Настоящая заявка относится к области конструирования взаимодействия антитело-FcRn. В данном документе описан способ получения и отбора антител с конструированным взаимодействием антитела с FcRn.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Неонатальный рецептор Fc (FcRn) важен для метаболической судьбы антител класса IgG in vivo. FcRn функционирует для спасения IgG от пути лизосомальной деградации, приводя к пониженному клиренсу и увеличенному времени полужизни. Он представляет собой гетеродимерный белок состоящий из двух полипептидов: 50 кДа белка, подобного главному комплексу гистосовместимости класса I (α-FcRn), и 15 кДа β2-микроглобулина (β2m). FcRn с высокой аффинностью связывается с СН2-СН3 частью области Fc антитела класса IgG. Взаимодействие между антителом класса IgG и FcRn является pH-зависимым и происходит в стехиометрии 1:2, т.е. одна молекула антитела IgG может взаимодействовать с двумя молекулами FcRn через его два полипептида области Fc тяжелых цепей (см., например, Huber, А.Н., et al., J. Mol. Biol. 230 (1993) 1077-1083).
Houde D., et al., описали характеризацию конформации IgG1 и конформационной динамики посредством масс-спектрометрии с водород/дейтериевым обменом (Anal. Chem. 81 (2009) 2644-2651). Важность неонатального FcR в регуляции сывороточного периода полужизни терапевтических белков, содержащих домен Fc человеческого IgG1: сравнительное исследование аффинности моноклональных антител и слитых с Fc белков с человеческим неонатальным FcR, была описана Suzuki, Т., et al. (J. Immunol. 184 (2010) 1968-1976). Wang, W., et al. описали то, что моноклональные антитела с идентичными последовательностями Fc могут дифференциально связываться с FcRn с фармакокинетическими последствиями (Drug Metabol. Dispos. 39 (2011) 1469-1477).
Аналитическая аффинная хроматография на основе FcRn для функциональной характеризации моноклональных антител была описана Schlothauer, Т., et al. (mAbs 5 (2013) 576-586).
Houde, D., et al., описали то, что посттрансляционные модификации дифференциально воздействуют на конформацию IgG1 и связывание с рецептором (Mol. Cell Proteom. 9 (2010) 1716-1728).
В ЕР 14165987.0 описано прогнозирование in vitro периода полужизни in vivo.
В ЕР 2233500 описаны оптимизированные варианты Fc. Антитела с модифицированными изоэлектрическими точками описаны в US 2012/028304.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Обнаружили, что во взаимодействии полноразмерного антитела с человеческим неонатальным рецептором Fc (FcRn) принимают участие области, включающие остатки 1-23, 145-174, 180-197 в тяжелой цепи, а также остатки 55-83 в легкой цепи (нумерация по Kabat (вариабельный домен) и индексу EU (Европейский Союз) (константная область) соответственно) моноклонального полноразмерного антитела.
Таким образом, одним аспектом, как описано в данном документе, является способ отбора полноразмерного антитела, включающий следующие этапы:
а) получение из родительского полноразмерного антитела множества вариантов полноразмерных антител посредством рандомизации одного или более чем одного аминокислотного остатка, выбранного из группы аминокислотных остатков, состоящей из остатков в положениях 1-23 в вариабельном домене тяжелой цепи (нумерация согласно Kabat), остатков в положениях 55-83 в вариабельном домене легкой цепи (нумерация согласно Kabat), остатков в положениях 145-174 в первом константном домене тяжелой цепи (нумерация согласно индексу EU) и остатков в положениях 180-197 в первом константном домене тяжелой цепи (нумерация согласно индексу EU),
б) определение силы связывания каждого полноразмерного антитела из множества антител с человеческим неонатальным рецептором (FcRn) и
в) отбор полноразмерного антитела из множества вариантов полноразмерных антител, которое имеет отличную силу связывания с FcRn, чем родительское полноразмерное антитело.
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 3-18 в вариабельном домене тяжелой цепи (нумерация согласно Kabat), в положениях 57-71 в вариабельном домене легкой цепи (нумерация согласно Kabat), в положениях 161-174 в первом константном домене тяжелой цепи (нумерация согласно индексу EU) и в положениях 182-197 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 5-18 в вариабельном домене тяжелой цепи (нумерация согласно Kabat), в положениях 57-70 в вариабельном домене легкой цепи (нумерация согласно Kabat), в положениях 161-174 в первом константном домене тяжелой цепи (нумерация согласно индексу EU) и в положениях 181-196 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 1-23 в вариабельном домене тяжелой цепи (нумерация согласно Kabat).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 3-18 в вариабельном домене тяжелой цепи (нумерация согласно Kabat).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 5-18 в вариабельном домене тяжелой цепи (нумерация согласно Kabat).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 145-174 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 161-174 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 180-197 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 181-196 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 182-197 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 55-83 в вариабельном домене легкой цепи (нумерация согласно Kabat).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 55-73 в вариабельном домене легкой цепи (нумерация согласно Kabat).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 57-71 в вариабельном домене легкой цепи (нумерация согласно Kabat).
В одном воплощении аминокислотные остатки выбраны из группы аминокислотных остатков, содержащей аминокислотные остатки 6, 162, 164, 165, 191, 194, 195 и 196 в тяжелой цепи и аминокислотные остатки 57 и 60 в легкой цепи (нумерация по Kabat (вариабельный домен) и индексу EU (константная область) соответственно).
В одном воплощении аминокислотный остаток в положении 6 вариабельного домена тяжелой цепи рандомизирован до Q.
В одном воплощении аминокислотные остатки в одном или более чем одном положении аминокислот 162, 164, 165, 191, 194, 195 и 196 первого константного домена тяжелой цепи (СН1) рандомизированы независимо друг от друга до кислотного аминокислотного остатка.
В одном воплощении аминокислотные остатки в одном или более чем одном положении аминокислот 57 и 60 вариабельного домена легкой цепи рандомизированы независимо друг от друга до основного аминокислотного остатка.
В одном воплощении всех аспектов, как описано в данном документе, определение силы связывания каждого полноразмерного антитела из множества антител с человеческим неонатальным рецептором Fc (FcRn) осуществляется в отсутствие соответствующего антигена антител.
В одном воплощении всех аспектов, как описано в данном документе, определение силы связывания каждого полноразмерного антитела из множества антител с человеческим неонатальным рецептором Fc (FcRn) осуществляется для свободного, т.е. не образующего комплекс с антигеном, антитела.
В одном воплощении антитело представляет собой полноразмерное антитело IgG. В одном воплощении антитело представляет собой полноразмерное антитело IgG1.
Полноразмерное антитело содержит четыре цепи антитела: две легкие цепи (первую легкую цепь и вторую легкую цепь) и две тяжелые цепи (первую тяжелую цепь и вторую тяжелую цепь). Первая и вторая тяжелая цепь, а также, независимо от них, первая и вторая легкая цепь могут быть либо идентичными, либо отличными в отношении их аминокислотной последовательности.
В одном воплощении антитело представляет собой одноплечее антитело.
Одноплечее антитело содержит i) одну полноразмерную легкую цепь, ii) одну полноразмерную тяжелую цепь, которая образует пару с полноразмерной легкой цепью, и iii) одну укороченную тяжелую цепь, содержащую по меньшей мере часть шарнирной области, домен СН2 и домен СН3, которая образует пару с полноразмерной тяжелой цепью.
В одном воплощении антитело представляет собой биспецифичное антитело.
В одном воплощении в одном или более чем одном из следующих положений аминокислот осуществляется замена (нумерация согласно нумерации вариабельного домена по Kabat и схеме нумерации индекса EU по Kabat соответственно) заряженного аминокислотного остатка нейтральным аминокислотным остатком или нейтрального аминокислотного остатка заряженным аминокислотным остатком, независимо друг от друга: в положении 6, 16, 19, 57, 66, 83, 162, 164, 165, 191, 194, 195, 196 тяжелой цепи, в положении 57, 60 легкой цепи.
В одном воплощении в одном или более чем одном из следующих положений аминокислот осуществляется замена (нумерация согласно нумерации вариабельного домена по Kabat и схеме нумерации индекса EU по Kabat соответственно) заряженного аминокислотного остатка нейтральным аминокислотным остатком или нейтрального аминокислотного остатка заряженным аминокислотным остатком, независимо друг от друга: в положении 6, 16, 19, 162, 164, 165, 191, 194, 195, 196 тяжелой цепи, в положении 57, 60 легкой цепи.
В одном воплощении вводится одна или более чем одна из следующих мутаций аминокислот (рандомизаций) (нумерация согласно нумерации вариабельного домена по Kabat и схеме нумерации индекса EU по Kabat соответственно), независимо друг от друга:
- E6Q тяжелой цепи и/или
- A162D тяжелой цепи, и/или
- А162Е тяжелой цепи, и/или
- T164D тяжелой цепи, и/или
- Т164Е тяжелой цепи, и/или
- S165D тяжелой цепи, и/или
- S165E тяжелой цепи, и/или
- S191D тяжелой цепи, и/или
- S191E тяжелой цепи, и/или
- G194D тяжелой цепи, и/или
- G194E тяжелой цепи, и/или
- T195D тяжелой цепи, и/или
- Т195Е тяжелой цепи, и/или
- Q196D тяжелой цепи, и/или
- Q196E тяжелой цепи, и/или
- G57K легкой цепи, и/или
- G57R легкой цепи, и/или
- S60K легкой цепи, и/или
- S60R легкой цепи.
В одном воплощении вводится следующая мутация аминокислоты (нумерация согласно нумерации вариабельного домена по Kabat и схеме нумерации индекса EU по Kabat соответственно):
i) E6Q тяжелой цепи.
В одном воплощении вводится следующая мутация аминокислоты (нумерация согласно нумерации вариабельного домена по Kabat и схеме нумерации индекса EU по Kabat соответственно):
i) Т164Е тяжелой цепи.
В одном воплощении вводятся следующие мутации аминокислот (нумерация согласно нумерации вариабельного домена по Kabat и схеме нумерации индекса EU по Kabat соответственно):
i) A162D и S165D тяжелой цепи.
В одном воплощении вводится следующая мутация аминокислоты (нумерация согласно нумерации вариабельного домена по Kabat и схеме нумерации индекса EU по Kabat соответственно):
i) G194E тяжелой цепи.
В одном воплощении вводятся следующие мутации аминокислот (нумерация согласно нумерации вариабельного домена по Kabat и схеме нумерации индекса EU по Kabat соответственно):
i) S191D и Q196D тяжелой цепи.
В одном воплощении вводится следующая мутация аминокислоты (нумерация согласно нумерации вариабельного домена по Kabat и схеме нумерации индекса EU по Kabat соответственно):
i) G57R легкой цепи.
В одном воплощении вводятся следующие мутации аминокислот (нумерация согласно нумерации вариабельного домена по Kabat и схеме нумерации индекса EU по Kabat соответственно):
i) G57K и S60K легкой цепи.
В одном воплощении вводятся следующие мутации аминокислот (нумерация согласно нумерации вариабельного домена по Kabat и схеме нумерации индекса EU по Kabat соответственно):
i) Т164Е тяжелой цепи и
ii) A162D и S165D тяжелой цепи, и
iii) G194E тяжелой цепи, и
iv) S191D и Q196D тяжелой цепи.
В одном воплощении вводятся следующие мутации аминокислот (нумерация согласно нумерации вариабельного домена по Kabat и схеме нумерации индекса EU по Kabat соответственно):
i) E6Q тяжелой цепи и
ii) Т164Е тяжелой цепи, и
iii) A162D и S165D тяжелой цепи, и
iv) G194E, S191D и Q196D тяжелой цепи, и
v) G57K и S60K легкой цепи.
В одном воплощении вводятся следующие мутации аминокислот (нумерация согласно нумерации вариабельного домена по Kabat и схеме нумерации индекса EU по Kabat соответственно):
i) E6Q тяжелой цепи и/или
ii) Т164Е тяжелой цепи, и/или
iii) A162D и S165D тяжелой цепи, и/или
iv) G194E тяжелой цепи, и/или
v) Т164Е, A162D и S165D тяжелой цепи, и/или
vi) G194E, S191D и Q196D тяжелой цепи, и/или
vii) G57R легкой цепи, и/или
viii) G57K и S60K легкой цепи.
В одном воплощении вводятся следующие мутации аминокислот (нумерация согласно нумерации вариабельного домена по Kabat и схеме нумерации индекса EU по Kabat соответственно):
i) E6Q в первой и/или второй тяжелой цепи, и/или
ii) а) Т164Е в первой тяжелой цепи, и б) A162D и S165D во второй тяжелой цепи, и/или
iii) a) G194E в первой тяжелой цепи, и б) S191D и Q196D во второй тяжелой цепи, и/или
iv) а) Т164Е, A162D и S165D в первой тяжелой цепи, и б) G194Q, S191D и Q196D во второй тяжелой цепи, и/или
v) a) E6Q в первой и/или второй тяжелой цепи, б) Т164Е, A162D и S165D в первой тяжелой цепи, в) G194E, S191D и Q196D во второй тяжелой цепи, и г) G57K и S60K в первой и/или второй легкой цепи.
В одном воплощении рандомизация осуществляется посредством мутирования аминокислотного остатка до другого аминокислотного остатка из той же самой группы аминокислотных остатков.
В одном воплощении рандомизация осуществляется посредством мутирования аминокислотного остатка до другого аминокислотного остатка из другой группы аминокислотных остатков.
В одном воплощении рандомизируются от одного до пятнадцати аминокислотных остатков. В одном воплощении рандомизируются от одного до десяти аминокислотных остатков. В одном воплощении рандомизируются от одного до пяти аминокислотных остатков.
В одном воплощении полноразмерное антитело выбрано из множества полноразмерных антител, которые имеют повышенную силу связывания с FcRn.
В одном воплощении полноразмерное антитело выбрано из множества полноразмерных антител, которые имеют пониженную силу связывания с FcRn.
В одном воплощении сила связывания определяется по значению KD.
В одном воплощении сила связывания определяется по времени удерживания на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH.
В одном воплощении сила связывания определяется по времени полужизни in vivo.
Одним аспектом, как описано в данном документе, является множество вариантов полноразмерных антител, полученных из одного родительского полноразмерного антитела посредством рандомизации одного или более чем одного аминокислотного остатка, выбранного из группы аминокислотных остатков, содержащей положения 1-23 в вариабельном домене тяжелой цепи (нумерация согласно Kabat), положения 55-83 в вариабельном домене легкой цепи (нумерация согласно Kabat), положения 145-174 в первом константном домене тяжелой цепи (нумерация согласно индексу EU) и положения 180-197 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 3-18 в вариабельном домене тяжелой цепи (нумерация согласно Kabat), в положениях 57-71 в вариабельном домене легкой цепи (нумерация согласно Kabat), в положениях 161-174 в первом константном домене тяжелой цепи (нумерация согласно индексу EU) и в положениях 182-197 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 5-18 в вариабельном домене тяжелой цепи (нумерация согласно Kabat), в положениях 57-70 в вариабельном домене легкой цепи (нумерация согласно Kabat), в положениях 161-174 в первом константном домене тяжелой цепи (нумерация согласно индексу EU) и в положениях 181-196 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 1-23 в вариабельном домене тяжелой цепи (нумерация согласно Kabat).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 3-18 в вариабельном домене тяжелой цепи (нумерация согласно Kabat).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 5-18 в вариабельном домене тяжелой цепи (нумерация согласно Kabat).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 145-174 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 161-174 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 180-197 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 181-196 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 182-197 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 55-83 в вариабельном домене легкой цепи (нумерация согласно Kabat).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 55-73 в вариабельном домене легкой цепи (нумерация согласно Kabat).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 57-71 в вариабельном домене легкой цепи (нумерация согласно Kabat).
В одном воплощении аминокислотные остатки выбраны из группы аминокислотных остатков, содержащей аминокислотные остатки 6, 162, 164, 165, 191, 194, 195 и 196 в тяжелой цепи и аминокислотные остатки 57 и 60 в легкой цепи (нумерация по Kabat (вариабельный домен) и индексу EU (константная область) соответственно).
В одном воплощении аминокислотный остаток в положении 6 вариабельного домена тяжелой цепи рандомизирован до Q.
В одном воплощении аминокислотный остаток в одном или более чем одном положении аминокислоты 162, 164, 165, 191, 194, 195 и 196 первого константного домена тяжелой цепи (СН1) рандомизирован до кислотного аминокислотного остатка.
В одном воплощении аминокислотный остаток в одном или более чем одном положении аминокислоты 57 и 60 вариабельного домена легкой цепи рандомизирован до основного аминокислотного остатка.
В одном воплощении независимо друг от друга вводится одна или более чем одна из следующих рандомизаций аминокислот
- E6Q тяжелой цепи и/или
- A162D тяжелой цепи, и/или
- А162Е тяжелой цепи, и/или
- T164D тяжелой цепи, и/или
- Т164Е тяжелой цепи, и/или
- S165D тяжелой цепи, и/или
- S165E тяжелой цепи, и/или
- S191D тяжелой цепи, и/или
- S191E тяжелой цепи, и/или
- G194D тяжелой цепи, и/или
- G194E тяжелой цепи, и/или
- T195D тяжелой цепи, и/или
- Т195Е тяжелой цепи, и/или
- Q196D тяжелой цепи, и/или
- Q196E тяжелой цепи, и/или
- G57K легкой цепи, и/или
- G57R легкой цепи, и/или
- S60K легкой цепи, и/или
- S60R легкой цепи.
В одном воплощении антитело представляет собой полноразмерное антитело IgG. В одном воплощении антитело представляет собой полноразмерное антитело IgG1.
В одном воплощении рандомизация осуществляется посредством мутирования аминокислотного остатка до другого аминокислотного остатка из той же самой группы аминокислотных остатков.
В одном воплощении рандомизация осуществляется посредством мутирования аминокислотного остатка до другого аминокислотного остатка из другой группы аминокислотных остатков.
В одном воплощении рандомизируются от одного до пятнадцати аминокислотных остатков. В одном воплощении рандомизируются от одного до десяти аминокислотных остатков. В одном воплощении рандомизируются от одного до пяти аминокислотных остатков.
В одном воплощении полноразмерное антитело выбрано из множества полноразмерных антител, которые имеют повышенную силу связывания с FcRn.
В одном воплощении полноразмерное антитело выбрано из множества полноразмерных антител, которые имеют пониженную силу связывания с FcRn.
В одном воплощении сила связывания определяется по значению KD.
В одном воплощении сила связывания определяется по времени удерживания на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH.
В одном воплощении сила связывания определяется по времени полужизни in vivo.
Другим аспектом, как описано в данном документе, является применение одной или более чем одной мутации аминокислоты в положениях, выбранных из группы положений, содержащей положения 1-23 в вариабельном домене тяжелой цепи (нумерация согласно Kabat), положения 55-83 в вариабельном домене легкой цепи (нумерация согласно Kabat), положения 145-174 в первом константном домене тяжелой цепи (нумерация согласно индексу EU) и положения 180-197 в первом константном домене тяжелой цепи (нумерация согласно индексу EU), для изменения времени полужизни полноразмерного антитела in vivo.
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 3-18 в вариабельном домене тяжелой цепи (нумерация согласно Kabat), в положениях 57-71 в вариабельном домене легкой цепи (нумерация согласно Kabat), в положениях 161-174 в первом константном домене тяжелой цепи (нумерация согласно индексу EU) и в положениях 182-197 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 5-18 в вариабельном домене тяжелой цепи (нумерация согласно Kabat), в положениях 57-70 в вариабельном домене легкой цепи (нумерация согласно Kabat), в положениях 161-174 в первом константном домене тяжелой цепи (нумерация согласно индексу EU) и в положениях 181-196 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 1-23 в вариабельном домене тяжелой цепи (нумерация согласно Kabat).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 3-18 в вариабельном домене тяжелой цепи (нумерация согласно Kabat).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 5-18 в вариабельном домене тяжелой цепи (нумерация согласно Kabat).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 145-174 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 161-174 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 180-197 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 181-196 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 182-197 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 55-83 в вариабельном домене легкой цепи (нумерация согласно Kabat).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 55-73 в вариабельном домене легкой цепи (нумерация согласно Kabat).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 57-71 в вариабельном домене легкой цепи (нумерация согласно Kabat).
В одном воплощении аминокислотные остатки выбраны из группы аминокислотных остатков, содержащей аминокислотные остатки 6, 162, 164, 165, 191, 194, 195 и 196 в тяжелой цепи и аминокислотные остатки 57 и 60 в легкой цепи (нумерация по Kabat (вариабельный домен) и индексу EU (константная область) соответственно).
В одном воплощении аминокислотный остаток в положении 6 вариабельного домена тяжелой цепи мутирован до Q.
В одном воплощении аминокислотный остаток в одном или более чем одном положении аминокислоты 162, 164, 165, 191, 194, 195 и 196 первого константного домена тяжелой цепи (СН1) мутирован, независимо друг от друга, до кислотного аминокислотного остатка.
В одном воплощении аминокислотный остаток в одном или более чем одном положении аминокислоты 57 и 60 вариабельного домена легкой цепи мутирован, независимо друг от друга, до основного аминокислотного остатка.
В одном воплощении, независимо друг от друга, вводится одна или более чем одна из следующих мутаций аминокислот:
- E6Q тяжелой цепи и/или
- A162D тяжелой цепи, и/или
- А162Е тяжелой цепи, и/или
- T164D тяжелой цепи, и/или
- Т164Е тяжелой цепи, и/или
- S165D тяжелой цепи, и/или
- S165E тяжелой цепи, и/или
- S191D тяжелой цепи, и/или
- S191E тяжелой цепи, и/или
- G194D тяжелой цепи, и/или
- G194E тяжелой цепи, и/или
- T195D тяжелой цепи, и/или
- Т195Е тяжелой цепи, и/или
- Q196D тяжелой цепи, и/или
- Q196E тяжелой цепи, и/или
- G57K легкой цепи, и/или
- G57R легкой цепи, и/или
- S60K легкой цепи, и/или
- S60R легкой цепи.
В одном воплощении антитело представляет собой полноразмерное антитело IgG. В одном воплощении антитело представляет собой полноразмерное антитело IgG1.
В одном воплощении рандомизация осуществляется посредством мутирования аминокислотного остатка до другого аминокислотного остатка из той же самой группы аминокислотных остатков.
В одном воплощении рандомизация осуществляется посредством мутирования аминокислотного остатка до другого аминокислотного остатка из другой группы аминокислотных остатков.
В одном воплощении рандомизируются от одного до пятнадцати аминокислотных остатков. В одном воплощении рандомизируются от одного до десяти аминокислотных остатков. В одном воплощении рандомизируются от одного до пяти аминокислотных остатков.
В одном воплощении полноразмерное антитело выбрано из множества полноразмерных антител, которые имеют увеличенную силу связывания с FcRn.
В одном воплощении полноразмерное антитело выбрано из множества полноразмерных антител, которые имеют уменьшенную силу связывания с FcRn.
В одном воплощении сила связывания определяется по значению KD.
В одном воплощении сила связывания определяется по времени удерживания на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH.
В одном воплощении сила связывания определяется по времени полужизни in vivo.
Другим аспектом, как описано в данном документе, является вариантное полноразмерное антитело, содержащее два полипептида легкой цепи и два полипептида тяжелой цепи, где этот вариант антитела получен из родительского полноразмерного антитела посредством введения мутаций аминокислот в одном или более чем одном положении, выбранном из группы положений, содержащей положения 1-23 в вариабельном домене тяжелой цепи (нумерация согласно Kabat), положения 55-83 в вариабельном домене легкой цепи (нумерация согласно Kabat), положения 145-174 в первом константном домене тяжелой цепи (нумерация согласно индексу EU) и положения 180-197 в первом константном домене тяжелой цепи (нумерация согласно индексу EU), и где этот вариант антитела имеет отличную аффинность в отношении человеческого неонатального рецептора Fc, чем родительское полноразмерное антитело.
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 3-18 в вариабельном домене тяжелой цепи (нумерация согласно Kabat), в положениях 57-71 в вариабельном домене легкой цепи (нумерация согласно Kabat), в положениях 161-174 в первом константном домене тяжелой цепи (нумерация согласно индексу EU) и в положениях 182-197 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 5-18 в вариабельном домене тяжелой цепи (нумерация согласно Kabat), в положениях 57-70 в вариабельном домене легкой цепи (нумерация согласно Kabat), в положениях 161-174 в первом константном домене тяжелой цепи (нумерация согласно индексу EU) и в положениях 181-196 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 1-23 в вариабельном домене тяжелой цепи (нумерация согласно Kabat).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 3-18 в вариабельном домене тяжелой цепи (нумерация согласно Kabat).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 5-18 в вариабельном домене тяжелой цепи (нумерация согласно Kabat).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 145-174 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 161-174 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 180-197 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 181-196 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 182-197 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 55-83 в вариабельном домене легкой цепи (нумерация согласно Kabat).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 55-73 в вариабельном домене легкой цепи (нумерация согласно Kabat).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 57-71 в вариабельном домене легкой цепи (нумерация согласно Kabat).
В одном воплощении аминокислотные остатки выбраны из группы аминокислотных остатков, содержащей аминокислотные остатки 6, 162, 164, 165, 191, 194, 195 и 196 в тяжелой цепи и аминокислотные остатки 57 и 60 в легкой цепи (нумерация по Kabat (вариабельный домен) и индексу EU (константная область) соответственно).
В одном воплощении аминокислотный остаток в положении 6 вариабельного домена тяжелой цепи мутирован до Q.
В одном воплощении аминокислотный остаток в одном или более чем одном положении аминокислоты 162, 164, 165, 191, 194, 195 и 196 первого константного домена тяжелой цепи (СН1) мутирован, независимо друг от друга, до кислотного аминокислотного остатка.
В одном воплощении аминокислотный остаток в одном или более чем одном положении аминокислоты 57 и 60 вариабельного домена легкой цепи мутирован, независимо друг от друга, до основного аминокислотного остатка.
В одном воплощении независимо друг от друга вводится одна или более чем одна из следующих мутаций аминокислот:
- E6Q тяжелой цепи и/или
- A162D тяжелой цепи, и/или
- А162Е тяжелой цепи, и/или
- T164D тяжелой цепи, и/или
- Т164Е тяжелой цепи, и/или
- S165D тяжелой цепи, и/или
- S165E тяжелой цепи, и/или
- S191D тяжелой цепи, и/или
- S191E тяжелой цепи, и/или
- G194D тяжелой цепи, и/или
- G194E тяжелой цепи, и/или
- T195D тяжелой цепи, и/или
- Т195Е тяжелой цепи, и/или
- Q196D тяжелой цепи, и/или
- Q196E тяжелой цепи, и/или
- G57K легкой цепи, и/или
- G57R легкой цепи, и/или
- S60K легкой цепи, и/или
- S60R легкой цепи.
В одном воплощении антитело представляет собой полноразмерное антитело IgG. В одном воплощении антитело представляет собой полноразмерное антитело IgG1.
В одном воплощении рандомизация осуществляется посредством мутирования аминокислотного остатка до другого аминокислотного остатка из той же самой группы аминокислотных остатков.
В одном воплощении рандомизация осуществляется посредством мутирования аминокислотного остатка до другого аминокислотного остатка из другой группы аминокислотных остатков.
В одном воплощении рандомизируются от одного до пятнадцати аминокислотных остатков. В одном воплощении рандомизируются от одного до десяти аминокислотных остатков. В одном воплощении рандомизируются от одного до пяти аминокислотных остатков.
В одном воплощении полноразмерное антитело выбрано из множества полноразмерных антител, которые имеют увеличенную силу связывания с FcRn.
В одном воплощении полноразмерное антитело выбрано из множества полноразмерных антител, которые имеют уменьшенную силу связывания с FcRn.
В одном воплощении сила связывания определяется по значению KD.
В одном воплощении сила связывания определяется по времени удерживания на колонке для аффинной хроматографии на основе FcRn с элюцией положительным линейным градиентом pH.
В одном воплощении сила связывания определяется по времени полужизни in vivo.
Одним аспектом, как описано в данном документе, является вариант антитела, который имеет мутацию одного или более чем одного из следующих аминокислотных остатков относительно его родительского антитела:
- остатки 6, 162, 164, 165, 191, 194, 195 и 196 в тяжелой цепи
- остатки 57 и 60 в легкой цепи
(нумерация согласно Kabat (вариабельный домен) и индексу EU (константная область) соответственно).
В одном воплощении вариант антитела имеет мутацию одного или более чем одного из следующих аминокислотных остатков относительно его родительского антитела:
- остатки 162, 164, 165, 191, 194, 195 и 196 в тяжелой цепи
- остатки 57 и 60 в легкой цепи
(нумерация согласно Kabat (вариабельный домен) и индексу EU (константная область) соответственно).
В одном воплощении вариант антитела имеет мутацию одного или более чем одного из следующих аминокислотных остатков относительно его родительского антитела:
- остатки 162, 164, 165, 191, 194, 195 и 196 в тяжелой цепи
(нумерация согласно Kabat (вариабельный домен) и индексу EU (константная область) соответственно).
В одном воплощении антитело представляет собой полноразмерное антитело IgG. В одном воплощении антитело представляет собой полноразмерное антитело IgG1.
В одном воплощении антитело представляет собой биспецифичное антитело.
В одном воплощении антитело имеет мутацию аминокислоты E6Q в вариабельном домене тяжелой цепи.
В одном воплощении антитело имеет кислотную аминокислоту в одном или более чем одном из положений аминокислот 162, 164, 165, 191, 194, 195 и 196 первого константного домена тяжелой цепи (СН1). В одном воплощении кислотная аминокислота представляет собой D или Е. В одном воплощении антитело имеет кислотный аминокислотный остаток в двух или более чем двух из положений аминокислот 162, 164, 165, 191, 194, 195 и 196 первого константного домена тяжелой цепи (СН1), при этом кислотные аминокислотные остатки выбраны независимо друг от друга.
В одном воплощении антитело имеет основной аминокислотный остаток в одном или в обоих положениях аминокислот 57 и 60 вариабельного домена легкой цепи. В одном воплощении основной аминокислотный остаток представляет собой K или R. В одном воплощении антитело имеет основной аминокислотный остаток в обоих положениях 57 и 60 вариабельного домена легкой цепи, при этом основные аминокислотные остатки выбраны независимо друг от друга.
В одном воплощении антитело имеет одну или более чем одну из следующих мутаций аминокислот, независимо друг от друга:
- E6Q тяжелой цепи и/или
- A162D тяжелой цепи, и/или
- А162Е тяжелой цепи, и/или
- T164D тяжелой цепи, и/или
- Т164Е тяжелой цепи, и/или
- S165D тяжелой цепи, и/или
- S165E тяжелой цепи, и/или
- S191D тяжелой цепи, и/или
- S191E тяжелой цепи, и/или
- G194D тяжелой цепи, и/или
- G194E тяжелой цепи, и/или
- T195D тяжелой цепи, и/или
- Т195Е тяжелой цепи, и/или
- Q196D тяжелой цепи, и/или
- Q196E тяжелой цепи, и/или
- G57K легкой цепи, и/или
- G57R легкой цепи, и/или
- S60K легкой цепи, и/или
- S60R легкой цепи.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Данное изобретение, по меньшей мере частично, основано на открытии того, что несколько областей моноклонального антитела как в области Fc, так и во фрагменте Fab демонстрируют уменьшение поглощения дейтерия при связывании с FcRn (см. Фиг. 1 и 2). Данные области включают остатки 1-23, 145-174 (аминокислотная последовательность GGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVL, SEQ ID NO: 11), 180-197 (аминокислотная последовательность YSLSSVVTVPSSSLGTQT, SEQ ID NO: 13) в тяжелой цепи, а также остатки 55-83 в легкой цепи (нумерация по Kabat (вариабельный домен) и индексу EU (константная область) соответственно).
I. ОПРЕДЕЛЕНИЯ
Положения аминокислот всех константных областей и доменов тяжелой и легкой цепи в том виде, в котором они используются в данном документе, пронумерованы согласно системе нумерации по Kabat, описанной в Kabat, et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991), и она в данном документе называется «нумерация согласно Kabat». Конкретно система нумерации по Kabat (см. страницы 647-660) Kabat, et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991) используется для константного домена легкой цепи CL изотипа каппа и лямбда, и система нумерации индекса EU по Kabat (см. страницы 661-723) используется для константных доменов тяжелой цепи (СН1, шарнирная область, СН2 и СН3).
Термин «примерно» обозначает интервал плюс/минус 20% от следующего далее числового значения. В одном воплощении термин «примерно» обозначает интервал плюс/минус 10% от следующего далее числового значения. В одном воплощении термин «примерно» обозначает интервал плюс/минус 5% от следующего далее числового значения.
Термин «акцепторный человеческий каркас» для целей данного документа представляет собой каркас, содержащий аминокислотную последовательность каркаса вариабельного домена легкой цепи (VL) или каркаса вариабельного домена тяжелой цепи (VH), происходящую из каркаса человеческого иммуноглобулина или человеческого консенсусного каркаса, как определено ниже. Акцепторный человеческий каркас, «происходящий из» каркаса человеческого иммуноглобулина или человеческого консенсусного каркаса, может содержать такую же аминокислотную последовательность, как и они, или он может содержать изменения аминокислотной последовательности. В некоторых воплощениях число изменений аминокислот составляет 10 или менее, 9 или менее, 8 или менее, 7 или менее, 6 или менее, 5 или менее, 4 или менее, 3 или менее, или 2 или менее. В некоторых воплощениях акцепторный человеческий каркас VL является идентичным по последовательности каркасной последовательности VL человеческого иммуноглобулина или человеческой консенсусной каркасной последовательности.
Термин антитело «с созревшей аффинностью» относится к антителу с одним или более чем одним изменением в одной или более чем одной гипервариабельной области (HVR) по сравнению с родительским антителом, которое не обладает такими изменениями, причем такие изменения приводят к улучшению аффинности антитела в отношении антигена.
Термин «изменение» обозначает мутацию (замену), вставку (присоединение) или делецию одного или более чем одного аминокислотного остатка в родительском антителе или слитом полипептиде, например, в слитом полипептиде, содержащем по меньшей мере часть области Fc, связывающуюся с FcRn, с получением модифицированного антитела или слитого полипептида. Термин «мутация» обозначает то, что определенный аминокислотный остаток заменен другим аминокислотным остатком. Например, мутация K234A обозначает то, что аминокислотный остаток лизин в положении 234 в области Fc антитела (полипептиде) заменен аминокислотным остатком аланином (замена лизина аланином) (нумерация согласно системе нумерации индекса EU по Kabat).
Фраза «встречающиеся в природе аминокислотные остатки» обозначает аминокислотный остаток из группы, состоящей из аланина (трехбуквенный код: Ala, однобуквенный код: А), аргинина (Arg, R), аспарагина (Asn, N), аспарагиновой кислоты (Asp, D), цистеина (Cys, С), глутамина (Gln, Q), глутаминовой кислоты (Glu, Е), глицина (Gly, G), гистидина (His, Н), изолейцина (lle, I), лейцина (Leu, L), лизина (Lys, K), метионина (Met, М), фенилаланина (Phe, F), пролина (Pro, Р), серина (Ser, S), треонина (Thr, Т), триптофана (Trp, W), тирозина (Tyr, Y) и валина (Val, V).
Термин «мутация аминокислоты» обозначает замену по меньшей мере одного существующего аминокислотного остатка другим отличным аминокислотным остатком (заменяющим аминокислотным остатком). Заменяющий аминокислотный остаток может представлять собой «встречающийся в природе аминокислотный остаток» и может быть выбран из группы, состоящей из аланина (трехбуквенный код: Ala, однобуквенный код: А), аргинина (Arg, R), аспарагина (Asn, N), аспарагиновой кислоты (Asp, D), цистеина (Cys, С), глутамина (Gln, Q), глутаминовой кислоты (Glu, Е), глицина (Gly, G), гистидина (His, Н), изолейцина (lle, I), лейцина (Leu, L), лизина (Lys, K), метионина (Met, М), фенилаланина (Phe, F), пролина (Pro, Р), серина (Ser, S), треонина (Thr, Т), триптофана (Trp, W), тирозина (Tyr, Y) и валина (Val, V). Заменяющий аминокислотный остаток может представлять собой «не встречающийся в природе аминокислотный остаток». См., например, US 6586207, WO 98/48032, WO 03/073238, US 2004/0214988, WO 2005/35727, WO 2005/74524, Chin, J.W., et al., J. Am. Chem. Soc. 124 (2002) 9026-9027; Chin, J.W. and Schultz, P.G., ChemBioChem 11 (2002) 1135-1137; Chin, J.W., et al., PICAS United States of America 99 (2002) 11020-11024 и Wang, L. and Schultz, P.G., Chem. (2002) 1-10 (все полностью включенные в данный документ посредством ссылки).
Термин «аминокислотная вставка» обозначает (дополнительное) включение по меньшей мере одного аминокислотного остатка в заданном положении в аминокислотной последовательности. В одном воплощении вставка будет представлять собой вставку одного или двух аминокислотных остатков. Вставленный(ные) аминокислотный(ные) остаток(тки) может(гут) представлять собой любой встречающийся в природе или не встречающийся в природе аминокислотный остаток.
Термин «делеция аминокислоты» обозначает удаление по меньшей мере одного аминокислотного остатка в заданном положении в аминокислотной последовательности.
Термин «антитело» в данном документе используется в самом широком смысле и охватывает разные структуры антитела, включающие моноклональные антитела, мультиспецифичные антитела (например, биспецифичные антитела, триспецифичные антитела) и фрагменты антител, при условии, что они демонстрируют желательную активность связывания в отношении антигена и/или белка А, и/или FcRn, но не ограничивающиеся ими.
Термин «гетеродимерная область Fc» обозначает область Fc, которая состоит из двух полипептидных цепей, которые имеют разные аминокислотные остатки в соответствующих положениях, при этом данные положения определяются согласно системе нумерации индекса EU по Kabat, при этом разные положения влияют на образование гетеродимеров. Примерами таких различий являются так называемые замены по типу «выступы во впадины» (см., например, US 7695936 и US 2003/0078385). Было обнаружено, что следующие замены по типу «выступы во впадины» в индивидуальных полипептидных цепях области Fc антитела IgG подкласса IgG1 увеличивают образование гетеродимера: 1) Y407T в одной цепи и T366Y в другой цепи; 2) Y407A в одной цепи и T366W в другой цепи; 3) F405A в одной цепи и T394W в другой цепи; 4) F405W в одной цепи и T394S в другой цепи; 5) Y407T в одной цепи и T366Y в другой цепи; 6) T366Y и F405A в одной цепи и T394W и Y407T в другой цепи; 7) T366W и F405W в одной цепи и T394S и Y407A в другой цепи; 8) F405W и Y407A в одной цепи и T366W и T394S в другой цепи; и 9) T366W в одной цепи и T366S, L368A и Y407V в другой цепи, при этом последние перечисленные являются особенно подходящими. Кроме того, образование гетеродимера облегчают замены, создающие новые дисульфидные мостики между двумя полипептидными цепями области Fc (см., например, US 2003/0078385). Обнаружили, что образование гетеродимера усиливают следующие замены, приводящие к подходящим образом расположенным в пространстве остаткам цистеина для образования новых внутрицепочечных дисульфидных связей в индивидуальных полипептидных цепях области Fc антитела IgG подкласса IgG1: Y349C в одной цепи и S354C в другой; Y349C в одной цепи и Е356С в другой; Y349C в одной цепи и Е357С в другой; L351C в одной цепи и S354C в другой; Т394С в одной цепи и Е397С в другой; или D399C в одной цепи и K392C в другой. Другие примеры аминокислотных замен, облегчающих гетеродимеризацию, представляют собой так называемые «замены пары зарядов» (см., например, WO 2009/089004). Обнаружили, что образование гетеродимера усиливают следующие замены пары зарядов в индивидуальных полипептидных цепях области Fc антитела IgG подкласса IgG1: 1) K409D или K409E в одной цепи и D399K или D399R в другой цепи; 2) K392D или K392E в одной цепи и D399K или D399R в другой цепи; 3) K439D или K439E в одной цепи и Е356К или E356R в другой цепи; 4) K370D или K370E в одной цепи и E357K или E357R в другой цепи; 5) K409D и K360D в одной цепи плюс D399K и Е356К в другой цепи; 6) K409D и K370D в одной цепи плюс D399K и E357K в другой цепи; 7) K409D и K392D в одной цепи плюс D399K, E356K и E357K в другой цепи; 8) K409D и K392D в одной цепи и D399K в другой цепи; 9) K409D и K392D в одной цепи и D399K и E356K в другой цепи; 10) K409D и K392D в одной цепи и D399K and D357K в другой цепи; 11) K409D и K370D в одной цепи и D399K и D357K в другой цепи; 12) D399K в одной цепи и K409D и K360D в другой цепи; и 13) K409D и K439D в одной цепи и D399K и E356K на другой.
Термин «химерное» антитело относится к антителу, в котором часть тяжелой и/или легкой цепи происходит из конкретного источника или вида, тогда как остальная тяжелая и/или легкая цепь происходит из другого источника или вида.
Термин «класс» антитела относится к типу константного домена или константной области, которыми обладает его тяжелая цепь. Существуют пять главных классов антител: IgA, IgD, IgE, IgG и IgM, и некоторые из них могут дополнительно подразделяться на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелой цепи, которые соответствуют разным классам иммуноглобулинов, называются α, δ, ε, γ и μ соответственно.
Термин «сравнимая длина» обозначает то, что два полипептида содержат идентичное число аминокислотных остатков или могут отличаться по длине на один или более чем один и, самое большее, вплоть до 10 аминокислотных остатков. В одном воплощении полипептиды (области Fc) содержат идентичное число аминокислотных остатков или отличаются числом от 1 до 10 аминокислотных остатков. В одном воплощении полипептиды (области Fc) содержат идентичное число аминокислотных остатков или отличаются числом от 1 до 5 аминокислотных остатков. В одном воплощении полипептиды (области Fc) содержат идентичное число аминокислотных остатков или отличаются числом от 1 до 3 аминокислотных остатков.
Термин «эффекторные функции» относится к тем биологическим активностям, приписываемым области Fc антитела, которые варьируют с классом антитела. Примеры эффекторных функций антитела включают: связывание с C1q и комплементзависимую цитотоксичность (CDC); связывание с рецептором Fc; антителозависимую клеточную цитотоксичность (ADCC); фагоцитоз; понижающую регуляцию рецепторов поверхности клетки (например, рецептора В-клеток) и активацию В-клеток.
Термин «эффективное количество» агента, например, фармацевтической композиции, относится к эффективному количеству в дозировках и в течение периодов времени, необходимых для достижения желательного терапевтического или профилактического результата.
Термин «область Fc человеческого происхождения» обозначает С-концевую область тяжелой цепи иммуноглобулина человеческого происхождения, которая содержит по меньшей мере часть шарнирной области, домен СН2 и домен СН3. В одном воплощении область Fc тяжелой цепи человеческого IgG простирается от Cys226 или от Pro230 до карбоксильного конца тяжелой цепи. В одном воплощении область Fc имеет аминокислотную последовательность SEQ ID NO: 02. Однако С-концевой лизин (Lys447) области Fc может присутствовать или может не присутствовать.
Термин «FcRn» обозначает человеческий неонатальный рецептор Fc. FcRn функционирует для спасения IgG от пути лизосомальной деградации, приводя к пониженному клиренсу и увеличенному времени полужизни. FcRn представляет собой гетеродимерный белок состоящий из двух полипептидов: 50 кДа белка, подобного главному комплексу гистосовместимости класса I (α-FcRn) и 15 кДа β2-микроглобулина (β2m). FcRn с высокой аффинностью связывается с частью СН2-СН3 области Fc IgG. Взаимодействие между IgG и FcRn является строго pH-зависимым и происходит в стехиометрии 1:2 - с одним IgG, связывающимся с двумя молекулами FcRn через его две тяжелые цепи (Huber, А.Н., et al., J. Mol. Biol. 230 (1993) 1077-1083). Связывание с FcRn происходит в эндосоме при кислотном pH (рН меньше 6,5), и IgG высвобождается на нейтральной поверхности клетки (pH примерно 7,4). pH-чувствительная природа взаимодействия облегчает опосредованную FcRn защиту IgG, пиноцитированных в клетки, от внутриклеточной деградации посредством связывания с рецептором в кислотной среде эндосом. FcRn затем облегчает рециклирование IgG к поверхности клетки и последующее высвобождение в кровоток при воздействии на комплекс FcRn-IgG среды с нейтральным pH вне клетки.
Термин «связывающаяся с FcRn часть области Fc» обозначает часть полипептида тяжелой цепи антитела, которая простирается приблизительно от положения 243 по EU до положения 261 по EU и приблизительно от положения 275 по EU до положения 293 по EU, и приблизительно от положения 302 по EU до положения 319 по EU, и приблизительно от положения 336 по EU до положения 348 по EU, и приблизительно от положения 367 по EU до положения 393 по EU, и положения 408 по EU, и приблизительно от положения 424 по EU до положения 440 по EU. В одном воплощении изменен один или более чем один из следующих аминокислотных остатков согласно нумерации EU по Kabat: F243, Р244, Р245 Р, K246, Р247, K248, D249, Т250, L251, М252, I253, S254, R255, Т256, Р257, Е258, V259, Т260, С261, F275, N276, W277, Y278, V279, D280, V282, Е283, V284, Н285, N286, А287, K288, Т289, K290, Р291, R292, Е293, V302, V303, S304, V305, L306, Т307, V308, L309, Н310, Q311, D312, W313, L314, N315, G316, K317, Е318, Y319, I336, S337, K338, А339, K340, G341, Q342, Р343, R344, Е345, Р346, Q347, V348, С367, V369, F372, Y373, Р374, S375, D376, I377, А378, V379, Е380, W381, Е382, S383, N384, G385, Q386, Р387, Е388, N389, Y391, Т393, S408, S424, С425, S426, V427, М428, Н429, Е430, А431, L432, Н433, N434, Н435, Y436, Т437, Q438, K439 и S440 (нумерация по EU).
Термин «каркас» или «FR» относится к остаткам вариабельного домена, отличным от остатков гипервариабельной области (HVR). FR вариабельного домена обычно состоит из четырех доменов FR: FR1, FR2, FR3 и FR4. Соответственно, последовательности HVR и FR обычно появляются в VH (или в VL) в следующей последовательности: FR1-H1(L1)-FR2-H2(L2)-FR3-H3(L3)-FR4.
Термин «полноразмерное антитело» обозначает антитело, имеющее структуру, по существу аналогичную структуре нативного антитела, содержащее четыре полипептида или имеющее тяжелые цепи, которые содержат область Fc, как определено в данном документе. Полноразмерное антитело может содержать дополнительные домены, такие как, например, scFv или scFab, конъюгированные с одной или более чем одной из цепей полноразмерного антитела. Данные конъюгаты также охватываются термином полноразмерное антитело.
Термин «димерный полипептид» обозначает комплекс, содержащий по меньшей мере два полипептида, которые ассоциированы ковалентно. Данный комплекс может содержать дополнительные полипептиды, которые также ковалентно или нековалентно ассоциированы с другими полипептидами. В одном воплощении димерный полипептид содержит два или четыре полипептида.
Термины «гетеродимер» или «гетеродимерный» обозначают молекулу, которая содержит два полипептида (например, сравнимой длины), где два данных полипептида имеют аминокислотную последовательность, которая имеет по меньшей мере один отличный аминокислотный остаток в соответствующем положении, при этом соответствующее положение определяется согласно системе нумерации индекса EU по Kabat.
Термины «гомодимер» или «гомодимерный» обозначают молекулу, которая содержит два полипептида сравнимой длины, где два данных полипептида имеют аминокислотную последовательность, которая является идентичной в соответствующих положениях, при этом соответствующие положения определяются согласно системе нумерации индекса EU по Kabat.
Термины «клетка-хозяин», «линия клетки-хозяина» и «культура клетки-хозяина» используются взаимозаменяемо и относятся к клеткам, в которые была введена экзогенная нуклеиновая кислота, включая потомство таких клеток. Клетки-хозяева включают «трансформантов» и «трансформированные клетки», которые включают первично трансформированную клетку и происходящее от нее потомство, не придавая значения числу пассажей. Потомство может не быть полностью идентичным родительской клетке по составу нуклеиновых кислот, но может содержать мутации. В данный документ включается мутантное потомство, которое имеет такую же функцию или биологическую активность, что и исходно трансформированная клетка, подвергнутая скринингу или отобранная на них.
«Человеческое антитело» представляет собой антитело, которое обладает аминокислотной последовательностью, которая соответствует аминокислотной последовательности антитела, продуцированного человеком или человеческой клеткой, или происходящее из источника, не являющегося человеческим, в котором используется человеческий репертуар антител или других последовательностей, кодирующих человеческое антитело. Данное определение человеческого антитела конкретно исключает гуманизированное антитело, содержащее антигенсвязывающие остатки, не являющиеся человеческими.
«Человеческий консенсусный каркас» представляет собой каркас, который представляет собой чаще всего встречающиеся аминокислотные остатки в наборе каркасных последовательностей VL или VH человеческого иммуноглобулина. Обычно набор последовательностей VL или VH человеческого иммуноглобулина происходит из подгруппы последовательностей вариабельного домена. Обычно данная подгруппа последовательностей представляет собой такую же подгруппу, как и в Kabat, Е.А. et al., Sequences of Proteins of Immunological Interest, 5th ed., Bethesda MD (1991), NIH Publication 91-3242, Vols. 1-3. В одном воплощении для VL данная подгруппа представляет собой подгруппу каппа I, такую же, как в Kabat et al., выше. В одном воплощении для VH данная подгруппа представляет собой подгруппу III, такую же, как в Kabat et al., выше.
Термин «происходящий от» обозначает то, что аминокислотная последовательность происходит от родительской аминокислотной последовательности посредством введения изменений по меньшей мере в одном положении. Таким образом, полученная аминокислотная последовательность отличается от соответствующей родительской аминокислотной последовательности по меньшей мере в одном соответствующем положении (нумерация согласно индексу EU по Kabat для областей Fc антител). В одном воплощении аминокислотная последовательность, происходящая от родительской аминокислотной последовательности, отличается по одному-пятнадцати аминокислотным остаткам в соответствующих положениях. В одном воплощении аминокислотная последовательность, происходящая от родительской аминокислотной последовательности, отличается по одному-десяти аминокислотным остаткам в соответствующих положениях. В одном воплощении аминокислотная последовательность, происходящая от родительской аминокислотной последовательности, отличается по одному-шести аминокислотным остаткам в соответствующих положениях. Подобным образом, полученная аминокислотная последовательность имеет высокую идентичность аминокислотной последовательности с ее родительской аминокислотной последовательностью. В одном воплощении аминокислотная последовательность, происходящая от родительской аминокислотной последовательности, имеет 80%-ную или большую идентичность аминокислотной последовательности. В одном воплощении аминокислотная последовательность, происходящая от родительской аминокислотной последовательности, имеет 90%-ную или большую идентичность аминокислотной последовательности. В одном воплощении аминокислотная последовательность, происходящая от родительской аминокислотной последовательности, имеет 95%-ную или большую идентичность аминокислотной последовательности.
Термин «полипептид человеческой области Fc» обозначает аминокислотную последовательность, которая является идентичной «нативному» полипептиду человеческой области Fc или полипептиду человеческой области Fc «дикого типа». Термин «полипептид варианта (человеческой) области Fc» обозначает аминокислотную последовательность, которая происходит от «нативного» полипептида человеческой области Fc или полипептида человеческой области Fc «дикого типа» посредством по меньшей мере одного «изменения аминокислоты». «Человеческая область Fc» состоит из двух полипептидов человеческой области Fc. «Вариант (человеческой) области Fc» состоит из двух полипептидов области Fc, при этом оба могут представлять собой полипептиды варианта (человеческой) области Fc, или один представляет собой полипептид человеческой области Fc, а другой представляет собой полипептид варианта (человеческой) области Fc.
Термин «гуманизированное» антитело относится к химерному антителу, содержащему аминокислотные остатки из HVR (гипервариабельная область), не являющихся человеческими, и аминокислотные остатки из человеческих FR (каркасная область). В некоторых воплощениях гуманизированное антитело будет содержать по существу все из по меньшей мере одного, и типично два вариабельных домена, в которых все или по существу все из HVR (например, CDR (область, определяющая комплементарность)) соответствуют HVR антитела, не являющегося человеческим, и все или по существу все из FR соответствуют FR человеческого антитела. Гуманизированное антитело возможно может содержать по меньшей мере часть константной области антитела, происходящую от человеческого антитела. Термин «гуманизированная форма» антитела, например, антитела, не являющегося человеческим, относится к антителу, которое подверглось гуманизации.
Термин «гипервариабельная область» или «HVR» в том виде, в котором он используется в данном документе, относится к каждой из областей вариабельного домена антитела, которые являются гипервариабельными по последовательности («области, определяющие комплементарность» или «CDR») и образуют структурно определенные петли («гипервариабельные петли») и/или содержат контактирующие с антигеном остатки («контакты с антигеном»). Обычно антитела содержат шесть HVR: три в VH (Н1, Н2 и Н3) и три в VL (L1, L2, L3). HVR в том виде, в котором они обозначены в данном документе, включают:
(а) гипервариабельные петли, встречающиеся в области аминокислотных остатков 26-32 (L1), 50-52 (L2), 91-96 (L3), 26-32 (Н1), 53-55 (Н2) и 96-101 (Н3) (Chothia, С. and Lesk, A.M., J. Mol. Biol. 196 (1987) 901-917);
(б) CDR, встречающиеся в области аминокислотных остатков 24-34 (L1), 50-56 (L2), 89-97 (L3), 31-35b (Н1), 50-65 (Н2) и 95-102 (Н3) (Kabat, Е.А. et al., Sequences of Proteins of Immunological Interest, 5th ed. Public Health Service, National Institutes of Health, Bethesda, MD (1991), NIH Publication 91-3242);
(в) контакты с антигеном, встречающиеся в области аминокислотных остатков 27c-36 (L1), 46-55 (L2), 89-96 (L3), 30-35b (Н1), 47-58 (Н2) и 93-101 (Н3) (MacCallum et al. J. Mol. Biol. 262: 732-745 (1996)); и
(г) комбинации (а), (б) и/или (в), включающие аминокислотные остатки HVR 46-56 (L2), 47-56 (L2), 48-56 (L2), 49-56 (L2), 26-35 (Н1), 26-35b (Н1), 49-65 (Н2), 93-102 (Н3) и 94-102 (Н3).
Если не указано иное, остатки HVR и другие остатки в вариабельном домене (например, остатки FR) пронумерованы в данном документе согласно системе нумерации индекса EU по Kabat (Kabat et al., выше).
«Выделенное» антитело представляет собой антитело, которое было отделено от компонента его природного окружения. В некоторых воплощениях антитело очищают до более чем 95%-ной или 99%-ной чистоты, например, при определении электрофоретическими (например, SDS-PAGE (электрофорез в полиакриламидном геле с додецилсульфатом натрия), изоэлектрофокусировка (IEF), капиллярный электрофорез) или хроматографическими (например, гель-фильтрация, ионообменная ВЭЖХ (высокоэффективная жидкостная хроматография) или ВЭЖХ с обращенной фазой) способами. Относительно обзора способов оценки чистоты антител, см., например, Flatman, S. et al., J. Chrom. В 848 (2007) 79-87.
Термин «выделенная» нуклеиновая кислота относится к молекуле нуклеиновой кислоты, которая была отделена от компонента ее природного окружения. Выделенная нуклеиновая кислота включает молекулу нуклеиновой кислоты, содержащуюся в клетках, которые обычно содержат данную молекулу нуклеиновой кислоты, но молекула нуклеиновой кислоты присутствует внехромосомно или в участке хромосомы, который отличается от его природного хромосомного участка.
Термин «моноклональное антитело» в том виде, как он используется в данном документе, относится к антителу, полученному из популяции по существу гомогенных антител, т.е. индивидуальные антитела, составляющие популяцию, являются идентичными и/или связываются с тем же самым эпитопом, за исключением возможных вариантов антител, например, содержащих встречающиеся в природе мутации или возникающие во время получения препарата моноклонального антитела, причем такие варианты обычно присутствуют в минорных количествах. В отличие от препаратов поликлональных антител, которые типично включают разные антитела, направленные против разных детерминант (эпитопов), каждое моноклональное антитело препарата моноклонального антитела направлено против одной детерминанты на антигене. Таким образом, модификатор «моноклональное» указывает на характер антитела, как полученного из по существу гомогенной популяции антител, и его не следует истолковывать как требующий получения антитела каким-либо конкретным способом. Например, моноклональные антитела, подлежащие применению согласно настоящему изобретению, можно получать целым рядом методик, включающих способ гибридомы, способы генной инженерии, способы фагового дисплея и способы с использованием трансгенных животных, содержащих все локусы человеческого иммуноглобулина или их часть, но не ограничивающихся ими, причем такие способы и другие типичные способы получения моноклональных антител описываются в данном документе.
Термин «нативные антитела» относится к встречающимся в природе молекулам иммуноглобулинов с варьирующими структурами. Например, нативные антитела IgG представляют собой гетеротетрамерные гликопротеины с массой примерно 150000 дальтон, состоящие из двух идентичных легких цепей и двух идентичных тяжелых цепей, которые связаны дисульфидными связями. От N- до С-конца каждая тяжелая цепь имеет вариабельную область (VH), также именуемую вариабельный тяжелый домен или вариабельный домен тяжелой цепи, с последующими тремя константными доменами (СН1, СН2 и СН3). Аналогичным образом, от N- до С-конца каждая легкая цепь имеет вариабельную область (VL), также именуемую вариабельный легкий домен или вариабельный домен легкой цепи, с последующим константным легким (CL) доменом. Легкая цепь антитела может быть приписана к одному из двух типов, именуемых каппа (κ) и лямбда (λ), на основе аминокислотной последовательности ее константного домена.
«Процент (%) идентичности аминокислотной последовательности» по отношению к эталонной полипептидной последовательности определяется как процентная доля аминокислотных остатков в последовательности-кандидате, которые являются идентичными аминокислотным остаткам в эталонной полипептидной последовательности, после выравнивания данных последовательностей и введения, при необходимости, пробелов для достижения максимального процента идентичности последовательностей, и не рассматривая какие-либо консервативные замены как часть идентичности последовательности. Выравнивание в целях определения процента идентичности аминокислотных последовательностей может достигаться разными способами, которые находятся в пределах квалификации в данной области, например, с использованием общедоступных компьютерных программ, таких как программы BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Специалисты в данной области могут определять подходящие параметры для выравнивания последовательностей, включая любые алгоритмы, необходимые для достижения максимального выравнивания по всей длине сравниваемых последовательностей. Однако для указанных в данном документе целей значения % идентичности аминокислотных последовательностей получают с использованием компьютерной программы для сравнения последовательностей ALIGN-2. Автором компьютерной программы для сравнения последовательностей ALIGN-2 является Genentech, Inc., и исходный код с пользовательской документацией был подан в Бюро регистрации авторских прав США, Washington D.C., 20559, где он зарегистрирован под регистрационным №авторских прав США TXU510087. Программа ALIGN-2 является общедоступной от Genentech, Inc., Южный Сан-Франциско, Калифорния, или может быть компилирована из исходного кода. Программа ALIGN-2 должна быть компилирована для применения на операционной системе UNIX, включая цифровую UNIX V4.0D. Все параметры сравнения последовательностей заданы программой ALIGN-2 и не изменяются.
В ситуациях, когда ALIGN-2 используется для сравнений аминокислотных последовательностей, % идентичности аминокислотной последовательности данной аминокислотной последовательности А относительно, при сравнении или против данной аминокислотной последовательности Б (что, в качестве альтернативы, может быть выражено как данная аминокислотная последовательность А, которая имеет или содержит определенный % идентичности аминокислотной последовательности относительно, при сравнении или против данной аминокислотной последовательности Б) рассчитывается следующим образом:
100 умножить на отношение X/Y,
где X равен числу аминокислотных остатков, подсчитанных программой для выравнивания последовательностей ALIGN-2 как идентичные соответствия, в выравнивании данной программой А и Б, и где Y равен общему числу аминокислотных остатков в Б. Будет понятно, что когда длина аминокислотной последовательности А не равна длине аминокислотной последовательности Б, % идентичности аминокислотной последовательности А с Б не будет равен % идентичности аминокислотной последовательности Б с А. Если конкретно не утверждается иное, все значения % идентичности аминокислотных последовательностей, использованные в данном документе, получают, как описано в непосредственно предшествующем абзаце, с использованием компьютерной программы ALIGN-2.
Термин «рекомбинантное антитело» в том виде, как он используется в данном документе, обозначает все антитела (химерные, гуманизированные и человеческие), которые получают, экспрессируют, создают или выделяют способами генной инженерии. Это включает антитела, выделенные из клетки-хозяина, такой как клетка NS0 или СНО (яичники китайского хомяка), или из животного (например, мыши), которое является трансгенным в отношении генов человеческих иммуноглобулинов, или антитела, экспрессируемые с использованием рекомбинантного экспрессионного вектора, трансфицированного в клетку-хозяина. Такие рекомбинантные антитела имеют вариабельные и константные области в реаранжированной форме. Рекомбинантные антитела могут подвергаться соматической гипермутации in vivo. Таким образом, аминокислотные последовательности областей VH и VL рекомбинантных антител представляют собой последовательности, которые, происходя от и будучи родственными последовательностям VH и VL человеческой зародышевой линии, могут не существовать в природе в репертуаре человеческих антител зародышевой линии in vivo.
Термин «валентный» в том виде, как он используется в настоящей заявке, обозначает присутствие определенного числа сайтов связывания в молекуле (антитела). Термины «двухвалентный», «четырехвалентный» и «шестивалентный», как таковые, обозначают присутствие в молекуле (антитела) двух сайтов связывания, четырех сайтов связывания и шести сайтов связывания соответственно. Биспецифичные антитела, как описано в данном документе, в одном предпочтительном воплощении являются «двухвалентными».
Термин «вариабельная область» или «вариабельный домен» относится к домену тяжелой или легкой цепи антитела, который участвует в связывании антитела с его антигеном. Вариабельные домены тяжелой цепи и легкой цепи (VH и VL соответственно) антитела обычно имеют аналогичные структуры, причем каждый домен содержит четыре каркасные области (FR) и три гипервариабельные области (HVR) (см., например, Kindt, T.J. et al. Kuby Immunology, 6th ed., W.H. Freeman and Co., N.Y. (2007), стр. 91). Один домен VH или VL может быть достаточным для придания специфичности связывания антигена. Кроме того, антитела, которые связываются с конкретным антигеном, могут быть выделены с использованием домена VH или VL из антитела, которое связывается с антигеном, для скрининга библиотеки комплементарных доменов VL или VH соответственно (см., например, Portolano, S. et al., J. Immunol. 150 (1993) 880-887; Clackson, T. et al., Nature 352 (1991) 624-628).
Термин «вектор» в том виде, в котором он используется в данном документе, относится к молекуле нуклеиновой кислоты, способной размножать другую нуклеиновую кислоту, с которой она связана. Данный термин включает вектор в виде самореплицируемой структуры нуклеиновой кислоты, а также вектор, включенный в геном клетки-хозяина, в которую он был введен. Некоторые векторы способны управлять экспрессией нуклеиновых кислот, с которыми они связаны функциональным образом. Такие векторы называются в данном документе «экспрессионные векторы».
II. НАСТОЯЩЕЕ ИЗОБРЕТЕНИЕ
Неонатальный рецептор Fc (FcRn) является важным для метаболической судьбы антител класса IgG in vivo. FcRn функционирует для спасения IgG дикого типа от пути лизосомальной деградации, приводя к пониженному клиренсу и увеличенному времени полужизни. Он представляет собой гетеродимерный белок, состоящий из двух полипептидов: 50 кДа белка, подобного главному комплексу гистосовместимости класса I (α-FcRn) и 15 кДа β2-микроглобулина (β2m). FcRn связывается с высокой аффинностью с СН2-СН3 частью области Fc антитела класса IgG. Взаимодействие между антителом класса IgG и FcRn является pH-зависимым и происходит в стехиометрии 1:2, т.е. одна молекула антитела IgG может взаимодействовать с двумя молекулами FcRn через ее два полипептида области Fc тяжелой цепи (см., например, Huber, А.Н., et al., J. Mol. Biol. 230 (1993) 1077-1083).
Таким образом, свойства/характеристики связывания IgG с FcRn in vitro указывают на его фармакокинетические свойства in vivo в системе кровообращения.
Во взаимодействии между FcRn и областью Fc антитела класса IgG участвуют разные аминокислотные остатки домена СН2 и СН3 тяжелой цепи. Аминокислотные остатки, взаимодействующие с FcRn, расположены приблизительно между положением 243 по EU и положением 261 по EU, приблизительно между положением 275 по EU и положением 293 по EU, приблизительно между положением 302 по EU и положением 319 по EU, приблизительно между положением 336 по EU и положением 348 по EU, приблизительно между положением 367 по EU и положением 393 по EU, в положении 408 по EU и приблизительно между положением 424 по EU и положением 440 по EU. Более конкретно, во взаимодействии между областью Fc и FcRn участвуют следующие аминокислотные остатки согласно нумерации EU по Kabat: F243, Р244, Р245 Р, K246, Р247, K248, D249, Т250, L251, М252, I253, S254, R255, Т256, Р257, Е258, V259, Т260, С261, F275, N276, W277, Y278, V279, D280, V282, Е283, V284, Н285, N286, А287, K288, Т289, К290, Р291, R292, Е293, V302, V303, S304, V305, L306, Т307, V308, L309, Н310, Q311, D312, W313, L314, N315, G316, K317, Е318, Y319, I336, S337, K338, А339, К340, G341, Q342, Р343, R344, Е345, Р346, Q347, V348, С367, V369, F372, Y373, Р374, S375, D376, I377, А378, V379, Е380, W381, Е382, S383, N384, G385, Q386, Р387, Е388, N389, Y391, Т393, S408, S424, С425, S426, V427, М428, Н429, Е430, А431, L432, Н433, N434, Н435, Y436, Т437, Q438, K439 и S440.
Исследования посредством сайт-направленного мутагенеза подтвердили то, что критически важными сайтами связывания в области Fc IgG для FcRn являются гистидин 310, гистидин 435 и изолейцин 253, и в меньшей степени - гистидин 433 и тирозин 436 (см., например, Kim, J.K., et al., Eur. J. Immunol. 29 (1999) 2819-2825; Raghavan, M., et al., Biochem. 34 (1995) 14649-146579; Medesan, C, et al., J Immunol. 158 (1997) 2211-2217).
Способы для увеличения связывания IgG с FcRn осуществляли посредством мутирования IgG по разным аминокислотным остаткам: треонину 250, метионину 252, серину 254, треонину 256, треонину 307, глутаминовой кислоте 380, метионину 428, гистидину 433 и аспарагину 434 (см. Kuo, Т.Т., et al., J. Clin. Immunol. 30 (2010) 777-789).
В некоторых случаях желательными являются антитела с уменьшенным временем полужизни в системе кровообращения. Например, лекарственные средства для интравитреального применения должны иметь продолжительное время полужизни в глазу и короткое время полужизни в системе кровообращения пациента. Такие антитела также имеют преимущество усиленного воздействия на сайт заболевания, например, в глазу.
Известны разные мутации, которые влияют на связывание с FcRn и, вместе с этим, на время полужизни в системе кровообращения. Посредством сайт-направленного мутагенеза были идентифицированы критически важные остатки области Fc для взаимодействия область Fc мыши-FcRn мыши (см., например, Dall'Acqua, W.F., et al. J. Immunol 169 (2002) 5171-5180). В данном взаимодействии участвуют остатки I253, Н310, Н433, N434 и Н435 (нумерация EU согласно Kabat) (Medesan, С., et al., Eur. J. Immunol. 26 (1996) 2533-2536; Firan, M., et al., Int.
Immunol. 13 (2001) 993-1002; Kim, J.K., et al., Eur. J. Immunol. 24 (1994) 542-548). Обнаружили, что остатки I253, H310 и Н435 являются критически важными для взаимодействия человеческого Fc с мышиным FcRn (Kim, J.K., et al., Eur. J. Immunol. 29 (1999) 2819-2825). Dall'Acqua et al. посредством исследований взаимодействия белок-белок описали, что остатки M252Y, S254T, Т256Е улучшают связывание с FcRn (Dall'Acqua, W.F., et al. J. Biol. Chem. 281 (2006) 23514-23524). Исследования комплекса человеческий Fc-человеческий FcRn показали, что остатки I253, S254, Н435 и Y436 являются решающими для данного взаимодействия (Firan, М., et al., Int. Immunol. 13 (2001) 993-1002; Shields, R.L., et al., J. Biol. Chem. 276 (2001) 6591-6604). У Yeung, Y.A., et al. (J. Immunol. 182 (2009) 7667-7671) были описаны и проверены разные мутанты по остаткам 248-259 и 301-317, и 376-382, и 424-437.
Теперь обнаружили, что несколько областей моноклонального антитела во фрагменте Fab демонстрируют уменьшение поглощения дейтерия при связывании с FcRn (см. Фиг. 1 и 2). Данные области включают остатки 1-23, 145-174, 180-197 в тяжелой цепи, а также остатки 55-83 в легкой цепи (нумерация по Kabat (вариабельный домен) и индексу EU (константная область) соответственно).
Таким образом, не только аминокислотные остатки области Fc способствуют силе и, всеете с тем, компактности взаимодействия антитело-FcRn, но также и остатки, расположенные в домене СН1 и в домене VH/VL.
На основе данного открытия теперь можно предложить новые мутации аминокислот и комбинации мутаций аминокислот для получения заданного времени полужизни антител in vivo.
Для картирования сайтов полноразмерного антитела IgG1, участвующих в связывании с человеческим FcRn, использовали HDX-MS (масс-спектрометрия с водород/дейтериевым обменом). Эффективное покрытие последовательности HDX-MS антитела против дигоксигенина с FcRn составляло 82% и для НС, и для LC, и оно свидетельствует о поглощении дейтерия 89 пептидами (см. Фиг. 1).
Несколько областей антитела и в домене Fc, и в домене Fab демонстрируют уменьшение поглощения дейтерия при связывании с FcRn (см. Фиг. 1 и 2). Данные области включают остатки 1-23, 145-174, 180-197 в тяжелой цепи, а также остатки 55-83 в легкой цепи (нумерация по Kabat (вариабельный домен) и индексу EU (константная область) соответственно).
В области 1-23 анализ HDX в перекрывающихся пептидах 5-18, 5-22 и 5-23 показывает то, что для С-конца удлиненных пептидов (19-23) не наблюдается дополнительная защита. Сравнение пептида 1-18 с другими пептидами (4-18, 5-18) не осуществляется, так как различие в обратном обмене полностью дейтерированного образца составляет больше, чем 15%. Таким образом, принимая во внимание то, что в экспериментах HDX-MS дейтерий на первых двух N-концевых остатках в пептидах теряется до выявления MS, уменьшенное поглощение дейтерия посредством связывания с FcRn может быть локализовано на остатках 3-18.
Сравнение HDX у перекрывающихся пептидов (145-174, 146-174, 156-174, 159-174) в области 145-174 в домене СН1 антитела и принятие во внимание того, что в экспериментах HDX-MS дейтерий на первых двух N-концевых остатках в пептидах теряется до выявления MS, показывает то, что защита HDX посредством связывания с FcRn может быть локализована на остатках 161-174.
Для области 180-197 перекрывающийся пептид 186-197 демонстрирует только примерно половину уменьшения поглощения дейтерия при связывании с FcRn, по сравнению с пептидом 180-197. Таким образом, принимая во внимание то, что в экспериментах HDX-MS дейтерий на первых двух N-концевых остатках в пептидах теряется до выявления MS, защита HDX посредством FcRn может быть локализована на остатках 182-197.
В LC несколько пептидов в области 55-83 в каркасной области 3 демонстрируют уменьшенное поглощение дейтерия при связывании с FcRn. Посредством анализа перекрывающихся пептидов (55-71, 55-73, 55-83, 71-82, 71-83) и принимая во внимание то, что в экспериментах HDX-MS дейтерий на первых двух N-концевых остатках в пептидах теряется до выявления MS, взаимодействие связывания, главным образом, локализуется на остатках 57-71.
Уменьшенный обмен HD данных областей, удаленных от области Fc, при связывании с FcRn показывает то, что данные области содержат аминокислотные остатки, которые участвуют во взаимодействии антитело-FcRn.
Одним аспектом, как описано в данном документе, является способ отбора полноразмерного антитела, включающий следующие этапы:
а) получение из родительского полноразмерного антитела множества (пула) полноразмерных антител посредством рандомизации одного или более чем одного аминокислотного остатка, выбранного из аминокислотных остатков в положениях 1-23 в вариабельном домене тяжелой цепи (нумерация согласно Kabat), в положениях 55-83 в вариабельном домене легкой цепи (нумерация согласно Kabat), в положениях 145-174 в первом константном домене тяжелой цепи (нумерация согласно индексу EU) и в положениях 180-197 в первом константном домене тяжелой цепи (нумерация согласно индексу EU),
б) определение силы связывания каждого полноразмерного антитела из множества антител с человеческим неонатальным рецептором (FcRn) и
в) отбор полноразмерного антитела из множества полноразмерных антител, которые имеют отличную силу связывания с FcRn, чем родительское полноразмерное антитело.
Не будучи связанными данной теорией, предполагается, что после установления первого взаимодействия между областью Fc антитела и FcRn формируется второе взаимодействие между Fab-частью антитела и FcRn. Это второе взаимодействие модифицирует силу первого взаимодействия, как продемонстрировано представленными здесь данными.
В представленных в данном документе данных по обменам HD обнаружили, что существует прямое взаимодействие Fab с FcRn, основанное на относительно малом количестве положительных зарядов в вариабельном домене.
Кроме того, как описано в данном документе, в устекинумаб в определенных участках области Fab были введены дополнительные положительные заряды, приводя к значительно улучшенной силе взаимодействия с FcRn. Было обнаружено, что данное усиление взаимодействия не следует характеристикам pH-зависимого связывания из-за более поздней элюции в градиенте pH на аффинной колонке на основе FcRn.
Во многих анализах времени удерживания на колонке на основе FcRn обнаружили, что все IgG демонстрируют индивидуальный профиль времени удерживания, в зависимости от их последовательности Fab и, посредством этого, их индивидуальной картины зарядов Fab. Это наблюдение можно модифицировать посредством нанесения высокой концентрации соли в подвижном буфере, выявляя дополнительное зависимое от заряда взаимодействие.
Обнаружили, что можно экранировать влияние Fab посредством предынкубации соответствующего антитела с мишенью.
Таким образом, в данном документе было обнаружено и представляется то, что каждое антитело имеет индивидуальную картину зарядов в области Fab, которая может влиять на связывание антитела с FcRn.
Если антитело уже имеет очень сильное или даже слишком сильное связывание с FcRn (что, например, иллюстрируется значением pH элюции на аффинной колонке на основе FcRn, близким к или даже превышающим pH 7,4, или неожиданно укороченным временем полужизни in vivo, несмотря на сильное связывание с FcRn), взаимодействие антитело-FcRn может быть модифицировано уменьшением числа зарядов в области Fab антитела. Например, в аффинной хроматографии на основе FcRn, как описано в примерах настоящей патентной заявки, антитело, имеющее pH элюции 7,4 или выше, представляет собой антитело со слишком сильным взаимодействием с FcRn. Посредством уменьшения числа зарядов в области Fab (в конечном счете, наряду с генетической модификацией области Fc, если генетическая модификация области Fab не является достаточной) время удерживания также может быть уменьшено, делая рН элюции меньшим, чем pH 7,4. Такое антитело выигрывает от взаимодействия с FcRn увеличенным временем полужизни in vivo.
Если антитело имеет слабое связывание с FcRn (что, например, иллюстрируется значением рН элюции на аффинной колонке на основе FcRn, заметно меньшим pH 7,4), взаимодействие антитело-FcRn может быть модифицировано увеличением числа зарядов в области Fab антитела. Например, в аффинной хроматографии на основе FcRn, как описано в примерах настоящей патентной заявки, антитело, имеющее pH элюции 6,5, представляет собой антитело со слабым взаимодействием с FcRn. Посредством увеличения числа зарядов в области Fab (в конечном счете, наряду с генетической модификацией области Fc, если генетическая модификация области Fab не является достаточной) время удерживания также может быть увеличено, делая рН элюции близким к pH 7,4. Такое антитело выигрывает от взаимодействия с FcRn увеличенным временем полужизни in vivo.
Обычно измененные заряды в области Fab влияют на общий положительный заряд, т.е. число положительных зарядов уменьшается или увеличивается, или, в качестве альтернативы, число отрицательных зарядов увеличивается или уменьшается.
Поскольку не все области Fab могут взаимодействовать с FcRn, который связан с областью Fc соответствующего антитела, обнаружили, что посредством модификации остатков в областях, как описано в данном документе, т.е. остатков в области, охватывающей положения 1-23 в вариабельном домене тяжелой цепи (нумерация согласно Kabat), в положениях в области, охватывающей положения 55-83 в вариабельном домене легкой цепи (нумерация согласно Kabat), в положениях в области, охватывающей положения 145-174 в первом константном домене тяжелой цепи (нумерация согласно индексу EU), и в положениях в области, охватывающей положения 180-197 в первом константном домене тяжелой цепи (нумерация согласно индексу EU), может быть модифицировано взаимодействие антитело-FcRn.
Кроме того, в случае биспецифичного антитела, каждая область Fab может быть генетически модифицирована индивидуально, так как одна область Fc антитела может взаимодействовать с двумя молекулами FcRn, приводя к более сильному взаимодействию модифицированных сайтов.
Таким образом, в способе, как описано в данном документе, введенные мутации в областях, как определено в данном документе, вводят, удаляют или модифицируют положительно заряженные участки. Такие мутации известны специалисту в данной области, такие как (в однобуквенном коде аминокислот), например, от Е до Q (введение положительного заряда), от Т до Е (уменьшение положительного заряда), от А до D (уменьшение положительного заряда), от S до D (уменьшение положительного заряда), от G до Е (уменьшение положительного заряда), от G до R (введение положительного заряда) или от S до К (введение положительного заряда). При физиологическом значении рН аминокислотные остатки аргинина (R), лизина (K) и гистидина (Н) преимущественно протонируются и, следовательно, заряжены положительно, тогда как аминокислотные остатки аспартата (D) и глутамата (Е) преимущественно депротонируются (т.е. заряжены отрицательно).
Далее изобретение было проиллюстрировано антителом против дигоксигенина.
В антитело против дигоксигенина были введены следующие мутации:
- HC-E6Q (вариант 1),
- НС-Т164Е (вариант 2),
- НС-A162D и HC-S165D (вариант 3),
- HC-G194E (вариант 4),
- HC-S191D и HC-Q196D (вариант 5),
- LC-G57R (вариант 6),
- LC-G57R и LC-S60K (вариант 7),
- НС164Е, HC-A162D, HC-S165D, HC-G194E, HC-S191D и HC-Q196D (вариант 8),
- HC-E6Q, HC-A162D, НС-Т164Е, HC-S165D, HC-S191D, HC-G194E, НС-Q196D, LC-G57K и LC-S60K (вариант 9).
Свойства данных мутантов на аффинной колонке на основе FcRn и в анализе связывания на основе SPR (поверхностный плазмонный резонанс) показаны в следующей Таблице (эффект на взаимодействие антитело-FcRn):
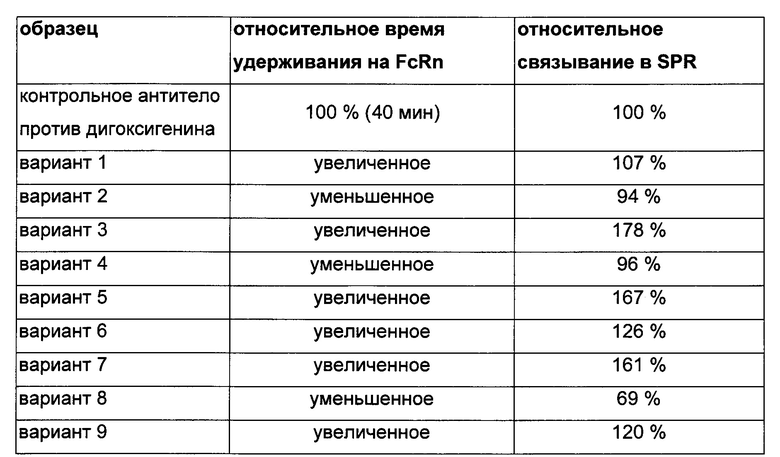
Таким образом, из данных, представленных выше, можно видеть, что посредством изменения картины зарядов в вариабельном домене и/или в первом константном домене (VH1 и CL) может осуществляться изменение в связывании с FcRn.
Одним аспектом, как описано в данном документе, является множество (пул) вариантов полноразмерных антител, полученных из одного родительского полноразмерного антитела посредством рандомизации одного или более чем одного аминокислотного остатка, выбранного из аминокислотных остатков в положениях 1-23 в вариабельном домене тяжелой цепи (нумерация согласно Kabat), в положениях 55-83 в вариабельном домене легкой цепи (нумерация согласно Kabat), в положениях 145-174 в первом константном домене тяжелой цепи (нумерация согласно индексу EU) и в положениях 180-197 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 3-18 в вариабельном домене тяжелой цепи (нумерация согласно Kabat), в положениях 57-71 в вариабельном домене легкой цепи (нумерация согласно Kabat), в положениях 161-174 в первом константном домене тяжелой цепи (нумерация согласно индексу EU) и в положениях 182-197 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 5-18 в вариабельном домене тяжелой цепи (нумерация согласно Kabat), в положениях 57-70 в вариабельном домене легкой цепи (нумерация согласно Kabat) и в положениях 181-196 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
Другим аспектом, как описано в данном документе, является применение одной или более чем одной мутации аминокислоты в положениях, выбранных из группы положений, состоящей из положений 1-23 в вариабельном домене тяжелой цепи (нумерация согласно Kabat), положений 55-83 в вариабельном домене легкой цепи (нумерация согласно Kabat), положений 145-174 в первом константном домене тяжелой цепи (нумерация согласно индексу EU) и положений 180-197 в первом константном домене тяжелой цепи (нумерация согласно индексу EU), для изменения времени полужизни полноразмерного антитела in vivo.
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 3-18 в вариабельном домене тяжелой цепи (нумерация согласно Kabat), в положениях 57-71 в вариабельном домене легкой цепи (нумерация согласно Kabat), в положениях 161-174 в первом константном домене тяжелой цепи (нумерация согласно индексу EU) и в положениях 182-197 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 5-18 в вариабельном домене тяжелой цепи (нумерация согласно Kabat), в положениях 57-70 в вариабельном домене легкой цепи (нумерация согласно Kabat) и в положениях 181-196 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
Другим аспектом, как описано в данном документе, является вариант полноразмерного антитела, содержащий два полипептида легкой цепи и два полипептида тяжелой цепи, где данный вариант антитела получен из родительского полноразмерного антитела посредством введения мутаций аминокислот в одном или более чем одном положении, выбранном из группы положений, состоящей из положений 1-23 в вариабельном домене тяжелой цепи (нумерация согласно Kabat), положений 55-83 в вариабельном домене легкой цепи (нумерация согласно Kabat), положений 145-174 в первом константном домене тяжелой цепи (нумерация согласно индексу EU) и положений 180-197 в первом константном домене тяжелой цепи (нумерация согласно индексу EU), и где вариант антитела имеет отличную аффинность в отношении человеческого неонатального рецептора Fc, чем родительское полноразмерное антитело.
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 3-18 в вариабельном домене тяжелой цепи (нумерация согласно Kabat), в положениях 57-71 в вариабельном домене легкой цепи (нумерация согласно Kabat), в положениях 161-174 в первом константном домене тяжелой цепи (нумерация согласно индексу EU) и в положениях 182-197 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
В одном воплощении один или более чем один аминокислотный остаток выбран из аминокислотных остатков в положениях 5-18 в вариабельном домене тяжелой цепи (нумерация согласно Kabat), в положениях 57-70 в вариабельном домене легкой цепи (нумерация согласно Kabat) и в положениях 181-196 в первом константном домене тяжелой цепи (нумерация согласно индексу EU).
Одним аспектом, как описано в данном документе, является антитело, которое имеет мутацию одного или более чем одного из следующих аминокислотных остатков:
-остатки 6, 162, 164, 165, 191, 194, 195 и 196 в тяжелой цепи
- остатки 57 и 60 в легкой цепи
(нумерация согласно Kabat (вариабельный домен) и индексу EU (константная область) соответственно).
В одном воплощении антитело представляет собой полноразмерное антитело IgG. В одном воплощении антитело представляет собой полноразмерное антитело IgG1.
В одном воплощении антитело представляет собой биспецифичное антитело.
Обнаружили, что для увеличения существующего участка поверхности с положительным зарядом, который может взаимодействовать с его отрицательно заряженным партнером на FcRn, может быть модифицирован отрезок пептида тяжелой цепи, содержащий аминокислотные остатки 3-18. Только мутация E6Q должно быть является приемлемой с энергетической точки зрения.
В одном воплощении антитело имеет мутацию аминокислоты E6Q в вариабельном домене тяжелой цепи.
Обнаружили, что отрезок пептида тяжелой цепи, содержащий аминокислотные остатки 159-174, разделяет два смежных отрицательно заряженных участка, которые располагаются напротив положительно заряженного участка на домене СН2. Таким образом, усиленное взаимодействие СН1-СН2 может ставить Fab ближе к FcRn, что, в свою очередь, может благоприятствовать связыванию с FcRn. Данный участок лучше всего модифицировать вместе с участком пептида тяжелой цепи, содержащим аминокислотные остатки 182-197, так как оба находятся рядом с тем же самым участком домена СН2. Таким образом, для того чтобы усилить связывание/взаимодействие антитело-FcRn один или более чем один из аминокислотных остатков в положениях 162 и/или 164, и/или 165, и/или 191, и/или 194, и/или 195, и/или 196 должен быть модифицирован до кислотного остатка, в одном предпочтительном воплощении - до D или Е.
В одном воплощении антитело имеет кислотную аминокислоту в одном или более чем одном из положений аминокислот 162, 164, 165, 191, 194, 195 и 196 первого константного домена тяжелой цепи (СН1). В одном воплощении кислотная аминокислота представляет собой D или Е. В одном воплощении антитело имеет кислотный аминокислотный остаток в двух или более чем двух положениях аминокислот 162, 164, 165, 191, 194, 195 и 196 первого константного домена тяжелой цепи (СН1), при этом кислотные аминокислотные остатки выбраны независимо друг от друга.
Обнаружили, что отрезок пептида, содержащий аминокислотные остатки 57-71 вариабельного домена легкой цепи, образует участок со слабым положительным зарядом напротив отрицательно заряженного участка на FcRn. Мутирование положительно заряженных остатков в данном отрезке, т.е. основных остатков, до отрицательно заряженных может усиливать взаимодействие антитело-FcRn. В одном предпочтительном воплощении аминокислотные остатки в положении 57 и/или 60 вариабельного домена легкой цепи, независимо друг от друга, представляют собой основные аминокислотные остатки. В одном воплощении основной аминокислотный остаток выбран из K и R.
В одном воплощении антитело имеет основной аминокислотный остаток в одном или в обоих положениях аминокислот 57 и 60 вариабельного домена легкой цепи. В одном воплощении основной аминокислотный остаток представляет собой K или R. В одном воплощении антитело имеет основной аминокислотный остаток в обоих положениях 57 и 60 вариабельного домена легкой цепи, при этом основные аминокислотные остатки выбраны независимо друг от друга.
В одном воплощении антитело имеет одну или более чем одну из следующих мутаций аминокислот:
- E6Q тяжелой цепи и/или
- A162D тяжелой цепи, и/или
- А162Е тяжелой цепи, и/или
- T164D тяжелой цепи, и/или
- Т164Е тяжелой цепи, и/или
- S165D тяжелой цепи, и/или
- S165E тяжелой цепи, и/или
- S191D тяжелой цепи, и/или
- S191E тяжелой цепи, и/или
- G194D тяжелой цепи, и/или
- G194E тяжелой цепи, и/или
- T195D тяжелой цепи, и/или
- Т195Е тяжелой цепи, и/или
- Q196D тяжелой цепи, и/или
- Q196E тяжелой цепи, и/или
- G57K легкой цепи, и/или
- G57R легкой цепи, и/или
- S60K легкой цепи, и/или
- S60R легкой цепи.
В одном аспекте, как описано в данном документе, антитело может быть отобрано посредством скрининга случайных библиотек антител с желательными взаимодействиями антитело-FcRn.
Например, в данной области известно множество способов получения библиотек фагового дисплея и скрининга таких библиотек на антитела, обладающие желательными характеристиками связывания. Обзор таких способов дается, например, в Hoogenboom, H.R. et al., Methods in Molecular Biology 178 (2001) 1-37, и они дополнительно описаны, например, в McCafferty, J. et al., Nature 348 (1990) 552-554; Clackson, T. et al., Nature 352 (1991) 624-628; Marks, J.D. et al., J. Mol. Biol. 222 (1992) 581-597; Marks, J.D. and Bradbury, A., Methods in Molecular Biology 248 (2003) 161-175; Sidhu, S.S. et al., J. Mol. Biol. 338 (2004) 299-310; Lee, C.V. et al., J. Mol. Biol. 340 (2004) 1073-1093; Fellouse, F.A., Proc. Natl. Acad. Sci. USA 101 (2004) 12467-12472 и Lee, C.V. et al., J. Immunol. Methods 284 (2004) 119-132.
В некоторых способах фагового дисплея репертуары генов VH и VL по отдельности клонируют полимеразной цепной реакцией (ПЦР) и случайным образом рекомбинируют в фаговые библиотеки, которые затем можно подвергать скринингу на антигенсвязывающий фаг, как описано в Winter, G. et al., Ann. Rev. Immunol. 12 (1994) 433-455. Фаг типично подвергает дисплею фрагменты антитела либо в виде одноцепочечных фрагментов Fv (scFv), либо в виде фрагментов Fab. Патентные публикации, описывающие фаговые библиотеки человеческих антител, включают, например: US 5750373 и US 2005/0079574, US 2005/0119455, US 2005/0266000, US 2007/0117126, US 2007/0160598, US 2007/0237764, US 2007/0292936 и US 2009/0002360.
Антитела или фрагменты антител, выделенные из библиотек человеческих антител, считаются в данном документе человеческими антителами или фрагментами человеческих антител.
Система скрининга, основанная на опосредованной вирусом осповакцины экспрессии полных антител в клетках млекопитающих, описана в US 2002/0123057. Другая система скрининга основана на экспрессии антител на поверхности клеток в клетках млекопитающих (Но, et al., Proc. Natl. Acad. Sci. USA 103 (2006) 9637-9642).
Клеточный дисплей описан Higuchi et al. в клетках COS (клетки почки обезьяны) (J. Immunol. Meth. 202 (1997) 193-204). Beerli et al. описывают библиотеку дисплея на поверхности клетки на основе вируса Синдбис, продуцированную из антигенспецифичных В-клеток, в клетках ВНК (клетки почки новорожденного хомяка) (Proc. Natl. Acad. Sci. USA 105 (2008) 14336-14341). Но и Pastan описывают способы с клетками НЕК293 (scFv) (Methods Mol. Biol. 562 (2009) 99-113). Лимфоцитарный дисплей описан Alonso-Camino et al. (PLoS One. 4 (2009) e7174). Zhou et al. описывают способы с использованием клеток HEK293 (Acta Biochim. Biophys. Sin. 42 (2010) 575-584). Zhou et al. описывают систему Flp-ln (MAbs. 2 (2010) 508-518).
Taube, R., et al описывают (PLOS One 3 (2008) e3181) стабильную экспрессию человеческих антител на поверхности человеческих клеток и частиц лентивируса.
В WO 2007/047578 описан клеточный дисплей библиотек антител.
Дисплей и/или секреция антител на эукариотических клетках представляет(ют) собой дополнительный(ные) способ(бы) выделения антител. Их получают посредством клонирования нуклеиновых кислот, кодирующих антитело, в лентивирусные векторы, которые используются для трансдукции клеток для обеспечения экспрессии антител на поверхности данных клеток. Использованные векторы несут легкую цепь антитела, тяжелую цепь антитела, мембранный якорь, образующийся в результате альтернативного сплайсинга, или метку, или флуоресцентный маркерный белок.
Sasaki-Haraguchi, N., et al. (Biochem. Biophys. Res. Commun. 423 (2012) 289-294) описывают механистическое понимание сплайсинга человеческих ультракоротких интронов человеческой пред-мРНК: потенциального необычного механизма, идентифицирующего интроны, обогащенные G.
В одном воплощении в способе, описанном в данном документе, используется способ/система дисплея, описанная в WO 2013/092720, с использованием лентивирусной экспрессионной библиотеки в комбинации с лентивирусным экспрессионным вектором, содержащим бицистронную экспрессионную кассету, связанную с EV71-IRES, для экспрессии легкой цепи полноразмерного антитела и тяжелой цепи полноразмерного антитела в растворимой, а также мембраносвязанной форме. Используемый лентивирусный вектор включает вирусные длинные концевые повторы 3'LTR и 5'LTR, легкую цепь антитела, EV71 IRES, тяжелую цепь антитела.
В одном воплощении используется способ/система дисплея, как описано в ЕР 13178581.8.
В некоторых воплощениях предложенное в данном документе антитело представляет собой мультиспецифичное антитело, например, биспецифичное антитело. Мультиспецифичные антитела представляют собой моноклональные антитела, которые имеют специфичности связывания в отношении по меньшей мере двух разных сайтов/антигенов.
Методики получения мультиспецифичных антител включают рекомбинантную соэкспрессию двух пар тяжелая цепь-легкая цепь иммуноглобулина, имеющих разные специфичности (см. Milstein, С. and Cuello, А.С., Nature 305 (1983) 537-540, WO 93/08829 и Traunecker, A. et al., EMBO J. 10 (1991) 3655-3659), и инженерию «выступ во впадину» (см., например, US 5731168), но не ограничиваются ими. Мультиспецифичные антитела также можно получать посредством инженерии эффектов электростатического наведения для получения гетеродимерных молекул на основе Fc антитела (WO 2009/089004); поперечного связывания двух или более чем двух антител или фрагментов (см., например, US 4676980 и Brennan, М. et al., Science 229 (1985) 81-83); с использованием лейциновых молний для получения биспецифичных антител (см., например, Kostelny, S.A. et al., J. Immunol. 148 (1992) 1547-1553; с использованием технологии «диатела» для получения биспецифичных фрагментов антитела (см., например, Holliger, P. et al., Proc. Natl. Acad. Sci. USA 90 (1993) 6444-6448); с использованием димеров на основе одноцепочечной Fv (scFv) (см., например, Gruber, М et al., J. Immunol. 152 (1994) 5368-5374) и получения триспецифичных антител, как описано, например, в Tutt, A. et al., J. Immunol. 147 (1991) 60-69).
В данный документ также включены генетически модифицированные антитела с тремя или более чем тремя функциональными антигенсвязывающими сайтами, включая «антитела-осьминоги» (см., например, US 2006/0025576).
Описанное в данном документе антитело также включает "Fab двойного действия» или «DAF» (см., например, US 2008/0069820).
Описанное в данном документе антитело также включает мультиспецифичные антитела, описанные в WO 2009/080251, WO 2009/080252, WO 2009/080253, WO 2009/080254, WO 2010/112193, WO 2010/115589, WO 2010/136172, WO 2010/145792 и WO 2010/145793.
В некоторых воплощениях предложены варианты/мутанты антител, имеющие одну или более чем одну мутацию аминокислоты. Консервативные мутации показаны в следующей Таблице под заголовком «предпочтительные мутации». Более существенные изменения предложены в следующей Таблице под заголовком «типичные мутации» и, как дополнительно описано ниже, в связи с классами боковых целей аминокислот. В интересующее антитело можно вводить мутации аминокислот и продукты подвергать скринингу на желательное связывание с FcRn.
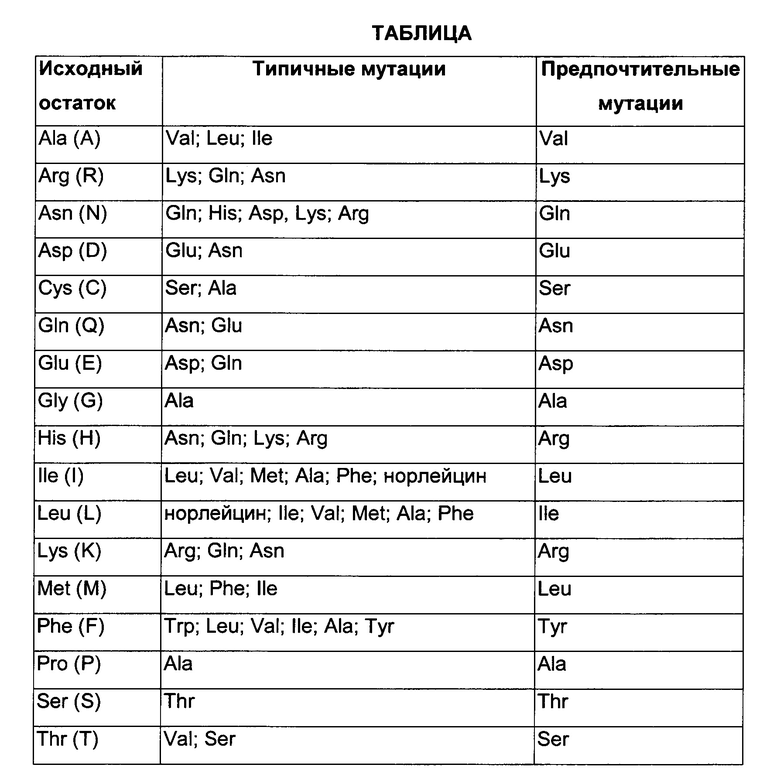

Аминокислоты можно группировать согласно общим свойствам боковой цепи:
(1) гидрофобные: норлейцин, Met, Ala, Val, Leu, lle;
(2) нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gln;
(3) кислотные: Asp, Glu;
(4) основные: His, Lys, Arg;
(5) остатки, которые влияют на ориентацию цепи: Gly, Pro;
(6) ароматические: Trp, Tyr, Phe.
Неконсервативные мутации будут влечь за собой замену члена одного из данных классов на другой класс.
Один тип варианта с мутацией включает мутирование одного или более чем одного остатка родительского антитела (например, гуманизированного или человеческого антитела). В общем, образующийся вариант(ты), отобранный для дальнейшего исследования, будет иметь модификации (например, улучшения) свойств связывания с FcRn относительно родительского антитела и будет иметь по существу сохраненные другие биологические свойства родительского антитела.
В некоторых воплощениях мутации вводятся в кодирующую нуклеиновую кислоту любым из множества способов (например, ПЦР, склонная к ошибкам, перетасовка цепи или мутагенез, направляемый олигонуклеотидами). Затем создается библиотека. Библиотека затем подвергается скринингу для идентификации любых вариантов антител с желательной аффинностью в отношении FcRn. Другой способ введения разнообразия включает подходы, направленные на положение, в которых рандомизируются несколько аминокислотных остатков (например, 4-6 остатков единовременно).
В одном воплощении мутация приводит к замене мутировавшего аминокислотного остатка на другой остаток той же самой группы.
В одном воплощении мутация приводит к замене мутировавшего аминокислотного остатка на другой остаток из той же другой группы.
В некоторых воплощениях в область Fc антитела, предложенного в данном документе, могут быть введены одна или более чем одна аминокислотная модификация, генерируя, посредством этого, вариант области Fc. Вариант области Fc может содержать последовательность человеческой области Fc (например, области Fc человеческого IgG1, IgG2, IgG3 или IgG4), содержащую аминокислотную модификацию (например, замену) в одном или более чем одном положении аминокислоты.
В некоторых воплощениях в данном изобретении рассматривается вариант антитела, который обладает некоторыми, но не всеми эффекторными функциями, что делает его желательным кандидатом для применений, в которых важным является время полужизни антитела in vivo, тем не менее, определенные эффекторные функции (такие как опосредованные комплементом и ADCC (антителозависимая клеточная цитотоксичность)) не требуются или являются вредными. Можно проводить анализы цитотоксичности in vitro и/или in vivo для подтверждения уменьшения/устранения активностей CDC (комплементзависимая клеточная цитотоксичность) и/или ADCC. Например, для того, чтобы убедиться, что антитело не имеет связывания с FcγR (следовательно, вероятно, не имеет активности ADCC), но сохраняет способность к связыванию с FcRn, можно проводить анализы связывания с рецептором Fc (FcR). Первичные клетки, опосредующие ADCC - клетки NK (природный киллер), экспрессируют только FcRIII, тогда как моноциты экспрессируют FcγRI, FcγRII и FcγRIII. Экспрессия FcR на гематопоэтических клетках обобщена в Таблице 3 на странице 464 Ravetch, J.V. and Kinet, J.P., Annu. Rev. Immunol. 9 (1991) 457-492. Неограничивающие примеры анализов in vitro для оценки активности ADCC интересующей молекулы описаны в US 5500362 (см., например, Hellstrom, I. et al., Proc. Natl. Acad. Sci. USA 83 (1986) 7059-7063 и Hellstrom, I. et al., Proc. Natl. Acad. Sci. USA 82 (1985) 1499-1502); US 5821337 (см. Bruggemann, M. et al., J. Exp. Med. 166 (1987) 1351-1361). В качестве альтернативы, можно использовать нерадиоактивные способы анализа (см., например, нерадиоактивный анализ цитотоксичности ACTI™ для проточной цитометрии (CellTechnology, Inc. Mountain View, CA) и нерадиоактивный анализ цитотоксичности CytoTox 96® (Promega, Madison, WI). Полезные эффекторные клетки для таких анализов включают одноядерные клетки периферической крови (РВМС) и клетки-природные киллеры (NK). В качестве альтернативы или дополнительно, активность ADCC интересующей молекулы можно оценивать in vivo, например, в такой животной модели, как модель, раскрытая в Clynes, R. et al., Proc. Natl. Acad. Sci. USA 95 (1998) 652-656. Также можно проводить анализы связывания C1q для подтверждения того, что антитело не способно связываться с C1q и, следовательно, не имеет активности CDC. См., например, ELISA связывания C1q и С3с в WO 2006/029879 и WO 2005/100402. Для оценки активации комплемента можно проводить анализ CDC (см., например, Gazzano-Santoro, Н. et al., J. Immunol. Methods 202 (1996) 163-171; Cragg, M.S. et al., Blood 101 (2003) 1045-1052 и Cragg, M.S. and M.J. Glennie, Blood 103 (2004) 2738-2743). Определения связывания с FcRn и клиренса/времени полужизни in vivo также можно проводить с использованием способов, известных в данной области (см., например, Petkova, S.B. et al., Int. Immunol. 18 (2006: 1759-1769).
Антитела с ослабленной эффекторной функцией включают антитела с заменой одного или более чем одного остатка области Fc 238, 265, 269, 270, 297, 327 и 329 (US 6737056). Такие мутанты Fc включают мутантов Fc с заменами в двух или более чем двух положениях аминокислот 265, 269, 270, 297 и 327, включая так называемого мутанта Fc «DANA» с заменой остатков 265 и 297 на аланин (US 7332581).
Описаны определенные варианты антител с улучшенным или ослабленным связыванием с FcR. (См., например, US 6737056; WO 2004/056312 и Shields, R.L. et al., J. Biol. Chem. 276 (2001) 6591-6604).
В некоторых воплощениях вариант антитела содержит область Fc с одной или более чем одной аминокислотной заменой, которая улучшает ADCC, например, с заменами в положениях 298, 333 и/или 334 области Fc (нумерация остатков ЕС).
В некоторых воплощениях в области Fc делают изменения, которые приводят к измененным (т.е. либо улучшенному, либо ослабленному) связыванию C1q и/или комплементзависимой цитотоксичности (CDC), например, как описано в US 6194551, WO 99/51642 и Idusogie, Е.Е. et al., J. Immunol. 164 (2000) 4178-4184.
Антитела с увеличенным временем полужизни и улучшенным связыванием с неонатальным рецептором Fc (FcRn), который отвечает за перенос материнских IgG в плод (Guyer, R.L. et al., J. Immunol. 117 (1976) 587-593 и Kim, J.K. et al., J. Immunol. 24 (1994) 2429-2434), описаны в US 2005/0014934. Данные антитела содержат область Fc с одной или более чем одной заменой в ней, которая улучшает связывание области Fc с FcRn. Такие варианты Fc включают варианты с заменами в одном или более чем одном остатке области Fc: 238, 256, 265, 272, 286, 303, 305, 307, 311, 312, 317, 340, 356, 360, 362, 376, 378, 380, 382, 413, 424 или 434, например, с заменой остатка 434 области Fc (US 7371826).
См. также Duncan, A.R. and Winter, G., Nature 322 (1988) 738-740; US 5648260; US 5624821 и WO 94/29351 относительно других примеров вариантов области Fc.
Идея настоящего изобретения
В данном изобретении обнаружили, что модификация определенных остатков в домене VL/CH1 антитела может использоваться для влияния на взаимодействия антитело-FcRn.
Способы генной инженерии
Антитела можно получать с использованием способов и композиций генной инженерии, например, как описано в US 4816567. В одном воплощении предложена выделенная нуклеиновая кислота, кодирующая антитело, описанное в данном документе. Такая нуклеиновая кислота может кодировать аминокислотную последовательность, содержащую VL, и/или аминокислотную последовательность, содержащую VH антитела (например, легкую и/или тяжелую цепи антитела). В другом воплощении предложен один или более чем один вектор (например, экспрессионные векторы), содержащий такую нуклеиновую кислоту. В другом воплощении предложена клетка-хозяин, содержащая такую нуклеиновую кислоту. В одном таком воплощении клетка-хозяин содержит (например, была трансформирована): (1) вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VL антитела, и аминокислотную последовательность, содержащую VH антитела, или (2) первый вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VL антитела, и второй вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VH антитела. В одном воплощении клетка-хозяин является эукариотической, например, клеткой яичника китайского хомяка (СНО) или лимфоидной клеткой (например, клетка Y0, NS0, Sp20). В одном воплощении предложен способ получения антитела, как описано в данном документе, где данный способ включает культивирование клетки-хозяина, содержащей нуклеиновую кислоту, кодирующую антитело, как приведено выше, при подходящих условиях для экспрессии антитела, и возможно выделение антитела из клетки-хозяина (или культуральной среды клетки-хозяина).
Для рекомбинантной продукции антитела выделяют нуклеиновую кислоту, кодирующую антитело, например, как описано выше, и вставляют в один или более чем один вектор для дальнейшего клонирования и/или экспрессии в клетке-хозяине. Такую нуклеиновую кислоту можно легко выделять и секвенировать с использованием традиционных методик (например, посредством применения олигонуклеотидных зондов, которые способны к специфичному связыванию с генами, кодирующими тяжелые и легкие цепи антитела).
Подходящие клетки-хозяева для клонирования или экспрессии векторов, кодирующих антитело, включают прокариотические или эукариотические клетки, описанные в данном документе. Например, антитела можно продуцировать в бактериях, в частности, когда не нужны гликозилирование и эффекторная функция Fc. Относительно экспрессии фрагментов и полипептидов антитела в бактериях см., например, US 5648237, US 5789199 и US 5840523. (См. также Charlton, K.А., In: Methods in Molecular Biology, Vol. 248, Lo, B.K.C. (ed.), Humana Press, Totowa, NJ (2003), pp. 245-254, описывающую экспрессию фрагментов антитела в E. coli.). После экспрессии антитело можно выделять из пасты бактериальных клеток в растворимой фракции и можно дополнительно очищать.
Помимо прокариот, подходящими хозяевами для клонирования или экспрессии векторов, кодирующих антитело, являются эукариотические микробы, такие как нитчатые грибки или дрожжи, включая штаммы грибков и дрожжей, пути гликозилирования которых были «гуманизированы», приводя к продукции антитела с частично или полностью человеческой картиной гликозилирования. См. Gerngross, T.U., Nat. Biotech. 22 (2004) 1409-1414 и Li, Н. et al., Nat. Biotech. 24 (2006) 210-215.
Подходящие клетки-хозяева для экспрессии гликозилированного антитела также происходят из многоклеточных организмов (беспозвоночных и позвоночных). Примеры клеток беспозвоночных включают клетки растений и насекомых. Были идентифицированы многочисленные бакуловирусные штаммы, которые можно использовать в сочетании с клетками насекомых, в частности, для трансфекции клеток Spodoptera frugiperda.
Культуры растительных клеток также можно использовать в качестве хозяев. См., например, US 5959177, US 6040498, US 6420548, US 7125978 и US 6417429 (описывающие технологию PLANTIBODIES™ для продуцирования антител в трансгенных растениях).
Клетки позвоночных также можно использовать в качестве хозяев. Например, могут быть полезными линии клеток млекопитающих, которые адаптированы для роста в суспензии. Другими примерами полезных линий клеток-хозяев млекопитающих являются линия CV1 почки обезьяны, трансформированная SV40 (COS-7); линия эмбриональной почки человека (293 или клетки 293, как описано, например, в Graham, F.L. et al., J. Gen Virol. 36 (1977) 59-74); клетки почки новорожденного хомяка (ВНК); мышиные клетки Сертоли (клетки ТМ4, как описано, например, в Mather, J.P., Biol. Reprod. 23 (1980) 243-252); клетки почки обезьяны (CV1); клетки почки африканской зеленой мартышки (VERO-76); клетки карциномы шейки матки человека (HELA); клетки почки собаки (MDCK); клетки печени серой крысы (BRL 3А); клетки легкого человека (W138); клетки печени человека (Hep G2); клетки опухоли молочной железы мыши (ММТ 060562); клетки TRI, как описано, например, в Mather, J.P. et al., Annals N.Y. Acad. Sci. 383 (1982) 44-68; клетки MRC 5 и клетки FS4. Другие полезные линии клеток-хозяев млекопитающих включают клетки яичника китайского хомяка (СНО), включая клетки СНО  (негативные по дегидрофолатредуктазе) (Urlaub, G. et al., Proc. Natl. Acad. Sci. USA 77 (1980) 4216-4220); и линии клеток миеломы, такие как Y0, NS0 и Sp2/0. Для обзора определенных линий клеток-хозяев млекопитающих, полезных для продукции антител, см., например, Yazaki, P. and Wu, A.M., Methods in Molecular Biology, Vol. 248, Lo, B.K.C. (ed.), Humana Press, Totowa, NJ (2004), pp. 255-268.
(негативные по дегидрофолатредуктазе) (Urlaub, G. et al., Proc. Natl. Acad. Sci. USA 77 (1980) 4216-4220); и линии клеток миеломы, такие как Y0, NS0 и Sp2/0. Для обзора определенных линий клеток-хозяев млекопитающих, полезных для продукции антител, см., например, Yazaki, P. and Wu, A.M., Methods in Molecular Biology, Vol. 248, Lo, B.K.C. (ed.), Humana Press, Totowa, NJ (2004), pp. 255-268.
ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг. 1. Пептидная карта HDX-MS антитела против дигоксигенина. Пептические пептиды, из которых могли быть получены данные HDX, показаны в виде белых полосок для НС (А) и LC (Б). Незакрашенные большие прямоугольники обозначают пептиды с пониженным содержанием дейтерия при связывании с FcRn. Закрашенные прямоугольники показывают целевые области, где использовали ETD (диссоциация с переносом электрона) дейтерированных пептических пептидов для разрешения дифференциального мечения дейтерием индивидуальных сайтов при связывании с FcRn. Остатки антитела, вовлеченные в связывание с FcRn в кристаллических структурах крысиная Fc-FcRn и человеческий FcRn-Fc-YTE, показаны звездочками (см. Martin, W.L., et al., Mol. Cell. 7 (2001) 867-877, Oganesyan, V., et al., J. Biol. Chem. 289 (2014) 7812-7824).
Фиг. 2. Профили HDX областей антитела против дигоксигенина в присутствии и в отсутствие человеческого FcRn. Графики HDX показаны для пептида 55-71 легкой цепи и пептидов 1-18, 57-79, 159-174 и 180-197 тяжелой цепи. Верхние кривые показывают HDX одного антитела, и нижние кривые показывают HDX пептидов НС и LC соответственно в присутствии FcRn. НС 57-79 с FcRn показан в виде пунктирной черной кривой. Включение дейтерия отслеживали в тройных повторностях в 1 мин, 1 ч, 2,5 ч и 5 ч. Полный уровень дейтерия, измеренный в контрольных экспериментах (при 90% D2O) показан черным в момент времени 5 ч.
Фиг. 3. Выравнивание аминокислотной последовательности вариабельных доменов тяжелой цепи антитела против дигоксигенина, бриакинумаба и устекинумаба. Отрезки аминокислотных остатков, в которых для всех трех антител была обнаружена защита от HDX, заключены в прямоугольник.
Фиг. 4. Выравнивание аминокислотной последовательности вариабельных доменов легкой цепи антитела против дигоксигенина, бриакинумаба и устекинумаба. Отрезки аминокислотных остатков, в которых для всех трех антител была обнаружена защита от HDX, заключены в прямоугольник.
Следующие примеры, последовательности и графические материалы приведены для помощи в понимании настоящего изобретения, истинный объем которого изложен в приложенной формуле изобретения. Понятно, что в изложенных методиках могут быть сделаны модификации без отступления от сущности изобретения.
ПРИМЕРЫ
Материалы и методы
Методики генной инженерии
Для манипуляции с ДНК использовали стандартные способы, как описано в Sambrook, J. et al., Molecular cloning: A laboratory manual; Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1989. Молекулярно-биологические реактивы использовали согласно инструкциям изготовителя.
Синтез генов и олигонуклеотидов
Желательные отрезки генов получали химическим синтезом в Geneart GmbH (Регенсбург, Германия). Синтезированные фрагменты генов клонировали в плазмиду Е. coli для размножения/амплификации. Последовательности ДНК субклонированных фрагментов генов подтверждали секвенированием ДНК. В качестве альтернативы, короткие синтетические фрагменты ДНК собирали отжигом химически синтезированных олигонуклеотидов или посредством ПЦР (полимеразная цепная реакция). Соответствующие олигонуклеотиды были получены Metabion GmbH (Планегг-Мартинсрид, Германия).
Реактивы
Все имеющиеся в продаже реактивы, антитела и наборы использовали в том виде, в котором они были предоставлены, согласно протоколу изготовителя, если не утверждается иное.
Пример 1
Получение рекомбинантных экспрессионных векторов для антитела против дигоксигенина
Получение вектора для экспрессии тяжелой цепи антитела против дигоксигенина
Слитый ген, кодирующий тяжелую цепь, содержащую константную область человеческого IgG1 (СН1, шарнирная область, СН2, СН3) и вариабельный домен тяжелой цепи антитела против дигоксигенина, собирали посредством слияния фрагмента ДНК, кодирующего соответствующий вариабельный домен тяжелой цепи антитела против дигоксигенина, с элементом последовательности, кодирующим константную область человеческого IgG1.
Вариабельный домен тяжелой цепи антитела против дигоксигенина имеет следующую аминокислотную последовательность:

(SEQ ID NO: 01).
Константная область человеческого IgG1 имеет следующую аминокислотную последовательность:

(SEQ ID NO: 02).
Экспрессионный вектор также содержал репликатор из вектора pUC18, который обеспечивает репликацию данной плазмиды в Е. coli, и ген бета-лактамазы, который придает устойчивость к ампициллину в Е. coli.
Транскрипционная единица тяжелой цепи антитела содержит следующие функциональные элементы в направлении 5' - 3':
- немедленный ранний энхансер и промотор из человеческого цитомегаловируса (P-CMV), включая интрон А,
- 5'-нетранслируемая область тяжелой цепи человеческого иммуноглобулина (5'UTR),
- сигнальная последовательность тяжелой цепи мышиного иммуноглобулина,
- нуклеиновая кислота, кодирующая вариабельный домен тяжелой цепи (VH),
- нуклеиновая кислота, кодирующая константную область человеческого IgG1, и
- последовательность полиаденилирования коровьего гормона роста (BGH pA).
Получение вектора для экспрессии легкой цепи антитела против дигоксигенина
Слитый ген, кодирующий легкую цепь каппа, содержащую константную область человеческого Ig-каппа (CL-каппа) и вариабельный домен легкой цепи антитела против дигоксигенина изотипа каппа, собирали посредством слияния фрагмента ДНК, кодирующего соответствующий вариабельный домен легкой цепи антитела против дигоксигенина, с элементом последовательности, кодирующим константную область человеческого Ig-каппа.
Вариабельный домен легкой цепи антитела против дигоксигенина имеет следующую аминокислотную последовательность:

(SEQ ID NO: 03).
Константная область человеческого lg-каппа имеет следующую аминокислотную последовательность:

(SEQ ID NO: 04).
Экспрессионный вектор также содержал репликатор из вектора pUC18, который обеспечивает репликацию данной плазмиды в E. coli, и ген бета-лактамазы, который придает устойчивость к ампициллину в Е. coli.
Транскрипционная единица легкой цепи каппа антитела содержит следующие функциональные элементы в направлении 5' - 3':
- немедленный ранний энхансер и промотор из человеческого цитомегаловируса (P-CMV), включая интрон А,
- 5'-нетранслируемая область тяжелой цепи человеческого иммуноглобулина (5'UTR),
- сигнальная последовательность тяжелой цепи мышиного иммуноглобулина,
- нуклеиновая кислота, кодирующая вариабельный домен легкой цепи (VL),
- нуклеиновая кислота, кодирующая константную область человеческого Ig-каппа, и
- последовательность полиаденилирования коровьего гормона роста (BGH рА).
Пример 2
Рекомбинантная продукция антитела против дигоксигенина
Антитело продуцировали во временно трансфицированных клетках НЕК293 (происходящих от линии клеток эмбриональной почки человека 293), культивируемых в среде F17 (Invitrogen Corp.). Для трансфекции соответствующих векторов, как описано в Примере 1, использовали трансфекционный реактив «293-free» (Novagen). Антитело экспрессировали из индивидуальных экспрессионных плазмид. Трансфекции проводили, как подробно описано в инструкциях изготовителя. Супернатанты культуры клеток, содержащие рекомбинантное антитело, отбирали через три-семь суток после трансфекции. Супернатанты хранили при пониженной температуре (например, -80°С) до очистки.
Общая информация относительно рекомбинантной экспрессии человеческих иммуноглобулинов, например, в клетках НЕК293, приведена в: Meissner, P. et al., Biotechnol. Bioeng. 75 (2001) 197-203.
Пример 3
Очистка рекомбинантного антитела против дигоксигенина
Супернатанты культуры, содержащие антитело, фильтровали и очищали посредством двух хроматографических стадий.
Антитело захватывали посредством аффинной хроматографии с использованием HiTrap MabSelectSuRe (GE Healthcare), уравновешенной PBS (фосфатно-солевой буферный раствор) (1 мМ KH2PO4, 10 мМ Na2HPO4, 137 мМ NaCl, 2,7 мМ KCl), pH 7,4. Несвязавшиеся белки удаляли посредством промывки уравновешивающим буфером, и антитело выделяли 25 мМ цитратным буфером, pH 3,1, который немедленно после элюции доводили до pH 6,0 с использованием 1 М основания Tris, pH 9,0.
В качестве второй стадии очистки использовали гель-фильтрацию на Superdex 200™ (GE Healthcare). Гель-фильтрацию проводили в 20 мМ гистидиновом буфере, 0,14 М NaCl, pH 6,0. Растворы, содержащие антитело, концентрировали с использованием блока центрифужного фильтра Ultrafree-CL, оснащенного мембраной Biomax-SK (Millipore, Billerica, MA, США) и хранили при -80°С.
Пример 4
Получение рекомбинантных экспрессионных векторов для устекинумаба и бриакинумаба
Устекинумаб: CNTO 1275, Stelara™, регистрационный номер CAS 815610-63-0.
Бриакинумаб: АВТ 874, J 695, Ozespa™, SEQ ID NO: 36, WO 2001/014162.
Для экспрессии вышеописанных антител использовали экспрессионные плазмиды для временной экспрессии (например, в клетках HEK293-F) на основе либо организации кДНК с промотором CMV-интроном А (цитомегаловирус) или без них, либо геномной организации с промотором CMV.
Помимо экспрессионной кассеты антитела плазмиды содержали:
- репликатор, который обеспечивает репликацию данной плазмиды в Е. coli,
- ген β-лактамазы, который придает устойчивость к ампициллину в Е. coli, и
- ген дигидрофолатредуктазы из Mus musculus в качестве селектируемого маркера в эукариотических клетках.
Транскрипционная единица гена антитела состояла из следующих элементов:
- уникальный(ные) сайт(ты) рестрикции на 5'-конце
- немедленный ранний энхансер и промотор из человеческого цитомегаловируса,
- с последующей последовательностью интрона А в случае организации кДНК,
- 5' нетранслируемая область гена человеческого антитела,
- сигнальная последовательность тяжелой цепи иммуноглобулина,
- цепь человеческого антитела либо в виде кДНК, либо в виде геномной организации с экзон-интронной организацией иммуноглобулина,
- 3' нетранслируемая область с сигнальной последовательностью полиаденилирования и
- уникальный(ные) сайт(ты) рестрикции на 3'-конце.
Слитые гены, содержащие цепи антитела, получали посредством ПЦР и/или синтеза гена и собирали известными способами и методиками генной инженерии соединением соответствующих отрезков нуклеиновой кислоты, например, с использованием уникальных рестрикционных сайтов в соответствующих плазмидах. Субклонированные последовательности нуклеиновой кислоты подтверждали секвенированием ДНК. Для временных трансфекций получали большие количества плазмид посредством получения плазмид из культур трансформированной Е. coli (Nucleobond АХ, Macherey-Nagel).
Пример 5
Рекомбинантная продукция устекинумаба и бриакинумаба
Использовали стандартные методики культуры клеток, как описано в Current Protocols in Cell Biology (2000), Bonifacino, J.S., Dasso, M., Harford, J.В., Lippincott-Schwartz, J. and Yamada, K.M. (eds.), John Wiley & Sons, Inc.
Антитела получали временной трансфекцией соответствующими плазмидами (например, кодирующими тяжелую цепь, а также соответствующую легкую цепь) с использованием системы HEK293-F (Invitrogen) согласно инструкции изготовителя. Вкратце, клетки HEK293-F (Invitrogen), растущие в суспензии либо во встряхиваемой колбе, либо в ферментере с перемешиванием в бессывороточной среде для экспрессии FreeStyle™ 293 (Invitrogen), трансфицировали смесью соответствующих экспрессионных плазмид и 293fectin™ или фектина (Invitrogen). Для 2 л встряхиваемой колбы (Corning) клетки HEK293-F высевали в плотности 1*106 клеток/мл в 600 мл и инкубировали при 120 об/мин, 8% CO2. Через одни сутки клетки трансфицировали при плотности клеток примерно 1,5*106 клеток/мл с использованием примерно 42 мл смеси А) 20 мл Opti-MEM (Invitrogen) с 600 мкг общей плазмидной ДНК (1 мкг/мл), кодирующей тяжелую цепь соответственно и соответствующую легкую цепь в эквимолярном соотношении, и Б) 20 мл Opti-MEM плюс 1,2 мл 293 фектина или фектина (2 мкл/мл). Согласно потреблению глюкозы по ходу ферментации добавляли раствор глюкозы. Супернатант, содержащий секретируемое антитело, отбирали через 5-10 суток, и антитела либо непосредственно очищали из супернатанта, либо супернатант замораживали и хранили.
Пример 6
Очистка рекомбинантного устекинумаба и бриакинумаба
Антитела очищали из супернатантов культуры клеток посредством аффинной хроматографии с использованием MabSelectSure-Sepharose™ (GE Healthcare, Швеция), хроматографии на основе гидрофобных взаимодействий с использованием бутил-сефарозы (GE Healthcare, Швеция) и гель-фильтрации на Superdex 200 (GE Healthcare, Швеция).
Вкратце, супернатанты культуры клеток после стерилизующей фильтрации захватывали на смоле MabSelectSuRe, уравновешенной буфером PBS (10 мМ Na2HPO4, 1 мМ KH2PO4, 137 мМ NaCl и 2,7 мМ KCl, pH 7,4), промытой уравновешивающим буфером, и элюировали 25 мМ цитратом натрия при pH 3,0. Элюированные фракции антител объединяли и нейтрализовали 2 М Tris, pH 9,0. Для хроматографии на основе гидрофобных взаимодействий получали пулы антител посредством добавления 1,6 М раствора сульфата аммония до конечной концентрации 0,8 М сульфата аммония, и pH доводили до рН 5,0 с использованием уксусной кислоты. После уравновешивания бутил-сефарозной смолы 35 мМ ацетатом натрия, 0,8 М сульфатом аммония, pH 5,0, антитела наносили на смолу, промывали уравновешивающим буфером и элюировали линейным градиентом до 35 мМ ацетата натрия, pH 5,0. Фракции, содержащие антитело, объединяли и дополнительно очищали гель-фильтрацией с использованием колонки Superdex 200 26/60 GL (GE Healthcare, Швеция), уравновешенной 20 мМ гистидином, 140 мМ NaCl, pH 6,0. Фракции, содержащие антитело, объединяли, концентрировали до требующейся концентрации с использованием устройств для ультрафильтрации Vivaspin (Sartorius Stedim Biotech S.A., Франция) и хранили при -80°С.
Чистоту и целостность антител анализировали после каждого этапа очистки посредством CE-SDS (капиллярный электрофорез с додецилсульфатом натрия) с использованием микрожидкостной технологии Labchip (Caliper Life Science, США). Для анализа CE-SDS готовили пять мкл раствора белка с использованием набора реактивов НТ Protein Express согласно инструкциям изготовителя и анализировали на системе LabChip GXII с использованием чипа НТ Protein Express. Данные анализировали с использованием программы LabChip GX.
Пример 7
Экспрессия FcRn в клетках НЕК293
FcRn временно экспрессировали посредством трансфекции клеток НЕК293 двумя плазмидами, содержащими кодирующую последовательность FcRn (SEQ ID NO: 09) и бета-2-микроглобулина (SEQ ID NO: 10). Трансфицированные клетки культивировали во встряхиваемых колбах при 36,5°С, 120 об/мин (амплитуда шейкера - 5 см), 80%-ной влажности и 7% CO2. Клетки разводили каждые 2-3 суток до плотности 3-4*105 клеток/мл.
Для временной экспрессии запускали 14 л биореактор из нержавеющей стали с объемом культуры 8 л при 36,5°С, рН 7,0 плюс/минус 0,2, pO2 35% (продувание N2 и воздухом, общий ток газа 200 мл мин-1) и скоростью мешалки 100-400 об/мин. При достижении плотности клеток 20*105 клеток/мл 10 мг плазмидной ДНК (эквимолярные количества обеих плазмид) разводили в 400 мл Opti-MEM (Invitrogen). В данную смесь добавляли 20 мл 293фектина (Invitrogen), которую затем инкубировали в течение 15 минут при комнатной температуре и затем переносили в ферментер. Начиная со следующих суток клеткам поставляли питательные вещества в непрерывном режиме: подпиточный раствор добавляли со скоростью 500 мл в сутки и глюкозу, по мере необходимости, для поддержания уровня более 2 г/л. Супернатант отбирали через 7 суток после трансфекции с использованием центрифуги с поворотными головками с 1 л бакетами: 4000 об/мин в течение 90 минут. Супернатант (13 л) осветляли посредством фильтра Sartobran Р (0,45 мкм плюс 0,2 мкм, Sartorius), и очищали из него комплекс FcRn-бета-2-м и кроглобулин.
Пример 8
Характеризация моноклональных антител и FcRn посредством MS
Масс-спектрометрию восстановленных интактных белков проводили на 50 мкг моноклонального антитела или FcRn, которые были восстановлены и денатурированы с использованием 0,5 М ТСЕР (трис-2-карбоксиэтилфосфин) (Perbio, Бонн, Германия) в 4 М растворе гуанидиния гидрохлорида при 37°С в течение 30 минут. Образцы обессоливали гель-фильтрацией (Sephadex G-25, изократическая элюция 40% ацетонитрилом с 2% муравьиной кислоты (об./об.). Масс спектры на основе ESI (электрораспылительная ионизация) записывали на установке Q-TOF (MaXis, Bruker, Германия), оснащенной Triversa NanoMate (Advion, Ithaca, США). Для оценки данных использовали программу, разработанную своими силами.
Пептидное картирование моноклонального антитела и FcRn (250 мкг) осуществляли денатурированием соединений посредством добавления 0,4 М Tris, 8 М гуанидиния гидрохлорида, рН 8, и 0,24 М DTT (дитиотрейтол) в течение одного часа при 37°С и алкилированием посредством добавления 0,6 М йодуксусной кислоты в воде в течение 15 минут при комнатной температуре в темноте. В образцах заменяли буфер на 50 мМ Tris/HCl, рН 7,5, с использованием колонок ДНК класса NAP 5 Sephadex G-25 (GE Healthcare, Мюнхен, Германия). Расщепление осуществляли трипсином (Promega, Мангейм, Германия) в течение 5 часов при 37°С (отношение фермента к субстрату 1:37). Полученную пептидную смесь инъецировали и разделяли без предобработки с использованием U-HPLC (сверхвысокоэффективная жидкостная хроматография) с обращенной фазой (NanoAcquity, Waters GmbH, Эшборн, Германия). Для разделения использовали колонку Acquity UPLC ВЕН С18 (1×150 мм, диаметр частиц 1,7 мкм, размер пор 300 А) от Waters. Растворителями были 0,1% (об./об.) муравьиная кислота в воде (А) и в ацетонитриле (Б) (Sigma-Aldrich, Мюнхен, Германия). Линейный градиент 60 мкл/мин от 1 до 40% Б прогоняли за 120 мин при 50°С. Анализ массы осуществляли посредством сочетания системы UPLC с тандемным масс-спектрометром LTQ Orbitrap XL (Thermo Fisher Scientific, Драйайх, Германия), работающим в режиме положительных ионов через интерфейс Triversa NanoMate (Advion, Ithaca, США).
Пример 9
Масс-спектрометрия с обменом водород/дейтерий (HDX-MS)
Эксперименты HDX-MS проводили с использованием следующего приготовления образца:
Моноклональное антитело (73 пмоль/мкл) с или без FcRn (112 пмоль/мкл) смешивали с и разводили D20 (99,9 атомных % дейтерия), содержащей 50 мМ фосфат натрия, 50 мМ NaCl, рН 6,5, до конечного содержания дейтерия 90% и концентраций антитела 1,2 пмоль/мкл и FcRn - 8,96 пмоль/мкл (84% связавшегося антитела, отношение связывания 1:2 IgG1:FcRn).
Теоретические расчеты были основаны на KD 0,6 мкМ для взаимодействия IgG1-FcRn и конечном объеме 25 мкл после разведения D2O. После 15 мин предынкубации образцов начинали мечение дейтерием при комнатной температуре в течение разных интервалов времени: 0 мин, 1 мин, 1 час, 2,5 часа и 5 часов. В каждый интервал времени из метящей смеси удаляли аликвоты (25 мкл) 30 пмоль белка-мишени и гасили доведением до конечного рН 2,5 в ледяной смеси 25 мкл 50 мМ фосфата Na, 50 мМ NaCl, рН 6,5, и 50 мкл 0,5 М ТСЕР, 6 М гуанидиния гидрохлорида в фосфорной кислоте, рН 2,3, и замораживали до -80°С до анализа LC-MS (жидкостная хроматография-масс-спектрометрия). Полностью дейтерированные образцы получали посредством инкубации в течение ночи в 6 М дейтерированном гуанидиния гидрохлориде (конечное содержание дейтерия 90 атомных %).
Дейтерированные белки после гашения (30 пмоль) загружали на охлаждаемую систему HDX-UPLC в сочетании с гибридным масс-спектрометром Q-TOF Synapt G2 (Waters, Милфорд, США). Система UPLC работала при 0°С и была оснащена упакованной собственными силами пепсиновой колонкой с внутренним объемом 60 мкл (IDEX, Oak Harbor, США), содержащей пепсин, иммобилизованный на агарозе (Thermo Scientific Pierce, Рокфорд, США), захватывающей С18-колонкой (ACQUITY UPLC ВЕН С18 1,7 мкм колонка VanGuard (Waters, Милфорд, США) и аналитической С18-колонкой (ACQUITY UPLC ВЕН С18 1,7 мкм, 1×100 мм колонка (Waters, Милфорд, США)). Белки расщепляли в линии при температуре 20°С и обессоливали на захватывающей колонке со скоростью тока 200 мкл подвижной фазы А (0,23% FA). Пептические пептиды элюировали из захватывающей колонки и через аналитическую колонку при токе 40 мкл/мин и 7 мин градиентом от 8% до 40% подвижной фазы Б (ACN, 0,23% FA) и в масс-спектрометр для анализа массы. Источник ESI работал в режиме положительных ионов, установка была подключена для анализа подвижности ионов. Для внутренней калибровки получали контрольный сигнал запертого спрея Glu-фибринопептида (Sigma-Aldrich, Сент-Луис, США). Идентификацию пептидов осуществляли посредством комбинации MSe, HDMSe и DDA MS/MS. Идентификации пептидов осуществляли посредством поиска в базе данных в PLGS, версия 2.5, и данные HDX-MS обрабатывали в DynamX, версия 2.2.1. Данные HDX-MS перекрывающихся пептидов использовали только для локализации поглощения дейтерия меньшими отрезками, если обратный обмен полностью дейтерированных пептидов антитела был аналогичным (меньше 7%) (Sheff, J., et al., J. Am. Soc. Mass Spectrom. 24 (2013) 1006-1015).
Пример 10
Масс-спектрометрия с водород/дейтериевым обменом с ETD
Дейтерированные образцы получали такой же методикой, что и в предыдущем Примере, за исключением того, что инъекционное количество было доведено до 100 пмоль антитела, и использовали пятикратное разведение в буфере D2O (до конечных 80 атомных % дейтерия), что приводило к концентрации антитела 4 пмоль/мкл и концентрации FcRn 14 пмоль/мкл и 85% связавшегося антитела во время мечения. HDX-ETD проводили целевым способом на отобранных пептидных фрагментах антитела с дифференциальным поглощением дейтерия между состоянием, связанным и не связанным с FcRn. Источник ESI и источник Т-волны работали при установках, оптимизированных для минимального преобразования H/D, как описано в Rand, K.D., et al. (J. Am. Soc. Mass Spectrom. 22 (2011) 1784-1793), со следующими параметрами: напряжение капилляра 2,8 кВ, ток десольватационного газа 800 л/ч, ток газа в конусе 0 л/ч, температура источника 90°С, температура десольватационного газа 300°С, напряжение пробоотборного конуса 20 В, напряжение экстракционного конуса 2 В, скорость волны в ловушке Т-волны 300 м/с, высота волны 0,2 В. ETD осуществляли в ловушке Т-волны с использованием 1,4-дицианобензола (Sigma-Aldrich, Сент-Луис, США) в качестве реактива ETD. 1,4-дицианобензол вводили в источник анионов с использованием азотного тока подпитки 20 мл/мин над кристаллами реактива, которые хранили в запечатанном контейнере. Анион-радикалы генерировали посредством тлеющего разряда тока 40 мкА и газового тока подпитки 20 мл/мин. Данные ETD анализировали посредством определения средних масс ETD фрагментов ионов с- и z-типа. Содержание дейтерия рассчитывали посредством вычитания содержания дейтерия ионов немеченого продукта из содержания дейтерия дейтерированных образцов. Отсутствие преобразования H/D подтверждали отслеживанием потери аммиака из заряженных восстановленных соединений в спектрах ETD, записанных на пептических пептидах из образца полностью дейтерированного антитела, как описано в Rand, K.D., et al. (Anal. Chem. 82 (2010) 9755-9762).
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Ф. ХОФФМАНН-ЛЯ РОШ АГ
<120> Способ отбора антител с модифицированным взаимодействием с FcRn
<130> P32178-WO
<150> EP14172180.3
<151> 2014-06-12
<160> 14
<170> PatentIn version 3.5
<210> 1
<211> 125
<212> PRT
<213> Искусственная последовательность
<220>
<223> VH гуманизированного антитела против дигоксигенина
<400> 1
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Ala Met Ser Trp Ile Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Asn Ile Gly Ala Thr Tyr Ile Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Pro Gly Ser Pro Tyr Glu Tyr Asp Lys Ala Tyr Tyr Ser Met
100 105 110
Ala Tyr Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser
115 120 125
<210> 2
<211> 330
<212> PRT
<213> Homo sapiens
<400> 2
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 3
<211> 107
<212> PRT
<213> Искусственная последовательность
<220>
<223> VL гуманизированного антитела против дигоксигенина
<400> 3
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Lys Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Ser Ser Thr Leu Leu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Ile Thr Leu Pro Pro
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 4
<211> 107
<212> PRT
<213> Homo sapiens
<400> 4
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 5
<211> 115
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность VH бриакинумаба
<400> 5
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Phe Ile Arg Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Lys Thr His Gly Ser His Asp Asn Trp Gly Gln Gly Thr Met Val Thr
100 105 110
Val Ser Ser
115
<210> 6
<211> 112
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность VL бриакинумаба
<400> 6
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Gly Ala Pro Gly Gln
1 5 10 15
Arg Val Thr Ile Ser Cys Ser Gly Ser Arg Ser Asn Ile Gly Ser Asn
20 25 30
Thr Val Lys Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Tyr Asn Asp Gln Arg Pro Ser Gly Val Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Ala Ser Leu Ala Ile Thr Gly Leu Gln
65 70 75 80
Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Gln Ser Tyr Asp Arg Tyr Thr
85 90 95
His Pro Ala Leu Leu Phe Gly Thr Gly Thr Lys Val Thr Val Leu Gly
100 105 110
<210> 7
<211> 119
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность VH устекинумаба
<400> 7
Glu Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Glu
1 5 10 15
Ser Leu Lys Ile Ser Cys Lys Gly Ser Gly Tyr Ser Phe Thr Thr Tyr
20 25 30
Trp Leu Gly Trp Val Arg Gln Met Pro Gly Lys Gly Leu Asp Trp Ile
35 40 45
Gly Ile Met Ser Pro Val Asp Ser Asp Ile Arg Tyr Ser Pro Ser Phe
50 55 60
Gln Gly Gln Val Thr Met Ser Val Asp Lys Ser Ile Thr Thr Ala Tyr
65 70 75 80
Leu Gln Trp Asn Ser Leu Lys Ala Ser Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Arg Arg Pro Gly Gln Gly Tyr Phe Asp Phe Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 8
<211> 107
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность VL устекинумаба
<400> 8
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Glu Lys Ala Pro Lys Ser Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Ile Tyr Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 9
<211> 274
<212> PRT
<213> Homo sapiens
<400> 9
Ala Glu Ser His Leu Ser Leu Leu Tyr His Leu Thr Ala Val Ser Ser
1 5 10 15
Pro Ala Pro Gly Thr Pro Ala Phe Trp Val Ser Gly Trp Leu Gly Pro
20 25 30
Gln Gln Tyr Leu Ser Tyr Asn Ser Leu Arg Gly Glu Ala Glu Pro Cys
35 40 45
Gly Ala Trp Val Trp Glu Asn Gln Val Ser Trp Tyr Trp Glu Lys Glu
50 55 60
Thr Thr Asp Leu Arg Ile Lys Glu Lys Leu Phe Leu Glu Ala Phe Lys
65 70 75 80
Ala Leu Gly Gly Lys Gly Pro Tyr Thr Leu Gln Gly Leu Leu Gly Cys
85 90 95
Glu Leu Gly Pro Asp Asn Thr Ser Val Pro Thr Ala Lys Phe Ala Leu
100 105 110
Asn Gly Glu Glu Phe Met Asn Phe Asp Leu Lys Gln Gly Thr Trp Gly
115 120 125
Gly Asp Trp Pro Glu Ala Leu Ala Ile Ser Gln Arg Trp Gln Gln Gln
130 135 140
Asp Lys Ala Ala Asn Lys Glu Leu Thr Phe Leu Leu Phe Ser Cys Pro
145 150 155 160
His Arg Leu Arg Glu His Leu Glu Arg Gly Arg Gly Asn Leu Glu Trp
165 170 175
Lys Glu Pro Pro Ser Met Arg Leu Lys Ala Arg Pro Ser Ser Pro Gly
180 185 190
Phe Ser Val Leu Thr Cys Ser Ala Phe Ser Phe Tyr Pro Pro Glu Leu
195 200 205
Gln Leu Arg Phe Leu Arg Asn Gly Leu Ala Ala Gly Thr Gly Gln Gly
210 215 220
Asp Phe Gly Pro Asn Ser Asp Gly Ser Phe His Ala Ser Ser Ser Leu
225 230 235 240
Thr Val Lys Ser Gly Asp Glu His His Tyr Cys Cys Ile Val Gln His
245 250 255
Ala Gly Leu Ala Gln Pro Leu Arg Val Glu Leu Glu Ser Pro Ala Lys
260 265 270
Ser Ser
<210> 10
<211> 99
<212> PRT
<213> Homo sapiens
<400> 10
Ile Gln Arg Thr Pro Lys Ile Gln Val Tyr Ser Arg His Pro Ala Glu
1 5 10 15
Asn Gly Lys Ser Asn Phe Leu Asn Cys Tyr Val Ser Gly Phe His Pro
20 25 30
Ser Asp Ile Glu Val Asp Leu Leu Lys Asn Gly Glu Arg Ile Glu Lys
35 40 45
Val Glu His Ser Asp Leu Ser Phe Ser Lys Asp Trp Ser Phe Tyr Leu
50 55 60
Leu Tyr Tyr Thr Glu Phe Thr Pro Thr Glu Lys Asp Glu Tyr Ala Cys
65 70 75 80
Arg Val Asn His Val Thr Leu Ser Gln Pro Lys Ile Val Lys Trp Asp
85 90 95
Arg Asp Met
<210> 11
<211> 38
<212> PRT
<213> Homo sapiens
<400> 11
Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu
1 5 10 15
Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His
20 25 30
Thr Phe Pro Ala Val Leu
35
<210> 12
<211> 14
<212> PRT
<213> Homo sapiens
<400> 12
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
1 5 10
<210> 13
<211> 18
<212> PRT
<213> Homo sapiens
<400> 13
Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr
1 5 10 15
Gln Thr
<210> 14
<211> 16
<212> PRT
<213> Homo sapiens
<400> 14
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
1 5 10 15
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ВАРИАНТЫ Fc-ОБЛАСТИ С МОДИФИЦИРОВАННЫМИ СВОЙСТВАМИ СВЯЗЫВАНИЯ FcRn И БЕЛКА А | 2015 |
|
RU2713131C1 |
| ВАРИАНТЫ Fc-ОБЛАСТИ С МОДИФИЦИРОВАННЫМ СВЯЗЫВАНИЕМ FcRn И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2714116C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА К CD19 ЧЕЛОВЕКА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2731156C1 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА К ЧЕЛОВЕЧЕСКОМУ БЕТА-АМИЛОИДУ/ЧЕЛОВЕЧЕСКОМУ РЕЦЕПТОРУ ТРАНСФЕРРИНА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2730682C1 |
| АНТИТЕЛА ПРОТИВ ТРАНСФЕРРИНОВОГО РЕЦЕПТОРА, ОБЛАДАЮЩИЕ ИНДИВИДУАЛИЗИРОВАННОЙ АФФИННОСТЬЮ | 2016 |
|
RU2831724C2 |
| АНТИТЕЛА ПРОТИВ ТРАНСФЕРРИНОВОГО РЕЦЕПТОРА, ОБЛАДАЮЩИЕ ИНДИВИДУАЛИЗИРОВАННОЙ АФФИННОСТЬЮ | 2016 |
|
RU2729416C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА К ЧЕЛОВЕЧЕСКОМУ CD20/ЧЕЛОВЕЧЕСКОМУ РЕЦЕПТОРУ ТРАНСФЕРРИНА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2753390C1 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С CD3 | 2019 |
|
RU2810924C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С GPRC5D | 2019 |
|
RU2797268C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С CD3 | 2019 |
|
RU2832669C2 |

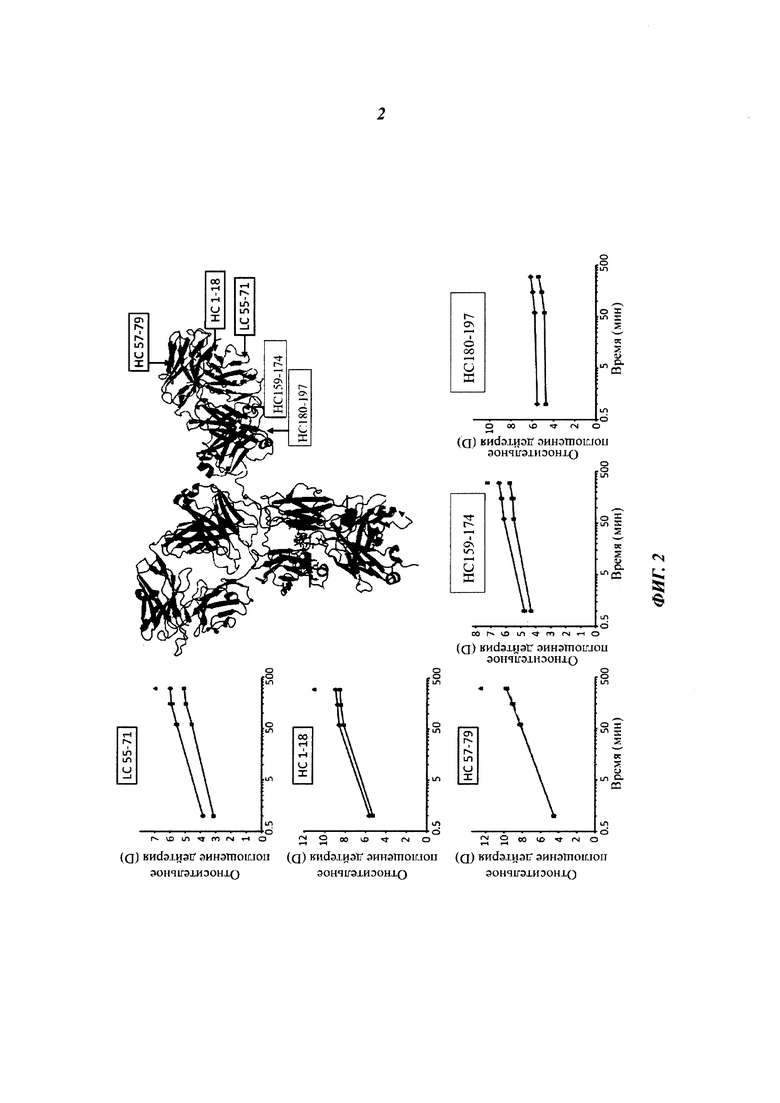
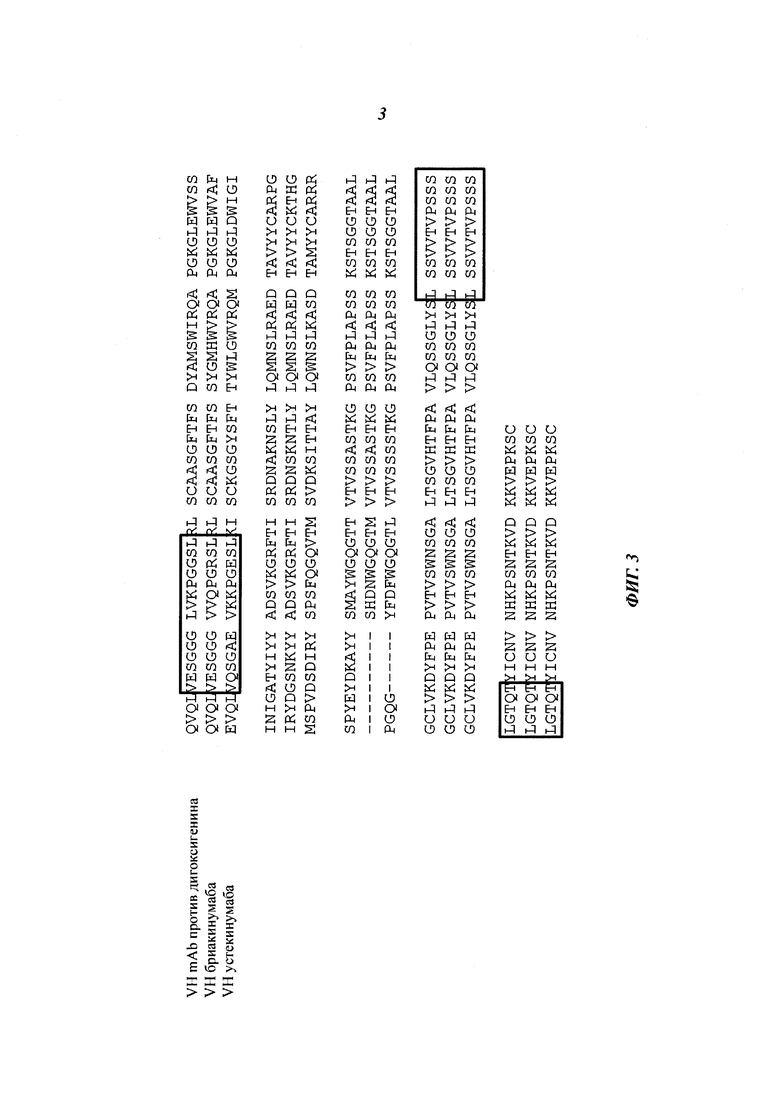

Изобретение относится к области биотехнологии, в частности к способу отбора полноразмерного антитела, которое имеет отличную силу связывания с FcRn, чем родительское полноразмерное антитело. 2 н.п. ф-лы, 4 ил., 10 пр.
1. Способ отбора полноразмерного антитела, которое имеет повышенную силу связывания с FcRn по сравнению с родительским полноразмерным антителом, при котором:
а) из родительского полноразмерного антитела посредством рандомизации одного или более чем одного аминокислотного остатка, выбранного из аминокислотных остатков в положениях 1-23 в вариабельном домене тяжелой цепи в нумерации согласно Kabat, в положениях 55-83 в вариабельном домене легкой цепи в нумерации согласно Kabat, в положениях 145-174 в первом константном домене тяжелой цепи в нумерации согласно индексу EU и в положениях 180-197 в первом константном домене тяжелой цепи в нумерации согласно индексу EU, получают множество полноразмерных антител,
б) определяют силу связывания каждого полноразмерного антитела из указанного множества антител с человеческим неонатальным рецептором Fc (FcRn) и
в) отбирают из указанного множества полноразмерных антител полноразмерное антитело, которое имеет повышенную силу связывания с FcRn по сравнению с родительским полноразмерным антителом, где родительское антитело представляет собой антитело IgG1.
2. Способ отбора полноразмерного антитела, которое имеет пониженную силу связывания с FcRn по сравнению с родительским полноразмерным антителом, при котором:
а) из родительского полноразмерного антитела посредством рандомизации одного или более чем одного аминокислотного остатка, выбранного из аминокислотных остатков в положениях 1-23 в вариабельном домене тяжелой цепи в нумерации согласно Kabat, в положениях 55-83 в вариабельном домене легкой цепи в нумерации согласно Kabat, в положениях 145-174 в первом константном домене тяжелой цепи в нумерации согласно индексу EU и в положениях 180-197 в первом константном домене тяжелой цепи в нумерации согласно индексу EU, получают множество полноразмерных антител,
б) определяют силу связывания каждого полноразмерного антитела из указанного множества антител с человеческим неонатальным рецептором Fc (FcRn) и
в) отбирают из указанного множества полноразмерных антител полноразмерное антитело, которое имеет пониженную силу связывания с FcRn по сравнению с родительским полноразмерным антителом, где родительское антитело представляет собой антитело IgG1.
3. Способ по п. 1 или 2, в котором один или более чем один аминокислотный остаток антитела выбран из аминокислотных остатков в положениях 5-18 в вариабельном домене тяжелой цепи в нумерации согласно Kabat.
4. Способ по п. 1 или 2, в котором один или более чем один аминокислотный остаток антитела выбран из аминокислотных остатков в положениях 145-174 в первом константном домене тяжелой цепи в нумерации согласно индексу EU.
5. Способ по п. 1 или 2, в котором один или более чем один аминокислотный остаток антитела выбран из аминокислотных остатков в положениях 161-174 в первом константном домене тяжелой цепи в нумерации согласно индексу EU.
6. Способ по п. 1 или 2, в котором один или более чем один аминокислотный остаток антитела выбран из аминокислотных остатков в положениях 181-196 в первом константном домене тяжелой цепи в нумерации согласно индексу EU.
7. Способ по п. 1 или 2, в котором один или более чем один аминокислотный остаток антитела выбран из аминокислотных остатков в положениях 182-197 в первом константном домене тяжелой цепи в нумерации согласно индексу EU.
8. Способ по п. 1 или 2, в котором один или более чем один аминокислотный остаток антитела выбран из аминокислотных остатков в положениях 55-83 в вариабельном домене легкой цепи в нумерации согласно Kabat.
9. Способ по п. 1 или 2, в котором один или более чем один аминокислотный остаток антитела выбран из аминокислотных остатков в положениях 55-73 в вариабельном домене легкой цепи в нумерации согласно Kabat.
10. Способ по п. 1 или 2, в котором один или более чем один аминокислотный остаток антитела выбран из аминокислотных остатков в положениях 57-70 в вариабельном домене легкой цепи в нумерации согласно Kabat.
11. Способ по п. 1 или 2, в котором мутация представляет собой мутацию от аминокислотного остатка антитела до другого аминокислотного остатка из той же самой группы аминокислотных остатков.
12. Способ по любому из пп. 1-11, где в антитело вводится одна или более чем одна из следующих мутаций в нумерации согласно нумерации вариабельного домена по Kabat и схеме нумерации индекса EU по Kabat соответственно:
- E6Q тяжелой цепи,
- A162D тяжелой цепи,
- A162E тяжелой цепи,
- T164D тяжелой цепи,
- T164E тяжелой цепи,
- S165D тяжелой цепи,
- S165E тяжелой цепи,
- S191D тяжелой цепи,
- S191E тяжелой цепи,
- G194D тяжелой цепи,
- G194E тяжелой цепи,
- T195D тяжелой цепи,
- T195E тяжелой цепи,
- Q196D тяжелой цепи,
- Q196E тяжелой цепи,
- G57K легкой цепи,
- G57R легкой цепи,
- S60K легкой цепи,
- S60R легкой цепи.
| US 7217797 B2, 15.05.2007 | |||
| IGAWA Т | |||
| et al | |||
| Reduced elimination of IgG antibodies by engineering the variable region, Protein Engineering, Design and Selection, 2010, Vol.23, N5, pp | |||
| Саморазгружающаяся платформа | 1922 |
|
SU385A1 |
| PYZIK M | |||
| et al | |||
| FcRn: The architect behind the immune and non-immune functions of IgG and albumin, The Journal of Immunology, 2015, Vol.194, N10, pp | |||

