в природе ассоциируются путем трехстороннего взаимодействия с образованием стабильного димера. Такие ассоциированные белки могут быть выделены в анализе связывания мишени. Преимущество этого способа заключается в том, что только связывающие домены, которые связываются с близкими соседними эпитопами, находящимися в правильной конформации, будут ассоциировать и, таким образом, будут выделены в результате их повышенной авидности в отношении мишени.
В альтернативном воплощении вышеприведенного аспекта второй конфигурации по изобретению, по меньшей мере один эпитоп-связывающий домен включает неиммуноглобулиновый "белковый каркас" или "белковый скелет", как определено в данном изобретении. Подходящие неиммуноглобулиновые белковые каркасы включают каркасы, выбранные из группы, состоящей из: SpA, фибронектина, GroEL и других шаперонов, липокаллина, CCTLA4 и аффител, как указано выше, но не ограничиваются ими.
В соответствии с вышеприведенным аспектом второй конфигурации по изобретению предпочтительно, чтобы эпитоп-связывающие домены были присоединены к белковому скелету. Предпочтительно, белковый скелет по изобретению представляет собой иммуноглобулиновый скелет.
Согласно настоящему изобретению, термин "иммуноглобулиновый скелет" относится к белку, который включает по меньшей мере одну иммуноглобулиновую складку, и который действует в качестве ядра для одного или более чем одного эпитоп-связывающего домена, как определено в данном изобретении.
Предпочтительные иммуноглобулиновые скелеты, как определено в данном изобретении, включают любой один или более чем один из следующих: молекула иммуноглобулина, включающая по меньшей мере (1) CL-домен антитела (подкласс каппа или лямбда); или (2) СН1-домен тяжелой цепи антитела; молекула иммуноглобулина, включающая СН1- и СН2-домены тяжелой цепи антитела; молекула иммуноглобулина, включающая СН1-, СН2- и СН3-домены тяжелой цепи антитела; или любой из подгруппы (2) в комбинации с CL-доменом (подкласс каппа или лямбда) антитела. Также может быть включен домен шарнирной области. Такие комбинации доменов могут, например, имитировать природные антитела, такие как IgG или IgM, или их фрагменты, такие как молекулы Fv, scFv, Fab или F(ab')2. Специалист в данной области понимает, что этот перечень не следует рассматривать как исчерпывающий.
Связывание скелета с определенными в данном изобретении эпитоп-связывающими доменами может быть достигнуто на полипептидном уровне, то есть после экспрессии нуклеиновой кислоты, кодирующей скелет и/или эпитоп-связывающие домены. Альтернативно, стадия связывания может быть осуществлена на уровне нуклеиновой кислоты. Способы связывания белкового скелета по настоящему изобретению с одним или более чем одним эпитоп-связывающим доменом включают применение способов белковой химии и/или молекулярной биологии, которые знакомы специалистам в данной области и описаны в данном изобретении.
Предпочтительно, полиспецифический лиганд с закрытой конформацией может включать первый домен, способный связывать молекулу-мишень, и второй домен, способный связывать молекулу или группу, увеличивающую период полувыведения лиганда. Например, молекула или группа может представлять собой объемный агент, такой как HSA или белок клеточного матрикса. Используемая в данном изобретении фраза "молекула или группа, увеличивающая период полувыведения лиганда" относится к молекуле или химической группе, которая при связывании с лигандом с двойной специфичностью, как описано в данном изобретении, увеличивает период полувыведения in vivo такого лиганда с двойной специфичностью при введении животному относительно лиганда, который не связывает данную молекулу или группу. Примеры молекул или групп, которые увеличивают период полувыведения лиганда, описаны ниже. В предпочтительном воплощении полиспецифический лиганд с закрытой конформацией может связывать молекулу-мишень только после замещения молекулы или группы, увеличивающей период полувыведения. Таким образом, например, полиспецифический лиганд с закрытой конформацией поддерживается в системе кровообращения субъекта с помощью объемной молекулы, такой как HSA. При столкновении с молекулой-мишенью конкуренция между связывающими доменами полиспецифического лиганда с закрытой конформацией приводит в результате к замещению HSA и связыванию мишени.
В еще одном аспекте второй конфигурации по изобретению в настоящем изобретении предложена одна или более чем одна молекула нуклеиновой кислоты, кодирующая по меньшей мере полиспецифический лиганд, как определено в данном изобретении. В одном из воплощений лиганд представляет собой лиганд с закрытой конформацией. В еще одном воплощении он представляет собой лиганд с открытой конформацией. Полиспецифический лиганд может кодироваться на одной молекуле нуклеиновой кислоты; альтернативно, каждый эпитоп-связывающий домен может кодироваться отдельной молекулой нуклеиновой кислоты. Когда лиганд кодируется одной молекулой нуклеиновой кислоты, тогда домены могут экспрессироваться в виде слитого полипептида или могут экспрессироваться раздельно и затем связываться вместе, например, с использованием химических связывающих агентов. Лиганды, экспрессирующиеся с различных нуклеиновых кислот, связаны вместе с использованием подходящих средств.
Нуклеиновая кислота может дополнительно кодировать сигнальную последовательность для экспорта полипептидов из клетки-хозяина после экспрессии и может быть слита с поверхностным компонентом частицы филаментного бактериофага (или другого компонента дисплейной системы для селекции) после экспрессии. Лидерные последовательности, которые могут быть использованы в бактериальной экспрессии и/или фаговом или фагмидном дисплее, включают pelB, stll, ompA, phoA, bla и pelA.
В еще одном аспекте второй конфигурации по изобретению в настоящем изобретении предложен вектор, содержащий нуклеиновую кислоту по настоящему изобретению.
В еще одном аспекте настоящего изобретения предложена клетка-хозяин, трансфицированная вектором по настоящему изобретению.
Экспрессия с такого вектора может быть сконфигурирована для получения, например, на поверхности частицы бактериофага эпитоп-связывающих доменов для селекции. Это дает возможность для селекции представленных доменов и таким образом для селекции 'полиспецифических лигандов' с использованием способа по настоящему изобретению.
В настоящем изобретении дополнительно предложен набор, включающий по меньшей мере полиспецифический лиганд по настоящему изобретению, который может представлять собой лиганд с открытой конформацией или закрытой конформацией. Наборы по изобретению могут представлять собой, например, диагностические наборы, терапевтические наборы, наборы для обнаружения химических или биологических видов и т.п.
В настоящем изобретении предложен способ получения полиспецифического лиганда, включающий стадии:
а) селекции первого эпитоп-связывающего домена по его способности связываться с первым эпитопом,
б) селекции второго эпитоп-связывающего домена по его способности связываться со вторым эпитопом,
в) комбинирования эпитоп-связывающих доменов; и
г) селекции полиспецифического лиганда с закрытой конформацией по его способности связываться с указанным первым вторым эпитопом и указанным вторым эпитопом.
В еще одном аспекте второй конфигурации в изобретении предложен способ получения полиспецифического лиганда с закрытой конформацией, включающего первый эпитоп-связывающий домен, обладающий первой эпитоп-связывающей специфичностью, и некомплементарный второй эпитоп-связывающий домен, обладающий второй эпитоп-связывающей специфичностью, где первая и вторая связывающие специфичности конкурируют за связывание с эпитопом, так что полиспецифический лиганд с закрытой конформацией не может связывать оба эпитопа одновременно, включающий стадии:
а) селекции первого эпитоп-связывающего домена по его способности связываться с первым эпитопом,
б) селекции второго эпитоп-связывающего домена по его способности связываться со вторым эпитопом,
в) комбинирования эпитоп-связывающих доменов таким образом, что домены находятся в закрытой конформации; и
г) селекции полиспецифического лиганда с закрытой конформацией по его способности связываться с указанным первым и вторым эпитопом и указанным вторым эпитопом, но не с обоими указанными первыми и вторыми эпитопами одновременно.
Альтернативное воплощение вышеуказанного аспекта второй конфигурации по изобретению возможно включает дополнительную стадию (61), включающую селекцию третьего или дополнительного эпитоп-связывающего домена. Таким образом, полиспецифический лиганд, полученный в открытой или закрытой конформации, включает более чем две эпитоп-связывающие специфичности. В предпочтительном аспекте второй конфигурации по изобретению, когда полиспецифический лиганд включает более чем два эпитоп-связывающих домена, по меньшей мере два из указанных доменов находятся в закрытой конформации и конкурируют за связывание; другие домены могут конкурировать за связывание или могут быть свободны для независимой ассоциации с их когнатным(и) эпитопом(ами).
Во второй конфигурации по изобретению первый и второй эпитопы предпочтительно разные. Они могут представлять собой или составлять часть полипептидов, белков или нуклеиновых кислот, которые могут быть природными или синтетическими. В этом отношении лиганд по изобретению может связывать эпитоп или антиген и действовать в качестве антагониста или агониста (например, агониста рецептора эритропоэтина (ЕРО)). Эпитоп-связывающие домены лиганда в одном из воплощений имеют одинаковую специфичность в отношении эпитопа и могут, например, одновременно связывать свой эпитоп, когда на одном и том же антигене представлено множество копий данного эпитопа. В еще одном воплощении эти эпитопы предложены на разных антигенах, так что лиганд может связывать эпитопы и соединять антигены. Специалисту в данной области понятно, что выбор эпитопов и антигенов огромен и разнообразен. Они могут представлять собой, например, человеческие или животные белки, цитокины, рецепторы цитокинов, ферменты, кофакторы ферментов или ДНК-связывающие белки. Подходящие цитокины и факторы роста включают в себя: АроЕ, Apo-SAA, BDNF, кардиотрофин-1, EGF, рецептор EGF, ENA-78, эотаксин, эотаксин-2, Exodus-2, EpoR, кислый FGF, основный FGF, фактор роста фибробластов-10, лиганд FLT3, фракталкин (СХ3С), GDNF, G-CSF, GM-CSF, GF-β1, инсулин, IFN-γ, IGF-I, IGF-II, интерлейкин IL-1α, IL-1β, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8 (72 аминокислоты), IL-8 (77 аминокислот), IL-9, IL-10, IL-11, IL-12, IL-13, IL-15, IL-16, IL-17, IL-18 (IGIF), ингибин α, ингибин β, IP-10, фактор роста кератиноцитов-2 (KGF-2), KGF, лептин, LIF, лимфотактин, ингибитор Мюллера, моноцитарный колониеингибирующий фактор, белок-аттрактант моноцитов, M-CSF, MDC (67 аминокислот), MDC (69 аминокислот), МСР-1 (MCAF), МСР-2, МСР-3, МСР-4, MDC (67 аминокислот), MDC (69 аминокислот), MIG, MIP-1α, MIP-1β, MIP-3α, MIP-3β, MDP-4, фактор-ингибитор миелоидного предшественника-1 (MPIF-1), NAP-2, нейртурин, фактор роста нервов, β-NGF, NT-3, NT-4, онкостатин М, PDGF-AA. PDGF-AB, PDGF-BB, PF-4, RANTES, SDF1α, SDF1β, SCF, SCGF, фактор стволовых клеток (SCF), TARC, TGF-α, TGF-β, TGF-β2, TGF-β3, фактор некроза опухоли (TNF), TNF-α, TNF-β, рецептор TNF I, рецептор TNF II, TNIL-1, TPO, VEGF, рецептор VEGF 1, рецептор VEGF 2, рецептор VEGF 3, GCP-2, GRO/MGSA, GRO-β, GRO-γ, HCC1, 1-309, HER 1, HER 2, HER 3 и HER 4, сайт распознавания ТАСЕ, TNF BP-I и TNF BP-II, а также любую мишень, раскрытую Приложении 2 или Приложении 3 к данному описанию, в комбинации, представленной в Приложениях, в разной комбинации или индивидуально. Рецепторы цитокинов включают рецепторы вышеуказанных цитокинов, например IL-1 R1; IL-6R; IL-10R; IL-18R, а также рецепторы цитокинов, представленных в Приложении 2 или Приложении 3, а также рецепторы, раскрытые в Приложении 2 и 3. Понятно, что этот перечень не следует рассматривать как исчерпывающий. Когда полиспецифический лиганд связывается с двумя эпитопами (на одном и том же или разных антигенах), антиген(ы) может быть выбран из этого перечня. В конкретных воплощениях лиганд включает dAb, которое связывает TNFR1, и второе dAb или эпитоп-связывающий домен, который связывает любой из этих антигенов. В таких воплощениях полиспецифический лиганд может включать любую комбинацию вариабельных доменов иммуноглобулина (например, VHVH, VHVL, VLVL). Получение полиспецифических иммуноглобулиновых лигандов Лиганды с двойной специфичностью по изобретению, имеющие открытую или закрытую конформацию в соответствии с желаемой конфигурацией по изобретению, могут быть получены в соответствии с ранее известными способами, используемыми в области инженерии антител, для получения scFv, "фаговых" антител и других сконструированных молекул антитела. Способы получения антител и, в частности, биспецифических антител описаны, например, в следующих обзорах и ссылках, процитированных в данном описании: Winter & Milstein, (1991) Nature 349:293-299; Pluckthun (1992) Immunological Reviews 130:151-188; Wright et al, (1992) Crti. Rev. lmmunol.12:125-168; Holliger. P. & Winter, G. (1993) Curr. Op. Biotechn. 4, 446-449; Carter, et al. (1995) J. Hematother. 4, 463-470; Chester, K.A. & Hawkins, R.E. (1995) Trends Biotechn. 13, 294-300; Hoogenboom, H.R. (1997) Nature Biotechnol. 15, 125-126; Fearon, D. (1997) Nature Biotechnol. 15, 618-619; Pluckthun, A. & Pack, P. (1997) Inumunotechnology 3, 83-105; Carter, P. & Merchant, A.M. (1997) Curr. Opin. Biotechnol. 8, 449-454; Holliger, P. & Winter, G. (1997) Cancer Immunol. Immunother. 45, 128-130.
В изобретении предложена селекция вариабельных доменов против двух разных антигенов или эпитопов, и последующая комбинация вариабельных доменов. В способах, используемых для селекции вариабельных доменов, используют библиотеки и способы селекции, которые известны в данной области. Природные библиотеки (Marks et al (1991) J. Mol. Biol, 222: 581; Vaughan et al (1996) Nature Biotech., 14: 309), в которых используются перегруппированные гены V, полученные из человеческих В-клеток, хорошо известны специалистам в данной области. Синтетические библиотеки (Hoogenboom & Winter (1992) J. Mol. Biol, 227: 381; Barbas et al. (1992) Proc. Natl. Acad. Sci. USA, 89: 4457; Nissim et al (1994) EMBO J., 13: 692; Griffiths et al. (1994) EMBO J, 13: 3245; De Kruif et al. (1995) J. Mol. Biol., 248: 97) получают путем клонирования иммуноглобулиновых V генов, обычно с использованием ПЦР. Ошибки в процессе ПЦР могут приводить к высокой степени рандомизации. Библиотеки VH и/или VL могут быть выбраны против целевых антигенов или эпитопов отдельно, в этом случае связывание единичного домена отбирается непосредственно или вместе.
В предпочтительном способе получения лиганда с двойной специфичностью по настоящему изобретению используют систему селекции, в которой репертуар вариабельных доменов выбран для связывания с первым антигеном или эпитопом, а репертуар вариабельных доменов выбран для связывания со вторым антигеном или эпитопом. Селектированные первый и второй вариабельные домены затем комбинируют и выбирают лиганд с двойной специфичностью для связывания с первым и вторым антигеном или эпитопом. Лиганды с закрытой конформацией отбирают для связывания с первым и вторым антигеном или эпитопом при выделении, но не одновременно.
А. Библиотечные векторные системы
В данной области известно множество систем селекции, которые подходят для применения в настоящем изобретении. Примеры таких систем описаны ниже.
Экспрессирующие системы на основе бактериофага лямбда могут быть подвергнуты скринингу непосредственно в виде бляшек бактериофагов или в виде колоний лизогенов, как описано ранее (Huse et al. (1989) Science, 246: 1275; Caton and Koprowski (1990) Proc. Natl. Acad. Sci. U.S.A., 87; Mullinax et al. (1990) Proc. Natl. Acad. Sci. U.S.A., 87: 8095; Persson et al. (1991) Proc. Natl Acad. Sci. U.S.A., 88: 2432), и находят применение в изобретении. Хотя такие экспресссирующие системы могут быть использованы для скрининга вплоть до 106 различных членов библиотеки, они реально не подходят для скрининга больших количеств (более 106 членов).
Особое применение в конструировании библиотек представляют селективные дисплейные системы, которые позволяют нуклеиновой кислоте связываться с полипептидом, который с нее экспрессируется. Используемая в данном изобретении селективная дисплейная система представляет собой систему, дающую возможность для селекции с помощью подходящих дисплейных средств индивидуальных членов библиотеки путем связывания с общими лигандами и/или лигандами-мишенями.
Протоколы селекции для выделения желаемых членов крупных библиотек известны в данной области, в качестве типичного примера выступают способы фагового дисплея. Такие системы, в которых отличающиеся пептидные последовательности представлены на поверхности филаментного бактериофага (Scott and Smith (1990) Science, 249: 386), оказались полезными для создания библиотек фрагментов антител (и нуклеотидных последовательностей, которые кодируют их) для селекции in vitro и амплификации специфических фрагментов антител, которые связываются с антигеном-мишенью (McCafferty et al., WO 92/01047). Нуклеотидные последовательности, кодирующие области VH и VL, связаны с генными фрагментами, которые кодируют лидерные сигналы, направляющие их в периплазматическое пространство Е. coli, и в результате фрагменты антител представлены на поверхности бактериофага, как правило, в виде слияний с белками оболочки бактериофага (например, pIII или pVIII). Альтернативно, фрагменты антитела представлены снаружи на капсидах фага лямбда (фаговые тела). Преимущество фаговых дисплейных систем заключается в том, что поскольку они представляют собой биологические системы, выбранные члены библиотеки могут быть амплифицированы просто путем выращивания фага, содержащего выбранный член библиотеки, в бактериальных клетках. Кроме того, поскольку нуклеотидная последовательность, которая кодирует член полипептидной библиотеки, содержится на фаге или фагмидном векторе, секвенирование, экспрессия и последующая генетическая манипуляция относительно просты.
Способы конструирования бактериофаговых дисплейных библиотек антител и библиотек экспрессии фага лямбда хорошо известны в данной области (McCafferty et al. (1990) Nature, 348: 552; Kang et al. (1991) Proc. Natl. Acad. Sci. U.S.A., 88: 4363; Clackson et al. (1991) Nature, 352: 624; Lowman et al. (1991) Biochemistry, 30: 10832; Burton et al (1991) Proc. Natl. Acad. Sci. U.S.A., 88: 10134; Hoogenboom et al. (1991) Nucleic Acids Res., 19: 4133; Chang et al. (1991) J. Immunol., 147: 3610; Breitling et al. (1991) Gene, 104: 147; Marks et al. (1991) выше; Baxbas et al. (1992) выше; Hawkins and Winter (1992) J. Immunol, 22: 867; Marks et at, 1992, J. Biol. Chem., 267: 16007; Lerner et al. (1992) Science, 258: 1313, включены в данное изобретение путем ссылки).
Один из особенно предпочтительных подходов представляет собой применение scFv фаговых библиотек (Huston et al, 1988, Proc. Natl. Acad. Sci. U.S.A., 85: 5879-5883; Chaudhary et al (1990) Proc. Natl. Acad. Sci U.S.A., 87: 1066-1070; McCafferty et al (1990) выше; Clackson et al (1991) Nature, 352: 624; Marks et al (1991) J. Mol. Biol., 222: 581; Chiswell et al (1992) Trends Biotech., 10: 80; Marks et al (1992) J. Biol. Chem., 267). Описаны различные воплощения библиотек scFv, представленных на белках оболочки бактериофагов. Также известны тонкости подходов фагового дисплея, например, как описано в WO96/06213 и WO92/01047 (Medical Research Council et al.) и WO97/08320 (Morphosys), которые включены в данное изобретение путем ссылки.
Другие системы создания библиотек полипептидов включают применение бесклеточной ферментативной системы для синтеза членов библиотеки in vitro. В одном из способов молекулы РНК выбраны с помощью чередующихся раундов селекции против лиганда-мишени и ПЦР-амплификации (Tuerk and Gold (1990) Science, 249: 505; Ellington and Szostak (1990) Nature, 346: 818). Похожий способ может быть использован для идентификации последовательностей ДНК, которые связывают заданный человеческий транскрипционный фактор (Thiesen and Bach (1990) Nucleic Acids Res., 18: 3203; Beaudry and Joyce (1992) Science, 257: 635; WO92/05258 и WO92/14843). Аналогично, трансляция in vitro может быть использована для синтеза полипептидов в качестве способа создания крупных библиотек. Эти способы, которые обычно включают стабилизированные полисомные комплексы, дополнительно описаны в WO88/08453, WO90/05785, WO90/07003, WO91/02076, WO91/05058 и WO92/02536. Альтернативные дисплейные системы, которые не являются фаговыми, такие как раскрытые в WO95/22625 и WO95/11922 (Affymax), используют полисомы для представления полипептидов для селекции.
Еще одна дополнительная категория способов включает селекцию репертуаров в искусственных компартментах, которые дают возможность для связи гена с его генным продуктом. Например, выбор системы, в которой нуклеиновые кислоты, кодирующие желаемые генные продукты, могут быть выбраны в микрокапсулах, образованных эмульсиями вода-в-масле, описан в WO99/02671, WO00/40712 и Tawfik & Griffiths (1998) Nature Biotechnol 16(7), 652-6. Генетические элементы, кодирующие генный продукт, имеющий желаемую активность, компартментализуются в микрокапсулы и затем транскрибируются и/или транслируются с получением соответствующих генных продуктов (РНК или белок) в микрокапсулах. Генетические элементы, которые продуцируют генный продукт, имеющий желаемую активность, затем сортируют. Этот подход позволяет отбирать интересующие генные продукты путем обнаружения желаемой активности с помощью различных способов.
Б. Конструирование библиотек
Библиотеки, предназначенные для селекции, могут быть сконструированы с использованием способов, известных в данной области, например, как изложено выше, или могут быть приобретены из коммерческих источников. Библиотеки, которые полезны в настоящем изобретении, описаны, например, в WO99/20749. После выбора векторной системы и клонирования одной или более чем одной последовательности нуклеиновой кислоты, кодирующей интересующие полипептиды, в библиотечном векторе, можно создавать разнообразие в клонируемой молекуле путем мутагенеза перед экспресиией; альтернативно, кодируемые белки могут быть экспрессированы и селектированы, как описано выше, перед мутагенезом и осуществлением дополнительных раундов селекции. Мутагенез последовательностей нуклеиновой кислоты, кодирующих структурно оптимизированные полипептиды, осуществляют с помощью стандартных молекулярных способов. Особенное применение представляет полимеразная цепная реакция или ПЦР (Mullis and Faloona (1987) Methods Enzymol, 155: 335, включенная в данное изобретение путем ссылки). ПЦР, в которой используются многочисленные циклы ДНК репликации, катализируемой термостабильной ДНК-зависимой ДНК-полимеразой для амплификации интересующей последовательности-мишени, хорошо известна в данной области. Конструкция различных библиотек антитела обсуждается в Winter et al. (1994) Ann. Rev. Immunology 12, 433-55, и процитированных в ней ссылках.
ПЦР осуществляют с использованием матричной ДНК (по меньшей мере 1 фг; более эффективно 1-1000 нг) и по меньшей мере 25 пмоль олигонуклеотидных праймеров; можно предпочтительно использовать большее количество праймера, когда праймерный набор весьма гетерогенен, поскольку каждая последовательность представлена только небольшой фракцией молекул в наборе, и количества становятся ограничивающими в более поздних циклах амплификации. Типичная реакционная смесь включает: 2 мкл ДНК, 25 пмоль олигонуклеотидного праймера, 2,5 мкл 10Х буфера 1 для ПЦР (Perkin-Elmer, Foster City, CA), 0,4 мкл 1,25 мкМ dNTP, 0,15 мкл (или 2,5 единицы) Taq ДНК-полимеразы (Perkin Elmer, Foster City, CA) и деионизированную воду до общего объема 25 мкл. Наслаивают минеральное масло, и ПЦР осуществляют с использованием программируемого термоциклера. Длительность и температуру каждой стадии цикла ПЦР, а также количество циклов, корректируют в соответствии с фактическими требованиями жесткости. Температуру отжига и время определяют по эффективности, с которой, как ожидают, праймер отжигается на матрице, и степени несоответствий, которая допускается; очевидно, когда молекулы нуклеиновой кислоты одновременно амплифицируются и подвергаются мутагенезу, несоответствие необходимо, по меньшей мере в первом раунде синтеза. Способность оптимизировать жесткость условий отжига праймера находится в пределах знаний специалиста в данной области. Используют температуру отжига от 30°С до 72°С. Начальная денатурация матричных молекул обычно происходит при температуре от 92°С до 99°С в течение 4 минут с последующими 20-40 циклами, состоящими из денатурации (94-99°С в течение от 15 секунд до 1 минуты), отжига (температура определяется, как обсуждается выше; 1-2 минуты) и удлинения (72°С в течение 1-5 минут, в зависимости от длины амплифицируемого продукта). Окончательное удлинение осуществляется, как правило, в течение 4 минут при 72°С, и за этим может следовать неопределенная (0-24 часа) стадия при 4°С.
В. Комбинирование единичных вариабельных доменов
Домены, полезные в изобретении, после селекции можно комбинировать множеством способов, известных в данной области, включая ковалентные и нековалентные способы.
Предпочтительные способы включают применение полипептидных линкеров, как описано, например, в связи с молекулами scFv (Bird et al, (1988) Science 242:423-426). Обсуждение подходящих линкеров предложено в Bird et al. Science 242, 423-426; Hudson et al, Journal Immunol Methods 231 (1999) 177-189; Hudson et al, Proc Nat Acad Sci USA 85, 5879-5883. Линкеры являются предпочтительно гибкими, обеспечивающими возможность для взаимодействия двух единичных доменов. Один из примеров линкера представляет собой линкер (Gly4Ser)n, где n=1-8, например, 2, 3, 4, 5 или 7. Также можно использовать линкеры, используемые в диа-антителах, которые являются менее гибкими (Holliger et al, (1993) PNAS (USA) 90:6444-6448).
В одном из воплощений используемый линкер не представляет собой шарнирную область иммуноглобулина.
Вариабельные домены можно комбинировать с использованием иных способов, чем линкеры. Например, применение дисульфидных мостиков, полученных с помощью встречающихся в природе или сконструированных цистеиновых остатков, может быть использовано для стабилизации димеров VH-VH, VL-VL или VH-VL (Reiter et al, (1994) Protein Eng. 7:697-704) или с помощью ремоделирования поверхности раздела между вариабельными доменами для улучшения "соответствия" и, следовательно, стабильности взаимодействия (Ridgeway et al, (1996) Protein Eng. 7:617-621; Zhu et al, (1997) Protein Science 6:781-788).
Другие способы соединения или стабилизации вариабельных доменов иммуноглобулинов, в частности VH-доменов антитела, могут быть использованы подходящим образом.
В соответствии с настоящим изобретением лиганды с двойной специфичностью могут находиться в растворе в "закрытых" конформациях. "Закрытая" конфигурация заключается в том, что два домена (например, VH и VL) присутствуют в ассоциированной форме, такой как ассоциированная пара VH-VL, которая образует сайт связывания антитела. Например, scFv может находиться в закрытой конформации, зависимой от структуры линкера, используемого для связи доменов VH и VL. Если он достаточно гибкий для того, чтобы обеспечить ассоциацию доменов, или жестко удерживает их в ассоциированном положении, то, вероятно, что домены адаптированы к закрытой конформации.
Аналогично, пары VH-доменов и пары VL-доменов могут существовать в закрытой конформации. Как правило, она будет представлять собой функцию закрытой ассоциации доменов, например с помощью жесткого линкера, в молекуле лиганда. Оба лиганда в закрытой конформации будут не способны связываться с молекулой, увеличивающей период полувыведения лиганда, и со второй молекулой-мишенью. Таким образом, лиганд будет связываться со второй молекулой-мишенью только после диссоциации от молекулы, которая увеличивает период полувыведения лиганда.
Кроме того, конструкция димеров VH/VH, VL/VL или VH/VL без линкеров предложена для конкуренции между доменами.
Лиганды по изобретению, кроме того, могут находиться в открытой конформации. В такой конформации оба лиганда способны одновременно связываться с молекулой, которая увеличивает период полувыведения лиганда, и со второй молекулой - мишенью. Как правило, вариабельные домены в открытой конфигурации (в случае пар VH-VL) достаточно далеко удерживаются от доменов для того, чтобы не взаимодействовать и образовывать сайт связывания антитела и не конкурировать за связывание с соответствующими эпитопами. В случае димеров VH/VH или VL/VL, домены не сжимаются вместе жесткими линкерами. В норме такие пары доменов не конкурируют за связывание с антигеном или образуют сайт, связывающий антитело.
Fab-Фрагменты и целые антитела существуют в основном в закрытой конформации, хотя понятно, что лиганды с двойной специфичностью в открытой и закрытой конформации, вероятно, существуют во множестве равновесий в различных условиях. Связывание лиганда с мишенью, вероятно, сдвигает баланс равновесия в направлении открытой конфигурации. Таким образом, некоторые лиганды по изобретению могут существовать в растворе в двух конформациях, одна из которых (открытая форма) может независимо связывать два антигена или эпитопа, тогда как альтернативная конформация (закрытая форма) может связываться лишь с одним антигеном или эпитопом; таким образом, антигены или эпитопы конкурируют за связывание с лигандом в этой конформации.
Хотя открытая форма лиганда с двойной специфичностью, таким образом, может существовать в равновесии с закрытой формой в растворе, предполагают, что равновесие будет способствовать закрытой форме; кроме того, открытая форма может быть ограничена связыванием мишени в закрытой конформации. Таким образом, предпочтительно, чтобы некоторые лиганды с двойной специфичностью по изобретению находились в равновесии между двумя (открытой и закрытой) конформациями.
Лиганды с двойной специфичностью по изобретению могут быть модифицированы для того, чтобы благоприятствовать открытой или закрытой конформации. Например, стабилизация взаимодействий VH-VL с дисульфидными связями стабилизирует закрытую конформацию. Кроме того, линкеры, используемые для связывания доменов, включая пары доменов VH и VL, могут быть сконструированы таким образом, чтобы благоприятствовать открытой форме; например, линкеры стерически могут препятствовать ассоциации доменов, например, путем включения крупных аминокислотных остатков в противоположных положениях, или разработки подходящей жесткой структуры, которая будет сохранять домены на физическом расстоянии друг от друга.
Г. Характеристика лиганда с двойной специфичностью
Связывание лиганда с двойной специфичностью с его специфическими антигенами или эпитопами можно протестировать с использованием способов, которые знакомы специалистам в данной области и включают ИФА. В предпочтительном воплощении изобретения связывание тестируют с использованием моноклонального фагового ИФА.
Фаговый ИФА может быть осуществлен в соответствии с любым подходящим способом: протокол примера представлен ниже.
Популяции фага, продуцируемые при каждом раунде селекции, могут быть подвергнуты скринингу в отношении связывания с помощью ИФА с выбранным антигеном или эпитопом для идентификации "поликлональных" фаговых антител. Фаг из единичных инфицированных бактериальных колоний из этих популяций затем может быть подвергнут скринингу с помощью ИФА для идентификации "моноклональных" фаговых антител. Также желательно подвергнуть скринингу растворимые фрагменты антител на связывание с антигеном или эпитопом, и это также может быть осуществлено с помощью ИФА с использованием реагентов, например против С- или N-концевой метки (смотри например Winter et al. (1994) Ann. Rev. Immunology 12, 433-55 и цитированные в ней ссылки.
Разнообразие выбранных фаговых моноклональных антител также может быть оценено с помощью гель-электрофореза ПЦР-продуктов (Marks et al. 1991, выше; Nissim et al. 1994 выше), зондирования (Tomlinson et al, 1992) J. Mol. Biol. 227, 776) или путем секвенирования векторной ДНК.
Д. Структура лигандов с двойной специфичностью
Как описано выше, антитело определено в данном изобретении как антитело (например, IgG, IgM, IgA, IgA, IgE) или фрагмент (Fab, Fv, дисульфид-связанный Fv, scFv, диа-антитела), которые включают по меньшей мере один вариабельный домен тяжелой и легкой цепи, по меньшей мере два вариабельных домена тяжелой цепи или по меньшей мере два вариабельных домена легкой цепи. Они по меньшей мере частично могут быть получены из любых видов, продуцирующих в природе антитело, или получен с помощью технологии рекомбинантной ДНК; выделенные из сыворотки крови, В-клеток, гибридом, трансфектом, дрожжей или бактерий).
В предпочтительном воплощении изобретения лиганд с двойной специфичностью включает по меньшей мере один единичный вариабельный домен тяжелой цепи антитела и один единичный вариабельный домен легкой цепи антитела, или два единичных вариабельных домена тяжелой или легкой цепей. Например, лиганд может включать пару VH/VL, пару VH-доменов или пару VH-доменов.
Первый и второй вариабельные домены такого лиганда могут находиться на одной и той же полипептидной цепи. Альтернативно, они могут располагаться на отдельных полипептидных цепях. В случае когда они находятся на одной и той же полипептидной цепи, они могут быть связаны линкером, который предпочтительно представляет собой пептидную последовательность, как описано выше.
Первый и второй вариабельные домены могут быть ковалентно или нековалентно ассоциированы. В случае когда они ковалентно ассоциированы, ковалентные связи могут представлять собой дисульфидные связи.
В случае когда вариабельные домены выбраны из репертуаров V-генов, например, с использованием описанной в данном изобретении технологии фагового дисплея, тогда эти вариабельные домены включают универсальную каркасную область, так что они могут распознаваться специфическим общим лигандом, как определено в данном изобретении. Применение универсальных каркасов, общих лигандов и т.п. описано в WO 99/20749.
Когда используют репертуары V-генов, тогда вариация полипептидной последовательности предпочтительно находится в структурных петлях вариабельных доменов. Полипептидные последовательности любого вариабельного домена могут быть изменены путем "перетасовки" ДНК или с помощью мутации для усиления взаимодействия каждого вариабельного домена с его комплементарной парой. "Перетасовка" ДНК известна в данной области и изложена, например, в Stemmer, 1994, Nature 370: 389-391, и патенте США №6297053, которые оба включены в данное изобретение путем ссылки. Другие способы мутагенеза хорошо известны специалистам в данной области.
В предпочтительном воплощении изобретения 'лиганд с двойной специфичностью' представляет собой одноцепочечный Fv-фрагмент. В альтернативном воплощении изобретения 'лиганд с двойной специфичностью' состоит из Fab-формата.
В еще одном аспекте настоящего изобретения предложена нуклеиновая кислота, кодирующая по меньшей мере 'лиганд с двойной специфичностью', как определено в данном изобретении.
Специалисту в данной области понятно, что в зависимости от аспекта изобретения, оба антигена или эпитопа могут связываться одновременно с одной и той же молекулой антитела. Альтернативно, они могут конкурировать за связывание с одной и той же молекулой антитела. Например, когда оба эпитопа связаны одновременно, тогда оба вариабельных домена лиганда с двойной специфичностью способны независимо связываться со своими эпитопами-мишенями. Когда домены конкурируют, тогда один вариабельный домен способен связывать свою мишень, но не одновременно с тем, когда другой вариабельный домен связывает свою когнатную мишень; или первый вариабельный домен способен связывать свою мишень, но не одновременно с тем, когда второй вариабельный домен связывает свою когнатную мишень.
Вариабельные области могут происходить из антител, направленных против целевых антигенов или эпитопов. Альтернативно, они могут происходить из репертуара единичных доменов антитела, например, экспрессирующихся на поверхности филаментного бактериофага. Селекцию можно осуществлять, как описано ниже.
Как правило, молекулы нуклеиновой кислоты и векторные конструкции, необходимые для осуществления настоящего изобретения, могут быть сконструированы и с ними можно манипулировать, как представлено в стандартных лабораторных руководствах, таких как Sambrook et al. (1989) Molecular Cloning: A Laboratory Manual, Cold Spring Harbor, USA.
Манипуляции с нуклеиновыми кислотами, полезными в настоящем изобретении, как правило, осуществляют в рекомбинантных векторах.
Таким образом, в еще одном аспекте настоящего изобретения предложен вектор, содержащий нуклеиновую кислоту, кодирующую по меньшей мере 'лиганд с двойной специфичностью', как определено в данном изобретении.
Используемый в данном изобретении вектор относится к дискретному элементу, который используют для введения гетерологичной ДНК в клетки для ее экспрессии и/или репликации. Способы, с помощью которых осуществляют селекцию или конструирование, и затем применение таких векторов, хорошо известны специалистам в данной области. Многочисленные векторы общественно доступны, включая бактериальные плазмиды, бактериофаг, искусственные хромосомы и эписомные векторы. Такие векторы могут быть использованы для простого клонирования и мутагенеза; альтернативно используют вектор, экспрессирующий ген. Используемый по изобретению вектор может быть выбран для того, чтобы вмещать полипептид-кодирующую последовательность желаемого размера, типично от 0,25 килобаз (кб) до 40 кб или более в длину. Подходящую клетку-хозяина трансформируют вектором после манипуляций клонирования in vitro. Каждый вектор содержит различные функциональные компоненты, которые обычно включают клонирующий (или "полилинкерный") сайт, точку начала репликации и по меньшей мере один селектируемый маркерный ген. Если заданный вектор представляет собой экспрессирующий вектор, то он дополнительно обладает одним или более чем одним из следующего: последовательностей энхансерного элемента, промотора, терминации трансляции и сигнальной последовательности, каждая из которых расположена по близости от клонирующего сайта, так что они функционально связаны с геном, кодирующим лиганд по изобретению.
И клонирующий, и экспрессирующий векторы, как правило, содержат последовательности нуклеиновых кислот, которые позволяют вектору реплицироваться в одной или более чем одной выбранной клетке-хозяине. Типично в клонирующих векторах эта последовательность представляет собой последовательность, которая позволяют вектору реплицироваться независимо от хозяйской хромосомной ДНК и включает точки начала репликации или автономно реплицирующиеся последовательности. Такие последовательности хорошо известны для множества бактерий, дрожжей и вирусов. Точка начала репликации из плазмидной pBR322 подходит для большинства грамотрицательных бактерий, 2-микронная плазмидная точка начала репликации подходит для дрожжей, а различные вирусные точки начала (например, аденовируса SV 40) полезны для клонирующих векторов в клетках млекопитающих. Как правило, точка начала репликации не обязательна для экспрессирующих векторов млекопитающих, если только они не используются в клетках млекопитающих, способных реплицировать высокие уровни ДНК, таких как клетки COS.
Предпочтительно, клонирующий или экспрессирующий вектор может содержать селективный ген, также известный как селективный маркер. Этот ген кодирует белок, необходимый для выживания или роста трансформированных клеток-хозяев, выращиваемых в селективной культуральной среде. Клетки-хозяева, не трансформированные вектором, содержащим селективный ген, таким образом, не будут выживать в данной культуральной среде. Типичные селективные гены кодируют белки, которые придают устойчивость к антибиотикам и другим токсинам, например ампициллину, неомицину, метотрексату или тетрациклину, дополнительные ауксотрофные дефициты, или обеспечивают критические питательные вещества, не содержащиеся в ростовых средах.
Поскольку репликацию векторов, кодирующих лиганд по настоящему изобретению, для максимального удобства осуществляют в Е. coli, используют E. coli-селективный маркер, например ген β-лактамазы, который придает устойчивость к антибиотику ампициллину. Он может быть получен из плазмид на основе Е. coli, такой как pBR322, или плазмиды pUC, такой как pUC18 или pUC19.
Экспрессирующие векторы обычно содержат промотор, который распознается организмом хозяина и функционально связан с интересующей кодирующей последовательностью. Такой промотор может быть индуцибельным или конститутивным. Термин "функционально связанный" относится к непосредственному соседству, где описанные компоненты находятся в связи, позволяющей им функционировать предполагаемым образом. Регуляторную последовательность, "функционально связанную" с кодирующей последовательностью, лигируют таким образом, чтобы экспрессия кодирующей последовательности достигалась в условиях, совместимых с регуляторными последовательностями.
Промоторы, подходящие для применения с прокариотическими хозяевами, включают, например β-лактамазную и лактозную промоторную системы, щелочную фосфатазу, триптофановую (trp) промоторную систему и гибридные промоторы, такие как tac промотор. Промоторы для применения в бактериальных системах также как обычно содержат последовательность Шайна-Далгарно, функционально связанную с кодирующей последовательностью.
Предпочтительные векторы представляют собой экспрессирующие векторы, которые позволяют экспрессироваться нуклеотидной последовательности, соответствующей члену полипептидной библиотеки. Таким образом, выбор первого и/или второго антигена или эпитопа может осуществляться путем раздельного размножения и экспрессии единичного клона, экспрессирующего член полипептидной библиотеки, или путем использования любой селективной дисплейной системы. Как описано выше, предпочтительная селективная дисплейная система представляет собой бактериофаговый дисплей. Таким образом, могут быть использованы фаговые или фагмидные векторы, например р1Т1 или р1Т2. Лидерные последовательности, полезные в изобретении, включают в себя peIB, stll, ompA, phoA, bla и peIA. Один из примеров являются фагмидные векторы, которые имеют точку начала репликации Е. coli (для двухцепочечной репликации), а также точку начала репликации фага (для получения одноцепочечной ДНК). Манипуляция с такими векторами и экспрессия таких векторов хорошо известна в данной области (Hoogenboom and Winter (1992) выше; Nissim et al. (1994) выше). Кратко, вектор содержит р-лактамазный ген для придания селективности на фагмиде и lac промотор выше экспрессирующей кассеты, которая состоит (от N- к С-концу) из pelB лидерной последовательности (которая направляет экспрессирующийся полипептид в периплазматическое пространство), сайт множественного клонирования (для клонирования нуклеотидного варианта члена библиотеки), возможно одну или более чем одну пептидную метку (для обнаружения), возможно один или более чем один TAG стоп-кодон и фаговый белок pIII. Таким образом, с использованием различных супрессорных и несупрессорных штаммов Е. coli и с добавлением глюкозы, изопропилтио-β-D-галактозида (IPTG) или хелперного фага, такого как VCS М13, вектор способен реплицироваться в виде плазмиды при отсутствии экспрессии, продуцировать значительные количества только члена полипептидной библиотеки или продуцировать фаг, некоторые из которых содержат по меньшей мере одну копию слияния полипептид-pIII на своей поверхности.
При конструировании векторов, кодирующих лиганды по изобретению, используют стандартные способы лигирования. Выделенные векторы или фрагменты ДНК расщепляют, подгоняют и повторно лигируют в форме, желательной для получения требуемого вектора. При желании, может быть осуществлен известным образом анализ для подтверждения того, что в конструируемом векторе находятся правильные последовательности. Подходящие способы для конструирования экспрессирующих векторов, получения транскриптов in vitro, введения ДНК в клетки-хозяева и проведения анализов для оценки экспрессии и функции известны специалистам в данной области. Обнаруживают присутствие генной последовательности в образце, или ее амплификацию и/или экспрессию количественно определяют стандартными способами, такими как Саузерн- или Нозерн-анализ, Вестерн-блоттинг, дот-блоттинг ДНК, РНК или белка, гибридизация in situ, иммуноцитохимия или анализ последовательнсти нуклеиновой кислоты или белковых молекул. Специалист в данной области легко представит, каким образом эти способы, при желании, могут быть модифицированы.
Структура полиспецифических лигандов с закрытой конформацией В соответствии с одним из аспектов второй конфигурации по настоящему изобретению, два или более некомплементарных эпитоп-связывающих домена связывают таким образом, что они находятся в закрытой конформации, как определено в данном изобретении. Предпочтительно, они могут быть дополнительно присоединены к скелету, который, в качестве альтернативы или дополнительно к описанному в данном изобретении линкеру, облегчает образование и/или поддержание закрытой конформации эпитоп-связывающих сайтов в отношении друг друга.
(I) Скелеты
Скелеты могут быть основаны на молекулах иммуноглобулина или могут быть неиммуноглобулиновыми по происхождению, как изложено выше. Предпочтительные иммуноглобулиновые скелеты, как определено в данном описании, включают любой один или более из следующего: молекула иммуноглобулина, включающая по меньшей мере (1) CL-домен (подкласс каппа или лямбда) антитела; или (2) СН1-домен тяжелой цепи антитела; молекула иммуноглобулина, включающая СН1- и СН2-домены тяжелой цепи антитела; молекула иммуноглобулина, включающая СН1-, СН2- и СН3-домены тяжелой цепи антитела; или любая из подгрупп (2) в сочетании с CL-доменом (подкласс каппа или лямбда) антитела. Также может быть включен домен шарнирной области. Такие комбинации доменов могут, например, имитировать природные антитела, такие как IgG или IgM, или их фрагменты, такие как молекулы Fv, scFv, Fab или F(ab')2. Специалист в данной области понимает, что этот перечень не следует рассматривать как исчерпывающий.
(II) Белковые каркасы
Каждый эпитоп-связывающий домен включает белковый каркас и одну или более CDR, которые вовлечены в специфическое взаимодействие домена с одним или более эпитопами. Предпочтительно, эпитоп-связывающий домен по настоящему изобретению включает три CDR. Подходящие белковые каркасы включают любые из белковых каркасов, выбранных из группы, состоящей из следующего: белковые каркасы на основе иммуноглобулиновых доменов, на основе фибронектина, на основе аффител, на основе CTLA4, на основе шаперонов, таких как GroEL, на основе липокаллина и на основе бактериальных Fc-рецепторов SpA и SpD. Специалисту в данной области понятно, что этот перечень не следует рассматривать как исчерпывающий.
Е: Каркасы для применения при конструировании лигандов с двойной специфичностью
1. Выбор конформации основной цепи
Все члены суперсемейства иммуноглобулинов имеют похожую укладку своей полипептидной цепи. Например, хотя антитела весьма разнообразны с точки зрения своей первичной последовательности, сравнение последовательностей и кристаллографических структур выявило, что в противоположность ожидаемому, пять из шести антиген-связывающих петель антитела (H1, Н2, L1, L2, L3) используют ограниченное количество конформации основной цепи, или канонических структур (Chothia and Lesk (1987) J. Mol. Biol, 196: 901; Chothia et al (1989) Nature, 342: 877). Таким образом, анализ длин петель и ключевых остатков дает возможность для предсказания конформации основной цепи Н1, Н2, L1, L2 и L3, обнаруженных в большинстве человеческих антител (Chothia et al. (1992) J Mol. Biol, 227: 799; Tomlinson et al. (1995) EMBO J., 14: 4628; Williams et al. (1996) J. Mol. Biol, 264: 220). Хотя область Н3 гораздо более разнообразна с точки зрения последовательности, длины и структуры (благодаря применению D-сегментов), она также образует ограниченное количество конформации основной цепи для коротких длин петель, которые зависят от длины и присутствия конкретных остатков или типов остатков, находящихся в ключевых положениях в петле и каркасе антитела (Martin et al (1996) J. Mol Biol, 263: 800; Shirai et al. (1996) FEBS Letters, 399: 1).
Лиганды с двойной специфичностью по настоящему изобретению предпочтительно собраны из библиотек доменов, таких как библиотеки VH-доменов и/или библиотек VL-доменов. Кроме того, лиганды с двойной специфичностью по изобретению сами могут быть предложены в форме библиотек. В одном из аспектов настоящего изобретения разработаны библиотеки лигандов с двойной специфичностью и/или домены, в которых длины некоторых петель и ключевые остатки были выбраны для обеспечения того, что конформация основной цепи членов известна. Предпочтительно, они представляют собой реальные конформации молекул суперсемейства иммуноглобулинов, обнаруженных в природе, для минимизации шансов того, что они не функциональны, как обсуждается выше. Сегменты герминального V-гена служат в качестве одного из подходящих основных скелетов для конструирования библиотек антитела или Т-клеточного рецептора; также используют другие последовательности. С низкой частотой могут возникать вариации, так что небольшое количество функциональных членов может иметь измененную конформацию основной цепи, которая не влияет на ее функцию.
Теорию канонической структуры также используют для оценки количества различных конформации основной цепи, кодируемых лигандами, для предсказания конформации основной цепи на основе лигандных последовательностей и для выбора остатков для диверсификации, которая не влияет на каноническую структуру. Известно, что в человеческом домене VK петля L1 может использовать одну из четырех канонических структур, петля L2 имеет единичную каноническую структуру и что 90% человеческих доменов VK используют одну из четырех или пяти канонических структур для петли L3 (Tomlinson et al. (1995) выше); таким образом, в одном домене VK, различные канонические структуры могут быть комбинированы с получением диапазона различных конформации основной цепи. Считая, что Vλ-домен кодирует различный диапазон канонических структур для петель L1, L2 и L3, и что домены VK и Vλ могут образовывать пару с доменом VH, который может кодировать несколько канонических структур для петель H1 и Н2, количество комбинаций канонических структур, обнаруженных для этих пяти петель, очень велико. Это означает, что получение разнообразия в конформации основной цепи может быть важным для продукции широкого диапазона связывающих специфичностей. Однако при конструировании библиотеки антител на основе единичной известной конформации основной цепи обнаружено, в противоположность ожидаемому, что разнообразие в конформации основной цепи не требуется для создания достаточного разнообразия для нацеливания по существу на все антигены. Еще более неожиданно, единичная конформация основной цепи не обязательно должна представлять собой консенсусную структуру - единичная природная конформация может быть использована в качестве основы для всей библиотеки. Таким образом, в предпочтительном аспекте лиганды с двойной специфичностью по изобретению обладают единичной известной конформацией основной цепи.
Единичная выбранная конформация основной цепи предпочтительно распространена среди интересующего типа молекул суперсемейства иммуноглобулинов. Конформация является распространенной, когда обнаружено, что значительное количество встречающихся в природе молекул используют ее. Соответственно, в предпочтительном аспекте изобретения обнаружение в природе различных конформаций основной цепи для каждой связывающей петли домена иммуноглобулина рассматривают раздельно и затем выбирают встречающийся в природе вариабельный домен, который обладает желаемой комбинацией конформаций основной цепи для различных петель. Если ни один из доменов не присутствует, то может быть выбран самый близкий эквивалент.Предпочтительно, чтобы желаемая комбинация конформаций основной цепи для различных петель была создана путем отбора герминальных генных сегментов, которые кодируют желаемые конформаций основной цепи. Более предпочтительно, чтобы выбранные герминальные генные сегменты часто экспрессировались в природе, и наиболее предпочтительно, чтобы они чаще всего экспрессировались из всех природных герминальных генных сегментов.
При конструировании лигандов с двойной специфичностью или их библиотек частота различных конформаций основной цепи для каждого из шести антиген-связывающих петель может быть рассмотрена по отдельности. Для Н1, Н2, L1, L2 и L3 выбирают заданную конформацию, которую используют от 20% до 100% антиген-связывающих петель природных молекул. Как правило, обнаруженная частота составляет более 35% (т.е. между 35% и 100%) и в идеале более 50% или даже более 65%. Поскольку подавляющее большинство петель Н3 не обладает канонической структурой, предпочтительно выбирать конформацию основной цепи, которая распространена среди петель, которые не демонстрируют канонические структуры. Для каждой из петель, таким образом, выбирают конформацию, которую чаще всего обнаруживают в природном репертуаре. В человеческих антителах наиболее популярными каноническими структурами (КС) для каждой петли являются следующие: Н1 - КС 1 (79% экспрессируемого репертуара), Н2 -КС 3 (46%), L1 - КС 2 в VK (39%), L2 - КС 1 (100%), L3 - КС 1 в VK (36%) (расчет предполагает отношение К:λ 70:30, Hood et al. (1967) Cold Spring Harbor Symp.Quant. Biol, 48: 133). Для петель Н3, имеющих канонические структуры, длина CDR3 (Kabat et al. (1991) Sequences of proteins of immunological interest, U.S. Department of Health and Human Services) из семи остатков с солевым мостиком от остатка 94 к остатку 101, по-видимому, является наиболее распространенной. Существует по меньшей мере 16 последовательностей человеческого антитела в библиотеке данных EMBL, имеющих требуемую длину Н3 и ключевые остатки для образования этой конформации и по меньшей мере две кристаллографические структуры в белковом банке данных, которые могут быть использованы в качестве основы для моделирования антител (2 cgr и 1tet). Наиболее часто экспрессируемые герминальные генные сегменты этой комбинации канонических структур представляют собой VH-сегмент 3-23 (DP-47), JH-сегмент JH4b, VK-сегмент 02/012 (DPK9) и JK-сегмент JK1. VH-Сегменты DP45 и DP38 также подходят. Таким образом, эти сегменты могут быть использованы в комбинации в качестве основы для конструирования библиотеки с желаемой единичной конформацией основной цепи.
Альтернативно, вместо выбора единичной конформации основной цепи на основе появления в природе различных конформации основной цепи для каждой из связывающих петель по отдельности, появление в природе комбинаций конформации основной цепи используют в качестве основы для выбора единичной конформации основной цепи. В случае антител, например, может быть определено появление в природе комбинаций канонической структуры для любых двух, трех, четырех, пяти или всех шести антиген-связывающих петель. Следовательно, предпочтительно, чтобы выбранная конформация была распространена в природных антителах и наиболее предпочтительно, чтобы она чаще всего встречалась в природном репертуаре. Таким образом, в человеческих антителах, например, когда рассматривают природные комбинации пяти антиген-связывающих петель, Н1, Н2, L1, L2 и L3, определяют наиболее частую комбинацию канонических структур и затем комбинируют с наиболее популярной конформацией для Н3-петли в качестве основы для выбора единичной конформации основной цепи.
2. Диверсификация канонической последовательности
Выбрав несколько известных конформации основной цепи или предпочтительно единичную известную конформацию основной цепи, лиганды с двойной специфичностью по изобретению или библиотеки для применения в изобретении могут быть созданы путем варьирования связывающего сайта молекулы для создания репертуара со структурным и/или функциональным разнообразием. Это означает, что создают такие варианты, которые обладают достаточным разнообразием в своей структуре и/или в функции, так что они способны обеспечить диапазон активности.
Желаемое разнообразие типично получают путем варьирования выбранной молекулы в одной или более положениях. Изменяемые положения могут быть выбраны случайно или предпочтительно. Вариация затем может быть достигнута либо путем рандомизации, во время которой резидентная аминокислота замещается любой аминокислотой или ее аналогом, природным или синтетическим, с получением очень большого количества вариантов, либо путем замещения резидентной аминокислоты одной или более чем одной из определенного набора аминокислот с получением более ограниченного количества вариантов.
Сообщали о различных способах введения такого разнообразия. Подверженная ошибкам ПЦР (Hawkins et al. (1992) J. Mol. Biol, 226: 889), химический мутагенез (Deng et al. (1994) J. Biol. Chem., 269; 9533) или бактериальные штаммы-мутаторы (Low et al (1996) J. Mol. Biol, 260: 359) могут быть использованы для введения случайных мутаций в гены, которые кодируют молекулу. Способы мутации выбранных положений также хорошо известны в данной области и включают применение несоответствующих олигонуклеотидов или вырожденных олигонуклеотидов, с использованием ПЦР или без нее. Например, несколько синтетических библиотек антител создано путем нацеливания мутаций на антиген-связывающие петли. Область Н3 человеческого Fab, который связывает столбнячный токсин, случайным образом рандомизирована для создания диапазона новых связывающих специфичностей (Barbas et al (1992) Proc. Natl. Acad. Sci USA, 89: 4457).
Случайные или наполовину случайные области Н3 и L3 прикреплены к герминальным генным сегментам V с получением больших библиотек с немутантными каркасными областями (Hoogenboom & Winter (1992) J. Mol Biol, 227: 381; Barbas et al (1992) Proc. Natl. Acad. Sci. USA, 89: 4457; Nissim et al (1994) EMBO J, 13: 692; Griffiths et al. (1994) EMBO J., 13: 3245; De Kruif et al (1995) J Mol Biol, 248: 97). Такая диверсификация была расширена для включения некоторых или всех других антиген-связываних петель (Crameri et al. (1996) Nature Med., 2: 100; Riechmann et al. (1995) Bio/Technology, 13: 475; Morphosys, WO97/08320, выше).
Поскольку рандомизация петли обладает потенциалом создания приблизительно более чем 1015 структур для одного Н3 и аналогично значительного количества вариантов для других пяти петель, не возможно использование современной технологии трансформации или даже использование бесклеточных систем для получения библиотеки, представляющей все возможные комбинации. Например, в одной из самых крупных библиотек, созданных к настоящему моменту, получили 6×1010 различных антител, что представляет собой единственную фракцию из возможного разнообразия для библиотеки этой конструкции (Griffiths et al. (1994) выше).
В предпочтительном воплощении диверсифицированы только те остатки, которые непосредственно вовлечены в создание или модификацию желаемой функции молекулы. Для множества молекул функция заключается в связывании мишени и таким образом разнообразие должно быть сконцентрировано в сайте связывания мишени, в то же время избегая изменений остатков, которые являются критическими для общей упаковки молекулы или для поддержания выбранной конформации основной цепи.
Диверсификация канонической последовательности в применении к доменам антитела
В случае лигандов антител с двойной специфичностью сайт связывания мишени чаще всего представляет собой антиген-связывающий сайт. Таким образом, в высоко предпочтительном аспекте в изобретении предложены библиотеки или для сборки лигандов антител с двойной специфичностью, в которых только эти остатки в антиген-связывающем сайте варьируют. Эти остатки чрезвычайно разнообразны в репертуаре человеческих антител и, как известно, образуют контакты в комплексах антитело/антиген с высоким разрешением. Например, в L2 известно, что положения 50 и 53 отличаются в природных антителах и обнаружено, что они образуют контакт с антигеном. В отличие от этого, стандартный подход заключается в диверсификации всех остатков в соответствующей области, определяющей комплементарность (CDR1), как определено Kabat et al. (1991, выше), некоторые семь остатков сравнивают с двумя диверсифицированными в библиотеке для применения в изобретении. Это представляет собой значительное улучшение с точки зрения функционального разнообразия, необходимого для создания диапазона антиген-связывающих специфичностей.
В природе, разнообразие антител является результатом двух процессов: соматической рекомбинации герминальных генных сегментов V, D и J для создания наивного первоначального репертуара (так называемого герминального разнообразия и множественности J-сегментов) и соматической гипермутации полученных в результате перестроенных V-генов. Анализ последовательностей человеческого антитела показал, что разнообразие первоначального репертуара сфокусировано в центре антиген-связывающего сайта, тогда как соматическая гипермутация расширяет разнообразие на области, находящиеся на периферии антиген-связывающего сайта, которые высококонсервативны в первоначальном репертуаре (смотри Tomlinson et al. (1996) J. Mol. Biol, 256: 813). Эта комплементарность, возможно, разрабатывается как эффективная стратегия для поиска пространства последовательностей и, хотя, по-видимому, уникально для антител, она легко может быть использована для других полипептидных репертуаров. Остатки, которые варьируют, представляют собой подгруппу остатков, которые образуют сайт связывания мишени. Различные (включая перекрывающиеся) подгруппы остатков в сайте связывания мишени, при желании, диверсифицируются на различных стадиях во время селекции.
В случае репертуара антитела, первоначальный 'наивный' репертуар создают, когда некоторые, а не все, остатки в сайте связывания антигена диверсифицируются. Используемый в данном изобретении в этом контексте термин "наивный" относится к молекуле антитела, которая не имеет предопределенной мишени. Эти молекулы напоминают молекулы, которые кодируются генами иммуноглобулинов индивидуума, которые не подвергались иммунной диверсификации, как в случае с эмбриональными и новорожденными индивидуумами, иммунная система которых еще не сенсибилизирована широким разнообразием антигенных стимулов. Этот репертуар затем отбирается против диапазона антигенов или эпитопов. При необходимости, дополнительное разнообразие может быть затем введено за пределами области, диверсифицированной в первоначальном репертуаре. Этот зрелый репертуар может быть выбран для модифицированной функции, специфичности или аффинности.
В изобретении предложены два различных наивных репертуара связывающих доменов для конструкции лигандов с двойной специфичностью, или наивной библиотеки лигандов с двойной специфичностью, в которых некоторые или все из остатков в антиген-связывающем сайте варьируют."Первичная" библиотека имитирует природный первоначальный репертуар, с разнообразием, ограниченным остатками в центре антиген-связыващего сайта, которые различны в герминальных генных сегментах V (герминальное разнообразие) или диверсифицированы в процессе рекомбинации (множественность J-сегментов). Остатки, которые диверсифицированы, включают в себя Н50, Н52, Н52а, Н53, Н55, Н56, Н58, Н95, Н96, Н97, Н98, L50, L53, L91, L92, L93, L94 и L96, но не ограничиваются ими. В "соматической" библиотеке разнообразие ограничено остатками, которые диверсифицированы в процессе рекомбинации (множественность J-сегментов) или в высокой степени подвергнуты соматическим мутациям). Остатки, которые диверсифицированы, включают в себя 1-131, Н33, Н35, Н95, Н96, Н97, Н98, L30, L31, L32, L34 и L96, но не ограничиваются ими. Все остатки, перечисленные выше, как подходящие для диверсификации в этих библиотеках, как известно, образуют контакты в одном или более комплексах антитело-антиген. Поскольку в обеих библиотеках не все остатки в антиген-связывающем сайте варьируют, дополнительное разнообразие вводят во время селекции путем варьирования оставшихся остатков, если желательно это сделать. Специалисту в данной области должно быть понятно, что любая подгруппа любых из этих остатков (или дополнительных остатков, которые включают антиген-связывающий сайт) может быть использована для первоначальной и/или последующей диверсификации антиген-связывающего сайта.
В конструкции библиотек для применения в изобретении диверсификации выбранных положений обычно достигают на уровне нуклеиновых кислот путем изменения кодирующей последовательности, которая устанавливает последовательность полипептида, так что целый ряд возможных аминокислот (всех 20 или их подгруппы) может быть включен по этому положению. С использованием номенклатуры IUPAC, наиболее изменчивый кодон представляет собой NNK, который кодирует все аминокислоты, а также стоп-кодон TAG. Кодон NNK предпочтительно используют для введения требуемого разнообразия. Другие кодоны, которые достигают тех же концов, также используют, включая кодон NNN, который приводит к получению дополнительных стоп-кодонов TGA и ТАА.
Особенность разнообразия боковой цепи в антиген-связывающем сайте человеческих антител представляет собой явное отклонение, которое благоприятствует определенным аминокислотным остаткам. Если суммировать аминокислотную композицию десяти наиболее разнообразных положений в каждой из областей VH, VK и Vλ, то более 76% разнообразия боковой цепи происходит только из семи различных остатков, представляющих собой серин (24%), тирозин (14%), аспрагин (11%), глицин (9%), аланин (7%), аспартат (6%) и треонин (6%). Это отклонение в сторону гидрофильных остатков и небольших остатков, которые могут обеспечить гибкость основной цепи, возможно отражает эволюцию поверхностей, которые предрасположены к связыванию широкого диапазона антигенов или эпитопов и может способствовать объяснению требуемой разнородности антител в первоначальном репертуаре.
Поскольку предпочтительно имитировать это распределение аминокислот, распределение аминокислот в варьирующих положениях предпочтительно имитирует распределение, наблюдаемое в антиген-связывающем сайте антител. Такое отклонение в замещении аминокислот, которое обеспечивает возможность для выбора некоторых полипептидов (не только полипептидов антитела) против диапазона антигенов-мишеней, легко используют для любого полипептидного репертуара. Существуют различные способы отклонения распределения аминокислот по положению, которое подлежит варьированию (включая применение трехнуклеотидного мутагенеза, смотри WO97/08320), из которых предпочтительным способом, благодаря легкости синтеза, является применение стандартных вырожденых кодонов. Путем сравнения аминокислотного профиля, кодируемого всеми комбинациями вырожденных кодонов (с единичной, двойной, тройной и четырехкратной вырожденностью в равных отношениях в каждом положении) с применением природных аминокислот, можно рассчитать наиболее типичный кодон. Кодоны (AGT)(AGC)T, (AGT)(AGC)C и (AGT)(AGC)(CT) - то есть DVT, DVC и DVY, соответственно с использованием номенклатуры IUPAC - представляют собой кодоны, которые наиболее близки к желаемому аминокислотному профилю: они кодируют 22% серина и 11% тирозина, аспарагина, глицина, аланина, аспартата, треонина и цистеина. Предпочтительно, таким образом, конструировать библиотеки с использованием кодона DVT, DVC или DVY в каждом из диверсифицированных положений.
Ж: Антигены, способные увеличивать период полувыведения лиганда
Лиганды с двойной специфичностью по изобретению в одной из его конфигураций способны связываться с одной или более молекулами, которые могут увеличивать период полувыведения лиганда in vivo. Типично, такие молекулы представляют собой полипептиды, которые в природе возникают in vivo и которые устойчивы к деградации или удалению эндогенными механизмами, которые удаляют нежелательный материал из организма. Например, молекула, которая увеличивает период полувыведения из организма, может быть выбрана из следующего:
Белки внеклеточного матрикса; например коллаген, ламинины, интегрины и фибронектин. Коллагены представляют собой основные белки внеклеточного матрикса. Примерно 15 типов коллагеновых молекул известно в настоящее время, обнаружено в различных частях организма, например, коллаген I типа (составляющий 90% коллагена в организме), обнаруженный в костях, коже, сухожилиях, связках, роговице, внутренних органах, или коллаген II типа, обнаруженный в хряще, межпозвоночных дисках, хорде, стекловидном теле глаза.
Белки, обнаруженные в крови, включая:
Белки плазмы, такие как фибрин, α-2 макроглобулин, сывороточный альбумин, фибриноген А, фибриноген В, сывороточный амилоидный белок А, гептаглобин, профилин, убиквитин, утероглобулин и β-2-микроглобулин;
Ферменты и ингибиторы, такие как плазминоген, лизоцим, цистатин С, альфа-1-антитрипсин и ингибитор панкреатического трипсина. Плазминоген представляет собой неактивный предшественник трипсиноподобной сериновой протеазы плазмина. Он в норме обнаруживается в системе кровообращения. Когда плазминоген активируется и превращается в плазмин, от разворачивает активный ферментативный домен, который расщепляет волокна фибриногена, которые запутывают клетки крови в кровяном сгустке. Это называется фибринолизом.
Белки иммунной системы, такие как IgE, IgG, IgM.
Транспортные белки, такие как ретинол-связывающий белок, α-1 микроглобулин.
Дефензины, такие как бета-дефензин 1, дефензины нейтрофилов 1, 2 и 3.
Белки, обнаруженные в гематоэнцефалическом барьере или нервных тканях, такие как меланокортиновый рецептор, миелин, переносчик аскорбата.
Специфические для рецептора трансферрина слитые белки лиганд-нейрофармацевтический агент (смотри US5977307);
Рецептор эндотелиальных клеток капилляров головного мозга, трансферрин, рецептор трансферрина, инсулин, рецептор инсулиноподобного фактора роста 1 (IGF 1), рецептор инсулиноподобного фактора роста 2 (IGF 2), инсулиновый рецептор.
Белки, локализованные в почках, такие как полицистин, коллаген IV типа, переносчик органических анионов К1, антиген Хейманна.
Белки, локализованные в печени, например алкогольдегидрогеназа G250.
Фактор свертывания крови Х
α1 антитрипсин
HNF 1α
Белки, локализованные в легких, такие как секреторный компонент (связывает IgA).
Белки, локализованные в сердце, например HSP 27. Они ассоциированы с дилатационной кардиомиопатией.
Белки, локализованные в коже, например кератин.
Белки, специфические для костей, такие как костные морфогенные белки (BMP), которые представляют собой подгруппу суперсемейства трансформирующего фактора роста β, которые проявляют остеогенную активность. Примеры включают ВМР-2, -4, -5, -6, -7 (также названные как остеогенный белок (ОР-1) и -8 (ОР-2).
Белки, специфические для опухолей, включая человеческий трофобластный антиген, герцептиновый рецептор, эстрогеновый рецептор, катепсины, например катепсин В (обнаруженный в печени и селезенке).
Белки, специфические для заболевания, такие как антигены, экспрессирующиеся только на активированных Т-клетках, включая:
LAG-3 (ген активации лимфоцитов), остеопротегериновый лиганд (OPGL) смотри Nature 402, 304-309; 1999, OX40 (член семейства рецепторов TNF, экспресирующихся на активированных Т-клетках и единственная костимулирующая Т-клеточная молекула, которая, как известно, подвергается специфической повышающей регуляции в клетках, продуцирующих человеческий вирус Т-клеточной лейкемии I типа (HTLV-I)) Смотри J Immunol. 2000 Jul I; 165(I):263-70; металлопротеазы (ассоциированные с артритом/раком), включая CG6512 Drosophila, человеческий параплегии, человеческий ftsH, человеческий AFG3L2, мышиный ftsH; ангиогенные факторы роста, включая кислый фактор роста фибробластов (FGF-1), основный фактор роста фибробластов (FGF-2), фактор роста эндотелия сосудов/фактор проницаемости сосудов (VE-GF/VPF), трансформирующий фактор роста-α (TGF α), фактор некроза опухоли-альфа (TNF-α), ангиогенин, интерлейкин-3 (IL-3), интерлейкин-8 (IL-8), тромбоцитарный фактор роста эндотелия (PD-ECGF), плацентарный фактор роста (PIGF), midkine тромбоцитарный фактор роста ВВ (PDGF), фракталкин.
Стресс-белки (белки теплового шока)
HSP обычно обнаруживают внутри клеток. Когда их обнаруживают вне клеток, они представляют собой показатель того, что клетка погибла и выплеснула наружу свое содержимое. Эта незапрограммированная гибель клеток (некроз) возникает только тогда, когда в результате травмы, заболевания или повреждения и, следовательно, in vivo, внеклеточные HSP запускают реакцию иммунной системы, которая борется с инфекцией и заболеванием. Двойная специфичность, которая связывается с внеклеточными HSP, может быть локализована в месте заболевания.
Белки, вовлеченные в транспорт Fc
Рецептор Брамбелла (также известный как FcRB).
Этот рецептор Fc имеет две функции, обе из которых потенциально полезны для доставки
Эти функции представляют собой:
Транспорт IgG от матери к ребенку через плаценту
защита IgG от деградации, пролонгирование таким образом периода полувыведения IgG из сыворотки крови.
Полагают, что рецептор рециркулирует IgG из эндосомы.
Смотри Holliger et al, Nat Biotechnol 1997 Jul; 15(7):632-6.
Лиганды по изобретению могут быть сконструированы таким образом, чтобы быть специфичными в отношении вышеуказанных мишеней без необходимости увеличения периода полувыведения in vivo. Например, лиганды по изобретению могут быть специфичными в отношении мишеней, выбранных из вышеуказанных, которые являются тканеспецифическими, таким образом обеспечивая возможность тканеспецифического нацеливания на лиганд с двойной специфичностью или мономер dAb, который связывает тканеспецифическую терапевтически подходящую мишень, независимо от какого-либо любого увеличения периода полувыведения, хотя это может быть результатом. Кроме того, когда лиганд или мономер dAb нацелен на почку или печень, тогда можно перенаправить лиганд или мономер dAb на альтернативный путь клиренса in vivo (например, лиганд может быть направлен из печеночного клиренса к почечному клиренсу).
3: Применение полиспецифических лигандов в соответствии со второй конфигурацией изобретения
Полиспецифические лиганды в соответствии со способом второй конфигурации настоящего изобретения могут быть использованы в терапевтических и профилактических применениях in vivo, диагностических применениях in vitro и in vivo, анализе in vitro и реагентных применениях и т.п. Например, молекулы антител могут быть использованы в способах анализа на основе антител, таких как способы ИФА, в соответствии со способами, известными специалистам в данной области.
Как упомянуто выше, полиспецифические лиганды по изобретению используют в диагностических, профилактических и терапевтических способах. Полиспецифические антитела по изобретению диагностически используют в Вестерн-анализе и в обнаружении белка in situ с помощью стандартных иммуногистохимических способов; для использования в этих применениях лиганды могут быть мечены в соответствии со способами, известными в данной области. Кроме того, такие полипептиды антитела могут быть использованы препаративно в способах аффинной хроматографии, когда они образуют комплекс с хроматографическим носителем, таким как смола. Все такие способы хорошо известны специалистам в данной области.
Диагностические применения полиспецифических лигандов с закрытой конформацией по изобретению включают гомогенные анализы аналитов, в которых используют способность полиспецифических лигандов с закрытой конформацией связываться две конкурирующие мишени, так что две мишени не могут связываться одновременно (закрытая конформация), или альтернативно, их способность связывать две мишени одновременно (открытая конформация).
В еще одном аспекте второй конфигурации по изобретению в настоящем изобретении предложен гомогенный иммуноанализ с использованием лиганда по настоящему изобретению.
Действительный гомогенный формат иммуноанализа пристально рассматривался производителями диагностических средств и исследовательских аналитических систем, используемых при открытии и разработке лекарств. Основные диагностические рынки включают тестирование людей в больницах, врачебных кабинетах и клиниках, коммерческих справочных лабораториях, банках крови и дома, не предназначенные для применения у людей диагностические средства (например, тестирование продуктов питания, тестирование воды, тестирование окружающей среды, биозащита и ветеринарное тестирование), и наконец исследования (включая разработку лекарств; основное исследование и академическое исследование).
В настоящее время на всех этих рынках используют системы иммуноанализа на основе хемилюминисценции, ИФА, флуоресценции или в редких случаях технологий радиоиммуноанализа. Каждый из этих форматов анализа требует стадии разделения (отделения связанных реагентов от несвязанных). В некоторых случаях требуется несколько стадий разделения. Добавление этих дополнительных стадий приводит к добавлению реагентов и автоматизации, занимает время и влияет на конечный выход анализов. В диагностике людей стадия разделения может быть автоматизирована, что маскирует проблемы, но не решает их. Автоматы, дополнительные реагенты, дополнительное время инкубации и т.п. добавляет значительную стоимость и сложность. При разработке лекарств, например, скрининг с высоким выходом, когда буквально миллионы образцов тестируются одновременно, с очень низкими уровнями тестируемой молекулы, добавление дополнительных стадий разделения может устранить возможность проведения скрининга. Однако избегая этого разделения, получают слишком значительный шум при считывании. Таким образом, существует необходимость в действительном гомогенном формате, который обеспечивает чувствительности, находящиеся в диапазоне, получаемом из настоящих форматов анализа. Предпочтительно, анализ обладает полностью количественными считываниями с высокой чувствительностью и значительным динамическим диапазоном. Чувствительность представляет собой важное требование, поскольку она уменьшает требуемое количество образца. Обе эти особенности представляют собой особенности, которые предлагает гомогенная система. Это очень важно с точки зрения тщательного тестирования и при разработке лекарств, когда образцы драгоценны. Гетерогенные системы, доступные в данной области в настоящее время, требуют значительных количеств образца и дорогих реагентов.
Применения для гомогенных анализов включают тестирование рака, где наибольший анализ представляет собой анализ простат-специфического антигена, используемого при скрининге людей на присутствие рака простаты. Другие применения включают тестирование фертильности, которое обеспечивает серию тестов для женщин, пытающихся забеременеть, включая бета-хорионический гонадотропин человека (бета-ХГТ) на беременность. Тесты на инфекционные заболевания, включая гепатит, ВИЧ, краснуху и другие вирусы и микроорганизмы, и заболевания, передаваемые половым путем. Тесты используются в банках крови, в особенности тесты на ВИЧ, гепатит А, В, С, ни А ни В. Тесты для терапевтического лекарственного мониторинга включают мониторинг уровней прописанных лекарств у пациентов на эффективность и во избежание токсичности, например дигоксин при аритмии, и уровни фенобарбитала в психотических случаях; теофиллин при астме. Диагностические тесты также полезны при тестировании наркотических средств, таких как тестирование кокаина, марихуаны и т.п. Метаболические тесты используются для измерения тиреоидной функции, анемии и других физиологических расстройств и функций.
Гомогенный формат иммуноанализа также полезен при изготовлении стандартных клинических химических анализов. Включение иммуноанализов и химических анализов в одном и том же приборе весьма предпочтительно при диагностическом тестировании. Подходящие химические анализы включают тесты на глюкозу, холестерин, калий и т.п.
Дополнительное основное применение гомогенных иммуноанализов представляет собой открытие и разработку лекарства: высокопроизводительный скрининг включает тестирование комбинаторных химических библиотек против мишеней в крупном масштабе. Сигнал обнаруживают, и положительные группы затем разделяют на более мелкие группы, и в конечном счете тестируют в клетках и затем в животных. Гомогенный анализ может быть использован во всех этих типах теста. При разработке лекарства, в особенности в исследованиях на животных и клинических испытаниях осуществляют интенсивное использование иммуноанализов. Гомогенные анализы значительно ускоряют и упрощают эти способы. Другие применения включают тестирование продуктов питания и напитков: тестирование мяса и других продуктов питания на E. coli, сальмонеллу и т.д.; тестирование воды, включая тестирование на водоочистительных станциях на все типы примесей, включающих Е. coli; и ветеринарное тестирование.
В широком воплощении в изобретении предложен анализ связывания, включающий детектируемый агент, который связан с полиспецифическим лигандом с закрытой конформацией по изобретению, и детектируемые свойства которого изменяются путем связывания аналита с указанным полиспецифическим лигандом с закрытой конформацией. Такой анализ может быть конфигурирован несколькими различными путями, каждый из которых использует вышеуказанные свойства полиспецифических лигандов с закрытой конформацией.
Анализ основан на прямом или опосредованном замещении агента аналитом, приводя в результате к изменению детектируемых свойств агента. Например, когда агент представляет собой фермент, способный катализировать реакцию, которая имеет детектируемую конечную точку, тогда указанный фермент может быть связан лигандом таким образом, чтобы блокировать его активный сайт, при этом инактивируя фермент. Аналит, который также связан полиспецифическим лиганом с закрытой конформацией, замещает фермент, делая его активным за счет высвобождения активного сайта. Фермент затем способен реагировать с субстратом, приводя к детектируемому событию. В альтернативном воплощении лиганд может связывать фермент вне активного сайта, влияя на конформацию фермента и таким образом изменяя его активность. Например, структура активного сайта может быть ограничена путем связывания лиганда, или можно предотвратить связывание кофакторов, необходимых для активности.
Физическое воплощение анализа может принимать любую форму, известную в данной области. Например, комплекс полиспецифического лиганда с закрытой конформацией и фермента может быть предложен на тестируемой полоске; субстрат может быть предложен в разной области тестируемой полоски, и растворитель, содержащий аналит, которому разрешено мигрировать через комплекс лиганд/фермент, замещая фермент, и перенос его в область субстрата с получением сигнала. Альтернативно, комплекс лиганд/фермент может быть представлен на тестируемой палочке или другой твердой фазе и погружен в раствор аналита/субстрата, высвобождая фермент в раствор в ответ на присутствие аналита.
Поскольку каждая молекула аналита потенциально высвобождает одну молекулу фермента, анализ является количественным, причем сила сигнала, генерируемого в заданный момент времени, зависит от концентрации аналита в растворе.
Возможны дополнительные конфигурации с использованием аналита в закрытой конформации. Например, полиспецифический лиганд с закрытой конформацией может быть конфигурирован для связывания фермента в аллостерическом сайте, таким образом активируя данный фермент. В таком воплощении фермент является активным в отсутствие аналита. Добавление аналита замещает фермент и устраняет аллостерическую активацию, таким образом инактивируя данный фермент.
В контексте вышеуказанных воплощений, в которых используется активность фермента как показатель концентрации аналита, активация или инактивация фермента относится к увеличению или уменьшению активности фермента, измеренной как способность фермента катализировать реакцию, генерирующую сигнал. Например, фермент может катализировать превращение недетектируемого субстрата в его детектируемую форму. Например, пероксидаза хрена широко используется в данной области вместе с хромогенными или хемилюминисцентными субстратами, которые имеются в продаже. Уровень увеличения или уменьшения активности фермента может находиться между 10% и 100%, такой как 20%, 30%, 40%, 50%, 60%, 70%, 80% или 90%; в случае увеличения активности, данное увеличение может составлять более 100%, т.е. 200%, 300%, 500% или более чем 500%, или не может быть измерено в виде процента, если базовая активность ингибируемого фермента не выявлена.
В еще одной конфигурации полиспецифический лиганд с закрытой конформацией может связывать субстрат пары фермент/субстрат, а не фермент. Таким образом, субстрат недоступен для фермента до высвобождения из полиспецифического лиганда с закрытой конформацией через связывание аналита. Реализации этой конфигурации такие же, как для конфигураций, которые связывают фермент.
Кроме того, анализ может быть конфигурирован для связывания флуоресцентной молекулы, такой как флуоресцеин или другой хромофор, в такой конформации, что флуоресценция гасится при связывании с лиганом. В этом случае связывание аналита с лигандом замещает флуоресцентную молекулу, генерируя таким образом сигнал. Альтернативы флуоресцентных молекул, которые полезны в настоящем изобретении, включают люминесцентные агенты, такие как люциферин/люцифераза, и хромогенные агенты, включая агенты, обычно используемые в иммуноанализах, такие как HRP.
Терапевтические и диагностические композиции и применения
В изобретении предложены композиции, содержащие антагонист TNFR1 (например, лиганд) по изобретению (например, лиганд с двойной специфичностью, полиспецифический лиганд, мономер dAb) и фармацевтически приелемый носитель, разбавитель или эксципиент, и терапевтические и диагностические способы, в которых используются лиганды или композиции по изобретению. Антагонисты и лиганды (например, лиганды с двойной специфичностью, полиспецифические лиганды, мономеры dAb) в соответствии со способом по настоящему изобретению могут быть использованы в терапевтических и профилактических применениях in vivo, диагностических применениях in vivo и т.п.
Терапевтические и профилактические применения антагонистов и лигандов (например, полиспецифических лигандов, лигандов с двойной специфичностью, мономеров dAb) по изобретению включают введение антагонистов и/или лигандов по изобретению млекопитающему реципиенту, такому как человек. Лиганды с двойной специфичностью и полиспецифические лиганды (например, форматы антител с двойной специфичностью) связываются с мультимерным антигеном со значительной авидностью. Лиганды с двойной специфичностью или полиспецифические лиганды могут перекрестно связывать два антигена, например в рекрутирующих цитотоксических Т-клетках для того, чтобы опосредовать килинг клеток опухолевых линий.
По существу чистые лиганды или связывающие их белки, например мономеры dAb, с по меньшей мере 90-95% гомогенностью предпочтительны для введения млекопитающему, и 98-99% или большая гомогенность наиболее предпочтительна для фармацевтических применений, особенно когда млекопитающее представляет собой человека. После очистки, частичной или до гомогенности, по желанию, лиганды могут быть использованы дагностически или терапевтически (включая экстракорпорально) или в разработке и осуществлении аналитических процедур, иммунофлуоресцентных окрашиваний и т.п. (Lefkovite and Pernis, (1979 и 1981) Immunological Methods, Volumes I и II, Academic Press, NY).
Например, лиганды или связывающие их белки, например мономеры dAb, по настоящему изобретению будут типично использовать в предупреждении, подавлении или лечении воспалительных состояний, включающих острые и хронические воспалительные заболевания. Например, антагонисты и/или лиганды могут быть введены для лечения, подавления или предупреждения хронического воспалительного заболевания, аллергической гиперчувствительности, рака, бактериальной или вирусной инфекции, аутоиммунных расстройств (которые включают, но не ограничиваются этим, диабет I типа, астму, рассеянный склероз, ревматоидный артрит, ювенильный ревматоидный артрит, псориатический артрит, спондилоартропатию (например, анкилозирующий спондилит), системную красную волчанку, воспалительное заболевание кишечника (например, болезнь Крона, неспецифический язвенный колит), астенический бульбарный паралич и болезнь Бехчета), псориаз, эндометриоз и абдоминальные адгезии (например, после абдоминальной хирургии).
Антагонисты (например, лиганды) по изобретению (например, лиганды с двойной специфичностью, полиспецифические лиганды, мономеры dAb), которые способны связываться с внеклеточными мишенями, вовлеченными в эндоцитоз (например, клатрин), могут быть подвергнуты эндоцитозу, обеспечивая доступ к внутриклеточным мишеням. Кроме того, лиганды с двойной специфичностью или полиспецифические лиганды обеспечивают способы, с помощью которых связывающий домен (например, мономер dAb), который способен специфически связываться с внеклеточной мишенью, может быть доставлен во внеклеточную среду. Эта стратегия требует, например лиганда с двойной специфичностью, с физическими свойствами, которые позволяют ему оставаться функциональным внутри клетки. Альтернативно, если конечный заданный внеклеточный компартмент является окисляющим, хорошо свернутому лиганду может быть не обязательно быть свободным от дисульфидных связей.
В настоящей заявке термин "предупреждение" включает введение защитной композиция до индукции заболевания. "Подавление" относится к введению композиции после индуктивного события, но перед клиническим проявлением заболевания. "Лечение" включает введение защитной композиции после того, как появились сиптомы заболевания.
Предпочтительно, лиганды с двойной специфичностью или полиспецифические лиганды могут быть использованы для нацеливания на цитокины и другие молекулы, которые синергически кооперируются в терапевтических ситуациях в теле организма. Таким образом, в изобретении предложен способ суммирования активности двух или более связывающих доменов (например, dAb), которые связывают цитокины или другие молекулы, включающий введение лиганда с двойной специфичностью или полиспецифического лиганда, способного связываться с указанными двумя или более молекулами (например, цитокинами). В этом аспекте изобретения лиганд с двойной специфичностью или полиспецифический лиганд может представлять собой любой лиганд с двойной специфичностью или полиспецифический лиганд, включая лиганд, состоящий из комплементарных и/или некомплементарных доменов, лиганд в открытой конформации и лиганд в закрытой конформации. Например, этот аспект изобретения относится к комбинации VH-доменов и VL-доменов, только VH-доменов и только VL-домснов.
Синергизм в терапевтическом контексте может быть достигнут многими путями. Например, целевые комбинации могут быть терапевтически активными, только если лиганд нацелен на обе мишени, тогда как нацеливание только на одну мишень терапевтически не эффективно. В еще одном воплощении только одна мишень может обеспечить некоторый низкий или минимальный терапевтический эффект, но вместе со второй мишенью комбинация обеспечивает синергетическое повышение терапевтического эффекта. Предпочтительно, цитокины, связанные лигандом с двойной специфичностью или полиспецифическими лигандами в соответствии с этим аспектом изобретения, выбраны из перечня, представленного в Приложении 2.
Кроме того, лиганды с двойной специфичностью или полиспецифические лиганды могут быть использованы в онкологических применениях, где одна специфичность нацелена на CD89, который экспрессируется цитотоксическими клетками, а другие являются опухолеспецифическими. Примеры опухолевых антигенов, на которые может быть осуществлено нацеливание, приведены в Приложении 3.
Имеются животные модельные системы, которые могут быть использованы для скрининга эффективности антагонистов TNFR1 (например, лиганды, антитела или связывающие их белки) при защите от заболевания или лечении заболевания. Способы тестирования системной красной волчанки (СКВ) у чувствительных мышей известны в данной области (Knight et al. (1978) J. Exp. Med., 147: 1653; Reinersten et al. (1978) New Eng. J. Med., 299: 515). Астенический бульбарный паралич (MG) тестируют у самок мышей SJL/J путем индукции заболевания растворимым белком AchR от других видов (Lindstrom et al. (1988) Adv. Immunol., 42: 233). Артрит индуцируют в чувствительной линии мышей путем инъекции коллагена II типа (Stuart et al. (1984) Ann. Rev. Immunol, 42: 233). Описана модель, с помощью которой адъювантный артрит вызывают у чувствительных мышей путем инъекции микобактериального белка теплового шока (Van Eden et al. (1988) Nature, 331: 171). Тиреоидит вызывают у мышей путем введения тиреоглобулина, как описано (Maron et al. (1980) J. Exp. Med., 152: 1115). Инсулинозависимый сахарный диабет (IDDM) наблюдается в природе или может быть индуцирован в некоторых линиях мышей, таких как описаны у Kanasawa et al. (1984) Diabetologia, 27: 113. ЕАЕ (аутоиммунный энцефаломиелит) у мышей и крыс служит в качестве модели рассеянного склероза (PC) у людей. В этой модели демиелинизирующее заболевание индуцируют путем введения основного белка миелина (смотри Paterson (1986) Textbook of Immunopathology, Mischer et al, eds., Grune and Stratton, New York, pp.179-213; McFarlin et al (1973) Science, 179: 478: и Satoh et al (1987) J. Immunol. 138: 179).
Как правило, антагонисты по настоящему изобретению (например, лиганды) используют в очищенной форме вместе с фармацевтически приемлемыми носителями. Типично, эти носители включают водные или спиртовые/водные растворы, эмульсии или суспензии, любые из которых включают физиологический раствор и/или забуференные среды. Растворы для парентерального введения включают раствор хлорида натрия, декстрозу Рингера, декстрозу и хлорид натрия и раствор Рингера с лактозой. Подходящие физиологически приемлемые адъюванты, при необходимости удерживать полипептидный комплекс в суспензии, могут быть выбраны из загустителей, таких как карбоксиметилцеллюлоза, поливинилпирролидон, желатин и альгинаты.
Внутривенные разбавители включают жидкие и питательные наполнители и электролитные наполнители, такие как наполнители на основе декстрозы Рингера. Также могут быть представлены консерванты и другие добавки, такие как антимикробные агенты, антиоксиданты, хелатообразующие агенты и инертные газы (Mack (1982) Remington's Pharmaceutical Sciences, 16th Edition). Может быть использовано множество подходящих препаратов, включая препараты с пролонгированным высвобождением.
Антагонисты (например, лиганды) по настоящему изобретению могут быть использованы в виде раздельно вводимых композиций или в сочетании с другими агентами. Они могут включать различные иммунотерапевтические лекарства, такие как циклоспорин, метотрексат, адриамицин или цисплатин и иммунотоксины. Фармацевтические композиции могут включать "коктейли" различных цитотоксических или других агентов в сочетании с антагонистами (например, лигандами) по настоящему изобретению, или даже комбинации лигандов по настоящему изобретению, имеющими различные специфичности, такие как лиганды, выбранные с использованием различных целевых антигенов или эпитопов, где их объединяют или не объединяют перед введением.
Путь введения фармацевтических композиций по изобретению может представлять собой любой путь, обычно известный специалистам в данной области. Для лечения, включая без ограничения иммунотерапию, выбранные лиганды по изобретению могут быть введены любому пациенту в соответствии со стандартными способами. Введение может представлять собой любой подходящий способ, включая парентеральное, внутривенное, внутримышечное, интраперитонеальное, трансдермальное, через легкие, или также соответственно путем прямой инфузии через катетер. Доза и частота введения зависит от возраста, пола и состояния пациента, одновременного введения других лекарств, противопоказаний и других параметров, принятых во внимание клиническими врачами. Введение может быть локальным (например, локальная доставка в легкое путем легочного введения, например интраназального введения) или системным, как указано.
Лиганды по данному изобретению могут быть лиофилизированы для хранения и ресуспендированы в подходящем носителе перед применением. Эта методика, как показано, может быть эффективной со стандартными иммуноглобулинами, и могут быть использованы известные в данной области способы лиофилизации и разведения. Специалисту в данной области известно, что лиофилизация и разведение могут привести к различным степеням потери активности антитела (например, что касается стандартных иммуноглобулинов, то антитела IgM имеют тенденцию больше терять активность по сравнению с антителами IgG) и что уровни применения могут быть увеличены для компенсации.
Композиции, содержащие антагонисты по настоящему изобретению (например, лиганды) или их коктейль, могут быть введены для профилактических и/или терапевтических лечений. В некоторых терапевтических применениях, соответствующее количество для осуществления по меньшей мере частичного ингибирования, подавления, модуляции, киллинга, или некоторого другого измеряемого параметра, популяции выбранных клеток определено как "терапевтически эффективная доза". Количества, необходимые для достижения этой дозы, зависят от тяжести заболевания и общего состояния иммунной системы пациента, но как правило находятся в диапазоне от 0,005 до 5,0 мг лиганда, например антитела, рецептора (например, Т-клеточного рецептора) или его связывающего белка на килограмм массы тела, причем более предпочтительно используют дозы от 0,05 до 2,0 мг/кг/дозу. Для профилактических применений композиции, содержащие лиганды по настоящему изобретению или их коктейли, также могут быть введены в аналогичных или слегка меньших дозах, для предупреждения, ингибирования или замедления начала заболевания (например, для поддержания ремиссии или покоя, или для предотвращения острой фазы). Опытный лечащий врач способен определить подходящий дозовый интервал для лечения, подавления или предупреждения заболевания. Когда антагонист TNFR1 (например, лиганд) вводят для лечения, подавления или предупреждения хронического воспалительного заболевания, тогда его можно вводить до четырех раз в сутки, два раза в неделю, один раз в неделю, один раз каждые две недели, один раз в месяц или один раз каждые два месяца, в дозе, составляющей, например, от примерно 10 мкг/кг до примерно 80 мг/кг, от примерно 100 мкг/кг до примерно 80 мг/кг, от примерно 1 мг/кг до примерно 80 мг/кг, от примерно 1 мг/кг до примерно 70 мг/кг, от примерно 1 мг/кг до примерно 60 мг/кг, от примерно 1 мг/кг до примерно 50 мг/кг, от примерно 1 мг/кг до примерно 40 мг/кг, от примерно 1 мг/кг до примерно 30 мг/кг, от примерно 1 мг/кг до примерно 20 мг/кг, от примерно 1 мг/кг до примерно 10 мг/кг, от примерно 10 мкг/кг до примерно 10 мг/кг, от примерно 10 мкг/кг до примерно 5 мг/кг, от примерно 10 мкг/кг до примерно 2,5 мг/кг, от примерно 1 мг/кг, примерно 2 мг/кг, примерно 3 мг/кг, примерно 4 мг/кг, примерно 5 мг/кг, примерно 6 мг/кг, примерно 7 мг/кг, примерно 8 мг/кг, примерно 9 мг/кг или примерно 10 мг/кг. В конкретных воплощениях антагонист TNFR1 (например, лиганд) вводят для лечения, подавления или предупреждения хронического воспалительного заболевания один раз каждые две недели или один раз в месяц в дозе от примерно 10 мкг/кг до примерно 10 мг/кг (например, примерно 10 мкг/кг, примерно 100 мкг/кг, примерно 1 мг/кг, примерно 2 мг/кг, примерно 3 мг/кг, примерно 4 мг/кг, примерно 5 мг/кг, примерно 6 мг/кг, примерно 7 мг/кг, примерно 8 мг/кг, примерно 9 мг/кг или примерно 10 мг/кг.)
Лечение или терапию, осуществляемую с использованием композиций, описанных в данном изобретении, рассматривают как "эффективные", если один или более симптомов уменьшаются (например, по меньшей мере на 10% или по меньшей мере на одно значение по шкале клинической оценки) относительно таких симптомов, присутствующих до лечения, или относительно таких симптомов у индивидуума (человека или модельного животного), которого не лечили с использованием такой композиции или другого подходящего контроля. Симптомы обычно варьируют в зависимости от предполагаемого заболевания или расстройства, но могут быть измерены клиническим врачом, обладающим обычными знаниями, или лаборантом. Такие симптомы могут быть измерены, например путем мониторинга уровня одного или более биохимических показателей заболевания или расстройства (например, уровней фермента или метаболита, коррелирующего с заболеванием, влияющего на количество клеток и т.д.), путем мониторинга физических проявлений (например, воспаления, размера опухоли и т.д.), или при помощи приемлемой шкалы клинической оценки, например Расширенной шкалы нетрудоспособности (для рассеянного склероза), Опросника Ирвина по определению качества жизни больных воспалительным заболеванием кишечника (32-балльная оценка качества жизни в отношении функции кишечника, системных симптомов, социальной функции и эмоционального статуса - оценка варьирует от 32 до 224, причем более высокие баллы указывают на лучшее качество жизни), Шкалы качества жизни при ревматоидном артрите, или другой приемлемой шкалы клинической оценки, известной в данной области. Длительное (например, на одни сутки или более, предпочтительно более) уменьшение симптомов заболевания или расстройства по меньшей мере на 10% или на одну или более точек на заданной клинической шкале указывает на "эффективное" лечение. Аналогично, профилактика, осуществляемая с использованием описанной в данном изобретении композиции, является "эффективной", если начало или тяжесть одного или более симптомов замедлено, уменьшено или отменено относительно таких симптомов у аналогичного индивидуума (человека или животной модели), которого не лечили данной композицией.
Композиция, содержащая антагонисты (например, лиганд) или их коктейль по настоящему изобретению, может быть использована в профилактических и терапевтических целях для облегчения изменения, инактивации, килинга или удаления выбранной популяции клеток-мишеней у млекопитающего. Кроме того, выбранные репертуары описанных в данном изобретении полипептидов могут быть использованы экстракорпорально или избирательно in vitro для уничтожения, истощения или другого эффективного удаления популяции клеток-мишеней из гетерогенной коллекции клеток. Кровь из млекопитающего может быть объединена экстракорпорально с лигандами, например антителами, рецепторами клеточной поверхности или их связывающими белками, в результате чего нежелательные клетки уничтожаются или иначе удаляются из крови для возвращения млекопитающему в соответствии со стандартными способами.
Композиция, содержащая антагонист (например, лиганд) по настоящему изобретению, может быть использована в профилактических и терапевтических целях для облегчения изменения, инактивации, уничтожения или удаления популяции выбранных клеток-мишеней у млекопитающего.
Антагонисты TNFR1 (например, лиганды, мономеры dAb) могут быть введены и/или приготовлены вместе с одним или более чем одним дополнительным терапевтическим или активным агентом. Когда антагонист TNFR1 (например, лиганд, мономер dAb) вводят с дополнительным терапевтическим агентом, тогда антагонист TNFR1 можно вводить до, одновременно или после введения дополнительного агента. Как правило, антагонист TNFR1 (например, лиганд, мономер dAb) и дополнительный агент вводят способом, который обеспечивает перекрывание терапевтического эффекта.
В одном из воплощений изобретение представляет собой способ лечения, подавления или предупреждения хронического воспалительного заболевания, включающий введение млекопитающему, нуждающемуся в этом, терапевтически эффективной дозы или количества антагониста TNFR1 (например, лиганда, который включает мономер dAb, который связывает TNFR1).
В одном из воплощений изобретение представляет собой способ лечения, подавления или предупреждения артрита (например, ревматоидного артрита, ювенильного ревматоидного артрита, анкилозирующего спондилита, псориатического артрита), включающий введение терапевтически эффективной дозы или количества антагониста TNFR1 (например, лиганда, включающего мономер dAb, который связывает TNFR1) млекопитающему, нуждающемуся в этом.
В еще одном воплощении изобретение представляет собой способ лечения, подавления или предупреждения псориаза, включающий введение терапевтически эффективной дозы или количества антагониста TNFR1 (например, лиганда, включающего мономер dAb, который связывает TNFR1) млекопитающему, нуждающемуся в этом.
В еще одном воплощении изобретение представляет собой способ лечения, подавления или предупреждения воспалительного заболевания кишечника (например, болезни Крона, неспецифического язвенного колита), включающий введение терапевтически эффективной дозы или количества антагониста TNFR1 (например, лиганда, включающего мономер dAb, который связывает TNFR1) млекопитающему, нуждающемуся в этом.
В еще одном воплощении изобретение представляет собой способ лечения, подавления или предупреждения хронического обструктивного заболевания легких (например, хронического бронхита, хронического обструктивного бронхита, эмфиземы), включающий введение терапевтически эффективной дозы или количества антагониста TNFR1 (например, лиганда, включающего мономер dAb, который связывает TNFR1) млекопитающему, нуждающемуся в этом.
В еще одном воплощении изобретение представляет собой способ лечения, подавления или предупреждения пневмонии (например, бактериальной пневмонии, такой как стафилококковая пневмония), включающий введение терапевтически эффективной дозы или количества антагониста TNFR1 (например, лиганда, включающего мономер dAb, который связывает TNFR1) млекопитающему, нуждающемуся в этом.
В изобретении предложен способ лечения, подавления или предупреждения других легочных заболеваний в дополнение к хроническому обструктивному заболеванию легких и пневмонии. Другие легочные заболевания, которые можно лечить, подавлять или предотвращать в соответствии с изобретением, включают, например, муковисцидоз и астму (например, стероидорезистентную астму). Таким образом, в еще одном воплощении изобретение представляет собой способ лечения, подавления или предупреждения легочного заболевания (например, муковисцидоза, астмы), включающий введение терапевтически эффективной дозы или количества антагониста TNFR1 (например, лиганда, включающего мономер dAb, который связывает TNFR1) млекопитающему, нуждающемуся в этом.
В конкретных воплощениях антагонист TNFR1 вводят через легкие, например с помощью ингаляции (например, с помощью внурибронхиальной, интраназальной или пероральной ингаляции, с помощью интраназальных капель) или с помощью системной доставки (например, парентерально, внутривенно, внутримышечно, интраперитонеально, подкожно).
В еще одном воплощении изобретение представляет собой способ лечения, подавления или предупреждения септического шока, включающий введение терапевтически эффективной дозы или количества антагониста TNFR1 (например, лиганда, включающего мономер dAb, который связывает TNFR1) млекопитающему, нуждающемуся в этом.
В еще одном аспекте второй конфигурации изобретения в настоящем изобретении предложена композиция, содержащая полиспецифический лиганд с закрытой конформацией, полученный с использованием способа по настоящему изобретению, и фармацевтически приемлемый носитель, разбавитель или эксципиент.
Кроме того, в настоящем изобретении предложен способ лечения заболевания с использованием "полиспецифического лиганда с закрытой конформацией" или композиции по настоящему изобретению.
В предпочтительном воплощении изобретения заболевание представляет собой рак или воспалительное заболевание, например ревматоидный артрит, асму или болезнь Крона.
В еще одном аспекте второй конфигурации по изобретению, в настоящем изобретении предложен способ диагностики, включая диагностику заболевания с использованием полиспецифического лиганда с закрытой конформацией или композиции по настоящему изобретению. Таким образом, в основном связывание аналита с полиспецифическим лигандом с закрытой конформацией может быть осуществлено для замещения агента, который приводит к образованию сигнала при замещении. Например, связывание аналита (второго антигена) может замещать фермент (первый антиген), связанный с антителом, обеспечивающим основу для иммуноанализа, особенно если фермент удерживается на антителе через его активный сайт.
Таким образом, в настоящем изобретении предложен способ обнаружения присутствия молекулы-мишени, включающий:
(а) обеспечение полиспецифического лиганда с закрытой конформацией, связанного с агентом, причем указанный лиганд является специфичным в отношении молекулы-мишени и данного агента, где агент, который связан с лигандом, приводит к образованию детектируемого сигнала при замещении из лиганда;
(б) приведение полиспецифического лиганда с закрытой конформацией в контакт с молекулой-мишенью; и
(в) обнаружение сигнала, генерируемого в результате замещения агента.
В соответствии с вышеуказанным аспектом второй конфигурации по изобретению, предпочтительно, агент представляет собой фермент, который является неактивным, когда связан полиспецифическим лигандом с закрытой конформацией. Альтернативно, агент может представлять собой один или более чем один, выбранный из группы, состоящей из следующего: субстрат для фермента, и флуоресцентная, люминисцентная или хромогенная молекула, которая неактивна или гасится после связывания с лигандом.
Кроме того, описанные в данном изобретении выбранные репертуары полипептидов могут быть использованы экстракорпорально или селективно in vitro для уничтожения, истощения или иным способом эффективного удаления популяции клеток-мишеней из гетерогенной коллекции клеток. Кровь от млекопитающего можно объединять экстракорпорально с лигандом, например антителами, рецепторами клеточной поверхности или связывающими их белками, в результате чего нежелательные клетки уничтожаются или иным образом удаляются из крови для возврата млекопитающему в соответствии со стандартными способами.
ПРИМЕРЫ
Изобретение дополнительно описано в следующих примерах только для целей иллюстрации. Как использовано в данном изобретении, для целей номенклатуры dAb, человеческий TNFa называют как TAR1, а человеческий рецептор TNFα 1 (рецептор р55) называют как TAR2.
Пример 1. Выбор scFv антитела (К8) с двойной специфичностью, направленного против человеческого сывороточного альбумина (HSA) и β-галактозидазы (β-gal)
Этот пример объясняет способ получения антитела с двойной специфичностью, направленного против β-gal и HSA, в котором репертуар вариабельных доменов VK, связанных с герминальным (пустым) VH-доменом выбран для связывания с β-gal, а репертуар вариабельных VH-доменов связан с герминальным (пустым) VK-доменом, выбранным для связывания с HSA. Затем комбинируют выбранные вариабельные домены VH HSA и VK β-gal и антитела, выбранные для связывания с β-gal и HSA. HSA представляет собой белок, увеличивающий период полувыведения, обнаруженный в человеческой крови.
В этом эксперименте используют четыре фаговые библиотеки человеческих антител.
Библиотека 1 Герминальное VK/DVT VH 8,46×107
Библиотека 2 Герминальное VK/NNK VH 9,64×107
Библиотека 3 Герминальное VH/DVT VK 1,47×108
Библиотека 4 Герминальное VH/NNK VK 1,45×108
Все библиотеки основаны на единичном каркасе человеческого антитела для VH (V3-23/DP47 и ин4b) и VK (012/02/DPK9 и JK1) с разнообразием боковой цепи, включенной в области, определяющие комплементарность (CDR2 и CDR3).
Библиотека 1 и Библиотека 2 содержат пустую последовательность VK, тогда как последовательность VH диверсифицирована в положениях Н50, Н52, Н52а, Н53, Н55, Н56, Н58, Н95, Н96, Н97 и Н98 (соответственно DVT или NNK кодируемые) (Фиг.1). Библиотека 3 и Библиотека 4 содержат пустую последовательность VH, тогда как последовательность VK диверсифицирована в положениях L50, L53, L91, L92, L93, L94 и L96 (соответственно DVT или NNK кодируемые) (Фиг.1). Библиотеки находятся в формате фагмиды pIT2/ScFv (Фиг.2) и подвергнуты предварительной селекции на связывание с общими лигандами Белком А и Белком L, так что большая часть клонов в нежелательных библиотеках являются функциональными. Эти размеры представленных выше библиотек соответствуют размерам после предварительной селекции. Библиотеку 1 и Библиотеку 2 смешивали перед селекцией на антигене с получением едининой библиотеки VH/пустой VK, и Библиотеку 3 и Библиотеку 4 смешивали с образованием единичной библиотеки VK/пустой VH.
Три раунда селекции осуществляли на β-gal с использованием библиотеки VK/пустой VH, и три раунда селекции осуществляли на HSA с использованием библиотеки VH/пустой VK. В случае β-gal, титры фага увеличиваются с 1,1×106 в первом раунде до 2,0×108 в третьем раунде. В случае HSA, титры фагов увеличиваются с 2×104 в первом раунде до 1,4×109 в третьем раунде. Селекцию осуществляли, как описано Griffith et al., (1993), за исключением того, что использовали хелперный фаг КМ13 (который содержит белок pIII с сайтом расщепления протеазой между доменами D2 и D3), и фаг элюировали 1 мг/мл трипсина в PBS. Добавление трипсина расщепляет белки pIII, полученные из хелперного фага (но не белки из фагмиды), и элюирует связанные слияния scFv-фаг путем расщепления в с-myc метке (Фиг.2), таким образом обеспечивая дополнительное обогащение в отношении фагов, экспрессирующих функциональные scFv, и соответствующее уменьшение скелета (Kristensen & Winter, Folding & Design 3: 321-328, Jul 9, 1998). Селекцию осуществляли с использованием иммунопробирок, покрытых HSA или β-gal в концентрации 100 мкг/мл.
Для контроля связывания 24 колонии из третьего раунда каждой селекции подвергали скринингу с помощью моноклонального фагового ИФА. Фаговые частицы получали, как описано Harrison et al. Methods Enzymol. 1996;267:83-109. 96-луночные планшеты для ИФА покрывали 100 мкл HSA или β-gal в концентрации 10 мкг/мл в PBS в течение ночи при 4°С. Следовали стандартному протоколу ИФА (Hoogenboom et al., 1991), используя детекцию связанного фага конъюгатом анти-М13-НКР. Селекция клонов позволила получить сигналы ИФА более чем 1,0 с 50 мкл супернатанта.
Затем препараты ДНК приготавливали из библиотеки VH/пустой VK, селектированной на HSA, и из VK/пустой VH, селектированной на β-gal с использованием набора QIAprep Spin Miniprep (Qiagen). Для оценки наибольшего разнообразия, препараты ДНК приготавливали из каждого из трех раундов селекции и затем объединяли вместе для каждого из антигенов. Препараты ДНК затем переваривали с помощью SaI//Not/ в течение ночи при 37°С. После очистки фрагментов на геле цепи VK из библиотеки VK/пустой VH, селектированные на β-gal, лигировали вместо пустой цепи VK библиотеки VH/пустой VK, селектированной на HSA, с получением библиотеки, состоящей из 3,3×109 клонов.
Эту библиотеку затем подвергали селекции на HSA (первый раунд) и β-gal (второй раунд), селекции HSA/β-gal, или на β-gal (первый раунд) и HSA (второй раунд), селекции β-gal/HSA. Селекцию осуществляли, как описано выше. В каждом случае после второго раунда тестировали 48 клонов на связывание с HSA и β-gal с помощью моноклонального фагового ИФА (как описано выше) и с помощью ИФА растворимых фрагментов scFv. Растворимые фрагменты антитела получали, как описано Harrison et al., (1996), и следовали стандартному протоколу ИФА в соответствии с Hoogenboom et al. (1991) Nucleic Acids Res., 19: 4133, за исключением того, что в качестве блокирующего буфера использовали 2% Tween/PBS, и связанные scFv обнаруживали с помощью конъюгата Белок L-HRP. Три клона (Е4, Е5 и Е8) из селекции HSA/β-gal и два клона (К8 и К10) из селекции β-gal/HSA оказались способными связываться с обоими антигенами. scFv из этих клонов амплифицировали с помощью ПЦР и секвенировали, как описано Ignatovich et al, (1999) J Mol Biol 1999 Nov 26; 294(2):457-65, с использованием прамеров LMB3 и pHENseq. Анализ последовательности выявил, что все клоны были идентичны. Таким образом, только один клон, кодирующий антитело с двойной специфичностью (К8), клонировали для дальнейшей работы (Фиг.3).
Пример 2. Характеристика связывающих свойств антитела К8
Во-первых, связывающие свойства антитела К8 характеризовали с помощью моноклональной фаговой ИФА. 96-луночный планшет покрывали 100 мкл HSA и β-gal наряду с щелочной фосфатазой (APS), бычьим сывороточным альбумином (BSA), агглютинином арахиса, лизоцимом и цитохромом с (для проверки перекрестной реактивности) в концентрации 10 мкг/мл в PBS в течение ночи при 4°С. Фагмиду из клона К8 высвобождали с помощью КМ13, как описано Harrison et al, (1996), и супернатант (50 мкл), содержащий фаг, непосредственно анализировали. Следовали стандартному протоколу ИФА (Hoogenboom et al, 1991), используя обнаружение связанного фага с помощью конъюгата анти-М13-НРР. Обнаружено, что антитело К8 с двойной специфичностью связывается с HSA и β-gal, когда присутствует на поверхности фага, с сигналами абсорбции больше 1,0 (Фиг.4). Также обнаружено сильное связывание с BSA (Фиг.4). Поскольку HSA и BSA на 76% гомологичны на аминокислотном уровне, то не является неожиданным, что антитело К8 распознает оба этих структурно родственных белка. Не обнаружили перекрестной реактивности с другими белками (Фиг.4).
Во-вторых, связывающие свойства антитела К8 тестировали в ИФА растворимого scFv. Продукцию растворимого фрагмента scFv индуцировали с помощью IPTG, как описано у Harrison et al. (1996). Для определения уровней экспрессии К8 scFv, растворимые фрагменты антитела очищали из супернатанта 50 мл индукций с использованием колонок протеин А-сефароза, как описано Harlow and Lane, Antibodies: a Laboratory Manual, (1988) Cold Spring Harbor. Затем измеряли OD280 и рассчитывали концентрацию белка, как описано Sambrook et al, (1989). К8 scFv продуцировался в супернатанте в концентрации 19 мг/л.
ИФА растворимого scFv затем осуществляли с использованием известных концентраций фрагмента антитела К8. 96-луночный планшет покрывали 100 мкл HSA, BSA и β-gal в концентрации 10 мкг/мл и 100 мкл Белка А в концентрации 1 мкг/мл. Наносили 50 мкл серийных разведении К8 scFv, и связанные фрагменты антитела обнаруживали с помощью конъюгата Белок L-HRP. Результаты ИФА подтвердили двойную специфичность антитела К8 (Фиг.5).
Для подтверждения того, что связывание с β-gal определяется доменом VK, а связывание с HSA/BSA - VH-доменом анитела К8 scFv, домен VK вырезали из ДНК К8 scFv путем переваривания рестриктазами Sal//Not/ и лигировали в вектор р1Т2, переваренный Sal//Not/, содержащий пустую цепь VH (Фиг.1 и 2). Связывающие характеристики полученного клона К8VK/пустой VH анализировали с помощью ИФА растворимого scFv. Продукцию фрагментов растворимого scFv индуцировали с помощью IPTG, как описано Harrison et al., (1996), и супернатант (50 мкл), содержащий scFv, непосредственно анализировали. ИФА растворимого scFv осуществляли, как описано в Примере 1, и связанные scFv обнаруживали с помощью конъюгата Белок L-HRP. Результаты ИФА выявили, что этот клон способен связывать β-gal, тогда как связывание с BSA отменяется (Фиг.6).
Пример 3. Селекция антител с единичным доменом VH, направленных против антигенов А и В, и антител с единичным доменом VK, направленных против антигенов С и D.
Этот пример описывает способ получения антител с единичным доменом VH, направленных против антигенов А и В, и антител с единичным доменом VK, направленных против антигенов С и D, путем селекции репертуаров первичных антител с единичным вариабельным доменом для связывания с этими антигенами в отсутствие комплементарных вариабельных доменов.
Селекцию и характеристику связывающих клонов осуществляют, как описано ранее (смотри Пример 5, PCT/GB02/003014). Четыре клона выбирают для дальнейшей работы:
VH1 - Анти-А VH
VH2 - Анти-В VH
VK1 - Анти-С VK
VK2 - Анти-D VK
Эти способы, описанные выше в Примерах 1-3, могут быть использованы по аналогии с описанным для получения молекул димеров, включающих комбинации доменов VH (т.е. лиганды VH-VH) и комбинации доменов VL (лиганды VL-VL.
Пример 4. Получение и характеристика ScFv антител с двойной специфичностью (VH1/VH2, направленных против антигенов А и В, и VK1/VK2, направленных против антигенов С и D).
Этот пример демонстрирует, что ScFv антитела с двойной специфичностью (VH1/VH2, направленные против антигенов А и В, и VK1/VK2, направленные против антигенов С и D) могут быть получены путем комбинирования единичных доменов VK и VH, направленных против соответствующих антигенов в векторе ScFv.
Для создания антитела VH1/VH2 с двойной специфичностью, единичный домен VH1 вырезают из вектора 1 с вариабельным доменом (Фиг.7) путем переваривания рестриктазами NcoI/XhoI и лигируют в переваренный рестриктазами NcoI/XhoI вектор 2 с вариабельным доменом (Фиг.7) с получением вектора 2 VHI/вариабельный домен. Единичный домен VH2 амплифицруют с помощью ПЦР из вектора 1 с вариабельным доменом с использованием праймеров для введения рестрикционного сайта SalI в 5'-конец и рестрикционного сайта Notl в 3'-конец. Продукт ПЦР затем переваривают рестриктазами SalI/Notl и лигируют в расщепленный рестриктазами SalI/Notl вектор 2 VHI/вариабельный домен с получением вектора 2 VH1/VH2/вариабельный домен.
Вектор 2 VК1/VK2/вариабельный домен получают аналогичным образом. Двойную специфичность полученных VH1/VH2 ScFv и VK1/VK2 ScFv тестируют в ИФА растворенного ScFv, как описано ранее (смотри Пример 6, PCT/GB 02/003014). Конкурентный ИФА проводят, как описано ранее (смотри Пример 8, PCT/GB 02/003014).
Возможные результаты:
VH1/VH2 ScFv способен одновременно связывать антигены А и В VK1/VK2 ScFv способен одновременно связывать антигены С и D VH1/VH2 ScFv связывание является конкурентным (при связывании с антигеном А, VH1/VH2 ScFv не может связываться с антигеном В) VK1/VK2 ScFv связывание является конкурентным (при связывании с антигеном С, VK1/VK2 ScFv не может связываться с антигеном D)
Пример 5. Конструирование VH1/VH2 Fab и VK1/VK2 Fab с двойной специфичностью и анализ их связывающих свойств.
Для создания VH1/VH2 Fab, единичный домен VH1 лигируют в переваренный рестриктазами NcoI/XhoI вектор СН (Фиг.8) для получения VH1/CH, и единичный домен VH2 лигируют в переваренный рестриктазами Sal/I/Notl вектор СК (Фиг.9) для получения VH2/CK. Плазмидную ДНК из VH1/CH и VH2/CK используют для котрансформации компетентных клеток E. coli, как описано ранее (смотри Пример 8, PCT/GB02/003014).
Клон, содержащий плазмиды VH1/CH и VH2/CK, затем индуцируют с помощью IPTG для продукции растворимого VH1/VH2 Fab, как описано ранее (смотри Пример 8, PCT/GB02/003014).
VK1/VK2 Fab получают аналогично.
Связывающие свойства полученных Fab тестируют с помощью конкурентного ИФА, как описано ранее (смотри Пример 8, PCT/GB 02/003014).
Возможные результаты:
- VH1/VH2 Fab способен одновременно связывать антигены А и В
- VK1/VK2 Fab способен одновременно связывать антигены С и D
- VH1/VH2 Fab связывание является конкурентным (при связывании с антигеном А,
- VH1/VH2 Fab не может связываться с антигеном В)
- VK1/VK2 Fab связывание является конкурентным (при связывании с антигеном С,
- VK1/VK2 Fab не может связываться с антигеном D)
Пример 6. Хелатообразующие димеры dAb
Краткое изложение
Гомодимеры VH и VK получают в формате dAb-линкер-dAb с использованием гибких полипептидных линкеров. Векторы создавали в формате dAb-линкер-dAb, содержащем глицин-сериновые линкеры разной длины SU:(Glu4Ser)3 (SEQ ID NO: 199), 5U:(Gly4Ser)5 (SEQ ID NO:629), 7U:(Gly4Ser)7 (SEQ ID NO:630). Библиотеки димеров создавали с использованием направляющих dAb выше линкера: TAR1-5 (VK), TAR1-27 (VK), TAR2-5 (VH) или TAR2-6 (VK) и библиотеки соответствующих вторых dAb после линкера. С использованием этого способа были выбраны новые димерные dAb. Влияние димеризации на связывание антигена определяли с помощью ИФА и BIAcore исследований и в анализах клеточной нейтрализации и связывания рецепторов. Димеризация TAR1-5 и TAR1-27 привела к значительному улучшению связывающей аффинности и уровней нейтрализации.
1.0 Способы
1.1 Получение библиотеки
1.1.1 Векторы pEDA3U, pEDA5U и pEDA7U разработаны для введения линкеров разной длины, совместимых с форматом dAb-линкер-dAb. Для pEDA3U смысловые и антисмысловые олигонуклеотидные линкеры длиной 73 пары оснований отжигали с использованием медленной программы отжига (95°С - 5 мин, 80°С - 10 мин, 70°С - 15 мин, 56°С - 15 мин, 42°С до использования) в буфере, содержащем 0,1 М NaCl, 10 мМ Трис-HCl рН 7,4, и клонировали с использованием рестрикционных сайтов Xho1 и Not1. Линкеры, охватывающие 3 единицы (Gly4Ser) и область вставки, расположенную между клонирующими сайтами Sail и Not1 (Схема 1). Для уменьшения возможности мономерных dAb селектироваться с помощью фагового дисплея, вставочную область конструировали таким образом, чтобы включить 3 стоп-кодона, рестрикционный сайт Sac1 и мутацию сдвига рамки считывания для того, чтобы исключить область из рамки, когда отсутствует второе dAb. Для pEDA5U и 7U, в зависимости от длины требуемых линкеров, перекрывание олигонуклеотидных линкеров было разработано для каждого вектора, отжигаемого и удлиняемого с использованием фрагмента Кленова. Фрагмент затем очищали и переваривали с использованием подходящих ферментов перед клонированном, используя рестрикционные сайты Xho1 и Not1.

1.1.2 Получение библиотеки
N-концевой V ген, соответствующий направляющему dAb, клонировали выше линкера с использованием рестрикционных сайтов Nco1 и Xho1. Гены VH имеют существующие совместимые сайты, однако для клонирования генов VK требуется введение подходящих рестрикционных сайтов. Этого достигали с использованием модифицирующих праймеров ПЦР (VK-DLIBF: 5' cggccatggcgtcaacggacat (SEQ ID NO:377); VKXhoIR: 5' atgtgcgctcgagcgtttgattt 3' (SEQ ID NO:378)) в 30 циклах ПЦР-амплификации с использованием смеси 2:1 SuperTaq (HTBiotechnology Ltd) и pfu turbo (Stratagene). Это поддерживало сайт Nco1 у 5'-конца, в то же время разрушая соседний сайт Sail и вводили сайт Xho1 у 3'-конца. 5 направляющих dAb клонировали в каждом из 3 димерных векторов: TAR1-5 (VK), TAR1-27 (VK), TAR2-5 (VH), TAR2-6 (VK) и TAR2-7 (VK). Все конструкции подтверждали с помощью анализа последовательностей.
Имея клонированные направляющие dAb выше линкера в каждом из векторов (pEDA3U, 5U и 7U): TAR1-5 (VK), TAR1-27(VK), TAR2-5(VH) или TAR2-6(VK) библиотеку соответствующих вторых dAb клонировали после линкера. Для достижения этого комплементарные библиотеки dAb амплифицировали с помощью ПЦР из фага, полученного после 1 раунда селекции библиотеки VK против человеческого TNFα (при приблизительном разнообразии 1×106 после 1 раунда), когда TAR1-5 или TAR1-27 представляют собой направляющие dAb, или библиотеки VH или VK против человеческого рецептора TNF р55 (при приблизительном разнообразии 1×105 после 1 раунда), когда TAR2-5 или TAR2-6 соответственно представляют собой направляющие dAb. Для библиотек VK ПЦР-амплификацию осуществляли с использованием праймеров в 30 циклах ПЦР-амплификации с использованием смеси 2:1 SuperTaq и pfu turbo. Библиотеки VH амплифицировали с помощью ПЦР с использованием праймеров для введения рестрикционного сайта Sail в 5'-конец гена. Продукты ПЦР библиотеки dAb расщепляли с использованием подходящих рестриктаз, лигировали в соответствующие векторы ниже линкера, используя рестрикционные сайты Sal1/Not1, и подвергали электропорации в свежеприготовленные компетентные клетки TG1.
Достигнутые для каждой библиотеки титры были следующими:
TAR1-5: pEDA3U=4×108, pEDA5U=8×107, pEDA7U=1×108
TAR1-27: pEDA3U=6,2×108, pEDA5U=1×108, pEDA7U=1×109
TAR2h-5: pEDA3U=4×107, pEDA5U=2×108, pEDA7U=8×107
TAR2h-6: pEDA3U=7,4×108, pEDA5U=1,2×108, pEDA7U=2,2×108
1.2 Селекции
1.2.1 TNFα
Селекции проводили с использованием человеческого TNFα, пассивно нанесенного на иммунопробирки. Кратко, иммунопробирки покрывали в течение ночи 1-4 мл требуемого антигена. Иммунопробирки затем 3 раза промывали PBS и блокировали с помощью 2% сухого молока в PBS в течение 1-2 ч и дополнительно промывали 3 раза PBS. Раствор фага разбавляют в 2% сухого молока в PBS и инкубируют при комнатной температуре в течение 2 ч. Пробирки затем промывали PBS, и фаг элюировали 1 мг/мл трипсин-PBS. Три стратегии селекции исследовали для библиотек димеров TAR1-5. Первый раунд селекции осуществляли в иммунопробирках, покрытых человеческим TNFα в концентрации 1 мкг/мл или 20 мкг/мл, с 20 промывками в PBS 0,1% Tween. Клетки TG1 инфицируют элюированным фагом и определяют титры (например, Marks et al J Mol Biol. 1991 Dec 5; 222(3):581-97, Richmann et al Biochemistry. 1993 Aug 31; 32(34):8848-55).
Полученные титры представляли собой:
pEDA3U=2,8×107 (1 мкг/мл TNF) 1,5×108 (20 мкг/мл TNF),
pEDA5U=1,8×107 (1 мкг/мл TNF), 1,6×108 (20 мкг/мл TNF)
pEDA7U=8×106 (1 мкг/мл TNF), 7×107 (20 мкг/мл TNF).
Второй раунд селекции осуществляли с использованием 3 различных способов.
1. В иммунопробирках, 20 промывок с инкубацией в течение ночи с последующими дополнительными 10 промывками.
2. В иммунопробирках, 20 промывок с последующей инкубации в течение 1 ч при КГ в промывочном буфере с (1 мкг/мл TNFα) и дополнительными 10 промывками.
3. Селекцию на стрептавидиновых шариках с использованием 33 пмоль биотинилированного человеческого TNFα (Henderikx et ah, 2002, Selection of antibodies against biotinylated antigens. Antibody Phage Display: Methods and protocols, Ed. O'Brien and Atkin, Humana Press). Единичные клоны из 2 раунда селекции переносили в 96-луночные планшеты, и неочищенные препараты супернатанта приготавливали в 2 мл формате 96-луночных планшетов.
Для TAR1-27, селекции осуществляли, как описано ранее со следующими модификациями. Первый раунд селекции осуществляли в иммунопробирках, покрытых человеческим TNFα в концентрации 1 мкг/мл или 20 мкг/мл, с 20 промывками в PBS 0,1% Tween. Второй раунд селекции осуществляли в иммунопробирках с использованием 20 промывок с ночной инкубацией с последующими дополнительными 20 промывками. Единичные клоны из раунда 2 селекции переносили в 96-луночные планшеты и неочищенные препараты супернатанта приготавливали в 2 мл формате 96-луночных планшетов.
Титры TAR1-27 представляют собой следующее:
1.2.2 РЕЦЕПТОР TNF 1 (РЕЦЕПТОР р55; TAR2)
Селекцию осуществляли, как описано ранее только для библиотек TAR2h-5. 3 раунда селекции осуществляли в иммунопробирках с использованием 1 мкг/мл человеческого рецептора TNF р55 или 10 мкг/мл человеческого рецептора TNF р55 с 20 промывками в PBS 0,1% Tween с ночной инкубацией с последующими дополнительными 20 промывками. Единичные клоны после раундов 2 и 3 селекции переносили в 96-луночные планшеты, и неочищенные препараты супернатантов приготавливали в 2 мл формате 96-луночных планшетов.
Титры TAR2h-5 представляют собой следующее:
1.3 Скрининг
Единичные клоны из раундов 2 или 3 селекции переносили из каждой из библиотек 3U, 5U и 7U из различных способов селекции, когда это целесообразно. Клоны выращивали в 2xTY с 100 мкг/мл ампициллина и 1% глюкозы в течение ночи при 37°С. Разведение 1/100 этой культуры инокулировали в 2 мл 2xTY с 100 мкг/мл ампициллина и 0,1% глюкозы в 2 мл формате 96-луночных планшетов и выращивали при 37°С, встряхивая до тех пор, пока OD600 не составит примерно 0,9. Культуру затем индуцировали 1 мМ IPTG в течение ночи при 30°С. Супернатанты осветляли путем центрифугирования при 4000 об/мин в течение 15 мин в центрифуге для планшетов sorval. Препараты супернатанта использовали для первоначального скрининга.
1.3.1 ИФА
Связывающую активность димерных рекомбинантных белков сравнивали с мономером с помощью белкового A/L ИФА или с помощью антигенного ИФА. Кратко, 96-луночный планшет покрывают антигеном или белком A/L в течение ночи при 4°С. Планшет промывают 0,05% Tween-PBS, блокируют в течение 2 ч 2% Tween-PBS. Образец добавляют к планшету, инкубированному в течение 1 ч при комнатной температуре. Планшет промывают и инкубируют вторичным реагентом в течение 1 ч при комнатной температуре. Планшет промывают и проявляют с использованием субстрата ТМВ (тетраметилбензидина). Белок A/L-HRP или India-HRP использовали в качестве вторичного реагента. Для антигенного ИФА используемые концентрации антигена составляли 1 мкг/мл в PBS для человеческого TNFα и человеческого рецептора THF 1. Благодаря присутствию направляющего dAb в большинстве случаев димеры давали положительный сигнал ИФА, поэтому определение скорости диссоциации антитела и антигена осуществляли с помощью BIAcore.
1.3.2 BIAcore
Анализ BIAcore проводили для клонов TAR1-5 и TAR2h-5. Для скрининга человеческий TNFα присоединяли к чипу СМ5 с высокой плотностью (приблизительно 10000 ОЕ (относительных единиц)). 50 мкл человеческого TNFα (50 мкг/мл) присоединяли к чипу при 5 мкл/мин в ацетатном буфере, рН 5,5. Регенерация чипа после анализа с использованием стандартных способов невозможна из-за нестабильности человеческого TNFα, таким образом после анализа каждого образца чип промывали в течение 10 мин буфером. Для TAR1-5 супернатанты клонов после 2 раунда селекции подвергали скринингу с помощью BIAcore.
48 клонов подвергали скринингу из каждой из библиотек 3U, 5U и 7U, полученных с использованием следующих способов селекции:
R1: иммунопробирка с 1 мкг/мл человеческого TNFα, R2 иммунопробирка с 1 мкг/мл человеческого TNFα, промывка в течение ночи.
R1: иммунопробирка с 20 мкг/мл человеческого TNFα, R2 иммунопробирка с 20 мкг/мл человеческого TNFα, промывка в течение ночи.
R1: иммунопробирка с 1 мкг/мл человеческого TNFα, R2 33 пмоль биотинилированного человеческого TNFα на шариках.
R1: иммунопробирка с 20 мкг/мл человеческого TNFα, R2 33 пмоль биотинилированного человеческого TNFα на шариках.
Для скрининга человеческий рецептор TNF р55 связывали с чипом СМ5 с с высокой плотностью (примерно 4000 ОЕ). 100 мкл человеческого рецептора TNF р55 (10 мкг/мл) связывали с чипом в концентрации 5 мкл/мин в ацетатном буфере, рН 5,5. Исследовали стандартные условия регенерации (глицин рН 2 или рН 3), но в каждом случае антиген удаляли с поверхности чипа, как в случае TNFα, таким образом после анализа каждого образца чип промывали в течение 10 мин буфером.
Для TAR2-5, супернатанты клонов после 2 раунда селекции подвергали скринингу. 48 клонов подвергали скринингу из каждой из библиотек 3U, 5U и 7U с использованием следующих способов селекции:
R1: иммунопробирка с 1 мкг/мл человеческого рецептора TNF р55, R2 иммунопробирка с 1 мкг/мл человеческого рецептора TNF р55, промывка в течение ночи.
R1: иммунопробирка с 10 мкг/мл человеческого рецептора TNF р55, R2 иммунопробирка с 10 мкг/мл человеческого рецептора TNF р55, промывка в течение ночи.
1.3.3 Анализы рецептора и клеточные анализы
Способность димеров нейтрализоваться в анализе рецептора определяли следующим образом:
Связывание рецептора
dAb против TNF тестировали в отношении способности ингибировать связывание TNF с рекомбинантным рецептором TNF 1 (р55). Кратко, планшеты Maxisorp инкубировали в течение ночи с 30 мг/мл мышиного моноклонального антитела против Fc человека (Zymed, San Francisco, USA). The лунки промывали физиологическим раствором, забуференным фосфатом (PBS), содержащим 0,05% Tween-20, и затем блокировали с помощью 1% BSA в PBS перед инкубацией с 100 нг/мл слитого белка, представляющего собой рецептор TNF 1 - Fc (R&D Systems, Minneapolis, USA). dAb против TNF смешивали с TNF, который добавляли к промытым лункам в конечной концентрации 10 нг/мл. Связывание TNF обнаруживали с помощью 0,2 мг/мл биотинилированного антитела против TNF (HyCult biotechnology, Uben, Netherlands), затем разведения (1:500) пероксидазы хрена, меченной стрептавидином (Amersham Biosciences, UK), и затем инкубации с субстратом ТМВ (KPL, Gaithersburg, USA). Реакцию останавливали путем добавления HCl, и абсорбцию регистрировали при 450 нм. Активность dAb против TNF приводила к уменьшению связывания TNF и, следовательно, уменьшению абсорбции по сравнению только с контролем TNF.
Анализ цитотоксичности в отношении L929
dAb против TNF также тестировали на мышиных фибробластах L929 в отношении способности нейтрализовать цитотоксическую активность TNF (Evans, Т. (2000) Molecular Biotechnology 15, 243-248). Кратко, клетки L929, посеянные в микротитрационные планшеты, инкубировали в течение ночи с dAb против TNF, 100 пг/мл TNF и 1 мг/мл актиномицина D (Sigma, Poole, UK). Жизнеспособность клеток измеряли путем считывания абсорбции при 490 нм с последующей инкубацией с [3-(4,5-диметилтиазол-2-ил)-5-(3-карбоксиметокси-фенил)-2-(4-сульфофенил)-2Н-тетразолием (Promega, Madison, USA). Активность dAb против TNF приводит к уменьшению цитотоксичности TNF и, следовательно, к увеличению абсорбции по сравнению только с контролем TNF.
В первоначальном скрининге супернатанты, приготовленные для описанного выше анализа BIAcore, использовали также в анализе рецептора. Дополнительный анализ выбранных димеров также проводили в анализе рецептора и клеточном анализе с использованием очищенных белков.
Анализ IL-8 в HeLa
Антитела против TNFR1 или dAb против TNF-альфа тестировали в отношении способности нейтрализовать индукцию секреции IL-8 с помощью TNF в клетках HeLa (способ, адаптированный из способа Akeson, L. et al (1996) Journal of Biological Chemistry 271, 30517-30523, описывающий индукцию IL-8 с помощью IL-I в HUVEC; в данном изобретении авторы изобретения рассматривают при индукции TNF-альфа человека и используют клетки HeLa вместо клеточной линии HUVEC). Кратко, клетки HeLa, посеянные в микротитрационные планшеты, инкубировали в течение ночи с dAb и 300 пг/мл TNF. После инкубации супернатант отсасывали из клеток и измеряли концентрацию IL-8 с помощью "сэндвич"-ИФА (R&D Systems). Активность dAb против TNFR1 приводила к уменьшению секреции IL-8 в супернатант по сравнению только с контролем TNF.
Анализ L929 используют в следующих экспериментах; однако применение анализа IL-8 в HeLa является предпочтительным для измерения лигандов против рецептора TNF 1 (р55); присутствие мышиного р55 в анализе L929 накладывает некоторые ограничения на его использование.
1.4 Анализ последовательностей
Димеры, которые, как доказано, имеют интересующие свойства в BIAcore и анализе рецептора, секвенировали. Последовательности подробно приведены в перечне последовательностей.
1.5 Задание формата
1.5.1 Димеры TAR 1-5-19
Димеры TAR1-5, которые, как показано, имеют хорошие нейтрализующие свойства, переформатировали и анализировали в клеточных анализах и анализах рецепторов. TAR1-5, направляющие dAb, замещали аффинно зрелым клоном TAR1-5-19. Для достижения этого TAR1-5 клонировали из индивидуальных димерных пар и замещали TAR1-5-19, который был амплифицирован с помощью ПЦР. Кроме того, гомодимеры TAR1-5-19 также конструировали в векторах 3D, 5U и TU. N-коцевую копию гена амплифицировали с помощью ПЦР и клонировали, как описано выше, и С-концевой генный фрагмент клонировали, используя рестрикционные сайты Sal1 и Not1.
1.5.2 Мутагены
Амбер стоп-кодон, представленный в dAb2, один из С-концевых dAb в димерных парах TAR1-5 подвергали мутации до глутамина с помощью сайт-направленного мутагенеза.
1.5.3 Fab
Димеры, содержащие TAR1-5 или TAR1-5-19, реформатировали в экспрессирующие векторы Fab. dAb клонировали в экспрессирующие векторы, содержащие либо СК, либо СН гены, с использованием рестрикционных сайтов Sfi1 и Not1 и подтверждали с помощью анализа последовательностей. Вектор СК получен на основе ампициллин-устойчивого вектора pUC, а вектор СН получен из хлорамфеникол-устойчивого вектора pACYC. Для экспрессии Fab, конструкции dAb-CH и dAb-CK котрансформировали в клетки НВ2151 и выращивали в 2xTY, содержащей 0,1% глюкозы, 100 мкг/мл ампициллина и 10 мкг/мл хлорамфеникола.
1.5.3 Димеризация шарниров
Димеризацию dAb оценивали через образование цистеиновой связи. Короткую последовательность аминокислот EPKSGDKTHTCPPCP (SEQ ID NO:379), представляющую собой модифицированную форму человеческого шарнира IgGd, конструировали в С-концевой области на dAb. Олиголинкер, кодирующий эту последовательность, синтезировали и отжигали, как описано ранее. Линкер клонировали в вектор pEDA, содержащий TAR1-5-19, используя рестрикционные сайты Xho1 и Not1. Димеризация происходит in situ в периплазме.
1.6 Экспрессия и очистка
1.6.1 Экспрессия
Супернатанты получали в 2 мл формате в 96-луночном планшете для первоначального скрининга, как описано ранее. После способа первоначального скрининга выбранные димеры дополнительно анализировали. Димерные конструкции экспрессировали в клетках TOP10F' или НВ2151 как супернатанты. Кратко, индивидуальную колонию из свежезасеянной чашки выращивали в течение ночи при 37°С в 2xTY с 100 мкг/мл ампициллина и 1% глюкозы. Разведение 1/100 этой культуры инокулировали в 2xTY с 100 мкг/мл ампициллина и 0,1% глюкозы и выращивали при 37°С, встряхивая до тех пор, пока OD600 не достигала примерно 0,9. Культуру затем индуцировали с помощью 1 мМ IPTG в течение ночи при 30°С. Клетки удаляли центрифугированием и супернатант очищали с помощью белок А или L агарозы.
Fab и димеры цистеинового шарнира экспрессировали в виде периплазматичских белков в клетках НВ2152. Разбавление 1/100 ночной культуры инокулировали в 2xTY с 0,1% глюкозы и подходящими антибиотиками и выращивали при 30°С со встряхиванием до тех пор, пока OD600 не достигала примерно 0,9. Культуру затем индуцировали с помощью 1 мМ IPTG в течение 3-4 часов при 25°С. Клетки собирали центрифугированием и остаток ресуспендировали в буфере для периплазматического препарата (30 мМ Трис-HCl, рН 8,0, 1 мМ ЭДТА, 20% сахарозы). После центрифугирования супернатант сохраняли и осадок ресуспендировали в 5 мМ MgSO4. Супернатант вновь собирали центрифугированием, объединяли и очищали.
1.6.2 Очистка белка A/L
Оценивали оптимизацию очистки димерных белков из белок L агарозы (Affitech, Norway) или белок А агарозы (Sigma, UK). Белок элюировали партиями или с помощью колоночной элюции с использованием перистальтического насоса. Исследовали три буфера 0,1 М фосфат-цитратный буфер, рН 2,6, 0,2 М глицин, рН 2,5 и 0,1 М глицин, рН 2,5. Оптимальные условия определяли как условия с перистальтическим насосом с использованием 0,1 М глицина, рН 2,5 в 10 объемах колонки. Очистку из белка А осуществляли в условиях с использованием перистальтического насоса и 0,1 М глицина, рН 2,5.
1.6.3 Очистка с помощью FPLC
Дополнительную очистку осуществляли с помощью FPLC в системе АКТА Explorer 100 (Amersham Biosciences Ltd). Димеры TAR1-5 и TAR1-5-19 фракционировали с помощью катионообменной хроматографии (1 мл Resource 5 - Amersham Biosciences Ltd), элюируя градиентом 0-1 М NaCl в 50 мМ ацетатном буфере, рН 4. Шарнирные димеры очищали с помощью ионного обмена (1 мл Resource Q Amersham Biosciences Ltd), элюируя 0,1 М градиентом NaCl в 25 мМ Трис HCl рН 8,0. Fab очищали с помощью гель-фильтрации с использованием колонки с superose 12 (Amersham Biosciences Ltd) при скорости потока 0,5 мл/мин в PBS с 0,05% tween. После очистки образцы концентрировали с использованием отсекающих концентраторов vivaspin 5K (Vivascience Ltd).
2.0 Результаты
2.1.1. Димеры TAR1-5
6×96 клонов отбирали из раунда 2 селекции, охватывающего все библиотеки и условия селекции. Приготавливали препараты супернатанта и анализировали с помощью антигенного и белок L-ИФА, BIAcore и в анализах рецептора. В ИФА положительные связывающие клоны идентифицировали из каждого способа селекции и распределяли между библиотеками 3U, 5U и 7U. Однако, поскольку всегда присутствует направляющее dAb, с помощью этого способа невозможно было разделить связывающие вещества с высокой и низкой аффинностью, таким образом проводили анализ BIAcore.
Анализ BIAcore осуществляли с использованием 2 мл супернатантов. Анализ BIAcore выявил, что скорости Koff для димера в значительной степени улучшались по сравнению с мономерным TAR1-5. Koff для мономера находилась на уровне 10-1 М по сравнению с Koff для димера, которая находилась в диапазоне 10-3-10-4 М. Отбирали 16 клонов, которые, по-видимому, имеют очень низкие скорости диссоциации, они были получены из библиотек 3U, 5U и 7U и подвергнуты секвенированию. Кроме того, супернатанты анализировали в отношении способности нейтрализовать TNFα человека в анализе рецептора.
6 ведущих клонов (d1-d6 ниже), нейтрализующих в этих анализах, были подвергнуты секвенированию. Результаты демонстрируют, что из 6 полученных клонов существует только 3 различных вторых dAb (dAb1, dAb2 и dAb3), однако когда второе dAb обнаружено более чем один раз, тогда они связаны линкерами различной длины.
TAR1-5d1: 3U линкер 2е dAb=dAb1 - 1 мкг/мл Ag, отмывка иммунопробирки в течение ночи
TAR1-5d2: 3U линкер 2е dAb=dAb2 - 1 мкг/мл Ag, отмывка иммунопробирки в течение ночи
TAR1-5d3: 5U линкер 2e dAb=dAb2 - 1 мкг/мл Ag, отмывка иммунопробирки в течение ночи
TAR1-5d4: 5U линкер 2е dAb=dAb3 - 20 мкг/мл Ag, отмывка иммунопробирки в течение ночи
TARI-5d5: 5U линкер 2е dAb=dAb1 - 20 мкг/мл Ag, отмывка иммунопробирки в течение ночи
TAR1-5d6: 7U линкер 2е dAb=dAb1 - R1: 1 мкг/мл Ag, отмывка иммунопробирки в течение ночи, Р2:шарики
Дополнительно анализировали 6 ведущих клонов. Белок получали из периплазмы и супернатанта, очищенного на агарозе с белком L, и оценивали в клеточном анализе и анализе рецептора. Уровни нейтрализации варьировали (Таблица 1). Определяли оптимальные условия для получения белка. Белок, продуцируемый из клеток НВ2151 в виде супернатантов, давал самый высокий выход (примерно 10 мкг/л культуры). Супернатанты инкубировали с агарозой с белком L в течение 2 ч при комнатной температуре или в течение ночи при 4°С. Шарики промывали PBS/NaCl и набивали в колонку FPLC, используя перистальтический насос. Шарики промывали 10 колоночными объемами PBS/NaCl и элюировали 0,1 М глицином, рН 2,5. Обычно димерный белок элюируется после мономера.
Димеры TAR1-5d1-6 очищали с помощью FPLC. Три вида получали путем очистки на FPLC и идентифицировали с помощью электрофореза в ПААГ-ДСН (полиакриламидный гель с додецилсульфатом натрия). Один из видов соответствует мономеру, два других соответствуют димерам разных размеров. Более крупный из двух видов, возможно, является следствием присутствия С-концевых меток. Эти белки исследовали в анализе рецептора. Данные, представленные в Таблице 1, представляют собой оптимальные результаты, полученные для двух видов димеров (Фиг.11).
Три вторых dAb из димерных пар (т.е. dAb1, dAb2 и dAb3) клонировали в виде мономеров и исследовали с помощью ИФА и в клеточном анализе и анализе рецептора. Все три dAb специфически связываются с TNF в антигенном ИФА и не реагируют перекрестно с пластиком или BSA. В виде мономеров ни один из dAb не нейтрализует в клеточном анализе или анализе рецептора.
2.1.2 Димеры TAR1-5-19
TAR1-5-19 замещали на TAR1-5 в 6 ведущих клонах. Если не указано иначе, то анализ всех димеров TAR1-5-19 в клеточном анализе и анализе рецепторов проводили с использованием общего белка (белок L, только очищенный) (Таблица 2). TAR1-5-19d4 и TAR1-5-19d3 имели наилучшую ND50 (примерно 5 нМ) в клеточном анализе, что согласуется с результатами анализа рецептора и является улучшением по сравнению с мономером TAR1-5-19 (ND50 примерно 30 нМ). Хотя очищенные димеры TAR1-5 дают различные результаты в анализе рецептора и клеточном анализе, димеры TAR1-5-19 были более согласованными. Вариабельность была продемонстрирована при использовании различных элюирующих буферов в процессе очистки белка. Элюция с использованием 0,1 М фосфат-цитратного буфера, рН 2,6 или 0,2 М глицина, рН 2,5, хотя и удаляла весь белок из агарозы с белком L, в большинстве случаев делала его менее функционален.
TAR1-5-19d4 экспрессировали в ферментере и очищали на катионнообменной FPLC с получением полностью очищенного димера. Как с TAR1-5d4, путем очистки на FPLC получали три вида, соответствующие мономерному и двум димерным видам. Этот димер подвергали аминокислотному секвенированию. Мономер TAR1-5-19 и TAR1-5-19d4 затем исследовали в анализе рецептора, и полученные IC50 для мономера составляли 30 нМ, а для димера 8 нМ. Результаты анализа рецептора, в котором сравнивается мономер TAR1-5-19, TAR1-5-19d4 и TAR1-5d4, представлены на Фиг.10.
Гомодимеры TAR1-5-19 получали в векторах 3U, 5U и 7U, экспрессировали и очищали на белке L. Белки исследовали в клеточном анализе и анализе рецептора и определяли полученные в результате IC50 (для анализа рецептора) и ND50 (для клеточного анализа) (Таблица 3, Фиг.12).
2.2 Fab
Димеры TAR1-5 и TAR1-5-19 также клонировали в формате Fab, экспрессировали и очищали на агарозе с белком L. Fab оценивали в анализах рецептора (Таблица 4). Результаты показали, что для димеров TAR1-5-19 и TAR1-5 уровни нейтрализации близки к первоначальным димерам с линкером Gly4Ser, из которых они получены. TAR1-5-19 Fab, где TAR1-5-19 представлен на СН и СК, экспрессировали, очищали на белке L и оценивали в анализе рецептора. Полученные IC50 составляли примерно 1 нМ.
2.3 Димеры TAR1-27
3×96 клонов отбирали из 2 раунда селекции, охватывающей все библиотеки и условия селекции. 2 мл препараты супернатантов приготавливали для анализа в ИФА и биоанализов. Антигенный ИФА дал 71 положительный клон. Анализ рецептора в неочищенных супернатантах дал 42 клона с ингибиторными свойствами (связывание TNF на 0-60%). В большинстве случаев ингибиторные свойства коррелировали с сильным сигналом ИФА. 42 клона секвенировали, 39 из них имеют уникальные последовательности второго dAb. Дополнительно анализировали 12 димеров, которые имели наилучшие ингибирующие свойства.
12 нейтрализующих клонов экспрессировали в виде препаратов супернатанта объемом 200 мл и очищали на белке L. Их оценивали с помощью белок L- и антигенного ИФА, BIAcore и в анализе рецептора. Во всех случаях получали сильные положительные сигналы ИФА. BIAcore анализ выявил все клоны, обладающие быстрыми скоростями ассоциации и диссоциации. Скорости диссоциации улучшались по сравнению с мономерным TAR1-27, однако скорость диссоциации для димеров TAR1-27 была быстрее (Koff находится приблизительно в диапазоне от 10-1 до 10-2 М) по сравнению с ранее исследованными димерами TAR1-5 (Koff находится приблизительно в диапазоне от 10-3 до 10-4 М). Выясняли стабильность очищенных димеров и поэтому для улучшения стабильности включили добавление 5% глицерина, 0,5% Triton Х100 или 0,5% NP40 (Sigma) в очистку 2 димеров TAR1-27 (d2 и d16). Добавление NP40 или Triton X100 улучшило выход очищенного продукта приблизительно в два раза. Димеры оценивали в анализе рецептора. TAR1-27d2 дал IC50 примерно 30 нМ при всех условиях очистки. TAR1-27d16 продемонстрировал отсутствие нейтрализующего эффекта при очистке без использования стабилизирующих агентов, но дал IC50 примерно 50 нМ при очистке в стабилизирующих условиях. Дополнительный анализ не проводили.
2.4 Димеры TAR2-5
3×96 клонов отбирали из второго раунда селекции, охватывающей все библиотеки и условия селекции. Для анализа приготавливали 2 мл препараты супернатанта. ИФА с белком А и антигеном осуществляли для каждого планшета. 30 интересующих клонов были идентифицированы с помощью BIAcore как имеющие хорошие скорости диссоциации (Koff находится в диапазоне от 10-2 до 10-3 М). Клоны секвенировали, и 13 уникальных димеров идентифицировали с помощью анализа последовательностей.
Димеры TAR1-5
* димер 2 и димер 3 имеют одинаковое второе dAb (названное dAb2), но имеют линкеры разной длины (d2=(Gly4Ser)3, d3=(Gly4Ser)3). dAb1 является партнером для димеров 1, 5 и 6. dAb3 является партнером для димера 4. Ни один из партнеров dAb не нейтрализует самостоятельно. Очистку с помощью FPLC, если не указано иначе, осуществляли с использованием катионного обмена. Оптимальные димерные частицы для каждого димера, полученного с помощью FPLC, определены в этих анализах.
Димеры TAR1-5-19
КА~12нМ
КА~10 нМ
Гомодимеры TAR1-5-19
КА~30 нМ
КА~3 нМ
КА~15нМ
TAR1-5/TAR1-5-19 Fab
КА~60 нМ
Пример 7. Димеризация dAb с помощью концевой цистеиновой связи
Краткое изложение
Для димеризации dAb свободный цистеин сконструировали на С-конце белка. При экспрессии белка образуется димер, который может быть очищен с помощью двухстадийного способа очистки.
Конструирование димера TAR1-5-19CYS с помощью ПЦР
Смотри пример 8, описывающий тример dAb. Протокол для тримера приводит к смеси мономера, димера и тримера.
Экспрессия и очистка димера TAR1-5-19CYS
Димер очищали из супернатанта культуры путем захвата на агарозе с белком L, как изложено в примере 8.
Отделение мономера TAR1-5-19CYS от димера TAR1-5-19CYS
Перед разделением с помощью катионного обмена смешанный образец мономера/димера путем замены буфера переводили в 50 мМ натрий-ацетатный буфер, рН 4,0, используя колонку PD-10 (Amersham Pharmacia), в соответствии с указаниями производителя. Затем наносили образец на 1 мл катионообменную колонку Resource S (Amersham Pharmacia), которую предварительно уравновешивали с помощью 50 мМ ацетата натрия, рН 4,0. Мономер и димер разделяли с помощью следующего солевого градиента в 50 мМ ацетате натрия, рН 4,0:
150-200 мМ хлорид натрия в 15 объемах колонки;
200-450 мМ хлорид натрия в 10 объемах колонки;
450-1000 мМ хлорид натрия в 15 объемах колонки.
Фракции, содержащие только димер, идентифицировали, используя ПААГ-ДСН, и затем объединяли, и рН увеличивали до 8 путем добавления 1/5 объема 1М Трис рН 8,0.
Функциональный анализ связывания in vitro: анализ рецептора TNF и клеточный анализ
Аффинность димера в отношении человеческого TNFα определяли, используя анализ рецептора TNF и клеточный анализ. IC50 в анализе рецептора составляла приблизительно 0,3-0,8 нМ; ND50 в клеточном анализе составляла приблизительно 3-8 нМ.
Другие возможные форматы димера TAR1-5-19CYS
ПЭГ (полиэтиленгликолевые) димеры и синтезированные на заказ малеимидные димеры
Nektar (Shearwater) предлагает диапазон бималеимидных ПЭГ [mPEG2-(MAL)2 или mPEG-(MAL)2], которые позволяют получить формат мономера в виде димера с небольшим линкером, разделяющим dAb, которые оба связаны с ПЭГ, размер которого находится в диапазоне от 5 до 40 кДа. Показано, что 5 кДа mPEG-(MAL)2 (т.е. [TAR1-5-19]-Cys-малеимид-PEGx2, где малеимиды связаны вместе в димере) имеет аффинность в анализе рецептора TNF примерно 1-3 нМ. Кроме того, димер также может быть получен с использованием ТМЕА (Трис[2-малеимидоэтил]амин) (Pierce Biotechnology) или других бифункциональных линкеров.
Также можно получить дисульфидный димер, используя метод химического сочетания с использованием 2,2'-дитиодипиридина (Sigma Aldrich) и укороченного мономера.
Добавление полипептидного линкера или шарнира к С-концу dAb.
Небольшой линкер (Gly4Ser)n, где п=1-10, например, 1, 2, 3, 4, 5, 6 или 7, иммуноглобулин (например, шарнирная область IgG или случайая пептидная последовательность (например, выбранная из библиотеки случайных пептидных последовательностей) может быть сконструирована между dAb и концевым цистеиновым остатком. Это можно затем использовать для получения димеров, как изложено выше.
Пример 8. Тримеризация dAb
Краткое изложение
Для тримеризации dAb требуется свободный цистеин на С-конце белка. Цистеиновый остаток, после восстановления с получением свободного тиола, затем может быть использован для специфического связывания белка с тримерной малеимидной молекулой, например ТМЕА (Трис[2-малеимидоэтил] амин).
Конструирование TAR1-5-19CYS с помощью ПЦР
Следующие олигонуклеотиды использовали для специфической ПЦР TAR1-5-19 с сайтами для клонирования SalI и BamHI, а также для введения С-концевого цистеинового остатка;

(*начало последовательности TAR1-5-19CYS; аминокислотная последовательность TAR1-5-19CYS (SEQ ID NO:293; нуклеотидные последовательности TAR1-5-19CYS (SEQ ID NO:294, кодирующая цепь; SEQ ID NO:295, некодирующая цепь))
Прямой праймер
5'-TGGAGCGCGTCGACGGACATCCAGATGACCCAGTCTCCA-3' (SEQ ID NO:296)
Обратный праймер
5'-TTAGCAGCCGGATCCTTATTAGCACCGTTTGATTTCCAC-3' (SEQ ID NO:297)
Реакцию ПЦР (объем 50 мкл) осуществляли в соответствии со следующим: брали 200 мкМ dNTP, 0.4 мкМ каждого праймера, 5 мкл 10х PfuTurbo буфера (Stratagene), 100 нг матричной плазмиды (кодирующей TAR1-5-19), 1 мкл фермента PfuTurbo (Stratagene) и объем доводили до 50 мкл с помощью стерильной воды. Использовали следующие условия ПЦР: первоначальная денатурирующая стадия при 94°С в течение 2 минут, затем 25 циклов при 94°С в течение 30 секунд, 64°С в течение 30 сек и 72°С в течение 30 сек. Конечная стадия также включала 72°С в течение 5 мин. Продукт ПЦР очищали и переваривали с помощью SalI и BamHI и лигировали в вектор, который разрезали теми же рестриктазами. Правильные клоны подтверждали секвенированием ДНК.
Экспрессия и очистка TAR1-5-19CYS
Вектор TAR1-5-19CYS трансформировали в химически компетентные клетки BL21 (DE3) pLysS (Novagen) в соответствии с протоколом производителя. Клетки, несущие плазмиду dAb, подвергали селекции с использованием 100 мкг/мл карбенициллина и 37 мкг/мл хлорамфеникола. Культуры приготавливали во встряхиваемых колбах объемом 2 л, содержащих 500 мл terrific бульона (Sigma-Aldrich), 100 мкг/мл карбенициллина и 37 мкг/мл хлорамфеникола. Культуры выращивали при 30°С при 200 об/мин до OD600 1-1,5 и затем индуцировали с помощью 1 мМ IPTG (изопропил-бета-D-тиогалактопиранозид, из Melford Laboratories). Экспрессию dAb продолжали в течение 12-16 ч при 30°С. Обнаружено, что большая часть dAb находилась в культуральных средах. Поэтому клетки отделяли от сред центрифугированием (8000xg в течение 30 минут), и супернатант использовали для очистки dAb. На литр супернатанта добавляли 30 мл агарозы с белком L (Affitech), и dAb давали возможность связываться в партии при перемешивании в течение 2 часов. Смолу затем оставляли осаждаться под действием силы тяжести в течение еще часа перед откачиванием супернатанта. Агарозу затем упаковывали в колонку ХК 50 (Amersham Phamacia) и промывали 10 колоночными объемами PBS. Связанный dAb элюировали 100 мМ глицином, рН 2,0, и фракции, содержащие белок, затем нейтрализовали путем добавления 1/5 объема 1 М Трис, рН 8,0. На литр культурального супернатанта выделяли 20 мг чистого белка, который содержал соотношение 50:50 мономера к димеру.
Тримеризация TAR1-5-19CYS
2,5 мл 100 мкМ TAR1-5-19CYS восстанавливали с помощью 5 мМ дитиотреитола и оставляли при комнатной температуре на 20 минут. Затем в обраце замещали буфер с помощью колонки PD-10 (Amersham Pharmacia). Колонку предварительно уравновешивали 5 мМ ЭДТА (этилендиамитетрауксусной кислотой), 50 мМ фосфатом натрия рН 6,5, и образец наносили и элюировали в соответствии с указаниями производителя. Образец помещали на лед до тех пор, пока не понадобится. ТМЕА (Трис[2-малеимидоэтил]амин) был приобретен в Pierce Biotechnology. Концентрированный раствор 20 мМ ТМЕА приготавливали в 100% ДМСО (диметилсульфоксид). Обнаружено, что концентрация ТМЕА более чем 3:1 (молярное соотношение dAb:TMEA) вызывала быстрое осаждение и перекрестное связывание белка. Кроме того, скорость осаждения и перекрестного связывания была еще выше при увеличении рН. Таким образом, используя 100 мкМ восстановленного TAR1-5-19CYS, добавляли 25 мкМ ТМЕА для тримеризации белка и реакции давали возможность протекать при комнатной температуре в течение двух часов. Обнаружено, что добавление дополнительных компонентов, таких как глицерин или этиленгликоль до 20% (об./об.), значительно уменьшало осаждение тримера по мере прохождения реакции сочетания. После сочетания анализ с ПААГ-ДСН показал присутствие мономера, димера и тримера в растворе.
Очистка тримерного TAR1-5-19CYS
40 мкл 40% ледяной уксусной кислоты добавляли на 1 мл реакционной смеси TMEA-TAR1-5-19cys для уменьшения рН до примерно 4. Образец затем наносили на 1 мл катионообменную колонку Resource S (Amersham Pharmacia), которую предварительно уравновешивали 50 мМ ацетата натрия, рН 4,0. Димер и тример частично разделяли, используя солевой градиент от 340 до 450 мМ хлорида натрия, 50 мМ ацетата натрия рН 4,0 в 30 объемах колонки. Фракции, содержащие только тример, идентифицировали с помощью ПААГ-ДСН и затем объединяли и рН увеличивали до 8 путем добавления 1/5 объема 1 М Трис рН 8,0. Для предотвращения осаждения тримера на стадиях концентрирования (с использованием 5К отсекающих концентраторов Viva spin;
Vivascience) к образцу добавляли 10% глицерина.
Функциональный анализ связывания in vitro: анализ рецептора TNF и клеточный анализ
Аффинность тримера в отношении человеческого TNFα определяли с использованием анализа рецептора TNF и клеточного анализа. IC50 в анализе рецептора составлял 0,3 нМ; ND50 в клеточном анализе находился в диапазоне от 3 до 10 нМ (например, 3 нМ).
Другие возможные форматы тримеров TAR1-5-19CYS
TAR1-5-19CYS также может быть переведен в формат тримера с использованием следующих реагентов:
ПЭГ тримеры и синтезированные на заказ малеимидные тримеры
Nektar (Shearwater) предлагает диапазон мультиплечевых ПЭГ, которые могут быть химически модифицированы на конце ПЭГ. Таким образом, использование ПЭГ тримера с малеимидной функциональной группой на конце каждого плеча дает возможность для тримеризации dAb по аналогии с изложенным выше с использованием ТМЕА. ПЭГ также может иметь преимущество при увеличении растворимости тримера, таким образом предотвращая проблему агрегации. Таким образом, можно получать тример dAb, в котором каждый dAb имеет С-концевой цистеин, который связан с малеимидной функциональной группой, причем малеимидные функциональные группы связаны с ПЭГ тримером.
Добавление полипептидного линкера или шарнира к С-концу dAb
Небольшой линкер, (Gly4Ser)n, где n=1-10, например 1, 2, 3, 4, 5, 6 или 7, иммуноглобулин (например, шарнирная область IgG или произвольная пептидная последовательность (например, выбранная из библиотеки случайных пептидных последовательностей) может быть сконструирована между dAb и концевым цистеиновым остатком. Когда линкер используют при получении мультимеров (например, димеров или тримеров), это снова может вносить большую степень гибкости и расстояние между индивидуальными мономерами, которые могут улучшить связывающие характеристики в отношении мишени, например мультисубъединичной мишени, такой как человеческий TNFα.
Пример 9. Селекция коллекции однодоменных антител (dAb), направленных против человеческого сывороточного альбумина (HSA) и мышиного сывороточного альбумина (MSA)
Этот пример объясняет способ получения однодоменного антитела (dAb), направленного против сывороточного альбумина. Описана селекция dAb против мышиного сывороточного альбумина (MSA) и человеческого сывороточного альбумина (HSA). В этом эксперименте использовали три фаговые дисплейные библиотеки человеческих антител, каждая из которых основана на единичном каркасе человеческого антитела для VH (смотри Фиг.13: последовательность пустого VH, основана на V3-23/DP47 и JH4b) или VK (смотри Фиг.15: последовательность пустого VK, основана на o12/o2/DPK9 и Jic1) с разнообразием боковой цепи, кодируемой кодонами NNK, включенными в области, определяющие комплементарность (CDR1, CDR2 и CDR3).
Библиотка 1 (VH):
Разнообразие в положениях: Н30, Н31, НЗЗ, Н35, Н50, Н52, Н52а, Н53, Н55, Н56, Н58, Н95, Н97, Н98.
Размер библиотеки: 6,2×109
Библиотка 2 (VH):
Разнообразие в положениях: Н30, Н31, Н33, Н35, Н50, Н52, Н52а, Н53, Н55, Н56, Н58, Н95, Н97, Н98, Н99, Н100, Н100а, H100b.
Размер библиотеки: 4,3×109
Библиотка 3 (VK):
Разнообразие в положениях: L30, L31, L32, L34, L50, L53, L91, L92, L93, L94, L96
Размер библиотеки: 2×109
Библиотеки VH и VK предварительно подвергали селекции в отношении связывания с общими лигандами белком А и белком L, соответственно, так что большинство клонов в неселектированных библиотеках являются функциональными. Размеры библиотек, представленных выше, соответствуют размерам после предварительной селекции.
Два раунда селекции осуществляли на сывороточном альбумине с использованием каждой из библиотек раздельно. Для каждой селекции антиген наносили на иммунопробирку (nunc) в 4 мл PBS в концентрации 100 мкг/мл. В первом раунде селекции каждую из трех библиотек разделяли методом пэннинга отдельно против HSA (Sigma) и MSA (Sigma). Во втором раунде селекции фаг из каждого из шести первых раундов селекции разделяли методом пэннинга против (1) того же антигена снова (например, 1й раунд MSA, 2й раунд MSA) и (2) против реципрокного антигена (например, 1й раунд MSA, 2й раунд HSA), что привело в результате к двенадцати 2м раундам селекции. В каждом случае после второго раунда селекции 48 клонов тестировали в отношении связывания с HSA и MSA. Растворимые фрагменты dAb получали, как описано для фрагментов scFv у Harrison etal, Methods Enzymol. 1996; 267:83-109, и следовали стандартному протоколу ИФА (Hoogenboom et al. (1991) Nucleic Acids Res., 19: 4133), за исключением того, что 2% tween PBS использовали в качестве блокирующего буфера, и связанные dAb обнаруживали с использованием белок L-HRP (Sigma) (для VK) и белок A-HRP (Amersham Pharmacia Biotech) (для VH).
dAb, которые дали сигнал больше фонового, указывающие на связывание с MSA, HSA или обоими, тестировали в нерастворимой форме ИФА на связывание только с пластиком, но все были специфичными в отношении сывороточного альбумина. Клоны затем секвенировали (смотри таблицу ниже), обнаружив, что идентифицирована 21 уникальная последовательность dAb. Минимальная близость (на аминокислотном уровне) между выбранными клонами VK dAb составляла 86,25% ((69/80)×100; результат, когда все диверсифицированные остатки различны, например клоны 24 и 34). Минимальная близость между выбранными клонами VH dAb составляла 94% ((127/136)×100).
Затем dAb, связывающие сывороточный альбумин, тестировали в отношении их способности захватывать биотинилированный антиген из раствора. Следовали протоколу ИФА (как выше), за исключением того, что планшет для ИФА покрывали 1 мкг/мл белка L (для клонов VK) и 1 мкг/мл белка А (для клонов VH). Растворимое dAb захватывали из раствора, как в протоколе, и обнаружение осуществляли с использованием биотинилированного MSA или HSA и стрептавидин-HRP. Биотинилированный MSA и HSA приготавливали в соответствии с инструкциями производителя с целью достижения в среднем 2 биотинов на молекулу сывороточного альбумина. Идентифицированы двадцать четыре клона, которые захватывали биотинилированный MSA из раствора в ИФА. Два из них (клоны 2 и 38 ниже) также захватывали 10 биотинилированных HSA. Затем dAb тестировали в отношении их способности связываться с MSA, нанесенными на biacore чип СМ5. Обнаружены восемь клонов, которые связывают MSA на biacore.
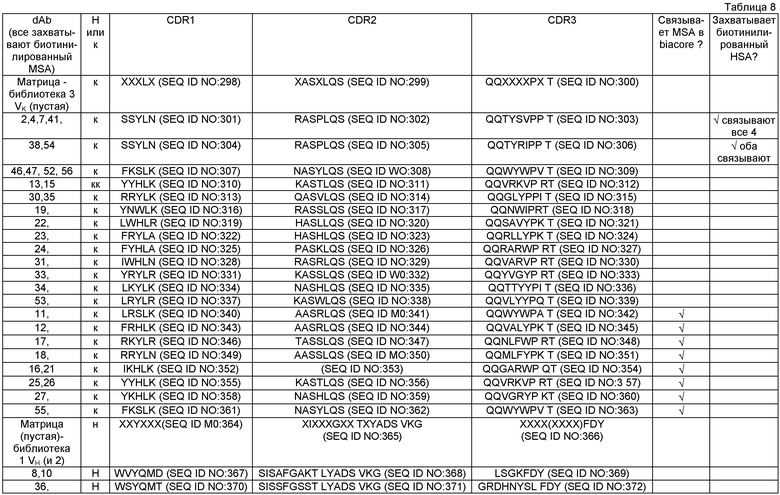
Во всех случаях каркасы были идентичны каркасам в соответствующей пустой последовательности, с различием в CDR, как указано в таблице выше.
Из восьми клонов, которые связывают MSA на biacore, два клона, которые высоко экспрессированы в E. coli (клоны MSA16 и MSA26), выбрали для дальнейшего исследования (смотри Пример 10). Полные нуклеотидные и аминокислотные последовательности для MSA16 и 26 приведены на Фиг.16.
Пример 10. Определение аффинности и периода полувыведения из сыворотки крови мыши dAb, связывающих MSA, MCA16 и МСА26
dAb MSA16 и MSA26 экспрессировали в периплазме Е. coli и очищали, используя абсорбцию партии на аффинной смоле белок L-агароза (Affitech, Norway) с последующей элюцией глицином при рН 2,2. Очищенные dAb затем тестировали с помощью biacore ингибирования для определения Kd. Кратко, очищенные MCA16 и МСА26 тестировали для определения концентрации dAb, необходимой для достижения 200 ОЕ ответа на biacore чипе СМ5, покрытом высокой плотностью MSA. После определения необходимых концентраций dAb, антиген MSA в диапазоне концентраций около ожидаемой Kd предварительно смешивали с dAb и инкубировали в течение ночи. Связывание с MSA-покрытым biacore чипом dAb в каждой из предварительных смесей затем измеряли при высокой скорости потока 30 мкл/минуту. Полученные кривые использовали для построения графиков Клотца, которые дали оцениваемую Кd 200 нМ для MSA16 и 70 нМ для MSA26 (Фиг.17 А и В),
Затем клоны MSA16 и MSA26 клонировали в экспрессирующий вектор с НА меткой (последовательность нуклеиновой кислоты:
TATCCTTATGATGTTCCTGATTATGCA (SEQ ID NO: 373) и аминокислотная последовательность: YPYDVPDYA (SEQ ID NO:374)) и количества 2-10 мг экспрессировали в Е. coli и очищали из супернатанта с аффинной смолой белок L-агарозой (Affitech, Norway) и элюировали глицином при рН 2,2. Период полувыведения dAb из сыворотки крови определяли у мышей. МСА26 и MCA16 дозировали в виде одноразовых внутривенных (в/в) инъекций при примерно 1,5 мг/кг мышам CD1. Анализ сывороточных уровней осуществляли с помощью захвата козьих антител против НА (Abeam, UK) и ИФА с обнаружением белка L-HRP (invitrogen), который блокировали с помощью 4% Marvel. Промывку осуществляли 0,05% tween PBS. Стандартные кривые с известными концентрациями dAb строили в присутствии 1х мышиной сыворотки крови для обеспечения совместимости с тестируемыми образцами. Моделирование с 2-компартментной моделью показало, что MSA-26 имел 11/2α 0,98 ч, t1/2β 14,5 ч и площадь под кривой (AUC) 465 ч.мг/мл (данные не представлены), и MSA-16 имел t1/2α 0,98 ч, t1/2α 36,5 ч и AUC 913 ч.мг/мл (Фиг.18). Оба клона против MSA имели значительно удлиненный период полувыведения по сравнению с HEL4 (dAb против лизоцима белка куриного яйца), который имел t1/2α 0,06 ч и t1/2β 0,34 ч.
Пример 11. Создание Fab-подобных фрагментов VH-VH и VK-VK с двойной специфичностью
Этот пример описывает способ получения двойных специфичностей VH-VH и VK-VK в виде Fab-подобных фрагментов. Перед конструированием каждого из описанных Fab-подобных фрагментов, dAb, которые связываются с выбранными мишенями, сначала селектировали из библиотек dAb, похожих на те, которые описаны в Примере 9. Выделяли VH dAb, HEL4, который связывает лизоцимом куриного яйца (Sigma), а также выделяли второй VH dAb (TAR2h-5), который связывает рецептор TNFα (R and D systems). Их последовательности приведены в перечне последовательностей. VK dAb, которое связывает TNFα (TAR1-5-19), выделяли путем селекции и аффинного созревания, и данная последовательность также приведена в перечне последовательности. Второе VK dAb (MCA26), описанное в Примере 9, последовательность которого приведена на Фиг.17 В, также использовали в этих экспериментах.
ДНК из экспрессирующих векторов, содержащих четыре dAb, описанных выше, переваривали ферментами SaiI и NotI для вырезания ДНК, кодирующей dAb. Полосу ожидаемого размера (300-400 п.о.) очищали путем разгонки перевара на агарозном геле и вырезания полосы с последующей очисткой на геле с использованием набора для очистки на геле Qiagen (Qiagen, UK). ДНК, кодирующую dAb, затем встраивали в векторы CH или CK (Фиг.8 и 9), как указано в таблице ниже.
Конструкции VH CH и VH CK котрансформировали в клетки НВ2151. Отдельно конструкции VK CH и VK CK котрансформировали в клетки НВ2151. Культуры каждой из котрансформированных клеточных линий выращивали в течение ночи (в 2xTy, содержащей 5% глюкозы, 10 мкг/мл хлорамфеникола и 100 мкг/мл ампициллина для поддержания антибиотической селекции для обеих плазмид CH и CK). Ночные культуры использовали для инокуляции свежих сред (2xTy, 10 мкг/мл хлорамфеникола и 100 мкг/мл ампициллина) и выращивали до OD 0,7-0,9 перед индукцией путем добавления IPTG для экспрессии их конструкций CH и CK. Экспрессирующийся Fab-подобный фрагмент затем очищали из периплазмы путем очистки на белке А (для котрансформированных VH CH и VH CK) и очистки на MSA аффинной смоле (для котрансформированных VK CH и VK CK).
VH-VH с двойной специфичностью
Экспрессию VH CH и VH CK с двойной специфичностью тестировали путем разгонки белка на геле. Гель подвергали блоттингу, и полосу ожидаемого размера для Fab-фрагмента можно было обнаружить на Вестерн-блоте посредством тус-метки и flag-метки, указывающих на то, что присутствовали обе части VH CH и VH CK Fab-подобного фрагмента. Затем для определения того, присутствуют ли две половины с двойной специфичностью в одном и том же Fab-подобном фрагменте, планшет для ИФА покрывали в течение ночи при 4°С 100 мкл на лунку лизоцима куриного яйца (HEL) в концентрации 3 мг/мл в натрий-бикарбонатном буфере. Планшет затем блокировали (как описано в Примере 1) с помощью 2% tween PBS с последующей инкубацией с Fab-подобным фрагментом VH CH/Vн CK с двойной специфичностью. Обнаружение связывания двойной специфичности с HEL осуществляли через некогнатную цепь, используя 9е10 (моноклональное антитело, которое связывает myc-метку, Roche) и антимышиный IgG-HRP (Amersham Pharmacia Biotech). Сигнал для Fab-подобного фрагмента VH CH/VH CK с двойной специфичностью составлял 0,154, по сравнению с фоновым сигналом 0,069 только для экспрессирующейся цепи VH CK. Это демонстрирует, что Fab-подобный фрагмент обладает связывающей специфичностью в отношении антигена-мишени.
VK-VK с двойной специфичностью
После очистки котрансформированного Fab-подобного фрагмента VK CH и VK CK с двойной специфичностью на MSA аффинной смоле, полученный белок использовали для зондирования планшета для ИФА, покрытого 1 мкг/мл TNFα, и планшета для ИФА, покрытого 10 мкг/мл MSA. Как предсказано, существовал сигнал выше фонового при детекции белком L-HRP на обоих планшетах для ИФА (данные не представлены). Это показало, что фракция белка, способная связываться с MSA (и таким образом очищенная на MSA аффинной колонке), также способна связываться с TNFα в последующем ИФА, подтверждая двойную специфичность фрагмента антитела. Эту фракцию белка затем использовали для двух последующих экспериментов. Во-первых, планшет для ИФА, покрытый 1 мкг/мл TNFα, зондировали Fab-подобным фрагментом VK CH и VK CK с двойной специфичностью, а также контрольным TNFα-связывающим dAb в концентрации, рассчитанной на получение похожего сигнала на ИФА. dAb с двойной специфичностью и контрольное dAb использовали для зондирования планшета для ИФА в присутствии и в отсутствие 2 мг/мл MSA. Сигнал в лунке с двойной специфичностью уменьшался более чем на 50%, а сигнал в лунке с dAb не уменьшался вовсе (смотри Фиг.19а). Тот же самый белок использовали также в анализе рецептора с MSA и без него, и также представлена конкуренция с MSA (смотри Фиг.19с). Это демонстрирует, что связывание MSA с двойной специфичностью конкурирует со связыванием с TNFα/
Пример 12. Создание cys-связанного VK-VK с двойной специфичностью, имеющего специфичность в отношении мышиного сывороточного альбумина и TNFα
Этот пример описывает способ получения фрагмента антитела с двойной специфичностью, специфичного в отношении как мышиного сывороточного альбумина, так и TNFα, с помощью химического сочетания через дисульфидную связь. MSA16 (из примера 1) и TAR1-5-19 dAb повторно клонировали в вектор на основе рЕТ с С-концевым цистеином и без меток. Два dAb экспрессировали при уровнях 4-10 мг и очищали из супернатанта, используя аффинную смолу белок L-агароза (Affitiech, Norway). dAb с цистеиновой меткой затем восстанавливали дитиотреитолом. TAR1-5-19 dAb затем подвергали сочетанию с дитиопиридином для блокирования преобразования дисульфидных связей, приводя к образованию гомодимеров PEP 1-5-19. Два различных dAb затем смешивали при рН 6,5, чтобы обеспечить образование дисульфидных связей и образование TAR1-5-19, MSA16 cys-связанных гетеродимеров. Этот способ получения конъюгатов двух непохожих белков первоначально описан Кингом с соавт.(King TP, Li Y Kochoumian L Biochemistry. 1978 vol 17:1499-506 Preparation of protein conjugates via intermolecular disulfide bond formation). Гетеродимеры отделяли от мономерных частиц посредством катионного обмена. Разделение подтверждали наличием полосы ожидаемого размера на геле с ДСН. Полученные гетерод и мерные частицы тестировали в анализе рецептора TNF и обнаружили, что они имеют IC50 для нейтрализации TNF приблизительно 18 нМ. Затем анализ рецептора повторяли с постоянной концентрацией гетеродимера (18 нМ) и сериями разведении MSA и HSA. Присутствие HSA в диапазоне концентраций (до 2 мг/мл) не вызывало уменьшения способности димера ингибировать TNFα. Однако добавление MSA вызывало дозозависимое уменьшение способности димера ингибировать TNFα (Фиг.20). Это демонстрирует, что MSA и TNFα конкурируют за связывание с cys-связанным димером TAR1-5-19, MSA16.
Сводка данных
Сводка данных, полученных в экспериментах, изложенных в следующих примерах, представлена в Приложении 4.
Пример 13. Активность dAb против мышиного TNFR1 и dAb против человеческого TNFR1
Активность dAb против мышиного TNFR1
Активность dAb против TNFR1
Анализ высвобождения IL-8 клетками MRC-5
Активности некоторых dAb, которые связывают человеческий TNFR1, оценивали в следующем анализе на клетках MRC-5. Анализ основан на индукции секреции IL-8 с помощью TNF в клетках MRC-5 и адаптирован из способа, описанного в Alceson, L. et al. Journal of Biological Chemistry 277:30517'-30523 (1996), описывающего индукцию IL-8 с помощью IL-1 в HUVEC. Активность dAb оценивали путем измерения индукции IL-8 человеческим TNFα с использованием клеток MRC-5 вместо клеточной линии HUVEC. Кратко, клетки MRC-5 высевали в микротитрационные планшеты, и данные планшеты инкубировали в течение ночи с dAb и человеческим TNFα (300 пг/мл). После инкубации культуральный супернатант отбирали и измеряли концентрацию IL-8 в супернатанте с помощью "сэндвич"-ИФА (R&D Systems). Активность dAb против TNFR1 приводила к уменьшению секреции IL-8 в супернатант, по сравнению с контрольными лунками, которые инкубировали только с TNFα.
Пример 14. Модель септического шока у мышей
Эффективность dAb против TNFR1 in vivo оценивали в луночной экспериментальной модели синдрома септического шока (Rothe etal, Circulatory Shock 44:51 -56, (1995)). ЛПС-индуцированная смерть в этой модели зависит от активации TNFR-1 (р55). В этой модели мышей сенситизировали в отношении токсичности ЛПС, используя D-галактозамин (D-GaIN). Летальная доза ЛПС для животных дикого типа в этом исследовании составляла примерно 10 нг.
ЛПС (Salmonella enteritidis, Sigma, USA) и D-галактозамин (D-GaIN, Sigma, USA) инъецировали внутрибрюшинно. D-GaIN-сенситизированных (10 мг/мышь) контрольных мышей умерщвляли в течение 18 часов после иммунизации ЛПС (10 нг). Смертность несенситизированных мышей регистрировали в течение периода 1 сутки после иммунизации.
Мышам вводили лиганд с двойной специфичностью, который связывает мышиный TNFR1 и мышиный сывороточный альбумин (TAR2m-21-23 3D TAR7m-16; TAR7m-16 также назван в данном изобретении как MSA16) или ENBREL® (entarecept; Immunex Corporation) с помощью внутрибрюшинных инъекций за 4 часа до введения ЛПС.(Смотри Таблицу 12). Выживание контролировали с 4-6 часовыми интервалов в течение 48 часов. Эффективность dAb против мышиного TNFR1 была продемонстрирована с помощью выживания.
TAR2m-21-23 3U TAR7m-16 представляет собой лиганд с двойной специфичностью, который содержит dAb, связывающее мышиный TNFR1, который соединен через пептидный линкер с dAb, связывающим мышиный сывороточный альбумин. Нуклеотидная последовательность, кодирующая TAR2m-21-23 3U TAR7m-16, и аминокислотная последовательность лиганда с двойной специфичностью представлены ниже соответственно как SEQ ID NO:375 и SEQ ID NO:376.

Присутствие выживших в группах, которые обрабатывали TAR2m-21-23 3U TAR7m-16, демонстрирует, что dAb против TNFR1 эффективны в ингибировании активности рецептора in vivo, и результаты демонстрируют, что эффект был дозозависимым. Кроме того, эффективность обработки TAR2m-21-23 3U TAR7m-16 предпочтительно сравнивали с эффективностью ENBREL® (entarecept; Immunex Corporation). Выживание животных, которых обрабатывали только TAR2m-21-23 3U TAR7m-16 (Группа 5, без ЛПС иммунизации) также демонстрирует, что TAR2m-21-23 3U TAR7m-16 не был токсичным и не оказывал агонистического действия на рецептор in vivo путем перекрестного связывания с рецептором.
Дополнительные исследования подтвердили, что dAb против TNFR1 не оказывало агонистического действия против TNFR1 (действует как агонист TNFR1) в отсутствие TNFα. Клетки L929 культивировали в средах, содержащих диапазон концентраций либо мономера TAR2m-21-23, либо мономера TAR2m-21-23, перекрестно связанного с коммерчески доступным антителом против туе (9Е10), TAR2m-21-23 3U TAR7m-16 или TAR2m-21-23 40К ПЭГ. В случае с мономером TAR2m-21-23, перекрестно связанным с антителом против myc, dAb и антитело смешивали в соотношении 2:1 и предварительно инкубировали в течение одного часа при комнатной температуре для стимуляции эффектов иммунного перекрестного связывания in vivo перед культивированием. Мономер TAR2m-21-23 инкубировали с клетками L929 в концентрации 3000 нМ. Мономер TAR2m-21-23 и антитело против Myc инкубировали при концентрации dAb 3000 нМ. TAR2m-21-23 3U TAR7m-16 инкубировали с клетками в концентрациях 25 нМ, 83,3 нМ, 250 нМ, 833 нМ и 2500 нМ. TAR2m-21-23 40К ПЭГ инкубировали с клетками в концентрациях 158,25 нМ, 527,5 нМ, 1582,5 нМ, 5275 нМ и 15825 нМ. После инкубации в течение ночи жизнеспособность клеток оценивали, как описано для анализа цитотоксичности в отношении клеток L929. Результаты выявили, что инкубация с различными количествами dAb не приводила к увеличению количества нежизнеспособных клеток в культурах. Инкубация клеток L929 с 10 нМ, 1 нМ и 0,1 нМ имеющегося в продаже IgG антитела против TNFR1 приводила к дозозависимому увеличению нежизнеспособных клеток, демонстрируя таким образом чувствительность этих клеток к TNFR1-опосредован ному агонизму (Фиг.26).
Пример 15. Модели хронических воспалительных заболеваний
А. Модель коллаген-индуцированного артрита у мышей
Мышам DBA/1 однократно инъецировали эмульсию адъюванта Arthrogen-CIA и коллагена Arthrogen-CIA (MD-biosciences). На 21 сутки животных с высокими оценками артрита удаляли из исследования, а оставшихся животных разделяли на группы по 10 с равными количествами самцов и самок. На 21 сутки обработки начинали с внутрибрюшинных инъекций физиологичекого раствора, ENBREL® (entarecept; Immunex Corporation) или TAR2m-21-23 40k ПЭГ и продолжали в течение 28 суток. Клинические оценки артрита по шкале от О до 4 измеряли для каждой из 4 конечностей животных, оценку 0 присваивали нормальной конечности, а оценку 4 присваивали максимально воспаленной конечности с вовлечением множества суставов.
Уменьшение суммирования оценок артрита для четырех конечностей от максимальной 16 до (а) 14-15, (б) 12-15, (в) 10-15. (г) 9-15, (д) 7-15, (е) 5-15, (ж), 3-15 или (з) 1-15 представляет собой благоприятный эффект в этой модели. Благоприятный эффект может приводить к суммированию оценок артрита для четырех конечностей 0, 1,2, 3, 4, 5, 6, 7, 8, 9, 1 0, 11, 12, 13, 14 или 15. Задержку начала артрита по сравнению с необработанной контрольной группой также представляет собой благоприятный эффект в этой модели.
Клинические оценки ясно продемонстрировали, что обработка TAR2m-21-23 40К ПЭГ оказывали очень благоприятное влияние на ингибирование развития артрита по сравнению с физиологическим раствором в качестве контроля, и, кроме того, обработку TAR2m-21-23 40K ПЭГ предпочтительно сравнивали с ENBREL® (entarecept; Immunex Corporation). Это представляет собой первую демонстрацию того, что ингибирование TNFR1 является эффективным в модели хронического воспалительного заболевания.
Б. Модель IBD и артрита у мышей AARE
Мыши, которые несли целевую делецию в 3' AU-обогащенных элементах (ARE) мpHК TNF (названных мышами TnfΔARE), сверхпродуцировали TNF, и у них развивалось воспалительное заболевание кишечника, которое гистопатологически напоминает болезнь Крона (Kontoyiannis et al, J Exp Med 196:1563-74 (2002). У этих мышей заболевание, подобное болезни Крона, развивается в возрасте между 4 и 8 неделями, и у животных также развивались клинические признаки ревматоидного артрита.
dAb, которые связывают мышиные TNFR-1, оценивали в отношении эффективности в ингибировании патологии, подобной болезни Крона, и артрита у мышей TnfΔARE ENBREL® (entarecept; Immunex Corporation) использовали в качестве положительного контроля. Агенты вводили с помощью внутрибрюшинных инъекций в соответствии с введением и дозовым режимом, представленными в Таблице 13.
dAb, которые исследовали, включают:
1) TAR2m-21-23, пэгилированные одной группировкой 40 кДа ПЭГ;
2) TAR2m-21-23 3U TAR7m-16 с двойной специфичностью, который связывают мышиный TNFR-1 и мышиный сывороточный альбумин.
По завершении дозового периода мышей умерщвляли и для анализа удаляли концевые отделы подвздошной кишки и проксимальные отделы толстой кишки.
Для гистологического анализа делали срезы тканевых образцов и окрашивали гематоксилином и эозином, и острое и хроническое воспаление оценивали с использованием полуколичественной системы оценки. Оценку осуществляли в соответствии со следующим: оценка острого воспаления 0=0-1 полиморфно-ядерная клетка (PMN) в высокоэнергетическом поле (PMN/hpf);
1=2-10 PMN/hpf в слизистой оболочке; 2=11-20 PMN/hpf в слизистой оболочке;
3=21-30 PMN/hpf в слизистой оболочке или 11-20 PMN/hpf с распространением ниже muscularis mucosae (мышечной пластинки слизистой оболочки); 4 не меньше 30 PMN/hpf в слизистой оболочке или более 20 PMN/hpf с распространением ниже muscularis mucosae; оценка хронического воспаления 0=0-10 мононуклеарные лейкоциты (ML) на hpf (ML/hpf) в слизистой оболочке; 1=11-20 ML/hpf в слизистой оболочке; 2=21-30 ML/hpf в слизистой оболочке или 11-20 ML/hpf с распространением ниже muscularis mucosae; 3=31-40 ML/hpf в слизистой оболочке или фолликулярной гиперплазии; и 4 не меньше 40 ML/hpf в слизистой оболочке или более 30 ML/hpf с распространением ниже musculan's mucosae или фолликулярной гиперплазии.
Макрофенотипические симптомы артрита оценивали еженедельно в соответствии со следующей системой: 0=отсутствие артрита (нормальный внешний вид и гибкость); 1=умеренный артрит (деформация сустава); 2=умеренный артрит (опухание, деформация суставов); 3=тяжелый артрит (сильно нарушенное движение).
TAR2m-21-23 dAb продемонстрировали хорошую эффективность in vivo в модели артрита у мышей дельта ARE в виде 40 кДа пэгилированного мономера (TAR2m21-23 40 кДа ПЭГ) и в виде обладающего двойной специфичностью формата антитела против TNFRI/антитела против SA (TAR2m-21-23 3U TAR7m-16). На 9 неделю средние оценки артрита для групп, обработанных TAR2m-21-23 и TAR2m-21-23 3U TAR7m-16, были меньше 0,4. В отличие от этого, контрольная группа, обработанная физиологическим раствором, имела от умеренного до тяжелого артрита со средней оценкой артрита, составляющей более 1,0. Группа, обработанная ENBREL® (entarecept; Immunex Corporation), который вводили три раза в неделю, по сравнению с TAR2m-21-23 и TAR2m-21-23 3U TAR7m-16, которые вводили два раза в неделю, имела среднюю оценку 0,5-1,0. Эти результаты показывают, что лечение форматами dAb, которые связывают TNFR1, представляет собой высокоэффективную антиартритную терапию, и что исследованные пэгилированный и обладающие двойной специфичностью форматы dAb представляют собой высокоэффективные лекарства при хроническом воспалительном заболевании. Кроме того, эти результаты дополнительно демонстрируют, что dAb, которые связывают TNFR1, соединяются с рецептором только антагонистическим образом.
В. Модель IBD у мышей DSS
IBD индуцируют у мышей путем введения декстрансульфата натрия (DSS) в питьевой воде (смотри, например Okayasu I. et al, Gastroenterology 98:694-702 (1990); Podolsky K., J Gasteruenterol. 38 suppi XV.-63-66 (2003).) Взрослых мышей BDF1, у которых отсутствовала Н. pylori, содержали в течение 2 недель для стабилизации их суточных ритмов. Всех мышей держали в отдельных вентилируемых контейнерах в специальном блоке, имеющем барьер против патогенов (SPF), с 12-часовым циклом день/ночь. Животных обеспечивали кормом и водой ad libitum.
Исследование продолжали в течение 7 суток. Питьевая вода содержала 5% DSS для продолжения исследования. Всех животных обрабатывали на 1-7 сутки утром (0900-1000) и вечером (1600-1700). (Смотри Таблицу 14.) 1 сутки эквивалентны единичной профилактической дозе. Всех животных ежедневно взвешивали и регистрировали любой симптом диареи. Всех животных умерщвляли через 24 часа после последней обработки и однократно вводили бромдезоксиуридин за 40 минут до умерщвления. Дистальные отделы толстой кишки извлекали из животных. Небольшой образец дистального отдела толстой кишки помещали в "RNAIater", а оставшуюся часть фиксировали в фиксаторе Карноя, заливали в парафин, делали срезы (несерийные срезы на слайд) и окрашивали гематоксилином и эозином. Срезы визуально оценивали в отношении тяжести IBD и присваивали оценку тяжести. Осуществляли гистометрические анализы и определяли среднюю площадь поражения (площадь язвы), среднюю площадь эпителия и среднюю интрамуральную площадь воспаления.
Поперечные срезы Н&Е толстой кишки использовали для регистрации серии размеров ткани с использованием микроскопа Zeiss Axiohome, который позволяет точно количественно оценить площади. Для каждого поперечного среза измеряли площадь эпителия вместе с собственной пластинкой (lamina ргорпа) и площадь соединительной ткани. Площадь эпителия затем измеряли отдельно, разница относится к площади lamina ргорпа. В нормальной ткани относительный вклад этой ткани в площадь составляет примерно 10%, но она увеличивается по мере увеличения воспаления. Поэтому изменяется относительная пропорция эпителия:lamina ргорпа.
При увеличении тяжести глубина этой области уменьшается (внося вклад в изъязвление) и длина толстой кишки сокращается. Вместе эти явления вызывают увеличение площади просвета на поперечном срезе. Поэтому этот параметр также может быть полезным измерением тяжести заболевания.
Образцы ткани наблюдали под микроскопом и присваивали оценку тяжести, где 0=нет воспаления; 1=умеренное воспаление вокруг основания крипты; 2=массивная воспалительная инфильтрация и нарушенная архитектура слизистой оболочки; 3=массивная воспалительная инфильтрация и нарушенная архитектура слизистой оболочки плюс изъязвление.
Эффективность указывается в этот модели, когда лечение приводит к уменьшению оценки тяжести по сравнению с оценкой тяжести в контрольной группе, обработанной физиологическим раствором. Например, оценка тяжести обработанной группы может быть уменьшена от примерно 0,1 до примерно 1, от 1 до примерно 2, или от 2 до примерно 3. Эффективность может быть указана при оценке примерно 2 или меньше, от 1 до примерно 2, или 1 или меньше.
9 групп из 6 животных обрабатывают в соответствии со следующим:
Пероральные зонды вводят с ZANTAC® (ранитидина гидрохлорид; GlaxoSmithKline).
Г. Модель хронического обструктивного заболевания легких (ХОЗЛ) у мышей
Оценивают эффективность dAb против TNFR1 в развитии заболевания в модели субхронического отравления табачным дымом (ТД) у мышей (смотри, например Wright JL and Churg A., Chest 122:306S-309S (2002). dAb против мышиного TNFR1 вводят с помощью внутрибрюшинных инъекций каждые 48 часов (начиная за 24 часа до первого воздействия ТД) и вводят в виде формата, беспечивающего удлиненный период полувыведения из сыворотки крови (например, пэгилированный лиганд с двойной специфичностью, включающий dAb против SA).
Альтернативно, dAb против TNFR1 вводят посредством интраназальной доставки каждые 24 часа (начиная с момента 4 часа до первого воздействия ТД) и в виде мономера dAb. ENBREL® (entarecept; Immunex Corporation) используют в качестве положительного контроля. Воздействие ТД осуществляют ежедневно, и исследование длится в течение 1-2 недель. (Смотри, например Vitalis et al, Eur. Respir. J, 77:664-669 (1998). После последнего воздействия ТД бронхоальвеолярные смывы анализируют в отношении общего и дифференцированного подсчета клеток для включения подгрупп нейтрофилов, эозинофилов, макрофагов и Т-лимфоцитов. Легочные доли фиксируют в 10%-ном забуференном формалине, и тканевые срезы анализируют в отношении увеличения альвеол и альвеолярных ходов, утолщения стенок малых дыхательных путей и подсчета клеток для включения подгрупп нейтрофилов, эозинофилов, макрофагов и Т-лимфоцитов. Эффективность будет очевидна по уменьшению количества подгрупп нейтрофилов, эозинофилов, макрофагов и Т- увеличения альвеол и альвеолярных ходов и утолщения стенок лимфоцитов, которые увеличивались при воздействии ТД, и уменьшения ТД-индуцированного малых дыхательных путей.
Пример 16. Конструирование и экспрессия рекомбинантной химерной молекулы TNFR1
Этот пример поясняет способ получения молекулы, состоящей из различных доменов мышиных TNFR1 и доменов человеческих TNFR1 (Banner DW, et al. Cell, 75(3):431-45 (1993), так что молекула содержит четыре определенных внеклеточных домена TNFR1, и что они варьируют по происхождению между мышиными и человеческими белками TNFR1. Продуцируемые химерные рецепторы разделяют свойства как человеческого TNFR1, так и мышиного TNFR1 в соответствии с различными ролями доменов и функциональностью. Молекулы обеспечили средства для оценки доменной специфичности dAb, антител и их антиген-связывающих фрагментов и других молекул (например, органические химические соединения, NCE; или белковые домены, такие как аффитела, домены рецептора LDL или домены EGF), которые связывают человеческий или мышиный TNFR1.
Способы
Последовательности человеческих и мышиных TNFR1 ранее клонировали в экспрессирующем векторе pPicZalpha (hivitrogen) на основе Pichia через рестрикционные сайты эндонуклеаз EcoRI и Notl. Матричная ДНК мышиного TNFR1 (и, следовательно, химерные рецепторные конструкции, заканчивающиеся мышиным доменом 4) содержала 3' 6х гистидиновую метку. Человеческий TNFR1 (и, следовательно, химерные рецепторные конструкции, заканчивающиеся человеческим доменом 4) содержал Myc и 6х гистидиновые метки последовательности на 3'-конце.
Первоначальные ПЦР осуществляли в соответствии со стандартными условиями ПЦР с использованием RubyTaq ДНК-полимеразы (USB Corporation, Cleveland, Ohio), 100 нг матричной ДНК (включающий соответствующую ДНК минипреп матрицу полноразмерной ДНК mTNFRI или hTNFRI).
Типичные реакции ПЦР были следующими: 25 мкл 10Х RubyTaq буфера для ПЦР, содержащего полимеразу; 2 мкл первого праймера (из 10 мкМ исходного раствора); 2 мкл второго праймера (из 10 мкМ исходного раствора); 1 мкл (100 нг) полноразмерной матрицы ДНК TNFR1; 20 мкл деионизированной H2O (до конечного объема 50 мкл). Реакции проводили в тонкостенных пробирках в термоциклере в соответствии со следующими параметрами.
Сводка по начальным реакциям ПЦР, используемым при получении химерных конструкций
Обозначения: Н=человеческий домен; М=мышиный домен; например, МННН=мышиный домен 1, человеческие домены 2-4.
Продукты ПЦР, полученные в этих первоначальных ПЦР, вырезали из 1%-ного агарозного геля и очищали с использованием набора для очистки в геле (Qiagen) перед элюцией в 50 мкл dH2O.
Праймеры, используемые для получения химерной конструкции TNFR1
SOE PCR
Сборочная ПЦР (assembly PCR) (также известная как "протягивание" или сплайсинг путем удлинения перекрывающихся областей (SOE, Splicing by Overlap Extension) смотри Gen, 15:77(1):61-8 (1989)) позволяет объединять первичные ПЦР-продукты без расщепления или лигирования с использованием комплементарных концов первичных ПЦР-продуктов. В этом способе первичные продукты объединяют и денатурируют перед тем, как позволяют их комплементарным концам отжигаться вместе в присутствии Taq ДНК-полимеразы и dNTP. Несколько циклов повторного отжига и удлинения приводят к заполнению комплементарных цепей и получению полноразмерной матрицы. Добавляют праймеры, которые фланкируют полноразмерную конструкцию кассеты, и стандартную ПЦР проводят для амплификации собранного продукта. SOE PCR проводили для отжига вместе и амплификации различных доменов TNFR1. происходящих из описанных выше первоначальных ПЦР. Сборочные SOE PCR представляли собой следующее: 40 мкл 10х буфера для ПЦР, содержащего MgCl2; примерно 2 мкл (100 нг) очищенного продукта первоначальной ПЦР 1; примерно 2 мкл (100 нг) очищенного продукта первоначальной ПЦР 2; 36 мкл дистиллированной H2O (до конечного объема 80 мкл). Смесь праймеров SOE добавляли после стадии сборки в соответствии со следующим: 2 мкл 5'-фланкирующего праймера (Праймер 1); 2 мкл 3'-фланкирующего праймера (Праймер 2); 10 мкл 10х буфера для ПЦР; 6 мкл дистиллированной H2O (до конечного объема 20 мкл).
Реакции ПЦР осуществляли с использованием описанной ниже программы. Первоначальные сборочные циклы требовали приблизительно 45 минут, после чего термоциклер устанавливали на паузу при 94°С. 20 мкл смеси праймера добавляли к каждой реакционной смеси и смешивали.
Стадия 1. Сборка
Стадия 2. Амплификация (пауза при 94°С, затем добавляли смесь праймеров)
Продукты ПЦР исследовали путем прогона 3-5 мкл каждой реакционной смеси на 1%-ном агарозном геле.
3) Клоны собранных химер TNFR1 в экспрессирующем векторе pPicZalpha на основе Pichia (Invitrogen) затем переваривали ферментами EcoRI и NotI перед очисткой на гель-фильтрационной колонке Chromaspin ТЕ-1000 (Clontech, Mountain View, CA).
4) Трансформация химерных конструкций TNFR1 в Е. соli
Лигированные химерные конструкции трансформировали в электрокомпетентные клетки НВ2151 E. coli и выделяли в течение одного часа в низкосолевых средах LB перед нанесением на низкосолевой агар LB с 0,25 мкг/мл ZEOCIN, представляющим собой антибиотический препарат, содержащий флеомицин D (Cayla, Toulouse, France), в течение 24 ч при 37°С. Индивидуальные колонии затем исследовали на обеспечение правильной последовательности химерной конструкции в экспрессирующем векторе и крупномасштабные плазмидные препараты Maxiprep приготавливали из каждого вектора химерной конструкции.
Нуклеотидные последовательности полученных химерных конструкций представлены ниже. Химерные конструкции названы в соответствии с происхождением их доменов (начиная с домена 1 слева до домена 4 справа). Например, НМММ содержит человеческий домен 1 и мышиные домены 2-4. Химерные белки, которые содержат мышиный домен 4, имеют только His метки и лишены спейсерной области между трансмембранной областью и доменом 4.



5) Получение химерной конструкции TNFR1 и трасформация в Pichia pastoris
Плазмидную ДНК, полученную в каждом максипрепарате, переваривали редко режущей рестрикционной эндонуклеазой Pmel для линеаризации ДНК перед трансформацией в pichia. Линеаризованную ДНК затем очищали с помощью экстракции в фенол/хлороформе и осаждения в этаноле перед суспендированием в 30 мкл дистиллированной Н20. 10 мкл раствора линеаризованной ДНК смешивали с 80 мкл электрокомпетентных клеток КМ71Н Pichia в течение 5 минут перед электропорацией при 1,5 кВ, 200Ω, 25 мкФ. Клетки немедленно выделяли с YPDS и инкубировали в течение 2 часов при 30°С перед посевом на YPDS агаровые чашки, содержащие 100 мкг/мл ZEOCIN, представляющего собой антибиотический препарат, содержащий флеомицин D (Cayla, Toulouse, France), в течение 2 суток.
6) Экспрессия конструкций в Pichia
Индивидуальную колонию трансформантов для каждой конструкции отбирали в 5 мл BMGY в качестве стартовой культуры и выращивали в течение 24 ч при 30°С.Эту культуру использовали для инокуляции 500 мл сред BMGY, которые выращивали в течение 24 ч при 30°С до того, как клетки собирали путем цнтрифугирования при 1500-3000g в течение 5 минут при комнатной температуре. Клетки затем ресуспендировали в 100 мл BMMY и выращивали в течение 4 суток с постепенным увеличением концентрации метанола (0,5% в сутки 1, 1% в сутки 2, 1,5% в сутки 3 и 2% в сутки 4). После экспрессии супернатант отделяли после центрифугирования культур при 3300g в течение 15 минут.
7) Очистка химерных конструкций TNFR1 с использованием никелевой смолы
Культуральные супернатанты сначала забуферивали путем добавления 10 мМ конечной концентрации имидазола и 2х PBS. His-меченый белок абсорбировали в партии в течение 4 часов (встряхивание) при комнатной температуре путем добавления смолы Nickel-NTA. Смесь супернатант/смола затем пропускали через препаративную колонку poly-prep (Biorad). Смолу затем промывали 10 объемами колонки 2xPBS перед элюцией с использованием 250 мМ имидазола 1xPBS. После замены буфера химерную экспрессирующую конструкцию дегликозилировали с использованием дегликозилазы EndoH перед подтверждением с помощью ПААГ-ДСН.
Матричные последовательности ДНК, используемые для ПЦР
Человеческий (Homo sapiens) TNFR1 (внеклеточный участок, per. №в Genbank 33991418)

Кодируемый внеклеточный участок человеческого TNFR1 имел следующую аминокислотную последовательность.

Мышиный (Mus musculus) TNFR1 (внеклеточный участок, per. № в Genbank №31560798)

Кодируемый внеклеточный участок мышиного (Mus musculus) TNFR1 имел следующую аминокислотную последовательность.

Пример 17. Доменная специфичность dAb против TNFR1
Этот пример описывает способ, который использовали для определения доменной специфичности dAb, которые связывают TNFR1. В способе использовали поверхностный плазменный резонанс (SPR) (Detection of immuno-complex formation via surface plasmon resonance on gold-coated diffraction gratings.' Biosensors. 1987-88; 3(4):211-25.) для определения способности антител связываться с полноразмерным человеческим или мышиным биотинилированным TNFR1, который иммобилизовали на поверхности чипа SPR, после этого антитела инкубировали и уравновешивали с избытком химерных молекул, описанных в Примере 16. В этом анализе поток dAb против TNFR1 над поверхностью TNFR1 вызывает сигнал SPR, указывающий на количество dAb, которое связывает TNFR1, иммобилизованный на чипе SPR. Если dAb предварительно инкубировали и уравновешивали с химерной молекулой, которая включает домен(ы) TNFR1, который связывает конкретное dAb, то поток этой смеси над поверхностью TNFR1 приведет к меньшему сигналу SPR по сравнению только с dAb. Однако если dAb предварительно инкубируют и уравновешивают с химерной молекулой, которая не содержит домен(ы) TNFR1, который связывает конкретное dAb, то поток этой смеси над поверхностью TNFR1 приведет к сигналу SPR, который является примерно таким же, как сигнал, полученный с использованием только dAb.
Способ
1) Создание TNFR1 поверхности на чипе SPR
Выбор поверхности TNFR1 определяется видоспецифичностью тестируемого dAb против TNFR1. Таким образом, dAb против человеческого TNFR1 оценивали с использованием поверхности, покрытой человеческим TNFR1, и dAb против мышиного TNFR1 оценивали с использованием чипа, покрытого мышиным TNFR1.
Биотинилированный TNFR1 разводили в подходящем буфере SPR и пропускали через сенсорный чип со стрептавидином (SA) в приборе BIACORE 3000 SPR (Biacore International AB, Uppsala, Sweden). Ннизкую скорость потока (5-10 мкл/минуту) использовали для максимизации времени контакта между биотинилированным TNFR1 и стрептавидиновой поверхностью. Поток продолжали до насыщения стрептавидиновой поверхности биотинилированным материалом для получения чипа с максимальной поверхностью TNFR1. Чип, как правило, связывал от нескольких сотен до нескольких тысяч единиц ответа биотинилированного материала на SPR.
2) Титрование ответа против TNFR1 на чипе SPR
Успешный конкурентный эксперимент требует первоначальной оптимизации концентрации dAb против TNFR1, так что минимальное количество dAb протекает над поверхностью, что обеспечивает значительный сигнал SPR. В некотором диапазоне концентраций dAb будет связываться с поверхностью дозозависимым образом, так что количество ОЕ связанного dAb отражает концентрацию dAb, протекающего через поверхность чипа.
Для установления концентрационного диапазона этой дозозависимости dAb против TNFR1 титровали в сериях 10-кратных разведении буфера Biacore в диапазоне разведении от 1 в 10 до 1 в 1000000. Разведения затем индивидуально и последовательно вводили на поверхность чипа TNFR1, начиная с наиболее разведенного образца. Измеряли максимальное количество ОЕ, достигаемое при каждом разведении. После каждой инъекции поверхность TNFR1 регенерировали для удаления связанного dAb против TNFR1, для чего необходимо использовать подходящий SPR буфер для регенерации. При использовании этого способа определяли минимальную концентрацию dAb против TNFR1, необходимую для получения сигнала, представляющего приблизительно 100 ОЕ.
3) Предварительное уравновешивание анти-TNFRI dAb/химер
После определения оптимальной концентрации dAb против TNFR1, приготавливали смеси анти-TNFRI dAb/химерных TNFR1. Смеси приготавливали таким образом, что конечная концентрация dAb против TNFR1 была идентична оптимальной концентрации, определенной ранее. Реакционные смеси обычно приготавливали в объеме 100 мкл, содержащем 50 микролитров 2х концентрации dAb против TNFR1, 40 микролитров буфера Biacore и 10 микролитров неразбавленного очищенного химерного белка. Типичные концентрации для конечной смеси составляли примерно 10-100 мкМ химерного белка и примерно 10-100 нМ dAb против TNFR1. Смеси оставляли уравновешиваться в течение 30 минут при комнатной температуре.
4) Конкурентный эксперимент Biacore
После уравновешивания каждую смесь анти-TNFRI dAb/химерный TNFR1 последовательно пропускали над поверхностью TNFR1 SPR и измеряли количество ОЕ. После инъекции каждой смеси, поверхность регенерировали для удаления связанного dAb против TNFR1 перед инъекцией следующей смеси. Разные ответы получали при использовании разных химер, позволяющих определять домены TNFR1, связанные конкретными dAb.
Эти исследования выявили, что TAR2m-21-23 связывает домен 1 мышиного TNFR1, TAR2h-205 связывает домен 1 человеческого TNFR1, и что TAR2h-10-27, TAR2h-131-8, TAR2h-15-8, TAR2h-35-4, TAR2D-154-7, TAR2h-154-10 и TAR2h-185-25 связывают домен 3 человеческого TNFR1.
Пример 18. Способы скрининга
Эти химерные рецепторные белки, описанные в Примере 16, могут быть использованы в анализах или способах скрининга для выделения агентов (например, антител, dAb, химических соединений), которые связываются с конкретными доменами в TNFR1. Кратко, эти способы описывают добавление химерных белков к неочищенным препаратам антител перед их скринингом в отношении TNFR1-связывания с помощью ИФА или поверхностного плазменного резонанса. Кроме того, они описывают применение химерных белков, нанесенных на поверхность (например, планшет для ИФА или чип SPR), и скрининга антител путем тестирования их связывания с химерными белками на этой поверхности.
1) Скрининг с помощью растворимого ИФА
Этот способ может быть использован для быстрого выделения антитела или фрагментов антитела (например, dAb), которые связывают специфические домены TNFR1 из большого репертуара антител или фрагментов антител, обладающих неизвестной специфичностью.
96-Луночный планшет для анализа покрывают в течение ночи при 4°С 100 мкл на лунку химерного TNFR1. Лунки трижды промывают 0,1% TPBS (забуференный фосфатом физиологический раствор, содержащий Tween-20 в концентрации 0,1%). Добавляют 200 мкл 1% TPBS на лунку для блокирования планшета и планшет инкубируют в течение 1-2 часов при комнатной температуре. Лунки затем трижды промывают PBS перед добавлением 50 мкл бактериального супернатанта или periprep, содержащего растворимое антитело или фрагмент антитела (которое содержит эпитопную метку с-Myc), в 50 мкл 0,2% TPBS. Планшет затем в течение 1 часа инкубируют при комнатной температуре. После этого планшет 5 раз промывают 0,1% TPBS (0,1% Tween-20 в PBS). Затем в каждую лунку добавляют 100 мкл первичных мышиных моноклональных антител против с-Myc в 0,1% TPBS и планшет инкубируют в течение 1 часа при комнатной температуре. Этот раствор первичного антитела удаляют и планшет затем промывают 5 раз 0,1% TPBS. Затем добавляют 100 мкл предварительно разведенного конъюгата козьих антител против мышиного IgG (специфичного в отношении Fc) с HRP (Sigma, кат. №: А0168) и планшет инкубируют в течение 1 часа при комнатной температуре. Вторичное антитело затем удаляют и планшет промывают 6 раз 0,1% TPBS с последующими 2 отмывками PBS. Затем в каждую лунку добавляют 50 мкл раствора ТМВ пероксидазы и планшет оставляют при комнатной температуре в течение 2-60 минут. Реакцию останавливают путем добавления 50 мкл 1 М соляной кислоты. OD в планшете считывают при 450 нм в аппарате для считывания 96-луночных планшетов в течение 30 минут после добавления кислоты. Антитела, которые представлены в неочищенном бактериальном супернатанте, или peripreps, которые связывают домены TNFR1, представленные в химерном белке, приводят к более сильному сигналу ИФА.
2) Скрининг с помощью конкурентного ИФА
Этот способ может быть использован для быстрого скрининга разнообразных наборов препаратов неочищенного антитела или фрагмента антитела, которое связывает TNFR1, для определения их домен-связывающей специфичности.
96-Луночный планшет для анализа покрывают в течение ночи при 4°С 100 мкл на лунку мышиного или человеческого TNFR1 (либо человеческого, либо мышиного). Лунки трижды промывают 0,1% TPBS (забуференный фосфатом физиологический раствор, содержащий Tween-20 в концентрации 0,1%). Добавляют 200 мкл 1% TPBS на лунку для блокирования планшета и планшет инкубируют в течение 1-2 часов при комнатной температуре. Лунки затем трижды промывают PBS. В то же самое время бактериальные супернатанты или periprep предварительно уравновешивают предварительно оптимизированной концентрацией химерного белка TNFR1 в растворе. 50 мкл этой смеси неочищенный бактериальный препарат/химерный белок, содержащей растворимое антитело или фрагмент антитела затем добавляют к планшету для ИФА. Планшет инкубируют в течение 1 часа при комнатной температуре. Затем планшет промывают 5 раз 0,1% TPBS (0,1% Tween-20 в PBS) и 100 мкл первично детектируемого антитела (Белок A-HRP или Белок L-HRP) добавляют в 0,1% TPBS в каждую лунку, планшет инкубируют в течение 1 часа при комнатной температуре. Этот раствор первичного антитела удаляют и планшет 5 раз промывают 0,1% TPBS. При необходимости затем добавляют 100 мкл предварительно разведенного конъюгата вторичного козьего антитела/HRP и планшет инкубируют в течение 1 часа при комнатной температуре. Вторичное антитело удаляют и планшет 6 раз промывают 0,1% TPBS с последующими 2 отмывками в PBS. В каждую лунку добавляют 50 мкл раствора ТМВ пероксидазы и планшет оставляют при комнатной температуре на 2-60 минут. Реакцию останавливают путем добавления 50 мкл 1 М соляной кислоты. OD на планшете при 450 нм считывают в аппарате для считывания 96-луночных планшетов в течение 30 минут после добавления кислоты. Уменьшение сигнала ИФА является показателем связывания антитела с химерными доменами TNFR1, а не с полноразмерным TNFR1, нанесенным на планшет, и, следовательно, того антитела, которое связывает один из доменов в химерном белке.
3) Скрининг с помощью конкурентного ИФА в отношении антител и фрагментов антител, конкурирующих со ссылочным антителом или фрагментом антитела за связывание с TNFR1
Этот способ может быть использован для быстрого скрининга разнообразных наборов препаратов неочищенного антитела или фрагмента антитела, которые связывают TNFR1, для антител или фрагментов антител, конкурирующих со ссылочным антителом или фрагментом антитела (например, TAR2m-21-23) за связывание с TNFR1 или связывание желаемого домена TNFR1 (например, домена 1). В способе используют ссылочное антитело или фрагмент антитела и тестируемое антитело или фрагмент антитела (например, популяцию антител, подлежащих скринингу), которые содержат различные детектируемые метки (эпитопные метки).
96-Луночный планшет для анализа в течение ночи при 4°С покрывают 100 мкл на лунку мышиного или человеческого TNFR1. Лунки трижды промывают 0,1% TPBS (физиологический раствор, забуференный фосфатом, содержащий Tween-20 в концентрации 0,1%). 200 мкл на лунку 1% TPBS (1% Tween-20 в PBS) добавляют для блокирования планшета и планшет инкубируют в течение 1-2 часов при комнатной температуре. Затем лунки трижды промывают PBS. В то же время тестируемые препараты неочищенного антитела смешивают с предварительно оптимизированной концентрацией ссылочного антитела или фрагмента антитела (например, антитело, связывающее домен 1; TAR2m-21-23) в растворе. Как уже указано, важно, чтобы это антитело не включало одинаковые метки для обнаружения, которые присутствуют в антителах, подвергнутых скринингу в отношении домен-связывающей специфичности. 50 мкл этой неочищенной смеси, содержащей антитело/ссылочное антитело, добавляют в планшет для ИФА. Планшет затем в течение 1 часа инкубируют при комнатной температуре. После этого планшет 5 раз промывают 0,1% TPBS, в каждую лунку добавляют 100 мкл первичного детектирующего антитела (которое связывает метку, представленную только в популяции антител, подвергаемых скринингу) в 0,1% TPBS и планшет инкубируют в течение 1 часа при комнатной температуре. Этот раствор первичного антитела удаляют и планшет затем 5 раз промывают 0,1% TPBS. Затем добавляют 100 мкл предварительно разведенного конъюгата вторичного антитела-HRP, распознающего первичное детектирующее антитело и планшет инкубируют в течение 1 часа при комнатной температуре. Раствор вторичного антитела затем удаляют и планшет 6 раз отмывают 0,1% TPBS с последующими 2 отмывками PBS. Затем в каждую лунку добавляют 50 мкл раствора ТМВ пероксидазы и планшет оставляют при комнатной температуре на 2-60 минут. Реакцию останавливают путем добавления 50 мкл 1 М соляной кислоты. OD в планшете при 450 нМ считывают в аппарате для считывания 96-луночных планшетов в течение 30 минут после добавления кислоты. Должен быть проведен отдельный и параллельный ИФА с использованием этого способа, но без добавления ссылочного антитела или фрагмента антитела. Уменьшение сигнала ИФА в присутствии ссылочного антитела или фрагмента антитела по сравнению с сигналом ИФА для препарата того же самого антитела без конкурентного ссылочного антитела или фрагмента антитела является показателем того, что конкретное антитело или фрагмент антитела конкурирует с ссылочным антителом или фрагментом антитела за связывание с TNFR1 и связывает тот же самый домен TNFR1, что и ссылочное антитело или фрагмент антитела.
4) Скрининг методом поверхностного плазменного резонанса (SPR)
Описанные выше способы ИФА могут быть легко адаптированы для формата, который использет поверхностный плазменный резонанс, например с использованием прибора BIACORE 3000 SPR (Biacore International AB, Uppsala, Sweden). Как правило, химерный белок иммобилизован на чипе для SPR, или химерный белок уравновешивают в неочищенном бактериальном супернатанте, содержащем антитела или фрагменты антител против TNFR1, и полученную смесь пропускают через чип SPR, покрытый полноразмерным человеческим TNFR1 или мышиным TNFR1.
Пример 19. Димеры TAR2m21-23 представляют собой высокоавидные антагонисты TNFR1
Димеры TAR2m21-23 получали путем продуцирования формы TAR2m21-23, содержащей остаток cys на карбоксильном конце, используя способы, описанные в Примере 8. Белок (TAR2m21-23 CYS) экспрессировали в Pichia и очищали, используя Streamline Protein А. Анализ с помощью ПААГ-ДСН в невосстанавливающих условиях продемонстрировал, что 40-50% белка представлено в растворе в виде димера. Димер дополнительно очищали с использованием гель-фильтрации.
2 мг белка концентрировали до примерно 250 мкл и наносили на колонку для гель-фильтрации Superdex 75 HR (Amersham Bioscience), ранее уравновешенную PBS. Пропускание через колонку осуществляли при скорости потока 0,5 мл/мин и собирали фракции объемом 0,5 мл. За элюцией белка из колонки наблюдали при 280 нм, и фракции, содержащие димер, идентифицировали с помощью ПААГ-ДСН в невосстанавливающих условиях. Фракции, содержащие димеры, но не мономерные TAR2m-21-23 CYS, объединяли. Объединенные фракции концентрировали и определяли эффективность димерного dAb в анализе цитотоксичности TNF в отношении клеток L929 (Пример 6).
Биологическую эффективность димера TAR2m21-23 сравнивали с мономерным TAR2m-21-23 в анализе цитотоксичности в отношении клеток L929. В этом анализе оценивали ингибирование TNF-индуцированной цитотоксичности в отношении мышиных клеток L929 мономером TAR2m21-23 и димером TAR2m21-23, и результаты выражали в виде концентрации мономера dAb или димера, которая на 50% ингибировала цитотоксичность в анализе (50% нейтрализующая доза, ND50).
Мономерное dAb имело NDso в анализе примерно 600 пМ. ND50 димеризованного dAb (димер TAR2m21-23) в анализе было примерно в 10 раз меньше (NDso примерно 60-70 пМ). Эти результаты показывают, что димер TAR2m21-23 обладает значительно улучшенной аффинностью в отношении TNFR1 клеточной поверхности по сравнению с мономером dAb. Результаты показывают, что димер TAR2m21-23 одновременно связывается с двумя отдельными молекулами TNFR1 на клеточной поверхности, и возникающий в результате мультимеризации эффект авидности приводит к улучшенному ингибированию TNFR1.
Затем тестировали димерный формат (димер TAR2m21-23) для того, чтобы определить, вызывает ли он какие-либо признаки агонистического действия на TNFR1, т.е. способности вызывать перекрестное связывание TNFR1 и инициировать внутриклеточную передачу сигнала и клеточную смерть. Этого достигали с использованием модифицированного анализа цитотоксичности в отношении клеток L929, к которым не добавляли TNF-α, но антитела против TNFR1 или форматы dAb тестировали для того, чтобы определить, оказывают ли они ангонистическое действие на TNFR1 и вызывают ли клеточную смерть. В анализе тестировали анти-TNFRI антитело AF-425-PB (R&D Systems), которое представляет собой известный агонист TNFR1, и анти-TNFR1 антитело МАВ430 (R&D Systems), о котором сообщалось, что оно представляет собой антагонист TNFR1, TAR2m-21-23 и димер TAR2m21-23.
Антитело AF-425-PB активировало TNFR1 и индуцировало цитотоксичность в анализе с ND50 примерно 100 пМ. Даже антитело МАВ430, которое, как сообщалось, представляет собой антитело-антагонист, вызывало перекрестное связывание рецептора и клеточную смерть в анализе с ND50 примерно 10 нМ. В отличие от этого, димер TAR2m-21-23 не вызывал клеточную гибель в анализе, даже когда присутствовал в очень высоких концентрациях (более 1 мкМ). Эти результаты демонстрируют, что димер TAR2m21-23 не является агонистом TNFR1.
Данные результаты показывают, что димеры, тримеры или другие мультимеры dAb, которые связывают TNFR1, обладают высокой авидностью в отношении TNFR1, экспрессируемого на поверхности клеток, и представляют собой эффективные антагонисты TNFR1. Кроме того, результаты этого исследования демонстрируют, что мультимеры dAb, которые связывают домен 1 TNFR1, такие как димер TAR2m21-23, могут связывать две молекулы TNFR1 (как могут антитела, которые действуют в качестве агонистов в анализе), и что связывание домена или эпитопа мишени на TNFR1 (Домен 1) предотвращало тесную ассоциацию рецепторных цепей, которая необходима для инициации передачи сигнала через TNFR1. Это свойство является уникальным для двухвалентной молекулы тем, что она способна перекрестно связываться с TNFR1 на клеточной поверхности и не вызывать передачу сигнала через TNFR1 и клеточную смерть.
Пример 20. Выделение dAb, которое связывает человеческий TNFR1 и мышиный TNFR1
dAb, обладающие известной последовательностью, экспрессировали в Е. coli и очищали с помощью смолы Protein A streamline. После элюции в Трис-глицине dAb пропускали через чип SPR, на котором иммобилизован биотинилированный человеческий TNFR1 (проточная кювета 2) и биотинилированный мышиный TNFR1 (проточная кювета 4). (Чип SPR покрывали человеческим TNFR1 и мышиным TNFR1 с близкими плотностями). Проточные кюветы 1 и 3 оставляли чистыми, и они действовали в качестве ссылочных поверхностей без антигена для обнаружения и вычитания неспецифического связывания.
dAb пропускали через каждую из 4 проточных кювет последовательно (т.е. проточная кювета 1, затем 2, 3 и наконец 4) с измерением различий в ответе между проточными кюветами 2 и 1, а также измерением различий в ответе между проточными кюветами 4 и 3. Первое представляет собой меру связывания с человеческим TNFR1, а последнее - связывание с мышиным TNFR1. Кривые специфического связывания строили для связывания как с человеческим TNFR1, так и с мышиным TNFR1, причем природа кривых такова, что отмечалась более быстрая скорость ассоциации для человеческого TNFR1, чем для мышиного TNFR1. Скорости диссоциации были похожими. Оценка перекрестной реактивности дается по количеству единиц ответа (RU), максимально достигаемых при каждом событии связывания. В этом примере поверхность с человеческим биотинилированным TNFR1 включала приблизительно 900 RU TNFR1 на проточной кювете 2, тогда как проточная кювета 4 включала примерно 1400 RU мышиного TNFR1. В качестве контроля человеческое специфическое dAb TAR2D-154-7 в концентрации 2 микромоль связанное с поверхностью, покрытой человеческим TNFR1, с максимальным ответом 385 RU, давало ответ на поверхности, покрытой мышиным TNFR1, только 4,5 RU. 2 микромоль TAR2h-205 давало ответ на поверхности, покрытой человеческим TNFR1, с ответом 435 RU, и ответом на поверхности, покрытой мышиным TNFR1, 266 RU.
Все публикации и ссылки, цитированные в указанных публикациях, включены в данное изобретение путем ссылки. Различные модификации и вариации описанных способов и систем по изобретению будут понятны специалистам в данной области без отклонения от объема и сущности изобретения. Хотя изобретение описано в связи со специфическими предпочтительными воплощениями, понятно, что заявленное изобретение не должно чрезмерно ограничиваться такими конкретными воплощениями. Действительно, предполагается, что различные модификации описанных способов реализации изобретения, которые очевидны специалистам в молекулярной биологии или родственных областях, находятся в объеме следующей формулы изобретения.
Приложение 1: полипептиды, увеличивающие период полувыведения in vivo
Альфа-1 гликопротеин (орозомукоид) (AAG)
Альфа-1 антихиромотрипсин (ACT)
Альфа-1 антитрипсин (ААТ)
Альфа-1 микроглобулин (белок НС) (AIM)
Альфа-2 макроглобулин (А2М)
Антитромбин III (AT III)
Аполипопротеин А-1 (АроА-1)
Аполипопротеин В (Аро В)
Бета-2-микроглобулин (В2М)
Церулоплазмин (Ср)
Компонент комплемента (С3)
Компонент комплемента (С4)
Ингибитор С1 эстеразы (С1 INH)
С-реактивный белок (CRP)
Цистатин С (Cys С)
Ферритин (FER)
Фибриноген (FIB)
Фибронектин (FN)
Гаптоглобин (Нр)
Гемопексин (НРХ)
Иммуноглобулин A (IgA)
Иммуноглобулин D (IgD)
Иммуноглобулин Е (IgE)
Иммуноглобулин G (IgG)
Иммуноглобулин М (IgM)
Легкие цепи иммуноглобулина (каппа/лямбда)
Липопротеин (a)[Lp(a)]
Манноза-связывающий белок (МВР)
Миоглобин (Myo)
Плазминоген (PSM)
Преальбумин (Транстиретин) (PAL)
Ретинол-связывающий белок (RBP)
Ревматоидный фактор (RF)
Сывороточный амилоид A (SAA)
Растворимый рецептор трансферрина (sTfR)
Трансферрин (Tf)
Приложение 2
- Синергизм TNF и IL-1 в патологии малярии (гипогликемия, NO).
- Синергизм TNF и IL-1 при индукции миграции полиморфно-ядерных (PMN) клеток при воспалении.
- Синергизм IL-1 и TNF при индукции инфильтрации PMN в брюшную полость.
- Синергизм IL-1 и TNF при индукции секреции IL-1 эндотелиальными клетками. Важно при воспалении.
- IL-1 или TNF самостоятельно вызывали некоторую клеточную инфильтрацию в коленную синовиальную оболочку. IL-1 индуцировал PMN, TNF-моноциты. Вместе они индуцировали более сильную инфильтрацию вследствие увеличения PMN.
- Циркулирующее в системе кровообращения вещество-депрессант миокарда (представленное при сепсисе) представляет собой низкие уровни действующих синергично IL-1 и TNF.
- Cancer Res. 1992 Apr 15; 52(8):2197-201.
- Синергизм TNF и IL-4 при индукции экспрессии IL-6 в кераноцитах.
- Sustained elevated levels of VCAM-1 in cultured fibroblast-like synoviocytes can be achieved by TNF-Alpha in combination with either IL-4 or IL-13 through increased mRNA stability. Am J Pathol. 1999 Apr; 154(4); 1149-58
human myeloma cell line. Eur J Haematol. 1994 Jul;53(l):31-7.
Вовлечена в острый респираторный дистресс-синдром. - Смотри IL-5/TNF (астма). Synergism between interleukin-8 and tumor necrosis factor-alpha for neutrophil-mediated platelet activation. Eur Cytokine Netw. 1994 Sep-0ct;5(5):455-60. (респираторный дистресс-синдром у взрослых (ARDS))
- lnterleukin-13 and tumour necrosis factor-alpha synergistically induce eotaxin production in human nasal fibroblasts. Clin Exp Allergy. 2000 Mar; 30(3):348-55.
- lnterleukin-13 and tumour necrosis factor-alpha synergistically induce eotaxin production in human nasal fibroblasts. Clin Exp Allergy. 2000 Mer; 30(3):348-55 (аллергическое воспаление)
- Implications of serum TNF-beta and IL-13 in the treatment response of childhood nephrotic syndrome. Cytokine. 2003 Feb 7; 21(3):155-9.
- Correlation of circulating interleukin 16 with proinflammatory cytokines in patients with rheumatoid arthritis. Rheumatology (Oxford). 2001 Apr; 40(4):474-5. Реферат отсутствует.
- Interleukin 16 is up-regulated in Crohn's disease and participates in TNBS colitis in mice. Gastroenterology. 2000 Oct; 119(4):972-82.
- Interleukin 17 synergises with tumour necrosis factor alpha to induce cartilage destruction in vitro. Ann Rheum Dis. 2002
Oct;61(10):870-6.
- Реферат: lnterleukin-1, tumor necrosis factor alpha, and interleukin-17 synergistically up-regulate nitric oxide and prostaglandin E2 production in explants of human osteoarthritic knee menisci. Arthritis Rheum. 2001 Sep; 44(9):2078-83.
- Реферат: Elevated levels of interieukin-18 and tumor necrosis factor-alpha in serum of patients with type 2 diabetes mellitus: relationship with diabetic nephropathy. Metabolism. 2003 May; 52(5):605-8.
- Синергизм при противовирусном ответе/индукции IFN-β.
- Активация нейтрофилов/окислительного взрыва.
- Активация эндотелиальных клеток
- Токсичность обнаружена при лечении пациентов с помощью TNF/IFN-γ в качестве противовирусной терапии
- Экспрессия фракталкина человеческими астроцитами.
- Множество статей по воспалительным ответам - т.е. ЛПС, также активация макрофагов.
- Антитела против TNF и антитела против IFN-y оказывают синергетическое действие для защиты мышей от летального эндотоксикоза.
(lymphotoxin). Cytokine. 1992 Nov; 4(6):479-87.
- IL-4 индуцирует экспрессию IL-1 при активации эндотелиальных клеток.
- Т-клеточная активация (может заменять вспомогательные клетки)
- IL-1 индуцирует экспрессию IL-6
- Экспрессия С3 и сывороточного амилоида (ответ острой фазы)
- Экспрессия ВИЧ
- Разрушение коллагена хряща.
- В-клеточная пролиферация
- Т-клеточная пролиферация
- (селективно индуцирует активацию CD8 и NK-лимфоцитов) IL-2R beta agonist P1-30 acts in synergy with IL-2, IL-4, IL-9, and IL-15: biological and molecular effects. J Immunol. 2000 Oct 15; 165(8): 4312-8.
- IL-5 индуцирует рецепторы IL-2 на В-клетках
- (Т-клеточная активация и пролиферация) IL-15 and IL-2: а matter of life and death for Т cells in vivo. Nat Med. 2001 Jan; 7(1): 114-8.
- IL-2 индуцирует экспрессию IFN-γ Т-клетками
- Synergistic effects of IL-4 and either GM-CSF or IL-3 on the induction of CD23 expression by human monocytes: regulatory effects of IFN-alpha and IFN-gamma. Cytokine. 1994 Jul; 6(4):407-13.
- A Th2-like cytokine response is involved in bullous pempnigoid. the role of IL-4 and IL-5 in the pathogenesis of the disease. Int J Immunopathol Pharmacol. 1999 May-Aug; 12(2):55-61.
- IL-4 synergistically enhances both IL-2- and IL-12-induced IFN-{gamma} expression in murine NK cells. Blood. 2003 Mar 13 [Электронная публикация перед печатью]
- Inhibition of the IL-4/IL-13 receptor system prevents allergic sensitization without affecting established allergy in a mouse model for allergic asthma. J Allergy Clin Immunol. 2003 Jun; 111(6):1361-1369.
- Реферат: Mast cells produce interleukin-25 upon Fcepsilon R1-mediated activation. Blood. 2003 May 1;101(9):3594-6. Электронная публикация 2003 Jan 02, (аллергическое воспаление)
- Реферат: Mast cells produce interleukm-25 upon Fcepsilon R1-mediated activation. Blood. 2003 May 1; 101(9):3594-6. Электронная публикация 2003 Jan 02. (аллергическое воспаление)
2002 Jan; 15(1): 17-23. (артрит)
- Реферат: lnterleukin-17 stimulates the expression of interleukin-8, growth-related oncogene-alpha, and granulocyte-colony-stimulating factor by human airway epithelial cells. Am J Respir Cell Mol Biol. 2002 Jun; 26(6):748-53. (воспаление дыхательных путей)
- Th2 cytokines and asthma, lnterleukin-9 as a therapeutic target for asthma. Respir Res. 2001; 2(2):80-4. Электронная публикация 2001 Feb 15. Обзор.
- Реферат: lnterleukin-9 enhances interleukin-5 receptor expression, differentiation, and survival of human eosinophils. Blood. 2000 Sep 15; 96(6):2163-71 (астма)
- Direct effects of interleukin-13 on epithelial cells cause airway hyperreactivity and mucus overproduction in asthma. Nat Med. 2002 Aug; 8(8):885-9.
- IL-17 promotes bone erosion in murine collagen-induced arthritis through loss of the receptor activator of NF-kappa В ligand/osteoprotegerin balance. J Immunol. 2003 Mar 1; 170(5):2655-62.
- Inflammatory Liver Steatosis Caused by IL-12 and IL-18. J Interferon Cytokine Res. 2003 Mar;23(3): 155-62.
- Реферат: A unique role for IL-23 in promoting cellular immunity. J Leukoc Biol. 2003 Jan; 73(1):49-56. Обзор.
- Реферат: Mast cells produce interleukin-25 upon Fcepsilon R1-mediated activation. Blood. 2003 May 1; 101(9):3594-6. Epub 2003 Jan 02. (аллергическое воспаление)
- Реферат: Inhibition of in vitro immunoglobulin production by IL-12 in murine chronic graft-vs.-host disease: synergism with IL-18. Eur J Immunol. 1998 Jun; 28(6):2017-24.
Приложение 3: Онкологические комбинации
IL-5
Эотаксин(ы)
MDC
TARC
TNFα
IL-9
EGFR
CD40L
IL-25
МСР-1
TGFP
Приложение 4: Сводные данные
димеры
TAR1-5-
19
(Gly4Ser)n
TAR1-5-19d2
=2нМ TAR1-5-19d3=8 нМ TAR1-5-19d4=2-5 нМ TAR1-5-19d5=8 нМ 3 формате Fab TAR1-5-19CH d1CK=
6 нМ TAR1-5-
19CK d1CH=
6 нМ TAR1-5-
19CHd2CK=
8 нМ TAR1-5-
19CHd3CK=
3 нМ
=10 нМ
=12 нМ
двойной специфичностью аффинность в отношении мишени от 1
до 100000х относительно аффинности SA dAb,
например 100 пМ (мишень) и 10 мкМ аффинность SA.
двойной специфичностью аффинность в отношении
мишени от 1 до 100000 х относительно
аффинности SA dAb, например 100 пМ (мишень) и 10 мкМ аффинность SA.
| название | год | авторы | номер документа |
|---|---|---|---|
| ХИМЕРНЫЙ БЕЛОК, СОСТАВЛЕННЫЙ ИЗ ДОМЕНА АНТАГОНИСТА NGF И ДОМЕНА АНТАГОНИСТА TNFα | 2015 |
|
RU2678810C2 |
| Антитела, специфичные в отношении лиганда 1А, подобного фактору некроза опухолей, и их композиции и применения | 2014 |
|
RU2708140C1 |
| ЛЕЧЕНИЕ ИНФАРКТА МИОКАРДА С ИСПОЛЬЗОВАНИЕМ АНТАГОНИСТОВ TGF-БЕТА | 2011 |
|
RU2637088C2 |
| АНТАГОНИСТИЧЕСКИЕ АНТИТЕЛА ПРОТИВ РЕЦЕПТОРА IL-7 И СПОСОБЫ | 2011 |
|
RU2533809C9 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА К Toll-ПОДОБНЫМ РЕЦЕПТОРАМ 2 И ИХ ПРИМЕНЕНИЯ | 2010 |
|
RU2563344C2 |
| СЛИТЫЕ КОНСТРУКЦИИ ЛЕКАРСТВЕННОГО СРЕДСТВА И КОНЪЮГАТЫ | 2005 |
|
RU2428431C2 |
| ЧЛЕНЫ СВЯЗЫВАНИЯ ПРОТИВ IL-1R1 | 2009 |
|
RU2555532C2 |
| Антагонистические антитела против рецептора IL-7 и способы | 2014 |
|
RU2653430C2 |
| СОЕДИНЕНИЯ | 2007 |
|
RU2466139C2 |
| НОВЫЕ МОЛЕКУЛЫ АНТАГОНИСТИЧЕСКИХ АНТИ-TNFR2-АНТИТЕЛ | 2019 |
|
RU2829587C2 |
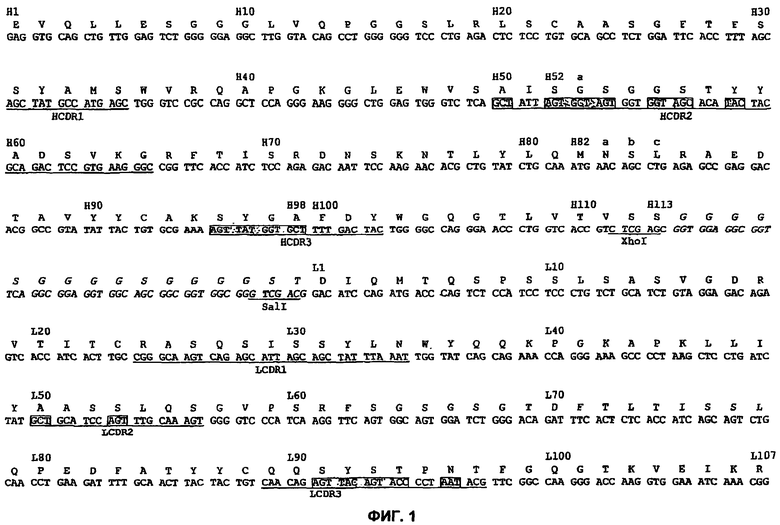
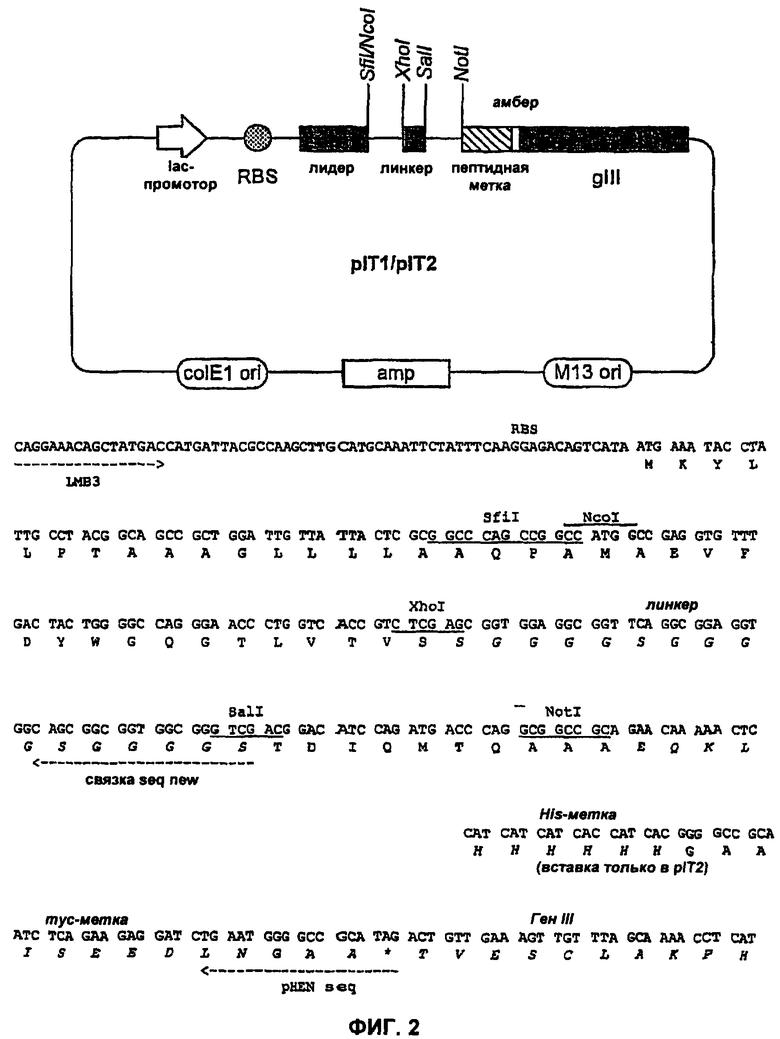
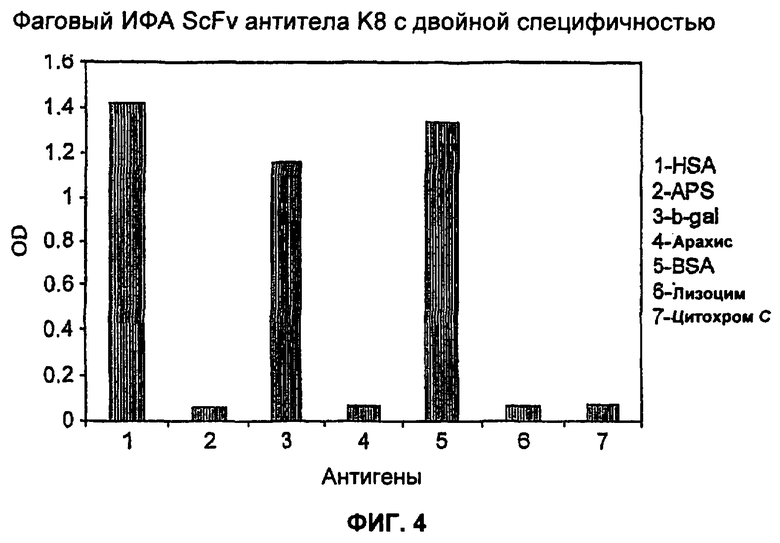
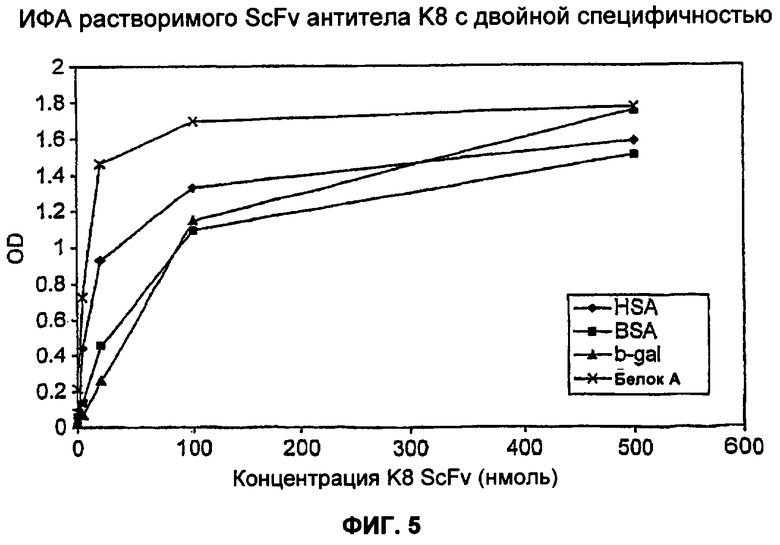
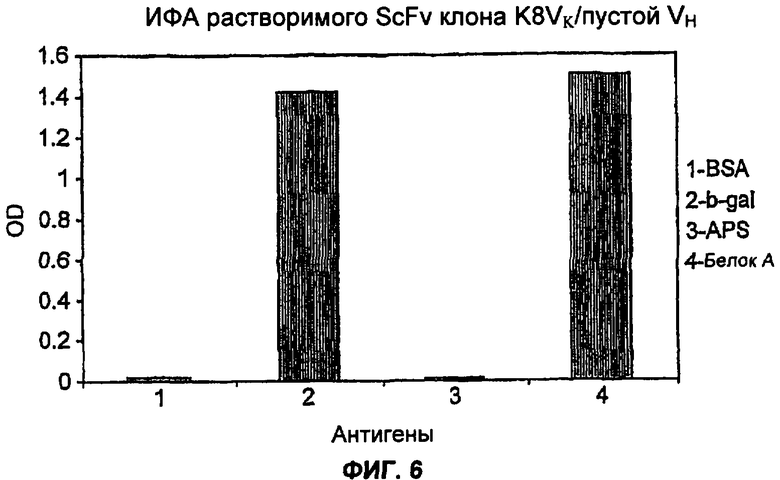
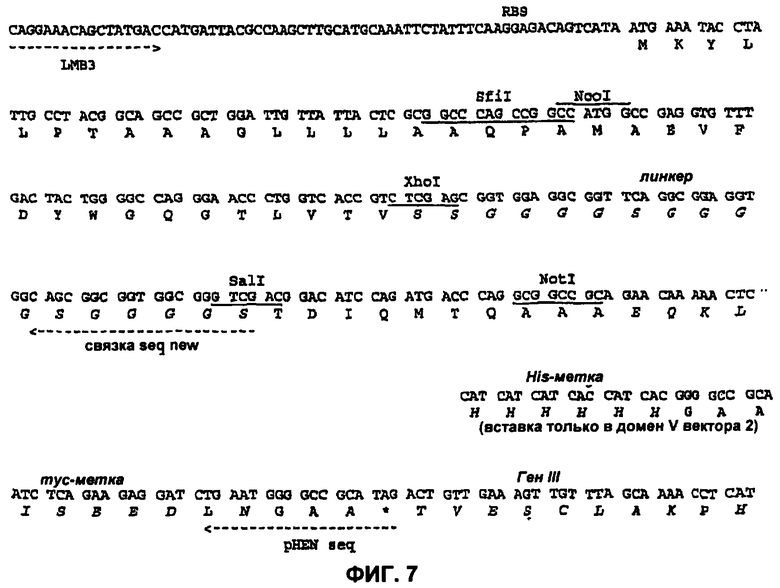
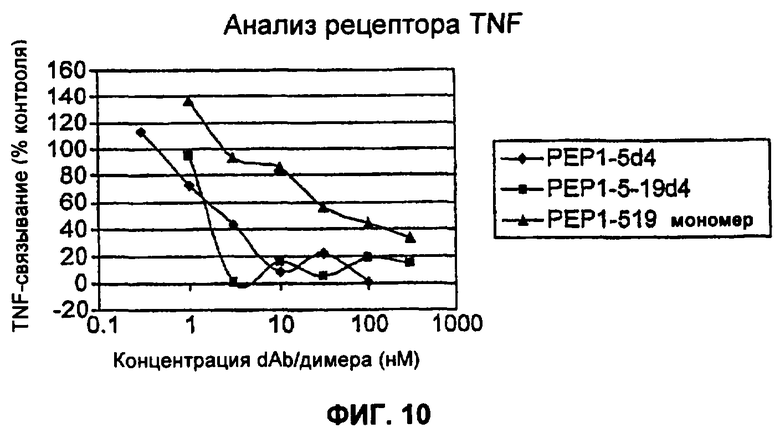
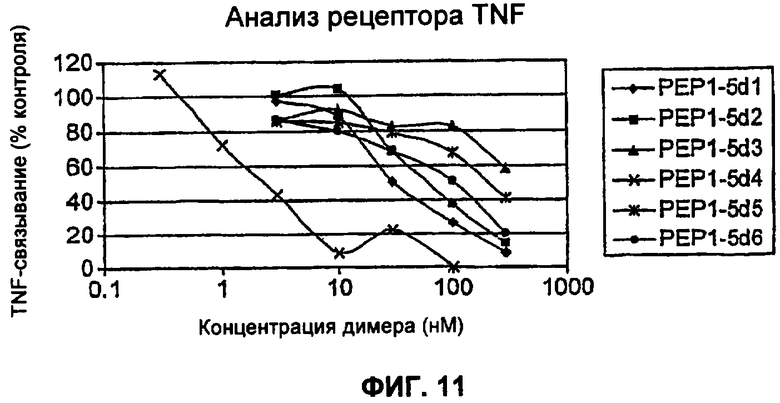
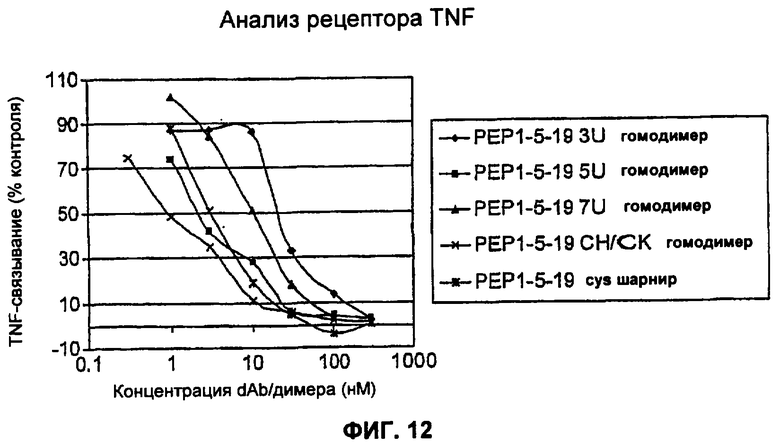




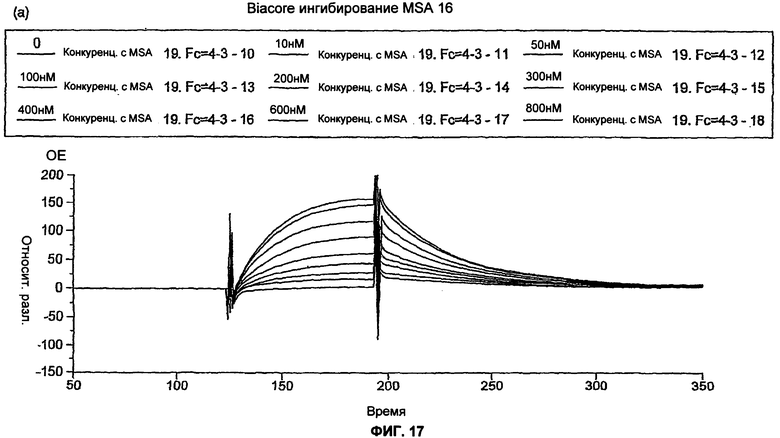
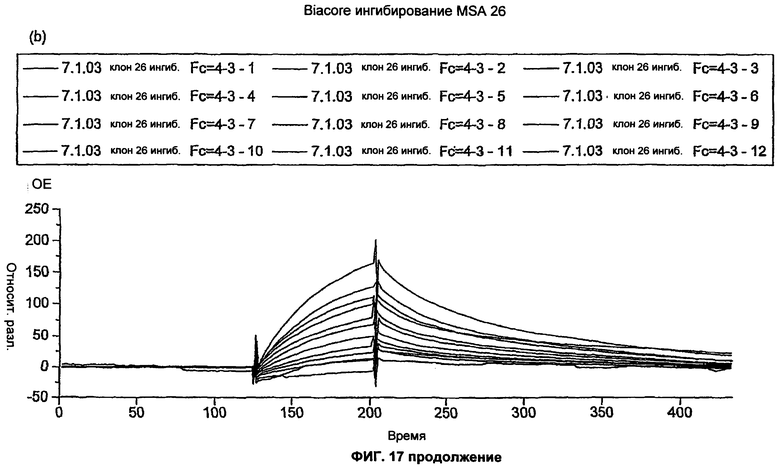
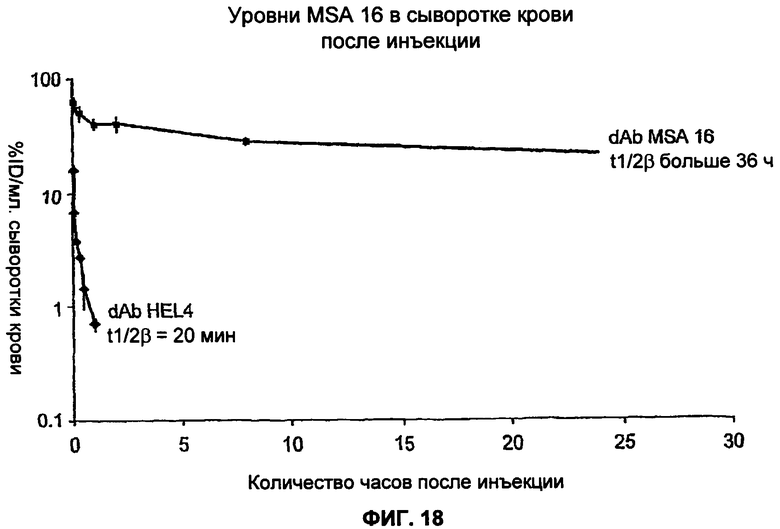
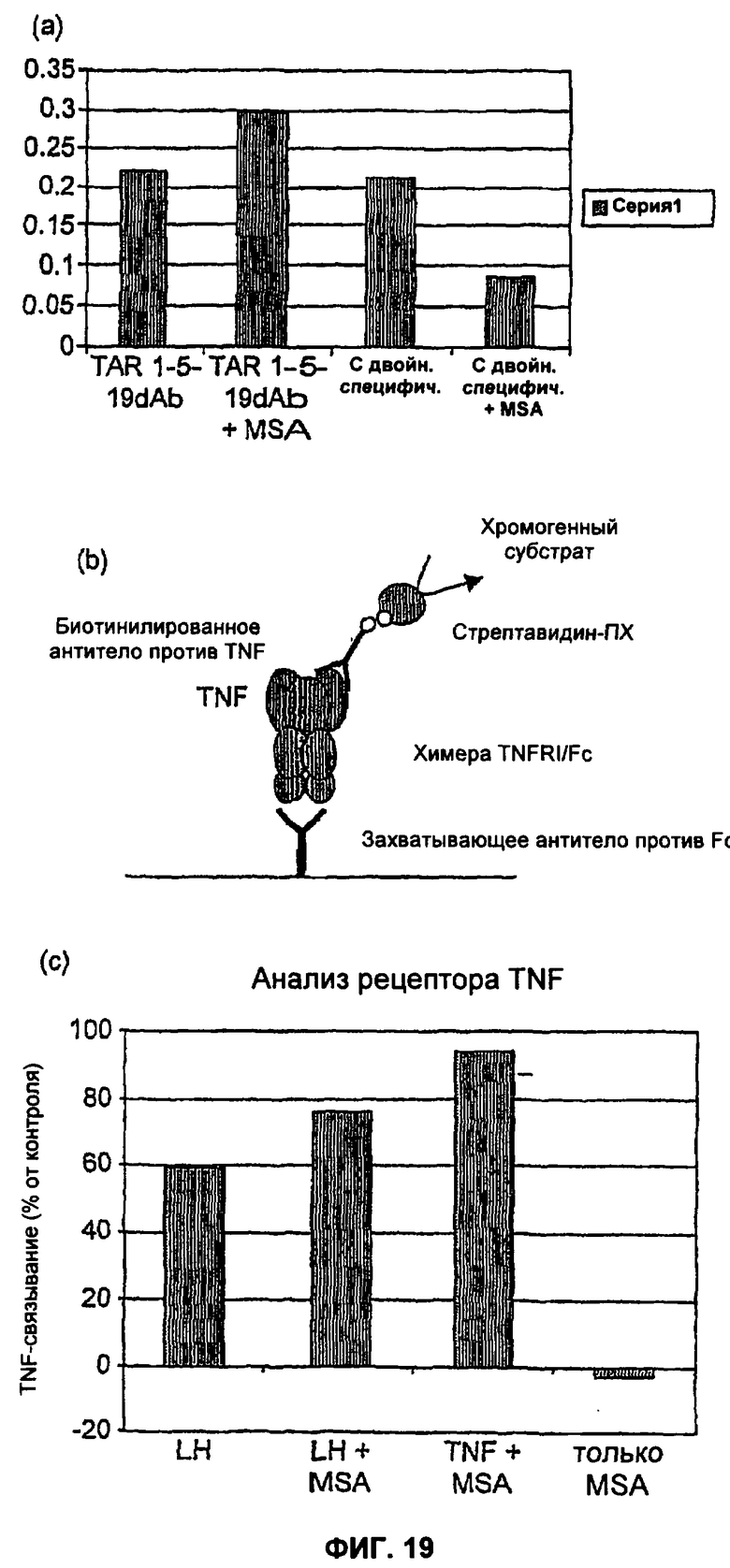
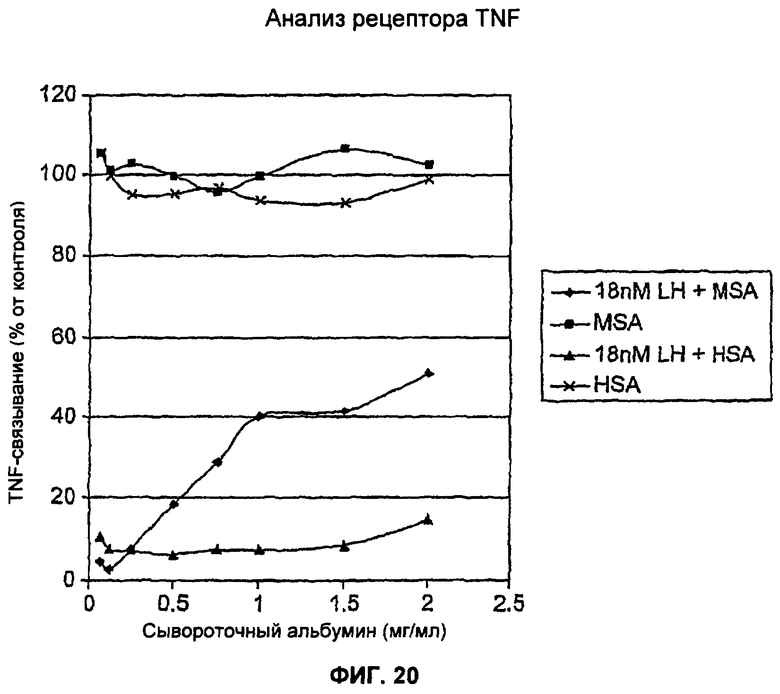





































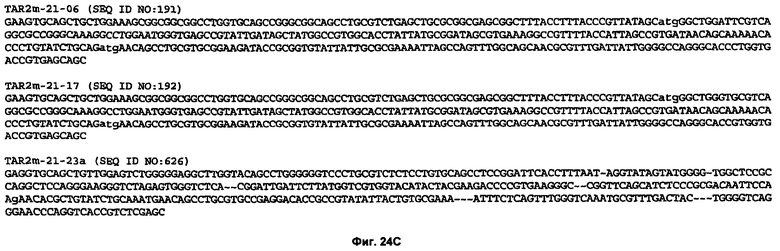





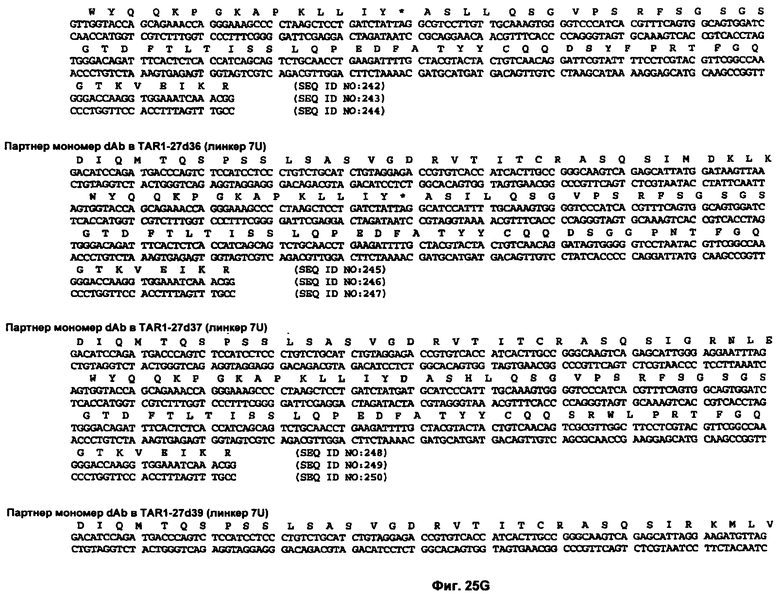


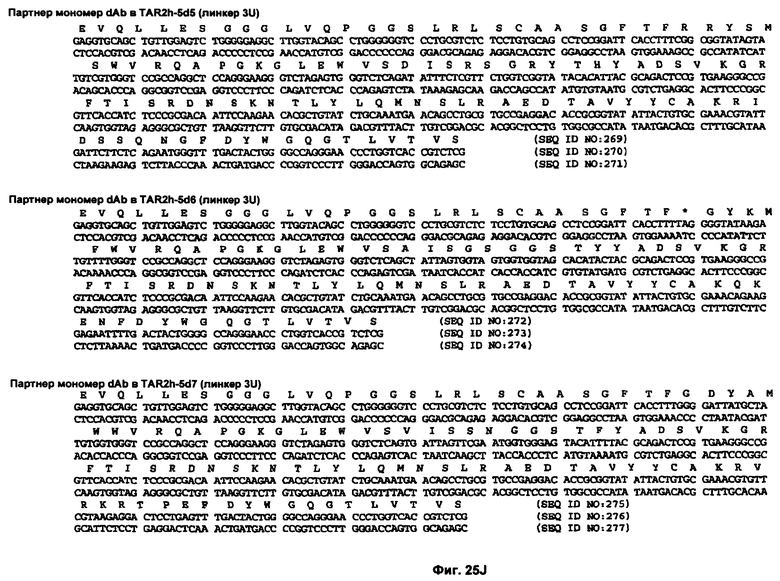


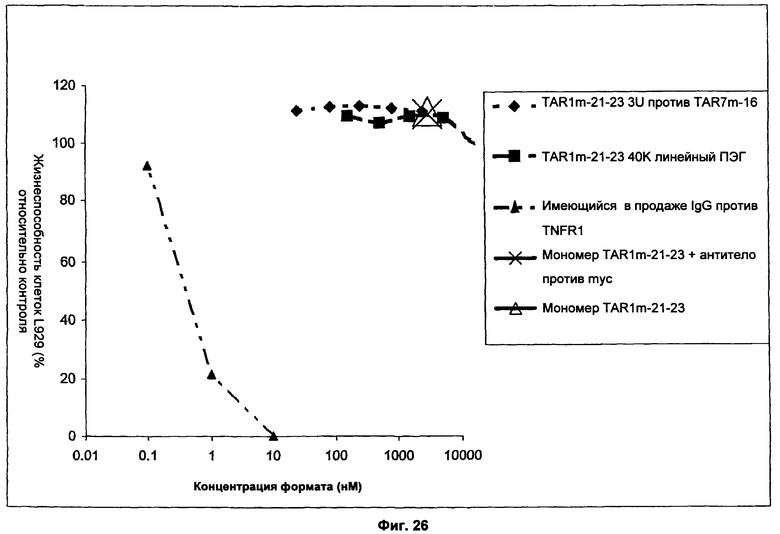











































































































































































































Изобретение относится к области биотехнологии и представляет собой антагонист рецептора фактора некроза опухоли 1, представляющий собой доменное антитело против TNFR1 (dAb). Также изобретение представляет собой к мономеру доменного антитела (dAb) против TNFR1, которое связывает рецептор фактора некроза опухоли 1, а также к лиганду, содержащему такой мономер. Кроме того, изобретение относится к способу ингибирования передачи сигнала через рецептор TNFR1 с использованием данных антагонистов. Также предложены нуклеиновая кислота, кодирующая указанный антагонист, вектор, содержащий указанную нуклеиновую кислоту, клетку-хозяина, содержащую указанный вектор, фармацевтическую композицию, содержащую указанный антагонист или указанный мономер dAb. Изобретение можно эффективно использовать при лечении воспалительных заболеваний (например, хронических воспалительных заболеваний). 23 н. и 34 з.п. ф-лы, 28 ил., 14 табл.
1. Антагонист рецептора фактора некроза опухоли I (TNFR1), где указанный антагонист представляет собой доменное антитело против TNFR1 (dAb), которое связывает домен 1 мышиного и/или человеческого TNFR1 и конкурирует с TAR2m-21-23 (SEQ ID NO:173) за связывание с мышиным TNFR1 или конкурирует с TAR2h-205 (SEQ ID NO:627) за связывание с человеческим TNFR1.
2. Антагонист по п.1, где указанный антагонист ингибирует TNFα-индуцированную гибель клеток в стандартном анализе цитотоксичности в отношении L929 или TNFα-индуцированную секрецию IL-8 в стандартном анализе IL-8 в HeLa.
3. Антагонист по п.1 или 2, где указанные антагонисты связывают домен 1 человеческого TNFR1.
4. Антагонист по п.1, где указанный антагонист связывает домен 1 указанного TNFR1 и конкурирует с TAR2m-21-23 (SEQ ID NO:173) за связывание с мышиным TNFR1.
5. Антагонист по п.1, где указанный антагонист связывает домен 1 указанного TNFR1 и конкурирует с TAR2h-205 (SEQ ID NO:627) за связывание с человеческим TNFR1.
6. Антагонист по п.1, где указанный антагонист связывает человеческий и мышиный TNFR1.
7. Антагонист по п.1, где указанный антагонист, по существу, не ингибирует шеддинг указанного TNFR1.
8. Антагонист рецептора фактора некроза опухоли I (TNFR1), который связывает мышиный или человеческий TNFR1 и ингибирует передачу сигнала через указанный TNFR1, причем указанный антагонист, по существу, не ингибирует связывание TNFα с указанным TNFR1, где указанный антагонист представляет собой доменное антитело против TNFR1, которое конкурирует с TAR2m-21-23 (SEQ ID NO:173) за связывание с мышиным TNFR1 или конкурирует с TAR2h-205 (SEQ ID NO:627) за связывание с человеческим TNFR1.
9. Антагонист по п.8, где указанный антагонист ингибирует TNFα-индуцированную гибель клеток в стандартном анализе цитотоксичности в отношении L929 или TNFa-индуцированную секрецию IL-8 в стандартном анализе IL-8 в HeLa.
10. Антагонист по п.8 или 9, где указанный антагонист связывает человеческий TNFR1.
11. Антагонист по п.8, где указанный антагонист связывает домен 1 указанного TNFR1.
12. Мономер доменного антитела (dAb) против TNFR1, который специфически связывает человеческий рецептор фактора некроза опухоли I (TNFR1), причем указанный мономер dAb связывает TNFR1 с Кd от 300 нМ до 5 пМ и содержит аминокислотную последовательность, которая по меньшей мере примерно на 95% гомологична аминокислотной последовательности dAb, выбранной из группы, состоящей из TAR2h-14 (SEQ ID NO:34), TAR2h-16 (SEQ ID NO:35), TAR2h-17 (SEQ ID NO:36), TAR2h-22 (SEQ ID NO:41), TAR2h-27 (SEQ ID NO:46), TAR2h-29 (SEQ ID NO:47), TAR2h-10-1 (SEQ ID NO:51), TAR2h-10-2 (SEQ ID NO:52), TAR2h-10-3 (SEQ ID NO:53), TAR2h-10-4 (SEQ ID NO:54), TAR2h-10-5 (SEQ ID NO:55), TAR2h-34 (SEQ ID NO:373), TAR2h-41 (SEQ ID NO:380), TAR2h-42 (SEQ ID NO:381), TAR2h-44 (SEQ ID NO:383), TAR2h-47 (SEQ ID NO:385), TAR2h-51 (SEQ ID NO:388) и TAR2h-15 (SEQ ID NO:431).
13. Лиганд для связывания мышиного или человеческого TNFR1, содержащий антагонист по п.1 или 8 или мономер dAb по п.12.
14. Лиганд по п.13, содержащий TNFR1-связывающую группировку и группировку, увеличивающую период полувыведения.
15. Лиганд по п.14, где группировка, увеличивающая период полувыведения, представляет собой полиэтиленгликолевую группировку, сывороточный альбумин или его фрагмент, рецептор трансферрина или его трансферрин-связывающий участок, или антитело или фрагмент антитела, содержащий сайт связывания полипептида, увеличивающего период полувыведения in vivo.
16. Лиганд по п.15, где группировка, увеличивающая период полувыведения, представляет собой антитело или фрагмент антитела, содержащий сайт связывания сывороточного альбумина или неонатального рецептора Fc.
17. Лиганд по п.16, где группировка, увеличивающая период полувыведения, представляет собой dAb.
18. Выделенная нуклеиновая кислота, кодирующая антагонист по п.1 или 8 или мономер dAb по п.12.
19. Рекомбинантная нуклеиновая кислота, кодирующая антагонист по п.1 или 8 или мономер dAb по п.12.
20. Вектор для экспрессии, содержащий рекомбинантную нуклеиновую кислоту, кодирующую антагонист по п.1 или 8 или мономер dAb по п.12.
21. Вектор по п.20, дополнительно содержащий регулирующую экспрессию последовательность, функционально связанную с указанной рекомбинантной нуклеиновой кислотой.
22. Клетка-хозяин для экспрессии, включающая вектор по п.20 или рекомбинантную нуклеиновую кислоту по п.19.
23. Способ получения полипептида, при котором клетку-хозяина по п.22 поддерживают в условиях, подходящих для экспрессии рекомбинантной нуклеиновой кислоты или вектора, в результате чего продуцируется полипептид.
24. Фармацевтическая композиция для связывания мышиного или человеческого TNFR1, содержащая эффективное количество антагониста по любому из пп.1-8, лиганда по любому из пп.13-17, или мономера dAb по п.12 и фармакологически приемлемый носитель.
25. Способ ингибирования передачи сигнала через мышиный или человеческий TNFR1 у млекопитающего, где связывание TNFα с указанным TNFR1, по существу, не ингибируется, при котором субъекту, нуждающемуся в этом, вводят терапевтически эффективную дозу антагониста по любому из пп.8-11.
26. Способ по п.25, где указанный антагонист связывает домен 1 указанного TNFR1.
27. Способ по п.26, где указанный антагонист конкурирует с TAR2m-21-23 (SEQ ID NO:173) за связывание с мышиным TNFR1.
28. Способ по п.26, где указанный антагонист конкурирует с TAR2h-205 (SEQ ID NO:627) за связывание с человеческим TNFR1.
29. Способ ингибирования передачи сигнала через мышиный или человеческий TNFR1 у млекопитающего, где шеддинг указанного TNFR1, по существу, не ингибируется, при котором субъекту, нуждающемуся в этом, вводят терапевтически эффективную дозу антагониста по любому из пп.1-11.
30. Способ по п.29, где указанный антагонист связывает домен 1 указанного TNFR1.
31. Способ по п.30, где указанный антагонист конкурирует с TAR2m-21-23 (SEQ ID NO:173) за связывание с мышиным TNFR1.
32. Способ по п.30, где указанный антагонист конкурирует с TAR2h-205 (SEQ ID NO: 627) c человеческим TNFR1.
33. Способ по п.29, где указанный антагонист ингибирует связывание TNFα с указанным TNFR1.
34. Способ по любому из пп.29 и 33, где указанный антагонист dAb связывает домен 3 указанного TNFR1.
35. Способ по любому из пп.29 и 33, где указанный антагонист конкурирует с TAR-2h-131-8, TAR2h-15-8, TAR2h-35-4, TAR2h-154-7, TAR2h-154-10 или TAR2h-185-25 за связывание с человеческим TNFR1.
36. Способ связывания с доменом 1 мышиного или человеческого TNFR1 и ингибирования передачи сигнала через указанный TNFR1 у млекопитающего, при котором субъекту, нуждающемуся в этом, вводят терапевтически эффективную дозу антагониста по любому из пп.1-7.
37. Способ по п.36, где указанный антагонист конкурирует с TAR2m-21-23 (SEQ ID NO:173) за связывание с мышиным TNFR1.
38. Способ по п.36, где указанный антагонист конкурирует с TAR2h-205 (SEQ ID NO:627) за связывание с человеческим TNFR1.
39. Способ по п.36, где указанный антагонист, по существу, не ингибирует связывание TNFα с указанным TNFR1.
40. Способ по любому из пп.36-39, где указанный антагонист, по существу, не ингибирует шеддинг указанного TNFR1.
41. Способ связывания с доменом 3 мышиного или человеческого TNFR1 и ингибирования передачи сигнала через указанный TNFR1 у млекопитающего, при котором субъекту, нуждающемуся в этом, вводят терапевтически эффективную дозу антагониста по любому из пп.1-11.
42. Способ по п.41, где указанный антагонист конкурирует с TAR2h-131-8, TAR2h-15-8, TAR2h-35-4, TAR2h-154-7, TAR2h-154-10 или TAR2h-185-25 за связывание с человеческим TNFR1.
43. Антагонист рецептора фактора некроза опухоли I (TNFR1), который связывает человеческий TNFR1 и ингибирует передачу сигнала через TNFR1, причем связывание указанного антагониста с указанным человеческим TNFR1 ингибируется полипептидом, состоящим из аминокислотной последовательности, кодируемой SEQ ID NO:619, и где указанный антагонист представляет собой доменное антитело против TNFR1 (dAb).
44. Антагонист по п.43, где связывание указанного антагониста с указанным человеческим TNFR1 также ингибируется полипептидом, состоящим из аминокислотной последовательности, кодируемой SEQ ID NO:623.
45. Антагонист по п.43, где связывание указанного антагониста с указанным человеческим TNFR1 также ингибируется полипептидом, состоящим из аминокислотной последовательности, кодируемой нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NO:620, SEQ ID NO:621 и SEQ ID NO:622.
46. Антагонист по п.43, где связывание указанного антагониста с указанным человеческим TNFR1 также ингибируется полипептидом, состоящим из аминокислотной последовательности, кодируемой нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NO:619, SEQ ID NO:620, SEQ ID NO:621 и SEQ ID NO:622, но не ингибируется полипептидом, состоящим из аминокислотной последовательности, кодируемой SEQ ID NO:623.
47. Способ ингибирования передачи сигнала через человеческий рецептор фактора роста некроза опухоли I (TNFR1), при котором субъекту, нуждающемуся в этом, вводят терапевтически эффективную дозу антагониста по п.43.
48. Выделенная нуклеиновая кислота, кодирующая dAb, как оно определено в п.43.
49. Антагонист рецептора фактора некроза опухоли I (TNFR1), который связывает человеческий TNFR1 и ингибирует передачу сигнала через TNFR1, причем связывание указанного антагониста с указанным человеческим TNFR1 ингибируется полипептидом, состоящим из аминокислотной последовательности, кодируемой нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NO:620, SEQ ID NO:622 и SEQ ID NO:623, и где указанный антагонист представляет собой доменное антитело против TNFR1 (dAb).
50. Антагонист по п.49, где связывание указанного антагониста с указанным человеческим TNFR1 не ингибируется полипептидом, состоящим из аминокислотной последовательности, кодируемой нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NO:619 и SEQ ID NO:621.
51. Антагонист по п.49, где связывание указанного антагониста с указанным человеческим TNFR1 ингибируется полипептидом, состоящим из аминокислотной последовательности, кодируемой нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NO:620, SEQ ID NO:622 и SEQ ID NO:623, но не ингибируется полипептидом, состоящим из аминокислотной последовательности, кодируемой нуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NO:619 и SEQ ID NO:621.
52. Способ ингибирования передачи сигнала через человеческий рецептор фактора роста некроза опухоли I (TNFR1), при котором субъекту, нуждающемуся в этом, вводят терапевтически эффективную дозу антагониста по п.49.
53. Выделенная нуклеиновая кислота, кодирующая dAb, как оно определено в п.49.
54. Антагонист рецептора фактора некроза опухоли I (TNFR1), который связывает человеческий TNFR1 и ингибирует передачу сигнала через TNFR1, причем связывание указанного антагониста с указанным человеческим TNFR1 ингибируется полипептидом, состоящим из аминокислотной последовательности, кодируемой SEQ ID NO:620, и где указанный антагонист представляет собой доменное антитело против TNFR1 (dAb).
55. Антагонист по п.54, где связывание указанного антагониста с указанным человеческим TNFR1 ингибируется полипептидом, состоящим из аминокислотной последовательности, кодируемой последовательностью нуклеиновой кислоты, выбранной из группы, состоящей из SEQ ID NO:621 и SEQ ID NO:623.
56. Способ ингибирования передачи сигнала через человеческий рецептор фактора роста некроза опухоли I (TNFR1), где шеддинг указанного TNFR1, по существу, не ингибируется, при котором субъекту, нуждающемуся в этом, вводят терапевтически эффективную дозу антагониста по п.54.
57. Выделенная нуклеиновая кислота, кодирующая dAb, как оно определено в п.54.
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| OHNO ET AL | |||
| Antigen-binding specificities of antibodies are primarily determined by seven residues of VH | |||
| Proc | |||
| Natl | |||
| Acad | |||
| Sci | |||
| USA | |||
| Приспособление для установки двигателя в топках с получающими возвратно-поступательное перемещение колосниками | 1917 |
|
SU1985A1 |
| Машина для разделения сыпучих материалов и размещения их в приемники | 0 |
|
SU82A1 |
| НАСОНОВ Е.Л | |||
| Перспективы фармакотерапии воспалительных ревматических заболеваний: моноклональные антитела к фактору некроза опухоли | |||
| РМЖ, 2001, 9, с | |||
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |

