ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к антителам и слитым конструкциям на основе полипептидов Fc-области, которые асимметрично модифицированы применительно к взаимодействию со своим Fc-рецептором, в частности своим неонатальным Fc-рецептором (FcRn), и к способам их применения.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Неонатальный Fc-рецептор (FcRn) важен для метаболического пути антител класса IgG in vivo. Функция FcRn заключается в спасении IgG от процесса лизосомальной деградации, что приводит к уменьшению клиренса и увеличению полупериода существования. Он представляет собой гетеродимерный белок, состоящий из двух полипептидов: белка, подобного белку главного комплекса гистосовместимости класса I с молекулярной массой (ММ) 50 кДа (α-FcRn) и β2-микроглобулина (β2m) с ММ 15 кДа. FcRn связывается с высокой аффинностью с CH2-CH3-частью (CH означает константный домен тяжелой цепи) Fc-области антитела класса IgG. Взаимодействие между антителом класса IgG и FcRn зависит от pH и осуществляется в стехиометрическом соотношении 1:2, т.е. одна молекула антитела IgG может взаимодействовать с двумя молекулами FcRn посредством двух своих полипептидов Fc-области тяжелой цепи (см., например, Huber A.H., et al., J. Mol. Biol., 230 (1993) 1077-1083).
Таким образом, свойства/характеристики связывания молекул IgG с FcRn in vitro являются показателями его фармакокинетических свойств in vivo в кровотоке.
Во взаимодействии между FcRn и Fc-областью антитела класса IgG принимают участие разные аминокислотные остатки CH2- и CH3-домена тяжелой цепи.
Известны различные мутации, оказывающие влияние на связывание с FcRn и тем самым на полупериод существования в кровотоке. С использованием сайт-специфического мутагенеза идентифицированы аминокислотные остатки Fc-области, критичные для взаимодействия Fc-области IgG мыши и мышиного FcRn (см., например, Dall'Acqua W.F., et al., J. Immunol., 169 (2002) 5171-5180). В это взаимодействие вовлечены остатки I253, H310, H433, N434 и H435 (нумерация соответствует системе нумерации EU-индекс по Kabat) (Medesan C., et al., Eur. J. Immunol., 26 (1996) 2533-2536; Firan M., et al., Int. Immunol., 13 (2001) 993-1002; Kim J.K., et al., Eur. J. Immunol., 24 (1994) 542-548). Обнаружено, что присутствие остатков I253, H310 и H435 критично для взаимодействия Fc-области IgG человека с мышиным FcRn (Kim J.K., et al., Eur. J. Immunol., 29 (1999) 2819-2885).
Способы усиления связывания Fc-области (и также IgG) с FcRn были осуществлены путем внесения мутаций по различным аминокислотным остаткам в Fc-области: Thr 250, Met 252, Ser 254, Thr 256, Thr 307, Glu 380, Met 428, His 433 и Asn 434 (см. Kuo T.T., et al., J. Clin. Immunol., 30 (2010) 777-789; Ropeenian D.C., et al., Nat. Rev. Immunol., 7 (2007) 715-725).
В результате исследований белок-белковых взаимодействий с целью улучшения связывания с FcRn Dall'Acqua и др. была описана комбинация мутаций M252Y, S254T, T256E (Dall'Acqua W.F., et al. J. Biol. Chem., 281 (2006) 23514-23524). Исследования комплекса (Fc-область IgG человека)-(FcRn человека) показали, что остатки I253, S254, H435 и Y436 являются важнейшими остатками для данного взаимодействия (Firan M., et al., Int. Immunol., 13 (2001) 993-1002; Shields R.L., et al., J. Biol. Chem., 276 (2001) 6591-6604). В работе Yeung Y.A. и др. (J. Immunol., 182 (2009) 7667-7671) приведены различные мутанты по остаткам 248-259, и 301-317, и 376-382, и 424-437 и проведено их изучение.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ Изобретения
Обнаружено, что связывание антитела или слитой конструкции на основе полипептидов Fc-области с FcRn можно модифицировать посредством изменения аминокислотных остатков в не соответствующих этому связыванию положениях в индивидуальных полипептидах Fc-области, поскольку совместное действие этих изменений модифицирует связывание с FcRn. Антитела и слитые конструкции на основе полипептидов Fc-области, описанные в данной заявке, полезны, например, для лечения заболеваний, при которых необходимы специально выбранные для системного введения времена удерживания.
Согласно данному изобретению предложены вариантные Fc-области, которые имеют модифицированные свойства в отношении связывания с FcRn по сравнению с соответствующей Fc-областью дикого типа. Эти вариантные Fc-области содержат специфические аминокислотные мутации в CH2- и/или CH3-домене. Обнаружено, что эти мутации, в случае их использования либо по отдельности, либо в комбинации, в одной и той же или в обеих тяжелых цепях в Fc-области, позволяют получить вариантную Fc-область со специально разработанным полупериодом существования in vivo.
Согласно одному из аспектов, описанных в данной заявке, предложена вариантная Fc-область класса IgG (человека), содержащая первый полипептид Fc-области и второй полипептид Fc-области, при этом
a) первый полипептид Fc-области и второй полипептид Fc-области происходят из одного и того же полипептида родительской Fc-области класса IgG (человека), и
b) первый полипептид Fc-области имеет аминокислотную последовательность, которая отличается от аминокислотной последовательности второго полипептида Fc-области по меньшей мере в одном соответствующем положении согласно системе нумерации EU-индекс по Kabat,
на основании чего вариантная Fc-область класса IgG (человека) имеет другую аффинность к Fc-рецептору человека по сравнению с Fc-областью класса IgG (человека), которая имеет те же аминокислотные остатки (что и полипептид родительской Fc-области IgG человека по пункту (a)) в соответствующих положениях согласно системе нумерации EU-индекс по Kabat в первом полипептиде Fc-области и втором полипептиде Fc-области.
Согласно одному из аспектов, описанных в данной заявке, предложена вариантная Fc-область класса IgG (человека), содержащая первый полипептид Fc-области и второй полипептид Fc-области,
при этом
a) первый полипептид Fc-области имеет аминокислотную последовательность, которая отличается от аминокислотной последовательности второго полипептида Fc-области по меньшей мере в одном соответствующем положении согласно системе нумерации EU-индекс по Kabat,
на основании чего вариантная Fc-область класса IgG (человека) имеет другую аффинность к Fc-рецептору человека по сравнению с Fc-областью класса IgG, которая имеет тот же аминокислотный остаток (что и в соответствующей Fc-области IgG человека) в первом и втором полипептиде Fc-области в соответствующем положении.
Согласно одному из аспектов, описанных в данной заявке, предложена вариантная Fc-область класса IgG (человека), содержащая первый полипептид Fc-области и второй полипептид Fc-области,
при этом
a) аминокислотная последовательность первого полипептида Fc-области отличается от аминокислотной последовательности первого полипептида родительской Fc-области класса IgG по одному или более аминокислотным остаткам,
и
аминокислотная последовательность второго полипептида Fc-области отличается от аминокислотной последовательности второго полипептида родительской Fc-области класса IgG по одному или более аминокислотным остаткам, и
b) первый полипептид Fc-области имеет аминокислотную последовательность, которая отличается от аминокислотной последовательности второго полипептида Fc-области по меньшей мере в одном соответствующем положении согласно системе нумерации EU-индекс по Kabat,
на основании чего вариантная Fc-область класса IgG (человека) имеет другую аффинность к Fc-рецептору человека по сравнению с родительской Fc-областью класса IgG, содержащей первый и второй полипептид родительской Fc-области класса IgG по пункту (a).
Согласно одному из аспектов, описанных в данной заявке, предложена вариантная Fc-область класса IgG (человека), содержащая первый полипептид Fc-области и второй полипептид Fc-области,
при этом
a) аминокислотная последовательность первого полипептида Fc-области происходит из первого полипептида родительской Fc-области класса IgG, а аминокислотная последовательность второго полипептида Fc-области происходит из второго полипептида родительской Fc-области класса IgG, и
b) в первый полипептид Fc-области и/или во второй полипептид Fc-области вносят одну или более мутаций, с тем чтобы первый полипептид Fc-области имел аминокислотную последовательность, которая отличается от аминокислотной последовательности второго полипептида Fc-области по меньшей мере в одном соответствующем положении согласно системе нумерации EU-индекс по Kabat,
на основании чего вариантная Fc-область класса IgG (человека) имеет другую аффинность к Fc-рецептору человека по сравнению с Fc-областью класса IgG, содержащей первый и второй полипептид родительской Fc-области класса IgG по пункту (a)
В одном из воплощений всех аспектов вариантная Fc-область класса IgG (человека) представляет собой вариантную гетеродимерную Fc-область класса IgG (человека).
В одном из воплощений всех аспектов первый полипептид родительской Fc-области класса IgG и второй полипептид родительской Fc-области класса IgG представляют собой полипептиды Fc-области класса IgG, не относящегося к человеку.
В одном из воплощений всех аспектов первый полипептид родительской Fc-области класса IgG и второй полипептид родительской Fc-области класса IgG представляют собой один и тот же полипептид Fc-области класса IgG.
В одном из воплощений всех аспектов в результате образования пары с участием первого полипептида Fc-области и второго полипептида Fc-области с получением димерной (функциональной) Fc-области образуется гетеродимер.
В одном из воплощений всех аспектов первый и второй полипептид Fc-области отличаются независимо друг от друга по меньшей мере по одному аминокислотному остатку от соответствующего полипептида родительской Fc-области класса IgG.
В одном из воплощений всех аспектов класс IgG выбран из подклассов IgG1, IgG2, IgG3 и IgG4.
В одном из воплощений всех аспектов Fc-рецептор человека выбран из неонатального Fc-рецептора человека и Fcγ-рецептора человека.
В одном из воплощений всех аспектов первый полипептид Fc-области отличается по 1 аминокислотному остатку, или 2, или 3, или 4, или 5, или 6, или 7, или 8, или 9, или 10, или 11, или 12 аминокислотным остаткам в соответствующем(им) положении(ям) согласно системе нумерации EU-индекс по Kabat от второго полипептида Fc-области.
В одном из воплощений всех аспектов, описанных в данной заявке, либо первый полипептид Fc-области, либо второй полипептид Fc-области, либо оба полипептида Fc-области содержат одну из приведенных ниже мутаций или комбинаций мутаций:
- T307H, или
- Q311H, или
- E430H, или
- N434H, или
- T307H и Q311H, или
- T307H и E430H, или
- T307H и N434A, или
- T307H и N434H, или
- T307Q и Q311H, или
- T307Q и E430H, или
- T307Q и N434H, или
- T307H, и Q311H, и E430H, и N434A, или
- T307H, и Q311H, и E430H, и N434H, или
- T307H, и Q311H, и E430H, и N434Y, или
- T307Q, и Q311H, и E430H, и N434A, или
- T307Q, и Q311H, и E430H, и N434H, или
- T307Q, и Q311H, и E430H, и N434Y, или
- T307Q, и V308P, и N434Y, и Y436H, или
- T307H, и M252Y, и S254T, и T256E, или
- T307Q, и M252Y, и S254T, и T256E, или
- Q311H, и M252Y, и S254T, и T256E, или
- E430H, и M252Y, и S254T, и T256E, или
- N434H, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и M252Y, и S254T, и T256E, или
- T307H, и E430H, и M252Y, и S254T, и T256E, или
- T307H, и N434A, и M252Y, и S254T, и T256E, или
- T307H, и N434H, и M252Y, и S254T, и T256E, или
- T307Q, и Q311H, и M252Y, и S254T, и T256E, или
- T307Q, и E430H, и M252Y, и S254T, и T256E, или
- T307Q, и N434H, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и E430H, и N434A, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и E430H, и N434H, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и E430H, и N434Y, и M252Y, и S254T, и T256E, или
- T307Q, и Q311H, и E430H, и N434A, и M252Y, и S254T, и T256E, или
- T307Q, и Q311H, и E430H, и N434H, и M252Y, и S254T, и T256E, или
- T307Q, и Q311H, и E430H, и N434Y, и M252Y, и S254T, и T256E, или
- T307Q, и V308P, и N434Y, и Y436H, и M252Y, и S254T, и T256E.
В одном из воплощений всех аспектов, описанных в данной заявке, либо первый полипептид Fc-области, либо второй полипептид Fc-области, либо оба полипептида Fc-области содержат одну из приведенных ниже мутаций или комбинаций мутаций:
- T307H, или
- E430H, или
- T307H и Q311H, или
- T307H и E430H, или
- T307H и N434A, или
- T307H и N434H, или
- T307Q и Q311H, или
- T307Q и E430H, или
- T307Q и N434H, или
- T307H, и Q311H, и E430H, и N434A, или
- T307H, и Q311H, и E430H, и N434H, или
- T307H, и Q311H, и E430H, и N434Y, или
- T307Q, и Q311H, и E430H, и N434A, или
- T307Q, и Q311H, и E430H, и N434H, или
- T307Q, и Q311H, и E430H, и N434Y, или
- T307Q, и V308P, и N434Y, и Y436H, или
- T307H, и M252Y, и S254T, и T256E или
- T307Q, и M252Y, и S254T, и T256E, или
- Q311H, и M252Y, и S254T, и T256E, или
- E430H, и M252Y, и S254T, и T256E, или
- N434H, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и M252Y, и S254T, и T256E, или
- T307H, и E430H, и M252Y, и S254T, и T256E, или
- T307H, и N434A, и M252Y, и S254T, и T256E, или
- T307H, и N434H, и M252Y, и S254T, и T256E, или
- T307Q, и Q311H, и M252Y, и S254T, и T256E, или
- T307Q, и E430H, и M252Y, и S254T, и T256E, или
- T307Q, и N434H, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и E430H, и N434A, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и E430H, и N434H, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и E430H, и N434Y, и M252Y, и S254T, и T256E, или
- T307Q, и Q311H, и E430H, и N434A, и M252Y, и S254T, и T256E, или
- T307Q, и Q311H, и E430H, и N434H, и M252Y, и S254T, и T256E, или
- T307Q, и Q311H, и E430H, и N434Y, и M252Y, и S254T, и T256E, или
- T307Q, и V308P, и N434Y, и Y436H, и M252Y, и S254T, и T256E.
В одном из воплощений всех аспектов, описанных в данной заявке,
первый полипептид Fc-области содержит, независимо от второго полипептида Fc-области, одну из приведенных ниже мутаций или комбинаций мутаций:
- T307H, или
- Q311H, или
- E430H, или
- N434H, или
- T307H и Q311H, или
- T307H и E430H, или
- T307H и N434A, или
- T307H и N434H, или
- T307Q и Q311H, или
- T307Q и E430H, или
- T307Q и N434H, или
- M252Y, и S254T, и T256E, или
- I253A, и H310A, и H435A, или
- H310A, и H433A, и Y436A, или
- T307H, и Q311H, и E430H, и N434A, или
- T307H, и Q311H, и E430H, и N434H, или
- T307H, и Q311H, и E430H, и N434Y, или
- T307Q, и Q311H, и E430H, и N434A, или
- T307Q, и Q311H, и E430H, и N434H, или
- T307Q, и Q311H, и E430H, и N434Y, или
- T307Q, и V308P, и N434Y, и Y436H, или
- T307H, и M252Y, и S254T, и T256E или
- Q311H, и M252Y, и S254T, и T256E, или
- E430H, и M252Y, и S254T, и T256E, или
- N434H, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и M252Y, и S254T, и T256E, или
- T307H, и E430H, и M252Y, и S254T, и T256E, или
- T307H, и N434A, и M252Y, и S254T, и T256E, или
- T307H, и N434H, и M252Y, и S254T, и T256E, или
- T307Q, и Q311H, и M252Y, и S254T, и T256E, или
- T307Q, и E430H, и M252Y, и S254T, и T256E, или
- T307Q, и N434H, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и E430H, и N434A, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и E430H, и N434H, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и E430H, и N434Y, и M252Y, и S254T, и T256E, или
- T307Q, и V308P, и N434Y, и Y436H, и M252Y, и S254T, и T256E, или
- T307Q, и V308P, и N434Y, и Y436H, и M252Y, и S254T, и T256E,
и
второй полипептид Fc-области содержит, независимо от первого полипептида Fc-области, одну из приведенных ниже мутаций или комбинаций мутаций:
- T307H, или
- T307Q, или
- Q311H, или
- E430H, или
- N434H, или
- T307H и Q311H, или
- T307H и E430H, или
- T307H и N434A, или
- T307H и N434H, или
- T307Q и Q311H, или
- T307Q и E430H, или
- T307Q и N434H, или
- T307H, и Q311H, и E430H, и N434A, или
- T307H, и Q311H, и E430H, и N434H, или
- T307H, и Q311H, и E430H, и N434Y, или
- T307Q, и Q311H, и E430H, и N434A, или
- T307Q, и Q311H, и E430H, и N434H, или
- T307Q, и Q311H, и E430H, и N434Y, или
- T307Q, и V308P, и N434Y, и Y436H, или
- T307H, и M252Y, и S254T, и T256E, или
- Q311H, и M252Y, и S254T, и T256E, или
- E430H, и M252Y, и S254T, и T256E, или
- N434H, и M252Y, и S254T, и T256E.
В одном из воплощений всех аспектов, описанных в данной заявке,
первый полипептид Fc-области содержит, независимо от второго полипептида Fc-области, одну из приведенных ниже мутаций или комбинаций мутаций:
- T307H, или
- E430H, или
- T307H и Q311H, или
- T307H и E430H, или
- T307H и N434A, или
- T307H и N434H, или
- T307Q и Q311H, или
- T307Q и E430H, или
- T307Q и N434H, или
- M252Y, и S254T, и T256E, или
- I253A, и H310A, и H435A, или
- H310A, и H433A, и Y436A, или
- T307H, и Q311H, и E430H, и N434A, или
- T307H, и Q311H, и E430H, и N434H, или
- T307H, и Q311H, и E430H, и N434Y, или
- T307Q, и Q311H, и E430H, и N434A, или
- T307Q, и Q311H, и E430H, и N434H, или
- T307Q, и Q311H, и E430H, и N434Y, или
- T307Q, и V308P, и N434Y, и Y436H, или
- T307H, и M252Y, и S254T, и T256E, или
- Q311H, и M252Y, и S254T, и T256E, или
- E430H, и M252Y, и S254T, и T256E, или
- N434H, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и M252Y, и S254T, и T256E, или
- T307H, и E430H, и M252Y, и S254T, и T256E, или
- T307H, и N434A, и M252Y, и S254T, и T256E, или
- T307H, и N434H, и M252Y, и S254T, и T256E, или
- T307Q, и Q311H, и M252Y, и S254T, и T256E, или
- T307Q, и E430H, и M252Y, и S254T, и T256E, или
- T307Q, и N434H, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и E430H, и N434A, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и E430H, и N434H, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и E430H, и N434Y, и M252Y, и S254T, и T256E, или
- T307Q, и V308P, и N434Y, и Y436H, и M252Y, и S254T, и T256E, или
- T307Q, и V308P, и N434Y, и Y436H, и M252Y, и S254T, и T256E,
и
второй полипептид Fc-области содержит, независимо от первого полипептида Fc-области, одну из приведенных ниже мутаций или комбинаций мутаций:
- T307H, или
- T307Q, или
- E430H, или
- T307H и Q311H, или
- T307H и E430H, или
- T307H и N434A, или
- T307H и N434H, или
- T307Q и Q311H, или
- T307Q и E430H, или
- T307Q и N434H, или
- T307H, и Q311H, и E430H, и N434A, или
- T307H, и Q311H, и E430H, и N434H, или
- T307H, и Q311H, и E430H, и N434Y, или
- T307Q, и Q311H, и E430H, и N434A, или
- T307Q, и Q311H, и E430H, и N434H, или
- T307Q, и Q311H, и E430H, и N434Y, или
- T307Q, и V308P, и N434Y, и Y436H, или
- T307H, и M252Y, и S254T, и T256E или
- Q311H, и M252Y, и S254T, и T256E, или
- E430H, и M252Y, и S254T, и T256E, или
- N434H, и M252Y, и S254, и T256E.
В одном из воплощений всех аспектов, описанных в данной заявке, первый полипептид Fc-области содержит
одну из приведенных ниже комбинаций мутаций:
- ни одной, или
- M252Y, и S254T, и T256E, или
- I253A, и H310A, и H435A, или
- H310A, и H433A, и Y436A,
и
одну из приведенных ниже мутаций или комбинаций мутаций:
- ни одной,
- T307H, или
- T307Q, или
- Q311H, или
- E430H, или
- N434H, или
- T307H и Q311H, или
- T307H и E430H, или
- T307H и N434A, или
- T307H и N434H, или
- T307Q и Q311H, или
- T307Q и E430H, или
- T307Q и N434H, или
- T307Q и N434A, или
- T307H, и Q311H, и E430H, и N434A, или
- T307H, и Q311H, и E430H, и N434H, или
- T307H, и Q311H, и E430H, и N434Y, или
- T307Q, и Q311H, и E430H, и N434A, или
- T307Q, и Q311H, и E430H, и N434H, или
- T307Q, и Q311H, и E430H, и N434Y, или
- T307Q, и V308P, и N434Y, и Y436H,
и второй полипептид Fc-области содержит
одну из приведенных ниже мутаций или комбинаций мутаций:
- ни одной, если первый полипептид Fc-области содержит по меньшей мере одну мутацию, или
- T307H, или
- T307Q, если первый полипептид Fc-области не содержит исключительно мутацию T307Q, или
- Q311H, или
- E430H, или
- N434H, или
- T307H и Q311H, или
- T307H и E430H, или
- T307H и N434A, или
- T307H и N434H, или
- T307Q и Q311H, или
- T307Q и E430H, или
- T307Q и N434H, или
- T307Q и N434A, или
- M252Y, и S254T, и T256E, если первый полипептид Fc-области не содержит исключительно комбинацию мутаций M252Y, и S254T, и T256E, или
- I253A, и H310A, и H435A, если первый полипептид Fc-области не содержит исключительно комбинацию мутаций I253A, и H310A, и H435A, или
- H310A, и H433A, и Y436A, если первый полипептид Fc-области не содержит исключительно комбинацию мутаций H310A, и H433A, и Y436A, или
- T307H, и Q311H, и E430H, и N434A, или
- T307H, и Q311H, и E430H, и N434H, или
- T307H, и Q311H, и E430H, и N434Y, или
- T307Q, и Q311H, и E430H, и N434A, или
- T307Q, и Q311H, и E430H, и N434H, или
- T307Q, и Q311H, и E430H, и N434Y, или
- T307Q, и V308P, и N434Y, и Y436H.
В одном из воплощений всех аспектов, описанных в данной заявке, первый полипептид Fc-области содержит
одну из приведенных ниже комбинаций мутаций:
- ни одной, или
- M252Y, и S254T, и T256E, или
- I253A, и H310A, и H435A, или
- H310A, и H433A, и Y436A,
и
одну из приведенных ниже мутаций или комбинаций мутаций:
- ни одной,
- T307H, или
- T307Q, или
- E430H, или
- T307H и Q311H, или
- T307H и E430H, или
- T307H и N434A, или
- T307H и N434H, или
- T307Q и Q311H, или
- T307Q и E430H, или
- T307Q и N434H, или
- T307Q и N434A, или
- T307H, и Q311H, и E430H, и N434A, или
- T307H, и Q311H, и E430H, и N434H, или
- T307H, и Q311H, и E430H, и N434Y, или
- T307Q, и Q311H, и E430H, и N434A, или
- T307Q, и Q311H, и E430H, и N434H, или
- T307Q, и Q311H, и E430H, и N434Y, или
- T307Q, и V308P, и N434Y, и Y436H,
и второй полипептид Fc-области содержит
одну из приведенных ниже мутаций или комбинаций мутаций:
- ни одной, если первый полипептид Fc-области содержит по меньшей мере одну мутацию, или
- T307H, или
- T307Q, если первый полипептид Fc-области не содержит исключительно мутацию T307Q, или
- E430H, или
- T307H и Q311H, или
- T307H и E430H, или
- T307H и N434A, или
- T307H и N434H, или
- T307Q и Q311H, или
- T307Q и E430H, или
- T307Q и N434H, или
- T307Q и N434A, или
- M252Y, и S254T, и T256E, если первый полипептид Fc-области не содержит исключительно комбинацию мутаций M252Y, и S254T, и T256E, или
- I253A, и H310A, и H435A, если первый полипептид Fc-области не содержит исключительно комбинацию мутаций I253A, и H310A, и H435A, или
- H310A, и H433A, и Y436A, если первый полипептид Fc-области не содержит исключительно комбинацию мутаций H310A, и H433A, и Y436A, или
- T307H, и Q311H, и E430H, и N434A, или
- T307H, и Q311H, и E430H, и N434H, или
- T307H, и Q311H, и E430H, и N434Y, или
- T307Q, и Q311H, и E430H, и N434A, или
- T307Q, и Q311H, и E430H, и N434H, или
- T307Q, и Q311H, и E430H, и N434Y, или
- T307Q, и V308P, и N434Y, и Y436H.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутации I253A, и H310A, и H435A, а второй полипептид Fc-области содержит мутации M252Y, и S254T, и T256E.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутации I253A, и H310A, и H435A, а второй полипептид Fc-области содержит мутации M252Y, и S254T, и T256E, и T307Q, и N434Y.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутации I253A, и H310A, и H435A, а второй полипептид Fc-области содержит мутации M252Y, и S254T, и T256E, и T307Q, и V308P, и N434Y, и Y436H.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутации T307H, и Q311H, и E430H, и N434H, а второй полипептид Fc-области содержит мутации M252Y, и S254T, и T256E.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутации T307H и N434H, а второй полипептид Fc-области содержит мутации M252Y, и S254T, и T256E.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутации T307Q и N434A, а второй полипептид Fc-области содержит мутации M252Y, и S254T, и T256E.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутации T250Q и M428L, а второй полипептид Fc-области содержит мутации M252Y, и S254T, и T256E.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутации T307Q и N434H, а второй полипептид Fc-области содержит мутации M252Y, и S254T, и T256E, и T307Q, и N434H.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутации T307H, и Q311H, и E430H, и N434H, и второй полипептид Fc-области содержит мутации T307H, и Q311H, и E430H, и N434H.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутации T307H и N434H, и второй полипептид Fc-области содержит мутации T307H и N434H.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутации T307H, и N434H, и M252Y, и S254T, и T256E, и второй полипептид Fc-области содержит мутации T307H, и N434H, и M252Y, и S254T, и T256E.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутацию N434H, и второй полипептид Fc-области содержит мутацию N434H.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутации T307Q и N434A.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутации T307H и N434H.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутации T307H, и N434H, и M252Y, и S254T, и T256E.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутацию N434H.
В одном из воплощений первый полипептид Fc-области дополнительно содержит мутации Y349C, T366S, L368A и Y407V (“полипептид Fc-области, формирующий впадину”), а второй полипептид Fc-области дополнительно содержит мутации S354C и T366W (“полипептид Fc-области, формирующий выступ”).
В одном из воплощений всех аспектов вариантная Fc-область класса IgG (человека) содержит первый и второй полипептид Fc-области подкласса IgG1 человека, где
a) и первый, и второй полипептид Fc-области оба дополнительно содержат мутации L234A и L235A (нумерация соответствует системе нумерации EU-индекс по Kabat), или
b) и первый, и второй полипептид Fc-области оба дополнительно содержат мутацию P329G (нумерация соответствует системе нумерации EU-индекс по Kabat), или
c) и первый, и второй полипептид Fc-области оба дополнительно содержат мутации L234A, и L235A, и P329G (нумерация соответствует системе нумерации EU-индекс по Kabat), или
d) и первый, и второй полипептид Fc-области оба дополнительно содержат мутации L234A и L235A (нумерация соответствует системе нумерации EU-индекс по Kabat), и первый полипептид Fc-области дополнительно содержит мутацию Y349C или S354C и мутацию T366W, а второй полипептид Fc-области дополнительно содержит мутацию Y349C или S354C и мутации T366S, L368A и Y407V, или
e) и первый, и второй полипептид Fc-области оба дополнительно содержат мутации L234A, и L235A, и P329G (нумерация соответствует системе нумерации EU-индекс по Kabat), и первый полипептид Fc-области дополнительно содержит мутацию Y349C или S354C и мутацию T366W, а второй полипептид Fc-области дополнительно содержит мутацию Y349C или S354C и мутации T366S, L368A и Y407V.
В одном из воплощений вариантная Fc-область класса IgG (человека) содержит первый и второй полипептид Fc-области подкласса IgG4 человека, где
a) и первый, и второй полипептид Fc-области оба дополнительно содержат мутации S228P и L235E (нумерация соответствует системе нумерации EU-индекс по Kabat), или
b) и первый, и второй полипептид Fc-области оба дополнительно содержат мутацию P329G (нумерация соответствует системе нумерации EU-индекс по Kabat), или
c) и первый, и второй полипептид Fc-области оба дополнительно содержат мутации S228P, и L235E, и P329G (нумерация соответствует системе нумерации EU-индекс по Kabat), или
d) и первый, и второй полипептид Fc-области оба дополнительно содержат мутации S228P и L235E (нумерация соответствует системе нумерации EU-индекс по Kabat), и первый полипептид Fc-области дополнительно содержит мутацию Y349C или S354C и мутацию T366W, а второй полипептид Fc-области дополнительно содержит мутацию Y349C или S354C и мутации T366S, L368A и Y407V,
e) и первый, и второй полипептид Fc-области оба дополнительно содержат мутации S228P, и L235E, и P329G (нумерация соответствует системе нумерации EU-индекс по Kabat), и первый полипептид Fc-области дополнительно содержит мутацию Y349C или S354C и мутацию T366W, а второй полипептид Fc-области дополнительно содержит мутацию Y349C или S354C и мутации T366S, L368A и Y407V.
Согласно одному из аспектов, описанных в данной заявке, предложены антитело или слитая конструкция на основе полипептидов Fc-области, содержащие вариантную Fc-область класса IgG (человека), описанную в данной заявке.
В одном из воплощений антитело представляет собой моноклональное антитело.
В одном из воплощений антитело представляет собой человеческое, гуманизированное или химерное антитело.
Согласно одному из аспектов, описанных в данной заявке, предложена нуклеиновая кислота, кодирующая вариантную Fc-область класса IgG (человека), описанную в данной заявке.
Согласно одному из аспектов, описанных в данной заявке, предложена нуклеиновая кислота, кодирующая антитело, описанное в данной заявке.
Согласно одному из аспектов, описанных в данной заявке, предложена нуклеиновая кислота, кодирующая слитую конструкцию на основе полипептидов Fc-области, описанную в данной заявке.
Согласно одному из аспектов, описанных в данной заявке, предложена клетка-хозяин, содержащая нуклеиновую кислоту, описанную в данной заявке.
Согласно одному из аспектов, описанных в данной заявке, предложен способ получения вариантной Fc-области класса IgG (человека), описанной в данной заявке, включающий культивирование клетки хозяина, описанной в данной заявке, с целью получения вариантной Fc-области класса IgG (человека).
Согласно одному из аспектов, описанных в данной заявке, предложен способ получения антитела, описанного в данной заявке, включающий культивирование клетки хозяина, описанной в данной заявке, с целью получения данного антитела.
Согласно одному из аспектов, описанных в данной заявке, предложен способ получения слитой конструкции на основе полипептидов Fc-области, описанной в данной заявке, включающий культивирование клетки хозяина, описанной в данной заявке, с целью получения слитой конструкции на основе полипептидов Fc-области.
Согласно одному из аспектов, описанных в данной заявке, предложена фармацевтическая композиция, содержащая вариантную Fc-область класса IgG (человека), описанную в данной заявке, или антитело, описанное в данной заявке, или слитую конструкцию на основе полипептидов Fc-области, описанную в данной заявке.
Согласно одному из аспектов, описанных в данной заявке, предложена вариантная Fc-область класса IgG (человека), описанная в данной заявке, или антитело, описанное в данной заявке, или слитая конструкция на основе полипептидов Fc-области, описанная в данной заявке, для применения в качестве лекарственного средства.
Согласно одному из аспектов, описанных в данной заявке, предложено применение вариантной Fc-области класса IgG (человека), описанной в данной заявке, или антитела, описанного в данной заявке, или слитой конструкции на основе полипептидов Fc-области, описанной в данной заявке, для изготовления лекарственного средства.
Антитела, описанные в данной заявке, можно использовать в качестве, например, рекрутеров T-клеток, в качестве связывающегося с Fc-гамма-рецептором вещества с высокой биологической активностью (эффективностью) и быстрым выведением из кровотока (из сыворотки крови), в виде конъюгатов антитело-лекарственное средство с быстрым выведением для снижения системных побочных эффектов или в качестве антител с предварительно определенной направленной доставкой.
КРАТКОЕ ОПИСАНИЕ ФИГУР
Фиг. 1. Полученный при проведении FcRn-аффинной хроматографии илллюстративный схематический график элюирования антитела, содержащего разные (вариантные) Fc-области: 1: с мутациями I253A/H310A/H435A; 2: Fc-область дикого типа; 3: M252Y/S254T/T256E в одном полипептиде Fc-области; другая Fc-область дикого типа; 4: M252Y/S254T/T256E в обоих полипептидах Fc-области; 5: цепь, формирующая выступ: M252Y/S254T/T256E; цепь, формирующая впадину: T307Q/N434A.
Фиг. 2. Полученный при проведении FcRn-аффинной хроматографии график элюирования антитела, содержащего разные (вариантные) Fc-области: 1: Fc-область дикого типа; 2: подвергнутая инженерии гликозилирования Fc-область; 3: T307Q/N434A; 4: T307H/N434H; 5: T307H/N434H/M252Y/S254T/T256E; 6: N434H; 7: M252Y/S254T/T256E.
Фиг. 3. Полученный при проведении FcRn-аффинной хроматографии график элюирования антитела, содержащего разные (вариантные) Fc-области: 1: Fc-область по типу (цепь, формирующая впадину)-(цепь, формирующая выступ); 2: цепь, формирующая впадину: T307Q/N434A; цепь, формирующая выступ: M252Y/S254T/T256E; 3: цепь, формирующая впадину: T307H/N434H; цепь, формирующая выступ: M252Y/S254T/T256E; 4: цепь, формирующая впадину: T250Q/M428L; цепь, формирующая выступ: M252Y/S254T/T256E; 5: цепь, формирующая впадину: T307Q, N434H; цепь, формирующая выступ: M252Y/S254T/T256E/T307Q/N434H; 6: цепь, формирующая впадину: T307H/Q311H/E430H/N434H; цепь, формирующая выступ: M252Y/S254T/T256E; 7: цепь, формирующая впадину: T307H/Q311H/E430H/N434H; цепь, формирующая выступ: M252Y/S254T/T256E/T307H/Q311H/E430H/N434H; 8: цепь, формирующая впадину/выступ: M252Y/S254T/T256E.
ПОДРОБНОЕ ОПИСАНИЕ ВОПЛОЩЕНИЙ ИЗОБРЕТЕНИЯ
I. ОПРЕДЕЛЕНИЯ
Термин “примерно” означает диапазон, составляющий +/- 20% от следующего далее численного значения. В одном из воплощений термин “примерно” означает диапазон, составляющий +/- 10% от следующего далее численного значения. В одном из воплощений термин “примерно” означает диапазон, составляющий +/- 5% от следующего далее численного значения.
“Акцепторный каркас IgG человека” для целей данного изобретения представляет собой каркас, содержащий аминокислотную последовательность каркаса вариабельного домена легкой цепи (VL) или каркаса вариабельного домена тяжелой цепи (VH), происходящего из каркаса иммуноглобулина человека или консенсусного каркаса IgG человека, который определен ниже. Акцепторный каркас IgG человека, “происходящий из” каркаса иммуноглобулина человека или консенсусного каркаса IgG человека, может содержать такую же, как у них, аминокислотную последовательность, или он может содержать изменения в аминокислотной последовательности. В некоторых воплощениях количество изменений аминокислот составляет 10 или меньше, 9 или меньше, 8 или меньше, 7 или меньше, 6 или меньше, 5 или меньше, 4 или меньше, 3 или меньше либо 2 или меньше. В некоторых воплощениях последовательность акцепторного каркаса VL иммуноглобулина человека идентична последовательности каркаса VL иммуноглобулина человека или последовательности консенсусного каркаса IgG человека.
Антитело с “созревшей аффинностью” относится к антителу с одним или более изменениями в одном или более чем одном гипервариабельном участке (HVR) по сравнению с родительским антителом, которое не претерпело таких изменений, приводящих к улучшению аффинности антитела к антигену.
Термин “изменение” означает мутацию (замену), вставку (добавление) или делецию одного или более аминокислотных остатков в родительском антителе или слитом полипептиде, например слитом полипептиде, содержащем по меньшей мере FcRn-связывающую часть Fc-области, приводящих к получению модифицированного антитела или слитого полипептида. Термин “мутация” означает, что определенный аминокислотный остаток замемен на другой аминокислотный остаток. Например, мутация L234A означает, что остаток аминокислоты лизина в положении 234 в (полипептиде) Fc-области антитела заменен на остаток аминокислоты аланина (замена лизина на аланин) (нумерация соответствует EU-индексу).
Термин “аминокислотная мутация” означает замену по меньшей мере одного имеющегося аминокислотного остатка на другой отличающийся от него аминокислотный остаток (т.е. заменяющий аминокислотный остаток). Заменяющий аминокислотный остаток может представлять собой “природный аминокислотный остаток” и быть выбран из группы, состоящей из аланина (трехбуквенный код: ala, однобуквенный код: A), аргинина (arg, R), аспарагина (asn, N), аспарагиновой кислоты (asp, D), цистеина (cys, C), глутамина (gln, Q), глутаминовой кислоты (glu, E), глицина (gly, G), гистидина (his, H), изолейцина (ile, I), лейцина (leu, L), лизина (lys, K), метионина (met, M), фенилаланина (phe, F), пролина (pro, P), серина (ser, S), треонина (thr, T), триптофана (trp, W), тирозина (tyr, Y) и валина (val, V). Заменяющим аминокислотным остатком может быть “неприродный аминокислотный остаток”. См., например, US 6586207, WO 98/48032, WO 03/073238, US 2004/0214988, WO 2005/35727, WO 2005/74524, Chin J.W., et al., J. Am. Chem. Soc., 124 (2002) 9026-9027; Chin J.W. and Schultz P.G., ChemBioChem., 11 (2002) 1135-1137; Chin J.W. et al., PICAS United States of America, 99 (2002) 11020-11024; и Wang L. and Schultz P.G., Chem. (2002) 1-10 (все они полностью включены в данное описание посредством ссылки).
Термин “аминокислотная вставка” означает (дополнительное) встраивание по меньшей мере одного аминокислотного остатка в предварительно заданное положение в аминокислотной последовательности. В одном из воплощений вставка будет представлять собой вставку одного или двух аминокислотных остатков. Встроенный(ые) аминокислотный(ые) остаток(ки) может(гут) представлять собой любой(ые) природный(ые) или неприродный(ые) аминокислотный(ые) остаток(ки).
Термин “аминокислотная делеция” означает удаление по меньшей мере одного аминокислотного остатка из предварительно заданного положения в аминокислотной последовательности.
Термин “антитело” используется в данном описании в самом широком смысле и охватывает различные структуры антител, включая, но не ограничиваясь этим, моноклональные антитела (mAb), поликлональные антитела, мультиспецифичные антитела (например, биспецифичные антитела, триспецифичные антитела) и фрагменты антител, при условии, что они демонстрируют желаемую антиген-, и/или белок A-, и/или FcRn-связывающую активность.
Термин “асимметричная Fc-область” означает пару полипептидов Fc-области, которые имеют разные аминокислотные остатки в соответствующих положениях согласно системе нумерации EU-индекс по Kabat.
Термин “Fc-область, асимметричная применительно к связыванию с FcRn”, означает Fc-область, состоящую из двух полипептидных цепей, которые имеют разные аминокислотные остатки в соответствующих положениях, причем данные положения определены согласно системе нумерации EU-индекс по Kabat, при этом присутствие разных аминокислот в этих положениях влияет на связывание Fc-области с неонатальным Fc-рецептором (FcRn) человека. В рамках данного описания различия между двумя полипептидными цепями Fc-области в “Fc-области, асимметричной применительно к связыванию с FcRn”, не включают различий, которые внесены с целью облегчения образования гетеродимерных Fc-областей, например, для получения биспецифичных антител. Эти различия также могут быть асимметричными, т.е. когда две цепи отличаются аминокислотными остатками в не соответствующих друг другу положениях согласно системе нумерации EU-индекс по Kabat. Присутствие таких различий облегчает осуществление гетеродимеризации и уменьшает вероятность гомодимеризации. Примерами таких различий являются так называемые замены по типу “выступы во впадины” (см., например, US 7695936 и US 2003/0078385). Обнаружено, что вероятность образования гетеродимеров повышают следующие замены по типу “выступы” и “впадины” в индивидуальных полипептидных цепях Fc-области антитела IgG подкласса IgG1: 1) в одной цепи Y407T, а в другой цепи T366Y; 2) в одной цепи Y407A, а в другой цепи T366W; 3) в одной цепи F405A, а в другой цепи T394W; 4) в одной цепи F405W, а в другой цепи T394S; 5) в одной цепи Y407T, а в другой цепи T366Y; 6) в одной цепи T366Y и F405A, а в другой цепи T394W и Y407T; 7) в одной цепи T366W и F405W, а в другой цепи T394S и Y407A; 8) в одной цепи F405W и Y407A, а в другой цепи T366W и T394S; и 9) в одной цепи T366W, а в другой цепи T366S, L368A и Y407V, при этом из перечисленного особенно подходит последний вариант. Кроме того, образованию гетеродимеров способствуют изменения, в результате которых создаются новые дисульфидные мостики между двумя полипептидными цепями Fc-области (см., например, US 2003/0078385). Обнаружено, что вероятность образования гетеродимеров повышают следующие замены, которые приводят к появлению соответствующим образом расположенных в пространстве остатков цистеина для образования новых внутрицепочечных дисульфидных связей в индивидуальных полипептидных цепях Fc-области антитела IgG подкласса IgG1: в одной цепи Y349C, а в другой S354C; в одной цепи Y349C, а в другой E356C; в одной цепи Y349C, а в другой E357C; в одной цепи L351C, а в другой S354C; в одной цепи T394C, а в другой E397C; или в одной цепи D399C, а в другой K392C. Следующими примерами изменений аминокислот, способствующих гетеродимеризации, являются так называемые “парные замены с изменением заряда” (см., например, WO 2009/089004). Обнаружено, что вероятность образования гетеродимеров повышают следующие парные замены с изменением заряда в индивидуальных полипептидных цепях Fc-области антитела IgG подкласса IgG1: 1) в одной цепи K409D или K409E, а в другой цепи D399K или D399R; 2) в одной цепи K392D или K392E, а в другой цепи D399K или D399R; 3) в одной цепи K439D или K439E, а в другой цепи E356K или E356R; 4) в одной цепи K370D или K370E, а в другой цепи E357K или E357R; 5) в одной цепи K409D и K360D, а в другой цепи D399K и E356K; 6) в одной цепи K409D и K370D, а в другой цепи D399K и E357K; 7) в одной цепи K409D и K392D, а в другой цепи D399K, E356K и E357K; 8) в одной цепи K409D и K392D, а в другой цепи D399K; 9) в одной цепи K409D и K392D, а в другой цепи D399K и E356K; 10) в одной цепи K409D и K392D, а в другой цепи D399K и D357K; 11) в одной цепи K409D и K370D, а в другой цепи D399K и D357K; 12) в одной цепи D399K, а в другой цепи K409D и K360D; и 13) в одной цепи K409D и K439D, а в другой D399K и E356K.
Термин “связывание (с антигеном)” означает связывание антитела со своим антигеном в анализе in vitro, в одном из воплощений в анализе связывания, в котором антитело связывают с поверхностью и связывание антигена с антителом измеряют с использованием поверхностного плазмонного резонанса (SPR). “Связывание” означает, что аффинность связывания (KD) составляет 10-8 М или меньше, в некоторых воплощениях от 10-13 до 10-8 М, в некоторых воплощениях от 10-13 до 10-9 М.
Связывание можно изучать в анализе с использованием BIAcore (биосенсор AB от GE Healthcare, Uppsala, Sweden). Аффинность связывания определяют в терминах ka (константа скорости ассоциации для антитела при образовании комплекса антитело/антиген), kd (константа диссоциации) и KD (kd/ka).
Термин “химерное” антитело относится к антителу, в котором часть тяжелой и/или легкой цепи происходит из определенного источника или вида, в то время как остальная часть тяжелой и/или легкой цепи происходит из другого источника или вида.
Термин “CH2-домен” означает часть полипептида тяжелой цепи антитела, которая простирается приблизительно от положения 231 согласно EU до положения 340 согласно EU (EU система нумерации по Kabat). В одном из воплощений CH2-домен имеет аминокислотную последовательность с SEQ ID NO: 01: APELLGG PSVFLFPPKP KDTLMISRTP EVTCVWDVS HEDPEVKFNW YVDGVEVHNA KTKPREEQ E STYRWSVLT VLHQDWLNGK EYKCKVSNKA LPAPIEKTIS KAK.
Термин “CH3-домен” означает часть полипептида тяжелой цепи антитела, которая простирается приблизительно от положения 341 согласно EU до положения 446 согласно EU. В одном из воплощений CH3-домен имеет аминокислотную последовательность с SEQ ID NO: 2: GQPREPQ VYTLPPSRDE LTKNQVSLTC LVKGFYPSDI AVEWESNGQP ENNYKTTPPV LDSDGSFFLY SKLTVDKSRW QQGNVFSCSV MHEALHNHYT QKSLSLSPG.
“Класс” антитела относится к типу константного домена или константной области его тяжелой цепи. Существуют пять основных классов антител: IgA, IgD, IgE, IgG и IgM, и некоторые из них можно дополнительно подразделить на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные домены тяжелой цепи, которые соответствуют разным классам иммуноглобулинов, называются α, δ, ε, γ и μ, соответственно.
Термин “сопоставимая длина” означает, что два полипептида содержат одинаковое число аминокислотных остатков или могут отличаться по длине на один или более и вплоть до, самое большее, 10 аминокислотных остатков. В одном из воплощений полипептиды Fc-области содержат одинаковое число аминокислотных остатков или отличаются числом аминокислотных остатков от 1 до 10. В одном из воплощений полипептиды Fc-области содержат одинаковое число аминокислотных остатков или отличаются числом аминокислотных остатков от 1 до 5. В одном из воплощений полипептиды Fc-области содержат одинаковое число аминокислотных остатков или отличаются числом аминокислотных остатков от 1 до 3.
Термин “происходящий из” означает, что аминокислотную последовательность получают из родительской аминокислотной последовательности путем внесения изменений по меньшей мере в одно положение. Такая измененная аминокислотная последовательность отличается от соответствующей родительской аминокислотной последовательности по меньшей мере в одном соответствующем положении (нумерация для Fc-областей антител согласно системе нумерации EU-индекс по Kabat). В одном из воплощений аминокислотная последовательность, происходящая из родительской аминокислотной последовательности, отличается по одному-пятнадцати аминокислотным остаткам в соответствующих положениях. В одном из воплощений аминокислотная последовательность, происходящая из родительской аминокислотной последовательности, отличается по одному-десяти аминокислотным остаткам в соответствующих положениях. В одном из воплощений аминокислотная последовательность, происходящая из родительской аминокислотной последовательности, отличается по одному-шести аминокислотным остаткам в соответствующих положениях. Аналогичным образом, измененная аминокислотная последовательность имеет высокую идентичность аминокислотной последовательности со своей родительской аминокислотной последовательностью. В одном из воплощений аминокислотная последовательность, происходящая из родительской аминокислотной последовательности, имеет идентичность аминокислотной последовательности, составляющую 80% или более. В одном из воплощений аминокислотная последовательность, происходящая из родительской аминокислотной последовательности, имеет идентичность аминокислотной последовательности, составляющую 90% или более. В одном из воплощений аминокислотная последовательность, происходящая из родительской аминокислотной последовательности, имеет идентичность аминокислотной последовательности, составляющую 95% или более.
“Эффекторные функции” относятся к тем биологическим активностям, присущим Fc-области антитела, которые варьируют в зависимости от класса антитела. Примеры эффекторных функций антител включают: связывание с C1q и комплемент-зависимую цитотоксичность (CDC); связывание с Fc-рецепторами; антитело-зависимую клеточно-опосредованную цитотоксичность (ADCC); фагоцитоз; отрицательную регуляцию рецепторов клеточной поверхности (например, B-клеточного рецептора); и активацию B-клеток.
“Эффективное количество” агента, например, фармацевтической композиции, относится к количеству, эффективному в необходимых дозировках и в течение необходимого периода времени для достижения желаемого терапевтического или профилактического результата.
Термин “слитая конструкция на основе полипептидов Fc-области” означает слитую конструкцию связывающего домена (например, антигенсвязывающего домена, такого как одноцепочечное антитело или полипептид, как например, лиганд рецептора) с Fc-областью антитела, которая демонстрирует желаемую активность в отношении связывания с мишенью, и/или белком A, и/или FcRn.
Термин “Fc-область человеческого происхождения” означает C-концевую область тяжелой цепи иммуноглобулина человеческого происхождения, которая содержит по меньшей мере часть шарнирной области, CH2-домен и CH3-домен. В одном из воплощений Fc-область тяжелой цепи IgG человека простирается от Cys226 или от Pro230 до карбоксильного конца тяжелой цепи. В одном из воплощений Fc-область имеет аминокислотную последовательность с SEQ ID NO: 60. Тем не менее, C-концевой остаток лизина (Lys447) Fc-области может присутствовать или может не присутствовать. Если в данном описании не указано иное, то нумерация аминокислотных остатков в Fc-области или константной области соответствует системе нумерации EU, также называемой EU-индексом, как описано в Kabat E.A., et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991), публикация NIH (Национальный институт здравоохранения) 91-3242. Fc-область состоит двух полипептидов Fc-области тяжелой цепи, которые могут быть ковалентно соединены друг с другом через остатки цистеина шарнирной области, участвующие в образовании дисульфидных связей между полипептидами.
Термин “FcRn” означает неонатальный Fc-рецептор человека. Функция FcRn заключается в спасении IgG от процесса лизосомальной деградации, что приводит к уменьшению клиренса и увеличению полупериода существования. FcRn представляет собой гетеродимерный белок, состоящий из двух полипептидов: белка, подобного белку главного комплекса гистосовместимости класса I с ММ 50 кДа (α-FcRn) и β2-микроглобулина (β2m) с ММ 15 кДа. FcRn связывается с высокой аффинностью с CH2-CH3-частью Fc-области IgG. Взаимодействие между IgG и FcRn строго зависит от pH и осуществляется в стехиометрическом соотношении 1:2, при этом одна молекула IgG связывается с двумя молекулами FcRn посредством двух своих тяжелых цепей (Huber A.H., et al., J. Mol. Biol., 230 (1993) 1077-1083). Связывание с FcRn происходит в эндосоме при кислотных значениях pH (pH меньше 6,5), а IgG высвобождается на клеточной поверхности при нейтральных значениях pH (pH примерно 7,4). Зависимость характера этого взаимодействия от pH способствует FcRn-опосредуемой защите молекул IgG, которые попадают в клетки посредством пиноцитоза, от внутриклеточной деградации, благодаря связыванию с рецептором в кислотных условиях внутри эндосом. Кроме того, FcRn способствует возвращению IgG на поверхность клеток и последующему высвобождению в кровоток после воздействия на комплекс FcRn-IgG среды с нейтральным значением pH вне клетки.
Термин “FcRn-связывающая часть Fc-области” означает часть полипептида тяжелой цепи антитела, которая простирается, начиная приблизительно от положения 243 согласно EU до положения 261 согласно EU, и приблизительно от положения 275 согласно EU до положения 293 согласно EU, и приблизительно от положения 302 согласно EU до положения 319 согласно EU, и приблизительно от положения 336 согласно EU до положения 348 согласно EU, и приблизительно от положения 367 согласно EU до положения 393 согласно EU и положения 408 согласно EU, и приблизительно от положения 424 согласно EU до положения 440 согласно EU. В одном из воплощений изменен один или более чем один из следующих аминокислотных остатков, соответствующих EU нумерации по Kabat: F243, P244, P245 P, K246, P247, K248, D249, T250, L251, M252, I253, S254, R255, T256, P257, E258, V259, T260, C261, F275, N276, W277, Y278, V279, D280, V282, E283, V284, H285, N286, A287, K288, T289, K290, P291, R292, E293, V302, V303, S304, V305, L306, T307, V308, L309, H310, Q311, D312, W313, L314, N315, G316, K317, E318, Y319, I336, S337, K338, A339, K340, G341, Q342, P343, R344, E345, P346, Q347, V348, C367, V369, F372, Y373, P374, S375, D376, I377, A378, V379, E380, W381, E382, S383, N384, G385, Q386, P387, E388, N389, Y391, T393, S408, S424, C425, S426, V427, M428, H429, E430, A431, L432, H433, N434, H435, Y436, T437, Q438, K439 и S440 (нумерация согласно EU).
Термин “каркас” или “каркасные области (FR)” относится к остаткам вариабельного домена, не являющимся остатками гипервариабельных участков (HVR). FR вариабельного домена обычно состоит из четырех доменов FR: FR1, FR2, FR3 и FR4. Соответственно, последовательности HVR и FR обычно располагаются в VH (или VL) в следующем порядке: FR1-H1(L1)-FR2-H2(L2)-FR3-H3(L3)-FR4.
Термин “полноразмерное антитело” означает антитело, имеющее структуру, по существу аналогичную структуре нативного антитела, или имеющее тяжелые цепи, содержащие Fc-область, определенную в данном описании. Полноразмерное антитело может содержать дополнительные домены, такие как, например, одноцепочечный Fv (scFv) или scFab, конъюгированные с одной или более чем одной из цепей полноразмерного антитела. Эти конъюгаты также охватываются термином “полноразмерное антитело”.
Термины “гетеродимер” или “гетеродимерный” означают молекулу, которая содержит две полипептидные цепи (например, сопоставимой длины), причем эти две полипептидные цепи имеют аминокислотную последовательность, в которой по меньшей мере один аминокислотный остаток в соответствующем положении будет другим, при этом данное соответствующее положение определено согласно EU-индексу по Kabat.
Термины “гомодимер” и “гомодимерный” означают молекулу, которая содержит две полипептидные цепи сопоставимой длины, причем эти две полипептидные цепи имеют аминокислотную последовательность, идентичную в соответствующих положениях, при этом данные соответствующие положения определены согласно EU-индексу по Kabat.
Антитело или слитая конструкция на основе полипептидов Fc-области, описанные в данной заявке, могут быть гомодимерными или гетеродимерными применительно к своей Fc-области, что определяется в плане рассматриваемых мутаций или свойств. Например, что касается связывания с FcRn и/или белком A (т.е. при обращении внимания на эти свойства), то Fc-область (антитело) является гомодимерной(ым) (т.е. оба полипептида Fc-области тяжелой цепи содержат эти мутации) в плане наличия мутаций H310A, H433A и Y436A (эти мутации рассматриваются в отношении свойства слитой конструкции на основе полипептидов Fc-области или антитела связываться с FcRn и/или белком A), но в то же самое время является гетеродимерной(ым) в плане наличия мутаций Y349C, T366S, L368A и Y407V (эти мутации не учитываются, поскольку эти мутации касаются гетеродимеризации тяжелых цепей и не затрагивают свойств, относящихся к связыванию с FcRn/белком A), а также в плане наличия мутаций S354C и T366W, соответственно (первый набор содержится только в первом полипептиде Fc-области, в то время как второй набор содержится только во втором полипептиде Fc-области). Кроме того, например, слитая конструкция на основе полипептидов Fc-области или антитело, описанные в данной заявке, могут быть гетеродимерными в плане наличия мутаций I253A, H310A, H433A, H435A и Y436A (т.е. все эти мутации направлены на свойства димерного полипептида связываться с FcRn и/или белком A), т.е. один полипептид Fc-области содержит мутации I253A, H310A и H435A, в то время как другой полипептид Fc-области содержит мутации H310A, H433A и Y436A.
Термины “клетка-хозяин”, “линия клеток хозяина” и “культура клеток хозяина” используются взаимозаменяемо и относятся к клеткам, в которые введена экзогенная нуклеиновая кислота, включая потомство таких клеток. Клетки хозяина включают “трансформанты” и “трансфицированные клетки”, которые включают первичную трансфицированную клетку и ее потомство безотносительно числа пересевов. Потомство может быть не полностью идентичным родительской клетке по составу нуклеиновых кислот, а может содержать мутации. Мутантное потомство, которое обладает той же функцией или биологической активностью, по которой проводят скрининг или отбирают первоначально трансфицированную клетку, включено в данное изобретение.
“Человеческое антитело” представляет собой антитело, имеющее аминокислотную последовательность, которая соответствует последовательности антитела, вырабатываемого человеком или продуцируемого человеческой клеткой, или происходящее из источника, не являющегося человеком, в котором используются репертуары человеческих антител или другие последовательности, кодирующие человеческие антитела. Такое определение человеческого антитела, в частности, исключает гуманизированное антитело, содержащее антигенсвязывающие остатки, не относящиеся к человеческому антителу.
“Консенсусным каркасом IgG человека” является каркас, который представляет собой наиболее часто встречающиеся аминокислотные остатки при выборе каркасных последовательностей VL или VH иммуноглобулина человека. Обычно, выбор последовательностей VL или VH иммуноглобулина человека осуществляют из подгруппы последовательностей вариабельных доменов. Как правило, эта подгруппа последовательностей представляет собой ту же подгруппу, что и в Kabat E.A. et al., Sequences of Proteins of Immunological Interest, 5th ed., Bethesda, MD (1991), NIH Publication 91-3242, vol. 1-3. В одном из воплощений для VL данная подгруппа представляет собой подгруппу каппа I, как и у Kabat и др., выше. В одном из воплощений для VH данная подгруппа представляет собой подгруппу III, как и у Kabat и др., выше.
Термин “полипептид Fc-области IgG человека” относится к аминокислотной последовательности, которая идентична “нативному” полипептиду Fc-области IgG человека или полипептиду Fc-области человеческого IgG “дикого типа”. Термин “полипептид вариантной Fc-области (IgG человека)” относится к аминокислотной последовательности, получающейся из полипептида “нативной” Fc-области человеческого IgG или полипептида Fc-области человеческого IgG “дикого типа” в результате по меньшей мере “изменения одной аминокислоты”. “Fc-область IgG человека” состоит из двух полипептидов Fc-области IgG человека. “Вариантная Fc-область (IgG человека)” состоит из двух полипептидов Fc-области, при этом оба могут быть полипептидами вариантной Fc-области (IgG человека), или один представляет собой полипептид Fc-области IgG человека, а другой представляет собой полипептид вариантной Fc-области.
В одном из воплощений полипептид Fc-области IgG человека имеет аминокислотную последовательность полипептида Fc-области человеческого IgG1 с SEQ ID NO: 03, или полипептида Fc-области человеческого IgG2 с SEQ ID NO: 04, или полипептида Fc-области человеческого IgG3 с SEQ ID NO: 05, или полипептида Fc-области человеческого IgG4 с SEQ ID NO: 06. В одном из воплощений полипептид вариантной Fc-области IgG (человека) происходит из полипептида Fc-области с SEQ ID NO: 03, или 04, или 05, или 06 и имеет по меньшей мере одну аминокислотную мутацию по сравнению с полипептидом Fc-области человеческого IgG, имеющим SEQ ID NO: 03, или 04, или 05, или 06. В одном из воплощений полипептид вариантной Fc-области IgG (человека) содержит/имеет от примерно одной до примерно двенадцати аминокислотных мутаций, а в одном из воплощений от примерно одной до примерно восьми аминокислотных мутаций. В одном из воплощений полипептид вариантной Fc-области IgG (человека) имеет по меньшей мере примерно 80% гомологии с полипептидом Fc-области IgG человека с SEQ ID NO: 03, или 04, или 05, или 06. В одном из воплощений полипептид вариантной Fc-области IgG (человека) имеет по меньшей мере примерно 90% гомологии с полипептидом Fc-области IgG человека с SEQ ID NO: 03, или 04, или 05, или 06. В одном из воплощений полипептид вариантной Fc-области IgG (человека) имеет по меньшей мере примерно 95% гомологии с полипептидом Fc-области IgG человека с SEQ ID NO: 03, или 04, или 05, или 06.
Полипептид вариантной Fc-области IgG (человека), происходящий из полипептида Fc-области человеческого IgG с SEQ ID NO: 03, или 04, или 05, или 06, определяется теми аминокислотными изменениями, которые в нем содержатся. Так, например, термин P329G относится к происходящему из полипептида вариантной Fc-области IgG (человека) полипептиду Fc-области человеческого IgG с мутацией пролин→глицин в аминокислотном положении 329 по сравнению с полипептидом Fc-области человеческого IgG с SEQ ID NO: 03, или 04, или 05, или 06.
Как использовано в данном описании, аминокислотные положения во всех константных областях и доменах тяжелой и легкой цепи нумеруют согласно системе нумерации по Kabat, описанной в Kabat et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991), и в данном описании в этом отношении используют фразу “нумерация согласно Kabat”. Как правило, систему нумерации по Kabat (см. стр. 647-660) в Kabat et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991) используют для константных доменов легкой цепи (CL) изотипа каппа и лямбда, а систему нумерации EU-индекс по Kabat (см. стр. 661-723) используют для константных доменов тяжелой цепи (CH1, шарнирной области, CH2 и CH3).
Полипептид Fc-области IgG1 человека имеет следующую аминокислотную последовательность:
DKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 03).
Происходящий из Fc-области IgG1 человека полипептид Fc-области с мутациями L234A, L235A имеет следующую аминокислотную последовательность:
DKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 07).
Происходящий из Fc-области IgG1 человека полипептид Fc-области с мутациями Y349C, T366S, L368A и Y407V имеет следующую аминокислотную последовательность:
DKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVCTLPPSRDELTKNQVSLSCAVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLVSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 08).
Происходящий из Fc-области IgG1 человека полипептид Fc-области с мутациями S354C, T366W имеет следующую аминокислотную последовательность:
DKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPCRDELTKNQVSLWCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 09).
Происходящий из Fc-области IgG1 человека полипептид Fc-области с мутациями L234A, L235A и мутациями Y349C, T366S, L368A, Y407V имеет следующую аминокислотную последовательность:
DKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVCTLPPSRDELTKNQVSLSCAVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLVSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 10).
Происходящий из Fc-области IgG1 человека полипептид Fc-области с мутациями L234A, L235A и S354C, T366W имеет следующую аминокислотную последовательность:
DKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPCRDELTKNQVSLWCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 11).
Происходящий из Fc-области IgG1 человека полипептид Fc-области с мутацией P329G имеет следующую аминокислотную последовательность:
DKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALGAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 12).
Происходящий из Fc-области IgG1 человека полипептид Fc-области с мутациями L234A, L235A и мутацией P329G имеет следующую аминокислотную последовательность:
DKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALGAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 13).
Происходящий из Fc-области IgG1 человека полипептид Fc-области с мутацией P239G и мутациями Y349C, T366S, L368A, Y407V имеет следующую аминокислотную последовательность:
DKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALGAPIEKTISKAKGQPREPQVCTLPPSRDELTKNQVSLSCAVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLVSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 14).
Происходящий из Fc-области IgG1 человека полипептид Fc-области с мутацией P329G и мутациями S354C, T366W имеет следующую аминокислотную последовательность:
DKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALGAPIEKTISKAKGQPREPQVYTLPPCRDELTKNQVSLWCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 15).
Происходящий из Fc-области IgG1 человека полипептид Fc-области с мутациями L234A, L235A, P329G и Y349C, T366S, L368A, Y407V имеет следующую аминокислотную последовательность:
DKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALGAPIEKTISKAKGQPREPQVCTLPPSRDELTKNQVSLSCAVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLVSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 16).
Происходящий из Fc-области IgG1 человека полипептид Fc-области с мутациями L234A, L235A, P329G и мутациями S354C, T366W имеет следующую аминокислотную последовательность:
DKTHTCPPCPAPEAAGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALGAPIEKTISKAKGQPREPQVYTLPPCRDELTKNQVSLWCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 17).
Полипептид Fc-области IgG4 человека имеет следующую аминокислотную последовательность:
ESKYGPPCPSCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK (SEQ ID NO: 06).
Происходящий из Fc-области IgG4 человека полипептид Fc-области с мутациями S228P и L235E имеет следующую аминокислотную последовательность:
ESKYGPPCPPCPAPEFEGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK (SEQ ID NO: 18).
Происходящий из Fc-области IgG4 человека полипептид Fc-области с мутациями S228P, L235E и мутацией P329G имеет следующую аминокислотную последовательность:
ESKYGPPCPPCPAPEFEGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLGSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK (SEQ ID NO: 19).
Происходящий из Fc-области IgG4 человека полипептид Fc-области с мутациями S354C, T366W имеет следующую аминокислотную последовательность:
ESKYGPPCPSCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPCQEEMTKNQVSLWCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK (SEQ ID NO: 20).
Происходящий из Fc-области IgG4 человека полипептид Fc-области с мутациями Y349C, T366S, L368A, Y407V имеет следующую аминокислотную последовательность:
ESKYGPPCPSCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVCTLPPSQEEMTKNQVSLSCAVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLVSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK (SEQ ID NO: 21).
Происходящий из Fc-области IgG4 человека полипептид Fc-области с мутациями S228P, L235E и S354C, T366W имеет следующую аминокислотную последовательность:
ESKYGPPCPPCPAPEFEGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPCQEEMTKNQVSLWCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK (SEQ ID NO: 22).
Происходящий из Fc-области IgG4 человека полипептид Fc-области с мутациями S228P, L235E и Y349C, T366S, L368A, Y407V имеет следующую аминокислотную последовательность:
ESKYGPPCPPCPAPEFEGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVCTLPPSQEEMTKNQVSLSCAVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLVSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK (SEQ ID NO: 23).
Происходящий из Fc-области IgG4 человека полипептид Fc-области с мутацией P329G имеет следующую аминокислотную последовательность:
ESKYGPPCPSCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLGSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK (SEQ ID NO: 24).
Происходящий из Fc-области IgG4 человека полипептид Fc-области с мутацией P239G и мутациями Y349C, T366S, L368A, Y407V имеет следующую аминокислотную последовательность:
ESKYGPPCPSCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLGSSIEKTISKAKGQPREPQVCTLPPSQEEMTKNQVSLSCAVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLVSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK (SEQ ID NO: 25).
Происходящий из Fc-области IgG4 человека полипептид Fc-области с мутацией P329G и мутациями S354C, T366W имеет следующую аминокислотную последовательность:
ESKYGPPCPSCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLGSSIEKTISKAKGQPREPQVYTLPPCQEEMTKNQVSLWCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK (SEQ ID NO: 26).
Происходящий из Fc-области IgG4 человека полипептид Fc-области с мутациями S228P, L235E, P329G и Y349C, T366S, L368A, Y407V имеет следующую аминокислотную последовательность:
ESKYGPPCPPCPAPEFEGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLGSSIEKTISKAKGQPREPQVCTLPPSQEEMTKNQVSLSCAVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLVSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK (SEQ ID NO: 27).
Происходящий из Fc-области IgG4 человека полипептид Fc-области с мутациями S228P, L235E, P329G и S354C, T366W имеет следующую аминокислотную последовательность:
ESKYGPPCPPCPAPEFEGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLGSSIEKTISKAKGQPREPQVYTLPPCQEEMTKNQVSLWCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK (SEQ ID NO: 28).
“Гуманизированное” антитело относится к химерному антителу, содержащему аминокислотные остатки из HVR антител, не являющихся человеческими, и аминокислотные остатки из FR человеческих антител. В некоторых воплощениях гуманизированное антитело будет содержать по существу все или по меньшей мере один и в типичном случае два вариабельных домена, в которых все или по существу все HVR (например, определяющие комплементарность участки (CDR)) соответствуют HVR антитела, не являющегося человеческим, а все или по существу все FR соответствуют FR человеческого антитела. Гуманизированное антитело возможно может содержать по меньшей мере часть константной области антитела, происходящей из человеческого антитела. “Гуманизированный вариант” антитела, например, антитела, не являющегося человеческим, относится к антителу, которое подвергнуто гуманизации.
Термин “гипервариабельный участок” или “HVR”, использованный в данном описании, относится к каждому из участков вариабельного домена антитела, которые являются гипервариабельными по последовательности (“определяющими комплементарность участками” или “CDR”), и образуют петли определенной структуры (“гипервариабельные петли”), и/или содержат контактирующие с антигеном остатки (“места контакта с антигеном”). Как правило, антитела содержат шесть HVR: три в VH (H1, H2, H3) и три в VL (L1, L2, L3). HVR, указанные в данном описании, включают:
(a) гипервариабельные петли, образуемые аминокислотными остатками 26-32 (L1), 50-52 (L2), 91-96 (L3), 26-32 (H1), 53-55 (H2) и 96-101 (H3) (Chothia C. and Lesk A.M., J. Mol. Biol., 196 (1987) 901-917;
(b) CDR образуемые аминокислотными остатками 24-34 (L1), 50-56 (L2), 89-97 (L3), 31-35b (H1), 50-65 (H2) и 95-102 (H3) (Kabat E.A. et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991), NIH Publication 91-3242);
(c) места контакта с антигеном, образуемые аминокислотными остатками 27c-36 (L1), 46-55 (L2), 89-96 (L3), 30-35b (H1), 47-58 (H2) и 93-101 (H3) (MacCallum et al., J. Mol. Biol., 262: 732-745 (1996)); и
(d) комбинации (a), (b) и/или (c), включающие аминокислотные остатки HVR 46-56 (L2), 47-56 (L2), 48-56 (L2), 49-56 (L2), 26-35 (H1), 26-35b (H1), 49-65 (H2), 93-102 (H3) и 94-102 (H3).
Если не указано иное, остатки HVR и другие остатки в вариабельном домене (например, остатки FR) нумеруются в данном описании согласно системе нумерации EU-индекс по Kabat (Kabat и др., выше).
“Индивидуумом” или “субъектом” является млекопитающее. Млекопитающие включают, но не ограничиваются этим, одомашенных животных (например, крупный рогатый скот, овец, кошек, собак и лошадей), приматов (например, людей и приматов, не являющихся людьми, таких как обезьяны), кроликов и грызунов (например, мышей и крыс). В некоторых воплощениях индивидуумом или субъектом является человек.
“Выделенное” антитело представляет собой антитело, которое отделено от компонентов своего природного окружения. В некоторых воплощениях антитело очищено до чистоты более 95% или 99%, как определено, например, электрофоретически (например, по данным электрофореза в полиакриамидном геле в присутствии додецилсульфата натрия (SDS-PAGE), изоэлектрофокусирования (IEF), капиллярного электрофореза) или хроматографически (например, посредством гель-проникающей хроматографии или ионообменной либо обращенно-фазовой высокоэффективной жидкостной хроматографии (HPLC)). В качестве обзора методов оценки чистоты антител см., например, Flatman S. et al., J. Chromatogr. B, 848 (2007) 79-87.
“Выделенная” нуклеиновая кислота относится к молекуле нуклеиновой кислоты, которая отделена от компонентов своего природного окружения. Выделенная нуклеиновая кислота включает молекулу нуклеиновой кислоты, содержащуюся в клетках, которые обычно содержат молекулу нуклеиновой кислоты, но молекула данной нуклеиновой кислоты присутствует вне хромосомы или в таком месте хромосомы, которое отличается от ее природного расположения в хромосоме.
Термин “моноклональное антитело”, использованный в данном описании, относится к антителу, полученному из популяции по существу гомогенных антител, т.е. индивидуальные антитела, составляющие популяцию, являются идентичными и/или связываются с одним и тем же эпитопом, за исключением возможных вариантных антител, например, содержащих встречающиеся в природе мутации или появляющихся в процессе приготовления препарата на основе моноклональных антител, при этом такие варианты обычно присутствуют в незначительных количествах. В противоположность препаратам на основе поликлональных антител, которые обычно включают различные антитела, направленные против разных детерминант (эпитопов), каждое моноклональное антитело в препарате на основе моноклональных антител направлено против одной детерминанты на антигене. Таким образом, определение “моноклональное” указывает на характер антитела, полученного по существу из гомогенной популяции антител, и его не следует истолковывать в смысле требования получения антитела каким-либо конкретным методом. Например, моноклональные антитела, предназначенные для применения в соответствии с настоящим изобретением, могут быть получены различными методами, включая, но не ограничиваясь этим, гибридомный метод, методы рекомбинантных ДНК, методы фагового дисплея и методы с использованием трансгенных животных, содержащих весь локус или часть локуса иммуноглобулина человека, при этом такие методы и другие типичные методы получения моноклональных антител изложены в данном описании.
“Нативные антитела” относятся к молекулам природных иммуноглобулинов различной структуры. Например, нативные антитела IgG представляют собой гетеротетрамерные гликопротеины с ММ примерно 150000 дальтон, состоящие из двух идентичных легких цепей и двух идентичных тяжелых цепей, соединенных дисульфидными связями. В направлении от N-конца к C-концу каждая тяжелая цепь имеет вариабельную область (VH), также называемую вариабельным тяжелым доменом или вариабельным доменом тяжелой цепи, за которым следуют три константных домена (CH1, CH2 и CH3). Аналогично, в направлении от N-конца к C-концу каждая легкая цепь имеет вариабельную область (VL), также называемую вариабельным легким доменом или вариабельным доменом легкой цепи, за которым следует константный домен легкой цепи (CL). Легкая цепь антитела может быть отнесена к одному из двух типов, называемых каппа (κ) и лямбда (λ), на основании аминокислотной последовательности ее константного домена.
Термин “инструкция по применению” используется для обозначения инструкций, обычно вкладываемых в коммерческие упаковки терапевтических продуктов, которые содержат информацию о показаниях, применении, дозировке, введении, комбинированной терапии, противопоказаниях и/или предупреждениях, касающихся применения таких терапевтических продуктов.
“Процент (%) идентичности аминокислотной последовательности” применительно к референсной полипептидной последовательности определяют как процентную долю аминокислотных остатков в последовательности-кандидате, которые идентичны аминокислотным остаткам в референсной полипептидной последовательности после выравнивания последовательностей и внесения, в случае необходимости, разрывов, для достижения максимального процента идентичности последовательности, и без учета любых консервативных замен как части идентичности последовательностей. Выравнивания, необходимого для определения процента идентичности аминокислотных последовательностей, можно достичь различными способами, известными специалисту в данной области техники, например, используя общедоступное компьютерное программное обеспечение, такое как программное обеспечение BLAST (средство поиска основного локального выравнивания), BLAST-2, ALIGN или Megalign (DNASTAR). Специалисты в данной области техники могут определить соответствующие параметры для выравнивания последовательностей, включая любые алгоритмы, необходимые для достижения максимального выравнивания по всей длине сравниваемых последовательностей. Однако же, для целей данного изобретения значения % идентичности аминокислотных последовательностей получают, используя компьютерную программу для сравнения последовательностей ALIGN-2. Компьютерная программа для сравнения последовательностей ALIGN-2 разработана Genentech, Inc., и исходная программа с документацией для пользователя подана в бюро по охране авторских прав США (U.S. Copyright Office), Вашингтон, округ Колумбия (D.C.), 20559, где она зарегистрирована под № TXU510087 в бюро регистрации авторского права США. Программа ALIGN-2 общедоступна через Genentech, Inc., Южный Сан-Франциско, Калифорния, или ее можно компилировать с исходной программы. Программу ALIGN-2 следует компилировать для использования в операционной системе UNIX, включая Digital UNIX V4.0D. Все параметры сравнения последовательностей устанавливаются программой ALIGN-2 и не изменяются.
В тех ситуациях, когда для сравнений аминокислотных последовательностей используют ALIGN-2, % идентичности аминокислотных последовательностей для заданной аминокислотной последовательности A относительно, по сравнению с или против заданной аминокислотной последовательности B (что можно альтернативно перефразировать как “заданная аминокислотная последовательность A, которая имеет или содержит определенный % аминокислотной последовательности, идентичной относительно, по сравнению с или против заданной аминокислотной последовательности B”) вычисляют так, как приведено ниже:
100 умножить на отношение X/Y,
где X представляет собой количество аминокислотных остатков, отмечаемых как идентичные совпадения программой для выравнивания последовательностей ALIGN-2 при выравнивании A и B этой программой, и где Y представляет собой общее количество аминокислотных остатков в B. Будет очевидно, что там, где длина аминокислотной последовательности A не равна длине аминокислотной последовательности B, % идентичности аминокислотной последовательности A относительно B не будет равен % идентичности аминокислотной последовательности B относительно A. Если конкретно не указано иное, то все используемые в данном описании значения % идентичности аминокислотных последовательностей получают так, как описано в непосредственно предшествующем абзаце, с использованием компьютерной программы ALIGN-2.
Термин “фармацевтическая композиция” относится к препарату, форма которого способствует эффективному проявлению биологической активности активного ингредиента, содержащегося в нем, и который не содержит никаких дополнительных компонентов, являющихся неприемлемо токсичными для субъекта, которому будет введена данная композиция.
“Фармацевтически приемлемый носитель” относится к ингредиенту в фармацевтической композиции, отличающемуся от активного ингредиента и являющемуся нетоксичным для субъекта. Фармацевтически приемлемый носитель включает, но не ограничивается этим, буфер, эксципиент, стабилизатор или консервант.
Термин “пептидный линкер”, использованный в данном описании, означает пептид с разными аминокислотными последовательностями, который в одном из воплощений имеет синтетическое происхождение. В одном из воплощений пептидный линкер представляет собой пептид, имеющий аминокислотную последовательность длиной по меньшей мере 30 аминокислот, в одном из воплощений длиной 32-50 аминокислот. В одном из воплощений пептидный линкер представляет собой пептид, имеющий аминокислотную последовательность длиной 32-40 аминокислот. В одном из воплощений пептидный линкер представляет собой (GxS)n, где G означает глицин, S означает серин (× равно 3, n равно 8, 9 или 10, либо × равно 4, и n равно 6, 7 или 8), в одном из воплощений × равно 4, n равно 6 или 7, в одном из воплощений × равно 4, n равно 7. В одном из воплощений пептидный линкер представляет собой (G4S)6G2.
Термин “рекомбинантное антитело” означает все антитела (химерное, гуманизированное и человеческое), которые получают, экспрессируют, создают или выделяют с использованием рекомбинантных методов. Термин включает в себя антитела, выделенные из клетки хозяина, такой как клетка NS0 или клетка яичников китайского хомячка (CHO), или из животного (например, мыши), генетически модифицированного генами иммуноглобулина человека, или антитела, экспрессируемые с использованием рекомбинантного экспрессирующего вектора, которым трансфицирована клетка хозяина. Такие рекомбинантные антитела имеют вариабельную и константную области в реаранжированной форме). Рекомбинантные антитела, описанные в данной заявке, могут быть подвергнуты in vivo соматической гипермутации. Таким образом, аминокислотные последовательности VH- и VL-областей рекомбинантных антител представляют собой последовательности, которые, будучи происходящими из последовательностей и родственными последовательностям VH и VL антител зародышевой линии человека, не могут естественным образом существовать в репертуаре антител зародышевой линии человека in vivo.
Использованный в данном описании термин “лечение” (и такие его грамматические варианты, как “лечить” или “подвергание лечению”) относится к клиническому воздействию в попытке изменить естественное течение болезни у подвергаемого лечению индивидуума, и оно может быть проведено либо с целью профилактики, либо в процессе курса лечения клинической патологии. Желательные эффекты лечения включают, но не ограничиваются этим, предупреждение возникновения или рецидива заболевания, облегчение симптомов, сведение к минимуму любых прямых или косвенных патологических последствий заболевания, предотвращение метастазирования, уменьшение скорости прогрессирования заболевания, уменьшение интенсивности симптомов или временное облегчение болезненного состояния и ремиссию или улучшенный прогноз. В некоторых воплощениях, антитела или слитые конструкции на основе полипептидов Fc-области, описанные в данной заявке, используют для задержки развития заболевания или для замедления прогрессирования заболевания.
Термин “валентный”, который используется в рамках настоящей заявки, относится к наличию конкретного количества сайтов связывания в молекуле (антитела). По существу, термины “бивалентный”, “тетравалентный” и “гексавалентный” относятся к наличию двух сайтов связывания, четырех сайтов связывания и шести сайтов связывания, соответственно, в молекуле (антитела). Биспецифичные антитела, описанные в данной заявке, в одном из предпочтительных воплощений являются “бивалентными”.
Термин “вариабельная область” или “вариабельный домен” относится к домену тяжелой или легкой цепи антитела, который вовлечен в связывание антитела с антигеном. Вариабельные домены тяжелой цепи и легкой цепи (VH и VL, соответственно) антитела обычно имеют аналогичные структуры, при этом каждый домен содержит четыре каркасные области (FR) и три гипервариабельных участка (HVR) (см., например, Kindt T.J. et al., Kuby Immunology, 6th ed., W.H. Freeman and Co., N.Y. (2007), page 91). Одного VH- или VL-домена может быть достаточно для придания антигенсвязывающей специфичности. Кроме того, для скрининга библиотеки комплементарных VL- или VH-доменов можно выделить антитела, связывающиеся с конкретным антигеном, используя VH- или VL-домен из антитела, которое связывается с данным антигеном, соответственно. См., например, Portolano S. et al., J. Immunol., 150 (1993) 880-887; Clarkson T. et al., Nature, 352 (1991) 624-628.
Термин “сосудистое заболевание глаз” включает, но не ограничивается этим, интраокулярные неоваскулярные синдромы, такие как диабетическая ретинопатия, диабетический макулярный отек, ретинопатия недоношенных, неоваскулярная глаукома, окклюзии вен сетчатки, окклюзии центральной вены сетчатки, макулярная дегенерация, возрастная макулярная дегенерация, пигментный ретинит, ретинальная ангиоматозная пролиферация, макулярная телеангиэктазия, ишемическая ретинопатия, неоваскуляризация радужной оболочки глаза, интраокулярная неоваскуляризация, роговичная неоваскуляризация, неоваскуляризация сетчатки, хориоидальная неоваскуляризация и дегенеративные изменения сетчатки (см., например, Garner A., “Vascular diseases” в: Pathobiology of ocular disease, A dynamic approach, Garner A. and Klintworth G.K., (eds.), 2nd edition, Marcel Dekker, New York (1994), pp. 1625-1710).
Термин “вектор”, использованный в данном описании, относится к молекуле нуклеиновой кислоты, обладающей способностью репродуцировать другую нуклеиновую кислоту, с которой она соединена. Термин включает в себя вектор как самореплицирующуюся нуклеиновокислотную структуру, а также вектор, встроенный в геном клетки хозяина, в которую он введен. Некоторые векторы способны направлять экспрессию нуклеиновых кислот, с которыми они функционально связаны. Такие векторы называются в данном описании “экспрессирующими векторами”.
Термин “с мутацией IHH-AAA”, использованный в данном описании, относится к комбинации мутаций I253A (Ile253Ala), H310A (His310Ala) и H435A (His435Ala), термин “с мутацией HHY-AAA”, использованный в данном описании, относится к комбинации мутаций H310A (His310Ala), H433A (His433Ala) и Y436A (Tyr436Ala), а термин “с мутацией YTE”, использованный в данном описании, относится к комбинации мутаций M252Y (Met252Tyr), S254T (Ser254Thr) и T256E (Thr256Glu) в константной области тяжелой цепи подкласса IgG1 или IgG4, при этом нумерация соответствует EU-индексу по Kabat.
Термин “с мутациями P329G LALA”, использованный в данном описании, относится к комбинации мутаций L234A (Leu234Ala), L235A (Leu235Ala) и P329G (Pro329Gly) в константной области тяжелой цепи подкласса IgG1, при этом нумерация соответствует EU-индексу по Kabat. Термин “с мутацией SPLE”, использованный в данном описании, относится к комбинации мутаций S228P (Ser228Pro) и L235E (Leu235Glu) в константной области тяжелой цепи подкласса IgG4, при этом нумерация соответствует EU-индексу по Kabat. Термин “с мутацией SPLE и P239G”, использованный в данном описании, относится к комбинации мутаций S228P (Ser228Pro), L235E (Leu235Glu) и P329G (Pro329Gly) в константной области тяжелой цепи подкласса IgG4, при этом нумерация соответствует EU-индексу по Kabat.
II. НАСТОЯЩЕЕ ИЗОБРЕТЕНИЕ
В основе данного изобретения лежит, по меньшей мере и помимо всего прочего, обнаружение того, что связывание антитела или слитой конструкции на основе полипептидов Fc-области с FcRn можно модифицировать посредством изменения аминокислотных остатков в не соответствующих этому связыванию положениях в индивидуальных полипептидах Fc-области, поскольку совместное действие этих изменений модифицирует связывание с FcRn. Fc-области, антитела и слитые конструкции на основе полипептидов Fc-области, описанные в данной заявке, полезны, например, для лечения заболеваний, при которых необходимы специально выбранные для системного введения времена удерживания.
Согласно данному изобретению предложены вариантные Fc-области, которые имеют модифицированные свойства в отношении связывания с FcRn по сравнению с соответствующей Fc-областью дикого типа. Эти вариантные Fc-области содержат специфические аминокислотные мутации в CH2- и/или CH3-домене. Обнаружено, что эти мутации, в случае их использования либо по отдельности, либо в комбинации, в одной и той же или в обеих тяжелых цепях в Fc-области, позволяют получить вариантную Fc-область со специально разработанным полупериодом существования i vivo.
A. Неонатальный Fc-рецептор (FcRn)
Неонатальный Fc-рецептор (FcRn) важен для метаболического пути антител класса IgG in vivo. Функция FcRn заключается в спасении IgG дикого типа от процесса лизосомальной деградации, что приводит к уменьшению клиренса и увеличению полупериода существования. Он представляет собой гетеродимерный белок, состоящий из двух полипептидов: белка, подобного белку главного комплекса гистосовместимости класса I с ММ 50 кДа (α-FcRn) и β2-микроглобулина (β2m) с ММ 15 кДа. FcRn связывается с высокой аффинностью с CH2-CH3-частью Fc-области антитела класса IgG. Взаимодействие между антителом класса IgG и FcRn зависит от pH и осуществляется в стехиометрическом соотношении 1:2, т.е. одна молекула IgG антитела может взаимодействовать с двумя молекулами FcRn посредством своих двух полипептидов Fc-области тяжелой цепи (см., например, Huber A.H., et al., J. Mol. Biol., 230 (1993) 1077-1083).
Таким образом, свойства/характеристики связывания молекул IgG с FcRn in vitro являются показателями его фармакокинетических свойств in vivo в кровотоке.
Во взаимодействии между FcRn и Fc-областью антитела класса IgG принимают участие разные аминокислотные остатки CH2- и CH3-домена тяжелой цепи. Аминокислотные остатки, участвующие во взаимодействии с FcRn, локализованы, начиная приблизительно от положения 243 согласно EU до положения 261 согласно EU, приблизительно от положения 275 согласно EU до положения 293 согласно EU, приблизительно от положения 302 согласно EU до положения 319 согласно EU, приблизительно от положения 336 согласно EU до положения 348 согласно EU, в положении 408 согласно EU, и приблизительно от положения 424 согласно EU до положения 440 согласно EU. Более конкретно, во взаимодействие между Fc-областью и FcRn вовлечены следующие аминокислотные остатки, соответствующие EU нумерации по Kabat: F243, P244, P245 P, K246, P247, K248, D249, T250, L251, M252, I253, S254, R255, T256, P257, E258, V259, T260, C261, F275, N276, W277, Y278, V279, D280, V282, E283, V284, H285, N286, A287, K288, T289, K290, P291, R292, E293, V302, V303, S304, V305, L306, T307, V308, L309, H310, Q311, D312, W313, L314, N315, G316, K317, E318, Y319, I336, S337, K338, A339, K340, G341, Q342, P343, R344, E345, P346, Q347, V348, C367, V369, F372, Y373, P374, S375, D376, I377, A378, V379, E380, W381, E382, S383, N384, G385, Q386, P387, E388, N389, Y391, T393, S408, S424, C425, S426, V427, M428, H429, E430, A431, L432, H433, N434, H435, Y436, T437, Q438, K439 и S440.
Исследованиями с использованием сайт-специфического мутагенеза доказано, что в Fc-области IgG критичными сайтами для связывания с FcRn, являются гистидин 310, гистидин 435 и изолейцин 253 и в меньшей степени гистидин 433 и тирозин 436 (см., например, Kim J.K., et al., Eur. J. Immunol., 29 (1999) 2819-2825; Raghavan M., et al., Biochem., 34 (1995) 14649-14657; Medesan C., et al., J. Immunol., 158 (1997) 2211-2217).
Способы усиления связывания IgG с FcRn были осуществлены путем внесения мутаций в различные аминокислотные остатки в IgG: треонин 250, метионин 252, серин 254, треонин 256, треонин 307, глутаминовую кислоту 380, метионин 428, гистидин 433 и аспарагин 434 (см. Kuo T.T., et al., J. Clin. Immunol., 30 (2010) 777-789).
В некоторых случаях желательны антитела с более низким полупериодом существования в кровотоке. Например, лекарственные средства для интравитреального введения должны иметь продолжительный полупериод существования в глазу и короткий полупериод существования в кровотоке пациента. Кроме того, преимущество таких антител заключается в их усиленном воздействии в месте заболевания, например в глазу.
Известны различные мутации, оказывающие влияние на связывание с FcRn и тем самым на полупериод существования в кровотоке. С использованием сайт-специфического мутагенеза идентифицированы аминокислотные остатки Fc-области, критичные для взаимодействия Fc-области IgG мыши и мышиного FcRn (см., например, Dall'Acqua W.F., et al., J. Immunol., 169 (2002) 5171-5180). В это взаимодействие вовлечены остатки I253, H310, H433, N434 и H435 (согласно EU нумерации по Kabat) (Medesan C., et al., Eur. J. Immunol., 26 (1996) 2533-2536; Firan M., et al., Int. Immunol., 13 (2001) 993-1002; Kim J.K., et al., Eur. J. Immunol., 24 (1994) 542-548). Обнаружено, что присутствие остатков I253, H310 и H435 критично для взаимодействия Fc-области IgG человека с мышиным FcRn (Kim J.K., et al., Eur. J. Immunol., 29 (1999) 2819-2825). В результате исследований белок-белковых взаимодействий с целью улучшения связывания с FcRn Dall'Acqua и др. были описаны мутации по остаткам M252Y, S254T, T256E (Dall'Acqua W.F., et al. J. Biol. Chem., 281 (2006) 23514-23524). Исследования комплекса Fc-область IgG человека-FcRn человека показали, что остатки I253, S254, H435 и Y436 являются важнейшими для данного взаимодействия остатками (Firan M., et al., Int. Immunol., 13 (2001) 993-1002; Shields R.L., et al., J. Biol. Chem., 276 (2001) 6591-6604). В работе Yeung Y.A. и др. (J. Immunol., 182 (2009) 7667-7671) описаны различные мутанты по остаткам 248-259, и 301-317, и 376-382, и 424-437 и проведено их изучение. Типичные мутации и их влияние на связывание с FcRn приведены в следующей таблице.
Таблица
H310Q/H433N
(мышиный IgG1)
(с FcRn мыши)
(у мыши)
H310A
H435A
H436A
(мышиный IgG1)
(с FcRn мыши)
(у мыши)
T252A/T254S/T256A
(мышиный IgG1)
(с FcRn мыши)
(у мыши)
H310A
H435A
H436A
H433A/N434Q
(мышиный IgG1)
(с FcRn мыши)
(у мыши)
H310A
H435A
H435R
(человеческий IgG1)
H310A: <0,1 отн. связывания с muFcRn
(FcRn мыши)
(у мыши)
(человеческий IgG1)
(мыши)
S254A
H435A
Y436A
(человеческий IgG1)
<0,1 относительно связывания с FcRn человека
K288A
L309A
S415A
H433A
(человеческий IgG1)
(с FcRn человека)
T256A
E272A
V305A
T307A
Q311A
D312A
K317A
D376A
A378Q
E380A
E382A
S424A
N434A
K288A/N434A
E380A/N434A
T307A/E380A/N434A
(человеческий IgG1)
(с FcRn человека)
(гуманизированный IgG1)
<0,1 относительно связывания с FcRn человека
M252W
M252Y
M252Y/T256Q
M252F/T256D
N434F/Y436H
M252Y/S254T/T256E
G385A/Q386P/N389S
H433K/N434F/Y436H
H433R/N434Y/Y436H
G385R/Q386T/P387R/N389P
M252Y/S254T/T256E/H433K/N434F/Y436H
M252Y/S254T/T256E/G385R /Q386T/P387R/N389P
(человеческий IgG1)
(с FcRn мыши и человека)
(у мыши)
T250Q/M428L
(человеческий IgG2)
(с FcRn человека)
(у макака)
(человеческий IgG)
(с FcRn человека)
(у мыши)
(химерный IgG1)
E380A
M428L
N434A
K288A/N434A
E380A/N434A
T307A/E380A/N434A
(человеческий IgG1)
(с FcRn человека)
(человеческий IgG1)
(с FcRn человека)
M252Y/S254T/T256E
(гуманизированный)
(с FcRn человека и яванского макака)
M428L
T250Q/M428L
(человеческий IgG1)
(с FcRn человека)
P257I/Q311I
(гуманизированный IgG1)
(с FcRn мыши и яванского макака)
увеличивали у мыши
P257I/N434H
D376V/N434H
(гуманизированный IgG1)
при pH 6
(с FcRn человека, яванского макака, мыши)
P257I/N434H: уменьшали у яванского макака
I253,
H310,
H433,
H435;
ослабляли связывание с FcRn:
Y436;
усиливали связывание с FcRn:
T250,
N252,
S254,
T256,
T307,
M428,
N434
T307Q/N434A
T307Q/N434S
V308P/N434A
T307Q/E380A/N434A
(человеческий IgG1)
(с FcRn яванского макака)
280K
339T
385H
428L
434W/Y/F/A/H
(человеческий IgG)
Обнаружено, что для существенного ослабления связывания с Fc-рецептором достаточно одной односторонней мутации в одном полипептиде Fc-области. Чем больше мутаций внесено в Fc-область, тем слабее становится связывание с FcRn. Однако, односторонних асимметричных мутаций не достаточно для полного ингибирования связывания с FcRn. Для полного ингибирования связывания с FcRn необходимы мутации, располагающиеся на обеих сторонах.
Таким образом, вариантная Fc-область класса IgG (человека) представляет собой гетеродимер, и в результате образования пары с участием первого полипептида Fc-области (тяжелой цепи) и второго полипептида Fc-области (тяжелой цепи) с получением функциональной Fc-области образуется гетеродимер.
Результаты симметричного конструирования Fc-области IgG1 с целью оказания влияния на связывание с FcRn показаны в следующей далее таблице (выравнивание мутаций и время удерживания на колонке для хроматографии на основе аффинности к FcRn).
Таблица
Времена удерживания меньше 3 минут соответствуют отсутствию связывания, поскольку вещество находится в проходящем потоке (“проскоке”) (в пике, выходящем со “свободным” объемом).
Одиночная мутация H310A является “самой сильной” молчащей симметричной мутацией с точки зрения ликвидации любого связывания с FcRn.
Симметричные одиночные мутации I253A и H435A приводят к относительному смещению времени удерживания на 0,3-0,4 мин. В общем случае, это можно рассматривать как недетектируемое связывание.
Внесение одиночной мутации Y436A приводит к детектированию сильного взаимодействия с колонкой при хроматографии на основе аффинности к FcRn. Не вдаваясь в теоретические рассуждения, отметим, что эта мутация может оказать влияние на FcRn-опосредуемый полупериод существования, в отличие от случая отсутствия взаимодействия, как например, при внесении комбинации мутаций I253A, H310A и H435A (мутация IHH-AAA).
Результаты, полученные с использованием симметрично модифицированного антитела к HER2, приведены в следующей далее таблице (в качестве ссылки см. WO 2006/031370).
Таблица
[мин]
Fc-область в слитой конструкции на основе полипептидов Fc-области придает своему партнеру слияния описанные выше характеристики. Партнером слияния могут быть любые молекулы, обладающие биологической активностью, полупериод существования которых in vivo должен быть уменьшен или увеличен, т.е. полупериод существования которых in vivo должен быть четко определен и специально выбран для его предполагаемого применения.
Слитые конструкции на основе полипептидов Fc-области могут содержать, например, вариантную Fc-область класса IgG (человека), описанную в данной заявке, и рецепторный белок, который связывается с мишенью, включая лиганд, такие как, например, слитая с TNFR конструкция на основе полипептидов Fc-области (TNFR означает рецептор фактора некроза опухоли человека), или слитая с IL-1R конструкция на основе полипептидов Fc-области (IL-1R означает рецептор интерлейкина-1 человека), или слитая с VEGFR конструкция на основе полипептидов Fc-области (VEGFR означает рецептор сосудистого эндотелиального ростового фактора человека), или слитые с ANG2R конструкции на основе полипептидов Fc-области (ANG2R означает рецептор 2 ангиопоэтина человека).
Слитые конструкции на основе полипептидов Fc-области могут содержать, например, вариантную Fc-область класса IgG (человека), описанную в данной заявке, и фрагмент антитела, который связывается с мишенью, в том числе, такой как, например, Fab фрагмент антитела, scFγ фрагменты (см., например, Nat. Biotechnol., 23 (2005) 1126-1136) или однодоменные антитела (dAb) (см., например, WO 2004/058821, WO 2003/002609).
Слитые конструкции на основе полипептидов Fc-области могут содержать, например, вариантную Fc-область класса IgG (человека), описанную в данной заявке, и лиганд рецептора (или природный, или искусственный).
B. Иллюстративные Fc-области и антитела, содержащие эти Fc-области
Согласно одному из аспектов изобретения предложены Fc-области, обладающие модифицированными свойствами в отношении связывания с FcRn, т.е. эти Fc-области связываются с FcRn человека с более высокой или более низкой аффинностью, чем у Fc-области, не имеющей никаких мутаций, влияющих на связывание с FcRn.
Согласно одному из аспектов изобретения предложены выделенные антитела, обладающие модифицированными свойствами в отношении связывания с FcRn, т.е. эти антитела связываются с FcRn человека с более высокой или более низкой аффинностью, чем у антитела, не имеющего никаких мутаций, влияющих на связывание с FcRn.
Согласно одному из аспектов, описанных в данной заявке, предложено антитело, содержащее (вариантную) Fc-область, содержащую первый полипептид Fc-области и второй полипептид Fc-области,
при этом
a) первый полипептид Fc-области и второй полипептид Fc-области происходят из полипептида одной и той же Fc-области IgG человека, и
b) первый полипептид Fc-области модифицирован таким образом, что его аминокислотная последовательность отличается от аминокислотной последовательности второго полипептида Fc-области по меньшей мере в одном соответствующем положении согласно системе нумерации EU-индекс по Kabat, и второй полипептид Fc-области модифицирован таким образом, что его аминокислотная последовательность отличается от аминокислотной последовательности первого полипептида Fc-области по меньшей мере в одном соответствующем положении согласно системе нумерации EU-индекс по Kabat, в результате чего модифицированное положение в первом полипептиде Fc-области и модифицированное положение во втором полипептиде Fc-области отличаются, и
c) данная Fc-область имеет другую аффинность к Fc-рецептору человека по сравнению с Fc-областью, которая содержит в качестве первого и второго полипептида Fc-области полипептид Fc-области IgG человека по пункту (a) (т.е. которая имеет те же аминокислотные остатки, что и полипептид Fc-области IgG человека по пункту (a) в соответствующих положениях согласно системе нумерации EU-индекс по Kabat).
В одном из воплощений всех аспектов, описанных в данной заявке, либо первый полипептид Fc-области, либо второй полипептид Fc-области, либо оба полипептида Fc-области содержат одну из приведенных ниже мутаций или комбинаций мутаций:
- T307H, или
- Q311H, или
- E430H, или
- N434H, или
- T307H и Q311H, или
- T307H и E430H, или
- T307H и N434A, или
- T307H и N434H, или
- T307Q и Q311H, или
- T307Q и E430H, или
- T307Q и N434H, или
- T307H, и Q311H, и E430H, и N434A, или
- T307H, и Q311H, и E430H, и N434H, или
- T307H, и Q311H, и E430H, и N434Y, или
- T307Q, и Q311H, и E430H, и N434A, или
- T307Q, и Q311H, и E430H, и N434H, или
- T307Q, и Q311H, и E430H, и N434Y, или
- T307Q, и V308P, и N434Y, и Y436H, или
- T307H, и M252Y, и S254T, и T256E, или
- T307Q, и M252Y, и S254T, и T256E, или
- Q311H, и M252Y, и S254T, и T256E, или
- E430H, и M252Y, и S254T, и T256E, или
- N434H, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и M252Y, и S254T, и T256E, или
- T307H, и E430H, и M252Y, и S254T, и T256E, или
- T307H, и N434A, и M252Y, и S254T, и T256E, или
- T307H, и N434H, и M252Y, и S254T, и T256E, или
- T307Q, и Q311H, и M252Y, и S254T, и T256E, или
- T307Q, и E430H, и M252Y, и S254T, и T256E, или
- T307Q, и N434H, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и E430H, и N434A, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и E430H, и N434H, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и E430H, и N434Y, и M252Y, и S254T, и T256E, или
- T307Q, и Q311H, и E430H, и N434A, и M252Y, и S254T, и T256E, или
- T307Q, и Q311H, и E430H, и N434H, и M252Y, и S254T, и T256E, или
- T307Q, и Q311H, и E430H, и N434Y, и M252Y, и S254T, и T256E, или
- T307Q, и V308P, и N434Y, и Y436H, и M252Y, и S254T, и T256E.
В одном из воплощений всех аспектов, описанных в данной заявке,
первый полипептид Fc-области содержит, независимо от второго полипептида Fc-области, одну из приведенных ниже мутаций или комбинаций мутаций:
- T307H, или
- Q311H, или
- E430H, или
- N434H, или
- T307H и Q311H, или
- T307H и E430H, или
- T307H и N434A, или
- T307H и N434H, или
- T307Q и Q311H, или
- T307Q и E430H, или
- T307Q и N434H, или
- M252Y, и S254T, и T256E, или
- I253A, и H310A, и H435A, или
- H310A, и H433A, и Y436A или
- T307H, и Q311H, и E430H, и N434A, или
- T307H, и Q311H, и E430H, и N434H, или
- T307H, и Q311H, и E430H, и N434Y, или
- T307Q, и Q311H, и E430H, и N434A, или
- T307Q, и Q311H, и E430H, и N434H, или
- T307Q, и Q311H, и E430H, и N434Y, или
- T307Q, и V308P, и N434Y, и Y436H, или
- T307H, и M252Y, и S254T, и T256E, или
- Q311H, и M252Y, и S254T, и T256E, или
- E430H, и M252Y, и S254T, и T256E, или
- N434H, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и M252Y, и S254T, и T256E, или
- T307H, и E430H, и M252Y, и S254T, и T256E, или
- T307H, и N434A, и M252Y, и S254T, и T256E, или
- T307H, и N434H, и M252Y, и S254T, и T256E, или
- T307Q, и Q311H, и M252Y, и S254T, и T256E, или
- T307Q, и E430H, и M252Y, и S254T, и T256E, или
- T307Q, и N434H, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и E430H, и N434A, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и E430H, и N434H, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и E430H, и N434Y, и M252Y, и S254T, и T256E, или
- T307Q, и V308P, и N434Y, и Y436H, и M252Y, и S254T, и T256E, или
- T307Q, и V308P, и N434Y, и Y436H, и M252Y, и S254T, и T256E,
и
второй полипептид Fc-области содержит, независимо от первого полипептида Fc-области, одну из приведенных ниже мутаций или комбинаций мутаций:
- T307H, или
- T307Q, или
- Q311H, или
- E430H, или
- N434H, или
- T307H и Q311H, или
- T307H и E430H, или
- T307H и N434A, или
- T307H и N434H, или
- T307Q и Q311H, или
- T307Q и E430H, или
- T307Q и N434H, или
- T307H, и Q311H, и E430H, и N434A, или
- T307H, и Q311H, и E430H, и N434H, или
- T307H, и Q311H, и E430H, и N434Y, или
- T307Q, и Q311H, и E430H, и N434A, или
- T307Q, и Q311H, и E430H, и N434H, или
- T307Q, и Q311H, и E430H, и N434Y, или
- T307Q, и V308P, и N434Y, и Y436H, или
- T307H, и M252Y, и S254T, и T256E, или
- Q311H, и M252Y, и S254T, и T256E, или
- E430H, и M252Y, и S254T, и T256E, или
- N434H, и M252Y, и S254T, и T256E.
В одном из воплощений всех аспектов, описанных в данной заявке, первый полипептид Fc-области содержит
одну из приведенных ниже комбинаций мутаций:
- ни одной, или
- M252Y, и S254T, и T256E, или
- I253A, и H310A, и H435A, или
- H310A, и H433A, и Y436A,
и
одну из приведенных ниже мутаций или комбинаций мутаций:
- ни одной,
- T307H, или
- T307Q, или
- Q311H, или
- E430H, или
- N434H, или
- T307H и Q311H, или
- T307H и E430H, или
- T307H и N434A, или
- T307H и N434H, или
- T307Q и Q311H, или
- T307Q и E430H, или
- T307Q и N434H, или
- T307Q и N434A, или
- T307H, и Q311H, и E430H, и N434A, или
- T307H, и Q311H, и E430H, и N434H, или
- T307H, и Q311H, и E430H, и N434Y, или
- T307Q, и Q311H, и E430H, и N434A, или
- T307Q, и Q311H, и E430H, и N434H, или
- T307Q, и Q311H, и E430H, и N434Y, или
- T307Q, и V308P, и N434Y, и Y436H,
и второй полипептид Fc-области содержит
одну из приведенных ниже мутаций или комбинаций мутаций:
- ни одной, если первый полипептид Fc-области содержит по меньшей мере одну мутацию, или
- T307H, или
- T307Q, если первый полипептид Fc-области не содержит исключительно мутацию T307Q, или
- Q311H, или
- E430H, или
- N434H, или
- T307H и Q311H, или
- T307H и E430H, или
- T307H и N434A, или
- T307H и N434H, или
- T307Q и Q311H, или
- T307Q и E430H, или
- T307Q и N434H, или
- T307Q и N434A, или
- M252Y, и S254T, и T256E, если первый полипептид Fc-области не содержит исключительно комбинацию мутаций M252Y, и S254T, и T256E, или
- I253A, и H310A, и H435A, если первый полипептид Fc-области не содержит исключительно комбинацию мутаций I253A, и H310A, и H435A, или
- H310A, и H433A, и Y436A, если первый полипептид Fc-области не содержит исключительно комбинацию мутаций H310A, и H433A, и Y436A, или
- T307H, и Q311H, и E430H, и N434A, или
- T307H, и Q311H, и E430H, и N434H, или
- T307H, и Q311H, и E430H, и N434Y, или
- T307Q, и Q311H, и E430H, и N434A, или
- T307Q, и Q311H, и E430H, и N434H, или
- T307Q, и Q311H, и E430H, и N434Y, или
- T307Q, и V308P, и N434Y, и Y436H.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутации I253A, и H310A, и H435A, а второй полипептид Fc-области содержит мутации M252Y, и S254T, и T256E.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутации I253A, и H310A, и H435A, а второй полипептид Fc-области содержит мутации M252Y, и S254T, и T256E и T307Q и N434Y.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутации I253A, и H310A, и H435A, а второй полипептид Fc-области содержит мутации M252Y, и S254T, и T256E и T307Q, и V308P, и N434Y, и Y436H.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутации T307H, и Q311H, и E430H, и N434H, а второй полипептид Fc-области содержит мутации M252Y, и S254T, и T256E.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутации T307H и N434H, а второй полипептид Fc-области содержит мутации M252Y, и S254T, и T256E.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутации T307Q и N434A, а второй полипептид Fc-области содержит мутации M252Y, и S254T, и T256E.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутации T250Q и M428L, а второй полипептид Fc-области содержит мутации M252Y, и S254T, и T256E.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутации T307Q и N434H, а второй полипептид Fc-области содержит мутации M252Y, и S254T, и T256E и T307Q и N434H.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутации T307H, и Q311H, и E430H, и N434H, и второй полипептид Fc-области содержит мутации T307H, и Q311H, и E430H, и N434H.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутации T307H и N434H, и второй полипептид Fc-области содержит мутации T307H и N434H.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутации T307H, и N434H, и M252Y, и S254T, и T256E, и второй полипептид Fc-области содержит мутации T307H, и N434H, и M252Y, и S254T, и T256E.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутацию N434H, и второй полипептид Fc-области содержит мутацию N434H.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутации T307Q и N434A.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутации T307H и N434H.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутации T307H, и N434H, и M252Y, и S254T, и T256E.
В одном из предпочтительных воплощений первый полипептид Fc-области содержит мутацию N434H.
В одном из воплощений всех аспектов Fc-область представляет собой вариантную Fc-область класса IgG (человека). В одном из воплощений вариантная Fc-область класса IgG (человека) представляет собой гетеродимерную Fc-область класса IgG.
В одном из воплощений всех аспектов образование пар с участием первого полипептида Fc-области и второго полипептида Fc-области с получением (функциональной) Fc-области приводит к образованию гетеродимера.
В одном из воплощений полипептид Fc-области IgG человека представляет собой полипептид Fc-области IgG человека подкласса IgG1 или подкласса IgG4.
В одном из воплощений полипептид Fc-области IgG человека представляет собой полипептид Fc-области IgG человека подкласса IgG1, который дополнительно содержит мутации L234A, L235A и P329G.
В одном из воплощений полипептид Fc-области IgG человека представляет собой полипептид Fc-области IgG человека подкласса IgG4, который дополнительно содержит мутации S228P и L235E.
В одном из воплощений первый полипептид Fc-области дополнительно содержит мутации S354C и T366W, а второй полипептид Fc-области дополнительно содержит мутации Y349C, T366S, L368A и Y407V.
В одном из воплощений биспецифичное антитело характеризуется тем, что Fc-область по пункту (3) происходит из подкласса IgG1 человека. В одном из воплощений биспецифичное антитело характеризуется тем, что Fc-область подкласса IgG1 человека дополнительно содержит мутации L234A, L235A и P329G (нумерация соответствует EU-индексу по Kabat).
В одном из воплощений биспецифичное антитело характеризуется тем, что Fc-область по пункту (3) происходит из подкласса IgG4 человека. В одном из воплощений биспецифичное антитело характеризуется тем, что Fc-область подкласса IgG4 человека дополнительно содержит мутации S228P и L235E (нумерация соответствует EU-индексу по Kabat). В одном из воплощений биспецифичное антитело характеризуется тем, что Fc-область подкласса IgG4 человека дополнительно содержит мутации S228P, L235E и P329G (нумерация соответствует EU-индексу по Kabat).
Следующие дополнительные аспекты, описанные в данной заявке, относятся к фармацевтической композиции, содержащей биспецифичное антитело, к фармацевтической композиции для применения в лечении сосудистых заболеваний глаз, к применению биспецифичного антитела для изготовления лекарственного средства для лечения сосудистых заболеваний глаз, к способу лечения пациента, страдающего сосудистыми заболеваниями глаз, путем введения биспецифичного антитела пациенту, нуждающемуся в таком лечении. В одном из воплощений биспецифичное антитело или фармацевтическую композицию, содержащую биспецифичное антитело, применяют посредством интравитреального введения.
Согласно другому аспекту настоящего изобретения предложена молекула нуклеиновой кислоты, кодирующей тяжелую и/или легкую цепь биспецифичного антитела, описанного в данной заявке.
Согласно изобретению дополнительно предложены экспрессирующие векторы, содержащие нуклеиновую кислоту, описанную в данной заявке, способные экспрессировать эту нуклеиновую кислоту в прокариотической или эукариотической клетке хозяина, и клетки хозяина, содержащие такие векторы для получения рекомбинантными методами биспецифичного антитела, описанного в данной заявке.
Изобретение дополнительно включает прокариотическую или эукариотическую клетку хозяина, содержащую вектор, описанный в данной заявке.
Изобретение дополнительно включает способ получения биспецифичного антитела, описанного в данной заявке, характеризующийся экспрессированием нуклеиновой кислоты, описанной в данной заявке, в прокариотической или эукариотической клетке хозяина и извлечением биспецифичного антитела из клетки или супернатанта клеточной культуры. Одно из воплощений относится к способу получения биспецифичного антитела, описанного в данной заявке, включающему стадии:
a) трансфекции клетки хозяина векторами, содержащими молекулы нуклеиновой кислоты, кодирующей данное антитело;
b) культивирования клетки хозяина в условиях, которые позволяют осуществлять синтез антитела; и
c) извлечения антитела из культуры.
Изобретение дополнительно включает антитело, полученное согласно такому способу получения биспецифичного антитела.
Антитела, описанные в данной заявке, обладают в высокой степени полезными свойствами, обусловленными специфическими модификациями в своей Fc-области, приносящими пользу пациенту, страдающему сосудистыми заболеваниями глаз. Они демонстрируют высокую стабильность в интравитреальном пространстве и медленную диффузию из глаза (по сравнению с более мелкими фрагментами антител, не содержащими константной области тяжелой цепи), т.е. из места, где фактически локализовано заболевание и куда должно быть направлено лечение (поэтому существует возможность улучшения схемы лечения по сравнению с не-IgG-подобными антителами типа, например, фрагментов Fab и (Fab)2). С другой стороны, антитела, описанные в данной заявке, достаточно быстро выводятся из сыворотки крови (что крайне желательно для снижения возможных побочных эффектов, возникающих при системном воздействии). Кроме того, неожиданным образом они демонстрируют более низкую вязкость (по сравнению с вариантами без комбинации мутаций I253A, H310A и H435A в константной области) и поэтому особенно полезны для интравитреального введения через тонкие иглы в процессе лечения заболеваний глаз (для такого введения обычно используют тонкие иглы, и высокая вязкость сильно затрудняет соответствующий способ применения). Кроме того, более низкая вязкость способствует введению композиций в более высоких концентрациях.
Кроме того, неожиданным образом антитела, описанные в данной заявке, демонстрируют более слабую тенденцию к агрегации в процессе хранения (по сравнению с вариантами без комбинации мутаций I253A, H310A и H435A в Fc-области), что является критичным в случае интравитреального введения в глаз (поскольку в ходе такого лечения агрегация в глазу может вызывать осложнения).
Биспецифичные антитела, описанные в данной заявке, демонстрируют хорошую эффективность в подавлении сосудистых заболеваний.
В некоторых воплощениях биспецифичные антитела, описанные в данной заявке, благодаря наличию у них специфических модификаций в константной области (например, P329G LALA), демонстрируют полезные свойства, такие как отсутствие связывания с Fc-гамма-рецепторами/связывания Fc-гамма-рецепторов, что снижает риск побочных эффектов типа тромбоза и/или нежелательной гибели клеток (вследствие, например, ADCC).
В одном из воплощений, описанных в данной заявке, биспецифичное антитело, описанное в данной заявке, является бивалентным.
В одном из воплощений вариабельный домен тяжелой цепи антитела (VH) и вариабельный домен легкой цепи антитела (VL) тяжелой и легкой цепи второго полноразмерного антитела дополнительно стабилизированы путем введения дисульфидной связи между следующими положениями: положением 44 вариабельного домена тяжелой цепи и положением 100 вариабельного домена легкой цепи (нумерация согласно Kabat (Kabat E.A., et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991))). Методы введения дисульфидных мостиков для стабилизации описаны, например, в WO 94/029350, Rajagopal V., et al., Prot. Eng., 10 (1997) 1453-1459, Kobayashi et al., Nuclear Medicine & Biology, 25 (1998) 387-393 и Schmidt M., et al., Oncogene, 18 (1999) 1711-1721.
В одном из воплощений CH3-домены биспецифичного, бивалентного антитела, описанного в данной заявке, изменены с использованием технологии “выступ-во-впадину”, которая описана подробно на нескольких примерах, например, в WO 96/027011, Ridgway J.B., et al., Protein Eng., 9 (1996) 617-621 и Merchant A.M., et al., Nat. Biotechnol., 16 (1998) 677-681. В этом методе взаимодействующие поверхности двух CH3-доменов изменяют с целью усиления гетеродимеризации обеих тяжелых цепей, содержащих эти два CH3-домена. Каждый из двух CH3-доменов (двух тяжелых цепей) может представлять собой “цепь, формирующую выступ”, в то время как другой представляет собой “цепь, формирующую впадину”. Кроме того, введение дисульфидного мостика стабилизирует гетеродимеры (Merchant A.M, et al., Nature Biotech., 16 (1998) 677-681, Atwell S., et al. J. Mol. Biol., 270 (1997) 26-35) и повышает выход.
В одном из предпочтительных воплощений всех аспектов, описанных в данной заявке, биспецифичные антитела характеризуются тем, что
CH3-домен одной тяжелой цепи и CH3-домен другой тяжелой цепи оба встречаются на поверхности контакта, которая представляет собой исходную поверхность контакта между CH3-доменами антитела,
причем поверхность контакта изменена, чтобы стимулировать образование биспецифичного антитела, при этом данное изменение характеризуется тем, что:
a) CH3-домен одной тяжелой цепи изменен
так, что в исходной поверхности контакта CH3-домена одной тяжелой цепи, которая встречается c исходной поверхностью контакта CH3-домена другой тяжелой цепи в биспецифичном антителе,
аминокислотный остаток заменен на аминокислотный остаток, имеющий боковую цепь большего объема, тем самым создавая выступающую часть в поверхности контакта CH3-домена одной тяжелой цепи, которая располагается в полости поверхности контакта CH3-домена другой тяжелой цепи,
и
b) CH3-домен другой тяжелой цепи изменен
так, что в исходной поверхности контакта второго CH3-домена, которая встречается c исходной поверхностью контакта первого CH3-домена в биспецифичном антителе,
аминокислотный остаток заменен на аминокислотный остаток, имеющий боковую цепь меньшего объема, тем самым создавая полость в поверхности контакта второго CH3-домена, в которой располагается выступающая часть поверхности контакта первого CH3-домена.
Таким образом, антитело по изобретению в одном из предпочтительных воплощений характеризуется тем, что
CH3-домен тяжелой цепи полноразмерного антитела по пункту (a) и CH3-домен тяжелой цепи полноразмерного антитела по пункту b) оба встречаются на поверхности контакта, которая изменена по сравнению с исходной поверхностью контакта между CH3-доменами антитела,
при этом
1) в CH3-домене одной тяжелой цепи
аминокислотный остаток заменен на аминокислотный остаток, имеющий боковую цепь большего объема, тем самым создавая выступающую часть в поверхности контакта CH3-домена одной тяжелой цепи, которая располагается в полости поверхности контакта CH3-домена другой тяжелой цепи,
и при этом
2) в CH3-домене другой тяжелой цепи
аминокислотный остаток заменен на аминокислотный остаток, имеющий боковую цепь меньшего объема, тем самым создавая полость в поверхности контакта второго CH3-домена, в которой располагается выступающая часть поверхности контакта первого CH3-домена.
В одном из предпочтительных воплощений аминокислотный остаток, имеющий боковую цепь большего объема, выбран из группы, состоящей из аргинина (R), фенилаланина (F), тирозина (Y), триптофана (W).
В одном из предпочтительных воплощений аминокислотный остаток, имеющий боковую цепь меньшего объема, выбран из группы, состоящей из аланина (A), серина (S), треонина (T), валина (V).
В одном из воплощений оба CH3-домена дополнительно изменены посредством введения остатка цистеина (C) в соответствующие положения каждого CH3-домена для того, чтобы мог образоваться дисульфидный мостик между обоими CH3-доменами.
В одном из воплощений биспецифичное антитело содержит мутацию T366W в CH3-домене “цепи, формирующей выступы” и мутации T366S, L368A и Y407V в CH3-домене “цепи, формирующей впадину”. Также можно использовать дополнительный межцепочечный дисульфидный мостик между CH3-доменами (Merchant A.M, et al., Nature Biotech., 16 (1998) 677-681), например, путем внесения мутации Y349C или S354C в CH3-домен “цепи, формирующей выступы” и мутации Y439C, или E356C, или S354C в CH3-домен “цепи, формирующей впадину”.
В одном из воплощений биспецифичное антитело, описанное в данной заявке, содержит мутацию Y349C или S354C и мутацию T366W в одном из двух CH3-доменов и мутации S354C, или E356C, или Y349C и мутации T366S, L368A и Y407V в другом из двух CH3-доменов. В одном из предпочтительных воплощений биспецифичное антитело содержит мутации Y349C, T366W в одном из двух CH3-доменов и мутации S354C, T366S, L368A, Y407V в другом из двух CH3-доменов (дополнительную мутацию Y349C в одном CH3-домене и дополнительную мутацию S354C в другом CH3-домене, формирующие межцепочечный дисульфидный мостик) (нумерация соответствует EU-индексу по Kabat (Kabat E.A., et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991)). В одном из предпочтительных воплощений биспецифичное антитело содержит мутации S354C, T366W в одном из двух CH3-доменов и мутации Y349C, T366S, L368A, Y407V в другом из двух CH3-доменов (дополнительную мутацию Y349C в одном CH3-домене и дополнительную мутацию S354C в другом CH3-домене, формирующие межцепочечный дисульфидный мостик) (нумерация соответствует EU-индексу по Kabat (Kabat E.A., et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991)).
Тем не менее, альтернативно или помимо этого, также можно использовать другие технологии “выступы-во-впадины”, описанные в EP 1870459A1. Так, другим примером для биспецифичного антитела являются мутации R409D и K370E в CH3-домене “цепи, формирующей выступы” и мутации D399K и E357K в CH3-домене “цепи, формирующей впадину” (нумерация соответствует EU-индексу по Kabat (Kabat E.A., et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991)).
В другом воплощении биспецифичное антитело содержит мутацию T366W в CH3-домене “цепи, формирующей выступы” и мутации T366S, L368A и Y407V в CH3-домене “цепи, формирующей впадину” и помимо этого мутации R409D, K370E в CH3-домене “цепи, формирующей выступы” и мутации D399K, E357K в CH3-домене “цепи, формирующей впадину”.
В одном из воплощений биспецифичное антитело содержит мутации Y349C, T366W в одном из двух CH3-доменов и мутации S354C, T366S, L368A и Y407V в другом из двух CH3-доменов, или биспецифичное антитело содержит мутации Y349C, T366W в одном из двух CH3-доменов и мутации S354C, T366S, L368A и Y407V в другом из двух CH3-доменов и кроме того мутации R409D, K370E в CH3-домене “цепи, формирующей выступы” и мутации D399K, E357K в CH3-домене “цепи, формирующей впадину”.
В одном из воплощений биспецифичное антитело содержит мутации S354C, T366W в одном из двух CH3-доменов и мутации Y349C, T366S, L368A, Y407V в другом из двух CH3-доменов, или биспецифичное антитело содержит мутации S354C, T366W в одном из двух CH3-доменов и мутации Y349C, T366S, L368A и Y407V в другом из двух CH3-доменов и кроме того мутации R409D, K370E в CH3-домене “цепи, формирующей выступы” и мутации D399K, E357K в CH3-домене “цепи, формирующей впадину”.
Антиген-связывающий сайт биспецифичного антитела, описанного в данной заявке, содержит шесть определяющих комплементарность участков (CDR), которые в разной степени вносят вклад в аффинность сайта связывания со своим антигеном. Ими являются три CDR вариабельного домена тяжелой цепи (CDRH1, CDRH2 и CDRH3) и три CDR вариабельного домена легкой цепи (CDRL1, CDRL2 и CDRL3). Протяженность CDR и каркасных областей (FR) определяют путем сравнения с компилированной базой данных аминокислотных последовательностей, где эти области определены в соответствии с вариабельностью среди последовательностей.
В одном из воплощений всех аспектов антитело не связывается специфически с FcRn человека. В одном из воплощений всех аспектов антитело, помимо этого, связывается специфически со стафилококковым белком A.
В одном из воплощений всех аспектов антитело не связывается специфически с FcRn человека. В одном из воплощений всех аспектов антитело, помимо этого, не связывается специфически со стафилококковым белком A.
В одном из воплощений всех аспектов первый полипептид дополнительно содержит мутации Y349C, T366S, L368A и Y407V („впадина“), а второй полипептид содержит мутации S354C и T366W („выступ“).
В одном из воплощений всех аспектов первый полипептид дополнительно содержит мутации S354C, T366S, L368A и Y407V („впадина“), а второй полипептид содержит мутации Y349C и T366W („выступ“).
В одном из воплощений всех аспектов полипептиды Fc-области происходят из подкласса IgG1 человека. В одном из воплощений первый полипептид Fc-области и второй полипептид Fc-области дополнительно содержат мутации L234A и L235A. В одном из воплощений первый полипептид Fc-области и второй полипептид Fc-области дополнительно содержат мутацию P329G.
В одном из воплощений всех аспектов Полипептиды Fc-области происходят из подкласса IgG4 человека. В одном из воплощений первый полипептид Fc-области и второй полипептид Fc-области дополнительно содержат мутации S228P и L235E. В одном из воплощений первый полипептид Fc-области и второй полипептид Fc-области дополнительно содержат мутацию P329G.
Антитело, описанное в данной заявке, получают с использованием рекомбинантных методов. Так, один из аспектов, описанных в данной заявке, относится к нуклеиновой кислоте, кодирующей антитело, описанное в данной заявке, а другой аспект относится к клетке, содержащей нуклеиновую кислоту, кодирующую антитело, описанное в данной заявке. Способы получения рекомбинантными методами широко известны в документах данной области техники и включают экспрессию белка в прокариотических и эукариотических клетках с последующим выделением антитела и обычно очистку до фармацевтически приемлемой чистоты. Чтобы осуществить экспрессию антител, которые упомянуты выше, в клетке хозяина, нуклеиновые кислоты, кодирующие соответствующие (модифицированные) легкую и тяжелую цепи, встраивают в экспрессирующие векторы стандартными методами. Экспрессию проводят в соответствующих прокариотических или эукариотических клетках хозяина типа клеток CHO, клеток NS0, клеток SP2/0, клеток HEK293, клеток COS, клеток PER.C6, клеток дрожжей или клеток E. coli, и антитело извлекают из клеток (из культурального супернатанта или клеток после разрушения). Общие способы получения антител рекомбинантными методами хорошо известны в документах данной области техники и описаны, например, в обзорных статьях Makrides S.C., Protein Expr. Purif., 17 (1999) 183-202; Geisse S., et al., Protein Expr. Purif., 8 (1996) 271-282; Kaufman R.J., Mol. Biotechnol., 16 (2000) 151-160; и Werner R.G., Drug Res., 48 (1998) 870-880.
Соответственно, один из аспектов, описанных в данной заявке, относится к способу получения биспецифичного антитела, описанного в данной заявке, включающему стадии:
a) трансфекции клетки хозяина векторами, содержащими молекулы нуклеиновой кислоты, кодирующей антитело,
b) культивирования клетки хозяина в условиях, которые позволяют осуществлять синтез антитела, и
c) извлечения антитела из культивируемой смеси.
В одном из воплощений стадия извлечения по пункту (c) включает применение реагента специфичного захвата константного домена легкой цепи (который обладает специфичностью, например, к константной области легкой цепи каппа или лямбда, в зависимости от того какая из легких цепей, каппа или лямбда, содержится в биспецифичном антителе). В одном из воплощений этот реагент специфичного захвата легкой цепи используется в режиме связывания-элюирования. Примерами таких реагентов специфичного захвата константного домена легкой цепи являются, например, KappaSelect™ и LambdaFabSelect™ (поставляемые GE Healthcare/BAC), изготовленные на базе особо прочной матрицы на основе агарозы, которая позволяет работать при высоких скоростях потока и низком противодавленим в случае получения крупномасштабных партий. Эти носители содержат лиганд, который связывается с константной областью легкой цепи каппа или лямбда, соответственно (т.е. фрагменты, у которых отсутствует константная область легкой цепи, связываться не будут). Таким образом, они оба могут связывать другие целевые молекулы, содержащие константную область легкой цепи, например, IgG, IgA и IgM. Лиганды присоединены к матрице через длинную гидрофильную спейсерную ножку, что делает их легко доступными для связывания с целевой молекулой. В качестве них используют одноцепочечный фрагмент антитела, скрининг которого осуществляют либо по каппа-, либо по лямбда-цепи Ig человека.
В одном из воплощений стадия извлечения по пункту (c) включает применение реагента специфичного захвата Fc-области. В одном из воплощений реагент специфичного захвата Fc-области используют в режиме связывания-элюирования. Примерами таких реагентов специфичного захвата Fc-области являются, например, носители для аффинной хроматографии на основе стафилококкового белка A.
Биспецифичные антитела подходящим образом отделяют от культуральной среды, используя традиционные методики очистки иммуноглобулинов, такие как, например, аффинная хроматография (на сефарозе с иммобилизованным на ней белком A или на KappaSelect™, LambdaFabSelect™), хроматография на гидроксилапатите, гель-электрофорез или диализ.
ДНК и РНК, кодирующие моноклональные антитела, легко выделяют и секвенируют с использованием традиционных методик. B-клетки или гибридомные клетки могут служить в качестве источника таких ДНК и РНК. После выделения ДНК может быть встроена в экспрессирующие векторы, которыми затем трансфицируют клетки хозяина, такие как клетки HEK 293, клетки CHO или миеломные клетки, которые в отсутствие этого не продуцируют иммуноглобулиновый белок, для синтеза рекомбинантных моноклональных антител в клетках хозяина.
Простота выделения/очистки некоторых из молекул, описанных в данной заявке, обеспечивается присутствием Fc-областей, которые модифицированы различным образом, при этом по меньшей мере один из результатов модификации заключается в достижении 1) различной аффинности данной молекулы к (стафилококковому) белку A и 2) различной аффинности данной молекулы к FcRn человека, и эту молекулу можно выделить из разрушенной клетки, из среды или из смеси молекул на основе ее аффинности к белку A.
Очистку антител осуществляют с целью устранения клеточных компонентов или других примесей, например, других нуклеиновых кислот или белков клетки, стандартными методами, включая обработку щелочью/SDS, разделение полос при центрифугировании в градиенте CsCl, колоночную хроматографию, электрофорез в агарозном геле и другие хорошо известные в данной области методы (см., например, Ausubel F. et al., ed., Current Ptotocos in Molecular Biology, Greene Publishing and Wiley Interscience, New York (1987)). Для очистки белков общепризнанными и широко используемыми считаются различные методы, такие как аффинная хроматография с использованием микробных белков (например, аффинная хроматография на носителях с иммобилизованным белком A или белком G), ионообменная хроматография (например, катионообменная (на карбоксиметил-содержащих носителях), анионообменная (на аминоэтил-содержащих носителях) и хроматография на смешанных ионообменниках), тиофильная адсорбционная хроматография (например, с использованием бета-меркаптоэтанола и других SH-содержащих лигандов), адсорбционная хроматография гидрофобных или ароматических взаимодействий (например, с использованием фенил-сефарозы, аза-аренофильных носителей или м-аминофенилбороновой кислоты), металл-хелатирующая аффинная хроматография (например, с использованием Ni(II)- и Cu(II)-аффинного носителя), гель-проникающая хроматография и электрофоретические методы (такие как гель-электрофорез, капиллярный электрофорез) (Vijayalakshmi M.A., Appl. Biochem. Biotech., 75 (1998) 93-102).
Один из аспектов, описанных в данной заявке, относится к фармацевтической композиции, содержащей антитело, описанное в данной заявке. Другой аспект, описанный в данной заявке, относится к применению антитела, описанного в данной заявке, для изготовления фармацевтической композиции. Следующий аспект, описанный в данной заявке, относится к способу изготовления фармацевтической композиции, содержащей антитело, описанное в данной заявке. Согласно другому аспекту предложена композиция, например фармацевтическая композиция, содержащая антитело, описанное в данной заявке, при изготовлении которой вместе с ним использован фармацевтический носитель.
Композицию по настоящему изобретению можно вводить разнообразными способами, известными в данной области техники. Специалисту в данной области будет очевидно, что путь и/или способ введения будет варьировать в зависимости от желаемых результатов. Чтобы ввести соединение, описанное в данной заявке, с использованием определенных путей введения, может потребоваться нанесение на данное соединение покрытия из материала, предотвращающего его инактивацию, или совместное с этим материалом введение соединения. Например, соединение можно вводить субъекту в соответствующем носителе, например, в составе липосом, или разбавителе. Фармацевтически приемлемые разбавители включают физиологический раствор и водные буферные растворы. Фармацевтически приемлемые носители включают стерильные водные растворы или дисперсии и стерильные порошки для экстемпорального приготовления стерильных инъекционных растворов или дисперсий. Применение таких сред и агентов для фармацевтически активных веществ известно в данной области техники.
Можно использовать много возможных способов доставки, включая, но не ограничиваясь этим, интраокулярное введение или местное нанесение. В одном из воплощений введение осуществляют интраокулярно, и оно включает, но не ограничивается этим, субконъюнктивальную инъекцию, интракамеральную инъекцию, инъекцию в переднюю камеру глаза через височную сторону лимба, интрастромальную инъекцию, внутрироговичную инъекцию, субретинальную инъекцию, инъекцию во внутриглазную жидкость, субтеноновую инъекцию или использование устройства непрерывной доставки, интравитреальную инъекцию (например, инъекцию в переднюю, среднюю или заднюю часть стекловидного тела). В одном из воплощений введение выполняют местно, и оно включает, но не ограничивается этим, нанесение глазных капель на роговицу.
В одном из воплощений биспецифичное антитело или фармацевтическую композицию, описанные в данной заявке, вводят, используя способ интравитреального введения, например, посредством интравитреальной инъекции. Это может быть осуществлено в соответствии со стандартными методиками, известными в данной области техники (см., например, Ritter et al., J. Clin. Invest., 116 (2006) 3266-3276, Russelakis-Carneiro et al., Neuropathol. Appl. Neurobiol., 25 (1999) 196-206 и Wray et al., Arch. Neurol., 33 (1976) 183-185).
В некоторых воплощениях терапевтические наборы, описанные в данной заявке, могут содержать одну или более доз (биспецифичного) антитела, присутствующего в фармацевтической композиции, описанной в данной заявке, устройство, подходящее для интравитреальной инъекции фармацевтической композиции, и инструкцию с подробным описанием подходящих субъектов и протоколов для выполнения такой инъекции. Обычно в этих воплощениях композиции вводят субъекту, нуждающемуся в лечении, посредством интравитреальной инъекции. Это может быть осуществлено в соответствии со стандартными методиками, известными в данной области техники (см., например, Ritter et al., J. Clin. Invest., 116 (2006) 3266-3276, Russelakis-Carneiro et al., Neuropathol. Appl. Neurobiol., 25 (1999) 196-206 и Wray et al., Arch. Neurol., 33 (1976) 183-185).
Эти композиции также могут содержать такие адъюванты, как консерванты, увлажняющие агенты, эмульгирующие агенты и диспергирующие агенты. Предотвращение действия микроорганизмов можно обеспечить как посредством описанных выше процедур стерилизации, так и посредством включения различных антибактеральных и противогрибковых агентов, например, парабена, хлорбутанола, фенола, сорбиновой кислоты и тому подобного. Кроме этого, желательным может быть включение в композиции изотонических агентов, таких как сахара, хлорид натрия и тому подобное. Помимо этого, посредством включения агентов, замедляющих всасывание, таких как моностеарат алюминия и желатин, можно способствовать пролонгированному всасыванию инъекционной фармацевтической формы.
Независимо от выбранного пути введения, на основе соединений, описанных в данной заявке, которые могут быть использованы в подходящей гидратированной форме, и/или фармацевтических композиций, описанных в данной заявке, готовят фармацевтически приемлемые лекарственные формы традиционными способами, известными специалистам в данной области техники.
Фактические уровни дозировок активных ингредиентов в фармацевтических композициях, описанных в данной заявке, можно варьировать, чтобы получить количество активного ингредиента, которое будет эффективным с точки зрения достижения желаемого терапевтического ответа для конкретного пациента, с точки зрения композиции и способа введения, причем без оказания токсичного действия пациенту. Выбранный уровень дозировок будет зависеть от ряда фармакокинетических факторов, включая активность конкретных используемых композиций, путь введения, время введения, скорость выделения конкретного используемого соединения, продолжительность лечения, другие лекарственные средства, соединения и/или материалы, применяемые в комбинации с конкретными используемыми композициями, возраст, пол, массу, состояние, общее состояние здоровья и предшествующая история болезни подвергаемого лечению пациента, и подобных факторов, хорошо известных в области медицины.
Композиция должна быть стерильной и текучей до такой степени, чтобы ее можно было доставлять с помощью шприца. Помимо воды, носителем предпочтительно является изотонический забуференный физиологический раствор.
Надлежащую текучесть можно поддерживать, например, путем использования покрытия, такого как лецитин, путем поддержания требуемого размера частиц в случае дисперсии и путем использования поверхностно-активных веществ. Во многих случаях, предпочтительно включать в композицию изотонические агенты, например, сахара, полиспирты, такие как маннит или сорбит, и хлорид натрия.
Композиция может представлять собой офтальмическую депо-композицию, содержащую активный агент для субконъюнктивального введения. Такая офтальмическая депо-композиция содержит микрочастицы по-существу чистого активного агента, например, биспецифичного антитела, описанного в данной заявке. Микрочастицы, содержащие биспецифичное антитело, описанное в данной заявке, могут быть встроены в биосовместимый фармацевтически приемлемый полимер или липидный инкапсулирующий агент. Такие депо-композиции могут быть приспособлены для того, чтобы осуществлять высвобождения всего или по существу всего активного вещества в течение длительного периода времени. Полимерная или липидная матрица, если она присутствует, может быть приспособлена для того, чтобы в достаточной степени быть подвергнутой разрушению при транспортировке от места введения после высвобождения всего или по существу всего активного агента. Депо-композиция может представлять собой композицию в жидкой форме, содержащую фармацевтически приемлемый полимер и растворенный или диспергированный активный агент. После инъекции полимер образует депо в месте инъекции, например, посредством гелеобразования или осаждения.
Другой аспект, описанный в данной заявке, относится к биспецифичному антителу, описанному в данной заявке, для применения в лечении сосудистых заболеваний глаз.
Другой аспект, описанный в данной заявке, относится к фармацевтической композиции, описанной в данной заявке, для применения в лечении сосудистых заболеваний глаз.
Другой аспект, описанный в данной заявке, относится к применению антитела, описанного в данной заявке, для изготовления лекарственного средства для лечения сосудистого заболевания глаз.
Другой аспект, описанный в данной заявке, относится к способу лечения пациента, страдающего сосудистыми заболеваниями глаз, путем введения антитела, описанного в данной заявке, пациенту, нуждающемуся в таком лечении.
В данном описании особо указано, что термин “содержащий”, использованный в данном описании, включает в себя термин “состоящий из”. Таким образом, все аспекты и воплощения, которые содержат термин “содержащий”, подобным же образом описываются термином “состоящий из”.
Модификации
В следующем аспекте Fc-область или антитело, описанное в данной заявке, а также антитело по любому из вышеупомянутых воплощений может включать любой из признаков, по отдельности или в комбинации, которые описаны в приведенных ниже разделах 1-6.
1. Аффинность антител
В одном из воплощений KD измеряют в анализах с применением поверхностного плазмонного резонанса, используя BIACORE®. Например, анализ с использованием BIACORE®-2000 или BIACORE®-3000 (GE Healthcare Inc., Piscataway, NJ) проводят при 25°C с CM5-чипами с иммобилизованным антигеном, на уровне приблизительно 10 единиц ответного сигнала (RU). В одном из воплощений биосенсорные чипы с карбоксиметилированным декстраном (CM5, GE Healthcare Inc.) активируют N-этил-N'-(3-диметиламинопропил)-карбодиимида гидрохлоридом (EDC) и N-гидроксисукцинимидом (NHS) в соответствии с инструкциями поставщика. Антиген разбавляют 10 мМ ацетатом натрия, pH 4,8, до концентрации 5 мкг/мл (примерно 0,2 мкМ), после чего вводят со скоростью потока 5 мкл/минута до момента достижения приблизительно 10 единиц ответного сигнала (RU) для связанного белка. После введения антигена вводят 1 М этаноламин, чтобы блокировать непрореагировавшие группы. Для кинетических измерений вводят Fab в двукратных серийных разведениях (от 0,78 нМ до 500 нМ) в забуференном фосфатом физиологическом растворе (PBS) с 0,05% полисорбата 20 (TWEEN-20™) в качестве поверхностно-активного вещества (PBST - PBS/твин) при 25°C и скорости потока приблизительно 25 мкл/мин. Константы скорости ассоциации (kon) и константы скорости диссоциации (koff) рассчитывают, используя простую модель связывания Ленгмюра “один к одному” (аналитическое программное обеспечение BIACORE®, версия 3.2), посредством совместной аппроксимации сенсограмм ассоциации и диссоциации. Равновесную константу диссоциации (KD) рассчитывают в виде соотношения koff/kon (см., например, Chen et al., J. Mol. Biol., 293 (1999), 865-881). Если константа скорости ассоциации превышает 106 М-1с-1 по данным вышеупомянутого анализа с использованием поверхностного плазмонного резонанса, то ее можно определить, применяя метод гашения флуоресценции, в котором измеряют при 25°C увеличение или уменьшение интенсивности флуоресценции (возбуждение соответствует 295 нм; излучение соответствует 340 нм, полоса пропускания 16 нм) для 20 нМ антитела к антигену (в форме Fab) в PBS, pH 7,2, в присутствии возрастающих концентраций антигена по результатам измерений на спектрометре, таком как спектрофотометр, приспособленный для измерения методом остановленной струи, (Aviv Instruments) или спектрофотометр SLM-AMINCO™ серии 8000 (ThermoSpectronic), снабженный кюветой с перемешиванием.
2. Химерные и гуманизированные антитела
В некоторых воплощениях антитело, предложенное в данном изобретении, представляет собой химерное антитело. Некоторые химерные антитела описаны, например, US 4816567 и в Morrison S.L. et al., Proc. Natl. Acad. Sci. USA, 81 (1984) 6851-6855. В одном из примеров химерное антитело содержит вариабельную область антитела, не являющегося человеческим, (например, вариабельную область, происходящую из антитела мыши, крысы, хомяка, кролика или примата, не являющегося человеком, такого как обезьяна) и константную область антитела человека. В другом примере химерное антитело представляет собой антитело с переключенным изотипом, в котором класс или подкласс был изменен по сравнению с родительским антителом. Химерные антитела включают свои антигенсвязывающие фрагменты.
В некоторых воплощениях химерное антитело представляет собой гуманизированное антитело. Обычно антитело, не являющееся человеческим, подвергают гуманизации с целью снижения иммуногенности в отношении людей, с сохранением при этом специфичности и аффинности родительского антитела, не являющегося человеческим. Как правило, гуманизированное антитело содержит один или более вариабельных доменов, в которых HVR, например, CDR, (или их части) происходят из антитела, не являющегося человеческим, а FR (или их части) происходят из последовательностей человеческого антитела. Гуманизированное антитело возможно также будет содержать по меньшей мере часть константной области антитела человека. В некоторых воплощениях несколько остатков FR в гуманизированном антителе заменены на соответствующие остатки из антитела, не являющегося человеческим (например, антитела, из которого данные остатки HVR происходят), например, для восстановления или улучшения специфичности или аффинности антитела.
Обзор по гуманизированным антителам и способам их получения приведен, например, в Almagro J.C. and Fransson J., Front. Biosci., 13 (2008) 1619-1633, и они также описаны, например, в Riechmann I. et al., Nature, 332 (1988) 323-329; Queen C. et al., Proc. Nat'l. Acad. Sci. USA, 86 (1989) 10029-10033; US 5821337, US 7527791, US 6982321 и US 7087409; Kashmiri S.V. et al., Methods, 36 (2005) 25-34 (где описывается привитие “определяющих специфичность участков” (SDR); Padlan E.A., Mol. Immunol., 28 (1991) 489-498 (где описывается “изменение поверхности”); Dall'Acqua W.F. et al., Methods, 36 (2005) 43-60 (где описывается “перетасовка FR”); Osbourn J. et al., Methods, 36 (2005) 61-68 и Klimka A. et al., Br. J. Cancer, 83 (2000) 252-260 (где описывается подход “направленной селекции” для перетасовки FR).
Каркасные области антител человека, которые могут быть использованы для гуманизации, включают, но не ограничиваются этим: каркасные области, отобранные с использованием метода “наилучшего соответствия” (см., например, Sims M.J. et al., J. Immunol., 151 (1993) 2296-2308); каркасные области, происходящие из консенсусной последовательности конкретной подгруппы вариабельных областей легкой или тяжелой цепей человеческих антител (см., например, Carter P. et al., Proc. Natl. Acad. Sci. USA, 89 (1992) 4285-4289 и Presta L.G. et al., J. Immunol., 151 (1993) 2623-2632); каркасные области зрелых человеческих антител (претерпевших соматические мутации) или каркасные области антител зародышевой линии человека (см., например, Almagro J.C. and Fransson J., Front. Biosci., 13 (2008) 1619-1633); и каркасные области, полученные в результате скрининга библиотек FR (см., например, Baca M. et al., J. Biol. Chem., 272 (1997) 10678-10684 и Rosok M.J. et al., J. Biol. Chem., 271 (1996) 22611-22618).
3. Человеческие антитела
В некоторых воплощениях антитело, предложенное в данном изобретении, представляет собой человеческое антитело. Человеческие антитела могут быть получены с использованием различных методов, известных в данной области техники. Человеческие антитела в целом описаны в van Dijk M.A. and van de Winkel J.G., Curr. Opin. Pharmacol., 5 (2001) 368-374 и Lonberg N., Curr. Opin. Immunol., 20 (2008) 450-459.
Человеческие антитела могут быть получены путем введения иммуногена трансгенному животному, которое было модифицировано с целью получения интактных человеческих антител или интактных антител с вариабельными областями антител человека в ответ на антигенную стимуляцию. Такие животные обычно содержат все иммуноглобулиновые локусы человека или их часть, которые заменяют эндогенные иммуноглобулиновые локусы или которые присутствуют вне хромосомы либо случайным образом интегрированы в хромосомы животных. В таких трансгенных мышах эндогенные иммуноглобулиновые локусы, как правило, инактивированы. Обзор способов получения человеческих антител с использованием трансгенных животных см. в Lonberg N., Nat. Biotech., 23 (2005) 1117-1125. Также см., например, US 6075181 и US 6150584, где описывается технология XENOMOUSE™; US 5770429, где описывается технология HuMab®; US 7041870, где описывается технология K-M MOUSE®, и US 2007/0061900, где описывается технология VELOCIMOUSE®. Вариабельные области антител человека из интактных антител, производимых такими животными, могут быть дополнительно модифицированы, например, путем комбинирования с другой константной областью антитела человека.
Человеческие антитела также могут быть получены гибридомными методами. Описаны клеточные линии миеломы человека и гетеромиеломы мышь-человек для получения человеческих моноклональных антител (см., например, Kozbor D., J. Immunol., 133 (1984) 3001-3005; Brodeur B.R. et al., Monoclonal Antibody Production Techniques and Applications, Marcel Dekker, Inc., New York (1987), pp. 51-63; и Boerner P. et al., J. Immunol., 147 (1991) 86-95). Человеческие антитела, созданные посредством гибридомной технологии с использованием B-клеток человека, также описаны в Li J. et al., Proc. Natl. Acad. Sci. USA, 103 (2006) 3557-3562. Дополнительные способы включают способы, описанные, например, в US 7189826 (где описывается получение моноклональных антител IgM человека из гибридомных клеточных линий) и в Ni J., Xiandai Mianyixue, 26 (2006) 265-268 (где описываются гибридомы человек-человек). Технология гибридом человека (триомная технология) также описана в Vollmers H.P. and Brandlein S., Histology and Histopathology, 20 (2005) 927-937 и Vollmers H.P. and Brandlein S., Methods and Findings in Experimental and Clinical Pharmacology, 27 (2005) 185-191.
Человеческие антитела также могут быть созданы посредством выделения последовательностей вариабельных доменов Fv-клонов, отобранных из библиотек фагового дисплея, полученных на основе антител человека. Такие последовательности вариабельных доменов далее могут быть объединены с желаемым константным доменом антитела человека. Методы отбора человеческих антител из библиотек антител описываются ниже.
4. Антитела, полученные с использованием библиотек
Антитела, описанные в данной заявке, могут быть выделены посредством скрининга комбинаторных библиотек на антитела с желаемой(ыми) активностью или активностями. Например, в данной области техники известен ряд способов создания библиотек фагового дисплея и скрининга таких библиотек на антитела, обладающие желаемыми характеристиками в отношении связывания. Обзор таких способов приведен, например, в Hoogenboom H.R. et al., Methods in Molecular Biology, 178 (2001) 1-37, и помимо этого они описаны, например, в McCafferty J. et al., Nature, 348 (1990) 552-554; Clackson T. et al., Nature, 352 (1991) 624-628; Marks J.D. et al., J. Mol. Biol., 222 (1992) 581-597; Marks J.D. and Bradbury A., Methods in Molecular Biology, 248 (2003) 161-175; Sidhu S.S. et al., J. Mol. Biol., 338 (2004) 299-310; Lee C.V. et al., J. Mol. Biol., 340 (2004) 1073-1093; Fellouse F.A., Proc. Natl. Acad. Sci. USA, 101 (2004) 12467-12472; и Lee C.V. et al., J. Immunol. Methods, 284 (2004) 119-132.
В некоторых методах фагового дисплея репертуары генов VH и VL клонируют по отдельности, используя полимеразную цепную реакцию (ПЦР), и повторно объединяют случайным образом в фаговых библиотеках, которые затем могут быть подвергнуты скринингу в отношении антигенсвязывающего фага, как описано в Winter G. et al., Ann. Rev. Immunol., 12 (1994) 433-455. Обычно фаг проявляет фрагменты антител либо в виде одноцепочечных Fv (scFv) фрагментов, либо в виде Fab фрагментов. Библиотеки на основе иммунизированных источников обеспечивают получение высокоаффинных в отношении иммуногена антител без необходимости конструирования гибридом. Альтернативно, можно клонировать репертуар интактных антител (например, человека) для обеспечения единого источника антител к разнообразному ассортименту как чужих, так и собственных антигенов, и без какой-либо иммунизации, как описано в Griffiths A.D. et al., EMBO J., 12 (1993) 725-734. И наконец, наивные библиотеки также могут быть созданы синтетическим путем посредством клонирования нереаранжированных сегментов V-генов из стволовых клеток и использования ПЦР-праймеров, содержащих случайную последовательность для кодирования высоко вариабельных участков CDR3 и для завершения реаранжировки in vitro, как описано в Hoogenboom H.R. and Winter G., J. Mol. Biol., 227 (1992) 381-388. Патентные публикации, описывающие фаговые библиотеки человеческих антител, включают, например: US 5750373 и US 2005/0079574, US 2005/0119455, US 2005/0266000, US 2007/0117126, US 2007/0160598, US 2007/0237764, US 2007/0292936 и US 2009/0002360.
Антитела или фрагменты антител, выделенные из библиотек антител человека, считаются в данном описании человеческими антителами или фрагментами человеческих антител.
5. Мультиспецифичные антитела
В некоторых воплощениях антитело, предложенное в данном изобретении, представляет собой мультиспецифичное антитело, например биспецифичное антитело. Мультиспецифичные антитела представляют собой моноклональные антитела, обладающие специфичностями связывания по меньшей мере к двум разным сайтам. Биспецифичные антитела также могут быть использованы для локализации цитотоксических агентов в клетках, которые экспрессируют один или более чем один из целевых антигенов. Биспецифичные антитела могут быть получены в виде полноразмерных антител или фрагментов антител.
Методы получения мультиспецифичных антител включают, но не ограничиваются этим, рекомбинантную коэкспрессию двух пар тяжелая цепь-легкая цепь иммуноглобулина, имеющих разные специфичности (см. Milstein C. and Cuello A.C., Nature, 305 (1983) 537-540, WO 93/08829 и Traunecker A. et al., EMBO J., 10 (1991) 3655-3659), и создание конструкций типа “выступ-во-впадину” (см., например, US 5731168). Мультиспецифичные антитела также могут быть получены путем конструирования электростатических направляющих взаимодействий для создания Fc-гетеродимерных молекул антител (WO 2009/089004); перекрестного сшивания двух или более антител или фрагментов (см., например, US 4676980 и Brennan M. et al., Science, 229 (1985) 81-83); использования лейциновых “застежек” для получения биспецифичных антител (см., например, Kostelny S.A. et al., J. Immunol., 148 (1992) 1547-1553); использования технологии “диател” для получения биспецифичных фрагментов антител (см., например, Hollinger P. et al., Proc. Natl. Acad. Sci. USA, 90 (1993) 6444-6448); использования димеров одноцепочечных Fv (scFv) (см., например Gruber M. et al., J. Immunol., 152 (1994) 5368-5374) и получения триспецифичных антител, как описано, например, в Tutt A. et al., J. Immunol., 147 (1991) 60-69.
Сконструированные антитела с тремя или более функциональными антигенсвязывающими сайтами, включая “антитела-осьминоги (octopus antibodies)”, также включены в данное описание (см., например, US 2006/0025576).
Антитело или фрагмент, упомянутые в данном описании, также включают “Fab двойного действия” или “DAF”, содержащий антигенсвязывающий сайт, который связывается с первым антигеном, а также с другим отличным от него антигеном (см., например, US 2008/0069820).
Антитело или фрагмент, упомянутые в данном описании, также включают мультиспецифичные антитела, описанные в WO 2009/080251, WO 2009/080252, WO 2009/080253, WO 2009/080254, WO 2010/112193, WO 2010/115589, WO 2010/136172, WO 2010/145792 и WO 2010/145793.
6. Варианты Fc-областей и антител
В некоторых воплощениях предусмотрены варианты аминокислотных последовательностей Fc-областей или антител, предложенных согласно данному изобретению. Например, может оказаться желательным улучшить аффинность связывания с антигеном и/или другие биологические свойства антитела. Варианты аминокислотной последовательности Fc-области или антитела могут быть получены посредством внесения соответствующих модификаций в нуклеотидную последовательность, кодирующую антитело, или в результате пептидного синтеза. Такие модификации включают, например, делеции из, и/или вставки в, и/или замены остатков в аминокислотных последовательностях Fc-области или антитела. Для получения окончательной конструкции, при условии, что такая окончательная конструкция будет обладать желаемыми характеристиками, например, антигенсвязывающими свойствами, можно выполнить любую комбинацию делеции, вставки и замены.
a) Варианты с заменами, вставками и делециями
В некоторых воплощениях предложены варианты Fc-областей или антител, имеющие одну или более аминокислотных замен. Сайты, представляющие интерес с точки зрения мутагенеза с введением замен, включают HVR и FR. Консервативные замены показаны в приведенной ниже таблице под заголовком “предпочтительные замены”. Более существенные изменения приведены в следующей далее таблице под заголовком “типичные замены” и, как дополнительно описано ниже, они имеют отношение к классам боковых цепей аминокислот. Аминокислотные замены могут быть внесены в антитело, представляющее интерес, и продукты могут быть подвергнуты скринингу в отношении желаемой активности, например, сохраненного/улучшенного связывания с антигеном, ослабленной иммуногенности или улучшенной ADCC или CDC.
ТАБЛИЦА
Аминокислоты могут быть классифицированы в соответствии с общими свойствами боковой цепи на:
(1) гидрофобные: норлейцин, Met, Ala, Val, Leu, Ile;
(2) нейтральные гидрофильные: Cys, Ser, Thr, Asn, Gln;
(3) кислые: Asp, Glu;
(4) основные: His, Lys, Arg;
(5) содержащие остатки, влияющие на ориентацию цепи: Gly, Pro;
(6) ароматические: Trp, Tyr, Phe.
Неконсервативные замены повлекут за собой замену представителя одного из этих классов на представителя другого класса.
Один из типов варианта с заменами включает замену одного или более остатков в гипервариабельных участках родительской Fc-области или родительского антитела (например, гуманизированного или человеческого антитела). Как правило, полученный(ые) вариант(ы), отобранный(ые) для последующего изучения, будет(ут) иметь модификации (например, улучшения) некоторых биологических свойств (например, повышенную аффинность, сниженную иммуногенность) по сравнению с родительской Fc-областью или родительским антителом и/или будет(ут) иметь существенно сохраненные некоторые биологические свойства родительской Fc-области или родительского антитела. Типичным вариантом с заменами является антитело с “созревшей аффинностью”, которое подходящим образом может быть создано, например, с использованием методов “созревания аффинности”, основанных на применении фагового дисплея, например, изложенных в данном описании. Кратко, вносят мутацию по одному или более остаткам HVR, проявляют вариантные антитела на поверхности фага и проводят скрининг в отношении конкретной биологической активности (например, аффинности связывания).
Изменения (например, замены) могут быть выполнены в HVR, например, для улучшения аффинности антитела. Такие изменения могут быть выполнены в “горячих точках” HVR, т.е. в остатках, кодируемых кодонами, которые с высокой частотой подвержены мутации в процессе соматического созревания (см., например, Chowdhury P.S., Methods Mol. Biol., 207 (2008) 179-196), и/или в остатках, которые контактируют с антигеном, при этом полученный вариант VH или VL тестируют в отношении аффинности связывания. Метод созревания аффинности путем конструирования вторичных библиотек и повторного отбора из них описан, например, Hoogenboom H.R. и др. в Methods in Molecular Biology, 178 (2002) 1-37. В некоторых воплощениях метода созревания аффинности разнообразие в гены вариабельной области, выбранные для созревания, вносят любым из ряда способов (например, посредством допускающей ошибки ПЦР, перетасовки цепей или олигонуклеотид-направленного мутагенеза). Затем создают вторичную библиотеку. После этого проводят скрининг данной библиотеки с целью идентификации любых вариантов антитела с желаемой аффинностью. Другой способ внесения разнообразия включает HVR-направленные подходы, при которых рандомизируют несколько остатков HVR (например, 4-6 остатков одновременно). При этом остатки HVR, вовлеченные в связывание антигена, можно специфически идентифицировать, например, используя аланин-сканирующий мутагенез или моделирование. Особенно часто мишенью становятся CDR-H3 и CDR-L3.
В некоторых воплощениях замены, вставки или делеции могут осуществляться в пределах одного или более HVR при условии, что такие изменения по существу не снижают способности антитела связываться с антигеном. Например, в HVR могут быть выполнены консервативные изменения (например, консервативные замены, предложенные в данном изобретении), которые по существу не снижают аффинности связывания. Такие изменения могут быть, например, вне контактирующих с антигеном остатков в HVR. В некоторых воплощениях предложенных выше последовательностей вариантов VH и VL каждый HVR либо не изменен, либо содержит не более одной аминокислотной замены, двух или трех аминокислотных замен.
Полезный метод идентификации остатков или участков Fc-области или антитела, которые могут быть мишенью для мутагенеза, называется “аланин-сканирующим мутагенезом”, описанным в Cunningham B.C. and Wells J.A., Science, 244 (1989) 1081-1085. В этом методе идентифицируют остаток или группу целевых остатков (например, заряженных остатков, таких как остатки arg, asp, his, lys и glu) и заменяют на остатки нейтральной или отрицательно заряженной аминокислоты (например, аланина или полиаланина), чтобы определить, повлияет ли это на желаемое биологическое свойство, такое как, например, взаимодействие антитела с антигеном. В положениях аминокислот, демонстрирующих функциональную чувствительность к первоначальным заменам, могут быть выполнены дальнейшие замены. Альтернативно или помимо этого, для идентификации точек контакта можно использовать кристаллическую структуру комплекса, содержащего Fc-область или антитело. Такие контактирующие остатки и соседние с ними остатки могут быть выявлены или устранены как кандидаты на замену. Варианты можно подвергнуть скринингу, чтобы определить, имеют ли они желаемые свойства.
Вставки в аминокислотную последовательность включают присоединяемые к амино- и/или карбоксильному концу вставки длиной от одного остатка до полипептидов, содержащих сто или более остатков, а также вставки одного или нескольких аминокислотных остатков внутрь последовательности. Примеры концевых вставок включают Fc-область или антитело с N-концевым метионильным остатком. Другие варианты молекулы Fc-области или антитела со вставками включают слитую конструкцию, в которой N- или C-конец Fc-области или антитела соединен с ферментом (например, для антитело-опосредованной терапии с использованием ферментов и пролекарств (ADEPT)) или полипептидом, что увеличивает полупериод существования Fc-области или антитела в сыворотке крови.
b) Варианты гликозилирования
В некоторых воплощениях Fc-область или антитело, предложенные согласно данному изобретению, изменяют с целью увеличения или уменьшения степени гликозилирования Fc-области или антитела. Добавление или удаление сайтов гликозилирования Fc-области или антитела можно подходящим образом осуществлять путем изменения аминокислотной последовательности таким образом, чтобы создавался или удалялся один или более чем один сайт гликозилирования.
В тех случаях, когда антитело содержит Fc-область, присоединенный к ней углевод может быть изменен. Нативные Fc-области или антитела, продуцируемые клетками млекопитающих, обычно содержат разветвленный, биантенный олигосахарид, как правило, присоединенный посредством N-гликозидной связи к Asn297 CH2-домена Fc-области. См., например, Wright A. and Morrison S.L., TIBTECH, 15 (1997) 26-32. Олигосахарид может включать различные углеводы, например, маннозу, N-ацетилглюкозамин (GlcNAc), галактозу и сиаловую кислоту, а также фукозу, присоединенную к GlcNAc в “стволе” биантенной структуры олигосахарида. В некоторых воплощениях могут быть выполнены модификации олигосахарида в Fc-области или антителе, описанных в данной заявке, с целью создания вариантов антитела с определенными улучшенными свойствами.
В одном из воплощений предложены варианты антитела, содержащие углеводную структуру с недостаточным числом остатков фукозы, присоединенных (непосредственно или опосредованно) к Fc-области. Например, количество остатков фукозы в таком антителе может составлять от 1% до 80%, от 1% до 65%, от 5% до 65% или от 20% до 40%. Количество остатков фукозы определяют, подсчитывая среднее количество остатков фукозы в углеводной цепи при Asn297 относительно общего количества всех гликоструктур, присоединенных к Asn 297 (например, комплексных, гибридных и высокоманнозных структур) по результатам измерений с использованием время-пролетной масс-спектрометрии с ионизацией посредством лазерной десорбции ионов из матрицы (MALDI-TOF масс-спектрометрии), как описано, например в WO 2008/077546. Asn297 относится к остатку аспарагина, локализованному примерно в положении 297 в Fc-области (нумерация остатков Fc-области соответствует EU-индексу); однако, Asn297 также может быть локализован на расстоянии примерно ± 3 аминокислоты вверх по течению или вниз по течению относительно положения 297, т.е. между положениями 294 и 300, вследствие незначительных вариаций последовательности антител. Такие фукозилированные варианты могут иметь улучшенную ADCC-функцию. См., например, US 2003/0157108, US 2004/0093621. Примеры публикаций, касающихся “дефукозилированных” или “дефицитных по фукозе” вариантов антител, включают: US 2003/0157108; WO 2000/61739; WO 2001/29246; US 2003/0115614; US 2002/0164328; US 2004/0093621; US 2004/0132140; US 2004/0110704; US 2004/0110282; US 2004/0109865; WO 2003/085119; WO 2003/084570; WO 2005/035586; WO 2005/035778; WO 2005/053742; WO 2002/031140; Okazaki A. et al., J. Mol. Biol., 336 (2004) 1239-1249; Yamane-Ohnuki N. et al., Biotech. Bioeng., 87 (2004) 614-622. Примеры клеточных линий, способных продуцировать дефукозилированные антитела, включают клетки CHO линии Lec13, дефицитные по фукозилированию белка (Ripka J., et al., Arch. Biochem. Biophys., 249 (1986) 533-545; US 2003/0157108 и WO 2004/056312, в частности, пример 11), и нокаутные клеточные линии, как например, клетки CHO, нокаутные по гену альфа-1,6-фукозилтрансферазы, FUT8 (см., например, Yamane-Ohnuki N., et al., Biotech. Bioeng., 87 (2004) 614-622; Kanda Y. et al., Biotechnol. Bioeng., 94 (2006) 680-688 и WO 2003/085107).
Также предложены варианты Fc-области или антитела с раздвоенными олигосахаридами, например, в которых биантенный олигосахарид, присоединенный к Fc-области антитела, имеет точки ветвления при GlcNAc. Такие варианты Fc-области или антитела могут иметь более низкую степень фукозилирования и/или улучшенную ADCC-функцию. Примеры таких вариантов антитела описаны, например, в WO 2003/011878, US 6602684 и US 2005/0123546. Также предложены варианты Fc-области или антитела по меньшей мере с одним остатком галактозы в олигосахариде, присоединенном к Fc-области. Такие варианты антитела могут иметь улучшенную CDC-функцию. Такие варианты антитела описаны, например, в WO 1997/30087, WO 1998/58964 и WO 1999/22764.
c) Варианты Fc-области
В некоторых воплощениях в Fc-область, предложенную в данном изобретении, может быть введена одна или более чем одна аминокислотная модификация с получением тем самым варианта Fc-области. Вариант Fc-области может содержать последовательность Fc-области антитела человека (например, Fc-область IgG1, IgG2, IgG3 или IgG4 человека), содержащую аминокислотную модификацию (например, замену/мутацию) в одном или более положениях аминокислот.
В некоторых воплощениях изобретения предусматривается вариант Fc-области, обладающий некоторыми, но не всеми эффекторными функциями, делающими его желаемым кандидатом для практических применений, при которых полупериод существования Fc-области in vivo является важным, в то время как некоторые эффекторные функции (такие как CDC и ADCC) не являются необходимыми или являются вредными. Для подтверждения снижения/истощения CDC- и/или ADCC-активностей можно провести анализы цитотоксичности in vitro и/или in vivo. Например, можно провести анализы связывания с Fc-рецепторами (FcR), чтобы убедиться, что у Fc-области или антитела отсутствует способность связываться с FcγR (поэтому, вероятнее всего, у них отсутствует ADCC-активность), но сохраняется способность связываться с FcRn. Первичные клетки для опосредования ADCC, NK-клетки (природные киллерные клетки), экспрессируют только FcγRIII, в то время как моноциты экспрессируют FcγRI, FcγRII и FcγRIII. Данные по экспрессии FcR на гематопоэтических клетках суммированы в Таблице 3 на странице 464 в Ravetch J.V. and Kinet J.P., Annu. Rev. Immunol., 9 (1991) 457-492. Неограничивающие примеры анализов in vitro для оценки ADCC-активности молекулы, представляющей интерес, описаны в US 5500362 (см., например, Hellstrom I. et al., Proc. Nat'l Acad. Sci. USA, 83 (1986) 7059-7063, и Hellstrom I. et al., Proc. Nat'l Acad. Sci. USA, 82 (1985) 1499-1502); US 5821337 (см. Bruggemann M. et al., J. Exp. Med., 166 (1987) 1351-1361). Альтернативно, можно применять нерадиоактивные методы анализа (см., например, анализ цитотоксичности ACTI™ для проточной цитометрии без использования радиоактивности (CellTechnology, Inc., Mountain View, CA) и анализ цитотоксичности CytoTox 96® без использования радиоактивности (Promega, Madison, WI)). Полезные для таких анализов эффекторные клетки включают мононуклеарные клетки периферической крови (PBMC) и природные киллерные (NK) клетки. Альтернативно или помимо этого, ADCC-активность молекулы, представляющей интерес, можно оценить in vivo, например, в животной модели, такой как описанная в Clynes et al., Proc. Nat'l Acad. Sci. USA, 95 (1998) 652-656. Также можно провести анализы связывания с C1q для подтверждения того, что Fc-область или антитело не обладает способностью связываться с C1q и, следовательно, не имеет CDC-активности. См., например, данные иммуноферментного твердофазного анализа (ELISA) для связывания с C1q и C3c в WO 2006/029879 и WO 2005/100402. Для оценки активации комплемента можно провести анализ CDC (см., например, Gazzano-Santoro H. et al., J. Immunol. Methods, 202 (1996) 163-171; Cragg M.S. et al., Blood, 101 (2003) 1045-1052; и Cragg M.S. and M.J. Glennie, Blood, 103 (2004) 2738-2743). Определения связывания с FcRn и клиренса/полупериода существования in vivo также могут быть выполнены с использованием методов, известных в данной области техники (см., например, Petkova S.B. et al., Int. Immunol. 18 (2006) 1759-1769).
Fc-области или антитела с пониженной эффекторной функцией включают антитела с заменой одного или более остатков в Fc-области в положениях 238, 265, 269, 270, 297, 327 и 329 (US 6737056). Такие варианты Fc-области включают Fc-области с заменами аминокислот в двух или более положениях 265, 269, 270, 297 и 327, включая так называемый мутант Fc-области “DANA” с заменой остатков 265 и 297 на аланин (US 7332581).
Описаны некоторые варианты Fc-области или антитела с улучшенным или ослабленным связыванием с FcR (см., например, US 6737056, WO 2004/056312 и Shields R.L. et al., J. Biol. Chem., 276 (2001) 6591-6604).
В некоторых воплощениях вариант Fc-области или антитела содержит Fc-область с одной или более аминокислотными заменами, которые улучшают ADCC, например, заменами в положениях 298, 333 и/или 334 в Fc-области (нумерация остатков соответствует EU-индексу).
В некоторых воплощениях в Fc-области выполнены изменения, которые приводят к изменению (т.е. или к улучшению, или к ослаблению) связывания с C1q и/или комплемент-зависимой цитотоксичности (CDC), например, как описано в US 6194551, WO 99/51642 и в Idusogie E.E. et al., J. Immunol., 164 (2000) 4178-4184.
Антитела с увеличенными полупериодами существования и улучшенным связыванием с неонатальным Fc-рецептором (FcRn), который отвечает за перенос материнских IgG к плоду (Guyer R.L. et al., J. Immunol., 117 (1976) 587-593; и Kim J.K. et al., J. Immunol., 24 (1994) 2429-2434), описаны в US 2005/0014934. Эти антитела содержат Fc-область с одной или более заменами в ней, которые улучшают связывание Fc-области с FcRn. Такие варианты Fc-области включают варианты с заменами одного или более остатков Fc-области в положениях: 238, 256, 265, 272, 286, 303, 305, 307, 311, 312, 317, 340, 356, 360, 362, 376, 378, 380, 382, 413, 424 или 434, например, с заменой остатка 434 Fc-области (US 7371826).
Относительно других примеров вариантов Fc-области также см. Duncan A.R. and Winter G., Nature, 322 (1988) 738-740, US 5648260, US 5624821 и WO 94/29351.
d) Варианты антител, сконструированных с цистеином
В некоторых воплощениях может быть желательно создать сконструированные с цистеином антитела, например, “тиоMAb”, в которых один или более остатков аминокислот в антителе заменены на остатки цистеина. В конкретных воплощениях заменяемые остатки находятся в доступных сайтах антитела. В результате замены этих остатков на цистеин реакционно-способные тиоловые группы будут тем самым располагаться в доступных сайтах антитела и могут быть использованы для конъюгирования антитела с другими группировками, такими как группировки лекарственного средства или группировки соединенного с линкером лекарственного средства, с целью создания иммуноконъюгата, как описано далее в данной заявке. В некоторых воплощениях могут быть заменены на цистеин какой-либо один или более из следующих остатков: V205 (нумерация по Kabat) в легкой цепи; A118 (нумерация согласно EU) в тяжелой цепи и S400 (нумерация согласно EU) в Fc-области тяжелой цепи. Сконструированные с цистеином антитела могут быть созданы так, как описано, например, в US 7521541.
e) Производные Fc-области и антител
В некоторых воплощениях Fc-область или антитело, предложенные в данном изобретении, могут быть дополнительно модифицированы с тем, чтобы они содержали дополнительные небелковые группировки, которые известны в данной области техники и легко доступны. Группировки, подходящие для получения производных Fc-области или антитела, включают, но не ограничиваются этим, водорастворимые полимеры. Неограничивающие примеры водорастворимых полимеров включают, но не ограничиваются этим, полиэтиленгликоль (ПЭГ), coполимеры этиленгликоля/пропиленгликоля, карбоксиметилцеллюлозу, декстран, поливиниловый спирт, поливинилпирролидон, поли-1,3-диоксолан, поли-1,3,6-триоксан, coполимер этилена и малеинового ангидрида, полиаминокислоты (или гомополимеры, или статистические coполимеры), поли(н-винилпирролидон)полиэтиленгликоль, гомополимеры пропиленгликоля, сополимеры полипропиленоксида/этиленоксида, полиоксиэтилированные полиолы (например, глицерин), и их смеси. Полиэтиленгликоль-пропиональдегид может иметь преимущества при получении ввиду его стабильности в воде. Полимер может иметь любую молекулярную массу и может быть разветвленным или неразветвленным. Число полимеров, присоединенных к Fc-области или антителу, может варьировать и, если присоединено более одного полимера, они могут представлять собой одинаковые или разные молекулы. В общем случае число и/или тип полимеров, используемых для получения производных, можно определить исходя из соображений, включающих, но не ограничивающихся этим, конкретные свойства или функции Fc-области или антитела, подлежащие улучшению, а также будет ли производное Fc-области или антитела использоваться в терапии определенных состояний и т.д.
В другом воплощении предложены конъюгаты Fc-области или антитела и одной или более чем одной небелковой группировки, которая может избирательно нагреваться под действием излучения. В одном из воплощений небелковая группировка представляет собой углеродную нанотрубку (Kam N.W. et al., Proc. Natl. Acad. Sci. USA, 102 (2005) 11600-11605). Излучение может иметь любую длину волны и включает, но не ограничивается этим, длины волн, которые не вредят обычным клеткам, но которые нагревают небелковую группировку до температуры, при которой клетки, расположенные в непосредственной близости от конъюгата антитела и небелковой группировки, погибают.
f) Гетеродимеризация
Имеется несколько подходов к CH3-модификациям для должного обеспечения гетеродимеризации, которые хорошо описаны, например, в WO 96/27011, WO 98/050431, EP 1870459, WO 2007/110205, WO 2007/147901, WO 2009/089004, WO 2010/129304, WO 2011/90754, WO 2011/143545, WO 2012058768, WO 2013157954, WO 2013096291. Обычно во всех таких подходах первый CH3-домен и второй CH3-домен оба сконструированы по комплементарному типу с тем, чтобы каждый CH3-домен (или каждая содержащая его тяжелая цепь) больше не могли гомодимеризоваться сами с собой, а инициировали гетеродимеризацию с комплементарно сконструированным другим CH3-доменом (для того, чтобы первый и второй CH3-домен гетеродимеризовались, и никаких гомодимеров между двумя первыми или двумя вторыми CH3-доменами не образовывалось). Применение этих разных подходов с целью улучшения гетеродимеризации тяжелых цепей в мультиспецифичных антителах по изобретению предусматривается в виде разных альтернативных вариантов в комбинации с модификациями тяжелой-легкой цепи (обмен/замена в VH и VL в одном связывающемся плече и внесение в качестве замен заряженных аминокислот с противоположными зарядами в поверхность контакта CH1/CL), что снижает вероятность ненадлежащего образования пар легких цепей как побочных продуктов типа белка Бенс-Джонса.
В одном из предпочтительных воплощений изобретения (в случае, если мультиспецифичное антитело содержит CH3-домены в тяжелых цепях) CH3-домены указанного мультиспецифичного антитела по изобретению могут быть изменены с использованием технологии “выступ-во-впадину”, которая описана подробно на нескольких примерах, например, в WO 96/027011, Ridgway J.B., et al., Protein Eng., 9 (1996) 617-621; и Merchant A.M., et al., Nat. Biotechnol., 16 (1998) 677-681; WO 98/050431. В этом методе взаимодействующие поверхности двух CH3-доменов изменяют с целью усиления гетеродимеризации обеих тяжелых цепей, содержащих эти два CH3-домена. Каждый из двух CH3-доменов (двух тяжелых цепей) может представлять собой “выступ”, в то время как другой представляет собой “впадину”. Кроме того, введение дисульфидного мостика стабилизирует гетеродимеры (Merchant A.M, et al., Nature Biotech., 16 (1998) 677-681, Atwell S., et al. J. Mol. Biol., 270 (1997) 26-35) и повышает выход.
Так, в одном из воплощений изобретения указанное мультиспецифичное антитело (содержит CH3-домен в каждой тяжелой цепи и) дополнительно характеризуется тем, что:
первый CH3-домен первой тяжелой цепи антитела по пункту (a) и второй CH3-домен второй тяжелой цепи антитела по пункту (b) оба встречаются на поверхности контакта, которая представляет собой исходную поверхность контакта между CH3-доменами антитела,
причем указанная поверхность контакта изменена, чтобы стимулировать образование мультиспецифичного антитела, при этом данное изменение характеризуется тем, что:
1) CH3-домен одной тяжелой цепи изменен
так, что в исходной поверхности контакта CH3-домена одной тяжелой цепи, которая встречается c исходной поверхностью контакта CH3-домена другой тяжелой цепи в мультиспецифичном антителе,
аминокислотный остаток заменен на аминокислотный остаток, имеющий боковую цепь большего объема, тем самым создавая выступающую часть в поверхности контакта CH3-домена одной тяжелой цепи, которая располагается в полости поверхности контакта CH3-домена другой тяжелой цепи,
и
2) CH3-домен другой тяжелой цепи изменен
так, что в исходной поверхности контакта второго CH3-домена, которая встречается c исходной поверхностью контакта первого CH3-домена в мультиспецифичном антителе,
аминокислотный остаток заменен на аминокислотный остаток, имеющий боковую цепь меньшего объема, тем самым создавая выступающую часть в поверхности контакта второго CH3-домена, которая располагается в полости поверхности контакта первого CH3-домена.
Предпочтительно, чтобы указанный аминокислотный остаток, имеющий боковую цепь большего объема, был выбран из группы, состоящей из аргинина (R), фенилаланина (F), тирозина (Y), триптофана (W).
Предпочтительно, чтобы указанный аминокислотный остаток, имеющий боковую цепь меньшего объема, был выбран из группы, состоящей из аланина (A), серина (S), треонина (T), валина (V).
Согласно одному из аспектов изобретения оба CH3-домена дополнительно изменены посредством введения остатка аминокислоты цистеин (C) в соответствующие положения каждого CH3-домена для того, чтобы мог образовываться дисульфидный мостик между обоими CH3-доменами.
В одном из предпочтительных воплощений изобретения указанное мультиспецифичное антитело содержит аминокислотную мутацию T366W в первом CH3-домене “цепи, формирующей выступ” и аминокислотные мутации T366S, L368A, Y407V во втором CH3-домене “цепи, формирующей впадину”. Также можно использовать дополнительный межцепочечный дисульфидный мостик между CH3-доменами (Merchant A.M., et al., Nature Biotech., 16 (1998) 677-681), например, путем внесения аминокислотной мутации Y349C в CH3-домен “цепи, формирующей впадину” и аминокислотной мутации E356C или аминокислотной мутации S354C в CH3-домен “цепи, формирующей выступы”.
В одном из предпочтительных воплощений указанное мультиспецифичное антитело (которое содержит CH3-домен в каждой тяжелой цепи) содержит аминокислотные мутации S354C, T366W в одном из двух CH3-доменов и аминокислотные мутации Y349C, T366S, L368A, Y407V в другом из двух CH3-доменов (дополнительную аминокислотную мутацию S354C в одном CH3-домене и дополнительную аминокислотную мутацию Y349C в другом CH3-домене, формирующие межцепочечный дисульфидный мостик) (нумерация согласно Kabat).
Другие способы CH3-модификаций для усиления гетеродимеризации включены в качестве альтернативных вариантов изобретения и описаны, например, в WO 96/27011, WO 98/050431, EP 1870459, WO 2007/110205, WO 2007/147901, WO 2009/089004, WO 2010/129304, WO 2011/90754, WO 2011/143545, WO 2012/058768, WO 2013/157954, WO 2013/096291.
В одном из воплощений, альтернативно, можно использовать связанный с гетеродимеризацией подход, описанный в EP 1870459 A1. Этот подход основан на внесении замен/мутаций заряженных аминокислот с противоположным зарядом в конкретные аминокислотные положения на поверхности контакта CH3/CH3-доменов между обеими тяжелыми цепями. Одно из предпочтительных воплощений для указанного мультиспецифичного антитела относится к аминокислотным мутациям R409D, K370E в первом CH3-домене (мультиспецифичного антитела) и аминокислотным мутациям D399K, E357K во втором CH3-домене мультиспецифичного антитела (нумерация согласно Kabat).
В другом воплощении указанное мультиспецифичное антитело содержит аминокислотную мутацию T366W в CH3-домене “цепи, формирующей выступы” и аминокислотные мутации T366S, L368A, Y407V в CH3-домене “цепи, формирующей впадину” и помимо этого аминокислотные мутации R409D, K370E в CH3-домене “цепи, формирующей выступы” и аминокислотные мутации D399K, E357K в CH3-домене “цепи, формирующей впадину”.
В другом воплощении указанное мультиспецифичное антитело содержит аминокислотные мутации S354C, T366W в одном из двух CH3-доменов и аминокислотные мутации Y349C, T366S, L368A, Y407V в другом из двух CH3-доменов или указанное мультиспецифичное антитело содержит аминокислотные мутации Y349C, T366W в одном из двух CH3-доменов и аминокислотные мутации S354C, T366S, L368A, Y407V в другом из двух CH3-доменов и помимо этого аминокислотные мутации R409D, K370E в CH3-домене “цепи, формирующей выступы” и аминокислотные мутации D399K, E357K в CH3-домене “цепи, формирующей впадину”.
В одном из воплощений, альтернативно, можно использовать подход с применением гетеродимеризации, описанный в WO2013/157953. В одном из воплощений первый CH3-домен содержит аминокислотную мутацию T366K, а полипептид второго CH3-домена содержит аминокислотную мутацию L351D. В следующем воплощении первый CH3-домен содержит другую аминокислотную мутацию L351K. В следующем воплощении второй CH3-домен содержит другую аминокислотную мутацию, выбранную из Y349E, Y349D и L368E (предпочтительно L368E).
В одном из воплощений, альтернативно, можно использовать подход с применением гетеродимеризации, описанный в WO2012/058768. В одном из воплощений первый CH3-домен содержит аминокислотные мутации L351Y, Y407A, а второй CH3-домен содержит аминокислотные мутации T366A, K409F. В следующем воплощении второй CH3-домен содержит другую аминокислотную мутацию в положении T411, D399, S400, F405, N390 или K392, например, выбранную из a) T411N, T411R, T411Q, T411K, T411D, T411E или T411W, b) D399R, D399W, D399Y или D399K, c) S400E, S400D, S400R или S400K, d) F405I, F405M, F405T, F405S, F405V или F405W, e) N390R, N390K или N390D, f) K392V, K392M, K392R, K392L, K392F или K392E. В следующем воплощении первый CH3-домен содержит аминокислотные мутации L351Y, Y407A, а второй CH3-домен содержит аминокислотные мутации T366V, K409F. В следующем воплощении первый CH3-домен содержит аминокислотную мутацию Y407A, а второй CH3-домен содержит аминокислотные мутации T366A, K409F. В следующем воплощении второй CH3-домен содержит другие аминокислотные мутации K392E, T411E, D399R и S400R.
В одном из воплощений, альтернативно, можно использовать подход с применением гетеродимеризации, описанный в WO2011/143545, например, проводя модификацию аминокислот в положении, выбранном из группы, состоящей из положений 368 и 409.
В одном из воплощений, альтернативно, можно использовать подход с применением гетеродимеризации, описанный в WO2011/090762, где также используется изложенная выше технология “выступы-во-впадины”. В одном из воплощений первый CH3-домен содержит аминокислотную мутацию T366W, а второй CH3-домен содержит аминокислотную мутацию Y407A. В одном из воплощений первый CH3-домен содержит аминокислотную мутацию T366Y, а второй CH3-домен содержит аминокислотную мутацию Y407T.
В одном из воплощений мультиспецифичное антитело происходит из изотипа IgG2, и, альтернативно, можно использовать подход с применением гетеродимеризации, описанный в WO2010/129304.
В одном из воплощений, альтернативно, можно использовать подход с применением гетеродимеризации, описанный в WO2009/089004. В одном из воплощений первый CH3-домен содержит замену аминокислоты K392 или N392 на отрицательно заряженную аминокислоту (например, на глутаминовую кислоту (E) или аспарагиновую кислоту (D), предпочтительно замену K392D или N392D), а второй CH3-домен содержит замену аминокислоты D399, E356, D356 или E357 на положительно заряженную аминокислоту (например, на лизин (K) или аргинин (R), предпочтительно замену D399K, E356K, D356K или E357K и более предпочтительно D399K и E356K. В следующем воплощении первый CH3-домен дополнительно содержит замену аминокислоты K409 или R409 на отрицательно заряженную аминокислоту (например, на глутаминовую кислоту (E) или аспарагиновую кислоту (D), предпочтительно замену K409D или R409D). В следующем воплощении первый CH3-домен дополнительно или альтернативно содержит замену аминокислоты K439 и/или K370 на отрицательно заряженную аминокислоту (например, на глутаминовую кислоту (E) или аспарагиновую кислоту (D)).
В одном из воплощений, альтернативно, можно использовать подход с применением гетеродимеризации, описанный в WO2007/147901. В одном из воплощений первый CH3-домен содержит аминокислотные мутации K253E, D282K и K322D, а второй CH3-домен содержит аминокислотные мутации D239K, E240K и K292D.
В одном из воплощений, альтернативно, можно использовать подход с применением гетеродимеризации, описанный в WO2007/110205.
Рекомбинантные методы и технологии изготовления препаратов
Fc-области и антитела могут быть получены с использованием рекомбинантных методов и технологий изготовления препаратов, например, как описано в US 4816567. В одном из воплощений предложена(ы) выделенная(ые) нуклеиновая(ые) кислота(ы), кодирующая(ие) Fc-область или антитело, описанные в данной заявке. Такая нуклеиновая кислота может кодировать аминокислотную последовательность, содержащую последовательность VL, и/или аминокислотную последовательность, содержащую последовательность VH антитела (например, легкой и/или тяжелой цепей антитела). В следующем воплощении предложены один или более векторов (например, экспрессирующих векторов), содержащих такую нуклеиновую кислоту. В следующем воплощении предложена клетка-хозяин, содержащая такую нуклеиновую кислоту. В одном из таких воплощений клетка-хозяин содержит (например, трансфицирована нижеперечисленным): (1) вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VL антитела, и аминокислотную последовательность, содержащую VH антитела, или (2) первый вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VL антитела, и второй вектор, содержащий нуклеиновую кислоту, которая кодирует аминокислотную последовательность, содержащую VH антитела. В одном из воплощений клетка-хозяин представляет собой эукариотическую клетку, например клетку яичников китайского хомячка (CHO) или лимфоидную клетку (например, клетку Y0, NS0, Sp20). В одном из воплощений предложен способ получения антитела, описанного в данной заявке, включающий культивирование клетки хозяина, содержащей нуклеиновую кислоту, кодирующую антитело, предложенное выше, в условиях, подходящих для экспрессии данного антитела, и возможно извлечение антитела из клетки хозяина (или среды культивирования клетки хозяина).
Чтобы получить рекомбинантными методами вариантную Fc-область, нуклеиновую кислоту, кодирующую вариантную Fc-область, например, которая описана выше, выделяют и встраивают в один или более векторов для дальнейшего клонирования и/или экспрессии в клетке хозяина. Такую нуклеиновую кислоту можно легко выделить и секвенировать, используя традиционные методики (например, используя олигонуклеотидные зонды, которые способны специфически связываться с генами, кодирующими полипептиды вариантной Fc-области или тяжелую и легкую цепи антитела).
Клетки хозяина, подходящие для клонирования или экспрессии векторов, кодирующих антитела, включают прокариотические или эукариотические клетки, описанные в данной заявке. Например, антитела могут продуцироваться бактериями, в частности, когда нет необходимости в гликозилировании и Fc-эффекторной функции. Для экспрессии фрагментов антител и полипептидов в бактериях см., например, US 5648237, US 5789199 и US 5840523 (см. также Charlton K.A. в Methods in Molecular Biology, Vol. 248, Lo B.K.C. (ed.), Humana Press, Totowa, NJ (2003), pp. 245-254, где описывается экспрессия фрагментов антител в E. coli). После экспрессии антитело можно выделять из бактериальной клеточной массы в растворимой фракции и можно очищать далее.
Помимо прокариотов подходящими хозяевами для клонирования или экспрессии векторов, кодирующих антитела, являются эукариотические микроорганизмы, такие как мицелиальные грибы или дрожжи, включая штаммы грибов и дрожжей, пути гликозилирования в которых “гуманизированы”, что приводит к получению антитела, частично или полностью воспроизводящего картину гликозилирования у человека. См. Gerngross T.U., Nat. Biotech., 22 (2004), 1409-1414 и Li H. et al., Nat. Biotech., 24 (2006), 210-215.
Кроме того, подходящие клетки хозяина для экспрессии гликозилированной Fc-области или гликозилированного антитела происходят из многоклеточных организмов (беспозвоночных и позвоночных). Примеры клеток беспозвоночных включают клетки растений и насекомых. Идентифицированы многочисленные бакуловирусные штаммы, которые могут быть использованы вместе с клетками насекомых, в частности, для трансфекции клеток Spodoptera frugiperda.
В качестве хозяев также можно использовать культуры растительных клеток. См., например, US 5959177, US 6040498, US 6420548, US 7125978 и US 6417429 (описывающие технологию PLANTIBODIES™ для получения антител в трансгенных растениях).
В качестве хозяев также можно использовать клетки позвоночных. Например, могут быть полезны линии клеток млекопитающих, адаптированные к росту в суспензии. Другими примерами полезных в качестве хозяев линий клеток млекопитающих являются линия клеток CV1 почки обезьяны, трансфицированная SV40 (обезьяний вирус 40) (COS-7); линия клеток почки эмбриона человека (клетки HEK293 или 293, которые описаны, например, в Graham F.L. et al., J. Gen. Virol., 36 (1977) 59-74); клетки почки новорожденного хомячка (BHK); клетки Сертоли у мышей (клетки TM4, которые описаны, например, в Mather J.P., Biol. Reprod., 23 (1980) 243-251); клетки почки обезьяны (CV1); клетки почки африканской зеленой мартышки (VERO-76); клетки карциномы шейки матки человека (HELA); клетки почки собаки (MDCK); клетки печени крысы линии Buffalo (BRL 3A); клетки легкого человека (W138); клетки печени человека (Hep G2); клетки опухоли молочной железы мышей (MMT 060562); клетки TRI, которые описаны, например, в Mather J.P. et al., Annals N.Y. Acad. Sci., 383 (1982) 44-68; клетки MRC 5 и клетки FS4. Другие полезные линии клеток млекопитающих в качестве клеток хозяина включают клетки яичников китайского хомячка (CHO), в том числе DHFR- (дефицитные по дигидрофолатредуктазе) клетки CHO (Urlaub G. et al., Proc. Natl. Acad. Sci. USA, 77 (1980) 4216-4220), и клеточные линии миеломы, такие как Y0, NS0 и Sp2/0. Обзор некоторых линий клеток млекопитающих в качестве клеток хозяина, подходящих для получения антител, см., например, в Yazaki P. and Wu A.M., Methods in Molecular Biology, Vol. 248, Lo B.K.C. (ed.), Humana Press, Totowa, NJ (2004), pp. 255-268.
Анализы
В отношении антител, предложенных в данном изобретении, можно провести идентификацию, скрининг или охарактеризовать их по физико-химическим свойствам и/или биологическим активностям, используя различные виды анализов, известных в данной области техники.
Согласно одному из аспектов антитело, описанное в данной заявке, тестируют по его антигенсвязывающей активности, например, известными методами, такими как ELISA, вестерн-блоттинг и т.д.
Иммуноконъюгаты
Согласно изобретению также предложены иммуноконъюгаты, содержащие антитело, описанное в данной заявке, конъюгированное с одним или более цитотоксическими агентами, такими как химиотерапевтические агенты или лекарственные средства, агенты, подавляющие рост, токсины (например, белковые токсины, обладающие ферментативной активностью токсины бактериального, грибного, растительного или животного происхождения или их фрагменты) или радиоактивные изотопы.
В одном из воплощений иммуноконъюгат представляет собой конъюгат антитело-лекарственное средство (ADC), в котором антитело конъюгировано с одним или более чем одним из лекарственных средств, включая, но не ограничиваясь этим, майтанзиноид (см. US 5208020, US 5416064 и EP 0425235 B1); ауристатин, такой как лекарственные группировки DE и DF монометилауристатина (MMAE и MMAF) (см. US 5635483, US 5780588 и US 7498298); доластатин; калихемицин или его производное (см. US 5712374, US 5714586, US 5739116, US 5767285, US 5770701, US 5770710, US 5773001 и US 5877296; Hinman L.M. et al., Cancer Res., 53 (1993) 3336-3342; и Lode H.N. et al., Cancer Res., 58 (1998) 2925-2928); антрациклин, такой как дауномицин или доксорубицин (см. Kratz F. et al., Current Med. Chem., 13 (2006) 477-523; Jeffrey S.C. et al., Bioorg. Med. Chem. Lett., 16 (2006) 358-362; Torgov M.Y. et al., Bioconjug. Chem., 16 (2005) 717-721; Nagy A. et al., Proc. Natl. Acad. Sci. USA, 97 (2000) 829-834; Dubowchik G.M. et al., Bioorg. & Med. Chem. Letters, 12 (2002) 1529-1532; King H.D. et al., J. Med. Chem., 45 (2002) 4336-4343; и US 6630579); метотрексат; виндезин; таксан, такой как доцетаксел, паклитаксел, ларотаксел, тезетаксел и ортатаксел; трихотецен и CC1065.
В другом воплощении иммуноконъюгат содержит антитело, описанное в данной заявке, конъюгированное с обладающим ферментативной активностью токсином или его фрагментом, включая, но не ограничиваясь этим, A-цепь дифтерийного токсина, несвязывающие активные фрагменты дифтерийного токсина, A-цепь экзотоксина (из Pseudomonas aeruginosa), A-цепь рицина, A-цепь абрина, A-цепь модессина, альфа-сарцин, белки китайского тунгового дерева, диантиновые белки, белки лаконоса американского (PAPI, PAPII и PAP-S), ингибитор из момордики харантской, курцин, кротин, ингибитор из мыльнянки лекарственной, гелонин, митогелин, рестриктоцин, феномицин, эномицин и трихотецены.
В другом воплощении иммуноконъюгат содержит антитело, описанное в данной заявке, конъюгированное с радиоактивным атомом с образованием радиоактивного конъюгата. Для получения радиоактивных конъюгатов доступны различные радиоактивные изотопы. Примеры включают At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb212 и радиоактивные изотопы Lu. В том случае, когда для детекции используют радиоактивный конъюгат, он может содержать радиоактивный атом для сцинтиграфических исследований, например Tc99m или I123, или спиновую метку для визуализации с применением ядерного магнитного резонанса (ЯМР) (также известной как магнитно-резонансная визуализация, MRI), такую как йод-123, йод-131, индий-111, фтор-19, углерод-13, азот-15, кислород-17, гадолиний, марганец или железо.
Конъюгаты антитела и цитотоксического агента могут быть получены с использованием целого ряда бифункциональных агентов для сочетания белков, таких как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные сложных имидоэфиров (такие как диметиладипимидат HCl), активированные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаровый альдегид), бис-азидо-соединения (такие как бис(пара-азидобензоил)гександиамин), производные на основе бис-диазония (такие как бис-(пара-диазонийбензоил)этилендиамин), диизоцианаты (такие как толуол-2,6-диизоцианат) и активные бис-фторсодержащие соединения (такие как 1,5-дифтор-2,4-динитробензол). Например, иммунотоксин на основе рицина может быть получен так, как описано в Vitetta E.S. et al., Science, 238 (1987) 1098-1104. Меченная углеродом-14 1-изотиоцианатобензил-3-метилдиэтилентриаминпентауксусная кислота (МХ-DTPA) является типичным хелатирующим агентом для конъюгирования радионуклида с антителом. См. WO 94/11026. Линкер может быть “расщепляемым линкером”, способствующим высвобождению цитотоксического лекарственного средства в клетке. Например, можно использовать кислотолабильный линкер, чувствительный к пептидазам линкер, фотолабильный линкер, диметил- или дисульфид-содержащий линкер (Chari R.V. et al., Cancer Res., 52 (1992) 127-131; US 5208020).
Иммуноконъюгаты или ADC, упомянутые в данном описании, прямо предусматривают, но не ограничиваются этим, такие конъюгаты, которые получены с использованием реагентов для перекрестного сшивания, включая, но не ограничиваясь этим, BMPS (N-(β-малеимидо-пропилокси)сукцинимидный эфир), EMCS (N-(ε-малеимидокапроилокси)сукцинимидный эфир), GMBS (N-[γ-малеимидобутирилокси]-сукцинимидный эфир), HBVS (1,6-гексан-бис-винилсульфон), LC-SMCC (длинноцепочечный аналог SMCC), MBS (м-малеинимидобензоил-N-гидроксисукцинимидный эфир), MPBH (4-(4-N-малеимидофенил)масляной кислоты гидразид), SBAP (N-сукцинимидил-3-(бромацетатамидо)пропионат), SIA (N-сукцинимидил-иодацетат), SIAB (N-сукцинимидил-(4-иодацетат)аминобензоат), SMCC, SMPB (сукцинимидил-4-(п-малеимидо-фенил)бутират), SMPH (сукцинимидил-6-[β-малеимидопропионамидо]гексаноат), сульфо-EMCS, сульфо-GMBS, сульфо-KMUS (N-сукцинимидный эфир κ-малеинимидоундекановой кислоты), сульфо-MBS, сульфо-SIAB, сульфо-SMCC и сульфо-SMPB и SVSB (сукцинимидил-(4-винилсульфон)бензоат), которые имеются в продаже (например, у Pierce Biotechnology, Inc., Rockford, IL., USA).
Способы и композиции для диагностики и детекции
В некоторых воплощениях любое из антител, предложенных согласно данному изобретению, является полезным для детекции присутствия родственного(ых) ему антигена(ов) в биологическом образце. Термин “детекция”, использованный в данном описании, охватывает количественную или качественную детекцию. В некоторых воплощениях биологический образец содержит клетку или ткань.
В одном из воплощений предложено антитело, описанное в данной заявке, для применения в способе диагностики или детекции.
В некоторых воплощениях предложены меченые антитела, описанные в данной заявке. Метки включают, но не ограничиваются этим, метки или группировки, которые детектируют напрямую (как например, флуоресцентные, хромофорные, электронно-плотные, хемилюминесцентные и радиоактивные метки), а также такие группировки, как ферменты или лиганды, детекцию которых осуществляют опосредованно, например, с использованием ферментативной реакции или молекулярного взаимодействия. Типичные метки включают, но не ограничиваются этим, радиоактивные изотопы 32P, 14C, 125I, 3H и 131I, флуорофоры, такие как редкоземельные хелаты или флуоресцеин и его производные, родамин и его производные, данзил, умбеллиферон, люциферазы, например, люциферазу светлячков и бактериальную люциферазу (US 4737456), люциферин, 2,3-дигидрофталазиндионы, пероксидазу хрена (HRP), щелочную фосфатазу, β-галактозидазу, глюкоамилазу, лизоцим, оксидазы сахаров, например, глюкозооксидазу, галактозооксидазу и глюкозо-6-фосфатдегидрогеназу, оксидазы гетероциклических соединений, такие как уриказа и ксантиноксидаза, в сочетании с ферментом, который использует перекись водорода для окисления предшественника красителя, таким как HRP, лактопероксидаза или микропероксидаза; биотин/авидин, спиновые метки, метки на основе бактериофагов, стабильные свободные радикалы и тому подобное.
Фармацевтические композиции
Фармацевтические композиции на основе антитела, описанного в данной заявке, готовят путем смешивания такого антитела, имеющего желаемую степень чистоты, с одним или более чем одним возможным фармацевтически приемлемым носителем (Remington's Pharmaceutical Sciences, 16th edition, Osol, A. (ed.) (1980)) в форме лиофилизированных композиций или водных растворов. Фармацевтически приемлемые носители, как правило, нетоксичны для реципиентов в используемых дозировках и концентрациях и включают, но не ограничиваются этим: буферы, например, на основе фосфата, цитрата и других органических кислот; антиоксиданты, в том числе аскорбиновую кислоту и метионин; консерванты (такие как октадецилдиметилбензилхлорид аммония; гексаметонийхлорид; бензалконийхлорид; бензетонийхлорид; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцин; циклогексанол; 3-пентанол; и м-крезол); низкомолекулярные (менее чем примерно 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поли(винилпирролидон); аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, в том числе глюкозу, маннозу или декстрины; хелатирующие агенты, такие как EDTA (этилендиаминтетрауксусная кислота); сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как ион натрия; комплексы с металлами (например, комплексы Zn-белок); и/или неионные поверхностно-активные вещества, такие как полиэтиленгликоль (ПЭГ). Типичные фармацевтически приемлемые носители, упомянутые в данном описании, также включают агенты диспергирования лекарственных средств в интерстициальном пространстве (interstitial drug dispersion agents), такие как активные растворимые в нейтральных условиях гликопротеины гиалуронидазы (sHASEGP), например, растворимые гликопротеины гиалуронидазы PH-20 человека, такие как рекомбинантная гиалуронидаза человека PH-20 (rHuPH20) (HYLENEX®, Baxter International, Inc.). Некоторые типичные sHASEGP и способы их применения, включая rHuPH20, описаны в US 2005/0260186 и US 2006/0104968. Согласно одному из аспектов sHASEGP комбинируют с одной или более чем одной дополнительной гликозаминогликаназой, такой как хондроитиназа.
Типичные лиофилизированные композиции на основе антител описаны в US 6267958. Водные композиции на основе антител включают композиции, описанные в US 6171586 и WO2006/044908, причем последние композиции включают в себя гистидин-ацетатный буфер.
Композиция, описанная в данной заявке, также может содержать более одного из активных ингредиентов, которые необходимы для конкретного подвергаемого лечению показания, предпочтительно, ингредиентов с дополняющими активностями, которые не оказывают неблагоприятного действия друг на друга. Такие активные ингредиенты соответственно представлены в комбинации в количествах, которые эффективны для предполаемой цели.
Активные ингредиенты могут быть заключены в микрокапсулы, приготовленные, например, методами коацервации или межфазной полимеризации, например, в гидроксиметилцеллюлозные или желатиновые микрокапсулы и микрокапсулы из поли(метилметакрилата), соответственно, в коллоидные системы доставки лекарственных средств (например, липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсии. Такие методы описаны в Remington's Pharmaceutical Sciences, 16th edition, Osol, A. (ed.) (1980).
Можно изготовить препараты с длительным высвобождением. Подходящие примеры препаратов с длительным высвобождением включают полупроницаемые формы из твердых гидрофобных полимеров, содержащие антитело, которые представлены в форме штампованных изделий, например, пленок или микрокапсул.
Композиции, предназначенные для введения in vivo, как правило, должны быть стерильными. Стерильности легко добиться, например, с помощью фильтрования через стерилизующие фильтрационные мембраны.
Способы терапии и композиции
Любое из антител, предложенных в данном изобретении, можно использовать в способах терапии.
Согласно одному из аспектов предложено антитело, описанное в данной заявке, для применения в качестве лекарственного средства.
В некоторых воплощениях предложено антитело для применения в способе лечения. В одном из таких воплощений способ дополнительно включает введение данному индивидууму эффективного количества по меньшей мере одного дополнительного терапевтического агента, например, описанного ниже. “Индивидуумом”, соответствующим любому из упомянутых выше воплощений, в одном из предпочтительных воплощений является человек.
Согласно следующему аспекту изобретения предложено применение антитела для изготовления или получения лекарственного средства. “Индивидуумом”, соответствующим любому из упомянутых выше воплощений, может быть человек.
Согласно следующему аспекту изобретения предложены фармацевтические композиции, содержащие любое из антител, предложенных в данном изобретении, например, для применения в любом из указанных выше способов терапии. В одном из воплощений фармацевтическая композиция содержит любое из антител, предложенных в данном изобретении, и фармацевтически приемлемый носитель. В другом воплощении фармацевтическая композиция содержит любое из антител, предложенных в данном изобретении, и по меньшей мере один дополнительный терапевтический агент, например, описанный ниже.
Антитела, описанные в данной заявке, можно использовать либо по отдельности, либо в комбинации с другими агентами в терапии. Например, антитело, описанное в данной заявке, может быть введено по меньшей мере вместе с одним дополнительным терапевтическим агентом.
Антитело, описанное в данной заявке, (и какой-либо дополнительный терапевтический агент) можно вводить любым подходящим способом, включая парентеральное, внутрилегочное и интраназальное введение, и, при необходимости местного лечения, введение в область поражения. Парентеральные инфузии включают внутримышечное, внутривенное, интраартериальное, внутрибрюшинное или подкожное введение. Введение может быть осуществлено любым подходящим путем, например посредством инъекций, таких как внутривенные или подкожные инъекции, зависимым в некоторой степени от того, является ли это введение кратковременным или продолжительным. Данным изобретением охвачены различные режимы введения, включая, но не ограничиваясь этим, однократные или многократные введения в разные моменты времени, струйное введение и прерывистую инфузию.
На основе антител, описанных в данной заявке, будут изготовлены лекарственные формы, которые могут быть разделены на дозы и введены способом, соответствующим надлежащей медицинской практике. Факторы, рассматриваемые в этом контексте, включают конкретное подвергаемое лечению расстройство, конкретное подвергаемое лечению млекопитающее, клиническое состояние отдельного пациента, причину расстройства, место доставки агента, способ введения, схему введения и другие факторы, известные врачам. Необходимости в этом нет, но на основе такого антитела можно изготовить лекарственную форму, содержащую один или более агентов, используемых в настоящее время для предупреждения или лечения рассматриваемого расстройства. Эффективное количество таких других агентов зависит от количества антитела, присутствующего в композиции, типа расстройства или лечения и других факторов, рассмотренных выше. Они, как правило, используются в тех же дозировках и вводятся теми же путями, какие изложены в данном описании, или в количестве, составляющем примерно от 1 до 99% от дозировок, изложенных в данном описании, или в любой дозировке и любым путем, которые эмпирически/клинически определены как соответствующие.
Для предупреждения или лечения заболевания соответствующая дозировка описанного в данной заявке антитела (при использовании по отдельности или в комбинации с одним или более чем одним другим дополнительным терапевтическим агентом) будет зависеть от типа подлежащего лечению заболевания, типа антитела, тяжести и течения заболевания, от того, вводят ли антитело в профилактических или терапевтических целях, от предыдущей терапии, от клинической истории пациента и восприимчивости к антителу и от усмотрения лечащего врача. Соответственно, антитело вводят пациенту за один раз или в ходе курса лечений. В зависимости от типа и тяжести заболевания первоначальной возможной дозировкой для введения пациенту может быть дозировка антитела, составляющая от примерно 1 мкг/кг до 15 мг/кг (например, 0,5 мг/кг - 10 мг/кг), будь то выполнено, например, в виде одного или более отдельных введений, или в виде непрерывной инфузии. Одна из типичных суточных дозировок может находиться в диапазоне от примерно 1 мкг/кг до 100 мг/кг или более в зависимости от вышеупомянутых факторов. При повторных введениях в течение нескольких суток или больше, в зависимости от состояния, лечение, как правило, будет продолжено до тех пор, пока не наступит желаемое подавление симптомов заболевания. Одна из типичных дозировок антитела может находиться в диапазоне от примерно 0,05 мг/кг до примерно 10 мг/кг. Таким образом, пациенту можно вводить одну дозу или большее число доз, составляющих примерно 0,5 мг/кг, 2,0 мг/кг, 4,0 мг/кг или 10 мг/кг (или любую их комбинацию). Такие дозы можно вводить периодически, например каждую неделю или каждые три недели (например, таким образом, чтобы пациент получал от примерно двух до примерно двадцати доз или, например, приблизительно шесть доз антитела). Сначала можно вводить более высокую ударную дозу, затем одну или несколько более низких доз. Ход такого лечения легко отслеживать посредством традиционных методов и анализов.
Очевидно, что любая из указанных выше (технологий приготовления) композиций или любой из приведенных выше способов терапии могут быть осуществлены с использованием иммуноконъюгата, описанного в данной заявке, вместо или помимо антитела, описанного в данной заявке.
Изделия производства
Согласно другому аспекту, описанному в данной заявке, предложено изделие производства, содержащее материалы, полезные для лечения, предупреждения и/или диагностики описанных выше расстройств. Изделие производства содержит контейнер и этикетку или инструкцию по применению на контейнере или в соединении с ним. Подходящие контейнеры включают, например, бутылки, флаконы, шприцы, пакеты для внутривенных (в./в.) растворов и т.д. Контейнеры могут быть выполнены из разнообразных материалов, таких как стекло или пластик. Контейнер содержит в себе композицию, которая отдельно или в комбинации с другой композицией эффективна для лечения, предупреждения и/или диагностики данного состояния, и может иметь стерильное входное отверстие (например, контейнер может представлять собой пакет для внутривенного раствора или флакон, снабженный пробкой, поддающейся прокалыванию иглой для подкожных инъекций). По меньшей мере один активный агент в композиции представляет собой антитело, описанное в данной заявке. Этикетка или инструкция по применению указывает на то, что композицию применяют для лечения выбранного состояния. Кроме того, изделие производства может содержать (a) первый контейнер с содержащейся в нем композицией, при этом композиция содержит антитело, описанное в данной заявке; и (b) второй контейнер с содержащейся в нем композицией, при этом композиция содержит дополнительный цитотоксический или иной терапевтический агент. Изделие производства по этому воплощению, как описанно в данной заявке, может дополнительно содержать инструкцию по применению, указывающую, что данные композиции можно использовать для лечения конкретного состояния. Альтернативно или помимо этого, изделие производства может дополнительно содержать второй (или третий) контейнер, включающий в себя фармацевтически приемлемый буфер, такой как бактериостатическая вода для инъекций (BWFI), забуференный фосфатом физиологический раствор, раствор Рингера и раствор декстрозы. Кроме того, оно может содержать другие необходимые с коммерческой и потребительской точки зрения материалы, включая другие буферы, разбавители, фильтры, иглы и шприцы.
Очевидно, что любое из упомянутых выше изделий производства может включать в себя иммуноконъюгат по изобретению вместо или помимо антитела, описанного в данной заявке.
III. КОНКРЕТНЫЕ ВОПЛОЩЕНИЯ
1. Fc-область класса IgG, содержащая первый полипептид вариантной Fc-области и второй полипептид вариантной Fc-области,
при этом
a) первый полипептид вариантной Fc-области происходит из первого полипептида родительской Fc-области класса IgG, а второй полипептид вариантной Fc-области происходит из второго полипептида родительской Fc-области класса IgG, вследствие чего первый полипептид родительской Fc-области класса IgG идентичен второму полипептиду или отличается от второго полипептида родительской Fc-области класса IgG, и
b) первый полипептид вариантной Fc-области отличается от второго полипептида вариантной Fc-области по одному или более аминокислотным остаткам, отличающимся от тех аминокислотных остатков, по которым первый полипептид родительской Fc-области класса IgG отличается от второго полипептида родительской Fc-области класса IgG, и
c) Fc-область класса IgG, содержащая первый полипептид вариантной Fc-области и второй полипептид вариантной Fc-области, обладает аффинностью к Fc-рецептору человека, которая отличается от таковой у Fc-области класса IgG, содержащей первый полипептид родительской Fc-области класса IgG по пункту (a) и второй полипептид родительской Fc-области класса IgG по пункту (a).
2. Fc-область класса IgG по воплощению 1, где Fc-рецептор человека представляет собой неонатальный Fc-рецептор (FcRn) человека или Fc-гамма-III рецептор (FcγRIII) человека.
3. Fc-область класса IgG по любому из воплощений 1-2, где Fc-рецептор человека представляет собой неонатальный Fc-рецептор человека.
4. Fc-область класса IgG по любому из воплощений 1-3, где аффинность Fc-области класса IgG, содержащей первый полипептид вариантной Fc-области и второй полипептид вариантной Fc-области, к Fc-рецептору человека повышена или снижена на 10% или более, что определено с использованием поверхностного плазмонного резонанса (SPR), по сравнению с аффинностью Fc-области класса IgG, содержащей первый полипептид родительской Fc-области класса IgG по пункту (a) и второй полипептид родительской Fc-области класса IgG по пункту (a).
5. Fc-область класса IgG по любому из воплощений 1-4, где по меньшей мере некоторые из тех аминокислотных остатков, по которым первый полипептид родительской Fc-области класса IgG отличается от второго полипептида родительской Fc-области класса IgG, стимулируют образование гетеродимерной Fc-области класса IgG.
6. Fc-область класса IgG по любому из воплощений 1-5, где
1) первый полипептид родительской Fc-области класса IgG выбран из группы, содержащей
- полипептид Fc-области человеческого IgG1,
- полипептид Fc-области человеческого IgG2,
- полипептид Fc-области человеческого IgG3,
- полипептид Fc-области человеческого IgG4,
- полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A,
- полипептид Fc-области человеческого IgG1 с мутациями Y349C, T366S, L368A, Y407V,
- полипептид Fc-области человеческого IgG1 с мутациями S354C, T366S, L368A, Y407V,
- полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, Y349C, T366S, L368A, Y407V,
- полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, S354C, T366S, L368A, Y407V,
- полипептид Fc-области человеческого IgG1 с мутацией P329G,
- полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, P329G,
- полипептид Fc-области человеческого IgG1 с мутациями P329G, Y349C, T366S, L368A, Y407V,
- полипептид Fc-области человеческого IgG1 с мутациями P329G, S354C, T366S, L368A, Y407V,
- полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, P329G, Y349C, T366S, L368A, Y407V,
- полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, P329G, S354C, T366S, L368A, Y407V,
- полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E,
- полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E, P329G,
- полипептид Fc-области человеческого IgG4 с мутациями Y349C, T366S, L368A, Y407V,
- полипептид Fc-области человеческого IgG4 с мутациями S354C, T366S, L368A, Y407V,
- полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E, Y349C, T366S, L368A, Y407V,
- полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E, S354C, T366S, L368A, Y407V,
- полипептид Fc-области человеческого IgG4 с мутацией P329G,
- полипептид Fc-области человеческого IgG4 с мутациями P329G, Y349C, T366S, L368A, Y407V,
- полипептид Fc-области человеческого IgG4 с мутациями P329G, S354C, T366S, L368A, Y407V,
- полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E, P329G, Y349C, T366S, L368A, Y407V,
- полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E, P329G, S354C, T366S, L368A, Y407V,
- человеческий IgG1, IgG2 или IgG4 с мутацией K392D и
- человеческий IgG3 с мутацией N392D,
и
2) второй полипептид родительской Fc-области класса IgG выбран из группы, содержащей
- полипептид Fc-области человеческого IgG1,
- полипептид Fc-области человеческого IgG2,
- полипептид Fc-области человеческого IgG3,
- полипептид Fc-области человеческого IgG4,
- полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A,
- полипептид Fc-области человеческого IgG1 с мутациями S354C, T366W,
- полипептид Fc-области человеческого IgG1 с мутациями Y349C, T366W,
- полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, S354C, T366W,
- полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, Y349C, T366W,
- полипептид Fc-области человеческого IgG1 с мутацией P329G,
- полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, P329G,
- полипептид Fc-области человеческого IgG1 с мутациями P329G, S354C, T366W,
- полипептид Fc-области человеческого IgG1 с мутациями P329G, Y349C, T366W,
- полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, P329G, S354C, T366W,
- полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, P329G, Y349C, T366W,
- полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E,
- полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E, P329G,
- полипептид Fc-области человеческого IgG4 с мутациями S354C, T366W,
- полипептид Fc-области человеческого IgG4 с мутациями Y349C, T366W,
- полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E, S354C, T366W,
- полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E, Y349C, T366W,
- полипептид Fc-области человеческого IgG4 с мутацией P329G,
- полипептид Fc-области человеческого IgG4 с мутациями P329G, S354C, T366W,
- полипептид Fc-области человеческого IgG4 с мутациями P329G, Y349C, T366W,
- полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E, P329G, S354C, T366W,
- полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E, P329G, Y349C, T366W,
- человеческий IgG1 с мутациями D399K, D356K и/или E357K и
- человеческий IgG2, IgG3 или IgG4 с мутациями D399K, E356K и/или E357K.
7. Fc-область класса IgG по любому из воплощений 1-6, где
1) первый полипептид родительской Fc-области класса IgG представляет собой полипептид Fc-области человеческого IgG1, и второй полипептид родительской Fc-области класса IgG представляет собойполипептид Fc-области человеческого IgG1, или
2) первый полипептид родительской Fc-области класса IgG представляет собой полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, и второй полипептид родительской Fc-области класса IgG представляет собой полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, или
3) первый полипептид родительской Fc-области класса IgG представляет собой полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, P329G, и второй полипептид родительской Fc-области класса IgG представляет собой полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, P329G, или
4) первый полипептид родительской Fc-области класса IgG представляет собой полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, S354C, T366W, а второй полипептид родительской Fc-области класса IgG представляет собой полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, Y349C, T366S, L368A, Y407V, или
5) первый полипептид родительской Fc-области класса IgG представляет собой полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, P329G, S354C, T366W, а второй полипептид родительской Fc-области класса IgG представляет собой полипептид Fc-области человеческого IgG1 с мутациями L234A, L235A, P329G, Y349C, T366S, L368A, Y407V, или
6) первый полипептид родительской Fc-области класса IgG представляет собой полипептид Fc-области человеческого IgG4, и второй полипептид родительской Fc-области класса IgG представляет собой полипептид Fc-области человеческого IgG4, или
7) первый полипептид родительской Fc-области класса IgG представляет собой полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E, и второй полипептид родительской Fc-области класса IgG представляет собой полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E, или
8) первый полипептид родительской Fc-области класса IgG представляет собой полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E, P329G, и второй полипептид родительской Fc-области класса IgG представляет собой полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E, P329G, или
9) первый полипептид родительской Fc-области класса IgG представляет собой полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E, S354C, T366W, а второй полипептид родительской Fc-области класса IgG представляет собой полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E, Y349C, T366S, L368A, Y407V, или
10) первый полипептид родительской Fc-области класса IgG представляет собой полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E, P329G, S354C, T366W, а второй полипептид родительской Fc-области класса IgG представляет собой полипептид Fc-области человеческого IgG4 с мутациями S228P, L235E, P329G, Y349C, T366S, L368A, Y407V.
8. Fc-область класса IgG по любому из воплощений 1-7, где первый полипептид Fc-области, или второй полипептид Fc-области, или оба полипептида Fc-области содержат одну из приведенных ниже мутаций или комбинаций мутаций:
- T307H, или
- Q311H, или
- E430H, или
- N434H, или
- T307H и Q311H, или
- T307H и E430H, или
- T307H и N434A, или
- T307H и N434H, или
- T307Q и Q311H, или
- T307Q и E430H, или
- T307Q и N434H, или
- T307H, и Q311H, и E430H, и N434A, или
- T307H, и Q311H, и E430H, и N434H, или
- T307H, и Q311H, и E430H, и N434Y, или
- T307Q, и Q311H, и E430H, и N434A, или
- T307Q, и Q311H, и E430H, и N434H, или
- T307Q, и Q311H, и E430H, и N434Y, или
- T307Q, и V308P, и N434Y, и Y436H, или
- T307H, и M252Y, и S254T, и T256E, или
- T307Q, и M252Y, и S254T, и T256E, или
- Q311H, и M252Y, и S254T, и T256E, или
- E430H, и M252Y, и S254T, и T256E, или
- N434H, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и M252Y, и S254T, и T256E, или
- T307H, и E430H, и M252Y, и S254T, и T256E, или
- T307H, и N434A, и M252Y, и S254T, и T256E, или
- T307H, и N434H, и M252Y, и S254T, и T256E, или
- T307Q, и Q311H, и M252Y, и S254T, и T256E, или
- T307Q, и E430H, и M252Y, и S254T, и T256E, или
- T307Q, и N434H, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и E430H, и N434A, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и E430H, и N434H, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и E430H, и N434Y, и M252Y, и S254T, и T256E, или
- T307Q, и Q311H, и E430H, и N434A, и M252Y, и S254T, и T256E, или
- T307Q, и Q311H, и E430H, и N434H, и M252Y, и S254T, и T256E, или
- T307Q, и Q311H, и E430H, и N434Y, и M252Y, и S254T, и T256E, или
- T307Q, и V308P, и N434Y, и Y436H, и M252Y, и S254T, и T256E.
9. Fc-область класса IgG по любому из воплощений 1-8, где первый полипептид Fc-области содержит, независимо от второго полипептида Fc-области, одну из приведенных ниже мутаций или комбинаций мутаций:
- T307H, или
- Q311H, или
- E430H, или
- N434H, или
- T307H и Q311H, или
- T307H и E430H, или
- T307H и N434A, или
- T307H и N434H, или
- T307Q и Q311H, или
- T307Q и E430H, или
- T307Q и N434H, или
- M252Y, и S254T, и T256E, или
- I253A, и H310A, и H435A, или
- H310A, и H433A, и Y436A, или
- T307H, и Q311H, и E430H, и N434A, или
- T307H, и Q311H, и E430H, и N434H, или
- T307H, и Q311H, и E430H, и N434Y, или
- T307Q, и Q311H, и E430H, и N434A, или
- T307Q, и Q311H, и E430H, и N434H, или
- T307Q, и Q311H, и E430H, и N434Y, или
- T307Q, и V308P, и N434Y, и Y436H, или
- T307H, и M252Y, и S254T, и T256E, или
- Q311H, и M252Y, и S254T, и T256E, или
- E430H, и M252Y, и S254T, и T256E, ил
- N434H, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и M252Y, и S254T, и T256E, или
- T307H, и E430H, и M252Y, и S254T, и T256E, или
- T307H, и N434A, и M252Y, и S254T, и T256E, или
- T307H, и N434H, и M252Y, и S254T, и T256E, или
- T307Q, и Q311H, и M252Y, и S254T, и T256E, или
- T307Q, и E430H, и M252Y, и S254T, и T256E, или
- T307Q, и N434H, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и E430H, и N434A, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и E430H, и N434H, и M252Y, и S254T, и T256E, или
- T307H, и Q311H, и E430H, и N434Y, и M252Y, и S254T, и T256E, или
- T307Q, и V308P, и N434Y, и Y436H, и M252Y, и S254T, и T256E, или
- T307Q, и V308P, и N434Y, и Y436H, и M252Y, и S254T, и T256E,
и
второй полипептид Fc-области содержит, независимо от первого полипептида Fc-области, одну из приведенных ниже мутаций или комбинаций мутаций:
- T307H, или
- T307Q, или
- Q311H, или
- E430H, или
- N434H, или
- T307H и Q311H, или
- T307H и E430H, или
- T307H и N434A, или
- T307H и N434H, или
- T307Q и Q311H, или
- T307Q и E430H, или
- T307Q и N434H, или
- T307H, и Q311H, и E430H, и N434A, или
- T307H, и Q311H, и E430H, и N434H, или
- T307H, и Q311H, и E430H, и N434Y, или
- T307Q, и Q311H, и E430H, и N434A, или
- T307Q, и Q311H, и E430H, и N434H, или
- T307Q, и Q311H, и E430H, и N434Y, или
- T307Q, и V308P, и N434Y, и Y436H, или
- T307H, и M252Y, и S254T, и T256E или
- Q311H, и M252Y, и S254T, и T256E, или
- E430H, и M252Y, и S254T, и T256E, или
- N434H, и M252Y, и S254T, и T256E.
10. Fc-область класса IgG по любому из воплощений 1-9, где первый полипептид Fc-области содержит
одну из приведенных ниже комбинаций мутаций:
- ни одной, или
- M252Y, и S254T, и T256E, или
- I253A, и H310A, и H435A, или
- H310A, и H433A, и Y436A,
и
одну из приведенных ниже мутаций или комбинаций мутаций:
- ни одной,
- T307H, или
- T307Q, или
- Q311H, или
- E430H, или
- N434H, или
- T307H и Q311H, или
- T307H и E430H, или
- T307H и N434A, или
- T307H и N434H, или
- T307Q и Q311H, или
- T307Q и E430H, или
- T307Q и N434H, или
- T307Q и N434A, или
- T307H, и Q311H, и E430H, и N434A, или
- T307H, и Q311H, и E430H, и N434H, или
- T307H, и Q311H, и E430H, и N434Y, или
- T307Q, и Q311H, и E430H, и N434A, или
- T307Q, и Q311H, и E430H, и N434H, или
- T307Q, и Q311H, и E430H, и N434Y, или
- T307Q, и V308P, и N434Y, и Y436H,
и второй полипептид Fc-области содержит
одну из приведенных ниже мутаций или комбинаций мутаций:
- ни одной, если первый полипептид Fc-области содержит по меньшей мере одну мутацию, или
- T307H, или
- T307Q, если первый полипептид Fc-области не содержит исключительно мутацию T307Q, или
- Q311H, или
- E430H, или
- N434H, или
- T307H и Q311H, или
- T307H и E430H, или
- T307H и N434A, или
- T307H и N434H, или
- T307Q и Q311H, или
- T307Q и E430H, или
- T307Q и N434H, или
- T307Q и N434A, или
- M252Y, и S254T, и T256E, если первый полипептид Fc-области не содержит исключительно комбинацию мутаций M252Y, и S254T, и T256E, или
- I253A, и H310A, и H435A, если первый полипептид Fc-области не содержит исключительно комбинацию мутаций I253A, и H310A, и H435A, или
- H310A, и H433A, и Y436A, если первый полипептид Fc-области не содержит исключительно комбинацию мутаций H310A, и H433A, и Y436A, или
- T307H, и Q311H, и E430H, и N434A, или
- T307H, и Q311H, и E430H, и N434H, или
- T307H, и Q311H, и E430H, и N434Y, или
- T307Q, и Q311H, и E430H, и N434A, или
- T307Q, и Q311H, и E430H, и N434H, или
- T307Q, и Q311H, и E430H, и N434Y, или
- T307Q, и V308P, и N434Y, и Y436H.
11. Fc-область класса IgG по любому из воплощений 1-10, где первый полипептид Fc-области содержит мутации I253A, и H310A, и H435A, а второй полипептид Fc-области содержит мутации M252Y, и S254T, и T256E.
12. Fc-область класса IgG по любому из воплощений 1-10, где первый полипептид Fc-области содержит мутации I253A, и H310A, и H435A, а второй полипептид Fc-области содержит мутации M252Y, и S254T, и T256E, и T307Q, и N434Y.
13. Fc-область класса IgG по любому из воплощений 1-10, где первый полипептид Fc-области содержит мутации I253A, и H310A, и H435A, а второй полипептид Fc-области содержит мутации M252Y, и S254T, и T256E, и T307Q, и V308P, и N434Y, и Y436H.
14. Fc-область класса IgG по любому из воплощений 1-10, где первый полипептид Fc-области содержит мутации T307H, и Q311H, и E430H, и N434H, а второй полипептид Fc-области содержит мутации M252Y, и S254T, и T256E.
15. Fc-область класса IgG по любому из воплощений 1-10, где первый полипептид Fc-области содержит мутации T307H и N434H, а второй полипептид Fc-области содержит мутации M252Y, и S254T, и T256E.
16. Fc-область класса IgG по любому из воплощений 1-10, где первый полипептид Fc-области содержит мутации T307Q и N434A, а второй полипептид Fc-области содержит мутации M252Y, и S254T, и T256E.
17. Fc-область класса IgG по любому из воплощений 1-10, где первый полипептид Fc-области содержит мутации T250Q и M428L, а второй полипептид Fc-области содержит мутации M252Y, и S254T, и T256E.
18. Fc-область класса IgG по любому из воплощений 1-10, где первый полипептид Fc-области содержит мутации T307Q и N434H, а второй полипептид Fc-области содержит мутации M252Y, и S254T, и T256E, и T307Q, и N434H.
19. Fc-область класса IgG по любому из воплощений 1-10, где первый полипептид Fc-области содержит мутации T307H, и Q311H, и E430H, и N434H, и второй полипептид Fc-области содержит мутации T307H, и Q311H, и E430H, и N434H.
20. Fc-область класса IgG по любому из воплощений 1-10, где первый полипептид Fc-области содержит мутации T307H и N434H, и второй полипептид Fc-области содержит мутации T307H и N434H.
21. Fc-область класса IgG по любому из воплощений 1-10, где первый полипептид Fc-области содержит мутации T307H, и N434H, и M252Y, и S254T, и T256E, и второй полипептид Fc-области содержит мутации T307H, и N434H, и M252Y, и S254T, и T256E.
22. Fc-область класса IgG по любому из воплощений 1-10, где первый полипептид Fc-области содержит мутацию N434H, и второй полипептид Fc-области содержит мутацию N434H.
23. Fc-область класса IgG по любому из воплощений 1-10, где первый полипептид Fc-области содержит мутации T307Q и N434A.
24. Fc-область класса IgG по любому из воплощений 1-10, где первый полипептид Fc-области содержит мутации T307H и N434H.
25. Fc-область класса IgG по любому из воплощений 1-10, где первый полипептид Fc-области содержит мутации T307H, и N434H, и M252Y, и S254T, и T256E.
26. Fc-область класса IgG по любому из воплощений 1-10, где первый полипептид Fc-области содержит мутацию N434H.
27. Антитело, содержащее Fc-область класса IgG по любому из воплощений 1-26.
28. Антитело по воплощению 27, где антитело представляет собой моноклональное антитело.
29. Антитело по любому из воплощений 27-28, где антитело представляет собой человеческое, гуманизированное или химерное антитело.
30. Антитело по любому из воплощений 27-29, где антитело представляет собой биспецифичное антитело.
31. Антитело по любому из воплощений 27-30, где антитело представляет собой бивалентное антитело.
32. Слитая конструкция на основе полипептидов Fc-области, содержащая Fc-область класса IgG по любому из воплощений 1-26.
33. Фармацевтическая композиция, содержащая антитело по любому из воплощений 27-31 или слитую конструкцию на основе полипептидов Fc-области по воплощению 32.
34. Фармацевтическая композиция по воплощению 33, где фармацевтическая композиция предназначена для применения при лечении сосудистых заболеваний глаз.
35. Антитело по любому из воплощений 27-31 или слитая конструкция на основе полипептидов Fc-области по воплощению 32 для применения в качестве лекарственного средства.
36. Применение по воплощению 35, где применение предполагается для лечения сосудистых заболеваний глаз.
37. Применение антитела по любому из воплощений 27-31 или слитой конструкции на основе полипептидов Fc-области по воплощению 32 для изготовления лекарственного средства.
38. Применение по воплощению 37, где применение предполагается для изготовления лекарственного средства для лечения сосудистого заболевания глаз.
39. Антитело по любому из воплощений 27-31 или слитая конструкция на основе полипептидов Fc-области по воплощению 32 для применения в лечении сосудистого заболевания глаз.
40. Способ лечения пациента, страдающего сосудистыми заболеваниями глаз, путем введения антитела по любому из воплощений 27-31 или слитой конструкции на основе полипептидов Fc-области по воплощению 32 пациенту, нуждающемуся в таком лечении.
IV. ПРИМЕРЫ
Далее приведены примеры способов и композиций, описанных в данной заявке. Очевидно, что с учетом приведенного выше общего описания на практике могут быть осуществлены различные другие воплощения.
Несмотря на то, что для ясности понимания вышеизложенное изобретение описано довольно подробно посредством иллюстрации и примеров, эти описания и примеры не следует истолковывать как ограничивающие объем, описанный в данной заявке. Описания всей патентной и научной литературы, приведенные в данном документе, явным образом включены во всей своей полноте посредством ссылки.
Методы
Масс-спектрометрия с ионизацией электрораспылением (ESI-MS)
Аликвоты белков (50 мкг) подвергали дегликозилированию путем добавления 0,5 мкл N-гликаназы плюс (Roche) и буферного раствора на основе фосфата натрия (0,1 М; pH 7,1) с получением конечного объема образца 115 мкл. Смесь инкубировали при 37°C в течение 18 ч. После этого добавляли для восстановления и денатурации по 60 мкл 0,5 М раствора TCEP (трис(2-карбоксиэтил)фосфин; Pierce) в 4 М растворе гуанидина⋅HCl (Pierce) и по 50 мкл 8 М раствора гуанидина⋅HCl. Смесь инкубировали при 37°C в течение 30 мин. Образцы обессоливали посредством гель-проникающей хроматографии (Сефароза G-25, изократический режим, 40%-ый ацетонитрил с 2% муравьиной кислоты). ESI-масс-спектры (+ve) регистрировали на квадрупольном времяпролетном (Q-TOF) приборе (maXis, Bruker), оборудованном источником нано-ESI (TriVersa NanoMate, Advion). Использовали приведенные ниже параметры настройки для MS: параметры переноса ионов: радиочастота (RF) ионной воронки, 400 Vpp; энергия индуцированной столкновениями диссоциации в источнике (ISCID), 0 эВ; RF мультиполя, 400 Vpp; квадруполь: энергия ионов, 4,0 эВ; нижний предел для массы, 600 m/z; источник: сухой газ, 8 л/мин; температура сухого газа, 160°C; столкновительная ячейка: энергия столкновений, 10 эВ; RF столкновений: 2000 Vpp; система охлаждения ионов: RF системы охлаждения ионов, 300 Vpp; время переноса: 120 мкс; накопление предимпульсов, 10 мкс; диапазон сканирования m/z 600-2000. Для оценки данных использовали программное обеспечение собственной разработки (MassAnalyzer).
Анализ связывания с FcRn с использованием поверхностного плазмонного резонанса (SPR)
Анализ свойств антитела дикого типа и мутантов в отношении связывания с FcRn проводили, применяя технологию поверхностного плазмонного резонанса (SPR) с использованием прибора BIAcore T100 (BIAcore AB, Uppsala, Sweden). Эта система является общепризнанной системой для исследования молекулярных взаимодействий. Она позволяет проводить непрерывный мониторинг связывания лиганд/аналит в режиме реального времени и, таким образом, определение кинетических параметров в различных условиях анализа. SPR-технология основана на измерении показателя преломления вблизи поверхности покрытого золотом биосенсорного чипа. Изменения показателя преломления указывают на изменения массы на поверхности, вызванные взаимодействием иммобилизованного лиганда с введенным в раствор аналитом. Если молекулы связаны с иммобилизованным на поверхности лигандом, то масса увеличивается, в случае же диссоциации масса уменьшается. В применяемом в настоящем изобретении анализе рецептор FcRn иммобилизовали на поверхности CM5-биосенсорного чипа BIAcore (GE Healthcare Bioscience, Uppsala, Sweden) посредством аминного сочетания на уровне 400 единиц ответного сигнала (RU). Анализ проводили при комнатной температуре, используя PBS, содержащий 0,05% Tween-20™, pH 6,0 (GE Healthcare Bioscience), в качестве рабочего буфера и буфера для разведения. 200 нМ растворы образцов антитела вводили при скорости потока 50 мкл/мин при комнатной температуре. Время ассоциации составляло 180 секунд, фаза диссоциации занимала 360 секунд. Регенерацию поверхности чипа осуществляли посредством непродолжительной инъекции HBS-P, pH 8,0. Оценку SPR-данных выполняли путем сравнения высоты сигнала биологического ответа через 180 секунд после инъекции и через 300 секунд после инъекции. Соответствующими параметрами являются максимальный уровень RU (через 180 секунд после инъекции) и стабильность на более поздний момент времени (через 300 секунд после окончания инъекции).
Анализ связывания с белком A с использованием поверхностного плазмонного резонанса (SPR)
Данный анализ основан на спектроскопии поверхностного плазмонного резонанса. Белок A иммобилизуют на поверхности биосенсора для SPR. После введения образца в проточные ячейки SPR-спектрометра происходит образование его комплекса с иммобилизованным белком A, что приводит к увеличению массы на поверхности сенсорного чипа и, следовательно, к возрастанию ответного сигнала (поскольку 1 RU определяется как 1 пг/мм²). После этого сенсорный чип регенерируют путем растворения комплекса образец-белок A. Затем полученные ответные сигналы оценивают по высоте сигнала в единицах ответа (RU) и по показателям диссоциации.
Проводили реакцию сочетания белка A (20 мкг/мл) на уровне приблизительно 3500 единиц ответного сигнала (RU) на поверхности CM5-чипа (GE Healthcare) при pH 4,0, используя набор для аминного сочетания от GE Healthcare.
В качестве буфера для образцов и системы использовали буфер HBS-P+ (состава: 0,01 М 4-(2-гидроксиэтил)-1-пиперазинэтансульфоновая кислота (HEPES), 0,15 М NaCl, 0,005% поверхностно-активного вещества P20; стерилизованный фильтрованием, pH 7,4). Температуру проточной ячейки устанавливали на 25°C, а температуру в отделении для образца на 12°C. Систему заполняли рабочим буфером. Затем в течение 120 секунд со скоростью потока 30 мкл/мин вводили 5 нМ растворы конструкций образца, после чего проводили фазу диссоциации продолжительностью 300 секунд. Затем поверхность сенсорного чипа регенерировали, выполняя две 30-секундные инъекции раствора глицина-HCl, pH 1,5, при скорости потока 30 мкл/мин. Каждый образец измеряли в трех повторах.
Термин “с мутацией IHH-AAA”, использованный в данном описании, относится к комбинациям мутаций I253A (Ile253Ala), H310A (His310Ala) и H435A (His435Ala) в константной области тяжелой цепи подкласса IgG1 или IgG4 (нумерация соответствует EU-индексу по Kabat), термин “с мутацией HHY-AAA”, использованный в данном описании, относится к комбинациям мутаций H310A (His310Ala), H433A (His433Ala) и Y436A (Tyr436Ala) в константной области тяжелой цепи подкласса IgG1 или IgG4 (нумерация соответствует EU-индексу по Kabat), термин “с мутацией P329G LALA”, использованный в данном описании, относится к комбинациям мутаций L234A (Leu234Ala), L235A (Leu235Ala) и P329G (Pro329Gly) в константной области тяжелой цепи подкласса IgG1 (нумерация соответствует EU-индексу по Kabat), а термин “с мутацией SPLE”, использованный в данном описании, относится к комбинациям мутаций S228P (Ser228Pro) и L235E (Leu235Glu) в константной области тяжелой цепи подкласса IgG4 (нумерация соответствует EU-индексу по Kabat).
Общая информация
Общая информация, касающаяся нуклеотидных последовательностей легкой и тяжелой цепей иммуноглобулина человека, приведена в: Kabat E.A. et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991). Аминокислотные остатки цепей антитела нумеруют и обозначают согласно EU нумерации (Edelman G.M., et al., Proc. Natl. Acad. Sci. USA, 63 (1969) 78-85; Kabat E.A., et al., Sequences of Proteins of Immunological Interest, 5th ed., Public Health Service, National Institutes of Health, Bethesda, MD (1991)).
Методы рекомбинантной ДНК
Для работы с ДНК использовали стандартные методы, описанные в Sambrook J. et al., Molecular Cloning: A laboratory manual; Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York (1989). Реагенты для молекулярной биологии использовали в соответствии с инструкциями производителя.
Синтез генов
Желаемые сегменты генов нумеровали согласно заданным спецификациям в Geneart (Regensburg, Germany).
Определение последовательности ДНК
Последовательности ДНК определяли посредством секвенирования двухцепочечных молекул, выполненного в MediGenomix GmbH (Martinsried, Germany) или SequiServe GmbH (Vaterstetten, Germany).
Анализ ДНК и белковых последовательностей и обработка полученных для последовательностей данных
Для создания, картирования, анализа, аннотирования и иллюстрирования последовательностей использовали пакет программ от GCG (Genetics Computer Group, Madison, Wisconsin), версию 10.2, и пакет программ Vector NT1 Advance, версию 8.0, от Infomax.
Экспрессирующие векторы
Для экспрессии описанных антител применяли экспрессирующие плазмиды для временной экспрессии (например, в клетках HEK293-F) либо на основе организации кДНК с промотором CMV и следующим за ним интроном A или без него, либо на основе геномной организации с цитомегаловирусным (CMV) промотором.
В состав транскрипционной единицы гена, кодирующего антитело, входили следующие элементы:
- уникальный(ые) сайт(ы) рестрикции на 5'-конце,
- энхансер и промотор непосредственно ранних генов цитомегаловируса человека,
- последовательность интрона A, в случае организации на основе кДНК,
- 5'-нетранслируемый участок гена, кодирующего иммуноглобулин человека,
- нуклеиновая кислота, кодирующая сигнальную последовательность тяжелой цепи иммуноглобулина,
- нуклеиновая кислота, кодирующая цепь человеческого антитела (дикого типа или с заменой в домене) либо в виде кДНК, либо в виде геномной организации с экзон-интронным построением для иммуноглобулина,
- 3'-нетранслируемый участок с последовательностью сигнала полиаденилирования и
- уникальный(ые) сайт(ы) рестрикции на 3'-конце.
Помимо экспрессирующей антитело кассеты такие плазмиды содержали:
- ориджин репликации, обеспечивающий осуществление репликации этой плазмиды в E. coli,
- ген β-лактамазы, придающий E. coli устойчивость к ампициллину, и
- ген дигидрофолатредуктазы из Mus. musculus в качестве селектируемого маркера в эукариотических клетках.
Нуклеиновые кислоты, кодирующие цепи антител, создавали посредством ПЦР и/или синтеза генов и сборку осуществляли с применением известных рекомбинантных методов и технологий, соединяя сообразные сегменты нуклеиновых кислот, например, с использованием уникальных сайтов рестрикции в соответствующих векторах. Последовательности субклонированных нуклеиновых кислот подтверждали посредством секвенирования ДНК. Для осуществления процедур временной трансфекции готовили большие количества плазмид, используя для их получения трансформированные культуры E. coli (Nucleobond AX, Macherey-Nagel).
Методы культивирования клеток
Использовали стандартные методы культивирования клеток, описанные в Current Protocols in Cell Biology (2000), Bonifacino J.S., Dasso M., Harford J.B., Lippincott-Schwartz J. and Yamada, K.M. (eds.), John Wiley & Sons, Inc.
Экспрессию биспецифичных антител осуществляли посредством временной котрансфекции клеток HEK293-F, выращиваемых в суспензии, соответствующими экспрессирующими плазмидами, как описано ниже.
Пример 1
Экспрессия и очистка
Процедуры временной трансфекции в системе HEK293-F
Моноспецифичные и биспецифичные антитела создавали посредством временной трансфекции соответствующими плазмидами (например, кодирующими тяжелую и модифицированную тяжелую цепь, а также соответствующую легкую и модифицированную легкую цепь) с использованием системы HEK293-F (Invitrogen) в соответствии с инструкцией производителя. Кратко, клетки HEK293-F (Invitrogen), выращиваемые в суспензии либо во встряхиваемой колбе, либо в ферментере с перемешиванием в бессывороточной среде FreeStyle™ для экспрессии в клетках 293 (Invitrogen), трансфицировали смесью, состоящей из соответствующих экспрессирующих плазмид и 293Fectin™ или Fectin™ (Invitrogen). В случае использования встряхиваемой колбы емкостью 2 л (Corning) клетки HEK293-F рассевали при плотности 106 клеток/мл в объеме 600 мл и инкубировали в условиях 120 об./мин, 8% CO2. На следующий день клетки трансфицировали при плотности клеток приблизительно 1,5.106 клеток/мл, используя приблизительно 42 мл смеси, состоящей из A) 20 мл среды Opti-MEM (Invitrogen) с содержанием 600 мкг суммарной плазмидной ДНК (1 мкг/мл), кодирующей тяжелую или модифицированную тяжелую цепь, соответственно, и соответствующую легкую цепь, в эквимолярном соотношении, и B) 20 мл Opti-MEM с 1,2 мл 293Fectin™ или Fectin™ (2 мкл/мл). По мере расходования глюкозы в ходе процесса ферментации добавляли раствор глюкозы. Супернатант, содержащий секретированное антитело, собирали через 5-10 суток, и антитела либо незамедлительно очищали из супернатанта, либо супернатант замораживали и оставляли на хранение.
Очистка
Биспецифичные антитела очищали из супернатантов клеточных культур аффинной хроматографией с использованием MabSelectSureSepharose™ (для мутантов, не являющихся IHH-AAA мутантами) (GE Healthcare, Sweden) или CappaSelect-Agarose (для IHH-AAA мутантов) (GE Healthcare, Sweden), гидрофобной хроматографией с использованием butyl-Sepharose (GE Healthcare, Sweden) и гель-проникающей (GE Healthcare, Sweden) хроматографией на Superdex 200.
Кратко, стерильные отфильтрованные супернатанты клеточных культур сорбировали на носителе MabSelectSure, уравновешенном (для антител, не содержащих мутаций IHH-AAA и антител дикого типа) буфером на основе PBS (10 мМ Na2HPO4, 1 мМ KH2PO4, 137 мМ NaCl и 2,7 мМ KCl, pH 7,4), промывали уравновешивающим буфером и элюировали 25 мМ раствором цитрата натрия при pH 3,0. IHH-AAA мутанты сорбировали на носителе CappaSelect, уравновешенном 25 мМ раствором Трис, с 50 мМ NaCl, pH 7,2, промывали уравновешивающим буфером и элюировали 25 мМ раствором цитрата натрия, pH 2,9. Фракции антител после элюирования объединяли и нейтрализовали, используя 2 М Трис, pH 9,0. Проводили подготовку пулов антител для гидрофобной хроматографии, добавляя 1,6 М раствор сульфата аммония до конечной концентрации 0,8 М по сульфату аммония и подводя pH до значения 5,0 с использованием уксусной кислоты. После уравновешивания на носителе butyl-Sepharose с использованием 35 мМ раствора ацетата натрия, 0,8 М раствора сульфата аммония, pH 5,0, на носитель наносили антитела, промывали уравновешивающим буфером и элюировали линейным градиентом до 35 мМ раствора ацетата натрия, pH 5,0. Фракции, содержащие (моноспецифичные или биспецифичные) антитела, объединяли и далее очищали гель-проникающей хроматографией, используя колонку Superdex 200 26/60 GL (GE Healthcare, Sweden), уравновешенную 20 мМ раствором гистидина с 140 мМ NaCl, pH 6,0. Фракции, содержащие (моноспецифичные или биспецифичные) антитела, объединяли, концентрировали до необходимой концентрации, используя устройства для ультрафильтрации Vivaspin (Sartorius Stedim Biotech S.A., France) и хранили при -80°C.
Анализ чистоты и целостности антител проводили после каждой стадии очистки посредством капиллярного электрофореза в присутствии додецилсульфата натрия (CE-SDS) с применением микрожидкостной технологии Labchip (Caliper Life Science, USA). Для CE-SDS-анализа белка готовили пять мкл раствора, используя набор реагентов для высокопроизводительного экспресс-анализа белков (HT Protein Express) в соответствии с инструкциями производителя, и анализ на системе LabChip GXII проводили, используя чип HT Protein Express. Анализ данных проводили, используя программное обеспечение LabChip GX.
Анализ содержания агрегатов в образцах антител проводили посредством высокоэффективной гель-проникающей хроматографии (SEC) с использованием аналитической колонки для гель-проникающей хроматографии, заполненной Superdex 200 (GE Healthcare, Sweden), в рабочем буфере 2xPBS (20 мМ Na2HPO4, 2 мМ KH2PO4, 274 мМ NaCl и 5,4 мМ KCl, pH 7,4) при 25°C. Раствор, содержащий 25 мкг белка, вводили в колонку со скоростью потока 0,75 мл/мин и элюировали в изократическом режиме в течение 50 минут.
Пример 2
Хроматография с использованием иммобилизованного FcRn
Связывание со стрептавидин-сефарозой
Один грамм стрептавидин-сефарозы (GE Healthcare) добавляли к раствору биотинилированного и диализованного рецептора и инкубировали в течение двух часов со встряхиванием. Сефарозой с иммобилизованным на ней рецептором заполняли XK колонку емкостью 1 мл (GE Healthcare).
Колоночная хроматография на основе аффинности к FcRn
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> F. Hoffmann-La Roche AG
<120> ВАРИАНТЫ FC-ОБЛАСТИ С МОДИФИЦИРОВАННЫМ СВЯЗЫВАНИЕМ С FCRN
И СПОСОБЫ ИХ ПРИМЕНЕНИЯ
<130> P32408-WO
<150> EP14192052.0
<151> 2014-11-06
<160> 30
<170> PatentIn version 3.5
<210> 1
<211> 107
<212> ПРТ
<213> Homo sapiens
<400> 1
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Trp Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
35 40 45
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
50 55 60
Glu Ser Thr Tyr Arg Trp Ser Val Leu Thr Val Leu His Gln Asp Trp
65 70 75 80
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
85 90 95
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
100 105
<210> 2
<211> 106
<212> ПРТ
<213> Homo sapiens
<400> 2
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1 5 10 15
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
20 25 30
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
35 40 45
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
50 55 60
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
65 70 75 80
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
85 90 95
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
100 105
<210> 3
<211> 227
<212> ПРТ
<213> Homo sapiens
<400> 3
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys
225
<210> 4
<211> 223
<212> ПРТ
<213> Homo sapiens
<400> 4
Val Glu Cys Pro Pro Cys Pro Ala Pro Pro Val Ala Gly Pro Ser Val
1 5 10 15
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
20 25 30
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
35 40 45
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
50 55 60
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Phe Arg Val Val Ser
65 70 75 80
Val Leu Thr Val Val His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
85 90 95
Cys Lys Val Ser Asn Lys Gly Leu Pro Ala Pro Ile Glu Lys Thr Ile
100 105 110
Ser Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
115 120 125
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
130 135 140
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ser Val Glu Trp Glu Ser Asn
145 150 155 160
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Met Leu Asp Ser
165 170 175
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
180 185 190
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
195 200 205
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
210 215 220
<210> 5
<211> 227
<212> ПРТ
<213> Homo sapiens
<400> 5
Asp Thr Pro Pro Pro Cys Pro Arg Cys Pro Ala Pro Glu Leu Leu Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Gln Phe Lys Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Phe
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
130 135 140
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Ser Gly Gln Pro Glu Asn Asn Tyr Asn Thr Thr Pro Pro
165 170 175
Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Ile Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn Arg Phe Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys
225
<210> 6
<211> 229
<212> ПРТ
<213> Homo sapiens
<400> 6
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro Glu Phe
1 5 10 15
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
20 25 30
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
35 40 45
Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val
50 55 60
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser
65 70 75 80
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
85 90 95
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser
100 105 110
Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
115 120 125
Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln
130 135 140
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
145 150 155 160
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
165 170 175
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu
180 185 190
Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser
195 200 205
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
210 215 220
Leu Ser Leu Gly Lys
225
<210> 7
<211> 227
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> полипептид Fc-области, происходящей из Fc-области человеческого
IgG1, с мутациями L234A, L235A
<400> 7
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys
225
<210> 8
<211> 227
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> полипептид Fc-области, происходящей из Fc-области человеческого
IgG1, с мутациями Y349C, T366S, L368A и Y407V
<400> 8
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys
225
<210> 9
<211> 227
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> полипептид Fc-области, происходящей из Fc-области человеческого
IgG1, с мутациями S354C, T366W
<400> 9
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys
225
<210> 10
<211> 227
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> полипептид Fc-области, происходящей из Fc-области человеческого
IgG1, с мутациями L234A, L235A и мутациями Y349C, T366S, L368A, Y407V
<400> 10
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys
225
<210> 11
<211> 227
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> полипептид Fc-области, происходящей из Fc-области человеческого
IgG1, с мутациями L234A, L235A и S354C, T366W
<400> 11
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys
225
<210> 12
<211> 227
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> полипептид Fc-области, происходящей из Fc-области человеческого
IgG1, с мутацией P329G
<400> 12
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys
225
<210> 13
<211> 227
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> полипептид Fc-области, происходящей из Fc-области человеческого
IgG1, с мутациями L234A, L235A и мутацией P329G
<400> 13
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys
225
<210> 14
<211> 227
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> полипептид Fc-области, происходящей из Fc-области человеческого
IgG1, с мутацией P239G и мутациями Y349C, T366S, L368A, Y407V
<400> 14
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys
225
<210> 15
<211> 227
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> полипептид Fc-области, происходящей из Fc-области человеческого
IgG1, с мутацией P329G и мутациями S354C, T366W
<400> 15
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys
225
<210> 16
<211> 227
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> полипептид Fc-области, происходящей из Fc-области человеческого
IgG1, с мутациями L234A, L235A, P329G и Y349C, T366S, L368A, Y407V
<400> 16
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys
225
<210> 17
<211> 227
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> полипептид Fc-области, происходящей из Fc-области человеческого
IgG1, с мутациями L234A, L235A, P329G и мутациями S354C, T366W
<400> 17
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Gly Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Cys Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys
225
<210> 18
<211> 229
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> полипептид Fc-области, происходящей из Fc-области человеческого
IgG4, с мутациями S228P и L235E
<400> 18
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe
1 5 10 15
Glu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
20 25 30
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
35 40 45
Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val
50 55 60
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser
65 70 75 80
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
85 90 95
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser
100 105 110
Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
115 120 125
Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln
130 135 140
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
145 150 155 160
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
165 170 175
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu
180 185 190
Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser
195 200 205
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
210 215 220
Leu Ser Leu Gly Lys
225
<210> 19
<211> 229
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> полипептид Fc-области, происходящей из Fc-области человеческого
IgG4, с мутациями S228P, L235E и мутацией P329G
<400> 19
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe
1 5 10 15
Glu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
20 25 30
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
35 40 45
Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val
50 55 60
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser
65 70 75 80
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
85 90 95
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Gly Ser
100 105 110
Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
115 120 125
Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln
130 135 140
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
145 150 155 160
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
165 170 175
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu
180 185 190
Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser
195 200 205
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
210 215 220
Leu Ser Leu Gly Lys
225
<210> 20
<211> 229
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> полипептид Fc-области, происходящей из Fc-области человеческого
IgG4, с мутациями S354C, T366W
<400> 20
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro Glu Phe
1 5 10 15
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
20 25 30
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
35 40 45
Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val
50 55 60
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser
65 70 75 80
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
85 90 95
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser
100 105 110
Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
115 120 125
Gln Val Tyr Thr Leu Pro Pro Cys Gln Glu Glu Met Thr Lys Asn Gln
130 135 140
Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
145 150 155 160
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
165 170 175
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu
180 185 190
Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser
195 200 205
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
210 215 220
Leu Ser Leu Gly Lys
225
<210> 21
<211> 229
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> полипептид Fc-области, происходящей из Fc-области человеческого
IgG4, с мутациями Y349C, T366S, L368A, Y407V
<400> 21
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro Glu Phe
1 5 10 15
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
20 25 30
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
35 40 45
Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val
50 55 60
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser
65 70 75 80
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
85 90 95
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser
100 105 110
Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
115 120 125
Gln Val Cys Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln
130 135 140
Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
145 150 155 160
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
165 170 175
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Arg Leu
180 185 190
Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser
195 200 205
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
210 215 220
Leu Ser Leu Gly Lys
225
<210> 22
<211> 229
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> полипептид Fc-области, происходящей из Fc-области человеческого
IgG4, с мутациями S228P, L235E и S354C, T366W
<400> 22
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe
1 5 10 15
Glu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
20 25 30
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
35 40 45
Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val
50 55 60
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser
65 70 75 80
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
85 90 95
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser
100 105 110
Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
115 120 125
Gln Val Tyr Thr Leu Pro Pro Cys Gln Glu Glu Met Thr Lys Asn Gln
130 135 140
Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
145 150 155 160
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
165 170 175
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu
180 185 190
Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser
195 200 205
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
210 215 220
Leu Ser Leu Gly Lys
225
<210> 23
<211> 229
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> полипептид Fc-области, происходящей из Fc-области человеческого
IgG4, с мутациями S228P, L235E и Y349C, T366S, L368A, Y407V
<400> 23
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe
1 5 10 15
Glu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
20 25 30
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
35 40 45
Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val
50 55 60
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser
65 70 75 80
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
85 90 95
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser
100 105 110
Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
115 120 125
Gln Val Cys Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln
130 135 140
Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
145 150 155 160
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
165 170 175
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Arg Leu
180 185 190
Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser
195 200 205
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
210 215 220
Leu Ser Leu Gly Lys
225
<210> 24
<211> 229
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> полипептид Fc-области, происходящей из Fc-области человеческого
IgG4, с мутацией P329G
<400> 24
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro Glu Phe
1 5 10 15
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
20 25 30
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
35 40 45
Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val
50 55 60
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser
65 70 75 80
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
85 90 95
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Gly Ser
100 105 110
Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
115 120 125
Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln
130 135 140
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
145 150 155 160
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
165 170 175
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu
180 185 190
Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser
195 200 205
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
210 215 220
Leu Ser Leu Gly Lys
225
<210> 25
<211> 229
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> полипептид Fc-области, происходящей из Fc-области человеческого
IgG4, с мутациями P239G и Y349C, T366S, L368A, Y407V
<400> 25
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro Glu Phe
1 5 10 15
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
20 25 30
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
35 40 45
Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val
50 55 60
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser
65 70 75 80
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
85 90 95
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Gly Ser
100 105 110
Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
115 120 125
Gln Val Cys Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln
130 135 140
Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
145 150 155 160
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
165 170 175
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Arg Leu
180 185 190
Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser
195 200 205
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
210 215 220
Leu Ser Leu Gly Lys
225
<210> 26
<211> 229
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> полипептид Fc-области, происходящей из Fc-области человеческого
IgG4, с мутациями P329G и S354C, T366W
<400> 26
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Ser Cys Pro Ala Pro Glu Phe
1 5 10 15
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
20 25 30
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
35 40 45
Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val
50 55 60
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser
65 70 75 80
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
85 90 95
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Gly Ser
100 105 110
Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
115 120 125
Gln Val Tyr Thr Leu Pro Pro Cys Gln Glu Glu Met Thr Lys Asn Gln
130 135 140
Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
145 150 155 160
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
165 170 175
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu
180 185 190
Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser
195 200 205
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
210 215 220
Leu Ser Leu Gly Lys
225
<210> 27
<211> 229
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> полипептид Fc-области, происходящей из Fc-области человеческого
IgG4, с мутациями S228P, L235E, P329G и Y349C, T366S, L368A, Y407V
<400> 27
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe
1 5 10 15
Glu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
20 25 30
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
35 40 45
Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val
50 55 60
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser
65 70 75 80
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
85 90 95
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Gly Ser
100 105 110
Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
115 120 125
Gln Val Cys Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln
130 135 140
Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
145 150 155 160
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
165 170 175
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Arg Leu
180 185 190
Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser
195 200 205
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
210 215 220
Leu Ser Leu Gly Lys
225
<210> 28
<211> 229
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> полипептид Fc-области, происходящей из Fc-области человеческого
IgG4, с мутациями S228P, L235E, P329G и S354C, T366W
<400> 28
Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe
1 5 10 15
Glu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
20 25 30
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
35 40 45
Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val
50 55 60
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser
65 70 75 80
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
85 90 95
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Gly Ser
100 105 110
Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
115 120 125
Gln Val Tyr Thr Leu Pro Pro Cys Gln Glu Glu Met Thr Lys Asn Gln
130 135 140
Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
145 150 155 160
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
165 170 175
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu
180 185 190
Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser
195 200 205
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
210 215 220
Leu Ser Leu Gly Lys
225
<210> 29
<211> 105
<212> ПРТ
<213> Homo sapiens
<400> 29
Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu
1 5 10 15
Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe
20 25 30
Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val
35 40 45
Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys
50 55 60
Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser
65 70 75 80
His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu
85 90 95
Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 30
<211> 107
<212> ПРТ
<213> Homo sapiens
<400> 30
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА С МОДИФИЦИРОВАННОЙ АФФИННОСТЬЮ К FcRn, КОТОРЫЕ УВЕЛИЧИВАЮТ КЛИРЕНС АНТИГЕНОВ | 2011 |
|
RU2745989C2 |
| ВАРИАНТЫ Fc-ОБЛАСТИ С МОДИФИЦИРОВАННЫМИ СВОЙСТВАМИ СВЯЗЫВАНИЯ FcRn И БЕЛКА А | 2015 |
|
RU2713131C1 |
| СПОСОБ ИЗМЕНЕНИЯ УДЕРЖАНИЯ В ПЛАЗМЕ И ИММУНОГЕННОСТИ АНТИГЕНСВЯЗЫВАЮЩЕЙ МОЛЕКУЛЫ | 2012 |
|
RU2799423C1 |
| ВАРИАНТЫ С FC-ФРАГМЕНТОМ, ОБЛАДАЮЩИЕ ПОВЫШЕННОЙ АФФИННОСТЬЮ К FCRN И ПОВЫШЕННОЙ АФФИННОСТЬЮ ПО МЕНЬШЕЙ МЕРЕ К ОДНОМУ РЕЦЕПТОРУ FC-ФРАГМЕНТА | 2018 |
|
RU2820162C2 |
| МОЛЕКУЛЫ, СВЯЗЫВАЮЩИЕ ADAM9, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2783619C2 |
| СКОНСТРУИРОВАННЫЕ МУЛЬТИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА И ДРУГИЕ МУЛЬТИМЕРНЫЕ БЕЛКИ С АСИММЕТРИЧНЫМИ МУТАЦИЯМИ В ОБЛАСТИ CH2-CH3 | 2018 |
|
RU2804031C2 |
| АНТИ-IL-5 АНТИТЕЛА | 2017 |
|
RU2758008C2 |
| АНТИТЕЛА ПРОТИВ SARS-COV-2 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2021 |
|
RU2832025C1 |
| ВАРИАНТ Fc И ЕГО ПОЛУЧЕНИЕ | 2021 |
|
RU2828852C1 |
| ВАРИАНТЫ FC-ОБЛАСТИ С МОДИФИЦИРОВАННОЙ СПОСОБНОСТЬЮ СВЯЗЫВАТЬСЯ С FCRN И С СОХРАНЕННОЙ СПОСОБНОСТЬЮ СВЯЗЫВАТЬСЯ С БЕЛКОМ А | 2015 |
|
RU2727639C2 |

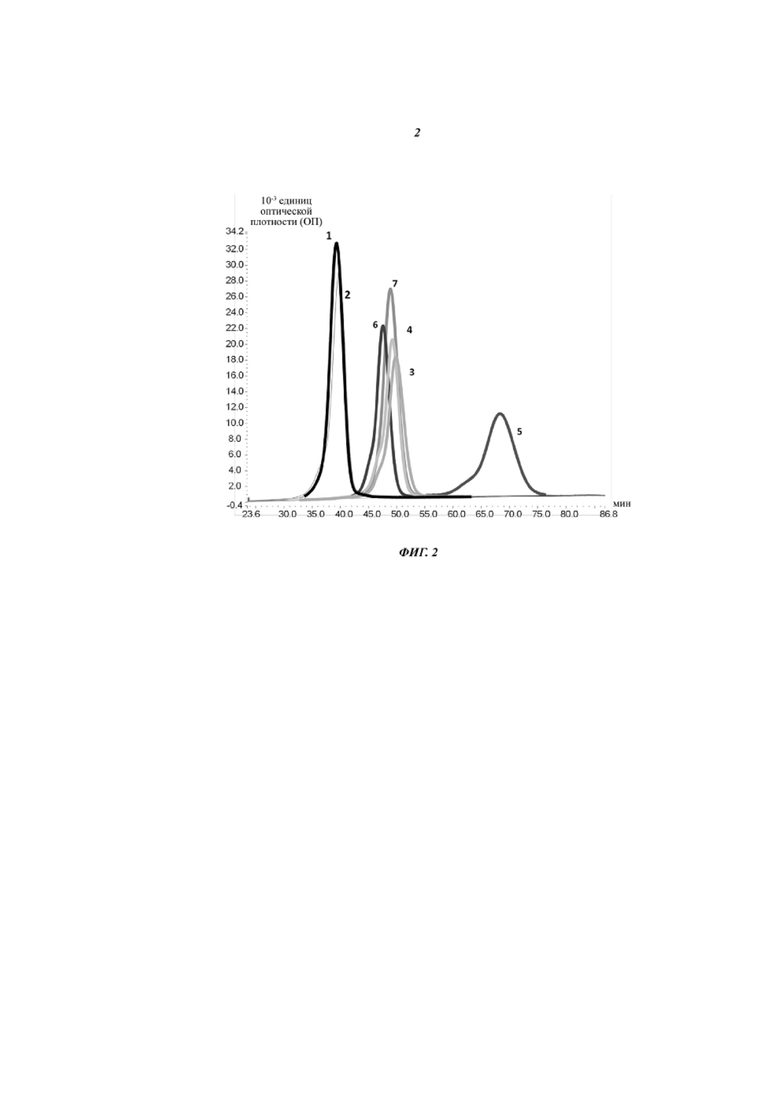
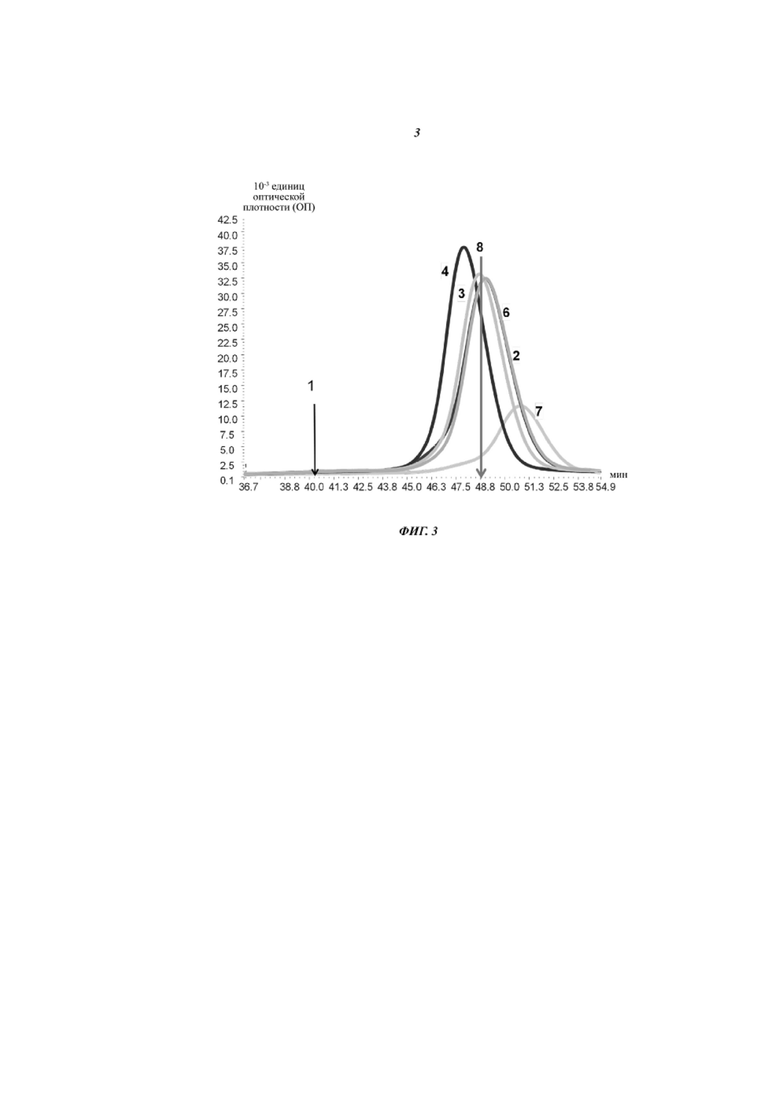
Изобретение относится к области биотехнологии и иммунологии. Предложен способ получения Fc-области класса IgG человека, содержащей первый вариантный полипептид Fc-области и второй вариантный полипептид Fc-области. Первый вариантный полипептид Fc-области происходит из первого родительского полипептида Fc-области класса IgG, а второй вариантный полипептид Fc-области происходит из второго родительского полипептида Fc-области класса IgG. Причем первый родительский полипептид идентичен второму родительскому полипептиду или отличается от второго родительского полипептида. В родительские полипептиды вносят мутации, включающие комбинацию: T307H и N434H, где нумерация соответствует EU-индексу по Kabat. В результате первый вариантный полипептид Fc-области отличается от второго вариантного полипептида Fc-области по одному или более аминокислотным остаткам. Причем если первый родительский полипептид отличался от второго родительского полипептида, то первый вариантный полипептид отличается от второго вариантного полипептида по остаткам, отличающимся от тех аминокислотных остатков, по которым первый родительский полипептид отличался от второго родительского полипептида. При этом полученная Fc-область класса IgG человека обладает аффинностью к неонатальному Fc-рецептору человека, которая выше таковой у Fc-области класса IgG без внесенных мутаций, что может найти применение в лечении заболеваний, требующих увеличенного времени удержания антител. 2 з.п. ф-лы, 3 ил., 4 табл., 2 пр.
1. Способ получения Fc-области класса IgG человека, содержащей первый вариантный полипептид Fc-области и второй вариантный полипептид Fc-области, где первый вариантный полипептид Fc-области происходит из первого родительского полипептида Fc-области класса IgG, а второй вариантный полипептид Fc-области происходит из второго родительского полипептида Fc-области класса IgG, причем первый родительский полипептид идентичен второму родительскому полипептиду или отличается от второго родительского полипептида,
при котором в родительские полипептиды вносят мутации, включающие комбинацию:
- T307H и N434H, где нумерация соответствует EU-индексу по Kabat, в результате чего
а) первый вариантный полипептид Fc-области отличается от второго вариантного полипептида Fc-области по одному или более аминокислотным остаткам, причем если первый родительской полипептид отличался от второго родительского полипептида, то первый вариантный полипептид отличается от второго вариантного полипептида по остаткам, отличающимся от тех аминокислотных остатков, по которым первый родительский полипептид отличался от второго родительского полипептида, и
б) Fc-область класса IgG, содержащая первый вариантный полипептид Fc-области и второй вариантный полипептид Fc-области, обладает аффинностью к неонатальному Fc-рецептору человека, которая выше таковой у Fc-области класса IgG, содержащей первый родительский полипептид и второй родительский полипептид.
2. Способ по п. 1, где те аминокислотные остатки, по которым первый родительский полипептид Fc-области класса IgG отличается от второго родительского полипептида Fc-области класса IgG, стимулируют образование гетеродимерной Fc-области класса IgG.
3. Способ по любому из пп. 1, 2, где либо в первый полипептид Fc-области, либо во второй полипептид Fc-области, либо в оба полипептида Fc-области независимо друг от друга дополнительно вносят одну или обе из приведенных ниже комбинаций мутаций:
- Q311H, и E430H, или
- M252Y, и S254T, и T256E.
| WO 2012125850 А1, 20.09.2012 | |||
| WO 2009086320 А1, 09.07.2009 | |||
| DALL’ ACQUA et al., "Increasing the affinity of a human IgG1 for the neonatal Fc receptor: biological consequences", J | |||
| Immunol., 2002, 169: pp.5171-5180 | |||
| YEUNG et al., "Engineering human IgG1 affinity to human neonatal Fc receptor: impact of affinity improvement on pharmacokinetics in |

