В данном документе описан способ улучшения стабильности антител путем обнаружения в аминокислотной последовательности антитела сайтов деградации аспарагина и аспартата и их удаления, основанный на моделировании структуры.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Моноклональные антитела и молекулы на основе доменов антител составляют основную часть белков терапевтического назначения, проходящих клинические испытания (1, 2). Моноклональные антитела эффективны при различных показаниях к применению и удобны в получении. Специфичность антитела в первую очередь определяется последовательностями CDR-участков вариабельных доменов. Процесс отбора возможных кандидатов для клинических исследований начинается со скрининга функциональных свойств большого числа моноклональных антител. После скрининга сотни молекул антител подробно анализируют in vitro с целью выявления моноклонального антитела, соответствующего всем желаемым функциональным критериям и помимо этого обладающего химической и биофизической стабильностью. Таким образом, чтобы избежать нежелательной деградации в ходе получения, хранения и после применения in vivo, необходимо выявлять и отбраковывать моноклональные антитела, обладающие недостаточной стабильностью, которая может сказаться на их структуре и биологической функции.
Одной из реакций разложения, протекающей в белках, является химическое разложение остатков аспарагина (Asn) (3) и аспартата (Asp) (4, 5). Указанные реакции возможно держать под контролем при надлежащих условиях (6-9). Если остатки Asn и Asp задействованы в распознавании антигена, их химическая модификация может привести к снижению активности антитела (10-14).
Изучали различные параметры, которые способны повлиять на предрасположенность к деградации остатков Asn и Asp, входящих в состав пептида (52, 53, 57-59) и белка (7, 10, 35, 60), например, такие как первичная последовательность (3, 5, 16, 33, 40, 51-56), диэлектрическая постоянная растворителя, температура и pH. Уже в 1980-х было сделано предположение, что определяющими факторами для дезаминирования белков (5, 61) являются определенные структурные характеристики, впоследствии нашедшее свое подтверждение и развитие (34, 37, 38, 40, 46, 51, 62-64).
Несмотря на накопленные знания о механизме деградации и условиях его осуществления, спонтанные реакции дезаминирования и изомеризации, протекающие в моноклональных антителах, по-прежнему остаются нерешенной проблемой.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В данном документе описана корреляция между деградацией остатков Asp и Asn в полипептидах и рядом структурных параметров. Обнаружили, что т.н. «горячие точки» деградации можно отличить по (i) их конформационной подвижности, (ii) размеру аминокислотного остатка, примыкающего со стороны С-конца, и (iii) параметрам вторичной структуры. Основываясь на данных характеристиках, разработали способ предсказания предрасположенности остатков Asn и Asp к деградации.
Один аспект согласно данному описанию представляет собой способ отбора антитела, стабильного к деградации (= модификации), включающий следующие стадии
а) определение для каждого остатка Asn и Asp в Fv домене антитела конформационной подвижности Cα-атома с помощью ансамбля/в ансамбле/используя ансамбль моделей, основанных на гомологии,
б) определение для каждого остатка Asn и Asp в Fv домене антитела размера аминокислотного остатка, непосредственно примыкающего к остатку Asn и Asp со стороны С-конца,
в) выбора антитела, в котором Cα-атом является конформационно неподвижным и/или в котором к Asn или Asp с С-конца примыкает аминокислотный остаток большого размера.
Один аспект согласно данному описанию представляет собой способ отбора одного или нескольких антител, стабильных к деградации (= модификации), который включает стадии, при которых
а) берут два или более антител,
б) определяют для каждого остатка Asn и Asp в Fv домене антитела конформационную подвижность Cα-атома с помощью ансамбля/в ансамбле/используя ансамбль моделей, основанных на гомологии,
в) определяют для каждого остатка Asn и Asp в Fv домене антитела размер аминокислотного остатка, непосредственно примыкающего к остатку Asn и Asp со стороны С-конца,
г) выбирают одно или несколько антител, у которых Cα-атом Asn или Asp является конформационно неподвижным и/или у которых к Asn или Asp со стороны С-конца примыкает аминокислотный остаток большого размера,
и таким образом отбирают одно или несколько антител, стабильных к деградации (модификации).
Один аспект согласно данному описанию представляет собой способ отбраковки антитела (отбора для удаления из множества антител), включающий следующие стадии
а) определение для каждого остатка Asn и Asp в Fv домене антитела (Fv доменах множества антител) конформационной подвижности Cα-атома с помощью ансамбля/в ансамбле/используя ансамбль моделей, основанных на гомологии,
б) определение для каждого остатка Asn и Asp в Fv домене антитела размера аминокислотного остатка, непосредственно примыкающего к остатку Asn и Asp со стороны С-конца,
в) отбраковку/удаление (из Fv доменов множества антител) антитела, в котором Cα-атом Asn или Asp обладает умеренной или высокой конформационной подвижностью и/или у которого к остатку Asn или Asp с С-конца примыкает аминокислотный остаток малого размера.
Один аспект согласно данному описанию представляет собой способ отбраковки антитела (отбора для удаления из множества антител) одного или нескольких антител, включающий стадии, при которых
а) берут два или более антител,
б) определяют для каждого остатка Asn и Asp в Fv домене антитела (Fv доменах множества антител) конформационную подвижность Cα-атома с помощью ансамбля/в ансамбле/используя ансамбль моделей, основанных на гомологии,
в) определяют для каждого остатка Asn и Asp в Fv домене антитела размер аминокислотного остатка, непосредственно примыкающего к остатку Asn и Asp со стороны С-конца,
г) отбраковывают/удаляют (из Fv доменов множества антител) одно или несколько антител, у которых Cα-атом Asn или Asp обладает умеренной или высокой конформационной подвижностью и/или у которых к остатку Asn или Asp с С-конца примыкает аминокислотный остаток малого размера.
Один аспект согласно данному описанию представляет собой способ выявления или отбраковки антитела, предрасположенного к деградации аспарагина (Asn) (дезаминированию и/или образованию сукцинимида), включающий стадии
а) определения для каждого остатка Asn в Fv домене антитела конформационной подвижности Cα-атома с помощью ансамбля/в ансамбле/используя ансамбль моделей, основанных на гомологии,
б) определения для каждого остатка Asn в Fv домене антитела, имеющего с С-конца аминокислотный остаток малого размера, торсионного (двугранного) угла фи остова,
в) определения для каждого остатка Asn в Fv домене антитела, имеющего с С-конца аминокислотный остаток малого размера и торсионный угол фи остова, превышающий -75,2 градуса, доступности для растворителя и возникновения/наличия изменения вторичной структуры, расположенной в направлении N-конца относительно остатка Asn, и расстояния между этим возникшим изменением и остатком Asn, выраженного в аминокислотных остатках, и
г) выявления или отбраковки антитела, подверженного деградации аспарагина (Asn) (дезаминированию и/или образованию сукцинимида), если
i) остаток Asn находится в петле CDR 1, его Cα-атом обладает конформационной подвижностью и прилегающий со стороны С-конца аминокислотный остаток представляет собой Asp, Pro, Thr или Asn,
ii) Cα-атом Asn является конформационно подвижным, имеет торсионный угол фи остова менее -75,2 градусов, а аминокислотный остаток, расположенный по отношению к остатку Asn в направлении С-конца имеет малый размер, или
iii) Cα-атом остатка Asn обладает конформационной подвижностью, имеет торсионный угол фи остова более -75,2 градусов, имеет высокую доступность для растворителя, а изменение вторичной структуры, расположенной в направлении амино-конца относительно остатка Asn, происходит внутри участка, превышающего 3 аминокислотных остатка.
Один аспект согласно данному описанию представляет собой способ выявления или отбраковки одного или нескольких антител, подверженных деградации аспарагина (Asn) (дезаминированию и/или образованию сукцинимида, включающий стадии, при которых
а) берут два или более антител,
б) у каждого остатка Asn в Fv домене антитела определяют конформационную подвижность Cα-атома с помощью ансамбля/в ансамбле/используя ансамбль моделей, основанных на гомологии,
в) у каждого остатка Asn в Fv домене антитела, к которому со стороны С-конца прилегает аминокислотный остаток малого размера, определяют торсионный угол фи остова,
г) у каждого остатка Asn в Fv домене антитела, к которому со стороны С-конца прилегает аминокислотный остаток малого размера, и у которого торсионный угол фи остова превышает -75,2 градуса, определяют доступность для растворителя и возникновение/наличие изменения вторичной структуры, расположенной в направлении N-конца относительно остатка Asn, а также расстояние, выраженное в аминокислотных остатках, между этим возникшим изменением и остатком Asn, и
д) выявляют или отбраковывают одно или несколько антител, подверженных деградации аспарагина (Asn) (дезаминированию и/или образованию сукцинимида), если
i) остаток Asn находится в петле CDR 1, его Cα-атом обладает конформационной подвижностью и прилегающий со стороны С-конца аминокислотный остаток представляет собой Asp, Pro, Thr или Asn,
ii) Cα-атом Asn является конформационно подвижным, имеет торсионный угол фи остова менее -75,2 градусов, и аминокислотный остаток, расположенный по отношению к остатку Asn в направлении С-конца имеет малый размер, или
iii) Cα-атом остатка Asn обладает конформационной подвижностью, имеет торсионный угол фи остова более -75,2 градусов, имеет высокую доступность для растворителя, а изменение вторичной структуры, расположенной в направлении амино-конца относительно остатка Asn, происходит внутри участка, превышающего 3 аминокислотных остатка.
Один аспект согласно данному описанию представляет собой способ отбора одного или нескольких антител с улучшенной стабильностью аспарагина (Asn) (пониженным дезаминированием и/или образованием сукцинимида), который включает стадии, при которых
а) берут два или более антител,
б) у каждого остатка Asn в Fv домене каждого антитела определяют конформационную подвижность Cα-атома с помощью ансамбля/в ансамбле/используя ансамбль моделей, основанных на гомологии,
в) у каждого остатка Asn в Fv домене каждого антитела, к которому со стороны С-конца прилегает аминокислотный остаток малого размера, определяют торсионный угол фи остова,
г) у каждого остатка Asn в Fv домене каждого антитела, у которого со стороны С-конца расположен аминокислотный остаток малого размера и у которого торсионный угол фи остова составляет более -75,2 градусов, определяют доступность для растворителя и возникновение/наличие изменения вторичной структуры, расположенной в направлении N-конца относительно остатка Asn, а также расстояние между этим возникшим изменением и остатком Asn, выраженное в аминокислотных остатках, и
д) удаляют/исключают/отбраковывают из двух или более антител те антитела, которые имеют по меньшей мере одно из следующего:
i) остаток Asn в петле CDR 1, у которого Cα-атом обладает конформационной подвижностью и у которого примыкающий с С-конца аминокислотный остаток представляет собой Asp, Pro, Thr или Asn,
ii) остаток Asn, у которого Cα-атом является конформационно подвижным, торсионный угол фи остова составляет менее -75,2 градусов, а примыкающий со стороны С-конца аминокислотный остаток имеет малый размер, или
iii) остаток Asn, у которого Cα-атом является конформационно подвижным, торсионный угол фи остова составляет более -75,2 градусов, который имеет высокую доступность для растворителя, а изменение вторичной структуры, расположенной в направлении амино-конца относительно остатка Asn, происходит внутри участка, превышающего 3 аминокислотных остатка,
и таким образом отбирают одно или несколько антител с улучшенной стабильностью аспарагина (Asn).
Один аспект согласно данному описанию представляет собой способ получения антитела с пониженной деградацией аспарагина (Asn) (дезаминированием и/или образованием сукцинимида), включающий одну или несколько следующих стадий
- снижение конформационной подвижности Cα-атома,
- увеличение размера аминокислотного остатка, примыкающего к остатку Asn со стороны С-конца.
Один аспект согласно данному описанию представляет собой способ выявления или отбраковки антитела, подверженного деградации аспартата (Asp) (изомеризации и/или образованию сукцинимида), включающий стадии:
а) определения для каждого остатка Asp в Fv домене антитела конформационной подвижности Cα-атома с помощью ансамбля/в ансамбле/используя ансамбль моделей, основанных на гомологии,
б) определения для каждого остатка Asp в Fv домене антитела размера аминокислотного остатка, прилегающего к остатку Asp со стороны С-конца,
в) определения для каждого остатка Asp в Fv домене антитела возникновения/наличия изменения вторичной структуры, расположенной в направлении С-конца относительно остатка Asp, а также расстояния между возникшим изменением и остатком Asp, выраженного в аминокислотных остатках, и
г) выявления или отбраковки антитела, подверженного деградации Asp (изомеризации и/или образованию сукцинимида), если
i) Cα-атом остатка Asp обладает высокой конформационной подвижностью, и аминокислотный остаток, прилегающий к остатку Asp со стороны С-конца, имеет малый размер, или
ii) Cα-атом остатка Asp обладает умеренной конформационной подвижностью, соседний с Asp аминокислотный остаток имеет малый размер, или изменение вторичной структуры, расположенной в направлении С-конца, происходит на расстоянии менее 3 аминокислотных остатков от остатка Asp, или
iii) Cα-атом остатка Asp обладает умеренной конформационной подвижностью, аминокислотный остаток, прилегающий к остатку Asp со стороны С-конца, представляет собой остаток Gly, и изменение вторичной структуры, расположенной в направлении С-конца, происходит на расстоянии более 3 аминокислотных остатков от остатка Asp.
Один аспект согласно данному описанию представляет собой способ выявления или отбраковки одного или нескольких антител, подверженных деградации аспартата (Asp) (изомеризации и/или образованию сукцинимида), включающий стадии, при которых
а) берут два или более антител,
б) у каждого остатка Asp в Fv домене антитела определяют конформационную подвижность Cα-атома с помощью ансамбля/в ансамбле/используя ансамбль моделей, основанных на гомологии,
в) у каждого остатка Asp в Fv домене антитела определяют размер аминокислотного остатка, прилегающего к остатку Asp со стороны С-конца,
г) у каждого остатка Asp в Fv домене антитела определяют возникновение/наличие изменения вторичной структуры, расположенной в направлении С-конца относительно остатка Asp, а также расстояние между возникшим изменением и остатком Asp, выраженное в аминокислотных остатках, и
д) выявляют или отбраковывают одно или несколько антител, подверженных деградации Asp (изомеризации и/или образованию сукцинимида), если
i) Cα-атом остатка Asp обладает высокой конформационной подвижностью, и аминокислотный остаток, прилегающий к остатку Asp со стороны С-конца, имеет малый размер, или
ii) Cα-атом остатка Asp обладает умеренной конформационной подвижностью, соседний с Asp аминокислотный остаток имеет малый размер, или изменение вторичной структуры, расположенной в направлении карбокси-конца относительно остатка Asp, происходит внутри участка менее чем из 3 аминокислотных остатков, или
iii) Cα-атом остатка Asp обладает умеренной конформационной подвижностью, аминокислотный остаток, прилегающий к остатку Asp со стороны С-конца, представляет собой остаток Gly, и изменение вторичной структуры, расположенной в направлении карбокси-конца относительно остатка Asp, происходит на расстоянии более 3 аминокислотных остатков.
Один аспект согласно данному описанию представляет собой способ отбора одного или нескольких антител с улучшенной стабильностью аспартата (Asp) (пониженной изомеризацией и/или образованием сукцинимида), который включает стадии, при которых:
а) берут два или более антител,
б) у каждого остатка Asp в Fv домене каждого антитела определяют конформационную подвижность Cα-атома с помощью ансамбля/в ансамбле/используя ансамбль моделей, основанных на гомологии,
в) у каждого остатка Asp в Fv домене каждого антитела определяют размер аминокислотного остатка, прилегающего к остатку Asp со стороны С-конца,
г) у каждого остатка Asp в Fv домене каждого антитела определяют возникновение/наличие изменения вторичной структуры, расположенной в направлении С-конца относительно остатка Asp, а также расстояние между этим возникшим изменением и остатком Asp, выраженное в аминокислотных остатках, и
д) удаляют/исключают/отбраковывают из двух или более антител те антитела, которые имеют по меньшей мере одно из следующего:
i) остаток Asp, у которого Cα-атом является конформационно подвижным и прилегающий с С-конца аминокислотный остаток имеет малый размер,
ii) остаток Asp, у которого Cα-атом обладает умеренной конформационной подвижностью, прилегающий с С-конца аминокислотный остаток имеет малый размер, и изменение вторичной структуры, расположенной в направлении С-конца относительно остатка Asp, происходит внутри участка менее чем из 3 аминокислотных остатков, или
iii) остаток Asp, у которого Cα-атом обладает умеренной конформационной подвижностью, аминокислотный остаток, прилегающий с С-конца представляет собой Gly, и изменение вторичной структуры, расположенной в направлении карбокси-конца, происходит на расстоянии более 3 аминокислотных остатков от остатка Asp,
и таким образом отбирают одно или несколько антител с улучшенной стабильностью аспартата (Asp).
Один аспект согласно данному описанию представляет собой способ получения антитела с пониженной деградацией аспартата (Asp) (изомеризацией и/или образованием сукцинимида), включающий одну или несколько следующих стадий:
- снижение конформационной подвижности Cα-атома,
- увеличение размера аминокислотного остатка, прилегающего к остатку Asp со стороны С-конца.
В одном воплощении аминокислотный остаток, прилегающий к остатку Asp со стороны С-конца, заменяют на аминокислотный остаток большого размера.
Один аспект согласно данному описанию представляет собой способ отбора антитела, стабильного при длительном хранении, включающий следующие стадии
а) определение для каждого остатка Asn и Asp в Fv домене антитела конформационной подвижности Cα-атома с помощью ансамбля/в ансамбле/используя ансамбль моделей, основанных на гомологии,
б) определение для каждого остатка Asn и Asp в Fv домене антитела размера аминокислотного остатка, прилегающего к остатку Asn или Asp со стороны С-конца,
в) выбор антитела, в котором Cα-атом является конформационно неподвижным и/или к Asn или Asp со стороны С-конца прилегает аминокислотный остаток большого размера.
Один аспект согласно данному описанию представляет собой способ отбора одного или нескольких антител, стабильных при длительном хранении, включающий стадии, при которых
а) берут два или более антител,
б) у каждого остатка Asn и Asp в Fv домене антитела определяют конформационную подвижность Cα-атома с помощью ансамбля/в ансамбле/используя ансамбль моделей, основанных на гомологии,
в) у каждого остатка Asn и Asp в Fv домене антитела определяют размер аминокислотного остатка, прилегающего к остатку Asn или Asp с С-конца,
г) выбирают антитело, в котором Cα-атом является конформационно неподвижным и/или к Asn или Asp со стороны С-конца примыкает аминокислотный остаток большого размера,
и таким образом отбирают одно или несколько антител с улучшенной стабильностью при хранении.
Один аспект согласно данному описанию представляет собой способ получения антитела, включающий следующие стадии:
а) культивирование клеток млекопитающего, содержащих нуклеиновую кислоту, кодирующую антитело, отобранное способом, описанным в данном документе, или полученное способом, описанным в данном документе,
б) выделение антитела из клеток или среды культивирования и таким образом, получение антитела.
Один аспект согласно данному описанию представляет собой способ получения антитела, включающий следующие стадии:
а) культивирование клеток млекопитающего, содержащих нуклеиновую кислоту, кодирующую антитело, отобранное способом, описанным в данном документе,
б) выделение антитела из клеток или среды культивирования и таким образом, получение антитела.
Ниже представлены воплощения всех аспектов данного изобретения. Любая комбинация отдельных воплощений также входит в объем изобретения.
В одном воплощении конформационная подвижность представляет собой среднеквадратичное отклонение (RMSD, от англ. root mean square deviation) соответствующих Cα-атомов остатков Asn/Asp в ансамбле моделей, основанных на гомологии. В одном воплощении ансамбль моделей, основанных на гомологии, представляет собой ансамбль из 5 моделей.
В одном воплощении
i) конформационно неподвижным является RMSD 0,01  или менее,
или менее,
ii) конформационно подвижным является RMSD более чем 0,01 ,
iii) умеренной конформационной подвижностью является RMSD между 0,145 и 0,485 , и
iv) высокой конформационной подвижностью является RMSD более 0,485 .
В одном воплощении ансамбль моделей, основанных на гомологии, получают для Fv фрагмента антитела.
В одном воплощении аминокислотный остаток малого размера представляет собой Gly, Ala, Ser, Cys или Asp. В одном воплощении аминокислотный остаток малого размера представляет собой Gly, Ala, Ser или Cys.
В одном воплощении способ включает этап, при котором:
- берут аминокислотную последовательность Fv домена антитела.
В одном воплощении высокая доступность для растворителя представляет собой значение SASA, превышающее 89,4 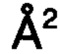 .
.
В одном воплощении для получения ансамбля моделей, основанных на гомологии, используют стандартный набор экспериментальных масс-спектрометрических данных.
В одном воплощении в процессе моделирования петель задействованы все CDR, в результате чего получают ансамбль из 5 моделей, основанных на гомологии.
В одном воплощении способ не требует симуляции молекулярной динамики.
В одном воплощении остаток считают точкой деградации, если он классифицирован данным образом по меньшей мере в одной из 5 построенных по гомологии моделей.
В одном воплощении используют алгоритм рекурсивного разбиения с возможностью предварительного просмотра, реализованный в программе Pipeline Pilot.
В одном воплощении доступную для растворителя площадь поверхности следующего атома азота остова определяют путем компьютерного моделирования и подсчитывают число водородных связей.
В одном воплощении переходную конформацию исследуют путем измерения расстояния между Cγ-атомом боковой цепи и Nn+1-атомом, торсионного угла боковой цепи χ1, и торсионного угла CGONC, который определяют как угол между атомами Cγ, O, Nn+1 и С.
В одном воплощении определяют доступную для растворителя площадь поверхности каждого Asp или Asn.
В одном воплощении определяют размер прилегающей со стороны С-конца (следующей) аминокислоты и торсионные углы остова φ (C'n-1N-Cα-C') и ψ (N-Cα-C''-Nn+1).
В одном воплощении среднеквадратичное отклонение (RMSD) соответствующих Cα-атомов остатков Asn/Asp в ансамбле моделей, основанных на гомологии (ансамбле из 5 моделей), отражает структурное разнообразие внутри ансамбля и расценивается как показатель возможной конформационной подвижности.
В одном воплощении в качестве параметров включают элемент вторичной структуры (остаток, находящийся в спирали, слое, изгибе или случайной спирали) и расстояние до следующего отличающегося элемента вторичной структуры, расположенного в направлении N- и С-конца.
В одном воплощении определяют расстояние между Cα-атомами остатков n-1 и n+1.
СВЕДЕНИЯ, ПОДТВЕРЖДАЮЩИЕ ВОЗМОЖНОСТЬ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Моноклональные антитела являются наиболее перспективными белками, используемыми в терапевтических целях по различным показаниям. Стандартные подходы к получению антител всегда приводят к получению нескольких подходящих кандидатов. Из этих кандидатов необходимо отобрать моноклональные антитела с высокой терапевтической активностью и химической стабильностью, чтобы избежать деградации в процессе получения, хранения и in vivo. Антитела часто подвержены деградации из-за дезаминирования аспарагина (Asn) и изомеризации аспаратата (Asp).
У остатков Asn и Asp общий путь деградации, который протекает с образованием циклического промежуточного соединения сукцинимида (Фиг. 1) (3, 5, 33). Образование сукцинимида происходит в результате внутримолекулярной перестройки после дезаминирования Asn или дегидратации Asp при нуклеофильной атаке атома азота, принадлежащего следующей аминокислоте, на γ-карбонильную группу боковой цепи Asn/Asp. Любая из двух карбонильных групп метастабильного циклического имида может подвергаться гидролизу с образованием аспартильных или изо-аспартильных связей в различном соотношении, в зависимости от условий гидролиза и конформационных ограничений (3, 5, 20, 34-36). Кроме этого, предполагают существование альтернативных механизмов деградации (10), таких как нуклеофильная атака атома кислорода карбонила остова с образованием циклического изоимида (5, 37, 38) или непосредственный гидролиз Asn в Asp при участии молекулы воды (39, 40). Описано несколько аналитических методов для определения любого из продуктов деградации, т.е. сукцинимида, аспартата или изо-аспартата, в основном, это основанные на различии зарядов методы, такие как ионообменная хроматография или изоэлектрическое фокусирование. Наиболее подходящим для количественного определения и определения местоположения сайтов деградации в белках является анализ с помощью жидкостной хроматографии с тандемной масс-спектрометрией (LC-MS/MS) (12, 13, 38, 43-50).
На основании масс-спектрометрических данных по сайт-специфической деградации 37 моноклональных антител, полученных экспериментально в стандартных условиях, в сочетании со структурными параметрами, полученными на основании построенных по гомологии моделей, определили параметры, способствующие деградации, и их соответствующую роль в пути деградации. Разработали способ выявления и отбора химически стабильных моноклональных антител.
Термины «модель, основанная на гомологии» обозначает трехмерную модель аминокислотной последовательности, полученную путем конструирования трехмерной модели, в одном воплощении трехмерная модель указанной аминокислотной последовательности с атомным разрешением основана на экспериментально полученной референтной структуре родственной гомологичной аминокислотной последовательности. Построение модели по гомологии основано на определении (общих) элементов последовательности в аминокислотной последовательности, представляющей интерес, и референтной аминокислотной последовательности, которая предположительно имеет аналогичную структуру, и (трехмерном) выравнивании данных аминокислотных последовательностей.
Поскольку белковые структуры являются высоко консервативными, высокая степень сходства последовательностей обычно предполагает существенное структурное сходство (Marti-Renom, М.А., et al. (2000) Annu. Rev. Biophys. Biomol. Struct. 29: 291-325).
В одном воплощении способ построения моделей на основе гомологии включает следующие стадии:
- выбор референтной структуры,
выравнивание последовательности, представляющей интерес, с референтной структурой,
- построение модели и
- оценку модели.
Выравнивание последовательностей можно осуществлять с помощью любого протокола выравнивания, например, такого как FASTA, BLAST, PSI-BLAST.
Модель, основанная на гомологии, используемая в способах согласно данному изобретению, может представлять собой любую модель, построенную по гомологии, например, модель, основанную на гомологии, полученную с использованием SWISS-model, CPHmodels, MODELER или LOOPER. В одном воплощении модель на основе гомологии построена с помощью алгоритмов MODELER и LOOPER.
Модели на основе гомологии строили с использованием автоматизированного сценария выполнения (скрипта), написанного для программы MODELER 9v7 (83). Матрицы для моделирования выбирали из базы референтных структур, состоящей из кристаллических структур Fab фрагментов человеческих, мышиных и химерных антител, имеющих минимальное разрешение 2,8, учитывая консервативность последовательностей и не пропуская остатки, расположенные во внутренней части их вариабельных областей. Наилучшую итоговую модель каждого моноклонального антитела брали за основу для процедуры уточнения структуры петель (LOOPER, Discovery Studio, Accelrys Inc., San Diego, USA) (84). После уточнения структуры петель отбирали пять наиболее правдоподобных моделей, которые использовали в качестве ансамбля структур для каждого моноклонального антитела. Ансамбли моделей, основанных на гомологии, использовали для расчета параметров (Таблица 2). Параметры «следующая отличающаяся вторичная структура, расположенная в направлении N-конца», «следующая отличающаяся вторичная структура, расположенная в направлении С-конца» и «местоположение в случайной спирали» устанавливали на основании информации о вторичной структуре, используя булевы функции, реализованные в Pipeline Pilot (Accelrys Inc., San Diego, USA). Термин «размер аминокислотного остатка, прилегающего со стороны С-конца» обозначает доступную для растворителя площадь поверхности (SASA, от англ. solvent accessible surface area, 85), выраженную в , и определяется следующим образом: Ala, 64.78; Cys, 95.24; Asp, 110.21; Glu, 143.92; Phe, 186.7; Gly, 23.13; His, 146.45; He, 151.24; Lys, 177.37; Leu, 139.52; Met, 164.67; Asn, 113.19; Pro, 111.53; Gin, 147.86; Arg, 210.02; Ser, 81.22; Thr, 111.6; Val, 124.24; Trp, 229.62; Tyr, 200.31. Аминокислотный остаток малого размера, прилегающий со стороны С-конца, имеет SASA менее чем 111 . Аминокислотный остаток большого размера, прилегающий со стороны С-конца, имеет SASA более чем 111 .
Аминокислотная последовательность антител указана для каждой полипептидной цепи в направлении от N-конца к С-концу. В аминокислотной последовательности каждый аминокислотный остаток (за исключением N-концевого аминокислотного остатка) имеет предшествующий аминокислотный остаток. Указанный предшествующий аминокислотный остаток расположен в направлении N-конца относительно аминокислотного остатка, представляющего интерес. Кроме того, в аминокислотной последовательности каждый аминокислотный остаток (за исключением C-концевого аминокислотного остатка) имеет последующий аминокислотный остаток. Указанный последующий аминокислотный остаток расположен в направлении С-конца относительно аминокислотного остатка, представляющего интерес. Таким образом, термин «аминокислотный остаток, прилегающий со стороны С-конца» обозначает аминокислотный остаток, который непосредственно примыкает к Asn или Asp, представляющему интерес, со стороны С-конца, т.е. аминокислотный остаток, образующий N-концевую амидную связь с соответствующим остатком Asn или Asp.
Термин Fv домен обозначает пару вариабельных доменов в соседних легкой цепи антитела и тяжелой цепи антитела.
Термин «изменение вторичной структуры в направлении карбокси-конца» обозначает переход от первой вторичной структуры ко второй отличающейся вторичной структуре. Термин «вторичная структура» обозначает вторичную структуру (альфа)-спираль, (бета-слой) поворот и случайную спираль. Таким образом, изменение вторичной структуры, представляет собой, например, переход от спирали к слою, повороту или случайной спирали, или от слоя к спирали, повороту или случайной спирали, или от случайной спирали к спирали, слою или повороту.
Экспериментальное исследование сайтов деградации и скорости деградации антител
Исследовали набор 37 различных моноклональных антител lgG1 и lgG4, используемых в терапевтических целях (Таблица 1).

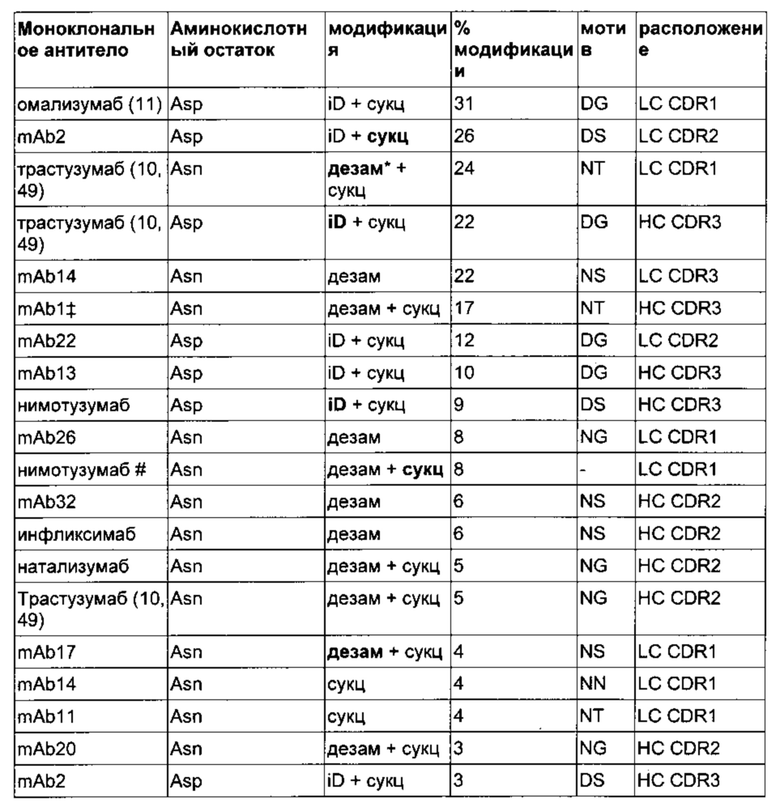
Основные модификации указаны жирным шрифтом. 15 из 37 исследованных моноклональных антител содержали по меньшей мере одну горячую точку Asn/Asp в одном из CDR.
iD = изомеризация, сукц = сукцинимид, дезам = дезаминирование
* только образцы с дезаминированным Asp
 исключили из набора горячих точек из-за взаимодействия с сайтом гликозилирования CDR, который не представлен в моделях, основанных на гомологии
исключили из набора горячих точек из-за взаимодействия с сайтом гликозилирования CDR, который не представлен в моделях, основанных на гомологии
# невозможно подтвердить наличие модификации сайта доступными методами (триптический пептид, AspN пептид, фрагментация методом столкновительной диссоциации, фрагментация методом высокоэнергетической столкновительной диссоциации)
Данные антитела подвергали контролируемому тепловому стрессу в стандартных условиях при pH 6,0 и температуре 40°C в течение 2 недель (подвергнутые стрессовому воздействию образцы) и затем проводили масс-спектрометрический анализ произошедших событий деградации, что позволило установить местоположение поврежденных остатков и определить степень их модификации в образцах, подвергнутых стрессовому воздействию, и в соответствующих референтных образцах.
Из всех 559 остатков Asn и Asp в Fv доменах 37 моноклональных антител, у 60 остатков (11%) степень модификации поддавалась количественному определению. Их подразделили на 19 горячих точек, 13 слабых точек и 28 реакционно-способных точек. Термин горячая точка соответствует степени модификации в образцах, подвергнутых стрессовому воздействию, 3% или более, термин слабая точка - степени модификации от 1% до менее 3% и термин реакционно-способная точка - степени модификации менее 1%.
Расположение сайтов деградации
Обнаружили, что горячие точки деградации, у которых степень модификации составляла 3% или более, располагались в петлях CDR (см. Табл. 1). Большинство горячих точек расположены в CDR 1 легкой цепи и CDR 3 тяжелой цепи, тогда как CDR 1 тяжелой цепи не содержит горячих точек. 15 из 37 исследованных моноклональных антител содержали по меньшей мере одну горячую точку Asn/Asp в одном из CDR. В составе Fv доменов моноклональных антител mAb3, mAb4, mAb9, mAb10, mAb12, mAb16, mAb18, mAb19, mAb21, mAb27, mAb28, mAb29, mAb31, mAb33, бевацизумаб, цетуксимаб, адалимумаб, деносумаб, эфализумаб, базиликсимаб, паливизумаб и панитумаб горячих точек не обнаружили.
В одном воплощении всех аспектов описан способ определения горячих точек дезаминирования/изомеризации/образования сукцинимида (или деградации Asn/Asp) в CDR 1 легкой цепи и/или CDR 3 тяжелой цепи.
В предыдущих исследованиях было показано, что аминокислотный остаток, следующий за Asn и Asp, влияет на скорость образования сукцинимида в белках (40, 51). Так, описано 18 различных мотивов последовательности в составе Fv доменов терапевтически активных антител, которые затрагивает химическая деградация (Asn, за которым следует Gly, Ser или Thr, и Asp, за которым следует Gly, Ser, Thr, Asp или His) (10-14, 35, 46, 65-73). Согласно предыдущим наблюдениям, мотивы Asn-Gly и Asp-Gly значительно больше подвержены модификации, что соответствует 67% и 36% горячих точек в составе CDR (Фиг. 2).
Системный анализ структуры сайтов деградации
Структурное окружение остатков Asn и Asp в Fv фрагментах антител (т.е. подверженных и не подверженных деградации) охарактеризовали набором из 20 параметров, потенциально играющих роль в механизме деградации. Модели Fab фрагментов строили с использованием современной программы моделирования на основе гомологии и оценивали полученные разрешения по баллам, выставленным программой. Модели, основанные на гомологии, использовали для нахождения параметров in silico с использованием автоматизированной процедуры. Как правило, высокая степень гомологии со структурами шаблона позволяет получить точные модели каркасных и коротких CDR участков. Однако, моделирование длинных CDR петель отличается высокой степенью неопределенности, отчасти из-за большой подвижности, присущей таким петлям (74-77). Поэтому в процедуре моделирования петель были задействованы все CDR, и в результате получали ансамбль из 5 моделей, основанных на гомологии. Таким образом, получали дополнительную информацию о возможных конформациях CDR, не прибегая к трудоемкой симуляции молекулярной динамики. Корреляцию между структурными параметрами и деградацией in vitro исследовали с использованием алгоритмов машинного обучения. Обнаружили, что предсказанная модель достаточно точная, а ошибка прогнозирования низкая по сравнению со стандартными способами предсказания по мотивам последовательности.
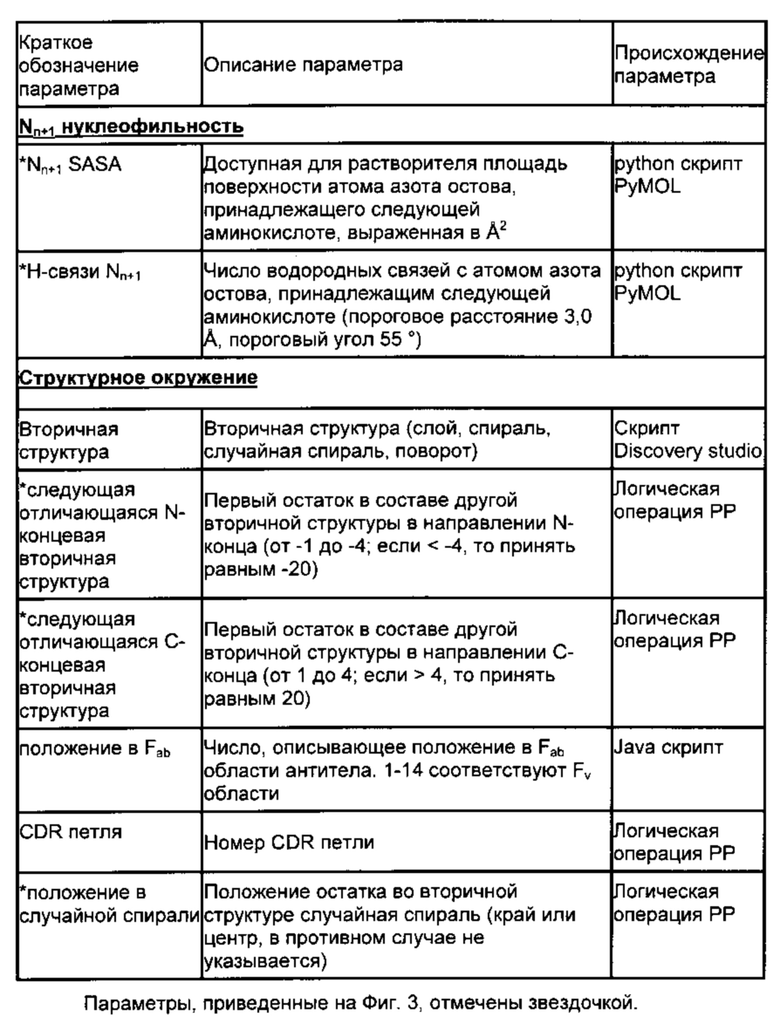
Поскольку при различении горячих точек деградации Asn/Asp и стабильных остатков Asn/Asp только на основании первичной последовательности возможна завышенная оценка значимости признака (51), выбрали набор из 20 структурных параметров, характеризующих трехмерное окружение этих аминокислот. Указанные параметры описаны ниже. Их выбрали с учетом их возможной роли в механизме деградации (см. Фиг. 1 и 3, Табл. 2) и рассчитали на основе моделей, построенных по гомологии.
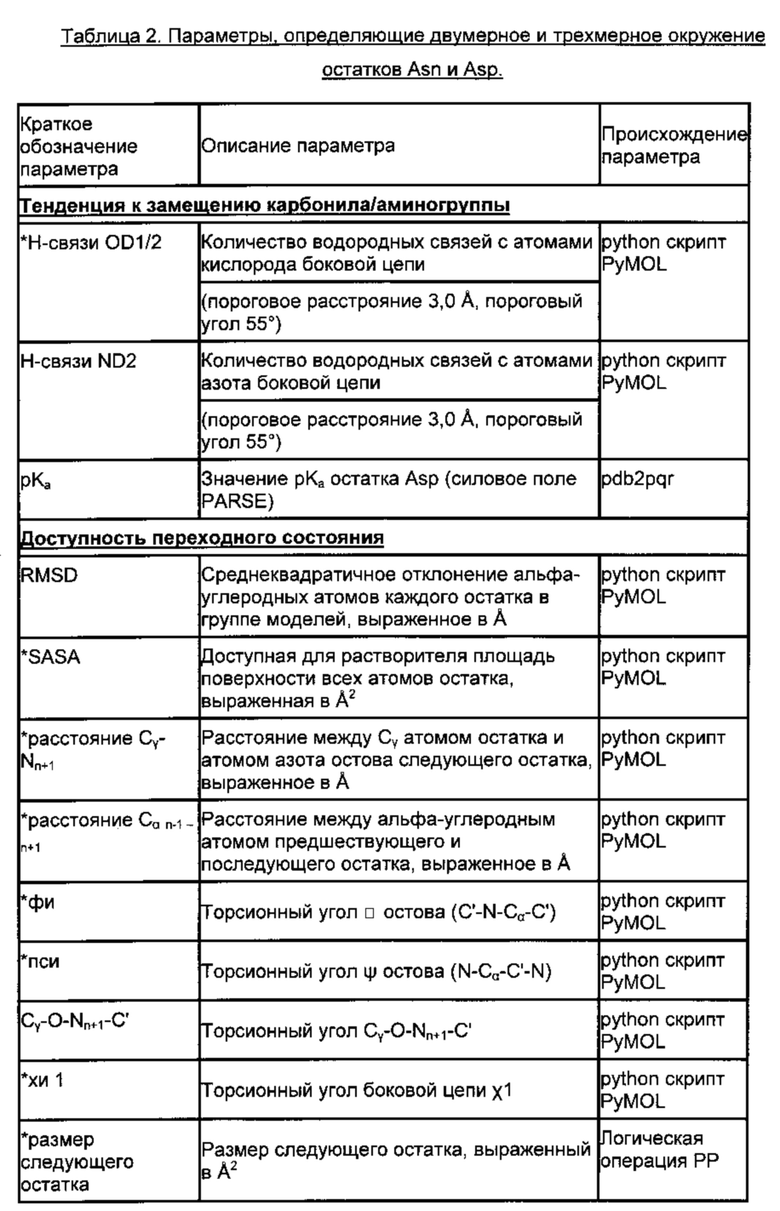
Предпосылкой для образования циклического имида является тенденция к отщеплению гидроксила или аминогруппы боковой цепи Asp или Asn, соответственно. Для оценки данной тенденции подсчитывали количество водородных связей, образованных с атомами кислорода боковой цепи или атомом азота боковой цепи. Для образования сукцинимида карбоксильная группа боковой цепи Asp должна быть протонирована (33, 78). Возможное состояние протонирования оценивали путем расчета значений рКа Asp, зависящих от структуры, используя общепринятый алгоритм PROPKA (79). Другими возможными предпосылками для образования сукцинимида являются доступность и высокая нуклеофильность следующего атома азота остова (см. Фиг. 1). Таким образом, доступную для растворителя площадь поверхности следующего атома азота остова определяли путем компьютерного моделирования и подсчитывали число водородных связей.
Переходное состояние реакции образования сукцинимида требует сближения концевой группы Asp или Asn с атомом азота остова, принадлежащего следующему остатку. Переходную конформацию исследовали путем измерения расстояния между Cγ-атомом боковой цепи и Nn+1-атомом (Фиг. 1, 3 (61)), торсионного угла χ1 боковой цепи и торсионного угла CGONC, который определяли как угол между атомами Cγ, O, Nn+1 и C. Кроме того, рассчитывали доступную для растворителя площадь каждого Asp или Asn. Показано, что боковая цепь п+1 влияет на скорость образования сукцинимида (3, 5, 16, 29, 33, 51, 52, 54). Таким образом, определяли размер следующей аминокислоты, а также торсионные углы каркаса φ (C'n-1N-Cα-C') и ψ (N-Cα-C'-Nn+1) и получали основные трехмерные характеристики локальной структурной конформации и, следовательно, возможной доступности переходного состояния.
Следующие параметры описывают более обширное структурное окружение. Среднеквадратичное отклонение (RMSD, от англ. root mean square deviation) соответствующих Cα-атомов остатков Asn/Asp в ансамбле моделей, основанных на гомологии, отражает структурное разнообразие внутри ансамбля и расценивается как показатель возможной конформационной подвижности. В качестве дополнительных параметров включали элемент вторичной структуры (остаток, находящийся в спирали, слое, изгибе или случайной спирали) (34, 62) и расстояние до следующего отличающегося элемента вторичной структуры в направлении N- и С-конца. Если остаток располагался в такой вторичной структуре, как случайная спираль, описывали его положение в случайной спирали (на краю или в центре). Для количественной характеристики «изгиба» конца случайной спирали, измеряли расстояние между Cα-атомами n-1 и n+1 остатков. Наконец, устанавливали местоположение каждого остатка в составе Fab фрагмента, а именно в одном из CDR, в каркасной области или домене CH1/CL.
Для классификации использовали только остатки в Fv области антител, поскольку в составе CH1/CL горячих точек не наблюдалось. Для статистического анализа использовали 2460 остатков Asn и Asp (492 остатка × 5 моделей) из 185 построенных по гомологии моделей (37 × 5 моделей), включавших 95 горячих точек (19 × 5 моделей) с уровнем модификации в подвергнутом стрессовому воздействию образце 3% или более, а также все 397 не-горячих точек. Обучение классификатора проводили на случайной обучающей 75% выборке (ансамбли из 5 моделей всегда рассматривали вместе), исключая концевые остатки, а также слабые точки и реакционно-способные точки во избежание неверной классификации. Классификацию по Байесу, рекурсивное разбиение, метод опорных векторов, случайный лес, регуляризованный дискриминантный анализ и нейронные сети оценивали на случайных обучающих выборках, в 40 повторах, используя все 20 параметров (Фиг. 4 и 5).
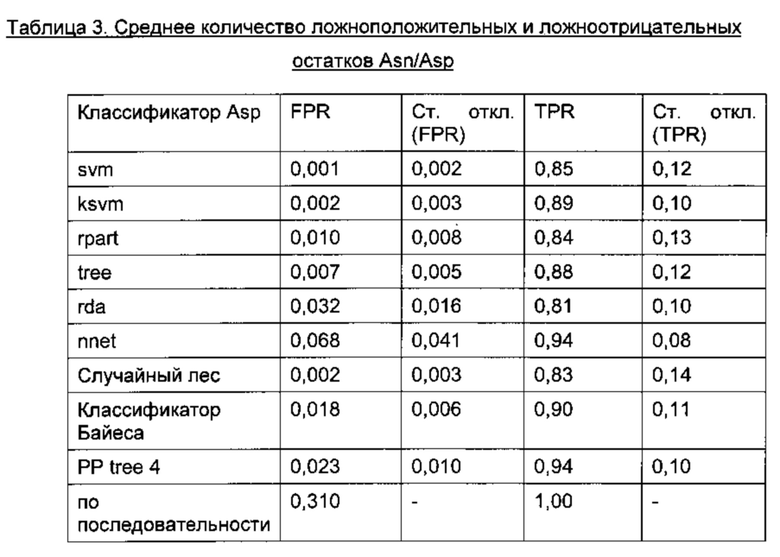
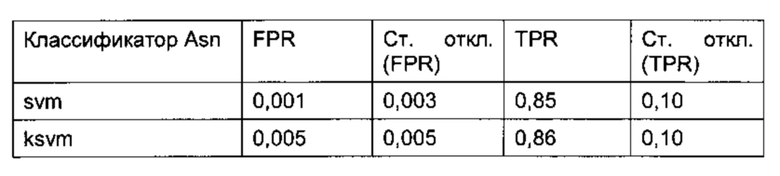
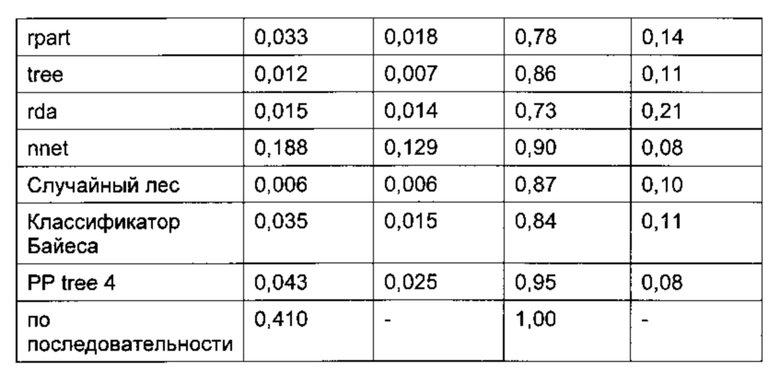
TPR (доля истинно положительных результатов, от англ. true positive rate) = число истинно положительных результатов, поделенное на число положительных результатов; FPR (доля ложноположительных результатов, от англ. false positive rate) = число ложноположительных результатов, поделенное на число отрицательных результатов.
Классификацию осуществляли отдельно для Asn и Asp, поскольку деградация Asn могла протекать по различным механизмам (5, 37-40), (Фиг. 1). Такое разделение позволило улучшить схему классификации. В одном воплощении остаток считают предсказанной точкой деградации, если он классифицирован данным образом по меньшей мере в одной из 5 моделей ансамбля. Для выбора наиболее подходящего классификатора из 9 различных моделей классификации использовали ROC-анализ (от англ. receiver operating characteristic), который обычно применяют для оценки качества систем бинарной классификации. При этом долю истинно положительных результатов от количества положительных результатов (TPR) откладывают против доли ложноположительных результатов от количества отрицательных результатов (FPR). Считая наиболее значимым критерием высокую долю истинно положительных результатов, в качестве наиболее подходящего классификатора выбрали алгоритм рекурсивного разбиения single-tree («одиночное дерево») с возможностью предварительного просмотра, реализованный в программе Pipeline Pilot (Фиг. 4 и 5). На каждом этапе алгоритм рекурсивного разбиения выбирает параметр, позволяющий наилучшим образом разделить массив данных на гомогенные подмножества, принадлежащие к одному классу (горячие точки или не-горячие точки), при этом точку разделения называют узлом, а класс - листом. Встроенная функция предварительного просмотра позволяет убедиться, что выбранный параметр разделения и значение являются оптимальными не только для данного этапа, но также и для последующих этапов. Таким образом, модель позволяет выбрать ключевые параметры, чтобы отличить горячие точки от не-горячих точек. Данный классификатор, предсказывающий предрасположенность Asn и Asp к деградации, отличается наилучшей комбинацией высокой TPR и приемлемого уровня FPR, а также хорошей интерпретируемостью алгоритма даже после применения процедуры оптимизации в целях улучшения предсказания.
Алгоритмы рекурсивного разбиения single-tree для Asn и Asp с предварительным просмотром оптимизировали, чтобы улучшить классификацию новых данных и избежать чрезмерно близкой подгонки. С этой целью, выполняли усечение деревьев классификации для Asn и Asp, т.е. систематически удаляли ветви для формирования деревьев меньшего размера. Для проверки предсказательной силы модели с «отсечением ветвей» их валидацию проводили на 25% тестовой выборке в 40 независимых прогонах (Фиг. 6). Полученные алгоритмы для Asn и Asp обучали на 100% данных и отбирали на основании соответствующих ROC-кривых (Фиг. 6) и интерпретируемости дерева. Алгоритмы принятия решений представлены в виде деревьев решений на Фиг. 7 и 8.
После валидации в 40 прогонах на тестовой выборке с использованием модели, обученной на рандомизированной обучающей 75% выборке, в среднем 0,5 из 8 горячих точек Asp не были распознаны, и в среднем 6,6 из 285 не-горячих точек Asp оказались ложноположительными. Это соответствует TPR 0,94, рассчитанному как количество истинно положительных результатов (7,5), поделенное на количество положительных результатов (8), и FPR 0,02, рассчитанному как количество ложноположительных результатов (6,6), поделенное на количество отрицательных результатов (285) (Фиг. 4а). В случае Asn ложноотрицательными оказались в среднем 0,6 из 11 горячих точек Asn (TPR = 0,95) и ложноположительными оказались 8,1 из 188 не-горячих точек (FPR = 0,04) (Фиг. 46). Этот результат существенно лучше, чем при классификации, основанной исключительно на информации о первичной последовательности и ведущей к завышенной оценке значимости признака (Asp TPR = 1,0, FPR = 0,31; Asn TPR = 0,91, FPR = 0,43) (Фиг 4).
Предрасположенность Asp и Asn к деградации зависит от подвижности остатков, размера следующего остатка и вторичной структуры
В случае Asp массив данных содержит только 2,7% горячих точек, которые необходимо отличить от остатков Asp, не являющихся горячими точками. Первые два расщепления дерева решений позволяют отделить 93% всех не-горячих точек (1105 при первом расщеплении; 260 при втором расщеплении). He-горячие точки являются неподвижными или в направлении карбокси-конца за ними следует аминокислота большого размера, например, такая как Pro, Thr, Asn, Val, Leu, Glu, His, Gin, He, Met, Lys, Phe, Tyr, Arg или Trp. Таким образом, оставшиеся Asp, которые необходимо классифицировать, являются подвижными и за ними следует аминокислота малого размера, которой может быть Gly, Ala, Ser, Cys или Asp. Из их числа отделяется первый и самый большой класс горячих точек Asp, который характеризуется высокой конформационной подвижностью (RMSD > 0,485 ) и наличием Asp, Cys, Ser, Ala или Gly на месте следующего остатка. Он содержит 5 горячих точек (классифицированных данным образом в 5 моделях), а также 2 ложноположительных остатка Asp (классифицированных данным образом в 5 моделях).
В следующем узле происходит отделение 2 класса горячих точек. 3 из них (1 классифицирована данным образом в 5 моделях, 1 в 2 и 1 только в 1 модели) обладают умеренной конформационной подвижностью (RMSD между 0,145 и 0,485 ), следующим аминокислотным остатком для них является Asp, Cys, Ser, Ala или Gly, и изменение вторичной структуры происходит в пределах участка менее чем из 3 аминокислот в направлении карбокси-конца.
Третий класс горячих точек отличается наличием остатка Asp в мотиве Asp-Gly. Кроме того, он характеризуется умеренной конформационной подвижностью (RMSD 0,145 - 0,485 ), как и класс 2, а также изменением вторичной структуры в пределах участка более чем из 3 аминокислот в направлении карбокси-конца. Он содержит 2 горячих точки (1 классифицирована данным образом в 4 моделях и 1 в 3 моделях) и 1 ложноположительный Asp (в 5 моделях).
Что касается классификации горячих точек деградации Asn, основными критериями здесь также являются размер прилегающей со стороны карбокси-конца аминокислоты и конформационная подвижность (Фиг. 8). По сравнению с массивом данных Asp, доля горячих точек Asn относительно количества не-горячих точек, в два раза выше и составляет 5,5%. При этом, первые два расщепления дерева решений также позволяют отделить большую часть всех не-горячих точек (72%; 395 в первом расщеплении; 320 во втором расщеплении). За не-горячими точками в направлении карбокси-конца следует аминокислота большого размера (Val, Leu, Glu, His, Gin, He, Met, Lys, Phe, Tyr, Arg или Trp) или они являются неподвижными (RMSD < 0,01 ). Другим критерием разделения является размер следующего остатка, при этом получают 2 ветви, где размер остатка, следующего за Asn, составляет менее или более 102,7 . Последние также классифицируются по положению CDR петли. Так, первый класс остатков Asn, представляющих собой горячие точки, включает остатки, находящиеся в петле CDR 1, а также характеризуется наличием Asp, Pro, Thr или Asn, прилегающих со стороны карбокси-конца, и отсутствием подвижности (RMSD > 0,01 ). Он содержит 3 представителя горячих точек (классифицированы данным образом в 5 моделях).
Остатки, у которых размер следующего остатка составляет менее 102,7 далее классифицируют в зависимости от величины торсионного угла фи остова. Остатки Asn, за которыми следуют Gly, Ala, Ser или Cys (< 102,7 ), не являющиеся неподвижными, и у которых угол фи составляет менее чем -75,2 градуса, составляют второй и самый большой 2 класс горячих точек. Он содержит 6 представителей горячих точек (4 классифицированы данным образом в 5 моделях, 1 в 4 и 1 в 2 моделях) и 4 ложноположительных точки (1 классифицирована данным образом в 5 моделях, 2 в 3 и 1 в 1 модели).
3 класс горячих точек отличается такой же подвижностью и характеристиками последующей аминокислоты, как и 2 класс, но 4 его представителя (2 классифицированы данным образом в 5 моделях, 1 в 3 и 1 только в 1 модели) имеют угол фи, превышающий -75,2 градуса, высокую доступность для растворителя (SASA > 89,4 ) (рассчитанную, например, с использованием PyMOL) и изменение вторичной структуры в направлении карбокси-конца в пределах участка более чем из 3 аминокислот. В этот класс также входят два ложноположительных остатка Asn (классифицированы данным образом в 1 и 2 моделях).
Выводы
Спонтанная деградация остатков Asn и Asp в составе терапевтически активных белков может произойти в процессе получения, хранения, а также in vivo. Если они задействованы в связывании мишени, то образование продуктов деградации - сукцинимида, isoAsp и Asp, находящихся в CDR, может привести к снижению эффективности связывания с мишенью и снижению эффективности лекарства.
Для облегчения отбора стабильных антител-кандидатов был разработан способ предсказания in silico. С этой целью вначале был получен однородный массив данных, содержащий качественную и количественную информацию о продуктах деградации антител. Обнаруженные модификации согласуются с известными данными о горячих точках.
Для возникновения деградации карбоксильная группа боковой цепи остатков аспартила должна быть протонирована, поскольку гидроксильная группа карбоновой кислоты является лучшей замещаемой группой, чем соответствующий анион. Повышение pH способствует ионизации атома азота остова, принадлежащего следующему остатку, делая его более нуклеофильным. Когда pH достигает значений выше 6, эти противоположные движущие силы стремятся компенсировать друг друга и зависимость от pH отчетливо не проявляется (81).
Наиболее предпочтительно детектировать деградацию остатков Asn при слегка кислых значениях pH, поскольку увеличение концентрации гидроксид-ионов искусственным образом завышает скорость дезаминирования, при этом невозможно отличить артефакты, обусловленные изменением pH при осуществлении способа, от искомых сайтов деградации (49).
Обнаружили, что информация, полученная при исследовании стабильности при щелочных значениях pH, не утрачивается в условиях слегка кислой среды. Чувствительные к щелочным значениям pH горячие точки подвергаются модификации в ходе ферментации (pH 7,4) в результате чего скорость их деградации в референтных и подвергнутых стрессовому воздействию образцах будет одинаковой, а следовательно, не будет существенно повышаться при индукции деградации при pH 6. Как правило, после гидролиза сукцинимида получают смесь Asp и iso-Asp в различных пропорциях (3, 53, 57). Образование только одного продукта, которым является Asp, предположительно свидетельствует о сукцинимид-независимом пути деградации, вследствие альтернативного механизма нуклеофильной атаки, в результате чего образуется изоимид (37), или вследствие непосредственного гидролиза боковой цепи Asn (39). Данный феномен наблюдался для мотива Asn-Thr трастузумаба.
Поразительным был тот факт, что все обнаруженные горячие точки располагались в CDR петлях исследуемых антител (Таблица 1). Таким образом, Fab фрагмент и каркасные участки Fv являются стабильными. Чтобы оценить соответствие нашей коллекции терапевтически активных моноклональных антител антителам естественного происхождения, сравнивали частоту известных мотивов последовательности, содержащих Asn и Asp, подвергающиеся деградации (NG, NN, NS, NT, DG, DS, DT, DD, DH), в CDR моноклональных антител нашей коллекции (для определения CDR комбинировали системы нумерации Kabat и Chothia (82)) и в последовательностях 16286 человеческих моноклональных антител естественного происхождения (последовательности 9990 V-D-J и 6296 V-J) из международной информационной системы ImMunoGeneTics (IMGT) базы данных моноклональных антител (www.IMGT.org). Несмотря на огромную разницу в размерах сравниваемых баз данных, частота, с которой наблюдались мотивы, содержащие Asn и Asp, была относительно одинаковой, состав последовательностей исследуемых молекул антител был сбалансированным (Фиг. 2). Единственным исключением являлся мотив NT, который встречался в два раза чаще в терапевтически активных моноклональных антителах, чем в IMGT. Очевидно, что частота наиболее значимых в отношении деградации мотивов Asn-Gly и Asp-Gly по сравнению с другими мотивами последовательности невысока в обоих массивах данных.
Как известно из уровня техники, предрасположенность Asn/Asp к деградации можно предсказывать на основе сведений о первичной последовательности и информации о пространственной структуре (5, 34, 37, 38, 40, 46, 51, 61-64). Способ предсказания дезаминирования Asn в белках был предложен Robinson & Robinson в 2001 (51). Авторы использовали известные скорости дезаминирования 198 остатков Asn в 23 различных белках и 70 остатков Asn в 61 вариантах гемоглобина человека, которые исследовали в сильно различающихся экспериментальных условиях. Основные отличия от данного исследования заключаются в том, что (i) предсказание возможно только для Asn, (ii) экспериментальные условия, в которых была получена коллекция горячих точек, на основе которой осуществляли предсказание, не были одинаковыми, (iii) информация о трехмерной структуре получена с использованием рентгеноструктурного анализа, а не моделирования по гомологии, (iv) обычные пользователи могут использовать для предсказания информацию о белках, которая была занесена в базу данных PDB до 2001, и (v) для новых белков предсказание возможно только при наличии данных рентгеноструктурного анализа. В отличие от указанного способа, способ, предложенный в данной заявке, предназначен для вариабельных областей терапевтически активных антител и основан на расчетах, выполненных с помощью компьютерного моделирования, без необходимости получения рентгеноструктурных данных. Для осуществления способа, описанного в данном документе, необходимы: (i) аминокислотная последовательность тяжелой и легкой цепей антитела, (ii) средство для моделирования гомологии, (iii) программное обеспечение для визуализации молекул и (iv) статистическая модель, описанная в данном документе. Сокращение количества неверно классифицированных горячих точек (2,3% Asp, 4,3% Asn) по сравнению с предсказанием только на основании последовательности (31% Asp, 43% Asn) позволяет сберечь время и ресурсы. Соотношение не-горячих точек к горячим точкам удалось сократить благодаря работе только с Fv областью Fab фрагмента, поскольку только вариабельная область содержала предрасположенные к деградации Asn и Asp. Проведение классификации только по остаткам, расположенным в петле CDR, приводило к получению статистических значений меньшей предсказательной силы.
В данном документе описан способ предсказания сайтов деградации антитела и приведены основные характеристики, отличающие нестабильные и стабильные аминокислотные остатки Asn и Asp в вариабельных областях моноклональных антител. Остатки Asn и Asp, обладающие высокой подвижностью и расположенные перед остатком малого размера, подвержены деградации. Для дополнительной характеристики используют элементы вторичной структуры. Неожиданно было обнаружено, что параметры, наиболее точно описывающие механизм реакции (Фиг. 1), такие как расстояние между Сγ атомом и атомом азота остова, принадлежащим аминокислоте, прилегающей со стороны карбокси-конца, значение pKa Asp или торсионный угол χ1 боковой цепи, не важны для классификации.
Описанный здесь способ позволяет проводить более эффективный отбор моноклональных антител. Он позволит избежать возможных неудач на завершающем этапе процесса отбора наиболее стабильных и в то же время наиболее эффективных молекул-кандидатов, которые можно совершенствовать дальше и внедрять в клиническую практику, и обеспечить максимум пользы для пациента.
Правило, указывающее на возможное существование горячей точки, можно сформулировать следующим образом: если Asn/Asp согласно предсказанию является горячей точкой по меньшей мере в одной из пяти моделей на основе гомологии, то остаток классифицируют как горячую точку. В каждой из моделей ансамбля вероятность отнесения к горячей точке варьирует от минимального значения 0,5 до максимального значения 1,0. Таким образом, результаты предсказания являются не только качественными, но и количественными, и выражаются в виде среднего арифметического вероятности отнесения остатка к горячим точкам в каждой модели и стандартного отклонения. Следовательно, результаты предсказания содержат информацию о том, находится ли остаток в конформации горячей точки в одной, двух, трех, четырех или пяти моделях ансамбля.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг. 1. Пути деградации аспарагина и аспартата. Дезаминирование аспарагина или дегидратация аспарагиновой кислоты при нуклеофильной атаке α-аминогруппы аминокислоты, примыкающей со стороны С-конца. Это приводит к образованию метастабильного сукцинимида (циклического имида), промежуточного соединения, которое гидролизуется с образованием смеси аспартила и изо-аспартила. В альтернативном случае, нуклеофильная атака кислорода карбонила остова приводит к образованию циклического изоимида, промежуточного соединения, которое в результате гидролиза приводит только к образованию остатков аспартила, независимо от места атаки молекулы воды. Остатки аспарагина могут подвергаться дезаминированию в Asp непосредственно в результате гидролиза, протекающего при участии воды. Стандартные аминокислоты (Asn, Asp) выделены черными рамками.
Фиг. 2. Встречаемость в CDR аминокислотных мотивов, содержащих Asn и Asp, в коллекции моноклональных терапевтически активных антител и в группе антител естественного происхождения (IMGT). Черные треугольники показывают процентное содержание горячих точек в составе мотивов, содержащих Asn и Asp, в экспериментальной коллекции из 37 моноклональных антител. Столбики показывают процентное содержание указанных мотивов последовательности среди всех остатков Asn и Asp в вариабельной области (верхняя панель) или только в участках CDR (нижняя панель). Процентное содержание, изображенное в виде темных столбиков, относится к неизбыточной коллекции 37 терапевтически активных моноклональных антител, исследованных аналитически, светло-серые столбики относятся к коллекции 9990 V-D-J- и 6296 V-J сегментов антител естественного происхождения из базы данных IMGT; а) мотивы последовательности, содержащие Asn, б) мотивы последовательности, содержащие Asp.
Фиг. 3. Параметры, характеризующие остатки Asn и Asp в их структурном окружении, показанные на примере остатка Asp. Параметры, характеризующие тенденцию к замещению карбоксила/аминогруппы, доступность переходного состояния, нуклеофильность Nn+1 и структурное окружение, показаны розовым, голубым, лиловым и синим, соответственно. Наименования параметров соответствуют указанным в Таблице 2.
Фиг. 4. ROC-кривая, позволяющая сравнить классификацию на основе трехмерной структуры и предсказания на основе последовательности, показывает существенное снижение доли ложноположительных результатов. Сравнивали различные статистические методы и предсказание только на основе последовательности. При классификации статистическими методами среднее количество ложноположительных и ложноотрицательных остатков Asn/Asp получили в результате 40 раундов валидации на тестовых выборках. TPR (доля истинно положительных результатов) = число истинно положительных результатов, поделенное на число положительных результатов. FPR (доля ложноположительных результатов) = число ложноположительных результатов, поделенное на число отрицательных результатов. Tree  , rpart (◊), PP (Pipeline Pilot) Tree (темные ○) и случайный лес (○) представляют собой алгоритмы рекурсивного разбиения; svm (Δ), ksvm
, rpart (◊), PP (Pipeline Pilot) Tree (темные ○) и случайный лес (○) представляют собой алгоритмы рекурсивного разбиения; svm (Δ), ksvm  представляют собой алгоритмы метода опорных векторов; rda
представляют собой алгоритмы метода опорных векторов; rda  представляет собой алгоритм регуляризованного дискриминантного анализа; nnet (+) представляет собой нейронную сеть; «на основе последовательности» означает предсказание на основе мотивов последовательности NG, NS, NT и DG, DS, DT, DD, DH. Для предсказания был выбран алгоритм дерево Pipeline Pilot, с отсечением ветвей на 4 уровне; а) валидация для аспартата, б) валидация для аспарагина.
представляет собой алгоритм регуляризованного дискриминантного анализа; nnet (+) представляет собой нейронную сеть; «на основе последовательности» означает предсказание на основе мотивов последовательности NG, NS, NT и DG, DS, DT, DD, DH. Для предсказания был выбран алгоритм дерево Pipeline Pilot, с отсечением ветвей на 4 уровне; а) валидация для аспартата, б) валидация для аспарагина.
Фиг. 5. ROC-кривая в увеличенном масштабе (Фиг. 4) и таблица для сравнения трехмерных классификаторов. Среднее количество ложноположительных и ложноотрицательных остатков Asn/Asp получено в результате 40 раундов валидации на тестовых выборках. TPR (доля истинно положительных результатов) = число истинно положительных результатов, поделенное на число положительных результатов. FPR (доля ложноположительных результатов) = число ложноположительных результатов, поделенное на число отрицательных результатов. Tree, rpart, РР (Pipeline Pilot) Tree и случайный лес представляют собой алгоритмы рекурсивного разбиения; svm, ksvm представляют собой алгоритмы метода опорных векторов; rda представляет собой алгоритм регуляризованного дискриминантного анализа; nnet представляет собой нейронную сеть. Для предсказания был выбран алгоритм дерево Pipeline Pilot (заштрихованные круги), с отсечением ветвей на 4 уровне; а) валидация для аспартата, б) валидация для аспарагина.
Фиг. 6. ROC-кривая для сравнения различных уровней отсечения ветвей дерева решений. Усечение деревьев решений осуществляли автоматически, как предусмотрено в программе Pipeline Pilot (Accelrys Inc., San Diego, USA). Среднее количество ложноположительных и ложноотрицательных остатков Asn/Asp получено в результате 40 раундов валидации на тестовых выборках (25%). TPR (доля истинно положительных результатов)=число истинно положительных результатов, поделенное на число положительных результатов. FPR (доля ложноположительных результатов)=число ложноположительных результатов, поделенное на число отрицательных результатов. Деревья 1-3 и 5-6 обозначены сферами, дерево 4 обозначено черным треугольником. Дерево 1 представляет собой модель без отсечения ветвей. Дерево 4 выбрали для предсказания. Валидацию уровня отсечения проводили на основании ROC-кривой, под увеличением показано, как перекрываются стандартные отклонения при отсечении на разных уровнях.
Фиг. 7. Дерево решений для аспартата, уровень отсечения 4 при глубине предварительного просмотра 4 и возможности предварительно просматривать 7 альтернативных вариантов. Модель обучали на 1425 не-горячих точках (белый) и 40 горячих точках (черный). Контуры узлов и листьев окрашены в соответствии со взвешенным большинством класса. Высота закрашивания столбцов в правой части каждого узла/листа соответствует доле в выборке. Доля каждого класса в узле/листе изображена в виде закрашенного сегмента круга. Основными критериями для принятия решений являются конформационная подвижность (RMSD) и размер аминокислоты, прилегающей со стороны С-конца (размер следующей аминокислоты); белый = не-горячая точка; черный = горячая точка.
Фиг. 8. Дерево решений для аспарагина, уровень отсечения 4 при глубине предварительного просмотра 4 и возможности предварительно просматривать 7 альтернативных вариантов. Модель обучали на 940 не-горячих точках (белый) и 55 горячих точках (черный). Контуры узлов и листьев окрашены в соответствии со взвешенным большинством класса. Высота закрашивания столбцов в правой части каждого узла/листа соответствует доле в выборке. Доля каждого класса в узле/листе изображена в виде закрашенного сегмента круга. Основными критериями для принятия решений являются размер следующей аминокислоты и конформационная подвижность (RMSD); белый = не-горячая точка; черный = горячая точка.
ОПИСАНИЕ ПРИМЕРОВ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Приведенные ниже примеры и графические материалы дают возможность лучше понять изобретение, объем которого ограничен прилагаемой формулой. Следует понимать, что указанные методики могут быть модифицированы, но при этом суть изобретения сохранится.
Материалы и методы
Происхождение моноклональных антител
24 моноклональных антитела представляли собой человеческие или гуманизированные lgG1 или lgG4 антитела. 13 моноклональных антител представляли собой коммерциализированные продукты, включая авастин (бевацизумаб, Genentech/Roche); CYT387 (нимотузумаб, Oncoscience, Ch.B.: 911017W002); эрбитукс (цетуксимаб, Bristol-Myers Squibb and Eli Lilly and Company, лот: 7666001); герцептин (трастузумаб, RO-45-2317/000, лот: HER401-4, Genentech); хумира (адалимумаб, Abbott, Ch.B.: 90054XD10); пролиа (деносумаб, Amgen, Ch.B.: 1021509); раптива (эфализумаб, Genentech, Merck Serono, лот: Y11A6845); ремикад (инфликсимаб, Centocor, Ch.B.: 0RMA66104); симулект (базиликсимаб, Novartis, Ch.B.: S0014); синагис (паливизумаб, Medimmune, лот: 122-389-12); тизабри (натализумаб, Biogen Idee and Elan, LotA: 080475); вектибикс (панитумумаб, Amgen, Ch.B.: 1023731); и ксолар (омализумаб, Genentech/Novartis, Ch.B.: S0053).
Пример 1. Получение образцов при индукции деградации
Все терапевтически активные моноклональные антитела подвергали индуцированной деградации (стрессовому воздействию). 2 мг каждого антитела диализовали в течение ночи при 4°C в буфере для разведения (20 мМ гистидин-хлорид, pH 6,0) в пробирках для диализа D-Tube Dialyzers (Novagen, MWCO 6-8 кДа). Определяли концентрации и доводили до 5 мг/мл буфером для разведения. После стерилизации фильтрованием (Pall Nanosep MF, 0,2 мкм) и переноса в стерильные пробирки с закручивающимися крышками, все образцы моноклональных антител инкубировали в статическом режиме при 40°C в течение 2 недель.
Пример 2. Подготовка образцов моноклональных антител для триптического пептидного картирования
80 мкг референтного и подвергнутого стрессовому воздействию образцов моноклональных антител денатурировали и восстанавливали в течение 1 ч в конечном объеме 124,5 мкл 100 мМ Tris, 5,6 М гуанидина гидрохлорид, 10 мМ ТСЕР (трис(2-карбоксиэтил)фосфин), Pierce Protein Biology Products, Thermo Fisher Scientific, Waltham, MA, USA), pH 6,0 при 37°C. Буфер заменяли на 20 мМ гистидина хлорид, 0,5 мМ ТСЕР, pH 6,0 на обессоливающих колонках Zeba Spin объемом 0,5 мл (Pierce Protein Biology Products, Thermo Fisher Scientific, Waltham, MA, USA). Моноклональные антитела расщепляли путем добавления 0,05 мкг трипсина (Promega, Madison) на мкг антитела в конечном объеме 140 мкл и инкубации в течение ночи при 37°C. Реакцию останавливали путем добавления 7 мкл 10% раствора муравьиной кислоты и замораживали образцы при -80°C до последующего анализа.
Пример 3. Определение модифицированных пептидов при помощи жидкостной хроматографии с тандемной масс-спектрометрией
14 мкг расщепленного антитела наносили на колонку Varian Polaris 3 С18 -Ether (1×250 мм; диаметр частиц 3 мкм, размер пор 180 ) компании Varian (Darmstadt, Germany) для разделения при помощи обращенно-фазовой высокоэффективной жидкостной хроматографии (ВЭЖХ, Agilent 1100 Cap LC, Agilent Technologies, Boeblingen, Germany). MAb2, mAM4 и нимотузумаб после расщепления дополнительно разделяли при помощи обращенно-фазовой сверхпроизводительной жидкостной хроматографии (СВЭЖХ, колонка ACQUITY ВЕН300 С18, 1×150 мм, размер гранул 1,7 мкм, размер пор 300 A, Waters, Manchester, UK). Элюат ВЭЖХ или СВЭЖХ разделяли с использованием Triversa NanoMate (Advion, Ithaca, NY, USA) и вводили 380 нл/мин в классический тандемный масс-спектрометр LTQ Orbitrap (Thermo Fisher Scientific, Waltham, MA, USA) в режиме регистрации положительно заряженных ионов. Подвижные фазы обращенно-фазовой ВЭЖХ представляли собой 0,1% раствор муравьиной кислоты в воде (растворитель А) и 0,1% раствор муравьиной кислоты в ацетонитриле (растворитель Б). При ВЭЖХ использовали ступенчатый градиент, начиная от 2% растворителя Б, с увеличением до 15% с 5 мин до 15 мин, до 32% с 15 мин до 70 мин, до 38% с 70 мин до 80 мин, до 100% с 80 мин до 90 мин, и наконец снижением до 2% с 92 мин до 110 мин при скорости тока 60 мкл/мин. При СВЭЖХ использовали линейный градиент от 1 до 40% растворителя Б с 0 до 130 мин. Поглощение УФ излучения измеряли при длинах волн 220 и 280 нм. Регистрацию данных контролировали при помощи программы Xcalibur™ (Thermo Fisher Scientific, Waltham, MA, USA). При тандемной масс-спектрометрии фрагментацию осуществляли методом столкновительной диссоциации при низкой энергии, в качестве газа для соударений использовали гелий, энергия соударений в линейной квадрупольной ловушке составляла 35%. Для получения более высокого разрешения фрагментов ионов mAb14 и нимотузумаба при фрагментации в Orbitrap использовали список масс ионов-предшественников, ширину изолируемого пика 3, масс-фильтр ионов-предшественников 0,2 Да, значение автоматического регулирования усиления 400000 и время накопления фрагментов 5000 мс.
Пример 4. Подготовка образцов mAb14 и нимотузумаба для анализа с помощью тандемной масс-спектрометрии
Для дальнейшей характеризации подвергнутые стрессовому воздействию образцы mAb14 и нимотузумаба обрабатывали следующим образом. 250 мкг моноклонального антитела подвергали денатурации путем добавления денатурирующего буфера (0,4 М Трис (Sigma-Aldrich, Taufkirchen, Germany), 8 М гуанидина гидрохлорид (Sigma-Aldrich, Taufkirchen, Germany), pH 8) до конечного объема 240 мкл. Для восстановления добавляли 20 мкл 0,24 М дитиотрейтола (ДТТ) (Roche Diagnostics GmbH, Mannheim, Germany) свежеприготовленного в денатурирующем буфере и инкубировали при 37°C в течение 60 мин. Затем образец алкилировали путем добавления 20 мкл 0,6 М йодуксусной кислоты (Merck KgaA, Darmstadt, Germany) в воде и инкубации в течение 15 мин при комнатной температуре в темноте. Избыток алкилирующего реагента инактивировали путем добавления 30 мкл раствора ДТТ. Затем в образцах производили замену буфера приблизительно на 480 мкл 50 мМ Трис/HCl, pH 7,5 с использованием колонок NAP5 Sephadex G-25, предназначенных для работы с ДНК (GE Healthcare, Germany). Моноклональные антитела расщепляли путем добавления 0,03 мкг трипсина (Promega, Madison) на мкг белка в конечном объеме 500 мкл и инкубации в течение 5 ч при 37°C. Расщепление останавливали путем добавления 20 мкл 10% раствора муравьиной кислоты и замораживали образцы при -80°C для последующего анализа.
Пример 5. Анализ данных для количественной оценки уровней модификации
Для предварительного отбора данных, выявивших различия между подвергнутыми стрессовому воздействию и референтными образцами, использовали программу SIEVE версии 2.0 (VAST Scientific Inc., Cambridge, MA). Ключевыми параметрами для программы SIEVE были интервал времени 1,0 мин, разброс m/z 8,0 ppm и пороговое значение интенсивности 50 ООО ионов. После отсева по моноизотопной массе в программе SIEVE (prelement = 0), данные импортировали в файл EXCEL с поддержкой макросов наряду с данными о расщеплении тяжелых и легких цепей моноклональных антител трипсином, полученными in silico и отражающими теоретически расчитанные значения отношения массы к заряду модифицированных и немодифицированных пептидов. Различия в интенсивности сигналов или времени удерживания (для референтных и подвергнутыми стрессовому воздействию образцов) определяли полуавтоматизированным способом с использованием файла EXCEL с поддержкой макросов (Microsoft, Redmond, WA, USA). Полученные в результате предварительного отсева пептиды из 76 пептидных карт проверяли вручную на наличие модификаций Asn и Asp по значениям m/z в масс-спектре, полученном экспериментально. Для количественной оценки строили экстрагированные (извлеченные) ионные хроматограммы (XIC, от англ. extracted ion chromatograms) на основе моноизотопных масс пептидов, представляющих интерес, и определяли зарядовое состояние с помощью программы Xcalibur (Thermo Fisher Scientific, Waltham, MA, USA). Относительное количество модифицированных и немодифицированных пептидов рассчитывали после интегрирования площади соответствующих пиков вручную. Кроме того, для полноты информации анализировали все пептиды, расположенные в CDR областях, содержащие мотивы, предположительно содержащие горячие точки (Asn-Gly, Asn-Thr, Asn-Ser, Asn-Asn, Asp-Gly, Asp-Thr, Asp-Ser, Asp-Asp, Asp-His), даже если они не были отобраны по результатам анализа в программе SIEVE.
Пример 6. Моделирование по гомологии и расчет двух- и трехмерных параметров
Модели на основе гомологии строили с использованием автоматизированного сценария выполнения (скрипта), написанного для программы MODELER 9v7 (83). Матрицы для моделирования выбирали из базы референтных структур, состоящей из кристаллических структур Fab фрагментов человеческих, мышиных и химерных антител, имеющих минимальное разрешение 2,8 , учитывая консервативность последовательностей и не пропуская остатки, расположенные во внутренней части их вариабельных областей. Наилучшую итоговую модель каждого моноклонального антитела брали за основу для процедуры уточнения структуры петель (LOOPER, Discovery Studio, Accelrys Inc., San Diego, USA) (84). После уточнения структуры петель отбирали пять наиболее правдоподобных моделей, которые использовали в качестве ансамбля структур для каждого моноклонального антитела. Ансамбли моделей, основанных на гомологии, использовали для определения параметров (Таблица 2). Значения pKa рассчитывали с помощью программы propka в составе пакета pdb2pqr (79). Элементы вторичной структуры (слой, спираль, поворот, случайная спираль) были найдены с помощью специально написанного скрипта с использованием пакета Discovery Studio (Accelrys Inc., San Diego, USA). Параметры «следующая отличающаяся вторичная структура в направлении N-конца», «следующая отличающаяся вторичная структура в направлении С-конца» и «местоположение в случайной спирали» получали на основании информации о вторичной структуре окружающих остатков, используя логические правила, реализованные в Pipeline Pilot (Accelrys Inc., San Diego, USA). «Местоположение в случайной спирали» обозначали «краевым», если элемент следующей отличающейся вторичной структуры находился на расстоянии одного или двух остатков в направлении N- или С-конца. «Местоположение в случайной спирали» обозначали «центральным», если вторичная структура была одинаковой для 4 остатков в направлении N- и C-конца или для более чем 4 остатков в обоих направлениях. Параметр «положение в Fab» представлял собой значение, которое определяли в соответствии с системами нумерации антител, предложенными Chothia и Kabat (82) (Kabat). «Положение в Fab» со значением 1 соответствует 1 каркасному участку тяжелой цепи (FR Н), 2 соответствует CDR Н 1, 3 соответствует FR Н 2, 4 соответствует CDR Н 2, 5 соответствует FR Н 3, 6 соответствует CDR Н 3, 7 соответствует FR Н 4, 8 соответствует 1 каркасному участку легкой цепи (FR L), 9 соответствует CDR L 1, 10 соответствует FR L 2, 11 соответствует CDR L 2, 12 соответствует FR L 3, 13 соответствует CDR L 3, а 14 соответствует FR L 4. «Петля CDR» представляет собой значение от 1 до 3, как для легкой, так и для тяжелой цепей. «Размер следующего аминокислотного остатка» обозначает доступную для растворителя площадь поверхности (85), выраженную в , и определяется следующим образом: Ala, 64.78; Cys, 95.24; Asp, 110.21; Glu, 143.92; Phe, 186.7; Gly, 23.13; His, 146.45; He, 151.24; Lys, 177.37; Leu, 139.52; Met, 164.67; Asn, 113.19; Pro, 111.53; Gin, 147.86; Arg, 210.02; Ser, 81.22; Thr, 111.6; Val, 124.24; Trp, 229.62; Tyr, 200.31. Концевые остатки (не имеющие углов фи и пси) отмечены в коллекции данных. Все остальные параметры находили с использованием PDB файлов с помощью скриптов, самостоятельно написанных на языке python для PyMOL (5) (Таблица 2).
Пример 7. Алгоритмы машинного обучения, использованные для оценки классификации
Для того, чтобы выбрать наилучший из возможных классификаторов, были протестированы различные способы, которые были наиболее подходящими для решения проблемы классификации данного типа, а именно алгоритмы метода опорных векторов, рекурсивного разбиения, регуляризованный дискриминантный анализ и нейронные сети. Они были реализованы в пакете статистических программ R или в Pipeline Pilot (Accelrys Inc., San Diego, USA). Метод опорных векторов позволяет различными способами преобразовать имеющийся массив данных в пространство более высокой размерности с помощью так называемой функции ядра. В данном случае использовали метод опорных векторов (86) из пакета е1071 и метод ksvm из пакета kernlab (87). Методы рекурсивного разбиения выявляют параметры пошаговым способом для разбиения заданного массива данных на подмножества, таким образом получая дерево решений. Различие между алгоритмами в основном обусловлено различными способами принятия решений о наилучшем разделяющем параметре на заданном этапе. Использовали методы «tree» (88) и «rpart» (89) пакета программ R, при этом тестировали различные способы разбиения. Более общую форму классификтора можно получить путем объединения деревьев решений, построенных на подмножествах исходных обучающих выборок, в так называемый случайный лес. Регуляризованный дискриминантный анализ позволяет построить классификатор путем объединения подмножества имеющихся параметров с использованием регуляризованных групповых ковариационных матриц, чтобы добиться наилучшего разделения. Данный метод реализован в пакете программ klaR как функция "rda" (90). Нейронная сеть имитирует базовые функции одного или нескольких соединенных между собой слоев нейронов. Использовали так называемую нейронную сеть с одним скрытым слоем, реализованную в методе "nnet" пакета программ R (91). Наконец, был испытан наивный классификатор Байеса, реализованный в R как метод "NaiveBayes", вероятностный метод, в котором вероятность принадлежности данных к определенному классу рассчитывают на основе обучающих примеров с использованием теоремы Байеса.
Поскольку массив данных был очень несбалансированным, включал очень мало горячих точек и много не-горячих точек, для придания большей весомости минорному классу вводили весовые коэффициенты классов. Разработали стандартную схему присвоения весов, используя значение, обратное частоте класса, что позволило минимизировать ошибку классификации и, что особенно важно, уменьшить долю ложноотрицательных результатов.
Пример 8. Рекурсивное разбиение и предсказание
Как показала сравнительная оценка методов, описанных в Примере 7, наилучшим алгоритмом классификации оказался алгоритм рекурсивного разбиения single-tree с возможностью предварительного просмотра, реализованный в Pipeline Pilot (Accelrys Inc., San Diego, USA). Данную модель обучали отдельно для Asn и Asp, используя для предсказания исключительно остатки, находящиеся в Fv доменах моделей, основанных на гомологии. Таким образом, обучение проводили на 1045 остатках Asn и 1520 остатках Asp, из которых 60 и 35, соответственно, представляли собой горячие точки, и выученный признак обозначали горячей точкой. Концевые остатки, а также остатки, у которых степень модификации в подвергнутых стрессовому воздействию образцах составляла менее 3% (слабые точки и реакционно-способные точки), исключали из обучения. Все 20 описанных параметров были включены в обучающую выборку. Главной характеристикой алгоритма классификации на основе рекурсивного разбиения single-tree, реализованного в Pipeline Pilot, является возможность устанавливать определенную глубину «предварительного просмотра», что позволяет улучшить классификацию благодаря тестированию большего количества альтернативных разбиений.
Две полученные предсказывающие модели используют на новых данных. Правило, указывающее на возможное существование горячей точки, можно сформулировать следующим образом: если Asn/Asp согласно предсказанию является горячей точкой по меньшей мере в одной из пяти моделей, основанных на гомологии, то остаток классифицируют как горячую точку. В каждой из моделей ансамбля вероятность отнесения к горячей точке варьирует от минимального значения 0,5 до максимального значения 1,0. Таким образом, результаты предсказания являются не только качественными, но и количественными, выражаются в виде среднего арифметического вероятностей отнесения остатка к горячим точкам в каждой модели и стандартного отклонения. Следовательно, результаты предсказания содержат информацию о том, находится ли остаток в конформации горячей точки в одной, двух, трех, четырех или пяти моделях ансамбля.
Список литературы
(1) Reichert, J.М., et al. (2005) Nat. Biotechnol. 23, 1073-1078.
(2) Swann, P.G., et al. (2008) Curr. Opin. Immunol. 20, 493-499.
(3) Geiger, T. & Clarke, S. (1987) J. Biol. Chem. 262, 785-794.
(4) Joshi, А.В., et al. (2005) J. Pharm. Sci. 94, 1912-1927.
(5) Clarke, S. (1987) Int. J. Pept. Protein Res. 30, 808-821.
(6) Manning, M.C., et al. (1989) Pharm. Res. 6, 903-918.
(7) Wakankar, A.A. & Borchardt, R.T. (2006) J. Pharm. Sci. 95, 2321-2336.
(8) Simpson, R.J. (2010) Cold Spring Harb. Protoc. 2010.
(9) Wakankar, A.A, et al. (2007) J. Pharm. Sci. 96, 1708-1718.
(10) Harris, R.J., et al. (2001) J. Chromatogr. В Biomed. Sci. Appl. 752, 233-245.
(11) Cacia, J., et al. (1996) Biochemistry35, 1897-1903.
(12) Huang, L, et al. (2005) Anal. Chem. 77, 1432-1439.
(13) Yan, В., et al. (2009) J. Pharm. Sci. 98, 3509-3521.
(14) Rehder, D.S., et al. (2008) Biochemistry 47, 2518-2530.
(15) Weintraub, S.J. & Manson, S.R. (2004) Mech. Ageing Dev. 125, 255-257.
(16) Robinson, N.E. & Robinson, A.B. (2001) Proc. Natl. Acad. Sci. USA 98, 944-949.
(17) Robinson, N.E. & Robinson, A.B. (2001) Proc. Natl. Acad. Sci. USA 98, 12409-12413.
(18) Robinson, N.E. (2002) Proc. Natl. Acad. Sci. U.S.A 99, 5283-5288.
(19) Robinson, А.В., et al. (1970) Proc. Natl. Acad. Sci. U.S.A 66, 753-757.
(20) Wright, H.T. (1991) Crit. Rev. Biochem. Mol. Biol. 26, 1-52.
(21) Harding, J.J., et al. (1989) Mech. Ageing Dev. 50, 7-16.
(22) Zhao, R., et al. (2004) Cancer Cell 5, 37-49.
(23) Zhao, R., et al. (2007) PLoS. Biol. 5, e1. doi:10.1371.
(24) Deverman, В.E., et al. (2002) Cell 111, 51-62.
(25) Weintraub, S.J. & Deverman, В.E. (2007) Sci. STKE. 2007..
(26) Takata, Т., et al. (2008) Protein Sci. 17, 1565-1575.
(27) Takata, Т., et al. (2007) Biochemistry 46, 8861-8871.
(28) Kosugi, S., et al. (2008) Biochem. Biophys. Res. Commun. 371, 22-27.
(29) Tomizawa, H., et al. (1994) Biochemistry 33, 8770-8774.
(30) Shimizu, Т., et al. (2005) Biol. Pharm. Bull. 28, 1590-1596.
(31) Bohme, L., et al. (2008) Biol. Chem. 389, 1055-1066.
(32) Bohme, L., et al. (2008) Biol. Chem. 389, 1043-1053.
(33) Stephenson, R.C. & Clarke, S. (1989) J. Biol. Chem. 264, 6164-6170.
(34) Xie, M., et al. (2000) J. Pept. Res. 56, 165-171.
(35) Chu, G.C., et al. (2007) Pharm. Res. 24, 1145-1156.
(36) Oliyai, C. & Borchardt, R.T. (1993) Pharm. Res. 10, 95-102.
(37) Athmer, L., et al. (2002) J. Biol. Chem. 277, 30502-30507.
(38) Sinha, S., et al. (2009) Protein Sci. 18, 1573-1584.
(39) Catak, S., et al. (2009) J. Phys. Chem. A 113, 1111-1120.
(40) Wright, H.T. (1991) Protein Eng. 4, 283-294.
(41) Vlasak, J. & lonescu, R. (2008) Curr. Pharm. Biotechnol. 9, 468-481.
(42) Zhang, W. & Czupryn, M.J. (2003) J. Pharm. Biomed. Anal. 30, 1479-1490.
(43) Kroon, D.J., et al. (1992) Pharm. Res. 9, 1386-1393.
(44) Zabrouskov, V., et al. (2006) Biochemistry 45, 987-992.
(45) Liu, H., et al. (2008) J. Chromatogr. A 1210, 76-83.
(46) Sreedhara, A., et al. (2012) Pharm. Res. 29, 187-197.
(47) Chelius, D., et al. (2005) Anal. Chem. 77, 6004-6011.
(48) Liu, H., et al. (2006) J. Chromatogr. В Analyt. Technol. Biomed. Life Sci. 837, 35-43.
(49) Diepold, K., et al. (2012) PLoS. One. 7, e30295. doi: 10.1371.
(50) Yu, X.C., et al. (2011) Anal. Chem. 83, 5912-5919.
(51) Robinson, N.E. & Robinson, A.B. (2001) Proc. Natl. Acad. Sci. USA 98, 4367-4372.
(52) Brennan, Т.V. & Clarke, S. (1995) Int. J. Pept. Protein Res. 45, 547-553.
(53) Tyler-Cross, R. & Schirch, V. (1991) J. Biol. Chem. 266, 22549-22556.
(54) Oliyai, C. & Borchardt, R.T. (1994) Pharm. Res. 11, 751-758.
(55) Kosky, A.A., et al. (2009) Pharm. Res. 26, 2417-2428.
(56)Capasso, S. (2000) J. Pept. Res. 55, 224-229.
(57) Patel, K. & Borchardt, R.T. (1990) Pharm. Res. 7, 703-711.
(58) Oliyai, C., et al. (1994) Pharm. Res. 11, 901-908.
(59) Brennan, Т.V. & Clarke, S. (1993) Protein Sci. 2, 331-338.
(60) Zheng, J.Y. & Janis, L.J. (2006) Int. J. Pharm. 308, 46-51.
(61) Kossiakoff, A.A. (1988) Science 240, 191-194.
(62) Xie, M. & Schowen, R.L. (1999) J. Pharm. Sci. 88, 8-13.
(63) Kosky, A.A., et al. (1999) Protein Sci. 8, 2519-2523.
(64) Bischoff, R. & Kolbe, H.V. (1994) J. Chromatogr. В Biomed. Appl. 662, 261-278.
(65) Wakankar, A.A., et al. (2007) Biochemistry 46, 1534-1544.
(66) Yi, L., et al. (2012) J. Pharm. Sci. 102, 947-959.
(67) Zhang, J., et al. (2011) Anal. Biochem. 410, 234-243.
(68) Xiao, G., et al. (2007) Anal. Chem. 79, 2714-2721.
(69) Timm, V., et al. (2010) J. Chromatogr. В Analyt. Technol. Biomed. Life Sci. 878, 777-784.
(70) Perkins, M., et al. (2000) Pharm. Res. 17, 1110-1117.
(71) Vlasak, J., et al. (2009) Anal. Biochem. 392, 145-154.
(72) Xiao, G. & Bondarenko, P.V. (2008) J. Pharm. Biomed. Anal. 47, 23-30.
(73) Valliere-Douglass, J., et al. (2008) Anal. Chem. 80, 3168-3174.
(74) Al-Lazikani, В., et al. (1997) J. Mol. Biol. 273, 927-948.
(75) Morea, V., et al. (1998) J. Mol. Biol. 275, 269-294.
(76) Martin, A.C. & Thornton, J.M. (1996) J. Mol. Biol. 263, 800-815.
(77) Whitelegg, N. & Rees, A.R. (2004) Methods Mol. Biol. 248, 51-91.
(78) Capasso, S., et al. (1992) Pept. Res. 5, 325-330.
(79) Li, H., et al. (2005) Proteins 61, 704-721.
(80) Hambly, D.M., et al. (2009) Anal. Chem. 81, 7454-7459.
(81) Zhigiang, A. (2009) in Therapeutic monoclonal antibodies: from bench to clinic. (Wiley & Sons, Inc., Hoboken, New Jersey).
(82) Chothia, C., et al. (1989) Nature 342, 877-883.
(83) Sali, A. & Blundell, T.L. (1993) J. Mol. Biol. 234, 779-815.
(84) Spassov, V.Z., et al. (2008) Protein Eng. Des. Sel. 21, 91-100.
(85) Chennamsetty, N., et al. (2009) Proc. Natl. Acad. Sci. USA 106, 11937-11942.
(86) Dimitriadou, E., et al. (2011), http://CRAN.R-project.org/package=e1071.
(87) Karatzoglou, A., et al. (2004) Journal of Statistical Software 11, 1-20.
(88) Ripley, B. (2011) http://CRAN.R-project.org/package=tree).
(89) Therneau, R.M., et al. (2010) http://CRAN.R-project.org/package=rpart).
(90) Weihs, C., et al. (2005) in Data Analysis and Decision, eds. Baier, D., Decker, R., & Schmidt-Thieme, L. (Springer-Verlag, Berlin), pp.335-343.
(91) Venables, W.N. & Ripley, B.D. (2002) Modern Applied Statistics with S. (Springer, New York.)
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕ IL-4 И/ИЛИ IL-13, И ИХ ПРИМЕНЕНИЕ | 2008 |
|
RU2488595C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕ IL-4 И/ИЛИ IL-13, И ИХ ПРИМЕНЕНИЕ | 2015 |
|
RU2721236C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕ IL-4 И/ИЛИ IL-13, И ИХ ПРИМЕНЕНИЕ | 2015 |
|
RU2705551C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕ IL-4 И/ИЛИ IL-13, И ИХ ПРИМЕНЕНИЕ | 2008 |
|
RU2580049C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА К CXCR5, ИХ ПРОИЗВОДНЫЕ И ИХ ПРИМЕНЕНИЕ | 2008 |
|
RU2571515C2 |
| АНТИТЕЛА ПРОТИВ TFR И ИХ ПРИМЕНЕНИЕ ПРИ ЛЕЧЕНИИ ПРОЛИФЕРАТИВНЫХ И ВОСПАЛИТЕЛЬНЫХ РАССТРОЙСТВ | 2016 |
|
RU2737637C2 |
| АНТИ-IgE АНТИТЕЛА | 2017 |
|
RU2816207C2 |
| Белок, связанный с гликопротеином человека, композиция, фармацевтическая композиция, вектор экспрессии | 2016 |
|
RU2773819C2 |
| Выделенный гуманизированный белок, связывающийся с гликопротеином VI человека, выделенное антитело, связывающееся с гликопротеином VI человека | 2018 |
|
RU2778884C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА, СПЕЦИФИЧНЫЕ К АЛЬФА-ЭНОЛАЗЕ, И СПОСОБЫ ПРИМЕНЕНИЯ В ПРОТИВООПУХОЛЕВОЙ ТЕРАПИИ | 2014 |
|
RU2761662C2 |
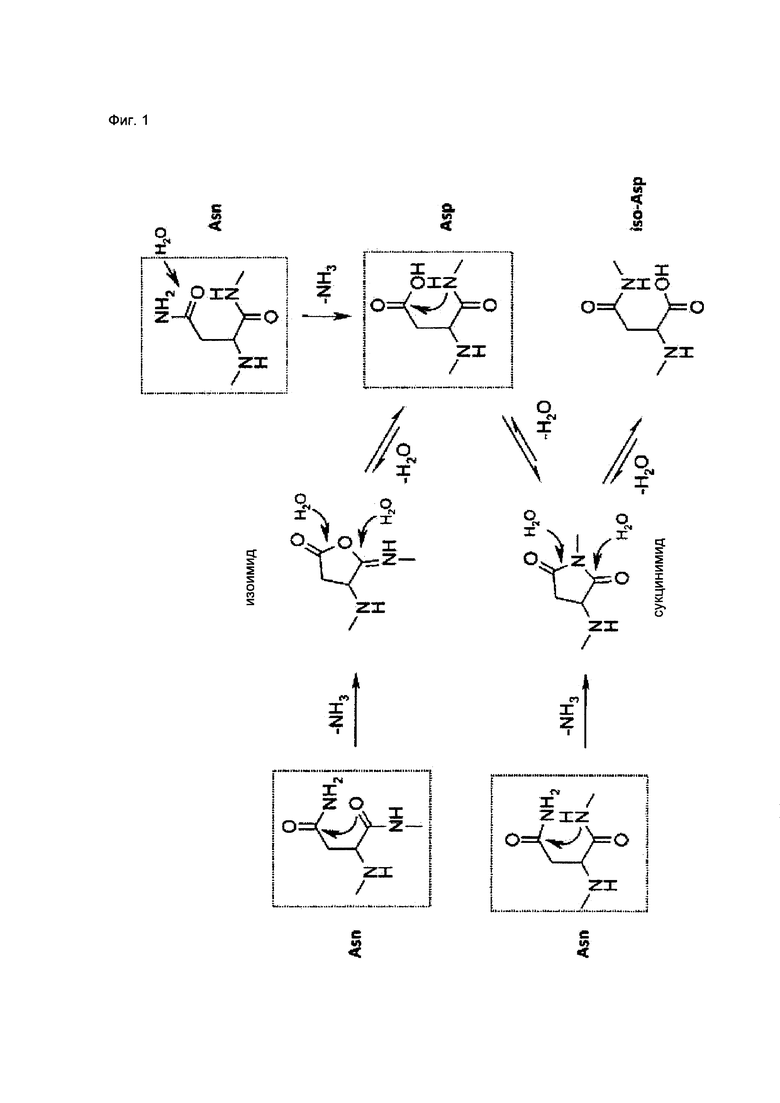
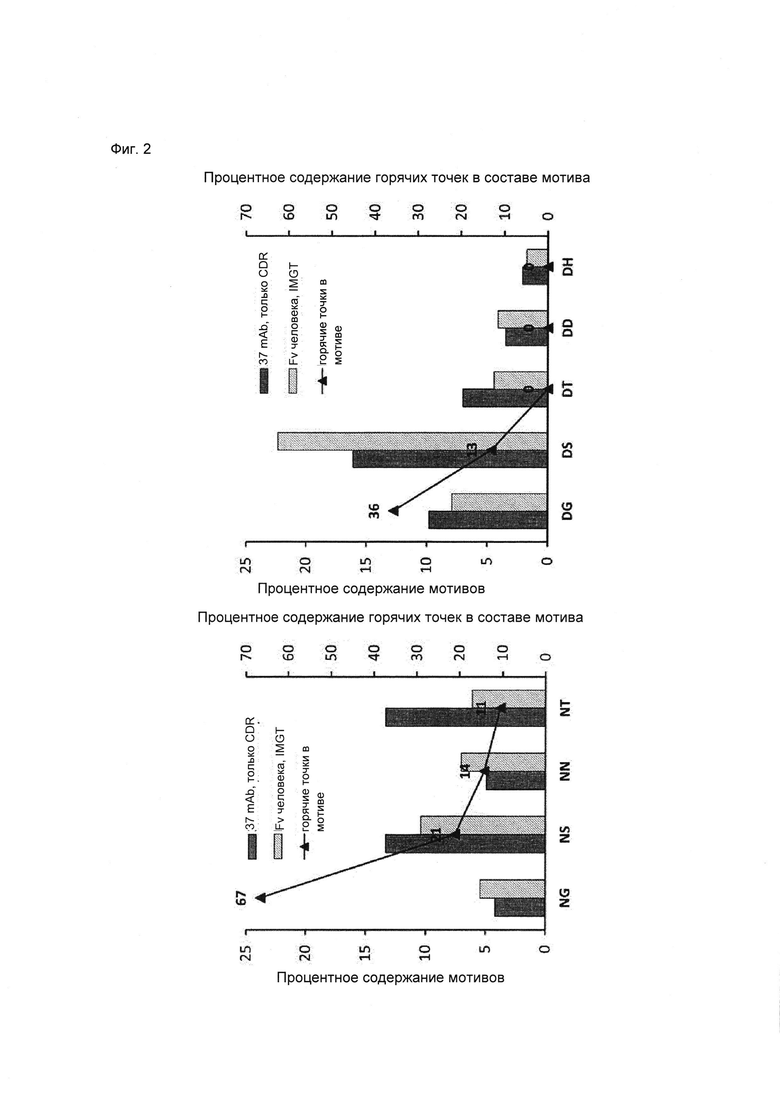
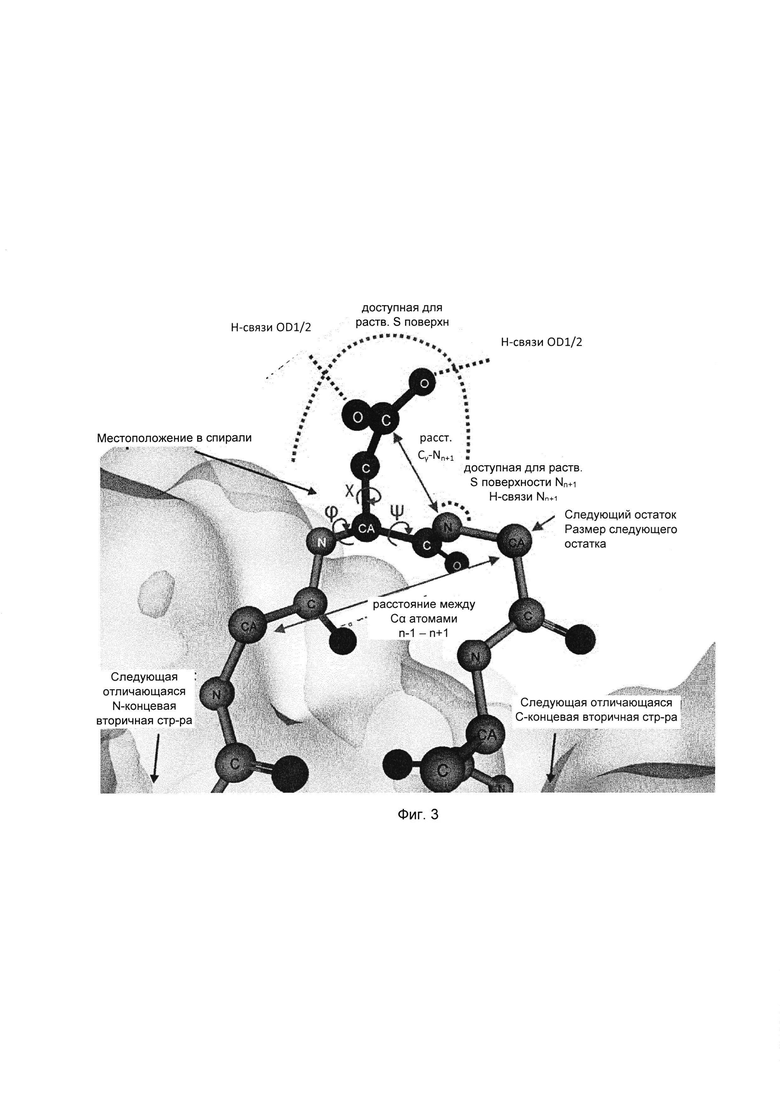
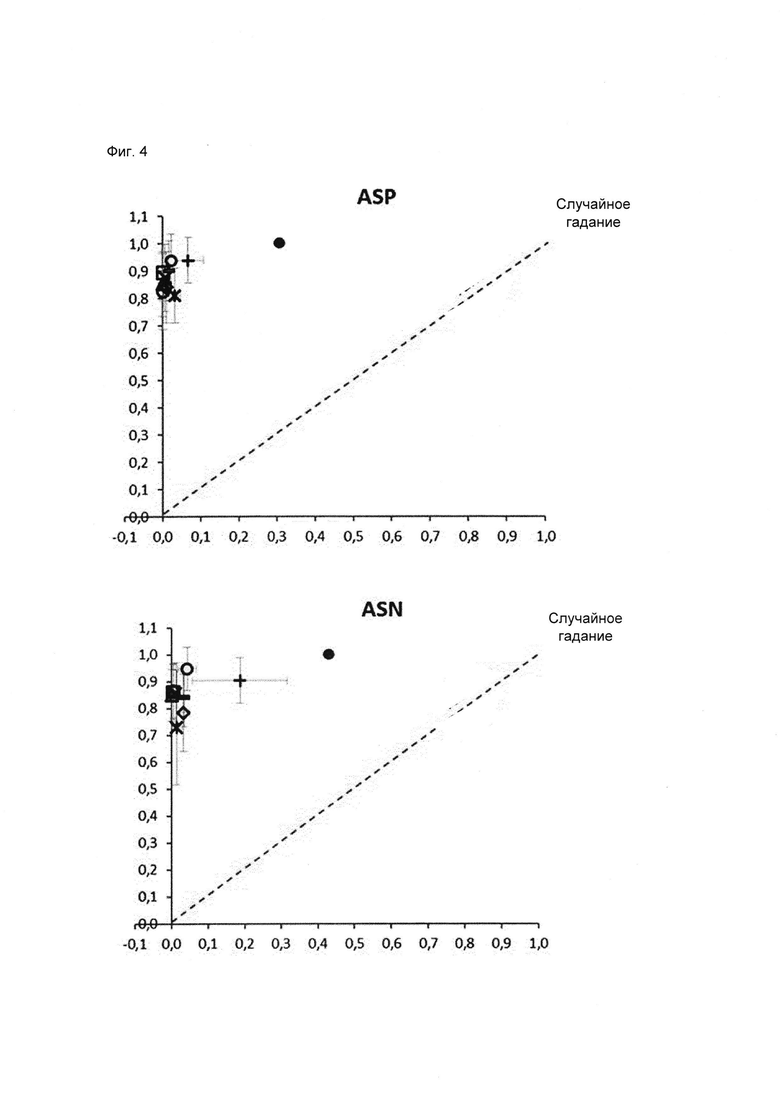
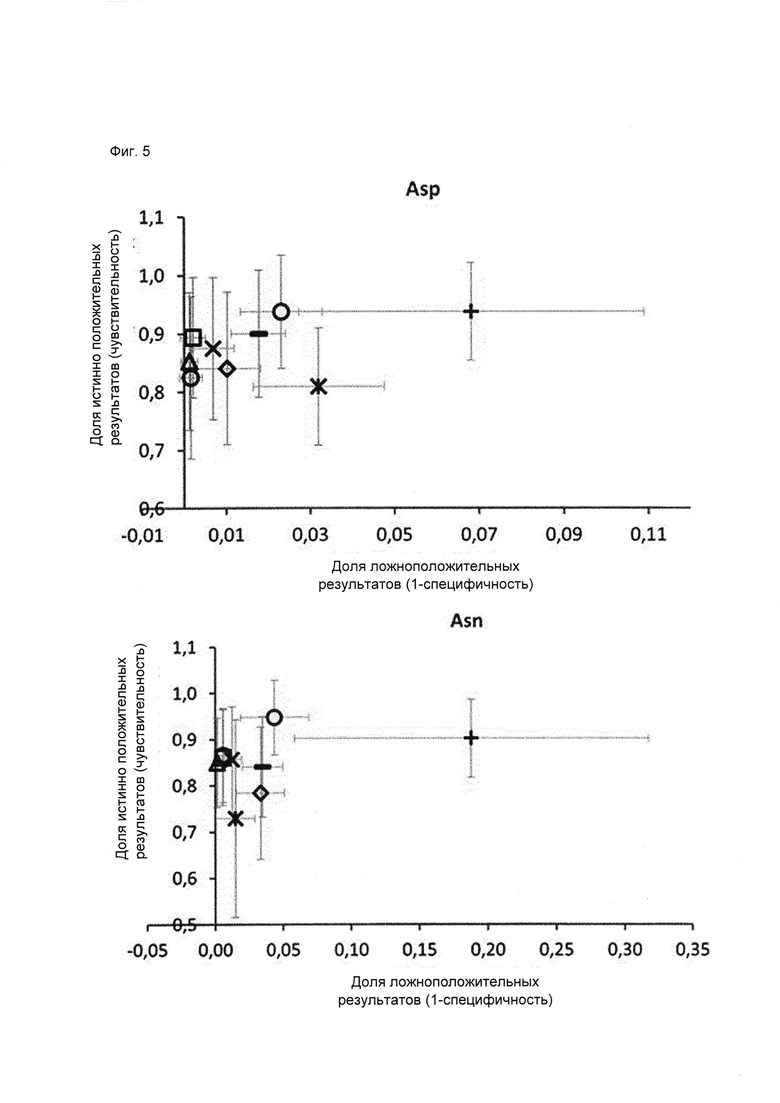
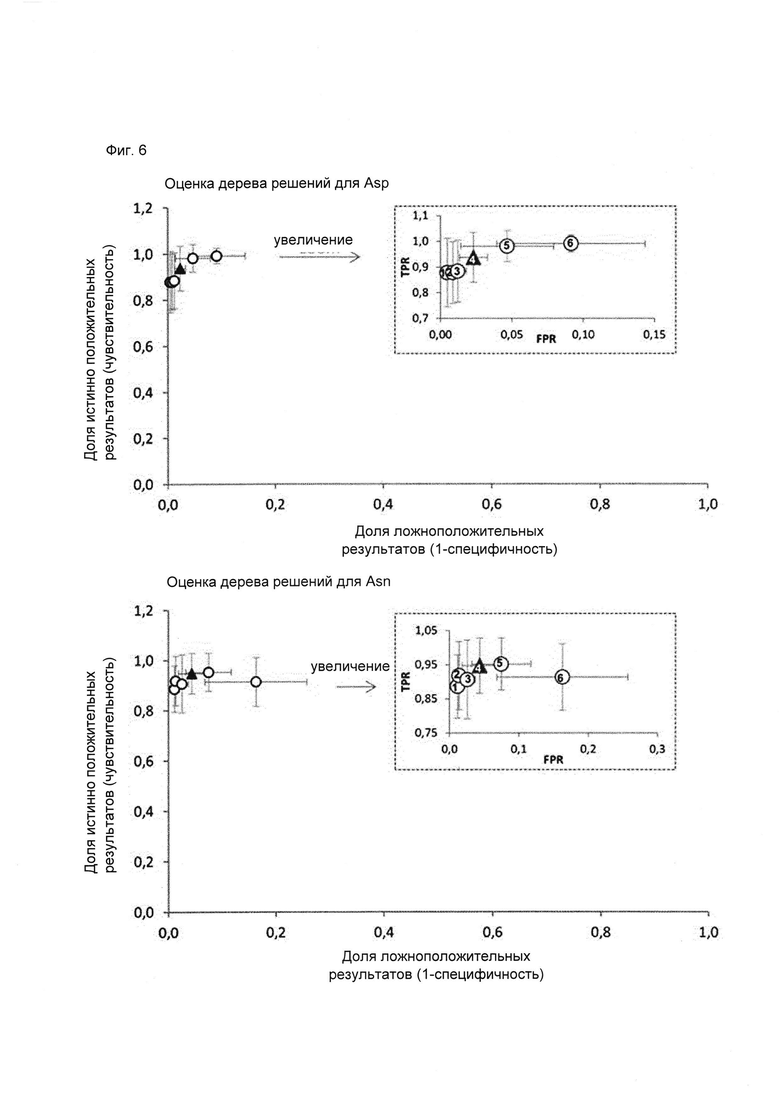
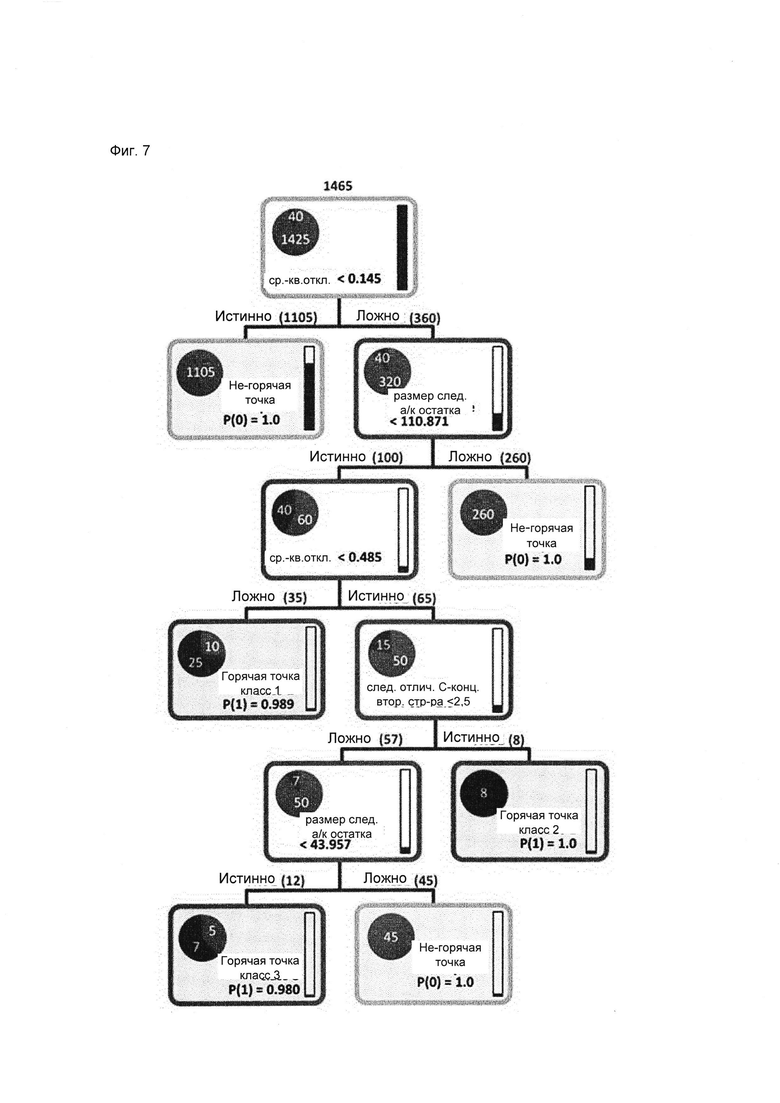
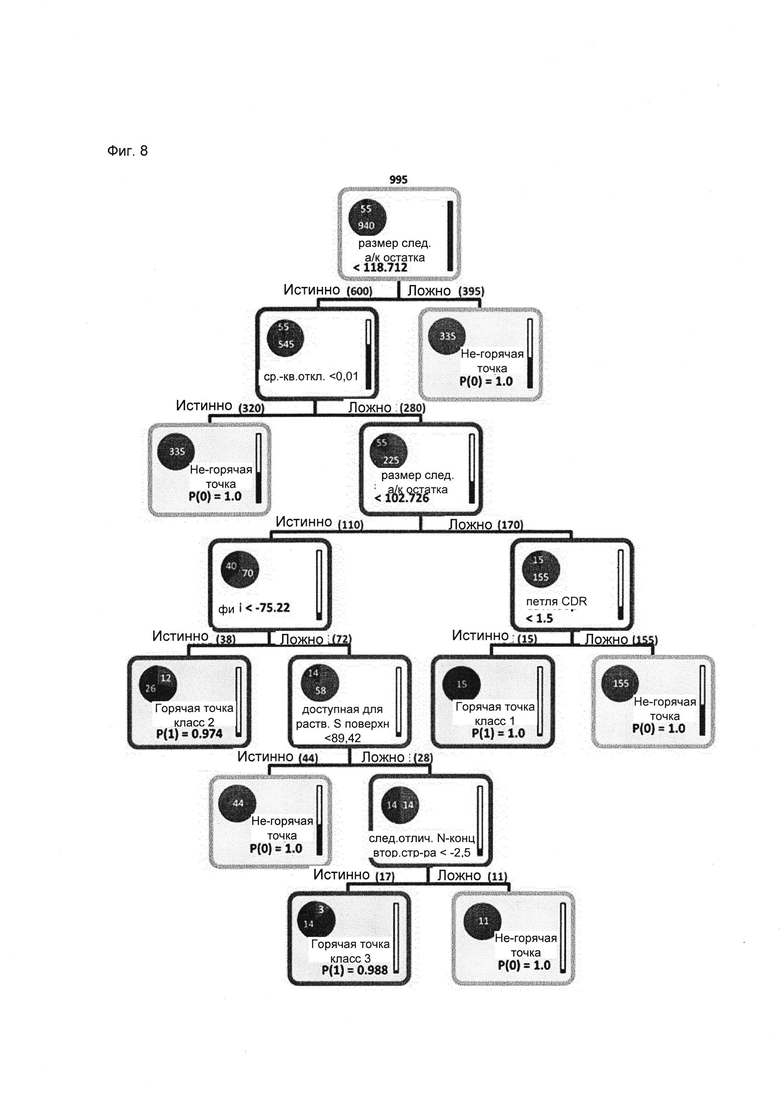
Изобретение относится к области биохимии, в частности к способу отбора одного или нескольких антител, стабильных к деградации. Указанный способ включает стадии, при которых: а) берут два или более антител, б) у каждого остатка Asn и Asp в Fv домене антитела определяют конформационную подвижность Сα-атома с использованием ансамбля моделей, основанных на гомологии, в) у каждого остатка Asn и Asp в Fv домене антитела определяют размер аминокислотного остатка, прилегающего к остатку Asn или Asp со стороны С-конца, г) выбирают одно или несколько антител, у которых Сα-атом является конформационно неподвижным и у которого со стороны С-конца к Asn или Asp прилегает аминокислотный остаток с доступной для растворителя площадью поверхности, составляющей 111  или более, где конформационная подвижность представляет собой среднеквадратичное отклонение (RMSD) соответствующих Сα-атомов остатков Asn/Asp в ансамбле моделей, основанных на гомологии. Изобретение позволяет эффективно отбирать одно или нескольких антител, стабильных к деградации. 1 з.п. ф-лы, 8 ил., 3 табл., 8 пр.
или более, где конформационная подвижность представляет собой среднеквадратичное отклонение (RMSD) соответствующих Сα-атомов остатков Asn/Asp в ансамбле моделей, основанных на гомологии. Изобретение позволяет эффективно отбирать одно или нескольких антител, стабильных к деградации. 1 з.п. ф-лы, 8 ил., 3 табл., 8 пр.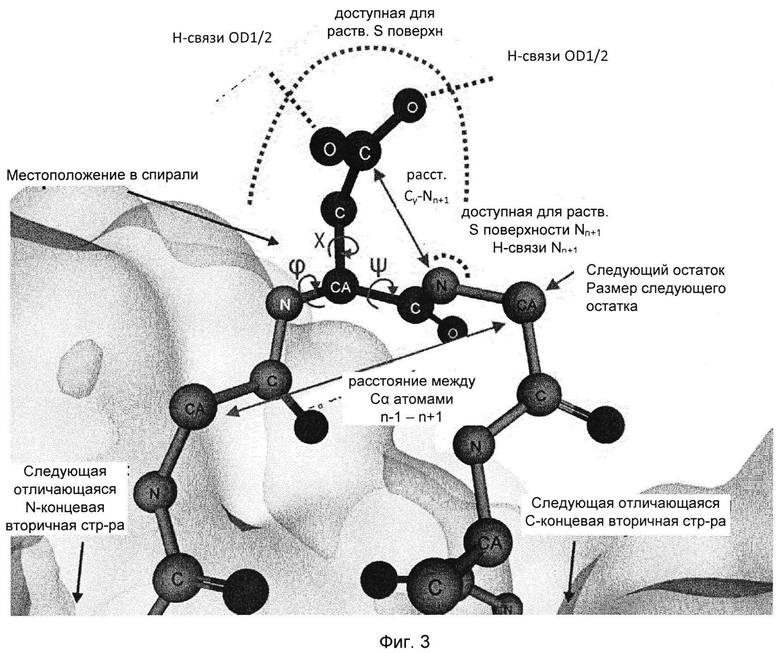
1. Способ отбора одного или нескольких антител, стабильных к деградации, включающий стадии, при которых
а) берут два или более антител,
б) у каждого остатка Asn и Asp в Fv домене антитела определяют конформационную подвижность Сα-атома с использованием ансамбля моделей, основанных на гомологии,
в) у каждого остатка Asn и Asp в Fv домене антитела определяют размер аминокислотного остатка, прилегающего к остатку Asn или Asp со стороны С-конца,
г) выбирают одно или несколько антител, у которых Сα-атом является конформационно неподвижным и у которого со стороны С-конца к Asn или Asp прилегает аминокислотный остаток с доступной для растворителя площадью поверхности, составляющей 111  или более,
или более,
где конформационная подвижность представляет собой среднеквадратичное отклонение (RMSD) соответствующих Сα-атомов остатков Asn/Asp в ансамбле моделей, основанных на гомологии и где
i) конформационно неподвижным является RMSD в 0,01  или менее,
или менее,
ii) конформационно подвижным является RMSD более чем 0,01 ,
iii) умеренной конформационной подвижностью является RMSD между 0,145 и 0,485 и
iv) высокой конформационной подвижностью является RMSD более чем 0,485 ,
и таким образом отбирают одно или несколько антител, стабильных к деградации.
2. Способ по п. 1, отличающийся тем, что ансамбль моделей, основанных на гомологии, строят для Fv фрагмента антитела.
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО И СПОСОБ ЕГО ПОЛУЧЕНИЯ, ПОЛИНУКЛЕОТИД (ВАРИАНТЫ), ГИБРИДОМНАЯ КЛЕТОЧНАЯ ЛИНИЯ (ВАРИАНТЫ) | 1993 |
|
RU2134724C1 |
| ROBINSON N.E | |||
| et al | |||
| Prediction of protein deamidation rates from primary and three-dimensional structure | |||
| Proceedings of the National Academy of Sciences, 2001, т | |||
| Дорожная спиртовая кухня | 1918 |
|
SU98A1 |
| WAKANKAR A.A | |||
| et al | |||
| Formulation considerations for proteins susceptible to asparagine deamidation and aspartate isomerization | |||
| Journal of | |||

