Область техники
По настоящей заявке испрашивается приоритет корейской патентной заявки № 10-2015-0045684, поданной 31 марта 2015 г., которая настоящим включена посредством ссылки во всей полноте.
Настоящее изобретение относится к фармацевтической композиции для предотвращения и лечения заболевания глаз, причем данная композиция содержит в качестве активного ингредиента белок слияния, в котором проникающий в ткань пептид слит со средством против фактора роста эндотелия сосудов (анти-VEGF). Более конкретно, настоящее изобретение относится к фармацевтической композиции для предотвращения и лечения заболевания глаз, причем данная композиция содержит в качестве активного ингредиента белок слияния, в котором проникающий в ткань пептид слит с анти-VEGF средством. Настоящее изобретение также относится к способу получения анти-VEGF средства с улучшенной эффективностью и способностью преодолевать резистентность, причем данный способ содержит: трансформацию клеток-хозяев с помощью рекомбинантного вектора, причем данный рекомбинантный вектор содержит последовательность нуклеиновой кислоты, кодирующую белок слияния, в котором проникающий в ткань пептид слит с анти-VEGF средством; культивирование данных клеток; и получение белка слияния из данных клеток. Настоящее изобретение также относится к способу лечения заболевания глаз, причем данный способ содержит введение эффективного количества белка слияния в соответствии с настоящим изобретением нуждающемуся в этом субъекту; и к применению белка слияния в соответствии с настоящим изобретением для получения терапевтического средства для заболевания глаз, содержащего данный белок слияния в качестве активного ингредиента.
Уровень техники
В то время как желтое пятно является частью сетчатки глаза, в которой сконцентрированы зрительные клетки для наиболее четкого и точного приема света, заболевание, которое вызывает нарушение зрения из-за дегенерации желтого пятна, вызываемой различными причинами, называется макулярной дегенерацией. Макулярная дегенерация является одной из трех причин слепоты, а также глаукомы и диабетической ретинопатии. Главной причиной макулярной дегенерации является увеличение возраста, но семейная история, раса, курение и т.п. также известны как причины макулярной дегенерации. Повреждение желтого пятна вызывает потерю способности распознавать детали, такие как мелкая печать, черты лица или мелкие предметы. Макулярная дегенерация может быть двух типов: неэкссудативная (сухая) макулярная дегенерация и экссудативная (влажная) макулярная дегенерация, причем распространенность сухой макулярной дегенерации составляет 90%. При сухой макулярной дегенерации выделения образуют желтые отложения, называемые друзами, которые могут накапливаться в ткани ниже макулярной ткани. Присутствие друз воздействует на кровоток в сетчатку, особенно в желтое пятно, а снижение кровотока уменьшает подачу питательных веществ в желтое пятно, прекращая или ограничивая эффективную работу фоточувствительных клеток. В случае влажной макулярной дегенерации новые слабые кровеносные сосуды растут в сетчатке или под ней, вызывая подтекание жидкости крови в пространство ниже желтого пятна. Влажную макулярную дегенерацию иногда описывают как хориоидальную неоваскуляризацию. Хориоидеа представляет собой сосудистый участок ниже желтого пятна, и неоваскуляризация относится к росту новых кровеносных сосудов в ткани. Как можно понять из названия хориоидальной неоваскуляризации, при влажной макулярной дегенерации кровеносные сосуды вновь образуются для прорастания из хориоидеи в желтое пятно. Макулярную дегенерацию считают заболеванием пожилых людей, но известно, что в последние годы быстро растет число пациентов в возрасте от 40 до 50 лет. Основными причинами снижения возраста начала макулярной дегенерации являются вестернизация привычек питания, такая как увеличение потребления жиров, и вредные привычки, такие как курение, употребление алкоголя и открытость для воздействия ультрафиолетовых лучей.
Диабетический макулярный отек (DME) объясняется утолщением сетчатки и/или твердым экссудатом в пределах одного диаметра диска от центра сетчатки. DME и диабетическая ретинопатия (DR) являются микрососудистыми осложнениями, возникающими у пациентов с диабетом, они ослабляют зрение и в конечном итоге приводят к слепоте. Пациенты с DR демонстрируют прогрессирование DME, а расширенные гиперпроницаемые капилляры и просачивание из микроаневризмы могут вызвать DME после повреждения гематоретинальных барьеров. Как и DR, DME связан с хориоидальной неоваскуляризацией, которая проникает в поврежденные или с разрушенной тканью мембраны Бруха.
При этом примеры типичных лекарственных средств, которые используются для предупреждения или лечения различных заболеваний глаз, связанных с неоваскуляризацией в глазах (таких как макулярная дегенерация и диабетический макулярный отек) включают ранибизумаб, бевацизумаб, афлиберцепт, конберцепт и т.п. Разработанные в настоящее время биофармацевтические препараты, включая упомянутые выше лекарственные средства, для лечения основных заболеваний глаз, таких как макулярная дегенерация и отек при диабетическаой ретинопатии, в основном используются для лечения заболеваний задней части глазных яблок, включая сетчатку, в форме внутриглазной инъекции. В последнее время были предприняты попытки разработки для ранибизумаба, афлиберцепта и т.п. формы глазных капель для решения таких проблем, как снижение удобства для пациента, увеличение числа побочных эффектов и психологический страх перед инъекциями. Однако, как показали результаты тестов, в которых у кроликов ранибизумаб достигал ткани сетчатки через 3-7 дней после закапывания половинной дозы (250 мкг) ранибизумаба в глаза 6 раз с интервалами 2 часа (Chen et al., 2011. Eye), ранибизумаб обладает плохой проницаемостью глаз и с трудом достигает задней части глазного яблока, где находится повреждение, причем большое количество лекарственного средства теряется из-за оттока воды при мигании глаз при нанесении глазных капель, и, следовательно, его фармацевтический эффект в форме глазных капель труднодостижим в глазах. Кроме того, конберцепт (Chengdu Kanghong pharm.), который был одобрен в качестве терапевтического средства для макулярной дегенерации в 2013 г., как и афлиберцепт, представляет собой белок слияния, в котором второй домен рецептора-1 фактора роста эндотелия сосудов (VEGFR-1) связан с третьим и четвертым доменами VEGFR-2 посредством Fc, и его лекарственная форма глазных капель также разрабатывается в настоящее время. Однако, судя по сообщениям, конберцепт в момент нанесения его глазных капель обладает биодоступностью менее приблизительно 5% (Wang et al., 2013. PLOS ONE).
Кроме того, внимание к ранибизумабу, который продается под торговым наименованием Lucentis, обусловлено тем, что к моменту разработки приблизительно 90% пациентов с макулярной дегенерацией продемонстрировали ответ. Однако только 30% реагирующих пациентов продемонстрировали терапевтический эффект, такой как улучшение зрения, и его постоянное введение вызывало резистентность к лекарственным средствам (Syed et al., 2012. Nature Rev. Drug Discov.). Для устранения таких недостатков в 2011 г. был выпущен афлиберцепт, который представляет собой белок слияния с Fc антитела, разработанный для 100-кратного повышения способности к связыванию с VEGF-A и для возможности ингибирования даже VEGF-B и PIGF, и он продемонстрировал значительный коммерческий рост, но, фактически, было подтверждено, что у этих двух продуктов нет различий в клинической эффективности.
Приблизительно 10% пациентов с макулярной дегенерацией не отвечают на анти-VEGF средства, и, следовательно, у таких пациентов нельзя ожидать от них терапевтического эффекта. Полагают, что это обусловлено другим зависимым от факторов роста каналом неоваскуляризации, отличным от VEGF-A. Кроме того, было подтверждено, что повторное введение вызывает резистентность приблизительно у 45% пациентов, и ответ на лекарственное средство снижается при увеличении числа введений (Lux et al., 2007. Br. J. Ophthalmol.). Известно, что эта резистентность вызвана укреплением сосудов, происходящим из-за увеличения покрытия перицитами, которое вносит вклад в стабилизацию эндотелиальных клеток во время повторных введений анти-VEGF средства и перицит-зависимое продуцирование VEGF.
Поэтому последние тенденции разработки терапевтических средств для заболеваний глаз заключаются в разработке комбинационной терапии с помощью ингибитора PDGF для вызывания диссоциации перицитов для преодоления резистентности и повышения эффективности анти-VEGF средств. Поэтому терапевтическое средство для заболевания глаз может: (i) иметь дополнительную функцию блокирования VEGF-A и других связанных с неоваскуляризацией лигандов; (ii) преодолевать резистентность к средству против фактора роста эндотелия сосудов; (iii) разрушать покрытие перицитами для улучшения эффективности лекарственного средства; (iv) иметь повышенную частоту введения; и (v) быть выпущено в виде глазных капель.
Подробное описание изобретения
Техническая задача
Авторы настоящего изобретения обнаружили, что слияние средства против фактора роста эндотелия сосудов (анти-VEGF) и проникающего в ткань пептида увеличивает проникновение лекарственного средства в ткань, разрушает покрытие перицитами для воздействия даже на пациентов, демонстрирующих резистентность к лекарственному средству, уменьшает дозу или повышает частоту введения и делает возможным выпуск в виде глазных капель, и этим завершили настоящее изобретение.
Таким образом, аспектом настоящего изобретения является предоставление фармацевтической композиции для предотвращения и лечения заболевания глаз, причем данная композиция содержит в качестве активного ингредиента белок слияния, в котором проникающий в ткань пептид слит со средством против фактора роста эндотелия сосудов (анти-VEGF).
Другим аспектом настоящего изобретения является предоставление способа получения средства против фактора роста эндотелия сосудов (анти-VEGF) с улучшенной эффективностью и способностью преодолевать резистентность, причем данный способ содержит: (a) трансформацию клеток-хозяев с помощью рекомбинантного вектора, причем данный рекомбинантный вектор содержит последовательность нуклеиновой кислоты, кодирующую белок слияния, в котором проникающий в ткань пептид слит со средством против фактора роста эндотелия сосудов (анти-VEGF); (b) культивирование данных клеток; и (c) получение белка слияния из данных клеток.
Еще одним аспектом настоящего изобретения является предоставление способа лечения заболевания глаз, причем данный способ содержит введение эффективного количества белка слияния, в котором проникающий в ткань пептид слит со средством против фактора роста эндотелия сосудов (анти-VEGF), нуждающемуся в этом субъекту.
Еще одним аспектом настоящего изобретения является предоставление применения белка слияния, в котором проникающий в ткань пептид слит со средством против фактора роста эндотелия сосудов (анти-VEGF), для получения терапевтического средства для заболевания глаз, содержащего данный белок слияния в качестве активного ингредиента.
Техническое решение
В соответствии с аспектом настоящего изобретения предлагается фармацевтическая композиция для предотвращения и лечения заболевания глаз, причем данная композиция содержит в качестве активного ингредиента белок слияния, в котором проникающий в ткань пептид слит со средством против фактора роста эндотелия сосудов (анти-VEGF).
В соответствии с другим аспектом настоящего изобретения предлагается способ получения средства против фактора роста эндотелия сосудов (анти-VEGF) с улучшенной эффективностью и способностью преодолевать резистентность, причем данный способ содержит: (a) трансформацию клеток-хозяев с помощью рекомбинантного вектора, причем данный рекомбинантный вектор содержит последовательность нуклеиновой кислоты, кодирующую белок слияния, в котором проникающий в ткань пептид слит со средством против фактора роста эндотелия сосудов (анти-VEGF); (b) культивирование данных клеток; и (c) получение белка слияния из данных клеток.
В соответствии с еще одним аспектом настоящего изобретения предлагается способ лечения заболевания глаз, причем данный способ содержит введение эффективного количества белка слияния, в котором проникающий в ткань пептид слит со средством против фактора роста эндотелия сосудов (анти-VEGF), нуждающемуся в этом субъекту.
В соответствии с еще одним аспектом настоящего изобретения предлагается применение белка слияния, в котором проникающий в ткань пептид слит со средством против фактора роста эндотелия сосудов (анти-VEGF), для получения терапевтического средства для заболевания глаз, содержащего данный белок слияния в качестве активного ингредиента.
Далее настоящее изобретение будет описано подробно.
В соответствии с аспектом настоящего изобретения предлагается фармацевтическая композиция для предотвращения и лечения заболевания глаз, причем данная композиция содержит в качестве активного ингредиента белок слияния, в котором проникающий в ткань пептид слит со средством против фактора роста эндотелия сосудов (анти-VEGF).
Фактор роста эндотелия сосудов-A (VEGF-A) хорошо известен среди белков, присутствующих в природе, индуцированием экстравазации крови. Его также называют фактором проницаемости сосудов. Известно, что это воздействие обусловлено его соединением с рецептором фактора роста эндотелия сосудов (VEGFR2), но интересно, что эксперимент по мутации фактора роста эндотелия сосудов-A продемонстрировал, что проникновение фактора роста эндотелия сосудов-A в сосуды возрастало, даже если фактор роста эндотелия сосудов-A не связывался с рецептором фактора роста эндотелия сосудов. Это позволило предположить, что существует еще один рецептор для фактора роста эндотелия сосудов-A (Stacker et al., 1999. J. Biol. Chem.). Другие современные исследователи установили, что этим рецептором является нейропилин (NRP) (Makinen et al., 1999. J. Biol. Chem.).
Нейропилин был впервые обнаружен в нервной системе Xenopus. Нейропилин представляет собой трансмембранный гликопротеин и имеет два типа: NRP1 и NRP2. Нейропилин действует как корецептор рецепторов VEGF (VEGFR), связывая лиганды семейства VEGF. В частности, NRP1 связывается с различными лигандами VEGF, действуя как корецептор для VEGFR1, VEGFR2 и VEGFR3. С другой стороны, NRP2 вносит вклад в лимфангиогенез и клеточную адгезию, действуя как корецептор для VEGFR2 и VEGFR3. Кроме того, NRP1/NRP2 (NRP1/2) действуют как корецептор для рецепторов семейства плексинов и связываются как с лигандами с секретируемыми семафоринами класса 3 (Sema3A, Sema3B, Sema3C, Sema3D, Sema3E, Sema3F и Sema3G).
Как использовано в настоящем документе, термин "проникающий в ткань" или "проникновение в ткань" говорит о наличии любой характеристики из: специфического распознавания ткани, сверхэкспрессирующей нейропилин для накопления в тканях; расширения клеточного зазора между клетками эндотелия сосудов для облегчения экстравазации лекарственного средства; или способствования распределению лекарственного средства в глазу посредством коррекции зазора между клетками роговицы, которые представляют собой ткань, действующую как барьер для водорастворимых молекул.
Как использовано в настоящем документе, термин "нейропилин (NRP)" относится к трансмембранному гликопротеину, и имеет два типа: NRP1 и NRP2. Нейропилин в основном состоит из пяти доменов. Домены a1/a2 с N-конца классифицируют как домены CUB, с которыми связывается Ig-подобный домен типа C2 семафорина. В частности, эти домены образуют комплекс с плексином для увеличения связывающей способности семафорин-плексин. Домены b1 и b2 нейропилина классифицируют как домены FV/VIII, с которыми связываются C-концы лигандов семейства VEGF или лигандов секретируемых семафоринов класса 3 (Sema3). Лиганды VEGF и лиганды семафорины класса 3 имеют сайт распознавания (RXRR, Arg-X-Arg-Arg) протеазы фурина, и поэтому вследствие обработки фурином лиганды обычно заканчиваются аминокислотным остатком аргинина (Arg) на C-конце (Adams et al., 1997. EMBO J.). Сообщалось, что остаток Arg на C-конце лигандов VEGF и Sema3 очень важен при взаимодействии нейропилиновых доменов b1b2 (Teesalu et al., 2009. Proc. Natl. Acad. Sci. USA). Была раскрыта третичная структура комплекса между лигандом VEGF и нейропилиновым доменом b1b2 (Parker et al., 2012. J. Biol. Chem.), и, соответственно, может быть определена аминокислотная последовательность VEGF, которая важна при связывании с нейропилиновым доменом b1b2. Однако еще не было установлено, какой участок C-конца Sema3A, связывающегося с NRP1, специфически связывается с NRP.
Средство против фактора роста эндотелия сосудов включает в себя молекулу, которая вмешивается во взаимодействие между VEGF и природным рецептором VEGF, например молекулу, которая связывается с VEGF или рецептором VEGF для предотвращения или препятствования взаимодействию между VEGF и рецептором VEGF. Примеры антагонистов VEGF включают антитела против VEGF, антитела против рецептора VEGF и химерные молекулы на основе рецептора VEGF.
Как использовано в настоящем документе, термин "слияние" относится к интеграции двух молекул, которые обладают одинаковыми или различными функциями или структурами, и может представлять собой слияние с помощью любого физического, химического или биологического способа, посредством которого проникающий в ткань пептид может связываться со средством против фактора роста эндотелия сосудов. Слияние может осуществляться, предпочтительно, с помощью линкерного пептида, причем этот линкерный пептид может связываться, например, с C-концом Fab (антиген-связывающего фрагмента) или Fc-фрагмента в антителе.
Заболевание глаз настоящего изобретения, предпочтительно, означает заболевание глаз из-за неоваскуляризации. Как использовано в настоящем документе, выражение "заболевание глаз из-за неоваскуляризации" относится к любому заболеванию глаз из-за сосудистого роста или пролиферации, просачивания из сосудов или связанному с ними заболеванию.
Белок слияния в соответствии с настоящим изобретением повышает проникновение лекарственного средства в ткань посредством связывания с нейропилиновым рецептором, разрушает покрытие перицитами для воздействия даже на пациентов, демонстрирующих резистентность к лекарственному средству, повышает частоту введения и делает возможным выпуск в виде глазных капель.
Точнее говоря, в соответствии с примером настоящего изобретения, производительность мутантного ранибизумаба, имеющего точечную мутацию, в которой цистеин (C) в аминокислотной последовательности ранибизумаба замещен серином (S), значительно увеличивается, причем повышенная производительность модифицированной формы, в которой проникающий в ткань пептид (TPP) слит с мутантным ранибизумабом, также сохраняется (пример 1).
В соответствии с другим примером настоящего изобретения в результате сравнения аффинности к нейропилиновому рецептору и способности к разрушению плотного соединения между эндотелиальными клетками белка слияния, в котором проникающий в ткань пептид слит с C-концом мутантного ранибизумаба, было подтверждено, что белок слияния очень хорошо на одном уровне связывается с нейропилиновым рецептором NRP1 и лигандом Sema3A, и заметно ингибирует VE-кадгерин, что говорит о его превосходной способности к разрушению плотных соединений между эндотелиальными клетками (пример <1-3>).
В соответствии с другим примером настоящего изобретения для исследования того, может ли белок слияния, в котором проникающий в ткань пептид слит с C-концом мутантного ранибизумаба, улучшать проникновение посредством связывания с нейропилиновыми рецепторами, широко распространенными в клетках глазного эндотелия, извлеченные глазные яблоки погружали в раствор, содержащий белок слияния, и раствор, содержащий ранибизумаб, и затем сравнивали их по проникновению с течением времени (пример 4). Результаты экспериментов подтвердили, что связывание средства против фактора роста эндотелия сосудов с проникающим в ткань пептидом инициирует проникновение из корнеальных эндотелиальных тканей от одного часа после эксперимента и демонстрирует очень высокое распределение лекарственного средства в глазу в течение 2 часов по сравнению с ранибизумабом, и, таким образом, подтвердили возможность колнтроля дозы и увеличения интервала введения посредством улучшения проникновения.
То есть можно подтвердить, что по сравнению с предыдущим исследованием, в котором глазное проникновение ранибизумаба оказалось очень низким, белок слияния в соответствии с настоящим изобретением проникает в роговицу, которая служит барьером для водорастворимых молекул, быстрее, чем ранибизумаб, и достигает внутренней части глазного яблока. Таким образом, белок слияния в соответствии с настоящим изобретением может быть выпущен в виде глазных капель за счет улучшения состава в будущем. Эффект увеличения проникновения в ткань глаза благодаря слиянию с проникающим в ткань пептидом аналогичным образом наблюдали для формы цельного антитела с большим молекулярным весом и сложной белковой структурой, а также модифицированной формы ранибизумаба, имеющего Fab-тип. Белок слияния, в котором проникающий в ткань пептид слит с Fc-концом бевацизумаба, имеющего тип иммуноглобулина G (IgG), также продемонстрировал значительно увеличенную способность к проникновению в глаз по сравнению с бевацизумабом, что говорит о том, что модифицированная форма бевацизумаба в соответствии с настоящим изобретением также может быть выпущена в виде лекарственного средства с увеличенным терапевтическим эффектом.
По результатам настоящих примеров можно предсказать, что, аналогично бевацизумабу, афлиберцепт, конберцепт и т.п. в форме белка слияния, в котором рецепторы VEGF (VEGFR1 и VEGFR2) слиты с Fc-фрагментом, если проникающий в ткань пептид слит с Fc-концом, также могут быть выпущены в виде лекарственных средств с заметно увеличенным по сравнению с существующими белками слияния с Fc терапевтическим эффектом.
В другом примере настоящего изобретения эффект ингибирования неоваскуляризации и эффект ингибирования неоваскуляризации в моделях резистентности к лекарственным средствам сравнивали и оценивали с использованием различных моделей заболеваний животных для белка слияния настоящего изобретения и ранибизумаба. В результате эксперимента с использованием моделей неоваскуляризации роговицы белок слияния настоящего изобретения продемонстрировал значительный эффект ингибирования неоваскуляризации, составляющий 50% или более, по сравнению с контрольной группой, и эквивалентный ранибизумабу эффект, что подтверждает улучшение эффективности из-за проникающего в ткань пептида, слитого с C-концом (пример <5-1>). Кроме того, в отношении эффекта ингибирования неоваскуляризации в моделях резистентности к лекарственным средствам, белок слияния настоящего изобретения продемонстрировал отличную эффективность в два или более раза превосходящую ранибизумаб, причем это значение аналогично результатам, приведенным в литературе, раскрывающей совместное введение анти-VEGF аптамера и средства, являющегося анти-PDGF антителом (Jo et al., 2006. Am. J. Pathol. 168), и, таким образом, можно предсказать, что белок слияния настоящего изобретения будет обладать способностью улучшения зрения у приблизительно 70% пациентов, которым вводили ранибизумаб, и у которых зрение только сохранялось без улучшения зрения (пример <5-2>). Аналогично, отличный эффект ингибирования неоваскуляризации при заболевании глаз с помощью белка слияния в соответствии с настоящим изобретением также наблюдали в модели хориоидальной неоваскуляризации (CNV), используемой в качестве модели фактической эффективности макулярной дегенерации, и при кислород-индуцированной ретинопатии (OIR), используемой в качестве модели эффективности ретинопатии, и, таким образом, можно предсказать, что белок слияния настоящего изобретения будет обладать клинически заметно увеличенным терапевтическим эффектом по сравнению с ранибизумабом (пример 6 и пример 7).
Как использовано в настоящем документе, термин "предупреждение" относится ко всем действиям по подавлению заболевания глаз или задержке прогрессирования заболевания глаз посредством введения композиции настоящего изобретения.
Как использовано в настоящем документе, термин "лечение" относится ко всем действиям по улучшению или изменению в лучшую сторону заболевания глаз посредством введения композиции настоящего изобретения. Более конкретно, термин "лечение" полностью относится к облегчению симптомов заболевания глаз, что может охватывать излечение, по существу предотвращение или облегчение состояния заболевания глаз и может охватывать облегчение, излечение или предотвращение одного симптома или большинства симптомов, возникающих из-за заболевания глаз, но не ограничивается этим.
При реализации настоящего изобретения на практике специалист в данной области техники может определять эффективную дозу (эффективное количество), количество введений и способ введения, для того чтобы предотвращать или лечить заболевание глаз, при надлежащем учете различных факторов, таких как тип и тяжесть соответствующего заболевания глаз, возраст, вес, состояние здоровья, пол, питание и интенсивность экскреции у нуждающегося во введении субъекта. Термин "эффективное количество" полностью относится к количеству для улучшения симптомов заболевания глаз при введении субъекту и охватывает количество для излечения или по существу предотвращения заболевания глаз или облегчения состояния заболевания глаз. Термин "субъект" может представлять собой животное, предпочтительно млекопитающее, и особенно животное, включая человека, и может включать клетки, ткани, органы или т.п., которые происходят от животного. Субъект может представлять собой пациента, нуждающегося в лечении. Композиция настоящего изобретения может быть введена млекопитающим, включая людей, любыми способами. Например, композиция настоящего изобретения может быть введена перорально или парентерально. Парентеральное введение может представлять собой, но без ограничения, внутривенное, внутримышечное, внутриартериальное, интрамедуллярное, внутрижелудочное, внутрисердечное, трансдермальное, подкожное, внутрибрюшинное, интраназальное, кишечное, местное, подъязычное или ректальное введение. Иммунногенный белковый комплекс в соответствии с настоящим изобретением может быть введен отдельно или в комбинации с известным соединением, имеющим эффект предотвращения и лечения целевого заболевания.
Например, доза фармацевтической композиции настоящего изобретения для человеческого тела может изменяться в зависимости от возраста, веса, пола, лекарственной формы, состояния здоровья и тяжести заболевания пациента. Для глазной инъекции доза может обычно составлять 0,001-10 мг/день на глазное яблоко и, предпочтительно, 0,1-2 мг/день на глазное яблоко. Для глазных капель доза может составлять 0,001-100 мг/день и, предпочтительно, 0,01-10 мг/день на 1 мл раствора глазных капель. Композицию можно также вводить дробно с предварительно определенными интервалами в соответствии с определением, сделанным врачом или фармацевтом.
Композиция настоящего изобретения может дополнительно содержать фармацевтически приемлемую добавку. Примеры фармацевтически приемлемых добавок могут включать крахмал, желатинизированный крахмал, микрокристаллическую целлюлозу, лактозу, повидон, коллоидный диоксид кремния, гидрофосфат кальция, лактозу, маннит, карамель, аравийский каучук, прежелатинизированный крахмал, кукурузный крахмал, порошок целлюлозы, гидроксипропилцеллюлозу, опадрай, натрийкарбоксиметилкрахмал, карнаубский воск, синтетический алюмосиликат, стеариновую кислоту, стеарат магния, стеарат алюминия, стеарат кальция, белый сахар, декстрозу, сорбит, тальк и т.д. Фармацевтически приемлемая добавка в соответствии с настоящим изобретением, предпочтительно, составляет 0,1-90 весовых частей относительно фармацевтической композиции.
Кроме того, фармацевтическая композиция может быть в различных формах, подходящих для любого способа введения, включая, но без ограничения, инъекцию, глазные капли, глазную мазь и внутриглазную лекарственную форму. При составлении фармацевтической композиции можно использовать разбавители или вспомогательные вещества, такие как наполнитель, расширитель, связующее вещество, смачивающее средство, дезинтегратор или поверхностно-активное вещество. Инъекция может включать все варианты интравитреальных инъекций и внутриглазных местных инъекций, включая инъекцию конъюнктивы, но без ограничения ими. Инъекция может содержать обычные добавки, такие как солюбилизатор, изотоническое средство, суспендирующее средство, эмульгатор, стабилизатор и консервант.
Подходящие носители для инъекции настоящего изобретения включают физиологический раствор, бактериостатическую воду, Cremophor EL (BASF, Parsippany, NJ, США) или фосфатный солевой буфер (ФСБ). Во всех случаях композиция должна быть стерильной и должна быть текучей до такой степени, чтобы имела место легкая проходимость через иглу. Композиция должна быть стабильной в условиях производства и хранения и должна быть защищена от загрязняющего воздействия микроорганизмов, таких как бактерии и грибы. Носитель может представлять собой растворитель или дисперсионную среду, содержащие, например, воду, этанол, полиол (например, глицерин, пропиленгликоль и жидкий полиэтиленгликоль и т.п.) и их подходящие смеси. Соответствующую текучесть можно поддерживать, например, с помощью покрытия, такого как лецитин, с помощью поддержания требуемого размера частиц в случае дисперсии и с помощью поверхностно-активных веществ. Воздействие микроорганизмов может быть предотвращено с помощью различных антибактериальных и противогрибковых средств, например парабенов, хлорбутанола, фенола, аскорбиновой кислоты, тимеросала и т.п. Во многих случаях композиция будет, предпочтительно, содержать изотонические средства, например сахара, полиспирты, такие как маннит или сорбит, или хлорид натрия. Длительная абсорбция инъекционных композиций может быть достигнута путем включения в композицию средства, которое замедляет абсорбцию, например моностеарата алюминия, гиалуроновой кислоты и желатина.
Глазные капли могут представлять собой водорастворимый офтальмологический раствор, водонерастворимый офтальмологический раствор или офтальмологическую эмульсию. Глазные капли настоящего изобретения могут содержать в дополнение к белку слияния, в котором вышеуказанный существенный компонент, проникающий в ткань пептид, слит со средством против фактора роста эндотелиальных клеток сосудов (анти-VEGF), любой обычно используемый компонент глазных капель, например буфер, средство для регулирования вязкости, стабилизатор, изотоническое средство, консервант и т.п., смешанный с белком слияния. В том числе, примеры буферов могут включать известные буферы цитратных, фосфатных, ацетатных и аминокислотных солей. Кроме того, примеры средств для регулирования вязкости могут включать поливиниловый спирт, гидроксипропилцеллюлозу, метилцеллюлозу, повидон, гидроксипропилметилцеллюлозу, этилцеллюлозу, гидроксиэтилцеллюлозу, кармелозу, полиэтиленгликоль, хондроитин или их соли. Примеры стабилизаторов могут включать: антиоксиданты, такие как нитрит натрия, гидросульфит натрия и метабисульфит натрия; и хелатирующие средства, такие как эдетат натрия, циклодекстрин, лимонная кислота и цитрат. Кроме того, примеры изотонизирующих средств могут включать: соли, такие как хлорид натрия и хлорид калия; многоатомные спирты, такие как глицерин и пропиленгликоль; сахара, такие как глюкоза, сахароза и трегалоза; сахарные спирты, такие как ксилит и сорбит; полиэфиры, такие как полиэтиленгликоль; и амидосульфоновые кислоты, такие как таурин. Кроме того, примеры консервантов могут включать хлорид бензалкония, хлорид бензетония, хлорбутанол, параоксибензойный эфир, тимеросал, сорбиновую кислоту, сорбат, хлоргексидина глюконат и т.п. В глазных каплях настоящего изобретения pH при комнатной температуре составляет, предпочтительно, 4,5-8,5, более предпочтительно 5,5-8 и, особенно предпочтительно, 6-8. pH измеряют при комнатной температуре с использованием pH-метра (например, pH-метра/иономера Accumet модель 25, производства Fisher Scientific).
Некоторые офтальмологические лекарственные средства не могут быть введены в виде глазных капель, поскольку они имеют низкую проницаемость через глазной барьер. Поэтому для продления времени контакта и увеличения количества абсорбируемого лекарственного средства может быть использована мазь. Примеры водонерастворимых полимеров в качестве компонента носителя, которые можно использовать в глазных каплях, могут включать этилцеллюлозу, сополимер этилена-винилацетата, полиметилметакрилат, сополимер этилакрилата-метилметакрилата-метакрилата этилхлорида триметиламмония и сополимер метилметакрилата-бутилметакрилата-диметиламиноэтилметакрилата и т.п. Примеры биоразлагаемых полимеров могут включать полимолочную кислоту, сополимер полимолочной кислоты-гликолевой кислоты, полицианоакрилат, полиалкилцианоакрилат, поли-ε-капролактон и т.п. Примеры водорастворимых полимеров могут включать: производные целлюлозы, такие как фталат гидроксипропилметилцеллюлозы, карбоксиметилэтилцеллюлоза и гидроксипропилцеллюлоза; альгинат кальция, хитозан, альбумин, желатин, сополимер метакриловой кислоты-метилметакрилата и т.п. Примеры маслянистых компонентов могут включать трипальмитин, цетиловый спирт, холестерин, различные фосфолипиды, цетилпальмитат, холестеринпальмитат и т.п. Эти компоненты носителя обычно обладают способностью к замедленному высвобождению активного компонента, содержащего активное вещество для офтальмологической терапии. Компонент носителя с высокой относительной плотностью, который может использоваться без добавления какой-либо формы для модификации относительной плотности, может представлять собой, например, фталат гидроксипропилметилцеллюлозы 200731 (относительная плотность: 1,65), фталат гидроксипропилметилцеллюлозы 220824 (относительная плотность: 1,82), карбоксиметилэтилцеллюлозу (относительная плотность: 1,59) и т.п. Тем не менее, даже при использовании такого компонента носителя предпочтительно добавлять форму для модификации относительной плотности для дополнительного увеличения относительной плотности. В настоящем изобретении примеры формы для модификации относительной плотности, используемые для коррекции относительной плотности частиц носителя, могут включать, но без ограничения: нерастворимые компоненты, такие как оксид титана (относительная плотность: 4,17); плохо растворимые компоненты, такие как фосфат трикальция (относительная плотность: 3,14), безводный гидрофосфат кальция (относительная плотность: 2,89) и гидрофосфат кальция дегидрат (относительная плотность: 2,30); водорастворимые компоненты, такие как хлорид натрия (относительная плотность: 2,17), хлорид калия (относительная плотность: 1,98), хлорид кальция (относительная плотность: 2,0), хлорид магния (относительная плотность: 2,41), карбонат натрия (относительная плотность: 2,53), дигидрофосфат натрия (относительная плотность: 1,95), моногидрофосфат натрия (относительная плотность: 1,7) и дигидрофосфат калия (относительная плотность: 2,34).
Вставка обычно схожа с мягкой контактной линзой, помещенной в роговице, за исключением того, что вставку размещают в верхний мешок, а не прикрепляют к открытой роговице, или, реже, в нижний конъюнктивальный мешок. Вставку обычно изготавливают из биологически растворимого материала, который растворяется или разрушается в слезной жидкости при высвобождении лекарственного средства.
Твердые препараты для перорального введения могут включать таблетку, пилюлю, порошок, гранулу, капсулу и т.п. Эти твердые препараты могут быть получены посредством смешивания тирозола с по меньшей мере одним вспомогательным веществом, например крахмалом, карбонатом кальция, сахарозой, лактозой, желатином и т.п. Помимо простых вспомогательных веществ можно использовать смазывающие вещества, такие как стеарат магния и тальк. Жидкие препараты для перорального введения включают суспензию, жидкость для внутреннего применения, масло, сироп и т.п., и могут также включать, помимо к простых разбавителей, таких как вода и жидкий парафин, несколько вспомогательных веществ, например увлажняющее средство, подсластитель, ароматизатор, консервант и т.п.
Кроме того, терапевтическая композиция настоящего изобретения может дополнительно содержать любой физиологически приемлемый носитель, вспомогательное вещество или стабилизатор (Remington: The Science and Practice of Pharmacy, l9th Edition, Alfonso, R., ed, Mack Publishing Co. (Easton, PA: 1995)). Приемлемый носитель, вспомогательное вещество или стабилизатор не токсичны для пользователя в используемой дозе и концентрации, и их примеры включают: буферы, например фосфорную кислоту, лимонную кислоту и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту; полипептиды с низким молекулярным весом (менее приблизительно 10 остатков); белки, например сывороточный альбумин, желатин или иммуноглобулин; гидрофильные полимеры, например поливинилпирролидон; аминокислоты, например, глицин, глутамин, аспарагин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрин; хелатирующие средства, например EDT; сахарные спирты, например маннит или сорбит; солеобразующие противоионы, например натрий; и (или) неионные поверхностно-активные вещества, например Tween, плюроники или полиэтиленгликоль (PEG).
В соответствии с другим аспектом настоящего изобретения предлагается фармацевтическая композиция, причем проникающий в ткань пептид содержит любую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-7.
Проникающий в ткань пептид, представленный аминокислотной последовательностью, выбранной из SEQ ID NO: 1-7, конструируют исходя из анализа аминокислотной последовательности связывающей области лиганда VEGFA, который связывается с доменом b1b2 нейропилина, и длины данной аминокислотной последовательности, и анализа нуклеотидных последовательностей фуриновых C-концевых последовательностей семафорина 3A и семафорина 3F, о которых известно, что они связываются с нейропилином, и, таким образом, последовательности их C-концов оказываются схожими друг с другом.
В соответствии с другим аспектом настоящего изобретения настоящее изобретение предлагает фармацевтическую композицию, в которой средство против фактора роста эндотелия сосудов включает: средство, используемое для предотвращения или лечения заболевания глаз и выбранное из группы, состоящей из бевацизумаба, ранибизумаба, r84 (PLoS One. 2010 Aug 6;5(8)), афлиберцепта, конберцепта, CT01 (WO2005056764A2), DOM15-10-11 (WO2008149147A2), DOM15-26-593 (WO2008149143A2), PRS-050 (Mross et al., 2013. PLoS One), CT-322 (Dineen et al., 2008. BMC Cancer), ESBA903 (Asmus et al., 2015. Eur J Pharm Biopharm.) и EPI-0030(WO2011023130A1); их биоаналогов и их мутантов. Более предпочтительно, средство против фактора роста эндотелия сосудов может представлять собой ранибизумаб, бевацизумаб, афлиберцепт или конберцепт, но без ограничения ими.
Как использовано в настоящем документе, термин "биоаналог" относится к копии медицинского продукта, для которой подтверждена эквивалентность в отношении качества, эффективности и безопасности, имитирующей оригинальный биологический медицинский продукт с истекшим сроком действия патента, который уже был разработан/выведен на рынок с использованием биотехнологии (такой как технология рекомбинации генов и культивирования клеток).
В соответствии с другим аспектом настоящего изобретения предлагается фармацевтическая композиция, в которой мутант может представлять собой мутант, в котором цистеин удален или замещен другим аминокислотным остатком, включая серин, исключая цистеин, в константном домене тяжелой цепи и константном домене легкой цепи.
В примере настоящего изобретения мутантный ранибизумаб, в котором последний остаток цистеина замещен серином в нуклеотидной последовательности ранибизумаба, был сконструирован с помощью метода белковой инженерии с использованием конверсии аминокислотной последовательности ранибизумаба (пример <1-1>).
В соответствии с другим аспектом настоящего изобретения предлагается фармацевтическая композиция, в которой мутант представляет собой мутантный ранибизумаб, состоящий из легкой цепи, представленной SEQ ID NO: 8, и тяжелой цепи, представленной SEQ ID NO: 10.
Проникающий в ткань пептид в соответствии с аспектом настоящего изобретения может дополнительно включать в себя линкерный пептид. Линкерный пептид может состоять из 1-50 аминокислот, предпочтительно 4-20 аминокислот, более предпочтительно 4-15 аминокислот. Кроме того, линкерный пептид может состоять из глицина (G), серина (S) или аланина (A), тогда как последовательность линкерного пептида может представлять собой, предпочтительно, аминокислотную последовательность (GA)n или (GGGGS)m (при условии, что n и m независимо представляют собой целые числа от 1 до 20 и означают количество повторений последовательности в скобках) и, более предпочтительно, аминокислотную последовательность GAGA или (GGGGS)3.
В примере настоящего изобретения конструировали белок слияния, в котором проникающий в ткань пептид (TPP) слит с мутантным ранибизумабом через линкер, и исследовали его эффект. Белок слияния может включать белок слияния (IDB0062), имеющий форму, в которой TPP#2 связан с мутантным ранибизумабом (IDB0061) в соответствии с настоящим изобретением с помощью линкера ((GGGGS)3), и состоящий из аминокислотных последовательностей, представленных SEQ ID NO: 12 и SEQ ID NO: 14; белок слияния (IDB0064), имеющий форму, в которой TPP#5 связан с мутантным ранибизумабом (IDB0061) в соответствии с настоящим изобретением с помощью линкера ((GGGGS)3), и состоящий из аминокислотных последовательностей, представленных SEQ ID NO: 16 и SEQ ID NO: 18; и белок слияния (IDB0072), имеющий форму, в которой TPP#2 связан с тяжелой цепью бевацизумаба с помощью линкера ((GGGGS)3), и состоящий из аминокислотных последовательностей, представленных SEQ ID NO: 20 и SEQ ID NO: 22.
Фармацевтическую композиция настоящего изобретения можно использовать для лечения любого заболевания глаз из-за неоваскуляризации. Как использовано в настоящем документе, выражение "заболевание глаз из-за неоваскуляризации" относится к любому заболеванию глаз из-за сосудистого роста или пролиферации, просачивания из сосудов или связанному с ними заболеванию.
Заболевание глаз из-за неоваскуляризация может быть выбрано из группы, состоящей из пролиферативной витреоретинопатии, макулярной дегенерации, пигментной ретинопатии, диабетической ретинопатии, хориоидальной неоваскуляризации, неоваскулярной глаукомы, ишемической оптической нейропатии, ретинопатии недоношенных, ретинопатии при незрелости, эпидемического конъюнктивита, неоваскулярного заболевания радужной оболочки, ретролентальной фиброплазии, атопического кератита, верхнего лимбального кератита, птеригия, сухого кератита, фликтенулезного кератоконъюнктивита, склерита и диабетического макулярного отека, и, более предпочтительно, его примерами могут быть макулярная дегенерация и диабетический макулярный отек, но без ограничения ими.
В соответствии с другим аспектом настоящего изобретения предлагается способ получения средства против фактора роста эндотелия сосудов (анти-VEGF) с улучшенной эффективностью и способностью преодолевать резистентность, причем данный способ содержит: (a) трансформацию клеток-хозяев с помощью рекомбинантного вектора, причем данный рекомбинантный вектор содержит последовательность нуклеиновой кислоты, кодирующую белок слияния, в котором проникающий в ткань пептид слит со средством против фактора роста эндотелия сосудов (анти-VEGF); (b) культивирование данных клеток; и (c) получение белка слияния из данных клеток.
Последовательность нуклеиновой кислоты, кодирующая белок слияния, может быть выбрана из группы, состоящей из SEQ ID NO: 13, SEQ ID NO: 15, SEQ ID NO: 17, SEQ ID NO: 19, SEQ ID NO: 21 и SEQ ID NO: 23. Точнее говоря, нуклеиновые кислоты, кодирующие легкие и тяжелые цепи модифицированной формы IDB0062 ранибизумаба в соответствии с настоящим изобретением, описаны нуклеотидными последовательностями SEQ ID NO: 13 и SEQ ID NO: 15; нуклеиновые кислоты, кодирующие легкие и тяжелые цепи модифицированной формы IDB0064 ранибизумаба в соответствии с настоящим изобретением, описаны нуклеотидными последовательностями SEQ ID NO: 17 и SEQ ID NO: 19; и нуклеиновые кислоты, кодирующие легкие и тяжелые цепи модифицированной формы IDB0072 бевацизумаба в соответствии с настоящим изобретением, описаны нуклеотидными последовательностями SEQ ID NO: 21 и SEQ ID NO: 23.
Как использовано в настоящем документе, вектор относится к вектору экспрессии, который получают путем введения полинуклеотида настоящего изобретения в вектор с помощью способа, хорошо известного в данной области техники, для экспрессии белка слияния настоящего изобретения с использованием подходящих регуляторных последовательностей транскрипции/трансляции.
Полинуклеотидная последовательность, клонированная в соответствии с настоящим изобретением, может быть функционально связана с подходящей последовательностью контроля экспрессии, причем функционально связанные последовательность гена и последовательность контроля экспрессии могут содержаться в одном векторе экспрессии, содержащем как селективный маркер, так и точку начала репликации. Термин "функционально связанный" означает, что последовательность полинуклеотида (нуклеиновой кислоты) связана таким образом, что последовательность контроля экспрессии допускает экспрессию гена. Термин "последовательность контроля экспрессии" относится к последовательности ДНК, которая управляет экспрессией функционально связанной полинуклеотидной последовательностью в конкретной клетке-хозяине. Такая последовательность контроля экспрессии может включать по меньшей мере одну последовательность, выбранную из группы, состоящей из промотора для осуществления транскрипции, операторной последовательности для управления транскрипцией, последовательности для кодирования подходящего сайта связывания рибосомы на мРНК, последовательности для управления терминацией транскрипции и трансляции и т.п.
Вектор, используемый в качестве родительского вектора для вектора экспрессии, конкретно не ограничен, причем можно использовать любые плазмиду, вирус или другой носитель, которые обычно используют для экспрессии в микроорганизме, используемом в качестве клетки-хозяина в области техники, к которой относится настоящее изобретение. Примеры плазмид могут включать плазмиды, получаемые из Escherichia coli (pBR322, pBR325, pUC118, pUC119 и pET-22b (+)), плазмиды, получаемые из Bacillus subtilis (pUB110 и pTP5), и плазмиды, получаемые из дрожжей (YEp13, YEp24 и YCp50), но без ограничения ими. Примеры вирусов могут включать вирусы животных (такие как ретровирус, аденовирус и вирус осповакцины), вирусы насекомых (такие как акуловирус) и т.п., но без ограничения ими.
Клетки-хозяева могут быть выбраны из клеток, которые могут управлять экспрессией вставленной последовательности или производить целевой продукт из гена определенным предпочтительным образом. Различные клетки-хозяева имеют свои собственные характерные и специфические механизмы трансляции, посттрансляционного процессинга и трансформации белка. Подходящая система клеточной линии или хозяина может быть выбрана из тех, которые обеспечивают предпочтительную трансформацию и процессинг экспрессируемых гетерологичных белков. Экспрессия в дрожжах может давать биологически активные продукты. Экспрессия в эукариотических клетках может увеличивать вероятность "природного" фолдинга.
В качестве клетки-хозяина можно использовать любую клетку-хозяина, известную в данной области техники, при условии, что она способна к непрерывным клонированию и экспрессии при стабилизации вектора настоящего изобретения. Например, E. coli JM109, E. coli BL21DE, E. coli DH5, E. coli RR1, E.coli LE392, E. coli B, E. coli X 1776 и E. coli W3110. Кроме того, в качестве клетки-хозяева можно использовать штаммы Agrobacterium spp. (такой как Agrobacterium A4), штаммы Bacilli spp. (такие как Bacillus subtilis), другие кишечные бактерии, такие как Salmonella typhimurium или Serratia marcescens, и различные штаммы Pseudomonas spp.
Кроме того, в случаях, когда вектор настоящего изобретения трансфицируют в эукариотические клетки, в качестве клеток-хозяев можно использовать дрожжи (Saccharomyces cerevisiae) и клетки насекомых и человеческие клетки (например, клеточную линию CHO (яичника китайского хомячка), клеточные линии W138, BHK, COS-7, 293, HepG2, 3T3, RIN и MDCK).
Можно использовать любой известный способ, с помощью которого вектор доставляют в клетку-хозяина для трансформации клетки-хозяина, но без конкретного ограничения. Например, E. coli может быть трансформирована с помощью теплового шока или электропорации. Если продуцирующую клеточную линию конструируют с использованием клеток животных, клетки могут быть трансфицированы с помощью преципитации фосфатом кальция, метода с ДЭАЭ-декстраном, электропорации, прямой микроинъекции, метода с ДНК-нагруженными липосомами, метода с комплексом липофектамин-ДНК, обработки клеток ультразвуком, генной бомбардировки с использованием высокоскоростных микрочастиц, поликатионного метода и рецептор-опосредованной трансфекции. Некоторые из этих методов могут быть модифицированы для использования in vivo или ex vivo.
Трансгенные клетки культивируют в подходящих условиях, допускающих экспрессию белков слияния, причем эти условия могут быть реализованы в соответствии со способами, хорошо известными специалисту в данной области техники. Трансгенные клетки можно культивировать в больших количествах с помощью обычного способа культивирования. В качестве культуральной среды можно использовать среду, содержащую источники углерода, источники азота, витамины и минералы, одним примером которой является среда 2xYT. Клетки можно культивировать в обычных условиях культивирования клеток. Например, клетки можно культивировать при диапазоне температуры 15-45°C в течение 10-40 часов. Для удаления клеток в культуральной жидкости и сбора только культуральной среды можно проводить центрифугирование или фильтрацию, и такой этап при необходимости может быть проведен специалистом в данной области техники. Культуральную среду (фильтрат) с удаленными клетками охлаждают обычным способом, так что культуральная среда может быть сохранена на короткое время без потери своей активности.
Белки слияния, экспрессируемые в трансгенных клетках (или трансформантах), могут быть очищены обычным способом, и, например, белки слияния настоящего изобретения могут быть очищены с использованием высаливания (например, преципитации сульфатом аммония или преципитации фосфатом натрия), преципитации растворителем (например, преципитации белковой фракции с использованием ацетона, этанола и т.п.), диализа, гель-фильтрации, ионного обмена, колоночной хроматографии (такой как колоночная хроматография с обращенной фазой и аффинная колоночная хроматография) и ультрафильтрации по-отдельности или в комбинации (Maniatis et al, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.(1982); Sambrook et al, Molecular Cloning: A Laboratory Manual, 2d Ed., Cold Spring Harbor Laboratory Press(1989); Deutscher, M., Guide to Protein Purification Methods Enzymology, vol. 182. Academic Press. Inc., San Diego, CA(1990)).
Благоприятные эффекты
Фармацевтическая композиция настоящего изобретения, содержащая в качестве активного ингредиента белок слияния, в котором проникающий в ткань пептид слит со средством против фактора роста эндотелия сосудов (анти-VEGF), увеличивает проникновение средства против фактора роста эндотелия сосудов в ткань и улучшает способность лекарственного средства проходить в хориоидальные ткани во время внутриглазной инъекции, что позволяет ее выпуск в форме глазных капель, а также приводит к лечению пациентов с резистентностью к лекарственному средству, уменьшению дозы и повышению частоты введения.
Краткое описание чертежей
Фиг. 1A представляет собой схему белка слияния, в котором проникающий в ткань пептид слит с антиген-связывающим фрагментом (Fab) против фактора роста эндотелия сосудов. Фиг. 1B представляет собой схему, демонстрирующую связывание белка слияния, в котором проникающий в ткань пептид слит со средством против фактора роста эндотелия сосудов, с VEGF и нейропилиновым рецептором-1. Фиг. 1C представляет собой схему модифицированных форм, в которых проникающий в ткань пептид связан с различными формами ингибиторов фактора роста эндотелия сосудов (белок слияния с Fab, цельным IgG и Fc).
Фиг. 2 показывает результаты ВЭЖХ-анализа основных продуктов производства ранибизумаба и белка слияния настоящего изобретения (IDB0062: белок слияния с Fab, в котором проникающий в ткань пептид TPP#2 слит с мутантным ранибизумабом). AU обозначает относительные единицы.
Фиг. 3 показывает результаты анализа в 12% невосстанавливающем ДСН-ПААГ геле первичных очищенных продуктов белков слияния в соответствии с настоящим изобретением (IDB0062: белок слияния с Fab, в котором проникающий в ткань пептид TPP#2 слит с мутантным ранибизумабом; IDB0064: белок слияния с Fab, в котором проникающий в ткань пептид TPP#5 слит с мутантным ранибизумабом).
Фиг. 4A показывает результаты анализа аффинности связывания VEGF для ранибизумаба, мутантного ранибизумаба и белка слияния в соответствии с настоящим изобретением. Фиг. 4B показывает результаты оценки анализа связывающей способности нейропилинового рецептора-1 для проникающего в ткань пептида и белка слияния в соответствии с настоящим изобретением (IDB0061: мутантный ранибизумаб; IDB0062: белок слияния с Fab, в котором проникающий в ткань пептид TPP#2 слит с мутантным ранибизумабом; Fc-TPP#2: белок слияния с Fc, в котором проникающий в ткань пептид TPP#2 слит с Fc-концом IgG1).
Фиг. 5A показывает результаты оценки стабильности при различных условиях хранения белка слияния в соответствии с настоящим изобретением. Фиг. 5B показывает результаты оценки стабильности при повторном замораживании/оттаивании белка слияния.
Фиг. 6A показывает результаты оценки эффекта ингибирования васкуляризации для ранибизумаба, мутантного ранибизумаба и белка слияния в соответствии с настоящим изобретением в моделях предупреждения неоваскуляризации роговицы. Фиг. 6B представляет собой график, полученный с помощью количественной оценки результатов, показанных на фиг. 6A.
Фиг. 7A показывает результаты оценки эффекта ингибирования васкуляризации для ранибизумаба и белка слияния в соответствии с настоящим изобретением в моделях резистентности к неоваскуляризации роговицы. Фиг. 7B представляет собой график, полученный с помощью количественной оценки результатов, показанных на фиг. 7A. Фиг. 7C показывает результаты оценки эффекта уменьшения покрытия перицитами для ранибизумаба и белка слияния в соответствии с настоящим изобретением.
Фиг. 8A показывает результаты оценки с помощью оптической микроскопии глазного проникновения модифицированной формы IDB0062 ранибизумаба, который представляет собой белок слияния в соответствии с настоящим изобретением. Фиг. 8B представляет собой график, полученный с помощью количественной оценки результатов, показанных на фиг. 8A. Фиг. 8C показывает результаты анализа распределения FITC-конъюгированного белка в срезе глазной ткани после реакции в течение 2 часов с использованием флуоресцентного микроскопа при окрашивании DAPI. Фиг. 8D представляет собой график, полученный с помощью количественной оценки результатов, показанных на фиг. 8C. Фиг. 8E показывает результат анализа распределения IDB0062 в соответствии с его способностью к ретинальному проникновению с помощью анализа фрагмента ткани сетчатки с помощью конфокального микроскопа.
Фиг. 9A показывает результаты оценки способности к проникновению в глаз модифицированной формы IDB0072 бевацизумаба, который представляет собой белок слияния в соответствии с настоящим изобретением, с использованием флуоресцентного микроскопа. Фиг. 9B представляет собой график, полученный с помощью количественной оценки результатов, показанных на фиг. 9A.
Фиг. 10A показывает аспект ингибирования хориоидальной неоваскулярной пролиферации с помощью лекарственного лечения ранибизумабом или белком слияния в соответствии с настоящим изобретением в моделях хориоидальной неоваскуляризации (CNV). Фиг. 10B представляет собой график, полученный с помощью количественной оценки результатов, показанных на фиг. 10A.
Фиг. 11A показывает результаты анализа ингибирования просачивания на концах сосудов с помощью лекарственного лечения ранибизумабом или белком слияния в соответствии с настоящим изобретением в моделях кислород-индуцированной ретинопатии (OIR). Фиг. 11B представляет собой график, полученный с помощью количественной оценки результатов, показанных на фиг. 11A.
Фиг. 12A показывает результаты анализа ингибирования образования неоваскулярных и сосудистых пучков с помощью лекарственного лечения ранибизумабом или белком слияния в соответствии с настоящим изобретением в моделях кислород-индуцированной ретинопатии (OIR) с использованием флуоресцентного микроскопа после окрашивания изолектином B4. Фиг. 12B представляет собой график, полученный с помощью количественной оценки результатов, показанных на фиг. 12A.
Фиг. 13 показывает результаты анализа с помощью вестерн-блота количества лекарственного средства, присутствующего в тканях сетчатки, для различного времени после внутриглазной инъекции IDB0062 и ранибизумаба в крысах.
Способ осуществления изобретения
Далее настоящее изобретение будет описано подробно.
Однако следующие примеры служат только для иллюстрации настоящего изобретения и не предназначены для ограничения объема настоящего изобретения.
<Пример 1>
Конструирование модифицированной формы ранибизумаба, содержащей слитый проникающий в ткань пептид
<1-1> Конструирование мутантного ранибизумаба IDB0061 с увеличенной производительностью
Низкий выход ранибизумаба обусловлен низким выходом фрагментов Fab в культуре и сложным процессом после лечения из-за полиморфизма получаемых с помощью периплазматической экспрессии белков. Для преодоления этой проблемы конкретную аминокислотную последовательность ранибизумаба преобразовали с помощью белковой инженерии (например, с помощью введения точечных мутаций) таким образом, что продуцирующая клеточная линия была оптимизирована для экспорта большинства производимых белков из клеток для повышения уровня экспрессии и для получения белков только однородной формы, тем самым получив мутантный ранибизумаб IDB0061. Соответственно, было подтверждено, что производительность культуры была улучшена и процесс очистки был упрощен, что привело к заметному улучшению выхода продукта.
Точнее говоря, в соответствии с белковой инженерией для конверсии аминокислотной последовательности ранибизумаба был синтезирован ген посредством замены цистеина на серин на конце нуклеотидной последовательности ранибизумаба. После этого проводили тест экспрессии после трансформации в продуцирующую клеточную линию SUPEX5 (KCTC 12657BP).
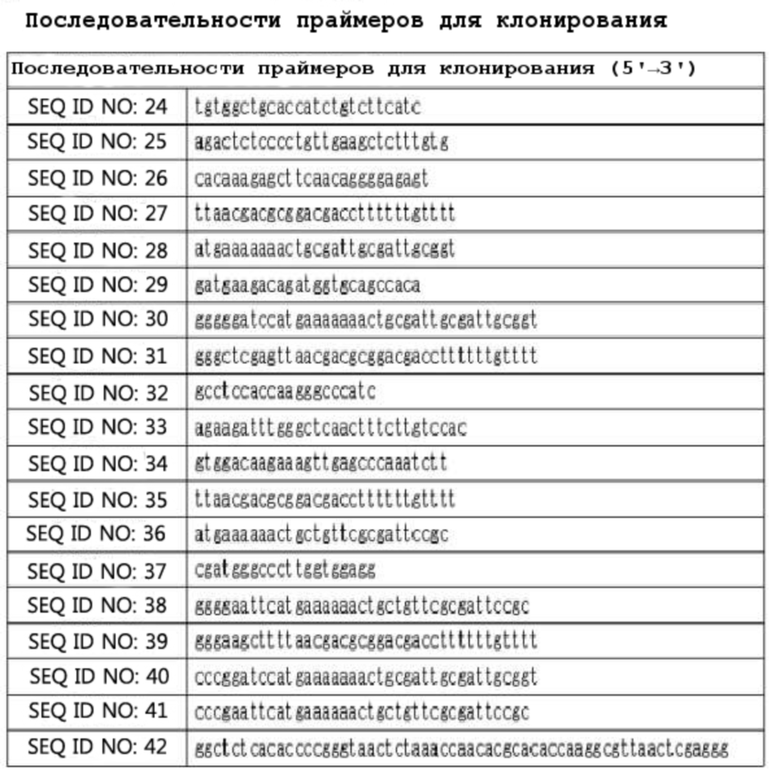
Процедура конструирования области легкой цепи мутантного ранибизумаба IDB0061 была следующей: Последовательность нуклеиновой кислоты ранибизумаба получали из соответствующего патента и подвергали оптимизации кодонов, на ее основе синтезировали нуклеиновую кислоту вариабельной области легкой цепи (VL) Fab, содержащую сигнальную последовательность. Область CL Fab конструировали с помощью ПЦР с использованием вектора с антиальбуминовым Fab SL335 в качестве матрицы и праймеров (SEQ ID NO: 24 и 25). Процедура конструирования области тяжелой цепи мутантного ранибизумаба IDB0061 была следующей: Последовательность нуклеиновой кислоты ранибизумаба получали из соответствующего патента и подвергали оптимизации кодонов, на ее основе синтезировали нуклеиновую кислоту вариабельной области тяжелой цепи (VH) Fab, содержащую сигнальную последовательность (gIII). Область CH1 Fab конструировали с помощью ПЦР с использованием вектора с антиальбуминовым Fab SL335 в качестве праймера и праймеров (SEQ ID NO: 32 и 33).
[Таблица 1]
Аминокислотную последовательность ранибизумаба подвергали точечной мутации с использованием способа, обычно используемого в области белковой инженерии. Точнее говоря, цистеин замещали серином в 214-м аминокислотном остатке аминокислотной последовательности легкой цепи ранибизумаба, а цистеин замещали серином в 226-м аминокислотном остатке аминокислотной последовательности тяжелой цепи ранибизумаба. Аминокислотные последовательности легкой цепи и тяжелой цепи мутантного ранибизумаба IDB0061 с точечными мутациями обозначены SEQ ID NO: 8 и SEQ ID NO: 10, соответственно, тогда как нуклеиновокислотные последовательности, кодирующие эти аминокислотные последовательности, обозначены SEQ ID NO: 9 и SEQ ID NO: 11, соответственно.
Последовательность мутантного ранибизумаба IDB0061, окончательно завершенную с помощью ПЦР, клонировали в вектор pHEKA и снова трансформировали в штамм SUPEX5. Точнее говоря, после добавления 100 мкл компетентных клеток в кювету толщиной 1 мм, к ним добавляли 3 мкл ДНК, а затем выполняли введение генов с помощью электрического шока при 1800 В, завершая тем самым получение продуцирующей клеточной линии.
Клеточные линии, продуцирующие ранибизумаб и IDB0061, соответственно, инокулировали при 1% (об/об) в 50 мл среды 2×YT (100 мМ фосфатно-калиевого буфера, pH 7,2, 50 мкг/мл канамицина) и предварительно культивировали при 220 об/мин в течение 18 часов при 28°C. Затем в колбу с дефлекторами объемом 1 л добавляли 180 мл среды 2×YT, и добавляли к стерилизованной основной культуральной среде 20 мл 1 М фосфата калия (pH 7,2) и 50 мкг/мл канамицина, которые ранее были бактериостатическими, а затем жидкость предварительного культивирования инокулировали до значения OD600 приблизительно 0,15. После этого клетки культивировали при 28°C и 220 об/мин до значения OD600 0,5-0,7, затем добавляли 0,1 мМ IPTG, и затем температуру понижали до 20°C для индуцирования экспрессии. После 21 часа индуцирования экспрессии культуральную жидкость центрифугировали для сбора культурального супернатанта, который затем фильтровали с помощью глубинного фильтра и мембранного стерильного фильтра и загружали на колонку с белком L, тем самым очищая целевые белки.
В результате приведенного в таблице 2 сравнения между мутантным ранибизумабом IDB0061 в соответствии с настоящим изобретением и оригинальным ранибизумабом по производительности было подтверждено, что в одной периодической культуре с использованием базальной среды IDB0061 имеет более высокую производительность, чем ранибизумаб, и посредством одностадийного способа с использованием колонки адсорбции может быть достигнута очистка с чистотой 95% или выше из культуральной жидкости без разрушения клеток, что приводит к упрощению способа.
[Таблица 2]
Сравнение между ранибизумабом, мутантным ранибизумабом и модифицированной формой ранибизумаба в соответствии с настоящим изобретением по производительности
1. Анти-VEGF антитело, патент (PCT/US1998/006604, Genetech)
2. Для ранибизумаба Novartis указана чистота финального очищенного продукта, тогда как для IDB0061 и IDB0062 указана чистота первичного очищенного продукта с помощью колонки с белком L.
<1-2> Слияние мутантного ранибизумаба и проникающего в ткань пептида
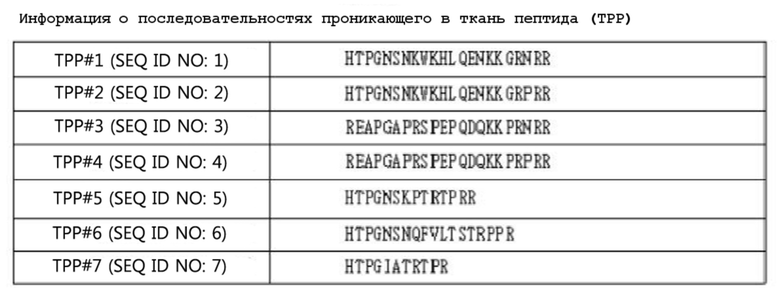
Для улучшения эффективности и преодоления резистентности к ранибизумабу как анти-VEGF средству авторы настоящего изобретения попытались слить проникающий в ткань пептид (TPP), который способен к связыванию как с нейропилином 1 (NRP1), так и с нейропилином 2 или только с нейропилином 1, с C-концом ранибизумаба. Аминокислотные последовательности TPP показаны в таблице 3 ниже. В списках последовательностей в таблице 3 TPP, имеющие аминокислотные последовательности с SEQ ID NO: 1 до 4, могут связываться с обоими нейропилинами 1 и 2, тогда как TPP, имеющие аминокислотные последовательности с SEQ ID NO: 5 до 7, могут специфически связываться с нейропилином 1. Схема белка слияния, в котором TPP слит с ранибизумабом, показана на фиг. 1. Точнее говоря, белки слияния, в которых различные TPP (пептиды, специфически связывающиеся с нейропилиновыми рецепторами), описанные в таблице 3, слиты с C-концом мутантного ранибизумаба IDB0061, сконструированы в примере <1-1>, а клеточные линии, продуцирующие белки слияния, сконструированы APRILBIO.
В то же время, из TPP, показанных в таблице 3, TPP#2, имеющий аминокислотную последовательность SEQ ID NO: 2, получали путем модификации C-концевых областей VEGF165 как собственного лиганда нейропилина и лигандов семафоринов класса 3; а TPP#5 получали путем выделения и идентификации пептида, получаемого из клона, селективно связывающегося с доменом b1 нейропилина 1 при использовании как белка домена b1b2 нейропилина 1, так и белка домена b1b2 нейропилина 2 в качестве конкурентов. В данном случае с ними был слит AVASTIN с использованием линкера для двухвалентного действия в нейропилиновом рецепторе, так что эти пептиды были сконструированы так, чтобы они проникали в ткань, имея при этом схожую аффинность к лигандам VEGF и Sema3A.
[Таблица 3]
Процедура конструирования IDB0062, имеющего форму, в которой пептид TPP#2 связан с мутантным ранибизумабом IDB0061 без связанного TPP (форма, в которой в ранибизумаб введена только точечная мутация) с помощью линкера, была следующей.
На этот раз область легкой цепи мутантного ранибизумаба IDB0062, содержащую слитый TPP, конструировали путем получения последовательности нуклеиновой кислоты ранибизумаба из соответствующего патента и выполнения оптимизации кодонов, и на ее основе синтезировали нуклеиновую кислоту вариабельной области легкой цепи (VL), содержащую сигнальную последовательность. Затем синтезировали нуклеиновую кислоту (CL'-линкер-TPP#2), содержащую линкер и последовательность TPP#2 помимо частичной последовательности, в которой цистеин замещен серином на C-конце константной области легкой цепи (CL). Область CL Fab конструировали с помощью ПЦР с использованием антиальбуминового Fab SL335 в качестве матрицы и праймеров (SEQ ID NO: 24 и 25), а CL'-линкер-TPP#2 конструировали с помощью ПЦР с использованием синтезированной нуклеиновой кислоты CL'-линкер-TPP#2 в качестве матрицы и праймеров (SEQ ID NO: 26 и 27). Эти два продукта связывали посредством связывающей ПЦР для конструирования олигонуклеотида CL-линкер-TPP#2. VL конструировали с использованием синтезированной нуклеиновой кислоты VL в качестве матрицы и праймеров (SEQ ID NO: 28 и 29), и затем проводили ПЦР-сборку с использованием праймеров (SEQ ID NO: 30 и 31, включая последовательности BamHI и XhoI) вместе с CL-линкер-TPP#2 для завершения создания финальной последовательности VL-CL-линкер-TPP#2.
Область тяжелой цепи мутантного ранибизумаба IDB0062, содержащую слитый TPP#2, конструировали путем получения последовательности нуклеиновой кислоты ранибизумаба из соответствующего патента и выполнения оптимизации кодонов, и на ее основе синтезировали нуклеиновую кислоту вариабельной области тяжелой цепи (VH), содержащую сигнальную последовательность (gIII). Затем синтезировали нуклеиновую кислоту (CH1'-линкер-TPP#2), содержащую линкер и последовательность TPP#2 помимо частичной последовательности, в которой цистеин замещен серином на C-конце константной области тяжелой цепи (CH1). Область CH1 Fab конструировали с помощью ПЦР с использованием антиальбуминового Fab SL335 в качестве матрицы и праймеров (SEQ ID NO: 32 и 33), а CH1'-линкер-TPP#2 конструировали с помощью ПЦР с использованием синтезированной нуклеиновой кислоты CH1'-линкер-TPP в качестве матрицы и праймеров (SEQ ID NO: 34 и 35). Эти два продукта связывали посредством связывающей ПЦР для конструирования олигонуклеотида CH1-линкер-TPP#2. VH конструировали с использованием синтезированной нуклеиновой кислоты VH в качестве матрицы и праймеров (SEQ ID NO: 36 и 37), и затем проводили ПЦР-сборку с использованием праймеров (SEQ ID NO: 38 и 39, включая последовательности EcoRI и HindIII) вместе с CL-линкер-TPP#2 для завершения создания финальной последовательности VH-CH1-линкер-TPP#2. Легкую цепь фрагмента, окончательно завершенную посредством ПЦР, расщепляли BamHI и XhoI, а его тяжелую цепь расщепляли EcoRI и HindIII, и клонировали их в вектор pHEKA, расщепленный теми же ферментами рестрикции. Вектор pHEKA, содержащий последовательность IDB0062, который был окончательно создан с помощью описанного выше способа, трансформировали в штамм SUPEX5. Точнее говоря, после добавления 100 мкл компетентных клеток в кювету толщиной 1 мм, к ним добавляли 3 мкл ДНК, а затем выполняли введение генов с помощью электрического шока при 1800 В, завершая тем самым получение продуцирующей клеточной линии.
Аминокислотные последовательности легкой цепи и тяжелой цепи сконструированного IDB0062 обозначены SEQ ID NO: 12 и SEQ ID NO: 13, соответственно, а нуклеиновокислотные последовательности, кодирующие данные аминокислотные последовательности, обозначены SEQ ID NO: 13 и SEQ ID NO: 15, соответственно.
<1-3> Селекция модифицированной формы IDB0062 ранибизумаба
Аффинность к нейропилиновому рецептору и способность к разрушению плотных соединений между эндотелиальными клетками сравнивали для различных белков-кандидатов для модифицированных форм ранибизумаба, сконструированных в примере <1-2>.
Во-первых, проводили аффинность к нейропилиновому рецептору на нейропилине 1 (NRP 1). Точнее говоря, для исследования способности TPP к связыванию с доменом нейропилина 1 проводили поверхностный плазмонный резонанс (SPR) с использованием Biacore 2000 (GE Healthcare). Точнее говоря, каждый домен нейропилина 1 разбавляли в 10 мМ Na-ацетатном буфере (pH 4,0), и фиксировали на сенсорном чипе CM5 (GE Healthcare, США) при приблизительно 1000 единиц ответа (RU). Для анализа использовали буфер HBS-EP (10 мМ HEPES, 2 мМ этилендиаминтетрауксусной кислоты и 0,005% поверхностно-активного вещества P20, pH 7,4, GE Healthcare) при скорости потока 30 мкл/мин, и использовали для анализа концентрации VEGF165 от 80 нМ до 5 нМ, семафорина 3A от 1 мкМ до 62,5 нМ и TPP от 25 мкМ до 1,5625 мкМ. После анализа ассоциации/диссоциации чип CM5 регенерировали, позволяя буферу (20 мМ NaOH, 1 М NaCl, pH 10,0) течь со скоростью потока 30 мкл/мин в течение 1 минуты. Сенсограммы, полученные при ассоциации в течение 3 минут и диссоциации в течение 3 минут, подвергали нормализации и субтракции по сравнению с необработанными клетками для вычисления аффинности.
Кроме того, оценивали способность к разрушению плотных соединений между эндотелиальными клетками путем измерения степени ингибирования белками VE-кадгерина и E-кадгерина. Известно, что уменьшение экспрессии VE-кадгерина и E-кадгерина в эндотелиальных клетках приводит к разрушению плотного соединения между эндотелиальными клетками, и в результате возрастает способность к доставке лекарственного средства в хориоидальную ткань (или способность к проникновению в ткань) при внутриглазном введении лекарственного средства. Точнее говоря, для экспериментального способа косвенного подтверждения увеличения сосудистого проникновения TPP исследовали изменение VE-кадгерина с помощью вестерн-блота. Точнее говоря, для подтверждения увеличения сосудистого проникновения клетки HUVEC высевали с плотностью 3×105 клеток на лунку в 6-луночном планшете, культивировали в течение 24 часов и обрабатывали 1 мкМ TPP в течение 10 минут с последующим вестерн-блотом. Гели, подвергавшиеся ДСН-ПААГ, переносили на мембраны из PVDF, и осуществляли детектирование с использованием первичных антител (SantaCruz), распознающих VE-кадгерин и β-актин, и HRC-конъюгированных вторичных антител (SantaCruz). Анализ проводили с использованием ImageQuant LAS4000 mini (GE Healthcare).
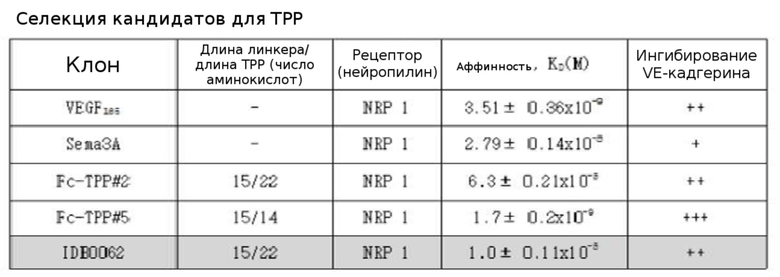
Результаты экспериментов, приведенные в таблице 4, подтвердили, что слитый с TPP#2 Fc (Fc-TPP#2) на высоком уровне связывается с нейропилиновым рецептором (NRP 1) (уровень, аналогичный лиганду Sema3A) и заметно ингибирует VE-кадгерин. Было показано, что слитый с TPP#5 Fc (Fc-TPP#5) обладает более высокой способностью связывания с NRP 1, чем Sema3A, а также оказывает лучший эффект ингибирования на VE-кадгерин. Было подтверждено, что IDB0062 на высоком уровне связывается с нейропилиновым рецептором (NRP 1) и ингибирует VE-кадгерин на уровне, аналогичном Fc-TPP#2.
[Таблица 4]
<1-4> Подтверждение производительности модифицированной формы IDB0062 ранибизумаба
Производительность IDB0062 исследовали с помощью такого же способа, как в примере <1-1>. Как показано в таблице 1, результаты подтверждают, что IDB0062, в котором TPP#2 слит с IDB0061, также демонстрирует приблизительно в 5 или более раз более высокую производительность, чем ранибизумаб.
Кроме того, чистоту первого очищенного продукта, полученного на колонке адсорбции, анализировали с помощью ВЭЖХ. ВЭЖХ анализ проводили следующим образом. Первый очищенный продукт, полученный на колонке адсорбции, концентрировали с помощью Amicon (Millipore, 10K), а затем разбавляли до конечной концентрации 0,5 мг/мл путем обмена с буфером для составления (10 мМ гистидина, 0,1% Tween20, 10% трегалозы). Использовали Waters Alliance e2695 в качестве аналитического прибора, BioSuite 250 UHR SEC (4,6 × 300 мм, 4 мкм, Waters) в качестве колонки и 20 мМ фосфатно-калиевый буфер (250 мМ KCl, pH 6,2) в качестве мобильной фазы. Для анализа вводили 20 мкл концентрированного образца, и анализировали его при скорости потока 0,35 мл/мин в течение 20 минут. Белковые пики анализировали на длине волны УФ 280 нм.
Результаты эксперимента подтвердили, что ранибизумаб является смесью трех основных компонентов, из которых только третий компонент является активным ингредиентом, тогда как IDB0062 был получен в одной форме (фиг. 2).
<1-5> Конструирование модифицированной формы IDB0064 ранибизумаба и подтверждение ее производительности
После модифицированной формы IDB0062 ранибизумаба конструировали модифицированную форму IDB0064 ранибизумаба путем связывания и слияния с ней TPP#5, для которого эффект ингибирования VE-кадгерина был подтвержден в примере <1-3>, с использованием линкера, а затем исследовали ее производительность.
Подробные процедуры конструирования IDB0064 были следующими. Для связывание TPP#5 в качестве нового TPP с IDB0061 осуществляли клонирование генов. Для использования IDB0062 в качестве матрицы и замены TPP на C-конце с TPP#2 на TPP#5 использовали специфические праймеры (SEQ ID NO: 40, 41 и 42). Праймеры были получены путем установки области отжига в начале последовательностей части, общей для TPP#2 и TPP#5, и затем продолжения от нее последовательностей TPP#5, а клонирование осуществляли с использованием данных праймеров. Выполняли 30 циклов ПЦР в порядке денатурация (95°C, 40 сек), отжиг (65°C, 40 сек), удлинение (72°C, 1 мин) для получения генов легкая цепь Fab-TPP#5 и тяжелая цепь Fab-TPP#5. Легкую цепь продукта ПЦР обрабатывали BamHI и XhoI, а тяжелую цепь обрабатывали EcoRI и HindIII с последующим лигированием с вектором pHEKA, который затем трансформировали в продуцирующую клеточную линию SUPEX5.
Аминокислотные последовательности легкой цепи и тяжелой цепи сконструированного IDB0064 обозначены SEQ ID NO: 16 и SEQ ID NO: 18, соответственно, а нуклеиновокислотные последовательности, кодирующие данные аминокислотные последовательности, обозначены SEQ ID NO: 17 и SEQ ID NO: 19, соответственно.
Исследовали производительность сконструированного IDB0064 в продуцирующей клеточной линии. Продуцирующую IDB0064 клеточную линию инокулировали при 1% (об/об) в 50 мл среды 2×YT (100 мМ фосфатно-калиевого буфера, pH 7,2, 50 мкг/мл канамицина), и предварительно культивировали при 220 об/мин в течение 18 часов при 28°C. Затем в колбу с дефлекторами объемом 1 л добавляли 180 мл среды 2×YT, и добавляли к стерилизованной основной культуральной среде 20 мл 1 М фосфата калия (pH 6,4) и 50 мкг/мл канамицина, которые ранее были бактериостатическими, а затем жидкость предварительного культивирования инокулировали до значения OD600 приблизительно 0,15. После этого клетки культивировали при 28°C и 220 об/мин до значения OD600 0,5-0,7, затем добавляли 0,05 мМ IPTG, и затем температуру понижали до 20°C для индуцирования экспрессии. После 21 часа индуцирования экспрессии культуральную жидкость центрифугировали для сбора культурального супернатанта, который затем фильтровали с помощью глубинного фильтра и мембранного стерильного фильтра и загружали на колонку с белком L, тем самым очищая целевые белки. Как показано в таблице 2 и на фиг. 3, результаты очистки подтвердили, что даже связь последовательности TPP#5, сконструированной как искусственная нуклеотидная последовательность, с Fab IDB0061 положительно индуцировала экспрессию, так что целевой белок с относительно высокой чистотой мог быть получен с производительностью приблизительно 3-4 мг/л посредством только первичной очистки.
<Пример 2>
Подтверждение двухвалентных характеристик модифицированной формы IDB0062 ранибизумаба
Исследовали способность модифицированной формы IDB0062 ранибизумаба к связыванию с VEGF-A и рецептором нейропилина 1.
<2-1> Анализ с помощью SPR (Biacore2000) способности к связыванию с рецептором нейропилина 1
Для исследования способности IDB0062 и IDB0072 к связыванию с NRP1 выполняли анализ с помощью SPR. После активации чипа Biacore CM5 с помощью смеси EDC/NHS целевой белок NRP1 разбавляли в фиксирующем буфере (10 мМ ацетата натрия, pH 5,5), и фиксировали до финальных 79 Ru с помощью расчета при Rmax:200. Затем перед анализом образцы IDB0062 и IDB0072 разбавляли вплоть до от 12,5 нМ до 400 нМ в буфере HBSEP. Скорость потока при анализе составляла 30 мкл/мин, и в результате на основе итоговых графиков были получены сенсограммы для вычисления значений Kd.
Как показано в таблице 4, было подтверждено, что способность IDB0062 к связыванию с рецептором нейропилина 1 сохраняется на аналогичном уровне с активностью связывания контрольного препарата (Fc-TPP#2). Эти результаты указывают на то, что пептид TPP#2, слитый с C-концом ранибизумаба, успешно связывается с рецептором нейропилина 1.
<2-2> ELISA-анализ способности к связыванию с рецептором нейропилина 1
NRP1 (собственного производства) разбавляли в карбонатном покрывающем буфере (0,1 М NaHCO3, pH 9,6) до конечной концентрации 10 мкг/мл, и разбавленный NRP1 добавляли в планшет для ELISA по 100 мкл на лунку (SPL, Immunoplate Maxi binding) с последующим покрытием при 37°C в течение 2 часов. Затем планшет промывали три раза, блокировали (4% обезжиренного молока, pH 7,4) при 37°C в течение 1 часа и промывали три раза. Затем каждый образец разбавляли в подходящей кратности и использовали для обработки по 100 мкл на лунку с последующей реакцией при 37°C в течение 1 часа. После завершения реакции образцов планшет промывали три раза, а затем козью античеловеческую легкую каппа-цепь Ab-HRP (Sigma Aldrich, A7164) разбавляли в 5000 раз в блокирующем буфере и использовали для обработки по 100 мкл на лунку с последующей реакцией при 37°C в течение 1 часа. Планшет промывали пять раз и обрабатывали субстратом ™B (Bethyl, E102) по 100 мкл на лунку с последующей реакцией в течение 2-3 минут. После этого реакцию останавливали останавливающим раствором (1 н HCl) по 100 мкл на лунку, и измеряли поглощение на 450 нм с использованием планшет-ридера для ELISA.
ELISA-анализ также подтвердил, что способность IDB0062 к связыванию с рецептором нейропилина 1 эквивалентна связывающей способности контрольного лекарственного средства (Fc-TPP#2) (фиг. 4B). Таким образом, было снова подтверждено, что характеристики пептида TPP#2 сохраняются при слиянии с C-концом ранибизумаба.
<2-3> ELISA-анализ способности связывания с VEGF
VEGF (R&D system, 293-VE-500/CF) разбавляли в карбонатном покрывающем буфере (0,1 М NaHCO3, pH 9,6) до конечной концентрации 3 мкг/мл, и разбавленный VEGF добавляли в планшет для ELISA по 100 мкл на лунку (SPL, Immunoplate Maxi binding) с последующим покрытием при 37°C в течение 2 часов. Затем планшет промывали три раза, блокировали (4% обезжиренного молока, pH 7,4) при 37°C в течение 1 часа и промывали три раза, а затем каждый образец разбавляли в подходящей кратности и использовали для обработки по 100 мкл на лунку, а затем оставляли при 37°C на 1 час. После завершения реакции образцов планшет промывали три раза, а затем козье антитело против человеческой легкой каппа-цепи Ab-HRP (Sigma Aldrich, A7164) разбавляли в 5000 раз в блокирующем буфере и использовали для обработки по 100 мкл на лунку, и затем оставляли при 37°C на 30 минут. Планшет промывали семь раз и обрабатывали субстратом ™B (Bethyl, E102) по 100 мкл на лунку с последующей реакцией в течение 2-3 минут. После этого реакцию останавливали останавливающим раствором (1 н HCl) по 100 мкл на лунку, и измеряли поглощение на 450 нм с использованием планшет-ридера для ELISA.
Как показано на фиг. 4A, результаты ELISA-анализа IDB0062 подтвердили, что способность IDB0062 к связыванию с VEGF-A несколько уменьшилась по сравнению с ранибизумабом. Предполагаемой причиной является то, что связывание области определения комплементарности (CDR) с VEGF-A частично изменилось при изменении третичной структуры IDB0062 из-за изменения определенной аминокислоты в последовательности. Однако, поскольку связывающая способность ранибизумаба значительно выше, чем у обычных антител, можно считать, что такое уменьшение связывающей способности не приведет к значительному снижению его клинической эффективности.
<Пример 3>
Оценка стабильности модифицированной формы IDB0062 ранибизумаба
Было подтверждено, что изменение определенной аминокислоты в последовательность IDB0062 вызывает его структурную модификацию, и поэтому способность IDB0062 к связыванию с VEGF-A несколько уменьшается по сравнению с ранибизумабом. Поэтому, для того чтобы исследовать, как это изменение влияет на стабильность IDB0062, анализировали его стабильность в зависимости от условий хранения и повторения замораживания/оттаивания.
Первый очищенный продукт после замены буфера на буфер для составления концентрировали до концентрации (5 мг/мл), используемой в эксперименте на животных, и затем проводили эксперимент по определению стабильности. Для эксперимента по определению стабильности в зависимости от условий хранения образец дозировали на аликвоты по 10 мкл, и во время хранения при 4°C и -80°C в течение 5 недель образец еженедельно вынимали, оттаивали на льду и анализировали с помощью ELISA-анализа связывания VEGF. Во время повторенной пять раз процедуры, в которой образец замораживали при -80°C и оттаивали на льду, часть образца отбирали и оценивали в отношении стабильности при повторном замораживании/оттаивании с помощью ELISA-анализа связывания VEGF.
В результате, как показано на фиг. 5, способность IDB0062 к связыванию с VEGF-A стабильно сохранялась при 4°C и -80°C после пяти недель хранения, и даже физическое воздействие пятикратного повторного замораживания/оттаивания не повлияло на его активность, и, таким образом, было определено, что отсутствует ухудшение его стабильности из-за изменения аминокислотной последовательности.
<Пример 4>
Оценка проникновения модифицированной формы анти-VEGF средства в ткань глаза
Для анализа глазного проникновения модифицированной формы ранибизумаба ex vivo конструировали модель глазного проникновения, и оценивали его эффективность. Кроме того, для того чтобы оценить, улучшилось ли проникновение в ткань благодаря слиянию проникающего в ткань пептида с антителом с большим молекулярным весом и более сложной третичной структурой (включая белок слияния с Fc), в то же время анализировали глазное проникновение бевацизумаба и его модифицированной формы IDB0072.
<4-1> Оценка проникновения в ткань модифицированной формы IDB0062 ранибизумаба
Для того чтобы исследовать, может ли TPP, слитый с C-концом IDB0062, фактически улучшать проникновение в ткань посредством связывания с нейропилиновым рецептором, широко распространенным в глазных эндотелиальных клетках; извлеченные глазные яблоки погружали в растворы FITC-конъюгированного ранибизумаба и IDB0062 для сравнения степени их проникновения с течением времени.
Для того чтобы увеличить эффективность конъюгирования белка и FITC, образец белка доводили до концентрации 1 мг/мл посредством обмена с 100 мМ натрий-карбонатным буфером (pH 9,0) и смешивали с FITC (1 мг/мл в DMSO) с последующей реакцией при комнатной температуре в течение 2 часов. Поскольку пептид TPP#2 содержит много свободных аминогрупп, способных к связыванию FITC, для реакции использовали различные количества молей FITC по отношению к белку. После этого, когда неконъюгировавший FITC удаляли с использованием обессоливающей колонки PD-10, буфер заменяли на ФСБ, а затем количественно оценивали очищенный продукт, и использовали его в качестве образца для оценки эффективности. Результаты конъюгирования подтвердили, что отношения F/P ранибизумаба и IDB0062 составляли 1,127 и 1,133, соответственно, что указывает на то, что с одной молекулой белка связывается почти одинаковое число молей FITC. После разбавления растворов FITC-конъюгированного ранибизумаба и IDB0062 с помощью ФСБ до 0,3 мг/мл глазные яблоки мышей C57BL/6 извлекали, погружали в них при 37°C на 1 час и 2 часа и промывали ФСБ два раза, каждый раз в течение 10 минут, а затем получали парафиновые стекла. Глазные яблоки фиксировали при 4°C в течение 4 часов с использованием раствора Дэвидсона (ледяная уксусная кислота: этиловый спирт: нейтрализованный формалин: дистиллированная вода=1:3:2:3), затем переднюю часть ткани конъюнктивы частично надрезали ножницами, и минимизировали ретинальную метаплазию на этапе получения парафиновых стекол. После фиксации в течение ночи при 4°C с использованием 10% (об/об) раствора формалина влагу из тканей удаляли, повышая концентрацию спирта от низкой до высокой концентрации с использованием машины для автоматической инфильтрации, с последующим просветлением ксилолом, а затем пропитыванием парафином. После завершения пропитывания парафином ткань помещали в форму для получения парафинового блока, и получали 4 среза с использованием микротома. Парафиновые срезы разгибали для прикрепления к стеклам, предварительно покрытым альбумином, поли-L-лизиномом и солевым раствором в плавающей водяной бане с постоянной температурой. Стекла с тканью депарафинизировали и затем монтировали непосредственно, и затем наблюдали FITC с использованием конфокального микроскопа для исследования глазного проникновения в ткань и распределения.
Как показано на фиг. 8A и 8B, было подтверждено, что IDB0062 быстро начинает проникать в глазное яблоко через роговицу и заднюю часть глазного яблока от 1 часа после эксперимента, и в пределах 2 часов количественное распределение в глазной ткани и стекловидном теле значительно возрастает приблизительно в четыре раза по сравнению с ранибизумабом как контрольным лекарственным средством. Поскольку глазная ткань может быть четко отделена и проанализирована посредством окрашивания DAPI, было подтверждено, что, в отличие от ранибизумаба, IDB0062 сильно распространен даже внутри роговицы (фиг. 8C, белая стрелка), и распределение лекарственного средства IDB0062 в глазу возрастает приблизительно в 10 раз по сравнению с ранибизумабом (фиг. 8D). Поскольку данные фиг. 8E показывают поперечный разрез ткани сетчатки глаза, было подтверждено, что контрольное лекарственное средство ранибизумаб по большей части присутствует около склеры, тогда как IDB0062 достигает сетчатки через слой пигментных эндотелиальных клеток сетчатки (RPE). В результате, связь проникающего в ткань пептида с ранибизумабом, который обладает плохим относительным внутриглазным проникновением в ткань, может повышать относительное глазное проникновение ранибизумаба приблизительно в 4 раза. Было подтверждено, что такая связь приводит к возможности контроля дозы лекарственного средства, увеличения интервала введения лекарственного средства и выпуска в виде глазных капель путем улучшения лекарственной формы из-за улучшения проникновения лекарственного средства.
<4-2> Оценка проникновения в ткань модифицированной формы IDB0072 бевацизумаба
Цельные белки антител, такие как бевацизумаб, имеют более высокие молекулярные массы и сложные третичные структуры по сравнению с фрагментами антител (Fab) и, таким образом, имеют неблагоприятные характеристики с точки зрения проникновения в ткань. Несмотря на это, для того, чтобы исследовать, преодолевает ли слияние проникающего в ткань пептида с C-концом бевацизумаба эти физические ограничения и улучшает его проникновение по сравнению с контрольным лекарственным средством бевацизумаб, проводили такой же эксперимент.
Процедуры конструирования модифицированной формы IDB0072 бевацизумаба были следующими. IDB0072 является белком слияния антитела такого типа, в котором TPP слит с C-концом бевацизумаба с помощью линкера, причем аминокислотная последовательность TPP показана в таблице 3 выше. В списках последовательностей в таблице 3 TPP, имеющие аминокислотные последовательности с SEQ ID NO: 1 до 4, могут связываться с обоими нейропилинами 1 и 2, а TPP, имеющие аминокислотные последовательности с SEQ ID NO: 5 до 7, могут специфически связываться с нейропилином 1. В то же время, из TPP, показанных в таблице 3, TPP#2 получали путем модификации C-концевых областей VEGF165 как собственного лиганда нейропилина и лиганда семафоринов класса 3; а TPP#5 получали путем выделения и идентификации пептида, получаемого из клона, селективно связывающегося с доменом b1b2 нейропилина 1. Из этих TPP TPP#2 сливали с бевацизумабом с использованием линкера для двухвалентного действия на нейропилиновый рецептор, так что IDB0072 был сконструирован так, чтобы он проникал в ткань, имея схожую аффинность к VEGF и Sema3A.
Точнее говоря, конструировали клеточную линию, продуцирующую IDB0072, в которой TPP #2, имеющий аминокислотную последовательность SEQ ID NO: 2, сливали с C-концом бевацизумаба. IDB0072 клонировали в вектор pcDNA3.4. Аминокислотные последовательности легкой цепи и тяжелой цепи сконструированного IDB0072 обозначены SEQ ID NO: 20 и SEQ ID NO: 22, соответственно, а нуклеиновокислотные последовательности, кодирующие данные аминокислотные последовательности, обозначены SEQ ID NO: 21 и SEQ ID NO: 23, соответственно. Плазмиду, кодирующую белок, в котором связывающий NRP1 пептид был слит с константной областью тяжелой цепи сконструированного антитела, и плазмиду, кодирующую белок легкой цепи, трансфицировали в клетки CHO DG44 с использованием электрофореза Neon™, и затем клетки высевали по 3×106 клеток в колбу T25 и культивировали при 37°C. Стабильные клетки соединяли с селективным маркером и затем культивировали в плавающем состоянии в течение 7 дней в условиях 100 об/мин, 37°C, pH 7,2, 50% DO2 с использованием бессывороточного SFM4CHO (Hyclone) в биореакторе. Супернатант отделяли от клеток центрифугированием и стерилизовали с помощью фильтра 0,22.
Культуральную жидкость IDB0072 собирали, и каждый белок очищали в соответствии со стандартным протоколом. Культуральную жидкость наносили на колонку с белком A (смола MabselectSure, GE healthcare) с последующим промыванием ФСБ (pH 7,4). Антитело элюировали при pH 3,0 с использованием 0,1 М глицинового буфера, и затем образец нейтрализовали до pH 7,0 с использованием 1 М Tris-буфера. Элюированные фракции антител концентрировали с использованием центробежного концентратора MILLIPORE Amicon Ultra (30 MWCO) с последующим обменом с буфером ФСБ (pH 7,4). Очищенный белок слияния, в котором пептид, специфически связывающийся с выбранным NRP1, слит с константной областью тяжелой цепи антитела, количественно оценивали с использованием коэффициента поглощения и абсорбции на скорректированной длине волны 280 нм.
Для повышения эффективности конъюгирования белка и FITC, бевацизумаб и IDB0072 доводили до концентрации 3 мг/мл после обмена с 100 мМ натрий-карбонатным буфером (pH 9,0) и смешивали с FITC (1 мг/мл в DMSO) с последующей реакцией при комнатной температуре в течение 2 часов. Для уменьшения различия в конъюгировании из-за слияния TPP#2 для реакции применяли другое число молей FITC по отношению к белку для IDB0072, чем для контрольного лекарственного средства бевацизумаб. После этого, когда неконъюгировавший FITC удаляли с использованием обессоливающей колонки PD-10, буфер заменяли на ФСБ, а затем количественно оценивали очищенный продукт, и использовали его в качестве образца для оценки эффективности. Результаты конъюгирования подтвердили, что отношения F/P бевацизумаба и IDB0072 составляли 2,18 и 1,98, соответственно, что указывает на то, что с одной молекулой белка связывается почти одинаковое число молей FITC. После разбавления растворов FITC-конъюгированного бевацизумаба и IDB0072 с помощью ФСБ до 0,9 мг/мл глазные яблоки мышей C57BL/6 извлекали, погружали в них при 37°C на 1 час и 2 часа и промывали ФСБ пять раз в течение 10 минут каждый раз,а затем получали парафиновые стекла. Последующие процедуры проводили также, как в примере <4-1>.
Для оценки активности плучаемой модифицированной формы IDB0072 бевацизумаба исследовали способность IDB0072 к связыванию с рецептором нейропилина 1 с помощью анализа SPR. Как показано в таблице 5, было подтверждено, что способность IDB0062 к связыванию с рецептором нейропилина 1 демонстрирует тот же уровень, что и контрольное лекарственное средство (Fc-TPP#2) и модифицированная форма IDB0062 ранибизумаба. Эти результаты указывают на то, что пептид TPP#2, слитый с C-концом бевацизумаба, успешно связывается с рецептором нейропилина 1.
[Таблица 5]
Оценка активности слитой с TPP формы по сравнению с NRP1
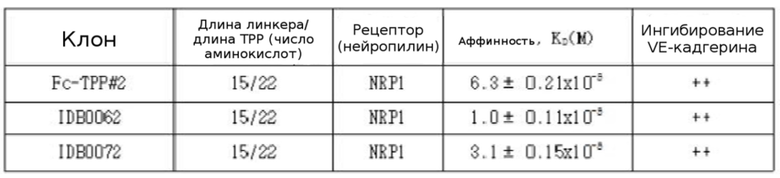
Как показано на фиг. 9A, было подтверждено, что IDB0072 также улучшает глазное проникновение, по сравнению с контрольным лекарственным средством бевацизумаб. Было подтверждено, что внутриглазное распределение IDB0072 значительно возрастает, превосходя приблизительно в 1,5 раза распределение бевацизумаба через 1 час и 2 часа (фиг. 9B). Было также подтверждено, что несмотря на свою большую молекулярную массу и структуру IDB0072 диффундирует в роговицу через конъюнктиву лучше бевацизумаба. Таким образом, было подтверждено, что даже для антитела, имеющего высокую молекулярную массу и сложную структуру, проникновение в ткань может значительно увеличиваться благодаря связыванию проникающего в ткань пептид с C-концом Fc. Эти результаты указывают на то, что глазное проникновение в ткань различных белков слияния с Fc, включая белки антител, можно также улучшать с помощью связывания с проникающим в ткань пептидом.
<Пример 5>
Оценка эффективности модифицированной формы ранибизумаба с использованием моделей неоваскуляризации роговицы
Для оценки эффективности IDB0062 конструировали модели неоваскуляризации роговицы. После разделения на модели предупреждения и модели резистентности их использовали для сравнения IDB0062 и ранибизумаба по ингибированию васкуляризации и снижению количества новых сосудов.
<5-1> Модели предупреждения
Для сравнения IDB0062 и ранибизумаба по эффекту ингибирования неоваскуляризации модели неоваскуляризации роговицы, индуцированной ожогом щелочью, конструировали следующим образом: Из целлюлозной фильтровальной бумаги вырезали круг с диаметром 2 мм, и затем погружали его в 1 М раствор NaOH. Использовали самок мышей C57BL/6 возрастом 6 недель. После анестезии (золетил 40 мг/кг+ромпун 5 мг/кг, и/п) фильтровальную бумагу с NaOH помещали на 30 секунд на роговицу левого глаза для индуцирования ожога щелочью с последующим достаточным промыванием 40 мл ФСБ, конструируя таким образом модели неоваскуляризации роговицы на животном. В отношении моделей предупреждения лекарственное лечение осуществляли в тот же день, в который индуцировали ожог щелочью. Каждое лекарственное средство вводили в глазных каплях в концентрации 5 мг/мл каждый раз по 5 мкл, четыре раза в день в течение 5 дней. После завершения введения глазные яблоки извлекали, погружали в 4% раствор параформальдегида и фиксировали в течение одного часа и промывали ФСБ, а затем отделяли роговицу с использованием препаровального микроскопа. Отделенную роговицу дополнительно фиксировали в 4% растворе параформальдегида в течение 12 часов. Фиксированную роговицу промывали ФСБ и подвергали реакции в блокирующем буфере (ФСБ, 0,3% БСА, 0,1% Triton X100) при комнатной температуре в течение 2 часов. Первичное антитело (BD Pharmingen), специфичное к PECAM-1 (CD31) в качестве сосудистого эндотелиального маркера, и первичное антитело (Millipore), специфичное к NG-2 в качестве маркера перицитов, реагировали в течение ночи в холодильнике, а вторичные флуоресцентные антитела (Alexa Fluor488, Alexa Fluor594, Life Technologies) реагировали при комнатной температуре в течение 4 часов для окрашивания ткани. После завершения процедур окрашивания роговицу переносили на стекло и монтировали на стекле, причем делали четыре линейных надреза в четырех направлениях к центру роговицы с использованием препаровального микроскопа. После завершения процедур монтирования на стеклах с роговицей наблюдали вид сосудов и перицитов с помощью флуоресцентной микроскопии/конфокальной микроскопии.
Как показано на фиг. 6, результаты эксперимента на моделях предупреждения подтвердили, что IDB0062 дополнительно ингибирует неоваскуляризацию на 25-30% по сравнению с ранибизумабом, демонстрируя значительный профилактический эффект, составляющий 50% или более по сравнению с носителем, причем IDB0061 также продемонстрировал эквивалентную эффективность по сравнению с ранибизумабом, что указывает на то, что уменьшение способности к связыванию VEGF из-за структурных изменений ранибизумаба по существу не влияет на его эффективность при ингибировании неоваскуляризации, и что улучшение его эффективности может быть достигнуто с помощью TPP, слитого с его C-концом.
<5-2> Модели резистентности
Модели резистентности конструировали путем индуцирования ожога щелочью таким же образом, как и в моделях предупреждения, и оставляли на 10 дней, так что достаточно проступала васкуляризация роговицы. Концентрация и цикл введения были такими же, как и в моделях предупреждения. Введение осуществляли в течение 10 дней от 10 дней после ожога щелочью, и сравнивали IDB0062 и ранибизумаб по степени уменьшения генерации новых сосудов (последующее получение корнеальных стекол было таким же, как и в моделях предупреждения).
Как показано на фиг. 7A, в группах обработки носителем и ранибизумабом сосуды развивались вплоть до роговицы, тогда как в группе обработки IDB0062 сосуды были локально распространены только в глазном лимбе, что говорит об уменьшении количества новых сосудов в группе обработки IDB0062. В результате, IDB0062 продемонстрировал значительную активность по уменьшению количества новых сосудов, составляющую 30% или более по сравнению с носителем, и продемонстрировал улучшение эффективности в два или более раза по сравнению с ранибизумабом, который не продемонстрировал значительной эффективности (фиг. 7B). Эти результаты аналогичны результатам в литературе, полученным при совместном введении анти-VEGF аптамера и средства, представляющего собой анти-PDGF антитело (Jo et al., 2006. Am. J. Pathol.). Таким образом, ожидается, что IDB0062, вероятно, будет индуцировать улучшение зрения у приблизительно 70% пациентов, которым вводят ранибизумаб, и у которых зрение сохраняется без улучшения зрения.
Кроме того, как показано на фиг. 7C, было подтверждено, что группа обработки IDB0062 показывает значительное уменьшение покрытия перицитами почти на 40% по сравнению с группой обработки носителем, и эти результаты аналогичны результатам для ингибитора PDGF, который разрабатывают в качестве средства для совместного введения с целью лечения пациентов, имеющих резистентность к существующим лекарственных средствам. Таким образом, предполагается, что IDB0062 может быть применим в качестве монотерапии для лечения резистентности, которая, как известно, имеет место у 45% пациентов, которым вводят анти-VEGF ингибитор.
<Пример 6>
Оценка эффективности модифицированной формы ранибизумаба с использованием моделей хориоидальной неоваскуляризации
Хориоидальную неоваскуляризацию (CNV), которая соответствует модели возрастной макулярной дегенерации, индуцировали с помощью индуцированной лазером хориоидальной неоваскуляризации, и исследовали терапевтический эффект модифицированной формы IDB0062 ранибизумаба.
Самцов крыс Brown Norway (BN) возрастом семь недель (SLC Japan, Токио, Япоия) акклиматизировали в течение одной недели, а затем анестезировали путем перитонеальной инъекции пентобарбитала натрия (Hanlim Pharm, 25 мг/кг). После этого зрачок расширяли с помощью глазных капель с 1% тропикамидом, и затем формировали шесть фотокоагуляционных пятен вокруг головки зрительного нерва с использованием диодного лазера (длина волны 532 нм; диаметр 100 мкм; мощность 150 мВт; период времени 0,1 сек). Разрушение мембраны Бруха подтверждали по образованию характерных пузырьков. Глазные яблоки с кровотечением или без пузырьков исключали из последующих экспериментальных процедур. После фотокоагуляционной обработки крыс случайным образом разделяли по пяти группам (по 10 крыс на группу), как показано в таблице 6. Лекарственные средства вводили интраокулярно в концентрациях, подходящих для соответствующих групп, с использованием шприца Hamilton (Hamilton, США). Такое же количество носителя вводили группе CNV, и в качестве контрольного лекарственного средства вводили 100 мкг ранибизумаба на глаз. Субъектов с хирургическим повреждением, вызванным введением лекарственного средства, или субъектов с помутнением хрусталика и т.п. исключали из последующих экспериментальных процедур.
[Таблица 6]
Экспериментальные группы для модели хориоидальной неоваскуляризации
Степень хориоидальной неоваскуляризации оценивали через 10 дней после введения лекарственного средства. Глазное яблоко каждой крысы извлекали, а затем рассекали в области, примыкающей к роговице и склере, под микроскопом. После этого сетчатку снимали с помощью тканей задней части глазного яблока, а затем отделяли ткань конъюнктивы, содержащую субретинальную область. Отделенную ткань фиксировали в 4% параформальдегиде в течение 1 часа, промывали ФСБ и затем перемешивали в ФСБ, содержащем 5% Triton X-100 и 1% БСА, в течение 3 часов. После повторного промывания изолектин B4 (Sigma) в качестве маркера эндотелиальных клеток, растворенный в количестве 1 мг/мл в ФСБ, разбавляли 1:50 с последующей реакцией при 4°C в течение ночи. После промывания ФСБ, содержащим 0,05% Tween 20, в течение 2 часов, стрептавидин-TRITC разбавляли 1:500 с последующей реакцией при 37°C в течение 4 часов, и затем после промывания ФСБ в течение 30 минут ткань наблюдали под флуоресцентным микроскопом (BX51, Olympus, Япония). Размеры субретинальных неоваскулярных областей анализировали с использованием программного обеспечения Image J (NIH, США).
Как показано на фиг. 10A, было обнаружено, что в группе носителя было много новых сосудов, отмеченных черным цветом, вытянутых с внутренней стороны поврежденной лазером области (хориоидеа) к наружней (сетчатка). В группе, которой вводили по 100 мкг контрольного лекарственного средства ранибизумаб, неоваскуляризация имела тенденцию к некоторому ингибированию по сравнению с группой носителя, но у некоторых субъектов все же наблюдались вытянутые сосуды. При этом было подтверждено, что IDB0062 имеет тенденцию к ингибированию неоваскуляризации в зависимости от концентрации. Было подтверждено, что группа, которой вводили по 50 мкг IDB0062, обладает эффективностью ингибирования неоваскуляризации на уровне, аналогичном группе, которой вводили по 100 мкг контрольного лекарственного средства ранибизумаб. В группе, которой вводили по 100 мкг IDB0062, на сетчатке оставались только индуцированные лазером рубцы, а неоваскуляризация в основном была ингибирована. Как показано на фиг. 10B, полученной количественной обработкой приведенных выше результатов, было подтверждено, что васкуляризация была ингибирована приблизительно на 31% по сравнению с группой носителя в группе, которой вводили по 100 мкг ранибизумаба, и в группе, которой вводили по 50 мкг IDB0062, и неоваскуляризация была ингибирована приблизительно на 36% в группе, которой вводили по 100 мкг IDB0062. В результате, было подтверждено, что IDB0062 проявляет такую же эффективность ингибирования неоваскуляризации, как и ранибизумаб, при использовании половины от дозы ранибизумаба.
<Пример 7>
Оценка эффективности модифицированной формы ранибизумаба с использованием модели кислород-индуцированной ретинопатии
Для моделей ретинопатии недоношенных индуцировали кислород-индуцированную ретинопатию (OIR), и исследовали эффект модифицированной формы IDB0062 ранибизумаба.
В качестве экспериментальных животных использовали выведенных скрещиванием мышей C57BL/6 возрастом 7-8 недель, приобретенных у Koatech. На день 7 после рождения мышей (постнатальный день 7, P7) помещали в кислородную камеру, и концентрацию кислорода в камере доводили до 75% и поддерживали такой (гипероксия) в течение 5 дней (P7-P11). Освещение в лаборатории включали и выключали каждые 12 часов, и поддерживали температуру 24±2°C. Мышей кормили со свободным доступом к пище и питьевой воде. Через 5 дней мышей подвергали воздействию внутреннего воздуха (нормоксия) вне камеры в течение 5 дней (P12-P17) для индуцирования ретинального ангиогенеза. Для введения лекарственного средства мышей случайным образом разделяли на пять групп (по 10 мышей в группе) непосредственно после воздействия нормоксии на P12, как показано в таблице 7. Лекарственные средства вводили интраокулярно в концентрациях, подходящий для соответствующих групп, с использованием шприца Hamilton (Hamilton, США). Такое же количество носителя вводили группе OIR, и в качестве контрольного лекарственного средства вводили 10 мкг ранибизумаба на глаз. Субъектов с хирургическим повреждением, вызванным введением лекарственного средства, или субъектов с помутнением хрусталика и т.п. исключали из последующих экспериментальных процедур.
[Таблица 7]
Экспериментальные группы для модели кислород-индуцированной ретинопатии (OIR)
Степень ретинального отека оценивали на день 17 после рождения. Мышей анестезировали с помощью перитонеальной инъекции пентобарбитала натрия (Hanlim Pharm, 25 мг/кг). После того, как открывали брюшную полость, в сердце инъецировали 10 мкл декстрана (FD40S-1G, Sigma; 50 мг/мл в ФСБ). Через 10 минут глазные яблоки извлекали и фиксировали в 4% параформальдегиде в течение 10 минут, а затем отделяли сетчатку. После этого получали стекла с монтированной на плоскости сетчаткой, и наблюдали их под флуоресцентным микроскопом (BX51, Olympus, Япония). Количество флуоресценции, выходящей из сосудов, количественно анализировали с использованием Image J.
Как показано на фиг. 11A, было обнаружено, что в группе носителя, не получавшей лекарственное средство, экстравазация флуоресценции из-за просачивания увеличивалась в центральной части области диска зрительного нерва и в области окончаний сосудов снаружи сетчатки. В группе, получавшей по 10 мкг контрольного лекарственного средства ранибизумаб, просачивание из сосудов по сравнению с группой носителя было несколько снижено, но экстравазация флуоресценции из-за просачивания из сосудов сохранялась на некотором уровне в области окончаний сосудов. При этом было обнаружено, что группа, получавшая IDB0062, демонстрировала тенденцию к ингибированию просачивания из сосудов в зависимости от концентрации, и группы, которым вводили IDB0062 (по меньшей мере по 10 мкг, что соответствует такой же дозе ранибизумаба), обладали, хотя и не статистически значимо, возможностью более эффективного ингибирования просачивания из сосудов по сравнению с ранибизумабом (фиг. 11B). В частности, в группе, которой вводили по 15 мкг IDB0062, форма сосудов имеет плотную и стабильную структуру, и область просачивания вовсе не показана.
Кроме того, также оценивали степень неоваскуляризации в сетчатке. Отделенную сетчатку фиксировали в 4% параформальдегиде в течение 3 часов, промывали ФСБ, и затем перемешивали в ФСБ, содержащем 5% Triton X-100 и 1% БСА, в течение 3 часов. После повторного промывания изолектин B4(L2140, sigma), растворенный в количестве 1 мг/мл в ФСБ, разбавляли 1:50 с последующей реакцией при 4°C в течение ночи. После промывания ФСБ, содержащим 0,05% Tween 20, в течение 2 часов стрептавидин-TRITC разбавляли 1:500 с последующей реакцией при 37°C в течение 4 часов, и затем после промывания ФСБ в течение 30 минут ткань наблюдали под флуоресцентным микроскопом (BX51, Olympus, Япония).
Когда мыши P7 были в условиях гипероксии в течение 5 дней, а затем были перенесены в нормоксию, мыши P7 оказывались в состоянии относительной ишемии. При этом в сетчатке росли лишние новые сосуды, и особенностью новых образующихся сосудов оказалось то, что сосуды росли из сетчатки в направлении стекловидного тела, что соответствует аваскулярной области, и образовывалась специфическая структура, называемая пучками, имеющая форму неупорядоченных сосудов, которая была легко подвержена просачиванию. Образование таких пучков можно использовать в качестве меры количественной оценки неоваскуляризации. Фиг. 12A подтверждает, что в окрашенных сосудах в группе носителя границы между сосудами четко различаются одна от другой, и сосуды проявляют характеристики нормальных сосудов сетчатки с относительно тонкой и стабильной структурой, но при этом наблюдают множество новых сосудов, которые образуют пучки, в которые собраны неупорядоченные и толстые сосуды. Было подтверждено, что, как и в группе носителя, множество новых сосудов в форме пучков были распространены в группе, получавшей по 10 мкг контрольного лекарственного средства ранибизумаб. В то время как IDB0062 значительно уменьшал распределение пучков, определяемых как новые сосуды, в зависимости от концентрации, и в группах, в которых вводили по 10 мкг или более, большая часть сосудистых структур четко отличались одна от другой, и наблюдались плотные и твердые нормальные формы сосудов, а не форма пучка. График, полученный с помощью количественной оценки результатов, подтверждает, что все группы из группы носителя, группы, получавшей по 10 мкг ранибизумаба, и группы, которой вводили по 5 мкг IDB0062, демонстрировали одинаковую степень неоваскуляризации, но в группе, которой вводили по 10 мкг IDB0062, неоваскуляризация была ингибирована приблизительно на 40% по сравнению с вышеупомянутыми группами (фиг. 12B). В результате было подтверждено, что в моделях OIR лечение с помощью ранибизумаба не демонстрировало эффективности ингибирования ретинальной неоваскуляризации, тогда как IDB0062 мог значительно ингибировать индуцированную OIR неоваскуляризацию в той же дозе, что и ранибизумаб.
<Пример 8>
Анализ распределения лекарственного средства после внутриглазной инъекции
Для исследования распределения лекарственного средства в ткани при интраокулярном инъецировании ранибизумаба и IDB0062, соответственно, белки из ткани сетчатки экстрагировали белки, и выполняли анализ с помощью вестерн-блота.
Извлекали глаз у каждой крысы, и затем отделяли сетчатку через 4 часа и 72 часов после внутриглазной инъекции. Отделенную сетчатку промывали три раза ФСБ, затем переносили в буфер для лизиса (20 мМ Tris-Cl, 150 мМ NaCl, 1 мМ ЭДТА, 0,1% triton X-100), и разрушали ткани с использованием гомогенизатора. После этого собирали супернатант с помощью центрифугирования (14000 об/мин, 4°C, 10 мин), и содержание белка в лизате измеряли с помощью анализа Бредфорда. Для анализа с помощью вестерн-блота использовали 12% невосстанавливающий гель ДСН-ПААГ. Образец белка загружали по 40 мкг на лунку и прогоняли при 25 мА в течение 1 часа. После переноса на мембрану из PVDF белки детектировали с использованием антитела HRP против каппа (Sigma, F3761), и каждую полосу белка количественно оценивали с использованием системы Bio-Rad Chemidoc.
Поскольку тяжелая цепь и легкая цепь ранибизумаба связаны дисульфидной связью, в невосстанавливающем ДСН-ПААГ наблюдали одну полосу при приблизительно 48 кДа. При этом IDB0062 не имеет дисульфидной связи между тяжелой цепью и легкой цепью, и поэтому в анализе с помощью ДСН-ПААГ наблюдали две полосы (тяжелой цепи и легкой цепи) при 28 кДа. Как показано на фиг. 13, было подтверждено, что количество лекарственного средства, присутствующего в сетчатке через 4 часов после внутриглазной инъекции для IDB0062 оказалось приблизительно в 3,6 раз больше, чем для ранибизумаба. Эти результаты указывают на то, что IDB0062 специфически связывается с NRP1-экспрессирующей тканью посредством TPP, демонстрируя, что IDB0062 может воздействовать на ткань сетчатки, которая является участком заболевания для различных заболеваний, вызываемых ретинальной неоваскуляризацией. При внутриглазной инъекции лекарственного средства лекарственное средство, связывающееся с поврежденной тканью, а не лекарственное средство, распределенное в стекловидном теле, имеет повышенную возможность для удаления VEGF, так что лекарственное средство, вероятно, будет проявлять такую же эффективность в меньшей дозе. Можно считать, что эти результаты подтверждают высокую эффективность IDB0062 при оценке эффективности с использованием моделей хориоидальной неоваскуляризации и моделей ретинопатии.
Промышленная применимость
По сравнению с существующими средствами против фактора роста эндотелия сосудов фармацевтические композиции в соответствии с настоящим изобретением, содержащие в качестве активного ингредиента белок слияния, в котором проникающий в ткань пептид слит со средством против фактора роста эндотелия сосудов (анти-VEGF), могут ингибировать различные факторы роста, связанные с неоваскуляризацией, а также эндотелиальные факторы роста, уменьшать покрытие перицитами для улучшения эффективности и лечить даже резистентных к лекарственному средству пациентов. Кроме того, высокая промышленная применимость фармацевтических композиций настоящего изобретения заключается в том, что улучшена способность к доставке лекарственного средства в хориоидальную ткань при внутриглазной инъекции, так что лекарственное средство с большой вероятностью может быть выпущено в виде глазных капель благодаря снижению дозы, повышению частоты введения и улучшению глазного проникновения.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ILDONG PHARM CO.,LTD.
<120> Фармацевтическая композиция для предотвращения и лечения
заболеваний глаз, содержащая в качестве активного ингредиента
белок слияния, в котором слиты проникающий в ткань пептид и
препарат против фактора роста эндотелия сосудов
<130> OP17-0118/PCT/RU
<150> KR 10-2015-0045684
<151> 2015-03-31
<160> 42
<170> KopatentIn 2.0
<210> 1
<211> 22
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Проникающий в ткань пептид (TPP) #1
<400> 1
His Thr Pro Gly Asn Ser Asn Lys Trp Lys His Leu Gln Glu Asn Lys
1 5 10 15
Lys Gly Arg Asn Arg Arg
20
<210> 2
<211> 22
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Проникающий в ткань пептид (TPP) #2
<400> 2
His Thr Pro Gly Asn Ser Asn Lys Trp Lys His Leu Gln Glu Asn Lys
1 5 10 15
Lys Gly Arg Pro Arg Arg
20
<210> 3
<211> 22
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Проникающий в ткань пептид (TPP) #3
<400> 3
Arg Glu Ala Pro Gly Ala Pro Arg Ser Pro Glu Pro Gln Asp Gln Lys
1 5 10 15
Lys Pro Arg Asn Arg Arg
20
<210> 4
<211> 22
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Проникающий в ткань пептид (TPP) #4
<400> 4
Arg Glu Ala Pro Gly Ala Pro Arg Ser Pro Glu Pro Gln Asp Gln Lys
1 5 10 15
Lys Pro Arg Pro Arg Arg
20
<210> 5
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Проникающий в ткань пептид (TPP) #5
<400> 5
His Thr Pro Gly Asn Ser Lys Pro Thr Arg Thr Pro Arg Arg
1 5 10
<210> 6
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Проникающий в ткань пептид (TPP) #6
<400> 6
His Thr Pro Gly Asn Ser Asn Gln Phe Val Leu Thr Ser Thr Arg Pro
1 5 10 15
Pro Arg
<210> 7
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Проникающий в ткань пептид (TPP) #7
<400> 7
His Thr Pro Gly Ile Ala Thr Arg Thr Pro Arg
1 5 10
<210> 8
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь мутантного ранибизумаба (IDB0061)
<400> 8
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Val Leu Ile
35 40 45
Tyr Phe Thr Ser Ser Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Thr Val Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Ser
210
<210> 9
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь мутантного ранибизумаба (IDB0061)
<400> 9
gatatccagt tgacgcagtc tccaagctcc ctgtctgcct ctgtggggga tcgtgtgacc 60
atcacctgca gcgccagcca ggatattagc aactatttaa actggtacca gcagaaacct 120
ggcaaagctc ccaaagttct catctatttc acctcctctc tgcactctgg tgtcccatct 180
cgcttcagtg gcagtggttc tgggacagac ttcactctca ctatcagcag cctgcagcct 240
gaagattttg caacttacta ttgtcaacag tatagcaccg tgccgtggac gttcggccaa 300
gggaccaagg tggaaatcaa acgtactgtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagttcgc ccgtcacaaa gagcttcaac aggggagagt ct 642
<210> 10
<211> 231
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь мутантного ранибизумаба (IDB0061)
<400> 10
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Asp Phe Thr His Tyr
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Trp Ile Asn Thr Tyr Thr Gly Glu Pro Thr Tyr Ala Ala Asp Phe
50 55 60
Lys Arg Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Tyr Pro Tyr Tyr Tyr Gly Thr Ser His Trp Tyr Phe Asp Val
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Glu Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Ser Asp Lys Thr His Leu
225 230
<210> 11
<211> 693
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь мутантного ранибизумаба (IDB0061)
<400> 11
gaagtgcagc tggtggagtc tgggggaggc ctggtccagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggata tgatttcacc cattatggta tgaattgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtcggctgg attaacacct ataccggtga accgacctac 180
gcagcggatt tcaagcgtcg attcaccttt tccctggaca ccagcaagtc aacggcgtat 240
ctacaaatga acagcctgag agccgaggac acggccgtct attactgtgc gaaatacccg 300
tattactatg gtaccagcca ctggtatttc gacgtctggg gccaaggaac cctggtcacc 360
gtctcctcag cctccaccaa gggcccatcg gtcttccccc tggcaccctc ctccaagagc 420
acctctgagg gcacagcggc cctgggctgc ctggtcaagg actacttccc cgaaccggtg 480
acggtgtcgt ggaactcagg cgccctgacc agcggcgtgc acaccttccc ggctgtccta 540
cagtcctcag gactctactc cctcagcagc gtggtgaccg tgccctccag cagcttgggc 600
acccagacct acatctgcaa cgtgaatcac aagcccagca acaccaaggt ggacaagaaa 660
gttgagccca aatcttctga caaaacccac ctg 693
<210> 12
<211> 251
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь белка слияния (IDB0062)
<400> 12
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Val Leu Ile
35 40 45
Tyr Phe Thr Ser Ser Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Thr Val Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
210 215 220
Gly Gly Gly Gly Ser His Thr Pro Gly Asn Ser Asn Lys Trp Lys His
225 230 235 240
Leu Gln Glu Asn Lys Lys Gly Arg Pro Arg Arg
245 250
<210> 13
<211> 753
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь белка слияния (IDB0062)
<400> 13
gatatccagt tgacgcagtc tccaagctcc ctgtctgcct ctgtggggga tcgtgtgacc 60
atcacctgca gcgccagcca ggatattagc aactatttaa actggtacca gcagaaacct 120
ggcaaagctc ccaaagttct catctatttc acctcctctc tgcactctgg tgtcccatct 180
cgcttcagtg gcagtggttc tgggacagac ttcactctca ctatcagcag cctgcagcct 240
gaagattttg caacttacta ttgtcaacag tatagcaccg tgccgtggac gttcggccaa 300
gggaccaagg tggaaatcaa acgtactgtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagttcgc ccgtcacaaa gagcttcaac aggggagagt ctggtggcgg tggctctggt 660
ggcggtggca gcggtggcgg tggctctcac accccgggta actctaacaa atggaaacac 720
ctgcaggaaa acaaaaaagg tcgtccgcgt cgt 753
<210> 14
<211> 268
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь белка слияния (IDB0062)
<400> 14
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Asp Phe Thr His Tyr
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Trp Ile Asn Thr Tyr Thr Gly Glu Pro Thr Tyr Ala Ala Asp Phe
50 55 60
Lys Arg Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Tyr Pro Tyr Tyr Tyr Gly Thr Ser His Trp Tyr Phe Asp Val
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Glu Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Ser Asp Lys Thr His Leu Gly Gly Gly Gly Ser Gly Gly Gly Gly
225 230 235 240
Ser Gly Gly Gly Gly Ser His Thr Pro Gly Asn Ser Asn Lys Trp Lys
245 250 255
His Leu Gln Glu Asn Lys Lys Gly Arg Pro Arg Arg
260 265
<210> 15
<211> 804
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь белка слияния (IDB0062)
<400> 15
gaagtgcagc tggtggagtc tgggggaggc ctggtccagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggata tgatttcacc cattatggta tgaattgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtcggctgg attaacacct ataccggtga accgacctac 180
gcagcggatt tcaagcgtcg attcaccttt tccctggaca ccagcaagtc aacggcgtat 240
ctacaaatga acagcctgag agccgaggac acggccgtct attactgtgc gaaatacccg 300
tattactatg gtaccagcca ctggtatttc gacgtctggg gccaaggaac cctggtcacc 360
gtctcctcag cctccaccaa gggcccatcg gtcttccccc tggcaccctc ctccaagagc 420
acctctgagg gcacagcggc cctgggctgc ctggtcaagg actacttccc cgaaccggtg 480
acggtgtcgt ggaactcagg cgccctgacc agcggcgtgc acaccttccc ggctgtccta 540
cagtcctcag gactctactc cctcagcagc gtggtgaccg tgccctccag cagcttgggc 600
acccagacct acatctgcaa cgtgaatcac aagcccagca acaccaaggt ggacaagaaa 660
gttgagccca aatcttctga caaaacccac ctgggtggcg gtggctctgg tggcggtggc 720
agcggtggcg gtggctctca caccccgggt aactctaaca aatggaaaca cctgcaggaa 780
aacaaaaaag gtcgtccgcg tcgt 804
<210> 16
<211> 243
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь белка слияния (IDB0064)
<400> 16
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Gln Asp Ile Ser Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Val Leu Ile
35 40 45
Tyr Phe Thr Ser Ser Leu His Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Thr Val Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
210 215 220
Gly Gly Gly Gly Ser His Thr Pro Gly Asn Ser Lys Pro Thr Arg Thr
225 230 235 240
Pro Arg Arg
<210> 17
<211> 729
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь белка слияния (IDB0064)
<400> 17
gatatccagt tgacgcagtc tccaagctcc ctgtctgcct ctgtggggga tcgtgtgacc 60
atcacctgca gcgccagcca ggatattagc aactatttaa actggtacca gcagaaacct 120
ggcaaagctc ccaaagttct catctatttc acctcctctc tgcactctgg tgtcccatct 180
cgcttcagtg gcagtggttc tgggacagac ttcactctca ctatcagcag cctgcagcct 240
gaagattttg caacttacta ttgtcaacag tatagcaccg tgccgtggac gttcggccaa 300
gggaccaagg tggaaatcaa acgtactgtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagttcgc ccgtcacaaa gagcttcaac aggggagagt ctggtggcgg tggctctggt 660
ggcggtggca gcggtggcgg tggctctcat actcctggaa atagcaaacc aacacgcaca 720
ccaaggcgt 729
<210> 18
<211> 260
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь белка слияния (IDB0064)
<400> 18
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Asp Phe Thr His Tyr
20 25 30
Gly Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Trp Ile Asn Thr Tyr Thr Gly Glu Pro Thr Tyr Ala Ala Asp Phe
50 55 60
Lys Arg Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Tyr Pro Tyr Tyr Tyr Gly Thr Ser His Trp Tyr Phe Asp Val
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Glu Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Ser Asp Lys Thr His Leu Gly Gly Gly Gly Ser Gly Gly Gly Gly
225 230 235 240
Ser Gly Gly Gly Gly Ser His Thr Pro Gly Asn Ser Lys Pro Thr Arg
245 250 255
Thr Pro Arg Arg
260
<210> 19
<211> 780
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь белка слияния (IDB0064)
<400> 19
gaagtgcagc tggtggagtc tgggggaggc ctggtccagc ctggggggtc cctgagactc 60
tcctgtgcag cctctggata tgatttcacc cattatggta tgaattgggt ccgccaggct 120
ccagggaagg ggctggagtg ggtcggctgg attaacacct ataccggtga accgacctac 180
gcagcggatt tcaagcgtcg attcaccttt tccctggaca ccagcaagtc aacggcgtat 240
ctacaaatga acagcctgag agccgaggac acggccgtct attactgtgc gaaatacccg 300
tattactatg gtaccagcca ctggtatttc gacgtctggg gccaaggaac cctggtcacc 360
gtctcctcag cctccaccaa gggcccatcg gtcttccccc tggcaccctc ctccaagagc 420
acctctgagg gcacagcggc cctgggctgc ctggtcaagg actacttccc cgaaccggtg 480
acggtgtcgt ggaactcagg cgccctgacc agcggcgtgc acaccttccc ggctgtccta 540
cagtcctcag gactctactc cctcagcagc gtggtgaccg tgccctccag cagcttgggc 600
acccagacct acatctgcaa cgtgaatcac aagcccagca acaccaaggt ggacaagaaa 660
gttgagccca aatcttctga caaaacccac ctgggtggcg gtggctctgg tggcggtggc 720
agcggtggcg gtggctctca tactcctgga aatagcaaac caacacgcac accaaggcgt 780
780
<210> 20
<211> 233
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь белка слияния (IDB0072)
<400> 20
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala
20 25 30
Ser Val Gly Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Gln Asp Ile
35 40 45
Ser Asn Tyr Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys
50 55 60
Val Leu Ile Tyr Phe Thr Ser Ser Leu His Ser Gly Val Pro Ser Arg
65 70 75 80
Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
85 90 95
Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Ser Thr
100 105 110
Val Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr
115 120 125
Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu
130 135 140
Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro
145 150 155 160
Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly
165 170 175
Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr
180 185 190
Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His
195 200 205
Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val
210 215 220
Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230
<210> 21
<211> 699
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Легкая цепь белка слияния (IDB0072)
<400> 21
atgggctggt cctgcatcat cctgttcctg gtggccaccg ccaccggcgt gcactccgac 60
atccagatga cccagtcccc ctcctccctg tccgcctccg tgggcgaccg ggtgaccatc 120
acctgctccg cctcccagga catctccaac tacctgaact ggtaccagca gaagcccggc 180
aaggccccca aggtgctgat ctacttcacc tcctccctgc actccggcgt gccctcccgg 240
ttctccggct ccggctccgg caccgacttc accctgacca tctcctccct gcagcccgag 300
gacttcgcca cctactactg ccagcagtac tccaccgtgc cctggacctt cggccagggc 360
accaaggtgg agatcaagcg gaccgtggcc gccccctccg tgttcatctt ccccccctcc 420
gacgagcagc tgaagtccgg caccgcctcc gtggtgtgcc tgctgaacaa cttctacccc 480
cgggaggcca aggtgcagtg gaaggtggac aacgccctgc agtccggcaa ctcccaggag 540
tccgtgaccg agcaggactc caaggactcc acctactccc tgtcctccac cctgaccctg 600
tccaaggccg actacgagaa gcacaaggtg tacgcctgcg aggtgaccca ccagggcctg 660
tcctcccccg tgaccaagtc cttcaaccgg ggcgagtgc 699
<210> 22
<211> 508
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь белка слияния (IDB0072)
<400> 22
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Asn Tyr Gly Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Gly Trp Ile Asn Thr Tyr Thr Gly Glu Pro Thr Tyr Ala
65 70 75 80
Ala Asp Phe Lys Arg Arg Phe Thr Phe Ser Leu Asp Thr Ser Lys Ser
85 90 95
Thr Ala Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Lys Tyr Pro His Tyr Tyr Gly Ser Ser His Trp Tyr
115 120 125
Phe Asp Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser
130 135 140
Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr
145 150 155 160
Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro
165 170 175
Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val
180 185 190
His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser
195 200 205
Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile
210 215 220
Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val
225 230 235 240
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
245 250 255
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
260 265 270
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
275 280 285
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
290 295 300
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
305 310 315 320
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
325 330 335
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
340 345 350
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
355 360 365
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
370 375 380
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
385 390 395 400
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
405 410 415
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
420 425 430
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
435 440 445
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
450 455 460
Ser Leu Ser Leu Ser Pro Gly Lys Gly Gly Gly Gly Ser Gly Gly Gly
465 470 475 480
Gly Ser Gly Gly Gly Gly Ser His Thr Pro Gly Asn Ser Asn Gln His
485 490 495
Ala His Gln Gly Val Glu Tyr Pro Glu Lys Asp Gln
500 505
<210> 23
<211> 1524
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь белка слияния (IDB0072)
<400> 23
atgggctggt cctgcatcat cctgttcctg gtggccaccg ccaccggcgt gcactccgag 60
gtgcagctgg tggagtccgg cggcggcctg gtgcagcccg gcggctccct gcggctgtcc 120
tgcgccgcct ccggctacac cttcaccaac tacggcatga actgggtgcg gcaggccccc 180
ggcaagggcc tggagtgggt gggctggatc aacacctaca ccggcgagcc cacctacgcc 240
gccgacttca agcggcggtt caccttctcc ctggacacct ccaagtccac cgcctacctg 300
cagatgaact ccctgcgggc cgaggacacc gccgtgtact actgcgccaa gtacccccac 360
tactacggct cctcccactg gtacttcgac gtgtggggcc agggcaccct ggtgaccgtg 420
tcctccgcct ccaccaaggg cccatcggtc ttccccctgg caccctcctc caagagcacc 480
tctgggggca cagcggccct gggctgcctg gtcaaggact acttccccga accggtgacg 540
gtgtcgtgga actcaggcgc cctgaccagc ggcgtgcaca ccttcccggc tgtcctacag 600
tcctcaggac tctactccct cagcagcgtg gtgaccgtgc cctccagcag cttgggcacc 660
cagacctaca tctgcaacgt gaatcacaag cccagcaaca ccaaggtgga caagaaagtt 720
gagcccaaat cttgtgacaa aactcacaca tgcccaccgt gcccagcacc tgaactcctg 780
gggggaccgt cagtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 840
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 900
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 960
tacaacagca cgtaccgtgt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 1020
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc 1080
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 1140
gatgagctga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 1200
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 1260
cccgtgctgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 1320
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 1380
tacacgcaga agagcctctc cctgtccccg ggtaaaggtg gaggaggatc tggaggagga 1440
ggaagtggag gtggaggatc acatactcct ggaaatagca accaacacgc acaccaaggc 1500
gttgaatatc ccgagaagga tcaa 1524
<210> 24
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер для клонирования #1
<400> 24
tgtggctgca ccatctgtct tcatc 25
<210> 25
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер для клонирования #2
<400> 25
agactctccc ctgttgaagc tctttgtg 28
<210> 26
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер для клонирования #3
<400> 26
cacaaagagc ttcaacaggg gagagt 26
<210> 27
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер для клонирования #4
<400> 27
ttaacgacgc ggacgacctt ttttgtttt 29
<210> 28
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер для клонирования #5
<400> 28
atgaaaaaaa ctgcgattgc gattgcggt 29
<210> 29
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер для клонирования #6
<400> 29
gatgaagaca gatggtgcag ccaca 25
<210> 30
<211> 38
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер для клонирования #7
<400> 30
gggggatcca tgaaaaaaac tgcgattgcg attgcggt 38
<210> 31
<211> 38
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер для клонирования #8
<400> 31
gggctcgagt taacgacgcg gacgaccttt tttgtttt 38
<210> 32
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер для клонирования #9
<400> 32
gcctccacca agggcccatc 20
<210> 33
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер для клонирования #10
<400> 33
agaagatttg ggctcaactt tcttgtccac 30
<210> 34
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер для клонирования #11
<400> 34
gtggacaaga aagttgagcc caaatctt 28
<210> 35
<211> 29
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер для клонирования #12
<400> 35
ttaacgacgc ggacgacctt ttttgtttt 29
<210> 36
<211> 28
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер для клонирования #13
<400> 36
atgaaaaaac tgctgttcgc gattccgc 28
<210> 37
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер для клонирования #14
<400> 37
cgatgggccc ttggtggagg 20
<210> 38
<211> 37
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер для клонирования #15
<400> 38
ggggaattca tgaaaaaact gctgttcgcg attccgc 37
<210> 39
<211> 38
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер для клонирования #16
<400> 39
gggaagcttt taacgacgcg gacgaccttt tttgtttt 38
<210> 40
<211> 38
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер для клонирования #17
<400> 40
cccggatcca tgaaaaaaac tgcgattgcg attgcggt 38
<210> 41
<211> 37
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер для клонирования #18
<400> 41
cccgaattca tgaaaaaact gctgttcgcg attccgc 37
<210> 42
<211> 59
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Праймер для клонирования #19
<400> 42
ggctctcaca ccccgggtaa ctctaaacca acacgcacac caaggcgtta actcgaggg 59
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ В КАЧЕСТВЕ АКТИВНОГО ИНГРЕДИЕНТА СЛИТЫЙ БЕЛОК, В КОТОРОМ СЛИТЫ ПРОНИКАЮЩИЙ В ОПУХОЛЬ ПЕПТИД И АНТИАНГИОГЕННОЕ СРЕДСТВО, ДЛЯ ПРЕДУПРЕЖДЕНИЯ И ЛЕЧЕНИЯ РАКА ИЛИ СВЯЗАННЫХ С АНГИОГЕНЕЗОМ ЗАБОЛЕВАНИЙ | 2016 |
|
RU2727238C2 |
| Fab ФРАГМЕНТ ГУМАНИЗИРОВАННОГО АНТИТЕЛА ПРОТИВ VEGF И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2802960C2 |
| БЕЛКОВЫЕ КОМПОЗИЦИИ ПРОТИВ VEGF И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2020 |
|
RU2795973C1 |
| АНТИТЕЛА С ПРИВИТЫМ МОЗГОВЫМ НАТРИЙУРЕТИЧЕСКИМ ПЕПТИДОМ | 2019 |
|
RU2839970C2 |
| АНТИТЕЛА С ПРИВИТЫМ ПРЕДСЕРДНЫМ НАТРИЙУРЕТИЧЕСКИМ ПЕПТИДОМ | 2019 |
|
RU2822433C2 |
| БЕЛКОВЫЕ КОМПОЗИЦИИ ПРОТИВ VEGF И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2020 |
|
RU2785994C1 |
| БЕЛКОВЫЕ КОМПОЗИЦИИ ПРОТИВ VEGF И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2020 |
|
RU2778325C1 |
| БЕЛКОВЫЕ КОМПОЗИЦИИ ПРОТИВ VEGF И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2020 |
|
RU2788949C1 |
| АНТИ-СЕАСАМ6 АНТИТЕЛА И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2739163C2 |
| ПОЛИФУНКЦИОНАЛЬНЫЕ БЕЛКОВЫЕ МОЛЕКУЛЫ, СОДЕРЖАЩИЕ ДЕКОРИН, И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2823357C2 |

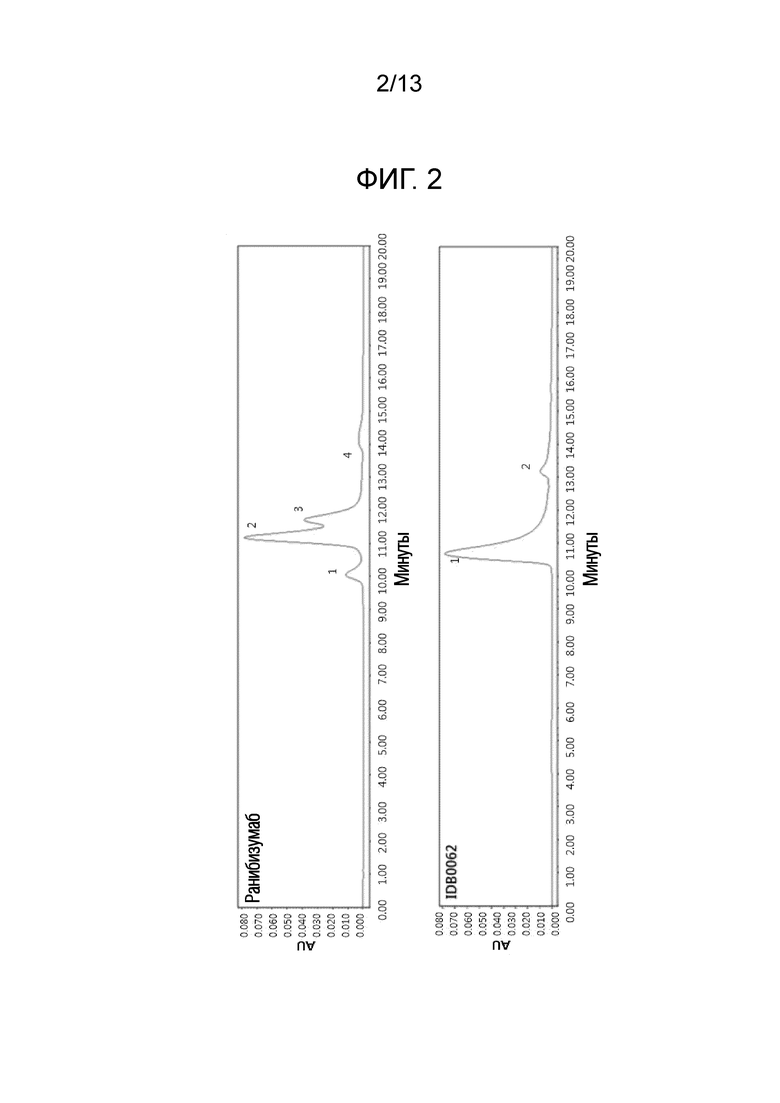
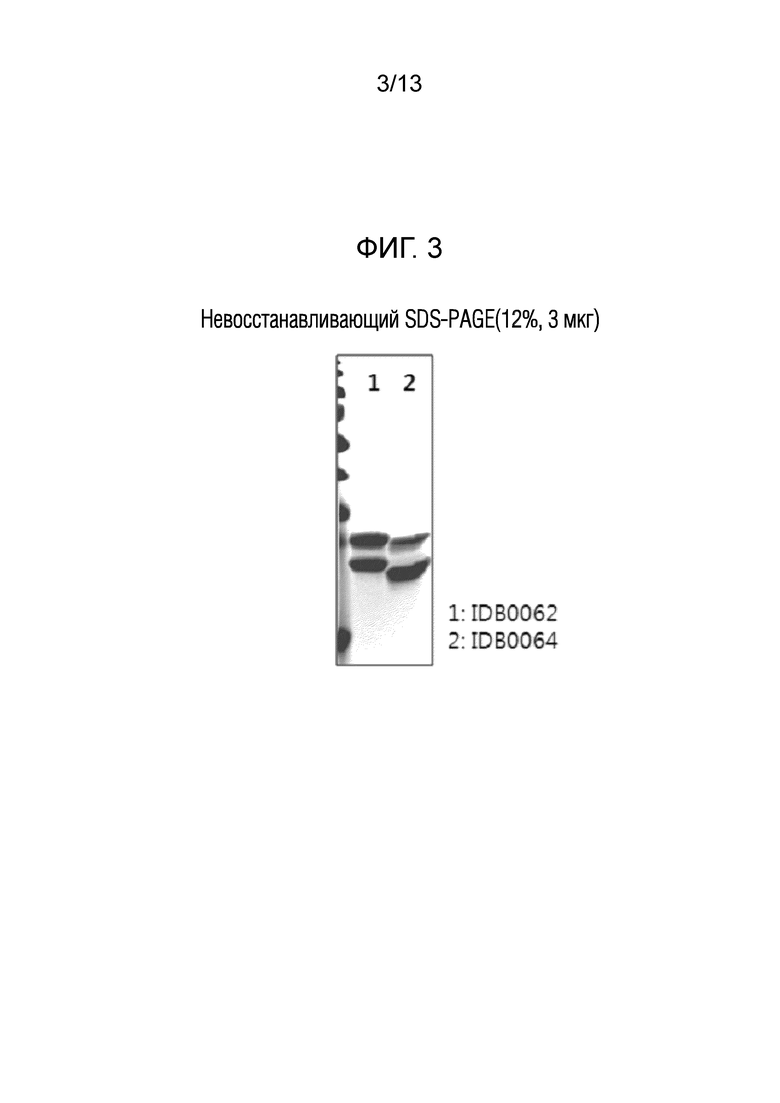


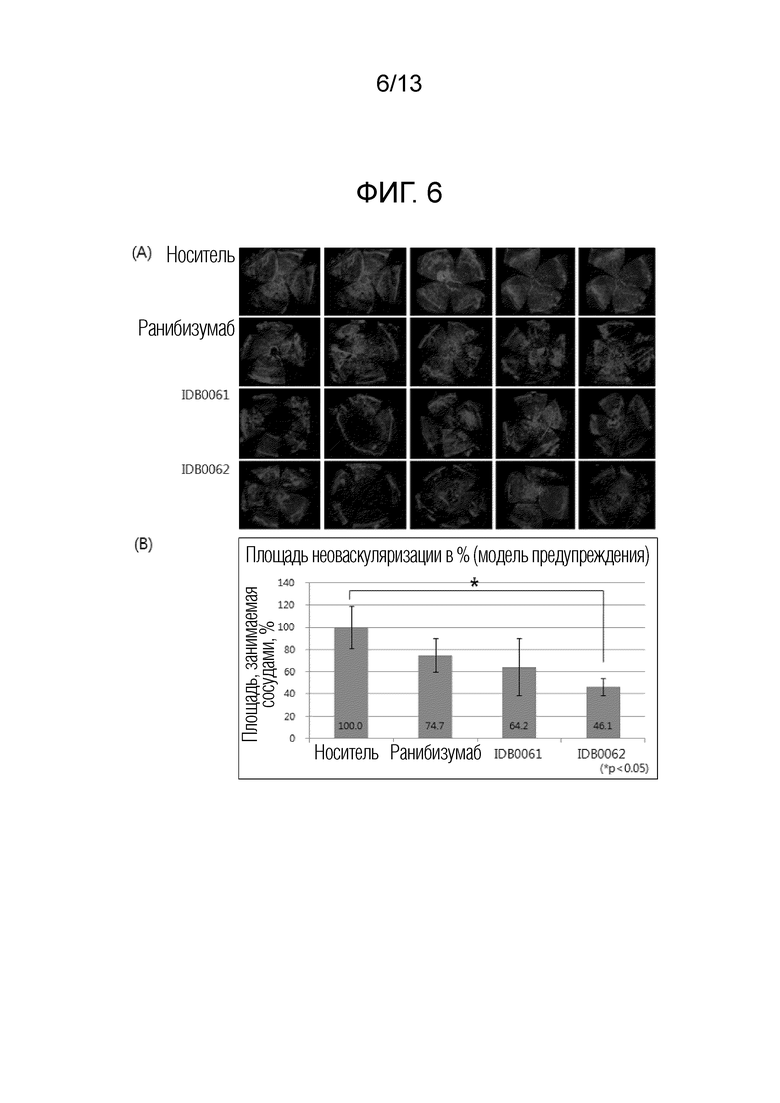
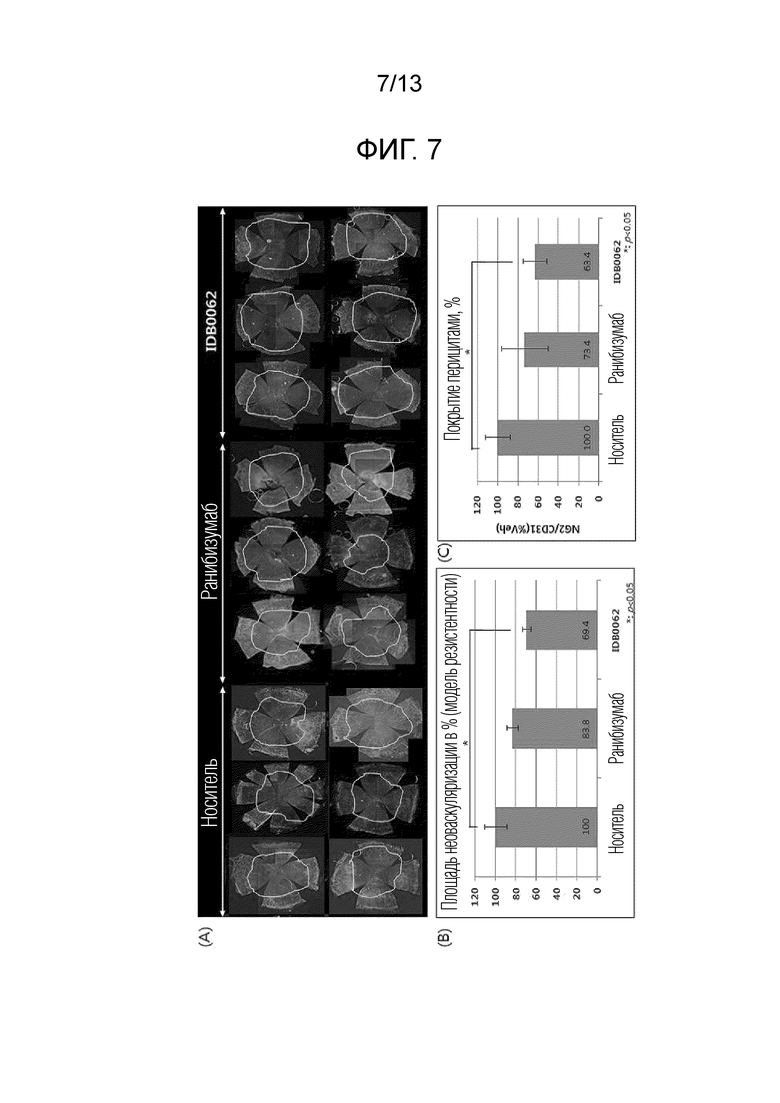
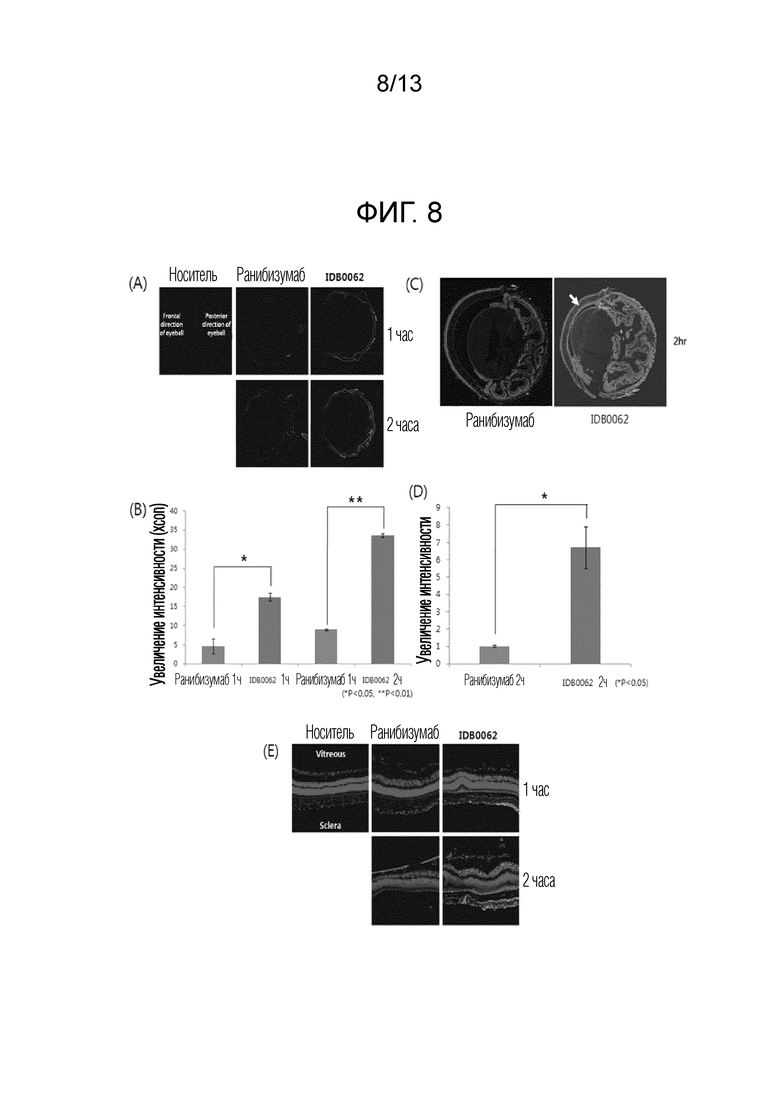
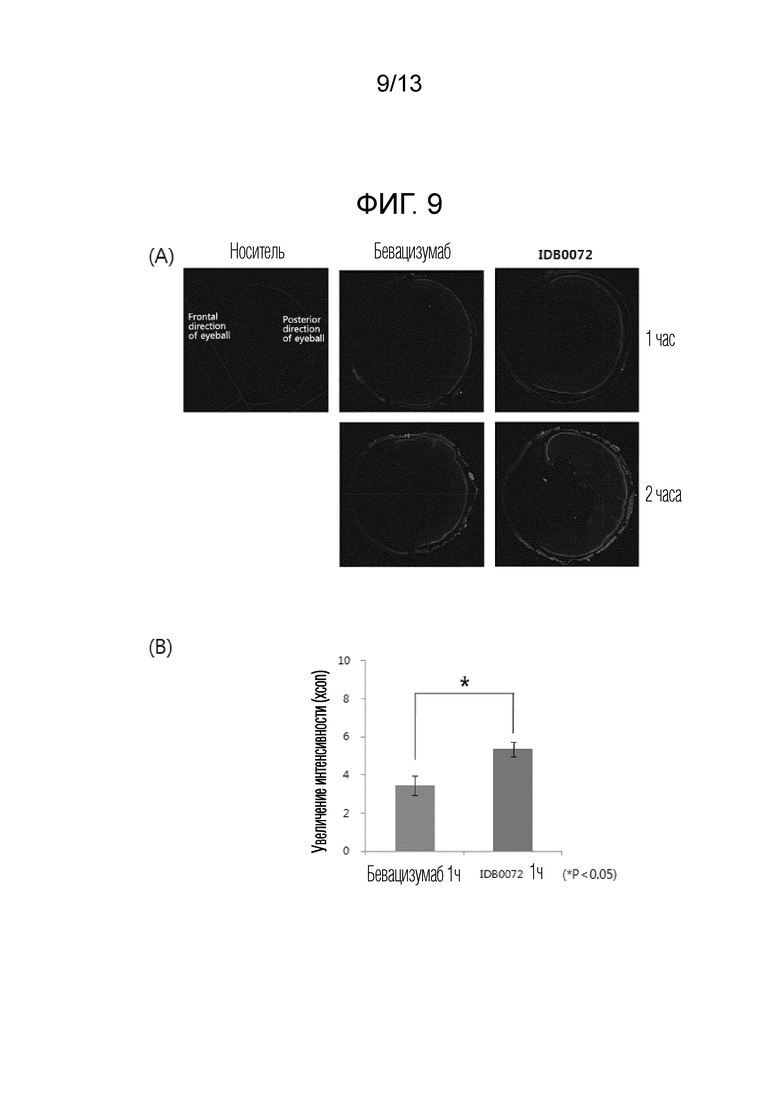
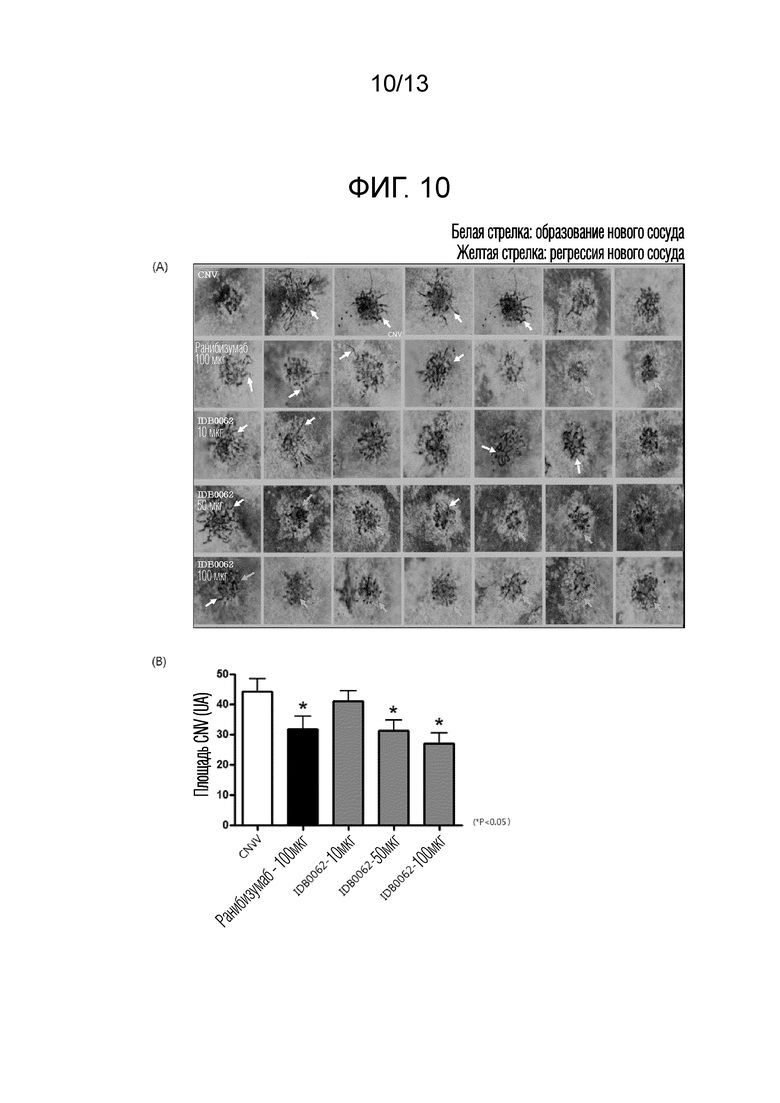

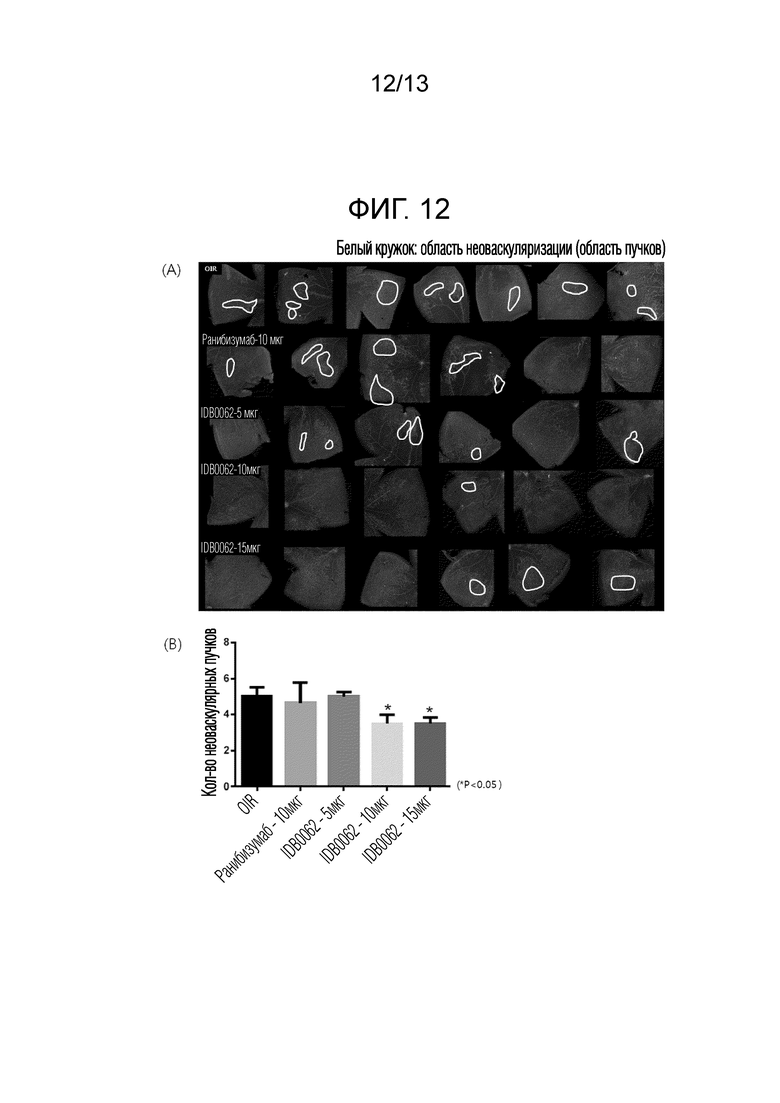
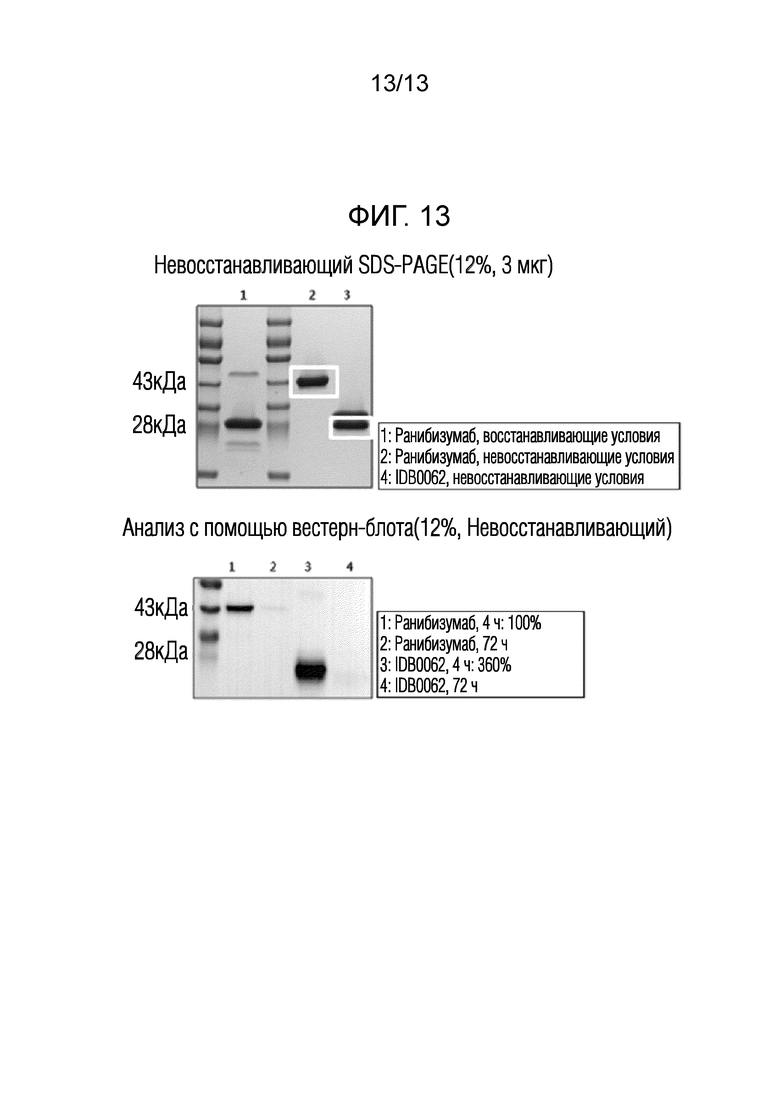
Предложенная группа изобретений относится к области медицины. Предложена фармацевтическая композиция для предотвращения и лечения вызванного неоваскуляризацией заболевания глаз, содержащая белок слияния, в котором проникающий в ткань пептид слит с анти-VEGF. Предложен способ получения слитого белка, его применение для получения терапевтического средства для вызванного неоваскуляризацией заболевания глаз и способ лечения такого заболевания. Предложенная группа изобретений обеспечивает эффективное лечение вызванного неоваскуляризацией заболевания глаз. 4 н. и 7 з.п. ф-лы, 13 ил., 7 табл., 8 пр.
1. Фармацевтическая композиция для предотвращения и лечения вызванного неоваскуляризацией заболевания глаз, причем данная фармацевтическая композиция содержит в качестве активного ингредиента белок слияния, в котором проникающий в ткань пептид слит со средством против фактора роста эндотелия сосудов (анти-VEGF), где проникающий в ткань пептид содержит любую одну аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-7.
2. Фармацевтическая композиция по п. 1, в которой средство против фактора роста эндотелия сосудов (анти-VEGF) представляет собой любое средство, выбранное из группы, состоящей из ранибизумаба, бевацизумаба, афлиберцепта, конберцепта, r84, CT01, DOM15-10-11, DOM15-26-593, PRS-050, CT-322, ESBA903, EPI-0030 и мутанта ранибизумаба, состоящего из легкой цепи, представленной SEQ ID NO: 8, и тяжелой цепи, представленной SEQ ID NO: 10.
3. Фармацевтическая композиция по п. 1, в которой белок слияния состоит из аминокислотной последовательности, представленной SEQ ID NO: 12 или 16, и аминокислотной последовательности, представленной SEQ ID NO: 14 или 18.
4. Фармацевтическая композиция по п. 1, в которой белок слияния состоит из аминокислотной последовательности, представленной SEQ ID NO: 20, и аминокислотной последовательности, представленной SEQ ID NO: 22.
5. Фармацевтическая композиция по п. 1, в которой слияние достигается с помощью линкерного пептида.
6. Фармацевтическая композиция по любому из пп. 1-5, в которой вызванное неоваскуляризацией заболевание глаз выбирают из группы, состоящей из пролиферативной витреоретинопатии, макулярной дегенерации, пигментной ретинопатии, диабетической ретинопатии, хориоидальной неоваскуляризации, неоваскулярной глаукомы, ишемической оптической нейропатии, ретинопатии недоношенных, ретинопатии при незрелости, эпидемического конъюнктивита, неоваскулярного заболевания радужной оболочки, ретролентальной фиброплазии, атопического кератита, верхнего лимбального кератита, птеригия, сухого кератита, фликтенулезного кератоконъюнктивита, склерита и диабетического макулярного отека.
7. Способ получения слитого белка, в котором проникающий в ткани пептид слит с агентом против васкулярного фактора роста эндотелия (анти-VEGF) для профилактики и лечения вызванного неоваскуляризацией заболевания глаз, причем данный способ содержит:
(a) трансформацию клеток-хозяев с помощью рекомбинантного вектора, причем данный рекомбинантный вектор содержит последовательность нуклеиновой кислоты, кодирующую белок слияния, в котором проникающий в ткань пептид слит со средством против фактора роста эндотелия сосудов (анти-VEGF);
(b) культивирование данных клеток и
(c) получение белка слияния из данных клеток, где проникающий в ткань пептид содержит любую одну аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-7.
8. Способ по п. 7, в котором средство против фактора роста эндотелия сосудов (анти-VEGF) представляет собой любое средство, выбранное из группы, состоящей из ранибизумаба, бевацизумаба, афлиберцепта, конберцепта, r84, CT01, DOM15-10-11, DOM15-26-593, PRS-050, CT-322, ESBA903, EPI-0030 и мутанта ранибизумаба, состоящего из легкой цепи, представленной SEQ ID NO: 8, и тяжелой цепи, представленной SEQ ID NO: 10.
9. Способ по п. 7, в котором слияние достигается с помощью линкерного пептида.
10. Способ лечения вызванного неоваскуляризацией заболевания глаз, причем данный способ содержит введение эффективного количества белка слияния, в котором проникающий в ткань пептид слит со средством против фактора роста эндотелия сосудов (анти-VEGF), нуждающемуся в этом субъекту, где проникающий в ткань пептид содержит любую одну аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-7.
11. Применение белка слияния, в котором проникающий в ткань пептид слит со средством против фактора роста эндотелия сосудов (анти-VEGF), для получения терапевтического средства для вызванного неоваскуляризацией заболевания глаз, содержащего данный белок слияния в качестве активного ингредиента, где проникающий в ткань пептид содержит любую одну аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1-7.
| US 20140193402 A1, 10.07.2014 | |||
| JOHNSON L.N | |||
| et al | |||
| Cell penetrating peptide POD mediates delivery of recombinant proteins to retina, cornea and skin | |||
| Vision Res | |||
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| Способ очистки нефти и нефтяных продуктов и уничтожения их флюоресценции | 1921 |
|
SU31A1 |
| WO 2009105669 A2, 27.08.2009 | |||
| БЕЛИКОВ В.Г | |||
| Фармацевтическая химия | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Общая фармацевтическая химия: Учеб | |||
| для фармац | |||
| ин-тов и фак | |||
| мед | |||
| ин-тов | |||
| - | |||

