Изобретение относится к области биотехнологии, средствам молекулярной диагностики и производству противоящурных вакцин, а именно к способу опосредованного определения полноты инактивации антигена вируса ящура в сырье для вакцины с применением обратно-транскриптазной полимеразной цепной реакции в режиме реального времени (ОТ-ПЦР-РВ) при амплификации большеразмерного фрагмента.
Ящур является высококонтагиозным инфекционным заболеванием парнокопытных животных, возбудитель которого - вирус порядка Picornavirales семейства Picornaviridae рода Aphthovirus [1]. Характерной особенностью вируса ящура является наличие 7 типов: А, О, С, Asia-1, SAT-1, SAT-2, SAT-3. В пределах каждого типа существует множество генетических вариантов вируса [2].
Геном вируса ящура представлен одноцепочечной позитивной молекулой РНК (ssRNA(+)) длиной около 8400 нуклеотидов. РНК одновременно является матрицей для репликации генома и трансляции вирусных белков. РНК вируса ящура включает три отдельные части, а именно относительно длинную 5'-нетранслируемую область (5-UTR), длинную кодирующую область с одной открытой рамкой считывания и 3'-нетранслируемую область (3'-UTR) (Фиг. 1).
РНК окружена белковой оболочкой (капсидом), состоящей из 60 капсомеров. Каждый капсомер представлен четырьмя структурными полипептидами VP1, VP2, VP3 и VP4. Белки VP1, VP2 и VP3 находятся на поверхности вируса и вносят вклад в антигенные свойства вируса, a VP4 располагается внутри вириона.
5'-UTR имеет длину около 1300 нуклеотидов [3]. Небольшой белок (длиной 23-24 н.о.), называемый VPg, который кодируется частью 3В области вирусного генома, ковалентно связан с 5'-концом генома. Пептид VPg производится в 3 различных формах (кодируемых генами 3В1-3), которые играют роль праймеров для синтеза РНК. 5'-UTR-область включает несколько различных структурных элементов: S-фрагмент, поли(С) тракт (Cn), 3 или 4 псевдоузла (PK), элемент cre/bus, внутренний сайт для рибосомы (IRES).
S-фрагмент находится на 5'-конце, имеет длину 360-370 нуклеотидов и сворачивается с образованием большой шпилечной структуры. Тракт poly С имеет переменную длину (150-250 нуклеотидов), но состоит из более чем 90% остатков С. Окончательно роль тракта poly С не известна. За трактом poly С расположена нетранслируемая область длиной около 720 н.о., в которую входят 3-4 структуры псевдокнот. Их функция неизвестна. Cre/bus - участок представляет собой стабильный элемент типа «стебель-петля» из примерно 55 нуклеотидов и содержит консервативный мотив (АААСА), который действует как матрица для уридилилирования белка VPg вирусной РНК-полимеразой. Таким образом, cre/bus участвует в инициации репликации РНК. Внутренний сайт связывания с рибосомой IRES имеет длину около 450 нуклеотидов и отвечает за инициирование синтеза вирусного белка. Данный участок характеризуется сложной пространственной структурой. Трансляцию инициируют два AUG, расположенные друг от друга на расстоянии 84 н.о.
Транслируемая область следует за 5'-UTR. РНК транслируется в виде единой длинной открытой рамки считывания (ORF) в полипротеин, после чего следует серия посттрансляционных протеолитических расщеплений с образованием как промежуточных, так и зрелых структурных и неструктурных белков [4, 5]. Это основная часть вирусного генома, длина которой составляет около 7000 нуклеотидов и включает 14 генов. Она кодирует большой полипротеин (около 2330 а.о.), который быстро расщепляется вирусными протеазами с образованием четырех различных структурных и одиннадцати неструктурных белков, которые выполняют разные функции. После трансляции первоначально образуются четыре первичных продукта, а именно Lpro (лидерная протеаза), Р1-2А, Р2 и Р3. Lpro представляет собой N-концевой компонент полипротеина. Лидерная протеаза отщепляется от полипротеина автолитически. L-кодирующая область содержит два отдельных инициирующих кодона AUG (обычно 84 нуклеотида друг от друга), которые приводят к образованию двух разных L-протеаз, названных Lab и Lb и отличающихся друг от друга по длине на 28 а.о. Lpro отвечает за ингибирование синтеза белка в клетке-хозяине, индуцируя расщепление белка-хозяина eIF4G, который является фактором инициации трансляции. В результате РНК вируса ящура может свободно использовать аппарат синтеза белка клетки-хозяина для собственного синтеза белка, поскольку IRES ящура может функционировать с остаточным С-концевым фрагментом eIF4G белка. Полипептид Р1-2А расщепляется протеазой 3С (3Cpro) с образованием 1АВ (VP0), 1С (VP3) и ID (VP1), 2А белков. 2А - очень короткий пептид, размер около 20 нуклеотидных остатков. Во время инкапсидации генома VP0 расщепляется с образованием белков VP4 и VP2 автолитически. Протеин VP4 полностью находится внутри вирусной частицы, тогда как VP1, VP2 и VP3 располагаются на поверхности и вносят свой вклад в антигенные свойства вируса [2, 3]. VP1 содержит, по крайней мере, два важных иммуногенных сайта, петлю G-H (в положениях аминокислот 141-160) и С-конец (остатки 200-213). Петля G-H включает мотив аргинин-глицин-аспарагиновая кислота (RGD), который необходим для прикрепления вируса к клетке-хозяину через рецептор интегрина [4, 5]. Интегрины представляют собой группу α-β-гетеродимерных гликопротеинов, которые расположены на поверхности клетки; около 15 а и 8 Р субъединиц объединяются с образованием 20 различных α-β гетеродимеров. Гетеродимер αvβ6 является рецептором белков внеклеточного матрикса, экспрессия которых ограничена эпителиальными клетками, и он также связывается с вирусом ящура посредством взаимодействия с мотивом RGD [6].
Белок VP1 является наиболее вариабельным, поскольку на него приходится около 90% мутаций всех структурных генов. Самыми вариабельными областями являются участки 40-60, 130-160 и 190-213 а.о. [7]. Участок поверхностного белка VP1 в регионе 130-160 а.о. отличается высокой вариабельностью, поскольку участвует в процессе связывания с рецепторами клетки-хозяина [8]. Изменчивость данного региона дает возможность вирусу ящура взаимодействовать с рецепторами клеток разных типов и облегчает переход от одного вида хозяина к другому. Нуклеотидные последовательности кодирующей области VP1 используются для генетической характеристики штаммов ящура из-за их значимости для прикрепления и проникновения вируса в клетку, защитного иммунитета и специфичности серотипа. Филогенетический анализ на основе последовательности VP1 широко используется для вывода эволюционной динамики, эпидемиологических отношений между генетическими линиями и для отслеживания происхождения и перемещения штаммов вируса ящура [9].
Белок 2А является вирусной протеазой и по функциям напоминает лидерную протеазу. По всей видимости, данный белок отвечает за расщепление сайта 2А/2В, которое является вторым событием процессинга у вируса ящура [10].
Области Р2 и Р3 полипротеина процессируются в неструктурные белки (NSP) вируса ящура. Область протеина Р2 расщепляется на белки 2В и 2С. Функция белка 2В не известна. Белок 2С определяет устойчивость вирусной частицы к гуанидину [11]. Протеин Р3 расщепляется с образованием 6 белков: 3А, трех отдельных копий VPg (3B1, 3В2, 3В3), 3Cpro и 3Dpol. Функция белка 3А не известна. Ген 3В кодирует три различные формы белка VPg. Протеаза 3Cpro отвечает за расщепление Р1-2А на белки VP0, VP1, VP3, а также за образование различных неструктурных белков. Белок также осуществляет расщепление клеточного гистона Н3 и, тем самым, ингибирует синтез белков клетки на этапе транскрипции матричной РНК (мРНК) [12]. 3Dpol кодируется высококонсервативным 3D-геном и является РНК-зависимой РНК-полимеразой для репликации вирусной РНК.
В трансляции РНК вируса ящура определенная роль отводится клеточным белкам. Luz и Beck в пределах участка IRES вируса ящура определил 2 области, которые взаимодействуют с белком клетки р57, необходимым для сплайсинга ядерных мРНК [13]. В инфицированной клетке-хозяине под действием лидерной протеазы происходит разрушение компонента кеп р220-связывающих комплексов. В результате прекращается синтез клеточных белков и включается трансляция вирусных составляющих [14].
3'-UTR-область намного короче 5' UTR-области, ее длина составляет около 90-100 нуклеотидов. Данный участок сворачивается, образуя специфическую структуру «стебель-петля», за которой следует поли(А)-фрагмент переменной длины [13, 14, 15, 17]. 3'-UTR-область играет важную роль в репликации вирусного генома. На конце 3'-UTR-области расположен poly (А)-участок, длина которого варьирует от 50 до 100 н.о. (Фиг. 1).
Ящур как заболевание причиняет серьезный экономический ущерб в связи с затратами на ликвидацию болезни и введением строгих ограничений, налагаемых на внутреннюю и международную торговлю продукцией животноводства. Система мер для борьбы с данным заболеванием и его профилактики предусматривает стемпинг-аут, массовую иммунизацию восприимчивых животных, а также контроль уровня напряженности поствакцинального иммунитета [1, 2, 16].
В процессе производства противоящурных инактивированных сорбированных и эмульсионных вакцин после инактивации антигена вируса ящура требуется проводить оценку отсутствия вирулентного возбудителя данного заболевания для подтверждения безопасности вакцинного препарата.
Инактивация вируса ящура может достигаться за счет влияния различных физических и химических факторов. Так, при увеличении температуры прогревания до 45°С уменьшается относительное процентное содержание 146S компонента при сохранении титра инфекционной активности вируса ящура на уровне контрольного образца, хранящегося при температуре 4°С, что свидетельствует о разрушении неинфекционных 146S частиц. Температура 55-60°С вызывает спонтанный распад полных вирионов и резкое снижение титра инфекционной активности вируса ящура. Воздействие ультрафиолетовых лучей инактивирует вирус через 5-10 минут.Солнечный свет при температуре 5-6°С обезвреживает вирус через 5-7 дней, при 16-18°С - через 3-4 дня, при 37°С - через 40 часов [1,18].
Вирус инактивируют различными химическими агентами: 0,5-1,0% раствор фенола при температуре плюс 4°С через 8 недель, при температуре плюс 18-20°С - через 10-14 дней, при температуре плюс 37°С - через 3 суток; 2% раствор формалина при температуре плюс 4°С - через 2 часа; 45-70% раствор этилового спирта при температуре плюс 4°С - через 1-2 ч [1].
В настоящее время в соответствии с требованиями МЭБ (OIE) [2] при изготовлении противоящурных вакцин при достижении достаточной концентрации 146S компонента и титра инфекционной активности вируса ящура, вируссодержащую суспензию осветляют посредством ультрафильтрации. Вирус ящура впоследствии инактивируют, добавляя инактивант первого порядка этиленимин (EI) в форме бинарного этиленимина (BEI), а именно 1,2-аминоэтилэтиленимин [19, 20]. После завершения инактивации любые остатки EI/BEI в инактивированной вирусной суспензии подвергают нейтрализации, например, посредством добавления раствора тиосульфата натрия до получения конечной концентрации 2%.
Инактивант 1,2-аминоэтилэтиленимин (АЭЭИ) - азотсодержащее гетероциклическое соединение, которое относится к циклическим аминам. АЭЭИ вступает в реакцию замещения водорода в N-H-связи вирусной молекулой РНК [20]. Данный процесс приводит к деградации генома вируса ящура, в результате этого утрачивается вирулентность возбудителя, но при этом сохраняется структура сформированных 146S иммуногенных частиц и их антигенные свойства.
За счет воздействия 1,2-аминоэтилэтиленимина вирус ящура теряет свою инфекционную активность за счет многочисленных делеций и разрывов вирусной РНК, разрушения вирусного генома до низкомолекулярных фрагментов [19, 20, 21], что блокирует процессы обратной транскрипции, трансляции и процессинга белковых составляющих вируса ящура, тем самым, репродукция вирусной частицы в клеточных системах in vivo и in vitro становится невозможной. Повреждения в 1А, 1В, 1С, ID-генах приводят к прекращению процесса синтеза белков VP4, VP2, VP1, VP3 вируса ящура, соответственно. Разрушения РНК в 3D-гене делает невозможным синтез РНК-зависимой-ДНК-полимеразы, следовательно, и формирование перечисленных выше структурных белков возбудителя ящура. Иными словами, при любом повреждений РНК антиген вируса ящура становится авирулентным. Данный факт можно обнаружить с помощью проведения молекулярно-биологического исследования с оригинальной системой олигонуклеотидных праймеров, зонда и подобранными условиями проведения анализа.
В соответствии с требованиями МЭБ (OIE) [2] полноту инактивации верифицируют с использованием теста на остаточное содержание вирулентного вируса ящура. С этой целью суспензией антигена инактивированного вируса ящура инокулируют чувствительную клеточную линию, в частности первичную монослойную культуру клеток почки свиньи СП, с последующим микроскопированием раз в сутки в течение 72 ч. Суспензия инактивированного антигена считается соответствующего качества, если вирулентный вирус в ней не обнаружен. Данный анализ проводят на протяжении 3 последовательных пассажей в чувствительной клеточной линии СП по 72 ч [22].
Существенными недостатками данного метода являются: 1) длительная процедура анализа, связанная с развитием цитопатического действия, 2) определенная степень субъективности при оценке результатов анализа, 3) высокая стоимость первичной монослойной клеточной линии как тест-системы и затраты на ее поддержание. В связи с этим целесообразно провести поиск способа определения полноты инактивации антигена вируса ящура с применением более быстрого и менее затратного метода исследования, который отличается при этом высокой диагностической чувствительностью, специфичностью и общей точностью.
Проблемой является отсутствие чувствительного и специфичного способа опосредованного определения полноты инактивации антигена вируса ящура с целью устранения вышеуказанных недостатков.
Данная проблема была решена благодаря разработке нового способа опосредованного определения полноты инактивации антигена вируса ящура с применением обратно-транскриптазной полимеразной цепной реакции в режиме реального времени при амплификации болыиеразмерного фрагмента, с его помощью возможно выявлять неповрежденный участок генома вируса ящура размером 7279 п.н. в диапазоне от 5'-NTR до 3D-гена включительно и тем самым определять наличие вирулентного вируса ящура, либо обнаруживать отсутствие ампликонов и, тем самым, доказывать отсутствие вирулентного вируса ящура в сырье для изготовления противоящурных инактивированных вакцин. Данная возможность позволит предварительно определять полноту инактивации антигена вируса ящура, который используется для изготовления противоящурных инактивированных сорбированных и эмульсионных вакцин.
Сущность изобретения заключается в новом подходе по опосредованному определению полноты инактивации антигена вируса ящура с применением обратно-транскриптазной полимеразной цепной реакции в режиме реального времени при амплификации болыиеразмерного фрагмента. Заявляемый способ основан на проведении экстрагирования молекул РНК возбудителя ящура до и после процесса инактивации с помощью 5 М гуанидинизотиоцианата (ГТЦ), 0,5% раствора меркаптоэтанола и 80% раствора изопропанола, оценке степени чистоты полученного элюата РНК и ее концентрации методом спектрального сканирования, синтезе комплементарной ДНК (кДНК) и ПЦР в режиме реального времени, предполагающей использование одной системы оригинальных олигонуклеотидных праймеров и зонда, рассчитанных на участки от 5'-NTR до 3D-гена включительно, размером ампликонов 7279 п.н. с применением высокоточных ферментов (ревертазы Maxima Н Minus Reverse Transcriptase и Taq-ДНК-полимеразы Platinum Polymerase High Fidelity). По итогам анализа оценивают наличие или отсутствие продуктов ПЦР-РВ в виде сформированных сигмоидных графиков (экспонент). На основании полученных экспоненциальных функций и значений пороговых циклов амплификации (Ct) делают заключение о целостности анализируемого участка генома вируса ящура, кодирующего структурные и неструктурные протеины возбудителя ящура и, как следствие, о полноте инактивации его антигена.
В настоящее время метод ОТ-ПЦР-РВ применяют для проведения молекулярно-биологической диагностики ящура (Reid et al., 2003; Vangrysperre & De Clerq, 1996; Callahan et al., 2002 и др.) [23-27].
Для оценки полноты инактивации антигена вируса ящура для вакцин используют в качестве биологической тест-системы первичную монослойную клеточную линию почки свиньи СП (прототип). По сравнению с прототипом метод ОТ-ПЦР-РВ с амплификацией большеразмерного участка генома вируса ящура отличается высокой чувствительностью и специфичностью, является более экономичным, позволяет одновременно исследовать несколько десятков проб инактивированного вируссодержащего материала для вакцин, а время проведения анализа сократить до 9-10 часов. Исходя из этого, актуально применять метод ОТ-ПЦР-РВ с амплификацией большеразмерного участка генома вируса ящура для опосредованного определения полноты инактивации антигена возбудителя ящура при изготовлении противоящурных инактивированных сорбированных и эмульсионных вакцин.
Ключевым элементом заявляемого способа является опосредованное определение полноты инактивации антигена вируса ящура благодаря проведению обратно-транскриптазной и полимеразной цепной реакции в режиме реального времени с использованием одной оригинальной системы олигонуклеотидных праймеров и зонда, позволяющих амплифицировать болыперазмерный участок генома вируса ящура длиной 7279 п.н., который включает в себя гены, кодирующие все структурные и неструктурные протеины вируса ящура (составляет около 7279 п.н./8400 п.н.=86,65% от полной длины РНК вируса ящура, что является высоким значением) (участок от 5'-NTR-участка до 3D-гена включительно). Оценка значения порогового цикла амплификации позволит сделать вывод о появлении повреждений амплифицируемого большеразмерного участка генома вируса ящура и, как следствие, о полноте инактивации антигена возбудителя ящура.
Сопоставительный анализ с прототипом позволяет сделать вывод, что новизна и изобретательский уровень заявляемого изобретения заключаются в применении способа элюирования молекул РНК вируса ящура, оценке степени чистоты полученного экстракта и концентрации РНК методом спектрального сканирования, проведении ОТ-ПЦР-РВ с использованием оригинальной системы олигонуклеотидных праймеров и зонда для синтеза большеразмерного ампликона генома вируса ящура и опосредованного определения полноты инактивации антигена возбудителя, применяемого для изготовления вакцин.
Сведений об аналогах предлагаемого способа опосредованного определения полноты инактивации антигена вируса ящура в сырье для противоящурных вакцин авторами не обнаружено. Возможность применения метода ОТ-ПЦР-РВ в иной модификации и с другими системами олигонуклеотидных праймеров и зондом в литературе не представлена. Найдены сведения о применении метода обратно-транскриптазной и полимеразной цепной реакции для определения полноты инактивации вируса классической чумы свиней (КЧС) [28].
Сущность изобретения отражена на графических изображениях:
Фиг. 1 - Структура генома вируса ящура с указанием структурных и неструктурных протеинов.
Фиг. 2 - Дизайн оптимальных олигонуклеотидных прямого праймера (А), обратного праймера и зонда (Б) для опосредованного определения полноты инактивации антигена вируса ящура в сырье для вакцины с применением обратно-транскриптазной полимеразной цепной реакции в режиме реального времени при амплификации большеразмерного фрагмента.
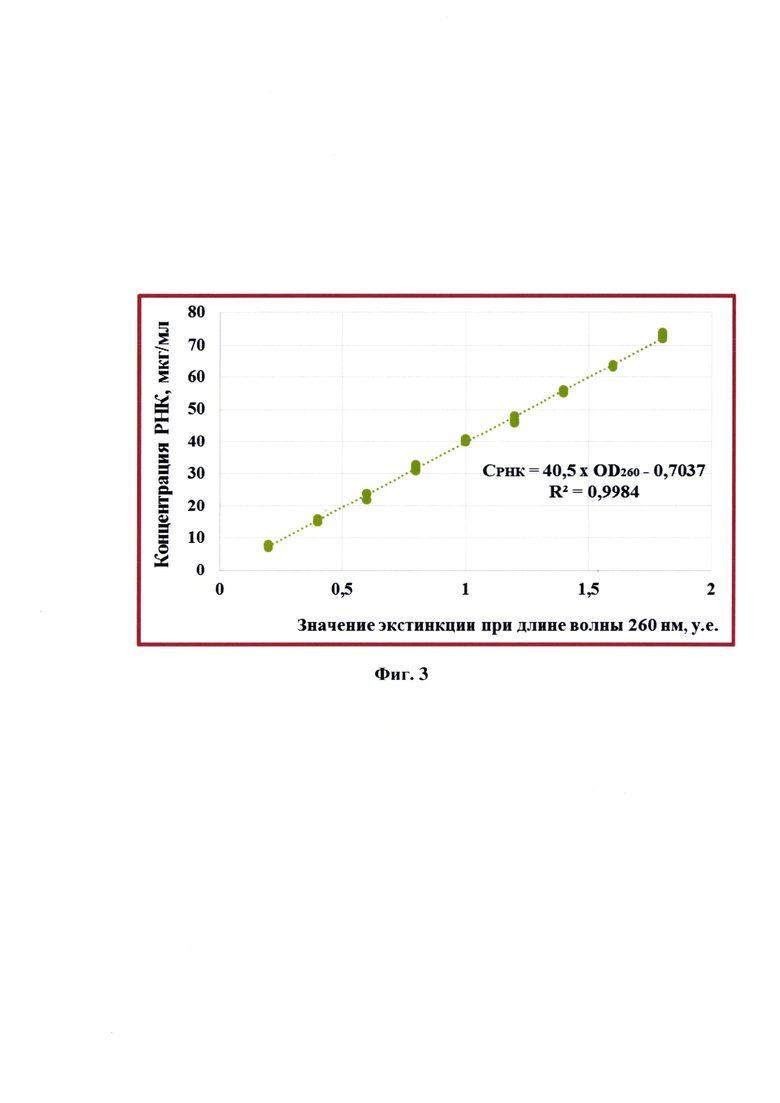
Фиг. 3 - Калибровочная кривая для определения концентрации РНК в очищенных элюатах, полученная с применением 9 стандартов в трех повторностях каждый.
Фиг. 4 - Дизайн оригинальных шести олигонуклеотидных праймеров и трех зондов (изначально предложенных для анализа) (А - прямые праймеры, Б - обратные праймеры и зонды) для опосредованного определения полноты инактивации антигена вируса ящура в сырье для вакцины с применением обратно-транскриптазной полимеразной цепной реакции в режиме реального времени при амплификации большеразмерного фрагмента.
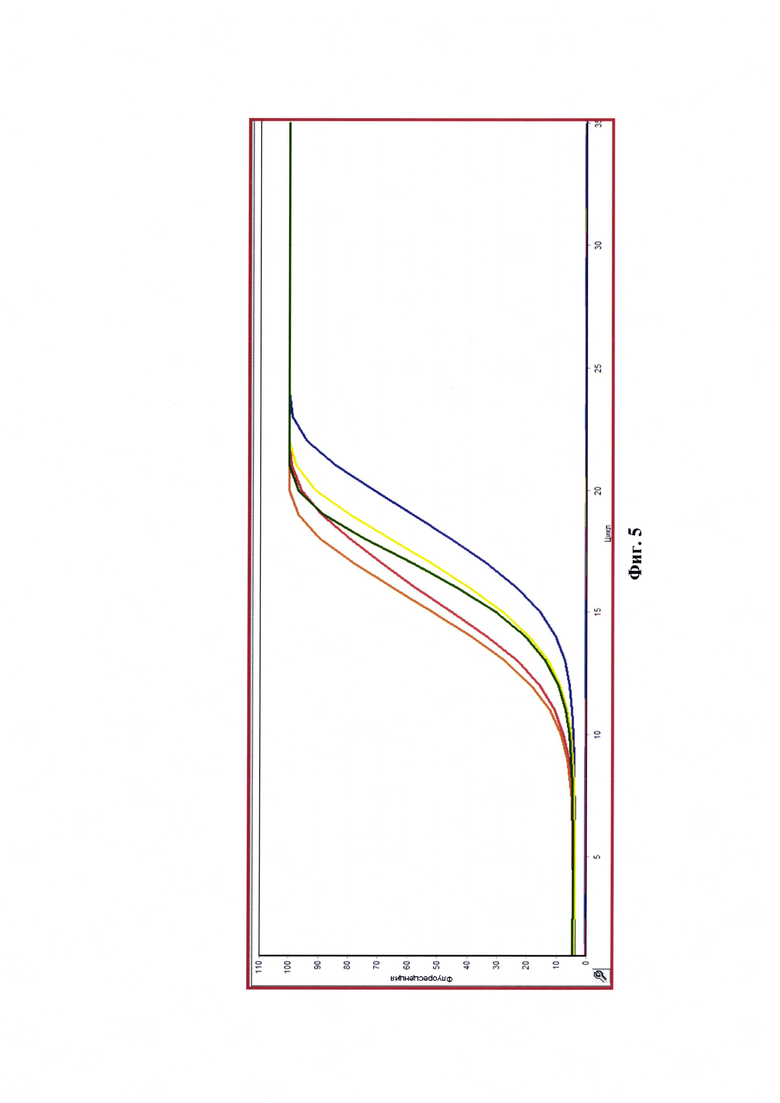
Фиг. 5 - Экспоненты реакции амплификации в режиме реального времени при тестировании пробы и контролей (ПКОвыд-е РНК - положительный контрольный образец на этапе выделения РНК (исходный не инактивированный вирус ящура) - рыжий цвет графика; ПКОПЦР - положительный контрольный образец на этапе ПЦР-РВ (проверенная кДНК не инактивированного вируса ящура) - зеленый цвет графика; ВКОвыд-е РНК - внутренний контрольный образец на этапе выделения РНК (исходный не инактивированный вирус бешенства) - желтый цвет графика; ВКОПЦР - внутренний контрольный образец на этапе ПЦР-РВ (проверенная кДНК не инактивированного вируса бешенства) - фиолетовый цвет графика; ОКО - отрицательный контрольный образец, контроль фона (деионизированная вода); ОИКОвыд-е РНК - отрицательный инактивированный контрольный образец на этапе выделения РНК (инактивированная суспензия вируса ящура); ОИКОПЦР - отрицательный инактивированный контрольный образец на этапе ПЦР-РВ (проверенная к ДНК вируса ящура, выделенная из инактивированной суспензии)). Неинактивированный контрольный образец - красный цвет графика. Данные представлены до 35 цикла, так как характер графиков с 35 по 45 циклы не изменялся.
Фиг. 6 - Монослойная перевиваемая клеточная линия почки свиньи СП без цитопатического действия (А) и с цитопатическим действием (Б).
При проведении определения полноты инактивации антигена вируса ящура требуется применять ряд контролей:
1) ПКОвыд-е РНК- положительный контрольный образец на этапе выделения РНК (исходный не инактивированный вирус ящура);
2) ПКОПЦР - положительный контрольный образец на этапе ПЦР-РВ (проверенная кДНК не инактивированного вируса ящура);
3) ВКОвыд-е РНК - внутренний контрольный образец на этапе выделения РНК (исходный не инактивированный вирус бешенства штамма РВ-97);
4) ВКОПЦР - внутренний контрольный образец на этапе ПЦР-РВ (проверенная кДНК не инактивированного вируса бешенства штамма РВ-97);
5) ОКО - отрицательный контрольный образец, контроль фона (деионизированная вода);
6) ОИКОвыд-е РНК - отрицательный инактивированный контрольный образец на этапе выделения РНК (инактивированная суспензия вируса ящура);
7) ОИКОПЦР - отрицательный инактивированный контрольный образец на этапе ПЦР-РВ (проверенная кДНК вируса ящура, выделенная из инактивированной суспензии).
Из суспензии исследуемого образца, а также из ПКОвыд-е РНК, BKOвыд-e РНК, ОКО и ОИКОвыд-е РНК выделяют РНК в процессе лизиса белков и липопротеинов, очистки нуклеиновой кислоты от примесей, ее концентрирования на стекловолокнистом фильтре и получения готового элюата. Данный метод выделения РНК позволяет достичь высокой стандартизации в процедуре экстракции и получить из образцов высокоочищенные препараты нуклеиновой кислоты. Для работы используют 5 М гуанидинтиоцианат (ГТЦ), сильный денатурирующий агент, который обеспечивает одновременный лизис клеток и денатурации всех клеточных белков (в том числе РНКаз). Усиление эффекта обеспечивается применением 0,5% меркаптоэтанола. Для очистки РНК от примесей дважды применяют 80% раствор изопропанола. Суммарную РНК после экстрагирования на стекловолокнистом фильтре элюируют в 0,2 см3 деионизированной воды. Таким образом, получают по 0,2 см3 элюатов суммарной РНК.
Для оценки качества и количества РНК в полученных экстрактах проводят спектральное сканирование, определяя поглощение аналитом монохроматического ультрафиолетового света, что позволяет оценить степень чистоты и концентрации продукта, который в последующем используют в ОТ-ПЦР-РВ. Измерения спектральной поглощающей способности образцов осуществляют при длинах волны в диапазоне 205-325 нм и температуре 23-25°С в кювете с длиной оптического пути 10 мм. При измерениях экстинкции нужно учитывать коэффициент разведения раствора исследуемого образца и длину оптического пути (это, чаще всего, компенсируется спектрофотометром).
Индикатором остатков фосфолипидов, полисахаридов и ГТЦ, полипептидов и крупных взвешенных частиц являются значения экстинкции при длинах волны 230, 280 и 320 нм, соответственно [29]. Элюат РНК считают свободным от примесей полипептидов, если OD260/OD280 (коэффициент экстинкции R1) ≥2,0 [29, 30]. Более низкие значения R1 указывают на наличие пептидных связей составляющих в элюате. Более высокие значения коэффициента R1 свидетельствуют о деградации РНК и наличии свободных рибонуклеотидов. Экстракт нуклеиновой кислоты считают незагрязненным полисахаридами, если OD260/OD230 (коэффициент экстинкции R2) ≥2,000 [29, 30]. Более низкие значения указывают, как правило, на загрязнение ГТЦ и углеводными составляющими. При замещении 1% РНК на полисахаридные составляющие R2 снижается на 0,002. Значения коэффициента R2 большие 2,200 могут указывать на деградацию молекул РНК. Отсутствие взвеси крупных частиц в элюате подтверждается, если OD320 приближено к нулевому значению, т.е. OD260/OD320 (коэффициент экстинкции R3) стремится к OD260 [29, 30]. При несоответствии требованиям чистоты повторно проводят этап выделения РНК из исходного материала.
Для определения концентрации молекул РНК в растворе учитывают фактор пересчета (F), равный 40. При этом величина оптической плотности 1,000 о.е. соответствует концентрации примерно 40 мкг/см3 раствора РНК, что обусловлено первичной структурой молекулы. Концентрацию РНК определяли, пользуясь разработанной формулой СРНК=40,5 × OD260 - 0,7037. График по данной формуле представлен на фиг. 3 и выведен эмпирически.
На следующем этапе исследования полученные экстракты вирусной РНК исследуемого образца и контролей исследуют в ОТ-ПЦР-РВ. Для постановки реакции готовят ОТ-ПЦР-РВ-смесь, рецептура приготовления которой представлена в таблице 1. В качестве гомологичного 5'-NTR-гену участка вируса ящура используют олигонуклеотидный прямой праймер 5'-NTR-F-fov2 (TTTCCAGGTCTAGAGGGGTGA), олигонуклеотидный обратный праймер 3D-R1-rev (GCGAGTCCTGCCACGGA) и олигонуклеотидный 3DP-rev-зонд (ROX-GTCCCACGGCGTGCAAAGGA-BHQ2), меченый красителем карбокси-Х-родамином (ROX) и гасителем флуоресценции BHQ 2 в концентрации 10 пМ на реакцию с внесением в ОТ-ПЦР-РВ-смесь по 0,1 мкл. Для формирования нуклеотидных цепей продуктов реакции применяют дезоксирибонуклеозидтрифосфаты с концентрацией в ОТ-ПЦР-РВ-смеси по 2,00 мМ. В качестве основы используют Thermo Scientific™ 10Х DreamTaq™ Buffer, содержание которого составляет 10% от общего объема реакционной смеси. DreamTaq™ Buffer специально оптимизирован для ПНР с использованием ДНК-полимеразы DreamTaq™. Буфер содержит как KCl, так и (NH4)2SO4 для обеспечения высокой специфичности отжига праймеров в широком диапазоне концентраций MgCl2. В реакционную смесь добавляют 3,0 мМ хлорида магния. В качестве ферментов применяют ревертазу Maxima Н Minus (Thermo Scientific) (10 е.а.) и ДНК-полимеразу Platinum High Fidelity (Invitrogene) (10 е.a.).
Ревертаза обладает РНК- и ДНК-зависимой полимеразной активностью, но при этом отсутствует активность РНКазы Н, характеризуется высокой степенью эффективности. Ревертаза Maxima Н Minus (Thermo Scientific) хранится в буфере следующего состава: 50 мМ трис-HCl (рН 7,5), 0,1 М NaCl, 1 мМ ЭДТА, 5 мМ ДТТ, 0,1% Triton Х-100 и 50% глицерина.
ДНК-полимераза Platinum High Fidelity (Invitrogene) была выбрана, поскольку данный энзим идеально подходит для амплификации фрагментов ДНК, когда требуются высокий выход ампликонов с большим размером и надежное усиление процесса. Точность данного фермента более чем в шесть раз выше, чем у ДНК-полимераза Taq. Благодаря этому обеспечивается высокой выход полноразмерных продуктов кДНК размером до 20000 п.н. В случае разработанного способа требуется амплифицировать фрагмент размером 7279 п.н. Полимераза Platinum® Taq хранится в буферном растворе (600 мМ Tris-S04 (рН 8,9), 180 мМ (NH4)2SO4).
Объем ОТ-ПЦР-РВ-смеси компонентов для проведения одной реакции составляет 20 мкл. Элюаты РНК каждого образца добавляют к ОТ-ПЦР-РВ-смеси по 5 мкл. Объем реакционной смеси составляет 25 мкл.
Постановку реакции осуществляют при температурных и временных параметрах, сведения о которых представлены в таблице 1. Обратную транскрипцию проводят при температуре 50°С в течение 30 мин. за 1 цикл, активацию ДНК-полимеразы Platinum High Fidelity - при температуре 95°С за 5 мин. в течение 1 цикла. Реакцию амплификации в режиме реального времени осуществляют в течение 45 циклов, каждый из которых складывается из 3 под этапов: «денатурации», проводимой при температуре 95°С в течение 30 с, этапа «отжига праймеров и зонда» - при температуре 57°С в течение 30 с, этапа «элонгации и аккумулирования флуоресцентного сигнала», осуществляемых при температуре 63°С за 8 минут. Время элонгации увеличено до 8 минут, исходя из ряда проведенных исследований (данные представлены в примере 1). Общее время проведения ОТ-ПЦР-РВ в данном случае составляет 440 мин. или 7 ч 20 мин.
Процесс основан на использовании 5'-экзонуклеазной активности ДНК-полимеразу Platinum High Fidelity [31]. В отсутствии мишени флуорофор ROX и гаситель флуоресценции BHQ2 в составе 3DP-rev-зонда сближены за счет максимального использования водородных связей между атомами Н, О и N олигонуклеотидов. Благодаря механизму флуоресцентно-резонансного переноса энергии свечение подавлено. За счет 5'-экзонуклеазной активности полимеразы после отжига оригинальных праймеров и зонда происходит разрушение гибридизованного зонда и ампликона, наблюдается их пространственное разделение, что приводит к росту детектируемого сигнала. Увеличение уровня флуоресценции (F1) пропорционально количеству образующихся продуктов реакции. Мониторинг сигнала в течение 45 циклов (С) ПЦР в режиме реального времени позволяет построить кинетическую кривую флуоресценции, которая задана функцией Fl=f (С).
Осуществляют интерпретацию полученных данных. Важным условием качественно проведенных исследований являются результаты испытания контрольных образцов. Если положительный контрольный образец на этапе выделения РНК (исходный не инактивированный вирус ящура) (ПКОвыд-е РНК); положительный контрольный образец на этапе ПЦР-РВ (проверенная кДНК не инактивированного вируса ящура) (ПКОПЦР); внутренний контрольный образец на этапе выделения РНК (исходный не инактивированный вирус бешенства) (ВКОвыд-е РНК); внутренний контрольный образец на этапе ПЦР-РВ (проверенная кДНК не инактивированного вируса бешенства) (ВКОПЦР) после всех стадий исследования дают положительный результат (наличие экспонент со значением порогового цикла амплификации <40,0 ед.), отрицательный контрольный образец (деионизированная вода) (ОКО); отрицательный инактивированный контрольный образец на этапе выделения РНК (инактивированная суспензия вируса ящура) (ОИКОвыд-е РНК); отрицательный инактивированный контрольный образец на этапе ПЦР-РВ (проверенная кДНК вируса ящура, выделенная из инактивированной суспензии) (ОИКОПЦР) характеризуются в качестве отрицательных образцов (отсутствие экспонент), то в таком случае стадии выделения РНК, получения кДНК и реакции амплификации проведены правильно, и можно учитывать результаты исследуемых проб.
Если для исследуемой пробы обнаружены графики ПЦР-РВ с пороговым циклом <40,00 ед., то анализируемый участок нуклеиновой кислоты вируса ящура размером 7279 п.н. не поврежден и поврежден только у части вирусных частиц, следовательно, данный образец считается вирулентным. Если экспонента и, как следствие, накопление ампликона отсутствует, то данный участок генома вируса ящура поврежден, тем самым, репликация вируса не возможна и антиген считается авирулентным. Образцы с показателями порогового значения, находящимися в пределах 40-45 ед., обозначаются как «пограничные» и должны быть протестированы повторно. Иными словами, на основании отсутствия графиков ПЦР-РВ тестируемого образца по сравнению с положительными контролями позволяет сделать заключение о разрушении генома вируса ящура всех вирионов в инактивированных суспензиях и, как следствие, об отсутствии вирулентного вируса.
Таким образом, неинактивированные тестируемые образцы и положительные (неинактивированные) контроли должны иметь показатель порогового цикла (Ct) <40,00 ед. Образцы с показателями порогового значения, находящимися в пределах 40-45 ед., обозначаются как «пограничные» и должны быть протестированы повторно. В том случае, если обнаружены несоответствия в результатах анализа контролей, требуется повторно провести исследование именно того этапа (выделение РНК и ОТ-ПЦР-РВ), на котором выявлено отклонение от нормы.
Пример 1. Подбор оптимальных олигонуклеотидных праймеров и зонда для синтеза болъшеразмерного фрагмента генома вируса ящура при исследовании полноты инактивации антигена вируса с помощью ОТ-ПЦР-РВ.
Оптимизация параметров ОТ-ПЦР-РВ для определения полноты инактивации антигена вируса ящура проведена с использованием часто применяемого в производстве вакцин штамма Азия-1/Шамир/89 (JF739177.1 - адрес последовательности нуклеотидов на сайте https://www.ncbi.nlm.nih.gov/search/all/?term=FMDV).
С целью подбора олигонуклеотидной системы праймеров и зонда для выявления участка генома вируса ящура длиной 7279 п.н., который включает в себя гены, кодирующие все структурные и неструктурные протеины вируса, были проанализированы полные нуклеотидные последовательности 5'-NTR-участка и 3D-гена, выделенных в период с 1999 по 2021 гг. в странах Африки и Азии и принадлежащих к различным генетическим группам, опубликованные в базе данных GeneBank электронного ресурса NCBI [15]. Множественное выравнивание нуклеотидных последовательностей с использованием алгоритма Clustal W и поиск наиболее консервативных участков проводили с помощью программы BioEdit 7.0. Специфичность праймеров и зондов была оценена с помощью on-line-pecypca Blast (NCBI) (http://blast.ncbi.nlm.nih.gov) [15], с использованием производственных штаммов вируса ящура, отраженных в таблице 2.
Для сравнительного анализа была сформирована первоначальная выборка из 11 912 нуклеотидных последовательностей 5'-NTR-участка и 3D-гена изолятов и штаммов вируса ящура. После пошаговой оптимизации, удаления из выборки наиболее генетически схожих последовательностей для анализа было выбрано 100 наиболее отличающихся между собой. Для всех последовательностей характерно наличие относительно консервативных участков, расположенных в середине 5'-NTR-участка и в конце 3D-гена. Оптимальной для отжига праймеров и зонда выбраны именно эти области, поскольку количество замен в этих областях наименьшее по сравнению с другими участками гена для всех последовательностей в указанной выборке.
В результате проведенной работы были выбраны и синтезированы 6 олигонуклеотидных праймеров и 3 флуоресцентных зонда для последующего их испытания в ОТ-ПЦР-РВ с целью поиска системы с высокой специфичностью и чувствительностью для производственных штаммов вируса ящура по данным пороговых циклов амплификации (Ct). Дизайн оптимальных оригинальных прямого, обратного праймеров и зонда осуществлен в лаборатории профилактики ящура ФГБУ «ВНИИЗЖ» и отражен в таблице 3 и на фиг. 4. Для тестирования предложены 3 системы олигонуклеотидных праймеров и зондов: 1) 5'-NTR-F-fovl-праймер, 3DP-2 rev-зонд, 3D-R2-rev-npafiMep; 2) 5'-NTR-F2-fov2-праймер, 3DP-rev-зонд, 3D-R1-rev-праймер; 3) 5'-NTR-F3-fov-праймер, 3DP-3 rev-зонд, 3D-R3-rev-праймер (таблица 3).
Проведена постановка ОТ-ПЦР-РВ с указанными системами праймеров и зондов для анализа производственных штаммов вируса ящура, отраженных в таблице 2. Титр инфекционной активности вируса во всех образцах составлял 7,00-8,00 lg ТЦД50/см3. Исследования с каждым штаммом и системой олигонуклеотидов в ОТ-ПЦР-РВ проводили в пяти повторностях. Полученные данные пороговых циклов амплификации представлены в таблице 4, из которой видно, что вторая система праймеров и зонда (5'-NTR-F2-fov2-праймер, SDP-rev-зонд, 3D-R1-rev-праймер) давала стабильно более высокие значения Ct, при этом были выявлены все исследуемые штаммы вируса ящура.
В ходе экспериментального проведения ОТ-ПЦР-РВ было выявлено, что из указанных олигонуклеотидных систем выбрана наиболее подходящая вторая система праймеров и зонда (в таблице 3 указаны полужирным шрифтом), с которой в дальнейшем проводилась оптимизация реакции для повышения степени ее чувствительности.
Пример 2. Подбор условий ОТ-ПЦР-РВ для синтеза большеразмерного фрагмента генома вируса ящура при оценке полноты инактивации антигена вируса.
Для достижения высокой чувствительности, специфичности и скорости ОТ-ПЦР-РВ для амплификации большеразмерного участка генома вируса ящура требовалось подобрать оптимальную концентрацию компонентов реакционной смеси и температурно-временной режим реакции. Непосредственно перед осуществлением процесса оптимизации условий постановки ОТ-ПЦР-РВ на участок длиной 7279 п.н. (5'-NTR-участок - 3D-ген) были приготовлены десятикратные разведения (10-1-10-8) выделенной суммарной РНК штамма вируса ящура Азия-1/Шамир 3/89. В целях повышения чувствительности и специфичности ОТ-ПЦР-РВ для амплификации большеразмерного участка генома вируса ящура и определения на основе результатов полноты инактивации антигена вируса основной задачей был подбор концентраций компонентов реакционной смеси. Для этого проведены постановки ОТ-ПЦР-РВ с использованием цельного и десятикратных разведений (10-1-10-8) вакцинного штамма Азия-1/Шамир 3/89. Для постановки реакции использовали тот же температурно-временной режим, что и в ОТ-ПЦР-РВ на ген 3D по данным OIE [2]. В таблицах 5-8 представлены средние значения порогового цикла (Ct) реакции по цельной РНК вируса и ее 8 разведениям (10-1-10-8) (n=3).
Подбор оптимальной концентрации хлорида магния в ОТ-ПЦР-РВ для определения полноты инактивации антигена вируса ящура. Для функционирования ДНК-полимеразу Platinum High Fidelity, являются катионы Mg2+, которые влияют на специфичность гибридизации олигонуклеотидов. Кроме того, катионы Mg2+ формируют растворимые комплексы с дезоксирибонуклеозидтрифосфатами, образуя субстрат для фермента. Оптимальная концентрация катионов Mg2+ может находиться в широком диапазоне в зависимости от используемых систем олигонуклеотидных праймеров и зондов, а также энзимов, и, чаще всего, составляет 1-5 мМ [32]. Как правило, для достижения наилучших результатов концентрацию Mg2+ подбираются эмпирическим путем для применяемой системы олигонуклеотидов и ферментов. Для анализа применяли концентрации хлорида магния 1,0, 2,0, 3,0, 4,0, 5,0 мМ. Результаты исследований представлены в таблице 5. Из представленных в таблице 5 данных видно, что изменение концентрации хлорида магния оказывает значительное влияние на результат реакции амплификации. Выявлено, что оптимальная концентрация MgCl2 для второй системы олигонуклеотидных праймеров и зонда, а также для ДНК-полимеразы Platinum High Fidelity составляет 3 мМ. При указанной концентрации значения пороговых циклов амплификации били наименьшими. При содержании хлорида магния 1,0 мМ выявить продукты реакции амплификации не удалось, при концентрациях 2,0, 4,0 и 5,0 мМ значения пороговых циклов амплификации для элюатов РНК были выше по сравнению с выбранным оптимальным количеством 3,0 мМ MgCl2. Увеличение концентрации соли магния может приводить к уменьшению специфичности реакции, поскольку свободные катионы магния существенно повышают температуру гибридизации двуцепочечной ДНК [33-35].
Подбор оптимальной концентрации олигонуклеотидных праймеров в ОТ-ПЦР-РВ для определения полноты инактивации антигена вируса ящура. Оптимальную концентрацию олигонуклеотидных праймеров 5'-NTR-F2-fov2, 3D-R1-rev подбирали экспериментально. С представленными в таблице 6 разведениями РНК вируса ящура штамма Азия-1/Шамир 3/89 готовили реакционные смеси, добавляя в каждую из них разный объем праймеров, а именно 0,1, 0,2, 0,3 и 0,4 мкл. Концентрации исходных растворов олигонуклеотидов составляли по 50 пмоль/мкл. В результате проведенных исследований выявлено, что увеличение концентрации олигонуклеотидов приводит к уменьшению чувствительности реакции, что отражено в таблице 6. Средние пороговые значения отличались на 1,10-2,38 цикла. Наименьшие значения пороговых циклов получены в реакциях, в которых к реакционной смеси добавляли 0,5 мкл каждого из олигонуклеотидов. По литературным данным повышение концентрации праймеров вызывает увеличение вероятности формирования неспецифических продуктов реакции амплификации, в том числе образование димеров праймеров. Уменьшение количества олигонуклеотидов может привести к снижению эффективности реакции и образованию меньшего количества ампликонов [34-38].
Подбор оптимальной концентрации флуоресцентного зонда для ОТ-ПЦР-РВ при определении полноты инактивации антигена вируса ящура. С целью повышения эффективности реакции амплификации и увеличения уровня флуоресценции сигмоидных кривых подобран объем вносимого на одну реакцию зонда. Анализ проводили с применением зонда 3DP-rev с вносимыми объемами на одну реакцию 0,10, 0,15, 0,20, 0,30 0,40 мкл с концентрацией исходного раствора зонда 50 пмоль/мкл. Объем праймеров составлял 0,10 мкл на реакцию с концентрацией исходного раствора 50 пмоль/мкл. Исследовали ранее полученные разведения выделенной РНК вируса ящура штамма Азия-1/Шамир 3/89. Значения полученных пороговых циклов реакции амплификации представлены в таблице 7. Выявлено, что при добавлении в реакционную смесь от 0,10 до 0,40 мкл флуоресцентного зонда регистрировали незначительные изменения чувствительности ОТ-ПЦР-РВ. Средние пороговые значения отличались на 0,29-1,63 цикла. Наименьшие значения пороговых циклов амплификации получены при использовании 1,0 мкл 3DP-rev-зонда. При использовании остальных компонентов реакции придерживались рекомендаций производителей.
Подбор температурно-временных параметров ОТ-ПЦР-РВ для определения полноты инактивации антигена вируса ящура. Проводили подбор температурно-временного режима, а именно ПЦР-РВ, поскольку температурный режим обратной транскрипции определяется обратной транскриптазой Maxima Н Minus (Thermo Scientific) [39]. Как правило, данный фермент эффективный синтез кДНК проводит за 15-30 минут [40].
В работе проводили определении оптимальной температуры отжига для олигонуклеотидов (5'-NTR-F2-fov2-праймер, 3DP-rev-зонд, 3D-R1-rev-праймер). Остальные этапы ПЦР-РВ - денатурация и элонгация -осуществляются в достаточно узком температурном диапазоне. Денатурацию обычно проводят при 90-95°С, а синтез цепи ДНК - при 68-72°С (при этом считается, что скорость работы полимеразы составляет около 50-100 нуклеотидов за секунду) [33].
Температура отжига олигонуклеотидов (Та) является одним из важнейших факторов в разработке высокоспецифичной ПЦР-РВ-системе. При занижении температуры могут образовываться неспецифические продукты реакции амплификации, а при завышении - может значительно снижаться эффективность ПЦР. Отжиг праймеров и зонда для амплификации большеразмерного фрагмента в ОТ-ПЦР-РВ проводили при следующих температурах: 53, 55, 57, 60, 65°С. Результаты подбора температуры отжига олигонуклеотидов при постановке ПЦР-РВ представлены в таблице 8, из которой следует, что сопоставимые значения пороговых циклов амплификации наблюдаются при температуре отжига праймеров и зонда 57°С, что подходит для большинства олигонуклеотидных систем, используемых в ОТ-ПЦР-РВ для молекулярной диагностики ящура. Представляется вполне логичным использование этой же температуры отжига для амплификации фрагмента генома вируса ящура размером 7279 п.н. В разработанном варианте для постановки ОТ-ПЦР-РВ и определения полноты инактивации антигена вируса ящура данным методом устанавливали следующие температурно-временные параметры: 30 мин при 50°С (обратная транскрипция), 5 мин при 95°С (активация ДНК-полимеразы Platinum High Fidelity), далее 45 циклов ПЦР, состоящие из денатурации ДНК в течение 0,5 мин при 95°С, отжига праймеров и зонда - 0,5 мин при 57°С и элонгации кДНК - 8 мин при 72°С. Большое время элонгации в каждом цикле обусловлено длиной амплифицирумого фрагмента (примерно по 1 минуте на 1000 п.н., в данном случае 7279 п.н. - около 8 минут).
Пример 3. Инактивация антигена вируса ящура и исследование полученной суспензии в клеточной линии и разработанным способом в ОТ-ПЦР-РВ.
Суспензию вируса ящура штамма Азия-1/Шамир 3/89 с титром инфекционной активности 7,75 lg ТЦИД50/см3, по данным титрования в перевиваемой монослойной клеточной линии IB-RS-2 [2], сразу после репродукции в суспензионной культуре клеток ВНК-21 разделяли контроль неинактивированный и исследуемый образец. Тестируемый образец подвергали процессу инактивации с применением аминоэтилэтиленимина в концентрации 0,025%, температура - плюс 37±0,5°С.время экспозиции - 18 ч. Водородный показатель вирусной суспензии поддерживали в диапазоне 7,4-7,6. Суспензию подвергали тщательному перемешиванию в течение 3-5 минут через каждый час. Условия инактивации соответствовали требованиям МЭБ (OIE) [2].
Параллельно исследовали вирус ящура штамма Азия-1/Шамир 3/89, не подвергнутый инактивации, экспозиция которого проводилась при той же температуре и в тот период времени.
Контрольный (неинактивированный) и опытный (инактивированный) образцы исследовали на наличие инфекционной активности вируса ящура в перевиваемой монослойной культуре клеток почки свиньи IB-RS-2 в соответствии с требованиями МЭБ (OIE) [2]. Исследование каждого образца проводили в 4 повторностях. Анализ монослоя на наличие/отсутствие цитопатического действия осуществляли раз в сутки в течение 3 дней. Изображение монослоя на третьи сутки после инокуляции контрольным и опытным образцами представлено на фиг. 6, на котором видно, что наличие ЦПД характерно для неинактивированного материала и его отсутствие - для инактивированной суспензии вируса ящура.
Параллельно данные суспензии анализировали с помощью ОТ-ПЦР-РВ для исследования полноты инактивации антигена вируса ящура при амплификации большеразмерного ампликона.
При проведении оценки полноты инактивации антигена вируса ящура разработанным способом применяли следующие контроли: ПКОвыд-е РНК - положительный контрольный образец на этапе выделения РНК (исходный не инактивированный вирус ящура); ПКОПЦР - положительный контрольный образец на этапе ПЦР-РВ (проверенная кДНК не инактивированного вируса ящура); ВКОвыд-я РНК - внутренний контрольный образец на этапе выделения РНК (исходный не инактивированный вирус бешенства штамма РВ-97); ВКОПЦР - внутренний контрольный образец на этапе ПЦР-РВ (проверенная кДНК не инактивированного вируса бешенства штамма РВ-97); ОКО -отрицательный контрольный образец, контроль фона (деионизированная вода); ОИКОвыд-е РНК- отрицательный инактивированный контрольный образец на этапе выделения РНК (инактивированная суспензия вируса ящура); ОИКОПЦР - отрицательный инактивированный контрольный образец на этапе ПЦР-РВ (проверенная кДНК вируса ящура, выделенная из инактивированной суспензии).
Из суспензии контрольного (инактивированного) и исследуемого (инактивированного) образца, а также из ПКОвыд-е РНК, ВКОвыд-е РНК, ОКО и ОИКОвыд-е РНК выделяли РНК вируса ящура. К 100 мл образца добавляли 400 мкл смеси 5 М гуанидинтиоцианата (ГТЦ) и 0,5% меркаптоэтанола (1:1), инкубировали при комнатной температуре в течение 5 минут. Лизат помещали в колонки на стекловолокнистое волокно. Процесс выполняли на системе Manifold, к которой подключали вакуумный насос Vacuum Pump 2С (Vacuubrand). Для очистки РНК от примесей белков дважды применяли 80% раствор изопропанола, внося его по 800 мкл. Суммарную РНК после экстрагирования на стекловолокнистом фильтре элюировали в 0,2 см3 деионизированной воды. Таким образом, получили по 0,2 см3 элюатов РНК.
Для оценки качества и количества РНК в полученных экстрактах проводили спектральное сканирование при длинах волны в диапазоне 205-325 нм и температуре 23-25°С в кювете с длиной оптического пути 10 мм. Индикатором остатков фосфолипидов, полисахаридов и ГТЦ, полипептидов и крупных взвешенных частиц являются значения экстинкции при длинах волны 230, 280 и 320 нм, соответственно [29]. Коэффициент экстинкции R1 (OD260/OD280) для полученных элюатов РНК был свободным от примесей полипептидов, поскольку R1 находился в диапазоне 2,002-2,009 о.е. (норма ≥2,000 о.е.) [29, 30]. Экстракты нуклеиновой кислоты считались незагрязненными полисахаридами и ГТЦ так как коэффициенты экстинкции R2 (OD260/OD230) составляли 2,001-2,007 о.е. (норма ≥2,000 о.е.) [29, 30]. В полученных элюатах не обнаружено крупных взвешенных частиц, поскольку OD320 приближено к нулевому значению (0,001-0,004 о.е.) [29, 30].
Для определения концентрации молекул РНК в элюатах учитывали фактор пересчета (F), равный 40. При этом величина оптической плотности 1,000 о.е. соответствует концентрации примерно 40 мкг/см3 раствора РНК, что обусловлено первичной структурой молекулы.
Изначально эмпирическим путем получили формулу расчета концентрации РНК в элюате. Для этого брали стандарты очищенных элюатов РНК в количестве 9 штук с концентрациями: 0,2, 0,4, 0,6, 0,8, 1,0, 1,2, 1,4, 1,6, 1,8 мкг/мл. Определяли их значения оптических плотностей при длине волны 260 нм. Полученные данные отразили на фиг. 3, на которой изображен линейный график, позволяющий определять по результатам спектрального сканирования концентрацию РНК в экстракте. График представлен формулой: СРНК=40,5 × OD260 - 0,7037. Пользуясь выведенной формулой для исследуемых образцов рассчитали концентрации РНК, которые составили 50,33-57,78 мкг/мл. Все данные по спектральному сканированию отражены в таблице 8, из которой следует, что полученные элюаты очищены и содержат в своем составе РНК.
На следующем этапе исследования полученные экстракты РНК исследуемого образца и контролей исследовали в ОТ-ПЦР-РВ. Для постановки реакции готовили ОТ-ПЦР-РВ-смесь, рецептура приготовления которой представлена в таблице 1. Использовали олигонуклеотидный прямой праймер 5'-NTR-F-fov2 (TTTCCAGGTCTAGAGGGGTGA), олигонуклеотидный обратный праймер 3D-R1-rev (GCGAGTCCTGCCACGGA) и олигонуклеотидный 3DP-rev-зонд (ROX-GTCCCACGGCGTGCAAAGGA-В HQ2), меченый красителем карбокси-Х-родамином (ROX) и гасителем флуоресценции BHQ2 в концентрации 10 пМ на реакцию с внесением в ОТ-ПЦР-РВ-смесь по 0,1 мкл. Для формирования нуклеотидных цепей продуктов реакции применяют дезоксирибонуклеозидтрифосфаты с концентрацией в ОТ-ПЦР-РВ-смеси по 2,00 мМ. В качестве основы используют Thermo Scientific™ 10Х DreamTaq™ Buffer, содержание которого составляет 10% от общего объема реакционной смеси. DreamTaq™ Buffer специально оптимизирован для ПЦР с использованием ДНК-полимеразы DreamTaq™. В реакционную смесь добавляли 3,0 мМ хлорида магния. В качестве ферментов применяли ревертазу Maxima Н Minus (Thermo Scientific) (10 е.а.) и ДНК-полимеразу Platinum High Fidelity (Invitrogene) (10 е.а.). Объем ОТ-ПЦР-РВ-смеси компонентов для проведения одной реакции составлял 20 мкл. Элюаты РНК вируса ящура каждого образца добавляли к ОТ-ПЦР-РВ-смеси по 5 мкл. Объем реакционной смеси составляет 25 мкл.
Обратную транскрипцию проводили при температуре 50°С в течение 30 мин. за 1 цикл, активацию ДНК-полимеразы Platinum High Fidelity - при температуре 95°С за 5 мин. в течение 1 цикла. Реакцию амплификации в режиме реального времени осуществляли в течение 45 циклов, каждый из которых складывается из 3 под этапов: «денатурации», проводимой при температуре 95°С в течение 30 с, этапа «отжига праймеров и зонда» - при температуре 57°С в течение 30 с, этапа «элонгации и аккумулирования флуоресцентного сигнала», осуществляемых при температуре 63°С за 8 минут.
Осуществляли интерпретацию полученных данных, учитывая изначально результаты испытания контрольных образцов. Положительный контрольный образец на этапе выделения РНК (исходный не инактивированный вирус ящура) (ПКОвыд-е РНК), положительный контрольный образец на этапе ПЦР (проверенная кДНК не инактивированного вируса ящура) (ПКОПЦР), внутренний контрольный образец на этапе выделения РНК (исходный не инактивированный вирус бешенства) (ВКОвыд-е РНК), внутренний контрольный образец на этапе ПЦР (проверенная кДНК не инактивированного вируса бешенства) (ВКОПЦР) после всех стадий исследования дали положительный результат (наличие экспонент со значением порогового цикла амплификации <40,0 у.е.); отрицательный контрольный образец (деионизированная вода) (ОКО), отрицательный инактивированный контрольный образец на этапе выделения РНК (инактивированная суспензия вируса ящура) (ОИКОвыд-е РНК), отрицательный инактивированный контрольный образец на этапе ПЦР (проверенная кДНК вируса ящура, выделенная из инактивированной суспензии) (ОИКОПЦР) дали отрицательный результат (отсутствие экспонент). Таким образом, этапы выделения РНК, получения кДНК и реакции амплификации проведены правильно, и можно учитывать результаты исследуемой инактивированной пробы суспензии вируса ящура штамма Азия-1/Шамир 3/89. Результаты исследования продемонстрированы на фиг. 5.
Для исследуемой инактивированной пробы не обнаружено наличия графика ПЦР-РВ, иными словами, накопление ампликона отсутствует, данный участок генома вируса ящура (размером 7279 п.н., диапазон от 5'-NTR-участка до 3D-гена включительно) поврежден, тем самым, репликация вируса не возможна и антиген считается авирулентным. Полученные результаты исследования контрольного (неинактивированного) и опытного (инактивированного) образца разработанным способом и в клеточной линии почки свиньи IB-RS-2 (прототип) согласовывались.
Таким образом, способ опосредованного определения полноты инактивации антигена вируса ящура в сырье для вакцины с применением ОТ-ПЦР-РВ при амплификации большеразмерного фрагмента не уступает по своей эффективности традиционному биологическому методу и позволяет значительно сократить время проведения исследования.
Пример 4. Определение диагностической чувствительности способа опосредованного определения полноты инактивации антигена вируса ящура в сырье для вакцины с применением ОТ-ПЦР-РВ при амплификации болъшеразмерного фрагмента.
Каждый метод анализа имеет свои пределы диагностических возможностей, которые отражены в их основных рабочих характеристиках: чувствительность, специфичность и общая точность [41]. Точность метода определяется, во-первых, его диагностической чувствительностью, то есть выявляется, что проба инактивирована в том случае, если это истина (ДП - достоверно положительный результат), при этом процент выявленных ложноинактивированных проб (ЛП - ложноположительный результат) должен стремиться к нулю; во-вторых, специфичностью, то есть полнота инактивации не выявляется, когда проба вирулентна (неинактивирована) (ДО - достоверно отрицательный результат), при этом процент ложно вирулентных проб (ЛО - ложноотрицательный результат) должен стремиться к нулю.
Чувствительность (истинно положительная пропорция) отражает долю положительных результатов анализа, которые правильно идентифицированы как таковые. Иными словами, чувствительность диагностического теста показывает вероятность того, что инактивированная суспензия вируса ящура будет определена с помощью разработанного способа именно как инактивированная [41]. Чувствительность разработанного теста определяли по формуле: Ч=ДП/(ДП+ЛО), где Ч - диагностическая чувствительность, ДП - достоверноположительный результат, ЛО - ложноотрицательный результат.
Для определения чувствительности разработанного способа опосредованного определения полноты инактивации антигена вируса ящура исследовали 360 суспензий, которые являлись истинно инактивированными по данным исследования в монослойной перевиваемой клеточной линии почки свиньи IB-RS-2 (прототип) [2]. Постановку теста проводили, как отражено в примере 3. В результате анализа в ОТ-ПЦР-РВ при амплификации большеразмерного фрагмента генома вируса ящура определили, что из 360 истинно инактивированных суспензий 358 определены в качестве инактивированных, а 2 - в качестве неинактивированных, т.е. количество ДП составило 358, а ЛО - 2 (таблица 9). Таким образом, диагностическая чувствительность разработанного способа опосредованного определения полноты инактивации антигена вируса ящура в сырье для вакцины с применением ОТ-ПЦР-РВ при амплификации большеразмерного фрагмента составила 99,44%.
Пример 5. Определение диагностической специфичности способа опосредованного определения полноты инактивации антигена вируса ящура в сырье для вакцины с применением ОТ-ПЦР-РВ при амплификации болъшеразмерного фрагмента.
Специфичность (истинно отрицательная пропорция) отражает долю отрицательных результатов, которые правильно идентифицированы как таковые, то есть вероятность того, что неинактивированные суспензии будут классифицированы именно как авирулентные [41]. Диагностическую специфичность разработанного теста определяли по формуле: Сп=ДО/(ДО+ЛП), где Сп - диагностическая специфичность, ДО - достоверноотрицательный результат, ЛП - ложноположительный результат.
Для исследования специфичности разработанного способа опосредованного определения полноты инактивации антигена вируса ящура тестировали 360 суспензий, которые являлись истинно неинактивированными по данным исследования в монослойной перевиваемой клеточной линии почки свиньи IB-RS-2 (прототип) [2]. Анализ проводили, как отражено в примере 3. В результате исследования в ОТ-ПЦР-РВ при амплификации большеразмерного фрагмента генома вируса ящура определили, что из 360 истинно неинактивированных суспензий 360 определены в качестве неинактивированных, статус инактивированных суспензий присвоен не был, т.е. количество ДО составило 360 (таблица 9). Таким образом, диагностическая специфичность разработанного способа опосредованного определения полноты инактивации антигена вируса ящура в сырье для вакцины с применением ОТ-ПЦР-РВ при амплификации большеразмерного фрагмента составила 100,00%.
Пример 6. Определение общей точности способа опосредованного определения полноты инактивации антигена вируса ящура в сырье для вакцины с применением ОТ-ПЦР-РВ при амплификации болъшеразмерного фрагмента.
Исследовали общую точность способа опосредованного определения полноты инактивации антигена вируса ящура в сырье для вакцины с применением ОТ-ПЦР-РВ при амплификации большеразмерного фрагмента.
Общая точность показывает соотношение числа достоверноположительных (ДП) и достоверноотрицательных (ДО) результатов анализа к общему числу окончательных результатов, заключенных с помощью разработанного способа. Понятие «общая точность теста» является объективным отражением процесса выдачи ложноотрицательных (ЛО) и ложноположительных (ЛП) результатов исследования с помощью разработанного способа [41].
Обитую точность разработанного способа определяли по формуле: Т=(ДП+ДО)/(ДП+ДО+ЛП+ЛО), где Т - общая точность, ДП - достоверноположительный результат, ДО - достоверноотрицательный результат, ЛП - ложноположительный результат, ЛО - ложноотрицательный результат.Данные для расчета общей точности представлены в примерах 4 и 5. ДП составляло 358, ДО - 360, ЛП - 0, ЛО - 2. Иными словами, общая точность разработанного способа опосредованного определения полноты инактивации антигена вируса ящура в сырье для вакцины с применением ОТ-ПЦР-РВ при амплификации большеразмерного фрагмента составила 99,72%.
Основным преимуществом предлагаемого изобретения является возможность одновременного исследования большого количества проб для опосредованного определения полноты инактивации антигена вируса ящура в сырье для вакцин с применением обратно-транскриптазной полимеразной цепной реакции в режиме реального времени при амплификации большеразмерного фрагмента в течение 9-10 часов. В предлагаемом изобретении для исследования полноты инактивации вируса ящура проводят выделение его нуклеиновой кислоты, анализ полученного элюата на степень чистоты и количество РНК с помощью спектрального сканирования, применяется оптимизированной по компонентам и температурно-временным параметрам ОТ-ПЦР-РВ с использованием системы оригинальных праймеров и зонда, позволяющих амплифицировать участок генома вируса ящура размером 7279 п.н. (от диапазон от 5'-NTR-участка до 3D-гена включительно). Данный подход дает возможность выявить поврежден ли геном вируса ящура в амплифицируемой зоне и, тем самым, опосредованно определить полноту инактивации антигена вируса ящура в сырье для вакцины.
Предлагаемый способ опосредованного определения полноты инактивации антигена вируса ящура в сырье для вакцины с применением обратно-транскриптазной полимеразной цепной реакции в режиме реального времени при амплификации большеразмерного фрагмента характеризуется высокими показаниями диагностической чувствительности, специфичности и общей точности.
Источники информации, принятые во внимание при составлении описания изобретения к заявке на выдачу патента РФ на изобретение «Способ опосредованного определения полноты инактивации антигена вируса ящура в сырье для вакцины с применением обратно-транскриптазной полимеразной цепной реакции в режиме реального времени при амплификации большеразмерного фрагмента»:
1. Пономарев А.П., Узюмов В.Л., Груздев К.Н. Вирус ящура: структура, биологические и физико-химические свойства. Владимир: Фолиант, 2006. - 250 с.
2. OIE. Manual of diagnostic tests and vaccines for terrestrial animals. - 7th ed. - Paris, 2018. - Vol.1, Chap. 2.1.8.
3. Acharia R., Fry E., Stuard D et al. The three-dimensional structure of FMDV at 2.9 A resolution // Nature (London). - 1989. - V. 337. - P. 709-716.
4. Grubman, M.J. Biochemical map of polypeptides specified by foot-and-mouth disease virus / M.J. Grubman, В.H. Robertson, D.O. Morgan // J. Virol. - 1984. - V. 50. - P. 579-586.
5. Rueckert, R.R. Systematic nomenclature of picornavirus proteins / R.R. Rueckert, E. Wimmer // J. Virol. - 1984. - V. 50. - P. 957-959.
6. Cottam E. M. Transmission pathways of foot-and-mouth disease virus in the United Kingdom in 2007 / E.M. Cottam, J. Wadsworth, A.E. Shaw, R.J. Rowlands // PLoS Pathog. - 2008. - V. 4: e100005010.1371/journal.ppat.1000050.
7. Cheung A. Comparision of the major antigenic determinants of different serotypes of FMDV / A. Cheung, J. DeLamarter, S. Weiss // J. Virol. -1983. - V. 48. - P. 451-459.
8. Marquardt O. Sequences of capsid protein VP1 of two type A FMDV / O. Marquardt, K.-H. Adam // Virus genes. - 1988. - V. 2. - P. 283-291.
9. Jamal S.M. Evolutionary analysis of serotype A foot-and-mouth disease viruses circulating in Pakistan and Afghanistan during 2002-2009 / S.M. Jamal, G. Ferrari, S. Ahmed // J. Gen. Virol. - 2011. - V. 92. - P.2 849-2864.
10. Ryan M.D. Cleavage of FMD polyprotein is mediated by residues located withig a 19 amino acid sequence / M.D. Ryan, A.M.Q. King, G.P. Thomas // J. Gen. Virol. - 1991. - V. 72. - P. 2727-2732.
11. Saunders K. Recombination and oligonucleotide analisis of guanidine-resistant FMDV mutants / K. Saunders, A.M.Q. King, D. McCahon // J. Virol. - 1985. - V. 56. - P. 921-929.
12. Falk M.M. FMDV protease 3C inducing specific proteolitic cleavage of host cell histon H3 / M.M. Falk, P.R. Crigera, J.E. Bergmann // J. Virol. - 1990. - V. 64, N. 2. - P. 748-756.
13. Luz N. A cellular 57 kD protein binds to two region of the internal translation initiation site of FMDV / N. Luz, E. Beck // FEBS. - 1990. - V. 269, N. 2. - P. 311-314.
14. Belsham G.J. Translation and replication of FMDV RNA // Curr Top Microbiol Immunol. - 2005. - V.288. - P. 43-70.
15. National Library of Medicine. FMDV genome. [Электронный ресурс]. - Режим доступа: https://www.ncbi.nlm.nih.gov/search/all/?term=FMDV (Дата обращения: 13 декабря 2020 г.).
16. Alexandersen, S. The pathogenesis and diagnosis of foot and mouth disease / S. Alexandersen, Z. Zhang, A.L. Donaldson // J. Compr. Pathol. - 2003 - V. 129. - P. 268-282.
17. Shaw A. Implementation of a one-step real-time RT-PCR protocol for diagnosis of foot-and-mouth disease / A. Shaw, S. M. Reid, K. Ebert [et al.] // J. Virol. Methods. - 2007. - Vol. 143, N 1. - P. 81-85.
18. Lubroth, J., Rodriguez, L. and Dekker, A. Vesicular diseases. In: Straw, B.E., Zimmerman, J.J., D'Allaire, S. and Taylor, D.J., editors. Diseases of Swine. // Blackwell Publishing Professional, Ames, Iowa, USA. - 2006. - 9th ed. - P. 517-536.
19. Bahnemann H.G. Inactivation of viral antigens for vaccine preparation with particular reference to the application of binary ethylenimine // Vaccine. - 1990. - V. 8(4). - P. 299-303.
20. Ethyleneimine [Электронный ресурс]. - Режим доступа: http://www.cdc.gov/niosh/npg/npgd0274.html (Дата обращения: 21 декабря 2020 г.).
21. Курашова С.С. Сравнительная характеристика инактивирующих агентов для создания вакцин против геморрагичексой лифхорадки с печечным синдромом / С.С. Курашова, А.А. Ишмухаметов, М.С. Егорова [и др.] // Эпидемиология и вакцинопрофилактика. - 2018. - №17(4). - С. 26-29.
22. Жильцова М.В. Биологические свойства эпизоотических изолятов вируса ящура типов А, О и Азия-1: Дис.… канд. ветер, наук: 16.00.03. - Владимир, 2008. - 146 с.
23. Reid S. Primary diagnosis of foot-and-mouth disease by reverse transcription polymerase chain reaction / S. Reid, N.P. Ferris, G.H. Hutchings // J. Virol. Methods. - 2000. - V. 89. - P. 167-176.
24. Vangrysperre W., De Clerco K. Rapid and sensitive polymerase chain reaction based detection and typing of foot-and-mouth disease virus in clinical samples and cell culture isolates, combined with a simultaneous differentiation with other genomically and/or symptomatically related viruses / Vangrysperre W., De Clerco K. // Arch. Virol. - 1996. - V. 141. - P. 331-344.
25. Callahan J.D. Use of a portable real-time reverse transcriptase-polymerase chain reaction assay for rapid detection of foot-and-mouth disease virus / J.D. Callahan, F. Brown, F.A. Csorio // J. Am. Vet. Med. Assoc. - 2002. - V. 220. - P. 1636-1642.
26. Калмыкова М.С, Калмыков M.B., Белоусова P.B. Основы полимеразной цепной реакции с разными формами детекции: Учебное пособие. - СПб.: Издательство «Лань». - 2009. - 80 с.
27. Shaw A.E. Enhanced laboratory diagnosis of foot-and-mouth disease by real-time polymerase chain reaction / A.E. Shaw, S.M. Reid, D.P. King [et al.] // Rev. Sci. Techn. OIE. - 2004. - Vol. 23, №3. - P. 1003-1009.
28. Федоров Д.Г. Усовершенствование технологии изготовления и контроля инактивированной вакцины против классической чумы свиней // Автореферат.… канд-та ветер, наук. - 1999. - Покров. - 24 с.
29. The Analysis of DNA or RNA using Its Wavelengths: 230 nm, 260 nm, 280 nm. Bioteachnology.com [Электронный ресурс]. - Режим доступа: http://bioteachnology.com/dna/analysis-dna-rna-wavelengths-230-260-280-nm. (Дата обращения 01 ноября 2020 г.).
30. WPA BioChrom. [Электронный ресурс]. - Режим доступа: https://www.biochemmack.ru/upload/uf/f4d/f4daefea6a77965d02ddbe780a813aad.pdf (Дата обращения 14 ноября 2020 г.).
31. ДНК-полимераза Taq Platinum High Fidelity [Электронный ресурс]. - Режим доступа: https://www.laboratorii.com/reaktivy/reaktivy-invitrogen/1241/ (Дата обращения 184 ноября 2020 г.).
32. Burgers P. Eukaryotic DNA polymerases: proposal for a revised nomenclature (англ.) / P. Burgers, E. Koonin, E. Bruford [et al.] // J. Biol. Chem.: Journal. - 2001. - Vol. 276, no. 47. - P. 43487-43490.
33. Rebrikov D.V., Samatov G.A., Trofimov D. Yu. Real-time PCR. Ed. by D.V. Rebrikov. M.: BINOM. Laboratoriya znaniy. 2009. 223 p.(in Russian).
34. Bogdanova E.A. Normalization of full-length enriched cDNA / E.A. Bogdanova, D.A. Shagin, S.A. Lukyanov // Mol. Biosyst. - 2008. - V. 4(3). - P. 205-212.
35. Shcheglov A., Zhulidov P., Bogdanova E. Generation of normalized cDNA libraries. In: Nucleic Acids Hybridization: Modern Applications. Buzdin, Anton; Lukyanov, Sergey (Eds.) Hardcover. - 2007. - P. 97-124.
36. Lukyanov S., Lukyanov K., Gurskaya N. Selective suppression of polymerase chain reaction and its most popular applications. In: Nucleic Acids Hybridization: Modern Applications. Buzdin, Anton; Lukyanov, Sergey (Eds.) Hardcover. - 2007. - P. 29-51.
37. Zhulidov P.A. A method for the preparation of normalized cDNA libraries enriched with full-length sequences / P.A. Zhulidov, E.A. Bogdanova, A.S. Shcheglov // Bioorg Khim. - 2005. - V. 31(2). - P. 186-194.
38. Bogdanova E.A. A family of genes of multidomain free lectins from a planarian: structure, expression, and use as markers for regeneration monitoring / E.A. Bogdanova, E.V. Barsova, N.I. Pounkova // Bioorg. Khim. - 2004. - V. 30(6). - P. 626-637.
39. Wiame I. Irreversible heat inactivation of DNase I without RNA degradation // BioTechniques. - 2000. - V. 29. - P. 252-256.
40. Ревертаза Maxima H Minus (Thermo Scientific) [Электронный ресурс]. - Режим доступа: https://www.thermofisher.com/document-connect/document-connect.html?url=https%3A%2F%2Fassets.thermofisher. com%2FTFS-Assets%2FLSG%2Fmanuals%2FMAN0012047_TS_Maxima_H_Minus_Reverse_Transcriptase_2000U_UG.pdf&title=VXNlciBHdWlkZTogVF MgTWF4aWlhIEggTWludXMgUmV2ZXJzZSBUcmFuc2NyaXB0YXNlLCAyMDAwIFU=(Дата обращения: 06 октября 2020 г.).
41. Сиделев, С.И. Математические методы в биологии и экологии: введение в элементарную биометрию: учебное пособие / С.И. Сиделев; Яросл. гос. ун-т им. П.Г. Демидова. - Ярославль: ЯрГУ, 2012. - 140 с.





Изобретение относится к области биотехнологии, средствам молекулярной диагностики и производству противоящурных вакцин. Описан способ опосредованного определения полноты инактивации антигена вируса ящура в сырье для вакцины с применением обратно-транскриптазной полимеразной цепной реакции в режиме реального времени (ОТ-ПЦР-РВ) при амплификации большеразмерного фрагмента. Исследование полноты инактивации антигена вируса ящура в сырье для инактивированных противоящурных вакцин определяют при проведении элюирования вирусной РНК, оценки степени чистоты и концентрации РНК с помощью спектрального сканирования и формулы СРНК = 40,5 × OD260 - 0,7037, обратно-транскриптазной полимеразной цепной реакции в режиме реального времени, с помощью которой выявляют неповрежденный участок генома вируса ящура размером 7279 н.о. в диапазоне от 5'-NTR до 3D-гена включительно, наличие экспоненты и, тем самым, определяют наличие вирулентного вируса ящура, либо выявляют отсутствие ампликона нужного размера, отсутствие экспоненты и, тем самым, доказывать отсутствие вирулентного вируса ящура в сырье для изготовления противоящурных инактивированных вакцин. Разработанный способ является экономичным, позволяет одновременно исследовать несколько десятков проб, а время проведения анализа сократить до 9-10 часов, характеризуется высокими показателями диагностической чувствительности - 99,44%, специфичности - 100% и общей точности - 99,72%. 7 з.п. ф-лы, 6 ил., 9 табл., 6 пр.
1. Способ опосредованного определения полноты инактивации антигена вируса ящура в сырье для вакцины с применением обратно-транскриптазной полимеразной цепной реакции в режиме реального времени при амплификации большеразмерного фрагмента, отличающийся тем, что с его помощью возможно выявлять неповрежденный участок генома вируса ящура размером 7279 п.н., наличие экспоненты с пороговым циклом амплификации Ct<40,0 ед. в диапазоне от 5'-NTR до 3D-гена включительно с помощью оригинальных специфических олигонуклеотидов 5'-NTR-F-fov2-праймера, 3D-R1-rev-праймера и 3DP-rev-зонда со следующими последовательностями:
5'-ТТТСС AGGTCTAGAGGGGTGA-3',
5'-GCGAGTCCTGCC ACGGA-3',
ROX-GTCCCACGGCGTGCAAAGGA-BHQ2, соответственно,
и, тем самым, определять наличие вирулентного вируса ящура, либо выявлять отсутствие ампликона или отсутствие экспоненты - отсутствие вирулентного вируса ящура в сырье при изготовлении противоящурных инактивированных вакцин.
2. Способ по п. 1, отличающийся тем, что основан на проведении экстрагирования молекул РНК возбудителя ящура до и после процесса инактивации с помощью 5 М гуанидинизотиоцианата (ГТЦ), 0,5% раствора меркаптоэтанола и 80% раствора изопропанола.
3. Способ по п. 1, отличающийся тем, что степень чистоты элюата вирусной РНК определяют методом спектрального сканирования, а концентрацию РНК, пользуясь разработанной формулой СРНК = 40,5 × OD260 - 0,7037.
4. Способ по п. 1, отличающийся тем, что смесь компонентов для проведения реакции включает в свой состав следующие компоненты: олигонуклеотидный прямой праймер 5'NTR-F-fov2, обратный праймер 3D-R1-rev и олигонуклеотидный 3DP-rev-зонд - 10 пМ, дезоксирибонуклеозидтрифосфаты - 2,00 мМ, Thermo Scientific™ 10Х DreamTaq™ Buffer - 10% от общего объема реакционной смеси, хлорид магния - 3,0 мМ, ревертаза Maxima Н Minus - 10 е.а., ДНК-полимераза Platinum High Fidelity - 10 е.а.
5. Способ по п. 1, отличающийся тем, что применяются высокоточные ферменты: ревертаза Maxima Н Minus, которая обладает РНК- и ДНК-зависимой полимеразной активностью с отсутствием активности РНКазы Н и характеризуется высокой степенью эффективности; ДНК-полимераза Platinum High Fidelity, позволяющая амплифицировать фрагмент генома вируса ящура размером 7279 п.н. с надежным усилением процесса.
6. Способ по п. 1, отличающийся тем, что реакцию проводят с соблюдением следующих режимов и количеством циклов амплификации - 45:
- обратная транскрипция: температура 50°С в течение 30 мин;
- активация ДНК-полимеразы Platinum High Fidelity, температура 95°С в течение 1 мин;
- ОТ-ПЦР-РВ: денатурация: температура 95°С в течение 30 с, отжиг олигонуклеотидов: температура 57°С в течение 30 с, элонгация: температура 63°С в течение 8 мин.
7. Способ по п. 1, отличающийся тем, что является экономичным, позволяет одновременно исследовать несколько десятков проб инактивированного вируссодержащего материала для вакцин, а время проведения анализа сократить до 9-10 часов.
8. Способ по п. 1, отличающийся тем, что его диагностическая чувствительность составляет 99,44%, диагностическая специфичность - 100%, общая точность - 99,72%.
| СПОСОБ КОНТРОЛЯ ПОЛНОТЫ ИНАКТИВАЦИИ АНТИРАБИЧЕСКОЙ ИНАКТИВИРОВАННОЙ ВАКЦИНЫ | 2012 |
|
RU2492452C1 |
| СПОСОБ ТЕХНОЛОГИЧЕСКОГО КОНТРОЛЯ ПОЛНОТЫ ИНАКТИВАЦИИ УБИТЫХ ВАКЦИН ПРИ ИСПОЛЬЗОВАНИИ В КАЧЕСТВЕ ИНАКТИВАТОРА СЕРНОКИСЛОЙ МЕДИ | 1995 |
|
RU2089893C1 |
| CN 102230029 A, 02.11.2011 | |||
| КУДАШЕВА Э.Ю., СОВРЕМЕННЫЕ ТЕХНОЛОГИЧЕСКИЕ ПОДХОДЫ К ОБЕСПЕЧЕНИЮ ВИРУСНОЙ БЕЗОПАСНОСТИ ПРЕПАРАТОВ ИММУНОГЛОБУЛИНОВ ЧЕЛОВЕКА, Успехи современного естествознания, 2015, N 5, С | |||
| Способ получения нерастворимых лаков основных красителей в субстанции и на волокнах | 1923 |
|
SU132A1 |

