Предпосылки создания изобретения
Настоящее изобретение относится в основном к материалам и методам для предупреждения и лечения инфекций, вызванных вирусом иммунодефицита человека (ВИЧ-1). Более конкретно, изобретние относится к моноклональным антителам, пригодным для пассивной иммунизации восприимчивых к или инфицированных ВИЧ-1 животных, особенно людей.
Инфекционный процесс, вызванный ВИЧ-1 in vivo недавно был темой обзорной статьи McCune, Cell 64, pp.351-363 (1991). Вкратце, Вич-1 инфицирует ряд линий клеток, таких как Т-клетки, моноциты/макрофаги и нейроны, которые экспрессируют рецептор CD4. Поскольку подавляющее большинство клеток CD4+ в организме являются "отдыхающими" или покоящимися и делятся только в ответ на специфические сигналы, инфицирование ВИЧ-1 приводит к тому, что клетки CD4+ включают транскрипционно неактивный вирус. Стимуляция иммунной системы инфицированных животных, включая активную иммунизацию, может приводить к поликлональной активации и сигналу для покоящихся клеток CD4+ перейти в S фазу клеточного цикла. Реплицирующиеся клетки затем активно продуцируют вирусные частицы, вызывая распространенные инфекции. Принимая во внимание этот отрицательный эффект стимуляции иммунной системы инфицированного ВИЧ-1 животного, возможно, что наиболее эффективным методом предупреждения и лечения инфекции ВИЧ-1 является пассивная иммунизация, то есть введение анти-ВИЧ-1 антител восприимчивому или инфицированному животному.
Jackson et al., Lancet, 2, pp. 647-652 (1988) сообщает, что однократное введение анти-ВИЧ-1 антител в форме плазмы больным людям, страдающим развитием синдрома приобретенного иммунодефицита (СПИД, синдром прогрессирующей деградации иммунной системы, связанной с инфекцией ВИЧ-1), временно приводило к уменьшению симптомов, кратковременному повышению числа Т-лимфоцитов, снижению частоты инфекций, вызванных условно-патогенными микроорганизмами и к снижению степени, с которой ВИЧ-1 мог выделяться из плазмы или лимфоцитов больных. См. также Karpas et al., Proc. Natl. Acad. Sci. USA,85, pp. 9234-9237 (1988). Кроме того, Emini et al., Nature, 355, pp. 728-730 (1992) сообщает, что введение шимпанзе антител, специфически реагирующих с ВИЧ-1, перед заражением животного ВИЧ-1 приводило к тому, что у шимпанзе не проявлялись признаки вирусной инфекции. Эти исследования показывают, что антитела, способные к нейтрализации ВИЧ-1, могут быть полезны при предупреждении/лечении инфекции ВИЧ-1.
Главный гликопротеин оболочки ВИЧ-1, gp 120, связывается с клеточным рецептором CD4 и облегчает внедрение вируса. С разработкой нейтрализующих антител связано несколько эпитопов гликопротеина. Но et al., Science, 239, pp. 1021-1023 (1988) сообщает, что аминокислоты 254 - 274 gp 120 вызывают образование поликлональной антисыворотки, способной к группоспецифической нейтрализации трех различных штаммов ВИЧ-1. Конформационно зависимые эпитопы, составляемые не первичными последовательностями аминокислот, на gp 120 также включены в индукцию синтеза антител, которые нейтрализуют разные штаммы вируса, согласно Haigwood et al., Vaccines 90, pp. 313-320 (1990) и Ho et al., J. Virol, 65(1), pp, 489-493 (1991). Так называемая "главная нейтрализующая детерминанта" (PND) gp 120 ВИЧ-1 локализована на "V3 петле" gp 120. См. Puthey et al., Science, 234, pp 1392-1395 (1986), Pusche et al., Proc. Natl. Acad. Sci. USA, 85, pp. 3198-3202 (1988), Coudsmit et al., Proc. Natl. Acad. Sci. USA, 85, pp. 4478-4482 (1988), Palker et al., Proc. Natl. Acad. Sci USA, 85, pp. 1932-1936 (1988), Holley et al., Froc. Natl. Acad. Sci. USA, 85, pp. 6800-6804 (1991). Петля V3 состоит из гипервариабельной области, которая формируется с помощью дисульфидной связи между цистеиновыми остатками, фланкирующими домен. Петля V3 ВИЧ-1MN, например, образуется с помощью дисульфидной связи между цистеиновыми остатками в позициям 320 и 336 gp 120.
Рекомбинантные и синтетические протеиновые фрагменты, включающие ряд аминокислотных остатков петли V3 из различных изолятов ВИЧ, как сообщалось, вызывают образование изолят- и типоспецифических нейтрализующих антител у грызунов, согласно Lasky et al., Science, 233, pp. 209-212 (1986), Palker et al., выше Matsushita et al., J. Virol., 62, pp. 2107-2114 (1988), Javaherian et al., Proc., Natl. Acad. Sci. USA 86, pp 6768-6772 (1989). Более недавние исследования (Puthey et al., выше и La Posa et al., Science 249, pp. 932-935 (1990) показали, что структура β- поворота петли V3 - это сайт, распознаваемый изолят-специфическими антителами. Scott et al., Proc. Natl. Acad. Sci. USA, 87, pp. 8597-8601 (1990) сообщают, что PND может также индуцировать образование типоспецифических антител у людей. Гипервариабельность PND может объяснить типоспецифическую нейтрализующую активность, генерируемую эпитопом.
Несколько исследований позволяют предположить, что антитела, полученные против рекомбинантного gp 120, очищенного gp 120 или синтетических пептидов из области V3, могут нейтрализовать различные изоляты ВИЧ-1. Javarian et al. , Science, 250 pp. 1590-1593 (1990) и Weiss et al., Natuve, 324, pp. 572-575 (1986), каждый, описываемый нейтрализацию как MN, так и IIIB изолятов поликлональной сывороткой, полученной от кроликов, соответственно иммунизированных пептидом, соответствующим PND из MN изоляторов и рекомбинантным gp 120, полученным из изолята IIIB. См. также Haynes et al., US Letters Patent 5019387.
Akerblom et al., AIDS, 4, pp. 953-960 (1990) описывает препараты моноклональных антител, которые нейтрализуют IIIB и одиннадцать первичных изолятов ВИЧ-1. См. также Patent Cooperation Treaty (PCT) Publication No. WO 91/11198 of Wahren et al., опубликованную 8 августа 1991 г. Гомология штаммов первичных изолятов Akerblom не определена, однако одиннадцать изолятов могут также быть IIIB . Durda et al., AIDS Res. Hum. Retrov., 6, pp. 1115-1123 (1990) сообщают о моноклональных антителах, которые блокируют образование синцития клетками, инфицированными как MN, так и IIIB, но не нейтрализуют инфекционности MN, что установлено с помощью "LAV улавливающего иммуноанализа" ("LAV capture immunoassay"), анализ, который имеет целью дать результаты, которые коррелировали бы с активностью обратной транскриптазы. Патентная заявка по договору о патентной кооперации N WO 90/15078 of Scott et al., опубликованная 13 декабря 1990 г., описывает моноклональные антитела, которые подавляют образование синцития клетками, инфицированными вирусом осповакцины, экспрессирующим PND, изолятов MN или "подобных MN". С помощью стандартных исследований на обратную транскриптазу, p24 или МТ-2 показано, что ни одно из антител с подтвержденным "широким нейтрализующим" действием не нейтрализует многочисленные штаммы живых ВИЧ-1. См. также PCТ Publication Nos. WO 88/09181, WO 90/12868, WO 91/09625 of Tanox Biosystems, Inc., опубликованных 1 декабря 1988 г., 1 ноября 1990 г. и 11 июля 1991 г. соответственно, PCT Publication N WO 91/19797 Нью-йоркского университета, опубликованную 26 декабря 1991 г. и Liou et al., J. Immunol., 143 (12), pp. 3967-3975 (1989).
Вышеприведенные публикации показывают, что моноклональные антитела, реагирующие с PND ВИЧ-1, полученные к настоящему времени, проявляют различные уровни групповой реактивности, но могут не обладать широким нейтрализующим действием. Различные примеры типо- и группоспецифической реактивности, показанные при этих исследованиях, могут быть связаны как с аминокислотной последовательностью, так и с конформацией области петли gp 120.
Несколько исследований наводят на мысль, что рецептор CD4 не может представлять собой единственный клеточный рецептор, ответственный за инвазию вируса. Результаты этих исследований увеличивают вероятность того, что введение описанных здесь ранее антител, которые блокируют инфицирование клеток CD4+, больному может обеспечить только ограниченную защиту против инфекции ВИЧ-1. Cheng. Mayer et al., Proc. Natl. Acad. Sci USA, 84, pp. 3526-3539 (1987) сообщают об инфицировании ВИЧ-1 глиальных клеток, включающих другой рецептор, а не молекулу CD4. Кроме того, Takeda et al., Science, 242, pp. 580-583 (1988) показали, что комплексы антитело/ВИЧ-1 могут инфицировать моноциты путем опосредуемого рецептором эндоцитоза и усиливать репликацию вируса. Сходное зависимое от антител усиление инфекции было описано у Halsted et al., Nature, 265, pp. 739-741 (1977), Peiris et al., Narure, 289, pp. 189-191 (1981) и Schlesihger et al., J. Immunol., 127, pp. 659-665 (1981),
Предшествующая работа показала, что определенные вирусы животных инактивируются комплементом, особенно Clg, через антителозависимый механизм. См. Weiss, in Molecular Biology of Tumor Viruses, RNA Tumor Viruses, Wliss et al. , Eds. , Cold Spring Harbor Laboratory, New York, pp. 1219-1220 (1982), Welsh et al., Virology, 74, pp. 432-440 (1976), Bartholomew et al., J. Exp. Med., 147, pp. 844-853 (1978), Cooper et al., J. Exp. Med., 144, pp. 970-984 (1976) и Sherwin et al., Int. J. Cancer, 21, pp. 6-11 (1978). В то время как Banapour et al., Virology. 152, pp. 268-271 (1986) описывают препараты непрогретой сыворотки, которые не действуют на плотность ВИЧ-1 или его способность инфицировать мононуклеарные клетки периферической крови, Spear et al., J. Virol. , 64 (12), pp. 5869-5873 (1990) сообщают, что ВИЧ-1, обработанный комбинацией комплемента и цельной сывороткой от больных, серопозитивных на ВИЧ-1, проявляет сниженную инфекционность.
Таким образом сохраняется потребность в новых препаратах моноклональных антител (включая, например, мышиные антитела, "очеловеченные" антитела и иммунологически активные фрагменты антител), которые являются специфически иммунореактивными с ВИЧ-1. В идеале такие антитела должны характеризоваться способностью осуществлять нейтрализацию многочисленных штаммов ВИЧ-1 (например, IIIB и MN), что определяется с помощью стандартных исследований на обратную транскриптазу, p24, MT-2 и образование синцития, включающих подходящие культивируемые клетки-хозяева (например, клетки H9). Ввиду предполагаемого применения для пассивной иммунизации инфицированных и неинфицированных пациентов, такие моноклональные антитела должны оптимально быть способны к участию в (т.е. опосредовании) комплемент-зависимости виролизе частиц ВИЧ-1 и антитело-зависимом цитолизе клеток, инфицированных ВИЧ-1.
Краткое изложение изобретения
Настоящее изобретение представляет моноклональные антитела, которые специфически реактивы с той частью белка gp 120 или gp 160 ВИЧ-1, которая включает аминокислотную последовательность глицинпролин-глицин-аргинин (G-P-G-R), показанную в ПОСЛ ИД N 1, и характеризуются способностью нейтрализовать инфицирование клеток H9 в культуре штаммами MN и IIIB живого ВИЧ-1, что определяется с помощью исследований на обратную транскриптазу, p24, MT-2 и образование синцития. Продукты этого изобретения могут, кроме того, характеризоваться своей способностью опосредовать комплемент-зависимый виролиз частиц ВИЧ-1 и/или антитело-зависимую клеточную цитотоксичность инфицированных ВИЧ-1 клеток.
Моноклональные антитела настоящего изобретения могут использоваться в диагностических методах и/или наборах для определения наличия ВИЧ-1 в жидкости (например, в крови). Моноклональные антитела по настоящему изобретению, предпочтительно IgG-антитела, также особенно пригодны для использования при противо-ВИЧ-1 лечении животных, особенно людей, восприимчивых к или инфицированных ВИЧ-1. Иммунологически эффективные количества моноклональных антител вводятся пациенту, инфицированному ВИЧ-1, или при риске инфицирования этим вирусом для создания пассивного иммунитета к инфекции ВИЧ-1 и предпочтительно осуществляют комплемент-зависимый виролиз частиц ВИЧ-1 и/или антитело-зависимую цитотоксичность для инфицированных ВИЧ-1 клеток у больного.
Химерные или "очеловеченные" антитела (включая CDR-трансплантированные антитела), фрагменты антител и особенно биспецифические антитела на основе заявляемых моноклональных антител включены в рассмотрение настоящего изобретения так же, как и рекомбинантные, имеющие отношение к антителам продукты, продуцируемые в прокариотических и эукариотических клетках. Например, фрагменты антител, такие как Fab и F(ab')2 фрагменты, могут продуцировать в культуре клеток-хозяев, таких как E.coli, дрожжи, клетки насекомых и клетки млекопитающих на основе определения информации о структуре (последовательности) для вариабельных областей антител этого изобретения. Информация о последовательности вариабельных областей также дает возможным получение CDR-трансплантированных антител. Кроме того, химерные антитела (например, мышино/человеческие антитела) могут быть получены при использовании трансформированных клеток мышиной миеломы или гибридных клеток, и биспецифические антитела могут продуцироваться гибридными гибридомными клетками. Особому рассмотрению подлежат антитела, которые состоят по существу из вариабельных областей человеческих антител, включающих последовательность аминокислот из по крайней мере одной определяющей комплементарность области антител, характеризующегося способностью специфически связываться с последовательностью аминокислот gp 120 или gp 160 ВИЧ-1, состоящую по существу из последовательности, показанной в последовательности ПОСЛ ИД N 1, и способностью нейтрализовать in vitro инфицирование клеток H9 живыми штаммами MN и IIIB ВИЧ-1 при определении по обратной транскриптазе, p24, MT-2 и образованию синцития. Подлежат рассмотрению последовательности ДНК, кодирующие такие антитела, клетки-хозяева, продуцирующие такие антитела, и рекомбинантные методы для получения таких антител.
Также в рассмотрение настоящего изобретения включается применение, при противо-ВИЧ-1 лечении, комбинации продуктов настоящего изобретения и других иммунологических средств и/или химиотерапевтических средств. Потенциальные средства для комбинированного применения включают комплемент, антитела, которые связываются с различными нейтрализующими и ненейтрализующими областями белков ВИЧ-1 и химические средства, такие как AZT.
Как излагается в последующем детальном описании, моноклональные антитела настоящего изобретения получали путем иммунизации подходящего хозяина живыми ВИЧ-1, представляя таким образом gp120 в его природной конформации.
Конкретно иллюстрируют настоящее изобретение мышиные моноклональные антитела (обозначенные NM-01), продуцируемые линией гибридомных клеток HB 10726, которая была получена на хранение American Type Culture Collection, 12301 Parklawn Drive, Rockville, Maryland 20852, 9 апреля 1991 и обозначена АТСС, поступление N HB 10726, и очеловеченные варианты антител NM-01, обозначенные NM-01 HuVH/HuVK, MN-01 HuVH/HuVKF, NM-01 HuVHМ/HuVK, NM-01 HuVHS/HuVK, NM-0,1 HuVHS/HuVKF и NМ-01 HuVHM/HuVKF, продуцируемые линиями клеток, которые приняты на хранение European Collection of Animal Cell Cultures (ЕСАСС) 20 августа 1993 г., PHLS Centre for Applied Microbiology & Research, Porten Down, Salisbury, Creat Britain SP4 PJG и были обозначены ЕСАСС поступления NN 93082022, 93082019, 93082020, 93082023, 93082018 и 93082021 соответственно.
Многочисленные аспекты и преимущества настоящего изобретения будут очевидны при рассмотрении иллюстрирующих примеров и описаний практических осуществлений настоящего изобретения в последующем его детальном описании, причем ссылки даются на фигуры, где фиг. 1 является сложной ауторадиограммой неинфицированных клеток H9, белков ВИЧ-1MN и 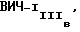 подвергнутых иммуноблоттингу с моноклональными антителами этого изобретения и иммунной сыворотки от сероположительных больных СПИД.
подвергнутых иммуноблоттингу с моноклональными антителами этого изобретения и иммунной сыворотки от сероположительных больных СПИД.
Фиг. 2 графически представляет результаты определения иммунореактивности антител этого изобретения с пептидами, соответствующими области петли V3 из разных штаммов ВИЧ-1.
Фиг. 3 является "столбчатой" диаграммой, показывающей действие пептидов, соответствующих области петли V3, на связывание антител этого изобретения и двух других антител анти-ВИЧ с gp120.
Фиг. 4A-4C, 5, 6A по 6B, 7A по 7B 8, 9A, 9B графически представляют результаты скрининга по исследованиям на обратную транскриптазу, p24, MT-2 и образование синцития соответственно, моноклональных антител этого изобретения на способность нейтрализовать инфицирование клеток H9 живыми штаммами ВИЧ-1.
Фиг. 10 графически представляет результат исследования по определению пептидной блокады нейтрализации инфекционности антителами этого изобретения.
Фиг. 11A, 11B, 12A-12F и 13A-13F являются электронными микрофотографиями частиц ВИЧ-1, которые были обработаны комбинацией моноклональных антител этого изобретения и комплемента.
Фиг. 14 и 15 - это выравнивание аминокислотных последовательностей вариабельных областей легкой и тяжелой цепей соответственно, моноклональных антител этого изобретения, NM-01, с аминокислотными последовательностями легких и тяжелых цепей трех различных анти-ВИЧ-1 моноклональных антител.
Фиг. 16 и 17 представляют выравнивание аминокислотных последовательностей вариабельных областей легкой и тяжелой цепей соответственно, мышиных моноклональных антител NM-01 с аминокислотными последовательностями легкой и тяжелой цепей гуманизированных антител NM-01 этого изобретения, обозначенных HuVH/HuVKF.
Фиг. 18, 19, 20, 21 и 22 графически представляют результаты проверки по исследованиям на обратную транскриптазу, p24, MT-2 и образование синцития соответственно, биологической активности химерных и гуманизированных антител этого изобретения.
Примеры
Следующие примеры иллюстрируют практическое осуществление этого изобретения при получении линии клеток гибридомы HB 10726, выделение из нее моноклональных антител, иммунореактивных с белками gp120 (или его предшественников gp160) ВИЧ-1 так же, как и с пептидами, включающими аминокислотную последовательность G-P-G-R, представленную в ПОСЛ ИД N 1, определение характеристик таких моноклональных антител.
Более конкретно, пример 1 предназначен для демонстрации получения линии гибридомных клеток HB 10726 и выделение из нее моноклональных антител NM-01. Пример 2 имеет отношение к картированию вирусного этипота, распознаваемого антителом NM-01. Пример 3 описывает характеристики реактивности моноклональных антител с разными изолятами ВИЧ-1. Пример 4 связан с проверкой антител NM-01 на способность нейтрализовать инфицирование клеток H9 различными живыми штаммами ВИЧ-1, что демонстрируется анализами обратной транскрипазы и p24. Пример 5 посвящен дальнейшей проверке антител на их способность нейтрализовать инфекционность живых изолятов ВИЧ-1, что демонстрируется определением МТ-2 и образования синцития. Пример 6 относится к пептидному блокированию свойств нейтрализации инфекционности ВИЧ-1 моноклональных антител NM-01. Пример 7 описывает анализ способности моноклональных антител NM-01 опосредовать комплект-зависимый лизис ВИЧ-1. Пример 8 относится к определению действия комбинации моноклональных антител NM-01 и комплемента на инфицирование ВИЧ-1 восприимчивых клеток в культуре. Пример 9 описывает ДНК и выведенные аминокислотные последовательности вариабельных областей тяжелых и легких цепей моноклональных антител NM-01. Пример 10 относится к получению химерных или гуманизированных вариантов моноклональных антител NM-01 и определению им иммунологической и биологической активности.
Пример 1.
Линия клеток гибридом HB 10726 была получена с использованием стандартных иммунологических методик, таких, которые описаны у Oi и Herzenberg, Selected Methods Cell Immunology, pp. 351-372 (1979) и Godding, J. Immunol. Meth., 39, pp. 285-308 (1980) и конкретно представлены ниже.
A. Очистка живых ВИЧ-1MN
300 мл культуры клеток H9, инфицированных ВИЧ-1MN, центрифугировали при 1500 об/мин в течение 5 минут, чтобы осадить клетки. Содержащий вирус супернатант удаляли и сохраняли, тогда как осадок повторно центрифугировали при 2100 об/мин в течение 20 минут. Второй супернатант собирали и объединяли с первым и этот супернатант подвергали ультрацентрифугированию в роторе SW 27 при 25000 об/мин в течение 90 минут при 4oC, чтобы осадить вирусные частицы. Полученный в результате супернатант отбрасывали. Осадок вирусов ресуспендировали в примерно 10 мл буфера ТНЭ (100 мМ NaCl, 10 мМ Трис-HCl, pH 7,7, 1 мМ ЭДТА). Готовили ультрацентрифужную пробирку, содержащую нижний слой из 10 мл ТРЭ с 50% сахарозы, средний слой на 10 мл ТРЭ с 25% сахарозы и верхний слой из 10 мл образца вирусов, и центрифугировали в ультрацентрифуге при 25000 об/мин при 4oC в течение 90 минут. Вирус осаждался в виде белой полосы между слоями ТРЭ с сахарозой и собирался пастеровской пипеткой. К вирусам добавляли двадцать мл ТРЭ/15 мМ ЭДТА (100 мМ NaCl, 10 мМ Трис-HCl, pH 7,7, 15 мМ ЭДТА) и образец вирусов снова центрифугировали при 25000 об/мин при 4oC в течение 90 минут. Полученный в результате осадок содержал очищенный живой ВИЧ-1NM.
В. Иммунизация и получение гибридомы
Для иммунизации каждой из трех двухмесячных мышей Balb/ с путем внутрибрюшинной инъекции использовали 100 мкг живого вируса ВИЧ-INM. Вторичную инъекцию 30 мкг вируса мышам делали через 3 недели и еще через 3 недели опять, 100 мкг вирусного препарата. Мышей забивали через 3 дня после второй реиммунизации и получали линии клеток гибридомы путем слияния спленоцитов с клетками РЗ-Х63-Ag8-U1 (АТСС CRL 1597). Линии гибридомных клеток получали также из селезенок мышей, иммунизированных хронически инфицированными клетками H9 (10 мышей), остро инфицированными клетками H9 (9 мышей) и мембранами инфицированных клеток H9 (3 мыши). Хронически инфицированные клетки H9 - это клетки спустя 2-3 недели после инфицирования, имеющие значения при исследовании на обратную транскриптазу (ОТ) от 100 000 до 150 000 имп/мин, в то время как остро инфицированные клетки H9 - это клетки спустя 10-12 дней после инфицирования, имеющие значения ОТ от 200 000 до 250 000 имп/мин.
Линии гибридомных клеток получали следующим способом.
Смесь клеток селезенки от иммунизированных мышей центрифугировали при 800 g в течение 5 минут. Супернатант отсасывали с осадка клеток и добавляли 1 мл теплого (37oC) 50% PEG-1500 на 108 клеток к осадку в течение периода в 1 минуту (добавить 0,25 мл, осторожно перемешать кончиком пипетки в течение 15 секунд и повторить). Смесь перемешивали дополнительно в течение минуты концом той же самой пипетки, не разбивая комочки клеток. В течение 1 минуты таким же путем (0,25 мл каждые 15 секунд) затем добавляли один мл "неполной среды" (RPMI 1640 (JRH Biosciences), дополненной 25 мМ HEPES (Sigma Co.), 10 000 ЕД/мл пенициллина и 10 000 мг/мл стрептомицина) и еще 1 мл добавлялся в течение еще 1 минуты. Затем 7 мл неполной среды подмешивали в течение 2-3 минут (1 мл каждые 20 секунд), что давало в результате суспензию с мелкими комочками клеток. Окончательную суспензию центрифугировали при 500 g на клинической центрифуге в течение 5 минут и супернатант удаляли. Осадок ресуспендировали вращением пробирки (не взвихривая или пепетируя раствор вверх и вниз) в "полной среде" ("неполная среда", которая описана выше дополненная 15% плодной телячьей сывороткой (FBS) до концентрации 2•106 клеток на мл среды. Затем 0,1 мл этой суспензии (2•105 клеток всего) на ячейку заполняли 96-ячеичные платы. Платы инкубировали при 37oC в атмосфере с 7% CO2. День слияния считался днем 0.
C. Селекция в НАТ и первоначальный скрининг гибридом
Через двадцать четыре часа после слияния (день 1) в каждую ячейку добавляли 0,1 мл среды НАТ (10-4 M гипоксантина, 5•10-7 M аминоптерина и 1,6•10-5 M тимидина). В дни 2, 3, 5, 8, 11, 14, 17 и 21 0,1 мл среды удаляли из каждой ячейки и заменяли 0,1 мл свежей среды НАТ. В дни со 2 по 5 ячейки, по-видимому, содержат только мертвые клетки. Гибридомы начинают появляться между днем 5 и 10. Гибридомы были легко заметны в виде колоний сильно преломляющих клеток, окруженных клеточными остатками.
D. Скрининг гибридом
Для отбора гибридомных супернатантов использовалось несколько исследований. Гибридомы, секретирующие антитела, реагирующие с ВИЧ-1, первоначально идентифицировали путем отбора мембран, полученных из неинфицированных и инфицированных MN клеток H9 с помощью EL ISA с супернатантами культур гибридом. За этим первоначальным отбором следовал отбор по имммунофлюоресцентному и радиоиммунному исследования в дополнение к данным EL ISA с данными по связыванию антител с живыми инфицированными клетками.
Клеточные мембраны для EL ISA получали из инфицированных или неинфицированных клеток H9. Клетки суспендировали в 250 мМ сахарозы/Трис-HCl буфере при pH 7,4, содержащем 1 мМ ЭДТА. Суспензию гомогенизировали в гомогенизаторе Dounce, помещенном на ледяную баню, до тех пор пока не обнаруживалось живых клеток исключением по трипановому синему. Смесь центрифугировали в течение 2 минут при 50 g. Полученный в результате осадок повторно гомогенизировали и центрифугировали. Два супернатанта соединяли и центрифугировали при 20 000 g в течение 20 минут. Осадок опять гомогенизировали в том же самом буфере и центрифугировали в течение 20 минут, и осадок ресуспендировали в 7 мл первоначального буфера 250 мМ сахарозы-ЭДТА. Этот раствор затем наслаивали поверх буфера 2 М сахарозы/10 мМ Трис-HCl, содержащего 1 мМ ЭДТА и центрифугировали в течение 1 часа при 80 000 g. В результате получалась пушистая белая промежуточная поверхность, которая собиралась и ресуспендировалась в 250 мМ сахарозном буфере. Содержание белка определяли методом BCA (Pierce Chemical Company). Суспензию делили на равные образцы и хранили при -70oC.
Для EL ISA в 96-ячеичную плату добавляли клеточные мембраны при концентрации 400 нг/ячейку и высушивали в течение ночи при 25oC. Платы промывали 0,5% Тритон X® фосфатно-буферным физраствором (PBS), блокировали 5% плодной бычьей сывороткой (FBS)/PBS и снова промывали. Супернатант гибридом (40 мкл) разводили в 50 мкл PBS и добавляли в ячейки, оставляя на ночь при 4oC. После промывания в ячейки добавляли кроличий антимышиный IgG (H+L), конъюгированный с пероксидазой хрена (HRP) (Zymed) на 2 часа при 25oC. Ячейки промывали 0,5% Тритон X® /PBS и затем инкубировали в присутствии ABTS (Bio-Rad субстратный набор) в течение 20 минут перед контролем оптической плотности при 405 и 650 нм.
Супернанты гибридом, полученных из клеток селезенки мышей, иммунизированных хронически инфицированными клетками и остро инфицированными клетками, отбирали на положительную реакцию как на мембраны неинфицированных клеток, так и на мембраны инфицированных клеток при EL ISA, указывающую на то, что антитела, продуцируемые гибридомами, не являются ВИЧ-1-специфичными. Из 1039 гибридом, полученным из клеток селезенки мышей, иммунизированных мембранами инфицированных клеток, 5 их супернатантов сильно реагировали с мембранами инфицированных клеток и очень слабо реагировали с мембранами неинфицированных клеток. Проводился Вестерн-блоттинг с супернатантами от этих линий клеток гибридом, и было установлено, что три из этих продуцируемых моноклональных антител связывались с p55 ВИЧ-1, одно связывалось с p55 и p24 ВИЧ-1, и последнее не давало полосы при Вестерн-блоттинге (данные не показаны). Результаты EL ISA представлены в таблице 1 в виде отношений значений, полученных для мембран инфицированных клеток в сравнении с мембранами неинфицированных клеток.
Одна тысяча сто восемьдесят семь гибридов были получены из клеток селезенки мышей, иммунизированных живым ВИЧ-1MN. Четыре линии гибридомных клеток были выбраны для дальнейшего отбора, основанного на результатах EL ISA, показывающих, что антитела в четырех супернатантах сильно реагировали с мембранами инфицированных клеток и очень слабо с мембранами неинфицированных клеток.
Супернатанты четырех гибридом подвергали клонированию методом серийных разведений и производили отбор по радиоиммуноисследованию (РИА). Кроличий против - мышиного IgG меченный 125I (R α M IgG125I) очищали на колонке Sephadex G-50 (NEN-Du Pont). Неинфицированные клетки H9 или клетки H9 (7,5•105 клеток в 150 мкл), инфицированные ВИЧ-1MN, помещали в 15 мл пробирки. Пятьдесят мкл супернатанта от каждой гибридомы добавляли в каждую из пробирок, содержащих неинфицированные и инфицированные клетки, и смеси инкубировали в течение ночи при 4oC. Клетки промывали 2 раза в 2 мл PBS/50% Твин-20® с взбалтыванием между промываниями. Пятьдесят мкл R α M IgG125I (750 000 имп/мин) в PBS/5% FBS добавляли и смесь снова инкубировали в течение ночи при 4oC. После инкубации клетки 3 раза промывали PBS/50% Твин-20oC. Сто мкл PBS/5% Тритон-X® добавляли для дезинфекции клеток и 100 мкл 1 M NaOH добавляли, чтобы способствовать перенесению метки в сосуды для сцинтилляции. Образцы были просчитаны, и результаты РИА представлены в таблице 1, как отношения значений имп/мин, полученных для инфицированных клеток в сравнении со значениями для неинфицированных клеток.
Затем четыре линии гибридомных клеток отбирали по иммунофлюоресценции (ИФА). Два мл или неинфицированных или инфицированных ВИЧ-1 клеток H9 (примерно 1•106 клеток/мл) помещали в 10 мл стерильные центрифужные пробирки с 10 мл PBS (без Ca++ или Mg++). Клетки один раз промывали 10 мл PBS путем заполнения пробирки, взбалтывания, центрифугирования при 100 об/мин в течение 5 минут и отсасывания всего супернатанта, кроме примерно 100 мкл, оставляя "молочную" клеточную суспензию. При работе под колпаком с ламинарным потоком 51 мм предметные стекла с 10 ячейками (Cel I Line Association) покрывали суспензией клеток, путем заполнения каждой ячейки до краев и затем втягивания суспензии обратно в кончик пипетки. Покрытым суспензией предметным стеклам давали высохнуть на воздухе и затем фиксировали в метаноле при комнатной температуре в течение 10 минут. Супернатант от каждой из четырех гибридом испытывали неразведенными и при титре 1:50 (супернатант разводили в 0,02% снятом молоке) на реактивность с препаратами неинфицированных и инфицированных клеток на предметном стекле. В каждую ячейку предметного стекла добавляли пятнадцать мкл неразведенных или разведенных супернатантов. Предметные стекла инкубировали при 37oC в течение 30 минут и погружали в PBS при покачивании в течение 5 минут. Предметные стекла затем быстро смывали дистиллированной водой и высушивали на воздухе под колпаком с ламинарным потоком. Шестнадцать мкл фрагмента F(ab)2(H+L)козьезо-антимышенного IgG (Cappel Biomedical), разбавленного 1: 80 0,02% снятым молоком, добавляли в каждую ячейку. Предметные стекла снова инкубировали при 37oC в течение 30 минут и затем погружали в PBS. Предметные стекла промывали в 0,01% раствор Эванса-синего в PBS в течение 5 секунд и 2 раза ополаскивали дистиллированной водой. Предметные стекла просматривали на иммунофлюоресценцию, и результаты анализа представлены в таблице 2, причем мышиный IgG (MIgG), антитела 5C6 (анти-IIIB) и супернатант грп. 5 (от гибридомы, полученной из селезенок мышей, иммунизированных мембранами инфицированных клеток) являются контрольными антителами.
Гибридомная клеточная линия HB 10726 была выбрана как наиболее обещающая в отношении антител на основании данных РИА и иммунофлюоресценции. Эта линия клеток не обладала наибольшей степенью связывания в EL ISA, но так как результаты РИА и иммунофлюоресценции представляют связывание с живыми инфицированными клетками, тогда как EL ISA представляет связывание с высушенными мембранами клеток, данные РИА более значимы. Эту клеточную линию дважды субклонировали и продуцируемые ей моноклональные антитела обозначили NM-01. Мышам внутрибрюшинно стандартным способом вводили эту клеточную линию и моноклональные антитела из асцитной жидкости концентрировали с помощью очистки на аффинной колонке с протеином А. (Pierce). С помощью специфических антисывороток (Bi O-Rad) было определено, что по изотипу антитела NM-01 являются IgG2b. Антитела (1,8 мг/мл) разводили в среде RPM1 1640 с 15% FBS и использовали в последующих примерах.
Пример 2.
Чтобы охарактеризовать вирусный эпитоп, распознаваемый моноклональным антителом NM-01, сначала проверяли с помощью Вестерн-блот-анализа реактивность антител с очищенными белками вирионов MN и IIIB, а затем с помощью EL ISA реактивность с перекрывающими пептидами, соответствующими аминокислотной последовательности области петли V3 gp120 ВИЧ-1.
A. Вестерн-блот-анализ
Вирион MN и IIIB, очищенные из супернатантов культуры инфицированных клеток H9, разрушали в 1,3% додецилсульфате натрия (ДСН)/3% β- меркаптоэтаноле и затем подвергали электрофорезу в 0,1% ДСН-10% полиакриламидном геле. После перенесения белков на нитроцеллюлозную бумагу полоски инкубировали в течение ночи с моноклональными антителами NM-01 в блокирующем буфере (0,02 М Трис-HCl, pH 7,4, 0,1 M NaCl, 0,05% нормальной козлиной сыворотки и 5% нежирного сухого молока) при 4oC и затем промывали в 0,02 M Трис-HCl, pH 7,4, 0,1 M NaCl и 0,3% Твин®. Полоски затем инкубировали с биотинилированным козьим анти-мышиным IgG (Zymed) в течение 1 часа, промывали и давали прореагировать с 125I-стрептавидином (Amersham, Arlington Heights, IL) в течение дополнительного часа при 4oC, реакционную способность моноклональных антител контролировали с помощью ауторадиографии.
Результаты ауторадиографии представлены на фиг. 1, причем полосы геля 1 и 3 содержали мембраны неинфицированных клеток H9; полоса 4 содержала мембраны инфицированных ВИЧ-1MN клеток H9; полосы 2 и 5 содержали вирус ВИЧ-1MN; и полосы 6 и 7 содержали вирус  Антитела NM-01 реагировали с белками в полосах 1,2 и 6, тогда как сыворотка больного сероположительного на ВИЧ-1 реагировала с белками в полосах 3-5 и 7.
Антитела NM-01 реагировали с белками в полосах 1,2 и 6, тогда как сыворотка больного сероположительного на ВИЧ-1 реагировала с белками в полосах 3-5 и 7.
Моноклональные антитела NM-01 проявляли реактивность с вирусными белками MN и IIIB, имеющими примерный молекулярный вес 120 kD, но не реагировали с любыми другими вирусными антигенами, показывая, что антитела распознают эпитоп gp120.
Для сравнения, моноклональные антитела F58/H3 и P4/D10, описанные у Wahren et al. PCT Publication N 91/11198, были получены их ECACC (поступления NN 90011607 и 90011608 соответственно) и были испытаны на связывание с рекомбинантным gp120 ВИЧ-1MN (Agmed Inc., Bedford, MA), рекомбинантным gp120 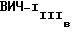 (DuPont-NEN, Boston, MA), природным gp120 ВИЧ-1MN и природным gp120
(DuPont-NEN, Boston, MA), природным gp120 ВИЧ-1MN и природным gp120 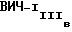 в Вестерн-блоттинге наряду с моноклональными антителами NM-01. Вестерн-блот выполнялся по существу так же, как описано выше, за исключением того, что кроличьи анти-мышиные вторичные антитела использовались в колориметрическом исследовании для определения связывания антител. Моноклональные антитела NM-01 реагировали с природным gp120 MN и IIIB и с рекомбинантными gp120 как MN, так и IIIB-производными. Моноклональные антитела F58/H3 и P4/D10, однако, реагировали только с природным gp120
в Вестерн-блоттинге наряду с моноклональными антителами NM-01. Вестерн-блот выполнялся по существу так же, как описано выше, за исключением того, что кроличьи анти-мышиные вторичные антитела использовались в колориметрическом исследовании для определения связывания антител. Моноклональные антитела NM-01 реагировали с природным gp120 MN и IIIB и с рекомбинантными gp120 как MN, так и IIIB-производными. Моноклональные антитела F58/H3 и P4/D10, однако, реагировали только с природным gp120 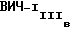 и рекомбинантным gp120, производным от
и рекомбинантным gp120, производным от 
B. Картирование эпитопа с помощью EL ISA
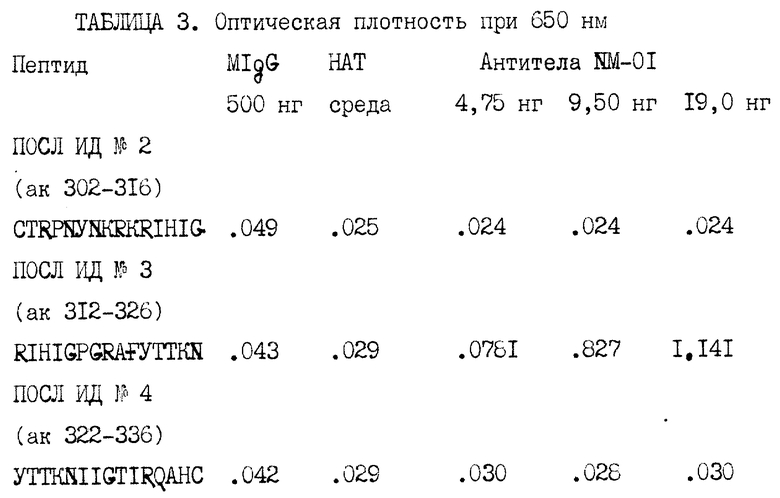
Чтобы идентифицировать специфический эпитоп gp120, распознаваемый антителами NM-01, антитела проверяли с помощью EL ISA на реактивность с перекрывающими пептидами, соответствующими области петли V3 gp120. Пептиды, синтезированные с помощью Multiple Peptide Systems, San Diego, CA, соответствовали аминокислотам 302-316, 312-326 и 322-336 gp120 ВИЧ-1MN.
Три пептида (250 нг/50 мкл 0,1 M боратного буфера, pH 8,0 на ячейку) инкубировали в течение ночи при 37oC в платах Immulon 2 (Dynatech). Платы промывали PBS и блокировали PBS/0,1% Твин® (0,1%) бычьим сывороточным альбумином (БСА) в течение 1 часа при комнатной температуре. Блокирующий агент удалялся, и различные количества антител NM-01 или мышиного IgG (MIgG), разведенных 100 мкл среды HAT, добавляли в платы. Антителам позволяли прореагировать в течение 2 часов при комнатной температуре. Затем платы 10 раз промывали водопроводной водой. Конъюгированные с пероксидазой хрена кроличьи анти-мышиные вторичные антитела, разведенные до 1:1000, доводили в PBS/0,05% Твин® /0,5% БСА и добавляли по 100 мкл на ячейку. Платы инкубировали в течение 1 часа при комнатной температуре и затем промывали 10 раз водопроводной водой. Субстрат ABTS (Bio-Rad) добавлялся на 20 минут, и платы анализировали при 650 нм. ПОСЛ ИД N 2-4 показывают аминокислотные последовательности пептидов, и таблица 3 представляет результаты исследования с использованием перекрывающих пептидов, причем антитела MIgG и среда HAT были отрицательными контролями.
В то время как не обнаружено реактивности на основе моноклональных антител NV-01 с пептидами, соответствующими аминокислотам 302-316 или 322-336 петли V3, наблюдалось связывание антител с пептидом, представляющим аминокислоты 312-326. Контрольные антитела, мышиный IgG не связывались с пептидами.
Пример 3.
Демонстрация того, что моноклональные антитела NM-01 связываются с областью петли V3 gp120 ВИЧ-1MN побудили дальнейшие исследования по определению степени этой реактивности с другими изолятами ВИЧ-1. Антитела проверяли с помощью EL ISA на реактивность с пептидами, соответствующими области петли V3 ВИЧ-1 изолятов IIIB, RF, CDC4, NY/5, Z6, Z2 и EL I. Аминокислотные последовательности пептидов показаны ниже в таблице 4 и в списке последовательности, как ПОСЛ ИД N 5-12 соответственно.
Пептиды (250 нг/0,1 M боратного буфера, pH 8,0, синтезированные American Biotechnologies, Cambridge, MA) инкубировали в течение ночи при 4oC в платах Immulon 2 (Dynatech). Платы промывали PBS, блокировали 0,1% Твин® /0,1% БСА/PBS в течение 2 часов при 25oC и затем инкубировали с моноклональными антителами NM-01 в течение 1 часа при 37oC. После промывания водопроводной водой платы инкубировали с конъюгированными с пероксидазой хрена кроличьими против мышиных вторичными антителами в течение 1 часа при 25oC и затем с субстратом ABTS (Bio-Rad) в течение 20 минут. Реактивность определяли путем контроля оптической плотности при 650-405 нм. Результаты исследования представлены на фиг. 2.
Моноклональные антитела NM-01 реагировали с пептидами петли из МN (закрашенный кружок), IIIB (пустой кружок), RF (пустой треугольник) и CDC4 (закрашенный треугольник) изолятов. Связывание антител с пептидами IIIB, RF и CDC4 было сравнимо со связыванием, полученным с пептидом MN. Антитела также показали меньшее сродство к пептиду NY/5 (звездочки). Моноклональные антитела NM-01 также ложно реактивны с RF-подобным пептидом, показанном в ПОСЛ ИД N 13. И наоборот, реактивность была слабой, если присутствовала, с пептидами петли из Z6 (закрашенный квадрат), Z2 (перевернутый пустой треугольник) и EL 1 (пустой квадрат) изолятов. Эти результаты показывают, что моноклональные антитела NM-01 распознают, в частности, эпитоп петли V3 gp120 (ВИЧ-1-изолятов, имеющих аминокислотную последовательность, представленную в ПОСЛ ИД N 1, G-P-G-R.
Моноклональные антитела F58/H3 и P4/D10 были также использованы на реактивность с пептидами петли V3 MN, IIIB, RF-подобными, CDC4, NY/5, Z2, Z6 и EL 1. В противоположность моноклональным антителам NM-01 как моноклональные антитела F58/H3, так и P4/D10 реагировали только с пептидом IIIB и в меньшей степени с RF-подобным пептидом, показанным в ПОСЛ ИД N 14.
Другие моноклональные антитела против gp120 ВИЧ-1, моноклональные антитела BAT123 описаны у Liou et al. выше, как реактивные с пептидами петли V3 MN-подобных и IIIB-подобных штаммов, и нереактивные с пептидом RF-подобных (ПОСЛ ИД N 13) (смотрите фиг. 5A на странице 3972 работы Liou et. al., выше). Эта сообщаемая реактивность отлична от реактивности моноклональных антител NM-01, как описано в предшествующем абзаце. В то время как антитела NM-01 и BAT123 связываются относительно хорошо с IIIB пептидом, необходимо примерно пятидесятикратное повышение концентрации BAT 123, чтобы получить связывание с пептидом MN, которое сходно со связыванием NM-01. Кроме того, NM-01 реагирует с пептидом RF, показанным в таблице 4 и ПОСЛ ИД N 7, и RF-подобным пептидом, показанным в ПОСЛ ИД N 14, тогда как BAT123 не связывается с RF-подобным пептидом ПОСЛ ИД N 13 даже при концентрациях антител 10000 мкг/мл.
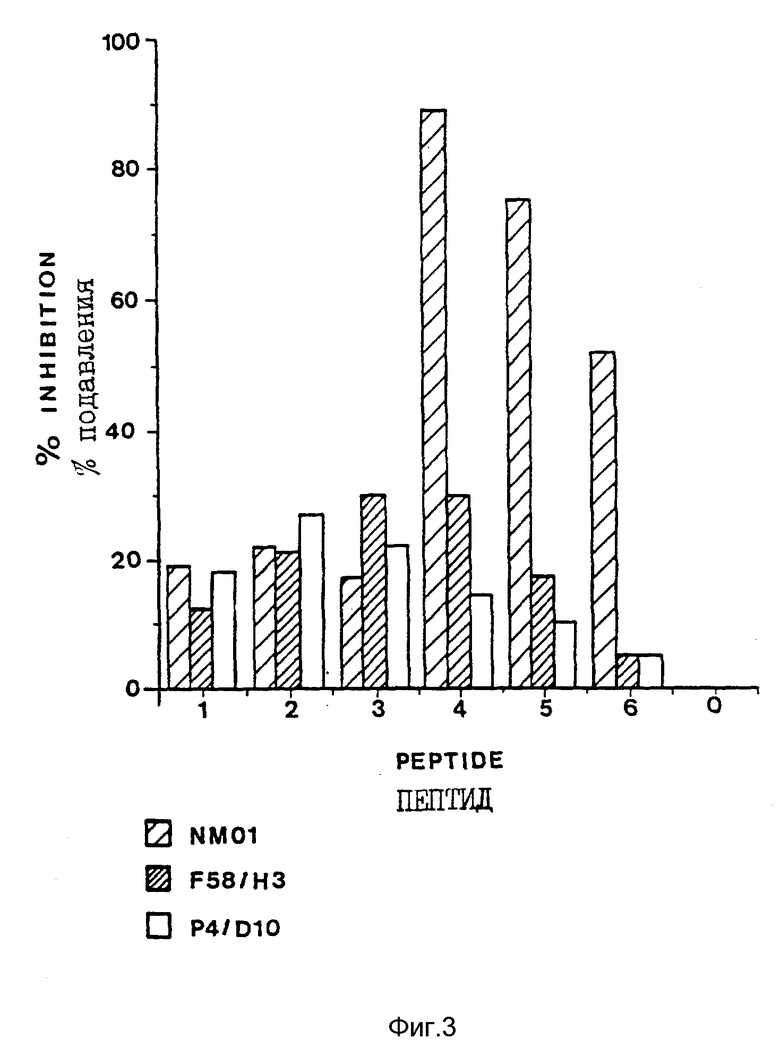
В конкурентном анализе, связывающем моноклональные антитела NM-01, F58/H3 и P4/D10 определялось в присутствии каждого из перекрывающих пептидов петли IIIB (которые являются частями ПОСЛ ИД N 7): IRIQRGPG (пептид 1), RIQRGPGR (пептид N 2), IQRGPGRA (пептид N 3), ORGPGRAF (пептид N 4), RGPGRAFV (пептид N 5) и GPGRAFVT (пептид N 6). Исследование выполнялось следующим образом. 100 мкл рекомбинантного gp120 IIIB (0,5 мгк/мл в PBS) покрывали плату Immuno 4 (Dynatech) и инкубировали при комнатной температуре в течение ночи. Затем плату блокировали 250 мкл блокирующего буфера (5% нормальной кроличьей сыворотки в PBS) в течение 1 часа при 37oC. Моноклональные антитела NM-01, F58/H3 и P4/D10 разводили до концентрации 10 мкг/мл блокирующим буфером, а каждый пептид из шести пептидов петли IIIB разбавляли до концентрации 100 мкг/мл блокирующим буфером. Затем каждое из антител отдельно смешивали с каждым пептидом 1:1 по объему с получением окончательной концентрации антител 5 мкг/мл и концентрации пептида 50 мкг/мл. Смеси антител и пептидов инкубировали при комнатной температуре в течение 40 минут и затем переносили в ячейки (100 мкл/ячейку) в блокированную, покрытую gp120 плату для исследования. Контрольные ячейки содержали 5 мкг/мл антитела и не содержали пептида. Платы инкубировали в течение 40 минут при 37oC и затем промывали четыре раза отмывающим буфером (0,005% Твин-20 в PBS). Связанные с пероксидазой хрена (ПХ) кроличьи анти-мышиные антитела (100 мкл/ячейку) использовали в качестве вторичного антитела в разведении 1:1000 в блокирующем буфере и инкубировали в течение 1 часа при 37oC. Затем плату опять промывали и проявляли, используя 100 мкл/ячейку ТМБ (тетраметилбензодина). Проявление останавливали 100 мкл/ячейку H2SO4 (0,36 N) и плату считывали при 450 нм - 650 нм.
Результаты исследования конкуренции представлены на фиг. 3. В этом исследовании пептид 4 был самым сильным ингибитором связывания моноклонального антитела NM-01 с рекомбинантным gp120 IIIB, тогда как пептиды 3 и 4 были самыми сильными ингибиторами связывания моноклонального антитела F58/H3 и пептид 2 был самым сильным ингибитором связывания моноклонального антитела P4/D10.
Пример 4.
Моноклональные антитела NM-01 испытывали на способность нейтрализовать инфицирование клеток H9 живыми штаммами MN, IIIB и RF, что оценивалось путем определения обратной транскриптазы, а штаммами MN и IIIB ВИЧ-1 - оценивалось путем исследования на p24.
Анализ обратный транскриптазы и p24
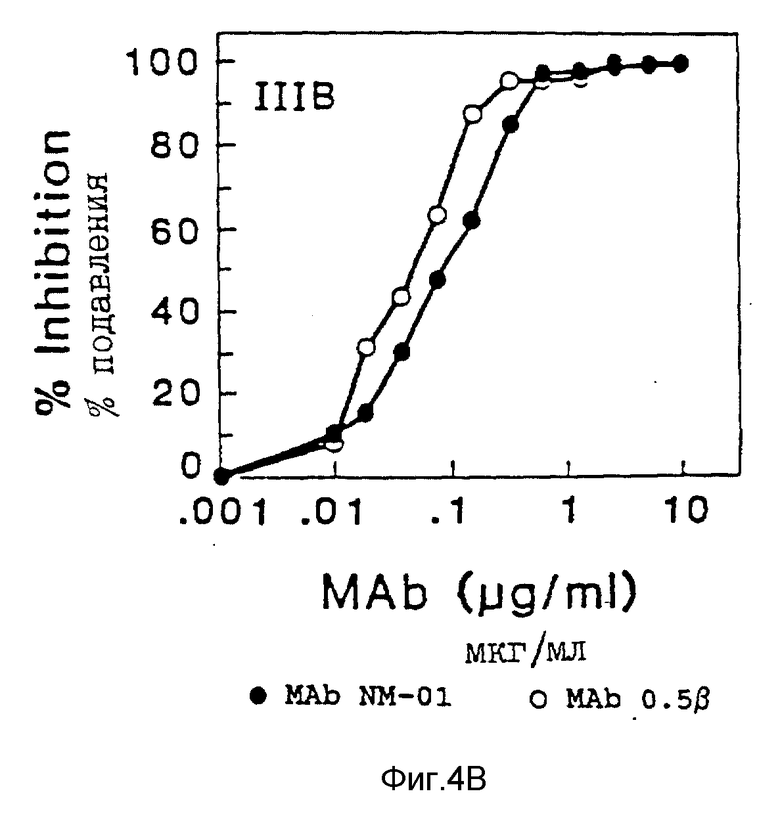
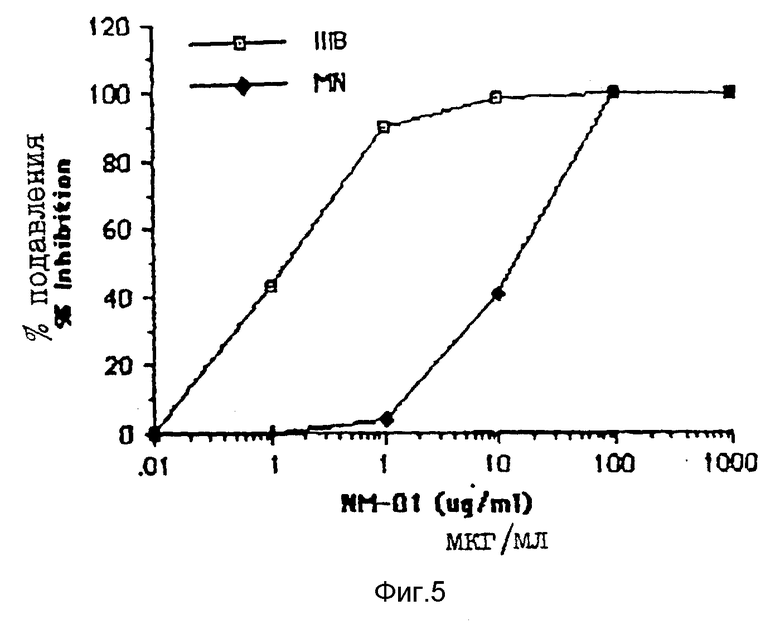
Разведения моноклональных антител NM-01 инкубировали с 40 TCID50 MN или 100 TCID50 IIIB живого вируса в 96-ячеечных платах в течение 1,5 часов при 37oC, моноклональные антитела 0,5 β (AIDS Reseаrch and Reference Reagent Program Catalog National Institute of Allergy and Indectious Diseases) использовались в качестве как положительного, так и отрицательного контроля в исследованиях на обратную транскриптазу; они связываются с gp120 ВИЧ-1. Затем в каждую ячейку добавляли клетки H9 (2,5 • 104) и платы инкубировали в течение еще одного часа при 37oC. Затем суспензию клеток H9 разводили в RPMI 1640/15% FBS и инкубировали в 24-ячеечной плате при 37oC. Продукцию вируса определяли анализом обратной транскриптазы (ОТ), выполняемым на 7 день, как описано у Poiesz et al. , Proc. Natl. Acad. Sci. USA. 77, pp. 7415-7419 (1980) и анализом p24, выполняемым на 5 день (Dupont HIV-1 p24 Core Profile EL ISA). Результаты этих двух исследований представлены на фиг. 4A по 4B и 5 соответственно.
Моноклональные антитела NM-01 (закрашенные кружочки на фиг. 4A) полностью нейтрализовали инфекционность живого вируса MN, что определено исследованием на ОТ, при концентрациях 10-100 мкг/мл. Кроме того, применение антител в концентрациях < 1 мкг/мл приводило к подавлению инфекционности вируса (ID50) на 50%. Эти результаты контрастировали с отсутствием обнаруживаемой нейтрализации моноклональными антителами 0,5β (пустые кружочки, фиг. 4A). Моноклональные антитела NM-01 также нейтрализовали живой вирус IIIB при ID50 примерно 0,1 мкг/мл (фиг. 4B). Моноклональные антитела 0,5β нейтрализовали IIIB немного более эффективно, чем моноклональные антитела NM-01 (фиг. 4B). Сходные результаты получены для MN и IIIB ВИЧ-1 при исследовании на р24. См. фиг. 5. При исследовании на обратную транскриптазу моноклональные антитела NM-01 также подавляли живой вирус RF при ID50 примерно 0,05 мкг/мл (фиг. 4C).
Эти данные показывают, что моноклональные антитела NM-01 нейтрализуют инфекционность по крайней мере трех разных штаммов ВИЧ-1.
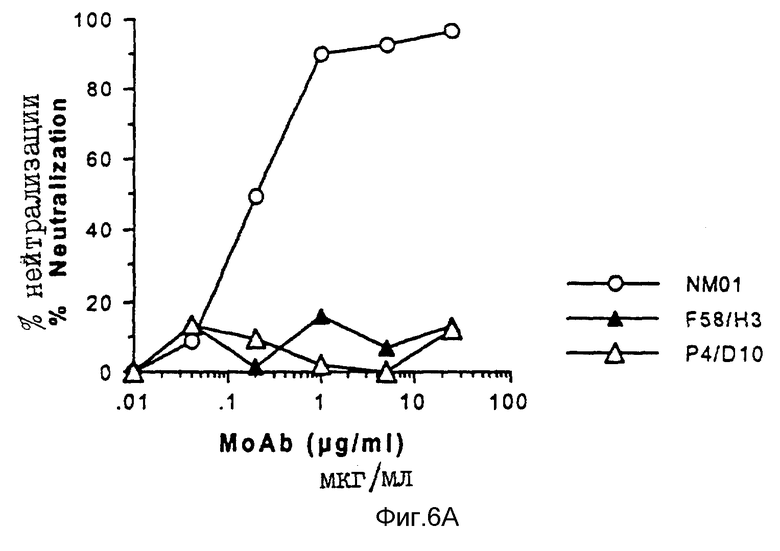
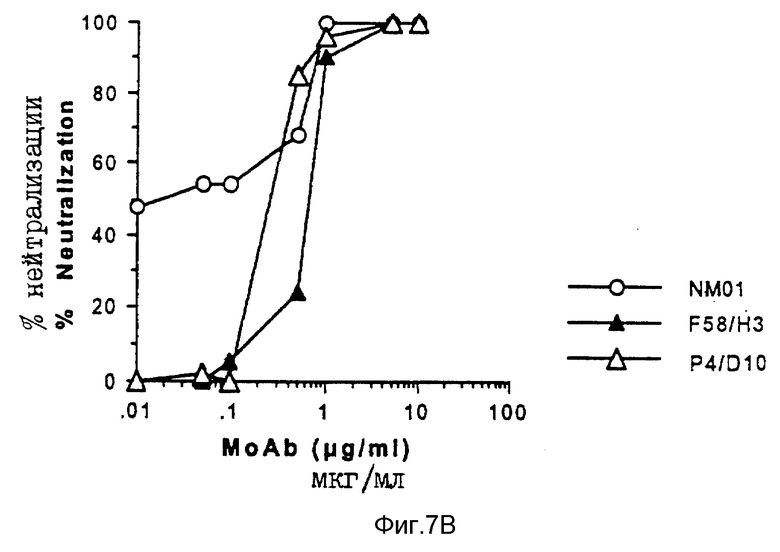
Способность моноклональных антител F58/H3 и P4/D10 нейтрализовать инфицирование клеток H9 живыми штаммами MN и IIIB ВИЧ-1 также оценивалось по определению обратной транскриптазы и по исследованию на p24, как описано выше для моноклональных антител NM-01. Результаты исследований представлены на фиг. 6A по 6B и 7A по 7B. При исследовании от ОТ снова обнаружено, что моноклональные антитела NM-01 полностью нейтрализуют инфекционность живого вируса MN при концентрациях 10-100 мкг/мл, и применение антител при концентрации < 1 мкг/мл приводило к 50% снижению инфекционности вируса (ID50) (см. пустые кружочки на фиг. 6A). Эти результаты контрастировали с отсутствием обнаруживаемой нейтрализации моноклональными антителами F58/H3 и P4/D10 (см. закрашенные и пустые треугольники на фиг. 6A). При исследовании на ОТ с использованием живого вируса IIIB, моноклональные антитела NM-01 нейтрализовали вирус при ID50 примерно 0,1 мкг/мл (пустые кружочки на фиг. 6B). Моноклональные антитела F58/H3 и P4/D10 нейтрализовали IIIB менее эффективно, чем моноклональные антитела NM-01 (см. фиг. 6B) с IC50S примерно 1,1 и 1,2 мкг/мл соответственно. Сходные результаты были получены для трех моноклональных антител при использовании MN и IIIB ВИЧ-1 исследовании на p24 (см. фиг. 7A и 7B).
Пример 5.
Изучение нейтрализации инфекционности живого ВИЧ-1, которая демонстрировалась анализом обратной транскриптазы и p24, были расширены с помощью исследования действия моноклональных антител NM-01 при анализах на MT-2 с использованием живых вирусов MN и IIIB и по образованию синцития с использованием живых вирусов MN, IIIB и RF.
A. Анализ с MT-2
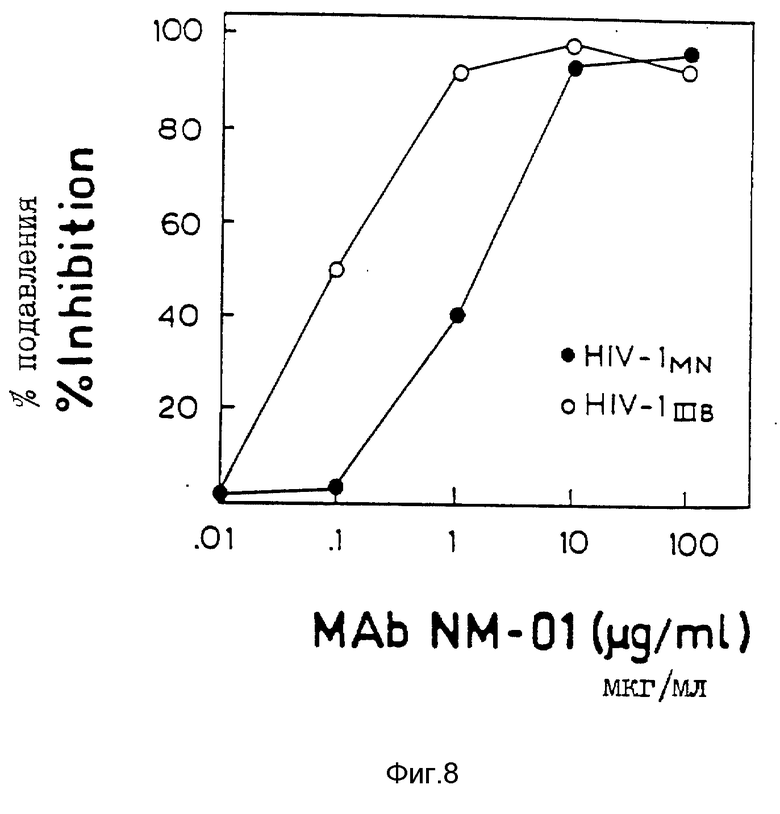
Исследование с МТ-2 выполнялось, как описано у Richman, AIDS Research and Reference Realent Program, Courier N 90-01, pp. 6-9 (1990) с некоторыми модификациями. Живые вирусы MN и IIIB инкубировали с разведениями моноклональных антител NM-01 в течение 1,5 часов при 4oC в 96-ячеечных платах. Клетки MT-2 (8 • 105) добавляли в ячейки и платы инкубировали в течение 3 дней при 37oC. Затем выполнялось исследование на снижение окрашивания MTT по Mosmann, J. Immunol. Meth., 65, pp 55-63 (1983) и Pauwels et al., J.Virol. Meth. , 20, pp. 55-63 (1983), чтобы определить жизнеспособность клеток. Результаты исследования с МТ-2 подтверждают результаты исследований ОТ и p24 и представлены на фиг. 8, причем пустые кружочки представляют значения для IIIB (100 TCID50) и закрашенные кружочки представляют значение для MN (40 TCID50).
Моноклональные антитела NM-01 нейтрализовали инфекционность живых изолятов MN и IIIB при ID50 2,0 и 0,1 мкг/мл соответственно.
В. Определение образования синцития
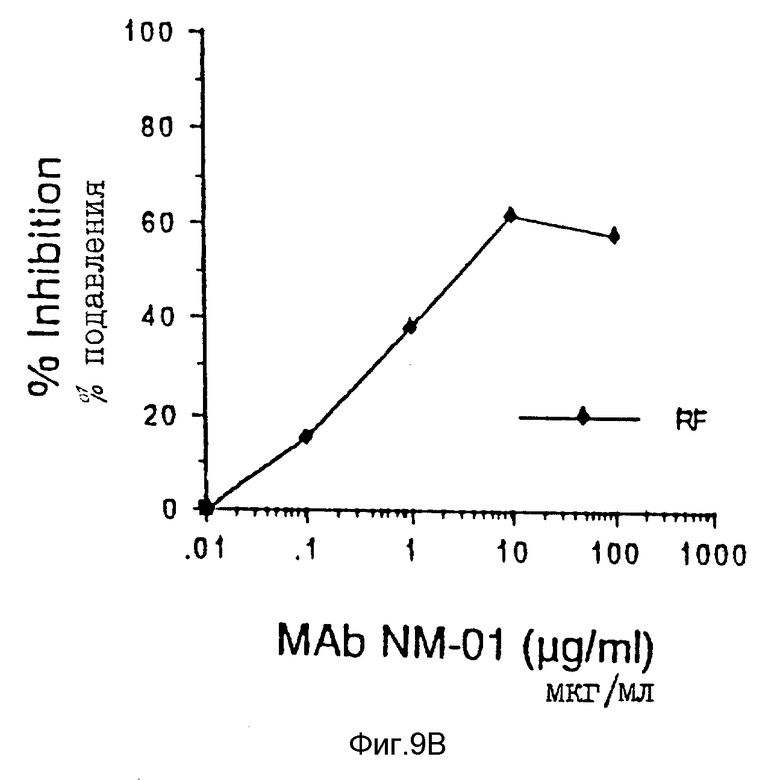
Исследование подавления связывания являлось модификацией метода, описанного ранее у Johnson and Walker, Eds., Techniques in HIV-1 Research, Stockton Press, New York, NY, pp. 92-97, (1990). Вкратце, клетки H9, хронически инцицированные или MN, или IIIB вирусом, инкубировали с разведениями моноклональных антител NM-01 в течение 1 часа при 37oC. Клетки C8166 добавляли затем в каждую ячейку и инкубировали в течение 2 часов при 37oC. Синцитии более чем трех лимфоцитных клеток в диаметре подсчитывали и сравнивали с синцитиями, полученными с контрольными инфицированными клетками H9, обрабатываемыми в отсутствие антител. Результаты исследования образования синцития также подтверждают результаты исследований на OT и p24 и представлены на фиг. 9A, причем пустые кружочки представляют значения для IIIB (100 TCID50)и закрашенные кружочки представляют значения для MN (40 TCID50). Моноклональные антитела NM-01 подавляли образование синцития инфицированными MN клетками H9 при ID50 равной 2 мкг/мл и инфицированными IIIB клетками H9 при ID50 равной 3 мкг/мл.
Соответствующие результаты подавления образования синцития представлены для моноклональных антител ВАТ 123 в таблице III WO 88/09181. Тогда как 25 мкг моноклональных антител NM-01 подавляют примерно 85% образования синцития клетками, инфицированными MN, сообщается, что 25 мкг ВАТ123 подавляют 51%, и в то время как 25 мкг NM-01 подавляют примерно 85% образования синцития инфицированными IIIB клетками, ВАТ123, как сообщается, подавляет 77,8% 25 мкг ВАТ 123, как также сообщается, подавляют образование синцития инфицированными RF клетками на 51%. Моноклональные антитела NM-01 также подавляют образование синцития инфицированными RF клетками (см. фиг. 9B). В исследовании, описанном в предшествующем абзаце, концентрация 25 мкг NM-01 подавляет примерно 59% образования синцития инфицированными RF клетками. Моноклональные антитела NM-01 подавляли образование синцития инфицированными RF клетками при ID50 равной 4 мкг/мл.
Взятые все вместе результаты исследований обратной транскриптазы, p24, МТ-2 и образования синцития примеров 4 и 5 показывают, что моноклональные антитела NM-01 нейтрализуют связывание и инфекционность различных штаммов ВИЧ-1 при концентрациях меньше 10 мкг/мл.
Пример 6.
Чтобы подтвердить, что моноклональные антитела NM-01 блокируют инфекционность MN и IIIB ВИЧ-1 путем связывания с частью V3 петли gp120, пептиды петли V3 испытывали на способность блокировать нейтрализацию антителами.
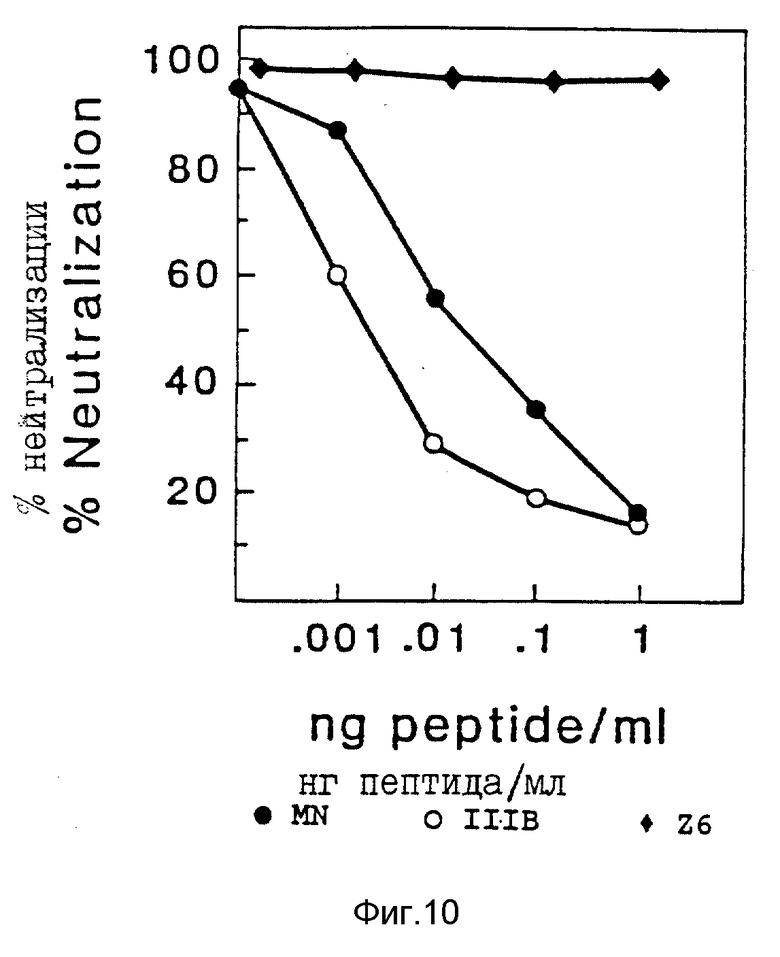
Моноклональные антитела NM-01 инкубировали с различными концентрациями пептидов, соответствующих петлям V3 штаммов MN, IIIB и Z6 (последовательности пептидов даны в таблице 4) в течение 30 минут при 37oC перед добавлением 100 TCID50 живого вируса IIIB. Затем клетки H9 добавляли на 1 час и определяли активность ОТ после подращивания клеток в полной среде в течение 7 дней, как описано в примере 4. Результаты исследования представлены на фиг. 10.
В то время как моноклональные антитела NM-01 полностью нейтрализовали инфекционность IIIB при самых низких концентрациях пептида, этот эффект прогрессивно блокировался преинкубацией с увеличивающимися концентрациями пептидов петли MN (закрашенные кружочки) и IIIB (пустые кружочки). При сходных концентрациях пептида, соответствующего петле V3 штамма Z6 (закрашенные ромбики), который не обладает последовательностью аминокислот, распознаваемой моноклональными антителами NM-01, регистрируемый эффект отсутствовал. Эти результаты показывают, что моноклональные антитела NM-01 блокируют инфекционность ВИЧ-1 путем реакции со специфической частью домена V3 gp120.
Пример 7.
Проводились также исследования по определению, могут ли моноклональные антитела NM-01 активировать метаболические пути комплемента и усиленно разрушать вирионы ВИЧ-1. В качестве источника комплемента использовалась кроличья сыворотка.
Лизис ВИЧ-1 моноклональными антителами NM-01 и комплементом
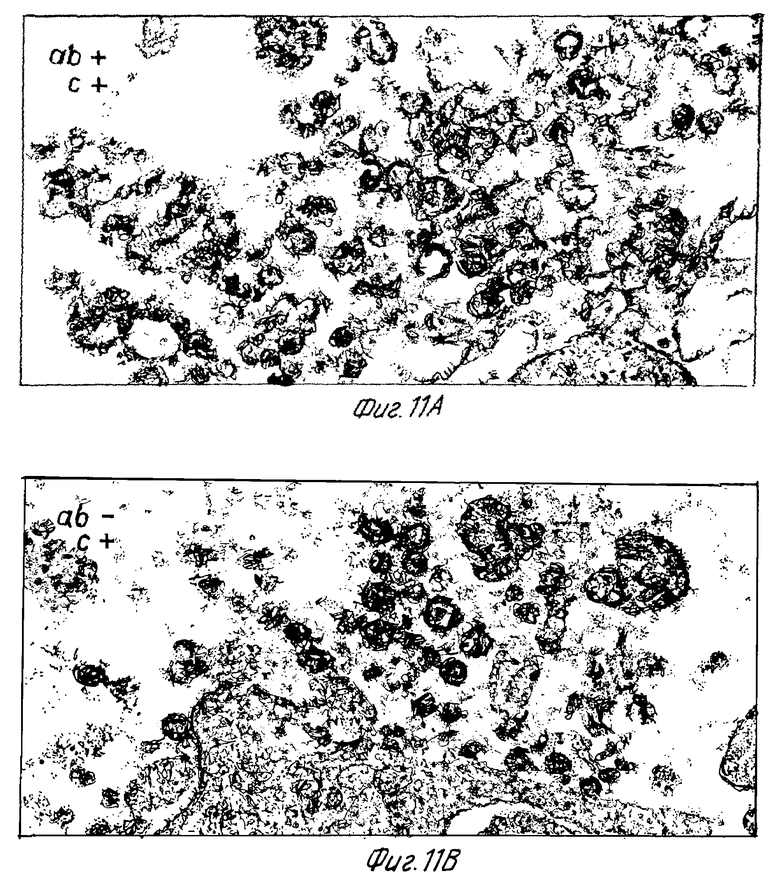
Клетки H9, инфицированные штаммом IIIB ВИЧ-1, отмывали в среде для определения цитотоксичности (Cedarlane Lab. Ltd.). Клетки ресуспендировали в среде для определения цитотоксичности или в отсутствие или в присутствии 40 мкг/мл моноклональных антител NM-01. После инкубации в течение 2 часов при 4oC добавляли кроличий комплемент (Low-tox-MA; Cedarlane Lab. Ltd.) в разведении 1: 6. Клеточную суспензию инкубировали при 4oC в течение 20 минут и затем в течение 45 минут при 37oC. Клетки дважды фиксировали 2% глютаральдегидом (0,1 М фосфатным буфером и 1% тетроксидом осмия) 0,1 М фосфатным буфером. После помещения в эпоксидную смолу делались тонкие срезы и дважды окрашивались уранилацетатом и цитратом свинца. Фиг. 11A по 11B, 12A по 12F и 13A по 13F представляют типичные электронные микрофотографии тонких срезов.
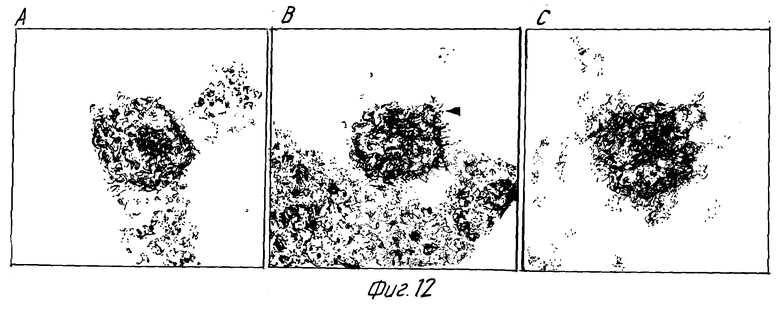
Одни - кроличья сыворотка (фиг. 11B) и моноклональные антитела NM-01 - не обладали заметным эффектом на морфологию ВИЧ-1. Экспозиция ВИЧ-1 с моноклональными антителами NM-01 и с комплементом была связана с появлением многочисленных вирусных частиц с разрушенными оболочками и потерей электронно плотной сердцевины (фиг. 11A). Типичные препараты показали примерно 90% разрушенных вирионов, большинство из которых имели потерю внутренней сердцевины. Остальные 10% вирионов были интактными или имели частично нарушенные внешние оболочки. Более высокое увеличение выявило, что разрушение ВИЧ-1 происходит путем прямого лизиса, как показано в сериях микрофотографий лизиса зрелых и неполных вирусных частиц на фиг. 12A по 12F и 13A по 13F соответственно.
Пример 8.
Комбинацию моноклональных антител NM-01 и комплемента затем подвергали анализу для определения ее воздействия на инфекционность ВИЧ-1.
Определение инфицирующей дозы для культуры ткани
Инфицированные  клетки H9 дважды промывали в среде для определения цитотоксичности (Cedarlane Lab. Ltd.) и затем ресуспендировали в среде для определения цитотоксичности, содержащей 2 мкг/мл моноклональных антител NM-01 или контрольного IgG2b. После инкубации при 4oC в течение 2 часов образцы делили на равные части и добавляли или кроличий комплемент, или инактивированную прогреванием кроличью сыворотку (Cedarlane Lab. Ltd.) в разведении 1: 6. Клетки инкубировали при 4oC в течение 20 минут и затем при 37oC в течение 45 минут промывали средой, ресуспендировали в среде 50% FBS/RPMI 1640 и встряхивали. Супернатант или вирусный изолят разводили десятикратно и затем серийно разводили в 2 раза перед добавлением 25 мкл клеток Н9(1 • 105/25 мкл). После инкубации в течение 3 часов при 37oC прошедшие экспозицию клетки разводили 10% средой FBS/RPMI 1640 и выдерживали при 37oC. Вирусная инфекция определялась через 6 дней путем исследования на обратную транскриптазу. Инфицирующая доза для культуры ткани для 50% образцов клеток H9 (TCID50) определялась по разведению, которое давало 50% инфицирование. Таблица 5 представляет результаты экспериментов.
клетки H9 дважды промывали в среде для определения цитотоксичности (Cedarlane Lab. Ltd.) и затем ресуспендировали в среде для определения цитотоксичности, содержащей 2 мкг/мл моноклональных антител NM-01 или контрольного IgG2b. После инкубации при 4oC в течение 2 часов образцы делили на равные части и добавляли или кроличий комплемент, или инактивированную прогреванием кроличью сыворотку (Cedarlane Lab. Ltd.) в разведении 1: 6. Клетки инкубировали при 4oC в течение 20 минут и затем при 37oC в течение 45 минут промывали средой, ресуспендировали в среде 50% FBS/RPMI 1640 и встряхивали. Супернатант или вирусный изолят разводили десятикратно и затем серийно разводили в 2 раза перед добавлением 25 мкл клеток Н9(1 • 105/25 мкл). После инкубации в течение 3 часов при 37oC прошедшие экспозицию клетки разводили 10% средой FBS/RPMI 1640 и выдерживали при 37oC. Вирусная инфекция определялась через 6 дней путем исследования на обратную транскриптазу. Инфицирующая доза для культуры ткани для 50% образцов клеток H9 (TCID50) определялась по разведению, которое давало 50% инфицирование. Таблица 5 представляет результаты экспериментов.
В то время как моноклональные антитела NM-01 сами по себе способны нейтрализовать инфекционность  обработка моноклональными антителами NM-01, вместе с комплементом снижала инфекционность
обработка моноклональными антителами NM-01, вместе с комплементом снижала инфекционность  больше, чем в 10 раз. Сходный эффект наблюдали также, когда применялся человеческий комплемент (в виде человеческой сыворотки) с моноклональными антителами NM-01. Эти результаты показывают, что экспозиция ВИЧ-1 с моноклональными антителами NM-01 и комплементом связана со значительным снижением вирусной инфекционности, и дополнительно подтверждают роль опосредуемого NM-01 зависимого от комплемента виролиза в терапии ВИЧ-1. См. также Nakamura et al. AIDS RESEARCH AND HYMAN RETROVIRUSES.
больше, чем в 10 раз. Сходный эффект наблюдали также, когда применялся человеческий комплемент (в виде человеческой сыворотки) с моноклональными антителами NM-01. Эти результаты показывают, что экспозиция ВИЧ-1 с моноклональными антителами NM-01 и комплементом связана со значительным снижением вирусной инфекционности, и дополнительно подтверждают роль опосредуемого NM-01 зависимого от комплемента виролиза в терапии ВИЧ-1. См. также Nakamura et al. AIDS RESEARCH AND HYMAN RETROVIRUSES.
9(7), pp. 619-626 (1993), которая приведена здесь в виде ссылки.
Пример 9.
Последовательности ДНК вариабельных областей тяжелой и легкой цепей моноклональных антител были клонированы с помощью PCR с использованием кДНК, произведенной из цитоплазматической PHK гибридом HB 10726 в качестве матрицы. ДНК вариабельных областей каждая затем встраивалась в M13mp18/mp19 (Pharmacia, Milton Keynes, UK) и секвенировалась. ДНК и выведенные аминокислотные последовательности вариабельных областей тяжелой и легкой цепей NM-01 представлены в ПОСЛ ИД NN 15 и 16 и ПОСЛ ИД NN 17 и 18 соответственно. Нуклеотиды 1-21 и 334-363 ПОСЛ ИД NN 15 соответствует праймерам PCR, использованным для амплификации последовательностей легкой цепи и нукленотиды 1-27 и 385-402 ПОСЛ ИД N 17 соответствуют праймерам PCR, использованным для амплификации последовательностей тяжелой цепи NM-01. Ресеквенирование вариабельных областей моноклональных антител NM-01 приводило к последовательностям, представленным в ПОСЛ ИД NN 19 и 20 и ПОСЛ ИД NN 21 и 22, которыми являются ДНК и выведенная аминокислотная последовательность вариабельной области тяжелой цепи и ДНК и выведенная аминокислотная последовательность вариабельной области легкой цепи соответственно. Аминокислотная последовательность вариабельной области легкой цепи (УК), как установлено, является наиболее гомологичной Kabat мышиной каппа подгруппе III, аминокислотная последовательность вариабельной области легкой цепи (VH), как установлено, является членом подгруппы IA Kabat мышиных тяжелых цепей.
Первые 120 остатков аминокислотных последовательностей вариабельных областей тяжелой (ПОСЛ ИД NN 20) и легкой (ПОСЛ ИД NN 22) цепей NM-01 также представлены на фиг. 14 и 15 соответственно, причем заключенные в них аминокислоты являются областями, определяющими комплементарность (CDR) антитела, которые определяют специфичность связывания антитела, CDR, идентифицированные на фиг. 14 и 15, сдвинуты в отношении CDR, показанных в предшествующей Международной патентной заявке N PCT/WO92/07111, чтобы привести их в соответствие с определениями CDR Kabat et al., Sequences of Proteins of Immunolоgical Interest, 5 th Edition, US Department of Health and Human Services, Government Printing. Office (1991).
На фиг. 14 и 15 каждая аминокислотная последовательность тяжелой или легкой цепи сравнивается с соответствующей аминокислотной последовательностью (ПОСЛ ИД N: 23 и 24) вариабельных областей тяжелой и легкой цепей моноклональных антител ВАТ123, с которых сообщено Liou et al., выше, и с соответствующими аминокислотными последовательностями (ПОСЛ ИД NN 25 и 26) вариабельных областей тяжелой и легкой цепей моноклональных антител F58/H3 и P4/D10, полученных из ECACC. Аминокислотные последовательности вариабельных областей моноклональных антител F58/H3 и P4/D10, как обнаружено, являются идентичными.
Вариабельная область тяжелой цепи NM-01 отличается от таковой ВАТ123 по сорока шести аминокислотам из общего числа в сто двадцать аминокислот. Вариабельные области легких цепей этих двух антител отличаются по двадцати трем аминокислотам. Важно, что три CDR в тяжелой цепи (V-H) NM-01 молекулы примерно на от 41 до 90% отличаются по последовательности от таковых в ВАТ-123, в то время как последовательности трех CDR в легкой цепи (V-L) отличаются примерно на от 29 до 47% по сравнению с NM-01.
Вариабельная область тяжелой цепи NM-01 отличается от таковой F58/H3 и P4/D10 по ста трем аминокислотам из общего количества сто двадцать, тогда как вариабельные области цепи отличаются по трем аминокислотам. Три CDR в тяжелой цепи (V-H) молекулы NM-01 различаются примерно на 86 до 100% последовательности от таковых F58/H3 и P4/D10, в то время как последовательности трех CDR в легкой цепи (V-L) различаются на от 13 до 19%.
Анализ первичной структуры NM-01 в сравнении с первичной структурой BAT123, F58/H3 и P4/D10 поэтому устанавливает, что NM-01 является новым антителом.
Пример 10.
На основании информации о последовательности ДНК, представленной в ПОСЛ ИД NN : 19 и 21 были получены химерные и очеловеченные/видоизмененные варианты антител NM-01. Для получения химерного варианта NM-01 применялись методы Orlandi et al., Proc.Natl. Acad. Sci. USA, 86, pp. 3833-3837 (1989). Очеловеченные варианты были получены методами, сходными с методами трансплантирования CDR Tempest et. al., BIO/TECHNOLOCY, 9, pp. 266-271, (1991) и Piechmann et al., Nature, 322, pp. 323-327 (1988).
A. Получение химерных антител
Вариабельные области NM-01 клонировали в две стадии в векторах экспрессии млекопитающих, чтобы иметь возможность получения химерных антител с мышиными вариабельными областями человека. В первую очередь, полностью секвенированные VH или VK амплифицировали из клонов M13mp19/mp19 NM-01, описанных в примере 9, используя праймеры, специфичные для концов 5' и 3' гена вариабельной области, и включая сайты рестрикции, позволяющие перенести полученный в результате амплифицированный фрагмент в вектор M13VHPCRI или M13VKPCRI (Orlandi et al., выше).
Таким образом вариабельная область помещалась за промотором и сигнальным пептидным геном в правильной ориентации для сшивания в ген константной области. На второй стадии вставки M13, включающие последовательности, кодирующие промотор, сигнальный пептид и вариабельную область (VH или VK), вырезали из ДНК RF и клонировали в вектор экспрессии млекопитающих, соответственно содержащий, в качестве подходящего, ген человеческого IgGl (вектор pSV-gpt) или константной области каппа (вектор pSV-hyg).
Плазмиды, кодирующие химерные легкие и тяжелые цепи NM-01 затем котрасфицировали в VB2/0 клетки миеломы крыс (ATCC CRL 1662), которые затем отсортировывали на присутствие ксантинфосфорибозилтрансферазного гена (gpt), обнаруживаемого на векторе экспрессии тяжелой цепи. Супернатант проверялся на присутствие человеческого IgG, и клетки, секретирующие антитела, размножались. Химерные антитела были обозначены NM-01 MuVH/MuVK.
B. Получение очеловеченных антител
CDR-трансплантация выполнялась путем сайт-направленного мутагенеза образцов вариабельной области человека. Гены вариабельной области человека, отобранные для CDR-трансплантации CDR NM-01, были NEWH VH (Saul et al., L. Biol. Chem. , 253, pp. 585-597 (1978) и REI VK (Epp et al., Eur. J. Biol. Chem., 45, pp. 513-524 (1974)).
В дополнение к мышиным CDR в очеловеченные MN-01 VH (обозначенные HuVH) включались четыре мышиных аминокислотных остатка перед первым CDR в положениях 27-30 с фиг. 16 и мышиный аргинин в положении 73 на фиг. 16 (положение 71 у Kabat). Четыре остатка перед первым CDR, хотя не гипервариабельны, как было показано, влияют на гипервариабельную конформацию петли по Chothia et al., J. Mol. Biol., 196, pp. 901-917 (1987). Остаток в положении 71 по Kabat, как было показано, помещается между петлями CDR 1 и 2, и важен в определении конформации CDR 2 (Tramontano et al., J. Mol. Biol., 215, pp. 175-182 (1990)).
Были получены два варианта HuVK NM-01, включая вариант с CDR-трансплантацией (HuVK) и вариант с CDR-трансплантацией (HuVKF), имеющий мышиный фенилаланин в положении 75 на фиг. 17. Боковая цепь аминокислоты в этом положении, как было показано, влияет на конформацию CDR 1 (Chothia et al., выше), и включение мышиного остатка положительно повлияло на связывающую способность других, очеловеченных антител. См. например, Foote et al., J. Mol. Biol., 224, pp. 487-499 (1992).
ДНК и производные аминокислотные последовательности HuVH, HuVK и HuVKF NM-01 соответственно показаны в ПОСЛ ИД N 27 и 28, 29 и 30, 31 и 32. Очеловеченные вариабельные области NM-01 были получены из фага M13, содержащего ген вариабельной области тяжелой или легкой цепи, следующим образом.
Фаг M13, содержащий гены вариабельных областей тяжелой или легкой цепей, выращивался в E. coli RZ1032 (dut ung) с целью получения однонитевых матричных ДНК, содержащих урацил вместо тимина. 1/2 мкг матричной ДНК смешивали с 1 пмоль олигонуклеотида, который ренатурирует матрицу M13 в прямом направлении от вставочной ДНК. Затем мутагенизирующие олигонуклеотиды, кодирующие мышиные остатки, ренатурировали матрицу в 20 мкл 40 мМ Трис-HCl pH 7,5, 20 мМ MgCl2, 50 мМ NaCl путем нагревания до 80oC в течение 5 минут и медленного охлаждения до комнатной температуры.
Для начальной реакции PCR вариабельной области тяжелой цепи использовали мутагенизирующие олигонуклеотиды:
VH олиго CDR 1 (ПОСЛ ИД N 33)
5' CTGTCTCACC CAGTGCCAGC AATAACTACT
ACTTGTGATG GAGAAGCCA ACAC 3'
причем олигонуклеотид является обратнокомплементарным ДНК, кодирующей аминокислоты  (аминокислоты 24-41 ПОСЛ ИД N 28 и последовательность HuVH на фиг. 16), и подчеркнутые аминокислоты являются мышиными остатками, введенными в матричную последовательность вариабельной области;
(аминокислоты 24-41 ПОСЛ ИД N 28 и последовательность HuVH на фиг. 16), и подчеркнутые аминокислоты являются мышиными остатками, введенными в матричную последовательность вариабельной области;
VH Олиго CDR 2 (ПОСД ИД N 34)
5' CATTGTCACT CTGCTTTTGA TGGATGGACT ATAGТCTATT
AACCTTCAT AACATATGCG TCC(A/C)ATCCAC TCAAGA 3'
причем олигонуклеотид является обратнокомплементарным ДНК, кодирующей аминокислоты L EW(1/M)  (аминокислоты 47-71 ПОСЛ ИД N 28 и последовательности HuVH на фиг. 16) и подчеркнутые аминокислоты являются мышиными остатками, введенными в матричную последовательность вариабельной области, и
(аминокислоты 47-71 ПОСЛ ИД N 28 и последовательности HuVH на фиг. 16) и подчеркнутые аминокислоты являются мышиными остатками, введенными в матричную последовательность вариабельной области, и
VH олиго CDR 3 (ПОСЛ ИД N 35)
5' CCAGTAGTCC ATAGAGGTCG TAGTACCATG GTTTTCTCTT
G(C/A)ACAATAAT AGAC 3'
причем олигонуклеотид является обратнокомплементарным ДНК, кодирующей аминокислоты  (аминокислоты 94-III ПОСЛ ИД N 28 и последовательность HuVH в фиг. 16) и подчеркнутые аминокислоты являются мышиными остатками, введенными в матричную последовательность вариабельной области.
(аминокислоты 94-III ПОСЛ ИД N 28 и последовательность HuVH в фиг. 16) и подчеркнутые аминокислоты являются мышиными остатками, введенными в матричную последовательность вариабельной области.
Человеческая матрица, использованная для мутагенеза вариабельной области легкой цепи, фактически кодирует структурные области, которые были родственны, но не идентичны REI и реакция мутагенеза устраняла эти несоответствия (используя неописанные нуклеотиды) так же, как и введение CDR NM-01. Единственное расхождение, которое нужно здесь обсудить особо, находится в положении 71 матрицы, которое кодирует фенилаланиновый остаток, отсутствующий в последовательности REI. Этот остаток сохранялся в HuVKF NM-01 (см. аминокислоту 75 HuVKF в фиг. 17), но изменялся на остаток REI в HuVK NM-01 (см. аминокислоту 75 HuVKF в фиг. 17) при использовании олигонуклеотида REI Y71, последовательность которого представлена ниже.
VK олиго REI Y71 (ПОСЛ ИД N 36)
5' ATGGTGAAGG TGTAGTCGGT ACCGC 3'
причем олигонуклеотид является обратнокомплементарным ДНК, кодирующей аминокислоты GTDYTFT (аминокислоты 72-78 ПОСЛ ИД N 32 и последовательность HuVK в фиг. 17). Этот праймер не включался в реакцию мутагенеза, которая производила HuVKF NM-01.
Для обоих вариабельных областей легких цепей HuVK и HuVKF мышиная CDR 1 NM-01 и матрица CDR 1 были идентичными, так что изменение CDR 1 было не нужно. Ограниченные различия между мышиными и матричными CDR 2 и 3 требовали изменения матричных CDR 2 и 3, и мутагенизированными олигонуклеотидами, использованными здесь, были
VK олиго CDR 2 (ПОСЛ ИД N 37)
5' AGGTTGGATG CAACGTAGAT CAGCAG 3'
причем олигонуклеотид является обратнокомплементарным ДНК, кодирующей аминокислоты (аминокислоты 50-57 ПОСЛ ИД N 30 и последовательность HuVK в фиг. 17), и подчеркнутые аминокислоты являются мышиными остатками, вводимыми в матричную последовательность вариабельной области, и
VK олиго CDR 3 (ПОСЛ ИД N 38)
5' CCGAACGTGA GCGGATCTTC ATTATTTTGC TGGCAGTA 3'
причем олигонуклеотид является обратнокомплементарным ДНК, кодирующей аминокислоты 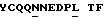 (аминокислоты 91-102 ПОСЛ ИД N 30 и последовательность HuVK в фиг. 17), и подчеркнутые аминокислоты являются мышиными остатками, введенными в матричную последовательность вариабельной области.
(аминокислоты 91-102 ПОСЛ ИД N 30 и последовательность HuVK в фиг. 17), и подчеркнутые аминокислоты являются мышиными остатками, введенными в матричную последовательность вариабельной области.
Чтобы произвести HuVH NM-01, VN олиго CDR 1, CDR 2 и CDR 3 ренатурировали в человеческую матрицу NEWH. Чтобы произвести HuVK NM-01, VK олиго REI Y71, VK олиго CDR2 и VK олиго CDR 3 ренатурировали в человеческую матричную ДНК. Чтобы получить HuVKF NM-01 ренатурировали VK олиго CDR 2 и VK олиго CDR 3. Затем в том же самом буфере добавляли dATP, dCTP, dGTP и dTTP до конечной концентрации 250 мкМ, DTT добавлялся до 7 мМ, ATP добавлялся до 1 мМ и добавляли 0,5 единицы T7 ДНК-полимеразы (United States Biochemical, Cleveland, OH) и 0,5 единицы T4 ДНК-лигазы (Life Technologies, Paisley, UK). Реакционную смесь в 30 мкл инкубировали при комнатной температуре в течение 1 часа и затем ДНК осаждали этанолом. Чтобы произвести однонитевой разрыв родительской матричной цепи, ДНК растворяли в 50 мкл 60 мМ Трис-HCl pH 8,0, 1 мМ DTT, 0,1 мг/мл БСА, содержащих 1 единицу урацил-ДНК-гликозилазы (Boehringer Mannheim, Kewis, Sussex, UK) и инкубировали при 37oC в течение 1 часа перед тем, как добавляли NaOH до 0,2 М, и инкубация продолжалась при комнатной температуре в течение 5 минут. ДНК опять осаждали этанолом, чтобы удалить фрагментированную родительскую ДНК. Мутантную ДНК затем растворяли в 20 мкл TE и вставку вариабельной области амплифицировали с помощью PCR, используя прямой и обратный праймеры M13. Реакционная смесь PCR содержала 2 мкл мутантной ДНК, 0,5 мкМ каждого праймера, 250 мкМ каждого dATP, dCTP, dGTP и dTTP, 10 мМ Трис HCl pH 8,3, 50 мМ KCl, 1,5 мМ MgCl2, 0,01% Твин-20, 0,01% желатина, 0,01% NP40 и 2 единицы термалазы (Thermalase) (IBI, Canbridge, UK) в 50 мкл. Амплификация достигалась с помощью 15 циклов, 94oC, 30 секунд, 50oC, 30 секунд, 72oC, 1 минуту, окончания с 72oC, 5 минут. Произведенные ДНК клонировали с M13 mp19 в виде Hind III-BamHI фрагментов и характерные клоны секвенировали. Сначала были получены только частичные мутанты с мышиными CDR 1 и 3 для тяжелой цепи. Чтобы получить мутанты с мышиными CDR 2, вышеприведенные реакции повторяли, используя частично мутированную ДНК в качестве матрицы и VH олиго CDR2. Полученный в результате продукт ДНК клонировали в M13mp19 в виде Hind III-BamHI фрагментов и характерные клоны секвенировали.
Фрагменты Hind III-BamHI, кодирующие правильные HuVH, HuVK и HVKF NM-01 затем соответственно клонировали в векторы экспрессии перед последовательностью кодирующей человеческой IgGI (вектор pSV-gpt) или каппа (вектор pSV-hyg) ген константной области как соответствующие. Полученные векторы коэлектропорировали в YB2/0 или NSO клетки (ECACC 85110503, чтобы получить клеточные линии, продуцирующие полностью очеловеченные антитела NM-01 (HuVH/HuVK), продуцируемые клеточной линией YB2/0, помещенных на хранение как ECACC 93082022, HuVH/HuVKF, продуцируемые клеточной линией, помещенной на хранение как ECACC 93082019) или их электропорировали отдельно вместе с векторами, кодирующими соответствующие тяжелые или легкие цепи химерных NM-01, описанных выше, чтобы получить линии клеток, продуцирующих смешенно-сочетанные (mix-and-match) антитела, где одна из цепей была химерной (например, MuVH/HuVKF). Антитела очищали путем аффинной хроматографии на агарозе с протеином A.
Четыре другие варианта очеловеченных антител NM-01 были получены методами, описанными выше. Первое HuVHM/HuVK (продуцируемое линией клеток YB2/0 ECACC 93082020) и второе HuVHM/HuVKF (продуцируемое линией клеток YB2/0, помещенной на хранение как 93082021) включают метионин в положении 48 HuVH. Третий вариант HuVHS/HuVK (продуцируемый линией клеток YB2/0, помещенной на хранение как ECACC 93082023) и четвертый вариант HuVHS/HuVKF (продуцируемый линией клеток YB2/0, помещенной на хранение как ECACC 93082018) включают серин в положении 93 HuVH. Эти очеловеченные антитела имели сходные свойства связывания с антигеном с таковыми антител NM-01 HuVH/HuVK или NM-01 HuVH/HuKF, в зависимости от включенной легкой цепи. Свойства связывания антигена NM-01 HuVH/HuVK и NM-01•HuVH/HuVKF описаны ниже.
C. Активность химерных и очеловеченных антител
Связывание очеловеченных антител NM-01 с gp120 оценивали в сравнении со связыванием мышиных NM-01 в конкурентном исследовании. Платы покрывали рекомбинантным gp120 (American Biotechnologies Inc. , Cambridge, MA) (5 нг/ячейку) и блокировали 5% нормальной козьей сывороткой (Life Technologies). Разведения (10-1000 нг/100 мкл) очеловеченных антител NM-01, химерных антител NM-01, мышиных антител NM-01 или отрицательный контроль очеловеченных антител добавляли в ячейки и платы инкубировали при 37oC в течение 30 минут. Добавляли биотинилированные мышиные антитела NM 01 (500 нг/50 мкл PBS на ячейку), и инкубация продолжалась в течение 1 часа. Платы промывали PBS - 0,05% Твин 20. Добавляли HRPO-стрептавидин (Sera-Lab Limited, Crawley Down, Sussek, UK; 40 нг/100 мкл PBS на ячейку), и платы инкубировали в течение 30 минут. Платы затем промывали и инкубировали в присутствии о-фенилдиамина в течение 5 минут или пока не резовьется окраска. Показания поглощения снимали при 492 нм.
Очеловеченные антитела NM-01 HuVH/HuVK были также эффективны /активным, как мышиные антитела NM-01 при блокировании связывания меченных мышиных антител NM-01 c gp120, в то время как очеловеченные NM-01 антитела HuVH/HuVKF были примерно в четыре раза более активными, чем мышиные антитела. Химерные антитела NM-01 были менее активны, чем мышиные антитела NM-01.
Химерные и очеловеченные антитела NM-01 также оценивали на активность по нейтрализации ВИЧ-1 с помощью анализа ОТ, p24 и образования синцития. Исследования, выполняемые по существу, как описано в предшествующих примерах, состояли в следующем.
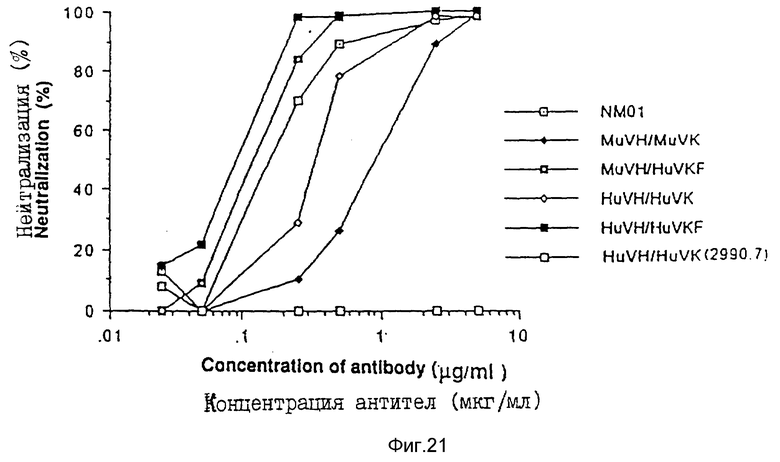
В анализе ОТ антитела разводили серийно в среде RPM1 1640 с 15% плодной бычьей сыворотки. Разведения антител инкубировали со 100 дозами, инфекционными до 50% тканевых культур (TCID50) вируса MN или IIIB в 96-ячеичных платах в течение 2 часов при 4oC. Затем в каждую ячейку добавляли клетки H9 (2,5•105 клеток) и плату инкубировали в течение еще одного часа при 37oC. Суспензию клеток H9 затем разводили в 2 мл среды RPMI 1640/15% плодной бычьей сыворотки и инкубировали в 24-ячеечной плате при 37oC. Продукцию вируса определяли с помощью исследования на ОТ на 7 день. Результаты исследования представлены на фиг. 18 (MN) и 19 (IIIB).
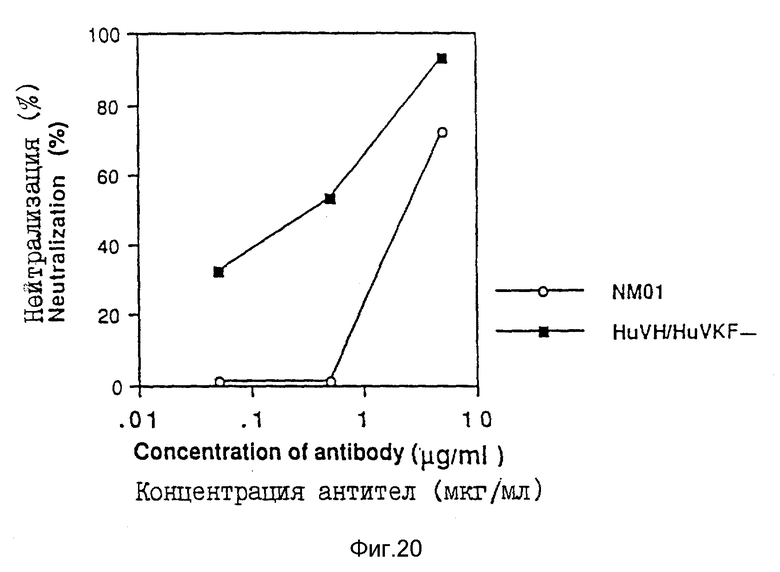
При исследовании на p24 клетки H9 инкубировали в течение от 6 до 8 дней с вирусом MN или IIIB (100•TCID50) и моноклональными антителами. Наличие антигена p24 в супернатанте культуры ткани затем определяли количественно с помощью профилированного для p24 ядра ВИЧ-I твердофазного иммуноферментного анализа (ELISA), с использованием методики, описанной производителем (DuPont-NEN). Вкратце, комплекс антиген-антитело определяли с помощью конъюгата с пероксидазой хрена (HRP). Конечный продукт количественно определялся по интенсивности желтого окрашивания, которая прямо пропорциональна количеству захваченного антигена p24 ядра ВИЧ-I. Проявление окрашивания регистрировалось при 450 нм, с использованием считывающего устройства для микроплат для EL ISA, и результаты исследования представлены на фиг. 20 (MN) и 21 (IIIB), причем моноклональные антитела 2990.7 служили отрицательным контролем.
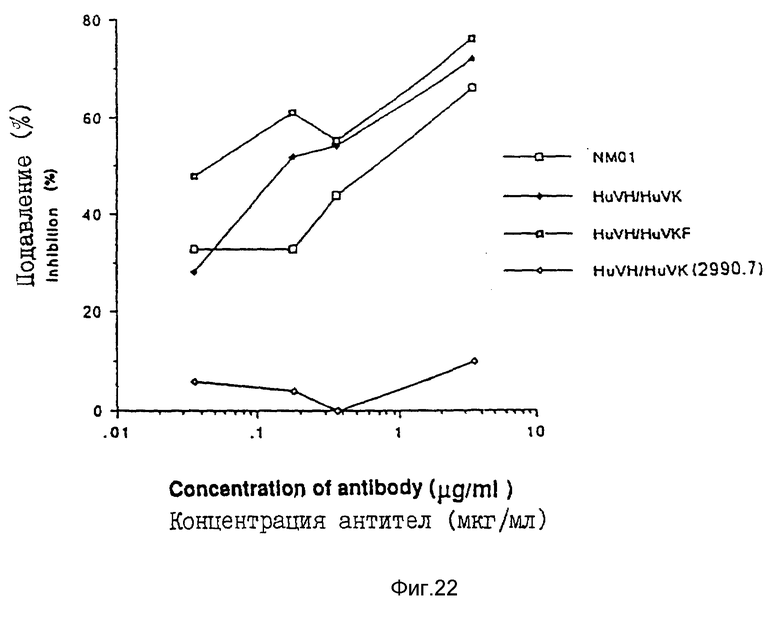
И наконец, в исследовании синцития клетки H9, хронически инфицированные любым вирусом MN, инкубировали с разведениями моноклональных антител NM-01 в течение часа при 37oC. Затем добавляли клетки из индикаторной клеточной линии С8166 (3•104 клеток/ячейку) и плату инкубировали дополнительно в течение от 2 до 12 часов при 37oC. Синцитии более трех лимфоцитных клеток диаметром подсчитывали и сравнивали с данными, полученными для контрольных инфицированных клеток H9 в отсутствие антител. Фиг. 22 представляет результаты исследования, причем антитела 2990.7 служат отрицательным контролем.
Результаты трех исследований показывают, что очеловеченные антитела NM-01 HuVH/HuVKF были одинаково эффективны или более эффективны, чем мышиные NM-01 моноклональные антитела при нейтрализации изолятов MN и IIIB ВИЧ-I.
Хотя настоящее изобретение было описано на примерах предпочтительных осуществлений, понятно, что изменения и улучшения будут проходить на ум специалистам. Поэтому имеется в виду, что прилагаемая форма изобретения охватывает все подобные эквивалентные изменения, которые входят в объем изобретения, которое заявляется.
Перечень последовательностей приведен в конце описанияа







































Изобретение представляет моноклональное антитело, которое специфически иммунореактивно с протеином ВИЧ-1 gpl20 или gpl60 и характеризуется способностью нейтрализовать in vitro инфицирование клеток Н9 живыми штаммами ВИЧ-1. Антитела выделяются гибридомными клеточными линиями. Фармацевтические композиции этого изобретения будут применяться при лечении людей, инфицированных ВИЧ-1, с помощью пассивной иммунизации. 10 с. и 6 з.п.ф-лы, 30 ил, 5 табл.
DIQMTQSPSS LSASVGDRVT ITCRASESVD SYGNSFMHWY QQTPGKAPKL LIYVASNLES GVPSRFSGSG SGTDFTFTIS SLQPEDIATY YCQQNNEDPL TFGOGTKLOI T
9. Полинуклеотид, кодирующий антитело NM-01 Hu VH/HuVKF, продуцируемое гибридомной клеточной линией ЕСАСС 93082019, причем тяжелая цепь указанного антитела NM-01 Hu VH/HuVKF имеет последовательность аминокислот:
DIQMTQSPSS LSASVGDRVT ITCRASESVD SYGNSFMHWY QQTPGKAPKL LIYVASNLES GVPSRFSGSG SGTDYTFTIS SLQPEDIATY YCQQNNEDPL TFGQGTKLQI T
10. Гибридомная клеточная линия ЕСААС 93082022, кодирующая антитело NM-01 Hu VH/HuVK.
| PCT 8809181 A1, 01.12.88 | |||
| PCT 9119797 A1, 26.12.91 | |||
| RU 2070926 C1, 27.12.96. |
