ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к области иммунотерапии опухолей, в частности относится к Fab фрагментам гуманизированного антитела против VEGF.
УРОВЕНЬ ТЕХНИКИ
Развитие сосудистой системы лежит в основе многих физиологических и патологических процессов. Фактор роста эндотелия сосудов (VEGF) представляет собой группу факторов роста, обладающих важной проангиогенной активностью, которые способствуют митозу эндотелиальных клеток и антиапоптозу, увеличивают проницаемость сосудов и способствуют миграции клеток. Ген VEGF человека локализован на хромосоме 6р21.3 и принадлежит к семейству супергенов VEGF/PDGF, которое кодирует VEGF, связанные дисульфидными связями в форме димера. У людей семейство VEGF включает несколько членов с разными функциями: VEGF A (VEGF, с несколькими различными вариантами сплайсинга), VEGFB, VEGFC, VEGFD, VEGFE, VEGFF, и плацентарный фактор роста (PIGF). Недавно в это семейство был также включен сосудистый эндотелиальный фактор роста эндокринных желез (EG-VEGF). (Samson М и др., J Clin Endocrinol Metab. 2004; 89(8):4078-4088). VEGF широко распространен в тканях и органах человека, среди которых клетки пигментного эпителия сетчатки глаза, эндотелиальные клетки сосудов, нервные клетки и т.д. (Goel Н L и др., Nat Rev Cancer. 2013; 13(12): 871). Существует три типа рецепторов VEGF: VEGFR1, VEGFR2 и VEGFR3. Связывание VEGF с внеклеточным доменом рецептора запускает димеризацию рецептора и способствует аутофосфорилированию остатков тирозина во внутриклеточном домене, тем самым активируя нижестоящие сигналы, которые способствуют клеточной пролиферации, миграции, антиапоптозу и повышению проницаемости сосудов. VEGFR1 и VEGFR2 экспрессируются главным образом в сосудистых эндотелиальных клетках, a VEGFR3 в основном экспрессируется в лимфатических эндотелиальных клетках.
Было подтверждено, что VEGF играет важную роль в регуляции нормального и патологического ангиогенеза (Melincovici С S и др., Rom J Morphol Embryol. 2018; 59(2): 455-467). VEGF сверхэкспрессируется в различных опухолях, которые могут вызывать злокачественный асцит, и экспрессия VEGF в опухолях коррелирует с миграционной способностью опухолевых клеток. Концентрация VEGF у пациентов с солидными опухолями с более низкой выживаемостью, такими как рак желудочно-кишечного тракта, яичников, молочной железы и легких, положительно коррелирует со стадией заболевания (Sebastian, K и др., Oncologist. 2009; 14(12): 1242-1251). Развитие некоторых поражений при заболеваниях заднего сегмента, таких как возрастная дегенерация желтого пятна (AMD), диабетический макулярный отек (DME), отек сетчатки, дегенеративная миопия и хориоидальная неоваскуляризация (CNV), также тесно связаны с уровнями экспрессии VEGF (Patel J R и др., Curr opin ophthalmol. 2016; 27(5): 387-392; Tan G S и др., Lancet Диабет Endo. 2017; 5(2): 143-155; Mitchell P и др., Lancet. 2018; 392(10153): 1147-1159).
Препараты VEGF моноклональное антитело ингибируют экспансию эндотелиальных клеток и неоваскуляризацию путем ингибирования взаимодействия VEGF с рецепторами поверхности эндотелиальных клеток VEGFR2 и VEGFR1 и последующей блокировкой нижестоящего сигнального пути. Управление по контролю за качеством пищевых продуктов и медикаментов (FDA) одобрило препараты против VEGF для лечения офтальмологических заболеваний, включая Луцентис (ранимизумаб, одобрен в 2006 г.), EYLEA (афлиберцепт, одобрен в 2004 г.) и конберцепт, который также одобрен к использованию в Китае. Луцентис представляет собой Fab фрагмент антитело человеческого происхождения против VEGFA, который связывает все активные формы VEGFA и ингибирует его связывание с VEGFR1 и VEGFR2, тем самым подавляя пролиферацию и миграцию клеток эндотелия сосудов и снижая проницаемость сосудов, тем самым подавляя образование хориоидальной неоваскуляризации. В отличие от полноразмерных антител, фрагменты антител в виде Fab-фрагментов могут легко проникать через сетчатку в субретинальное пространство и достигать ткани-мишени для связывания с VEGF, ингибируя, таким образом, формирование хориоидальной неоваскуляризации. Fab фрагменты антитела, попадающие в общую систему через кровоток, элиминируются всего за 0,09 дня или примерно за 2 ч, минимизируя влияние на физиологические функции нормального VEGF и уменьшая токсические эффекты, такие как желудочно-кишечные перфорации, гипертензия и кровотечения. (Ferrara, N и др., Retina. 2006; 26(8): 859-870; Van Wijngaarden и др., Clin Exp Optom. 2008; 91(5): 427-437). Исследования показали взаимосвязь между AMD и воспалительной реакцией, вызванной эффектами комплемента, a Fab фрагмент антитела, который не содержит Fc-фрагмент, не стимулирует каскад комплемента, что снижает риск эндофтальмита и аутоиммунных воспалительных реакций. (Ferrara, N и др., Retina. 2006; 26(8): 859-870). Луцентис был одобрен для лечения влажной формы AMD, CNV, DME и отека сетчатки. Бевацизумаб представляет собой рекомбинантное моноклональное антитело человека, одобренное FDA для лечения солидных опухолей, таких как метастатический рак толстой кишки и немелкоклеточный рак легких, и в настоящее время используется в качестве лекарственного средства не по прямому назначению для лечения AMD. Афлиберцепт и гонберцепт представляют собой гуманизированный рекомбинантный слитой белок, который содержит специфический домен VEGFR, который связывается с лигандами, и может связываться с VEGF со специфическим высоким сродством и блокировать связывание VEGF с рецепторами. Афлиберцепт имеет более высокое сродство к VEGF 165, чем Бевацизумаб и Луцентис, и продемонстрировал более высокую эффективность в отношении DME. Афлиберцепт одобрен для лечения влажной формы AMD, окклюзии ветвей вен сетчатки, окклюзии центральной вены сетчатки, CNV, DME и диабетической ретинопатии. Конберцепт одобрен в Китае для лечения влажной формы AMD.
Поскольку эти лекарственные средства вводят интравитреально местно, частое введение с высокой вероятностью может вызвать поражение глаз и периокулярных инфекций. Таким образом, оптимизация лекарственных средств представляет собой необходимость повышения их эффективности и уменьшения частоты введения для обеспечения большей терапевтической пользы для пациентов.
КРАТКОЕ ОПИСАНИЕ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
Настоящее изобретение обеспечивает новые Fab фрагменты гуманизированного антитела против VEGF, которые могут применяться для лечения болезней глаз, характеризующихся хориоидальной неоваскуляризацией, включая возникновение возрастной дегенерации желтого пятна (AMD), диабетического макулярного отека (DME), отек сетчатки, дегенеративную миопию и хориоидальную неоваскуляризацию (CNV), но не ограничиваются ими.
В одном аспекте настоящее изобретение обеспечивает изолированное антитело против VEGF или его антиген-связывающий фрагмент, содержащий вариабельный участок тяжелой цепи, содержащий участок CDR1 тяжелой цепи, содержащий последовательность аминокислот, указанную в SEQ ID NO: 13 и участок CDR2 тяжелой цепи, содержащий последовательность аминокислот, указанную в SEQ ID NO: 14 и участок CDR3 тяжелой цепи, содержащий последовательность аминокислот, указанную в SEQ ID NO: 15; и вариабельный участок легкой цепи, содержащий участок CDR1 легкой цепи, содержащий последовательность аминокислот, указанную в SEQ ID NO: 10, участок CDR2 легкой цепи, содержащий последовательность аминокислот, указанную в SEQ ID NO: 11, и участок CDR3 легкой цепи, содержащий последовательность аминокислот, указанную в SEQ ID NO: 12.
В некоторых вариантах реализации указанное антитело против VEGF или его антиген-связывающий фрагмент содержит вариабельный участок тяжелой цепи, содержащий последовательность аминокислот, указанную в SEQ ID NO: 22, или последовательность аминокислот, по меньшей мере на 90%, 92%, 95%, 98% или 99% идентичную последовательности, указанной в SEQ ID NO: 22; и вариабельный участок легкой цепи, содержащий последовательность аминокислот, указанную в SEQ ID NO: 23, или последовательность аминокислот, имеющую последовательность, по меньшей мере на 90%, 92%, 95%, 98% или 99% идентичную последовательности, указанной в SEQ ID NO: 23.
В некоторых вариантах реализации указанное антитело против VEGF или его антиген-связывающий фрагмент дополнительно включает константный участок тяжелой цепи и константный участок легкой цепи, предпочтительно указанный константный участок тяжелой цепи является константным участком тяжелой цепи IgG1, содержащим последовательность аминокислот, указанную в SEQ ID NO: 38, или последовательность аминокислот, по меньшей мере на 90%, 92%, 95%, 98%, или 99% идентичную последовательности, указанной в SEQ ID NO: 38; и/или указанный константный участок легкой цепи представляет собой константный участок легкой цепи, содержащий последовательность аминокислот, указанную в SEQ ID NO: 39, или последовательность аминокислот, по меньшей мере на 90%, 92%, 95%, 98%, или 99% идентичную последовательности, указанной в SEQ ID NO: 39.
В некоторых вариантах реализации указанное антитело против VEGF или его антиген-связывающий фрагмент представляет собой гуманизированное антитело или химерное антитело.
В другом аспекте настоящее изобретение обеспечивает изолированное антитело против VEGF или его антиген-связывающий фрагмент, содержащий вариабельный участок тяжелой цепи, содержащий участок CDR1 тяжелой цепи, содержащий последовательность аминокислот, указанную в SEQ ID NO: 27 и участок CDR2 тяжелой цепи, содержащий последовательность аминокислот, указанную в SEQ ID NO: 28 и участок CDR3 тяжелой цепи, содержащий последовательность аминокислот, указанную в SEQ ID NO: 29; и вариабельный участок легкой цепи, содержащий участок CDR1 легкой цепи, содержащий последовательность аминокислот, указанную в SEQ ID NO: 24, участок CDR2 легкой цепи, содержащий последовательность аминокислот, указанную в SEQ ID NO: 25, и участок CDR3 легкой цепи, содержащий последовательность аминокислот, указанную в SEQ ID NO: 26.
В некоторых вариантах реализации указанное антитело против VEGF или его антиген-связывающий фрагмент включает константный участок тяжелой цепи, содержащий последовательность аминокислот, указанную в SEQ ID NO: 36, или последовательность аминокислот, по меньшей мере на 90%, 92%, 95%, 98%, или 99% идентичную последовательности, указанной в SEQ ID NO: 36; и вариабельный участок легкой цепи, содержащий последовательность аминокислот, указанную в SEQ ID NO: 37, или последовательность аминокислот, по меньшей мере на 90%, 92%, 95%, 98%, или 99% идентичную последовательности, указанной в SEQ ID NO: 37.
В некоторых вариантах реализации указанное антитело против VEGF или его антиген-связывающий фрагмент представляет собой Fab фрагмент, при этом указанный Fab фрагмент дополнительно включает константный участок тяжелой цепи CH1 и константный участок легкой цепи, предпочтительно константный участок тяжелой цепи СН1, представляющий собой константный участок тяжелой цепи IgG1, содержащий последовательность аминокислот, указанную в SEQ ID NO: 40, или последовательность аминокислот, по меньшей мере на 90%, 92%, 95%, 98%, или 99% идентичную последовательности, указанной в SEQ ID NO: 40; и/или указанный константный участок легкой цепи представляет собой константный участок легкой цепи, содержащий последовательность аминокислот, указанную в SEQ ID NO: 39, или последовательность аминокислот, по меньшей мере на 90%, 92%, 95%, 98%, или 99% идентичную последовательности, указанной в SEQ ID NO: 39.
В некоторых вариантах реализации указанное антитело против VEGF или его антиген-связывающий фрагмент дополнительно включает сигнальный пептид тяжелой цепи и сигнальный пептид легкой цепи, предпочтительно указанный сигнальный пептид тяжелой цепи имеет последовательность аминокислот, указанную в SEQ ID NO: 34, или последовательность аминокислот, по меньшей мере на 90%, 92%, 95%, 98% или 99% идентичную последовательности, указанной в SEQ ID NO: 34, и/или указанный сигнальный пептид легкой цепи имеет последовательность аминокислот, указанную в SEQ ID NO: 35 или последовательность аминокислот, по меньшей мере на 90%, 92%, 95%, 98% или 99% идентичную последовательности, указанной в SEQ ID NO: 35.
В некоторых вариантах реализации указанное антитело против VEGF или его антиген-связывающий фрагмент представляет собой Fab фрагмент антитела.
В некоторых вариантах реализации указанное антитело против VEGF или его антиген-связывающий фрагмент представляет собой антитело IgG, предпочтительно антитело IgG1.
В некоторых вариантах реализации указанный Fab фрагмент антитела против VEGF представляет собой Fab фрагмент, связанный с антителом IgG, предпочтительно Fab фрагмент, связанный с антителом IgG1.
В некоторых вариантах реализации указанное антитело против VEGF или его антиген-связывающий фрагмент представляет собой моноклональное антитело.
В некоторых вариантах реализации указанный Fab фрагмент антитела против VEGF является моноклональным.
В некоторых вариантах реализации аффинность связывания KD указанного антитела против VEGF или его антиген-связывающий фрагмент с рекомбинантным белком VEGF 165 человека составляет 0,01-8 Е-10 М, предпочтительно 0,1-5 Е-10 М, и более предпочтительно 0,5-3 Е-10 М, наиболее предпочтительно 1,54 Е-10 М.
В некоторых вариантах реализации указанный антиген-связывающий фрагмент представляет собой фрагмент Fv, Fab, Fab', Fab'-SH, F(ab')2, Fd, Fd', молекулу одноцепочечного антитела или однодоменное антитело; при этом указанная молекула одноцепочечного антитела представляет собой предпочтительно scFv, ди-scFv, три-scFv, диатело или scFab.
В еще одном аспекте, настоящее изобретение обеспечивает конъюгат антитела и лекарственного средства, содержащий указанное антитело против VEGF или его антиген-связывающий фрагмент согласно настоящему изобретению и дополнительный терапевтический агент, предпочтительно указанное антитело против VEGF или его антиген-связывающий фрагмент связан с указанным дополнительным терапевтическим агентом посредством линкера.
В еще одном аспекте, настоящее изобретение обеспечивает нуклеиновую кислоту, кодирующую указанное антитело против VEGF или его антиген-связывающий фрагмент согласно настоящему изобретению.
В некоторых вариантах реализации указанная нуклеиновая кислота включает последовательность нуклеотидов, указанную в SEQ ID NO: 4 и/или последовательность нуклеотидов, указанную в SEQ ID NO: 5; или включает последовательность нуклеотидов, указанную в SEQ ID NO: 20 и/или последовательность нуклеотидов, указанную в SEQ ID NO: 21; или включает последовательность нуклеотидов, указанную в SEQ ID NO: 45 и/или последовательность нуклеотидов, указанную в SEQ ID NO: 46. В предпочтительном варианте указанная нуклеиновая кислота дополнительно включает последовательность нуклеотидов, указанную в SEQ ID NO: 49 и/или последовательность нуклеотидов, указанную в SEQ ID NO: 48. Более предпочтительно, указанная нуклеиновая кислота включает последовательность нуклеотидов, указанную в SEQ ID NO: 41 и/или последовательность нуклеотидов, указанную в SEQ ID NO: 42.
В еще одном аспекте, настоящее изобретение обеспечивает вектор экспрессии, содержащий указанную нуклеиновую кислоту согласно настоящему изобретению.
В еще одном аспекте, настоящее изобретение обеспечивает клетку-хозяина, содержащую указанную нуклеиновую кислоту согласно настоящему изобретению или указанный вектор экспрессии согласно настоящему изобретению.
В еще одном аспекте, настоящее изобретение обеспечивает способ получения антитела против VEGF или его антиген-связывающего фрагмента согласно настоящему изобретению, включающий выращивание указанной клетки-хозяина согласно настоящему изобретению в условиях, подходящих для экспрессии антитела, и сбора экспрессированного антитела из культуральной жидкости.
В еще одном аспекте, настоящее изобретение обеспечивает способ получения антитела против VEGF или его антиген-связывающего фрагмента согласно настоящему изобретению, включающий выращивание указанной клетки-хозяин согласно настоящему изобретению в условиях, подходящих для экспрессии Fab фрагмента антитела, и сбора экспрессированного Fab фрагмента антитела из культуральной жидкости.
В еще одном аспекте, настоящее изобретение обеспечивает фармацевтическую композицию, содержащую указанное антитело против VEGF или его антиген-связывающий фрагмент согласно настоящему изобретению, или конъюгат антитела и лекарственного средства согласно настоящему изобретению, или указанную нуклеиновую кислоту согласно настоящему изобретению, или указанный вектор экспрессии согласно настоящему изобретению, и фармацевтически приемлемый носитель.
В еще одном аспекте, настоящее изобретение обеспечивает указанное антитело против VEGF или его антиген-связывающий фрагмент согласно настоящему изобретению или конъюгат антитела и лекарственного средства согласно настоящему изобретению или указанную фармацевтическую композицию согласно настоящему изобретению, для применения в лечении заболеваний, связанных с ангиогенезом.
В некоторых вариантах реализации указанное заболевание, связанное с ангиогенезом, представляет собой заболевание глаза.
В некоторых вариантах реализации указанное заболевание глаза представляет собой заболевание глаза, характеризующееся хориоидальной неоваскуляризацией, включая вызванную возрастом дегенерацию желтого пятна (AMD), диабетический макулярный отек (DME), отек сетчатки, дегенеративную миопию, хориоидальную неоваскуляризацию (CNV).
В еще одном аспекте, настоящее изобретение обеспечивает способ для лечения заболеваний, связанных с ангиогенезом, включающий введение субъекту, нуждающемуся в этом, терапевтически эффективного количества указанного антитела против VEGF или антиген-связывающего фрагмента согласно настоящему изобретению или конъюгата антитела и лекарственного средства согласно настоящему изобретению или указанной фармацевтической композиции согласно настоящему изобретению, излечивая тем самым заболевание, связанное с ангиогенезом.
В некоторых вариантах реализации указанное заболевание, связанное с ангиогенезом представляет собой заболевание глаза.
В некоторых вариантах реализации указанное заболевание глаза представляет собой заболевание глаза, характеризующееся хориоидальной неоваскуляризацией, включая вызванную возрастом дегенерация желтого пятна (AMD), диабетический макулярный отек (DME), отек сетчатки, дегенеративную близорукость, хориоидальную неоваскуляризацию (CNV).
В еще одном аспекте, настоящее изобретение обеспечивает антитело против VEGF или его антиген-связывающий фрагмент согласно настоящему изобретению, или конъюгат антитела и лекарственного средства согласно настоящему изобретению, или фармацевтическую композицию согласно настоящему изобретению, для применения в получении лекарственного средства для лечения заболеваний, связанных с ангиогенезом.
В некоторых вариантах реализации указанное заболевание, связанное с ангиогенезом представляет собой заболевание глаза.
В некоторых вариантах реализации указанное заболевание глаза представляет собой заболевание глаза, характеризующееся хориоидальной неоваскуляризацией, включая вызванную возрастом дегенерацию желтого пятна (AMD), диабетический макулярный отек (DME), отек сетчатки, дегенеративную близорукость, хориоидальную неоваскуляризацию (CNV).
В еще одном аспекте, настоящее изобретение обеспечивает фармацевтическую комбинацию, содержащую антитело против VEGF или его антиген-связывающий фрагмент согласно настоящему изобретению, или конъюгат антитела и лекарственного средства согласно настоящему изобретению, или указанная фармацевтическая композиция согласно настоящему изобретению, и один или более дополнительный терапевтический агент.
В еще одном аспекте, настоящее изобретение обеспечивает набор, содержащий антитело против VEGF или его антиген-связывающий фрагмент согласно настоящему изобретению, или конъюгат антитела и лекарственного средства согласно настоящему изобретению, или указанную фармацевтическую композицию согласно настоящему изобретению, предпочтительно, дополнительно содержащую устройство для введения.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Настоящее изобретение проиллюстрировано в комбинации с приложенными чертежами, в которых:
На Фиг. 1 показано, что антитело VEGF-R988 кролика против VEGF165 блокирует связывание VEGF 165 с белком VEGFR2.
На Фиг. 2 показано, что антитело VEGF-R988 кролика против VEGF 165 нейтрализует эффект пролиферации VEGF165-HUVEC.
На Фиг. 3 показано связывание Fab гуманизированного антитела VEGF-H988 с VEGF165, обнаруженное с помощью ИФА.
На Фиг. 4 показано перекрестное межвидовое связывание Fab VEGF-H988 с mVEGF164, обнаруженное с помощью ИФА.
На Фиг. 5 показано, что Fab антитела VEGF-H988 блокирует связывание VEGF 165 с белком VEGFR2, обнаруженное с помощью ИФА.
На Фиг. 6 показано влияние Fab VEGF-H988 в нейтрализации VEGF 165 при различных концентрациях в сравнении с лекарственным средством Луцентис.
На Фиг. 7 показано влияние Fab VEGF-H988 в нейтрализации VEGF 165 при различных концентрациях в сравнении с лекарственным средством EYLEA.
На Фиг. 8 показано влияние Fab VEGF-H988 в нейтрализации VEGF 165 при различных концентрациях в сравнении с лекарственным средством Авастин.
На Фиг. 9 показано влияние Fab VEGF-H988 в нейтрализации VEGF165 при различных концентрациях в сравнении с лекарственным средством Конберцепт.
На Фиг. 10 показано влияние Fab VEGF-H988 в нейтрализации VEGF 165 при различных концентрациях в сравнении с лекарственным средством Бролуцизумаб.
ПОДРОБНОЕ ОПИСАНИЕ
Различные аспекты согласно настоящему изобретению относятся к изолированному Fab фрагменту антитела против VEGF, конъюгату антитела и лекарственного средства, содержащему указанный фрагмент антитела или его антиген-связывающий фрагмент, нуклеиновой кислоте и вектору экспрессии, кодирующим указанный Fab антитела, и клетке-хозяину, содержащей указанную нуклеиновую кислоту или вектор экспрессии, способу получения указанного Fab антитела против VEGF, фармацевтической композиции, содержащей указанный Fab антитела против VEGF, и способ применения Fab антитела против VEGF для лечения заболеваний, ассоциированных с ангиогенезом.
Определения
Если не указано иное, все используемые здесь технические и научные термины имеют значения, обычно понятные специалистам в данной области техники, к которой относится настоящее изобретение. Для целей настоящего изобретения следующие термины определяются в соответствии со значениями, обычно понимаемыми в данной области техники.
При использовании в настоящем описании и в прилагаемой формуле изобретения формы единственного числа «один», «другой» и «указанный» включают обозначение объекта во множественном числе, если контекст явно не указывает на иное.
Термин «антитело» относится к молекуле иммуноглобулина и относится к любой форме антитела, которая проявляет желаемую биологическую активность. К ним относятся, моноклональные антитела (включая полноразмерные моноклональные антитела), поликлональные антитела и мультиспецифические антитела (например, биспецифические антитела), и даже фрагменты антитела, но не ограничиваются ими. Обычно, структуры полноразмерных антител предпочтительно включают четыре цепи полипептидов, две тяжелых (Н) цепи и две легких (L) цепи, обычно взаимосвязанных дисульфидными связями. Каждая тяжелая цепь содержит вариабельный участок тяжелой цепи и константный участок тяжелой цепи. Каждая легкая цепь содержит вариабельный участок легкой цепи и константный участок легкой цепи. В дополнение к этой обычной структуре полноразмерного антитела, указанная структура также включает другие производные формы.
Указанные вариабельный участок тяжелой цепи и вариабельный участок легкой цепи могут быть дополнительно подразделяться на более консервативные участки (называемые каркасными участками (FR)) и гипервариабельные участки (называемые участками, определяющими комплементарность (CDR)), разделенные промежутками.
Термин «участок, определяющий комплементарность» (CDR, например, CDR1, CDR2 и CDR3) относится к таким остаткам аминокислот в указанном вариабельном участке антитела, чье присутствие необходимо для связывания антигена. Каждый вариабельный участок обычно содержит три участка CDR, обозначенные как CDR1, CDR2 и CDR3. Каждый участок, определяющий комплементарность, может содержать остатки аминокислот из «участка, определяющего комплементарность» в соответствии с нумерацией Kabat (Kabat и др., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Бетесда, Мэриленд, 1991) и/или остатки из «высоковариабельной петли» (Chothia и Lesk; J MolBiol 196:901-917 (1987)).
Термин остатки «каркаса» или «FR» означает остатки в указанном вариабельном участке, отличающиеся от остатков CDR, как определено в настоящем документе.
Каждый вариабельный участок тяжелой цепи и вариабельный участок легкой цепи обычно содержит 3 CDR и до 4 FR, при этом указанные CDR и FR расположены от аминоконца к карбоксильному концу в следующем порядке, например: FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4.
Участок, определяющий комплементарность, (CDR) и каркасный участок (FR) указанного антитела могут быть определены с использованием системы Kabat (Kabat и др.: Sequences of Proteins of Immunological Interest, 5th edition, US Department of Health and Human Services, PHS, NIH, NIH Publication No. 91- 3242, 1991).
Термин «константный участок» относится к такой последовательности аминокислот в легкой и тяжелой цепях антитела, которая впрямую не вовлечена в связывание указанного антитела с антигеном, но влияет на различные эффекторные функции, такие как вызываемая антителом цитотоксичность.
В соответствии с антигенными различиями последовательности аминокислот константного участка, указанная тяжелая цепь антитела может быть отнесена к одному из пяти классов: а, 5, е, у, и а. Когда она образует полноразмерное антитело с легкой цепью, оно может быть отнесено к одному из пяти классов: IgA, IgD, IgE, IgG и IgM, которые могут быть дополнительно отнесены к пяти подклассам (изотипам), таким как IgG1, IgG2, IgG3, IgG4, IgAn IgA2. На основании последовательности аминокислот его константного участка, легкая цепь антитела может быть классифицирована как κ и λ.
«Антиген-связывающий фрагмент антитела» означает часть интактной молекулы антитела, у которой остается по меньшей мере некоторая специфичность связывания исходного антитела и обычно включает по меньшей мере часть антиген-связывающего участка или вариабельный участок (например, один или более CDR) исходного антитело. Примеры антиген-связывающих фрагментов включают фрагмент Fv, Fab, Fab', Fab'-SH, F(ab')2, Fd, Fd', молекулы одноцепочечных антител (например, scFv, ди-scFv или три-scFv, диатело или scFab), однодоменные антитела, но не ограничиваются ими. Fab фрагмент обычно содержит вариабельный участок тяжелой цепи (VH) и константный участок тяжелой цепи 1 (CH1), и вариабельный участок легкой цепи (VL) и константный участок легкой цепи (CL).
Термин «фрагмент антитела» относится к молекуле неполноразмерного антитела, у которой остаются по меньшей мере некоторые биологические свойства исходного антитела, включая фрагмент Fc, в дополнение к фрагментам, описанным выше как «антиген-связывающие фрагменты», но не ограничиваются ими.
Термин «конъюгат антитела и лекарственного средства» или «ADC» относится к связывающему белку, такому как антитело или его антиген-связывающий фрагмент, который химически связан с одним или более химическим средством (также обозначенном в настоящем документе как «агент»), которое необязательно может быть терапевтическим агентом или цитотоксическим агентом. В одном из предпочтительных вариантов реализации ADC включает антитело, цитотоксическое или терапевтическое лекарственное средство, и линкер, с помощью которого указанное лекарственное средство может быть связано указанным антителом или может образовывать с ним конъюгат. ADC обычно могут содержать от 1 до 8 лекарственных средств, образующих конъюгат с указанным антителом, включая 2, 4, 6 или 8 лекарственных средств. Неограничивающими примерами лекарственных средств, которые могут быть включены в ADC, являются митотические ингибиторы, противоопухолевые антибиотики, иммуномодуляторы, векторы для генной терапии, алкилирующие агенты, антиангиогенные агенты, антиметаболиты, борсодержащие агенты, химиотерапевтические защитные агенты, гормоны, антигормональные агенты, кортикостероиды, фотоактивные терапевтические агенты, олигонуклеотиды, радионуклидные агенты, ингибиторы топоизомеразы, ингибиторы тирозинкиназы и радиосенсибилизаторы.
Термин «химерные антитело» относится к антителу, в котором часть указанной тяжелой цепи и/или легкой цепи получена из одного конкретного источника или вида, а оставшаяся часть получена из другого источника или вида. Указанное «химерное антитело» также может быть функциональным фрагментом, описанным выше. «Гуманизированное антитела» являются подмножеством «химерных антител».
Термин «гуманизированное антитело» или «гуманизированный антиген-связывающий фрагмент» в настоящем документе означает антитело или фрагмент антитела, который: (i) получен из источника, не являющегося человеком, (например, трансгенной мыши, имеющей гетерологичную иммунную систему) и основан на последовательности зародышевой линии человека; или (ii) является химерным антителом, в котором вариабельный участок имеет происхождение не из человека, а константный участок происходит из человека; или (iii) трансплантат CDR, в котором указанный CDR вариабельного участка имеет происхождение не из человека, один или более каркасных участков вариабельного участка происходят из человека и константный участок, если есть, происходит из человека. Целью «гуманизации» является устранение иммуногенности антител нечеловеческого происхождения в организме человека при сохранении максимально возможной аффинности. В качестве шаблона для гуманизации целесообразно выбрать последовательность каркаса человека, которая наиболее похожа на последовательность каркаса антитела, происходящего их источника, не являющегося человеком. В некоторых случаях может оказаться необходимым заменить одну или несколько аминокислот в каркасной последовательности человека соответствующими остатками в конструкции не из человека, чтобы избежать потери аффинности.
Термин «моноклональное антитело» относится к антителу, полученному из по существу гомогенной популяции антител, т.е. каждое отдельное антитело, входящее в указанную популяцию, является идентичным, за исключением возможных мутаций (например, природных мутаций), которые могут присутствовать в очень маленьких количествах. Таким образом, термин «моноклональное» указывает на природу рассматриваемого указанного антитела, т.е., а не на смесь неродственных антител. В отличие от препаратов поликлональных антител, которые обычно включают разные антитела против разных эпитопов, все моноклональные антитела в препаратах моноклональных антител направлены против одного эпитопа антигена. В дополнение к его специфичности, препараты моноклональных антител имеют то преимущество, что они обычно не загрязнены другими антителами. Термин «моноклональные» не следует понимать как требующее производства указанного антитела каким-либо конкретным способом.
Указанное антитело «специфически связывается» с целевым антигеном, таким как ассоциированный с опухолью пептидный антиген-мишень (в данном случае, VEGF), т.е. связывает указанный антиген с достаточной аффинностью, чтобы сделать возможным использование указанного антитело в качестве терапевтического агента, нацеливаясь на клетку или ткань, экспрессирующую указанный антиген, и не дает значительной перекрестной реакции с другими белками или не дает значительной перекрестной реакции с белками, отличными от гомологов и вариантов белков-мишеней, упомянутых выше (например, мутантные формы, варианты сплайсинга, или укороченные формы белка, полученные в результате гидролиза).
Термин «аффинность связывания» относится к силе суммы нековалентных взаимодействий между отдельными сайтами связывания молекулы и ее партнерами по связыванию. Если не указано иное, «аффинность связывания» при использовании в настоящем документе относится к внутренней аффинности связывания, которая отражает взаимодействие 1:1 между членами связывающей пары (например, антитело и антиген). В настоящем тексте термин «KD» относится к равновесной константе диссоциации взаимодействия антитело-антиген. В настоящем тексте термин «kon» относится к константе скорости, при которой антитело связывается с антигеном. В настоящем тексте термин «koff» относится к константе скорости, при которой антитело диссоциирует из комплекса антитело/антиген. «KD», «константа скорости связывания kon» и «константа скорости диссоциации koff» обычно используются для описания сродства между молекулой (например, антителом) и ее партнером по связыванию (например, антигеном). Аффинность, т.е., степень силы, при которой лиганд связывается с определенным белком. На аффинность связывания влияют нековалентные межмолекулярные взаимодействия, такие как водородные связи, электростатические взаимодействия, гидрофобные и ван-дер-ваальсовы силы между двумя молекулами. Кроме того, на аффинность связывания между лигандом и его целевой молекулой может влиять присутствие других молекул. Аффинность может быть проанализирована традиционными методами, известными в данной области техники, включая ИФА, описанный в настоящем документе.
Термин «эпитоп» включает любой кластер белковой детерминанты, который специфически связывается с антителом или Т-клеточным рецептором. Кластеры эпитопной детерминанты обычно состоят из химически активных поверхностных групп молекулы (например, боковых цепей аминокислот или Сахаров или их комбинации) и часто имеют специфические трехмерные структурные признаки, а также специфические характеристики заряда.
Термин «изолированное» антитело представляет собой антитело, которое было идентифицировано и выделено из компонентов клетки, где экспрессируется указанное антитело. Однако, как правило, изолированное антитело получают посредством по меньшей мере одной стадии очистки.
«Идентичность последовательностей» между двумя последовательностями полипептидов или нуклеиновых кислот указывает количество остатков, которые идентичны между указанными последовательностями в процентах от общего числа остатков, и рассчитывается на основе размера меньшей из сравниваемых молекул. При вычислении идентичности в процентах сравниваемые последовательности сопоставляются таким образом, чтобы получить максимальное совпадение между указанными последовательностями, при этом пробелы при сравнении (если они есть) разрешаются с помощью определенного алгоритма. Предпочтительные компьютерные программные методы для определения идентичности между двумя последовательностями включают наборы программ GCG, включая GAP, BLASTP, BLASTN и FASTA (Altschul и др., 1990, J. Mol. Biol. 215: 403-410), но не ограничиваются ими. Вышеупомянутые процедуры общедоступны в Международном центре биотехнологической информации (NCBI) и других источниках. Известный алгоритм Смита-Уотермана также может быть использован для определения идентичности.
Термин «Fc рецептор» или «FcR» относится к рецептору, который связывается с участком Fc антитела. Предпочтительны природные последовательности FcR человека и предпочтительно рецепторы, которые связываются с IgG антителами (гамма-рецепторы), которые включают FcγRI, FcγRII и изоформы FcγRIII, а также варианты из этих рецепторов. Все остальные FcR включены в термин «FcR». Термин также включает неонатальный рецептор (FcRn), который отвечает за транспорт материнского IgG к плоду (Guyer и др., Journal of Immunology 117: 587 (1976) и Kim и др., Journal of Immunology 24: 249 (1994)).
Термин «неонатальный Fc-рецептор», сокращенно «FcRn», связывается с участком Fc IgG-антитела. Неонатальный Fc-рецептор (FcRn) играет важную роль в метаболической судьбе IgG-подобных антител in vivo. FcRn функционирует для спасения IgG от пути лизосомной деградации, тем самым снижая его клиренс в сыворотке и удлиняя период его полужизни. Таким образом, свойства связывания FcRn /характеристики IgG in vivo указывают на его фармакокинетические свойства в кровотоке in vivo.
Термин «эффекторная функция» относится к той биологической активности, которая приписывается участку Fc антитела, которая варьирует от изотипа к изотипу. Примеры эффекторной функции антитела включают связывание C1q и комплемент-зависимую цитотоксичность (CDC), связывание рецептора Fc, антитело-зависимую клеточно-опосредованная цитотоксичность (ADCC), антителозависимый клеточный фагоцитоз (ADCP), секреция цитокинов, опосредованное иммунными комплексами поглощение антигена антигенпрезентирующими клетками, подавление рецепторов клеточной поверхности (например, рецепторов В-клеток) и активация В-клеток.
Термин «эффекторная клетка» относится к клетке, которая экспрессирует один или несколько FcR и выполняет эффекторные функции. В одном аспекте указанные эффекторные клетки экспрессируют в меньшей степени FcγRIII и выполняют эффекторные функции ADCC. Примеры клеток человека, которые опосредуют ADCC, включают мононуклеарные клетки периферической крови, (РВМС), натуральные киллеры (NK), моноциты, цитотоксические Т-клетки и нейтрофилы. Эффекторные клетки могут быть выделены из природных источников, например, крови. Эффекторные клетки обычно представляют собой лимфоциты, ассоциированные с эффекторной фазой и функционирующие для продукции цитокинов (хелперные Т-клетки), уничтожения клеток, инфицированных патогенами (цитотоксические Т-клетки) или секреции антитела (дифференцированные В-клетки).
«Иммунные клетки» включают клетки, которые имеют гемопоэтическое происхождение и играют роль в иммунном ответе. Иммунные клетки включают: лимфоциты, такие как В-клетки и Т-клетки; естественные клетки-киллеры; и миелоидные клетки, такие как моноциты, макрофаги, эозинофилы, тучные клетки, базофилы и гранулоциты.
«Антитело-зависимая клеточно-опосредованная цитотоксичность» или «ADCC» относится к форме цитотоксичности, при которой секретируемый Ig связывается с рецепторами Fcγ, представленными на определенных цитотоксических клетках (например, NK-клетках, нейтрофилах и макрофагах), что позволяет этим цитотоксическим эффекторным клеткам специфически связываться с клетками-мишенями, несущими антигены, и последующего уничтожения указанных клеток-мишеней с использованием, например, цитотоксина. Для оценки ADCC-активности целевого антитела могут быть выполнены анализы ADCC in vitro, такие как анализы ADCC in vitro, задокументированные в патенте США №5,500,362 или 5,821,337 или патенте США №6,737,056 (Presta). Полезные эффекторные клетки для применения в таких случаях включают РВМС и NK-клетки.
«Комплемент-зависимая цитотоксичность» или «CDC» относится к лизису клеток-мишеней в присутствии комплемента. Классический путь активации комплемента инициируется связыванием первого компонента системы комплемента (C1q) с антителом (соответствующего подкласса), которое связывается с соответствующим антигеном. Для оценки активации комплемента проводят анализ CDC, такой как анализ CDC, описанный в Gazzano-Santoro и др., J. Immunol Methods 202: 163 (1996). Например, в патенте США №6,194,551 В1 и WO 1999/51642, описаны варианты полипептидов, содержащие измененную последовательность аминокислот участка Fc (полипептиды, содержащие вариант участка Fc) и варианты полипептидов, имеющие повышенное или пониженное связывание C1q.
«Эндотелиальные клетки пупочной вены человека (HUVEC)» выделяют из вен пуповины и обычно используют для физиологических и фармакологических исследований, например, для изучения макромолекулярного транспорта, свертывания крови, ангиогенеза и фибринолиза. В частности, они могут быть использованы в качестве модели для исследований в отношении ангиогенеза и других исследований, касающихся VEGF-зависимого сигнального пути (связанных эндотелиальных факторов роста).
Последовательность аминокислот указанного антитела согласно настоящему изобретению
В настоящем изобретении использовали рекомбинантный белок VEGF 165 человека для иммунизации кролика, а затем получали клоны антитела VEGF165-R988, которые специфически связываются с рекомбинантным белком VEGF165 человека, посредством скрининга библиотеки фагового дисплея. Указанную последовательность нуклеотидов, кодирующую вариабельные участки тяжелой и легкой цепи антитела scFv VEGF165-R988, затем вводили посредством ПЦР в вектор pSTEP2, содержащий последовательность нуклеотидов, кодирующую константный участок IgG1 кролика или константный участок каппа кролика, и выращивали для экспрессии. Антитела высокой степени очистки получали посредством очистки на колонке с белком А. ИФА показал, что указанное антитело кролика VEGF165-R988 было способно блокировать связывание белка VEGF165 с белком VEGFR2, a VEGF165-R988 было способно эффективно нейтрализовать способность VEGF165 к стимулированию пролиферации HUVEC.
Затем, используя классический метод гуманизированной трансплантации CDR, в качестве матрицы были выбраны вариабельные участки легкой цепи или тяжелой цепей антитела человека, последовательность которых ближе к указанной последовательности вариабельных участков легкой или тяжелой цепей кролика, гуманизированный вариабельный участок легкой цепи (VL) и вариабельный участок тяжелой цепи (VH) по очереди путем вставки каждого из трех CDR (Таблица 1) легкой цепи или тяжелой цепи антитела кролика в вариабельные участки указанного антитела человека. Поскольку ключевые сайты каркасного участка антитела кролика необходимы для поддержания стабильности активности CDR, ключевые сайты были мутированы обратно в соответствующую последовательность антитела кролика. Вектор экспрессии легкой/тяжелой цепей VEGF-H988-10 получали путем полного синтеза гена, трансфицировали его в клетки HEK-293 и культивировали для экспрессии, культуральный супернатант очищали с использованием колонки с белком А, в результате чего получали антитело VEGF-H988-10 высокой степени очистки. Для улучшения аффинности VEGF-H988-10 конструировали библиотеки SDM участков CDR вариабельных участков тяжелой и легкой цепей (включая LCDR1, LCDR3, HCDR2 и HCDR3), и библиотеки четырех мутантов конструировали в форме scFv и клонировали в фаговые векторы в виде слитого белка svFv-gIII; для каждого CDR проводили скрининг клонов CDR, обладающих оптимальной способностью к связыванию с растворимым антигеном VEGF, и получали конечное антитело VEGF-Н988, обладающее оптимизированной аффинностью CDR и стабильностью.
По сравнению с полноразмерным антителом, антитела в виде Fab фрагментов обладают большей проникающей способностью и менее токсичны в отношении перфорации желудочно-кишечного тракта, гипертензии и кровоизлияния и не стимулируют реакцию каскада комплемента, что снижает риск развития эндофтальмита и аутоиммунных воспалительных реакций.
Нуклеиноные кислоты согласно настоящего изобретению
Настоящее изобретение также относится к молекулам нуклеиновой кислоты, кодирующей антитела или их фрагменты согласно настоящему изобретению. Последовательности этих молекул нуклеиновый кислоты включают SEQ ID NO: 2-3, 4-7, 16-17, 20-21, 41-49 и 52-53, но не ограничиваются ими.
Указанные молекулы нуклеиновой кислоты согласно настоящему изобретению не ограничены указанными последовательностями, описанными в настоящем документе, но также включают их варианты. Варианты в настоящем изобретении могут быть описаны со ссылкой на их физические свойства при гибридизации. Специалисту в данной области техники понятно, что с использованием методов гибридизации нуклеиновых кислот, нуклеиновые кислоты может применяться для идентификации их комплементов, а также их эквивалентов и гомологов. Также понятно, что гибридизация может происходить при комплементарности, меньшей 100%. Однако при соответствующем выборе условий методы гибридизации могут применяться для различения указанных последовательностей ДНК на основе структурной релевантности к последовательности ДНК конкретного зонда. Руководство по таким условиям смотри Sambrook и др., Molecular Cloning: A Laboratory Manual, 2nd Ed. Cold Spring Harbor Press, Cold Spring Harbor, N.Y., 1989 и Ausubel, F.M., Brent, R., Kingston, R.E., Moore, D.D., Sedman, J.G., Smith, J.A., &Struhl, K. eds. (1995). Current Protocols in Molecular Biology. New York: John Wiley and Sons.
Рекомбинантные нектары и экспрессия
Согласно настоящему изобретению также предложены рекомбинантные конструкции, содержащие одну или более последовательностей нуклеотидов согласно настоящему изобретению. Указанную рекомбинантную конструкцию согласно настоящему изобретению получают посредством вставки указанной молекулы нуклеиновой кислоты, кодирующей указанное антитело согласно настоящему изобретению, в вектор, такой как плазмида, фагмида, фаг или вирусный вектор.
Антитела, предлагаемые в настоящем документе, могут быть получены посредством рекомбинантной экспрессии последовательности нуклеотидов, кодирующей легкую и тяжелую цепи или их части в клетке-хозяине. Для рекомбинантной экспрессии указанного антитела, указанная клетка-хозяин может быть трансфицирована одним или более рекомбинантными векторами экспрессии, несущими последовательность нуклеотидов, кодирующую легкую и/или тяжелую цепи или их части, так, что указанная легкая и тяжелая цепи экспрессируются в указанной клетке-хозяине. Стандартную методологию рекомбинантных ДНК используют для получения нуклеиновых кислот, кодирующих тяжелую и легкую цепи, для включения указанных нуклеиновых кислот в рекомбинантные векторы экспрессии и для введения указанных векторов в клетки-хозяева, например, Sambrook, Fritsch и Maniatis (eds.), Molecular Cloning; A Laboratory Manual, Second Edition, Cold Spring Harbor, N.Y., (1989), Ausubel, F.M. и др. (eds.) Current Protocols in Molecular Biology, Greene Publishing Associates, (1989) и описанную в патенте США №4,816,397 Boss и др.
Подходящими клетками-хозяевами являются прокариотические и эукариотические клетки. Примерами прокариотических клеток-хозяев являются бактерии, а примерами эукариотических клеток-хозяев являются клетки дрожжей, насекомых или млекопитающего. Следует понимать, что дизайн векторной экспрессии, включая выбор регуляторной последовательности, определяется рядом факторов, таких как выбор клетки-хозяина, уровень экспрессии желаемого белка, и является ли экспрессия конститутивной или индуцируемой.
Бактериальная экспрессия
Вектор экспрессии для применения в бактерии конструируют путем вставки структурной последовательности ДНК, кодирующей желаемое антитело вместе с соответствующими сигналами инициации и терминации трансляции и функциональными промоторами в действующую рамку считывания. Вектор содержит один или несколько маркеров фенотипической селекции и точку начала репликации, чтобы обеспечить сохранение вектора и обеспечить амплификацию в хозяине по мере необходимости. Подходящие прокариотические хозяева для трансформации включают несколько видов Е. coli, Bacillus subtilis, Salmonella typhimurium, а также Pseudomonas, Streptomyces и Staphylococcus.
Указанный бактериальный вектор может быть на основе, например, фага, плазмиды или фагмиды. Эти векторы могут содержать маркеры селекции и бактериальные точки начала репликации, которые получены из коммерчески доступных плазмид, которые обычно содержат элементы хорошо известного вектора клонирования pBR322 (АТСС 37017). после трансформации соответствующего штамма-хозяина и выращивания штамма-хозяина до соответствующей плотности клеток выбранный промотор дерепрессируют/индуцируют соответствующим методом (например, изменением температуры или химической индукцией), и клетки культивируют в течение еще некоторого времени. Клетки обычно собирают центрифугированием, разрушают физическими или химическими методами, а полученный неочищенный экстракт сохраняют для дополнительной очистки.
В бактериальной системе множество векторов экспрессии могут быть выгодно подобранными в соответствии с предполагаемым использованием экспрессированного белка. Например, когда необходимо произвести большое количество таких белков для производства антител или для скрининга библиотеки пептидов, например, может потребоваться вектор, который позволяет осуществить высокоуровневую экспрессию слитого белка продукта для легкой очистки.
Экспрессия в клетках млекопитающих и очистка
Предпочтительные регуляторные последовательности для экспрессии в клетках-хозяевах млекопитающих включают вирусные элементы, которые позволяют осуществить высокоуровневую экспрессию белка в клетках млекопитающего, такие как промоторы и/или энхансеры, полученные из цитомегаловируса (CMV) (например, промотор/энхансер CMV), промоторы и/или энхансеры вируса обезьян 40 (SV40) (например, промотор/энхансер SV40), промоторы и/или энхансеры аденовируса (например, главный поздний промотор аденовируса (AdMLP)) и промоторы и/или энхансеры вируса полиомы. Для дополнительного описания вирусных регуляторных элементов и их последовательностей, см., например, патент США №5,168,062, Stinski, патент США №4,510,245, Bell и др., и патента США №4,968,615, Schaffner и др. Указанные рекомбинантные векторы экспрессии также могут включать точку начала репликации и маркер селекции (см., например, патенты США №4,399,216, №4,634,665 и №5,179,017, Axel и др.), подходящие маркеры селекции включают гены, которые обеспечивают устойчивость к лекарственным средствам, таким как G418, гигромицин или метотрексат, клеткам-хозяевам, в которые введен указанный вектор. Например, ген дигидрофолатредуктазы (DHFR) обеспечивает устойчивость к метотрексату, в то время как ген neo обеспечивает устойчивость к G418.
Трансфекцию указанного вектора экспрессии в клетки-хозяева можно проводить с использованием стандартных методов, таких как электропорация, осаждение с фосфатом кальция и трансфекция DEAE-декстраном.
Подходящие клетки-хозяева млекопитающих для экспрессии представленного в настоящем документе антитела включают клетки яичника китайского хомячка (клетки СНО) [включая клетки dhfr-CHO, описанные Urlaub и Chasin, (1980) Proc. Natl. Acad. Sci. USA 77:4216-4220, с применением маркера селекции DHFR, как описано, например, в R.J. Kaufman и Р.А. Sharp (1982) Mol. Biol. 159:601-621], клетки миеломы NSO, клетки COS и клетки SP2.
Антитела согласно настоящему изобретению могут быть выделены и очищены от культуры рекомбинантных клеток известными способами, включая осаждение сульфатом аммония или этанолом, кислотную экстракцию, аффинную хроматографию на белке А, аффинную хроматографию на белке G, анионо- или катионообменную хроматография, хроматографию на фосфоцеллюлозе, хроматографию гидрофобного взаимодействия, аффинную хроматографию, хроматографию на гидроксиапатите, хроматографию на лектинах, но не ограничиваются ими. Высокоэффективная жидкостная хроматография (ВЭЖХ) также может применяться для очистки. См., например, Colligan, Current Protocols in Immunology, или Current Protocols in Protein Science, John Wiley & Sons, NY, N.Y., (1997-2001), например, главы 1, 4, 6, 8, 9, и 10, каждая из которых полностью включена в настоящий документ посредством ссылки.
Характеристики и функции антитела согласно настоящему изобретению
Осуществляли анализ характеристик и функциональный анализ Fab гуманизированного антитела VEGF-H988 согласно настоящему изобретению. Анализ показал, что указанное антитело согласно настоящему изобретению обладает следующими преимуществами:
(1) способность Fab VEGF-H988 связывать белок VEGF165 сходна с Луцентисом;
(2) аффинность связывания Fab VEGF165-H988 с белком VEGF 165 выше, чем у Луцентиса, а именно примерно в 3,75 раза выше, чем у Луцентиса;
(3) Fab VEGF165-H988 специфически связывается с рекомбинантным белком VEGF165 человека и не связывается перекрестно с рекомбинантным белком mVEGF164 мыши;
(4) Fab VEGF165-H988 может эффективно ингибировать связывание белка VEGFR2 с белком VEGF165, и его ингибирующая способность выше, чем у Луцентиса;
(5) нейтрализующая активность Fab VEGF-H988 сильнее, чем у Луцентиса при различных концентрациях рекомбинантного VEGF165 человека; его нейтрализующая активность выше, чем у EYLEA при высоких концентрациях VEGF165; его нейтрализующая активность выше, чем у Бевацизумаба и Бролуцизумаба при разных концентрациях VEGF165, но сравнима с активностью Конберцепта.
Применения
Антитела согласно настоящему изобретению могут применяться для лечения заболеваний, ассоциированных с ангиогенезом, включая болезни глаз, характеризующиеся хориоидальной неоваскуляризацией, включая возникновение возрастной дегенерации желтого пятна (AMD), диабетический макулярный отек (DME), отек сетчатки, дегенеративную близорукость и хориоидальную неоваскуляризацию (CNV), но не ограничиваются ими.
Фармацевтические композиции
Антитела согласно настоящему изобретению могут быть получены с по меньшей мере одним другим агентом (например, стабильным соединением) с образованием фармацевтической композиции, содержащий антитело согласно настоящему изобретению и один или более фармацевтически приемлемые носители, разбавители или вспомогательные вещества. Указанная фармацевтическая композиция необязательно может содержать дополнительные терапевтические агенты.
Наборы
Настоящее изобретение также относится к фармацевтическому набору и набору, содержащему один или более контейнеров, где указанные контейнеры содержат указанные выше фармацевтические композиции согласно настоящему изобретению. К таким контейнерам могут прилагаться спецификации по форме, установленной государственным органом, регулирующим производство, использование или распространение лекарственного средства или биологического продукта, которые отражают одобрение его введения человеку указанным органом страны, в которой указанный продукт производится, используется или распространяется.
Получение и хранение
Указанная фармацевтическая композиция согласно настоящему изобретению может быть получена способом, известным в данной области техники, например, традиционными методами смешивания, растворения, грануляции, приготовления пастилок, измельчения, эмульгирования, инкапсуляции, заливки или лиофилизации.
Будучи приготовленными, фармацевтические композиции, содержащие соединения согласно настоящему изобретению, изготовленные в виде лекарственной формы в приемлемом носителе, они могут быть помещены в соответствующие контейнеры и помечены для лечения указанного состояния. Такая маркировка должна включать количество, частоту и способы введения препарата.
Комбинации
Указанная фармацевтическая композиция, содержащая антитела согласно настоящему изобретению, описанные выше, также может быть скомбинирована с одним или более другими терапевтическими агентами, такими как противоопухолевые агенты, если при этом полученная комбинация не вызывает неприемлемых побочных эффектов.
Последующие примеры способствуют лучшему пониманию настоящего изобретения, но не предназначены для ограничения рамок настоящего изобретения. Все экспериментальные методы в последующих примерах, если не указано иное, являются обычными методами. Экспериментальные материалы, использованные в последующих примерах, если не указано иное, были приобретены у обычных дистрибьюторов биохимических реагентов.
ПРИМЕРЫ
Пример 1: Скрининг антител кролика, которые блокируют связывание VEGF165 с VEGFR1/VEGFR2 с использованием библиотека фагового дисплея антител
1.1 Иммунизация кроликов
Рекомбинантный белок VEGF165 человека (от Sino Biological, Inc, кат. №11066-HNAH) использовали для иммунизации кроликов. Последовательность аминокислот внеклеточного участка Metl-Arg191 белка VEGF165 человека (UniProt Р15692-4) приведена в SEQ ID NO: 1.
Подробное описание метода следующее: рекомбинантный белок VEGF165 человека смешивали с адъювантом Фрейнда, кроликов подкожно иммунизировали указанной смесью 4 раза с интервалами 3 недели, 2 недели и 2 недели, соответственно, дозировкой 500 мкг каждый раз. После четвертой иммунизации кровь собирали через 4 дня после иммунизации через медиальное кантальное сплетение глаза. Титр антитела против VEGF165 в сыворотке кролика измеряли методом ИФА с использованием покрытия рекомбинантным белком VEGF165 человека. Титр в сыворотке после четвертой иммунизации достигал 1:250000, и кроликам вводили бустер внутривенно с 25 мкг рекомбинантного белка VEGF165 человека через 9 недель после четвертой иммунизации. Через 7 дней, кроликов умерщвляли, а ткань селезенки удаляли и замораживали в жидком азоте.
1.2 Скрининг библиотеки фагового дисплея антител
РНК экстрагировали из ткани селезенки кролика с использованием TriPure Isolation Reagent (от Roche, кат. №11 667 165 001), и получали кДНК обратной транскрипцией РНК с использованием набора для обратной транскрипции (от Invitrogen кат. №18080-051). Конструировали 10 пар праймеров для амплификации последовательности вариабельного участка легкой цепи антитела кролика и 4 пары праймеров для амплификации последовательности вариабельного участка тяжелой цепи (Barbas С F и др., CSHL Press. 2004). Указанные последовательности, кодирующую вариабельные участки легкой и тяжелой цепей антитела кролика объединяли с последовательностью нуклеотидов, кодирующей scFv, с помощью ПНР с удлинением перекрывающихся участков, вариабельные участки легкой и тяжелой цепей соединяли (Jones S Т и др., Bio/technology. 1991; 9(1): 88) с помощью следующего линкера:
TCTAGTGGTGGCGGTGGTTCGGGCGGTGGTGGAGGTGGTAGTTCTAGATCTTCC
(SSGGGGSGGGGGGSSRSS) (SEQ ID NO: 2);
затем лигировали в фаговый вектор pComb3x (от Sino Biological, Inc.) с помощью эндонуклеазы рестрикции SfiI (от Fermentas), и электротрансформировали в компетентые клетки Х-Blue для получения библиотеки фагового дисплея антител scFv кролика. Рекомбинантный белок VEGF165 человека наносили в качестве покрытия на чашку для ИФА, и фаговую библиотеку, обогащенную положительными антителами против VEGF165, скринировали в соответствии с указанным фаговым способом сортировки антител (O'Brien, РМ, & Aitken, R. (Eds.), Springer Science & Business Media. 2002; ISBN: 9780896037113). Для экспрессии из обогащенной библиотеки были отобраны одиночные колонии фагов, а их связывание с рекомбинантным белком VEGF165 человека определяли с помощью ИФА. Клон антитела, которое специфически связывается с рекомбинантным VEGF165 человека, отбирали и направляли в компанию, осуществляющую услуги по секвенированию, в результате чего получали последовательность нуклеотидов антител (SEQ ID NO: 3) антитела scFv VEGF165-R988.
1.3 Получение антитела кролика, связывающегося с VEGF165
указанную последовательность нуклеотидов, кодирующую вариабельный участок тяжелой цепи scFv антитела VEGF165-R988, амплифицировали с помощью ПЦР и вставляли в вектор pSTEP2, разрезанный ScaI + KpnI (Fermentas), несущий последовательность нуклеотидов, кодирующую сигнальный пептид тяжелой цепи (SEQ ID NO: 43) и константный участок тяжелой цепи IgG1 кролика (SEQ ID NO: 6), методом слияния с получением вектора экспрессии тяжелой цепи (SEQ ID NO: 52). Указанную последовательность нуклеотидов, кодирующую вариабельный участок легкой цепи scFv антитела VEGF165-R988, амплифицировали с помощью ПЦР и вставляли в pSTEP2 вектор, разрезанный ScanI + BamHI (Fermentas), несущий последовательность нуклеотидов, кодирующую сигнальный пептид легкой цепи (SEQ ID NO: 44) и константный участок каппа легкой цепи кролика (SEQ ID NO: 7), методом слияния с получением вектора экспрессии легкой цепи (SEQ ID NO: 53). Указанные рекомбинантные плазмиды экстрагировали, трансфицировали ими клетки HEK-293 и выращивали для экспрессии в течение 7 суток, супернатант культуры очищали на колонке с белком А в результате чего получали высокоочищенные антитела.
Праймеры для амплификации вариабельного участка тяжелой цепи:

Праймеры для амплификации вариабельного участка легкой цепи:

1.4 Функциональный анализ антитела кролика, связывающегося с VEGF165
1.4.1 Антитело кролика блокирует связывание VEGF165 с VEGFR2-his
Белок VEGF165 (от SinoBiological, Inc.) при концентрации 1 мкг/мл наносили в качестве покрытия на 96-луночный планшет в количестве 100 мкл/на лунку в течение ночи при 4°С. Планшет промывали на следующий день и блокировали при комнатной температуре в течение 1 ч. Добавляли 100 мкл раствора белка VEGFR2-биотин концентрации 5 мкг/мл (от SinoBiological, Inc.) и указанного антитела VEGF-R988 при различных концентрациях и совместно инкубировали. Планшет промывали для удаления несвязавшихся антител, инкубировали со стрептавидином/HRP (от Beijing ZSGB-Bio Co., Ltd.), а затем несколько промывали, и раствор хромогенного субстрата добавляли для появления окраски. Измеряли OD450 после появления окраски. Принимая концентрацию антитела кролика, связывающегося с VEGF165, в качестве горизонтальной координаты и степень ингибирования PI% в качестве вертикальной координаты, использовали программное обеспечение graphPad Prism 6.0 для анализа данных и строили график кривой зависимости. Степень ингибирования (%) = (ODkoнтpoль - ODобразец)/ODконтроль× 100%, где ODконтроль обозначает значение OD для лунок только с добавленным VEGFR2-биотин, но без антитела кролика, а ODобразец обозначает значение OD лунки с добавленными VEGFR2-биотин и антителом кролика.
Как видно на Фиг. 1, антитело VEGF-R988 может эффективно связываться с покрытием из белка VEGF165 и может эффективно ингибировать связывание белка VEGFR165 с белком VEGFR2.
1.4.2 Антитело кролика ингибирует пролиферацию HUVEC, вызванную VEGF165
Эффект антитела, нейтрализующего пролиферацию эндотелиальных клеток пупочной вены, индуцированную VEGF165, определяли с использованием метода WST-8. Эндотелиальные клетки HUVEC пупочной вены человека инокулировали в 96-луночный планшет в количестве 4×103 клеток на лунку, выращивали в среде M199, содержащей 10% ФБС и 5% L-Gln в течение 4 часов, а затем различные концентрации антитела VEGF-R988 добавляли в количестве 50 мкл на лунку, затем VEGF-165 в конечной концентрации 10 нг/мл добавляли в количестве 10 мкл на лунку, 96-луночный планшет инкубировали при 37°С, 5% СО2 в инкубаторе клеток в течение 3 суток, и использовали пустую лунку В (без клеток), отрицательный контроль М (клетки инокулировали, без образца антитела, добавлен VEGF-165) и М' (клетки инокулировали, без образца антитела и без VEGF-165). После инкубации добавляли 10 мкл на лунку хромогенного раствора WST-8, и указанный 96-луночный планшет инкубировали в инкубаторе СО2 до появления окраски, измеряли OD450 и OD630 с помощью считывателя микропланшетов после стабилизации появления окраски. Для каждой лунки считанное значение составляло (OD450 - OD630), и степень нейтрализации для указанного антитела, рассчитанное как значение OD для каждой группы, определяли как считанное значение группы минус считанное значение пустой лунки В, степень нейтрализации % = (значение OD отрицательного контроля М - значение OD образца) / (значение OD отрицательного контроля М - значение OD М') × 100%. Стандартную кривую рассчитывали с использованием функции автоматического анализа статистического программного обеспечения GraphPad Prism, выбирая концентрацию образца указанного антитела в качестве горизонтальной координаты и степень нейтрализации в качестве вертикальной координаты, и уравнение логистической регрессии с четырьмя параметрами использовали для соответствия стандартной кривой «S» для расчета половины максимальной эффективной концентрации (ЕС50) образца указанного антитела.
Результаты, показанные на Фиг. 2 демонстрируют, что антитело VEGF-R988 эффективно снижает способность VEGF165 инициировать пролиферацию HUVEC.
Пример 2: Гуманизация, модификация антитела VEGF-R988 кролика и получение его Fab фрагмента
2.1 Определение последовательностей CDR легкой и тяжелой цепей антитела VEGF-R988 кролика
На основании указанной последовательности нуклеотидов scFv антитело VEGF-R988, определенной в Примере 1.2, получали последовательность аминокислот вариабельных участков тяжелой цепи и легкой цепи VEGF-R988 scFv, смотри SEQ ID NO: 8/9.
Принимая во внимание индекс Кабат и систему нумерации IMGT, определяли последовательность аминокислот каждого трех CDR легкой и тяжелой цепей антитела кролика VEGF-R988-scFv, см. Таблицу 1. Указанные выше соответствующие три CDR указанной легкой цепи и тяжелой цепи переносили в гуманизированное антитело VEGF-R988-scFv в последующих стадиях, см. Пример 2.2.

2.2 Гуманизация антитела VEGF-R988 кролика посредством трансплантации CDR Гуманизацию антитела кролика осуществляли с использованием классического способа гуманизации трансплантацией CDR. Вариабельные участки легкой или тяжелой цепи антитела, человека, чьи последовательности являются наиболее близкими указанным последовательностям вариабельных участков легкой или тяжелой цепи кролика, выбирали в качестве матрицы, и каждый из трех CDR (Таблица 1) легкой или тяжелая цепи кролика вставляли в указанные вариабельные участки антитела человека, в результате чего получали последовательности гуманизированного вариабельного участка легкой цепи (VL) или вариабельного участка тяжелой цепи (VH), соответственно. Матрицей человека для вариабельного участка легкой цепи VEGF-R988 является IGKV1-27*01, который на 65,30% гомологичен легкой цепи VEGF-R988, и матрицей человека для вариабельного участка тяжелой цепи является IGHV4-4*08, который на 53,20% гомологичен тяжелой цепи VEGF-R988.
2.3 Обратные мутации в каркасном участке гуманизированного вариабельного участка
Поскольку некоторые ключевые аминокислоты в каркасном участке кролика необходимы для поддержания активности CDR, ключевые аминокислоты были обратно мутированы в соответствующие последовательности аминокислот антитела кролика, следующие сайты были обратно мутированы: в легкой цепи, позиция 1 была обратно мутировала в Е, позиция 2 была обратно мутирована в L, позиция 4 была обратно мутирована в L, и позиция 63 была обратно мутирована в К; в то время как в тяжелой цепи позиция 3 была обратно мутирована в V, позиция 37 была обратно мутирована в V, позиция 47 была обратно мутирована в Y, позиция 78 была обратно мутирована в V, позиция 79 была обратно мутирована в D, и позиция 91 была обратно мутирована в F; все вышеперечисленные сайты были пронумерованы по схеме нумерации Кабата. Гуманизированное антитело VEGF-H988-10 получали гуманизированной трансплантацией CDR и обратными мутациями каркасного участка.
2.4 Получение гуманизированного моноклонального антитела VEGF-H988-10 и аффинная модификация CDR
Вариабельный участок тяжелой цепи VEGF-H988-10 (SEQ IN NO: 20) получали методом полногенного синтеза, а затем вводили инфузионным методом в предварительно разрезанный ScaI + NheI (Fermentas) вектор pSTEP2 вектор, несущий последовательность нуклеотидов, кодирующую сигнальный пептид тяжелой цепи (SEQ ID NO: 43) и последовательность нуклеотидов, кодирующую константный участок IgGl человека (SEQ ID NO: 47), в результате чего получали вектор экспрессии тяжелой цепи VEGF-H988-10 (SEQ ID NO: 16). Вариабельный участок легкой цепи VEGF-H988-10 (SEQ ID NO: 21) получали методом полногенного синтеза, а затем вводили инфузионным методом в предварительно разрезанный ScaI + BsiWI (Fermentas) вектор pSTEP2 вектор, несущий последовательность нуклеотидов, кодирующую сигнальный пептид легкой цепи (SEQ ID NO: 44) и последовательность нуклеотидов, кодирующую константный участок каппа человека (SEQ ID NO: 48), в результате чего получали вектор экспрессии легкой цепи VEGF-H988-10 (SEQ ID NO: 17). Плазмиды экстрагировали и совместно трансфицировали в клетки HEK-293, указанный клетки выращивали в течение 7 суток. Супернатант культуры очищали на колонке с белком А, в результате чего получали высокоочищенные антитела.
Праймеры для синтеза полного гена вариабельного участка тяжелой цепи

Праймеры для синтеза полного гена вариабельного участка легкой цепи

Для улучшения аффинности VEGF-H988-10, конструировали библиотеки SDM участков CDR вариабельных участков тяжелой и легкой цепи (включая три библиотеки, насыщенные мутациями LCDR1, LCDR3, и HCDR2); тем временем, для улучшения химической стабильности указанного антитела, остатки аминокислот, способные подвергаться дезамидированию или изомеризации, должны быть модифицированы другими остатками аминокислот. Дезаминирование аспарагина может происходить, такого как NG, NS, NA, NT и т.д., что приводит к образованию остатков изоаспарагиновой кислоты, влияющих на стабильность или биологическую функцию указанного антитела. Вариабельный участок HCDR3 VEGF-H988 содержит (а) участок(ки), восприимчивый(е) к дезамидированию, поэтому конструировали библиотеки SDM для улучшения химической стабильности и биологической функции указанного антитела. Вышеупомянутые четыре библиотеки мутантов сконструировали в форме scFv и клонировали в фаговый вектор в виде слитого белка scFv-gIII; для каждого CDR, скринировали клоны CDR, имеющие оптимальную способность к связыванию с растворимым антигеном VEGF, и наконец получали указанное антитело VEGF-H988, содержащее оптимизированную аффинность и стабильность CDR. Указанные последовательности CDR легкой и тяжелой цепи VEGF-H988 показаны в Таблице 2.

2.5 Получение гуманизированного Fab фрагмента антитела VEGF-H988
Последовательность нуклеотидов (SEQ ID NO: 42), кодирующую легкую цепь и сигнальный пептид указанного Fab фрагмента VEGF-H988, которая в следующем порядке содержит последовательность нуклеотидов, кодирующую сигнальный пептид легкой цепи (SEQ ID NO: 44), вариабельный участок легкой цепи гуманизированного антитела (SEQ IN NO: 46) и константный участок каппа легкой цепи антитела человека (SEQ IN NO: 48), амплифицировали с помощью ПЦР и вставляли в саморазвитый вектор pGS (KpnI+XbaI) инфузионным методом, и правильность последовательности плазмиды проверяли секвенированием. Последовательность нуклеотидов (SEQ ID NO: 41), кодирующую тяжелую цепь и сигнальный пептид указанного выше Fab фрагмента антитела VEGF-H988, которая в следующем порядке содержит последовательность нуклеотидов, кодирующую сигнальный пептид тяжелой цепи (SEQ ID NO: 43), вариабельный участок тяжелой цепи гуманизированного антитела (SEQ ID NO: 45) и константный участок СН1 тяжелой цепи IgGl человека (SEQ ID NO: 49), амплифицировали с помощью ПЦР и инфузионным методом вставляли в вектор pGS (NheI+NotI), который содержит легкую цепь, правильность последовательности которой была проверена, и правильность последовательностей векторов, экспрессирующих как легкую, так и тяжелую цепи Fab фрагмента VEGF-R988, проверяли секвенированием. Эти векторы экспрессии являются эукариотическими векторами экспрессии, содержащими гены GS в качестве маркеров селекции и элементы экспрессии легкой и тяжелой цепи указанного антитела. Эти векторы экспрессии трансфицировали в CHO-K1-GS-дефицитные клетки и линии клеток с высокой экспрессией Fab фрагмента VEGF-H988 получали в результате скрининга MSX. Клоны с высокой экспрессией антитела экспрессия отбирали методом ИФА, и указанные линии клеток с высокой экспрессией выбирали, принимая во внимание как статус роста клеток, так и ключевые характеристики качества для лекарственного препарата антитела. Суспензионные культуры без сыворотки использовали для выращивания линии клеток СНО, продуцирующей Fab фрагмент VEGF-H988, в результате чего получали Fab фрагменты VEGF-H988 высокой чистоты и качества.
Пример 3: Характеристический анализ Fab фрагмента гуманизированного антитела VEGF-H988
3.1 Характеристический анализ Fab фрагмента гуманизированного антитела VEGF-H988, связывающегося с VEGF165
3.1.1 Fab фрагмент VEGF-H988 специфически связывается с VEGF 165 Рекомбинантный белок VEGF 165 человека (от SinoBiological, Inc.) при различных концентрациях (0,15 нг/мл, 0,46 нг/мл, 1,37 нг/мл, 4,12 нг/мл, 12,35 нг/мл, 37,04 нг/мл, 111,11 нг/мл, 333,33 нг/мл, 1000 нг/мл и 3000 нг/мл) наносили на 96-луночный планшет в течение ночи при 4°С в количестве 100 мкл на лунку. Планшет промывали на следующий день и блокировали при комнатной температуре в течение 1 часа. После инкубации с 100 мкл 1 мг/мл Fab фрагмента VEGF165-H988, Луцентис (от Norvatis) или Fab фрагментом H7N9-R1 в качестве отрицательного контроля (от SinoBiological, Inc.) соответственно, планшет промывали для удаления несвязавшихся антител, затем инкубировали с F(ab')2 козы против IgG F(ab')2/HRP человека (от Jackson ImmunoResearch Laboratories, Inc.) и промывали несколько раз, а затем добавляли раствор хромогенного субстрата до появления окраски. OD450 измеряли после завершения образования окраски, человека в качестве горизонтальной координаты и значение OD450 в качестве вертикальной координаты, использовали программное обеспечение graphPad Prism 6.0 для получения кривой графика "S" и анализировали связывание фрагмента антитела с рекомбинантным белком VEGF165 человека.
Результаты, показанные на Фиг. 3, демонстрируют, что значение EC50 Fab фрагмента гуманизированного VEGF165-H988, специфически связывающегося с рекомбинантным VEGF165 человека, составляет 18,75 нг/мл, R2=0,993; значение ЕС50 для Луцентиса, связывающегося с рекомбинантным белком VEGF165 человека составляет 12,87 нг/мл, R2=0,989. Это указывает на то, что способность Fab фрагмента VEGF165-H988 связываться с рекомбинантным белком VEGF165 человека сходна со способностью к связыванию Луцентиса. Fab фрагмент H7N9-R1, использованный в качестве отрицательного контроля, не обладает способностью к связыванию с рекомбинантным белком VEGF165 человека.
3.1.2 Тест на аффинность связывания Fab фрагмента VEGF-H988 с белком VEGF 165
Аффинности Fab фрагмента VEGF165-H988 и Луцентиса измеряли при различных концентрациях с использованием сенсора, покрытого стрептавидином, и иммобилизованного белка VEGF165, меченного биотином.
Сначала рекомбинантный белок VEGF165 человека метили биотином в молярном соотношении 1:2 в ходе следующего процесса: буфер (20 мМ Трис, 150 мМ NaCl, рН 8,0) рекомбинантного белка VEGF замещали ФБС методом ультрафильтрации в пробирке для ультрафильтрации центрифугированием 5000 MW и получали 567,57 мкг белка, измеренного методом количественной оценки с помощью УФ, полученные белки смешивали с 20 мМ раствором биотина в молярном соотношении 1:2 для инкубации в течение 30 мин при комнатной температуре в темноте, затем снова фильтровали в пробирке для ультрафильтрации центрифугированием 5000 MW для удаления несвязавшегося биотина. После оценки количества методом УФ, меченые биотином белки получали путем добавления равного объема глицерина и БСА до финальной концентрации 0.1%. Концентрация белка VEGF165, определенная с помощью УФ, составляла 2,08 мг/мл.
Затем измеряли аффинности Fab фрагмента VEGF165-H988 и Луцентиса при различных концентрациях с биотинилированными рекомбинантными белками VEGF человека, и полученные значения KD означали финальные аффинности.
Результаты, показанные в Таблице 3 демонстрируют, что, значение аффинности связывания KD Fab фрагмента VEGF-H988 с рекомбинантным белком VEGF165 человека составило 1,54 Е-10 М, значение константы связывания kon составило 2,74 Е+05 М-1с-1, и значение константы диссоциации kdis составило 4,21 Е-05 с-1; значение аффинности связывания KD Луцентиса с белком VEGF165 составило 5,78 Е-11 М, значение константы связывания kon составило 5,36 Е+04 М-1с-1, а значение константы диссоциации kdis составило 3,10 Е-05 с-1, как показано в Таблице 3. На основании этих результатов можно сделать вывод, что аффинность Fab фрагмента VEGF-H988 была выше, чем у Луцентиса, а именно примерно в 3,75 раз выше, чем у Луцентиса. Таким образом, Fab фрагмент VEGF-H988 обладает более сильной способностью к связыванию белка VEGF165, чем Луцентис.

3.1.3 Определение видоспецифичной кросс-реактивности Fab фрагмента VEGF165-H988
Рекомбинантный белок VEGF 165 человека или рекомбинантный белок mVEGF164 мыши (от Sino Biological., Inc.) разбавляли до 0,1 мкг/мл, 1 мкг/мл и 10 мкг/мл, соответственно, и наносили на 96-луночный планшет в течение ночи при 4°С в количестве 100 мкл на лунку. Планшет промывали на следующий день, блокировали при комнатной температуре в течение 1 ч. 100 мкл Fab фрагмента VEGF165-H988, Луцентис или отрицательный контроль H7N9-R-Fab добавляли соответственно в концентрации 2 мкг/мл и инкубировали в течение 1 ч. Планшет промывали для удаления несвязавшихся антител. Планшет инкубировали с F(ab')2 козы против IgG F(ab')2/HRP человека (Jackson ImmunoResearch Laboratories, Inc.) и затем несколько раз промывали, и добавляли раствор хромогенного субстрата до появления окраски. После завершения образования окраски измеряли OD450. Выбирая концентрацию белка в качестве горизонтальной координаты и значение OD450 в качестве вертикальной координаты, использовали программное обеспечение graphPad Prism 6.0 для построения гистограммы.
Результаты, показанные на Фиг. 4 демонстрируют, что Fab фрагмент VEGF165-Н988 специфически связывается с рекомбинантным белком VEGF165 человека обладает перекрестным связыванием с рекомбинантным белком mVEGF164 мыши; в то время как Луцентис не демонстрирует перекрестного связывания с рекомбинантным белком mVEGF164 мыши.
3.2 Свойства Fab фрагмента VEGF-H988 по блокированию рецептора
Белок VEGF165 в концентрации 1 мкг/мл наносили на 96-луночный планшет в количестве 100 мкл/на лунку в течение ночи при 4°С. На следующий день планшет промывали и блокировали при комнатной температуре в течение 1 ч. В каждую лунку добавляли 100 мкл 2 мкг/мл белка VEGFR2-his (от SinoBiological, Inc.), различные концентрации Fab фрагмента гуманизированного VEGF-H988, Луцентиса или H7N9-R1-Fab в качестве отрицательного контроля (от SinoBiological, Inc.), соответственно и совместно инкубировали. Планшет промывали для удаления несвязавшихся антител. Планшет инкубировали с C-his-R023/HRP и затем несколько раз промывали, и добавляли раствор хромогенного субстрата до появления окраски. Для каждой исследованной группы измеряли OD450 в двух повторах после стабилизации появления окраски. Используя концентрацию указанного антитела в качестве горизонтальной координаты и степень ингибирования Р1% в качестве вертикальной координаты, использовали программное обеспечение graphPad Prism 6.0 для анализа данных и строили график кривой зависимости для расчета значения IC50. Степень ингибирования (%)=(ODконтроль-ODобразец)/ODконтроль×100%, где ODконтроль обозначает значение OD лунки только с добавленным VEGFR2-his, но без добавки гуманизированного антитела, а ODобразец обозначает значение OD лунки как с добавленным VEGFR2-his, так и с гуманизированным антителом.
Как видно на Фиг. 5, белок VEGFR2 может эффективно связываться с белком VEGF165 на подложке, a Fab фрагмент VEGF-H988 может эффективно ингибировать связывание белка VEGFR2 с белком VEGFR165, с заметно более высокой ингибирующей способностью, чем Луцентис, при это указанный отрицательный контроль не демонстрировал какой-либо ингибирующий эффект.
3.3 Fab фрагмент VEGF-H988 блокирует активность VEGF165 при различных концентрациях
Эффект гуманизированного Fab фрагмента по нейтрализации пролиферации клеток HUVEC, вызванной VEGF 165, определяли с использованием метода WST-8. Клетки HUVEC инокулировали в 96-луночный планшет в количестве 4×103 клеток на лунку, выращивали в среде М199, содержащей 10% ФБС и 5% L-Gln в течение 4 ч, а затем добавляли гуманизированный Fab фрагмент при различных концентрациях в количестве 50 мкл на лунку, затем добавляли VEGF-165 в конечных концентрациях 1000 нг/мл, 100 нг/мл или 10 нг/мл в количестве 10 мкл на лунку, указанный 96-луночный планшет инкубировали при 37°С, 5% СО2 в инкубаторе клеток в течение 3 суток, и использовали контрольную лунку В (без клеток), отрицательный контроль М (с добавлением клеток, без добавления образца антитела, с добавлением VEGF-165) и М' (с добавлением клеток, без добавления образца антитела и без добавления VEGF-165). После инкубации добавляли 10 мкл на лунку хромогенного раствора WST-8 и указанный 96-луночный планшет инкубировали в инкубаторе с СО2 до появления окраски, OD450 и OD630 измеряли с помощью считывателя микропланшетов после стабилизации появления окраски. Для каждой лунки считывали значение (OD450-OD630), и степень нейтрализации указанного антитела рассчитывали как значение OD для каждой группы, определенное как считанное значение указанной группы минус считанное значение пустой лунки В, степень нейтрализации, % = (значение OD отрицательного контроля М - значение OD образца) / (значение OD отрицательного контроля М - значение OD М') × 100%. Стандартную кривую рассчитывали с использованием функции автоматического анализа статистического программного обеспечения GraphPad Prism, используя концентрацию образца указанного антитела в качестве горизонтальной координаты и степень нейтрализации в качестве вертикальной координаты, и использовали уравнение логистической регрессии с 4 параметрами для построения стандартной кривой «S» для расчета половины максимальной эффективной концентрации (ЕС50) образца указанного антитела.
Как видно из Фиг. 6-10 и Таблицы 4, нейтрализующая активность Fab фрагмента VEGF-H988 была сильнее при всех различных концентрациях рекомбинантного белка VEGF165 человека, чем Луцентис. Когда концентрации VEGF165 была 1000 нг/мл или 100 нг/мл, нейтрализующая активность Fab фрагмента VEGF-H988 была сильнее, чем активность EYLEA, но когда концентрация VEGF165 была 10 нг/мл, нейтрализующая активность Fab фрагмента VEGF-H988 была немного слабее, чем EYLEA. Нейтрализующая активность Fab фрагмента VEGF-H988 была превосходной при различных концентрациях рекомбинантного белка VEGF165 человека по сравнению с Бевацизумабом и Бролуцизумаб, и сравнимой с активностью Конберцепта. При повышении концентрации VEGF165, Fab фрагмент VEGF-H988 сохранял высокую степень максимальной нейтрализации, в то время как EYLEA и Авастин демонстрировали снижение активности.

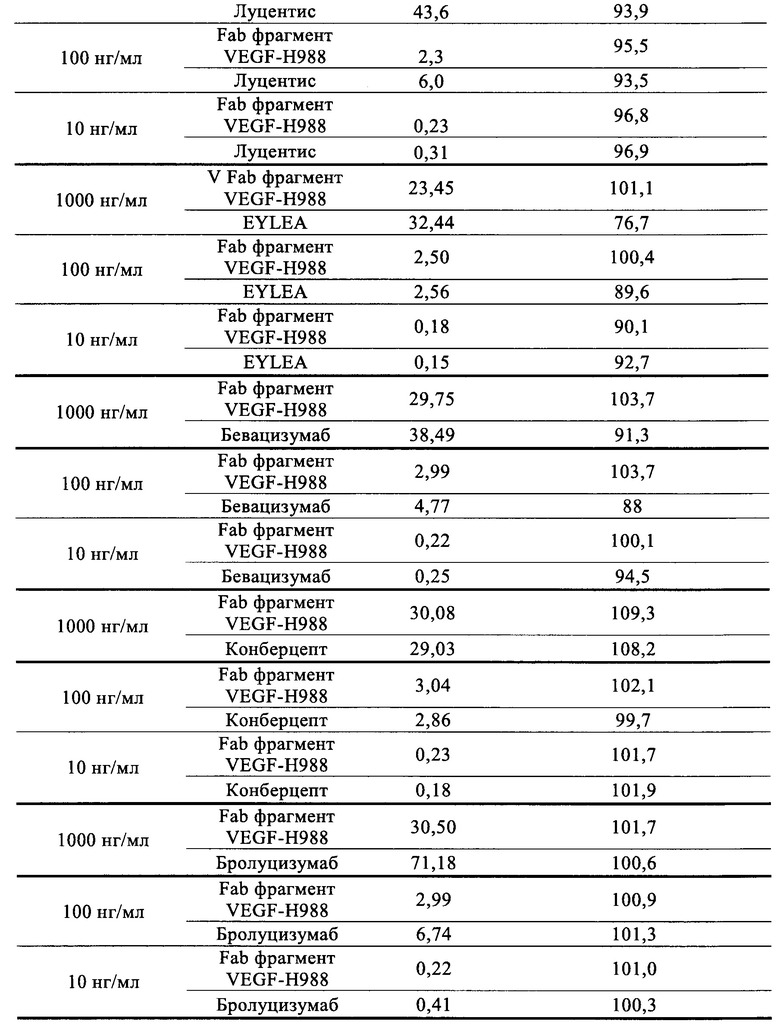
Перечень последовательностей


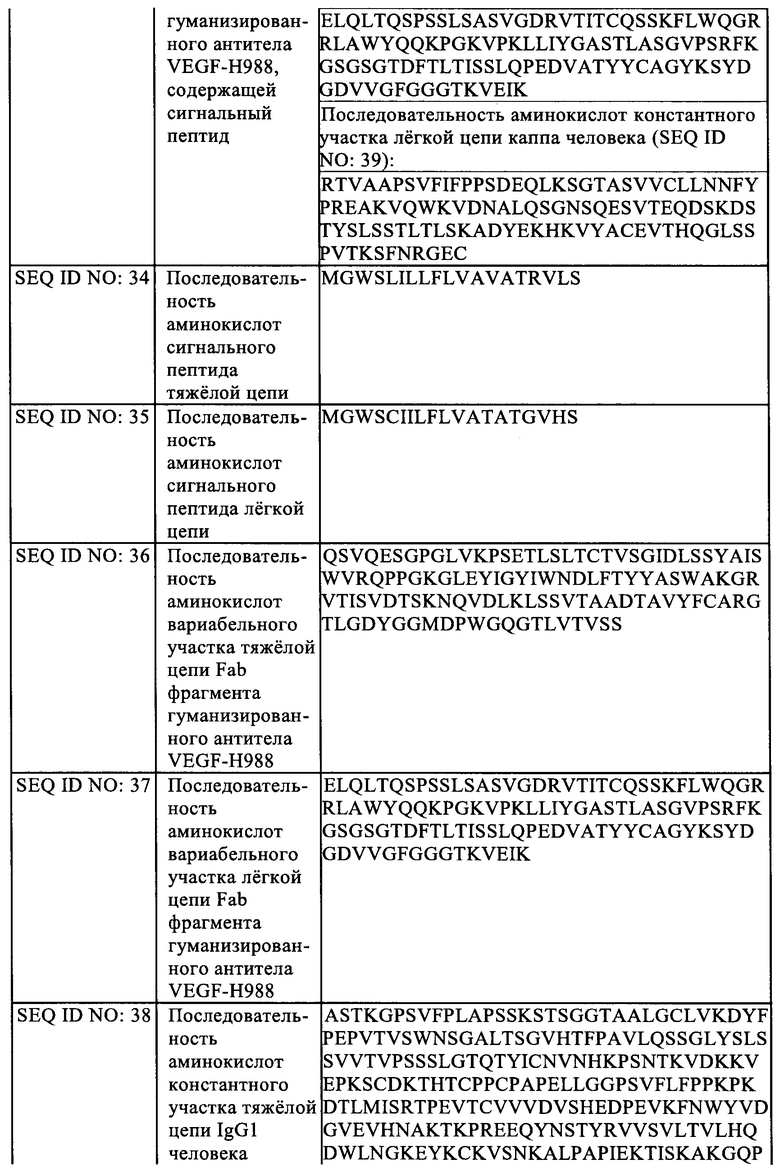
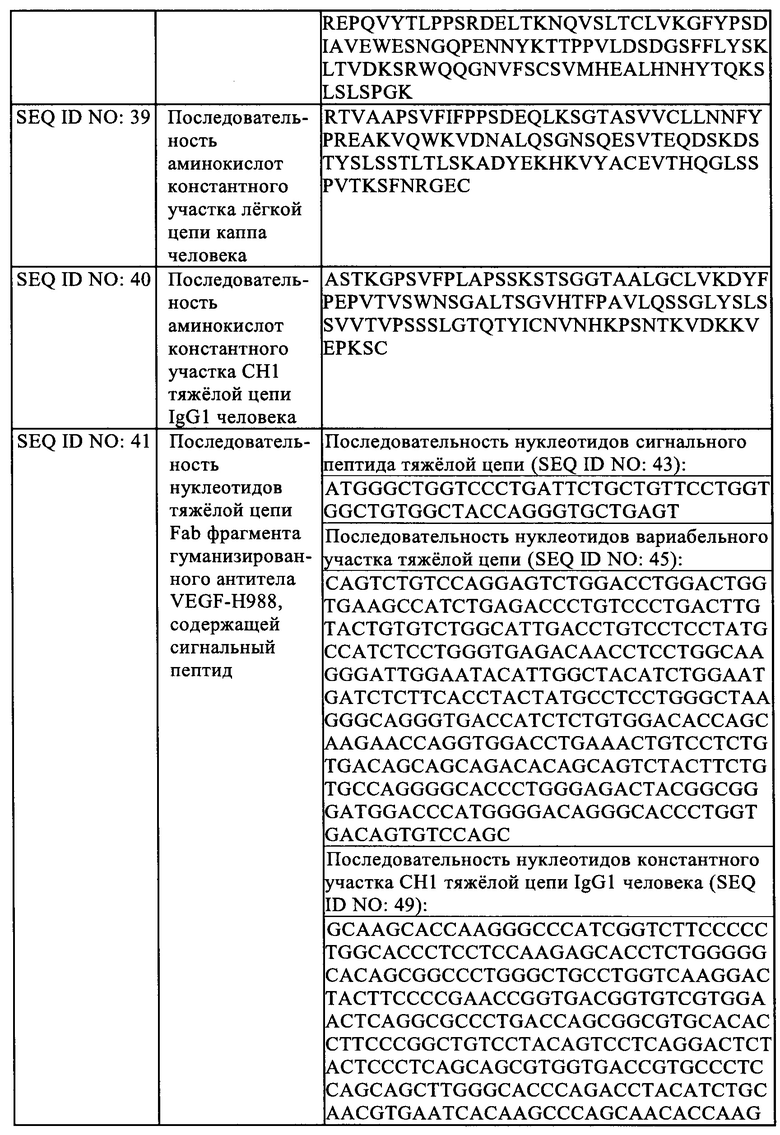


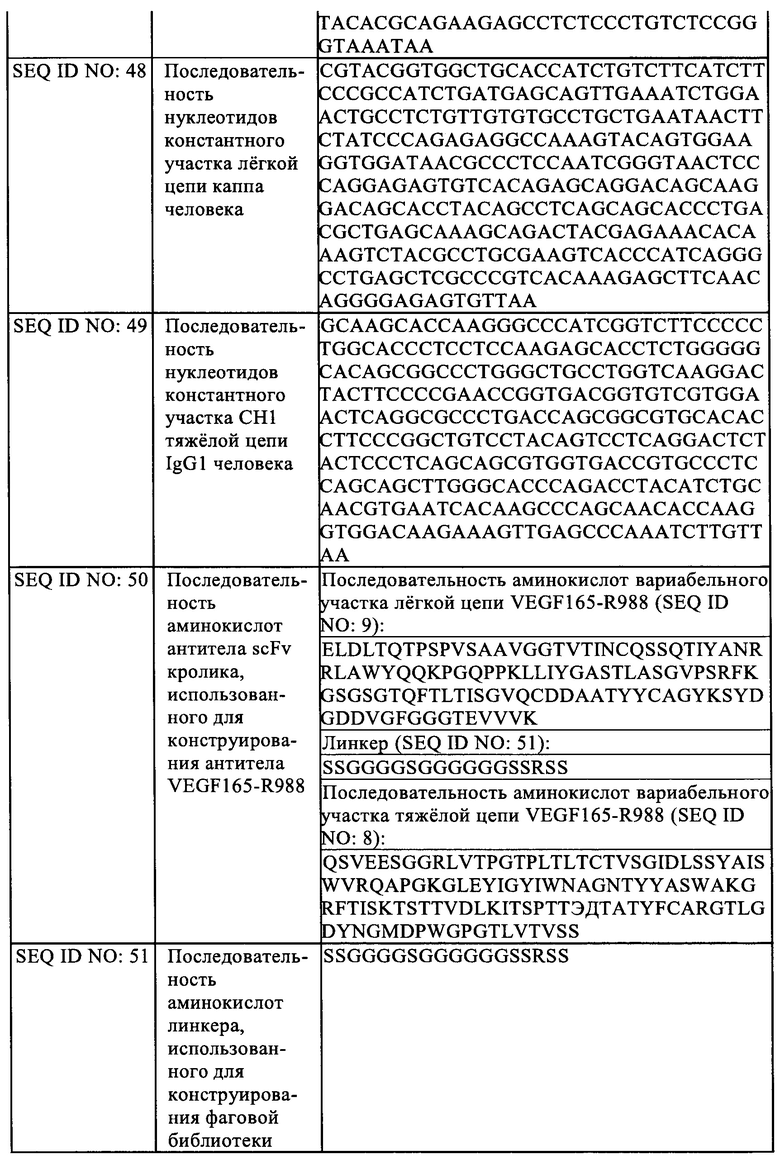

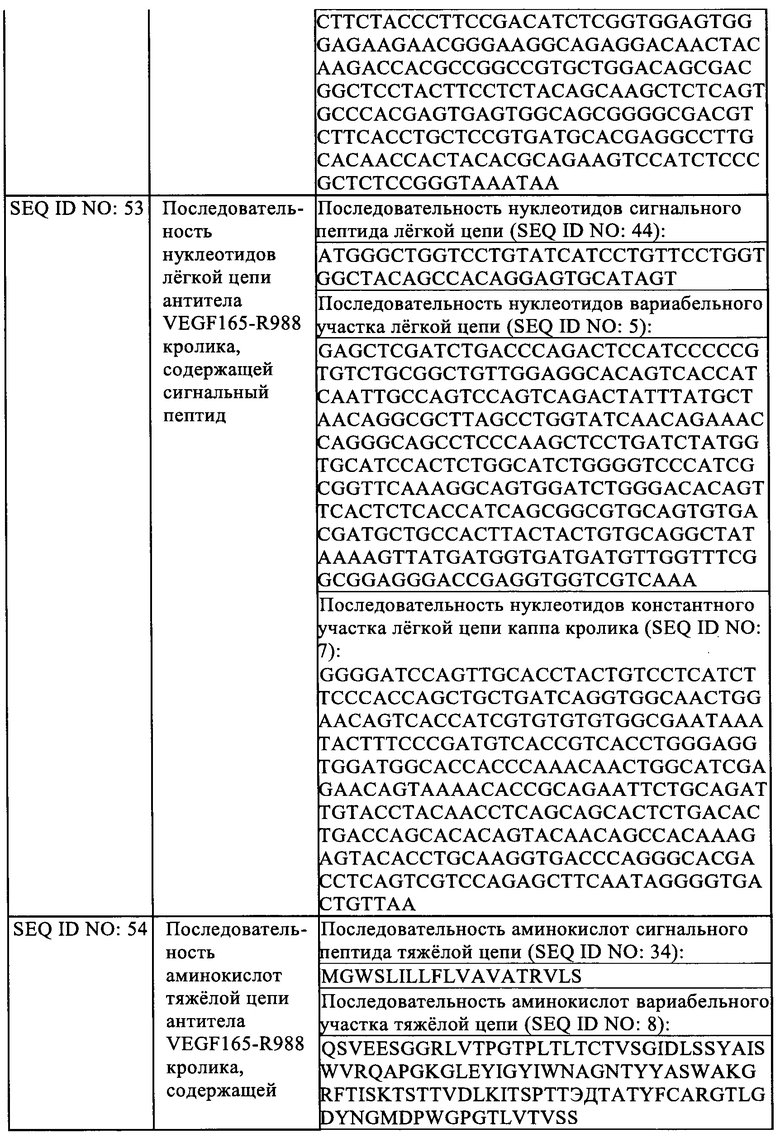

--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> SinoCellTech Ltd.
<120> Fab ФРАГМЕНТ ГУМАНИЗИРОВАННОГО АНТИТЕЛА ПРОТИВ VEGF И ЕГО
ПРИМЕНЕНИЕ
<130> CIE200066PCT
<160> 55
<170> BiSSAP 1.3.6
<210> 1
<211> 191
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот Met1 - Arg191 белка VEGF
человека
(UniProtKB P15692)
<400> 1
Met Asn Phe Leu Leu Ser Trp Val His Trp Ser Leu Ala Leu Leu Leu
1 5 10 15
Tyr Leu His His Ala Lys Trp Ser Gln Ala Ala Pro Met Ala Glu Gly
20 25 30
Gly Gly Gln Asn His His Glu Val Val Lys Phe Met Asp Val Tyr Gln
35 40 45
Arg Ser Tyr Cys His Pro Ile Glu Thr Leu Val Asp Ile Phe Gln Glu
50 55 60
Tyr Pro Asp Glu Ile Glu Tyr Ile Phe Lys Pro Ser Cys Val Pro Leu
65 70 75 80
Met Arg Cys Gly Gly Cys Cys Asn Asp Glu Gly Leu Glu Cys Val Pro
85 90 95
Thr Glu Glu Ser Asn Ile Thr Met Gln Ile Met Arg Ile Lys Pro His
100 105 110
Gln Gly Gln His Ile Gly Glu Met Ser Phe Leu Gln His Asn Lys Cys
115 120 125
Glu Cys Arg Pro Lys Lys Asp Arg Ala Arg Gln Glu Lys Lys Ser Val
130 135 140
Arg Gly Lys Gly Lys Gly Gln Lys Arg Lys Arg Lys Lys Ser Arg Tyr
145 150 155 160
Lys Ser Trp Ser Val Tyr Val Gly Ala Arg Cys Cys Leu Met Pro Trp
165 170 175
Ser Leu Pro Gly Pro His Pro Cys Gly Pro Cys Ser Glu Arg Arg
180 185 190
<210> 2
<211> 54
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов линкера, использованного для
конструирования фаговой библиотеки антител, для связывания антитела
scFv
кролика
<400> 2
tctagtggtg gcggtggttc gggcggtggt ggaggtggta gttctagatc ttcc
54
<210> 3
<211> 738
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов антитела scFv кролика,
использованного
для конструирования антитела VEGF165-R988
<400> 3
gagctcgatc tgacccagac tccatccccc gtgtctgcgg ctgttggagg cacagtcacc
60
atcaattgcc agtccagtca gactatttat gctaacaggc gcttagcctg gtatcaacag
120
aaaccagggc agcctcccaa gctcctgatc tatggtgcat ccactctggc atctggggtc
180
ccatcgcggt tcaaaggcag tggatctggg acacagttca ctctcaccat cagcggcgtg
240
cagtgtgacg atgctgccac ttactactgt gcaggctata aaagttatga tggtgatgat
300
gttggtttcg gcggagggac cgaggtggtc gtcaaatcta gtggtggcgg tggttcgggc
360
ggtggtggag gtggtagttc tagatcttcc cagtcggtgg aggagtccgg gggtcgcctg
420
gtaacgcctg ggacacccct gacactcacc tgcacagtct ctggaatcga cctcagtagc
480
tatgcaataa gctgggtccg ccaggctcca gggaaggggc tggaatacat cggatacatt
540
tggaatgctg gtaacaccta ctacgcgagc tgggcaaaag gccgattcac catctccaaa
600
acctcgacca cggtggatct gaaaatcacc agtccgacaa ccgaggacac ggccacctat
660
ttctgtgcca gaggaacatt aggggactac aatggcatgg acccctgggg cccagggacc
720
ctcgtcaccg tctcttca
738
<210> 4
<211> 348
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов вариабельного участка тяжёлой
цепи
антитела VEGF165-R988 кролика
<400> 4
cagtcggtgg aggagtccgg gggtcgcctg gtaacgcctg ggacacccct gacactcacc
60
tgcacagtct ctggaatcga cctcagtagc tatgcaataa gctgggtccg ccaggctcca
120
gggaaggggc tggaatacat cggatacatt tggaatgctg gtaacaccta ctacgcgagc
180
tgggcaaaag gccgattcac catctccaaa acctcgacca cggtggatct gaaaatcacc
240
agtccgacaa ccgaggacac ggccacctat ttctgtgcca gaggaacatt aggggactac
300
aatggcatgg acccctgggg cccagggacc ctcgtcaccg tctcttca
348
<210> 5
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов вариабельного участка лёгкой
цепи
антитела VEGF165-R988 кролика
<400> 5
gagctcgatc tgacccagac tccatccccc gtgtctgcgg ctgttggagg cacagtcacc
60
atcaattgcc agtccagtca gactatttat gctaacaggc gcttagcctg gtatcaacag
120
aaaccagggc agcctcccaa gctcctgatc tatggtgcat ccactctggc atctggggtc
180
ccatcgcggt tcaaaggcag tggatctggg acacagttca ctctcaccat cagcggcgtg
240
cagtgtgacg atgctgccac ttactactgt gcaggctata aaagttatga tggtgatgat
300
gttggtttcg gcggagggac cgaggtggtc gtcaaa
336
<210> 6
<211> 972
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов константного участка тяжёлой
цепи IgG1
кролика
<400> 6
ggtcaaccta aggctccgtc agtcttccca ctggccccct gctgcgggga cacacccagc
60
tccacggtga ccctgggctg cctggtcaaa ggctacctcc cggagccagt gaccgtgacc
120
tggaactcgg gcaccctcac caatggggta cgcaccttcc cgtccgtccg gcagtcctca
180
ggcctctact cgctgagcag cgtggtgagc gtgacctcaa gcagccagcc cgtcacctgc
240
aacgtggccc acccagccac caacaccaaa gtggacaaga ccgttgcgcc ctcgacatgc
300
agcaagccca cgtgcccacc ccctgaactc ctggggggac cgtctgtctt catcttcccc
360
ccaaaaccca aggacaccct catgatctca cgcacccccg aggtcacatg cgtggtggtg
420
gacgtgagcc aggatgaccc cgaggtgcag ttcacatggt acataaacaa cgagcaggtg
480
cgcaccgccc ggccgccgct acgggagcag cagttcaaca gcacgatccg cgtggtcagc
540
accctcccca tcgcgcacca ggactggctg aggggcaagg agttcaagtg caaagtccac
600
aacaaggcac tcccggcccc catcgagaaa accatctcca aagccagagg gcagcccctg
660
gagccgaagg tctacaccat gggccctccc cgggaggagc tgagcagcag gtcggtcagc
720
ctgacctgca tgatcaacgg cttctaccct tccgacatct cggtggagtg ggagaagaac
780
gggaaggcag aggacaacta caagaccacg ccggccgtgc tggacagcga cggctcctac
840
ttcctctaca gcaagctctc agtgcccacg agtgagtggc agcggggcga cgtcttcacc
900
tgctccgtga tgcacgaggc cttgcacaac cactacacgc agaagtccat ctcccgctct
960
ccgggtaaat aa
972
<210> 7
<211> 315
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов константного участка лёгкой цепи
каппа
кролика
<400> 7
ggggatccag ttgcacctac tgtcctcatc ttcccaccag ctgctgatca ggtggcaact
60
ggaacagtca ccatcgtgtg tgtggcgaat aaatactttc ccgatgtcac cgtcacctgg
120
gaggtggatg gcaccaccca aacaactggc atcgagaaca gtaaaacacc gcagaattct
180
gcagattgta cctacaacct cagcagcact ctgacactga ccagcacaca gtacaacagc
240
cacaaagagt acacctgcaa ggtgacccag ggcacgacct cagtcgtcca gagcttcaat
300
aggggtgact gttaa
315
<210> 8
<211> 116
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот вариабельного участка тяжёлой
цепи
антитела VEGF165-R988 кролика
<400> 8
Gln Ser Val Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly Thr Pro
1 5 10 15
Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser Ser Tyr Ala
20 25 30
Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr Ile Gly
35 40 45
Tyr Ile Trp Asn Ala Gly Asn Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Phe Thr Ile Ser Lys Thr Ser Thr Thr Val Asp Leu Lys Ile Thr
65 70 75 80
Ser Pro Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Gly Thr
85 90 95
Leu Gly Asp Tyr Asn Gly Met Asp Pro Trp Gly Pro Gly Thr Leu Val
100 105 110
Thr Val Ser Ser
115
<210> 9
<211> 112
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот вариабельного участка лёгкой
цепи
антитела VEGF165-R988 кролика
<400> 9
Glu Leu Asp Leu Thr Gln Thr Pro Ser Pro Val Ser Ala Ala Val Gly
1 5 10 15
Gly Thr Val Thr Ile Asn Cys Gln Ser Ser Gln Thr Ile Tyr Ala Asn
20 25 30
Arg Arg Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Lys Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Gly Val
65 70 75 80
Gln Cys Asp Asp Ala Ala Thr Tyr Tyr Cys Ala Gly Tyr Lys Ser Tyr
85 90 95
Asp Gly Asp Asp Val Gly Phe Gly Gly Gly Thr Glu Val Val Val Lys
100 105 110
<210> 10
<211> 13
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот CDR1 лёгкой цепи антитела
VEGF165-R988
кролика
<400> 10
Gln Ser Ser Gln Thr Ile Tyr Ala Asn Arg Arg Leu Ala
1 5 10
<210> 11
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот CDR2 лёгкой цепи антитела
VEGF165-R988
кролика
<400> 11
Gly Ala Ser Thr Leu Ala Ser
1 5
<210> 12
<211> 12
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот CDR3 лёгкой цепи антитела
VEGF165-R988
кролика
<400> 12
Ala Gly Tyr Lys Ser Tyr Asp Gly Asp Asp Val Gly
1 5 10
<210> 13
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот CDR1 тяжёлой цепи антитела
VEGF165-R988
кролика
<400> 13
Gly Ile Asp Leu Ser Ser Tyr Ala Ile Ser
1 5 10
<210> 14
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот CDR2 тяжёлой цепи антитела
VEGF165-R988
кролика
<400> 14
Tyr Ile Trp Asn Ala Gly Asn Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
1 5 10 15
<210> 15
<211> 13
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот CDR3 тяжёлой цепи антитела
VEGF165-R988
кролика
<400> 15
Ala Arg Gly Thr Leu Gly Asp Tyr Asn Gly Met Asp Pro
1 5 10
<210> 16
<211> 1404
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов тяжёлой цепи гуманизированного
антитела
VEGF165-H988-10, содержащей сигнальный пептид
<400> 16
atgggctggt ccctgattct gctgttcctg gtggctgtgg ctaccagggt gctgagtcag
60
tctgtccagg agtctggacc tggactggtg aagccatctg agaccctgtc cctgacttgt
120
actgtgtctg gcattgacct gtcctcctat gccatctcct gggtgagaca acctcctggc
180
aagggattgg aatacattgg ctacatctgg aatgctggca acacctacta tgcctcctgg
240
gctaagggca gggtgaccat ctctgtggac accagcaaga accaggtgga cctgaaactg
300
tcctctgtga cagcagcaga cacagcagtc tacttctgtg ccaggggcac cctgggagac
360
tacaatggga tggacccatg gggacctggc accctggtga cagtgtccag cgctagcacc
420
aagggcccat cggtcttccc cctggcaccc tcctccaaga gcacctctgg gggcacagcg
480
gccctgggct gcctggtcaa ggactacttc cccgaaccgg tgacggtgtc gtggaactca
540
ggcgccctga ccagcggcgt gcacaccttc ccggctgtcc tacagtcctc aggactctac
600
tccctcagca gcgtggtgac cgtgccctcc agcagcttgg gcacccagac ctacatctgc
660
aacgtgaatc acaagcccag caacaccaag gtggacaaga aagttgagcc caaatcttgt
720
gacaaaactc acacatgccc accgtgccca gcacctgaac tcctgggggg accgtcagtc
780
ttcctcttcc ccccaaaacc caaggacacc ctcatgatct cccggacccc tgaggtcacg
840
tgcgtggtgg tggacgtgag ccacgaagac cccgaggtca agttcaactg gtacgtggac
900
ggcgtggagg tgcataatgc caagacaaag ccgcgggagg agcagtacaa cagcacgtac
960
cgtgtggtca gcgtcctcac cgtcctgcac caggactggc tgaatggcaa ggagtacaag
1020
tgcaaggtct ccaacaaagc cctcccagcc cccatcgaga aaaccatctc caaagccaaa
1080
gggcagcccc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gctgaccaag
1140
aaccaggtca gcctgacctg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag
1200
tgggagagca atgggcagcc ggagaacaac tacaagacca cgcctcccgt gctggactcc
1260
gacggctcct tcttcctcta cagcaagctc accgtggaca agagcaggtg gcagcagggg
1320
aacgtcttct catgctccgt gatgcatgag gctctgcaca accactacac gcagaagagc
1380
ctctccctgt ctccgggtaa atga
1404
<210> 17
<211> 717
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов лёгкой цепи гуманизированного
антитела
VEGF165-H988-10, содержащей сигнальный пептид
<400> 17
atgggctggt cctgtatcat cctgttcctg gtggctacag ccacaggagt gcatagtgaa
60
ctccaactta cccagagccc atcctccctg tctgcctctg tgggagacag ggtgaccatc
120
acttgtcagt ccagccagac catctatgcc aacaggagac tggcttggta tcaacagaag
180
cctggcaagg tgccaaaact gctgatttat ggagccagca ccctggcatc tggagtgcca
240
agcaggttca agggctctgg ctctggcaca gacttcaccc tgaccatctc ctccctccaa
300
cctgaggatg tggctaccta ctactgtgct ggctacaagt cctatgatgg agatgatgtg
360
ggctttggag gaggcaccaa ggtggagatt aagcgtacgg tggctgcacc atctgtcttc
420
atcttcccgc catctgatga gcagttgaaa tctggaactg cctctgttgt gtgcctgctg
480
aataacttct atcccagaga ggccaaagta cagtggaagg tggataacgc cctccaatcg
540
ggtaactccc aggagagtgt cacagagcag gacagcaagg acagcaccta cagcctcagc
600
agcaccctga cgctgagcaa agcagactac gagaaacaca aagtctacgc ctgcgaagtc
660
acccatcagg gcctgagctc gcccgtcaca aagagcttca acaggggaga gtgttag
717
<210> 18
<211> 467
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот тяжёлой цепи гуманизированного
антитела
VEGF165-H988-10, содержащей сигнальный пептид
<400> 18
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Gln Ser Val Gln Glu Ser Gly Pro Gly Leu Val Lys Pro
20 25 30
Ser Glu Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser
35 40 45
Ser Tyr Ala Ile Ser Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu
50 55 60
Tyr Ile Gly Tyr Ile Trp Asn Ala Gly Asn Thr Tyr Tyr Ala Ser Trp
65 70 75 80
Ala Lys Gly Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Val
85 90 95
Asp Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe
100 105 110
Cys Ala Arg Gly Thr Leu Gly Asp Tyr Asn Gly Met Asp Pro Trp Gly
115 120 125
Pro Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
130 135 140
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
145 150 155 160
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
165 170 175
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
180 185 190
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
195 200 205
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
210 215 220
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
225 230 235 240
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
245 250 255
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
260 265 270
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
275 280 285
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
290 295 300
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
305 310 315 320
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
325 330 335
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
340 345 350
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
355 360 365
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
370 375 380
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
385 390 395 400
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
405 410 415
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
420 425 430
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
435 440 445
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
450 455 460
Pro Gly Lys
465
<210> 19
<211> 238
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот лёгкой цепи гуманизированного
антитела
VEGF165-H988-10, содержащей сигнальный пептид
<400> 19
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Glu Leu Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala
20 25 30
Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ser Ser Gln Thr Ile
35 40 45
Tyr Ala Asn Arg Arg Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Val
50 55 60
Pro Lys Leu Leu Ile Tyr Gly Ala Ser Thr Leu Ala Ser Gly Val Pro
65 70 75 80
Ser Arg Phe Lys Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
85 90 95
Ser Ser Leu Gln Pro Glu Asp Val Ala Thr Tyr Tyr Cys Ala Gly Tyr
100 105 110
Lys Ser Tyr Asp Gly Asp Asp Val Gly Phe Gly Gly Gly Thr Lys Val
115 120 125
Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro
130 135 140
Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu
145 150 155 160
Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn
165 170 175
Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser
180 185 190
Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala
195 200 205
Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly
210 215 220
Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230 235
<210> 20
<211> 354
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов вариабельного участка тяжёлой
цепи
гуманизированного антитела VEGF165-H988-10
<400> 20
cagtctgtcc aggagtctgg acctggactg gtgaagccat ctgagaccct gtccctgact
60
tgtactgtgt ctggcattga cctgtcctcc tatgccatct cctgggtgag acaacctcct
120
ggcaagggat tggaatacat tggctacatc tggaatgctg gcaacaccta ctatgcctcc
180
tgggctaagg gcagggtgac catctctgtg gacaccagca agaaccaggt ggacctgaaa
240
ctgtcctctg tgacagcagc agacacagca gtctacttct gtgccagggg caccctggga
300
gactacaatg ggatggaccc atggggacct ggcaccctgg tgacagtgtc cagc
354
<210> 21
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов вариабельного участка лёгкой
цепи
гуманизированного антитела VEGF165-H988-10
<400> 21
gaactccaac ttacccagag cccatcctcc ctgtctgcct ctgtgggaga cagggtgacc
60
atcacttgtc agtccagcca gaccatctat gccaacagga gactggcttg gtatcaacag
120
aagcctggca aggtgccaaa actgctgatt tatggagcca gcaccctggc atctggagtg
180
ccaagcaggt tcaagggctc tggctctggc acagacttca ccctgaccat ctcctccctc
240
caacctgagg atgtggctac ctactactgt gctggctaca agtcctatga tggagatgat
300
gtgggctttg gaggaggcac caaggtggag attaag
336
<210> 22
<211> 118
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот вариабельного участка тяжёлой
цепи
антитела VEGF165-H988-10
<400> 22
Gln Ser Val Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu Thr
1 5 10 15
Leu Ser Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser Ser Tyr Ala
20 25 30
Ile Ser Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Tyr Ile Gly
35 40 45
Tyr Ile Trp Asn Ala Gly Asn Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Val Asp Leu Lys
65 70 75 80
Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala Arg
85 90 95
Gly Thr Leu Gly Asp Tyr Asn Gly Met Asp Pro Trp Gly Pro Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 23
<211> 112
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот вариабельного участка лёгкой
цепи
антитела VEGF165-H988-10
<400> 23
Glu Leu Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ser Ser Gln Thr Ile Tyr Ala Asn
20 25 30
Arg Arg Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Lys Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Pro Glu Asp Val Ala Thr Tyr Tyr Cys Ala Gly Tyr Lys Ser Tyr
85 90 95
Asp Gly Asp Asp Val Gly Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 24
<211> 13
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот CDR1 лёгкой цепи Fab фрагмента
гуманизированного антитела VEGF-H988
<400> 24
Gln Ser Ser Lys Phe Leu Trp Gln Gly Arg Arg Leu Ala
1 5 10
<210> 25
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот CDR2 лёгкой цепи Fab фрагмента
гуманизированного антитела VEGF-H988
<400> 25
Gly Ala Ser Thr Leu Ala Ser
1 5
<210> 26
<211> 12
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот CDR3 лёгкой цепи Fab фрагмента
гуманизированного антитела VEGF-H988
<400> 26
Ala Gly Tyr Lys Ser Tyr Asp Gly Asp Val Val Gly
1 5 10
<210> 27
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот CDR1 тяжёлой цепи Fab фрагмента
гуманизированного антитела VEGF-H988
<400> 27
Gly Ile Asp Leu Ser Ser Tyr Ala Ile Ser
1 5 10
<210> 28
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот CDR2 тяжёлой цепи Fab фрагмента
гуманизированного антитела VEGF-H988
<400> 28
Tyr Ile Trp Asn Asp Leu Phe Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
1 5 10 15
<210> 29
<211> 13
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот CDR3 тяжёлой цепи Fab фрагмента
гуманизированного антитела VEGF-H988
<400> 29
Ala Arg Gly Thr Leu Gly Asp Tyr Gly Gly Met Asp Pro
1 5 10
<210> 30
<211> 221
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот тяжёлой цепи Fab фрагмента
гуманизированного антитела VEGF-H988
<400> 30
Gln Ser Val Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu Thr
1 5 10 15
Leu Ser Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser Ser Tyr Ala
20 25 30
Ile Ser Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Tyr Ile Gly
35 40 45
Tyr Ile Trp Asn Asp Leu Phe Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Val Asp Leu Lys
65 70 75 80
Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala Arg
85 90 95
Gly Thr Leu Gly Asp Tyr Gly Gly Met Asp Pro Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
<210> 31
<211> 219
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот лёгкой цепи Fab фрагмента
гуманизированного антитела VEGF-H988
<400> 31
Glu Leu Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ser Ser Lys Phe Leu Trp Gln Gly
20 25 30
Arg Arg Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Lys Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Pro Glu Asp Val Ala Thr Tyr Tyr Cys Ala Gly Tyr Lys Ser Tyr
85 90 95
Asp Gly Asp Val Val Gly Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 32
<211> 240
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот тяжёлой цепи Fab фрагмента
гуманизированного антитела VEGF-H988, содержащей сигнальный пептид
<400> 32
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Gln Ser Val Gln Glu Ser Gly Pro Gly Leu Val Lys Pro
20 25 30
Ser Glu Thr Leu Ser Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser
35 40 45
Ser Tyr Ala Ile Ser Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu
50 55 60
Tyr Ile Gly Tyr Ile Trp Asn Asp Leu Phe Thr Tyr Tyr Ala Ser Trp
65 70 75 80
Ala Lys Gly Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Val
85 90 95
Asp Leu Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe
100 105 110
Cys Ala Arg Gly Thr Leu Gly Asp Tyr Gly Gly Met Asp Pro Trp Gly
115 120 125
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
130 135 140
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
145 150 155 160
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
165 170 175
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
180 185 190
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
195 200 205
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
210 215 220
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
225 230 235 240
<210> 33
<211> 238
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот лёгкой цепи Fab фрагмента
гуманизированного антитела VEGF-H988, содержащей сигнальный пептид
<400> 33
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Glu Leu Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala
20 25 30
Ser Val Gly Asp Arg Val Thr Ile Thr Cys Gln Ser Ser Lys Phe Leu
35 40 45
Trp Gln Gly Arg Arg Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Val
50 55 60
Pro Lys Leu Leu Ile Tyr Gly Ala Ser Thr Leu Ala Ser Gly Val Pro
65 70 75 80
Ser Arg Phe Lys Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile
85 90 95
Ser Ser Leu Gln Pro Glu Asp Val Ala Thr Tyr Tyr Cys Ala Gly Tyr
100 105 110
Lys Ser Tyr Asp Gly Asp Val Val Gly Phe Gly Gly Gly Thr Lys Val
115 120 125
Glu Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro
130 135 140
Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu
145 150 155 160
Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn
165 170 175
Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser
180 185 190
Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala
195 200 205
Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly
210 215 220
Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230 235
<210> 34
<211> 19
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот сигнального пептида тяжёлой цепи
<400> 34
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser
<210> 35
<211> 19
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот сигнального пептида лёгкой цепи
<400> 35
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser
<210> 36
<211> 118
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот вариабельного участка тяжёлой
цепи Fab
фрагмента гуманизированного антитела VEGF-H988
<400> 36
Gln Ser Val Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Glu Thr
1 5 10 15
Leu Ser Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser Ser Tyr Ala
20 25 30
Ile Ser Trp Val Arg Gln Pro Pro Gly Lys Gly Leu Glu Tyr Ile Gly
35 40 45
Tyr Ile Trp Asn Asp Leu Phe Thr Tyr Tyr Ala Ser Trp Ala Lys Gly
50 55 60
Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Val Asp Leu Lys
65 70 75 80
Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Phe Cys Ala Arg
85 90 95
Gly Thr Leu Gly Asp Tyr Gly Gly Met Asp Pro Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 37
<211> 112
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот вариабельного участка лёгкой
цепи Fab
фрагмента гуманизированного антитела VEGF-H988
<400> 37
Glu Leu Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Gln Ser Ser Lys Phe Leu Trp Gln Gly
20 25 30
Arg Arg Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Lys Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu
65 70 75 80
Gln Pro Glu Asp Val Ala Thr Tyr Tyr Cys Ala Gly Tyr Lys Ser Tyr
85 90 95
Asp Gly Asp Val Val Gly Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 38
<211> 330
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот константного участка тяжёлой
цепи IgG1
человека
<400> 38
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 39
<211> 107
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот константного участка лёгкой цепи
каппа
человека
<400> 39
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 40
<211> 103
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот константного участка CH1 тяжёлой
цепи
IgG1 человека
<400> 40
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys
100
<210> 41
<211> 723
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов тяжёлой цепи Fab фрагмента
гуманизированного антитела VEGF-H988, содержащей сигнальный пептид
<400> 41
atgggctggt ccctgattct gctgttcctg gtggctgtgg ctaccagggt gctgagtcag
60
tctgtccagg agtctggacc tggactggtg aagccatctg agaccctgtc cctgacttgt
120
actgtgtctg gcattgacct gtcctcctat gccatctcct gggtgagaca acctcctggc
180
aagggattgg aatacattgg ctacatctgg aatgatctct tcacctacta tgcctcctgg
240
gctaagggca gggtgaccat ctctgtggac accagcaaga accaggtgga cctgaaactg
300
tcctctgtga cagcagcaga cacagcagtc tacttctgtg ccaggggcac cctgggagac
360
tacggcggga tggacccatg gggacagggc accctggtga cagtgtccag cgcaagcacc
420
aagggcccat cggtcttccc cctggcaccc tcctccaaga gcacctctgg gggcacagcg
480
gccctgggct gcctggtcaa ggactacttc cccgaaccgg tgacggtgtc gtggaactca
540
ggcgccctga ccagcggcgt gcacaccttc ccggctgtcc tacagtcctc aggactctac
600
tccctcagca gcgtggtgac cgtgccctcc agcagcttgg gcacccagac ctacatctgc
660
aacgtgaatc acaagcccag caacaccaag gtggacaaga aagttgagcc caaatcttgt
720
taa
723
<210> 42
<211> 717
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов лёгкой цепи Fab фрагмента
гуманизированного антитела VEGF-H988, содержащей сигнальный пептид
<400> 42
atgggctggt cctgtatcat cctgttcctg gtggctacag ccacaggagt gcatagtgaa
60
ctccaactta cccagagccc atcctccctg tctgcctctg tgggagacag ggtgaccatc
120
acttgtcagt ccagcaagtt cctctggcag ggcaggagac tggcttggta tcaacagaag
180
cctggcaagg tgccaaaact gctgatttat ggagccagca ccctggcatc tggagtgcca
240
agcaggttca agggctctgg ctctggcaca gacttcaccc tgaccatctc ctccctccaa
300
cctgaggatg tggctaccta ctactgtgct ggctacaagt cctatgatgg agatgttgtg
360
ggctttggag gaggcaccaa ggtggagatt aagcgtacgg tggctgcacc atctgtcttc
420
atcttcccgc catctgatga gcagttgaaa tctggaactg cctctgttgt gtgcctgctg
480
aataacttct atcccagaga ggccaaagta cagtggaagg tggataacgc cctccaatcg
540
ggtaactccc aggagagtgt cacagagcag gacagcaagg acagcaccta cagcctcagc
600
agcaccctga cgctgagcaa agcagactac gagaaacaca aagtctacgc ctgcgaagtc
660
acccatcagg gcctgagctc gcccgtcaca aagagcttca acaggggaga gtgttaa
717
<210> 43
<211> 57
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов сигнального пептида тяжёлой цепи
<400> 43
atgggctggt ccctgattct gctgttcctg gtggctgtgg ctaccagggt gctgagt
57
<210> 44
<211> 57
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов сигнального пептида лёгкой цепи
<400> 44
atgggctggt cctgtatcat cctgttcctg gtggctacag ccacaggagt gcatagt
57
<210> 45
<211> 354
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов вариабельного участка тяжёлой
цепи Fab
фрагмента гуманизированного антитела VEGF-H988
<400> 45
cagtctgtcc aggagtctgg acctggactg gtgaagccat ctgagaccct gtccctgact
60
tgtactgtgt ctggcattga cctgtcctcc tatgccatct cctgggtgag acaacctcct
120
ggcaagggat tggaatacat tggctacatc tggaatgatc tcttcaccta ctatgcctcc
180
tgggctaagg gcagggtgac catctctgtg gacaccagca agaaccaggt ggacctgaaa
240
ctgtcctctg tgacagcagc agacacagca gtctacttct gtgccagggg caccctggga
300
gactacggcg ggatggaccc atggggacag ggcaccctgg tgacagtgtc cagc
354
<210> 46
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов вариабельного участка лёгкой
цепи Fab
фрагмента гуманизированного антитела VEGF-H988
<400> 46
gaactccaac ttacccagag cccatcctcc ctgtctgcct ctgtgggaga cagggtgacc
60
atcacttgtc agtccagcaa gttcctctgg cagggcagga gactggcttg gtatcaacag
120
aagcctggca aggtgccaaa actgctgatt tatggagcca gcaccctggc atctggagtg
180
ccaagcaggt tcaagggctc tggctctggc acagacttca ccctgaccat ctcctccctc
240
caacctgagg atgtggctac ctactactgt gctggctaca agtcctatga tggagatgtt
300
gtgggctttg gaggaggcac caaggtggag attaag
336
<210> 47
<211> 993
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов константного участка тяжёлой
цепи IgG1
человека
<400> 47
gcaagcacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg
60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg
120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca
180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc
240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc
300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga
360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct
420
gaggtcacgt gcgtggtggt ggacgtgagc cacgaagacc ccgaggtcaa gttcaactgg
480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac
540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag
600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc
660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggatgag
720
ctgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc
780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg
840
ctggactccg acggctcctt cttcctctac agcaagctca ccgtggacaa gagcaggtgg
900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg
960
cagaagagcc tctccctgtc tccgggtaaa taa
993
<210> 48
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов константного участка лёгкой цепи
каппа
человека
<400> 48
cgtacggtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct
60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag
120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac
180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag
240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagctcgcc cgtcacaaag
300
agcttcaaca ggggagagtg ttaa
324
<210> 49
<211> 312
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов константного участка CH1 тяжёлой
цепи
IgG1 человека
<400> 49
gcaagcacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg
60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg
120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca
180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc
240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc
300
aaatcttgtt aa
312
<210> 50
<211> 246
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот антитела scFv кролика,
использованного
для конструирования антитела VEGF165-R988
<400> 50
Glu Leu Asp Leu Thr Gln Thr Pro Ser Pro Val Ser Ala Ala Val Gly
1 5 10 15
Gly Thr Val Thr Ile Asn Cys Gln Ser Ser Gln Thr Ile Tyr Ala Asn
20 25 30
Arg Arg Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu
35 40 45
Leu Ile Tyr Gly Ala Ser Thr Leu Ala Ser Gly Val Pro Ser Arg Phe
50 55 60
Lys Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile Ser Gly Val
65 70 75 80
Gln Cys Asp Asp Ala Ala Thr Tyr Tyr Cys Ala Gly Tyr Lys Ser Tyr
85 90 95
Asp Gly Asp Asp Val Gly Phe Gly Gly Gly Thr Glu Val Val Val Lys
100 105 110
Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Gly Gly Ser Ser Arg
115 120 125
Ser Ser Gln Ser Val Glu Glu Ser Gly Gly Arg Leu Val Thr Pro Gly
130 135 140
Thr Pro Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser Ser
145 150 155 160
Tyr Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Tyr
165 170 175
Ile Gly Tyr Ile Trp Asn Ala Gly Asn Thr Tyr Tyr Ala Ser Trp Ala
180 185 190
Lys Gly Arg Phe Thr Ile Ser Lys Thr Ser Thr Thr Val Asp Leu Lys
195 200 205
Ile Thr Ser Pro Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg
210 215 220
Gly Thr Leu Gly Asp Tyr Asn Gly Met Asp Pro Trp Gly Pro Gly Thr
225 230 235 240
Leu Val Thr Val Ser Ser
245
<210> 51
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот линкера, использованного для
конструирования фаговой библиотеки антител для связывания антитела
scFv
кролика
<400> 51
Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Gly Gly Ser Ser Arg
1 5 10 15
Ser Ser
<210> 52
<211> 1377
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов тяжёлой цепи антитела
VEGF165-R988
кролика, содержащей сигнальный пептид
<400> 52
atgggctggt ccctgattct gctgttcctg gtggctgtgg ctaccagggt gctgagtcag
60
tcggtggagg agtccggggg tcgcctggta acgcctggga cacccctgac actcacctgc
120
acagtctctg gaatcgacct cagtagctat gcaataagct gggtccgcca ggctccaggg
180
aaggggctgg aatacatcgg atacatttgg aatgctggta acacctacta cgcgagctgg
240
gcaaaaggcc gattcaccat ctccaaaacc tcgaccacgg tggatctgaa aatcaccagt
300
ccgacaaccg aggacacggc cacctatttc tgtgccagag gaacattagg ggactacaat
360
ggcatggacc cctggggccc agggaccctc gtcaccgtct cttcaggtca acctaaggct
420
ccgtcagtct tcccactggc cccctgctgc ggggacacac ccagctccac ggtgaccctg
480
ggctgcctgg tcaaaggcta cctcccggag ccagtgaccg tgacctggaa ctcgggcacc
540
ctcaccaatg gggtacgcac cttcccgtcc gtccggcagt cctcaggcct ctactcgctg
600
agcagcgtgg tgagcgtgac ctcaagcagc cagcccgtca cctgcaacgt ggcccaccca
660
gccaccaaca ccaaagtgga caagaccgtt gcgccctcga catgcagcaa gcccacgtgc
720
ccaccccctg aactcctggg gggaccgtct gtcttcatct tccccccaaa acccaaggac
780
accctcatga tctcacgcac ccccgaggtc acatgcgtgg tggtggacgt gagccaggat
840
gaccccgagg tgcagttcac atggtacata aacaacgagc aggtgcgcac cgcccggccg
900
ccgctacggg agcagcagtt caacagcacg atccgcgtgg tcagcaccct ccccatcgcg
960
caccaggact ggctgagggg caaggagttc aagtgcaaag tccacaacaa ggcactcccg
1020
gcccccatcg agaaaaccat ctccaaagcc agagggcagc ccctggagcc gaaggtctac
1080
accatgggcc ctccccggga ggagctgagc agcaggtcgg tcagcctgac ctgcatgatc
1140
aacggcttct acccttccga catctcggtg gagtgggaga agaacgggaa ggcagaggac
1200
aactacaaga ccacgccggc cgtgctggac agcgacggct cctacttcct ctacagcaag
1260
ctctcagtgc ccacgagtga gtggcagcgg ggcgacgtct tcacctgctc cgtgatgcac
1320
gaggccttgc acaaccacta cacgcagaag tccatctccc gctctccggg taaataa
1377
<210> 53
<211> 708
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность нуклеотидов лёгкой цепи антитела
VEGF165-R988
кролика, содержащей сигнальный пептид
<400> 53
atgggctggt cctgtatcat cctgttcctg gtggctacag ccacaggagt gcatagtgag
60
ctcgatctga cccagactcc atcccccgtg tctgcggctg ttggaggcac agtcaccatc
120
aattgccagt ccagtcagac tatttatgct aacaggcgct tagcctggta tcaacagaaa
180
ccagggcagc ctcccaagct cctgatctat ggtgcatcca ctctggcatc tggggtccca
240
tcgcggttca aaggcagtgg atctgggaca cagttcactc tcaccatcag cggcgtgcag
300
tgtgacgatg ctgccactta ctactgtgca ggctataaaa gttatgatgg tgatgatgtt
360
ggtttcggcg gagggaccga ggtggtcgtc aaaggggatc cagttgcacc tactgtcctc
420
atcttcccac cagctgctga tcaggtggca actggaacag tcaccatcgt gtgtgtggcg
480
aataaatact ttcccgatgt caccgtcacc tgggaggtgg atggcaccac ccaaacaact
540
ggcatcgaga acagtaaaac accgcagaat tctgcagatt gtacctacaa cctcagcagc
600
actctgacac tgaccagcac acagtacaac agccacaaag agtacacctg caaggtgacc
660
cagggcacga cctcagtcgt ccagagcttc aataggggtg actgttaa
708
<210> 54
<211> 458
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот тяжёлой цепи антитела
VEGF165-R988
кролика, содержащей сигнальный пептид
<400> 54
Met Gly Trp Ser Leu Ile Leu Leu Phe Leu Val Ala Val Ala Thr Arg
1 5 10 15
Val Leu Ser Gln Ser Val Glu Glu Ser Gly Gly Arg Leu Val Thr Pro
20 25 30
Gly Thr Pro Leu Thr Leu Thr Cys Thr Val Ser Gly Ile Asp Leu Ser
35 40 45
Ser Tyr Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu
50 55 60
Tyr Ile Gly Tyr Ile Trp Asn Ala Gly Asn Thr Tyr Tyr Ala Ser Trp
65 70 75 80
Ala Lys Gly Arg Phe Thr Ile Ser Lys Thr Ser Thr Thr Val Asp Leu
85 90 95
Lys Ile Thr Ser Pro Thr Thr Glu Asp Thr Ala Thr Tyr Phe Cys Ala
100 105 110
Arg Gly Thr Leu Gly Asp Tyr Asn Gly Met Asp Pro Trp Gly Pro Gly
115 120 125
Thr Leu Val Thr Val Ser Ser Gly Gln Pro Lys Ala Pro Ser Val Phe
130 135 140
Pro Leu Ala Pro Cys Cys Gly Asp Thr Pro Ser Ser Thr Val Thr Leu
145 150 155 160
Gly Cys Leu Val Lys Gly Tyr Leu Pro Glu Pro Val Thr Val Thr Trp
165 170 175
Asn Ser Gly Thr Leu Thr Asn Gly Val Arg Thr Phe Pro Ser Val Arg
180 185 190
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Ser Val Thr Ser
195 200 205
Ser Ser Gln Pro Val Thr Cys Asn Val Ala His Pro Ala Thr Asn Thr
210 215 220
Lys Val Asp Lys Thr Val Ala Pro Ser Thr Cys Ser Lys Pro Thr Cys
225 230 235 240
Pro Pro Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Ile Phe Pro Pro
245 250 255
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
260 265 270
Val Val Val Asp Val Ser Gln Asp Asp Pro Glu Val Gln Phe Thr Trp
275 280 285
Tyr Ile Asn Asn Glu Gln Val Arg Thr Ala Arg Pro Pro Leu Arg Glu
290 295 300
Gln Gln Phe Asn Ser Thr Ile Arg Val Val Ser Thr Leu Pro Ile Ala
305 310 315 320
His Gln Asp Trp Leu Arg Gly Lys Glu Phe Lys Cys Lys Val His Asn
325 330 335
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Arg Gly
340 345 350
Gln Pro Leu Glu Pro Lys Val Tyr Thr Met Gly Pro Pro Arg Glu Glu
355 360 365
Leu Ser Ser Arg Ser Val Ser Leu Thr Cys Met Ile Asn Gly Phe Tyr
370 375 380
Pro Ser Asp Ile Ser Val Glu Trp Glu Lys Asn Gly Lys Ala Glu Asp
385 390 395 400
Asn Tyr Lys Thr Thr Pro Ala Val Leu Asp Ser Asp Gly Ser Tyr Phe
405 410 415
Leu Tyr Ser Lys Leu Ser Val Pro Thr Ser Glu Trp Gln Arg Gly Asp
420 425 430
Val Phe Thr Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
435 440 445
Gln Lys Ser Ile Ser Arg Ser Pro Gly Lys
450 455
<210> 55
<211> 235
<212> PRT
<213> Искусственная последовательность
<220>
<223> Последовательность аминокислот лёгкой цепи антитела
VEGF165-R988
кролика, содержащей сигнальный пептид
<400> 55
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Glu Leu Asp Leu Thr Gln Thr Pro Ser Pro Val Ser Ala
20 25 30
Ala Val Gly Gly Thr Val Thr Ile Asn Cys Gln Ser Ser Gln Thr Ile
35 40 45
Tyr Ala Asn Arg Arg Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Pro
50 55 60
Pro Lys Leu Leu Ile Tyr Gly Ala Ser Thr Leu Ala Ser Gly Val Pro
65 70 75 80
Ser Arg Phe Lys Gly Ser Gly Ser Gly Thr Gln Phe Thr Leu Thr Ile
85 90 95
Ser Gly Val Gln Cys Asp Asp Ala Ala Thr Tyr Tyr Cys Ala Gly Tyr
100 105 110
Lys Ser Tyr Asp Gly Asp Asp Val Gly Phe Gly Gly Gly Thr Glu Val
115 120 125
Val Val Lys Gly Asp Pro Val Ala Pro Thr Val Leu Ile Phe Pro Pro
130 135 140
Ala Ala Asp Gln Val Ala Thr Gly Thr Val Thr Ile Val Cys Val Ala
145 150 155 160
Asn Lys Tyr Phe Pro Asp Val Thr Val Thr Trp Glu Val Asp Gly Thr
165 170 175
Thr Gln Thr Thr Gly Ile Glu Asn Ser Lys Thr Pro Gln Asn Ser Ala
180 185 190
Asp Cys Thr Tyr Asn Leu Ser Ser Thr Leu Thr Leu Thr Ser Thr Gln
195 200 205
Tyr Asn Ser His Lys Glu Tyr Thr Cys Lys Val Thr Gln Gly Thr Thr
210 215 220
Ser Val Val Gln Ser Phe Asn Arg Gly Asp Cys
225 230 235
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| БИФУНКЦИОНАЛЬНЫЕ БЕЛКИ, КОМБИНИРУЮЩИЕ БЛОКАДУ КОНТРОЛЬНОЙ ТОЧКИ, ДЛЯ ТАРГЕТНОЙ ТЕРАПИИ | 2019 |
|
RU2756899C1 |
| СТАБИЛЬНЫЕ И РАСТВОРИМЫЕ АНТИТЕЛА, ИНГИБИРУЮЩИЕ VEGF | 2019 |
|
RU2747735C2 |
| БИСПЕЦИФИЧНЫЙ СЛИТЫЙ БЕЛОК И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2801528C2 |
| БЕЛКОВЫЕ КОМПОЗИЦИИ ПРОТИВ VEGF И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2020 |
|
RU2795973C1 |
| АНТИТЕЛА ПРОТИВ НtrА1 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2750285C2 |
| АНТИТЕЛА К IL-6 И ИХ СЛИТЫЕ КОНСТРУКЦИИ И КОНЪЮГАТЫ | 2019 |
|
RU2810777C2 |
| АНТИТЕЛА, СОДЕРЖАЩИЕ ПОЛИПЕПТИД, ВСТРОЕННЫЙ В УЧАСТОК КАРКАСНОЙ ОБЛАСТИ 3 | 2019 |
|
RU2796254C2 |
| ПОЛИПЕПТИДНЫЙ ЛИНКЕР ДЛЯ ПОЛУЧЕНИЯ МУЛЬТИСПЕЦИФИЧЕСКИХ АНТИТЕЛ | 2018 |
|
RU2776302C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С GPRC5D | 2020 |
|
RU2841523C2 |
| АНТИ-PD-1 АНТИТЕЛА | 2019 |
|
RU2788095C2 |
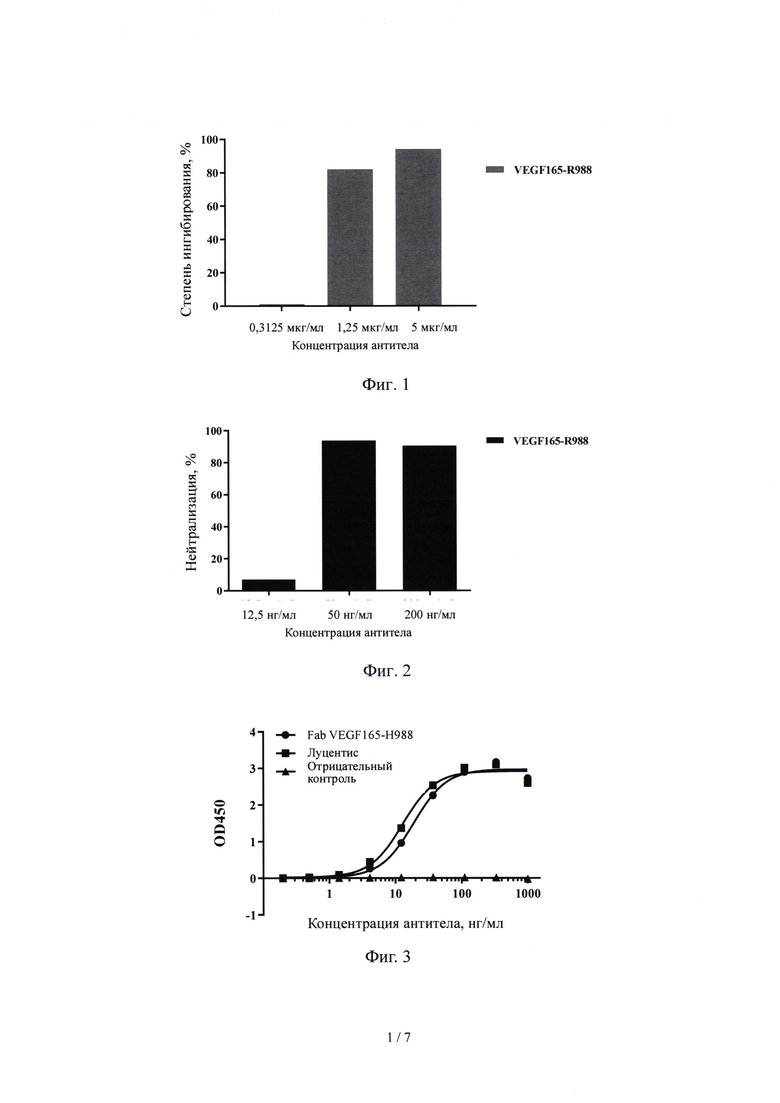
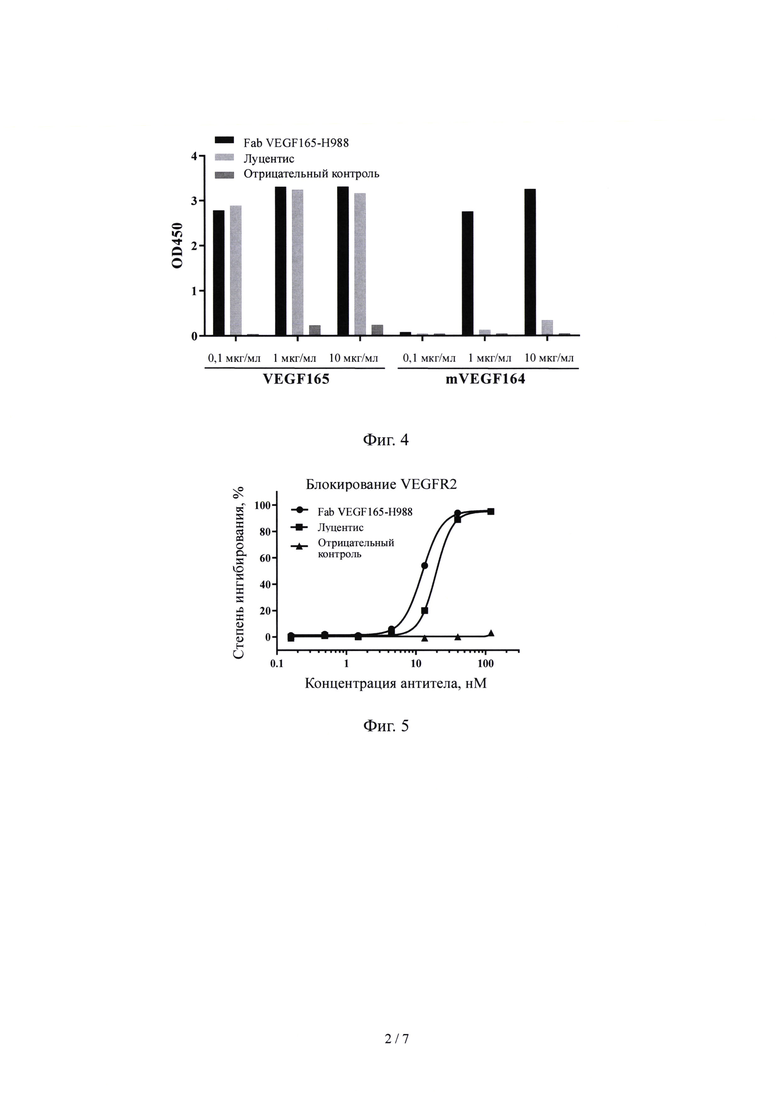
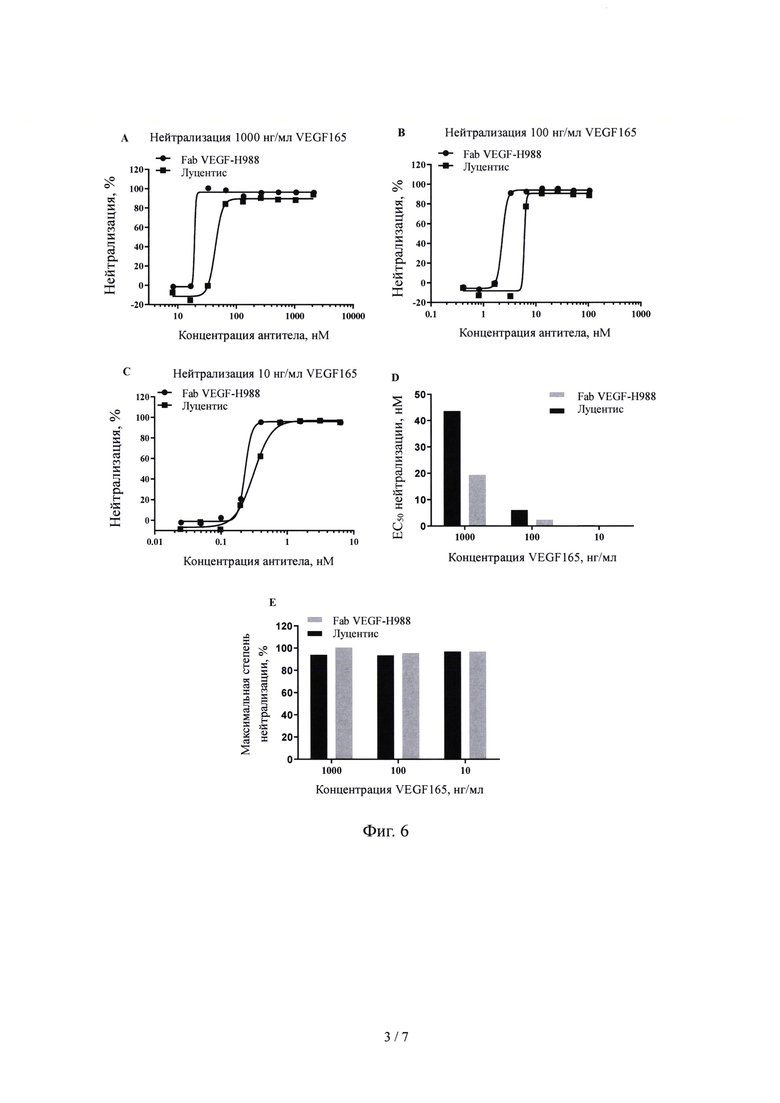
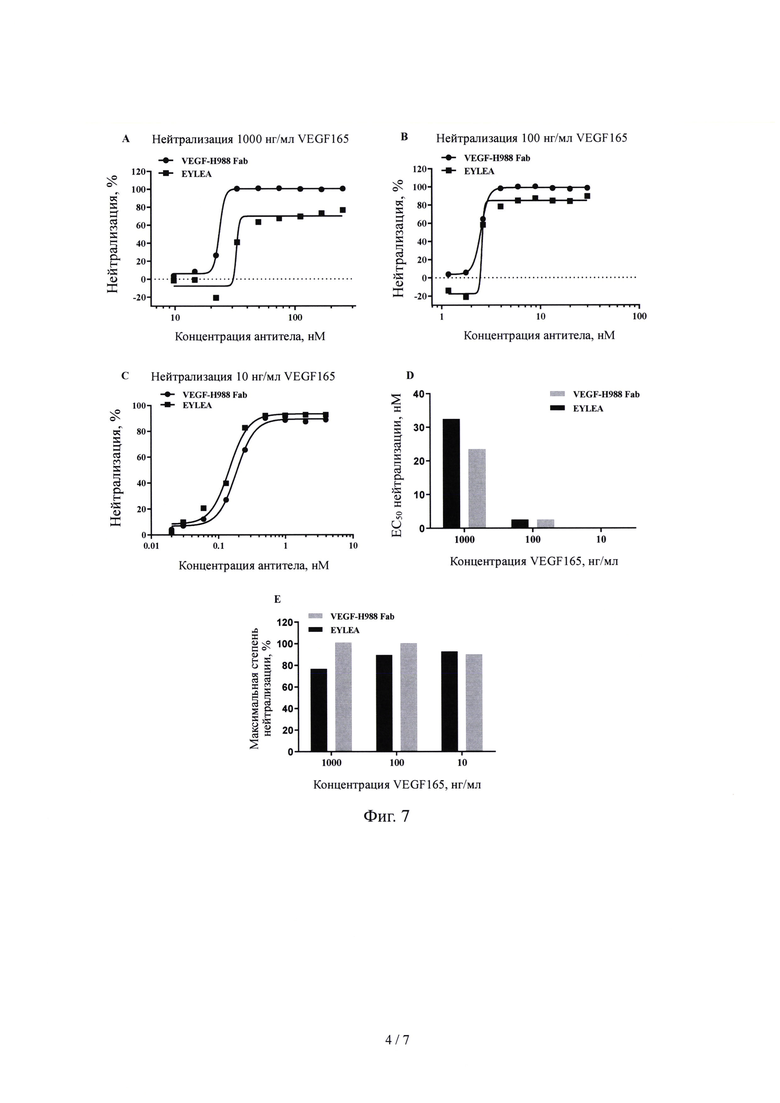
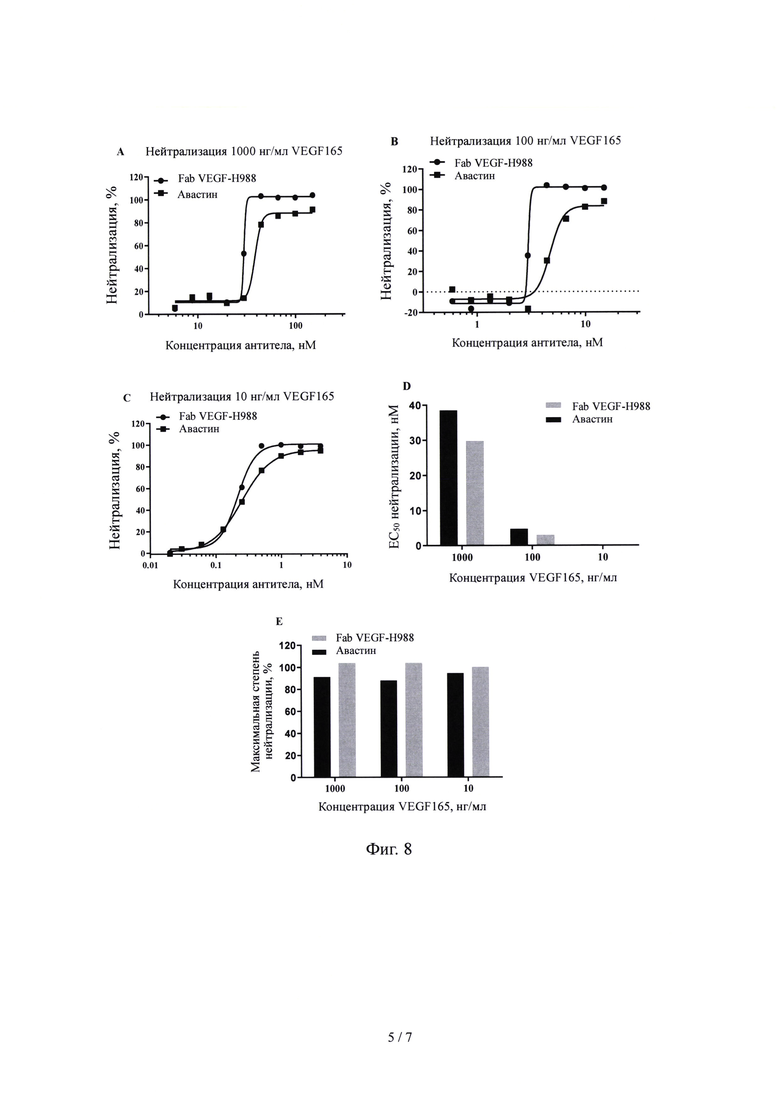
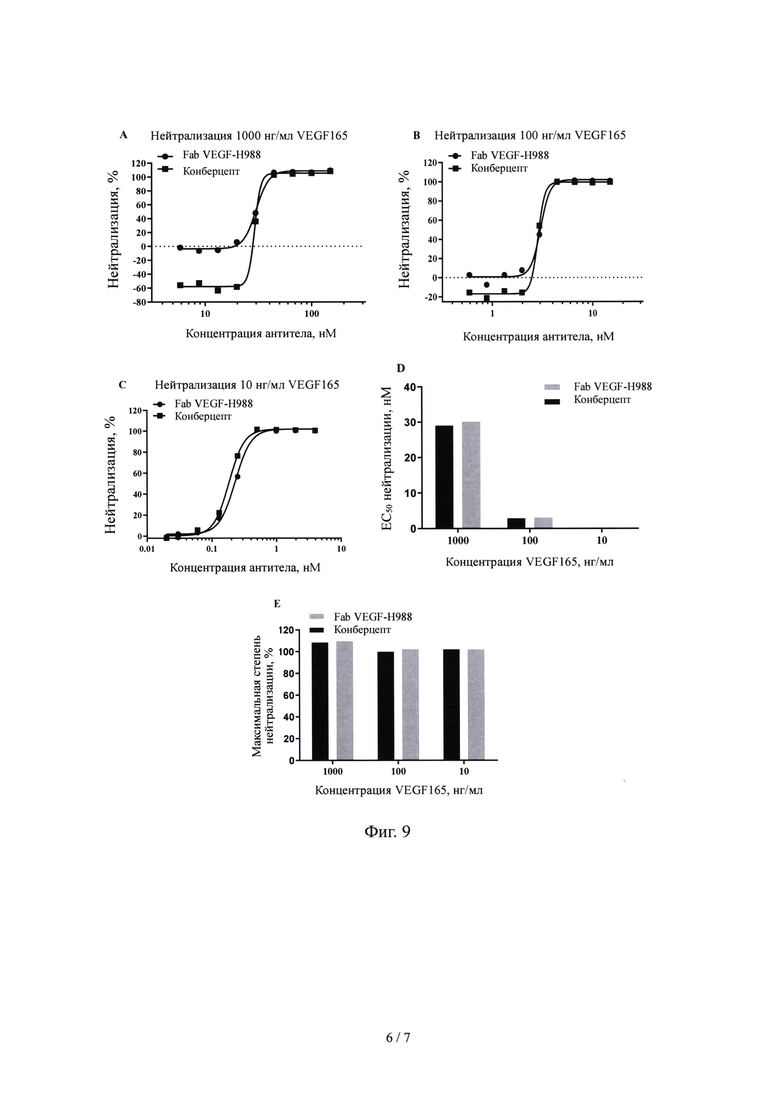
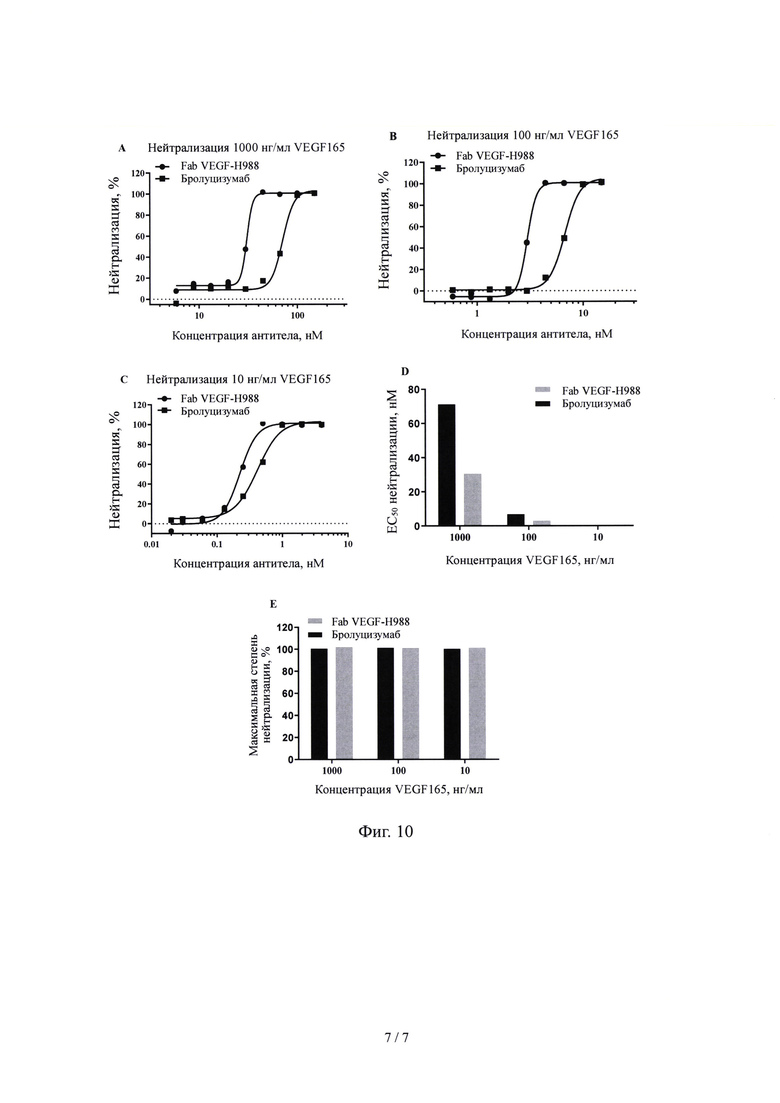
Группа изобретений относится к области иммунотерапии опухолей. Предложен Fab фрагмент антитела против VEGF. Изолированный Fab фрагмент антитела против VEGF содержит вариабельный участок тяжелой цепи, содержащий участок CDR1 с SEQ ID NO: 27, CDR2 с SEQ ID NO: 28 и CDR3 с SEQ ID NO: 29; и вариабельный участок легкой цепи, содержащий участок CDR1 с SEQ ID NO: 24, CDR2 с SEQ ID NO: 25 и CDR3 с SEQ ID NO: 26. Также предложены конъюгат, содержащий Fab фрагмент антитела и терапевтический агент, нуклеиновая кислота, кодирующая Fab фрагмент антитела, вектор экспрессии, клетка-хозяин, способ получения Fab фрагмента антитела, фармацевтические композиция и комбинация и способ лечения заболевания глаза, связанного с ангиогенезом. Fab фрагмент антитела эффективно блокирует активность VEGF165. Изобретения можно использовать для лечения заболеваний глаз, характеризующихся хориоидальной неоваскуляризацией, включая вызванную возрастом дегенерацию желтого пятна (AMD), диабетический макулярный отек (DME), отек сетчатки, дегенеративную близорукость, хориоидальную неоваскуляризацию (CNV). 12 н. и 13 з.п. ф-лы, 10 ил., 4 табл., 3 пр.
1. Изолированный Fab фрагмент антитела против VEGF, содержащий вариабельный участок тяжелой цепи, содержащий участок CDR1 тяжелой цепи, содержащий последовательность аминокислот, указанную в SEQ ID NO: 27, и участок CDR2 тяжелой цепи, содержащий последовательность аминокислот, указанную в SEQ ID NO: 28, и участок CDR3 тяжелой цепи, содержащий последовательность аминокислот, указанную в SEQ ID NO: 29; и
вариабельный участок легкой цепи, содержащий участок CDR1 легкой цепи, содержащий последовательность аминокислот, указанную в SEQ ID NO: 24, участок CDR2 легкой цепи, содержащий последовательность аминокислот, указанную в SEQ ID NO: 25, и участок CDR3 легкой цепи, содержащий последовательность аминокислот, указанную в SEQ ID NO: 26.
2. Fab фрагмент антитела против VEGF по п. 1, содержащий вариабельный участок тяжелой цепи, содержащий последовательность аминокислот, указанную в SEQ ID NO: 36, или последовательность аминокислот, по меньшей мере на 90%, 92%, 95%, 98% или 99% идентичную последовательности, указанной в SEQ ID NO: 36; и вариабельный участок легкой цепи, содержащий последовательность аминокислот, указанную в SEQ ID NO: 37, или последовательность аминокислот, по меньшей мере на 90%, 92%, 95%, 98% или 99% идентичную последовательности, указанной в SEQ ID NO: 37.
3. Fab фрагмент антитела против VEGF по п. 1 или 2, который представляет собой Fab фрагмент, дополнительно содержащий константный участок тяжелой цепи СН1 и константный участок легкой цепи, предпочтительно константный участок тяжелой цепи СН1, являющийся константным участком тяжелой цепи IgG1, содержащим последовательность аминокислот, указанную в SEQ ID NO: 40, или последовательность аминокислот, по меньшей мере на 90%, 92%, 95%, 98% или 99% идентичную последовательности, указанной в SEQ ID NO: 40; и/или константный участок легкой цепи, являющийся константным участком легкой цепи, содержащим последовательность аминокислот, указанную в SEQ ID NO: 39, или последовательность аминокислот, по меньшей мере на 90%, 92%, 95%, 98% или 99% идентичную последовательности, указанной в SEQ ID NO: 39.
4. Fab фрагмент антитела против VEGF по любому из пп. 1-3, дополнительно содержащий сигнальный пептид тяжелой цепи и сигнальный пептид легкой цепи, где предпочтительно указанный сигнальный пептид тяжелой цепи представляет собой последовательность аминокислот, указанную в SEQ ID NO: 34, или последовательность аминокислот, по меньшей мере на 90%, 92%, 95%, 98% или 99% идентичную последовательности, указанной в SEQ ID NO: 34, и/или указанный сигнальный пептид легкой цепи представляет собой последовательность аминокислот, указанную в SEQ ID NO: 35, или последовательность аминокислот, по меньшей мере на 90%, 92%, 95%, 98% или 99% идентичную последовательности, указанной в SEQ ID NO: 35.
5. Fab фрагмент антитела против VEGF по любому из пп. 1-4, отличающийся тем, что указанный Fab фрагмент антитела против VEGF представляет собой Fab фрагмент, связанный с антителом IgG, предпочтительно Fab фрагмент, связанный с антителом IgG1.
6. Fab фрагмент антитела против VEGF по любому из пп. 1-5, отличающийся тем, что указанный Fab фрагмент антитела против VEGF является моноклональным.
7. Fab фрагмент антитела против VEGF по любому из пп. 1-6, отличающийся тем, что аффинность связывания KD указанного антитела против VEGF или его антиген-связывающего фрагмента с рекомбинантным белком VEGF 165 человека составляет 0,01-8 Е-10 М, предпочтительно 0,1-5 Е-10 М и более предпочтительно 0,5-3 Е-10 М, наиболее предпочтительно 1,54 Е-10 М.
8. Конъюгат антитела и лекарственного средства для использования при лечении заболеваний глаза, связанных с ангиогенезом, содержащий Fab фрагмент антитела против VEGF по любому из пп. 1-7 и дополнительный терапевтический агент.
9. Конъюгат антитела и лекарственного средства по п. 8, отличающийся тем, что указанный Fab фрагмент антитела против VEGF связан с указанным дополнительным терапевтическим агентом посредством линкера.
10. Нуклеиновая кислота, кодирующая Fab фрагмент антитела против VEGF по любому из пп. 1-7.
11. Нуклеиновая кислота по п. 10, содержащая последовательность нуклеотидов, указанную в SEQ ID NO: 45, и последовательность нуклеотидов, указанную в SEQ ID NO: 46.
12. Нуклеиновая кислота по п. 10, дополнительно содержащая последовательность нуклеотидов, указанную в SEQ ID NO: 49, и/или последовательность нуклеотидов, указанную в SEQ ID NO: 48.
13. Вектор экспрессии, содержащий нуклеиновую кислоту по любому из пп. 10-12.
14. Клетка-хозяин для получения Fab фрагмента по любому из пп. 1-7, содержащая нуклеиновую кислоту по любому из пп. 10-12 или вектор экспрессии по п. 13.
15. Способ получения Fab фрагмента антитела против VEGF по любому из пп. 1-7, включающий выращивание клетки-хозяина по п. 14 в условиях, подходящих для экспрессии Fab фрагмента антитела и сбора экспрессированного Fab фрагмента антитела из культуральной жидкости.
16. Фармацевтическая композиция для лечения заболеваний глаза, связанных с ангиогенезом, содержащая Fab фрагмент антитела против VEGF по любому из пп. 1-7 или конъюгат антитела и лекарственного средства по п. 8 или 9, и фармацевтически приемлемый носитель.
17. Применение Fab фрагмента антитела против VEGF по любому из пп. 1-7, или конъюгата антитела и лекарственного средства по п. 8 или 9, или фармацевтической композиции по п. 16 для лечения заболеваний глаза, связанных с ангиогенезом.
18. Применение Fab фрагмента антитела против VEGF, или конъюгата антитела и лекарственного средства, или фармацевтической композиции по п. 17, отличающееся тем, что указанное заболевание глаза представляет собой хориоидальную неоваскуляризацию, включая вызванную возрастом дегенерацию желтого пятна (AMD), диабетический макулярный отек (DME), отек сетчатки, дегенеративную близорукость, хориоидальную неоваскуляризацию (CNV).
19. Способ лечения заболевания глаза, связанного с ангиогенезом, включающий введение субъекту, нуждающемуся в этом, терапевтически эффективного количества Fab фрагмента антитела против VEGF по любому из пп. 1-7, или конъюгата антитела и лекарственного средства по п. 8 или 9, или фармацевтической композиции по п. 16, и лечения таким образом заболевания глаза, связанного с ангиогенезом.
20. Способ по п. 19, отличающийся тем, что указанное заболевание глаза, связанное с ангиогенезом, представляет собой хориоидальную неоваскуляризацию, включая вызванную возрастом дегенерацию желтого пятна (AMD), диабетический макулярный отек (DME), отек сетчатки, дегенеративную близорукость, хориоидальную неоваскуляризацию (CNV).
21. Применение Fab фрагмента антитела против VEGF по любому из пп. 1-7, или конъюгата антитела и лекарственного средства по п. 8 или 9, или фармацевтической композиции по п. 16 в получении лекарственного средства для лечения заболеваний глаза, связанных с ангиогенезом.
22. Применение по п. 21, отличающееся тем, что указанное заболевание глаза, связанное с ангиогенезом, представляет собой хориоидальную неоваскуляризацию, включая вызванную возрастом дегенерацию желтого пятна (AMD), диабетический макулярный отек (DME), отек сетчатки, дегенеративную близорукость, хориоидальную неоваскуляризацию (CNV).
23. Фармацевтическая комбинация для лечения заболеваний глаза, связанных с ангиогенезом, содержащая Fab фрагмент антитела против VEGF по любому из пп. 1-7, или конъюгат антитела и лекарственного средства по п. 8 или 9, или фармацевтическую композицию по п. 16, и один или более дополнительный терапевтический агент.
24. Набор для лечения заболеваний глаза, связанных с ангиогенезом, содержащий Fab фрагмент антитела против VEGF по любому из пп. 1-7, или конъюгат антитела и лекарственного средства по п. 8 или 9, или фармацевтическую композицию по п. 16.
25. Набор по п. 24, дополнительно содержащий устройство для введения.
| Машина для послойного фрезирования и размешивания торфа в залежи | 1928 |
|
SU12835A1 |
| Зажимное приспособление к динамометрам для испытания пряжи пасмами | 1932 |
|
SU29277A1 |
| СТАБИЛЬНЫЕ И РАСТВОРИМЫЕ АНТИТЕЛА, ИНГИБИРУЮЩИЕ VEGF | 2016 |
|
RU2648152C2 |
| CN 102286101 B, 08.05.2013 | |||
| US 7667004 B2, 23.02.2010. | |||
