ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Настоящая заявка заявляет приоритет в соответствии с 35 USC 119(e) по предварительной заявке США №62/898651, поданной 11 сентября 2019 г., которая включена в данный документ посредством ссылки в полном объеме.
ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к векторам на основе рекомбинантных вирусов для вставки и экспрессии чужеродных генов для применения в безопасных иммунизациях для защиты от различных патогенов. Это также относится к поливалентным композициям или вакцинам, содержащим один или несколько векторов на основе рекомбинантных вирусов для защиты от различных патогенов. Настоящее изобретение связано со способами получения и применения указанных векторов на основе рекомбинантных вирусов.
УРОВЕНЬ ТЕХНИКИ
Болезнь Марека, высококонтагиозное лимфопролиферативное заболевание, представляет собой одно из наиболее распространенных инфекций птиц, которое поражает преимущественно молодых цыплят.Болезнь Марека вызвана вирусом болезни Марека. Вирус болезни Марека (MDV), представляющий собой вирус герпеса, является представителем рода Mardivirus, который содержит три серотипа (вида): MDV-1 (вирус герпеса Gallid 2), MDV-2 (вирус герпеса Gallid 3) и MDV-3 (вирус герпеса Meleagrid 1, вирус герпеса индеек (HVT)). MDV-1 представляет собой наиболее вирулентный из трех серотипов, вызывающих широко распространенное заболевание у невакцинированной домашней птицы. У птиц, инфицированных MDV-1, проявляются неврологические, висцеральные и кожные клинические симптомы, такие как паралич нижних конечностей, крыльев и шеи; поражения глаз и нарушение зрения, потеря веса, раковые опухоли во многих органах, таких как тимус, сердце, легкие, гонады, мышцы и перьевые фолликулы. Заболеваемость пораженных птиц составляет 10-50%, а смертность может достигать до 100%. Несмотря на то, что болезнь Марека может поражать птиц в любом возрасте, острая форма болезни Марека вызывает гибель большого количества не вакцинированных птиц в раннем возрасте от четырех до восьми недель. Болезнь Марека распространяется при непосредственном или опосредованном контакте с перхотью инфицированных кур, а вирус попадает в организм при вдыхании. MDV-2 и MDV-3 представляют собой авирулентные вирусные штаммы и их использовали при подготовке вакцинации против родственного и вирулентного MDV-1.
Кроме болезни Марека существует несколько патогенов, которые поражают домашнюю птицу и представляют угрозу для птицеводства. Производители должны полагаться на иммунитет, обеспечиваемый вакцинами, для защиты стад от вирусных, бактериальных и других патогенов. Для вакцинации птиц применяли живые, убитые и рекомбинантные вакцины. Преимущество живых вакцин заключается в сильном и длительном иммунитете, однако они требуют осторожного обращения, поскольку они могут вызвать легкую или тяжелую реакцию. С другой стороны, убитые вакцины являются более стабильными и безопасными, чем живые вакцины, но вызывают более слабый иммунный ответ, что требует многократных введений. Как живые, так и убитые вакцины доказали свою безопасность и эффективность, однако остается потребность в разработке и постоянном совершенствовании поливалентных вакцин для обеспечения защиты от более чем одного патогена при одной вакцинации.
Рекомбинантные векторные вакцины были разработаны для обеспечения иммунитета к нескольким патогенам одновременно. Эти вакцины изготавливаются посредством удаления некоторых несущественных участков гена в геноме хозяина непатогенного организма и замены их одним или несколькими генами, кодирующими антигены, которые отвечают за выработку иммунного ответа против патогенного организма. Затем вновь полученный вектор используется для инфицирования хозяина, в котором он будет реплицироваться и экспрессировать антигены вирулентного (вирулентных) организма (организмов), индуцируя иммунный ответ. Рекомбинантные векторные вакцины сочетают в себе преимущества живых и убитых вакцин. Рекомбинантные векторы, аналогично живой вакцине, обеспечивают более длительный иммунитет и в то же время вызывают более легкую реакцию после вакцинации, чем убитые вакцины. Кроме того, как вектор, так и вставленный (вставленные) ген (гены) могут обеспечить иммунитет, защищающий птиц от двух или более заболеваний.
Вирусы болезни Марека представляют собой одни из наиболее эффективных векторов поливалентных вакцин для иммунизации против болезней домашней птицы, поскольку эти вирусы обеспечивают прижизненную защиту всего после одной вакцинации. Кроме того, эти вирусы ограничены хозяевами-птицами, поэтому опасность заражения других животных и людей, работающих на птицефабриках, отсутствует. Среди вирусов болезни Марека вирус герпеса индеек (HVT) более широко использовался как в качестве живой вакцины, так и в качестве вектора на основе рекомбинантной вакцины против более вирулентного MDV-1. HVT был впервые выделен от индеек в 1969-1970 годах, и вскоре было обнаружено, что он защищает от MDV, после чего он был получил разрешение на применение в качестве вакцины в 1971 году. Вирус герпеса индеек (HVT) характеризуется аналогичными антигенными свойствами с вирусом болезни Марека (MDV-1), но не является патогенным для кур. Кроме того, HVT не чувствителен к материнским антителам против MDV или HVT, поэтому живая вакцина HVT использовалась для эффективной вакцинации против MDV-1 in ovo или в раннем возрасте до вылупления. Кроме того, геном HVT использовался в качестве вакцинного вектора для содержания чужеродных последовательностей ДНК других патогенов птиц.
КРАТКАЯ СУЩНОСТЬ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
В настоящем изобретении предусмотрены векторы на основе рекомбинантных вирусов для вставки и экспрессии чужеродных генов для применения в безопасных иммунизациях для защиты птиц от различных патогенов. В настоящем изобретении также предусмотрены поливалентные композиции или вакцины, содержащие один или несколько векторов на основе рекомбинантного вируса HVT для защиты от различных патогенов. Кроме того, в настоящем изобретении также предусмотрены способы получения и применения векторов на основе рекомбинантных вирусов отдельно или в комбинации с другими вакцинами или фармацевтическими композициями.
В одном аспекте в настоящем изобретении предусмотрен геном на основе рекомбинантного вируса герпеса индеек (HVT), содержащий одну или несколько нуклеотидных последовательностей, кодирующих один или несколько гетерологичных антигенов, вставленных в межгенные локусы UL 35/UL 36 в уникальной длинной (UL) области генома HVT.
В одном аспекте в настоящем изобретении предусмотрен геном рекомбинантного вируса герпеса индеек (HVT), содержащий одну или несколько нуклеотидных последовательностей, кодирующих один или несколько гетерологичных антигенов, вставленных в межгенные локусы UL 35/UL 36 в уникальной длинной области генома HVT, и одну или несколько нуклеотидных последовательностей, кодирующих один или несколько гетерологичных антигенов, вставленных в сайт UL55/ген 3 уникальной длинной области (UL) генома HVT.
В одном или нескольких вариантах осуществления в настоящем изобретении предусмотрен рекомбинантный HVT, в котором один или несколько гетерологичных антигенов характеризуются защитным действием от патогенов птиц или патогенов, выбранных из группы, состоящей из: вируса инфекционного бурсита (IBDV); вируса болезни Ньюкасла (NDV); вируса инфекционного бронхита (IBV); вируса инфекционного ларинготрахеита (ILTV); вируса анемии цыплят (CAV); и вируса птичьего гриппа (AIV).
В одном или нескольких вариантах осуществления в настоящем изобретении предусмотрен рекомбинантный HVT, в котором один или один или несколько гетерологичных антигенов выбраны из группы, состоящей из белков VP2, VP3 или VP4 вируса инфекционного бурсита (IBDV); белков VP1 или VP2 вируса анемии цыплят (CAV); химерного белка F/HN или белков F, NP, Р, М, HN или L вируса болезни Ньюкасла (NDV); белков S1, S2 или М вируса инфекционного бронхита (IBV); белков gB, gC, gD, gE, gH, gI или gL вируса инфекционного ларинготрахеита (ILTV); и любого из белков НА, NA, NP или М вируса птичьего гриппа (AIV).
В одном или нескольких вариантах осуществления рекомбинантный HVT по настоящему изобретению предусматривает, что один или несколько гетерологичных антигенов характеризуются защитным действием от IBDV. В одном варианте осуществления рекомбинантный HVT по настоящему изобретению предусматривает, что один или несколько гетерологичных антигенов представляют собой белок VP2 из IBDV. В одном варианте осуществления рекомбинантный HVT по настоящему изобретению предусматривает, что белок VP2 кодируется нуклеотидной последовательностью, на по меньшей мере 80% идентичной нуклеотидной последовательности, содержащей SEQ ID NO. 5 или SEQ ID NO. 10. В одном варианте осуществления рекомбинантный HVT по настоящему изобретению предусматривает, что белок VP2 кодируется нуклеотидной последовательностью, содержащей либо SEQ ID NO. 5, либо SEQ ID NO. 10.
В одном или нескольких вариантах осуществления рекомбинантный HVT по настоящему изобретению предусматривает, что один или несколько гетерологичных антигенов характеризуются защитным действием от вируса болезни Ньюкасла (NDV). В одном варианте осуществления рекомбинантный HVT по настоящему изобретению предусматривает, что один или несколько гетерологичных антигенов представляют собой белок F NDV. В одном варианте осуществления рекомбинантный HVT по настоящему изобретению предусматривает, что белок F из NDV кодируется нуклеотидной последовательностью, на по меньшей мере 80% идентичной нуклеотидной последовательности, содержащей SEQ ID NO. 3. В одном варианте осуществления рекомбинантный HVT по настоящему изобретению предусматривает, что белок F из NDV кодируется нуклеотидной последовательностью, содержащей SEQ ID NO. 3.
В одном или нескольких вариантах осуществления рекомбинантный HVT по настоящему изобретению предусматривает, что один или несколько гетерологичных антигенов характеризуются защитным действием от NDV и IBDV. В одном или нескольких вариантах осуществления рекомбинантный HVT по настоящему изобретению предусматривает, что по меньшей мере один гетерологичный антиген представляет собой белок F из NDV и белок VP2 из IBDV.
В одном или нескольких вариантах осуществления рекомбинантный HVT по настоящему изобретению предусматривает, что белок F из NDV, кодируемый нуклеотидной последовательностью, на по меньшей мере 80% идентичной нуклеотидной последовательности, содержащей SEQ ID NO. 3, и белок VP2 из IBDV, кодируемый нуклеотидной последовательностью, на по меньшей мере 80% идентичной нуклеотидной последовательности, содержащей SEQ ID NO. 5 или SEQ ID NO. 10.
В одном или нескольких вариантах осуществления рекомбинантный HVT по настоящему изобретению предусматривает белок F из NDV, кодируемый нуклеотидной последовательностью, содержащей SEQ ID NO. 3, а белок VP2 из IBDV кодируется нуклеотидной последовательностью, содержащей SEQ ID NO. 5 или SEQ ID NO. 10.
В одном или нескольких вариантах осуществления рекомбинантный HVT по настоящему изобретению содержит геном, содержащий одну или несколько кассет экспрессии, содержащих одну или несколько нуклеотидных последовательностей, которые кодируют один или несколько гетерологичных антигенов. В одном варианте осуществления рекомбинантный HVT содержит геном на основе рекомбинантного HVT, кассету экспрессии, которая содержит нуклеотидную последовательность, кодирующую промоторы, которые функционально связаны с одним или несколькими нуклеотидами, которые кодируют антигены, подлежащие экспрессии. В одном варианте осуществления антиген, подлежащий экспрессии, содержит белок F из NDV. В одном варианте осуществления антиген, подлежащий экспрессии, содержит белок VP2 из IBDV. В одном варианте осуществления антигены, подлежащие экспрессии, содержат как белок F из NDV, так и белок VP2 из IBDV.
В одном варианте осуществления рекомбинантный HVT по настоящему изобретению предусматривает один или несколько промоторов, выбранных из группы, состоящей из: немедленно раннего промотора цитомегаловируса человека (hCMV); немедленно раннего промотора CMV морской свинки; немедленно раннего промотора CMV мыши; промотора Рес; промотора β-актина кур; промотора SV40; промоторов гликопротеина X вируса псевдобешенства; промотора вируса простого герпеса-1 альфа 4; промоторов гликопротеинов gA, gC, gB, gE или gI вируса болезни Марека; промоторов гликопротеина gB, gE, gI, gD вирусов инфекционного ларинготрахеита; и промотора VP8 вируса герпеса крупного рогатого скота 1/1. В одном варианте осуществления рекомбинантный HVT содержит промотор CMV человека. В одном варианте осуществления рекомбинантный HVT содержит промотор CMV мыши. В одном варианте осуществления рекомбинантный HVT содержит промотор hCMV и mCMV.
В одном или нескольких вариантах осуществления рекомбинантный HVT содержит нуклеотидную последовательность, кодирующую сигнальную последовательность полиаденилирования (poly А). В одном или нескольких вариантах осуществления рекомбинантный HVT содержит нуклеотидную последовательность, кодирующую сигнальную последовательность poly А, и выбран из последовательности poly A BGH (SEQ ID NO.6) или последовательности poly A SV40 (SEQ ID NO. 12). В одном варианте осуществления сигнальная последовательность poly А представляет собой сигнальную последовательность poly A BGH. В одном варианте осуществления сигнальная последовательность poly А представляет собой сигнальную последовательность poly A SV40.
В одном аспекте рекомбинантный HVT по настоящему изобретению содержит промотор CMV, функционально связанный с нуклеотидной последовательностью, кодирующей белок VP2 из IBDV, дополнительно содержащий нуклеотидную последовательность, кодирующую сигнальную последовательность полиаденилирования, при этом вся кассета экспрессии на основе VP2 вставлена в некодирующую область генома HVT. В одном варианте осуществления промотор CMV содержит промотор hCMV (SEQ ID NO. 1). В одном варианте осуществления нуклеотидная последовательность, кодирующая белок VP2 IBDV, выбрана из SEQ ID NO.5 или SEQ ID NO. 10. В одном варианте осуществления нуклеотидная последовательность, кодирующая белок VP2, содержит SEQ ID NO.5. В одном варианте осуществления нуклеотидная последовательность, кодирующая белок VP2, содержит SEQ ID NO. 10. В одном варианте осуществления сигнальная последовательность полиаденилирования содержит SEQ ID NO. 6. В одном варианте осуществления сигнальная последовательность полиаденилирования содержит SEQ ID NO. 12. В одном варианте осуществления промотор, нуклеотидная последовательность, кодирующая белок VP2, и сигнальная последовательность poly А содержат кассету экспрессии. В одном варианте осуществления кассета экспрессии вставлена в геном HVT в сайте UL55/ген 3. В одном варианте осуществления кассета экспрессии вставлена в геном HVT в сайте UL35/36 в геноме. В одном варианте осуществления кассета экспрессии содержит по порядку SEQ ID NO.1, SEQ ID NO.5 или SEQ ID NO. 10 и SEQ ID NO.6, вставленные в геном HVT в сайте UL55/ген 3.
В одном аспекте рекомбинантный HVT по настоящему изобретению содержит промотор CMV, функционально связанный с нуклеотидной последовательностью, кодирующей белок F из NDV, дополнительно содержащий нуклеотидную последовательность, кодирующую сигнальную последовательность полиаденилирования, при этом вся часть кассеты на основе белка F из NDV вставлена в некодирующую область в геноме HVT. В одном варианте осуществления промотор CMV содержит промотор mCMV (SEQ ID NO.2). В одном варианте осуществления нуклеотидная последовательность кодирует белок F из NDV, который содержит SEQ ID NO.3. В одном варианте осуществления сигнальная последовательность полиаденилирования кодируется нуклеотидной последовательностью, содержащей SEQ ID NO. 12. В одном варианте осуществления промотор, нуклеотидная последовательность, кодирующая белок F, и сигнальная последовательность poly А содержат кассету экспрессии. В одном варианте осуществления кассета экспрессии вставлена в геном HVT в сайте UL55/ген 3. В одном варианте осуществления кассета экспрессии вставлена в геном HVT в сайте UL35/36 в геноме. В одном варианте осуществления кассета экспрессии содержит по порядку SEQ ID NO.2, SEQ ID NO.3 и SEQ ID NO. 12, вставленные в геном HVT в сайте UL55/ген 3.
В одном аспекте рекомбинантный HVT по настоящему изобретению содержит промотор CMV, функционально связанный с нуклеотидной последовательностью, кодирующей белок VP2 из IBDV, дополнительно содержащий нуклеотидную последовательность, кодирующую сигнальную последовательность полиаденилирования, при этом все содержат кассету экспрессии VP2, вставленную в некодирующую область в геноме HVT. В одном варианте осуществления рекомбинантный HVT по настоящему изобретению дополнительно содержит промотор CMV, функционально связанный с нуклеотидной последовательностью, кодирующей белок F из NDV, дополнительно содержащий нуклеотидную последовательность, кодирующую сигнальную последовательность полиаденилирования, в виде части кассеты экспрессии на основе F из NDV, вставленной в тот же сайт вставки, что и кассета на основе VP2. В одном варианте осуществления рекомбинантный HVT по настоящему изобретению дополнительно содержит промотор CMV, функционально связанный с нуклеотидной последовательностью, кодирующей белок F из NDV, дополнительно содержащий нуклеотидную последовательность, кодирующую сигнальную последовательность полиаденилирования, в виде части кассеты экспрессии на основе F из NDV, вставленной в другой сайт по сравнению с кассетой на основе VP2.
В одном варианте осуществления рекомбинантный HVT по настоящему изобретению предусматривает, что кассета экспрессии на основе VP2 содержит по порядку нуклеотидную последовательность, кодирующую промотор hCMV (SEQ ID NO. 1), нуклеотидную последовательность, кодирующую VP2 из IBDV (выбранный из SEQ ID NO. 5 или SEQ ID NO. 10), и нуклеотидную последовательность, кодирующую сигнальную последовательность полиаденилирования BGH (SEQ ID NO.6), вставленную в геном HVT в некодирующей области UL35/36, и по порядку нуклеотидную последовательность, кодирующую промотор mCMV (SEQ ID NO.2), нуклеотидную последовательность, кодирующую белок F из NDV (SEQ ID NO.3), и нуклеотидную последовательность, кодирующую сигнальную последовательность полиаденилирования SV40 (SEQ ID NO. 12), вставленную в геном HVT в некодирующей области UL55/ген 3. В одном аспекте рекомбинантный HVT по настоящему изобретению содержит промотор, функционально связанный с нуклеотидной последовательностью, кодирующей антиген вируса инфекционного ларинготрахеита, дополнительно содержащей нуклеотидную последовательность, кодирующую сигнальную последовательность полиаденилирования. В одном варианте осуществления антиген ILT содержит один или несколько антигенов, выбранных из группы, состоящей из: gB, gC, gD, gE, gH, gI, gL или химерных белков одного или нескольких антигенов ILT вируса инфекционного ларинготрахеита (ILTV). В одном варианте осуществления рекомбинантный HVT по настоящему изобретению дополнительно содержит нуклеотидную последовательность, кодирующую один или несколько антигенов, выбранных из группы, состоящей из вируса инфекционного бурсита, вируса анемии цыплят, вируса болезни Ньюкасла, вируса инфекционного бронхита и вируса птичьего гриппа. В одном варианте осуществления рекомбинантный HVT по настоящему изобретению дополнительно предусматривает промотор, функционально связанный с нуклеотидной последовательностью, кодирующей антигены, выбранные из группы, состоящей из: антигена VP1, VP2, VP3 или VP4 вируса инфекционного бурсита (IBDV); белков VP1 или VP2 вируса анемии цыплят (CAV); химерного белка F/HN или белков F, NP, Р, М, HN или L вируса болезни Ньюкасла (NDV); белков S1, S2 или М вируса инфекционного бронхита (IBV); и любого из белков НА, NA, NP или М вируса птичьего гриппа (AIV).
В одном варианте осуществления рекомбинантный HVT по настоящему изобретению содержит один или несколько антигенов ILT в качестве части кассеты экспрессии, содержащей промотор, функционально связанный с нуклеотидом, кодирующим антиген ILT, и дополнительно содержащий нуклеотидную последовательность, кодирующую сигнальную последовательность полиаденилирования. В одном варианте осуществления рекомбинантный HVT по настоящему изобретению содержит вторую и третью кассету экспрессии, каждая из которых содержит нуклеотидные последовательности, кодирующие промотор, функционально связанный с нуклеотидной последовательностью, кодирующей антигены птиц, выбранные из группы, состоящей из антигена VP1, VP2, VP3 или VP4 вируса инфекционного бурсита (IBDV); белков VP1 или VP2 вируса анемии цыплят (CAV); химерного белка F/HN или белков F, NP, Р, М, HN или L вируса болезни Ньюкасла (NDV); белков S1, S2 или М вируса инфекционного бронхита (IBV); и любого из белков НА, NA, NP или М вируса птичьего гриппа (AIV), и дополнительно содержит нуклеотидную последовательность, кодирующую сигнальную последовательность полиаденилирования.
В одном или нескольких аспектах в настоящем изобретении предусмотрена рекомбинантная ДНК, кодирующая рекомбинантный геном HVT по настоящему изобретению.
В одном или нескольких аспектах в настоящем изобретении предусмотрена иммуногенная композиция, содержащая рекомбинантный HVT по настоящему изобретению и дополнительно содержащая фармацевтически приемлемый носитель, вспомогательное вещество или адъювант.
В одном или нескольких аспектах в настоящем изобретении предусмотрена вакцинная композиция, содержащая рекомбинантный HVT по настоящему изобретению и дополнительно содержащая фармацевтически приемлемый носитель, вспомогательное вещество или адъювант.
В одном варианте осуществления вакцина по настоящему изобретению дополнительно содержит дополнительный вирус болезни Марека (MDV), выбранный из группы, состоящей из: естественно аттенуированного штамма MDV-1 Rispens (CVI-988); или вируса герпеса Gallid 3 штамма SB-1. В одном варианте осуществления вакцина по настоящему изобретению предусматривает, что дополнительный MDV содержит рекомбинантный геном. В одном варианте осуществления вакцина по настоящему изобретению предусматривает, что дополнительный геном рекомбинантного MDV содержит одну или несколько нуклеотидных последовательностей, кодирующих один или несколько гетерологичных антигенов, которые характеризуются защитным действием от одного или нескольких патогенов птиц.
В одном варианте осуществления вакцина по настоящему изобретению предусмотрена для вакцинации птицы против одного или нескольких заболеваний, вызванных одним или несколькими патогенами птиц. В одном или нескольких вариантах осуществления вакцина по настоящему изобретению предусмотрена для применения в защите птицы от клинических симптомов, вызванных одним или несколькими патогенами птиц. В одном или нескольких вариантах осуществления вакцина по настоящему изобретению предусмотрена для применения в защите птицы от клинических симптомов, вызванных вирусом болезни Марека, и клинических симптомов, вызванных одним или несколькими патогенами птиц. В одном или нескольких вариантах осуществления вакцина по настоящему изобретению содержит один или несколько патогенов птиц, выбранных из группы, состоящей из: вируса инфекционного бурсита (IBDV); вируса болезни Ньюкасла (NDV); вируса инфекционного бронхита (IBV); вируса инфекционного ларинготрахеита (ILTV); вируса анемии цыплят (CAV); и вируса птичьего гриппа (AIV). В одном варианте осуществления вакцина по настоящему изобретению предусматривает, что один или несколько патогенов птиц включают вирус болезни Ньюкасла. В одном варианте осуществления вакцина по настоящему изобретению предусматривает, что один или несколько патогенов птиц включают вирус инфекционного бурсита (IBDV). В одном варианте осуществления вакцина по настоящему изобретению предусматривает, что один или несколько патогенов птиц включают вирус болезни Ньюкасла и вирус инфекционного бурсита.
В одном или нескольких вариантах осуществления вакцина по настоящему изобретению предусмотрена для применения в вакцинации птиц, при этом вакцину вводят посредством по меньшей мере одного введения вакцины путем распыления, введения in ovo, подкожного введения, внутримышечного введения, перорального введения, назального введения или их комбинации. В одном варианте осуществления вакцина по настоящему изобретению предусматривает, что вакцину вводят путем введения in ovo. В одном варианте осуществления вакцина по настоящему изобретению предусматривает, что введение in ovo происходит в яйцеклетку с развивающимся эмбрионом в период между от около 16 до 22 дней развития. В одном или нескольких вариантах осуществления вакцина по настоящему изобретению предусматривает, что введение in ovo происходит в яйцеклетку с развивающимся эмбрионом в период около 18 дней развития. В одном варианте осуществления вакцина по настоящему изобретению предусматривает, что введение вакцины включает введение in ovo с последующим введением спрея. В одном варианте осуществления вакцина по настоящему изобретению предусматривает, что введение вакцины включает введение спрея.
В одном аспекте в настоящем изобретении предусмотрен способ вакцинации птицы для лечения или предупреждения болезни Марека и одного или нескольких заболеваний птиц, вызванных одним или несколькими патогенами птиц, включающий стадию введения эффективного количества вакцинной композиции по настоящему изобретению. В одном варианте осуществления способ по настоящему изобретению предусматривает, что один или несколько патогенов птиц выбраны из группы, состоящей из вируса инфекционного бурсита (IBDV); вируса болезни Ньюкасла (NDV); вируса инфекционного бронхита (IBV); вируса инфекционного ларинготрахеита (ILTV); вируса анемии цыплят (CAV); и вируса птичьего гриппа (AIV). В одном варианте осуществления способ по настоящему изобретению предусматривает, что один или несколько патогенов птиц включают вирус инфекционного бурсита (IBDV). В одном варианте осуществления способ по настоящему изобретению предусматривает, что один или несколько патогенов птиц включают вирус болезни Ньюкасла (NDV). В одном варианте осуществления способ по настоящему изобретению предусматривает, что один или несколько патогенов птиц включают вирус инфекционного бурсита (IBDV) и вирус болезни Ньюкасла (NDV).
В одном аспекте настоящего изобретения предусмотрен способ индукции иммунного ответа у животного-птицы на вирус болезни Марека и один или несколько патогенов птиц, включающий стадию введения птице эффективного количества иммуногенной или вакцинной композиции по настоящему изобретению. В одном варианте осуществления способ по настоящему изобретению предусматривает, что один или несколько патогенов птиц выбраны из группы, состоящей из вируса инфекционного бурсита (IBDV); вируса болезни Ньюкасла (NDV); вируса инфекционного бронхита (IBV); вируса инфекционного ларинготрахеита (ILTV); вируса анемии цыплят (CAV); и вируса птичьего гриппа (AIV). В одном варианте осуществления способ по настоящему изобретению предусматривает, что один или несколько патогенов птиц включают вирус инфекционного бурсита (IBDV). В одном варианте осуществления способ по настоящему изобретению предусматривает, что один или несколько патогенов птиц включают вирус болезни Ньюкасла (NDV). В одном варианте осуществления способ по настоящему изобретению предусматривает, что один или несколько патогенов птиц включают вирус инфекционного бурсита (IBDV) и вирус болезни Ньюкасла (NDV). В одном или нескольких вариантах осуществления способ по настоящему изобретению предусматривает, что введение осуществляется путем распыления, введения in ovo, подкожного введения, внутримышечного введения, перорального введения или назального введения. В одном варианте осуществления способ предусматривает введение in ovo. В одном варианте осуществления способ предусматривает, что введение in ovo происходит в яйцеклетку с развивающимся эмбрионом в период между от около 16 до 22 дней развития. В одном или нескольких вариантах осуществления способ предусматривает, что введение in ovo происходит в яйцеклетку с развивающимся эмбрионом в период около 18 дней развития. В одном или нескольких вариантах осуществления способ предусматривает, что путь введения включает введение in ovo с последующим введением спрея. В одном варианте осуществления способ предусматривает, что способ введения включает введение спрея. В одном или нескольких вариантах осуществления способ предусматривает, что птица выбрана из группы, состоящей из курицы, индейки, гуся, утки, фазана, страуса, голубя и перепела. В одном варианте осуществления способ предусматривает, что птица представляет собой курицу.
В одном аспекте настоящего изобретения предусмотрена вакцинная композиция, содержащая рекомбинантный HVT по настоящему изобретению, который содержит нуклеотидную последовательность, кодирующую белок F вируса болезни Ньюкасла, дополнительно содержащая композицию, содержащую аттенуированный вирус инфекционного бурсита и антитело, которое специфически связывается с вирусом инфекционного бурсита. В одном или нескольких вариантах осуществления композиция, содержащая IBDV, представляет собой аттенуированный штамм I2512 BD и содержит вакцину Bursaplex™. В одном или нескольких вариантах осуществления композиция, содержащая IBDV, представляет собой аттенуированный штамм V877 IBD и содержит вакцину Magniplex™.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На Фиг. 1 представлена реакция ПЦР, демонстрирующая надлежащую вставку гена gfp в сайт UL55/ген 3 генома HVT.
На Фиг. 2 представлена реакция ПЦР, демонстрирующая сайт интеграции гена gfp в сайте интеграции UL35/36 генома HVT.
На Фиг. 3А, 3В, 3С представлены реакции ПЦР, демонстрирующие надлежащую интеграцию гена VP2 в геном HVT для HVT IBD 1.
На Фиг. 4А и 4В представлены изображения трансфицированных/инфицированных клеток JBJ-1, окрашенных для IBDV VP2 (панель А) и инфекции HVT (панель В) для HVT IBD5.
На Фиг. 5 представлено изображение реакций ПЦР, осуществляемых для подтверждения надлежащей ориентации вставки VP2 в сайт интеграции UL35/36 генома HVT для HVT IBD 5.
На Фиг. 6 представлен вестерн-блоттинг лизата инфицированных клеток с использованием моноклонального антитела против IBDVR63, демонстрирующий полосу белка около 50 кДа для HVT IBD 6а.
На Фиг. 7А и 7В представлено изображение реакций ПЦР, демонстрирующие надлежащую интеграцию гена VP2 в сайт UL35/36 в геноме HVT для HVT IBD 6а.
На Фиг. 8 представлено изображение реакций ПЦР, демонстрирующие надлежащую интеграцию гена VP2 в сайт UL55/ген 3 в геноме HVT для HVT IBD 9.
На Фиг. 9 представлено изображение реакций ПЦР, демонстрирующие надлежащую интеграцию гена VP2 в сайт UL55/ген 3 в геноме HVT для HVT IBD 30.
На Фиг. 10А и 10В представлены изображения на основе вестерн-блоттинга трансфицированных/инфицированных клеточных лизатов с использованием моноклонального антитела против IBDVR63 для HVT IBD 31.
На Фиг. 11 представлены изображения реакций ПНР, демонстрирующие надлежащую интеграцию гена VP2 в сайт интеграции UL35/36 в геноме HVT для HVT IBD 31.
На Фиг. 12 представлены изображения реакций ПЦР, демонстрирующие надлежащую интеграцию гена VP2 в сайт интеграции ИЬ55/ген 3 в геноме HVT для HVT IBD 34.
На Фиг. 13 представлено графическое изображение IBDV серологических ответов HVT-IBD 1, 5, 9 и 15.
На Фиг. 14 представлено графическое изображение IBDV серологических ответов HVT-IBD 6а, 30, 31.
На Фиг. 15А и 15В представлены изображения реакций ПЦР, демонстрирующих надлежащую ориентацию вставки NDVF для HVT ND №38.
На Фиг. 16А и 16В представлены изображения реакций ПЦР, демонстрирующих надлежащую ориентацию вставки NDVF для HVT ND №39.
На Фиг. 17А, 17В, 17С представлены изображения реакций ПЦР, демонстрирующих надлежащую ориентацию вставки NDV F для HVT ND №40.
На Фиг. 18 представлено изображение нескольких реакций ПЦР, демонстрирующих надлежащую ориентацию вставки NDVF для HVT NDV 42.
На Фиг. 19 представлено изображение реакций ПЦР, демонстрирующих надлежащую ориентацию вставки NDVF для HVT NDV 45.
На Фиг. 20А, 20В, 20С представлено изображение реакций ПЦР, демонстрирующих надлежащую ориентацию вставки NDVF для HVT NDV 46.
На Фиг. 21 представлено линейное изображение генома HVT, обозначающее сайт вставки A, UL35 (HVT043)-UL36 (HVT044).
На Фиг. 22 представлено линейное изображение генома HVT, обозначающее сайт вставки В, UL55 (HVT065)-ген 3 (HVT066).
На Фиг. 23 представлено изображение штамма Faragher IBDV VP2 F52/70.
На Фиг. 24 представлено изображение синтеза плазмиды pHVT-IBD №30.
На Фиг. 25 представлено изображение синтеза плазмиды pHVT-ND №42.
На Фиг. 26 представлено изображение круговой карты pSiteA №30 трансферной плазмиды.
На Фиг. 27 представлено изображение круговой карты pSiteB №42 трансферной плазмиды.
На Фиг. 28 представлено изображение продуцирования промежуточного рекомбинантного HVT-ND №42.
На Фиг. 29 представлено изображение продуцирования HVT-IBD №30-ND №42.
На Фиг. 30 представлено изображение характеристики конструкции HVT-IBD №30-ND №42 на основе ПЦР и расщепления сайта А рестрикционными эндонуклеазами.
На Фиг. 31 представлено изображение характеристики конструкции HVT-IBD №30-ND №42 на основе ПЦР и расщепления сайта В рестрикционными эндонуклеазами.
На Фиг. 32 представлено изображение вестерн-блоттинга экспрессии целевого белка HVT-IBD №30-ND №42 IBD VP2.
На Фиг. 33 представлено изображение вестерн-блоттинга экспрессии целевого белка HVT-IBD №30-ND №42 NDV F.
КРАТКОЕ ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ







ПОДРОБНОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Ниже приводится подробное описание, предусмотренное для содействия специалистам в данной области техники. Специалисты в данной области техники могут выполнять модификации и вариации в вариантах осуществления, описанных в данном документе, без отклонения от идеи или объема настоящего изобретения.
Настоящее изобретение связано с вакцинами для применения у птиц на основе живых рекомбинантных вирусов герпеса птиц, а именно, в частности, вируса болезни Марека (MDV) и более конкретно на основе вируса HVT (вируса герпеса индеек), в который была вставлена одна или несколько нуклеотидных последовательностей, кодирующих и экспрессирующих антигенный полипептид патогенного агента в условиях, обеспечивающих иммунизацию, приводящую к эффективной защите вакцинированного животного от указанного патогенного агента или агентов. Болезнь Марека (MD) представляет собой распространенное лимфопролиферативное заболевание кур, вызываемое вирусом болезни Марека (MDV), которое может привести к значительным потерям в птицеводстве. В настоящее время MD контролируется у домашней птицы с помощью вакцин с использованием серотипа 3 MDV, который является родственным вирусом герпеса индеек (HVT). В результате введения генов вирусов домашней птицы, отличных от MDV, в геном HVT в определенных генетических положениях, авторы настоящего изобретения смогли разработать новые вакцины на основе рекомбинантных вирусов, которые обеспечивают одновременную защиту домашней птицы от MD и одного или нескольких дополнительных заболеваний посредством введения одной вирусной вакцины.
В настоящем изобретении предусмотрены векторы на основе рекомбинантных вирусов для вставки и экспрессии чужеродных генов для применения в безопасных иммунизациях для защиты птиц от различных патогенов. В настоящем изобретении также предусмотрены поливалентные композиции или вакцины, содержащие один или несколько векторов на основе рекомбинантного вируса HVT для защиты от различных патогенов. Кроме того, в настоящем изобретении также предусмотрены способы получения и применения векторов на основе рекомбинантных вирусов отдельно или в комбинации с другими вакцинами или фармацевтическими композициями.
В одном аспекте в настоящем изобретении предусмотрен геном на основе рекомбинантного вируса герпеса индеек (HVT), содержащий одну или несколько нуклеотидных последовательностей, кодирующих один или несколько гетерологичных антигенов, вставленных в межгенные локусы UL 35/UL 36 в уникальной длинной (UL) области генома HVT.
В одном аспекте в настоящем изобретении предусмотрен геном рекомбинантного вируса герпеса индеек (HVT), содержащий одну или несколько нуклеотидных последовательностей, кодирующих один или несколько гетерологичных антигенов, вставленных в межгенные локусы UL 35/UL 36 в уникальной длинной области генома HVT, и одну или несколько нуклеотидных последовательностей, кодирующих один или несколько гетерологичных антигенов, вставленных в сайт UL55/ген 3 уникальной длинной области (UL) генома HVT.
В одном или нескольких вариантах осуществления в настоящем изобретении предусмотрен рекомбинантный HVT, в котором один или несколько гетерологичных антигенов характеризуются защитным действием от патогенов птиц или патогенов, выбранных из группы, состоящей из: вируса инфекционного бурсита (IBDV); вируса болезни Ньюкасла (NDV); вируса инфекционного бронхита (IBV); вируса инфекционного ларинготрахеита (ILTV); вируса анемии цыплят (CAV); и вируса птичьего гриппа (AIV).
В одном или нескольких вариантах осуществления в настоящем изобретении предусмотрен рекомбинантный HVT, в котором один или один или несколько гетерологичных антигенов выбраны из группы, состоящей из белков VP2, VP3 или VP4 вируса инфекционного бурсита (IBDV); белков VP1 или VP2 вируса анемии цыплят (CAV); химерного белка F/HN или белков F, NP, Р, М, HN или L вируса болезни Ньюкасла (NDV); белков S1, S2 или М вируса инфекционного бронхита (IBV); белков gB, gC, gD, gE, gH, gI или gL вируса инфекционного ларинготрахеита (ILTV); и любого из белков НА, NA, NP или М вируса птичьего гриппа (AIV).
В одном или нескольких вариантах осуществления рекомбинантный HVT по настоящему изобретению предусматривает, что один или несколько гетерологичных антигенов характеризуются защитным действием от IBDV. В одном варианте осуществления рекомбинантный HVT по настоящему изобретению предусматривает, что один или несколько гетерологичных антигенов представляют собой белок VP2 из IBDV. В одном варианте осуществления рекомбинантный HVT по настоящему изобретению предусматривает, что белок VP2 кодируется нуклеотидной последовательностью, на по меньшей мере 80% идентичной нуклеотидной последовательности, содержащей SEQ ID NO. 5 или SEQ ID NO. 10. В одном варианте осуществления рекомбинантный HVT по настоящему изобретению предусматривает, что белок VP2 кодируется нуклеотидной последовательностью, содержащей либо SEQ ID NO. 5 или SEQ ID NO. 10.
В одном или нескольких вариантах осуществления рекомбинантный HVT по настоящему изобретению предусматривает, что один или несколько гетерологичных антигенов характеризуются защитным действием от вируса болезни Ньюкасла (NDV). В одном варианте осуществления рекомбинантный HVT по настоящему изобретению предусматривает, что один или несколько гетерологичных антигенов представляют собой белок F NDV. В одном варианте осуществления рекомбинантный HVT по настоящему изобретению предусматривает, что белок F из NDV кодируется нуклеотидной последовательностью, на по меньшей мере 80% идентичной нуклеотидной последовательности, содержащей SEQ ID NO. 3. В одном варианте осуществления рекомбинантный HVT по настоящему изобретению предусматривает, что белок F из NDV кодируется нуклеотидной последовательностью, содержащей SEQ ID NO. 3.
В одном или нескольких вариантах осуществления рекомбинантный HVT по настоящему изобретению предусматривает, что один или несколько гетерологичных антигенов характеризуются защитным действием от NDV и IBDV. В одном или нескольких вариантах осуществления рекомбинантный HVT по настоящему изобретению предусматривает, что по меньшей мере один гетерологичный антиген представляет собой белок F из NDV и белок VP2 из IBDV.
В одном или нескольких вариантах осуществления рекомбинантный HVT по настоящему изобретению предусматривает, что белок F из NDV, кодируемый нуклеотидной последовательностью, на по меньшей мере 80% идентичной нуклеотидной последовательности, содержащей SEQ ID NO. 3, и белок VP2 из IBDV, кодируемый нуклеотидной последовательностью, на по меньшей мере 80% идентичной нуклеотидной последовательности, содержащей SEQ ID NO. 5 или SEQ ID NO. 10.
В одном или нескольких вариантах осуществления рекомбинантный HVT по настоящему изобретению предусматривает белок F из NDV, кодируемый нуклеотидной последовательностью, содержащей SEQ ID NO. 3, а белок VP2 из IBDV кодируется нуклеотидной последовательностью, содержащей SEQ ID NO. 5 или SEQ ID NO. 10.
В одном или нескольких вариантах осуществления рекомбинантный HVT по настоящему изобретению содержит геном, содержащий одну или несколько кассет экспрессии, содержащих одну или несколько нуклеотидных последовательностей, которые кодируют один или несколько гетерологичных антигенов. В одном варианте осуществления рекомбинантный HVT содержит геном на основе рекомбинантного HVT, кассету экспрессии, которая содержит нуклеотидную последовательность, кодирующую промоторы, которые функционально связаны с одним или несколькими нуклеотидами, которые кодируют антигены, подлежащие экспрессии. В одном варианте осуществления антиген, подлежащий экспрессии, содержит белок F из NDV. В одном варианте осуществления антиген, подлежащий экспрессии, содержит белок VP2 из IBDV. В одном варианте осуществления антигены, подлежащие экспрессии, содержат как белок F из NDV, так и белок VP2 из IBDV.
В одном варианте осуществления рекомбинантный HVT по настоящему изобретению предусматривает один или несколько промоторов, выбранных из группы, состоящей из: немедленно раннего промотора цитомегаловируса человека (hCMV); немедленно раннего промотора CMV морской свинки; немедленно раннего промотора CMV мыши; промотора Рес; промотора р-актина кур; промотора SV40; промоторов гликопротеина X вируса псевдобешенства; промотора вируса простого герпеса-1 альфа 4; промоторов гликопротеинов gA, gC, gB, gE или gI вируса болезни Марека; промоторов гликопротеина gB, gE, gI, gD вирусов инфекционного ларинготрахеита; и промотора VP8 вируса герпеса крупного рогатого скота 1/1. В одном варианте осуществления рекомбинантный HVT содержит промотор CMV человека. В одном варианте осуществления рекомбинантный HVT содержит промотор CMV мыши. В одном варианте осуществления рекомбинантный HVT содержит промотор hCMV и mCMV.
В одном или нескольких вариантах осуществления рекомбинантный HVT содержит нуклеотидную последовательность, кодирующую сигнал полиаденилирования (poly А). В одном или нескольких вариантах осуществления рекомбинантный HVT содержит нуклеотидную последовательность, кодирующую сигнал poly А, и выбран из последовательности poly A BGH (SEQ ID NO.6) или последовательности poly A SV40 (SEQ ID NO. 12). В одном варианте осуществления сигнальная последовательность poly А представляет собой сигнальную последовательность poly A BGH. В одном варианте осуществления сигнальная последовательность poly А представляет собой сигнальную последовательность poly A SV40.
В одном аспекте рекомбинантный HVT по настоящему изобретению содержит промотор CMV, функционально связанный с нуклеотидной последовательностью, кодирующей белок VP2 из IBDV, дополнительно содержащий нуклеотидную последовательность, кодирующую сигнальную последовательность полиаденилирования, при этом вся кассета экспрессии на основе VP2 вставлена в некодирующую область генома HVT. В одном варианте осуществления промотор CMV содержит промотор hCMV (SEQ ID NO. 1). В одном варианте осуществления нуклеотидная последовательность, кодирующая белок VP2 IBDV, выбрана из SEQ ID NO.5 или SEQ ID NO. 10. В одном варианте осуществления нуклеотидная последовательность, кодирующая белок VP2, содержит SEQ ID NO. 5. В одном варианте осуществления нуклеотидная последовательность, кодирующая белок VP2, содержит SEQ ID NO. 10. В одном варианте осуществления сигнальная последовательность полиаденилирования содержит SEQ ID NO. 6. В одном варианте осуществления сигнальная последовательность полиаденилирования содержит SEQ ID NO. 12. В одном варианте осуществления промотор, нуклеотидная последовательность, кодирующая белок VP2, и сигнальная последовательность poly А содержат кассету экспрессии. В одном варианте осуществления кассета экспрессии вставлена в геном HVT в сайте UL55/ген 3. В одном варианте осуществления кассета экспрессии вставлена в геном HVT в сайте UL35/36 в геноме. В одном варианте осуществления кассета экспрессии содержит по порядку SEQ ID NO.l, SEQ ID NO.5 или SEQ ID NO. 10 и SEQ ID NO.6, вставленные в геном HVT в сайте UL55/ген 3.
В одном аспекте рекомбинантный HVT по настоящему изобретению содержит промотор CMV, функционально связанный с нуклеотидной последовательностью, кодирующей белок F из NDV, дополнительно содержащий нуклеотидную последовательность, кодирующую сигнальную последовательность полиаденилирования, при этом вся часть кассеты на основе белка F из NDV вставлена в некодирующую область в геноме HVT. В одном варианте осуществления промотор CMV содержит промотор mCMV (SEQ ID NO.2). В одном варианте осуществления нуклеотидная последовательность кодирует белок F из NDV, который содержит SEQ ID NO.3. В одном варианте осуществления сигнальная последовательность полиаденилирования кодируется нуклеотидной последовательностью, содержащей SEQ ID NO. 12. В одном варианте осуществления промотор, нуклеотидная последовательность, кодирующая белок F, и сигнальная последовательность poly А содержат кассету экспрессии. В одном варианте осуществления кассета экспрессии вставлена в геном HVT в сайте UL55/ген 3. В одном варианте осуществления кассета экспрессии вставлена в геном HVT в сайте UL35/36 в геноме. В одном варианте осуществления кассета экспрессии содержит по порядку SEQ ID NO.2, SEQ ID NO.3 и SEQ ID NO. 12, вставленные в геном HVT в сайте UL55/ген 3.
В одном аспекте рекомбинантный HVT по настоящему изобретению содержит промотор CMV, функционально связанный с нуклеотидной последовательностью, кодирующей белок VP2 из IBDV, дополнительно содержащий нуклеотидную последовательность, кодирующую сигнальную последовательность полиаденилирования, при этом все содержат кассету экспрессии VP2, вставленную в некодирующую область в геноме HVT. В одном варианте осуществления рекомбинантный HVT по настоящему изобретению дополнительно содержит промотор CMV, функционально связанный с нуклеотидной последовательностью, кодирующей белок F из NDV, дополнительно содержащий нуклеотидную последовательность, кодирующую сигнальную последовательность полиаденилирования, в виде части кассеты экспрессии на основе F из NDV, вставленной в тот же сайт вставки, что и кассета на основе VP2. В одном варианте осуществления рекомбинантный HVT по настоящему изобретению дополнительно содержит промотор CMV, функционально связанный с нуклеотидной последовательностью, кодирующей белок F из NDV, дополнительно содержащий нуклеотидную последовательность, кодирующую сигнальную последовательность полиаденилирования, в виде части кассеты экспрессии на основе F из NDV, вставленной в другой сайт по сравнению с кассетой на основе VP2.
В одном варианте осуществления рекомбинантный HVT по настоящему изобретению предусматривает, что кассета экспрессии на основе VP2 содержит по порядку нуклеотидную последовательность, кодирующую промотор hCMV (SEQ ID NO. 1), нуклеотидную последовательность, кодирующую VP2 из IBDV (выбранный из SEQ ID NO. 5 или SEQ ID NO. 10), и нуклеотидную последовательность, кодирующую сигнальную последовательность полиаденилирования BGH (SEQ ID NO.6), вставленную в геном HVT в некодирующей области UL35/36, и по порядку нуклеотидную последовательность, кодирующую промотор mCMV (SEQ ID NO.2), нуклеотидную последовательность, кодирующую белок F из NDV (SEQ ID NO.3), и нуклеотидную последовательность, кодирующую сигнальную последовательность полиаденилирования SV40 (SEQ ID NO. 12), вставленную в геном HVT в некодирующей области UL55/ген 3. В одном варианте осуществления рекомбинантный HVT по настоящему изобретению дополнительно содержит нуклеотидную последовательность, кодирующую один или несколько антигенов, выбранных из группы, состоящей из вируса инфекционного бурсита, вируса анемии цыплят, вируса болезни Ньюкасла, вируса инфекционного бронхита, вируса инфекционного ларинготрахеита и вируса птичьего гриппа. В одном варианте осуществления рекомбинантный HVT по настоящему изобретению дополнительно предусматривает промотор, функционально связанный с нуклеотидной последовательностью, кодирующей антигены, выбранные из группы, состоящей из: антигена VP1, VP2, VP3 или VP4 вируса инфекционного бурсита (IBDV); белков VP1 или VP2 вируса анемии цыплят (CAV); химерного белка F/HN или белков F, NP, Р, М, HN или L вируса болезни Ньюкасла (NDV); белков S1, S2 или М вируса инфекционного бронхита (IBV); и любого из белков НА, NA, NP или М вируса птичьего гриппа (AIV).
В одном варианте осуществления рекомбинантный HVT по настоящему изобретению содержит один или несколько антигенов ILT в качестве части кассеты экспрессии, содержащей промотор, функционально связанный с нуклеотидом, кодирующим антиген ILT, и дополнительно содержащий нуклеотидную последовательность, кодирующую сигнальную последовательность полиаденилирования. В одном варианте осуществления рекомбинантный HVT по настоящему изобретению содержит вторую и третью кассету экспрессии, каждая из которых содержит нуклеотидные последовательности, кодирующие промотор, функционально связанный с нуклеотидной последовательностью, кодирующей антигены птиц, выбранные из группы, состоящей из антигена VP1, VP2, VP3 или VP4 вируса инфекционного бурсита (IBDV); белков VP1 или VP2 вируса анемии цыплят (CAV); химерного белка F/HN или белков F, NP, Р, М, HN или L вируса болезни Ньюкасла (NDV); белков S1, S2 или М вируса инфекционного бронхита (IBV); и любого из белков НА, NA, NP или М вируса птичьего гриппа (AIV), и дополнительно содержит нуклеотидную последовательность, кодирующую сигнальную последовательность полиаденилирования.
В одном или нескольких аспектах в настоящем изобретении предусмотрена рекомбинантная ДНК, кодирующая рекомбинантный геном HVT по настоящему изобретению.
В одном или нескольких аспектах в настоящем изобретении предусмотрена иммуногенная композиция, содержащая рекомбинантный HVT по настоящему изобретению и дополнительно содержащая фармацевтически приемлемый носитель, вспомогательное вещество или адъювант.
В одном или нескольких аспектах в настоящем изобретении предусмотрена вакцинная композиция, содержащая рекомбинантный HVT по настоящему изобретению и дополнительно содержащая фармацевтически приемлемый носитель, вспомогательное вещество или адъювант.
В одном варианте осуществления вакцина по настоящему изобретению дополнительно содержит дополнительный вирус болезни Марека (MDV), выбранный из группы, состоящей из: естественно аттенуированного штамма MDV-1 Rispens (CVI-988); или вируса герпеса Gallid 3 штамма SB-1. В одном варианте осуществления вакцина по настоящему изобретению предусматривает, что дополнительный MDV содержит рекомбинантный геном. В одном варианте осуществления вакцина по настоящему изобретению предусматривает, что дополнительный геном рекомбинантного MDV содержит одну или несколько нуклеотидных последовательностей, кодирующих один или несколько гетерологичных антигенов, которые характеризуются защитным действием от одного или нескольких патогенов птиц.
В одном варианте осуществления вакцина по настоящему изобретению предусмотрена для вакцинации птицы против одного или нескольких заболеваний, вызванных одним или несколькими патогенами птиц. В одном или нескольких вариантах осуществления вакцина по настоящему изобретению предусмотрена для применения в защите птицы от клинических симптомов, вызванных одним или несколькими патогенами птиц. В одном или нескольких вариантах осуществления вакцина по настоящему изобретению предусмотрена для применения в защите птицы от клинических симптомов, вызванных вирусом болезни Марека, и клинических симптомов, вызванных одним или несколькими патогенами птиц. В одном или нескольких вариантах осуществления вакцина по настоящему изобретению содержит один или несколько патогенов птиц, выбранных из группы, состоящей из: вируса инфекционного бурсита (IBDV); вируса болезни Ньюкасла (NDV); вируса инфекционного бронхита (IBV); вируса инфекционного ларинготрахеита (ILTV); вируса анемии цыплят (CAV); и вируса птичьего гриппа (AIV). В одном варианте осуществления вакцина по настоящему изобретению предусматривает, что один или несколько патогенов птиц включают вирус болезни Ньюкасла. В одном варианте осуществления вакцина по настоящему изобретению предусматривает, что один или несколько патогенов птиц включают вирус инфекционного бурсита (IBDV). В одном варианте осуществления вакцина по настоящему изобретению предусматривает, что один или несколько патогенов птиц включают вирус болезни Ньюкасла и вирус инфекционного бурсита.
В одном или нескольких вариантах осуществления вакцина по настоящему изобретению предусмотрена для применения в вакцинации птиц, при этом вакцину вводят посредством по меньшей мере одного введения вакцины путем распыления, введения in ovo, подкожного введения, внутримышечного введения, перорального введения, назального введения или их комбинации. В одном варианте осуществления вакцина по настоящему изобретению предусматривает, что вакцину вводят путем введения in ovo. В одном варианте осуществления вакцина по настоящему изобретению предусматривает, что введение in ovo происходит в яйцеклетку с развивающимся эмбрионом в период между от около 16 до 22 дней развития. В одном или нескольких вариантах осуществления вакцина по настоящему изобретению предусматривает, что введение in ovo происходит в яйцеклетку с развивающимся эмбрионом в период около 18 дней развития. В одном варианте осуществления вакцина по настоящему изобретению предусматривает, что введение вакцины включает введение in ovo с последующим введением спрея. В одном варианте осуществления вакцина по настоящему изобретению предусматривает, что введение вакцины включает введение спрея.
В одном аспекте в настоящем изобретении предусмотрен способ вакцинации птицы для лечения или предупреждения болезни Марека и одного или нескольких заболеваний птиц, вызванных одним или несколькими патогенами птиц, включающий стадию введения эффективного количества вакцинной композиции по настоящему изобретению. В одном варианте осуществления способ по настоящему изобретению предусматривает, что один или несколько патогенов птиц выбраны из группы, состоящей из вируса инфекционного бурсита (IBDV); вируса болезни Ньюкасла (NDV); вируса инфекционного бронхита (IBV); вируса инфекционного ларинготрахеита (ILTV); вируса анемии цыплят (CAV); и вируса птичьего гриппа (AIV). В одном варианте осуществления способ по настоящему изобретению предусматривает, что один или несколько патогенов птиц включают вирус инфекционного бурсита (IBDV). В одном варианте осуществления способ по настоящему изобретению предусматривает, что один или несколько патогенов птиц включают вирус болезни Ньюкасла (NDV). В одном варианте осуществления способ по настоящему изобретению предусматривает, что один или несколько патогенов птиц включают вирус инфекционного бурсита (IBDV) и вирус болезни Ньюкасла (NDV).
В одном аспекте настоящего изобретения предусмотрен способ индукции иммунного ответа у животного-птицы на вирус болезни Марека и один или несколько патогенов птиц, включающий стадию введения птице эффективного количества иммуногенной или вакцинной композиции по настоящему изобретению. В одном варианте осуществления способ по настоящему изобретению предусматривает, что один или несколько патогенов птиц выбраны из группы, состоящей из вируса инфекционного бурсита (IBDV); вируса болезни Ньюкасла (NDV); вируса инфекционного бронхита (IBV); вируса инфекционного ларинготрахеита (ILTV); вируса анемии цыплят (CAV); и вируса птичьего гриппа (AIV). В одном варианте осуществления способ по настоящему изобретению предусматривает, что один или несколько патогенов птиц включают вирус инфекционного бурсита (IBDV). В одном варианте осуществления способ по настоящему изобретению предусматривает, что один или несколько патогенов птиц включают вирус болезни Ньюкасла (NDV). В одном варианте осуществления способ по настоящему изобретению предусматривает, что один или несколько патогенов птиц включают вирус инфекционного бурсита (IBDV) и вирус болезни Ньюкасла (NDV). В одном или нескольких вариантах осуществления способ по настоящему изобретению предусматривает, что введение осуществляют путем введения спрея, введения in ovo, подкожного введения, внутримышечного введения, перорального введения или назального введения. В одном варианте осуществления способ предусматривает введение in ovo. В одном варианте осуществления способ предусматривает, что введение in ovo происходит в яйцеклетку с развивающимся эмбрионом в период между от около 16 до 22 дней развития. В одном или нескольких вариантах осуществления способ предусматривает, что введение in ovo происходит в яйцеклетку с развивающимся эмбрионом в период около 18 дней развития. В одном или нескольких вариантах осуществления способ предусматривает, что путь введения включает введение in ovo с последующим введением спрея. В одном варианте осуществления способ предусматривает, что способ введения включает введение спрея. В одном или нескольких вариантах осуществления способ предусматривает, что птица выбрана из группы, состоящей из курицы, индейки, гуся, утки, фазана, страуса, голубя и перепела. В одном варианте осуществления способ предусматривает, что птица представляет собой курицу.
Общие методологии:
Следует понимать, что настоящее изобретение не ограничивается конкретной методологией, протоколами, реагентами и т.д., описанными в данном документе, и, таким образом, может варьироваться. Терминология, используемая в данном документе, предназначена только для целей описания конкретных вариантов осуществления и не предназначена для ограничения объема настоящего изобретения.
Если не указано иное, научные и технические термины, используемые в связи с настоящим изобретением, описанным в данном документе, должны иметь значения, обычно понятные специалистам в данной области техники. Кроме того, если контекстом не требуется иное, термины в единственном числе должны включать формы множественного числа, а термины во множественном числе должны включать форму единственного числа. Как правило, номенклатура, используемая в связи с культивированием клеток и тканей, молекулярной биологией, а также химией белков и олиго- или полинуклеотидов и гибридизацией, и методики, описанные в данном документе, хорошо известны и обычно используются в данной области техники.
Для рекомбинантной ДНК, синтеза олигонуклеотидов, культивирования тканей и трансфекции используются стандартные методики, которые хорошо известны специалистам в данной области техники. Ферментативные реакции и методики очистки осуществляют в соответствии с описаниями производителя или так, как это обычно делается в данной области техники, или как описано в данном документе. Вышеупомянутые методики и процедуры, как правило, осуществляют в соответствии с общепринятыми способами, хорошо известными из уровня техники, и как описано, но без ограничения различными общими и более конкретными ссылками, которые цитируются и обсуждаются в настоящем описании, см., например, Sambrook et al. MOLECULAR CLONING: LAB. MANUAL (3rd ed., Cold Spring Harbor Lab. Press, Cold Spring Harbor, N.Y., 2001), и Ausubel et al. Current Protocols in Molecular Biology (New York: Greene Publishing Association JWiley Interscience), Oligonucleotide Synthesis (M. J. Gait, ed.,1984); Methods in Molecular Biology, Humana Press; Cell Biology: A Laboratory Notebook (J. E. Cellis, ed., 1998) Academic Press; Animal Cell Culture (R. 1. Freshney, ed.1987); Introduction to Cell and Tissue Culture (1. P. Mather and P. E. Roberts, 1998) Plenum Press; Cell and Tissue Culture: Laboratory Procedures (A. Doyle, J. B. Griffiths, and D. G. Newell, eds., 1993-1998) J. Wiley and Sons; Methods in Enzymology (Academic Press, Inc.); Handbook of Experimental Immunology (D. M. Weir and С.C. Blackwell, eds.); Gene Transfer Vectors for Mammalian Cells (J. M. Miller and M. P. Calos, eds., 1987); Current Protocols in Molecular Biology (F. M. Ausubel et al., eds., 1987); PCR: The Polymerase Chain Reaction, (Mullis et al., eds., 1994); Current Protocols in Immunology (E. Coligan et al., eds., 1991); Short Protocols in Molecular Biology (Wiley and Sons, 1999); Immunobiology (C. A. Janeway and P. Travers, 1997); Antibodies (P. Finch, 1997); Antibodies: a practical approach (D. Catty., ed., IRL Press, 1988-1989); Monoclonal antibodies: a practical approach (P. Shepherd and C. Dean, eds., Oxford University Press, 2000); Using antibodies: a laboratory manual (E. Harlow and D. Lane (Cold Spring Harbor Laboratory Press, 1999); The Antibodies (M. Zanetti and J. D. Capra, eds., Harwood Academic Publishers, 1995); и Cancer: Principles and Practice of Oncology (Y. T. DeVita et al., eds., J.B. Lippincott Company, 1993).
За исключением того, что представлено в рабочих примерах, или в случае, если указано иное, все числа, выражающие количества ингредиентов или условия реакций, используемые в данном документе, необходимо понимать как модифицированные во всех случаях с помощью термина «около».
Все указанные патенты и другие публикации непосредственно включены в данный документ посредством ссылки с целью описания и раскрытия, например, методологий, описанных в таких публикациях, которые могут быть применены в связи с настоящим изобретением. Эти публикации предусмотрены исключительно для их раскрытия до даты подачи настоящей заявки.
Определения
Перед подробным описанием настоящего изобретения будут определены несколько терминов, используемых в контексте настоящего изобретения. В дополнение к этим терминам другие термины определяются в другом месте описания по мере необходимости. Если в данном документе явно не указано иное, термины из данной области техники, используемые в настоящем описании, будут иметь значения, признанные в данной области техники.
Следует отметить, что в настоящем изобретении такие термины, как «содержит», «содержал», «содержащий», «заключал», «заключающий», «состоящий», «состоял», «состоящий по сути из», «включает», «включал» и т.п.определяются в соответствии со стандартной практикой патентного права Соединенных Штатов Америки и международной практикой.
Термин «около» используется в данном документе для обозначения того, что значение включает в себя стандартное отклонение ошибки для устройства или способа, используемого для определения значения. Использование термина «или» в формуле изобретения используется для обозначения «и/или», если явно не указано, что он относится только к альтернативам или альтернативы являются взаимоисключающими, хотя настоящее изобретение поддерживает определение, которое относится только к альтернативам и к «и/или». Когда в формуле изобретения не используется в сочетании с закрытой формулировкой или особо не указано иное, формы единственного числа означают «один или несколько». Таким образом, термин «сообщенный трансгеном», например, охватывает один или несколько трансгенов.
Термин «аминокислота» относится к встречающимся в природе и синтетическим аминокислотам, а также к аналогам аминокислот и миметикам аминокислот, которые функционируют аналогично встречающимся в природе аминокислотам. Встречающиеся в природе аминокислоты представляют собой те аминокислоты, которые закодированы генетическим кодом, а также те аминокислоты, которые позже модифицированы, например, гидроксипролин, γ-карбоксиглутамат и О-фосфосерин. Аналоги аминокислот относятся к соединениям, которые характеризуются той же самой основной химической структурой, что и встречающаяся в природе аминокислота, т.е. α-углеродом, связанным с водородом, карбоксильной группой, аминогруппой и R-группой, например, гомосерину, норлейцину, метионину сульфоксиду, метионину метилсульфонию. Такие аналоги содержат модифицированные R-группы (например, норлейцин) или модифицированные пептидные остовы, однако сохраняют ту же самую основную химическую структуру, что и встречающаяся в природе аминокислота. Миметики аминокислот относятся к химическим соединениям, которые имеют структуру, которая отличается от общей химической структуры аминокислоты, однако которые функционируют аналогично встречающейся в природе аминокислоте.
Аминокислоты могут называться в данном документе или по их общеизвестным трехбуквенным символам, или по однобуквенным символам, рекомендованным комиссией по биохимической номенклатуре IUPAC-IUB. Аналогичным образом, нуклеотиды могут быть обозначены по их общепринятым однобуквенным кодам. Макромолекулярные структуры, такие как полипептидные структуры, могут быть описаны с точки зрения различных уровней организации. «Первичная структура» относится к аминокислотной последовательности определенного пептида. «Вторичная структура» относится к локально упорядоченным трехмерным структурам в пределах полипептида. Эти структуры обычно известны как домены, например, ферментативные домены, внеклеточные домены, трансмембранные домены, домены пор или домены цитоплазматического хвоста. Домены представляют собой части полипептида, которые образуют компактную единицу полипептида. Иллюстративные домены включают домены с ферментативной активностью. Домен может состоять из участков меньшей организации, таких как участки β-листа и α-спирали. «Третичная структура» относится к полной трехмерной структуре мономера полипептида. «Четверичная структура» относится к трехмерной структуре, образованной путем нековалентной ассоциации независимых третичных звеньев. Анизотропные термины также известны как энергетические термины.
Используемый в данном документе термин «антитело» относится к полипептиду, содержащему каркасную область гена иммуноглобулина или его фрагментов, которая специфически связывается и распознает антиген.
Общепризнанные гены иммуноглобулинов могут включать гены константных областей каппа, лямбда, альфа, гамма, дельта, эпсилон и мю, а также множество генов вариабельных областей иммуноглобулинов. Легкие цепи могут быть классифицированы либо как каппа, либо как лямбда. Тяжелые цепи могут быть классифицированы как гамма, мю, альфа, дельта или эпсилон, которые, в свою очередь, определяют классы иммуноглобулинов IgY, IgG, IgM, IgA, IgD и IgE соответственно.
Иллюстративная структурная единица иммуноглобулина (антитела) может содержать тетрамер, при этом каждый тетрамер состоит из двух идентичных пар полипептидных цепей, каждая пара содержит одну «легкую» (около 25 кДа) и одну «тяжелую» цепь (около 50-70 кДа). N-конец каждой цепи определяет вариабельную область от около 100 до 110 или более аминокислот, в первую очередь ответственную за распознавание антигенов. Термины вариабельная легкая цепь и вариабельная тяжелая цепь относятся к этим легким и тяжелым цепям. Антитела существуют, например, в виде интактных иммуноглобулинов или в виде нескольких хорошо охарактеризованных фрагментов, полученных при расщеплении различными пептидазами. В то время как различные фрагменты антител определяются с точки зрения расщепления интактного антитела, специалисту в данной области техники будет понятно, что такие фрагменты могут быть синтезированы de novo либо химически, либо с использованием методологии рекомбинантной ДНК. Таким образом, термин «антитело», используемый в данном документе, также включает фрагменты антител, либо полученные путем модификации целых антител, либо синтезированные de novo с использованием методологий рекомбинантной ДНК, либо идентифицированные с использованием других способов, известных из уровня техники.
Для получения антител, например, рекомбинантных, моноклональных или поликлональных антител, можно применять многие способы, известные из уровня техники. Гены, кодирующие тяжелую и легкую цепи антитела, представляющего интерес, можно клонировать из клетки и использовать для получения рекомбинантного моноклонального антитела. Также можно использовать генные библиотеки, кодирующие тяжелые и легкие цепи моноклональных антител. Случайные комбинации продуктов генов тяжелой и легкой цепей создают большой пул антител с различной антигенной специфичностью. Способы получения одноцепочечных антител или рекомбинантных антител известны из уровня техники и могут быть адаптированы для получения антител к полипептидам в соответствии с настоящим изобретением. Технология фагового дисплея также может быть использована для идентификации антител и гетеромерных фрагментов, которые специфически связываются с выбранными антигенами. Антитела также могут быть биспецифическими, т.е. способными распознавать два разных антигена, или представлять собой гетероконъюгаты, например, два ковалентно соединенных антитела или иммунотоксина.
Используемый в данном документе термин «антиген» относится к вирусному белку или полипептиду, такому как вирусный полипептид, а также к вирусным частицам. В некоторых вариантах осуществления антиген в соответствии с настоящим изобретением также может представлять собой вирусную нуклеиновую кислоту. Антиген представляет собой молекулу, которая распознается иммунной системой и способна индуцировать иммунный ответ в организме хозяина. Антиген может представлять собой целый, аттенуированный, убитый или живой организм или субъединицу или часть организма. Он также может представлять собой часть или фрагмент ДНК, полипептид, эпитоп, гаптен или любую их комбинацию, которая может индуцировать иммунный ответ.
Используемый в данном документе термин «птица» включает домашнюю птицу, такую как представители отряда Galliformes. Более конкретно, класс птиц, главным образом представляющих экономический и/или агрономический интерес, таких как куры, индейки, гуси, утки, фазаны, страусы, голуби и перепела и т.п.
Используемый в данном документе термин «биологический образец» или «образец» может включать кровь и части крови, в том числе без ограничения сыворотку крови, плазму крови, тромбоциты или эритроциты; мокроту, клоакальные мазки, слизистую оболочку, ткань, культивированные клетки, в том числе первичные культуры, эксплантаты и трансформированные клетки; биологические жидкости, кал и мочу. Биологический образец может также включать срезы тканей, такие как образцы, отбираемые после биопсии и вскрытия, а также замороженные срезы, взятые для гистологических целей. Биологический образец может быть получен из эукариотического организма, такого как птица, в том числе без ограничения птицы из отряда Galliformes, такой как куры, перепела и индейки. Можно использовать любую ткань, подходящую для применения в соответствии с настоящим изобретением, например, кожу, головной мозг, спинной мозг, надпочечники, грудные мышцы, легкое, сердце, печень, зоб, желудок, желудочек, двенадцатиперстную кишку, тонкий кишечник, толстый кишечник, клоаку, почку, сумку Фабрициуса, селезенку, поджелудочную железу, костный мозг, пояснично-крестцовый отдел спинного мозга или кровь.
Термин «консервативная аминокислотная замена» означает любую аминокислотную замену определенного аминокислотного остатка, где замещающий остаток настолько химически подобен остатку определенного остатка, что не приводит к существенному снижению функции полипептида (например, ферментативной активности). Консервативные аминокислотные замены широко известны из уровня техники, и их примеры описаны, например, в патентах США №№6790639, 6774107, 6194167 или 5350576. В предпочтительном варианте осуществления консервативная аминокислотная замена представляет собой любую замену, которая встречается в одной из следующих шести групп:
• 1. Небольшие алифатические, практически неполярные остатки: Ala, Gly, Pro, Ser и Thr;
• 2. Крупные алифатические, неполярные остатки: Не, Leu и Val; Met;
• 3. Полярные, отрицательно заряженные остатки и их амиды: Asp и Glu;
• 4. Амиды полярных отрицательно заряженных остатков: Asn и Gin; His;
• 5. Полярные, положительно заряженные остатки: Arg и Lys; His; и
• 6. Крупные ароматические остатки: Trp и Tyr; Phe.
В предпочтительном варианте осуществления консервативная аминокислотная замена представляет собой любую из следующих пар, перечисленных в виде пар нативных остатков (консервативные замены): Ala (Ser); Arg (Lys); Asn (Gin; His); Asp (Glu); Gin (Asn); Glu (Asp); Gly (Pro); His (Asn; Gin); He (Leu; Val); Leu (Ile; Val); Lys (Arg; Gin; Glu); Met (Leu; Ile); Phe (Met; Leu; Tyr); Ser (Thr); Thr (Ser); Trp (Tyr); Tyr (Trp; Phe); и Val (He; Leu).
Фраза «функциональные эффекты» в контексте анализов для тестирования соединений, которые модулируют активность вируса, как описано в данном документе, включает определение параметра, на который непосредственно или опосредованно влияет такой вирус, например, фенотипический или химический эффект.«Функциональные эффекты» могут включать активность in vitro, in vivo и ex vivo и могут быть измерены посредством любых способов, известных специалистам в данной области техники, таких как изменения спектроскопических характеристик, формы, хроматографических свойств или свойств растворимости белка, измерение индуцируемых маркеров или транскрипционной активации белка; измерение активности связывания или анализы связывания, т.е. связывание с антителами; измерение изменений активности связывания лиганда или субстрата, измерение вирусной репликации, измерение экспрессии маркера клеточной поверхности, измерение изменений уровней белка, измерение стабильности РНК, идентификацию экспрессии нижерасположенного или репортерного гена посредством, например, хемилюминесценции, флуоресценции, колориметрических реакций, связывания антител и/или индуцируемых маркеров.
Термин «ген» относится к компонентам, которые включают вирусную ДНК или РНК, cDNA, ДНК вирусных интронов и экзонов, искусственный полинуклеотид вирусной ДНК или другую ДНК, которая кодирует вирусный пептид, вирусный полипептид, вирусный белок или молекулу транскрипта вирусной РНК, и генетические элементы, которые могут фланкировать кодирующую последовательность, которые участвуют в регуляции экспрессии, такие как промоторные области, 5'-лидерные области, 3'-нетранслируемая область, которые могут существовать в виде нативных генов или трансгенов в вирусном геноме. Ген или его фрагмент можно подвергать способам секвенирования полинуклеотидов, которые определяют порядок нуклеотидов, которые составляют ген.
Термин «вирус герпеса индейки (HVT)» определяется как непатогенный вирус домашних индеек и классифицируется в качестве третьего серотипа в группе вирусов болезни Марека антигенно и генетически родственных лимфотропных вирусов герпеса птиц.
Термин «гетерологичный» при использовании в отношении частей нуклеиновой кислоты, указывает, что нуклеиновая кислота содержит две или более последовательностей, которые не встречаются в том же самом соотношении друг по отношению к другу в природе. Например, нуклеиновую кислоту, как правило, получают рекомбинантным путем, имея две или более последовательностей из неродственных генов, организованных таким образом, чтобы получить новую функциональную нуклеиновую кислоту, например, промотор из одного источника и кодирующую область из другого источника. Аналогично, гетерологичный белок указывает на то, что белок содержит две или более субпоследовательности, которые не обнаруживаются в одинаковом отношении друг по отношению к другу в природе (например, слитый белок). Гетерологичный может также относиться к вирусной последовательности, такой как ген или трансген, или их части, вставленной в вирусный геном, в котором она обычно не обнаруживается, или к гену, введенному в организм, в котором он обычно не обнаруживается.
Термин «клетка-хозяин» означает любую клетку любого организма, которая отобрана, модифицирована, трансформирована, выращена или использована или обработана каким-либо образом для продуцирования клеткой вещества, например, экспрессии клеткой гена, последовательности ДНК или РНК, белка или фермента. Предполагается, что клетка-хозяин включает любую отдельную клетку или клеточную культуру, которая может представлять собой или представляла собой реципиента для векторов или для включения экзогенных молекул нуклеиновой кислоты, полинуклеотидов и/или белков. Он также предназначен для включения потомства одной клетки. Потомство не обязательно может быть полностью идентичным (по морфологии или по составу геномной или общей ДНК) исходной родительской клетке в связи с естественной, случайной или преднамеренной мутацией. Клетки могут быть прокариотическими или эукариотическими.
Используемый в данном документе термин «хозяин», «субъект», «пациент» или «организм» может включать животных, в частности, птиц, особенно домашнюю птицу. Для ветеринарных вариантов применения птицы могут принадлежать к отряду Galliformes, который включает кур, перепелов, индеек и т.п.Термин «живой хозяин» относится к хозяину, как указано выше, или к другому организму, который является живым. Этот термин может также относиться ко всему хозяину или организму, а не только к части, оперативно удаленной (например, головному мозгу или другому органу) из живого хозяина. Эти термины также включают человека на всех стадиях развития, в том числе стадии эмбриона и плода.
Термины «идентичный» или «процент идентичности» в контексте двух или более последовательностей нуклеиновых кислот или полипептидов относятся к двум или более последовательностям или подпоследовательностям, которые являются одинаковыми или характеризуются определенным процентом аминокислотных остатков или нуклеотидов, которые являются одинаковыми (т.е. около 60% идентичность, предпочтительно 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более высокая идентичность по указанной области при сравнении и выравнивании для максимального соответствия по окну сравнения или обозначенной области) при измерении с применением алгоритмов сравнения последовательностей BLAST или BLAST 2.0 с параметрами по умолчанию, описанными ниже, или посредством выравнивания вручную и визуального осмотра (см., например, веб-сайт NCBI по адресу ncbi.nlm.nih.gov/BLAST/ или аналогичный). Такие последовательности затем называют «по сути идентичными». Это определение также относится или применяется к комплименту определенной последовательности. Определение может также включать последовательности, которые содержат делеции, добавления и/или замены.
При сравнении последовательностей одна последовательность, как правило, выступает в качестве референтной последовательности, с которой сравнивают другие последовательности. При использовании алгоритма сравнения последовательностей референтные последовательности и последовательности сравнения могут быть введены в компьютер, а параметры программы алгоритма последовательности выбраны по желанию. Затем генерируют процент идентичности последовательностей для последовательностей сравнения по отношению к референтной последовательности на основе выбранных параметров. Примером алгоритма, который может быть подходящим для определения процента идентичности последовательностей и сходства последовательностей, являются алгоритмы BLAST и BLAST 2.0, которые описаны в Altschul et al., (Nuc Acids Res 25:3389-3402, 1977) и Altschul et al, (JMol Biol 215:403-410, 1990) соответственно. BLAST и BLAST 2.0 хорошо известны из уровня техники и могут применяться для определения процента идентичности последовательностей любых нуклеиновых кислот или белков, таких как те, которые описаны в данном документе.
Используемые в данном документе термины «иммуногенная композиция», или «фармацевтическая композиция», или «вакцина» предполагают охватывать композицию, содержащую антиген, подходящий для введения субъекту, такому как субъект-птица. Указанная композиция обычно предназначена для индукции иммунного ответа у субъекта. Иммунный ответ может включать Т-клеточный ответ, В-клеточный ответ или как Т-клеточный, так и В-клеточный ответ.Композиция может выступать для сенсибилизации субъекта посредством презентации антигена в ассоциации с молекулами МНС на поверхности клетки. Кроме того, можно получить антигенспецифические Т-лимфоциты или антитела для обеспечения последующей защиты иммунизированного хозяина. «Иммуногенная композиция» может содержать живую, аттенуированную или убитую/инактивированную вакцину, содержащую целый микроорганизм или полученную из него иммуногенную часть, которая индуцирует либо клеточно-опосредованный (Т-клеточный) иммунный ответ, либо антителоопосредованный (В-клеточный) иммунный ответ, либо или то и другое, и может защитить животное от одного или нескольких симптомов, ассоциированных с инфекцией микроорганизмом, или может защитить животное от летального исхода в связи с инфекцией микроорганизмом. Как правило, «иммуногенная композиция» является стерильной и предпочтительно не содержит примесей, которые могут индуцировать нежелательный ответ у субъекта (например, соединение (соединения) в иммуногенной композиции характеризуется (характеризуются) фармацевтической чистотой). Иммуногенные композиции могут быть разработаны для введения субъектам, нуждающимся в этом, посредством ряда различных путей введения, в том числе in ovo, перорального, внутривенного, буккального, ректального, парентерального, внутрибрюшинного, внутрикожного, интратекального, внутримышечного, подкожного, ингаляционного и т.п.
Термин «иммуногенный» белок или пептид, используемый в данном документе, включает полипептиды, которые являются иммунологически активными в том смысле, что после введения хозяину они способны индуцировать иммунный ответ гуморального и/или клеточного типа, направленный против белка. Предпочтительно белковый фрагмент характеризуется такой же иммунологической активностью, что и полноразмерный белок. Таким образом, белковый фрагмент в соответствии с настоящим изобретением содержит по меньшей мере один эпитоп или антигенную детерминанту или по сути состоит из или состоит из них. Термин «иммуногенный» белок или полипептид, используемый в данном документе, включает полноразмерную последовательность белка, его аналоги или его иммуногенные фрагменты. Под «иммуногенным фрагментом» понимают фрагмент белка, который включает один или несколько эпитопов и, таким образом, индуцирует иммунологический ответ, описанный выше.
Термин «иммуногенный белок или пептид» дополнительно предполагает делеции, добавления и замены в последовательности до тех пор, пока полипептид функционирует, приводя к иммунологическому ответу, как определено в данном документе. Термин «консервативная вариация» означает замену аминокислотного остатка другим биологически подобным остатком или замену нуклеотида в последовательности нуклеиновой кислоты таким образом, что кодируемый аминокислотный остаток не изменяется или представляет собой другой биологически подобный остаток. В этом отношении особенно предпочтительные замены, как правило, будут носить консервативный характер, т.е. те замены, которые имеют место в семействе аминокислот. Например, аминокислоты обычно делятся на четыре семейства: (1) кислые - аспартат и глутамат; (2) основные - лизин, аргинин, гистидин; (3) неполярные аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан; и (4) незаряженные полярные глицин, аспарагин, глутамин, цистеин, серии, треонин, тирозин. Фенилаланин, триптофан и тирозин иногда классифицируют как ароматические аминокислоты. Примеры консервативных вариаций включают замену одного гидрофобного остатка, такого как изолейцин, валин, лейцин или метионин, на другой гидрофобный остаток, или замену одного полярного остатка на другой полярный остаток, например, замену аргинина на лизин, глутаминовой кислоты на аспарагиновую кислоту или глутамин на аспарагин и т.п.; или подобную консервативную замену аминокислоты структурно родственной аминокислотой, которая не будет оказывать существенного влияния на биологическую активность. Таким образом, белки, содержащие по сути такую же аминокислотную последовательность, как референтная молекула, но характеризующиеся минорными аминокислотными заменами, которые не оказывают существенного влияния на иммуногенность белка, подпадают под определение референтного полипептида. В данный документ включены все полипептиды, полученные посредством этих модификаций. Термин «консервативная вариация» также включает применение замещенной аминокислоты вместо незамещенной исходной аминокислоты при условии, что антитела, индуцированные замещенным полипептидом, также вступают в иммунологическую реакцию с незамещенным полипептидом.
Термин «иммунологически эффективное количество», используемый в данном документе, относится к количеству антигена или вакцины, достаточному для того, чтобы индуцировать иммунный ответ, либо клеточный (Т-клеточный), либо гуморальный (В-клеточный или антителогенез) ответ, измеряемый посредством стандартных анализов, известных специалисту в данной области техники. Например, в отношении настоящего изобретения термин «иммунологически эффективное количество» представляет собой минимальную защитную дозу (титр). Эффективность антигена в качестве иммуногена можно измерить либо посредством анализов пролиферации, либо посредством цитолитических анализов, таких как анализы высвобождения хрома, для измерения способности Т-клеток лизировать свою специфическую целевую клетку, либо посредством измерения уровней активности В-клеток посредством измерения уровней циркулирующих антител, специфичных к антигену, в сыворотке крови, или посредством других анализов, которые известны и используются специалистами в данной области техники. Кроме того, уровень защиты иммунного ответа может быть измерен посредством воздействия на иммунизированного хозяина антигена, который был инъецирован. Например, если антиген, к которому требуется иммунный ответ, является вирусом или опухолевой клеткой, уровень защиты, индуцируемый «иммунологически эффективным количеством» антигена, измеряется посредством определения процента выживаемости или процента смертности после воздействия вируса или опухолевых клеток у животных.
Специалист в данной области техники может определить, какое количество вакцины в соответствии с настоящим изобретением является иммунологически эффективным, например, посредством мониторинга иммунологического ответа после вакцинации или после воздействия инфекции, например, посредством повторного выделения возбудителя или посредством наблюдения за клиническими признаками заболевания или серологическими параметрами у мишеней и их сравнения с ответами, наблюдаемыми у вакцинированных имитационным контролем животных. Схема введения доз для применения вакцины по настоящему изобретению к целевому организму может быть представлена в виде однократной или многократной доз, которые можно вводить одновременно или последовательно, способом, совместимым с составом вакцины, и в таком количестве, которое будет иммунологически эффективным.
Термины «ингибиторы», активаторы и «модуляторы» последовательностей вирусных нуклеиновых кислот и полипептидов используются для обозначения активирующих, ингибирующих или модулирующих молекул, идентифицированных с использованием анализов последовательностей вирусных нуклеиновых кислот и полипептидов in vitro и in vivo. Ингибиторы представляют собой соединения, которые могут связываться, частично или полностью блокировать активность, снижать, предупреждать, задерживать активацию, инактивировать, снижать чувствительность или подавлять активность или экспрессию вируса. Активаторы относятся к соединениям, которые повышают, открывают, активируют, облегчают, усиливают активацию, сенсибилизируют, агонизируют или усиливают активность вируса. Ингибиторы, активаторы или модуляторы также включают генетически модифицированные версии вируса, как описано в данном документе, например, варианты с измененной активностью, а также встречающиеся в природе и синтетические лиганды, субстраты, антагонисты, агонисты, антитела, пептиды, циклические пептиды, нуклеиновые кислоты, антисмысловые молекулы, рибозимы, малые химические молекулы и т.п. Анализы в отношении ингибиторов и активаторов включают, например, экспрессию вируса по настоящему изобретению in vitro, в клетках или клеточных мембранах, применение предполагаемых соединений-модуляторов и затем определение функциональных эффектов в отношении активности, как описано в данном документе.
Тестируемые образцы или анализы, содержащие вирус по настоящему изобретению, которые обработанные потенциальным активатором, ингибитором или модулятором, можно сравнить с контрольным образцом, не содержащим ингибитора, активатора или модулятора, для определения степени ингибирования. Контрольным образцам, с которыми сравнивается тестируемый образец или анализ, может быть присвоено значение относительной активности белка, равное 100%. Ингибирование вируса достигается при значении активности тестируемого образца по отношению к контрольному образцу менее около 80%, в том числе около 75%, около 70%, около 65%, около 60%, около 55%, около 50%, около 45%, около 40%, около 35%, около 30%, около 25%, около 20%, около 15%, около 10%, около 5% и около 0%.
Межгенные локусы, как описано в данном документе, определяются как область последовательности ДНК, расположенная между генами, в том числе нетранслируемые области, 5'- и 3'-фланкирующие области, интроны и т.д. Межгенные области являются частью некодирующей ДНК, которая может содержать элементы контроля генов, такие как как промоторы и энхансеры. Термин «выделенный» означает вещество, которое было значительно отделено от других веществ, с которыми оно встречается в природе, или обогащено по сравнению с ними. Выделенные вещества обычно характеризуются чистотой, составляющей по меньшей мере около 80%, по меньшей мере 90%, по меньшей мере 98% или по меньшей мере около 99% по массе.
«Метка» или «определяемый фрагмент» представляет собой композицию, обнаруживаемую посредством спектроскопических, фотохимических, биохимических, иммунохимических, химических или других физических средств. Например, применимые метки включают 32Р, флуоресцентные красители, электронно-плотные реагенты, ферменты (например, те, которые обычно используются в ELISA), биотин, дигоксигенин или гаптены и белки, которые можно сделать обнаружимыми, например, посредством включения радиоактивной метки в пептид или использования для обнаружения антител, специфически реагирующих с пептидом.
Используемый в данном документе термин «вирус болезни Марека» или «MDV» относится к любому альфагерпесвирусу рода Mardivirus, который включает вирус герпеса индеек (HVT), как описано в данном документе. В конкретном варианте осуществления настоящее изобретение связано с вирусом болезни Марека, его генетическим компонентам, генами и белкам, продуцируемыми ими. Как используется в данном документе, такой вирус может включать генетические компоненты вируса, т.е. геном и его транскрипты, белки, кодируемые геномом (в том числе структурные и неструктурные белки), и функциональные или нефункциональные вирусные частицы. Полинуклеотидные и полипептидные последовательности, кодирующие такие вирусы, хорошо известны из уровня техники и могут быть легко обнаружены специалистом в данной области техники.
Термины «мутант» и «мутация» означают любое обнаружимое изменение в генетическом материале, например, ДНК или любой процесс, механизм или результат такого изменения. Они включают генные мутации, при которых структура (например, последовательность ДНК) гена изменена, любой ген или ДНК, возникшие в результате любого процесса мутации, и любой продукт экспрессии (например, белок или фермент), экспрессируемый модифицированным геном или последовательностью ДНК. Термин «вариант» может также использоваться для обозначения модифицированного или измененного гена, последовательности ДНК, фермента, клетки и т.д., т.е. любого вида мутанта.
Используемый в данном документе термин «нуклеиновая кислота» относится к одноцепочечному или двухцепочечному полимеру дезоксирибонуклеотидных оснований или рибонуклеотидных оснований, прочитанных от 5'-конца к 3'-концу. «Нуклеиновая кислота» может также необязательно содержать не встречающиеся в природе или измененные нуклеотидные основания, которые обеспечивают надлежащее считывание полимеразой и не снижают экспрессию полипептида, кодируемого этой нуклеиновой кислотой. Термин «нуклеотидная последовательность» или «последовательность нуклеиновой кислоты» относится как к смысловым, так и к антисмысловым цепям нуклеиновой кислоты либо в виде отдельных одиночных цепей, либо в виде дуплекса. Термин «рибонуклеиновая кислота» (РНК) включает RNAi (ингибиторную РНК), dsRNA (двухцепочечную РНК), siRNA (малую интерферирующую РНК), mRNA (информационную РНК), miRNA (микроРНК), tRNA (транспортную РНК, заряженную соответствующей ацилированной аминокислотой или освобожденную от нее) и cRNA (комплементарную РНК). Термины «сегмент нуклеиновой кислоты», «сегмент нуклеотидной последовательности» или в более общем смысле «сегмент» будут пониматься специалистами в данной области техники в качестве функционального термина, который включает геномные последовательности, последовательности рибосомной РНК, последовательности транспортной РНК, последовательности информационной РНК, последовательности оперонов и более мелкие сконструированные нуклеотидные последовательности, которые экспрессируют или могут быть адаптированы для экспрессии белков, полипептидов или пептидов. В данном документе используется номенклатура, требуемая разделом 37 Свода федеральных правил США §1.822 и изложенная в таблицах WIPO Standard ST.25 (1998 г. ), Приложение 2, Таблицы 1 и 3.
Термин «функционально связанный» используется в данном документе для обозначения расположения фланкирующих последовательностей, при этом фланкирующие последовательности, описанные таким образом, сконфигурированы или собраны для выполнения своей обычной функции. Таким образом, фланкирующая последовательность, функционально связанная с кодирующей последовательностью, может быть способна осуществлять репликацию, транскрипцию и/или трансляцию кодирующей последовательности. Например, промотор или энхансер функционально связаны с кодирующей последовательностью, если они влияют на транскрипцию последовательности. Фланкирующая последовательность не обязательно должна быть смежной с кодирующей последовательностью, если она функционирует надлежащим образом. Таким образом, например, между промоторной последовательностью и кодирующей последовательностью могут присутствовать промежуточные нетранслируемые, но транскрибируемые последовательности, и промоторная последовательность все еще может считаться «функционально связанной» с кодирующей последовательностью.
Термин «фармацевтически приемлемый носитель» относится к ингредиенту в фармацевтическом составе, отличном от активного ингредиента, который физиологически совместим для введения субъекту. Фармацевтически приемлемый носитель включает без ограничения буфер, вспомогательное вещество, стабилизатор, адъювант, консервант, разбавитель, водный или неводный носитель и другие добавки. Кроме того, этот термин относится к элементу иммуногенной композиции или вакцины, который обычно одобрен регулирующим органом федерального правительства, правительства штата или другого регулирующего органа, или указан в Фармакопее США или другой общепризнанной фармакопее для применения как у человека, так и у отличных от человека животных. Такие фармацевтические носители могут представлять собой стерильные жидкости, такие как вода, и масла, в том числе нефтяного, животного, растительного или синтетического происхождения, такие как арахисовое масло, соевое масло, минеральное масло, кунжутное масло и т.п. Вода является предпочтительным носителем, если фармацевтическая композиция вводится внутривенно. В качестве жидких носителей также могут использоваться физиологический раствор и водные растворы декстрозы и глицерина, особенно для инъекционных растворов. Подходящие фармацевтические вспомогательные вещества включают крахмал, глюкозу, лактозу, сахарозу, желатин, солод, рис, муку, мел, силикагель, натрия стеарат, глицеролмоностеарат, тальк, натрия хлорид, сухое обезжиренное молоко, глицерол, пропиленгликоль, воду, этанол и т.п. При необходимости композиция также может содержать незначительные количества увлажняющих или эмульгирующих средств, или рН-буферных средств. Эти композиции могут принимать форму растворов, суспензий, эмульсии, таблеток, пилюль, капсул, порошков, составов с замедленным высвобождением и т.п.Композицию также можно составить в виде суппозитория, с традиционными связывающими средствами и носителями, такими как триглицериды. Состав для перорального применения может включать стандартные носители, такие как маннит, лактозу, крахмал, стеарат магния, сахарин натрия, целлюлозу, карбонат магния фармацевтической степени чистоты и т.д. Примеры подходящих фармацевтических носителей описаны в "Remington's Pharmaceutical Sciences" E.W. Martin. Состав должен соответствовать способу введения.
Используемый в данном документе термин «домашняя птица» относится к домашней или коммерческой птице, содержащейся для производства яиц, а также их мяса и перьев. В некоторых вариантах осуществления домашняя птица может включать птиц из отряда Galliformes, который включает кур, перепелов и индеек, а также может включать гусей, уток, лебедей, цесарок, голубей и т.п.
Полинуклеотиды, описанные в данном документе, могут быть комплементарны всей или части последовательности вирусного гена, в том числе промотору, интрону, кодирующей последовательности, экзону, 5'-нетранслируемой области и З'-нетранслируемой области.
Определенная последовательность нуклеиновой кислоты может также включать «сплайс-варианты». Аналогично, конкретный белок, кодируемый нуклеиновой кислотой, неявно охватывает любой белок, кодируемый сплайс-вариантом этой нуклеиновой кислоты. Сплайс-варианты являются продуктами альтернативного сплайсинга гена. После транскрипции исходный транскрипт нуклеиновой кислоты может быть сплайсирован таким образом, что разные (альтернативные) продукты сплайсинга нуклеиновых кислот кодируют разные полипептиды. Механизмы получения сплайс-вариантов различаются, но включают альтернативный сплайсинг экзонов. Альтернативные полипептиды, полученные из одной и той же нуклеиновой кислоты посредством сквозной транскрипции, также охватываются настоящим определением. Любые продукты реакции сплайсинга, в том числе рекомбинантные формы продуктов сплайсинга, включены в настоящее определение.
Термины «полипептид», «пептид» и «белок» используются в данном документе взаимозаменяемо для обозначения полимера из аминокислотных остатков. Термины применимы к полимерам из аминокислот, в которых один или несколько аминокислотных остатков представляют собой искусственный химический миметик соответствующей встречающейся в природе аминокислоты, а также к полимерам из встречающихся в природе аминокислот и полимеру из не встречающихся в природе аминокислот.
Термины «поливалентная вакцина», «комбинированная или комбо-вакцина» и «поливалентная вакцина» используются взаимозаменяемо для обозначения вакцины, содержащей более одного антигена. Поливалентная вакцина может содержать два, три, четыре или более антигенов. Поливалентная вакцина может содержать векторы на основе рекомбинантных вирусов, активные, аттенуированные или убитые вирусы дикого типа или смесь векторов на основе рекомбинантных вирусов и вирусов дикого типа в активной, аттенуированной или убитой формах.
Термин «промоторы», используемые в данном документе, относятся к последовательностям ДНК, которые определяют положение, где начинается транскрипция гена с помощью РНК-полимеразы. Промоторы обычно располагаются выше сайта инициации транскрипции. Промотор также может содержать дистальные энхансерные или репрессорные элементы, которые могут располагаться на расстоянии до нескольких тысяч нуклеотидов от сайта начала транскрипции. Промоторы определяют направление транскрипции и указывают, какая цепь ДНК будет транскрибироваться. Промотор может быть получен из источников, в том числе, вирусов, бактерий, грибов, растений, насекомых и животных. Промотор может регулировать экспрессию генного компонента конститутивно или дифференциально по отношению к клетке, ткани или органу, в котором происходит экспрессия или по отношению к стадии развития, на которой происходит экспрессия, или в ответ на внешние раздражители, такие как физиологические стрессы, патогены, ионы металлов или индуцирующие средства. Типичные примеры промоторов включают промотор бактериофага Т7, промотор бактериофага Т3, промотор SP6, промотор-оператор lac, промотор tac, промотор RSV-LTR, промотор CMV IE, промотор CMV человека; промотор CMV мыши; промотор Рес; промотор р-актина кур; промотор CMV морской свинки, промотор вируса псевдобешенства; промотор гликопротеина X, промотор вируса простого герпеса 1; промотор вируса болезни Марека; и промотор SV40.
Используемые в данном документе термины «профилактически лечить» или «профилактически осуществлять лечение» относятся к полному или частичному предупреждению заболевания или его симптома и/или могут быть терапевтическими в смысле частичного или полного излечения заболевания и/или нежелательного эффекта, связанного с заболеванием.
Термин «рекомбинантный» при использовании со ссылкой, например, на клетку или нуклеиновую кислоту, белок или вектор, указывает, что клетка, нуклеиновая кислота, белок или вектор были модифицированы посредством введения гетерологичной нуклеиновой кислоты или белка или изменения нативной нуклеиновой кислоты или белка, или что клетка получена из клетки, модифицированной таким образом. Так, например, рекомбинантные клетки экспрессируют гены, которые не встречаются в нативной (нерекомбинантной) форме клетки, или экспрессируют нативные гены, которые иным образом экспрессируются аномально, экспрессируются недостаточно или вообще не экспрессируются. В некоторых вариантах осуществления рекомбинантные последовательности могут также включать нуклеиновые кислоты, белки или рекомбинантные геномы, такие как вирусные геномы. Векторы на основе рекомбинантных вирусов, как описано в данном документе, могут содержать трансгены, которые функционально связаны с гетерологичным промотором для осуществления транскрипции трансгена.
Фраза «жесткие условия гибридизации» относится к условиям, при которых зонд будет гибридизоваться со своей целевой последовательностью, обычно в сложной смеси нуклеиновых кислот, но не с другими последовательностями. Жесткие условия могут зависеть от последовательности и будут отличаться в разных обстоятельствах. Более длинные последовательности гибридизируются специфическим образом при более высоких температурах. Жесткие условия могут быть достигнуты при добавлении дестабилизирующих средств, таких как формамид.
Соответствующие жесткие условия, способствующие гибридизации ДНК, хорошо известны специалистам в данной области техники и могут включать, например, 6Х хлорид натрия/цитрат натрия (SSC) при температуре около 45°С с последующей промывкой 2Х SSC при 50°С. Концентрация соли на стадии промывки может быть выбрана от низкой жесткости примерно 2Х SSC при 50°С до высокой жесткости примерно 0,2Х SSC при 50°С. Температура на стадии промывки может быть повышена от условий низкой жесткости при комнатной температуре, около 22°С, до условий высокой жесткости, при около 65°С. Температуру и/или солевые условия можно варьировать в зависимости от обстоятельств для получения оптимальных результатов. В соответствии с настоящим изобретением нуклеиновая кислота может проявлять по меньшей мере от около 80% до около 100% идентичности последовательности с одной или несколькими молекулами нуклеиновой кислоты, как описано в данном документе, например, по меньшей мере от около 80%, около 85%, около 90%, около 95%, около 98%, около 99% или около 100% идентичности последовательности.
Нуклеиновые кислоты, которые не гибридизуются друг с другом в жестких условиях, по-прежнему являются по сути идентичными, если полипептиды, которые они кодируют, являются по сути идентичными. Это происходит, например, когда копия нуклеиновой кислоты создается с использованием максимальной вырожденности кодона, допускаемого генетическим кодом. В таких случаях нуклеиновые кислоты обычно гибридизуются в умеренно жестких условиях гибридизации.
Термин «терапевтически эффективное количество», «эффективное количество» или «терапевтически эффективная доза», используемый в данном документе, относится к дозе, которая вызывает эффект, по поводу которого ее вводят.Такая доза или количество может также относиться к количеству вводимого варианта осуществления средства, которое в некоторой степени облегчит один или несколько симптомов заболевания, т.е. инфекции, которую лечат, и/или тому количеству, которое предупреждает, в некоторой степени, один или несколько симптомов заболевания, т.е. инфекции, которая имеется у получающего лечение хозяина или угрожает риск развития у него. Точная доза будет зависеть от цели лечения, и специалист в данной области техники сможет определить такую дозу с применением методик, известных из уровня техники.
Используемый в данном документе термин «трансген» относится к сегменту ДНК, содержащему гетерологичную кодирующую последовательность или другой генетический материал для введения из одного организма в другой. Например, в некоторых вариантах осуществления трансген по настоящему изобретению может содержать последовательность, кодирующую антиген, такую как вирусный ген, или последовательность, кодирующую вирусный белок.
Используемые в данном документе термины «лечение», «осуществление лечения» и «лечить» определяются как воздействие на заболевание, нарушение или состояние с помощью средства для снижения или улучшения фармакологических и/или физиологических эффектов заболевания, нарушения или состояния и/или их симптомов. Термин «лечение», как используется в данном документе, охватывает любое лечение заболевания у субъекта или хозяина (например, у животного, представляющего ветеринарный интерес), и включает: (а) снижение риска возникновения заболевания у субъекта, который, как установлено, предрасположен к заболеванию, но который еще не диагностирован как инфицированный заболеванием, (b) предупреждение развития заболевания и (с) облегчение заболевания, т.е., путем индукции регрессии заболевания и/или облегчения одного или нескольких симптомов заболевания. Также подразумевается, что термин «лечение» включает доставку ингибирующего средства для обеспечения фармакологического эффекта даже в отсутствие заболевания или состояния. Например, термин «лечение» включает доставку средства, ингибирующего заболевание, или патогена, который обеспечивает усиленные или требуемые эффекты у субъекта (например, снижение патогенной нагрузки, уменьшение симптомов заболевания и т.д.).
Термин «единичная лекарственная форма», используемый в данном документе, относится к физически дискретным единицам, подходящим в качестве разовых доз для субъектов-животных, при этом каждая единица содержит предварительно определенное количество соединения (например, противовирусного соединения, как описано в данном документе), рассчитанное в достаточном количестве для получения требуемого эффекта в сочетании с фармацевтически приемлемым разбавителем, носителем или основой. Спецификации единичных лекарственных форм зависят от конкретного используемого соединения, пути и частоты введения, эффекта, который должен быть достигнут, и фармакодинамики, ассоциированной с каждым соединением у хозяина.
Термины «вакцина» или «вакцинная композиция», используемые в данном документе взаимозаменяемо, относятся к фармацевтическим композициям, содержащим по меньшей мере одну иммуногенную композицию по настоящему изобретению, которая индуцирует иммунный ответ у субъекта. Вакцина или вакцинная композиция могут защитить субъекта от заболевания или возможной смерти и могут содержать или не содержать один или несколько дополнительных компонентов, которые усиливают иммунологическую активность активного компонента. Композиция по настоящему изобретению, которая индуцирует защитный иммунный ответ, содержит рекомбинантный вирус HVT, содержащий один или несколько генов, кодирующих гетерологичный антиген, вставленных в геном HVT в межгенной области UL 35/36. В некоторых вариантах осуществления композиция по настоящему изобретению содержит рекомбинантный вирус HVT, содержащий один или несколько генов, кодирующих гетерологичный антиген, вставленных в геном HVT в UL 35/36, и один или несколько генов, кодирующих антиген, вставленных в геном HVT в UL55. В некоторых вариантах осуществления гены, кодирующие антиген, представляют собой антигены, полученные из патогенов домашней птицы, таких как вирус болезни Ньюкасла, вирус инфекционного бурсита, вирус инфекционного бронхита, вирус птичьего гриппа, вирус инфекционного ларинготрахеита и/или вирус анемии цыплят.В некоторых вариантах осуществления рекомбинантный HVT комбинируют с другой рекомбинантной вакциной против вируса болезни Марека, которая вызывает защитный иммунный ответ у домашней птицы. Вакцина или вакцинная композиция по настоящему изобретению могут дополнительно содержать дополнительные компоненты, типичные для вакцин или вакцинных композиций, в том числе, например, адъювант или иммуномодулятор. Вакцина может содержать один или одновременно несколько элементов, описанных выше.
Вакцина по настоящему изобретению может дополнительно содержать подходящий фармацевтический носитель. Предполагается, что термин «фармацевтически приемлемый носитель» включает любые и все растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые средства, изотонические средства и средства, замедляющие абсорбцию, в отношении хозяев. Термин «носитель» относится к разбавителю, адъюванту, вспомогательному веществу или наполнителю, с которым вводят фармацевтическую композицию. Такие фармацевтические носители могут представлять собой стерильные жидкости, такие как вода, и масла, в том числе нефтяного, животного, растительного или синтетического происхождения, такие как арахисовое масло, соевое масло, минеральное масло, кунжутное масло и т.п. Вода является предпочтительным носителем, если фармацевтическая композиция вводится внутривенно. В качестве жидких носителей также могут использоваться физиологический раствор и водные растворы декстрозы и глицерина, особенно для инъекционных растворов. Подходящие фармацевтические вспомогательные вещества включают крахмал, глюкозу, лактозу, сахарозу, желатин, солод, рис, муку, мел, силикагель, натрия стеарат, глицеролмоностеарат, тальк, натрия хлорид, сухое обезжиренное молоко, глицерол, пропиленгликоль, воду, этанол и т.п. При необходимости композиция также может содержать незначительные количества увлажняющих или эмульгирующих средств, или рН-буферных средств. Эти композиции могут принимать форму растворов, суспензий, эмульсии, таблеток, пилюль, капсул, порошков, составов с замедленным высвобождением и т.п. Композиция может быть составлена с традиционными связывающими веществами и носителями, такими как триглицериды, в зависимости от способа введения. Определенные составы могут включать стандартные носители, такие как маннит, лактозу, крахмал, стеарат магния, сахарин натрия, целлюлозу, карбонат магния фармацевтической степени чистоты и т.д. Примеры подходящих фармацевтических носителей описаны в "Remington's Pharmaceutical Sciences" E.W. Martin. Состав должен соответствовать способу введения. Подходящий носитель очевиден для специалистов в данной области техники и в значительной степени зависит от пути введения. Дополнительные компоненты, которые могут присутствовать в настоящем изобретении, представляют собой адъюванты, консерванты, поверхностно-активные средства, химические стабилизаторы, суспендирующие или диспергирующие средства. Как правило, стабилизаторы, адъюванты и консерванты оптимизируются для определения наилучшего состава для эффективности у целевого субъекта.
«Вариант» пептида относится в данном документе к пептиду, аминокислотная последовательность которого отличается от аминокислотной последовательности «исходного» вакцинного пептида в связи с добавлением, делецией и/или заменой одного или нескольких аминокислотных остатков в исходной пептидной последовательности, и сохраняет по меньшей мере одну требуемую активность исходного вакцинного пептида. Например, вариант может содержать по меньшей мере один, например, от около одной до примерно десяти, а предпочтительно от около двух до около пяти замен в одной или нескольких аминокислотных последовательностях пептида, применяемого в качестве части вакцины по настоящему изобретению. Обычно вариант будет содержать аминокислотную последовательность, характеризующуюся на по меньшей мере 50 % идентичностью аминокислотной последовательности с исходными аминокислотными последовательностями, предпочтительно по меньшей мере 65%, более предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 75%, более предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95% идентичностью последовательности. Идентичность или гомология по отношению к этой последовательности определяется в данном документе в виде процента аминокислотных остатков в кандидатной последовательности, которые идентичны остаткам исходного пептида, после выравнивания последовательностей и введения гэпов, при необходимости, для достижения максимального процента идентичности последовательности. Никакие N-концевые, С-концевые или внутренние удлинения, делеции или вставки в пептидную последовательность не должны рассматриваться как влияющие на идентичность или гомологию последовательности. Вариант сохраняет способность индуцировать иммунный ответ и предпочтительно характеризуется требуемой активностью, которая превосходит активность исходного пептида.
Варианты пептидов могут быть полностью функциональными или могут не выполнять одну или несколько функций. Полностью функциональные варианты обычно содержат только консервативные вариации или вариации в некритических остатках или в некритических областях. Функциональные варианты также могут содержать замену подобных аминокислот, что приводит к отсутствию изменения или незначительному изменению функции. В качестве альтернативы такие замены могут в некоторой степени оказывать положительное или отрицательное влияние на функцию. Нефункциональные варианты, как правило, содержат одну или несколько неконсервативных аминокислотных замен, делеций, вставок, инверсий или усечений, или замену, вставку, инверсию или делецию в критическом остатке или критической области.
Более того, полипептиды часто содержат аминокислоты, отличные от двадцати «встречающихся в природе» аминокислот.Кроме того, многие аминокислоты, в том числе концевые аминокислоты, могут быть модифицированы естественными процессами, такими как процессинг и другие посттрансляционные модификации, или посредствов способов химической модификации, хорошо известных в данной области техники. Известные модификации включают без ограничения ацетилирование, ацилирование, АДФ-рибозилирование, амидирование, ковалентное присоединение флавина, ковалентное присоединение гема, ковалентное присоединение нуклеотида или производного нуклеотида, ковалентное присоединение липида или производного липида, ковалентное присоединение фосфотидилинозитола, сшивание, циклизацию, образование дисульфидной связи, деметилирование, образование ковалентных поперечных связей, образование цистина, образование пироглутамата, формилирование, гамма-карбоксилирование, гликозилирование, образование GPI-якоря, гидроксилирование, йодирование, метилирование, миристоилирование, окисление, протеолитический процессинг, фосфорилирование, пренилирование, рацемизацию, селеноилирование, сульфатирование, опосредованное транспортной РНК добавление аминокислот к белкам, такое как аргинилирование и убиквитинирование. Такие модификации хорошо известны специалистам в данной области техники и очень подробно описаны в научной литературе. Несколько особенно распространенных модификаций, например, гликозилирование, присоединение липидов, сульфатирование, гамма-карбоксилирование остатков глутаминовой кислоты, гидроксилирование и рибозилирование АДФ, описаны в большинстве основных публикаций, таких как Proteins-Structure and Molecular Properties (2nd ed., Т. E. Creighton, W.H. Freeman & Co., NY, 1993). В отношении этого предмета имеется множество подробных обзоров, таких как Wold, Posttranslational Covalent Modification of proteins, 1-12 (Johnson, ed., Academic Press, NY, 1983); Seifter et al. 182 Meth. Enzymol. 626-46 (1990); и Rattan et al. 663 Ann. NY Acad. Sci. 48-62 (1992).
Соответственно, пептиды по настоящему изобретению также охватывают производные или аналоги, в которых замещенный аминокислотный остаток не кодируется генетическим кодом. Подобным образом добавления и замены в аминокислотной последовательности, а также только что описанные вариации и модификации могут быть в равной степени применимы к аминокислотной последовательности антигена и/или его эпитопа или пептидов и, таким образом, охватываются настоящим изобретением.
«Вариант» нуклеиновой кислоты относится в данном документе к молекуле, которая отличается по последовательности от «исходной» нуклеиновой кислоты. Дивергенция полинуклеотидной последовательности может быть результатом мутационных изменений, таких как делеции, замены или добавления одного или нескольких нуклеотидов. Каждое из этих изменений может происходить отдельно или в комбинации, один или несколько раз в определенной последовательности.
Точно так же, как полипептид может содержать консервативную (консервативные) аминокислотную (аминокислотные) замену (замены), его полинуклеотид может содержать консервативную (консервативные) замену (замены) кодона. Замена кодона считается консервативной, если при экспрессии она приводит к консервативной аминокислотной замене, как описано выше. Замена вырожденного кодона, которая не приводит к аминокислотной замене, также применима в полинуклеотидах по настоящему изобретению. Таким образом, например, полинуклеотид, кодирующий выбранный полипептид, применимый в варианте осуществления настоящего изобретения, может быть мутирован посредством замены вырожденного кодона, чтобы приблизиться к частоте использования кодона, демонстрируемой экспрессирующей клеткой-хозяином, которая будет трансформирована ею, или иным образом улучшить его экспрессию.
Используемый в данном документе термин «вектор» означает конструкцию, которая способна доставлять и предпочтительно экспрессировать один или несколько генов или последовательностей, представляющих интерес, в клетке-хозяине. Примеры векторов включают без ограничения вирусные векторы, векторы экспрессии «голой» ДНК или РНК, плазмидные, космидные или фаговые векторы, векторы экспрессии ДНК или РНК, ассоциированные с катионными конденсирующими средствами, векторы экспрессии ДНК или РНК, инкапсулированные в липосомы, и определенные эукариотические клетки, такие как клетки-продуценты. Векторы, как описано в данном документе, содержат последовательности контроля экспрессии, что означает последовательность нуклеиновой кислоты, которая управляет транскрипцией нуклеиновой кислоты. Последовательность контроля экспрессии может представлять собой промотор, такой как конститутивный или индуцируемый промотор, или энхансер. Последовательность контроля экспрессии «функционально связана» с последовательностью нуклеиновой кислоты, подлежащей транскрибированию. Нуклеиновая кислота является «функционально связанной», когда она помещена в функциональную взаимосвязь с другой последовательностью нуклеиновой кислоты. Например, ДНК препоследовательности или секреторной лидерной последовательности функционально связана с ДНК полипептида, если она экспрессируется как пре-белок, который принимает участие в секреции полипептида; промотор или энхансер функционально связан с кодирующей последовательностью, если он оказывает влияние на транскрипцию последовательности; или сайт связывания рибосомы функционально связан с кодирующей последовательностью, если он расположен так, чтобы облегчить трансляцию. Как правило, «функционально связанный» означает, что связываемые последовательности ДНК являются смежными, а в случае секреторной лидерной последовательности смежными и находятся в фазе чтения. Однако энхансеры не обязательно должны быть смежными. Связывание осуществляется посредством лигирования в удобных сайтах рестрикции. Если таких сайтов не существует, синтетические олигонуклеотидные адаптеры или линкеры используют в соответствии с общепринятой практикой.
Используемый в данном документе термин «вирусные белки» или «вирусные полипептиды» относится к белку, кодируемому вирусом, описанным в данном документе, в том числе структурным и неструктурным белкам. Такие белки могут включать встречающиеся в природе или не встречающиеся в природе вирусные белки MDV, NDV и/или IBDV, в том числе белки VP2, F и/или HN, NP, Р, М или L. Такие белки могут также включать встречающиеся в природе или не встречающиеся в природе вирусные белки ILTV, такие как gB, gC, gD, gE, gH, gI или gL, белки S1, S2 или M вируса инфекционного бронхита (IBV), белки VP1 или VP2 вируса анемии цыплят (CAV) и/или любой из белков НА, NA, NP или М вируса птичьего гриппа (AIV).
В соответствии с настоящим изобретением векторы на основе рекомбинантных вирусов, описанные в данном документе, могут обеспечить защиту домашней птицы от двух или более различных вирусных патогенов за счет введения векторов на основе рекомбинантных вирусов, которые экспрессируют гены таких вирусных патогенов. В некоторых вариантах осуществления векторы на основе рекомбинантных вирусов по настоящему изобретению могут быть введены домашней птице в иммуногенной композиции, как описано в данном документе. Можно использовать гены любого вирусного патогена, подходящего для применения с вектором на основе рекомбинантного вируса, как описано в данном документе. Например, в некоторых вариантах осуществления вектор на основе рекомбинантного вируса может экспрессировать гены вируса болезни Ньюкасла (NDV), вируса инфекционного бурсита (IBDV), вируса птичьего гриппа (AIV), вируса анемии цыплят (CAV), вируса инфекционного бронхита (IBV) и вируса инфекционного ларинготрахеита (ILTV) или т.п.
В соответствии с настоящим изобретением трансген, обеспечивающий защиту или устойчивость к определенному вирусу или вирусам, может быть вставлен в вирусный геном в определенном месте. Например, в некоторых вариантах осуществления трансген, описанный в данном документе, может быть вставлен в вирусный геном в межгенной области, фланкированной HVT UL 35/UL 36 в уникальной длинной области генома. В другом варианте осуществления настоящего изобретения трансген, описанный в данном документе, содержит один или несколько гетерологичных генов, вставленных в вирусный геном в межгенной области, фланкированной HVT UL 35/UL 36 генома HVT, в дополнение ко второму используемому сайту, при этом один или несколько гетерологичных генов вставлены в сайт UL55 в геноме HVT. В других вариантах осуществления в одну или обе эти области можно вставить более одного трансгена.
В некоторых вариантах осуществления вектор на основе рекомбинантного вируса может экспрессировать несколько генов одного вида вируса или может экспрессировать гены более чем одного вида вируса для получения устойчивости к множеству вирусов. Например, в одном варианте осуществления в настоящем изобретении предусмотрен вектор на основе рекомбинантного вируса, содержащий геном HVT и по меньшей мере один трансген другого вирусного патогена, таким образом обеспечивая защиту птицы, такой как домашняя птица, от болезни Марека и по меньшей мере от одного другого вирусного заболевания. Например, в одном варианте осуществления вектор на основе рекомбинантного вируса в соответствии с настоящим изобретением может обеспечивать защиту домашней птицы от MDV и NDV, или может обеспечивать защиту от MDV и IBDV, или может обеспечивать защиту от MDV, NDV и IBDV.
Вирусные антигены для экспрессии у домашней птицы с помощью вектора на основе рекомбинантного вируса по настоящему изобретению могут кодироваться вирусным геном, таким как вирусный ген, описанный в данном документе. Специалисту в данной области техники будет понятно в этом отношении, что может не потребоваться включение всего определенного вирусного гена для получения требуемой устойчивости к вирусам. Скорее можно использовать часть такого гена. Может быть желательным выбор определенной части требуемого гена, которая является специфичной для любого определенного целевого вируса или вирусов. Оптимизация требуемого вирусного белка или последовательности, кодирующей такой белок, независимо от длины белка, может быть легко осуществленас применением методологий, известных из уровня техники, которые являются подходящими для применения в настоящем изобретении. Специалисту в данной области техники будет понятно, что модификации могут быть выполнены в вирусном гене или генах или белках, кодируемых ими, для повышения активности вирусного белка при введении субъекту. Модификации, внесенные в вирусные гены или белки, могут усилить или ослабить реакцию хозяина в отношении специфического вируса.
В определенных вариантах осуществления рекомбинантный вирус болезни Марека или вектор на основе рекомбинантного вируса по настоящему изобретению могут содержать трансген, кодирующий вирусный белок IBDV или продукт гена, такой как белок или продукт гена VP2 IBDV. В другом варианте осуществления такой рекомбинантный вирус или вирусный вектор может содержать трансген, кодирующий вирусный белок или продукт гена NDV, такой как белок или продукт гена F или HN NDV. В другом варианте осуществления такой рекомбинантный вирус или вирусный вектор может содержать трансген, кодирующий вирусный белок или продукт гена вируса птичьего гриппа (AIV), такой как белок или продукт гена НА или N AIV. В другом варианте осуществления такой рекомбинантный вирус или вирусный вектор может содержать трансген, кодирующий вирусный белок или продукт гена вируса инфекционного бронхита (IBV), такой как белок или продукт гена S1 или S2 IBV. Трансген по настоящему изобретению может содержать более одного гена, включая слитый белок на основе гена или продукт гена, такой как слитый белок, химеру или продукт гена F-HN NDV. В некоторых вариантах осуществления можно использовать полную кодирующую последовательность такого гена для получения полноразмерного или полностью функционального белка или полипептида. В качестве альтернативы часть или фрагмента вирусного белка или полипептида может быть достаточным для обеспечения защиты от определенного вируса или вирусов или устойчивости к ним.
В определенных вариантах осуществления рекомбинантный вирус болезни Марека или вектор на основе рекомбинантного вируса по настоящему изобретению могут содержать трансген, кодирующий иммуномодулятор, такой как белок или продукт гена цитокина. В соответствии с настоящим изобретением цитокин может представлять собой интерлейкин (IL), в том числе без ограничения IL2, IL6, IL7, IL8, IL12, IL18 и т.п. Такой трансген, кодирующий цитокин, может быть вставлен в один или оба геномных сайта, как описано в данном документе. В некоторых вариантах осуществления трансген, кодирующий цитокин, может быть вставлен в один сайт, описанный в данном документе, а трансген, кодирующий вирусный белок, может быть вставлен в другой сайт.Могут быть использованы и другие иммуномодуляторы, такие как интерфероны, хемокины, глюканы, гранулоцитарные колониестимулирующие факторы, олигодезоксинуклеотиды, которые также могут быть использованы в соответствии с настоящим изобретением.
Выделение вирусных генов или белков
В вариантах осуществления настоящего изобретения вирусный ген, описанный в данном документе, может быть выделен с использованием зондов нуклеиновых кислот и/или олигонуклеотидов в жестких условиях гибридизации, ПЦР или микрочипов, скрининга библиотек ДНК или с применением любых других способов, известных из уровня техники. Специалисту в данной области техники будет легко понять, как выделить вирусные гены или белки для применения по настоящему изобретению. В качестве альтернативы библиотеки экспрессии можно использовать для клонирования вируса, его полиморфных вариантов, ортологов или аллелей путем иммунологической детекции гомологов с помощью антисыворотки или очищенных антител, направленных против вируса другого вида или его частей.
Способы создания и скрининга библиотек cDNA хорошо известны из уровня техники. Например, для создания библиотеки cDNA для клонирования вирусных генов, экспрессируемых геномом, mRNA можно подвергнуть обратной транскрипции в cDNA с использованием обратной транскриптазы. Затем cDNA может быть лигирована в вектор, такой как рекомбинантный вектор, и введена в клетку-хозяина или организм для размножения, скрининга и клонирования.
Для геномной библиотеки ДНК может быть извлечена из требуемой ткани и может быть расщеплена с использованием биологических ферментов или может быть подвергнута механическому расщеплению. Полученные фрагменты ДНК могут быть затем выделены из нежелательных фрагментов ДНК и сконструированы в соответствующий вектор, который затем может быть упакован in vitro. Рекомбинантные векторы можно анализировать посредством любого способа, известного из уровня техники.
Такие способы, как полимеразная цепная реакция (ПЦР и RT-PCR) и лигазная цепная реакция (LCR), могут быть использованы для амплификации последовательностей нуклеиновых кислот непосредственно из mRNA, из геномных библиотек или библиотек cDNA. Вырожденные олигонуклеотиды могут быть сконструированы для амплификации гомологов с использованием последовательностей, предусмотренных в данном документе. Сайты рестрикционных эндонуклеаз могут быть включены в праймеры. Полимеразная цепная реакция или другие способы амплификации in vitro также могут быть применимы, например, для клонирования последовательностей нуклеиновых кислот, которые кодируют белки, подлежащие экспрессии, для получения нуклеиновых кислот, которые можно использовать в качестве зондов для обнаружения наличия заболевания, подлежащего целенаправленному воздействию, таких как кодирующая mRNA MDV, NDV и/или IBDV, в биологических образцах, для секвенирования нуклеиновых кислот или для других целей. Гены, амплифицированные посредством ПЦР, могут быть очищены от агарозы и клонированы в соответствующий вектор.
Экспрессию вирусных генов также можно анализировать посредством методик, известных из уровня техники, например, обратной транскрипции и амплификации mRNA, выделения общей РНК или РНК polyA, нозерн-блоттинга, дот-блоттинга, гибридизации in situ, защиты от RNase, технологии полинуклеотидных чипов высокой плотности и т.п.
Нуклеиновые кислоты, кодирующие вирусный геном или белок, можно использовать посредством технологии олигонуклеотидных чипов высокой плотности (например, GeneChip™) для идентификации вирусных генов, ортологов, аллелей, их вариантов и полиморфных вариантов в настоящем изобретении. Выбранный ген может быть клонирован в промежуточный вектор перед трансформацией в прокариотические или эукариотические клетки для репликации и/или экспрессии. Эти промежуточные векторы могут представлять собой прокариотические векторы, например, плазмиды или челночные векторы.
Модификация нуклеиновых кислот
Любое количество способов, хорошо известных специалистам в данной области техники, может быть применено для выделения молекулы ДНК и манипулирования с ней. Например, технология полимеразной цепной реакции (ПЦР) может быть использована для амплификации конкретной исходной молекулы ДНК и/или для получения вариантов исходной молекулы ДНК. Молекулы ДНК или их фрагменты также могут быть получены посредством любых методик, известных из уровня техники, в том числе прямого синтеза фрагмента химическими средствами. Таким образом, может быть синтезирована вся нуклеиновая кислота или ее часть, как описано в данном документе.
Используемый в данном документе термин «комплементарные нуклеиновые кислоты» относится к двум молекулам нуклеиновых кислот, которые способны специфически гибрид изо ваться друг с другом, при этом две молекулы могут образовывать антипараллельную двухцепочечную структуру нуклеиновой кислоты. В связи с этим считается, что молекула нуклеиновой кислоты комплементарна другой молекуле нуклеиновой кислоты, если они проявляют полную комплементарность. Две молекулы называются «минимально комплементарными», если они могут гибридизоваться друг с другом с достаточной стабильностью, позволяющей им оставаться отожженными друг от друга по меньшей мере в обычных условиях с низкой жесткостью. Аналогичным образом считается, что молекулы комплементарны, если они могут гибридизоваться друг с другом с достаточной стабильностью, позволяющей им оставаться отожженными друг от друга в обычных условиях с высокой жесткостью. Обычные условия жесткости описаны Sambrook, et al. (1989) и Haymes et al. (1985).
Отклонения от полной комплементарности допустимы, пока сохраняется способность молекул образовывать двухцепочечную структуру. Таким образом, для того, чтобы молекула нуклеиновой кислоты или фрагмент молекулы нуклеиновой кислоты выступали в качестве праймера или зонда, такая молекула или фрагмент должны быть достаточно комплементарны в последовательности, чтобы иметь возможность образовывать стабильную двухцепочечную структуру при определенных условиях использования концентрации растворителя и соли.
Используемые в данном документе термины «идентичность последовательностей», «сходство последовательностей» или «гомология» используются для описания взаимосвязей последовательностей между двумя или более нуклеотидными последовательностями. Процент «идентичности последовательностей» между двумя последовательностями определяется посредством сравнения двух оптимально выровненных последовательностей по определенному количеству нуклеотидов, при этом часть последовательности в окне сравнения может содержать добавления или делеции (т.е. гэпы) по сравнению с референтной последовательностью. Две последовательности называются идентичными, если нуклеотиды в каждом положении являются одинаковыми. Нуклеотидная последовательность, наблюдаемая в направлении от 5' к 3', считается «комплементом» или комплементарной второй нуклеотидной последовательности, наблюдаемой в направлении от 3' к 5', если первая нуклеотидная последовательность демонстрирует полную комплементарность со второй или референтной последовательностью. Как используется в данном документе, считается, что молекулы последовательности нуклеиновой кислоты проявляют «полную комплементарность», когда каждый нуклеотид одной из последовательностей, прочитанной в направлении с 5' по 3', комплементарен каждому нуклеотиду другой последовательности, прочитанной в направлении с 3' по 5'. Нуклеотидная последовательность, которая является комплементарной референтной нуклеотидной последовательности, будет демонстрировать последовательность, идентичную обратной последовательности комплемента референтной нуклеотидной последовательности.
Рекомбинантные векторы и клетки-хозяева
Вектор на основе рекомбинантной ДНК может представлять собой, например, линейную или кольцевую плазмиду. Векторная система может представлять собой единственный вектор или плазмиду или два или более вектора или плазмиды, которые совместно содержат общую ДНК для введения в геном клетки-хозяина. Рекомбинантный вектор, описанный в данном документе, может представлять собой вектор экспрессии, например, для обеспечения продуцирования требуемого белка в клетке-хозяине, такой как бактериальная клетка. Молекулы нуклеиновой кислоты, описанные в данном документе, или их комплементы или фрагменты могут быть вставлены в вектор под контролем подходящего промотора, который функционирует в одном или нескольких хозяевах-микроорганизмах для управления экспрессией связанной кодирующей последовательности или другой последовательности ДНК. Многие векторы доступны и известны из уровня техники для этой цели, и выбор подходящего вектора зависит от размера нуклеиновой кислоты, которую нужно вставить в вектор, и клетки-хозяина, которая подлежит трансформации вектором. Каждый вектор может содержать различные компоненты в зависимости от его функции (например, амплификация ДНК или экспрессия ДНК) и конкретной клетки-хозяина, с которой он совместим. Компоненты вектора для бактериальной трансформации обычно включают без ограничения одно или несколько из следующего: сигнальную последовательность, точку начала репликации, один или несколько селектируемых маркерных генов и индуцируемый промотор, обеспечивающий экспрессию экзогенной ДНК.
Используемый в данном документе термин «рекомбинантный вирус болезни Марека» или «рекомбинантный HVT» или «рекомбинантный вирус» обозначает инфекционный вирус или вирусную частицу, которые были генетически модифицированы посредством включения в вирусный геном одной или нескольких последовательностей гетерологичных нуклеиновых кислот, т.е. ДНК, кодирующую вирусный ген или его фрагмент или часть, не идентичную последовательности нуклеиновой кислоты гена, естественным образом присутствующего в вирусе. При заражении клетки рекомбинантным вирусом болезни Марека рекомбинантный вирус экспрессирует гетерологичный ген в форме гетерологичного полипептида.
Термин «вектор на основе рекомбинантного вируса» или «вирусный вектор», используемый в данном документе, относится к рекомбинантной конструкции, которую вставляют в вирус для введения в клетку-хозяина. Такой вектор по настоящему изобретению может быть получен из любого штамма HVT. При необходимости вирусные гены или последовательности, кодирующие белок, могут быть включены в такой вектор на основе рекомбинантного вируса, как описано в данном документе, для введения курице или другой домашней птице для защиты от одного или нескольких вирусных заболеваний.
Используемый в данном документе термин «сайт вставки» относится к области вирусного генома, в которую вставлена трансгенная или экзогенная ДНК. Сайты вставки по настоящему изобретению могут представлять собой межгенные области. Межгенная область в соответствии с настоящим изобретением может быть фланкирована HVT UL35 и HVT UL36 в уникальной длинной области генома. В некоторых вариантах осуществления настоящего изобретения один или несколько гетерологичных нуклеотидов, кодирующих антигены, также могут быть вставлены в области, определяемые локусом UL55 генома HVT. В некоторых вариантах осуществления сайты вставки по настоящему изобретению могут включать весь фланкирующий ген или его часть с любой стороны межгенной области. Вставка одного или нескольких трансгенов в одну из этих областей позволяет получить вектор на основе рекомбинантного вируса, который затем можно ввести курице или другой домашней птице для защиты от одного или нескольких заболеваний.
Используемый в данном документе термин «функционально связанный» при использовании в отношении регуляторной последовательности и нуклеотидной последовательности означает, что регуляторная последовательность вызывает регулируемую экспрессию связанной структурной нуклеотидной последовательности. Термины «регуляторные последовательности», «регуляторные элементы» или «контрольные элементы» относятся к нуклеотидным последовательностям, расположенным выше (5' последовательности), в пределах или ниже (3' последовательности) структурной нуклеотидной последовательности. Такие последовательности оказывют влияние на время и уровень или количество транскрипции, процессинга или стабильности РНК или трансляции ассоциированной структурной нуклеотидной последовательности. Регуляторные последовательности могут включать без ограничения промоторы, лидерные последовательности, интроны, энхансеры, структуры «стебель-петля», последовательности связывания репрессора и последовательности распознавания полиаденилирования, в том числе без ограничения сигнальную последовательность polyA бычьего гормона роста, сигнальную последовательность polyA вируса 40 обезьян (SV40), сигнальную последовательность polyA 1629 ORF вируса ядерного nojm3ppo3a,Autographa californica (AcNPV) и сигнальную последовательность polyA тимидинкиназы (ТК) вируса простого герпеса (HSV). Специалисту в данной области техники будет понятно, что для повышения или снижения экспрессии трансгена, как описано в данном документе, можно использовать различные комбинации промоторов и/или регуляторных элементов.
Промоторы, функционирующие у разных видов, также хорошо известны из уровня техники. Промоторы, применимые для экспрессии полипептидов, включают промоторы, которые являются индуцируемыми, вирусными, синтетическими или конститутивными, и/или промоторы, которые являются тканеспецифичными, регулируемыми во времени, регулируемыми в пространстве и регулируемыми в пространстве и времени. Например, промоторы, применимые в соответствии с настоящим изобретением, могут включать без ограничения промотор немедленно раннего (IE) цитомегаловируса (CMV), промотор CMV морской свинки, промотор SV40, промоторы вируса псевдобешенства, такие как промотор гликопротеина X, промотор вируса простого герпеса 1, такой как промотор альфа-4, промоторы вирусов болезни Марека, в том числе любой изолят или штамм MDV, такой как MDV-1, MDV-2 и HVT, например, промотор, контролирующий экспрессию гликопротеинов, таких как как gC, gB, gE или gI, промоторы вируса инфекционного ларинготрахеита, такие как промоторы гликопротеина gB, gE, gI, gD, или любые другие подходящие промоторы. Специалисту в данной области техники должно быть известно, как идентифицировать промотор, применимый в соответствии с настоящим изобретением.
В соответствии с настоящим изобретением рекомбинантный вирус болезни Марека или вектор на основе рекомбинантного вируса, описанные в данном документе, могут содержать один или несколько трансгенов, функционально связанных с одним или несколькими промоторами для экспрессии одного или нескольких вирусных белков или пептидов, или их фрагментов или частей. В некоторых вариантах осуществления один трансген может быть функционально связан с одним промотором, или несколько трансгенов могут быть функционально связаны с одним промотором. В других вариантах осуществления в рекомбинантном векторе может присутствовать более одного трансгена, при этом первый трансген функционально связан с первым промотором, а второй трансген функционально связан со вторым промотором.
Конструирование и выбор векторов
Специалистам в данной области техники известно конструирование векторов, содержащих один или несколько компонентов, описанных в данном документе, применимых для вставки генов или трансгенов или их частей в целевой сайт, и в них можно применять стандартные методики рекомбинантных ДНК. Вектор или конструкция на основе рекомбинантной ДНК могут содержать селектируемый маркер, который придает клетке селектируемый фенотип.Селектируемые маркеры также можно использовать для отбора клеток, содержащих экзогенные нуклеиновые кислоты, кодирующие полипептиды или белки, как описано в данном документе. Такой маркер может кодировать, например, устойчивость к биоцидам или устойчивость к антибиотикам (например, канамицину, G418, блеомицину, гигромицину и т.д.). Селектируемые маркеры хорошо известны специалистам в данной области техники и могут включать любые маркеры, подходящие для применения в соответствии с настоящим изобретением.
Рекомбинантный вектор или конструкция могут также включать поддающийся скринингу маркер, который можно использовать для мониторинга экспрессии, но который может не приводить к гибели клетки. Подходящие маркеры для скрининга могут включать, например, ген β-глюкуронидазы или uidA (GUS), один или несколько различных генов флуоресцентных белков, таких как зеленый флуоресцентный белок (GFP), красный флуоресцентный белок (RFP), или любой из белков крупного семейства, которые флуоресцируют при характерных длинах волн, ген, который кодирует фермент, для которого известны различные хромогенные субстраты, ген люциферазы, ген xylE, который кодирует катехолдиоксигеназу, которая конвертирует хромогенные катехолы, ген β-амилазы, ген тирозиназы, который кодирует фермент, способный окислять тирозин до DOPA и допахинона, которые, в свою очередь, конденсируются в меланин, или α-галактозидазу, которая катализирует хромогенный субстрат α-галактозы.
Экспрессия белков в клетках-хозяевах
Для получения высокого уровня экспрессии клонированного вирусного гена, как описано в данном документе, нуклеиновую кислоту можно субклонировать в вектор экспрессии, который содержит сильный промотор для направления транскрипции и терминатор транскрипции/трансляции. Для кодируемых белков также может быть включен сайт связывания рибосомы для инициации трансляции. Подходящие промоторы для применения в векторах экспрессии хорошо известны из уровня техники, такие как бактериальный промотор, вирусный промотор и т.п. Системы экспрессии для экспрессии белка доступны у нескольких прокариотических и эукариотических видов, известных из уровня техники. Коммерческие наборы для таких систем экспрессии также легко доступны. Эукариотические системы экспрессии для клеток млекопитающих, дрожжей и клеток насекомых хорошо известны из уровня техники и также коммерчески доступны.
Выбор подходящего промотора для непосредственной экспрессии гетерологичной нуклеиновой кислоты будет зависеть от определенного применения. Такой промотор может быть расположен на расстоянии от гетерологичного сайта инициации транскрипции, аналогичном расстоянию в его естественных условиях, хотя специалисту в данной области техники будет понятно, что некоторое изменение этого расстояния может быть разрешено без потери функции промотора.
Кроме промотора, вектор экспрессии обычно содержит кассету транскрипции или экспрессии, которая содержит все элементы, необходимые для экспрессии нуклеиновой кислоты в клетке-хозяине. Любые стандартные векторы, известные из уровня техники, которые можно применять для экспрессии в эукариотических или прокариотических клетках, можно использовать для переноса генетической информации в клетку. Таким образом, типичная кассета экспрессии содержит промотор, функционально связанный с последовательностью нуклеиновой кислоты, кодирующей выбранную нуклеиновую кислоту, и соответствующие сигнальные последовательности, необходимые для эффективного процессинга, например, сайты связывания рибосом, полиаденилирования и терминации трансляции. Дополнительные элементы могут включать энхансеры и, в случае геномной ДНК в качестве структурного гена, интроны с функциональными донорными и акцепторными сайтами сплайсинга.
Кроме промоторной последовательности, такой как промотор, указанный в данном документе, кассета экспрессии может также содержать область терминации транскрипции ниже структурного гена, чтобы обеспечить эффективную терминацию транскрипции. Область терминации может происходить из того же гена, что и промоторная последовательность, или она может происходить из другого гена. Маркеры, такие как флуоресцентные белки, зеленый или красный флуоресцентный белок, β-gal, CAT и т.п., могут быть включены в векторы в качестве маркеров для векторной трансдукции. Метки эпитопов или метки последовательности также могут быть добавлены к рекомбинантным белкам для обеспечения удобных способов выделения.
Векторы экспрессии, содержащие регуляторные элементы из эукариотических вирусов, как правило, используют в эукариотических векторах экспрессии, например, векторах на основе SV40, векторах на основе папилломавируса, ретровирусных векторах и векторах, полученных из вируса Эпштейна-Барр. Другие иллюстративные эукариотические векторы включают pMSG, pAV009/A+, рМТО10/А+, pMAMneo-5, бакуловирус pDSVE и любой другой вектор, обеспечивающий экспрессию белков под управлением промотора CMV, раннего промотора SV40, позднего промотора SV40, металлотионеинового промотора, промотор вируса опухоли молочной железы мышей, промотора вируса саркомы Рауса, промотора полиэдрина или других промоторов, известных из уровня техники, которые могут быть эффективны для экспрессии в эукариотических клетках.
Экспрессию белков из эукариотических векторов также можно регулировать с применением индуцируемых промоторов. В случае индуцируемых промоторов уровни экспрессии связаны с концентрацией индуцирующих средств, таких как тетрациклин или экдизон, за счет включения в промотор элементов ответа для этих средств. Высокие уровни экспрессии могут быть получены из индуцируемых промоторов в присутствии индуцирующего средства. Некоторые системы экспрессии содержат маркеры, такие как тимидинкиназа и дигидрофолатредуктаза, которые обеспечивают амплификацию генов.
Вектор экспрессии может также содержать репликон, функционирующий в Е. coli, ген устойчивости к антибиотикам для селекции бактерий, содержащих рекомбинантные плазмиды, и уникальные сайты рестрикции в несущественных областях плазмиды, позволяющие вставлять эукариотические последовательности. Можно использовать любой ген устойчивости к антибиотикам, применимый для применения в настоящем изобретении.
Стандартные способы трансфекции, известные из уровня техники, могут быть применены для получения клеточных линий бактерий, млекопитающих, дрожжей или насекомых, которые экспрессируют значительные количества белка. Затем такие клеточные линии могут быть очищены с применением стандартных методик, известных из уровня техники, а прокариотические и/или эукариотические клетки могут быть трансформированы в соответствии с любым способом, известным в данной области техники, для введения клонированной геномной ДНК, cDNA, синтетической ДНК или другого чужеродного генетического материала в клетку-хозяина. Такие способы могут включать без ограничения плазмидные или вирусные векторы, трансфекцию фосфатом кальция, слияние протопластов, электропорацию, биолистику, липосомы, микроинъекцию или любые способы, доступные в данной области техники.
После введения вектора экспрессии или трансгена в клетку-хозяина, клетку затем можно культивировать в условиях, оптимальных для экспрессии требуемого белка, который может быть выделен с применением стандартных способов, известных из уровня техники. Вирусные патогены или вирусные белки, такие как описанные в данном документе, могут быть затем очищены для применения в диагностических анализах, для получения антител и иммуногенных композиций и для идентификации противовирусных соединений. Встречающиеся в природе белки могут быть очищены из биологических образцов, таких как образец ткани птицы, инфицированной вирусом, как описано в данном документе, в то время как рекомбинантные белки могут быть очищены с применением любых подходящих способов или систем экспрессии, известных из уровня техники.
В данной области техники доступен ряд процедур очистки рекомбинантного белка. Например, белки, характеризующиеся определенными свойствами молекулярной адгезии, могут быть обратимо слиты с другим белком. Кроме того, определенный белок может быть селективно адсорбирован на колонке для очистки, а затем выделен из колонки в относительно чистой форме с использованием соответствующих лигандов или субстратов. Затем слитый белок может быть удален посредством ферментативной активности. Белок также может быть очищен с использованием аффинных колонок. Рекомбинантный белок может быть очищен из любого подходящего источника.
Очистка белка от рекомбинантных бактерий
Рекомбинантные белки могут экспрессироваться бактериями в значительных количествах, например, с использованием индуцируемого или конститутивного промотора. Индукция промотора с использованием IPTG является примером индуцируемой промоторной системы. Бактерии можно выращивать из свежей или замороженной культуры в соответствии со стандартными процедурами, известными из уровня техники.
Белки, экспрессированные в бактериях, могут образовывать нерастворимые агрегаты, называемые тельцами включения. Подходящие протоколы очистки белковых телец включения известны из уровня техники. Лизирование бактерий для выделения экспрессированных белков можно проводить с применением любых способов, известных из уровня техники, которые могут включать введение химических буферов, обработку ультразвуком, механическое разрушение и т.п. Тельца включения также могут быть солюбилизированы, а суспензия лизированных клеток может быть центрифугирована для удаления нежелательных клеточных остатков. Белки телец включения можно ренатурировать посредством разведения или диализа соответствующим буфером.
Рекомбинантные белки также могут быть получены из периплазмы бактерий. После лизиса бактериальных клеток периплазматическая фракция бактерий может быть выделена посредством любых способов, известных из уровня техники. Рекомбинантные белки, присутствующие в супернатанте, могут быть отделены от белков-хозяев посредством стандартных методик разделения, хорошо известных специалистам в данной области техники.
Белки могут быть разделены с применением любых методик, известных из уровня техники, например, фракционирования на основе растворимости или дифференциальной фильтрации на основе размеров, при которой белок выделяют на основе молекулярной массы с использованием фильтрации через мембраны с различным размером пор. Колоночную хроматографию можно использовать для выделения белка из других белков на основе размера, суммарного поверхностного заряда, гидрофобности или аффинности в отношении лигандов или субстратов. Кроме того, антитела, выработанные против белка, представляющего интерес, могут быть конъюгированы с колонкой, а белки подвергнуты иммуноочистке. Все эти способы хорошо известны специалистам в данной области техники. Специалисту будет очевидно, что хроматографические методики можно применять в любом масштабе и с использованием любого подходящего коммерческого оборудования. Продуцирование антител
Способы получения поликлональных и моноклональных антител, которые специфически реагируют с вирусными белками, вирусными частицами и/или нуклеиновыми кислотами, известны из уровня техники. Такие способы могут включать получение антител посредством отбора антител из библиотек рекомбинантных антител в фаговых или других векторах, а также получение поликлональных и моноклональных антител посредством иммунизации кроликов или мышей.
Ряд антигенов или антигенных областей, содержащих вирусный белок или его части, вирусные частицы и/или нуклеиновые кислоты, можно применять для получения антител, специфически реагирующих с требуемым вирусным патогеном. Например, рекомбинантный вирусный белок или его антигенный фрагмент можно выделить с применением любых способов, описанных в данном документе или известных из уровня техники. Рекомбинантные белки могут быть экспрессированы в прокариотических или эукариотических клетках и очищены, как описано в данном документе. Моноклональные и/или поликлональные антитела могут быть получены с использованием встречающихся в природе (в чистой форме или с примесями) или рекомбинантных белков с применением способов, известных из уровня техники. Синтетические пептиды, полученные из вирусной последовательности, также можно использовать для получения антител, их можно конъюгировать с белком-носителем и вводить животному, способному продуцировать антитела (например, кролику).
Способы получения поликлональных антител известны специалистам в данной области техники. Например, инбредную линию мышей или кроликов можно иммунизировать белком с помощью стандартного адъюванта, такого как адъювант, описанный в данном документе, с использованием стандартного протокола иммунизации, известного из уровня техники. Когда получают достаточно высокие титры антител к белку, можно приготовить антисыворотку и провести обогащение для получения антител, реагирующих с белком.
Моноклональные антитела также могут быть получены посредством различных способов, известных из уровня техники. Например, клетки селезенки животного, иммунизированного требуемым антигеном, могут быть иммортализованы, обычно посредством слияния с клеткой миеломы или посредством трансформации вирусом Эпштейна-Барра (EBV), онкогенами или ретровирусами, или других способов, хорошо известных из уровня техники. Затем иммортализованные клетки могут быть подвергнуты скринингу в отношении продуцирования антител с требуемой специфичностью и аффинностью к антигену. Выход моноклональных антител, продуцируемых такими клетками, можно повысить посредством различных методик, известных из уровня техники, например, путем инъекции в брюшную полость позвоночного хозяина.
Моноклональные антитела и поликлональные сыворотки могут быть собраны и титрованы против требуемого антигена или белка в иммунологическом анализе, например, твердофазном иммунологическом анализе с белком, иммобилизованным на твердой подложке. Антитела, специфичные только в отношении конкретного вирусного белка, также могут быть получены путем удаления других перекрестно реагирующих белков. Таким образом могут быть получены антитела, которые связываются только с выбранным белком.
Когда специфические антитела против требуемого вирусного антигена, такого как белок, вирус и/или нуклеиновая кислота, становятся доступными, требуемый антиген может быть обнаружен с применением различных способов иммунологического анализа. Антитело также может быть использовано терапевтически.
Белок, либо ассоциированный с вирусной частицей, либо отдаленный от нее, как описано в данном документе, может быть обнаружен и/или количественно определен с применением любого из ряда хорошо известных иммунологических анализов связывания. Вирусные частицы могут быть обнаружены на основе эпитопа, определяемого вирусными белками, презентируемыми в вирусной частице, и/или эпитопа, определяемого вирусным белком, который отделен от вирусной частицы (например, такой, который может присутствовать в инфицированной клетке). В иммунологических анализах может использоваться антитело, которое специфически связывается с выбранным белком или антигеном. Антитело может быть получено посредством любого из ряда способов, хорошо известных специалистам в данной области техники. В иммунологических анализах также может использоваться метящее средство для специфического связывания комплекса, образованного антителом и антигеном, в целях обнаружения. Метящее средство само по себе может быть одним из фрагментов, содержащих комплекс антитело/антиген. Таким образом, метящее средство может представлять собой меченую нуклеиновую кислоту вирусного белка или меченое противовирусное антитело. В качестве альтернативы метящее средство может представлять собой третий фрагмент, такой как вторичное антитело, которое специфически связывается с комплексом антитело/антиген. Вторичное антитело может быть специфичным в отношении к антителам того вида, из которого получено первое антитело. Метящее средство может быть модифицировано с помощью обнаружимой части, такой как биотин, с которой может специфически связываться другая молекула, такая как стрептавидин. Специалистам в данной области техники хорошо известны различные обнаружимые фрагменты.
Иммунологические анализы для обнаружения вирусного белка, вируса и/или нуклеиновой кислоты в образцах хорошо известны из уровня техники. Такие анализы могут быть как конкурентными, так и неконкурентными, а также количественными или неколичественными. Неконкурентные иммунологические анализы представляют собой анализы, в которых антиген может быть обнаружен непосредственно, и в некоторых случаях непосредственно измерено количество антигена. В конкурентных анализах вирусный антиген, присутствующий в образце, выявляется опосредованно с помощью детектируемого сигнала, ассоциированного с известным добавленным (экзогенным) вирусным антигеном, вытесненным из антитела к противовирусному антигену вирусным антигеном, присутствующим в образце. Таким образом, такие анализы также можно адаптировать для опосредованного измерения количества вирусного антигена, присутствующего в образце. Иммунологические анализы конкурентного связывания также могут использоваться для определения перекрестной реактивности, при которой любые перекрестно реагирующие антитела могут быть удалены из объединенной антисыворотки. Дополнительные типы анализов, в том числе без ограничения вестерн-блоттинг или иммунологические анализы с применением липосом, также могут быть использованы в соответствии с настоящим изобретением.
Специалисту в данной области техники будет понятно, что часто желательно свести к минимуму неспецифическое связывание в иммунологических анализах. В частности, когда анализ включает антиген или антитело, иммобилизованное на твердом субстрате, желательно свести к минимуму степень неспецифического связывания с субстратом. Средства снижения такого неспецифического связывания хорошо известны специалистам в данной области техники.
Анализ, описанный в данном документе, может включать метку или обнаружимую группу, которые не оказывают существенного влияния на специфическое связывание антитела, используемого в анализе. Обнаружимой группой может быть любой материал, характеризующийся обнаружимым физическим или химическим свойством. Такие обнаружимые метки известны из уровня техники, и, как правило, любая метка, применяемая в таких способах, может применяться в настоящем изобретении. Таким образом, термин «метка», используемый в данном документе, может представлять собой любую композицию, определяемую спектроскопическими, фотохимическими, биохимическими, иммунохимическими, электрическими, оптическими или химическими средствами. Применимые метки в настоящем изобретении могут включать магнитные гранулы (например, DYNABEADS™), флуоресцентные красители (например, изотиоцианат флуоресцеина, техасский красный, родамин и т.п.), радиоактивные метки (например, 3Н, 125I, 35S, 14С или 32Р), ферменты (например, пероксидаза хрена, щелочная фосфатаза и/или любые другие, известные из уровня техники и используемые в ELISA), и колориметрические метки, такие как коллоидное золото, цветное стекло или пластмассовые гранулы (например, полистирол, полипропилен, латекс и т.д.).
Метка в соответствии с настоящим изобретением может быть связана непосредственно или опосредованно с требуемым компонентом анализа в соответствии со способами, хорошо известными из уровня техники. Как описано выше, можно использовать широкий спектр меток, при этом выбор метки зависит, среди прочего, от чувствительности, простоты конъюгации с соединением, требований к стабильности или доступного оборудования.
Нерадиоактивные метки могут быть нанесены опосредованными средствами. Как правило, молекула лиганда (например, биотина) ковалентно связана с молекулой. Затем лиганд может связываться с другой молекулой (например, стрептавидином), которая может обнаруживаться либо сама по себе, либо ковалентно связанной с сигнальной системой, такой как обнаружимый фермент, флуоресцентное соединение или хемилюминесцентное соединение. Лиганды и соответствующие им мишени можно использовать в любой подходящей комбинации с антителами, распознающими вирусный антиген, или вторичными антителами, распознающими противовирусный антиген. Молекулы также могут быть конъюгированы непосредственно с соединениями, генерирующими сигнал, например, посредством конъюгации с ферментом или флуорофором. Ферменты, представляющие интерес для применения в качестве меток, могут представлять собой гидролазы, например, фосфатазы, эстеразы и гликозидазы, или оксидотазы, такие как пероксидазы. Флуоресцентные соединения могут включать флуоресцеин и его производные, родамин и его производные, дансил, умбеллиферон и т.п. Хемилюминесцентные соединения могут включать люциферин, 2,3-дигидрофталазиндионы, например, люминол или другие соединения, известные из уровня техники.
Способы обнаружения меток хорошо известны специалистам в данной области техники и будут зависеть от типа используемой метки. Например, для обнаружения радиоактивной метки можно использовать авторадиографический анализ, а для обнаружения флуоресцентной метки можно использовать флуорохромы. Флуоресценцию можно обнаружить визуально, например, посредством электронных детекторов, таких как устройства с зарядовой связью (CCD) или фотоэлектрические умножители и т.п. Аналогичным образом ферментативные метки могут быть обнаружены посредством введения соответствующих субстратов для фермента и обнаружения полученного продукта реакции. Колориметрические или хемилюминесцентные метки можно обнаружить, наблюдая за цветом, ассоциированным с определенной меткой. В некоторых вариантах осуществления форматы анализа могут не требовать использования меченого компонента, а могут быть обнаружены посредством простого визуального осмотра.
Фармацевтические/иммупогенные композиции и их введение
В некоторых аспектах рекомбинантные векторы, содержащие один или несколько трансгенов, экспрессирующих один или несколько вирусных белков или пептидов или их фрагментов, как описано в данном документе, могут быть использованы в качестве фармацевтических композиций или иммуногенных композиций для введения субъекту, такому как курица или другая домашняя птица, для обеспечения защиты от одного или нескольких вирусов. Например, иммуногенная композиция, описанная в данном документе, содержит рекомбинантный вектор с одним или несколькими трансгенами, описанными в данном документе, которые вставлены в вирусный геном, например, в межгенную область, фланкированную межгенными локусами UL 35/UL 36 в уникальной длинной (UL) области генома HVT. В одном аспекте в настоящем изобретении предусмотрен геном рекомбинантного вируса герпеса индеек (HVT), содержащий одну или несколько нуклеотидных последовательностей, кодирующих один или несколько гетерологичных антигенов, вставленных в межгенные локусы UL 35/UL 36 в уникальной длинной области генома HVT и одну или несколько нуклеотидных последовательностей, кодирующих один или несколько гетерологичных антигенов, вставленных в сайт UL55 уникальной длинной области (UL) генома HVT.
В других аспектах белки или пептиды и их иммуногенные фрагменты и/или полинуклеотиды, а также противовирусные антитела и/или Т-клетки могут быть включены в фармацевтические композиции или иммуногенные композиции (например, вакцины). В другом варианте осуществления иммуногенная композиция по настоящему изобретению может содержать по меньшей мере третий трансген, четвертый трансген и т.п., которые могут кодировать дополнительные вирусные белки. Таким образом, можно ввести субъекту, такому как домашняя птица, иммуногенную композицию, которая обеспечивает защиту от любого требуемого количества вирусов. Цельно-вирусная вакцина (живая и аттенуированная, или неспособная к репликации, или убитая) или субъединичные вакцины, такие как структурные или неструктурные вирусные белки или их иммуногенные фрагменты, могут быть использованы для лечения или предупреждения вирусных инфекций посредством индукции иммунного ответа у субъекта. В качестве альтернативы фармацевтическая композиция может содержать антигенпрезентирующую клетку, транс фицированную вирусным полинуклеотид ом, так что антигенпрезентирующая клетка экспрессирует вирусный пептид.
Иммуногенные композиции в соответствии с настоящим изобретением могут быть разработаны для создания иммунитета к антителам и/или клеточного иммунитета у субъекта. Такие композиции могут содержать одно или несколько таких соединений совместно с не встречающимся в природе фармацевтически приемлемым носителем. В других вариантах осуществления иммуногенная композиция в соответствии с настоящим изобретением может включать более одного адъюванта или фармацевтически приемлемого носителя, из которых по меньшей мере один не встречается в природе. Фармацевтически приемлемый носитель или адъювант может представлять собой любое вещество, которое усиливает иммунный ответ у субъекта на экзогенный антиген, в том числе без ограничения адъюванты, липосомы, биоразлагаемые микросферы. Фармацевтически приемлемый носитель или адъювант может содержать вещество, предназначенное для защиты антигена от быстрого катаболизма, такое как гидроксид алюминия или минеральное масло, или стимулятор иммунных реакций, такой как белки, полученные из Bortadella pertussis или Mycobacterium tuberculosis. Коммерчески доступные адъюванты могут включать, например, неполный адъювант и полный адъювант Фрейнда, адъювант Merck 65, соли алюминия, такие как гель гидроксида алюминия (квасцы) или фосфат алюминия; олигонуклеотиды CpG, соли кальция, железа или цинка; нерастворимую суспензию ацилированного тирозина; ацилированные сахара; катионно или анионно дериватизированные полисахариды; полифосфазены; биоразлагаемые микросферы; и монофосфориллипид А. Специалист в данной области техники сможет идентифицировать подходящие фармацевтически приемлемые носители для применения в настоящем изобретении.
Фармацевтические или иммуногенные композиции и/или вакцины в объеме настоящего изобретения могут также содержать другие соединения, которые могут быть биологически активными или неактивными. Например, одна или несколько иммуногенных частей других антигенов могут присутствовать либо в составе слитого полипептида, либо в виде отдельного соединения в составе композиции или вакцины по настоящему изобретению. В некоторых вариантах осуществления полипептиды, используемые в настоящем изобретении, могут быть конъюгированы с другими макромолекулами. Фармацевтические или иммуногенные композиции и вакцины, как правило, можно использовать в профилактических и/или терапевтических целях. Например, в соответствии с настоящим изобретением композиция, описанная в данном документе, может быть введена субъекту, такому как птица, до инфицирования или воздействия вируса, чтобы обеспечить защиту от инфекции одним или несколькими вирусами или развития симптомов инфекции. В других вариантах осуществления такую композицию можно вводить субъекту, такому как птица, после инфицирования или воздействия одного или нескольких вирусов, чтобы обеспечить лечение вирусов у субъекта, например, посредством уменьшения или устранения инфекции в организме субъекта.
Вакцины на основе нуклеиновых кислот, кодирующие геном, структурный или неструктурный белок или его фрагмент вируса, описанного в данном документе, также могут быть использованы для индукции иммунного ответа для лечения или предупреждения вирусной инфекции. Из уровня техники хорошо известны многочисленные методики доставки генов. Подходящие системы экспрессии нуклеиновых кислот могут содержать необходимые последовательности ДНК для экспрессии у субъекта (такие как подходящий промотор и сигнальную последовательность терминации). В некоторых вариантах осуществления ДНК, описанная в данном документе, может быть введена с использованием системы вирусной экспрессии (например, вирус болезни Марека или HVT), что может включать использование непатогенного вируса, способного к репликации.
Фармацевтические или иммуногенные композиции могут поставляться в однодозовых или многодозовых контейнерах, таких как запаянные ампулы или флаконы. Такие контейнеры могут быть запаяны для сохранения стерильности композиции до ее применения. Как правило, композиции, описанные в данном документе, можно хранить в виде суспензий, растворов или эмульсий в масляных или водных носителях. В качестве альтернативы такую композицию можно хранить в лиофилизированном состоянии, требуя только добавления стерильного жидкого носителя непосредственно перед применением.
Как описано в данном документе, иммуногенная композиция может быть объединена с фармацевтически приемлемым носителем. Выбор подходящего носителя может частично определяться определенной вводимой композицией (например, нуклеиновой кислотой, белком, модуляторными соединениями или трансдуцированной клеткой), а также определенным способом, применяемым для введения композиции. Соответственно, доступно большое разнообразие подходящих составов фармацевтических или иммуногенных композиций, которые можно использовать в настоящем изобретении. Введение может осуществляться любым удобным способом, например, путем инъекции, перорального введения, ингаляции, трансдермального применения или ректального введения. Инъекция рекомбинантного вектора или иммуногенной композиции, как описано в данном документе, может быть предоставлена субъекту, такому как домашняя птица, в виде однократного введения или дозы, или может быть введена более одного раза, например, в виде повторных доз.
Составы, подходящие для парентерального введения, такого как, например, внутрисуставное (в суставы), внутривенное, внутримышечное, внутрикожное, внутрибрюшинное, in ovo и подкожное введение, включают водные и неводные изотонические стерильные растворы для инъекций, которые могут содержать антиоксиданты, буферы, бактериостатические средства и растворенные вещества, которые делают состав изотоническим с кровью предполагаемого субъекта, а также водные и неводные стерильные суспензии, которые могут включать суспендирующие средства, солюбилизаторы, загустители, стабилизаторы и консерванты. При осуществлении настоящего изобретения композиции можно вводить, например, путем внутривенной инфузии, перорально, местно, внутрибрюшинно, внутрипузырно или интратекально.
Такие композиции могут также содержать буферы (например, нейтральный забуференный физиологический раствор или фосфатно-солевой буфер), углеводы (например, глюкозу, маннозу, сахарозу или декстраны), маннит, белки, полипептиды или аминокислоты, такие как глицин, антиоксиданты, бактериостаты, хелатирующие агенты, такие как как EDTA или глутатион, адъюванты (например, гидроксид алюминия), растворенные вещества, которые делают состав изотоническим, гипотоническим или слабо гипертоническим по отношению к крови субъекта, суспендирующие средства, загустители и/или консерванты. В качестве альтернативы композиции по настоящему изобретению могут быть составлены в виде лиофилизата. Соединения также могут быть инкапсулированы в липосомы с применением способов, известных из уровня техники.
Растворы и суспензии для инъекций можно приготовить из стерильных порошков, гранул и таблеток, как описано в данном документе. Клетки, трансдуцированные нуклеиновыми кислотами для терапии ex vivo, можно также вводить внутривенно или парентерально, как описано выше. Инъекция, как описано в данном документе, может содержать суспензию одной или нескольких убитых, инактивированных, аттенуированных или иным образом невирулентных вирусных культур, очищенный или неочищенный раствор вирусного белка или нуклеиновую кислоту, как описано в данном документе. Раствор для инъекций может также содержать фармацевтически приемлемый носитель, как описано в данном документе.
Составы, подходящие для перорального введения, могут состоять из (а) жидких растворов, таких как эффективное количество упакованного вирусного белка или нуклеиновой кислоты, суспендированных в разбавителях, таких как вода, физиологический раствор или PEG 400; (b) капсулы или таблетки, каждая из которых содержит предварительно определенное количество активного ингредиента в виде жидкости, твердого вещества, гранул или желатина; (с) суспензии в соответствующей жидкости; или (d) подходящих эмульсий. Таблетированные формы могут включать лактозу, сахарозу, маннит, сорбит, фосфаты кальция, кукурузный крахмал, картофельный крахмал, микрокристаллическую целлюлозу, желатин, коллоидный диоксид кремния, тальк, стеарат магния, стеариновую кислоту и другие вспомогательные вещества, красители, наполнители, связующие вещества, разбавители, буферные средства, увлажняющие средства, консерванты, ароматизаторы, красители, разрыхлители и фармацевтически совместимые носители. Пастилки для рассасывания могут содержать активный ингредиент в ароматизаторе, например, сахарозе, а также пастилки, содержащие активный ингредиент в инертной основе, такой как желатин и глицерин или эмульсии сахарозы и акации, гели и т.п., содержащие, помимо активного ингредиента, носители, известные из уровня техники.
Выбранное соединение, отдельно или в комбинации с другими подходящими компонентами, может быть приготовлено в виде аэрозольных составов для введения путем ингаляции. Аэрозольные составы могут быть помещены в приемлемые пропелленты под давлением, такие как дихлордифторметан, пропан, азот и т.п.
Доза, вводимая субъекту в контексте настоящего изобретения, должна быть достаточной для оказания положительного терапевтического ответа у субъекта в динамике. Доза будет определяться эффективностью определенного используемого вектора и состоянием субъекта, а также массой тела и/или площадью поверхности пациента, подлежащего лечению. Размер дозы также может определяться наличием, характером и степенью любых неблагоприятных побочных эффектов, которые сопровождают введение определенного вектора или типа трансдуцированных клеток у определенного пациента. Для композиций, содержащих вектор, как описано в данном документе, эффективное количество вводимого вектора может быть определено частично на основании уровней вектора в циркулирующей плазме крови, токсичности вектора, состояния здоровья субъекта и продуцирования антител к вектору.
Что касается введения, то соединения и трансдуцированные клетки по настоящему изобретению можно вводить со скоростью, определяемой LD-50 ингибитора, вектора или типа трансдуцированных клеток, а также побочными эффектами ингибитора, вектора или типа клеток при различных концентрациях, применительно к массе и общему состоянию здоровья субъекта. Введение может осуществляться в виде однократных, многократных или разделенных доз.
Иммунологическое определение полипептидое и нуклеиновых кислот
Иммунологические анализы можно использовать для обнаружения вирусных белков, вирусных частиц и/или нуклеиновых кислот.Такие анализы могут быть применимы для терапевтических и/или диагностических вариантов применения, таких как описанные в данном документе. Иммунологические анализы хорошо известны из уровня техники и могут использоваться для качественного или количественного анализа белков, вирусных частиц и/или нуклеиновых кислот.
Анализы вирусных белков и антител к вирусным антигенам
В одном варианте осуществления настоящего изобретения присутствие вируса, описанного в данном документе, вирусной нуклеиновой кислоты или вирусного белка в образце можно определить посредством иммунологического анализа. Ферментно-опосредованные иммунологические анализы, такие как иммунофлуоресцентный анализ (IFA), твердофазный иммуноферментный анализ (ELISA), анализы захвата, тесты микроагглютинации и анализы иммуноблотинга (например, вестерн-блоттинг), могут быть легко адаптированы для обнаружения вируса или вирусных белков. Способ ELISA может быть эффективным для обнаружения вируса или вирусного белка, как описано в данном документе. Такой ELISA может, например, включать такие стадии, как: (1) связывание противовирусного антитела или антигена с субстратом; (2) приведение в контакт связанного рецептора с биологическим образцом, содержащим вирус, вирусный антиген, вирусный белок или антитела к вирусу; (3) приведение в контакт биологического образца с антителом, связанным с обнаружимым фрагментом (например, ферментом пероксидазой хрена или ферментом щелочной фосфатазой); (4) приведение в контакт биологического образца с субстратом для фермента; (5) приведение в контакт биологического образца с детектирующим реагентом, таким как окрашивающий реагент; (6) наблюдение за обнаружимым результатом. В некоторых вариантах осуществления биологический образец, подходящий для применения в таком ELISA, может представлять собой кровь или другие жидкости. В другом варианте осуществления ELISA, описанный в данном документе, может обнаруживать вирус или вирусный белок в образце ткани. Такие способы могут быть легко модифицированы специалистами для обнаружения присутствия противовирусного антитела в образце или специфического вирусного белка, а также самого вируса. В некоторых вариантах осуществления ELISA по настоящему изобретению может обнаруживать присутствие противовирусного антитела.
Анализы ELISA, как описано в данном документе, могут включать применение нитроцеллюлозной полоски, пропитанной вирусным белком, как описано в данном документе. Нитроцеллюлозная полоска может дать визуальный результат при контакте с тестируемым образцом, содержащим противовирусные нуклеопротеиновые антитела. Такой тест может идентифицировать субъекта, у которого уже имеются антитела против вирусного белка, и, таким образом, у субъекта может иметь место иммунитет к вирусу. Введение иммуногенной композиции для предупреждения вирусной инфекции, такой как описано в данном документе, может быть излишним такому субъекту, и, следовательно, идентификация субъектов, у которых уже имеются иммуногенные антитела, может предупредить излишнее введение иммуногенного соединения такому субъекту. В этом отношении вариант осуществления настоящего изобретения может включать идентификацию субъекта, у которого отсутствуют противовирусные антитела, с применением анализа, описанного в данном документе, такого как анализ ELISA, а затем введение иммуногенной композиции, описанной в данном документе, этому субъекту для предупреждения вирусной инфекции. В другом варианте осуществления нитроцеллюлозная полоска для применения в ELISA по настоящему изобретению может быть пропитана антителом, таким как противовирусное антитело, и может давать визуальный результат при контакте с тестируемым образцом, содержащим вирусный белок. Такой тест может идентифицировать субъекта, инфицированного вирусом, описанным в данном документе.
Еще одной иммунологической методикой, которая может быть применимы для обнаружения вируса, является анализ конкурентного ингибирования. В таком анализе используются моноклональные антитела (МАВ), реагирующие со специфическим вирусом. Биологическая жидкость (например, кровь) субъекта может быть приведена в контакт с первым антителом, связанным с субстратом, а меченое моноклональное антитело приведено в контакт с первым комплексом антитело-вирус. Величину ингибирования связывания моноклональных антител измеряют по отношению к контролю.
Как будет понятно специалисту в данной области техники, биологический образец для использования в вышеуказанных анализах может быть взят непосредственно у субъекта или может находитсья в частично очищенной форме. Антитело, специфичное в отношении определенного вируса, будет реагировать посредством связывания с вирусом в качестве первичной реакции. После этого также может быть добавлена вторичная реакция с антителом, связанным или меченым обнаружимым фрагментом, для усиления обнаружения первичной реакции. Как правило, во вторичной реакции антитело или другой лиганд, специфически или неспецифически реагирующий с другим сайтом связывания (эпитопом) вируса, выбирают по его способности реагировать с несколькими сайтами комплекса антитела и вируса. Так, например, несколько молекул антитела во вторичной реакции могут реагировать с каждым комплексом, образованным в результате первичной реакции, что делает первичную реакцию более обнаружимой.
Обнаружимый фрагмент может обеспечить визуальное обнаружение осадка или изменение цвета, визуальное обнаружение посредсвтом микроскопии или автоматическое обнаружение посредством спектрометрии, радиометрических измерений и т.п. Примеры обнаружимых фрагментов включают флуоресцеин и родамин (для флуоресцентной микроскопии), пероксидазу хрена (для световой или электронной микроскопии и биохимического обнаружения), биотин-стрептавидин (для световой или электронной микроскопии) и щелочную фосфатазу (для биохимического обнаружения по изменению цвета). Применяемые способы обнаружения и фрагменты могут быть выбраны, например, из любых раскрытых в данном документе или доступных из уровня техники.
Обнаружение присутствия вирусной нуклеиновой кислоты
В некоторых вариантах осуществления вирусная инфекция, описанная в данном документе, может быть обнаружена на основании уровня определенной РНК или ДНК в биологическом образце. Праймеры от определенного вируса или вирусного патогена можно использовать для обнаружения, диагностики и определения наличия вируса. Любой подходящий праймер можно использовать для обнаружения геномной ДНК или любой ее последовательности, открытой рамки считывания или гена или выбранного белка с применением любых подходящих способов, известных из уровня техники. Подходящая последовательность нуклеиновой кислоты может быть использована в качестве одноцепочечных или двухцепочечных зондов или праймеров для обнаружения вирусной mRNA или cDNA, полученной из нее, которые могут присутствовать в биологическом образце. Вирусные полинуклеотиды, описанные в данном документе, также могут быть использованы для получения дополнительных копий полинуклеотидов, для получения антисмысловых олигонуклеотидов или в качестве олигонуклеотидов, образующих тройную цепь. Например, два олигонуклеотидных праймера можно использовать в анализе на основе ПЦР для амплификации части вирусной cDNA, полученной из биологического образца, при этом по меньшей мере один из олигонуклеотидных праймеров является специфичным по отношению к вирусному полинуклеотиду (т.е. гибридизуется с ним). Такие праймеры могут характеризоваться любой длиной, достаточной для гибридизации и обеспечения возможности амплификации вирусной нуклеиновой кислоты, как описано в данном документе, в том числе по меньшей мере или около 10 нуклеотидами, 11 нуклеотидами, 12 нуклеотидами, 13 нуклеотидами, 14 нуклеотидами, 15 нуклеотидами, 16 нуклеотидами, 17 нуклеотидами, 18 нуклеотидами, 19 нуклеотидами, 20 нуклеотидами, 21 нуклеотидом, 22 нуклеотидами, 23 нуклеотидами, 24 нуклеотидами, 25 нуклеотидами, 26 нуклеотидами, 27 нуклеотидами, 28 нуклеотидами, 29 нуклеотидами, 30 нуклеотидами, 35 нуклеотидами, 40 нуклеотидами, 45 нуклеотидами или 50 нуклеотидами; или длиной от около 12 до около 50 нуклеотидов, длиной от 15 до 30 нуклеотидов, длиной от 15 до 25 нуклеотидов или длиной от 20 до 30 нуклеотидов. ДНК-праймеры, подходящие для применения в данном изобретении, могут представлять собой любые праймеры, описанные в данном документе, такие как указанные в виде SEQ ID NO: 40-157.
Затем амплифицированный нуклеотид, например, cDNA, может быть выделен и обнаружен с применением методик, хорошо известных из уровня техники, таких как гель-электрофорез. Аналогично, о л иго нуклеотид ные зонды, которые специфически гибридизуются с вирусным полинуклеотидом, можно использовать в анализе гибридизации для обнаружения присутствия вирусного полинуклеотида в биологическом образце.
Зонды нуклеиновой кислоты или праймеры, специфичные для вируса, как описано в данном документе, могут быть созданы с использованием полинуклеотидных последовательностей, раскрытых в данном документе. Зонды предпочтительно представляют собой по меньшей мере около 12, 15, 16, 18, 20, 22, 24 или 25 нуклеотидных фрагментов или другую полинуклеотидную последовательность, кодирующую вирусную нуклеиновую кислоту или полипептид. Длина зондов нуклеиновой кислоты может составлять менее около 200 п. о., 150 п. о., 100 п. о., 75 п. о., 50 п. о., 60 п. о., 40 п. о., 30 п. о., 25 п. о., 2 т.п.о., 1,5 т.п.о., 1 т.п.о., 0,5 т.п.о., 0,25 т.п.о., 0,1 т.п.о. или 0,05 т.п.о. Зонды могут быть получены, например, посредством химического синтеза, ПЦР-амплификации, получения из более длинных полинуклеотидов с использованием ферментов рестрикции или других способов, хорошо известных из уровня техники. Полинуклеотиды, описанные в данном документе, можно также применять в способах или анализах, включающих применение твердых субстратов, таких как чипы. Такой чип может содержать один или несколько различных полинуклеотидов, которые могут быть иммобилизованы на чипах с применением способов, известных из уровня техники.
В некоторых вариантах осуществления полинуклеотид по настоящему изобретению может быть помечен обнаружимой меткой. Обнаружимые метки могут включать без ограничения радиоактивные метки, флуорохромы, в том числе флуоресцеинизотиоцианат (FITC), родамин, техасский красный, фикоэритрин, аллофикоцианин, 6-карбоксифлуоресцеин (6-FAM), 2',7'-диметокси-4',5'-дихлор-6-карбоксифлуоресцеин, 6-карбокси-Х-родамин (ROX), 6-карбокси-2',4',7',4,7-гексахлорфлуоресцеин (HEX), 5-карбоксифлуоресцеин (5-FAM) или N,N,N',N'-тетраметил-6-карбоксиродамин (TAMRA); радиоактивные метки, такие как 32Р, 35S и 3Н, и т.п. В некоторых вариантах осуществления обнаружимая метка может включать несколько стадий (например, биотин-авидин, гаптен-антитело к гаптену и т.п.).
В соответствии с настоящим изобретением можно применять любые подходящие качественные или количественные способы, известные из уровня техники для обнаружения специфических вирусных нуклеиновых кислот (например, РНК или ДНК). Вирусная нуклеиновая кислота, как описано в данном документе, может быть обнаружена, например, посредством гибридизации in situ в срезах ткани с применением способов, которые обнаруживают различия в одной паре оснований между гибридизующимися нуклеиновыми кислотами, посредством ПЦР с обратной транскриптазой или в нозерн-блотах, содержащих mRNA poly А, или другие способы, хорошо известные из уровня техники. Для обнаружения вирусных полинуклеотидов в крови или образцах, полученных из крови, можно применять способы, которые позволяют обнаруживать ошибочно спаренные основания одиночных пар оснований.
Последовательность вирусной нуклеиновой кислоты может присутствовать в биологическом образце, полученном от инфицированного человека, в относительно низких количествах, и, таким образом, методики амплификации, известные из уровня техники (например, ПЦР) можно применять для амплификации последовательности перед проведением анализов гибридизации.
Зонды нуклеиновой кислоты могут быть получены с использованием вирусного генома, как описано в данном документе. Такой зонд может содержать по меньшей мере около 8 нуклеотидов или более и может быть получен синтетическим путем или путем вырезания из рекомбинантных полинуклеотидов. Зонд, описанный в данном документе, может гибридизоваться с вирусной нуклеиновой кислотой и, таким образом, такой зонд может быть применим для обнаружения определенного вируса в биологическом образце. Зонды, описанные в данном документе, также могут быть применимы для идентификации инфицированных субъектов, а также для дополнительной характеристики вирусных геномов. Зонд для обнаружения вирусных полинуклеотидов (природных или производных) может характеризоваться определенной длиной или содержать последовательность, которая позволяет обнаруживать уникальные вирусные последовательности посредством гибридизации. Хотя могут быть применимы около 6-8 нуклеотидов, более длинные последовательности могут быть предпочтительными, например, последовательности из около 10-12 нуклеотидов или около 20 нуклеотидов или больше. Специалисту в данной области техники будет известно, как изготавливать и применять зонд, как описано в данном документе.
Зонды нуклеиновой кислоты могут быть получены с применением обычных способов, в том числе без ограничения автоматизированных способов синтеза олигонуклеотидов. Последовательность, применимая для получения такого зонда, может включать комплемент любой уникальной части вирусного генома, например, части вирусного генома, которая позволяет отличать определенный вирус от других вирусов, которые могут присутствовать в образце. Зонд, описанный в данном документе, может характеризоваться полной комплементарностью в отношении целевой последовательности, представляющей интерес, или может содержать одно или несколько ошибочно спаренных оснований. Зонд, применяемый в соответствии с настоящим изобретением, содержащий одно или несколько ошибочно спаренных оснований, по-прежнему будет гибридизоваться с целевой последовательностью, представляющей интерес.Для использования таких зондов в качестве диагностики анализируемый биологический образец может быть обработан перед анализом, при необходимости, для извлечения нуклеиновых кислот, содержащихся в нем. Полученные нуклеиновые кислоты из образца могут быть подвергнуты гель-электрофорезу или другим методикам разделения на основе размеров. Зонд может быть помечен обнаружимой меткой, как описано в данном документе. Подходящие метки и способы мечения зондов известны из уровня техники и могут включать любые метки, описанные в данном документе, или другие метки, применимые в настоящем изобретении.
Зонд может быть полностью комплементарным вирусному геному или его части (например, всей или части последовательности, кодирующей вирусный белок, как описано в данном документе). Могут быть желательны условия высокой жесткости для предупреждения или по меньшей мере сведения к минимуму ложно положительных результатов. Жесткость гибридизации может определяться несколькими факторами во время гибридизации и промывки, в том числе температурой, ионной силой, продолжительностью времени и концентрацией реагентов. Зонд или нуклеиновая кислота из образца могут быть представлены в растворе для таких анализов или могут быть прикреплены к подложке (например, твердой или полутвердой подложке). Примеры подложек, которые можно применять, включают без ограничения нитроцеллюлозу (например, форму мембраны или лунки), поливинилхлорид (например, листы или микротитрационные лунки), латекс из полистирола (например, гранулы или микротитрационные планшеты, поливинилидинфторид, диазотиро ванную бумагу, нейлоновые мембраны, активированные гранулы и гранулы с белком А).
В одном варианте осуществления зонд или образец нуклеиновой кислоты могут быть представлены на чипе для обнаружения. Чипы могут быть созданы, например, путем нанесения полинуклеотидных зондов на подложку (например, стекло, нитроцеллюлозу и т.п.) в двумерной матрице или чипе. Зонды могут быть связаны с подложкой либо ковалентными связями, либо неспецифическими взаимодействиями, такими как гидрофобные взаимодействия. Образцы полинуклеотидов могут быть помечены обнаружимой меткой (например, с использованием радиоактивных или флуоресцентных меток), а затем гибридизированы с зондами. Двухцепочечные полинуклеотиды, содержащие меченые полинуклеотиды образца, связанные с полинуклеотидами зонда, могут быть обнаружены после удаления несвязанной части образца. Методики конструирования чипов и способы применения этих чипов известны из уровня техники. Чипы можно использовать для анализа одного образца в отношении наличия двух или более целевых областей нуклеиновой кислоты. В таком случае зонды для каждой из целевых областей, а также контроли (как положительные, так и отрицательные) могут быть представлены на одном чипе. Таким образом, чипы облегчают быстрый и удобный анализ.
Диагностические тесты и наборы
В настоящем изобретении дополнительно предусмотрены диагностические реагенты и наборы, содержащие один или несколько таких реагентов, для применения в различных диагностических анализах, в том числе, например, иммунологических анализах, таких как ELISA, и иммунологических анализах типа «сэндвич», а также анализах нуклеиновых кислот, например, анализах на основе ПЦР. В связанном варианте осуществления анализ можно проводить в формате проточного теста или тест-полоски, при этом связывающее средство иммобилизовано на мембране, такой как нитроцеллюлоза. Такие наборы могут предпочтительно содержать по меньшей мере первый пептид, или первое антитело, или антигенсвязывающий фрагмент по настоящему изобретению, их функциональный фрагмент, или их смесь, или первую пару олигонуклеотидов, а также средства для генерирования сигнала. В некоторых вариантах осуществления набор может содержать иммуногенную композицию, такую как рекомбинантный вирус, как описано в данном документе. В набор могут быть включены реагенты и другие соединения, такие как фармацевтически приемлемый носитель. Иммуногенная композиция, представленная в таком наборе, может находиться в растворе, например, в предварительно измеренной дозе или количестве, или может представлять собой сухую композицию, например, в высушенной или лиофилизированной форме, подходящей для регидратации или ресуспендирования. Компоненты набора могут быть предварительно прикреплены к твердой подложке или могут быть нанесены на поверхность твердой подложки при применении набора. Средство генерирования сигнала может быть предварительно ассоциировано с антителом или нуклеиновой кислотой по настоящему изобретению или может требоваться комбинация с одним или несколькими компонентами, например, буферами, нуклеиновыми кислотами, конъюгатами антитело-фермент, ферментными субстратами и т.п. перед применением.
Наборы могут также содержать дополнительные реагенты, например, блокирующие реагенты для снижения неспецифического связывания с поверхностью твердой фазы, промывочные реагенты, ферментные субстраты, ферменты и т.п. Поверхность твердой фазы может быть представлена в форме микротитрационных планшетов, микросфер или других материалов, подходящих для иммобилизации нуклеиновых кислот, белков, пептидов или полипептидов. Фермент, который катализирует образование хемилюминесцентного или хромогенного продукта или восстановление хемилюминесцентного или хромогенного субстрата, является одним из таких компонентов средств, генерирующих сигнал. Такие ферменты хорошо известны из уровня техники. Если в набор включена радиоактивная метка, хромогенная, флуорогенная или обнаружимая метка другого типа или средства обнаружения, метящее средство может быть предоставлено либо в том же контейнере, что и сама диагностическая или терапевтическая композиция, либо в качестве альтернативы может быть помещено во второй отдельный контейнер, в котором эта вторая композиция может быть помещена и подходящим образом разделена на аликвоты. В качестве альтернативы реагент для обнаружения и метка могут быть приготовлены в одном контейнере.
Все указанные патенты и другие публикации непосредственно включены в данный документ посредством ссылки с целью описания и раскрытия, например, методологий, описанных в таких публикациях, которые могут быть применены в связи с настоящим изобретением. Эти публикации предусмотрены исключительно для их раскрытия до даты подачи настоящей заявки.
Настоящее изобретение дополнительно иллюстрируется и поддерживается следующими примерами. Однако эти примеры никоим образом не следует рассматривать как дополнительные ограничения объема настоящего изобретения. В отличие от этого, специалисту в данной области техники будет легко понять, что существуют другие варианты осуществления, модификации и эквиваленты настоящего изобретения без отклонения от сущности настоящего изобретения и/или объема прилагаемой формулы изобретения.
ПРИМЕРЫ
Пример 1
Конструирование плазмид на основе HVT-gfp
Конструирование трансферной тазмиды на основе HVT-зеленого флуоресцентного белка (gfp)-B
Трансферную плазмиду HVT-gfp-B (SEQ ID NO. 18) химически синтезировали GeneArt, Thermo Fisher. 2,5 мкг плазмиды транс фицировали во вторичные клетки CEF с использованием реагента для трансфекции LTX (Invitrogen) в 6-луночном планшете. Через около 4-6 часов трансфицированные клетки инфицировали HVT при moi 0,006 (1,5×104 БОЕ/2,5×106 клеток). Через три дня клетки пассировали 1:15 в отношении Т75 со свежеприготовленными CEF (1×107 клеток/Т75). Затем через три дня клетки высевали в соотношении 1:50 на 24-луночные планшеты. Клетки из лунок, содержащих фокусы зеленого флуоресцентного белка, высевали на 96-луночный планшет в разведениях 1:200, 1:500 и 1:1000 со свежеприготовленными клетками (6×104 клеток/лунка). Лунки, содержащие одиночные фокусы зеленого цвета, очищали в течение 3 раундов посредством метода предельных разведений с использованием 96-луночных планшетов. Очищенный вирус экспандировали с использованием клеток CEF и получали из замороженного запасного раствора. Его обозначали в виде «HVT-gfp-В».
ПЦР-анализ трех очищенных клонов с использованием праймеров непосредственно за пределами сайта интеграции UL55-ген 3 (верхний праймер: SEQ ID NO. 49; нижний праймер: 5'-SEQ ID NO. 50) приводил к образованию полосы размером 1,893 т.п.н., как и предполагали. HVT приводил к образованию полосы размером 0,15 т.п.н., как и ожидали. См. Фиг. 1.
Конструирование модифицированной трансферной плазмиды HVT-gfp-A
Модифицированную трансферную плазмиду HVT-gfp-A (SEQ ID NO. 16) создавали путем применения сайт-специфического мутагенеза с использованием двух пар праймеров (верхние пары праймеров для получения SbfI выше гена gfp: SEQ ID NO.40 и SEQ ID NO.41; нижние праймеры для получения SbfI ниже гена gfp: - SEQ ID NO. 42 и SEQ ID NO.43 для исходной трансферной плазмиды HVT-gfp-A (SEQ ID NO. 17), которую химически синтезировали GeneArt, ThermoFisher. 0,01 мкг модифицированной трансферной плазмиды HVT-gfp-A котрансфицировали 2,5 мкг ДНК HVT с использованием 7,5 мкл PEI (полиэтиленимина) в отношении вторичных клеток CEF на 6-луночном планшете. Фокусы зеленого флуоресцентного белка становились заметными при пассаже 1. После трех раундов очистки методом предельных разведений 1 клон HVT-gfp-A дополнительно экспандировали и получали из замороженного запасного раствора.
ПЦР-анализ очищенного клона (левая полоса) с использованием праймеров непосредственно за пределами сайта интеграции UL35-UL36 (верхний праймер: SEQ ID NO. 44; нижний праймер: SEQ ID NO. 45) приводил к образованию полосы размером 1,922 т.п.н., как и предполагали (Фиг. 4). ДНК модифицированной трансферной плазмиды HVT-gfp-A использовали в качестве контроля (правая полоса). См. Фиг. 2.
Пример 2
Констурирование HVT-IBD
Конструировние HVT-IBD №1
Трансферную плазмиду HVT-IBD №1 (SEQ ID NO. 20) химически синтезировали GeneArt, ThermoFisher. 2,5 мкг плазмиды трансфицировали во вторичные клетки CEF с использованием реагента для трансфекции LTX (Invitrogen) в 6-луночном планшете. Через примерно 4-6 часов трансфицированные клетки инфицировали HVT при moi 0,055. Через три дня клетки пассировали 1:7,5 в отношении Т75 со свежеприготвленными CEF (1×107 клеток/Т75). Затем клетки высевали на 10 96-луночных планшетах и через три дня создавали планшеты в двух повторностях. Один набор планшетов фиксировали и окрашивали куриной антисывороткой к IBDV. Идентифицировали две лунки, содержащие фокусы, положительно окрашенные в отношении IBD. Соответствующие лунки, которые содержат фокусы положительного окрашивания, очищали в течение трех раундов методом предельных разведений с использованием 96-луночных планшетов.
Очищенный вирус экс панд ировали с использованием клеток CEF и получали из замороженного запасного раствора. Его обозначали в виде «HVT-IBD №1».
ГЩР-анализ различных клонов с использованием праймеров непосредственно за пределами сайта интеграции UL55-ген 3 (верхний праймер: SEQ ID NO. 46; нижний праймер: SEQ ID NO. 47, панель А) приводил к образованию полосы размером 2,414 т.п.н., в то время как полоса ПЦР исходного вектора составляет 1,922 т.п.о. Надлежающую интеграцию дополнительно подтверждали с использованием праймеров, окружающих расположенную ниже соединения вставку (верхний праймер SEQ ID NO. 48, который локализован в кодирующей области IBDV VP2; нижний праймер SEQ ID NO.49, локализованный ниже трансферной плазмиды, панель В). Как и ожидали, получали полосу ПЦР размером 1,118 т.п.н. Надлежащую интеграцию для расположенного выше сайта интеграции выполняли с использованием праймеров, окружающих расположенную выше соединения вставку (верхний праймер SEQ ID NO. 50, который локализован выше трансферной плазмиды; нижний праймер SEQ ID NO. 51, который находится в пределах кодирующей области IBDV VP2, панель С). Как и ожидали, получали полосу ПЦР размером 1,428 т.п.н. См. Фиг. 3А, В и С.
Конструировние HVT-IBD №5
Трансферную плазмиду HVT-IBD №5 (SEQ ID NO. 21) химически синтезировали Gene Art, ThermoFisher. Клетки JBJ-1 (клеточная линия фибробластов курицы) в 6-луночном планшете трансфицировали 2,5 мкг плазмиды с использованием реагента для трансфекции LTX (mvitrogen). Транс фи цированные клетки инфицировали HVT при moi 0,05 через примерно 5 часов после трансфекции. Трансфицированные/инфицированные клетки амплифицировали последовательным пассажем (1:4-1:10) и часть затем высевали в 96-луночные планшеты в предельных разведениях. Экспрессию антигена IBDV VP2 оценивали посредством окрашивания монослоев живых клеток антителом без фиксации. См. Фиг. 4А и 4В. Окрашенные фокусы собирали путем трипсинизации клеток с помощью клонирующих цилиндров, помещенных вокруг положительных фокусов. Это «живое окрашивание» с последующим пассажем клонирующего цилиндра повторяли 4 раза и получали чистые VP2 положительные культуры. Культуры амплифицировали посредством серийных пассажей на клетках JBJ-1 и перед окончательной амплификации на первичных клетках CEF в роллерных флаконах. Собранные клетки CEF использовали для приготовления замороженного раствора клеток и обозначали в виде «HVT-IBD №5».
ПЦР-анализ клона №7 выполняли с использованием 2 наборов праймеров для подтверждения интеграции вставки в оба сайта вставки. В ПЦР А верхний праймер (SEQ ID NO.52) связывается с кодирующей областью IBDV VP2, в то время как нижний праймер (SEQ ID NO.53) связывается ниже сайта интеграции UL35-UL36. Этот набор праймеров приводил к образованию полосы ПЦР размером 1,244 т.п.н., как и ожидали. В ПЦР В верхний праймер (SEQ ID NO. 54) связывается выше сайта вставки UL35-UL36, а нижний праймер (SEQ ID NO.55) связывается в пределах промотора CMV человека вставки и приводит к образованию полосы ПЦР размером 0,926 т.п.н., как и ожидали. См. Фиг. 5.
Конструирование HVT-IBD №6а
Трансферную плазмиду HVT-IBD №6а (SEQ ID NO.22) химически синтезировали BioBasic Inc. 0,1 мкг и 0,01 мкг линеаризованной трансферной плазмиды (посредством расщепления EcoR1 и HindIII) котрансфицировали 2,5 мкг HVT-gfp-A, которую расщепляли Sbfl во вторичных клетках CEF с использованием реагента для трансфекции PEI (полиэтиленимин) в 6-луночном планшете. Через 4 дня после трансфекции 4 фокуса, не характеризующиеся зеленым цветом, наблюдали при трансфекции 0,01 мкг трансферной плазмидой и 3 фокуса, не характеризующиеся зеленым цветом, наблюдали при трансфекции 0,1 мкг трансферной плазмидой, в то время как при использовании HVT-gfp-A, расщепленной только Sbf1, фокусов не наблюдали. 2 фокуса, не характеризующиеся зеленым цветом, очищали 3 раза методом предельных разведений с использованием 96-луночного планшета. Очищенный вирус экспандировали с использованием клеток CEF и получали из замороженного запасного раствора. Его обозначали в виде «HVT-IBD №6а».
Готовили инфицированный клеточный лизат и проводили вестерн-блоттинг с использованием моноклонального антитела к IBDV R63. Полоса белка около 50 кДа была заметна на всех полосах, за исключением полосы, содержащей лизат вектора HVT-gfp-A. См. Фиг. 6.
ПЦР-анализ клонов с использованием 2 наборов праймеров для подтверждения надлежащей интеграции. Первый набор праймеров, нацеленный на расположенный выше сайт интеграции: верхний праймер 5' SEQ ID NO. 56, локализованный выше сайта интеграции UL35-UL36; нижний праймер SEQ ID NO. 57, который локализован в пределах промотора Рес. Этот набор праймеров, как и ожидали, приводил к образованию полосы ПЦР размером 0,911 т.п.н. Второй набор праймеров, нацеленный на расположенный ниже сайт интеграции: верхний праймер 5' SEQ ID NO. 58, который локализован в кодирующей области IBDV VP2; нижний праймер 5' SEQ ID NO. 59, который локализован ниже сайта вставки UL35-UL36. Как и ожидали, получали полосу ПЦР размером 1,244 т.п.н. См. Фиг. 7А и 7В.
Конструироение HVT-IBD №9
Трансферную плазмиду HVT-IBD №9 (SEQ ID NO.23) химически синтезировали GeneArt, Thermo Fisher. 2,5 мкг плазмиды транс фицировали во вторичные клетки CEF с использованием реагента для трансфекции LTX (Invitrogen) в 6-луночном планшете. Через около 4-6 часов трансфицированные клетки инфицировали HVT-gfp-B при moi 0,075. Через три дня клетки пассировали 1:10 в отношении Т75 со свежеприготовленными CEF (1×107 клеток/Т75) 3 раза. Затем клетки высевали на 10 96-луночных планшетах и получали 90 фокусов, не характеризующихся зеленым цветом. Три из них были положительно окрашены куриной антисывороткой к IBDV. Два клона очищали в течение 3 раундов методом предельных разведений с использованием 96-луночных планшетов. Очищенный вирус экспандировали с использованием клеток CEF и получали из замороженного запасного раствора. Его обозначали в виде «HVT-IBD №9».
ПЦР-анализ различных клонов с использованием праймеров непосредственно за пределами сайта интеграции UL55-ген 3 (верхний праймер: SEQ ID NO.60; нижний праймер: SEQ ID NO. 61, панель А) приводил к образованию полосы размером 2,536 т.п.н., в то время как полоса ПЦР исходного вектора HVT-gfp-B составляет 1,922 т.п.о. Надлежающую интеграцию дополнительно подтверждали с использованием праймеров, окружающих расположенную выше соединения вставку (верхний праймер SEQ ID NO. 62, который локализован выше сайта вставки UL55-ген 3; нижний праймер SEQ ID NO. 63, локализованный в кодирующей области IBDV VP2. Как и ожидали, получали полосу ПЦР размером 1,482 т.п.н. Надлежающую интеграцию расположенного ниже сайта осуществляли с использованием праймеров, окружающих расположенную ниже соединение вставку (верхний праймер SEQ ID NO. 64, который локализован в кодирующей области IBDV VP2; нижний праймер SEQ ID NO. 65, который локализован ниже сайта вставки UL55-ген 3. Как и ожидали, получали полосу ПЦР размером 1,166 т.п.н. См. Фиг. 8А и В.
Трансферную плазмиду HVT-IBD №30 (SEQ ID NO.24) химически синтезировали GeneArt, ThermoFisher. Вторичные клетки CEF котрансфицировали ОД мкг плазмиды и 2,5 мкг HVT с использованием реагента для трансфекции PEI (полиэтиленимин) в 6-луночном планшете. Через три дня клетки пассировали в соотношении 1:12 со свежеприготовленными клетками CEF. Фокусы, экспрессирующие IBD VP2, визуализировали посредсвтом окрашивания нефиксированных культур куриной поликлональной антисывороткой к IBDV и маркировали эти фокусы с помощью флуоресцентного микроскопа. В общей сложности 16 положительных фокусов пассировали со свежеприготовленными клетками CEF посредством трипсинизации с использованием клонирующих цилиндров для отделения фокусов от фокусов, не экспрессирующих VP2. Четыре из этих культур клонировали три раза по одной и той же процедуре до амплифицикации на первичных клетках CEF в роллерных флаконах. Замороженный запасной раствор клеток откладывали и обозначали в виде «HVT-IBD №30».
ПЦР-анализ 4 различных клонов с использованием праймеров для расположенной выше области сайта интеграции UL55-ген 3 (верхний праймер: SEQ ID NO.66; нижний праймер: SEQ ID NO.67, панель А) приводил к образованию полосы размером 1,673 т.п.о. Надлежающую интеграцию дополнительно подтверждали с использованием праймеров, окружающих 3' соединение вставки (верхний праймер SEQ ID NO. 68, который локализован в кодирующей области IBDV VP2; нижний праймер SEQ ID NO. 69, который локализован ниже сайта вставки UL55-ген 3, панель В). Как и ожидали, получали полосу ПЦР размером 1,082 т.п.н. Надлежащую интеграцию расположенного ниже сайта дополнительно подтверждали с использованием праймеров за пределами кассеты экспрессии (верхний праймер SEQ ID NO.70; нижний праймер SEQ ID NO. 71 (панель С). Как и ожидали, получали полосу ПЦР размером 2,558 т.п.н. См. Фиг. 9А-С.
Копструировние HVT-IBD №31
Трансферную плазмиду HVT-IBD №31 (SEQ ID NO. 25) химически синтезировали GeneArt, ThermoFisher. 0,1 мкг и 0,01 мкг линеаризованной трансферной плазмиды (посредством расщепления EcoR1 и HindIII) котрансфицировали 2,5 мкг HVT-gfp-A, которую расщепляли Sbf1 во вторичных клетках CEF с использованием реагента для трансфекции PEI (полиэтиленимин) в 6-луночном планшете. Через 4 дня после трансфекции наблюдали 1 фокус, не характеризующийся зеленым цветом, в то время как при использовании HVT-gfp-A, расщепленного только Sbf1, фокусов не наблюдали. После пассажа 2 фокуса, не характеризующихся зеленым цветом, очищали 3 раза методом предельных разведений с использованием 96-луночного планшета. Очищенный вирус экспандировали с использованием клеток CEF и получали из замороженного запасного раствора. Его обозначали в виде «HVT-IBD №31».
Готовили инфицированный клеточный лизат и проводили вестерн-блоттинг с использованием моноклонального антитела к IBDV R63. Полоса белка около 50 кДа была заметна на всех полосах с куриной антисывороткой к IBDV только в качестве зонда, но не с mAb для IBDV R63. См. Фиг. 10А и В.
ПЦР-анализ клонов с использованием 2 наборов праймеров для подтверждения надлежащей интеграции. Первый набор праймеров, нацеленный на расположенный выше сайт интеграции: верхний праймер SEQ ID NO. 72, который локализован выше сайта интеграции UL35-UL36; нижний праймер SEQ ID NO. 73, который локализован в пределах промотора бета-актина кур. Этот набор праймеров, как и ожидали, приводил к образованию полосы ПЦР размером 0,835 т.п.н. Второй набор праймеров, нацеленный на расположенный ниже сайт интеграции: верхний праймер SEQ ID NO.76, который локализован в кодирующей области IBDV VP2; нижний праймер SEQ ID NO.77, который локализован ниже сайта вставки UL35-UL36. Как и ожидали, получали полосу ПЦР размером 1,248 т.п.н. См. Фиг. 11А и В.
Конструирование HVT-IBD №34
Трансферную плазмиду HVT-IBD №34 (SEQ ID NO. 28) химически синтезировали GeneArt, ThermoFisher. 2,5 мкг плазмиды трансфицировали во вторичные клетки CEF с использованием реагента для трансфекции LTX (Invitrogen) в 6-луночном планшете. Через около 4-6 часов трансфицированные клетки инфицировали HVT-gfp-B при moi 0,05. Через три дня клетки пассировали 1:15 в отношении Т75 со свежеприготовленными CEF (1×107 клеток/Т75). Инфицированные клетки высевали на 10 × 96-луночных планшетах, выращивали в течение 3 дней, затем пересевали в 96-луночные планшеты в двух повторностях. Одну повторность фиксировали и окрашивали куриной антисывороткой к IBDV, и идентифицировали 3 лунки, содержащие фокусы, которые окрашивались положительно в отношении IBDV. Соответствующие лунки из планшета с живыми клетками для выполнения повторности очищали посредством 3 раундов клонирования посредством предельного разведения с использованием 96-луночных планшетов. Один из очищенных вирусов экспандировали с использованием клеток CEF и получали из замороженного запасного раствора. Его обозначали в виде «HVT-IBD №34».
ГЩР-анализ 3 различных клонов с использованием праймеров для расположенной выше области сайта интеграции ген 3-UL55 (верхний праймер: SEQ ID NO.78; нижний праймер: SEQ ID NO.79, который локализован в промоторе бета-актина кур, SEQ ID NO:79, панель А) приводил к образованию полосы размером 0,815 т.п.н., как и ожидали. Надлежащую интеграцию дополнительно подтверждали с использованием праймеров, окружающих расположенную ниже соединения вставку (верхний праймер SEQ ID NO. 80, который локализован в кодирующей области IBDV VP2; нижний праймер SEQ ID NO.81, который локализован ниже сайта вставки ген 3-UL55, панель В). Как и ожидали, получали полосу ПЦР размером 1,296 т.п.н. Надлежащую конструкцию дополнительно подтверждали с использованием праймеров за пределами кассеты экспрессии (верхний праймер SEQ ID NO.82; нижний праймер SEQ ID NO.83 (панель С). Как и ожидали, получали полосу ПЦР размером 3,001 т.п.н. См. Фиг. 12А-С.
Конструирование HVT-ND №38
Трансферную плазмиду HVT-IBD №38 (SEQ ID NO.29) химически синтезировали BioBasic, Inc. Трансферную плазмиду HVT-ND №38, расщепленную HindIII и ApoI, котрансфицировали Sbf1, расщепленной ДНК HVT-gfp-A с использованием PEI (полиэтиленимин, 7,5 мкл) в 6-луночном планшете с вторичными клетками CEF. Через 6 дней после трансфекции трансфицированные клетки высевали на 96-луночный планшет и живыми окрашивали куриной антисывороткой к NDV. Семь лунок, содержащих фокусы с положительным окрашиванием, очищали 3 раза посредством предельного разведения. Один из очищенных вирусов экспандировали с использованием клеток CEF и получали из замороженного запасного раствора. Его обозначали в виде «HVT-ND №38».
ПЦР-анализ 5 различных клонов с использованием праймеров для расположенной выше области сайта интеграции UL35-UL36 (верхний праймер: SEQ ID NO. 84; нижний праймер: SEQ ID NO.85, который локализован в пределах кодирующей области NDV F, панель А) приводил к образованию полосы размером 2,122 т.п.о. Надлежащую интеграцию дополнительно подтверждали с использованием праймеров, окружающих расположенную 3' соединения вставку (верхний праймер SEQ ID NO. 86, который локализован в кодирующей области NDV F; нижний праймер SEQ ID NO.87, который локализован ниже сайта вставки UL35-UL36, панель В). Как и ожидали, получали полосу ПЦР размером 1,127 т.п.н. Надлежащую конструкцию дополнительно подтверждали с использованием праймеров за пределами кассеты экспрессии (верхний праймер SEQ ID NO. 88; нижний праймер SEQ ID NO. 89 (панель С). Как и ожидали, получали полосу ПЦР размером 3,657 т.п.н. См. Фиг. 15А и В.
Конструирование HVT-ND №39
Трансферную плазмиду HVT-IBD №39 (SEQ ID NO. 30) химически синтезировали BioBasic, Inc. Вторичные клетки CEF котрансфицировали 0,01 мкг плазмиды и 2,5 мкг HVT с использованием реагента для трансфекции PEI (полиэтиленимин) в 6-луночном планшете. Через шесть дней клетки пассировали в соотношении 1:24 со свежеприготовленными клетками CEF. Через три дня после пассажа фокусы, экспрессирующие белок NDV F, визуализировали посредством окрашивания нефиксированных культур поликлональной специфичной куриной антисывороткой к NDV, и эти фокусы маркировали с помощью флуоресцентного микроскопа. В общей сложности 4 положительных фокусов пассировали со свежеприготовленными клетками CEF посредством трипсинизации с использованием клонирующих цилиндров для отделения фокусов от фокусов, не экспрессирующих белок F. Четыре из этих культур клонировали три раза по одной и той же процедуре до амплифицикации на первичных клетках CEF в роллерных флаконах. Замороженный запасной раствор клеток (клон 2 на фигурах) откладывали и обозначали в виде «HVT-ND №39».
ПЦР-анализ 3 различных клонов с использованием праймеров для расположенной выше области сайта интеграции UL35-UL36 (верхний праймер: SEQ ID NO 90; нижний праймер: SEQ ID NO. 92, который локализован в промоторе бета-актина кур, SEQ ID NO:79, панель А) приводил к образованию полосы размером 0,835 т.п.н. Надлежащую интеграцию дополнительно подтверждали с использованием праймеров, окружающих расположенную ниже соединения вставку (верхний праймер SEQ ID NO.92, который локализован в кодирующей области poly А; нижний праймер SEQ ID NO.93, который локализован ниже сайта вставки UL35-UL36, панель В). Как и ожидали, получали полосу ПЦР размером 0,856 т.п.н. Надлежащую конструкцию дополнительно подтверждали с использованием праймеров за пределами кассеты экспрессии (верхний праймер SEQ ID NO. 94; нижний праймер SEQ ID NO. 95 (панель С). Как и ожидали, получали полосу ПЦР размером 3,449 т.п.н. См. Фиг. 16А и В.
Конструирование HVT-ND №40
Трансферную плазмиду HVT-IBD №40 (SEQ ID NO.31) химически синтезировали BioBasic, Inc. Трансферную плазмиду HVT-ND №40, расщепленную HindIII, котрансфицировали Sbf1, расщепленной ДНК HVT-gfp-A с использованием PEI (полиэтиленимин, 7,5 мкл) в 6-луночном планшете с вторичными клетками CEF. Через 7 дней после трансфекции трансфицированные клетки высевали на 24-луночные планшеты и живыми окрашивали куриной антисывороткой к NDV. Четыре лунки, содержащие фокусы с положительным окрашиванием, очищали 3 раза посредством предельного разведения. Один из очищенных вирусов экспандировали с использованием клеток CEF и получали из замороженного запасного раствора. Его обозначали в виде «HVT-ND №40».
ПЦР-анализ 4 различных клонов с использованием праймеров для расположенной выше области сайта интеграции UL35-UL36 (верхний праймер: SEQ ID NO.96; нижний праймер: SEQ ID NO.97, который локализован в промоторе бета-актина кур, SEQ ID NO:79, панель А) приводил к образованию полосы размером 0,835 т.п.н. Надлежащую интеграцию дополнительно подтверждали с использованием праймеров, окружающих расположенную ниже соединения вставку (верхний праймер SEQ ID NO.98, который локализован в кодирующей области NDV F; нижний праймер SEQ ID NO. 99, который локализован ниже сайта вставки UL35-UL36, панель В). Как и ожидали, получали полосу ПЦР размером 0,856 т.п.н. Надлежащую конструкцию дополнительно подтверждали с использованием праймеров за пределами кассеты экспрессии (верхний праймер SEQ ID NO. 100; нижний праймер SEQ ID NO. 101 (панель С). Как и ожидали, получали полосу ПЦР размером 3,449 т.п.н. См. Фиг. 17 А-С.
Конструирование HVT-ND №42
Исходную трансферную плазмиду HVT-ND №42 (SEQ ID NO. 33) химически синтезировали компанией BioBasic, Inc. Клонирующую плазмиду химически синтезировали посредством амплификации DNA2.0.PCR кассеты экспрессии гена NDV F трансферной плазмиды HVT-ND №42 с использованием следующих праймеров: верхний праймер, SEQ ID NO. 102; нижний праймер, 5' SEQ ID NO. 103. Амплифицированный фрагмент ПЦР клонировали в сайты AscI и NheI UL55/ген 3 для получения конечной трансферной плазмиды HVT ND №42 (SEQ ID NO.35).
Трансферную плазмиду трансфицировали в клетки CEF, инфицированные HVT, с использованием липофектамина LTX в 6-луночном планшете. Через 3 дня после трансфекции трансфицированные/инфицированные клетки высевали на 6-луночный планшет в двух повторностях, а затем на 96-луночный планшет для скрининга фокусов, экспрессирующих ND, посредством окрашивания антисывороткой к NDV. Лунки, соответствующие фокусам экспрессии ND, очищали 3 раза посредством предельного разведения. Один из очищенных вирусов экспандировали с использованием клеток CEF и получали из замороженного запасного раствора. Его обозначали в виде «HVT-ND №42».
ПЦР-анализ одного конечного клона с использованием праймеров за пределами кассеты экспрессии (набор праймеров 1: верхний праймер SEQ ID NO. 104; нижний праймер SEQ ID NO. 105) приводил к образованию полосы размером 3,597 т.п.н., как и ожидали. Четыре набора праймеров для расположенной выше области интеграции UL55-ген 3, при этом все верхние праймеры были расположены выше и за пределами кассеты экспрессии, а все нижние праймеры были расположены в кодирующей области ND F. Набор праймеров 2: верхний праймер: SEQ ID NO. 106, нижний праймер: SEQ ID NO. 107, который приводил к образованию полосы размером 2,243 т.п.о.; набор праймеров 3: верхний праймер: SEQ ID NO. 108; нижний праймер: SEQ ID NO. 109, который приводил к образованию полосы ПЦР размером 2,356 т.п.о. Набор праймеров 4: верхний праймер: SEQ ID NO. 110; нижний праймер: SEQ ID NO. 111, который приводил к образованию полосы ПЦР размером 2,424 т.п.о. Набор праймеров 5: верхний праймер: SEQ ID NO. 112; нижний праймер: SEQ ID NO. 113, который приводил к образованию полосы ПЦР размером 2,170 т.п.о. Надлежающую интеграцию дополнительно подтверждали с использованием праймеров, окружающих расположенную ниже соединения вставку (набор праймеров 6: верхний праймер SEQ ID NO. 114, который локализован в кодирующей последовательности гена NDV F; нижний праймер SEQ ID NO. 115, который локализован ниже сайта вставки UL55-ген 3, панель С). Как и ожидали, получали полосу ПЦР размером 0,971 т.п.н. См. Фиг. 18.
Трансферную плазмиду HVT-IBD №44 (SEQ ID NO.36) химически синтезировали BioBasic, Inc. Трансферную плазмиду №44 расщепляли рестрикционными ферментами EcoRI и HindIII для высвобождения вставки из плазмидных последовательностей, а полученную расщепленную ДНК (10 нг) использовали вместе с 2,5 мкг ДНК HVT-gfpB для котрансфекции вторичных клеток PEI (полиэтиленимином). Через четыре дня после трансфекции трансфицированные клетки пассировали в соотношении 1:6 со свежеприготовленными вторичными клетками и живыми окрашивали куриной поликлональной антисывороткой к NDV для идентификации фокусов, экспрессирующих NDV, через три или четыре дня после пассажа. Три положительно окрашенных фокуса собирали путем трипсинизации с использованием клонирующих цилиндров. Собранные клетки серийно разводили и высевали на свежеприготовленные вторичные клетки CEF. Этот процесс повторяли каждые три или четыре дня до тех пор, пока окрашивание NDV не демонстрировало гомогенность, а затем выполняли четыре последовательных клонирования. Затем клонированную культуру амплифицировали и готовили из замороженного запасного раствора. Замороженный запасной раствор обозначали в виде «HVT-ND №44».
ПЦР-анализ 1 конечного клона с использованием праймеров для расположенной выше области сайта интеграции UL55-ген 3 (верхний праймер: SEQ ID NO. 116, который локализован выше UL55; нижний праймер: SEQ ID NO. 117, который локализован в промоторе бета-актина кур, SEQ ID NO:79, панель А) приводил к образованию полосы размером 0,71 т.п.н.; аналогично локализованная пара праймеров: верхний праймер: SEQ ID NO. 118, нижний праймер: SEQ ID NO. 119 (панель В, которая приводила к образованию полосы ПЦР размером 0,965 т.п.о.). Надлежающую интеграцию дополнительно подтверждали с использованием праймеров, окружающих расположенную ниже соединения вставку (верхний праймер SEQ ID NO. 120, который локализован в кодирующей последовательности гена NDV F; нижний праймер SEQ ID NO. 121, который локализован ниже сайта вставки UL55-ген 3, панель С). Как и ожидали, получали полосу ПЦР размером 0,971 т.п.н. Надлежащую конструкцию дополнительно подтверждали с использованием праймеров за пределами кассеты экспрессии (верхний праймер SEQ ID NO. 122; нижний праймер SEQ ID NO. 123 (панель D). Как и ожидали, получали полосу ПЦР размером 3,438 т.п.н.
Конструирование HVT-ND №45
Трансферную плазмиду HVT-IBD №45 (SEQ ID) химически синтезировали BioBasic, Inc. Эту плазмиду трансфицировали в клетки CEF, которые инфицировали HVT-GFP-B, с использованием липофектамина LTX в 6-луночном планшете. Через 3 дня после трансфекции трансфицированные/инфицированные клетки высевали на 96-луночный планшет для скрининга GFP отрицательных фокусов. Лунки, содержащие GFP отрицательные фокусы, очищали 3 раза посредством предельного разведения. Очищенные вирусы окрашивали IFA куриной антисывороткой к NDV для подтверждения экспрессии гена NDV F. Один из очищенных вирусов экспандировали с использованием клеток CEF и получали из замороженного запасного раствора. Его обозначали в виде «HVT-ND №45».
ПЦР-анализ 1 конечного клона с использованием праймеров за пределами кассеты экспрессии (набор праймеров 1: верхний праймер SEQ ID NO. 124; нижний праймер SEQ ID NO. 125) приводил к образованию полосы размером 2,830 т.п.н., как и ожидали. Два набора праймеров для расположенной выше области интеграции ген 3-UL55, при этом оба верхних праймера были расположены выше и за пределами кассеты экспрессии, а оба нижних праймера были расположены в кодирующей области ND F.. Набор праймеров 2: верхний праймер: SEQ ID NO. 126; нижний праймер: SEQ ID NO. 127, который приводил к образованию полосы размером 1,635 т.п.о.; набор праймеров 3: верхний праймер SEQ ID NO. 128; нижний праймер: SEQ ID NO. 129, который приводил к образованию полосы ПЦР размером 1,588 т.п.о. Надлежающую интеграцию дополнительно подтверждали с использованием 2 наборов праймеров, окружающих расположенную ниже соединения вставку: набор праймеров 4: верхний праймер SEQ ID NO. 130, который локализован в кодирующей последовательности гена NDV F; нижний праймер SEQ ID NO. 131, который локализован ниже сайта вставки ген 3-UL55. Как и ожидали, получали полосу ПЦР размером 0,993 т.п.н. Набор праймеров 5: верхний праймер: SEQ ID NO. 132; нижний праймер: SEQ ID NO. 133, как и ожидали, получали полосу ПЦР размером 1,137 т.п.н. См. Фиг. 19.
Конструирование HVT-ND №46
Трансферную плазмиду HVT-ND №46 (SEQ ID NO.38) химически синтезировали BioBasic, Inc. Эту плазмиду трансфицировали вместе с вирусной ДНК HVT-GFP-B в клетки CEF с использованием PEI (полиэтиленимина, 7,5 мкл) в 6-луночном планшете. Через 4 дня после трансфекции трансфицированные клетки высевали на 96-луночный планшет для скрининга GFP отрицательных фокусов. Лунки, содержащие GFP отрицательные фокусы, очищали 3 раза посредством предельного разведения. Очищенные вирусы окрашивали IFA куриной антисывороткой к NDV для подтверждения экспрессии гена NDV F. Один из очищенных вирусов экспандировали с использованием клеток CEF и получали из замороженного запасного раствора. Его обозначали в виде «HVT-ND №46».
ПЦР-анализ 1 конечного клона с использованием праймеров за пределами кассеты экспрессии (верхний праймер SEQ ID NO. 134; нижний праймер SEQ ID NO. 135) приводил к образованию полосы размером 3,597 т.п.н., как и ожидали. Один набор праймеров для расположенной выше области интеграции ген 3-UL55, при этом верхние праймеры были расположены выше и за пределами кассеты экспрессии (SEQ ID NO. 136), а нижние праймеры были расположены в промоторе CMV мыши (верхний праймер SEQ ID NO. 137), который приводил к образованию полосы ПЦР размером 1,107 т.п.о., как и ожидали. Надлежащую интеграцию дополнительно подтверждали с использованием 4 наборов праймеров, окружающих расположенную ниже соединения вставку: Р1: верхний праймер SEQ ID NO. 138, который локализован в кодирующей последовательности гена NDV F; нижний праймер SEQ ID NO. 139, который локализован ниже и за пределами кассеты экспрессии. Как и ожидали, получали полосу ПЦР размером 1,003 т.п.н. Р2: верхний праймер: SEQ ID NO. 140, нижний праймер: SEQ ID NO. 141, как и ожидали, получали полосу ПЦР размером 1,147 т.п.н. РЗ: верхний праймер: SEQ ID NO. 142, нижний праймер: SEQ ID NO. 143, как и ожидали, получали полосу ПЦР размером 1,019 т.п.н. Р4: верхний праймер: SEQ ID NO. 144, нижний праймер: SEQ ID NO. 145, как и ожидали, получали полосу ПЦР размером 1,018 т.п.н. См. Фиг. 20 А-С.
Конструирование HVT-ND №48
Линеаризованную трансферную плазмиду для HVT-ND №48 (SEQ ID NO.39) котрансфицировали ДНК HVT-gfp-B с использованием PEI (полиэтиленимина, 7,5 мкл) в 6-луночном планшете с вторичными клетками CEF. Через 4 дней после трансфекции трансфицированные клетки высевали на 96-луночный планшет и живыми окрашивали куриной антисывороткой к NDV. Обнаруживали четыре фокуса, не характеризующиеся зеленым цветом, и 2 были положительно окрашены с использованием куриной антисыворотки к NDV. См. Фиг. 33А и В. Два клона очищали 3 раза посредством предельного разведения. Очищенный вирус экспандировали с использованием клеток CEF и получали из замороженного запасного раствора. Его обозначали в виде «HVT-ND №48».
Трансферную плазмиду HVT-IBD №48 химически синтезировали BioBasic, Inc. Трансферную плазмиду расщепляли EcoRI и HindIII для высвобождения вставки из плазмидных последовательностей, а полученную расщепленную ДНК (10 нг) использовали вместе с 2,5 мкг ДНК HVT-gfp-B для котрансфекции вторичных клеток PEI (полиэтиленимином). Через три дня после трансфекции транс фицированные клетки пассировали в соотношении 1:6 со свежеприготовленными вторичными клетками и живыми окрашивали куриной поликлональной антисывороткой к NDV для идентификации фокусов, экспрессирующих NDV, через четыре дня после пассажа. Три положительно окрашенных фокуса собирали путем трипсинизации с использованием клонирующих цилиндров. Собранные клетки серийно разводили и высевали на свежеприготовленные вторичные клетки CEF. Этот процесс повторяли каждые три или четыре дня до тех пор, пока окрашивание NDV не демонстрировало гомогенность, а затем выполняли четыре последовательных клонирования. Затем клонированную культуру амплифицировали из шестилуночного планшета в колбу объемом 75 см2, затем в колбу объемом 225 см2 перед окончательной амплификацией в роллерной колбе на 850 см2 с использованием первичных клеток CEF. Конечную культуру собирали и обозначали в виде «HVT-ND №48».
ПЦР-анализ 1 конечного клона с использованием праймеров для расположенной выше области сайта интеграции ген 3-UL55 (верхний праймер: SEQ ID NO. 146; нижний праймер: SEQ ID NO: 147, который локализован в промоторе бета-актина кур, панель А), как и ожидали, приводил к образованию полосы размером 0,815 т.п.н.; надлежащую интеграцию дополнительно подтверждали с использованием праймеров, окружающих расположенную ниже соединения вставку (верхний праймер SEQ ID NO. 148, который локализован в кодирующей области NDV F; нижний праймер SEQ ID NO. 149, локализованный ниже сайта вставки ген 3-UL55, панель В). Как и ожидали, получали полосу ПЦР размером 1,003 т.п.н. Другие аналогично локализованные праймеры: верхний праймер: SEQ ID NO. 150; нижний праймер: SEQ ID NO. 151 (панель С), которая приводила к образованию полосы ПЦР размером 1,147 т.п.н., как и ожидали. Надлежащую конструкцию дополнительно подтверждали с использованием праймеров за пределами кассеты экспрессии (верхний праймер SEQ ID NO. 152; нижний праймер SEQ ID NO. 153 (панель D). Как и ожидали, получали полосу ПЦР размером 3,430 т.п.н.
Пример 3
Тест в отношении эффективности против IBDV in vivo HVT-IBD
№9, №34 у SPF птиц против IBDV
Два рекомбинанта HVT-IBD, HVT-IBD №9, №34 тестировали в отношении их эффективности in vivo против заражения вирулентным IBDV (штамм STD, предоставленный USDA) у SPF птиц. В этом исследовании использовали положительный контроль в виде коммерческой вакцины Vaxxitek (Merial). 1500 БОЕ каждого рекомбинантного вируса вводили in ovo в El8. Обратный титр каждого вакцинного вируса также определяли для каждого рекомбинанта после вакцинации. В то время как 100% HVT-IBD №9 экспрессируют антиген VP2 IBDV, было обнаружено, что только 96% HVT-IBD №34 экспрессируют этот антиген. Заражение IBDV STD проводили в день 28 в соответствии с инструкцией USDA. Вскрытие всех птиц проводили через 5 дней после заражения. Наблюдали 100% защиту от HVT-IBD №9 и 90% защиту от HVT-IBD №34, в то время как положительный контроль Vaxxitek обеспечивал защиту 97%.

Пример 4
Тест в отношении эффективности против IBDV in vivo HVT-IBD
№1, №5, №6a, №9, №30, №34 SPF птиц
Шесть рекомбинантов HVT-IBD, HVT-IBD №1, №5, №6а, №9, №30, №34 тестировали в отношении их эффективности in vivo против заражения вирулентным IBDV (штамм STD, предоставленный USDA) у SPF птиц. В этом исследовании использовали положительный контроль в виде коммерческой вакцины Vaxxitek (Merial). 1500 БОЕ каждого рекомбинантного вируса вводили in ovo в E18. Обратный титр каждого вакцинного вируса также определяли для каждого рекомбинанта после вакцинации. Было обнаружено, что все рекомбинанты характеризуются 100% экспрессией антигена VP2 IBDV. Заражение IBDV STD проводили в день 28 в соответствии с инструкцией USDA. Вскрытие всех птиц проводили через 5 дней после заражения. Наблюдали 100% защиту от HVT-IBD №9 и 96% защиту от HVT-IBD №1, №30, №34, и 92% защиту от HVT-IBD №6а, в то время как положительный контроль Vaxxitek обеспечивал защиту 92%.

Пример 5:
Серологические ответы в отношении IBDV HVT-IBD
№1, №5, №9, №15 коммерческих птиц-бройлеров
Серологические ответы в отношении антигена IBDV измеряли с использованием коммерческого набора для ELISA ProFlok ND plus. 1500 БОЕ (0,2 мл) каждого рекомбинанта (HVT-IBD №1, №5, №9, №15) вводили подкожно (SC) цыплятам в возрасте 1 дня. Образцы сыворотки крови выделяли в дни 12, 19, 26, 33, 39, 47 и 54, и их можно обнаружить в Табл. 3 ниже. Процент положительных образцов для каждой конструкции в динамике показан на Фиг. 13.

Пример 6:
Серологические ответы в отношении IBDV HVT-IBD
№6а, №30, №31 коммерческих птиц-бройлеров
Серологические ответы в отношении антигена IBDV измеряли с использованием коммерческого набора для ELISA ProFlok IBD plus. 1500 БОЕ (0,2 мл) каждого рекомбинанта (HVT-IBD №6а, №30, №31) вводили подкожно (SC) цыплятам в возрасте 1 дня. Образцы сыворотки крови выделяли в дни 12, 19, 26, 33, 39, 47 и 54, и они представлены в Табл. 4 ниже. Процент положительных образцов для каждой конструкции в динамике показан на Фиг. 14.

Пример 7:
Тест в отношении эффективности in vivo HVT-ND
№38, №39, №44, №48 у SPF птиц
Четыре рекомбинанта HVT-IBD, HVT-IBD №38, №39, №44, №48 тестировали в отношении их эффективности in vivo против заражения вирулентным NDV (штамм Texas GB, предоставленный USDA) у SPF птиц. В этом исследовании использовали положительный контроль в виде коммерческой вакцины Vectormune ND (Ceva). 1500 БОЕ каждого рекомбинантного вируса вводили in ovo в Е18. Обратный титр вакцинного вируса также определяли для каждого рекомбинанта после вакцинации. В то время как 100% HVT-ND №38 и №48 экспрессируют антиген NDV F, было обнаружено, что только 95-96% HVT-IBD №39 и №44 экспрессируют этот антиген. Заражение NDV Texas GB проводили в день 28 в соответствии с инструкцией USDA. Наблюдение за всеми птицами проводили через 2 недели после заражения. Наблюдали 90% защиту от HVT-ND №38 и №48, и 50% защиту от HVT-ND №39, и 60% защиту от HVT-ND №44, в то время как положительный контроль Vectormune ND обеспечивал защиту 90%. См. Табл. 5 ниже.

Ответ антител на различные вакцины-кандидаты HVT-ND анализировали с использованием набора ProFlok ND plus (Zoetis LLC). Все титры включали без использования порогового значения (345), рекомендованного набором. Процент птиц с положительным титром ND показан в Табл. 6 ниже.
Пример 8:
Тест в отношении эффективности против NDV in vivo HVT-ND
№40, №42, №45, №46 у SPF птиц
Четыре рекомбинанта HVT-IBD, HVT-IBD №40, №42, №45, №46 тестировали в отношении их эффективности in vivo против заражения вирулентным NDV (штамм Texas GB, предоставленный USDA) у SPF птиц. В этом исследовании использовали положительный контроль в виде коммерческой вакцины Vectormune ND (Ceva). 1500 БОЕ каждого рекомбинантного вируса вводили in ovo в Е18. Обратный титр вакцинного вируса определяли для каждого рекомбинанта после вакцинации. В то время как 100% HVT-ND №42, №45, №46 экспрессируют антиген NDV F, было обнаружено, что только 94-99% HVT-IBD №40 экспрессируют этот антиген. Заражение NDV Texas GB проводили в день 28 в соответствии с инструкцией USDA. Наблюдение за всеми птицами проводили через 2 недели после заражения. Наблюдали 95% защиту от HVT-ND №42 и №45, и 80% защиту от HVT-ND №46, 55% защиту от HVT-ND №40, в то время как положительный контроль Vectormune ND обеспечивал защиту 90%. См. Табл. 7 ниже.

Ответ антител на различные вакцины-кандидаты HVT-ND анализировали с использованием набора ProFlok ND plus (Zoetis LLC). Все титры включали без использования порогового значения (345), рекомендованного набором. Процент птиц с положительными титрами ND показан в Табл. 8 ниже.

Пример 9
Тест в отношении эффективности против MDV in vivo HVT-ND
№38, №42, №45 у SPF птиц
Три рекомбинанта HVT-IBD, HVT-IBD №38, №42, №45 тестировали в отношении их эффективности in vivo против заражения вирулентным MDV (GA22) у SPF птиц. В этом исследовании использовали положительный контроль в виде коммерческой вакцины Vectormune ND (Ceva). 1500 БОЕ каждого рекомбинантного вируса вводили in ovo в Е18. Обратный титр вакцинного вируса определяли для каждого рекомбинанта после вакцинации. Заражение MDV GA22 проводили в день 5 в соответствии с инструкцией USDA. Наблюдение за всеми птицами проводили через 54 дня после заражения. Наблюдали 69% защиту от HVT-ND №42 и №45, и 46% защиту от HVT-ND №38, в то время как положительный контроль Vectormune ND обеспечивал защиту 62%. См. Табл. 9 ниже.

Пример 10
Тест в отношении эффективности против ND in vivo HVT-ND
№38, №42, №45 у птиц-бройлеров
Три рекомбинанта HVT-IBD, HVT-IBD №38, №42, №45 тестировали в отношении их эффективности in vivo против заражения вирулентным NDV (штамм Texas GB, предоставленный USDA) у птиц-бройлеров. В этом исследовании использовали положительный контроль в виде коммерческой вакцины Vectormune ND (Ceva). 4000 БОЕ каждого рекомбинантного вируса вводили in ovo в Е18. Обратный титр вакцинного вируса определяли для каждого рекомбинанта после вакцинации. Заражение NDV Texas GB проводили в день 28 в соответствии с инструкцией USDA. Наблюдение за всеми птицами проводили через 2 недели после заражения. Наблюдали 100% защиту от HVT-ND №42 и №45, и 37% защиту от HVT-ND №38, в то время как положительный контроль Vectormune ND обеспечивал защиту 50%. Обратный титр Vectormune ND составлял 0.

Ответ антител на различные вакцины-кандидаты HVT-ND анализировали с использованием набора ProFlok ND plus (Zoetis LLC). Все титры включали без использования порогового значения (345), рекомендованного набором.
Пример 11
Тест в отношении эффективности против NDV in vivo HVT-ND
№38, №42, №45 у SPF птиц при заражении в день 20
Три рекомбинанта HVT-IBD, HVT-IBD №38, №42, №45 тестировали в отношении их эффективности in vivo против заражения вирулентным NDV (штамм Texas GB, предоставленный USDA) у SPF птиц при заражении в день 20. В этом исследовании использовали положительный контроль в виде коммерческой вакцины Vectormune ND (Ceva). 2000 БОЕ каждого рекомбинантного вируса вводили in ovo в El8. Обратный титр вакцинного вируса определяли для каждого рекомбинанта после вакцинации. Заражение NDV Texas GB проводили в день 20 в соответствии с инструкцией USDA. Наблюдение за всеми птицами проводили через 2 недели после заражения. Наблюдали 87,5% защиту от HVT-ND №42, и 70% защиту от HVT-ND №45, 65% защиту от HVT-ND №38, в то время как положительный контроль Vectormune ND обеспечивал защиту 87,5%. См. Табл. 11 ниже.

Пример 12
Тест в отношении эффективности против NDV in vivo HVT-ND
(№42, MSV+5) у SPF птиц при заражении в день 17, 18 и 19
Три рекомбинанта HVT-ND, HVT-ND (№42, MSV+5) тестировали в отношении их эффективности in vivo против заражения вирулентным NDV (штамм Texas GB) у SPF птиц при заражении в день 17, 18 и 19. Наблюдение за всеми птицами проводили через 2 недели после заражения. Наблюдали 100% (40/40), 88% (35/40), 98% (39/40) защиты при заражениях NDV в день 17, 18, 19 соответственно для вакцинации in ovo. 75% (30/40), 88% (35/40), 93% (37/40) защиты наблюдали при подкожной вакцинации в день вылупления. См. Табл. 12 ниже.

Пример 13
Тест в отношении эффективности против NDV in vivo HVT-ND
(№42, MSV+5) у SPF птиц при заражении в день 16 и 19
Три рекомбинанта HVT-ND, HVT-ND (№42, MSV+5) тестировали в отношении их эффективности in vivo против заражения вирулентным NDV (штамм Texas GB) у SPF птиц при заражении в день 16 и 19. Наблюдение за всеми птицами проводили через 2 недели после заражения. Наблюдали 85% (37/40) и 93% (37/40) защиты в день 16 и день 19 при заражении NDV для вакцинации in ovo. 70% (28/40) и 95% (38/40) защиты наблюдали при подкожной вакцинации в день вылупления. См. Табл. 13 ниже.

Пример 14
Тест на продолжительность иммунитета при использовании HVT-ND
(№42, MSV+5) у SPF птиц при заражении в день 63
Три рекомбинанта HVT-ND, HVT-ND (№42, MSV+5) тестировали в отношении продолжительности иммунитета против заражения вирулентным NDV (штамм Texas GB) у SPF птиц при заражении в день 63. Наблюдение за всеми птицами проводили через 2 недели после заражения. Наблюдали 100% (30/30) защиту как при вакцинации in ovo, так и при подкожной вакцинации в день вылупления. См. Табл. 14 ниже.

Пример 15
Тест в отношении иммуногенности ND HVT-ND (№42, MSV+5) у SPF птиц
Три рекомбинанта HVT-ND, HVT-ND (№42, MSV+5) тестировали в отношении иммуногенности против заражения вирулентным NDV (штамм Texas GB, предоставленный USDA) в день 28 у SPF птиц. Наблюдение за всеми птицами проводили через 2 недели после заражения. Наблюдали 100% (30/30) защиту как при вакцинации in ovo, так и при подкожной вакцинации в день вылупления. См. Табл. 15 ниже.

Пример 16
Тест в отношении иммуногенности MD HVT-ND (№42, MSV+5) у SPF птиц
Три рекомбинанта HVT-ND, HVT-ND (№42, MSV+5) тестировали в отношении иммуногенности против заражения вирулентным MDV (штамм GA22) в день 5 у SPF птиц. Наблюдение за всеми птицами проводили через 54 дня после заражения. Наблюдали 100% (30/30) защиту как при вакцинации in ovo, так и при подкожной вакцинации в день вылупления. См. Табл. 16 ниже.

Пример 17
Эксперимент по росту in vitro
Эксперимент по росту in vitro проводили для HVT-ND №38, №42, №45. Роллерные флаконы площадью 490 см2 засевали 5×108 первичными клетками CEF. HVT-ND №38, №42, №45 инокулировали в каждую роллерную бутыль при трех различных MOI: 0,001, 0,003, 0,008. Инфицированные клетки собирали через 48 часов после инфицирования и титровали на клетках CEF. Как HVT-ND №42, так и №45 удовлетворительно растут и характеризуются титром 2,86 × 106 и 2,97 × 106 БОЕ/мл соответственно. HVT-ND №38 характеризовался титром 1,67 × 106 БОЕ/мл. См. Табл. 17 ниже.
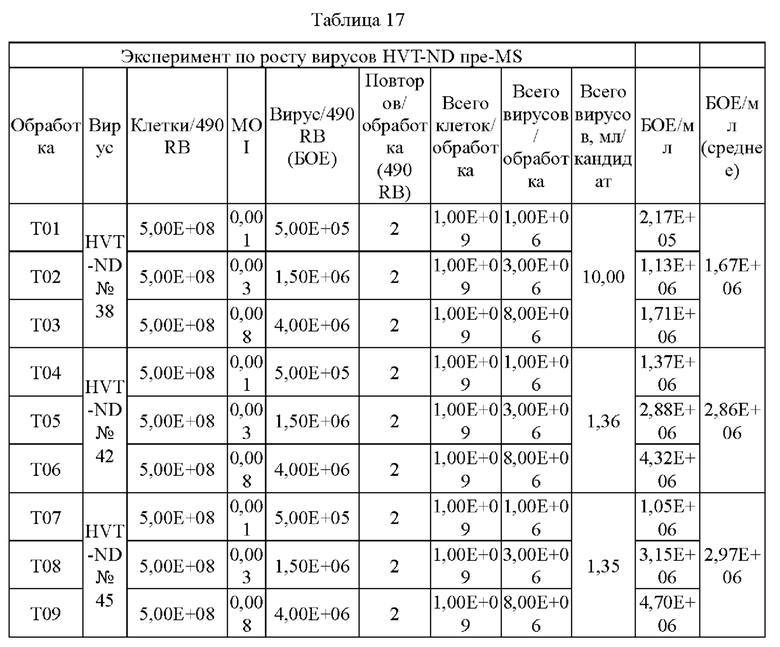
Пример 18
Конструирование HVT-IBD-ND №42-№30 LP С2
Создание трансферной плазмиды -№42:
Исходную трансферную плазмиду HVT-ND №42 химически синтезировали BioBasic, Inc. Клонирующую плазмиду UL55/ген 3 химически синтезировали посредством DNA2.0, как описано выше. ПЦР-амплификация кассеты экспрессии гена F NDV трансферной плазмиды HVT-ND №42 с использованием следующих праймеров: верхний праймер SEQ ID NO. 154; нижний праймер SEQ ID NO. 155.
Амплифицированный продукт ПЦР клонировали в сайты AscI и NheI UL55/ген 3 для получения конечной трансферной плазмиды №42. Эту плазмиду использовали для трансфекции/инфицирования для получения HVT-ND №42.
Создание трансферной плазмиды -№30:
Исходную трансферную плазмиду HVT-IBD №30 химически синтезировали BioBasic, Inc. Клонирующую плазмиду химически синтезировали посредством DNA2.0. ПЦР-амплификация кассеты экспрессии гена IBD плазмиды №30 с использованием следующих праймеров: верхний праймер SEQ ID NO. 156, нижний праймер SEQ ID NO. 157. Амплифицированный продукт ПЦР клонировали в сайты AgeI и KpnI UL35/36 для получения конечной трансферной плазмиды №30. Эту плазмиду использовали для трансфекции/инфицирования для получения HVT-IBD-ND №42-№30 LP С2.
Конструирование HVT-ND №42:
Коинфицирование/трансфекция: высевали клетки CEF в 6-луночный планшет, на следующий день проводили инфицирование рабочего посевного материала HVT (140 мкл) + трансфекцию плазмидой - №42 (линеаризованную посредством расщепления SpeI + SbfI) с использованием реагента Lipofectamine™ LTX (ThermoFisher). Собирали трансфицированные клетки в день 2 после трансфекции. Положительные фокусы подвергали скринингу в 6-луночном планшете посредством IFA с куриным поликлональным антителом к NDV (живое пятно, разведение ~1:250), затем дополнительно однократно очищали (живым пятном) в 96-луночном планшете посредством предельного разведения для получения отдельных клонов. Очищенный клон дважды пассировали в 6-луночном планшете в двух повторностях, и чистоту клона подтверждали посредством IFA (посредством фиксации и окрашивания). Для конструирования HVT-IBD-ND №42-№30 использовали сбор из 6-луночного планшета.
Конструирование HVT-IBD-ND №42-№30:
Коинфицирование/трансфекция: высевали клетки CEF в 6-луночный планшет, на следующий день проводили инфицирование HVT-ND №42 + трансфекцию плазмидой - №30 (линеаризованную посредством расщепления SbfI) с использованием реагента Lipofectamine™ LTX (ThermoFisher). Собирали трансфицированные клетки в день 3 после трансфекции. Положительные фокусы подвергали скринингу в 6-луночном планшете посредством IFA с куриным поликлональным антителом к IBD (живое пятно, разведение ~1:250), затем дополнительно однократно очищали (живым пятном) посредством предельного разведения для получения отдельных клонов. Два очищенных клона отбирали и пассировали в 6-луночном планшете в двух повторностях, и чистоту клонов подтверждали посредством IFA (посредством фиксации и окрашивания). Клоны воспроизводили последовательно в колбе Т-75, колбе Т-150, колбе Т-225, роллерной колбе объемом 850 мл. Рекомбинантный вирус собирали и разделяли на аликвоты по 1 мл/флакон, замораживали при -80°С в течение ночи, затем переносили в резервуар с LN.
Пример 19:
Тест в отношении эффективности против NDV in vivo HVT-IBD-ND
№42-№30, №42-№32, №104 у SPF птиц
Семь рекомбинантов HVT-IBD-ND, HVT-IBD-ND №42-№30 (3 клона), №42-№32 (2 клона), №104 (2 клона) тестировали в отношении их эффективности in vivo против заражения вирулентным NDV у SPF птиц. Заражение NDV Texas GB проводили в день 28. Около 1500 БОЕ каждого рекомбинантного вируса вводили in ovo в E18. Наблюдение за всеми птицами проводили через 2 недели после заражения. См. Табл. 18 ниже.

Пример 20:
Тест в отношении эффективности против IBD in vivo HVT-IBD-ND
№42-№30, №42-№32, №104 у SPF птиц
Семь рекомбинантов HVT-IBD-ND, HVT-IBD-ND №42-№30 (3 клона), №42-№32 (2 клона), №104 (2 клона) тестировали в отношении их эффективности in vivo против заражения вирулентным IBDV у SPF птиц в день 14 и день 21 соответственно. Около 2000 БОЕ каждого рекомбинантного вируса вводили in ovo в El8. Вскрытие всех птиц проводили через 5 дней после заражения. См. Табл. 19 ниже.

Пример 21
Тест в отношении эффективности против MDV in vivo HVT-IBD-ND №42 - №30
(4 клона)
у SPF птиц
Три рекомбинанта HVT-IBD-ND №42-№30 (4 клона) тестировали в отношении их эффективности in vivo против заражения вирулентным MDV (GA22) у SPF птиц. Около 1500 БОЕ каждого рекомбинантного вируса вводили in ovo в E18. Заражение MDV GA22 проводили в день 5. Наблюдение за всеми птицами проводили через 54 дня после заражения. См. Табл. 20 ниже.

Пример 22
Тест в отношении эффективности против wIBD in vivo HVT-IBD-ND
№42-№30, №42-№32, №04 у SPF птиц
Три рекомбинанта HVT-IBD-ND, №42-№30 (2 клона), №42-№32 (2 клона), №104 тестировали в отношении их эффективности in vivo против заражения крайне вирулентным IBDV у SPF птиц. Около 1500 БОЕ каждого рекомбинантного вируса вводили in ovo в Е18. Заражение vvIBDV проводили в день 14 и день 21. Наблюдение за всеми птицами проводили через 10 дня после заражения. Гистологическое исследование сумки Фабрициуса проводили для каждой птицы в конце исследования. См. Табл. 21 ниже.

Пример 23
Тест на продолжительность иммунитета против IBD при использовании
HVT-IBD-ND (№42-№30, Х + 5) у SPF птиц при заражении в день 63
Рекомбинант HVT-IBD-ND, №42-№30 (MSV+5) тестировали в отношении продолжительности иммунитета против заражения вирулентным классическим IBDV у SPF птиц при заражении в день 63. Наблюдение за всеми птицами проводили через четыре дня после заражения с последующим вскрытием. См. Табл. 22.
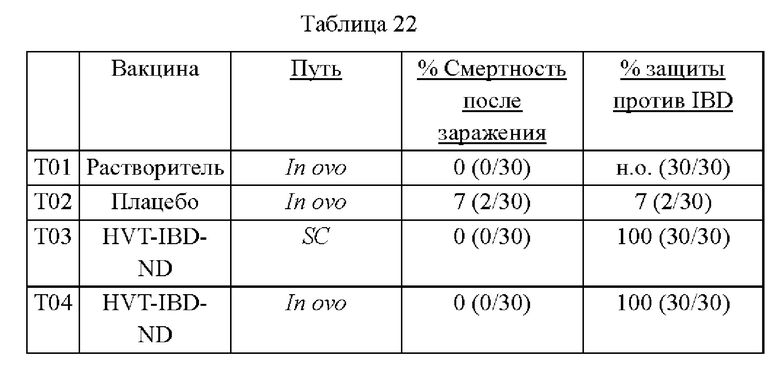
Пример 24
Тест в отношении иммуногенности ND HVT-IBD-ND (№42-№30, MSV+5) у SPF птиц
Рекомбинант HVT-IBD-ND, №42-№30 (MSV+5) тестировали в отношении иммуногенности против заражения вирулентным NDV (штамм Texas GB) в день 28 у SPF птиц. Наблюдение за всеми птицами проводили через 2 недели после заражения. См. Табл. 23.

Пример 25
Тест в отношении иммуногенности IBD HVT-IBD-ND (№42-№30, MSV+5) у SPF птиц
Рекомбинант HVT-IBD-ND, №42-№30 (MSV+5) тестировали в отношении иммуногенности против заражения вирулентным IBDV в день 34 у SPF птиц. Наблюдение за всеми птицами проводили через 4 дня после заражения с последующим вскрытием в отношении поражений сумки Фабрициуса. См. Табл. 24 ниже.

Пример 26
Тест в отношении иммуногенности MD HVT-IBD-ND (№42-№30, MSV+5) у SPF птиц
Три рекомбинанта HVT-IBD-ND, №42-№30 (MSV+5) тестировали в отношении иммуногенности против заражения вирулентным MDV (штамм GA22) в день 5 у SPF птиц. Наблюдение за всеми птицами проводили через 54 дня после заражения. См. Табл. 25 ниже.
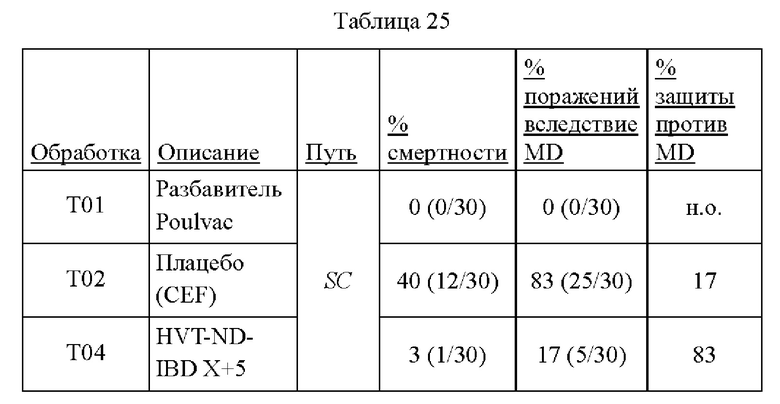
Пример 27
Тест в отношении иммуногенности ND HVT-IBD-ND
(№42-№30, MSV+5) у SPF птиц против заражения штаммом EU
Рекомбинант HVT-IBD-ND, №42-№30 (MSV+5) тестировали в отношении иммуногенности против заражения вирулентным NDV Europe (Herts Weybridge 33/56) в день 21 у SPF птиц. Наблюдение за всеми птицами проводили через 2 недели после заражения. См. Табл. 26 ниже.

Пример 28 Совместимость HVT-ND с Bursaplex
Эффективность против IBDV
BURSAPLEX™ (Zoetis, US 5871748, включенный в данный документ посредством ссылки) представляет собой вакцину против инфекционного бурсита (IBD), которая содержит вакцинный конъюгат, состоящий из живого аттенуированного штамма IBD 2512 и нейтрализующего антитела, BDA, связанного с вирусом. Bursaplex генерирует активный иммунитет против IBD у домашней птицы, в частности, у кур. В Е18 яйца инъецировали in ovo либо контрольной, либо тестируемой вакциной (HVT-ND с Bursaplex в соотношении 1:1) и переносили в отведенный для этого инкубатор, как указано Biometrics, вместе с яйцами, в которые не вводили инъекцию. В день вылупления птиц Т04 вакцинировали подкожно. Образцы крови собирали в дни 5, 12, 19, 26 и 33 для серологического исследования IBDV. В день 34 определенных птиц заражали классическим вирулентным IBDV, а в день 38 всех птиц вскрывали в отношении наличия поражений сумки Фабрициуса. Ни у одного цыпленка в отрицательной группе Т01 не было заметно заметных поражений, а у 100 процентов цыплят в контрольной группе заражения Т02 развивались хорошо заметные поражения. Как Т03 (HVT-ND + Bursaplex, in ovo), так и Т04 (HVT-ND + Bursaplex, SC) оказывали защитный эффект на 100%. Можно сделать вывод, что Poulvac Procerta HVT-ND и Poulvac Bursaplex совместимы при совместном введении и остаются эффективными против заражения IBDV при введении либо in ovo, либо подкожно.
Эффективность против NDV
В El8 яйца инъецировали in ovo либо контрольной, либо тестируемой вакциной (HVT-ND с Bursaplex в соотношении 1:1) и переносили в отведенный для этого инкубатор, как указано Biometrics, вместе с яйцами, в которые не вводили инъекцию. В день вылупления птиц Т04 вакцинировали подкожно. Образцы крови собирали в дни 6, 13, 20 и 27 для серологического исследования NDV. В день 28 определенных птиц заражали велогенным NDV, а в день 42 всех выживших птиц умерщвляли. Ни у одного цыпленка в отрицательной группе Т01 не развились клинические признаки, а у 100 процентов цыплят в контрольной группе заражения Т02 развились клинические признаки болезни Ньюкасла, в том числе смертность. Как Т03 (HVT-ND + Bursaplex, in ovo), так и Т04 (HVT-ND + Bursaplex, SC) оказывали защитный эффект на 92,5% и 95% соответственно. Можно сделать вывод, что Poulvac Procerta HVT-ND и Poulvac Bursaplex совместимы при совместном введении и остаются эффективными против заражения NDV при введении либо in ovo, либо подкожно.
Пример 29 Совместимость HVT-ND с Magniplex Эффективность против IBD
MAGNIPLEX™ (Zoetis) представляет собой вакцину против инфекционного бурсита (IBD), которая содержит вакцинный конъюгат, состоящий из живого аттенуированного штамма IBD V877 и нейтрализующего антитела, BDA, связанного с вирусом. Bursaplex генерирует активный иммунитет против IBD у домашней птицы, в частности, у кур. В Е18 яйца инъецировали in ovo либо контрольной, либо тестируемой вакциной (серия HVT-ND до получения разрешения с Magniplex в соотношении 1:1) и переносили в отведенный для этого инкубатор, как указано Biometrics, вместе с яйцами, в которые не вводили инъекцию. В день вылупления птиц Т04 вакцинировали подкожно. Образцы крови собирали в дни 5, 12, 19, 26 и 33 для серологического исследования IBDV. В день 34 определенных птиц заражали классическим вирулентным IBDV, а в день 38 всех птиц вскрывали в отношении наличия поражений сумки Фабрициуса. Ни у одного цыпленка в отрицательной группе Т01 не было заметно заметных поражений, а у 100 процентов цыплят в контрольной группе заражения Т02 развивались хорошо заметные поражения. Как Т03 (HVT-ND + Magniplex, in ovo), так и Т04 (HVT-ND + Magniplex, SC) оказывали защитный эффект на 100%. Можно сделать вывод, что Poulvac Procerta HVT-ND и Poulvac Magniplex совместимы при совместном введении и остаются эффективными против заражения IBDV при введении либо in ovo, либо подкожно.


Эффективность против ND
В Е18 яйца инъецировали in ovo либо контрольной, либо тестируемой вакциной (HVT-ND с Magniplex в соотношении 1:1) и переносили в отведенный для этого инкубатор, как указано Biometrics, вместе с яйцами, в которые не вводили инъекцию. В день вылупления птиц Т04 вакцинировали подкожно. Образцы крови собирали в дни 6, 13, 20 и 27 для серологического исследования NDV. В день 28 определенных птиц заражали велогенным NDV, а в день 42 всех выживших птиц умерщвляли. Ни у одного цыпленка в отрицательной группе Т01 не развились клинические признаки, а у 100 процентов цыплят в контрольной группе заражения Т02 развились клинические признаки болезни Ньюкасла, в том числе смертность. Как Т03 (HVT-ND + Magniplex, in ovo), так и Т04 (HVT-ND + Magniplex, SC) оказывали защитный эффект на 92,5% и 95% соответственно. Можно сделать вывод, что Poulvac Procerta HVT-ND и Poulvac Magniplex совместимы при совместном введении и остаются эффективными против заражения NDV при введении либо in ovo, либо подкожно.


--->
Перечень последовательностей
<110> Zoetis
Rong, Sing
Luo, Yugang
Brown, Tyler
<120> ВЕКТОРЫ НА ОСНОВЕ РЕКОМБИНАНТНОГО ВИРУСА ГЕРПЕСА ИНДЕЕК,
ЭКСПРЕССИРУЮЩИЕ АНТИГЕНЫ ПАТОГЕНОВ ПТИЦ,
И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ
<130> ZP000168
<140> 62/898651
<141> 4 сентября 2020 г.
<160> 158
<170> PatentIn версия 3.5
<210> 1
<211> 687
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК промотора CMV человека (hCMV)
<400> 1
ttgacattga ttattgacta gttattaata gtaatcaatt acggggtcat tagttcatag 60
cccatatatg gagttccgcg ttacataact tacggtaaat ggcccgcctg gctgaccgcc 120
caacgacccc cgcccattga cgtcaataat gacgtatgtt cccatagtaa cgccaatagg 180
gactttccat tgacgtcaat gggtggagta tttacggtaa actgcccact tggcagtaca 240
tcaagtgtat catatgccaa gtacgccccc tattgacgtc aatgacggta aatggcccgc 300
ctggcattat gcccagtaca tgaccttatg ggactttcct acttggcagt acatctacgt 360
attagtcatc gctattacca tggtgatgcg gttttggcag tacatcaatg ggcgtggata 420
gcggtttgac tcacggggat ttccaagtct ccaccccatt gacgtcaatg ggagtttgtt 480
ttggcaccaa aatcaacggg actttccaaa atgtcgtaac aactccgccc cattgacgca 540
aatgggcggt aggcgtgtac ggtgggaggt ctatataagc agagctctct ggctaactag 600
agaacccact gcttactggc ttatcgaaat taatacgact cactataggg agacccaagc 660
tggctagcgt ttaaacttaa gcttacc 687
<210> 2
<211> 1391
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК промотора CMV мыши (mCMV)
<400> 2
aactccgccc gttttatgac tagaaccaat agtttttaat gccaaatgca ctgaaatccc 60
ctaatttgca aagccaaacg ccccctatgt gagtaatacg gggacttttt acccaatttc 120
ccaagcggaa agccccctaa tacactcata tggcatatga atcagcacgg tcatgcactc 180
taatggcggc ccatagggac tttccacata gggggcgttc accatttccc agcatagggg 240
tggtgactca atggccttta cccaagtaca ttgggtcaat gggaggtaag ccaatgggtt 300
tttcccatta ctggcaagca cactgagtca aatgggactt tccactgggt tttgcccaag 360
tacattgggt caatgggagg tgagccaatg ggaaaaaccc attgctgcca agtacactga 420
ctcaataggg actttccaat gggtttttcc attgttggca agcatataag gtcaatgtgg 480
gtgagtcaat agggactttc cattgtattc tgcccagtac ataaggtcaa tagggggtga 540
atcaacagga aagtcccatt ggagccaagt acactgcgtc aatagggact ttccattggg 600
ttttgcccag tacataaggt caatagggga tgagtcaatg ggaaaaaccc attggagcca 660
agtacactga ctcaataggg actttccatt gggttttgcc cagtacataa ggtcaatagg 720
gggtgagtca acaggaaagt cccattggag ccaagtacat tgagtcaata gggactttcc 780
aatgggtttt gcccagtaca taaggtcaat gggaggtaag ccaatgggtt tttcccatta 840
ctggcacgta tactgagtca ttagggactt tccaatgggt tttgcccagt acataaggtc 900
aataggggtg aatcaacagg aaagtcccat tggagccaag tacactgagt caatagggac 960
tttccattgg gttttgccca gtacaaaagg tcaatagggg gtgagtcaat gggtttttcc 1020
cattattggc acgtacataa ggtcaatagg ggtgagtcat tgggtttttc cagccaattt 1080
aattaaaacg ccatgtactt tcccaccatt gacgtcaatg ggctattgaa actaatgcaa 1140
cgtgaccttt aaacggtact ttcccatagc tgattaatgg gaaagtaccg ttctcgagcc 1200
aatacacgtc aatgggaagt gaaagggcag ccaaaacgta acaccgcccc ggttttcccc 1260
tggaaattcc atattggcac gcattctatt ggctgagctg cgttctacgt gggtataaga 1320
ggcgcgacca gcgtcggtac cgtcgcagtc ttcggtctga ccaccgtaga acgcagagct 1380
cctcgctgca g 1391
<210> 3
<211> 1662
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК белка NDV F штамма D26-76
<400> 3
atgggctcca gatcttctac caggatccca gtacctctga tgctgaccgt ccgaatcatg 60
ttggcactga gttgcgtctg tccgaccagc tcccttgatg gcaggcctct tgcagctgca 120
gggattgtgg taacaggaga caaagcagtc aacatataca cctcatctca gacagggtca 180
atcataatca agttactccc aaatatgccc aaggataaag aggcgtgtgc aaaagcccca 240
ttggaagcat acaacaggac attgactact ttgctcaccc cccttggtga ttctatccgt 300
aggatacaag agtctgtgac cacatccgga ggagggaaac agggacgtct tataggcgcc 360
attatcggtg gtgtagctct cggggttgca accgctgcac agataacagc agcctcggct 420
ctgatacaag ccaatcaaaa tgctgccaac atcctccggc tcaaagagag cattgctgca 480
accaatgagg ctgtgcacga ggtcactgac ggattatcac aactagcagt ggcagttggg 540
aagatgcagc aatttgttaa tgaccagttt aataaaacag ctcaggaatt ggactgtata 600
aaaattacac agcaggttgg tgtagaactc aacctgtacc taactgaatt gactacagta 660
ttcgggccac aaatcacttc ccctgcctta actcagctga ctatccaggc gctttacaat 720
ctagctggtg ggaatatgga ttacttgttg actaagttag gtgtaggaaa caaccaactc 780
agctcattaa ttggtagtgg cctgattacc ggcaacccta tcctgtacga ctcacagact 840
caactcttgg gtatacaggt caccctaccc tcagtcggga atctaaataa tatgcgtgcc 900
acctacctgg aaaccttgtc tgtaagtaca accaaaggat ttgcctcagc acttgtccca 960
aaagtagtga cacaggttgg ttccgtgata gaagagcttg acacctcgta ctgtatcgag 1020
accgatttgg acctatattg tacaagaata gtgacattcc ctatgtctcc tggtatttat 1080
tcctgtttga gtggcaatac atctgcttgc atgtattcaa agactgaagg cgcactcact 1140
acgccgtata tgaccctcaa aggctcagtt attgccaact gtaagatgac aacatgtaga 1200
tgtgcagacc ccccgggtat catatcgcag aattatggag aagctgtgtc tctaatagat 1260
aggcaatcat gcaatatctt atccttagac gggataactt tgaggctcag tggggaattt 1320
gatgcaactt atcaaaagaa tatctcaata caagattctc aagtaatagt tacaggcaat 1380
cttgacatct cgactgagct tgggaatgtc aacaactcga taagtaatgc tttggataag 1440
ttagaggaaa gcaacagcaa actagacaag gtcaatgtta aactgaccag cacatccgct 1500
cttattacct atatcgtttt aactgtcata tctcttgtat gtggtatact tagcctggtt 1560
ctagcatgct acctgatgta caagcaaaag gcgcaacaga agaccttgtt gtggcttggg 1620
aataataccc tagaccagat gagggccact acaaaaatgt ag 1662
<210> 4
<211> 553
<212> Белок
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность ДНК белка NDV F штамма D26-76
<400> 4
Met Gly Ser Arg Ser Ser Thr Arg Ile Pro Val Pro Leu Met Leu Thr
1 5 10 15
Val Arg Ile Met Leu Ala Leu Ser Cys Val Cys Pro Thr Ser Ser Leu
20 25 30
Asp Gly Arg Pro Leu Ala Ala Ala Gly Ile Val Val Thr Gly Asp Lys
35 40 45
Ala Val Asn Ile Tyr Thr Ser Ser Gln Thr Gly Ser Ile Ile Ile Lys
50 55 60
Leu Leu Pro Asn Met Pro Lys Asp Lys Glu Ala Cys Ala Lys Ala Pro
65 70 75 80
Leu Glu Ala Tyr Asn Arg Thr Leu Thr Thr Leu Leu Thr Pro Leu Gly
85 90 95
Asp Ser Ile Arg Arg Ile Gln Glu Ser Val Thr Thr Ser Gly Gly Gly
100 105 110
Lys Gln Gly Arg Leu Ile Gly Ala Ile Ile Gly Gly Val Ala Leu Gly
115 120 125
Val Ala Thr Ala Ala Gln Ile Thr Ala Ala Ser Ala Leu Ile Gln Ala
130 135 140
Asn Gln Asn Ala Ala Asn Ile Leu Arg Leu Lys Glu Ser Ile Ala Ala
145 150 155 160
Thr Asn Glu Ala Val His Glu Val Thr Asp Gly Leu Ser Gln Leu Ala
165 170 175
Val Ala Val Gly Lys Met Gln Gln Phe Val Asn Asp Gln Phe Asn Lys
180 185 190
Thr Ala Gln Glu Leu Asp Cys Ile Lys Ile Thr Gln Gln Val Gly Val
195 200 205
Glu Leu Asn Leu Tyr Leu Thr Glu Leu Thr Thr Val Phe Gly Pro Gln
210 215 220
Ile Thr Ser Pro Ala Leu Thr Gln Leu Thr Ile Gln Ala Leu Tyr Asn
225 230 235 240
Leu Ala Gly Gly Asn Met Asp Tyr Leu Leu Thr Lys Leu Gly Val Gly
245 250 255
Asn Asn Gln Leu Ser Ser Leu Ile Gly Ser Gly Leu Ile Thr Gly Asn
260 265 270
Pro Ile Leu Tyr Asp Ser Gln Thr Gln Leu Leu Gly Ile Gln Val Thr
275 280 285
Leu Pro Ser Val Gly Asn Leu Asn Asn Met Arg Ala Thr Tyr Leu Glu
290 295 300
Thr Leu Ser Val Ser Thr Thr Lys Gly Phe Ala Ser Ala Leu Val Pro
305 310 315 320
Lys Val Val Thr Gln Val Gly Ser Val Ile Glu Glu Leu Asp Thr Ser
325 330 335
Tyr Cys Ile Glu Thr Asp Leu Asp Leu Tyr Cys Thr Arg Ile Val Thr
340 345 350
Phe Pro Met Ser Pro Gly Ile Tyr Ser Cys Leu Ser Gly Asn Thr Ser
355 360 365
Ala Cys Met Tyr Ser Lys Thr Glu Gly Ala Leu Thr Thr Pro Tyr Met
370 375 380
Thr Leu Lys Gly Ser Val Ile Ala Asn Cys Lys Met Thr Thr Cys Arg
385 390 395 400
Cys Ala Asp Pro Pro Gly Ile Ile Ser Gln Asn Tyr Gly Glu Ala Val
405 410 415
Ser Leu Ile Asp Arg Gln Ser Cys Asn Ile Leu Ser Leu Asp Gly Ile
420 425 430
Thr Leu Arg Leu Ser Gly Glu Phe Asp Ala Thr Tyr Gln Lys Asn Ile
435 440 445
Ser Ile Gln Asp Ser Gln Val Ile Val Thr Gly Asn Leu Asp Ile Ser
450 455 460
Thr Glu Leu Gly Asn Val Asn Asn Ser Ile Ser Asn Ala Leu Asp Lys
465 470 475 480
Leu Glu Glu Ser Asn Ser Lys Leu Asp Lys Val Asn Val Lys Leu Thr
485 490 495
Ser Thr Ser Ala Leu Ile Thr Tyr Ile Val Leu Thr Val Ile Ser Leu
500 505 510
Val Cys Gly Ile Leu Ser Leu Val Leu Ala Cys Tyr Leu Met Tyr Lys
515 520 525
Gln Lys Ala Gln Gln Lys Thr Leu Leu Trp Leu Gly Asn Asn Thr Leu
530 535 540
Asp Gln Met Arg Ala Thr Thr Lys Met
545 550
<210> 5
<211> 1362
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК vIBDV (F52/70) VP2
<400> 5
atgaccaatc tccaggacca gacccagcag attgtgcctt ttattaggag tctcttgatg 60
cctacaaccg gccccgccag catcccggac gacacactgg aaaaacatac actgagaagc 120
gagacatcta catacaattt gaccgtgggc gataccggct ccgggcttat cgtgttcttc 180
ccaggttttc ccggatctat cgtaggagcg cactacaccc tccaaagtaa cggcaattac 240
aaattcgacc agatgctcct gacagcccag aaccttcctg cttcttacaa ttactgtaga 300
cttgtgtcca ggtccctgac tgtgcggagt agcacgcttc caggaggcgt atacgccctg 360
aacggaacta taaacgccgt caccttccag ggctccttgt ccgaacttac cgacgtgtcc 420
tacaatggcc tcatgagcgc aacggccaac ataaacgata agatcggcaa tgttcttgtg 480
ggcgaggggg ttacagtcct ttctctgcca accagttatg atctgggata cgtgcggctt 540
ggcgatccca ttcccgctat cggtctcgac cctaaaatgg tggctacttg cgactcatct 600
gaccgcccaa gggtctatac aattactgca gccgatgact atcagttttc cagccaatac 660
cagccagggg gtgtgacaat cacacttttc agcgccaata ttgacgctat cacatccctc 720
tcaatcggag gtgagcttgt gttccagact tctgttcagg gcttggtatt gggcgccact 780
atttacttga tcgggttcga cgggaccgca gtgatcactc gggcagtggc tgcggataac 840
ggactcactg ccggaactga caaccttatg ccttttaatc tggtcatccc cactaacgag 900
atcacccagc ctattacctc cataaagctc gaaattgtga ccagcaagag cggagggcag 960
gcaggcgacc aaatgagttg gtctgcaagc gggtccctcg ccgtgaccat ccacggtggc 1020
aactatcctg gggcgctcag acccgtcacc ctggtagcct acgaaagggt tgccacaggc 1080
tcagttgtca cggtggctgg agtaagcaat ttcgagctca tcccgaatcc tgagctcgct 1140
aaaaatcttg tgaccgagta tggaaggttc gaccctggcg caatgaatta cacaaagctg 1200
attctgtccg aacgggatag gctgggtatc aagacagttt ggcccacgcg cgaatacaca 1260
gatttcaggg agtactttat ggaggtcgca gatttgaata gcccacttaa gatcgctgga 1320
gcatttggct ttaaggatat tatccgcgca atcagaaggt ag 1362
<210> 6
<211> 225
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК Poly A бычьего гормона роста
<400> 6
tgtgccttct agttgccagc catctgttgt ttgcccctcc cccgtgcctt ccttgaccct 60
ggaaggtgcc actcccactg tcctttccta ataaaatgag gaaattgcat cgcattgtct 120
gagtaggtgt cattctattc tggggggtgg ggtggggcag gacagcaagg gggaggattg 180
ggaagacaat agcaggcatg ctggggatgc ggtgggctct atggt 225
<210> 7
<211> 580
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для фланкирования HVT UL55
<400> 7
atggcagcga gcatgggata ttcatcctcg tcatcgttaa catctctacg ggttcagaat 60
gtttggcatg tcgtcgatcc tttgcccatc gttgcaaatt acaagtccga tcgccatgac 120
cgcgataagc ctgtaccatg tggcattagg gtgacatctc gatcatacat tataagacca 180
acgtgcgagt cttccaaaga cctgcacgcc ttcttcttcg gattgtcaac gggttcttca 240
gaatctatgc ccatatctgg cgttgagacc attgtgcgtt taatgaacaa taaagcggca 300
tgccatggaa aggagggctg cagatctcca ttttctcacg ccactatcct ggacgctgta 360
gacgataatt ataccatgaa tatagagggg gtatgtttcc actgccactg tgatgataag 420
ttttctccag attgttggat atctgcattt tctgctgccg aacaaacttc atcgctatgc 480
aaagagatgc gtgtgtacac gcgccgttga gtatacggga aactaaatgt tcatagaggt 540
ctttgggcta tatgttatta aataaaataa ttgaccagtg 580
<210> 8
<211> 678
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для фланкирования HVT гена 3
<400> 8
gtttaatgtt agtttattca atgcattggt tgcaaatatt cattacttct ccaatcccag 60
gtcattcttt agcgagatga tgttatgaca ttgctgtgaa aattactaca ggatatattt 120
ttaagatgca ggagtaacaa tgtgcatagt aggcgtagtt atcgcagacg tgcaacgctt 180
cgcatttgag ttaccgaagt gcccaacagt gctgcggtta tggtttatgc gcacagaatc 240
catgcatgtc ctaattgaac catccgattt ttcttttaat cgcgatcgtt gtttgggcaa 300
ctgcgttatt tcagatctaa aaaatttacc ctttatgacc atcacatctc tctggctcat 360
accccgcttg gataagatat catgtagatt ccgccctaag aaatgcaaac taacattatt 420
gtcggttcca tatacacttc catcttgtcc ttcgaaaata acaaactcgc gcaatagacc 480
gtccgtacat gcatggccga tgtgtgtcaa catcattggt ctgctagatc ccgatgggac 540
gaatcgtaca gtcgtcgctc cagcattggc aaaaatcccc agataccctc catgcggcaa 600
atctaaattg cgaccccgaa gagactgcac caaagtctta tcgacgcacg ctgatttttt 660
tgaacagcgg gagcccat 678
<210> 9
<211> 561
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК промотора Pec
<400> 9
agttattaat agtaatcaat tacggggtca ttagttcata gcccatatat ggagttccgc 60
gttacataac ttacggtaaa tggcccgccg gctgaccgcc caacgacccc cgcccattga 120
cgtcaataat gacgtatgtt cccatagtaa cgccaatagg gactttccat tgacgtcaat 180
gggtggagta tttacggtaa actgcccatt ggcagtacat caagtgtatc atatgccaag 240
tacgccccct attgacgtca atgacggtaa atggatgcag tattttgtgc agcgatgggg 300
gcgggggggg ggggggcgcg cgccaggcgg ggcggggcgg ggcgaggggc ggggcggggc 360
gaggcggaga ggtgcggcgg cagccaatca gagcggcgcg ctccgaaagt ttccttttat 420
ggcgaggcgg cggcggcggc ggccctataa aaagcgaagc gcgcggcggg cgggagtcgc 480
tgcgcgctgc cttcgccccg tgccccgctc cgccgccgcc tcgcgccgcc cgccccggct 540
ctgactgacc gcgtctagag g 561
<210> 10
<211> 1362
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК vvIBDV VP2
<400> 10
atgacaaatc ttcaggatca gacccagcag atagttccct ttattaggtc ccttctgatg 60
ccaaccacag ggcccgctag cattccggac gataccctgg agaaacacac tttgcggagt 120
gagacaagca cttacaatct gacggtggga gataccggct caggtcttat cgtgtttttt 180
cccggctttc ctggatccat tgttggcgcg cattacacgc tgcagagcaa cggcaactat 240
aaattcgatc agatgctcct gacggctcag aatctccccg ccagttacaa ttactgccgc 300
cttgtaagta ggtccttgac tgttagaagc tcaacgctgc caggcggagt atatgccctg 360
aatggaacca ttaatgctgt aacattccaa ggatcactgt ccgagctcac cgatgtgtct 420
tacaatggat tgatgtctgc cacggctaac attaacgaca agatcgggaa tgtgctcgtg 480
ggcgagggag tgaccgtttt gagcctgccg acaagctacg acctcggcta cgtaaggctc 540
ggggatccaa tccccgcgat cggcttggat cccaaaatgg ttgctacgtg cgacagcagc 600
gatagaccca gggtctatac catcaccgct gccgatgatt accagtttag ctcccagtac 660
caggcgggag gggtcacgat cacccttttt agcgccaaca tcgacgccat aacctcactt 720
tctatagggg gcgagttggt ttttcagacc agtgtccagg ggctcatcct cggtgcgaca 780
atctatctga tcggctttga cggaacagct gtcatcacga gggccgtagc tgcagataat 840
ggcctgactg ctgggacaga taatctgatg ccgttcaaca tagtgatccc caccagtgag 900
attacgcaac ccatcacgag catcaaactg gagatcgtga cgtcaaaatc cggcggtcag 960
gcaggtgacc agatgtcttg gtccgcaagc ggaagtttgg ccgtgacaat tcacgggggg 1020
aattaccccg gcgcactcag gcccgtgacc ctcgtcgcct acgaaagagt tgcaacggga 1080
agtgtagtga cagtcgctgg agtgagtaac ttcgaactca tccctaatcc cgagctcgcc 1140
aaaaatctcg tcacggagta tgggaggttt gatcccggcg ccatgaacta cacaaaactg 1200
atattgtccg aaagggatag gttgggcatt aaaaccgtgt ggcctactag ggaatacacc 1260
gatttccgcg aatattttat ggaggtcgcg gatctgaact ctcccctgaa gatagcaggc 1320
gcttttgggt tcaaggatat tatccgggcg ttgcggcggt ag 1362
<210> 11
<211> 197
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для промотора SV40
<400> 11
tgcatctcaa ttagtcagca accatagtcc cgcccctaac tccgcccatc ccgcccctaa 60
ctccgcccag ttccgcccat tctccgcccc atcgctgact aatttttttt atttatgcag 120
aggccgaggc cgcctcggcc tctgagctat tccagaagta gtgaggaggc ttttttggag 180
gcctaggctt ttgcaaa 197
<210> 12
<211> 122
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для последовательности polyA SV40
<400> 12
taagatacat tgatgagttt ggacaaacca caactagaat gcagtgaaaa aaatgcttta 60
tttgtgaaat ttgtgatgct attgctttat ttgtaaccat tataagctgc aataaacaag 120
tt 122
<210> 13
<211> 531
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для фланкирования HVT UL35
<400> 13
gccactgtat gggccattta tgtttatcga gtctaaaagt cgtggaagcg cctggccatc 60
gccagtctcg acctgccact gctgagttcg gcttcaattc cagcatccca gaaatgtcgc 120
agtcaccatc acgccgtgca tcgcttccta cgaggccttt tgacgcttct gatttgggca 180
catacaccct ggacatactc caccgctatt cgctcgtaga tttagtacaa ctactgaatg 240
acttgccgcg taacattacc tccacgcccg cttctaatgt agaaaccatg gcaaaaatta 300
atgttttaag ggccatttgc gtaggatttg ccgaggtccg tcgccacaac gacgcgcgaa 360
ctttacagcg aacggcaatg tttgccgccg acgacgtcgc atcacggatc agaccatcca 420
ttggattaaa gcgcacctac ccaccgggta tattttccac agctattacc gtatctaatt 480
ccgaggatga agagcgaaat tcgtgatcgt aaaaataaaa aatacaagat a 531
<210> 14
<211> 700
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для фланкирования HVT UL36
<400> 14
gtacataatt cttatttatc tttaatccat gaggagcatt tttattttaa aaatgtcagc 60
cgccagccct ataaccctag atcgcaactg atccctagtc tgcgttattt gtcttgcaat 120
cttttcgcac gcctttgtga gtgcatacaa tgcccccctg ctcgcttttc tgaaatcgcg 180
tcgggtcatt aatgtgtcgg ctatcacaat gcgagatgta ctcgacatgt ccgtgtctgt 240
actattggga ttgtaaatag tcgaccgcga atcatcagag tcggaatctg taaaggatac 300
agattccgac tctgagcgct tatgaatggg atccactcgg acgttgttga acttccgttc 360
ggattctgct tcagtcaaca ccggcccccg atagctacta aggttggggg gtttgtgggt 420
tgtttgtgaa actgctttgc ggtgtgcatt accacggggg gtgtggggaa gtatctgttt 480
ccacgatgcg ataacgttcg gtggcggagg gggcgattca ttctctagtg tacgcgtttc 540
aacttcagga acgtgattat ttctttcagg acactctttc caatttcctt cttccttcac 600
ttcgggtaca ggtatattct taatgtttac atacatgtcg tctgctcgtc tcaactgcgg 660
ggttatgatg ggtggtggtg acagtctctc cgaatgatcg 700
<210> 15
<211> 1278
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК промотора бета-актина кур
<400> 15
tcgaggtgag ccccacgttc tgcttcactc tccccatctc ccccccctcc ccacccccaa 60
ttttgtattt atttattttt taattatttt gtgcagcgat gggggcgggg gggggggggg 120
cgcgcgccag gcggggcggg gcggggcgag gggcggggcg gggcgaggcg gagaggtgcg 180
gcggcagcca atcagagcgg cgcgctccga aagtttcctt ttatggcgag gcggcggcgg 240
cggcggccct ataaaaagcg aagcgcgcgg cgggcgggag tcgctgcgtt gccttcgccc 300
cgtgccccgc tccgcgccgc ctcgcgccgc ccgccccggc tctgactgac cgcgttactc 360
ccacaggtga gcgggcggga cggcccttct cctccgggct gtaattagcg cttggtttaa 420
tgacggctcg tttcttttct gtggctgcgt gaaagcctta aagggctccg ggagggccct 480
ttgtgcgggg gggagcggct cggggggtgc gtgcgtgtgt gtgtgcgtgg ggagcgccgc 540
gtgcggcccg cgctgcccgg cggctgtgag cgctgcgggc gcggcgcggg gctttgtgcg 600
ctccgcgtgt gcgcgagggg agcgcggccg ggggcggtgc cccgcggtgc gggggggctg 660
cgaggggaac aaaggctgcg tgcggggtgt gtgcgtgggg gggtgagcag ggggtgtggg 720
cgcggcggtc gggctgtaac ccccccctgc acccccctcc ccgagttgct gagcacggcc 780
cggcttcggg tgcggggctc cgtgcggggc gtggcgcggg gctcgccgtg ccgggcgggg 840
ggtggcggca ggtgggggtg ccgggcgggg cggggccgcc tcgggccggg gagggctcgg 900
gggaggggcg cggcggcccc ggagcgccgg cggctgtcga ggcgcggcga gccgcagcca 960
ttgcctttta tggtaatcgt gcgagagggc gcagggactt cctttgtccc aaatctggcg 1020
gagccgaaat ctgggaggcg ccgccgcacc ccctctagcg ggcgcgggcg aagcggtgcg 1080
gcgccggcag gaaggaaatg ggcggggagg gccttcgtgc gtcgccgcgc cgccgtcccc 1140
ttctccatct ccagcctcgg ggctgccgca gggggacggc tgccttcggg ggggacgggg 1200
cagggcgggg ttcggcttct ggcgtgtgac cggcggggtt tatatcttcc cttctctgtt 1260
cctccgcagc cagccatg 1278
<210> 16
<211> 2882
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК модифицированной трансферной плазмиды для
HVT-gfp-A № 14*
<400> 16
gccactgtat gggccattta tgtttatcga gtctaaaagt cgtggaagcg cctggccatc 60
gccagtctcg acctgccact gctgagttcg gcttcaattc cagcatccca gaaatgtcgc 120
agtcaccatc acgccgtgca tcgcttccta cgaggccttt tgacgcttct gatttgggca 180
catacaccct ggacatactc caccgctatt cgctcgtaga tttagtacaa ctactgaatg 240
acttgccgcg taacattacc tccacgcccg cttctaatgt agaaaccatg gcaaaaatta 300
atgttttaag ggccatttgc gtaggatttg ccgaggtccg tcgccacaac gacgcgcgaa 360
ctttacagcg aacggcaatg tttgccgccg acgacgtcgc atcacggatc agaccatcca 420
ttggattaaa gcgcacctac ccaccgggta tattttccac agctattacc gtatctaatt 480
ccgaggatga agagcgaaat tcgtgatcgt aaaaataaaa aatacaagat attgacattg 540
attattgact agttattaat agtaatcaat tacggggtca ttagttcata gcccatatat 600
ggagttccgc gttacataac ttacggtaaa tggcccgcct ggctgaccgc ccaacgaccc 660
ccgcccattg acgtcaataa tgacgtatgt tcccatagta acgccaatag ggactttcca 720
ttgacgtcaa tgggtggagt atttacggta aactgcccac ttggcagtac atcaagtgta 780
tcatatgcca agtacgcccc ctattgacgt caatgacggt aaatggcccg cctggcatta 840
tgcccagtac atgaccttat gggactttcc tacttggcag tacatctacg tattagtcat 900
cgctattacc atggtgatgc ggttttggca gtacatcaat gggcgtggat agcggtttga 960
ctcacgggga tttccaagtc tccaccccat tgacgtcaat gggagtttgt tttggcacca 1020
aaatcaacgg gactttccaa aatgtcgtaa caactccgcc ccattgacgc aaatgggcgg 1080
taggcgtgta cggtgggagg tctatataag cagagctctc tggctaacta gagaacccac 1140
tgcttactgg cttatcgaaa ttaatacgac tcactatagg gagacccaag ctggctagcg 1200
tttaaactta agcttacctg caggccacca tggtgagcaa gggcgccgag ctgttcaccg 1260
gcatcgtgcc catcctgatc gagctgaatg gcgatgtgaa tggccacaag ttcagcgtga 1320
gcggcgaggg cgagggcgat gccacctacg gcaagctgac cctgaagttc atctgcacca 1380
ccggcaagct gcctgtgccc tggcccaccc tggtgaccac cctgagctac ggcgtgcagt 1440
gcttctcacg ctaccccgat cacatgaagc agcacgactt cttcaagagc gccatgcctg 1500
agggctacat ccaggagcgc accatcttct tcgaggatga cggcaactac aagtcgcgcg 1560
ccgaggtgaa gttcgagggc gataccctgg tgaatcgcat cgagctgacc ggcaccgatt 1620
tcaaggagga tggcaacatc ctgggcaata agatggagta caactacaac gcccacaatg 1680
tgtacatcat gaccgacaag gccaagaatg gcatcaaggt gaacttcaag atccgccaca 1740
acatcgagga tggcagcgtg cagctggccg accactacca gcagaatacc cccatcggcg 1800
atggccctgt gctgctgccc gataaccact acctgtccac ccagagcgcc ctgtccaagg 1860
accccaacga gaagcgcgat cacatgatct acttcggctt cgtgaccgcc gccgccatca 1920
cccacggcat ggatgagctg tacaagtgac ctgcaggtgt gccttctagt tgccagccat 1980
ctgttgtttg cccctccccc gtgccttcct tgaccctgga aggtgccact cccactgtcc 2040
tttcctaata aaatgaggaa attgcatcgc attgtctgag taggtgtcat tctattctgg 2100
ggggtggggt ggggcaggac agcaaggggg aggattggga agacaatagc aggcatgctg 2160
gggatgcggt gggctctatg gtgtacataa ttcttattta tctttaatcc atgaggagca 2220
tttttatttt aaaaatgtca gccgccagcc ctataaccct agatcgcaac tgatccctag 2280
tctgcgttat ttgtcttgca atcttttcgc acgcctttgt gagtgcatac aatgcccccc 2340
tgctcgcttt tctgaaatcg cgtcgggtca ttaatgtgtc ggctatcaca atgcgagatg 2400
tactcgacat gtccgtgtct gtactattgg gattgtaaat agtcgaccgc gaatcatcag 2460
agtcggaatc tgtaaaggat acagattccg actctgagcg cttatgaatg ggatccactc 2520
ggacgttgtt gaacttccgt tcggattctg cttcagtcaa caccggcccc cgatagctac 2580
taaggttggg gggtttgtgg gttgtttgtg aaactgcttt gcggtgtgca ttaccacggg 2640
gggtgtgggg aagtatctgt ttccacgatg cgataacgtt cggtggcgga gggggcgatt 2700
cattctctag tgtacgcgtt tcaacttcag gaacgtgatt atttctttca ggacactctt 2760
tccaatttcc ttcttccttc acttcgggta caggtatatt cttaatgttt acatacatgt 2820
cgtctgctcg tctcaactgc ggggttatga tgggtggtgg tgacagtctc tccgaatgat 2880
cg 2882
<210> 17
<211> 2869
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК исходной трансферной плазмиды для HVT-gfp-A № 14
<400> 17
gccactgtat gggccattta tgtttatcga gtctaaaagt cgtggaagcg cctggccatc 60
gccagtctcg acctgccact gctgagttcg gcttcaattc cagcatccca gaaatgtcgc 120
agtcaccatc acgccgtgca tcgcttccta cgaggccttt tgacgcttct gatttgggca 180
catacaccct ggacatactc caccgctatt cgctcgtaga tttagtacaa ctactgaatg 240
acttgccgcg taacattacc tccacgcccg cttctaatgt agaaaccatg gcaaaaatta 300
atgttttaag ggccatttgc gtaggatttg ccgaggtccg tcgccacaac gacgcgcgaa 360
ctttacagcg aacggcaatg tttgccgccg acgacgtcgc atcacggatc agaccatcca 420
ttggattaaa gcgcacctac ccaccgggta tattttccac agctattacc gtatctaatt 480
ccgaggatga agagcgaaat tcgtgatcgt aaaaataaaa aatacaagat attgacattg 540
attattgact agttattaat agtaatcaat tacggggtca ttagttcata gcccatatat 600
ggagttccgc gttacataac ttacggtaaa tggcccgcct ggctgaccgc ccaacgaccc 660
ccgcccattg acgtcaataa tgacgtatgt tcccatagta acgccaatag ggactttcca 720
ttgacgtcaa tgggtggagt atttacggta aactgcccac ttggcagtac atcaagtgta 780
tcatatgcca agtacgcccc ctattgacgt caatgacggt aaatggcccg cctggcatta 840
tgcccagtac atgaccttat gggactttcc tacttggcag tacatctacg tattagtcat 900
cgctattacc atggtgatgc ggttttggca gtacatcaat gggcgtggat agcggtttga 960
ctcacgggga tttccaagtc tccaccccat tgacgtcaat gggagtttgt tttggcacca 1020
aaatcaacgg gactttccaa aatgtcgtaa caactccgcc ccattgacgc aaatgggcgg 1080
taggcgtgta cggtgggagg tctatataag cagagctctc tggctaacta gagaacccac 1140
tgcttactgg cttatcgaaa ttaatacgac tcactatagg gagacccaag ctggctagcg 1200
tttaaactta agcttaccgc caccatggtg agcaagggcg ccgagctgtt caccggcatc 1260
gtgcccatcc tgatcgagct gaatggcgat gtgaatggcc acaagttcag cgtgagcggc 1320
gagggcgagg gcgatgccac ctacggcaag ctgaccctga agttcatctg caccaccggc 1380
aagctgcctg tgccctggcc caccctggtg accaccctga gctacggcgt gcagtgcttc 1440
tcacgctacc ccgatcacat gaagcagcac gacttcttca agagcgccat gcctgagggc 1500
tacatccagg agcgcaccat cttcttcgag gatgacggca actacaagtc gcgcgccgag 1560
gtgaagttcg agggcgatac cctggtgaat cgcatcgagc tgaccggcac cgatttcaag 1620
gaggatggca acatcctggg caataagatg gagtacaact acaacgccca caatgtgtac 1680
atcatgaccg acaaggccaa gaatggcatc aaggtgaact tcaagatccg ccacaacatc 1740
gaggatggca gcgtgcagct ggccgaccac taccagcaga atacccccat cggcgatggc 1800
cctgtgctgc tgcccgataa ccactacctg tccacccaga gcgccctgtc caaggacccc 1860
aacgagaagc gcgatcacat gatctacttc ggcttcgtga ccgccgccgc catcacccac 1920
ggcatggatg agctgtacaa gtgatgtgcc ttctagttgc cagccatctg ttgtttgccc 1980
ctcccccgtg ccttccttga ccctggaagg tgccactccc actgtccttt cctaataaaa 2040
tgaggaaatt gcatcgcatt gtctgagtag gtgtcattct attctggggg gtggggtggg 2100
gcaggacagc aagggggagg attgggaaga caatagcagg catgctgggg atgcggtggg 2160
ctctatggtg tacataattc ttatttatct ttaatccatg aggagcattt ttattttaaa 2220
aatgtcagcc gccagcccta taaccctaga tcgcaactga tccctagtct gcgttatttg 2280
tcttgcaatc ttttcgcacg cctttgtgag tgcatacaat gcccccctgc tcgcttttct 2340
gaaatcgcgt cgggtcatta atgtgtcggc tatcacaatg cgagatgtac tcgacatgtc 2400
cgtgtctgta ctattgggat tgtaaatagt cgaccgcgaa tcatcagagt cggaatctgt 2460
aaaggataca gattccgact ctgagcgctt atgaatggga tccactcgga cgttgttgaa 2520
cttccgttcg gattctgctt cagtcaacac cggcccccga tagctactaa ggttgggggg 2580
tttgtgggtt gtttgtgaaa ctgctttgcg gtgtgcatta ccacgggggg tgtggggaag 2640
tatctgtttc cacgatgcga taacgttcgg tggcggaggg ggcgattcat tctctagtgt 2700
acgcgtttca acttcaggaa cgtgattatt tctttcagga cactctttcc aatttccttc 2760
ttccttcact tcgggtacag gtatattctt aatgtttaca tacatgtcgt ctgctcgtct 2820
caactgcggg gttatgatgg gtggtggtga cagtctctcc gaatgatcg 2869
<210> 18
<211> 2895
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК трансферной плазмиды для HVT-gfp-B № 13
<400> 18
atggcagcga gcatgggata ttcatcctcg tcatcgttaa catctctacg ggttcagaat 60
gtttggcatg tcgtcgatcc tttgcccatc gttgcaaatt acaagtccga tcgccatgac 120
cgcgataagc ctgtaccatg tggcattagg gtgacatctc gatcatacat tataagacca 180
acgtgcgagt cttccaaaga cctgcacgcc ttcttcttcg gattgtcaac gggttcttca 240
gaatctatgc ccatatctgg cgttgagacc attgtgcgtt taatgaacaa taaagcggca 300
tgccatggaa aggagggctg cagatctcca ttttctcacg ccactatcct ggacgctgta 360
gacgataatt ataccatgaa tatagagggg gtatgtttcc actgccactg tgatgataag 420
ttttctccag attgttggat atctgcattt tctgctgccg aacaaacttc atcgctatgc 480
aaagagatgc gtgtgtacac gcgccgttga gtatacggga aactaaatgt tcatagaggt 540
ctttgggcta tatgttatta aataaaataa ttgaccagtg ttgacattga ttattgacta 600
gttattaata gtaatcaatt acggggtcat tagttcatag cccatatatg gagttccgcg 660
ttacataact tacggtaaat ggcccgcctg gctgaccgcc caacgacccc cgcccattga 720
cgtcaataat gacgtatgtt cccatagtaa cgccaatagg gactttccat tgacgtcaat 780
gggtggagta tttacggtaa actgcccact tggcagtaca tcaagtgtat catatgccaa 840
gtacgccccc tattgacgtc aatgacggta aatggcccgc ctggcattat gcccagtaca 900
tgaccttatg ggactttcct acttggcagt acatctacgt attagtcatc gctattacca 960
tggtgatgcg gttttggcag tacatcaatg ggcgtggata gcggtttgac tcacggggat 1020
ttccaagtct ccaccccatt gacgtcaatg ggagtttgtt ttggcaccaa aatcaacggg 1080
actttccaaa atgtcgtaac aactccgccc cattgacgca aatgggcggt aggcgtgtac 1140
ggtgggaggt ctatataagc agagctctct ggctaactag agaacccact gcttactggc 1200
ttatcgaaat taatacgact cactataggg agacccaagc tggctagcgt ttaaacttaa 1260
gcttaccgcc accatggtga gcaagggcgc cgagctgttc accggcatcg tgcccatcct 1320
gatcgagctg aatggcgatg tgaatggcca caagttcagc gtgagcggcg agggcgaggg 1380
cgatgccacc tacggcaagc tgaccctgaa gttcatctgc accaccggca agctgcctgt 1440
gccctggccc accctggtga ccaccctgag ctacggcgtg cagtgcttct cacgctaccc 1500
cgatcacatg aagcagcacg acttcttcaa gagcgccatg cctgagggct acatccagga 1560
gcgcaccatc ttcttcgagg atgacggcaa ctacaagtcg cgcgccgagg tgaagttcga 1620
gggcgatacc ctggtgaatc gcatcgagct gaccggcacc gatttcaagg aggatggcaa 1680
catcctgggc aataagatgg agtacaacta caacgcccac aatgtgtaca tcatgaccga 1740
caaggccaag aatggcatca aggtgaactt caagatccgc cacaacatcg aggatggcag 1800
cgtgcagctg gccgaccact accagcagaa tacccccatc ggcgatggcc ctgtgctgct 1860
gcccgataac cactacctgt ccacccagag cgccctgtcc aaggacccca acgagaagcg 1920
cgatcacatg atctacttcg gcttcgtgac cgccgccgcc atcacccacg gcatggatga 1980
gctgtacaag tgatgtgcct tctagttgcc agccatctgt tgtttgcccc tcccccgtgc 2040
cttccttgac cctggaaggt gccactccca ctgtcctttc ctaataaaat gaggaaattg 2100
catcgcattg tctgagtagg tgtcattcta ttctgggggg tggggtgggg caggacagca 2160
agggggagga ttgggaagac aatagcaggc atgctgggga tgcggtgggc tctatggttt 2220
taatgttagt ttattcaatg cattggttgc aaatattcat tacttctcca atcccaggtc 2280
attctttagc gagatgatgt tatgacattg ctgtgaaaat tactacagga tatattttta 2340
agatgcagga gtaacaatgt gcatagtagg cgtagttatc gcagacgtgc aacgcttcgc 2400
atttgagtta ccgaagtgcc caacagtgct gcggttatgg tttatgcgca cagaatccat 2460
gcatgtccta attgaaccat ccgatttttc ttttaatcgc gatcgttgtt tgggcaactg 2520
cgttatttca gatctaaaaa atttaccctt tatgaccatc acatctctct ggctcatacc 2580
ccgcttggat aagatatcat gtagattccg ccctaagaaa tgcaaactaa cattattgtc 2640
ggttccatat acacttccat cttgtccttc gaaaataaca aactcgcgca atagaccgtc 2700
cgtacatgca tggccgatgt gtgtcaacat cattggtctg ctagatcccg atgggacgaa 2760
tcgtacagtc gtcgctccag cattggcaaa aatccccaga taccctccat gcggcaaatc 2820
taaattgcga ccccgaagag actgcaccaa agtcttatcg acgcacgctg atttttttga 2880
acagcgggag cccat 2895
<210> 19
<211> 2908
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК трансферной плазмиды для HVT-gfp-B № 13
<400> 19
atggcagcga gcatgggata ttcatcctcg tcatcgttaa catctctacg ggttcagaat 60
gtttggcatg tcgtcgatcc tttgcccatc gttgcaaatt acaagtccga tcgccatgac 120
cgcgataagc ctgtaccatg tggcattagg gtgacatctc gatcatacat tataagacca 180
acgtgcgagt cttccaaaga cctgcacgcc ttcttcttcg gattgtcaac gggttcttca 240
gaatctatgc ccatatctgg cgttgagacc attgtgcgtt taatgaacaa taaagcggca 300
tgccatggaa aggagggctg cagatctcca ttttctcacg ccactatcct ggacgctgta 360
gacgataatt ataccatgaa tatagagggg gtatgtttcc actgccactg tgatgataag 420
ttttctccag attgttggat atctgcattt tctgctgccg aacaaacttc atcgctatgc 480
aaagagatgc gtgtgtacac gcgccgttga gtatacggga aactaaatgt tcatagaggt 540
ctttgggcta tatgttatta aataaaataa ttgaccagtg ttgacattga ttattgacta 600
gttattaata gtaatcaatt acggggtcat tagttcatag cccatatatg gagttccgcg 660
ttacataact tacggtaaat ggcccgcctg gctgaccgcc caacgacccc cgcccattga 720
cgtcaataat gacgtatgtt cccatagtaa cgccaatagg gactttccat tgacgtcaat 780
gggtggagta tttacggtaa actgcccact tggcagtaca tcaagtgtat catatgccaa 840
gtacgccccc tattgacgtc aatgacggta aatggcccgc ctggcattat gcccagtaca 900
tgaccttatg ggactttcct acttggcagt acatctacgt attagtcatc gctattacca 960
tggtgatgcg gttttggcag tacatcaatg ggcgtggata gcggtttgac tcacggggat 1020
ttccaagtct ccaccccatt gacgtcaatg ggagtttgtt ttggcaccaa aatcaacggg 1080
actttccaaa atgtcgtaac aactccgccc cattgacgca aatgggcggt aggcgtgtac 1140
ggtgggaggt ctatataagc agagctctct ggctaactag agaacccact gcttactggc 1200
ttatcgaaat taatacgact cactataggg agacccaagc tggctagcgt ttaaacttaa 1260
gcttacctgc aggccaccat ggtgagcaag ggcgccgagc tgttcaccgg catcgtgccc 1320
atcctgatcg agctgaatgg cgatgtgaat ggccacaagt tcagcgtgag cggcgagggc 1380
gagggcgatg ccacctacgg caagctgacc ctgaagttca tctgcaccac cggcaagctg 1440
cctgtgccct ggcccaccct ggtgaccacc ctgagctacg gcgtgcagtg cttctcacgc 1500
taccccgatc acatgaagca gcacgacttc ttcaagagcg ccatgcctga gggctacatc 1560
caggagcgca ccatcttctt cgaggatgac ggcaactaca agtcgcgcgc cgaggtgaag 1620
ttcgagggcg ataccctggt gaatcgcatc gagctgaccg gcaccgattt caaggaggat 1680
ggcaacatcc tgggcaataa gatggagtac aactacaacg cccacaatgt gtacatcatg 1740
accgacaagg ccaagaatgg catcaaggtg aacttcaaga tccgccacaa catcgaggat 1800
ggcagcgtgc agctggccga ccactaccag cagaataccc ccatcggcga tggccctgtg 1860
ctgctgcccg ataaccacta cctgtccacc cagagcgccc tgtccaagga ccccaacgag 1920
aagcgcgatc acatgatcta cttcggcttc gtgaccgccg ccgccatcac ccacggcatg 1980
gatgagctgt acaagtgacc tgcaggtgtg ccttctagtt gccagccatc tgttgtttgc 2040
ccctcccccg tgccttcctt gaccctggaa ggtgccactc ccactgtcct ttcctaataa 2100
aatgaggaaa ttgcatcgca ttgtctgagt aggtgtcatt ctattctggg gggtggggtg 2160
gggcaggaca gcaaggggga ggattgggaa gacaatagca ggcatgctgg ggatgcggtg 2220
ggctctatgg ttttaatgtt agtttattca atgcattggt tgcaaatatt cattacttct 2280
ccaatcccag gtcattcttt agcgagatga tgttatgaca ttgctgtgaa aattactaca 2340
ggatatattt ttaagatgca ggagtaacaa tgtgcatagt aggcgtagtt atcgcagacg 2400
tgcaacgctt cgcatttgag ttaccgaagt gcccaacagt gctgcggtta tggtttatgc 2460
gcacagaatc catgcatgtc ctaattgaac catccgattt ttcttttaat cgcgatcgtt 2520
gtttgggcaa ctgcgttatt tcagatctaa aaaatttacc ctttatgacc atcacatctc 2580
tctggctcat accccgcttg gataagatat catgtagatt ccgccctaag aaatgcaaac 2640
taacattatt gtcggttcca tatacacttc catcttgtcc ttcgaaaata acaaactcgc 2700
gcaatagacc gtccgtacat gcatggccga tgtgtgtcaa catcattggt ctgctagatc 2760
ccgatgggac gaatcgtaca gtcgtcgctc cagcattggc aaaaatcccc agataccctc 2820
catgcggcaa atctaaattg cgaccccgaa gagactgcac caaagtctta tcgacgcacg 2880
ctgatttttt tgaacagcgg gagcccat 2908
<210> 20
<211> 3538
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК трансферной плазмиды для HVT-IBD № 1
<400> 20
atggcagcga gcatgggata ttcatcctcg tcatcgttaa catctctacg ggttcagaat 60
gtttggcatg tcgtcgatcc tttgcccatc gttgcaaatt acaagtccga tcgccatgac 120
cgcgataagc ctgtaccatg tggcattagg gtgacatctc gatcatacat tataagacca 180
acgtgcgagt cttccaaaga cctgcacgcc ttcttcttcg gattgtcaac gggttcttca 240
gaatctatgc ccatatctgg cgttgagacc attgtgcgtt taatgaacaa taaagcggca 300
tgccatggaa aggagggctg cagatctcca ttttctcacg ccactatcct ggacgctgta 360
gacgataatt ataccatgaa tatagagggg gtatgtttcc actgccactg tgatgataag 420
ttttctccag attgttggat atctgcattt tctgctgccg aacaaacttc atcgctatgc 480
aaagagatgc gtgtgtacac gcgccgttga gtatacggga aactaaatgt tcatagaggt 540
ctttgggcta tatgttatta aataaaataa ttgaccagtg ttgacattga ttattgacta 600
gttattaata gtaatcaatt acggggtcat tagttcatag cccatatatg gagttccgcg 660
ttacataact tacggtaaat ggcccgcctg gctgaccgcc caacgacccc cgcccattga 720
cgtcaataat gacgtatgtt cccatagtaa cgccaatagg gactttccat tgacgtcaat 780
gggtggagta tttacggtaa actgcccact tggcagtaca tcaagtgtat catatgccaa 840
gtacgccccc tattgacgtc aatgacggta aatggcccgc ctggcattat gcccagtaca 900
tgaccttatg ggactttcct acttggcagt acatctacgt attagtcatc gctattacca 960
tggtgatgcg gttttggcag tacatcaatg ggcgtggata gcggtttgac tcacggggat 1020
ttccaagtct ccaccccatt gacgtcaatg ggagtttgtt ttggcaccaa aatcaacggg 1080
actttccaaa atgtcgtaac aactccgccc cattgacgca aatgggcggt aggcgtgtac 1140
ggtgggaggt ctatataagc agagctctct ggctaactag agaacccact gcttactggc 1200
ttatcgaaat taatacgact cactataggg agacccaagc tggctagcgt ttaaacttaa 1260
gcttaccgcc accatgacaa atcttcagga tcagacccag cagatagttc cctttattag 1320
gtcccttctg atgccaacca cagggcccgc tagcattccg gacgataccc tggagaaaca 1380
cactttgcgg agtgagacaa gcacttacaa tctgacggtg ggagataccg gctcaggtct 1440
tatcgtgttt tttcccggct ttcctggatc cattgttggc gcgcattaca cgctgcagag 1500
caacggcaac tataaattcg atcagatgct cctgacggct cagaatctcc ccgccagtta 1560
caattactgc cgccttgtaa gtaggtcctt gactgttaga agctcaacgc tgccaggcgg 1620
agtatatgcc ctgaatggaa ccattaatgc tgtaacattc caaggatcac tgtccgagct 1680
caccgatgtg tcttacaatg gattgatgtc tgccacggct aacattaacg acaagatcgg 1740
gaatgtgctc gtgggcgagg gagtgaccgt tttgagcctg ccgacaagct acgacctcgg 1800
ctacgtaagg ctcggggatc caatccccgc gatcggcttg gatcccaaaa tggttgctac 1860
gtgcgacagc agcgatagac ccagggtcta taccatcacc gctgccgatg attaccagtt 1920
tagctcccag taccaggcgg gaggggtcac gatcaccctt tttagcgcca acatcgacgc 1980
cataacctca ctttctatag ggggcgagtt ggtttttcag accagtgtcc aggggctcat 2040
cctcggtgcg acaatctatc tgatcggctt tgacggaaca gctgtcatca cgagggccgt 2100
agctgcagat aatggcctga ctgctgggac agataatctg atgccgttca acatagtgat 2160
ccccaccagt gagattacgc aacccatcac gagcatcaaa ctggagatcg tgacgtcaaa 2220
atccggcggt caggcaggtg accagatgtc ttggtccgca agcggaagtt tggccgtgac 2280
aattcacggg gggaattacc ccggcgcact caggcccgtg accctcgtcg cctacgaaag 2340
agttgcaacg ggaagtgtag tgacagtcgc tggagtgagt aacttcgaac tcatccctaa 2400
tcccgagctc gccaaaaatc tcgtcacgga gtatgggagg tttgatcccg gcgccatgaa 2460
ctacacaaaa ctgatattgt ccgaaaggga taggttgggc attaaaaccg tgtggcctac 2520
tagggaatac accgatttcc gcgaatattt tatggaggtc gcggatctga actctcccct 2580
gaagatagca ggcgcttttg ggttcaagga tattatccgg gcgttgcggc ggtagtgtgc 2640
cttctagttg ccagccatct gttgtttgcc cctcccccgt gccttccttg accctggaag 2700
gtgccactcc cactgtcctt tcctaataaa atgaggaaat tgcatcgcat tgtctgagta 2760
ggtgtcattc tattctgggg ggtggggtgg ggcaggacag caagggggag gattgggaag 2820
acaatagcag gcatgctggg gatgcggtgg gctctatggt gtttaatgtt agtttattca 2880
atgcattggt tgcaaatatt cattacttct ccaatcccag gtcattcttt agcgagatga 2940
tgttatgaca ttgctgtgaa aattactaca ggatatattt ttaagatgca ggagtaacaa 3000
tgtgcatagt aggcgtagtt atcgcagacg tgcaacgctt cgcatttgag ttaccgaagt 3060
gcccaacagt gctgcggtta tggtttatgc gcacagaatc catgcatgtc ctaattgaac 3120
catccgattt ttcttttaat cgcgatcgtt gtttgggcaa ctgcgttatt tcagatctaa 3180
aaaatttacc ctttatgacc atcacatctc tctggctcat accccgcttg gataagatat 3240
catgtagatt ccgccctaag aaatgcaaac taacattatt gtcggttcca tatacacttc 3300
catcttgtcc ttcgaaaata acaaactcgc gcaatagacc gtccgtacat gcatggccga 3360
tgtgtgtcaa catcattggt ctgctagatc ccgatgggac gaatcgtaca gtcgtcgctc 3420
cagcattggc aaaaatcccc agataccctc catgcggcaa atctaaattg cgaccccgaa 3480
gagactgcac caaagtctta tcgacgcacg ctgatttttt tgaacagcgg gagcccat 3538
<210> 21
<211> 3511
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК трансферной плазмиды для HVT-IBD № 5
<400> 21
gccactgtat gggccattta tgtttatcga gtctaaaagt cgtggaagcg cctggccatc 60
gccagtctcg acctgccact gctgagttcg gcttcaattc cagcatccca gaaatgtcgc 120
agtcaccatc acgccgtgca tcgcttccta cgaggccttt tgacgcttct gatttgggca 180
catacaccct ggacatactc caccgctatt cgctcgtaga tttagtacaa ctactgaatg 240
acttgccgcg taacattacc tccacgcccg cttctaatgt agaaaccatg gcaaaaatta 300
atgttttaag ggccatttgc gtaggatttg ccgaggtccg tcgccacaac gacgcgcgaa 360
ctttacagcg aacggcaatg tttgccgccg acgacgtcgc atcacggatc agaccatcca 420
ttggattaaa gcgcacctac ccaccgggta tattttccac agctattacc gtatctaatt 480
ccgaggatga agagcgaaat tcgtgatcgt aaaaataaaa aatacaagat attgacattg 540
attattgact agttattaat agtaatcaat tacggggtca ttagttcata gcccatatat 600
ggagttccgc gttacataac ttacggtaaa tggcccgcct ggctgaccgc ccaacgaccc 660
ccgcccattg acgtcaataa tgacgtatgt tcccatagta acgccaatag ggactttcca 720
ttgacgtcaa tgggtggagt atttacggta aactgcccac ttggcagtac atcaagtgta 780
tcatatgcca agtacgcccc ctattgacgt caatgacggt aaatggcccg cctggcatta 840
tgcccagtac atgaccttat gggactttcc tacttggcag tacatctacg tattagtcat 900
cgctattacc atggtgatgc ggttttggca gtacatcaat gggcgtggat agcggtttga 960
ctcacgggga tttccaagtc tccaccccat tgacgtcaat gggagtttgt tttggcacca 1020
aaatcaacgg gactttccaa aatgtcgtaa caactccgcc ccattgacgc aaatgggcgg 1080
taggcgtgta cggtgggagg tctatataag cagagctctc tggctaacta gagaacccac 1140
tgcttactgg cttatcgaaa ttaatacgac tcactatagg gagacccaag ctggctagcg 1200
tttaaactta agcttaccgc caccatgaca aatcttcagg atcagaccca gcagatagtt 1260
ccctttatta ggtcccttct gatgccaacc acagggcccg ctagcattcc ggacgatacc 1320
ctggagaaac acactttgcg gagtgagaca agcacttaca atctgacggt gggagatacc 1380
ggctcaggtc ttatcgtgtt ttttcccggc tttcctggat ccattgttgg cgcgcattac 1440
acgctgcaga gcaacggcaa ctataaattc gatcagatgc tcctgacggc tcagaatctc 1500
cccgccagtt acaattactg ccgccttgta agtaggtcct tgactgttag aagctcaacg 1560
ctgccaggcg gagtatatgc cctgaatgga accattaatg ctgtaacatt ccaaggatca 1620
ctgtccgagc tcaccgatgt gtcttacaat ggattgatgt ctgccacggc taacattaac 1680
gacaagatcg ggaatgtgct cgtgggcgag ggagtgaccg ttttgagcct gccgacaagc 1740
tacgacctcg gctacgtaag gctcggggat ccaatccccg cgatcggctt ggatcccaaa 1800
atggttgcta cgtgcgacag cagcgataga cccagggtct ataccatcac cgctgccgat 1860
gattaccagt ttagctccca gtaccaggcg ggaggggtca cgatcaccct ttttagcgcc 1920
aacatcgacg ccataacctc actttctata gggggcgagt tggtttttca gaccagtgtc 1980
caggggctca tcctcggtgc gacaatctat ctgatcggct ttgacggaac agctgtcatc 2040
acgagggccg tagctgcaga taatggcctg actgctggga cagataatct gatgccgttc 2100
aacatagtga tccccaccag tgagattacg caacccatca cgagcatcaa actggagatc 2160
gtgacgtcaa aatccggcgg tcaggcaggt gaccagatgt cttggtccgc aagcggaagt 2220
ttggccgtga caattcacgg ggggaattac cccggcgcac tcaggcccgt gaccctcgtc 2280
gcctacgaaa gagttgcaac gggaagtgta gtgacagtcg ctggagtgag taacttcgaa 2340
ctcatcccta atcccgagct cgccaaaaat ctcgtcacgg agtatgggag gtttgatccc 2400
ggcgccatga actacacaaa actgatattg tccgaaaggg ataggttggg cattaaaacc 2460
gtgtggccta ctagggaata caccgatttc cgcgaatatt ttatggaggt cgcggatctg 2520
aactctcccc tgaagatagc aggcgctttt gggttcaagg atattatccg ggcgttgcgg 2580
cggtagtgtg ccttctagtt gccagccatc tgttgtttgc ccctcccccg tgccttcctt 2640
gaccctggaa ggtgccactc ccactgtcct ttcctaataa aatgaggaaa ttgcatcgca 2700
ttgtctgagt aggtgtcatt ctattctggg gggtggggtg gggcaggaca gcaaggggga 2760
ggattgggaa gacaatagca ggcatgctgg ggatgcggtg ggctctatgg tgtacataat 2820
tcttatttat ctttaatcca tgaggagcat ttttatttta aaaatgtcag ccgccagccc 2880
tataacccta gatcgcaact gatccctagt ctgcgttatt tgtcttgcaa tcttttcgca 2940
cgcctttgtg agtgcataca atgcccccct gctcgctttt ctgaaatcgc gtcgggtcat 3000
taatgtgtcg gctatcacaa tgcgagatgt actcgacatg tccgtgtctg tactattggg 3060
attgtaaata gtcgaccgcg aatcatcaga gtcggaatct gtaaaggata cagattccga 3120
ctctgagcgc ttatgaatgg gatccactcg gacgttgttg aacttccgtt cggattctgc 3180
ttcagtcaac accggccccc gatagctact aaggttgggg ggtttgtggg ttgtttgtga 3240
aactgctttg cggtgtgcat taccacgggg ggtgtgggga agtatctgtt tccacgatgc 3300
gataacgttc ggtggcggag ggggcgattc attctctagt gtacgcgttt caacttcagg 3360
aacgtgatta tttctttcag gacactcttt ccaatttcct tcttccttca cttcgggtac 3420
aggtatattc ttaatgttta catacatgtc gtctgctcgt ctcaactgcg gggttatgat 3480
gggtggtggt gacagtctct ccgaatgatc g 3511
<210> 22
<211> 3385
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК трансферной плазмиды для HVT-IBD № 1
<400> 22
gccactgtat gggccattta tgtttatcga gtctaaaagt cgtggaagcg cctggccatc 60
gccagtctcg acctgccact gctgagttcg gcttcaattc cagcatccca gaaatgtcgc 120
agtcaccatc acgccgtgca tcgcttccta cgaggccttt tgacgcttct gatttgggca 180
catacaccct ggacatactc caccgctatt cgctcgtaga tttagtacaa ctactgaatg 240
acttgccgcg taacattacc tccacgcccg cttctaatgt agaaaccatg gcaaaaatta 300
atgttttaag ggccatttgc gtaggatttg ccgaggtccg tcgccacaac gacgcgcgaa 360
ctttacagcg aacggcaatg tttgccgccg acgacgtcgc atcacggatc agaccatcca 420
ttggattaaa gcgcacctac ccaccgggta tattttccac agctattacc gtatctaatt 480
ccgaggatga agagcgaaat tcgtgatcgt aaaaataaaa aatacaagat aagttattaa 540
tagtaatcaa ttacggggtc attagttcat agcccatata tggagttccg cgttacataa 600
cttacggtaa atggcccgcc ggctgaccgc ccaacgaccc ccgcccattg acgtcaataa 660
tgacgtatgt tcccatagta acgccaatag ggactttcca ttgacgtcaa tgggtggagt 720
atttacggta aactgcccat tggcagtaca tcaagtgtat catatgccaa gtacgccccc 780
tattgacgtc aatgacggta aatggatgca gtattttgtg cagcgatggg ggcggggggg 840
gggggggcgc gcgccaggcg gggcggggcg gggcgagggg cggggcgggg cgaggcggag 900
aggtgcggcg gcagccaatc agagcggcgc gctccgaaag tttcctttta tggcgaggcg 960
gcggcggcgg cggccctata aaaagcgaag cgcgcggcgg gcgggagtcg ctgcgcgctg 1020
ccttcgcccc gtgccccgct ccgccgccgc ctcgcgccgc ccgccccggc tctgactgac 1080
cgcgtctaga gggccaccat gacaaatctt caggatcaga cccagcagat agttcccttt 1140
attaggtccc ttctgatgcc aaccacaggg cccgctagca ttccggacga taccctggag 1200
aaacacactt tgcggagtga gacaagcact tacaatctga cggtgggaga taccggctca 1260
ggtcttatcg tgttttttcc cggctttcct ggatccattg ttggcgcgca ttacacgctg 1320
cagagcaacg gcaactataa attcgatcag atgctcctga cggctcagaa tctccccgcc 1380
agttacaatt actgccgcct tgtaagtagg tccttgactg ttagaagctc aacgctgcca 1440
ggcggagtat atgccctgaa tggaaccatt aatgctgtaa cattccaagg atcactgtcc 1500
gagctcaccg atgtgtctta caatggattg atgtctgcca cggctaacat taacgacaag 1560
atcgggaatg tgctcgtggg cgagggagtg accgttttga gcctgccgac aagctacgac 1620
ctcggctacg taaggctcgg ggatccaatc cccgcgatcg gcttggatcc caaaatggtt 1680
gctacgtgcg acagcagcga tagacccagg gtctatacca tcaccgctgc cgatgattac 1740
cagtttagct cccagtacca ggcgggaggg gtcacgatca ccctttttag cgccaacatc 1800
gacgccataa cctcactttc tatagggggc gagttggttt ttcagaccag tgtccagggg 1860
ctcatcctcg gtgcgacaat ctatctgatc ggctttgacg gaacagctgt catcacgagg 1920
gccgtagctg cagataatgg cctgactgct gggacagata atctgatgcc gttcaacata 1980
gtgatcccca ccagtgagat tacgcaaccc atcacgagca tcaaactgga gatcgtgacg 2040
tcaaaatccg gcggtcaggc aggtgaccag atgtcttggt ccgcaagcgg aagtttggcc 2100
gtgacaattc acggggggaa ttaccccggc gcactcaggc ccgtgaccct cgtcgcctac 2160
gaaagagttg caacgggaag tgtagtgaca gtcgctggag tgagtaactt cgaactcatc 2220
cctaatcccg agctcgccaa aaatctcgtc acggagtatg ggaggtttga tcccggcgcc 2280
atgaactaca caaaactgat attgtccgaa agggataggt tgggcattaa aaccgtgtgg 2340
cctactaggg aatacaccga tttccgcgaa tattttatgg aggtcgcgga tctgaactct 2400
cccctgaaga tagcaggcgc ttttgggttc aaggatatta tccgggcgtt gcggcggtag 2460
tgtgccttct agttgccagc catctgttgt ttgcccctcc cccgtgcctt ccttgaccct 2520
ggaaggtgcc actcccactg tcctttccta ataaaatgag gaaattgcat cgcattgtct 2580
gagtaggtgt cattctattc tggggggtgg ggtggggcag gacagcaagg gggaggattg 2640
ggaagacaat agcaggcatg ctggggatgc ggtgggctct atggtgtaca taattcttat 2700
ttatctttaa tccatgagga gcatttttat tttaaaaatg tcagccgcca gccctataac 2760
cctagatcgc aactgatccc tagtctgcgt tatttgtctt gcaatctttt cgcacgcctt 2820
tgtgagtgca tacaatgccc ccctgctcgc ttttctgaaa tcgcgtcggg tcattaatgt 2880
gtcggctatc acaatgcgag atgtactcga catgtccgtg tctgtactat tgggattgta 2940
aatagtcgac cgcgaatcat cagagtcgga atctgtaaag gatacagatt ccgactctga 3000
gcgcttatga atgggatcca ctcggacgtt gttgaacttc cgttcggatt ctgcttcagt 3060
caacaccggc ccccgatagc tactaaggtt ggggggtttg tgggttgttt gtgaaactgc 3120
tttgcggtgt gcattaccac ggggggtgtg gggaagtatc tgtttccacg atgcgataac 3180
gttcggtggc ggagggggcg attcattctc tagtgtacgc gtttcaactt caggaacgtg 3240
attatttctt tcaggacact ctttccaatt tccttcttcc ttcacttcgg gtacaggtat 3300
attcttaatg tttacataca tgtcgtctgc tcgtctcaac tgcggggtta tgatgggtgg 3360
tggtgacagt ctctccgaat gatcg 3385
<210> 23
<211> 3538
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК трансферной плазмиды для HVT-IBD № 9
<400> 23
atggcagcga gcatgggata ttcatcctcg tcatcgttaa catctctacg ggttcagaat 60
gtttggcatg tcgtcgatcc tttgcccatc gttgcaaatt acaagtccga tcgccatgac 120
cgcgataagc ctgtaccatg tggcattagg gtgacatctc gatcatacat tataagacca 180
acgtgcgagt cttccaaaga cctgcacgcc ttcttcttcg gattgtcaac gggttcttca 240
gaatctatgc ccatatctgg cgttgagacc attgtgcgtt taatgaacaa taaagcggca 300
tgccatggaa aggagggctg cagatctcca ttttctcacg ccactatcct ggacgctgta 360
gacgataatt ataccatgaa tatagagggg gtatgtttcc actgccactg tgatgataag 420
ttttctccag attgttggat atctgcattt tctgctgccg aacaaacttc atcgctatgc 480
aaagagatgc gtgtgtacac gcgccgttga gtatacggga aactaaatgt tcatagaggt 540
ctttgggcta tatgttatta aataaaataa ttgaccagtg ttgacattga ttattgacta 600
gttattaata gtaatcaatt acggggtcat tagttcatag cccatatatg gagttccgcg 660
ttacataact tacggtaaat ggcccgcctg gctgaccgcc caacgacccc cgcccattga 720
cgtcaataat gacgtatgtt cccatagtaa cgccaatagg gactttccat tgacgtcaat 780
gggtggagta tttacggtaa actgcccact tggcagtaca tcaagtgtat catatgccaa 840
gtacgccccc tattgacgtc aatgacggta aatggcccgc ctggcattat gcccagtaca 900
tgaccttatg ggactttcct acttggcagt acatctacgt attagtcatc gctattacca 960
tggtgatgcg gttttggcag tacatcaatg ggcgtggata gcggtttgac tcacggggat 1020
ttccaagtct ccaccccatt gacgtcaatg ggagtttgtt ttggcaccaa aatcaacggg 1080
actttccaaa atgtcgtaac aactccgccc cattgacgca aatgggcggt aggcgtgtac 1140
ggtgggaggt ctatataagc agagctctct ggctaactag agaacccact gcttactggc 1200
ttatcgaaat taatacgact cactataggg agacccaagc tggctagcgt ttaaacttaa 1260
gcttaccgcc accatgacca atctccagga ccagacccag cagattgtgc cttttattag 1320
gagtctcttg atgcctacaa ccggccccgc cagcatcccg gacgacacac tggaaaaaca 1380
tacactgaga agcgagacat ctacatacaa tttgaccgtg ggcgataccg gctccgggct 1440
tatcgtgttc ttcccaggtt ttcccggatc tatcgtagga gcgcactaca ccctccaaag 1500
taacggcaat tacaaattcg accagatgct cctgacagcc cagaaccttc ctgcttctta 1560
caattactgt agacttgtgt ccaggtccct gactgtgcgg agtagcacgc ttccaggagg 1620
cgtatacgcc ctgaacggaa ctataaacgc cgtcaccttc cagggctcct tgtccgaact 1680
taccgacgtg tcctacaatg gcctcatgag cgcaacggcc aacataaacg ataagatcgg 1740
caatgttctt gtgggcgagg gggttacagt cctttctctg ccaaccagtt atgatctggg 1800
atacgtgcgg cttggcgatc ccattcccgc tatcggtctc gaccctaaaa tggtggctac 1860
ttgcgactca tctgaccgcc caagggtcta tacaattact gcagccgatg actatcagtt 1920
ttccagccaa taccagccag ggggtgtgac aatcacactt ttcagcgcca atattgacgc 1980
tatcacatcc ctctcaatcg gaggtgagct tgtgttccag acttctgttc agggcttggt 2040
attgggcgcc actatttact tgatcgggtt cgacgggacc gcagtgatca ctcgggcagt 2100
ggctgcggat aacggactca ctgccggaac tgacaacctt atgcctttta atctggtcat 2160
ccccactaac gagatcaccc agcctattac ctccataaag ctcgaaattg tgaccagcaa 2220
gagcggaggg caggcaggcg accaaatgag ttggtctgca agcgggtccc tcgccgtgac 2280
catccacggt ggcaactatc ctggggcgct cagacccgtc accctggtag cctacgaaag 2340
ggttgccaca ggctcagttg tcacggtggc tggagtaagc aatttcgagc tcatcccgaa 2400
tcctgagctc gctaaaaatc ttgtgaccga gtatggaagg ttcgaccctg gcgcaatgaa 2460
ttacacaaag ctgattctgt ccgaacggga taggctgggt atcaagacag tttggcccac 2520
gcgcgaatac acagatttca gggagtactt tatggaggtc gcagatttga atagcccact 2580
taagatcgct ggagcatttg gctttaagga tattatccgc gcaatcagaa ggtagtgtgc 2640
cttctagttg ccagccatct gttgtttgcc cctcccccgt gccttccttg accctggaag 2700
gtgccactcc cactgtcctt tcctaataaa atgaggaaat tgcatcgcat tgtctgagta 2760
ggtgtcattc tattctgggg ggtggggtgg ggcaggacag caagggggag gattgggaag 2820
acaatagcag gcatgctggg gatgcggtgg gctctatggt gtttaatgtt agtttattca 2880
atgcattggt tgcaaatatt cattacttct ccaatcccag gtcattcttt agcgagatga 2940
tgttatgaca ttgctgtgaa aattactaca ggatatattt ttaagatgca ggagtaacaa 3000
tgtgcatagt aggcgtagtt atcgcagacg tgcaacgctt cgcatttgag ttaccgaagt 3060
gcccaacagt gctgcggtta tggtttatgc gcacagaatc catgcatgtc ctaattgaac 3120
catccgattt ttcttttaat cgcgatcgtt gtttgggcaa ctgcgttatt tcagatctaa 3180
aaaatttacc ctttatgacc atcacatctc tctggctcat accccgcttg gataagatat 3240
catgtagatt ccgccctaag aaatgcaaac taacattatt gtcggttcca tatacacttc 3300
catcttgtcc ttcgaaaata acaaactcgc gcaatagacc gtccgtacat gcatggccga 3360
tgtgtgtcaa catcattggt ctgctagatc ccgatgggac gaatcgtaca gtcgtcgctc 3420
cagcattggc aaaaatcccc agataccctc catgcggcaa atctaaattg cgaccccgaa 3480
gagactgcac caaagtctta tcgacgcacg ctgatttttt tgaacagcgg gagcccat 3538
<210> 24
<211> 3511
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК трансферной плазмиды для HVT-IBD № 30
<400> 24
gccactgtat gggccattta tgtttatcga gtctaaaagt cgtggaagcg cctggccatc 60
gccagtctcg acctgccact gctgagttcg gcttcaattc cagcatccca gaaatgtcgc 120
agtcaccatc acgccgtgca tcgcttccta cgaggccttt tgacgcttct gatttgggca 180
catacaccct ggacatactc caccgctatt cgctcgtaga tttagtacaa ctactgaatg 240
acttgccgcg taacattacc tccacgcccg cttctaatgt agaaaccatg gcaaaaatta 300
atgttttaag ggccatttgc gtaggatttg ccgaggtccg tcgccacaac gacgcgcgaa 360
ctttacagcg aacggcaatg tttgccgccg acgacgtcgc atcacggatc agaccatcca 420
ttggattaaa gcgcacctac ccaccgggta tattttccac agctattacc gtatctaatt 480
ccgaggatga agagcgaaat tcgtgatcgt aaaaataaaa aatacaagat attgacattg 540
attattgact agttattaat agtaatcaat tacggggtca ttagttcata gcccatatat 600
ggagttccgc gttacataac ttacggtaaa tggcccgcct ggctgaccgc ccaacgaccc 660
ccgcccattg acgtcaataa tgacgtatgt tcccatagta acgccaatag ggactttcca 720
ttgacgtcaa tgggtggagt atttacggta aactgcccac ttggcagtac atcaagtgta 780
tcatatgcca agtacgcccc ctattgacgt caatgacggt aaatggcccg cctggcatta 840
tgcccagtac atgaccttat gggactttcc tacttggcag tacatctacg tattagtcat 900
cgctattacc atggtgatgc ggttttggca gtacatcaat gggcgtggat agcggtttga 960
ctcacgggga tttccaagtc tccaccccat tgacgtcaat gggagtttgt tttggcacca 1020
aaatcaacgg gactttccaa aatgtcgtaa caactccgcc ccattgacgc aaatgggcgg 1080
taggcgtgta cggtgggagg tctatataag cagagctctc tggctaacta gagaacccac 1140
tgcttactgg cttatcgaaa ttaatacgac tcactatagg gagacccaag ctggctagcg 1200
tttaaactta agcttaccgc caccatgacc aatctccagg accagaccca gcagattgtg 1260
ccttttatta ggagtctctt gatgcctaca accggccccg ccagcatccc ggacgacaca 1320
ctggaaaaac atacactgag aagcgagaca tctacataca atttgaccgt gggcgatacc 1380
ggctccgggc ttatcgtgtt cttcccaggt tttcccggat ctatcgtagg agcgcactac 1440
accctccaaa gtaacggcaa ttacaaattc gaccagatgc tcctgacagc ccagaacctt 1500
cctgcttctt acaattactg tagacttgtg tccaggtccc tgactgtgcg gagtagcacg 1560
cttccaggag gcgtatacgc cctgaacgga actataaacg ccgtcacctt ccagggctcc 1620
ttgtccgaac ttaccgacgt gtcctacaat ggcctcatga gcgcaacggc caacataaac 1680
gataagatcg gcaatgttct tgtgggcgag ggggttacag tcctttctct gccaaccagt 1740
tatgatctgg gatacgtgcg gcttggcgat cccattcccg ctatcggtct cgaccctaaa 1800
atggtggcta cttgcgactc atctgaccgc ccaagggtct atacaattac tgcagccgat 1860
gactatcagt tttccagcca ataccagcca gggggtgtga caatcacact tttcagcgcc 1920
aatattgacg ctatcacatc cctctcaatc ggaggtgagc ttgtgttcca gacttctgtt 1980
cagggcttgg tattgggcgc cactatttac ttgatcgggt tcgacgggac cgcagtgatc 2040
actcgggcag tggctgcgga taacggactc actgccggaa ctgacaacct tatgcctttt 2100
aatctggtca tccccactaa cgagatcacc cagcctatta cctccataaa gctcgaaatt 2160
gtgaccagca agagcggagg gcaggcaggc gaccaaatga gttggtctgc aagcgggtcc 2220
ctcgccgtga ccatccacgg tggcaactat cctggggcgc tcagacccgt caccctggta 2280
gcctacgaaa gggttgccac aggctcagtt gtcacggtgg ctggagtaag caatttcgag 2340
ctcatcccga atcctgagct cgctaaaaat cttgtgaccg agtatggaag gttcgaccct 2400
ggcgcaatga attacacaaa gctgattctg tccgaacggg ataggctggg tatcaagaca 2460
gtttggccca cgcgcgaata cacagatttc agggagtact ttatggaggt cgcagatttg 2520
aatagcccac ttaagatcgc tggagcattt ggctttaagg atattatccg cgcaatcaga 2580
aggtagtgtg ccttctagtt gccagccatc tgttgtttgc ccctcccccg tgccttcctt 2640
gaccctggaa ggtgccactc ccactgtcct ttcctaataa aatgaggaaa ttgcatcgca 2700
ttgtctgagt aggtgtcatt ctattctggg gggtggggtg gggcaggaca gcaaggggga 2760
ggattgggaa gacaatagca ggcatgctgg ggatgcggtg ggctctatgg tgtacataat 2820
tcttatttat ctttaatcca tgaggagcat ttttatttta aaaatgtcag ccgccagccc 2880
tataacccta gatcgcaact gatccctagt ctgcgttatt tgtcttgcaa tcttttcgca 2940
cgcctttgtg agtgcataca atgcccccct gctcgctttt ctgaaatcgc gtcgggtcat 3000
taatgtgtcg gctatcacaa tgcgagatgt actcgacatg tccgtgtctg tactattggg 3060
attgtaaata gtcgaccgcg aatcatcaga gtcggaatct gtaaaggata cagattccga 3120
ctctgagcgc ttatgaatgg gatccactcg gacgttgttg aacttccgtt cggattctgc 3180
ttcagtcaac accggccccc gatagctact aaggttgggg ggtttgtggg ttgtttgtga 3240
aactgctttg cggtgtgcat taccacgggg ggtgtgggga agtatctgtt tccacgatgc 3300
gataacgttc ggtggcggag ggggcgattc attctctagt gtacgcgttt caacttcagg 3360
aacgtgatta tttctttcag gacactcttt ccaatttcct tcttccttca cttcgggtac 3420
aggtatattc ttaatgttta catacatgtc gtctgctcgt ctcaactgcg gggttatgat 3480
gggtggtggt gacagtctct ccgaatgatc g 3511
<210> 25
<211> 4102
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК трансферной плазмиды для HVT-IBD № 31
<400> 25
gccactgtat gggccattta tgtttatcga gtctaaaagt cgtggaagcg cctggccatc 60
gccagtctcg acctgccact gctgagttcg gcttcaattc cagcatccca gaaatgtcgc 120
agtcaccatc acgccgtgca tcgcttccta cgaggccttt tgacgcttct gatttgggca 180
catacaccct ggacatactc caccgctatt cgctcgtaga tttagtacaa ctactgaatg 240
acttgccgcg taacattacc tccacgcccg cttctaatgt agaaaccatg gcaaaaatta 300
atgttttaag ggccatttgc gtaggatttg ccgaggtccg tcgccacaac gacgcgcgaa 360
ctttacagcg aacggcaatg tttgccgccg acgacgtcgc atcacggatc agaccatcca 420
ttggattaaa gcgcacctac ccaccgggta tattttccac agctattacc gtatctaatt 480
ccgaggatga agagcgaaat tcgtgatcgt aaaaataaaa aatacaagat atcgaggtga 540
gccccacgtt ctgcttcact ctccccatct cccccccctc cccaccccca attttgtatt 600
tatttatttt ttaattattt tgtgcagcga tgggggcggg gggggggggg gcgcgcgcca 660
ggcggggcgg ggcggggcga ggggcggggc ggggcgaggc ggagaggtgc ggcggcagcc 720
aatcagagcg gcgcgctccg aaagtttcct tttatggcga ggcggcggcg gcggcggccc 780
tataaaaagc gaagcgcgcg gcgggcggga gtcgctgcgt tgccttcgcc ccgtgccccg 840
ctccgcgccg cctcgcgccg cccgccccgg ctctgactga ccgcgttact cccacaggtg 900
agcgggcggg acggcccttc tcctccgggc tgtaattagc gcttggttta atgacggctc 960
gtttcttttc tgtggctgcg tgaaagcctt aaagggctcc gggagggccc tttgtgcggg 1020
ggggagcggc tcggggggtg cgtgcgtgtg tgtgtgcgtg gggagcgccg cgtgcggccc 1080
gcgctgcccg gcggctgtga gcgctgcggg cgcggcgcgg ggctttgtgc gctccgcgtg 1140
tgcgcgaggg gagcgcggcc gggggcggtg ccccgcggtg cgggggggct gcgaggggaa 1200
caaaggctgc gtgcggggtg tgtgcgtggg ggggtgagca gggggtgtgg gcgcggcggt 1260
cgggctgtaa cccccccctg cacccccctc cccgagttgc tgagcacggc ccggcttcgg 1320
gtgcggggct ccgtgcgggg cgtggcgcgg ggctcgccgt gccgggcggg gggtggcggc 1380
aggtgggggt gccgggcggg gcggggccgc ctcgggccgg ggagggctcg ggggaggggc 1440
gcggcggccc cggagcgccg gcggctgtcg aggcgcggcg agccgcagcc attgcctttt 1500
atggtaatcg tgcgagaggg cgcagggact tcctttgtcc caaatctggc ggagccgaaa 1560
tctgggaggc gccgccgcac cccctctagc gggcgcgggc gaagcggtgc ggcgccggca 1620
ggaaggaaat gggcggggag ggccttcgtg cgtcgccgcg ccgccgtccc cttctccatc 1680
tccagcctcg gggctgccgc agggggacgg ctgccttcgg gggggacggg gcagggcggg 1740
gttcggcttc tggcgtgtga ccggcggggt ttatatcttc ccttctctgt tcctccgcag 1800
ccagccatgg ccaccatgac caatctccag gaccagaccc agcagattgt gccttttatt 1860
aggagtctct tgatgcctac aaccggcccc gccagcatcc cggacgacac actggaaaaa 1920
catacactga gaagcgagac atctacatac aatttgaccg tgggcgatac cggctccggg 1980
cttatcgtgt tcttcccagg ttttcccgga tctatcgtag gagcgcacta caccctccaa 2040
agtaacggca attacaaatt cgaccagatg ctcctgacag cccagaacct tcctgcttct 2100
tacaattact gtagacttgt gtccaggtcc ctgactgtgc ggagtagcac gcttccagga 2160
ggcgtatacg ccctgaacgg aactataaac gccgtcacct tccagggctc cttgtccgaa 2220
cttaccgacg tgtcctacaa tggcctcatg agcgcaacgg ccaacataaa cgataagatc 2280
ggcaatgttc ttgtgggcga gggggttaca gtcctttctc tgccaaccag ttatgatctg 2340
ggatacgtgc ggcttggcga tcccattccc gctatcggtc tcgaccctaa aatggtggct 2400
acttgcgact catctgaccg cccaagggtc tatacaatta ctgcagccga tgactatcag 2460
ttttccagcc aataccagcc agggggtgtg acaatcacac ttttcagcgc caatattgac 2520
gctatcacat ccctctcaat cggaggtgag cttgtgttcc agacttctgt tcagggcttg 2580
gtattgggcg ccactattta cttgatcggg ttcgacggga ccgcagtgat cactcgggca 2640
gtggctgcgg ataacggact cactgccgga actgacaacc ttatgccttt taatctggtc 2700
atccccacta acgagatcac ccagcctatt acctccataa agctcgaaat tgtgaccagc 2760
aagagcggag ggcaggcagg cgaccaaatg agttggtctg caagcgggtc cctcgccgtg 2820
accatccacg gtggcaacta tcctggggcg ctcagacccg tcaccctggt agcctacgaa 2880
agggttgcca caggctcagt tgtcacggtg gctggagtaa gcaatttcga gctcatcccg 2940
aatcctgagc tcgctaaaaa tcttgtgacc gagtatggaa ggttcgaccc tggcgcaatg 3000
aattacacaa agctgattct gtccgaacgg gataggctgg gtatcaagac agtttggccc 3060
acgcgcgaat acacagattt cagggagtac tttatggagg tcgcagattt gaatagccca 3120
cttaagatcg ctggagcatt tggctttaag gatattatcc gcgcaatcag aaggtagtgt 3180
gccttctagt tgccagccat ctgttgtttg cccctccccc gtgccttcct tgaccctgga 3240
aggtgccact cccactgtcc tttcctaata aaatgaggaa attgcatcgc attgtctgag 3300
taggtgtcat tctattctgg ggggtggggt ggggcaggac agcaaggggg aggattggga 3360
agacaatagc aggcatgctg gggatgcggt gggctctatg gtgtacataa ttcttattta 3420
tctttaatcc atgaggagca tttttatttt aaaaatgtca gccgccagcc ctataaccct 3480
agatcgcaac tgatccctag tctgcgttat ttgtcttgca atcttttcgc acgcctttgt 3540
gagtgcatac aatgcccccc tgctcgcttt tctgaaatcg cgtcgggtca ttaatgtgtc 3600
ggctatcaca atgcgagatg tactcgacat gtccgtgtct gtactattgg gattgtaaat 3660
agtcgaccgc gaatcatcag agtcggaatc tgtaaaggat acagattccg actctgagcg 3720
cttatgaatg ggatccactc ggacgttgtt gaacttccgt tcggattctg cttcagtcaa 3780
caccggcccc cgatagctac taaggttggg gggtttgtgg gttgtttgtg aaactgcttt 3840
gcggtgtgca ttaccacggg gggtgtgggg aagtatctgt ttccacgatg cgataacgtt 3900
cggtggcgga gggggcgatt cattctctag tgtacgcgtt tcaacttcag gaacgtgatt 3960
atttctttca ggacactctt tccaatttcc ttcttccttc acttcgggta caggtatatt 4020
cttaatgttt acatacatgt cgtctgctcg tctcaactgc ggggttatga tgggtggtgg 4080
tgacagtctc tccgaatgat cg 4102
<210> 26
<211> 3394
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК трансферной плазмиды для HVT-IBD № 32
<400> 26
gccactgtat gggccattta tgtttatcga gtctaaaagt cgtggaagcg cctggccatc 60
gccagtctcg acctgccact gctgagttcg gcttcaattc cagcatccca gaaatgtcgc 120
agtcaccatc acgccgtgca tcgcttccta cgaggccttt tgacgcttct gatttgggca 180
catacaccct ggacatactc caccgctatt cgctcgtaga tttagtacaa ctactgaatg 240
acttgccgcg taacattacc tccacgcccg cttctaatgt agaaaccatg gcaaaaatta 300
atgttttaag ggccatttgc gtaggatttg ccgaggtccg tcgccacaac gacgcgcgaa 360
ctttacagcg aacggcaatg tttgccgccg acgacgtcgc atcacggatc agaccatcca 420
ttggattaaa gcgcacctac ccaccgggta tattttccac agctattacc gtatctaatt 480
ccgaggatga agagcgaaat tcgtgatcgt aaaaataaaa aatacaagat aactctcagt 540
acaatctgct ctgatgccgc atagttaagc cagtatctgc tccctgcttg tgtgttggag 600
gtcgctgagt agtgcgcgag caaaatttaa gctacaacaa ggcaaggctt gaccgacaat 660
tgcatgaaga atctgcttag ggttaggcgt tttgcgctgc ttcgcgatgt acgggccaga 720
tatacgcgta tctgagggga ctagggtgtg tttaggcgaa aagcggggct tcggttgtac 780
gcggttagga gtcccctcag gatatagtag tttcgctttt gcatagggag ggggaaatgt 840
agtcttatgc aatactcttg tagtcttgca acatggtaac gatgagttag caacatgcct 900
tacaaggaga gaaaaagcac cgtgcatgcc gattggtgga agtaaggtgg tacgatcgtg 960
ccttattagg aaggcaacag acgggtctga catggattgg acgaaccact gaattccgca 1020
ttgcagagat attgtattta agtgcctagc tcgatacaat aaacgccatt tgaccattca 1080
ccacattggt gtgcacctcc agccaccatg accaatctcc aggaccagac ccagcagatt 1140
gtgcctttta ttaggagtct cttgatgcct acaaccggcc ccgccagcat cccggacgac 1200
acactggaaa aacatacact gagaagcgag acatctacat acaatttgac cgtgggcgat 1260
accggctccg ggcttatcgt gttcttccca ggttttcccg gatctatcgt aggagcgcac 1320
tacaccctcc aaagtaacgg caattacaaa ttcgaccaga tgctcctgac agcccagaac 1380
cttcctgctt cttacaatta ctgtagactt gtgtccaggt ccctgactgt gcggagtagc 1440
acgcttccag gaggcgtata cgccctgaac ggaactataa acgccgtcac cttccagggc 1500
tccttgtccg aacttaccga cgtgtcctac aatggcctca tgagcgcaac ggccaacata 1560
aacgataaga tcggcaatgt tcttgtgggc gagggggtta cagtcctttc tctgccaacc 1620
agttatgatc tgggatacgt gcggcttggc gatcccattc ccgctatcgg tctcgaccct 1680
aaaatggtgg ctacttgcga ctcatctgac cgcccaaggg tctatacaat tactgcagcc 1740
gatgactatc agttttccag ccaataccag ccagggggtg tgacaatcac acttttcagc 1800
gccaatattg acgctatcac atccctctca atcggaggtg agcttgtgtt ccagacttct 1860
gttcagggct tggtattggg cgccactatt tacttgatcg ggttcgacgg gaccgcagtg 1920
atcactcggg cagtggctgc ggataacgga ctcactgccg gaactgacaa ccttatgcct 1980
tttaatctgg tcatccccac taacgagatc acccagccta ttacctccat aaagctcgaa 2040
attgtgacca gcaagagcgg agggcaggca ggcgaccaaa tgagttggtc tgcaagcggg 2100
tccctcgccg tgaccatcca cggtggcaac tatcctgggg cgctcagacc cgtcaccctg 2160
gtagcctacg aaagggttgc cacaggctca gttgtcacgg tggctggagt aagcaatttc 2220
gagctcatcc cgaatcctga gctcgctaaa aatcttgtga ccgagtatgg aaggttcgac 2280
cctggcgcaa tgaattacac aaagctgatt ctgtccgaac gggataggct gggtatcaag 2340
acagtttggc ccacgcgcga atacacagat ttcagggagt actttatgga ggtcgcagat 2400
ttgaatagcc cacttaagat cgctggagca tttggcttta aggatattat ccgcgcaatc 2460
agaaggtagt gtgccttcta gttgccagcc atctgttgtt tgcccctccc ccgtgccttc 2520
cttgaccctg gaaggtgcca ctcccactgt cctttcctaa taaaatgagg aaattgcatc 2580
gcattgtctg agtaggtgtc attctattct ggggggtggg gtggggcagg acagcaaggg 2640
ggaggattgg gaagacaata gcaggcatgc tggggatgcg gtgggctcta tggtgtacat 2700
aattcttatt tatctttaat ccatgaggag catttttatt ttaaaaatgt cagccgccag 2760
ccctataacc ctagatcgca actgatccct agtctgcgtt atttgtcttg caatcttttc 2820
gcacgccttt gtgagtgcat acaatgcccc cctgctcgct tttctgaaat cgcgtcgggt 2880
cattaatgtg tcggctatca caatgcgaga tgtactcgac atgtccgtgt ctgtactatt 2940
gggattgtaa atagtcgacc gcgaatcatc agagtcggaa tctgtaaagg atacagattc 3000
cgactctgag cgcttatgaa tgggatccac tcggacgttg ttgaacttcc gttcggattc 3060
tgcttcagtc aacaccggcc cccgatagct actaaggttg gggggtttgt gggttgtttg 3120
tgaaactgct ttgcggtgtg cattaccacg gggggtgtgg ggaagtatct gtttccacga 3180
tgcgataacg ttcggtggcg gagggggcga ttcattctct agtgtacgcg tttcaacttc 3240
aggaacgtga ttatttcttt caggacactc tttccaattt ccttcttcct tcacttcggg 3300
tacaggtata ttcttaatgt ttacatacat gtcgtctgct cgtctcaact gcggggttat 3360
gatgggtggt ggtgacagtc tctccgaatg atcg 3394
<210> 27
<211> 3243
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК трансферной плазмиды для HVT-IBD № 33
<400> 27
gccactgtat gggccattta tgtttatcga gtctaaaagt cgtggaagcg cctggccatc 60
gccagtctcg acctgccact gctgagttcg gcttcaattc cagcatccca gaaatgtcgc 120
agtcaccatc acgccgtgca tcgcttccta cgaggccttt tgacgcttct gatttgggca 180
catacaccct ggacatactc caccgctatt cgctcgtaga tttagtacaa ctactgaatg 240
acttgccgcg taacattacc tccacgcccg cttctaatgt agaaaccatg gcaaaaatta 300
atgttttaag ggccatttgc gtaggatttg ccgaggtccg tcgccacaac gacgcgcgaa 360
ctttacagcg aacggcaatg tttgccgccg acgacgtcgc atcacggatc agaccatcca 420
ttggattaaa gcgcacctac ccaccgggta tattttccac agctattacc gtatctaatt 480
ccgaggatga agagcgaaat tcgtgatcgt aaaaataaaa aatacaagat agcgcagcac 540
catggcctga aataacctct gaaagaggaa cttggttagc taccttctga ggcggaaaga 600
accagctgtg gaatgtgtgt cagttagggt gtggaaagtc cccaggctcc ccagcaggca 660
gaagtatgca aagcatgcat ctcaattagt cagcaaccag gtgtggaaag tccccaggct 720
ccccagcagg cagaagtatg caaagcatgc atctcaatta gtcagcaacc atagtcccgc 780
ccctaactcc gcccatcccg cccctaactc cgcccagttc cgcccattct ccgccccatg 840
gctgactaat tttttttatt tatgcagagg ccgaggccgc ctctgcctct gagctattcc 900
agaagtagtg aggaggcttt tttggaggcc taggcttttg caaaaagctc gccaccatga 960
ccaatctcca ggaccagacc cagcagattg tgccttttat taggagtctc ttgatgccta 1020
caaccggccc cgccagcatc ccggacgaca cactggaaaa acatacactg agaagcgaga 1080
catctacata caatttgacc gtgggcgata ccggctccgg gcttatcgtg ttcttcccag 1140
gttttcccgg atctatcgta ggagcgcact acaccctcca aagtaacggc aattacaaat 1200
tcgaccagat gctcctgaca gcccagaacc ttcctgcttc ttacaattac tgtagacttg 1260
tgtccaggtc cctgactgtg cggagtagca cgcttccagg aggcgtatac gccctgaacg 1320
gaactataaa cgccgtcacc ttccagggct ccttgtccga acttaccgac gtgtcctaca 1380
atggcctcat gagcgcaacg gccaacataa acgataagat cggcaatgtt cttgtgggcg 1440
agggggttac agtcctttct ctgccaacca gttatgatct gggatacgtg cggcttggcg 1500
atcccattcc cgctatcggt ctcgacccta aaatggtggc tacttgcgac tcatctgacc 1560
gcccaagggt ctatacaatt actgcagccg atgactatca gttttccagc caataccagc 1620
cagggggtgt gacaatcaca cttttcagcg ccaatattga cgctatcaca tccctctcaa 1680
tcggaggtga gcttgtgttc cagacttctg ttcagggctt ggtattgggc gccactattt 1740
acttgatcgg gttcgacggg accgcagtga tcactcgggc agtggctgcg gataacggac 1800
tcactgccgg aactgacaac cttatgcctt ttaatctggt catccccact aacgagatca 1860
cccagcctat tacctccata aagctcgaaa ttgtgaccag caagagcgga gggcaggcag 1920
gcgaccaaat gagttggtct gcaagcgggt ccctcgccgt gaccatccac ggtggcaact 1980
atcctggggc gctcagaccc gtcaccctgg tagcctacga aagggttgcc acaggctcag 2040
ttgtcacggt ggctggagta agcaatttcg agctcatccc gaatcctgag ctcgctaaaa 2100
atcttgtgac cgagtatgga aggttcgacc ctggcgcaat gaattacaca aagctgattc 2160
tgtccgaacg ggataggctg ggtatcaaga cagtttggcc cacgcgcgaa tacacagatt 2220
tcagggagta ctttatggag gtcgcagatt tgaatagccc acttaagatc gctggagcat 2280
ttggctttaa ggatattatc cgcgcaatca gaaggtagtg tgccttctag ttgccagcca 2340
tctgttgttt gcccctcccc cgtgccttcc ttgaccctgg aaggtgccac tcccactgtc 2400
ctttcctaat aaaatgagga aattgcatcg cattgtctga gtaggtgtca ttctattctg 2460
gggggtgggg tggggcagga cagcaagggg gaggattggg aagacaatag caggcatgct 2520
ggggatgcgg tgggctctat ggtgtacata attcttattt atctttaatc catgaggagc 2580
atttttattt taaaaatgtc agccgccagc cctataaccc tagatcgcaa ctgatcccta 2640
gtctgcgtta tttgtcttgc aatcttttcg cacgcctttg tgagtgcata caatgccccc 2700
ctgctcgctt ttctgaaatc gcgtcgggtc attaatgtgt cggctatcac aatgcgagat 2760
gtactcgaca tgtccgtgtc tgtactattg ggattgtaaa tagtcgaccg cgaatcatca 2820
gagtcggaat ctgtaaagga tacagattcc gactctgagc gcttatgaat gggatccact 2880
cggacgttgt tgaacttccg ttcggattct gcttcagtca acaccggccc ccgatagcta 2940
ctaaggttgg ggggtttgtg ggttgtttgt gaaactgctt tgcggtgtgc attaccacgg 3000
ggggtgtggg gaagtatctg tttccacgat gcgataacgt tcggtggcgg agggggcgat 3060
tcattctcta gtgtacgcgt ttcaacttca ggaacgtgat tatttctttc aggacactct 3120
ttccaatttc cttcttcctt cacttcgggt acaggtatat tcttaatgtt tacatacatg 3180
tcgtctgctc gtctcaactg cggggttatg atgggtggtg gtgacagtct ctccgaatga 3240
tcg 3243
<210> 28
<211> 4124
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК трансферной плазмиды для HVT-IBD № 34
<400> 28
atgggctccc gctgttcaaa aaaatcagcg tgcgtcgata agactttggt gcagtctctt 60
cggggtcgca atttagattt gccgcatgga gggtatctgg ggatttttgc caatgctgga 120
gcgacgactg tacgattcgt cccatcggga tctagcagac caatgatgtt gacacacatc 180
ggccatgcat gtacggacgg tctattgcgc gagtttgtta ttttcgaagg acaagatgga 240
agtgtatatg gaaccgacaa taatgttagt ttgcatttct tagggcggaa tctacatgat 300
atcttatcca agcggggtat gagccagaga gatgtgatgg tcataaaggg taaatttttt 360
agatctgaaa taacgcagtt gcccaaacaa cgatcgcgat taaaagaaaa atcggatggt 420
tcaattagga catgcatgga ttctgtgcgc ataaaccata accgcagcac tgttgggcac 480
ttcggtaact caaatgcgaa gcgttgcacg tctgcgataa ctacgcctac tatgcacatt 540
gttactcctg catcttaaaa atatatcctg tagtaatttt cacagcaatg tcataacatc 600
atctcgctaa agaatgacct gggattggag aagtaatgaa tatttgcaac caatgcattg 660
aataaactaa cattaaactc gaggtgagcc ccacgttctg cttcactctc cccatctccc 720
ccccctcccc acccccaatt ttgtatttat ttatttttta attattttgt gcagcgatgg 780
gggcgggggg ggggggggcg cgcgccaggc ggggcggggc ggggcgaggg gcggggcggg 840
gcgaggcgga gaggtgcggc ggcagccaat cagagcggcg cgctccgaaa gtttcctttt 900
atggcgaggc ggcggcggcg gcggccctat aaaaagcgaa gcgcgcggcg ggcgggagtc 960
gctgcgttgc cttcgccccg tgccccgctc cgcgccgcct cgcgccgccc gccccggctc 1020
tgactgaccg cgttactccc acaggtgagc gggcgggacg gcccttctcc tccgggctgt 1080
aattagcgct tggtttaatg acggctcgtt tcttttctgt ggctgcgtga aagccttaaa 1140
gggctccggg agggcccttt gtgcgggggg gagcggctcg gggggtgcgt gcgtgtgtgt 1200
gtgcgtgggg agcgccgcgt gcggcccgcg ctgcccggcg gctgtgagcg ctgcgggcgc 1260
ggcgcggggc tttgtgcgct ccgcgtgtgc gcgaggggag cgcggccggg ggcggtgccc 1320
cgcggtgcgg gggggctgcg aggggaacaa aggctgcgtg cggggtgtgt gcgtgggggg 1380
gtgagcaggg ggtgtgggcg cggcggtcgg gctgtaaccc ccccctgcac ccccctcccc 1440
gagttgctga gcacggcccg gcttcgggtg cggggctccg tgcggggcgt ggcgcggggc 1500
tcgccgtgcc gggcgggggg tggcggcagg tgggggtgcc gggcggggcg gggccgcctc 1560
gggccgggga gggctcgggg gaggggcgcg gcggccccgg agcgccggcg gctgtcgagg 1620
cgcggcgagc cgcagccatt gccttttatg gtaatcgtgc gagagggcgc agggacttcc 1680
tttgtcccaa atctggcgga gccgaaatct gggaggcgcc gccgcacccc ctctagcggg 1740
cgcgggcgaa gcggtgcggc gccggcagga aggaaatggg cggggagggc cttcgtgcgt 1800
cgccgcgccg ccgtcccctt ctccatctcc agcctcgggg ctgccgcagg gggacggctg 1860
ccttcggggg ggacggggca gggcggggtt cggcttctgg cgtgtgaccg gcggggttta 1920
tatcttccct tctctgttcc tccgcagcca gccatggcca ccatgacaaa cctgcaagat 1980
caaacccaac agattgttcc gttcatacgg agccttctga tgccaacaac cggaccggcg 2040
tccattccgg acgacaccct ggagaagcac actctcaggt cagagacctc gacctacaat 2100
ttgactgtgg gggacacagg gtcagggcta attgtctttt tccctggatt ccctggctca 2160
attgtgggtg ctcactacac actgcagagc aatgggaact acaagttcga tcagatgctc 2220
ctgactgccc agaacctacc ggccagctac aactactgca gactagtgag tcggagtctc 2280
acagtgaggt caagcacact ccctggtggc gtttatgcac taaacggcac cataaacgcc 2340
gtgaccttcc aaggaagcct gagtgaactg acagatgtta gctacaatgg gttgatgtct 2400
gcaacagcca acatcaacga caaaattggg aatgtcctgg taggggaagg ggtcactgtc 2460
ctcagcctac ccacatcata tgatcttggg tatgtgaggc ttggtgaccc cattcccgct 2520
atagggcttg acccaaaaat ggtagctaca tgcgacagca gtgacaggcc cagagtctac 2580
accataactg cagccgatga ttaccaattc tcatcacagt accaaccagg tggggtaaca 2640
atcacactgt tctcagccaa cattgatgct atcacaagcc tcagcattgg gggagagctc 2700
gtgtttcaaa caagcgtcca aggccttgta ctgggcgcca ccatctacct tataggcttt 2760
gatgggactg cggtaatcac cagagctgta gccgcagata atgggctgac ggccggcacc 2820
gacaatctta tgccattcaa tcttgtcatt ccaaccaatg agataaccca gccaatcaca 2880
tccatcaaac tggagatagt gacctccaaa agtggtggtc aggcagggga tcagatgtca 2940
tggtcggcaa gtgggagcct agcagtgacg atccatggtg gcaactatcc aggggccctc 3000
cgtcccgtca cactagtagc ctacgaaaga gtggcaacag gatccgtcgt tacggtcgct 3060
ggggtgagta acttcgagct gattccaaat cctgaactag caaagaacct ggttacagaa 3120
tacggccgat ttgacccagg agccatgaac tacacaaaat tgatactgag tgagagggac 3180
cgtcttggca tcaagaccgt ctggccaaca agggagtaca ctgattttcg tgagtacttc 3240
atggaggtgg ccgacctcaa ctctcccctg aagattgcag gagcatttgg cttcaaagac 3300
ataatccggg ctataaggag gtaagctgtg ccttctagtt gccagccatc tgttgtttgc 3360
ccctcccccg tgccttcctt gaccctggaa ggtgccactc ccactgtcct ttcctaataa 3420
aatgaggaaa ttgcatcgca ttgtctgagt aggtgtcatt ctattctggg gggtggggtg 3480
gggcaggaca gcaaggggga ggattgggaa gacaatagca ggcatgctgg ggatgcggtg 3540
ggctctatgg tcaattattt tatttaataa catatagccc aaagacctct atgaacattt 3600
agtttcccgt atactcaacg gcgcgtgtac acacgcatct ctttgcatag cgatgaagtt 3660
tgttcggcag cagaaaatgc agatatccaa caatctggag aaaacttatc atcacagtgg 3720
cagtggaaac ataccccctc tatattcatg gtataattat cgtctacagc gtccaggata 3780
gtggcgtgag aaaatggaga tctgcagccc tcctttccat ggcatgccgc tttattgttc 3840
attaaacgca caatggtctc aacgccagat atgggcatag attctgaaga acccgttgac 3900
aatccgaaga agaaggcgtg caggtctttg gaagactcgc acgttggtct tataatgtat 3960
gatcgagatg tcaccctaat gccacatggt acaggcttat cgcggtcatg gcgatcggac 4020
ttgtaatttg caacgatggg caaaggatcg acgacatgcc aaacattctg aacccgtaga 4080
gatgttaacg atgacgagga tgaatatccc atgctcgctg ccat 4124
<210> 29
<211> 4572
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК трансферной плазмиды для HVT-ND № 38
<400> 29
gccactgtat gggccattta tgtttatcga gtctaaaagt cgtggaagcg cctggccatc 60
gccagtctcg acctgccact gctgagttcg gcttcaattc cagcatccca gaaatgtcgc 120
agtcaccatc acgccgtgca tcgcttccta cgaggccttt tgacgcttct gatttgggca 180
catacaccct ggacatactc caccgctatt cgctcgtaga tttagtacaa ctactgaatg 240
acttgccgcg taacattacc tccacgcccg cttctaatgt agaaaccatg gcaaaaatta 300
atgttttaag ggccatttgc gtaggatttg ccgaggtccg tcgccacaac gacgcgcgaa 360
ctttacagcg aacggcaatg tttgccgccg acgacgtcgc atcacggatc agaccatcca 420
ttggattaaa gcgcacctac ccaccgggta tattttccac agctattacc gtatctaatt 480
ccgaggatga agagcgaaat tcgtgatcgt aaaaataaaa aatacaagat agaattcact 540
agtggatccc ccaactccgc ccgttttatg actagaacca atagttttta atgccaaatg 600
cactgaaatc ccctaatttg caaagccaaa cgccccctat gtgagtaata cggggacttt 660
ttacccaatt tcccaagcgg aaagccccct aatacactca tatggcatat gaatcagcac 720
ggtcatgcac tctaatggcg gcccataggg actttccaca tagggggcgt tcaccatttc 780
ccagcatagg ggtggtgact caatggcctt tacccaagta cattgggtca atgggaggta 840
agccaatggg tttttcccat tactggcaag cacactgagt caaatgggac tttccactgg 900
gttttgccca agtacattgg gtcaatggga ggtgagccaa tgggaaaaac ccattgctgc 960
caagtacact gactcaatag ggactttcca atgggttttt ccattgttgg caagcatata 1020
aggtcaatgt gggtgagtca atagggactt tccattgtat tctgcccagt acataaggtc 1080
aatagggggt gaatcaacag gaaagtccca ttggagccaa gtacactgcg tcaataggga 1140
ctttccattg ggttttgccc agtacataag gtcaataggg gatgagtcaa tgggaaaaac 1200
ccattggagc caagtacact gactcaatag ggactttcca ttgggttttg cccagtacat 1260
aaggtcaata gggggtgagt caacaggaaa gtcccattgg agccaagtac attgagtcaa 1320
tagggacttt ccaatgggtt ttgcccagta cataaggtca atgggaggta agccaatggg 1380
tttttcccat tactggcacg tatactgagt cattagggac tttccaatgg gttttgccca 1440
gtacataagg tcaatagggg tgaatcaaca ggaaagtccc attggagcca agtacactga 1500
gtcaataggg actttccatt gggttttgcc cagtacaaaa ggtcaatagg gggtgagtca 1560
atgggttttt cccattattg gcacgtacat aaggtcaata ggggtgagtc attgggtttt 1620
tccagccaat ttaattaaaa cgccatgtac tttcccacca ttgacgtcaa tgggctattg 1680
aaactaatgc aacgtgacct ttaaacggta ctttcccata gctgattaat gggaaagtac 1740
cgttctcgag ccaatacacg tcaatgggaa gtgaaagggc agccaaaacg taacaccgcc 1800
ccggttttcc cctggaaatt ccatattggc acgcattcta ttggctgagc tgcgttctac 1860
gtgggtataa gaggcgcgac cagcgtcggt accgtcgcag tcttcggtct gaccaccgta 1920
gaacgcagag ctcctcgctg caggcggccg ctctagaact cgtcgatcgc agcgatgggc 1980
tccagatctt ctaccaggat cccagtacct ctgatgctga ccgtccgaat catgttggca 2040
ctgagttgcg tctgtccgac cagctccctt gatggcaggc ctcttgcagc tgcagggatt 2100
gtggtaacag gagacaaagc agtcaacata tacacctcat ctcagacagg gtcaatcata 2160
atcaagttac tcccaaatat gcccaaggat aaagaggcgt gtgcaaaagc cccattggaa 2220
gcatacaaca ggacattgac tactttgctc accccccttg gtgattctat ccgtaggata 2280
caagagtctg tgaccacatc cggaggaggg aaacagggac gtcttatagg cgccattatc 2340
ggtggtgtag ctctcggggt tgcaaccgct gcacagataa cagcagcctc ggctctgata 2400
caagccaatc aaaatgctgc caacatcctc cggctcaaag agagcattgc tgcaaccaat 2460
gaggctgtgc acgaggtcac tgacggatta tcacaactag cagtggcagt tgggaagatg 2520
cagcaatttg ttaatgacca gtttaataaa acagctcagg aattggactg tataaaaatt 2580
acacagcagg ttggtgtaga actcaacctg tacctaactg aattgactac agtattcggg 2640
ccacaaatca cttcccctgc cttaactcag ctgactatcc aggcgcttta caatctagct 2700
ggtgggaata tggattactt gttgactaag ttaggtgtag gaaacaacca actcagctca 2760
ttaattggta gtggcctgat taccggcaac cctatcctgt acgactcaca gactcaactc 2820
ttgggtatac aggtcaccct accctcagtc gggaatctaa ataatatgcg tgccacctac 2880
ctggaaacct tgtctgtaag tacaaccaaa ggatttgcct cagcacttgt cccaaaagta 2940
gtgacacagg ttggttccgt gatagaagag cttgacacct cgtactgtat cgagaccgat 3000
ttggacctat attgtacaag aatagtgaca ttccctatgt ctcctggtat ttattcctgt 3060
ttgagtggca atacatctgc ttgcatgtat tcaaagactg aaggcgcact cactacgccg 3120
tatatgaccc tcaaaggctc agttattgcc aactgtaaga tgacaacatg tagatgtgca 3180
gaccccccgg gtatcatatc gcagaattat ggagaagctg tgtctctaat agataggcaa 3240
tcatgcaata tcttatcctt agacgggata actttgaggc tcagtgggga atttgatgca 3300
acttatcaaa agaatatctc aatacaagat tctcaagtaa tagttacagg caatcttgac 3360
atctcgactg agcttgggaa tgtcaacaac tcgataagta atgctttgga taagttagag 3420
gaaagcaaca gcaaactaga caaggtcaat gttaaactga ccagcacatc cgctcttatt 3480
acctatatcg ttttaactgt catatctctt gtatgtggta tacttagcct ggttctagca 3540
tgctacctga tgtacaagca aaaggcgcaa cagaagacct tgttgtggct tgggaataat 3600
accctagacc agatgagggc cactacaaaa atgtagcttg atctagagcg gccgcgggga 3660
tccagacatg ataagataca ttgatgagtt tggacaaacc acaactagaa tgcagtgaaa 3720
aaaatgcttt atttgtgaaa tttgtgatgc tattgcttta tttgtaacca ttataagctg 3780
caataaacaa gttaacaaca acaattgcat tcattttatg tttcaggttc agggggaggt 3840
gtgggaggtt ttttcggatc ctctagagtc gagtacataa ttcttattta tctttaatcc 3900
atgaggagca tttttatttt aaaaatgtca gccgccagcc ctataaccct agatcgcaac 3960
tgatccctag tctgcgttat ttgtcttgca atcttttcgc acgcctttgt gagtgcatac 4020
aatgcccccc tgctcgcttt tctgaaatcg cgtcgggtca ttaatgtgtc ggctatcaca 4080
atgcgagatg tactcgacat gtccgtgtct gtactattgg gattgtaaat agtcgaccgc 4140
gaatcatcag agtcggaatc tgtaaaggat acagattccg actctgagcg cttatgaatg 4200
ggatccactc ggacgttgtt gaacttccgt tcggattctg cttcagtcaa caccggcccc 4260
cgatagctac taaggttggg gggtttgtgg gttgtttgtg aaactgcttt gcggtgtgca 4320
ttaccacggg gggtgtgggg aagtatctgt ttccacgatg cgataacgtt cggtggcgga 4380
gggggcgatt cattctctag tgtacgcgtt tcaacttcag gaacgtgatt atttctttca 4440
ggacactctt tccaatttcc ttcttccttc acttcgggta caggtatatt cttaatgttt 4500
acatacatgt cgtctgctcg tctcaactgc ggggttatga tgggtggtgg tgacagtctc 4560
tccgaatgat cg 4572
<210> 30
<211> 4402
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК трансферной плазмиды для HVT-ND № 39
<400> 30
gccactgtat gggccattta tgtttatcga gtctaaaagt cgtggaagcg cctggccatc 60
gccagtctcg acctgccact gctgagttcg gcttcaattc cagcatccca gaaatgtcgc 120
agtcaccatc acgccgtgca tcgcttccta cgaggccttt tgacgcttct gatttgggca 180
catacaccct ggacatactc caccgctatt cgctcgtaga tttagtacaa ctactgaatg 240
acttgccgcg taacattacc tccacgcccg cttctaatgt agaaaccatg gcaaaaatta 300
atgttttaag ggccatttgc gtaggatttg ccgaggtccg tcgccacaac gacgcgcgaa 360
ctttacagcg aacggcaatg tttgccgccg acgacgtcgc atcacggatc agaccatcca 420
ttggattaaa gcgcacctac ccaccgggta tattttccac agctattacc gtatctaatt 480
ccgaggatga agagcgaaat tcgtgatcgt aaaaataaaa aatacaagat atcgaggtga 540
gccccacgtt ctgcttcact ctccccatct cccccccctc cccaccccca attttgtatt 600
tatttatttt ttaattattt tgtgcagcga tgggggcggg gggggggggg gcgcgcgcca 660
ggcggggcgg ggcggggcga ggggcggggc ggggcgaggc ggagaggtgc ggcggcagcc 720
aatcagagcg gcgcgctccg aaagtttcct tttatggcga ggcggcggcg gcggcggccc 780
tataaaaagc gaagcgcgcg gcgggcggga gtcgctgcgt tgccttcgcc ccgtgccccg 840
ctccgcgccg cctcgcgccg cccgccccgg ctctgactga ccgcgttact cccacaggtg 900
agcgggcggg acggcccttc tcctccgggc tgtaattagc gcttggttta atgacggctc 960
gtttcttttc tgtggctgcg tgaaagcctt aaagggctcc gggagggccc tttgtgcggg 1020
ggggagcggc tcggggggtg cgtgcgtgtg tgtgtgcgtg gggagcgccg cgtgcggccc 1080
gcgctgcccg gcggctgtga gcgctgcggg cgcggcgcgg ggctttgtgc gctccgcgtg 1140
tgcgcgaggg gagcgcggcc gggggcggtg ccccgcggtg cgggggggct gcgaggggaa 1200
caaaggctgc gtgcggggtg tgtgcgtggg ggggtgagca gggggtgtgg gcgcggcggt 1260
cgggctgtaa cccccccctg cacccccctc cccgagttgc tgagcacggc ccggcttcgg 1320
gtgcggggct ccgtgcgggg cgtggcgcgg ggctcgccgt gccgggcggg gggtggcggc 1380
aggtgggggt gccgggcggg gcggggccgc ctcgggccgg ggagggctcg ggggaggggc 1440
gcggcggccc cggagcgccg gcggctgtcg aggcgcggcg agccgcagcc attgcctttt 1500
atggtaatcg tgcgagaggg cgcagggact tcctttgtcc caaatctggc ggagccgaaa 1560
tctgggaggc gccgccgcac cccctctagc gggcgcgggc gaagcggtgc ggcgccggca 1620
ggaaggaaat gggcggggag ggccttcgtg cgtcgccgcg ccgccgtccc cttctccatc 1680
tccagcctcg gggctgccgc agggggacgg ctgccttcgg gggggacggg gcagggcggg 1740
gttcggcttc tggcgtgtga ccggcggggt ttatatcttc ccttctctgt tcctccgcag 1800
ccagccatgg ccaccatggg ctccagatct tctaccagga tcccagtacc tctgatgctg 1860
accgtccgaa tcatgttggc actgagttgc gtctgtccga ccagctccct tgatggcagg 1920
cctcttgcag ctgcagggat tgtggtaaca ggagacaaag cagtcaacat atacacctca 1980
tctcagacag ggtcaatcat aatcaagtta ctcccaaata tgcccaagga taaagaggcg 2040
tgtgcaaaag ccccattgga agcatacaac aggacattga ctactttgct cacccccctt 2100
ggtgattcta tccgtaggat acaagagtct gtgaccacat ccggaggagg gaaacaggga 2160
cgtcttatag gcgccattat cggtggtgta gctctcgggg ttgcaaccgc tgcacagata 2220
acagcagcct cggctctgat acaagccaat caaaatgctg ccaacatcct ccggctcaaa 2280
gagagcattg ctgcaaccaa tgaggctgtg cacgaggtca ctgacggatt atcacaacta 2340
gcagtggcag ttgggaagat gcagcaattt gttaatgacc agtttaataa aacagctcag 2400
gaattggact gtataaaaat tacacagcag gttggtgtag aactcaacct gtacctaact 2460
gaattgacta cagtattcgg gccacaaatc acttcccctg ccttaactca gctgactatc 2520
caggcgcttt acaatctagc tggtgggaat atggattact tgttgactaa gttaggtgta 2580
ggaaacaacc aactcagctc attaattggt agtggcctga ttaccggcaa ccctatcctg 2640
tacgactcac agactcaact cttgggtata caggtcaccc taccctcagt cgggaatcta 2700
aataatatgc gtgccaccta cctggaaacc ttgtctgtaa gtacaaccaa aggatttgcc 2760
tcagcacttg tcccaaaagt agtgacacag gttggttccg tgatagaaga gcttgacacc 2820
tcgtactgta tcgagaccga tttggaccta tattgtacaa gaatagtgac attccctatg 2880
tctcctggta tttattcctg tttgagtggc aatacatctg cttgcatgta ttcaaagact 2940
gaaggcgcac tcactacgcc gtatatgacc ctcaaaggct cagttattgc caactgtaag 3000
atgacaacat gtagatgtgc agaccccccg ggtatcatat cgcagaatta tggagaagct 3060
gtgtctctaa tagataggca atcatgcaat atcttatcct tagacgggat aactttgagg 3120
ctcagtgggg aatttgatgc aacttatcaa aagaatatct caatacaaga ttctcaagta 3180
atagttacag gcaatcttga catctcgact gagcttggga atgtcaacaa ctcgataagt 3240
aatgctttgg ataagttaga ggaaagcaac agcaaactag acaaggtcaa tgttaaactg 3300
accagcacat ccgctcttat tacctatatc gttttaactg tcatatctct tgtatgtggt 3360
atacttagcc tggttctagc atgctacctg atgtacaagc aaaaggcgca acagaagacc 3420
ttgttgtggc ttgggaataa taccctagac cagatgaggg ccactacaaa aatgtagtgt 3480
gccttctagt tgccagccat ctgttgtttg cccctccccc gtgccttcct tgaccctgga 3540
aggtgccact cccactgtcc tttcctaata aaatgaggaa attgcatcgc attgtctgag 3600
taggtgtcat tctattctgg ggggtggggt ggggcaggac agcaaggggg aggattggga 3660
agacaatagc aggcatgctg gggatgcggt gggctctatg gtgtacataa ttcttattta 3720
tctttaatcc atgaggagca tttttatttt aaaaatgtca gccgccagcc ctataaccct 3780
agatcgcaac tgatccctag tctgcgttat ttgtcttgca atcttttcgc acgcctttgt 3840
gagtgcatac aatgcccccc tgctcgcttt tctgaaatcg cgtcgggtca ttaatgtgtc 3900
ggctatcaca atgcgagatg tactcgacat gtccgtgtct gtactattgg gattgtaaat 3960
agtcgaccgc gaatcatcag agtcggaatc tgtaaaggat acagattccg actctgagcg 4020
cttatgaatg ggatccactc ggacgttgtt gaacttccgt tcggattctg cttcagtcaa 4080
caccggcccc cgatagctac taaggttggg gggtttgtgg gttgtttgtg aaactgcttt 4140
gcggtgtgca ttaccacggg gggtgtgggg aagtatctgt ttccacgatg cgataacgtt 4200
cggtggcgga gggggcgatt cattctctag tgtacgcgtt tcaacttcag gaacgtgatt 4260
atttctttca ggacactctt tccaatttcc ttcttccttc acttcgggta caggtatatt 4320
cttaatgttt acatacatgt cgtctgctcg tctcaactgc ggggttatga tgggtggtgg 4380
tgacagtctc tccgaatgat cg 4402
<210> 31
<211> 4413
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК трансферной плазмиды для HVT-ND № 40
<400> 31
gccactgtat gggccattta tgtttatcga gtctaaaagt cgtggaagcg cctggccatc 60
gccagtctcg acctgccact gctgagttcg gcttcaattc cagcatccca gaaatgtcgc 120
agtcaccatc acgccgtgca tcgcttccta cgaggccttt tgacgcttct gatttgggca 180
catacaccct ggacatactc caccgctatt cgctcgtaga tttagtacaa ctactgaatg 240
acttgccgcg taacattacc tccacgcccg cttctaatgt agaaaccatg gcaaaaatta 300
atgttttaag ggccatttgc gtaggatttg ccgaggtccg tcgccacaac gacgcgcgaa 360
ctttacagcg aacggcaatg tttgccgccg acgacgtcgc atcacggatc agaccatcca 420
ttggattaaa gcgcacctac ccaccgggta tattttccac agctattacc gtatctaatt 480
ccgaggatga agagcgaaat tcgtgatcgt aaaaataaaa aatacaagat atcgaggtga 540
gccccacgtt ctgcttcact ctccccatct cccccccctc cccaccccca attttgtatt 600
tatttatttt ttaattattt tgtgcagcga tgggggcggg gggggggggg gcgcgcgcca 660
ggcggggcgg ggcggggcga ggggcggggc ggggcgaggc ggagaggtgc ggcggcagcc 720
aatcagagcg gcgcgctccg aaagtttcct tttatggcga ggcggcggcg gcggcggccc 780
tataaaaagc gaagcgcgcg gcgggcggga gtcgctgcgt tgccttcgcc ccgtgccccg 840
ctccgcgccg cctcgcgccg cccgccccgg ctctgactga ccgcgttact cccacaggtg 900
agcgggcggg acggcccttc tcctccgggc tgtaattagc gcttggttta atgacggctc 960
gtttcttttc tgtggctgcg tgaaagcctt aaagggctcc gggagggccc tttgtgcggg 1020
ggggagcggc tcggggggtg cgtgcgtgtg tgtgtgcgtg gggagcgccg cgtgcggccc 1080
gcgctgcccg gcggctgtga gcgctgcggg cgcggcgcgg ggctttgtgc gctccgcgtg 1140
tgcgcgaggg gagcgcggcc gggggcggtg ccccgcggtg cgggggggct gcgaggggaa 1200
caaaggctgc gtgcggggtg tgtgcgtggg ggggtgagca gggggtgtgg gcgcggcggt 1260
cgggctgtaa cccccccctg cacccccctc cccgagttgc tgagcacggc ccggcttcgg 1320
gtgcggggct ccgtgcgggg cgtggcgcgg ggctcgccgt gccgggcggg gggtggcggc 1380
aggtgggggt gccgggcggg gcggggccgc ctcgggccgg ggagggctcg ggggaggggc 1440
gcggcggccc cggagcgccg gcggctgtcg aggcgcggcg agccgcagcc attgcctttt 1500
atggtaatcg tgcgagaggg cgcagggact tcctttgtcc caaatctggc ggagccgaaa 1560
tctgggaggc gccgccgcac cccctctagc gggcgcgggc gaagcggtgc ggcgccggca 1620
ggaaggaaat gggcggggag ggccttcgtg cgtcgccgcg ccgccgtccc cttctccatc 1680
tccagcctcg gggctgccgc agggggacgg ctgccttcgg gggggacggg gcagggcggg 1740
gttcggcttc tggcgtgtga ccggcggggt ttatatcttc ccttctctgt tcctccgcag 1800
ccagccatgg ccaccatggg ctccagatct tctaccagga tcccagtacc tctgatgctg 1860
accgtccgaa tcatgttggc actgagttgc gtctgtccga ccagctccct tgatggcagg 1920
cctcttgcag ctgcagggat tgtggtaaca ggagacaaag cagtcaacat atacacctca 1980
tctcagacag ggtcaatcat aatcaagtta ctcccaaata tgcccaagga taaagaggcg 2040
tgtgcaaaag ccccattgga agcatacaac aggacattga ctactttgct cacccccctt 2100
ggtgattcta tccgtaggat acaagagtct gtgaccacat ccggaggagg gaaacaggga 2160
cgtcttatag gcgccattat cggtggtgta gctctcgggg ttgcaaccgc tgcacagata 2220
acagcagcct cggctctgat acaagccaat caaaatgctg ccaacatcct ccggctcaaa 2280
gagagcattg ctgcaaccaa tgaggctgtg cacgaggtca ctgacggatt atcacaacta 2340
gcagtggcag ttgggaagat gcagcaattt gttaatgacc agtttaataa aacagctcag 2400
gaattggact gtataaaaat tacacagcag gttggtgtag aactcaacct gtacctaact 2460
gaattgacta cagtattcgg gccacaaatc acttcccctg ccttaactca gctgactatc 2520
caggcgcttt acaatctagc tggtgggaat atggattact tgttgactaa gttaggtgta 2580
ggaaacaacc aactcagctc attaattggt agtggcctga ttaccggcaa ccctatcctg 2640
tacgactcac agactcaact cttgggtata caggtcaccc taccctcagt cgggaatcta 2700
aataatatgc gtgccaccta cctggaaacc ttgtctgtaa gtacaaccaa aggatttgcc 2760
tcagcacttg tcccaaaagt agtgacacag gttggttccg tgatagaaga gcttgacacc 2820
tcgtactgta tcgagaccga tttggaccta tattgtacaa gaatagtgac attccctatg 2880
tctcctggta tttattcctg tttgagtggc aatacatctg cttgcatgta ttcaaagact 2940
gaaggcgcac tcactacgcc gtatatgacc ctcaaaggct cagttattgc caactgtaag 3000
atgacaacat gtagatgtgc agaccccccg ggtatcatat cgcagaatta tggagaagct 3060
gtgtctctaa tagataggca atcatgcaat atcttatcct tagacgggat aactttgagg 3120
ctcagtgggg aatttgatgc aacttatcaa aagaatatct caatacaaga ttctcaagta 3180
atagttacag gcaatcttga catctcgact gagcttggga atgtcaacaa ctcgataagt 3240
aatgctttgg ataagttaga ggaaagcaac agcaaactag acaaggtcaa tgttaaactg 3300
accagcacat ccgctcttat tacctatatc gttttaactg tcatatctct tgtatgtggt 3360
atacttagcc tggttctagc atgctacctg atgtacaagc aaaaggcgca acagaagacc 3420
ttgttgtggc ttgggaataa taccctagac cagatgaggg ccactacaaa aatgtagctt 3480
gatctagagc ggccgcgggg atccagacat gataagatac attgatgagt ttggacaaac 3540
cacaactaga atgcagtgaa aaaaatgctt tatttgtgaa atttgtgatg ctattgcttt 3600
atttgtaacc attataagct gcaataaaca agttaacaac aacaattgca ttcattttat 3660
gtttcaggtt cagggggagg tgtgggaggt tttttcggat cctctagagt cgagtacata 3720
attcttattt atctttaatc catgaggagc atttttattt taaaaatgtc agccgccagc 3780
cctataaccc tagatcgcaa ctgatcccta gtctgcgtta tttgtcttgc aatcttttcg 3840
cacgcctttg tgagtgcata caatgccccc ctgctcgctt ttctgaaatc gcgtcgggtc 3900
attaatgtgt cggctatcac aatgcgagat gtactcgaca tgtccgtgtc tgtactattg 3960
ggattgtaaa tagtcgaccg cgaatcatca gagtcggaat ctgtaaagga tacagattcc 4020
gactctgagc gcttatgaat gggatccact cggacgttgt tgaacttccg ttcggattct 4080
gcttcagtca acaccggccc ccgatagcta ctaaggttgg ggggtttgtg ggttgtttgt 4140
gaaactgctt tgcggtgtgc attaccacgg ggggtgtggg gaagtatctg tttccacgat 4200
gcgataacgt tcggtggcgg agggggcgat tcattctcta gtgtacgcgt ttcaacttca 4260
ggaacgtgat tatttctttc aggacactct ttccaatttc cttcttcctt cacttcgggt 4320
acaggtatat tcttaatgtt tacatacatg tcgtctgctc gtctcaactg cggggttatg 4380
atgggtggtg gtgacagtct ctccgaatga tcg 4413
<210> 32
<211> 3849
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК трансферной плазмиды для HVT-ND № 41
<400> 32
atggcagcga gcatgggata ttcatcctcg tcatcgttaa catctctacg ggttcagaat 60
gtttggcatg tcgtcgatcc tttgcccatc gttgcaaatt acaagtccga tcgccatgac 120
cgcgataagc ctgtaccatg tggcattagg gtgacatctc gatcatacat tataagacca 180
acgtgcgagt cttccaaaga cctgcacgcc ttcttcttcg gattgtcaac gggttcttca 240
gaatctatgc ccatatctgg cgttgagacc attgtgcgtt taatgaacaa taaagcggca 300
tgccatggaa aggagggctg cagatctcca ttttctcacg ccactatcct ggacgctgta 360
gacgataatt ataccatgaa tatagagggg gtatgtttcc actgccactg tgatgataag 420
ttttctccag attgttggat atctgcattt tctgctgccg aacaaacttc atcgctatgc 480
aaagagatgc gtgtgtacac gcgccgttga gtatacggga aactaaatgt tcatagaggt 540
ctttgggcta tatgttatta aataaaataa ttgaccagtg ttgacattga ttattgacta 600
gttattaata gtaatcaatt acggggtcat tagttcatag cccatatatg gagttccgcg 660
ttacataact tacggtaaat ggcccgcctg gctgaccgcc caacgacccc cgcccattga 720
cgtcaataat gacgtatgtt cccatagtaa cgccaatagg gactttccat tgacgtcaat 780
gggtggagta tttacggtaa actgcccact tggcagtaca tcaagtgtat catatgccaa 840
gtacgccccc tattgacgtc aatgacggta aatggcccgc ctggcattat gcccagtaca 900
tgaccttatg ggactttcct acttggcagt acatctacgt attagtcatc gctattacca 960
tggtgatgcg gttttggcag tacatcaatg ggcgtggata gcggtttgac tcacggggat 1020
ttccaagtct ccaccccatt gacgtcaatg ggagtttgtt ttggcaccaa aatcaacggg 1080
actttccaaa atgtcgtaac aactccgccc cattgacgca aatgggcggt aggcgtgtac 1140
ggtgggaggt ctatataagc agagctctct ggctaactag agaacccact gcttactggc 1200
ttatcgaaat taatacgact cactataggg agacccaagc tggctagcgt ttaaacttaa 1260
gcttaccgcc accatgggct ccagatcttc taccaggatc ccagtacctc tgatgctgac 1320
cgtccgaatc atgttggcac tgagttgcgt ctgtccgacc agctcccttg atggcaggcc 1380
tcttgcagct gcagggattg tggtaacagg agacaaagca gtcaacatat acacctcatc 1440
tcagacaggg tcaatcataa tcaagttact cccaaatatg cccaaggata aagaggcgtg 1500
tgcaaaagcc ccattggaag catacaacag gacattgact actttgctca ccccccttgg 1560
tgattctatc cgtaggatac aagagtctgt gaccacatcc ggaggaggga aacagggacg 1620
tcttataggc gccattatcg gtggtgtagc tctcggggtt gcaaccgctg cacagataac 1680
agcagcctcg gctctgatac aagccaatca aaatgctgcc aacatcctcc ggctcaaaga 1740
gagcattgct gcaaccaatg aggctgtgca cgaggtcact gacggattat cacaactagc 1800
agtggcagtt gggaagatgc agcaatttgt taatgaccag tttaataaaa cagctcagga 1860
attggactgt ataaaaatta cacagcaggt tggtgtagaa ctcaacctgt acctaactga 1920
attgactaca gtattcgggc cacaaatcac ttcccctgcc ttaactcagc tgactatcca 1980
ggcgctttac aatctagctg gtgggaatat ggattacttg ttgactaagt taggtgtagg 2040
aaacaaccaa ctcagctcat taattggtag tggcctgatt accggcaacc ctatcctgta 2100
cgactcacag actcaactct tgggtataca ggtcacccta ccctcagtcg ggaatctaaa 2160
taatatgcgt gccacctacc tggaaacctt gtctgtaagt acaaccaaag gatttgcctc 2220
agcacttgtc ccaaaagtag tgacacaggt tggttccgtg atagaagagc ttgacacctc 2280
gtactgtatc gagaccgatt tggacctata ttgtacaaga atagtgacat tccctatgtc 2340
tcctggtatt tattcctgtt tgagtggcaa tacatctgct tgcatgtatt caaagactga 2400
aggcgcactc actacgccgt atatgaccct caaaggctca gttattgcca actgtaagat 2460
gacaacatgt agatgtgcag accccccggg tatcatatcg cagaattatg gagaagctgt 2520
gtctctaata gataggcaat catgcaatat cttatcctta gacgggataa ctttgaggct 2580
cagtggggaa tttgatgcaa cttatcaaaa gaatatctca atacaagatt ctcaagtaat 2640
agttacaggc aatcttgaca tctcgactga gcttgggaat gtcaacaact cgataagtaa 2700
tgctttggat aagttagagg aaagcaacag caaactagac aaggtcaatg ttaaactgac 2760
cagcacatcc gctcttatta cctatatcgt tttaactgtc atatctcttg tatgtggtat 2820
acttagcctg gttctagcat gctacctgat gtacaagcaa aaggcgcaac agaagacctt 2880
gttgtggctt gggaataata ccctagacca gatgagggcc actacaaaaa tgtagcttga 2940
tctagagcgg ccgcggggat ccagacatga taagatacat tgatgagttt ggacaaacca 3000
caactagaat gcagtgaaaa aaatgcttta tttgtgaaat ttgtgatgct attgctttat 3060
ttgtaaccat tataagctgc aataaacaag ttaacaacaa caattgcatt cattttatgt 3120
ttcaggttca gggggaggtg tgggaggttt tttcggatcc tctagagtcg agtttaatgt 3180
tagtttattc aatgcattgg ttgcaaatat tcattacttc tccaatccca ggtcattctt 3240
tagcgagatg atgttatgac attgctgtga aaattactac aggatatatt tttaagatgc 3300
aggagtaaca atgtgcatag taggcgtagt tatcgcagac gtgcaacgct tcgcatttga 3360
gttaccgaag tgcccaacag tgctgcggtt atggtttatg cgcacagaat ccatgcatgt 3420
cctaattgaa ccatccgatt tttcttttaa tcgcgatcgt tgtttgggca actgcgttat 3480
ttcagatcta aaaaatttac cctttatgac catcacatct ctctggctca taccccgctt 3540
ggataagata tcatgtagat tccgccctaa gaaatgcaaa ctaacattat tgtcggttcc 3600
atatacactt ccatcttgtc cttcgaaaat aacaaactcg cgcaatagac cgtccgtaca 3660
tgcatggccg atgtgtgtca acatcattgg tctgctagat cccgatggga cgaatcgtac 3720
agtcgtcgct ccagcattgg caaaaatccc cagataccct ccatgcggca aatctaaatt 3780
gcgaccccga agagactgca ccaaagtctt atcgacgcac gctgattttt ttgaacagcg 3840
ggagcccat 3849
<210> 33
<211> 4599
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК исходной трансферной плазмиды для HVT-ND № 42
<400> 33
atggcagcga gcatgggata ttcatcctcg tcatcgttaa catctctacg ggttcagaat 60
gtttggcatg tcgtcgatcc tttgcccatc gttgcaaatt acaagtccga tcgccatgac 120
cgcgataagc ctgtaccatg tggcattagg gtgacatctc gatcatacat tataagacca 180
acgtgcgagt cttccaaaga cctgcacgcc ttcttcttcg gattgtcaac gggttcttca 240
gaatctatgc ccatatctgg cgttgagacc attgtgcgtt taatgaacaa taaagcggca 300
tgccatggaa aggagggctg cagatctcca ttttctcacg ccactatcct ggacgctgta 360
gacgataatt ataccatgaa tatagagggg gtatgtttcc actgccactg tgatgataag 420
ttttctccag attgttggat atctgcattt tctgctgccg aacaaacttc atcgctatgc 480
aaagagatgc gtgtgtacac gcgccgttga gtatacggga aactaaatgt tcatagaggt 540
ctttgggcta tatgttatta aataaaataa ttgaccagtg gaattcacta gtggatcccc 600
caactccgcc cgttttatga ctagaaccaa tagtttttaa tgccaaatgc actgaaatcc 660
cctaatttgc aaagccaaac gccccctatg tgagtaatac ggggactttt tacccaattt 720
cccaagcgga aagcccccta atacactcat atggcatatg aatcagcacg gtcatgcact 780
ctaatggcgg cccataggga ctttccacat agggggcgtt caccatttcc cagcataggg 840
gtggtgactc aatggccttt acccaagtac attgggtcaa tgggaggtaa gccaatgggt 900
ttttcccatt actggcaagc acactgagtc aaatgggact ttccactggg ttttgcccaa 960
gtacattggg tcaatgggag gtgagccaat gggaaaaacc cattgctgcc aagtacactg 1020
actcaatagg gactttccaa tgggtttttc cattgttggc aagcatataa ggtcaatgtg 1080
ggtgagtcaa tagggacttt ccattgtatt ctgcccagta cataaggtca atagggggtg 1140
aatcaacagg aaagtcccat tggagccaag tacactgcgt caatagggac tttccattgg 1200
gttttgccca gtacataagg tcaatagggg atgagtcaat gggaaaaacc cattggagcc 1260
aagtacactg actcaatagg gactttccat tgggttttgc ccagtacata aggtcaatag 1320
ggggtgagtc aacaggaaag tcccattgga gccaagtaca ttgagtcaat agggactttc 1380
caatgggttt tgcccagtac ataaggtcaa tgggaggtaa gccaatgggt ttttcccatt 1440
actggcacgt atactgagtc attagggact ttccaatggg ttttgcccag tacataaggt 1500
caataggggt gaatcaacag gaaagtccca ttggagccaa gtacactgag tcaataggga 1560
ctttccattg ggttttgccc agtacaaaag gtcaataggg ggtgagtcaa tgggtttttc 1620
ccattattgg cacgtacata aggtcaatag gggtgagtca ttgggttttt ccagccaatt 1680
taattaaaac gccatgtact ttcccaccat tgacgtcaat gggctattga aactaatgca 1740
acgtgacctt taaacggtac tttcccatag ctgattaatg ggaaagtacc gttctcgagc 1800
caatacacgt caatgggaag tgaaagggca gccaaaacgt aacaccgccc cggttttccc 1860
ctggaaattc catattggca cgcattctat tggctgagct gcgttctacg tgggtataag 1920
aggcgcgacc agcgtcggta ccgtcgcagt cttcggtctg accaccgtag aacgcagagc 1980
tcctcgctgc aggcggccgc tctagaactc gtcgatcgca gcgatgggct ccagatcttc 2040
taccaggatc ccagtacctc tgatgctgac cgtccgaatc atgttggcac tgagttgcgt 2100
ctgtccgacc agctcccttg atggcaggcc tcttgcagct gcagggattg tggtaacagg 2160
agacaaagca gtcaacatat acacctcatc tcagacaggg tcaatcataa tcaagttact 2220
cccaaatatg cccaaggata aagaggcgtg tgcaaaagcc ccattggaag catacaacag 2280
gacattgact actttgctca ccccccttgg tgattctatc cgtaggatac aagagtctgt 2340
gaccacatcc ggaggaggga aacagggacg tcttataggc gccattatcg gtggtgtagc 2400
tctcggggtt gcaaccgctg cacagataac agcagcctcg gctctgatac aagccaatca 2460
aaatgctgcc aacatcctcc ggctcaaaga gagcattgct gcaaccaatg aggctgtgca 2520
cgaggtcact gacggattat cacaactagc agtggcagtt gggaagatgc agcaatttgt 2580
taatgaccag tttaataaaa cagctcagga attggactgt ataaaaatta cacagcaggt 2640
tggtgtagaa ctcaacctgt acctaactga attgactaca gtattcgggc cacaaatcac 2700
ttcccctgcc ttaactcagc tgactatcca ggcgctttac aatctagctg gtgggaatat 2760
ggattacttg ttgactaagt taggtgtagg aaacaaccaa ctcagctcat taattggtag 2820
tggcctgatt accggcaacc ctatcctgta cgactcacag actcaactct tgggtataca 2880
ggtcacccta ccctcagtcg ggaatctaaa taatatgcgt gccacctacc tggaaacctt 2940
gtctgtaagt acaaccaaag gatttgcctc agcacttgtc ccaaaagtag tgacacaggt 3000
tggttccgtg atagaagagc ttgacacctc gtactgtatc gagaccgatt tggacctata 3060
ttgtacaaga atagtgacat tccctatgtc tcctggtatt tattcctgtt tgagtggcaa 3120
tacatctgct tgcatgtatt caaagactga aggcgcactc actacgccgt atatgaccct 3180
caaaggctca gttattgcca actgtaagat gacaacatgt agatgtgcag accccccggg 3240
tatcatatcg cagaattatg gagaagctgt gtctctaata gataggcaat catgcaatat 3300
cttatcctta gacgggataa ctttgaggct cagtggggaa tttgatgcaa cttatcaaaa 3360
gaatatctca atacaagatt ctcaagtaat agttacaggc aatcttgaca tctcgactga 3420
gcttgggaat gtcaacaact cgataagtaa tgctttggat aagttagagg aaagcaacag 3480
caaactagac aaggtcaatg ttaaactgac cagcacatcc gctcttatta cctatatcgt 3540
tttaactgtc atatctcttg tatgtggtat acttagcctg gttctagcat gctacctgat 3600
gtacaagcaa aaggcgcaac agaagacctt gttgtggctt gggaataata ccctagacca 3660
gatgagggcc actacaaaaa tgtagcttga tctagagcgg ccgcggggat ccagacatga 3720
taagatacat tgatgagttt ggacaaacca caactagaat gcagtgaaaa aaatgcttta 3780
tttgtgaaat ttgtgatgct attgctttat ttgtaaccat tataagctgc aataaacaag 3840
ttaacaacaa caattgcatt cattttatgt ttcaggttca gggggaggtg tgggaggttt 3900
tttcggatcc tctagagtcg agtttaatgt tagtttattc aatgcattgg ttgcaaatat 3960
tcattacttc tccaatccca ggtcattctt tagcgagatg atgttatgac attgctgtga 4020
aaattactac aggatatatt tttaagatgc aggagtaaca atgtgcatag taggcgtagt 4080
tatcgcagac gtgcaacgct tcgcatttga gttaccgaag tgcccaacag tgctgcggtt 4140
atggtttatg cgcacagaat ccatgcatgt cctaattgaa ccatccgatt tttcttttaa 4200
tcgcgatcgt tgtttgggca actgcgttat ttcagatcta aaaaatttac cctttatgac 4260
catcacatct ctctggctca taccccgctt ggataagata tcatgtagat tccgccctaa 4320
gaaatgcaaa ctaacattat tgtcggttcc atatacactt ccatcttgtc cttcgaaaat 4380
aacaaactcg cgcaatagac cgtccgtaca tgcatggccg atgtgtgtca acatcattgg 4440
tctgctagat cccgatggga cgaatcgtac agtcgtcgct ccagcattgg caaaaatccc 4500
cagataccct ccatgcggca aatctaaatt gcgaccccga agagactgca ccaaagtctt 4560
atcgacgcac gctgattttt ttgaacagcg ggagcccat 4599
<210> 34
<211> 3974
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК клонирующей плазмиды для pSiteB
<400> 34
tctgcttaga aaaactcatc gagcatcaaa tgaaactgca atttattcat atcaggatta 60
tcaataccat atttttgaaa aagccgtttc tgtaatgaag gagaaaactc accgaggcag 120
ttccatagga tggcaagatc ctggtatcgg tctgcgattc cgactcgtcc aacatcaata 180
caacctatta atttcccctc gtcaaaaata aggttatcaa gtgagaaatc accatgagtg 240
acgactgaat ccggtgagaa tggcaaaagt ttatgcattt ctttccagac ttgttcaaca 300
ggccagccat tacgctcgtc atcaaaatca ctcgcatcaa ccaaaccgtt attcattcgt 360
gattgcgcct gagcgaggcg aaatacgcga tcgctgttaa aaggacaatt acaaacagga 420
atcgagtgca accggcgcag gaacactgcc agcgcatcaa caatattttc acctgaatca 480
ggatattctt ctaatacctg gaacgctgtt tttccgggga tcgcagtggt gagtaaccat 540
gcatcatcag gagtacggat aaaatgcttg atggtcggaa gtggcataaa ttccgtcagc 600
cagtttagtc tgaccatctc atctgtaaca tcattggcaa cgctaccttt gccatgtttc 660
agaaacaact ctggcgcatc gggcttccca tacaagcgat agattgtcgc acctgattgc 720
ccgacattat cgcgagccca tttataccca tataaatcag catccatgtt ggaatttaat 780
cgcggcctcg acgtttcccg ttgaatatgg ctcatattct tcctttttca atattattga 840
agcatttatc agggttattg tctcatgagc ggatacatat ttgaatgtat ttagaaaaat 900
aaacaaatag gggtcagtgt tacaaccaat taaccaattc tgaacattat cgcgagccca 960
tttatacctg aatatggctc ataacacccc ttgcagtgcg actaacggca tgaagctcgt 1020
cggggctgga tcgcttcgta ttgcgagctg tgcggctgag ttgacgtatc tgtgctggat 1080
gattactcat aacggcaccg ctatcaaacg tgccacgttc atgtccgtgt cgcccttcgc 1140
tgaaactagt ctctacactt cttgttaaat ggaaagtgca tttgcttgtt cttacaatcg 1200
gcccgagtct cgttcacagc gcctcgttca cacttaaacc acaaatagtc tacaggctat 1260
atgggagcca gactgaaact cacatatgac taatattcgg gggtgttagt cacgtgtagc 1320
ccattgtgtg catataacga tgttggacgc gtccttattc gcggtgtact tgatactatg 1380
gcagcgagca tgggatattc atcctcgtca tcgttaacat ctctacgggt tcagaatgtt 1440
tggcatgtcg tcgatccttt gcccatcgtt gcaaattaca agtccgatcg ccatgaccgc 1500
gataagcctg taccatgtgg cattagggtg acatctcgat catacattat aagaccaacg 1560
tgcgagtctt ccaaagacct gcacgccttc ttcttcggat tgtcaacggg ttcttcagaa 1620
tctatgccca tatctggcgt tgagaccatt gtgcgtttaa tgaacaataa agcggcatgc 1680
catggaaagg agggctgcag atctccattt tctcacgcca ctatcctgga cgctgtagac 1740
gataattata ccatgaatat agagggggta tgtttccact gccactgtga tgataagttt 1800
tctccagatt gttggatatc tgcattttct gctgccgaac aaacttcatc gctatgcaaa 1860
gagatgcgtg tgtacacgcg ccgttgagta tacgggaaac taaatgttca tagaggtctt 1920
tgggctatat gttattaaat aaaataattg ggcgcgccac cggtacgagt cactggatcc 1980
tctagtcagc ctcgagtgac tagcgtgcta gcagtggccg gccgtttaat gttagtttat 2040
tcaatgcatt ggttgcaaat attcattact tctccaatcc caggtcattc tttagcgaga 2100
tgatgttatg acattgctgt gaaaattact acaggatata tttttaagat gcaggagtaa 2160
caatgtgcat agtaggcgta gttatcgcag acgtgcaacg cttcgcattt gagttaccga 2220
agtgcccaac agtgctgcgg ttatggttta tgcgcacaga atccatgcat gtcctaattg 2280
aaccatccga tttttctttt aatcgcgatc gttgtttggg caactgcgtt atttcagatc 2340
taaaaaattt accctttatg accatcacat ctctctggct cataccccgc ttggataaga 2400
tatcatgtag attccgccct aagaaatgca aactaacatt attgtcggtt ccatatacac 2460
ttccatcttg tccttcgaaa ataacaaact cgcgcaatag accgtccgta catgcatggc 2520
cgatgtgtgt caacatcatt ggtctgctag atcccgatgg gacgaatcgt acagtcgtcg 2580
ctccagcatt ggcaaaaatc cccagatacc ctccatgcgg caaatctaaa ttgcgacccc 2640
gaagagactg caccaaagtc ttatcgacgc acgctgattt ttttgaacag cgggagccca 2700
ttatcttcag tggagcgtag acgggcgagg ctaattatgt gacatagcaa cactgcatgt 2760
atgtttttat aaatcaataa gagtacataa tttattacgt atcatttccg tttgtaatat 2820
actcctgcag gcgtcaaaag ggcgacacac tgtcattagc aactccttgt ccttcgatct 2880
cgtcaacaac agcttgcagt tcaaatacaa gacccagaag gcgactattc tggaagcgag 2940
cttgaagcca gacgctgagt acgaaaaggg gcccgagctt aagactggcc gtcgttttac 3000
aacacagaaa gagtttgtag aaacgcaaaa aggccatccg tcaggggcct tctgcttagt 3060
ttgatgcctg gcagttccct actctcgcct tccgcttcct cgctcactga ctcgctgcgc 3120
tcggtcgttc ggctgcggcg agcggtatca gctcactcaa aggcggtaat acggttatcc 3180
acagaatcag gggataacgc aggaaagaac atgtgagcaa aaggccagca aaaggccagg 3240
aaccgtaaaa aggccgcgtt gctggcgttt ttccataggc tccgcccccc tgacgagcat 3300
cacaaaaatc gacgctcaag tcagaggtgg cgaaacccga caggactata aagataccag 3360
gcgtttcccc ctggaagctc cctcgtgcgc tctcctgttc cgaccctgcc gcttaccgga 3420
tacctgtccg cctttctccc ttcgggaagc gtggcgcttt ctcatagctc acgctgtagg 3480
tatctcagtt cggtgtaggt cgttcgctcc aagctgggct gtgtgcacga accccccgtt 3540
cagcccgacc gctgcgcctt atccggtaac tatcgtcttg agtccaaccc ggtaagacac 3600
gacttatcgc cactggcagc agccactggt aacaggatta gcagagcgag gtatgtaggc 3660
ggtgctacag agttcttgaa gtggtgggct aactacggct acactagaag aacagtattt 3720
ggtatctgcg ctctgctgaa gccagttacc ttcggaaaaa gagttggtag ctcttgatcc 3780
ggcaaacaaa ccaccgctgg tagcggtggt ttttttgttt gcaagcagca gattacgcgc 3840
agaaaaaaag gatctcaaga agatcctttg atcttttcta cggggtctga cgctcagtgg 3900
aacgacgcgc gcgtaactca cgttaaggga ttttggtcat gagcttgcgc cgtcccgtca 3960
agtcagcgta atgc 3974
<210> 35
<211> 7246
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК конечной трансферной плазмиды для pSiteB № 42
<400> 35
tctgcttaga aaaactcatc gagcatcaaa tgaaactgca atttattcat atcaggatta 60
tcaataccat atttttgaaa aagccgtttc tgtaatgaag gagaaaactc accgaggcag 120
ttccatagga tggcaagatc ctggtatcgg tctgcgattc cgactcgtcc aacatcaata 180
caacctatta atttcccctc gtcaaaaata aggttatcaa gtgagaaatc accatgagtg 240
acgactgaat ccggtgagaa tggcaaaagt ttatgcattt ctttccagac ttgttcaaca 300
ggccagccat tacgctcgtc atcaaaatca ctcgcatcaa ccaaaccgtt attcattcgt 360
gattgcgcct gagcgaggcg aaatacgcga tcgctgttaa aaggacaatt acaaacagga 420
atcgagtgca accggcgcag gaacactgcc agcgcatcaa caatattttc acctgaatca 480
ggatattctt ctaatacctg gaacgctgtt tttccgggga tcgcagtggt gagtaaccat 540
gcatcatcag gagtacggat aaaatgcttg atggtcggaa gtggcataaa ttccgtcagc 600
cagtttagtc tgaccatctc atctgtaaca tcattggcaa cgctaccttt gccatgtttc 660
agaaacaact ctggcgcatc gggcttccca tacaagcgat agattgtcgc acctgattgc 720
ccgacattat cgcgagccca tttataccca tataaatcag catccatgtt ggaatttaat 780
cgcggcctcg acgtttcccg ttgaatatgg ctcatattct tcctttttca atattattga 840
agcatttatc agggttattg tctcatgagc ggatacatat ttgaatgtat ttagaaaaat 900
aaacaaatag gggtcagtgt tacaaccaat taaccaattc tgaacattat cgcgagccca 960
tttatacctg aatatggctc ataacacccc ttgcagtgcg actaacggca tgaagctcgt 1020
cggggctgga tcgcttcgta ttgcgagctg tgcggctgag ttgacgtatc tgtgctggat 1080
gattactcat aacggcaccg ctatcaaacg tgccacgttc atgtccgtgt cgcccttcgc 1140
tgaaactagt ctctacactt cttgttaaat ggaaagtgca tttgcttgtt cttacaatcg 1200
gcccgagtct cgttcacagc gcctcgttca cacttaaacc acaaatagtc tacaggctat 1260
atgggagcca gactgaaact cacatatgac taatattcgg gggtgttagt cacgtgtagc 1320
ccattgtgtg catataacga tgttggacgc gtccttattc gcggtgtact tgatactatg 1380
gcagcgagca tgggatattc atcctcgtca tcgttaacat ctctacgggt tcagaatgtt 1440
tggcatgtcg tcgatccttt gcccatcgtt gcaaattaca agtccgatcg ccatgaccgc 1500
gataagcctg taccatgtgg cattagggtg acatctcgat catacattat aagaccaacg 1560
tgcgagtctt ccaaagacct gcacgccttc ttcttcggat tgtcaacggg ttcttcagaa 1620
tctatgccca tatctggcgt tgagaccatt gtgcgtttaa tgaacaataa agcggcatgc 1680
catggaaagg agggctgcag atctccattt tctcacgcca ctatcctgga cgctgtagac 1740
gataattata ccatgaatat agagggggta tgtttccact gccactgtga tgataagttt 1800
tctccagatt gttggatatc tgcattttct gctgccgaac aaacttcatc gctatgcaaa 1860
gagatgcgtg tgtacacgcg ccgttgagta tacgggaaac taaatgttca tagaggtctt 1920
tgggctatat gttattaaat aaaataattg ggcgcgccaa ctccgcccgt tttatgacta 1980
gaaccaatag tttttaatgc caaatgcact gaaatcccct aatttgcaaa gccaaacgcc 2040
ccctatgtga gtaatacggg gactttttac ccaatttccc aagcggaaag ccccctaata 2100
cactcatatg gcatatgaat cagcacggtc atgcactcta atggcggccc atagggactt 2160
tccacatagg gggcgttcac catttcccag cataggggtg gtgactcaat ggcctttacc 2220
caagtacatt gggtcaatgg gaggtaagcc aatgggtttt tcccattact ggcaagcaca 2280
ctgagtcaaa tgggactttc cactgggttt tgcccaagta cattgggtca atgggaggtg 2340
agccaatggg aaaaacccat tgctgccaag tacactgact caatagggac tttccaatgg 2400
gtttttccat tgttggcaag catataaggt caatgtgggt gagtcaatag ggactttcca 2460
ttgtattctg cccagtacat aaggtcaata gggggtgaat caacaggaaa gtcccattgg 2520
agccaagtac actgcgtcaa tagggacttt ccattgggtt ttgcccagta cataaggtca 2580
ataggggatg agtcaatggg aaaaacccat tggagccaag tacactgact caatagggac 2640
tttccattgg gttttgccca gtacataagg tcaatagggg gtgagtcaac aggaaagtcc 2700
cattggagcc aagtacattg agtcaatagg gactttccaa tgggttttgc ccagtacata 2760
aggtcaatgg gaggtaagcc aatgggtttt tcccattact ggcacgtata ctgagtcatt 2820
agggactttc caatgggttt tgcccagtac ataaggtcaa taggggtgaa tcaacaggaa 2880
agtcccattg gagccaagta cactgagtca atagggactt tccattgggt tttgcccagt 2940
acaaaaggtc aatagggggt gagtcaatgg gtttttccca ttattggcac gtacataagg 3000
tcaatagggg tgagtcattg ggtttttcca gccaatttaa ttaaaacgcc atgtactttc 3060
ccaccattga cgtcaatggg ctattgaaac taatgcaacg tgacctttaa acggtacttt 3120
cccatagctg attaatggga aagtaccgtt ctcgagccaa tacacgtcaa tgggaagtga 3180
aagggcagcc aaaacgtaac accgccccgg ttttcccctg gaaattccat attggcacgc 3240
attctattgg ctgagctgcg ttctacgtgg gtataagagg cgcgaccagc gtcggtaccg 3300
tcgcagtctt cggtctgacc accgtagaac gcagagctcc tcgctgcagg cggccgctct 3360
agaactcgtc gatcgcagcg atgggctcca gatcttctac caggatccca gtacctctga 3420
tgctgaccgt ccgaatcatg ttggcactga gttgcgtctg tccgaccagc tcccttgatg 3480
gcaggcctct tgcagctgca gggattgtgg taacaggaga caaagcagtc aacatataca 3540
cctcatctca gacagggtca atcataatca agttactccc aaatatgccc aaggataaag 3600
aggcgtgtgc aaaagcccca ttggaagcat acaacaggac attgactact ttgctcaccc 3660
cccttggtga ttctatccgt aggatacaag agtctgtgac cacatccgga ggagggaaac 3720
agggacgtct tataggcgcc attatcggtg gtgtagctct cggggttgca accgctgcac 3780
agataacagc agcctcggct ctgatacaag ccaatcaaaa tgctgccaac atcctccggc 3840
tcaaagagag cattgctgca accaatgagg ctgtgcacga ggtcactgac ggattatcac 3900
aactagcagt ggcagttggg aagatgcagc aatttgttaa tgaccagttt aataaaacag 3960
ctcaggaatt ggactgtata aaaattacac agcaggttgg tgtagaactc aacctgtacc 4020
taactgaatt gactacagta ttcgggccac aaatcacttc ccctgcctta actcagctga 4080
ctatccaggc gctttacaat ctagctggtg ggaatatgga ttacttgttg actaagttag 4140
gtgtaggaaa caaccaactc agctcattaa ttggtagtgg cctgattacc ggcaacccta 4200
tcctgtacga ctcacagact caactcttgg gtatacaggt caccctaccc tcagtcggga 4260
atctaaataa tatgcgtgcc acctacctgg aaaccttgtc tgtaagtaca accaaaggat 4320
ttgcctcagc acttgtccca aaagtagtga cacaggttgg ttccgtgata gaagagcttg 4380
acacctcgta ctgtatcgag accgatttgg acctatattg tacaagaata gtgacattcc 4440
ctatgtctcc tggtatttat tcctgtttga gtggcaatac atctgcttgc atgtattcaa 4500
agactgaagg cgcactcact acgccgtata tgaccctcaa aggctcagtt attgccaact 4560
gtaagatgac aacatgtaga tgtgcagacc ccccgggtat catatcgcag aattatggag 4620
aagctgtgtc tctaatagat aggcaatcat gcaatatctt atccttagac gggataactt 4680
tgaggctcag tggggaattt gatgcaactt atcaaaagaa tatctcaata caagattctc 4740
aagtaatagt tacaggcaat cttgacatct cgactgagct tgggaatgtc aacaactcga 4800
taagtaatgc tttggataag ttagaggaaa gcaacagcaa actagacaag gtcaatgtta 4860
aactgaccag cacatccgct cttattacct atatcgtttt aactgtcata tctcttgtat 4920
gtggtatact tagcctggtt ctagcatgct acctgatgta caagcaaaag gcgcaacaga 4980
agaccttgtt gtggcttggg aataataccc tagaccagat gagggccact acaaaaatgt 5040
agcttgatct agagcggccg cggggatcca gacatgataa gatacattga tgagtttgga 5100
caaaccacaa ctagaatgca gtgaaaaaaa tgctttattt gtgaaatttg tgatgctatt 5160
gctttatttg taaccattat aagctgcaat aaacaagtta acaacaacaa ttgcattcat 5220
tttatgtttc aggttcaggg ggaggtgtgg gaggtttttt cggatcctct agagtcgagc 5280
tagcagtggc cggccgttta atgttagttt attcaatgca ttggttgcaa atattcatta 5340
cttctccaat cccaggtcat tctttagcga gatgatgtta tgacattgct gtgaaaatta 5400
ctacaggata tatttttaag atgcaggagt aacaatgtgc atagtaggcg tagttatcgc 5460
agacgtgcaa cgcttcgcat ttgagttacc gaagtgccca acagtgctgc ggttatggtt 5520
tatgcgcaca gaatccatgc atgtcctaat tgaaccatcc gatttttctt ttaatcgcga 5580
tcgttgtttg ggcaactgcg ttatttcaga tctaaaaaat ttacccttta tgaccatcac 5640
atctctctgg ctcatacccc gcttggataa gatatcatgt agattccgcc ctaagaaatg 5700
caaactaaca ttattgtcgg ttccatatac acttccatct tgtccttcga aaataacaaa 5760
ctcgcgcaat agaccgtccg tacatgcatg gccgatgtgt gtcaacatca ttggtctgct 5820
agatcccgat gggacgaatc gtacagtcgt cgctccagca ttggcaaaaa tccccagata 5880
ccctccatgc ggcaaatcta aattgcgacc ccgaagagac tgcaccaaag tcttatcgac 5940
gcacgctgat ttttttgaac agcgggagcc cattatcttc agtggagcgt agacgggcga 6000
ggctaattat gtgacatagc aacactgcat gtatgttttt ataaatcaat aagagtacat 6060
aatttattac gtatcatttc cgtttgtaat atactcctgc aggcgtcaaa agggcgacac 6120
actgtcatta gcaactcctt gtccttcgat ctcgtcaaca acagcttgca gttcaaatac 6180
aagacccaga aggcgactat tctggaagcg agcttgaagc cagacgctga gtacgaaaag 6240
gggcccgagc ttaagactgg ccgtcgtttt acaacacaga aagagtttgt agaaacgcaa 6300
aaaggccatc cgtcaggggc cttctgctta gtttgatgcc tggcagttcc ctactctcgc 6360
cttccgcttc ctcgctcact gactcgctgc gctcggtcgt tcggctgcgg cgagcggtat 6420
cagctcactc aaaggcggta atacggttat ccacagaatc aggggataac gcaggaaaga 6480
acatgtgagc aaaaggccag caaaaggcca ggaaccgtaa aaaggccgcg ttgctggcgt 6540
ttttccatag gctccgcccc cctgacgagc atcacaaaaa tcgacgctca agtcagaggt 6600
ggcgaaaccc gacaggacta taaagatacc aggcgtttcc ccctggaagc tccctcgtgc 6660
gctctcctgt tccgaccctg ccgcttaccg gatacctgtc cgcctttctc ccttcgggaa 6720
gcgtggcgct ttctcatagc tcacgctgta ggtatctcag ttcggtgtag gtcgttcgct 6780
ccaagctggg ctgtgtgcac gaaccccccg ttcagcccga ccgctgcgcc ttatccggta 6840
actatcgtct tgagtccaac ccggtaagac acgacttatc gccactggca gcagccactg 6900
gtaacaggat tagcagagcg aggtatgtag gcggtgctac agagttcttg aagtggtggg 6960
ctaactacgg ctacactaga agaacagtat ttggtatctg cgctctgctg aagccagtta 7020
ccttcggaaa aagagttggt agctcttgat ccggcaaaca aaccaccgct ggtagcggtg 7080
gtttttttgt ttgcaagcag cagattacgc gcagaaaaaa aggatctcaa gaagatcctt 7140
tgatcttttc tacggggtct gacgctcagt ggaacgacgc gcgcgtaact cacgttaagg 7200
gattttggtc atgagcttgc gccgtcccgt caagtcagcg taatgc 7246
<210> 36
<211> 4440
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК трансферной плазмиды для HVT-ND № 44
<400> 36
atggcagcga gcatgggata ttcatcctcg tcatcgttaa catctctacg ggttcagaat 60
gtttggcatg tcgtcgatcc tttgcccatc gttgcaaatt acaagtccga tcgccatgac 120
cgcgataagc ctgtaccatg tggcattagg gtgacatctc gatcatacat tataagacca 180
acgtgcgagt cttccaaaga cctgcacgcc ttcttcttcg gattgtcaac gggttcttca 240
gaatctatgc ccatatctgg cgttgagacc attgtgcgtt taatgaacaa taaagcggca 300
tgccatggaa aggagggctg cagatctcca ttttctcacg ccactatcct ggacgctgta 360
gacgataatt ataccatgaa tatagagggg gtatgtttcc actgccactg tgatgataag 420
ttttctccag attgttggat atctgcattt tctgctgccg aacaaacttc atcgctatgc 480
aaagagatgc gtgtgtacac gcgccgttga gtatacggga aactaaatgt tcatagaggt 540
ctttgggcta tatgttatta aataaaataa ttgaccagtg tcgaggtgag ccccacgttc 600
tgcttcactc tccccatctc ccccccctcc ccacccccaa ttttgtattt atttattttt 660
taattatttt gtgcagcgat gggggcgggg gggggggggg cgcgcgccag gcggggcggg 720
gcggggcgag gggcggggcg gggcgaggcg gagaggtgcg gcggcagcca atcagagcgg 780
cgcgctccga aagtttcctt ttatggcgag gcggcggcgg cggcggccct ataaaaagcg 840
aagcgcgcgg cgggcgggag tcgctgcgtt gccttcgccc cgtgccccgc tccgcgccgc 900
ctcgcgccgc ccgccccggc tctgactgac cgcgttactc ccacaggtga gcgggcggga 960
cggcccttct cctccgggct gtaattagcg cttggtttaa tgacggctcg tttcttttct 1020
gtggctgcgt gaaagcctta aagggctccg ggagggccct ttgtgcgggg gggagcggct 1080
cggggggtgc gtgcgtgtgt gtgtgcgtgg ggagcgccgc gtgcggcccg cgctgcccgg 1140
cggctgtgag cgctgcgggc gcggcgcggg gctttgtgcg ctccgcgtgt gcgcgagggg 1200
agcgcggccg ggggcggtgc cccgcggtgc gggggggctg cgaggggaac aaaggctgcg 1260
tgcggggtgt gtgcgtgggg gggtgagcag ggggtgtggg cgcggcggtc gggctgtaac 1320
ccccccctgc acccccctcc ccgagttgct gagcacggcc cggcttcggg tgcggggctc 1380
cgtgcggggc gtggcgcggg gctcgccgtg ccgggcgggg ggtggcggca ggtgggggtg 1440
ccgggcgggg cggggccgcc tcgggccggg gagggctcgg gggaggggcg cggcggcccc 1500
ggagcgccgg cggctgtcga ggcgcggcga gccgcagcca ttgcctttta tggtaatcgt 1560
gcgagagggc gcagggactt cctttgtccc aaatctggcg gagccgaaat ctgggaggcg 1620
ccgccgcacc ccctctagcg ggcgcgggcg aagcggtgcg gcgccggcag gaaggaaatg 1680
ggcggggagg gccttcgtgc gtcgccgcgc cgccgtcccc ttctccatct ccagcctcgg 1740
ggctgccgca gggggacggc tgccttcggg ggggacgggg cagggcgggg ttcggcttct 1800
ggcgtgtgac cggcggggtt tatatcttcc cttctctgtt cctccgcagc cagccatggc 1860
caccatgggc tccagatctt ctaccaggat cccagtacct ctgatgctga ccgtccgaat 1920
catgttggca ctgagttgcg tctgtccgac cagctccctt gatggcaggc ctcttgcagc 1980
tgcagggatt gtggtaacag gagacaaagc agtcaacata tacacctcat ctcagacagg 2040
gtcaatcata atcaagttac tcccaaatat gcccaaggat aaagaggcgt gtgcaaaagc 2100
cccattggaa gcatacaaca ggacattgac tactttgctc accccccttg gtgattctat 2160
ccgtaggata caagagtctg tgaccacatc cggaggaggg aaacagggac gtcttatagg 2220
cgccattatc ggtggtgtag ctctcggggt tgcaaccgct gcacagataa cagcagcctc 2280
ggctctgata caagccaatc aaaatgctgc caacatcctc cggctcaaag agagcattgc 2340
tgcaaccaat gaggctgtgc acgaggtcac tgacggatta tcacaactag cagtggcagt 2400
tgggaagatg cagcaatttg ttaatgacca gtttaataaa acagctcagg aattggactg 2460
tataaaaatt acacagcagg ttggtgtaga actcaacctg tacctaactg aattgactac 2520
agtattcggg ccacaaatca cttcccctgc cttaactcag ctgactatcc aggcgcttta 2580
caatctagct ggtgggaata tggattactt gttgactaag ttaggtgtag gaaacaacca 2640
actcagctca ttaattggta gtggcctgat taccggcaac cctatcctgt acgactcaca 2700
gactcaactc ttgggtatac aggtcaccct accctcagtc gggaatctaa ataatatgcg 2760
tgccacctac ctggaaacct tgtctgtaag tacaaccaaa ggatttgcct cagcacttgt 2820
cccaaaagta gtgacacagg ttggttccgt gatagaagag cttgacacct cgtactgtat 2880
cgagaccgat ttggacctat attgtacaag aatagtgaca ttccctatgt ctcctggtat 2940
ttattcctgt ttgagtggca atacatctgc ttgcatgtat tcaaagactg aaggcgcact 3000
cactacgccg tatatgaccc tcaaaggctc agttattgcc aactgtaaga tgacaacatg 3060
tagatgtgca gaccccccgg gtatcatatc gcagaattat ggagaagctg tgtctctaat 3120
agataggcaa tcatgcaata tcttatcctt agacgggata actttgaggc tcagtgggga 3180
atttgatgca acttatcaaa agaatatctc aatacaagat tctcaagtaa tagttacagg 3240
caatcttgac atctcgactg agcttgggaa tgtcaacaac tcgataagta atgctttgga 3300
taagttagag gaaagcaaca gcaaactaga caaggtcaat gttaaactga ccagcacatc 3360
cgctcttatt acctatatcg ttttaactgt catatctctt gtatgtggta tacttagcct 3420
ggttctagca tgctacctga tgtacaagca aaaggcgcaa cagaagacct tgttgtggct 3480
tgggaataat accctagacc agatgagggc cactacaaaa atgtagcttg atctagagcg 3540
gccgcgggga tccagacatg ataagataca ttgatgagtt tggacaaacc acaactagaa 3600
tgcagtgaaa aaaatgcttt atttgtgaaa tttgtgatgc tattgcttta tttgtaacca 3660
ttataagctg caataaacaa gttaacaaca acaattgcat tcattttatg tttcaggttc 3720
agggggaggt gtgggaggtt ttttcggatc ctctagagtc gagtttaatg ttagtttatt 3780
caatgcattg gttgcaaata ttcattactt ctccaatccc aggtcattct ttagcgagat 3840
gatgttatga cattgctgtg aaaattacta caggatatat ttttaagatg caggagtaac 3900
aatgtgcata gtaggcgtag ttatcgcaga cgtgcaacgc ttcgcatttg agttaccgaa 3960
gtgcccaaca gtgctgcggt tatggtttat gcgcacagaa tccatgcatg tcctaattga 4020
accatccgat ttttctttta atcgcgatcg ttgtttgggc aactgcgtta tttcagatct 4080
aaaaaattta ccctttatga ccatcacatc tctctggctc ataccccgct tggataagat 4140
atcatgtaga ttccgcccta agaaatgcaa actaacatta ttgtcggttc catatacact 4200
tccatcttgt ccttcgaaaa taacaaactc gcgcaataga ccgtccgtac atgcatggcc 4260
gatgtgtgtc aacatcattg gtctgctaga tcccgatggg acgaatcgta cagtcgtcgc 4320
tccagcattg gcaaaaatcc ccagataccc tccatgcggc aaatctaaat tgcgaccccg 4380
aagagactgc accaaagtct tatcgacgca cgctgatttt tttgaacagc gggagcccat 4440
<210> 37
<211> 3832
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК трансферной плазмиды для HVT-ND № 45
<400> 37
atgggctccc gctgttcaaa aaaatcagcg tgcgtcgata agactttggt gcagtctctt 60
cggggtcgca atttagattt gccgcatgga gggtatctgg ggatttttgc caatgctgga 120
gcgacgactg tacgattcgt cccatcggga tctagcagac caatgatgtt gacacacatc 180
ggccatgcat gtacggacgg tctattgcgc gagtttgtta ttttcgaagg acaagatgga 240
agtgtatatg gaaccgacaa taatgttagt ttgcatttct tagggcggaa tctacatgat 300
atcttatcca agcggggtat gagccagaga gatgtgatgg tcataaaggg taaatttttt 360
agatctgaaa taacgcagtt gcccaaacaa cgatcgcgat taaaagaaaa atcggatggt 420
tcaattagga catgcatgga ttctgtgcgc ataaaccata accgcagcac tgttgggcac 480
ttcggtaact caaatgcgaa gcgttgcacg tctgcgataa ctacgcctac tatgcacatt 540
gttactcctg catcttaaaa atatatcctg tagtaatttt cacagcaatg tcataacatc 600
atctcgctaa agaatgacct gggattggag aagtaatgaa tatttgcaac caatgcattg 660
aataaactaa cattaaactt gacattgatt attgactagt tattaatagt aatcaattac 720
ggggtcatta gttcatagcc catatatgga gttccgcgtt acataactta cggtaaatgg 780
cccgcctggc tgaccgccca acgacccccg cccattgacg tcaataatga cgtatgttcc 840
catagtaacg ccaataggga ctttccattg acgtcaatgg gtggagtatt tacggtaaac 900
tgcccacttg gcagtacatc aagtgtatca tatgccaagt acgcccccta ttgacgtcaa 960
tgacggtaaa tggcccgcct ggcattatgc ccagtacatg accttatggg actttcctac 1020
ttggcagtac atctacgtat tagtcatcgc tattaccatg gtgatgcggt tttggcagta 1080
catcaatggg cgtggatagc ggtttgactc acggggattt ccaagtctcc accccattga 1140
cgtcaatggg agtttgtttt ggcaccaaaa tcaacgggac tttccaaaat gtcgtaacaa 1200
ctccgcccca ttgacgcaaa tgggcggtag gcgtgtacgg tgggaggtct atataagcag 1260
agctctctgg ctaactagag aacccactgc ttactggctt atcgaaatta atacgactca 1320
ctatagggag acccaagctg gctagcgttt aaacttaagc ttaccgccac catgggctcc 1380
agatcttcta ccaggatccc agtacctctg atgctgaccg tccgaatcat gttggcactg 1440
agttgcgtct gtccgaccag ctcccttgat ggcaggcctc ttgcagctgc agggattgtg 1500
gtaacaggag acaaagcagt caacatatac acctcatctc agacagggtc aatcataatc 1560
aagttactcc caaatatgcc caaggataaa gaggcgtgtg caaaagcccc attggaagca 1620
tacaacagga cattgactac tttgctcacc ccccttggtg attctatccg taggatacaa 1680
gagtctgtga ccacatccgg aggagggaaa cagggacgtc ttataggcgc cattatcggt 1740
ggtgtagctc tcggggttgc aaccgctgca cagataacag cagcctcggc tctgatacaa 1800
gccaatcaaa atgctgccaa catcctccgg ctcaaagaga gcattgctgc aaccaatgag 1860
gctgtgcacg aggtcactga cggattatca caactagcag tggcagttgg gaagatgcag 1920
caatttgtta atgaccagtt taataaaaca gctcaggaat tggactgtat aaaaattaca 1980
cagcaggttg gtgtagaact caacctgtac ctaactgaat tgactacagt attcgggcca 2040
caaatcactt cccctgcctt aactcagctg actatccagg cgctttacaa tctagctggt 2100
gggaatatgg attacttgtt gactaagtta ggtgtaggaa acaaccaact cagctcatta 2160
attggtagtg gcctgattac cggcaaccct atcctgtacg actcacagac tcaactcttg 2220
ggtatacagg tcaccctacc ctcagtcggg aatctaaata atatgcgtgc cacctacctg 2280
gaaaccttgt ctgtaagtac aaccaaagga tttgcctcag cacttgtccc aaaagtagtg 2340
acacaggttg gttccgtgat agaagagctt gacacctcgt actgtatcga gaccgatttg 2400
gacctatatt gtacaagaat agtgacattc cctatgtctc ctggtattta ttcctgtttg 2460
agtggcaata catctgcttg catgtattca aagactgaag gcgcactcac tacgccgtat 2520
atgaccctca aaggctcagt tattgccaac tgtaagatga caacatgtag atgtgcagac 2580
cccccgggta tcatatcgca gaattatgga gaagctgtgt ctctaataga taggcaatca 2640
tgcaatatct tatccttaga cgggataact ttgaggctca gtggggaatt tgatgcaact 2700
tatcaaaaga atatctcaat acaagattct caagtaatag ttacaggcaa tcttgacatc 2760
tcgactgagc ttgggaatgt caacaactcg ataagtaatg ctttggataa gttagaggaa 2820
agcaacagca aactagacaa ggtcaatgtt aaactgacca gcacatccgc tcttattacc 2880
tatatcgttt taactgtcat atctcttgta tgtggtatac ttagcctggt tctagcatgc 2940
tacctgatgt acaagcaaaa ggcgcaacag aagaccttgt tgtggcttgg gaataatacc 3000
ctagaccaga tgagggccac tacaaaaatg tagctgtgcc ttctagttgc cagccatctg 3060
ttgtttgccc ctcccccgtg ccttccttga ccctggaagg tgccactccc actgtccttt 3120
cctaataaaa tgaggaaatt gcatcgcatt gtctgagtag gtgtcattct attctggggg 3180
gtggggtggg gcaggacagc aagggggagg attgggaaga caatagcagg catgctgggg 3240
atgcggtggg ctctatggtc aattatttta tttaataaca tatagcccaa agacctctat 3300
gaacatttag tttcccgtat actcaacggc gcgtgtacac acgcatctct ttgcatagcg 3360
atgaagtttg ttcggcagca gaaaatgcag atatccaaca atctggagaa aacttatcat 3420
cacagtggca gtggaaacat accccctcta tattcatggt ataattatcg tctacagcgt 3480
ccaggatagt ggcgtgagaa aatggagatc tgcagccctc ctttccatgg catgccgctt 3540
tattgttcat taaacgcaca atggtctcaa cgccagatat gggcatagat tctgaagaac 3600
ccgttgacaa tccgaagaag aaggcgtgca ggtctttgga agactcgcac gttggtctta 3660
taatgtatga tcgagatgtc accctaatgc cacatggtac aggcttatcg cggtcatggc 3720
gatcggactt gtaatttgca acgatgggca aaggatcgac gacatgccaa acattctgaa 3780
cccgtagaga tgttaacgat gacgaggatg aatatcccat gctcgctgcc at 3832
<210> 38
<211> 4592
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК трансферной плазмиды для HVT-ND № 46
<400> 38
atgggctccc gctgttcaaa aaaatcagcg tgcgtcgata agactttggt gcagtctctt 60
cggggtcgca atttagattt gccgcatgga gggtatctgg ggatttttgc caatgctgga 120
gcgacgactg tacgattcgt cccatcggga tctagcagac caatgatgtt gacacacatc 180
ggccatgcat gtacggacgg tctattgcgc gagtttgtta ttttcgaagg acaagatgga 240
agtgtatatg gaaccgacaa taatgttagt ttgcatttct tagggcggaa tctacatgat 300
atcttatcca agcggggtat gagccagaga gatgtgatgg tcataaaggg taaatttttt 360
agatctgaaa taacgcagtt gcccaaacaa cgatcgcgat taaaagaaaa atcggatggt 420
tcaattagga catgcatgga ttctgtgcgc ataaaccata accgcagcac tgttgggcac 480
ttcggtaact caaatgcgaa gcgttgcacg tctgcgataa ctacgcctac tatgcacatt 540
gttactcctg catcttaaaa atatatcctg tagtaatttt cacagcaatg tcataacatc 600
atctcgctaa agaatgacct gggattggag aagtaatgaa tatttgcaac caatgcattg 660
aataaactaa cattaaacga attcactagt ggatccccca actccgcccg ttttatgact 720
agaaccaata gtttttaatg ccaaatgcac tgaaatcccc taatttgcaa agccaaacgc 780
cccctatgtg agtaatacgg ggacttttta cccaatttcc caagcggaaa gccccctaat 840
acactcatat ggcatatgaa tcagcacggt catgcactct aatggcggcc catagggact 900
ttccacatag ggggcgttca ccatttccca gcataggggt ggtgactcaa tggcctttac 960
ccaagtacat tgggtcaatg ggaggtaagc caatgggttt ttcccattac tggcaagcac 1020
actgagtcaa atgggacttt ccactgggtt ttgcccaagt acattgggtc aatgggaggt 1080
gagccaatgg gaaaaaccca ttgctgccaa gtacactgac tcaataggga ctttccaatg 1140
ggtttttcca ttgttggcaa gcatataagg tcaatgtggg tgagtcaata gggactttcc 1200
attgtattct gcccagtaca taaggtcaat agggggtgaa tcaacaggaa agtcccattg 1260
gagccaagta cactgcgtca atagggactt tccattgggt tttgcccagt acataaggtc 1320
aataggggat gagtcaatgg gaaaaaccca ttggagccaa gtacactgac tcaataggga 1380
ctttccattg ggttttgccc agtacataag gtcaataggg ggtgagtcaa caggaaagtc 1440
ccattggagc caagtacatt gagtcaatag ggactttcca atgggttttg cccagtacat 1500
aaggtcaatg ggaggtaagc caatgggttt ttcccattac tggcacgtat actgagtcat 1560
tagggacttt ccaatgggtt ttgcccagta cataaggtca ataggggtga atcaacagga 1620
aagtcccatt ggagccaagt acactgagtc aatagggact ttccattggg ttttgcccag 1680
tacaaaaggt caataggggg tgagtcaatg ggtttttccc attattggca cgtacataag 1740
gtcaataggg gtgagtcatt gggtttttcc agccaattta attaaaacgc catgtacttt 1800
cccaccattg acgtcaatgg gctattgaaa ctaatgcaac gtgaccttta aacggtactt 1860
tcccatagct gattaatggg aaagtaccgt tctcgagcca atacacgtca atgggaagtg 1920
aaagggcagc caaaacgtaa caccgccccg gttttcccct ggaaattcca tattggcacg 1980
cattctattg gctgagctgc gttctacgtg ggtataagag gcgcgaccag cgtcggtacc 2040
gtcgcagtct tcggtctgac caccgtagaa cgcagagctc ctcgctgcag gcggccgctc 2100
tagaactcgt cgatcgcagc gatgggctcc agatcttcta ccaggatccc agtacctctg 2160
atgctgaccg tccgaatcat gttggcactg agttgcgtct gtccgaccag ctcccttgat 2220
ggcaggcctc ttgcagctgc agggattgtg gtaacaggag acaaagcagt caacatatac 2280
acctcatctc agacagggtc aatcataatc aagttactcc caaatatgcc caaggataaa 2340
gaggcgtgtg caaaagcccc attggaagca tacaacagga cattgactac tttgctcacc 2400
ccccttggtg attctatccg taggatacaa gagtctgtga ccacatccgg aggagggaaa 2460
cagggacgtc ttataggcgc cattatcggt ggtgtagctc tcggggttgc aaccgctgca 2520
cagataacag cagcctcggc tctgatacaa gccaatcaaa atgctgccaa catcctccgg 2580
ctcaaagaga gcattgctgc aaccaatgag gctgtgcacg aggtcactga cggattatca 2640
caactagcag tggcagttgg gaagatgcag caatttgtta atgaccagtt taataaaaca 2700
gctcaggaat tggactgtat aaaaattaca cagcaggttg gtgtagaact caacctgtac 2760
ctaactgaat tgactacagt attcgggcca caaatcactt cccctgcctt aactcagctg 2820
actatccagg cgctttacaa tctagctggt gggaatatgg attacttgtt gactaagtta 2880
ggtgtaggaa acaaccaact cagctcatta attggtagtg gcctgattac cggcaaccct 2940
atcctgtacg actcacagac tcaactcttg ggtatacagg tcaccctacc ctcagtcggg 3000
aatctaaata atatgcgtgc cacctacctg gaaaccttgt ctgtaagtac aaccaaagga 3060
tttgcctcag cacttgtccc aaaagtagtg acacaggttg gttccgtgat agaagagctt 3120
gacacctcgt actgtatcga gaccgatttg gacctatatt gtacaagaat agtgacattc 3180
cctatgtctc ctggtattta ttcctgtttg agtggcaata catctgcttg catgtattca 3240
aagactgaag gcgcactcac tacgccgtat atgaccctca aaggctcagt tattgccaac 3300
tgtaagatga caacatgtag atgtgcagac cccccgggta tcatatcgca gaattatgga 3360
gaagctgtgt ctctaataga taggcaatca tgcaatatct tatccttaga cgggataact 3420
ttgaggctca gtggggaatt tgatgcaact tatcaaaaga atatctcaat acaagattct 3480
caagtaatag ttacaggcaa tcttgacatc tcgactgagc ttgggaatgt caacaactcg 3540
ataagtaatg ctttggataa gttagaggaa agcaacagca aactagacaa ggtcaatgtt 3600
aaactgacca gcacatccgc tcttattacc tatatcgttt taactgtcat atctcttgta 3660
tgtggtatac ttagcctggt tctagcatgc tacctgatgt acaagcaaaa ggcgcaacag 3720
aagaccttgt tgtggcttgg gaataatacc ctagaccaga tgagggccac tacaaaaatg 3780
tagcttgatc tagagcggcc gcggggatcc agacatgata agatacattg atgagtttgg 3840
acaaaccaca actagaatgc agtgaaaaaa atgctttatt tgtgaaattt gtgatgctat 3900
tgctttattt gtaaccatta taagctgcaa taaacaagtt aacaacaaca attgcattca 3960
ttttatgttt caggttcagg gggaggtgtg ggaggttttt tcggatcctc tagagtcgac 4020
aattatttta tttaataaca tatagcccaa agacctctat gaacatttag tttcccgtat 4080
actcaacggc gcgtgtacac acgcatctct ttgcatagcg atgaagtttg ttcggcagca 4140
gaaaatgcag atatccaaca atctggagaa aacttatcat cacagtggca gtggaaacat 4200
accccctcta tattcatggt ataattatcg tctacagcgt ccaggatagt ggcgtgagaa 4260
aatggagatc tgcagccctc ctttccatgg catgccgctt tattgttcat taaacgcaca 4320
atggtctcaa cgccagatat gggcatagat tctgaagaac ccgttgacaa tccgaagaag 4380
aaggcgtgca ggtctttgga agactcgcac gttggtctta taatgtatga tcgagatgtc 4440
accctaatgc cacatggtac aggcttatcg cggtcatggc gatcggactt gtaatttgca 4500
acgatgggca aaggatcgac gacatgccaa acattctgaa cccgtagaga tgttaacgat 4560
gacgaggatg aatatcccat gctcgctgcc at 4592
<210> 39
<211> 4433
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК трансферной плазмиды для HVT-ND № 48
<400> 39
atgggctccc gctgttcaaa aaaatcagcg tgcgtcgata agactttggt gcagtctctt 60
cggggtcgca atttagattt gccgcatgga gggtatctgg ggatttttgc caatgctgga 120
gcgacgactg tacgattcgt cccatcggga tctagcagac caatgatgtt gacacacatc 180
ggccatgcat gtacggacgg tctattgcgc gagtttgtta ttttcgaagg acaagatgga 240
agtgtatatg gaaccgacaa taatgttagt ttgcatttct tagggcggaa tctacatgat 300
atcttatcca agcggggtat gagccagaga gatgtgatgg tcataaaggg taaatttttt 360
agatctgaaa taacgcagtt gcccaaacaa cgatcgcgat taaaagaaaa atcggatggt 420
tcaattagga catgcatgga ttctgtgcgc ataaaccata accgcagcac tgttgggcac 480
ttcggtaact caaatgcgaa gcgttgcacg tctgcgataa ctacgcctac tatgcacatt 540
gttactcctg catcttaaaa atatatcctg tagtaatttt cacagcaatg tcataacatc 600
atctcgctaa agaatgacct gggattggag aagtaatgaa tatttgcaac caatgcattg 660
aataaactaa cattaaactc gaggtgagcc ccacgttctg cttcactctc cccatctccc 720
ccccctcccc acccccaatt ttgtatttat ttatttttta attattttgt gcagcgatgg 780
gggcgggggg ggggggggcg cgcgccaggc ggggcggggc ggggcgaggg gcggggcggg 840
gcgaggcgga gaggtgcggc ggcagccaat cagagcggcg cgctccgaaa gtttcctttt 900
atggcgaggc ggcggcggcg gcggccctat aaaaagcgaa gcgcgcggcg ggcgggagtc 960
gctgcgttgc cttcgccccg tgccccgctc cgcgccgcct cgcgccgccc gccccggctc 1020
tgactgaccg cgttactccc acaggtgagc gggcgggacg gcccttctcc tccgggctgt 1080
aattagcgct tggtttaatg acggctcgtt tcttttctgt ggctgcgtga aagccttaaa 1140
gggctccggg agggcccttt gtgcgggggg gagcggctcg gggggtgcgt gcgtgtgtgt 1200
gtgcgtgggg agcgccgcgt gcggcccgcg ctgcccggcg gctgtgagcg ctgcgggcgc 1260
ggcgcggggc tttgtgcgct ccgcgtgtgc gcgaggggag cgcggccggg ggcggtgccc 1320
cgcggtgcgg gggggctgcg aggggaacaa aggctgcgtg cggggtgtgt gcgtgggggg 1380
gtgagcaggg ggtgtgggcg cggcggtcgg gctgtaaccc ccccctgcac ccccctcccc 1440
gagttgctga gcacggcccg gcttcgggtg cggggctccg tgcggggcgt ggcgcggggc 1500
tcgccgtgcc gggcgggggg tggcggcagg tgggggtgcc gggcggggcg gggccgcctc 1560
gggccgggga gggctcgggg gaggggcgcg gcggccccgg agcgccggcg gctgtcgagg 1620
cgcggcgagc cgcagccatt gccttttatg gtaatcgtgc gagagggcgc agggacttcc 1680
tttgtcccaa atctggcgga gccgaaatct gggaggcgcc gccgcacccc ctctagcggg 1740
cgcgggcgaa gcggtgcggc gccggcagga aggaaatggg cggggagggc cttcgtgcgt 1800
cgccgcgccg ccgtcccctt ctccatctcc agcctcgggg ctgccgcagg gggacggctg 1860
ccttcggggg ggacggggca gggcggggtt cggcttctgg cgtgtgaccg gcggggttta 1920
tatcttccct tctctgttcc tccgcagcca gccatggcca ccatgggctc cagatcttct 1980
accaggatcc cagtacctct gatgctgacc gtccgaatca tgttggcact gagttgcgtc 2040
tgtccgacca gctcccttga tggcaggcct cttgcagctg cagggattgt ggtaacagga 2100
gacaaagcag tcaacatata cacctcatct cagacagggt caatcataat caagttactc 2160
ccaaatatgc ccaaggataa agaggcgtgt gcaaaagccc cattggaagc atacaacagg 2220
acattgacta ctttgctcac cccccttggt gattctatcc gtaggataca agagtctgtg 2280
accacatccg gaggagggaa acagggacgt cttataggcg ccattatcgg tggtgtagct 2340
ctcggggttg caaccgctgc acagataaca gcagcctcgg ctctgataca agccaatcaa 2400
aatgctgcca acatcctccg gctcaaagag agcattgctg caaccaatga ggctgtgcac 2460
gaggtcactg acggattatc acaactagca gtggcagttg ggaagatgca gcaatttgtt 2520
aatgaccagt ttaataaaac agctcaggaa ttggactgta taaaaattac acagcaggtt 2580
ggtgtagaac tcaacctgta cctaactgaa ttgactacag tattcgggcc acaaatcact 2640
tcccctgcct taactcagct gactatccag gcgctttaca atctagctgg tgggaatatg 2700
gattacttgt tgactaagtt aggtgtagga aacaaccaac tcagctcatt aattggtagt 2760
ggcctgatta ccggcaaccc tatcctgtac gactcacaga ctcaactctt gggtatacag 2820
gtcaccctac cctcagtcgg gaatctaaat aatatgcgtg ccacctacct ggaaaccttg 2880
tctgtaagta caaccaaagg atttgcctca gcacttgtcc caaaagtagt gacacaggtt 2940
ggttccgtga tagaagagct tgacacctcg tactgtatcg agaccgattt ggacctatat 3000
tgtacaagaa tagtgacatt ccctatgtct cctggtattt attcctgttt gagtggcaat 3060
acatctgctt gcatgtattc aaagactgaa ggcgcactca ctacgccgta tatgaccctc 3120
aaaggctcag ttattgccaa ctgtaagatg acaacatgta gatgtgcaga ccccccgggt 3180
atcatatcgc agaattatgg agaagctgtg tctctaatag ataggcaatc atgcaatatc 3240
ttatccttag acgggataac tttgaggctc agtggggaat ttgatgcaac ttatcaaaag 3300
aatatctcaa tacaagattc tcaagtaata gttacaggca atcttgacat ctcgactgag 3360
cttgggaatg tcaacaactc gataagtaat gctttggata agttagagga aagcaacagc 3420
aaactagaca aggtcaatgt taaactgacc agcacatccg ctcttattac ctatatcgtt 3480
ttaactgtca tatctcttgt atgtggtata cttagcctgg ttctagcatg ctacctgatg 3540
tacaagcaaa aggcgcaaca gaagaccttg ttgtggcttg ggaataatac cctagaccag 3600
atgagggcca ctacaaaaat gtagcttgat ctagagcggc cgcggggatc cagacatgat 3660
aagatacatt gatgagtttg gacaaaccac aactagaatg cagtgaaaaa aatgctttat 3720
ttgtgaaatt tgtgatgcta ttgctttatt tgtaaccatt ataagctgca ataaacaagt 3780
taacaacaac aattgcattc attttatgtt tcaggttcag ggggaggtgt gggaggtttt 3840
ttcggatcct ctagagtcga caattatttt atttaataac atatagccca aagacctcta 3900
tgaacattta gtttcccgta tactcaacgg cgcgtgtaca cacgcatctc tttgcatagc 3960
gatgaagttt gttcggcagc agaaaatgca gatatccaac aatctggaga aaacttatca 4020
tcacagtggc agtggaaaca taccccctct atattcatgg tataattatc gtctacagcg 4080
tccaggatag tggcgtgaga aaatggagat ctgcagccct cctttccatg gcatgccgct 4140
ttattgttca ttaaacgcac aatggtctca acgccagata tgggcataga ttctgaagaa 4200
cccgttgaca atccgaagaa gaaggcgtgc aggtctttgg aagactcgca cgttggtctt 4260
ataatgtatg atcgagatgt caccctaatg ccacatggta caggcttatc gcggtcatgg 4320
cgatcggact tgtaatttgc aacgatgggc aaaggatcga cgacatgcca aacattctga 4380
acccgtagag atgttaacga tgacgaggat gaatatccca tgctcgctgc cat 4433
<210> 40
<211> 59
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК праймера 1 для мутагенеза расположенного
выше 5’ гена Sbfi gfp
<400> 40
ggctagcgtt taaacttaag cttacctgca ggccaccatg gtgagcaagg gcgccgagc 59
<210> 41
<211> 59
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК праймера 2 для мутагенеза расположенного
выше 5’ гена Sbfi gfp
<400> 41
gctcggcgcc cttgctcacc atggtggcct gcaggtaagc ttaagtttaa acgctagcc 59
<210> 42
<211> 61
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК праймера 1 для мутагенеза расположенного
ниже 3’ гена Sbfi gfp
<400> 42
cacggcatgg atgagctgta caagtgacct gcaggtgtgc cttctagttg ccagccatct 60
g 61
<210> 43
<211> 61
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК праймера 2 для мутагенеза расположенного
ниже 3’ гена Sbfi gfp
<400> 43
cagatggctg gcaactagaa ggcacacctg caggtcactt gtacagctca tccatgccgt 60
g 61
<210> 44
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК более верхнего праймера UL55-ген 3
<400> 44
ccaccgggta tattttccac agc 23
<210> 45
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК нижнего праймера UL55-ген 3
<400> 45
gacccgacgc gatttcaga 19
<210> 46
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК верхнего праймера для ПЦР UL55-ген 3
<400> 46
actgccactg tgatgataag 20
<210> 47
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК нижнего праймера для ПЦР UL55-ген 3
<400> 47
ctcgctaaag aatgacctg 19
<210> 48
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
в пределах кодирующей области IBD VP2
<400> 48
ggccgacctc aactctccac tcaa 24
<210> 49
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
ниже в пределах HVT IBD № 1
<400> 49
ttcacagcgc ctcgttcaca ctta 24
<210> 50
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
выше соединения
сайта вставки трансферной плазмиды HVT IBD № 1
<400> 50
aatattcggg ggtgttagtc 20
<210> 51
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера в пределах
кодирующей области IBDV VP2
HVT IBD № 1
<400> 51
tgtggttggc atcagaag 18
<210> 52
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера в пределах
кодирующей области IBDV VP2
<400> 52
cggcgccatg aactacacaa aact 24
<210> 53
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
ниже сайта вставки UL35/36
HVT-IBD № 5
<400> 53
ggggcggaaa caaataaact ctcg 24
<210> 54
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера сайта вставки
UL35/36
HVT IBD № 5
<400> 54
actcggcggt aggggcattt g 21
<210> 55
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера в пределах
промотора hCMV HVT IBD
№ 5
<400> 55
gggtcgttgg gcggtcagc 19
<210> 56
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего промотора, расположенного
выше сайта интеграции
HVT IBD № 6a
<400> 56
tagactcggc ggtaggggca tttg 24
<210> 57
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
в пределах промотора pec
HVT IBD № 6a
<400> 57
gggggtcgtt gggcggtcag c 21
<210> 58
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
в пределах кодирующей области IBD VP2
HVT IBD № 6a
<400> 58
cggcgccatg aactacacaa aact 24
<210> 59
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
ниже сайта вставки UL35/36
<400> 59
ggggcggaaa caaataaact ctcg 24
<210> 60
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего промотора, расположенного
выше сайта интеграции
UL55/ген 3 для HVT IBD № 9
<400> 60
actgccactg tgatgataag 20
<210> 61
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
ниже сайта интеграции
UL55/ген для HVT IBD № 9
<400> 61
ctcgctaaag aatgacct 18
<210> 62
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для вставки ориентации верхнего праймера,
окружающего расположенные выше соединение вставки гена HVT IBD № 9 VP2
<400> 62
aatattcggg ggtgttagtc 20
<210> 63
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для вставки ориентации нижнего праймера,
расположенного в пределах
кодирующей области IBDV VP2 для HVT IBD № 9
<400> 63
ctcgcttctc agtgtatgtt tttc 24
<210> 64
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
ниже сайта, определяющего надлежащую
вставку кодирующей области IBDV VP2 для HVT IBDV № 9
<400> 64
atcctgagct cgctaaaaat cttg 24
<210> 65
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
ниже сайта, определяющего надлежащую
вставку кодирующей области IBDV VP2 для HVT IBDV № 9
<400> 65
cctcgcccgt ctacgctcca ct 22
<210> 66
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера для расположенной
выше области сайта интеграции
UL55-ген для HVT IBD № 30
<400> 66
tagcgacccg agtagga 17
<210> 67
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера для расположенной
выше области сайта интеграции
UL55-ген для HVT IBD № 30
<400> 67
cgcccacggt caaattgtat gtag 24
<210> 68
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера для подтверждения
надлежащей ориентации
вставки VP2, окружающей 3’ соединение сайта вставки
HVT IBD № 30
<400> 68
ttatccgcgc aatcagaagg 20
<210> 69
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера для подтверждения
надлежащей ориентации
вставки VP2, окружающей 3’ соединение сайта вставки
HVT IBD № 30
<400> 69
atggggcgga aacaaataaa ct 22
<210> 70
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера для подтверждения
надлежащей ориентации интеграции вставки VP2 за пределами кассеты экпрессии
HVT IBD № 30
<400> 70
ccaccgggta tattttccac agc 23
<210> 71
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера для подтверждения
надлежащей ориентации интеграции вставки VP2
за пределами кассеты экпрессии HVT IBD № 30
<400> 71
gacccgacgc gatttcaga 19
<210> 72
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера для подтверждения
надлежащей ориентации
вставки VP2, расположенной выше сайта интеграции UL35/36 HVT IBD № 31
№ 31
<400> 72
ctcggcggta ggggcatttg ataa 24
<210> 73
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера для подтверждения
надлежащей ориентации
вставки VP2, расположенной в пределах промотора бета-актина кур
HVT IBD № 31
<400> 73
agatggggag agtgaagcag aacg 24
<210> 74
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера для подтверждения
вставки VP2,
расположенной в пределах кодирующей области IBDV VP2
<400> 74
ttatccgcgc aatcagaagg 20
<210> 75
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера для подтверждения
надлежащей ориентации
вставки VP2, расположенной ниже сайта интеграции UL35/36 HVT
IBD № 31
<400> 75
atggggcgga aacaaataaa ct 22
<210> 76
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, нацеленного на расположенный
ниже
сайт интеграции вставки VP2 HVT IBD № 31, расположенный в пределах
вставки VP2
<400> 76
ttatccgcgc aatcagaagg 20
<210> 77
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
ниже сайта UL35/36
HVT IBD № 31
<400> 77
atggggcgga aacaaataaa ct 22
<210> 78
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера для расположенной
выше области сайта интеграции
ген 3-UL55 для HVT-IBD № 34
<400> 78
cctcgcccgt ctacgctcca ctga 24
<210> 79
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
в пределах промотора бета-актина кур
для HVT-IBD № 34
<400> 79
ccgcccccat cgctgcacaa aata 24
<210> 80
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
в пределах кодирующей области IBDV VP2
для HVT-IBD № 34
<400> 80
ggccctccgt cccgtcac 18
<210> 81
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
ниже сайта вставки ген 3-UL55
для HVT IBD № 34
<400> 81
ttcacagcgc ctcgttcaca ctta 24
<210> 82
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
за пределами кассеты экспрессии VP2
HVT IBD № 34
<400> 82
ctcgctaaag aatgacctg 19
<210> 83
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
за пределами кассеты экспрессии VP2
HVT IBD № 34
<400> 83
actgccactg tgatgataag 20
<210> 84
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера для расположенной
выше области сайта интеграции
UL35-UL36 HVT ND № 38
<400> 84
tgtttaaggc attttcaagt 20
<210> 85
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, который расположен
в пределах кодирующей области NDV F
HVT ND № 38
<400> 85
attcggacgg tcagcatca 19
<210> 86
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, окружающего 3’ соединение
вставки, расположенной в пределах кодирующей области NDV F HVT ND № 38
<400> 86
accagatgag ggccactaca aaaa 24
<210> 87
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
ниже сайта вставки UL35-UL36
HVT ND № 38
<400> 87
atggggcgga aacaaataaa ct 22
<210> 88
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
за пределами кассеты экспрессии
HVT ND № 38
<400> 88
gaggtccgtc gccacaa 17
<210> 89
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
за пределами кассеты экспрессии
HVT ND № 38
<400> 89
gcaagacaaa taacgcagac ta 22
<210> 90
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
выше области сайта интеграции
UL35-UL36 для HVT-ND № 39
<400> 90
ctcggcggta ggggcatttg ataa 24
<210> 91
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
в пределах промотора бета-актина кур
HVT-ND № 39
<400> 91
agatggggag agtgaagcag aacg 24
<210> 92
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, окружающего расположенное
ниже соединение
вставки, расположенное в пределах области poly A HVT-ND № 39
<400> 92
tggggatgcg gtgggctcta 20
<210> 93
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного ниже
сайта вставки UL35-UL36
HVT-ND № 39
<400> 93
ggggcggaaa caaataaact ctcg 24
<210> 94
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
за пределами кассеты экспрессии
HVT-ND № 39
<400> 94
ccaccgggta tattttccac agc 23
<210> 95
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
за пределами кассеты экспрессии
HVT-ND № 39
<400> 95
gacccgacgc gatttcaga 19
<210> 96
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
выше сайта интеграции
UL35/36 для HVT ND № 40
<400> 96
ctcggcggta ggggcatttg ataa 24
<210> 97
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
в пределах промотора бета-актина кур
для HVT ND № 40
<400> 97
agatggggag agtgaagcag aacg 24
<210> 98
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
в пределах кодирующей области NDVF
для HVT ND № 40
<400> 98
accagatgag ggccactaca aaaa 24
<210> 99
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
ниже соединения
сайта вставки для HVT ND № 40
<400> 99
atggggcgga aacaaataaa ctct 24
<210> 100
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
за пределами кассеты экспрессии
для HVT ND № 40
<400> 100
ccaccgggta tattttccac agc 23
<210> 101
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
за пределами кассеты экспрессии
для HVT № 40
<400> 101
gacccgacgc gatttcaga 19
<210> 102
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера для ПЦР-амплификации кассеты
для HVT ND № 42
<400> 102
tacttagtca taggcgcgcc aactccgccc gttttatgac tagaacca 48
<210> 103
<211> 49
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера для ПЦР-амплификации кассеты
для HVT ND № 42
<400> 103
tgacagtatc tagctagctc gactctagag gatccgaaaa aacctccca 49
<210> 104
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
за пределами кассеты для HVT
ND № 42
<400> 104
actgccactg tgatgataag 20
<210> 105
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
за пределами кассеты для HVT
ND № 42
<400> 105
ctcgctaaag aatgacctg 19
<210> 106
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
выше и за пределами кассеты экспрессии
для HVT ND № 42
<400> 106
ttcacagcgc ctcgttcaca ctta 24
<210> 107
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
в пределах кодирующей области ND F
для HVT ND № 42
<400> 107
ttcggacggt cagcatcaga ggta 24
<210> 108
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
выше и за пределами кассеты экспрессии
для HVT ND № 42
<400> 108
gcctcggcgt ggtagttctc 20
<210> 109
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
в пределах кодирующей области ND F
для HVT ND № 42
<400> 109
attcggacgg tcagcatca 19
<210> 110
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
выше и за пределами кассеты экспрессии
для HVT ND № 42
<400> 110
ttcacagcgc ctcgttcaca ctta 24
<210> 111
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
в пределах кодирующей области NDV F
для HVT ND № 42
<400> 111
gcttttgcac acgcctcttt atcc 24
<210> 112
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
выше и за пределами кассеты экспрессии
<400> 112
ttcacagcgc ctcgttcaca ctta 24
<210> 113
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
в пределах кодирующей области ND F
для HVT ND № 42
<400> 113
ttcggacggt cagcatcaga ggta 24
<210> 114
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
выше и за пределами кассеты экспрессии
для HVT ND № 42
<400> 114
accagatgag ggccactaca aaaa 24
<210> 115
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
в пределах кодирующей области ND F
для HVT ND № 42
<400> 115
cgcccgtcta cgctccactg a 21
<210> 116
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
выше UL55 для HVT
ND № 44
<400> 116
aatattcggg ggtgttag 18
<210> 117
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
в пределах промотора бета-актина кур
для HVT ND № 44
<400> 117
gggggagatg gggagagtga 20
<210> 118
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
выше UL55 для HVT
ND № 44
<400> 118
gcctcggcgt ggtagttctc 20
<210> 119
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
в пределах промотора бета-актина кур
для HVT ND № 44
<400> 119
ccgcccccat cgctgcacaa aata 24
<210> 120
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
в пределах последовательности, кодирующей ген NDV F,
для HVT ND № 44
<400> 120
accagatgag ggccactaca aaaa 24
<210> 121
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
ниже сайта вставки UL55-ген 3
для HVT ND № 44
<400> 121
ctcgcccgtc tacgctccac tgaa 24
<210> 122
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
за пределами кассеты экспрессии
для HVT ND № 44
<400> 122
actgccactg tgatgataag 20
<210> 123
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
за пределами кассеты экспрессии
для HVT № 44
<400> 123
ctcgctaaag aatgacctg 19
<210> 124
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
за пределами кассеты экспрессии
для HVT № 45
<400> 124
actgccactg tgatgataag 20
<210> 125
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
за пределами кассеты экспрессии
для HVT № 45
<400> 125
ctcgctaaag aatgacctg 19
<210> 126
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
выше и за пределами кассеты экспрессии
для HVT ND № 45
<400> 126
ctcgcccgtc tacgctccac tgaa 24
<210> 127
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
в пределах кодирующей области ND F
для HVT ND № 45
<400> 127
gcttttgcac acgcctcttt atcc 24
<210> 128
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
выше и за пределами кассеты экспрессии
для HVT ND № 45
<400> 128
cacgctgcta ttgtaacg 18
<210> 129
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
в пределах кодирующей области ND F
для HVT ND № 45
<400> 129
actgggatcc tggtagaag 19
<210> 130
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, окружающего расположенное
ниже соединение
сайта вставки для HVT ND № 45
<400> 130
accagatgag ggccactaca aaaa 24
<210> 131
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
ниже сайта вставки ген 3-UL55
<400> 131
ttcacagcgc ctcgttcaca ctta 24
<210> 132
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, окружающего расположенное
ниже соединение
сайта вставки для HVT ND № 45
<400> 132
gcacatccgc tcttattacc tat 23
<210> 133
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
ниже сайта
вставки ген 3-UL55
<400> 133
ttcacagcgc ctcgttcaca ctta 24
<210> 134
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
за пределами кассеты экспрессии
для HVT ND № 46
<400> 134
tagagggggt atgtttccac tgc 23
<210> 135
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
за пределами кассеты экспрессии
для HVT ND № 46
<400> 135
gtcataacat catctcgcta aag 23
<210> 136
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
выше и за пределами
сайта интеграции для HVT ND № 46
<400> 136
cacgctgcta ttgtaacg 18
<210> 137
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
в пределах промотора mCMV
для HVT ND № 46
<400> 137
ggtgaacgcc ccctatgtgg a 21
<210> 138
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
в пределах последовательности,
кодирующей ген NDV F, для HVT ND № 46
<400> 138
accagatgag ggccactaca aaaa 24
<210> 139
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
ниже и за пределами
кассеты экспрессии для HVT ND № 46
<400> 139
ttcacagcgc ctcgttcaca ctta 24
<210> 140
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
в пределах последовательности,
кодирующей ген NDV F, для HVT ND № 46
<400> 140
gcacatccgc tcttattacc tat 23
<210> 141
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
ниже и за пределами
кассеты экспрессии для HVT ND № 46
<400> 141
ttcacagcgc ctcgttcaca ctta 24
<210> 142
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
в пределах последовательности,
кодирующей ген NDV F, для HVT ND № 46
<400> 142
atgtggtata cttagcctgg ttc 23
<210> 143
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
ниже и за пределами
кассеты экспрессии для HVT ND № 46
<400> 143
aatattcggg ggtgttag 18
<210> 144
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
в пределах последовательности,
кодирующей ген NDV F, для HVT ND № 46
<400> 144
ggaataatac cctagaccag atg 23
<210> 145
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
ниже и за пределами
кассеты экспрессии для HVT ND № 46
<400> 145
gtcataacat catctcgcta aag 23
<210> 146
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
выше области
сайта интеграции ген 3-UL55 для HVT ND № 48
<400> 146
cctcgcccgt ctacgctcca ctga 24
<210> 147
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
в пределах промотора бета-актина кур
для HVT ND № 48
<400> 147
ccgcccccat cgctgcacaa aata 24
<210> 148
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, окружающего расположенное
ниже соединение сайта вставки для HVT ND № 48
<400> 148
accagatgag ggccactaca aaaa 24
<210> 149
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
ниже сайта вставки ген 3-UL55
для HVT ND № 48
<400> 149
ttcacagcgc ctcgttcaca ctta 24
<210> 150
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, окружающего расположенное
ниже соединение сайта вставки для HVT ND № 48
<400> 150
gcacatccgc tcttattacc tat 23
<210> 151
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного ниже
сайта вставки ген 3-UL55
для HVT ND № 48
<400> 151
ttcacagcgc ctcgttcaca ctta 24
<210> 152
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера, расположенного
за пределами кассеты экспрессии
для HVT ND № 48
<400> 152
actgccactg tgatgataag 20
<210> 153
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера, расположенного
за пределами кассеты экспрессии
для HVT ND № 48
<400> 153
ctcgctaaag aatgacctg 19
<210> 154
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера для амплификации кассеты
экспрессии на основе NDV F
HVT ND № 42 для HVT-IBD-ND № 42-№ 30
<400> 154
tacttagtca taggcgcgcc aactccgccc gttttatgac tagaacca 48
<210> 155
<211> 49
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера для амплификации кассеты
экспрессии на основе NDV F
HVT ND № 42 для HVT-IBD-ND № 42-№ 30
<400> 155
tgacagtatc tagctagctc gactctagag gatccgaaaa aacctccca 49
<210> 156
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для верхнего праймера для амплификации кассеты
экспрессии на основе гена IBD
плазмиды № 30 для HVT-IBD-ND № 42-№ 30
<400> 156
ctgtcagacg agtaccggtt tgacattgat tattgactag ttattaatag ta 52
<210> 157
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для нижнего праймера для амплификации кассеты
экспрессии на основе гена IBD
плазмиды № 30 для HVT-IBD-ND № 42-№ 30
<400> 157
tctagtactg atggtaccac catagagccc accgcatccc ca 42
<210> 158
<211> 570
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность ДНК для промотора LTR RSV
<400> 158
actctcagta caatctgctc tgatgccgca tagttaagcc agtatctgct ccctgcttgt 60
gtgttggagg tcgctgagta gtgcgcgagc aaaatttaag ctacaacaag gcaaggcttg 120
accgacaatt gcatgaagaa tctgcttagg gttaggcgtt ttgcgctgct tcgcgatgta 180
cgggccagat atacgcgtat ctgaggggac tagggtgtgt ttaggcgaaa agcggggctt 240
cggttgtacg cggttaggag tcccctcagg atatagtagt ttcgcttttg catagggagg 300
gggaaatgta gtcttatgca atactcttgt agtcttgcaa catggtaacg atgagttagc 360
aacatgcctt acaaggagag aaaaagcacc gtgcatgccg attggtggaa gtaaggtggt 420
acgatcgtgc cttattagga aggcaacaga cgggtctgac atggattgga cgaaccactg 480
aattccgcat tgcagagata ttgtatttaa gtgcctagct cgatacaata aacgccattt 540
gaccattcac cacattggtg tgcacctcca 570
<---


































Настоящее изобретение относится к биотехнологии и вирусологии. Предложен геном рекомбинантного вируса герпеса индеек (HVT), кодирующий и экспрессирующий антигенный полипептид патогенного агента, где указанный геном содержит одну или несколько нуклеотидных последовательностей, кодирующих один или несколько гетерологичных антигенов, вставленных в межгенные локусы UL 35/UL 36 в уникальной длинной (UL) области генома HVT. Предложен геном рекомбинантного вируса герпеса индеек (HVT), кодирующий и экспрессирующий антигенный полипептид патогенного агента, где указанный геном содержит одну или несколько нуклеотидных последовательностей, кодирующих один или несколько гетерологичных антигенов, вставленных в межгенные локусы UL 35/UL 36 в уникальной длинной области генома HVT и одну или несколько нуклеотидных последовательностей, кодирующих один или несколько гетерологичных антигенов, вставленных в сайт UL55/ген 3 уникальной длинной области (UL) генома HVT. Предложены: выделенная ДНК, кодирующая рекомбинантный геном HVT; иммуногенная композиция; вакцинная композиция; способ вакцинации птицы для лечения или предупреждения болезни Марека и одного или нескольких заболеваний птиц, вызванных одним или несколькими патогенами птиц, и способ индукции иммунного ответа у животного-птицы на вирус болезни Марека и один или несколько патогенов птиц. Изобретение позволяет получить вакцины для обеспечения иммунитета к нескольким патогенам одновременно. 8 н. и 52 з.п. ф-лы, 33 ил., 30 табл., 29 пр.
1. Геном рекомбинантного вируса герпеса индеек (HVT), кодирующий и экспрессирующий антигенный полипептид патогенного агента, где указанный геном содержит одну или несколько нуклеотидных последовательностей, кодирующих один или несколько гетерологичных антигенов, вставленных в межгенные локусы UL 35/UL 36 в уникальной длинной (UL) области генома HVT.
2. Геном рекомбинантного вируса герпеса индеек (HVT), кодирующий и экспрессирующий антигенный полипептид патогенного агента, где указанный геном содержит одну или несколько нуклеотидных последовательностей, кодирующих один или несколько гетерологичных антигенов, вставленных в межгенные локусы UL 35/UL 36 в уникальной длинной области генома HVT, и одну или несколько нуклеотидных последовательностей, кодирующих один или несколько гетерологичных антигенов, вставленных в сайт UL55/ген 3 уникальной длинной области (UL) генома HVT.
3. Рекомбинантный геном HVT по п. 1 или 2, отличающийся тем, что один или несколько указанных гетерологичных антигенов характеризуются защитным эффектом в отношении патогена (патогенов) птиц, выбранного (выбранных) из группы, состоящей из: вируса инфекционного бурсита (IBDV); вируса болезни Ньюкасла (NDV); вируса инфекционного бронхита (IBV); вируса инфекционного ларинготрахеита (ILTV); вируса анемии цыплят (CAV); и вируса птичьего гриппа (AIV).
4. Рекомбинантный геном HVT по любому из предыдущих пунктов, отличающийся тем, что один или несколько указанных гетерологичных антигенов выбраны из группы, состоящей из:
a) белков VP2, VP3 или VP4 вируса инфекционного бурсита (IBDV);
b) белков VP1 или VP2 вируса анемии цыплят (CAV);
c) химерного белка F/HN или белка F, NP, Р, М, HN или L вируса болезни Ньюкасла (NDV);
d) белков S1, S2 или М вируса инфекционного бронхита (IBV);
e) белков gB, gC, gD, gE, gH, gl или gL вируса инфекционного ларинготрахеита (ILTV); и
f) любого из белков НА, NA, NP или М вируса птичьего гриппа (AIV).
5. Рекомбинантный геном HVT по любому из предыдущих пунктов, отличающийся тем, что один или несколько указанных гетерологичных антигенов характеризуются защитным эффектом в отношении IBDV.
6. Рекомбинантный геном HVT по п. 5, отличающийся тем, что один или несколько указанных гетерологичных антигенов представляют собой белок VP2 IBDV.
7. Рекомбинантный геном HVT по п. 6, отличающийся тем, что указанная последовательность белка VP2 кодируется нуклеотидной последовательностью, на по меньшей мере 80% идентичной нуклеотидной последовательности, содержащей SEQ ID NO. 5 или SEQ ID NO. 10.
8. Рекомбинантный геном HVT по п. 6, отличающийся тем, что указанный белок VP2 кодируется нуклеотидной последовательностью, содержащей SEQ ID NO. 5 или SEQ ID NO. 10.
9. Рекомбинантный геном HVT по любому из пп. 1-4, отличающийся тем, что один или несколько гетерологичных антигенов характеризуются защитным эффектом в отношении вируса болезни Ньюкасла (NDV).
10. Рекомбинантный геном HVT по п. 9, отличающийся тем, что один или несколько указанных гетерологичных антигенов представляют собой белок F NDV.
11. Рекомбинантный геном HVT по п. 10, отличающийся тем, что указанный белок F NDV кодируется нуклеотидной последовательностью, на по меньшей мере 80% идентичной последовательности, содержащей SEQ ID NO. 3.
12. Рекомбинантный геном HVT по п. 10, отличающийся тем, что указанный белок F NDV кодируется нуклеотидной последовательностью, содержащей SEQ ID NO. 3.
13. Рекомбинантный геном HVT по любому из предыдущих пунктов, отличающийся тем, что один или несколько указанных гетерологичных антигенов оказывают защитный эффект в отношении NDV и IBDV.
14. Рекомбинантный геном HVT по п. 13, отличающийся тем, что по меньшей мере один указанный гетерологичный антиген содержит белок F NDV и белок VP2 IBDV.
15. Рекомбинантный геном HVT по п. 14, отличающийся тем, что указанный белок F NDV кодируется нуклеотидной последовательностью, на по меньшей мере 80% идентичной нуклеотидной последовательности, содержащей SEQ ID NO. 3, и указанный белок VP2 IBDV кодируется нуклеотидной последовательностью, на по меньшей мере 80% идентичной нуклеотидным последовательностям, содержащим SEQ ID NO. 5 или SEQ ID NO. 10.
16. Рекомбинантный геном HVT по п. 14 или 15, отличающийся тем, что указанный белок F NDV кодируется нуклеотидной последовательностью, содержащей SEQ ID NO. 3, а белок VP2 из IBDV кодируется нуклеотидными последовательностями, содержащими SEQ ID NO. 5 или SEQ ID NO. 10.
17. Рекомбинантный геном HVT по любому из предыдущих пунктов, содержащий геном, содержащий одну или несколько кассет экспрессии, содержащих одну или несколько нуклеотидных последовательностей, которые кодируют один или несколько гетерологичных антигенов.
18. Рекомбинантный геном HVT по п. 17, отличающийся тем, что указанная кассета экспрессии содержит один или несколько промоторов.
19. Рекомбинантный геном HVT по п. 18, отличающийся тем, что один или несколько указанных промоторов выбраны из группы, состоящей из: немедленно раннего промотора цитомегаловируса человека (hCMV); немедленно раннего промотора CMV морской свинки; немедленно раннего промотора CMV мыши; промотора Рес; промотора β-актина кур; промотора SV40; промоторов гликопротеина X вируса псевдобешенства; промотора вируса простого герпеса-1 альфа 4; промоторов гликопротеинов gA, gC, gB, gE или gI вируса болезни Марека; промоторов гликопротеина gB, gE, gI, gD вирусов инфекционного ларинготрахеита; и промотора VP8 вируса герпеса крупного рогатого скота 1/1.
20. Рекомбинантный геном HVT по п. 19, отличающийся тем, что один или несколько указанных промоторов содержат промотор CMV человека.
21. Рекомбинантный геном HVT по п. 19, отличающийся тем, что один или несколько указанных промоторов содержат промотор CMV мыши.
22. Рекомбинантный геном HVT по п. 19, отличающийся тем, что один или несколько указанных промоторов содержат промотор hCMV и mCMV.
23. Выделенная ДНК, кодирующая рекомбинантный геном HVT по любому из пп. 1 и 3-22.
24. Выделенная ДНК, кодирующая рекомбинантный геном HVT по любому из пп. 2-22.
25. Иммуногенная композиция, содержащая рекомбинантный HVT по любому из пп. 1-22, дополнительно содержащая фармацевтически приемлемый носитель, вспомогательное вещество или адъювант.
26. Вакцинная композиция, содержащая рекомбинантный HVT по любому из пп. 1-22, дополнительно содержащая фармацевтически приемлемый носитель, вспомогательное вещество или адъювант.
27. Вакцинная композиция по п. 26, дополнительно содержащая дополнительный вирус болезни Марека (MDV), выбранный из группы, состоящей из: естественно аттенуированного штамма MDV-1 Rispens (CVI-988); или вируса герпеса Gallid 3 штамма SB-1.
28. Вакцинная композиция по п. 27, отличающаяся тем, что указанный дополнительный MDV содержит рекомбинантный геном.
29. Вакцинная композиция по п. 28, отличающаяся тем, что указанный геном рекомбинантного MDV содержит одну или несколько нуклеотидных последовательностей, кодирующих один или несколько гетерологичных антигенов, которые оказывают защитный эффект в отношении одного или нескольких патогенов птиц.
30. Вакцинная композиция по любому из пп. 26-29 для применения при вакцинации птицы против одного или нескольких заболеваний, вызванных одним или несколькими патогенами птиц.
31. Вакцинная композиция по любому из пп. 26-29 для применения для защиты птицы от клинических симптомов, вызванных одним или несколькими патогенами птиц.
32. Вакцинная композиция по любому из пп. 26-29 для применения для защиты птицы от клинических симптомов, вызванных вирусом болезни Марека, и клинических симптомов, вызванных одним или несколькими патогенами птиц.
33. Вакцинная композиция по любому из пп. 30-32, отличающаяся тем, что один или несколько указанных патогенов птиц выбраны из группы, состоящей из: вируса инфекционного бурсита (IBDV); вируса болезни Ньюкасла (NDV); вируса инфекционного бронхита (IBV); вируса инфекционного ларинготрахеита (ILTV); вируса анемии цыплят (CAV); и вируса птичьего гриппа (AIV).
34. Вакцинная композиция по п. 33, отличающаяся тем, что один или несколько указанных патогенов птиц включают вирус болезни Ньюкасла.
35. Вакцинная композиция по п. 33, отличающаяся тем, что один или несколько указанных патогенов птиц включают вирус инфекционного бурсита (IBDV).
36. Вакцинная композиция по п. 33, отличающаяся тем, что один или несколько указанных патогенов птиц включают вирус болезни Ньюкасла и вирус инфекционного бурсита (IBDV).
37. Вакцинная композиция по любому из пп. 26-36 для применения в вакцинации птиц, при этом вакцинную композицию вводят посредством по меньшей мере одного или нескольких введений вакцинной композиции путем распыления, введения in ovo, подкожного введения, внутримышечного введения, перорального введения, назального введения или их комбинации.
38. Вакцинная композиция по п. 37, отличающаяся тем, что указанную вакцинную композицию вводят путем введения in ovo.
39. Вакцинная композиция по п. 38, отличающаяся тем, что указанное введение in ovo происходит в яйцеклетку с развивающимся эмбрионом в период между от около 16 до 22 дней развития.
40. Вакцинная композиция по любому из пп. 37-39, отличающаяся тем, что указанное введение in ovo происходит в яйцеклетку с развивающимся эмбрионом в период около 18 дней развития.
41. Вакцинная композиция по любому из пп. 37-40, отличающаяся тем, что указанное введение вакцинной композиции включает введение in ovo с последующим введением спрея.
42. Вакцинная композиция по п. 37, отличающаяся тем, что указанное введение вакцинной композиции включает введение спрея.
43. Способ вакцинации птицы для лечения или предупреждения болезни Марека и одного или нескольких заболеваний птиц, вызванных одним или несколькими патогенами птиц, включающий стадию введения эффективного количества вакцинной композиции по пп. 26-36.
44. Способ по п. 43, отличающийся тем, что один или несколько указанных патогенов птиц выбраны из группы, состоящей из вируса инфекционного бурсита (IBDV); вируса болезни Ньюкасла (NDV); вируса инфекционного бронхита (IBV); вируса инфекционного ларинготрахеита (ILTV); вируса анемии цыплят (CAV); и вируса птичьего гриппа (AIV).
45. Способ по п. 44, отличающийся тем, что один или несколько указанных патогенов птиц включают вирус инфекционного бурсита (IBDV).
46. Способ по п. 44, отличающийся тем, что один или несколько указанных патогенов птиц включают вирус болезни Ньюкасла (NDV).
47. Способ по п. 44, отличающийся тем, что один или несколько указанных патогенов птиц включают вирус инфекционного бурсита (IBDV) и вирус болезни Ньюкасла (NDV).
48. Способ индукции иммунного ответа у животного-птицы на вирус болезни Марека и один или несколько патогенов птиц, включающий стадию введения птице эффективного количества иммуногенной композиции по п. 25 или вакцинной композиции по любому из пп. 26-36.
49. Способ по п. 48, отличающийся тем, что один или несколько указанных патогенов птиц выбраны из группы, состоящей из вируса инфекционного бурсита (IBDV); вируса болезни Ньюкасла (NDV); вируса инфекционного бронхита (IBV); вируса инфекционного ларинготрахеита (ILTV); вируса анемии цыплят (CAV); и вируса птичьего гриппа (AIV).
50. Способ по п. 49, отличающийся тем, что один или несколько указанных патогенов птиц включают вирус инфекционного бурсита (IBDV).
51. Способ по п. 49, отличающийся тем, что один или несколько указанных патогенов птиц включают вирус болезни Ньюкасла (NDV).
52. Способ по п. 49, отличающийся тем, что один или несколько указанных патогенов птиц включают вирус инфекционного бурсита (IBDV) и вирус болезни Ньюкасла (NDV).
53. Способ по любому из пп. 43-52, отличающийся тем, что указанное введение осуществляется путем распыления, введения in ovo, подкожного введения, внутримышечного введения, перорального введения или назального введения.
54. Способ по п. 53, отличающийся тем, что указанный путь введения включает введение in ovo.
55. Способ по п. 54, отличающийся тем, что указанное введение in ovo происходит в яйцеклетку с развивающимся эмбрионом в период между от около 16 до 22 дней развития.
56. Способ по любому из пп. 53-55, отличающийся тем, что указанное введение in ovo происходит в яйцеклетку с развивающимся эмбрионом в период около 18 дней развития.
57. Способ по пп. 53-56, отличающийся тем, что указанный путь введения включает введение in ovo с последующим введением спрея.
58. Способ по п. 53, отличающийся тем, что указанный путь введения включает введение спрея.
59. Способ по любому из пп. 45-58, отличающийся тем, что указанная птица выбрана из группы, состоящей из курицы, индейки, гуся, утки, фазана, страуса, голубя и перепела.
60. Способ по п. 59, отличающийся тем, что указанная птица представляет собой курицу.
| Токарный резец | 1924 |
|
SU2016A1 |
| Способ получения цианистых соединений | 1924 |
|
SU2018A1 |
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| Станок для придания концам круглых радиаторных трубок шестигранного сечения | 1924 |
|
SU2019A1 |
| Автомобиль-сани, движущиеся на полозьях посредством устанавливающихся по высоте колес с шинами | 1924 |
|
SU2017A1 |
| Способ получения цианистых соединений | 1924 |
|
SU2018A1 |
| РЕКОМБИНАНТНЫЕ ВЕКТОРЫ HVT, ЭКСПРЕССИРУЮЩИЕ АНТИГЕНЫ ПАТОГЕНОВ ПТИЦ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2620936C2 |

