ОБЛАСТЬ ТЕХНИКИ
Изобретение относится к области противораковой иммунотерапии. В частности, изобретение касается индукции врожденного или приобретенного иммунитета путем введения молекул IL-12 направленного действия, предпочтительно совместно с IL-2 и/или IL-7, пациенту, страдающему раком мышц, костей, нервной системы, хрящей, сухожилий, кровеносных сосудов и т.д., преимущественно саркомой.
Изобретение относится в частности к применению IL-12 в виде конкретного химерного белка под названием NHS-IL12, слитого из иммуноглобулина и цитокина, предпочтительно в комбинации с IL-2 и/или IL-7 в форме, демонстрирующей пролонгированные фармакокинетические свойства, для лечения указанных видов рака, особенно сарком.
УРОВЕНЬ ТЕХНИКИ
Противораковая иммунотерапия охватывает широкое разнообразие терапевтических подходов, включая "пассивное" введение опухолеспецифичных моноклональных антител и других компонентов иммунной системы, "активную" иммунизацию с целью вызвать или усилить специфический иммунный ответ против клеток опухоли, опосредованный T-клетками, адоптивный перенос модифицированных ex vivo T-клеток и неспецифическое усиление иммунологической реактивности с помощью иммуномодуляторов. Иммунотерапия уже оказала сильное влияние на тактику лечения широкого спектра раковых заболеваний, однако это влияние в основном ограничивается пассивной иммунотерапией моноклональными антителами. Область противораковой иммунотерапии является сложной и быстро развивающейся. Иммунотерапия отличается от традиционной химиотерапии как механизмами действия, так и типами вызванных ответов, и стандартные критерии оценки ответа на лечение могут не предоставлять достоверной характеристики болезнь-модифицирующей активности иммунотерапевтических агентов. Во многих видах активной иммунотерапии используют несколько компонентов (например, антигены, адъюванты и носители). К тому же растет понимание того, что для достижения устойчивого терапевтического эффекта необходимо комбинировать иммунизацию с другими иммуномодулирующими стратегиями, направленными на усиление общей иммунологической реактивности и преодоление механизмов подавления иммунитета, с помощью которых опухоли вызывают иммунологическую толерантность своего окружения.
Рабдомиосаркома (РМС) является наиболее часто встречающейся опухолью мягких тканей у детей и имеет крайне неблагоприятный прогноз на поздних стадиях (Oberlin et al., 2008, J. Clin. Oncol. 26: 2384-2389). Удаление хирургическим путем, химиотерапия и лучевая терапия часто не оказывают действия из-за локализации опухолей в хрупких анатомических структурах и их предрасположенности к распространению. Таким образом, существует большая неудовлетворенная потребность в альтернативных стратегиях лечения.
Недавние исследования наводят на мысль, что аллогенная трансплантация стволовых клеток (аллоТСК) является подходящей стратегией лечения плотных опухолей на поздней стадии, включая PMC (Koscielniak et al., 2005, J. Clin. Oncol. 23: 242-244). Однако механизмы контроля опухоли в таких условиях изучены недостаточно. Хотя ответы цитотоксичных Т-клеток и естественных киллеров на РМС могут быть воспроизведены in vitro (van den Broeke et al., 2006, Cancer Res. 66: 1818-1823), противоопухолевая активность иммунных эффекторных клеток является многогранной. Кроме киллерной активности, комбинированные сигналы по путям, зависящим от интерферона-гамма (IFN-γ)-и рецептора1 фактора некроза опухолей (TNFR1), могут вызвать переход опухоли в состояние покоя (Muller-Hermelink, et al 2008, Cancer Cell 13: 507-518), соответственно, индуцированная секреция этих цитокинов может быть важным способом локального контроля роста опухоли. Действительно, несколько исследований показали улучшенную выживаемость после аллоТСК пациентов с увеличенной частотой встречаемости T-клеток, производящих IFN-γ (Wiegering et al., 2011, Cancer Immunol. Immunother. 60: 693-703). Таким образом, CD4+ T-клетки могут быть так же важны, как и CD8+ ЦТЛ.
IL-12, основной участник этой сети, может вызывать регрессию опухоли и влияет на врожденный и приобретенный иммунитет (например, Trinchieri, G. 2003, Nat. Rev. Immiinol. 3: 133-146). Кроме роли в примировании Т-клеток, IL-12 восстанавливает фенотип TH1 у TH17-клеток, восстанавливает функцию макрофагов M1 и является посредником во взаимодействиях естественных киллеров и дендритных клеток (DC-NK).
Интерлейкин-12 (IL-12) является плейотропным провоспалительным цитокином, который вырабатывается в ответ на инфекцию несколькими разновидностями клеток иммунной системы, включая фагоцитарные клетки, B-клетки и активированные дендритные клетки (Colombo and Trinchieri (2002) Cytokine & Growth Factor Reviews, 13: 155-168). IL-12 играет важную роль в качестве промежуточного звена во взаимодействии врожденного и приобретенного иммунитета, воздействуя на Т-клетки и естественные киллеры (NK-клетки), улучшая пролиферацию и активность цитотоксичных лимфоцитов, а также выработку других воспалительных цитокинов, особенно интерферона-γ. IL-12 является гетеродимерной молекулой, состоящей из α-цепи (субъединица p35, IL-12p35) и β-цепи (субъединица p40, IL-12p40), соединенных ковалентно дисульфидным мостиком и образующих биологически активный гетеродимер массой 74 кДа.
Было показано, что присутствие эндогенного IL-12 необходимо для иммунологического сопротивления широкому спектру патогенов, а также трансплантированным и химически индуцированным опухолям (Gateley et al. (1998), Annu. Rev. Immunol., 16: 495-521). Было продемонстрировано, что IL-12 обладает сильным противоопухолевым действием, основанным на индукции IFN-γ и активации эффекторных клеток, таких как CD8+ T-клетки и NK-клетки (Brunda et al. (1993), J. Exp. Med., 178: 1223-30). В ответ на IL-12 T-клетки и NK-клетки вырабатывают большие количества IFN-гамма [Kobayashi et al., 1989, J Exp Med; 170: 827-45], приводя к усилению антиген-презентации за счет паракринного повышения экспрессии МНС класса I и класса II Wallach et al., 1982 Nature 1982; 299: 833-69]. Вследствие доказанной противоопухолевой активности IL-12 его испытывали в клинических исследованиях на людях в качестве иммунотерапевтического агента для лечения широкого спектра разновидностей рака (Atkins et al. (1997), Clin. Cancer Res., 3: 409-17; Gollob et al. (2000), Clin. Cancer Res., 6: 1678-92; и Hurteau et al. (2001), Gynecol. Oncol., 82: 7-10), в том числе рака почек, рака толстой кишки, рака яичников, меланомы и T-клеточной лимфомы, а также в качестве адъюванта для противораковых вакцин (Lee et al. (2001), J. Clin. Oncol. 19: 3836-47).
Системное введение IL-12 преимущественно демонстрирует эффективность против некоторых плотных опухолей, но его использование в терапии ограничено из-за его дозолимитирующей токсичности (Gollob et al., 2000, Clin. Cancer Res. 6: 1678-1692). После более чем десятилетних клинических исследований ранних фаз, задокументированные случаи сильной токсичности и в целом низкий процент положительного ответа на лечение rIL-12 воспрепятствовали его дальнейшим клиническим исследованиям.
Таким образом, целевая доставка IL-12 в микроокружение опухоли представляет собой многообещающее направление в иммунотерапии опухолей, поскольку она могла бы сделать цитокин более эффективным и менее токсичным. Поэтому основной задачей данного изобретения является предоставление эффективного и прежде всего могущего быть использованным подхода к противораковой иммунотерапии, используя для этого эффективный лекарственный противоопухолевый иммунитет.
КОРОТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Одна из стратегий повышения безопасности провоспалительных цитокинов, таких как IL-2 и IL-12, заключается в том, чтобы направить их доставку к опухолям через слияние с нацеленным на опухоль антителом. Такие белки, слитые из антитела и цитокина, или "иммуноцитокины", предварительно показали способность усиливать противоопухолевый иммунитет в доклинических моделях [Gillies SD. In Lustgarten J, Cui Y, Li S, eds. Targeted Cancer Immune Therapy. New York, New York, США Springer; 2009: 241-256]. Для максимизации переносимости иммуноцитокина антитело, выбранное в качестве носителя, должно специфически связываться с антигеном, встречающимся исключительно в опухолях. Привлекательным способом доставки представляются антитела против антигенов, ассоциированных с некрозом, которые в избытке присутствуют в опухолях, но не в нормальных тканях [например, Epstein et al., 1988, Cancer Res 1988; 48: 5842-48, 20].
Способность ДНК/гистон-связывающих антител селективно нацеливаться на области опухолевого некроза хорошо исследована как доклинически, так и клинически. С использованием этой концепции был создан некрозо-таргетный иммуноцитокин на основе IL-12, получивший название NHS-IL12, через генетическое слияние двоих гетеродимеров человеческого IL-12 с C-концами тяжелых цепей антитела NHS76 (WO 2000/001822). NHS76 - это полностью человеческое, полученное методом фаг-дисплея антитело IgG1, выбранное за его способность специфически связываться с ДНК/гистонами и таким образом нацеленность на опухоли in vivo [Sharifi et al., 2001, Hybrid Hybridomics; 20: 305-127.
Было обнаружено, что побуждаемая IL-12 человеческая иммунная система с помощью введения вышеозначенного таргетного препарата IL-12 не только убивает раковые клетки, но и использует альтернативные механизмы ослабления роста опухоли, т.е. через индуцирование старения и/или дифференциации раковых клеток, таким образом приводя к ремиссии опухолевых клеток или тканей до клеток или тканей происхождения. Предлагаемая таргетная терапия IL-12 вызывает специфически индуцируемую TH1 остановку роста и дифференциацию раковых клеток. Этот эффект особенно возрастает в случае введения таргетного IL-12 совместно или в комбинации с иммуномодуляторами, такими как интерлейкины, например, IL-2 и IL-7.
Предлагается использовать IL-12 в связанной таргетной форме, такой как химерный IL-12-белок, предпочтительно белок, слитый из иммуноглобулина, способного нацеливаться на антиген или специфическую экспрессируемую тканями пациента рецепторную молекулу, и IL-12, в котором предпочтительно C-конец иммуноглобулина связан с N-концом иммуноцитокина. Предлагаемый нацеленный на опухоль IL-12 является терапевтически эффективным моноклональным антителом или его биологически активным фрагментом (таким, как Fab фрагмент или scFv), связанным с IL-12.
В предпочтительном варианте осуществления изобретения терапевтически эффективное антитело или его таргетирующий фрагмент направлены на комплекс ДНК-гистон H1, представленный в опухолевом некрозе. В следующем конкретном варианте осуществления изобретения таргетная молекула IL-12 является соответствующим белком, слитым из IL-12 и известного полностью человеческого антитела IgG1 NHS76, в соответствии с выше- и нижеизложенным детальным описанием.
Предлагаемый таргетный IL-12 может быть использован в противораковой иммунотерапии. Предлагаемый таргетный IL-12 может быть использован в противораковой иммунотерапии, где рак представляет собой рак мышц, костей, нервной системы, хрящей, сухожилий, кровеносных сосудов и т.д., главным образом саркому, и более предпочтительно, рабдомиосаркому (РМС). Предлагаемый таргетный IL-12 может быть использован в монотерапии или в комбинации с иммуномодуляторами, такими как интерлейкины, интерфероны, CpG-островки, хемокины или глюканы.
В конкретном варианте осуществления изобретения иммуномодулятором является IL-2 и/или IL-7, предпочтительно IL-2. В дальнейшем конкретном и предпочтительном варианте осуществления изобретения иммуномодулятором является IL-2. Иммуномодулятор предпочтительно ковалентно связан или слит с более крупным белком или иммуноглобулином или его фрагментом (таким как Fab, scFv, Fc), или находится в комплексе с иммуноглобулином. В наиболее предпочтительном варианте осуществления изобретения IL-2 связан в комплексной форме с анти-IL-2 антителом, а IL-7 ковалентно слит с Fc-фрагментом антитела.
Таким образом, предлагается нацеленный на опухоль IL-12 для использования в противораковой терапии, отличающийся тем, что препарат индуцирует и/или стимулирует иммунный ответ пациента против рака, такого как саркома, но также и против других плотных опухолей, причем указанная индукция или стимуляция иммунного ответа инициирует старение раковых клеток, и/или неожиданную ремиссию раковых клеток или тканей до клеток или тканей происхождения, где нацеленный на опухоль IL-12 является терапевтически эффективным моноклональным антителом или его биологически активным фрагментом, направленным на человеческий комплекс ДНК-гистон H1, представленный в опухолевом некрозе, и слитым предпочтительно через С-конец с IL-12, предпочтительно с субъединицей р35 IL-12.
Кроме того, предлагается нацеленный на опухоль IL-12 для использования в комбинированной терапии с иммуномодулятором и/или иммунокомплементарным агентом, таким как интерлейкин, интерферон или хемокин, с целью индукции и/или стимуляции иммунного ответа против рака у пациента, больного указанным раком, причем указанная индукция или стимуляция иммунного ответа инициирует старение раковых клеток, а также ремиссию раковых клеток или тканей до клеток или тканей происхождения, где нацеленный на опухоль IL-12 является терапевтически эффективным моноклональным антителом или его биологически активным фрагментом, направленным на человеческий комплекс ДНК-гистон H1, представленный в опухолевом некрозе, и слитым через С-конец с IL-12.
Предлагаемая таргетная терапия опухолей с применением IL-12 способствует вызыванию старения раковых клеток, которое предпочтительно вызывается выработкой эндогенного IFNy и/или TNF вследствие указанной стимуляции или индукции иммунной системы пациента, запущенных указанной таргетной терапией с применением IL-12. Было обнаружено, что при применении предлагаемого лечения старение раковых клеток имело своим результатом стабильную блокировку роста. Более того, было обнаружено, что старение раковых клеток и ремиссия тканей не зависят от прямых иммуноспецифических цитотоксичных эффектов, таких как генерация и активация NK-клеток и макрофагов, хотя эти клетки и запускаются IL-12.
Предлагаемая таргетная терапия опухолей с применением IL-12 является применимой в противораковой иммунотерапии, где рак представляет собой плотные опухоли или опухоли мышц, костей, нервной системы, хрящей, сухожилий, кровеносных сосудов и жировых или волокнистых тканей. В конкретном варианте осуществления изобретения предлагаемую терапию используют для лечения саркомы, предпочтительно рабдомиосаркомы (РМС).
Было обнаружено, что предлагаемая противораковая иммунотерапия, где NHS-IL12 и IL-2 или IL-7, в соответствии с выше- и нижеизложенным, используются в комбинации, не только провоцирует старение раковых клеток, но и дополнительно инициирует индукцию миогенной дифференциации и ремиссии раковых клеток до клеток ткани происхождения, указывая на то, что побуждаемая IL-12 человеческая иммунная система не только убивает раковые клетки, но и использует альтернативные механизмы ослабления роста опухоли. Нацеленный на опухоль IL-12 в значительной степени подавляет рост опухоли по меньшей мере у мышей линии NSG, на которых воспроизведена полностью гуманизированная аллогенная иммунная система.
Предлагаемые IL-2 и IL-7 по отдельности не блокировали рост опухоли и не индуцировали эффективный противоопухолевый иммунный ответ. Стимуляция аллогенной человеческой иммунной системы IL-12 необходима, чтобы эффективно контролировать рост опухоли у гуманизированных мышей. Более эффективной в этих обстоятельствах является комбинация NHS-IL12 и IL-2 или IL-7, прежде всего, когда IL-2 или IL-7 находятся в форме, продлевающей срок их полураспада в организма. В одном из вариантов осуществления изобретения IL-2 или IL-7 соединены или связаны с иммуноглобулином, предпочтительно IL-2 или IL-7 слиты с С-концом Fc-фрагмента иммуноглобулина. В еще одном варианте осуществления изобретения IL-2 находится в комплексе с анти-IL-2 антителом, таким как MAb602.
Комбинация NHS-IL12/ILMAB602 показывает значительно более высокую эффективность в отношении общей выживаемости и уменьшения объема опухоли. Исходя из гистологических исследований, можно предположить, что IL-12 не требуется для инфильтрации опухоли NK-клетками или CD68+ макрофагами. Наоборот, предлагаемая конструкция IL-12 необходима, чтобы привести макрофаги к M1-типу, экспрессирующему МНС класса II, который эффективно вырабатывает молекулы-эффекторы и воспалительные цитокины (TNF, IL-12, IL-6, IL-1β).
Предлагаемые данные могут привести к предположению, что IL-12 стимулирует аллогенную иммунную систему убивать раковые клетки, такие как раковые клетки мышц, костей, нервной системы, хрящей, сухожилий, кровеносных сосудов и жировых или волокнистых тканей, предпочтительно клетки РМС по меньшей мере в данной конкретной системе при данном применении. Примечательно, что иммуногистологические исследования не дают подтверждения гибели, апоптоза или некроза раковых клеток, заставляя предположить, что таргетная противораковая иммунотерапия с применением IL-12 в комбинации с IL-2 или IL-7, предпочтительно в связанной или комплексной форме, приводит в действие критические механизмы контроля опухоли, не зависящие от гибели клеток. Но наиболее важно то, что опухоли, рост которых был блокирован введением предлагаемой таргетной терапии IL-12, начинают de novo экспрессировать десмин, мышечно-специфичный белок и ключевую субъединицу промежуточных филаментов сердечных, скелетных и гладких мышц, в стерической поперечноисчерченной конфигурации. Присутствие десмина в исчерченной форме особенно указывает на высокую степень дифференциации мышечных клеток (van, d., V, Schaart, et al., 1992, Cell Tissue Res. 270: 189-1981).
Подводя итог вышесказанному, предлагаемые изобретателями данные являются первым полученным in vivo доказательством индуцируемой TH1 блокировки роста и дифференциации раковых клеток. Это может быть достигнуто целевой доставкой IL-12, предпочтительно в комбинации с IL-2 или IL-7, в микроокружение опухоли с последующей активацией метаболического пути p16INK4a.
Детальное описание изобретения
Термин "старение" в предлагаемом изобретении означает программу блокировки роста, ограничивающую продолжительность жизни раковых клеток и предотвращающую неограниченную пролиферацию клеток.
Термин "прямые иммуноспецифические цитотоксичные эффекты" в предлагаемом изобретении означает активное вовлечение и действие во время иммунного ответа иммуноспецифических цитотоксичных клеток, таких как NK-клетки, макрофаги, T-цитотоксичные клетки или дендритные клетки.
Термин "противораковая иммунотерапия" в предлагаемом изобретении означает терапевтическое воздействие, стимулирующее или восстанавливающее способность иммунной системы бороться с раком с помощью индукции, усиления или подавления иммунного ответа. Результатом противораковой иммунотерапии является направленная иммунная активность против специфичного для конкретной болезни антигена, через повышение распознаваемости цели иммунными клетками или снижение связанной с болезнью супрессии иммунной системы.
Предпочтительно комбинируют введение предлагаемого таргетного IL-12, такого как NHS-IL-12, с IL-2 или IL-7, предпочтительно с IL-2 и IL-7 в связанной, слитой или комплексной форме, в соответствии с выше- и нижеизложенным детальным описанием. Совместное введение может осуществляться одновременно или последовательно, причем в последнем случае иммуномодулятор может быть введен за несколько часов или дней до или после введения таргетной молекулы IL-12 наподобие NHS-IL-12, следуя конкретной дозировке и протоколу лечения.
Принципиально возможно комбинировать введение предлагаемого таргетного IL-12, такого как NHS-IL-12, с лучевой терапией и/или химиотерапией.
Химиотерапевтическим агентом, используемым в комбинации с вышеуказанными таргетными химерными молекулами IL-12, согласно изобретению может быть, например, метотрексат, винкристин, адриамицин, цисплатин, не содержащие сахара хлорэтил-производные нитрозомочевины, 5-фторурацил, митомицин C, блеомицин, доксорубицин, дакарбазин, таксол, фраджилин, мегламин GLA, валрубицин, кармустин, UFT (Тегафур/Урацил), ZD 9331, Таксотер/Доцетаксел, Фторурацил (5-FU), винбластин, и другие хорошо известные соединения этого класса. Предлагается применять по меньшей мере два цикла химиотерапии, предпочтительно 2-8 циклов, более предпочтительно 2-5 циклов. Один цикл длится от 21 до 35 дней, предпочтительно от 21 до 28 дней. Дозировки и протоколы приема химиотерапевтических агентов зависят от различных возможных условий и свойств, зависящих от конкретного пациента и препарата.
Лучевую терапию предлагается проводить стандартным облучением, где общая доза облучения составляет 40-120 Гр, предпочтительно не менее, чем 50 Гр, более предпочтительно от 50 до 75 Гр. Лучевую терапию обычно разбивают на блоки, причем применяют дозы по 1,5-3,5 Гр в день на протяжении по меньшей мере четырех дней, предпочтительно 5-7 дней последовательно. Предлагается применить общую дозу радиации за время 21-35 дней, предпочтительно до 28 дней. В случае необходимости или благоприятности в начале облучения или посредине интервала могут быть применены очаговые дозы 3,5-15 Гр, предпочтительно 5-10 Гр. Предлагаемая лучевая терапия может быть проведена до или после введения молекул таргетного IL-12 и, опционально, предлагаемых иммуномодуляторов, или же одновременно с ними.
Химиотерапевтическое и лучевое лечение может сопровождаться введением агентов, способных модулировать иммунную систему. Например, с помощью применения относительно малой дозы циклофосфамида 100-400 мг/м2, предпочтительно 250 мг/м2 иммунная система пациента может быть активирована или усилена. Обычно для оказания эффекта достаточно одной дозы перед началом вакцинации, как правило, за 1-5 дней до последней, предпочтительно за 2-5 дней.
Предпочтительно предлагаемый таргетный IL-12 является вышеозначенным NHS-IL12. Как упоминалось выше, IL-12 является гетеродимерной молекулой, состоящей из α-цепи (субъединица р35, IL-12p35) и β-цепи (субъединица р40, IL-12p40), соединенных ковалентно дисульфидным мостиком. В предлагаемом слитом белке субъединица р35 связана с С-концом каждой из двух тяжелых цепей (димерного) антитела NHS76. Субъединица р40 связана с субъединицей р35 ковалентно. На практике, молекулу синтезируют методом рекомбинации, используя ДНК-конструкт, экспрессирующий тяжелую цепь и субъединицу р35 как слитый белок, и отдельно субъединицу р40, которая связывается in situ с экспрессованным белком, слитым из NHS и субъединицы р35.
SEQ ID No.1 отображает зрелую аминокислотную последовательность α-цепи, т.е., человеческую субъединицу р35, зрелого (дикого типа) человеческого IL-12:

SEQ ID No.2 отображает зрелую аминокислотную последовательность β-цепи, т.е., человеческую субъединицу р40, зрелого (дикого типа) человеческого IL-12:

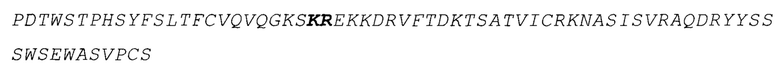
SEQ ID No.3 отображает аминокислотную последовательность лямбда-легкой цепи Mab NHS, которую предлагается использовать и модифицировать, включая сигнальную последовательность (выделена курсивом, первые 19 аминокислот), вариабельный домен (выделен подчеркиванием) и деиммунизирующую мутацию L103V (выделена жирным). Сигнальная последовательность не является частью зрелой полипептидной цепи:
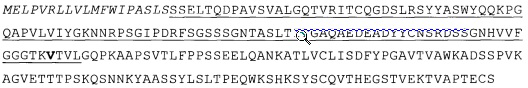
SEQ ID No.4 отображает аминокислотную последовательность слитых тяжелой цепи / человеческого р35, включая сигнальную последовательность (выделена курсивом, первые 19 аминокислот), вариабельный домен (выделен подчеркиванием) и замену R на A в первой аминокислоте hu р35 (выделена жирным), hu р35 (курсивом). Сигнальная последовательность не является частью зрелой полипептидной цепи:

SEQ ID No.5 отображает аминокислотную последовательность человеческого p40, включая сигнальную последовательность (выделена курсивом) и вариантную последовательность, выделенную жирным (KSKREKKDRV, мутировавшее в KDNTEGRV). Сигнальная последовательность не является частью зрелой полипептидной цепи:

Создание полнофункционалъной иммунной системы в мышах линии NSG
Для тестирования токсичности и потенциальной терапевтической эффективности конструкции NHS-IL-12 в человеческой системе in vivo, мышам линии NSG трансплантировали стволовые клетки huCD34+ и стимулировали приживление трансплантата с помощью FcIL-7 (Фигура 1А). Трансплантация на >99.9% чистых huCD34+ клеток мышам линии NSG воссоздала все ростки гемопоэза в течение 12 недель. К тому же, в костном мозге, тимусе и селезенке, а также эквивалентах тимуса и селезенки были обнаружены комплексные репертуары T-клеточных рецепторов (TCR). В совокупности данные демонстрируют, что мыши, которым была осуществлена трансплантация, развили иммунную систему человеческих Т-клеток, которая имеет сильное сходство с таковой в организме человека in vivo. У мышей также развились нормальные NK-клетки, что показано наличием рецепторов NKp и иммуноглобулиноподобных рецепторов клеток-киллеров (KIR), отображающих репертуар NK донора.
Инокуляция опухоли
Подкожное введение 106 аллогенных клеток A204 через 12 недель после трансплантации стволовых клеток за 3 недели привело к агрессивно растущим опухолям у всех мышей. Как и у пациентов с РМС, получивших трансплантацию стволовых клеток, иммунная система не отторгла аллогенный А204. Опухоли стремительно росли, несмотря на значительную экспрессию поверхностных HLA класса I и II, MICA/B, Нектина-2 (CD112) и полиовирусного рецептора (PVR, CD155), но UL16-связывающие белки (ULBP) 1-4 в них совершенно отсутствовали.
Лечение сарком гистон-таргетными слитыми белками IL-12
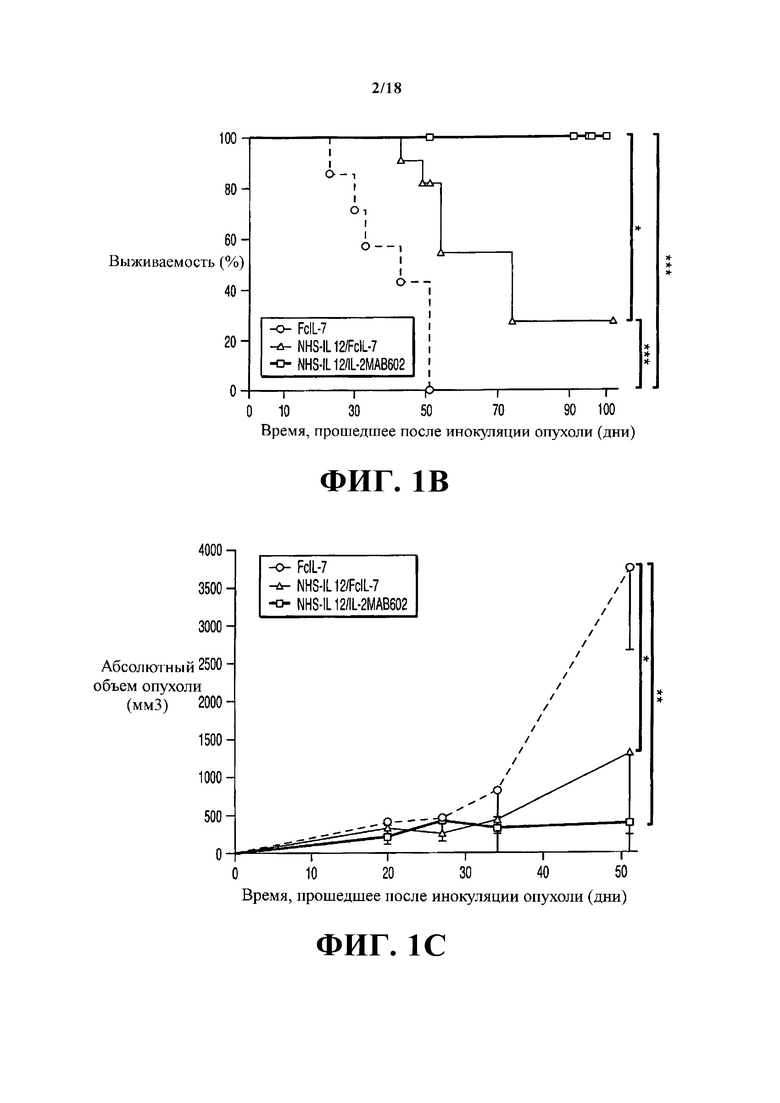
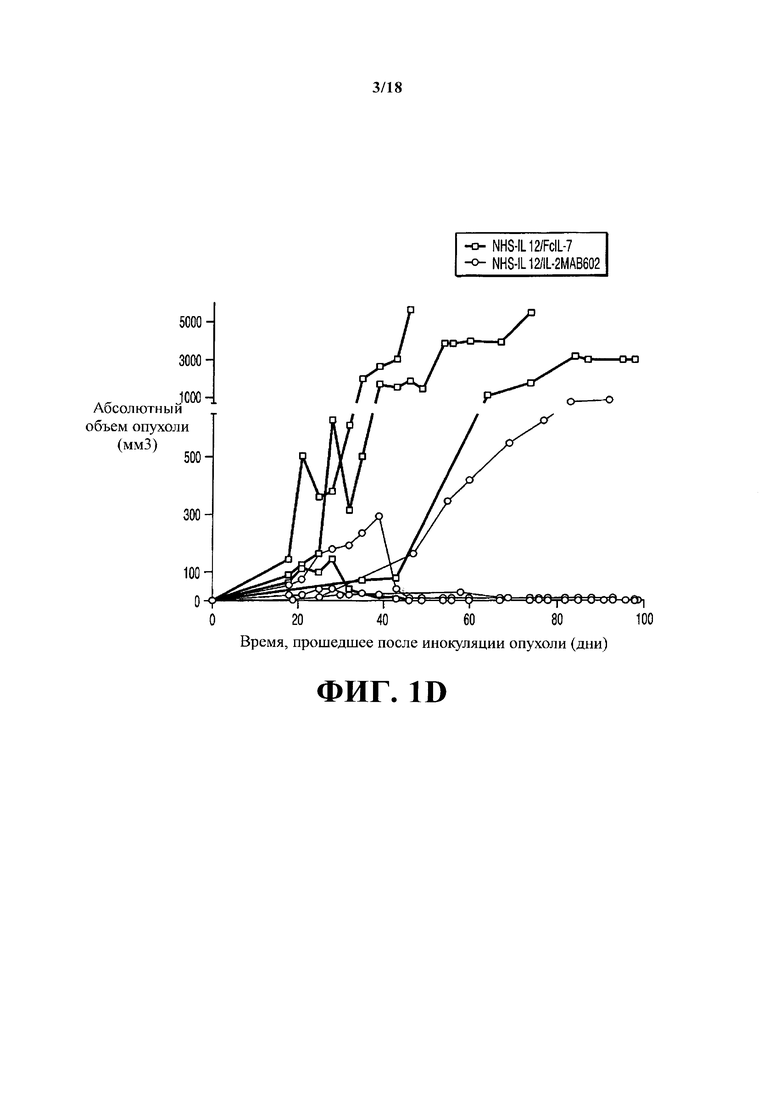
После инокуляции опухолей в течение 3-х недель развились плотные опухоли PMC А204. В дальнейшем мыши получали лечение раз в неделю на протяжении 5 недель только FcIL-7 (контрольная группа), NHS-IL12/FcIL-7 или NHS-IL12/IL-2MAB602 (Фигура 1А). Внутривенная инъекция конструкций не вызвала видимой системной токсичности, как острой, так и в течение продолжительного периода (Фигура 1B: 4 мыши/группа, 100 дней). У мышей, получавших лечение только FcIL-7, саркомы демонстрировали экспоненциальный рост. 4/7 мышей умерли до 5-й недели, а 3 мыши достигли конечных критериев из-за тяжести саркомы на 52-й день (Фигура 1C). В группе Fc-IL7 наблюдался рост опухоли в 6,5 раз за время от 25-го до 52-го дня, тогда как в группе NHS-IL12/IL-2MAB602 этот показатель был снижен до 1,8 раз (P≤0.05, односторонний t-критерий). Таким образом, рост сарком оставался в значительной степени блокированным у мышей, получающих NHS-IL12/IL-2МАВ602 (n=11) (Фигура 1C), приведя к выживанию всех мышей (Фигура 1В и 1D). NHS-IL12/FcIL-7 защищал 2/11 мышей на протяжении немногим более короткого периода времени (Фигуры 1B и 1C), и они умерли на 43-й и на 49-й день, соответственно. Чтобы получить результаты анализа от всех трех групп, мыши были умерщвлены на 52-й день (короткосрочное лечение), за исключением 4-х мышей из группы лечения NHS-IL12, которые оставались в живых и получали лечение до 100-го дня (долгосрочное лечение). Долгосрочное лечение NHS-IL12/FcIL-7 успешно остановило рост опухоли у 1/4, задержало рост опухоли у 2/4, и уничтожило опухоль у 1 мыши. Долгосрочное лечение NHS-IL12/IL-2MAB602 уничтожило опухоль у 3/4 мышей и задержало рост опухоли у оставшейся мыши (Фигура 1D).
Биораспределение конструкции NHS-IL12
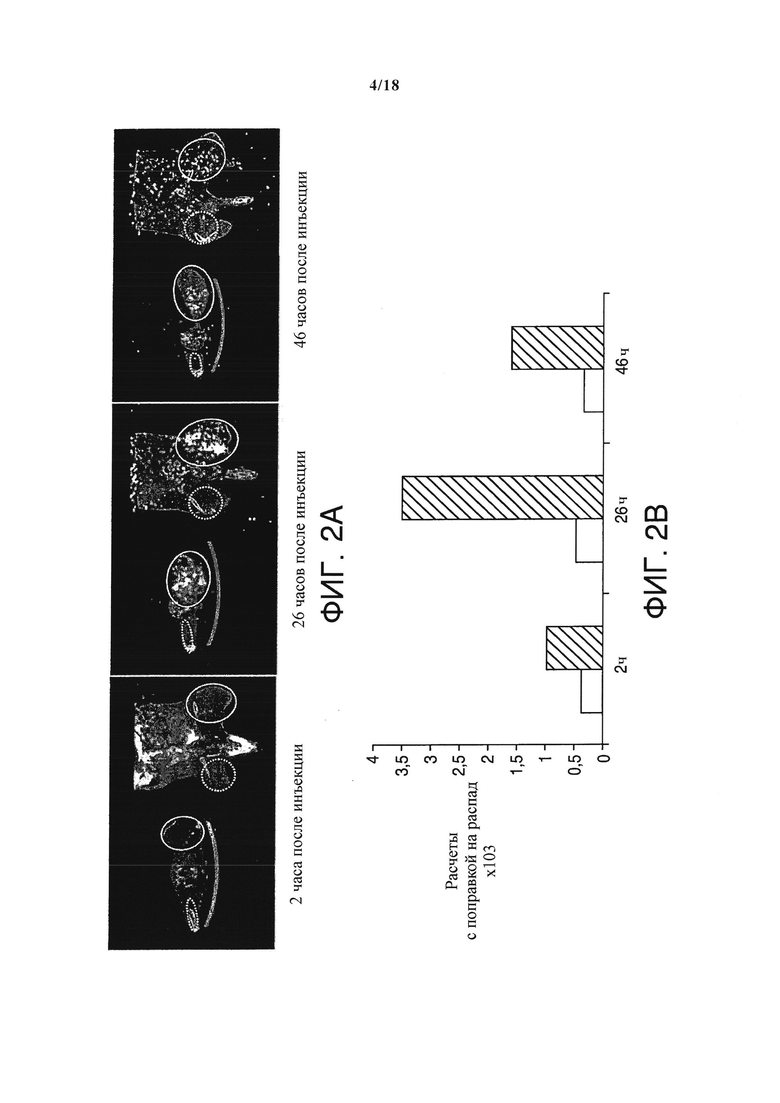
Моноклональное антитело NHS76 распознает внутриклеточный антиген в областях ракового некроза. Поэтому было исследовано, связывается ли NHS-IL12 в первую очередь с местами расположения сарком. Исследования биораспределения с помощью SPECT/CT 123I-меченого NHS-IL12 выявили значительное in vivo обогащение NHS-IL12 внутри микроокружения саркомы (Фигура 2А). Определение количества 123I-меченого NHS-IL12 показало от 4- до 6-кратное обогащение радионуклидов в опухолях по сравнению с контралатеральными мышцами. Количество 123I достигло пика в области опухоли через 26 часов после внутривенного введения NHS-IL12, тогда как в нормальной мышечной ткани количество 123I оставалось стабильным в течение продолжительного периода (Фигура 2B), подтверждая, что NHS-IL12 в первую очередь связывается с человеческими саркомами.
Опухолеспецифичные иммунные ответы
С целью понять различия, лежащие в основе терапевтической эффективности разных протоколов лечения, изобретатели провели гистологические, иммуногистохимические (ИГХ) исследования, а также дали обширную молекулярную и функциональную характеристику человеческим иммунным клеткам, инфильтрирующим саркомы А204.
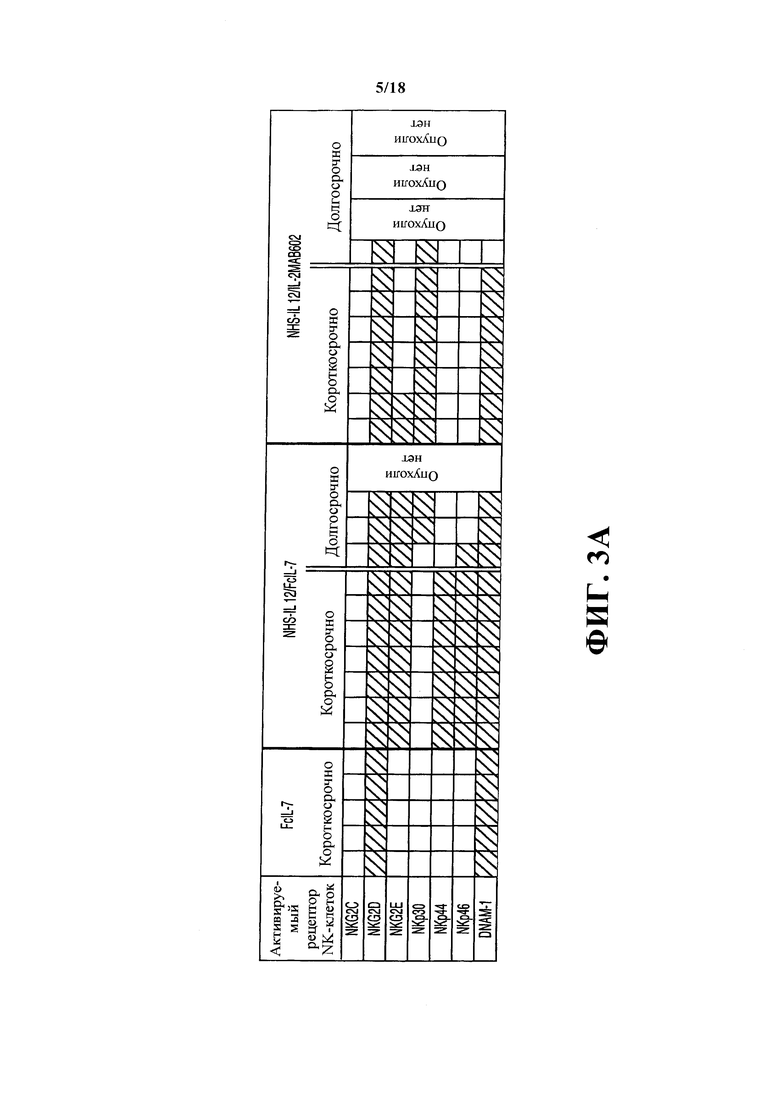
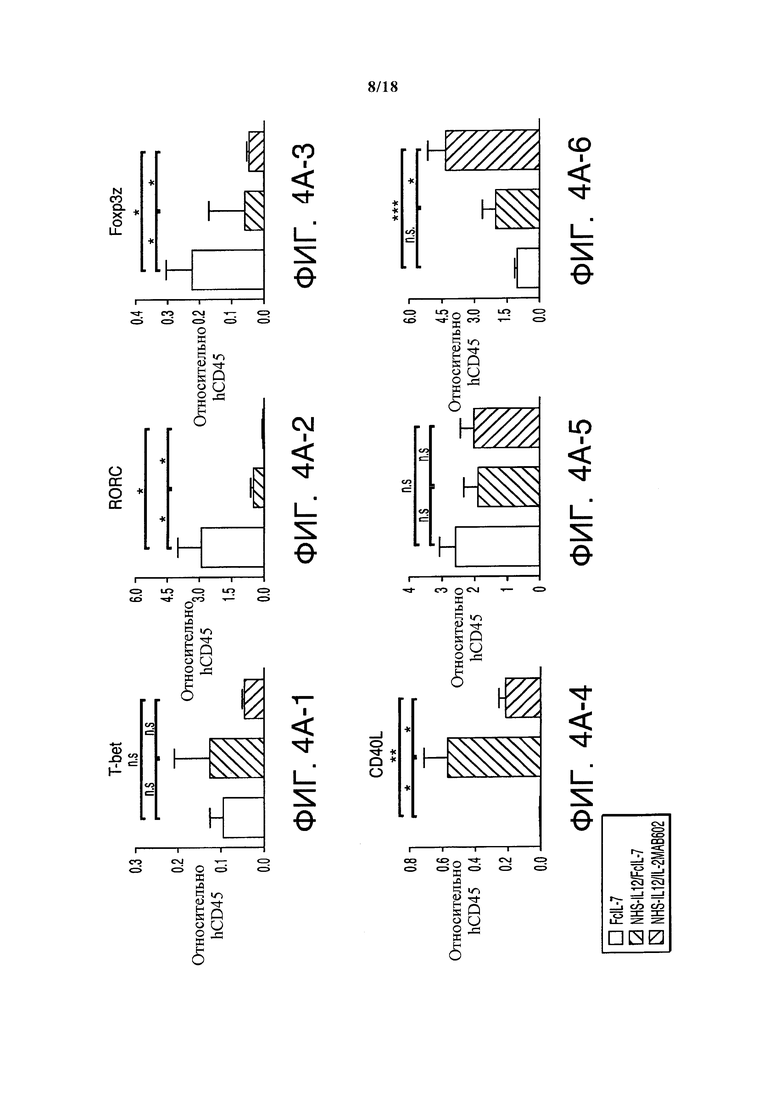
Удивительно, но саркомы мышей, получавших лечение FcIL-7, содержали лишь незначительное количество инфильтрата, содержащего исключительно макрофаги (CD68+) и NK-клетки (CD56+). Напротив, саркомы мышей, получавших лечение по любому из протоколов NHS-IL12, демонстрировали густой одноядерный инфильтрат NK-клетками, макрофагами и большим количеством CD4+ и CD8+ T-клеток. NK-клетки всех групп лечения экспрессовали мРНК NKG2D и DNAM-1 (Фигура 3A), лиганд саркомо-ассоциированных поверхностных молекул нектина-2 (CD112) и PVR (CD155). Экспрессия мРНК поверхностных молекул, управляющих дифференциацией и активацией NK-клеток, строго требовала наличия конструкции NHS-IL12. FcIL-7 или IL-2MAB602 затем модулировали эффект конструкции NHS-IL12 на популяцию инфильтрирующих NK-клеток. NKG2E, NKp44 и NKp46 были обнаружены только в опухолях мышей, получавших лечение NHS-IL12/FcIL-7, в то время как экспрессия NKp30 ограничивалась саркомами мышей, получавших лечение NHS-IL12/IL-2MAB602 и мышей, получавших долгосрочное лечение NHS-IL12/FcIL-7 (Фигура 3A). Саркомы мышей, получавших лечение FcIL-7, демонстрировали сильную экспрессию CD 161 (Фигура 3B) и RORC - мастер-фактора транскрипции TH17 (Фигура 4А), характеризуя IL-17-вырабатывающий фенотип (Billerbeck et al. (2010) Proc. Natl. Acad. Sci. USA. 107: 3006-11). Напротив, мыши, получавшие лечение NHS-IL12/FcIL-7, также показывали сильную экспрессию CD 161 (Фигура 4B), но значительно меньшую экспрессию RORC (Фигура 4А), характеризуя эффекторные Т-клетки и клетки центральной памяти как вырабатывающие высокий уровень IFN-γ и TNF, но недостаточно литически активные (Takahashi et al. (2006) J. Immunol 176: 211-216). У мышей, получавших лечение NHS-IL12/IL-2MAB602, отсутствовала мРНК CD161 (Фигура 3B) и мРНК RORC (Фигура 4А), что дает веские основания предполагать подавление IL-17-вырабатывающих фенотипов NK-клеток или Т-клеток у этих мышей (Laurence et al. (2007) Immunity 26: 371-381; Ghoreschi et al. (2010) Nature 467: 967-971).
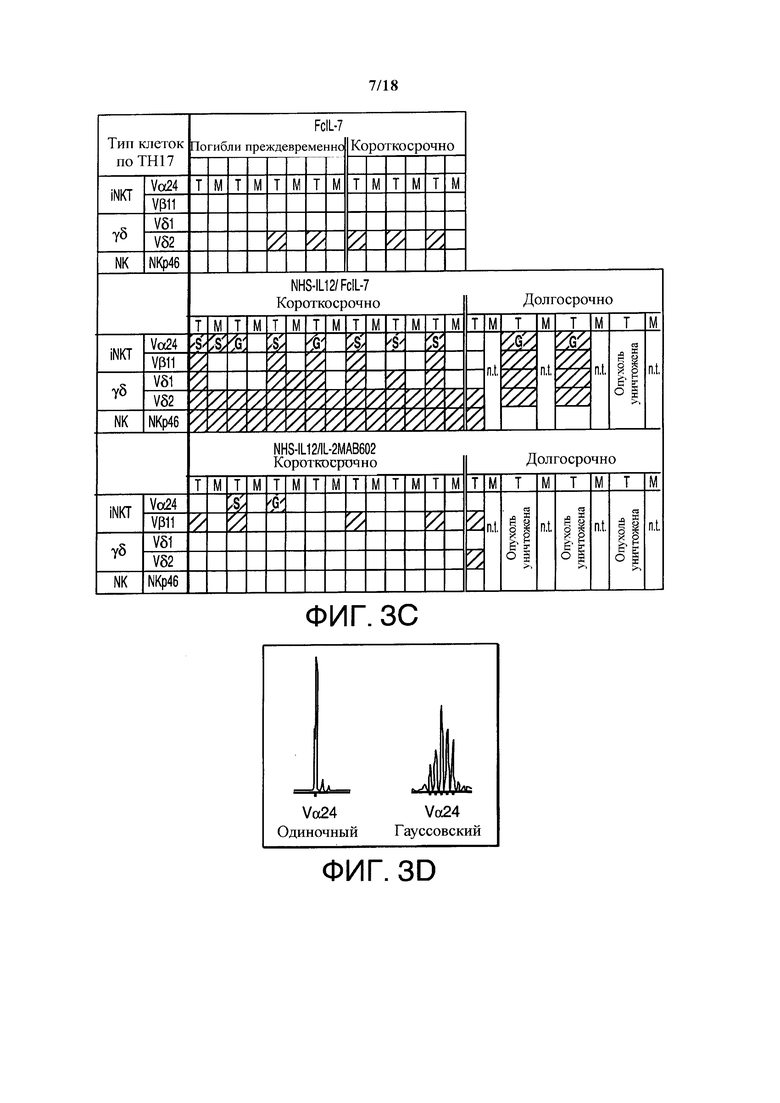
Поскольку молекулы KIR ухудшают функции NK-клеток даже в дефицитном по MHC-I окружении (27), то анализировали экспрессию KIR в саркомах мышей, получавших лечение FcIL-7- или NHS-IL12/FcIL-7.qRT-ПЦP всей саркомы выявила одинакомые уровни в обоих группах. Как и ожидалось, экспрессия KIR в NK-клетках нормальной мышечной ткани мышей и в лимфоцитах опухолевого инфильтрата ксенотрансплантата человеческой саркомы отличалась, так как в мышцах мыши были обнаружены KIR2DL3 и KIR2DL4, а в человеческих саркомах - дополнительно KIR2DL1 и KIR3DL1. Несмотря на экспрессию различных KIR, NK-клетки сохраняли функциональную активность, так как свежеизолированные NK-клетки тканей саркомы выделяли IFN-γ после in vitro стимуляции с NHS-IL12 (не показано). Было выявлено, что кроме NK-клеток, экспрессия мРНК характеризует популяцию Тн17 врожденных лимфоцитов, наподобие TCRVα24-экспрессирующих iNKT клеток, NKp46+ NK или γδ Т-клеток почти исключительно в саркомах мышей, получавших лечение NHS-IL12/FcIL-7 (Фигуры 3A, 3C, 3D).
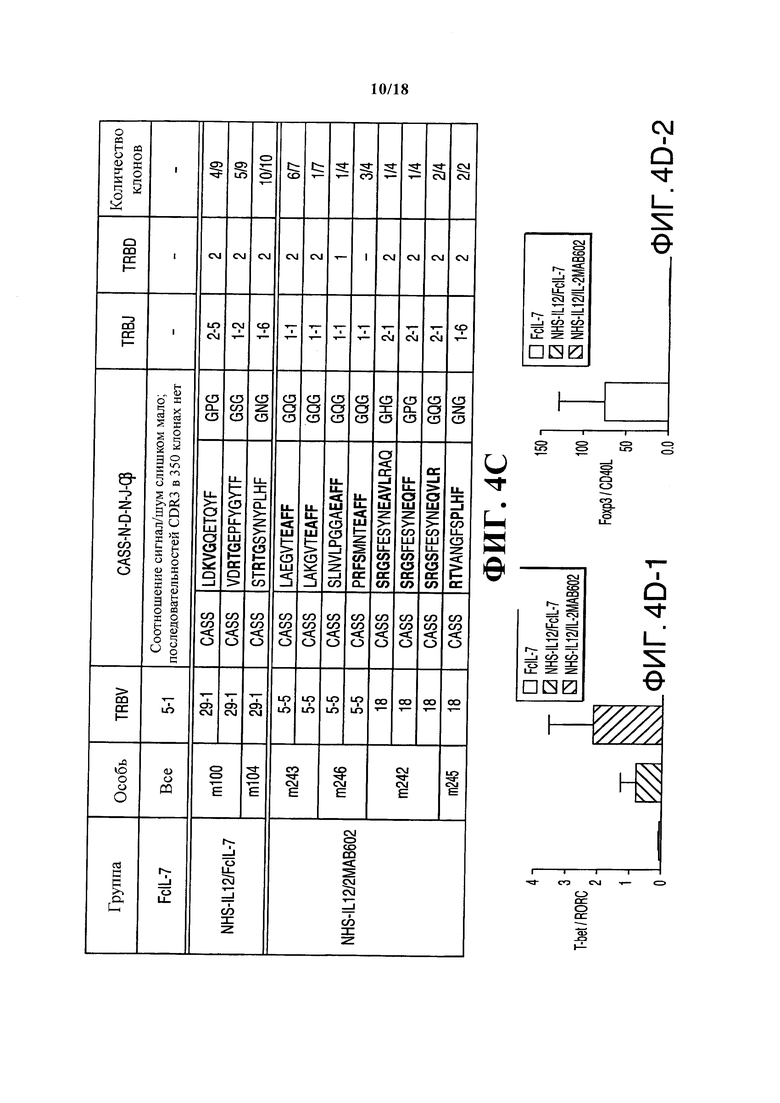
Опухоли всех групп, получавших лечение NHS-IL12, демонстрировали, помимо врожденных лимфоцитных популяций, широкий спектр CD3+ T-клеток (Фигуры 4B и 4C). Эти клетки отсутствовали в саркомах мышей, получавших лечение FcIL-7, и показывавших слабый сигнал при анализе Vβ-спектрального типа (Фигуры 4B и 4C), а также отсутствие инфильтрирующих CD8+ Т-клеток. Саркомы, лечение которых проводилось NHS-IL12, демонстрировали обширный репертуар TCR, при этом с олигоклональными или моноклональными пиками внутри различных Vβ-семейств (Фигура 4С), как это случается во время преференциальной экспансии рестриктированных клонов Т-клеток. Клонирование и секвенирование области CDR3 подтвердило, что пики содержали ограниченное число разных клонов Т-клеток с сильно увеличенными сегментами TRBV в двоих группах лечения, таких как TRBV29-1 у всех особей группы NHS-IL12/FcIL-7, или TRBV5-5 и TRBV18 в группе NHS-IL12/IL-2МАВ602 (Фигуры 4B и 4С). Относительная экспрессия факторов транскрипции T-bet и RORC, регулирующих IFN-γ и IL-17, соответственно отражала экспрессию мРНК цитокина в трех группах. Соотношение T-bet/RORC составляло <0,05 в группе лечения только FcIL-7, тогда как оно было в 19 раз выше (0,8) и в 44 раза выше (2,2) у мышей, получающих NHS-IL12 с IL-7 или IL-2 (Фигура 4D). Согласуясь с экспрессией T-bet/RORC, уровень Foxp3 был примерно в десять раз ниже в группах лечения NHS-IL12, чем в группе лечения только FcIL-7 (Фигура 4А). Соответственно этому, слабая экспрессия Foxp3 имеет обратную корреляцию с сильной экспрессией маркера активации Т-клеток CD40L (Фигура 4А и 4D).
Индукция старения сарком с помощью NHS-IL12
Конструкция NHS-IL12 сильно подавляла развитие саркомы у всех получавших ее мышей (Фигура 1). Удивительно, но только саркомы группы NHS-IL12/IL-2MAB602 содержали большие количества белка перфорина и мРНК гранзима K (Фигура 4А), в то время как мыши, получавшие лечение NHS-IL12/FcIL-7, были практически лишены белка перфорина и содержали малые количества мРНК гранзима K (Фигура 4А). Это дает веские основания предполагать, что контролирующий саркому иммунный ответ включал отличные от цитолиза механизмы. Более того, саркомы не содержали достаточных количеств CD4+ или CD8+ Т-клеток, чтобы объяснить контроль рака гибелью клеток или апоптозом.
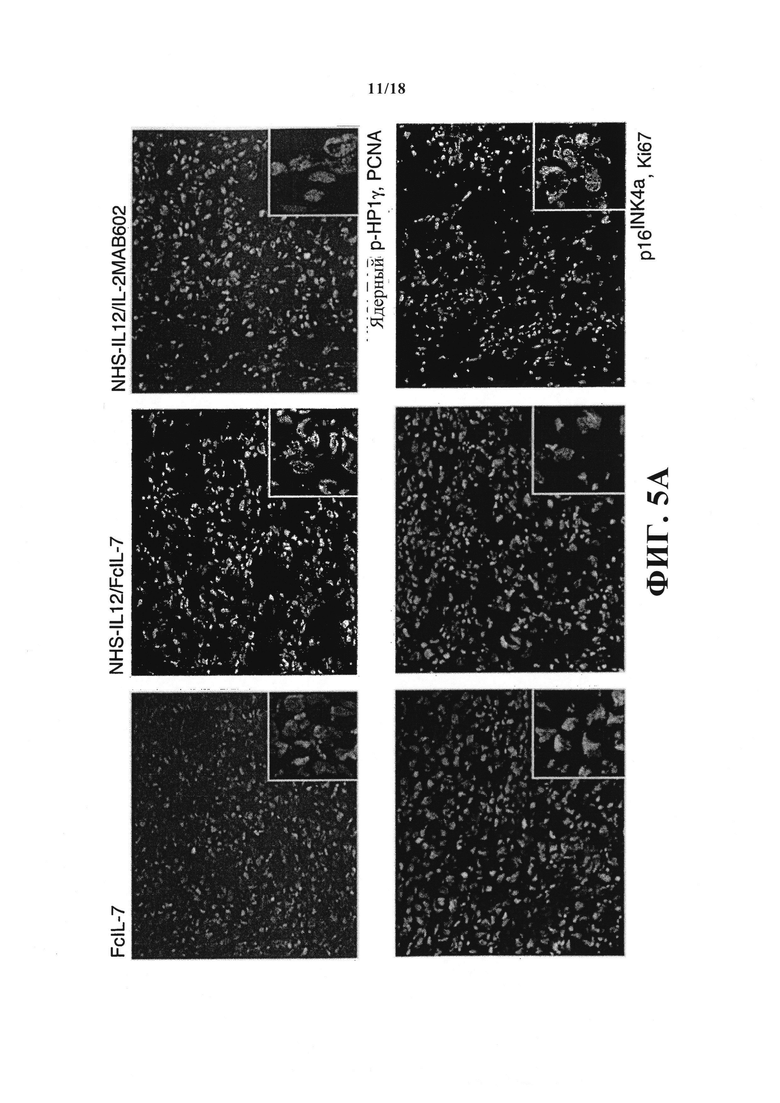
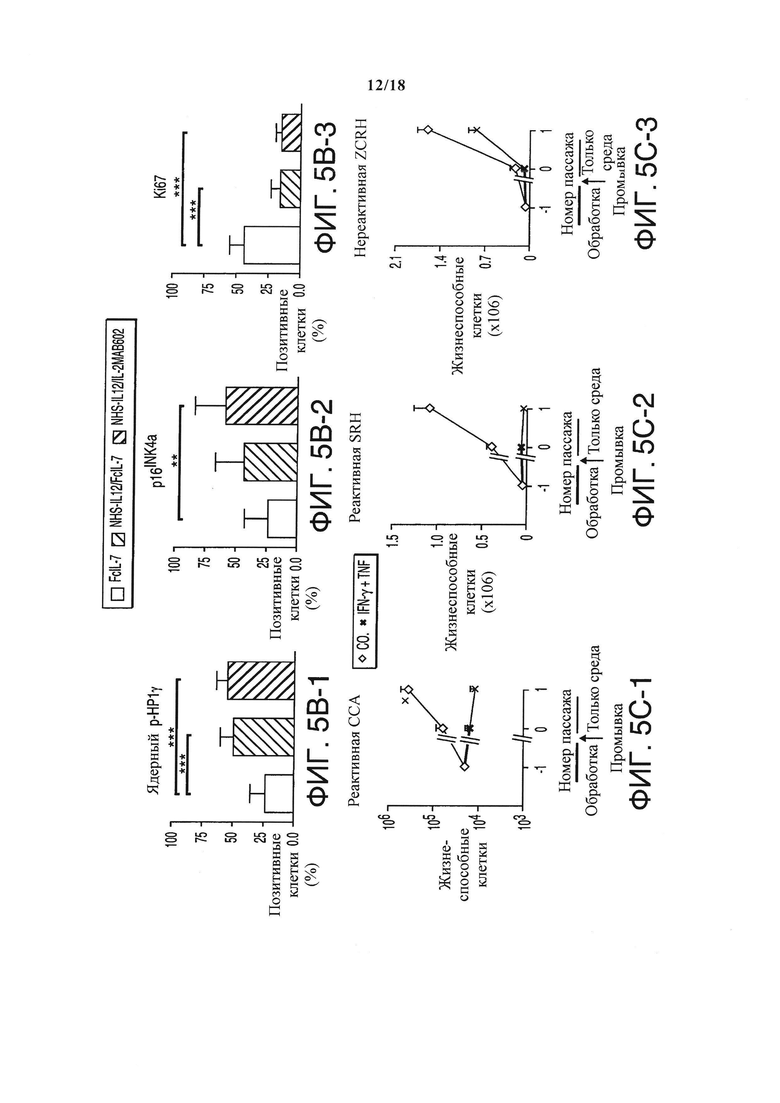
Поэтому был проанализирован эффект иммунного ответа на пролиферацию клеток саркомы путем определения маркера пролиферации - ядерного антигена пролиферирующих клеток (PCNA) или Ki67. В быстрорастущих опухолях, контролируемых с помощью FcIL-7, 50% клеток саркомы окрашивались позитивно на PCNA или Ki67, демонстрируя, что большинство клеток пролиферировали (Фигура 5A и 5B). В группах, получавших лечение NHS-IL12, как PCNA-окраска (Фигура 5А), так и Ki67+ окраска клеток саркомы были значительно ниже, чем в FcIL-7-контроле (Фигура 5А и 5B). Чтобы определить, остановил ли иммунный ответ только клеточный цикл или же индуцировал стабильную блокировку роста, как при цитокин-индуцированном старении, саркомы окрашивали дважды: на один маркер пролиферации и либо на ассоциированный со старением фосфорилированный белок гетерохроматина 1 (p-НР1γ), либо на p16INK4a, регулятор клеточного цикла, ингибитор циклин-зависимой киназы 2А (CDKN2A). Саркомы, лечение которых производилось только FcIL-7, показали сильную экспрессию PCNA/Ki67 и в то же время очень слабую экспрессию p16INK4a/ядерного p-НР1γ, подтверждая, что клетки саркомы быстро пролиферировали (Фигура 5А и 5B). И наоборот, до 70% клеток саркомы мышей, получавших лечение NHS-IL12/IL-2MAB602 или NHS-IL12/FcIL-7, экспрессовали маркер старения p-HP1γ или p16INK4a (Фигура 5A и 5B), при отсутствии PCNA (PCNA-/p-HP1γ+) (Фигура 5А, вставки на верхней линии) и Ki67 (Ki67-/p16INK4a) (Фигура 5А, вставки на нижней линии).
Полученные данные ясно показывают, что побуждаемая IL-12 стимуляция человеческой иммунной системы может использовать старение раковых клеток как важный механизм сдерживания роста опухоли.
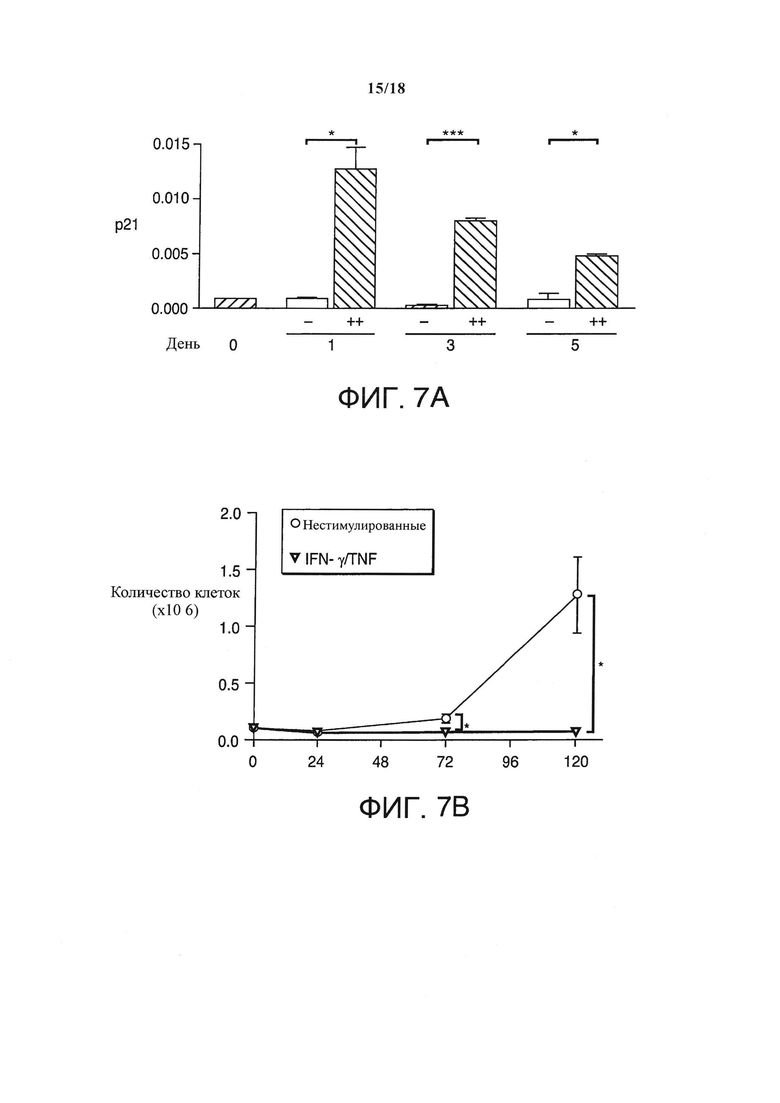
Поскольку IFN-γ и TNF являются двумя главными эффекторными цитокинами спровоцированного IL-12 TH1-иммунитета, и поскольку эти два цитокина могут вызывать старение, то различные происходящие от пациентов клеточные линии человеческой РМС на очень ранних пассажах инкубировали с возрастающими дозами IFN-γ и TNF. Каждый цитокин по отдельности не ингибировал рост или ингибировал его слабо. Однако в комбинации они вызывали постоянную, определяющуюся как старение блокировку роста в 2-х из 3-х сарком (Фигура 5С). Что важно, резистентная к старению саркома не экспрессировала регулятор клеточного цикла p16INK4a (не показан), подтверждая, что индуцированное IFN-γ и TNF старение требует строго активации p16INK4a.
Индукция дифференциации в рабдомиосаркомах A204
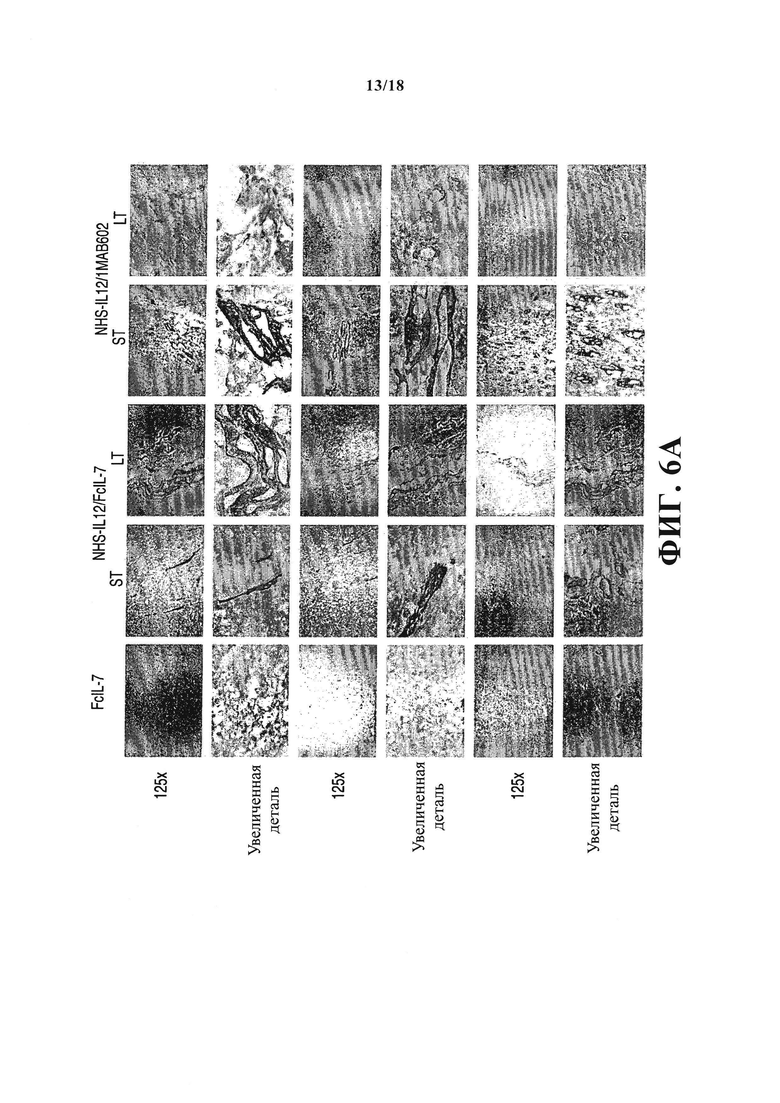
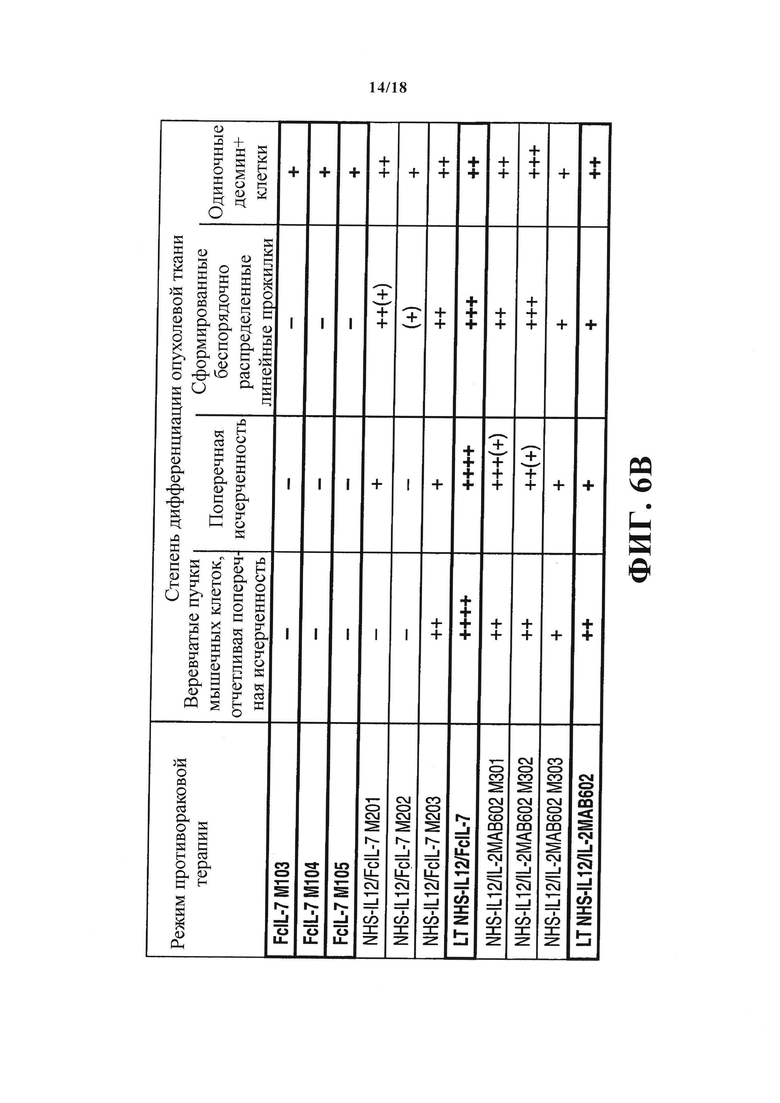
Поскольку определяемый IFN-γ и TNF иммунный ответ вызвал постоянную блокировку клеточного цикла в РМС, а дифференциация скелетных мышц зависит от раннего выхода миобластов из клеточного цикла, что позволяет начать экспрессию специфичных для мышц генов и сливание клеток в мультиядерные миотрубки, возникает вопрос, может ли эта блокировка роста повлиять на дифференциацию сарком A204. Десмин является надежным маркером рабдомиобластической дифференциации, так как он отсутствует в недифференциированных или слабо дифференциированных РМС. Вследствие этого саркомы А204 не демонстрируют ни поперечной расчерченности, характеризующей миоциты, ни экспрессии десмина до трансплантации (не показана). После трансплантации гуманизированным мышам пролиферирующие саркомы, пролечиваемые FcIL-7, оставались слабо дифференциированными, с малым количеством одиночных десмин+ клеток, диффузно распределенных в опухолях, но без поперечной исчерченности (Фигура 6A и 6B). И наоборот, лечение NHS-IL12 не только ингибировало рост опухоли и индуцировало старение, но также постепенно восстанавливало миогенную структуру и антиген-экспрессию в саркомах А204. NHS-IL12/FcIL-7 вызывали образование беспорядочно распределенных линейных зон зрелых клеток РМС, ясно выражающих наличие десмина и зон поперечной исчерченности. Такая дифференциация в направлении функциональной мышечной ткани была еще более выраженной в саркомах мышей, получавших лечение NHS-IL12/IL-2МАВ602. Такие саркомы с блокированным ростом показывали веревчатое распространение дифференциированных зон с трубчатыми или решетчатыми структурами, пронизывающими опухоль во всех направлениях (Фигура 6А и 6B). Таким образом, блокировка роста сарком in vivo вызывала старение и была ассоциирована с восстановлением специфических маркеров предопределенности клеток для миоцитов, клеток происхождения РМС А204.
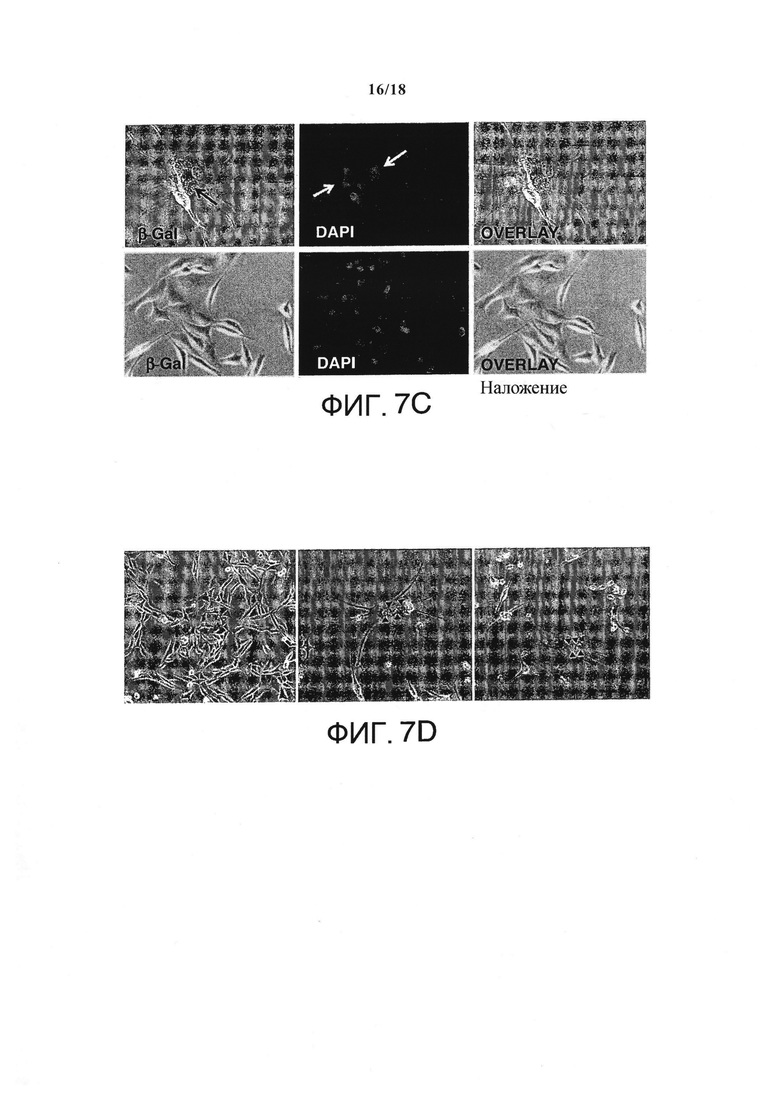
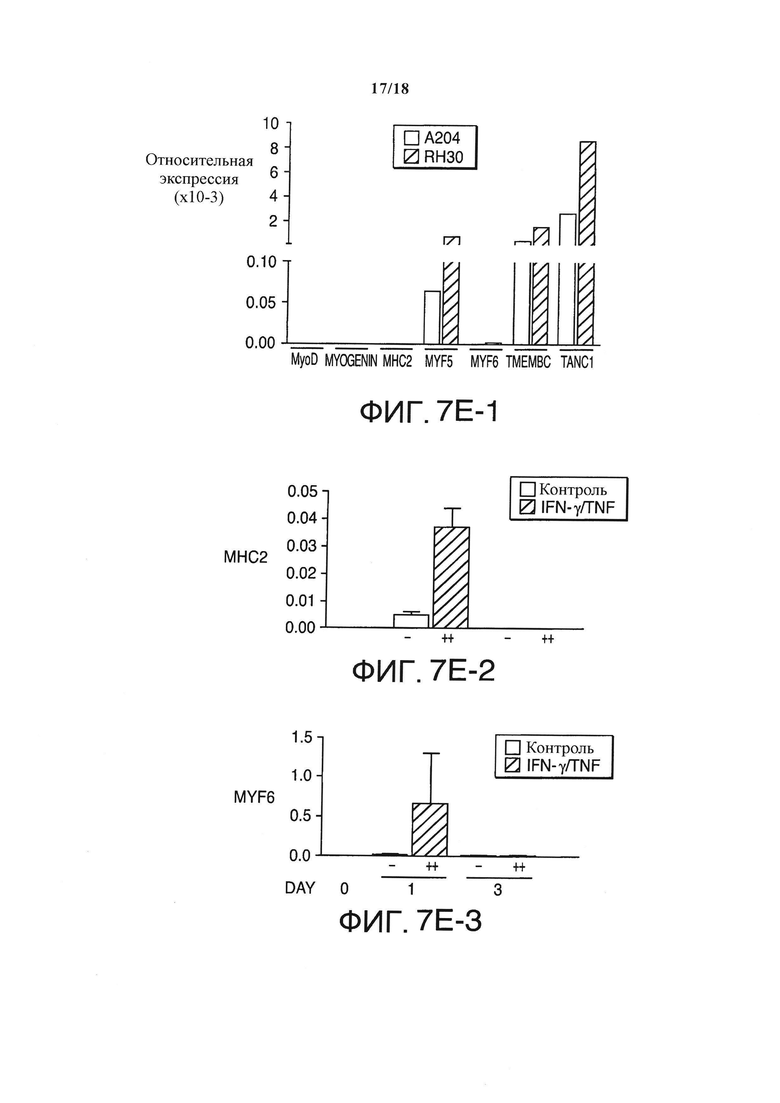
In vitro, единоразовое применение комбинированных IFN-γ/TNF безвозвратно блокировало рост клеток саркомы А204 и немедленно пробуждало мРНК ингибитора циклин-зависимой киназы p21CIP1/WAF1 (семикратно на день +1, десятикратно на 5 день, по сравнению с непролеченным контролем), который в скелетных мышцах естественно прерывает клеточный цикл (Фигура 7А и 7B). Неожиданно в течение 5-ти дней IFN-γ/TNF также массово индуцировали появление би- и полиядерных клеток в культурах саркомы (>80% клеток - полиядерные) (Фигура 7С и 7D), с ближайшими ядрами как в поздней анафазе, заставляя предположить наличие опосредованной p16INK4a необратимой блокировки цитокинеза. К тому же, клетки саркомы de novo экспрессировали еще два маркера регулятора дифференциации мышечных клеток (Myf-6 и MyosinHeavyChain-II) (Фигура 7Е), и трансформировались в SA-β-галактозидаза+, удлиненные мультиядерные клетки, формируя миогенные гигантские клетки (Фигура 7С и 7D), этап в дальнейшей дифференциации мышечных клеток. Единоразовое применение IFN-γ/TNF in vitro также необратимо блокировало рост клеточной линии РМС RH30 и полученной от пациентов линии РМС SRH (пассаж 14) (Фигура 8), но не ZCRH (дефицитная по p16, пассаж 9).
Индукция дифференциации в панели опухолевых клеточных линий, представляющих ткани трех зародышевых слоев
С целью проверить, могут ли блокировка роста и дифференциация, вызванные IFN-γ/TNF, обладать более широкой клинической значимостью, две комбинации цитокинов TH1 IFN-γ/TNF были протестированы in vitro на панели опухолевых клеточных линий, представляющих ткани трех зародышевых слоев 3/3 глиобластомные клеточные линии (T98G, Ln229, А172), 2/2 нейробластомные клеточные линии (LS and LAN-1) и линии раковых клеток MCF-7 (рак груди), HCT-116 (колоректальный рак) и Hep3b (печеночно-клеточная карцинома) прекратили пролиферацию, в унисон экспрессировали SA-β-Gal и демонстрировали клеточную и ядерную атипию дегенеративного типа (Фигура 8) Гистопатологические характеристики включали большие гиперхроматиновые плеоморфные ядра и морфологию, напоминающую реактивные/не-неопластичные мультиядерные миогенные гигантские клетки в скелетных мышцах, вызванные повреждением клеток. В глиобластомах ядерная атипия имела сходство с типом, встречающимся в доброкачественных "давних шванномах" периферических нервов (Фигура 8). Развитие мультиядерных гигантских клеток отмечалось во многих органах и связано с инфекцией, повреждением, аутоиммунитетом и опухолями.
Фармацевтические композиции
Таргетные слитые белки IL-12 согласно изобретению могут быть включены в фармацевтическую композицию, пригодную для введения. Подобные композиции обычно включают вариабельные области антител и фармацевтически приемлемый носитель. Здесь и далее термин "фармацевтически приемлемый носитель" включает все возможные растворители, диспергенты, покрытия, антибактериальные и антигрибковые агенты, изотонические и задерживающие абсорбцию агенты и прочие подобные вещества, совместимые с фармацевтическим применением. Использование подобных сред и агентов для фармацевтически активных субстанций хорошо известно из уровня техники.
Предлагаемую фармацевтическую композицию формулируют так, чтобы она была совместимой с намеченным путем ее введения. Пути введения, например, включают парэнтеральный, напр., внутривенный, внутрикожный, подкожный, оральный (напр., ингаляции), трансдермальный (местный), трансмукозальный и ректальный. Растворы или суспензии, используемые для парэнтерального, внутрикожного или подкожного применения, могут включать следующие компоненты: стерильный разбавитель, такой как инъекционная вода, солевой раствор, нелетучие масла, полиэтиленгликоли, глицерин, пропиленгликоль или другие синтетические растворители; антибактериальные агенты, такие как бензиловый спирт или метилпарабены; антиоксиданты, такие как аскорбиновая кислота или бисульфит натрия; хелатирующие агенты, такие как этилендиаминтетрауксусная кислота; буферы, такие как ацетатный, цитратный или фосфатный, и регулирующие тоничность агенты, такие как хлорид натрия или декстроза. pH может регулироваться кислотами или основаниями, такими как соляная кислота или гидроксид натрия. Препарат для парэнтерального введения может быть упакован в ампулы, одноразовые шприцы или флаконы с несколькими дозами, сделанные из стекла либо пластика.
Медикаменты, содержащие предлагаемые таргетные слитые белки IL-12 и молекулы IL-7/IL-2, могут иметь концентрацию от 0,01 до 100% масс., хотя конкретное количество изменяется в зависимости от лекарственной формы медикамента.
Введение осуществляют предпочтительно один раз в две недели или один раз в месяц, но частота введения может изменяться в большую или меньшую стороны в зависимости от фармакокинетического поведения организма конкретного больного. Дозировка слитого с антителом белка, предлагаемого в данной заявке, для взрослого массой тела около 70 килограмм, находится в пределах от около 50 до 1000 миллиграмм на дозу, предпочтительный диапазон - от около 100 до 500 миллиграмм на дозу. Наиболее предпочтительная доза - около 200-400 миллиграмм для взрослого массой тела около 70 килограмм, введение которому препарата осуществляют раз в месяц.
КОРОТКОЕ ОПИСАНИЕ ФИГУР
Фигура 1
План клинического исследования, рост опухоли и выживаемость после заражения РМС и ее лечения. (А) Мыши линии NSG возрастом 4-6 недель получали сублетальную дозу облучения и были гуманизированы трансплантатами CD34+, CD3-. Мышам с полностью прижившимся трансплантатом вводили 1×106 клеток А204 на 12-й неделе. Иммунотерапию начинали через 18 дней, когда объем опухоли составлял 500-200 мм3. Мышей умерщвляли после 5-недельного лечения, когда опухоли у группы FcIL-7 достигли 20% массы тела. Четырех мышей из групп NHS-IL12/FcIL-7 и NHS-IL12/IL-2MAB602 оставили в живых и продолжали лечение по меньшей мере до 95-го дня после инокуляции опухоли. (В) Влияние FcIL-7, NHS-IL12/FcIL-7 и NHS-IL12/IL-2MAB602 на выживаемость. Кривые выживаемости сравнивали, используя логарифмический ранговый критерий. Выживаемость была значительно лучше у группы NHS-IL12 по сравнению с группой FcIL-7. В контрольной группе FcIL-7 4 животных умерло до 52-го дня, а 3 были умерщвлены на 52-й день опыта из-за чрезмерного роста опухолей. В группе лечения NHS-IL12/FcIL-7 2 мыши умерло до 52-го дня, и в группе долгосрочного лечения - преждевременно умерло по одной на 56-й и 74-й день. (С) Влияние FcIL-7, NHS-IL12/FcIL-7 и NHS-IL12/IL-2MAB602 на рост опухолей. Мыши-носители человеческой рабдомиосаркомы А204 раз в неделю получали внутривенно FcIL-7 (20 мкг) (кружки), NHS-IL12 (20 мкг)/FcIL-7 (20 мкг) (треугольники) или NHS-IL12 (20 мкг) и IL-2 (1.5 мкг) в комплексе с МАВ602 (15 мкг) (квадраты). Объем опухолей в мм3 обозначен как средняя величина ± статистическая ошибка среднего 7 мышей на группу, получавших короткосрочное лечение [5 недель]. (D) Индивидуальные объемы опухолей 4-х мышей из группы NHS-IL12 (NHS-IL12 /FcIL-7: квадраты; NHS-IL12/IL-2МАВ602: кружки) на протяжении долгосрочного лечения [14 недель, >95 дней].
Фигура 2
Накопление 123I-меченого NHS-IL12 в опухолевом поражении ксенотрансплантата человеческой опухоли А204.(А) Результаты in vivo SPECT-сканирования, сделанные через 2, 26 и 46 часов после инъекции терапевтической дозы (30 мкг) 123I-меченого NHS-IL12, показывают специфическое накопление NHS-IL12 в опухоли (обведенные сплошной линией кружки) по сравнению с мышечной тканью (пунктирные кружки). (В) Поглощение 123I-NHS-IL12 опухолевым поражением достигло максимума через 26 часов после введения, в то время как в мышцах за все время сканирования не был выявлен ни один специфический сигнал. В расчетах была сделана поправка на распад с целью учесть радиоактивный распад 123I между моментами времени проведения измерений (n=2). *P<0.05.
Фигура 3
Влияние FcIL-7, NHS-IL12/FcIL-7 и NHS-IL12/IL-2MAB602 на врожденный иммунитет. (A) Гомогенаты опухолей особей из каждой группы были протестированы на наличие основных рецепторов запуска NKG2C, -D и -E, DNAM-1 и рецепторов естественных киллеров NKp30, -44 и -46 с помощью основанного на RT-ПЦР анализа длины фрагмента. В группе отмечена высокая согласованность. (В) Экспрессия CD161 в гомогенатах опухолей и мышц. Количество представлено как средняя интенсивность флюоресценции. Каждая точка представляет одну отдельную опухоль. **P≤0.01, ***Р≤0.001, оцененные с помощью одностороннего t-критерия Стьюдента. (С) TCR-транскрипты, указывающие на iNKT-клетки (инвариантные Vα24 и Vβ11), Vδ1 и -2 цепи, а также NKp46 на 52-й день. T = ткань опухоли, M = мышечная ткань, LT = мышь, получавшая долгосрочное лечение, на 100-й день, S - отдельный пик, G - нормальное распределение Гаусса для экспрессии цепи Vα24. (D) Экспрессия мРНК TCRVα24 в опухолях А204, определенная как отдельный пик или в нормальном распределении Гаусса. В человеческих iNKT TCRVα24 главным образом ассоциируется с TCRVβ11 цепью.
Фигура 4
Анализ клональных свойств αβ Т-клеток под влиянием FcIL-7, NHS-IL12/FcIL-7 и NHS-IL12/IL-2MAB602. (А) Обнаружение с помощью ПЦР в реальном времени различных иммунных маркеров в опухолевых гомогенатах. (В) Экспрессия целевого гена, нормализованная до экспрессии человеческого CD45. Экспрессия 25 сегментов TRBV, определенная анализом спектрального типа по размеру CDR3. Закрашенные квадраты обозначают экспрессию до 12 фрагментов. Каждый вертикальный ряд представляет одну мышь, темные квадраты обозначают сегменты TRBV, выбранные для анализа последовательности CDR3. (С) Региональные белковые последовательности CDR3 выбранных сегментов TRBV, выделенные жирным обозначения аминокислот отмечают гомологические последовательности. 

 . (D) Степень экспрессии T-bet/RORC и Foxp3/CD40L в гомогенатах опухолей (n=4). *P≤0.05, **Р≤0.01, ***P≤0.001.
. (D) Степень экспрессии T-bet/RORC и Foxp3/CD40L в гомогенатах опухолей (n=4). *P≤0.05, **Р≤0.01, ***P≤0.001.
Фигура 5
Индукция маркеров старения и антипролиферативного эффекта лечением NHS-IL12/FcIL-7 и NHS-IL12/IL-2MAB602. (А) Клеточное старение и пролиферацию на опухолевых срезах определяли двойной иммунофлюоресцентной окраской на ядерный p-НР1γ в комбинации с PCNA (верхние панели) или p16INK4a в комбинации с Ki67 (нижние панели) (1:100). Вставки показывают увеличенные области изображения (1:300) для визуализации ядерных точек окраски р-НР1γ или p16INK4a. (В) Среднее процентное содержание р-НР1γ-позитивных клеток (т.е., клеток с более чем пятью ядерными точками), p16INK4a-позитивных клеток (как определено при большем увеличении (1:300)) или Ki67-позитивных клеток после лечения FcIL-7, NHS-IL12/FcIL-7 или NHS-IL12/IL-2MAB602 (n=3). **P≤0.01, ***P≤0.001, оцененные с помощью одностороннего t-критерия Стьюдента. (C) Вызванная цитокинами блокировка роста в препаратах клеток первичной человеческой РМС. Клетки линий ССА (эРМС, пассаж 7), SRH (эРМС, пассаж 8) или ZCRH (аРМС, пассаж >9) были посеяны с плотностью 20000 клеток/9.6 см2. На 3-й и 4-й дни клетки подвергали воздействию 100 нг/мл IFN-γ и 10 нг/мл TNF или простой среды (контроль). На 7-й день цитокины убирали, клетки трипсинизировали, считали и пересевали на 20000 клеток/9.6 см2. После инкубации на протяжении еще 4-х дней (ZCRH и SRH) или 10-ти дней (ССА), живые клетки считали. Кривые роста реактивных клеток ССА и SRH или нереактивных клеток ZCRH в отсутствие (Со.) или присутствие IFN-γ плюс TNF. Показано среднее количество клеток ± стандартная ошибка среднего (n=3).
Фигура 6
Экспрессия и организация десмина_in vivo как маркер миогенной дифференциации в РМС А204. (А) Гистологические препараты опухолей (n=3/группа) всех групп (получавших лечение FcIL-7, NHS-IL12/FcIL-7, или NHS-IL12/IL-2MAB602) окрашивали на десмин, и (В) патоморфолог анализировал их в слепом исследовании. LT: долгосрочное лечение; ST: короткосрочное лечение.
Фигура 7
Мультиядерные, стареющие клетки А204 и экспрессия р21 и миогенных маркеров в природных и обработанных цитокинами клетках А204. (А) Относительная экспрессия р21 до и после обработки IFN-γ и TNF (++) или средой (-) в качестве контроля, измерялась количественной ПЦР в реальном времени (n=3). (В) Обработка IFN-γ и TNF (обернутые треугольники) прекращает пролиферацию раковых клеток, но не убивает клетки саркомы (n=3). Для сравнения, культура нормальных клеток (кружки) показывает пролиферацию без задержек (n=3). (С) верхний ряд: Обработанные цитокином раковые клетки А204 стареют (черная стрелка: голубовато-серая окраска) и мультиядерны (белые стрелки, окраска DAPI). Нижний ряд: Клетки саркомы А204, обработанные средой в качестве негативного контроля, негативны по SA-β-Gal и моноядерны (окраска DAPI). (D) Индуцированная цитокинами удлиненная, мультиядерная и синцитиальная морфология клеток А204 (посередине и справа) по сравнению с клетками А204 стандартной культуры (слева), изображение получено с помощью микроскопии просвечивающего типа. (Е) Относительная экспрессия миогенных факторов транскрипции в клеточных линиях сарком А204 и RH30. Индукция экспрессии мРНК МНСП и MYF-6в клетках А204 с помощью 100 нг/мл IFN-γ и 10 нг/мл TNF (++) по сравнению со средой (-) на протяжении 3-х дней (n=3).
Фигура 8
Клеточные линии, представляющие мезодермное, эндодермное и эктодермное происхождение, показывали мультиядерный фенотип и блокировку роста после единократной обработки дозой 100 нг/мл IFN-γ в комбинации с 10 нг/мл TNF (день 1), по сравнению с контрольной средой (не обрабатывалась). Изображения глиобластомы были получены на 37-й день, все остальные - на 6-й день после начала обработки. Клетки всех обработанных клеточных линий выжили до 43-го дня, до момента времени заключительного обследования клеток.
RMS: рабдомиосаркома, GBM: глиобластома, BC: рак груди, CRC: колоректальный рак, HCC: печеночно-клеточная карцинома, NB: нейробластома.
ПРИМЕРЫ:
(1) Гуманизация мышей линии NSG. Человеческие стволовые клетки CD34+ (huCD34+) получали от избытка G-CSF мобилизированных стволовых клеток периферической крови родительских доноров, истощенной по Т-клеткам селекцией CD34+ (CliniMACS, Miltenyi, Германия). Клетки суспендировали 1:2 в 20% ДМСО / 80% 5%-HSA растворе и затем криоконсервировали с использованием устройства кубикового льда Sylab и контролируемой скорости замораживания. После размораживания клетки окрашивали трипановым синим и считали в счетной камере Нойбауэра. В соответствии с Хельсинкской Декларацией, все доноры дали информированное согласие на использование избыточных клеток в научных целях. Чистота популяции CD34+ была повышена до >99.99% вторым раундом истощения CD3+ после размораживания (LS MACS, Miltenyi, Германия). Все доноры стволовых клеток были HLA-несовместимы с клеточной линией РМС А204. 1×106 huCD34+ клеток в 100 мкл подогретого натрий-фосфатного буфера вводили в хвостовую вену мышей линии NSG, получивших сублетальную дозу облучения (250 сГр). Трансплантацию поддерживали еженедельным применением 20 мкг FcIL-7 (Merck, Германия). В каждой из групп лечения NHS-IL12 4 животных получали долгосрочное лечение цитокином NHS-IL12 с FcIL-7 или IL-2MAB602 на протяжении максимум 15-ти недель (100 дней).
(2) Имплантация и измерение опухоли. Клетки эмбриональной детской рабдомиосаркомы А204 получали из АТСС (НТВ 82), размораживали, субкультивировали 1 пассаж в среде RPMI 1640+10% сыворотки крови эмбрионов коров, трижды промывали стерильным солевым раствором, проверяли на микоплазмы с помощью PCR Mycoplasma Test Kit AppliChem (Германия) и имплантировали мышам линии NSG с трансплантатами. Одной мыши имплантировали 1×106 опухолевых клеток подкожно в правый бок. Объем опухоли определяли по следующей формуле: VT=a×b×d×π/6, где a, b и d - соответственно длина, ширина и глубина опухоли. Мышей умерщвляли через 52 дня после инокуляции опухоли, в момент времени, в который опухоли животных из группы FcIL-7 достигали ≥20% массы тела. Четырех мышей из группы NHS-IL12 оставили в живых для долгосрочного лечения.
(3) Введение комплексов человеческого анти-гистона/слитого белка человеческого IL-12 и IL-2/анти-IL-2 антител. Раз в неделю 20 мкг слитого белка NHS-IL12 вместе с 20 мкг FcIL-7 (оба произведены Merck, Германия) вводили уколом в хвостовую вену гуманизированных мышей линии NSG с трансплантатами. Лечение начинали по достижении опухолью объема ≥150 мкл. Как изложено выше, одна дополнительная группа мышей получала смесь 1.5 мкг IL-2 плюс 15 мкг анти-IL-2 mAB МАВ602 в неделю. Рекомбинантный человеческий IL-2 (PROLEUKIN, Aldesleukin, Chiron, США) и МАВ602 (анти-hIL-2 mABCD122, клон 5355, R&D Systems) совместно инкубировали на протяжении 15 минут при комнатной температуре перед инъекцией.
(4) SPECT/CT визуализация In Vivo. Визуализацию инокулированных мышей in vivo выполняли с использованием Inveon Multimodality SPECT/CT (Siemens Healthcare, Ноксвилл, TN, США). Свободный от носителя йодид натрия (123I) приобрели у GE Healthcare, а радиойодирование NHS-IL12 выполняли с помощью Pierce® Iodination Reagent (Thermo Scientific). Мышам вводили 30 мкг NHS-IL12, меченого 18 МБк 123I, через хвостовую вену и получали in vivo SPECT/CT изображения через 2, 26 и 46 часов после введения метки. Во время инъекции и измерения мышей анестезировали 1,5% изофлураном в кислороде (0,5 л/мин). Исследуемые области (ИО) обводили на реконструированных изображениях SPECT, основываясь на информации CT, поверх нескольких слоев, чтобы покрыть всю опухоль. Для сравнения ИО эквивалентного размера размещали на незатронутой мышечной ткани на левой задней лапе того же животного. Для оценки поглощения 123I-NHS-IL12 в расчетах делали поправку на распад.
(5) Шестицветная проточная цитометрия. Восстановление иммунитета оценивали у нескольких особей через 10-12 недель после трансплантации, анализируя периферическую кровь, селезенку, костный мозг и тимус с использованием следующих мышиных моноклональных антител, не реагирующих перекрестно с мышиными эпитопами, но специфичных для человеческого эпитопа: CD62L(Dreg56)-FITC, CD25(2A3)-APC, CD3(SK7)-PerCP, CD8(SK1)-PerCP, CD8(SK1)-APC-H7, CD4(SK3)-FITC, CD4(SK3)-PerCP, CD14(M5E2)-PE, CD56(My31)-PE, HLA-DR(L243)-PerCp, NKp30(P30-15)-PE, NKp44(P44-8)-APC, NKp46(9E2)-PE, а также соответствующие им изотипы IgG (все произведены BD Pharmingen, Германия). CD45(MEM-28)-PacifIc Blue, CD19(HIB19)-PerCP, CD3(MEM-57)-Alexa Fluor 700, CD4(MEM-241)-Alexa Fluor 700 (Exbio, Чехословакия). CD45(HI30)-PE и соответствующий изотип IgG (Biolegend, Германия). Приживление трансплантата регулярно контролировали через 12-14 недель после трансплантации ретроорбитальным забором крови и окраской FACS. Клеточную линию А204 характеризовали с ULBP-1(Z-9), ULBP-2(2F9), ULBP-3(F16), ULBP-4(6E6) (all Santa Cruz, США), MICA/B(6D4)APC CD112(R2.525)PE, CD155(SKII.4)PE (все - Biolegend, Германия), HLA-ABC(W6/32)PE (DAKO Cytomation, Германия), вторичным антителом RAM-PE(X56) (BD Pharmingen, Германия), и изотипными контролями (Beckman Coulter and R&D Systems, Германия). Проточную цитометрию выполняли на LSR II (BD Biosciences) с использованием программного обеспечения  .
.
(6) Иммуногистологическая окраска. Замороженные срезы тканей (толщиной 5 мкм) инкубировали в 4% буферного раствора формалина (2 минуты), промывали дистиллированной водой., кипятили в цитратном буфере pH 6,0 под давлением (4 минуты) и промывали трис-NaCl-твином. Образцы срезов перемещали во влажную камеру и окрашивали Zytochem-Plus AP Polymer-Kit (Zytomed Systems, Германия). Основными антителами являлись: CD3 (клон SP7, 1:50; DCS Innovative Diagnostic Systeme GmBH, Германия), моноклональное кроличье антитело против человеческого CD4 (клон SP35, 1:50, Zytomed Systems), CD8 (клон С8/144B, 1:100), CD56 (клон 123C3-D5, 1:20), CD68 (клон PG-M1, 1:150), HLA-DR-α (клон TAL.1B5, 1:200), десмина (клон D33, 1:100, все - DAKO, Германия), перфорина (клон 5810, 1:200, Novocastra/Leica, Германия). Финальное окрашивание выполняли Permanent AP Red Kit (Zytomed Systems, Германия). Анализ иммуногистологических срезов выполняли одинарным слепым методом.
(7) Иммунофлюоресценция. Свежезамороженные криостатные срезы ксенотрансплантированных человеческих опухолей А204 окрашивали как описано выше (Zhang and Adams (2007) Cell Cycle 6: 784-789). Вкратце, криостатные срезы фиксировали перйодат-лизин-параформальдегидом и блокировали сывороткой ослиной крови (1:20) на протяжении 30 минут при комнатной температуре (КТ). Затем срезы инкубировали с кроличьим антителом против р-НР1γ (1:80; Abeam, UK), мышиным антителом против PCNA (1:50; Cell Signaling Technology, США), мышиным антителом против p16INK4a (1:50; Santa Cruz Biotechnology, Германия), или кроличьим антителом против Ki67 (1:100; Abeam, США) на протяжении 1 часа, при KT. После трех промываний срезы инкубировали с Cy3-конъюгированным ослиным-анти-кроличьим антителом, Cy3-конъюгированным ослиным-анти-мышиным антителом, Cy5-конъюгированным ослиным-анти-кроличьим антителом, Cy5-конъюгированным ослиным-анти-мышиным антителом (все - Dianova, Германия). Перед заливкой срезов средой Mowiol (Hoechst, Германия), ядра окрашивали Yopro (1:2000; Invitrogen, Германия) (5 минут), и срезы анализировали на конфокальном лазерном сканирующем микроскопе Leica TCS-Sp/Leica DM RB (Leica Microsystems, Германия). Изображения обрабатывали Leica Confocal Software LCS (версия 2.61).
(8) Лечение происходящих от пациентов первичных клеточных линий РМС и раковых клеточных линий с помощью TNF and IFN-γ. Постоянную блокировку роста различных раковых клеточных линий (рабдомиосаркомы RH30, SRH; глиобластомы T98G, Ln229, A172; рака груди MCF-7; колоректального рака HCT-116; печеночно-клеточной карциномы Hep3b и нейробластомы LAN-1, LS) после обработки цитокинами определяли с использованием процедуры, детально описанной ранее (Braumuller et al., 2013, Nature 494: 361-365). Молодые культуры для изучения событий ранней дифференциации в РМС получали посевом опухолевых клеток в малых количествах (4.000 или 8.000 клеток/см2) и инкубацией со средой, содержащей 10 нг/мл TNF и 100 нг/мл IFN-γ, а также с обычной средой.
(8) Анализ экспрессии KIR. Анализ экспрессии проводили как описано выше (77). Транскрипты NKp30, -44, -46, DNAM-1, CD161 определяли с помощью специфических праймеров в ПЦР "по конечной точке", используя для этого 5`FAM-меченые обратные праймеры. Продукты ПЦР анализировали на длину фрагмента в секвенаторе ABI GeneScan-600 LIZ для получения стандарта длины и программным обеспечением GeneMapper (AppliedBiosystems, Германия); для полуколичественного анализа использовали MFI.
(9) Экспрессия T-bet, RORC, Foxp3, CD40L, гранзима B, гранзима K. Внутриопухолевую экспрессию генов определяли с помощью ПЦР в реальном времени на термоциклере BioRad С1000 / системе реального времени CFX96 (Мюнхен, Германия), используя кДНК-специфичные праймеры (все последовательности могут быть получены по запросу) и iQ SYBR Green Supermix BioRad (Мюнхен, Германия). Экспрессию нормализовали до экспрессии человеческого CD45. Экспрессию RORC выявляли набором специфических праймеров (Search LC, Германия) и FastStart DNA SYBR Green I (Roche, Германия).
(10) Идентификация цепи Vα-24. Для этого использовали протокол, опубликованный Han et al. (Han et al., 1999, J. Immunol. 163: 301-31).
(11) Анализ Vβ-спектрального типа. Разнообразие экспрессии Vβ-цепи TCR и сложность репертуара TCR анализировали согласно Gorski et al. (Gorsk et al., 1994, J. Immunol. 152: 5109-511) с небольшими модификациями. Использовали 5`FAM-меченый Cβ-праймер, а для определения ампликонов ПЦР использовали секвенатор ABI с программным обеспечением Gene Scan-600 LIZ и GeneMapper (оба - Applied Biosystems, Германия).
(12) γδ иммуноскопия. Иммуноскопию проводили согласно прежней публикации (Dechanet et al., ,J., 1999, Clin. Invest 103: 1437-1449).
(13) Идентификация областей TCR-CDR3. Продукты Vβ ПЦР клонировали в pGEMTeasy (Promega, Германия) и амплифицировали в XL1-Blue компетентных клетках (Stratagene, США), используя стандартные процедуры. Клоны, содержащие вставки, передавали напрямую на реамплификацию в Vβ ПЦР. Аликвоты ПЦР по 5 мкл анализировали на 2.5% агарозном геле, и продукты ПЦР подходящей длины отправляли в Seqlab, Германия, на анализ последовательностей. Трансляцию кДНК в белковую последовательность осуществляли на свободном программном обеспечении EMBOSS Transeq.




Группа изобретений относится к противораковой иммунотерапии. Предложено применение нацеленного на опухоль IL-12 для комбинированной терапии с IL-7 для индукции и/или стимуляции иммунного ответа против рака у пациента, где рак - плотные опухоли или опухоли мышц, костей, нервной системы, хрящей, сухожилий, кровеносных сосудов и жировых или волокнистых тканей, и указанная индукция (стимуляция) вызывает старение раковых клеток или блокировку их роста, и/или ремиссию раковых клеток или раковой ткани до клеток или ткани происхождения, где IL-12 слит через N-конец своих субъединиц р40 и/или р35 с С-концом полностью человеческого антитела NHS76 или его биологически активного фрагмента, и где IL-7 ковалентно слит с тяжелой цепью иммуноглобулина или Fc-фрагментом тяжелой цепи иммуноглобулина; применение нацеленного на опухоль IL-12 для комбинированной терапии с IL-2 саркомы (вариант); набор того же назначения, включающий (а) полностью человеческое антитело NHS76, направленное на человеческий комплекс ДНК-гистон HI, представленный в опухолевом некрозе, и слитое через С-конец с N-концом субъединиц р40 и/или р35 IL-12, и (б) иммуномодулятор и/или иммунокомплементарный агент, представляющий собой IL-7, ковалентно слитый с тяжелой цепью иммуноглобулина или Fc-фрагментом тяжелой цепи иммуноглобулина, или представляющий собой IL-2 в комплексе с анти-IL-2-антителом (варианты). Технический результат: побуждаемая IL-12 иммунная система пациента не только убивает раковые клетки, но и использует альтернативные механизмы ослабления роста опухоли. 4 н. и 6 з.п. ф-лы, 8 ил., 13 пр.
1. Применение нацеленного на опухоль IL-12 для комбинированной терапии с IL-7 для индукции и/или стимуляции иммунного ответа против рака у пациента, страдающего указанным раком, где рак представляет собой плотные опухоли или опухоли мышц, костей, нервной системы, хрящей, сухожилий, кровеносных сосудов и жировых или волокнистых тканей,
где указанная индукция или стимуляция вызывает:
(I) старение раковых клеток или блокировку их роста, и/или
(II) ремиссию раковых клеток или раковой ткани до клеток или ткани происхождения,
вызванные указанным препаратом,
где IL-12 слит через N-конец своих субъединиц р40 и/или р35 с С-концом полностью человеческого антитела NHS76 или его биологически активного фрагмента,
и где IL-7 ковалентно слит с тяжелой цепью иммуноглобулина или Fc-фрагментом тяжелой цепи иммуноглобулина.
2. Применение нацеленного на опухоль IL-12 для комбинированной терапии с IL-2, который находится в комплексе с анти-IL-2 антителом, для индукции и/или стимуляции иммунного ответа против рака у пациента, страдающего указанным раком, где рак представляет собой саркому,
где указанная индукция или стимуляция вызывает:
(I) старение раковых клеток или блокировку их роста, и/или
(II) ремиссию раковых клеток или раковой ткани до клеток или ткани происхождения,
вызванные указанным препаратом,
где IL-12 слит через N-конец своих субъединиц р40 и/или р35 с С-концом полностью человеческого антитела NHS76 или его биологически активного фрагмента.
3. Применение по п. 1 или 2, отличающееся тем, что указанное старение раковых клеток вызвано выработкой эндогенного IFNγ и/или TNF вследствие указанной стимуляции или индукции иммунной системы пациента, запущенных указанным препаратом.
4. Применение по п. 1 или 2, отличающееся тем, что старение раковых клеток приводит к стабильной блокировке роста.
5. Применение по п. 1 или 2, отличающееся тем, что старение раковых клеток не зависит от прямых иммуноспецифических цитотоксичных эффектов.
6. Применение по п. 1 или 2, отличающееся тем, что терапия инициирует индукцию миогенной дифференциации.
7. Применение по п. 1 или 2, скомбинированное с лучевой терапией или радио-химиотерапией.
8. Применение по п. 1 или 2, отличающееся тем, что антитело NHS76 представляет собой NHS-IL12.
9. Фармацевтический набор для лечения рака, который представляет собой плотные опухоли или опухоли мышц, костей, нервной системы, хрящей, сухожилий, кровеносных сосудов и жировых или волокнистых тканей, причем указанный набор включает:
(I) первый пакет, включающий полностью человеческое антитело NHS76, направленное на человеческий комплекс ДНК-гистон HI, представленный в опухолевом некрозе, и слитое через С-конец с N-концом субъединиц р40 и/или р35 IL-12, и
(II) второй пакет, включающий иммуномодулятор и/или иммунокомплементарный агент для индукции и/или стимуляции иммунного ответа против рака, указанный иммуномодулятор и/или иммунокомплементарный агент представляет собой IL-7, ковалентно слитый с тяжелой цепью иммуноглобулина или Fc-фрагментом тяжелой цепи иммуноглобулина.
10. Фармацевтический набор для лечения саркомы, причем указанный набор включает:
(I) первый пакет, включающий полностью человеческое антитело NHS76, направленное на человеческий комплекс ДНК-гистон H1, представленный в опухолевом некрозе, и слитое через С-конец с N-концом субъединиц р40 и/или р35 IL-12, и
(II) второй пакет, включающий иммуномодулятор и/или
иммунокомплементарный агент для индукции и/или стимуляции иммунного ответа против саркомы, где указанный иммуномодулятор и/или иммунокомплементарный агент представляет собой IL-2, который находится в комплексе с анти-IL-2 антителом.
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| TEVS G | |||
| et al | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| The Journal of Gene Medicine | |||
| Колосоуборка | 1923 |
|
SU2009A1 |
| Вилка для выдергивания сорных трав | 1929 |
|
SU16038A1 |
| DAVIS C.B | |||
| et al | |||
| Immunocytokines: amplification of anti-cancer immunity | |||
| Cancer Immunol | |||
| Immunother | |||
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |
| JOHNSON E.E | |||
| et al | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Clin.Cancer Res | |||
| Колосоуборка | 1923 |
|
SU2009A1 |
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| LAURENT J | |||
| et al | |||
| Устройство для формования изделий с начинкой | 1974 |
|
SU521873A1 |
| J.Transl | |||
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |

