ОБЛАСТЬ ТЕХНИКИ
В настоящем изобретении предложены антигенсвязывающие белки, способные связываться с полипептидами KIR3DL2. Указанные антитела обладают повышенной активностью в лечении нарушений, характеризующихся экспрессирующими KIR3DL2 клетками, в частности, Т-клетками CD4+, в том числе злокачественных новообразований, таких как фунгоидный микоз и синдром Сезари, и аутоиммунных расстройств, при которых экспрессируется KIR3DL2.
ПЕРЕКРЕСТНЫЕ ССЫЛКИ НА РОДСТВЕННЫЕ ЗАЯВКИ
Настоящая заявка испрашивает приоритет по предварительной заявке на патент США №61/702834, поданной 19 сентября 2012 г.; описание которой полностью включено в настоящий документ посредством ссылки; в том числе включены все чертежи.
ССЫЛКИ НА ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
При подаче к настоящей заявке прилагается перечень последовательностей в электронном формате. Указанный перечень последовательностей представлен файлом с названием «KIR-3 PCT_ST25», созданным 13 сентября 2013 г., и имеющим размер 91 кб. Информация, содержащаяся в электронном формате в указанном перечне последовательностей, полностью включена в настоящий документ посредством ссылки.
уровень ТЕХНИКИ
Иммуноглобулин-подобные рецепторы киллеров (KIR) представляют собой семейство рецепторов, которые, наряду с рецепторами пектинов С-типа (CD94-NKG2), используются NK-клетками человека и субпопуляциями Т-лимфоцитов для специфического распознавания молекул ГКГС класса I. Определенные ингибирующие и активирующие KIR имеют очень сходные внеклеточные домены и распознаются одним и тем же моноклональным антителом, например, как KIR2DL1, так и KIR2DS1 распознаются ЕВ6, a 2DL2 и 2DS2 распознаются GL183. Три критерия (количество внеклеточных Ig-подобных доменов (доменов D0, D1, D2), длина цитоплазматического сегмента и сходство последовательностей) были использованы для разделения белков KIR на 13 групп, а именно, KIR3DL1-2, KIR3DS1, KIR2DL1-5 и KIR2DS1-5. Номенклатура 2D для 2 доменов или 3D для 3 доменов отражает количество Ig-подобных доменов; рецепторы с длинными или с короткими цитоплазматическими доменами дополнительно классифицируют как L или S. (Pascal V. et al., 2007 J. Immunol. 179: 1625-1633) Ингибирующие рецепторы обладают длинными (L) цитоплазматическими сегментами (т.е. KIR2DL или KIR3DL), содержащими классический иммунорецепторный тирозиновый ингибирующий мотив (ITIM), который подвергается тирозиновому фосфорилированию при взаимодействии их лигандов HLA класса I с KIR. Фосфорилированный ITIM активирует содержащую гомологичный Src домен 2 протеинтирозинфосфатазу, содержащую гомологичный Src домен 2 фосфатазу 1 и/или содержащую гомологичный Src домен 2 фосфатазу 2, которые дефосфорилируют клеточные субстраты, устраняя, такие образом, сигнал активации NK, соответственно, сохраняя клетки-мишени с надлежащей экспрессией ауто-ГКГС класса I. В рецепторах с короткими (S) цитоплазматическими сегментами отсутствуют ITIM (т.е. KIR2DS или KIR3DS). Указанные активирующие KIR содержат заряженный остаток в составе трансмембранного домена, что облегчает взаимодействие с сигнальной цепью KARAP/DAP12. Было показано, что активация рецепторов семейства KIR2DS вызывает каскад опосредованных KARAP/DAP12 сигналов, что обуславливает повышение цитолитической активности NK-клеток и продуцирование провоспалительных цитокинов, таких как ИФН-7 (Pascal et al. 2007) J. Immunol. 179: 1625-1633). Зрелые NK-клетки предположительно приобретают по меньшей мере один ингибирующий рецептор, специфичный в отношении молекулы ауто-ГКГС класса I, который, как правило, функционально превалирует над потенциально аутореактивными активирующими молекулами. Предполагается, что ответ NK-клеток представляет собой комплексный результат и активирующей, и ингибирующей сигнализации KIR и других рецепторов.
KIR3DL2 исследовали в качестве мишени для лечения злокачественных новообразований, в которых задействованы Т-клетки CD4+, экспрессирующие рецепторы KIR3DL2, в частности, Т-клетки CD4+, включая такие злокачественные новообразования, как фунгоидный микоз и синдром Сезари (см., например, РСТ-публикации WO 2010/081890 и WO 02/50122).
Лиганд KIR3DL2, HLA-B27, тесно связан со спондилоартритами (СпА), группой инвалидизирующих воспалительных артритических расстройств, примером которых является анкилозирующий спондилит (АС). Полногеномные исследования связей показали глубокую вовлеченность генов, участвующих в регуляции ИЛ-17, продуцируемого клетками Th17 при СпА (Reveille, et al. (2011) Nat Genet 43: 761-767.). Показано, что IL17 играет роль в разнообразных аутоиммунных расстройствах, в том числе в СпА (Shen, et al. (2009) Arthritis Rheum 60: 1647-1656; Wendling, et al. (2007) Joint Bone Spine 74:304-305). HLA-B27 (B27) экспрессируется на поверхности антиген-экспрессирующих клеток (АРС, АПК) при заболевании как в виде классических связанных с β2m гетеротримеров, так и в виде неканонических, не содержащих β2m, связанных дисульфидными мостиками димеров тяжелых цепей (называемых В272) (Bird, et al. (2003) Eur J Immunol 33: 748-759; Kollnberger, et al. (2002) Arthritis Rheum 46: 2972-2982). Димеры B27, но не гетеротримеры B27, являются лигандами иммуноглобулин-подобного рецептора KIR3DL2 киллерных клеток (Kollnberger et al. (2002)). Три иммуноглобулин-подобных домена KIR3DL2, D0, D1 и D2, задействованы в связывании лиганда. Связывание KIR3DL2 димерами B27 способствует выживанию субпопуляций Th17 и NK-клеток (Bowness, et al. (2011) Journal of immunology 186: 2672-2680; Chan, et al. (2005) Arthritis Rheum 52: 3586-3595). Было показано, что у пациентов с СпА повышено содержание патогенных субпопуляций Th17 и NK-клеток, экспрессирующих KIR3DL2, (Bowness et al. (2011) и Chan et al. (2005). Исследования убедительно демонстрируют, что взаимодействия KIR3DL2-B27 играют центральную роль в СпА, и что KIR3DL2 представляет собой перспективную терапевтическую мишень.
Сообщалось о существовании реакционноспособных антител против различных полипептидов KIR3D. Сообщалось о существовании двух антител против KIR3DL2: Q241 и Q66 (Pende, et al. (1996) J Exp Med 184: 505-518). Однако указанные два антитела принадлежат к изотипу IgM (пентамеры) и их фармацевтическое применение затруднено; кроме того, их аффинность при помещении их вариабельных областей в бивалентное антитело типа IgG, предположительно, будет низкой. Были описаны клетки, называемые «AZ158» и продуцирующие другое антитело (Parolini, S., et al. (2002) In Leucocyte typing VII. D. Mason, editor. Oxford University Press, Oxford. 415-417; PCT публикация WO 2010/081890). Антитело 5.133 поставляется Miltenty Biotech (Оберн, Калифорния). Оба антитела, AZ158 и 5.133, связывают KIR3DL2, а также KIR3DL1 (и также высокогомологичный KIR3DS1). KIR3DL2 и KIR3DL1 отличаются относительно высокой идентичностью последовательности аминокислот, и различные HLA-лиганды, связывающие KIR3DL2, распознает также и KIR3DL1. Несмотря на иммунизацию, обеспечивающую получение AZ158, Q241 и Q66, существует потребность в усовершенствованных антителах для терапевтических и других применений.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Согласно одному аспекту настоящее изобретение является результатом, в числе прочего, обнаружения того, что KIR3DL2 может интернализоваться при связывании с антителом. Авторы настоящего изобретения, в свою очередь, идентифицируют ряд моноклональных антител (mAb, MAT) против KIR3DL2, которые не интернализуются. Продемонстрировано, что интернализация KIR3DL2 сильно осложняет применение подходов на основе АЗКЦ. В настоящем описании авторы также предлагают антитела против KIR3DL2, которые подавляют взаимодействия димеров В27 с KIR3DL2. Примечательно, что блокада лиганда может быть достигнута без сопутствующей интернализации рецептора. Авторы также предлагают антитела, которые избирательно блокируют взаимодействия KIR3DL2-HLA В27, не блокируя взаимодействия KIR3DL2-HLA-A3.
Предложены антитела, которые связывают основные (в смысле частоты встречаемости в популяциях человека) аллели KIR3DL2, однако не связывают близкородственный полипептид KIR3DL1 (например, аллель *00101, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 169). Согласно одному варианту реализации указанные антитела связываются с 1, 2, 3, 4 или 5 или большим количеством полипептидов KIR3DL2 (например, аллелями *002, *003, *005, *007, и/или *008), соответствующими последовательностям SEQ ID NO: 1 и 159-168. Соответственно, предложены антитела, имеющие улучшенный функциональные характеристики, описанный в настоящем документе, и подходящие для введения для лечения заболевания по существу во всей человеческой популяции, например, без необходимости проведения диагностических тестов для оценки экспрессии аллеля KTR3DL2 у индивидуума.
Также предложены определенные путем исследования эпитопов антител области на KIR3DL2 (в доменах D0 и D2), которые могут являться мишенями для антител, для обеспечения улучшенных характеристик.
Согласно одному аспекту указанные антитела также обладают дополнительным преимуществом, заключающемся в связывании с несколькими аллелями KIR3DL2 человека при сохранении специфичности к KIR3DL2 относительно KIR3DL1.
Предложены антитела, обладающие преимуществом, заключающемся в блокировании природных лигандов KIR3DL2, и, соответственно, хорошо подходящие для лечения или предотвращения воспалительных расстройств, в формате истощающих или неистощающих mAb. Кроме того, разные эпитопы обеспечивают разную специфичность для блокирования лигандов.
Также предложены антитела, в том числе неинтернализующиеся антитела, которые не блокируют лиганды KIR3DL2 (HLA-A3 и HLA-B27); применение указанных антител может быть целесообразно в способах на основе АЗКЦ, где они могут помогать избежать конкуренции с лигандами.
Согласно одному варианту реализации предложено антитело, которое связывает полипептид KIR3DL2, отличающееся тем, что указанное антитело не связывается существенно с полипептидом KIR3DL1 (например, если указанный полипептид KIR3DL1 содержит аминокислотную последовательность SEQ ID NO: 169), и при этом указанное антитело не интернализируется в экспрессирующие KIR3DL2 клетки.
Согласно одному варианту реализации предложено антитело, которое связывает по меньшей мере два полипептида KIR3DL2 (аллели), причем указанное антитело не связывается существенно с полипептидом KIR3DL1 (например, аллелем KIR3DL1 *00101, содержащим аминокислотную последовательность, представленную в SEQ ID NO: 169).
Согласно одному варианту реализации указанные антитела связываются с 1, 2, 3, 4 или 5 полипептидами KIR3DL2 (аллели *002, *003, *005, *007 и/или *008), соответствующим последовательностям SEQ ID NO: 1, 161, 163, 165 и/или 166.
Согласно одному варианту реализации указанные антитела связываются с каждым из полипептидов KIR3DL2, имеющим аминокислотную последовательность, приведенную в последовательностях SEQ ID NO: 1, 171 и 176 (аллели_*002, *001 и *007, соответственно). Согласно одному варианту реализации указанные антитела связываются с каждым из полипептидов KIR3DL2, имеющим аминокислотную последовательность, приведенную в последовательностях SEQ ID NO: 171 и 178 (аллели_*001 и *009, соответственно). Согласно одному варианту реализации указанные антитела связываются с каждым из полипептидов KIR3DL2, имеющим аминокислотную последовательность, приведенную в последовательностях SEQ ID NO: 171, 1, 176 и 178 (аллели_*001, *002, *007 и *009, соответственно). Согласно одному варианту реализации указанные антитела связываются с каждым из полипептидов KIR3DL2, имеющим аминокислотную последовательность, приведенную в последовательностях SEQ ID NO: 171, 1, 172, 174 и 176 (аллели_*001, *002, *003, *005 и *007, соответственно). Согласно одному варианту реализации указанные антитела связываются с каждым из полипептидов KIR3DL2, имеющим аминокислотную последовательность, приведенную в последовательностях SEQ ID NO: 171. 1, 176 и 177 (аллели_*001, *002, *007 и *008, соответственно). Согласно одному варианту реализации указанные антитела связываются с каждым из полипептидов KIR3DL2, имеющим аминокислотную последовательность, приведенную в последовательностях SEQ ID NO: 171, 1, 172. 174, 176 и 177 (аллели_*001, *002, *003, *005, *007 и *008, соответственно). Согласно одному варианту любого описанного выше вариант указанные антитела также связывают полипептид KIR3DL2, имеющий аминокислотную последовательность, представленную в SEQ ID NO: 178 (аллель *09). Согласно одному варианту реализации любого описанного выше варианта указанные антитела также связывают полипептид KIR3DL2, имеющий аминокислотную последовательность, представленную в SEQ ID NO: 173 (аллель *004). Согласно одному варианту реализации любого описанного выше варианта указанные антитела также связывают аллель *010 полипептида KIR3DL2 (содержащий тот же внеклеточный домен последовательности SEQ ID NO: 171, что и *001). Согласно одному варианту реализации любого описанного выше варианта указанные антитела также связывают полипептид KIR3DL2 аллель *011 (содержащий тот же внеклеточный домен (последовательности SEQ ID NO: 179), что и *003). Согласно одному варианту реализации любого описанного выше варианта указанные антитела также связывают полипептид KIR3DL2 аллель *006. Необязательно в каждом случае антитело связывается с указанным полипептидом KIR3DL2, экспрессируемым на поверхности клетки (например, репортерной клеточной линии, при этом KIR3DL2 находится в природной конформации). Необязательно указанное антитело связывает конформационный эпитоп.
Необязательно в каждом случае антитело связывается с указанным полипептидом KIR3DL2, экспрессируемым на поверхности клетки, с аффинностью связывания (KD), необязательно бивалентным, к полипептиду KIR3DL2 человека, составляющие менее чем 10-8 М. Предпочтительно указанное антитело связывает конформационный эпитоп на KIR3DL2.
Согласно одному варианту реализации предложено антитело, которое связывается с остатком аминокислоты в домене D0 или D2 полипептида KIR3DL2, отличающееся тем, что указанное антитело не связывается существенно с полипептидом KIR3DL1.
Необязательно указанное антитело имеет аффинность связывания (KD), необязательно бивалентное, к полипептиду KIR3DL2 человека, составляющую менее (т.е. имеющее более высокую аффинность, чем) 10-8 М, предпочтительно менее чем 10-9 М, или предпочтительно менее чем 10-10 M.
Необязательно указанные антитела имеют ЕС50, не превышающую 5 мкг/мл, необязательно не более чем 3 мкг/мл, не более чем 2 мкг/мл, не более чем 1 мкг/мл или не более чем 0,5 мкг/мл, для связывания с клетками, адаптированными для экспрессии на поверхности конкретного аллеля KIR3DL2 (например, аллели_*001, *002, *003, *005, *007 и/или *008).
Согласно одному аспекту предложены антитела, которые связывают полипептид KIR3DL2 в лиганд-связывающей (HLA-связывающей) области (например, HLA-связывающем кармане) или по меньшей мере частично на HLA-связывающей поверхности белка KIR3DL2.
Предпочтительно, согласно любым представленным в настоящем документе вариантам реализации, предложено антитело, которое связывается с остатком аминокислоты в составе домена D0 (остатки 1-98 последовательности SEQ ID NO: 1) и/или домена D2 (остатки 193-292 последовательности SEQ ID NO: 1) полипептида KIR3DL2. Необязательно связывание указанного антитела с полипептидом KIR3DL2, содержащим мутацию в остатке в составе домена D0 и/или D2 существенно снижено по сравнению со связыванием с полипептидом KIR3DL2 дикого типа последовательности SEQ ID NO: 1.
Согласно одному аспекту указанные антитела связывают эпитоп, содержащий один, два, три, четыре, пять или большее количество остатков, выбранных из группы, состоящей из: R13, Р14, S15, Н23, А25, Q27, 160 и G62 (относительно SEQ ID NO: 1), и/или связывание указанных антител с полипептидом KIR3DL2, содержащим мутацию в остатке, выбранном из группы, состоящей из: R13, Р14, S15, Н23, А25. Q27,160 и G62 (относительно SEQ ID NO: 1), понижено.
Для мутаций в настоящем документе используются следующие сокращенные обозначения: остаток дикого типа: положение в полипептиде, с нумерацией остатков согласно заданной в последовательности SEQ ID NO: 1: мутантный остаток.
Согласно одному аспекту предложены антитела, которые связывают эпитоп, содержащий остатки R13, А25 и/или Q27 полипептида KIR3DL2, и/или связывание которых с полипептидом KIR3DL2, содержащим мутацию в остатках R13, А25 и/или Q27 (относительно SEQ ID NO: 1), снижено. Например, может быть снижено связывание антитела с полипептидом KIR3DL2, содержащим мутации R13W, А25Т и/или Q27R. Необязательно указанный эпитоп дополнительно содержит один или большее количество остатков 160 и/или G62 (относительно SEQ ID NO: 1), и/или связывание указанных антител с полипептидом KIR3DL2, содержащим мутацию в остатках 160 и/или G62 (относительно SEQ ID NO: 1, например, I60N, G62S), снижено. Необязательно указанный эпитоп дополнительно или как вариант содержит один или большее количество остатков Р14, S15 и/или Н23 (относительно SEQ ID NO: 1), и/или связывание указанных антител с полипептидом KIR3DL2, содержащим мутацию в остатках Р14, S15 и/или Н23 (относительно SEQ ID NO: 1, например, P14S, S15A, H23S), снижено. Необязательно указанный эпитоп не содержит остатков R32 и/или G33 (относительно SEQ ID NO: 1), и/или связывание указанных антител с полипептидом KIR3DL2, содержащим мутацию в остатках R32 и/или G33 (относительно SEQ ID NO: 1, например, R32H и/или G33R), не понижено. Необязательно указанный эпитоп не содержит остатков F50 и/или R53 (относительно SEQ ID NO: 1), и/или связывание указанных антител с полипептидом KIR3DL2, содержащим мутацию в остатках F50 и/или R53 (относительно SEQ ID NO: 1, например, F50A, R53S), не понижено. Указанное антитело может связывать (например, антитела, блокирующие взаимодействия KIR3DL2 с HLA B27 и HLA A3) или может не связывать (например, неинтернализующиеся антитела) остатки Q56 и/или Е57, и/или остатки F9 и/или S11; соответственно, согласно одному варианту реализации. Необязательно указанный эпитоп не содержит остатков F9, S11, Q56 и/или Е57 (относительно SEQ ID NO: 1), и/или связывание указанных антител с полипептидом KIR3DL2, содержащим мутацию в остатках F9, S11, Q56 и/или Е57 (относительно SEQ ID NO: 1, например, P9S и S11A, Q56S и Е57А), не понижено; согласно другому варианту реализации указанный эпитоп необязательно содержит остатки F9, S11, Q56 и/или Е57 (относительно SEQ ID NO: 1), и/или связывание указанных антител с полипептидом KIR3DL2, содержащим мутацию в остатках F9, S11, Q56 и/или Е57 (относительно SEQ ID NO: 1, например, F9S и S11A, Q56S и Е57А), снижено. Необязательно указанный эпитоп не содержит остатков Н29 и/или F34 (относительно SEQ ID NO: 1), и/или связывание указанных антител с полипептидом KIR3DL2, содержащим мутацию в остатках Н29 и/или F34 (относительно SEQ ID NO: 1, например, H29S, F34A), не понижено. Необязательно указанный эпитоп не содержит один или большее количество остатков F9 и/или S11 (относительно SEQ ID NO: 1), и/или связывание указанных антител с полипептидом KIR3DL2, содержащим мутацию в остатках F9 и/или S11 (относительно SEQ ID NO: 1, например, F9S, S11A), не понижено.
Согласно одному аспекту предложены антитела, которые связывают эпитоп, содержащий остатки 160 и/или G62 полипептида KIR3DL2 последовательности SEQ ID NO: 1, и/или связывание которых с полипептидом KIR3DL2, содержащим мутацию в остатках 160 и/или G62 (относительно SEQ ID NO: 1), снижено. Например, может быть снижено связывание антитела с полипептидом KIR3DL2, содержащим мутации I60N и/или G62S. Необязательно, указанный эпитоп дополнительно или как вариант содержит один или большее количество остатков Р14, S15 и/или Н23 (относительно SEQ ID NO: 1), и/или связывание указанных антител с полипептидом KIR3DL2, содержащим мутацию в остатках Р14, S15 и/или Н23 (относительно SEQ ID NO: 1, например, P14S, S15A, H23S), снижено. Необязательно указанные антитела не связывают остатки R13, А25 и/или Q27 полипептида KIR3DL2, и/или их связывание с полипептидом KIR3DL2, содержащим мутацию в остатках R13, А25 и/или Q27 (например, полипептидом KIR3DL2, содержащим мутации R13W, А25Т и/или Q27R), не понижено.
Согласно одному аспекту предложены антитела, которые связывают эпитоп, содержащий остатки Р14, S15 и/или Н23 полипептида KIR3DL2 последовательности SEQ ID NO: 1, и/или связывание которых с полипептидом KIR3DL2, содержащим мутацию в остатках Р14, S15 и/или Н23 (относительно SEQ ID NO: 1, например, P14S, S15A, H23S), снижено.
Согласно одному аспекту предложены антитела, связывание которых с (1) полипептидом KIR3DL2, содержащим мутацию в остатках 160 и/или G62 (относительно SEQ ID NO: 1, например, I60N, G62S), и (2) полипептидом KIR3DL2, содержащим мутацию в остатках Р14, S15 и/или Н23 (относительно SEQ ID NO: 1, например, P14S, S15A, H23S), снижено.
Согласно одному аспекту предложены антитела, которые связывают эпитоп, содержащий: (а) 1, 2 или 3 остатка R13, А25 и/или Q27 и (b) один из остатков или оба остатка 160 и/или G62 полипептида KIR3DL2. Согласно одному аспекту снижено связывание антител с полипептидом KIR3DL2, содержащим: (а) мутацию 1, 2 или 3 остатков R13, А25 и/или Q27, и (b) мутацию одного из остатков или обоих остатков 160 и/или G62.
Согласно одному аспекту предложены антитела, которые связывают эпитоп, содержащий остатки R78 и/или L82 полипептида KIR3DL2 последовательности SEQ ID NO: 1, и/или связывание которых с полипептидом KIR3DL2, содержащим мутацию в остатках R78 и/или L82 (относительно SEQ ID NO: 1), снижено. Например, может быть снижено связывание антитела с полипептидом KIR3DL2, содержащим мутации R78H и L82P. Необязательно указанный эпитоп дополнительно содержит или исключает один или большее количество остатков K7, Y30, R31, Р79, Н80, S81, Т83, G84, W85, S86 и/или А87 (относительно SEQ ID NO: 1), и/или указанные антитела характеризуются пониженным связыванием или не характеризуются пониженным связыванием с полипептидом KIR3DL2, содержащим мутацию в остатках K7, Y30, R31, Р79, Н80, S81, Т83, G84, W85, S86 и/или А87 (относительно SEQ ID NO: 1). Согласно одному варианту реализации указанные антитела связывают эпитоп, содержащий 1, 2, 3, 4, 5, 6, 7 или большее количество остатков в сегменте, соответствующем остаткам 1-98 полипептида KIR3DL2 (относительно SEQ ID NO: 1), необязательно при этом указанный эпитоп также содержит один или большее количество (например, 1, 2, 3, 4, 5) остатков K7, Y30, R31, R78, Р79, Н80, S81, L82, Т83, G84, W85, S86 и/или А87.
Согласно одному аспекту предложены антитела, которые связывают эпитоп, содержащий остатки W226 полипептида KIR3DL2 последовательности SEQ ID NO: 1, и/или связывание которых с полипептидом KIR3DL2, содержащим мутацию в остатках W226 (относительно SEQ ID NO: 1), понижено. Необязательно указанный эпитоп дополнительно содержит один или большее количество остатков I231 и/или R246 (относительно SEQ ID NO: 1), и/или связывание указанных антител с полипептидом KIR3DL2, содержащим мутацию в остатках I231 и/или R246 (относительно SEQ ID NO: 1, например, I231M, R246P), понижено. Необязательно указанный эпитоп дополнительно содержит остаток Е239 (относительно SEQ ID NO: 1), и/или связывание указанных антител с полипептидом KIR3DL2, содержащим мутацию в остатке Е239 (относительно SEQ ID NO: 1, например, E239G), понижено.
Согласно одному аспекту предложены антитела, которые связывают эпитоп, содержащий остатки I231 и/или R246 полипептида KIR3DL2 последовательности SEQ ID NO: 1, и/или связывание которых с полипептидом KIR3DL2, содержащим мутацию в остатках I231 и/или R246 (относительно SEQ ID NO: 1), понижено.
Согласно одному аспекту предложены антитела, которые связывают эпитоп, содержащий остаток W226 и один или оба из остатков I231 и/или R246 полипептида KIR3DL2.
Согласно одному аспекту связывание антител с полипептидом KIR3DL2, содержащим мутацию в остатках W226 и по одному или обоим остаткам I231 и/или R246, понижено.
Согласно любому варианту реализации описанного изобретения антитело необязательно не вызывает интернализации полипептидов KIR3DL2 в экспрессирующих KIR3DL2 клетках и/или не интернализируется в экспрессирующие KIR3DL2 клетки.
Согласно одному варианту реализации предложено антитело, которое связывает полипептид KIR3DL2, отличающееся тем, что указанное антитело детектируемо снижает (или устраняет) связывание между KIR3DL2 и первым природным HLA-лигандом KIR3DL2, но не приводит к детектируемому снижению (или не устраняет) связывания между KIR3DL2 и вторым природным HLA-лигандом KIR3DL2.
Согласно одному варианту реализации антитело необязательно детектируемо снижает связывание между KIR3DL2 и HLA класса 1-лигандом KIR3DL2 (например, HLA-B27, HLA-A3, HLA-B7, HLA-B35 и/или HLA-A2).
Согласно одному варианту реализации антитело необязательно детектируемо снижает связывание между KIR3DL2 и HLA-B27, но не приводит к детектируемому снижению связывания между KIR3DL2 и HLA-A3.
Согласно одному варианту реализации антитело необязательно детектируемо снижает связывание между KIR3DL2 и HLA-A3, но не приводит к детектируемому снижению связывания между KIR3DL2 и HLA-B27.
Согласно одному варианту реализации антитело необязательно не приводит к детектируемому снижению связывания между KIR3DL2 и HLA-B27, или между KIR3DL2 и HLA-A3.
Согласно одному варианту реализации антитело необязательно связывает по меньшей мере два полипептида (аллеля) KIR3DL2, имеющие разные аминокислотные последовательности.
Согласно одному варианту реализации антитело необязательно не связывается существенно с полипептидом KIR3DL1.
Согласно представленным в настоящем документе вариантам реализации блокирующих лиганд антител и/или антител, которые связывают эпитоп, содержащий остатки Н32 и/или G33 полипептида KIR3DL2, указанное антитело может необязательно вызывать интернализацию полипептидов KIR3DL2 в экспрессирующие KIR3DL2 клетки и/или интернализуется экспрессирующими KIR3DL2 клетками.
Антитело против KIR3DL2 может подходить для лечения раковых заболеваний, воспалительных расстройств и аутоиммунных расстройств, например, у субъектов-людей. Указанное антитело может применяться в связанном или не связанном с токсическим или другим агентом виде, в зависимости от требуемого эффекта или применения антител. Согласно одному варианту реализации антитело против KIR3DL2 представляет собой «голое антитело» («naked»), не связанное с токсическим агентом. Согласно одному варианту реализации голое или связанное антитело содержит тяжелую цепь, содержащую область Fc (например, IgG1), которая связывает рецепторы Fcγ (например, CD16). При этом необязательно такое антитело индуцирует комплементзависимую цитотоксичность (КЗЦ) и/или антителозависимую клеточную цитотоксичность (АЗКЦ) в отношении клетки, которая экспрессирует KIR3DL2.
Необязательно согласно любому варианту реализации указанное антитело (например, IgG4, IgG1, фрагмент антитела и т.д.) дополнительно содержит токсический агент (например, химиотерапевтический агент), токсичный для клетки при интернализации конъюгата антитело-токсин. Согласно одному варианту реализации указанное антитело конъюгировано с радиоактивным агентом.
В настоящем описании также предложены антитела, фрагменты и производные антител, которые специфически связывают KIR3DL2 человека. В настоящем описании предложены композиции таких антител, а также их применение согласно любым способам раскрытого в настоящем документе лечения, предотвращения и диагностики раковых заболеваний, воспалительных расстройств или аутоиммунных расстройств.
Согласно одному варианту реализации указанные антитела имеют аффинность связывания (KD) к полипептиду KIR3DL2 человека, составляющее менее чем 10-8 М, предпочтительно менее чем 10-9 М, или предпочтительно менее чем 10-10 M. Аффинность необязательно относится к бивалентному связыванию.
Согласно одному аспекту любого из описываемых в настоящем документе вариантов реализации антитело может содержать тяжелую и/или легкую цепь, содержащую одну, две или три участка CDR из соответствующих тяжелой и/или легкой цепи антитела, выбранного из группы, состоящей из антитела 10F6, 2В12, 18С6, 9Е10, 10G5, 13Н1, 5Н1, 1Е2, 1С3 и/или 20Е9.
Согласно одному аспекту любого из описываемых в настоящем документе вариантов реализации антитело конкурирует за связывание с полипептидом KIR3DL2 с любым одним или с любой комбинацией моноклональных антител 10F6, 2В12, 18С6, 9Е10, 10G5, 13Н1, 5Н1, 1Е2, 1С3 и/или 20Е9. Согласно одному варианту реализации антитело конкурирует за связывание с полипептидом KIR3DL2 с антителом, выбранным из группы, состоящей из:
Согласно одному аспекту в настоящем описании предложено моноклональное антитело, которое специфически связывает KIR3DL2, выбранный из группы, состоящей из:
(a) антитела, содержащего (i) тяжелую цепь, содержащую участки CDR 1, 2 и 3 (HCDR1, HCDR2, HCDR3), содержащие последовательность, соответствующую последовательностям SEQ ID NO: 4, 5 или 6 (HCDR1), последовательностям SEQ ID NO: 7 или 8 (HCDR2) и SEQ ID NO: 9 (HCDR3), соответственно, и (ii) легкую цепь, содержащую участки CDR 1, 2 и 3 (LCDR1, LCDR2, LCDR3), содержащие последовательность SEQ ID NO: 10, 11 или 12, соответственно, при этом каждый участок CDR может необязательно содержать 1, 2, 3 или 4 замены, удаления или вставки аминокислот;
(b) антитела, содержащего (i) тяжелую цепь, содержащую участки CDR 1, 2 и 3 (HCDR1, HCDR2, HCDR3), содержащие последовательность, соответствующую последовательностям SEQ ID NO: 15, 16 или 17 (HCDR1), последовательностям SEQ ID NO: 18 или 19 (HCDR2) и SEQ ID NO: 20 (HCDR3), соответственно, и (ii) легкую цепь, содержащую участки CDR 1, 2 и 3 (LCDR1, LCDR2, LCDR3), содержащие последовательность SEQ ID NO: 10, 21 или 22, соответственно, при этом каждый участок CDR может необязательно содержать 1, 2, 3 или 4 замены, удаления или вставки аминокислот;
(c) антитела, содержащего (i) тяжелую цепь, содержащую участки CDR 1, 2 и 3 (HCDR1, HCDR2, HCDR3), содержащие последовательность, соответствующую последовательностям SEQ ID NO: 25, 26 или 27 (HCDR1), последовательностям SEQ ID NO: 28 или 29 (HCDR2) и SEQ ID NO: 30 (HCDR3), соответственно, и (ii) легкую цепь, содержащую участки CDR 1, 2 и 3 (LCDR1, LCDR2, LCDR3), содержащие последовательность SEQ ID NO: 31, 32 или 33, соответственно, при этом каждый участок CDR может необязательно содержать 1, 2, 3 или 4 замены, удаления или вставки аминокислот;
(d) антитела, содержащего (i) тяжелую цепь, содержащую участки CDR 1, 2 и 3 (HCDR1, HCDR2, HCDR3), содержащие последовательность, соответствующую последовательностям SEQ ID NO: 36, 37 или 38 (HCDR1), последовательностям SEQ ID NO: 39 или 40 (HCDR2) и SEQ ID NO: 41 (HCDR3), соответственно, и (ii) легкую цепь, содержащую участки CDR 1, 2 и 3 (LCDR1, LCDR2, LCDR3), содержащие последовательность SEQ ID NO: 42, 43 или 44, соответственно, при этом каждый участок CDR может необязательно содержать 1, 2, 3 или 4 замены, удаления или вставки аминокислот;
(e) антитела, содержащего (i) тяжелую цепь, содержащую участки CDR 1, 2 и 3 (HCDR1, HCDR2, HCDR3), содержащие последовательность, соответствующую последовательностям SEQ ID NO: 47, 48 или 49 (HCDR1), последовательностям SEQ ID NO: 50 или 51 (HCDR2) и SEQ ID NO: 52 (HCDR3), соответственно, и (ii) легкую цепь, содержащую участки CDR 1, 2 и 3 (LCDR1, LCDR2, LCDR3), содержащие последовательность SEQ ID NO: 53, 54 или 55, соответственно, при этом каждый участок CDR может необязательно содержать 1, 2, 3 или 4 замены, удаления или вставки аминокислот;
(f) антитела, содержащего (i) тяжелую цепь, содержащую участки CDR 1, 2 и 3 (HCDR1, HCDR2, HCDR3), содержащие последовательность, соответствующую последовательностям SEQ ID NO: 58, 59 или 60 (HCDR1), последовательностям SEQ ID NO: 61 или 62 (HCDR2) и SEQ ID NO: 63 (HCDR3), соответственно, и (ii) легкую цепь, содержащую участки CDR 1, 2 и 3 (LCDR1, LCDR2, LCDR3), содержащие последовательность SEQ ID NO: 64, 65 или 66, соответственно, при этом каждый участок CDR может необязательно содержать 1, 2, 3 или 4 замены, удаления или вставки аминокислот;
(g) антитела, содержащего (i) тяжелую цепь, содержащую участки CDR 1, 2 и 3 (HCDR1, HCDR2, HCDR3), содержащие последовательность SEQ ID NO: 172, 173 или 174 (HCDR1), SEQ ID NO: 175 или 176 (HCDR2) и SEQ ID NO: 177 (HCDR3), соответственно, и (ii) легкую цепь, содержащую участки CDR 1, 2 и 3 (LCDR1, LCDR2, LCDR3), содержащие последовательность SEQ ID NO: 178, 179 или 180, соответственно, при этом каждый участок CDR может необязательно содержать 1, 2, 3 или 4 замены, удаления или вставки аминокислот; и
(h) антитела, содержащего (i) тяжелую цепь, содержащую участки CDR 1, 2 и 3 (HCDR1, HCDR2, HCDR3), содержащие последовательность SEQ ID NO: 183, 184 или 185 (HCDR1), SEQ ID NO: 186 или 187 (HCDR2) и SEQ ID NO: 188 (HCDR3), соответственно, и (ii) легкую цепь, содержащую участки CDR 1, 2 и 3 (LCDR1, LCDR2, LCDR3), содержащие последовательность SEQ ID NO: 189, 190 или 191, соответственно, при этом каждый участок CDR может необязательно содержать 1, 2, 3 или 4 замены, удаления или вставки аминокислот.
Согласно одному аспекту предложено антитело, которое специфически связывает KIR3DL2, при этом указанное антитело обладает одним или несколькими из следующих свойств (включая любую их комбинацию, при условии, что такая комбинация не является несовместимой):
(a) имеет Kd связывания с полипептидом KIR3DL2, составляющую менее чем 10-8 М, предпочтительно менее чем 10-9 М, или предпочтительно менее чем 10-10 M;
(b) связывается по меньшей мере с одним остатком в сегменте, соответствующем остаткам 1-98 или остаткам 193-292 полипептида KIR3DL2;
(c) конкурирует за связывание с полипептидом KIR3DL2 с антителом 10F6, 2В12, 18С6, 9Е10, 10G5, 13Н1, 5Н1, 1Е2, 1С3 и/или 20Е9;
(d) конкурирует или не конкурирует с природным лигандом KIR3DL2 (например, HLA-полипептидами HLA-A3, HLA-11 и/или HLA-B27) за связывание с полипептидом KIR3DL2 (например, в анализе взаимодействия полипептидов);
(e) не вызывает интернализации полипептидов KIR3DL2 в экспрессирующие KIR3DL2 клетки и/или не интернализируется в экспрессирующие KIR3DL2 клетки;
(f) подавляет или не подавляет сигнализацию KIR3DL2, индуцируемую природным лигандом KIR3DL2 (например, HLA-полипептидов HLA-A3, HLA-11 и/или HLA-B27);
(g) не связывается существенно с полипептидом KIR3DL1, KIR3DS1, KIR3DL3, KIR2DS1, KIR2DS2, KIR2DL3, KIR2DL1 и/или KIR2DS4;
(h) связывается с 1, 2, 3, 4, 5 или 6 полипептидами KIR3DL2 (например, аллели *001, *002, *003, *005, *007 и/или *008), соответствующими последовательностям SEQ ID NO: 160, 1, 161, 163, 165 и/или 166);
(i) связывается с эпитопом, содержащим любой один остаток или большее количество остатков аминокислот R13, Р14, S15, Н23, А25, Q27, Н32, G33, I60, G62, R78, L82, W226, I231 и/или R246 полипептида KIR3DL2; и
(j) связывание которого понижено с полипептидом KIR3DL2, содержащим мутацию в одном или большем числе остатков R13, Р14, S15, Н23, А25, Q27, Н32, G33, I60, G62, R78, L82, W226, I231 и/или R246 полипептида KIR3DL2.
Согласно любым представленным в настоящем документе вариантам реализации антитело может характеризоваться любым одним или большим количеством признаков (a)-(j), выше.
Согласно одному варианту реализации указанное антитело подходит для человека. Согласно одному варианту реализации указанное антитело является гибридным, например, содержит константную область, не принадлежащую мыши, необязательно принадлежащую человеку. Согласно одному варианту реализации указанное антитело представляет собой антитело человека или гуманизированное антитело. Согласно другому варианту реализации указанное антитело представляет собой антитело мыши.
Согласно одному аспекту любого из описываемых в настоящем документе вариантов реализации, изотип указанного антитела представлен IgG, необязательно IgG1, IgG2, IgG3 или IgG4. Согласно одному варианту реализации указанное антитело содержит домен Fс или относится к изотипу, связываемому FcγR (например, FcγRIIIA), например, представляет собой антитело изотипа IgG1 или IgG3.
Согласно одному аспекту любого из описываемых в настоящем документе вариантов реализации антитело представляет собой фрагмент антитела, выбранный из Fab, Fab', Fab'-SH, F(ab')2, Fv, диател, одноцепочечного фрагмента антитела или мультиспецифического антитела, содержащего несколько разных фрагментов антител. Согласно одному аспекту любого из описываемых в настоящем документе вариантов реализации антитело не содержит домен Fc или относится к изотипу, которое не связывается существенно FcγR. Согласно одному варианту реализации указанное антитело относится к изотипу IgG4 или IgG2.
Необязательно такие антитела также являются тетрамерными (две тяжелых и две легких цепи) и, соответственно, бивалентны (например, антитела IgG).
Согласно определенным вариантам реализации указанные антитела также содержат токсический агент. Согласно одному варианту реализации антитела, содержащие токсический агент, способны непосредственно вызывать гибель клеток, экспрессирующих KIR3DL2. Согласно одному варианту реализации указанные антитела способны непосредственно индуцировать (например, в отсутствие иммунных эффекторных клеток) по меньшей мере 20%, 30%, 40% или 50% гибель клеток, например, в анализе in vitro, экспрессирующих KIR3DL2 клеток.
Согласно одному варианту реализации указанные антитела способны индуцировать КЗЦ и/или АЗКЦ клеток, экспрессирующих KIR3DL2. Согласно одному варианту реализации указанные антитела способны индуцировать по меньшей мере 20%, 30, 40 или 50% лизис, в анализе цитотоксичности, экспрессирующих KIR3DL2 клеток (например, клеток Т-клеточной лимфомы, клеток пациентов с СС или клеточных линий СС).
Согласно одному варианту реализации предложен способ тестирования антитела против KIR3DL2, при этом указанный способ включает приведение антитела, которое связывает полипептид KIR3DL2, в контакт с клеткой, экспрессирующей полипептид KIR3DL2, и оценку того, интернализируется ли указанное антитело в экспрессирующие KIR3DL2 клетки и/или индуцирует и/или повышает ли указанное антитело внутриклеточную интернализацию полипептида KIR3DL2, и отбор антитела, если антитело не индуцирует и/или не повышает внутриклеточную интернализацию полипептида KIR3DL2.
Согласно другому варианту реализации предложен способ получения антитела, которое связывает полипептид KIR3DL2 у субъекта-млекопитающего, необязательно для лечения ракового заболевания, воспалительного расстройства или аутоиммунного расстройства, при этом указанный способ включает этапы: а) получения множества антител, необязательно иммунизации не являющегося человеком млекопитающего иммуногеном, содержащим полипептид KIR3DL2 человека; b) определение того, способно ли каждое из указанного множества антител связываться с 1, 2, 3, 4, 5, или большим количеством разных аллелей полипептидов KIR3DL2 (например, аллелями *001, *002, *003, *005, *007, *008, *009 и/или *011), необязательно в каждом случае, когда указанный полипептид KIR3DL2 экспрессируется на поверхности клетки, и с) выбора (например, для получения, разработки, применения в терапии и т.д.) из указанного множества антител, которые способны связываться с 1, 2, 3, 4, 5, или большим количеством разных аллелей полипептидов KIR3DL2 (например, аллелями *001, *002, *003, *005, *007, *008, *009 и/или *011), необязательно в каждом случае, когда указанный полипептид KIR3DL2 экспрессируется на поверхности клетки. Указанный способ необязательно также включает определение того, способно ли каждое из указанного множества антител связываться с полипептидом KIR3DL1, и выбор антитела из указанного множества, которое способно связываться с указанным полипептидом KIR3DL1.
Согласно другому варианту реализации предложен способ получения антитела, которое связывает полипептид KIR3DL2 у субъекта-млекопитающего, необязательно для лечения ракового заболевания, воспалительного расстройства или аутоиммунного расстройства, при этом указанный способ включает этапы: а) получения множества антител, необязательно иммунизация не являющегося человеком млекопитающего иммуногеном, содержащим полипептид KIR3DL2 человека; и b) выбора (например, для получения, разработки, применения в терапии и т.д.) из указанного множества антитела, которое:
(i) связывается с полипептидом KIR3DL2, но не с полипептидом KIR3DL1; и/или
(ii) (а) связывается по меньшей мере с одним остатком в сегменте, соответствующем остаткам 99-192 зрелого полипептида KIR3DL2 последовательности SEQ ID NO: 1, и/или с любым одним остатком или большим количеством (например, 2, 3, 4, 5 или более) остатков R13, Р14, S15, Н23, А25, Q27, Н32, G33, I60, G62, R78, L82, W226, I231 и/или R246, и/или связывание которого с полипептидом KIR3DL2 содержащим замену аминокислоты в положении указанного(ых) остатка(ов), понижено или
(b) связывается по меньшей мере с одним остатком в сегменте, соответствующем остаткам 1-98, зрелого полипептида KIR3DL2 последовательности SEQ ID NO: 1, и/или любому одному или большему количеству (например, 2, 3, 4, 5 или большее количество) остатков R13, Р14, S15, Н23, А25, Q27, Н32, G33, I60, G62, R78, L82, W226, I231 и/или R246, и/или связывание которого с полипептидом KIR3DL2 содержащим замену аминокислоты в положении указанного(ых) остатка(ов), понижено; и/или
(iii) не интернализируется в экспрессирующие KIR3DL2 клетки и/или не индуцирует и/или не увеличивает внутриклеточную интернализацию полипептида KIR3DL2.
Согласно одному аспекту предложены способы подавления биологической активности экспрессирующей KIR3DL2 клетки, включающий приведение указанной клетки в контакт с антителами против KIR3DL2, in vitro, ex vivo или in vivo. Необязательно указанное приведение в контакт происходит в присутствии лиганда (например, HLA) KIR3DL2, необязательно - клетки, экспрессирующей лиганд (например, HLA) KIR3DL2. Предпочтительно указанная экспрессирующая KIR3DL2 клетка представляет собой иммунную клетку, например, Т-клетку или NK-клетку, злокачественную Т-клетку или NK-клетку, клетку CD4 Th17 (например, провоспалительные Т-клетки CD4, которые экспрессируют ИЛ-23R и продуцируют ИЛ-17А) или провоспалительную NK-клетку, которая экспрессирует и продуцирует ИЛ-17А. Согласно одному варианту реализации предложены способы подавления биологической активности экспрессирующей KIR3DL2 Т-клетки или NK-клетки, которая продуцирует ИЛ-17А, включающие приведение указанной клетки в контакт с антителами против KIR3DL2, in vitro, ex vivo или in vivo. Предпочтительно указанная биологическая активность представлена активацией, литической активностью, продуцированном цитокинов (например, ИЛ-17А) и/или клеточной пролиферацией. Предпочтительно указанная биологическая активность представлена лиганд-индуцированной (например, HLA-индуцированной) сигнализацией. Согласно одному аспекту предложены способы подавления биологической активности экспрессирующей KIR3DL2 клетки, включающие приведение указанной клетки в контакт с антителами против KIR3DL2, in vitro, ex vivo или in vivo.
Согласно одному аспекту предложены способы устранения или истощения экспрессирующих KIR3DL2 клеток, включающие приведение указанной клетки в контакт с антителом против KIR3DL2, in vitro, ex vivo или in vivo. Указанная клетка может представлять собой, например, злокачественную Т-клетку или NK-клетку, Т-клетку или NK-клетку, клетку CD4 Th17 (например, провоспалительную Т-клетку CD4, которая экспрессирует ИЛ-23R и продуцирует ИЛ-17А) или провоспалительную NK-клетку, которая экспрессирует и продуцирует ИЛ-17А.
Согласно одному аспекту предложены способы лечения с применением антител против KIR3DL2 согласно настоящему описанию. Указанные антитела могут применяться в качестве профилактического или терапевтического лечения; согласно любым представленным в настоящем документе вариантам реализации терапевтически эффективное количество антитела может быть взаимозаменяемым с профилактически эффективным количеством антитела. Согласно одному аспекту предложен способ лечения пациента с раковым заболеванием, например, Т-клеточной лимфомой, CD4+ или CD8+ CTCL, синдромом Сезари (СС), фунгоидным микозом (ФМ), CD30+ Т-клеточной лимфомой, отличающийся тем, что указанный способ включает введение указанному пациенту фармацевтически эффективного количества антигенсвязывающего соединения, описанного в настоящем документе, которое специфически связывается с полипептидом KIR3DL2. Согласно другому варианту реализации предложен способ лечения пациента с аутоиммунным или воспалительным расстройством, опосредованным по меньшей мере частично экспрессирующими KIR3DL2 Т-клетками, отличающийся тем, что указанный способ включает введение указанному пациенту фармацевтически эффективного количества антигенсвязывающего соединения, описанного в настоящем документе, которое специфически связывается с полипептидом KIR3DL2.
Указанные способы лечения и антитело против KIR3DL2 могут применяться для лечения индивидуума в комбинации со вторым терапевтическим агентом, включая иммуномодуляторы (например, химиотерапевтические лекарственные средства, противовоспалительные лекарственные средства, противоопухолевые вакцины, антитела, которые связываются с опухолеспецифичными антигенами на опухолевых клетках, антитела, которые индуцируют АЗКЦ в отношении опухолевых клеток, антитела, которые усиливают иммунный ответ, модифицирующие течение болезни противоревматическое лекарственные средства (БПРП) и т.д.). Согласно одному варианту реализации второй терапевтический агент представляет собой антитело против CD4 или антитело против CD30.
Раскрытое в настоящем описании изобретение также относится к способу выбора субъектов с заболеванием, отвечающим на лечение с применением антагониста KIR3DL2 (например, антитела, которое связывается с полипептидом KIR3DL2), отличающийся тем, что указанный способ включает определение того, экспрессируют ли связанные с заболеванием клетки указанного субъекта рецептор KIR3DL2, при этом экспрессия рецептора KIR3DL2 указывает на то, что пациент отвечает на лечение. Необязательно указанный способ также включает введение отвечающему на лечение пациенту антитела (например, антитела против KIR3DL2 согласно настоящему изобретению), которое связывается с полипептидом KIR3DL2. Согласно одному варианту реализации указанный способ используют для выбора субъектов, имеющих раковое заболевание, и связанные с заболеванием клетки представляют собой раковые клетки. Согласно одному варианту реализации указанный способ используют для выбора субъектов, имеющих воспалительное или аутоиммунное расстройство, и связанные с заболеванием клетки представляют собой Т-клетки.
Экспрессия рецептора KIR3DL2 в указанной связанной с заболеванием клетке могут быть определены с применением KIR3DL2-специфического лиганда. Предпочтительно указанный лиганд представляет собой антитело или его фрагмент или производное. Согласно одному аспекту в настоящем изобретении предложены композиции, содержащие моноклональные антитела, и способы применения моноклональных антител, включая, но не ограничиваясь перечисленными, фрагменты и производные антител, которые специфически связывают KIR3DL2 человека.
Согласно другому аспекту предложен способ (например, способ выполнения диагностического анализа, анализа реактивности и т.д.), включающий оценку того, присутствуют ли у пациента связанные с заболеванием клетки, экспрессирующие полипептид KIR3DL2, например, полипептид KIR3DL2 (один или большее количество аллелей KIR3DL2), связываемый антителом, описанным в настоящем документе. Указанный способ может включать, например, получение биологического образца от пациента, содержащего связанные с заболеванием клетки, приведение указанных связанных с заболеванием клеток в контакт с таким антителом и оценка того, связывается ли указанное антитело со связанными с заболеванием клетками. Обнаружение того, что KIR3DL2 экспрессируется связанными с заболеванием клетками, указывает на то, что у указанного пациента имеется состояние, характеризующееся экспрессирующими KIR3DL2 клетками и/или для лечения которого подходит антитело против KIR3DL2, описанное в настоящем документе. После этого может быть проведено лечение пациента с применением лечения, подходящего для конкретного заболевания, характеризующегося экспрессирующими KIR3DL2 клетками. Необязательно проводят лечение указанного пациента антителом против KIR3DL2. Согласно одному варианту реализации указанный способ используют для выбора субъектов, имеющих раковое заболевание, и связанные с заболеванием клетки представляют собой раковые клетки. Согласно одному варианту реализации указанный способ используют для выбора субъектов, имеющих воспалительное или аутоиммунное расстройство, и связанные с заболеванием клетки представляют собой Т-клетки. Согласно одному варианту реализации антитело, которое приводят в контакт со связанными с заболеванием клетками для оценки того, связывается ли указанное антитело со связанными с заболеванием клетками, представляет собой антитело, описанное в настоящем документе.
Также предложен способ лечения пациента, отличающийся тем, что указанный способ включает:
a) определение того, имеются ли у пациента патогенные экспрессирующие KIR3DL2 клетки, и
b) в том случае, если установлено, что у пациента имеются патогенные экспрессирующие KIR3DL2 клетки, введение антигенсвязывающего соединения (например, антитело) согласно настоящему описанию.
Также предложен способ оценки уровня развития кожной Т-клеточной лимфомы CTCL (стадирование заболевания), позволяющий определить долю (например, процент) злокачественных клеток CTCL CD4+, присутствующих в определенной части организма пациента. В соответствии с указанным способом клетки из биологического образца, полученного из указанной части организма, приводят в контакт с антителом против KIR3DL2 согласно настоящему описанию, и измеряют долю клеток CD4+, экспрессирующих на поверхности полипептид KIR3DL2. Долю клеток CTCL CD4+, фактически присутствующих в указанной части организма, можно считать по существу равной указанной измеренной доле, например, в диапазоне ±10% от указанной измеренной доли.
Также предложен способ диагностики CTCL, включающий приведение клеток из биологического образца индивидуума в контакт с антителом против KIR3DL2 согласно настоящему описанию, и измеряют долю (например, процент) Т-клеток, экспрессирующих на поверхности полипептид KIR3DL2, и сравнение такой доли со средней долей (например, процентом) Т-клеток, экспрессирующих на поверхности полипептид KIR3DL2, наблюдаемой у людей, не страдающих CTCL (предпочтительно, у здоровых людей), при этом положительный по CTCL диагноз устанавливают в том случае, если измеренная доля существенно выше, чем указанная средняя доля.
Указанные и дополнительные благоприятные аспекты и признаки настоящего изобретения могут быть дополнительно описаны в других разделах настоящего описания.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг. 1 приведено изображение полипептида KIR3DL2, включая фрагменты в составе домена D0, демонстрирующее мутации остатков аминокислот, обозначенные как «Мутант 1», «Мутант 2», «Мутант 3» и «Мутант 6», которые приводили (в различных комбинациях) утрате связывания антителами.
На фиг. 2 приведено изображение полипептида KIR3DL2, включая фрагменты в составе домена D0, демонстрирующее мутации остатков аминокислот, обозначенные как «Мутант 1», «Мутант 2» и «Мутант 3»; мутанты 1, 2 и 6 обуславливают (в различных комбинациях) утрату связывания антителами 10F6, 2В12, 18С6, 9Е10, 10G5 и 13Н1, с выделением цветом остатков, смежных с остатками (F9, S11, Р14, F34 и/или S140, смежные с мутантом 2, и G21, G22, Н23, Е57, S58, F59, Р63 и/или Н68, смежные с мутантом 1).
На фиг. 3 приведено изображение каждой поверхности полипептида KIR3DL2, включая фрагменты в составе домена D0, демонстрирующее мутации остатков аминокислот, обозначенные как «Мутант 6», которые приводили к утрате связывания антителом 5Н1; показан «Мутант 3», не приводивший к утрате связывания. Также выделены цветом остатки, смежные с остатками, смежными с мутантом 6, которые также могут связываться антителами (K7, Y30, R31, Р79, Н80, S81, Т83, G84, W85, S86 и/или А87).
На фиг. 4 приведено изображение полипептида KIR3DL2, включающее фрагменты в составе домена D2 (граница D1/D2), демонстрирующее мутации остатков аминокислот обозначенные как «Мутант 14», с которыми утрачено связывание антител 1С3 и 20Е9, и «Мутант 12» и «Мутант 17», которые не приводили к утрате связывания антителами; также выделены цветом остатки, смежные с остатками (Q201, K202, Р203, S204, S224, S225, S227, S228, N252, R253 и/или Т254, смежные с мутантом 14).
На фиг. 5 приведено изображение полипептида KIR3DL2, включая фрагменты в составе домена D2 (граница D1/D2), демонстрирующее мутации остатков аминокислот, обозначенные как «Мутант 15», с которым у антитела 20Е9 утрачено связывание; также выделены цветом остатки, смежные с остатками (D230, I231, R244, L245, R246, А247, V248, S275, R277 и/или Р280), смежные с мутантом 14).
На фиг. 6 показана способность антител опосредовать КЗЦ; антитела mAb против KIR3DL2, связывающие домен D0, обозначены серым цветом, антитела, которые связывают домен D1 - черным цветом; видно, что в случае исходных антител mAb мыши наиболее заметное влияние на КЗЦ оказывает изотип указанных mAb.
На фиг. 7 показано, что интернализация KIR3DL2 при связывании полностью устраняет способность mo19H12 к киллингу B221-KIR3DL2 за счет активации комплемента, тогда как в температурных условиях, ограничивающих интернализацию, отчетливо наблюдается КЗЦ-активность mo19H12.
На фиг. 8 показана способность гибридных mAb против KIR3DL2 опосредовать КЗЦ против B221-KIR3DL2 in vitro.
На фиг. 9 показана способность ряда mAb против KIR3DL2, протестированных в одной конечной концентрации (10 мкг/мл), к киллингу прототипичных клеток Сезари клеточной линии HUT78 за счет опосредованного АЗКЦ механизма.
На фиг. 10 показана способность mAb против KIR3DL2 к АЗКЦ-опосредованному киллингу трансфицированных KIR3DL2 клеток В221. Антитела mAb, обозначенные серым цветом, индуцируют интернализацию рецептора и, судя по всему, менее эффективны, чем 4 других антитела mAb, которые не индуцируют интернализацию KIR3DL2.
На фиг. 11 представлено сравнение в эксперименте с подбором дозы способности гибридизованных антител mAb huIgG1 против KIR3DL2 опосредовать АЗКЦ против экспрессирующих KIR3DL2 целевых клеток В221.
На фиг. 12 представлены результаты эксперимента (n=6 мышей NOD-SCID на группу), в котором эффективность трех антител мыши изотипа IgG2b против KIR3DL2, 9Е10 и 19Н12, тестировали на п/к ксенотрансплантатах В221-KIR3DL2. Неинтернализующееся антитело против D0 9E10 обеспечивало повышенную выживаемость по сравнению как с ФСБ, так и с интернализующимся антителом против D1 19H12.
На фиг. 13 представлены результаты другого эксперимента (n=6 NOD-SCID мышей на группу) в котором тестировали эффективность антитела мыши против KIR3DL2 19H12 против п/к ксенотрансплантатов RAJI-KIR3DL2. hi vitro трансфицированные KIR3DL2 клетки RAJI демонстрировали меньшую интернализацию при связывании mAb, чем клеточные линии В221-KIR3DL2 или Сезари. В модели с ксенотрансплантатом RAJI-KIR3DL2 mAb mo 19H12 было более эффективным, чем в модели B221-KIR3DL2. Это обусловлено менее эффективной интернализацией мишени in vivo.
На фиг. 14 показано, что антитела к домену D0 KIR3DL2 подавляют связывание HLA-A3 и димера тяжелых цепей В27 (В272). Репрезентативное FACS-окрашивание демонстрирует эффект антител против D0 KIR3DL2 на связывание тетрамера HLA-A3 и B272 с трансдуцированными KIR3DL2 клетками Baf3. (Репрезентативное изображение из одного из трех независимых экспериментов).
На фиг. 15 показано, что антитела против доменов D1 и D2 (антитело 1С3) KIR3DL2 подавляют связывание HLA-A3, но не димера тяжелых цепей В27 (В272). Репрезентативное FACS-окрашивание демонстрирует эффект антител против D1/D2 KIR3DL2 на связывание тетрамера HLA-A3 и В272 с трансдуцированными KIR3DL2 клетками Baf3. (Репрезентативное изображение из одного из трех независимых экспериментов).
На фиг. 16А показано, что антитела к домену D0 KIR3DL2 подавляют связывание тетрамера HLA-A3 и димера тяжелых цепей В27. Фиг. 16В: mAb против домена D2 (1С3) подавляет связывание тетрамера HLA-A3, но не димера тяжелой цепи В27 (В272). Результаты выражены как % от окрашивания тетрамера в присутствии контрольного изотипа MAb.
На фиг. 17 показано, что антитела к домену D0 KIR3DL2, но не к домену D1/D2 подавляют секретирование ИЛ-2 репортерной клеткой KIR3DL2 CD3e, стимулированной экспрессирующими HLA-B27 В-клеточными линиями (221В27). Антитела к D0 подавляют синтез ИЛ-2 за счет репортерных клеток, стимулированных В-клеточными линиями, экспрессирующими контрольный HLA класса 1, в меньшей степени, чем клетки, стимулированные HLA-B27. Представлены репрезентативные ИФА ELISA для синтеза ИЛ-2 в одном из трех независимых экспериментов.
На фиг. 18 приведено изображение полипептида KIR3DL2 аллеля *001, включая сайт связывания антитела, соответствующий мутанту 2 с заменами I60N и G62S в составе домена D0 (например, сайт связывания антител 2В12, 10F6, 18С10, 10G5 и 13Н1). Также на фигуре показаны аминокислотные различия между аллелем *001 и аллелями *002, *004, *006/*007, *008 и *009 KIR3DL2.
На фиг. 19 приведено альтернативное изображение полипептида KIR3DL2 аллеля *001, включая сайт связывания антитела, соответствующий мутанту 2 с заменами I60N и G62S в составе домена D0. Также на фигуре показаны аминокислотные различия между аллелем *001 и аллелями *005 и *003/*011 KIR3DL2.
На фиг. 20 приведены два альтернативных изображения (вид спереди и сзади) полипептида KIR3DL2 аллеля *001, включая сайт связывания антитела, соответствующий мутанту 2 с заменами I60N и G62S в составе домена D0. Также на фигуре показаны аминокислотные различия между аллелем *001 и аллелем *004KIR3DL2.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Введение
Антитела согласно настоящему описанию способны к прямому специфическому направленному действию на экспрессирующие KIR3DL2 клетки, в частности, Т-клетки CD4+, KIR3DL2+, без направленного действия на другие клетки, такие как клетки KIR3DL1+ (или клетки KIR3DL2+ KIR3DL1+, клетки KIR3DS1+; или клетки KIR3DS1 KIR3DL2+), и не интернализуются в клетки KIR3DL2+. Также предложены антитела, которые подавляют или не подавляют связывание природных лигандов KIR3DL2 (или лиганд-индуцированную KIR3DL2-сигнализацию). В настоящем описании предложен ряд антител, обладающих такими свойствами, которые конкурируют между собой за связывание с областью KIR3DL2+, которая включает домены 0 и 2, ограниченные аминокислотными остатками 1-98 и 193-292, соответственно, зрелых полипептидов KIR3DL2 последовательности SEQ ID NO: 1.
KIR3DL2 (CD158k) представляет собой дисульфид-связанный гомодимер трехдоменных молекул Ig, размером приблизительно 140 кДа, описанный в источнике: Pende et al. (1996) J. Exp. Med. 184: 505-518, содержание которого включено в настоящий документ посредством ссылки. KIR3DL1 (CD158e1) представляет собой мономерную молекулу размером приблизительно 70 кДа, описанную в: Colonna and Samaridis (1995) Science 268 (5209), 405-408; HLA-связывающий карман был описан Vivian et al. (2011) Nature 479: 401-405. Природные лиганды KIR3DL2 включают в том числе полипептиды HLA-A и HLA-B, в частности, HLA-A3 и HLA-A11 (см. Hansasuta et al. (2004) Eur. J. Immunol. 34: 1673-1679 и HLA-B27. HLA-B27 (см., например, информацию относительно организации, последовательности и экспрессии гена HLA-B27 у Weiss et al. (1985), Immunobiology 170(5): 367-380, и информацию относительно мультимеров HLA-B27 и гомодимеров HLA-B272 у Alien et al. (1999), J. Immunol. 162: 5045-5048, и Kollnberger et al. (2007), Eur. J. Immunol. 37: 1313-1322. Содержание всех вышеуказанных источников включено в настоящий документ посредством ссылки. В настоящем документе «KIR3D» относится к любому рецептору KIR3D (например, KIR3DL1, KIR3DL2, KIR3DS1) по отдельности или в совокупности, и термин «KIR3D» может быть заменен на термин «KIR3DL1, KIR3DL2 и/или KIR3DS1». Аналогичным образом, «KIR3DL» относится к любому рецептору KIR3DL (например, KIR3DL1, KIR3DL2) по отдельности или в совокупности, и термин «KIR3DL» может быть заменен на термин «KIR3DL1 и/или KIR3DL2». Каждый из терминов «KIR3D», «KIR3DL», «KIR3DL1», «KIR3DL2», «KIR3DS1» также включает любой вариант, производное или изоформу гена KIR3D или кодируемого(ых) белка(ов), к которому(ым) они относятся. Было описано несколько аллельных вариантов для полипептидов KIR3D (например, KIR3DL2), каждый из которых охвачен соответствующими терминами. Аминокислотная последовательность зрелого KIR3DL2 человека (аллель *002) представлена в последовательности SEQ ID NO: 1, соответствующей № доступа в Genbank AAB52520, в которой отсутствует лидерная последовательность размером 21 аминокислотный остаток, и соответствующей № доступа в базе IPD KIR (публикуемой EMBL-EBI, European Bioinformatics Institute, Великобритания) KIR00066. кДНК KIR3DL2 (аллель *002) приведена в Genbank под № доступа U30272. Аминокислотная последовательность предшественника (включающая лидерную последовательность) KIR3DL2 человека аллеля *002 представлена в последовательности SEQ ID NO: 159, соответствующей № доступа в Genbank AAB52520. Аминокислотная последовательность KIR3DL2 человека аллеля *001 представлена в последовательности SEQ ID NO: 160, соответствующей № доступа в базе данных IPD KIR KIR00065. Аминокислотная последовательность KIR3DL2 человека аллеля *003 представлена в последовательности SEQ ID NO: 161, соответствующей № доступа в Genbank AAB36593 и № доступа в базе данных IPD KIR KIR00067. Аминокислотная последовательность KIR3DL2 человека аллеля *004 представлена в последовательности SEQ ID NO: 162, соответствующей № доступа в базе данных IPD KIR KIR00068. Аминокислотная последовательность KIR3DL2 человека аллеля *005 представлена в последовательности SEQ ID NO: 163, соответствующей № доступа в базе данных IPD KIR KIR00069. Аминокислотная последовательность KIR3DL2 человека аллеля *006 (зрелого) представлена в последовательности SEQ ID NO: 164, соответствующей № доступа в Genbank AAK30053 и № доступа в базе данных IPD KIR KIR00070. Аминокислотная последовательность KIR3DL2 человека аллеля *007 (зрелого) представлена в последовательности SEQ ID NO: 165, соответствующей № доступа в Genbank AAK30052 и № доступа в базе данных IPD KIR KIR00071. Аминокислотная последовательность KIR3DL2 человека аллеля *008 представлена в последовательности SEQ ID NO: 166, соответствующей № доступа в Genbank AAK30054 и № доступа в базе данных IPD KIR KIR00072. Аминокислотная последовательность KIR3DL2 человека аллеля *009 представлена в последовательности SEQ ID NO: 167, соответствующей № доступа в базе данных IPD KIR KIR00457. Аминокислотная последовательность KIR3DL2 человека аллеля *011 представлена в последовательности SEQ ID NO: 168, соответствующей № доступа в базе данных IPD KIR KIR00544. кДНК, кодирующая полипептид KIR3DL1 (CD158e2) (аллель *00101), приведена в Genbank под № доступа L41269; кодируемая аминокислотная последовательность представлена в последовательности SEQ ID NO: 169, соответствующей № доступа в Genbank AAA69870. Если в конкретной последовательности SEQ ID NO, описывающей последовательность полипептида KIR3DL2 (например, последовательностях SEQ ID NO: 1 и 159-168) присутствует лидерная последовательность, любое упоминание положений остатков аминокислот в настоящем документе относится к зрелому полипептиду KIR3DL.
Предложены способы применения антигенсвязывающих соединений; например, способ подавления клеточной пролиферации или активности, для доставки молекулы в клетку (например, токсической молекулы, детектируемого маркера и т.д.), для таргетинга, идентификации или очищения клетки, для истощения, киллинга или устранения клетки, для снижения клеточной пролиферации, при этом указанный способ включает воздействие на клетку, такую как Т-клетка, которая экспрессирует полипептид KIR3DL, антигенсвязывающим соединением согласно настоящему описанию, связывающим полипептид KIR3DL2. Следует понимать, что «клеточная пролиферация», для целей описываемого изобретения, может относиться к любому аспекту роста или пролиферации клеток, например, росту клетки, делению клетки или любому аспекту клеточного цикла. Указанная клетка может присутствовать в культуре клеток (in vitro) или в организме млекопитающего (in vivo), например, млекопитающего, страдающего связанной с экспрессией KIR3DL2 патологией. Также предложен способ индуцирования клеточной смерти или подавления пролиферации или активности клетки, которая экспрессирует полипептид KIR3DL2, включающий воздействие на указанную клетку антигенсвязывающим соединением, которое связывает полипептид KIR3DL2, соединенный с токсическим агентом, в количестве, эффективном для индуцирования смерти и/или подавления пролиферации указанной клетки. Соответственно, предложен способ лечения млекопитающего, страдающего пролиферативным заболеванием и любым состоянием, характеризующимся патогенным размножением или активацией клеток, экспрессирующих полипептид KIR3DL2, отличающийся тем, что указанный способ включает введение фармацевтически эффективного количества раскрытого в настоящем документе антигенсвязывающего соединения млекопитающему. Примеры таких состояний включают синдром Сезари, фунгоидный микоз, CTCL; и аутоиммунные или воспалительные состояния, например, артрит, заболевание сердечно-сосудистой системы. Предпочтительно такие патогенно распространяющиеся клетки экспрессируют KIR3DL2, однако выражение не экспрессируют KIR3DL1 (например, KIR3DL1 экспрессируют не более чем 20%, 40%, 50% или 60% патогенных клеток, поскольку при указанных состояниях благоприятным является, в частности, применение селективных антител.
Лечение и диагностика ряда связанных с экспрессией KIR3DL2 расстройств, в частности, опосредованных Т-клетками и NK-клетками расстройств, могут проводиться с применением способов и композиций согласно настоящему описанию. Указанные расстройств могут представлять собой, например, CD4+ Т-клеточные злокачественные новообразования, например, CTCL, ФМ или СС, или аутоиммунные или воспалительные расстройства, при которых благоприятным было бы устранение или подавление активности и/или пролиферации Т- и/или NK-клеток. Т-клетки СЕМ+ включают, например, активированные Т-клетки CD4+, Th17 Т-клетки, Т-клетки CD4+, экспрессирующие или не экспрессирующие один или большее количество других маркеров (например, CD2+, CD3+, CD5+, CD8-, CD28+, CD28-, CD45RO+ и TCRαβ+). Известно, например, что CD4+ CD28- Т-клетки способны экспрессировать KIR3DL2 и с высокой частотой встречаются в клонально пролиферирующих клетках при некоторых аутоиммунных и воспалительных расстройствах, но редко у здоровых индивидуумов. Указанные Т-клетки могут быть цитотоксическими, секретировать значительные количества ИФН-гамма, и пролиферировать при стимуляции аутологичными адгезивными мононуклеарными клетками.
Антитела согласно настоящему описанию обладают преимуществом, заключающимся в связывании разных аллелей KIR3DL2, что позволяет широко применять их для лечения, характеризации и диагностики заболеваний. Сообщалось, что клетки ФМ/СС в коже и кровотоке не экспрессируют какие-либо предпочтительные аллели из девяти протестированных аллелей KIR3DL2. На сегодняшний день описано еще 13 аллелей. Хотя рецептор KIR3DL2 р140 экспрессируется в минорной субпопуляции NK-клеток и на редко встречающихся у здоровых людей Т-клетках CD8+, он, очевидно, ограничен Т-клетками CD4+ опухоли CTCL у пациентов с ФМ/СС. Другие рецепторы, которые, как правило, наблюдаются на поверхности NK-клеток (например, р58.1, р58.2, p70KIRs, CD94/NKG2A), не обнаруживаются на поверхности злокачественных CD4+ Т-клеток (Bahler D.W. et аl., (2008) Cytometry In Clin Cytom. 74(3): 156-62). Клетки СС также, как правило, характеризуются, помимо CD4+, фенотипом зрелого Т-лимфоцита, CD2+, CD3+, CD5+, CD8-, CD28+, CD45RO+ и TCRap+.
Способы и композиции согласно настоящему описанию могут применяться при лечении аутоиммунных и воспалительных состояний, характеризующихся экспрессией KIR3DL2, посредством устранения экспрессирующих KIR3DL2 клеток и/или посредством подавления биологической активности экспрессирующих KIR3DL2 клеток (т.е. посредством блокирования KIR3DL2-сигнализации, индуцируемой его природными лигандами). Подавление биологической активности экспрессирующих KIR3DL2 клеток может включать, например, уменьшение пролиферации экспрессирующих KIR3DL2 клеток, уменьшение реакционноспособности или цитотоксичности экспрессирующих KIR3DL2 клеток в отношении целевых клеток, уменьшение активации, подавление маркеров активации (например, экспрессии CD 107) и/или синтеза цитокинов (например, синтеза ИФН-γ) экспрессирующими KIR3DL2 клетками, и/или уменьшение частоты встречаемости in vivo таких активированных, реакционноспособных, цитотоксических и/или активированных экспрессирующими KIR3DL2 клеток.
Например, было показано, что ряд таких расстройств опосредован по меньшей мере отчасти Т-клетками CD4+, включая специфические Т-клетки CD4+CD28null. Обычно считается, что активация CD4+ Т-клеток управляется за счет взаимодействия стимулирующих и ингибирующих рецепторов, при этом преобладание стимулирующих сигналов способствует аутоиммунным реакциям. Chan с коллегами (Chan et al. ((2005) Arthrit. Rheumatism 52(11): 3586-3595) сообщают о повышенных количествах CD4+ Т-клеток и NK-клеток, экспрессирующих KIR3DL2, в периферической крови и синовиальной жидкости при спондилоартрите. У пациентов с ревматоидным артритом часто прекращается экспрессия важной костимулирующей молекулы CD28. Вместо этого популяция Т-клеток CD4+, где отсутствует CD28 (Т-клетки CD4+CD28-), экспрессирует иммуноглобулин-подобные рецепторы киллеров (KIR). В частности, сообщалось, что CD4+CD28null Т-клетки экспрессируют полипептиды KIR3D. По сравнению со своими CD28+-аналогами клетки CD4+CD28 - продуцируют значительно более высокие уровни ИФН-γ, что придает им способность функционировать в качестве провоспалительных клеток. Клоны Т-клеток CD4+CD28null сохраняются в кровотоке в течение многих лет. Известно, что указанные Т-клетки отличаются от CD28+ Т-клеток устойчивостью к Fas-опосредованному апоптозу при перекрестном связывании CD3. CD28null Т-клетки проходят клеточный цикл, и на всех стадиях клеточного цикла устойчивы к апоптозу, в отличие от их аналогов CD28*. Дисрегуляция путей апоптоза в Т-клетках CD4+CD28null, как было показано, способствует клональной экспансии и стабилизации in vivo. Namekawa et al. ((2000) J. Immunol. 165: 1138-1145 сообщают, что KIR, включая KIR3DL2, присутствовали на Т-клетках CD4+CD28null, экспансия которых происходит при ревматоидном артрите. При ревматоидном артрите задействованы лимфоцитарные инфильтраты, медиаторы воспаления и наблюдается синовиальная гиперплазия, обусловленная агрессивной пролиферацией фибробластоподобных синовиоцитов и макрофагов. Прогноз суставных эрозий и тяжесть заболевания коррелируют с высокими частотами встречаемости клонально пролиферирующих Т-клеток CD4+CD28-. Lamprecht с коллегами (Lamprecht et al. (2001) Thorax 56: 751-757) описывают рекрутинг Т-клеток CD4+CD28- при гранулематозе Вегенера. Markovic-Plese с коллегами (Markovic-Plese et al. (2001) J Clin Invest. 108: 1185-1194) сообщают о присутствии независимых от костимуляции Т-клеток CD4+CD28- в ЦНС и их связи с рассеянным склерозом. Указанные способы и композиции могут, соответственно, использоваться при лечении или предотвращении гранулематоза Вегенера, рассеянного склероза или других воспалительных или аутоиммунных расстройств центральной нервной системы, артрита или других ревматических расстройств, характеризующихся воспалением.
Т-клетки CD4+CD28- были также ассоциированы с сердечно-сосудистыми расстройствами. Betjes с коллегами (Betjes et al. (2008) Kidney International 74, 760-767) сообщают, что повышенный риск атеросклеротических заболеваний у серопозитивных по цитомегаловирусу (ЦМВ) пациентов связан с возрастным увеличением количества Т-клеток CD4+CD28-, которые могут составлять более половины циркулирующих Т-клеток CD4 у индивидуумов. Сообщалось, что у пациентов в возрасте старше 50 лет процент CD4+CD28- Т-клеток в 50 раз выше по сравнению с ЦМВ-серонегативными пациентами и в 5 раз выше по сравнению с серопозитивными здоровыми контролями. Nakajima с коллегами (Nakajima et al. ((2003) Circ. Res. 93: 106-113)) сообщают о de novo экспрессии KIR при остром коронарном синдроме, когда Т-клетки CD4+ от пациентов с острым коронарным синдромом (ОКС) экспрессируют несколько KIR, тогда как нормальные Т-клетки CD4+CD28null от здоровых доноров не экспрессируют KIR. Yen с коллегами (Yen et al. Journal of Experimental Medicine, Volume 193, Number 10, May 21, 2001 1159-1168) исследовали CD4+CD28null Т-клеточные клоны, полученные от пациентов с ревматоидным васкулитом, для экспрессии ингибирующих и стимулирующих KIR с применением ОТ-ПЦР. У пациентов с ревматоидным артритом и пациента с ОКС паттерны экспрессии благоприятствовали ингибирующим KIR, включая KIR3DL2, тогда как экспрессия стимулирующих рецепторов была выражение ограничена KIR2DS2. Указанные способы и композиции могут, соответственно, использоваться при лечении или предотвращении сердечно-сосудистых расстройств, например, ОКС, атеросклеротического заболевания, ревматоидного васкулита, характеризующегося воспалением.
Bowness с коллегами (Bowness et al (2011) J. Immunol. 186: 2672-2680) сообщают, что Т-клетки KIR3DL2+ CD4, на которые приходится большая часть экспрессии ИЛ-23Р Т-клетками CD4 периферической крови, и что такие клетки KIR3DL2+ Th17-типа продуцируют больше ИЛ-17 в присутствии ИЛ-23. Несмотря на то, что клетки KIR3DL2+ содержат в среднем только 15% Т-клеток CD4 в периферической крови пациентов с СпА, указанная субпопуляция отвечает за 70% наблюдаемого увеличения количества Th17 у пациентов с СпА по сравнению с контрольными субъектами. Стимулированные TCR Т-клеточные линии периферической крови KIR3DL2+ CD4 от пациентов с СпА секретировали в 4 раза больше ИЛ-17 по сравнению с линиями KIR3DL2+ от контролей или Т-клетками KIR3DL2- CD4.
Предложены способы получения и применения антител и других соединений, подходящих для лечения расстройств (например, раковых заболеваний, воспалительных и аутоиммунных расстройств), при которых благоприятным было бы устранение экспрессирующих KIR3DL2 клеток. Включены антитела, производные антител, фрагменты антител и продуцирующие их клетки, а также способы их получения и способы лечения пациентов с применением указанный антителам и соединений.
Поскольку предлагаемые антитела являются специфическими в отношении KIR3DL2, они могут применяться для различных целей, включая очищение KIR3DL2 или экспрессирующих KIR3DL2 клеток, модулирование (например, активацию или подавление) рецепторов KIR3DL2 in vitro, ex vivo или in vivo, направленное воздействие на экспрессирующие KIR3DL2 клетки для разрушения in vivo, или специфическое мечение/связывание KIR3DL2 in vivo, ex vivo, или in vitro, в том числе для таких методов, как иммуноблоттинг, иммуногистохимический анализ, а именно, биопсия замороженных срезов, анализ FACS и иммунопреципитация.
Определения
В настоящем описании термины, приведенные в единственном числе, могут означать «один или большее количество». В пункте(ах) формулы изобретения при использовании в сочетании с выражением «содержащий», термины в единственном числе могут означать один или количество, большее чем один. В настоящем документе «другой/еще один» может означать по меньшей мере второй или более.
При использовании термина «содержащий» он может предпочтительно заменяться на «состоящий по существу из», более предпочтительно на «состоящий из».
«Лечение пролиферативного заболевания» или «лечение опухоли», или «лечение раковых заболеваний» или т.п. в отношении связывающего анти-KIR3DL2 агента (например, антитела), включает, не ограничиваясь перечисленными: (а) способ лечения пролиферативного заболевания, при этом указанный способ включает этап введения (для по меньшей мере одного эпизода лечения) связывающего анти-KIR3DL2 агента, (например, в материале фармацевтически приемлемого носителя) теплокровному животному, в частности, человеку, нуждающемуся в таком лечении, в дозе, обеспечивающей лечение указанного заболевания (в терапевтически эффективном количестве), например, в дозе (количестве) согласно описанию выше и ниже в настоящем документе; (b) применение анти-KIR3DL2 связывающего агента для лечения пролиферативного заболевания, или анти-KIR3DL2 связывающего агента, для применения в указанном лечении (в частности, у человека); (с) применение анти-KIR3DL2 связывающего агента для получения фармацевтического состава для лечения пролиферативного заболевания, способ применения анти-KIR3DL2 связывающего агента для получения фармацевтического состава для лечения пролиферативного заболевания, включающий смешивание анти-KIR3DL2 связывающего агента с фармацевтически приемлемым носителем, или фармацевтического состава, содержащего эффективную дозу анти-KIR3DL2 связывающего агента, подходящего для лечения пролиферативного заболевания; или (d) любая комбинация а), b) и с), в соответствии с патентоспособностью предмета изобретения в стране подачи настоящей заявки. В случаях, когда упоминается конкретное заболевание (например, воспалительное или аутоиммунное заболевание) или конкретная опухоль (например, CTCL), а не «пролиферативное заболевание», категории а) - е) также включены, то есть в пунктах а) - е) выше может быть указано соответствующее заболевание вместо «пролиферативного заболевания», в соответствии с патентоспособностью предмета изобретения.
Термины «рак/раковое заболевание» и «опухоль» в настоящем документе определены как новообразование клеток или ткани, включающее неконтролируемое и прогрессирующее размножение. Согласно конкретному варианту реализации при естественном развитии событий раковое заболевание приводит к летальному исходу. Согласно конкретным вариантам реализации раковое заболевание является инвазивным, метастатическим и/или анапластическим (утрата дифференцировки и ориентации друг относительно друга и относительно осевого каркаса).
«Аутоиммунные» расстройства включают любое расстройство, состояние или заболевание, при котором иммунная система запускает реакцию против собственных клеток или тканей по причине разрушения способности различать собственные клетки от чужих, или др. Примеры аутоиммунных расстройств включают ревматоидный артрит, ревматоидный васкулит, системную красную волчанку, рассеянный склероз, гранулематоз Вегенера, спондилоартрит и др. «Воспалительное расстройство» включает любое расстройство, характеризующееся нежелательным иммунным ответом. Аутоиммунные и воспалительные расстройства могут задействовать любой компонент иммунной системы, и их мишенью может быть любой тип клеток или тканей организма.
Термин «биопсия» в настоящем документе определен как извлечение ткани из органа (например, сустава) с целью исследования, например, установления диагноза. Примеры типов биопсии включают применение аспирации, например, через присоединенную к шприцу иглу; инструментальное извлечение фрагмента ткани; извлечение с применением подходящих инструментов через эндоскоп; хирургическое удаление, например, повреждения полностью; и т.п.
Термин «антитело» в настоящем документе относится к поликлональным и моноклональным антителам. В зависимости от типа константного домена в тяжелых цепях антитело распределяют в один из пяти основных классов: IgA, IgD, IgE, IgG и IgM. Некоторые из них дополнительно подразделяются на подклассы или изотопы, такие как IgG1, IgG2, IgG3, IgG4 и т.п. Типовая структурная единица иммуноглобулина (антитела) содержит тетрамер. Каждый тетрамер состоит из двух идентичных пар полипептидных цепей, каждая пара содержит одну «легкую» (приблизительно 25 кДа) и одну «тяжелую» цепь (приблизительно 50-70 кДа). N-конец каждой цепи определяет вариабельную область длиной приблизительно 100-110 или более аминокислот, которая несет основную ответственность за распознавание антигена. Термины «вариабельная легкая цепь» (VL) и «вариабельная тяжелая цепь» (VH) относятся к указанным легкой и тяжелой цепям, соответственно. Указанные константные домены тяжелых цепей соответствуют разным классам иммуноглобулинов называют «альфа», «дельта», «эпсилон», «гамма» и «мю», соответственно. Структуры и трехмерные конфигурации субъединиц разных классов иммуноглобулинов хорошо известны. IgG и/или IgM представляют собой предпочтительные классы антител для применения в настоящем изобретении, в частности, предпочтительным являются IgG, поскольку они являются наиболее распространенными в физиологических условиях антителами, и поскольку их очень легко получить в лабораторных условиях. Предпочтительно, антитело представляет собой моноклональное антитело. В частности, предпочтительными являются гуманизированные антитела, гибридные антитела, антитела человека или другие подходящие для применения у человека антитела. «Антитела» также включают любой фрагмент или производное любого из описанных в настоящем документе антител.
Термин «специфически связывается» означает, что антитело может предпочтительно связываться в конкурентном анализе связывания с партнером для связывания, например, KIR3DL2, по оценке с применением любых рекомбинантных форм белков, их эпитопов или нативных белков, присутствующих на поверхности выделенных целевых клеток. Конкурентный анализ связывания и другие способы определения специфического связывания дополнительно описаны ниже и хорошо известны в данной области техники.
Если говорится, что антитело «конкурирует с» конкретным моноклональным антителом (например, 10F6, 2В12, 18С6, 9Е10, 10G5, 13Н1, 5Н1, 1Е2, 1С3 или 20Е9), это означает, что указанное антитело конкурирует с указанным моноклональным антителом в анализе связывания с применением либо рекомбинантных молекул KIR3DL2, либо экспрессируемых на поверхности молекул KIR3DL2. Например, если экспериментальное антитело снижает связывание 10F6, 2В12, 18С6, 9Е10, 10G5, 13Н1, 5Н1, 1Е2, 1С3 или 20Е9 с полипептидом KIR3DL2 или экспрессирующей KIR3DL2 клеткой в анализе связывания, говорят, что указанное антитело «конкурирует», соответственно, с 10F6, 2В12, 18С6, 9Е10, 10G5, 13Н1, 5Н1, 1Е2, 1С3 или 20Е9.
Термин «сродство/аффинность» в настоящем документе означает силу связывания антитела с эпитопом. Аффинность антитела задается константой диссоциации Kd, определяемой как [Ab]×[Ag]/[Ab-Ag], где [Ab-Ag] - молярная концентрация комплекса антитело-антиген, [Ab] - молярная концентрация несвязанного антитела, и [Ag] - молярная концентрация несвязанного антигена. Константа аффинности Kа определена как 1/Kd. Примеры способов определения аффинности моноклональных антител приведены в источнике: Harlow, et al., Antibodies: A Laboratory Manual («Антитела: лабораторное руководство»). Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1988), Coligan et al., eds., Current Protocols in Immunology, Greene Publishing Assoc. and Wiley biterscience, N.Y., (1992, 1993), и Muller, Mem. Enzymol. 92: 589-601 (1983), полностью включенных в настоящий документ посредством ссылки. Одним из стандартных хорошо известных в данной области техники способов определения аффинности mAb является применение скрининга на основе поверхностного плазменного резонанса (ППР) (например, путем анализа с применением аналитического устройства для ППР BIAcore™).
В настоящем документе «детерминанта» означает сайт взаимодействия или связывания на полипептиде.
Термин «эпитоп» относится к антигенной детерминанте и представляет собой участок или область на антигене, с которым(ой) связывается антитело. Белковый эпитоп может содержать остатки аминокислот, прямо вовлеченные в связывание, а также остатки аминокислот, эффективно блокируемые специфическим антигенсвязывающим антителом или пептидом, т.е., остатки аминокислот в составе «области узнавания» антитела. Это простейшая форма или минимальный структурный участок комплексной молекулы антигена, способный соединяться, например, с антителом или рецептором. Эпитопы могут быть линейными или конформационными/структурными. Термин «линейный эпитоп» определен как эпитоп, состоящий из смежных остатков аминокислот в линейной последовательности аминокислот (первичная структура). Термин «конформационный или структурный эпитоп» определен как эпитоп, состоящий из остатков аминокислот, не все из которых являются смежными и, соответственно, представляют собой отдельные фрагменты линейной последовательности аминокислот, приводимые в непосредственную близость друг относительно друга за счет укладки молекулы (вторичные, третичные и/или четвертичные структуры). Конформационный эпитоп зависит от 3-мерной структуры. Термин «конформационный», соответственно, часто используются взаимозаменяемо с термином «структурный».
Термин «внутриклеточная интернализация», или «интернализация», в отношении полипептида KIR3DL2 и/или антитела, которое его связывает, относится к молекулярным, биохимическим и клеточным событиям, ассоциированным с процессом перемещения молекулы с внеклеточной поверхности клетки на внутриклеточную поверхность клетки. Процессы, отвечающие за внутриклеточную интернализацию молекул, хорошо известны и могут включать, в том числе, интернализацию внеклеточных молекул (таких как гормоны, антитела и малые органические молекулы); мембраносвязанных молекул (таких как рецепторы клеточной поверхности); и комплексов мембраносвязанных молекул, связанных с внеклеточными молекулами (например, лиганд, связанный с трансмембранным рецептором, или антитело, связанное с мембраносвязанной молекулой). Соответственно, «индуцирование и/или увеличение внутриклеточной интернализации» включает события, при которых инициируется внутриклеточная интернализация и/или увеличивается скорость и/или степень внутриклеточной интернализации.
Термин «истощение» в отношении экспрессирующих KIR3DL2 клеток означает процесс, способ или соединение, способное к киллингу, элиминированию, лизису, или способное индуцировать такой киллинг, элиминацию или лизис, с негативным влиянием на количество экспрессирующих KIR3DL2 клеток, присутствующих в образце или у субъекта.
Термин «агент» используется в настоящем документе для обозначения химического соединения, смеси химических соединений, биологической макромолекулы или экстракта, полученного из биологических материалов. Термин «терапевтический агент» относится к агенту, обладающему биологической активностью.
Термины «токсический агент» и «цитотоксический агент» охватывают любое соединение, способное замедлять, останавливать или обращать пролиферацию клеток, снижать их активность любым детектируемым образом, или прямо или непрямо обеспечивать их гибель. Предпочтительно, цитотоксические агенты вызывают клеточную смерть главным образом за счет непосредственного действия на функционирование клетки, и включают, не ограничиваясь перечисленными, алкилирующие агенты, ингибиторы фактора некроза опухоли, интеркаляторы, ингибиторы микротрубочек, ингибиторы киназ, ингибиторы протеасомы и ингибиторы топоизомеразы. «Токсический груз» в настоящем документе относится к достаточному количеству цитотоксического агента, который при введении в клетку приводит к клеточной смерти. Доставка токсического груза может быть реализована путем введения достаточного количества иммуноконъюгата, содержащего антитело или антигенсвязывающий фрагмент, и цитотоксический агент. Доставка токсического груза может также быть реализована путем введения достаточного количества иммуноконъюгата, содержащего цитотоксический агент, при этом указанный иммуноконъюгат содержит вторичное антитело или его антигенсвязывающий фрагмент, который распознает и связывает антитело или антигенсвязывающий фрагмент.
Для целей настоящего изобретения «гуманизированное» антитело или антитело «человека» относится к антителу, в котором константная и вариабельная каркасная область одного или большего количества иммуноглобулинов человека соединена с участком связывания, например, CDR, животного иммуноглобулина. Такие антитела сконструированы таким образом, чтобы сохранить специфичность связывания не принадлежащего человеку антитела, из которого получены области связывания, но при этом избежать иммунной реакции против указанного не принадлежащего человеку антитела. Такие антитела могут быть получены от трансгенных мышей или других животных, «сконструированных» для продуцирования специфических антител человека в ответ на антигенную стимуляцию (см., например, источник Green et al. (1994) Nature Genet 7: 13; Lonberg et al. (1994) Nature 368: 856; Taylor et al. (1994) fat Immun 6: 579, содержание которого полностью включено в настоящий документ посредством ссылки). Полностью человеческое антитело также может быть сконструировано с применением методов генной или хромосомной трансфекции, а также технологии фагового дисплея, известных в данной области техники (см., например, McCafferty et al. (1990) Nature 348: 552-553). Антитела человека могут также быть синтезированы in vitro активированными В-клетками (см., например, патенты США №5567610 и 5229275, которые полностью включены в настоящий документ посредством ссылки).
«Гибридное антитело» представляет собой молекулу антитела, в которой(а) константная область или ее фрагмент изменен(а), заменен(а) или рекомбинирован(а) таким образом, что антигенсвязывающий сайг (вариабельная область) соединен с константной областью другого(ой) или измененного(ой) класса, эффекторной функции и/или вида, или с совершенно отличной молекулой, придающей новые свойства гибридному антителу, например, ферментом, токсином, гормоном, фактором роста, лекарственным веществом и т.д.; или (b) вариабельная область или ее фрагмент изменен или заменен на вариабельную область, обладающую другой или измененной антигеноспецифичностью.
Термины «домен Fc», «фрагмент Fc» и «область Fc» относятся к С-концевому фрагменту тяжелой цепи антитела, например, приблизительно от аминокислоты (АК) 230 до приблизительно 450 АК. тяжелой цепи γ (гамма) человека или аналогичной последовательности других типов тяжелых цепей антител (например, α, δ, ε и μ для антител человека) или их встречающемуся в природе аллотипу. Если не указано иное, для иммуноглобулинов с настоящем описании используют общепринятую нумерацию аминокислот по Kabat (см. Kabat et al. (1991). Sequences of Protein of Immunological Interest, 5th ed., United States Public Health Service, National Institute of Health, Bethesda, MD)
Термин «антителозависимая клеточноопосредованная цитотоксичность», или «АЗКЦ», представляет собой хорошо известный в данной области техники термин, и относится к клеточноопосредованной реакции, в ходе которой неспецифические цитотоксические клетки, экспрессирующие рецепторы Fc (FcR), распознают связанное антитело на целевой клетке, и затем вызывают лизис целевой клетки. Неспецифические цитотоксические клетки, опосредующие АЗКЦ, включают естественные киллерные клетки (NK), макрофаги, моноциты, нейтрофилы и эозинофилы.
Термины «выделенный», «очищенный» или «биологически чистый» относятся к материалу, который по существу не содержит компонентов, которые обычно присутствуют в нем в естественном состоянии. Чистоту и гомогенность, как правило, определяют с применением техник аналитической химии, таких как электрофорез в полиакриламидном геле или высокоэффективная жидкостная хроматография. Белок, представляющий собой основное присутствующее в составе вещество, по существу очищен.
Термины «полипептид», «пептид» и «белок» используются в настоящем документе взаимозаменяемо и относятся к полимеру остатков аминокислот. Указанные термины относятся к полимерам аминокислот, в которых один или большее количество остатков аминокислот представляют собой искусственный химический миметик соответствующей встречающейся в природе аминокислоты, а также к встречающимся в природе полимерам аминокислот и не встречающемуся в природе полимеру аминокислот.
Термин «рекомбинантные» при использовании в отношении, например, клетки или нуклеиновой кислоты, белка или вектора указывает на то, что указанная(ый) клетка, нуклеиновая кислота, белок или вектор был(а) модифицирован(а) путем введения гетерологичной нуклеиновой кислоты или белка, или изменения нативной(ого) нуклеиновой кислоты или белка, или что указанная клетка получена из таким образом модифицированной клетки. Соответственно, например, рекомбинантные клетки экспрессируют гены, отсутствующие в нативной (нерекомбинантной) форме клетки или экспрессируют нативные гены, которые в ином случае экспрессируются аномально, зкспрессируются недостаточно или не экспрессируются.
Термин «модификация» в отношении последовательности аминокислот (например, «модификация аминокислоты») подразумевает замену аминокислоты, вставку и/или удаление в последовательности полипептида. Под «модификацией» или «модификацией аминокислоты» подразумевается замена аминокислоты, вставка и/или удаление в последовательности полипептидов. Под «заменой аминокислоты» или «заменой» в настоящем документе подразумевается замена аминокислоты в определенном положении в последовательности белка другой аминокислотой. Например, замена P14S относится к варианту исходного полипептида, в котором пролин в положении 14 заменен на серии. «Вариант» полипептида относится к полипептиду, имеющему аминокислотную последовательность, по существу идентичную референтному полипептиду, как правило, нативному или «исходному» полипептиду. Вариант полипептида может содержать одну или большее количество замен, удалений и/или вставок аминокислот в определенных положениях в составе нативной последовательности аминокислот.
В настоящем документе «Т-клетки» относится к субпопуляции лимфоцитов, созревающих в вилочковой железе и экспонирующих на поверхности, помимо других молекул, Т-клеточные рецепторы. Т-клетки могут быть идентифицированы по определенным характеристикам и биологическим свойствам, таким как экспрессия конкретных поверхностных антигенов, включая TCR, CD4 или CD8, необязательно CD4 и ИЛ-23R, способность определенных Т-клеток к киллингу опухолевых или инфицированных клеток, способность определенных Т-клеток активировать другие клетки иммунной системы и способность высвобождать молекулы белков, называемых цитокинами, которые стимулируют или подавляют иммунный ответ. Любые из указанных характеристик и активностей могут применяться для идентификации Т-клеток, с применением способов, хорошо известных в данной области техники. В настоящем документе «активными» или «активированными» Т-клетками называются биологически активные Т-клетки, конкретнее, Т-клетки, способные к цитолизу или стимуляции иммунного ответа посредством, например, секреции цитокинов. Активные клетки могут быть детектированы с применением любого из множества хорошо известных способов, включая функциональный анализ и способы анализа, основанные на экспрессии, например, экспрессии цитокинов, таких как ФНО-альфа или ИЛ-17А.
В настоящем документе термин «антитело, связывающее» полипептид или эпитоп, означает антитело, которое связывает указанную детерминанту со специфичностью и/или аффинностью.
Антитела и эпитопы
Раскрытые антитела представляют собой антитела, которые связывают KIR3DL2 человека. Согласно варианту реализации указанные антитела селективно связывают KIR3DL2 (например, 1, 2, 3, 4 или большее количество наиболее распространенных аллелей KIR3DL2) и не связывают KIR3DL1 (например, 1, 2, 3, 4 или большее количество наиболее распространенных аллелей KIR3DL1). Согласно одному варианту реализации указанные антитела связывают домен D0 KIR3DL2, соответствующий остаткам аминокислот 1-98 полипептида KIR3DL2 последовательности SEQ ID NO: 1. Согласно одному варианту реализации указанные антитела связывают домен D2 KIR3DL2, или область, расположенную в обоих доменах Din D2 (на границе доменов D1 и D2) полипептида KIR3DL2, последовательности SEQ ID NO: 1. Согласно одному варианту реализации указанные антитела имеют аффинность к KIR3DL2 человека, характеризующуюся KD, составляющей менее чем 10-9 М, предпочтительно менее чем 10-10 M.
Согласно другому варианту реализации указанные антитела связывают по существу тот же самый эпитоп, что и антитело 10F6, 2В12, 18С6, 9Е10, 10G5, 13Н1, 5Н1, 1Е2, 1С3 или 20Е9. Согласно другому варианту реализации антитела по меньшей мере частично перекрывается с сегментом, или включает по меньшей мере один остаток из сегмента, соответствующего остаткам 1-98 или остаткам 193-292 полипептида KIR3DL2 последовательности SEQ ID NO: 1 (или ее субпоследовательности). Согласно одному варианту реализации все ключевые остатки эпитопа расположены в сегменте, соответствующем остаткам 1-98. Согласно одному варианту реализации указанное антитело связывает остаток, присутствующий в домене D1, а также остаток, присутствующий в домене D2; необязательно один или большее количество ключевых остатков расположено на границе доменов D1 (остатки 99-192) и D2 (остатки 193-292). Согласно одному варианту реализации указанные антитела связывают эпитоп, содержащий 1, 2, 3, 4, 5, 6, 7 или большее количество остатков в сегменте, соответствующем остаткам 1-98, 99-292, 99-192 или 193-292 полипептида KIR3DL2 последовательности SEQ ID NO: 1. Предпочтительно указанные остатки, связанные антителом, располагаются на поверхности полипептида KIR3DL2.
Согласно одному варианту реализации указанные антитела связывают эпитоп, содержащий остатки R13, А25, и/или Q27. Необязательно указанные антитела связывают эпитоп, содержащий остатки R13, А25, и/или Q27, а также остатки 160 и/или G62. Необязательно указанные антитела не связывают остатки Н32 и/или Н33. Необязательно указанные антитела также связывают остатки Q56 и/или Е57.
Согласно одному варианту реализации указанные антитела связывают эпитоп, содержащий остатки 160 и/или G62. Необязательно указанные антитела связывают эпитоп, содержащий один или большее количество остатков 160 и/или G62, но не остатки R13, А25, и/или Q27.
Согласно одному варианту реализации указанные антитела связывают эпитоп, содержащий один или большее количество остатков 160 и/или G62, а также один или большее количество остатков Р14, S15 и/или Н23. Необязательно указанные антитела связывают эпитоп, содержащий 1, 2, 3, 4, 5, 6 или 7 остатков G21, G22, Н23, Е57, S58, F59, Р63 и/или Н68.
Необязательно указанные антитела связывают эпитоп, содержащий один или большее количество остатков R78 и/или L82. Необязательно указанные антитела связывают эпитоп, содержащий 1, 2, 3, 4, 5, 6 или 7 остатков K7, Y30, R31, Р79, Н80, S81, Т83, G84, W85, S86 и/или А87.
Необязательно указанные антитела связывают эпитоп, содержащий остаток W226. Необязательно указанные антитела связывают эпитоп, содержащий 1, 2, 3, 4, 5, 6 или 7 остатков Q201, K202, Р203, S204, S224, S225, S227, S228, N252, R253 и/или Т254.
Необязательно указанные антитела связывают эпитоп, содержащий один или большее количество остатков I231 и/или R246. Необязательно указанные антитела связывают эпитоп, содержащий остатки I231 и/или R246, а также эпитоп, содержащий остаток W226. Необязательно указанные антитела связывают эпитоп, содержащий 1, 2, 3, 4, 5, 6 или 7 остатков D230, I231, R244, L245, R246, А247, V248, S275, R277 и/или Р280.
Необязательно указанные антитела связывают эпитоп, содержащий остаток Е239. Необязательно указанные антитела также связывают один или большее количество остатков I231 и/или R246. Необязательно указанные антитела также связывают остаток W226.
В разделе «Примеры» настоящего документа описано конструирование серии мутантных полипептидов KIR3DL2 человека. Измеряли связывание антитела против KIR3DL2 с клетками, трансфицированными мутантными KIR3DL2, и сравнивали со способностью антитела против KIR3DL2 связывать полипептид KIR3DL2 дикого типа (SEQ ID NO: 1). Снижение связывания между антителом против KIR3DL2 и мутантным полипептидом KIR3DL2 в настоящем документе означает, что имеется снижение аффинности связывания (например, согласно оценке с применением известных способов, таких как тестирование клеток, экспрессирующих конкретный мутант, с применением FACS, или с помощью теста Biacore на связывание с мутантными полипептидами) и/или снижение общей связывающей способности указанного антитела против KIR3DL2 (например, по уменьшению Вшах на графике зависимости концентрации антитела против KIR3DL2 от концентрации полипептида). Значительное снижение связывания указывает на то, что мутантный остаток прямо вовлечен в связывание с антителом против KIR3DL2 или находится в непосредственной близости к связывающему белку при связывании антитела против KIR3DL2 с KIR3DL2. Эпитоп антитела, соответственно, предпочтительно включает такой остаток и может включать дополнительные остатки, смежные с таким остатком.
Согласно некоторым вариантам реализации значительное снижение связывания означает, что аффинность и/или способность к связыванию антитела против KIR3DL2 и мутантного полипептида KIR3DL2 снижается более чем на 40%, более чем на 50%, более чем на 55%, более чем на 60%, более чем на 65%, более чем на 70%, более чем на 75%, более чем на 80%, более чем на 85%, более чем на 90% или более чем на 95% относительно связывания между указанным антителом и полипептидом KIR3DL2 дикого типа (например, полипептидом, представленным в последовательности SEQ ID NO: 1). Согласно определенным вариантам реализации связывание снижается до уровня ниже порога детекции. Согласно некоторым вариантам реализации значительное снижение связывания наблюдается, когда связывание антитела против KIR3DL2 с мутантным полипептидом KIR3DL2 составляет менее чем 50% (например, менее чем 45%, 40%, 35%, 30%, 25%, 20%, 15% или 10%) от связывания, наблюдаемого между антителом против KIR3DL2 и полипептидом KIR3DL2 дикого типа (например, внеклеточным доменом, представленным в последовательности SEQ ID NO: 1). Такие измерения связывания могут быть выполнены с применением разнообразных анализов связывания, известных в данной области техники. Конкретный пример одного такого анализа описан в разделе «Примеры».
Согласно некоторым вариантам реализации предложены антитела против KIR3DL2, демонстрирующие значительно более слабое связывание с мутантным полипептидом KIR3DL2, в котором заменен остаток полипептида KIR3DL2 дикого типа (например, SEQ ID NO: 1). В настоящем документе используются сокращенные обозначения следующего формата: остаток дикого типа: положение в полипептиде: мутантный остаток, с нумерацией остатков согласно приведенной в последовательности SEQ ID NO: 1.
Согласно некоторым вариантам реализации антитело против KIR3DL2 связывает полипептид KIR3DL2 дикого типа, однако отличается сниженным связыванием с мутантным полипептидом KIR3DL2, содержащим любую одну или несколько следующих мутаций (относительно SEQ ID NO: 1):
R13W, А25Т и/или G25R;
I60N и/или G62S;
P14S, S15A и/или H23S;
что-либо одно или более из R13W, А25Т и/или G25R, и что-либо одно или более из I60N и/или G62S;
что-либо одно или более из P14S, S15A и/или H23S, и что-либо одно или более из I60N и/или G62S;
один или большее количество R13W, А25Т и/или G25R, что-либо одно или более из I60N и/или G62S; и что-либо одно или более из P14S, S15A и/или H23S;
что-либо одно или более из P14S. S15A и/или H23S, и что-либо одно или более из I60N и/или G62S;
R78H и/или L82P;
W226A;
I231M и/или R246P;
что-либо одно или более из I231M и/или R246P, и дополнительно W226A; или
что-либо одно или более из I231M и/или R246P, но при этом связывание указанного антитела с мутантом, содержащим мутацию W226A, не понижено.
Предпочтительно связывание с конкретным(и) мутантом(ами) KIR3DL2 значительно понижено по сравнению со связыванием с KIR3DL2 дикого типа.
Получение антител против KIR3DL2
Антитела могут быть получены при помощи разнообразных техник, известных в данной области техники. Как правило, их получают путем иммунизации не являющегося человеком животного, предпочтительно, мыши, иммуногеном, содержащим полипептид KIR3DL2, предпочтительно, полипептид KIR3DL2 человека. Полипептид KIR3DL2 может включать последовательность полноразмерного полипептида KIR3DL2 человека или его фрагмент или производное, как правило, иммуногенный фрагмент, т.е. фрагмент полипептида, содержащий эпитоп, экспонируемый на поверхности клеток, экспрессирующих полипептид KIR3DL2, предпочтительно, эпитоп, распознаваемый антителом 10F6, 2В12, 18С6, 9Е10, 10G5, 13Н1, 5Н1, 1Е2, 1С3 или 20Е9. Такие фрагменты, как правило, содержат по меньшей мере приблизительно 7 последовательных аминокислот последовательности зрелого полипептида, еще более предпочтительно - по меньшей мере приблизительно 10 последовательных аминокислот последовательности зрелого полипептида. Фрагменты, как правило, получены по существу из внеклеточного домена рецептора. Согласно одному варианту реализации иммуноген содержит полипептид KIR3DL2 человека дикого типа в липидной мембране, как правило, на поверхности клетки. Согласно конкретному варианту реализации иммуноген содержит интактные клетки, в частности, интактные клетки человека, необязательно обработанные или лизированные. Согласно другому варианту реализации полипептид представляет собой рекомбинантный полипептид KIR3DL2. Согласно конкретному варианту реализации иммуноген содержит интактные клетки СС или ФМ, в частности интактные злокачественную Т-клетки CD4+ или CD4+ CD28- Т-клетки человека, необязательно обработанные или лизированные. Согласно другому варианту реализации полипептид представляет собой рекомбинантный димерный полипептид KIR3DL2.
Этап иммунизации не являющегося человеком млекопитающего антигеном может проводиться любым хорошо известным в данной области техники способом стимуляции продуцирования антител у мыши (см., например, Е. Harlow and D. Lane, Antibodies: A Laboratory Manual., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (1988), полностью включенный в настоящий документ посредством ссылки). Указанный иммуноген суспендируют или растворяют в буфере, необязательно с адъювантом, таким как полный или неполный адъювант Фрейнда. Способы определения количества иммуногена, типов буферов и количеств адъюванта хорошо известны специалистам в данной области техники. Указанные параметры могут быть разными для разных иммуногенов, но могут быть легко установлены.
Аналогичным образом, локализация и частота иммунизации, достаточные для стимуляции продуцирования антител, также хорошо известны в данной области техники. Согласно типичному протоколу иммунизации не являющимся человеком животным внутрибрюшинно инъецируют антиген на 1 день и повторно приблизительно через неделю. После этого проводят вторичные инъекции антигена приблизительно на 20 день, необязательно с адъювантом, таким как неполный адъювант Фрейнда. Вторичные инъекции выполняются внутривенно и могут повторяться в течение нескольких последовательных дней. После этого проводят бустер-инъекцию на 40 день, внутривенно или внутрибрюшинно, как правило, без адъюванта. Указанный протокол обеспечивает образование продуцирующих антиген-специфическое антитело В-клеток приблизительно через 40 дней. Могут использоваться и другие протоколы, при условии, что они обеспечивают образование В-клеток, экспрессирующих антитело, направленное на антиген, используемый для иммунизации.
Для получения поликлональных антител получают сыворотку от иммунизированного не являющегося человеком животного, и выделяют присутствующие в ней антитела с применением общеизвестных техник. Сыворотка может быть очищена аффинным методом с применением любого из представленных выше иммуногенов, связанных с твердой подложкой, для получения антител, которые вступают в реакцию с полипептидами KIR3DL2.
Согласно альтернативному варианту реализации лимфоциты неиммунизированного не являющегося человеком млекопитающего выделяют, культивируют in vitro и затем подвергают воздействию иммуногена в культуре клеток. Затем лимфоциты собирают и проводят описанный ниже этап слияния.
В случае моноклональных антител следующий этап заключается в выделении спленоцитов иммунизированного не являющегося человеком млекопитающего и последующем слиянии указанных спленоцитов с иммортализированной клеткой для создания продуцирующей антитело гибридомы. Выделение спленоцитов не являющегося человеком млекопитающего общеизвестно в данной области техники и, как правило, включает извлечение селезенки анестезированного не являющегося человеком млекопитающего, измельчение на небольшие фрагменты и продавливание спленоцитов из капсулы селезенки через нейлоновое сито клеточного фильтра в подходящий буфер для получения суспензии отдельных клеток. Клетки промывают, центрифугируют и ресуспендируют в буфере, лизирующем все эритроциты. Раствор повторно центрифугируют и затем оставшиеся лимфоциты в осадке ресуспендируют в свежем буфере.
После выделения и получения в виде суспензии отдельных клеток лимфоциты могут быть слиты с клетками иммортализированной линии. Как правило, это клеточная линия миеломы мыши, хотя в данной области техники известны и многие другие подходящие для создания гибридом иммортализированные клеточные линии. Линии миеломы мыши включают, не ограничиваясь перечисленными, полученные из опухолей мышей МОРС-21 и МРС-11 доступны от Salk Institute Cell Distribution Center, Сан-Диего, США, клетки Х63 Ag8653 и SP-2 доступны от American Type Culture Collection, Роквилл, Мэриленд, США. Слияние реализуют с применением полиэтиленгликоля или т.п. Затем полученные гибридомы культивируют на селективной среде, которая содержит одно или большее количество веществ, подавляющих рост или выживание неслитых исходных клеток миеломы. Например, если в исходных клетках миеломы отсутствует фермент гипоксантингуанинфосфорибозилтрансфераза (HGPRT, или ГФРТ), культуральная среда для гибридом, как правило, включают гипоксантин, аминоптерин и тимидин (среда HAT), вещества, предотвращающие рост дефицитных по HGPRT клеток.
Гибридомы, как правило, культивируют на фидерном слое макрофагов. Макрофаги предпочтительно получены от однопометных особей не являющихся человеком млекопитающих, которых использовали для выделения спленоцитов, и, как правило, примируют неполным адъювантом Фрейнда или т.п. за несколько дней до высевания гибридом. Способы слияния описаны в источнике: Goding, «Monoclonal Antibodies: Principles and Practice» («Моноклинальные антитела: теория и практика»), pp. 59-103 (Academic Press, 1986), содержание которого включено в настоящий документ посредством ссылки.
Клетки культивируют на селективной среде в течение времени, достаточного для образования колоний и продуцирования антитела. Обычно указанный период составляет от приблизительно 7 до приблизительно 14 дней.
Затем в колониях гибридом анализируют продуцирование антител, которые специфически связываются с полипептидными генными продуктами KIR3DL2, необязательно эпитопом, специфически распознаваемым антителом 10F6, 2В12, 18С6, 9Е10, 10G5, 13Н1, 5Н1, 1Е2, 1С3 или 20Е9. Указанный анализ представлен, как правило, колориметрическим ИФА-анализом ELISA, хотя может применяться любой анализ, который может быть адаптирован для лунок, в которых культивируют гибридомы. Другие анализы включают радиоиммуноанализ или сортировку клеток с активацией флуоресценции. Лунки, положительные по продукции нужного антитела, исследуют для определения присутствия одной или большего количества отдельных колоний. При наличии более чем одной колонии клетки могут быть повторно клонированы и культивированы, чтобы гарантировать, что колония, продуцирующая нужное антитело, образована единственной клеткой.
Гибридомы, синтез подходящего моноклонального антитела в которых подтвержден, могут культивироваться в больших количествах на подходящей среде, такой как DMEM или RPMI-1640. Как вариант, клетки гибридомы могут культивироваться in vivo в виде асцитных опухолей у животного.
После культивирования, достаточного для продуцирования нужного моноклонального антитела, ростовую среду, содержащую моноклональное антитело (или асцитную жидкость), отделяют от клеток, и присутствующее в ней моноклональное антитело очищают. Очищение, как правило, проводят с применением гель-электрофореза, диализа, хроматографии на сефарозе с белком А или белком G, или антимышиного Ig, связанного с твердой подложкой, например, агарозными или сефарозными гранулами (все указанные методы описаны, например, в пособии: Antibody Purification Handbook («Руководство по очистке антител»), Biosciences, publication No. 18-1037-46, Edition AC, содержание которого включено в настоящий документ посредством ссылки). Связанное антитело, как правило, элюируют из колонок с белком А/белком G с применением буферов с низкими значениями рН (глициновый или ацетатный буферы с рН 3,0 или ниже) с немедленной нейтрализацией содержащей антитело фракции. Указанные фракции объединяют, диализируют и концентрируют подходящим образом.
Положительные лунки с одиночной видимой колонией, как правило, повторно клонируют и анализируют, чтобы гарантировать детекцию и продукцию единственного моноклонального антитела.
Антитела могут также получены отбором из комбинаторных библиотек иммуноглобулинов, согласно описанию, например, в (Ward et al. Nature, 341 (1989) p. 544, все содержание которого полностью включено в настоящий документ посредством ссылки).
Идентификация одного или большего количества антител, которые связываются с KIR3DL2, в частности, связывают по существу тот же эпитоп, что и моноклональное антитело 10F6, 2В12, 18С6, 9Е10, 10G5, 13Н1, 5Н1, 1Е2, 1С3 или 20Е9, может быть легко осуществлена с применением любого из разнообразных иммунологических методов скрининга, с помощью которых можно оценить конкуренцию антител. Многие из таких методов анализа являются общеупотребительными и хорошо известны в данной области техники (см., например, патент США №5660827, который прямо включен в настоящий документ посредством ссылки). Следует понимать, что фактически определение эпитопа, с которым связывается описанное в настоящем документе антитело, ни в коей мере не является необходимым для идентификации антитела, которое связывается с тем же или по существу тем же самым эпигоном, что и моноклональное антитело, описанное в настоящем документе.
Например, если экспериментальные антитела для анализа получены от различных животных, или даже если они принадлежат к разным изотипам Ig, может применяться простой конкурентный анализ, в ходе которого контрольное антитело (например, в целях иллюстрации, 10F6 или любое другое антитело, такое как 2В12, 18С6, 9Е10, 10G5, 13Н1, 5Н1, 1Е2, 1С3 или 20Е9) и экспериментальные антитела смешивают (или предадсорбируют) и наносят на образец, содержащий полипептиды KIR3DL2. Протоколы, основанные на вестерн-блоттинге и применении анализа BIACORE, подходят для применения в таких методах конкурентного анализа.
Согласно определенным вариантам реализации смешивают контрольные антитела (например, 10F6, хотя подходят и любые другие антитела) с варьирующими количествами экспериментальных антител (например, приблизительно 1:10 или приблизительно 1:100) на протяжении некоторого периода времени до нанесения на образец с антигеном KIR3DL2. Согласно другим вариантам реализации контрольные и варьирующие количества экспериментальных антител могут просто смешиваться при контакте с образцом антигена KIR3DL2. При условии, что связанные антитела могут быть отличены от свободных (например, с применением техник разделения или отмывки для устранения несвязанных антител) и (10F6 от экспериментальных антител (например, с применением видоспецифических или изотип-специфических вторичных антител или специфического мечения 10F6 детектируемой меткой), можно определить, снижают ли экспериментальные антитела связывание 10F6 с антигенами, что указывает на то, что экспериментальное антитело распознает по существу тот же самый эпитоп, что и 10F6. Связывание (меченых) контрольных антител в отсутствие полностью нерелевантного антитела может служить в качестве контрольного высокого значения. Контрольное низкое значение может быть получено путем инкубирования меченых (10F6) антител с немечеными антителами точно такого же типа (10F6), при котором наблюдается конкуренция и снижение связывания меченых антител. Значительное снижение реактивности меченого антитела в присутствии экспериментального антитела в экспериментальном анализе указывает на то, что экспериментальное антитело распознает по существу тот же самый эпитоп, т.е. «вступает в перекрестную реакцию» или конкурирует с меченым (10F6) антителом. Любое экспериментальное антитело, которое снижает связывание 10F6 с антигенами KIR3DL2 по меньшей мере приблизительно на 50%, например, по меньшей мере приблизительно на 60% или более, предпочтительно, по меньшей мере приблизительно на 80% или 90% (например, приблизительно на 65-100%), при любой пропорции 10F6: экспериментальное антитело приблизительно от 1:10 и приблизительно до 1:100, считают антителом, которое связывается по существу с тем же самым эпитопом или детерминантой, что и 10F6. Предпочтительно, такое экспериментальное антитело снижает связывание 10F6 с антигеном KIR3DL2 по меньшей мере приблизительно на 90% (например, приблизительно на 95%).
Конкуренция также может оцениваться, например, с применением проточно-цитометрического теста. В ходе такого теста клетки, несущие определенный полипептид KIR3DL2, могут инкубироваться сначала, например, с 10F6, а затем с экспериментальным антителом, меченым флуорохромом или биотином. Говорят, что антитело конкурирует с 10F6, если связывание, наблюдаемое при преинкубации с насыщающим количеством 10F6, составляет приблизительно 80%, предпочтительно приблизительно 50%, приблизительно 40% или менее (например, приблизительно 30%, 20% или 10%) от связывания (согласно оценке среднего флуоресценции), наблюдаемого для указанного антитела без преинкубации с 10F6. Как вариант, говорят, что антитело конкурирует с 10F6, если связывание, наблюдаемое с меченым антителом 10F6 (флуорохромом или биотином) на клетках, проинкубированных с насыщающим количеством экспериментального антитела, составляет приблизительно 80%, предпочтительно приблизительно 50%, приблизительно 40%, или менее (например, приблизительно 30%, 20% или 10%) от связывания, наблюдаемого без преинкубации с экспериментальным антителом.
Может также применяться простой конкурентный анализ, в ходе которого экспериментальное антитело предадсорбируют и наносят в насыщающей концентрации на поверхность, на которой иммобилизован антиген KIR3DL2. Поверхность при простом конкурентном анализе представлена предпочтительно чипом BIACORE (или другой средой, подходящей для анализа методом поверхностного плазменного резонанса). Затем контрольное антитело (например, 10F6) приводят в контакт с поверхностью в насыщающей KIR3DL2 концентрации и измеряют связывание с KIR3DL2 и поверхностное связывание контрольного антитела. Указанное связывание контрольного антитела сравнивают со связыванием контрольного антитела с содержащей KIR3DL2 поверхностью в отсутствие экспериментального антитела. В экспериментальном анализе значительное снижение связывания содержащей KIR3DL2 поверхности контрольным антителом в присутствии экспериментального антитела указывает на то, что указанное экспериментальное антитело распознает по существу тот же самый эпитоп, что и контрольное антитело, так что экспериментальное антитело «вступает в перекрестную реакцию» с контрольным антителом. Любое экспериментальное антитело, которое снижает связывание контрольного (например, 10F6) антитела с антигеном KIR3DL2 по меньшей мере приблизительно на 30% или более, предпочтительно приблизительно на 40%, может считаться антителом, которое связывается по существу с тем же эпитопом или детерминантой, что и контрольное (например, 10F6). Предпочтительно, такое экспериментальное антитело снижает связывание контрольного антитела (например, 10F6) с антигеном KIR3DL2 по меньшей мере приблизительно на 50% (например, по меньшей мере приблизительно на 60%, по меньшей мере приблизительно на 70%, или более). Следует понимать, что очередность контрольного и экспериментального антител может быть обратной: то есть сначала с поверхностью может быть связано контрольное антитело, а экспериментальное антитело приводят в контакт с поверхностью позже в ходе конкурентного анализа. Предпочтительно, антитело, имеющее более высокое аффинность к антигену KIR3DL2, связывается с поверхностью первым, поскольку можно ожидать, что наблюдаемое снижение связывания второго антитела (при предположении, что антитела вступают в перекрестную реакцию) будет иметь большую величину. Дополнительные примеры таких анализов приведены, например, в Saunal (1995) J. Immunol. Methods 183: 33-41, содержание которого включено в настоящий документ посредством ссылки.
Предпочтительно, моноклональные антитела, которые распознают эпитоп KIR3DL2, вступают в реакцию с эпитопом, который присутствует на существенном проценте или даже на всех релевантных клетках, например, на злокачественных Т-клетках CD4+, клетках пациента с СС или ФМ, однако не вступают в выраженную реакцию с другими клетками, т.е. клетками, которые не экспрессируют KIR3DL2. Согласно одному аспекту антитела против KIR3DL2 связывают KIR3DL2, но не связывают KIR3DL1 и/или KIR3DS1.
Согласно некоторым вариантам реализации антитела связываются с экспрессирующими KIR3DL2 клетками индивидуума или индивидуумов с заболеванием, характеризующимся экспрессированием положительных по KIR3DL2 клеток, т.е. индивидуума, являющегося кандидатом для лечения одним из описанных в настоящем документе способов с применением антитела против KIR3DL2. Соответственно, после получения антитела, которое специфически распознает KIR3DL2 на клетках, оно может быть протестировано на способность связываться с положительными по KIR3DL2 клетками (например, злокачественными Т-клетками CD4+), полученными от пациента с таким расстройством, как СС или ФМ. В частности, перед лечением пациента одним из предложенных антител целесообразно проведение тестирования способности указанного антитела связываться со злокачественными клетками, полученными от указанного пациента, например, в образце крови, чтобы максимизировать вероятность того, что терапия у указанного пациента будет благоприятной.
Согласно одному варианту реализации антитела валидируют с применением иммуноанализа для тестирования их способности связываться с экспрессирующими KIR3DL2 клетками, например, злокачественными Т-клетками CD4+, провоспалительными клетками CD4+. Например, получают от группы пациентов лимфоциты периферической крови (ЛПК), и указанные ЛПК обогащают по Т-клеткам CD4+, например, посредством проточной цитометрии с применением релевантных антител (информацию о злокачественных клетках CD4+ можно найти, например, в источнике Bagot et al. (2001) Blood 97: 1388-1391, содержание которого включено в настоящий документ посредством ссылки), или выделяют фракции клеток CD4+CD28- с применением магнитной сепарации на колонке MACS (Miltenyi Biotec). Затем способность определенного антитела связываться с клетками оценивают с применением стандартных способов, хорошо известных специалистам в данной области техники. Антитела, которые, как обнаружено, связываются с существенной долей (например, 20%, 30%, 40%, 50%, 60%, 70%, 80% или более) клеток, которые, как известно, экспрессируют KIR3DL2, например, Т-клеток, у значительного процента индивидуумов или пациентов (например, 5%, 10%, 20%, 30%, 40%, 50% или более), подходят для применения в настоящем документе, как для целей диагностики, для определения присутствия или уровня злокачественных Т-клеток у пациента, так и для применения описанных в настоящем документе терапевтических способов, например, для увеличения или уменьшения числа или активности злокачественных Т-клеток. Для оценки связывания антител с указанными клетками может проводиться прямое или непрямое мечение указанных антител. При непрямом мечении, как правило, добавляют вторичное меченое антитело. Связывание антител с указанными клетками после этого можно детектировать с применением, например, цитофлуориметрического анализа (например, FACScan). Такие способы хорошо известны специалистам в данной области техники.
Определение того, связывается ли антитело в области эпитопа, может быть проведено способами, известными специалисту в данной области техники. Согласно одному из примеров таких способов картирования/характеризации область эпитопа для антитела против KIR3DL2 может быть определена путем «футпринтинга» эпитопа с применением химической модификации экспонируемых аминов/карбоксилов белка KIR3DL2. Один из конкретных примеров такой техники футпринтинга представлен применением HXMS (детектируемый с помощью масс-спектрометрии водородно-дейтериевый обмен), при котором происходит водородно-дейтериевый обмен, связывание и обратный обмен амидных протонов между белками рецептора и лиганда, при этом амидные группы остова, участвующие в связывании белка, защищены от обратного обмена и, соответственно, остаются дейтерированными. На указанном этапе релевантные области могут быть идентифицированы посредством пептического протеолиза, быстрым разделением с применением высокоэффективной жидкостной хроматографии на микроколонке и/или масс-спектрометрии с ионизацией электрораспылением. См., например, Ehring H, Analytical Biochemistry, Vol. 267 (2) pp. 252-259 (1999), Engen, J.R. and Smith, D. L. (2001) Anal. Chem. 73, 256A - 265A. Другой пример подходящей техники идентификации эпитопов представлен картированием эпитопов с применением ядерного магнитного резонанса (ЯМР), при котором, как правило, сравнивают расположение сигналов в двумерном спектре ЯМР свободного антигена и антигена в комплексе с антигенсвязывающим пептидом, таким как антитело. Антиген, как правило, селективно метят изотопом 15N, так что в спектре ЯМР видны только сигналы, соответствующие указанному антигену, и не видны сигналы от антигенсвязывающего пептида. Расположение в спектре комплекса сигналов антигена, происходящих от аминокислот, вовлеченных во взаимодействие с антигенсвязывающим пептидом, как правило, сдвинуто относительно расположения в спектре свободного антигена, и таким образом могут быть идентифицированы аминокислоты, вовлеченные в связывание. См., например, Ernst Sobering Res Found Workshop. 2004; (44): 149-67; Huang et al. Journal of Molecular Biology, Vol. 281 (1) pp. 61-67 (1998); и Saito and Patterson, Methods. 1996 Jun; 9 (3): 516-24.
Картирование/характеризация эпитопов также может быть проведено с применением методов масс-спектрометрии. См., например. Downward, J Mass Spectrom. 2000 Apr; 35 (4): 493-503 и Kiselar and Downard, Anal Chem. 1999 May 1; 71 (9): 1792-801. В контексте картирования и идентификации эпитопов также может быть целесообразным использование техник расщепления протеазами. Релевантные области/последовательности антигенных детерминант могут быть определены с помощью расщепления протеазами, например, с применением трипсина в пропорции, составляющей приблизительно 1:50 к KIR3DL2, или расщеплением в течение ночи при рН 7-8, с последующим масс-спектрометрическим (МС) анализом для идентификации пептидов. Пептиды, защищенные анти-KIR3DL2 связывающим агентом от расщепления трипсином могут затем быть идентифицированы путем сравнения образцов, подвергаемых расщеплению трипсином, и образцов, инкубированных с антителом, а затем обработаны, например, трипсином для расщепления (выявляя таким образом область узнавания для связывающего агента). Дополнительно или в качестве альтернативного варианта в аналогичных способах характеризации эпитопов могут применяться другие ферменты, такие как химотрипсин, пепсин и т.д. Кроме того, ферментативное расщепление может обеспечить способ быстрого анализа для определения того, входит ли потенциальная последовательность антигенной детерминанты в состав области полипептида KIR3DL2, которая не экспонируется на поверхности и, соответственно, скорее всего, не является значимой в отношении иммуногенности/антигенности. См., например, Manca, Ann 1st Super Sanita. 1991; 27: 15-9, где обсуждаются аналогичные техники.
Сайт-специфический мутагенез представляет собой другую подходящую технику для исследования связывания эпитопов. Например, при «аланиновом сканировании» каждый остаток в пределах сегмента белка заменяется на остаток аланина, и оцениваются последствия в отношении аффинности связывания. Если мутация приводит к значительному снижению аффинности связывания, остаток, скорее всего, участвует в связывании. Могут применяться моноклональные антитела, специфические в отношении структурных эпитопов (т.е. антитела, которые не связывают белок без укладки) для проверки того, что замена на аланины не влияет на общую укладку белка. См., например, Clackson and Wells, Science 1995; 267: 383-386; и Wells, Proc Nati Acad Sci USA 1996; 93: 1-6.
Для «футпринтинга» эпитопов может также использоваться электронная микроскопия. Например, Wang et al., Nature 1992; 355: 275-278 скоординированным образом применяли криоэлектронную микроскопию, трехмерную реконструкцию изображений и рентгеновскую кристаллографию для определения физической области узнавания Fab-фрагмента на поверхности капсида природного вируса мозаики коровьего гороха.
Другие формы анализа «без меток» для исследования эпитопов включают поверхностный плазменный резонанс (ППР, BIACORE) и рефлектометрическую интерференционную спектроскопию (RifS). См., например, Fagerstam et al., Journal Of Molecular Recognition 1990; 3: 208-14; Nice et al., J. Chromatogr. 1993; 646: 159-168; Leipert et al., Angew. Chem. Int. Ed. 1998; 37: 3308-3311; Kroger et al., Biosensors and Bioelectronics 2002; 17: 937-944.
Следует также отметить, что антитело, связывающее тот же или по существу тот же самый эпитоп, что и антитело, описанное в настоящем документе, могут быть идентифицированы в ходе одного или нескольких видов конкурентного анализа, примеры которых описаны в настоящем документе.
Необязательно оценивают поглощение клетками или локализацию в клетках для выбора антитела, которое легко поглощается клеткой и/или попадает в клеточный компартмент, где присутствует KIR3DL2. Поглощение клетками или локализацию в клетках, как правило, измеряют в клетках, в которых требуется или предполагается наличие активности антитела. Поглощение клетками или локализация в клетках могут оцениваться с использованием стандартных способов, например, конфокальной микроскопии с окрашиванием с применением антитела, меченого детектируемым фрагментом (например, флуоресцентным фрагментом).
После иммунизации и получения антител у позвоночного животного или в клетке могут быть реализованы конкретные этапы отбора для выделения заявленных антител. Соответственно, согласно конкретному варианту реализации предложены способы получения таких антител, включающие: (а) иммунизацию не являющегося человеком млекопитающего иммуногеном, содержащим полипептид KIR3DL2; и (b) получение антител от указанного иммунизированного животного; и (с) выбор антител из полученных на этапе (b), которые способны связывать KIR3DL2.
Как правило, антитело против KIR3DL2 в настоящем документе обладает аффинностью к полипептиду KIR3DL2 в диапазоне от приблизительно 104 до приблизительно 1011 М-1 (например, от приблизительно 108 до приблизительно 1010 М-1). Например, антитело может иметь среднюю константу диссоциации (Kd), составляющую менее чем 1×10-9 М в отношении KIR3DL2, согласно оценке, полученной, например, путем скрининга с применением поверхностного плазменного резонанса (ППР) (например, анализа на аналитическом устройстве для ППР BIAcore™). Согласно примеру, соответствующему более конкретному аспекту, антитело может иметь Kd в отношении KIR3DL2, составляющую от приблизительно 1×10-8 М до приблизительно 1×10-10 М, или от приблизительно 1×10-9 М до приблизительно 1×10-11 М.
Антитела могут характеризоваться, например, средним значением Kd, не превышающим приблизительно 100, 60, 10, 5, или 1 нМ, предпочтительно в субнаномолярном диапазоне, или, необязательно, не более чем (т.е. обладать аффинностью, более высокой, чем) приблизительно 500, 200, 100 или 10 пМ. Kd может быть определена, например, путем иммобилизации полученных рекомбинантным образом белков KIR3DL2 человека на поверхности чипа, с последующим добавлением указанного подлежащего тестированию антитела в раствор. Согласно одному варианту реализации указанный способ также включает этап (d) выбора антител из полученных на этапе (b), способных конкурировать за связывание KIR3DL2 с антителом 10Р6, 2В12, 18С6, 9Е10, 10G5, 13Н1, 5Н1, 1Е2, 1С3 или 20Е9.
Согласно одному аспекту любого из вариантов реализации антитела, получаемые в соответствии с представленными способами, представляют собой моноклональные антитела. Согласно другому аспекту не являющееся человеком животное, используемое для продуцирования антител в соответствии со способами, предложенными в настоящем изобретении, представляет собой млекопитающее, например, грызуна, крупный рогатый скот, свинью, птицу, лошадь, кролика, козу или овцу. Антитела включают 10F6, 2В12, 18С6, 9Е10, 10G5, 13Н1, 5Н1, 1Е2, 1С3 или 20Е9. Дополнительно может быть указано, что антитела необязательно представлены отличными от любых из антител Q241 и Q66 (Pende et al. (1996) J Exp Med 184: 505-518), клона 5.133 (Miltenyi Biotec). «AZ158» (Parolini, S., et al. (2002), Leucocyte typing VII. D. Mason, editor. Oxford University Press, Oxford. 415-417, и WO 2010/081890 (например, антител, содержащих вариабельную область тяжелой и легкой цепи, соответствующую последовательностям SEQ ID NO: 8 и 10 из WO 2010/081890), или их производных, например, включающих антигенсвязывающую область полностью или частично.
В соответствии с альтернативным вариантом реализации ДНК, кодирующую антитело, которое связывает эпитоп, присутствующий на полипептидах KIR3DL2, выделяют из гибридомы и помещают в подходящий экспрессионный вектор для трансфицирования подходящего хозяина. Затем указанный хозяин используется для рекомбинантного получения антитела или его вариантов, таких как гуманизированная версия указанного моноклонального антитела, активные фрагменты указанного антитела, гибридные антитела, содержащие распознающий антиген фрагмент указанного антитела, или версии, содержащие детектируемый фрагмент.
ДНК, кодирующая моноклональные антитела, например, антитело 10F6, 2В12, 18С6, 9Е10, 10G5, 13Н1, 5Н1, 1Е2, 1С3 или 20Е9, может быть легко выделена и секвенирована с применением стандартных процедур (например, с применением олигонуклеотидных зондов, способных специфически связываться с генами, кодирующими тяжелую и легкую цепи антител мыши). После выделения ДНК может быть помещена в экспрессионные векторы, которыми затем трансфицируют клетки-хозяева, такие как клетки Е.coli, клетки COS обезьяны, клетки яичника китайского хомячка (СНО) или клетки миеломы, которые в иных случаях не продуцируют иммуноглобулиновый белок, чтобы обеспечить синтез моноклональных антител в указанных рекомбинантных клетках-хозяевах, согласно описанию в других разделах настоящего документа такие последовательности ДНК могут быть модифицированы для любых из широкого ряда возможных целей, например, для гуманизации антител, получения фрагментов или производных, или для модификации последовательности указанного антитела, например, в антигенсвязывающем сайте, для оптимизации специфичности связывания указанного антитела.
Рекомбинантная экспрессия в бактериях ДНК, кодирующей антитело, хорошо известна в данной области техники (см., например, Skerra et al., Curr. Opinion in Immunol, 5, pp. 256 (1993); и Pluckthun, Immunol. 130, p. 151 (1992).
Оценка активности
После получения антигенсвязывающего соединения, как правило, проводят оценку его способности интернализоваться в экспрессирующие KIR3DL2 целевые клетки или вызывать интернализацию KIR3DL2 экспрессирующими KIR3DL2 целевыми клетками для индуцирован™ АЗКЦ или КЗЦ, направленной на подавление провоспалительной активности и/или пролиферации, и/или вызывать элиминацию экспрессирующих KIR3DL2 целевых клеток. Оценка способности антигенсвязывающего соединения интернализоваться или индуцировать АЗКЦ, КЗЦ, или в целом приводить к элиминации или подавлению активности экспрессирующих KIR3DL2 целевых клеток, может проводиться на любом подходящем этапе реализации способа, например, согласно примерам, приведенным в настоящем документе. Проведение указанной оценки может быть целесообразным на одном или нескольких различных этапах, задействованных при идентификации, получении и/или усовершенствовании антитела (или другого соединения), предназначенного для терапевтического применения. Например, активность может оцениваться в контексте способа скрининга для идентификации кандидатных антигенсвязывающих соединений, или в способах, где выбирают антигенсвязывающее соединение и модифицируют для обеспечения возможности применения у человека (например, в случае антитела, получают гибридное или гуманизированное антитело), когда получают клетку, экспрессирующую антигенсвязывающее соединение (например, клетку-хозяина, экспрессирующую рекомбинантное антигенсвязывающее соединение), и оценивают ее способность продуцировать функциональные антитела (или другие соединения), и/или когда получено некоторое количество антигенсвязывающего соединения и необходимо оценить его активность (например, для тестирования партий или серий продукта). Как правило, известно, что указанное антигенсвязывающее соединение специфически связывается с полипептидом KIR3DL2. Указанный этап может включать тестирование множества (например, очень значительного количества, с применением высокопроизводительных методов скрининга, или меньшего количества) антигенсвязывающих соединений.
В настоящем документе антитело против KIR3DL2, которое не «интернализуется», представляет собой соединение, которое по существу не поглощается клеткой (т.е. не попадает в клетку) при связывании с KIR3DL2 на клетке млекопитающего (т.е. KIR3DL2 клеточной поверхности). Неинтернализующиеся антитела, разумеется, включают фрагменты антител, антитело человека или гуманизированное антитело, и конъюгаты антител.
Установить, интернализируется ли антитело против KIR3DL2 при связывании KIR3DL2 на клетке млекопитающего, или подвергается ли внутриклеточной интернализации полипептид KIR3DL2 (например, при связывании антителом) можно с помощью различных анализов, включая описанные в настоящем документе в экспериментальных примерах. Например, для тестирования интернализации in vivo экспериментальное антитело метят и вводят животному, у которого, как известно, происходит экспрессирование KIR3DL2 на поверхности определенных клеток. Указанное антитело может, например, содержать радиоактивную метку, флуоресцентные частицы или частицы золота. Животные, подходящие для указанного анализа, включают таких млекопитающих, как мыши «nude» с экспрессирующим KIR3DL2 трансплантатом или ксенотрансплантатом опухоли человека, или мыши, которым были введены клетки, трансфицированные KIR3DL2 человека, или трансгенные мыши, экспрессирующие трансген KIR3DL2 человека. Подходящие контроли включают животных, не получавших экспериментальное антитело или получавших постороннее антитело, и животных, получавших антитело к другому антигену на представляющих интерес клетках, которое, как известно, интернализируется при связывании с указанным антигеном. Указанное антитело может вводиться животному, например, путем внутривенной инъекции. Через подходящие интервалы времени могут быть получены срезы тканей животного с применением известных способов или согласно описанию ниже в экспериментальных примерах, и проведен анализ интернализации, а также локализации интернализованного антитела в клетке с применением световой микроскопии или электронной микроскопии. Для интернализации in vitro клетки могут инкубироваться в чашках для тканевых культур в присутствии или в отсутствие релевантных антител, добавляемых в культуральную среду, и подготовлены для микроскопического анализа в требуемые моменты времени. Присутствие интернализованного меченого антитела в клетках может быть прямо визуализировано посредством микроскопии или ауторадиографии при использовании радиоактивно меченого антитела. Необязательно при микроскопии может оцениваться колокализация с известным полипептидом или другим клеточным компонентом; например, колокализация с эндосомальным/лизосомальным маркером LAMP-1 (CD107a) может обеспечить получение информации относительно субклеточной локализации интернализованного антитела. Как вариант, при количественном биохимическом анализе популяцию клеток, содержащую экспрессирующие KIR3DL2 клетки, приводят в контакт in vitro или in vivo с радиоактивно меченым экспериментальным антителом, и указанные клетки (при контактировании in vivo клетки выделяют впоследствии, через подходящий промежуток времени) обрабатывают протеазой или подвергают кислотной промывке для удаления неинтернализованного антитела с клеточной поверхности. Клетки измельчают и, пропуская гомогенат через сцинтилляционный счетчик, измеряют количество нечувствительных к протеазе радиоактивных импульсов в минуту (имп./мин), связанное с каждой партией клеток. На основании известной специфической активности радиоактивно меченого антитела по полученному для измельченных клеток числу сцинцилляций может быть выведено число интернализованных молекул антитела на клетку. Клетки «приводят в контакт» с антителом in vitro предпочтительно в растворе, например, добавляя указанные клетки в клеточную культуральную среду в чашке или флаконе для культивирования, и тщательно перемешивая антитело со средой, чтобы обеспечить равномерное воздействие антитела на клетки.
Тестирование на КЗЦ и АЗКЦ может проводиться с применением различных методов анализа, включая описанные в экспериментальных примерах в настоящем документе (см. примеры 4 и 5). Тестирование на АЗКЦ, как правило, включает оценку клеточноопосредованной цитотоксичности, при которой экспрессирующая KIR3DL2 целевая клетка (например, клетка Cou-L, пораженная синдромом Сезари клетка или другие экспрессирующие KIR3DL2 клетки) со связанным антителом против KIR3DL2 распознается эффекторной клеткой, несущей рецепторы Fc, без участия комплемента. Клетка, которая не экспрессирует антиген KIR3DL2, может необязательно использоваться в качестве контроля. Активацию цитотоксичность NK-клеток оценивают путем измерения увеличения синтеза цитокинов (например, синтеза ИФН-D) или маркеров цитотоксичности (например, мобилизации CD 107). Предпочтительно, в присутствии целевых клеток указанное антитело индуцирует увеличение синтеза цитокинов, экспрессии маркеров цитотоксичности или лизиса целевых клеток по меньшей мере на 20%, 50%, 80%, 100%, 200% или 500% по сравнению с контрольным антителом (например, антителом, не связывающимся с KIR3DL2, антителом к KIR3DL2, содержащим константные области мыши). Согласно другому примеру проводят детекцию лизиса целевых клеток, например, в анализе высвобождения хрома; предпочтительно, указанное антитело индуцирует лизис по меньшей мере 10%, 20%, 30%, 40% или 50% целевых клеток. При тестировании антигенсвязывающего соединения как на способность (а) и индуцировать АЗКЦ, и (b) интернализоваться в экспрессирующие KIR3DL2 клетки, так и/или на способность индуцировать интернализацию KIR3DL2, анализы согласно (а) и (b) могут проводиться в любом порядке. При этом, как правило, предполагается, что большие степень и скорость интернализации ассоциированы с уменьшением степени активности КЗЦ и АЗКЦ.
Антитело 10F6
Аминокислотная последовательность вариабельной области тяжелой цепи антитела 10F6 приведена в последовательности SEQ ID NO: 2, аминокислотная последовательность вариабельной области легкой цепи приведена в последовательности SEQ ID NO: 3. Согласно конкретному варианту реализации предложено антитело, которое связывает по существу тот же эпитоп или детерминанту, что и моноклональные антитела 10F6; необязательно указанное антитело содержит антигенсвязывающую область антитела 10F6. Согласно любым представленным в настоящем документе вариантам реализации антитело 10F6 может характеризоваться его аминокислотной последовательностью и/или кодирующей его последовательностью нуклеиновых кислот. Согласно одному варианту реализации указанное моноклональное антитело содержит фрагмент Fab или F(ab')2 10F6. Также предложено моноклональное антитело, которое содержит вариабельную область тяжелой цепи 10F6. В соответствии с одним вариантом реализации указанное моноклональное антитело содержит три участка CDR вариабельной области тяжелой цепи 10F6. Также предложено моноклональное антитело, которое дополнительно содержит вариабельную область легкой цепи 10F6 или одну, две или три области из участков CDR вариабельной области легкой цепи 10F6. Необязательно любая одна или большее количество указанных участков CDR легкой или тяжелой цепи могут содержать одну, две, три, четыре или пять или большее количество модификаций аминокислот (например, замен, вставок или удалений). Необязательно, предложено антитело, в котором любые из вариабельных областей легкой и/или тяжелой цепи, содержащие часть антигенсвязывающей области или всю антигенсвязывающую область антитела 10F6 соединены с константной областью иммуноглобулина человека типа IgG, необязательно константной областью человека, необязательно изотипа IgG1 или IgG3 человека.
Согласно другому аспекту предложен очищенный полипептид, кодирующий антитело, при этом указанное антитело содержит: участок HCDR1, содержащий аминокислотную последовательность GYTFTIAGMQ, приведенную в последовательности SEQ ID NO: 6, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот (например, IAGMQ (SEQ ID NO: 4), GYTFTI (SEQ ID NO: 5)), при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; участок HCDR2, содержащий аминокислотную последовательность WINTHSGVPKYAEDFKG, приведенную в последовательности SEQ ID NO: 7, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот (например, WINTHSGVPK (SEQ ID NO: 8)), при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; участок HCDR3, содержащий аминокислотную последовательность GGDEGVMDY, приведенную в последовательности SEQ ID NO: 9, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот, при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; участок LCDR1, содержащую аминокислотную последовательность KASQDVSTAVA, приведенную в последовательности SEQ ID NO: 10, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот, при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; участок LCDR2, содержащую аминокислотную последовательность WASTRHT, приведенную в последовательности SEQ ID NO: 11, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот, при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; участок LCDR3, содержащую аминокислотную последовательность QQHYNTPWT, приведенную в последовательности SEQ ID NO: 12, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот, при этом одна или большее количество указанных аминокислот могут быть удалены или заменены на другую аминокислоту.
Согласно другому аспекту предложено антитело, которое связывает KIR3DL2 человека, содержащее:
(a) вариабельную область тяжелой цепи последовательности SEQ ID NO: 2, при этом необязательно один, два, три или большее количество остатков могут быть заменены на другую аминокислоту; и/или
(b) вариабельную область легкой цепи последовательности SEQ ID NO: 3, при этом необязательно один, два, три или большее количество остатков могут быть заменены на другую аминокислоту; и/или
(c) вариабельную область тяжелой цепи последовательности SEQ ID NO: 2, при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; и вариабельную область легкой цепи последовательности SEQ ID NO: 3, при этом необязательно один, два, три или большее количество остатков могут быть заменены на другую аминокислоту; и/или
(d) аминокислотные последовательности участков CDR 1, 2 и 3 тяжелой цепи (HCDR1, HCDR2, HCDR3) согласно представленным в последовательностях SEQ ID NO: 4-6, 7-8 и 9, соответственно, при этом необязательно один, два, три или большее количество остатков любого участка CDR могут быть заменены на другую аминокислоту; и/или
(e) аминокислотные последовательности участков CDR 1, 2 и 3 легкой цепи (LCDR1, LCDR2, LCDR3) согласно представленным в последовательностях SEQ ID NO: 10, 11 и 12, при этом необязательно один, два, три или большее количество остатков любого участка CDR могут быть заменены на другую аминокислоту; и/или
(f) аминокислотные последовательности участков CDR 1, 2 и 3 тяжелой цепи (HCDR1, HCDR2, HCDR3) согласно представленным в последовательностях SEQ ID NO: 4, 7 и 9, соответственно, при этом необязательно один, два, три или большее количество остатков любого участка CDR могут быть заменены на другую аминокислоту; и аминокислотные последовательности и участков CDR 1, 2 и 3 легкой цепи (LCDR1, LCDR2, LCDR3) согласно представленным в последовательностях SEQ ID NO: 10, 11 и 12, при этом необязательно один, два, три или большее количество остатков любого участка CDR могут быть заменены на другую аминокислоту.
Согласно другому аспекту любого из описываемых в настоящем документе вариантов реализации, любые из участков CDR 1, 2 и 3 тяжелой и легкой цепей могут характеризоваться последовательностью, состоящей по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот, и/или как имеющие аминокислотную последовательность, идентичную по меньшей мере на 50%, 60%, 70%, 80%, 85%, 90% или 95% конкретной области или набору участков CDR, приведенных в соответствующей последовательности SEQ ID NO.
Согласно другому аспекту предложено антитело, которое конкурирует за связывание KIR3DL2 с моноклональным антителом согласно пунктам (a)-(f), приведенным выше.
Антитело 2В12
Аминокислотная последовательность вариабельной области тяжелой цепи антитела 2В12 приведена в последовательности SEQ ID NO: 13, аминокислотная последовательность вариабельной области легкой цепи приведена в последовательности SEQ ID NO: 14. Согласно одному варианту реализации предложено антитело, которое связывает по существу тот же эпитоп или детерминанту, что и моноклональные антитела 2В12; необязательно указанное антитело содержит антигенсвязывающую область антитела 2В12. Согласно любым представленным в настоящем документе вариантам реализации антитело 2В12 может характеризоваться его аминокислотной последовательностью и/или кодирующей его последовательностью нуклеиновых кислот. Согласно одному варианту реализации указанное моноклональное антитело содержит фрагмент Fab или F(ab')2 2B12. Также предложено моноклональное антитело, которое содержит вариабельную область тяжелой цепи 2В12. В соответствии с одним вариантом реализации указанное моноклональное антитело содержит три участка CDR вариабельной области тяжелой цепи 2В12. Также предложено моноклональное антитело, которое дополнительно содержит вариабельную область легкой цепи 2В12 или одну, две или три области из участков CDR вариабельной области легкой цепи 2В12. Необязательно любая одна или большее количество указанных участков CDR легкой или тяжелой цепи могут содержать одну, две, три, четыре или пять модификаций аминокислот (например, замен, вставок или удалений). Необязательно, предложено антитело, в котором любые из вариабельных областей легкой и/или тяжелой цепи, содержащие часть антигенсвязывающей области или всю антигенсвязывающую область антитела 2В12, соединены с константной областью иммуноглобулина типа IgG, необязательно константной область человека, необязательно изотипа IgG1 или IgG4.
Согласно другому аспекту предложен очищенный полипептид, кодирующий антитело, при этом указанное антитело содержит: участок HCDR1, содержащий аминокислотную последовательность GYTFTTAGMQ, приведенную в последовательности SEQ ID NO: 17, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот (например, TAGMQ (SEQ ID NO: 15), GYTFTT (SEQ ID NO: 16)), при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; участок HCDR2, содержащий аминокислотную последовательность WINSHSGVPKYAEDFK, приведенную в последовательности SEQ ID NO: 18, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот (например, WINSHSGVP (SEQ ID NO: 19)), при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; участок HCDR3, содержащий аминокислотную последовательность GGDEGVMDYW, приведенную в последовательности SEQ ID NO: 20, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот, при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; участок LCDR1, содержащую аминокислотную последовательность KASQDVSTAVA, приведенную в последовательности SEQ ID NO: 10, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот, при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; участок LCDR2, содержащую аминокислотную последовательность WTSTRHT, приведенную в последовательности SEQ ID NO: 21, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот, при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; и/или участок LCDR3, содержащая аминокислотную последовательность QQHYSTPWT, приведенную в последовательности SEQ ID NO: 22, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот, при этом одна или большее количество указанных аминокислот могут быть удалены или заменены на другую аминокислоту, или при этом указанная последовательность может содержать вставку одной или большего количества аминокислот.
Согласно другому аспекту предложено антитело, которое связывает KIR3DL2 человека, содержащее:
(a) вариабельную область тяжелой цепи последовательности SEQ ID NO: 13, при этом необязательно один, два, три или большее количество остатков аминокислот могут быть заменены на другую аминокислоту; и/или
(b) вариабельную область легкой цепи последовательности SEQ ID NO: 14, при этом необязательно один, два, три или большее количество остатков аминокислот могут быть заменены на другую аминокислоту; и/или
(c) вариабельную область тяжелой цепи последовательности SEQ ID NO: 13, при этом необязательно один, два, три или большее количество остатков аминокислот могут быть заменены на другую аминокислоту; и вариабельную область легкой цепи последовательности SEQ ID NO: 14, при этом необязательно один или большее количество остатков аминокислот могут быть заменены на другую аминокислоту; и/или
(d) аминокислотные последовательности участков CDR 1, 2 и 3 тяжелой цепи (HCDR1, HCDR2, HCDR3) согласно представленным в последовательностях SEQ ID NO: 15-17, 18-19 и 20, соответственно, при этом необязательно один, два, три или большее количество остатков любого участка CDR могут быть заменены на другую аминокислоту; и/или
(e) аминокислотные последовательности участков CDR 1, 2 и 3 легкой цепи (LCDR1, LCDR2, LCDR3) согласно представленным в последовательностях SEQ ID NO: 10, 21 и 22, соответственно, при этом необязательно один, два, три или большее количество остатков любого участка CDR могут быть заменены на другую аминокислоту; и/или
(f) аминокислотные последовательности участков CDR 1, 2 и 3 тяжелой цепи (HCDR1, HCDR2, HCDR3) согласно представленным в последовательностях SEQ ID NO: 15, 18 и 20, соответственно, при этом необязательно один, два, три или большее количество остатков любого участка CDR могут быть заменены на другую аминокислоту; и аминокислотные последовательности участков CDR 1, 2 и 3 легкой цепи (LCDR1, LCDR2, LCDR3) согласно представленным в последовательностях SEQ ID NO: 10, 21 и 22, при этом необязательно один, два, три или большее количество остатков любого участка CDR могут быть заменены на другую аминокислоту.
Согласно другому аспекту любого из описываемых в настоящем документе вариантов реализации, любые из участков CDR 1, 2 и 3 тяжелой и легкой цепей могут характеризоваться последовательностью, состоящей по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот, и/или как имеющие аминокислотную последовательность, идентичную по меньшей мере на 50%, 60%, 70%, 80%, 85%, 90% или 95% конкретного участка CDR или набору участков CDR, приведенных в соответствующей последовательности SEQ ID NO.
Согласно другому аспекту предложено антитело, которое конкурирует за связывание KIR3DL2 с моноклональным антителом согласно пунктам (a)-(f), приведенным выше.
Антитело 10G5
Аминокислотная последовательность вариабельной области тяжелой цепи антитела 10G5 приведена в последовательности SEQ ID NO: 23, аминокислотная последовательность вариабельной области легкой цепи приведена в последовательности SEQ ID NO: 24. Согласно конкретному варианту реализации предложено антитело, которое связывает по существу тот же эпитоп или детерминанту, что и моноклональные антитела 10G5; необязательно указанное антитело содержит антигенсвязывающую область антитела 10G5. Согласно любым представленным в настоящем документе вариантам реализации антитело 10G5 может характеризоваться его аминокислотной последовательностью и/или кодирующей его последовательностью нуклеиновых кислот. Согласно одному варианту реализации указанное моноклональное антитело содержит фрагмент Fab или F(ab')2 10G5. Также предложено моноклональное антитело, которое содержит вариабельную область тяжелой цепи 10G5. В соответствии с одним вариантом реализации указанное моноклональное антитело содержит три участка CDR вариабельной области тяжелой цепи 10G5. Также предложено моноклональное антитело, которое дополнительно содержит вариабельную область легкой цепи 10G5 или одну, две или три области из участков CDR вариабельной области легкой цепи 10G5. Необязательно любая одна или большее количество указанных участков CDR легкой или тяжелой цепи могут содержать одну, две, три, четыре или пять или большее количество модификаций аминокислот (например, замен, вставок или удалений). Необязательно предложено антитело, в котором любые из вариабельных областей легкой и/или тяжелой цепи, содержащие часть антигенсвязывающей области или всю антигенсвязывающую область антитела 10G5 соединены с константной областью иммуноглобулина человека типа IgG, необязательно константной областью человека, необязательно изотипа IgG1 или IgG3 человека.
Согласно другому аспекту предложен очищенный полипептид, кодирующий антитело, при этом указанное антитело содержит: участок HCDR1, содержащий аминокислотную последовательность GYTFTSYTMH, приведенную в последовательности SEQ ID NO: 27, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот (например, SYTMH (SEQ ID NO: 25), GYTFTS (SEQ ID NO: 26)), при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; участок HCDR2, содержащий аминокислотную последовательность YINPSSGYTENNRKF, приведенную в последовательности SEQ ID NO: 28, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот (например, YINPSSGY (SEQ ID NO: 29)), при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; участок HCDR3, содержащий аминокислотную последовательность RLGKGLLPPFDY, приведенную в последовательности SEQ ID NO: 30, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот, при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; участок LCDR1, содержащую аминокислотную последовательность RASENIYSNLA, приведенную в последовательности SEQ ID NO: 31, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот, при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; участок LCDR2, содержащую аминокислотную последовательность AATNLAD, приведенную в последовательности SEQ ID NO: 32, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот, при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; участок LCDR3, содержащую аминокислотную последовательность QHFWGTPYT, приведенную в последовательности SEQ ID NO: 33, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот, при этом одна или большее количество указанных аминокислот могут быть удалены или заменены на другую аминокислоту.
Согласно другому аспекту предложено антитело, которое связывает KIR3DL2 человека, содержащее:
(a) вариабельную область тяжелой цепи последовательности SEQ ID NO: 23, при этом необязательно один, два, три или большее количество остатков могут быть заменены на другую аминокислоту; и/или
(b) вариабельную область легкой цепи последовательности SEQ ID NO: 24, при этом необязательно один, два, три или большее количество остатков могут быть заменены на другую аминокислоту; и/или
(c) вариабельную область тяжелой цепи последовательности SEQ ID NO: 23, при этом необязательно один или большее количество остатков могут быть заменены на другую аминокислоту; и вариабельную область легкой цепи последовательности SEQ ID NO: 24, при этом один, два, три или большее количество остатков могут быть заменены на другую аминокислоту; и/или
(d) аминокислотные последовательности участков CDR 1, 2 и 3 тяжелой цепи (HCDR1, HCDR2, HCDR3) согласно представленным в последовательностях SEQ ID NO: 25-27, 28-29 и 30, соответственно, при этом необязательно один, два, три или большее количество остатков любого участка CDR могут быть заменены на другую аминокислоту; и/или
(e) аминокислотные последовательности участков CDR 1, 2 и 3 легкой цепи (LCDR1, LCDR2, LCDR3) согласно представленным в последовательностях SEQ ID NO: 31, 32, и 33, при этом необязательно один, два, три или большее количество остатков любого участка CDR могут быть заменены на другую аминокислоту; и/или
(f) аминокислотные последовательности участков CDR 1, 2 и 3 тяжелой цепи (HCDR1, HCDR2, HCDR3) согласно представленным в последовательностях SEQ ID NO: 25, 28 и 30, соответственно, при этом необязательно один или большее количество остатки любого участка CDR могут быть заменены на другую аминокислоту; и аминокислотные последовательности и участков CDR 1, 2 и 3 легкой цепи (LCDR1, LCDR2, LCDR3) согласно представленным в последовательностях SEQ ID NO: 31, 32, и 33, при этом необязательно один, два, три или большее количество остатков любого участка CDR могут быть заменены на другую аминокислоту.
Согласно другому аспекту любого из описываемых в настоящем документе вариантов реализации, любые из участков CDR 1, 2 и 3 тяжелой и легкой цепей могут характеризоваться последовательностью, состоящей по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот, и/или аминокислотной последовательностью, идентичной по меньшей мере на 50%, 60%, 70%, 80%, 85%, 90% или 95% конкретной области или набору участков CDR, приведенных в соответствующей последовательности SEQ ID NO.
Согласно другому аспекту предложено антитело, которое конкурирует за связывание KIR3DL2 с моноклональным антителом согласно пунктам (a)-(f), приведенным выше.
Антитело 13Н1
Аминокислотная последовательность вариабельной области тяжелой цепи антитела 13Н1 приведена в последовательности SEQ ID NO: 34, аминокислотная последовательность вариабельной области легкой цепи приведена в последовательности SEQ ID NO: 35. Согласно конкретному варианту реализации предложено антитело, которое связывает по существу тот же эпитоп или детерминанту, что и моноклональное антитело 13Н1; необязательно указанное антитело содержит антигенсвязывающую область антитела 13Н1. Согласно любым представленным в настоящем документе вариантам реализации антитело 13Н1 может характеризоваться его аминокислотной последовательностью и/или кодирующей его последовательностью нуклеиновых кислот. Согласно одному варианту реализации указанное моноклональное антитело содержит фрагмент Fab или F(ab')2 13Н1. Также предложено моноклональное антитело, которое содержит вариабельную область тяжелой цепи 13Н1. В соответствии с одним вариантом реализации указанное моноклональное антитело содержит три участка CDR вариабельной области тяжелой цепи 13Н1. Также предложено моноклональное антитело, которое дополнительно содержит вариабельную область легкой цепи 13Н1 или одну, две или три области из участков CDR вариабельной области легкой цепи 13Н1. Необязательно любая одна или большее количество указанных участков CDR легкой или тяжелой цепи могут содержать одну, две, три, четыре или пять или большее количество модификаций аминокислот (например, замен, вставок или удалений). Необязательно, предложено антитело, в котором любые из вариабельных областей легкой и/или тяжелой цепи, содержащие часть антигенсвязывающей области или всю антигенсвязывающую область антитела 13Н1 соединены с константной областью иммуноглобулина человека типа IgG, необязательно константной областью человека, необязательно изотипа IgG1 или IgG3 человека.
Согласно другому аспекту предложен очищенный полипептид, кодирующий антитело, при этом указанное антитело содержит: участок HCDR1, содержащий аминокислотную последовательность HYSFIGYTM, приведенную в последовательности SEQ ID NO: 38, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот (например, GYTMN (SEQ ID NO: 36), HYSFIG (SEQ ID NO: 37)), при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; участок HCDR2, содержащий аминокислотную последовательность LINPYNGDTTYNQKFKG, приведенную в последовательности SEQ ID NO: 39, или последовательность, состоящую по меньшей мере из 4,5,6, 7, 8, 9 или 10 ее смежных аминокислот (например, UNPYNGDTT (SEQ ID NO: 40)), при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; участок HCDR3, содержащий аминокислотную последовательность ENWGYPYAMDY, приведенную в последовательности SEQ ID NO: 41, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот, при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; участок LCDR1, содержащую аминокислотную последовательность RASES VDNFGISFMN, приведенную в последовательности SEQ ID NO: 42, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот, при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; участок LCDR2, содержащую аминокислотную последовательность AASNQGS, приведенную в последовательности SEQ ID NO: 43, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот, при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; участок LCDR3, содержащую аминокислотную последовательность QQSKEVPYT, приведенную в последовательности SEQ ID NO: 44, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот, при этом одна или большее количество указанных аминокислот могут быть удалены или заменены на другую аминокислоту.
Согласно другому аспекту предложено антитело, которое связывает KIR3DL2 человека, содержащее:
(а) вариабельную область тяжелой цепи последовательности SEQ ID NO: 34, при этом необязательно один, два, три или большее количество остатков аминокислот могут быть заменены на другую аминокислоту; и/или
(b) вариабельную область легкой цепи последовательности SEQ ID NO: 35, при этом необязательно один, два, три или большее количество остатков аминокислот могут быть заменены на другую аминокислоту; и/или
(c) вариабельную область тяжелой цепи последовательности SEQ ID NO: 34, при этом необязательно один или большее количество остатков аминокислот могут быть заменены на другую аминокислоту; и вариабельную область легкой цепи последовательности SEQ ID NO: 35, при этом одна, две, три или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; и/или
(d) аминокислотные последовательности участков CDR 1, 2 и 3 тяжелой цепи (HCDR1, HCDR2, HCDR3) согласно представленным в последовательностях SEQ ID NO: 36-38, 39-40 и 41, соответственно, при этом необязательно один, два, три или большее количество остатков аминокислот любого участка CDR могут быть заменены на другую аминокислоту; и/или
(e) аминокислотные последовательности участков CDR 1, 2 и 3 легкой цепи (LCDR1, LCDR2, LCDR3) согласно представленным в последовательностях SEQ ID NO: 42, 43 и 44, при этом необязательно один, два, три или большее количество остатков аминокислот любого участка CDR могут быть заменены на другую аминокислоту; и/или
(f) аминокислотные последовательности участков CDR 1, 2 и 3 тяжелой цепи (HCDR1, HCDR2, HCDR3) согласно представленным в последовательностях SEQ ID NO: 36, 39 и 41, соответственно, при этом необязательно один или большее количество остатков аминокислот любого участка CDR могут быть заменены на другую аминокислоту; и аминокислотные последовательности и участков CDR 1, 2 и 3 легкой цепи (LCDR1, LCDR2, LCDR3) согласно представленным в последовательностях SEQ ID NO: 42, 43 и 44, при этом необязательно один, два, три или большее количество остатков аминокислот любого участка CDR могут быть заменены на другую аминокислоту.
Согласно другому аспекту любого из описываемых в настоящем документе вариантов реализации любые из участков CDR 1, 2 и 3 тяжелой и легкой цепей могут характеризоваться последовательностью, состоящей по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот, и/или как имеющие аминокислотную последовательность, идентичную по меньшей мере на 50%, 60%, 70%, 80%, 85%, 90% или 95% конкретной области или набору участков CDR, приведенных в соответствующей последовательности SEQ ID NO.
Согласно другому аспекту предложено антитело, которое конкурирует за связывание KIR3DL2 с моноклональным антителом согласно пунктам (a)-(f), приведенным выше.
Антитело 1Е2
Аминокислотная последовательность вариабельной области тяжелой цепи антитела 1Е2 приведена в последовательности SEQ ID NO: 45, аминокислотная последовательность вариабельной области легкой цепи приведена в последовательности SEQ ID NO: 46. Согласно конкретному варианту реализации предложено антитело, которое связывает по существу тот же эпитоп или детерминанту, что и моноклональные антитела 1Е2; необязательно указанное антитело содержит антигенсвязывающую область антитела 1Е2. Согласно любым представленным в настоящем документе вариантам реализации антитело 1Е2 может характеризоваться его аминокислотной последовательностью и/или кодирующей его последовательностью нуклеиновых кислот. Согласно одному варианту реализации указанное моноклональное антитело содержит фрагмент Fab или F(ab')2 1Е2. Также предложено моноклональное антитело, которое содержит вариабельную область тяжелой цепи 1Е2. В соответствии с одним вариантом реализации указанное моноклональное антитело содержит три участка CDR вариабельной области тяжелой цепи 1Е2. Также предложено моноклональное антитело, которое дополнительно содержит вариабельную область легкой цепи 1Е2 или одну, две или три области из участков CDR вариабельной области легкой цепи 1Е2. Необязательно любая одна или большее количество указанных участков CDR легкой или тяжелой цепи могут содержать одну, две, три, четыре или пять или большее количество модификаций аминокислот (например, замен, вставок или удалений). Необязательно, предложено антитело, в котором любые из вариабельных областей легкой и/или тяжелой цепи, содержащие часть антигенсвязывающей области или всю антигенсвязывающую область антитела 1Е2, соединены с константной областью иммуноглобулина человека типа IgG, необязательно константной областью человека, необязательно изотипа IgG1 или IgG3 человека.
Согласно другому аспекту предложен очищенный полипептид, кодирующий антитело, при этом указанное антитело содержит: участок HCDR1, содержащий аминокислотную последовательность GYTPTDYAMN, приведенную в последовательности SEQ ID NO: 49, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот (например, DYAMN (SEQ ID NO: 47), GYTFTD (SEQ ID NO: 48)), при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; участок HCDR2, содержащий аминокислотную последовательность VISTYYGDANYNQKFKG, приведенную в последовательности SEQ ID NO: 50, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот (например, VISTYYGDAN (SEQ ID NO: 51)), при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; участок HCDR3, содержащий аминокислотную последовательность IYYDYDGSY, приведенную в последовательности SEQ ID NO: 52, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот, при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; участок LCDR1, содержащую аминокислотную последовательность RSSQSLVHSNGNTYLH, приведенную в последовательности SEQ ID NO: 53, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот, при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; участок LCDR2, содержащую аминокислотную последовательность KVSNRFS, приведенную в последовательности SEQ ID NO: 54, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот, при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; участок LCDR3, содержащую аминокислотную последовательность SQSTHVPPYT, приведенную в последовательности SEQ ID NO: 55, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот, при этом одна или большее количество указанных аминокислот могут быть удалены или заменены на другую аминокислоту.
Согласно другому аспекту предложено антитело, которое связывает KIR3DL2 человека, содержащее:
(a) вариабельную область тяжелой цепи последовательности SEQ ID NO: 42, при этом необязательно один, два, три или большее количество остатков аминокислот могут быть заменены на другую аминокислоту; и/или
(b) вариабельную область легкой цепи последовательности SEQ ID NO: 43, при этом необязательно один, два, три или большее количество остатков аминокислот могут быть заменены на другую аминокислоту; и/или
(c) вариабельную область тяжелой цепи последовательности SEQ ID NO: 42, при этом необязательно один или большее количество остатков аминокислот могут быть заменены на другую аминокислоту; и вариабельную область легкой цепи последовательности SEQ ID NO: 43, при этом необязательно один, два, три или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; и/или
(d) аминокислотные последовательности участков CDR 1, 2 и 3 тяжелой цепи (HCDR1, HCDR2, HCDR3) согласно представленным в последовательностях SEQ ID NO: 47-49, 50-51 и 52, соответственно, при этом необязательно один, два, три или большее количество остатков аминокислот любого участка CDR могут быть заменены на другую аминокислоту; и/или
(e) аминокислотные последовательности участков CDR 1, 2 и 3 легкой цепи (LCDR1, LCDR2, LCDR3) согласно представленным в последовательностях SEQ ID NO: 53, 54 и 55, при этом необязательно один, два, три остатка аминокислот любого участка CDR могут быть заменены на другую аминокислоту; и/или
(f) аминокислотные последовательности участков CDR 1, 2 и 3 тяжелой цепи (HCDR1, HCDR2, HCDR3) согласно представленным в последовательностях SEQ ID NO: 47, 50 и 52, при этом необязательно один или большее количество остатков аминокислот любого участка CDR могут быть заменены на другую аминокислоту; и аминокислотные последовательности и участков CDR 1, 2 и 3 легкой цепи (LCDR1, LCDR2, LCDR3) согласно представленным в последовательностях SEQ ID NO: 53, 54 и 55, при этом необязательно один, два, три или большее количество остатков аминокислот любого участка CDR могут быть заменены на другую аминокислоту.
Согласно другому аспекту любого из описываемых в настоящем документе вариантов реализации, любые из участков CDR 1, 2 и 3 тяжелой и легкой цепей могут характеризоваться последовательностью, состоящей по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот, и/или как имеющие аминокислотную последовательность, идентичную по меньшей мере на 50%, 60%, 70%, 80%, 85%, 90% или 95% конкретной области или набору участков CDR, приведенных в соответствующей последовательности SEQ ID NO.
Согласно другому аспекту предложено антитело, которое конкурирует за связывание KIR3DL2 с моноклональным антителом согласно пунктам (a)-(f), приведенным выше.
Антитело 9Е10
Аминокислотная последовательность вариабельной области тяжелой цепи 9Е10 приведена в последовательности SEQ ID NO: 56, аминокислотная последовательность вариабельных областей легкой цепи (доступны два варианта легкой цепи) 9Е10 приведены в последовательностях SEQ ID NO: 57 и 67. Согласно конкретному варианту реализации предложено антитело, которое связывает по существу тот же эпитоп или детерминанту, что и моноклональные антитела 9Е10; необязательно указанное антитело содержит антигенсвязывающую область антитела 9Е10. Согласно любым представленным в настоящем документе вариантам реализации антитело 9Е10 может характеризоваться его аминокислотной последовательностью и/или кодирующей его последовательностью нуклеиновых кислот. Согласно одному варианту реализации указанное моноклональное антитело содержит фрагмент Fab или F(ab')2 9Е10. Также предложено моноклональное антитело, которое содержит вариабельную область тяжелой цепи 9Е10. В соответствии с одним вариантом реализации указанное моноклональное антитело содержит три участка CDR вариабельной области тяжелой цепи 9Е10. Также предложено моноклональное антитело, которое дополнительно содержит вариабельную область легкой цепи 9Е10 или одну, две или три области из участков CDR вариабельной области легкой цепи 9Е10. Необязательно любая одна или большее количество указанных участков CDR легкой или тяжелой цепи могут содержать одну, две, три, четыре или пять модификаций аминокислот (например, замен, вставок или удалений). Необязательно, предложено антитело, в котором любые из вариабельных областей легкой и/или тяжелой цепи, содержащие часть антигенсвязывающей области или всю антигенсвязывающую область антитела 9Е10 соединены с константной областью иммуноглобулина типа IgG, необязательно константной областью человека, необязательно изотипа IgG1 или IgG4 человека.
Согласно другому аспекту предложен очищенный полипептид, кодирующий антитело, при этом указанное антитело содержит: участок HCDR1, содержащий аминокислотную последовательность GYTFTSYTMH, приведенную в последовательности SEQ ID NO: 60, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот (например, SYTMH (SEQ ID NO: 58), GYTFTS (SEQ ID NO: 59)), при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; участок HCDR2, содержащий аминокислотную последовательность YINPSSGYTDYNQKPKD, приведенную в последовательности SEQ ID NO: 61, или последовательность, состоящую по меньшей мере из 4, 5, 6,7, 8, 9 или 10 ее смежных аминокислот (например, YINPSSGYTD (SEQ ID NO: 62)), при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; участок HCDR3, содержащий аминокислотную последовательность LGKGLLPPFDY, приведенную в последовательности SEQ ID NO: 63, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот, при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; участок LCDR1, содержащую аминокислотную последовательность KSNQNLLWSGNQRYCLV, приведенную в последовательности SEQ ID NO: 64, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот, при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; участок LCDR2, содержащую аминокислотную последовательность WTSDRYS, приведенную в последовательности SEQ ID NO: 65, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот, при этом одна или большее количество указанных аминокислот могут быть заменены на другую аминокислоту; участок LCDR3, содержащую аминокислотную последовательность QQHLHIPYT, приведенную в последовательности SEQ ID NO: 66, или последовательность, состоящую по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот, при этом одна или большее количество указанных аминокислот могут быть удалены или заменены на другую аминокислоту, или при этом указанная последовательность может содержать вставку из одной или большего количества аминокислот.
Согласно другому аспекту предложено антитело, которое связывает KIR3DL2 человека, содержащее:
(a) вариабельную область тяжелой цепи последовательности SEQ ID NO: 56, при этом необязательно один, два или три остатка аминокислот могут быть заменены на другую аминокислоту; и/или
(b) вариабельную область легкой цепи, соответствующую последовательности SEQ ID NO: 57 или 67, при этом необязательно один, два, три или большее количество остатков аминокислот могут быть заменены на другую аминокислоту; и/или
(c) вариабельную область тяжелой цепи последовательности SEQ ID NO: 56, при этом необязательно один, два, три или большее количество остатков аминокислот могут быть заменены на другую аминокислоту; и вариабельную область легкой цепи, соответствующую последовательности SEQ ID NO: 57 или 67, при этом необязательно один, два, три или большее количество остатков аминокислот могут быть заменены на другую аминокислоту; и/или
(d) аминокислотные последовательности участков CDR 1 и 2 тяжелой цепи (HCDR1, HCDR2) согласно представленным в последовательностях SEQ ID NO: 58, 59 или 60, 61-62 и 63, при этом необязательно один, два, три или большее количество остатков любого участка CDR могут быть заменены на другую аминокислоту; и/или
(e) аминокислотные последовательности участков CDR 1, 2 и 3 легкой цепи (LCDR1, LCDR2, LCDR3) согласно представленным в последовательностях SEQ ID NO: 64, 65 и 66, при этом необязательно один, два, три или большее количество остатков любого участка CDR могут быть заменены на другую аминокислоту; и/или
(f) аминокислотные последовательности участков CDR 1, 2 и 3 тяжелой цепи (HCDR1, HCDR2, HCDR3) согласно представленным в последовательностях SEQ ID NO: 58, 61 и 63, при этом необязательно один, два, три остатка любого участка CDR могут быть заменены на другую аминокислоту; и аминокислотные последовательности участков CDR 1, 2 и 3 легкой цепи (LCDR1, LCDR2, LCDR3) согласно представленным в последовательностях SEQ ID NO: 64, 65 и 66, при этом необязательно один, два, три или большее количество остатков любого участка CDR могут быть заменены на другую аминокислоту.
Согласно другому варианту реализации предложены антитело 1С3 (против домена D2), его вариабельная область и области CDR. Согласно одному варианту реализации предложено антитело, содержащее, соответственно, области VH и VL, соответствующие последовательностям SEQ ID NO: 170 и 171 (1С3). Согласно одному варианту реализации предложено антитело, содержащее тяжелую цепь, содержащую участки CDR 1, 2 и 3 (HCDR1, HCDR2, HCDR3), содержащие последовательность SEQ ID NO: 172, 173 или 174 (HCDR1), SEQ ID NO: 175 или 176 (HCDR2) и SEQ ID NO: 177 (HCDR3), соответственно, при этом каждый участок CDR может необязательно содержать 1, 2, 3 или 4 замены, удаления или вставки аминокислот. Согласно одному варианту реализации предложено антитело, содержащее (i) тяжелую цепь, содержащую участки CDR 1, 2 и 3 (HCDR1, HCDR2, HCDR3), содержащие последовательность SEQ ID NO: 172, 173 или 174 (HCDR1), SEQ ID NO: 175 или 176 (HCDR2) и SEQ ID NO: 177 (HCDR3), соответственно, и (ii) легкую цепь, содержащую участки CDR 1, 2 и 3 (LCDR1, LCDR2, LCDR3), содержащие последовательность SEQ ID NO: 178, 179 или 180, соответственно, при этом каждый участок CDR может необязательно содержать 1, 2, 3 или 4 замены, удаления или вставки аминокислот.
Согласно другому варианту реализации предложено антитело 20Е9 (против домена D2), его вариабельная область и области CDR. Согласно одному варианту реализации предложено антитело, содержащее, соответственно, области VH и VL, соответствующие последовательностям SEQ ID NO: 181 и 182 (20Е9). Согласно одному варианту реализации предложено антитело, содержащее тяжелую цепь, содержащую участки CDR 1, 2 и 3 (HCDR1, HCDR2, HCDR3), содержащие последовательность SEQ ID NO: 183, 184 или 185 (HCDR1), SEQ ID NO: 186 или 187 (HCDR2) и SEQ ID NO: 188 (HCDR3), соответственно, при этом каждый участок CDR может необязательно содержать 1, 2, 3 или 4 замены, удаления или вставки аминокислот. Согласно одному варианту реализации предложено антитело, содержащее (i) тяжелую цепь, содержащую участки CDR 1, 2 и 3 (HCDR1, HCDR2, HCDR3), содержащие последовательность SEQ ID NO: 183, 184 или 185 (HCDR1), SEQ ID NO: 186 или 187 (HCDR2) и SEQ ID NO: 188 (HCDR3), соответственно, и (ii) легкую цепь, содержащую участки CDR 1, 2 и 3 (LCDR1, LCDR2, LCDR3), содержащие последовательность SEQ ID NO: 189, 190 или 191, соответственно, при этом каждый участок CDR может необязательно содержать 1, 2, 3 или 4 замены, удаления или вставки аминокислот.
Согласно другому аспекту любого из описываемых в настоящем документе вариантов реализации, любые из участков CDR 1, 2 и 3 тяжелой и легкой цепей могут характеризоваться последовательностью, состоящей по меньшей мере из 4, 5, 6, 7, 8, 9 или 10 ее смежных аминокислот, и/или как имеющие аминокислотную последовательность, идентичную по меньшей мере на 50%, 60%, 70%, 80%, 85%, 90% или 95% конкретной области или набору участков CDR, приведенных в соответствующей последовательности SEQ ID NO.
Согласно другому аспекту предложено антитело, которое конкурирует за связывание KIR3DL2 с моноклональным антителом, описанным выше.
В любых из указанных антител конкретные последовательности вариабельных областей и CDR могут содержать одну, два, три, четыре, пять или большее количество консервативных модификаций последовательности. Консервативные модификации последовательностей относится к модификациям аминокислот, значительно не влияющим или не изменяющим связывающие характеристики антитела, содержащего указанную последовательность аминокислот. Такие консервативные модификации включают замены, добавления и удаления аминокислот. Модификации могут быть введены в антитело с помощью стандартных техник, известных в данной области техники, таких как сайт-специфический мутагенез и ПЦР-опосредованный мутагенез. Консервативные замены аминокислот, как правило, представляют собой замены, при которых остаток аминокислоты заменен на остаток аминокислоты с боковой цепью, имеющей аналогичные физико-химические свойства. Конкретные последовательности вариабельных областей и CDR могут содержать одну, две, три, четыре или большее количество вставок, удалений или замен аминокислот. При наличии замен указанные замены могут необязательно представлять собой консервативные модификации. Семейства остатков аминокислот, содержащие аналогичные боковые цепи, определены в данной области техники. Указанные семейства включают аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислотными боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин, триптофан), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин), бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Соответственно, один или большее количество остатков аминокислот с составе участков CDR антитела могут быть заменены на другие остатки аминокислот, принадлежащие к тому же семейству боковых цепей, и измененное антитело может быть протестировано для проверки сохранности функции (т.е., описанных в настоящем документе свойств) с применением описанных в настоящем документе анализов.
Термин «идентичность» или «идентичный», при использовании в отношении последовательностей двух или большего количества полипептидов, относится к степени родства последовательностей полипептидов, в соответствии с оценкой числа совпадений между линейными последовательностями двух или большего количества остатков аминокислот. «Идентичность» является процентной мерой идентичных совпадений между меньшим из двух или большего количества последовательностей при выравнивании с разрывами (при их наличии), определяемая с применением конкретной математической модели или компьютерной программы (т.е. «алгоритмов»). Идентичность родственных полипептидов может быть легко рассчитана с помощью известных способов. Такие способы включают, не ограничиваясь перечисленными, описанные в источниках: Computational Molecular Biology, Lesk, A.M., ed., Oxford University Press, New York, 1988; Biocomputing: Informatics and Genome Projects, Smith, D.W., ed., Academic Press, New York, 1993; Computer Analysis of Sequence Data, Part 1, Griffin, A.M., and Griffin, H.G., eds., Humana Press, New Jersey, 1994; Sequence Analysis in Molecular Biology, von Heinje, G., Academic Press, 1987; Sequence Analysis Primer, Gribskov, M. and Devereux, J., eds., M. Stockton Press, New York, 1991; и Carillo et al., SIAM J. Applied Math. 48, 1073 (1988).
Предпочтительные способы определения идентичности разработаны таким образом, чтобы отражать максимальный уровень совпадений между тестируемыми последовательностями. Способы определения идентичности описаны в общедоступных компьютерных программах. Предпочтительные способы определения идентичности двух последовательностей на основе компьютерных программ включают программный пакет GCG, в том числе GAP (Devereux et al., Nucl. Acid. Res. 12, 387 (1984); Genetics Computer Group, University of Wisconsin, Madison, Wis.), BLASTP, BLASTN, и PASTA (Altschul et al., J. Mol. Biol. 215, 403-410 (1990)). Открытый доступ к программе BLASTX предоставляет Национальный центр биотехнологической информации (National Center for Biotechnology Information, NCBI) и другие источники (BLAST Manual, Altschul et al. NCB/NLM/NIH Bethesda, Md. 20894; Altschul et al., выше). Хорошо известный алгоритм Смита-Уотермана также может быть использован для определения идентичности.
Последовательности участков CDR антител, в соответствии с системами определения AbM (определение согласно ПО для моделирования антител AbM от Oxford Molecular), Kabat и Chothia, обобщены в таблицах 1 для участков CDR тяжелых цепей, и в таблице 2 ниже для участков CDR легких цепей (определения для участков CDR легких цепей идентичны согласно AbM, Kabat и Chothia). Последовательности аминокислот, описанные в настоящем документе, пронумерованы в соответствии с системами нумерации Abm, Kabat и Chothia. Хотя для определяемых участков CDR может использоваться любая подходящая система нумерации, в отсутствие каких-либо других указаний, может применяться нумерация Abm. Такая нумерация была установлена с применением следующих обозначений: CDR-L1: начало: приблизительно на остатке 24, остаток перед участком: всегда Cys, остаток после: всегда Trp (как правило, Trp-Tyr-GIn, но также Trp-Leu-GIn, Trp-Phe-Gin, Trp-Tyr-Leu), длина: 10-17 остатков; CDR-L2: начало: всегда через 16 остатков за концом L1, остатки перед участком: как правило, Ile-Tyr (но также Val-Tyr, Ile-Lys, Ile-Phe), Длина: всегда 7 остатков; CDR-L3, начало: всегда через 33 остатка после конца L2, остаток перед участком: всегда Cys, остатки после: всегда Phe-Gly-Xaa-Gly, Длина: от 7 до 11 остатков; CDR-H1, Начало: приблизительно на остатке 26 (всегда 4 после Cys) (определение по Chothia / AbM, согласно определению по Kabat начало расположено на 5 остатков дальше), остатки перед участком: всегда Cys-Xaa-Xaa-Xaa, остатки после: всегда Trp (как правило, Trp-Val, но также Trp-Ile, Trp-Ala), Длина: от 10 до 12 остатков (определение по АЬМ, при определении по Chothia исключены последние 4 остатка); CDR-H2, Начало: всегда через 15 остатков после конца CDR-H1 по Kabat / AbM, остатки перед участком: как правило, Leu-Glu-Trp-De-Gly (хотя имеется ряд вариаций, остатки после: Lys/Arg-Leu/Ile/Val/Phe/Thr/Ala-Thr/Ser/De/Ala), Длина: согласно определению по Kabat от 16 до 19 остатков; согласно определению по AbM (и Chothia) заканчивается на 7 остатков раньше; CDR-H3, Начало: всегда через 33 остатка за концом CDR-H2 (всегда 2 после Cys), остатки перед участком: всегда Cys-Xaa-Xaa (как правило, Cys-Ala-Arg), остатки после: всегда Trp-Gly-Xaa-Gly, Длина: 3-25 остатков.
Согласно одному варианту реализации указанные антитела принадлежат к изотипу IgG1 человека или мыши. Согласно другому варианту реализации указанные антитела принадлежат к изотипу IgG1 человека. Согласно варианту реализации указанные антитела представлены фрагментами антител, сохраняющими связывающие и/или функциональные характеристики.

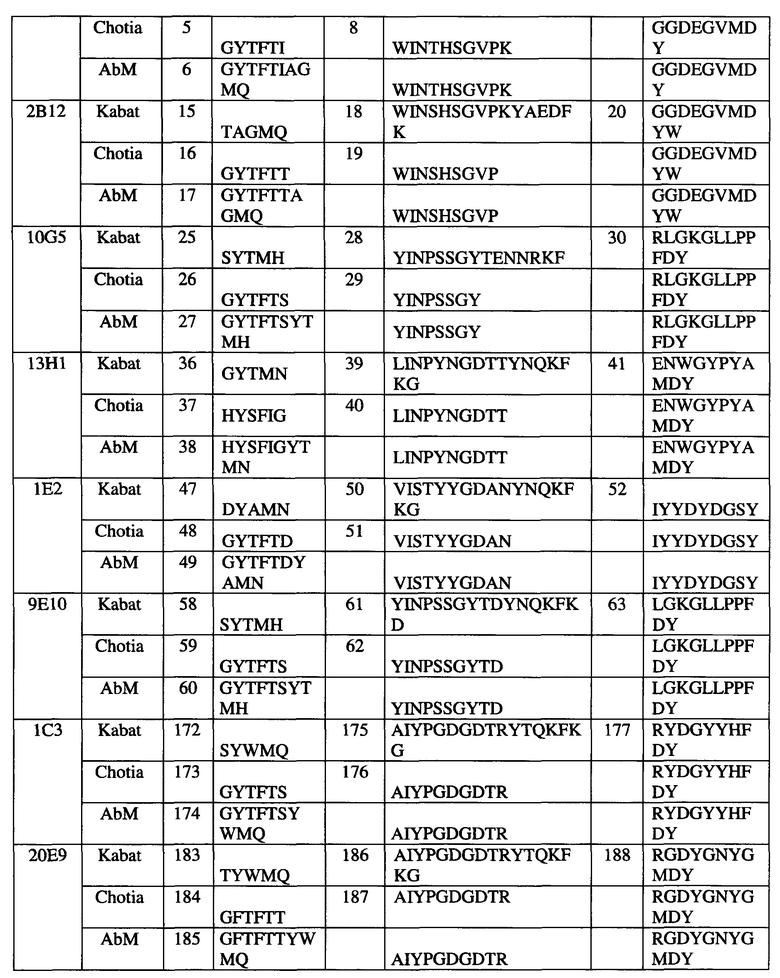
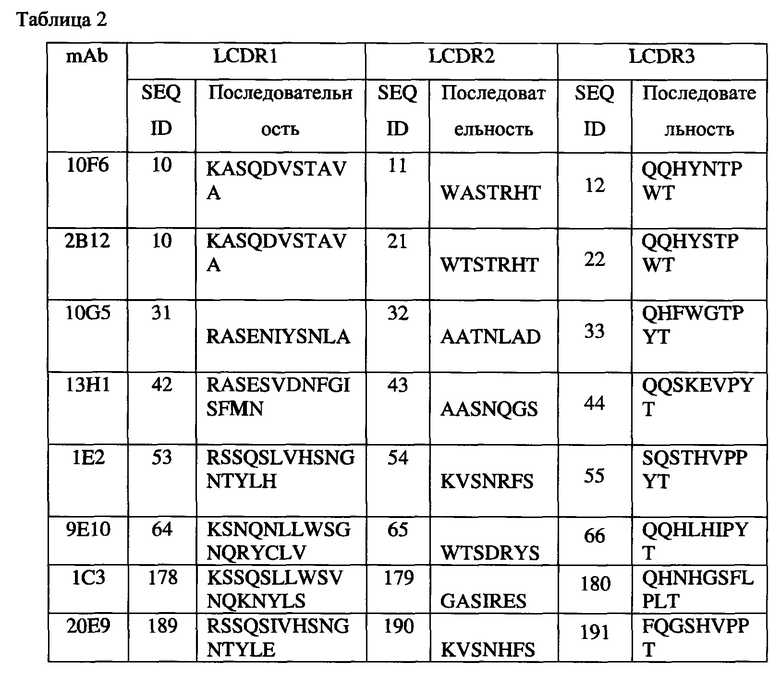
Последовательности вариабельных цепей указанных антител приведены в таблице 3 ниже; области CDR подчеркнуты. Согласно любому варианту реализации описанного изобретения последовательность VL или VH может быть задана или пронумерована так, чтобы содержать или не содержать сигнальный пептид или любую его часть.

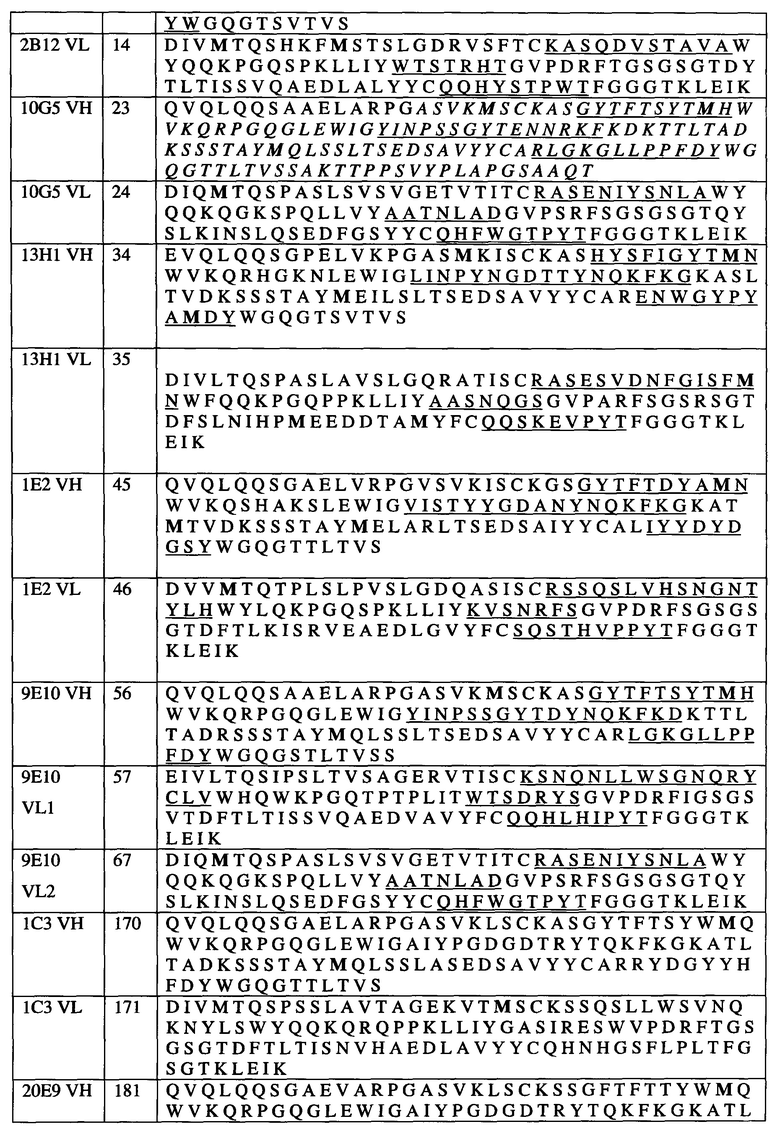

Фрагменты и производные
В настоящей заявке фрагменты и производные антител (охваченные термином «антитело» или «антитела», если не указано иное или это явным образом не противоречит контексту), предпочтительно, 10F6, 2В12, 18С6, 9Е10, 10G5, 13Н1, 5Н1, 1Е2, 1С3 или 20Е9-подобное антитело, могут быть получены с применением техник, известных в данной области техники. «Фрагменты» содержат фрагмент интактного антитела, как правило, антигенсвязывающий сайт или вариабельную область. Примеры фрагментов антител включают фрагменты Fab, Fab', Fab'-SH, F(ab')2, и Fv; диатела; любой фрагмент антитела, который представляет собой полипептид, имеющий первичную структуру, состоящую из одной непрерывной последовательности смежных остатков аминокислот (называемый в настоящем документе «одноцепочечным фрагментом антитела» или «одноцепочечным полипептидом»), в том числе, без ограничения, (1) одноцепочечные молекулы Fv (2) одноцепочечные полипептиды, содержащие только один вариабельный домен легкой цепи или его фрагмент, который содержит три участка CDR указанного вариабельного домена легкой цепи, без связанного фрагмента тяжелой цепи, и (3) одноцепочечные полипептиды, содержащие только одну вариабельную область тяжелой цепи или ее фрагмент, содержащий три участка CDR вариабельной области тяжелой цепи, без связанного фрагмента легкой цепи; и мультиспецифические антитела, образованные фрагментами антител. Включены, в том числе, нанотело, доменное антитело, однодоменное антитело, или «dAb».
Фрагменты предложенных антител могут быть получены с применением стандартных способов. Например, фрагменты Fab или F(ab')2 могут быть получены путем расщепления выделенных антител протеазами в соответствии с общепринятыми техниками. Следует понимать, что иммунореактивные фрагменты могут быть модифицированы с применением известных способов, например, для замедления клиренса in vivo и получения более благоприятного фармакокинетического профиля фрагмент может быть модифицирован полиэтиленгликолем (ПЭГ). Способы связывания и сайт-специфической конъюгации ПЭГ с фрагментом Fab' описаны, например, в источниках: Leong et al, 16 (3): 106-119 (2001) и Delgado et al., Br. J. Cancer 73 (2): 175-182 (1996), содержание которых включено в настоящий документ посредством ссылки.
Как вариант, ДНК гибридомы, продуцирующей антитело, предпочтительно, 10F6, 2В12, 18С6, 9Е10, 10G5, 13Н1, 5Н1, 1Е2, 1С3 или 20Е9-подобное антитело, может быть модифицирована таким образом, чтобы кодировать фрагмент. Модифицированную ДНК затем встраивают в экспрессионный вектор и используют для трансформации или трансфекции подходящей клетки, которая затем экспрессирует нужный фрагмент.
Согласно определенным вариантам реализации ДНК гибридомы, продуцирующей антитело, предпочтительно, 10F6, 2В12, 18С6, 9Е10, 10G5, 13Н1, 5Н1, 1Е2, 1С3 или 20Е9-подобное антитело, может быть модифицирована до встраивания в экспрессионный вектор, например, путем замены на кодирующую последовательность константных доменов тяжелой и легкой цепей человека гомологичных не принадлежащих человеку последовательностей (например, Morrison et al., PNAS pp. 6851 (1984)), или путем ковалентного связывания с кодирующей последовательностью иммуноглобулина всей или части кодирующей последовательности не являющегося иммуноглобулином полипептида. Таким образом получают «химерные» или «гибридные» антитела, обладающие специфичностью связывания исходного антитела. Как правило, такими не являющимися иммуноглобулинами полипептидами заменяют константные домены антитела.
Соответственно, согласно другому варианту реализации антитело, предпочтительно, 10F6, 2В12, 18С6, 9Е10, 10G5, 13Н1, 5Н1, 1Е2, 1С3 или 20Е9-подобное антитело, гуманизировано. «Гуманизированные» формы антител представляют собой специфические гибридные иммуноглобулины, иммуноглобулиновые цепи или их фрагменты (такие как Fv, Fab, Fab', F(ab')2, или другие антигенсвязывающие субпоследовательности антител), которые содержат минимальную последовательность, полученную из иммуноглобулина мыши. Гуманизированные антитела в общем являются иммуноглобулинами человека (реципиентное антитело), в которых остатки определяющей комплементарность области (CDR) реципиента заменены остатками CDR исходного антитела (донорного антитела), при этом сохраняются нужные специфичность, аффинность и активность исходного антитела.
В некоторых случаях каркасные остатки Fv иммуноглобулина человека могут быть заменены на соответствующие не принадлежащие человеку остатки. Кроме того, Гуманизированные антитела могут содержать остатки, которые отсутствуют как в реципиентном антителе, так и во встраиваемых CDR или каркасных последовательностях. Указанные модификации осуществляют для дополнительной коррекции и оптимизации характеристик антитела. В целом, гуманизированное антитело содержит по существу все из по меньшей мере одного, но обычно - двух вариабельных доменов, в которых все или по существу все области CDR соответствуют областям CDR исходного антитела, и все или по существу все области FR представляют собой области FR консенсусной последовательности иммуноглобулина человека. В оптимальном варианте указанное гуманизированное антитело также содержит по меньшей мере фрагмент константной области (Fc) иммуноглобулина, как правило, иммуноглобулина человека. Более подробную информацию см. в источниках: Jones et al., Nature, 321. pp. 522 (1986); Reichmann et al, Nature, 332, pp.323 (1988); Presta, Curr. Op. Struct. BioL, 2, pp. 593 (1992); Verhoeyen et Science, 239, pp. 1534; и в патенте США №4816567, содержание которых полностью включены в настоящий документ посредством ссылки.)
Выбор вариабельных доменов человека, как легкой, так и тяжелой цепи, для применения при получении гуманизированных антител очень важен для снижения антигенности. В соответствии с так называемым методом «наилучшего соответствия» проводят скрининг последовательности вариабельного домена антитела, сравнивая ее со всей библиотекой известных последовательностей вариабельных доменов человека. Затем последовательность человека, наиболее близкая последовательности мыши, выбирается в качестве каркасной области (FR) человека для гуманизированного антитела (Sims et al., J. Immunol. 151, pp. 2296 (1993); Chothia and Lesk, J. Mol. 196, 1987, pp. 901). В другом способе используется конкретная каркасная область консенсусной последовательности всех антител человека конкретной подгруппы легких или тяжелых цепей. Одна и та же каркасная область может использоваться для нескольких разных гуманизированных антител (Carter et al., PNAS 89, pp. 4285 (1992); Presta et al., J. Immunol., 151, p. 2623 (1993)).
Важно также, чтобы при гуманизации антител сохранялась высокая аффинность к рецепторам KIR3DL2 и другие благоприятные биологические свойства. С этой целью, в соответствии с иллюстративным способом, гуманизированные антитела получают путем анализа исходных последовательностей и различных концептуальных гуманизированных продуктов с применением трехмерных моделей исходных и гуманизированных последовательностей. Трехмерные модели иммуноглобулина общедоступны и знакомы специалистам в данной области техники. Доступны компьютерные программы для отображения и вывода возможных трехмерных структур выбранных кандидатных последовательностей иммуноглобулинов. Рассмотрение указанных изображений позволяет проанализировать вероятную роль остатков в функционировании кандидатной последовательности иммуноглобулина, т.е. проанализировать остатки, которые влияют на способность кандидатного иммуноглобулина к связыванию с его антигеном. Таким образом могут быть выбраны и скомбинированы остатки FR консенсусной и встраиваемой последовательностей, так что обеспечивается нужная характеристика антитела, такая как повышенная аффинность к целевому(ым) антигену(ам). В общем случае остатки CDR прямо и наиболее существенно влияют на связывание антигена.
Другой способ получения «гуманизированных» моноклональных антител заключается в использовании мыши в качестве хозяина; в соответствии с указанным способом гены иммуноглобулина мыши заменены функциональными генами иммуноглобулинов человека (см., например, патент США №6162963, полностью включенный в настоящий документ посредством ссылки).
Антитела человека могут также быть получены в соответствии с различными другими техниками, например, путем применения для иммунизации других трансгенных животных, сконструированных для экспрессии репертуара антител человека (Jakobovitz et al., Nature 362 (1993) 255), или путем отбора репертуаров антител с применением методов фагового дисплея. Такие техники известны специалистам и их можно реализовать, отталкиваясь от моноклональных антител, соответствующих описанию в настоящем документе.
Указанные антитела, необязательно 10F6, 2В12, 18С6, 9Е10, 10G5, 13Н1, 5Н1, 1Е2, 1С3 или 20Е9-подобное антитело, могут также быть дериватизированы с получением «гибридных» антител (иммуноглобулинов), в которых фрагмент тяжелой/легкой цепи(ей) идентичен или гомологичен соответствующим последовательностям исходного антитела, а остальная часть цепи(ей) идентична или гомологична соответствующим последовательностям антител, полученных от других видов или принадлежащих к другому классу или подклассу антител, а также фрагментов таких антител, при условии, что они проявляют нужную биологическую активность и специфичность связывания (Cabilly et al., выше; Morrison et al., Proc. Natl. Acad. Sci. U.S.A., pp. 6851 (1984)).
Предусмотрены различные формы гуманизированного антитела или антитела с созревшей аффинностью. Например, гуманизированное антитело или антитело с созревшей аффинностью может представлять собой фрагмент антитела, такой как Fab. Как вариант, гуманизированное антитело или антитело с созревшей аффинностью может представлять собой полноразмерное или интактное антитело, например, полноразмерное или интактное антитело IgG1 или IgG4. Согласно одному варианту реализации гуманизированное антитело представляет собой полноразмерное антитело IgG4 или его фрагмент. Для получения таких антител гуманизированные области VH и VL или их варианты могут быть клонированы в экспрессионные векторы, кодирующие полноразмерные или усеченные константные области антитела человека в соответствии со стандартными рекомбинантными методами (см., например, Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd Ed., Cold Spring Harbor Laboratory, Cold Spring Harbor, New York, 1989). В результате получают трансфицированную клеточную линию, которая экспрессирует и секретирует представляющую интерес молекулу гуманизированного антитела, содержащую выбранные области VH и VL и константные области. Последовательности кДНК, кодирующие константные области антител человека, известны.
Константная область может быть дополнительно модифицирована в соответствии с известными способами. Например, в константной области IgG4 остаток S241 может быть изменен на остаток пролина (Р), что обеспечивает образование полного дисульфидного мостика в шарнире (см., например, Angal et al., Mol Inununol. 1993; 30: 105-8).
Модифицированные константные области
Принимая во внимание способность антител против KIR3DL2 (в частности, неинтернализующихся антител) индуцировать АЗКЦ и КЗЦ, могут также быть получены указанные антитела, содержащие модификации, которые повышают их способность к связыванию рецепторов Fc, что может влиять на эффекторные функции, такие как антителозависимая цитотоксичность, дегрануляцию тучных клеток и фагоцитоз, а также иммуномодулирующие сигналы, например, регуляцию пролиферации лимфоцитов и секрецию антител. Типичные модификации включают модификации константных областей IgG1 человека, включающие по меньшей мере одну модификацию аминокислоты (например, замену, удаления, вставки) и/или изменение типа гликозилирования, например, гипофукозилирование. Такие модификации могут влиять на взаимодействие с рецепторами Fc: FcγRI (CD64), FcγRII (CD32) и FcγRIII (CD16). FcγRI (CD64), FcγRIIA (CD32A) и FcγRIII (CD16) являются активирующими (т.е. стимулирующими иммунную систему) рецепторами, a FcγRIIB (CD32B) представляет собой ингибирующий (т.е. подавляющий иммунную систему) рецептор. Модификация может, например, усиливать связывание домена Fc с FcγRIIIa на эффекторных клетках (например, NK-клетках).
Антитела против KIR3DL2 предпочтительно содержат домен Fc (или его фрагмент) изотипа IgG1 или IgG3 человека, необязательно модифицированный. Остатки 230-341 (Kabat EU) составляют область CH2Fc. Остатки 342-447 (Kabat EU) составляют область СН3 Fc. Антитела против KIR3DL2 могут содержать вариант области Fc, содержащий одну или большее количество модификаций аминокислот (например, замен, удалений, вставок) в одном или нескольких фрагментах; указанные модификации увеличивают аффинность и авидность варианта области Fc в отношении FcγR (включая активирующие и ингибирующие FcγR). Согласно некоторым вариантам реализации указанные одна или большее количество модификаций аминокислот увеличивают аффинность варианта области Fc FcγRIIIA и/или FcγRIIA. Согласно другому варианту реализации вариант области Fc также специфически связывает FcγRIIB с более низкой аффинностью по сравнению с областью Fc аналогичного исходного антитела (например, антитела, содержащего ту же аминокислотную последовательность, что и указанное антитело, за исключением одной или нескольких модификаций аминокислот в области Fc). Например, один или оба остатка гистидина в положениях АК 310 и 435 могут быть заменены, например, на лизин, аланин, глицин, валин, лейцин, изолейцин, пролин, метионин, триптофан, фенилаланин, серии или треонин (см., например, РСТ-публикацию, № WO 2007/080277); такие содержащие замены константные области обеспечивают сниженной связывание с ингибирующим FcγRIIB без снижения связывания с активирующим FcγRIIIA. Согласно некоторым вариантам реализации такие модификации увеличивают аффинность области Fc к FcγRDIA и/или FcγRIIA, а также повышают аффинность варианта области Fc к FcγyRIIB относительно исходного антитела. Согласно другим вариантам реализации указанные одна или несколько модификаций аминокислот увеличивают аффинность варианта области Fc к FcγRIIIA и/или FcγRIIA, но не изменяют аффинность варианта области Fс к FcγRIIB относительно области Fc исходного антитела. Согласно другому варианту реализации указанные одна или большее количество модификаций аминокислот повышают аффинность варианта области Fc к FcγRIIIA и FcγRIIA, но уменьшают аффинность к FcγRIIB относительно исходного антитела. Повышенная аффинность и/или авидность приводит к детектируемому связыванию с FcγR или связанной с FcγR активности в клетках, экспрессирующих незначительные количества FcγR, когда невозможно детектировать связывающую активность исходной молекулы (без модифицированной области Fc) в клетках.
Аффинность и связывающие свойства антител в отношении FcγR могут быть определены с применением известных в данной области техники in vitro анализов (биологических или иммунологических) для определения взаимодействий антитело-антиген или Fc-FcγR, т.е. специфического связывания антигена с антителом, или специфического связывания области Fc с FcγR, соответственно, включая, но не ограничиваясь перечисленными, ИФА ELISA, анализ с применением поверхностного плазмонного резонанса, иммунопреципитацию.
Согласно некоторым вариантам реализации антитела, содержащие вариант области Fc, содержат по меньшей мере одну модификацию аминокислоты (например, содержат 1, 2, 3, 4, 5, 6, 7, 8, 9, или большее количество модификаций аминокислот) в домене СН3 области Fc. Согласно другим вариантам реализации антитела, содержащие вариант области Fc, содержат по меньшей мере одну модификацию аминокислоты (например, содержат 1, 2, 3, 4, 5, 6, 7, 8, 9, или большее количество модификаций аминокислот) в домене области СН2 Fc, который определен как захватывающий аминокислоты 231-341. Согласно некоторым вариантам реализации антитела содержат по меньшей мере две модификации аминокислот (например, содержат 2, 3, 4, 5, 6, 7, 8, 9 или большее количество модификаций аминокислот), при этом по меньшей мере одна такая модификация находится в области СН3 и по меньшей мере одна такая модификация находится в области СН2. Предусмотрена также модификация аминокислоты в шарнирной области. Согласно одному варианту реализации предусмотрена модификация аминокислоты в области Fc домена СН1, который определен как захватывающий аминокислоты 216-230.
Может быть реализована любая комбинация модификаций Fc, например, любая комбинация различных модификаций, описанных в патентах США №№ US 7632497; 7521542; 7425619; 7416727; 7371826; 7355008; 7335742; 7332581; 7183387; 7122637; 6821505 и 6737056; в РСТ-публикациях №№ WO 2011/109400; WO 2008/105886; WO 2008/002933; WO 2007/021841; WO 2007/106707; WO 06/088494; WO 05/115452; WO 05/110474; WO 04/1032269; WO 00/42072; WO 06/088494; WO 07/024249; WO 05/047327; WO 04/099249 и WO 04/063351; и у Presta, L.G. et al. (2002) Biochem. Soc. Trans. 30(4): 487-490; Shields, R.L. et al. (2002) J. Biol. Chem. 26; 277(30): 26733-26740, и Shields, R.L. et al. (2001) J. Biol. Chem. 276(9): 6591-6604).
Антитела против KIR3DL2 могут содержать вариант области Fc, отличающийся тем, что указанный вариант области Fc содержит по меньшей мере одну модификацию аминокислоты (например, содержат 1, 2, 3, 4, 5, 6, 7, 8, 9, или большее количество модификаций аминокислот) относительно области Fc дикого типа, таким образом, что эффекторная функция указанной молекулы усилена по сравнению с молекулой, содержащей область Fc дикого типа, при этом необязательно указанный вариант области Fc содержит замену в любом одном положении или большем количестве положений из 221, 239, 243, 247, 255, 256, 258, 267, 268, 269, 270, 272, 276, 278, 280, 283, 285, 286, 289, 290, 292, 293, 294, 295, 296, 298, 300, 301, 303, 305, 307,308,309, 310, 311, 312, 316, 320, 322, 326, 329, 330, 332, 331, 332, 333, 334, 335, 337, 338, 339, 340, 359, 360, 370, 373, 376, 378, 392, 396, 399, 402, 404, 416, 419, 421, 430, 434, 435, 437, 438 и/или 439. Согласно одному варианту реализации антитела против KIR3DL2 могут содержать вариант области Fc, отличающийся тем, что. указанный вариант области Fc содержит по меньшей мере одну модификацию аминокислоты (например, содержит 1, 2, 3, 4, 5, 6, 7, 8, 9 или большее количество модификаций аминокислот) относительно области Fc дикого типа, таким образом, что эффекторная функция указанной молекулы усилена по сравнению с молекулой, содержащей область Fc дикого типа, при этом необязательно указанный вариант области Fc содержит замену в любом одном положении или большем количестве положений 329, 298, 330, 332, 333 и/или 334 (например, замены S239D, S298A, A330L, I332E, Е333А и/или K334A).
Согласно одному варианту реализации антитела, содержащие варианты области Fc или области Fc дикого типа, могут иметь измененные паттерны гликозилирования, которые повышают связывающую способность антител в отношении рецептора Fc. Такие модификации углеводов могут быть реализованы путем, например, экспрессии антитела в клетке-хозяине с измененными механизмами гликозилирования. Клетки с измененными механизмами гликозилирования были описаны в данной области техники и могут применяться в качестве клеток-хозяев для экспрессии рекомбинантных антител, с получением таким образом антитела с измененным гликозилированием. См., например, источники: Shields, R.L. et al. (2002) J. Biol. Chem. 277: 26733-26740; Umana et al. (1999) Nat. Biotech. 17: 176-1, а также европейский патент №: EP 1176195; PCT-публикации WO 06/133148; WO 03/035835; WO 99/54342, каждый из которых полностью включен в настоящий документ посредством ссылки.
Как правило, такие антитела с измененным гликозилированием «гликооптимизированы» таким образом, что антитело содержит конкретную N-гликановую структуру, которая обеспечивает определенные требуемые свойства, включая, но не ограничиваясь перечисленными, повышенную АЗКЦ и связывающую активность в отношении рецепторов эффекторных клеток по сравнению с немодифицированными антителами или антителами, содержащими встречающуюся в природе константную область, и продуцируемыми клетками NSO миеломы мыши и клетками яичников китайского хомячка (СНО) (Chu and Robinson, Current Opinion Biotechnol. 2001, 12: 180-7), антителами, экспрессируемыми НЕК293Т согласно описанию в настоящем документе в разделе «Примеры», или другими линиями клеток-хозяев млекопитающих, часто используемыми для получения рекомбинантных терапевтических антител.
Моноклональные антитела, продуцируемые в клетках-хозяевах млекопитающих, содержат сайг N-связанного гликозилирования по Asn297 каждой тяжелой цепи. Гликаны на антителах, как правило, представляют собой сложные биантеннарные структуры с очень низким содержанием или отсутствием N-ацетилглюкозамина (GlcNAc в точке ветвления) и высокими уровнями корового фукозилирования. Концы гликанов содержат очень незначительное количество или не содержат концевой сиаловой кислоты, и вариабельные количества галактозы. Обзор эффектов гликозилирования на функции антител приведен, например, у Wright & Morrison, Trend Biotechnol. 15: 26-31(1997). Существенный объем исследований демонстрирует, что изменения в составе Сахаров гликановой структуры антитела могут изменять эффекторные функции Fc. Предположительно, важными углеводными структурами, вносящими вклад в активность антитела, являются остатки фукозы, соединенные альфа-1,6-связью с наиболее глубоко расположенными остатками N-ацетилглюкозамина (GlacNAc) N-связанных олигосахаридов области Fc (Shields et al., 2002). Соответственно, могут быть получены антитела, содержащие меньшие уровни фукозы на N-связанных гликанах (гипофукозилированные N-связанные гликаны).
Для связывания FcγR требуется присутствие олигосахаридов, ковалентно связанных с консервативным Asn297 в области Fc иммуноглобулина человека типа IgG1, IgG2 или IgG3. Недавно была обнаружена связь между нефукозилированными олигосахаридными структурами и резким увеличением активности АЗКЦ in vitro. «Asn297» относится к аминокислоте аспарагину, расположенной приблизительно в положении 297 в области Fc; с учетом минорных вариаций последовательностей антител Asn297 также может быть расположена на несколько положений (обычно не более чем +3 аминокислоты) выше или ниже.
Как показывает опыт, приблизительно 2-6% популяции антител, продуцируемых клетками СНО, нефукозилировано. Было показано, что клеточные линии YB2/0 (миелома крысы) и Lecl3 (мутантная по пектину линия СНО с дефицитом СПР-маннозо-4,6-дегидратазы, приводящему к дефициту GDP-фукозы или промежуточных соединений GDP-сахаров, которые являются субстратом альфа-6-фукозилтрансферазы) продуцируют антитела, от 78 до 98% которых нефукозилировано. Согласно другим примерам может использоваться РНК-интерференция (RNAi) или нокаутные техники для конструирования клеток, либо для уменьшения уровней транскриптов мРНК FUT8, либо для полного нокаута генной экспрессии, и такие антитела, как сообщалось, содержат до 70% нефукозилированных гликанов.
Антитело, связывающееся с KIR3DL2, могут быть гликозилированы цепью сахара по Asn297; указанное антитело демонстрирует повышенное аффинность связывания с FcγRUI посредством фрагмента Fc. Согласно одному варианту реализации настоящего изобретения антитело содержит константную область, содержащую по меньшей мере одно изменение аминокислоты в области Fс, улучшающее связывание антитела с FcγRIIIa и/или АЗКЦ.
Согласно одному аспекту указанные антитела гипофукозилированы в константной области. Такие антитела могут содержать или не содержать модификацию аминокислоты, но быть получены или обработаны в условиях, обеспечивающих такое гипофукозилирование. Согласно одному аспекту композиция с антителом содержит гибридное антитело, антитело человека или гуманизированное антитело согласно описанию в настоящем документе, при этом по меньшей мере 20, 30, 40, 50, 60, 75, 85, 90, 95% или по существу все молекулы указанного антитела в композиции содержат константную область, содержащую коревую углеводную структуру (например, комплексные, гибридные и высокоманнозные структуры), в которой отсутствует фукоза. Согласно одному варианту реализации предложена композиция с антителом, где отсутствуют антитела, содержащие коровую углеводную структуру, содержащую фукозу. Коровые углеводы предпочтительно представляют собой цепь сахара на Asn297.
Согласно одному варианту реализации предложена композиция с антителом, например, композиция, содержащая антитела, которые связываются с KIR3DL2, гликозилированные цепью сахара по Asn297, при этом указанные антитела частично фукозилированы. Частично фукозилированные антитела характеризуются тем, что доля в композиции антител против KIR3DL2, в которых отсутствует фукоза в цепи сахара на Asn297, составляет от 20% до 90%, предпочтительно от 20% до 80%, предпочтительно от 20% до 50%, 55%, 60%, 70% или 75%, от 35% до 50%, 55%, 60%, 70% или 75%, или от 45% до 50%, 55%, 60%, 70% или 75%. Предпочтительно, указанное антитело относится к Ig человека типа IgG1 или IgG3.
Цепь сахара может дополнительно иметь любые характеристики (например, присутствие и доля комплексных, гибридных и высокоманнозных структур), включая характеристики N-связанных гликанов, присоединенных к Asn297 антитела, полученного из клетки человека, или антитела, рекомбинантно экспрессируемого в клетке грызуна, клетке мышеобразных (например, клетке СНО) или в клетке птицы.
Согласно одному варианту реализации антитело экспрессируется в клетке, в которой отсутствует фермент фукозилтрансфераза, так что указанная клеточная линия продуцирует белки, в которых отсутствует фукоза в коровых углеводах. Например, в клеточных линиях Ms704, Ms 705, и Ms709 отсутствует ген фукозилтрансферазы, FUT8 (альфа-(1,6)- фукозилтрансфераза), так что в антителах, экспрессируемых клеточными линиями Ms704, Ms705 и Ms709, отсутствует фукоза в коровых углеводах. Указанные клеточные линии были созданы путем направленного разрушения гена FUT8 в клетках CHO/DG44 с применением двух векторов замещения (см. патентную публикацию США №20040110704, Yamane et al.; и Yamane-Ohnuki et al. (2004) Biotechnol Bioeng 87: 614-22, содержание которых включено в настоящий документ посредством ссылки). Другие примеры включали применение антисмысловой супрессии, РНК-интерференции двуцепочечной РНК (dsRNA), РНК-интерференции шпилечной РНК (hpRNA) или РНК-интерференции интрон-содержащей шпилечной РНК (ihpRNA) для функционального разрушения гена FUT8. Согласно одному варианту реализации антитело экспрессируется в клеточной линии с функционально разрушенным геном FUT8, кодирующим фукозилтрансферазу, так что антитела, экспрессируемые в такой клеточной линии, демонстрируют гипофукозилирование за счет уменьшения количества или устранения относящегося к альфа-1,6-связям фермента.
Согласно одному варианту реализации антитело экспрессируется в клеточных линиях, сконструированных для экспрессии модифицирующих гликопротеины гликозилтрансфераз (например, бета(1,4)-N-ацетилглюкозаминил-трансферазы III (GnTHI)), так что антитела, экспрессируемые в указанных сконструированных клеточных линиях, демонстрируют увеличение количества GlcNac-структур в точках ветвления, что приводит к усилению АЗКЦ-активности антител (РСТ-публикация WO 99/54342, Umana et al.; и Umana et al. (1999) Nat. Biotech. 17: 176-180, содержание которых включено в настоящий документ посредством ссылки).
Согласно другому варианту реализации экспрессируется антитело и остаток(тки) фукозила расщепляют с применением фермента фукозидазы. Например, альфа-L-фукозидаза удаляет остатки фукозила из антител (Tarentino, et al. (1975) Biochem. 14: 5516-5523). Согласно другим примерам клеточная линия, продуцирующая антитело, может быть обработана ингибитором гликозилирования; у Zhou et al. Biotech. and Bioengin. 99: 652-665 (2008) описана обработка клеток СНО ингибитором альфа-маннозидазы I, кифунензином, приводящая к синтезу антител с нефукозилированными N-глюканами олигоманнозного типа.
Согласно одному варианту реализации антитело экспрессируется в клеточной линии, в которой наблюдается естественная низкая ферментативная активность в отношении добавления фукозила к N-ацетилглюкозамину, который связывается с областью Fc указанного антитела, либо указанная ферментативная активность отсутствует, например, в клеточной линии миеломы крысы YB2/0 (АТСС CRL 1662). Другие примеры клеточных линий включают вариант клеточной линии СНО, клетки Led 3 с пониженной способностью к присоединению фукозила к Asn(297)-связанным углеводам, что также приводит к гипофукозилированию антител, экспрессируемых такой клеткой-хозяином (WO 03/035835 (Presta et al); и Shields, RX. et al. (2002) J. Biol. Chem. 277: 26733-26740, содержание которых включено в настоящий документ посредством ссылки). Согласно другому варианту реализации антитело экспрессируется в клетке птиц, предпочтительно, в клетке ЕВх® (Vivalis, Франция), естественным образом продуцирующей антитела с низким содержанием фукозы, см., например, WO 2008/142124. Гипофукозилированные гликаны могут также продуцироваться в клеточных линиях растительного происхождения, см., например, WO 07/084926А2 (Biolex Inc.), WO 08/006554 (Greenovation Biotech GMBH), содержание которых включено в настоящий документ посредством ссылки.
Применение в диагностике и терапии
Согласно определенным вариантам реализации предложенные антитела используются для очищения или идентификации положительных по KIR3DL2 клеток из биологического образца. Биологические образцы могут быть получены от пациента, например, для диагностических или терапевтических целей ex vivo, или от индивидуумов или не являющихся человеком приматов для получения источника таких клеток для целей исследования.
Положительные по KIR3DL2 клетки могут быть очищены или идентифицированы с применением предложенных антител с помощью любых стандартных способов. Например, клетки периферической крови могут быть отсортированы с помощью FACS-сканера с применением меченых антител, специфических для KIR3DL2, и необязательно других молекул клеточной поверхности, как правило, присутствующих на клетках, например, CD4, CD8 или CD30 для Т-клеток; CD4 CD2+, CD3+, CD5+, CD8-, CD28+, CD45RO+ и/или TCRαβ+ для злокачественных клеток при синдроме Сезари; CD4+ (необязательно CD4+ и CD28-) при воспалительных, аутоиммунных или сердечно-сосудистых заболеваниях.
Кроме того, антитела могут быть конъюгированы или ковалентно связаны с твердой подложкой и использованы для очищения или идентификации положительных по KIR3DL2 клеток или любых клеток, экспрессирующих KIR3DL2, из биологического образца, например, образца крови или биоптата тканей слизистых оболочек пациента или другого индивидуума. В частности, указанный биологический образца приводят в контакт с антителами в условиях, обеспечивающих связывание клеток образца с указанным антителом, и затем указанные клетки отмывают от связанного с твердой подложкой антитела.
Независимо от способа, применяемого для выделения, очищения или идентификации положительные по KIR3DL2 клеток, возможность реализации таких выделения, очищения или идентификации полезна для применения в разнообразных целях, например, для диагностики расстройства, характеризующегося патогенным размножением экспрессирующих KIR3DL2 клеток, путем оценки количества или активности, или других характеристик положительных по KIR3DL2 клеток, полученных от пациента, или для оценки способности указанных антител или их фрагментов или производных модулировать активность или поведение клеток пациента, например, перед реализацией одного из вариантов описанного в настоящем документе лечения с применением указанных антител. Далее, очищенные положительные по KIR3DL2 клетки подходят для применения в контексте исследования, например, для лучшей характеризации клеток и различных их свойств и поведения, а также для идентификации соединений или способов, которые могут применяться для модулирования их поведения, активности или пролиферации. Указанные антитела могут также подходить для применения в способах диагностики, например, способах детекции полипептидов KIR на клетках, например, пораженных заболеванием клетках пациента.
В настоящем описании также предложены фармацевтические композиции, содержащие антитело, которое специфически связывается с полипептидами KIR3DL2 на поверхности клеток. Указанное антитело предпочтительно подавляет рост или активность (например, синтез цитокинов) клеток и/или приводит к элиминации положительных по KIR3DL2 клеток, предпочтительно, за счет индуцирован™ КЗЦ и/или АЗКЦ. Указанная композиция дополнительно содержит фармацевтически приемлемый носитель.
В описываемом изобретении также предложен способ подавления роста или активности, и/или истощения положительных по KIR3DL2 клеток у нуждающегося в этом пациента, включающий этап введения указанному пациенту композиции, описанной в настоящем документе. Такие способы лечения могут применяться для ряда расстройств, включая, но не ограничиваясь перечисленными, CTCL, СС и ФМ, воспалительные, аутоиммунные и сердечно-сосудистые расстройства.
Злокачественные Т-клетки CD4+, экспрессирующие на поверхности KIR3DL2, присутствуют независимо от формы CTCL CD4+. Соответственно, KIR3DL2 присутствует при различных вариантах CTCL CD4+, и, в частности, при синдроме Сезари («СС»), трансформированном фунгоидном микозе («трансформированный ФМ»), лимфоматоидном папулезе («ЛП») и лимфомах CD30+.
Диагноз (например, диагноз CTCL) может быть основан на анализе присутствия KIR3DL2 на поверхности клеток CD4+, полученных из исследуемой области организма (например, в образце кожной эритродермии при подозрении на трансформированный ФМ, или в образце периферической крови при подозрении на более агрессивную форму CTCL, такую как СС). Как правило, можно заключить, что Т-клетка CD4+ является опухолевой, если на поверхности указанных Т-клеток CD4+ детектированы полипептиды KIR3DL2. Можно измерить процент Т-клеток CD4+ KIR3DL2+ в образце периферической крови, полученном от пациента, у которого подозревают наличие СС, и такой процент по существу будет соответствовать проценту злокачественных клеток СС, фактически присутствующего в периферической крови указанного пациента (как правило, в диапазоне ±10% или даже ±5% для клеток KIR3DL2+ CD4+). KIR3DL2 и антитела против KIR3DL2, описанные в настоящем документе, соответственно, могут применяться для стадирования заболевания, в частности, СС.
Поскольку KIR3DL2 представляет собой универсальный маркер CTCL, указанные антитела могут применяться в комбинации с другим лечением или диагностическими маркерами CTCL, например, CD30, присутствие которого на поверхности злокачественных Т-клеток CD4+ указывает на то, что у указанного пациента имеется конкретная форма CTCL CD4+, называемая в данной области техники лимфомой CD30+. CD30, соответственно, является маркером конкретной формы CTCL (лимфомы CD30+), однако не охватывает все формы CTCL CD4+, поскольку при CTCL CD4+, например, при СС, трансформированном ФМ или ЛП, необязательно присутствуют злокачественные Т-клетки CD4+, которые экспрессировали бы CD30 на поверхности. Соответственно, CD30 может применяться дополнительно к KIR3DL2 в качестве маркера в диагностике и терапии CTCL. Кроме того, обнаружение того, что у пациента имеется CTCL CD4+, экспрессирующая CD30, может указывать на то, что для указанного пациента подходит лечение антителом против KIR3DL2 и антителом против CD30; необязательно указанный пациент может после этого получать лечение антителом против KIR3DL2 и антителом против CD30.
Согласно некоторым вариантам реализации перед введением указанного антитела против KIR3DL2 или airra-KIR3DL2 композиции анализируют присутствие CD2, CD3, CD4, CD5, CD8, CD28, CD30, CD45RO и/или TCRαβ на клетках (например, патогенных клетках) пациента. Пациент, клетки которого экспрессируют (или не экспрессируют, в соответствии с конкретным целевым расстройством и целевыми клетками) маркер, затем может получать лечение антителом против KIR3DL2 или анти-KIR3DL2 композицией. Согласно некоторым вариантам реализации перед введением указанного антитела против KIR3DL2 или -KIR3DL2 композиции анализируют присутствие KIR3DL2 на клетках пациента, например, для определения относительного содержания и активности положительных по KIR3DL2 клеток у указанного пациента, а также для подтверждения эффективности связывания антител с указанными клетками пациента. Пациент, клетки которого экспрессируют KIR3DL2, может затем получать лечение антителом против KIR3DL2 или анти-KIR3DL2 композицией. Это может быть осуществлено путем получения образца ЛПК или клеток из области, пораженной расстройством, и тестирования, например, с применением методов иммуноанализа, для определения относительной выраженности таких маркеров, как CD4, CD8, CD30 или KIR3DL2, на клетках.
Согласно одному варианту реализации, если требуется подавление активности или роста, либо истощение положительных по KIR3DL2 клеток у пациента, оценивают способность указанного антитела против KIR3DL2 подавлять пролиферацию или истощать положительные по KIR3DL2 клетки у пациента. В том случае, если происходит истощение по положительным по KIR3DL2 клеткам при применении антитела против KIR3DL2 или анти-KIR3DL2 композиции, указанный пациент определяется как отвечающий на терапию антителом против KIR3DL2 или анти-KIR3DL2 композицией, и необязательно указанный пациент получает лечение антителом против KIR3DL2 или анти-KIR3DL2 композицией.
Согласно некоторым вариантам реализации указанный способ может включать дополнительный этап введения указанному пациенту подходящего дополнительного (второго) терапевтического агента, выбранного из иммуномодулирующего агента, иммуносупрессивного агента, гормонального агента, химиотерапевтического агента, второго антитела (например, истощающего антитела), который связывается с полипептидом, присутствующим на экспрессирующей KIR3DL2 клетке. Такие дополнительные агенты могут вводиться указанному пациенту в составе одной лекарственной формы вместе с указанным антителом, или в виде отдельной лекарственной формы. Дозировка указанного антитела (или дозировка антитела и дозировка дополнительного терапевтического агента в совокупности) достаточна для того, чтобы детектируемо индуцировать, способствовать и/или усиливать терапевтический ответ у пациента. При раздельном введении антитело, фрагмент или производное и дополнительный терапевтический агент предпочтительно вводят в условиях (например, в отношении времени, количества доз и т.д.), которые приводят к детектируемому комбинированному благоприятному терапевтическому эффекту у указанного пациента.
Фунгоидный микоз и более агрессивный синдром Сезари представляют собой наиболее распространенные формы CTCL. Клиническое течение ФМ/СС, как правило, индолентное, с медленно развивающимися в течение длительных периодов зудящими эритематозными областями. Однако в конечном итоге эритематозные пятна становятся прогрессивно инфильтрующими, превращаются в бляшки и, наконец, в язвенные опухоли. Прогноз при ФМ/СС зависит от распространения заболевания на момент постановки диагноза. Медиана выживаемости пациентов с заболеванием на I стадии составляет 20 лет или более, в отличие от медианы выживаемости пациентов с заболеванием III/IV стадии, составляющей приблизительно 3-4 года.
Описанные в настоящем документе композиции может применяться для лечения в комбинации с любым известным агентом, подходящим для лечения конкретного злокачественного Т-клеточного заболевания. Применяются различные способы лечения CTCL, включая кортикостероиды, мустарген, кармустин, такролимус для местного применения (Protopic®), имиквимод (Aldara®; 3М Inc.), ретиноиды для местного применения, и рексиноиды (бексаротен; Таргретин®; Ligand Pharmaceuticals, Сан-Диего, Калифорния)), а также терапию ультрафиолетовым излучением (псорален + УФА (ГГУВА-терапия), узкополосный УФВ, и УФВ), фотодинамическая терапия (ФДТ) и облучение тела. Способы лечения также включают ингибиторы гистондеацетилазы, такие как вориностат (субероиланилид гидроксамовой кислоты, Zolinza®) и Ромидепсин (депсипептид, FK-228, Istodax®), циклический пептид, который селективно подавляет гистондеацетилазу изотопов 1, 2, 4 и 6. Также используется химиотерапия или комбинированная химиотерапия. Примеры включают гемцитабин, аналоги антифолатов, такие как пралатрексат (Folotyn®). Другие методы терапии включают IMiD (иммуномодулирующие лекарственные средства), аналоги, полученные из талидомида, обладающие широким спектром действия, включая как связанные, так и не связанные с иммунной системой эффекты. Представители класса IMiD включают СС-5013 (леналидомид; Ревлимид®), СС-4047 (Актимид), и ENMD-0995. Другие методы терапии включают ингибиторы протеосомы, такие как бортезомиб (Velcade®), обратимый ингибитор 26S-протеасомы. Также применяется трансплантация стволовых клеток.
Несмотря на то, что действующего стандарта медицинской помощи при ФМ/СС нет, имеется общая тенденция к использованию местных методов вмешательства на ранних стадиях заболевания, с откладыванием более токсичной системной терапии до развития обширных симптомов. Терапия псораленом и УФА-излучением (ПУВА-терапия), скомбинированная или не скомбинированная с низкими дозами интерферона-α, эффективна на ранних стадиях ФМ/СС, обеспечивая полную ремиссию (CR) у большинства пациентов. Местная лучевая терапия или тотальное облучение кожи электронным пучком (TSEB) успешно применялись для контроля распространенного кожного заболевания. Также может успешно применяться экстракорпоральный фотоферез, но он не является общедоступным. Если заболевание становится рефрактерным к местной терапии, интерферону-α, рексиноиду бексаротену (Таргретин®, Ligand Pharmaceuticals, Сан-Диего, Калифорния), могут применяться синтетический ретиноидный аналог, нацеленный на ретиноидный рецептор X, монохимиотерапия или комбинированная химиотерапия. Способы лечения, в частности, направленной терапии кожи, включают, например, кортикостероиды, мустарген, кармустин, такролимус для местного применения (Protopic®) и имиквимод (Aldara®; 3М Inc.). Однако часто длительность ответа составляет менее 1 года, в конечном итоге у всех пациентов наблюдается рецидив и заболевание становится рефракторным. Рекомбинантный ИЛ2-дифтерийный токсин денилейкин дифтитокс (Denileukin diftitox) (DAB389IL-2, ONTAK®, Ligand Pharmaceuticals, Сан-Диего, Калифорния) эффективен у пациентов с CTCL на стадиях Ib-IV, рефракторными к предшествующему лечению (суммарный объективный ответ у 30% из 71 пациентов с медианной длительностью ответа 7 месяцев) и, по всей видимости, оказывает благоприятный эффект в отношении купирования симптомов и качества жизни. Недавно денилейкин дифтитокс (Denileukin diftitox) был протестирован в исследовании фазы I в комбинации с бексаротеном, поскольку он индуцирует повышающую регуляцию CD25 in vitro. Комбинация хорошо переносилась и индуцировала объективный ответ у 67% из 14 пациентов. Наиболее значительными были нежелательные явления, о которых уже сообщалось при отдельном применении бексаротена (гипертриглицеридемия и подавление функции щитовидной железы из-за снижения синтеза ТТГ), и лимфопения 3-й или 4-й степени, однако они исчезали в течение одного месяца после прекращения терапии. Информация о продолжительности времени до констатации отсутствия эффекта лечения для указанного исследования не указана. В других исследованиях разрабатывались антитела против CD4, истощающие экспрессирующие CD4 клетки. Примеры включают полностью человеческое IgG1-антитело против CD4 занолимумаб (HuMax-CD4; Genmab A/S и TenX BioPharma hie.) и гибридное моноклональное антитело против CD4 (сМ-Т412, Centocor, Мальверн, Пенсильвания) вводили 8 пациентам с ФМ и индуцировали объективный ответ у 7 из них, однако медианная длительность ответа составляла всего 5 месяцев. Увадекс® (метоксален, Therakos Inc. Экстон, Пенсильвания) при экстракорпоральном фотоферезе также демонстрировал признаки эффективности. Гуманизированное моноклональное антитело алемтузумаб (моноклональное антитело человека IgG1 против CD52, Campath®, Millennium Pharmaceuticals, Inc. и ILEX Oncology, Inc., предлагаемое и распространяемое в США Berlex Laboratories, Inc., Монтвилл, Нью-Джерси) показано для лечения В-клеточного хронического лимфоцитарного лейкоза (B-CLL) у пациентов, получавших лечение алкилирующими агентами и не отвечавших на терапию флударабином. Его тестировали на пациентах с распространенным ФМ/СС (заболевание III или IV стадии) и оно вызывало объективный ответ по меньшей мере в половине случаев (55% из 22 пациентов). Профиль его побочных эффектов преимущественно представлен иммуносупрессией и инфузионными реакциями. В независимом ретроспективном исследовании описывается также значительная кардиотоксичность у 4 из 8 пациентов. С учетом долгосрочности наблюдаемой ремиссии (медиана времени до констатации отсутствия эффекта лечения составляет 12 месяцев, диапазон - от 5 до 32+ месяцев), терапия алемтузумабом, по-видимому, представляет собой лечение с более благоприятной медианной длительностью ответа из всех описанных на сегодняшний день методов лечения. Другие подходящие агенты могут включать антитела против CCR4 (С-С хемокиновый рецептор 4; CD194). Одним из примеров является могамулизумаб (KW-0761; AMG-761; торговое наименование Poteligeo, Kyowa Hakko Kirin Ltd., Япония и Amgen, США) и гуманизированное антитело против CCR4. Другие подходящие агенты могут включать антитела против CD30. Одним из примеров является SGN-35, который представляет собой конъюгат антитела с лекарственным препаратом (ADC), содержащий мощное антимитотическое лекарственное вещество, монометилауристатин Е (ММАЕ), соединенный с моноклональным антителом против CD30, сАС10 (Okeley et al. (2010) Clin. Cancer Res. 16(3): 888-897); другим примером является иммуноглобулин (Ig) 01к человека, моноклональное антитело против CD30 MDX-060 (Medarex Inc. и Bristol Myers Squibb; Ansell et al. (2007) J. Clin. Oncol. 25: 2767-2769). Каждый из указанных способов лечения может применяться в комбинации с антителами согласно настоящему описанию.
Антитела, полученные с применением предложенных способов эффективны, в частности, при лечении аутоиммунных и воспалительных расстройств, а также сердечно-сосудистых расстройств, в особенности, острого коронарного синдрома, артрита, ревматоидного артрита, ревматоидного васкулита, системной красной волчанки, рассеянного склероза и гранулематоза Вегенера, спондилоартрита. В целом, предложенные способы могут применяться для лечения любого расстройства, вызванного по меньшей мере отчасти присутствием или активностью экспрессирующих KIR3DL клеток, например, NK-клеток или Т-клеток, провоспалительных Т- или NK-клеток, продуцирующих ИЛ-17А, таких Т-клеток, как клетки Th17 или клетки CD4+CD28-, экспрессирующие KIR3DL2, и для лечения которого может, соответственно, быть эффективным селективный киллинг или подавление пролиферации или активации экспрессирующих KIR3DL2 клеток.
Согласно некоторым вариантам реализации перед введением указанного антитела против KIR3DL2 анализируется экспрессия KIR3DL2 на клетках, лежащих в основе конкретного расстройства. Это может осуществляться путем получения образца ЛПК или клеток из очага расстройства (например, из синовиальной оболочки пациентов с РА), и тестирования, например, с применением методов иммуноанализа, для определения относительной выраженности таких маркеров, как CD4, CD28 и т.д., а также KIR3DL2 на указанных клетках. Могут также использоваться другие способы детекции экспрессии KIR3DL2 и других генов, такие как способы на основе РНК, например, ОТ-ПЦР или нозерн-блоттинг.
Лечение может включать несколько циклов введения антитела или соединения. Например, после первого цикла введения уровень и/или активность экспрессирующих KIR3DL Т-клеток или NK-клеток (например, Т-клеток CD4+CD28-, злокачественных CD4+ Т-клеток) у пациента, как правило, повторно измеряют, и в том случае, если указанные значения все еще повышены, может проводиться дополнительный цикл введения. Таким образом могут быть проведены несколько циклов детекции рецептора и введения антитело или соединения, например, до тех пор, пока не будет установлен контроль над указанным расстройством.
При применении для лечения аутоиммунных или воспалительных расстройств антитела против KIR3DL2 согласно настоящему описанию могут применяться для лечения в комбинации с любым агентом, который, как известно, подходит при лечении конкретного воспалительного расстройства, аутоиммунного расстройства или сердечно-сосудистого расстройства. Антитела против KIR3DL2 могут быть скомбинированы, например, со стероидными противовоспалительными агентами, нестероидными противовоспалительными агентами, антиметаболитами и другими агентами, применяемыми при лечении сердечно-сосудистых, воспалительных или аутоиммунных заболеваний. Согласно некоторым вариантам реализации противовоспалительные агенты содержат стероидные противовоспалительные агенты, которые включают глюкокортикостероиды и минералокортикостероиды. Они могут вводиться любыми способами, подходящими для лечения воспалительных расстройств, включая, в числе прочих, пероральный, внутривенный, внутримышечный, дермальный или назальный пути введения. Согласно некоторым вариантам реализации противовоспалительные агенты содержат нестероидные противовоспалительные агенты. Указанные агенты, как правило, действуют за счет подавления действия ферментов циклооксигеназы и липоксигеназы, или рецепторов к медиаторам, синтезируемым указанными ферментами. Нестероидные противовоспалительные соединения включают неселективные ингибиторы СОХ, селективные ингибиторы СОХ, а также антагонисты FLAP и антагонисты 5- липоксигеназы. Согласно некоторым вариантам реализации противовоспалительные агенты могут содержать антиметаболиты, влияющие на пролиферацию клеток, вовлеченных в иммунный ответ. Подходящие антиметаболиты включают аналоги фолатов, такие как метотрексат; ингибиторы инозин-монофосфат-дегидрогеназы (IMPDH), такие как микофенолата мофетил; и азатиопурин. Соединения указанной группы, как правило, влияют на синтез субстратов, необходимых для репликации ДНК, подавляя таким образом пролиферацию клеток, вовлеченных или активируемых в ответ на воспалительную реакцию. Согласно некоторым вариантам реализации противовоспалительный агент представляет собой агент, блокирующий действие ФНО-альфа, основной цитокин, вовлеченный в воспалительные расстройства. Согласно некоторым вариантам реализации анти-ФНО агент представляет собой антитело, которое блокирует действие ФНО-альфа. Примером антитела против ФНО является инфликсимаб (Remicade®). Согласно другим вариантам реализации агент против ФНО-альфа представляет собой рецепторную конструкцию, которая связывает ФНО-альфа и предотвращает его взаимодействие с рецепторами ФНО, присутствующими на клетках, например, этанерцепт (Enbrel®). Согласно другим вариантам реализации противовоспалительный агент представляет собой любой другой агент (например, антитело) с иммуносупрессивными свойствами, подходящий при лечении расстройства, лечение которого проводят с применением антитела к KIR3DL2 согласно описанию в настоящем документе.
Фармацевтические составы
Фармацевтически приемлемые носители, которые могут использоваться в указанных композициях, включают, не ограничиваясь перечисленными, ионообменники, глинозем, стеарат алюминия, лецитин, сывороточные белки, такие как сывороточный альбумин человека, буферные вещества, такие как фосфаты, глицин, сорбиновая кислота, сорбат калия, смеси неполных глицеридов насыщенных растительных жирных кислот, вода, соли или электролиты, такие как протаминсульфат, вторичный кислый фосфат натрия, кислый фосфат калия, хлорид натрия, соли цинка, коллоидный диоксид кремния, трисиликат магния, поливинилпирролидон, вещества на основе целлюлозы, полиэтиленгликоль, карбоксиметилцеллюлоза натрия, полиакрилаты, воски, полиэтилен-полиоксипропилен-блокполимеры, полиэтиленгликоль и ланолин. Антитела, описанные в настоящем документе, могут применяться в способе модулирования, например, подавления, активности экспрессирующих KIR3DL2 клеток у пациента. Указанный способ включает этап приведения в контакт указанной композиции с указанным пациентом. Такой способ подходит как для профилактических, так и для терапевтических целей.
При применении для введения пациенту композицию вводят в состав для введения пациенту. Композиции, описанные в настоящем документе, могут вводиться перорально, парентерально, путем ингаляции, местно, ректально, назально, буккально, вагинально или с помощью имплантированного резервуара. Согласно настоящему описанию используются техники подкожного, внутривенного, внутримышечного, внутрисуставного, внутрисиновиального, внутригрудинного, внутриоболочечного, внутрипеченочного, внутриочагового и внутричерепного инъецирования или инфузии. Антитело может присутствовать в разовой дозе в количестве, например, от приблизительно 25 мг до 500 мг.
Стерильные инъецируемые формы описанных в настоящем документе композиций могут представлять собой водную или масляную суспензию. Составы с указанными суспензиями могут быть получены в соответствии с известными в данной области техниками с применением подходящих диспергирующих или смачивающих агентов и суспендирующих агентов. Стерильный инъецируемый состав может также представлять собой стерильный инъецируемый раствор или суспензию в нетоксичном приемлемом для парентерального применения разбавителе или растворителе, например, раствор в 1,3-бутандиоле. Подходящие для применения приемлемые основы и растворители включают воду, раствор Рингера и изотонический раствор хлорида натрия. Кроме того, обычно используются стерильные нелетучие масла в качестве растворителя или суспендирующей среды. Для указанной цели может использоваться любое мягкое нелетучее масло, включая синтетические моно- или диглицериды. Жирные кислоты, такие как олеиновая кислота и ее глицеридные производные, подходят для получения инъецируемых составов, как и природные фармацевтически приемлемые масла, такие как оливковое масло или касторовое масло, в частности, их полиоксиэтилированные варианты. Указанные масляные растворы или суспензии могут также содержать в качестве разбавителя или диспергирующего агента длинноцепочечный спирт, такой как карбоксиметилцеллюлоза, или аналогичные диспергирующие агенты, обычно используемые при получении фармацевтически приемлемых лекарственных форм, включая эмульсии и суспензии. Другие широко используемые поверхностно-активные вещества, такие как твины, спаны и другие эмульгирующие агенты или повышающие биодоступность вещества, обычно используемые при производстве фармацевтически приемлемых твердых, жидких или других лекарственных форм могут также быть использованы для получения составов.
Описанные в настоящем документе композиции могут вводиться перорально в виде любой приемлемой для перорального применения лекарственной форме, включая, но не ограничиваясь перечисленными, капсулы, таблетки, водные суспензии или растворы, В случае таблеток для перорального применения широко используемые носители включают лактозу и кукурузный крахмал. Также, как правило, добавляют смазывающие агенты, такие как стеарат магния. Для перорального введения в форме капсул включают подходящие разбавители, например, лактозу. Если необходимо получить водные суспензии для перорального применения, активный ингредиент комбинируют с эмульгирующими и суспендирующими агентами. При необходимости могут также быть добавлены определенные подсластители, вкусоароматические или подкрашивающие агенты.
Как вариант, описанные в настоящем документе композиции могут вводиться в форме суппозиториев для ректального введения. Указанные суппозитории могут быть получены путем смешивания агента с подходящим нераздражающим вспомогательным веществом, твердым при комнатной температуре, но жидким при ректальной температуре и, таким образом, правящимся в прямой кишке с высвобождением лекарственного средства. Такие материалы включают масло какао, пчелиный воск и полиэтиленгликоли.
Описанные в настоящем документе композиции могут также вводиться местно, в частности, если мишень лечения включает области или органы, легко доступные для местного нанесения, в том числе заболевания глаз, кожи или нижних отделов кишечника. Подходящее составы для местного применения могут быть легко получены для каждой(ого) из указанных областей или органов. Для местного применения композиции могут быть введены в состав подходящей мази, содержащей активный компонент, суспендированный или растворенный в одном или нескольких носителях. Носители для местного введения описанных в настоящем документе соединений включают, не ограничиваясь перечисленными, минеральное масло, жидкий вазелин, белый вазелин, пропиленгликоль, полиоксиэтилен, соединение полиоксипропилена, эмульгирующий воск и воду. Как вариант, указанные композиции могут входить в состав подходящего лосьона или крема, содержащего активные компоненты, суспендированные или растворенные в одном или нескольких фармацевтически приемлемых носителей. Подходящие носители включают, не ограничиваясь перечисленными, минеральное масло, сорбитан моностеарат, полисорбат 60, воск цетиловых эфиров, цетеариловый спирт, 2-октилдодеканол, бензиловый спирт и воду.
Представленные антитела могут входить в состав наборов. Наборы могут необязательно дополнительно содержать любое количество антител и/или других соединений, например, 1, 2, 3, 4, или любое другое количество терапевтических антител и/или соединений. Следует иметь в виду, что указанное описание содержимого наборов не является в каком-либо смысле ограничивающим. Например, набор может содержать другие типы терапевтических соединений. Предпочтительно указанные наборы также содержат инструкции по применению антител, например, с подробным описанием представленных в настоящем документе способов.
Лекарственные формы
Терапевтические составы с антителами подготавливают для хранения путем смешивания антител нужной степени чистоты с необязательными фармацевтически приемлемыми носителями, вспомогательными веществами или стабилизаторами в форме лиофилизированных составов или водных растворов. Общую информацию по составам можно найти, например, в источниках: Gilman et al. (eds.), The Pharmacological Bases of Therapeutics, 8th Ed. (Pergamon Press, 1990); Gennaro (ed.), Remington's Pharmaceutical Sciences, 18th Edition (Mack Publishing Co., Easton, Pa., 1990); Avis et al. (eds.), Pharmaceutical Dosage Forms: Parenteral Medications (Dekker, New York, 1993); Lieberman et al. (eds.). Pharmaceutical Dosage Forms: Tablets (Dekker, New York, 1990); Lieberman et al. (eds.) Pharmaceutical Dosage Forms: Disperse Systems (Dekker, New York, 1990); и Walters (ed.), Dermatological and Transdermal Formulations (Drugs and the Pharmaceutical Sciences), Vol 119 (Dekker, New York, 2002).
Приемлемые носители, вспомогательные вещества или стабилизаторы нетоксичны для реципиентов в используемых дозировках и концентрациях, и включают буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония, хлорид бензетония; феноловый, бутиловый или бензиловый спирт; алкилпарабены, таких как метил- или пропилпарабен; катехин; резорцин; циклогексанол; 3-пентанол; и м-крезол); полипептиды с низкой молекулярной массой (менее чем приблизительно 10 остатков); белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, таких как глицин, глутамин, аспарагин, гистидин, аргинин, или лизин; моносахариды, дисахариды, и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как этилендиаминтетрауксусная кислота (ЭДТК); сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; металлокомплексы (например, комплексы белков с Zn); и/или неионные поверхностно-активные вещества, такие как твины (TWEEN™), плюроники (PLURONICS™) или ПЭГ.
Примеры составов с антителами описаны, например, в WO 1998/56418, где описывается жидкий многодозовый состав с антителом против CD20, содержащий 40 мг/мл ритуксимаба, 25 мМ ацетат, 150 мМ трегалозу, 0,9% бензилового спирта и 0,02% полисорбата 20™ с рН 5,0, имеющий минимальный срок хранения 2 года при хранении при 2-8°С. Другой представляющий интерес анти-CD20 состав содержит 10 мг/мл ритуксимаба в 9,0 мг/мл хлорида натрия, 7,35 мг/мл дигидрата цитрата натрия, 0,7 мг/мл полисорбата 80™ и стерильную воду для инъекций, рН 6,5.
Лиофилизированные составы, предназначенные для подкожного введения, описаны, например, в патенте США №6267958 (Andya et al.). Такие лиофилизированные составы могут быть восстановлены подходящим разбавителем с получением высокой концентрации белка, и указанный восстановленный состав может вводиться подкожно млекопитающему, подлежащему лечению согласно настоящему документу.
Состав согласно настоящему описанию может также содержать более чем одно активное соединение (второй медикамент, как отмечалось выше), предпочтительно такие, дополнительные типы активности которых не влияют друг на друга неблагоприятным образом. Тип и эффективные количества таких медикаментов зависят, например, от количества и типа В-клеточного антагониста, присутствующего в составе, и клинических показателей субъектов. Примеры вторых медикаментов приведены выше.
Активные ингредиенты могут также быть заключены в микрокапсулы, полученные, например, с применением техник коацервации или межфазной полимеризации, например, гидроксиметилцеллюлозные или желатиновые микрокапсулы и поли-(метилметацилатные) микрокапсулы, соответственно, в коллоидных системах доставки лекарственных веществ (например, липосомах, альбуминовых микросферах, микроэмульсиях, наночастицах и нанокапсулах), или в макроэмульсиях. Такие техники описаны, например, в руководстве Remington's Pharmaceutical Sciences, см. выше.
Могут быть получены составы для длительного высвобождения. Подходящие примеры составов для длительного высвобождения включают полупроницаемые матрицы твердых гидрофобных полимеров, содержащих антагонист, в виде объектов определенной формы, например, пленок или микрокапсул. Примеры матриц для длительного высвобождения включают полиэфиры, гидрогели (например, поли(2-гидроксиэтил-метакрилат), или поли(виниловый спирт)), полилактиды (патент США №3773919), сополимеры L-глутаминовой кислоты и γ-этил-L-глутамата, неразлагаемый этиленвинилацетат, разлагаемые сополимеры молочной и гликолевой кислот, такие как Lupron Depot™ (инъецируемые микросферы, состоящие из сополимера 1 молочной и гликолевой кислот и лейпролида ацетата), и поли-D-(-)-3- гидроксимасляную кислоту.
Составы для применения для введения in vivo должны быть стерильными. Этого можно легко добиться с помощью фильтрации через стерильные фильтрующие мембраны. Фармацевтически приемлемые носители, которые могут использоваться в указанных композициях, включают, не ограничиваясь перечисленными, ионообменники, глинозем, стеарат алюминия, лецитин, сывороточные белки, такие как сывороточный альбумин человека, буферные вещества, такие как фосфаты, глицин, сорбиновая кислота, сорбат калия, смеси неполных глицеридов насыщенных растительных жирных кислот, воду, соли или электролиты, такие как протаминсульфат, вторичный кислый фосфат натрия, кислый фосфат калия, хлорид натрия, соли цинка, коллоидный диоксид кремния, трисиликат магния, поливинилпирролидон, вещества на основе целлюлозы, полиэтиленгликоль, карбоксиметилцеллюлоза натрия, полиакрилаты, воски, полиэтилен-полиоксипропилен-блокполимеры, полиэтиленгликоль и ланолин.
Дополнительные аспекты и преимущества, которые следует рассматривать как иллюстративные и не ограничивающие объем настоящей заявки, раскрыты в приведенном ниже экспериментальном разделе.
ПРИМЕРЫ
Пример 1 - Получение антител против KIR3DL2
Материалы и методы
Первичный и вторичный скрининг с применением проточной цитометрии
Скрининг антител mAb против KIR3DL2 проводили главным образом с применением проточной цитометрии для связывания с экспрессирующими KIR3DL2 клеточными линиями Сезари (HUT78 и COU-L) и с трансфицированными KIR3DL2 опухолевыми клеточными линиями (HEK-293Т). Устройства для проточной цитометрии включают: PACSarray (BD Biosciences, первичный скрининг), FACSCanto II n°1 и n°2 (BD Biosciences) (вторичный скрининг) и FC500 (Beckman Coulter) (вторичный скрининг). KIR3DL2+и другие использованные опухолевые клеточные линии включали:
- HUT-78 (положительная по KIR3DL2 клеточная линия Сезари), культивируемые на полной модифицированной по Исков среде Дульбекко (IMDM);
- Клеточные линии HEK-293Т (рак почки человека)/KIR3DL2 и HEK-293Т/ Домен 0 KIR3DL2 - eGFP (культивируемые на полной DMEM);
- COU-L (положительная по KIR3DL2 клеточная линия Сезари) (культивируемые на полной RPMI с добавлением 10% АВ-сыворотки человека);
- Клеточные линии HEK-293T/KIR3DL1 и HEK-293T/KIR3DL1-eGFP (культивируемые на полной DMEM);
- Клеточная линия В221 (В-лимфобластоидная положительная по CD20 клеточная линия человека)/KIR3DL2 (культивируемые на полной RPMI, содержащей фетальную бычью сыворотку); и
- Клеточная линия RAJI (положительная по CD20 клеточная линия лимфомы Беркитта человека)/KIR3DL2 (культивируемые на полной RPMI, содержащей фетальную бычью сыворотку).
В то время как ни одна из использованных клеточных линий Сезари не росла после в/в или п/к введения мышам с иммунной недостаточностью, трансфицированные KIR3DL2 клетки В221 или RAJI росли в виде диссеминированных (в/в) или солидных (п/к) опухолей после инъецирования мышам.
На основе информации, представленной Gardiner et al. Journal of Immunology 2001 (vol 166, p 2992-3001), определяли аллели гена KIR3DL2, присутствующего в используемых опухолевых клеточных линиях. Авторы настоящего изобретения установили, что клеточная линия Сезари COU-L гетерозиготна по аллелям 3DL2*003 и 3DL2*008, a HUT-78 гетерозиготна по аллелям 3DL2*002 и 3DL2*007. Все 4 аллеля 3DL2*003, 3DL2*008, 3DL2*002 и 3DL2*007 кодируют варианты белка KIR3DL2 с различиями во внеклеточных доменах. Отметим, что рекомбинантный гибридный белок KIR3DL2-Fc, который использовали для иммунизации мышей, кодируется разными аллелями гена KIR3DL2 3DL2*006 и 3DL2*007 (клон 1.1, оба аллеля кодируют одну и ту же последовательность внеклеточного домена белка).
Домены 0, 1 и 2 KIR3DL2 клеточных линий
Клетки НЕК293Т/17 культивировали на среде DMEM (Gibco) с добавлением пирувата натрия (1 мМ), пенициллина (100 Ед/мл), стрептомицина (100 мкг/мл) и 10% инактивированной нагреванием ФБС (PAN Biotech). Реагенты липофектамин 2000, Тризол, обратная транскриптаза Superscript П, вектор pcDNA3.1 и ФИТЦ-меченые антитела против V5 приобретали у Invitrogen. ФЭ-меченое антитело козы против тяжелых и легких цепей мыши приобретали у Beckman Coulter. МКПК (5×106 клеток) Homo Sapiens ресуспендировали в 1 мл реагента тризола. Выделение РНК осуществляли путем добавления 200 мкл хлороформа. После центрифугирования (15 мин, 13000 об/мин) РНК осаждали из водной фазы 500 мкл изопропанола. После инкубирования (10 мин, КТ) и центрифугирования (10 мин, 13 000) РНК промывали 70% этанолом и повторно центрифугировали (5 мин, 13,000 об/мин). РНК ресуспендировали в не содержащей РНКаз H2O (RNase-free Water). Получали кДНК с применением обратной транскриптазы Superscript II из 2 мкг специфической РНК, следуя инструкциям производителя. Последовательности домена 0, домена 1 и домена 2 KIR3DL2 человека (номер доступа U30272, аллель *002 KIR3DL2) приведены в таблице 4.

Последовательности домена 0, домена 1 и домена 2 KIR3DL2 Homo Sapiens (номер доступа U30272) амплифицировали с применением ПЦР-реакции из кДНК с применением олигонуклеотидов 5' АА GCT AGC GGT AAG ССТ АТС ССТ ААС ССТ СТС СТС GGT CTC GAT ТСТ ACG СТС ATG GGT GGT CAG GAC ААА С (SEQ ID NO: 71) (прямой) и 3' АА GGA ТСС СТС ТСС TGA ТТТ CAG CAG GGT (SEQ ID NO: 72) (обратный); 5' АА GCT AGC GGT AAG ССТ АТС ССТ ААС ССТ СТС СТС GGT СТС GAT ТСТ ACG АСА GTC АТС CTG САА TGT TGG (SEQ ID NO: 73) (прямой) и 3' АА GGA ТСС СТС ТСС TGC CTG ААС CGT GGG (SEQ ID NO: 74) (обратный); 5' АА GCT AGC GGT AAG ССТ АТС ССТ ААС ССТ СТС СТС GGT СТС GAT ТСТ ACG ААС GTG АСС TTG ТСС TGT AGC (SEQ ID NO: 75) (прямой) и 3' АА GGA ТСС ATG CAG GTG ТСТ GCA GAT АСС (SEQ ID NO: 76) (обратный), соответственно. После ТА-клонирования и секвенирования последовательности клонировали в вектор pcDNA3.1 между сайтами рестрикции Nhel и BamHI. Указанные конструкции встраивали между последовательностями лидерного пептида CD33 и ГФИ-якоря CD24 (ДНК и аминокислотные последовательности ГФИ-якоря CD24 приведены в последовательностях SEQ ID NO: 77 и 78, соответственно), синтезированными MWG Biotech (встраивали между сайтами рестрикции BamHI и HindIII).
Клетки HEK-293Т/17 высевали за 24 часа до трансфекции в 6-луночные планшеты (5,105 клеток на лунку) в DMEM без антибиотиков. Трансфекцию проводили с применением 5 мкг разных конструкций, pcDNA3.1/ домен 0 KIR3DL2, pcDNA3.1/ домен 1 KIR3DL2 или pcDNA3.1/ домен 2 KIR3DL2, с применением липофектамина 2000 в соответствии с инструкциями производителя. Для проверки чистоты ДНК для трансфекции использовали Maxi-prep набор без эндотоксинов от Qiagen. Использовали фиксированную пропорцию липофектамина/ДНК 2/1. Клетки собирали через 48 часов после трансфекции для проведения экспериментов с применением проточной цитометрии.
Иммунизация
Мышей иммунизировали рекомбинантным гибридным белком KIR3DL2-Fc (аллель *006). Супернатант (SN) от растущих гибридом тестировали с применением проточной цитометрии на клетках HUT78, COU-L и HEK-293Т/ Домен 0 KIR3DL2 - eGFP. Представляющие потенциальный интерес гибридомы, выбранные при первичном скрининге, клонировали с применением техник серийных разведении в 96-луночных планшетах. Вторичный скрининг включал отбор представляющих интерес гибридом путем тестирования супернатантов субклонов с применением проточной цитометрии на клетках HUT78, COU-L, HEK-293Т/ Домен 0 KIR3DL1 - eGFP и HEK-293Т/ Домен 0 KIR3DL2 - eGFP. Положительные субклоны инъецировали мышам для формирования асцита, и представляющие интерес антитела очищали до тестирования в анализе Biacore с применением чипов с рекомбинантным KIR3DL2, с последующими анализами в различном формате, основанными на связывании с экспрессирующими KIR3DL2 клетками человека. Среди выбранных клонов были супернатанты для антител 10F6, 2В12, 18С6, 9Е10, 10G5, 13Н1, 4В5, 5Н1, 1Е2, 1С3 и 20Е9. На основании скрининга, позволяющего выбирать между связыванием с доменом D0 и связыванием с доменами D1/2, установлено, что антитела 10F6, 2В12, 18С6, 9Е10, 10G5, 13Н1, 4В5, 5Н1 и 1Е2 связываются с KIR3DL2, присутствующим на внеклеточном домене 0 (D0), a 1C3 и 20Е9 связываются с эпитопом, присутствующем на домене 1/2 (D2).
Последовательности вариабельных доменов тяжелой (VH) и легкой (VL) цепей выбранных антител амплифицировали с применением ПЦР из кДНК каждого антитела. Амплифицированные последовательности проводили через агарозный гель, затем очищали с применением набора для экстракции Qiagen Gel Extraction kit. Затем последовательности VH и VL субклонировали в экспрессионные двойные («double-gene») векторы от Lonza (с применением системы biFusion (Clontech) в соответствии с инструкциями производителя. После секвенирования векторы, содержащие последовательности VH и VL, получали в виде Maxiprep с применением системы для выделения плазмид Promega PureYield™ Plasmid Maxiprep System. Затем векторы использовали для трансфекции клеток HEK-293Т с применением липофектамина 2000 от hivitrogen в соответствии с инструкциями производителя.
Пример 2 - Антитела, не индуцирующие интернализацию KIR3DL2
Вкратце, на протяжении 24 часов при 37°С свежевыделенные от 5 разных доноров-людей клетки, пораженные синдромом Сезари, инкубировали либо с 20 мкг/мл антитела против домена 0 KIR3DL2 или антитела 10F6, 2В12, 18С6, 9Е10, 10G5, 13Н1, 4В5, 5Н1, 1Е2, 1C3 или 20Е9, либо инкубировали без антитела. Затем клетки промывали, фиксировали и пермеабилизировали с применением реагента для пермеабилизации bitraPrep от Beckman Coulter. Присутствие связанного с KIR3DL2 10F6, 2В12, 18С6, 9Е10, 10G5, 13Н1, 4В5, 5Н1, 1Е2, 1C3 или 20Е9 антитела обнаруживали с применением меченого ФЭ антитела козы против антитела мыши (GAM-PE). В таблице 6 представлен пример антитела против домена 0 KIR3DL213H1 через 24 часа инкубирования, соответственно. В таблице 5 показано значительное уменьшение флуоресценции 13Н1 для нескольких разных доноров, подтверждающее, что связывание указанного антитела понижающе модулирует экспрессию KIR3DL2 на клетках СС. Аналогичные результаты были получены для антитела против D0 4B5, а также ряда антител против D1. И напротив, антитела против домена 0 или домена 2 KIR3DL2 10F6, 2В12, 18С6, 9Е10, 10G5, 5Н1, 1Е2, 1С3 и 20Е9 не вызывают уменьшения флуоресценции, что указывает на то, что указанное антитело не модулирует понижающе экспрессию KIR3DL2 на клетках СС. В таблице 6 представлен репрезентативный пример антитела 10G5.

Пример 3 - Антитела, которые не интернализируются клеточной линией с синдромом Сезари
Интернализацию антител 10F6, 2В12, 18С6, 9Е10, 10G5, 13Н1, 4B5, 5Н1, 1Е2, 1С3 и 20Е9, а также антитела AZ158 (mAb против домена 0) и других антител против D1 оценивали с помощью флуоресцентной микроскопии с применением клеточной линии HUT78 СС.
Материалы и методы:
Клетки Hut-78 инкубировали в течение 1 ч при 4°С с 10 мкг/мл разных антител. После указанного инкубирования клетки либо фиксировали (t=ОН) или инкубировали в течение 2 ч при 37°С. Затем клетки, инкубированные в течение 2 ч, фиксировали и окрашивали. Антитела окрашивали с применением антител козы против мыши, соединенных с Alexa594 (bivitrogen, А11032). Компартменты LAMP-1 окрашивали с применением антител кролика против LAMP-1 (Abeam, ab24170), обнаруживаемые с применением поликлональных антител козы против кролика, конъюгированных с ФИТЦ (Abeam ab6717). Изображения получали с применением устройства Apotome (Zeiss) и анализировали с применением программного обеспечения Axiovision.
Результаты:
Антитела mAb против KIR3DL2 окрашивались в красный цвет, а компартменты LAMP-1 окрашивались в зеленый цвет. В момент добавления антител окрашивание KIR3DL2 в красный цвет было заметно на клеточной поверхности, тогда как LAMP-1 зеленого цвета были видны внутри клеток. При этом через 2 часа после добавления антител каждое из антител AZ158, 13Н1 и 4В5, и антитела против D1 вызывали колокализацию окрашивания в красный цвет и окрашивания в зеленый цвет, а также уменьшение окрашивания в красный цвет на клеточной поверхности, что указывает на быструю интернализацию AZ158, 13Н1 и 4В5, и антител против D1. Однако антитела 10F6, 2В12, 18С6, 9Е10, 10G5, 5Н1, 1Е2, 1С3 и 20Е9 не интернализовались, и через 2 часа после добавления антитела окрашивание в красный цвет оставалось полностью на клеточной поверхности.
Пример 4 - Антитела способны к киллингу экспрессирующих KIR3DL2 мишеней за счет комплементзависимого механизма (КЗЦ)
Вкратце, 50 мкл 20 мкг/мл разбавленных антител (концентрация 2х) помещали в стандартные белые с прозрачным дном 96-луночные планшеты (кат. №655098 - Greiner), с которым добавляли 50 мкл суспензии клеток в концентрации 2 млн на мл (100000 клеток на лунку) в стандартной среде, и инкубировали в течение 30 мин при 4°С. Добавляли по 5 мкл на лунку свежевосстановленного комплемента (кат. № CL3441 - Cedarlan) с последующим инкубированием 1Н при 37°С. Добавляли по 100 мкл на лунку Cell Titer Glo (кат. № G7572 - Promega) с последующим инкубированием в течение 10 мин при комнатной температуре, предохраняя от света. Результаты считывали с применением люминометра (VICTOR).
С применением очищенного комплемента из крови кролика оценивали способность предложенных авторами антител mAb против KIR3DL2 к рекрутингу комплемента и лизису трансфицированных KIR3DL2 клеток В221 in vitro.
На фиг. 6 показана способность антител опосредовать КЗЦ; антитела mAb против KIR3DL2, которые связывают домен D0, обозначены серым цветом, антитела, которые связывают домен D1, обозначены черным цветом. В случае исходных mAb мыши наиболее выражение влияет на результат указанного анализа изотип mAb, поскольку антитела mAb IgG2b мыши связывают комплемент более эффективно, чем любой другой изотип (IgG1 мыши вообще не связывают комплемент).
Для оценки влияния интернализации KIR3DL2 на комплемент-опосредованную клеточную смерть мишеней при связывании с антителами mAb против KIR3DL2, авторы настоящего изобретения использовали mo19H12, антитело против KIR3DL2, которое индуцирует быструю интернализацию KIR3DL2 в клеточной линии Сезари HUT78 и B221-KIR3DL2. Перед инкубированием с комплементом авторы предварительно инкубировали целевые мишени B221-KIR3DL2 с mo19H12, либо при 4°С (ингернализация блокируется) либо при 37°С (что обеспечивает оптимальную интернализацию). Затем добавляли комплемент, инкубировали и измеряли КЗЦ согласно описанию выше.
В настоящем эксперименте интернализация KIR3DL2 при связывании полностью устраняет способность mo19H12 к киллингу B221-KIR3DL2 за счет активации комплемента, тогда как в температурных условиях ограничивающих интернализацию, КЗЦ активность mo19H12 отчетливо наблюдается (Фиг. 7). Ритуксимаб против CD20 используют в качестве контроля, который опосредует КЗЦ против мишеней CD20+, но не индуцирует интернализацию CD20.
Выбранные mAb гибридизовали в IgG1 человека для придания им способности опосредовать эффекторные функции (АЗКЦ и КЗЦ). На фиг. 8 показана способность гибридных антител mAb против KIR3DL2 опосредовать КЗЦ против B221-KIR3DL2 in vitro.
Определенные клоны mAb, такие как 1Е2 и 10G5, после гибридизации приобретали способность к киллингу положительных по KIR3DL2 мишеней за счет механизма КЗЦ. В настоящем эксперименте активная интернализация антител mAb против D0 (например, индуцируемая 13Н1, обозначено черным цветом), может предотвращать оптимальную эффективность, как наблюдалось, в частности, для 1Е2 и 10G5.
Пример 5 - Антитела способны к киллингу экспрессирующих KIR3DL2 мишеней за счет антителозависимой клеточной цитотоксичности (АЗКЦ)
Мониторинг лизиса клеток за счет механизма АЗКЦ осуществляли в эксперименте, основанном на высвобождении радиоактивного 51Cr (уровень радиоактивности, высвобождаемой предварительно нагруженными целевыми клетками, пропорционален клеточной смерти). 1 млн целевых клеток нагружали 51Cr на протяжении 1 часа при 37°С и троекратно промывали. Высевали по 3000 клеток на лунку (в U-образные 96-луночные планшеты) и добавляли экспериментальные антитела mAb в конечной концентрации 10 или 20 мкг/мл (или в возрастающих концентрациях, если исследуется зависимость доза-ответ). Добавляли эффекторные клетки до достижения определенной пропорции эффекторных: целевых клеток (как правило, 10:1) и смесь инкубировали при 37°С в течение 4 ч. Супернатант анализируют на аппарате Lumaplate. При применении гибридных антител mAb hulgGI в качестве эффекторных клеток использовались аллогенные NK-клетки человека от здорового донора-добровольца, очищенные от МКПК.
Для оптимального проведения оценки эксперименты с АЗКЦ проводили как правило, с применением гибридизованных антител mAb hulgG1, полученных из различных исходных mAb мыши против KIR3DL2. На фиг. 9 показана способность ряда антител mAb против KIR3DL2, протестированных в одинаковой конечной концентрации (10 мкг/мл), к киллингу клеток прототипичной клеточной линии синдрома Сезари HUT78 за счет опосредованного АЗКЦ механизма.
На фиг. 10 представлен аналогичный эксперимент, в котором в качестве целевых использовались трансфицированные KIR3DL2 клетки В221, в целом более чувствительные к опосредованному АЗКЦ киллингу антителами mAb против KIR3DL2. Выделенные серым цветом антитела mAb индуцируют интернализацию рецептора и, по-видимому, менее эффективны, чем 4 других антитела mAb, которые не индуцируют интернализацию KIR3DL2.
На фиг. 11 представлено сравнение в эксперименте с подбором дозы способности гибридизованных антител mAb hulgG1 против KIR3DL2 опосредовать АЗКЦ против экспрессирующих KIR3DL2 целевых клеток В221; профиль эффективности антител mAb, которые не индуцируют интернализацию мишеней (10F6, 2В12 и 10G5) благоприятнее, чем профиль антител mAb, индуцирующих интернализацию KIR3DL2 (13Н1 и mAb против D0 15C11 и 18В10).
Пример 6 - Активность экспрессирующих KIR3DL2 опухолей человека в моделях ксенотрансплантатов на мышах
Материалы и методы
Мышей NOD-SCID с иммунной недостаточностью для использования в моделях В221-KIR3DL2 и RAJI-KIR3DL2 приобретали у Charles River Laboratories. В описанных ниже моделях 5 млн опухолевых клеток человека (в 100 мкл ФСБ в качестве основы) трансплантировали в/в на 0 день (D0), т.е. за 1 день до начала лечения (D1). Начиная с D1 мыши получали в/в лечение разными дозами антител mAb (дозы подбирали в соответствии с массой тела мышей), разведенными в ФСБ, по 2 инъекции в неделю на протяжении всего эксперимента.
Контрольные группы включали, в зависимости от эксперимента:
- Мышей, получавших ФСБ/плацебо, в качестве контроля обычного/не подвергающегося воздействию роста опухоли;
- Мышей, которым инъецировали идентичные дозы совпадающих по изотипу контрольных антител mAb против нерелевантного антигена.
Мышей взвешивали и осматривали для выявления клинических признаков каждые 2-5 дней в зависимости от модели. Рассчитывали процент изменения массы тела относительно массы тела в день D0 до имплантирования опухоли или относительно максимальной массы тела, наблюдавшейся во время эксперимента. Гибель или существенную потерю веса у мышей регистрировали и использовали для построения кривых выживаемости Каплана-Майера и расчета увеличения выживаемости по сравнению с контрольными группами мышей.
Результаты
На фиг. 12 представлены результаты эксперимента (n=6 NOD-SCID мышей на группу) в котором тестировали эффективность 3 антител мыши изотипа IgG2b против KIR3DL2, 9Е10 и 19Н12 (оба вводили в дозе 300 мкг/ мышь, два раза в неделю) в отношении п/к ксенотрансплантатов В221-KIR3DL2. Неинтернализующееся антитело против D0 9E10 демонстрировало повышенную выживаемость по сравнению как с ФСБ, так и с интернализующимся антителом 19Н12 против D1/D2.
На фиг. 13 представлены результаты другого эксперимента (n=6 мышей NOD-SCID на группу), в котором тестировали эффективность антитела мыши 19Н12 против KIR3DL2 (вводимого в дозе 300 мкг/мышь, два раза в неделю) в отношении п/к ксенотрансплантатов RAJI-KIR3DL2. In vitro трансфицированные KIR3DL2 клетки RAJI демонстрировали меньшую интернализацию при связывании mAb, чем клеточные линии В221-KIR3DL2 или синдрома Сезари. В модели с ксенотрансплантатом RAJI-KIR3DL2 mAb mo19H12 было более эффективным, чем в модели с B221-KIR3DL2. Это обусловлено менее выраженной интернализацией мишени in vivo.
Пример 7 - Блокада лигандов
Материалы и способы
Подавление окрашивания тетрамера антителами
Состав с димером В27 и тетрамером HLA-A3 и FACS-окрашивание описывались ранее (Kollnberger, et al. (2007) Eur J Immunol 37: 1313-1322). Получали тетрамеры HLA-A3. Для экспериментов с подавлением антителами трансдуцированные KIR3DL2 клетки Baf3 окрашивали 5 мкг антитела или контрольным изотипом IgGl/IgG2a (Biolegend UK Ltd) при 4°С в течение 20 минут перед окрашиванием с тетрамером при комнатной температуре в течение 20 минут. Затем окрашенные клетки промывали и фиксировали согласно существующему описанию перед FACS-анализов на аппарате для FACS BD Fortessa. FACS-анализ осуществляли с применением программного обеспечения Flowjo (Tree star Inc., США).
Подавление антителами репортерных клеток Jurkat KIR3DL2 CD3ε
Репортерные клетки Jurkat, трансдуцированные для экспрессии гибридного белка KIR3DL2CD3ε, были описаны ранее (Payeli, et al. (2012) Arthritis Rheum.). Для экспериментов с подавлением антителами репортерные клетки в концентрации 100000 клеток на лунку в среде RPM1640 (Sigma, с добавлением 10% ФБС, пенициллина и стрептомицина) сначала окрашивали при 4°С с 10 мкг антитела или контрольного изотипа в течение 20 минут. Затем репортерные клетки стимулировали 200000 исходных клеток LCL.LBL.721.221 (далее называемых в настоящем документе клетками 221) или клеток 221, трансфицированных HLA-B27 или контрольным HLA класса 1 в конечном объеме 200 мкл. Супернатанты собирали после стимуляции на протяжении ночи для анализа на ИЛ-2 с применением ИФА ELISA (Ebiosciences UK Ltd), проводимого в соответствии с инструкциями производителя.
Результаты
Специфические антитела к домену D0 подавляют окрашивание тетрамера HLA-A3 и димера В27 (В272) трансдуцированных KIR3DL2 клеток
Сначала авторы исследовали способность специфических антител против доменов D0 и D1/D2 KIR3DL2 подавлять окрашивание тетрамера HLA-A3 и димера В27 KIR3DL2. Специфические антитела к домену D0 KIR3DL2 1Е2 и 13Н1 стабильно подавляли окрашивание HLA-A3 и тяжелой цепи В27 (В272) трансдуцированных KIR3DL2 клеток Baf3 (фиг. 14 и 16А). 2В12 подавляли В272, оказывая при этом пренебрежимый эффект на окрашивание тетрамера HLA-A3 KIR3DL2. Напротив, 10G5 не подавлял окрашивание KIR3DL2 чем-либо из тетрамеров HLA-A3 или В272.
Специфическое антитело к домену D2 1С3 подавляет окрашивание тетрамера HLA-A3, но не димера В27 (В272) KIR3DL2
Специфическое антитело к D2 1С3 стабильно подавляло окрашивание тетрамера HLA-A3 трансдуцированных KIR3DL2 клеток (фиг. 15 и 16В). Напротив, 1С3 MAb не влияло на окрашивание В272 клеток KIR3DL2 Baf3 и специфические антитела к D1 не оказывали значительного эффекта на окрашивание тетрамера HLA-A3 или B272 KIR3DL2 (фиг. 15 и 16В).
Специфические антитела mAb к домену D0, но не D1/D2 KIR3DL2 подавляют взаимодействие репортерных KIR3DL2-клеток с HLA класса 1.
Затем авторы настоящего изобретения определили эффект специфических антител к KIR3DL2 на распознавание KIR3DL2 HLA-B27 и других HLA класса 1 путем исследования эффекта антител на синтез ИЛ-2 трансдуцированными KIR3DL2 CD3ε репортерными клетками Jurkat, стимулированными трансфектантами 221. В соответствии с полученными авторами ранее предыдущими данными клетки 221, экспрессирующие, HLA-B27 стабильно стимулировали синтез ИЛ-2 в репортерных KIR3DL2-клетках, в 6 раз более высокий по сравнению с синтезом при стимуляции клетками, экспрессирующими контрольный HLA класса 1 (фиг. 17).
Все специфические антитела к домену D0, 2В12, 1Е2, 10G5 и 13Н1, в некоторой степени подавляли синтез ИЛ-2 репортерными KIR3DL2-клетками, стимулированными трансфицированными HLA-B27 клетками (фиг. 4), при этом для 2В12 и 1Е2 наблюдался максимальный ингибирующий эффект. Напротив, специфическое антитело к D2, 1С3, не оказывало значительного эффекта на распознавание HLA-B27 репортерными клетками.
Специфические антитела к домену D0 также подавляли синтез ИЛ-2 репортерными KIR3DL2-клетками, стимулированными клетками 221, трансфицированными HLA-B7, HLA-B35 и HLA-A2 и HLA-A3, хотя эффект был менее выраженным, чем при стимуляции клеток HLA-B27. Напротив, специфическое антитело к D2, 1С3, не оказывало значительного эффекта на синтез ИЛ-2 репортерными KIR3DL2-клетками, стимулированными клетками 221, трансфицированными HLA-B7, HLA-B35 и HLA-A2 и HLA-A3.
Выводы
В настоящем документе авторы показали, что моноклональные антитела против домена D0 KIR3DL2 (2В12, 1Е2, 13Н1) подавляют связывание с не содержащими β2m B27 димерами тяжелой цепи (В272) и связанными с β2m лигандами, такими как HLA-A3. Напротив, антитела против домена D2 KIR3DL2 (1С3) подавляют исключительно взаимодействие с HLA-A3 и оказывают незначительный эффект на связывание тетрамера В272.
Хотя KIR3DL2 репортерные Т-клетки продуцируют ИЛ-2 при стимуляции трансфицированными HLA-B27, HLA-B7, HLA-B35, HLA-A2 и HLA-A3 В-клетками LBL.721.221, репортерные клетки стабильно продуцируют в 6 раз более высокие уровни ИЛ-2 в ответ на HLA-B27. Взаимодействие KIR3DL2 с В-клетками, экспрессирующими HLA-B27 и другие HLA класса 1, стабильно подавляется специфическими антителами к домену D0 2B12 и 1Е2 и, в меньшей степени, 13Н1. Это свидетельствует о том, что домен D0 KIR3DL2 может обладать некоторой аффинностью к общим признакам разных HLA класса 1. Домен D0 KIR3DL2 может связываться по меньшей мере отчасти с областью HLA-B27, общей для разных HLA класса 1. Повышенная авидность KIR3DL2 в отношении HLA-B27 может быть обусловлена димеризацией тяжелых цепей B27.
Сообщалось, что три иммуноглобулин-подобных домена D0, D1 и D2 KIR3DL2 задействованы в связывающем лиганде. Результаты исследования подавления с применением антител указывают на то, что основной контакт KIR3DL2 с HLA класса 1 происходит в домене D0. Примечательно, что антитело к D2 1С3 подавляло исключительно связывание с KIR3DL2 HLA-A3, но не димера B27. Соответственно, авторы предполагают, что домен D0 контактирует с остатками, консервативными для разных HLA класса 1, а контактные области доменов D1 и D2 контактируют с полиморфными областями и пептидом в пептидном комплексе ГКГС. Идентифицированные антитела могут применяться в терапевтических, диагностических и других исследовательских приложениях, в зависимости от конкретного применения - для избирательной блокады различных лигандов, или блокады нескольких лигандов, или отсутствия конкуренции с лигандами за связывание с KIR3DL2.
Пример 8 - Картирование эпитопов
Для идентификации специфических в отношении KIR3DL2 антител, которые обладают нужными связывающими свойствами, были разработаны мутанты KIR3DL2. Антитела предпочтительно связываются с большинством или со всеми основными аллелями KIR3DL2 в популяции (в смысле частоты аллелей), не связываясь при этом с основными аллелями KIR3DL1 (например, аллелем *00101).
Были получены мутации, соответствующие остаткам, отличающимся у KIR3DL1 и у KIR3DL2. Был получен первый набор мутаций, характеризующихся тем, что аминокислоту KIR3DL1 встраивали в KIR3DL2. Тем не менее, многие из указанных мутированных белков не экспрессировались на клеточной поверхности, что указывает на то, что включение АK из KIR3DL1 оказывает глубокое влияние на укладку всей молекулы KIR3DL2. В частности, мутанты в кластерах D21, D22, D23, D26 и D27, представленные ниже в таблице 7А, не экспрессируются на клеточной поверхности.

Были повторно разработаны и исследованы дополнительные мутанты; окончательный набор мутаций представлен ниже в таблице 7В. Антитела тестировали на связывание KIR3DL2 с различными мутантами KIR3DL2. Мутанты KIR3DL2 были получены с применением ПЦР (см. таблицу 7 В ниже). Все праймеры Mx-R использовали со следующим 5'-праймером: ACCCAAGCTGGCTAGCATGTCGCTCACGGTCGTCAGCATG (SEQ ID NO: 79). Все праймеры Mx-F использовали со следующим 3'-праймером: AGCACAGTGGCGGCCGCCTAGAAAA CCCCCTCAAGACC (SEQ ID NO: 80). Амплифицированные последовательности проводили через агарозный гель и затем очищали с применением набора для экстракции Qiagen Gel Extraction kit.
Для получения мутантов 12 и 21 необходимо было провести третью реакцию ПЦР. Для указанной ПЦР использовали следующие праймеры: праймер M12a-F (5'-GCCACAGGTGCATATGAGAAACCTTCTCTCTCAGCC-3') (SEQ ID NO: 81) с праймером M12b-R (5'-TGGGTCACTTGCGGCTGACCACACGCAGGGCAGGG-3') (SEQ ID NO: 82) и праймером M21a-F (5'-CGTGCCCTGCCCTACGTGTGGTCAAACTCAAGTGAC-3') (SEQ ID NO: 83) с праймером M21b-R (5'-ATG CAGGTGTCTGGGGATACCAGATTTGGAGCTTGGTTC-3') (SEQ ID NO: 84).
Два или три продукта ПЦР, синтезированные для каждого мутанта, затем дотировали в вектор pcDNA3.1, расщепляли рестрикционным ферментом NheI и NotI с применением системы biFusion (Clontech) в соответствии с инструкциями производителя.
После секвенирования векторы, содержащие мутированные последовательности, получали в виде Maxiprep с применением системы для выделения плазмид Promega PureYield™ Plasmid Maxiprep System. Затем векторы использовали для трансфекции клеток HEK-293Т с применением липофектамина 2000 от bivitrogen в соответствии с инструкциями производителя.

Каждое антитело тестировали на связывание с KIR3DL2 дикого типа и с каждым из мутантов доменов D0, D1 и D2. Не наблюдалось какого-либо уменьшения связывания антител с немутированным KIR3DL2 дикого типа (WTaKIR3DL2), однако утрачивалось связывание с одним или несколькими мутантами; таким образом было идентифицировано несколько эпитопов.
Обзор результатов, приведенных в таблицах 7С и 7D («+», указывает на отсутствие значительного уменьшения связывания, «+/-» указывает на уменьшение связывания (или частичную утрату связывания) и «-» указывает на по существу полную утрату связывания). Большинство неинтернализующихся D0 антител по существу полностью утрачивали связывание с мутантом 2 (четыре антитела: 10F6, 2В12, 18С6, 10G5). Наблюдалась также по меньшей мере частичная утрата связывания всех указанных антител также с мутантом 2А3. Одно неинтернализующееся антитело к D0 демонстрировало утрату связывания с единственным мутантом 1 (9Е10). Одно неинтернализующееся антитело к D0 (1E2) утрачивало связывание исключительно с мутантом 2А3. Одно антитело (5Н1) утрачивало связывание с мутантом 6.
Наблюдалось сниженное связывание природного блокирующего лиганды интернализующегося антитела 13Н1 также с мутантом 2А2 и MD0/HLA1, помимо мутантов 1 и 2.
Что касается антител, которые связывали домен D2 KIR3DL2 (оба антитела не интернализуются), утрачивалось связывание антител 1С3 и 20Е9 с мутантом 14, а также происходила частичная утрата связывания с мутантом 15 и мутантом 16.
Наблюдалась утрата связывания антител 10F6, 2В12, 18С6 и 10G5 с мутантом 2, содержащим замены I60N и G62S, и уменьшение связывания с мутантом 2А3, содержащим замены P14S, S15A и H23S, но связывание с любыми другими мутантами не было утрачено. Основной эпитоп указанных антител, соответственно, включает остатки 160 и/или G62 (также указанный эпитоп необязательно включает один или большее количество остатков из Р14, S15, и Н23). Остатки 60 и 62 входят в состав домена D0 KIR3DL2.
Наблюдается утрата связывания антитела 13Н1 как с мутантом 1, содержащим замены R13W, А25Т и Q27R, так и с мутантом 2, содержащим замены I60N и G62S. Также понижено связывание 13Н1 с мутантом 2А2 (Q56S, Е57А) и мутантом MDO/HLA1 (F9S, S11A). Эпитоп 13Н1, соответственно, включает остатки F9, S11, Q56 и/или Е57. Указанные остатки входят в состав домена D0.
Наблюдается сниженное связывание антитела 9Е10 с мутантом 1, содержащим замены R13W, А25Т и Q27R, но не с какими-либо другими мутантами. Эпитоп 9Е10 и 10G5, соответственно, включает остатки R13, А25 и/или Q27.
На фиг. 1 приведено изображение полипептида KIR3DL2, включающее фрагменты в составе домена D0, демонстрирующее мутированные остатки аминокислот, обозначенные как «Мутант 1», «Мутант 2», «Мутант 3» и «Мутант 6», которые приводили (в различных комбинациях) к утрате связывания антителами. На фиг. 2 приведено изображение полипептида KIR3DL2, включающее фрагменты в составе домена D0, демонстрирующее модификации остатков аминокислот, обозначенные как «Мутант 1», «Мутант 2» и «Мутант 3», которые приводили (в различных комбинациях) к утрате связывания антителами, с выделением цветом остатков, смежных с остатками (F9, S11, Р14, F34 и/или S140, смежные с мутантом 2, и G21, G22, Н23, Е57, S58, F59, Р63 и/или Н68, смежные с мутантом 1).
Наблюдается утрата связывания антитела 5Н1 с мутантом 6, содержащим замены R78H и L82P, но не связывания с любыми другими мутантами. Основной эпитоп 5Н1, соответственно, включает остатки R78 и/или L82. Остатки R78 и L82 входят в состав домена D0 KIR3DL2. Экспонируемые на поверхности остатки, смежные с указанными мутированными остатками, могут также вносить вклад в эпитопы указанных антител. На фиг. 3 приведено изображение каждой поверхности полипептида KIR3DL2, включая фрагменты в составе домена D0, демонстрирующее мутированные остатки аминокислот, обозначенные как «Мутант 6», которые приводили (в различных комбинациях) к утрате связывания антителами; показан «Мутант 3», не приводивший к утрате связывания. Также выделены цветом остатки, смежные с остатками, смежными с мутантом 6, которые могут также связываться антителами (K7, Y30, R31, Р79, Н80, S81, Т83, G84, W85, S86 и/или А87).
Наблюдалась утрата связывания антител 1С3 и 20Е9 с мутантом 14, содержащим замену W226A. Дополнительно наблюдалось сниженное связывание указанных антител с мутантом 15, содержащим замены I231M и R246P, и с мутантом 16, содержащим замену E239G. Основной эпитоп 1С3, соответственно, включает остаток W226. Основной эпитоп 20Е9 может включать остатки I231M и/или R246P, и/или может дополнительно включать Е239. Остатки W226, I231 и R246 входят в состав области на границе доменов D1 и D2 KIR3DL2. Экспонируемые на поверхности остатки, смежные с мутированными остатками, также могут вносить вклад в эпитопы указанных антител, в том числе, например, остатки Q201, K202, Р203, S204, S224, S225, S227, S228, N252, R253 и/или Т254 (см. SEQ ID NO: 1), расположенные на поверхности KIR3DL2 в области эпитопа W226, но не в составе области мутаций KIR3DL2, которые не приводили к утрате связывания указанных антител (например, мутантов 12 и 17). Экспонируемые на поверхности остатки, смежные с мутированными остатками I231 и R246, также могут вносить вклад в эпитопы указанных антител, в том числе, например, остатки D230, I231, R244, L245, R246, А247, V248, S275, R277 и/или Р280 (см. SEQ ID NO: 1), расположенные на поверхности KIR3DL2 в области эпитопа I231/R246 но не в составе области модификаций KIR3DL2, которые не приводили к утрате связывания указанных антител (например, мутантов 12 и 17).
На фиг. 4 приведено изображение полипептида KIR3DL2, включая фрагменты в составе домена D2 (граница D1/D2), демонстрирующее модификации остатков аминокислот, обозначенные как «Мутант 14», связывание антител с которым утрачено, и «Мутант 12» и «Мутант 17», которые не вызывали утраты связывания антителами; также выделены цветом остатки, смежные с остатками (Q201, K202, Р203, S204, S224, S225, S227, S228, N252, R253 и/или Т254, смежные с мутантом 14), которые также могут связываться.
На фиг. 5 приведено изображение полипептида KIR3DL2, включая фрагменты в составе домена D2 (граница D1/D2), демонстрирующее модификации остатков аминокислот обозначенные как «Мутант 15», связывание антител с которым утрачено; также выделены цветом остатки, смежные с остатками (D230, I231, R244, L245, R246, А247, V248, S275, R277 и/или Р280) смежные с мутантом 14), которые также могут связываться.
На фиг. 18, 19 и 20 приведены изображения полипептида KIR3DL2, аллель *001, на которых показаны сайт связывания в мутанте 2 и аминокислотные различия, наблюдаемые в KIR3DL2 разных аллелей, имеющих максимальную частоту (согласно популяционным исследованиям в Соединенных Штатах). Можно видеть, что сайт связывания антитела располагается в участке, консервативном у всех аллелей KIR3DL2, что согласуется со способностью указанного антитела связывать клетки всех протестированных индивидуумов.
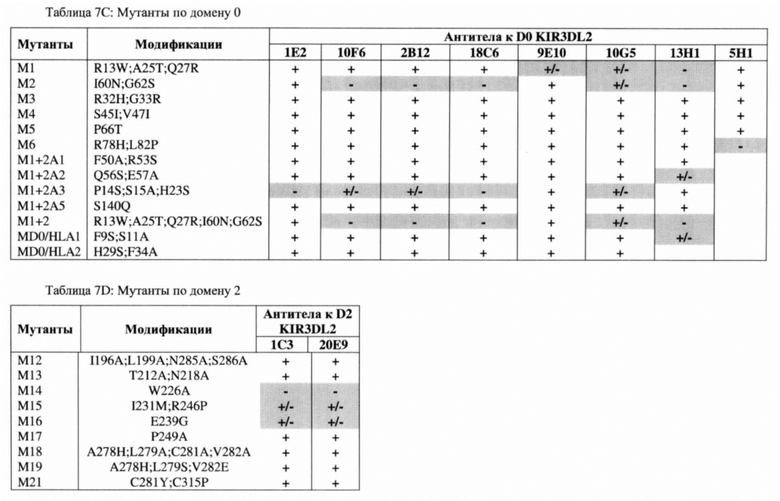
Все заголовки и подзаголовки используются в настоящем документе исключительно для удобства и не должны рассматриваться как каким-либо образом ограничивающие настоящее изобретение. Настоящим изобретением охвачена любая комбинация вышеописанных элементов во всех возможных вариантах, если иное не указано в настоящем документе или это иным образом явно не противоречит контексту. Указание диапазонов значений в настоящем документе используется исключительно в качестве способа сокращенного индивидуального обозначения каждого отдельного значения, входящего в диапазон, если иное не указано в настоящем документе, и каждое отдельное значение включено в описание в той же степени, как если бы оно было индивидуально указано в настоящем документе. Если не указано иное, все точные значения, приведенные в настоящем документе, представляют и соответствующие приблизительные значения (например, можно считать, что все приведенные примеры точных значений, относящихся к конкретному фактору или измерению, также предусматривают соответствующие приблизительные значения, модифицированные при необходимости термином «приблизительно»).
Все описанные в настоящем документе способы могут быть реализованы в любом подходящем порядке, если иное не указано в настоящем документе или это иным образом явно не противоречит контексту.
Использование каких бы то ни было примеров или связанной с примерами терминологии (например, «такой как») в настоящем документе предназначено исключительно для лучшего описания настоящего изобретения и не накладывает ограничений на объем настоящего изобретения, если не указано иное. Никакие выражения в настоящем описании не должны толковаться как указание на то, что какой-либо элемент является неотъемлемым для практической реализации настоящего изобретения, если это не указано явным образом.
Цитирование и включение патентных документов в настоящий документ осуществляется исключительно для удобства и не отражает какое-либо представление о действительности, патентоспособности и/или правового обеспечения таких патентных документов. Описание в настоящем документе какого-либо аспекта или варианта реализации согласно настоящему изобретению с применением каких-либо терминов, например, со ссылкой на элемент или элементы, предназначено для обоснования аналогичного аспекта или варианта реализации настоящего изобретения, который «состоит из», «состоит по существу из» или «по существу содержит» указанный конкретный элемент или элементы, если не указано иное или если это явным образом не противоречит контексту (например, следует понимать, что композиция, описанная в настоящем документе как содержащая конкретный элемент, также описывает композицию, состоящую из указанного элемента, если не указано иное или если это явным образом не противоречит контексту).
Указанное изобретение включает все модификации и эквиваленты предмета изобретения согласно представленным в настоящем документе аспектам или формуле изобретения, в максимальной степени, допускаемой применимым законодательством.
Все публикации и патентные заявки, цитируемые в настоящем описании, включены в настоящий документ посредством ссылки полностью, в той же мере, как если бы каждая индивидуальная публикация или патентная заявка была конкретным образом индивидуально включена посредством ссылки.
Хотя вышеизложенное изобретение было подробно описано с применением иллюстративного материала и примеров для ясности понимания, любому специалисту в данной области техники будет ясно, в свете идей настоящего изобретения, что определенные изменения и модификации могут осуществляться без отхода от сущности или объема прилагаемой формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА С ПОВЫШЕННОЙ СТАБИЛЬНОСТЬЮ | 2015 |
|
RU2684703C2 |
| ЛЕЧЕНИЕ Т-КЛЕТОЧНОЙ ЛИМФОМЫ | 2020 |
|
RU2835929C2 |
| СОСТАВЫ И СПОСОБЫ ДЛЯ ЛЕЧЕНИЯ РАКА | 2018 |
|
RU2781116C1 |
| СВЯЗЫВАЮЩИЕ MICA АГЕНТЫ | 2013 |
|
RU2656183C2 |
| СРЕДСТВА ДЛЯ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЯ | 2010 |
|
RU2598719C2 |
| ПЕПТИД ИЛИ ПЕПТИДНЫЙ КОМПЛЕКС, СВЯЗЫВАЮЩИЙСЯ С альфа2-ИНТЕГРИНОМ, СПОСОБЫ ЕГО ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ УКАЗАННОГО ВЕЩЕСТВА | 2011 |
|
RU2588668C2 |
| ЛЕЧЕНИЕ РАКОВЫХ ЗАБОЛЕВАНИЙ С ПРИМЕНЕНИЕМ АНТИ-NKG2A СРЕДСТВ | 2015 |
|
RU2721271C2 |
| АНТИТЕЛА И ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ, СПЕЦИФИЧНЫЕ К CD19 | 2015 |
|
RU2741105C2 |
| МОДИФИЦИРОВАННЫЕ ЭПИТОПЫ ДЛЯ УСИЛЕНИЯ ОТВЕТОВ CD4+ Т-КЛЕТОК | 2013 |
|
RU2724994C2 |
| НЕЙТРАЛИЗАЦИЯ ИНГИБИТОРНЫХ ПУТЕЙ В ЛИМФОЦИТАХ | 2015 |
|
RU2734771C2 |

























































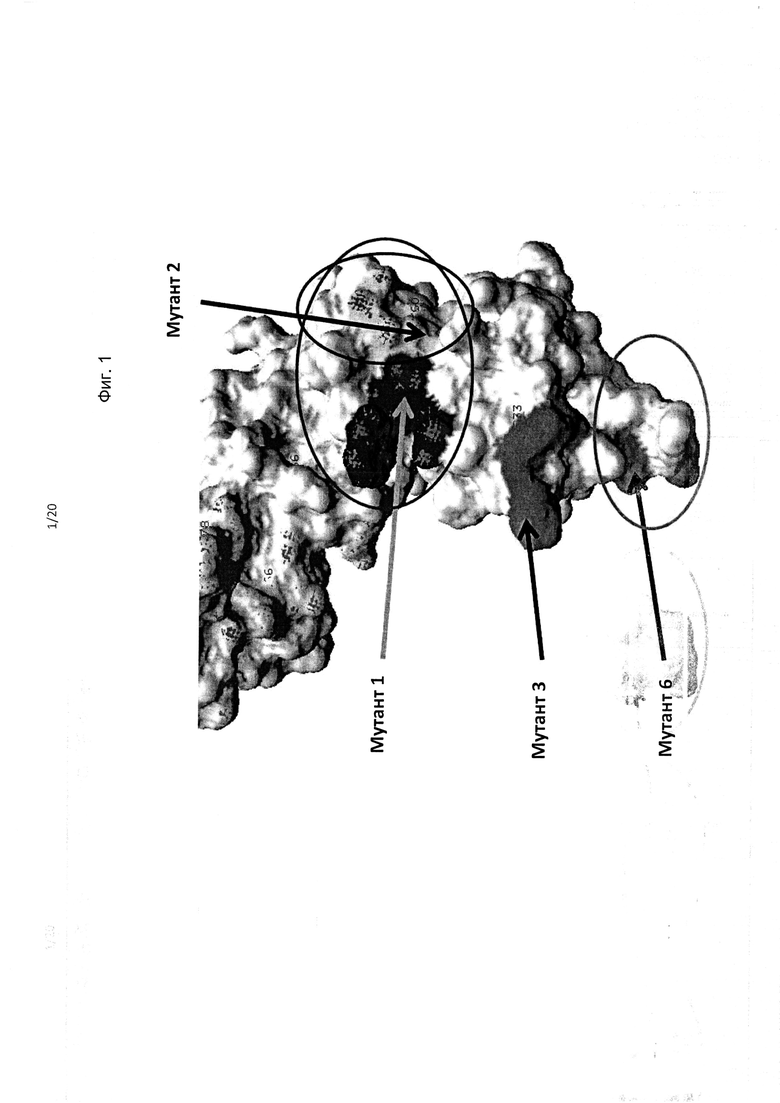
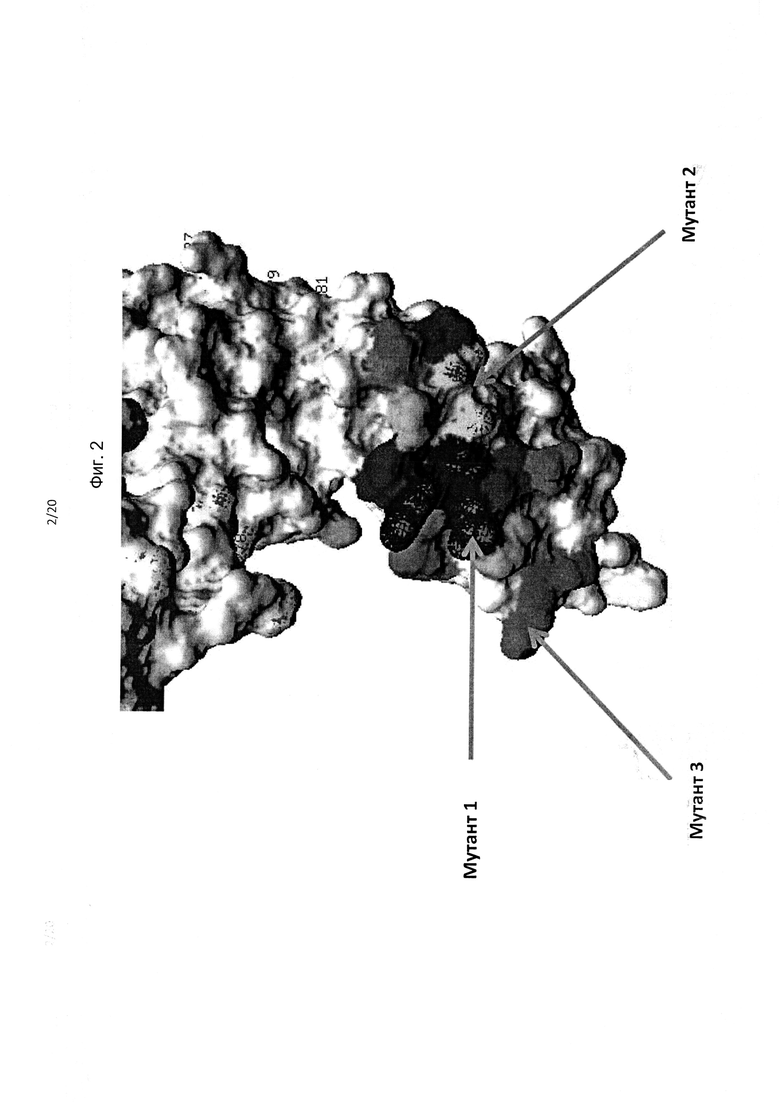
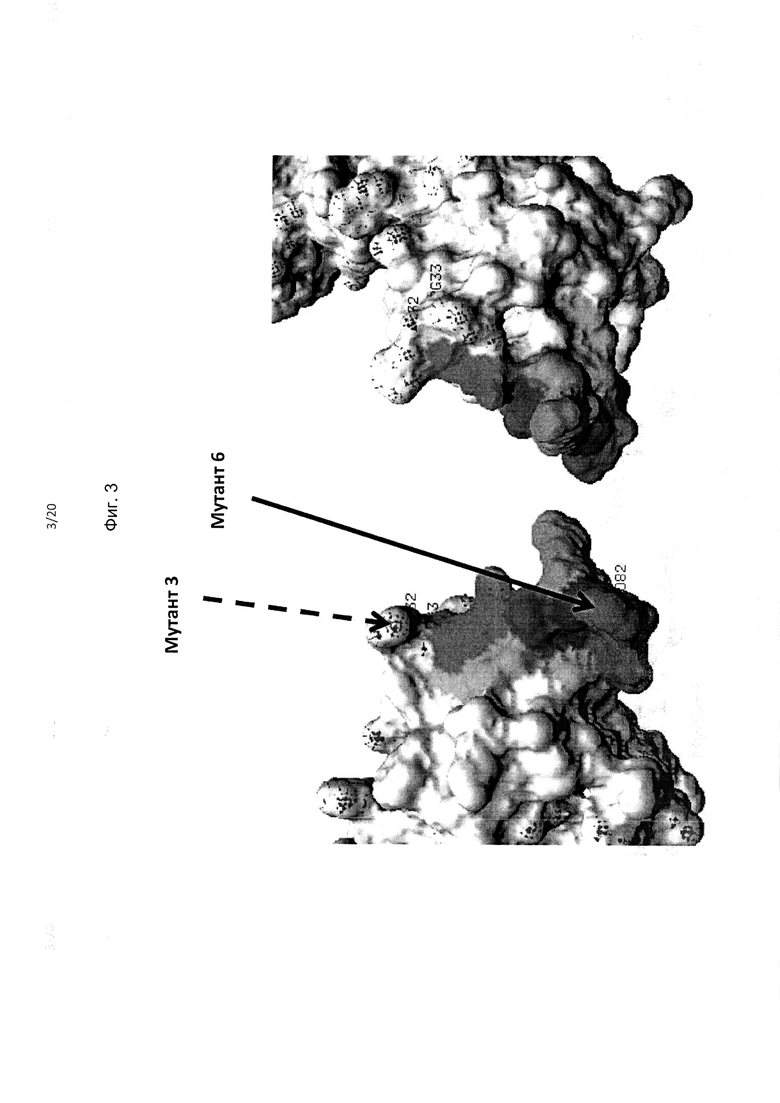
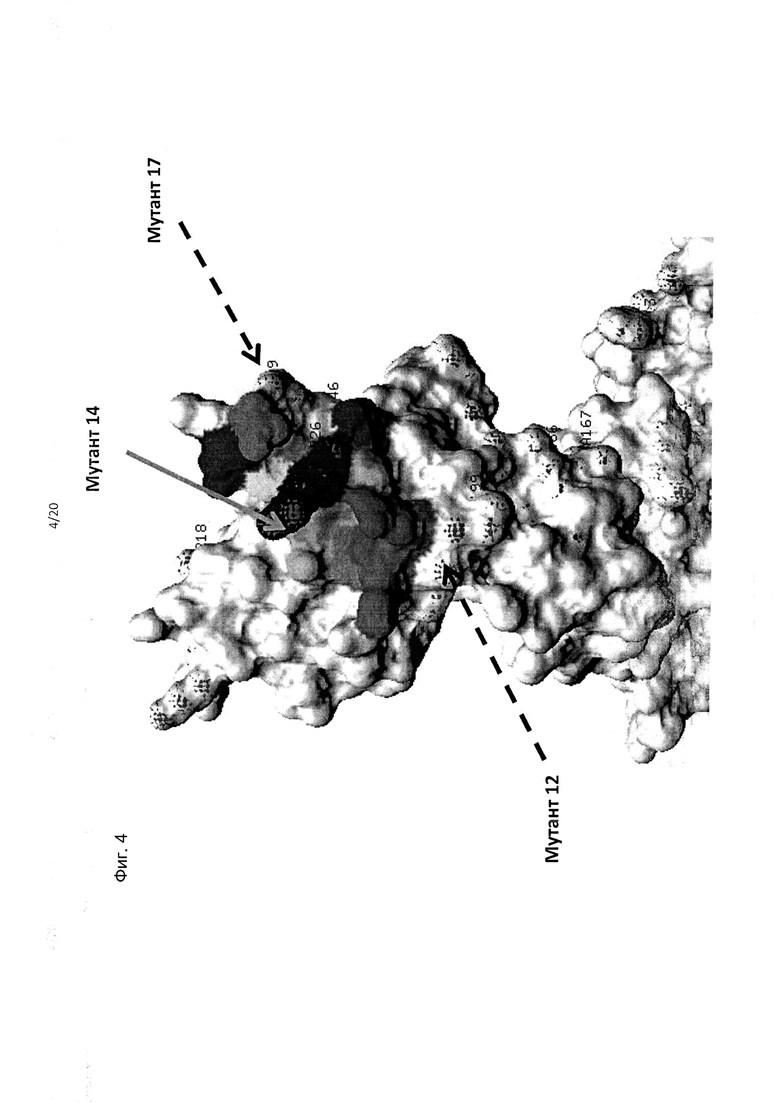

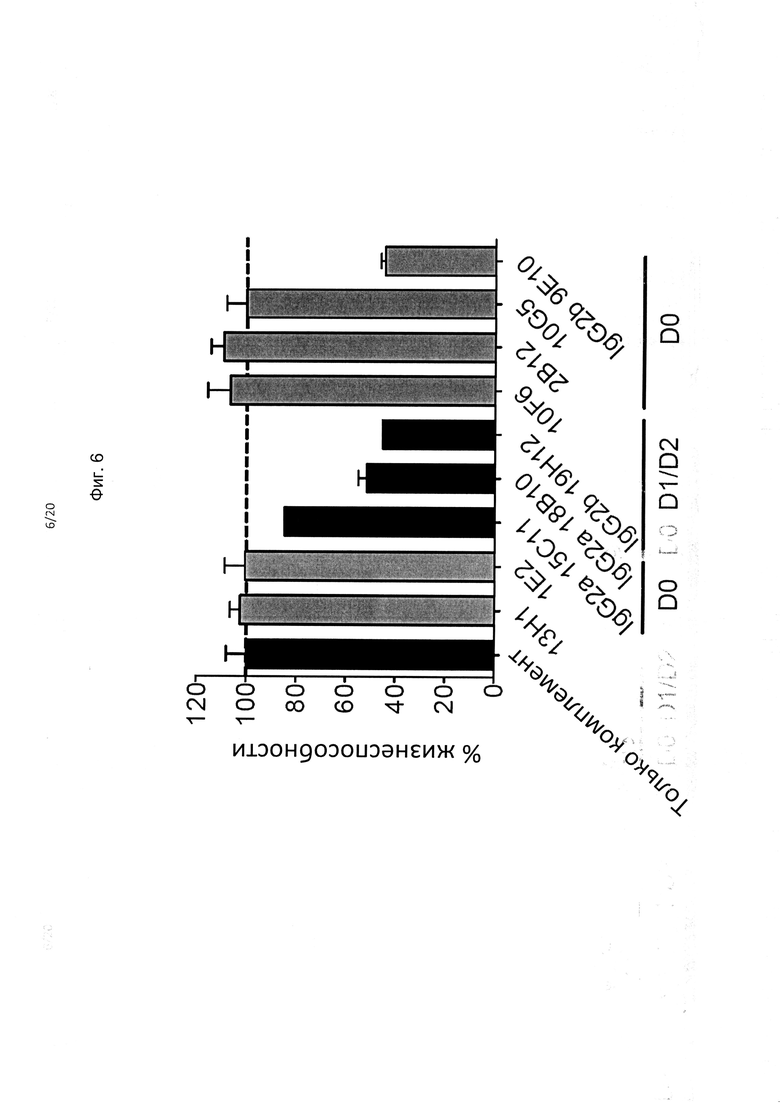
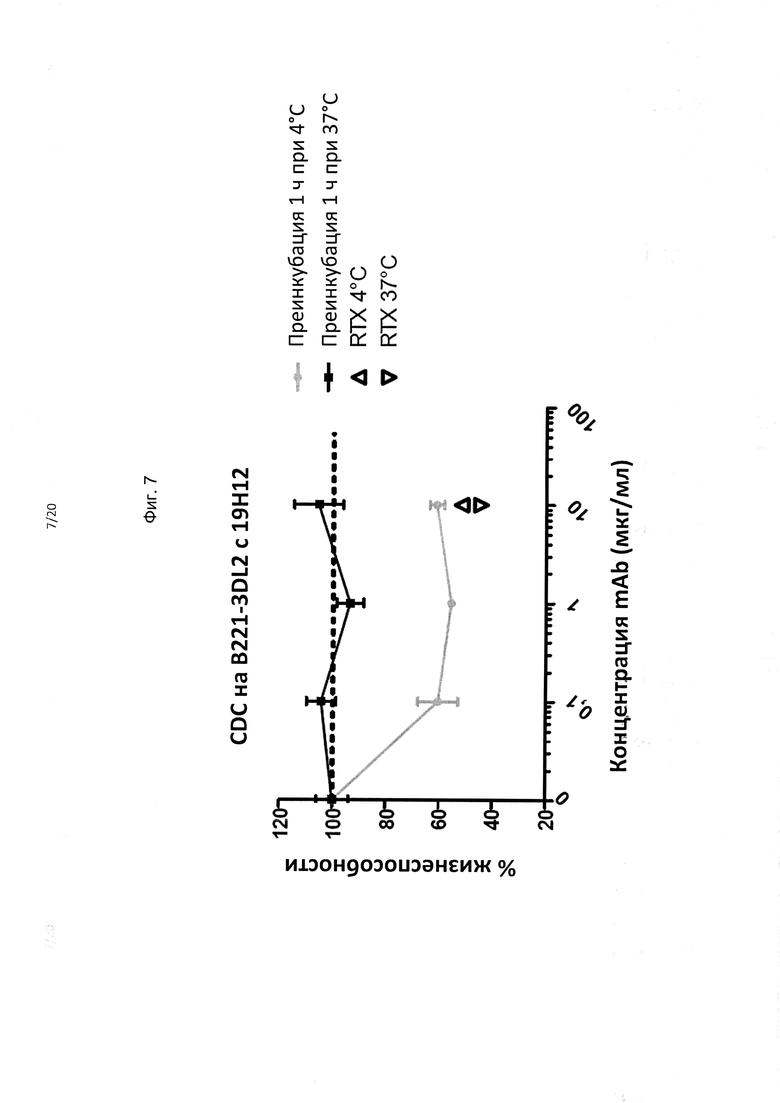
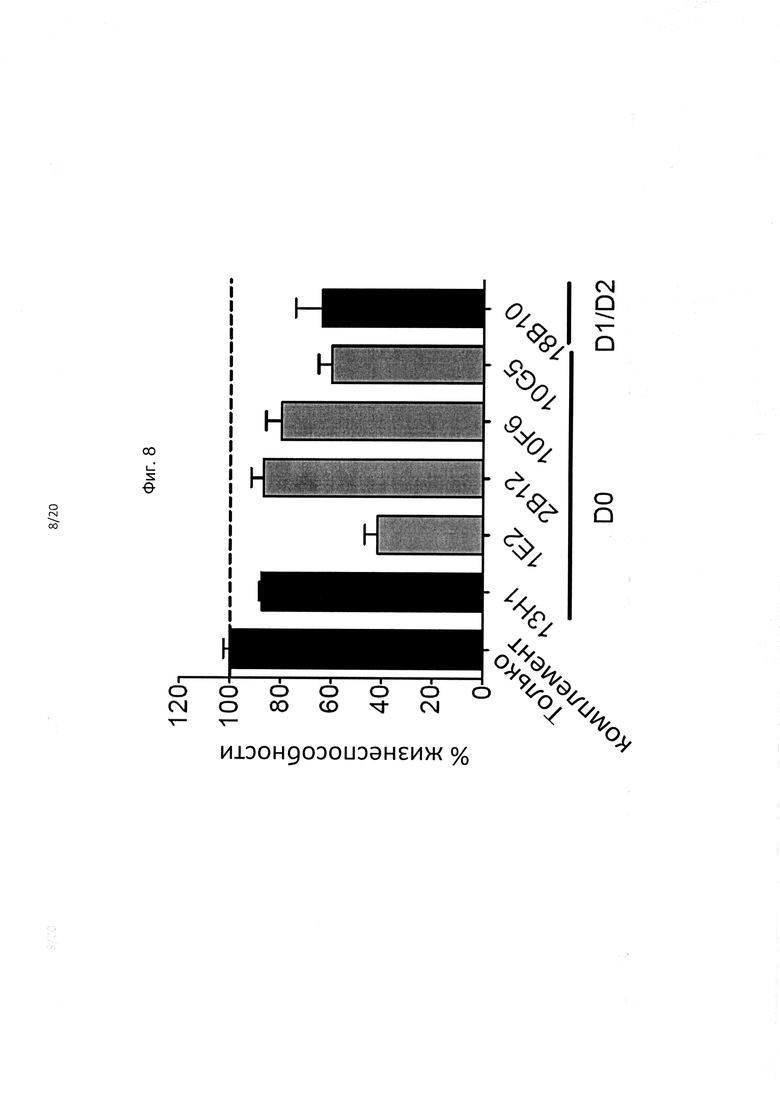
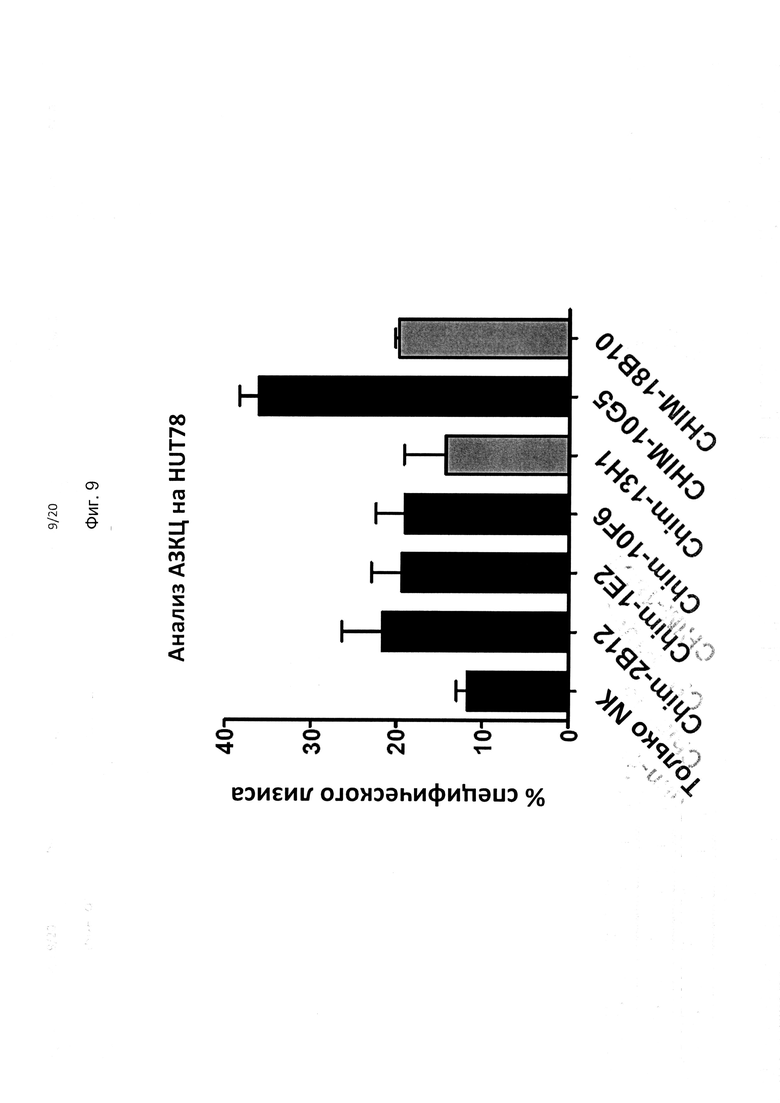
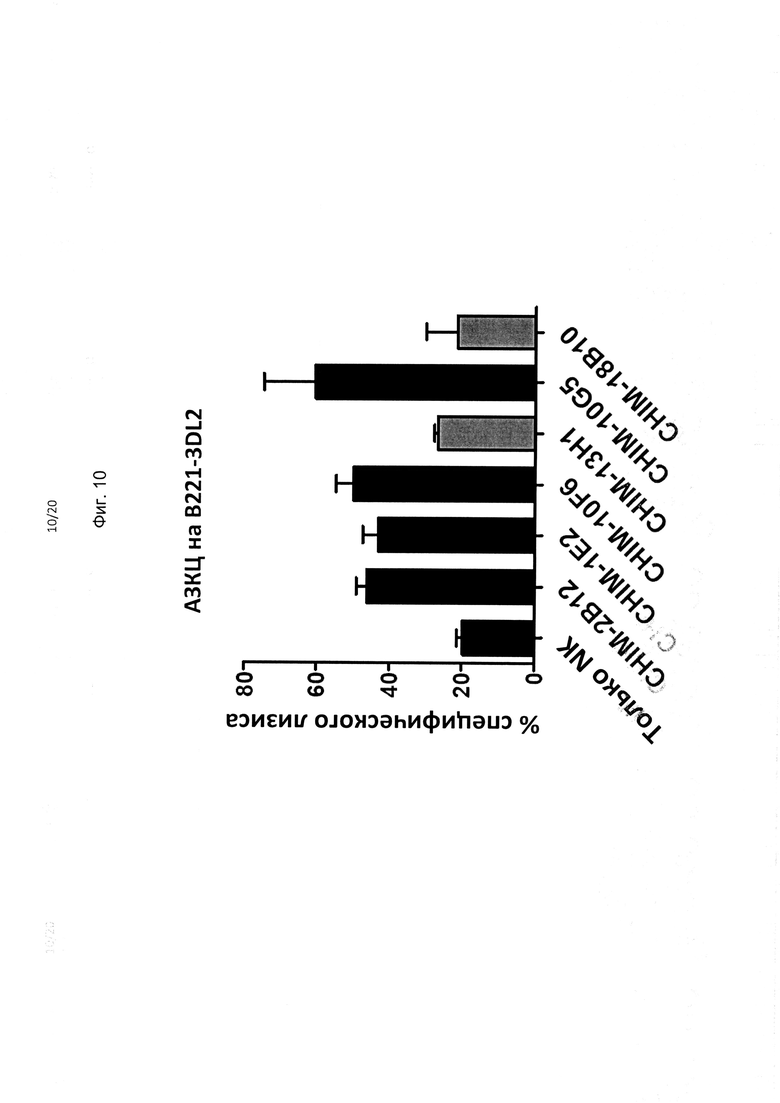
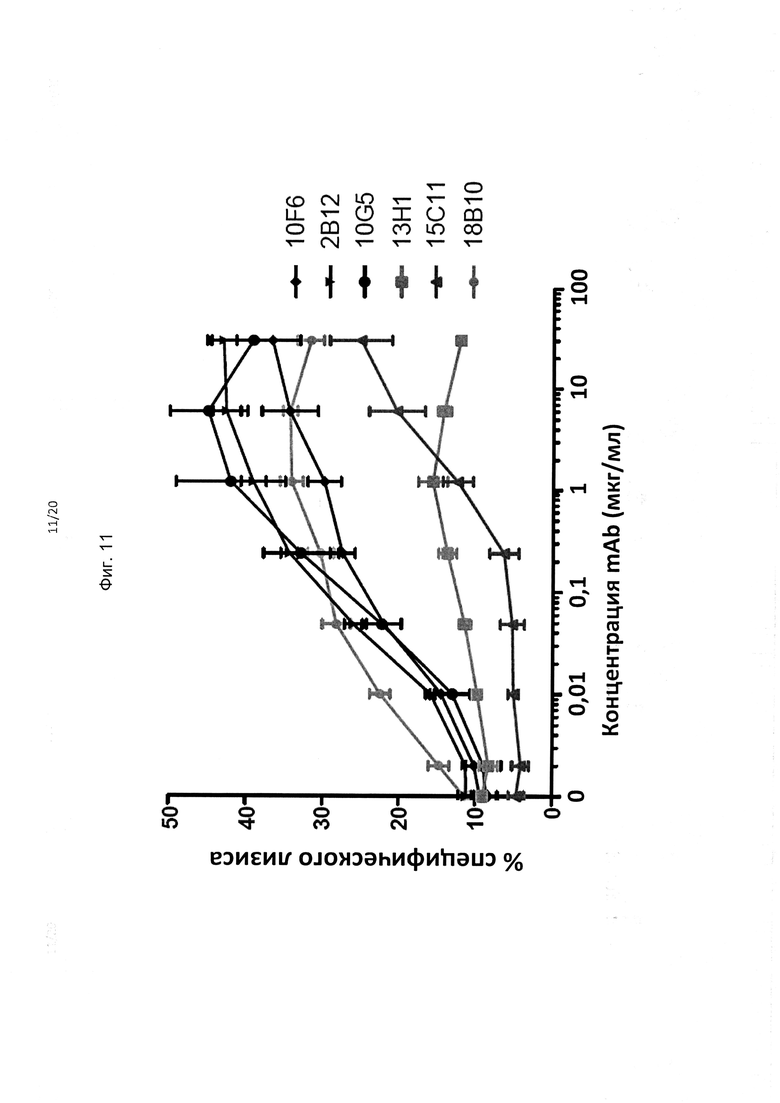
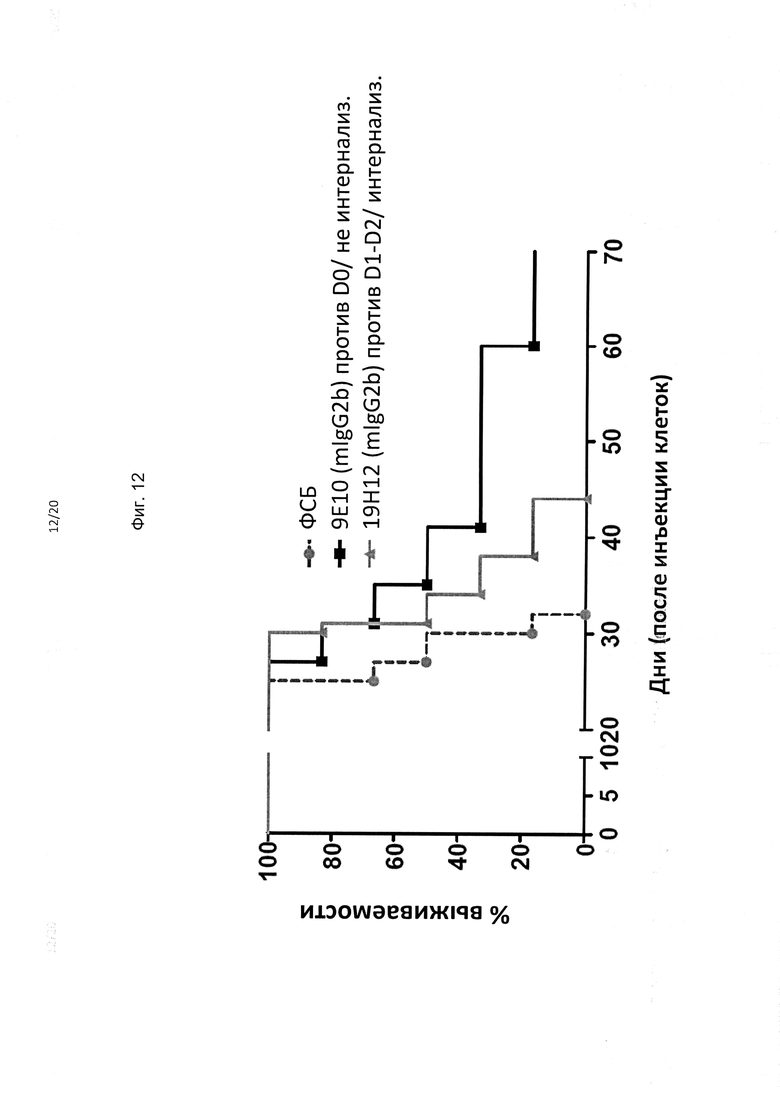
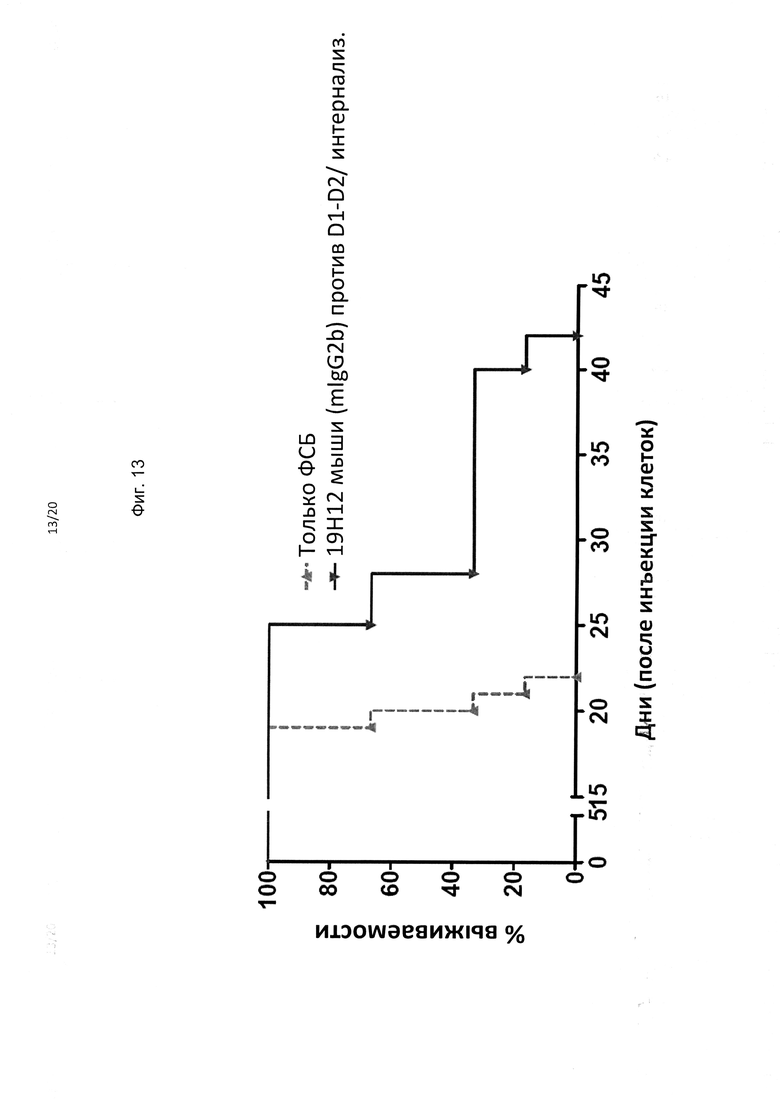
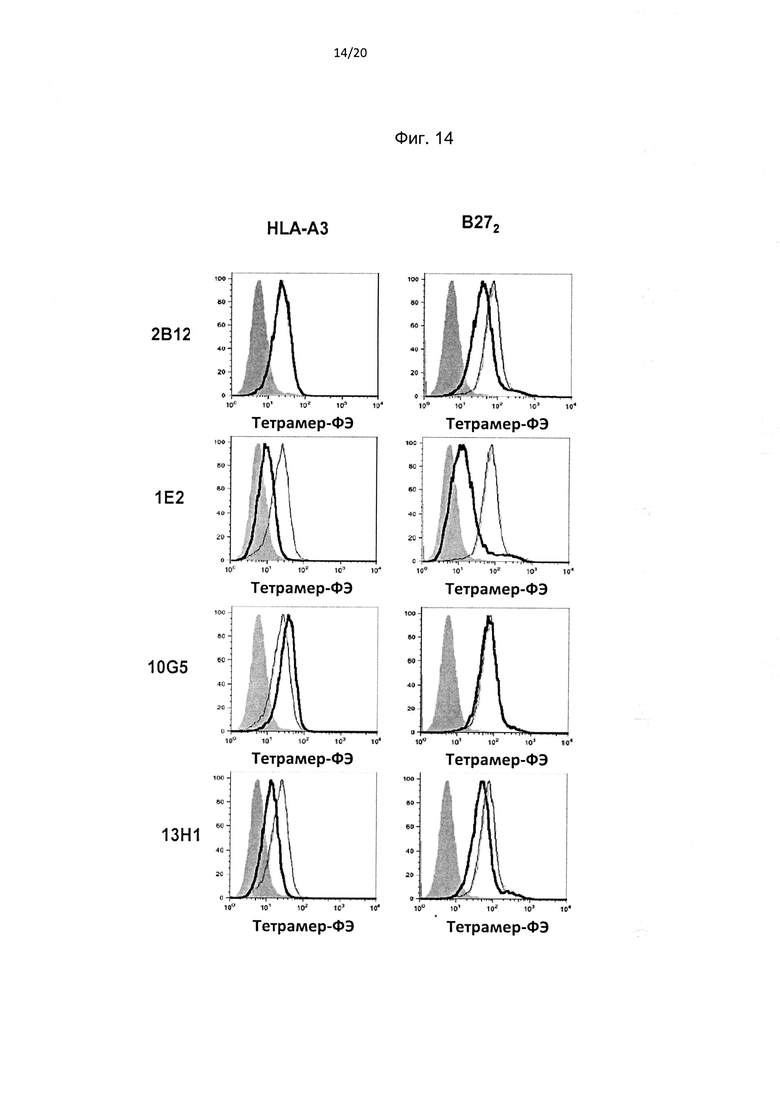
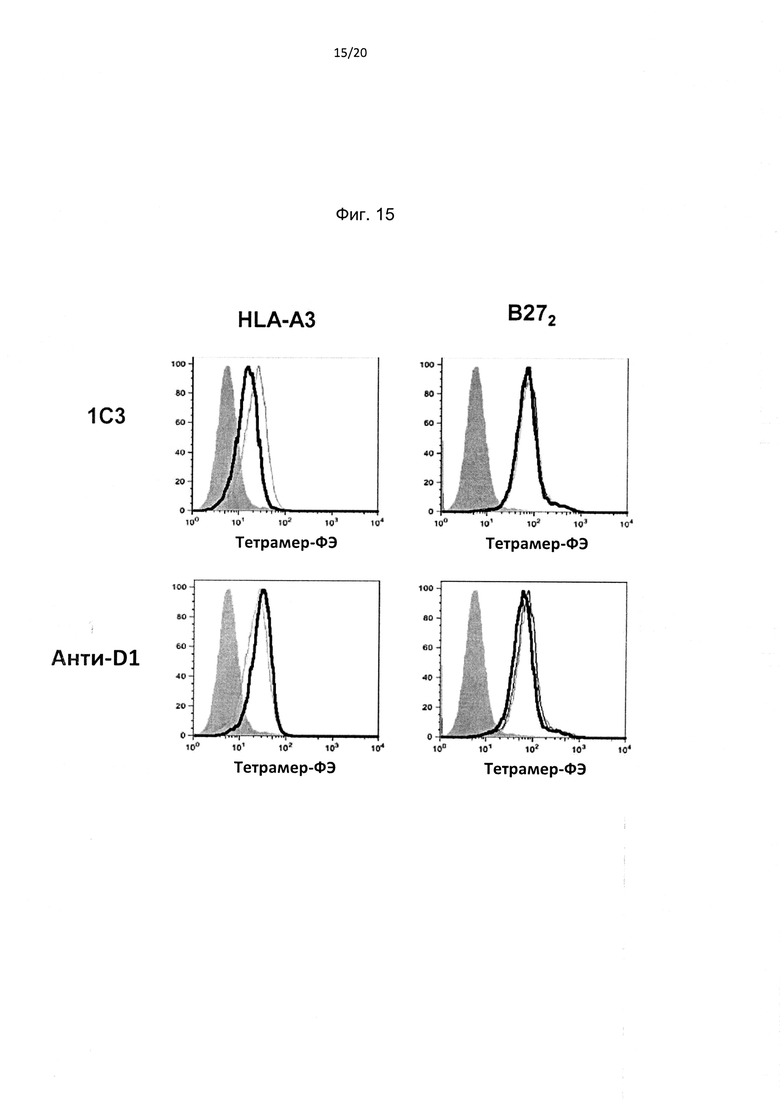
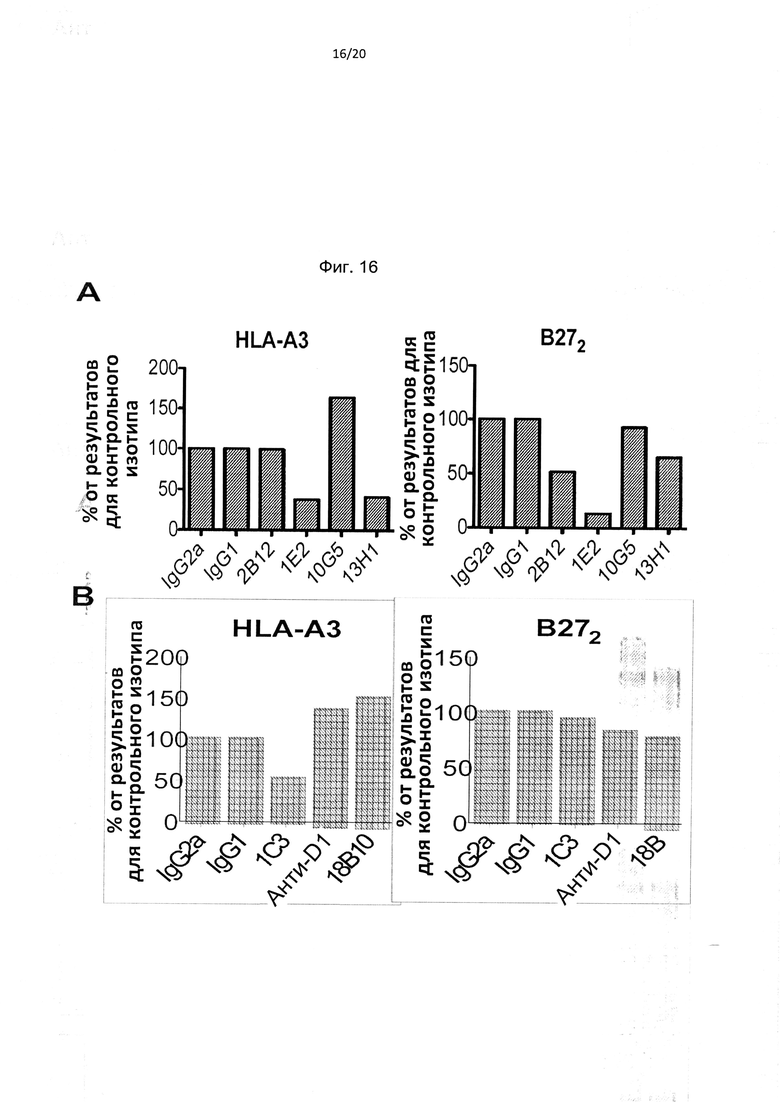
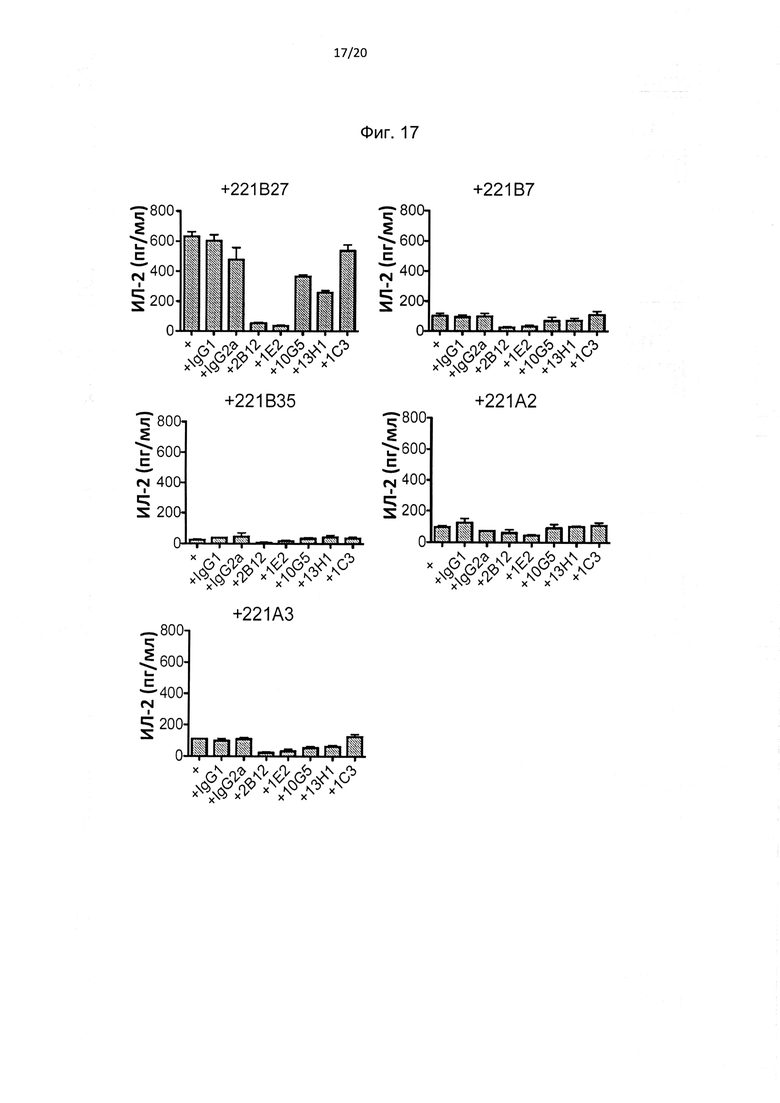
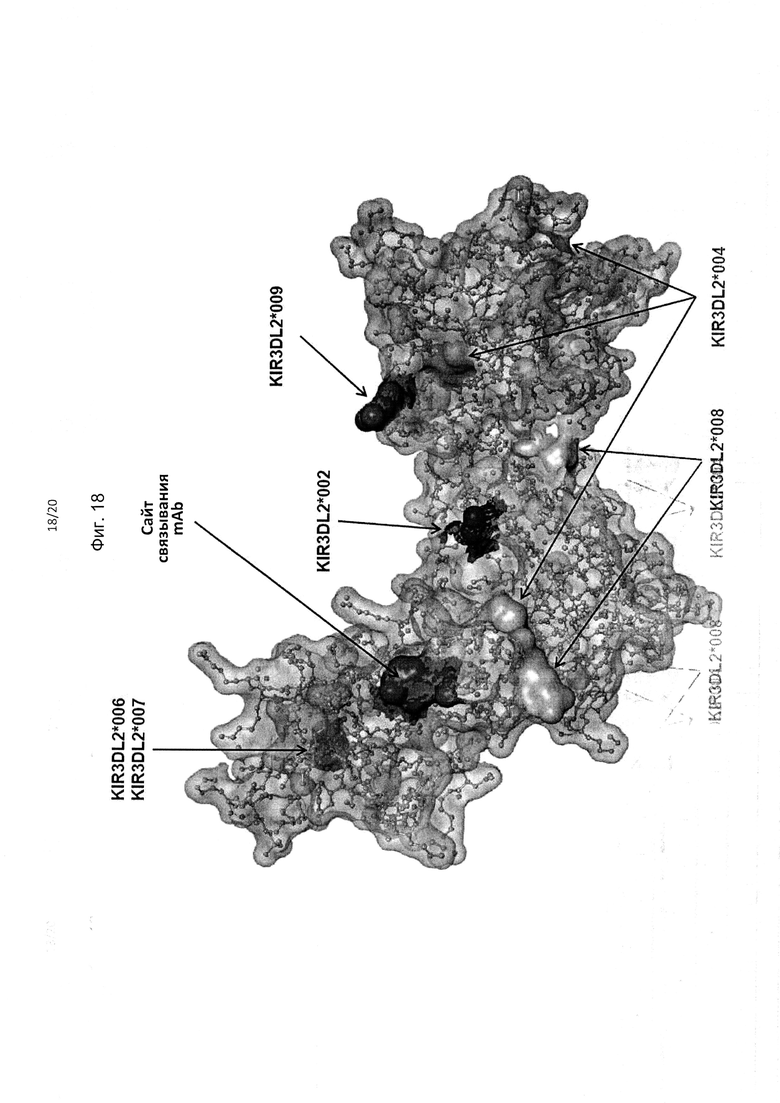
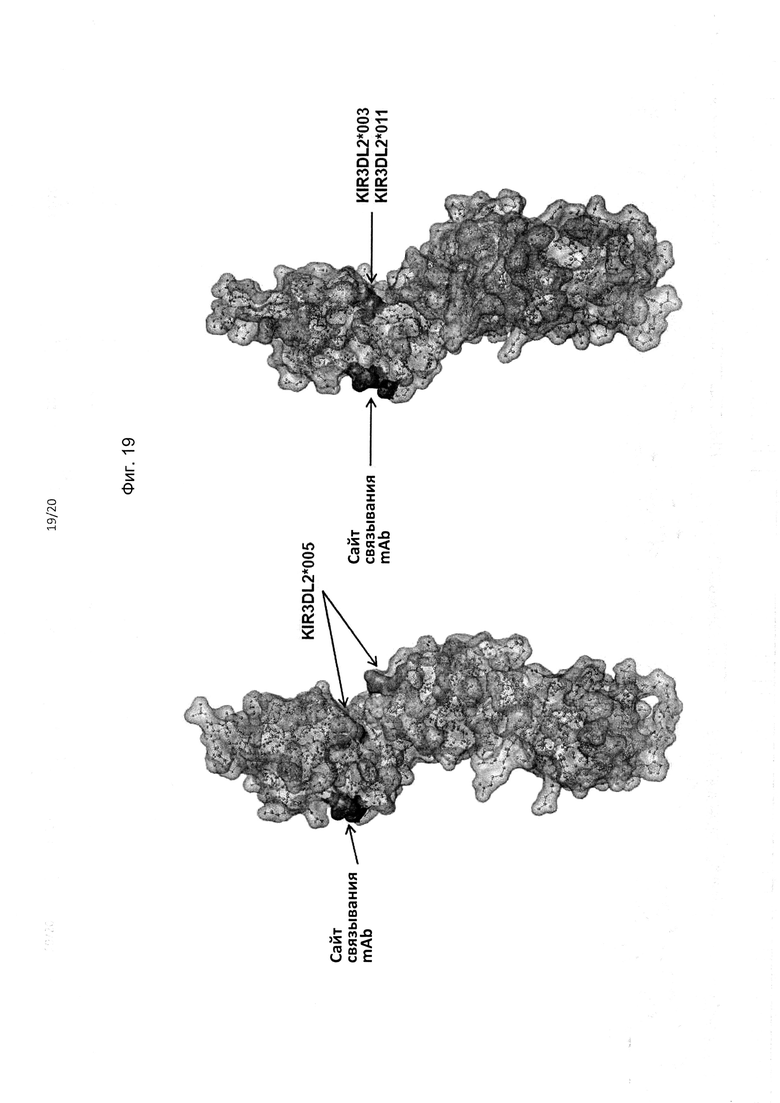
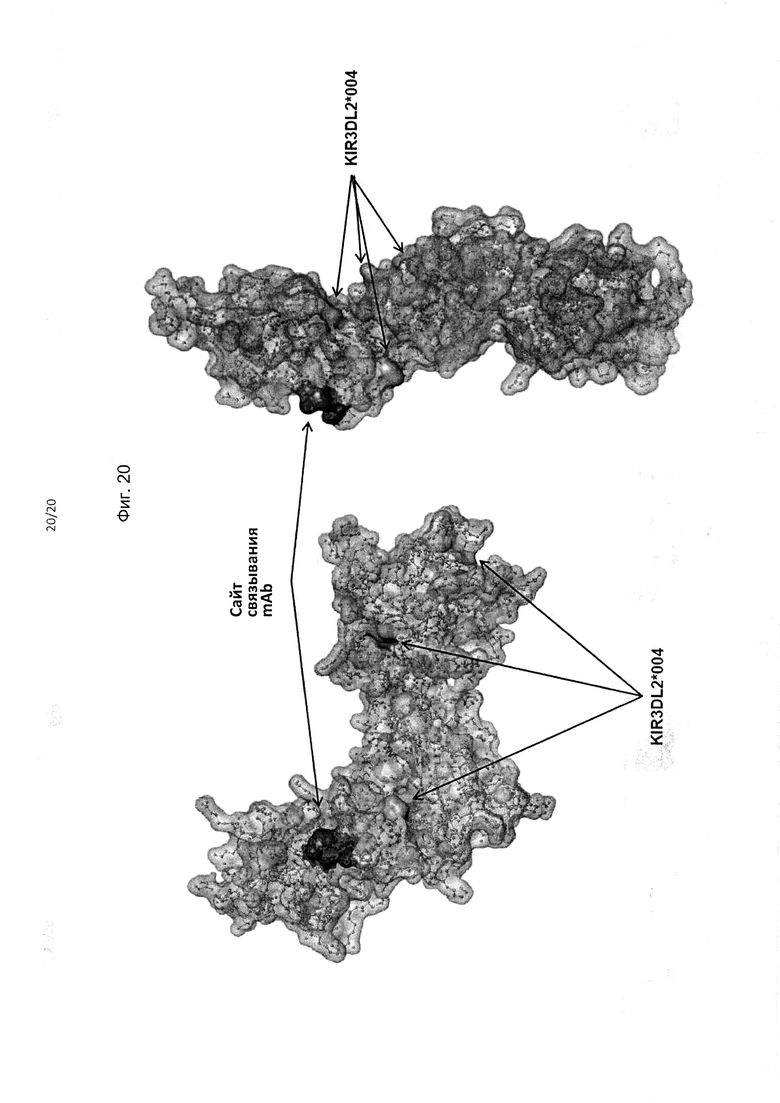
Изобретение относится к области биохимии, в частности к моноклональному антителу, которое специфически связывает полипептид KIR3DL2, а также к фармацевтической композиции для лечения рака или воспалительного или аутоиммунного нарушения, его содержащей. Также раскрыта клетка-хозяин, модифицированная гетерологичной ДНК, кодирующей вышеуказанное антитело. Изобретение также относится к способу лечения или предотвращения заболевания, при котором благоприятным было бы устранение экспрессирующих KIR3DL2 клеток, способу идентификации экспрессирующей KIR3DL2 клетки в организме субъекта, способу идентификации экспрессирующей KIR3DL2 клетки Т-клеточной лимфомы у субъекта, а также к способу отбора субъектов с Т-клеточной лимфомой, отвечающей на лечение антителом, которое специфически связывает полипептид KIR3DL2, с использованием вышеуказанного антитела. Изобретение позволяет эффективно осуществлять лечение рака или воспалительного или аутоиммунного нарушения, при котором благоприятным было бы устранение экспрессирующих KIR3DL2 клеток. 7 н. и 22 з.п. ф-лы, 20 ил., 7 табл., 8 пр.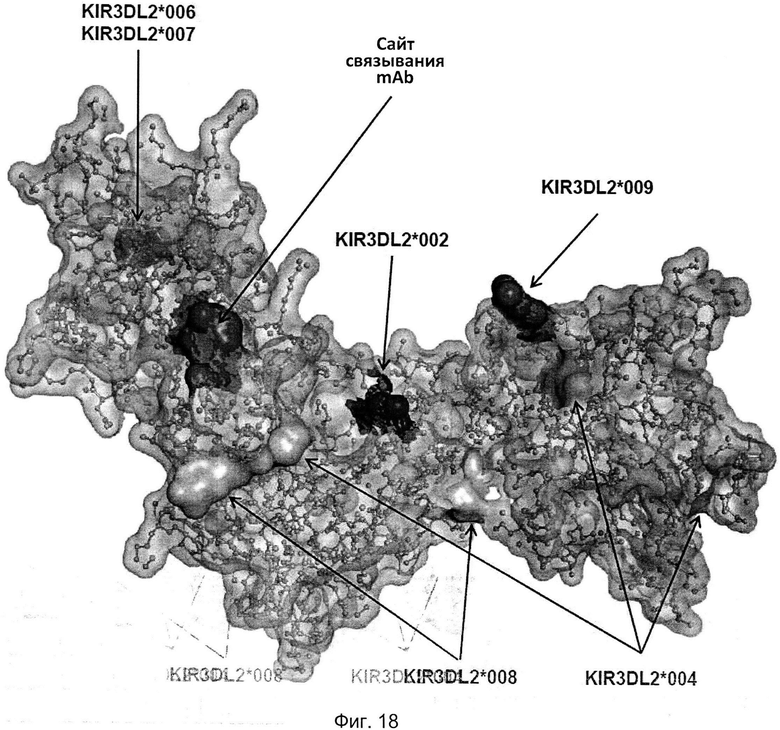
1. Моноклональное антитело, которое специфически связывает полипептид KIR3DL2, содержащий аминокислотную последовательность SEQ ID NO: 1, отличающееся тем, что указанное антитело выбрано из группы, состоящей из:
(a) антитела, содержащего (i) CDR 1, 2 и 3 тяжелой цепи (HCDR1, HCDR2, HCDR3), содержащие последовательность SEQ ID NO: 15 (HCDR1), SEQ ID NO: 18 (HCDR2) и SEQ ID NO: 20 (HCDR3) соответственно, и (ii) CDR 1, 2 и 3 легкой цепи (LCDR1, LCDR2, LCDR3), содержащие последовательности SEQ ID NO: 10, 21 и 22 соответственно;
(b) антитела, содержащего (i) CDR 1, 2 и 3 тяжелой цепи (HCDR1, HCDR2, HCDR3), содержащие последовательности SEQ ID NO: 4 (HCDR1), SEQ ID NO: 7 (HCDR2) и SEQ ID NO: 9 (HCDR3) соответственно, и (ii) CDR 1, 2 и 3 легкой цепи (LCDR1, LCDR2, LCDR3), содержащие последовательность SEQ ID NO: 10, 11 и 12 соответственно; и
(с) антитела, содержащего (i) CDR 1, 2 и 3 тяжелой цепи (HCDR1, HCDR2, HCDR3), содержащие последовательность SEQ ID NO: 25 (HCDR1), SEQ ID NO: 28 (HCDR2) и SEQ ID NO: 30 (HCDR3) соответственно, и (ii) CDR 1, 2 и 3 легкой цепи (LCDR1, LCDR2, LCDR3), содержащие последовательности SEQ ID NO: 31, 32 и 33 соответственно.
2. Антитело по п. 1, причем указанное антитело характеризуется пониженным связыванием с мутантным полипептидом KIR3DL2, содержащим аминокислотные мутации P14S, S15A и H23S, относительно связывания между указанным антителом и полипептидом KIR3DL2 дикого типа последовательности SEQ ID NO: 1.
3. Антитело по любому из пп. 1, 2, причем указанное антитело характеризуется пониженным связыванием с (а) мутантным полипептидом KIR3DL2, содержащим аминокислотные мутации I60N и G62, и (b) мутантным полипептидом KIR3DL2, содержащим аминокислотные мутации P14S, S15A и H23S, относительно связывания между указанным антителом и полипептидом KIR3DL2 дикого типа последовательности SEQ ID NO: 1.
4. Антитело по любому из пп. 1, 2, отличающееся тем, что указанное антитело детектируемо снижает связывание между KIR3DL2 и HLA класса I-лигандом KIR3DL2.
5. Антитело по любому из пп. 1, 2, отличающееся тем, что указанное антитело детектируемо снижает связывание между KIR3DL2 и HLA-B27.
6. Антитело по любому из пп. 1, 2, отличающееся тем, что указанное антитело детектируемо снижает связывание между KIR3DL2 и HLA-B27, но не приводит к детектируемому снижению связывания между KIR3DL2 и HLA-A3.
7. Антитело по любому из пп. 1, 2, отличающееся тем, что указанное антитело способно индуцировать, через антителозависимую клеточную цитотоксичность, лизис экспрессирующей KIR3DL2 клетки.
8. Антитело по любому из пп. 1, 2, отличающееся тем, что указанное антитело подавляет лиганд-индуцированную KIR3DL2-сигнализацию.
9. Антитело по любому из пп. 1, 2, отличающееся тем, что указанное антитело содержит (i) участки CDR 1, 2 и 3 тяжелой цепи (HCDR1, HCDR2, HCDR3), содержащие последовательность SEQ ID NO: 15 (HCDR1), SEQ ID NO: 18 (HCDR2) и SEQ ID NO: 20 (HCDR3) соответственно, и (ii) участки CDR 1, 2 и 3 легкой цепи (LCDR1, LCDR2, LCDR3), содержащие последовательность SEQ ID NO: 10, 21 или 22 соответственно.
10. Антитело по любому из пп. 1, 2, отличающееся тем, что указанное антитело содержит содержащую участки CDR 1, 2 и 3 тяжелой цепи (HCDR1, HCDR2, HCDR3), содержащие последовательность SEQ ID NO: 4 (HCDR1), SEQ ID NO: 7 (HCDR2) и SEQ ID NO: 9 (HCDR3) соответственно, и (ii) участки CDR 1, 2 и 3 легкой цепи (LCDR1, LCDR2, LCDR3), содержащие последовательность SEQ ID NO: 10, 11 или 12 соответственно.
11. Антитело по п. 1, отличающееся тем, что указанное антитело обладает бивалентной аффинностью связывания (KD) полипептида KIR3DL2 человека, составляющей менее 10-8 М.
12. Антитело по п. 1, отличающееся тем, что указанное антитело содержит константную область тяжелой цепи IgG человека.
13. Антитело по п. 1, отличающееся тем, что указанное антитело содержит константную область тяжелой цепи человека, которая связывает рецептор FcγIIIA.
14. Антитело по п. 1, отличающееся тем, что указанное антитело представляет собой химерное антитело, антитело человека или гуманизированное антитело.
15. Антитело по п. 13, отличающееся тем, что указанное антитело содержит модифицированную константную область тяжелой цепи человека с по меньшей мере одной заменой аминокислоты, при этом аффинность связывания указанной модифицированной константной области к рецептору FcγIIIA повышена по сравнению с константной областью, не содержащей указанной замены аминокислоты.
16. Антитело по п. 13, отличающееся тем, что указанное антитело содержит константную область тяжелой цепи человека, содержащую гипофукозилированные N-связанные гликаны, при этом аффинность связывания указанной модифицированной константной области к рецептору FcγIIIA повышена по сравнению с константной областью, не содержащей указанных гипофукозилированных N-связанных гликанов.
17. Фармацевтическая композиция для лечения рака или воспалительного или аутоиммунного нарушения, при котором благоприятным было бы устранение экспрессирующих KIR3DL2 клеток, содержащая эффективное количество антитела по любому из предшествующих пунктов и фармацевтически приемлемый носитель.
18. Композиция по п. 17, отличающаяся тем, что указанное антитело присутствует в количестве от приблизительно 25 мг до 500 мг.
19. Рекомбинантная клетка-хозяин для продуцирования антитела по пп. 1-16, модифицированная гетерологичной ДНК, кодирующей антитело по любому из пп. 1-16.
20. Способ лечения или предотвращения заболевания, при котором благоприятным было бы устранение экспрессирующих KIR3DL2 клеток, у нуждающегося в этом пациента, который включает введение указанному пациенту эффективного количества антитела по пп. 1-16 или композиции по пп. 17-18.
21. Способ по п. 20, отличающийся тем, что указанное заболевание представляет собой CD4+ Т-клеточную лимфому.
22. Способ по п. 21, отличающийся тем, что указанная CD4+ Т-клеточная лимфома выбрана из фунгоидного микоза и синдрома Сезари.
23. Способ по п. 20, отличающийся тем, что указанное заболевание представляет собой воспалительное или аутоиммунное нарушение.
24. Способ идентификации экспрессирующей KIR3DL2 клетки в организме субъекта, причем указанный способ включает получение биологического образца от субъекта, содержащего клетки, приведение указанных клеток в контакт с антителом по пп. 1-16 и оценку того, связывается ли указанное антитело с клетками, причем определение того, что указанное антитело связывается с клетками, указывает на то, что эти клетки являются экспрессирующими KIR3DL2 клетками.
25. Способ идентификации экспрессирующей KIR3DL2 клетки Т-клеточной лимфомы у субъекта, отличающийся тем, что указанный способ включает получение биологического образца от субъекта, содержащего связанные с заболеванием клетки, приведение указанных клеток Т-клеточной лимфомы в контакт с антителом по пп. 1-16 и оценку того, связывается ли указанное антитело с клетками Т-клеточной лимфомы, при этом определение того, что указанное антитело связывается с клетками Т-клеточной лимфомы, указывает на то, что указанная клетка Т-клеточной лимфомы экспрессирует KIR3DL2.
26. Способ отбора субъектов с Т-клеточной лимфомой, отвечающей на лечение антителом по пп. 1-16, причем указанный способ включает определение того, экспрессируют ли клетки Т-клеточной лимфомы указанного субъекта рецептор KIR3DL2, при этом экспрессия рецептора KIR3DL2 указывает на субъекта с Т-клеточной лимфомой, отвечающей на лечение антителом по пп. 1-16.
27. Способ по п. 26, включающий дополнительно введение отвечающему на лечение пациенту антитела по пп. 1-16.
28. Способ по пп. 26, 27, отличающийся тем, что указанная лимфома представляет собой CD4+ Т-клеточную лимфому.
29. Способ по п. 28, отличающийся тем, что указанная лимфома выбрана из фунгоидного микоза и синдрома Сезари.
| WO2010081890 A1, 22.07.2010 | |||
| RUDIKOFF S | |||
| et al., Single amino acid substitution altering antigen-binding specificity, Proc Natl Acad Sci U S A, 1982, Vol.79, N.6, pp | |||
| Дверной замок, автоматически запирающийся на ригель, удерживаемый в крайних своих положениях помощью серии парных, симметрично расположенных цугальт | 1914 |
|
SU1979A1 |
| WU Н | |||
| et al., Humanization of a Murine Monoclonal Antibody by Simultaneous Optimization of Framework and CDR Residues, J | |||
| Mol | |||
| Biol, 1999, Vol | |||
| ДИФФЕРЕНЦИАЛЬНАЯ ТЕРМИОННАЯ ЛАМПА | 1920 |
|
SU294A1 |
| Двухколейная подвесная дорога | 1919 |
|
SU151A1 |
| КОМПОЗИЦИЯ И СПОСОБ РЕГУЛИРОВАНИЯ АКТИВНОСТИ ЕСТЕСТВЕННЫХ КЛЕТОК-КИЛЛЕРОВ | 2004 |
|
RU2376315C2 |

