Сокращения и условные обозначения
БСА – бычий сывороточный альбумин;
Готовые реакционные смеси – заранее подготовленные смеси компонентов, необходимых для проведения стадии амплификации нуклеиновых кислот, обычно предназначенные для решения рутинных исследовательских задач;
ДTT – дитиотреитол;
ДНК – дезоксирибонуклеиновая кислота;
дц – двуцепочечная;
мяРНК – малая ядерная РНК;
НК – нуклеиновая кислота (ДНК или РНК);
ОТ – обратная транскрипция;
ОТ-ПЦР – ПЦР в сочетании с обратной транскрипцией;
п.н. – пары нуклеотидов;
Праймер – олигомер, состоящий из звеньев частично или полностью нуклеотидной природы, содержащий фрагменты структуры, обеспечивающие узнавание ДНК-матрицы по принципу комплементарного взаимодействия и начало полимеразной реакции, катализируемой ДНК-полимеразой, и удлиняемый с 3’-конца хотя бы на одно дополнительное мономерное звено;
ПЦР – полимеразная цепная реакция;
ревертаза – РНК-зависимая ДНК-полимераза;
РНК – рибонуклеиновая кислота;
Трис – трис(гидроксиметил)аминометан;
ФГ – фосфорилгуанидиновая группа;
ФГ-олигонуклеотиды – производные олигонуклеотидов, содержащие одну или более фосфатных групп, в которых на атоме фосфора введен остаток гуанидина или замещенного гуанидина;
Фрагмент Кленова – часть бактериальной ДНК-полимеразы I E.coli с полимеразной и 3'-5'-экзонуклеазной активностями и отсутствующей 5'-3'-экзонуклеазной активностью;
Ct – пороговый цикл;
GFP – зеленый флуоресцентный белок (от англ. green fluorescent protein);
keff – коэффициент амплификации;
LNA – «замкнутая» нуклеиновая кислота (от англ. «locked nucleic acid»);
ROX, BHQ2, FAM, TAMRA – флуоресцентные красители;
RCA – реакция амплификации катящегося кольца;
SD – стандартное отклонение;
SYBR Green I, SYTO-13 – интеркалирующие флуоресцентные красители;
KRAS - протоонкоген, представитель семейства белков Ras.
Изобретение относится к области молекулярной биологии и молекулярной диагностики. Предметом изобретения является использование новых производных олигонуклеотидов, а именно фосфорилгуанидинов, содержащих одну или более фосфатных групп, несущих на атоме фосфора остаток гуанидина или замещенного гуанидина, в качестве праймеров-затравок при проведении полимеразной цепной реакции (ПЦР) и ПЦР в сочетании с обратной транскрипцией.
1. Олигонуклеотиды широко используют в качестве праймеров для проведения ПЦР, позволяющей увеличивать копийность фрагмента ДНК, границы которого определяются нуклеотидной последовательностью праймеров-затравок (Saiki R.K., Scharf S., Faloona F., Mullis K.B., Horn G.T., Erlich H.A., Arnheim N. Enzymatic amplification of beta-globin genomic sequences and restriction site analysis for diagnosis of sickle cell anemia // Science. – 1985. – V. 230. – P. 1350-1354), (Mullis K.B., Faloona F.A. Specific synthesis of DNA in vitro via a polymerase-catalyzed chain reaction // Methods Enzymol. – 1987. – V. 155. – Р. 335-350). Структура праймеров в значительной степени определяет эффективность протекания ПЦР, что заставляет выбирать их последовательность в рамках рационального дизайна и в строгом соответствии с набором установленных критериев. Наиболее часто в качестве праймеров-затравок используют нативные олигодезоксирибонуклеотиды. Помимо нативных олигонуклеотидов в качестве праймеров предложен ряд олигонуклеотидных производных, в состав которых модифицированные фрагменты вводят с целью повышения эффективности ПЦР. Например, к числу таких олигонуклеотидных производных относят производные, содержащие нуклеотидные звенья на основе «замкнутых» нуклеиновых кислот (LNA) (Ballantyne K.N., van Oorschot R.A., Mitchell R.J. Locked nucleic acids in PCR primers increase sensitivity and performance // Genomics. – 2008. – V. 91. – P. 301-305.) или фосфодиэфира олигоэтиленгликоля (Brent C. Satterfield cooperative primers : 2.5 million–fold improvement in the reduction of nonspecific amplification // J. Mol. Diagn. – 2014. – V. 16. – P. 163-173) в составе так называемых «кооперативных» праймеров. Кроме того, наличие указанных модификаций значительно влияет на температуру плавления формируемых комплементарных комплексов между ДНК-матрицей и модифицированным олигонуклеотидом-праймером: LNA-звенья повышают, в то время как гексаэтиленгликоль снижает. Помимо изменения структуры углеводно-фосфатного остова, альтернативными вариантами, повышающими эффективность ПЦР, являются модификации гетероциклических азотистых оснований (Bodepudi V., Schoenbrunner N.J., Will S. Methods and reagents for reducing non-specific amplification // WO 2013091835 A1, 27 June 2013), введение группировок ненуклеотидной природы, повышающих стабильность комплексов праймера с амплифицируемой ДНК (Schneider U.V., Mikkelsen N.D., Lindqvist A., Okkels L.M., Jøhnk N., Lisby G. Improved efficiency and robustness in qPCR and multiplex end-point PCR by twisted intercalating nucleic acid modified primers // PLoS One. – 2012. – V. 7. - e38451). Другим способом повышения точности ПЦР является использование в качестве праймеров олигонуклеотидных производных с временной защитой 3′-концевого фрагмента, удаляемой под действием химических агентов или физических стимулов – света или температуры (Wanli Bi. Nucleic acid amplification using a reversibly modified oligonucleotide // US 8,334,099 B2. 18 December 2012), (Lebedev A.V., Paul N., Yee J., Timoshchuk V.A., Shum J., Miyagi K., Kellum J., Hogrefe R.I., Zon G. Hot start PCR with heat-activatable primers: a novel approach for improved PCR performance // Nucleic Acids Res. – 2008. – V. 36. - e131). Недостатком таких вышеописанных типов праймеров на основе производных олигонуклеотидов является необходимость предварительного получения дополнительных мономеров–модификаторов, которые затем должны быть использованы в ходе автоматического олигонуклеотидного синтеза. Полностью незаряженные производные олигонуклеотидов такие, как морфолиновые и пептидилнуклеотидные не используют в качестве праймеров, поскольку их углеводофосфатный остов не узнается ферментами (Summerton J., Stein D., Huang S.B., Matthews P., Weller D., Partridge M. Morpholino and phosphorothioate antisense oligomers compared in cell-free and in-cell systems // Antisense Nucleic Acid Drug Dev. – 1997. – V. 7. – P. 63-70.) и их синтез достаточно трудоемок (время- и ресурсозатратен). Частично незаряженные олигонуклеотиды, например, содержащие остатки фосфотриэфиров, то есть несущих остаткок алифатического спирта вместо одного из атомов кислорода, могут быть использованы в качестве праймеров при амплификации ДНК, однако их получение требует использования специальных фосфороамидных нуклеотидных мономеров и нестандартных условий пост-синтетического деблокирования и выделения синтезированных олигонуклеотидов (Chan H.W.-H., Yang Y.-S., Chen W.-Y. Partially neutral single-stranded oligonucleotide // US 20170015699 A1, January 2017). До сих пор не описано примера широкого применения частично незаряженных олигонуклеотидов в реакционных смесях с различным температурным режимом удлинения праймеров и амплификации фрагмента ДНК с применением широкого набора ДНК-полимераз.
Наиболее близким к предлагаемому способу – прототипом, является способ амплификации в присутствии модифицированных праймеров олигонуклеотидов, содержащих заместители при атоме фосфора в составе межнуклеотидных фосфатных группировок, вводимых вместо атома кислорода, а именно олигонуклеотиды с фосфоротиоатными остатками (Robinson P.S., Holme J., Jain N. Polymerase chain reaction detection system using oligonucleotides comprising a phosphorothioate group // WO2013140107 A1, 26 September 2013). Праймеры с таким типом модификации остова могут быть получены с использованием стандартных нуклеотидных мономеров, но специальных окисляющих агентов. Недостатком прототипа является ограниченность условий использования такой системы, заключающаяся в следующем:
1. фосфоротиоатсодержащие праймеры используют только при выявлении ДНК;
2. в прототипе заявлена полимераза, лишенная 3′→5′ экзонуклеазной активности, т.е. применение полимеразы с корректирующей активностью не будет приводить к уменьшению неспецифической амплификации;
3. полимераза прототипа относится к термофильному классу и температурные режимы удлинения лежат в диапазоне 55-61°C;
4. условия проведения реакции амплифицирования термоциклические;
5. необходимость введения 5’-концевой метки в модифицированный олигонуклеотид, используемый для детекции ДНК-продукта.
Ключевым отличием от аналогов представленного изобретения является включение в структуру олигонуклеотидного праймера одной или более фосфорилгуанидиновых группировок. Введение гуанидиновой группы вместо одного из атомов кислорода значительно увеличивает объем заместителя при атоме фосфора и делает ФГ незаряженной в физиологических условиях. Размер используемой модификации является существенным отличием от упомянутых фосфоротиоатсодержащих олигонуклеотидов.
Использование праймеров с фосфорилгуанидиновыми группировками (Stetsenko D., Kupryushkin M., Pyshnyi D. Modified oligonucleotides and methods for their synthesis // WO 2016028187 A1, publication date 25 February 2016), Купрюшкин М.С., Пышный Д.В., Стеценко Д.А.. Фосфорилгуанидины. Новый класс аналогов нуклеиновых кислот // Acta naturae. – 2014. – Т. 6. - № 4 (23). – С. 53-55), (Kuznetsov N.A., Kupryushkin M.S., Abramova T.V., Kuznetsova A.A., Miroshnikova A.D., Stetsenko D.A., Pyshnyi D.V, Fedorova O.S. New Oligonucleotide Derivatives as Unreactive Substrate Analogues and Potential Inhibitors of Human Apurinic/Apyrimidinic Endonuclease APE1 // Molecular BioSystems. – 2016. – V. 12. –N 1. – P. 67-75) по сравнению с аналогами сочетает в себе ряд преимуществ, комбинация которых обеспечивает достижение технического результата:
1. их синтез не требует включения в синтетический регламент дополнительных амидофосфитных мономеров предшественников и проводится на стандартном автоматическом ДНК/РНК синтезаторе;
2. число и положение фосфорилгуанидиновых групп (ФГ) в праймере задается и строго контролируется, что позволяет получать как полностью модифицированные производные, так и производные с частично замещенными фосфодиэфирными остатками;
3. наличие ФГ не приводит к значительному изменению температуры плавления ДНК/ДНК комплексов в условиях умеренной ионной силы растворов;
4. введение гуанидиновой группы вместо одного из атомов кислорода значительно увеличивает объем заместителя при атоме фосфора и делает ФГ незаряженной в физиологических условиях;
5. введение ФГ в состав олигонуклеотида приводит к его химической устойчивости в широком диапазоне рН и устойчивости к действию нуклеаз;
6. фосфорилгуанидины выступают субстратами для широкого набора ДНК-полимераз как мезофильных, так и термофильных вне зависимости от наличия 3′→5′/5′→3′ экзонуклеазной активности;
7. праймеры с фосфорилгуанидиновыми группировками могут быть использованы при различных температурах (температурных протоколах) проведения полимеразной реакции;
8. применение праймера с фосфорилгуанидиновыми группировками предполагает простой алгоритм дизайна структуры праймера, не требующий дополнительных критериев, кроме положения и числа модификаций, по сравнению со стандартными олигонуклеотидными праймерами;
9. изобретение обеспечивает возможность выявления как ДНК в реакции ПЦР, так и РНК в реакции ОТ с последующей ПЦР с повышенной специфичностью и чувствительностью за счет снижения выхода неспецифичных продуктов реакции.
Технической задачей изобретения является повышение достоверности, чувствительности и специфичности выявления анализируемых последовательностей НК, а также упрощение способа матричного синтеза ДНК. Поставленная техническая задача достигается тем, что при амплификации нуклеиновых кислот применяют праймер, содержащий в своем составе от одной до 100 % межзвенных фосфорилгуанидиновых групп. Общая структура праймера представлена на Фиг. 1А, где Z – фосфорилгуанидиновая группа. Каждый из заместителей R1, R2, R3 и R4 может быть атомом водорода Н или опционально замещенным органическим радикалом. Каждый из заместителей R1, R2, R3 и R4 независимо выбирается из группы, включающей H, C1-10 алкил, C2 10 алкенил, C2-10 алкинил, –C6-10 арил или –C5-10 гетероарил; где каждый алкил, алкенил, алкинил, арил, гетероарил, алкилен или гетероалкилен может быть дополнительно замещён; где дополнительно R1 и R2, вместе с атомом, с которым они связаны, образуют 5-8-членный гетероцикл; В некоторых применениях R1 и R2 вместе с атомом, с которым они связаны, образуют 5-8-членный гетероцикл, предпочтительно пирролидин, пиперидин, пиперазин, морфолин. Предпочтительно, R1 и R2 вместе с атомом, с которым они связаны, образуют 5-членный гетероцикл, предпочтительно пирролидин. В некоторых применениях R2 и R3 вместе образуют алкиленовую или гетероалкиленовую цепь длиной 2-4 атома, а R1 и R4 каждый независимо выбираются из группы, включающей H и C1 4 алкил. В некоторых применениях R2 и R3 вместе составляют цепь CH2 CH2 , а R1 и R4 есть –H или метил. Часть предпочтительных соединений, охватываемых настоящим изобретением, соответствуют формулам ФГ, несущих остаток гуанидина (g”), N,N,N’,N’-тетраметилгуанидина (g’), 1,3-диметилимидазолидин-2-имина (g), N,N’-бис(тетраметилен)гуанидина (g”’), приведённым на Фиг. 1Б. В качестве ферментов амплификации использованы полимеразы как лишенные, так и имеющие 3′→5′/5′→3′ нуклеазную активность, в условиях термоциклического и изотемпературного режима. Настоящее изобретение основано на следующих критериях пригодности олигонуклеотидов в качестве праймеров-затравок: замена отрицательно заряженных фосфатных групп нейтральными группами должна влиять на способность олигонуклеотида удлиняться ДНК- или РНК-зависимыми ДНК-полимеразами, а также на его способность выступать в качестве фрагмента ДНК-матричной цепи при синтезе комплементарной цепи, катализируемом ДНК-зависимой ДНК-полимеразой; структурное разнообразие, позволяющее варьировать набор дополнительных модификаций праймера при необходимости.
В данном изобретении раскрыты свойства фосфорилгуанидиновых олигонуклеотидов, которые соответствуют вышеизложенным критериям и могут быть использованы для создания улучшенных систем детекции и количественного определения нуклеиновых кислот, основанных на ПЦР и ОТ-ПЦР.
Изобретение подробнее проиллюстрировано ниже следующими примерами конкретного выполнения, которые не ограничивают объем изобретения. Многочисленные варианты осуществления изобретения в объеме формулы изобретения, которые вытекают из примеров, должны быть очевидны специалистам в данной области техники на основе вышеприведенного описания и последующих примеров. Пригодность конкретной группы, их комбинации и положение в составе олигонуклеотидов, используемых в качестве праймеров в реакциях матричного синтеза ДНК, определяют эмпирически самостоятельно специалистом в данной области техники.
Пример. 1 (описание используемых ниже систем)
ФГ-олигонуклеотиды и немодифицированные олигодезоксирибонуклеотиды представлены на Фиг. 2. Олигонуклеотиды содержали от 8 до 40 нуклеотидных звеньев. Фосфорилгуанидиновые группы (ФГ) представлены в виде немодифицированного (g”), разветвленного (g’), замкнутого (g) остатка и гуанидинового остатка с объемными заместителями (g”’) (Фиг. 1Б). Количество ФГ в составе олигонуклеотидов варьировали от одной группы до 100 % (Фиг. 2).
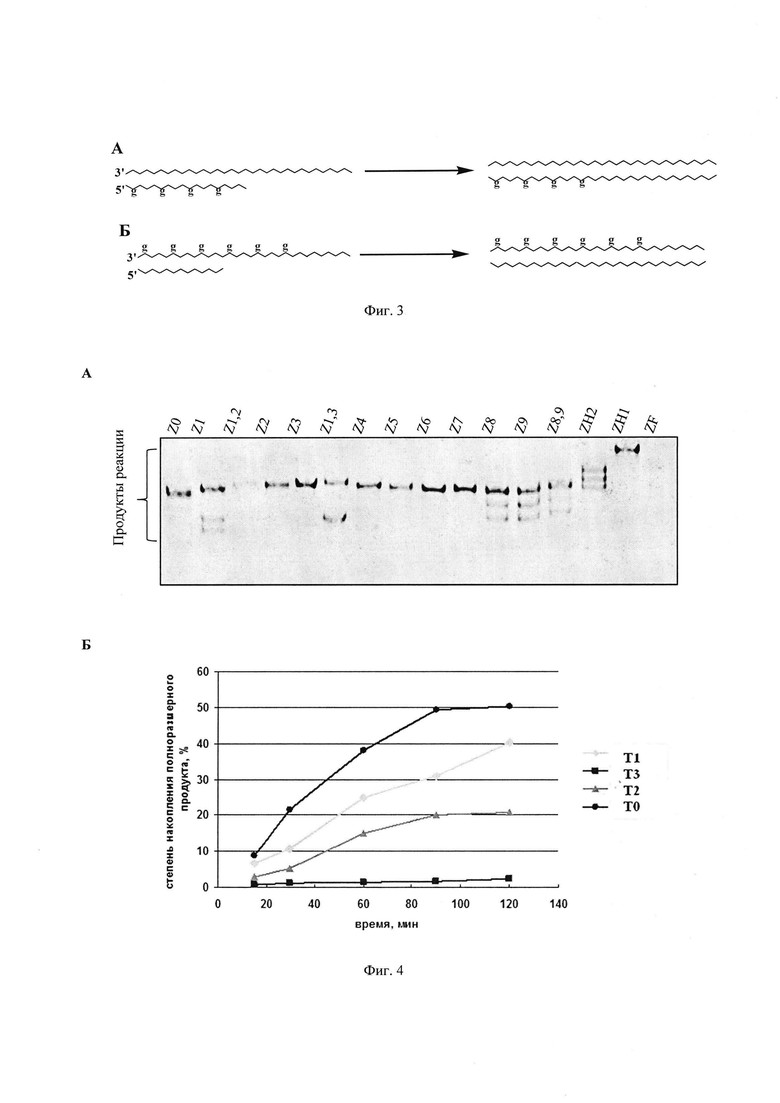
ФГ-олигонуклеотиды выступали в качестве олигонуклеотидов-праймеров (Фиг. 3А) или матриц (Фиг. 3Б), содержали флуоресцентную (* – FAM или TAMRA) или радиоактивную метку ([32P]).
В качестве матриц использовали нативные и ФГ-олигонуклеотиды (Фиг. 2), плазмидную ДНК, содержащую ген eGFP; препараты цельной крови и плазмы крови человека, содержащие предварительно добавленную плазмидную ДНК; РНК вируса гепатита С (ВГС); суммарную РНК клеток аденокарциномы молочной железы человека MCF-7; плазмиду pUC19; геномную ДНК-человека.
В качестве ДНК-полимераз использовали мезофильные и термофильные ферменты с и без 3′→5′/5′→3′ экзонуклеазной активности: Taq ДНК-полимеразу, ДНК-полимеразу фага Т5, ДНК полимераза I E. coli (Фрагмент Кленова), Pfu ДНК-полимеразу, РНК-зависимые ДНК-полимеразы (ревертазы) MMLV и HIV-p66; ДНК полимеразу phi29.
Пример. 2 (ФГ-олигонуклеотид как праймер в реакции, катализируемой термостабильной полимеразой, в термоциклическом (1) и изотермическом (2) режимах)
(1) Для демонстрации применения ФГ-олигонуклеотидов как праймеров (система представлена на Фиг. 3А) реакционные смеси (10 мкл) содержали: нативный олигонуклеотид М30 (10 мкM); немодифицированный олигонуклеотид Z0 либо ФГ-олигонуклеотид Z1чZН1 (10 мкM, последовательности праймеров Z0 ч ZF представлены на Фиг. 2); набор трифосфатов (по 0,1 мM каждый) dATP, dCTP, dGTP и флуоресцентно-меченный dUTP (флуоресцеин-6-аминотиокарбонил-[5-(3-аминоаллил)-2′-дезоксиуридин-5′–трифосфат]); 1,8 мМ MgCl2, Трис-HCl (10 мМ) pH 8,8, KCl (50 мМ); 0,1 % Tween-20, Taq ДНК-полимеразу (2 ед.акт.). Реакцию проводили в термоциклическом режиме: 95°C - 10 с, 61°C - 10 с, 72°C - 30 с (32 цикла). Через 25 минут реакцию останавливали добавлением 2% раствора перхлората лития в ацетоне. Все образцы разделяли с помощью электрофоретического анализа в 20% денатурирующем полиакриламидном геле, результат электрофоретического анализа фиксировали флуоресцентным сканером (Фиг. 4А). Эффективность удлинения ФГ-олигонуклеотидов на матрице ДНК (дорожки Z1 – ZF на Фиг. 4А) сравнивали с таковой для нативного олигонуклеотида Z0 (дорожка Z0 на Фиг. 4А).
(2) Для демонстрации применения ФГ-олигонуклеотидов как праймеров (система представлена на Фиг. 3А) реакционные смеси (10 мкл) содержали: нативный олигонуклеотид М40 (30 мкM), радиоактивномеченный олигонуклеотид Т0чТ3 (10 мкM, последовательности праймеров Т0 ч Т3 представлены на Фиг. 2); набор трифосфатов (по 0,2 мM каждый), 1,8 мМ MgCl2, Трис-HCl (10 мМ) pH 8,8, KCl (50 мМ); 0,1 % Tween-20, Taq ДНК- полимеразу (1 ед.акт.). Реакцию проводили в изотермическом режиме (Фиг. 4Б) течение 20-120 минут при 37°C и останавливали добавлением 2% раствора перхлората лития в ацетоне. Все образцы разделяли с помощью электрофоретического анализа в 15% денатурирующем полиакриламидном геле. Степень накопления полноразмерного продукта реакции рассчитывали как отношение интенсивности в полосе, соответствующей продукту реакции, к суммарной интенсивности всех полос в дорожке. Относительная погрешность определения во всех экспериментах не превышала 15% (Фиг. 4Б).
Видно (Фиг. 4А), что ФГ-олигонуклеотиды способны выступать в качестве праймеров в присутствии термостабильной Taq ДНК-полимеразы и в условиях термоциклического режима температур достраиваться до полноразмерного продукта, как и нативный олигонуклеотидный праймер Z0. Снижение эффективности удлинения наблюдается для олигонуклеотида Z1,2, содержащего вблизи 3′-конца две подряд ФГ. В изотермических условиях (Фиг. 4Б) наблюдался аналогичный результат: степень накопления полноразмерного продукта реакции для олигонуклеотидов Т1 и Т2 близка к таковому для Т0. Снижение степени накопления полноразмерного продукта реакции наблюдается для олигонуклеотида Т3, содержащего ФГ вблизи 3′-конца.
Пример. 3 (ФГ-олигонуклеотид как праймер в реакции, катализируемой мезофильной полимеразой)
Для демонстрации применения ФГ-олигонуклеотидов как праймеров (система представлена на Фиг. 3А) реакционные смеси (5 мкл) содержали: нативный олигонуклеотид М40 (30 мкM), радиоактивномеченный олигонуклеотид Т0÷Т3 (10 мкM, последовательности праймеров Т0 ч Т3 представлены на Фиг. 2); набор трифосфатов (по 0,2 мM каждый), MgCl2 (20 мM), Трис-HCl (50 мМ) pH 8,0, NaCl (100 мкМ), ДТТ (5 мМ), 0,1 мг/мл БСА; ДНК-полимеразу фага Т5 (500 мкМ). Реакцию проводили в течение 6-60 минут и останавливали добавлением раствора для нанесения проб при электрофорезе, содержащего 7 мМ ЭДТА. Все образцы разделяли и анализировали аналогично примеру 2 (2). Относительная погрешность определения во всех экспериментах не превышала 15% (Фиг. 5).
Видно (Фиг. 5), что все ФГ-олигонуклеотиды способны выступать в качестве праймеров в присутствии мезофильной ДНК-полимеразы фага Т5 в условиях изотермического режима, хотя эффективность достраивания ФГ-олигонуклеотидов до полноразмерного продукта меньше, чем в случае нативного олигонуклеотида Т0.
Пример. 4 (ФГ-олигонуклеотид как матрица в реакции, катализируемой термостабильной полимеразой)
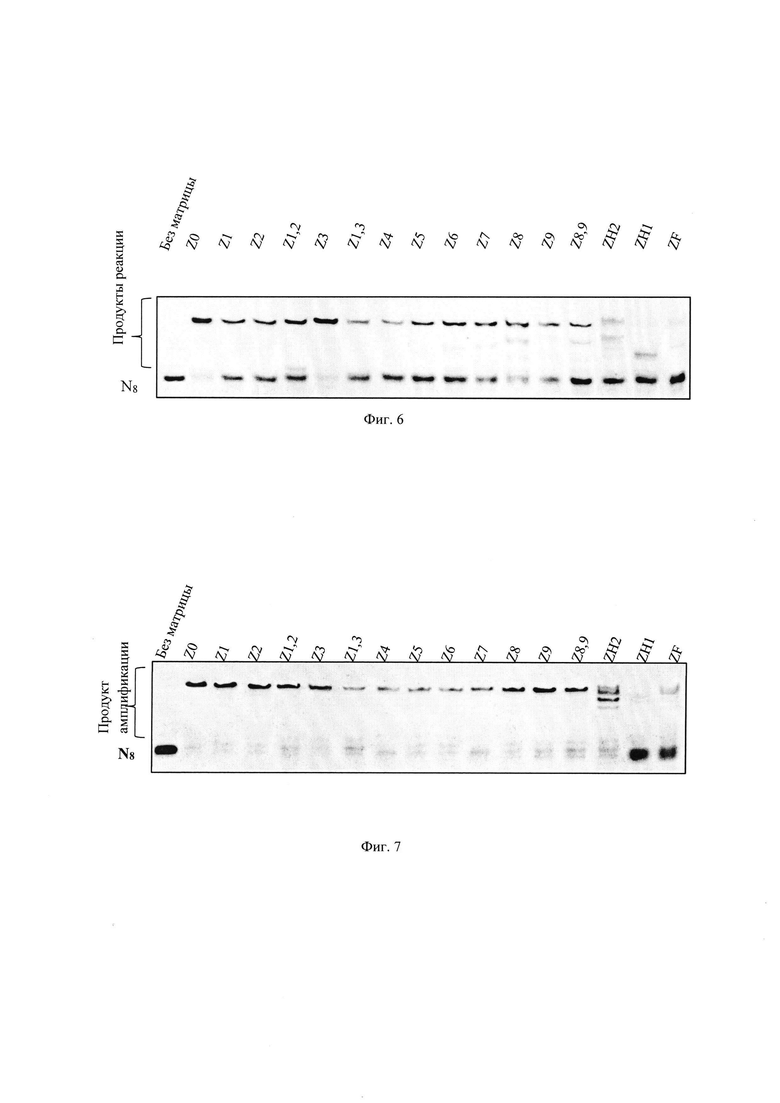
Для демонстрации применения ФГ-олигонуклеотидов как матриц (система представлена на Фиг. 3Б) реакционные смеси (10 мкл) содержали: немодифицированный олигонуклеотид Z0 либо ФГ-олигонуклеотид Z1чZF (10 мкM, последовательности праймеров Z0 ч ZF представлены на Фиг. 2), флуоресцентно-меченный нативный праймер FAM-N8 (10 мкM, последовательность представлена на Фиг. 2), полный набор трифосфатов dNTP (10 мM), Taq ДНК- полимеразу (2 ед.акт.), 1,8 мМ MgCl2, Трис-HCl (10 мМ) pH 8,8, KCl (50 мМ); 0,1 % Tween-20. Реакцию проводили в термоциклическом режиме: 95°C - 10 с, 61°C - 10 с, 72°C – 30 с (32 цикла). Реакцию останавливали и анализировали аналогично примеру 2(1).
Видно (Фиг. 6), что все ФГ-олигонуклеотиды способны выступать в качестве матриц в присутствии термостабильной Taq ДНК-полимеразы в условиях термоциклического режима. Эффективность достраивания нативного олигонуклеотида до полноразмерного продукта увеличивается при удалении ФГЗ от 3′-конца ФГ-олигонуклеотида. Максимальное влияние на выход полноразмерного продукта оказывает ФГ-олигонуклеотид, содержащий подряд (ZH1) и чередующиеся модифицированные ФГ (ZH2, ZF).
Пример. 5 (ФГ-олигонуклеотиды как матрица в реакции, катализируемой мезофильной полимеразой)
Для демонстрации применения ФГ-олигонуклеотидов как матриц (система представлена на Фиг. 3Б) реакционные смеси (10 мкл) содержали: 10 мкM немодифицированный олигонуклеотид Z0 либо ФГ-олигонуклеотид Z1чZF (последовательности праймеров Z0 ч ZF представлены на Фиг. 2), 10 мкM TAMRA-меченный нативный праймер *N8 (Фиг. 2), полный набор трифосфатов dNTP (10 мM), Фрагмент Кленова (2 ед.акт.), Трис-HCl (50 мМ), pH 7,6, MgCl2 (10 мМ), ДТТ (5 мМ). Реакцию проводили 30 минут в изотемпературном режиме при 37°C. Реакцию останавливали и анализировали аналогично примеру 2.
Было получено (Фиг. 7), что все ФГ-олигонуклеотиды способны выступать в качестве матриц в присутствии мезофильной полимеразы - Фрагмента Кленова, в условиях изотемпературного режима. Эффективность достраивания нативного олигонуклеотидов до полноразмерного продукта увеличивается при удалении ФГЗ от 3′-конца нативного олигонуклеотида. Максимальное влияние на выход полноразмерного продукта оказывает ФГ-олигонуклеотид ZH1, содержащий подряд фосфорилгуанидиновые группы.
Пример. 6 (эффективность ПЦР с использованием ФГ-олигодезоксирибонуклеотидов в качестве праймеров)
Амплификацию фрагмента гена eGFP проводили в реакционном буфере, содержащем 50 мM Трис-НCl, рН 8,5, 50 мM KCl, 0,2 мМ каждого дезоксинуклеозидтрифосфата, 2 мМ MgCl2, 0,03 ед.акт./мкл Taq ДНК-полимеразы.
Режим амплификации 95оС 5 мин, 47 циклов: 95°C - 10 с, 61°C - 10 с, 72оС - 10 с.
В качестве матрицы использовали последовательное 10-кратное разведение плазмидной ДНК, содержащей ген eGFP, от 10-9 г до 10-17 г.
В качестве праймеров использовали пары, представленные на Фиг. 8, (прямой + обратный), на основе стандартных нативных олигонуклеотидов и ФГ-олигонуклеотидов. ФГ-олигонуклеотиды отличались по количеству и расположению фосфорилгуанидиновых звеньев в олигонуклеотидной цепи. Последовательности олигонуклеотидов представлены на Фиг. 2. Концентрация каждого праймера составляла 500 нМ.
Эффективность амплификации определяли методом ПЦР в режиме реального времени в присутствии интеркалирующего красителя SYBR Green I на приборе LightCycler 96 (Roche, Швейцария).
Сравнение эффективности амплификации проводили по значениям коэффициентов амплификации (keff), используя подход линеаризующих координат зависимости Ct(lg C0) (где Ct – пороговый цикл, C0 – исходная концентрация матрицы), реализованный в программе LightCycler 96 Software версии 1.1.0.1320. Значения коэффициентов амплификации представлены на Фиг. 8 (приведены значения коэффициентов эффективности ПЦР, полученных по результатам трех или более независимых экспериментов; SD – стандартное отклонение). Видно, что единичные ФГ в структуре праймера (см. праймеры Z1÷Z9) несущественно влияют на эффективность амплификации. Влияние фосфорилгуанидиновых групп определяется их положением в цепи праймера-затравки. Подряд-расположенные ФГ (праймер ZH1) значительно уменьшают keff до 1,44. Введение модификаций в положения «через один» (чередующиеся модифицированные фосфорилгуанидиновые звенья) обеспечивает накопление полноразмерного продукта.
Пример. 7 (определение длины растущей цепи ДНК)
Амплификацию фрагмента гена eGFP проводили в реакционном буфере как описано ранее в примере 6.
Режим амплификации 95оС 5 мин, 28 циклов: 95°C - 10 с, 61°C - 10 с, 72°C - 10 с.
В качестве матрицы использовали плазмидную ДНК, содержащую ген eGFP, в количестве 10-10 г на реакцию.
В качестве праймеров использовали пары (*Q0 - Z0), (*Q0 - ZH1), (*Q0 - ZH2), где * - флуоресцентная метка FAM. Последовательности ФГ-олигонуклеотидов представлены на Фиг. 2. Концентрация каждого праймера составляла 500 нМ.
Определение точной длины продуктов ПЦР с использованием фрагментного анализа проводили на автоматическом капиллярном анализаторе. Из представленных на Фиг. 9 данных видно, что применение олигонуклеотидов Z0 и ZH2 позволяет получать полноразмерные ПЦР-продукты длиной 228 п.н. В то время как использование олигонуклеотида ZH1 приводит к терминации синтеза комплементарной цепи за 8 нуклеотидов до 5′-конца, что, очевидно, приводит к формированию ДНК-молекул с «липкими» концами.
Пример. 8 (сравнение накопления специфичных и неспецифичных продуктов ПЦР, катализируемой Taq-ДНК полимеразой при использовании стандартных и ФГ-модифицированных олигодезоксирибонуклеотидов в качестве праймеров)
Амплификацию фрагмента гена eGFP проводили в реакционном буфере как описано ранее в примере 6.
Режим амплификации описан ранее в примере 7.
Используемая матрица описана ранее в примере 6.
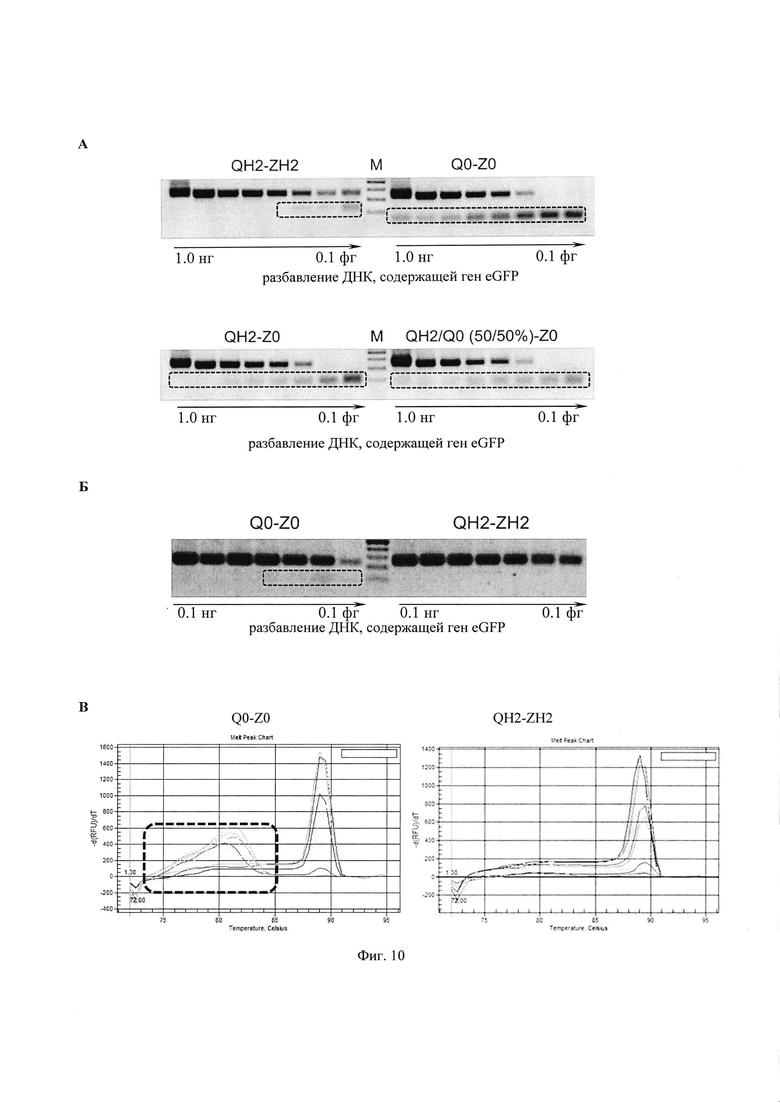
В качестве праймеров (прямой - обратный) использовали пары из стандартных нативных олигонуклеотидов (Q0 - Z0,) модифицированных ФГ-олигонуклеотидов (QH2 - ZH2), стандартного и модифицированного олигонуклеотидов (QH2 - Z0), а также смесь трех олигонуклеотидов (Z0 - QH2/Q0, последние в соотношении 50/50 %). Последовательности олигонуклеотидов представлены на Фиг. 2. Концентрация праймеров составляла 500 нМ.
Анализ продуктов ПЦР проводили методом электрофореза в агарозном геле (Фиг. 10А и Б, М – маркер длин ДНК от 100 до 400 п.н.) и методом термической денатурации продуктов ПЦР на приборе iQ5 (Bio-Rad, США) в присутствии интеркалирующего красителя SYBR Green I (Фиг. 10В).
Видно (Фиг. 10А и В), что применение пары модифицированных ФГ-олигонуклеотидов (QH2 - ZH2) увеличивает достоверность и позволяет значительно уменьшить образование неспецифических продуктов, по сравнению с использованием как нативных олигонуклеотидов (Q0 - Z0), так и смесей из нативного и модифицированного олигонуклеотидов (QH2 - Z0) и (Z0 - QH2/Q0).
Полное отсутствие неспецифических продуктов было достигнуто при использовании ПЦР-систем с отложенным «горячим» стартом на модифицированных ФГ-олигонуклеотидных праймерах, по сравнению с нативными праймерами (Фиг. 10Б).
Пример. 9 (анализ мутаций в растущей цепи ДНК)
Амплификацию фрагмента гена eGFP проводили в реакционном буфере как описано ранее в примере 6.
Режим амплификации описан ранее в примере 7.
Используемая матрица описана ранее в примере 6.
В качестве праймеров (прямой - обратный) использовали пары (Q0 - Z0), (Q0 - ZH1), (QН2 - ZH2). Последовательности олигонуклеотидов представлены на Фиг. 2.
По завершении амплификации осуществляли очистку продуктов ПЦР и их секвенирование по Сэнгеру по стандартной методике с применением флуоресцентно меченых терминаторов BigDye 3.1. При секвенировании использовали нативные олигонуклеотидные праймеры Q0 и Z0. На Фиг. 11 представлены результаты секвенирования ДНК-продуктов по Сэнгеру после амплификации в присутствии пар праймеров, приведенных по центру с помощью нативных олигонуклеотидных праймеров Q0 (колонка справа) и Z0 (колонка слева). Рамкой выделена область исходных праймеров.
Видно (Фиг. 11), что нуклеотидный состав растущей цепи ДНК в присутствии пар нативных праймеров соответствует составу в присутствии пар, один или оба из которых содержит ФГ. Наличие ФГ в исходном праймере не вызывает появления мутаций в растущей цепи ДНК при построении комплементарной цепи в области праймера. В то же время наличие ФГ подряд в соседних положениях терминирует удлинение растущей цепи на модифицированной матрице и позволяет контролировать длину конечного продукта, формируя одноцепочечные «липкие концы», как и описано в примере 7.
Пример. 10 (использование ФГ-олигонуклеотидов как праймеров для ДНК-полимераз, используемым в коммерческих ПЦР-системах)
Для демонстрации использования ФГ-олигонуклеотидов как праймеров были выбраны три коммерческие системы. Система I содержала ДНК-полимеразу Pfu (ЗАО «Силекс», Россия); система II - смесь Taq и Pfu полимераз для проведения ПЦР длинных фрагментов (ПЦР типа “Long Range”) (ООО «Биолабмикс», Россия); система III - для проведения ПЦР на цельной крови «InBlood PCR kit» (ЗАО «Евроген», Россия) с полимеразой “InBlood” (ЗАО «Евроген», Россия).
Для систем I и II в качестве матрицы использовали 1 нг плазмидной ДНК, содержащей ген eGFP, амплификацию проводили в режиме: предварительная денатурация 95°C - 5 мин, 32 цикла: 9°C - 10 с, 61°C - 10 с, 72°C.
Для системы III в качестве матрицы использовали препараты цельной крови и плазмы крови человека, содержащие предварительно добавленные 0,2 нг плазмидной ДНК на 1 мкл физиологической жидкости, в отношении 2% (только для плазмы крови), 5%, 10%, 20%, 25% от общего объема реакционной смеси в 25 мкл. Амплификацию проводили в режиме 95°C - 5 мин, 26 циклов: 95°C - 10 с, 61°C - 10 с, 72°C - 10 с.
В качестве пар праймеров использовали (Q0 – Z0), (Q0 – ZH2), (QH2 – Z0) , (QH2 – ZH2). Последовательности олигонуклеотидов представлены на Фиг. 2.
Результаты анализа продуктов ПЦР методом электрофореза в агарозном геле представлены на Фиг. 12 (где М – маркер длин ДНК от 100 до 500 п.н.). Видно (Фиг. 12 А и Б), что при использовании ФГ-олигонуклеотидов как праймеров в присутствии различных коммерческих полимераз и буферных смесей специфично образуется продукт ПЦР заданной длины. ФГ-олигонуклеотиды пригодны для проведения ПЦР в физиологических жидкостях. Представленные результаты позволяют сделать вывод о возможности эффективного применения ФГ-модифицированных олигодезоксирибонуклеотидов в качестве праймеров-затравок в ПЦР, катализируемых различными ДНК-полимеразами, используемыми в коммерческих ПЦР-системах.
Пример. 11 (ФГ-олигонуклеотиды как праймеры для РНК-зависимых ДНК-полимераз (ревертаз))
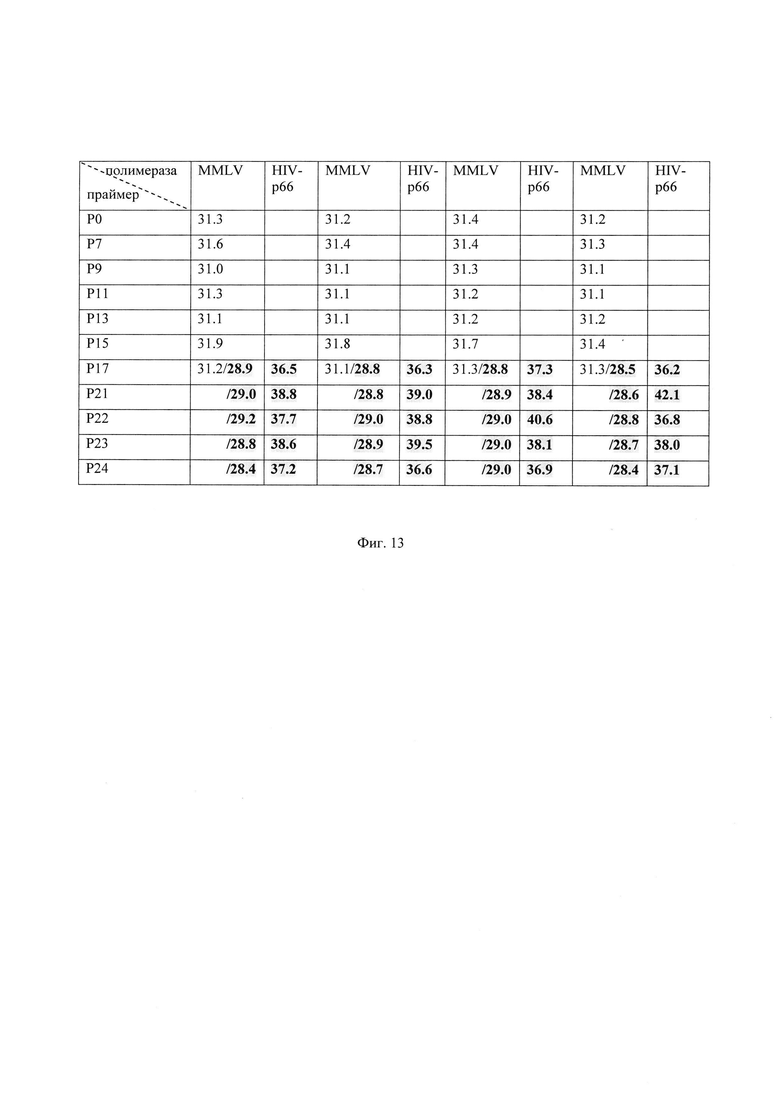
В данном примере продемонстрирована возможность использования ФГ-олигонуклеотидов Р7 ÷ Р24, меченных флуоресцеином, как праймеров при выявлении РНК вируса гепатита С (ВГС) по сравнению с нативным олигонуклеотидом Р0. Последовательности олигонуклеотидов представлены на Фиг. 2. Модифицированные ФГ-праймеры содержали от 7 (со стороны 5′-конца олигонуклеотидов) до 21 ФГ подряд.
В качестве исходного материала для анализа использовали свежую сыворотку крови ВГС-инфицированных доноров. РНК ВГС выделяли с использованием набора «РеалБест» Экстракция 100 (Вектор-Бест) и вносили ее в рамках описанной процедуры в реакцию ОТ-ПЦР, используя на этапе обратной транскрипции либо нативный олигонуклеотид Р0, либо ФГ-олигонуклеотиды Р7 ÷ Р21 и ревертазу (MMLV или HIV-p66). Реакцию обратной транскрипции (50 мкл) проводили в смеси следующего состава: ФГ-олигонуклеотиды (0,5 мкМ), MgCl2 (3 мМ), Трис-НСl (50 мМ) pH 8,0, (NH4)2SO4 (10 мМ), KCl (30 мМ), 0,01% Tween-20, набор трифосфатов dNTP (по 0,4 мM каждый), БСА 100 мкг/мл, MMLV или HIV-p66 (10 ед.акт.). Реакцию обратной транскрипции проводили не менее чем в двух повторах в температурном режиме: 45°C – 30 мин, 95°C – 3 мин.
Далее полученную кДНК использовали для ПЦР ВГС-специфичного фрагмента. ПЦР проводили в смеси следующего состава: праймеры CTCCCGGGAGAGCCATAG и TCCAAGAAAGGACCCGGTC (по 0,5 мкМ каждый), буфер (MgCl2 (3 мМ), Трис-SO4 (50 мМ) pH 8,0, (NH4)2SO4 (10 мМ), KCl (30 мМ), 0,01% Tween-20), Taq ДНК-полимеразу (1 ед.акт.), зонд флуоресцентно меченный гидролизующийся зонд зонд 5'-ROX-TCTGCGGAACCGGTGAGTACACCG-(BHQ2) (0,25 мкМ), SYBR Green I (в разведении 1/10000) или SYTO-13 (в разведении 1/2500). Амплификацию осуществляли в режиме 50°C – 2 мин, 49 циклов: 94°C – 10 с, 60°C – 20 с, 5°C – 5 с, 95°C – 1 мин. Процесс выявления ВГС с использованием прибора CFX96 (BioRad, США) и набора РеалБест РНК ВГС осуществляли в режиме реального времени по двум каналам: ROX–специфическая детекция флуоресцентно меченного гидролизующегося зонда и FAM–неспецифическая детекция дцДНК с помощью красителя SYBR Green I (Фиг. 13, обычное начертание) или SYTO-13 (Фиг. 13, выделено полужирным и цветом). В результате амплификации нарабатывали фрагмент ДНК длиной 79 п.н., имеющий в буферных условиях для ПЦР температуру плавления 86-88°C.
Сравнение субстратных свойств ФГ-олигонуклеотидов и нативного олигонуклеотида как праймеров затравок проводили с помощью определения значения порогового цикла реакции – Ct (cycle threshold), представленного на Фиг. 13. Значение Ct рассчитывали как описано ранее в примере 6. Видно, что ФГ-олигонуклеотиды могут выступать как праймеры-затравки при получении кДНК на матрице анализируемой РНК с помощью РНК-зависимых ДНК-полимераз (ревертаз MMLV и HIV-p66).
Пример 12 (ФГ-олигонуклеотиды как праймеры для ДНК-полимераз в одношаговых системах ОТ-ПЦР)
В системе для одношаговой ОТ-ПЦР «БиоМастер ОТ-ПЦР SYBR Blue (2×)» (ООО «Биолабмикс»), содержащей ревертазу MMLV и термостабильную ДНК-зависимую Taq ДНК-полимеразу, использовали специфичные к последовательности U12 мяРНК человека праймеры.
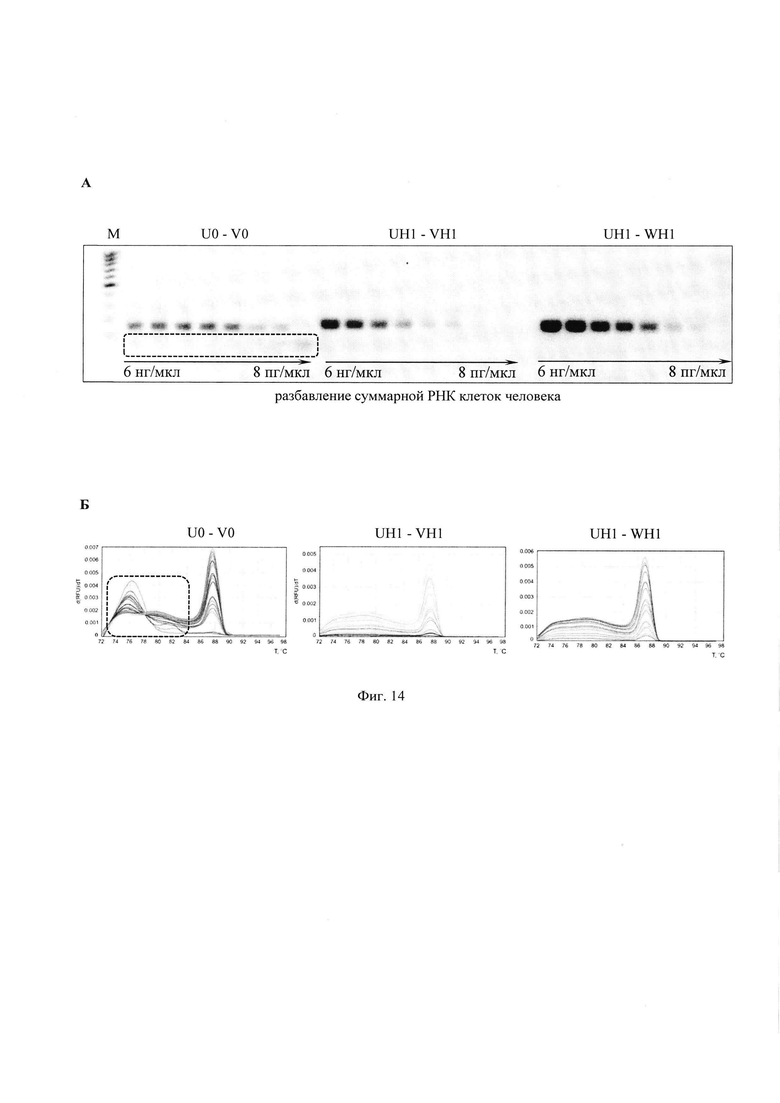
В качестве праймеров использовали нативные олигонуклеотиды (U0, V0) и ФГ-олигонуклеотиды (UH1, VH1, WH1). Последовательности олигонуклеотидов представлены на Фиг. 2. Концентрация каждого праймера составляла 480–700 нМ.
ОТ-ПЦР проводили на препарате суммарной РНК клеток аденокарциномы молочной железы человека MCF-7 в диапазоне концентрации от 6 нг/мкл до 8 пг/мкл.
Реакцию ревертирования проводили при 45°C – 45 мин
Режим амплификации 95°C 5 мин, 48 циклов: 95°C - 10 с, 60°C - 10 с, 72°C - 10 с.
Анализ продуктов ОТ-ПЦР проводили методами термической денатурации и горизонтального гель-электрофореза в 1,5% геле с последующей визуализацией нуклеотидного материала бромистым этидием (Фиг. 14А, где маркер длин ДНК от 100 до 400 п.н.). Для анализируемых пар праймеров определяли кажущиеся коэффициенты эффективности: 1,7 для пары (U0 - V0), 1,63 для (UH1 - VH1) и 1,56 для (UH1 - WH1).
Видно (Фиг. 14Б), что при использовании ФГ-модифицированных праймеров в значительно меньшей степени происходит образование неспецифических продуктов амплификации с пониженной температурой плавления. Полученные результаты демонстрируют принципиальную возможность использования ФГ-олигонуклеотидов в качестве праймеров в одношаговых системах для ОТ-ПЦР, т.е. ФГ-модифицированные праймеры способны выполнять роль затравок на стадии ОТ (катализируемой РНК-зависимой ДНК-полимеразой), а затем на стадии амплификации (катализируемой ДНК-зависимой ДНК-полимеразой).
Пример 13 (ФГ-олигонуклеотиды как праймеры в реакции амплификации по механизму катящегося кольца (RCA))
В данном примере продемонстрировано сравнение эффективности реакции амплификации ДНК по механизму катящегося кольца (RCA) при использовании ФГ-олигонуклеотидов и нативного олигонуклеотида.
Амплификацию ДНК плазмиды pUC19 (0,2 нг) проводили в реакционном буфере, содержащем Tris-HCl (50 мM) pH 7.5, MgCl2 (10 мМ), (NH4)2SO4 (10 мМ), ДТТ (4 мМ), в присутствии набора дезоксинуклеозидтрифосфатов (по 0,2 мМ каждый), БСА (200 нг/мкл), интеркалирующего красителя SYBR Green I, ДНК-полимеразы phi29 (0,5 ед.акт./мкл), 1мкМ одного из праймеров.
Режим амплификации 14 часов при 30°С.
В качестве праймеров использовали ФГ-олигонуклеотиды D2 и D3 и нативный олигонуклеотид D0. Последовательности олигонуклеотидов представлены на Фиг. 2. Модифицированные ФГ-праймеры содержали 2 и до 3 ФГ подряд со стороны 3′-конца олигонуклеотида.
Анализ продуктов амплификации проводили агарозным гель-электрофорезом в 0,8% агарозе, используя в качестве маркера (М) 200 нг плазмиды pUC19. Результаты анализа представлены на Фиг. 15 А, «K-» соответствует результатам контрольной реакции без праймеров. Видно (Фиг. 15 А), что в случае ФГ-олигонуклеотидов наблюдается накопление продукта амплификации. Одновременно продукт амплификации (для каждого праймера D2, D3 и D0) разводили в 3х10-6 раз и использовали по 50 мкл полученного раствора для оценки коэффициента амплификации методом ПЦР в девять повторениях в режиме реального времени в присутствии флуоресцентно меченного гидролизующегося зонда на участок pUC9 длиной 105 нуклеотидов. Далее рассчитывали среднее значение порогового цикла реакции (Ct) и разницу (∆Ct) между Ct конкретного праймера и разведения исходной матрицы, соответствующим начальным условиям реакции амплификации (амплификации ДНК плазмиды pUC19 в количестве 0,2 нг).
Полученное значение ∆Ct использовали для расчёта коэффициента амплификации RCA с допущением, что эффективность начальных условий составляет 1 (Фиг. 11 Б). При сравнении данных, представленных в Таблице, видно, что коэффициент амплификации при использовании ФГ-олигонуклеотидов D2 и D3 выше, чем у исходного нативного праймера D0, аименно, 247,3 и 29,9, соответсвенно, по сравнению с 9,4.
Пример 14 (повышение селективности выявления мутации с помощью ФГ-олигонуклеотида как праймера аллель-специфичной ПЦР)
Амплификацию матрицы проводили в буфере, содержащем Трис-НCl (65 мM) рН 8,9; (NH4)2SO4 (24 мM); MgSO4 (3 мM), 0,05% Tween-20, набора дезоксинуклеозидтрифосфатов (по 0,2 мМ каждого), Taq ДНК-полимеразы (0,03 ед.акт./мкл), флуоресцентно меченный гидролизующийся зонд 5’-HEX-CTGTATCGTCAAGGCACTCTTGC-BHQ2-3’ (100 нМ).
Режим амплификации: 95оС 3 мин, 50 циклов: 95°C - 10 с, 60°C - 40 с.
В качестве матриц использовали 2 нг геномной ДНК человека, выделенной из гистологического блока с тканью колоректального рака, без или с добавлением контрольной плазмиды, содержащей мутацию c.38G>A (G13D) в фрагменте гена KRAS (gactgaatataaacttgtggtagttggagctggtg(g/A)cgtaggcaagagtgccttgacgatacagctaattcagaatcattttgtggacgaatatg), в количестве 1,1% относительно общего количества фрагмента гена KRAS в геномной ДНК.
В качестве праймеров использовали пары олигонуклеотидов (прямой + обратный): S0+S и S1+S, где S и S0 нативные олигонуклеотиды, S1 – ФГ-олигонуклеотид. Каждый праймер вносили в реакционную смесь в концентрации 300 нМ. Структуры праймеров представлены на Фиг. 2. Схематическое изображение выявления однонуклеотидной мутации Т/С в составе матрицы методом аллель-специфичной ПЦР в присутствии пар праймеров S0+S и S1+S представлено на Фиг. 16.
Селективность выявления мутации определяли методом ПЦР в режиме реального времени на приборе LightCycler 96 (Roche, Швейцария).
Для каждой пары праймеров рассчитывали среднее значение порогового цикла реакции (Ct) и разницу (∆Ct) между образцом, содержащим 1,1% мутации и образцом без мутации. Было получено, что ∆Ct пары (S1+S) составляет 9,27, в то время как (S0+S) 4,42 (Фиг. 16, значение ∆Ct указано рядом с обозначениями соответствующих праймеров). Таким образом, можно заключить, что селективность пары, содержащей ФГ-олигонуклеотид, выше, чем пары на основе нативных олигонуклеотидов.
| название | год | авторы | номер документа |
|---|---|---|---|
| Фосфорамидные азольные олигонуклеотиды, способ синтеза фосфорамидных азольных олигонуклеотидов и способ матричного ферментативного синтеза ДНК с их использованием | 2023 |
|
RU2823099C1 |
| Способ оценки активности системы эксцизионной репарации нуклеотидов млекопитающих | 2024 |
|
RU2831226C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ УРОВНЯ ТРАНСКРИПЦИИ ГЕНА, КОДИРУЮЩЕГО ХЕМОКИН CCL2 (МСР-1) ЧЕЛОВЕКА И НАБОР ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 2012 |
|
RU2522801C2 |
| СУХАЯ СМЕСЬ ДЛЯ ПРИГОТОВЛЕНИЯ РЕАКЦИОННОЙ СМЕСИ ДЛЯ АМПЛИФИКАЦИИ НУКЛЕИНОВОЙ КИСЛОТЫ И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2012 |
|
RU2535995C2 |
| Тест-система "miR-M-SCREEN" для прогнозирования развития метастазов у больных колоректальным раком на основании уровня микро-РНК miR-26a и miR-143 в плазме крови | 2022 |
|
RU2786386C1 |
| Способ опосредованного определения титра инфекционной активности вируса бешенства штамма ВНИИЗЖ в сырье для инактивированной антирабической вакцины методом ПЦР в режиме реального времени | 2020 |
|
RU2755925C1 |
| СПОСОБ ИДЕНТИФИКАЦИИ НАТИВНЫХ ПАР ФРАГМЕНТОВ ДНК ИЛИ РНК, ПРИСУТСТВУЮЩИХ В ОДНИХ И ТЕХ ЖЕ ЖИВЫХ КЛЕТКАХ | 2015 |
|
RU2600873C1 |
| СПОСОБ ИДЕНТИФИКАЦИИ НАТИВНЫХ ПАР ФРАГМЕНТОВ ДНК ИЛИ РНК, ПРИСУТСТВУЮЩИХ В ОДНИХ И ТЕХ ЖЕ ЖИВЫХ КЛЕТКАХ | 2013 |
|
RU2578009C2 |
| Способ опосредованного определения полноты инактивации антигена вируса ящура в сырье для вакцины с применением обратно-транскриптазной полимеразной цепной реакции в режиме реального времени при амплификации большеразмерного фрагмента | 2021 |
|
RU2753969C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ СУБСТРАТНОЙ СОВМЕСТИМОСТИ МОДИФИЦИРОВАННЫХ ТРИФОСФАТОВ ДЕЗОКСИУРИДИНА И ДНК-ПОЛИМЕРАЗ СОЧЕТАНИЕМ МЕТОДОВ КОЛИЧЕСТВЕННОЙ ПЦР И ЭЛЕКТРОФОРЕЗА | 2019 |
|
RU2736969C1 |








Изобретение относится к биотехнологии. Изобретение может быть использовано в разработке и оптимизации ПЦР и ОТ-ПЦР систем, применяемых для выявления нуклеиновых кислот, в том числе при диагностике генетических, вирусных и других заболеваний. Предложен способ матричного ферментативного синтеза ДНК. Способ отличается тем, что в качестве праймеров при проведении полимеразной цепной реакции (ПЦР) и ПЦР в сочетании с обратной транскрипцией (ОТ-ПЦР) используют нейтральные производные олигонуклеотидов, а именно фосфорилгуанидины. Фосфорилгуанидины содержат одну или более фосфатных групп, в которых на атоме фосфора введен остаток гуанидина или замещенного гуанидина. Техническим результатом является повышение достоверности, чувствительности и специфичности выявления анализируемых последовательностей нуклеиновых кислот, а также упрощение способа матричного синтеза ДНК. 6 з.п. ф-лы, 16 ил., 14 пр.
1. Способ матричного ферментативного синтеза ДНК, отличающийся тем, что для инициации реакции используют праймер, содержащий по крайней мере одну межзвенную фосфорилгуанидиновую группу, соответствующую общей формуле (I):

X - звено или цепь звеньев олигомера (праймера) с 5'-стороны от межзвенной фосфорилгуанидиновой группы,
Y - звено или цепь звеньев олигомера (праймера) с 3'-стороны от межзвенной фосфорилгуанидиновой группы,
где каждый из заместителей R1, R2, R3 и R4 независимо выбирается из группы, включающей H, C1-10 алкил; R1 и R2 вместе с атомом, с которым они связаны, могут образовывать 5-членный N-гетероцикл, R2 и R3 вместе могут образовывать алкиленовую цепь длиной 2-4 атома.
2. Способ матричного ферментативного синтеза ДНК по п. 1, применяемый для амплификации нуклеиновых кислот.
3. Способ матричного ферментативного синтеза ДНК по п. 1, применяемый для проведения полимеразной цепной реакции.
4. Способ матричного ферментативного синтеза ДНК по п. 1, используемый для проведения аллель-специфичной полимеразной цепной реакции.
5. Способ матричного ферментативного синтеза ДНК по п. 1, применяемый для обратной транскрипции.
6. Способ матричного ферментативного синтеза ДНК по п. 1, применяемый на обеих стадиях обратной транскрипции и последующей полимеразной цепной реакции.
7. Способ матричного ферментативного синтеза ДНК по п. 1, применяемый для амплификации нуклеиновых кислот по механизму катящегося кольца.
| WO 2016028187, 25.02.2016 | |||
| Купрюшкин М.С., Пышный Д.В., Стеценко Д.А | |||
| Фосфорилгуанидины | |||
| Новый класс аналогов нуклеиновых кислот // Acta Naturae (русскоязычная версия) | |||
| Способ защиты переносных электрических установок от опасностей, связанных с заземлением одной из фаз | 1924 |
|
SU2014A1 |
| - Т | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| - n | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |

